Данная заявка испрашивает приоритет предварительной заявки США на патент №61/477.440, поданной 20 апреля 2011 г., полное содержание каждой из которых включено в данный документ посредством ссылки. В данной заявке даются ссылки на различные публикации. Раскрытие этих публикаций во всей их полноте включено в данную заявку посредством ссылки для более полного описания уровня техники, к которому относится настоящее изобретение.
ЗАЯВЛЕНИЕ ОТНОСИТЕЛЬНО ФИНАНСИРУЕМЫХ ИЗ ФЕДЕРАЛЬНОГО БЮДЖЕТА ИССЛЕДОВАНИЙ
Настоящее изобретение было сделано при поддержке правительства в виде гранта № AI 067769, выданного Национальными институтами здоровья. Правительство имеет определенные права на настоящее изобретение.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
Трансплантация гемопоэтических стволовых клеток (HSCT) является основой лечения многих наследственных заболеваний и лимфогемопоэтических злокачественных новообразований (1). Кроме того, гемопоэтические стволовые клетки (HSC) в целом представляют собой важную цель для генной терапии ex vivo. Перенос генов в HSC предоставляет потенциальную стратегию коррекции моногенных дефектов и изменения чувствительности нормального ВМ к цитотоксическим препаратам. Эти применения имеют значительный терапевтический потенциал, но были ограничены низким уровнем переноса генов в HSC. Последние достижения, такие как улучшенные цитокины для минимизации коммитирования в ходе процедуры ex vivo, перенос генов с помощью фибронектина и обогащение HSC перед переносом генов, улучшили эффективность переноса генов в клетки человека и усовершенствовали испытания генной терапии человека (2). Тем не менее, эффективность переноса генов в HSC и пересадка большого числа трансдуцированных клеток остается серьезной проблемой для расширения применения этой технологии для успешного лечения рака и моногенетических заболеваний.
Для улучшения пересадки генномодифицированных HSC и уменьшения времени, необходимого для лимфогемопоэтического восстановления после HSCT, исследовали стратегии селекции in vivo, использующие гены лекарственной устойчивости, такие как дигидрофолатредуктаза (DHFR) (3) или ген множественной лекарственной устойчивости 1 (MDR1) (4, US 1996/017660), но, как правило, они потерпели неудачу из-за неприемлемой токсичности (5) или недостаточной эффективности селекции (6). В настоящее время, мутантные формы O-6-метилгуанин-ДНК-метилтрансферазы (MGMT) исследуют на их способность придавать хемопротекцию против BCNU или темозоломида в сочетании с O-6-бензилгуанином (2,7, US 1997/004917), но эти средства представляют значительный риск токсичности, а последние наблюдения показывают, что мутантный MGMT может придавать селективный недостаток при высоком уровне экспрессии (8). В US 2003032003 AA описана стратегия селекции HPRT-дефицитных клеток in vivo с помощью 6TG. Однако в этой заявке на патент для предварительного кондиционирования перед селекцией in vivo по-прежнему используется облучение, или селекцию in vivo совершают циклами с периодами восстановления между введением 6TG.
Кроме того, предложенная доза 6TG является высокой и вводится в течение длительного периода времени (200 мг/кг общей дозы в течение 55 дней). Кроме того, Porter и DiGregori описали подход к инактивации экспрессии HPRT в ВМ и последующей селекции донорских клеток с 6TG in vivo как «опосредованную интерферирующей РНК резистентность к аналогам пурина» («iPAR»). Этот отчет показал возможность инактивации HPRT в HSC с помощью лентивирусного вектора, экспрессирующего shRNA, которая воздействует на HPRT, и обогащение этих измененных гемопоэтических клеток 6TG у мышей in vivo. Тем не менее, в данном отчете, предварительную подготовку осуществляли все еще путем облучения всего тела, а хемоселекцию in vivo не начинали до, по меньшей мере, 4 недель после трансплантации. Кроме того, 6TG вводили в виде короткого импульса или при дозировках, которые выбрали только для умеренной миелосупрессии, и не ясно, были ли достигнуты достаточные уровни трансдукции HSC лентивирусными векторами второго поколения, которые использовали в их исследовании. В целом, описанные уровни пересадки были переменными и относительно незначительными.
Остается необходимость более эффективных способов HSCT, чтобы избежать токсичности во время восстановления клеток костного мозга.
Литература:
(1) Bhattacharya D, Ehrlich LI, Weissman IL. Eur J Immunol. 2008; 38:2060-2067.
(2) Milsom MD, Williams DA. DNA Repair (Amst). 2007; 6:1210-1221.
(3) Williams DA, Hsieh K, DeSilva A, Mulligan RC. J Exp Med. 1987; 166:210-218.
(4) Sorrentino BP, Brandt SJ, Bodine D, et al. Science. 1992; 257:99-103.
(5) Zaboikin M, Srinivasakumar N, Schuening F. Cancer Gene Ther. 2006; 13:335-345.
(6) Southgate T, Fairbairn LJ. Expert Rev Mol Med. 2004; 6:1-24.
(7) Neff T, Beard ВС, Kiem HP. Blood. 2006; 107:1751-1760.
(8) Schambach A, Baum C. DNA Repair (Amst). 2007; 6:1187-1196.
(9) Porter CC, DeGregori J. Blood. 2008; 112:4466-4474.
Селекция in vivo примитивных гемопоэтических клеток (публикация патента WO /1998/ 019540) US 1996 /017660
Использование мутантных алкилтрансфераз для генной терапии для защиты от токсичности терапевтических алкилирующих средств (публикация патента WO /1997/ 035477) US 1997 /004917
Селекция in vivo (публикация патента WO /1997/ 043900) US 2003032003 АА
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение предлагает способ трансплантации гемопоэтических стволовых клеток (HSC) без облучения. Как правило, способ включает введение млекопитающему субъекту одной или двух доз от 2 до 10 мг/кг массы тела аналога пуринового основания в качестве стадии предварительного кондиционирования. Способ дополнительно включает пересадку субъекту гипоксантин-гуанин-фосфорибозилтрансфераз (HPRT)-дефицитных HSC донора в течение от 48 до 72 часов стадии предварительного кондиционирования и введение субъекту приблизительно от 1 до 5 мг/кг аналога пуриновых оснований каждые 2-4 дня в течение 2-8 недель после стадии пересадки. Способ осуществляют в отсутствие предварительного кондиционирования путем облучения. Поэтому, при подготовке к трансплантации субъект не подвергают миелоаблативному облучению, и, таким образом, субъект лишен токсичности, индуцированной миелоаблативным облучением.
Характерные примеры аналогов пуриновых оснований включают: 6-тиогуанин (6TG), 6-меркаптопурин (6-МР) и азатиопурин (AZA). В одном варианте осуществления аналогом пуриновых оснований является 6TG. В некоторых вариантах осуществления общая доза 6TG, которую вводят субъекту, не превышает 105 мг; обычно, общая доза 6TG, которую вводят субъекту, не превышает 75 мг. В одном варианте осуществления введение аналога пуринового основания осуществляют каждые 3 дня и на протяжении не более четырех недель после этапа пересадки.
У субъектов, которых лечили в соответствии с данным способом, будет наблюдаться более 75% генетически модифицированных гемопоэтических клеток. В некоторых вариантах осуществления у субъекта наблюдается более 95% генетически модифицированных гемопоэтических клеток.
HPRT-дефицитные HSC для трансплантации могут быть превращены в HPRT-дефицитные с применением традиционных способов, известных специалистам в данной области техники. Характерные способы включают, без ограничения, введение последовательностей, кодирующих нуклеазы ''цинковые пальцы'' (ZFN), эффекторные нуклеазы, подобные активаторам транскрипции (TALEN), матричные цепи небольшого фрагмента гомологичной рекомбинации (SFHR), ингибирующие РНК (siRNA) или микроРНК (miRNA), антисмысловые РНК, транс-сплайсинговые РНК, рибозимы, внутриклеточные антитела или доминантно-негативные, или конкурентные белки-ингибиторы. Трансплантированные HSC могут быть аутологичными, изогенными или аллогенными.
В некоторых вариантах осуществления HPRT-дефицитные HSC для трансплантации были генетически модифицированы. Субъект может иметь наследственное или генетическое заболевание, приобретенное заболевание, поражающее лимфогемопоэтические клетки, такое как инфекция, вызванная вирусом иммунодефицита человека (ВИЧ), или синдром приобретенного иммунодефицита (СПИД) или лимфогемопоэтические злокачественные новообразования. Генетическая модификация донорских HSC может выходить за пределы превращения клеток в HPRT-дефицитные, а также служить для лечения или исправления состояния.
КРАТКОЕ ОПИСАНИЕ ФИГУР
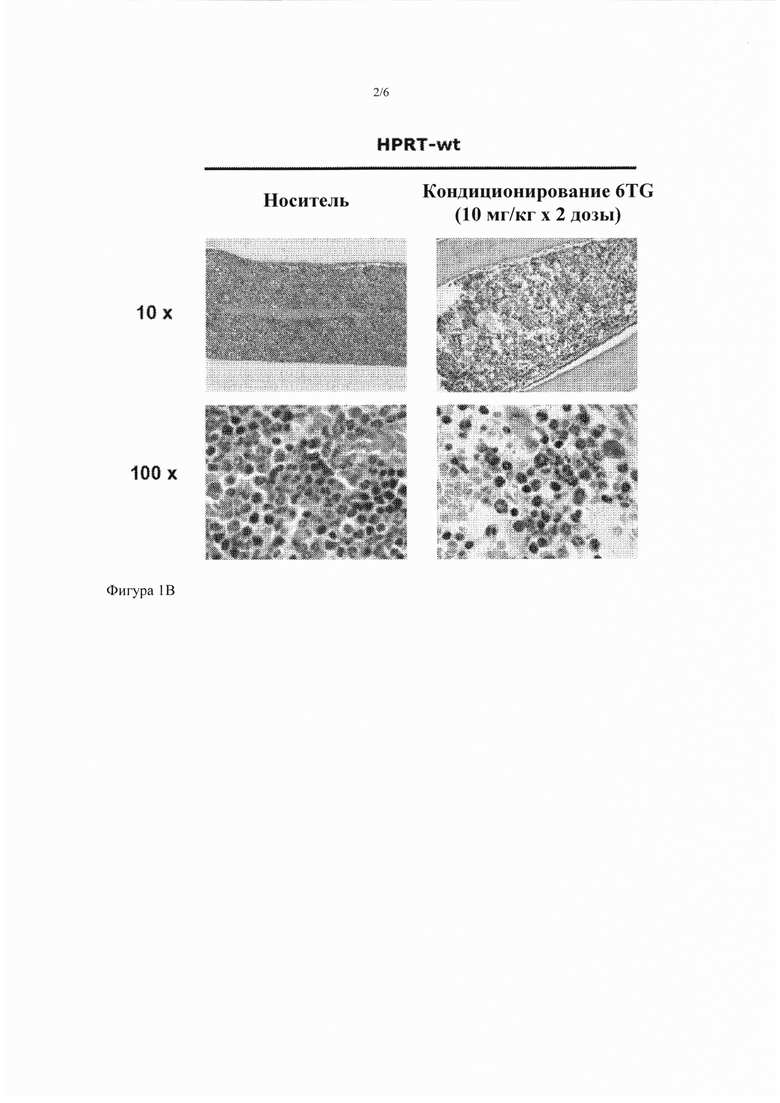
Фигура 1А-1В. Оптимизация режима кондиционирования 6TG. (Фиг. 1А) В качестве исследования подбора дозы мышам HPRT-wt интраперитонеально инъецировали носитель или изменяющиеся дозы 6TG от 2,5 до 10 мг/кг, как указано, в день 1 (n = 3 на группу) или две дозы 10 мг/кг в день 1 и день 3 (n = 3), соответственно. В день 4 после первой дозы 6TG фиксированные параформальдегидом срезы кости окрасили Н&Е и исследовали гистологию ВМ. Характерные микрофотографии (40-кратное увеличение) показаны для каждого режима кондиционирования 6TG. (Фиг. 1В) Характерные микрофотографии показывают при низких (10х) и высоких (100х) увеличениях общую и детальную гистологию ВМ мышей HPRT-wt, которые получили носитель, или с оптимизированным режимом кондиционирования, состоящим из двух доз 10 мг/кг 6TG в день 1 и день 3. Гистологические анализы провели в день 4, как указано выше.
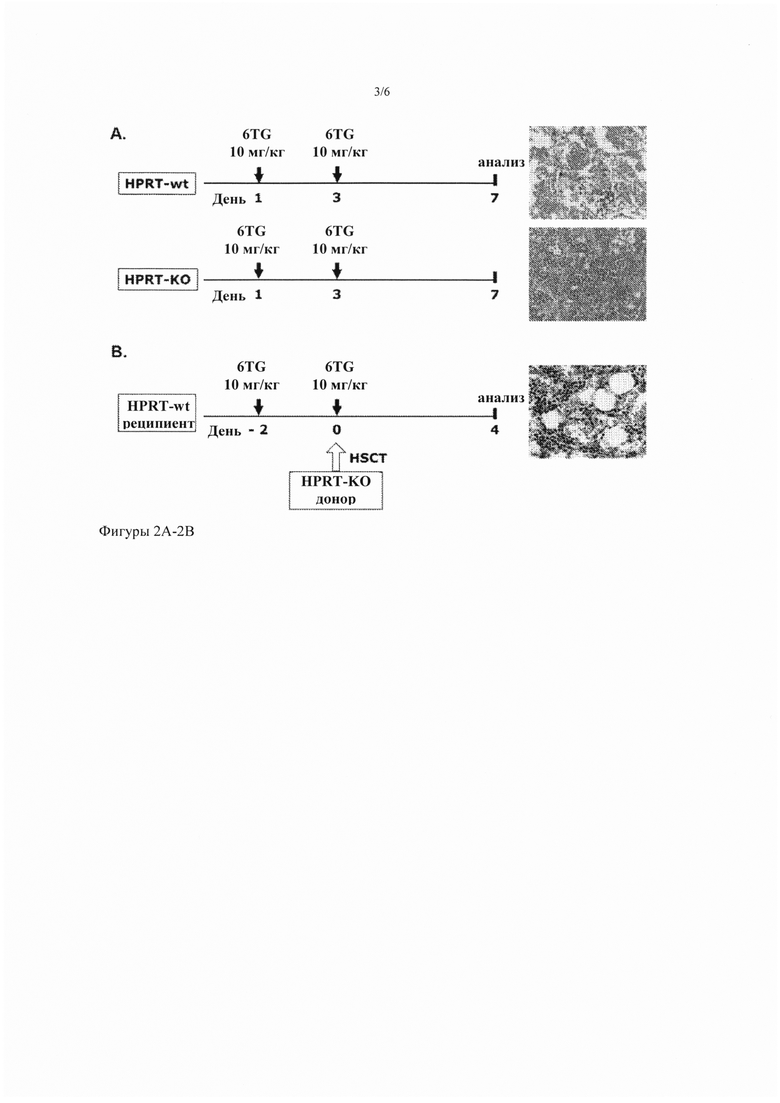
Фигура 2А-2В. Отсутствие прогрессивной миелотоксичности после инъекции кондиционирующих доз 6TG HPRT-дефицитным мышам и низкий уровень пересадки только с кондиционированием 6TG. (Фиг. 2А) HPRT-wt и HPRT-дефицитным мышам интраперитонеально инъецировали 10 мг/кг 6TG в дни 1 и 3. В день 7 после первой дозы 6TG, фиксированные параформальдегидом срезы кости окрашивали Н&Е и исследовали гистологию ВМ. Показаны характерные микрофотографии (40-кратное увеличение). (Фиг. 2В) График лечения: самки мышей-реципиентов HPRT-wt (n = 4) получили первую кондиционирующую дозу 6TG (10 мг/кг, интраперитонеально) в день -2, затем им пересадили ВМ HPRT-дефицитных самцов, а после этого - вторую кондиционирующую дозу 6TG (10 мг/кг, интраперитонеально) в день 0. ВМ проанализировали в день 7 после первой дозы 6TG. Показаны фиксированные параформальдегидом срезы кости, окрашенные Н&Е (40-кратное увеличение).
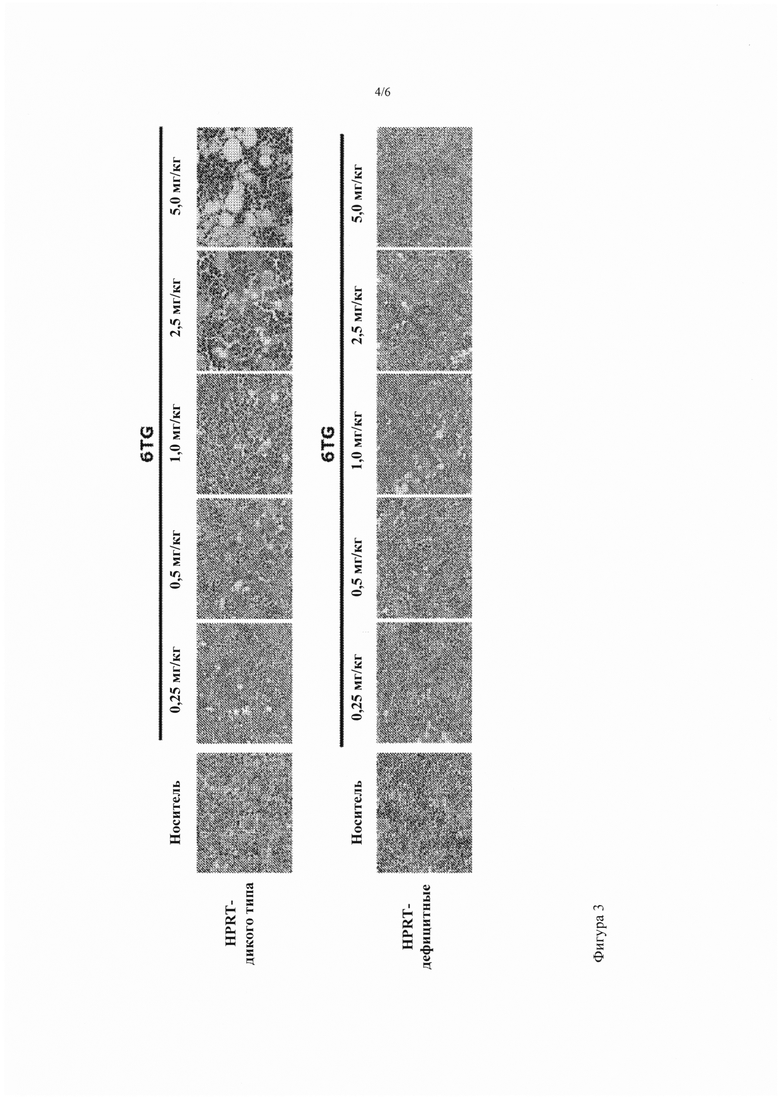
Фигура 3. Эффект дозы и период действия хронической миелотоксичности низких доз 6TG у HPRT-wt по сравнению с HPRT-дефицитными мышами. HPRT-мыши дикого типа и HPRT-дефицитные мыши получали каждые 3 дня различные дозировки 6TG или носитель (n = 3 на группу), как показано над каждой панелью. Для мышей HPRT дикого типа гистологию исследовали в следующие временные точки вплоть до 60 дней после начала лечения: носитель (день 60), 6TG 0,25 мг/кг (день 60), 6TG 0,5 мг/кг (день 60), 6TG 1,0 мг/кг (день 38), 6TG 2,5 мг/кг (день 28), 6TG 5,0 мг/кг (день 22). Для HPRT-дефицитных мышей гистологию исследовали в день 60 для всех животных. Показаны характерные микрофотографии фиксированных параформальдегидом срезов кости, окрашенных Н&Е (40-кратное исходное увеличение).
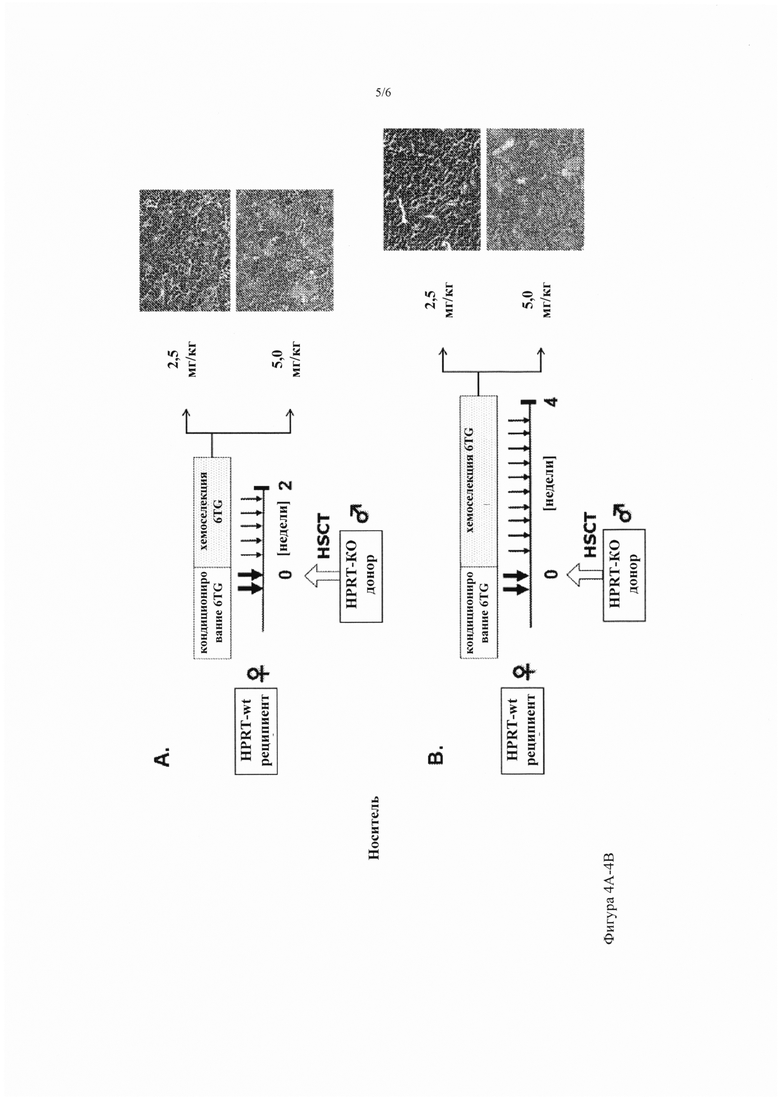
Фигура 4А-4В. Оптимизация совместного кондиционирования 6TG и стратегии хемоселекции in vivo. График лечения: самки мышей-реципиентов HPRT-wt получили первую кондиционирующую дозу 6TG (10 мг/кг, интраперитонеально) в день -2, затем им пересадили ВМ HPRT-дефицитных самцов вместе со второй кондиционирующей дозой 6TG (10 мг/кг, интраперитонеально) в день 0. Затем провели хемоселекцию in vivo с помощью многократных инъекций 2,5 мг/кг или 5,0 мг/кг 6TG каждые 3 дня в течение 2 недель (фиг. 4А) или 4 недель (фиг. 4В), как указано. Показаны характерные микрофотографии костного мозга из фиксированных параформальдегидом срезов, окрашенных Н&Е (40-кратное исходное увеличение).
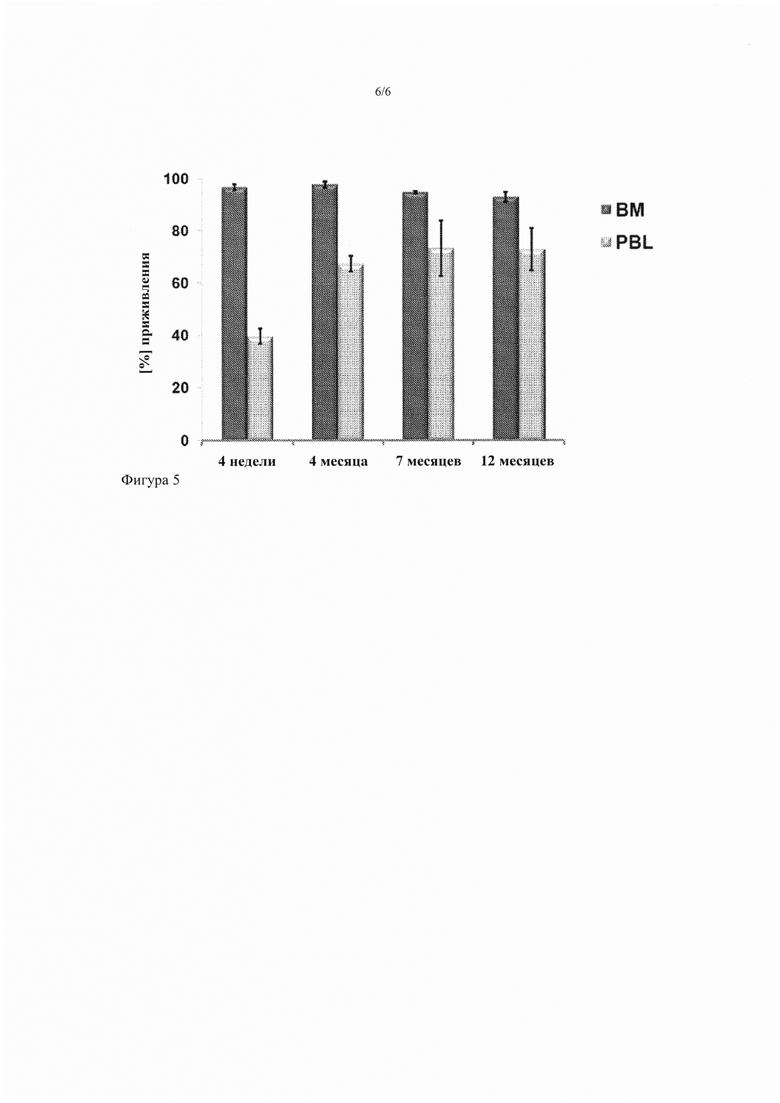
Фигура 5. Долгосрочное гемопоэтическое восстановление после трансплантации реципиентам HPRT-wt ВМ, полученного из HPRT-дефицитных доноров с помощью совместного кондиционирования 6TG и хемоселекции. Гистограммы показывают процентное отношение клеток костного мозга (ВМ), полученных от донора, и лейкоцитов периферической крови (PBL), определенное с помощью FISH анализа XY хромосом через 4 недели (т.е. сразу после окончания периода хемоселекции) и через 4 месяца, 7 месяцев и 12 месяцев после трансплантации, как указано.
ПОДРОБНОЕ ОПИСАНИЕ
Настоящее изобретение предлагает новую стратегию хемоселекции in vivo, которая применяет ключевую роль опосредованного гипоксантин-гуанин фосфорибозилтрансферазой (HPRT) преобразования 6-тиогуанина (6TG) в нуклеотид тиогуанина с миелотоксичностью 6TG. Поскольку дефицит HPRT сам по себе не ухудшает развитие или функцию гемопоэтических клеток, его можно удалить из гемопоэтических клеток, применяемых для трансплантации. Стратегия хемоселекции in vivo включает HSCT, которую осуществляют с применением HSC HPRT-дефицитных доноров, используя 6TG как для миелоаблативного кондиционирования реципиентов HPRT-дикого типа, так и процесса хемоселекции in vivo донорских клеток в один цикл. Настоящее изобретение основано на разработке и выявлении режима дозирования, при котором 6TG вызывает селективную миелоаблацию без каких-либо негативных последствий для экстра-гемопоэтических тканей, в то время как пересаженные HSC с недостаточной HPRT активностью обладают высокой устойчивостью к цитотоксическим эффектам 6TG. С помощью этой стратегии совместного кондиционирования 6TG и хемоселекции можно достигнуть эффективной и высокой пересадки HSC HPRT-дефицитных доноров с низкой общей токсичностью. 6TG в хемоселекции in vivo обеспечивает долгосрочное восстановление иммунофенотипически нормального костного мозга (ВМ), увеличивая популяцию самообновляющихся плюрипотентных HSC из ВМ HPRT-дефицитного донора или пересаженного ВМ.
Описанный в данном документе способ высокоэффективного и полностью нетоксичного кондиционирования, и хемоселекции in vivo в одном цикле, в общем применим в качестве стратегии улучшения эффективности пересадки HSCT и исход трансплантации, и придать селективное преимущество генетически модифицированным клеткам после генной терапии ex vivo. В стратегии хемоселекции in vivo используется исключительно 6TG или другой аналог пуриновых оснований, как для предварительного кондиционирования, так и хемоселекции HSC HPRT-дефицитных доноров в одном цикле, и она способна обеспечить высокоэффективную пересадку и долгосрочное восстановление с заменой >95% ВМ реципиента. Данная стратегия применима для улучшения пересадки большого числа HSC, манипулированных ex vivo, чтобы расширить применение генной терапии в целом.
Определения
Все научные и технические термины, используемые в данной заявке, имеют общеупотребимые в данной области техники значения, если не указано иное. Следующие слова или фразы, используемые в данной заявке, имеют указанные значения.
Используемый в данном документе термин ''трансплантация гемопоэтических стволовых клеток (HSC) без облучения" означает, что реципиент не подвергается миелоаблативному кондиционированию путем облучения. Вместо этого, кондиционирование осуществляют за счет введения 6TG, который обычно вводят в течение 48 часов до (и включая день) пересадки донорских HSC.
Используемый в данном документе термин ''HPRT-дефицитные'' включает как клетки, которые HPRT-дефицитные от природы, так и те, которые сделали HPRT-дефицитными путем генетической модификации.
Используемый в данном документе термин ''донорские HSC'' или ''донорские клетки'' относится к клеткам, которые будут пересажены, независимо от того, получены ли HSC от реципиента трансплантата или от другого субъекта. Таким образом, клетки, полученные из субъекта, могут быть модифицированы и пересажены обратно тому же субъекту, став ''донорскими клетками''. Они могут упоминаться в данном документе как ''донорские клетки'', ''пересаженные клетки'' или ''трансплантированные клетки''.
Используемые в данном документе артикли ''а'' или ''an'' означают, по меньшей мере, один, если явно не указано иное.
Совместное кондиционирование и хемоселекция для гемопоэтической трансплантации
Настоящее изобретение предоставляет способ трансплантации гемопоэтических стволовых клеток (HSC) без облучения. Как правило, способ включает введение млекопитающему субъекту одной или двух доз от 2 до 10 мг/кг массы тела аналога пуринового основания в качестве стадии предварительного кондиционирования. Способ дополнительно содержит пересадку субъекту доноров HSC с нехваткой гипоксантин-гуанин-фосфорибозилтрансферазы (HPRT) в пределах от 48 до 72 часов стадии предварительного кондиционирования и введение субъекту приблизительно от 1 до 5 мг/кг аналога пуриновых оснований каждые 2-4 дня в течение 2-8 недель после стадии пересадки.
Способ осуществляют в отсутствие предварительного кондиционирования путем облучения. Следовательно, субъект не подвергают миелоаблативному облучению при подготовке к трансплантации, и, таким образом, субъект не подвергается индуцированной миелоаблативным облучением токсичности. Способ рассматривается для применения с различными субъектами, в том числе с субъектами, которые никогда не подвергались лучевой терапии любого рода, субъектами, которые никогда не подвергались миелоаблативной лучевой терапии, а также субъектами, которых подвергали миелоаблативному лечению в прошлом, но не в сроки и/или с дозой, которая была бы предварительно кондиционирующей для способа, описанного в данном документе. Например, как правило, субъект не был бы подвержен миелоаблативному облучению в течение 2 недель, или даже в течение 8 недель, способа совместного кондиционирования и хемоселекции, описанного в данном документе.
Характерные примеры аналогов пуриновых оснований включают: 6-тиогуанин (6TG), 6-меркаптопурин (6-МР) и азатиопурин (AZA). В одном варианте осуществления аналогом пуриновых оснований является 6TG. В некоторых вариантах осуществления общая доза 6TG, которую вводят субъекту, не превышает 105 мг; обычно, общая доза 6TG, которую вводят субъекту, не превышает 75 мг.В одном варианте осуществления аналог пуринового основания вводят каждые 3 дня и на протяжении не более чем четырех недель после этапа пересадки.
Когда в качестве аналога пуринового основания применяют альтернативу 6TG, для инструктирования квалифицированного клинициста в определении подходящей дозы для применения в способе настоящего изобретения доступны известные онлайн (например, rxlist.com) и другие ресурсы. Например, обычная пероральная доза для однокомпонентной химиотерапии 6TG у детей и взрослых составляет 2 мг/кг массы тела в день; если после 4 недель эффект лечения не наблюдается, дозу можно увеличить до 3 мг/кг. В одной пероральной дозе давали вплоть до 35 мг/кг, наблюдая реверсивную миелосуппрессию.
В случае острой лимфатической лейкемии обычная начальная доза для пациентов педиатрического профиля и взрослых составляет 2,5 мг/кг массы тела 6-МР в день (от 100 до 200 мг для среднестатистического взрослого и 50 мг для среднестатистического 5-летнего ребенка). Пациенты педиатрического профиля с острой лейкемией в большинстве случаев легко переносили эту дозу; некоторым пациентам ее можно продолжать каждый день в течение нескольких недель или более. Если после 4 недель при такой дозировке нет клинического улучшения и точных признаков подавления лейкоцитов или тромбоцитов, дозу можно увеличить до 5 мг/кг в день. Дозировка 2,5 мг/кг/день может привести к быстрому падению количества лейкоцитов в течение от 1 до 2 недель у некоторых взрослых с острой лимфатической лейкемией и высоким общим количеством лейкоцитов. После достижения полной гематологической ремиссии считается необходимой поддерживающая терапия. Поддерживающие дозы будет изменяться от пациента к пациенту. Обычная суточная поддерживающая доза 6-МР составляет от 1,5 до 2,5 мг/кг/день в виде разовой дозы.
Дозировка для 6-TG и 6-МР в некоторой степени сопоставима, в то время как дозировку для AZA более трудно сравнивать, потому что его нужно сначала биоактивировать до 6-МР, и его, как правило, не применяют для лечения лейкемии. Для пациентов, которые получают трансплантацию твердого органа, доза AZA, которая требуется для предотвращения отторжения и минимизации токсичности, будет изменяться у отдельных пациентов, что требует тщательного управления. Начальная доза составляет обычно от 3 до 5 мг/кг ежедневно, начиная с момента трансплантации. AZA обычно дают в виде разовой ежедневной дозы в день, а в редких случаях от 1 до 3 дней до трансплантации. Снижение дозы до поддерживающих уровней от 1 до 3 мг/кг ежедневно обычно является возможным. Дозу AZA нельзя увеличить до токсичных уровней из-за угрозы отторжения.
HPRT-дефицитные доноры HSC могут быть HPRT-дефицитными от природы или их можно сделать HPRT-дефицитными путем генетической модификации. В этом контексте ''донорские HSC'' обозначает клетки, которые будут пересажены, независимо от того, получены HSC из реципиента трансплантата или другого субъекта. Трансплантированные HSC могут быть аутологичными, сингенными или аллогенными.
Генетическую модификацию можно осуществить с помощью любого из различных средств, известных специалистам в данной области техники. Примеры подходящих средств генетической модификации включают, без ограничения, введение последовательностей, кодирующих нуклеазы ''цинковые пальцы'' (ZFN), эффекторные нуклеазы, подобные активаторам транскрипции (TALEN), матричные цепи небольшого фрагмента гомологичной рекомбинации (SFHR), ингибирующие РНК (siRNA) или микроРНК (miRNA), антисмысловые РНК, транс-сплайсинговые РНК, рибозимы, внутриклеточные антитела или доминантно-негативные или конкурентные белки-ингибиторы. Модификация может быть применена непосредственно с донорскими HSC или клетками-предшественниками. Данные технологии могут быть применены для генетической модификации различных типов клеток, включая, без ограничения, гемопоэтических клеток-предшественников или непосредственно гемопоэтических стволовых клеток, а также других типов зрелых или эмбриональных стволовых клеток или индуцированных плюрипотентных стволовых клеток, которые могут быть дифференцированы или транс-дифференцированы в гемопоэтические клетки-предшественники или гемопоэтические стволовые клетки.
В некоторых вариантах осуществления HPRT-дефицитные HSC для трансплантации были генетически модифицированы для соответствия определенной терапевтической цели. Например, донорские HSC могут быть модифицированы для того, чтобы исправить наследственный генетический дефект для изменения чувствительности нормального костного мозга к цитотоксическим лекарственным средствам, чтобы придать устойчивость к инфекционным микроорганизмам, которые поражают лимфогематопоэтические клетки, чтобы заменить или возвратить в исходное состояние эндогенную иммунную систему, или для борьбы с лимфогематопоэтическими злокачественными новообразованиями путем замены эндогенного костного мозга и индукции эффекта трансплантат против лейкемии/лимфомы.
Более конкретно, наследственные генетические дефекты могут включать, без ограничения, расстройства гемопоэза, в том числе гемоглобинопатии, такие как серповидно-клеточная анемия, талассемия, наследственный сфероцитоз, нехватка G6PD и т.д., нарушения иммунологической или антимикробной функции, такие как тяжелый комбинированный иммунодефицит (SCID), хронический гранулематоз (CGD), расстройства тромбопоэза, ведущие к дефектам коагуляции, таким как синдром Вискотта-Олдрича (WAS), а также другие генетические структурные или метаболические расстройства, которые можно облегчить с помощью генной инженерии гемопоэтических клеток, которые передвигаются в места повреждения тканей, такие как различные формы буллезного эпидермолиза (ЕВ) и мукополисахаридоза.
Заболевания, при которых модификация чувствительности костного мозга к хемотоксическим лекарственным препаратам была бы полезной, включают, без ограничения, злокачественные заболевания, которые лечат химиотерапевтическими средствами, максимально переносимая доза которых ограничена миелотоксичностью. К ним относятся рак легких, рак толстой и прямой кишок, рак молочной железы, рак простаты, рак поджелудочной железы, рак желудка, рак печени, рак головы и шеи, почечно-клеточный рак, рак мочевого пузыря, рак шейки матки, рак яичников, рак кожи, саркомы и глиома.
Заболевания, при которых используют трансплантацию костного мозга или гемопоэтических стволовых клеток для замены или возвращения в исходное состояние эндогенной иммунной системы, включают, без ограничения, воспалительное заболевание кишечника, склеродермию и красную волчанку.
Заболевания, при которых было бы полезным придание устойчивости к инфекционным микроорганизмам, включают, без ограничения, ВИЧ-инфекцию и СПИД, HTLV-инфекцию и парвовирусную В19 инфекцию.
Злокачественные или предопухолевые заболевания лимфогеиопоэза, которые лечат трансплантацией костного мозга или гемопоэтических стволовых клеток, включают, без ограничения, острую миелоцитарную лейкемию, острый лимфоцитарный лейкоз, лимфому и миелодиспластические синдромы.
Другой пример терапевтического применения этой технологии будет заключаться в улучшении результата трансплантации костного мозга или гемопоэтических стволовых клеток после приобретенного нарушения эндогенного лимфогемопоэза, вызванного лучевым поражением и хемотоксинами.
Нетерапевтическим, но коммерчески полезным применением этой технологии будет его применение для создания гуманизированных животных моделей, в которых их эндогенный лимфогемопоэз почти полностью заменен клетками из человека-донора. После продуцирования, такие животные могут быть применены, например, для проверки новых лекарственных препаратов на миелотоксичность, которые рассматривают для применения к заболеваниям человека. Это является полезным, потому что чувствительность гемопоэза к различным лекарственным средствам может различаться в зависимости от вида животного, поэтому наиболее желательной является исследование таких препаратов на гуманизированной животной модели.
Как правило, субъектом является млекопитающее. Млекопитающий субъект может быть мышью, собакой, кошкой, крупным рогатым скотом, лошадью, овцой, приматом или человеком. В одном варианте осуществления субъектом является человек.
Введение и дозировка
Композиции вводят любым подходящим способом, часто с фармацевтически приемлемыми носителями. В контексте настоящего изобретения доступны подходящие способы введения лекарственного средства субъекту, и, хотя для введения конкретной композиции можно использовать более чем один путь, конкретный путь может часто обеспечить более быструю и более эффективную реакцию, чем другой путь.
Доза, которую вводят пациенту, в контексте настоящего изобретения должна быть достаточной, чтобы обеспечить благоприятный терапевтический ответ у пациента с течением времени, или чтобы ингибировать развитие заболевания. Таким образом, композицию вводят субъекту в количестве, достаточном, чтобы вызвать эффективный ответ и/или облегчить, уменьшить, вылечить или, по меньшей мере, частично остановить симптомы и/или осложнения болезни. Количество, необходимое для достижения этой цели, определяется как ''терапевтически эффективная доза''.
Пути и частота введения терапевтических композиций, раскрытых в данном документе, а также дозировка, будут изменяться от индивидуума к индивидууму, а также в зависимости от выбранного лекарственного средства, и могут быть легко установлены с помощью стандартных методов. В общем, фармацевтические композиции можно вводить путем инъекции (например, внутрикожной, внутриопухолевой, внутримышечной, внутривенной или подкожной), интраназально (например, путем аспирации) или перорально. Для отдельных пациентов могут быть подходящими альтернативные протоколы.
Как понятно специалистам в данной области техники, дозы могут быть преобразованы из мг/кг массы тела в мг/площадь поверхности тела, причем последнее подходит для применения с более крупными млекопитающими субъектами, включая человека. Калькуляторы для аллометрического вычисления известны в данной области техники и широко доступны в Интернете. Как правило, при аллометрическом вычислении используют экспоненту 0,75-0,80. Для получения дополнительной информации см. West & Brown, J Exp Bio 208, 1575-1592, 2005. Кроме того, Управление США по контролю пищевых продуктов и лекарственных средств публикует ''Guidance for Industry: Estimating the Maximum Safe Starting Dose in Initial Clinical Trials for Therapeutics in Adult Healthy Volunteers'', которое доступно от: Управления обучения и коммуникаций Отдела информации по лекарственным препаратам, HFD-240 Центр по оценке лекарственных препаратов и исследованиям пищевых продуктов и лекарственных препаратов, переулок Фишере, 5600, Роквилл, Мэриленд 20857.
Например, 5 мг/кг 6TG соответствует дозе 15,08 мг/м2 для мыши массой 20 г. Это соответствует 0,4 мг/кг для человека массой 65 кг.Абсорбция после перорального введения 6TG оценивается в 30%, поэтому эта интраперитонеальная доза у мышей соответствует абсорбированной дозе после перорального введения приблизительно 1,3 мг/кг у человека. Обычная пероральная доза для монокомпонентной химиотерапии 6TG у пациентов педиатрического профиля и взрослых составляет 2 мг/кг массы тела в день; если эффекта лечения не наблюдают после 4 недель, дозу можно увеличить до 3 мг/кг.
Способ настоящего изобретения предлагает неожиданное преимущество предотвращения токсичности, являющейся результатом или чрезмерного облучения субъекта, или чрезмерной дозировки 6TG. К удивлению, эффективное кондиционирование и восстановление костного мозга можно осуществить с помощью менее чем 105 мг общей дозы 6TG в течение курса лечения и в течение от двух до восьми недель. Эффективную пересадку наблюдали с общей дозой 6TG менее 65 мг и в течение всего лишь двух недель. Кроме того, заявленный способ делает возможным мониторинг токсичности у отдельного субъекта и регулировку дозирования для оптимизации эффективной пересадки с минимальной токсичностью для каждого субъекта. В некоторых вариантах осуществления субъекту вводят 1 или 2,5 мг/кг массы тела 6TG в ходе лечения после пересадки.
У субъектов, которых лечили в соответствии с данным способом, будет наблюдаться более 75% генетически модифицированных гемопоэтических клеток. В некоторых вариантах осуществления у субъекта наблюдали более 95% генетически модифицированных гемопоэтических клеток. Успешная пересадка у субъекта может быть подтверждена путем отбора проб периферической крови или костного мозга в различных интервалах после трансплантации и хемоселекции. Мононуклеарные клетки периферической крови могут быть изучены путем мониторинга уровней разрывов гена HPRT, нокдауна или снижения функциональной активности, используя различные стандартные методики, которые обычно известны любому специалисту в данной области техники, включая, без ограничения, полимеразную цепную реакцию (ПЦР), количественную ПЦР в реальном времени (Q-ПЦР), анализ с помощью нуклеазы surveyor (также называемый ''анализ Cel-I''), Саузерн-Блоттинг, Вестерн-Блоттинг/иммуноблот-анализ, иммуногистохимию или иммуноцитохимию, анализ проточной цитометрии с внутриклеточным окрашиванием, HPRT анализ ферментативной активности, HPLC, масс-спектрометрию и тому подобное.
ПРИМЕР
Следующий пример представлен для иллюстрации настоящего изобретения и для того, чтобы помочь специалистам при его получении и применении. Эти примеры никоим образом не предназначены для ограничения объема настоящего изобретения.
Пример 1: Совместное предварительное кондиционирование и хемоселекция in vivo только с 6-тиогуанином обеспечивают высокоэффективное восстановление нормального гемопоэза с помощью HPRT-дефицитного костного мозга
Пуриновые аналоги, такие как 6-тиогуанин (6TG), вызывают миелотоксичность при преобразовании в нуклеотиды гипоксантин-гуанин фосфорибозилтрансферазой (HPRT). Этот пример показывает разработку новой и высокоэффективной стратегии, использующей 6TG в качестве монотерапии как для кондиционирования, так и хемоселекции in vivo HPRT-дефицитных HSC. Эффект дозы и период действия миелотоксичности 6TG сначала сравнили в мышах HPRT дикого типа и HPRT-дефицитных трансгенных мышах. Параметры дозировки и режима оптимизировали, чтобы использовать 6TG для миелосупрессивного кондиционирования и сразу следующей за ним хемоселекции in vivo костного мозга (ВМ) HPRT-дефицитных трансгенных доноров, пересаженного сингенным реципиентам HPRT дикого типа.
В соответствующих дозах 6TG индуцировал избирательную миелотоксичность без какого-либо негативного воздействия на экстра-гемопоэтические ткани у мышей HPRT дикого типа, в то время как HSC с недостатком HPRT активности были весьма устойчивы к его цитотоксическому воздействию. Совместное кондиционирование 6TG и хемоселекция после трансплантации последовательно позволяли достичь -95% пересадки ВМ HPRT-дефицитных доноров, с низкой общей токсичностью. Долгосрочное восстановление иммунофенотипически нормального ВМ было достигнуто как у первичных, так и у вторичных реципиентов. Эти результаты предоставляют доказательство правильности концепции того, что монотерапия 6TG может быть применена как для миелосупрессивного кондиционирования, не требующего облучения, так и для хемоселекции in vivo HPRT-дефицитных донорских клеток. Результаты показывают, что путем применения миелосупрессивного воздействия 6TG как до (в качестве кондиционирования), так и после трансплантации (хемоселекция), можно достичь высокоэффективной пересадки HPRT-дефицитных гемопоэтических стволовых клеток.
Клиническая эффективность генной терапии ex vivo с использованием гемопоэтических стволовых клеток остается зависимой от придания селективного преимущества трансплантированным клеткам [1, 2]. В целях повышения пересадки и уменьшения времени, необходимого для лимфогемопоэтического восстановления, проверили стратегии селекции in vivo, использующие гены лекарственной устойчивости, такие как дигидрофолатредуктазу (DHFR) [3] или ген множественной лекарственной устойчивости 1 (MDR1) [4, 5], но, как правило, они потерпели неудачу из-за неприемлемой токсичности [6] или недостаточной эффективности селекции [7]. В настоящее время мутантные формы O-6-метилгуанин-ДНК-метилтрансферазы (MGMT) проходят испытания на их способность придавать хемопротекцию против BCNU или темозоломида в комбинации с O-6-бензилгуанином [8, 9], но эти средства также представляют собой значительный риск токсичности, и последние наблюдения показывают, что мутантная MGMT может придавать селективный недостаток при экспрессии на высоких уровнях [10].
Следует отметить, что эти подходы, как правило, основаны на трансплантации гемопоэтических клеток-предшественников, которые сверхэкспрессируют экзогенный ген лекарственной устойчивости, реципиентам, предварительно кондиционированным миелоаблативным облучением; однако хеморезистентность также можно придать понижением уровня эндогенных ферментов, которые обычно необходимы для активации цитотоксических лекарственных препаратов. В этом контексте мы ранее отметили, что высокие уровни фермента пути реутилизации пуриновых нуклеотидов гипоксантин-гуанин фосфорибозилтрансферазы (HPRT) приводят к повышенной чувствительности к пуриновому аналогу 6-тиогуанину (6TG) [11]. Первый этап метаболического преобразования 6TG катализируется HPRT [12], которая опосредует добавление рибозо-5-фосфата с получением тиогуанозин-монофосфата (TGMP). Таким образом, цитотоксичность 6TG, по существу, основывается на его опосредованном HPRT преобразовании в тио-дГТФ, которые затем включаются в ДНК, вызывая безрезультатное исправление ошибок спаривания и последующий апоптоз.
Чтобы придать миелопротекцию путем уменьшения активности, эндогенный фермент, активирующий лекарственные препараты, как правило, должен высоко экспрессироваться в гемопоэтических клетках-предшественниках, но быть несущественным для нормального гемопоэза. В действительности, гемопоэтические клетки-предшественники обычно экспрессируют высокие уровни HPRT [13-16], что делает их чрезвычайно чувствительными к 6TG. Действительно, на протяжении полувека для лечения лейкемии клинически применяли пуриновые аналоги, такие как 6-меркаптопурин (6МР), азатиопурин (Aza) и 6TG,, особенно у пациентов педиатрического профиля [17], а также для иммуносупрессии у пациентов с пересадкой органов, а в последнее время - для аутоиммунных заболеваний. При более высоких дозах, миелотоксичность является наиболее частым и постоянным побочным эффектом при клиническом применении 6TG, и при введении в соответствующих концентрациях в течение коротких периодов времени, 6TG является сильно миелосупрессивным с небольшой токсичностью для других тканей животных дикого типа с нормальной HPRT [11].
В отличие от этого, костный мозг (ВМ) HPRT-дефицитных животных обладает высокой устойчивостью к 6TG [11]. Примечательно, однако, что мы [11] и другие [18] обнаружили, что гемопоэз является фенотипически и функционально нормальным у животных с Hprt-иокаутом, и хотя случаи мегалобластной анемии связывают с наследственной нехваткой HPRT (синдром Леша-Нихена) у человека [19], сообщают, что это хорошо поддается лечению пероральным введением аденина [20]. Кроме того, нехватка HPRT не выглядит связанной с каким-либо тяжелым поражением иммунной системы у людей или у животных [21].
Эти наблюдения позволяют предположить, что HPRT-дефицитные гемопоэтические клетки-предшественники, которые являются устойчивыми к 6TG, но в остальном нормальными, должны иметь селективное преимущество после трансплантации реципиентам HPRT дикого типа, проходящих лечение 6TG, и что эту стратегию можно использовать для улучшения результата генной терапии ex vivo. В действительности, Porter и DeGregori [22] ранее показали возможность трансдукции HSC лентивирусным вектором, экспрессирующим shRNA с Hprt-мишенью, и обогащения этих сконструированных гемопоэтических клеток in vivo путем хемоселекции 6TG у мышей. Однако в этом предыдущем докладе 6TG применяли либо в дозах, которые выбрали только для умеренной миелосупрессии, или только в течение коротких периодов селекции, которые начали через 4 недели или более после трансплантации, и, несмотря на предварительное кондиционирование облучением всего тела, результаты пересадки были относительно незначительными и сильно различались, изменяясь в пределах 5 - 50% [22].
Теперь мы систематически изучили влияние изменения дозы, времени и продолжительности введения 6TG на пересадку и гемопоэтическое восстановление после трансплантации HPRT-дефицитного костного мозга. Чтобы исключить эффективность трансдукции вектора как переменную, мы использовали ВМ животных с Hprt - нокаутом в качестве "идеальных" доноров клеток, тем самым позволяя нам сосредоточиться на последствиях изменения дозировки 6TG и планировании параметров (i) предварительной трансплантации для миелосупрессивного кондиционирования реципиентов HPRT дикого типа, (ii) посттрансплантации для хемоселективного увеличения количества популяций клеток HPRT-дефицитных доноров. Следовательно, здесь мы опишем разработку нового режима, который последовательно использует 6TG в качестве монотерапии как для предварительного кондиционирования, так и хемоселекции in vivo, и покажем, что этот совместный режим быстро и регулярно обеспечивает высокоэффективную пересадку и долгосрочное восстановление.
Материалы и методы
Мыши
HPRT-дефицитных мышей B6.129P2-Hprt 1b-m3/J (CD45.2), мышей HPRT дикого типа (wt) C57BL/6J и B6.SJL-PtprcaPepcb/BoyJ (CD45.1) первоначально получили от лаборатории Джексона (Бар Харбор, Мэн). Мыши B6.129P2-Hprt 1b-m3/J содержат 55 kb делецию, которая охватывает промотор и первые 2 экзона гена Hprt [23]. Мышей выводили и содержали в институционном виварии, свободном от особых патогенов, при стандартных условиях в соответствии с нормами учреждения.
Лечение 6TG
Мышам C57BL/6J и B6.129P2-Hprt 1b-m3/J инъецировали интраперитонеально (i.p.) в различные временные точки 200 мкл 6TG в различных дозах (Sigma-Aldrich, Сент-Луис, Миссури), как описано в условных обозначениях к фигурам. Контрольным животным интраперитонеально инъецировали 200 мкл стерильной Н2О.
Трансплантация костного мозга и хемоселекция in vivo 6TG
Реципиентов-самок мышей C57BL/6J (HPRT-wt) или B6.SJL-PtprcaPepcb/BoyJ (HPRT-wt, CD45.1) лечили 10 мг/кг 6TG путем инъекции интраперитонеально за 48 часов до HSCT. Для HSCT 0,8-1 × 107 ядросодержащие клетки ВМ, взятые от мышей-самцов B6.129P2-Hprt 1b-m3/J (CD45.2), внутривенно инъецировали реципиентам HPRT-wt. 6TG (10 мг/кг) снова ввели путем инъекции интраперитонеально через 2 часа и каждые 3 дня после этого в дозе 5 мг/кг в течение 4 недель. Последовательную трансплантацию вторичным мышам-реципиентам провели, используя ту же дозу клеток и режим предварительного кондиционирования/хемоселекции 6TG, как описано выше, но с использованием ВМ первичных мышей-реципиентов, которые перенесли трансплантацию с хемоселекцией 6TG in vivo 6 месяцев ранее.
Флуоресцентная гибридизация in situ, специфическая для X и Y хромосом мыши FISH проводили на клетках ВМ и PBL, используя смесь специфических для мыши зонда окрашивания всей хромосомы Y/ДНК-зонда RAB9 (XqF1) (Kreatech, Амстердам, Нидерланды) в соответствии с протоколом производителя. Чтобы определить процент клеток самцов-доноров HPRT-wt в реципиентах-самках мышей C57BL/6J, подсчитали 200 ядер на слайд с помощью флуоресцентного микроскопа (Zeiss), снабженного соответствующими двойными и тройными цветными фильтрами. Для анализа FISH применяли следующие критерии: (1) качество ядер оценивали с помощью окрашивания DAPI, (2) для Y-хромосомы оценивали зеленый флуоресцентный сигнал, (3) для X-хромосомы оценивали красный флуоресцентный сигнал, (4) при отсутствии зеленого флуоресцентного сигнала для Y-хромосомы в ядре регистрировали самку, даже если обнаружили только одну Х-хромосому (дополнительная фиг. S1; см. дополнительные фигуры в Experimental Hematology 2012, 40:3-13).
Иммунофенотипический анализ гемопоэтических тканей
После блокирования с помощью Mouse BD Fc Block (BD Biosciences, Сан-Хосе, Калифорния), ВМ, PBL, тимус или клетки селезенки окрашивали FITC-, РЕ-, PerCP- или АРС-конъюгированными антимышиными антителами крысы против CD45, CD45.2, CD4, CD8, Mac1/Gr1, В220, Sca-1, c-kit мыши и коктейлем антител линии. Антитела получили от Biolegend (Сан Диего, Калифорния) или BD Biosciences. Данные проточной цитометрии получили на работающем на BD LSRII BD FACSDiva (BD Biosciences) и проанализировали с использованием программного обеспечения FlowJo (TreeStar, Ашленд, Орегон) (дополнительная фиг. S2).
Гистопатологический анализ
Вскрытие всех мышей, использованных в данном исследовании, и гистологическое исследование всех грудных органов и органов брюшной полости и костного мозга провели в отделе UCLA лаборатории ветеринарной службы диагностики. Ткани регулярно обрабатывали или, если необходимо, декальцинировали, делали парафиновые срезы и окрашивали гематоксилином и эозином (Н&Е).
Статистический анализ
Данные проанализировали с помощью статистического программного обеспечения QuickCalcs (GraphPad Software Inc.). Для расчета величины р использовали непарные t-тесты, причем р<0,05 рассматривалось как статистически значимое.
Результаты
Острая миелотоксичность 6TG у мышей HPRT-wt
Прогнозируют, что высокие уровни экспрессии HPRT в гемопоэтических клетках-предшественниках опосредуют селективный миелотоксический эффект 6TG и делают возможным его применение в качестве режима кондиционирования. Соответственно, мы исследовали краткосрочные влияния на ВМ после разовой инъекции 6TG в различных дозировках в мышей HPRT-wt. Провели внутрибрюшинную инъекцию однократной дозы 2,5 мг/кг, 5,0 мг/кг или 10 мг/кг 6TG в день 1 или инъекцию двух доз 10 мг/кг в дни 1 и 3 и исследовали гистологию ВМ в день 4. При увеличении общей дозировки 6TG наблюдали увеличение миелотоксичности (фиг. 1А). Сосудистые структуры ставали все более выраженными, а клетки как эритроидных, так и миелоидных линий были исчерпаны. При самых высоких исследованных дозах 6TG остались только эндотелий сосудов, мезенхимальные клетки, некоторые зрелые гранулоциты и макрофаги и единичные гемопоэтические клетки-предшественники (фиг. 1В).
Никаких очевидных клинических признаков не наблюдали в день 4 с любым из указанных выше режимов, но когда период наблюдения продлили, в день 7 наблюдали потерю веса и бледность конечностей. Гистологическое исследование ВМ показало увеличение интенсивности между днем 4 и днем 7 даже без введения дополнительных доз 6TG (фиг. 1А, 2А). В соответствии с клиническими и гистологическими данными ВМ мышей HPRT-wt, которые получили две дозы 10 мг/кг 6TG и которых анализировали на день 7, показал значительное количественное уменьшение ядросодержащих клеток ВМ, извлеченных из одного бедра и голени (4,7×106±1,0×106, n = 3) по сравнению с мышами, получавшими носитель HPRT-wt (1,1×107±0,2×107, n = 3) (p<0,01).
Иммунофенотипический анализ остальных гемопоэтичных клеток ВМ методом проточной цитометрии (таблица 1) показал, что относительная доля клеток-предшественников KLS (lin-/c-kit+/sca-1+), которая включает HSC, с долгосрочной мультилинейной восстанавливающей активностью [24], статистически достоверно уменьшилась в 3 раза ко дню 4 (р<0,01) и в 10 раз ко дню 7 (р<0,001) (таблица 1 - iii,-iv). Относительная доля зрелых Т-клеток CD8+ и CD4+ постепенно увеличивалась с течением времени после введения 6TG, доходя до увеличения в 7 раз по сравнению с контролями ко дню 7 (р<0,001), вероятно в связи с чрезвычайно расширенными кровеносными сосудами и притоком периферической крови/кровоизлиянием, как можно также видеть на гистологии ВМ. Относительный процент клеток В220+ не показал существенных изменений ко дню 4, но увеличился в два раза по сравнению с контролями ко дню 7 после введения 6TG.
Таблица 1. Иммунофенотипический анализ гемопоэтических клеток ВМ после режима кондиционирования 6TG.
ВМ клетки окрасили с помощью следующих антимышиных антител крысы: CD45-FITC, CD4-PE, CD8-APC, B220-PerCP, Macl/Grl-PE, Sea-1-РЕ и c-kit-FITC и исследовали с помощью проточной цитометрии. Режимы лечения и анализа обозначены малыми римскими цифрами и являются такими же, как на фигуре 1. (i): Носитель в день 1, анализ в день 4. (ii) Разовая доза 10 мг/кг 6TG в день 1, анализ в день 4. (iii): Две дозы 10 мг/кг 6TG в дни 1 и 3, анализ в день 4. (iv) и (v): Две дозы 6TG 10 мг/кг в дни 1 и 3, анализ в день 7. Последние две колонки показывают результаты той же дозировки 6TG и режима (10 мг/кг 6TG × 2 дозы) у HPRT-wt (iv) и HPRT-дефицитных (v) мышей, соответственно. Проценты указанных субпопуляций гемопоэтических клеток выражены как средний % ± SD от общего числа клеток CD45 + (n = 3 на группу).
Следует отметить, что ферменты печени не были повышены после этого режима кондиционирования у мышей HPRT-wt, что указывает на селективность цитотоксичности 6TG в этой дозировке для гемопоэтических клеток-предшественников, и предполагает его потенциал для применения в качестве режима миелосупрессивного кондиционирования.
Отсутствие миелотоксичности при кондиционирующей дозировке 6TG в HPRT-дефицитных мышей
В отличие от вышеуказанных полученных данных, когда HPRT-дефицитным мышам ввели максимальную дозу (две дозы 10 мг/кг 6TG в дни 1 и 3), гистология костного мозга осталась полностью неизмененной в день 7 (фиг. 2А) и была сравнимой с таковой в группе контрольного носителя (фиг. 1А). Важно отметить, что общее количество ядросодержащих клеток ВМ, полученных из одного бедра и голени обработанных 6TG HPRT-дефицитных мышей (1,5×107±0,3×107, n = 3) также было сопоставимо с таковым мышей HPRT-wt, которые получили носитель (1,1×107±0,2×107, n = 3), и было значительно выше, чем у мышей HPRT-wt, которые получили 6TG по тому же режиму (4,7×106±1,0×106, n = 3) (р<0,005).
Кроме того, соотношения субпопуляций гемопоэтических клеток в ВМ HPRT-дефицитных мышей, которые получили 6TG (таблица 1-v), были сопоставимы с таковыми мышей HPRT-wt, которые получили только носитель (таблица 1-i), и не получавших терапию мышей HPRT-wt (таблица 2, колонка ВМ%), а также не получавших терапию HPRT-дефицитных мышей (таблица 3, колонка ВМ%).
Таблица 2. Иммунофенотипический анализ гемопоэтических клеток мышей HPRT-wt, не получавших терапию.
ВМ, PBL, селезенку (S) и тимус (Т) окрасили с помощью следующих антимышиных антител крысы: CD45-FITC (ВМ, PBL, Т, S), CD4-PE (ВМ, PBL, Т, S), CD8-APC (ВМ, PBL, Т, S), Macl/Grl-PE (ВМ, PBL, S), B220-PerCP (ВМ, PBL, S), Sca-1-РЕ (ВМ) и c-kit-FITC (ВМ) и исследовали методом проточной цитометрии. Проценты указанных субпопуляций гемопоэтических клеток выражены как среднее % ±SD от общего количества клеток CD45+ (n = 3 на группу).
Таблица 3. Иммунофенотипический анализ гемопоэтических клеток в необработанных HPRT-дефицитных мышах.
ВМ, PBL, селезенку (S) и тимус (Т) окрасили с помощью следующих антител крысы против мышиных антител: CD45-FITC (ВМ, PBL, Т, S), CD4-PE (ВМ, PBL, Т, S), CD8-APC (ВМ, PBL, Т, S), Macl/Grl-PE (ВМ, PBL, S), B220-PerCP (ВМ, PBL, S), Sca-1-РЕ (ВМ) и c-kit-FITC (ВМ) и исследовали методом проточной цитометрии. Проценты указанных субпопуляций гемопоэтических клеток выражены как среднее % ±SD от общего количества клеток CD45+ (n = 3 на группу).
Вместе взятые, эти результаты свидетельствуют о том, что можно использовать до двух доз 10 мг/кг 6TG в качестве эффективного режима кондиционирования, который будет хорошо переносится, в течение до 3 дней до HSCT, причем постоянно возрастающая миелотоксичность происходит в течение 7 дней.
Трансплантация HPRT-дефицитного ВМ у реципиентов HPRT-wt после кондиционирования 6TG
На основании графика, установленного выше, мы использовали 6TG (10 мг/кг интраперитонеально) в качестве режима кондиционирования в реципиентов HPRT-wt (CD45.1), причем одну дозу ввели за 48 часов до (теперь обозначаемым днем -2, а не днем 1) и одну дозу ввели в день трансплантации (теперь обозначаемым днем 0, а не днем 3) в соответствии с режимом, установленным выше. После кондиционирования реципиентам HPRT-wt затем трансплантировали ВМ от HPRT-дефицитных конгенных доноров (CD45.2).
На день 4 в костном мозге обнаружили снижение насыщенности клетками с меньшим количеством ранних клеток-предшественников и увеличением васкуляризации (фиг. 2В), а общее количество содержащих ядро клеток ВМ, выделенных из этих кондиционированных 6TG мышей HPRT-wt, которым пересадили HPRT-дефицитный ВМ (3,8×106±0,5×106, n = 4), все еще было значительно уменьшенным (р<0,001) по сравнению с группой HPRT-wt (1,1×107±0,2×107, n = 3), которая получала носитель. Анализ ВМ проточной цитометрией показал, что в день 4 только 17,8% ± 4,4% от общей популяции клеток получили от доноров HPRT-дефицитных гемопоэтических клеток CD45.2+.
Порог миелотоксичности после хронического лечения низкими дозами 6TG
Поскольку наши результаты выше показали, что предварительное кондиционирование 6TG само по себе может быть недостаточным для достижения высоких уровней пересадки, мы затем провели исследование подбора дозы на мышах, не подвергнутых трансплантации, получавших длительное лечение более низкими дозами 6TG. HPRT-wt и HPRT-дефицитным мышам (n = 3 на группу) инъецировали интраперитонеально только носитель, 0,25 мг/кг, 0,5 мг/кг, 1,0 мг/кг, 2,5 мг/кг или 5,0 мг/кг 6TG каждые 3 дня в течение до 60 дней. Мыши HPRT-wt, контрольная группа, получавшая носитель, а также группы, получавшие 0,25 мг/кг и 0,5 мг/кг 6TG, показали 100% выживаемость в течение периода 60 дней, и гистологическое исследование ВМ в группах, получавшие 0,25 мг/кг и 0,5 мг/кг 6TG, показало нормальную насыщенность клетками в день 60 (фиг. 3).
В отличие от этого, в группе HPRT-wt, которая получала 1,0 мг/кг 6TG, смертельные случаи произошли в день 38 (общая доза 13 мг/кг), день 42 (общая доза 14 мг/кг) и день 51 (общая доза 17 мг/кг) (таблица S1). При более высоких дозах мыши HPRT-wt, получающие повторные инъекции 2,5 мг/кг или 5 мг/кг 6TG, систематически проявляли прогрессирующие клинические признаки недомогания (бездействие, выгибание спины, отсутствие ухода за поверхностью тела, анорексия), анемию (бледность конечностей) и потерю >10% веса, что, согласно нормам учреждения, влечет за собой необходимость умерщвления в день 28 (группа, которая получала 2,5 мг/кг 6TG; общая доза 22,5 мг/кг) и в день 22 (группа, которая получала 5,0 мг/кг 6TG; общая доза 35 мг/кг), соответственно. Гистологическое исследование ВМ группы, которая получила 1,0 мг/кг 6TG, показало несколько апоптических фигур и больше бластных клеток, чем в группах с более низкой дозировкой, вероятно отражая первоначальный активирующий ответ на травму (фиг. 3; день 38). Группы, которые получали более высокие дозы, также показали значительное снижение насыщенности клетками и расширение сосудистых структур, с тяжестью поражений, пропорциональной совокупной дозе 6TG в каждой группе. Мыши, получавшие повторные дозы 2,5 мг/кг 6TG, показали снижение насыщенности клетками и обширный апоптоз (день 28). При наивысшей хронической дозировке 5 мг/кг ВМ заметно истощился, причем большинство выживших клеток являются миелоидного происхождения (день 22).
В отличие от мышей HPRT-wt, все HPRT-дефицитные мыши выжили в течение эксперимента (60 дней), независимо от дозы инъецированного 6TG (до 105 мг/кг максимальной общей дозы, вводимой в соответствии с теми же режимами дозировок, как указано выше) (таблица S1). Никакой существенной патологии ВМ не наблюдали ни у одной из HPRT-дефицитных мышей, независимо от дозы 6TG, в последнюю временную точку эксперимента в день 60 (фиг. 3). Никаких значительных отклонений, связанных с лечением, не обнаружили ни в одной другой исследованной ткани, в том числе в сердце, легких, печени, поджелудочной железе, почках и селезенке. Таким образом, у HPRT-дефицитных мышей не обнаружили никаких токсических действий длительного лечения 6TG в дозах 1,0 мг/кг, 2,5 мг/кг и 5,0 мг/кг, которые вызвали смертельную миелотоксичность у мышей HPRT-wt.
Совместное кондиционирование и хемоселекция in vivo 6TG обеспечивают систематическое и высокоэффективную пересадку HPRT-дефицитного ВМ
На основе вышеизложенного исследования подбора дозы мы затем поинтересовались, может ли предварительное кондиционирование 6TG в сочетании с дальнейшим введением низких доз 6TG, начинаясь сразу после трансплантации HPRT-дефицитного ВМ реципиентам HPRT-wt, обеспечить дальнейшее хемоселективное увеличение количества пеерсаженных донорских клеток. Соответственно, самок мышей HPRT-wt предварительно кондиционировали 6TG (10 мг/кг IP), причем одну дозу ввели за 48 часов до (день -2), а одну дозу ввели в день трансплантации (день 0) согласно режима кондиционирования, установленного ранее. Реципиентам-самкам HPRT-wt затем трансплантировали ВМ HPRT-дефицитных самцов, и на основе вышеописанных результатов хронической миелотоксичности реципиентам затем дополнительно вводили повторные дозы 2,5 мг/кг 6TG каждые 3 дня в течение 2 недель (общая доза 30 мг/кг) или 4 недель (42,5 мг/кг) или повторные дозы 5,0 мг/кг 6TG каждые 3 дня в течение 2 недель (общая доза 40 мг/кг) или 4 недель (общая доза 65 мг/кг). Анализ провели непосредственно после периода хемоселекции in vivo через 2 или 4 недели, соответственно (фиг. 4).
Как и ожидалось, совместное кондиционирование 6TG, HSCT и хемоселекция in vivo 6TG хорошо переносились, и не наблюдалось никаких признаков недомогания. Во всех группах, которые получили 6TG, выжили 100% животных, подверженных трансплантации, (2 недели лечения: n = 3 и 4 недели лечения: n = 8). Массы тела животных, которые получали 6TG, первоначально снизилась в течение первой недели после трансплантации, но стабилизировалась, и все животные после этого заново набрали нормальный вес. Гистопатологический анализ показал, что общая насыщенность клетками и гемопоэз у животных, подвергнутых трансплантации, были неотличимы от контрольной группы HPRT-wt, не получавшей терапию, независимо от дозировки или продолжительности хемоселекции 6TG (фиг. 4).
FISH XY-хромосом [25, 26] показал, что в группах, получавших хемоселекцию 6TG в течение 2 недель, ВМ в этой временной точке уже был в значительной мере восстановлен полученным из донора костным мозгом на уровнях 89,3% ± 1,7 (группа 2,5 мг/кг) и 95,5% ± 1,2 (группа 5 мг/кг). Проценты полученных из донора лейкоцитов периферической крови (PBL) в 2 недели составляли 13,0% ± 4,6 (группа 2,5 мг/кг) и 12,7% ± 2,9 (группа 5,0 мг/кг), соответственно (таблица 4). Когда хемоселекцию in vivo продолжали вплоть до 4 недель после HSCT, процент полученных от донора клеток ВМ снова оказался чрезвычайно высоким, как в группе, которая получала 2,5 мг/кг (95,3% ± 0,9), так и в группе, которая получала 5,0 мг/кг (96,7% ± 1,2). Примечательно, что процент полученных от донора PBL был значительно выше в группе, которая получала 5,0 мг/кг (39,7% ± 3), по сравнению с группой, которая получала 2,5 мг/кг (29,9% ± 1,9), через 4 недели (р<0,005), а также значительно выше (р<0,0002), чем в группах, получавших хемоселекцию в любой дозе в течение 2 недель (таблица 4). Таким образом, предварительное кондиционирование 10 мг/кг 6TG в день -2 и день 0, в сочетании с продолжающейся хемоселекцией in vivo с 5 мг/кг 6TG каждые 3 дня в течение 4 недель, дало максимальные уровни пересадки ВМ из клеток HPRT-дефицитных доноров, а также самые высокие уровни полученных из доноров PBL; этот режим использовали в дальнейших исследованиях.
Таблица 4. Выживание и пересадка после совместного кондиционирования и хемоселекции in vivo 6TG.
Схемы лечения для хемоселекции in vivo являются, как указано: хемоселекция in vivo с 2,5 мг/кг 6TG в течение 2 недель, 5,0 мг/кг 6TG в течение 2 недель, 2,5 мг/кг 6TG в течение 4 недель или 5,0 мг/кг в течение 4 недель. Все группы, которые получали лечение, показали 100% выживаемость. Пересадку гемопоэтических клеток HPRT-дефицитных самцов в ВМ и PBL самок-реципиентов HPRT-wt определяли с помощью FISH XY-хромосом (среднее % ±SD).
Совместное кондиционирование и хемоселекция in vivo 6TG приводит к долгосрочному восстановлению HPRT-дефицитного ВМ
Надежность пересадки ВМ HPRT-дефицитного донора с помощью совместного кондиционирования и режима хемоселекции in vivo 6TG, установленного выше, исследовали через 4 месяца, 7 месяцев или 12 месяцев после трансплантации (т.е. 3 месяца, 6 месяцев и 11 месяцев после окончания 4-недельного периода хемоселекции in vivo, соответственно). Все животные, подверженные трансплантации, (4 месяца: n = 8; 7 месяцев: n = 6; 12 месяцев: n = 5) остались живы и здоровы, не показывая никаких признаков заболевания или дискомфорта, во всех обследованных временных точках. Макроскопическое патологическое и гистологическое исследование данных животных не выявило существенных нарушений.
Мультилинейное восстановление лимфогемопоэза посредством полученных от донора клеток-предшественников оценили через 4 месяца после HSCT с совметным предварительным кондиционированием и хемоселекцией in vivo 6TG, т.е. через 3 месяца после окончания 4-недельного периода хемоселекции. Иммунофенотипирование ВМ, PBL, тимуса и селезенки проводили в конгенном трансплантате CD45.1/CD45.2, применяя HPRT-дефицитных мышей CD45.2 в качестве доноров и мышей HPRT-wt CD45.1 в качестве реципиентов (n = 5). Все гемопоэтические ткани показали высокую пересадку донорских клеток CD45.2+ на уровнях, превышающих 75% общего объема костного мозга (таблица 5). Иммунофенотипирование полученной от доноров популяции CD45.2+ показало, что относительное процентное содержание Т-клеток (CD4/CD8), В220+ клеток и макрофагов/гранулоцитов (Mac-1/Gr1) было сопоставимо с таковым для контрольных групп, не получавших терапии (таблицы 2 и 3).
Таблица 5. Иммунофенотипический анализ гемопоэтических тканей через 4 месяца после трансплантации, применяя кондиционирование и режим хемоселекции in vivo 6TG в модели конгенного трансплантата CD45.1/CD45.2. Кондиционирование и хемоселекцию in vivo 6TG проводили, как описано согласно тексту. ВМ и PBL реципиента окрасили с помощью следующих антимышиных антител крысы: CD45.2-FITC (ВМ, PBL, Т, S), CD4-PE (ВМ, PBL, Т, S), CD8-APC (ВМ, PBL, Т, S), Mac1/Gr1-PE (ВМ, PBL), В220-PerCP (ВМ, PBL, S) и исследовали методом проточной цитометрии. Проценты указанных субпопуляций гемопоэтических клеток выражены как средний % ± SD от общего количества клеток CD45.2 + (n = 5 на группу).
клеток
Уровни пересадки также оценили с помощью FISH XY-хромосом через 4 месяца, 7 месяцев и 12 месяцев после трансплантации. Стабильное долгосрочное восстановление с помощью полученного от донора ВМ на высоких уровнях наблюдали во все моменты времени, с уровнями пересадки 97,7% ± 0,5% (4 месяца), 94,7% ± 1,9% (7 месяцев) и 93,0% ± 0,8% (12 месяцев), соответственно, после трансплантации (фиг. 5). Кроме того, процент полученных от донора PBL был значительно увеличен (67,4% ± 10,6% через 4 месяца, р=0,01; 73,3% ± 7,9% через 7 месяцев, р<0,01; 73,0% ± 10,7% через 12 месяцев, р<0,01) в сравнении с процентом PBL сразу же после 4-недельного периода селекции 6TG (39,7% ± 3,0%).
Относительные процентные содержания клеток CD4+ и CD8+, клеток В220+ и клеток Mac-1+/Gr1+, а также HSC KLS (lin-/sca-1+/c-kit+) определили в ВМ путем иммунофенотипирования через 4 месяца, 7 месяцев и 12 месяцев (таблица 6) и сравнили с контрольными мышами-самками HPRT-wt, которые не получали терапию (таблица 2) и HPRT-дефицитными самцами, которые не получали терапию (таблица 3). Во все три временнные точки относительное процентное содержание каждой клеточной популяции было сопоставима с таковым в контролях, хотя количество клеток KLS было повышено через 4 месяца и 7 месяцев и в пределах нормального диапазона через 12 месяцев после HSCT. Таким образом, с помощью полученного от доноров HPRT-дефицитного костного мозга удалось осуществить долгосрочное воссоздание нормального гемопоэза, по меньшей мере через 12 месяцев после трансплантации.
Таблица 6. Иммунофенотипический анализ ВМ через 4 месяца, 7 месяцев и 12 месяцев после трансплантации HPRT-дефицитного ВМ, применяя кондиционирование и режим хемоселекции in vivo 6TG. В указанные временные точки после HSCT с кондиционированием и хемоселекцией in vivo 6TG клетки ВМ реципиента окрасили следующими антимышиными антителами крысы: CD45-FITC, CD4-PE, CD8-АРС, Mac1/Gr1-PE, В220-PerCP, Sca-1-PE и c-kit-FITC и исследовали методом проточной цитометрии. Проценты указанных субпопуляций гемопоэтических клеток выражены как средний % ±SD от общего количества клеток CD45+.
клеток
ВМ (%)
ВМ (%)
ВМ (%)
Гемопоэтическое восстановление вторичных реципиентов после последовательной трансплантации ВМ HPRT-дефицитных доноров с совместным кондиционированием и хемоселекцией 6TG
Для дальнейшей оценки, селектирует ли оптимизированный режим совместного кондиционирования и хемоселекции 6TG долгосрочно репопулирующие HSC, мы затем трансплантировали ВМ от первичных реципиентов через 7 месяцев после трансплантации вторичным реципиентам [27], применяя тот же режим. Вторичных реципиентов, подверженных трансплантации, затем содержали в течение 3 месяцев после окончания их 4-недельного курса хемоселекции in vivo 6TG (дополнительная фигура S3). После кондиционирования и хемоселекции 6TG HPRT-дефицитные клетки самцов-доноров, которые приживили первичным реципиентам, были способны к последовательной репопуляции вторичных самок-реципиентов на высоких уровнях (95,5% ± 1,1%, как определили с помощью XY-FISH). Иммунофенотипический анализ показал, что проценты всех клеточных популяций, исследованных в ВМ и PBL вторичных реципиентов (таблица 7), снова были сопоставимы с таковыми контрольных групп, не получавших терапию (таблицы 2 и 3).
Таблица 7. Иммунофенотипический анализ гемопоэтических клеток вторичных реципиентов после последовательной трансплантации HPRT-дефицитного ВМ с кондиционированием и хемоселекцией in vivo 6TG.
Последовательную трансплантацию с кондиционированием и режим хемоселекции in vivo 6TG провели, как описано на фигуре S3. ВМ и PBL вторичного реципиента окрасили с помощью следующих антимышиных антител крысы: CD45-FITC (ВМ, PBL), CD4-PE (ВМ, PBL), CD8-APC (ВМ, PBL), Mac1/Gr1-PE (ВМ, PBL) и B220-PerCP (ВМ, PBL) и исследовали методом проточной цитометрии. Проценты указанных субпопуляций гемопоэтических клеток выражены как средний % ±SD от общего количества клеток CD45 + (n = 6 на группу).
Мы разработали оптимизированный режим, который применяет 6TG в качестве монотерапии для предтрансплантационного кондиционирования, а также продолжающейся после трансплантации хемоселекции in vivo HSC HPRT-дефицитных доноров. Эта стратегия совместного кондиционирования и хемоселекции 6TG обеспечила эффективную пересадку HSC с низкой общей токсичностью, через прогрессирующий и одновременный процесс замены, в котором реципиенты показали небольшое или не показали вообще никакого недомогания и 100% выживаемость, в то время как их ВМ был быстро и почти полностью заменен клетками HPRT-дефицитного донора, систематически достигая -95% пересадки по XY-FISH и >75% по иммунофенотипированию CD45.2. Процент полученных от доноров PBL значительно увеличился с течением времени (через 4 и 7 месяцев после трансплантации) по сравнению с процентом PBL сразу же после 4-недельного периода селекции 6TG. Остаточные клетки реципиента в PBL вероятно отражают более низкий оборот неделящихся зрелых клеток в PBL. Стабильное долгосрочное восстановление ВМ было достигнуто как у первичных, так и у вторичных реципиентов. Иммунофенотипический анализ ВМ, PBL, селезенки и тимуса показал, что после длительного восстановления гемопоэтическая дифференциация не была подвержена влиянию со стороны хемоселекции in vivo 6TG.
Эти результаты также подтверждают наше предыдущее наблюдение того, что 6TG в соответствующих концентрациях, по-видимому, вызывает селективную миелотоксичность без каких-либо негативных последствий для экстра-гемопоэтических тканей. HPRT экспрессируется на низких уровнях во всех соматических клетках [30], и унаследованная потеря HPRT вызывает синдром Леша-Нихена [31], который проявляется в виде тяжелой умственной отсталости и поведенческих отклонений, поскольку более высокие уровни экспрессии и активности HPRT были найдены в центральной нервной системе, в частности во время развития нервной системы [32]. Однако, поскольку полностью дифференцированные нейроны не подвергаются репликации, эти более высокие уровни не преобразовываются в более высокую нейротоксичность 6TG у зрелых особей.
В предыдущей работе Porter и DeGregori [22] для достижения миелоаблации использовали общее облучение тела величиной 4,5 Гр, a 6TG использовали только для хемоселекции в более поздние периоды времени. В отличие от этого, в нашем режиме 6TG в качестве монотерапии выполняет двойную роль кондиционирования (циторедукции ВМ хозяина) и хемоселективного лекарственного препарата (увеличение ВМ донора). Известно, что для токсичности 6TG требуются два цикла репликации ДНК, чтобы привести к апоптозу [33], и, следовательно, ее эффект замедленный. Наши результаты также указывают на зависимость миелотоксического эффекта 6TG от дозы и времени. Таким образом, кондиционирование с 10 мг/кг 6TG до трансплантации может улучшить результат хемоселекции in vivo 6TG путем компенсации за задержку в миелотоксичности 6TG и обеспечения адекватной ниши для HSC во время трансплантации.
Кроме того, Porter et al. использовали 6TG в гораздо более низких дозах 0,25-2 мг/кг в течение коротких периодов времени, начиная более чем через месяц после трансплантации, вызывая переменные уровни пересадки от 5 до 50%. В нашем исследовании мы подтвердили, что мыши, которым трансплантировали Hprt-дефицитный ВМ, могут переносить в 5-40 раз более высокие дозы 6TG, причем хемоселективный препарат начинают вводить сразу после трансплантации и продолжают в течение значительно более длительных периодов селекции.
Для того чтобы преобразовать применение стратегии хемоселекции in vivo 6TG в клинически осуществимый подход, необходимо разработать способы генной инженерии нормальных HSC, чтобы сделать их HPRT-дефицитными и, таким образом, устойчивыми к 6TG. Кроме того, следует подчеркнуть, что наше нынешнее исследование было ограничено применением этой стратегии для трансплантации костного мозга у сингенных мышей, что моделирует ситуацию в условиях аутотрансплантации. Остается выяснить, будет ли данная стратегия одинаково полезной в аллогенном контексте, поскольку режимы кондиционирования сниженной интенсивности регулярно используют, чтобы обойти токсичность традиционных миелоаблативных режимов. Потенциальные проблемы, которые, возможно, нужно будет решить, включают возможность возникновения спонтанной устойчивости к 6TG в лейкозных клетках после аллогенной трансплантации в случае гемопоэтических злокачественных новообразований, а также возможные обострения болезни трансплантат против хозяина, когда количество аллогенных донорских клеток избирательно увеличивается in vivo.
Литература
[1] Biffi A, Cesani М. Curr Gene Ther. 2008; 8:135-146.
[2] Persons DA, Nienhuis AW. Curr Opin Mol Ther. 2002; 4:491-498.
[3] Williams DA, Hsieh K, DeSilva A, Mulligan RC. J Exp Med. 1987; 166:210-218.
[4] Mickisch GH, Aksentijevich I, Schoenlein PV, et al. Blood. 1992; 79:1087-1093.
[5] Sorrentino BP, Brandt SJ, Bodine D, et al. Science. 1992; 257:99-103.
[6] Zaboikin M, Srinivasakumar N, Schuening F. Cancer Gene Ther. 2006; 13:335-345.
[7] Southgate T, Fairbairn LJ. Expert Rev Mol Med. 2004; 6:1 -24.
[8] Milsom MD, Williams DA. DNA Repair (Amst). 2007; 6:1210-1221.
[9] Neff T, Beard ВС, Kiem HP. Blood. 2006; 107:1751-1760.
[10] Schambach A, Baum C. DNA Repair (Amst). 2007; 6:1187-1196.
[11] Aubrecht J, GoadME, Schiestl RH. J Pharmacol Exp Ther. 1997; 282:1102-1108.
[12] Stout JT, Caskey CT. Annu Rev Genet. 1985; 19:127-148.
[13] Fontenelle LJ, Henderson JF. Biochim Biophys Acta. 1969; 177:175-176.
[14] Holmsen H, Rozenberg MC. Biochim Biophys Acta. 1968; 157:266-279.
[15] Lajtha LG, Vane JR. Nature. 1958; 182:191-192.
[16] Lowy BA, Ramot B, London IM. J Biol Chem. 1960; 235:2920-2923.
[17] Elion GB. Science. 1989; 244:41-47.
[18] Ansell JD, Samuel K, Whittingham DG, et al. Development. 1991; 112:489-498.
[19] Torres RJ, Puig JG. Orphanet J Rare Dis. 2007; 2:48.
[20] van der Zee SP, Schretlen ED, Monnens LA. Lancet. 1968; 1:1427.
[21] Seegmiller JE, etal.. Adv Exp Med Biol. 1977; 76A:412-433.
[22] Porter CC, DeGregori J. Blood. 2008; 112:4466-4474.
[23] Hooper M, Hardy K, Handyside A, Hunter S, Monk M. Nature. 1987; 326:292-295.
[24] Bryder D, Rossi DJ, Weissman IL. Am J Pathol. 2006; 169:338-346.
[25] Rondon G, Giralt S, Pereira M, et al. Leuk Lymphoma. 1997; 25:463-467.
[26] Brazelton TR, Blau HM. Stem Cells. 2005; 23:1251-1265.
[27] Krause DS, Theise ND, Collector Ml, et al. Cell. 2001; 105:369-377.
[28] Olsen NJ, Kovacs WJ. Immunol Res. 2001; 23:281-288.
[29] Olsen NJ, Gu X, Kovacs WJ. J Clin Invest. 2001; 108:1697-1704.
[30] Kelley WN, Wyngaarden, J.B. In: J.B. Stanbury JBW, et al., ed. The Metabolic Basis of Inherited Disease, 5th ed. New York: McGraw Hill; 1983. p. 1115-1143.
[31] Lesch M, Nyhan WL. Am J Med. 1964; 36:561-570.
[32] Lo YF, Palmour RM. Biochem Genet. 1979; 17:737-746.
[33] Mojas N, Lopes M, Jiricny J. Genes Dev. 2007; 21:3342-3355.
[34] An DS, Qin FX, Auyeung VC, et al. Mol Ther. 2006; 14:494-504.
[35] Grimm D, Streetz KL, Jopling CL, et al. Nature. 2006; 441:537-541.
Пример 2: Создание HPRT-дефицитных клеток
Этот пример направлен на переносное применение к генной терапии ex vivo в аутологичном контексте, используя как лентивирусные векторы третьего поколения, экспрессирующие различные вероятные последовательности shRNA, нацеленные на HPRT, так и нуклеофекцию нуклеаз ''цинковые пальцы'' (ZFN), нацеленные на HPRT. Последний подход имеет преимущество, поскольку только кратковременная экспрессия конструкта ZFN необходима для достижения постоянного нокаута гена-мишени, тем самым уменьшая потенциал инсерционного мутагенеза в результате этой процедуры генной инженерии. В этом контексте эта стратегия является уникальной в придании селективного преимущества трансплантированным клеткам через дефицит фермента, а не вставку нового трансгена для достижения химической устойчивости. shRNA, нацеленная на HPRT, успешно даунрегулировала HPRT до необнаруживаемого уровня. Кроме того, с помощью ZFN успешно делали нокаут HPRT.
Из вышесказанного будет понятно, что, хотя в данном документе были описаны конкретные варианты осуществления настоящего изобретения в целях иллюстрации, можно сделать различные модификации без отклонения от сущности и объема настоящего изобретения. Соответственно, настоящее изобретение не ограничено ничем, кроме как описано в прилагаемой формуле изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| Способ совместного кондиционирования и хемоселекции за один цикл | 2012 |
|
RU2611396C2 |
| КОМБИНИРОВАННАЯ ТЕРАПИЯ ДЛЯ СТАБИЛЬНОГО И ДОЛГОВРЕМЕННОГО ПРИЖИВЛЕНИЯ ТРАНСПЛАНТАТА | 2012 |
|
RU2657758C2 |
| КОМБИНИРОВАННАЯ ТЕРАПИЯ ДЛЯ СТАБИЛЬНОГО И ДОЛГОВРЕМЕННОГО ПРИЖИВЛЕНИЯ ТРАНСПЛАНТАТА С ИСПОЛЬЗОВАНИЕМ КОНКРЕТНЫХ ПРОТОКОЛОВ ДЛЯ Т/В-КЛЕТОЧНОЙ ДЕПЛЕЦИИ | 2012 |
|
RU2648354C2 |
| Т-КЛЕТКИ ЦЕНТРАЛЬНОЙ ПАМЯТИ ПРОТИВ ТРЕТЬЕЙ СТОРОНЫ, СПОСОБЫ ИХ ПОЛУЧЕНИЯ И ИХ ПРИМЕНЕНИЕ В ТРАНСПЛАНТАЦИИ И ЛЕЧЕНИИ ЗАБОЛЕВАНИЙ | 2009 |
|
RU2506311C2 |
| ПРОТИВОВИРУСНЫЕ КЛЕТКИ ВЕТО ЦЕНТРАЛЬНОЙ ПАМЯТИ CD8+ ПРИ ГАПЛОИДЕНТИЧНОЙ ТРАНСПЛАНТАЦИИ СТВОЛОВЫХ КЛЕТОК | 2020 |
|
RU2834894C2 |
| СПОСОБЫ ИНДУЦИРОВАНИЯ Т-КЛЕТОЧНОЙ ТОЛЕРАНТНОСТИ К ТКАНЕВОМУ ИЛИ ОРГАННОМУ ТРАНСПЛАНТАТУ | 1995 |
|
RU2169009C2 |
| СПОСОБЫ ИНГИБИРОВАНИЯ МИГРАЦИИ РАКОВЫХ КЛЕТОК | 2006 |
|
RU2404805C2 |
| ВЕТО-КЛЕТКИ, ПОЛУЧЕННЫЕ ИЗ Т-КЛЕТОК ПАМЯТИ | 2017 |
|
RU2779844C2 |
| КОМПОЗИЦИИ И СПОСОБЫ УСИЛЕНИЯ ЭКСПРЕССИИ ГЕНА PKLR | 2017 |
|
RU2773358C2 |
| Т-КЛЕТКИ ЦЕНТРАЛЬНОЙ ПАМЯТИ ПРОТИВ ТРЕТЬЕЙ СТОРОНЫ, СПОСОБЫ ИХ ПОЛУЧЕНИЯ И ИХ ПРИМЕНЕНИЕ В ТРАНСПЛАНТАЦИИ И ЛЕЧЕНИИ ЗАБОЛЕВАНИЙ | 2012 |
|
RU2636503C2 |

Группа изобретений относится к трансплантации гемопоэтических стволовых клеток (HSC). Способ трансплантации HSC без предварительного кондиционирования путем облучения включает: (a) введение млекопитающему субъекту одной или двух доз от 2 до 10 мг/кг массы тела аналога пуринового основания, выбранного из 6-тиогуанина (6TG), 6-меркаптопурина (6-МР) или азатиопурина (AZA) в качестве этапа предварительного кондиционирования; (b) пересадку субъекту HSC доноров с нехваткой гипоксантин-гуанин фосфорибозилтрансферазы (HPRT) в течение 48-72 часов после этапа предварительного кондиционирования; и (c) немедленное введение субъекту приблизительно от 1 до 5 мг/кг аналога пуринового основания и продолжающееся введение субъекту приблизительно от 1 до 5 мг/кг аналога пуринового основания каждые 2-4 дня в течение 2-8 недель без предусмотренного периода восстановления; где у субъекта предполагается наличие заболевания или расстройства, поражающего лимфогемопоэтические клетки. Также раскрыт способ лечения симптомов заболевания или расстройства, поражающего лимфогемопоэтические клетки, у субъекта. Группа изобретений обеспечивает отсутствие облучения при подготовке к трансплантации, поэтому субъект избавлен от токсичности, индуцированной миелоаблативным облучением. 2 н. и 42 з.п. ф-лы, 8 ил., 7 табл., 1 пр.
1. Способ трансплантации гемопоэтических стволовых клеток (HSC) без облучения, включающий:
(a) введение млекопитающему субъекту одной или двух доз от 2 до 10 мг/кг массы тела аналога пуринового основания, выбранного из 6-тиогуанина (6TG), 6-меркаптопурина (6-МР) или азатиопурина (AZA) в качестве этапа предварительного кондиционирования;
(b) пересадку субъекту HSC доноров с нехваткой гипоксантин-гуанин фосфорибозилтрансферазы (HPRT) в течение 48-72 часов после этапа предварительного кондиционирования; и
(c) немедленное введение субъекту приблизительно от 1 до 5 мг/кг аналога пуринового основания и продолжающееся введение субъекту приблизительно от 1 до 5 мг/кг аналога пуринового основания каждые 2-4 дня в течение 2-8 недель без предусмотренного периода восстановления,
где способ осуществляют в отсутствие предварительного кондиционирования путем облучения и где у субъекта предполагается наличие заболевания или расстройства, поражающего лимфогемопоэтические клетки.
2. Способ по п. 1, где субъект представляет собой человека.
3. Способ по п. 1, где субъект имеет наследственное или генетическое расстройство.
4. Способ по п. 1, где субъект имеет приобретенное заболевание, поражающее лимфогемопоэтические клетки.
5. Способ по п. 4, где заболевание или расстройство представляет собой инфекцию, вызванную вирусом иммунодефицита человека (ВИЧ), или синдром приобретенного иммунодефицита (СПИД).
6. Способ по п. 4, где расстройство или заболевание представляет собой лимфогемопоэтическое злокачественное новообразование.
7. Способ по п. 1, где расстройство или заболевание представляет собой расстройство или заболевание гемопоэтической, или тромбопоэтической, или лимфопоэтической системы.
8. Способ по п. 7, где расстройство или заболевание гемопоэтической системы представляет собой гемоглобинопатию.
9. Способ по п. 1, дополнительно включающий трансплантацию костного мозга.
10. Способ по п. 1, где аналогом пуринового основания является 6TG.
11. Способ по п. 1, отличающийся тем, что общая дозировка 6TG, которую вводят субъекту, не превышает 105 мг.
12. Способ по п. 1, отличающийся тем, что общая дозировка 6TG, которую вводят субъекту, не превышает 75 мг.
13. Способ по п. 1, отличающийся тем, что введение на этапе (с) выполняют каждые 3 дня и в течение от двух до четырех недель после этапа пересадки.
14. Способ по п. 1, где между стадиями (а) и (с) проходит не более 5 суток.
15. Способ по п. 1, отличающийся тем, что у субъекта наблюдают более 75% генетически модифицированных гемопоэтических клеток.
16. Способ по п. 1, отличающийся тем, что у субъекта наблюдают более 95% генетически модифицированных гемопоэтических клеток.
17. Способ по п. 1, отличающийся тем, что HPRT-дефицитные HSC для трансплантации сделали HPRT-дефицитными путем введения последовательностей, кодирующих нуклеазы "цинковые пальцы" (ZFN), эффекторные нуклеазы, подобные активаторам транскрипции (TALEN), матричные цепи небольшого фрагмента гомологичной рекомбинации (SFHR), ингибирующие РНК (siRNA) или микроРНК (miRNA), антисмысловые РНК, транс-сплайсинговые РНК, рибозимы, внутриклеточные антитела или доминантно-негативные или конкурентные белки-ингибиторы.
18. Способ по п. 1, где HPRT-дефицитные HSC для трансплантации сделаны HPRT-дефицитными путем введения последовательностей, кодирующих нуклеазы "цинковые пальцы" (ZFN).
19. Способ по п. 1, где HPRT-дефицитные HSC для трансплантации сделаны HPRT-дефицитными путем введения последовательностей, кодирующих эффекторные нуклеазы, подобные активаторам транскрипции (TALEN).
20. Способ по п. 1, где HPRT-дефицитные HSC для трансплантации сделаны HPRT-дефицитными путем введения последовательностей, кодирующих ингибирующие РНК (siRNA).
21. Способ по п. 1, отличающийся тем, что HPRT-дефицитные HSC для трансплантации были генетически модифицированы.
22. Способ по п. 1, отличающийся тем, что трансплантированные HSC являются аутологичными или сингенными.
23. Способ по п. 1, отличающийся тем, что трансплантированные HSC являются аллогенными.
24. Способ по п. 1, отличающийся тем, что субъекта не обрабатывают миелоаблативным облучением.
25. Способ лечения симптомов заболевания или расстройства, поражающего лимфогемопоэтические клетки, у субъекта, включающий:
(a) введение млекопитающему субъекту одной или двух доз от 2 до 10 мг/кг массы тела аналога пуринового основания, выбранного из 6-тиогуанина (6TG), 6-меркаптопурина (6-МР) или азатиопурина (AZA) в качестве этапа предварительного кондиционирования;
(b) пересадку субъекту HSC доноров с нехваткой гипоксантин-гуанин фосфорибозилтрансферазы (HPRT) в течение 48-72 часов после этапа предварительного кондиционирования; и
(c) немедленное введение субъекту приблизительно от 1 до 5 мг/кг аналога пуринового основания и продолжающееся введение субъекту приблизительно от 1 до 5 мг/кг аналога пуринового основания каждые 2-4 дня в течение 2-8 недель после этапа пересадки без предусмотренного периода восстановления,
где способ осуществляют в отсутствие предварительного кондиционирования путем облучения.
26. Способ по п. 25, где заболевание или расстройство представляет собой инфекцию, вызванную вирусом иммунодефицита человека (ВИЧ), или синдром приобретенного иммунодефицита (СПИД).
27. Способ по п. 25, где расстройство или заболевание представляет собой лимфогемопоэтическое злокачественное новообразование.
28. Способ по п. 25, где расстройство или заболевание представляет собой расстройство или заболевание гемопоэтической, или тромбопоэтической, или лимфопоэтической системы.
29. Способ по п. 28, где расстройство или заболевание гемопоэтической системы представляет собой гемоглобинопатию.
30. Способ по п. 25, дополнительно включающий трансплантацию костного мозга или гемопоэтических стволовых клеток.
31. Способ по п. 25, где субъект представляет собой человека.
32. Способ по п. 25, где аналогом пуринового основания является 6TG.
33. Способ по п. 25, отличающийся тем, что введение на этапе (с) выполняют каждые 3 дня и в течение от двух до четырех недель после этапа пересадки.
34. Способ по п. 25, где между введением доз аналога пуринового основания на стадиях (а) и (с) проходит не более 5 суток.
35. Способ по п. 25, где у субъекта наблюдают более 75% генетически
модифицированных гемопоэтических клеток.
36. Способ по п. 25, где у субъекта наблюдают более 95% генетически модифицированных гемопоэтических клеток.
37. Способ по п. 25, где HPRT-дефицитные HSC для трансплантации сделали HPRT-дефицитными путем введения последовательностей, кодирующих нуклеазы "цинковые пальцы" (ZFN), эффекторные нуклеазы, подобные активаторам транскрипции (TALEN), матричные цепи небольшого фрагмента гомологичной рекомбинации (SFHR), ингибирующие РНК (siRNA) или микроРНК (miRNA), антисмысловые РНК, транс-сплайсинговые РНК, рибозимы, внутриклеточные антитела или доминантно-негативные или конкурентные белки-ингибиторы.
38. Способ по п. 25, где HPRT-дефицитные HSC для трансплантации сделаны HPRT-дефицитными путем введения последовательностей, кодирующих нуклеазы "цинковые пальцы" (ZFN).
39. Способ по п. 25, где HPRT-дефицитные HSC для трансплантации сделаны HPRT-дефицитными путем введения последовательностей, кодирующих эффекторные нуклеазы, подобные активаторам транскрипции (TALEN).
40. Способ по п. 25, где HPRT-дефицитные HSC для трансплантации сделаны HPRT-дефицитными путем введения последовательностей, кодирующих ингибирующие РНК (siRNA).
41. Способ по п. 25, где HPRT-дефицитные HSC для трансплантации были генетически модифицированы.
42. Способ по п. 1, где трансплантированные HSC являются аутологичными или сингенными.
43. Способ по п. 1, где трансплантированные HSC являются аллогенными.
44. Способ по п. 1, где субъекта не обрабатывают миелоаблативным облучением.
| US 2003032003 A1, 13.02.2003 | |||
| US 2003073649 A1, 17.04.2003 | |||
| US 7094885 B2, 22.08.2006 | |||
| ТРАНСПЛАНТАЦИЯ ГЕМОПОЭТИЧЕСКИХ КЛЕТОК | 2000 |
|
RU2216336C2 |

