ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Настоящее изобретение, в целом, относится к новым анти-PD-1 антителам.
ПРЕДПОСЫЛКИ СОЗДАНИЯ ИЗОБРЕТЕНИЯ
Все большее число результатов доклинических и клинических исследований свидетельствуют в пользу того, что терапия, направленная на иммунные контрольные точки, становится наиболее многообещающим подходом для лечения пациентов, страдающих раком. Белок 1 программируемой клеточной гибели, один из белков иммунных контрольных точек, играет важную роль в ограничении активности T-клеток, обеспечивающих основной иммунный защитный механизм, за счет чего клетки опухолей избегают контроля иммунной системы. Взаимодействие PD-1, экспрессированного на активированных T-клетках, и PD-L1, экспрессированного на клетках опухолей, приводит к отрицательной регуляции иммунного ответа и ослаблению противоопухолевого иммунитета. Экспрессия PD-L1 на опухолях коррелирует с пониженной выживаемостью при раке пищевода, поджелудочной железы и других видах рака, свидетельствуя в пользу того, что данный путь является новой перспективной мишенью для противоопухолевой иммунотерапии. Множество средств, направленных на путь PD-1, было разработано фармацевтическими компаниями, такими как Bristol-Myers Squibb (BMS), Merck, Roche и GlaxoSmithKline (GSK). Результаты клинических исследований продемонстрировали начальные свидетельства устойчивой клинической активности и обнадеживающий профиль безопасности у пациентов с опухолями различных видов. Ниволумаб, направленное на PD-1 лекарственное средство, разработанное BMS, занимает центральное положение среди препаратов следующего поколения. В настоящее время в 6 исследованиях рака на поздней стадии лечение привело к уменьшению размеров опухолей в трех из 5 групп онкологических пациентов, включая 18% из 72 пациентов с раком легкого, около трети из 98 пациентов с меланомой и 27% из 33 пациентов с раком почки. Разработанный компанией Merck ламбролизумаб представляет собой полностью человеческое моноклональное IgG4 антитело, направленное против PD-1, которое получило одобрение FDA после впечатляющих данных фазы IB исследования, полученных при лечении рака кожи. Результаты фазы IB исследования показали наличие объективного противоопухолевого ответа у 51% из 85 онкологических пациентов, и полного ответа у 9% пациентов. Экспериментальный препарат MPDL3280A компании Roche продемонстрировал способность вызывать уменьшение размеров опухолей у 29 из 140 (21%) пациентов с раком на поздней стадии, имеющих опухоли разных размеров.
Однако не все существующие методы лечения могут приводить к удовлетворительным результатам и, следовательно, сохраняется потребность в новых анти-PD-1 антителах.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к новым моноклональным анти-PD-1 антителам (в частности, полностью человеческим антителам), кодирующим их полинуклеотидам и способам их применения.
В одном аспекте настоящее изобретение относится к выделенным моноклональным антителам, или их антигенсвязывающим фрагментам, которые способны специфически связывать человеческий PD-1 с величиной Kd не более 10-8 M (например, ≤9×10-9 M, ≤8×10-9 M, ≤7×10-9 M, ≤6×10-9 M, ≤5×10-9 M, ≤4×10-9 M, ≤3×10-9 M, ≤2×10-9 M или ≤10-9 M) при измерении в анализе связывания методом плазмонного резонанса.
В конкретных вариантах осуществления антитела, или их антигенсвязывающие фрагменты, связывают обезьяний PD-1 с величиной EC50 не более 100 нМ или не более 10 нМ (например, не более 50 нМ, 40 нМ, 30 нМ, 20 нМ, 10 нМ, 9 нМ, 8 нМ, 7 нМ, 6 нМ, 5 нМ, 4 нМ, 3 нМ, 2 нМ или 1 нМ). В конкретных вариантах осуществления антитела и их антигенсвязывающие фрагменты не связывают мышиный PD-1, но связывают обезьяний PD-1 с аффинностью связывания, аналогичной аффинности для человеческого PD-1. В конкретных вариантах осуществления антитела, или их антигенсвязывающие фрагменты, сильно ингибируют связывание человеческого или обезьяньего PD-1 с его лигандом (например, PD-L1 или PD-L2) с величиной IC50 не более 100 нМ (например, не более 50 нМ, 40 нМ, 30 нМ, 20 нМ, 10 нМ, 9 нМ, 8 нМ, 7 нМ, 6 нМ, 5 нМ, 4 нМ, 3 нМ, 2 нМ, 1 нМ, 0,9 нМ, 0,8 нМ, 0,7 нМ, 0,6 нМ, 0,5 нМ, 0,4 нМ, 0,3 нМ, 0,2 нМ или 0,1 нМ). В конкретных вариантах осуществления величину EC50 или IC50 измеряют в анализе активированной флуоресценцией сортировки клеток (FACS).
В конкретных вариантах осуществления антитела, или их антигенсвязывающие фрагменты, имеют существенно сниженную эффекторную функцию. В конкретных вариантах осуществления антитела, или их антигенсвязывающие фрагменты, не опосредуют ADCC или CDC, или обе из них.
В конкретных вариантах осуществления антитела, или их антигенсвязывающие фрагменты, предложенные в настоящем документе, содержат последовательности CDR тяжелой цепи, выбранные из группы, состоящей из SEQ ID NOs: 1, 3, 5, 13, 15, 21, 23, 25, 33, 35 и 37.
В одном аспекте антитела, или их антигенсвязывающие фрагменты, предложенные в настоящем документе, содержат последовательности CDR легкой цепи, выбранные из группы, состоящей из SEQ ID NOs: 7, 9, 11, 17, 19, 27, 29, 31, 39, 41, 43 и 65.
В конкретных вариантах осуществления антитела, или их антигенсвязывающие фрагменты, предложенные в настоящем документе, содержат по меньшей мере одну, две, три, четыре, пять или шесть областей CDR, выбранных из группы, состоящей из SEQ ID NOs: 1, 3, 5, 7, 9 и 11; или выбранных из группы, состоящей из SEQ ID NOs: 13, 15, 5, 7, 17 и 11; или выбранных из группы, состоящей из SEQ ID NOs: 1, 15, 5, 7, 17 и 19; или выбранных из группы, состоящей из SEQ ID NOs: 1, 15, 5, 7, 17 и 65; или выбранных из группы, состоящей из SEQ ID NOs: 21, 23, 25, 27, 29 и 31; или выбранных из группы, состоящей из SEQ ID NOs: 33, 35, 37, 39, 41 и 43.
В конкретных вариантах осуществления антитела, или их антигенсвязывающие фрагменты, предложенные в настоящем документе, содержат вариабельную область тяжелой цепи, выбранную из группы, состоящей из:
a) вариабельной области тяжелой цепи, содержащей SEQ ID NO: 1, SEQ ID NO: 3 и/или SEQ ID NO: 5;
b) вариабельной области тяжелой цепи, содержащей SEQ ID NO: 13, SEQ ID NO: 15 и/или SEQ ID NO: 5;
c) вариабельной области тяжелой цепи, содержащей SEQ ID NO: 1, SEQ ID NO: 15 и/или SEQ ID NO: 5;
d) вариабельной области тяжелой цепи, содержащей SEQ ID NO: 21, SEQ ID NO: 23 и/или SEQ ID NO: 25; и
e) вариабельной области тяжелой цепи, содержащей SEQ ID NO: 33, SEQ ID NO: 35 и/или SEQ ID NO: 37.
В конкретных вариантах осуществления антитела, или их антигенсвязывающие фрагменты, предложенные в настоящем документе, содержат вариабельную область легкой цепи, выбранную из группы, состоящей из:
a) вариабельной области легкой цепи, содержащей SEQ ID NO: 7, SEQ ID NO: 9 и/или SEQ ID NO: 11;
b) вариабельной области легкой цепи, содержащей SEQ ID NO: 7, SEQ ID NO: 17 и/или SEQ ID NO: 11;
c) вариабельной области легкой цепи, содержащей SEQ ID NO: 7, SEQ ID NO: 17 и/или SEQ ID NO: 19;
d) вариабельной области легкой цепи, содержащей SEQ ID NO: 27, SEQ ID NO: 29 и/или SEQ ID NO: 31;
e) вариабельной области легкой цепи, содержащей SEQ ID NO: 39, SEQ ID NO: 41 и/или SEQ ID NO: 43; и
f) вариабельной области легкой цепи, содержащей SEQ ID NO: 7, SEQ ID NO: 17 и/или SEQ ID NO: 65.
В конкретных вариантах осуществления антитела, или их антигенсвязывающие фрагменты, предложенные в настоящем документе, содержат:
a) вариабельную область тяжелой цепи, содержащую SEQ ID NO: 1, SEQ ID NO: 3 и/или SEQ ID NO: 5; и вариабельную область легкой цепи, содержащую SEQ ID NO: 7, SEQ ID NO: 9 и/или SEQ ID NO: 11;
b) вариабельную область тяжелой цепи, содержащую SEQ ID NO: 13, SEQ ID NO: 15 и/или SEQ ID NO: 5; и вариабельную область легкой цепи, содержащую SEQ ID NO: 7, SEQ ID NO: 17 и/или SEQ ID NO: 11;
c) вариабельную область тяжелой цепи, содержащую SEQ ID NO: 1, SEQ ID NO: 15 и/или SEQ ID NO: 5; и вариабельную область легкой цепи, содержащую SEQ ID NO: 7, SEQ ID NO: 17 и/или SEQ ID NO: 19;
d) вариабельную область тяжелой цепи, содержащую SEQ ID NO: 21, SEQ ID NO: 23 и/или SEQ ID NO: 25; и вариабельную область легкой цепи, содержащую SEQ ID NO: 27, SEQ ID NO: 29 и/или SEQ ID NO: 31;
e) вариабельную область тяжелой цепи, содержащую SEQ ID NO: 33, SEQ ID NO: 35 и/или SEQ ID NO: 37; и вариабельную область легкой цепи, содержащую SEQ ID NO: 39, SEQ ID NO: 41 и/или SEQ ID NO: 43; или
f) вариабельную область тяжелой цепи, содержащую SEQ ID NO: 1, SEQ ID NO: 15 и/или SEQ ID NO: 5; и вариабельную область легкой цепи, содержащую SEQ ID NO: 7, SEQ ID NO: 17 и/или SEQ ID NO: 65.
В конкретных вариантах осуществления антитела, или их антигенсвязывающие фрагменты, предложенные в настоящем документе, содержат вариабельную область тяжелой цепи, выбранную из группы, состоящей из SEQ ID NO: 45, SEQ ID NO: 49, SEQ ID NO: 53, SEQ ID NO: 57 и SEQ ID NO: 61.
В конкретных вариантах осуществления антитела, или антигенсвязывающие фрагменты, предложенные в настоящем документе, содержат вариабельную область легкой цепи, выбранную из группы, состоящей из SEQ ID NO: 47, SEQ ID NO: 51, SEQ ID NO: 55, SEQ ID NO: 59, SEQ ID NO: 63 и SEQ ID NO: 67.
В конкретных вариантах осуществления антитела, или их антигенсвязывающие фрагменты, предложенные в настоящем документе, содержат:
a) вариабельную область тяжелой цепи, содержащую SEQ ID NO: 45; и вариабельную область легкой цепи, содержащую SEQ ID NO: 47;
b) вариабельную область тяжелой цепи, содержащую SEQ ID NO: 49; и вариабельную область легкой цепи, содержащую SEQ ID NO: 51;
c) вариабельную область тяжелой цепи, содержащую SEQ ID NO: 53; и вариабельную область легкой цепи, содержащую SEQ ID NO: 55;
d) вариабельную область тяжелой цепи, содержащую SEQ ID NO: 57; и вариабельную область легкой цепи, содержащую SEQ ID NO: 59;
e) вариабельную область тяжелой цепи, содержащую SEQ ID NO: 61; и вариабельную область легкой цепи, содержащую SEQ ID NO: 63; или
f) вариабельную область тяжелой цепи, содержащую SEQ ID NO: 53; и вариабельную область легкой цепи, содержащую SEQ ID NO: 67.
В конкретных вариантах осуществления антитела, предложенные в настоящем документе, включают, например, 1.7.3 чАт, 1.49.9 чАт, 1.103.11 чАт, 1.103.11-v2 чАт, 1.139.15 чАт и 1.153.7 чАт.
В конкретных вариантах осуществления антитела, или их антигенсвязывающие фрагменты, предложенные в настоящем документе, конкурируют за один и тот же эпитоп с антителами 1.7.3 чАт, 1.49.9 чАт, 1.103.11 чАт, 1.103.11-v2 чАт, 1.139.15 чАт или 1.153.7 чАт. В конкретных вариантах осуществления антитела, или их антигенсвязывающие фрагменты, предложенные в настоящем документе, связывают эпитоп, содержащий по меньшей мере один из следующих аминокислотных остатков в PD-1: V64, P83, D85, L128, A129, P130, K131, A132 и Q133.
В конкретных вариантах осуществления антитела, или их антигенсвязывающие фрагменты, способны блокировать связывание человеческого PD-1 с его лигандом и за счет этого обеспечивать по меньшей мере одну из следующих активностей:
a) индукцию продуцирования IL-2 в CD4+ T-клетках;
b) индукцию продуцирования IFNγ в CD4+ T-клетках;
c) индукцию пролиферации CD4+ T-клеток и
d) реверсию супрессорной функции Treg.
В конкретных вариантах осуществления антитела, предложенные в настоящем документе, представляют собой моноклональное антитело, полностью человеческое антитело, гуманизированное антитело, химерное антитело, рекомбинантное антитело, биспецифическое антитело, меченое антитело, двухвалентное антитело или антиидиотипическое антитело. В конкретных вариантах осуществления антитела, или их антигенсвязывающие фрагменты, представляют собой полностью человеческие моноклональные антитела, необязательно, продуцируемые в организме трансгенной крысы, например, трансгенной крысы с инактивированной эндогенной экспрессией генов крысиных иммуноглобулинов, несущей локусы рекомбинантных человеческих иммуноглобулинов с делецией J-локуса и мутацией C-каппа.
В конкретных вариантах осуществления антигенсвязывающие фрагменты антител, предложенные в настоящем документе, представляют собой однодоменное антитело верблюдовых, диатело, scFv, димер scFv, BsFv, dsFv, (dsFv)2, dsFv-dsFv', Fv-фрагмент, Fab, Fab', F(ab')2, ds-диатело, нанотело, доменное антитело или двухвалентное доменное антитело.
В конкретных вариантах осуществления антитела, или их антигенсвязывающие фрагменты, предложенные в настоящем документе, дополнительно содержат константную область иммуноглобулина.
В конкретных вариантах осуществления антитела, или их антигенсвязывающие фрагменты, предложенные в настоящем документе, дополнительно содержат конъюгат.
В конкретных вариантах осуществления конъюгат может представлять собой детектируемую метку, модифицирующий фармакокинетику фрагмент или облегчающий очистку фрагмент.
В другом аспекте настоящее изобретение относится к выделенным полинуклеотидам, кодирующим антитела, или их антигенсвязывающие фрагменты, предложенные в настоящем документе. В конкретных вариантах осуществления предложены полинуклеотиды, кодирующие аминокислотные последовательности антител, или антигенсвязывающих фрагментов, раскрытых в настоящем документе. В других конкретных вариантах осуществления предложены векторы, содержащие эти полинуклеотиды, и в других конкретных вариантах осуществления предложены клетки-хозяева, содержащие такие векторы. В конкретных вариантах осуществления предложены способы экспрессии одного или более антител, или антигенсвязывающих фрагментов, раскрытых в настоящем документе, путем культивирования таких клеток-хозяев в условиях, в которых антитела, или антигенсвязывающие фрагменты, кодируемые полинуклеотидами, экспрессируются с вектора. В конкретных вариантах осуществления полинуклеотиды, предложенные в настоящем документе, функционально связаны в векторе с промотором, таким как промотор SV40. В конкретных вариантах осуществления клетки-хозяева, содержащие векторы, предложенные в настоящем документе, представляют собой клетки яичника китайского хомяка или клетки 293F.
В другом аспекте настоящее изобретение относится к наборам, включающим антитело, или его антигенсвязывающий фрагмент.
В другом аспекте анти-PD-1 антитела, предложенные в настоящем документе, такие как 1.7.3 чАт, 1.49.9 чАт, 1.103.11 чАт, 1.103.11-v2 чАт, 1.139.15 чАт и 1.153.7 чАт, характеризуются хорошей переносимостью и высокой in vivo противоопухолевой активностью у животного. В конкретных вариантах осуществления у животного, имеющего клетки опухолей, которому вводили анти-PD-1 антитела, предложенные в настоящем документе, происходит уменьшение объема опухоли на по меньшей мере 20%, по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 90% или по меньшей мере 95% по сравнению с контрольным животным, имеющим такой же исходный размер опухоли, но которому вводили растворитель.
В другом аспекте настоящее изобретение относится к способам лечения состояния, связанного с PD-1, у индивидуума, включающим введение индивидууму терапевтически эффективного количества антитела, или его антигенсвязывающего фрагмента, предложенного в настоящем документе. В конкретных вариантах осуществления индивидуум был идентифицирован, как имеющий заболевание или состояние, на которое может оказывать влияние антагонист PD-1. В конкретных вариантах осуществления индивидуума идентифицируют, как положительного в отношении наличия, или повышенных уровней, PD-L1 в тестируемом биологическом от индивидуума.
В другом аспекте настоящее изобретение относится к фармацевтическим композициям, содержащим антитело, или его антигенсвязывающий фрагмент, предложенное в настоящем документе, и один или более фармацевтически приемлемых носителей. В некоторых из этих вариантов осуществления фармацевтические носители могут представлять собой, например, разбавители, антиоксиданты, адъюванты, эксципиенты или нетоксичные вспомогательные вещества.
В другом аспекте настоящее изобретение относится к способам лечения состояния у субъекта, при котором будет полезно усиление иммунного ответа, включающим введение субъекту эффективного количества антитела, или его антигенсвязывающего фрагмента, предложенного в настоящем документе. В конкретных вариантах осуществления субъект имеет повышенную экспрессию PD-L1 или был положительно идентифицирован в отношении экспрессии PD-L1.
Предложено применение антитела, или его антигенсвязывающего фрагмента, предложенного в настоящем документе, в производстве лекарственного средства для лечения состояния, при котором может быть полено усиление иммунного ответа. В конкретных вариантах осуществления состояние представляет собой рак или хроническую вирусную инфекцию.
КРАТКОЕ ОПИСАНИЕ ФИГУР
На фигуре 1 показано связывание полностью человеческих анти-PD-1 антител с экспрессирующими PD-1 клетками CHO при определении в FACS-анализе.
На фигуре 2 показано связывание полностью человеческих анти-PD-1 антител с экспрессирующими PD-1 клетками CHO, с величиной EC50 примерно 2 нМ при определении в FACS-анализе.
На фигуре 3 показано связывание полностью человеческого анти-PD-1 антитела с PD-1, экспрессированным на активированных CD4+ T-клетках, при определении в FACS-анализе.
На фигуре 4 показано, что полностью человеческие анти-PD-1 антитела блокировали связывание PD-L1 с трансфицированными PD-1 клетками CHO, с величиной IC50 примерно 3-8 нМ при определении в FACS-анализе.
На фигуре 5 показано, что полностью человеческие анти-PD-1 антитела специфически связывают PD-1, но не связывают представителей семейства CD28 и CTLA4, при определении в FACS-анализе.
На фигуре 6 показано, что полностью человеческие анти-PD-1 антитела против PD-1 связывают PD-1 яванского макака, но не мышиный PD-1.
На фигуре 7 приведена полная кинетика аффинного связывания анти-PD-1 антител с человеческим PD-1 в диапазоне от 3,76 E-9 до 1,76 E-10 моль/л при определении методом поверхностного плазмонного резонанса.
На фигуре 8 показан эффект полностью человеческих анти-PD-1 антител на продуцирование IL-2 в реакции смешанных лимфоцитов (РСЛ).
На фигуре 9 показан эффект полностью человеческих анти-PD-1 антител на продуцирование IFNγ в РСЛ.
На фигуре 10 показано, что полностью человеческие анти-PD-1 антитела стимулируют пролиферацию T-клеток в РСЛ.
На фигуре 11 показано, что полностью человеческие анти-PD-1 антитела стимулируют пролиферацию T-клеток в специфическом T-клеточном ответе.
На фигуре 12 показано, что анти-PD-1 антитела вызывают реверсию супрессорной функции Treg.
На фигуре 13 показано, что анти-PD-1 антитела не опосредовали ADCC активированных T-клеток.
На фигуре 14 показано, что анти-PD-1 антитела не опосредовали CDC активированных T-клеток.
На фигуре 15 показано, что 1.103.11-v2 чАт в разных буферах связывают внеклеточный домен человеческого PD-1 со сходными значениями аффинности при измерении методом ELISA. «1.103.11-v2 чАт в буфере» означает антитело в буфере формулирования, и «1.103.11-v2 чАт в PBS» означает антитело в 1x буфере PBS, pH 7,4.
На фигуре 16 показано, что 1.103.11-v2 чАт в разных буферах связывают экспрессирующие PD-1 клетки CHO со сходными значениями аффинности при измерении методом FACS. «1.103.11-v2 чАт в буфере» означает антитело в буфере формулирования, и «1.103.11-v2 чАт в PBS» означает антитело в 1x буфере PBS, pH 7,4.
На фигуре 17 показаны остатки «горячей зоны» (затемненная область) на кристаллической структуре человеческого PD-L1, с которым связываются антитела. На (A) показаны общие остатки «горячей зоны»; на (B)-(D) показаны остатки «горячей зоны» для 1.103.11 чАт, Keytruda и 11.148.10 чАт, соответственно.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Следующее далее описание изобретения предназначено лишь для иллюстрации различных вариантов осуществления данного изобретения. Следовательно, конкретные описанные модификации не следует воспринимать как ограничение объема изобретения. Для специалиста в данной области будет очевидно, что различные эквиваленты, изменения и модификации могут быть использованы без отклонения от объема изобретения, и понятно, что такие эквивалентные варианты осуществления включены в настоящий документ. Все литературные источники, цитируемые в настоящем документе, включая публикации, патенты и патентные заявки, включены в настоящий документ посредством ссылки в полном объеме.
Определения
Используемый в настоящем документе термин «антитело» включает любой иммуноглобулин, моноклональное антитело, поликлональное антитело, мультиспецифическое антитело или биспецифическое (двухвалентное) антитело, которое связывает конкретный антиген. Природное интактное антитело содержит две тяжелые цепи и две легкие цепи. Каждая тяжелая цепь состоит из вариабельной области, а также первой, второй и третьей константных областей, в то время как легкая цепь состоит из вариабельной области и константной области. Тяжелые цепи антител млекопитающих классифицируют как α, δ, ε, γ и μ, и легкие цепи антител млекопитающих классифицируют как λ или κ. Антитело имеет форму «Y», при этом ствол «Y» состоит из второй и третьей константных областей двух тяжелых цепей, связанных вместе дисульфидными связями. Каждое плечо «Y» включает вариабельную область и первую константную область одной тяжелой цепи, связанные с вариабельной и константной областями одной легкой цепи. Вариабельные области легкой и тяжелой цепей отвечают за связывание антигена. Вариабельная область в обеих цепях, как правило, содержит три высоко вариабельные петли, называемые определяющими комплементарность областями (CDR) (области CDR легкой (L) цепи включают LCDR1, LCDR2 и LCDR3, области CDR тяжелой (H) цепи включают HCDR1, HCDR2, HCDR3). Границы CDR для антител и антигенсвязывающих фрагментов, раскрытых в настоящем документе, могут быть определены или идентифицированы в соответствии с системами Kabat, Chothia или Al-Lazikani (Al-Lazikani, B., Chothia, C., Lesk, A. M., J. Mol. Biol., 273(4), 927 (1997); Chothia, C. et al., J Mol Biol. Dec 5; 186(3): 651-63 (1985); Chothia, C. and Lesk, A.M., J. Mol. Biol., 196, 901 (1987); Chothia, C. et al., Nature. Dec 21-28; 342(6252): 877-83 (1989); Kabat E.A. et al., National Institutes of Health, Bethesda, Md. (1991)). Три области CDR располагаются между фланкирующими последовательностями, известными как каркасные области (FR), которые более консервативны, чем области CDR и образуют каркас для поддержки гипервариабельных петель. Константные области тяжелой и легкой цепей не участвуют в связывании антигена, однако обладают разными эффекторными функциями. Антитела подразделяют на классы на основании аминокислотной последовательности константной области их тяжелой цепи. Пять основных классов или изотипов антител включают IgA, IgD, IgE, IgG и IgM, которые характеризуются наличием тяжелых цепей α, δ, ε, γ и μ, соответственно. Несколько основных классов антител делят на подклассы, такие как IgG1 (тяжелая цепь γ1), IgG2 (тяжелая цепь γ2), IgG3 (тяжелая цепь γ3), IgG4 (тяжелая цепь γ4), IgA1 (тяжелая цепь α1) или IgA2 (тяжелая цепь α2).
Используемый в настоящем документе термин «антигенсвязывающий фрагмент» означает фрагмент антитела, образованный из части антитела, содержащей одну или более CDR, или любой другой фрагмент антитела, который связывает антиген, но не имеет структуру интактного природного антитела. Примеры антигенсвязывающих фрагментов включают, без ограничения, диатело, Fab, Fab', F(ab')2, Fv-фрагмент, дисульфид-стабилизированный Fv-фрагмент (dsFv), (dsFv)2, биспецифический dsFv (dsFv-dsFv'), дисульфид-стабилизированное диатело (ds-диатело), одноцепочечную молекулу антитела (scFv), димер scFv (двухвалентное антитело), мультиспецифическое антитело, однодоменное антитело верблюдовых, нанотело, доменное антитело и двухвалентное доменное антитело. Антигенсвязывающий фрагмент способен связывать тот же антиген, который связывает исходное антитело. В конкретных вариантах осуществления антигенсвязывающий фрагмент может содержать одну или более областей CDR из конкретного человеческого антитела, привитые на каркасную область из одного или более других человеческих антител.
«Fab» применительно к антителу означает фрагмент антитела, состоящий из одной легкой цепи (как вариабельной, так и константной областей), связанной дисульфидной связью с вариабельной областью и первой константной областью одной тяжелой цепи.
«Fab'» означает Fab-фрагмент, включающий часть шарнирной области.
«F(ab')2» означает димер Fab'.
«Fc» применительно к антителу означает фрагмент антитела, состоящий из второй и третьей константных областей первой тяжелой цепи, связанных дисульфидными связями со второй и третьей константными областями второй тяжелой цепи. Fc-фрагмент антитела отвечает за разные эффекторные функции, такие как ADCC и CDC, но не участвует в связывании антигена.
«Fv» применительно к антителу означает наименьший фрагмент антитела, содержащий полный антигенсвязывающий сайт. Fv-фрагмент состоит из вариабельной области одной легкой цепи, связанной с вариабельной областью одной тяжелой цепи.
«Одноцепочечное Fv-антитело» или «scFv» означает сконструированное антитело, состоящее из вариабельной области легкой цепи и вариабельной области тяжелой цепи, связанных друг с другом напрямую или через последовательность пептидного линкера (Huston J.S. et al. Proc Natl Acad Sci USA, 85: 5879 (1988)).
«Одноцепочечное Fv-Fc антитело» или «scFv-Fc» означает сконструированное антитело, состоящее из scFv, связанного с Fc-областью антитела.
«Однодоменное антитело верблюдовых», «антитело из тяжелой цепи» или «HCAb» означает антитело, содержащее два домена VH и не содержащее легкие цепи (Riechmann L. and Muyldermans S., J Immunol Methods. Dec 10; 231(1-2): 25-38 (1999); Muyldermans S., J Biotechnol. Jun; 74(4): 277-302 (2001); WO 94/04678; WO 94/25591; патент США № 6005079). Антитела из тяжелой цепи исходно были обнаружены у представителей семейства Camelidae (двугорбых верблюдов, одногорбых верблюдов и лам). Хотя и не содержащие легкие цепи, антитела верблюдовых имеют аутентичный антигенсвязывающий репертуар (Hamers-Casterman C. et al., Nature. Jun 3; 363(6428): 446-8 (1993); Nguyen VK. et al. «Heavy-chain antibodies in Camelidae; a case of evolutionary innovation,ʺ Immunogenetics. Apr; 54(1): 39-47 (2002); Nguyen VK. et al. Immunology. May; 109(1): 93-101 (2003)). Вариабельный домен антитела из тяжелой цепи (домен VHH) представляет собой наименьший известный антигенсвязывающий фрагмент, вырабатывающийся при адаптивных иммунных ответах (Koch-Nolte F. et al., FASEB J. Nov; 21(13): 3490-8. Epub 2007 Jun 15 (2007)).
«Нанотело» означает фрагмент антитела, состоящий из домена VHH из тяжелой цепи антитела и двух константных доменов, CH2 и CH3.
«Диатела» включают небольшие фрагменты антитела с двумя антигенсвязывающими сайтами, при этом фрагменты содержат домен VH, связанный с доменом VL в одной и той же полипептидной цепи (VH-VL или VL-VH) (смотри, например, Holliger P. et al., Proc Natl Acad Sci U S A. Jul 15; 90(14): 6444-8 (1993); EP404097; WO 93/11161). При использовании линкера, который является слишком коротким, чтобы допустить спаривание между двумя доменами на одной и той же цепи, домены вынужденно образуют пары с комплементарными доменами другой цепи, при этом образуются два антигенсвязывающих сайта. Антигенсвязывающие сайты могут связывать одинаковые или разные антигены (или эпитопы).
«Доменное антитело» означает фрагмент антитела, содержащий только вариабельную область тяжелой цепи или вариабельную область легкой цепи. В некоторых случаях два или более доменов VH ковалентно связаны через пептидный линкер, образуя двухвалентное или мультивалентное доменное антитело. Два домена VH двухвалентного доменного антитела могут связывать одинаковые или разные антигены.
В конкретных вариантах осуществления «(dsFv)2» фрагменты содержат три пептидные цепи: два VH-фрагмента, соединенные пептидным линкером и связанные дисульфидными связями с двумя VL-фрагментами.
В конкретных вариантах осуществления «биспецифическое ds-диатело» содержит области VH1-VL2 (соединенные пептидным линкером), связанные с областями VL1-VH2 (также соединенными пептидным линкером) дисульфидной связью между VH1 и VL1.
В конкретных вариантах осуществления «биспецифические dsFv» или «dsFv-dsFv'» содержат три пептидные цепи: фрагмент VH1-VH2, в котором тяжелые цепи соединены пептидным линкером (например, длинным гибким линкером) и связаны с фрагментами VL1 и VL2, соответственно, дисульфидными связями, при этом каждая связанная дисульфидной связью пара тяжелой и легкой цепей имеет отличающуюся антигенную специфичность.
В конкретных вариантах осуществления «димер scFv» представляет собой двухвалентное диатело или двухвалентный фрагмент scFv (BsFv), содержащий фрагмент VH-VL (соединенные пептидным линкером), димеризованный с другим фрагментом VH-VL, так что области VH одного фрагмента взаимодействуют с областями VL другого фрагмента и образуют два связывающих сайта, которые могут связывать одинаковые антигены (или эпитопы) или разные антигены (или эпитопы). В других вариантах осуществления «димер scFv» представляет собой биспецифическое диатело, содержащее VH1-VL2 (соединенные пептидным линкером), связанные с VL1-VH2 (также соединенными пептидным линкером) таким образом, что VH1 и VL1 взаимодействуют, и VH2 и VL2 взаимодействуют, и все взаимодействующие пары имеет разные антигенные специфичности.
Используемый в настоящем документе термин «полностью человеческие» применительно к антителу или антигенсвязывающему фрагменту означает, что антитело или антигенсвязывающий фрагмент имеет, или состоит из, аминокислотную последовательность(и), соответствующую последовательности антитела, продуцируемого в организме человека или человеческой иммунной клеткой, или полученного от источника, иного, нежели человек, такого как трансгенное отличное от человека животное, в организме которого используются репертуары человеческих антител или другие человеческие кодирующие антитела последовательности. В конкретных вариантах осуществления полностью человеческое антитело не содержит аминокислотные остатки (в частности, антигенсвязывающие остатки), происходящие из антитела, отличного от человеческого.
Используемый в настоящем документе термин «гуманизированное» применительно к антителу, или антигенсвязывающему фрагменту, означает, что антитело, или антигенсвязывающий фрагмент, содержит области CDR, происходящие из антител животных, отличных от человека, области FR, происходящие из антитела человека, и, когда это применимо, константные области, происходящие из антитела человека. Гуманизированное антитело, или антигенсвязывающий фрагмент, в конкретных вариантах осуществления полезно в качестве терапевтического средства для человека, поскольку оно имеет пониженную иммуногенность в организме человека. В некоторых вариантах осуществления отличное от человека животное представляет собой млекопитающее, например, мышь, крысу, кролика, козу, овцу, морскую свинку или хомяка. В некоторых вариантах осуществления гуманизированное антитело, или антигенсвязывающий фрагмент, состоит из по существу полностью человеческих последовательностей, за исключением последовательностей CDR, которые не принадлежат человеку. В некоторых вариантах осуществления области FR, происходящие из антитела человека, могут содержать ту же аминокислотную последовательность, что и человеческое антитело, из которого они получены, или могут иметь несколько аминокислотных изменений, например, не более 10, 9, 8, 7, 6, 5, 4, 3, 2 или 1 измененной аминокислоты. В некоторых вариантах осуществления такое изменение аминокислоты может иметь место только в областях FR тяжелой цепи, только в областях FR легкой цепи или в обеих цепях. В некоторых предпочтительных вариантах осуществления гуманизированные антитела содержат человеческие области FR1-3 и человеческие области JH и Jκ.
Используемый в настоящем документе термин «химерное» означает антитело, или антигенсвязывающий фрагмент, содержащее часть тяжелой и/или легкой цепи, происходящую из антитела одного биологического вида, и остальную часть тяжелой и/или легкой цепи, происходящую из антитела другого биологического вида. В иллюстративном примере химерное антитело может содержать константную область, происходящую из антитела человека, и вариабельную область из антитела биологического вида, отличного от человека, например, из антитела мыши.
Используемый в настоящем документе термин «PD-1» означает белок программируемой клеточной гибели, который принадлежит к суперсемейству иммуноглобулинов и действует в качестве коингибирующего рецептора, отрицательно регулируя функции иммунной системы. PD-1 является представителем семейства CD28/CTLA-4 и имеет два известных лиганда, включая PD-L1 и PD-L2. Репрезентативная аминокислотная последовательность человеческого PD-1 раскрыта под регистрационным номером NCBI: NP_005009.2, и репрезентативная нуклеотидная последовательность, кодирующая человеческий PD-1, раскрыта под регистрационным номером NCBI: NM_005018.2.
Используемый в настоящем документе термин «PD-L1» означает лиганд 1 программируемой клеточной гибели (PD-L1, смотри, например, Freeman et al. (2000) J. Exp. Med. 192: 1027). Репрезентативная аминокислотная последовательность человеческого PD-L1 раскрыта под регистрационным номером NCBI: NP_054862.1, и репрезентативная нуклеотидная последовательность, кодирующая человеческий PD-L1, раскрыта под регистрационным номером NCBI: NM_014143.3. PD-L1 экспрессируется в плаценте, селезенке, лимфатических узлах, тимусе, сердце, эмбриональной печени и также встречается на многих опухолевых или раковых клетках. PD-L1 связывается с его рецептором PD-1 или B7-1, который экспрессируется на активированных T-клетках, B-клетках и миелоидных клетках. Связывание PD-L1 и его рецептора индуцирует передачу сигнала для подавления опосредованной TCR активации продуцирования цитокинов и пролиферации T-клеток. Соответственно, PD-L1 играет важную роль в подавлении функций иммунной системы во время конкретных событий, таких как беременность, аутоиммунные заболевания, аллотрансплантация тканей, и считается, что он позволяет опухолевым или раковым клеткам обходить систему иммунологического контроля и избегать иммунного ответа.
Используемый в настоящем документе термин «анти-PD-1 антитело» означает антитело, способное специфически связывать PD-1 (например, человеческий или обезьяний PD-1) с аффинностью, которая достаточна для его диагностического и/или терапевтического применения.
Используемый в настоящем документе термин «специфическое связывание» или «специфически связывает» относится к неслучайной реакции связывания между двумя молекулами, например, между антителом и антигеном. В конкретных вариантах осуществления антитела, или антигенсвязывающие фрагменты, предложенные в настоящем документе, специфически связывают человеческий и/или обезьяний PD-1 с аффинностью связывания (KD), составляющей ≤10-6 M (например, ≤5×10-7 M, ≤2×10-7 M, ≤10-7 M, ≤5×10-8 M, ≤2×10-8 M, ≤10-8 M, ≤5×10-9 M, ≤2×10-9 M, ≤10-9 M, 10-10 M). Используемый в настоящем документе термин «KD» означает отношение скорости диссоциации к скорости ассоциации (koff/kon), этот показатель можно определять методом поверхностного плазмонного резонанса, например, с использованием такого инструмента, как Biacore.
При использовании в настоящем документе, способность «блокировать связывание» или «конкурировать за один и тот же эпитоп» означает способность антитела, или антигенсвязывающего фрагмента, ингибировать взаимодействие связывания между двумя молекулами (например, человеческим PD-1 и анти-PD-1 антителом) в поддающейся определению степени. В конкретных вариантах осуществления антитело, или антигенсвязывающий фрагмент, которое блокирует связывание между двумя молекулами, ингибирует взаимодействие связывания между двумя молекулами на по меньшей мере 50%. В конкретных вариантах осуществления степень этого ингибирования может составлять более 60%, более 70%, более 80% или более 90%.
Используемый в настоящем документе термин «эпитоп» означает конкретную группу атомов или аминокислот на антигене, с которым связывается антитело. Два антитела могут связывать один и тот же эпитоп в составе антигена, если они конкурируют за связывание с антигеном. Например, если антитело, или антигенсвязывающий фрагмент, раскрытое в настоящем документе, блокирует связывание иллюстративных антител, таких как 1.7.3 чАт, 1.49.9 чАт, 1.103.11 чАт, 1.103.11-v2 чАт, 1.139.15 чАт и 1.153.7 чАт, с человеческим PD-1, то можно считать, что антитело, или антигенсвязывающий фрагмент, связывает тот же эпитоп, что и эти иллюстративные антитела.
Конкретный аминокислотный остаток в составе эпитопа можно подвергать мутации, например, методом аланин-сканирующего мутагенеза, и выявлять мутации, которые приводят к уменьшению или предотвращению связывания белка. «Аланин-сканирующий мутагенез» представляет собой метод, который можно использовать для идентификации определенных остатков или областей белка, влияющих на взаимодействие эпитопа с другим соединением или белком, который с ним связывается. Остаток или группу целевых остатков в белке заменяют нейтральной или отрицательно заряженной аминокислотой (наиболее предпочтительно аланином или полиаланином, или путем консервативной аминокислотной замены). Любая мутация аминокислотных остатков или кодонов, кодирующих их, которая приводит к уменьшению связывания белка на величину более порогового значения или приводит к уменьшению связывания белка в максимальной степени относительно других мутаций, вероятно, имеет место в эпитопе, связываемом данным белком. В конкретных вариантах осуществления настоящего изобретения эпитоп, который является ключевым для анти-PD-1 антитела, содержит по меньшей мере один из аминокислотных остатков V64, P83, D85, L128, A129, P130, K131, A132 и Q133.
«1.7.3 чАт» при использовании в настоящем документе означает полностью человеческое моноклональное антитело, имеющее вариабельную область тяжелой цепи SEQ ID NO: 45, вариабельную область легкой цепи SEQ ID NO: 47 и константную область человеческого антитела изотипа IgG4.
«1.49.9 чАт» при использовании в настоящем документе означает полностью человеческое моноклональное антитело, имеющее вариабельную область тяжелой цепи SEQ ID NO: 49, вариабельную область легкой цепи SEQ ID NO: 51 и константную область человеческого антитела изотипа IgG4.
«1.103.11 чАт» при использовании в настоящем документе означает полностью человеческое моноклональное антитело, имеющее вариабельную область тяжелой цепи SEQ ID NO: 53, вариабельную область легкой цепи SEQ ID NO: 55 и константную область человеческого антитела изотипа IgG4.
«1.103.11-v2 чАт» при использовании в настоящем документе означает полностью человеческое моноклональное антитело, имеющее вариабельную область тяжелой цепи SEQ ID NO: 53, вариабельную область легкой цепи SEQ ID NO: 67 и константную область человеческого антитела изотипа IgG4.
«1.139.15 чАт» при использовании в настоящем документе означает полностью человеческое моноклональное антитело, имеющее вариабельную область тяжелой цепи SEQ ID NO: 57, вариабельную область легкой цепи SEQ ID NO: 59 и константную область человеческого антитела изотипа IgG4.
«1.153.7 чАт» при использовании в настоящем документе означает полностью человеческое моноклональное антитело, имеющее вариабельную область тяжелой цепи SEQ ID NO: 61, вариабельную область легкой цепи SEQ ID NO: 63 и константную область человеческого антитела изотипа IgG4.
«Консервативная замена» применительно к аминокислотной последовательности означает замену аминокислотного остатка другим аминокислотным остатком, имеющим боковую цепь со сходными физико-химическими свойствами. Например, консервативные замены можно выполнять между аминокислотными остатками с гидрофобными боковыми цепями (например, Met, Ala, Val, Leu и Ile), между остатками с нейтральными гидрофильными боковыми цепями (например, Cys, Ser, Thr, Asn и Gln), между остатками с кислыми боковыми цепями (например, Asp, Glu), между аминокислотами с основными боковыми цепями (например, His, Lys и Arg) или между остатками с ароматическими боковыми цепями (например, Trp, Tyr, и Phe). Как известно в данной области, консервативная замена, как правило, не вызывает существенного изменения в конформационной структуре белка и, таким образом, биологическая активность белка может сохраняться.
«Процент (%) идентичности последовательности» применительно к аминокислотной последовательности (или нуклеотидной последовательности) определяют, как процент аминокислотных остатков (или нуклеотидов) в последовательности-кандидате, которые идентичны аминокислотным остаткам (или нуклеотидам) в эталонной последовательности после выравнивания последовательностей и, при необходимости, внесения делеций для достижения максимального числа идентичных аминокислот (или нуклеотидов). Консервативно замененные аминокислотные остатки могут считаться или не считаться идентичными остатками. Выравнивание для целей определения процента идентичности аминокислотных (или нуклеотидных) последовательностей можно производить, например, с использованием общедоступных программных инструментов, таких как BLASTN, BLASTp (доступно на вебсайте Национального центра биотехнологической информации (NCBI) США, смотри также: Altschul S.F. et al., J. Mol. Biol., 215: 403-410 (1990); Stephen F. et al., Nucleic Acids Res., 25: 3389-3402 (1997)), ClustalW2 (доступно на вебсайте Европейского института биоинформатики, смотри также: Higgins D.G. et al., Methods in Enzymology, 266: 383-402 (1996); Larkin M.A. et al., Bioinformatics (Oxford, England), 23(21): 2947-8 (2007)), а также ALIGN или Megalign (DNASTAR). Специалисты в данной области могут использовать параметры по умолчанию, предоставляемые конкретной программой, или могут изменять параметры по мере необходимости для выравнивания, например, выбирая подходящий алгоритм.
Используемый в настоящем документе термин «T-клетка» охватывает CD4+ T-клетки, CD8+ T-клетки, T-хелперы 1 типа, T-хелперы 2 типа, T-хелперы 17 типа и ингибиторные T-клетки.
Используемый в настоящем документе термин «эффекторные функции» относится к видам биологической активности, которые вызваны связыванием Fc-области антитела с его эффекторами, такими как комплекс C1 и Fc-рецептор. Иллюстративные эффекторные функции включают комплементзависимую цитотоксичность (CDC), вызываемую взаимодействием антител и C1q в комплексе C1; антителозависимую опосредуемую клетками цитотоксичность (ADCC), вызываемую связыванием Fc-области антитела с Fc-рецептором на эффекторной клетке; и фагоцитоз.
Используемый в настоящем документе термин «рак» или «онкологическое заболевание» означает любое медицинское состояние, опосредованное ростом, пролиферацией или метастазированием неопластических, или злокачественных, клеток, и включает солидные формы рака и не солидные формы рака, такие как лейкоз. Используемый в настоящем документе термин «опухоль» означает солидную массу из неопластических и/или злокачественных клеток.
Используемый в настоящем документе термин «лечение», или «терапия», состояния включает предотвращение или ослабление состояния, отсрочку начала или замедление развития состояния, уменьшение риска развития состояния, предотвращение или отсрочку развития симптомов, ассоциированных с состоянием, уменьшение или устранение симптомов, ассоциированных с состоянием, достижение полной или частичной регрессии состояния, излечение состояния или некие комбинации перечисленного. Применительно к раку термин «лечение», или «терапия», может означать ингибирование или замедление роста, пролиферации или метастазирования неопластических, или злокачественных, клеток, предотвращение или замедление возникновения, роста, пролиферации или метастазирования неопластических, или злокачественных, клеток, или некие комбинации перечисленного. Применительно к опухоли термин «лечение», или «терапия», включает ликвидацию всей, или части, опухоли, ингибирование или замедление роста и метастазирования опухоли, предотвращение или задержку развития опухоли, или некие комбинации перечисленного.
«Выделенное» вещество было изменено рукой человека по сравнению с естественным состоянием. Если «выделенные» композиции или вещества существуют в природе, то они были изменены или удалены из их естественного окружения, либо и то, и другое. Например, полинуклеотид или полипептид, естественным образом присутствующий в организме живого животного, не является «выделенным», однако тот же полинуклеотид или полипептид является «выделенным», если он был в достаточной степени отделен от материалов, присутствующих совместно с ним в его естественном состоянии, чтобы существовать в по существу чистой форме. В конкретных вариантах осуществления антитела и антигенсвязывающие фрагменты имеют чистоту по меньшей мере 90%, 93%, 95%, 96%, 97%, 98%, 99%, что определяют электрофоретическими методами (такими как SDS-ПААГ, изоэлектрическое фокусирование, капиллярный электрофорез) или хроматографическими методами (такими как ионообменная хроматография или ВЭЖХ с обращенной фазой).
Используемый в настоящем документе термин «вектор» означает носитель, в который полинуклеотид, кодирующий белок, может быть функционально встроен для осуществления экспрессии данного белка. Вектор может быть использован для трансформации, трансдукции или трансфекции клетки-хозяина с целью осуществления экспрессии генетического элемента, который он несет, в клетке-хозяине. Примеры векторов включают плазмиды, фагмиды, космиды, искусственные хромосомы, такие как дрожжевая искусственная хромосома (YAC), бактериальная искусственная хромосома (BAC), или полученная из P1 искусственная хромосома (PAC), бактериофаги, такие как фаг лямбда или фаг M13, и вирусы животных. Категории вирусов животных, используемых в качестве векторов, включают ретровирус (в том числе лентивирус), аденовирус, аденоассоциированный вирус, вирус герпеса (например, вирус простого герпеса), поксвирус, бакуловирус, папилломавирус и паповавирус (например, SV40). Вектор может содержать различные элементы для контроля экспрессии, включая последовательности промотора, последовательности инициации транскрипции, последовательности энхансера, селективные элементы и гены репортеров. Кроме того, вектор может содержать точку начала репликации. Вектор также может содержать материалы, способствующие проникновению в клетку, включая, но без ограничения, вирусную частицу, липосому или белковое покрытие.
Используемый в настоящем документе термин «клетка-хозяин» означает клетку, в которую был введен экзогенный полинуклеотид и/или вектор.
Используемый в настоящем документе термин «заболевание, ассоциированное с, или связанное с, PD-1» относится к любому состоянию, которое вызывается, усугубляется или иным образом связано с увеличением, или уменьшением, экспрессии или активности PD-1 (например, человеческого PD-1).
Используемые в настоящем документе термины «терапевтически эффективное количество» или «эффективная доза» означают дозу или концентрацию лекарственного средства, эффективную для лечения заболевания или состояния, ассоциированного с человеческим PD-1. Например, применительно к использованию антител, или антигенсвязывающих фрагментов, раскрытых в настоящем документе, для лечения рака терапевтически эффективное количество представляет собой дозу или концентрацию антитела, или антигенсвязывающего фрагмента, способную приводить к уничтожению всей, или части, опухоли, ингибированию или замедлению роста опухоли, ингибированию роста или пролиферации клеток, опосредующих онкологическое заболевание, ингибированию метастазирования клеток опухоли, уменьшению проявления какого-либо симптома или содержания маркера, ассоциированного с опухолью или онкологическим заболеванием, предотвращению или замедлению развития опухоли или онкологического заболевания, или некую комбинацию перечисленного.
Термин «фармацевтически приемлемые» означает, что указанные: носитель, среда, разбавитель, эксципиент(ы) и/или соль, как правило, являются химически и/или физически совместимыми с другими ингредиентами, составляющими препарат, и физиологически совместимыми с их реципиентом.
Анти-PD-1 антитело
В одном аспекте настоящее изобретение относится к анти-PD-1 антителам и их антигенсвязывающим фрагментам. PD-1, также называемый CD279, известен как ключевой рецептор иммунологических контрольных точек, экспрессируемый активированными T-клетками, который опосредует иммуносупрессию. Лиганд 1 PD-1 (PD-L1) представляет собой 40-кДа трансмембранный белок, который экспрессируется на клетках различных опухолей, клетках стромы, или и тех, и других, и связывается с PD-1. Ингибирование взаимодействия между PD-1 и PD-L1 способно приводить к усилению T-клеточных ответов и, таким образом, опосредовать противораковую активность.
В конкретных вариантах осуществления настоящее изобретение относится к иллюстративным полностью человеческим моноклональным антителам 1.7.3 чАт, 1.49.9 чАт, 1.103.11 чАт, 1.103.11-v2 чАт, 1.139.15 чАт и 1.153.7 чАт, последовательности CDR которых приведены ниже в таблице 1; последовательности вариабельных областей тяжелой или легкой цепей также приведены ниже.
Таблица 1
STTYYWV
SISYSGNTYYNPSLKS
HLGYNGRYLPFDY
AGT ACT ACT TAC TAC TGG GTC
AGT ATC TCT TAT AGT GGG AAC ACC TAC TAC AAT CCG TCC CTC AAG AGT
CAT CTA GGG TAT AAT GGG AGG TAC CTC CCC TTT GAC TAC
TGTSSDVGFYNYVS
DVTNRPS
SSYTSISTWV
ACT GGA ACC AGC AGT GAC GTT GGT TTT TAT AAC TAT GTC TCC
GAT GTC ACT AAT CGG CCC TCA
AGC TCA TAT ACA AGC ATC AGC ACT TGG GTG
SSTYYWG
SISYSGSTYYNPSLKS
HLGYNGRYLPFDY
AGT AGT ACT TAC TAC TGG GGC
AGT ATC TCT TAT AGT GGG AGC ACC TAC TAC AAT CCG TCC CTC AAG AGT
CAT CTA GGG TAT AAT GGG AGG TAC CTC CCC TTT GAC TAC
TGTSSDVGFYNYVS
DVSNRPS
SSYTSISTWV
ACT GGA ACC AGC AGT GAC GTT GGT TTT TAT AAC TAT GTC TCC
GAT GTC AGT AAT CGG CCC TCA
AGC TCA TAT ACA AGC ATC AGC ACT TGG GTG
STTYYWV
SISYSGSTYYNPSLKS
HLGYNGRYLPFDY
AGT ACT ACT TAC TAC TGG GTC
AGT ATC TCT TAT AGT GGG AGC ACC TAC TAC AAT CCG TCC CTC AAG AGT
CAT CTA GGG TAT AAT GGG AGG TAC CTC CCC TTT GAC TAC
TGTSSDVGFYNYVS
DVSNRPS
SSYTNISTWV
ACT GGA ACC AGC AGT GAC GTT GGT TTT TAT AAC TAT GTC TCC
GAT GTC AGT AAT CGG CCC TCA
AGC TCA TAT ACA AAC ATC AGC ACT TGG GTG
STTYYWG
SISYSGTTYYNPSLKS
HLGYNSNWYPFDY
AGT ACT ACT TAC TAC TGG GGC
AGT ATC TCT TAT AGT GGG ACC ACC TAC TAC AAC CCG TCC CTC AAG AGT
CAT CTC GGG TAT AAC AGC AAC TGG TAC CCT TTT GAC TAC
TGTSSDVGSYNRVS
EVSNRPS
SSYTSSSTWV
ACT GGA ACC AGC AGT GAC GTT GGT AGT TAT AAC CGT GTC TCC
GAG GTC AGT AAT CGG CCC TCA
AGC TCA TAT ACA AGC AGC AGC ACT TGG GTG
SHAMS
TITGGGGSIYYADSVKG
NRAGEGYFDY
AGC CAT GCC ATG AGC
ACT ATT ACT GGT GGT GGT GGT AGC ATA TAC TAC GCA GAC TCC GTG AAG GGC
AAC CGC GCT GGG GAG GGT TAC TTT GAC TAC
GGDNIGNKDVH
RDSNRPS
QVWDSIWV
GGG GGA GAC AAC ATT GGA AAT AAA GAT GTG CAC
AGG GAT AGC AAC CGG CCC TCT
CAG GTG TGG GAC AGC ATT TGG GTG
STTYYWV
SISYSGSTYYNPSLKS
HLGYNGRYLPFDY
AGT ACT ACT TAC TAC TGG GTC
AGT ATC TCT TAT AGT GGG AGC ACC TAC TAC AAT CCG TCC CTC AAG AGT
CAT CTA GGG TAT AAT GGG AGG TAC CTC CCC TTT GAC TAC
TGTSSDVGFYNYVS
DVSNRPS
SSYTSISTWV
ACT GGA ACC AGC AGT GAC GTT GGT TTT TAT AAC TAT GTC TCC
GAT GTC AGT AAT CGG CCC TCA
AGC TCA TAT ACA AGC ATC AGC ACT TGG GTG
1.7.3 чАт -VH(23466-VH): (SEQ ID NO: 45 для аминокислотной последовательности и SEQ ID NO: 46 для нуклеотидной последовательности) с CDR 1-3 тяжелой цепи: SEQ ID NOs: 1, 3, 5 представляют собой аминокислотные последовательности и SEQ ID NOs: 2, 4, 6 представляют собой нуклеотидные последовательности, соответственно:
V-сегмент: IGHV4-39*01
D-сегмент: IGHD1-26*01
J-сегмент: IGHJ4*02
Q L Q L Q E S G P G L V K P S
1 CAG CTG CAG CTG CAG GAG TCG GGC CCA GGA CTG GTG AAG CCT TCG
E T L T L T C T V S G D S I S
46 GAG ACC CTG ACC CTC ACC TGC ACT GTC TCT GGT GAC TCC ATC AGC
CDR1
~~~~~~~~~~~~~~~~~~~~~~~~~
S T T Y Y W V W I R Q P P G K
91 AGT ACT ACT TAC TAC TGG GTC TGG ATC CGC CAG CCC CCA GGG AAG
CDR2
~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~
G L E W I G S I S Y S G N T Y
136 GGA CTG GAG TGG ATT GGG AGT ATC TCT TAT AGT GGG AAC ACC TAC
CDR2
~~~~~~~~~~~~~~~~~~~~~~~~~
Y N P S L K S R V T I S V D T
181 TAC AAT CCG TCC CTC AAG AGT CGA GTC ACC ATA TCC GTA GAC ACG
S K N H F S L K L S S V A A T
226 TCC AAG AAC CAC TTC TCC CTG AAG CTG AGT TCT GTG GCC GCC ACA
CDR3
~~~~~~~~~~~~~~~~~~~~~
D T A L Y Y C A R H L G Y N G
271 GAC ACG GCT CTA TAT TAC TGT GCG AGA CAT CTA GGG TAT AAT GGG
CDR3
~~~~~~~~~~~~~~~~~~~~~~~~~
R Y L P F D Y W G Q G T L V T
316 AGG TAC CTC CCC TTT GAC TAC TGG GGC CAG GGA ACC CTG GTC ACC
V S S (SEQ ID NO: 45)
361 GTC TCC TCC (SEQ ID NO: 46)
1.7.3 чАт -VL(23195-VL): (SEQ ID NO: 47 для аминокислотной последовательности и SEQ ID NO: 48 для нуклеотидной последовательности) с CDR 1-3 легкой цепи: SEQ ID NOs: 7, 9, 11 представляют собой аминокислотные последовательности и SEQ ID NOs: 8, 10, 12 представляют собой нуклеотидные последовательности, соответственно:
V-сегмент: IGLV2-14*01
J-сегмент: IGLJ3*02
Q S A L T Q P A S V S G S P G
1 CAG TCT GCC CTG ACT CAG CCT GCC TCC GTG TCT GGG TCT CCT GGA
CDR1
~~~~~~~~~~~~~~~~~~~~~~~~~~~~
Q S I T I S C T G T S S D V G
46 CAG TCG ATC ACC ATC TCC TGC ACT GGA ACC AGC AGT GAC GTT GGT
CDR1
~~~~~~~~~~~~~~~~~~~~~
F Y N Y V S W Y Q Q H P G K A
91 TTT TAT AAC TAT GTC TCC TGG TAC CAA CAG CAC CCA GGC AAA GCC
CDR2
~~~~~~~~~~~~~~~~~~~~~~~~~
P E L M I Y D V T N R P S G V
136 CCC GAA CTC ATG ATT TAT GAT GTC ACT AAT CGG CCC TCA GGG GTT
S D R F S G S K S G N T A S L
181 TCT GAT CGC TTC TCT GGC TCC AAG TCT GGC AAC ACG GCC TCC CTG
T I S G L Q A E D E A D Y Y C
226 ACC ATC TCT GGG CTC CAG GCT GAG GAC GAG GCT GAT TAT TAC TGC
CDR3
~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~
S S Y T S I S T W V F G G G T
261 AGC TCA TAT ACA AGC ATC AGC ACT TGG GTG TTC GGC GGA GGG ACC
K L T V L (SEQ ID NO: 47)
316 AAG CTG ACC GTC CTA (SEQ ID NO: 48)
1.49.9 чАт -VH(20951-VH): (SEQ ID NO: 49 для аминокислотной последовательности и SEQ ID NO: 50 для нуклеотидной последовательности) с CDR 1-3 тяжелой цепи: SEQ ID NOs: 13, 15, 5 представляют собой аминокислотные последовательности и SEQ ID NOs: 14, 16, 6 представляют собой нуклеотидные последовательности, соответственно:
V-сегмент: IGHV4-39*01
D-сегмент: IGHD1-26*01
J-сегмент: IGHJ4*02
Q L Q L Q E S G P G L V K P S
1 CAG CTG CAG CTG CAG GAG TCG GGC CCA GGA CTG GTG AAG CCT TCG
E T L S L T C T V S G G S I S
46 GAG ACC CTG TCC CTC ACC TGC ACT GTC TCT GGT GGC TCC ATC AGC
CDR1
~~~~~~~~~~~~~~~~~~~~~~~~~
S S T Y Y W G W I R Q P P G K
91 AGT AGT ACT TAC TAC TGG GGC TGG ATC CGC CAG CCC CCA GGG AAG
CDR2
~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~
G L E W I G S I S Y S G S T Y
136 GGA CTG GAG TGG ATT GGG AGT ATC TCT TAT AGT GGG AGC ACC TAC
CDR2
~~~~~~~~~~~~~~~~~~~~~~~~~
Y N P S L K S R V T I S V D T
181 TAC AAT CCG TCC CTC AAG AGT CGA GTC ACC ATA TCC GTA GAC ACG
S K N Q F S L K L S S V T D A
226 TCC AAG AAC CAG TTC TCC CTG AAG CTG AGC TCT GTG ACC GAC GCA
CDR3
~~~~~~~~~~~~~~~~~~~~~
D T A V Y Y C A R H L G Y N G
261 GAC ACG GCT GTG TAT TAC TGT GCG AGA CAT CTA GGG TAT AAT GGG
CDR3
~~~~~~~~~~~~~~~~~~~~~~~~~
R Y L P F D Y W G Q G T L V T
316 AGG TAC CTC CCC TTT GAC TAC TGG GGC CAG GGA ACC CTG GTC ACC
V S S (SEQ ID NO: 49)
361 GTC TCC TCC (SEQ ID NO: 50)
1.49.9 чАт -VL(21526-VL): (SEQ ID NO: 51 для аминокислотной последовательности и SEQ ID NO: 52 для нуклеотидной последовательности) с CDR 1-3 легкой цепи: SEQ ID NOs: 7, 17, 11 представляют собой аминокислотные последовательности и SEQ ID NOs: 8, 18, 12 представляют собой нуклеотидные последовательности, соответственно:
V-сегмент: IGLV2-14*01
J-сегмент: IGLJ3*02
Q S A L T Q P A S V S G S P G
1 CAG TCT GCC CTG ACT CAG CCT GCC TCC GTG TCT GGG TCT CCT GGA
CDR1
~~~~~~~~~~~~~~~~~~~~~~~~~~~~~
Q S I T I S C T G T S S D V G
46 CAG TCG ATC ACC ATC TCC TGC ACT GGA ACC AGC AGT GAC GTT GGT
CDR1
~~~~~~~~~~~~~~~~~~~~~
F Y N Y V S W Y Q Q H P G K A
91 TTT TAT AAC TAT GTC TCC TGG TAC CAA CAG CAC CCA GGC AAA GCC
CDR2
~~~~~~~~~~~~~~~~~~~~~~~~~
P E V M I Y D V S N R P S G V
136 CCC GAA GTC ATG ATT TAT GAT GTC AGT AAT CGG CCC TCA GGG GTT
S D R F S G S K S G N T A S L
181 TCT GAT CGC TTC TCT GGC TCC AAG TCT GGC AAC ACG GCC TCC CTG
T I S G L Q A E D E A D Y Y C
226 ACT ATC TCT GGG CTC CAG GCT GAG GAC GAG GCT GAT TAT TAC TGC
CDR3
~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~
S S Y T S I S T W V F G G G T
261 AGC TCA TAT ACA AGC ATC AGC ACT TGG GTG TTC GGC GGA GGG ACC
K L T V L (SEQ ID NO: 51)
316 AAG CTG ACT GTC CTA (SEQ ID NO: 52)
1.103.11 чАт -VH(20975-VH): (SEQ ID NO: 53 для аминокислотной последовательности и SEQ ID NO: 54 для нуклеотидной последовательности) с CDR 1-3 тяжелой цепи: SEQ ID NOs: 1, 15, 5 представляют собой аминокислотные последовательности и SEQ ID NOs: 2, 16, 6 представляют собой нуклеотидные последовательности, соответственно:
V-сегмент: IGHV4-39*01
D-сегмент: IGHD1-26*01
J-сегмент: IGHJ4*02
Q L Q L Q E S G P G L V K P S
1 CAG CTG CAG CTG CAG GAG TCG GGC CCA GGA CTG GTG AAG CCT TCG
E T L T L T C T V S A D S I S
46 GAG ACC CTG ACC CTC ACC TGC ACT GTC TCT GCT GAC TCC ATC AGC
CDR1
~~~~~~~~~~~~~~~~~~~~~~~~~
S T T Y Y W V W I R Q P P G K
91 AGT ACT ACT TAC TAC TGG GTC TGG ATC CGC CAG CCC CCA GGG AAG
CDR2
~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~
G L E W I G S I S Y S G S T Y
136 GGA CTG GAG TGG ATT GGG AGT ATC TCT TAT AGT GGG AGC ACC TAC
CDR2
~~~~~~~~~~~~~~~~~~~~~~~~~
Y N P S L K S R V T V S V D T
181 TAC AAT CCG TCC CTC AAG AGT CGA GTC ACC GTA TCC GTA GAC ACG
S K N Q F S L K L N S V A A T
226 TCC AAG AAC CAG TTC TCC CTG AAG CTG AAC TCT GTG GCC GCC ACA
CDR3
~~~~~~~~~~~~~~~~~~~~~
D T A L Y Y C A R H L G Y N G
261 GAC ACG GCT CTA TAT TAC TGT GCG AGA CAT CTA GGG TAT AAT GGG
CDR3
~~~~~~~~~~~~~~~~~~~~~~~~~
R Y L P F D Y W G Q G T L V T
316 AGG TAC CTC CCC TTT GAC TAC TGG GGC CAG GGA ACC CTG GTC ACC
V S S (SEQ ID NO: 53)
361 GTC TCC TCC (SEQ ID NO: 54)
1.103.11 чАт -VL(21038-VL): (SEQ ID NO: 55 для аминокислотной последовательности и SEQ ID NO: 56 для нуклеотидной последовательности) с CDR 1-3 легкой цепи: SEQ ID NOs: 7, 17, 19 представляют собой аминокислотные последовательности и SEQ ID NOs: 8, 18, 20 представляют собой нуклеотидные последовательности, соответственно:
V-сегмент: IGLV2-14*01
J-сегмент: IGLJ3*02
Q S A L T Q P A S V S G S P G
1 CAG TCT GCC CTG ACT CAG CCT GCC TCC GTG TCT GGG TCT CCT GGA
CDR1
~~~~~~~~~~~~~~~~~~~~~~~~~~~~~
Q S I T I S C T G T S S D V G
46 CAG TCG ATC ACC ATC TCC TGC ACT GGA ACC AGC AGT GAC GTT GGT
CDR1
~~~~~~~~~~~~~~~~~~~~~
F Y N Y V S W Y Q Q H P G K A
91 TTT TAT AAC TAT GTC TCC TGG TAC CAA CAG CAC CCA GGC AAA GCC
CDR2
~~~~~~~~~~~~~~~~~~~~~~~~~
P E L M I Y D V S N R P S G V
136 CCC GAA CTC ATG ATT TAT GAT GTC AGT AAT CGG CCC TCA GGG GTT
S D R F S G S K S G N T A S L
181 TCT GAT CGC TTC TCT GGC TCC AAG TCT GGC AAC ACG GCC TCC CTG
T I S G L Q A E D E A D Y Y C
226 ACC ATC TCT GGG CTC CAG GCT GAG GAC GAG GCT GAT TAT TAC TGC
CDR3
~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~
S S Y T N I S T W V F G G G T
261 AGC TCA TAT ACA AAC ATC AGC ACT TGG GTG TTC GGC GGA GGG ACC
K L T V L (SEQ ID NO: 55)
316 AAG CTG ACC GTC CTA (SEQ ID NO: 56)
1.139.15 чАт -VH(23521-VH) (SEQ ID NO: 57 для аминокислотной последовательности и SEQ ID NO: 58 для нуклеотидной последовательности) с CDR 1-3 тяжелой цепи: SEQ ID NOs: 21, 23, 25 представляют собой аминокислотные последовательности и SEQ ID NOs: 22, 24, 26 представляют собой нуклеотидные последовательности, соответственно:
V-сегмент: IGHV4-39*01
D-сегмент: IGHD6-13*01
J-сегмент: IGHJ4*02
Q L Q L Q E S G P G L V K P S
1 CAG CTG CAG CTG CAG GAG TCG GGC CCA GGA CTG GTG AAG CCC TCG
E T L S L T C T V S G G S I S
46 GAG ACC CTG TCC CTC ACC TGC ACT GTC TCT GGT GGC TCC ATC AGC
CDR1
~~~~~~~~~~~~~~~~~~~~~~~~~
S T T Y Y W G W I R Q P P G K
91 AGT ACT ACT TAC TAC TGG GGC TGG ATC CGC CAG CCC CCA GGG AAG
CDR2
~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~
G L E W I G S I S Y S G T T Y
136 GGG CTG GAG TGG ATT GGG AGT ATC TCT TAT AGT GGG ACC ACC TAC
CDR2
~~~~~~~~~~~~~~~~~~~~~~~~~
Y N P S L K S R V T I P V D T
181 TAC AAC CCG TCC CTC AAG AGT CGA GTC ACC ATC CCC GTA GAC ACG
S K N Q I S L K L S S V T A A
226 TCC AAG AAC CAG ATC TCC CTG AAA CTG AGC TCT GTG ACC GCC GCA
CDR3
~~~~~~~~~~~~~~~~~~~~~
D T S L Y Y C A R H L G Y N S
261 GAC ACG TCT TTG TAT TAT TGT GCG AGA CAT CTC GGG TAT AAC AGC
CDR3
~~~~~~~~~~~~~~~~~~~~~~~~~
N W Y P F D Y W G Q G T L V T
316 AAC TGG TAC CCT TTT GAC TAC TGG GGC CAG GGA ACC CTG GTC ACC
V S S (SEQ ID NO: 57)
361 GTC TCC TCA (SEQ ID NO: 58)
1.139.15 чАт -VL(22895-VL) (SEQ ID NO: 59 для аминокислотной последовательности и SEQ ID NO: 60 для нуклеотидной последовательности) с CDR 1-3 легкой цепи: SEQ ID NOs: 27, 29, 31 представляют собой аминокислотные последовательности и SEQ ID NOs: 28, 30, 32 представляют собой нуклеотидные последовательности, соответственно:
V-сегмент: IGLV2-18*02
J-сегмент: IGLJ3*02
Q S A L T Q P P S V S G S P G
1 CAG TCG GCC CTG ACT CAG CCT CCC TCC GTG TCC GGG TCT CCT GGA
CDR1
~~~~~~~~~~~~~~~~~~~~~~~~~~~~~
Q S V T I S C T G T S S D V G
46 CAG TCA GTC ACC ATC TCC TGC ACT GGA ACC AGC AGT GAC GTT GGT
CDR1
~~~~~~~~~~~~~~~~~~~~~
S Y N R V S W Y Q Q P P G T A
91 AGT TAT AAC CGT GTC TCC TGG TAC CAG CAG CCC CCA GGC ACA GCC
CDR2
~~~~~~~~~~~~~~~~~~~~~~~~~
P E V I I Y E V S N R P S G V
136 CCC GAA GTC ATT ATT TAT GAG GTC AGT AAT CGG CCC TCA GGG GTC
P D R F S G S K S G N T A S L
181 CCT GAT CGC TTC TCT GGG TCC AAG TCT GGC AAC ACG GCC TCC CTG
T I S G L Q A E D E A D Y Y C
226 ACC ATC TCT GGG CTC CAG GCT GAG GAC GAG GCT GAT TAT TAC TGC
CDR3
~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~
S S Y T S S S T W V F G G G T
261 AGC TCA TAT ACA AGC AGC AGC ACT TGG GTG TTC GGC GGA GGG ACC
K L T V L (SEQ ID NO: 59)
316 AAG CTG ACC GTC CTA (SEQ ID NO: 60)
1.153.7 чАт -VH(20942-VH): (SEQ ID NO: 61 для аминокислотной последовательности и SEQ ID NO: 62 для нуклеотидной последовательности) с CDR 1-3 тяжелой цепи: SEQ ID NOs: 33, 35, 37 представляют собой аминокислотные последовательности и SEQ ID NOs: 34, 36, 38 представляют собой нуклеотидные последовательности, соответственно:
V-сегмент: IGHV3-23*01
D-сегмент: IGHD7-27*01
J-сегмент: IGHJ4*02
E V Q L L E S G G G L V Q P G
1 GAG GTG CAG CTG TTG GAG TCT GGG GGA GGC TTG GTA CAG CCT GGG
G S L R L S C A A S G F T F S
46 GGG TCC CTG AGA CTG TCC TGC GCA GCC TCT GGA TTC ACC TTT AGC
CDR1
~~~~~~~~~~~~~~~~~
S H A M S W V R Q A P G K G L
91 AGC CAT GCC ATG AGC TGG GTC CGC CAG GCT CCA GGG AAG GGG CTG
CDR2
~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~
E W V S T I T G G G G S I Y Y
136 GAG TGG GTC TCA ACT ATT ACT GGT GGT GGT GGT AGC ATA TAC TAC
CDR2
~~~~~~~~~~~~~~~~~~~~~
A D S V K G R F T I S R D N S
181 GCA GAC TCC GTG AAG GGC CGG TTC ACC ATC TCC AGA GAC AAT TCC
K N T L Y L Q M N S L R A E D
226 AAG AAC ACG CTG TAT CTG CAA ATG AAC AGC CTG AGA GCC GAG GAC
CDR3
~~~~~~~~~~~~~~~~~~~~~~~~~
T A V Y Y C A K N R A G E G Y
261 ACG GCC GTA TAT TAT TGT GCG AAA AAC CGC GCT GGG GAG GGT TAC
CDR3
~~~~~~~~~
F D Y W G Q G T L V T V S S (SEQ ID NO: 61)
316 TTT GAC TAC TGG GGC CAG GGA ACC CTG GTC ACC GTC TCC TCA(SEQ ID NO: 62)
1.153.7 чАт -VL(21110-VL) (SEQ ID NO: 63 для аминокислотной последовательности и SEQ ID NO: 64 для нуклеотидной последовательности) с CDR 1-3 легкой цепи: SEQ ID NOs: 39, 41, 43 представляют собой аминокислотные последовательности и SEQ ID NOs: 40, 42, 44 представляют собой нуклеотидные последовательности, соответственно:
V-сегмент: IGLV3-9*01
J-сегмент: IGLJ3*02
S Y E L T Q P L S V S V A L G
1 TCC TAT GAG CTG ACT CAG CCA CTC TCA GTG TCA GTG GCC CTG GGA
CDR1
~~~~~~~~~~~~~~~~~~~~~~~~~~~~~
Q T A R I T C G G D N I G N K
46 CAG ACG GCC AGG ATT ACC TGT GGG GGA GAC AAC ATT GGA AAT AAA
CDR1
~~~~~~~~~
D V H W Y Q Q K P G Q A P V L
91 GAT GTG CAC TGG TAC CAG CAG AAG CCA GGC CAG GCC CCT GTG CTG
CDR2
~~~~~~~~~~~~~~~~~~~~~~~~~
V I Y R D S N R P S G I P E G
136 GTC ATC TAT AGG GAT AGC AAC CGG CCC TCT GGG ATC CCT GAG GGA
F S G S N S G N T A T L T I S
181 TTC TCT GGC TCC AAC TCG GGG AAC ACG GCC ACC CTG ACC ATC AGC
CDR3
~~~~~~~~~
R A Q A G D E A D Y Y C Q V W
226 AGA GCC CAA GCC GGG GAT GAG GCT GAC TAT TAC TGT CAG GTG TGG
CDR3
~~~~~~~~~~~~~~~~~
D S I W V F G G G T K L T V L (SEQ ID NO: 63)
261 GAC AGC ATT TGG GTG TTC GGC GGA GGG ACC AAG CTG ACC GTC CTA (SEQ ID NO: 64)
1.103.11-v2 чАт -VH(20975-VH): (SEQ ID NO: 53 для аминокислотной последовательности и SEQ ID NO: 54 для нуклеотидной последовательности) с CDR 1-3 тяжелой цепи: SEQ ID NOs: 1, 15, 5 представляют собой аминокислотные последовательности и SEQ ID NOs: 2, 16, 6 представляют собой нуклеотидные последовательности, соответственно:
V-сегмент: IGHV4-39*01
D-сегмент: IGHD1-26*01
J-сегмент: IGHJ4*02
Q L Q L Q E S G P G L V K P S
1 CAG CTG CAG CTG CAG GAG TCG GGC CCA GGA CTG GTG AAG CCT TCG
E T L T L T C T V S A D S I S
46 GAG ACC CTG ACC CTC ACC TGC ACT GTC TCT GCT GAC TCC ATC AGC
CDR1
~~~~~~~~~~~~~~~~~~~~~~~~~
S T T Y Y W V W I R Q P P G K
91 AGT ACT ACT TAC TAC TGG GTC TGG ATC CGC CAG CCC CCA GGG AAG
CDR2
~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~
G L E W I G S I S Y S G S T Y
136 GGA CTG GAG TGG ATT GGG AGT ATC TCT TAT AGT GGG AGC ACC TAC
CDR2
~~~~~~~~~~~~~~~~~~~~~~~~~
Y N P S L K S R V T V S V D T
181 TAC AAT CCG TCC CTC AAG AGT CGA GTC ACC GTA TCC GTA GAC ACG
S K N Q F S L K L N S V A A T
226 TCC AAG AAC CAG TTC TCC CTG AAG CTG AAC TCT GTG GCC GCC ACA
CDR3
~~~~~~~~~~~~~~~~~~~~~
D T A L Y Y C A R H L G Y N G
261 GAC ACG GCT CTA TAT TAC TGT GCG AGA CAT CTA GGG TAT AAT GGG
CDR3
~~~~~~~~~~~~~~~~~~~~~~~~~
R Y L P F D Y W G Q G T L V T
316 AGG TAC CTC CCC TTT GAC TAC TGG GGC CAG GGA ACC CTG GTC ACC
V S S (SEQ ID NO: 53)
361 GTC TCC TCC (SEQ ID NO: 54)
1.103.11-v2 чАт -VL(21038-2-VL): (SEQ ID NO: 67 для аминокислотной последовательности и SEQ ID NO: 68 для нуклеотидной последовательности) с CDR 1-3 легкой цепи: SEQ ID NOs: 7, 17, 65 представляют собой аминокислотные последовательности и SEQ ID NOs: 8, 18, 66 представляют собой нуклеотидные последовательности, соответственно:
V-сегмент: IGLV2-14*01
J-сегмент: IGLJ3*02
Q S A L T Q P A S V S G S P G
1 CAG TCT GCC CTG ACT CAG CCT GCC TCC GTG TCT GGG TCT CCT GGA
CDR1
~~~~~~~~~~~~~~~~~~~~~~~~~~~~~
Q S I T I S C T G T S S D V G
46 CAG TCG ATC ACC ATC TCC TGC ACT GGA ACC AGC AGT GAC GTT GGT
CDR1
~~~~~~~~~~~~~~~~~~~~~
F Y N Y V S W Y Q Q H P G K A
91 TTT TAT AAC TAT GTC TCC TGG TAC CAA CAG CAC CCA GGC AAA GCC
CDR2
~~~~~~~~~~~~~~~~~~~~~~~~~
P E L M I Y D V S N R P S G V
136 CCC GAA CTC ATG ATT TAT GAT GTC AGT AAT CGG CCC TCA GGG GTT
S D R F S G S K S G N T A S L
181 TCT GAT CGC TTC TCT GGC TCC AAG TCT GGC AAC ACG GCC TCC CTG
T I S G L Q A E D E A D Y Y C
226 ACC ATC TCT GGG CTC CAG GCT GAG GAC GAG GCT GAT TAT TAC TGC
CDR3
~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~
S S Y T S I S T W V F G G G T
261 AGC TCA TAT ACA AGC ATC AGC ACT TGG GTG TTC GGC GGA GGG ACC
K L T V L (SEQ ID NO: 67)
316 AAG CTG ACC GTC CTA (SEQ ID NO: 68)
В некоторых вариантах осуществления анти-PD-1 антитела и их антигенсвязывающие фрагменты содержат последовательности CDR тяжелой цепи, выбранные из группы, состоящей из SEQ ID NOs: 1, 3, 5, 13, 15, 21, 23, 25, 33, 35 и 37. В некоторых вариантах осуществления анти-PD-1 антитела и их антигенсвязывающие фрагменты содержат последовательности CDR легкой цепи, выбранные из группы, состоящей из SEQ ID NOs: 7, 9, 11, 17, 19, 27, 29, 31, 39, 41, 43 и 65. В конкретных вариантах осуществления одна или более последовательностей CDR, предложенных в настоящем документе, могут быть модифицированы или изменены так, что у полученного антитела по сравнению с исходным антителом улучшено одно или более свойств (например, улучшено связывание антигена, улучшена схема гликозилирования, снижен риск гликозилирования на остатке CDR, уменьшено дезаминирование на остатке CDR, увеличен фармакокинетический период полувыведения, pH-чувствительность и совместимость с конъюгацией), а в остальном оно сравнимо с исходным антителом (то есть, антителом, в остальном имеющим тот же набор последовательностей CDR за исключением вышеупомянутых модификаций или изменений) или по меньшей мере по существу сохраняет антигенсвязывающую способность исходного антитела.
В некоторых вариантах осуществления анти-PD-1 антитела и их антигенсвязывающие фрагменты содержат вариабельную область тяжелой цепи, выбранную из группы, состоящей из вариабельной области тяжелой цепи, содержащей SEQ ID NO: 1, SEQ ID NO: 3, и/или SEQ ID NO: 5; вариабельной области тяжелой цепи, содержащей SEQ ID NO: 13, SEQ ID NO: 15 и/или SEQ ID NO: 5; вариабельной области тяжелой цепи, содержащей SEQ ID NO: 1, SEQ ID NO: 15 и/или SEQ ID NO: 5; вариабельной области тяжелой цепи, содержащей SEQ ID NO: 21, SEQ ID NO: 23 и/или SEQ ID NO: 25; и вариабельной области тяжелой цепи, содержащей SEQ ID NO: 33, SEQ ID NO: 35 и/или SEQ ID NO: 37.
В некоторых вариантах осуществления анти-PD-1 антитела и их антигенсвязывающие фрагменты содержат вариабельную область легкой цепи, выбранную из группы, состоящей из вариабельной области легкой цепи, содержащей SEQ ID NO: 7, SEQ ID NO: 9 и/или SEQ ID NO: 11; вариабельной области легкой цепи, содержащей SEQ ID NO: 7, SEQ ID NO: 17 и/или SEQ ID NO: 11; вариабельной области легкой цепи, содержащей SEQ ID NO: 7, SEQ ID NO: 17 и/или SEQ ID NO: 19; вариабельной области легкой цепи, содержащей SEQ ID NO: 27, SEQ ID NO: 29 и/или SEQ ID NO: 31; вариабельной области легкой цепи, содержащей SEQ ID NO: 39, SEQ ID NO: 41 и/или SEQ ID NO: 43; и вариабельной области легкой цепи, содержащей SEQ ID NO: 7, SEQ ID NO: 17 и/или SEQ ID NO: 65.
В некоторых вариантах осуществления анти-PD-1 антитела и их антигенсвязывающие фрагменты содержат: a) вариабельную область тяжелой цепи, содержащую SEQ ID NO: 1, SEQ ID NO: 3 и/или SEQ ID NO: 5; и вариабельную область легкой цепи, содержащую SEQ ID NO: 7, SEQ ID NO: 9 и/или SEQ ID NO: 11; b) вариабельную область тяжелой цепи, содержащую SEQ ID NO: 13, SEQ ID NO: 15 и/или SEQ ID NO: 5; и вариабельную область легкой цепи, содержащую SEQ ID NO: 7, SEQ ID NO: 17 и/или SEQ ID NO: 11; c) вариабельную область тяжелой цепи, содержащую SEQ ID NO: 1, SEQ ID NO: 15 и/или SEQ ID NO: 5; и вариабельную область легкой цепи, содержащую SEQ ID NO: 7, SEQ ID NO: 17 и/или SEQ ID NO: 19; d) вариабельную область тяжелой цепи, содержащую SEQ ID NO: 21, SEQ ID NO: 23 и/или SEQ ID NO: 25; и вариабельную область легкой цепи, содержащую SEQ ID NO: 27, SEQ ID NO: 29 и/или SEQ ID NO: 31; e) вариабельную область тяжелой цепи, содержащую SEQ ID NO: 33, SEQ ID NO: 35 и/или SEQ ID NO: 37; и вариабельную область легкой цепи, содержащую SEQ ID NO: 39, SEQ ID NO: 41 и/или SEQ ID NO: 43; или f) вариабельную область тяжелой цепи, содержащую SEQ ID NO: 1, SEQ ID NO: 15 и/или SEQ ID NO: 5; и вариабельную область легкой цепи, содержащую SEQ ID NO: 7, SEQ ID NO: 17 и/или SEQ ID NO: 65.
Специалист в данной области понимает, что последовательности CDR, приведенные в таблице 1, могут быть модифицированы для включения одной или более аминокислотных замен с целью обеспечения улучшенной биологической активности, например, повышенной аффинности связывания с человеческим PD-1. Например, можно получать библиотеку вариантов антител (например, вариантов Fab или scFv) и осуществлять экспрессию с использованием технологии фагового дисплея, а затем проводить скрининг на аффинность связывания с человеческим PD-1. В качестве другого примера, можно использовать компьютерную программу для виртуальной имитации связывания антител с человеческим PD-1 и идентификации аминокислотных остатков в антителах, которые образуют поверхность связывания. Такие остатки можно либо не использовать для замен, чтобы предотвратить уменьшение аффинности связывания, либо выбирать мишенью для замен, чтобы обеспечить более сильное связывание. В конкретных вариантах осуществления по меньшей мере одна (или все) из замен в последовательностях CDR являются консервативными заменами.
В конкретных вариантах осуществления антитела и их антигенсвязывающие фрагменты содержат одну или более последовательностей CDR, имеющих по меньшей мере 80% (например, по меньшей мере 85%, 88%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%) идентичности последовательности с последовательностью (или последовательностями) в таблице 1, и при этом сохраняют аффинность связывания с человеческим PD-1 на уровне, аналогичном или даже более высоком, чем у исходного антитела, имеющего по существу ту же последовательность, за исключением того, что соответствующая последовательность CDR имеет 100% идентичности последовательности с последовательностью (или последовательностями) в таблице 1.
В конкретных вариантах осуществления анти-PD-1 антитела и их антигенсвязывающие фрагменты являются полностью человеческими. Полностью человеческие антитела помогают избежать проблем с иммуногенностью для человека и/или сниженной аффинностью связывания, что часто наблюдается в случае гуманизированных антител.
В некоторых вариантах осуществления полностью человеческие анти-PD-1 антитела и их антигенсвязывающие фрагменты содержат вариабельную область тяжелой цепи, выбранную из группы, состоящей из SEQ ID NO: 45, SEQ ID NO: 49, SEQ ID NO: 53, SEQ ID NO: 57, SEQ ID NO: 61 и их гомологичных последовательностей, имеющих по меньшей мере 80% (например, по меньшей мере 85%, 88%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%) идентичности последовательности; и/или вариабельную область легкой цепи, выбранную из группы, состоящей из SEQ ID NO: 47, SEQ ID NO: 51, SEQ ID NO: 55, SEQ ID NO: 59, SEQ ID NO: 63, SEQ ID NO: 67 и их гомологичных последовательностей, имеющих по меньшей мере 80% (например, по меньшей мере 85%, 88%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%) идентичности последовательности. Эти полностью человеческие антитела сохраняют аффинность связывания с человеческим PD-1, предпочтительно, на уровне, сходном с уровнем одного из иллюстративных антител: 1.7.3 чАт, 1.49.9 чАт, 1.103.11 чАт, 1.103.11-v2 чАт, 1.139.15 чАт и 1.153.7 чАт.
В некоторых вариантах осуществления полностью человеческие анти-PD-1 антитела и их антигенсвязывающие фрагменты содержат: a) вариабельную область тяжелой цепи, содержащую SEQ ID NO: 45, и вариабельную область легкой цепи, содержащую SEQ ID NO: 47; b) вариабельную область тяжелой цепи, содержащую SEQ ID NO: 49, и вариабельную область легкой цепи, содержащую SEQ ID NO: 51; c) вариабельную область тяжелой цепи, содержащую SEQ ID NO: 53, и вариабельную область легкой цепи, содержащую SEQ ID NO: 55; d) вариабельную область тяжелой цепи, содержащую SEQ ID NO: 57, и вариабельную область легкой цепи, содержащую SEQ ID NO: 59; e) вариабельную область тяжелой цепи, содержащую SEQ ID NO: 61, и вариабельную область легкой цепи, содержащую SEQ ID NO: 63, или f) вариабельную область тяжелой цепи, содержащую SEQ ID NO: 53, и вариабельную область легкой цепи, содержащую SEQ ID NO: 67.
В настоящем документе также предусмотрены антитела и антигенсвязывающие фрагменты, которые конкурируют за один и тот же эпитоп с анти-PD-1 антителами и их антигенсвязывающими фрагментами, предложенными в настоящем документе. В конкретных вариантах осуществления антитела блокируют связывание 1.7.3 чАт, 1.49.9 чАт, 1.103.11 чАт, 1.103.11-v2 чАт, 1.139.15 чАт или 1.153.7 чАт с человеческим или обезьяньим PD-1, например, с величиной IC50 (то есть, концентрация 50% ингибирования) ниже 10-6 M, ниже 10-7 M, ниже 10-7,5 M, ниже 10-8 M, ниже 10-8,5 M, ниже 10-9 M или ниже 10-10 M. Значения IC50 определяют на основании конкурентного анализа, такого как анализы ELISA, конкурентные анализы связывания радиоактивного лиганда и FACS-анализ.
В некоторых вариантах осуществления анти-PD-1 антитела и их антигенсвязывающие фрагменты, предложенные в настоящем документе, способны специфически связывать человеческий PD-1 с аффинностью связывания (Kd) ≤10-6 M (например, ≤5×10-7 M, ≤2×10-7 M, ≤10-7 M, ≤5×10-8 M, ≤2×10-8 M, ≤10-8 M, ≤5×10-9 M, ≤2×10-9 M, ≤10-9 M, 10-10 M) при измерении в анализе связывания методом плазмонного резонанса. Аффинность связывания может быть представлена значением KD, которое рассчитывают, как отношение скорости диссоциации к скорости ассоциации (koff/kon), когда связывание между антигеном и антигенсвязывающей молекулой достигает равновесия. Аффинность связывания антигена (например, KD) можно надлежащим образом определять соответствующими способами, известными в данной области, включая, например, анализ связывания методом плазмонного резонанса с использованием таких инструментов, как Biacore (смотри, например, Murphy, M. et al., Current protocols in protein science, Chapter 19, unit 19.14, 2006).
В конкретных вариантах осуществления антитела и их фрагменты, предложенные в настоящем документе, связывают человеческий PD-1 с величиной EC50 (то есть, концентрация 50% связывания), составляющей 0,1 нМ - 100 нМ (например, 0,1 нМ - 50 нМ, 0,1 нМ - 30 нМ, 0,1 нМ - 20 нМ, 0,1 нМ - 10 нМ или 0,1 нМ - 1 нМ). Связывание антител с человеческим PD-1 можно измерять методами, известными в данной области, например, с использованием сэндвич-анализа ELISA, вестерн-блоттинга, FACS или другого анализа связывания. В иллюстративном примере тестируемому антителу (то есть, первому антителу) дают возможность связываться с иммобилизованным человеческим PD-1 или клетками, экспрессирующими человеческий PD-1, после отмывания не связавшегося антитела добавляют меченое вторичное антитело, которое может связываться со связанным первым антителом и, таким образом, обеспечивать его обнаружение. Это обнаружение можно осуществлять на планшетном ридере, если используют иммобилизованный PD-1, или в FACS-анализе, если используют клетки, экспрессирующие человеческий PD-1. В конкретных вариантах осуществления антитела и их фрагменты, предложенные в настоящем документе, связывают человеческий PD-1 с величиной EC50 (то есть, 50% эффективная концентрация), составляющей 1 нМ - 10 нМ, или 1 нМ - 5 нМ при измерении в FACS-анализе.
В конкретных вариантах осуществления антитела и их фрагменты, предложенные в настоящем документе, ингибируют связывание человеческого PD-1 с его лигандом с величиной IC50, составляющей 0,2 нМ - 100 нМ (например, 0,2 нМ - 50 нМ, 0,2 нМ - 30 нМ, 0,2 нМ - 20 нМ, 0,2 нМ - 10 нМ или 1 нМ - 10 нМ) при измерении в конкурентном анализе.
В конкретных вариантах осуществления антитела и их фрагменты, предложенные в настоящем документе, блокируют связывание человеческого PD-1 с его лигандом и, тем самым, обеспечивают биологическую активность, включая, например, продуцирование цитокинов активированными T-клетками (такими как CD4+ T-клетки и CD8+ T-клетки), индукцию пролиферации активированных T-клеток (таких как CD4+ T-клетки и CD8+ T-клетки), а также реверсию супрессорной функции Treg. Иллюстративные цитокины включают IL-2 и IFNγ. Термин «IL-2» означает интерлейкин 2, вид сигнальной молекулы цитокина в иммунной системе, которая регулирует активность белых клеток крови (например, лейкоцитов). Термин «интерферон-гамма (IFNγ)» означает цитокин, который продуцируется клетками-естественными киллерами (NK), NK T-клетками, CD4+ и CD8+ T-клетками, и является важным активатором макрофагов и индуктором экспрессии молекул главного комплекса гистосовместимости (MHC). Продуцирование цитокинов можно определять методами, известными в данной области, например, методом ELISA. Можно также использовать методы определения пролиферации T-клеток, включая анализ с включением [3H] тимидина.
Анти-PD-1 антитела и их антигенсвязывающие фрагменты являются специфичными для PD-1. В конкретных вариантах осуществления антитела и их антигенсвязывающие фрагменты не связывают CD28 и/или CTLA-4. Например, аффинность связывания с CD28 и/или CTLA-4 составляет менее 15%, 10%, 9%, 8%, 7%, 6%, 5%, 4%, 3%, 2% или 1% от аффинности для PD-1.
В конкретных вариантах осуществления антитела и их антигенсвязывающие фрагменты связывают обезьяний PD-1 с величиной EC50 не более 100 нМ, например, не более или примерно 10 нМ, 9 нМ, 8 нМ, 7 нМ, 6 нМ, 5 нМ, 4 нМ, 3 нМ, 2 нМ, 1 нМ, 0,9 нМ, 0,8 нМ, 0,7 нМ, 0,6 нМ, 0,5 нМ, 0,4 нМ, 0,3 нМ, 0,2 нМ, 0,1 нМ, 0,09 нМ, 0,08 нМ, 0,07 нМ, 0,06 нМ, 0,05 нМ, 0,04 нМ, 0,03 нМ, 0,02 нМ или 0,01 нМ, при измерении методом ELISA. В конкретных вариантах осуществления антитела и их антигенсвязывающие фрагменты связывают обезьяний PD-1 с величиной EC50 примерно 1 нМ - 10 нМ.
В конкретных вариантах осуществления антитела и их антигенсвязывающие фрагменты не связывают мышиный PD-1, но связывают обезьяний PD-1 с аффинностью связывания, аналогичной аффинности для человеческого PD-1. Например, связывание иллюстративных антител 1.7.3 чАт, 1.49.9 чАт, 1.103.11 чАт, 1.103.11-v2 чАт, 1.139.15 чАт и 1.153.7 чАт с мышиным PD-1 невозможно обнаруживать в общепринятых анализах связывания, таких как ELISA или FACS-анализ, в то время как связывание этих антител с обезьяньим PD-1 имеет место с аффинностью или величиной EC50, сходной с величиной для человеческого PD-1, при измерении в анализе ELISA или FACS.
В некоторых вариантах осуществления у анти-PD-1 антител и их антигенсвязывающих фрагментов эффекторная функция ослаблена или отсутствует. В некоторых вариантах осуществления анти-PD-1 антитела и их антигенсвязывающие фрагменты имеют константную область изотипа IgG4, которая отличается ослабленной эффекторной функцией или ее отсутствием. Эффекторные функции, такие как ADCC и CDC, могут приводить к цитотоксичности в отношении клеток, экспрессирующих PD-1. Многие клетки, такие как T-клетки, обычно экспрессируют PD-1. Чтобы избежать потенциальной нежелательной токсичности для этих нормальных клеток, в конкретных вариантах осуществления у антител и антигенсвязывающих фрагментов, предложенных в настоящем документе, эффекторные функции могут быть ослаблены или отсутствовать. Различные анализы для оценки активностей ADCC или CDC, например, анализ связывания Fc-рецептора, анализ связывания C1q и анализ лизиса клеток, известны и могут быть с легкостью использованы специалистами в данной области. Без привязки к конкретной теории, считается, что антитела с ослабленными или отсутствующими эффекторными функциями, такими как ADCC или CDC, не будут вызывать, или будут вызывать минимальную, цитотоксичность в отношении PD-1-экспрессирующих клеток, например, T-клеток, и, таким образом, позволят избегать нежелательных побочных эффектов, при этом блокирование PD-1 приведет к стимуляции иммунной системы для лечения таких состояний, как рак или хроническая инфекция.
В конкретных вариантах осуществления анти-PD-1 антитела и их антигенсвязывающие фрагменты, предложенные в настоящем документе, вызывают меньше побочных эффектов. Например, антитела и их антигенсвязывающие фрагменты, предложенные в настоящем документе, могут иметь полностью человеческие последовательности IgG и, таким образом, сниженную иммуногенность по сравнению с гуманизированным аналогом антитела. В качестве другого примера, антитела и их антигенсвязывающие фрагменты, предложенные в настоящем документе, могут иметь формат IgG4 для элиминации ADCC и CDC.
В конкретных вариантах осуществления анти-PD-1 антитела и их антигенсвязывающие фрагменты, предложенные в настоящем документе, обладают тем преимуществом, что их можно использовать в сочетании с иммуногенными средствами, такими как клетки опухолей, очищенный опухолевый антиген и клетки, трансфицированные генами, кодирующими иммуностимулирующие цитокины, противоопухолевые вакцины. Кроме того, анти-PD-1 антитела и их антигенсвязывающие фрагменты можно использовать в комбинированной терапии, включающей стандартную химиотерапию и лучевую терапию, направленную терапию низкомолекулярными соединениями, развивающиеся другие виды терапии модуляторами иммунных контрольных точек. В конкретных вариантах осуществления антитела и их антигенсвязывающие фрагменты можно использовать в качестве основы для конъюгатов антитело-лекарственное средство, биспецифических или мультивалентных антител.
Анти-PD-1 антитела, или их антигенсвязывающие фрагменты, предложенные в настоящем документе, могут представлять собой моноклональное антитело, поликлональное антитело, полностью человеческое антитело, гуманизированное антитело, химерное антитело, рекомбинантное антитело, биспецифическое антитело, меченое антитело, двухвалентное антитело или антиидиотипическое антитело. Рекомбинантное антитело представляет собой антитело, полученное in vitro с использованием рекомбинантных методов, а не от животных. Биспецифическое или двухвалентное антитело представляет собой искусственное антитело, имеющее фрагменты двух разных моноклональных антител, и может связывать два разных антигена. Антитело, или его антигенсвязывающий фрагмент, которое является «двухвалентным», содержит два антигенсвязывающих сайта. Два антигенсвязывающих сайта могут связывать один и тот же антиген, или они могут связывать разные антигены, в этом случае антитело, или антигенсвязывающий фрагмент, называют «биспецифическим».
В некоторых вариантах осуществления анти-PD-1 антитела, или их антигенсвязывающие фрагменты, предложенные в настоящем документе, представляют собой полностью человеческие антитела. В конкретных вариантах осуществления полностью человеческие антитела получают с использованием рекомбинантных методов. Например, можно создавать трансгенное животное, такое как мышь, которое несет трансгены или трансхромосомы генов человеческих иммуноглобулинов и, вследствие этого, способно продуцировать полностью человеческие антитела после иммунизации нужным антигеном, таким как человеческий PD-1. Полностью человеческие антитела можно выделять из организма такого трансгенного животного или, альтернативно, можно получать с помощью технологии гибридом путем слияния клеток селезенки трансгенного животного с иммортализованными клетками для создания клеток гибридом, секретирующих полностью человеческие антитела. Иллюстративные трансгенные животные включают, но не ограничиваются ими, крыс OmniRat, у которых эндогенная экспрессия крысиных генов иммуноглобулинов инактивирована и которые в то же время генетически модифицированы для содержания локусов функциональных рекомбинантных человеческих иммуноглобулинов; мышей OmniMouse, у которых эндогенная экспрессия мышиных генов иммуноглобулинов инактивирована и которые в то же время генетически модифицированы для содержания локусов рекомбинантных человеческих иммуноглобулинов с делецией J-локуса и мутацией C-каппа; крыс OmniFlic, которые представляют собой трансгенных крыс, у которых эндогенная экспрессия крысиных генов иммуноглобулинов инактивирована и которые в то же время генетически модифицированы для содержания локусов рекомбинантных человеческих иммуноглобулинов, имеющих одну общую, перегруппированную легкую цепь VkJk и функциональную тяжелую цепь. Подробную информацию также можно получить в: Osborn M. et al., Journal of Immunology, 2013, 190: 1481-90; Ma B. et al., Journal of Immunological Methods 400-401 (2013) 78-86; Geurts A. et al., Science, 2009, 325: 433; патенте США 8907157; EP патенте 2152880B1; EP патенте 2336329B1, содержание всех из которых включено в настоящий документ посредством ссылки в полном объеме. Также можно использовать других подходящих трансгенных животных, например, мышь HuMab (для получение подробной информации смотри Lonberg, N. et al. Nature 368(6474): 856-859 (1994)), Xeno-Mouse (Mendez et al. Nat Genet., 1997, 15: 146-156), TransChromo Mouse (Ishida et al. Cloning Stem Cells, 2002, 4: 91-102) и VelocImmune Mouse (Murphy et al. Proc Natl Acad Sci USA, 2014, 111: 5153-5158), Kymouse (Lee et al. Nat Biotechnol, 2014, 32: 356-363) и трансгенный кролик (Flisikowska et al. PLoS One, 2011, 6: e21045).
В некоторых вариантах осуществления анти-PD-1 антитела и их антигенсвязывающие фрагменты представляют собой однодоменное антитело верблюдовых, диатело, scFv, димер scFv, BsFv, dsFv, (dsFv)2, dsFv-dsFv', Fv-фрагмент, Fab, Fab', F(ab')2, ds-диатело, нанотело, доменное антитело или двухвалентное доменное антитело.
В некоторых вариантах осуществления анти-PD-1 антитела и их антигенсвязывающие фрагменты также содержат константную область иммуноглобулина. В некоторых вариантах осуществления константная область иммуноглобулина содержит константную область тяжелой цепи и/или легкой цепи. Константная область тяжелой цепи содержит области CH1, CH1-CH2 или CH1-CH3. В некоторых вариантах осуществления константная область может дополнительно иметь одну или более модификаций для придания желательных свойств. Например, константная область может быть модифицирована для ослабления, или устранения, одной или более эффекторных функций, для улучшения связывания FcRn-рецептора, либо для введения одного или более остатков цистеина.
В некоторых вариантах осуществления анти-PD-1 антитела и их антигенсвязывающие фрагменты также содержат конъюгат. Предусмотрено, что различные конъюгаты могут быть связаны с антителами, или антигенсвязывающими фрагментами, предложенными в настоящем документе (смотри, например, «Conjugate Vaccines», Contributions to Microbiology and Immunology, J. M. Cruse and R. E. Lewis, Jr. (eds.), Carger Press, New York, (1989)). Эти конъюгаты могут быть связаны с антителами, или антигенсвязывающими фрагментами, за счет ковалентного связывания, аффинного связывания, интеркаляции, координационного связывания, образования комплекса, ассоциации, смешивания или присоединения, в числе прочих способов. В конкретных вариантах осуществления антитела и антигенсвязывающие фрагменты, раскрытые в настоящем документе, могут быть сконструированы для содержания определенных сайтов за пределами эпитоп-связывающей части, которые могут быть использованы для связывания одного или более конъюгатов. Например, такой сайт может содержать один или более реакционноспособных аминокислотных остатков, таких как, например, остатки цистеина или гистидина, для облегчения ковалентного связывания с конъюгатом. В конкретных вариантах осуществления антитела могут быть связаны с конъюгатом опосредованно, или через другой конъюгат. Например, антитело, или антигенсвязывающие фрагменты, может быть конъюгировано с биотином, затем опосредованно конъюгировано с другим конъюгатом, связанным с авидином. Конъюгат может представлять собой детектируемую метку, модифицирующий фармакокинетику фрагмент, облегчающий очистку фрагмент или цитотоксический фрагмент. Примеры детектируемой метки могут включать флуоресцентные метки (например, флуоресцеин, родамин, дансил, фикоэритрин или техасский красный), фермент-субстратные метки (например, пероксидазу хрена, щелочную фосфатазу, люциферазы, глюкоамилазу, лизоцим, сахарид оксидазы или β-D-галактозидазу), радиоактивные изотопы (например, 123I, 124I, 125I, 131I, 35S, 3H, 111In, 112In, 14C, 64Cu, 67Cu, 86Y, 88Y, 90Y, 177Lu, 211At, 186Re, 188Re, 153Sm, 212Bi и 32P, другие лантаниды, люминесцентные метки), хромофорный фрагмент, дигоксигенин, биотин/авидин, молекулу ДНК или золото для детектирования. В конкретных вариантах осуществления конъюгат может представлять собой модифицирующий фармакокинетику фрагмент, такой как ПЭГ, который помогает увеличивать период полувыведения антитела. Другие подходящие полимеры включают, например, карбоксиметилцеллюлозу, декстран, поливиниловый спирт, поливинилпирролидон, сополимеры этиленгликоля/пропиленгликоля и тому подобное. В конкретных вариантах осуществления конъюгат может представлять собой облегчающий очистку фрагмент, такой как магнитная гранула. «Цитотоксический фрагмент» может представлять собой любое средство, которое наносит вред клеткам, либо которое может повреждать или убивать клетки. Примеры цитотоксических фрагментов включают, без ограничения, таксол, цитохалазин B, грамицидин D, бромид этидия, эметин, митомицин, этопозид, тенопозид, винкристин, винбластин, колхицин, доксорубицин, даунорубицин, дигидроксиантрацин дион, митоксантрон, митрамицин, актиномицин D, 1-дегидротестостерон, глюкокортикоиды, прокаин, тетракаин, лидокаин, пропранолол, пуромицин и их аналоги, антиметаболиты (например, метотрексат, 6-меркаптопурин, 6-тиогуанин, цитарабин, 5-фторурацил декарбазин), алкилирующие средства (например, мехлорэтамин, тиотепу, хлорамбуцил, мелфалан, кармустин (BSNU) и ломустин (CCNU), циклофосфамид, бусульфан, дибромманнит, стрептозотоцин, митомицин C и цис-дихлордиамин платину (II) (DDP) цисплатин), антрациклины (например, даунорубицин (бывший дауномицин) и доксорубицин), антибиотики (например, дактиномицин (бывший актиномицин), блеомицин, митрамицин и антрамицин (AMC)), а также антимитотические средства (например, винкристин и винбластин).
Полинуклеотиды и рекомбинантные методы
Настоящее изобретение относится к выделенным полинуклеотидам, кодирующим анти-PD-1 антитела и их антигенсвязывающие фрагменты. В конкретных вариантах осуществления выделенные полинуклеотиды содержат одну или более из нуклеотидных последовательностей, приведенных в таблице 1, которые кодируют последовательности CDR, приведенные в таблице 1.
В некоторых вариантах осуществления выделенный полинуклеотид кодирует вариабельную область тяжелой цепи и содержит последовательность, выбранную из группы, состоящей из SEQ ID NO: 46, SEQ ID NO: 50, SEQ ID NO: 54, SEQ ID NO: 58, SEQ ID NO: 62, а также их гомологичных последовательностей, имеющих по меньшей мере 80% (например, по меньшей мере 85%, 88%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%) идентичности последовательности. В некоторых вариантах осуществления выделенный полинуклеотид кодирует вариабельную область легкой цепи и содержит последовательность, выбранную из группы, состоящей из: SEQ ID NO: 48, SEQ ID NO: 52, SEQ ID NO: 56, SEQ ID NO: 60, SEQ ID NO: 64, SEQ ID NO: 68, а также их гомологичных последовательностей, имеющих по меньшей мере 80% (например, по меньшей мере 85%, 88%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%) идентичности последовательности. В конкретных вариантах осуществления процент идентичности обусловлен вырожденностью генетического кода, при этом закодированная белковая последовательность остается неизменной.
Выделенные полинуклеотиды, кодирующие анти-PD-1 антитела и их антигенсвязывающие фрагменты (например, включая последовательности в таблице 1), можно включать в вектор для дальнейшего клонирования (амплификации ДНК) или для экспрессии с использованием рекомбинантных методов, известных в данной области. В другом варианте осуществления антитело можно получать путем гомологичной рекомбинации, известной в данной области. ДНК, кодирующую моноклональное антитело, можно с легкостью выделять и секвенировать с использованием общепринятых методов (например, с использованием олигонуклеотидных зондов, которые способны специфически связываться с генами, кодирующими тяжелую и легкую цепи антитела). Доступно множество векторов. Компоненты вектора, как правило, включают, но не ограничиваются ими, одно или более из следующего: сигнальную последовательность, точку начала репликации, один или более генов маркеров, элемент энхансера, промотор (например, SV40, CMV, EF-1α) и последовательность терминации транскрипции.
В некоторых вариантах осуществления векторная система включает систему для клеток млекопитающих, бактерий, дрожжей, и так далее, и включает плазмиды, такие как, но без ограничения, pALTER, pBAD, pcDNA, pCal, pL, pET, pGEMEX, pGEX, pCI, pCMV, pEGFP, pEGFT, pSV2, pFUSE, pVITRO, pVIVO, pMAL, pMONO, pSELECT, pUNO, pDUO, Psg5L, pBABE, pWPXL, pBI, p15TV-L, pPro18, pTD, pRS420, pLexA, pACT2.2 и так далее, а также другие лабораторные и коммерчески доступные векторы. Соответствующие векторы могут включать плазмидные или вирусные векторы (например, дефектные по репликации ретровирусы, аденовирусы и аденоассоциированные вирусы).
Векторы, содержащие полинуклеотидную последовательность, кодирующую антитело, или антигенсвязывающий фрагмент, можно вводить в клетку-хозяина для клонирования или экспрессии гена. Подходящими клетками-хозяевами для клонирования или экспрессии ДНК в векторах по настоящему изобретению являются клетки прокариот, дрожжей или высших эукариот, описанные выше. Соответствующие прокариоты для этой цели включают эубактерии, такие как грамотрицательные или грамположительные микроорганизмы, например, Enterobacteriaceae, такие как Escherichia, например, E. coli, Enterobacter, Erwinia, Klebsiella, Proteus, Salmonella, например, Salmonella typhimurium, Serratia, например, Serratia marcescans, и Shigella, а также Bacilli, такие как B. subtilis и B. licheniformis, Pseudomonas, такие как P. aeruginosa, и Streptomyces.
Помимо прокариот подходящими хозяевами для клонирования или экспрессии векторов, кодирующих анти-PD-1 антитела, являются эукариотические микроорганизмы, такие как нитчатые грибы или дрожжи. Среди низших эукариотических микроорганизмов-хозяев наиболее часто используемыми являются Saccharomyces cerevisiae, или пекарские дрожжи. Однако представители и других родов, видов и штаммов являются общедоступными и применимыми для настоящего изобретения, например, хозяева Schizosaccharomyces pombe; Kluyveromyces, такие как, например, K. lactis, K. fragilis (ATCC 12424), K. bulgaricus (ATCC 16045), K. wickeramii (ATCC 24178), K. waltii (ATCC 56500), K. drosophilarum (ATCC 36906), K. thermotolerans и K. marxianus; Yarrowia (EP 402226); Pichia pastoris (EP 183070); Candida; Trichoderma reesia (EP 244234); Neurospora crassa; Schwanniomyces, например, Schwanniomyces occidentalis; а также хозяева-нитчатые грибы, такие как, например, Neurospora, Penicillium, Tolypocladium и Aspergillus, например, A. nidulans и A. niger.
Подходящие клетки-хозяева для экспрессии гликозилированных антител, или антигенсвязывающих фрагментов, предложенных по настоящему изобретению, происходят из многоклеточных организмов. Примеры клеток беспозвоночных включают клетки растений и клетки насекомых. Известно множество бакуловирусных штаммов и вариантов, а также соответствующих пермиссивных клеток-хозяев насекомых из таких хозяев, как Spodoptera frugiperda (гусеница), Aedes aegypti (комар), Aedes albopictus (комар), Drosophila melanogaster (плодовая муха) и Bombyx mori. Различные вирусные штаммы для трансфекции являются общедоступными, например, вариант L-1 Autographa californica NPV и штамм Bm-5 Bombyx mori NPV, и такие вирусы могут быть использованы в качестве вирусов по настоящему изобретению, в частности, для трансфекции клеток Spodoptera frugiperda. В качестве хозяев также можно использовать культуры клеток растений, например, хлопчатника, кукурузы, картофеля, сои, петунии, томатов и табака.
Однако наибольший интерес представляют клетки позвоночных, и размножение клеток позвоночных в культуре (тканевой культуре) стало рутинной процедурой. Примерами полезных линий клеток-хозяев млекопитающих являются: линия клеток CV1 почки обезьяны, трансформированных SV40 (COS-7, ATCC CRL 1651); линия клеток эмбриональной почки человека (клетки 293 или 293, субклонированные для роста в суспензионной культуре, Graham et al., J. Gen Virol. 36: 59 (1977)); клетки почки новорожденного хомяка (BHK, ATCC CCL 10); клетки яичника китайского хомяка/-DHFR (CHO, Urlaub et al., Proc. Natl. Acad. Sci. USA 77: 4216 (1980)); мышиные клетки Сертоли (TM4, Mather, Biol. Reprod. 23: 243-251 (1980)); клетки почки обезьяны (CV1 ATCC CCL 70); клетки почки африканской зеленой мартышки (VERO-76, ATCC CRL-1587); клетки цервикальной карциномы человека (HELA, ATCC CCL 2); клетки почки собаки (MDCK, ATCC CCL 34); клетки печени крысы Buffalo (BRL 3A, ATCC CRL 1442); человеческие клетки легкого (W138, ATCC CCL 75); человеческие клетки печени (Hep G2, HB 8065); мышиные клетки опухоли молочной железы (MMT 060562, ATCC CCL51); клетки TRI (Mather et al., Annals N.Y. Acad. Sci. 383: 44-68 (1982)); клетки MRC 5; клетки FS4 и линия клеток гепатомы человека (Hep G2). В некоторых предпочтительных вариантах осуществления клетка-хозяин представляет собой клетку 293F.
Клетки-хозяева трансформируют вышеописанными экспрессионными или клонирующими векторами для продуцирования анти-PD-1 антитела и культивируют в общепринятых питательных средах, модифицированных соответствующим образом для индукции промоторов, отбора трансформантов или амплификации генов, кодирующих нужные последовательности.
Клетки-хозяева, используемые для продуцирования антител, или антигенсвязывающих фрагментов, предложенных в настоящем документе, можно культивировать в разных средах. Для культивирования клеток-хозяев подходят коммерчески доступные среды, такие как среда Хэма F10 (Sigma), минимальная поддерживающая среда (MEM) (Sigma), RPMI-1640 (Sigma) и среда Игла в модификации Дульбекко (DMEM) Sigma). Кроме того, в качестве сред для культивирования клеток-хозяев можно использовать любую из сред, описанных в Ham et al., Meth. Enz. 58: 44 (1979), Barnes et al., Anal. Biochem. 102: 255 (1980), патентах США №№ 4767704, 4657866, 4927762, 4560655 или 5122469, WO 90/03430, WO 87/00195 или патенте США Re. 30985. Любую из этих сред можно дополнять по мере необходимости гормонами и/или другими факторами роста (такими как инсулин, трансферрин или эпидермальный фактор роста), солями (такими как хлорид и фосфат натрия, кальция, магния), буферами (такими как HEPES), нуклеотидами (такими как аденозин и тимидин), антибиотиками (такими как лекарственное средство гентамицин™), микроэлементами (определяемыми, как неорганические соединения, обычно присутствующие в конечных концентрациях микромолярного диапазона), а также глюкозой или эквивалентным источником энергии. Также можно включать в соответствующих концентрациях любые другие необходимые добавки, известные специалистам в данной области. Условия культивирования, такие как температура, pH и тому подобное, являются такими, которые были использованы ранее в случае клеток-хозяев, выбранных для экспрессии, и которые известны специалистам в данной области.
При использовании рекомбинантных методов антитело может продуцироваться внутри клетки, в периплазматическом пространстве, или непосредственно секретироваться в среду. Если антитело продуцируется внутри клетки, в качестве первого этапа удаляют твердый детрит, либо клетки-хозяева, либо лизированные фрагменты, например, центрифугированием или ультрафильтрацией. В публикации Carter et al., Bio/Technology 10: 163-167 (1992) описан метод выделения антител, которые секретируются в периплазматическое пространство E. coli. Вкратце, пасту из клеток размораживают в присутствии ацетата натрия (pH 3,5), ЭДТА и фенилметилсульфонилфторида (PMSF) в течение примерно 30 мин. Клеточный детрит можно удалять центрифугированием. Если антитело секретируется в среду, супернатанты от таких систем экспрессии, как правило, сначала концентрируют с использованием коммерчески доступного фильтра для концентрирования белков, например, в ячейке для ультрафильтрации Amicon или Millipore Pellicon. На любом из вышеперечисленных этапов можно включать ингибитор протеаз, такой как PMSF, для ингибирования протеолиза и антибиотики для предотвращения роста случайных загрязняющих микроорганизмов.
Антитело, полученное из клеток, можно очищать с использованием, например, хроматографии на гидроксилапатите, электрофореза в геле, диализа, ионообменной хроматографии на DEAE-целлюлозе, осаждения сульфатом аммония, высаливания и аффинной хроматографии, при этом аффинная хроматография является предпочтительным методом очистки. Пригодность белка A в качестве аффинного лиганда зависит от вида и изотипа какого-либо Fc-домена иммуноглобулина, который имеется у антитела. Белок A можно использовать для очистки антител на основе тяжелых цепей.гамма.1,.гамма.2 или.гамма.4 человека (Lindmark et al., J. Immunol. Meth. 62: 1-13 (1983)). Белок G рекомендован для очистки всех изотипов мышиных антител и антител человека с тяжелой цепью.гамма.3 (Guss et al., EMBO J. 5: 1567-1575 (1986)). Матрицей, к которой присоединен аффинный лиганд, чаще всего является агароза, однако доступны и другие матрицы. Механически стабильные матрицы, такие как стекло с контролируемым размером пор или поли(стиролдивинил)бензол, допускают более высокие скорости потока и более короткое время процесса по сравнению с агарозными матрицами. Если антитело содержит домен CH3, для очистки полезно использовать смолу Bakerbond ABX.TM. (J. T. Baker, Phillipsburg, N.J.). Другие методы очистки белков, такие как фракционирование на ионообменной колонке, осаждение этанолом, ВЭЖХ с обращенной фазой, хроматография на силикагеле, хроматография на гепарин-сефарозе™, хроматография на анионо- или катионообменной смоле (например, на колонке с полиаспарагиновой кислотой), хроматофокусирование, SDS-ПААГ и осаждение сульфатом аммония, также можно использовать в зависимости от антитела, которое предстоит очищать.
После любого предварительного этапа(ов) очистки смесь, содержащую интересующее антитело и примеси, можно подвергать хроматографии гидрофобного взаимодействия при низких значениях pH с использованием буфера элюции при значениях pH примерно 2,5-4,5, предпочтительно выполняемой при низкой концентрации соли (например, примерно 0-0,25 M соли).
Наборы
Настоящее изобретение относится к наборам, включающим анти-PD-1 антитела, или их антигенсвязывающие фрагменты. В некоторых вариантах осуществления наборы полезны для определения наличия или уровня PD-1 в биологическом образце. Биологический образец может содержать клетку или ткань.
В некоторых вариантах осуществления набор включает анти-PD-1 антитело, или его антигенсвязывающий фрагмент, конъюгированное с детектируемой меткой. В других конкретных вариантах осуществления набор включает немеченое анти-PD-1 антитело, или антигенсвязывающий фрагмент, и дополнительно включает вторичное меченое антитело, которое способно связываться с немеченым анти-PD-1 антителом. Набор может дополнительно включать инструкции по применению и упаковку, которая разделяет каждый из компонентов в наборе.
В конкретных вариантах осуществления анти-PD-1 антитело, или его антигенсвязывающий фрагмент, связано с субстратом или устройством, полезным при выполнении сэндвич-анализа, такого как ELISA, или иммунографического анализа. Полезным субстратом или устройством может быть, например, микропланшет для титрования и тест-полоска.
Фармацевтическая композиция и способ лечения
Настоящее изобретение также относится к фармацевтическим композициям, содержащим анти-PD-1 антитела, или их антигенсвязывающие фрагменты, и один или более фармацевтически приемлемых носителей.
Фармацевтически приемлемые носители для использования в фармацевтических композициях, раскрытых в настоящем документе, могут включать, например, фармацевтически приемлемую жидкость, гель, или твердые носители, водные среды, неводные среды, противомикробные средства, изотонические средства, буферы, антиоксиданты, анестетики, суспендирующие/диспергирующие средства, секвестрирующие или хелатирующие средства, разбавители, адъюванты, эксципиенты или нетоксичные вспомогательные вещества, другие компоненты, известные в данной области, или различные их сочетания.
Подходящие компоненты могут включать, например, антиоксиданты, наполнители, связывающие вещества, дезинтегрирующие средства, буферы, консерванты, смазывающие средства, ароматизаторы, загустители, красители, эмульгаторы или стабилизаторы, такие как сахара и циклодекстрины. Подходящие антиоксиданты могут включать, например, метионин, аскорбиновую кислоту, ЭДТА, тиосульфат натрия, платину, каталазу, лимонную кислоту, цистеин, тиоглицерин, тиогликолевую кислоту, тиосорбит, бутилированный гидроксанизол, бутилированный гидрокситолуол, и/или пропилгаллат. Как описано в настоящем документе, включение одного или более антиоксидантов, таких как метионин, в композицию, содержащую антитело, или антигенсвязывающий фрагмент, и конъюгаты, предложенные в настоящем документе, позволяет уменьшить окисление антитела, или антигенсвязывающего фрагмента. Это уменьшение окисления позволяет предотвращать или уменьшать снижение аффинности связывания, тем самым способствуя повышению стабильности антитела и максимальному увеличению срока хранения. Вследствие этого, в конкретных вариантах осуществления предложены композиции, которые содержат одно или более антител, или антигенсвязывающих фрагментов, описанных в настоящем документе, и один или более антиоксидантов, таких как метионин. Также предложены способы предотвращения окисления, продления срока хранения и/или повышения эффективности антитела, или антигенсвязывающего фрагмента, предложенного в настоящем документе, путем смешивания антитела, или антигенсвязывающего фрагмента, с одним или более антиоксидантами, такими как метионин.
В качестве дополнительной иллюстрации, фармацевтически приемлемые носители могут включать, например, водные среды, такие как хлорид натрия для инъекций, раствор Рингера для инъекций, изотонический раствор декстрозы для инъекций, стерильная вода для инъекций или раствор декстрозы и раствор Рингера лактат для инъекций, неводные среды, такие как нелетучие масла растительного происхождения, хлопковое масло, кукурузное масло, кунжутное масло или арахисовое масло, противомикробные средства в бактериостатических или фунгистатических концентрациях, изотонические средства, такие как хлорид натрия или декстроза, буферы, такие как фосфатные или цитратные буферы, антиоксиданты, такие как бисульфат натрия, местные анестетики, такие как прокаина гидрохлорид, суспендирующие и диспергирующие средства, такие как натрий-карбоксиметилцеллюлоза, гидроксипропилметилцеллюлоза или поливинилпирролидон, эмульгаторы, такие как полисорбат 80 (TWEEN-80), секвестрирующие или хелатирующие средства, такие как ЭДТА (этилендиаминтетрауксусная кислота) или ЭГТА (этиленгликольтетрауксусная кислота), этиловый спирт, полиэтиленгликоль, пропиленгликоль, гидроксид натрия, соляную кислоту, лимонную кислоту или молочную кислоту. В фармацевтические композиции в многодозовых контейнерах можно добавлять противомикробные средства, используемые в качестве носителей, включая фенолы или крезолы, соединения ртути, бензиловый спирт, хлорбутанол, метиловые и пропиловые эфиры п-гидроксибензойной кислоты, тимеросал, бензалкония хлорид и бензэтония хлорид. Подходящие эксципиенты могут включать, например, воду, солевой раствор, декстрозу, глицерин или этанол. Подходящие нетоксичные вспомогательные вещества могут включать, например, увлажняющие вещества или эмульгаторы, поддерживающие pH буферные средства, стабилизаторы, усилители растворения или такие средства, как ацетат натрия, сорбитан монолаурат, триэтаноламин олеат или циклодекстрин.
Фармацевтические композиции могут представлять собой жидкий раствор, суспензию, эмульсию, пилюлю, капсулу, таблетку, препарат с замедленным высвобождением или порошок. Пероральные препараты могут содержать стандартные носители, такие как маннит, лактоза, крахмал, стеарат магния, поливинилпирролидон, сахарин натрия, целлюлоза, карбонат магния, и так далее, фармацевтической степени чистоты.
В вариантах осуществления фармацевтические композиции сформулированы в виде инъекционных композиций. Инъекционные фармацевтические композиции могут быть получены в любой общепринятой форме, такой как, например, жидкий раствор, суспензия, эмульсия, или твердых формах, подходящих для получения жидкого раствора, суспензии или эмульсии. Препараты для инъекций могут включать стерильные и/или непирогенные растворы, готовые для инъекций, стерильные сухие растворимые препараты, такие как лиофилизированные порошки, готовые для объединения с растворителем непосредственно перед использованием, включая таблетки для приготовления растворов для гиподермальных инъекций, стерильные суспензии, готовые для инъекций, стерильные сухие нерастворимые препараты, готовые для объединения со средой непосредственно перед использованием, а также стерильные и/или непирогенные эмульсии. Растворы могут быть либо водными, либо неводными.
В конкретных вариантах осуществления однократные дозы парентеральных препаратов упакованы в ампулы, флаконы или шприцы с иглами. Все препараты для парентерального введения должны быть стерильными и непирогенными, как известно и как осуществляется на практике в данной области.
В конкретных вариантах осуществления стерильный лиофилизированный порошок готовят путем растворения антитела, или антигенсвязывающего фрагмента, раскрытого в настоящем документе, в соответствующем растворителе. Растворитель может содержать эксципиент, который способствует улучшению стабильности или других фармакологических свойств порошка или восстановленного раствора, приготовленного из порошка. Эксципиенты, которые можно использовать, включают, но не ограничиваются ими, воду, декстрозу, сорбит, фруктозу, кукурузный сироп, ксилит, глицерин, глюкозу, сахарозу или другое подходящее средство. Растворитель может содержать буфер, такой как цитрат или фосфат натрия, или калия, или другой такой буфер, известный специалистам в данной области, в одном варианте осуществления имеющий примерно нейтральное значение pH. Последующая стерилизация раствора фильтрованием, с последующей лиофилизацией в стандартных условиях, известных специалистам в данной области, позволяет получать нужный препарат. В одном варианте осуществления полученный раствор будет разделен на порции во флаконы для лиофилизации. Каждый флакон может содержать одну дозу или несколько доз анти-PD-1 антитела, или его антигенсвязывающего фрагмента, или его композиции. Во флаконы можно вносить количество, немного превышающее необходимое количество для дозы или набора доз (например, примерно на 10%), для облегчения точного отбора образца и точного дозирования. Лиофилизированный порошок можно хранить в соответствующих условиях, например, при температуре от 4°C до комнатной температуры.
Восстановление лиофилизированного порошка в воде для инъекций позволяет получать препарат, используемый для парентерального введения. В одном варианте осуществления для восстановления к лиофилизированному порошку добавляют стерильную и/или непирогенную воду или другой подходящий жидкий носитель. Точное количество зависит от выбранного метода лечения и может быть определено эмпирически.
Также предложены терапевтические способы, включающие введение терапевтически эффективного количества антитела, или антигенсвязывающего фрагмента, предложенного в настоящем документе, субъекту, который нуждается в этом, тем самым осуществляя лечение, или предотвращение, состояния или заболевания, ассоциированного или связанного с PD-1. В другом аспекте предложены способы лечения состояния у субъекта, которому принесет пользу усиление иммунного ответа, включающие введение терапевтически эффективного количества антитела, или антигенсвязывающего фрагмента, предложенного в настоящем документе, субъекту, который нуждается в этом.
Терапевтически эффективное количество антитела, или антигенсвязывающего фрагмента, предложенного в настоящем документе, будет зависеть от разных факторов, известных в данной области, таких как, например, масса тела, возраст, прошлая медицинская история, принимаемые препараты, состояние здоровья субъекта и возможные перекрестные реакции, аллергия, чувствительность и неблагоприятные побочные эффекты, а также путь введения и степень развития опухоли. Специалист в данной области (например, врач или ветеринар) может пропорционально уменьшать или увеличивать дозы в зависимости от этих и других обстоятельств или потребностей.
В конкретных вариантах осуществления антитело, или антигенсвязывающий фрагмент, предложенное в настоящем документе, можно вводить в терапевтически эффективной дозе от примерно 0,01 мг/кг до примерно 100 мг/кг (например, примерно 0,01 мг/кг, примерно 0,5 мг/кг, примерно 1 мг/кг, примерно 2 мг/кг, примерно 5 мг/кг, примерно 10 мг/кг, примерно 15 мг/кг, примерно 20 мг/кг, примерно 25 мг/кг, примерно 30 мг/кг, примерно 35 мг/кг, примерно 40 мг/кг, примерно 45 мг/кг, примерно 50 мг/кг, примерно 55 мг/кг, примерно 60 мг/кг, примерно 65 мг/кг, примерно 70 мг/кг, примерно 75 мг/кг, примерно 80 мг/кг, примерно 85 мг/кг, примерно 90 мг/кг, примерно 95 мг/кг или примерно 100 мг/кг). В некоторых из этих вариантов осуществления антитело, или антигенсвязывающий фрагмент, вводят в дозе примерно 50 мг/кг или менее, и в некоторых из этих вариантов осуществления доза составляет 10 мг/кг или менее, 5 мг/кг или менее, 1 мг/кг или менее, 0,5 мг/кг или менее, или 0,1 мг/кг или менее. В конкретных вариантах осуществления вводимую дозу можно изменять в зависимости от курса лечения. Например, в конкретных вариантах осуществления начальная вводимая доза может быть выше, чем вводимые впоследствии дозы. В конкретных вариантах осуществления вводимая доза может варьироваться на протяжении курса лечения в зависимости от реакции субъекта.
Режим введения доз можно корректировать для обеспечения оптимального желательного ответа (например, терапевтического ответа). Например, можно вводить однократную дозу или несколько разделенных доз на протяжении некоторого времени.
Антитела и антигенсвязывающие фрагменты, раскрытые в настоящем документе, можно вводить любым путем введения, известным в данной области, таким как, например, парентеральный (например, подкожный, внутрибрюшинный, внутривенный, включая внутривенную инфузию, внутримышечную или внутрикожную инъекцию) или не парентеральный (например, пероральный, интраназальный, внутриглазной, подъязычный, ректальный или топический) пути введения.
Состояния и заболевания, ассоциированные с PD-1, могут представлять собой связанные с иммунитетом заболевания или нарушения. В конкретных вариантах осуществления ассоциированные с PD-1 состояния и заболевания включают опухоли и рак, например, немелкоклеточный рак легкого, мелкоклеточный рак легкого, почечно-клеточный рак, колоректальный рак, рак яичника, рак молочной железы, рак поджелудочной железы, карциному желудка, рак мочевого пузыря, рак пищевода, мезотелиому, меланому, рак головы и шеи, рак щитовидной железы, саркому, рак предстательной железы, глиобластому, рак шейки матки, карциному тимуса, лейкоз, лимфомы, миеломы, фунгоидную гранулему, рак из клеток Меркеля и другие гематологические злокачественные новообразования, такие как классическая лимфома Ходжкина (CHL), первичная медиастинальная B-крупноклеточная лимфома, богатая T-клетками/гистиоцитами B-клеточная лимфома, EBV-положительное и -отрицательное PTLD, а также EBV-ассоциированная диффузная B-крупноклеточная лимфома (DLBCL), плазмобластная лимфома, экстранодальная NK/T-клеточная лимфома, назофарингеальную карциному и HHV8-ассоциированную первичную эффузионную лимфому, лимфому Ходжкина, новообразование в центральной нервной системе (ЦНС), такое как первичная лимфома ЦНС, опухоль спинного мозга, глиома ствола головного мозга. В конкретных вариантах осуществления опухоли и рак являются метастатическими, в частности, метастатическими опухолями, экспрессирующими PD-L1. В конкретных вариантах осуществления ассоциированные с PD-1 состояния и заболевания включают аутоиммунные заболевания, такие как системная красная волчанка (СКВ), псориаз, системная склеродермия, аутоиммунный диабет и тому подобное. В конкретных вариантах осуществления ассоциированные с PD-1 состояния и заболевания включают инфекционные заболевания, такие как хроническая вирусная инфекция, например, инфекция вируса гепатита B, гепатита C, вируса герпеса, вируса Эпштейна-Барр, ВИЧ, цитомегаловируса, вируса простого герпеса I типа, вируса простого герпеса 2 типа, вируса папилломы человека, аденовируса, эпидемическое заболевание, вызванное вирусом герпеса саркомы Капоши, инфекция, вызванная тонким кольцевым вирусом (вирусом гепатита TTV), вирусом JC или вирусом BK.
Способы применения
Настоящее изобретение также относится к способам применения анти-PD-1 антител или их антигенсвязывающих фрагментов.
В некоторых вариантах осуществления настоящее изобретение относится к способам лечения состояния или заболевания, ассоциированного или связанного с PD-1, у индивидуума, включающим введение терапевтически эффективного количества анти-PD-1 антитела или его антигенсвязывающего фрагмента. В конкретных вариантах осуществления индивидуум был идентифицирован, как имеющий заболевание или состояние, на которое может оказывать влияние антагонист PD-1.
Наличие или уровень PD-L1 в интересующем биологическом образце может указывать на то, может ли индивидуум, от которого был получен биологический образец, реагировать на введение антагониста PD-1. Различные методы можно использовать для определения наличия или уровня PD-L1 в тестируемом биологическом образце от индивидуума. Например, тестируемый биологический образец можно подвергать воздействию анти-PD-L1 антитела, или его антигенсвязывающего фрагмента, которое связывает и позволяет обнаруживать экспрессированный белок PD-L1. Альтернативно, экспрессию PD-L1 также можно обнаруживать на уровне нуклеиновой кислоты с использованием таких методов, как кПЦР, ПЦР с обратной транскриптазой, микрочипы, SAGE, FISH и тому подобное. В некоторых вариантах осуществления тестируемый образец получают из раковой клетки или ткани, или инфильтрирующих опухоль иммунных клеток. В конкретных вариантах осуществления наличие или повышенный уровень PD-L1 в тестируемом биологическом образце указывает на вероятность ответной реакции. Используемый в настоящем документе термин «повышенный» означает, в целом, увеличенный на не менее чем 10%, 15%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80% или более, уровень белка PD-L1 в тестируемом образце, определенный с использованием антител, или антигенсвязывающих фрагментов, предложенных в настоящем документе, по сравнению с уровнем белка PD-L1 в эталонном образце, определенным с использованием того же антитела. Эталонным образцом может быть контрольный образец, полученный от здорового или не страдающего заболеванием индивидуума, либо здоровый или не затронутый заболеванием образец, полученный от того же индивидуума, от которого получен тестируемый образец. Например, эталонный образец может представлять собой не затронутый заболеванием образец, расположенный вблизи или рядом с тестируемым образцом (например, из опухоли).
Антитела, или антигенсвязывающие фрагменты, раскрытые в настоящем документе, можно вводить отдельно или в сочетании с одним или более дополнительными терапевтическими средствами или агентами. Например, антитела, или антигенсвязывающие фрагменты, раскрытые в настоящем документе, можно вводить в сочетании с химиотерапией, лучевой терапией, хирургической операцией для лечения рака (например, извлечением опухоли), одним или более средствами против рвоты или другими средствами против осложнений, вызванных применением химиотерапии, либо любым другим лекарственным средством, используемым для лечения рака или любого заболевания, опосредованного PD-1. В некоторых из этих вариантов осуществления антитело, или антигенсвязывающий фрагмент, раскрытое в настоящем документе, которое вводят в сочетании с одним или более дополнительными терапевтическими средствами, можно вводить одновременно с одним или более дополнительными терапевтическими средствами, и в некоторых из этих вариантов осуществления антитело, или антигенсвязывающий фрагмент, и дополнительное терапевтическое средство(а) можно вводить в составе одной и той же фармацевтической композиции. Однако антитело, или антигенсвязывающий фрагмент, которое вводят «в сочетании» с другим терапевтическим средством, не обязательно должно быть введено одновременно или в одной и той же композиции, что и средство. Антитело, или антигенсвязывающий фрагмент, которое вводят до или после введения другого средства, считают вводимым «в сочетании» с данным средством, в соответствии с выражением, используемым в настоящем документе, даже если антитело, или антигенсвязывающий фрагмент, и второе средство вводят разными путями введения. Где это возможно, дополнительные терапевтические средства, вводимые в сочетании с антителами, или антигенсвязывающими фрагментами, раскрытыми в настоящем документе, вводят в соответствии со схемой, изложенной в инструкции по применению дополнительного терапевтического средства, либо в соответствии с Physicians' Desk Reference 2003 (Physicians' Desk Reference, 57-е издание; Medical Economics Company; ISBN: 1563634457; 57-е издание (ноябрь 2002 г.)) или протоколами, хорошо известными в данной области.
В конкретных вариантах осуществления терапевтические средства могут индуцировать или усиливать иммунный ответ против рака. Например, можно использовать противоопухолевую вакцину для индукции иммунного ответа на некоторые виды опухолей или рака. Также можно использовать терапию цитокинами для усиления представления опухолевого антигена клеткам иммунной системы. Примеры цитокинов для терапии включают, без ограничения, интерфероны, такие как интерферон-α, -β и -γ, колониестимулирующие факторы, такие как макрофагальный CSF, гранулоцитарно-макрофагальный CSF и гранулоцитарный CSF, интерлейкины, такие как IL-1, IL-1α, IL-2, IL-3, IL-4, IL-5, IL-6, IL-7, IL-8, IL-9, IL-10, IL-11 и IL-12, факторы некроза опухолей, такие как TNF-α и TNF-β. Также можно использовать средства, инактивирующие иммуносупрессивные мишени, например, ингибиторы TGF-бета, ингибиторы IL-10 и ингибиторы Fas-лиганда. Другая группа средств включает средства, которые активируют иммунную реакцию на опухолевые или раковые клетки, например, те, которые повышают активацию T-клеток (например, агонисты рецепторов T-клеток, костимулирующие молекулы, такие как CTLA-4, ICOS и OX-40), и те, которые усиливают функцию дендритных клеток и представление антигенов.
Следующие далее примеры приведены для лучшей иллюстрации заявленного изобретения и не должны быть истолкованы, как ограничивающие объем изобретения. Все конкретные композиции, материалы и способы, описанные ниже, полностью и частично, входят в объем настоящего изобретения. Эти конкретные композиции, материалы и способы не предназначены для ограничения изобретения, а лишь для иллюстрации конкретных вариантов осуществления, входящих в объем изобретения. Специалист в данной области может создавать эквивалентные композиции, материалы и способы, не обладающие признаками изобретения и без отклонения от объема изобретения. Следует понимать, что можно использовать множество вариаций способов, описанных в настоящем документе, которые остаются в пределах объема настоящего изобретения. Авторы изобретения считают, что такие вариации входят в объем изобретения.
ПРИМЕР 1: Получение антител с помощью гибридом
1.1 Получение иммуногена: молекулы ДНК, кодирующие ECD или полноразмерные PD-1 и PD-L1, были синтезированы и вставлены в экспрессионный вектор pcDNA3.3. ДНК плазмид Maxprep и вставленные последовательности ДНК подтверждали секвенированием. Слитые белки PD-1 ECD и PD-L1 ECD, содержащие различные метки, включая Fc человека, Fc мыши и His-метки, получали путем трансфекции человеческого гена PD-1 ECD в клетки CHO-S или HEK293. Через 5 дней супернатанты, собранные с культур временно трансфицированных клеток, использовали для очистки белков. Слитые белки очищали и оценивали количественно с целью использования для иммунизации и скрининга.
1.2 Создание стабильной линии клеток. С целью создания инструментов для скрининга и валидации антител авторы изобретения создали линии клеток, трансфицированных PD-1 и PD-L1. Вкратце, клетки CHO-K1, 293F или Ba/F3 были трансфицированы экспрессионным вектором pCND3.3, содержащим последовательности для полноразмерного PD-1 или PD-L1, с использованием набора для трансфекции Lipofectamine 2000 Transfection kit в соответствии с протоколом производителя. В течение 48-72 часов после трансфекции трансфицированные клетки культивировали в среде, содержащей бластицидин или G418, для селекции. С течением времени это позволяет отбирать клетки со стабильно встроенными генами PD-1 или PD-L1 в их геномных ДНК. В то же время клетки проверяли на экспрессию интересующих генов PD-1 и PD-L1. После подтверждения экспрессии отдельные представляющие интерес клоны отбирали методом лимитирующего разведения и проводили масштабирование на большие объемы. Полученные моноклональные линии клеток затем поддерживали в среде, содержащей меньшую дозу антибиотиков бластицидина или G418.
1.3 Получение антител при помощи гибридом
1.3.1 Иммунизация и слияние клеток: крыс OMT (полученных от компании Open Monoclonal Technology, Inc., Palo Alto, US) в возрасте 8-10 недель иммунизировали 10 мкг человеческого белка PD-1 ECD в TiterMax в подушечки лап при первой инъекции и повторно иммунизировали каждые 3 дня белком PD-1 ECD с адъювантом на основе алюминия. У крыс отбирали кровь каждые две недели для сбора сыворотки, и титры антител измеряли методом ELISA или FACS-анализа. Когда титр антител становился достаточно высоким, крысам выполняли последнюю бустерную инъекцию без адъюванта (вместо этого добавляя 100 мкл 1X PBS) и проводили слияние клеток следующим образом: B-лимфоциты, выделенные из лимфатических узлов иммунизированных крыс OMT, объединяли с клетками миеломы (в соотношении 1:1). Смесь клеток промывали и суспендировали в 5-10 мл раствора ECF. Добавляли раствор ECF для доведения концентрации до 2×106 клеток/мл. После стимулированного электрическими импульсами слияния клеток суспензию клеток из камеры для слияния немедленно переносили в стерильную пробирку, содержащую больший объем среды. После инкубации в течение более 24 часов при 37°C суспензию клеток смешивали и пипеткой переносили в лунки 96-луночных планшетов (0,5×106 клеток/планшет). Клетки инкубировали в условиях 37°C, 5% CO2. Когда размеры клонов становились достаточно большими, отбирали 100 мкл супернатанта из 96-луночных планшетов для проведения скрининга антител.
1.3.2 Первичный и подтверждающий скрининг супернатантов гибридом: анализ методом ELISA использовали в качестве первого метода скрининга для тестирования связывания супернатантов гибридом с белком PD-1. Вкратце, планшеты (Nunc) покрывали растворимым белком внеклеточного домена человеческого PD-1 в концентрации 1 мкг/мл в течение ночи при 4°C. После блокирования и промывания супернатанты гибридом переносили на покрытые планшеты и инкубировали при комнатной температуре в течение 1 ч. Затем планшеты промывали и инкубировали со вторичным антителом козы против IgG1 крысы, конъюгированным с HRP (Bethyl), и антителом козы против IgG2b крысы, конъюгированным с HRP (Bethyl), в течение 45 мин. После промывания добавляли субстрат TMB и реакцию останавливали 2 M HCl. Поглощение при длине волны 450 нм измеряли на планшетном ридере (Molecular Device). Для подтверждения естественного связывания анти-PD-1 антител с конформационными молекулами PD-1, экспрессированными на клеточной мембране, проводили FACS-анализ на линии трансфицированных PD-1 клеток CHO-S. Клетки CHO-S, экспрессирующие человеческий PD-1, переносили в 96-луночные планшеты с U-образным дном (BD) при плотности 1×106 клеток/мл. Затем супернатанты гибридом переносили в планшеты и инкубировали в течение 1 ч при 4°C. После промывания 1X PBS/1% БСА добавляли вторичное антитело козы против иммуноглобулинов крысы, меченое FITC (Jackson Immunoresearch Lab), и инкубировали с клетками при 4°C в темноте в течение 1 ч. Затем клетки промывали и ресуспендировали в 1X PBS/1% БСА или фиксировали 4% параформальдегидом и анализировали методом проточной цитометрии (BD). Связывание антител клетками исходной линии CHO-S выполняли таким же образом. На фигуре 1 показано связывание антител против человеческого PD-1 с экспрессирующими PD-1 клетками CHO. Клетки CHO, трансфицированные полноразмерным человеческим PD-1, окрашивали антителами против человеческого PD-1 из крысиной гибридомы, а затем окрашивали конъюгированным с FITC антителом козы против IgG Fc крысы в качестве 2-го антитела и анализировали методом FACS. Данные показывают, что антитела специфически связывают PD-1, экспрессированный на клетках CHO.
Для тестирования аффинности связывания антител с нативным PD-1, экспрессированным на человеческих CD4+ T-клетках, человеческие CD4+ T-клетки получали из МКПК, культивировали с IL-2 и OKT3 в течение 3 дней и окрашивали антителами против человеческого PD-1. Связывание антител с PD-1 на T-клетках анализировали методом FACS. Как показано на фигуре 3, FACS-анализ продемонстрировал, что антитела специфически связывают нативный PD-1, экспрессированный на CD4+ T-клетках.
Тестирование блокирующей активности антител использовали в качестве подтверждающего скрининга для отбора потенциальных хитов - удачно выбранных антител. Выбранные антитела тестировали на способность блокировать связывание лиганда PD-L1 с трансфицированными PD-1 клетками CHO-S методом FACS-анализа. Клетки CHO-S, экспрессирующие человеческий PD-1, переносили в 96-луночные планшеты с U-образным дном (BD) при плотности 1×106 клеток/мл. Антитела серийно разводили в промывочном буфере (1X PBS/1% БСА) и инкубировали с клетками при 4°C в течение 1 ч. После промывания добавляли слитый белок человеческий Fc - человеческий PD-L1 и инкубировали при 4°C в течение 1 ч. Антитело козы против IgG Fc человека, меченое FITC, в качестве вторичного антитела (без перекрестной реактивности с IgG Fc крысы, Jackson Immunoresearch Lab) инкубировали с клетками при 4°C в темноте в течение 1 ч. Затем клетки промывали и ресуспендировали в 1X PBS/1% БСА или фиксировали 4% параформальдегидом и анализировали методом проточной цитометрии (BD).
1.3.3 Субклонирование гибридом: после подтверждения специфического связывания и блокирования при первичном и подтверждающем скрининге положительные линии клеток гибридомы можно использовать для субклонирования. Вкратце, для каждой линии клеток гибридомы клетки подсчитывали и разбавляли до плотности 5 клеток/лунку, 1 клетка/лунку и 0,5 клеток/лунку в среде для клонирования. По 200 мкл/лунку 96-луночных планшетов вносили: в один планшет по 5 клеток/лунку, в один планшет по 1 клетке/лунку и в четыре планшета по 0,5 клеток/лунку. Помещали все планшеты в инкубатор с условиями 37°C, 5% CO2. Инкубировали до тех пор, когда все линии клеток стало возможно проверять в анализе ELISA.
ПРИМЕР 2: Последовательности антител из клеток гибридом и характеризация полностью человеческих антител
2.1 Последовательности антител из клеток гибридом: РНК выделяли из моноклональных клеток гибридом с использованием реагента Trizol. Последовательности VH и VL анти-PD-1 антител амплифицировали в соответствии со следующим протоколом: вкратце, сначала РНК обратно транскрибировали в кДНК с использованием обратной транскриптазы, как описано в настоящем документе, реакционная система (20 мкл) содержала:
Условия реакции
Полученную кДНК использовали в качестве матрицы для последующей ПЦР-амплификации с использованием праймеров, специфических для интересующих генов. Реакцию ПЦР проводили следующим образом:
Условия реакции:
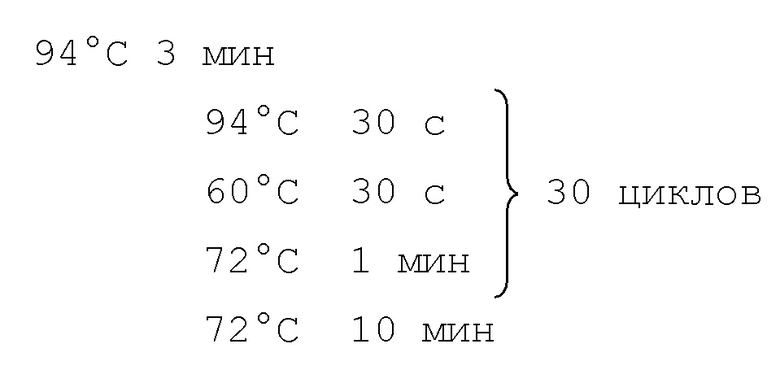
Отбирали 10 мкл реакционной смеси ПЦР для проведения лигирования с вектором pMD18-T. Выполняли трансформацию компетентных клеток Top10, используя 10 мкл продуктов лигирования, и переносили смесь на предварительно нагретые планшеты 2-YT+Cab, следуя стандартному протоколу, инкубировали в течение ночи. Положительные клоны проверяли ПЦР, используя праймеры M13-48 и M13-47, с последующим секвенированием.
2.2 Конструирование молекул полностью человеческих антител: Последовательности VH и VL анти-PD-1 антител амплифицировали, как описано выше. Продукты реакции ПЦР очищали при помощи набора PCR clean-up kit, и VL и вектор pCI расщепляли ферментами рестрикции Pme I и BssH II при 37°C в течение 2 часов. Проводили реакции в 1% агарозе и выполняли экстракцию из геля, используя набор в соответствии с инструкциями производителя. Проводили лигирование расщепленных VL и вектора pCI следующим образом:
Смесь инкубировали при 16°C в течение 30 минут. 10 мкл реакционной смеси использовали для трансформации и выращивания клонов. Подтвержденные клоны использовали для экстрагирования ДНК плазмиды pCI-VL. Затем вектор pCI-VL и фрагмент VH расщепляли Xbal и Sal I, и очищенные расщепленные VH и вектор лигировали T4 ДНК-лигазой в течение 30 минут при 16°C. После того, как последовательности вставленных VL и VH были подтверждены секвенированием, экспрессионный вектор, содержащий целый IgG полностью человеческого анти-PD-1 антитела, использовали для временной трансфекции и получения стабильной линии клеток.
ПРИМЕР 3: Характеризация полностью человеческих антител
3.1 Аффинность связывания анти-PD-1 антител с молекулами PD-1 на клеточной поверхности тестировали методом проточной цитометрии (FACS): Аффинность связывания антител с PD-1 на клеточной поверхности определяли методом FACS. Клетки CHO-S, экспрессирующие человеческий PD-1, переносили в 96-луночный планшет с U-образным дном (BD) при плотности 5×105 клеток/мл. Тестируемые антитела серийно разводили 1:2 в промывочном буфере (1X PBS/1% БСА) и инкубировали с клетками при 4°C в течение 1 ч. В качестве вторичного антитела добавляли антитело козы против IgG Fc человека, меченое FITC (3,0 моля FITC на моль IgG, (Jackson Immunoresearch Lab)), и инкубировали при 4°C в темноте в течение 1 ч. Затем клетки промывали один раз, ресуспендировали в 1X PBS/1% БСА и анализировали методом проточной цитометрии (BD). Показатели интенсивности флуоресценции были преобразованы в показатели количества связанных молекул/клетку на основании результатов, полученных с микрогранулами набора Quantum™ MESF, Bangs Laboratories, Inc.). Величину KD рассчитывали с использованием программы Graphpad Prism 5. На фигуре 2 показано связывание полностью человеческих анти-PD-1 антител (то есть, 1.7.3 чАт, 1.49.9 чАт, 1.103.11 чАт, 1.139.15 чАт и 1.153.7 чАт) с экспрессирующими PD-1 клетками CHO. Полностью человеческие антитела против человеческого PD-1 использовали для окрашивания трансфицированных PD-1 клеток CHO, и FACS-анализ показал, что полностью человеческие анти-PD-1 антитела специфически связывают PD-1 с величиной EC50 примерно 2 нмоль/л.
Антитело 1.103.11-v2 чАт было получено в результате мутации одной аминокислоты Asn93 (нумерация Kabat) в исходном антителе 1.103.11 чАт -VH(20975-VL) на остаток серина с целью уменьшения риска гликозилирования на остатке CDR. Хотя Asn93 расположен в CDR3 легкой цепи, модель комплекса антитело-антиген, созданная при компьютерном моделировании стыковки, указывала на то, что Asn93 не имеет прямого контакта с каким-либо остатком антигена человеческого PD-1. На связывающую функцию CDR3 легкой цепи, судя по всему, наибольшее влияние оказывает соседний остаток Tyr91, который взаимодействует с некоторыми остатками в FG-петле PD-1. Функциональные анализы 1.103.11-v2 чАт с клетками подтвердили, что мутация никак не затронула способность к связыванию (смотри экспериментальные результаты ниже и фигуры 15 и 16).
Аффинность связывания 1.103.11-v2 чАт с человеческим PD-1 измеряли методами FACS и ELISA. На фигуре 16 показано связывание 1.103.11-v2 чАт в разных буферах с экспрессирующими PD-1 клетками CHO, и связывание также тестировали в тех же условиях, что и в FACS-анализе, описанном выше, за исключением того, что антитело находилось либо в буфере формулирования, либо в 1x PBS (pH 7,4), и клетки CHO-S, экспрессирующие человеческий PD-1, переносили в 96-луночные планшеты с U-образным дном (BD) при плотности 2×105 клеток/мл. Результаты были сопоставимы с результатами в случае 1.103.11 чАт. «1.103.11-v2 чАт в буфере» означает антитело в буфере формулирования и «1.103.11-v2 чАт в PBS» означает антитело в 1x PBS, pH 7,4. Антитела в обоих растворах связывали PD-1 клеточной поверхности на клетках CHO, и не было существенных отличий в аффинности к человеческому PD-1 между этими двумя условиями (в случае 1x PBS величина EC50 составляла примерно 2,52 нмоль/л и в случае буфера формулирования она составляла примерно 3,12 нмоль/л).
На фигуре 15 показано связывание антитела 1.103.11-v2 чАт с белком PD-1 в разных растворах при измерении методом ELISA. Следовали протоколу ELISA, описанному выше, время инкубации для 1.103.11-v2 чАт составляло 2 ч, и время инкубации для вторичного антитела козы против IgG Fc человека, конъюгированного с HRP (1:5000, Abcam), составляло 1 ч. «1.103.11-v2 чАт в буфере» означает антитело в буфере формулирования и «1.103.11-v2 чАт в PBS» означает антитело в 1x PBS, pH 7,4. Аффинное связывание с человеческим PD-1 было продемонстрировано в обоих условиях.
Клетки CHO, экспрессирующие человеческий PD-1, инкубировали с антителами против PD-1 в разных концентрациях, затем к клеткам добавляли маркированный мышиным Fc человеческий PD-L1. Связывание человеческого PD-L1 с экспрессирующими PD-1 клетками обнаруживали при помощи конъюгированного с FITC антитела козы против IgG мыши, с последующим FACS-анализом. Как показано на фигуре 4, антитела против PD-1 блокировали связывание PD-L1 с трансфицированными PD-1 клетками CHO. Антитело 1.103.11-v2 чАт также тестировали на блокирование связывания PD-L1 с трансфицированными PD-1 клетками CHO, и результаты были сопоставимы с результатами для 1.103.11 чАт.
3.2 Полная кинетическая аффинность связывания, протестированная методом поверхностного плазмонного резонанса (ППР): Антитела характеризовали в отношении аффинности связывания и кинетики связывания с PD-1 методом ППР с использованием ProteOn XPR36 (Bio-Rad). Белок A (Sigma) иммобилизовали на сенсорном чипе GLM (Bio-Rad) связыванием через аминогруппы. Очищенные антитела были пропущены в потоке над сенсорным чипом и захвачены белком A. Чип поворачивали на 90° и промывали проточным буфером (1X PBS/0,01% Tween20, Bio-Rad) до достижения стабильной базовой линии. Человеческий PD-1 в пяти концентрациях и проточный буфер пропускали через проточную ячейку с антителом со скоростью потока 100 мкл/мин на протяжении фазы ассоциации длительностью 240 с, с последующей фазой диссоциации в течение 600 с. После каждого прогона чип восстанавливали при помощи H3PO4, pH 1,7. Кривую ассоциации и диссоциации подгоняли к модели связывания 1:1 Ленгмюра с использованием программы ProteOn.
Как показано на фигуре 7, при определении методом поверхностного плазмонного резонанса значения аффинности антител против PD-1 в отношении рекомбинантного человеческого PD-1 составляли от 3,76E-9 до 1,76E-10 моль/л. Ожидается, что аффинность 1.103.11-v2 чАт будет сопоставима с таковой для 1.103.11 чАт.
3.3 Скрининг ортологов (перекрестно-видовых) и гомологов (перекрестно-семейственных):
3.3.1 Перекрестная реактивность в отношении PD-1 яванского макака и PD-1 мыши: перекрестную реактивность определяли методом ELISA. Планшеты (Nunc) покрывали PD-1 яванского макака (Sino Biological) и PD-1 мыши (Sino Biological) в концентрации 1 мкг/мл в течение ночи при 4°C. После блокирования и промывания в планшеты добавляли антитела в концентрации 1 мкг/мл и инкубировали при комнатной температуре в течение 1 ч. Затем планшеты промывали, после чего инкубировали со вторичным антителом козы против IgG1 крысы, конъюгированным с HRP (Bethyl), и антителом козы против IgG2b крысы, конъюгированным с HRP (Bethyl), в течение 45 мин. После промывания добавляли субстрат TMB и реакцию останавливали 2 M HCl. Поглощение при длине волны 450 нм измеряли на планшетном ридере (Molecular Device).
Результаты эксперимента с белками из разных биологических видов показали, что антитела против PD-1 связывают PD-1 яванского макака, но не связывают мышиный PD-1 (фигура 6). Ожидается, что результат для 1.103.11-v2 чАт в таком же эксперименте будет сопоставимым с результатом для 1.103.11 чАт.
3.3.2 Перекрестная реактивность в отношении представителей семейства PD-1 CD28, CTLA4 и ICOS: для изучения перекрестно-семейственной активности связывания полностью человеческих антител линейные клетки, экспрессирующие PD-1, CD28, CTLA4 или ICOS, окрашивали антителами, с последующим использованием в качестве 2-го антитела конъюгированного с FITC антитела козы против IgG Fc человека. Экспрессирующие PD-1 клетки использовали в качестве положительного контроля. Соответствующие исходные линии клеток использовали в качестве отрицательного контроля. Окрашенные клетки анализировали с использованием BD Biosciences FACSCanto II и версии программы FlowJo.
На фигуре 5 показано, что клетки CHO, трансфицированные PD-1, CD28, и 293F, трансфицированные CTLA4, были окрашены антителами против PD-1 и проанализированы методом FACS. Результаты демонстрируют, что анти-PD-1 антитела специфически связывают PD-1, но не CD28 и CTLA4 из семейства PD-1. Ожидается, что результат для 1.103.11-v2 чАт в таком же эксперименте будет сопоставимым с результатом для 1.103.11 чАт.
3.4 Тест эпитоп-специфической сортировки:
3.4.1 Проводили сортировку эпитопов, связываемых анти-PD-1 антителами, против эталонных антител A и B методом ППР с использованием ProteOn XPR36 (Bio-Rad). Эталонные антитела A и B иммобилизовали на сенсорном чипе GLC (Bio-Rad) связыванием через аминогруппы. Человеческий PD-1 в растворе был пропущен в потоке по каналам с иммобилизованными антителами и захвачен эталонными антителами. Затем чип поворачивали на 90° и промывали проточным буфером до достижения стабильной базовой линии. Выбранные антитела пропускали в потоке через сенсорный чип.
3.4.2 Проводили сортировку эпитопов, связываемых анти-PD-1 антителами, против эталонных антител A и B методом FACS. Клетки CHO, экспрессирующие человеческий PD-1 на клеточной поверхности, инкубировали с эталонным антителом A или B в концентрации 10 мкг/мл в течение 1 часа. Клетки промывали, добавляли анти-PD-1 антитела по изобретению и инкубировали в течение 1 часа. В качестве второго антитела добавляли антитело против IgG крысы, конъюгированное с FITC, и инкубировали в течение 1 часа при 4°C. Затем клетки промывали один раз, ресуспендировали в 1X PBS/1% БСА и анализировали методом проточной цитометрии (BD).
Результаты измерения методами ППР и FACS в тесте на связывание показали, что эпитоп на человеческом PD-1, связываемый полностью человеческими анти-PD-1 антителами (то есть, 1.7.3 чАт, 1.49.9 чАт, 1.103.11 чАт, 1.139.15 чАт и 1.153.7 чАт), отличается от эпитопов для существующих анти-PD-1 антител (то есть, эталонных антител A и B). Ожидается, что результат для 1.103.11-v2 чАт в таком же эксперименте будет сопоставимым с результатом для 1.103.11 чАт.
3.5 In vitro функция анти-PD-1 антител, протестированная в анализах с использованием клеток:
3.5.1 Эффекты человеческих анти-PD-1 антител на пролиферацию T-клеток. Для тестирования эффектов анти-PD-1 антител на пролиферацию T-лимфоцитов использовали аллогенный ответ. Стимулированную первичными дендритными клетками (ДК) РСЛ проводили в 96-луночном планшете с U-образным дном для культивирования тканей в 200 мкл среды RPMI 1640, содержащей 10% ЭТС и антибиотики. ДК смешивали с аллогенными суммарными CD4+ T-клетками в количестве 1×105 при соотношении ДК:T-клетки, составляющем 1:10 и 1:100. Культивирование также проводили в присутствии или в отсутствие нейтрализующих мАт: человеческих анти-PD-1 антител и эталонных антител A и B, используемых в концентрации 10 мкг/мл. Инкубацию проводили в течение 5 дней, и на последние 16 ч добавляли [3H]тимидин по 1 мкКи/лунку. Включение [3H]тимидина измеряли в сцинтилляционном счетчике, и пролиферативные ответы выражали в виде среднего значения включения [3H]тимидина (импульсы в минуту) в троекратно продублированных лунках. Импульсы за счет одних только ДК, как правило, составляли <1000 имп/мин. Представленные результаты являются репрезентативными примерами минимум пяти проведенных экспериментов.
Человеческие дендритные клетки (ДК), а также CD4+T, CD8+T и суммарные клетки, используемые в вышеописанной алло-РСЛ, были получены из МКПК следующим методом: человеческие моноциты очищали из МКПК путем отрицательной селекции с использованием набора с коктейлем для обогащения человеческих моноцитов в соответствии с инструкциями производителя (StemCell Meylan). Вкратце, МКПК выделяли из крови здорового донора с использованием градиента Ficoll-Paque. Клетки два раза промывали PBS, затем ресуспендировали в концентрации 1×108 клеток/мл в буфере выделения и инкубировали со смесью Ат для обогащения моноцитов при 4°C в течение 30 мин. Клетки промывали, а затем инкубировали с магнитным коллоидом при 4°C в течение 30 мин. Немеченые моноциты пропускали через колонку MACS и собирали. Для получения нДК моноциты культивировали в среде RPMI 1640, содержащей 10% ЭТС, антибиотики с GM-CSF (PeproTech, Rocky Hill, NJ; 800 Ед/мл) и IL-4 (PeproTech; 500 Ед/мл), в концентрации 2×106 клеток/мл. Половину среды заменяли через день средой, содержащей GM-CSF и IL-4. Зрелые ДК получали путем стимуляции нДК при помощи LPS (026: B6; Sigma-Aldrich, St. Louis, MO; 1 мкг/мл) в день 5 в течение дополнительных 24 ч. CD4+T, CD8+T и суммарные T-клетки очищали путем отрицательной селекции, инкубируя МКПК со средой для обогащения человеческих CD4+T, CD8+T и суммарных T-клеток и магнитным коллоидом в соответствии с инструкциями производителя (Stemsep).
Человеческие CD4+ T-клетки стимулировали аллогенными ДК в присутствии или в отсутствие анти-PD-1 антител 1.7.3 чАт, 1.49.9 чАт, 1.103.11 чАт, 1.139.15 чАт и 1.153.7 чАт. Пролиферацию CD4+ T-клеток оценивали по включению [3H]тимидина. На фигуре 10 показано, что 1.7.3 чАт, 1.49.9 чАт, 1.103.11 чАт, 1.139.15 чАт и 1.153.7 чАт приводили к усилению пролиферации T-клеток зависимым от концентрации образом. Ожидается, что результат для 1.103.11-v2 чАт в таком же эксперименте будет сопоставимым с результатом для 1.103.11 чАт.
3.5.2 Эффекты человеческих анти-PD-1 антител на секрецию цитокина IFNγ in vitro: для прямой оценки эффекта блокады человеческими анти-PD-1 антителами на продуцирование цитокина IFNγ авторы изобретения проводили эксперименты по оценке продуцирования IFNγ в алло-РСЛ. Вкратце, человеческие CD4+ T-клетки очищали из МКПК отрицательной селекцией с использованием набора с коктейлем для обогащения CD4+ T-клеток в соответствии с инструкциями производителя. Незрелые ДК получали из моноцитов путем культивирования с GM-CSF и IL-4 в течение 5 дней, и дифференцировку в зрелые ДК проводили путем стимуляции LPS в концентрации 1 мкг/мл в течение ночи. CD4+ T-клетки смешивали с нДК/зДК в соотношении T:ДК от 10:1 до 100:1. Культивирование проводили в присутствии или в отсутствие человеческих анти-PD-1 антител и эталонных антител. Через 5 дней супернатанты с каждой культуры собирали для измерения количества цитокина IFNγ. Уровень IFNγ в супернатантах измеряли методом ELISA. Вкратце, планшеты Maxisorp покрывали мАт против IFN-гамма человека, разведенными в буфере для нанесения (0,75 мкг/мл; то есть, разведение 1/1360), по 50 мкл/лунку (то есть, для целого 96-луночного планшета добавляли 3,7 мкл антитела в 5 мл буфера для нанесения) и инкубировали в течение ночи при 4°C. Блокировали свободные участки связывания белка, добавляя по 200 мкл/лунку блокирующего буфера на 2 часа. Готовили разведения рекомбинантного IFN-гамма в качестве стандартов, выполняя двукратные разведения, начиная от концентрации 8000 пг/мл до 125 пг/мл, в полной среде, плюс отдельно пробу полной среды. Промывали планшеты, добавляли стандарты и тестируемые супернатанты (100 мкл/лунку) и инкубировали в течение 2-4 часов. Добавляли биотинилированное мАт против IFN-гамма, разведенное в блокирующем буфере (1/1333), с последующим добавлением конъюгата экстравидин-пероксидаза. Окраска развивалась при добавлении субстрата TMB, реакцию останавливали добавлением 2 M HCl. Измеряли поглощение при длине волны 450 нм.
На фигуре 9 показана стимуляция человеческих CD4+ T-клеток аллогенными ДК в присутствии или в отсутствие антител 1.7.3 чАт, 1.49.9 чАт, 1.103.11 чАт, 1.139.15 чАт и 1.153.7 чАт. Уровень IFNγ измеряли методом ELISA. Результаты показали, что полностью человеческие анти-PD-1 антитела вызывали увеличение секреции IFNγ зависимым от дозы образом. Ожидается, что результат для 1.103.11-v2 чАт в таком же эксперименте будет сопоставимым с результатом для 1.103.11 чАт.
3.5.3 Эффекты человеческих анти-PD-1 антител на продуцирование интерлейкина 2 (IL-2) in vitro: CD4+ T-клетки смешивали с нДК/зДК в соотношении T:ДК от 10:1 до 100:1. Культивирование проводили в присутствии или в отсутствие человеческих анти-PD-1 антител и эталонных антител. Через 5 дней супернатанты с каждой культуры собирали для измерения количества цитокина. Уровень IL-2 в супернатантах измеряли методом ELISA.
На фигуре 8 показана стимуляция человеческих CD4+ T-клеток аллогенными ДК в присутствии или в отсутствие тестируемых антител или контрольных Ат. Уровень IL-2 измеряли методом ELISA. Результаты показали, что антитела против PD-1 вызывали увеличение секреции IL-2 зависимым от дозы образом. Ожидается, что результат для 1.103.11-v2 чАт в таком же эксперименте будет сопоставимым с результатом для 1.103.11 чАт.
3.5.4 Эффект человеческих анти-PD-1 антител на пролиферацию клеток и продуцирование цитокинов при аутологичном антиген-специфическом иммунном ответе: в данном анализе T-клетки и ДК были от одного и того же донора. Вкратце, CD4+ T-клетки очищали из МКПК и культивировали в присутствии пептида pp65 из CMV и низкой дозы IL2 (20 Ед/мл), в то же время получали ДК, культивируя моноциты из МКПК того же донора с GM-CSF и IL-4. Через 5 дней обработанные пептидом pp65 из CMV CD4+ T-клетки совместно культивировали с ДК, добавляя пептид pp65 в отсутствие или в присутствии человеческих анти-PD-1 антител и эталонных антител (в качестве контроля). В день 5 по 100 мкл супернатанта каждой из культур отбирали для измерения количества цитокинов IFNγ и IL-2. Уровень продуцирования IFNγ и IL-2 определяли методом ELISA. Пролиферацию специфических T-клеток в присутствии нагруженных пептидом pp65 из CMV ДК оценивали по включению [3H]тимидина.
Как показано на фигуре 11, анти-PD-1 антитела зависимым от концентрации образом приводили к усилению пролиферации CMV+-CD4+ T-клеток, стимулированных нагруженными пептидом pp65 из CMV аутологичными ДК. Ожидается, что результат для 1.103.11-v2 чАт в таком же эксперименте будет сопоставимым с результатом для 1.103.11 чАт.
3.5.5 Эффект человеческих анти-PD-1 антител на супрессорную функцию регуляторных T-клеток (Treg): клетки Treg, представляющие собой субпопуляцию T-клеток, являются ключевым иммуномодулятором и играют важную роль в поддержании толерантности к собственным антигенам. CD4+CD25+ регуляторные T-клетки связаны с образованием опухолей, поскольку повышенные количества Treg были обнаружены у пациентов при различных формах рака, что также связано с плохим прогнозом. Для непосредственной оценки эффекта человеческих анти-PD-1 антител на иммуносупрессорный ответ авторы изобретения проводили эксперимент с клетками Treg. CD4+CD25+ и CD4+CD25- T-клетки разделяли с использованием специфических анти-CD25 микрогранул (Miltenyi Biotec, Auburn, CA) методами положительной и отрицательной селекции, соответственно. Сначала CD4+ T-клетки очищали отрицательной селекцией, инкубируя МКПК со смесью для обогащения человеческих CD4+ T-клеток и магнитным коллоидом в соответствии с инструкциями производителя (Stemsep). Затем CD4+ T-клетки ресуспендировали в буфере MACS, инкубировали с CD25+ микрогранулами на льду в течение 30 мин, промывали и загружали в колонку. CD4+CD25- T-клетки, которые не связывались на колонке, собирали в протекшем объеме и промывали перед использованием. CD4+CD25+ T-клетки впоследствии снимали с колонки и промывали перед использованием. Клетки Treg культивировали с CD4+CD25- T-клетками и ДК (соотношение Treg:Teff 1:1) в отсутствие или в присутствии человеческих анти-PD-1 антител в концентрации 10 мкг/мл. В качестве отрицательного контроля либо не добавляли антитело, либо добавляли контрольное по изотипу антитело. Супернатанты культур отбирали в день 5 для определения цитокинов методом ELISA, и пролиферацию клеток измеряли, добавляя [3H]тимидин в концентрации 1 мкКи/лунку и дополнительно инкубируя в течение 18 часов. Включение [3H]тимидина измеряли в сцинтилляционном счетчике. Как показано на фигуре 12, анти-PD-1 антитела приводили к исчезновению супрессорной функции Treg и восстановлению пролиферации отвечающих на стимул T-клеток и секреции IFNγ. Ожидается, что результат для 1.103.11-v2 чАт в таком же эксперименте будет сопоставимым с результатом для 1.103.11 чАт.
3.6 Анализ ADCC/CDC: для сведения к минимуму нежелательной токсичности в отношении здоровых PD-1+ клеток подтверждали, что выбранные полностью человеческие анти-PD-1 антитела не обладают функциями ADCC и CDC.
3.6.1 ADCC: активированные T-клетки, на высоком уровне экспрессирующие на клеточной поверхности PD-1, использовали в качестве клеток-мишеней и преинкубировали с полностью человеческими антителами в разных концентрациях в 96-луночных планшетах в течение 30 мин, затем добавляли IL-2-активированные МКПК (используемые в качестве источника клеток - естественных киллеров (NK), то есть, эффекторных клеток) в соотношении эффектор/мишень, составляющем 50:1. Планшеты инкубировали в течение 6 часов при 37°C в инкубаторе с 5% CO2. Лизис клеток-мишеней определяли при помощи набора для определения цитотоксичности (Roche). Оптическую плотность измеряли на планшетном ридере SpectraMax M5e от компании Molecular Devices. Результаты показали, что протестированные полностью человеческие антитела против PD-1 не опосредовали ADCC (фигура 13). Ожидается, что результат для 1.103.11-v2 чАт в таком же эксперименте будет сопоставимым с результатом для 1.103.11 чАт.
3.6.2 CDC: клетки-мишени (активированные T-клетки), разбавленный комплемент из человеческой сыворотки (Quidel-A112) и полностью человеческие анти-PD-1 антитела в разных концентрациях смешивали в 96-луночном планшете. Планшет инкубировали в течение 4 ч при 37°C в инкубаторе с 5% CO2. Лизис клеток-мишеней определяли при помощи CellTiter glo (Promega-G7573). Ритуксан (Roche) и линию клеток Raji человеческой B-клеточной лимфомы (CD20 положительную) использовали в качестве положительного контроля. Данные показали, что анти-PD-1 антитела не опосредовали CDC (фигура 14). Ожидается, что результат для 1.103.11-v2 чАт в таком же эксперименте будет сопоставимым с результатом для 1.103.11 чАт.
ПРИМЕР 4: Картирование эпитопов полностью человеческого антитела
Для определения различий в эпитопах между настоящим антителом 1.103.11 чАт, предложенным в настоящем документе, и Keytruda, известным анти-чPD-1 антителом, проводили эксперименты с аланин-сканирующим мутагенезом чPD-1, и эффект на связывание антител оценивали, используя 1.103.11 чАт, Keytruda и 11.148.10 (контрольное анти-чPD-1 антитело, которое связывает эпитоп, не перекрывающийся с эпитопом 1.103.11 чАт или эпитопом Keytruda).
Осуществляли мутацию кодонов для остатков аланина в чPD-1 в кодоны глицина, и кодонов всех других остатков в кодоны аланина. Для каждого остатка внеклеточного домена (ECD) чPD-1 выполняли точечные аминокислотные замены с использованием двух последовательных этапов ПЦР. Плазмиду pcDNA3.3-hPD-1_ECD.His, кодирующую ECD человеческого PD-1 и C-концевую His-метку, использовали в качестве матрицы, и набор мутагенных праймеров использовали для первого этапа ПЦР с применением набора для сайт-направленного мутагенеза QuikChange lightning multi-site-directed mutagenesis kit (Agilent technologies, Palo Alto, CA). Эндонуклеазу Dpn I использовали для расщепления исходной матрицы после реакции синтеза мутантной цепи. На втором этапе ПЦР линейную ДНК экспрессионную кассету, состоящую из промотора CMV, последовательности для внеклеточного домена (ECD) PD-1, His-метки и сигнала полиаденилирования тимидинкиназы (TK) вируса простого герпеса, амплифицировали и временно экспрессировали в клетках HEK293F (Life Technologies, Gaithersburg, MD).
Моноклональными антителами 1.103.11 чАт, Keytruda и 11.148.10 чАт покрывали лунки планшетов для анализа связывания методом ELISA. После взаимодействия с супернатантом, содержащим определенное количество мутанта PD-1, добавляли конъюгированное с HRP анти-His антитело (Rockland, каталожный № 200-303-382) в качестве обнаруживающего антитела. Поглощение нормировали на среднее поглощение контрольных мутантов. После установки дополнительного отсечения для кратности изменения связывания (<0,55) проводили окончательную идентификацию остатков эпитопа.
Основные 30 мутантов чPD-1 с точечными заменами, которые приводили к значительному ослаблению связывания антител, приведены в таблице 2. Проверка положений всех этих остатков на кристаллических структурах чPD-1 (PDB код 3RRQ и 4ZQK) выявила, что некоторые аминокислоты (например, Val144, Leu142, Val110, Met108, Cys123 и так далее) были полностью погружены в белок и вряд ли могли бы непосредственно контактировать с какими-либо антителами. Наблюдаемое ослабление связывания, наиболее вероятно, являлось следствием нестабильности или даже коллапса структуры чPD-1 после замен на остатки аланина. Во избежание неправильной интерпретации этих данных, как «горячих зон» эпитопов, авторы изобретения воспользовались контрольным антителом 11.148.10 чАт, которое связывается в совершенно противоположном участке антигена, но, как ожидается, будет реагировать на коллапс структуры чPD-1, если это действительно произойдет. Мутации, которые затрагивали взаимодействие с обоими антителами, считали ложными «горячими зонами» и удаляли из списка. Остатки эпитопа, окончательно определенные после установки дополнительного отсечения для кратности изменения связывания (<0,55), приведены в таблице 3. Они соответствуют 9 положениям в 1.103.11 чАт и 5 положениям в Keytruda, а также 10 остаткам в контрольном антителе 11.148.10 чАт.
Таблица 2. Эффект точечных мутаций PD-1 на связывание антител
Выделено жирным шрифтом: аминокислоты, перекрывающиеся с аминокислотами в случае контрольного 11.148.10 чАт, необходимые для поддержания структуры, которые были исключены из списка «горячих зон».
a Кратность изменения связывания относительно связывания при некоторых «молчащих» аланиновых заменах.
Таблица 3. Идентификация потенциальных эпитопов
Порог отсечения: кратность изменения <0,55
* C, C', C'', F, G, A' представляют собой пептидные цепи в кристаллической структуре чPD-1, представленной на фигуре 17. Цепь C'', наблюдаемая в мPD-1, не существует в структуре чPD-1. Этот β-слой заменен на бесструктурную петлю в чPD-1. Авторы изобретения все-еще использовали C'' для маркировки этой области, просто для облегчения сравнения с мPD-1.
Сравнение остатков эпитопов для 1.103.11 чАт и Keytruda в таблице 3 выявило только два перекрывающихся остатка «горячих зон». Остальные довольно сильно отличались, это указывало на то, что два антитела могут сильно отличаться по механизму связывания чPD-1 и блокирования чPD-L1. Идентификация остатков в таблице 3 недостаточна для интерпретации механизма действия. Вследствие этого, все данные из таблицы 3, а также сайт связывания чPD-L1, были картированы на кристаллическую структуру чPD-1 для лучшей визуализации и сравнения. (фигура 17).
Как показано на фигуре 17, все остатки «горячей зоны», ответственные за связывание чPD-L1, собраны в середине цепей C, F и G (фигура 17A). Два исследуемых антитела 1.103.11 чАт и Keytruda, хотя оба обладают функцией связывания чPD-1 и блокирования чPD-L1, безусловно, имеют разные эпитопы (Фигура 17B для 1.103.11 чАт, 17C для Keytruda). Эпитоп для Keytruda в основном состоит из остатков в петле C'D (соответствующей цепи C'' в мPD-1), которая совсем не пересекается с сайтом связывания PD-L1. Это свидетельствует о том, что функция блокирования чPD-L1 у антитела Keytruda больше зависит от эффектов стерического препятствия, создаваемых размером антитела. Напротив, эпитоп ведущего, по мнению авторов изобретения, антитела 1.103.11 чАт состоит из «горячих зон», находящихся в разных местах, и непосредственно перекрывается с сайтом связывания чPD-L1 (фигура 17A, 17B). Настоящее антитело 1.103.11 чАт блокирует чPD-L1 за счет большей конкурентоспособности по сравнению с чPD-L1 для связывания с их общим сайтом связывания. Вследствие этого, ожидается, что 1.103.11 чАт будет более функциональным при последующих разработках.
Антитело 11.148.10 чАт, однако, связывается в совершенно ином участке (фигура 17D), чем два функциональных антитела, являясь хорошим контрольным антителом для мониторинга функции чPD-1 при выполнении замен на остатки аланина.
Хотя изобретение представлено и описано со ссылкой на конкретные варианты осуществления (некоторые из которых являются предпочтительными вариантами осуществления), специалисты в данной области должны понимать, что различные изменения с точки зрения формы или деталей могут иметь место без отклонения от сущности и объема данного изобретения, описанного в настоящем документе.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> WuXi Biologics (Shanghai) Co. Ltd.
Open Monoclonal Technology, Inc
<120> НОВЫЕ АНТИ-PD-1 АНТИТЕЛА
<130> 053674-8008WO03
<160> 68
<170> PatentIn version 3.5
<210> 1
<211> 7
<212> БЕЛОК
<213> Homo sapiens
<400> 1
Ser Thr Thr Tyr Tyr Trp Val
1 5
<210> 2
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 2
agtactactt actactgggt c 21
<210> 3
<211> 16
<212> БЕЛОК
<213> Homo sapiens
<400> 3
Ser Ile Ser Tyr Ser Gly Asn Thr Tyr Tyr Asn Pro Ser Leu Lys Ser
1 5 10 15
<210> 4
<211> 48
<212> ДНК
<213> Homo sapiens
<400> 4
agtatctctt atagtgggaa cacctactac aatccgtccc tcaagagt 48
<210> 5
<211> 13
<212> БЕЛОК
<213> Homo sapiens
<400> 5
His Leu Gly Tyr Asn Gly Arg Tyr Leu Pro Phe Asp Tyr
1 5 10
<210> 6
<211> 39
<212> ДНК
<213> Homo sapiens
<400> 6
catctagggt ataatgggag gtacctcccc tttgactac 39
<210> 7
<211> 14
<212> БЕЛОК
<213> Homo sapiens
<400> 7
Thr Gly Thr Ser Ser Asp Val Gly Phe Tyr Asn Tyr Val Ser
1 5 10
<210> 8
<211> 42
<212> ДНК
<213> Homo sapiens
<400> 8
actggaacca gcagtgacgt tggtttttat aactatgtct cc 42
<210> 9
<211> 7
<212> БЕЛОК
<213> Homo sapiens
<400> 9
Asp Val Thr Asn Arg Pro Ser
1 5
<210> 10
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 10
gatgtcacta atcggccctc a 21
<210> 11
<211> 10
<212> БЕЛОК
<213> Homo sapiens
<400> 11
Ser Ser Tyr Thr Ser Ile Ser Thr Trp Val
1 5 10
<210> 12
<211> 30
<212> ДНК
<213> Homo sapiens
<400> 12
agctcatata caagcatcag cacttgggtg 30
<210> 13
<211> 7
<212> БЕЛОК
<213> Homo sapiens
<400> 13
Ser Ser Thr Tyr Tyr Trp Gly
1 5
<210> 14
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 14
agtagtactt actactgggg c 21
<210> 15
<211> 16
<212> БЕЛОК
<213> Homo sapiens
<400> 15
Ser Ile Ser Tyr Ser Gly Ser Thr Tyr Tyr Asn Pro Ser Leu Lys Ser
1 5 10 15
<210> 16
<211> 48
<212> ДНК
<213> Homo sapiens
<400> 16
agtatctctt atagtgggag cacctactac aatccgtccc tcaagagt 48
<210> 17
<211> 7
<212> БЕЛОК
<213> Homo sapiens
<400> 17
Asp Val Ser Asn Arg Pro Ser
1 5
<210> 18
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 18
gatgtcagta atcggccctc a 21
<210> 19
<211> 10
<212> БЕЛОК
<213> Homo sapiens
<400> 19
Ser Ser Tyr Thr Asn Ile Ser Thr Trp Val
1 5 10
<210> 20
<211> 30
<212> ДНК
<213> Homo sapiens
<400> 20
agctcatata caaacatcag cacttgggtg 30
<210> 21
<211> 7
<212> БЕЛОК
<213> Homo sapiens
<400> 21
Ser Thr Thr Tyr Tyr Trp Gly
1 5
<210> 22
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 22
agtactactt actactgggg c 21
<210> 23
<211> 16
<212> БЕЛОК
<213> Homo sapiens
<400> 23
Ser Ile Ser Tyr Ser Gly Thr Thr Tyr Tyr Asn Pro Ser Leu Lys Ser
1 5 10 15
<210> 24
<211> 48
<212> ДНК
<213> Homo sapiens
<400> 24
agtatctctt atagtgggac cacctactac aacccgtccc tcaagagt 48
<210> 25
<211> 13
<212> БЕЛОК
<213> Homo sapiens
<400> 25
His Leu Gly Tyr Asn Ser Asn Trp Tyr Pro Phe Asp Tyr
1 5 10
<210> 26
<211> 39
<212> ДНК
<213> Homo sapiens
<400> 26
catctcgggt ataacagcaa ctggtaccct tttgactac 39
<210> 27
<211> 14
<212> БЕЛОК
<213> Homo sapiens
<400> 27
Thr Gly Thr Ser Ser Asp Val Gly Ser Tyr Asn Arg Val Ser
1 5 10
<210> 28
<211> 42
<212> ДНК
<213> Homo sapiens
<400> 28
actggaacca gcagtgacgt tggtagttat aaccgtgtct cc 42
<210> 29
<211> 7
<212> БЕЛОК
<213> Homo sapiens
<400> 29
Glu Val Ser Asn Arg Pro Ser
1 5
<210> 30
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 30
gaggtcagta atcggccctc a 21
<210> 31
<211> 10
<212> БЕЛОК
<213> Homo sapiens
<400> 31
Ser Ser Tyr Thr Ser Ser Ser Thr Trp Val
1 5 10
<210> 32
<211> 30
<212> ДНК
<213> Homo sapiens
<400> 32
agctcatata caagcagcag cacttgggtg 30
<210> 33
<211> 5
<212> БЕЛОК
<213> Homo sapiens
<400> 33
Ser His Ala Met Ser
1 5
<210> 34
<211> 15
<212> ДНК
<213> Homo sapiens
<400> 34
agccatgcca tgagc 15
<210> 35
<211> 17
<212> БЕЛОК
<213> Homo sapiens
<400> 35
Thr Ile Thr Gly Gly Gly Gly Ser Ile Tyr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 36
<211> 51
<212> ДНК
<213> Homo sapiens
<400> 36
actattactg gtggtggtgg tagcatatac tacgcagact ccgtgaaggg c 51
<210> 37
<211> 10
<212> БЕЛОК
<213> Homo sapiens
<400> 37
Asn Arg Ala Gly Glu Gly Tyr Phe Asp Tyr
1 5 10
<210> 38
<211> 30
<212> ДНК
<213> Homo sapiens
<400> 38
aaccgcgctg gggagggtta ctttgactac 30
<210> 39
<211> 11
<212> БЕЛОК
<213> Homo sapiens
<400> 39
Gly Gly Asp Asn Ile Gly Asn Lys Asp Val His
1 5 10
<210> 40
<211> 33
<212> ДНК
<213> Homo sapiens
<400> 40
gggggagaca acattggaaa taaagatgtg cac 33
<210> 41
<211> 7
<212> БЕЛОК
<213> Homo sapiens
<400> 41
Arg Asp Ser Asn Arg Pro Ser
1 5
<210> 42
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 42
agggatagca accggccctc t 21
<210> 43
<211> 8
<212> БЕЛОК
<213> Homo sapiens
<400> 43
Gln Val Trp Asp Ser Ile Trp Val
1 5
<210> 44
<211> 24
<212> ДНК
<213> Homo sapiens
<400> 44
caggtgtggg acagcatttg ggtg 24
<210> 45
<211> 123
<212> БЕЛОК
<213> Homo sapiens
<400> 45
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Val Ser Gly Asp Ser Ile Ser Ser Thr
20 25 30
Thr Tyr Tyr Trp Val Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Ser Ile Ser Tyr Ser Gly Asn Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn His Phe
65 70 75 80
Ser Leu Lys Leu Ser Ser Val Ala Ala Thr Asp Thr Ala Leu Tyr Tyr
85 90 95
Cys Ala Arg His Leu Gly Tyr Asn Gly Arg Tyr Leu Pro Phe Asp Tyr
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 46
<211> 369
<212> ДНК
<213> Homo sapiens
<400> 46
cagctgcagc tgcaggagtc gggcccagga ctggtgaagc cttcggagac cctgaccctc 60
acctgcactg tctctggtga ctccatcagc agtactactt actactgggt ctggatccgc 120
cagcccccag ggaagggact ggagtggatt gggagtatct cttatagtgg gaacacctac 180
tacaatccgt ccctcaagag tcgagtcacc atatccgtag acacgtccaa gaaccacttc 240
tccctgaagc tgagttctgt ggccgccaca gacacggctc tatattactg tgcgagacat 300
ctagggtata atgggaggta cctccccttt gactactggg gccagggaac cctggtcacc 360
gtctcctcc 369
<210> 47
<211> 110
<212> БЕЛОК
<213> Homo sapiens
<400> 47
Gln Ser Ala Leu Thr Gln Pro Ala Ser Val Ser Gly Ser Pro Gly Gln
1 5 10 15
Ser Ile Thr Ile Ser Cys Thr Gly Thr Ser Ser Asp Val Gly Phe Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Glu Leu
35 40 45
Met Ile Tyr Asp Val Thr Asn Arg Pro Ser Gly Val Ser Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Ser Tyr Thr Ser Ile
85 90 95
Ser Thr Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 48
<211> 330
<212> ДНК
<213> Homo sapiens
<400> 48
cagtctgccc tgactcagcc tgcctccgtg tctgggtctc ctggacagtc gatcaccatc 60
tcctgcactg gaaccagcag tgacgttggt ttttataact atgtctcctg gtaccaacag 120
cacccaggca aagcccccga actcatgatt tatgatgtca ctaatcggcc ctcaggggtt 180
tctgatcgct tctctggctc caagtctggc aacacggcct ccctgaccat ctctgggctc 240
caggctgagg acgaggctga ttattactgc agctcatata caagcatcag cacttgggtg 300
ttcggcggag ggaccaagct gaccgtccta 330
<210> 49
<211> 123
<212> БЕЛОК
<213> Homo sapiens
<400> 49
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Ser
20 25 30
Thr Tyr Tyr Trp Gly Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Ser Ile Ser Tyr Ser Gly Ser Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Ser Ser Val Thr Asp Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg His Leu Gly Tyr Asn Gly Arg Tyr Leu Pro Phe Asp Tyr
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 50
<211> 369
<212> ДНК
<213> Homo sapiens
<400> 50
cagctgcagc tgcaggagtc gggcccagga ctggtgaagc cttcggagac cctgtccctc 60
acctgcactg tctctggtgg ctccatcagc agtagtactt actactgggg ctggatccgc 120
cagcccccag ggaagggact ggagtggatt gggagtatct cttatagtgg gagcacctac 180
tacaatccgt ccctcaagag tcgagtcacc atatccgtag acacgtccaa gaaccagttc 240
tccctgaagc tgagctctgt gaccgacgca gacacggctg tgtattactg tgcgagacat 300
ctagggtata atgggaggta cctccccttt gactactggg gccagggaac cctggtcacc 360
gtctcctcc 369
<210> 51
<211> 110
<212> БЕЛОК
<213> Homo sapiens
<400> 51
Gln Ser Ala Leu Thr Gln Pro Ala Ser Val Ser Gly Ser Pro Gly Gln
1 5 10 15
Ser Ile Thr Ile Ser Cys Thr Gly Thr Ser Ser Asp Val Gly Phe Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Glu Val
35 40 45
Met Ile Tyr Asp Val Ser Asn Arg Pro Ser Gly Val Ser Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Ser Tyr Thr Ser Ile
85 90 95
Ser Thr Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 52
<211> 330
<212> ДНК
<213> Homo sapiens
<400> 52
cagtctgccc tgactcagcc tgcctccgtg tctgggtctc ctggacagtc gatcaccatc 60
tcctgcactg gaaccagcag tgacgttggt ttttataact atgtctcctg gtaccaacag 120
cacccaggca aagcccccga agtcatgatt tatgatgtca gtaatcggcc ctcaggggtt 180
tctgatcgct tctctggctc caagtctggc aacacggcct ccctgactat ctctgggctc 240
caggctgagg acgaggctga ttattactgc agctcatata caagcatcag cacttgggtg 300
ttcggcggag ggaccaagct gactgtccta 330
<210> 53
<211> 123
<212> БЕЛОК
<213> Homo sapiens
<400> 53
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Val Ser Ala Asp Ser Ile Ser Ser Thr
20 25 30
Thr Tyr Tyr Trp Val Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Ser Ile Ser Tyr Ser Gly Ser Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Thr Val Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Asn Ser Val Ala Ala Thr Asp Thr Ala Leu Tyr Tyr
85 90 95
Cys Ala Arg His Leu Gly Tyr Asn Gly Arg Tyr Leu Pro Phe Asp Tyr
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 54
<211> 369
<212> ДНК
<213> Homo sapiens
<400> 54
cagctgcagc tgcaggagtc gggcccagga ctggtgaagc cttcggagac cctgaccctc 60
acctgcactg tctctgctga ctccatcagc agtactactt actactgggt ctggatccgc 120
cagcccccag ggaagggact ggagtggatt gggagtatct cttatagtgg gagcacctac 180
tacaatccgt ccctcaagag tcgagtcacc gtatccgtag acacgtccaa gaaccagttc 240
tccctgaagc tgaactctgt ggccgccaca gacacggctc tatattactg tgcgagacat 300
ctagggtata atgggaggta cctccccttt gactactggg gccagggaac cctggtcacc 360
gtctcctcc 369
<210> 55
<211> 110
<212> БЕЛОК
<213> Homo sapiens
<400> 55
Gln Ser Ala Leu Thr Gln Pro Ala Ser Val Ser Gly Ser Pro Gly Gln
1 5 10 15
Ser Ile Thr Ile Ser Cys Thr Gly Thr Ser Ser Asp Val Gly Phe Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Glu Leu
35 40 45
Met Ile Tyr Asp Val Ser Asn Arg Pro Ser Gly Val Ser Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Ser Tyr Thr Asn Ile
85 90 95
Ser Thr Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 56
<211> 330
<212> ДНК
<213> Homo sapiens
<400> 56
cagtctgccc tgactcagcc tgcctccgtg tctgggtctc ctggacagtc gatcaccatc 60
tcctgcactg gaaccagcag tgacgttggt ttttataact atgtctcctg gtaccaacag 120
cacccaggca aagcccccga actcatgatt tatgatgtca gtaatcggcc ctcaggggtt 180
tctgatcgct tctctggctc caagtctggc aacacggcct ccctgaccat ctctgggctc 240
caggctgagg acgaggctga ttattactgc agctcatata caaacatcag cacttgggtg 300
ttcggcggag ggaccaagct gaccgtccta 330
<210> 57
<211> 123
<212> БЕЛОК
<213> Homo sapiens
<400> 57
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Thr
20 25 30
Thr Tyr Tyr Trp Gly Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Ser Ile Ser Tyr Ser Gly Thr Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Thr Ile Pro Val Asp Thr Ser Lys Asn Gln Ile
65 70 75 80
Ser Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ser Leu Tyr Tyr
85 90 95
Cys Ala Arg His Leu Gly Tyr Asn Ser Asn Trp Tyr Pro Phe Asp Tyr
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 58
<211> 369
<212> ДНК
<213> Homo sapiens
<400> 58
cagctgcagc tgcaggagtc gggcccagga ctggtgaagc cctcggagac cctgtccctc 60
acctgcactg tctctggtgg ctccatcagc agtactactt actactgggg ctggatccgc 120
cagcccccag ggaaggggct ggagtggatt gggagtatct cttatagtgg gaccacctac 180
tacaacccgt ccctcaagag tcgagtcacc atccccgtag acacgtccaa gaaccagatc 240
tccctgaaac tgagctctgt gaccgccgca gacacgtctt tgtattattg tgcgagacat 300
ctcgggtata acagcaactg gtaccctttt gactactggg gccagggaac cctggtcacc 360
gtctcctca 369
<210> 59
<211> 110
<212> БЕЛОК
<213> Homo sapiens
<400> 59
Gln Ser Ala Leu Thr Gln Pro Pro Ser Val Ser Gly Ser Pro Gly Gln
1 5 10 15
Ser Val Thr Ile Ser Cys Thr Gly Thr Ser Ser Asp Val Gly Ser Tyr
20 25 30
Asn Arg Val Ser Trp Tyr Gln Gln Pro Pro Gly Thr Ala Pro Glu Val
35 40 45
Ile Ile Tyr Glu Val Ser Asn Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Ser Tyr Thr Ser Ser
85 90 95
Ser Thr Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 60
<211> 330
<212> ДНК
<213> Homo sapiens
<400> 60
cagtcggccc tgactcagcc tccctccgtg tccgggtctc ctggacagtc agtcaccatc 60
tcctgcactg gaaccagcag tgacgttggt agttataacc gtgtctcctg gtaccagcag 120
cccccaggca cagcccccga agtcattatt tatgaggtca gtaatcggcc ctcaggggtc 180
cctgatcgct tctctgggtc caagtctggc aacacggcct ccctgaccat ctctgggctc 240
caggctgagg acgaggctga ttattactgc agctcatata caagcagcag cacttgggtg 300
ttcggcggag ggaccaagct gaccgtccta 330
<210> 61
<211> 119
<212> БЕЛОК
<213> Homo sapiens
<400> 61
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser His
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Thr Ile Thr Gly Gly Gly Gly Ser Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Asn Arg Ala Gly Glu Gly Tyr Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 62
<211> 357
<212> ДНК
<213> Homo sapiens
<400> 62
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactg 60
tcctgcgcag cctctggatt cacctttagc agccatgcca tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcaact attactggtg gtggtggtag catatactac 180
gcagactccg tgaagggccg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtat attattgtgc gaaaaaccgc 300
gctggggagg gttactttga ctactggggc cagggaaccc tggtcaccgt ctcctca 357
<210> 63
<211> 105
<212> БЕЛОК
<213> Homo sapiens
<400> 63
Ser Tyr Glu Leu Thr Gln Pro Leu Ser Val Ser Val Ala Leu Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys Gly Gly Asp Asn Ile Gly Asn Lys Asp Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Arg Asp Ser Asn Arg Pro Ser Gly Ile Pro Glu Gly Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Arg Ala Gln Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Ser Ile Trp Val Phe
85 90 95
Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 64
<211> 315
<212> ДНК
<213> Homo sapiens
<400> 64
tcctatgagc tgactcagcc actctcagtg tcagtggccc tgggacagac ggccaggatt 60
acctgtgggg gagacaacat tggaaataaa gatgtgcact ggtaccagca gaagccaggc 120
caggcccctg tgctggtcat ctatagggat agcaaccggc cctctgggat ccctgaggga 180
ttctctggct ccaactcggg gaacacggcc accctgacca tcagcagagc ccaagccggg 240
gatgaggctg actattactg tcaggtgtgg gacagcattt gggtgttcgg cggagggacc 300
aagctgaccg tccta 315
<210> 65
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 65
Ser Ser Tyr Thr Ser Ile Ser Thr Trp Val
1 5 10
<210> 66
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 66
agctcatata caagcatcag cacttgggtg 30
<210> 67
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 67
Gln Ser Ala Leu Thr Gln Pro Ala Ser Val Ser Gly Ser Pro Gly Gln
1 5 10 15
Ser Ile Thr Ile Ser Cys Thr Gly Thr Ser Ser Asp Val Gly Phe Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Glu Leu
35 40 45
Met Ile Tyr Asp Val Ser Asn Arg Pro Ser Gly Val Ser Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Ser Tyr Thr Ser Ile
85 90 95
Ser Thr Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 68
<211> 330
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 68
cagtctgccc tgactcagcc tgcctccgtg tctgggtctc ctggacagtc gatcaccatc 60
tcctgcactg gaaccagcag tgacgttggt ttttataact atgtctcctg gtaccaacag 120
cacccaggca aagcccccga actcatgatt tatgatgtca gtaatcggcc ctcaggggtt 180
tctgatcgct tctctggctc caagtctggc aacacggcct ccctgaccat ctctgggctc 240
caggctgagg acgaggctga ttattactgc agctcatata caagcatcag cacttgggtg 300
ttcggcggag ggaccaagct gaccgtccta 330
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛО ПРОТИВ Fn14 ЧЕЛОВЕКА | 2019 |
|
RU2787044C2 |
| БИСПЕЦИФИЧЕСКОЕ АНТИТЕЛО | 2020 |
|
RU2814713C2 |
| МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА ЧЕЛОВЕКА К БЕЛКУ ПРОГРАММИРУЕМОЙ СМЕРТИ 1 (PD-1) И СПОСОБЫ ЛЕЧЕНИЯ РАКА С ИСПОЛЬЗОВАНИЕМ АНТИ-PD-1-АНТИТЕЛ САМОСТОЯТЕЛЬНО ИЛИ В КОМБИНАЦИИ С ДРУГИМИ ИММУНОТЕРАПЕВТИЧЕСКИМИ СРЕДСТВАМИ | 2016 |
|
RU2732924C2 |
| АНТИТЕЛО К CD73 ЧЕЛОВЕКА | 2017 |
|
RU2754058C2 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ ЛЕЧЕНИЯ И/ИЛИ ПРОФИЛАКТИКИ ЗЛОКАЧЕСТВЕННОЙ ОПУХОЛИ | 2019 |
|
RU2781542C2 |
| АНТИТЕЛО ДЛЯ ЛЕЧЕНИЯ АУТОИММУННЫХ ЗАБОЛЕВАНИЙ | 2017 |
|
RU2781148C2 |
| ИНГИБИТОР ИММУНОСУПРЕССИИ, СВЯЗАННЫЙ С РАКОМ | 2018 |
|
RU2805232C2 |
| БИСПЕЦИФИЧЕСКОЕ АНТИТЕЛО | 2019 |
|
RU2796019C2 |
| Соединения, связывающие AMHRII, для профилактики или лечения раковых заболеваний легкого | 2018 |
|
RU2797506C2 |
| НОВЫЕ АНТИТЕЛА ПРОТИВ PD-L1 | 2016 |
|
RU2736151C2 |
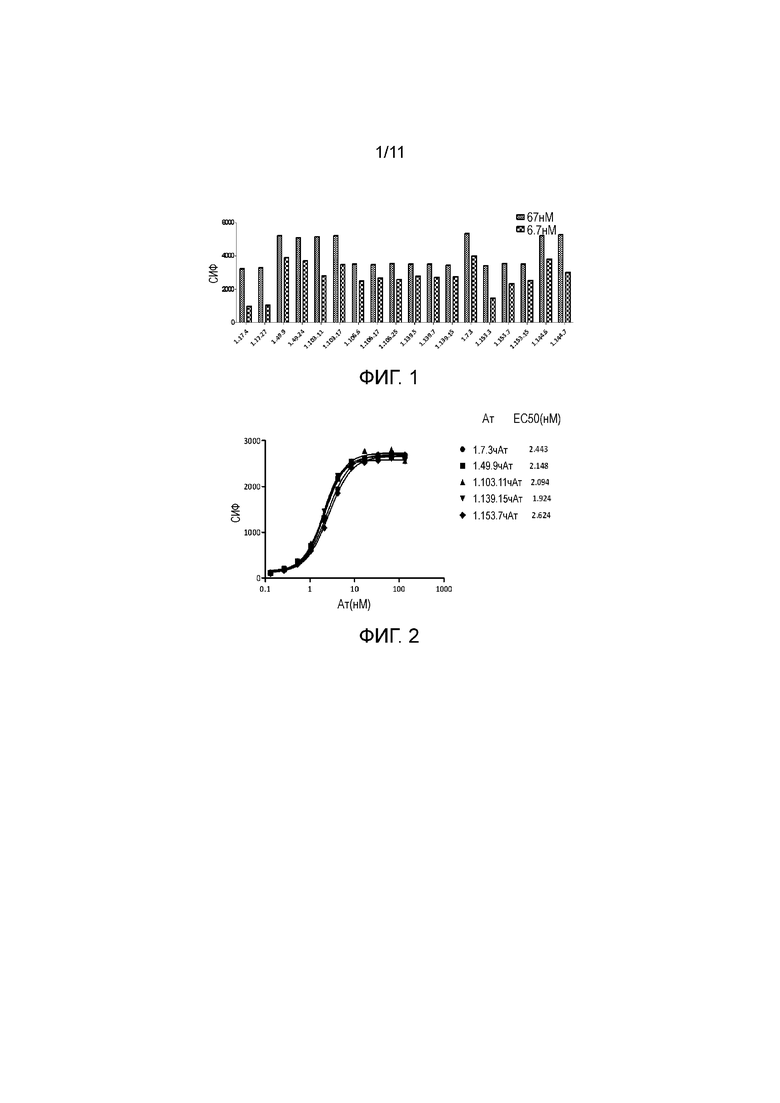
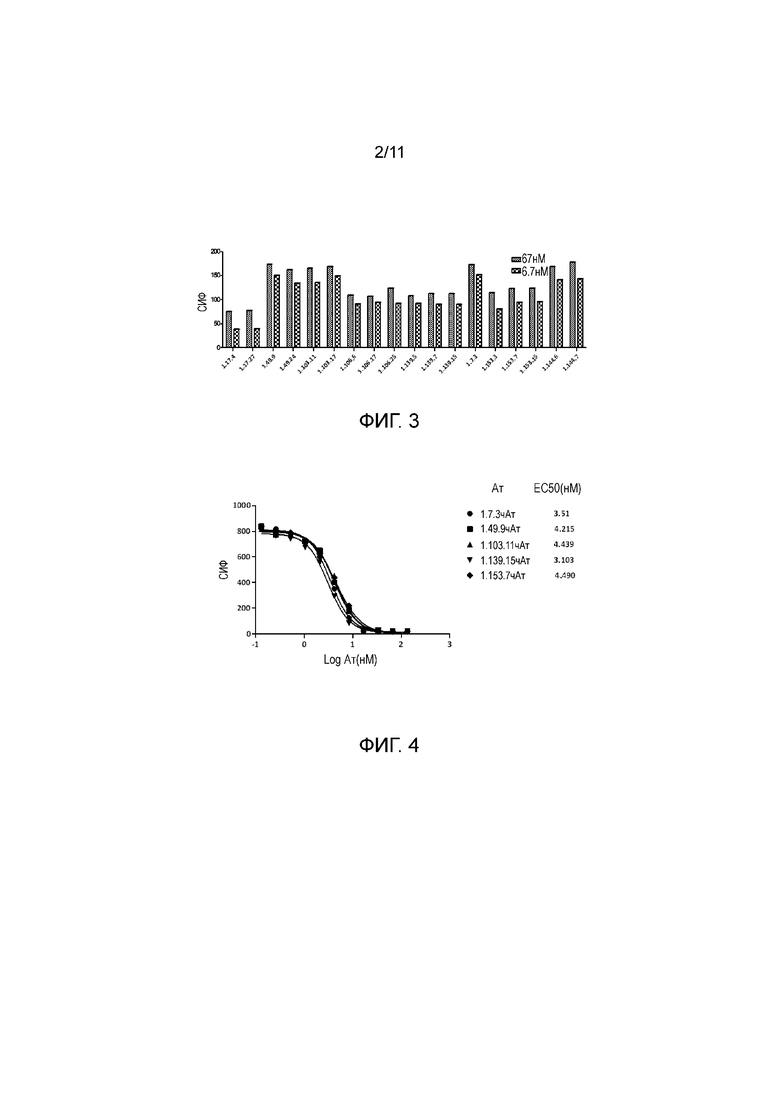
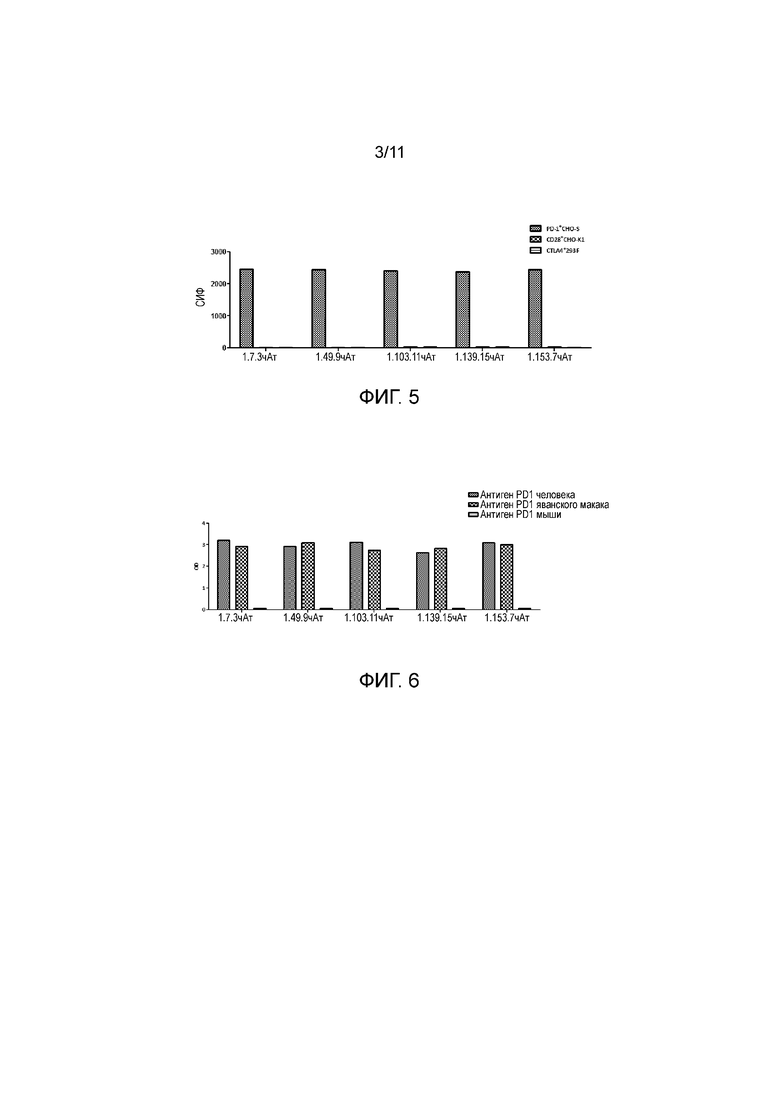

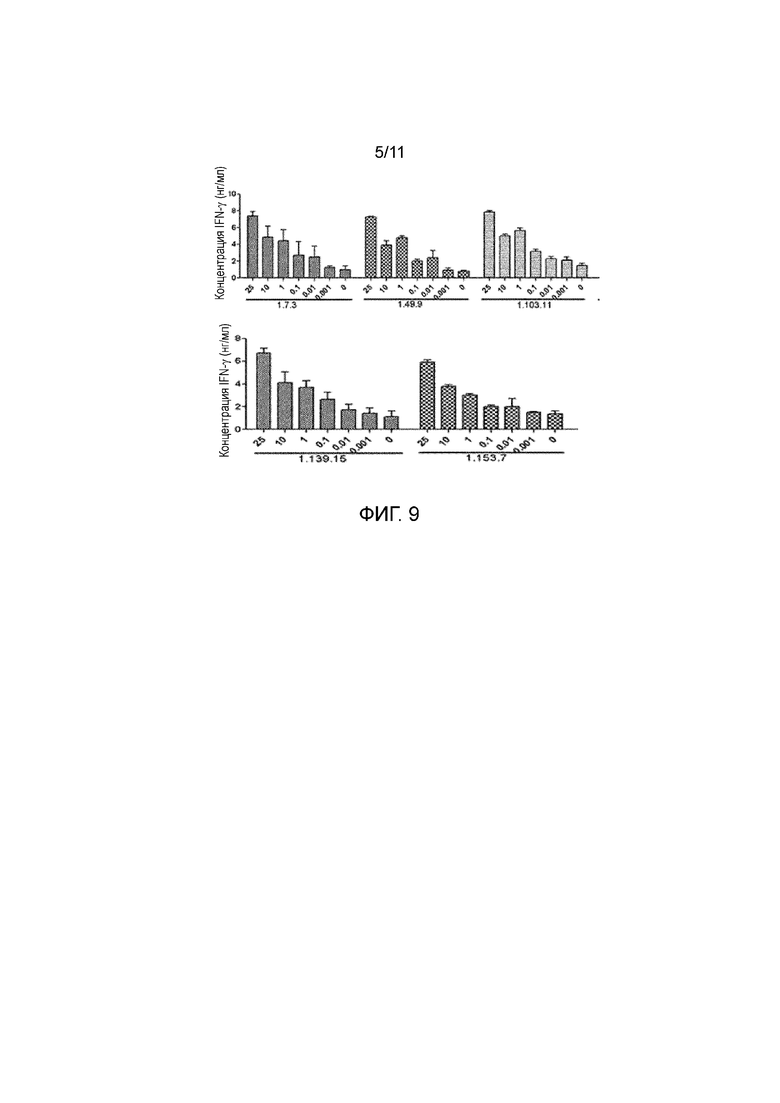
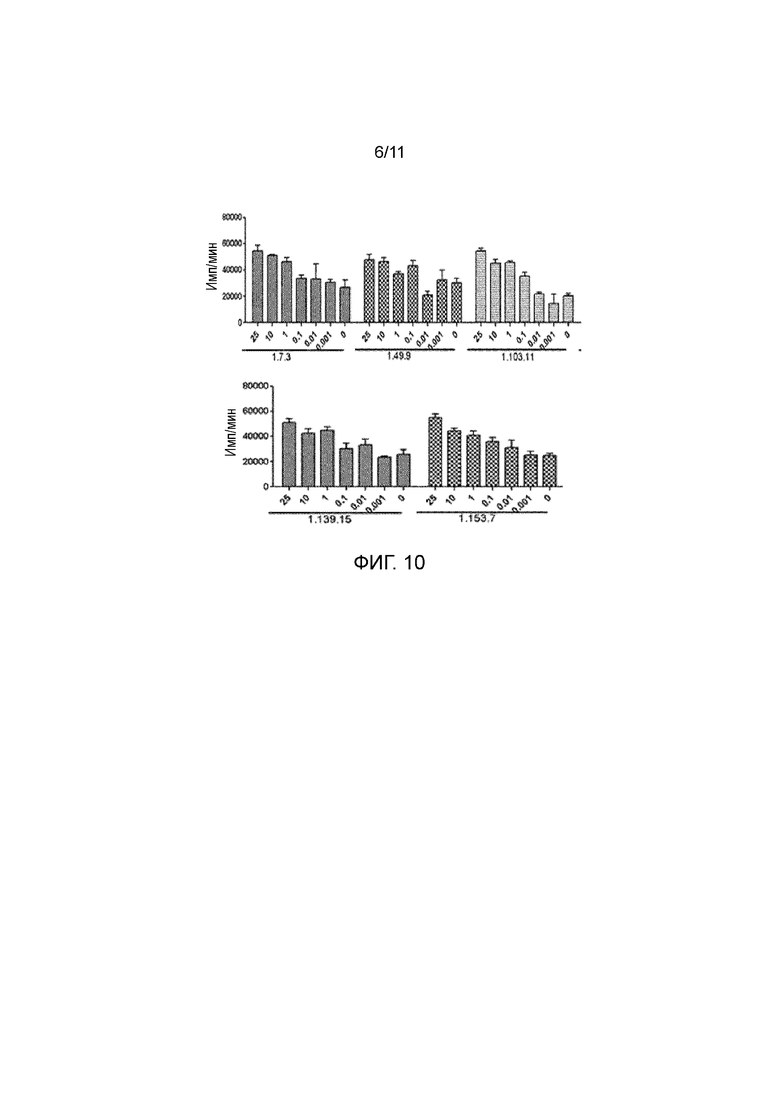
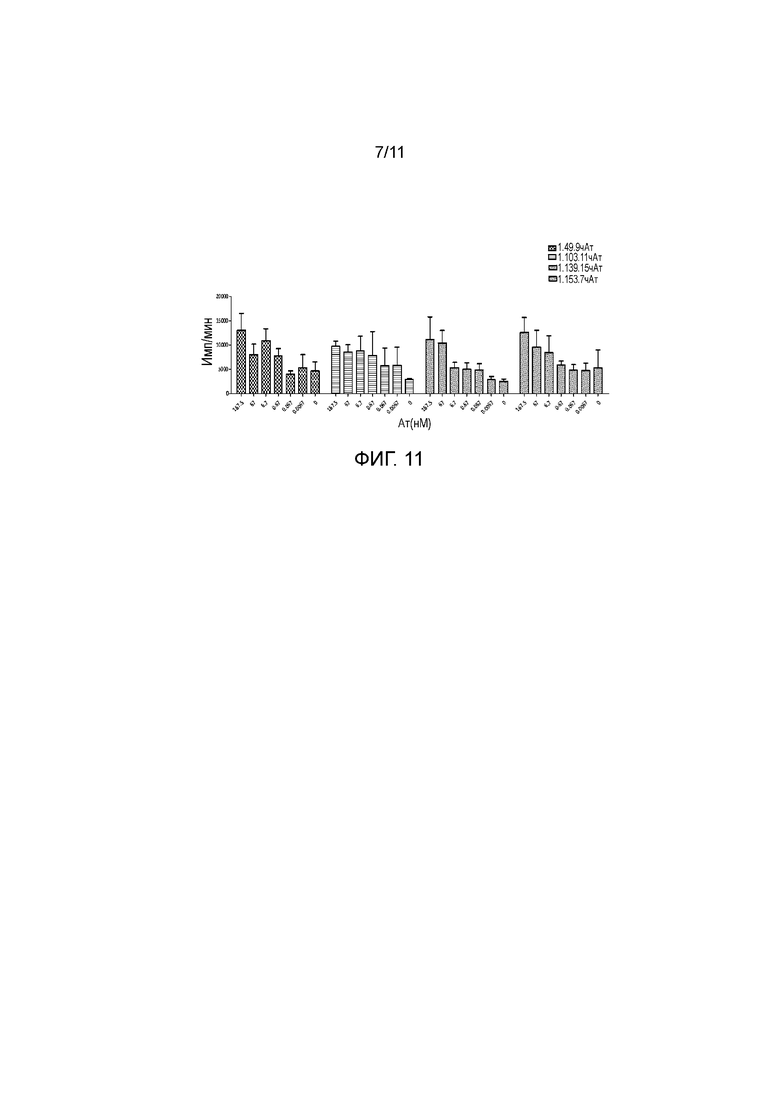

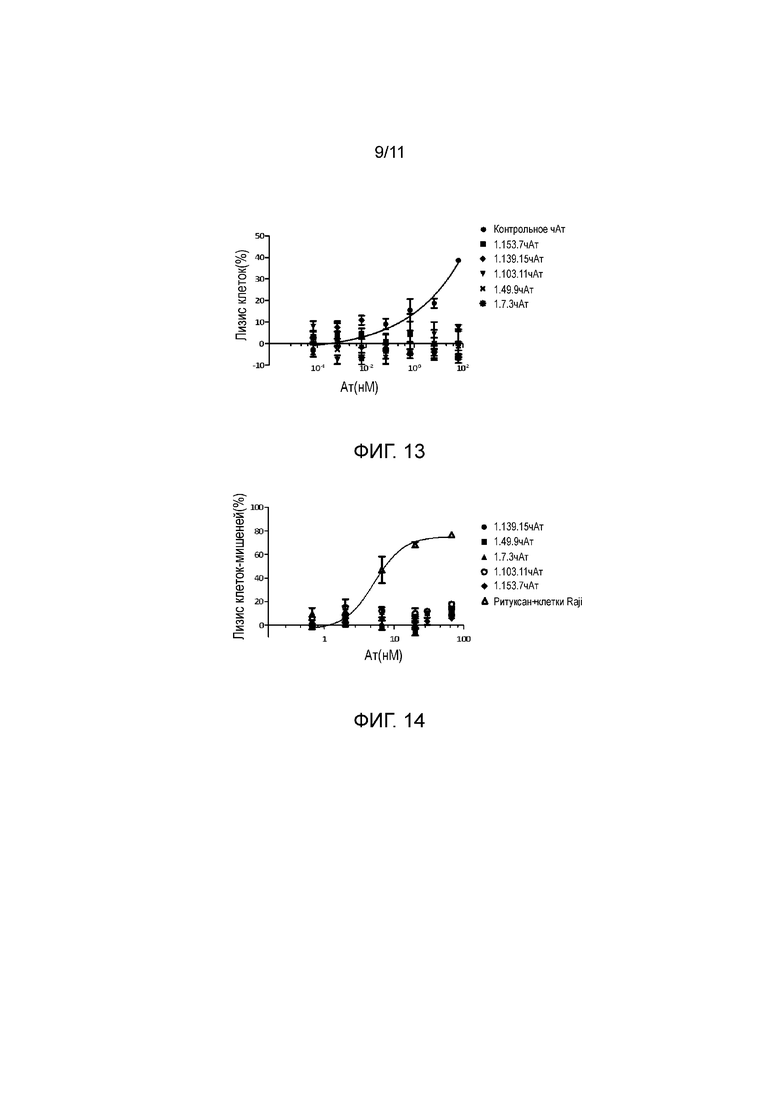
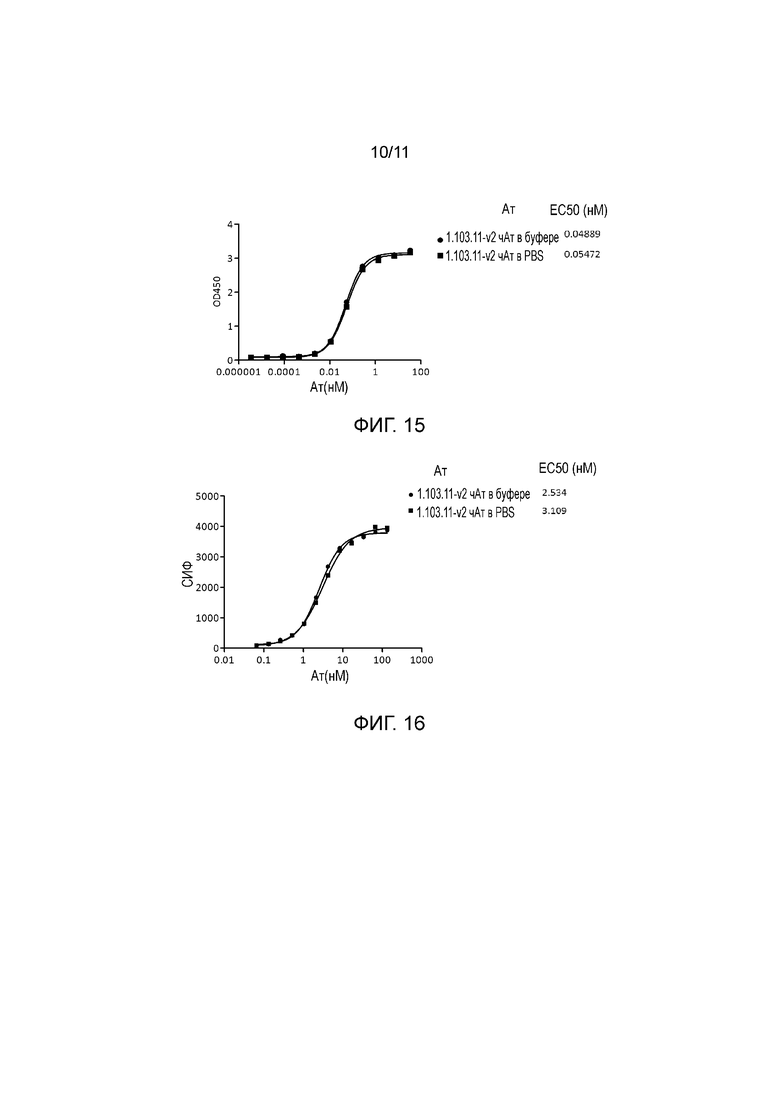

Изобретение относится к области биотехнологии. Предложены моноклональные антитела против белка 1 программируемой клеточной гибели (PD-1). Антитела могут блокировать связывание лигандов PD-1 с PD-1 и, таким образом, блокировать ингибирующую функцию лигандов PD-1 на экспрессирующие PD-1 T-клетки. Также изобретение относится к получению и применению указанных антител. Изобретение обеспечивает эффективные средства для лечения множества видов рака за счет модулирования иммунных функций человека. 11 н. и 16 з.п. ф-лы, 17 ил., 3 табл., 4 пр.
1. Выделенное анти-PD-1 антитело или его антигенсвязывающий фрагмент, содержащее:
a) вариабельную область тяжелой цепи, содержащую
область, определяющую комплементарность (CDR) 1 тяжелой цепи (H) с последовательностью SEQ ID NO:1,
HCDR2 с последовательностью SEQ ID NO:3 и
HCDR3 c последовательностью SEQ ID NO:5; и
вариабельную область легкой цепи, содержащую
CDR1 легкой цепи (L) с последовательностью SEQ ID NO:7,
LCDR2 с последовательностью SEQ ID NO:9 и
LCDR3 c последовательностью SEQ ID NO:11;
b) вариабельную область тяжелой цепи, содержащую
HCDR1 с последовательностью SEQ ID NO:13,
HCDR2 с последовательностью SEQ ID NO:15 и
HCDR3 c последовательностью SEQ ID NO:5; и
вариабельную область легкой цепи, содержащую
LCDR1 с последовательностью SEQ ID NO:7,
LCDR2 с последовательностью SEQ ID NO:17 и
LCDR3 c последовательностью SEQ ID NO:19;
c) вариабельную область тяжелой цепи, содержащую
HCDR1 с последовательностью SEQ ID NO:1,
HCDR2 с последовательностью SEQ ID NO:15 и
HCDR3 c последовательностью SEQ ID NO:5; и
вариабельную область легкой цепи, содержащую
LCDR1 с последовательностью SEQ ID NO:7,
LCDR2 с последовательностью SEQ ID NO:17 и
LCDR3 c последовательностью SEQ ID NO:19;
d) вариабельную область тяжелой цепи, содержащую
HCDR1 с последовательностью SEQ ID NO:21,
HCDR2 с последовательностью SEQ ID NO:23 и
HCDR3 c последовательностью SEQ ID NO:25; и
вариабельную область легкой цепи, содержащую
LCDR1 с последовательностью SEQ ID NO:27,
LCDR2 с последовательностью SEQ ID NO:29 и
LCDR3 c последовательностью SEQ ID NO:31;
e) вариабельную область тяжелой цепи, содержащую
HCDR1 с последовательностью SEQ ID NO:33,
HCDR2 с последовательностью SEQ ID NO:35 и
HCDR3 c последовательностью SEQ ID NO:37; и
вариабельную область легкой цепи, содержащую
LCDR1 с последовательностью SEQ ID NO:39,
LCDR2 с последовательностью SEQ ID NO:41 и
LCDR3 c последовательностью SEQ ID NO:43;
f) вариабельную область тяжелой цепи, содержащую
HCDR1 с последовательностью SEQ ID NO:1,
HCDR2 с последовательностью SEQ ID NO:15 и
HCDR3 c последовательностью SEQ ID NO:5; и
вариабельную область легкой цепи, содержащую
LCDR1 с последовательностью SEQ ID NO:7,
LCDR2 с последовательностью SEQ ID NO:17 и
LCDR3 c последовательностью SEQ ID NO:65.
2. Антитело или его антигенсвязывающий фрагмент по п.1, содержащее:
a) вариабельную область тяжелой цепи, последовательность которой по меньшей мере на 80% идентична SEQ ID NO:45; и
вариабельную область легкой цепи, последовательность которой по меньшей мере на 80% идентична SEQ ID NO:47;
b) вариабельную область тяжелой цепи, последовательность которой по меньшей мере на 80% идентична SEQ ID NO:49; и
вариабельную область легкой цепи, последовательность которой по меньшей мере на 80% идентична SEQ ID NO:51;
c) вариабельную область тяжелой цепи, последовательность которой по меньшей мере на 80% идентична SEQ ID NO:53; и
вариабельную область легкой цепи, последовательность которой по меньшей мере на 80% идентична SEQ ID NO:55;
d) вариабельную область тяжелой цепи, последовательность которой по меньшей мере на 80% идентична SEQ ID NO:57; и
вариабельную область легкой цепи, последовательность которой по меньшей мере на 80% идентична SEQ ID NO:59;
e) вариабельную область тяжелой цепи, последовательность которой по меньшей мере на 80% идентична SEQ ID NO:61; и
вариабельную область легкой цепи, последовательность которой по меньшей мере на 80% идентична SEQ ID NO:63;
f) вариабельную область тяжелой цепи, последовательность которой по меньшей мере на 80% идентична SEQ ID NO:53; и
вариабельную область легкой цепи, последовательность которой по меньшей мере на 80% идентична SEQ ID NO:63.
3. Антитело или его антигенсвязывающий фрагмент по п.1 или 2, содержащее:
a) вариабельную область тяжелой цепи, содержащую SEQ ID NO:45; и вариабельную область легкой цепи, содержащую SEQ ID NO:47;
b) вариабельную область тяжелой цепи, содержащую SEQ ID NO:49; и вариабельную область легкой цепи, содержащую SEQ ID NO:51;
c) вариабельную область тяжелой цепи, содержащую SEQ ID NO:53; и вариабельную область легкой цепи, содержащую SEQ ID NO:55;
d) вариабельную область тяжелой цепи, содержащую SEQ ID NO:57; и вариабельную область легкой цепи, содержащую SEQ ID NO:59;
e) вариабельную область тяжелой цепи, содержащую SEQ ID NO:61; и вариабельную область легкой цепи, содержащую SEQ ID NO:63; или
f) вариабельную область тяжелой цепи, содержащую SEQ ID NO:53; и вариабельную область легкой цепи, содержащую SEQ ID NO:67.
4. Антитело или его антигенсвязывающий фрагмент по любому из предшествующих пунктов, способное к специфическому связыванию человеческого PD-1 с величиной KD не более 10-8 M при измерении в анализе связывания методом поверхностного плазмонного резонанса.
5. Антитело или его антигенсвязывающий фрагмент по любому из предшествующих пунктов, которое связывает обезьяний PD-1 с величиной EC50 не более 100 нМ или не более 10 нМ и/или не связывает мышиный PD-1.
6. Антитело или его антигенсвязывающий фрагмент по любому из предшествующих пунктов, способное ингибировать связывание человеческим или обезьяньим PD-1 его лиганда с величиной IC50 не более 100 нМ.
7. Антитело или его антигенсвязывающий фрагмент по любому из предшествующих пунктов, которое по существу не связывает CD28 или CTLA4.
8. Антитело или его антигенсвязывающий фрагмент по любому из предшествующих пунктов, которое не опосредует ADCC, или CDC, или обе из них.
9. Антитело или его антигенсвязывающий фрагмент по любому из предшествующих пунктов, представляющее собой полностью человеческое моноклональное антитело.
10. Антитело или его антигенсвязывающий фрагмент по п.9, отличающееся тем, что полностью человеческое моноклональное антитело продуцируется в организме трансгенной крысы.
11. Антитело или его антигенсвязывающий фрагмент по любому из предшествующих пунктов, способное блокировать связывание человеческого PD-1 с его лигандом и за счет этого обеспечивать по меньшей мере одну из следующих активностей:
a) индукцию продуцирования IL-2 в CD4+ T-клетках;
b) индукцию продуцирования IFNγ в CD4+ T-клетках;
c) индукцию пролиферации CD4+ T-клеток и
d) реверсию супрессорной функции Treg.
12. Антитело или его антигенсвязывающий фрагмент по любому из предшествующих пунктов, представляющее собой однодоменное антитело верблюдовых, диатело, scFv, димер scFv, BsFv, dsFv, (dsFv)2, dsFv-dsFv', Fv-фрагмент, Fab, Fab', F(ab')2, ds-диатело, нанотело, доменное антитело или двухвалентное доменное антитело.
13. Антитело или его антигенсвязывающий фрагмент по любому из предшествующих пунктов, дополнительно содержащее константную область иммуноглобулина.
14. Конъюгат, содержащий антитело или его антигенсвязывающий фрагмент по любому из предшествующих пунктов и дектируемую метку, фармакокинетическую модифицирующую группу или группу для очистки.
15. Выделенный полинуклеотид, кодирующий антитело или его антигенсвязывающий фрагмент, по любому из пп.1-13.
16. Вектор экспрессии, содержащий выделенный полинуклеотид по п.15.
17. Клетка-хозяин для экспрессии антитела или его антигенсвязывающего фрагмента по любому из пп.1-13, содержащая вектор по п.16.
18. Способ экспрессии антитела или его антигенсвязывающего фрагмента по любому из пп.1-13, включающий культивирование клетки-хозяина по п.17 в условиях, при которых экспрессируется антитело или его антигенсвязывающий фрагмент.
19. Набор для детекции присутствия или уровня PD-1 в биологическом образце, содержащий антитело или его антигенсвязывающий фрагмент по любому из пп.1-13 или конъюгат по п.14 и инструкцию по применению.
20. Способ лечения состояния, ассоциированного с PD-1, у индивидуума, включающий введение индивидууму терапевтически эффективного количества антитела или его антигенсвязывающего фрагмента по любому из пп.1-13 или конъюгата по п.14.
21. Способ по п.25, где индивидуума идентифицируют как имеющего заболевание или состояние, на которое, вероятно, будет оказывать влияние антагонист PD-1.
22. Способ по п.21, где индивидуума идентифицируют как положительного в отношении наличия или повышенных уровней, PD-L1 в тестируемом биологическом образце от индивидуума.
23. Фармацевтическая композиция для лечения заболевания или профилактики состояния или заболевания, связанного с PD-1, или усиление иммунного ответа при котором будет иметь положительный эффект, содержащая терапевтически эффективное количество антитела или его антигенсвязывающего фрагмента по любому из пп.1-13 или конъюгата по п.14 и один или более фармацевтически приемлемых носителей.
24. Способ лечения состояния у индивида, усиление иммунного ответа при котором будет иметь положительный эффект, включающий введение индивиду терапевтически эффективного количества антитела или его антигенсвязывающего фрагмента по любому из пп.1-13 или конъюгата по п.14.
25. Способ по п.24, где индивид имеет повышенную экспрессию PD-L1.
26. Применение антитела или его антигенсвязывающего фрагмента по любому из пп.1-13 или конъюгата по п.14 в производстве лекарственного средства для лечения состояния у индивида, усиление иммунного ответа при котором будет иметь положительный эффект.
27. Применение по п.26, где состояние представляет собой рак или хроническую вирусную инфекцию.
| WO 2013059524 A2, 25.04.2013 | |||
| WO 2004056875 A1, 08.07.2004 | |||
| US 8460886 B2, 11.06.2013 | |||
| WO 2014194302 A2, 04.12.2014 | |||
| МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА ЧЕЛОВЕКА К БЕЛКУ ПРОГРАММИРУЕМОЙ СМЕРТИ 1 (PD-1) И СПОСОБЫ ЛЕЧЕНИЯ РАКА С ИСПОЛЬЗОВАНИЕМ АНТИ-PD-1-АНТИТЕЛ САМОСТОЯТЕЛЬНО ИЛИ В КОМБИНАЦИИ С ДРУГИМИ ИММУНОТЕРАПЕВТИЧЕСКИМИ СРЕДСТВАМИ | 2006 |
|
RU2494107C2 |

