Область техники
Изобретение относится к медицинской диагностике анеуплоидий и других аномалий генома молекулярно-генетическими методами, в частности высокопроизводительным секвенированием ДНК.
Уровень техники
Неинвазивная пренатальная диагностика анеуплоидий рекомендована беременным женщинам для получения более полной информации о здоровье будущего ребенка. Список наиболее часто встечающихся и тяжелых состояний, вызванных анеуплоидиями, включает в себя на сегодняшний день: синдром Дауна (трисомия по 21 паре хромосом), синдром Эдвардса (трисомия по 18 паре хромосом), синдром Патау (трисомия по 13 паре хромосом), а также синдромы Клайнфельтера и Шершевского-Тернера (полисомии по половым хромосомам и моносомия по хромосоме X, соответственно). На сегодняшний день выявление беременных, плод которых имеет одну из этих патологий начинается скринингом I триместра, который используется для оценки риска анеуплоидии. Согласно [Wapner R., et al. "First-trimesterscreeningfortrisomies 21 and 18", NewEnglandJournalofMedicine, 2003, Т. 349. №. 15. стр. 1405-1413], значения чувствительности и специфичности для трисомии по 21 хромосоме составляют 85,2 и 90,6%. После скрининга I триместра формируется группа с высоким риском патологий плода, входящие в нее пациенты направляются на процедуру инвазивной диагностики (ИД). ИД представляет собой забор биоматериала плода (как правило, на таких сроках беременности это амниотическая жидкость - амниоцентез) с помощью иглы под контролем УЗИ и дальнейший цитогенетический анализ фетальных клеток для определения их кариотипа. С учетом чувствительности, 9,4% всех женщин получают ложноположительный результат по результатам скрининга и тем не менее делают ИД. До 1% процедур ИД приводит к выкидышу, также повышаются риски развития других осложнений (повышение температуры и лихорадка, обильные кровотечения, отслойка плаценты, аллоиммунизация).
Во внеклеточной фракции крови человека присутствуют нуклеиновые кислоты. Внеклеточная ДНК (внДНК) является побочным продуктом апоптоза клеток и представляет собой короткие фрагменты геномной ДНК длиной 166 пар оснований (медианное значение). Во внДНК беременных женщин, начиная с самых ранних сроков беременности, определяется фетальная (плодная) фракция (вкфДНК, англ. - cell-free fetal DNA, cffDNA), причем с увеличением срока беременности, доля фетальной внДНК растет. У фракции вкфДНК наблюдается снижение среднего размера с 166 пар оснований (п.о.) до 143 п.о. ВнДНК выделяется из плазмы периферической крови человека и может быть использована для оценки состояния генома плода. Диагностика генетических нарушений плода по внеклеточной фетальной ДНК называется неинвазивным пренатальным тестированием (НИПТ, англ. - non-invasive prenatal testing, NIPT).
NIPT - это статистическое исследование, проводимое для оценки доли представленности каждой из хромосом в исследуемом образце. В норме на каждую из хромосом генома небеременной женщины будет приходиться пропорциональное ее длине количество прочтений (ридов), полученных методами высокопроизводительного секвенирования. Если женщина беременна ребенком с нормальным кариотипом, картина не изменится. Но если у ребенка имеется трисомия по, например, 21 хромосоме, то ее относительное покрытие вырастет. Длина 21 хромосомы составляет примерно 1,5% генома. Если доля фетальной ДНК у образца 10%, то третья фетальная 21 хромосома даст примерно 0,08% прибавку к общему покрытию 21 хромосомы. Чтобы оценить достоверность полученных данных результатов, используются различные статистические методики оценки. Наиболее распространенной является методика Z-критерия Фишера. Z-тест проверяет, не является ли увеличение покрытия хромосомы случайным, сравнивая ее значение с математическим ожиданием покрытия с учетом его стандартной ошибки. Математическое ожидание вычисляется с помощью проведенного заранее анализа выборки образцов внДНК беременных женщин с известным диагнозом у ребенка. Таким образом, чем больше выборка, тем ниже значение стандартной ошибки, а значит, больше точность теста.
Важнейшим критерием, влияющим на NIPT, является доля фетальной внДНК. Чем она больше, тем большее значение Z покажет анализ в случае анеуплоидии. Существует несколько общепринятых способов оценки доли вкфДНК. Их объединяет принцип поиска значимых отличий фетальной фракции внДНК от материнской. Наиболее очевидной является оценка по доле Y-хромосомы, значение которой составляет половину значения вкфДНК. Этот подход надежен, но имеет существенный недостаток - он применим только в случае вынашивания мальчика.
Универсальным и наиболее популярным в настоящее время подходом является оценка доли фетальной внДНК по однонуклеотидным полиморфизмам (single nucleotide polymorphism, SNP). В геноме можно отобрать биоинформатически те диаллельные SNP, которые имеют частоту минорного аллеля (minor allele frequency, MAF), близкую к 50%, входят в разные группы сцепления, и не подвергаются отбору. Так как половину своего генома ребенок наследует от отца, то при достаточной выборке SNP (80-150 шт.) обнаружатся те точки, в которых генотип матери будет гомозиготен, а плода - гетерозиготен (за счет другого аллеля, унаследованного от отца). Доля ридов, выравненных на альтернативные материнскому генотипу аллели, подсчитывается напрямую и равна половине доли фетальной внДНК. В пределе, увеличивая количество анализируемых SNP, можно определять анеуполидию, сравнивая долю покрытия фетальных и материнских хромосом по соотношению ридов соответствующих SNP. Подобным образом поступила компания Natera, тест Panorama которой основан на анализе около 20000 SNP, распределенных по геному [Zimmermann, В., Hill, М., Gemelos, G., Demko, Z., Banjevic, M., Baner, J., … & Levy, B, (2012) Noninvasive prenatal aneuploidy testing of chromosomes 13, 18, 21, X, and Y, using targeted sequencing of polymorphic loci. Prenataldiagnosis, 32(13), 1233-1241]. Для создания таких SNP-библиотек применяется ряд дополнительных лабораторных методик, направленных на обогащение библиотеки NGS соответствующими участками генома.
Недостатками существующих тестов типа NIPT является их высокая себестоимость по сравнению со стандартным биохимическим скринингом и даже инвазивной диагностикой. Таким образом, несмотря на существование ряда способов оценки наличия анеуплоидии, на сегодняшний день на рынке сохраняется потребность в более эффективном методе.
Сущность изобретения
Задачей настоящего изобретения является создание нового, более эффективного способа определения анеуплоидий плода по внеклеточной ДНК, циркулирующей в крови матери. Для решения этой задачи авторами предложен способ создания библиотек внеклеточной ДНК беременных женщин для проведения NIPT, путем «упаковки» в один рид нескольких различных фрагментов исходных внДНК с медианой длины 35 п.о., получая таким образом химерные линейные молекулы ДНК. Тем самым достигается снижение необходимого для анализа количества сырых данных как минимум в 4 раза, что существенно повышает эффективность идентификации анеуплоидий по внДНК для последующего определения анеуплоидий плода. Разработанный способ основан на том, что для картирования рида на геном достаточно 30-40 п.о., то есть не более четверти длины нативной внДНК длиной 166 п.о. Существующие тесты NIPT делают полное прочтение внДНК, либо два коротких (до 40 п.о.) прочтения с концов молекул внДНК. В первом случае информация оказывается избыточной. Однако реактивы для высокопроизводительных секвенаторов позволяют проводить анализ ридов большей длины, поэтому второй подход нерационально расходует реактивы для секвенирования. Таким образом, если создать химерные молекулы внДНК, состоящие из случайно соединенных фрагментов внДНК в одном риде, можно получить в несколько раз больше информации с одного прочтения, и использовать ее для определения анеуплоидий плода.
Указанная задача решается путем создания способа подготовки внеклеточной ДНК из образца крови беременной женщины к секвенированию для диагностики анеуплоидии плода, включающего следующие стадии: а) получают образец материнской крови, состоящий из цельной крови, или плазмы крови, или сыворотки крови, при этом для образца, состоящего из цельной крови, отделяют плазму крови и используют ее для последующих стадий; b) выделяют внеклеточную ДНК, присутствующую в указанном образце крови; с) осуществляют фрагментацию очищенной внеклеточной ДНК, получая в результате размер ДНК-фрагментов от 30 до 100 пар оснований со средним размером 40 пар оснований; d) отбирают фрагменты ДНК длиной между 35 и 45 пар оснований; е) осуществляют случайное лигирование отобранных на стадии d) фрагментов ДНК друг с другом для формирования химерных линейных молекул ДНК размером более 100 п.о.; f) осуществляют лигирование адаптерных последовательностей, специфических для используемого в дальнейшем метода секвенирования, с химерными линейными молекулами ДНК для создания библиотек химерных линейных молекул ДНК; g) проводят полимеразную цепную реакцию (ПЦР) для индексирования библиотек химерных линейных молекул ДНК уникальными последовательностями баркодов.
В некоторых вариантах изобретения данный способ характеризуется тем, что фрагментацию очищенной внеклеточной ДНК на стадии с) осуществляют с помощью ферментов и без обработки ДНК ультразвуком.
Указанная задача также решается путем создания способа определения анеуплоидии плода с помощью анализа внеклеточной ДНК из крови беременной женщины методом секвенирования, включающего следующие стадии: а) подготавливают внеклеточную ДНК из образца крови беременной женщины согласно описанному выше способу; б) осуществляют секвенирование образованных библиотек химерных линейных молекул ДНК одним из методов массового параллельного секвенирования, получая множество чтений; в) осуществляют картирование полученных чтений на референсный геном человека для определения присутствия в чтениях участков хромосом человека; г) определяют количество участков последовательностей ДНК в чтениях, соответствующих разным хромосомам человека, при этом исключают участки, которые являются неуникальными для отдельных хромосом генома человека; д) определяют наличие анеуплоидии по определенной хромосоме при обнаружении отличий в нормализованной на длину хромосомы представленности этой хромосомы от среднего значения по геному.
В некоторых вариантах изобретения данный способ характеризуется тем, что классификация представленности отдельных хромосом производится на основании z-статистики, вычисленной в регрессионной модели, и порогового значения z_0. Пороговое значение выбирается для каждой хромосомы отдельно в соответствии с критерием максимизации специфичности.
Техническим результатом настоящего изобретения является повышение эффективности процесса определения анеуплоидии плода путем анализа внеклеточной ДНК из крови беременной женщины методом массового параллельного секвенирования. Технический результат достигается тем, что за одно прочтение образованных библиотек ДНК потенциально получают информацию о нескольких различных хромосомах, и используют эту информацию для определения анеуплоидии плода.
Краткое описание рисунков
Фиг. 1. Схема строения рида, применяемого компаниями, занимающимися проведением NIPT (А), а также схема строения рида для проведения NIPT, разработанного авторами и состоящего из коротких фрагментов. Прямоугольники на концах - адаптерные последовательности, необходимые для закрепления библиотеки на подложке проточной ячейки высокопроизводительного секвенатора. Стрелки показывают прямые и обратные прочтения рида, проводящиеся в процессе секвенирования.
Фиг. 2. Схема картирования ридов для «классического» NIPT (А) и разработанного авторами (В) на геном человека. За счет химерного строения молекулы, картирование прочитанных фрагментов каждого из ридов (В) несет в себе больше информации для определения наличия анеуплоидии плода.
Фиг. 3. (а). Хроматограмма библиотеки химерных молекул ДНК высокого качества. Показана зависимость сигнала флуоресценции (ось Y) от длины ДНК (ось X). Цифрами 35 и 10380 обозначены нижний и верхний маркеры соответствующей длины в парах оснований.
Фиг. 3. (b). Хроматограмма библиотеки амплифицированных фрагментов ДНК высокого качества. Показана зависимость сигнала флуоресценции (ось Y) от длины ДНК (ось X). Цифрами 35 и 10380 обозначены нижний и верхний маркеры соответствующей длины в парах оснований.
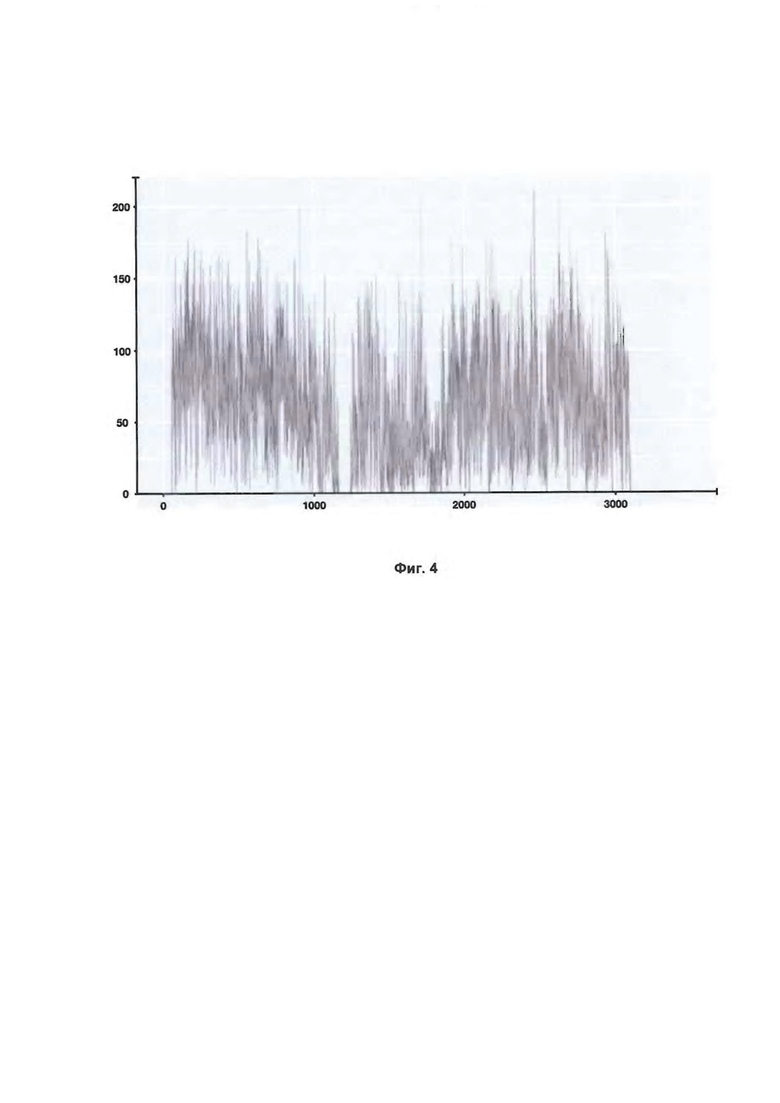
Фиг. 4. Пример покрытия хромосомы X с нормальным женским кариотипом для одного из образцов контрольной выборки. По оси X отложены окна (бины) размером 50 кб (тысяч пар оснований), на которые разбита хромосома, по оси Y - количество прочтений внутри бина.
Фиг. 5. (а). Значения Z-статистики по 13-й хромосоме для всех образцов контрольной выборки. Кругами обозначены средние значения Z для образцов с нормальным кариотипом, треугольниками - с трисомией по 13 хромосоме. «Усы» - диапазон значений Z для соответствующего образца, которые она принимала в рамках 200 симуляций. Жирная горизонтальная линия - значение Z, равное 3.
(b). Значения Z-статистики по 18-й хромосоме для всех образцов контрольной выборки. Кругами обозначены средние значения Z для образцов с нормальным кариотипом, треугольниками - с трисомией по 18 хромосоме. «Усы» - диапазон значений Z для соответствующего образца, которые она принимала в рамках 200 симуляций. Жирная горизонтальная линия - значение Z, равное 3.
(c). Значения Z-статистики по 21-й хромосоме для всех образцов контрольной выборки. Кругами обозначены средние значения Z для образцов с нормальным кариотипом, треугольниками - с трисомией по 21 хромосоме. «Усы» - диапазон значений Z для соответствующего образца, которые она принимала в рамках 200 симуляций. Жирная горизонтальная линия - значение Z, равное 3.
Фиг. 6. График зависимости значений характеристик модели, подсчитывающей z-статистику, от доли оставшихся данных, рассчитанный в одной из симуляций для трисомии по 21 хромосоме. По оси Y отложены значения от 0 до 1, по оси X отложены значения доли данных относительно исходно полученного количества прочтений, используемые для вычислений AUC, чувствительности и специфичности. Видно, что снижение количества данных в 2 раза от среднего не приводит к ухудшению показателей разработанного NIPT.
Фиг. 7. (a). ROC кривая для определения трисомии по 13 хромосоме.
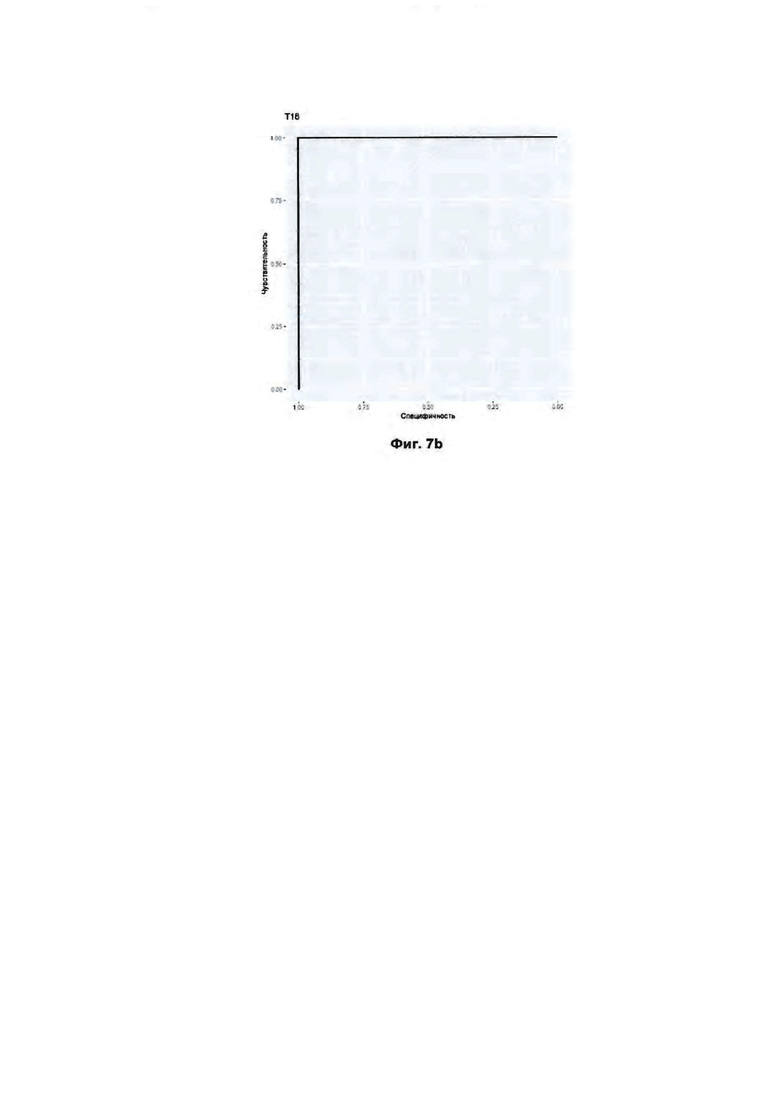
(b). ROC кривая для определения трисомии по 18 хромосоме.
(c). ROC кривая для определения трисомии по 21 хромосоме.
Фиг. 8. Значения доли фетальной ДНК, определенные для контрольной выборки с помощью анализа результатов секвенирования библиотек амплифицированных фрагментов ДНК. По оси Y отложено количество образцов, по оси X - значение доли фетальной ДНК в %.
Фиг. 9. Результаты определения пола плода по отношению нормированного покрытия Y хромосомы (значения отложены по оси Y) к отношению нормированного покрытия X хромосомы к аутосомам (значения отложены по оси X). Образцы делятся на две группы: группа А - беременные с плодами мужского пола, группа В - беременные с плодами женского пола. Стрелками обозначены два образца с плодами мужского пола, определенного как женский по причине низкой доли фетальной ДНК (менее 2%).
Подробное раскрытие изобретения
В описании данного изобретения термины «включает» и «включающий» интерпретируются как означающие «включает, помимо всего прочего». Указанные термины не предназначены для того, чтобы их истолковывали как «состоит только из». Если не определено отдельно, технические и научные термины в данной заявке имеют стандартные значения, общепринятые в научной и технической литературе.
Методы массового параллельного секвенирования (также называемыми методами секвенирования следующего поколения, NGS) - термин, относящийся ко всем высокопроизводительным технологиям секвенирования ДНК, которые способны расшифровать большое число различных последовательностей ДНК в одной реакции (то есть параллельно). Технологии массового параллельного секвенирования, в отличие от капиллярного секвенирования, основаны, как правило, на параллельном выявлении присоединяемых нуклеотидов при синтезе клона ДНК в каждой из миллионов микроскопических ячеек/участках матрицы. Каждая из наиболее распространенных платформ массового параллельного секвенирования (Illumina, Ion Torrent, MGI, Oxford Nanopore и др.) использует собственную стратегию для реализации этого принципа. Эти компании используют подходы, основанные на обратимой флуоресценции, выявлении изменений в уровне рН среды, протаскивании молекулы ДНК через белковую пору и измерении в ней электрического сопротивления.
В основе заявляемого способа определения анеуплоидии плода с помощью анализа внеклеточной ДНК из крови беременной женщины лежит разработанный авторами протокол пробоподготовки внеклеточной ДНК беременных женщин на ранних стадиях беременности (10-14 неделя), с помощью которого можно получать библиотеки ДНК, состоящие из коротких фрагментов внДНК, для высокопроизводительного секвенирования и последующего NIPT-анализа полученных данных. Ключевыми отличиями от существующих методов анализа внДНК для определения анеуплоидии являются:
1. Длительная фрагментация внДНК до сверхкоротких фрагментов;
2. Селективный отбор (size-select) фрагментов внДНК длиной около 40 п.о.;
3. Случайное лигирование коротких фрагментов внДНК (конкатемеров) друг с другом для формирования химерных линейных молекул ДНК размером более 200 пар оснований.
Комбинация перечисленных выше шагов не применяется ни в одной из описанных ранее методик пробоподготовки образцов для проведения NIPT. Основным преимуществом данного подхода является сниженное по меньшей мере вчетверо по сравнению с аналогичными подходами количество сырых данных, необходимое для проведения NIPT с такими же параметрами чувствительности и специфичности. На Фиг. 1, представлена схема строения рида, применяемого на сегодняшний день компаниями, занимающимися проведением NIPT, а также схема строения рида по настоящему изобретению (упаковка в один рид нескольких различных фрагментов внДНК). Прямоугольники на концах - адаптерные последовательности, необходимые для закрепления библиотеки на подложке проточной ячейки высокопроизводительного секвенатора. Стрелки показывают прямые и обратные прочтения рида, проводящиеся в процессе секвенирования. На Фиг. 2 представлена схема картирования ридов для «классического» NIPT (А) и разработанного авторами (В) на геном человека. Один рид, состоящий из коротких конкатемеров, содержит такое же количество клинически полезной информации, как и по меньшей мере 4 обычных рида.
Для осуществления заявляемого способа определения анеуплоидии плода авторами был разработан набор реагентов, который позволяет модифицировать внДНК с помощью ряда ферментативных реакций (фрагментация, фосфорилирование 5' концов, затупление 5' и 3' концов, ПЦР, лигирование), в результате чего из внДНК готовятся библиотеки химерных фрагментов ДНК и библиотеки амплифицированных фрагментов ДНК, необходимые для анализа одним из методов высокопроизводительного секвенирования, например, синтезом с использованием обратимых терминаторов.
Библиотеки ДНК позволяют провести молекулярное кариотипирование образца внДНК беременной женщины. Смесь экзонуклеаз обеспечивает внедрение случайных двунитевых разрывов в молекулах исходной ДНК. Смеси полинуклеотидкиназы и Т4 ДНК-полимеразы производит «полировку» концов разорванных молекул ДНК, в результате чего убираются выступающие одноцепочечные нити, а также 5' концы фрагментов фосфорилируются. Лигаза на первом этапе проводит случайное лигирование коротких фраментов внДНК, формируя цепочки химерных ДНК; на втором этапе лигаза пришивает к подготовленным концам фрагментов олигнуклеотидные адаптеры, необходимые для их индексирования специфическими последовательностями в процессе высокопроизводительного секвенирования. ДНК-полимераза осуществляет амплификацию библиотек ДНК, фланкированных адаптерами на втором этапе лигирования.
Библиотеки амплифицированных фрагментов ДНК позволяют проводить оценку доли фетальной ДНК у беременных женщин. Процедура пробоподготовки помимо описанных ранее этапов для подготовки химерных фрагментов ДНК содержит стадию амплификации целевых регионов генома, проводимую с помощью мультиплексной ПЦР.
Комплект реагентов для индексирования библиотек ДНК баркодами позволяет с помощью ПЦР-реакции присоединить к адаптерным концам фрагментов ДНК уникальные индексные нуклеотидные последовательности, благодаря которым возможно анализировать сразу несколько библиотек ДНК в одном запуске высокопроизводительного секвенатора.
Комплект реагентов для высокопроизводительного секвенирования позволяет провести анализ библиотек фрагментов ДНК и библиотек амплифицированных фрагментов ДНК по одной технологий массового параллельного секвенирования. В предпочтительных вариантах изобретения используют секвенирование синтезом с обратимыми терминаторами, разработанное компанией Illumina, США для платформы Illumina MiSeq или MiSeqDX.
Авторами также разработано программное обеспечение «НИПТ-Аналитика», которое позволяет проанализилировать данные секвенирования на IlluminaMiSeq/MiSeqDX и получить результаты о наличии или отсутствии анеуплоидий у плода по интересующим хромосомам.
Таким образом, разработанный авторами набор реагентов включает: смеси реагентов для конструирования библиотек химерных фрагментов ДНК, смеси реагентов для конструирования библиотек амплифицированных фрагментов ДНК, смеси реагентов для индексирования библиотек баркодами, реагенты для проведения высокопроизводительного секвенирования на платформе IlluminaMiSeq, программное обеспечение для анализа данных секвенирования, контрольные образцы, содержащие ДНК с нормальным кариотипом.
Нижеследующие примеры осуществления способа приведены в целях раскрытия характеристик настоящего изобретения и их не следует рассматривать как каким-либо образом ограничивающие объем изобретения.
Для осуществления способа предпочтительно использовать свежую цельную периферическую кровь беременных женщин. Также для осуществления способа пригодна свежеотобранная или замороженная плазма, или сыворотка периферической крови беременных женщин. Выделение геномной ДНК или внеклеточной ДНК можно проводить в соответствии с инструкцией производителя набора для выделения QIAamp DNA Blood mini kit, Qiagen, либо использовать аналогичные наборы.
Пример. Забор венозной крови общим объемом 20 мл осуществлялся квалифицированным персоналом в 2 пробирки StreckBCT. Режим хранения и транспортировки - только комнатная температура (+15 - +25°С) не более 7 дней с момента сбора крови.
Отделение плазмы крови для выделения внеклеточной ДНК.
1. Центрифугировать пробирки с цельной кровью 30 мин 1500 об/мин;
2. Промаркировать для каждого образца по 4-6 пробирок объемом 2 мл;
3. Отобрать верхнюю фракцию, не задевая раздела фаз (лейкоцитарное кольцо) по 2000 мкл в каждую пробирку;
4. Центрифугировать плазму крови 15 мин в микроцентрифуге на максимальных оборотах;
5. Промаркировать для каждого образца по 4-6 пробирок объемом 2 мл;
6. Отобрать в каждую пробирку по ~2000 мкл плазмы после центрифугирования, не задевая осадок;
7. Заморозить и хранить на -20°С; Срок хранения замороженной плазмы не более 4 месяцев.
Приготовление раствора протеиназы К. На 10 мл раствора необходимо смешать 100 мг протеиназы К, 200 мкл 0,5 М CaCl2, довести водой деионизированной до 10 мл.
Протокол выделения внДНКс помощью набора QIAampDNAMini от Qiagen.
1. Разморозить образцы плазмы по 4-5 мл плазмы.
2. В чистые 15 мл фальконы (по числу образцов) внести 4-5 (в зависимости от объема образца) мл буфера AL. К нему добавить один объем плазмы и 0,1 объема (от объема плазмы) протеиназы К. Тщательно перемешать на вортексе в течение 1 минуты и затем инкубировать 40 минут при температуре 56°С на водяной бане.
3. Добавить в каждую пробирку по одному объему [плазмы] 95% этанола, перемешать на вортексе в течение 1 минуты, оставить на 5 минут при комнатной температуре.
4. Подготовить манифолд QiaVac-24: вставить vac-переходники (по числу образцов), в них вставить подписанные (на крышке и сбоку) колонки. Открыть колонки, включить насос и проверить по манометру, что давление достигло -800-900 мбар.
5. Перенести 700 мкл образца на колонку. Подождать, пока жидкость пройдет через колонку.
6. Повторить пункт 5 необходимое количество раз (пока не закончится лизат).
7. После лизиса центрифугировать колонки на 7000 об/мин 3 минуты.
8. Внести на колонку 700 мкл буфера AW1, центрифугировать на 7000 об/мин 1 минуту. Вылить жидкость из пробирки и вставить колонку в новую пробирку.
9. Внести на колонку 700 мкл буфера AW2, центрифугировать на 7000 об/мин 1 минуту. Вылить жидкость из пробирки и вставить колонку в новую пробирку.
10. Внести на колонку 700 мкл 95% этанола, центрифугировать на 7000 об/мин 1 минуту. Вылить жидкость из пробирки и вставить колонку в новую пробирку.
11. Центрифугировать образцы на максимальной скорости в течение 3 минут, чтобы удалить остатки этанола. Вставить колонки в чистые 1,5 мл пробирки (LoBind).
12. Внести на колонку 41 мкл low ТЕ и инкубировать 30 минут.
13. Центрифугировать на максимальной скорости в течение 1 минуты. Выбросить колонку. ДНК выделена. Выделенную ДНК следует хранить на -20°С.
Для проведения пробоподготовки требуются следующие реагенты:

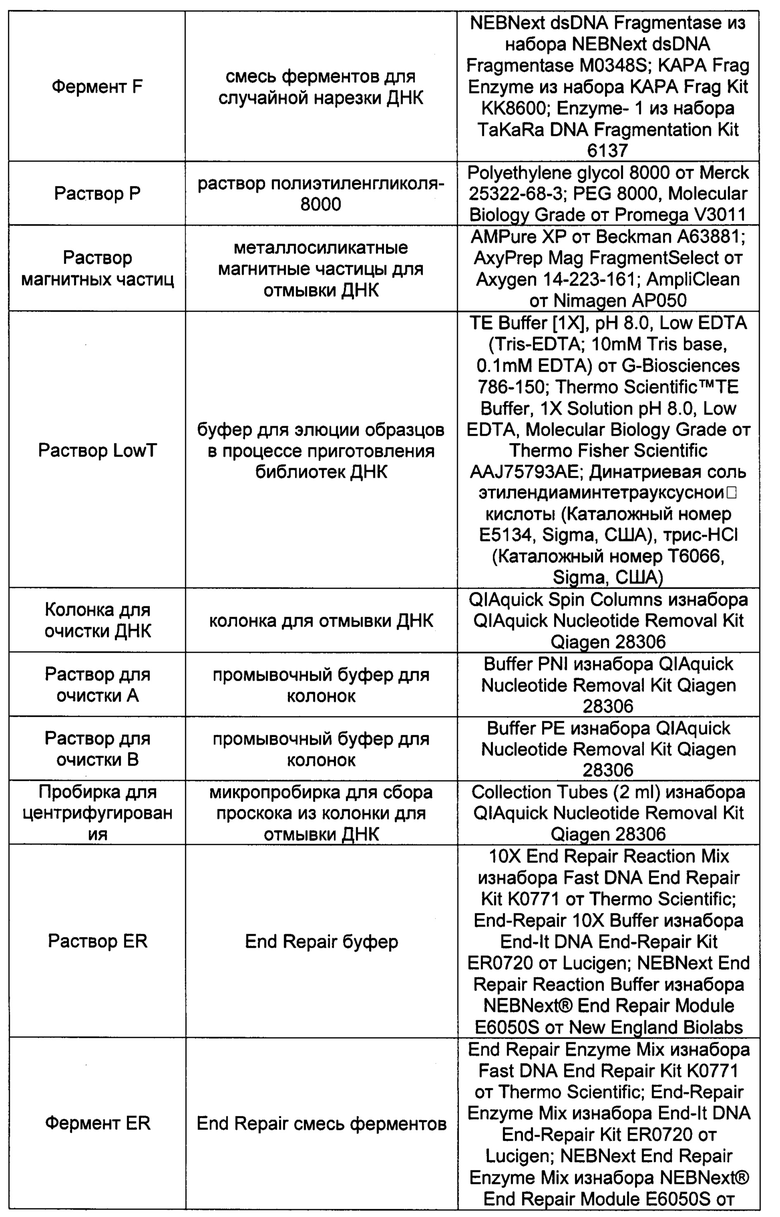

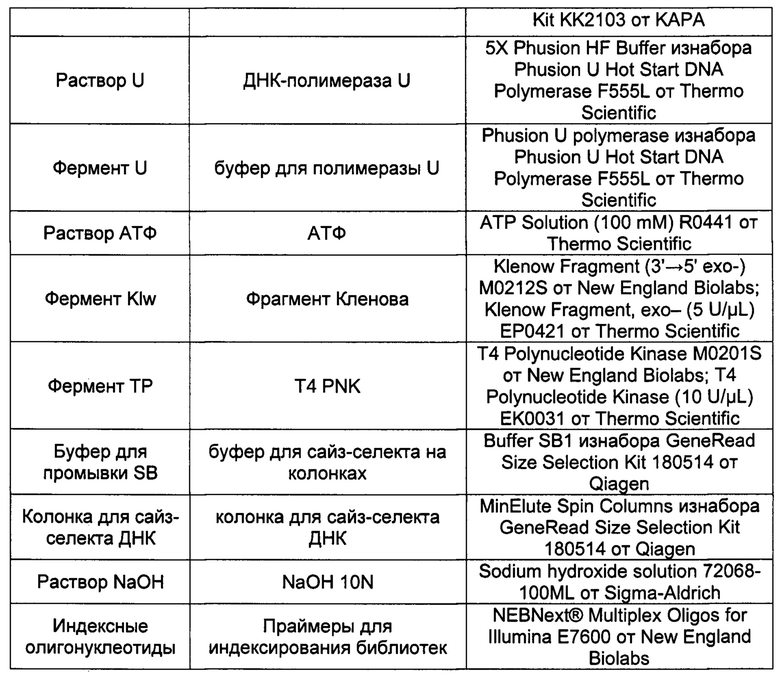
Протокол пробоподготовки рассчитан на образец внДНК общим количеством не менее 3 нг.
Протокол подготовки библиотек фрагментов ДНК
1. Модификация длин ДНК
Для подготовки библиотеки фрагментов ДНК из внеклеточной ДНК необходимо взять около 3-6 нг выделенной по описанному выше протоколу ДНК, пересчитав количество ДНК из значения ее концентрации и подставив его в таблицу ниже.
В микропробирку 1,5 мл необходимо внести следующие компоненты:


Тщательно перемешать пипетированием.
Инкубировать при следующих параметрах:

 После фрагментации добавить Раствор магнитных частиц (предварительно перемешав флакон с частицами на вортексе, чтобы получилась взвесь частиц равномерной концентрации) в количестве 60 мкл (×2 объема);
После фрагментации добавить Раствор магнитных частиц (предварительно перемешав флакон с частицами на вортексе, чтобы получилась взвесь частиц равномерной концентрации) в количестве 60 мкл (×2 объема);
Инкубировать смесь 5 минут;
Затем поставить на магнитный штатив пробирки со смесью на 5 минут;
Супернатант (надосадочную жидкость) перенести в новые пробирки на 1,5 мл;
Добавить 10 объемов Раствора для очистки А;
Инкубировать в течение 1 минуты;
Перенести 600 мкл раствора на Колонку для очистки ДНК;
Инкубировать 1 минуту;
Затем центрифугировать на 7000 об/мин в течение 1 минуты;
Поменять пробирку под колонкой на чистую Пробирку для центрифугирования;
Внести на колонку оставшийся объем Раствора для очистки А с разведенным в нем образцом;
Центрифугировать на 7000 об/мин в течение 1 минуты;
Промыть колонку Раствора для очистки В в количестве 500 мкл еще раз;
Центрифугируя на 7000 об/мин в течение 1 минуты;
Промыть колонку Раствора для очистки В в количестве 500 мкл еще раз;
Центрифугируя на 7000 об/мин в течение 1 минуты;
После отмывок центрифугировать колонки на 13400 об/мин в течение 1 минуты;
Внести на колонки по 20 мкл Раствора LowT;
Инкубировать 1 минуту;
Центрифугировать на 13400 об/мин в течение 1 минуты;
Перенести элюат на ту же колонку;
Снова центрифугировать на 13400 об/мин в течение 1 минуты;
Полученный элюат F использовать в следующей реакции модификации концов ДНК.
2. Модификация концов ДНК
В микропробирку 1,5 мл необходимо внести следующие компоненты:

Тщательно перемешать пипетированием.
Инкубировать при следующих параметрах:

3. Предварительное лигирование
В микропробирку 1,5 мл необходимо внести следующие компоненты:

Тщательно перемешать пипетированием.
Инкубировать при следующих параметрах:

4. Постлигатная модификация ДНК Смешать следующие компоненты:

Тщательно перемешать пипетированием.
Перенести подготовленную смесь в микропробирки 0,2 мл в стрипах и запустил амплификаторе следующую программу (режим нагрева крышки должен быть включен

5. Лигирование адаптера Y
В отдельной микропробирке смешать следующие компоненты:

Тщательно перемешать пипетированием. Полученную смесь добавить к 47,1 мкл продукта постлигатной модификации ДНК. Тщательно перемешать пипетированием.
Инкубировать в амплификаторе с нагреваемой крышкой при следующих параметрах:

6. Отмывка продуктов лигирования адаптера Y
Перенести продукт лигирования адаптера Y в микропробирку объемом 1,5 мл;
К продукту лигирования адаптера Y (65,7 мкл) добавить такой же объем воды деионизированной;
В полученный раствор добавить 0,4 объема раствора магнитных частиц. (предварительно перемешав флакон с частицами на вортексе, чтобы получилась взвесь частиц равномерной концентрации);
Инкубировать при комнатной температуре 5 минут;
Сбить капли мини-центрифугоий/вортексом;
Перенести микропробирки на магнитный штатив;
После того как смесь станет прозрачной (на это требуется до 5 минут), нужно осторожно удалить супернатант, не задевая магнитные частицы, которые содержат фрагменты ДНК;
Добавить 200 мкл 80% этилового спирта;
Проворачивать микропробирки в магнитном штативе на 180° таким образом, чтобы магнитные частицы пробежали через раствор спирта несколько раз;
Собрать все магнитные частицы на одной стенке микропробирки;
Осторожно удалить этиловый спирт, не задевая магнитные частицы;
Добавить 200 мкл 80% этилового спирта;
Проворачивать пробирки в магнитном штативе на 180° таким образом, чтобы магнитные частицы пробежали через раствор спирта несколько раз;
Собрать все магнитные частицы на одной стенке микропробирки;
Итого получилось две последовательные отмывки ДНК магнитными частицами;
Максимально отобрать микропипеткой остатки раствора спирта;
Оставить сушиться микропробирки с открытой крышкой прямо на магнитном штативе на 2-5 минут, следя за тем, чтобы магнитные частицы не пересушились, но при этом не осталось спирта.
7. ПЦР для индексирования библиотек ДНК баркодами
Смешать в отдельной микропробирке следующие компоненты:

Тщательно перемешать пипетированием.
Полученную смесь добавить к иммобилизированной на магнитных частицах ДНК. Снять микропробирки с магнитного штатива, тщательно перемешать пипетированием.
Перенести полученную смесь в пробирки 0,2 мл в стрипах.
Запустить индексную ПЦР в амплификаторе по следующей программе:
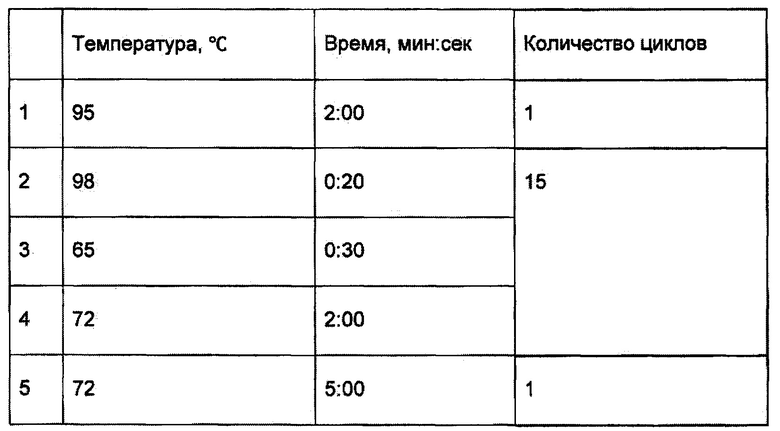
8. Отмывка продуктов ПЦР.
После окончания ПЦР необходимо провести очистку библиотек фрагментов ДНК.
Перенести продукт ПЦР в микропробирку объемом 1,5 мл;
К продукту ПЦР (~25 мкл) добавить 0,5 объема (13 мкл) раствора магнитных частиц (предварительно перемешав флакон с частицами на вортексе, чтобы получилась взвесь частиц равномерной концентрации);
Инкубировать при комнатной температуре 5 минут;
Сбить капли мини-центрифугой/вортексом;
Перенести микропробирки на магнитный штатив;
После того как смесь станет прозрачной (на это требуется до 5 минут), нужно осторожно удалить супернатант, не задевая магнитные частицы, которые содержат фрагменты ДНК;
Добавить 200 мкл 80% этилового спирта;
Проворачивать микропробирки в магнитном штативе на 180° таким образом, чтобы магнитные частицы пробежали через раствор спирта несколько раз;
Собрать все магнитные частицы на одной стенке микропробирки;
Осторожно удалить этиловый спирт, не задевая магнитные частицы;
Добавить 200 мкл 80% этилового спирта;
Проворачивать пробирки в магнитном штативе на 180° таким образом, чтобы магнитные частицы пробежали через раствор спирта несколько раз;
Собрать все магнитные частицы на одной стенке микропробирки;
Итого получилось две последовательные отмывки ДНК магнитными частицами;
Максимально отобрать микропипеткой остатки раствора спирта;
Оставить сушиться микропробирки с открытой крышкой прямо на магнитном штативе на 2-5 минут, следя за тем, чтобы магнитные частицы не пересушились, но при этом не осталось спирта;
Добавить к подсушенным магнитным частицам 20 мкл Раствора LowT;
Перемешать смесь на вортексе;
Сбить капли мини-центрифугой/вортексом;
Оставить инкубироваться на столе на 3-5 минут;
Перенести микропробирки на магнитный штатив;
Осторожно перенести супернатант в новую микропробирку, не задевая магнитных частиц.
Контроль качества библиотек фрагментов ДНК
Необходимо провести контроль качества полученных библиотек амплифицированных фрагментов ДНК на приборе Agilent Bioanalyzer 2100 с помощью набора High Sensitivity Kit по протоколу производителя (или аналогичном флуоресцентном анализаторе ДНК).
Правильным результатом является пик длин библиотек фрагментов в диапазоне 580-650 п.о. Молярность библиотек в диапазоне 200-1000 п.о. должна составлять не менее 2 наноМ (см. Фиг. 3а). В диапазоне 100-160 п.о. должны отсутствовать пики, свидетельствующие о недостаточной отмывке димеров адаптеров Y.
Протокол подготовки библиотек амплифицированных фрагментов ДНК
1. Постановка ПЦР №1.
В отдельной микропробирке 0,2 мл в стрипах смешать следующие компоненты:

Тщательно перемешать пипетированием.
Запустить ПЦР в амплификаторе по следующей программе:

2. Очистка продуктов ПЦР №1.
Перенести продукт ПЦР в микропробирку объемом 1,5 мл;
К продукту ПЦР (~35 мкл) добавить 3 объема (100 мкл) раствора магнитных частиц (предварительно перемешав флакон с частицами на вортексе, чтобы получилась взвесь частиц равномерной концентрации);
Инкубировать при комнатной температуре 5 минут;
Сбить капли мини-центрифугой/вортексом;
Перенести микропробирки на магнитный штатив;
После того как смесь станет прозрачной (на это требуется до 5 минут), нужно осторожно удалить супернатант, не задевая магнитные частицы, которые содержат фрагменты ДНК;
Добавить 200 мкл 80% этилового спирта;
Проворачивать микропробирки в магнитном штативе на 180° таким образом, чтобы магнитные частицы пробежали через раствор спирта несколько раз;
Собрать все магнитные частицы на одной стенке микропробирки;
Осторожно удалить этиловый спирт, не задевая магнитные частицы;
Добавить 200 мкл 80% этилового спирта;
Проворачивать пробирки в магнитном штативе на 180° таким образом, чтобы магнитные частицы пробежали через раствор спирта несколько раз;
Собрать все магнитные частицы на одной стенке микропробирки;
Итого получилось две последовательные отмывки ДНК магнитными частицами;
Максимально отобрать микропипеткой остатки раствора спирта;
Оставить сушиться микропробирки с открытой крышкой прямо на магнитном штативе на 2-5 минут, следя за тем, чтобы магнитные частицы не пересушились, но при этом не осталось спирта;
Добавить к подсушенным магнитным частицам 21 мкл Раствора LowT;
Перемешать смесь на вортексе;
Сбить капли мини-центрифугой/вортексом;
Оставить инкубироваться на столе на 3-5 минут;
Перенести микропробирки на магнитный штатив;
Осторожно перенести супернатант в новую микропробирку, не задевая магнитных частиц.
3. Лигирование адаптера Y.
В отдельной микропробирке смешать следующие компоненты:
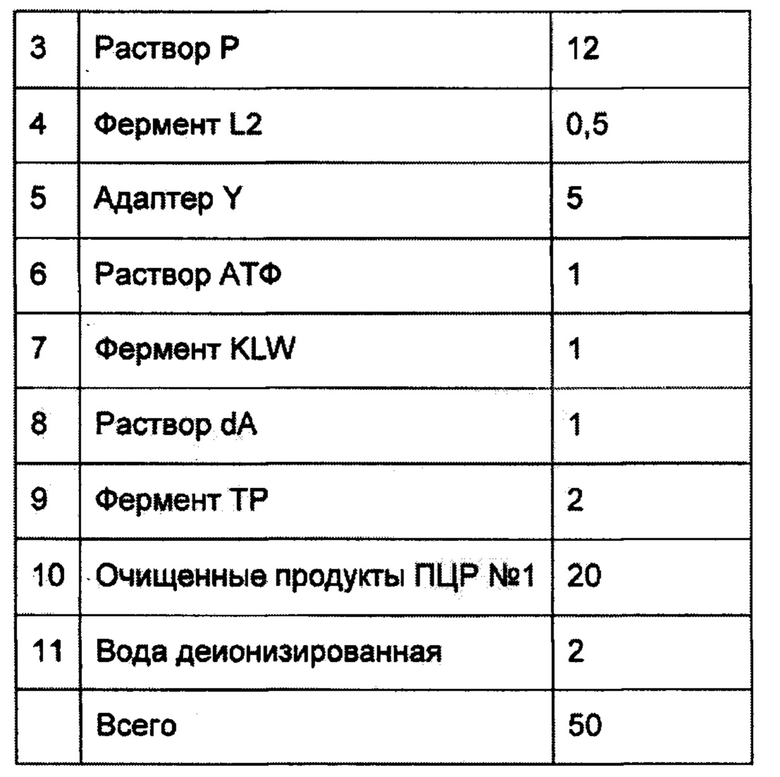
Тщательно перемешать пипетированием.
Инкубировать в амплификаторе с нагреваемой крышкой при следующих параметрах:

4. Отмывка продуктов лигирования адаптера Y.
Перенести продукт лигирования адаптера Y в микропробирку объемом 1,5 мл;
В раствор добавить 1,5 объема (75 мкл) раствора магнитных частиц (предварительно перемешав флакон с частицами на вортексе, чтобы получилась взвесь частиц равномерной концентрации);
Инкубировать при комнатной температуре 5 минут;
Сбить капли мини-центрифугой/вортексом;
Перенести микропробирки на магнитный штатив;
После того как смесь станет прозрачной (на это требуется до 5 минут), нужно осторожно удалить супернатант, не задевая магнитные частицы, которые содержат фрагменты ДНК;
Добавить 200 мкл 80% этилового спирта;
Проворачивать микропробирки в магнитном штативе на 180° таким образом, чтобы магнитные частицы пробежали через раствор спирта несколько раз;
Собрать все магнитные частицы на одной стенке микропробирки;
осторожно удалить этиловый спирт, не задевая магнитные частицы;
Добавить 200 мкл 80% этилового спирта;
Проворачивать пробирки в магнитном штативе на 180° таким образом, чтобы магнитные частицы пробежали через раствор спирта несколько раз;
Собрать все магнитные частицы на одной стенке микропробирки;
Итого получилось две последовательные отмывки ДНК магнитными частицами;
Максимально отобрать микропипеткой остатки раствора спирта;
Оставить сушиться микропробирки с открытой крышкой прямо на магнитном штативе на 2-5 минут, следя за тем, чтобы магнитные частицы не пересушились, но при этом не осталось спирта;
Добавить к подсушенным магнитным частицам 19 мкл Раствора LowT;
Перемешать смесь на вортексе;
Сбить капли мини-центрифугой/вортексом;
Оставить инкубироваться на столе на 3-5 минут;
Перенести микропробирки на магнитный штатив;
Осторожно перенести супернатант в новую микропробирку, не задевая магнитных частиц.
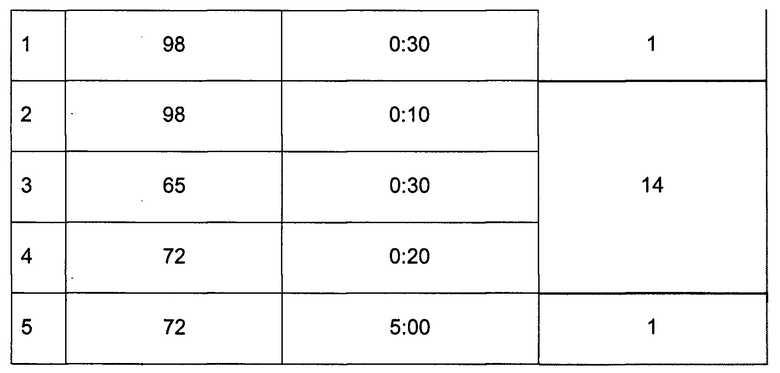
5. ПЦР для индексирования библиотек ДНК баркодами:

Тщательно перемешать пипетированием
Запустить индексную ПЦР в амплификаторе по следующей программе:

6. Отмывка продуктов индексной ПЦР с сайз-селектом
Добавить 4 объема (100 мкл) Буфера для промывки SB к 1 объему реакции;
Смешать пипетированием;
Перенести смесь на Колонка для сайз-селекта ДНК;
Центрифугировать 1 минуту на максимальной скорости;
Перенести колонку в новую Пробирку для центрифугирования;
Добавить 700 мкл 80% этилового спирта;
Центрифугировать 1 минуту;
Перенести колонку в новую Пробирку для центрифугирования;
Добавить 700 мкл 80% этилового спирта;
Центрифугировать 1 минуту;
Тем самым, отмывок 80% этиловым спиртом проводится две подряд;
Перенести колонку в новую Пробирку для центрифугирования;
Отцентрифугировать колонку на максимальной скорости минуту для удаления остатков этилового спирта;
Переместить колонку в чистую 1,5 мл микропробирку;
Добавить 20 мкл Раствора LowT прямо на мембрану колонки;
Инкубировать 5 минут;
Центрифугировать 1 мин на максимальной скорости;
Вынув колонку из пробирки, отобрать пипеткой весь объем элюата, поставить колонку в эту же микропробирку и вновь внести элюат на колонку;
Инкубировать при комнатной температуре в течение 5 минут;
Центрифугировать на максимальной скорости 1 минуту;
Выбросить колонку;
В микропробирке очищенная библиотека амплифицированных фрагментов ДНК.
Контроль качества библиотек фрагментов ДНК
Необходимо провести контроль качества полученных библиотек амплифицированных фрагментов ДНК на приборе Agilent Bioanalyzer 2100 с помощью набора High Sensitivity Kit по протоколу производителя (или аналогичном флуоресцентном анализаторе ДНК).
Правильным результатом является пик длин библиотек фрагментов в диапазоне 265-280 п.о. Молярность библиотек в диапазоне 200-1000 п.о. должна составлять не менее 2 наноМ (см. Фиг. 3b). В диапазоне 100-160 п.о. должны отсутствовать пики, свидетельствующие о недостаточной отмывке димеров адаптеров Y.
Секвенирование библиотек фрагментов ДНК и библиотек амплифицированных фрагментов ДНК на Illumina MiSeq.
Пулирование образцов. На основании результатов измерения концентрации библиотек ДНК и библиотек фрагментов ДНК необходимо провести три последовательных смешивания;
Пулирование библиотек фрагментов ДНК.
Пулирование библиотек амплифицированных фрагментов ДНК.
Смешивание пула библиотек фрагментов ДНК и пула библиотек амплифицированных фрагментов ДНК.
Для проведения первого и второго пулирования необходимо смешать все библиотеки соответствующего типа эквимолярно. Значения молярности можно определять на приборе Agilent Bioanalyzer 2100 с набором High Sensitivity DNA kit или аналогичном оборудовании и реактивах в диапазоне 200-1000 п.о. Предлагается следующая схема:
Определить библиотеку, имеющую наибольшее значение молярности [Xm наноМ];
Рассчитать, во сколько раз каждая из остальных библиотек того же типа меньше значения X. Обозначим это значение Yi;
Смешать по объемам все библиотеки одного типа в одной микропробирке. Объемная доля рассчитывается как Xm/Yi. На основании этой пропорции и проводится пулирование;
Итоговая концентрация пула определяется по формуле Xm/ΣYi;
Смешивание пула библиотек фрагментов ДНК и пула библиотек амплифицированных фрагментов ДНК проводится в пропорции 100/1 (на 100 объемов пула библиотеки фрагментов ДНК берется один объем пула библиотек амплифицированных фрагментов).
Разведение аликвот всех компонентов проводится Раствором LowT.
Оптимальную концентрацию конечной (денатурированной, готовой к секвенированию) смеси пулов можно определять для каждого прибора IlluminaMiSeq индивидуально. Практика показывает, что она лежит в диапазоне 8-13 пикоМ.
Денатурацию и разведение смеси пулов непосредственно перед запуском секвенирования рекомендуется проводить по инструкции производителя IlluminaMiSeq: "MiSeq System. DenatureandDiluteLibrariesGuide" под номером 15039740, расположенную по ссылке https://support.illumina.com/content/dam/illumina-support/documents/documentation/system_documentation/miseq/miseq-denature-dilute-libraries-guide-15039740-06.pdf.
Обслуживание Illumina MiSeq после и между запусками секвенирования рекомендуется проводить по инструкции производителя MiSeq System Guide for Local Run Manager под номером 15027617, расположенную по ссылке https://support.illumina.com/content/dam/illumina-support/documents/documentation/system_documentation/miseq/miseq-system-guide-for-local-run-manager-15027617-04.pdf.
Учет результатов реакции
1. Для всех образцов в зависимости от типа библиотеки рассчитывается ряд биоинформатических метрик (количество сырых ридов, количество картированных ридов, Q20, Q30, доля фетальной фракции, количество информативных для оценки доли фетальной фракции SNP, полезная эффективность данных библиотеки фрагментов ДНК), на основании которых программа принимает решение о качестве проведенного анализа. В случае обнаружения проблемы соответствующий образец имеет маркировку «BioinformaticQCfailed» в отчете;
2. Обработка выходных файлов с секвенатора в формате fastq.gz, а также учет и интерпретация результатов секвенирования проводится с помощью программного обеспечения «НИПТ-аналитика» версии не менее 1.0;
3. Для всех образцов программа рассчитывает долю фетальной ДНК во внДНК и выдает ее значение в отчете;
4. Для всех образцов программа рассчитывает наличие анеуплоидий по хромосомам, входящим в данный вариант исполнения набора (21 или 13, 18, 21 или 13, 18, 21, X, Y) и выдает ответ о наличии в отчете;
5. Для всех образцов, не прошедших контроль качества по доле фетальной внДНК, программа выдает отчет о непрохождении контроля качества в связи с низкой долей фетальной ДНК.
Пример осуществления предсказания хромосомных аномалий.
В данном примере авторами решались следующие задачи: 1) предсказать трисомию по хромосомам 13, 18, 21; 2) определить пол ребенка; 3) предсказать долю фетальной ДНК; 4) оценить минимально необходимое количество данных.
Предобработка данных: было проведено маскирование с помощью списка надежных регионов из базы 1000 Genomes.
Статистическая обработка данных:
Из контрольной выборки объемом 154 образца было взято 78 образцов с долей фетальной ДНК, превышающей 4%, а также имеющих не менее чем 2,5 млн фильтрованных фрагментов на образец (см ниже).
Доля внеклеточной фетальной ДНК была подсчитана методом, основанным на глубоком прочтении нескольких полиморфизмов (SNP), по которым мать гомозиготна, а ребенок гетерозиготен. Сравнение с работающим для мальчиков методом, основанным на количестве прочтений Y хромосомы, показано на Фиг. 8, на котором можно увидеть хорошую корреляцию результатов.
Для обработки данных использовалась библиотека NIPTeR языка R. Количество фрагментов генома подсчитали на участках в 50000 п.о. для каждого образца. Полученные суммы характеризуют интенсивность прочтений участков хромосом (Фиг. 4).
После этого была проведена GC-коррекция, то есть внесена поправка на регионы с повышенным количеством G-C связей, с которых удается считывать меньшее количество фрагментов. Следующим шагом проведена поправка на выбросы. Был использован метод, основанный на отбрасывании пиков с помощью квантилей, так и процедура уменьшения вариации, основанная на критерии хи-квадрат. При каждом запуске программы подсчета статистики контрольная выборка формировалась случайным образом из подвыборки не имеющих трисомии образцов. Алгоритм обучался на такой выборке и определял трисомию у оставшихся образцов. Многократный запуск программы позволяет судить о качестве выбранного метода. Всего было проделано 200 симуляций.
Результаты расчета чувствительности, специфичности и AUC для каждой анеуплоидии и всех вместе представлены в Таблице 1 (в скобках указано значение стандартного отклонения).

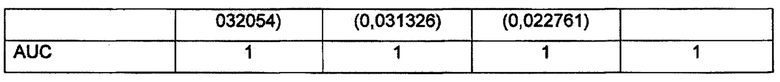
Результаты расчета Z статистики по 13, 18 и 21 хромосоме для контрольной выборки образцов представлены на Фиг. 5 (а, b, с) соответственно. Видно, что во всех трех вариантах образцы с нормальным и анеуплоидным кариотипом располагаются под и над пороговым значением (Z=3).
С целью выяснить, будет ли алгоритм работать при сокращении числа прочтений генома, авторы отбрасывали часть фрагментов и повторяли эксперимент на неполных данных. Результат не претерпевает серьезных ухудшений при уменьшении количества данных в 2 раза (Фиг. 6). Таким образом, порогом по количеству прошедших все фильтры и поправки значений авторы приняли цифру в 2,5 млн фрагментов на 1 образец.
На Фиг. 7 (а, b, с) представлены результаты построения ROC-кривой для определения анеуплоидий 13, 18 и 21 хромосом в контрольной выборке образцов.
Для определения пола ребенка авторы подсчитали у образцов нормализованное количество прочтений на Y хромосоме и нашли соотношение количества фрагментов с X хромосомы к количеству фрагментов аутосомных хромосом (Фиг. 9). Ошибка происходила на двух образцах - это мальчики, у которых нет достаточного количества прочтений на Y хромосоме.
Несмотря на то, что изобретение описано со ссылкой на раскрываемые варианты воплощения, для специалистов в данной области должно быть очевидно, что конкретные подробно описанные случаи приведены лишь в целях иллюстрирования настоящего изобретения, и их не следует рассматривать как каким-либо образом ограничивающие объем изобретения. Должно быть, понятно, что возможно осуществление различных модификаций без отступления от сути настоящего изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ НЕИНВАЗИВНОЙ ПРЕНАТАЛЬНОЙ ДИАГНОСТИКИ АНЕУПЛОИДИЙ ПЛОДА | 2015 |
|
RU2627673C2 |
| СПОСОБ НЕИНВАЗИВНОЙ ПРЕНАТАЛЬНОЙ ДИАГНОСТИКИ АНЕУПЛОИДИЙ ПЛОДА | 2014 |
|
RU2583830C2 |
| СПОСОБ ОПРЕДЕЛЕНИЯ АНЕУПЛОИДИИ ПЛОДА В ОБРАЗЦЕ КРОВИ БЕРЕМЕННОЙ ЖЕНЩИНЫ | 2021 |
|
RU2777072C1 |
| Способ неинвазивного пренатального скрининга анеуплоидий плода | 2019 |
|
RU2712175C1 |
| СПОСОБЫ И СИСТЕМЫ ДЛЯ ОПРЕДЕЛЕНИЯ ТОГО, ЯВЛЯЕТСЯ ЛИ ГЕНОМ АНОМАЛЬНЫМ | 2011 |
|
RU2599419C2 |
| СПОСОБ НЕИНВАЗИВНОЙ ПРЕНАТАЛЬНОЙ ДИАГНОСТИКИ ТРИСОМИИ И НАБОР ДЛЯ ЕЕ ПРОВЕДЕНИЯ | 2020 |
|
RU2734484C1 |
| СПОСОБ И УСТРОЙСТВО ДЛЯ ОПРЕДЕЛЕНИЯ ФРАКЦИИ ВНЕКЛЕТОЧНЫХ НУКЛЕИНОВЫХ КИСЛОТ В БИОЛОГИЧЕСКОМ ОБРАЗЦЕ И ИХ ПРИМЕНЕНИЕ | 2015 |
|
RU2699728C2 |
| СПОСОБ НЕИНВАЗИВНОЙ ДИАГНОСТИКИ АНЕУПЛОИДИЙ ПЛОДА МЕТОДОМ СЕКВЕНИРОВАНИЯ | 2014 |
|
RU2543155C1 |
| СПОСОБ ОПРЕДЕЛЕНИЯ ИСТОЧНИКА АНЕУПЛОИДНЫХ КЛЕТОК ПО КРОВИ БЕРЕМЕННОЙ ЖЕНЩИНЫ | 2016 |
|
RU2674700C2 |
| ТЕХНОЛОГИЯ ОПРЕДЕЛЕНИЯ АНЕУПЛОИДИИ МЕТОДОМ СЕКВЕНИРОВАНИЯ | 2012 |
|
RU2529784C2 |












Изобретение относится к биотехнологии и описывает способ неинвазивной диагностики анеуплоидий плода с помощью анализа внеклеточной ДНК из крови беременной женщины методом массового параллельного секвенирования ДНК. Описанный способ характеризуется новым способом пробоподготовки внеклеточной ДНК из образцов крови. Техническим результатом настоящего изобретения является повышение эффективности процесса определения анеуплоидии плода, достигаемое тем, что за одно прочтение образованных библиотек ДНК потенциально получают информацию о нескольких различных хромосомах и используют эту информацию для определения анеуплоидии плода. Данный способ может быть использован в клинико-диагностических лабораториях медицинских учреждений. 2 н. и 2 з.п. ф-лы, 9 ил., 1 табл., 1 пр.
1. Способ подготовки внеклеточной ДНК из образца крови беременной женщины к секвенированию для диагностики анеуплоидий плода, включающий следующие стадии:
a) получают образец материнской крови, состоящий из цельной крови, или плазмы крови, или сыворотки крови, при этом для образца, состоящего из цельной крови, отделяют плазму крови и используют ее для последующих стадий;
b) выделяют из указанного образца крови внеклеточную ДНК;
c) осуществляют фрагментацию выделенной внеклеточной ДНК, получая в результате размер ДНК-фрагментов от 30 до 100 пар оснований со средним размером около 40 пар оснований;
d) отбирают фрагменты ДНК длиной между 35 и 45 пар оснований;
e) осуществляют случайное лигирование отобранных на стадии d) фрагментов ДНК друг с другом для формирования химерных линейных молекул ДНК размером более 100 пар оснований;
f) осуществляют лигирование адаптерных последовательностей, специфических для используемого в дальнейшем метода секвенирования, к химерным линейным молекулам ДНК для создания библиотек химерных линейных молекул ДНК;
g) проводят ПЦР для индексирования библиотек химерных линейных молекул ДНК уникальными последовательностями баркодов.
2. Способ по п. 1, характеризующийся тем, что фрагментацию очищенной внеклеточной ДНК на стадии c) осуществляют с помощью ферментов и без обработки ДНК ультразвуком.
3. Способ определения анеуплоидии плода с помощью анализа внеклеточной ДНК из крови беременной женщины методом секвенирования, включающий следующие стадии:
а) подготавливают внеклеточную ДНК из образца крови беременной женщины по п. 1;
б) осуществляют секвенирование образованных библиотек химерных линейных молекул ДНК одним из методов массового параллельного секвенирования, получая множество чтений;
в) осуществляют картирование полученных чтений на референсный геном человека для определения присутствия в чтениях участков хромосом человека;
г) определяют количество участков последовательностей ДНК в чтениях, соответствующих разным хромосомам человека, при этом исключают участки, которые являются неуникальными для отдельных хромосом генома человека;
д) определяют наличие анеуплоидии по определенной хромосоме при обнаружении отличий в нормализованной на длину хромосомы представленности этой хромосомы от среднего значения по геному.
4. Способ по п. 3, характеризующийся тем, что классификация представленности отдельных хромосом производится на основании z-статистики, вычисленной в регрессионной модели, и порогового значения z_0, равного 3.
| EA 17966 B1, 30.04.2013 | |||
| EP 3026124 A1, 01.06.2016 | |||
| WO 2017009372 A2, 19.01.2017 | |||
| К.С | |||
| Пантюх, Е.С | |||
| Шубина, НЕИНВАЗИВНАЯ ПРЕНАТАЛЬНАЯ ДИАГНОСТИКА АНЕУПЛОИДИЙ ПЛОДА, ОСНОВАННАЯ НА СЕКВЕНИРОВАНИИ ВНЕКЛЕТОЧНОЙ ДНК КРОВИ БЕРЕМЕННОЙ ЖЕНЩИНЫ, акушерство и гинекология, номер 8, 2015, стр.5-11. |

