Изобретение относится к иммунологии и биотехнологии, а именно к способам индукции иммунологической толерантности на трансплантационные антигены с помощью трансфецированных дендритных клеток у млекопитающих, в том числе человека, и может быть использовано для снижения трансплантационных реакций, включая отторжение трансплантата и реакцию трансплантат против хозяина.
Проблемы отторжения пересаженных органов и реакции трансплантат против хозяина (РТПХ) остаются актуальными, несмотря на современные методы антигенного совмещения тканей донора и реципиента и эффективную медикаментозную терапию. Так, согласно литературным данным, 60 из 315 аллотрансплантатов почки подвергаются острой реакции отторжения [Understanding the Causes of Kidney Transplant Failure: The Dominant Role of Antibody-Mediated Rejection and Nonadherence J. Sellares et all, American Journal of Transplantation 2012; 12: 388-399], а в 1921 из 2813 случаев неродственной пересадки костного мозга возникает острая реакция трансплантат против хозяина [Graft-versus-host disease in recipients of male unrelated donor compared with parous female sibling donor transplants. Anita J. Kumar et. all Blood Advances 2018 2:1022-1031].
В основе отторжения трансплантатов и РТПХ лежит развитие иммунного ответа на трансплантационные антигены, представленные в пересаженных тканях и организме реципиента соответственно, главным образом на антигены основных и минорных молекул главного комплекса гистосовместимости (ГКГС). Принципиальная возможность индукции иммунологической толерантности, т.е. активной неотвечаемости иммунной системы, на трансплантационные антигены была показана в середине XX века П.Б. Медоваром, за что исследователь получил Нобелевскую премию [R.E. Billingham, L. Brent, Р.В. Medawar, Actively acquired tolerance of foreign cells, Nature 172 (1953) 603-606].
На данный момент для индукции иммунологической толерантности широко используются дендритные клетки (ДК). Дендритные клетки управляют иммунным ответом путем представления эпитопов антигена и ряда костимулирующих или ингибирующих сигналов лимфоцитам. В результате полученного сигнала лимфоциты приобретают способность игнорировать или уничтожать объекты, несущие представленные эпитопы антигена. Современный уровень биотехнологии позволяет модулировать активность дендритных клеток с целью получения иммунологической толерантности в ответ на определенный, в т.ч. трансплантационный, антиген.
Так, известно, что дендритные клетки, демонстрирующие незрелый фенотип, способствуют снижению иммунного ответа на трансплантационный антиген и продлевают выживание трансплантата, т.е. проявляют толерогенную функцию [Fu F., Li Y., Qian S., Lu L., Chambers F., Starzl Т.Е., et al. Costimulatory molecule-deficient dendritic cell progenitors (MHC class II+, CD80dim, CD86-) prolong cardiac allograft survival in nonimmunosuppressed recipients. Transplantation. 1996; 62: 659-65. Lutz M.B., Suri R.M., Niimi M., Ogilvie A.L., Kukutsch N.A., Rossner S. et al.].
Для получения таких незрелых-толерогенных ДК используют инкубирование клеток в среде, обедненной факторами созревания [Immature dendritic cells generated with low doses of GM-CSF in the absence of IL-4 are maturation resistant and prolong allograft survival in vivo. Eur. J. Immunol. 2000; 30: 1813-22.], а также культивирование клеток в среде с дексаметазоном, витамином D, рапамицином, циклоспорином, ретиноевой кислотой и др. Такие ДК обладают пониженной экспрессией молекул костимуляции (CD80 и CD86) и провоспалительных цитокинов (IL-1, IL-2, IL-12, INF-γ, TNF-α), повышенной экспрессией ингибиторных молекул (PD-L1, ICOSL) и противовоспалительных цитокинов (IL-10 и TGF-β), а также повышенной способностью активировать регуляторные Т-клетки, что и обеспечивает их толерогенную функцию [Haibin Li, Bingyi Shi, Tolerogenic dendritic cells and their applications in transplantation, Cell Mol Immunol. 2015 Jan; 12(1): 24-30.]. Однако, при попадании в условия in vivo, полученные таким образом клетки могут изменять свою функцию с толерогенной на иммунногенную в связи с провоспалительным влиянием их нового окружения [Generation, Characteristics and Clinical Trials of Ex Vivo Generated Tolerogenic Dendritic Cells, Sang-Hyun Kim, Ho-Hyun Jung, and Chong-Kil Lee, Yonsei Med J 2018 Sep; 59(7):807-815].
Для стабилизации толерогенных свойств ДК при переносе в организм реципиента используется метод трансфекции дендритных клеток ДНК-конструкциями, кодирующими различные иммунорегуляторные протеины.
Например, ДК мыши, трансфецированные лентивирусной конструкцией, кодирующей IL-10, вызывают снижение иммунного ответа на аллоантигены и трансплантационные антигены в реакции смешанной культуры лимфоцитов и реакции трансплантат против хозяина, соответственно [Wan J, Huang F, Hao S, Ни W, Liu С, Zhang W, Deng X, Chen L, Ma L, Tao R. lnterleukin-10 Gene-Modified Dendritic Cell-Induced Type 1 Regulatory T Cells Induce Transplant-Tolerance and Impede Graft Versus Host Disease After Allogeneic Stem Cell Transplantation. Cell Physiol Biochem. 2017; 43(1):353-366].
Для придания иммунологической толерантности антигенспецифичности дендритные клетки нагружают антигеном, против которого необходимо создать толерантность. На данный момент для нагрузки толерогенных ДК трансплантационными антигенами используют добавление к культуре клеток материала донора, например, клеточного или тканевого лизата донора [Alloantigen-specific T-cell hyporesponsiveness induced by dnIKK2 gene-transfected recipient immature dendritic cells, Fan C.B. et. all, Cell Immunol. 2015 Oct; 297(2):100-7] или везикул из донорской ткани [Renal Allograft Survival in Nonhuman Primates Infused with Donor Antigen-Pulsed Autologous Regulatory Dendritic Cells, M.B. Ezzelarab et. all, Am J Transplant. 2017 Jun; 17(6): 1476-1489]. Однако, такой способ не обеспечивает долговременного присутствия трансплантационного антигена в дендритных клетках, в результате чего свойство антигенспецифичности, индуцируемой ими толерантности, может пропадать при переносе в организм реципиента, что может приводить к индукции неспецифической толерантности в том числе и на потенциально опасные инфекционные и злокачественные агенты [Optimizing the process of nucleofection for professional antigen presenting cells, Mullins et al. BMC Res Notes (2015) 8:472; Dendritic cells as therapeutic vaccines against cancer, Jacques Banchereau, Nature Reviews (2005), Vol. 5, 296-306]. Для решения данной проблемы также используется метод трансфекции антигена в дендритные клетки для его длительной экспрессии в организме реципиента и формирования именно антигенспецифической толерантности.
Ряд способов индукции иммунологической толерантности на трансплантационные антигены с помощью дендритных клеток описаны в российских и зарубежных патентах.
В патенте [RU 2597976, А61К 48/00, 2011] толерогенные дендритные клетки для снижения реакции отторжения и РТПХ получают с помощью инкубирования их предшественников в присутствии (S)-(+)-1-(5-гидрокси-1,5-диметилгексил)-3-[7-(4-метоксифенил)-[1,2,4] триазоло[1,5-а]пиримидин-2-ил]мочевины. Нагрузка полученных толерогенных ДК антигеном в способе не предусмотрена.
В патентах [JP 2004298181, А61К 35/12, 2004; US 2005032210, А61К 35/12, 2005] для снижения реакции отторжения и РТПХ толерогенные дендритные клетки получают с помощью инкубирования их предшественников в присутствии иммунорегуляторных цитокинов IL-10 и TGF-β. Нагрузка ДК антигеном происходит также простым добавлением в среду трансплантационного антигена.
В патенте [US 2017340669, А61К 35/15, 2017] толерогенные дендритные клетки для снижения реакции отторжения и РТПХ получают с помощью инкубирования их предшественников в присутствии дексаметазона и витамина D2. Нагрузка ДК антигеном происходит также простым добавлением в среду трансплантационного антигена.
В патенте [WO 2013036300, А61К 39/00, 2013] толерогенные дендритные клетки для снижения реакции отторжения и РТПХ получают с помощью инкубирования их предшественников в присутствии рапамицина и TGF-β. Нагрузка ДК происходит добавлением к среде не самого трансплантационного антигена, а его рестриктированных по ГКГС 1го или 2го типа эпитопов.
Недостатком описанных способов является то, что индукцию толерогенности ДК и нагрузку их антигеном или эпитопами производят простым добавлением соответствующих факторов в среду к дендритным клеткам. После переноса таких ДК в организм реципиента, данные факторы будут отсутствовать, что приводит к нарушению индукции такими ДК антигенспецифической толерантности и, впоследствии, к нежелательным иммунным реакциям, таким как развитие иммунного ответа вместо его подавления (в случае отсутствия формирования толерантности), а также индукции толерантности на потенциально опасные инфекционные и опухолевые агенты (в случае отсутствия способности ДК индуцировать именно антигенспецифическую толерантность).
Данные недостатки частично устранены в способе снижения реакции отторжения, где для обеспечения толерогенности ДК предложена их генетическая модификация вирусным вектором, кодирующим IL-10, TGF-β или другие иммунорегуляторные цитокины. Для обеспечения антигенспецифичности индукции иммунологической толерантности предложено включение в тот же вирусный вектор последовательностей, кодирующих трансплантационный антиген, в частности ГКГС. Таким образом, предлагаемая в патенте трансфекция генетически модифицирует ДК и закрепляет в них экспрессию иммунорегуляторных цитокинов и антигенов, необходимых для индукции антигенспецифической толерантности в организме реципиента. Также в способе предложено сокультивирование полученных генетически модифицированных дендритных клеток и аутологичных лимфоцитов для получения регуляторных Т-клеток с последующим введением полученной культуры реципиенту для усиления толерогенного эффекта [US 2004132674, А61К 35/12, 2004].
Однако, способ предполагает обязательное включение в вирусный вектор патентуемую иммунорегуляторную последовательность из гена фасцина, а остальные описанные последовательности, кодирующие иммунорегуляторные протеины и антигены, используются лишь в качестве возможной модификации этого вирусного вектора. Таким образом, данный способ индукции иммунологической толерантности осложняется необходимостью кодирования иммунорегуляторной последовательности из гена фасцина, а может быть использован только для снижения реакции отторжения, но не РТПХ.
Наиболее близким к заявленному способу является способ индукции иммунологической толерантности на трансплантационные антигены у млекопитающих, включающий трансфекцию незрелых дендритных клеток ДНК-конструкцией, кодирующей иммунорегуляторный цитокин IL-10. В данном способе реципиенту вводят незрелые дендритные клетки, генетически модифицированные методом вирусной трансфекции для экспрессии минимум двух иммунорегуляторных белков в т.ч. IL-10, и трансплантационных антигенов, имеющих минимум 1 эпитоп. [US 2008311140, А61К 39/00, C12N 5/10, 2008].
Недостатком способа является сохраняющаяся возможность неэффективной презентации антигена дендритными клетками, поскольку целый белок антигена, трансфецированный в ДК, нуждается в дальнейшем процессинге и расщеплении до презентируемых эпитопов и, таким образом, сохраняет вероятность того, что: во-первых, антиген может вообще не расщепиться до требуемых эпитопов и не будет выполнять поставленную перед ним задачу обеспечения антигенспецифичности иммунологической толерантности; во-вторых, антиген может расщепиться до неиммуногенных эпитопов и тогда не будет выполнять данную задачу с должной эффективностью. Тем самым трансфекция целого белка антигена в ДК не гарантирует эффективной индукции антигенспецифической толерантности.
Кроме того, использование вирусного вектора для трансфекции ДК, которые далее вводятся реципиенту, требует дополнительных мер биологической безопасности, а сама вирусная трансфекция является генномодифицирующей и нарушает собственный геном ДК, что потенциально опасно при введении таких клеток реципиенту.
Задача изобретения заключается в создании более эффективного способа индукции антигенспецифической иммунологической толерантности, за счет доставки в ДК именно эпитопов трансплантационного антигена, а не целого антигена.
Поставленная задача решается в предложенном способе индукции иммунологической толерантности на трансплантационные антигены у млекопитающих, включающем трансфекцию незрелых дендритных клеток ДНК-конструкцией, кодирующей иммунорегуляторный цитокин IL-10, путем индукции толерантности на трансплантационные антигены с помощью котрансфекции незрелых дендритных клеток дополнительной ДНК-плазмидой, кодирующей специально подобранные иммуногенные различающиеся у донора и реципиента эпитопы белков главного комплекса гистосовместимости, при этом трансфекция является негенномодифицурующей, а полученные трансфецированные дендритные клетки сокультивируют с аутологичными лимфоцитами для индукции регуляторных Т-клеток, с последующим введением реципиенту полученных культур.
Изобретение, на наш взгляд, является новым.
Увеличение эффективности индукции иммунологической толерантности на трансплантационные антигены в заявленном способе обеспечивается котрансфекцией дендритных клеток дополнительной ДНК-плазмидой, кодирующей специально подобранные иммуногенные различающиеся у донора и реципиента эпитопы белков главного комплекса гистосовместимости, а не сам антиген, как в известном способе, что гарантирует презентацию дендритными клетками эпитопов, необходимых для эффективной индукции антигенспецифической иммунологической толерантности. Также эффективность индукции иммунологической толерантности на трансплантационные антигены увеличивается за счет этапов сокультивирования трансфецированных ДК и аутологичных лимфоцитов для индукции регуляторных Т-клеток. Полученные после сокультивирования клеточные культуры содержат регуляторные Т-клетки и обладают способностью к индукции толерантности к антигенам.
Изобретение осуществляется следующим образом
Дендритные клетки получают из костного мозга мышей или из мононуклеарных клеток периферической крови человека. Суспензия клеток костного мозга мышей или мононуклеарные клетки периферической крови человека дважды промываются средой RPMI-1640. Далее клетки высаживаются в культуральный флакон 75 см2 и инкубируются в атмосфере 5% CO2 при 37°С. Через 30 минут инкубирования неприлипшая фракция клеток сливается. Прилипшая фракция собирается скребком, отмывается в среде RPMI-1640 и высаживается во флакон в концентрации 1 млн. клеток на 1 мл среды для культивирования, содержащей среду RPMI-1640, с добавлением 10% FCS, 2 мМ L-глютамина, 10 мМ HEPES буфера, 5*10-4М 2-меркаптоэтанола, 80 мкг/мл гентамицина, 100 мкг/мл бензилпенициллина. Для дифференцировки клеток в незрелые дендритные клетки в среду добавляют 20 нг/мл rmGM-CSF и 20 нг/мл rmIL-4. Через 48 часов культивирования выполняется смена среды для культивирования с повторным добавлением 20 нг/мл rmGM-CSF и 20 нг/мл rmIL-4. Через 72 часа культивирования дифференцированные незрелые дендритные клетки снимаются с пластика скребком в собственной среде и центрифугируются для получения односуточной кондиционной среды и клеточного осадка. Односуточная кондиционная среда эвакуируется и сохраняется. Клеточный осадок, т.е. дифференцированные незрелые дендритные клетки, подвергаются трансфекции путем электропорации кольцевыми ДНК-плазмидами, кодирующими эпитопы белков ГКГС и кольцевыми ДНК-плазмидами, кодирующими эпитопы молекул ГКГС и кольцевыми ДНК-плазмидами, кодирующими мышиный или человеческий IL-10. Электропорация осуществляется в холодной среде OptiMem, содержащей 20 млн/мл дендритных клеток на электропораторе (например, ВТХ 830 square-wave) при следующих параметрах: сила импульса 260 V, длительность импульса 5 ms, концентрация плазмидной ДНК 60 мкг/мл. Далее, трансфецированные дендритные клетки ссаживаются с аутологичными спленоцитами мышей или аутологичными мононуклеарными клетками человека в соотношении 1:10 - 1:100 и культивируются в среде, состоящей из среды для культивирования и односуточной кондиционной среды в соотношении 1:1 без добавления факторов созревания. Через 72 часа полученная культура отмывается и используется для индукции иммунологической толерантности для снижения реакции отторжения или РТПХ. Причем для профилактики РТПХ готовят культуру толерогенных клеток донора а для профилактики отторжения культуру толерогенных клеток реципиента.
Способность изобретения индуцировать иммунологическую толерантность показана экспериментально в различных моделях, например, в смешанной культуре лимфоцитов, реакции отторжения кожного лоскута и в реакции трансплантат против хозяина.
Пример 1.
Индукция иммунологической толерантности у мышей линии C57BI/6 на белки ГКГС мышей линии СВА.
Незрелые дендритные клетки получали, как описано выше, из костного мозга мышей линии C57BI/6. Дендритные клетки трансфецировали кольцевыми ДНК-плазмидами, кодирующими эпитопы молекул Н-2K (pGVHD), т.е. аллеля ГКГС мышей линии СВА или кольцевыми ДНК-плазмидами, кодирующими эпитопы молекул Н-2К и кольцевыми ДНК-плазмидами, кодирующими мышиный IL-10 (pGVHD+pIL-10). Далее дендритные клетки сокультивировали с аутологичными спленоцитами как описано выше.
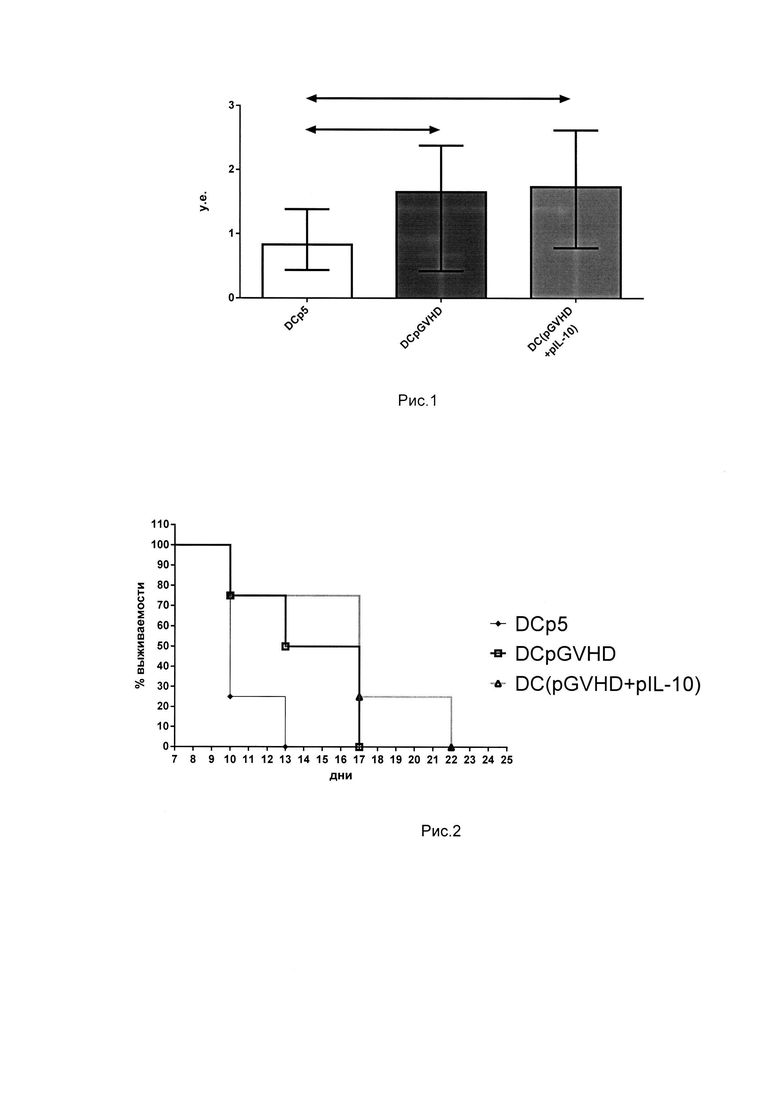
Для установления способности полученных толерогенных культур ограничивать активацию аутологичных спленоцитов в ответ на стимуляцию аллоантигенами была поставлена смешанная культура лимфоцитов. Полученные толерогенные культуры мышей C57BI/6 сокультивировались со спленоцитами мышей СВА, предварительно обработанных митомицином С. Через 72 часа совместного культивирования оценивался уровень пролиферации в культурах и высчитывался индекс подавления пролиферации (пролиферация толерогенной культуры/пролиферация толерогенной культуры при добавлении аллогенных спленоцитов мышей СВА, обработанных митомицином С). Таким образом, чем выше оказывался индекс подавления, тем меньше пролиферировала исследуемая культура в ответ на стимуляцию аллоантигенами. Результаты показали, что культуры, полученные с помощью толерогенных ДК, трансфецированных pGVHD и pGVHD+pIL-10 обладают статистически значимо большей способностью угнетать пролиферацию аутологичных спленоцитов в ответ на аллогенный стимул по сравнению с культурой, полученной с помощью ДК, трансфецированных контрольной плазмидой р5, кодирующей неиммунногенные эпитопы. На рис. 1 представлен индекс подавления пролиферации аутологичных спленоцитов в ответ на аллогенный стимул для полученных культур. Стрелками показаны статистически достоверные отличия р<0.05. Представленные данные говорят о выполнении толерогеннной функции культурами, полученными с помощью ДК, трансфецированными pGVHD и pGVHD+pIL-10 в условиях in vitro.
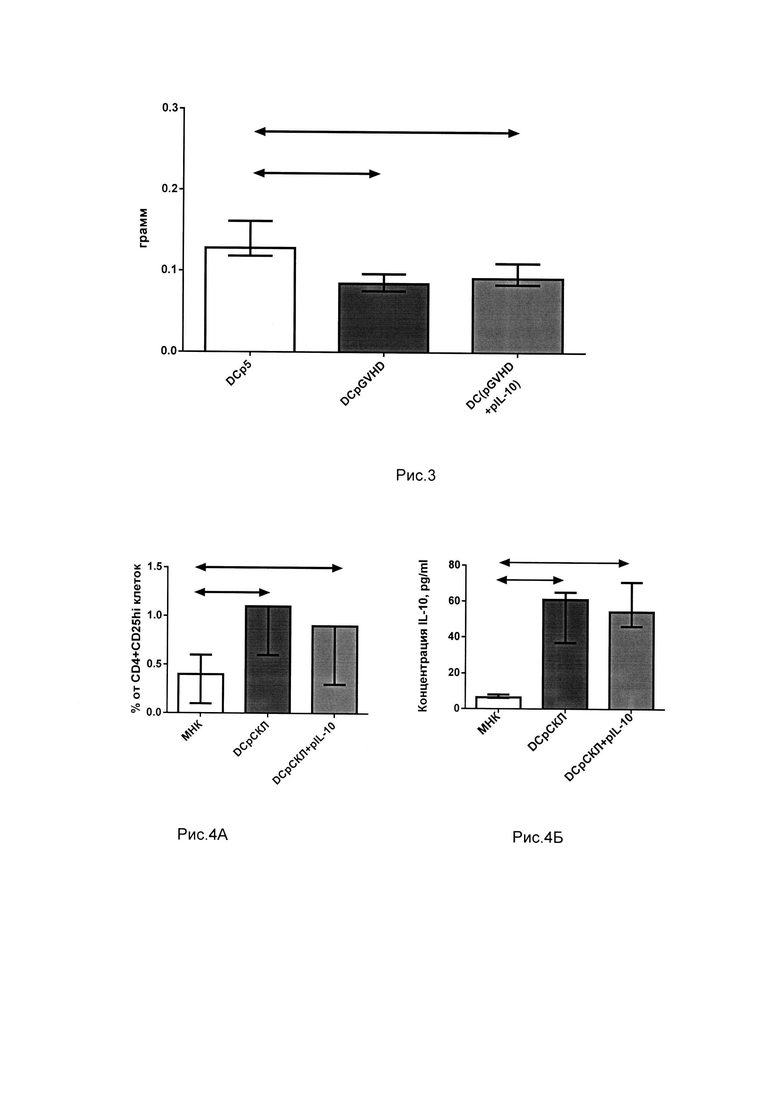
Для установления способности полученных культур снижать реакцию отторжения in vivo была использована модель отторжения кожного лоскута. До пересадки кожного лоскута мышам C57BI/6 подкожно в область будущей операции вводили 1 млн клеток из культур, полученных с помощью толерогенных дендритных клеток. Через 24 часа мышам линии C57BI/6 на спину трансплантировали кожный лоскут с хвоста мышей линии СВА. Через 72 часа после трансплантации осуществляли повторное введение клеток толерогенных культур в количестве 1 млн. Выживаемость кожного лоскута оценивалась по его некротическим изменениям. Медиана выживаемости кожного лоскута при введении мышам, культур, полученных с помощью толерогенных ДК, трансфецированных pGVHD и pGVHD+pIL-10 составила 15 и 17 суток соответственно, что статистически значимо больше по сравнению с группой введения мышам контрольных культур DCp5 (10 суток). На рис. 2, показаны кривые выживаемости кожных лоскутов от мышей СВА, трансплантированных мышам C57BI/6, которым были введены толерогенные культуры.
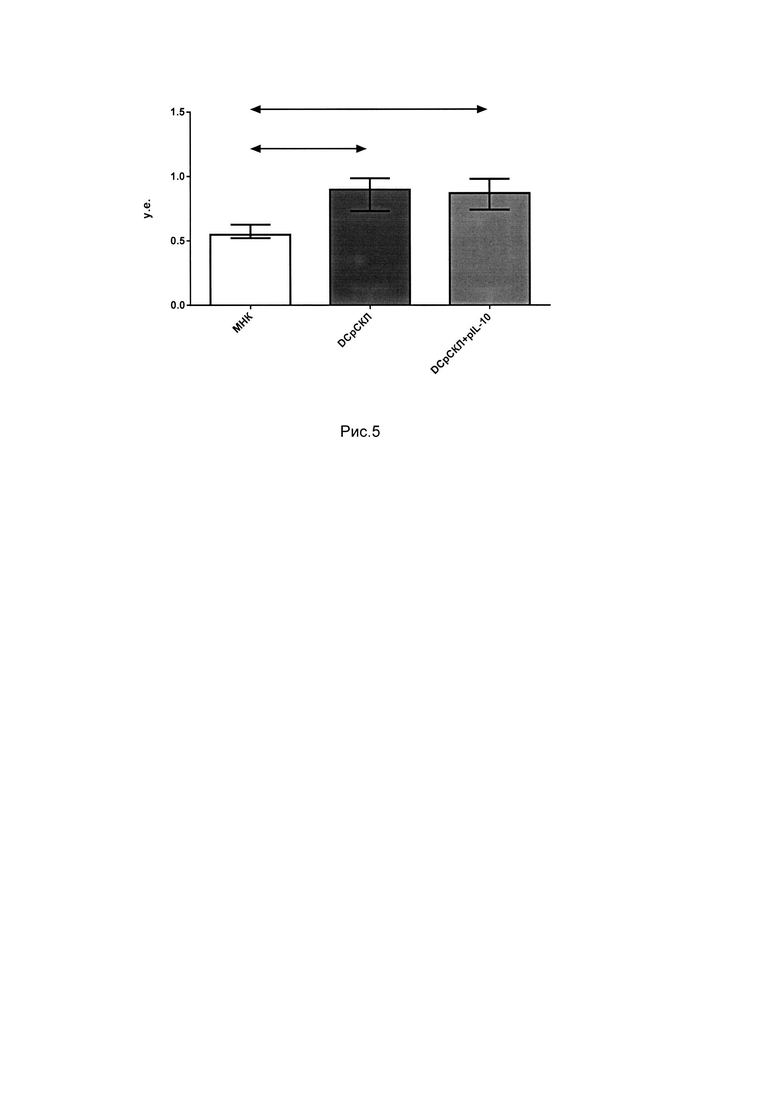
Для установления способности полученных толерогенных культур in vivo угнетать реакцию трансплантат против хозяина мышам линии CBF, т.е. первому поколению от скрещивания C57BI/6 и СВА, внутривенно вводили 100 млн. клеток из полученных толерогенных культур. Выраженность реакции РТПХ оценивали по весу селезенки. Результаты показали, что вес селезенок мышей, которым были введены культуры, полученные с помощью толерогенных ДК, трансфецированных pGVHD и pGVHD+pIL-10, оказался статистически значимо меньшим по сравнению с весом селезенок мышей, которым вводили контрольные культуры DCp5. На рис. 3 показан вес селезенки мышей CBF через 3 недели после введения им полученных толерогенных культур. Стрелками показаны статистически достоверные отличия р<0.05.
Полученные результаты говорят о том, что предлагаемый способ индукции иммунологической толерантности с помощью дендритных клеток, трансфецированных ДНК-конструкциями, кодирующими эпитопы белков главного комплекса гистосовместимости, действительно индуцирует толерантность на трансплантационные антигены и способен подавлять реакции отторжения и трансплантат против хозяина у мышей.
Пример 2.
Индукция иммунологической толерантности на аллоантигены ГКГС в смешанной культуре лимфоцитов человека.
Для установления способности ДК, трансфецированных эпитопами белков ГКГС, индуцировать иммунологическую толерантность в смешанной культуре лимфоцитов человека среди 10 условно-здоровых доноров были выбраны пары «стимулятор»-«ответчик», где МНК «ответчика» наиболее выражено пролиферировали на МНК «стимулятора» в условиях смешанной культуры лимфоцитов. Гены ГКГС выбранных «стимуляторов» и «ответчиков» были секвенированы. На основе данных секвенирования были разработаны и синтезированы кольцевые ДНК-плазмиды, кодирующие подобранные иммуногенные эпитопы белков ГКГС «стимуляторов», способные встраиваться в антигенсвязывающие желобки ГКГС I класса «ответчиков» (рСКЛ). Далее полученные плазмиды использовались в экспериментах с соответствующими парами «стимулятор»-«ответчик».
Незрелые дендритные клетки получали как описано выше из мононуклеарных клеток периферической крови человека. Дендритные клетки доноров-"ответчиков" трансфецировали кольцевыми ДНК-плазмидами, кодирующими эпитопы белков ГКГС доноров-"стимуляторов"(рСКЛ) или кольцевыми ДНК-плазмидами, кодирующими эпитопы ГКГС доноров-"стимуляторов" и кольцевыми ДНК-плазмидами, кодирующими человеческий IL-10 (рСКЛ+pIL-10). Далее дендритные клетки культивировали совместно с аутологичными МНК как описано выше. Полученные культуры использовали для дальнейшего анализа и постановки смешанной культуры лимфоцитов человека.
Через 96 часов в культурах, полученных с помощью ДК, трансфецированных рСКЛ и рСКЛ+pIL-10 была обнаружена значимо большая экспрессия FoxP3 среди CD4+CD25hi клеток по сравнению с исходной культурой мононуклеарных клеток (Рис. 4А). Также в кондиционных средах этих групп обнаружено значимо большее содержание IL-10 (Рис. 4Б). На рис. 4. показано содержание Т-регуляторных клеток (А) и продукция IL-10 (Б) в совместных культурах МНК и ДК, трансфицированных плазмидами, кодирующими антигены ГКГС и IL-10.
Полученные данные говорят о выполнении полученными дендритными клетками толерогенной функции через активацию регуляторных Т-клеток и секрецию IL-10.
Для оценки способности полученных толерогенных культур ограничивать активацию аутологичных МНК в ответ на стимуляцию аллоантигенами была поставлена смешанная культура лимфоцитов. Полученные с помощью трансфекции ДК рСКЛ или рСКЛ+pIL-10 культуры мононуклеарных клеток доноров-"ответчиков" культивировались совместно с МНК доноров-"стимуляторов", предварительно обработанных митомицином С. Через 96 часов совместного культивирования оценивался уровень пролиферации в культурах и высчитывался индекс подавления пролиферации (пролиферация толерогенной культуры доноров-«ответчиков»/пролиферация толерогенной культуры доноров-ответчиков» при добавлении аллогенных МНК доноров-"стимуляторов", обработанных митомицином С). Следовательно, чем выше оказывался индекс подавления, тем меньше пролиферировала исследуемая культура в ответ на стимуляцию аллоантигенами. Результаты показали, что культуры, полученные с помощью толерогенных ДК, трансфецированных рСКЛ или рСКЛ+pIL-10, обладают статистически значимо большей способностью угнетать пролиферацию аутологичных МНК в ответ на стимуляцию аллоантигенами по сравнению с исходной культурой мононуклеарных клеток (Рис. 5). На рис. 5 представлен индекс подавления пролиферации аутологичных МНК в ответ на аллогенный стимул для полученных культур. Стрелками показаны статически достоверные отличия р<0.05.
Таким образом, предлагаемый способ индукции иммунологической толерантности с помощью ДК, трансфецированных ДНК-плазмидами, кодирующими эпитопы ГКГС, действительно индуцирует иммунологическую толерантность и подавляет активацию аутологичных МНК в ответ на аллоантигенную стимуляцию в условиях смешанной культуры лимфоцитов человека in vitro.
Преимущества заявленного способа заключаются в увеличенной эффективности индукции именно антигенспецифической толерантности на трансплантационные антигены за счет котрансфекции в ДК дополнительной ДНК-плазмиды, кодирующей специально подобранные иммунногенные различающиеся у донора и реципиента эпитопы белков ГКГС, что гарантирует их презентацию дендритными клетками и индукцию толерантности именно на закодированные эпитопы трансплантационных антигенов. Вдобавок, эффективность индукции иммунологической толерантности повышается за счет этапов сокультивитрования трансфецированных ДК с аутологичными лимфоцитами для индукции регуляторных Т-клеток и последующего введения реципиенту полученных культур.
Предложенный способ является универсальным, поскольку может быть использован для снижения реакции отторжения органов и тканей в случае если иммунологическая толерантность индуцируется у реципиента на белки ГКГС трансплантата или для снижения РТПХ в случае если иммунологическая толерантность индуцируется у клеток трансплантата костного мозга донора на белки ГКГС реципиента.
Также, предложенный способ не требует дополнительных мер биологической безопасности, поскольку трансфекция ДК осуществляется методом электропорации с помощью невирусных кольцевых ДНК-плазмид, что, к тому же, не приводит к изменению собственного генома дендритных клеток.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ АНТИГЕН-СПЕЦИФИЧЕСКИХ ЦИТОТОКСИЧЕСКИХ КЛЕТОК, ОБЛАДАЮЩИХ АКТИВНОСТЬЮ ПРОТИВ КЛЕТОК РАКА МОЛОЧНОЙ ЖЕЛЕЗЫ | 2013 |
|
RU2596920C2 |
| ПОЛИЭПИТОПНЫЕ ИММУНОГЕННЫЕ ПОЛИПЕПТИДЫ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2013 |
|
RU2598265C2 |
| Способ увеличения миграции плазмоцитоидных дендритных клеток к тимоцитам мышей in vitro | 2020 |
|
RU2760215C1 |
| Способ получения антиген-специфических цитотоксических клеток, обладающих противоопухолевой цитотоксической активностью против клеток немелкоклеточного рака легкого | 2016 |
|
RU2639514C1 |
| СПОСОБ СТИМУЛЯЦИИ ЦИТОТОКСИЧЕСКОГО ИММУННОГО ОТВЕТА ПРОТИВ КЛЕТОК ОПУХОЛЕВОЙ ЛИНИИ АДЕНОКАРЦИНОМЫ МОЛОЧНОЙ ЖЕЛЕЗЫ, ЭКСПРЕССИРУЮЩИХ СПЕЦИФИЧЕСКИЕ АНТИГЕНЫ, С ПОМОЩЬЮ ДЕНДРИТНЫХ КЛЕТОК, ТРАНСФЕЦИРОВАННЫХ ПОЛИЭПИТОПНОЙ ДНК-КОНСТРУКЦИЕЙ | 2012 |
|
RU2520091C2 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pCI-UB-POLYEPI, СОДЕРЖАЩАЯ ЭПИТОПЫ ОПУХОЛЬ-АССОЦИИРОВАННЫХ АНТИГЕНОВ ДЛЯ КОЛОРЕКТАЛЬНОГО РАКА, И СПОСОБ ЕЕ ПРИМЕНЕНИЯ ДЛЯ СТИМУЛЯЦИИ СПЕЦИФИЧЕСКОГО ПРОТИВООПУХОЛЕВОГО ИММУННОГО ОТВЕТА ПРОТИВ КЛЕТОК КОЛОРЕКТАЛЬНОГО РАКА | 2012 |
|
RU2507265C2 |
| ИСКУССТВЕННЫЙ ГЕН MEL-TCI-A0201, КОДИРУЮЩИЙ ПОЛИЭПИТОПНЫЙ БЕЛОК-ИММУНОГЕН MEL-TCI-A0201, РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pMEL-TCI-A0201, ОБЕСПЕЧИВАЮЩАЯ ЭКСПРЕССИЮ ИСКУССТВЕННОГО ГЕНА MEL-TCI-A0201 И ИСКУССТВЕННЫЙ БЕЛОК-ИММУНОГЕН MEL-TCI-A0201, СОДЕРЖАЩИЙ МНОЖЕСТВЕННЫЕ CTL- И Th-ЭПИТОПЫ АНТИГЕНОВ МЕЛАНОМЫ | 2012 |
|
RU2522830C2 |
| СПОСОБ ИММУНОТЕРАПИИ ХРОНИЧЕСКОГО ВИРУСНОГО ГЕПАТИТА С | 2016 |
|
RU2637631C2 |
| ЛИГАНД NOTCH | 1997 |
|
RU2197262C2 |
| Искусственный ген MEL-TCI, кодирующий полиэпитопный белок-иммуноген MEL-TCI, рекомбинантная плазмидная ДНК pMEL-TCI, обеспечивающая экспрессию искусственного гена MEL-TCI и искусственный белок-иммуноген MEL-TCI, содержащий CTL- и Th-эпитопы антигенов меланомы, рестриктированные множественными аллелями HLA I и II класса | 2017 |
|
RU2650872C1 |
Настоящее изобретение относится к области иммунологии. Предложен способ индукции иммунологической толерантности на трансплантационные антигены у млекопитающих. Данное изобретение может найти дальнейшее применение в снижении трансплантационных реакций, включая отторжение трансплантата и реакцию трансплантата против хозяина. 6 ил., 2 пр.
Способ индукции иммунологической толерантности на трансплантационные антигены у млекопитающих, включающий трансфекцию незрелых дендритных клеток ДНК-конструкцией, кодирующей иммунорегуляторный цитокин IL-10, отличающийся тем, что индукцию толерантности на трансплантационные антигены обеспечивают котрансфекцией незрелых дендритных клеток дополнительной ДНК-плазмидой, кодирующей специально подобранные иммуногенные различающиеся у донора и реципиента эпитопы белков главного комплекса гистосовместимости, при этом трансфекция является негенномодифицурующей, а полученные трансфецированные дендритные клетки сокультивируют с аутологичными лимфоцитами для индукции регуляторных Т-клеток с последующим введением реципиенту полученных культур.
| Станок для изготовления деревянных ниточных катушек из цилиндрических, снабженных осевым отверстием, заготовок | 1923 |
|
SU2008A1 |
| Многоступенчатая активно-реактивная турбина | 1924 |
|
SU2013A1 |
| Способ приготовления мыла | 1923 |
|
SU2004A1 |
| Способ получения in vitro популяций активированных антигенспецифических противоопухолевых цитотоксических Т-лимфоцитов, специфичных к эпитопам опухоль-ассоциированного антигена | 2016 |
|
RU2619186C1 |
| СЕННИКОВ С.В | |||
| и др | |||
| "Молекулярно-клеточные механизмы, опосредуемые дендритными | |||

