Изобретение относится к использованию терапевтических соединений в модификации активации Т-клеток. В частности, оно имеет отношение к их использованию для модуляции взаимодействия между белками семейства Notch и их лигандами и использовании подобных соединений в терапии таких состояний как: реакция отторжения трансплантата, аутоиммунные реакции, аллергии, инфекционные болезни и опухоли.
Контролируемое взаимодействие между Т-клетками и между антиген-представляющими клетками (АП-клетками) и Т-клетками является жизненно важным для функционирования иммунной системы человека. Тем не менее при определенных патологиях может быть полезно модифицировать, положительно или отрицательно, подобные взаимодействия. Например, при таких состояниях как аутоиммунная реакция, аллергия или реакция отторжения трансплантата желательно индуцировать регуляцию по типу отрицательной обратной связи иммунного ответа путем стимуляции негативных Т-клеток или взаимодействия Т-клеток с АП-клетками. Модели "инфекционной толерантности" и "связанной супрессии" предполагают, что толерантность может быть индуцирована в небольшом количестве Т-клеток, и данные Т-клетки затем передают эту толерантность другим Т-клеткам, таким образом предупреждая эффективную иммунологическую атаку. При иных патологических состояниях, таких как индуцированная опухолью иммуносупрессия, паразитарные вирусные или бактериальные инфекции, иммуносупрессия является обычным явлением. В данных обстоятельствах было бы, следовательно, желательно ингибировать Т-клеточные взаимодействия, ведущие к инфекционной толерантности.
Однако до настоящего момента механизмы, поддерживающие подобные взаимодействия Т-клеток и Т-клеток с АП-клетками, не были изучены.
В заявке WO 92/19734 обнаружены нуклеотидные последовательности генов Notch и Delta и определены аминокислотные последовательности кодируемых ими белков. Показано, что семейство генов Notch играет существенную роль для правильного развития дифференцировки эмбриональных клеток насекомых, таких как Drosophila.
Белки, относящиеся к семейству Notch, являются трансмембранными рецепторами, которые содержат несколько консервативных пептидных мотивов. Каждый белок из этого семейства имеет характерные повторы подобно внеклеточному эпидермальному фактору роста (EGF) и околомембранный мотив Lin-12/Notch. Кроме того, каждый белок имеет 6-8 анкириновых повторяющихся мотивов в цитоплазматическом хвосте вместе с последовательностью PEST. Лиганды Notch имеют диагностический домен DSL (D, Delta, S, Serrate, L,Lag2), включающий 20-22 аминокислот на аминоконце белка и от 3 до 8 EGF-подобных повторов во внеклеточной области. Белки имеют короткий цитоплазматический хвост с несохраненными функциональными доменами.
Полученные данные свидетельствуют о том, что Notch способствует дифференцировочному коммитированию незрелых Т-клеток в тимусе, направляя тимоциты по пути дифференцировки в клетки CD8+, не зависящие от МНС (Robey E. с сотр. Cell, 1996, 87:483-492). В процессе созревания в тимусе Т-клетки приобретают способность распознавать свои собственные антигены и отличать их от чужих, данный процесс назван аутотолерантностью (von Boehmer Н. с.сотр. Ann Rev Immunol. 1990; 8:531). Механизмы индукции и поддержки толерантности существуют также и на периферии, и во многих отношениях их значение находится в стадии изучения. Существует много экспериментальных моделей реакции отторжения трансплантата, аутоиммунных заболеваний и специфических ответов на аллергены, которые четко иллюстрируют способность индуцировать состояние специфической ареактивности (толерантность или анергия) у реципиента при иммунизации определенным антигеном. В результате можно сделать два важных вывода. Во-первых, иммунизация пептидным фрагментом антигена в особых условиях может подавлять специфические ответы не только на себя самого, но и на другие области этой же молекулы в том случае, если интактный белок используется для провокационной иммунизации (связанная супрессия; Hoyne GF. с сотр. J. Exp Med. 1993; 178:183 и Metzler В., Wraith DC. Int. Immunol. 1993; 5: 1159). Во-вторых, наилучшим образом показано в экспериментальных моделях трансплантации в феномене "инфекционной толерантности", где постулировано, что иммунокомпетентные клетки, будучи невосприимчивыми к определенному антигену, способны подавлять клеточный ответ у других клеток и, более того, эта вторая популяция клеток становится регуляторной и толерантной (Qin SX. с сотр. Science, 1993; 258:974). Иммунологические механизмы, лежащие в основе этого явления пока не выяснены.
Стимулом для появления данного изобретения послужило открытие того, что рецепторы семейства Notch и их лиганды Delta и Serrate экспрессированы на поверхности нормальных зрелых клеток периферической иммунной системы.
Следовательно, в условиях согласно изобретению, Notch-лиганды могут быть использованы в производстве медицинских препаратов для иммунотерапии.
Характер экспрессии рецепторов семейства Notch и их лигандов в периферической иммунной системе здорового взрослого человека до сих пор не был описан, но в настоящем изобретении показано, что Т-клетки конститутивно экспрессируют Notch 1 мРНК, в то время как экспрессия Delta ограничена только субпопуляцией Т-клеток в периферических лимфоидных тканях. Экспрессия Serrate ограничена субпопуляцией антиген-представляющих клеток (АП-клеток) на периферии (фигура 1). Следовательно, семейство рецепторных лигандов может продолжать регулировать судьбу клеток в иммунной системе, как было показано на других тканях, после эмбрионального развития (Ellisen LW. с сотр. Cell, 1991; 66: 649). Сигнализация Notch может играть центральную роль в индукции периферической реакционной ареактивности (толерантности или анергии) и может обеспечивать физическое объяснение связанной супрессии и инфекционной толерантности.
Настоящее изобретение, кроме того, связано с применением лигандов Notch для воздействия на реакции, опосредованные Т-клетками. Так, было замечено, что при совместной экспозиции популяции нативных Т-клеток с лигандами Notch, экспрессированными на АП-клетках, в присутствии антигена или аллергена лиганды Notch способны вызвать толерантность Т-клеточной популяции к указанному антигену или аллергену. Более того, данная популяция Т-клеток при дальнейшей экспозиции со второй популяцией нативных Т-клеток, также способна передать второй популяции толерантность к указанным антигену или аллергену.
Таким образом, изобретение также связано с применением лигандов Notch для воздействия на связанную толерантность и/или перенесенную толерантность (также называемой инфекционной толерантностью).
Другой аспект изобретения связан с применением лигандов Notch для модуляции экспрессии функционального белка Notch или путей сигналирования Notch.
В приведенных выше аспектах согласно изобретению лиганд Notch может быть экспонирован Т-клеткам путем инкубации смешивания Т-клеток с АП-клетками или им подобными, которые экспрессируют или избыточно экспрессируют лиганды Notch в присутствии аллергена или антигена. Избыточная экспрессия гена лиганда Notch может быть вызвана АП-клетками при трансфекции их геном, способным вызывать экспрессию лиганда Notch, или АП-клетками, экспонированными с агентом, способным повышать экспрессию эндогенного гена (генов) лиганда Notch.
Соответствующие агенты, влияющие на экспрессию лигандов Notch, включают в себя агенты, воздействующие на экспрессию генов Delta и/или Serrate. Например, для экспрессии Delta связывание внеклеточных BMPs (морфогенетических белков кости, Wilson, P.А. и Hemmati-Brivanlou A., 1997, Neuron 18: 699-710, Hemmati-Brivanlou A. и Melton D. Cell 88:13-17) с их рецепторами приводит к обратной регуляции транскрипции Delta посредством ингибирования экспрессии фактора транскрипции комплекса Achaete/Scute. Есть основания полагать, что этот комплекс непосредственно вовлечен в регуляцию экспрессии Delta. Таким образом, любой агент, ингибирующий связывание BMPs с их рецепторами, способен вызвать увеличение экспрессии Delta и/или Serrate. К подобным агентам относятся: Noggin, Chordin, Follistatin, Xnr3, FGFs, Fringe, а также их производные и иные разновидности (см. ссылки на noggin-Smith W.C. и Наrland, R. M Cell 70:829-840, chordin-Sasai, Y. et al., 1994 Cell 79: 779-790). Таким образом, связь Noggin и Chordin BMPs препятствует активации их сигнального каскада, который приводит к подавлению транскрипции Delta.
При некоторых болезненных состояниях иммунитет оказывается ослабленным и может возникнуть необходимость подавить экспрессию Delta и/или Serrate с целью преодоления иммуносупрессии. При подобных обстоятельствах могут быть использованы агенты, способные ингибировать экспрессию Delta и/или Serrate. Примерами агентов, способных подавлять экспрессию Delta и/или Serrate, может служить белок Toll (Medzhitov R. с сотр., 1997 Nature 388: 394-397) BMPs и иные агенты, способные снижать или препятствовать образованию Noggin, Chordin, Follistatin, Xnr3, FGFs и Fringe. Таким образом, изобретение связано также с применением агента, способного по типу отрицательной обратной связи снижать экспрессию белков Delta или Serrate, в производстве медицинских препаратов для применения при лечении повторной бактериальной инфекции или иммуносупрессии, вызванной опухолью.
Как обсуждалось выше, изобретение также связано с модификацией экспрессии белка Notch, или презентации на клеточной мембране, либо путей сигнализации. Было показано, что все это вовлекается в опосредованные Т-клетками ответы, которые принимают участие в возникновении толерантности (связанной и/или инфекционной). Агенты, увеличивающие презентацию полностью функционального белка Notch на поверхности клетки, включают в себя металлопротеиназы матрикса, такие как продукт гена дрозофилы Kuzbanian (Dkuz, Pan, D и Rubin, G.M. 1997 Cell 90: 2.71-280) и других генов семейства ADAMALYSIN. Агентами, снижающими или препятствующими его презентации как полноценно функционирующего белка клеточной мембраны могут служить ингибиторы металлопротеиназ матрикса (ММР), такие как ингибиторы, основанные на гидроксиматах.
Понятие "антиген-представляющая клетка или ей подобная", здесь подразумевает не только АП-клетки. Специалисту понятно, что подразумевается любой наполнитель, способный представлять требуемый лиганд Notch популяции T-клеток; для обозначения всех подобных структур с целью упорядочивания терминологии используется понятие АП-клетки. Таким образом, понятие АП-клетки включает в себя также дендритные клетки, L-клетки, гибридомы, лимфомы, макрофаги, В-клетки или синтетические АП-клетки, такие как липидные мембраны.
Когда АП-клетки трансфицированы геном, способным к экспрессии лиганда Notch, трансфекция может быть осуществлена вирусом, таким как ретровирус или аденовирус, или другим носителем или методом, способным доставить ген клетке. Эффективность любых подобных носителей или методов показана в генной терапии, и к ним относятся ретровирусы, липосомы, метод электропорации, а также другие вирусы, такие как аденовирусы, адено-ассоциированные вирусы, вирус герпеса, вирус коровьей оспы, ДНК, преципитированная фосфатом кальция, трансфекция с помощью ДЕАЕ-декстрана, микроинъекции, полиэтиленгликоль, комплексы белок-ДНК.
Использование подобных носителей или методов по отдельности или в комбинации друг с другом делает возможным осуществлять сайтнаправленную передачу гена в определенную клеточную популяцию, делая возможным осуществление способа настоящего изобретения in vivo. Например, вирус может быть использован в комбинации с липосомами с целью повышения эффективности поглощения ДНК. Сайтспецифичность доставки может быть достигнута включением специфических белков (Например, одноцепочечного антитела, реагирующего с CD11c дендритных клеток/макрофагов) в вирусную оболочку или липосомы.
Предпочтительно использование конструкций, с помощью которых экспрессия гена (например, Serrate) была бы связана с экспрессией антигена. АП-клетки, избыточно экспрессирующие Serrate, имели бы, кроме того, высокий уровень экспрессии связанного антигена и предпочтительно реагировали бы с Т-клетками соответствующей специфичности.
Дальнейшее воплощение данного изобретения связано с молекулой, содержащей фрагмент лиганда Notch, оперативно связанный с Т-клеточным фрагментом аллергена или антигена, так что при экспозиции с Т-клетками оба фрагмента способны взаимодействовать своими соответствующими сайтами. Такая молекула способна превращать антиген/аллерген-специфичную Т-клетку, толерантную к аллергену или антигену, содержащему этот фрагмент, поскольку специфичность, требуемая от фрагмента лиганда Notch, обусловлена тесным контактом фрагмента аллергена или антигена.
Фрагментом антигена или аллергена может являться, например, синтетический МНС-пептидный комплекс. Это фрагмент молекулы МНС, несущий антигенный пептидсвязывающий участок, несущий элемент антигена. Подобные комплексы были описаны в Altman JD с сотр. Science 1996 274:94-6.
В приведенных далее пояснениях описываемое вещество является слитым белком, содержащим сегмент Notch или внеклеточный домен лиганда Notch и иммуноглобулин Fc, предпочтительно IgGFc или IgMFc.
Во всех описанных выше аспектах настоящего изобретения предпочтительно, чтобы лигандом Notch были белки семейства Serrate или белки семейства Delta, их производные, фрагменты или аналоги.
В список заболеваний или инфекций, которые могут быть охарактеризованы как опосредованные Т-клетками, можно включить одно или более из нижеперечисленного: астма, аллергия, отторжение трансплантата, аутоиммунные проявления, вызванные опухолью аберрации в системе Т-клеток, а также инфекционные заболевания, такие как вызванные видами плазмодия, микрофилярией, гельминтами, микобактериями, ВИЧ, цитомегаловирусом, Pseudomonas, Toxoplasma, Echinococcus, Haemophilus influenza типа В, возбудителями кори, гепатита С или Toxicara. Таким образом, при использовании подходящего аллергена или антигена, лиганд Notch может быть использован согласно изобретению для лечения указанных заболеваний или инфекций.
В изобретении также предусматривается способ, позволяющий обнаруживать иммуносупрессию, индуцированную поражающим организмом. Подобные организмы способны создавать растворимые формы из членов семейств Serrate, Notch и/или Delta или из их производных, или из белков, действующих на их экспрессию in vivo, индуцируя таким образом, инфекционную толерантность иммуносупрессии. Способ включает в себя анализ на присутствие in vivo немембраносвязанных Serrate, Delta, Notch или их производных и предпочтительно связан с антителами против Serrate, Delta или Notch или их производных.
Применяемые методы скрининговых анализов для регистрации сниженной или повышенной экспрессии и/или процессинга Notch, Delta/Serrate, включают в себя:
1. Экспрессию Delta/Serrate, Notch и Fringe, устанавливаемую после экспозиции изолированных клеток с исследуемыми соединениями в культуре, например,
a) на белковом уровне посредством специфического окрашивания антител с использованием методов иммуногистохимии или проточной цитометрии,
b) на уровне РНК посредством количественной полимеразной цепной реакции с обратной транскриптазой RT-PCR. RT-PCR может быть реализована при помощи контрольной плазмиды с заданными стандартами для измерения эндогенной экспрессии гена при помощи специфических для Notch 1 и Notch 2, Serrate 1 и Serrate 2, Delta 1 и Delta 2, Delta 3 и Fringe праймеров. Эта конструкция может быть модифицирована как только будут идентифицированы новые члены лигандной группы.
c) на функциональном уровне посредством анализа клеточной адгезии.
Повышенный уровень экспрессии Delta/Serrate или Notch должен приводить к усилению адгезии между клетками, экспрессирующими Notch, и его лигандами Delta/Serrate. Тестируемые клетки подвергаются частичной обработке в культуре, и выявляются меченные радиоактивными или флуоресцентными метками клетки-мишени (трансфицированные белком Notch/Delta/Serrate). Смеси клеток инкубируются при 37oС в течение 2 ч. Неприлипающие клетки смываются, и степень адгезии определяют по уровню, радиоактивности/иммунофлуоресценции на поверхности плашки.
Использование подобных методов позволяет выявлять соединения или лиганды Notch, нарушающие экспрессию или процессинг белка Notch или лиганда Notch. Изобретение также относится к соединениям или лигандам Notch, которые могут быть выявлены подобными аналитическими методами.
В сферу данного изобретения также входит способ анализа, включающий в себя контактирование (а) белка Notch и лиганда, способного связываться с белком Notch (b); и определение того, влияет ли соединение с на связывание лиганда с белком Notch, в особенности если белок Notch ассоциирован с Т-клеткой.
Лигандами Notch согласно изобретению предпочтительно являются белки - члены семейств Delta или Serrate или их полипептиды, или их производные. Предпочтительно, чтобы они были получены стандартными рекомбинантными методами, хорошо известными специалистам в данной области. Соответствующие последовательности генов для образования подобных соединений согласно изобретению могут быть получены из публикаций, таких как WO 97/01571, WO 96/27610 и WO 92/19734. Однако изобретение не ограничивается последовательностями Notch, Delta и Serrate, обнаруженными в данных публикациях. Еще более предпочтительно, чтобы члены семейств Notch, Delta или Serrate, белки или полипептиды, или их производные являлись фрагментами внеклеточных доменов Notch, Delta или Serrate или членами их семейств, или производными подобных фрагментов. Здесь понятие "лиганд Notch" дополнительно включает в себя любой лиганд или член семейства лигандов, взаимодействующий с членом семейства белков Notch и включает в себя группу белков, относящуюся как к "топоритмическим белкам", т.е. белкам - продуктам генов Delta, Serrate, Deltex и Enhancer of split (энхансер сублокуса), так и к другим членам этого семейства генов, идентифицируемых благодаря способности их генных последовательностей гибридизоваться с или их гомологии белками Notch, Delta и Serrate, или способности их генов проявлять фенотипические взаимодействия.
Notch, Delta или Serrate впервые были описаны у дрозофилы, и, следовательно, представляют собой белки-прототипы рецептора Notch и, соответственно, членов семейства лиганда Notch. Множественные белки и лиганды Notch в настоящий момент описаны у многих видов позвоночных и беспозвоночных, но их номенклатура может отличаться от таковой у мухи. Например, Notch является гомологом Lin 12 и Glp 1, Serrate/Delta являются гомологами Jagged, Apx1 и Lag-2.
Фармацевтические композиции согласно изобретению могут быть составлены в соответствии с общеизвестными специалистам в данной области принципами. Таким образом, природа наполнителя и уровень активности будут зависеть от соединения согласно изобретению, которое должно быть введено в состав композиции.
Предпочтительно, чтобы фармацевтические композиции были представлены в виде одноразовой дозировки.
Дозировки соединений настоящего изобретения, назначаемые пациенту в виде фармацевтической композиции, могут быть определены лечащим врачом.
Предпочтительным способом назначения композиции согласно изобретению является один из обычных способов, включающих внутримышечный и внутрибрюшинный, внутривенную инъекцию, интраназальную ингаляцию, легочную ингаляцию, введение подкожно, внутрикожно, внутрисуставно, внутриоболочковый, местный способы введения, а также способ введения через пищеварительный тракт (например, через пейеровы бляшки).
Понятие "производное", используемое здесь в отношении белков или полипептидов настоящего изобретения, может включать в себя любое замещение, вариацию, модификацию, делецию, замену или добавку одного (или более) аминокислотного остатка в последовательности, при условии, что полученный белок или полипептид обладает способностью модулировать взаимодействия лигандов Notch-Notch.
Понятие "вариант", используемое здесь в отношении белков или полипептидов настоящего изобретения, может включать в себя любое замещение, вариацию, модификацию, замену, делецию или добавку одного (или более) аминокислотного остатка в последовательности, при условии, что полученный белок или полипептид обладает способностью модулировать взаимодействия лигандов Notch-Notch.
Понятие "аналог", используемое здесь в отношении белков или полипептидов настоящего изобретения, включает в себя любой пептидомиметик, который является химическим соединением, обладающим способностью модулировать взаимодействия лигандов Notch-Notch аналогично родительским белку или полипептиду. К ним относятся соединения, которые могут положительно или отрицательно влиять на экспрессию или активность белка Notch или лиганда Notch.
Соединение может быть рассмотрено как модулирующее взаимодействие лигандов Notch-Notch, если оно способно либо подавлять, либо усиливать взаимодействие Notch с его лигандами, предпочтительно в достаточной для обеспечения терапевтической эффективности степени.
Под экспрессией "лиганда Notch-Notch" здесь подразумевается взаимодействие между членом семейства Notch и лигандом, способным связывать один или более подобных членов.
Понятие терапии, используемое здесь, охватывает диагностические и профилактические мероприятия.
Понятие "медицинский" включает в себя применение в ветеринарии и к человеку.
Здесь понятия белок и полипептид могут быть приняты как синонимы, при этом понятие белок обычно чаще используется для обозначения относительно более длинной аминокислотной последовательности, чем у полипептида.
Далее настоящее изобретение будет описано неограничивающими примерами со ссылкой на соответствующие чертежи, в которых:
на фигуре 1 показаны результаты гибридизации in situ, выполненной, как описано в примере 1;
на фигуре 2 показаны результаты эксперимента, описанного в примере 4;
на фигуре 3 показаны результаты эксперимента, описанного в примере 5;
на фигуре 4 показаны результаты эксперимента, описанного в примере 6;
на фигуре 5 показаны результаты эксперимента, описанного в примере 7;
на фигуре 6 показаны результаты эксперимента, описанного в примере 8;
на фигуре 7 показаны результаты эксперимента, описанного в примере 9;
на фигурах 8а и 8b показаны результаты эксперимента, описанного в примере 10;
на фигуре 9 показаны результаты эксперимента, описанного в примере 11.
Пример 1
Notch, Delta и Serrate экспрессируются в периферической иммунной системе.
Антисмысловые пробы РНК, специфичные для Notch 1, Delta 1 и Serrate 1, были синтезированы и включены с меченным дигоксигенином UTP. Каждая проба была растворена в гибридизационном буфере, нагретом до 70oС в течение 5-10 мин, и добавлена к покрытым TESPA микроскопическим препаратам, содержащим 10 мм среды селезенки и тимуса, которые предварительно были фиксированы в 4% р-ре параформальдегида + PBS. Микроскопические препараты были гибридизованы в течение ночи при 65oС. На следующий день эти препараты дважды промывали при 65oС и дважды - при комнатной температуре (КТ) 1хSSС/50%формамид/0,1%твин-20. Микроскопические препараты промывали дважды (МАВТ)-буфером 0,1М малеиновой кислотой /0,15М NaCl/0,1% твин-20, рН 7,5, при КТ, и затем блокированы на 2 ч МАВТ + 20% козьей сыворотки + 296-блокирующий реагент Берингера (BBR). Микропрепараты инкубировали в течение ночи при КТ с антидигоксигениновыми Fаb-фрагментами. После 4 промываний МАВТ микропрепараты промывали еще 2 раза в щелочном субстратном буфере. Наличие связанных проб антисмысловой РНК определяли посредством инкубации микропрепаратов в темноте в субстратном буфере, содержащем NBT + BCIP. Микропрепараты окрашивали гематоксилином и закрепляли в среднем закрепителе Depx.
Результаты
Результаты данных по гибридизации приведены на фигуре 1, где показано, что в селезенке трехмесячной мыши Delta и Serrate экспрессировались изолированными клетками в периартериолярном влагалище, но не в зародышевом центре (ЗЦ). Notch экспрессировался во множестве клеток, опять же в периартериолярном влагалище.
Пример 2
Получение слитого белка Delta-Fc
Вектор экспрессии pIG-1 [D.Simmons, "Cloning cell surface molecules by transient surface expression in mammalian cells", pp. 93-128, Cellular Interactions in Development Ed. D. Hartley, pub. Ox. Uni. Press (1993)] позволяет получать слитый белок, содержащий внеклеточную часть Delta 1, связанную с IgGl-Fc-доменом человека. Фрагмент фермента рестрикции, содержащий только внеклеточный домен белка Delta 1, клонирован в вектор pIG-1, Полученная плазмида была трансформирована в МС 1061 E. coli и выращена в SOB, содержащем 10 мкг/мл тетрациклина/ампициллина. Очищенный вектор использовали для трансфекции клеток COS in vitro. Клетки COS выращивали до 50-75% конфлюэнтности и были трансфицированы 10 мкг плазмидной ДНК на чашку методом DEAE-декстран. Через 24 ч после трансфекции культуральную среду заменяли на среду, содержащую 1% FCS, клетки культивировали еще 3-6 дней in vitro. Клетки центрифугировали в течение 5 мин при 5000 об/мин до осаждения клеточных обломков и образования клеточного осадка, супернатант удаляли и сохраняли для последующих опытов. Слияние Delta-Fc очищали от культуральных супернатантов добавлением 2 мл 50% смеси протеин-сефарозы (Pharmacia) и перемешивали в течение ночи при 4oС. Гранулы сефарозы удаляли, пропуская культуральные супернатанты через 0,45 мм фильтр, отмывали и переносили в 10 мл пластиковую колонку. Слитую конструкцию Delta-Fc элюировали 2 мл элюционного буфера, рН 4,0. Элюат дополнительно нейтрализовали добавлением 1М трис.
Пример 3
Примеры моделей, в которых можно исследовать сигналирование лиганда Notch
Периферическая толерантность к собственным антигенам может быть исследована в Т-клеточном рецепторе (TCR) трансгенных мышей, у которых лиганд TCR экспрессирован как собственный антиген только на периферии. Периферическая толерантность к трансплантационным антигенам может быть вызвана несколькими путями, включая преобработку реципиента антителами против Т-клеток или блокаду костимуляции. Посредством этого становится возможным продемонстрировать как связанную супрессию, так и инфекционную толерантность. Периферическая толерантность к аллергенам может быть индуцирована интраназальным введением выделенных из аллергена белков. Экспрессию лигандов Notch-Notch измеряли на клетках, взятых из воздухоносных путей и/или из лимфоидных тканей, прилежащих к областям ингаляции аллергена, и выявили изменения в толерантности. Более того, в экспериментальных моделях заражения инфекционными агентами экспрессия лигандов Notch-Notch может быть измерена в организме (патогенном) и в иммунокомпетентных клетках хозяина.
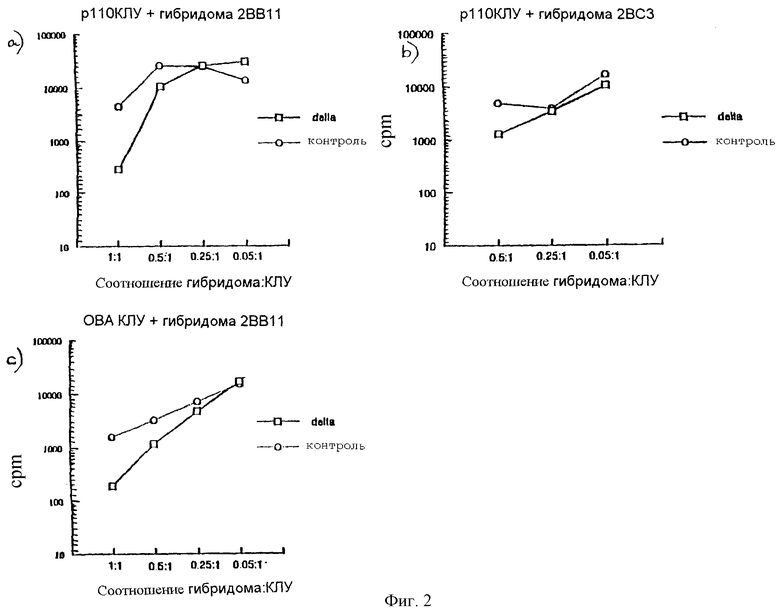
Пример 4
Экспрессирующие Delta гибридомы могут подавлять ответы примированных антигеном лимфоцитов
Мышей иммунизировали синтетическим пептидом, содержащим иммунодоминантный эпитоп аллергена домашнего пылевого клеща (НДМ), Der p1 (p110-131), или овальбумином (OVA, белок куриного яйца). Через неделю клетки лимфатических узлов (клетки ЛУ) выделяли и из них получали клеточные суспензии. Лимфатические узлы животных, иммунизированных другими антигенами, хранили отдельно. Эти клетки были названы примированными клетками ЛУ.
Т-клеточные гибридомы также транфицировали полноразмерной Delta или контрольной плазмидой, такой, чтобы Delta экспрессировалась как мембранный белок. Через два дня культивирования гибридомы облучали, чтобы предотвратить их пролиферацию или продукцию цитокинов. Таким образом, единственный ответ, который был измеряем в анализе, исходил только из клеток лимфатических узлов.
Облученные гибридомы добавляли в увеличенном количестве в культуры, содержащие примированные клетки ЛУ. Надлежащий антиген (напр., р110-131 или OVA) добавляли в клеточную культуру на 24 часа. Супернатанты затем собирали и определяли содержание IL-2 (главный фактор роста Т-клеток). Пролиферативный ответ клеток лимфатических узлов также измеряли через 72 часа.
Результаты: клетки лимфатических узлов, культивируемые в присутствии облученных гибридом, экспрессирующих контрольную плазмиду, продолжали пролиферировать, как показано на фигуре 2, и секретировали IL-2, когда стимулировались в культуре надлежащим антигеном. Их способность к ответу поддерживалась в соотношении 1:1 клетки ЛУ:гибридома. Напротив, пролиферативный ответ и продукция IL-2 клетками лимфоузлов снизилась на 88% при культивировании в присутствии гибридом, экспрессирующих полноразмерную Delta (в соотношении 1: 1), с соответствующим антигеном. Гибридомы трансфицировали контрольным вирусом (пустые кружки), Delta вирусом (пустые квадраты). На фигуре 2 приведены данные счета радиоактивности в минуту (cрm) метки 3H-Tdr, включенной за 72 часа с начала роста культуры. Счет/мин для клеток лимфатических узлов, выращиваемых в присутствии гибридом, экспрессирующих Delta или контрольные конструкции. Общее число клеток на лунку = 4•105 (т.е. количество клеток лимфоузла варьирует, в зависимости от отношения гибридом к клеткам ЛУ, т.о., cрm будет варьировать). Клетки ЛУ p110-131 являются клетками, примированными Der p1 (p110-131), OVA-клетки ЛУ являются клетками, примированными OVA. 2BB11 и 2ВСЗ являются двумя различными Der p1-гибридомами.
Эти данные показывают, что ингибирование ответов экспрессирующими Delta Т-клетками может быть отсрочено. Несмотря на то, что в этой культуральной системе Т-гибридомы, экспрессирующие Delta и специфичные в отношении Der р1, проявляли способность ингибировать ответ примированных OVA Т-клеток, эта кажущаяся потеря специфичности, по всей видимости, обусловлена непосредственной близостью с клетками, усиленными в культуральной системе. Действительно, данные представленные на фигурах 8а и 8b, показывают, что у животных гибридомы, экспрессирующие Delta, должны проявлять антигенную специфичность с иммуногеном, для которого имеет место модуляционный эффект на иммунный ответ на этот иммуноген. В этом случае вероятно, что Т-клетки, экспрессирующие Delta, могут оказаться в непосредственной близости только с отвечающими Т-клетками, если они распознают один и тот же антиген на одной и той же АП-клетке.
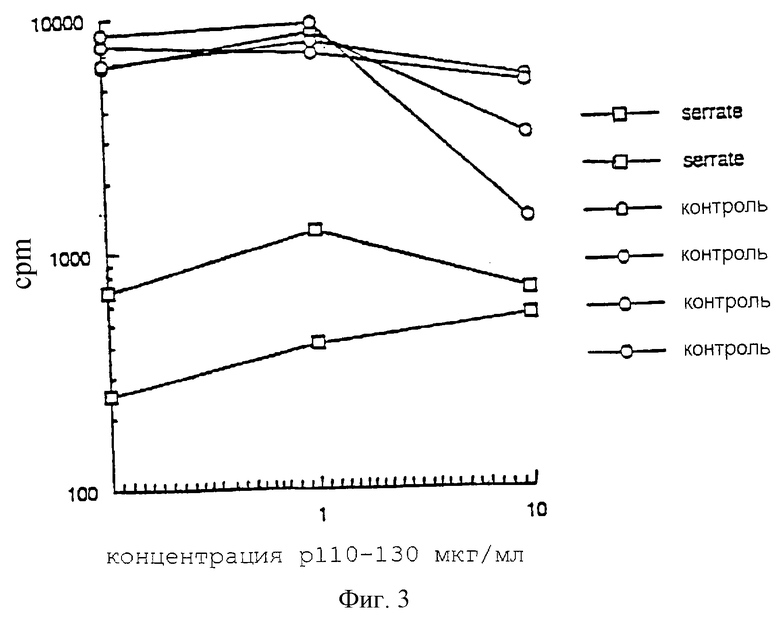
Пример 5
Дендритные клетки, экспрессирующие Serrate, предотвращают примирование антигеном Т-лимфоцитов
Дендритные клетки (ДК) являются основными антиген-представляющими клетками в иммунной системе и незаменимы при стимуляции Т-клеточного ответа. ДК получали из селезенки и трансфицировали либо ретровирусом, в результате чего происходит экспрессия полноразмерного белка Serrate, либо контрольным ретровирусом. Также ДК в течение 3 часов непрерывно взбалтывали с белком HDM p110-131 при температуре 37oС in vitro. Затем ДК промывали и использовали для иммунизации интактных мышей, вводя подкожно 105 клеток/мышь. Через 7 дней выходящие клетки ЛУ собирали и повторно стимулировали в культуре пептидом, 4•105 клеток/лунку. Поскольку мыши были иммунизированы только дендритными клетками, обработанными белком при взбалтывании, это дало нам возможность оценить способность этих клеток к примированию иммунного ответа.
Результаты: на фигуре 3 приведены данные счета радиоактивности в минуту в клетках ЛУ, выделенных через 72 часа из культуры; эти клетки были получены у животных, иммунизированных контрольными трансфицированными (пустые кружки) или трансфицированными Serrate (пустые квадраты) дендритными клетками (ДК).
Иммунизация мышей экспрессирующими Serrate дендритными клетками привела к 10-кратному уменьшению числа клеток, полученных из лимфатических узлов, по сравнению с иммунизацией контрольными ДК. Кроме того, мы обнаружили, что клетки ЛУ мышей, иммунизированных ДК+Serrate, не проявляли пролиферативной активности (93% снижение относительно контрольных значений, фигура 3) и не секретировали IL-2 по сравнению с мышами, иммунизированными контрольными ДК.
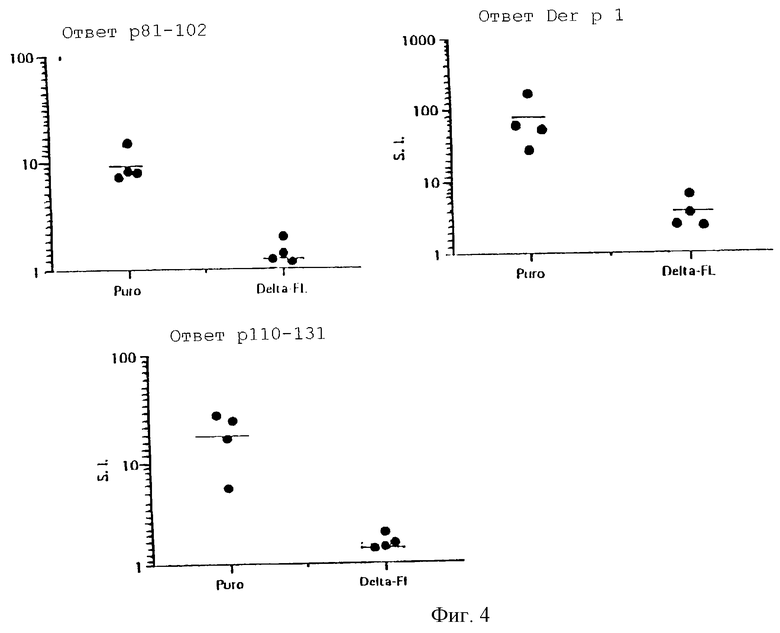
Пример 6
Экспрессирующие Delta Т-клеточные гибридомы способны ингибировать развитие иммунитета к Per p1-антигену у животных
Т-клеточные гибридомы (реактивные с Der p1) трансфицировали ретровирусом, содержащим Delta мыши, такой, что Delta был экспрессирован на поверхности клетки или с контрольным ретровирусом. С57 BL-мышам вводили внутрибрюшинно 10 млн. облученных гибридом и подкожно иммунизировали 50 мкг Der р1, эмульгированном в полном адъюванте Фрейнда (CFA). Через 7 дней собирали выходящие из лимфатических узлов клетки и выращивали их в количестве 4•105 клеток/лунку в присутствии Der р1 (10 мкг/мл), пептида 110-131 Der р1 (10 мкг/мл) или пептида 81-102 Der p1 (10 мкг/мл). Культуры инкубировали в течение 72 часов при 37oС и за 8 часов до конца инкубации добавляли меченный тритием тимидин. Результаты анализов пролиферации клеток животных, инъецированных контрольно трансфицированными (puro) или трансфицированными Delta (Delta-FL), показаны на фигуре 4.
Результаты: клетки ЛУ животных, инъецированных контрольными вирус-трансфицированными гибридомами, продуцировали высокие уровни IL-2 и пролиферировали в культуре в присутствии Der p1, пептида 110-131 или пептида 81-102. И наоборот, клетки животных, инъецированных гибридомами, экспрессирующими Delta, не отвечали ни на один из Der р1 антигенов (фиг.4).
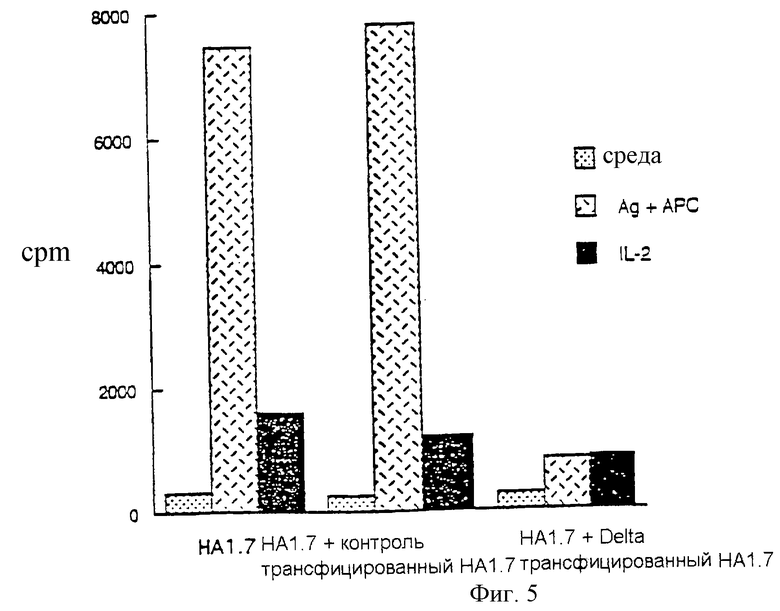
Пример 7
Экспрессирующие Delta Т-клетки человека могут блокировать ответ нормальных Т-клеток
Реактивный к гриппу клон Т-клеток (НА1.7) человека трансфицировали мышиным Delta, используя ретровирусную конструкцию, чтобы белок Delta экспрессировался на клеточной поверхности. Смешивание этой клеточной популяции с нормальными НА1.7 предотвращало дальнейшую реактивность этих нормальных НА1.7 с пептидом НА306-318 и с антиген-представляющими клетками. НА1.7 в количестве 5•105 смешали с 1•106 облученных DRB1*0101 мононуклеарных клеток периферической крови (РВМС) + 1 мкг НА306-318 и культивировали при 37oС. Через 6 часов добавляли 5% лимфокультуру (среду, содержащую IL-2) до общего объема 1 мл. Через 24 часа после инициации культуры добавляли Delta или контрольный ретровирус, или не добавляли ничего. Через 7 дней после начала культивирования клетки собирали, отмывали, и трансфицированные клетки облучали. Трансфицированные клетки смешивали в соотношении 2:1 с необработанными НА1.7 и культивировали в течение 2 дней. Затем смешанные культуры собирали, отмывали и переносили на плашки в количестве 2•104 жизнеспособных клеток на лунку вместе с:
a) 2,5•104 DRB1*0101 PBMCs (среда)
b) 2,5•104 DRB1*0101 PBMCs + пептид (Аg+АП-клетки)
c) 5% лимфокультурой (IL-2)
Клетки собирали через 68 часов, последние 8 часов клетки культивировали в присутствии меченного тритием тимидина.
Результаты показаны на фигуре 5.
Результаты
Описываемая культура как таковая или после трансфекции контрольным вирусом НА1.7; необработанные НА1.7 дают хорошую ответную реакцию на белок + антиген-представляющие клетки. Инкубация в присутствии Delta-трансфицированных облученных НА1.7 полностью препятствует ответу необработанных НА1.7 на антиген + АП-клетки. Однако такие клетки дают такой же хороший ответ на IL-2, как и необработанные или инкубированные с контрольным вирусом клетки НА1.7, что показывает, что они просто не способны к пролиферации (фиг.5).
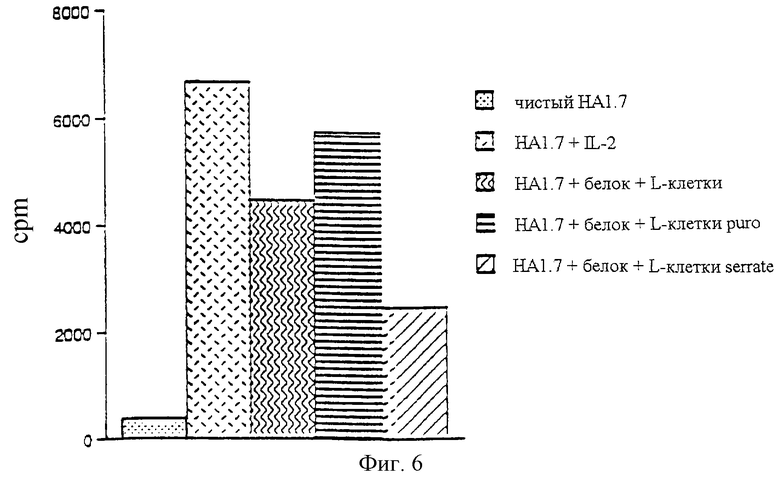
Пример 8
Экспрессия Serrate антиген-представляющими клетками препятствует Т-клеточным ответам
Клон НА1.7 смешивали с пептидом НА306-318 (1,0 мкг/мл) в присутствии L-клеток, экспрессирующих HLA-DRB1*0101 (как антиген-представляющие клетки), используя 2•104 клеток каждого типа. L-клетки трансфицировали либо контрольным (puro), либо Serrate (Serrate L-клетки)- экспрессирующим ретровирусом. Пролиферативный ответ измеряли через 72 часа, последние 8 часов культивирование проводили в присутствии меченного тритием тимидина. Результаты даны на фигуре 6 для культур НА1.7:
a) без добавок
b) HL-2
c) + пептид + DRВ1*0101-L-клетки
d) + пептид + DRВ1*0101-L-клетки, трансфицированные контрольным вирусом
е) + пептид + DRВ1*0101-L-клетки, трансфицированные вирусом Serrate
Результаты: НА1.7, стимулированные L-клетками, экспрессирующими Serrate, демонстрировал слабый ответ на антиген по сравнению с таковыми, стимулированными контрольно трансфицированными L-клетками (фиг.6).
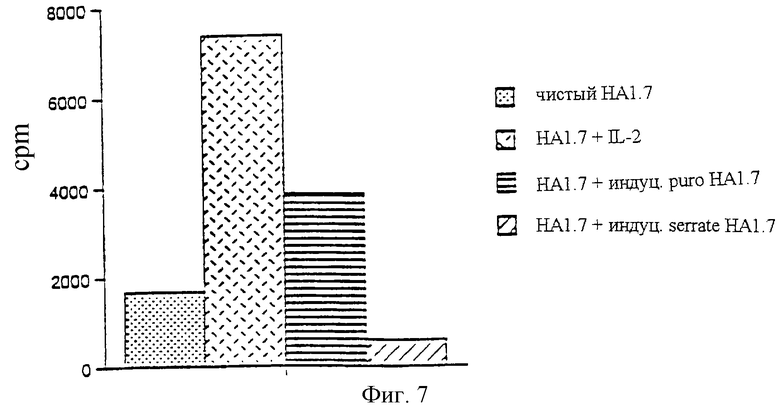
Пример 9
Экспрессирующие Serrate антиген-представляющие клетки индуцируют регуляторные Т-клетки
Клон НА1.7 смешивали с пептидом НА306-318 и L-клетками (экспрессирующими DRB1*0101 как антиген-представляющие клетки) в присутствии 2% IL-2. L-клетки трансфицировали либо контрольным, либо Serrate - экспрессирующим ретро-вирусом. Через 7 дней культивирования клетки НА1.7 собирали, промывали и облучали перед тем как смешать со свежими НА1.7 (используя по 2•104 клеток каждой популяции). Затем клетки культивировали еще 2 дня перед стимуляцией пептидом (10 мкг/мл) + нормальные антиген-представляющие клетки (DRB1*0101 PBMCs). Пролиферативный ответ оценивали через 72 часа после начала культивирования, причем за 8 часов до конца культивирования вводили 3H-тимидин. Результаты приведены на фиг. 7 для свежекультивированных клеток НА1.7:
a) без добавок
b) + IL-2
c) + контрольные вирус-индуцированные НА1.7, затем пептид + DRB1*1010 PBMC
d) + Serrate - вирус-индуцированные НА1.7, затем пептид + DRB1*0101 PBMC
Результаты: НА1.7, индуцированные Serrate-экспрессирующими L-клетками (Serrate-индуцированные НА1.7), утрачивали способность нормальных клеток НА1.7 отвечать на нормальный антигенный стимул (фиг.7). Это указывает на способность клеток, ставших толерантными под действием Serrate, преодолевать их толерантность, превращаясь в популяцию нативных клеток (инфекционная/перенесенная толерантность).
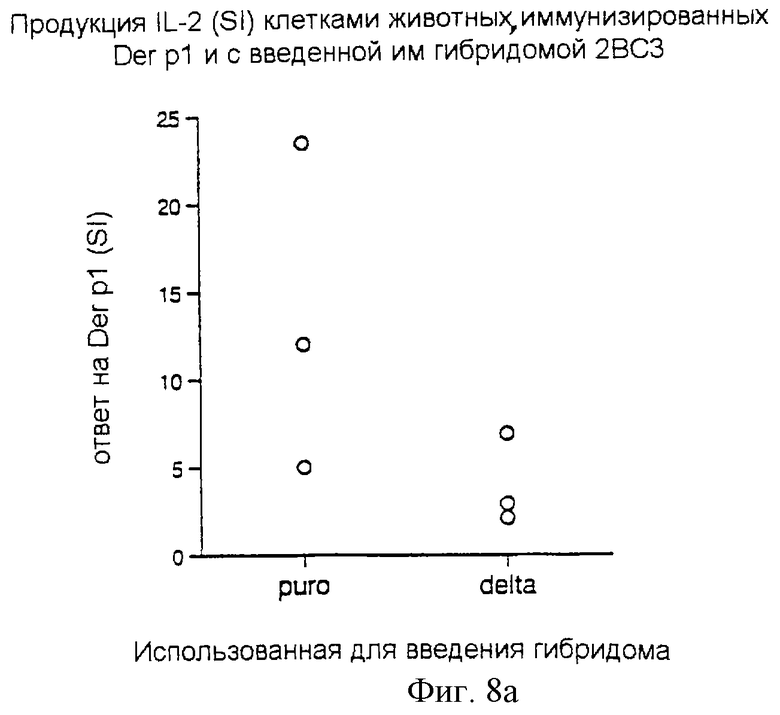
Пример 10
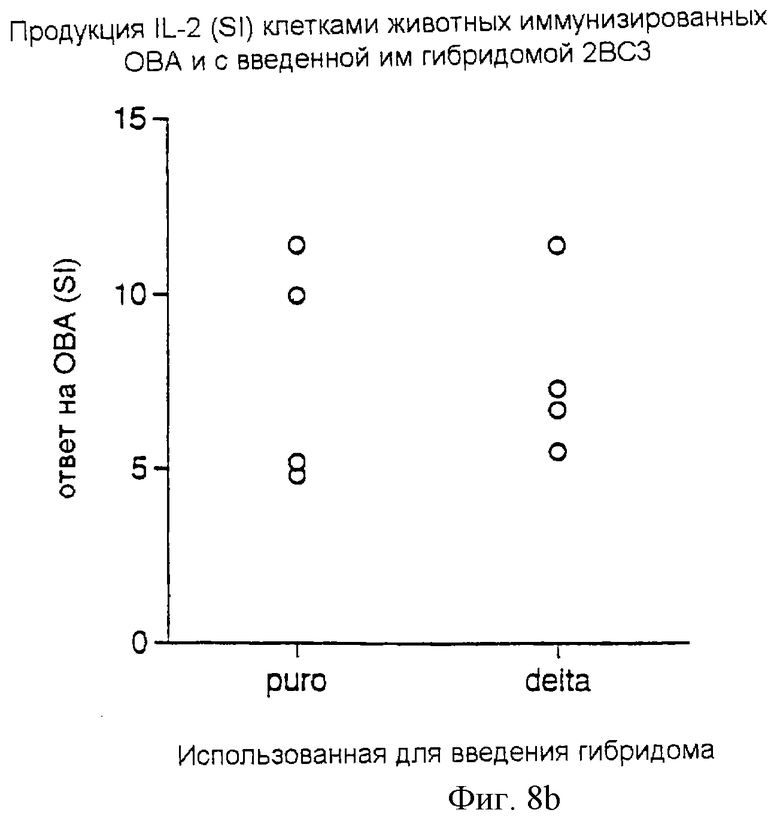
Регуляция, индуцируемая Delta-экспрессирующими Т-клетками, является антигеноспецифической
Мышам вводили 107 клеток Т-клеточной гибридомы, трансдуцированной Delta или контрольным ретровирусом. Эта гибридома 2ВС3 обладает специфичностью к белку 110-131 Der р1. В то же время животные получали либо Der р1, либо овальбумин, эмульгированный в полном адъюванте Фрейнда. Через 7 дней клетки лимфатических узлов удаляли, и 4•105 клеток культивировали в присутствии Der р1 или овальбумина (т.е. использовался тот же антиген, каким они уже были иммунизированы). Продукцию IL-2 измеряли через 24 часа после начала культивирования. Индексы стимуляции продукции IL-2 показаны на фигуре 8а (ответы животных, иммунизированных Der р1) и на фигуре 8b (ответы животных, иммунизированных ОBА) у животных, которым инъецировали гибридомы, трансфицированные контрольным (puro) или Delta (delta)-ретровирусами.
Результаты: ответ на Der р1 у животных, которым были введены гибридомы delta, экспрессирующие 2ВС3, был практически полностью подавлен, тогда как ответ на овальбумин был незатронут. Это показано на фиг.8а и 8b, где каждая точка представляет собой продукцию IL-2 у каждой отдельной мыши на антиген как индекс стимуляции (ИС) контрольных ответов (без добавления антигена).
Пример 11
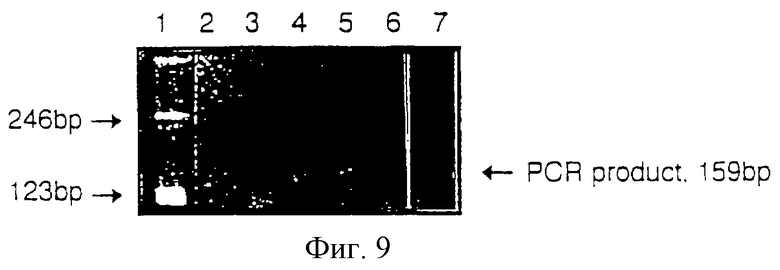
Т-клетки экспрессируют Delta в процессе индукции толерантности
НА1.7 культивировали в присутствии пептида НА306-318 (50 мкг/мл) в отсутствие АП-клеток. Подобные условия использовались с целью развития в НА1.7 толерантности. По прошествии 0, 30, 120, 240 или 360 минут клетки (1,5•106) собирали в капсулы и замораживали. РНК получали из клеточных осадков путем гомогенизации в растворе гуанидина тио-цианата и затем центрифугированием в плотности CsCl. 1 мкг РНК превращали в кДНК при помощи олигопраймера dT. Из полученной кДНК 1/20 подвергли PCR (40 циклов), используя специфические для delta-гомолога человека праймеры. Образцы PCR анализировали посредством гель-электрофореза (фигура 9), где: дорожка 1 - маркер, дорожка 2 - t=0 мин, дорожка 3 - t=30 мин, дорожка 4 - t=120 мин, дорожка 5 - t=240 мин, дорожка 6 - t=360 мин, дорожка 7 - негативный контроль.
Результаты
Покоящиеся НА1.7 не экспрессировали Delta-мРНК, но транскрипты появлялись по прошествии 2 часов после начала развития схемы по инициации толерантности.
Пример 12
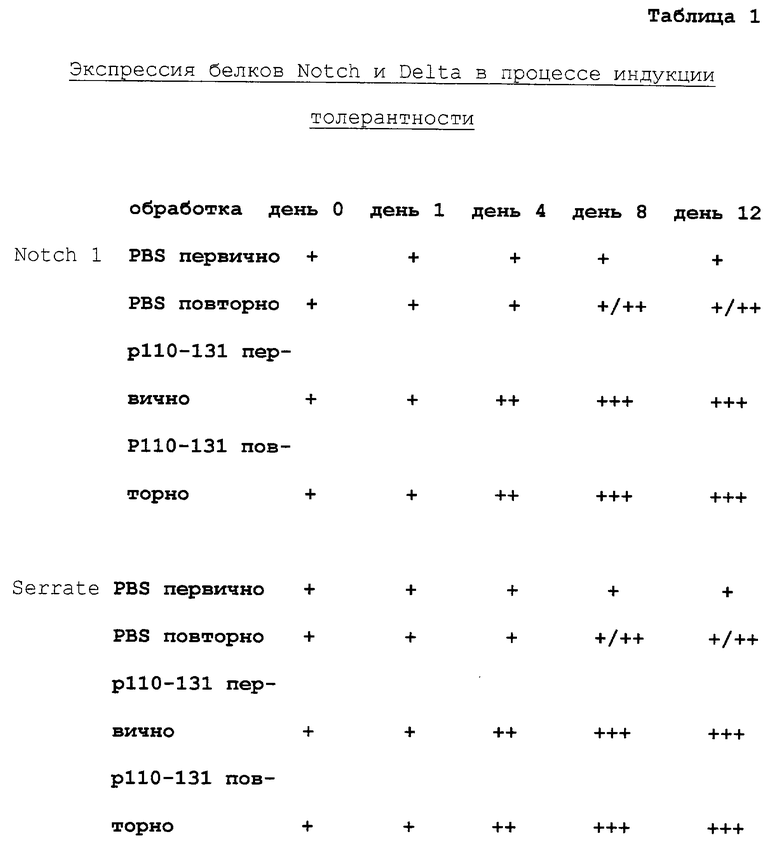
Анализы экспрессии Notch 1, Serrate 1 и Delta 1 в процессе индукции толерантности иммуногистологическим методом и методом гибридизации in situ
Мышам С57 BL/6J интраназально вводили либо 100 мкг 110-131 белка Der p1, либо контрольный раствор (солевой фосфатный буфер, PBS) в течение 3 дней последовательно. Показано, что интраназальное введение антигена подобным способом приводит к развитию невосприимчивости к данному антигену. Некоторую часть животных оставляли на две недели до повторного введения 50 мкг антигена Der p1 CFA подкожно в основание хвоста. Поверхностные лимфатические узлы и селезенки животных с этого времени выделяли через различные промежутки времени (d0 есть первый день введения антигена или повторной иммунизации) и использовали в иммуногистологических исследованиях или для гибридизации in situ. Для иммуногистологических исследований ткани замораживали и делали тонкие 3 мкм срезы, которые фиксировали в ледяном ацетоне и метили поликлональными антителами, специфичными к Notch 1 или Serrate 1. Связанные антитела определяли при помощи пероксидазы хрена, конъюгированной с козьими антикроличьими антителами, полученными в присутствии диаминобензидина в качестве субстрата. Антитела, специфичные к Delta 1, в исследованиях не применяли. Для гибридизации in situ делали срезы замороженных тканей и фиксировали их в 4% параформальдегиде. Срезы подвергали гибридизации с дигоксигенин-связанными пробами антисмысловой РНК, специфичными к Notch I, Serrate 1 и Delta 1, при 65oС. Связанную пробу определяли с помощью щелочной фосфатазы, конъюгированной с козьими антидигоксигениновыми антителами, полученными при использовании NBT и BCIP в качестве субстрата. Данные, показанные в таблицах 1 и 2, относятся к иммуногистологическим исследованиям и гибридизации in situ соответственно. Данные представляют собой анализ тканей 5 различных мышей после интраназального введения одного только пептида (первично PBS/p110-131) или интраназального и подкожного введения антигена (PBS/p110-131 повторно).
+ слабая окраска
++ умеренная окраска
+++ сильная окраска
Результаты
Начальные уровни Notch, Delta и Serrate экспрессировались у контрольных мышей, получавших только PBS. Мыши, получавшие PBS интраназально и антиген подкожно, демонстрировали умеренное повышение экспрессии всех трех молекул в течение 8 дней после повторной иммунизации. Животные, получавшие либо интраназально один пептид либо интраназально пептид и вслед за ним повторно антиген, демонстрировали одинаковую картину повышенной экспрессии Notch, Delta и Serrate, которая развивалась более быстро и до более высоких значений, чем у контрольных мышей.

Изобретение относится к применению терапевтических соединений в модификации Т-клеток взаимодействий, антиген-представляющих клеток с Т-клетками и взаимодействий между патогенными организмами и иммунокомпетентными клетками хозяина. В частности, оно относится к применению этих соединений в модификации взаимодействий между белками Notch и их лигандами и к применению таких соединений в производстве лекарственных средств для лечения состояний, таких как отторжение трансплантата, аутоиммунные процессы, аллергия и астма и инфекционные заболевания. Преимущество изобретения заключается в разработке средства, ингибирующего Т-клеточные взаимодействия, ведущие к инфекционной толерантности и которые могут применяться в производстве медицинских препаратов для иммунотерапии. 3 с. и 9 з.п. ф-лы, 2 табл., 9 ил.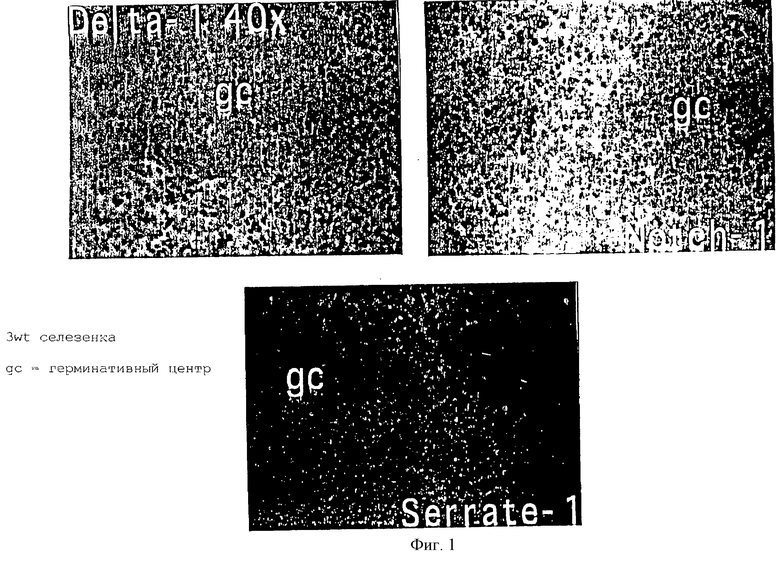
| Автоматический огнетушитель | 0 |
|
SU92A1 |
| Приспособление в пере для письма с целью увеличения на нем запаса чернил и уменьшения скорости их высыхания | 1917 |
|
SU96A1 |
| ARTAVANIS-TSAKOHAS et al., Notch Signallings, Science, 1995, v | |||
| Способ изготовления гибких труб для проведения жидкостей (пожарных рукавов и т.п.) | 1921 |
|
SU268A1 |

