ОБЛАСТЬ ИЗОБРЕТЕНИЯ
[0001] Предоставлена генетически модифицированная мышь, экспрессирующая антитела с общей легкой цепью с вариабельной областью человека/константной областью мыши, ассоциированной с различными тяжелыми цепями с вариабельной областью человека/константной областью мыши. Предоставлен способ получения биспецифического антитела человека на основании генных последовательностей вариабельных областей человека В-клеток мыши.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
[0002] Как правило, антитела содержат компонент гомодимерных тяжелых цепей, где каждый мономер тяжелой цепи ассоциирован с идентичной легкой цепью. В качестве терапевтических антител желательны антитела, содержащие компонент гетеродимерных тяжелых цепей (например, биспецифические антитела). Но получение биспецифических антител с подходящим компонентом легкой цепи, который может удовлетворительно ассоциировать с каждой из тяжелых цепей биспецифического антитела оказалось проблематичным.
[0003] В одном из подходов легкую цепь можно выбирать, изучая статистику использования всех вариабельных доменов легких цепей, идентифицируя наиболее часто используемую в антителах человека легкую цепь и спаривая эту легкую цепь in vitro с двумя тяжелыми цепями с различной специфичностью.
[0004] В другом подходе легкую цепь можно выбирать, исследуя последовательности легких цепей в библиотеке фагового дисплея (например, в библиотеке фагового дисплея, содержащей последовательности вариабельных областей легких цепей человека, например, библиотека ScFv человека) и выбирая из библиотеки наиболее часто используемую вариабельную область легкой цепи. Затем легкую цепь можно тестировать на двух представляющих интерес различных тяжелых цепях.
[0005] В другом подходе легкую цепь можно выбирать, анализируя библиотеку фагового дисплея последовательностей вариабельных областей легких цепей с использованием в качестве зондов последовательностей вариабельных областей тяжелых цепей обеих представляющих интерес тяжелых цепей. В качестве легкой цепи для тяжелых цепей можно выбирать легкую цепь, ассоциирующую с обеими последовательностями вариабельных областей тяжелых цепей.
[0006] В другом подходе легкую цепь-кандидата можно выравнивать с легкими цепями, родственными тяжелым цепям, и в легкой цепи производят модификации для более близкого соответствия характеристик последовательности, общих с легкими цепями, родственными с тяжелыми цепями. Если необходимо минимизировать вероятность иммуногенности, модификации предпочтительно имеют результатом последовательности, которые присутствуют в известных последовательности легких цепей человека, так, что протеолитический процессинг на основе известных в данной области параметров и способов оценки вероятности иммуногенности (т.е., анализов in silico, а также "мокрых" анализов) приводит к Т-клеточному эпитопу с минимальной вероятностью.
[0007] Все из указанных выше подходов основаны на способах in vitro, содержащих ряд априорных ограничений, например, идентичность последовательностей, способность к ассоциации с конкретными предварительно выбранными тяжелыми цепями и т.д. В данной области существует необходимость в композициях и способах, которые не основаны на регуляции условий in vitro, а в которых для получения связывающих эпитопы белков человека, содержащих общую легкую цепь, вместо этого используют более биологически разумные подходы.
КРАТКОЕ ОПИСАНИЕ ФИГУР
[0008] На фиг. 1 проиллюстрирована стратегия определения мишени для замены эндогенного участков гена вариабельной области легкой цепи иммуноглобулина мыши генной областью Vκ1-39Jκ5 человека.
[0009] На фиг. 2 проиллюстрирована стратегия определения мишени для замены эндогенного участков гена вариабельной области легкой цепи иммуноглобулина мыши генной областью Vκ3-20Jκ1 человека.
[0010] На фиг. 3 проиллюстрирована стратегия определения мишени для замены эндогенного участков гена вариабельной области легкой цепи иммуноглобулина мыши генной областью VpreB/Jλ5 человека.
СУЩНОСТЬ
[0011] Предоставлены генетически модифицированные мыши, экспрессирующие вариабельные домены тяжелых и легких цепей иммуноглобулинов человека, где у мышей существует ограниченный репертуар вариабельных областей легких цепей. Предоставлена биологическая система для получения вариабельного домена легкой цепи человека, ассоциирующего и экспрессирующегося с широким репертуаром вариабельных доменов тяжелых цепей человека с созревшей аффинностью. Предоставлены способы получения связывающих белков, содержащих вариабельные домены
иммуноглобулинов, включающие иммунизацию мышей с ограниченным репертуаром легких цепей иммуноглобулинов представляющим интерес антигеном, и использование генной последовательности вариабельной области иммуноглобулина мыши для связывающего белка, который специфически связывает представляющий интерес антиген. Способы включают способы получения вариабельных доменов тяжелых цепей иммуноглобулинов человека, пригодных для использования в получении полиспецифичных антигенсвязывающих белков.
[0012] Предоставлены полученные генно-инженерными способами мыши, у которых происходит отбор подходящих вариабельных доменов тяжелых цепей иммуноглобулинов человека с созревшей аффинностью, получаемых из репертуара неперегруппированных генных сегментов вариабельных областей тяжелых цепей человека, где вариабельные домены тяжелых цепей человека с созревшей аффинностью ассоциируют и экспрессируются с единственным вариабельным доменом легкой цепи человека, получаемым из единственного генного сегмента вариабельной области легкой цепи человека. Также предоставлены полученные генно-инженерными способами мыши, обеспечивающие выбор из двух генных сегментов вариабельных областей легких цепей человека.
[0013] Предоставлены полученные генно-инженерными способами мыши, экспрессирующие ограниченный репертуар вариабельных доменов легких цепей человека или единственный вариабельный домен легкой цепи человека из ограниченного репертуара генных сегментов вариабельных областей легких цепей человека. Мыши получены генно-инженерными способами так, чтобы содержать единственный неперегруппированный генный сегмент вариабельной области легкой цепи человека (или два генных сегмента вариабельных областей легких цепей человека), который перестраивается, формируя перегруппированный ген вариабельной области легкой цепи человека (или два перегруппированных гена вариабельных областей легких цепей), экспрессирующий единственную легкую цепь (или экспрессирующий любую или обе из двух легких цепей). Перегруппированные вариабельные домены легких цепей человека способны к спариванию со множеством тяжелых цепей человека с созревшей аффинностью, отобранных мышами, где вариабельные области тяжелых цепей специфически связывают различные эпитопы.
[0014] В одном из аспектов предоставлена генетически модифицированная мышь, экспрессирующая единственный генный сегмент вариабельной (VL) области легкой цепи иммуноглобулина человека, способный к перегруппировке и кодирующий домен VL легкой цепи иммуноглобулина человека. В другом аспекте мышь несет не более двух генных сегментов VL человека, способных к перегруппировке и кодирующих домен VL легкой цепи иммуноглобулина человека.
[0015] В одном из аспектов предоставлена генетически модифицированная мышь, несущая единственный перегруппированный (V/J) сегмент вариабельной (VL) области легкой цепи иммуноглобулина человека (т.е., сегмент V/J), кодирующий домен VL легкой цепи иммуноглобулина человека. В другом аспекте мышь несет не более двух перегруппированных генных сегментов VL человека, способных к кодированию домена VL легкой цепи иммуноглобулина человека.
[0016] В одном из вариантов осуществления генный сегмент VL представляет собой генный сегмент Vκ1-39Jκ5 человека или генный сегмент Vκ3-20Jκ1 человека. В одном из вариантов осуществления мышь несет генный сегмент Vκ1-39Jκ5 человека и генный сегмент Vκ3-20Jκ1 человека.
[0017] В одном из вариантов осуществления генный сегмент VL человека функционально связан с лидерной последовательностью человека или мыши. В одном из вариантов осуществления лидерная последовательность представляет собой лидерную
последовательность мыши. В конкретном варианте осуществления лидерная последовательность мыши представляет собой лидерную последовательность мыши Vκ3-7.
[0018] В одном из вариантов осуществления генный сегмент VL функционально связан с последовательность промотора иммуноглобулина. В одном из вариантов осуществления последовательность промотора представляет собой последовательность промотора человека. В конкретном варианте осуществления промотор иммуноглобулина человека представляет собой промотор Vκ3-15.
[0019] В одном из вариантов осуществления генетически модифицированная мышь несет локус VL, который не содержит эндогенный генный сегмент VL мыши, способный к перегруппировке с формированием гена легкой цепи иммуноглобулина, где локус VL содержит единственный генный сегмент VL человека, способный к перегруппировке с кодированием области VL гена легкой цепи. В конкретном варианте осуществления генный сегмент VL человека представляет собой генный сегмент Vκ1-39Jκ5 человека или генный сегмент Vκ3-20Jκ1 человека.
[0020] В одном из вариантов осуществления локус VL содержит лидерную последовательность, фланкированную на 5'-конце (относительно направления транскрипции генного сегмента VL) промотором иммуноглобулина человека, а на 3'-конце фланкированную генным сегментом VL человека, который перестраивается и кодирует домен VL обратной химерной легкой цепи, содержащей эндогенную константную область легкой цепи (CL) мыши. В конкретном варианте осуществления генный сегмент VL представляет собой локус VL каппа (к) мыши, a CL мыши представляет собой CL к мыши.
[0021] В одном из вариантов осуществления мышь несет нефункциональный локус легкой цепи иммуноглобулина лямбда (λ). В конкретном варианте осуществления локус λ содержит делецию одной или нескольких последовательностей из локуса, где одна или несколько делеций приводят локус λ в состояние неспособности к перегруппировке с формированием гена легкой цепи. В другом варианте осуществления удалены все или по существу все генные сегменты VL локуса λ.
[0022] В одном из вариантов осуществления локус VL модифицированной мыши представляет собой локус к, а локус к содержит интронный энхансер к мыши, 3'-энхансер к мыши или и интронный энхансер и 3'-энхансер.
[0023] В одном из вариантов осуществления у мыши образуется легкая цепь, содержащая соматически мутированный домен VL, образуемый из генного сегмента VL человека. В одном из вариантов осуществления легкая цепь содержит соматически мутированный домен VL, получаемый из генного сегмента VL человека, и область CL к мыши. В одном из вариантов осуществления мышь не экспрессирует легкую цепь λ.
[0024] В одном из вариантов осуществления генетически модифицированная мышь способна к соматической гипермутации последовательности области VL человека. В конкретном варианте осуществления у мыши присутствует клетка, содержащая перегруппированный ген легкой цепи иммуноглобулина, получаемый из генного сегмента VL человека, способный к перегруппировке и к кодированию домена VL, и перегруппированный ген легкой цепи иммуноглобулина содержит соматически мутированный домен VL.
[0025] В одном из вариантов осуществления у мыши присутствует клетка, экспрессирующая легкую цепь, содержащую соматически мутированный домен VL человека, связанный с CL к мыши, где легкая цепь ассоциирует с тяжелой цепью, содержащей соматически мутированный домен VH, получаемый из генного сегмента VH человека, и где тяжелая цепь содержит константную область тяжелой цепи мыши (СН).
[0026] В одном из вариантов осуществления у мыши присутствует замена эндогенных генных сегментов VH мыши одним или несколькими генными сегментами VH человека, где генные сегменты VH человека функционально связаны с геном области СН мыши так, что у мыши происходит перегруппировка генных сегментов VH человека и экспрессируется обратная химерная тяжелая цепь иммуноглобулина, содержащая домен VH человека и СН мыши. В одном из вариантов осуществления 90-100% неперегруппированных генных сегментов VH мыши замещены по меньшей мере одним неперегруппированным генным сегментом VH человека. В конкретном варианте осуществления все или по существу все эндогенные генные сегменты VH мыши замещены по меньшей мере одним неперегруппированным генным сегментом VH человека. В одном из вариантов осуществления замену производят по меньшей мере 19, по меньшей мере 39 или по меньшей мере 80 или 81 неперегруппированными генными сегментами VH человека. В одном из вариантов осуществления замену производят по меньшей мере 12 функциональными неперегруппированными генными сегментами VH человека, по меньшей мере 25 функциональными неперегруппированными генными сегментами VH человека или по меньшей мере 43 функциональными неперегруппированными генными сегментами VH человека. В одном из вариантов осуществления у мыши присутствует замена всех сегментов D и J мыши по меньшей мере одним неперегруппированным сегментом D человека и по меньшей мере одним неперегруппированным сегментом J человека. В одном из вариантов осуществления по меньшей мере один неперегруппированный сегмент D человека выбран из D1-7, D1-26, D3-3, D3-10, D3-16, D3-22, D5-5, D5-12, D6-6, D6-13, D7-27 и их сочетания. В одном из вариантов осуществления по меньшей мере один неперегруппированный сегмент J человека выбран из J1, J3, J4, J5, J6 и их сочетания. В конкретном варианте осуществления один или несколько генных сегментов VH человека выбраны из генных сегментов VH человека 1-2, 1-8, 1-24, 2-5, 3-7, 3-9, 3-11, 3-13, 3-15, 3-20, 3-23, 3-30, 3-33, 3-48, 4-31, 4-39, 4-59, 5-51, 6-1 и их сочетания.
[0027] В одном из вариантов осуществления у мыши присутствует В-клетка, экспрессирующая связывающий белок, специфически связывающийся с представляющим интерес антигеном, где связывающий белок содержит легкую цепь, получаемую на основе перегруппировки Vκ1-39/Jκ5 человека или перегруппировки Vκ3-20/JκI человека, и где клетка содержит перегруппированный ген тяжелой цепи иммуноглобулина, получаемый на основе перегруппировки генных сегментов человека, выбранных из генных сегментов VH2-5, VH3-23, VH3-30, VH4-39, VH4-59 и VH5-51. В одном из вариантов осуществления один или несколько генных сегментов VH человека подвергают перегруппировке с генным сегментом J тяжелой цепи человека, выбранным из J1, J3, J4, J5, и J6. В одном из вариантов осуществления один или несколько генных сегментов VH и J человека подвергают перегруппировке с генным сегментом D человека, выбранным из D1-7, D1-26, D3-3, D3-10, D3-16, D3-22, D5-5, D5-12, D6-6, D6-13 и D7-27. В конкретном варианте осуществления ген легкой цепи содержит 1, 2, 3, 4 или 5 или более соматических гипермутаций.
[0028] В одном из вариантов осуществления у мыши присутствует В-клетка, содержащая генную последовательность перегруппированной вариабельной области тяжелой цепи иммуноглобулина, содержащую генные сегменты VH, JH и DH, выбранные из VH2-5+JH1+D6-6, VH3-23+JH4+D3, VH3-23+JH4+D3-10, VH3-30+JH1+D6-6, VH3-30+JH3+D6-6, VH3-30+JH4+D1-7, VH3-30+JH4+D5-12, VH3-30+JH4+D6-13, VH3-30+JH4+D6-б, VH3-30+JH4+D7-27, VH3-3 0+JH5+D3-22, VH3-30+JH5+D6-6, VH3-30+JH5+D7-27, VH4-39+JH3+D1-26, VH4-59+JH3+D3-16, VH4-59+JH3+D3-22, VH4-59+JH4+D3-16, VH5-51+JH3+D5-5, VH5-51+JH5+D6-13 и VH5-51+JH6+D3-16. В конкретном варианте осуществления В-клетка экспрессирует связывающий белок, содержащий вариабельную область тяжелой цепи иммуноглобулина человека, слитую с константной областью тяжелой цепи мыши, и вариабельную область легкой цепи иммуноглобулина человека, слитую с константной областью легкой цепи мыши,
[0029] В одном из вариантов осуществления генный сегмент VL человека представляет собой генный сегмент Vκ1-39Jκ5 человека, и мышь экспрессирует обратную химерную легкую цепь, содержащую (i) домен VL, получаемый из генного сегмента VL человека, и (ii) CL мыши; где легкая цепь ассоциирована с обратной химерной тяжелой цепью, содержащей (i) СН мыши и (ii) соматически мутированный домен VH человека, получаемый из генного сегмента VH человека, выбранного из генных сегментов VH человека 1-2, 1-8, 1-24, 2-5, 3-7, 3-9, 3-11, 3-13, 3-15, 3-20, 3-23, 3-30, 3-33, 3-48, 4-31, 4-39, 4-59, 5-51 и 6-1 и их сочетания. В одном из вариантов осуществления мышь экспрессирует легкую цепь, являющуюся соматически мутированной. В одном из вариантов осуществления CL представляет собой CL к мыши.
[0030] В одном из вариантов осуществления генный сегмент VL человека представляет собой генный сегмент Vκ3-20Jκ1 человека, и мышь экспрессирует обратную химерную легкую цепь, содержащую (i) домен VL, получаемый из генного сегмента VL человека, и (ii) CL мыши; где легкая цепь ассоциирована с обратной химерной тяжелой цепью, содержащей (i) СН мыши и (ii) соматически мутированный VH человека, получаемый из генного сегмента VH человека, выбранного из генных сегментов VH человека 1-2, 2-5, 3-7, 3-9, 3-11, 3-20, 3-23, 3-30, 3-33, 4-59 и 5-51 и их сочетания. В одном из вариантов осуществления мышь экспрессирует легкую цепь, являющуюся соматически мутированной. В одном из вариантов осуществления CL представляет собой CL к мыши.
[0031] В одном из вариантов осуществления мышь несет генный сегмент Vκ1-39Jκ5 человека и генный сегмент Vκ3-20Jκ1 человека, и мышь экспрессирует обратную химерную легкую цепь, содержащую (i) домен VL, получаемый из генного сегмента Vκ1-39Jκ5 человека или генного сегмента Vκ3-20Jκ1 человека, и (ii) CL мыши; где легкая цепь ассоциирована с обратной химерной тяжелой цепью, содержащей (i) СН мыши и (ii) соматически мутированный VH человека, получаемый из генного сегмента VH человека, выбранного из генных сегментов VH человека 1-2, 1-8, 1-24, 2-5, 3-7, 3-9, 3-11, 3-13, 3-15, 3-20, 3-23, 3-30, 3-33, 3-48, 4-31, 4-39, 4-59, 5-51, 6-1 и их сочетания. В одном из вариантов осуществления мышь экспрессирует легкую цепь, являющуюся соматически мутированной. В одном из вариантов осуществления CL представляет собой CL к мыши.
[0032] В одном из вариантов осуществления 90-100% эндогенных неперегруппированных генных сегментов VH мыши замещены по меньшей мере одним неперегруппированным генным сегментом VH человека. В конкретном варианте осуществления все или по существу все эндогенные неперегруппированные генные сегменты VH мыши замещены по меньшей мере одним неперегруппированным генным сегментом VH человека. В одном из вариантов осуществления замену производят по меньшей мере 18, по меньшей мере 39, по меньшей мере 80 или 81 неперегруппированными генными сегментами VH человека. В одном из вариантов осуществления замену производят по меньшей мере 12 функциональными неперегруппированными генными сегментами VH человека, по меньшей мере 25 функциональными неперегруппированными генными сегментами VH человека или по меньшей мере 4 3 неперегруппированными генными сегментами VH человека.
[0033] В одном из вариантов осуществления генетически модифицированная мышь представляет собой линию C57BL, в конкретном варианте осуществления, выбранную из C57BL/A, C57BL/An, C57BL/GrFa, C57BL/KaLwN, C57BL/6, C57BL/6J, C57BL/6ByJ, C57BL/6NJ, C57BL/10, C57BL/10ScSn, C57BL/10Cr, C57BL/Ola. В конкретном варианте осуществления генетически модифицированная мышь представляет собой результат скрещивания указанных выше линий 129 и указанной выше линии C57BL/6. В другом конкретном варианте осуществления мышь представляет собой результат скрещивания указанных выше линий 12 9 или результат скрещивания указанных выше линий BL/6. В конкретном варианте осуществления линия 129 представляет собой результат скрещивания линии 129S6 (129/SvEvTac).
[0034] В одном из вариантов осуществления мышь экспрессирует обратное химерное антитело, содержащее легкую цепь, содержащую CL к мыши и соматически мутированный домен VL человека, получаемый из генного сегмента Vκ1-39Jκ5 человека или генного сегмента Vκ3-20Jκ1 человека, и тяжелую цепь, содержащую СН мыши и соматически мутированный домен VH человека, получаемый из генного сегмента VH человека, выбранного из генных сегментов VH человека 1-2, 1-8, 1-24, 2-5, 3-7, 3-9, 3-11, 3-13, 3-15, 3-20, 3-23, 3-30, 3-33, 3-48, 4-31, 4-39, 4-59, 5-51 и 6-1, где мышь не экспрессирует полностью мышиного антитела и не экспрессирует полностью человеческого антитела. В одном из вариантов осуществления мышь несет локус легкой цепи к, содержащий замену эндогенных генных сегментов VL к мыши генным сегментом Vκ1-39Jκ5 человека или генным сегментом Vκ3-20Jκ1 человека, и у нее произведена замена всех или по существу всех эндогенных генных сегментов VH мыши полным или по существу полным репертуаром генных сегментов VH человека.
[0035] В одном из аспектов предоставлена клетка мыши, выделенная у мыши, как описано в настоящем документе. В одном из вариантов осуществления клетка представляет собой ES клетку. В одном из вариантов осуществления клетка представляет собой лимфоцит. В одном из вариантов осуществления лимфоцит представляет собой В-клетку. В одном из вариантов осуществления В-клетка экспрессирует химерную тяжелую цепь, содержащую вариабельный домен, получаемый из генного сегмента человека; и легкую цепь, получаемую из перегруппированного сегмента Vκ1-39/J человека, перегруппированного сегмента Vκ3-20/J человека или их сочетания; где вариабельный домен тяжелой цепи слит с константной областью мыши, и вариабельный домен легкой цепи слит с константной областью мыши или человека.
[0036] В одном из аспектов предоставлена гибридома, где гибридому получают с использованием В-клетки мыши, как описано в настоящем документе. В конкретном варианте осуществления В-клетку получают у мыши, как описано в настоящем документе, которую иммунизировали иммуногеном, содержащим представляющий интерес эпитоп, и В-клетка экспрессирует связывающий белок, который связывает представляющий интерес эпитоп, где связывающий белок содержит соматически мутированный домен VH человека и СН мыши и содержит домен VL человека, получаемый из генных сегментов Vκ1-39Jκ5 человека или Vκ3-20Jκ1 человека, и CL мыши.
[0037] В одном из аспектов предоставлен эмбрион мыши, где эмбрион содержит донорскую ES клетку, получаемую у мыши, как описано в настоящем документе.
[0038] В одном из аспектов предоставлен направленный вектор, содержащий в направлении транскрипции от 5' к 3' относительно последовательностей 5' и 3' участков гомологии вектора с последовательностями мыши, 5' участок гомологии с последовательностью мыши, промотор иммуноглобулина человека или мыши, лидерная последовательность человека или мыши и генный сегмент LCVR человека, выбранный из генных сегментов Vκ1-3 9Jκ5 человека или Vκ3-20Jκ1 человека, и 3' участок гомологии с последовательностью мыши. В одном из вариантов осуществления 5' и 3' участки гомологии направляют вектор к последовательности 5' относительно энхансерной последовательности, находящейся с 5'-конца и проксимально к гену константной области к мыши. В одном из вариантов осуществления промотор представляет собой промотор генного сегмента вариабельной области иммуноглобулина человека. В конкретном варианте осуществления промотор представляет собой промотор Vκ3-15 человека. В одном из вариантов осуществления лидерная последовательность представляет собой лидерную последовательность мыши. В конкретном варианте осуществления лидерная последовательность мыши представляет собой лидерную последовательность Vκ3-7 мыши.
[0039] В одном из аспектов предоставлен направленный вектор, как описано выше, но вместо 5' участка гомологии с последовательностью мыши промотор человека или мыши на 5'-конце фланкирован участком распознавания сайт-специфической рекомбиназы (SRRS), и вместо 3' участка гомологии с последовательностью мыши генный сегмент LCVR человека на 3'-конце фланкирован SRRS.
[0040] В одном из аспектов обратное химерное антитело продуцирует мышь, как описано в настоящем документе, где обратное химерное антитело содержит легкую цепь, содержащую CL мыши и VL человека, и тяжелую цепь, содержащую VH человека и СН мыши.
[0041] В одном из аспектов предоставлен способ получения антитела, включающий экспрессию в одной клетке (а) первой генной последовательности VH иммунизированной мыши, как описано в настоящем документе, слитой с генной последовательностью СН человека; (b) генной последовательности VL иммунизированной мыши, как описано в настоящем документе, слитой с генной последовательностью CL человека; и (с) поддержание клетки в условиях, достаточных для экспрессии полностью человеческого антитела, и выделение антитела. В одном из вариантов осуществления клетка содержит вторую генную последовательность VH второй иммунизированной мыши, как описано в настоящем документе, слитую с генной последовательностью СН человека, где первая генная последовательность VH кодирует домен VH, распознающий первый эпитоп, а второй генная последовательность VH кодирует домен VH, распознающий второй эпитоп, где первый эпитоп и второй эпитоп не являются идентичными.
[0042] В одном из аспектов предоставлен способ получения эпитопсвязывающего белка, включающий воздействие на мышь, как описано в настоящем документе, иммуногеном, содержащим представляющий интерес эпитоп, содержание мыши в условиях, достаточных для мыши для получения молекулы иммуноглобулина, специфически связывающийся с представляющим интерес эпитопом, и выделение молекулы иммуноглобулина, специфически связывающейся с представляющим интерес эпитопом; где эпитопсвязывающий белок содержит тяжелую цепь, содержащую соматически мутированный VH человека и СН мыши, ассоциированную с легкой цепью, содержащей CL мыши и VL человека, получаемую из генных сегментов Vκ1-39Jκ5 человека или Vκ3-20Jκ1 человека.
[0043] В одном из аспектов предоставлена клетка, экспрессирующая эпитопсвязывающий белок, где клетка содержит: (а) нуклеотидную последовательность VL человека, кодирующую домен VL человека, получаемую из генных сегментов Vκ1-39Jκ5 человека или Vκ3-20Jκ1 человека, где нуклеотидная последовательность VL человека слита (непосредственно или через линкер) с последовательностью кДНК константного домена легкой цепи иммуноглобулина человека (например, с последовательностью ДНК константного домена к человека); и (b) первую нуклеотидную последовательность VH человека, кодирующую домен VH человека, получаемую из первой нуклеотидной последовательности VH человека, где первая нуклеотидная последовательность VH человека слита (непосредственно или через линкер) с последовательностью кДНК константного домена тяжелой цепи иммуноглобулина человека; где эпитопсвязывающий белок распознает первый эпитоп. В одном из вариантов осуществления эпитопсвязывающий белок связывает первый эпитоп с константой диссоциации меньшей 10-6 М, меньшей 10-8 М, меньшей 10-9 М, меньшей 10-10 М, меньшей 10-11 M или меньшей 10-12 М.
[0044] В одном из вариантов осуществления клетка содержит вторую нуклеотидную последовательность VH человека, кодирующую второй домен VH человека, где вторая последовательность VH человека слита (непосредственно или через линкер) с последовательностью кДНК константного домена тяжелой цепи иммуноглобулина человека, и где второй домен VH человека специфически не распознает первый эпитоп (например, демонстрирует константу диссоциации например, 10-6 М, 10-5 M, 10-4 M или более), и где эпитопсвязывающий белок распознает первый эпитоп и второй эпитоп, и где каждая из первой и второй тяжелых цепей иммуноглобулинов ассоциируют с идентичной легкой цепью (а).
[0045] В одном из вариантов осуществления второй домен VH связывает второй эпитоп с константой диссоциации меньшей 10-6 М, меньшей 10-7 М, меньшей 10-8 М, меньшей 10-9 М, меньшей 10-10 М, меньшей 10-11 M или меньшей 10-12 M.
[0046] В одном из вариантов осуществления эпитопсвязывающий белок содержит первую тяжелую цепь иммуноглобулина и вторую тяжелую цепь иммуноглобулина, где каждая ассоциирована с идентичной легкой цепью, получаемой из генного сегмента VL человека, выбранного из генных сегментов Vκ1-39Jκ5 человека или Vκ3-20Jκ1 человека, где первая тяжелая цепь иммуноглобулина связывает первый эпитоп с константой диссоциации в диапазоне от наномолярного до пикомолярного, вторая тяжелая цепь иммуноглобулина связывает второй эпитоп с константой диссоциации в диапазоне от наномолярного до пикомолярного, первый эпитоп и второй эпитоп являются неидентичными, первая тяжелая цепь иммуноглобулина не связывает второй эпитоп или связывает второй эпитоп с константой диссоциации, более слабой, чем в микромолярном диапазоне (например, в миллимолярном диапазоне), вторая тяжелая цепь иммуноглобулина не связывает первый эпитоп или связывает первый эпитоп с константой диссоциации, более слабой, чем в микромолярном диапазоне (например, в миллимолярном диапазоне) и один или несколько из VL, VH первой тяжелой цепи иммуноглобулина и VH второй тяжелой цепи иммуноглобулина являются соматически мутированными.
[0047] В одном из вариантов осуществления первая тяжелая цепь иммуноглобулина содержит связывающий белок А остаток, а во второй тяжелой цепи иммуноглобулина связывающий белок А остаток отсутствует.
[0048] В одном из вариантов осуществления клетка выбрана из СНО, COS, 293, HeLa и клетки сетчатки, экспрессирующей вирусную последовательность нуклеиновой кислоты (например, клетка PERC.6™).
[0049] В одном из аспектов предоставлено обратное химерное антитело, содержащее VH человека и константный домен тяжелой цепи мыши, VL человека и константный домен легкой цепи мыши, где антитело получают способом, включающим иммунизацию мыши, как описано в настоящем документе, иммуногеном, содержащим эпитоп, и антитело специфически связывается с эпитопом иммуногена, которым иммунизируют мышь. В одном из вариантов осуществления домен VL является соматически мутированным. В одном из вариантов осуществления домен VH является соматически мутированным. В одном из вариантов осуществления и домен VL, и домен VH являются соматически мутированными. В одном из вариантов осуществления VL связан с константным доменом к мыши.
[0050] В одном из аспектов предоставлена мышь, несущая генные сегменты вариабельных областей тяжелых цепей человека, замещающие в эндогенном локусе мыши все или по существу все генные сегменты вариабельных областей тяжелых цепей мыши; не более чем один или два генных сегмента вариабельных областей легких цепей человека, выбранных из перегруппированного сегмента Vκ1-39/J и перегруппированного сегмента Vκ3-20/J или их сочетания, замещающих все генные сегменты вариабельных областей легких цепей мыши; где генные сегменты вариабельных областей тяжелых цепей человека связаны с геном константной области мыши, и генный сегмент(ы) вариабельной области легкой цепи человека связан с геном константной области человека или мыши.
[0051] В одном из аспектов ES клетка мыши, содержащая замену всех или по существу всех генных сегментов вариабельных областей тяжелых цепей мыши генными сегментами вариабельных областей тяжелых цепей человека, и не более чем один или два перегруппированных сегментов V/J легких цепей человека, где генные сегменты вариабельных областей тяжелых цепей человека связаны с геном константной области тяжелой цепи иммуноглобулина мыши, и сегменты V/J легких цепей человека связаны с геном константной области легкой цепи иммуноглобулина мыши или человека. В конкретном варианте осуществления ген константной области легкой цепи представляет собой ген константной области мыши.
[0052] В одном из аспектов предоставлен антигенсвязывающий белок, продуцируемый мышью, как описано в настоящем документе. В конкретном варианте осуществления антигенсвязывающий белок содержит вариабельную область тяжелой цепи иммуноглобулина человека, слитую с константной областью мыши, и вариабельная область легкой цепи иммуноглобулина человека, получаемая из генного сегмента Vκ1-39 или генного сегмента Vκ3-20, где константная область легкой цепи представляет собой константную область мыши.
[0053] В одном из аспектов предоставлен полностью человеческий антигенсвязывающий белок, полученный из генной последовательности вариабельной области иммуноглобулина из мыши, как описано в настоящем документе, где антигенсвязывающий белок содержит полностью человеческую тяжелую цепь, содержащую вариабельную область человека, получаемую из последовательности мыши, как описано в настоящем документе, и полностью человеческую легкую цепь, содержащую вариабельную область Vκ1-39 или a Vκ3-20. В одном из вариантов осуществления вариабельная область легкой цепи содержит от одной до пяти соматических мутаций. В одном из вариантов осуществления вариабельная область легкой цепи представляет собой родственную вариабельную область легкой цепи, которая в В-клетке мыши спарена с вариабельной областью тяжелой цепи.
[0054] В одном из вариантов осуществления полностью человеческий антигенсвязывающий белок содержит первую тяжелую цепь и вторую тяжелую цепь, где первая тяжелая цепь и вторая тяжелая цепь независимо содержат неидентичные вариабельные области, получаемые у мыши, как описано в настоящем документе, и где каждая из первой и второй тяжелых цепей экспрессируется из клетки-хозяина в ассоциации с легкой цепью человека, получаемой из генного сегмента Vκ1-39 или генного сегмента Vκ3-20. В одном из вариантов осуществления первая тяжелая цепь содержит первую вариабельную область тяжелой цепи, специфически связывающуюся с первым эпитопом первого антигена, а вторая тяжелая цепь содержит вторую вариабельную область тяжелой цепи, специфически связывающуюся со вторым эпитопом второго антигена. В конкретном варианте осуществления первый антиген и второй антиген являются различными. В конкретном варианте осуществления первый антиген и второй антиген являются одинаковыми, а первый эпитоп и второй эпитоп не являются идентичными; в конкретном варианте осуществления связывание первого эпитопа первой молекулой связывающего белка не блокирует связывание второго эпитопа второй молекулой связывающего белка.
[0055] В одном из аспектов полностью человеческий связывающий белок, получаемый из последовательности иммуноглобулина человека у мыши, как описано в настоящем документе, содержит первую тяжелую цепь иммуноглобулина и вторую тяжелую цепь иммуноглобулина, где первая тяжелая цепь иммуноглобулина содержит первую вариабельную область, которая не идентична с вариабельной областью второй тяжелой цепи иммуноглобулина, и где первая тяжелая цепь иммуноглобулина содержит связывающую детерминанту белка А дикого типа, а во второй тяжелой цепи связывающая детерминанта белка А дикого типа отсутствует. В одном из вариантов осуществления первая тяжелая цепь иммуноглобулина в условиях выделения связывается с белком А, а вторая тяжел цепь иммуноглобулина в условиях выделения не связывается с белком А или связывается с белком А по меньшей мере в 10 раз, сто раз или тысячу раз слабее, чем с белком А связывается первая тяжелая цепь иммуноглобулина. В конкретном варианте осуществления первая и вторая тяжелые цепи представляют собой изотипы IgG1, где вторая тяжелая цепь содержит модификацию, выбранную из 95R (EU 435R), 96F (EU 436F) и их сочетания, и где у первой тяжелой цепи такая модификация отсутствует.
[0056] В одном из аспектов предоставлен способ получения биспецифического антигенсвязывающего белка, включающий воздействие на первую мышь, как описано в настоящем документе, первым представляющим интерес антигеном, содержащим первый эпитоп, воздействие на вторую мышь, как описано в настоящем документе, вторым представляющим интерес антигеном, содержащим второй эпитоп, обеспечение развития у каждой из первой и второй мышей иммунного ответа на представляющие интерес антигены, идентификацию у первой мыши первой вариабельной области тяжелой цепи человека, связывающей первый эпитоп первого представляющего интерес антигена, идентификацию у второй мыши второй вариабельной области тяжелой цепи человека, связывающей второй эпитоп второго представляющего интерес антигена, получение первого гена полностью человеческой тяжелой цепи, кодирующей первую тяжелую цепь, связывающую первый эпитоп первого представляющего интерес антигена, получение второго гена полностью человеческой тяжелой цепи, кодирующего вторую тяжелую цепь, связывающую второй эпитоп второго представляющего интерес антигена, экспрессию первой тяжелой цепи и второй тяжелой цепи в клетке, экспрессирующей единственную полностью человеческую легкую цепь, получаемую из генных сегментов Vκ1-39 человека или Vκ3-20 человека с формированием биспецифического антигенсвязывающего белка и выделение биспецифического антигенсвязывающего белка.
[0057] В одном из вариантов осуществления первый антиген и второй антиген являются неидентичными.
[0058] В одном из вариантов осуществления первый антиген и второй антиген являются идентичными, а первый эпитоп и второй эпитоп являются неидентичными. В одном из вариантов осуществления связывание первой вариабельной области тяжелой цепи с первым эпитопом не блокирует связывание второй вариабельной области тяжелой цепи со вторым эпитопом.
[0059] В одном из вариантов осуществления первый антиген выбран из растворимого антигена и клеточного поверхностного антигена (например, опухолевого антигена), а второй антиген включает клеточный поверхностный рецептор. В конкретном варианте осуществления клеточный поверхностный рецептор представляет собой рецептор иммуноглобулинов. В конкретном варианте осуществления рецептор иммуноглобулинов представляет собой рецептор Fc. В одном из вариантов осуществления первый антиген и второй антиген представляют собой один и тот же клеточный поверхностный рецептор, а связывание первой тяжелой цепи с первым эпитопом не блокирует связывание второй тяжелой цепи со вторым эпитопом.
[0060] В одном из вариантов осуществления вариабельный домен легкой цепи содержит от 2 до 5 соматических мутаций. В одном из вариантов осуществления вариабельный домен легкой цепи представляет собой соматически мутированную родственную легкую цепь, экспрессируемую в В-клетке первой или второй иммунизированной мыши с первым или вторым вариабельным доменом тяжелой цепи.
[0061] В одном из вариантов осуществления первая полностью человеческая тяжелая цепь содержит модификацию аминокислот, снижающую ее аффинность к белку А, а вторая полностью человеческая тяжелая цепь не содержит модификации, снижающей ее аффинность к белку А.
[0062] В одном из аспектов предоставлено антитело или биспецифическое антитело, содержащее вариабельный домен тяжелой цепи человека, получаемый по изобретению. В другом аспекте для получения полностью человеческого антитела или полностью человеческого биспецифического антитела предоставлено использование мышей, как описано в настоящем документе.
[0063] Если не указано иначе или не очевидно из контекста, любой из вариантов осуществления и аспектов, описываемых в настоящем документе, можно использовать в сочетании с любым другим. Другие варианты осуществления будут понятны специалистам в данной области из рассмотрения приведенного ниже описания.
ПОДРОБНОЕ ОПИСАНИЕ
[0064] Настоящее изобретение не ограничено конкретными описанными способами и экспериментальными условиями, по существу способы и условия могут варьировать. Также следует понимать, что терминологию, применяемую в настоящем документе, используют только с целью описания конкретных вариантов осуществления, и она не предназначена для ограничения, так как объем настоящего изобретения определен формулой изобретения.
[0065] Если не определено иначе, все термины и фразы, используемые в настоящем документе, включают значения, которые термины и фразы имеют в данной области, если обратное явно не указано или явно не очевидно из контекста, в котором используют термин или фразу. Хотя в практическом осуществлении или тестировании настоящего изобретения можно использовать любые способы и материалы, сходные или эквивалентные способам и материалам, описываемым в настоящем документе, в настоящем документе описаны конкретные способы и материалы. Все указанные публикации таким образом включены в качестве ссылки.
[0066] Как применяют в настоящем документе термин "антитело", включает молекулы иммуноглобулинов, содержащие четыре полипептидные цепи, две тяжелых (Н) цепи и две легких (L) цепи, связанные дисульфидными связями. Каждая тяжелая цепь содержит вариабельную область тяжелой цепи (VH) и константную область тяжелой цепи (СН). Константная область тяжелой цепи содержит три домена, CH1, СН2 и СНЗ. Каждая легкая цепь содержит вариабельную область легкой цепи (VL) и константную область легкой цепи (CL). Области VH и VL можно дополнительно подразделять на области гипервариабельности, называемые определяющими комплементарность областями (CDR), чередующиеся с областями, которые являются более консервативными, называемыми каркасными областями (FR). Каждая VH и VL содержит три CDR и четыре FR, расположенные, начиная с N-конца до С-конца, в следующем порядке: FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4 (CDR тяжелых цепей можно сокращать как HCDR1, HCDR2 и HCDR3; CDR легких цепей можно сокращать как LCDR1, LCDR2 и LCDR3). Термин "высокоаффинное" антитело относится к антителу с KD в отношении его эпитопа-мишени приблизительно 10-9 M или менее (например, приблизительно 1×10-9 M, 1×10-10 M, 1×10-11 M или приблизительно 1×10-12 M). В одном из вариантов осуществления KD измеряют посредством поверхностного плазмонного резонанса, например, BIACORE™; в другом варианте осуществления KD измеряют посредством ELISA.
[0067] Фраза "биспецифическое антитело" включает антитело, способное к селективному связыванию двух или более эпитопов. Биспецифические антитела, как правило, содержат две неидентичных тяжелых цепи, где каждая тяжелая цепь специфически связывает различные эпитопы - или на двух различных молекулах (например, различные эпитопы на двух различных иммуногенах) или на одной и той же молекуле (например, различные эпитопы на одном и том же иммуногене). Если биспецифическое антитело способно к селективному связыванию двух различных эпитопов (первого эпитопа и второго эпитопа), аффинность первой тяжелой цепи к первому эпитопу, как правило, по меньшей мере на один-два, или три, или четыре, или более порядков ниже аффинности первой тяжелой цепи ко второму эпитопу и наоборот.Эпитопы, специфически связываемые биспецифическим антителом, могут находиться на одной и той же или на различных мишенях (например, на одном и том же или на различных белках). Биспецифические антитела можно получать, например, комбинируя тяжелые цепи, распознающие различные эпитопы одного и того же иммуногена. Например, последовательности нуклеиновых кислот, кодирующие последовательности вариабельных областей тяжелых цепей, распознающих различные эпитопы одного и того же иммуногена, можно сливать с последовательностями нуклеиновых кислот, кодирующих одну и ту же или различные константные области тяжелых цепей, и такие последовательности можно экспрессировать в клетке, экспрессирующей легкую цепь иммуноглобулина. Типичное биспецифическое антитело содержит две тяжелых цепи, где каждая содержит три CDR тяжелой цепи с последующим (от N-конца к С-концу) доменом СН1, шарниром, доменом СН2 и доменом СН3, и легкую цепь иммуноглобулина, которая не обеспечивает специфичность связывания эпитопа, но которая может ассоциировать с каждой тяжелой цепью, или которая может ассоциировать с каждой тяжелой цепью и которая может связываться с одним или несколькими из эпитопов, связываемых эпитопсвязывающими областями тяжелых цепей, или которая может ассоциировать с каждой тяжелой цепью и обеспечивать связывание одной или обеих тяжелых цепей с одним или обоими эпитопами.
[0068] Термин "клетка" включает любую клетку, которая подходит для экспрессии рекомбинантной последовательности нуклеиновой кислоты. Клетки включают клетки прокариот и эукариот (одноклеточных или многоклеточных), бактериальные клетки (например, штаммы Е. coli, видов Bacillus, видов Streptomyces и т.д.), клетки микобактерий, клетки грибов, дрожжевые клетки (например, S. cerevisiae, S. pombe, P. pastoris, P. methanolica и т.д.), растительные клетки, клетки насекомых (например, SF-9, SF-21, инфицированные бакуловирусами клетки насекомых, Trichoplusia ni и т.д.), клетки не являющихся человеком животных, клетки человека или слитые клетки, такие как, например, гибридомы или квадромы. В некоторых вариантах осуществления клетка представляет собой клетку человека, нечеловекообразной обезьяны, человекообразной обезьяны, хомяка, крысы или мыши. В некоторых вариантах осуществления клетка является эукариотической и выбрана из следующих клеток: СНО (например, СНО K1, DXB-11 СНО, Veggie-СНО), COS (например, COS-7), клетка сетчатки, Vero, CV1, клетка почки (например, НЕК293, 293 EBNA, MSR 293, MDCK, НаК, ВНК), HeLa, HepG2, WI38, MRC 5, Colo205, НВ 8065, HL-60, (например, ВНК21), Jurkat, Daudi, А431 (клетка эпидермиса), CV-1, U937, 3Т3, клетка L, клетка С127, SP2/0, NS-0, ММТ 060562, клетка Сертоли, клетка BRL 3А, клетка НТ1080, миеломная клетка, опухолевая клетка и клеточные линии, получаемые из указанных выше клеток. В некоторых вариантах осуществления клетка содержит один или несколько вирусных генов, например, клетка сетчатки, экспрессирующая вирусный ген (например, клетка PER.C6™).
[0069] Фраза "определяющая комплементарность область" или термин "CDR", включает аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты генов иммуноглобулинов организма, которая в норме (т.е., у животного дикого типа) находится между двумя каркасными областями в вариабельной области легкой или тяжелой цепи молекулы иммуноглобулина (например, антитела или Т-клеточного рецептора). CDR могут кодировать, например, последовательность зародышевой линии или перегруппирования или неперегруппированная последовательность, и, например, наивная или зрелая В-клетка или Т-клетка. CDR может быть соматически мутированной (например, отличаться от последовательности, кодируемой зародышевой линией животного), гуманизированной и/или модифицированной заменами, добавлениями или делециями аминокислот.В некоторых случаях (например, для CDR3), CDR могут кодировать две или более последовательности (например, последовательности зародышевой линии), которые не являются смежными (например, в неперегруппированной последовательности нуклеиновой кислоты), но которые являются смежными в последовательности нуклеиновой кислоты В-клетки, например, в результате сплайсинга или соединения последовательностей (например, рекомбинации V-D-J с формированием CDR3 тяжелой цепи).
[0070] Термин "консервативная", когда его используют для описания консервативной замены аминокислот, включает замену остатка аминокислоты другим остатком аминокислоты с группой R боковой цепи со сходными химическими свойствами (например, зарядом или гидрофобностью). В основном, консервативная замена аминокислот существенно не изменяет представляющие интерес функциональные свойства белка, например, способность вариабельной области специфически связывать эпитоп-мишень с желаемой аффинностью. Примеры групп аминокислот с боковыми цепями со сходными химическими свойствами включают алифатические боковые цепи, такие как глицин, аланин, валин, лейцин и изолейцин; алифатическо-гидроксильные боковые цепи, такие как серии и треонин; амидсодержащие боковые цепи, такие как аспарагин и глутамин; ароматические боковые цепи, такие как фенилаланин, тирозин и триптофан; основные боковые цепи, такие как лизин, аргинин и гистидин; кислые боковые цепи, такие как аспарагиновая кислота и глутаминовая кислота; и серусодержащие боковые цепи, такие как цистеин и метионин. Группы консервативных замен аминокислот включают, например, валин/лейцин/изолейцин, фенилаланин/тирозин, лизин/аргинин, аланин/валин, глутаминат/аспартат и аспарагин/глутамин. В некоторых вариантах осуществления консервативная замена аминокислот может представлять собой замену любого природного остатка в белке на аланин, как используют, например, в мутагенезе при сканировании аланином. В некоторых вариантах осуществления производят консервативную замену, которая обладает положительным значением в матрице логарифмического правдоподобия РАМ250, описанной в Gonnet et at. (1992) Exhaustive Matching of the Entire Protein Sequence Database, Science 256:1443-45, включенной, таким образом, в качестве ссылки. В некоторых вариантах осуществления замена представляет собой умеренно консервативную замену, где замена обладает неотрицательным значением в матрице логарифмического правдоподобия РАМ250.
[0071] В некоторых вариантах осуществления положения остатков в легкой цепи иммуноглобулина или в тяжелой цепи отличаются по одной или нескольким консервативным заменам аминокислот. В некоторых вариантах осуществления положения остатков в легкой цепи иммуноглобулина или в ее функциональном фрагменте (например, фрагмент, позволяющий экспрессию и секрецию, например, из В-клетки) не идентичны с легкой цепью, аминокислотная последовательность которой приведена в настоящем документе, а отличаются на одну или несколько консервативных замен аминокислот.
[0072] Фраза "эпитопсвязывающий белок" включает белок, содержащий по меньшей мере одну CDR и способный к селективному распознаванию эпитопа, например, способный к связыванию эпитопа с KD, составляющей приблизительно один микромоль или менее (например, KD, которая приблизительно составляет 1×10-6 M, 1×10-7 M, 1×10-9 M, 1×10-9 M, 1×10-10 M, 1×10-11 M или приблизительно 1×10-12 M). Терапевтические эпитопсвязывающие белки (например, терапевтические антитела) часто требуют KD, находящуюся в наномолярном или пикомолярном диапазоне.
[0073] Фраза "функциональный фрагмент" включает фрагменты эпитопсвязывающих белков, способные к экспрессии, секреции и специфическому связыванию с эпитопом с KD в микромолярном, наномолярном или пикомолярном диапазонах. Специфическое распознавание включает наличие KD по меньшей мере в микромолярном диапазоне, наномолярном диапазоне или пикомолярном диапазоне.
[0074] Термин "зародышевая линия" включает указание на последовательность нуклеиновой кислоты иммуноглобулина в соматически немутированной клетка, например, соматически немутированной В-клетке или пре-В-клетке или в гематопоэтической клетке.
[0075] Фраза "тяжелая цепь" или "тяжелая цепь иммуноглобулина" включает последовательность константной области тяжелой цепи иммуноглобулина из любого организма. Если не указано иначе, вариабельные домены тяжелых цепей включают три CDR тяжелой цепи и четыре FR-области. Фрагменты тяжелой цепи включают CDR, CDR и FR и их сочетания. Типичная тяжелая цепь после вариабельного домена (от N-конца к С-концу) содержит домен СН1, шарнир, домен СН2 и домен СН3. Функциональный фрагмент тяжелой цепи включает фрагмент, способный к специфическому распознаванию эпитопа (например, распознаванию эпитопа с KD в микромолярном, наномолярном или пикомолярном диапазонах), способный к экспрессии и секреции из клетки и содержащий по меньшей мере одну CDR.
[0076] Термин "идентичность", когда его используют по отношению к последовательностям, включает идентичность, как ее определяют рядом различных алгоритмов, известных в данной области, которые можно использовать для определения идентичности последовательностей нуклеотидов и/или аминокислот. В некоторых вариантах осуществления, описываемых в настоящем документе, идентичность определяют с использованием выравнивания ClustalW v. 1.83 (медленно) с использованием штрафа за создание пропуска 10,0, штрафа за продление пропуска 0,1 и с использованием матрицы подобия Gonnet (MacVector™ 10.0.2, MacVector Inc., 2008). Длина сравниваемых последовательностей в отношении идентичности последовательностей зависит от конкретных последовательностей, но в случае константного домена легкой цепи, участок должен содержать последовательность достаточной длины для сворачивания в константный домен легкой цепи, способный к самоассоциации с формированием канонического константного домена легкой цепи, например, способного к формированию двух бета-слоев, содержащих бета-цепи, и способного к взаимодействию по меньшей мере с одним доменом СН1 человека или мыши. В случае домена СН1, участок последовательности должен содержать последовательность достаточной длины для сворачивания в домен СН1, способный к формированию двух бета-слоев, содержащих бета-цепи, и способный к взаимодействию по меньшей мере с одним константным доменом легкой цепи мыши или человека.
[0077] Фраза "молекула иммуноглобулина" включает две тяжелых цепи иммуноглобулина и две легких цепи иммуноглобулина. Тяжелые цепи могут быть идентичными или различными, и легкие цепи могут быть идентичными или различными.
[0078] Фраза "легкая цепь" включает последовательность легкой цепи иммуноглобулина из любого организма и, если не указано иначе, включает легкие цепи κ и λ человека и VpreB, а также суррогатные легкие цепи. Если не указано иначе, вариабельные домены легких цепей (VL), как правило, включают три CDR легких цепей и четыре каркасных (FR) области. Как правило, полноразмерная легкая цепь от N-конца до С-конца содержит домен VL, содержащий FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4, и константный домен легкой цепи. Например, легкие цепи включают легкие цепи, которые селективно не связываются с первым или вторым эпитопом, селективно связываемым эпитопсвязывающим белком, в котором они находятся. Легкие цепи также включают легкие цепи, которые связывают и распознают или помогают тяжелой цепи связывать и распознавать один или несколько эпитопов, селективно связываемых эпитопсвязывающим белком, в котором они находятся. Общие легкие цепи представляют собой легкие цепи, получаемые из генного сегмента Vκ1-39Jκ5 человека или генного сегмента Vκ3-20Jκ1 человека, и включают их соматически мутированные (например, аффинно зрелые) варианты.
[0079] Фраза "микромолярный диапазон" предназначена для обозначения 1-999 микромоль; фраза "наномолярный диапазон" предназначена для 1-999 наномоль; фраза "пикомолярный диапазон" предназначена для обозначения 1-999 пикомоль.
[0080] Фраза "соматически мутированный" включает указание на последовательность нуклеиновой кислоты из В-клетки, которая претерпела переключение класса, где последовательность нуклеиновой кислоты вариабельной области иммуноглобулина (например, вариабельный домен тяжелой цепи или включающая последовательность CDR или FR тяжелой цепи) в В-клетке с переключенным классом не идентична последовательности нуклеиновой кислоты в В-клетке до переключения класса, например, как различия в последовательностях нуклеиновых кислот CDR или каркаса между В-клеткой, не претерпевшей переключение класса, и В-клетка, претерпевшей переключение класса. "Соматически мутированный" включает указание на последовательности нуклеиновых кислот из В-клеток с созревшей аффинностью, которые не идентичны соответствующим последовательностям вариабельных областей иммуноглобулинов в В-клетках с непрошедшим созреванием аффинности (т.е., последовательностям в геноме клеток зародышевой линии). Фраза "соматически мутированный" также включает указание на последовательность нуклеиновой кислоты вариабельной области иммуноглобулина из В-клетки после воздействия на В-клетку представляющим интерес эпитопом, где последовательность нуклеиновой кислоты отличается от соответствующей последовательности нуклеиновой кислоты до воздействия на В-клетку представляющим интерес эпитопом. Фраза "соматически мутированный" относится к последовательностям из антител, полученным у животных, например, мыши, несущей последовательности нуклеиновых кислот вариабельных областей иммуноглобулинов человека, в ответ на стимуляцию иммуногеном, и которые являются результатом процесса отбора, врожденно функционирующего у такого животного.
[0081] Термин "неперегруппированная" относительно последовательности нуклеиновой кислоты, включает последовательности нуклеиновой кислоты, которые присутствуют в клетках зародышевой линии животного.
[0082] Фраза "вариабельный домен" включает аминокислотную последовательность легкой или тяжелой цепи иммуноглобулина (модифицированную по желанию), содержащую в последовательности от Ν-конца до С-конца следующие аминокислотные области (если не указано иначе): FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4. Общая легкая цепь
[0083] Предыдущим попыткам получения пригодных полиспецифичных эпитопсвязывающих белков, например, биспецифических антител, препятствовал ряд проблем, которые часто разделяли общий принцип: отбор или обработку последовательностей in vitro с рациональным конструированием или с конструированием методом проб и ошибок подходящего формата для спаривания гетеродимерного биспецифического иммуноглобулина человека. К сожалению, большинство, если не все, инженерных подходов in vitro в основном предусматривают подходящие поправки, которые подходят, если вообще подходят, для индивидуальных молекул. С другой стороны, способы in vivo с использованием сложных организмов для отбора подходящих пар, способных приводить к терапевтическим средствам для человека, не реализованы.
[0084] Как правило, природные последовательности мыши часто не являются хорошим источником для терапевтических последовательностей человека. По меньшей мере по этой причине получение вариабельных областей тяжелых цепей иммуноглобулинов мыши, спаривающихся с общей легкой цепью человека, имеет ограниченную практическую ценность. Большинство попыток конструирования in vitro израсходовалось бы в процессе проб и ошибок в попытке гуманизировать последовательности вариабельных областей тяжелых цепей мыши в надежде сохранить специфичность и аффинность к эпитопу при сохранении способности связываться с общей легкой цепью человека с неопределенным исходом. По окончании такого процесса конечный продукт мог бы сохранять некоторую специфичность и аффинность и ассоциировать с общей легкой цепью, но, в конечном счете, принципиальным риском, вероятно, осталась бы иммуногенность у человека.
[0085] Таким образом, подходящая мышь для получения терапевтических средств для человека должна была бы вместо эндогенных генных сегментов вариабельных областей тяжелых цепей мыши нести достаточно большой репертуар генных сегментов вариабельных областей тяжелых цепей человека. Генные сегменты вариабельных областей тяжелых цепей человека должны быть способными к перегруппировке и рекомбинации с эндогенным константным доменом тяжелой цепи мыши с формированием обратной химерной тяжелой цепи (т.е., тяжелой цепи, содержащей вариабельный домен человека и константную область мыши). Тяжелая цепь должна быть способна к переключению классов и соматическому гипермутированию так, что для мыши для выбора вариабельного домена тяжелой цепи, который может ассоциировать с ограниченным репертуаром вариабельных областей легких цепей человека, доступен достаточно большой репертуар вариабельных доменов тяжелых цепей.
[0086] Практическую ценность имеет мышь, у которой происходит отбор общей легкой цепи для множества тяжелых цепей. В различных вариантах осуществления антитела, экспрессируемые у мыши, которая экспрессирует только общую легкую цепь, содержат тяжелые цепи, которые могут ассоциировать и экспрессироваться с идентичными или по существу идентичными легкими цепями. Это особенно пригодно для получения биспецифических антител. Например, такую мышь можно иммунизировать первым иммуногеном с получением В-клетки, экспрессирующей антитело, специфически связывающееся с первым эпитопом. Мышь (или генетически одинаковую мышь) можно иммунизировать вторым иммуногеном с получением В-клетки, экспрессирующей антитело, специфически связывающееся со вторым эпитопом. Вариабельные области тяжелых цепей можно клонировать из В-клеток и экспрессировать с той же константной областью тяжелой цепи и той же легкой цепью, и экспрессировать в клетке с получением биспецифического антитела, где у мыши происходит отбор компонента легкой цепи биспецифического антитела для ассоциации и экспрессии с компонентом легкой цепи.
[0087] Авторы изобретения генно-инженерными способами получили мышь для получения легких цепей иммуноглобулинов, которые подходящим образом спариваются с достаточно разновидным семейством тяжелых цепей, включающим тяжелые цепи, вариабельные области которых отличаются от последовательностей зародышевой линии, например, аффинно зрелые или соматически мутированные вариабельные области. В различных вариантах осуществления мышь разработана для спаривания вариабельных доменов легких цепей человека с вариабельными доменами тяжелых цепей человека, содержащими соматические мутации, таким образом, обеспечивая путь к высокоаффинным связывающим белкам, пригодным для использования в качестве терапевтических средств для человека.
[0088] У полученной генно-инженерными способами мыши при длительном и сложном процессе отбора антител в организме происходит биологически соответствующий выбор спаривания разнообразного набора вариабельных доменов тяжелых цепей человека с ограниченным количеством вариантов легкой цепи человека. Для достижения этого генно-инженерными способами получают мышь, несущую ограниченное количество вариантов вариабельного домена легкой цепи человека в сочетании с широким разнообразием вариантов вариабельного домена тяжелой цепи человека. После стимуляции иммуногеном у мыши в ее репертуаре происходит максимизация количества решений для получения антитела к иммуногену, преимущественно ограниченных или ограниченных исключительно количеством вариантов легких цепей в ее репертуаре. В различных вариантах осуществления это включает обеспечение образования у мыши подходящих и сочетаемых соматических мутаций вариабельного домена легкой цепи, которые, тем не менее, совместимы с относительно большим разнообразием вариабельных доменов тяжелых цепей человека, включая, в частности, соматически мутированные вариабельные домены тяжелых цепей человека.
[0089] Для получения ограниченного репертуара вариантов легкой цепи генно-инженерными способами получают мышь, делая у нее нефункциональной или по существу нефункциональной возможность продукции или перегруппировки природного вариабельного домена легкой цепи мыши. Этого можно достигать, например, удаляя генные сегменты вариабельных областей легких цепей мыши. Затем эндогенный локус мыши можно модифицировать подходящим предпочтительным экзогенным генным сегментом вариабельной области легкой цепи человека, функционально связанным с эндогенным константным доменом легкой цепи мыши таким способом, что генные сегменты экзогенной вариабельной области человека могут перегруппировываться и рекомбинировать с геном эндогенной константной области легкой цепи мыши и формировать перегруппированный ген обратной химерной легкой цепи (вариабельная область человека, константная область мыши). В различных вариантах осуществления вариабельная область легкой цепи может являться соматически мутированной. В различных вариантах осуществления для максимизации возможности возникновения соматических мутаций в вариабельной области легкой цепи, у мыши сохраняют соответствующий энхансер(ы). Например, в модифицированном локусе мыши к для замены эндогенных генных сегментов вариабельных областей к мыши генными сегментами вариабельных областей к человека функционально сохраняют или не подвергают разрушению интронный энхансер к мыши и 3'-энхансер к мыши.
[0090] Предоставлена полученная генно-инженерными способами мышь, экспрессирующая ограниченный репертуар обратных химерных (вариабельная область человека, константная область мыши) легких цепей, ассоциированных с многообразием обратных химерных (вариабельная область человека, константная область мыши) тяжелых цепей. В различных вариантах осуществления эндогенные генные сегменты вариабельных областей легких цепей к мыши удалены и замещены единственным (или двумя) генными сегментами вариабельных областей легких цепей человека, функционально связанными с эндогенным геном константной области к мыши. В вариантах осуществления для максимизации соматического гипермутирования генных сегментов вариабельных областей легких цепей человека сохраняют интронный энхансер к мыши и 3'-энхансер к мыши. В различных вариантах осуществления у мышь также присутствует нефункциональный локус легкой цепи λ, или его делеция, или делеция, делающая локус неспособным к продукции легкой цепи λ.
[0091] Предоставлена полученная генно-инженерными способами мышь, которая в различных вариантах осуществления несет локус вариабельной области легкой цепи с отсутствием эндогенного генного сегмента вариабельной области легкой цепи мыши и содержащий генный сегмент вариабельной области легкой цепи человека, в одном из вариантов осуществления перегруппированную последовательность V/J человека, функционально связанную с константной областью мыши, где локус способен претерпевать соматическое гипермутирование, и где локус экспрессирует легкую цепь, содержащую последовательность V/J человека, связанную с константной областью мыши. Таким образом, в различных вариантах осуществления локус содержит 3'-энхансер к мыши, который коррелирует с нормальным, или дикого типа, уровнем соматического гипермутирования.
[0092] В различных вариантах осуществления у полученной генно-инженерными способами мыши при иммунизации представляющим интерес антигеном образуются В-клетки, демонстрирующие разнообразие перегруппировок вариабельных областей тяжелых цепей иммуноглобулинов человек, экспрессирующихся и функционирующих с одной или с двумя перегруппированными легкими цепями, включая варианты осуществления, где одна или две легкие цепи содержат вариабельные области легких цепей человека, например, содержащие от 1 до 5 соматических мутаций. В различных вариантах осуществления экспрессируемые таким образом легкие цепи человека способны к ассоциации и экспрессии с любой вариабельной областью тяжелой цепи иммуноглобулина человека, экспрессируемой у мыши. Эпитопсвязывающие белки, связывающие более одного эпитопа [0093] Композиции и способы, описываемые в настоящем документе, можно использовать для получения связывающих белков, которые с высокой аффинностью связывают более одного эпитопа, например, биспецифических антител. Преимущества изобретения включают возможность отбора подходящих сильно связывающихся (например, аффинно зрелых) тяжелых цепей иммуноглобулинов, каждая из которых ассоциирует с единственной легкой цепью.
[0094] Синтез и экспрессия биспецифических связывающих белков являлись проблемными, частично вследствие трудностей, связанных с определением подходящей легкой цепи, которая может ассоциировать и экспрессироваться с двумя различными тяжелыми цепями, а частично вследствие трудностей с выделением. Способы и композиции, описываемые в настоящем документе, предусматривают генетически модифицированную мышь для отбора посредством природных в остальном отношении процессов подходящей легкой цепи, которая может ассоциировать и экспрессироваться более чем с одной тяжелой цепью, включая тяжелые цепи, которые являются соматически мутированными (например, аффинно зрелыми). Последовательности VL и VH человека из подходящих В-клеток иммунизированных мышей, как описано в настоящем документе, экспрессирующих аффинно зрелые антитела с обратными химерными тяжелыми цепями (т.е., вариабельная область человека и константная область мыши) можно идентифицировать и клонировать в рамку в экспрессирующий вектор с подходящей генной последовательностью константной области человека (например, IgG1 человека). Можно получать две таких конструкции, где каждая конструкция кодирует вариабельный домен тяжелой цепи человека, связывающийся с отличным эпитопом. Одну из VL человека (например, Vκ1-39Jκ5 человека или Vκ3-20Jκ1 человека) в последовательности зародышевой линии или из В-клетки, где последовательность соматически мутирована, можно сливать в рамку с подходящим геном константной области человека (например, ген константной области к человека). Эти три полностью человеческие конструкции тяжелых и легких цепей можно вводить в подходящую клетку для экспрессии. Клетка будет экспрессировать две основных молекулы: гомодимерную тяжелую цепь с идентичной легкой цепью и гетеродимерную тяжелую цепь с идентичной легкой цепью. Для обеспечения легкого разделения этих основных молекул, одну из тяжелых цепей модифицируют, удаляя связывающую белок А детерминанту, что приводит к различной аффинности гомодимерного связывающего белка и гетеродимерного связывающего белка. Композиции и способы, решающие эту проблему, описаны в USSN 12/832838, поданной 25 июня 2010 года, озаглавленной "Readily Isolated Bispecific Antibodies with Native Immunoglobulin Format", опубликованной как US 2010/0331527A1, включенный, таким образом, в качестве ссылки.
[0095] В одном из аспектов предоставлен эпитопсвязывающий белок, как описано в настоящем документе, где последовательности VL и VH человека получают у мыши, описываемой в настоящем документе, которую иммунизировали антигеном, содержащим представляющий интерес эпитоп.
[0096] В одном из вариантов осуществления предоставлен эпитопсвязывающий белок, содержащий первый и второй полипептид, где первый полипептид от N-конца до С-конца содержит первую эпитопсвязывающую область, селективно связывающуюся с первым эпитопом, с последующей константной областью, содержащей первую область СН3 IgG человека, выбранного из IgG1, IgG2, IgG4 и их сочетания; и, второй полипептид, содержащий от N-конца до С-конца вторую эпитопсвязывающую область, селективно связывающуюся со вторым эпитопом, с последующей константной областью, содержащей вторую область СН3 IgG человека, выбранного из IgG1, IgG2, IgG4 и их сочетания, где вторая область СН3 содержит модификации, уменьшающие или устраняющие связывание второго домена СН3 с белком А.
[0097] В одном из вариантов осуществления вторая область СНЗ содержит модификацию H95R (по нумерации экзонов IMGT; H435R по нумерации EU). В другом варианте осуществления вторая область СНЗ дополнительно содержит модификацию Y96F (IMGT; Y436F по EU).
[0098] В одном из вариантов осуществления вторая область СНЗ происходит из модифицированного IgGl человека и дополнительно содержит модификацию, выбранную из группы, состоящей из D16E, L18M, N44S, K52N, V57M и V82I (IMGT; D356E, L358M, N384S, K392N, V397M и V422I по EU).
[0099] В одном из вариантов осуществления вторая область СН3 происходит из модифицированного IgG2 человека и дополнительно содержит модификацию, выбранную из группы, состоящей из N44S, K52N и V82I (IMGT; N384S, K392N и V422I по EU).
[0100] В одном из вариантов осуществления вторая область СН3 происходит из модифицированного IgG4 человека и дополнительно содержит модификацию, выбранную из группы, состоящей из Q15R, N44S, K52N, V57M, R69K, E79Q и V82I (IMGT; Q355R, N384S, K392N, V397M, R409K, E419Q и V422I по EU).
[0101] Один из способов получения эпитопсвязывающего белка, связывающего более одного эпитопа, представляет собой иммунизацию первой мыши по изобретению антигеном, содержащим первый представляющий интерес эпитоп, где мышь несет эндогенный локус вариабельной области легкой цепи иммуноглобулина, не содержащий эндогенной VL мыши, способный к перегруппировке и формированию легкой цепи, где в эндогенном локусе вариабельной области легкой цепи иммуноглобулина мыши находится единственный генный сегмент VL человека, функционально связанный с эндогенным геном легкой цепи константной области мыши, и генный сегмент VL человека выбран из Vκ1-39Jκ5 человека и Vκ3-20Jκ1 человека, и эндогенные генные сегменты VH мыши целиком или полностью замещены генными сегментами VH человека так, что тяжелые цепи иммуноглобулинов, продуцируемые мышью целиком или в значительной степени, представляют собой тяжелые цепи, которые содержат вариабельные домены человека и константные домены мыши. При иммунизации такая мышь продуцирует обратное химерное антитело, содержащее только один из двух вариабельных доменов легких цепей человека (например, один из Vκ1-39Jκ5 человека или Vκ3-20Jκ1 человека). После идентификации В-клетки, кодирующей VH, связывающую представляющий интерес эпитоп, можно выделять (например, посредством ПЦР) нуклеотидную последовательность VH (и, необязательно, VL) и клонировать в экспрессирующую конструкцию в рамку с подходящим константным доменом иммуноглобулина человека. Этот способ можно повторять для идентификации второго домена VH, связывающего второй эпитоп, и можно выделять вторую генную последовательность VH и клонировать в экспрессирующий вектор в рамку со вторым подходящим константным доменом иммуноглобулина. Первый и второй константные домены иммуноглобулинов могут иметь один и тот же или различные изотипы, а один из константных доменов иммуноглобулинов (но не другой) можно модифицировать, как описано в настоящем документе или в US 2010/0331527 А1, и эпитопсвязывающий белок можно экспрессировать в подходящей клетке и выделять на основе его отличающейся аффинности к белку А по сравнению с гомодимерным эпитопсвязывающим белком, например, как описано в US 2010/0331527 А1.
[0102] В одном из вариантов осуществления предоставлен способ получения биспецифического эпитопсвязывающего белка, включающий идентификацию у мыши, как описано в настоящем документе, первой нуклеотидной последовательности VH человека (VH1) с созревшей аффинностью (например, содержащей одну или несколько соматических гипермутаций), идентификации у мыши, как описано в настоящем документе, второй нуклеотидной последовательности VH человека (VH2) с созревшей аффинностью (например, содержащей одну или несколько соматических гипермутаций), клонирование VH1 в рамку с тяжелой цепью человека с модификацией отсутствия детерминанты белка А, как описано в US 2010/0331527 А1, с формированием тяжелой цепи 1 (НС1), клонирование VH2 в рамку с тяжелой цепью человека, содержащей детерминанту белка А, как описано в US 2010/0331527 А1, с формированием тяжелой цепи 2 (НС2), введения экспрессирующего вектора, содержащего НС1, и того же или другого экспрессирующего вектора, содержащего НС2, в клетку, где клетка также экспрессирует легкую цепь иммуноглобулина человека, содержащую Vκ1-39 человека/Jκ5 человека или Vκ3-20 человека/JK1 человека, слитую с константным доменом легкой цепи человека, обеспечение экспрессии клеткой биспецифического эпитопсвязывающего белка, содержащего домен VH, кодируемый VH1, и домен VH, кодируемый VH2, и выделение биспецифического эпитопсвязывающего белка на основе его отличающейся способности связывать белок А по сравнению с моноспецифичным гомодимерным эпитопсвязывающим белком. В конкретном варианте осуществления НС1 представляет собой IgG1, а НС2 представляет собой IgG1, содержащие модификацию H95R (IMGT; H435R по EU) и дополнительно содержащие модификацию Y96F (IMGT; Y436F по EU). В одном из вариантов осуществления домен VH, кодируемый VH1, домен VH, кодируемый VH2, или оба являются соматически мутированными.
Гены VH человека, экспрессирующиеся с общей VL человека [0103] Экспрессировали множество вариабельных областей человека из антител с созревшей аффинностью, индуцированных к четырем различным антигенам, с их родственной легкой цепью или по меньшей мере с одной из легких цепей человека, выбранных из Vκ1-39Jκ5 человека, Vκ3-20Jκ1 человека или VpreBJλ5 человека (см. пример 1). Для антител к каждому из антигенов соматически мутированные высокоаффинные тяжелые цепи из различных семейств генов успешно спаривались с перегруппированными областями Vκ1-39Jκ5 и Vκ3-20Jκ1 зародышевой линии человека и секретировались из клеток, экспрессирующих тяжелые и легкие цепи. Для Vκ1-39Jκ5 и Vκ3-20Jκ1, благоприятно экспрессировались домены VH, получаемые из следующих семейств VH человека: 1-2, 1-8, 1-24, 2-5, 3-7, 3-9, 3-11,3-13, 3-15, 3-20, 3-23, 3-30, 3-33, 3-48, 4-31, 4-39, 4-59, 5-51 и 6-1. Таким образом, мышь, которую генно-инженерными способами получали для экспрессии ограниченного репертуара доменов VL человека с одной или обеими Vκ1-39Jκ5 и Vκ3-20Jκ1, продуцирует с локуса VH, модифицированного с заменой генных сегментов VH мыши, генными сегментами VH человека, разнообразное множество соматически мутированных доменов VH человека.
[0104] Мыши, полученные генно-инженерными способами для экспрессии обратных химерных (вариабельная область человека, константная область мыши) тяжелых цепей иммуноглобулинов, ассоциированных с единственной перегруппированной легкой цепью (например, Vκ1-39/J или a Vκ3-20/J), при иммунизации представляющим интерес антигеном продуцировали В-клетки, содержащие множество перегруппировок сегмента V человека, и экспрессировали множество высокоаффинных антигенспецифичных антител с разнообразными свойствами в отношении их способности блокировать связывание антигена с его лигандом и в отношении их способности связывать варианты антигена (см. примеры 5-10).
[0105] Таким образом, мыши и способы, описываемые в настоящем документе, пригодны для получения и отбора вариабельных доменов тяжелых цепей иммуноглобулинов человека, включая соматически мутированные вариабельные домены тяжелых цепей человека, которые являются результатом многообразия перегруппировок, которые демонстрируют широкий спектр аффинности (включая демонстрацию KD приблизительно в наномолярном диапазоне или менее), широкий спектр специфичности (включая связывание с различными эпитопами одного и того же антигена) и которые ассоциируют и экспрессируются с одной и той же или по существу с одной и той же вариабельной областью легкой цепи иммуноглобулина человека.
[0106] Приведенные ниже примеры приведены для описания специалистам в данной области того, как получать и использовать способы и композиции по изобретению, и они не предназначены для ограничения объема того, что авторы изобретения рассматривают как свое изобретение. Были предприняты усилия для обеспечения точности в отношении используемых чисел (например, количеств, температуры и т.д.), но следует учитывать некоторые экспериментальные ошибки и отклонения. Если не указано иначе, части представляют собой массовые части, молекулярная масса представляет собой среднюю молекулярную массу, температура указана в градусах Цельсия, а давление является атмосферным или близким к нему.
ПРИМЕРЫ
Пример 1. Идентификация вариабельных областей тяжелых цепей человека, ассоциирующих с выбранными вариабельными областями легких цепей человека
[0107] Для определения единственной перегруппированной легкой цепи зародышевой линии человека, способной коэспрессироваться с тяжелыми цепями человека из антигенспецифичных антител человека, генно-инженерными способами получали экспрессирующую систему in vitro.
[0108] Способы получения антител человека у генетически модифицированной мыши известны (см. например, US 6596541, Regeneron Pharmaceuticals, VELOCIMMUNE®). Технология VELOCIMMUNE® включает получение генетически модифицированной мыши с геномом, содержащим вариабельные области тяжелых и легких цепей человека, функционально связанные с эндогенными локусами константных областей мыши так, что мышь в ответ на стимуляцию антигеном продуцирует антитела, содержащие вариабельную область человека и константную область мыши. ДНК, кодирующая вариабельные области тяжелых и легких цепей антител, продуцируемых у мыши VELOCIMMUNE®, является полностью человеческой. На начальном этапе выделяют высокоаффинные химерные антитела с вариабельной областью человека и константной областью мыши. Как описано ниже, антитела характеризуют и отбирают по желательным характеристикам, включая аффинность, селективность, эпитоп и т.д. Константные области мыши замещают желаемой константной областью человека с получением полностью человеческого антитела, включающего изотип не-IgM, например, IgG1, IgG2, IgG3 или IgG4 дикого типа или модифицированные IgG1, IgG2, IgG3 или IgG4. Хотя выбранная константная область может варьировать в соответствии с конкретным применением, у вариабельной области остаются характеристики высокоаффинного связывания антигена и специфичности к мишени.
[0109] Мышь VELOCIMMUNE® иммунизировали фактором роста, стимулирующим ангиогенез (антиген С), и выделяли антигенспецифические антитела человека и стандартными способами, известными в данной области, секвенировали для определения использования гена V. Выбранные антитела клонировали на константные области тяжелых и легких цепей человека и выбирали 69 тяжелых цепей для спаривания с одной из трех легких цепей человека: (1) родственная легкая цепь к, связанная с константной областью к человека, (2) перегруппированная Vκ1-39Jκ5 зародышевой линии человека, связанная с константной областью к человека, или (3) перегруппированная Vκ3-20Jκ1 зародышевой линии человека, связанная с константной областью к человека. Каждую пару тяжелых цепей и легких цепей стандартными способами котрансфицировали в клетки СНО-К1. Присутствие антитела в супернатанте детектировали посредством антител к IgG человека в анализе ELISA. Для каждой пары тяжелая цепь/легкая цепь определяли титр антител (нг/мл) и титры, получаемые с различными перегруппированными легкими цепями зародышевой линии, сравнивали с титрами, получаемыми с исходной молекулой антитела (т.е., тяжелая цепь, спаренная с родственной легкой цепью), и рассчитывали процент от титра природных антител (таблица 1).
VH: ген вариабельной области тяжелой цепи. Н.д.: экспрессию в текущих экспериментальных условиях не детектировали.
[0110]

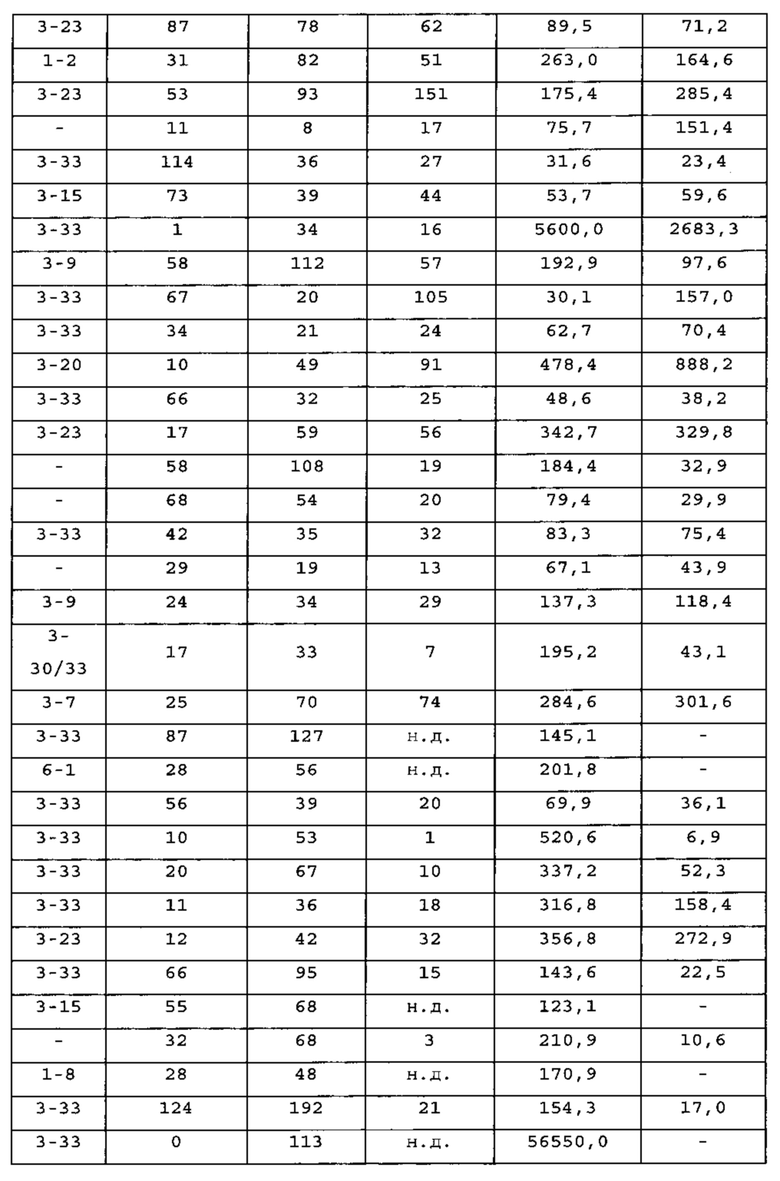
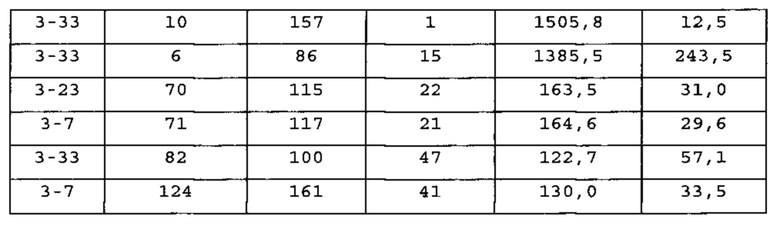
[0111] В сходном эксперименте мышей VELOCIMMUNE® иммунизировали несколькими различными антигенами, и выбранные тяжелые цепи антигенспецифичных антител человека тестировали на их способность спариваться с различными перегруппированными легкими цепями зародышевой линии человека (как описано выше). Антигены, используемые в этом эксперимент, включали фермент, вовлеченный в гомеостаз холестерина (антиген А), сывороточный гормон, вовлеченный в регуляцию гомеостаза глюкозы (антиген В), фактор роста, стимулирующий ангиогенез (антиген С) и клеточный поверхностный рецептор (антиген D). У мышей каждой иммунизированной группы выделяли антигенспецифичные антитела и вариабельные области тяжелых цепей и легких цепей клонировали и секвенировали. В последовательностях тяжелых и легких цепей определяли использование гена V и выбранные тяжелые цепи спаривали с их родственной легкой цепью или с перегруппированной областью Vκ1-39Jκ5 зародышевой линии человека. Каждую пару тяжелой/легкой цепей котрансфицировали в клетки СНО-К1 и присутствие антитела в супернатанте детектировали посредством антител к IgG человека в анализе ELISA. Для каждой пары тяжелой цепи/легкой цепи определяли титр антител (мкг/мл), и титры, получаемые с различными перегруппированными легкими цепями зародышевой линии человека, сравнивали с титрами, получаемыми с исходной молекулой антитела (т.е., тяжелой цепью, спаренной с родственной легкой цепью), и рассчитывали процент от титра природных антител (таблица 2). VH: ген вариабельной области тяжелой цепи. Vκ: ген вариабельной области легкой цепи к. Н.д.: экспрессию в текущих экспериментальных условиях не детектировали.
[0112]


[0113] Результаты, полученные в этих экспериментах, демонстрируют, что соматически мутированные, высокоаффинные тяжелые цепи из различных семейств генов способны спариваться с перегруппированными областями Vκ1-3 9Jκ5 и Vκ3-20Jκ1 зародышевой линии человека и секретироваться из клетки в виде нормальных молекул антител. Как продемонстрировано в таблице 1, приблизительно у 61% (42 из 69) тяжелых цепей при спаривании с перегруппированной Vκ1-39Jκ5 легкой цепи человека и приблизительно у 29% (20 из 69) тяжелых цепей при спаривании с перегруппированной Vκ3-20Jκ1 легкой цепи человека по сравнению с родственной легкой цепью исходного антитела увеличивался титр антител. Приблизительно у 20% (14 из 69) тяжелых цепей, обе перегруппированные легкие цепи зародышевой линии человека обеспечивают увеличение экспрессии по сравнению с родственной легкой цепью исходного антитела. Как продемонстрировано в таблице 2, перегруппированная область Vκ1-39Jκ5 зародышевой линии человека обеспечивала увеличение экспрессии нескольких тяжелых цепей, специфичных для диапазона различных классов антигенов, по сравнению с родственной легкой цепью исходных антител. Приблизительно у 35% (15/43) тяжелых цепей титр антител по сравнению с родственной легкой цепью исходных антител увеличивался более чем в два раза. У двух тяжелых цепей (315 и 316) увеличение по сравнению с исходным антителом было более чем в десять раз. Среди всех тяжелых цепей, продемонстрировавших увеличение экспрессии относительно родственной легкой цепи исходного антитела, третье семейство (VH3) тяжелых цепей по сравнению с другими семействами генов вариабельных областей тяжелых цепей является наиболее представленным. Это демонстрирует благоприятную зависимость VH3 тяжелых цепей человека в отношении спаривания с перегруппированными Vκ1-39Jκ5 и Vκ3-20Jκ1 легких цепей зародышевой линии человека.
Пример 2. Получение локуса перегруппированной легкой цепи зародышевой линии человека
[0114] С использованием технологии VELOCIGENE® получали различные направленные векторы с перегруппированными легкими цепями зародышевой линии человека (см., например, патент США №6586251 и Valenzuela et al. (2003) High-throughput engineering of the mouse genome coupled with high-resolution expression analysis, Nature Biotech. 21 (6): 652-659) для модификации клонов генома мыши в бактериальных искусственных хромосомах (ВАС) 302g12 и 254m04 (Invitrogen). С использованием этих двух клонов в ВАС получали геномные конструкции, содержащие одну область перегруппированной легкой цепи зародышевой линии человека, и встраивали в эндогенный локус легкой цепи κ, который предварительно модифицировали, удаляя эндогенные генные сегменты вариабельных областей и соединительные генные сегменты κ.
А. Конструирование направленных векторов с перегруппированными легкими цепями зародышевой линии человека
[0115] С использованием стандартных способов молекулярной биологии, известных в данной области, получали три различных перегруппированных области легких цепей зародышевой линии человека. Генные сегменты вариабельных областей человека, используемые для конструирования этих трех областей, включали последовательность перегруппированной Vκ1-39Jκ5 человека, последовательность перегруппированной Vκ3-20Jκ1 человека и последовательность перегруппированной VpreBJλ5 человека.
[0116] Посредством синтеза ДНК de novo (Integrated DNA Technologies) получали сегмент ДНК, содержащий экзон 1 (кодирующий лидерный пептид) и интрон 1 гена Vκ3-7 мыши. Включали часть 5'-нетранслируемой области до природного участка распознавания рестрикционного фермента BlpI. Из геномной библиотеки ВАС человека посредством ПЦР амплифицировали экзоны генов Vκ1-39 и Vκ3-20 человека. Прямые праймеры содержали дополнение 5'-конца, содержащее акцепторный участок сплайсинга интрона 1 гена Vκ3-7 мыши. Обратный праймер, используемый для ПЦР последовательности Vκ1-39 человека, содержал дополнение, кодирующее Jκ5 человека, тогда как обратный праймер, используемый для ПЦР последовательности Vκ3-20 человека, содержал дополнение, кодирующее Jκ1 человека. Посредством синтеза ДНК de novo (Integrated DNA Technologies) получали последовательность VpreBJA5 человека. Посредством ПЦР с плазмиды pBS-296-HA18-PISceI амплифицировали часть интрона Jk-Ck человека, включающую участок донора сплайсированного фрагмента. Прямой праймер ПЦР содержал дополнение, кодирующее часть последовательности Jκ5, Jκ1 или Jλ5 человека. Обратный праймер содержал участок PI-SceI, который предварительно посредством генно-инженерных способов встраивали в интрон.
[0117] Фрагменты экзона 1/интрона 1 Vκ3-7 мыши, экзонов вариабельной области легкой цепи человека и интрона Jk-Ck человека соединяли вместе посредством ПЦР с достройкой перекрывающихся участков, расщепляли BlpI и PI-SceI и лигировали в плазмиду pBS-296-HA18-PISceI, содержащую промотор из генного сегмента Vκ3-15 вариабельной области человека. Фланкированную участками lox кассету гигромицина в плазмиде pBS-296-HA18-PISceI замещали фланкированной FRT кассетой гигромицина, фланкированной участками NotI и AscI. Фрагмент NotI/PI-SceI этой плазмиды лигировали в модифицированную ВАС 254m04 мыши, содержащую часть интрона Jk-Ck мыши, экзон Ck мыши и приблизительно 75 т.п.н. геномной последовательности ниже локуса к мыши, обеспечивающих 3' участок гомологии для гомологической рекомбинации в ES клетки мыши. Затем фрагмент NotI/AscI этой ВАС лигировали в модифицированную ВАС 302g12 мыши, содержащую фланкированную FRT кассету неомицина и приблизительно 23 т.п.н. геномной последовательности выше эндогенного локуса к для гомологической рекомбинации в ES клетках мыши.
В. Направленный вектор с перегруппированной Vκ1-39Jκ5 зародышевой линии человека (фигура 1)
[0118] На 5'- и 3'-концах сконструированной вставки легкой цепи для клонирования в направленный вектор вводили участки распознавания рестрикционных ферментов: участок AscI на 5'-конце и участок PI-SceI на 3'-конце. В пределах 5' участка AscI и 3' участка PI-SceI направленная конструкция от 51 к 3' содержала 5' участок гомологии, содержащий последовательность с 5'-конца эндогенного локуса легкой цепи к мыши, полученную из клона ВАС 302g12 мыши, фланкированный FRT ген устойчивости к неомицину, геномную последовательность, содержащую промотор Vκ3-15 человека, лидерную последовательность генного сегмента вариабельной области Vκ3-7 мыши, интроную последовательность генного сегмента вариабельной области Vκ3-7 мыши, открытую рамку считывания перегруппированной области Vκ1-39Jκ5 зародышевой линии человека, геномную последовательность, содержащую часть интрона Jk-Ck человека и 3' участок гомологии, содержащий 3' последовательность эндогенного генного сегмента Jκ5 мыши, полученную из клона ВАС 254m04 мыши (фигура 1, середина). Гены и/или последовательности выше эндогенного локуса легкой цепи к мыши и ниже 3'-конца генного сегмента Jk (например, эндогенного 3' энхансера) в направленной конструкции оставляли немодифицированными (см. фигуру 1). Последовательность сконструированного локуса Vκ1-39Jκ5 человека представлена в SEQ ID NO: 1.
[0119] Направленную вставку перегруппированной области Vκ1-39Jκ5 зародышевой линии человека в ДНК ВАС подтверждали посредством полимеразной цепной реакции (ПЦР) с использованием праймеров, расположенных в последовательностях в перегруппированной области легкой цепи зародышевой линии человека. В кратком изложении последовательность интрона с 3'-конца лидерной последовательности Vκ3-7 мыши подтверждали с использованием праймеров ULC-m1F (AGGTGAGGGT ACAGATAAGT GTTATGAG; SEQ ID NO: 2) и ULC-m1R (TGACAAATGC CCTAATTATA GTGATCA; SEQ ID NO: 3). Открытую рамку считывания перегруппированной области Vκ1-39Jκ5 зародышевой линии человека подтверждали с использованием праймеров 1633-h2F (GGGCAAGTCA GAGCATTAGC A; SEQ ID NO: 4) и 1633-h2R (TGCAAACTGG ATGCAGCATA G; SEQ ID NO: 5). Кассету неомицина подтверждали с использованием праймеров neoF (GGTGGAGAGG CTATTCGGC; SEQ ID NO: 6) и neoR (GAACACGGCG GCATCAG; SEQ ID NO: 7). Затем ДНК ВАС со вставкой использовали для электропорации ES клеток мыши с получением модифицированных ES клеток для получения химерных мышей, экспрессирующих перегруппированные области Vκ1-39Jκ5 зародышевой линии человека.
[0120] Положительные клоны ES клеток подтверждали посредством скрининга TAQMAN™ и кариотипирования с использованием зондов, специфичных для сконструированной области Vκ1-39Jκ5 легкой цепи, встроенной в эндогенный локус. В кратком изложении зонд neoP (TGGGCACAAC AGACAATCGG CTG; SEQ ID NO: 8), связывающийся в области маркерного гена неомицина, зонд ULC-mlP (CCATTATGAT GCTCCATGCC TCTCTGTTC; SEQ ID NO: 9), связывающийся в области последовательности интрона с 3'-конца от лидерной последовательности Vκ3-7 мыши, и зонд 1633h2P (ATCAGCAGAA ACCAGGGAAA GCCCCT; SEQ ID NO: 10), связывающийся в области открытой рамки считывания перегруппированной Vκ1-39Jκ5 зародышевой линии человека. Затем положительные клоны ES клеток использовали для имплантации самкам мышей с получением выводка детенышей, экспрессирующих область Vκ1-39Jκ5 легкой цепи зародышевой линии.
[0121] Альтернативно, ES клетки, несущие перегруппированную область Vκ1-39Jκ5 легкой цепи зародышевой линии человека, трансфицируют конструкцией, экспрессирующей FLP для удаления фланкированной FRT кассеты неомицина, введенной направленной конструкцией. Необязательно, кассету неомицина удаляют, посредством скрещивания с мышами, экспрессирующими рекомбиназу FLP (например, US 6774279). Необязательно, кассету неомицина у мышей сохраняют.
С. Направленный вектор перегруппированной Vκ3-20Jκ1 зародышевой линии человека (фигура 2)
[0122] Подобным образом с использованием направленной конструкции, содержащей от 5'-конца к 3'-концу, 5' участок гомологии, содержащий последовательность с 5'-конца от эндогенного локуса легкой цепи к мыши, полученную из клона ВАС 302g12 мыши, фланкированный FRT ген устойчивости к неомицину, геномную последовательность, содержащую промотор Vκ3-15 человека, лидерную последовательность генного сегмента вариабельной области Vκ3-7 мыши, последовательность интрона генного сегмента вариабельной области Vκ3-7 мыши, открытую рамку считывания перегруппированной области Vκ3-20Jκ1 зародышевой линии человека, геномную последовательность, содержащую часть интрона Jk-Ck человека, и 3' участок гомологии, содержащий 3' последовательность эндогенного генного сегмента Jκ5 мыши, полученную из клона ВАС 254m04 мыши (фигура 2, середина), получали сконструированный локус легкой цепи, экспрессирующий перегруппированные области Vκ3-20Jκ1 зародышевой линии человека. Последовательность сконструированного локуса Vκ3-20Jκ1 человека представлена в SEQ ID NO: 11.
[0123] Направленную вставку перегруппированных областей Vκ3-20Jκ1 зародышевой линии человека в ДНК ВАС подтверждали посредством полимеразной цепной реакции (ПЦР) с использованием праймеров, расположенных в последовательностях в
перегруппированной области Vκ3-20Jκ1 легкой цепи зародышевой линии человека. В кратком изложении, последовательность интрона с 3'-конца от лидерной последовательности Vκ3-7 мыши подтверждали с использованием праймеров ULC-m1F (SEQ ID NO: 2) и ULC-m1R (SEQ ID NO: 3). Открытую рамку считывания перегруппированной области Vκ3-20Jκ1 зародышевой линии человека подтверждали с использованием праймеров 1635-h2F (TCCAGGCACC CTGTCTTTG; SEQ ID NO: 12) и 1635-h2R (AAGTAGCTGC TGCTAACACT CTGACT; SEQ ID NO: 13). Кассету неомицина подтверждали с использованием праймеров neoF (SEQ ID NO: 6) и neoR (SEQ ID NO: 7). Затем ДНК ВАС со вставкой использовали для электропорации ES клеток мыши с получением модифицированных ES клеток для получения химерных мышей, экспрессирующих перегруппированные области Vκ3-20Jκ1 легкой цепи зародышевой линии человека.
[0124] Положительные клоны ES клеток подтверждали посредством скрининга TAQMAN™ и кариотипирования с использованием зондов, специфичных для сконструированной области Vκ3-20Jκ1 легкой цепи, встроенной в эндогенный локус легкой цепи к. В кратком изложении зонд neoP (SEQ ID NO: 8), связывающийся в области маркерного гена неомицина, зонд ULC-m1P (SEQ ID NO: 9), связывающийся в области лидерной последовательности Vκ3-7 мыши, и зонд 1635h2P (AAAGAGCCAC CCTCTCCTGC AGGG; SEQ ID NO: 14), связывающийся в области открытой рамки считывания Vκ3-20Jκ1 человека. Затем положительные клоны ES клеток использовали для имплантации самкам мышей. Выводок детенышей экспрессирует область Vκ3-20Jκ1 легкой цепи зародышевой линии человека.
[0125] Альтернативно, ES клетки, несущие область Vκ3-20Jκ1 легкой цепи зародышевой линии человека, можно трансфицировать конструкцией, экспрессирующей FLP для удаления фланкированной FRT кассеты неомицина, введенной направленной конструкцией. Необязательно, кассету неомицина можно удалять, посредством скрещивания с мышами, экспрессирующими рекомбиназу FLP (например, US 6774279). Необязательно, кассету неомицина у мышей сохраняют.
D. Направленный вектор перегруппированной VpreBJX5 зародышевой линии человека (фигура 3)
[0126] Подобным образом с использованием направленной конструкции, содержащей от 5'-конца к 3'-концу, 5' участок гомологии, содержащий последовательность с 5'-конца от эндогенного локуса легкой цепи к мыши, полученную из клона ВАС 302g12 мыши, фланкированный FRT ген устойчивости к неомицину, геномную последовательность, содержащую промотор Vκ3-15 человека, лидерную последовательность генного сегмента вариабельной области Vκ3-7 мыши, последовательность интрона генного сегмента вариабельной области Vκ3-7 мыши, открытую рамку считывания перегруппированной области VpreBJλ5 зародышевой линии человека, геномную последовательность, содержащую часть интрона Jk-Ck человека, и 3' участок гомологии, содержащий 3' последовательность эндогенного генного сегмента Jκ5 мыши, полученную из клона ВАС 254m04 мыши (фигура 3, середина), получали сконструированный локус легкой цепи, экспрессирующий перегруппированные области VpreBJλ5 зародышевой линии человека. Последовательность сконструированного локуса VpreBJλ5 человека представлена в SEQ ID NO: 15.
[0127] Направленную вставку перегруппированных областей VpreBJλ5 зародышевой линии человека в ДНК ВАС подтверждали посредством полимеразной цепной реакции (ПЦР) с использованием праймеров, расположенных в последовательностях в перегруппированной области VpreBJλ5 легкой цепи зародышевой линии человека. В кратком изложении, последовательность интрона с 3'-конца от лидерной последовательности Vκ3-7 мыши подтверждали с использованием праймеров ULC-m1F (SEQ ID NO: 2) и ULC-m1R (SEQ ID NO: 3). Открытую рамку считывания перегруппированной области VpreBJλ5 зародышевой линии человека подтверждали с использованием праймеров 1616-h1F (TGTCCTCGGC CCTTGGA; SEQ ID NO: 16) и 1616-hlR (CCGATGTCAT GGTCGTTCCT; SEQ ID NO: 17). Кассету неомицина подтверждали с использованием праймеров neoF (SEQ ID NO: 6) и neoR (SEQ ID NO:7). Затем ДНК ВАС со вставкой использовали для электропорации ES клеток мыши с получением модифицированных ES клеток для получения химерных мышей, экспрессирующих перегруппированную VpreBJλ5 легкой цепи зародышевой линии человека.
[0128] Положительные клоны ES клеток подтверждали посредством скрининга TAQMAN™ и кариотипирования с использованием зондов, специфичных для сконструированной области VpreBJλ5 легкой цепи, встроенной в эндогенный локус легкой цепи κ. В кратком изложении зонд neoP (SEQ ID NO:8), связывающийся в области маркерного гена неомицина, зонд ULC-m1P (SEQ ID NO: 9), связывающийся в области лидерной последовательности IgVκ3-7 мыши, и зонд 1616h1P (ACAATCCGCC TCACCTGCAC ССТ; SEQ ID NO: 18), связывающийся в области открытой рамки считывания VpreBJλ5 человека. Затем положительные клоны ES клеток использовали для имплантации самкам мышей с получением выводка детенышей, экспрессирующих область легкой цепи зародышевой линии человека.
[0129] Альтернативно, ES клетки, несущие перегруппированную область VpreBJλ5 легкой цепи зародышевой линии человека трансфицируют конструкцией, экспрессирующей FLP для удаления фланкированной FRT кассеты неомицина, введенной направленной конструкцией. Необязательно, кассету неомицина удаляют, посредством скрещивания с мышами, экспрессирующими рекомбиназу FLP (например, US 6774279). Необязательно, кассету неомицина у мышей сохраняют.
Пример 3. Получение мышей, экспрессирующих единственную перегруппированную легкую цепь зародышевой линии человека
[0130] ES клетки-мишени, описанные выше, применяли в качестве ES клеток-доноров и вводили в эмбрион мыши на стадии 8-клеток способом VELOCIM0USE® (см., например, патент США №7294754 и Poueymirou et at. (2007) F0 generation mice that are essentially fully derived from the donor gene-targeted ES cells allowing immediate phenotypic analyses Nature Biotech. 25(1):91-99. Мышей VELOCIMICE®, независимо несущих сконструированную область Vκ1-39Jκ5 легкой цепи зародышевой линии человека, область Vκ3-20Jκ1 легкой цепи или область VpreBJλ5 легкой цепи, идентифицируют посредством генотипирования с использованием видоизмененного анализа аллелей (Valenzuela et al, выше), посредством которого детектируют присутствие уникальной перегруппированной области легкой цепи зародышевой линии человека.
[0131] Детенышей генотипируют, и детенышей, гетерозиготных по уникальной перегруппированной области легкой цепи зародышевой линии человека, выбирают для характеристики экспрессии перегруппированной области легкой цепи зародышевой линии человека.
Пример 4. Скрещивание мышей, экспрессирующих единственную перегруппированную легкую цепь зародышевой линии человека
А. Нокаут (КО) эндогенной Igλ
[0132] Для оптимизации использования сконструированного локуса легкой цепи мышей, несущих одну из перегруппированных областей легких цепей зародышевой линии человека, скрещивают с другими мышами, несущими делецию в эндогенном локусе легкой цепи λ. Таким образом, получаемое потомство в качестве своей единственной легкой цепи экспрессирует перегруппированную область легкой цепи зародышевой линии человека, как описано в примере 2. Скрещивание проводят стандартными способами, известными в данной области и, альтернативно, у коммерческих селекционеров (например, The Jackson Laboratory). Линии мышей, несущие сконструированный локус легкой цепи и делецию эндогенного локуса легкой цепи λ, подвергают скринингу на присутствие уникальной области легкой цепи и отсутствие эндогенных легких цепей λ мыши.
В. Гуманизированный локус эндогенной тяжелой цепи
[0133] Мышей, несущих сконструированный локус легкой цепи зародышевой линии человека, скрещивают с мышами, несущими замену эндогенного локуса гена вариабельной области тяжелой цепи мыши локусом гена вариабельной области тяжелой цепи человека (см. US 6596541; мышь VELOCIMMUNE®, Regeneron Pharmaceuticals, Inc.). Мышь VELOCIMMUNE® несет геном, содержащий вариабельные области тяжелых цепей человека, функционально связанные с эндогенными локусами константных областей мыши так, что мышь в ответ на стимуляцию антигеном продуцирует антитела, содержащие вариабельную область тяжелой цепи человека и константную область тяжелой цепи мыши. ДНК, кодирующую вариабельные области тяжелых цепей антител, выделяют и функционально связывают с ДНК, кодирующей константные области тяжелых цепей человека. Затем ДНК экспрессируют в клетке, способной экспрессировать полностью человеческую тяжелую цепь антитела.
[0134] Получают мышей, несущих замену эндогенного локуса VH мыши локусом VH человека и в эндогенном локусе легкой цепи к единственную перегруппированную область VL зародышевой линии человека. После иммунизации представляющим интерес антигеном получают обратные химерные антитела, содержащие соматически мутированные тяжелые цепи (VH человека и СН мыши) с единственной легкой цепью человека (VL человека и CL мыши). Идентифицируют нуклеотидные последовательности VH и VL В-клеток, экспрессирующих антитела, и получают полностью человеческие антитела посредством слияния нуклеотидных последовательностей VH и VL с нуклеотидными последовательностями СН и CL человека в подходящей экспрессирующей системе.
Пример 5. Получение антител у мышей, экспрессирующих тяжелые цепи человека и перегруппированную область легкой цепи зародышевой линии
[0135] После скрещивания мышей, несущих сконструированную область легкой цепи человека с различными желаемыми линиями, несущими модификации и делеции других эндогенных локусов Ig (как описано в примере 4), отобранных мышей можно иммунизировать представляющим интерес антигеном.
[0136] Как правило, мышей VELOCIMMUNE®, несущих одну из единственных перегруппированных областей легких цепей зародышевой линии человека, стимулируют антигеном, и из сыворотки животных выделяют лимфоциты (такие как В-клетки). Лимфоциты сливают с миеломной клеточной линией с получением бессмертных гибридомных клеточных линий, и такие гибридомные клеточные линии скринируют и отбирают для идентификации гибридомных клеточных линий, продуцирующих антитела, содержащие вариабельные области тяжелых цепей человека и перегруппированные легкие цепи зародышевой линии человека, которые специфичны к антигену, используемому для иммунизации. Выделяют ДНК, кодирующую вариабельные области тяжелых цепей и легких цепей, и связывают с желаемыми изотипическими константными областями тяжелой цепи и легкой цепи. Вследствие присутствия эндогенных последовательностей мыши и любых дополнительных действующих в цис-положении элементов, присутствующих в эндогенном локусе, единственная легкая цепь каждого антитела может быть соматически мутированной. Это добавляет дополнительное разнообразие антигенспецифического репертуара, включающего единственную легкую цепь и различные последовательности тяжелых цепей. Затем полученные клонированные последовательности антител экспрессируют в клетке, такой как клетка СНО. Альтернативно, ДНК, кодирующие антигенспецифические химерные антитела или вариабельные домены легких и тяжелых цепей, напрямую идентифицируют в антигенспецифических лимфоцитах.
[0137] Сначала выделяют высокоаффинные химерные антитела, содержащие вариабельную область человека и константную область мыши. Как описано выше, антитела характеризуют и отбирают по желаемым характеристикам, включающим аффинность, селективность, эпитоп и т.д. Константные области мыши замещают желаемой константной областью человека с получением полностью человеческого антитела, содержащего соматически мутированную тяжелую цепь человека и единственную легкую цепь, получаемую из перегруппированной области легкой цепи зародышевой линии человека по изобретению. Подходящие константные области человека включают, например, IgG1 или IgG4 дикого типа или модифицированные IgG1 или IgG4.
[0138] Отдельные группы мышей VELOCIMMUNE®, несущих замену эндогенного локуса тяжелой цепи мыши генными сегментами V, D и J человека и замену эндогенного локуса легкой цепи к мыши сконструированной областью Vκ1-39Jκ5 легкой цепи зародышевой линии человека или сконструированной областью Vκ3-20Jκ1 легкой цепи зародышевой линии человека (описанных выше), иммунизировали белком клеточного поверхностного рецептора человека (антиген Е). Антиген Ε вводят непосредственно в подушечку задней лапы мыши посредством шести последовательных инъекций каждые 3-4 суток. Перед инъекцией от двух до трех микрограмм антигена Ε смешивают с 10 мкг олигонуклеотида CpG (каталожный № tlrl-modn - олигонуклеотид ODN1826; InVivogen, San Diego, СА) и 25 мкг Adju-Phos (гелевый адъювант с фосфатом алюминия, кат. № Н-71639-250; Brenntag Biosector, Frederikssund, Denmark). Перед конечной стимуляцией антигеном, которую проводят за 3-5 суток перед умерщвлением, проводят всего шесть инъекций. После 4 и 6 инъекций получают образцы крови и иммунный ответ антителами подвергают мониторингу стандартным антигенспецифическим иммунологическим анализом.
[0139] Когда достигается желаемый иммунный ответ, собирают спленоциты и сливают с миеломными клетками мыши для сохранения их жизнеспособности и формирования гибридомных клеточных линий. Гибридомные клеточные линии подвергают скринингу и отбору с идентификацией клеточных линий, продуцирующих специфичные к антигену Ε антитела с общей легкой цепью. Этим способом получают несколько специфичных к антигену Ε антител с общей легкой цепью (т.е., антител, содержащих вариабельные домены тяжелых цепей человека, такой же вариабельный домен легкой цепи человека и константные домены мыши).
[0140] Альтернативно, антитела с общей легкой цепью к антигену Ε выделяют прямо из положительных к антигену В-клеток без слияния с миеломными клетками, как описано в U.S. 2007/0280945 А1, конкретно полностью включенном в настоящий документ в качестве ссылки. Этим способом получали несколько полностью человеческих антител к антигену Ε с общей легкой цепью (т.е., антитела, содержащие вариабельные домены тяжелых цепей человека, сконструированную область Vκ1-39Jκ5 легкой цепи человека или сконструированную область Vκ3-20Jκ1 легкой цепи человека и константные домены человека).
[0141] Биологические свойства иллюстративных антител с общей легкой цепью к антигену Е, получаемых способами этого примера, подробно описаны в разделах, приведенных ниже.
Пример 6. Использование генных сегментов тяжелых цепей в антигенспецифических антителах с общей леркой цепью
[0142] Для анализа структуры продуцируемых антител человека к антигену Ε с общей легкой цепью клонировали и секвенировали нуклеиновые кислоты, кодирующие вариабельные области тяжелых цепей антител. На основании последовательностей нуклеиновых кислот и теоретически рассчитанных последовательностей аминокислот антител для вариабельных областей тяжелых цепей (HCVR) выбранных антител с общей легкой цепью, получаемых у иммунизированных мышей VELOCIMMUNE®, несущих сконструированную область Vκ1-39Jκ5 легкой цепи человека или сконструированную область Vκ3-20Jκ1 легкой цепи человека, определяли использование генов. Результаты приведены в таблицах 3 и 4, в которых показано, что мыши по изобретению из множества генных сегментов тяжелых цепей человека, вследствие множества перегруппировок, при использовании мыши, экспрессирующей легкую цепь только из полученных из Vκ1-39 человека или Vκ3-20 человека легких цепей, продуцируют антигенспецифические антитела с общей легкой цепью. Генные сегменты VH человека семейств 2, 3, 4 и 5 перегруппировывали с множеством сегментов DH человека и сегментов JH человека с получением антигенспецифических антител.
[0143]
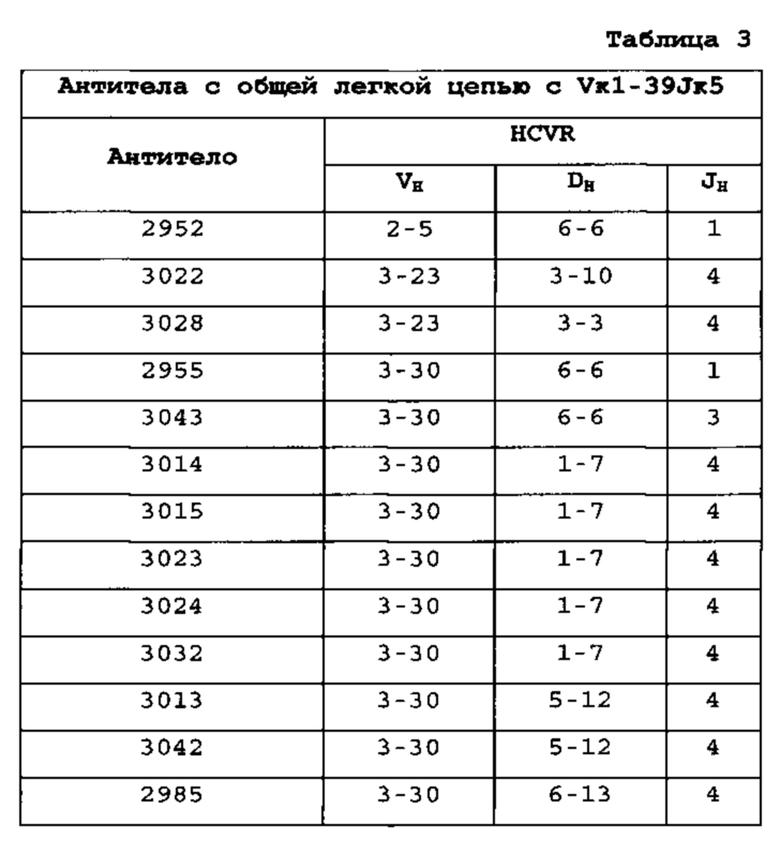
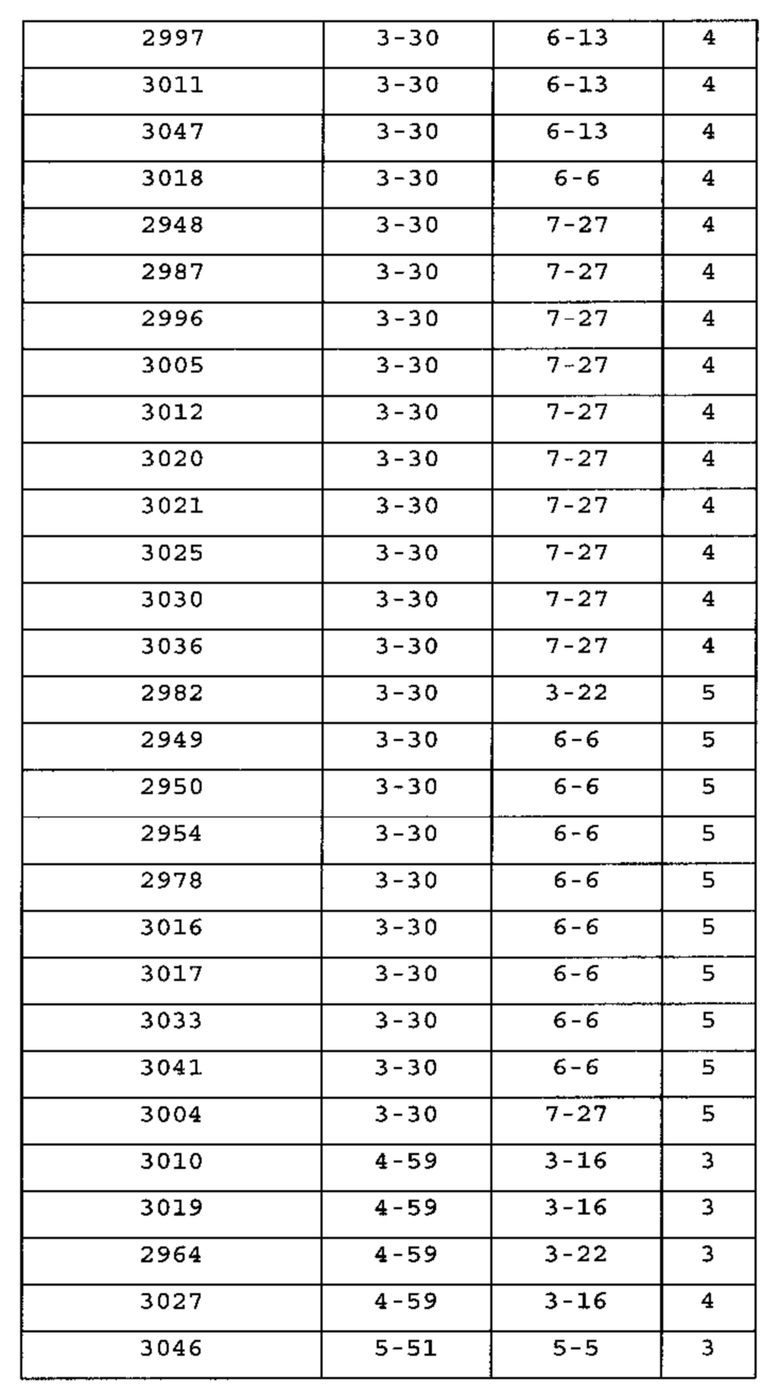
[0144]
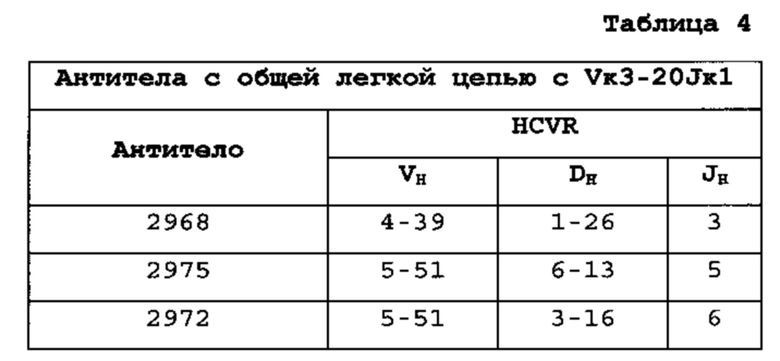
Пример 7. Определение блокирующей способности антигенспецифических антител с общей легкой цепью посредством анализа Luminex™
[0145] Девяносто восемь антител человека с общей легкой цепью, индуцированных антигеном Е, тестировали на их способность блокировать связывание природного лиганда антигена Е (лиганда Y) с антигеном Е в анализе с гранулами.
[0146] Внеклеточный домен (ECD) антигена Е конъюгировали с двумя эпитопными метками туе и меткой из 6Х гистидинов (антиген E-mmH) и в буфере MES посредством аминов связывали с карбоксилированными микросферами в концентрации 20 мкг/мл. Смесь инкубировали в течение двух часов при комнатной температуре с последующей дезактивацией гранул 1М Tris рН 8,0 с последующей отмывкой в PBS 0,05% (об./об.) Tween-20. Затем гранулы блокировали PBS (Irvine Scientific, Santa Ana, CA), содержащим 2% (масс./об.) BSA (Sigma-Aldrich Corp., St. Louis, МО). В 96-луночном планшете для фильтрования супернатанты, содержащие специфичные к антигену Е антитела с общей легкой цепью, разбавляли в буфере 1:15. Получали отрицательный контроль, содержащий пустой супернатант с теми же компонентами среды, что и супернатант с антителами. К супернатантам добавляли меченные антигеном Ε гранулы и инкубировали в течение ночи при 4°С. Добавляли биотинилированный белок лиганда Y до конечной концентрации 0,06 нМ и инкубировали в течение двух часов при комнатной температуре. Детекцию биотинилированного лиганда Y, связанного с меченными антигеном E-myc-myc-6His гранулами проводили с использованием R-фикоэритрина, конъюгированного со стрептавидином (Moss Inc, Pasadena, MD) с последующим измерением в анализаторе на основе проточной цитометрии Luminex™. Из всех образцов вычитали фоновую среднюю интенсивность флуоресценции (MFI) образца без лиганда Y. Процент блокирования рассчитывали, деля MFI каждого образца после вычитания фона на скорректированное значение отрицательного контроля, умножая на 100 и вычитая полученное значение из 100.
[0147] В сходном эксперименте те же 98 антител человека с общей легкой цепью, индуцированных антигеном Е, тестировали на их способность блокировать связывание антигена Ε с меченными лигандом Y гранулами.
[0148] В кратком изложении, лиганд Y посредством аминов амин связывали с карбоксилированными микросферами в концентрации 20 мкг/мл, разбавленной в буфере MES. Смесь инкубировали два часа при комнатной температуре с последующей дезактивацией гранул 1М Tris рН 8 с последующей отмывкой PBS с 0,05% (об./об.) Tween-20. Затем гранулы блокировали PBS (Irvine Scientific, Santa Ana, CA), содержащим 2% (масс./об.) BSA (Sigma-Aldrich Corp., St. Louis, MO). В 96-луночном планшете для фильтрования, супернатанты, содержащие специфичные к антигену Ε антитела с общей легкой цепью, разбавляли 1:15 в буфере. Получали отрицательный контроль, содержащий пустой супернатант с теми же компонентами среды, что и супернатант с антителами. Добавляли биотинилированный антиген E-mmH до конечной концентрации 0,42 нМ и инкубировали в течение ночи при 4°С. Затем к смеси антитело/антиген Ε добавляли меченные лигандом Y гранулы и инкубировали в течение двух часов при комнатной температуре. Детекцию биотинилированного антигена E-mmH, связанного с гранулами с лигандом Y, проводили с R-фикоэритрином, конъюгированным со стрептавидином (Moss Inc, Pasadena, MD), с последующим измерением в анализаторе на основе проточной цитометрии Luminex™. Из всех образцов вычитали фоновую среднюю интенсивность флуоресценции (MFI) образца без антигена Е. Процент блокирования рассчитывали, деля MFI каждого образца после вычитания фона на скорректированное значение отрицательного контроля, умножая на 100 и вычитая полученное значение из 100.
[0149] В таблицах 5 и 6 приведен процент блокирования для всех 98 антител к антигену Ε с общей легкой цепью, тестируемых в обоих анализах Luminex™. н.д.: в текущих экспериментальных условиях не детектировали.
[0150] В первом эксперименте Luminex™, описанном выше, 80 антител с общей легкой цепью, содержащих сконструированную с Vκ1-39Jκ5 легкую цепь, тестировали на их способность блокировать связывание лиганда Y с меченными антигеном Ε гранулами. Из этих 80 антител с общей легкой цепью, 68 продемонстрировали блокирование >50%, тогда как 12 продемонстрировали блокирование <50% (6 с блокированием 25-50% и 6 с блокированием <25%). Среди 18 антител с общей легкой цепью, содержащих сконструированную с Vκ3-20Jκ1 легкую цепь, 12 продемонстрировали блокирование связывания лиганда Y с меченными антигеном Ε гранулами >50%, тогда как 6 продемонстрировали блокирование связывания лиганда Y с меченными антигеном Ε гранулами <50% (3 с блокированием 25-50% и 3 с блокированием <25%).
[0151] Во втором эксперименте Luminex™, описанном выше, те же 80 антител с общей легкой цепью, содержащих сконструированную с Vκ1-39Jκ5 легкую цепь, тестировали на их способность блокировать связывание антигена Ε с меченными лигандом Y гранулами. Из этих 80 антител с общей легкой цепью, 36 продемонстрировали блокирование >50%, тогда как 44 продемонстрировали блокирование <50% (27 с блокированием 25-50% и 17 с блокированием <25%). Среди 18 антител с общей легкой цепью, содержащих сконструированную с Vκ3-20Jκ1 легкую цепь, 1 продемонстрировало блокирование связывания антигена Ε с меченными лигандом Y гранулами >50%, тогда как 17 продемонстрировали блокирование связывания антигена Ε с меченными лигандом Y гранулами <50% (5 с блокированием 25-50% и 12 с блокированием <25%).
[0152] Данные таблиц 5 и 6 подтверждают, что при перегруппировках, описанных в таблицах 3 и 4, получали специфичные к антигену Ε антитела с общей легкой цепью, которые с различными степенями эффективности блокировали связывание лиганда Y с узнающим его рецептором антигеном Е, что согласуется с тем, что антитела к антигену Ε с общей легкой цепью из таблиц 3 и 4 содержат антитела с перекрывающейся и неперекрывающейся специфичностью к эпитопу в отношении антигена Е.
[0153]


[0154]
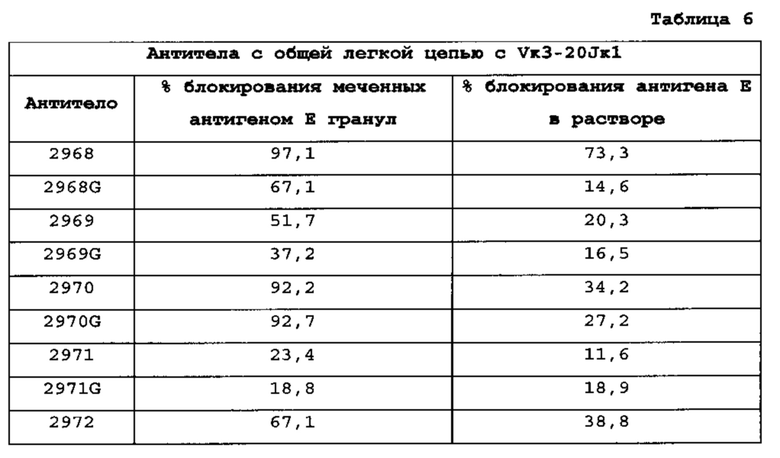

Пример 8. Определение блокирующей способности антигенаспецифических антител с общей легкой цепью посредством ELISA
[0155] Антитела человека с общей легкой цепью, индуцированные антигеном Е, тестировали на их способность блокировать связывание антигена Е с покрытой лигандом Y поверхностью в анализе ELISA.
[0156] Лиганд Y наносили на 96-луночные планшеты в концентрации 2 мкг/мл в PBS и инкубировали в течение ночи с последующей отмывкой четыре раза в PBS с 0,05% Tween-20. Затем планшет в течение одного часа при комнатной температуре блокировали PBS (Irvine Scientific, Santa Ana, CA), содержащим 0,5% (масс./об.) BSA (Sigma-Aldrich Corp., St. Louis, МО). В отдельном планшеты в буфере 1:10 разбавляли супернатанты, содержащие антитела к антигену Е с общей легкой цепью. В качестве отрицательного контроля использовали пустой супернатант с теми же компонентами антител. Добавляли антиген E-mmH (описанный выше) до конечной концентрации 0,150 нМ и инкубировали в течение одного часа при комнатной температуре. Затем к планшету, содержащему лиганд Y, добавляли смесь антитело/антиген E-mmH и инкубировали в течение одного часа при комнатной температуре. Детекцию антигена E-mmH, связанного с лигандом Y, проводили с использованием пероксидазы хрена (HRP), конъюгированной с антителом к Penta-His (Qiagen, Valencia, СА), и развитие окрашивания проводили посредством стандартного колориметрического ответа с использованием субстрата тетраметилбензидина (ТМВ) (BD Biosciences, San Jose, СА), нейтрализуемого серной кислотой. Оптическую плотность определяли при OD450 в течение 0,1 сек. Из всех образцов вычитали Фоновую оптическую плотность образца без антигена Е. Процент блокирования рассчитывали, деля MFI каждого образца после вычитания фона на скорректированное значение отрицательного контроля, умножая на 100 и вычитая полученное значение из 100.
[0157] В таблицах 7 и 8 приведен процент блокирования для всех 98 антител к антигену Ε с общей легкой цепью, тестируемых в анализе ELISA, н.д.: в текущих экспериментальных условиях не детектировали.
[0158] Как описано в этом примере, из 80 антител с общей легкой цепью, содержащих сконструированную с Vκ1-39Jκ5 легкую цепь, тестируемых на их способность блокировать связывание антигена Ε с покрытой лигандом Y поверхностью, 22 продемонстрировали блокирование >50%, тогда как 58 продемонстрировали блокирование <50% (20 с блокированием 25-50% и 38 с блокированием <25%). Для 18 антител с общей легкой цепью, содержащих сконструированную с Vκ3-20Jκ1 легкую цепь, 1 продемонстрировало блокирование связывания антигена Ε с покрытой лигандом Y поверхностью>50%, тогда как 17 продемонстрировали блокирование связывания антигена Е с покрытой лигандом Y поверхностью <50% (5 с блокированием 25-50% и 12 с блокированием <25%).
[0159] Эти результаты также согласуются с тем, что совокупность специфичных к антигену Е антител с общей легкой цепью содержит антитела с перекрывающейся и неперекрывающейся специфичностью к эпитопу в отношении антигена Е.
[0160]




Пример 9. Определение аффинности на BIAcore™ для антигенспецифических антител с общей легкой цепью
[0162] Для супернатантов выбранных антител посредством SPR (поверхностного плазмонного резонанса) с использованием устройства BIAcore™ Т100 (GE Healthcare) определяли равновесные константы диссоциации (KD). Все данные получали при 25°С с использованием HBS-EP (10 мМ Hepes, 150 мМ NaCl, 0,3 мМ ЭДТА, 0,05% поверхностно-активного вещества Р20, рН 7,4) в качестве буферов для запуска и образцов. Антитела захватывали из образцов неочищенных супернатантов на поверхности сенсорного чипа СМ5, предварительно дериватизированной с высокой плотностью антителами к Fc человека с использованием стандартной химической реакции связывания по аминам. На этапе захвата супернатанты впрыскивали параллельно поверхности с антителом к Fc человека при скорости потока 3 мкл/мин всего в течение 3 минут. После этапа захвата проводили впрыск буфера для запуска или анализируемого вещества при концентрации 100 нМ в течение 2 минут при скорости потока 35 мкл/мин. Диссоциацию антигена и захваченного антитела подвергали мониторингу в течение 6 минут. Захваченное антитело удаляли посредством кратковременного впрыска 10 мМ глицина, рН 1,5. Для всех сенсограмм проводили двойную коррекцию посредством вычитания сенсограмм после впрыска буфера из сенсограмм с анализируемым веществом, таким образом, удаляя артефакты, обусловленные диссоциацией антител с захватывающей поверхностью. Данные связывания для каждого антитела аппроксимировали по модели связывания 1:1 с переносом масс с использованием программного обеспечения BIAcore Т100 Evaluation software версии 2.1. Результаты представлены в таблицах 9 и 10.
[0163] Аффинности связывания антител с общей легкой цепью, содержащих перегруппировки, представленные в таблицах 3 и 4, варьируют, где почти все демонстрируют KD в наномолярном диапазоне. Данные аффинности соответствуют антителам с общей легкой цепью, являющихся результатом комбинаторной ассоциации перегруппированных вариабельных доменов, описанных в таблицах 3 и 4, являющихся высокоаффинными, клонально отобранными и соматически мутированными. В сочетании с ранее представленными данными антитела с общей легкой цепью, описанные в таблицах 3 и 4, составляют набор разнообразных, высокоаффинных антител, демонстрирующих специфичность к одному или нескольким эпитопам на антигене Е.



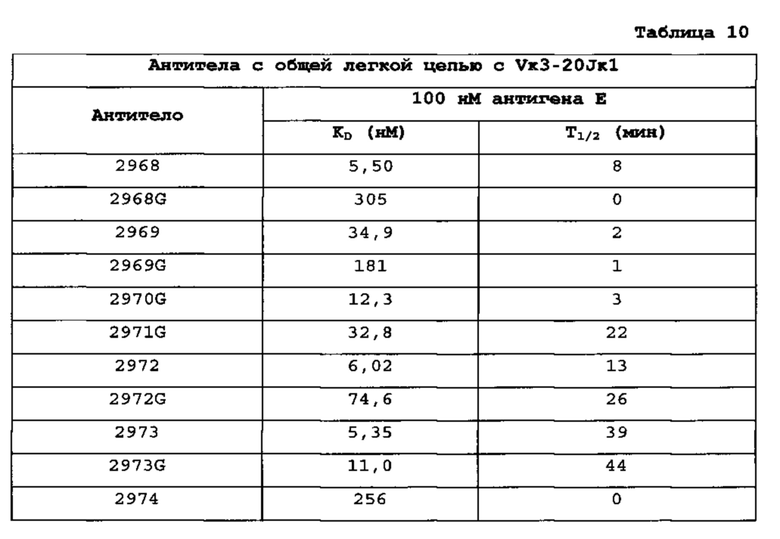

Пример 10. Определение специфичности связывания антигенаспецифических антител с общей легкой цепью посредством анализа Luminex™
[0166] Отобранные антитела к антигену Е с общей легкой цепью тестировали на их способность связываться с ECD антигена Е и вариантами ECD антигена Е, включая ортолог яванского макака (антиген Е Mf), отличающийся от белка человек приблизительно по 10% его аминокислотных остатков; делеционного мутанта антигена Е с отсутствием последних 10 аминокислот на С-конце ECD (антиген Е-ДСТ) и двух мутантов, содержащих замену предполагаемых положений взаимодействия с лигандом Y на аланин (антиген Е-Ala1 и антиген Е-Ala2). Белки антигена Е продуцировали в клетках СНО и каждый из них содержал С-концевую метку myc-myc-His.
[0167] Для исследований связывания белок ECD антигена Е или вариант белка (описанный выше) из 1 мл среды для культивирования захватывали посредством инкубации в течение 2 часов при комнатной температуре с 1×106 микросферических (Luminex™) гранул, ковалентно покрытых моноклональным антителом к myc (MAb 9Е10, гибридомная клеточная линия CRL-1729™; АТСС, Manassas, VA). Затем гранулы перед использованием отмывали PBS. Супернатанты, содержащие антитела к антигену Е с общей легкой цепью, разбавляли 1:4 в буфере и добавляли в 96-луночные планшеты для фильтрования. В качестве отрицательного контроля использовали пустой супернатант без антитела. Затем гранулы, содержащие захваченные белки антигена Е, добавляли в образцы антител (3000 гранул на лунку) и инкубировали в течение ночи при 4°С. На следующие сутки гранулы с образцами отмывали и связывание антител с общей легкой цепью детектировали с использованием конъюгированного с R-фикоэритрином антитела к IgG человека. Интенсивность флуоресценции гранул (для каждого связывания образца антитела с каждым белком антигена Е учитывали приблизительно 100 гранул) измеряли анализатором на основе проточной цитометрии Luminex™ и регистрировали среднюю интенсивность флуоресценции (MFI) по меньшей мере для 100 учитываемых гранул для взаимодействия гранула/антитело. Результаты приведены в таблицах 11 и 12.
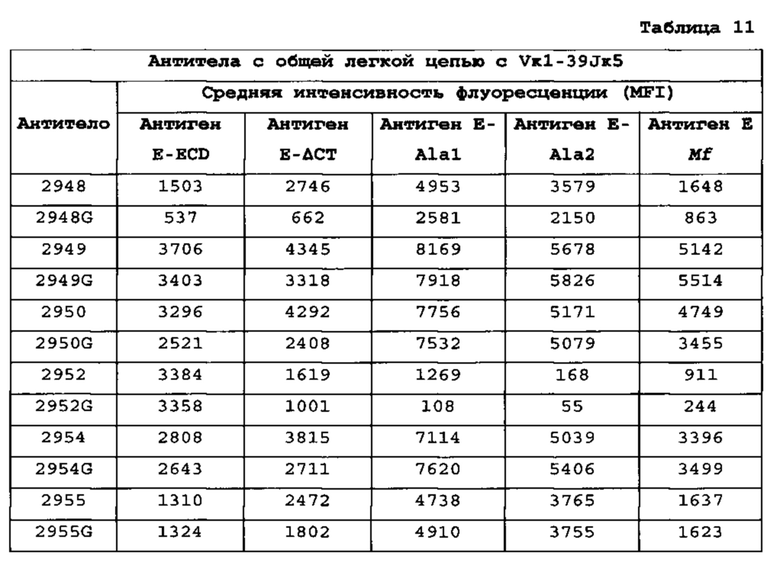
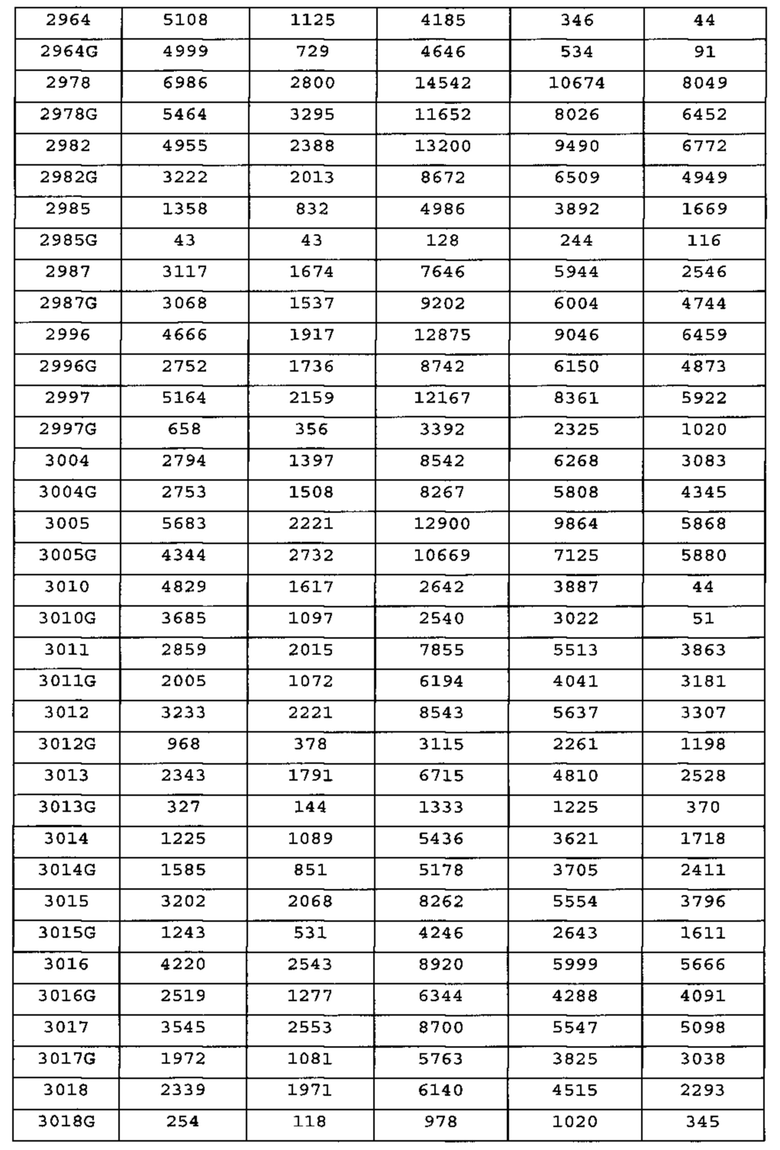

[0169]
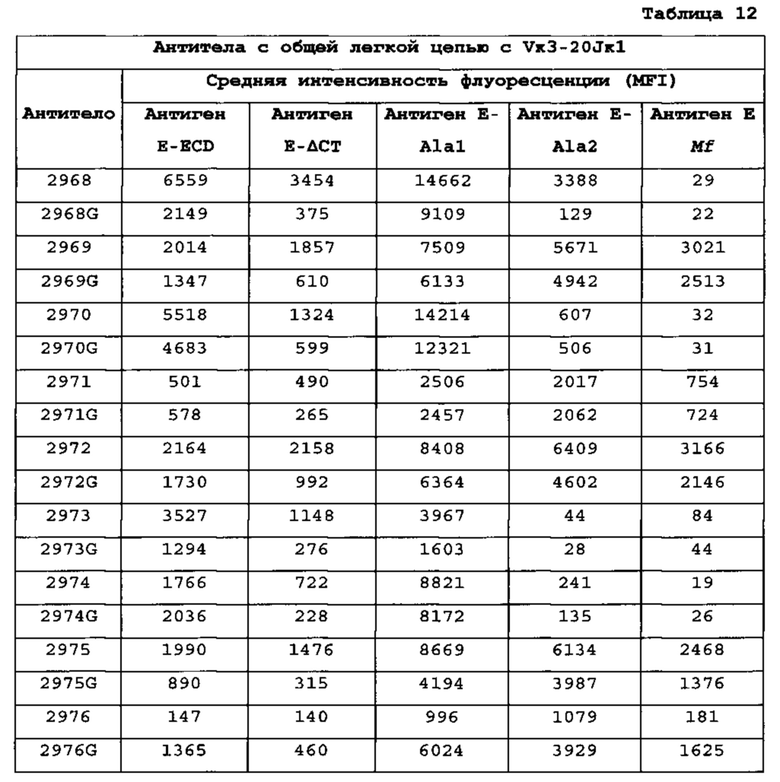
[0170] Супернатанты с антителом с общей легкой цепью к антигену Е демонстрировали высокоспецифическое связывание с гранулами, связанными с антигеном E-ECD. Для этих гранул пустой супернатант отрицательного контроля давал пренебрежимо малый сигнал (<10 MFI) при комбинировании с образцом гранул с антигеном E-ECD, тогда как супернатанты, содержащие антитела к антигену Е с общей легкой цепью, демонстрировали сильный сигнал связывания (средняя MFI для супернатантов 98 антител 2627; MFI для 91/98 образцов антител >500).
[0171] В качестве меры способности выбранных антител к антигену Ε с общей легкой цепью идентифицировать различные эпитопы на ECD антигена Е, определяли относительное связывание антител с вариантами. Все четыре варианта антигена Ε связывали с гранулами Luminex™ с антителом к туе, как описано выше для исследований связывания природного антигена E-ECD, и определяли относительные отношения связывания (MFIвариант/MFIантиген E-ECD). Для супернатантов 98 тестируемых антител с общей легкой цепью, представленных в таблицах 11 и 12, средние отношения (MFIвариант/MFIантиген E-ECD) отличились для каждого варианта, возможно отражая различные захватываемые на гранулах количества белков (средние отношения для антигена Е-ΔСТ, антигена Ε-Ala1, антигена Е-Ala2 и антигена Ε Mf 0,61, 2,9, 2,0 и 1,0, соответственно). Для каждого варианта белка, связывание с подмножеством 98 тестируемых антител с общей легкой цепью продемонстрировало существенно сниженное связывание, указывающее на чувствительность к мутации, характеризующей данный вариант. Например, 19 из образцов антител с общей легкой цепью связывались с антигеном Ε Mf с МFIвариант/МFIантиген E-ECD <8%. Так как значительное количество в этой группе включает очень или умеренно высокоаффинные антитела (5 с KD<5 нМ, 15 с KD<50 нМ), вероятно, что наименьший сигнал для этой группы является результатом чувствительности к различиям последовательностей (эпитопов) между природным антигеном E-ECD и данным вариантом, а не результатом меньшей аффинности.
[0172] Эти данные подтверждают, что антитела с общей легкой цепью, описанные в таблицах 3 и 4, фактически представляют группу разнообразных специфичных к антигену Е антител с общей легкой цепью, которые специфически распознают на антигене Е более одного эпитопа.
В частных вариантах осуществления настоящее изобретение может относиться к следующим объектам:
1. Мышь, несущая:
(a) замену в эндогенном локусе вариабельной области легкой цепи к иммуноглобулина мыши всех или по существу всех эндогенных генных сегментов вариабельной области легкой цепи к иммуноглобулина мыши генным сегментом Vκ1-39/J человека, генным сегментом Vκ3-20/J человека или их сочетанием, где генный сегмент человека функционально связан с эндогенным геном константной области к мыши; и,
(b) замену всего или по существу всего эндогенного локуса гена вариабельной области тяжелой цепи мыши множеством генных сегментов вариабельных областей тяжелых цепей человека, где генные сегменты вариабельных областей тяжелых цепей человека функционально связаны с эндогенным геном константной области тяжелой цепи мыши, и генные сегменты вариабельных областей тяжелых цепей человека способны к перегруппировке и формированию перегруппированного гена химерной тяжелой цепи человека/мыши.
2. Мышь по п. 1, где у мыши отсутствует эндогенный локус вариабельной области легкой цепи к иммуноглобулина мыши, способный к перегруппировке и формированию гена, кодирующего вариабельную область к мыши.
3. Мышь по п. 1, дополнительно несущая интронный энхансер к мыши с 5'-конца относительно константной область легкой цепи мыши.
4. Мышь по п. 1, дополнительно несущая 3'-энхансер к мыши.
5. Мышь по п. 1, где множество генных сегментов вариабельных областей тяжелых цепей человека содержит сегмент, выбранный из генных сегментов вариабельной области иммуноглобулина человека 1-2, 1-8, 1-24, 2-5, 3-7, 3-9, 3-11, 3-13, 3-15, 3-20, 3-23, 3-30, 3-33, 3-48, 4-31, 4-39, 4-59, 5-51, 6-1.
6. Мышь по п. 1, где множество генных сегментов вариабельных областей тяжелых цепей человека содержит сегмент, выбранный из генных сегментов D1-7, D1-26, D3-3, D3-10, D3-16, D3-22, D5-5, D5-12, D6-6, D6-13, D7-27 и их сочетания.
7. Мышь по п. 1, где мышь несет В-клетку, содержащую перегруппированную последовательность гена вариабельной области тяжелой цепи иммуноглобулина, содержащую вариабельную область гена тяжелой цепи человека, получаемую из генного сегмента VH, выбранного из VH2-5, VH3-23, VH3-30, VH4-39, VH4-59, VH5-51, и получаемую из генного сегмента D, выбранного из D1-7, D1-26, D3-3, D3-16, D3-10, D3-22, D5-5, D5-12, D6-6, D6-13 и D7-27.
8. Мышь по п. 1, у которой генный сегмент Vκ1-39 человека находится в перегруппировке с генным сегментом Jκ5 человека.
9. Мышь по п. 1, у которой генный сегмент Vκ3-20 человека находится в перегруппировке с генным сегментом Jκ1 человека.
10. Способ отбора вариабельной область человека для получения биспецифического антитела, включающий:
(а) иммунизацию генетически модифицированной мыши представляющим интерес антигеном, где мышь несет (i) замену в эндогенном локусе вариабельной области легкой цепи к иммуноглобулина мыши всех или по существу всех эндогенных генных сегментов вариабельной области легкой цепи к иммуноглобулина мыши на генный сегмент Vκ1-39/J человека, генный сегмент Vκ3-20/J человека или их сочетание, где генный сегмент человека функционально связан с эндогенным геном константной области к мыши; и (ii) замену всего или по существу всего эндогенного локуса гена вариабельной области тяжелой цепи мыши множеством генных сегментов вариабельных областей тяжелых цепей человека, где генные сегменты вариабельных областей тяжелых цепей человека функционально связаны с эндогенным геном константной области тяжелой цепи мыши, и генные сегменты вариабельных областей тяжелых цепей человека способны к перегруппировке и формированию перегруппированного гена химерной тяжелой цепи человека/мыши/
(b) обеспечение развития у мыши иммунного ответа на представляющий интерес антиген; и,
(c) идентификацию клонально отобранных лимфоцитов мыши, экспрессирующих антитела, специфически связывающиеся с представляющим интерес антигеном, и получение из лимфоцитов или антител нуклеотидной последовательности, кодирующей вариабельную область тяжелой цепи человека, специфически связывающуюся с представляющим интерес антигеном; и,
(d) применение нуклеотидной последовательности из (с) для получения биспецифического антитела.
11. Способ по п. 10, где у мыши отсутствует эндогенный локус вариабельной области легкой цепи к иммуноглобулина мыши, способный к перегруппировке и формированию гена, кодирующего вариабельную область к мыши.
12. Способ по п. 10, дополнительно включающий интронный энхансер к мыши с 5'-конца относительно константной области легкой цепи мыши.
13. Способ по п. 10, дополнительно включающий 3'-энхансер к мыши.
14. Способ по п. 10, где множество генных сегментов вариабельных областей тяжелых цепей человека содержит сегмент, выбранный из генных сегментов вариабельной области иммуноглобулина человека 1-2, 1-8, 1-24, 2-5, 3-7, 3-9, 3-11, 3-13, 3-15, 3-20, 3-23, 3-30, 3-33, 3-48, 4-31, 4-39, 4-59, 5-51, 6-1.
15. Способ по п. 10, где множество генных сегментов вариабельных областей тяжелых цепей человека содержит сегмент, выбранный из генных сегментов D1-7, D1-26, D3-3, D3-10, D3-16, D3-22, D5-5, D5-12, D6-6, D6-13, D7-27 и их сочетания.
16. Способ по п. 10, где мышь несет В-клетку, содержащую перегруппированную последовательность гена вариабельной области тяжелой цепи иммуноглобулина, содержащую ген вариабельной области тяжелой цепи человека, получаемую из генного сегмента VH, выбранного из VH2-5, VH3-23, VH3-30, VH4-39, VH4-59, VH5-51, и получаемую из генного сегмента D, выбранного из D1-7, D1-26, D3-3, D3-16, D3-10, D3-22, D5-5, D5-12, D6-6, D6-13 и D7-27.
17. Способ по п. 10, где генный сегмент Vκ1-39 человека находится в перегруппировке с генным сегментом Jκ5 человека.
18. Способ по п. 10, где генный сегмент Vκ3-20 человека находится в перегруппировке с генным сегментом Jκ1 человека.
19. Способ по п. 10, где этапы (а)-(d) первый раз проводят для первого представляющего интерес антигена с получением последовательности первой вариабельной области тяжелой цепи человека, и этапы (а)-(d) второй раз проводят для второго представляющего интерес антигена с получением последовательности второй вариабельной области тяжелой цепи человека, и где последовательность первой вариабельной области тяжелой цепи человека экспрессирована слитой с первой константной областью тяжелой цепи человека с формированием первой тяжелой цепи человека, последовательность второй вариабельной области тяжелой цепи человека экспрессирована слитой со второй константной областью тяжелой цепи человека с формированием второй тяжелой цепи человека, где первая и вторая тяжелые цепи человека экспрессированы в присутствии единственной легкой цепь человека, получаемой из генных сегментов Vκ1-39 или Vκ3-20.
20. Способ по п. 19, где первая тяжелая цепь человека содержит модификацию, устраняющую или в значительной степени снижающую аффинность первой тяжелой цепи человека к белку А, а вторая тяжелая цепь человека сохраняет способность связывать белок А.
21. Способ по п. 20, где модификация, устраняющая или в значительной степени снижающая аффинность первой тяжелой цепи человека к белку А, выбрана из 95R (EU 435R), 96F (EU 436F) и их сочетания.
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> MCWHIRTER, John
MACDONALD, Lynn
STEVENS, Sean
DAVIS, Samuel
MURPHY, Andrew J.
<120> Мышь с общей легкой цепью
<130> 0802A-WO
<140> To be assigned
<141>
<150> 61/302,282
<151> 2010-02-08
<160> 18
<170> FastSEQ for Windows Version 4.0
<210> 1
<211> 3155
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 1
ggcgcgccgt agctttgaat tttaaacatc tatttgacaa gaaatgcata gttccttctc 60
tttaaaataa tgtaatgttt ctttcaagaa taagcttggt ttgatgcctc tctccccaac 120
atgatagaag tgtagcataa atctatgaaa aattccattt ccctgtgcct acaacaacta 180
cctgggattg aaaacttctt cccttgctct agtcctttct tctacaccta cttccacatc 240
atctgtgact caaaacaata cttgtcagga aagatcccgg aaagagcaaa aaagacttcc 300
ttagaggtgt cagagattcc tatgccacta tctgtcatct ctagaagggg ttgtgagtat 360
gaggaagagc agagcttgta aattttctac ttgctttgac ttccactgta tttcctaaca 420
acaacaacca cagcaacacc cataacatca caggacaaac ttctagtact tccaaggctt 480
tagtctcagt aaatcttctc tacctccatc acagcagcta gaaggtttga tactcataca 540
aatagtactg tagctttctg ttcataattg gaaaaataga caagacccaa tgtaatacag 600
gctttccttc agccagttag cgttcagttt ttggatcacc attgcacaca tatacccagc 660
atatgtctaa tatatatgta gaaatccgtg aagcaagagt tataatagct tgtgttttct 720
attgtattgt attttcctct tatatcatct tcttcttcgt tcattaaaaa aaaaccgttc 780
aagtaggtct aaattaatta ttggatcata agtagataaa atattttatt tcataacaca 840
ttgacccgat gaatatgttt ctttgccaga catagtcctc atttccaagg taacaagcct 900
gaaaaaatta tactggagca agtcaacagg taatgatggt agcttttcct tattgtcctg 960
gggcaagaat aagacaaaag ataacagggt agaataaaga ttgtgtaaga aagaaggaca 1020
gcaacaggac atgggaacct tttatagagt aacattttga taatggatga tgagaattaa 1080
tgagttagac agggatgggt gggaatgatt gaaggtgtga gtactttagc acagattaag 1140
accaaatcat taggatttaa agagttgtgt agagttagtg aaggaaaagc cttagaatta 1200
aatttggctg cggataaaac attcttggat tagactgaag actcttttct gtgctaagta 1260
agtatattta tgataatgat gatgactgta gtgctgaata tttaataaat aaaaacaaaa 1320
ttaattgccg catacataat gtcctgaata ctattgtaaa tgttttatct tatttccttt 1380
aaactgtcta cagcactata aggtaggtac cagtattgtc acagttacac agatatggaa 1440
accgagacac agggaagtta agttacttga tcaatttcaa gcaatcggca agccatggag 1500
catctatgtc agggctgcca ggacatgtga ctgtaaacag aagtttttca ctttttaact 1560
caaagagggt atgtggctgg gttaatggaa agcttcagga ccctcagaaa acattactaa 1620
caagcaaatg aaaggtgtat ctggaagatt aagttttaac agactcttca tttccatcga 1680
tccaataatg cacttaggga gatgactggg catattgagg ataggaagag agaagtgaaa 1740
acacagcttt ttatattgtt cttaacaggc ttgtgccaaa catcttctgg gtggatttag 1800
gtgattgagg agaagaaaga cacaggagcg aaattctctg agcacaaggg aggagttcta 1860
cactcagact gagccaacag acttttctgg cctgacaacc agggcggcgc aggatgctca 1920
gtgcagagag gaagaagcag gtggtctttg cagctgaaag ctcagctgat ttgcatatgg 1980
agtcattata caacatccca gaattcttta agggcagctg ccaggaagct aagaagcatc 2040
ctctcttcta gctctcagag atggagacag acacactcct gctatgggtg ctgctgctct 2100
gggttccagg tgagggtaca gataagtgtt atgagcaacc tctgtggcca ttatgatgct 2160
ccatgcctct ctgttcttga tcactataat tagggcattt gtcactggtt ttaagtttcc 2220
ccagtcccct gaattttcca ttttctcaga gtgatgtcca aaattattct taaaaattta 2280
aatgaaaagg tcctctgctg tgaaggcttt taaagatata taaaaataat ctttgtgttt 2340
atcattccag gtgccagatg tgacatccag atgacccagt ctccatcctc cctgtctgca 2400
tctgtaggag acagagtcac catcacttgc cgggcaagtc agagcattag cagctattta 2460
aattggtatc agcagaaacc agggaaagcc cctaagctcc tgatctatgc tgcatccagt 2520
ttgcaaagtg gggtcccatc aaggttcagt ggcagtggat ctgggacaga tttcactctc 2580
accatcagca gtctgcaacc tgaagatttt gcaacttact actgtcaaca gagttacagt 2640
acccctccga tcaccttcgg ccaagggaca cgactggaga ttaaacgtaa gtaatttttc 2700
actattgtct tctgaaattt gggtctgatg gccagtattg acttttagag gcttaaatag 2760
gagtttggta aagattggta aatgagggca tttaagattt gccatgggtt gcaaaagtta 2820
aactcagctt caaaaatgga tttggagaaa aaaagattaa attgctctaa actgaatgac 2880
acaaagtaaa aaaaaaaagt gtaactaaaa aggaaccctt gtatttctaa ggagcaaaag 2940
taaatttatt tttgttcact cttgccaaat attgtattgg ttgttgctga ttatgcatga 3000
tacagaaaag tggaaaaata cattttttag tctttctccc ttttgtttga taaattattt 3060
tgtcagacaa caataaaaat caatagcacg ccctaagatc tagatgcatg ctcgagtgcc 3120
atttcattac ctctttctcc gcacccgaca tagat 3155
<210> 2
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 2
aggtgagggt acagataagt gttatgag 28
<210> 3
<211> 27
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 3
tgacaaatgc cctaattata gtgatca 27
<210> 4
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 4
gggcaagtca gagcattagc a 21
<210> 5
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 5
tgcaaactgg atgcagcata g 21
<210> 6
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 6
ggtggagagg ctattcggc 19
<210> 7
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 7
gaacacggcg gcatcag 17
<210> 8
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 8
tgggcacaac agacaatcgg ctg 23
<210> 9
<211> 29
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 9
ccattatgat gctccatgcc tctctgttc 29
<210> 10
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 10
atcagcagaa accagggaaa gcccct 26
<210> 11
<211> 3166
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 11
ggcgcgccgt agctttgaat tttaaacatc tatttgacaa gaaatgcata gttccttctc 60
tttaaaataa tgtaatgttt ctttcaagaa taagcttggt ttgatgcctc tctccccaac 120
atgatagaag tgtagcataa atctatgaaa aattccattt ccctgtgcct acaacaacta 180
cctgggattg aaaacttctt cccttgctct agtcctttct tctacaccta cttccacatc 240
atctgtgact caaaacaata cttgtcagga aagatcccgg aaagagcaaa aaagacttcc 300
ttagaggtgt cagagattcc tatgccacta tctgtcatct ctagaagggg ttgtgagtat 360
gaggaagagc agagcttgta aattttctac ttgctttgac ttccactgta tttcctaaca 420
acaacaacca cagcaacacc cataacatca caggacaaac ttctagtact tccaaggctt 480
tagtctcagt aaatcttctc tacctccatc acagcagcta gaaggtttga tactcataca 540
aatagtactg tagctttctg ttcataattg gaaaaataga caagacccaa tgtaatacag 600
gctttccttc agccagttag cgttcagttt ttggatcacc attgcacaca tatacccagc 660
atatgtctaa tatatatgta gaaatccgtg aagcaagagt tataatagct tgtgttttct 720
attgtattgt attttcctct tatatcatct tcttcttcgt tcattaaaaa aaaaccgttc 780
aagtaggtct aaattaatta ttggatcata agtagataaa atattttatt tcataacaca 840
ttgacccgat gaatatgttt ctttgccaga catagtcctc atttccaagg taacaagcct 900
gaaaaaatta tactggagca agtcaacagg taatgatggt agcttttcct tattgtcctg 960
gggcaagaat aagacaaaag ataacagggt agaataaaga ttgtgtaaga aagaaggaca 1020
gcaacaggac atgggaacct tttatagagt aacattttga taatggatga tgagaattaa 1080
tgagttagac agggatgggt gggaatgatt gaaggtgtga gtactttagc acagattaag 1140
accaaatcat taggatttaa agagttgtgt agagttagtg aaggaaaagc cttagaatta 1200
aatttggctg cggataaaac attcttggat tagactgaag actcttttct gtgctaagta 1260
agtatattta tgataatgat gatgactgta gtgctgaata tttaataaat aaaaacaaaa 1320
ttaattgccg catacataat gtcctgaata ctattgtaaa tgttttatct tatttccttt 1380
aaactgtcta cagcactata aggtaggtac cagtattgtc acagttacac agatatggaa 1440
accgagacac agggaagtta agttacttga tcaatttcaa gcaatcggca agccatggag 1500
catctatgtc agggctgcca ggacatgtga ctgtaaacag aagtttttca ctttttaact 1560
caaagagggt atgtggctgg gttaatggaa agcttcagga ccctcagaaa acattactaa 1620
caagcaaatg aaaggtgtat ctggaagatt aagttttaac agactcttca tttccatcga 1680
tccaataatg cacttaggga gatgactggg catattgagg ataggaagag agaagtgaaa 1740
acacagcttt ttatattgtt cttaacaggc ttgtgccaaa catcttctgg gtggatttag 1800
gtgattgagg agaagaaaga cacaggagcg aaattctctg agcacaaggg aggagttcta 1860
cactcagact gagccaacag acttttctgg cctgacaacc agggcggcgc aggatgctca 1920
gtgcagagag gaagaagcag gtggtctttg cagctgaaag ctcagctgat ttgcatatgg 1980
agtcattata caacatccca gaattcttta agggcagctg ccaggaagct aagaagcatc 2040
ctctcttcta gctctcagag atggagacag acacactcct gctatgggtg ctgctgctct 2100
gggttccagg tgagggtaca gataagtgtt atgagcaacc tctgtggcca ttatgatgct 2160
ccatgcctct ctgttcttga tcactataat tagggcattt gtcactggtt ttaagtttcc 2220
ccagtcccct gaattttcca ttttctcaga gtgatgtcca aaattattct taaaaattta 2280
aatgaaaagg tcctctgctg tgaaggcttt taaagatata taaaaataat ctttgtgttt 2340
atcattccag gtgccagatg tataccaccg gagaaattgt gttgacgcag tctccaggca 2400
ccctgtcttt gtctccaggg gaaagagcca ccctctcctg cagggccagt cagagtgtta 2460
gcagcagcta cttagcctgg taccagcaga aacctggcca ggctcccagg ctcctcatct 2520
atggtgcatc cagcagggcc actggcatcc cagacaggtt cagtggcagt gggtctggga 2580
cagacttcac tctcaccatc agcagactgg agcctgaaga ttttgcagtg tattactgtc 2640
agcagtatgg tagctcacct tggacgttcg gccaagggac caaggtggaa atcaaacgta 2700
agtaattttt cactattgtc ttctgaaatt tgggtctgat ggccagtatt gacttttaga 2760
ggcttaaata ggagtttggt aaagattggt aaatgagggc atttaagatt tgccatgggt 2820
tgcaaaagtt aaactcagct tcaaaaatgg atttggagaa aaaaagatta aattgctcta 2880
aactgaatga cacaaagtaa aaaaaaaaag tgtaactaaa aaggaaccct tgtatttcta 2940
aggagcaaaa gtaaatttat ttttgttcac tcttgccaaa tattgtattg gttgttgctg 3000
attatgcatg atacagaaaa gtggaaaaat acatttttta gtctttctcc cttttgtttg 3060
ataaattatt ttgtcagaca acaataaaaa tcaatagcac gccctaagat ctagatgcat 3120
gctcgagtgc catttcatta cctctttctc cgcacccgac atagat 3166
<210> 12
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 12
tccaggcacc ctgtctttg 19
<210> 13
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 13
aagtagctgc tgctaacact ctgact 26
<210> 14
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 14
aaagagccac cctctcctgc aggg 24
<210> 15
<211> 3187
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 15
ggcgcgccgt agctttgaat tttaaacatc tatttgacaa gaaatgcata gttccttctc 60
tttaaaataa tgtaatgttt ctttcaagaa taagcttggt ttgatgcctc tctccccaac 120
atgatagaag tgtagcataa atctatgaaa aattccattt ccctgtgcct acaacaacta 180
cctgggattg aaaacttctt cccttgctct agtcctttct tctacaccta cttccacatc 240
atctgtgact caaaacaata cttgtcagga aagatcccgg aaagagcaaa aaagacttcc 300
ttagaggtgt cagagattcc tatgccacta tctgtcatct ctagaagggg ttgtgagtat 360
gaggaagagc agagcttgta aattttctac ttgctttgac ttccactgta tttcctaaca 420
acaacaacca cagcaacacc cataacatca caggacaaac ttctagtact tccaaggctt 480
tagtctcagt aaatcttctc tacctccatc acagcagcta gaaggtttga tactcataca 540
aatagtactg tagctttctg ttcataattg gaaaaataga caagacccaa tgtaatacag 600
gctttccttc agccagttag cgttcagttt ttggatcacc attgcacaca tatacccagc 660
atatgtctaa tatatatgta gaaatccgtg aagcaagagt tataatagct tgtgttttct 720
attgtattgt attttcctct tatatcatct tcttcttcgt tcattaaaaa aaaaccgttc 780
aagtaggtct aaattaatta ttggatcata agtagataaa atattttatt tcataacaca 840
ttgacccgat gaatatgttt ctttgccaga catagtcctc atttccaagg taacaagcct 900
gaaaaaatta tactggagca agtcaacagg taatgatggt agcttttcct tattgtcctg 960
gggcaagaat aagacaaaag ataacagggt agaataaaga ttgtgtaaga aagaaggaca 1020
gcaacaggac atgggaacct tttatagagt aacattttga taatggatga tgagaattaa 1080
tgagttagac agggatgggt gggaatgatt gaaggtgtga gtactttagc acagattaag 1140
accaaatcat taggatttaa agagttgtgt agagttagtg aaggaaaagc cttagaatta 1200
aatttggctg cggataaaac attcttggat tagactgaag actcttttct gtgctaagta 1260
agtatattta tgataatgat gatgactgta gtgctgaata tttaataaat aaaaacaaaa 1320
ttaattgccg catacataat gtcctgaata ctattgtaaa tgttttatct tatttccttt 1380
aaactgtcta cagcactata aggtaggtac cagtattgtc acagttacac agatatggaa 1440
accgagacac agggaagtta agttacttga tcaatttcaa gcaatcggca agccatggag 1500
catctatgtc agggctgcca ggacatgtga ctgtaaacag aagtttttca ctttttaact 1560
caaagagggt atgtggctgg gttaatggaa agcttcagga ccctcagaaa acattactaa 1620
caagcaaatg aaaggtgtat ctggaagatt aagttttaac agactcttca tttccatcga 1680
tccaataatg cacttaggga gatgactggg catattgagg ataggaagag agaagtgaaa 1740
acacagcttt ttatattgtt cttaacaggc ttgtgccaaa catcttctgg gtggatttag 1800
gtgattgagg agaagaaaga cacaggagcg aaattctctg agcacaaggg aggagttcta 1860
cactcagact gagccaacag acttttctgg cctgacaacc agggcggcgc aggatgctca 1920
gtgcagagag gaagaagcag gtggtctttg cagctgaaag ctcagctgat ttgcatatgg 1980
agtcattata caacatccca gaattcttta agggcagctg ccaggaagct aagaagcatc 2040
ctctcttcta gctctcagag atggagacag acacactcct gctatgggtg ctgctgctct 2100
gggttccagg tgagggtaca gataagtgtt atgagcaacc tctgtggcca ttatgatgct 2160
ccatgcctct ctgttcttga tcactataat tagggcattt gtcactggtt ttaagtttcc 2220
ccagtcccct gaattttcca ttttctcaga gtgatgtcca aaattattct taaaaattta 2280
aatgaaaagg tcctctgctg tgaaggcttt taaagatata taaaaataat ctttgtgttt 2340
atcattccag gtgccagatg tgttgtggtc ctcagccggt gctgcatcag ccgccggcca 2400
tgtcctcggc ccttggaacc acaatccgcc tcacctgcac cctgaggaac gaccatgaca 2460
tcggtgtgta cagcgtctac tggtaccagc agaggccggg ccaccctccc aggttcctgc 2520
tgagatattt ctcacaatca gacaagagcc agggccccca ggtcccccct cgcttctctg 2580
gatccaaaga tgtggccagg aacagggggt atttgagcat ctctgagctg cagcctgagg 2640
acgaggctat gtattactgt gctatgcata actcagtgac gcatgtgttt ggcagcggga 2700
cccagctcac cgttttaagt aagtaatttt tcactattgt cttctgaaat ttgggtctga 2760
tggccagtat tgacttttag aggcttaaat aggagtttgg taaagattgg taaatgaggg 2820
catttaagat ttgccatggg ttgcaaaagt taaactcagc ttcaaaaatg gatttggaga 2880
aaaaaagatt aaattgctct aaactgaatg acacaaagta aaaaaaaaaa gtgtaactaa 2940
aaaggaaccc ttgtatttct aaggagcaaa agtaaattta tttttgttca ctcttgccaa 3000
atattgtatt ggttgttgct gattatgcat gatacagaaa agtggaaaaa tacatttttt 3060
agtctttctc ccttttgttt gataaattat tttgtcagac aacaataaaa atcaatagca 3120
cgccctaaga tctagatgca tgctcgagtg ccatttcatt acctctttct ccgcacccga 3180
catagat 3187
<210> 16
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 16
tgtcctcggc ccttgga 17
<210> 17
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 17
ccgatgtcat ggtcgttcct 20
<210> 18
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 18
acaatccgcc tcacctgcac cct 23
| название | год | авторы | номер документа |
|---|---|---|---|
| МЫШЬ С ОБЩЕЙ ЛЕГКОЙ ЦЕПЬЮ | 2011 |
|
RU2571205C2 |
| МЫШЬ С ОБЩЕЙ ЛЕГКОЙ ЦЕПЬЮ | 2020 |
|
RU2826121C2 |
| МЫШЬ С ОБЩЕЙ ЛЕГКОЙ ЦЕПЬЮ | 2013 |
|
RU2683514C2 |
| ОТЛИЧНЫЕ ОТ ЧЕЛОВЕКА ЖИВОТНЫЕ, ЭКСПРЕССИРУЮЩИЕ АНТИТЕЛА С ОБЩЕЙ ЛЕГКОЙ ЦЕПЬЮ | 2012 |
|
RU2614859C2 |
| МЫШИ, ЭКСПРЕССИРУЮЩИЕ ОГРАНИЧЕННЫЙ РЕПЕРТУАР ЛЕГКИХ ЦЕПЕЙ ИММУНОГЛОБУЛИНА | 2014 |
|
RU2689664C2 |
| МЫШИ, ЭКСПРЕССИРУЮЩИЕ ОГРАНИЧЕННЫЙ РЕПЕРТУАР ЛЕГКИХ ЦЕПЕЙ ИММУНОГЛОБУЛИНА | 2014 |
|
RU2812814C2 |
| МЫШИ С ГУМАНИЗИРОВАННОЙ УНИВЕРСАЛЬНОЙ ЛЕГКОЙ ЦЕПЬЮ | 2012 |
|
RU2664232C2 |
| АНТИТЕЛА СО ВСТРОЕННЫМ В ЛЕГКИЕ ЦЕПИ ГИСТИДИНОМ И ГЕНЕТИЧЕСКИ МОДИФИЦИРОВАННЫЕ ОТЛИЧНЫЕ ОТ ЧЕЛОВЕКА ЖИВОТНЫЕ ДЛЯ ИХ ПОЛУЧЕНИЯ | 2013 |
|
RU2644684C2 |
| МЫШИ С ГУМАНИЗИРОВАННОЙ УНИВЕРСАЛЬНОЙ ЛЕГКОЙ ЦЕПЬЮ | 2012 |
|
RU2787781C2 |
| АНТИТЕЛА СО ВСТРОЕННЫМ В ЛЕГКИЕ ЦЕПИ ГИСТИДИНОМ И ГЕНЕТИЧЕСКИ МОДИФИЦИРОВАННЫЕ ОТЛИЧНЫЕ ОТ ЧЕЛОВЕКА ЖИВОТНЫЕ ДЛЯ ИХ ПОЛУЧЕНИЯ | 2014 |
|
RU2694728C2 |
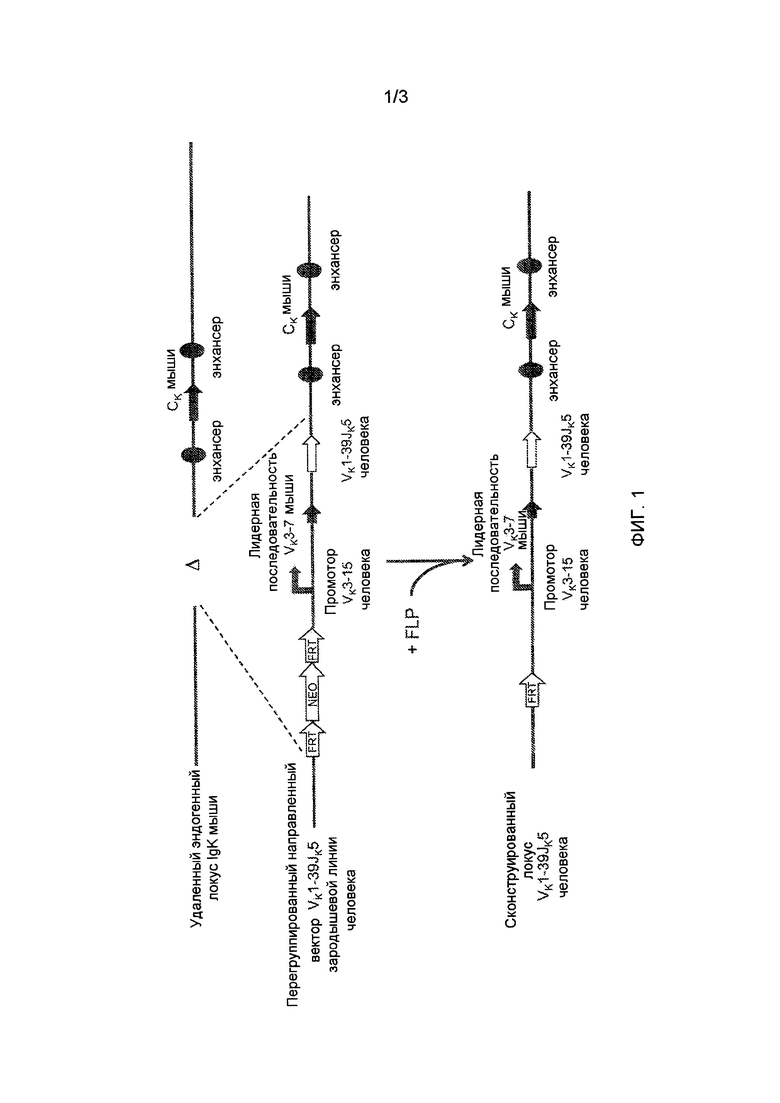

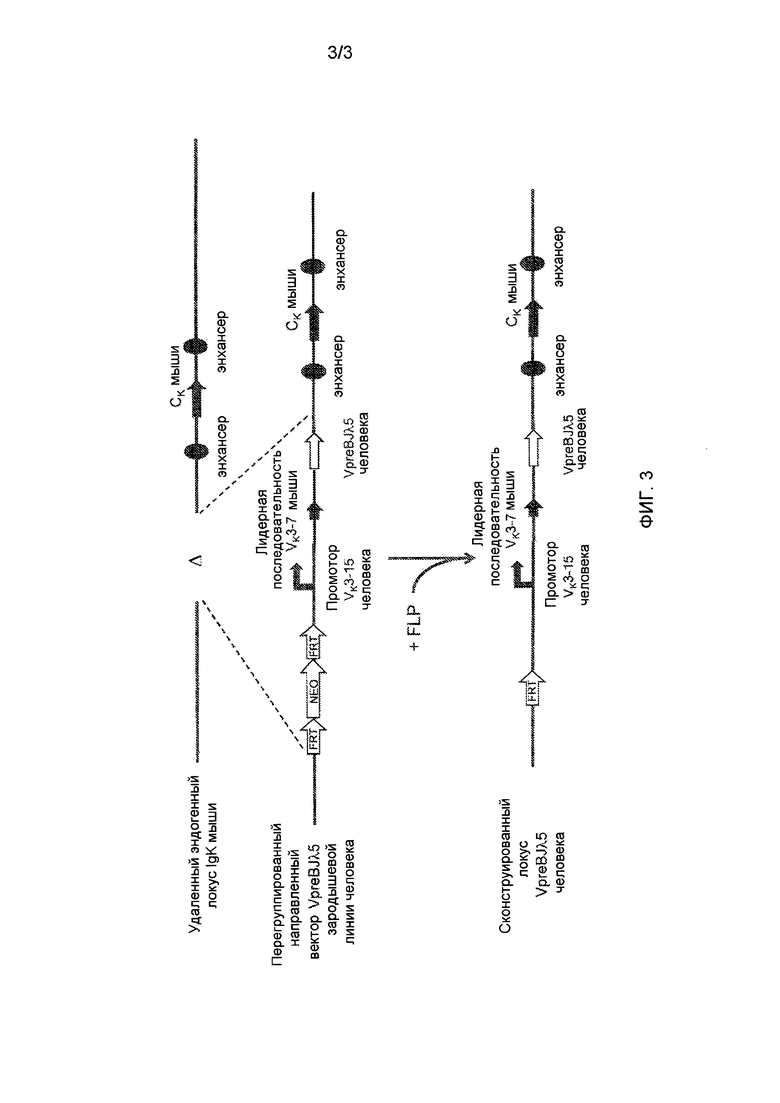
Изобретение относится к области биохимии, в частности к генетически модифицированной мыши, которая экспрессирует вариабельные домены тяжелой и легкой цепей иммуноглобулина человека, где вариабельные домены легкой цепи человека экспрессируются из единственной перегруппированной Vκ1-39/Jκ5 последовательности человека, присутствующей в геноме зародышевой линии мыши, а также к В-клетке, полученной из вышеуказанной мыши, и гибридоме, полученной из вышеуказанной клетки. Также раскрыты способ получения антитела и способ получения биспецифического антитела человека, которые предусматривают использование вышеуказанной мыши. Изобретение также относится к применению вышеуказанной мыши в получении антитела, для продукции клетки, которая экспрессирует биспецифическое антитело человека, а также для идентификации вариабельного домена тяжелой цепи иммуноглобулина человека, присутствующей в тяжелой цепи иммуноглобулина, которая ассоциирована с общей легкой цепью. Изобретение позволяет эффективно получать антитело. 8 н. и 11 з.п. ф-лы, 3 ил., 12 табл., 10 пр.
1. Генетически модифицированная мышь, которая экспрессирует вариабельные домены тяжелой и легкой цепей иммуноглобулина человека, где вариабельные домены легкой цепи человека экспрессируются из единственной перегруппированной Vκ1-39/Jκ5 последовательности человека, присутствующей в геноме зародышевой линии мыши, при этом указанная единственная перегруппированная Vκ1-39/Jκ5 последовательность человека вставлена в геном мыши в эндогенном локусе легкой цепи к иммуноглобулина мыши, и единственная перегруппированная Vκ1-39/Jκ5 последовательность человека функционально связана с эндогенной последовательностью константной области к мыши, где перегруппированная Vκ1-39/Jκ5 последовательность человека содержит генный сегмент Vκ1-39 зародышевой линии человека и генный сегмент Jκ5 зародышевой линии человека, при этом:
(a) вариабельные домены легкой цепи иммуноглобулина человека экспрессируются из последовательности, которая идентична или является соматически гипермутированным вариантом перегруппированной Vκ1-39/Jκ5 последовательности человека;
(b) у мыши отсутствует эндогенный локус вариабельной области легкой цепи к иммуноглобулина мыши, способный к перегруппировке и формированию гена, кодирующего вариабельную область к мыши;
(c) мышь экспрессирует общую легкую цепь иммуноглобулина человека для множества тяжелых цепей человека, и общая легкая цепь происходит из указанной единственной перегруппированной Vκ1-39/Jκ5 последовательности человека; и
(d) у мыши присутствует В-клетка, экспрессирующая антитело, которое специфически связывается с представляющим интерес антигеном, где антитело содержит указанную общую легкую цепь и тяжелую цепь иммуноглобулина, получаемую на основе перегруппировки генных сегментов вариабельной области тяжелой цепи иммуноглобулина человека, присутствующих в геноме указанной мыши, где перегруппировка генных сегментов вариабельной области тяжелой цепи иммуноглобулина человека содержит генный сегмент VH, выбранный из группы, состоящей из генных сегментов VH2-5, VH3-23, VH3-30, VH4-59 и VH5-51.
2. Генетически модифицированная мышь по п. 1, при этом 90-100% эндогенных неперегруппированных генных сегментов VH мыши замещены:
(i) по меньшей мере 12 функциональными неперегруппированными генными сегментами VH человека;
(ii) по меньшей мере 18 неперегруппированными генными сегментами VH человека;
(iii) по меньшей мере 39 неперегруппированными генными сегментами VH человека;
(iv) по меньшей мере 80 неперегруппированными сегментами VH человека; или
(v) по меньшей мере 81 неперегруппированными сегментами VH человека.
3. Генетически модифицированная мышь по п. 1, которая экспрессирует:
антитело, содержащее легкую цепь, включающую вариабельный домен легкой цепи иммуноглобулина человека и константный домен легкой цепи иммуноглобулина мыши, и тяжелую цепь, включающую вариабельный домен тяжелой цепи иммуноглобулина человека и константный домен тяжелой цепи иммуноглобулина мыши.
4. Генетически модифицированная мышь по п. 1, где генный сегмент Vκ человека функционально связан с последовательность промотора иммуноглобулина.
5. Генетически модифицированная мышь по п. 4, где последовательность промотора иммуноглобулина представляет собой последовательность промотора иммуноглобулина человека.
6. В-клетка, выделенная из генетически модифицированной мыши по любому из пп. 1-5, где выделенная В-клетка содержит в своем геноме:
единственную перегруппированную Vκ1-39/Jκ5 последовательность человека, функционально связанную с эндогенной последовательностью константной области к мыши, где перегруппированная Vκ1-39/Jκ5 последовательность человека происходит из генного сегмента Vk1-39 зародышевой линии человека и генного сегмента Jκ5 зародышевой линии человека, и
где В-клетка экспрессирует антитело, которое специфически связывается с представляющим интерес антигеном, где антитело содержит тяжелую цепь иммуноглобулина, получаемую на основе перегруппировки генных сегментов вариабельной области тяжелой цепи иммуноглобулина человека, присутствующих в геноме указанной генетически модифицированной мыши.
7. Гибридома, полученная из В-клетки по п. 6, где геном указанной гибридомы содержит:
единственную перегруппированную Vκ1-39/Jκ5 последовательность человека, функционально связанную с эндогенной последовательностью константной области к мыши, где перегруппированная Vκ1-39/Jκ5 последовательность человека происходит из генного сегмента Vκ1-39 зародышевой линии человека и генного сегмента Jκ5 зародышевой линии человека, и
где гибридома экспрессирует антитело, которое специфически связывается с представляющим интерес антигеном, где антитело содержит тяжелую цепь иммуноглобулина, получаемую на основе перегруппировки генных сегментов вариабельной области тяжелой цепи иммуноглобулина человека, где перегруппировка генных сегментов вариабельной области тяжелой цепи иммуноглобулина человека включает генный сегмент VH, выбранный из группы, состоящей из генных сегментов VH2-5, VH3-23, VH3-30, VH4-59 и VH5-51.
8. Способ получения антитела, включающий:
(a) иммунизацию мыши по любому из пунктов 1-5 представляющим интерес антигеном;
(b) обеспечение развития у мыши иммунного ответа на указанный представляющий интерес антиген, где мышь экспрессирует популяцию антител в ответ на представляющий интерес антиген, где популяция антиген-специфических антител содержит легкую цепь, которая является соматически мутированной, и множество тяжелых цепей иммуноглобулина, каждая из которых содержит вариабельную область тяжелой цепи иммуноглобулина, происходящую из одного из множества сегментов VH человека, в том числе по меньшей мере VH2-5, VH3-23, VH3-30 и VH4-59;
(c) идентификацию последовательности, которая кодирует вариабельную область тяжелой цепи человека, которая специфически связывается с представляющим интерес антигеном; и
(d) получение указанного антитела с применением одной или более генной(ых) последовательности(ей) вариабельной области иммуноглобулина человека из указанной мыши.
9. Способ по п. 8, в котором применение одной или более последовательности(ей) вариабельной области иммуноглобулина человека из мыши для получения антитела включает:
экспрессию в одной клетке:
(a) первой последовательности вариабельной области иммуноглобулина человека, полученной из мыши, при этом первая генная последовательность вариабельной области тяжелой цепи иммуноглобулина человека слита с генной последовательностью константной области иммуноглобулина человека; и
(b) генной последовательности вариабельной области легкой цепи иммуноглобулина человека, полученной из единственной перегруппированной Vκ1-39/Jκ5 последовательности человека, при этом генная последовательность вариабельной области легкой цепи иммуноглобулина человека слита с генной последовательностью константной области легкой цепи иммуноглобулина человека;
поддержание клетки в условиях, достаточных для экспрессии полностью человеческого антитела, и
выделение указанного антитела.
10. Способ по п. 9, где способ дополнительно включает экспрессию в той же одной клетке с указанной первой последовательностью вариабельной области иммуноглобулина человека, слитой с указанной генной последовательностью константной области иммуноглобулина человека, второй генной последовательности вариабельной области тяжелой цепи иммуноглобулина человека, полученной из мыши, при этом вторая генная последовательность вариабельной области тяжелой цепи иммуноглобулина человека слита с генной последовательностью константной области иммуноглобулина человека, где первая генная последовательность вариабельной области тяжелой цепи кодирует вариабельный домен тяжелой цепи иммуноглобулина человека, распознающий первый эпитоп, и вторая генная последовательность вариабельной области тяжелой цепи иммуноглобулина человека кодирует вариабельный домен тяжелой цепи иммуноглобулина человека, распознающий второй эпитоп, и при этом первый эпитоп и второй эпитоп не являются идентичными.
11. Способ по п. 10, где первый и второй эпитопы являются различными эпитопами одного и того же антигена; или
где первый эпитоп является эпитопом первого антигена и второй эпитоп является эпитопом второго антигена.
12. Применение мыши по любому из пп. 1-5 в получении антитела.
13. Применение по п. 12, где антитело представляет собой полностью человеческое антитело.
14. Применение по п. 12, где антитело представляет собой биспецифическое антитело.
15. Применение по п. 13, где антитело представляет собой биспецифическое антитело.
16. Способ получения биспецифического антитела человека, включающий
получение первой и второй генных последовательностей вариабельной области тяжелой цепи иммуноглобулина человека из одной или более В-клеток мыши по любому из пп. 1-5,
где указанные одна или более В-клеток указанной мыши содержат единственную перегруппированную Vκ1-39/Jκ5 последовательность человека, функционально связанную с эндогенной последовательностью константной области к мыши, где перегруппированная Vκ1-39/Jκ5 последовательность человека происходит из генного сегмента Vκ1-39 зародышевой линии человека и генного сегмента Jκ5 зародышевой линии человека; и
где каждая из одной или более В-клеток указанной мыши экспрессирует антитело, которое специфически связывается с представляющим интерес антигеном, где антитело содержит тяжелую цепь иммуноглобулина, получаемую на основе перегруппировки вариабельных (VH) генных сегментов тяжелой цепи иммуноглобулина человека, присутствующих в геноме указанной мыши, выбранных из группы, состоящей из генных сегментов VH2-5, VH3-23, VH3-30, VH4-59 и VH5-51; и
экспрессию в одной клетке легкой цепи, происходящей из указанной единственной перегруппированной Vκ1-39/Jκ5 последовательности человека, первой тяжелой цепи, получаемой на основе перегруппировки генных сегментов вариабельной области тяжелой цепи иммуноглобулина человека, где перегруппировка генных сегментов вариабельной области тяжелой цепи иммуноглобулина человека включает генный сегмент VH, выбранный из группы, состоящей из генных сегментов VH2-5, VH3-23, VH3-30, VH4-59 и VH5-51, и второй тяжелой цепи, получаемой на основе перегруппировки генных сегментов вариабельной области тяжелой цепи иммуноглобулина человека, где перегруппировка генных сегментов вариабельной области тяжелой цепи иммуноглобулина человека включает генный сегмент VH, выбранный из группы, состоящей из генных сегментов VH2-5, VH3-23, VH3-30, VH4-59 и VH5-51;
поддержание клетки в условиях, достаточных для экспрессии человеческого биспецифического антитела; и
выделение человеческого биспецифического антитела из указанной клетки.
17. Применение мыши по любому из пп. 1-5 для продукции клетки, которая экспрессирует биспецифическое антитело человека, где клетка содержит:
(a) единственную генную последовательность вариабельной области легкой цепи иммуноглобулина человека, кодирующую вариабельный домен легкой цепи иммуноглобулина человека, получаемую из перегруппированной Vκ1-39/Jκ5 последовательности человека, где генная последовательность вариабельной области легкой цепи иммуноглобулина человека слита непосредственно или через линкер с генной последовательностью константной области легкой цепи иммуноглобулина человека, с образованием гена, кодирующего легкую цепь иммуноглобулина человека;
(b) первую генную последовательность вариабельной области тяжелой цепи иммуноглобулина человека, получаемую из первой В-клетки мыши, кодирующую первый вариабельный домен тяжелой цепи иммуноглобулина человека, где первая генная последовательность вариабельной области тяжелой цепи иммуноглобулина человека слита непосредственно или через линкер с генной последовательностью константной области тяжелой цепи иммуноглобулина человека, с образованием гена, кодирующего первую тяжелую цепь иммуноглобулина человека, где первый вариабельный домен тяжелой цепи иммуноглобулина человека распознает первый эпитоп; и
(с) вторую генную последовательность вариабельной области тяжелой цепи иммуноглобулина человека, получаемую из второй В-клетки мыши, кодирующую второй вариабельный домен тяжелой цепи иммуноглобулина человека, где вторая генная последовательность вариабельной области тяжелой цепи иммуноглобулина человека слита непосредственно или через линкер с генной последовательностью константной области тяжелой цепи иммуноглобулина человека, с образованием гена, кодирующего вторую тяжелую цепь иммуноглобулина человека, где второй вариабельный домен тяжелой цепи иммуноглобулина человека распознает второй эпитоп и специфически не распознает первый эпитоп,
при этом каждую из первой и второй тяжелых цепей иммуноглобулина ассоциируют с легкой цепью иммуноглобулина человека из (а).
18. Применение по п. 17, где первый и второй эпитопы являются различными эпитопами одного и того же антигена; или
где первый эпитоп является эпитопом первого антигена и второй эпитоп является эпитопом второго антигена.
19. Применение мыши по любому из пп. 1-5 для идентификации вариабельного домена тяжелой цепи иммуноглобулина человека, присутствующей в тяжелой цепи иммуноглобулина, которая ассоциирована с общей легкой цепью.
| US2006015957 A1, 19.01.2006 | |||
| US2003078385 A1, 24.04.2003 | |||
| MERCHANT A.M | |||
| et al., An efficient route to human bispecific IgG, Nat Biotechnol, 1998, Vol.16, N.7, pp.677-681 | |||
| ЧЕЛОВЕЧЕСКОЕ МОНОКЛОНАЛЬНОЕ АНТИТЕЛО ПРОТИВ AILIM, КОСТИМУЛИРУЮЩЕЙ МОЛЕКУЛЫ ПЕРЕДАЧИ СИГНАЛА, И ЕГО ФАРМАЦЕВТИЧЕСКОЕ ПРИМЕНЕНИЕ | 2001 |
|
RU2262511C2 |

