Область техники
[0001] Настоящее изобретение относится, например, к способу получения клеточной популяции, произошедшей от единичной клетки, а именно клонированию, стволовых клеток или соматических клеток, состоящих из различных популяций, посредством использования репрограммирования соматических клеток.
Предшествующий уровень техники
[0002] Клонирование является важным этапом в развитии клеточных линий, и это клонирование стандартно проводили с использованием метода серийных разведений.
[0003] Хотя клетки превращают в желаемый тип клеток посредством введения экзогенного гена внутрь клеток (патентный документ 1, непатентные документы 1 и 2), полученные клетки не являются единообразными вследствие таких факторов, как различия в числе копий введенного экзогенного гена или различия во введенном сайте в хромосоме. Таким образом, хотя считается, что клонирование возможно произвести в соответствии с методом предельных разведений, этот метод предельных разведений необязательно может быть применен ко всем клеткам.
[0004] Кроме того, поскольку клетки, полученные посредством введения экзогенного гена, с низкой вероятностью обеспечивают возможность получить желаемые клетки снова, даже если предпринимаются попытки получить клетки тем же способом, в случае, когда клетки, требуют манипуляций с генами, таких как гомологичная рекомбинация, существуют ограничения клеточных типов, позволяющих произвести эту манипуляцию.
[0005] Хотя была предложена заместительная терапия, которая включает индуцированное превращение в T-лимфоциты из iPS клеток посредством репрограммирования T-лимфоцитов, сохраняющих желаемый TCR тип (патентный документ 2 или непатентный документ 3), эту терапию не проводят в целях клонирования или манипуляций с генами.
Список ссылок
Патентные документы
[0006] Патентный документ 1: WO 2014/148646
Патентный документ 2: WO 2011/096482
Непатентные документы
[0007] Непатентный документ 1: Nakamura S, et al, Cell Stem Cell. 14:535-548, 2014
Непатентный документ 2: Tanaka A, et al, PLoS One. 8:e61540, 2013
Непатентный документ 3: Nishimura T, et al., Cell Stem Cell. 12(1):114-126, 2013
Сущность изобретения
Техническая проблема
[0008] Целью настоящего изобретения является получить клеточную популяцию, происходящую из единичной клетки, а именно клонирование, стволовых клеток или соматических клеток, состоящих из различных популяций.
Решение проблемы
[0009] Когда изобретатели настоящего изобретения выделили стволовые клетки, имеющие ген, связанный с индуцированием дифференцировки в соматические клетки, входящий в состав их хромосомы, из колонии стволовых клеток, полученных посредством одного или множества раундов дедифференцировки, было обнаружено, что выделенные стволовые клетки пригодны для индуцирования дифференцировки в соматические клетки, таким образом, приводя к исполнению настоящего изобретения.
[0010] А именно, настоящее изобретение относится к изобретениям, указанным ниже.
[A1] Способ получения клона стволовых клеток, который содержит этапы:
(i) введения в стволовые клетки экзогенного гена, связанного с индукцией дифференцировки в соматические клетки;
(ii) индукция дифференцировки стволовых клеток, в которые введен экзогенный ген, в соматические клетки;
(iii) дедифференцировка индуцированных к дифференцировке соматических клеток; и
(iv) выделение стволовых клеток, несущих экзогенный ген, введенный в их хромосому, из колонии стволовых клеток, образованных на этапе (iii).
[A2] Способ, описанный в [A1], где эффективность индукции дифференцировки изолированных клонов стволовых клеток в соматические клетки выше по сравнению с индукцией стволовых клеток перед клонированиемм.
[A3] Способ, описанный в [A1] или [A2], где соматическими клетками являются гемопоэтические клетки-предшественники, мегакариоцитарные клетки-предшественники, эритробласты, нервные клетки, нейральные стволовые клетки, клетки нервного гребня, миокардиальные клетки, клетки скелетных мышц, хондроциты, гепатоциты или меланоциты.
[A4] Способ, описанный в [A3], где соматические клетки являются клетками-предшественниками мегакариоцитов, и экзогенный ген, связанный с индукцией дифференцировки, является, по меньшей мере, одним экзогенным геном, выбранным из группы, состоящей из онкогенов, включая гены семейства MYC, гены (поликомб гены), ингибирующие экспрессию гена p16 или гена p19, включая Bmi1, и гены, подавляющие апоптоз, включая ген BCL-XL.
[A5] Способ, описанный в любом из пунктов от [A1] до [A4], где выделяют стволовые клетки, экспрессирующие MEG3.
[A6] Способ, описанный в любом из пунктов от [A1] до [A5], где экзогенный ген, связанный с индукцией дифференцировки, функционально связан с отвечающим на препарат промотором.
[A7] Способ, описанный в любом из пунктов от [A1] до [A6], где дедифференцировку на этапе (iii) проводят посредством введения репрограммирующего фактора, выбранного из группы, состоящей из OCT3/4, SOX2 и KLF4.
[A8] Способ получения соматических клеток, который содержит этап:
индукции дифференцировки клонов стволовых клеток, полученных в соответствии со способом, описанным в любом из пунктов от [A1] до [A7], в соматические клетки.
[A9] Способ получения тромбоцитов, который содержит этапы:
индукции дифференцировки клонов стволовых клеток, полученных в соответствии со способом, описанным в любом из пунктов от [A1] до [A6] в клетки-предшественники мегакариоцитов; и
обеспечения созревания индуцированных к дифференцировке клеток-предшественников мегакариоцитов в мегакариоциты и высвобождения тромбоцитов.
[A10] Способ, описанный в [A9], где у полученных тромбоцитов отсутствует HLA.
[0011] [B1] Способ клонирования соматических клеток, который содержит этапы, указанные ниже, где стволовые клетки получают посредством экспрессии экзогенного гена:
(i) образование колонии стволовых клеток посредством введения репрограммирующего фактора в соматические клетки, несущие экзогенный ген, функционально связанный с отвечающим на препарат промотором, встроенным в их хромосому;
(ii) выделение колонии стволовых клеток, полученных на этапе (i); и
(iii) индуцирование стволовых клеток, содержащихся в колонии стволовых клеток, изолированной на этапе (ii), к дифференцировке в соматические клетки посредством контактирования клеток на любой стадии дифференцировки из стволовых клеток в соматические клетки с соответствующим лекарством.
[B2] Способ, описанный в [B1], где соматические клетки являются клетками-предшественниками мегакариоцитов, и экзогенный ген является, по меньшей мере, одним геном, выбранным из группы, состоящей из генов семейства MYC, генов поликомб и генов, подавляющих апоптоз.
[B3] Способ, описанный в [B2], где этап (iii) содержит этапы:
(a) индуцирования стволовых клеток, содержащихся в колонии стволовых клеток, изолированных на этапе (ii), к дифференцировке в гемопоэтические клетки-предшественники; и
(b) контактирования гемопоэтических клеток-предшественников, полученных на этапе (a), с соответствующим препаратом.
[B4] Способ, описанный в любом из пунктов от [B1] до [B3], где репрограммирующий фактор включает OCT3/4, SOX2 и KLF4.
[B5] Способ, описанный в любом из пунктов от [B1] до [B4], где отвечающий на препарат промотор является промотором, имеющим TRE последовательность, и дополнительно экспрессирует обратный tetR слитый белок, по меньшей мере, в клетках этапа (iii).
[B6] Способ, описанный в любом из пунктов от [B2] до [B5], где этап (iii) дополнительно содержит этап отбора стволовых клеток, экспрессирующих MEG3, среди стволовых клеток, содержащихся в колонии стволовых клеток, изолированной на этапе (ii).
[B7] Способ, описанный в любом из пунктов от [B1] до [B6], где этап (iii) дополнительно содержит этап, приводящий к тому, что у стволовых клеток, содержащихся в колонии стволовых клеток, изолированных на этапе (ii), отсутствует HLA.
[B8] Способ, описанный в [B7], где HLA является антигеном класса I.
[B9] Способ, описанный в [B8], где антигеном класса I является β2-микроглобулин.
[B10] Способ получения тромбоцитов, который содержит этапы:
Клонирования клеток-предшественников мегакариоцитов с применением способа, описанного в любом одном из пунктов от [B2] до [B9]; и
Обеспечения созревания клонированных клеток-предшественников мегакариоцитов в мегакариоциты и высвобождения тромбоцитов.
[B11] Способ получения HLA-дефицитных соматических клеток, который содержит этапы:
(i) образования плюрипотентных стволовых клеток посредством введения репрограммирующего фактора в соматические клетки;
(ii) инициирования того, чтобы плюрипотентные стволовые клетки, полученные на этапе (i), были дефицитными по HLA; и
(iii) индуцирование дифференцировки HLA-дефицитных плюрипотентных стволовых клеток, полученных на этапе (ii), в соматические клетки.
[B12] Способ, описанный в [B11], где репрограммирующий фактор включает OCT3/4, SOX2 и KLF4.
[B13] Способ, описанный в [B11] или [B12], где соматические клетки, применяемые на этапе (i), являются клетками-предшественниками мегакариоцитов, а клетки-предшественники мегакариоцитов являются клетками-предшественниками мегакариоцитов, полученными посредством встраивания, по меньшей мере, одного гена, который функционально связан с отвечающим на препарат промотором и выбран из группы, состоящей из генов семейства MYC, генов поликомб, и генов, подавляющих апоптоз, в их хромосоме.
[B14] Способ, описанный в [B13], где этап (iii) содержит этапы:
(A) индуцирования HLA-дефицитных плюрипотентных стволовых клеток, полученных на этапе (ii), к дифференцировке в гемопоэтические клетки-предшественники; и
(B) контактирования гемопоэтических клеток-предшественников, полученных на этапе (A), с соответствующим препаратом.
[B15] Способ, описанный в любом из пунктов от [B11] до [B14], где HLA является антигеном класса I.
[B16] Способ, описанный в [B15], где антигеном класса I является β2-микроглобулин.
[B17] Способ получения HLA-дефицитных тромбоцитов, который содержит этапы:
получения HLA-дефицитных клеток-предшественников мегакариоцитов с применением способа, описанного в любом из пунктов от [B13] до [B16], и
обеспечения созревания HLA-дефицитных клеток-предшественников мегакариоцитов в мегакариоциты и высвобождения тромбоцитов.
[B18] Клетки iPS, содержащие экзогенный онкоген, и экзогенный ген, который подавляет экспрессию гена p16 или гена p19, где соотношение содержания экзогенного гена, который подавляет экспрессию гена p16 или гена p19, и экзогенного онкогена, составляет от 2-кратного до 7-кратного.
[B19] Клетка iPS, описанная в [B18], где онкогеном является c-Myc, и геном, который подавляет экспрессию гена p16 или гена p19, является Bmi1.
[B20] Клетка-предшественник мегакариоцита, содержащая экзогенный онкоген и экзогенный ген, который подавляет экспрессию гена p16 или гена p19, где соотношение содержания экзогенного гена, который подавляет экспрессию гена p16 или гена p19, и экзогенного онкогена составляет от 2-кратного до 7-кратного.
[B21] Клетка-предшественник мегакариоцита, описанная в [B20], где онкогеном является c-Myc, и геном, который подавляет экспрессию гена p16 или гена p19, является Bmi1.
[B22] Способ отбора плюрипотентных стволовых клеток или гемопоэтических клеток-предшественников, пригодных для индукции дифференцировки клеток-предшественников мегакариоцитов, который содержит этап: отбора плюрипотентных стволовых клеток или гемопоэтических клеток-предшественников, которые экспрсессируют MEG3.
Полезные эффекты по изобретению
[0012] По настоящему изобретению, можно получать клон стволовых клеток, который является пригодным для индукции дифференцировки в соматические клетки. Кроме того, стволовые клетки, клонированные по настоящему изобретению, обладают превосходным пролиферативным потенциалом по сравнению со стволовыми клетками, клонированными в соответствии с общепринятым методом предельных разведений. Например, не только вторичные клоны клеток-предшественников мегакариоцитов, полученные в соответствии с настоящим изобретением, обладают превосходным потенциалом к клеточной пролиферации и способностью созревать в мегакариоциты по сравнению с общепринятыми клонами, мегакариоциты, полученные из этих клонов, обладают высокой способностью образовывать тромбоциты.
Краткое описание рисунков
[0013] Фиг. 1 является графиком, на котором показано число образовавшихся тромбоцитов, полученных из 22 кандидатных клонов клеток-предшественников мегакариоцитов, полученных посредством метода предельных разведений наряду с клетками-предшественниками мегакариоцитов (H+), состоящими из различных исходных популяций (фиг. 1A), и интенсивность флуоресценции вместо количества комплекса GPIIb/IIIa, образованного посредством PMA стимуляции тромбоцитов (фиг. 1B).
Фиг. 2 является графиком, на котором показаны результаты повторного измерения числа образовавшихся тромбоцитов, полученных из пяти из 22 кандидатных клонов клеток-предшественников мегакариоцитов на фиг. 1 наряду с клетками-предшественниками мегакариоцитов (H), состоящими из различных исходных популяций (фиг. 2A), и интенсивность флуоресценции вместо количества комплекса GPIIb/IIIa, образованного посредством PMA стимуляции тромбоцитов (фиг. 2B).
Фиг. 3 является графиком, на котором показаны результаты повторного измерения количеств образовавшихся тромбоцитов, полученных из 5 клонов клеток-предшественников мегакариоцитов на фиг. 2, наряду с исходными клетками-предшественниками мегакариоцитов (H) (фиг. 3A), и интенсивность флуоресценции вместо количества комплекса GPIIb/IIIa, образовавшегося посредством PMA стимуляции тромбоцитов (фиг. 3B).
Фиг. 4A является схематической диаграммой получения клеток-предшественников мегакариоцитов, клонирования посредством репрограммирующих факторов, и получения вторичных клеток-предшественников мегакариоцитов из вторичных iPS клеток. Фиг. 4B является графиком, на котором показано число копий на c-MYC или BMI1 клетку, содержащееся в хромосомах вторичных iPS клеточных клонов, полученных посредством репрограммирования клеток-предшественников мегакариоцитов.
На фиг. 5 показаны результаты применения проточного цитометра для измерения распределения клеток, экспрессирующих CD42b и CD41a, и распределения клеток, экспрессирующих CD235 и CD41a, среди клеток-предшественников мегакариоцитов, полученных из каждого вторичного клона iPS клеток.
Фиг. 6A является схематической диаграммой гомологичной рекомбинации для удаления Экзона1 β2-микроглобулина. На фиг. 6B представлены результаты PCR для подтверждения гомологичной рекомбинации во вторичных клонах iPS клеток после введения таргетного вектора.
На фиг. 7A показаны результаты применения проточного цитометра для измерения распределения клеток, экспрессирующих β2-микроглобулин и HLA в клетках-предшественников мегакариоцитов (imMKCL), полученных из вторичных клонов iPS клеток с удаленным Экзоном1 β2-микроглобулина (представлены слева), и тромбоциты, образовавшиеся из клеток-предшественников мегакариоцитов (представлены справа). На фиг. 7B показано соотношение интенсивности флуоресценции PAC1 после PMA стимуляции в тромбоцитах, образовавшихся из мегакариоцитов, полученных из вторичных клонов iPS клеток (HLA(-)) с удаленным Экзоном1 β2-микроглобулина или вторичных клонов iPS клеток (HLA(+)). Интенсивность флуоресценции представлена на основе значения 1 для тромбоцитов, полученных из вторичных клонов iPS клеток (HLA(+)).
На фиг. 8 указаны результаты сравнения уровней экспрессии гена с применением микрочипа, имеющегося в продаже в Illumina (фиг. 8A) или в Affymetrix (фиг. 8B) для вторичных клонов iPS клеток (хорошие iPS), обладающих способностью быть индуцированными к дифференцировке в клетки-предшественники мегакариоцитов, и вторичных клонов iPS клеток (плохие iPS), не обладающих способностью быть индуцированными к дифференцировке в клетки-предшественники мегакариоцитов, или гемопоэтических клеток-предшественников, полученных из вторичных клонов iPS клеток (хорошие HPC), обладающих способностью быть индуцированными к дифференцировке в клетки-предшественники мегакариоцитов, и вторичных клонов iPS клеток (плохие HPC), не способных быть индуцированными к дифференцировке в клетки-предшественники мегакариоцитов.
На фиг. 9A и 9B, соответственно, показаны кривые роста мегакариоцитов (клон 2), полученных из вторичного клона iPS клеток, и клеток-предшественников мегакариоцитов (родительские) перед клонированием, и время удвоения, рассчитанное на основе кривых роста (**: P<0.01).
На фиг. 10A и 10B, соответственно, показаны внешний вид клеток и изменения размера клеток до и после созревания мегакариоцитов (клон 2), полученных из вторичного клона iPS клеток, и клеток-предшественников мегакариоцитов (родительские) перед клонированием.
Фиг. 11 является графиком, сравнивающим количества протромбоцитов, которые являются прогениторами образовавшихся тромбоцитов.
Описание вариантов осуществления
[0014] Способ получения клона стволовых клеток по настоящему изобретению содержит этапы:
(i) введения в стволовые клетки экзогенного гена, связанного с индукцией дифференцировки в соматические клетки;
(ii) индукции дифференцировки стволовых клеток, в которые введен экзогенный ген, в соматические клетки;
(iii) дедифференцировки индуцированных к дифференцировке соматических клеток; и
(iv) выделения стволовых клеток, несущих экзогенный ген, встроенный в их хромосому, из колонии стволовых клеток, образовавшихся на этапе (iii).
[0015] Изолированные клоны стволовых клеток являются более пригодными для того, чтобы быть индуцированными к дифференцировке в соматические клетки по сравнению со стволовыми клетками перед клонированием, и имеют высокую эффективность, чтобы быть индуцированными к дифференцировке в соматические клетки, в пересчете на клетку.
[0016] В одном из вариантов осуществления, способ получения клона стволовых клеток по настоящему изобретению может включать этапы:
(i) образования колонии стволовых клеток посредством введения репрограммирующего фактора в соматические клетки, несущие экзогенный ген, функционально связанный с отвечающим на препарат промотором, встроенным в их хромосому; и
(ii) выделения колонии стволовых клеток, полученных на этапе (i).
[0017] Полученный клон стволовых клеток может быть дополнительно индуцирован к дифференцировке в соматическую клетку. Индукцию дифференцировки может проводить специалист в данной области посредством подходящего отбора способа, пригодного для индукции дифференцировки в желаемую соматическую клетку, и она может не быть ограничена конкретным способом, и может дополнительно включать этап:
(iii) индуцирования стволовых клеток, содержащихся в колонии стволовых клеток, изолированных на этапе (ii), к дифференцировке в соматические клетки посредством контакта клеток на любой стадии дифференцировки из стволовых клеток в соматические клетки с применением соответствующего препарата.
[0018] В настоящем изобретении клонирование относится к клонированию клеточной популяции в смысле выделения клеточной популяции, имеющей однообразную генетическую информацию, из клеточной популяции, имеющей не однообразную генетическую информацию.
[0019] Соматические клетки, представленные для клонирования
В настоящем изобретении нет никаких конкретных ограничений в отношении соматических клеток, представленных для клонирования (обозначаемых как первичные соматические клетки), в случаях, когда клетки получены посредством встраивания гена, функционально связанного с отвечающим на препарат промотором, в их хромосому, и их примеры включают нервные клетки (WO 2014/148646, Wapinski OL et al, Cell. 155:621-635, 2013), нейральные стволовые клетки (Han DW et al, Cell Stem Cell. 10:465-472, 2012), клетки нервного гребня (Kim YJ, et al, Cell Stem Cell. 15:497-506, 2014), миокардиальные клетки (Ieda M et al, Cell. 142:375-386, 2010), клетки скелетных мышц (Tanaka A, et al, PLoS One. 8:e61540, 2013), хондроциты (Outani H, et al, PLoS One. 8:e77365, 2013), гепатоциты (Huang P, et al, Cell Stem Cell. 14:370-384, 2014), меланоциты (Yang R, et al, Nat Commun. 5:5807, 2014), гемопоэтические клетки-предшественники (Batta K, Cell Rep. 9:1871-84, 2014), эритробласты (Hirose S, et al, Stem Cell Reports. 1:499-508, 2013) и клетки-предшественники мегакариоцитов (Nakamura S, et al, Cell Stem Cell. 14:535-548, 2014).
[0020] В настоящем изобретении, клетки-предшественники мегакариоцитов являются пригодными в качестве соматических клеток, клонированных в соответствии со способом по настоящему изобретению, ввиду того, что они не могут быть клонированными посредством метода предельных разведений. Эритробласты также являются пригодными в качестве соматических клеток в настоящем изобретении. Однако, ввиду того, что соматические клетки, отличные от этих клеток, также позволяют получить клоны стволовых клеток, несущие экзогенный ген, связанный с индукцией дифференцировки в желаемые соматические клетки, введенный в их хромосому, соматические клетки не ограничены клетками-предшественниками мегакариоцитов и эритробластами.
[0021] Экзогенный ген, связанный с индукцией дифференцировки в соматические клетки, в настоящем изобретении относится в широком смысле к гену, введенному в клетку, при индукции дифференцировки из стволовой клетки в соматическую клетку. Объясняя это, ген, при использовании в качестве примера случая соматических клеток, являющихся клетками-предшественниками мегакариоцитов, ген, связанный с индукцией дифференцировки, может быть, по меньшей мере, одним геном, выбранным из группы, состоящей из онкогенов, предпочтительно членом семейства генов MYC и более предпочтительно c-Myc, генами, подавляющими экспрессию гена p16 или гена p19 (гены поликомб) и предпочтительно Bmi1, и генами, подавляющими апоптоз, и предпочтительно геном BCL-XL. При использовании в качестве примера случая соматических клеток, являющихся эритробластами, ген, связанный с индукцией дифференцировки, может быть, по меньшей мере, одним геном, выбранным из группы, состоящей из онкогенов, предпочтительно членом семейства генов MYC и более предпочтительно c-Myc, и генами, подавляющими апоптоз, и предпочтительно геном BCL-XL. Экзогенный ген, связанный с индукцией дифференцировки в соматические клетки, может быть функционально связанным с отвечающим на препарат промотором.
[0022] В настоящем изобретении, отвечающий на препарат промотор относится к промотору, который экспрессирует ген в присутствии или отсутствии соответствующего препарата. Примером промотора, который экспрессирует ген в присутствии соответствующего препарата, является TRE промотор (CMV минимальный промотор, несущий Tet отвечающую последовательность, включая семь повторов tet0 последовательности). В случае использования TRE промотора, предпочтительно применяют систему, в которой экспрессия гена индуцирована в присутствии соответствующего препарата (такого как тетрациклин или доксициклин) посредством одновременно экспрессирующегося слитого белка (обратный tetR слитый белок) обратного tetR (rtetR) и VP16AD в тех же клетках. В случае использования обратного tetR слитого белка, необходимо, чтобы слитый белок, по меньшей мере, экспрессировался на "этапе индукции дифференцировки из стволовых клеток во вторичные соматические клетки", которые будут описаны далее. Например, посредством функционально связывающего гена, кодирующего обратный tetR слитый белок к отвечающему на препарат промотору и введения этого гена при получении первичных соматических клеток, обратный tetR слитый белок можно экспрессировать посредством добавления или удаления соответствующего препарата на "этапе индукции дифференцировки из стволовых клеток во вторичные соматические клетки". В случае функционально связанного отвечающего на препарат промотора с двумя или более типами генов, тот же тип отвечающего на препарат промотора можно использовать для всех генов или можно использовать два или более типов отвечающих на препарат промоторов.
[0023] Способ получения первичных клеток-предшественников мегакариоцитов
[0024] В настоящем изобретении, "клетки-предшественники мегакариоцитов" относятся к клеткам, которые становятся мегакариоцитами в результате созревания. Эти клетки не являются многоядерными, и включают клетки, характеризующиеся как CD41a-положительные/CD42a-положительные/CD42b-слабо положительные. Клетки-предшественники мегакариоцитов по настоящему изобретению являются предпочтительно клетками, которые можно выращивать посредством экспансивного культивирования, такими, как клетки, способные претерпевать экспансивное культивирование в течение, по меньшей мере, 60 дней при подходящих условиях. В настоящем изобретении, клетки-предшественники мегакариоцитов могут или могут не быть клонированными, и хотя нет конкретных ограничений относительно этого, те, которые были клонированы, обозначаются как клеточная линия предшественников мегакариоцитов. Клетки-предшественники мегакариоцитов в настоящем изобретении могут быть получены из гемопоэтических клеток-предшественников.
[0025] В настоящем изобретении, "мегакариоциты", которые также обозначают как клетки-предшественники и мегакариоцитарные клетки, являются клетками, которые продуцируют тромбоциты посредством отделения от их цитоплазмы, могут быть мультиядросодержащими клетками, и включают клетки, характеризующиеся как, например, CD41a-положительные/CD42a-положительные/CD42b-положительные. Кроме того, мегакариоциты могут также быть охарактеризованы как клетки, экспрессирующие GATA1, FOG1, NF-E2 и β1-тубулин. Многоядерные мегакариоциты относятся к клеткам или группе клеток, в которых число ядер претерпело относительное увеличение по сравнению с клетками-предшественниками мегакариоцитов. Например, в случае когда ядра клеток-предшественников мегакариоцитов, к которым применяют способ по настоящему изобретению, представляют собой 2N, клетки, в которых ядра представляют собой 4N или более, являются многоядерными мегакариоцитами. Кроме того, в настоящем изобретении, мегакариоциты могут быть иммортализованы в форме мегакариоцитарной клеточной линии или могут быть клонированной группой клеток.
[0026] В настоящем изобретении, гемопоэтические клетки-предшественники (HPC) относятся к клеткам, способным дифференцироваться в клетки крови, такие как лимфоциты, эозинофилы, нейтрофилы, базофилы, эритроциты или мегакариоциты, и в настоящем изобретении, нет разделения между гемопоэтическими клетками-предшественниками и гемопоэтическими стволовыми клетками, и это относится к одним и тем же клеткам, если не указано иначе. Гемопоэтические стволовые клетки/клетки-предшественники могут быть распознаны посредством, например, положительного признака для поверхностных антигенов, CD34 и/или CD43. В настоящем изобретении, гемопоэтические стволовые клетки также могут быть применены к гемопоэтическим клеткам-предшественникам, которые были индуцированы к дифференцировке из плюрипотентных стволовых клеток или гемопоэтических стволовых клеток, а также клеток-предшественников, полученных из плацентарной крови, крови костного мозга или периферической крови. Например, в случае применения плюрипотентных стволовых клеток, гемопоэтические клетки-предшественники можно получать из сетеподобной структуры (ES-мешок или iPS-мешок), полученной посредством культивирования плюрипотентных стволовых клеток на C3H10T1/2 в присутствии VEGF в соответствии со способом, описанным в Takayama N., et al. J Exp Med. 2817-2830 (2010). Здесь, "сетеподобная структура" относится к трехмерной мешкоподобной структуре (имеющей пространство внутри), полученной из плюрипотентных стволовых клеток, которая образована посредством популяции эндотелиальных клеток или подобных клеток и содержит гемопоэтические клетки-предшественники в своей внутренней части. Другие примеры способов, применяемых для индукции дифференцировки из плюрипотентных стволовых клеток в гемопоэтические клетки-предшественники, включают способ, в котором применяют образование эмбриоидного тела и добавление цитокина (Chadwick et al. Blood 2003, 102: 906-15, Vijayaragavan et al. Cell Stem Cell 2009, 4: 248-62, Saeki et al. Stem Cells 2009, 27: 59-67), и способ, включающий со-культивирование со стромальными клетками, полученными из различных видов (Niwa et al. J Cell Physiol. 2009 Nov;221(2):367-77.).
[0027] Примеры плюрипотентных стволовых клеток включают оплодотворенные яйцеклетки и такие клетки, как эмбриональные стволовые клетки (ES клетки), индуцированные плюрипотентные стволовые клетки (iPS клетки) или эмбриональные половые клетки (EG клетки). Клетки-предшественники мегакариоцитов, служащие в качестве соматических клеток, по настоящему изобретению являются предпочтительно теми, которые были индуцированы посредством этапа культивирования клеток посредством сверхэкспрессии онкогена, гена, подавляющего экспрессию гена p16 или ген p19 (поликомб ген), и/или гена, подавляющего апоптоз в гемопоэтических клетках-предшественниках.
[0028] В настоящем изобретении, "онкоген" относится к гену, который вызывает злокачественную трансформацию нормальных клеток в результате экспрессии, их структура или функция, являющиеся отличными от структуры и функции нормальных клеток, и их примеры включают гены семейства MYC, гены семейства Src, гены семейства Ras, гены семейства Raf, гены семейства c-Kit и протеинкиназ, такие как PDGFR или Abl. Примеры генов семейства MYC включают c-MYC, N-MYC и L-MYC. Гены c-MYC относятся, например, к генам, состоящим из последовательности нуклеиновой кислоты, представленной в NCBI, номер доступа. NM 002467. Кроме того, c-MYC гены включают их гомологи, и гомологи генов c-MYC относятся к генам, для которых, например, их cDNA последовательность состоит из последовательности, которая является по существу идентичной последовательности нуклеиновой кислоты, представленной в NCBI, номер доступа NM 002467. cDNA, состоящая из последовательности, по существу идентичной последовательности нуклеиновой кислоты, представленной в NCBI, номер доступа NM 002467, относится к DNA, состоящей из последовательности, имеющей идентификацию приблизительно 60% или более, предпочтительно приблизительно 70% или более, более предпочтительно приблизительно 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97% или 98%, и наиболее предпочтительно приблизительно 99%, с DNA, состоящей из последовательности, представленной в NCBI, номер доступа NM 002467, или DNA, способной гибридизоваться при жестких условиях с DNA, состоящей из последовательности, комплементарной последовательности нуклеиновой кислоты, представленной в NCBI, номер доступа NM 002467, с белком, кодируемым этими DNA, способствуя экспансии клеток, таких как гемопоэтические клетки-предшественники, которые находятся на стадии дифференцировки.
[0029] Здесь, жесткие условия относятся к условиям гибридизации, легко определяемым специалистом в данной области, которые, как правило, являются эмпирическими экспериментальными условиями, зависящими от длины зонда, температуры отмывки и концентрации соли. В основном, температура для надлежащего отжига становится выше с увеличением длины зонда, и температура становится ниже с уменьшением длины зонда. Образование гибрида, как правило, зависит от способности комплементарной цепи претерпевать повторный отжиг во внешней среде при температуре немного ниже, чем температура их плавления.
[0030] Например, пример в незначительной степени жестких условий включает отмывку при 0,1 х SSC в 0,1% растворе SDS при температурных условиях от 37°C до 42°C во время стадии отмывки фильтра после гибридизации. Кроме того, пример в высокой степени жестких условий включает отмывку при 5 х SSC в 0,1% растворе SDS при 65°C во время стадии отмывки. Полинуклеотиды более высокой гомологии можно получать посредством использования в более высокой степени жестких условий.
[0031] В настоящем изобретении, поскольку является предпочтительным подавлять уровень экспрессии c-MYC, c-MYC может быть таким, который кодирует белок, слитый с дестабилизирующим доменом. Можно использовать дестабилизирующий домен, приобретенный в ProteoTuner или Clontech Laboratories, Inc.
[0032] В настоящем изобретении, примеры «генов, подавляющих экспрессию гена p16 или ген p19» включают BMI1, Id1, Mel18, Ring1a/b, Phc1/2/3, Cbx2/4/6/7/8, Ezh2, Eed, Suz12, HDAC и Dnmt1/3a/3b. Ген BMI1 относится, например, к гену, состоящему из последовательности нуклеиновой кислоты, представленной в NCBI, номер доступа NM 005180. Кроме того, ген BMI1 включает его гомологи, и гомологи гена BMI1 относятся к генам, для которых их последовательность cDNA состоит из последовательности по существу идентичной последовательности нуклеиновой кислоты, представленной в NCBI, номер доступа NM 005180. cDNA, состоящая из последовательности, по существу идентичной последовательности нуклеиновой кислоты, представленной в NCBI, номер доступа NM 005180, относится к DNA, состоящей из последовательности, имеющей идентификацию приблизительно 60% или более, предпочтительно приблизительно 70% или более, более предпочтительно приблизительно 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97% или 98%, и наиболее предпочтительно приблизительно 99%, с DNA, состоящей из последовательности, представленной в NCBI, номер доступа NM 005180, или DNA, способной гибридизоваться при жестких условиях с DNA, состоящей из последовательности, комплементарной последовательности нуклеиновой кислоты, представленной в NCBI, номер доступа NM 005180, с белком, кодируемым DNA, стимулирующим экспансию клеток посредством подавления старения клеток, способных к индуцированию онкогенов, присутствующих в клетках, которые экспрессируются онкогенами, такими как гены семейства MYC.
[0033] В настоящем изобретении, "гены, подавляющие апоптоз" относятся к генам, которые подавляют апоптоз, в отношении них нет никаких ограничений, и их примеры включают ген BCL2 ген, ген BCL-XL, сурвивин и MCL1. Ген BCL-XL относится к гену, состоящему из последовательности нуклеиновой кислоты, представленной в NCBI, номер доступа NM 001191 или NM 138578. Кроме того, ген BCL-XL включает их гомологи, и гомологи гена BCL-XL относятся к генам, для которых, например, их cDNA последовательность состоит из последовательности, которая по существу идентична последовательности нуклеиновой кислоты, представленной в NCBI, номер доступа NM 001191 или NM 138578. cDNA, состоящая из последовательности, по существу идентичной последовательности нуклеиновой кислоты, представленной в NCBI, номер доступа NM 001191 или NM 138578 относится к DNA, состоящей из последовательности, имеющей идентификацию приблизительно 60% или более, предпочтительно приблизительно 70% или более, более предпочтительно приблизительно 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97% или 98%, и наиболее предпочтительно приблизительно 99%, с DNA, состоящей из последовательности, представленной в NCBI, номер доступа NM 001191 или NM 138578, или DNA, способной гибридизоваться при жестких условиях с DNA, состоящей из последовательности, комплементарной последовательности нуклеиновой кислоты, представленной в NCBI, номер доступа NM 001191 или NM 138578, с белком, кодируемым этой DNA, обладающим эффектом подавления апоптоза.
[0034] В настоящем изобретении, способ для сверхэкспрессирующих указанных выше генов в гемопоэтических клетках-предшественниках предпочтительно реализован посредством встраивания гена, функционально связанного с отвечающим на препарат промотором в его хромосоме, и может быть достигнут посредством, например, введения экспрессирующего вектора, содержащего ген, функционально связанный с отвечающим на препарат промотором, в гемопоэтическую клетку-предшественник. Примеры векторов, экспрессирующих эти гены, которые можно использовать, включают ретровирус, лентивирус и другие вирусные векторы, а также экспрессионные плазмиды животных клеток (такие как pA1-11, pXT1, pRc/смV, pRc/RSV или pcDNAI/Neo). Ретровирусные векторы или лентивирусные векторы применяют предпочтительно с точки зрения встраивания в хромосому.
[0035] В дополнение к промотору, экспрессирующий вектор может содержать энхансер, дополнительный сигнал поли(A), селективный маркерный ген или SV40 источник репликации и т.п. Примеры пригодных селективных маркерных генов включают ген дигидрофолат редуктазы, ген устойчивости к неомицину и ген устойчивости к пуромицину.
[0036] В настоящем изобретении, полицистронный вектор, в котором гены соединены в продольном направлении, можно получать, чтобы ввести множество генов одновременно. Чтобы обеспечить полицистронную экспрессию, последовательность 2A саморасщепляющегося пептида вируса ног и рта (ссылка, например, на Science, 322, 949-953, 2008), или IRES (внутренний участок связывания рибосомы), может быть пришит между множеством сверхэкспрессированных генов.
[0037] В настоящем изобретении, в случае вирусного вектора, способ введения экспрессирующего вектора в гемопоэтические клетки-предшественники можно производить посредством введения плазмиды, содержащей нуклеиновую кислоту, в подходящие упаковывающие клетки (такие как Plat-E клетки) или дополняющую клеточную линию (такую как клетки 293) с последующим восстановлением вируса, продуцированного в супернатант клеточной культуры, и инфицированием гемопоэтических клеток-предшественников посредством контакта с вирусом. В случае невирусного вектора, плазмидный вектор может быть введен в клетки посредством использования такого способа, как липофекция, липосомальный способ, электропорация, co-осаждение фосфата кальция, DEAE-декстран способ, микроинъекция или генная пушка.
[0038] В одном из аспектов способа для индуцирования клеток-предшественников мегакариоцитов по настоящему изобретению, ген, подавляющий апоптоз, может быть сверхэкспрессирован после сверхэкспрессирования онкогена или гена, подавляющего экспрессию гена p16 или гена p19 в гемопоэтических клетках-предшественниках. Сверхэкспрессию гена, подавляющего апоптоз, можно проводить таким же образом, как описано выше, посредством введения экспрессионного вектора, белка, кодируемого этими генами, или RNA, кодирующей эти гены, в гемопоэтические клетки-предшественники. В случае последующей экспрессии гена, подавляющего апоптоз, хотя в отношении него нет никаких ограничений, сверхэкспрессию гена, подавляющего апоптоз, предпочтительно производят после сверхэкспрессии онкогена или гена, подавляющего экспрессию гена p16 или гена p19 в течение, по меньшей мере, 14 дней.
[0039] В настоящем изобретении, ингибитор каспазы можно приводить в контакт с гемопоэтическими клетками-предшественниками вместо сверхэкспрессирующего гена, подавляющего апоптоз, в клетках. В настоящем изобретении, ингибитор каспазы может быть любым пептидным соединением, непептидным соединением или биологическим белком. Примеры пептидных соединений включают искусственные химически синтезированные пептидные соединения, указанные в пунктах от (1) до (10) ниже.
(1) Z-Asp-CH2-DCB (молекулярная масса: 454.26)
(2) Boc-Asp(OMe)-FMK (молекулярная масса: 263.3)
(3) Boc-Asp(OBzl)-CMK (молекулярная масса: 355.8)
(4) Ac-AAVALLPAVLLALLAP-YVAD-CHO (молекулярная масса: 1990.5) (SEQ ID NO: 1)
(5) Ac-AAVALLPAVLLALLAP-DEVD-CHO (молекулярная масса: 2000.4) (SEQ ID NO: 2)
(6) Ac-AAVALLPAVLLALLAP-LEVD-CHO (молекулярная масса: 1998.5) (SEQ ID NO: 3)
(7) Ac-AAVALLPAVLLALLAP-IETD-CHO (молекулярная масса: 2000.5) (SEQ ID NO: 4)
(8) Ac-AAVALLPAVLLALLAP-LEHD-CHO (молекулярная масса: 2036.5) (SEQ ID NO: 5)
(9) Z-DEVD-FMK (Z-Asp-Glu-Val-Asp-фторметилкетон) (SEQ ID NO: 6)
(10) Z-VAD FMK
[0040] Примеры ингибиторов каспаз пептидных соединений включают: (1) VX-740 - Vertex Pharmaceuticals (Leung-Toung et al., Curr. Med. Chem. 9, 979-1002 (2002)) и (2) HMR-3480 - Aventis Pharma AG (Randle et al., Expert Opin. Investig. Drugs 10, 1207-1209 (2001)).
[0041] Примеры ингибиторов каспаз непептидных соединений включают: (1) анилинохиназолины (AQZs), AstraZeneca Pharmaceuticals (Scott et al., J. Pharmacol. Exp. Ther. 304, 433-440 (2003)), (2) M826 - Merck Frosst (Han et al., J. Biol. Chem. 277, 30128-30136 (2002)), (3) M867 - Merck Frosst (Methot et al., J. Exp. Med. 199, 199-207 (2004)), и (4) никотинил аспартил кетоны - Merck Frosst (Isabel et al., Bioorg. Med. Chem. Lett. 13, 2137-2140 (2003)).
[0042] Кроме того, примеры ингибиторов каспаз других непептидных соединений включают: (1) IDN-6556 - Idun Pharmaceuticals (Hoglen et al., J. Pharmacol. Exp. Ther. 309, 634-640 (2004)), (2) MF-286 и MF-867 - Merck Frosst (Los et al., Drug Discov. Today 8, 67-77 (2003)), (3) IDN-5370 - Idun Pharmaceuticals (Deckwerth et al., Препарат Dev. Res. 52, 579-586 (2001)), (4) IDN-1965 - Idun Pharmaceuticals (Hoglen et al., J. Pharmacol. Exp. Ther. 297, 811-818 (2001)), и (5) VX-799 - Vertex Pharmaceuticals (Los et al., Drug Discov. Today 8, 67-77 (2003)). Другие примеры ингибиторов каспаз включают M-920 и M-791 - Merck Frosst (Hotchkiss et al., Nat. Immunol. 1, 496-501 (2000)).
[0043] В настоящем изобретении, ингибитором каспазы является предпочтительно Z-VAD FMK. В случае применения Z-VAD FMK для ингибитора каспазы, Z-VAD FMK добавляют к среде, в которой культивируют гемопоэтические клетки-предшественники. Предпочтительная концентрация Z-VAD FMK в среде составляет, например, 10 мкM или более, 20 мкM или более, 30 мкM или более, 40 мкM или более или 50 мкM или более, и составляет предпочтительно 30 мкM или более.
[0044] Хотя в этом отношении нет каких-либо ограничений, среду, применяемую для получения клеток-предшественников мегакариоцитов из гемопоэтических клеток-предшественников, можно получать посредством использования среды, применяемой для культивирования животных клеток, в качестве минимальной среды. Примеры минимальных сред включают среду IMDM, среду 199, минимальную поддерживающую среду Игла (EMEM), среду αMEM, модифицированную Дульбекко среду Игла (DMEM), среду Хэма F12, среду RPMI 1640, среду Фишера, нейроминимальную среду (Life Technologies) и их смешенную среду. Среда может содержать сыворотку или может быть свободной от сыворотки. Среда также может содержать одно или несколько веществ, таких как альбумин, инсулин, трансферрин, селен, жирные кислоты, микроэлементы, 2-меркаптоэтанол, тиол глицерин, липид, аминокислоты, L-глутамин, заменимые аминокислоты, витамины, факторы роста, соединения с низкой молекулярной массой, антибиотики, антиоксиданты, пировиноградную кислоту, буферы, неорганические соли или цитокины при необходимости. Цитокины относятся к белкам, которые стимулируют гемопоэтическую дифференцировку, и их примеры включают VEGF, TPO и SСМ. Предпочтительной средой в настоящем изобретении является среда IMDM, содержащая сыворотку, инсулин, трансферрин, серин, тиол глицерин, аскорбиновую кислоту и TPO. Среда более предпочтительно дополнительно содержит SСМ. Кроме того, в случае применения экспрессионного вектора, содержащего отвечающий на препарат промотор, соответствующий препарат, такой как тетрациклин или доксициклин, предпочтительно содержится в среде на этапе сверхэкспрессии.
[0045] В настоящем изобретении, хотя в данном отношении нет каких-либо ограничений, температурные условия для получения клеток-предшественников мегакариоцитов из гемопоэтических клеток-предшественников являются такими, что стимуляция дифференцировки в клетки-предшественники мегакариоцитов поддерживается посредством культивирования гемопоэтических клеток-предшественников при температуре 37°C или выше. Здесь, поскольку температура, которая не оказывает повреждающего действия на клетки является подходящей, температура 37°C или выше относится, например, к температуре от приблизительно 37°C до приблизительно 42°C и предпочтительно к температуре от приблизительно 37°C до приблизительно 39°C. Продолжительность культивирования при температуре 37°C или выше может быть подходящим образом определена специалистом в данной области во время мониторинга таких факторов как число клеток-предшественников мегакариоцитов. Хотя нет каких-либо конкретных ограничений на эту продолжительность, получают желаемое количество клеток-предшественников мегакариоцитов, ее примеры включают продолжительность, по меньшей мере, 6 дней или более, 12 дней или более, 18 дней или более, 24 дня или более, 30 дней или более, 42 дня или более, 48 дней или более, 54 дня или более или 60 дней или более, и предпочтительно 60 дней или более. Длинный период культивирования не представляет проблемы в отношении индукции клеток-предшественников мегакариоцитов. Кроме того, предпочтительным является подходящим образом производить субкультивирование во время периода культивирования.
[0046] Способ репрограммирования соматических клеток
В настоящем изобретении, введение репрограммирующего фактора в соматические клетки можно проводить для способа, применяемого для репрограммирования соматических клеток. Здесь, примеры репрограммирующих факторов включают гены или продукты генов, такие как Oct3/4, Sox2, Sox1, Sox3, Sox15, Sox17, Klf4, Klf2, c-Myc, N-Myc, L-Myc, Nanog, Lin28, Fbx15, ERas, ECAT15-2, Tcl1, бета-катенин, Lin28b, Sall1, Sall4, Esrrb, Nr5a2, Tbx3 или Glis1, и эти репрограммирующие факторы можно использовать в отдельности или в комбинации. Примеры комбинаций репрограммирующих факторов включают комбинации, описанные в WO 2007/069666, WO 2008/118820, WO 2009/007852, WO 2009/032194, WO 2009/058413, WO 2009/057831, WO 2009/075119, WO 2009/079007, WO 2009/091659, WO 2009/101084, WO 2009/101407, WO 2009/102983, WO 2009/114949, WO 2009/117439, WO 2009/126250, WO 2009/126251, WO 2009/126655, WO 2009/157593, WO 2010/009015, WO 2010/033906, WO 2010/033920, WO 2010/042800, WO 2010/050626, WO 2010/056831, WO 2010/068955, WO 2010/098419, WO 2010/102267, WO 2010/111409, WO 2010/111422, WO 2010/115050, WO 2010/124290, WO 2010/147395, WO 2010/147612, Huangfu D, et al. (2008), Nat. Biotechnol., 26: 795-797, Shi Y, et al. (2008), Cell Stem Cell, 2: 525-528, Eminli S, et al. (2008), Stem Cells. 26:2467-2474, Huangfu D, et al. (2008), Nat. Biotechnol. 26:1269-1275, Shi Y, et al. (2008), Cell Stem Cell, 3, 568-574, Zhao Y, et al. (2008), Cell Stem Cell, 3:475-479, Marson A, (2008), Cell Stem Cell, 3, 132-135, Feng B, et al. (2009), Nat. Cell Biol. 11:197-203, R. L. Judson et al., (2009), Nat. Biotechnol., 27:459-461, Lyssiotis CA, et al. (2009), Proc Natl Acad Sci USA. 106:8912-8917, Kim JB, et al. (2009), Nature. 461:649-643, Ichida JK, et al. (2009), Cell Stem Cell. 5:491-503, Heng JC, et al. (2010), Cell Stem Cell. 6:167-74, Han J, et al. (2010), Nature. 463:1096-100, Mali P, et al. (2010), Stem Cells. 28:713-720 и Maekawa M, et al. (2011), Nature. 474:225-9. Более предпочтительная комбинация репрограммирующих факторов включает Oct3/4, Sox2 и Klf4.
[0047] Указанные выше репрограммирующие факторы содержат факторы, применяемые с целью усилить установление эффективности, такие как ингибиторы гистон деацетилазы (HDAC) (такие как низкомолекулярные ингибиторы наподобие вальпроевой кислоты (VPA), трихостатина A, бутирата натрия, MC 1293 или M344, ингибиторы экспрессии нуклеиновых кислот, такие как siRNA и кшРНК против HDAC (например, HDAC1 siRNA Smartpool® (Millipore), HuSH 29mer кшРНК конструкты против HDAC1 (Ori ген)), ингибиторы MEK (такие как PD184352, PD98059, U0126, SL327 или PD0325901), ингибиторы гликоген синтаза киназы-3 (такие как Bio или CHIR99021), ингибиторы DNA метил трансферазы (такие как 5-азацитидин), ингибиторы гистон метил трансферазы (такие как низкомолекулярные ингибиторы наподобие BIX-01294 или ингибиторы экспрессии нуклеиновых кислот наподобие siRNA и кшРНК против Suv39hl, Suv39h2, SetDBl или G9a), агонисты кальциевых L-каналов (такие как Bayk8644), масляная кислота, ингибиторы TGFβ или ингибиторы ALK5 (такие как LY364947, SB431542, 616453 или A-83-01), ингибиторы p53 (такие как siRNA и кшРНК против p53), ингибиторы ARID3A (такие как siRNA и кшРНК против ARID3A), miRNA, такие как miR-291-3p, miR-294, miR-295 или miR-302), Wnt сигналинг (такой как растворимые Wnt3a), нейропептид Y, простагландины (такие как простагландин E2 или простагландин J2), hTERT, SV40LT, UTF1, IRX6, GLISl, PITX2 или DMRTBl, и в настоящем описании не делают конкретных различий между репрограммирующими факторами, и эти факторы, применяемые с целью улучшить установленную эффективность.
[0048] В случае, когда репрограммирующий фактор представляет собой белок, репрограммирующий фактор может быть введен в соматические клетки посредством такого метода, как липофекция, слияние с проникающим в клетку пептидом (таким как TAT или полиаргинин, полученные из HIV), или микроинъекции.
[0049] С другой стороны, в случае, когда репрограммирующий фактор представляет собой DNA, DNA может быть введено в соматические клетки посредством вектора наподобие вируса, плазмиды или искусственной хромосомы, и посредством таких методов, как липофекция, липосомы или микроинъекция. Примеры вирусные векторы включают ретровирусный вектор, лентивирусный вектор (описан в Cell, 126, pp. 663-676, 2006; Cell, 131, pp. 861-872, 2007; Science, 318, pp. 1917-1920, 2007), аденовирусный вектор (Science, 322, 945-949, 2008), аденоассоциированный вирусный вектор и вирусный вектор Сендай (WO 2010/008054). Кроме того, примеры искусственных хромосомных векторов включают искусственные хромосомы человека (HAC), искусственные дрожжевые хромосомы (YAC) и бактериальные искусственные хромосомы (BAC, PAC). В качестве плазмид можно использовать плазмиды клеток млекопитающих (Science, 322:949-953, 2008). Векторы могут содержать контрольную последовательность, такую как промотор, энхансер, последовательность связывания рибосом, терминатор или сайт полиаденилирования, чтобы обеспечить экспрессию ядерного репрограммирующего вещества, и могут дополнительно содержать ген устойчивости к препарату (такой как ген устойчивости к канамицину, ген устойчивости к ампициллину или ген устойчивости к пуромицину), последовательность селективного маркера, такую как ген тимидин киназы или ген дифтерийного токсина, или ген репортерной последовательности, такие как зеленый флуоресцентный белок (GFP), β-глюкуронидаза (GUS) или FLAG при необходимости. Кроме того, указанные выше векторы могут иметь LoxP последовательность до или после вектора, чтобы удалить гены, кодирующие репрограммирующие факторы, или и промотор, и ген, кодирующий репрограммирующий фактор, связанный с ним, после введения в соматические клетки.
[0050] В случае, когда репрограммирующий фактор находится в форме RNA, репрограммирующий фактор может быть введен в соматические клетки посредством метода, такого как липофекция или микроинъекция, и можно использовать RNA с встроенными 5-метилцитидином и псевдоуридином (TriLink Biotechnologies), чтобы ингибировать деградацию (Warren L, (2010) Cell Stem Cell. 7:618-630).
[0051] Примеры культуральной жидкости для клеток после репрограммирования включают DMEM, содержащую от 10% до 15% FBS, DMEM/F12 и DME культуральную жидкость (и эти культуральные жидкости могут соответствующим образом дополнительно содержать LIF, пенициллин/стрептомицин, пуромицин, L-глутамин, заменимые аминокислоты или β-меркаптоэтанол и т.п.), а также коммерчески доступные культуральные жидкости (такие как культуральная жидкость для культивирования ES клеток мыши (TX-WES культуральная жидкость, Thrombo-X), культуральная жидкость для культивирования ES клеток приматов (культуральная жидкость ES приматов/iPS клеток, ReproCELL Inc.), или среда, свободная от сыворотки (mTeSR, Stemcell Technology)).
[0052] Пример способа для культивирования клеток после репрограммирования содержит контактирование соматических клеток с репрограммирующим фактором в DMEM, содержащей 10% FBS или DMEM/F12 культуральной жидкости при 37°C в присутствии 5% CO2 и культивирование в течение приблизительно от 4 дней до 7 дней, с последующим пересевом клеток на фидерные клетки (такие как STO клетки или SNL клетки, обработанные митомицином C), и культивирование в культуральной жидкости для культивирования ES клеток приматов, содержащей bFGF, начиная через приблизительно 10 дней после контактирования соматических клеток с репрограммирующим фактором, чтобы обеспечить образование iPS-подобных колоний после от приблизительно 30 дней до приблизительно 45 дней или более от времени контакта.
[0053] Альтернативно, соматические клетки культивируют в DMEM среде, содержащей 10% FBS (которая может также подходящим образом содержать LIF, пенициллин/стрептомицин, пуромицин, L-глутамин, заменимые аминокислоты или β-меркаптоэтанол и т.п.) на фидерных клетках (таких как STO клетки или SNL клетки, обработанные митомицином C) при 37°C в присутствии 5% CO2, чтобы обеспечить образование ES-подобных колоний после от приблизительно 25 дней до приблизительно 30 дней. Репрограммированные соматические клетки предпочтительно применяют как есть вместо фидерных клеток (Takahashi K, et al. (2009), PLoS One. 4:e8067 или WO 2010/137746), или применяют внеклеточный матрикс (такой как ламинин-5 (WO 2009/123349) или матригель (Becton, Dickinson and Company)).
[0054] Кроме того, примеры способов культивирования включают способы с применением среды, которая не содержит сыворотку (Sun N, et al. Proc Natl Acad Sci USA. 106:15720-15725, 2009 или Nakagawa M, et al, Sci Rep. 4:3594, 2014). Кроме того, iPS клетки могут содержаться в условиях гипоксии (концентрация кислорода от 0,1% до 15%), чтобы повысить эффективность содержания (Yoshida Y, et al. (2009), Cell Stem Cell. 5:237-241 или WO 2010/013845).
[0055] Культуральную жидкость замещают свежей культуральной жидкостью один раз в день, начиная со 2 дня после начала культивирования во время указанного выше культивирования. Кроме того, хотя нет каких-либо конкретных ограничений в данном отношении, пример количества соматических клеток, применяемых при репрограммировании находится в диапазоне от приблизительно 5×103 до приблизительно 5×106 клеток на 10 см2 области культуральной чашки.
[0056] Этап выделения колоний стволовых клеток, полученных посредством репрограммирования соматических клеток
В настоящем изобретении, колонии стволовых клеток можно получать посредством введения репрограммирующего фактора в соматические клетки и культивирования клеток, как описано выше. В настоящем изобретении, стволовые клетки относятся к клеткам, обладающим способностью к саморепликации, которая позволяет клеткам образовывать клетки, идентичные тем клеткам посредством клеточного деления, и способностью дифференцироваться в различные типы клеток, в то же время обладая способностью пролиферировать без ограничения. Хотя нет каких-либо конкретных ограничений для стволовых клеток по настоящему изобретению, они образуют колонии, эти стволовые клетки являются плюрипотентными стволовыми клетками, обладающими способностью дифференцироваться в клетки ткани, исключая плацентарные клетки.
[0057] В настоящем изобретении, колония относится к клеточной массе, полученной из единичной клетки.
[0058] Способ клонирования по настоящему изобретению содержит этап выделения полученной колонии стволовых клеток. Это выделение можно проводить посредством подходящего сбора единичной колонии, а затем переноса в другую культуральную посуду.
[0059] iPS клетки для индуцирования клеток-предшественников мегакариоцитов
В настоящем изобретении, в случае, если соматические клетки являются клетками-предшественниками мегакариоцитов, экзогенный онкоген и экзогенный ген, подавляющий экспрессию гена p16 или ген p19, функционально связанный с отвечающим на препарат промотором, может содержаться в хромосомах клеток-предшественников мегакариоцитов, как описано ранее. В этом случае, вторичные iPS клетки, полученные посредством репрограммирования клеток-предшественников мегакариоцитов в соответствии со способом, описанным выше, сходным образом содержат экзогенный онкоген и экзогенный ген, подавляющий экспрессию гена p16 или гена p19, функционально связанный с отвечающим на препарат промотором в их хромосомах. На данный момент, соотношение содержания экзогенного гена, подавляющего экспрессию гена p16 или гена p19, функционально связанного с отвечающим на препарат промотором, и экзогенного онкогена, функционально связанного с отвечающим на препарат промотором в iPS клетках для индуцирования клеток-предшественников мегакариоцитов, составляет предпочтительно от 2-кратного до 7-кратного и более предпочтительно от 3-кратного до 5-кратного. Сходным образом, соотношение содержания экзогенного гена, подавляющего экспрессию гена p16 или гена p19, функционально связанного с отвечающим на препарат промотором, и экзогенного онкогена, функционально связанного с отвечающим на препарат промотором в клетках-предшественниках мегакариоцитов, составляет предпочтительно от 2-кратного до 7-кратного и более предпочтительно от 3-кратного до 5-кратного. Кроме того, онкоген и ген, который подавляет экспрессию гена p16 или гена p19, подходящим образом выбирают из указанных выше генов.
[0060] Этап индуцирования дифференцировки из стволовых клеток во вторичные соматические клетки
Способ клонирования по настоящему изобретению содержит этап индукции дифференцировки стволовых клеток, полученных в соответствии со способом, описанным выше, во вторичные соматические клетки. В настоящем изобретении, вторичные соматические клетки относятся к соматическим клеткам, полученным посредством репрограммирования первичных соматических клеток в стволовые клетки с последующим индуцированием к дифференцировке во вторичные соматические клетки, и первичные соматические клетки, и вторичные соматические клетки предпочтительно являются одними и теми же клетками. Настоящий этап индуцирования можно проводить посредством реэкспрессии встроенного гена. Реэкспрессию гена можно проводить посредством контактирования клеток на любой стадии дифференцировки из стволовых клеток, полученных посредством репрограммирования первичных соматических клеток, во вторичные соматические клетки с применением соответствующего препарата (в случае промотора, который экспрессирует ген в присутствии соответствующего препарата), или посредством прерывания контакта между клетками на любой стадии дифференцировки из стволовых клеток, полученных посредством репрограммирования первичных соматических клеток, во вторичные соматические клетки и соответствующего препарата (в случае промотора, который экспрессирует ген, когда соответствующий препарат удален). Например, в случае применения гибридного гена (обратный tetR) rtetR и VP16AD, ген может быть повторно экспрессирован посредством введения соответствующего препарата. "Клетки на любой стадии дифференцировки из стволовых клеток, полученные посредством репрограммирования первичных соматических клеток, во вторичные соматические клетки" могут быть любыми клетками, в которых стволовые клетки прошли через дифференцировку во вторичные соматические клетки. Таким образом, в настоящем изобретении, стволовые клетки могут быть индуцированы к дифференцировке в другие клетки перед повторной экспрессией гена, и примеры клеток, подверженных этой индукции дифференцировки, включают фибробласты и гемопоэтические клетки-предшественники. Индукцию дифференцировки в "другие клетки" можно проводить в соответствии с известными способами. В случае применения клеток-предшественников мегакариоцитов в качестве соматических клеток, описанный ранее способ индукции дифференцировки в гемопоэтические клетки-предшественники является примером способа, применяемого для индукции дифференцировки. А именно, посредством введения препарата, соответствующего среде, применяемого для индукции дифференцировки из стволовых клеток в гемопоэтические клетки-предшественники и индукции дифференцировки из ранее описанных гемопоэтических клеток-предшественников в клетки-предшественники мегакариоцитов, онкоген, ген, подавляющий экспрессию гена p16 или гена p19, и/или ген, подавляющий апоптоз, могут быть сверхэкспрессированы.
[0061] Этап отбора стволовых клеток или гемопоэтических клеток-предшественников
В настоящем изобретении, поскольку все стволовые клетки необязательно способны быть индуцированы к дифференцировке в клетки-предшественники мегакариоцитов в случае применения клеток-предшественников мегакариоцитов в качестве соматических клеток, предпочтительно соответствующим образом выбирают стволовые клетки, способные быть индуцированными к дифференцировке в клетки-предшественники мегакариоцитов, и пример способа проведения этой процедуры содержит отбор тех стволовых клеток, которые экспрессируют MEG3. В настоящем изобретении, также могут быть отобраны гемопоэтические клетки-предшественники, полученные из стволовых клеток, экспрессирующих MEG3. В настоящем изобретении, в случае людей, MEG3 относится к некодирующей RNA, состоящей из последовательности нуклеиновой кислоты, представленной в NCBI, номер доступа NR 002766, NR 003530, NR 003531, NR 033358, NR 033359, NR 033360, NR 046464, NR 046465, NR 046466, NR 046467, NR 046468, NR 046469, NR 046470, NR 046471, NR 046472 или NR 046473. Хотя нет каких-либо конкретных ограничений в данном отношении, стволовые клетки, к которым применяют способ по настоящему изобретению, могут быть первичными плюрипотентными стволовыми клетками и более предпочтительно являются клонами стволовых клеток, полученными в соответствии со способом описанным выше. Высокий уровень экспрессии может относиться к экспрессии на уровне, который выше, чем среднее значение в множестве одновременно измеренных стволовых клеток или гемопоэтических клеток-предшественников, или может относиться к уровню экспрессии, который выше по сравнению с экспрессией известных стволовых клеток или гемопоэтических клеток-предшественников, которые не могут быть индуцированы к дифференцировке в клетки-предшественники мегакариоцитов.
[0062] В настоящем изобретении, способ известный среди специалистов в данной области можно использовать в качестве способа подтверждения экспрессии MEG3, и его примеры включают PCR анализ с обратной транскрипцией, количественный PCR анализ с обратной транскрипцией, «нозерн»-блоттинг анализ, иммуногистохимический анализ, анализ на микрочипе и их сочетания.
[0063] Способ инициирования соматических клеток быть дефицитными по HLA и способ получения HLA-дефицитных соматических клеток
В одном из вариантов осуществления, настоящее изобретение относится к способу инициирования соматических клеток быть дефицитными по HLA, что включает следующие этапы, или способ получения HLA-дефицитных соматических клеток:
(i) образование плюрипотентных стволовых клеток посредством введения репрограммирующего фактора в соматические клетки;
(ii) инициирование плюрипотентных стволовых клеток, полученных на этапе (i) быть дефицитными по HLA; и
(iii) индуцирование HLA-дефицитных плюрипотентных стволовых клеток, полученных на этапе (ii) к дифференцировке в соматические клетки.
[0064] Соматические клетки, предоставленные для применения в способе для инициирования дефицита HLA по настоящему изобретению, применяемый репрограммирующий фактор, способ образования плюрипотентных стволовых клеток, и способ индукции плюрипотентных стволовых клеток к дифференцировке в соматические клетки являются теми же, что в случае соматических клеток, предоставленных для применения при указанном выше клонировании. Таким образом, настоящее изобретение также относится к способу клонирования соматических клеток и дополнительно инициированию этих клеток быть дефицитными по HLA.
[0065] В настоящем изобретении, HLA относится к человеческому лимфоцитарному антигену, и относится к антигену класса I, состоящему из α цепи и L цепи, антигену класса II, состоящему из β цепи, кодируемой геном DRB1 и α цепи, кодируемой геном DRA, и антигену класса III. Поскольку экспрессируемый HLA отличается в соответствии с соматической клеткой, HLA подлежащий удалению может подходящим образом быть выбран, и в случае применения клеток-предшественников мегакариоцитов в качестве соматических клеток, в качестве HLA предпочтительно выбирают и удаляют антиген класса I. Удаление HLA относится к удалению α цепи, L цепи или β цепи, и в случае удаления антигена класса I, предпочтительно удаляют L цепь, а именно β2-микроглобулин.
[0066] Способ инициирования хромосомы быть дефицитной по HLA в плюрипотентных стволовых клетках по настоящему изобретению можно проводить посредством подходящим образом выбранного известного способа, такого как гомологичная рекомбинация.
[0067] Способ получения тромбоцитов
Способ получения тромбоцитов по настоящему изобретению содержит этап клонирования клеток-предшественников мегакариоцитов с применением способа клонирования по настоящему изобретению, и этап обеспечения созревания клонированных клеток-предшественников мегакариоцитов в мегакариоциты и высвобождения тромбоцитов. Этап обеспечения созревания клонированных клеток-предшественников мегакариоцитов и высвобождения тромбоцитов можно проводить в соответствии с известным способом или способом, соответствующим ему. Например, в случае если клетки-предшественники мегакариоцитов содержат, по меньшей мере, один ген, выбранный из группы, состоящей из генов семейства MYC, поликомб генов и генов, подавляющих апоптоз, созревание мегакариоцитов можно проводить посредством подавления экспрессии генов семейства MYC, поликомб генов и/или генов, подавляющих апоптоз, посредством удаления соответствующего препарата из среды после указанного выше этапа (iii). Созревшие мегакариоциты становятся многоядерными и высвобождают тромбоциты.
[0068] Тромбоциты могут быть в форме препарата тромбоцитов посредством комбинирования с раствором ACD-A, FFP, цитратом натрия, лимонной кислотой или глюкозой и т.п., или могут быть в форме препарата крови посредством комбинирования с эритроцитами.
[0069] В случае получения клонов мегакариоцитов, дефицитных по HLA, в соответствии со способом, описанным выше, тромбоциты, дефицитные по HLA можно получать посредством обеспечения созревания клонов мегакариоцитов и высвобождения тромбоцитов. HLA-дефицитные тромбоциты пригодны, поскольку они могут быть перелиты безотносительно HLA типа реципиента.
[0070] Способ улучшения пролиферативной способности клеток-предшественников мегакариоцитов
Настоящее изобретение относится к способу улучшения пролиферативной способности клеток-предшественников мегакариоцитов посредством получения стволовых клеток посредством репрограммирования клеток-предшественников мегакариоцитов и последующего превращения в клетки-предшественники мегакариоцитов. Таким образом, в одном из его вариантов осуществления, способ улучшения пролиферативной способности клеток-предшественников мегакариоцитов по настоящему изобретению содержит этапы:
(i) образования колонии стволовых клеток посредством введения репрограммирующего фактора в клетки-предшественники мегакариоцитов, несущие экзогенный ген, экспрессирующийся в ответ на препарат;
(ii) выделения колонии стволовых клеток, полученных на этапе (i); и
(iii) индуцирования стволовых клеток, содержащихся в колонии стволовых клеток, изолированных на этапе (ii), к дифференцировке в клетки-предшественники мегакариоцитов, где индукция в соматические клетки содержит этап контактирования с соответствующим препаратом.
[0071] В настоящем изобретении, улучшение пролиферативной способности относится к увеличению длины теломерной последовательности в хромосоме. В настоящем изобретении, теломерная последовательность относится к повторяющейся последовательности, включая TTAGGG, и увеличение длины теломерной последовательности означает, что количество повторов увеличилось.
Примеры
[0072] Хотя далее представлено более подробное объяснение настоящего изобретения на основе примеров и тестовых примеров, настоящее изобретение не ограничено следующими примерами.
[0073] Получение клеток-предшественников мегакариоцитов
Гемопоэтические клетки-предшественники (HPC) получали посредством iPS-мешка из iPS клеток (SeV2: полученные посредством введения c-MYC, OCT3/4, SOX2 и KLF4 в неонатальные фибробласты человека с применением вирусного вектора Сендай в соответствии со способом, описанным в WO 2010/134526) в полуконфлюэнтном состоянии и поддерживали в 6 см чашке, в которой MEF рассевали в концентрации 3×105 клеток/чашку. Более конкретно, iPS клетки разделяли с применением раствора трипсина человека, и приблизительно от 1/30 до 1/50 клеток рассевали на C3H10T1/2 (доступных из Riken, Japan.), обработанных митомицином C (MMC) в форме массы колонии. Кроме того, обработанные MMC C3H10T1/2 получали посредством рассевания в 10 см чашке при концентрации 8×105 клеток/чашку в день накануне рассева iPS клеток. После рассева, культивирование начинали в минимальной среде Игла (EBM), содержащей 20 нг/мл VEGF в атмосфере 5% O2 и 5% CO2 при 37°C (день 0). Среду замещали той же средой на 3 день и 6 день.
[0074] На 7 день, культивирование продолжали в атмосфере 20% O2 и 5% CO2 при 37°C. Среду замещали той же средой на 9 день, 11 день и 13 день. На 14 день, клетки физически снимали с применением скребка для клеток или наконечника пипетки, и клетки однообразного размера вылавливали посредством пропускания через 40 микрометровое клеточное сито. Выловленные клетки подтверждали в качестве гемопоэтических клеток-предшественников (HPC) на основе клеточного размера.
[0075] На 14 день, выловленные HPC рассевали в обработанных MMC C3H10T1/2 в концентрации от 3×104 до 1×105 клеток/лунку. Для среды применяли EBM, содержащую SCF в концентрации 50 нг/мл, TPO в концентрации 50 нг/мл и доксициклин в концентрации 0,5 мкг/мл. Далее, c-MYC и BMI1 вводили в HPC с применением лентивирусного вектора. Применяемым лентивирусным вектором был индуцируемый вектор, контролируемый тетрациклином, и его получали посредством рекомбинирования mOKS кассеты LV-TRE-mOKS-Ubc-tTA-I2G с c-MYC или BMI1 (LV-TRE-c-MYC-xL-Ubc-tTA-I2G или LV-TRE-BMI1-Ubc-tTA-I2G, соответственно) (Nakamura S, et al, Cell Stem Cell. 14:535-548, 2014). Вирусные частицы, применяемые для инфицирования, получали посредством инфицирования 293T клеток с применением лентивирусного вектора (MOI 300). Протамин добавляли только во время инфицирования. Далее, среду заменяли через сутки, и C3H10T1/2 и среду заменяли один или два раза в неделю.
[0076] BCL-xl вводили при MOI 10 с применением лентивирусного вектора две недели после введения c-MYC и BMI1. Лентивирусным вектором, применяемым для введения BCL-xl, был индуцируемый вектор, контролируемый тетрациклином, и его получали посредством рекомбинации mOKS кассеты, чтобы содержать BCL-xl таки же образом, как описано выше (LV-TRE-BCL-xL-Ubc-tTA-I2G) (Nakamura S, et al, Cell Stem Cell. 14:535-548, 2014). Протамин добавляли только во время инфицирования. Далее, культивирование производили в EBM, содержащей SCF в концентрации 50 нг/мл, TPO в концентрации 50 нг/мл и доксициклин в концентрации 0,5 мкг/мл в 10T1/2 фидерных клетках на 10 см чашке, чтобы получить клетки-предшественники мегакариоцитов (также обозначаемые как imMKCL).
Справочный пример 1
[0077] Метод предельных разведений
Клетки-предшественники мегакариоцитов (imMKCL) рассевали в 96-луночный планшет при плотности 1,5 клетки/300 мкл/лунку с последующим культивированием в течение от 10 дней до 14 дней в среде Дульбекко, модифицированной по методу Исков (IMDM), содержащей 15% эмбриональной телячьей сыворотки (FBS), SCF человека (R&D Systems) в концентрации 50 нг/мл, TPO в концентрации 50 нг/мл, доксициклин (Clontech) в концентрации 5 мг/мл и пуромицин (Sigma-Aldrich) в концентрации 2 мг/мл в атмосфере 5% CO2 при 37°C. Культивирование продолжали тем же образом после переноса содержимого каждой лунки в 24-луночный планшет и 6-луночный планшет с целью увеличить масштаб культивирования. Клетки в каждой лунке были обозначены как клоны клеток-предшественников мегакариоцитов.
[0078] Анализ клонов клеток-предшественников мегакариоцитов (первый раунд)
Каждый из клонов клеток-предшественников мегакариоцитов, полученных в соответствии со способом, описанным выше отмывали дважды с применением PBS, рассевали в 6-луночный планшет в концентрации 4×105 клеток/3 мл, и культивировали в IMDM, содержащей SCF человека в концентрации 50 нг/мл, TPO человека в концентрации 50 нг/мл, SR1 (Calbiochem) в концентрации 750 нМ и 15% FBS. Супернатант собирали 7 дней спустя с последующей оценкой количества образовавшихся тромбоцитов и функции тромбоцитов. Оценку количества образовавшихся тромбоцитов проводили образом, описанным ниже. А именно, антитела, связанные с флуоресцентным красителем, CD41 (BioLegend), CD42a (eBioscience) и CD42b (BioLegend) и йодид пропидия (Sigma-Aldrich) добавляли к культуральному супернатанту и инкубировали в течение 30 минут с последующим анализом с использованием FACSVerseO (BD Biosciences). Анализ, включая исключение клеток-предшественников мегакариоцитов на основе размера, с последующим подсчетом клеток, положительных по CD41, CD42a и CD42b и подсчет количества тромбоцитов на клетку-предшественник мегакариоцита. Оценку функции тромбоцитов проводили образом, описанным ниже. А именно, антитела, связанные с флуоресцентным красителем, CD41, CD42b и активированный гликопротеин (GP) IIb/IIIa (PAC-1; BD Biosciences) и 0,4 мМ форбол 12-миристат 13-ацетат (PMA) (Sigma-Aldrich) добавляли в культуральный супернатант и инкубировали в течение 30 минут с последующим анализом с применением FACSVerseO. В анализе, уровень экспрессии активированного GP IIb/IIIa применяли для оценки посредством измерения в качестве интенсивности флуоресценции (MFI).
[0079] В результате оценки объема образования тромбоцитов и функции тромбоцитов для 22 клонов клеток-предшественников мегакариоцитов в соответствии со способом, описанным выше, подтверждали, что объем образования тромбоцитов в от 1,6 раз до 1,9 раз выше, чем контрольный (клетки-предшественники мегакариоцитов перед клонированием) для клона 1, клона 4 и клона 13 (фиг. 1A). В отношении функции тромбоцитов, наиболее высокий уровень активности был продемонстрирован клоном, 13 и было показано, что он хорошо реагирует на PMA стимуляцию по сравнению с контролем (фиг. 1B).
[0080] Анализ клонов клеток-предшественников мегакариоцитов (второй раунд)
Пять клонов (1, 2, 4, 11 и 13), которые продемонстрировали высокие объемы образования тромбоцитов и функцию тромбоцитов в результаты для первого раунда анализа были повторно подвергнуты анализу тем же образом. Только для клона 4 было подтверждено, что объем образования тромбоцитов приблизительно в 1,3 раза выше, чем контрольный (фиг. 2A). Кроме того, хотя было подтверждено, что функция тромбоцитов приблизительно в 3,7 раз выше, чем контрольная, для клона 13, результаты по функции тромбоцитов из клонов 1, 2, 4 и 11 отличались от результатов первого раунда анализа, в котором не было изменений по сравнению с контролем (фиг. 2B).
[0081] Анализ клонов клеток-предшественников мегакариоцитов (третий раунд)
Анализы проводили снова тем же образом, что для результатов второго раунда анализа. Не было изменений в объеме образования тромбоцитов в отношении контроля (фиг. 3A). Кроме того, хотя было подтверждено, что функция тромбоцитов в 1,7 раз и 1,6 раз выше, соответственно, чем в контроле для клонов 4 и 13, различия были небольшими (фиг. 3B).
[0082] В соответствии с этими результатами, было подтверждено, что клоны клеток-предшественников мегакариоцитов, полученные посредством метода предельных разведений, не демонстрируют стабильного функционирования в отношении способности образовывать тромбоциты и образованных тромбоцитов. Таким образом, было сделано предположение, что применение метода предельных разведений является непригодным для клонирования клеток-предшественников мегакариоцитов.
Пример 1
[0083] Клонирование клеток-предшественников мегакариоцитов посредством репрограммирования
iPS клетки (вторичные iPS клетки) получали посредством репрограммирования клеток-предшественников мегакариоцитов, полученных с применением описанного ранее способа с последующим произведением клонирования с применением вторичных iPS клеток и снова индуцирования дифференцировки клеток в клетки-предшественники мегакариоцитов для клонирования этих клеток (вторичные клетки-предшественники мегакариоцитов) (фиг. 4A). Далее представлено его подробное описание.
[0084] После введения четырех типов эписомальных плазмидных векторов (pCXLE-hOCT3/4-shp53-F, pCXLE-hSK, pCXLE-hUL и pCXWB-EBNA1; Okita K, et al., Stem Cells. 31:458-66, 2013 и Okita K, et al., Nat Methods. 8:409-12, 2011) в 1×106 клеток первично культивируемой клеточной линии клеток-предшественников мегакариоцитов посредством электропорации с использованием Amaxa Nucleofector, клетки рассевали в концентрации 3×105 клеток/чашку на фидерные клетки в форме MEF в концентрации 1-2×105 клеток/6 см чашку. Полученные колонии извлекали из чашки 14 дней спустя и подвергали экспансивному культивированию, после которого 10 вторичных клонов iPS клеток применяли в анализе.
[0085] Анализ вторичных клонов iPS клеток
Геномную ДНК экстрагировали из 10 вторичных клонов iPS клеток, полученных в соответствии со способом, описанным выше, и количественную PCR проводили на c-MYC и BMI1 внутри экзонов с применением праймеров, совместимых с PCR реакцией. В результате, проверку количества вставок экзогенных c-MYC и BMI1, отличных от двух характерных копий, выявили, что количество вставок экзогенного BMI1 10 типов вторичных клонов iPS клеток составяло приблизительно от 7 копий до 26 копий, в то время как количество вставок экзогенного c-MYC составляло приблизительно от 1 копии до 6 копий, и все были различными (фиг. 4B).
[0086] Геномную ДНК экстрагировали из 10 вторичных клонов iPS клеток (с применением некоторых клонов, которые отличались от описанных ранее) полученных в соответствии со способом, описанным выше с последующим анализом всей геномной последовательности, и когда количество вставок экзогенных c-MYC и BMI1 было проверено, было подтверждено, что они встроились в хромосомы в соотношениях, указанных в следующей Таблице 1. Было подтверждено, что вторичные клоны iPS клеток содержат приблизительно от 3-кратного до 5-кратного количества вставок BMI1 по сравнению с количеством вставок c-MYC.
[Таблица 1]
[0087] В соответствии с представленными выше результатами, хотя количество вставок экзогенных генов, введенных на этапе получения клеток-предшественников мегакариоцитов, отличалось для каждой клетки-предшественника мегакариоцитов, в случае если они были встроены при постоянном соотношении, предполагали, что образовывались клетки-предшественники мегакариоцитов.
[0088] Индукция вторичных клонов клеток-предшественников мегакариоцитов из вторичных клонов iPS клеток
HPC клоны были соответственно получены посредством iPS-мешка через 14 дней после трех вторичных клонов iPS клеток (#4, #11 и #12) в соответствии с описанным ранее способом. Полученные HPC клоны рассевали в 6-луночные планшеты в концентрации 1×105 клеток/лунку с последующим культивированием в течение 27 дней в EBM, содержащей SCF в концентрации 50 нг/мл, TPO в концентрации 50 нг/мл и доксициклин в концентрации 0,5 мкг/мл для получения вторичных клонов клеток-предшественников мегакариоцитов. Анализ полученных трех клонов клеток-предшественников мегакариоцитов выявил, что клоны были положительны по CD41a и CD41b, отрицательны по CD235, и были получены в форме единообразной клеточной группы (фиг. 5A и 5B).
[0089] Получение HLA-дефицитных (HLA-null) вторичных клонов iPS клеток
Таргетный вектор, показанный на фиг. 6A, был введен во вторичный клон iPS клеток (#11), полученный в соответствии со способом, описанным выше для удаления экзона 1 β2-Микроглобулина (β2m) посредством гомологичной рекомбинации. Была ли проведена гомологичная рекомбинация надлежащим образом или нет, определяли посредством подтверждения PCR продуктов с применением праймеров, сконструированных в сайтах, указанных на фиг. 6A для дикого типа и мутанта (фиг. 6B). В результате, β2m-дефицитные вторичные клоны iPS клеток были установлены для двух клонов (клоны 3 и 4).
[0090] Далее, β2m-дефицитные вторичные клоны клеток-предшественников мегакариоцитов индуцировали в соответствии с описанным ранее способом. Кроме того, тромбоциты получали из супернатанта в соответствии с ранее описанным способом. Когда проточную цитометрию проводили на β2m-дефицитных вторичных клонах клеток-предшественников мегакариоцитов и тромбоцитов с применением антител к β2m (BD Pharmingen) и HLA (BD Pharmingen), было подтверждено, что все являются дефицитными по β2m и HLA (фиг. 7A).
[0091] Подтверждение функции тромбоцитов β2m-дефицитных вторичных клонов клеток-предшественников мегакариоцитов
После стимуляции тромбоцитов, присутствующих в культуре, супернатант вторичного клона iPS клеток дикого типа (#11) и β2m-дефицитные вторичные клоны клеток-предшественников мегакариоцитов с PMA таким же образом, как описано ранее, клоны контактировали с CD42a антителом, CD42b антителом и PAC1, и когда положительные уровни CD42a, CD42b и PAC1 измеряли и оценивали на основе интенсивности флуоресценции (MFI), не было наблюдаемых различий между диким типом и β2m-дефицитным типом (фиг. 7B). На основе сказанного выше, было показано, что тромбоциты, дефицитные по β2m, имеют ту же функцию, что и тромбоциты дикого типа.
Пример 2
[0092] Поиск маркера iPS клеток, пригодного для индукции клеток-предшественников мегакариоцитов
mRNA экстрагировали из двух типов вторичных iPS клеток (обозначаемых как хорошие-iPSC), способных образовывать imMKCL, полученный в соответствии с ранее описанным способом и из двух типов iPS клеток (обозначаемых как плохие-iPSC), не способных образовывать imMKCL, полученные в соответствии с ранее описанным способом с последующим усреднением и анализом уровней экспрессии двух типов хороших-iPSC и плохих-iPSC с применением микрочипа с использованием Illumina HumanHT-12 v4.0 или Affymetrix Gene Chip Human Gene 2.0 ST Array в соответствии с руководствами, предоставленными производителями (фиг. 8A и 8B).
[0093] Кроме того, гемопоэтические клетки-предшественники (HPC) были индуцированы из хороших-iPSC и плохих-iPSC с применением того же способа, как описано ранее (соответственно, обозначаемые как хорошие-HPC и плохие-HPC) с последующим анализом с применением микрочипа с использованием Illumina HumanHT-12 v4.0 или Affymetrix Gene Chip Human Gene 2.0 ST Array (Figs. 8A и 8B).
[0094] В результате анализа с применением указанных выше микрочипов, было подтверждено, что MEG3to высоко экспрессируется и хорошими-iPSC и хорошими-HPC. Таким образом, во вторичных клонах iPS клеток и HPC, индуцированных из них, предполагалось, что подтверждение экспрессии MEG3 обеспечит отбор вторичных клонов iPS клеток, пригодных для индукции клеток-предшественников мегакариоцитов.
Пример 3
[0095] Пролиферативная способность клеток-предшественников мегакариоцитов, полученных из вторичных клонов iPS клеток
Клеточную пролиферативную способность клеток-предшественников мегакариоцитов, полученных из вторичного клона iPS клеток, сравнивали с клеточной пролиферативной способностью линии клеток-предшественников мегакариоцитов перед клонированием, чтобы проверить эффект клонирования по настоящему изобретению. В данном эксперименте, клон, полученный из 2nd-iPS_11, перечисленного в таблицу 1, применяли для вторичного клона iPS клеток. Более конкретно, HPC клон, полученный из 2nd-iPS_11 посредством iPS-мешка через 14 дней в соответствии с ранее описанным способом, рассевали в 6-луночную чашку в концентрации 1×105 клеток/лунку с последующим культивированием в течение 27 дней в EBM, содержащей SCF в концентрации 50 нг/мл, TPO в концентрации 50 нг/мл и доксициклин в концентрации 0,5 мкг/мл, чтобы получить вторичный клон клеток-предшественников мегакариоцитов (клон 2). Клетки-предшественники мегакариоцитов (родительские) перед клонированием, полученные в разделе, озаглавленном "Получение клеток-предшественников мегакариоцитов" применяли в качестве сравнительного примера. Эти клетки рассевали в 6-луночную чашку в концентрации 1×105 клеток/лунку и культивировали в течение 15 дней в EBM, содержащей SCF в концентрации 50 нг/мл, TPO в концентрации 50 нг/мл и доксициклин в концентрации 0,5 мкг/мл.
[0096] В результате периодического подсчета числа клеток во время 15-дневного периода культивирования, клетки-предшественники мегакариоцитов, полученные из вторичных клонов iPS клеток, четко продемонстрировали более быстрый рост, чем клетки-предшественники мегакариоцитов перед клонированием. Результаты показаны на фиг. 9A и 9B.
[0097] Способность к созреванию клеток-предшественников мегакариоцитов, полученных из вторичного клона iPS клеток
Вторичный клон клеток-предшественников мегакариоцитов (клон 2), культивируемый в течение 15 дней, культивировали в условиях содействия созреванию мегакариоцитов (включая добавление SCF в концентрации 50 нг/мл и TPO в концентрации 50 нг/мл к EBM с последующим добавлением SR-1 (StemRegenin1) (Selleckchem) в концентрации 750 нМ и Y27632 (Wako) в концентрации 10 мкМ). Клеткам-предшественникам мегакариоцитов (родительские) также позволяли созревать до мегакариоцитов при тех же условиях культивирования, что и вторичный клон клеток-предшественников мегакариоцитов. Изображения обеих клеток, сфотографированных посредством замедленной фотосъемки, представлены на фиг. 10A, в то время как результаты анализа полученных изображений с применением ImageJ представлены на фиг. 10B. На основе этих результатов, мегакариоциты, клонированные в соответствии с настоящим изобретением, были представлены, чтобы продемонстрировать более высокую способность созревания (клеточная гипертрофия), чем мегакариоциты, полученные из исходных клонов клеток.
[0098] Способность клеток-предшественников мегакариоцитов, полученных из вторичного клона iPS клеток, к образованию тромбоцитов
Число клеток, которые образовывали тромбоциты, измеряли визуально на изображениях, представленных на фиг. 10A, чтобы сравнить способность образования тромбоцитов из полученных мегакариоцитов. В результате, мегакариоциты, полученные из клеток, клонированных в соответствии с настоящим изобретением, четко продемонстрировали более высокую способность к образованию тромбоцитов (число образовавшихся протромбоцитов) (**: p<0.01).
| название | год | авторы | номер документа |
|---|---|---|---|
| НОВЫЙ СПОСОБ ПОЛУЧЕНИЯ ДИФФЕРЕНЦИРОВАННЫХ КЛЕТОК | 2010 |
|
RU2661107C1 |
| НОВЫЙ СПОСОБ ПОЛУЧЕНИЯ ДИФФЕРЕНЦИРОВАННЫХ КЛЕТОК | 2010 |
|
RU2603094C2 |
| СПОСОБ ПОЛУЧЕНИЯ КУЛЬТУРЫ, СОДЕРЖАЩЕЙ МЕГАКАРИОЦИТЫ, И СПОСОБ ПОЛУЧЕНИЯ ТРОМБОЦИТОВ С ЕЕ ПРИМЕНЕНИЕМ | 2016 |
|
RU2750740C2 |
| СПОСОБ ПОЛУЧЕНИЯ ВЫСОКОФУНКЦИОНАЛЬНЫХ ТРОМБОЦИТОВ | 2016 |
|
RU2724533C2 |
| СПОСОБ ПРОИЗВОДСТВА ТРОМБОЦИТОВ СПОСОБОМ РОТАЦИОННОГО ПЕРЕМЕШИВАНИЯ КУЛЬТУРЫ | 2016 |
|
RU2756000C2 |
| СПОСОБ ПОЛУЧЕНИЯ ТРОМБОЦИТОВ С ПОМОЩЬЮ УСТРОЙСТВА ДЛЯ ВОЗВРАТНО-ПОСТУПАТЕЛЬНОГО ПЕРЕМЕШИВАНИЯ | 2016 |
|
RU2741871C2 |
| СПОСОБ ПОЛУЧЕНИЯ ПОЛИПЛОИДНЫХ МЕГАКАРИОЦИТОВ И ТРОМБОЦИТОВ | 2012 |
|
RU2606764C2 |
| СПОСОБ ПОЛУЧЕНИЯ ИНДУЦИРОВАННЫХ ПЛЮРИПОТЕНТНЫХ СТВОЛОВЫХ КЛЕТОК ПАЦИЕНТОВ С СИНДРОМОМ ДАУНА | 2012 |
|
RU2492233C1 |
| СПОСОБ ПОЛУЧЕНИЯ ОЧИЩЕННЫХ ТРОМБОЦИТОВ | 2016 |
|
RU2766177C2 |
| КОНВЕРСИЯ СОМАТИЧЕСКИХ КЛЕТОК В ИНДУЦИРОВАННЫЕ РЕПРОГРАМИРОВАННЫЕ НЕЙРОНАЛЬНЫЕ СТВОЛОВЫЕ КЛЕТКИ (ИРНСК) | 2011 |
|
RU2562111C2 |
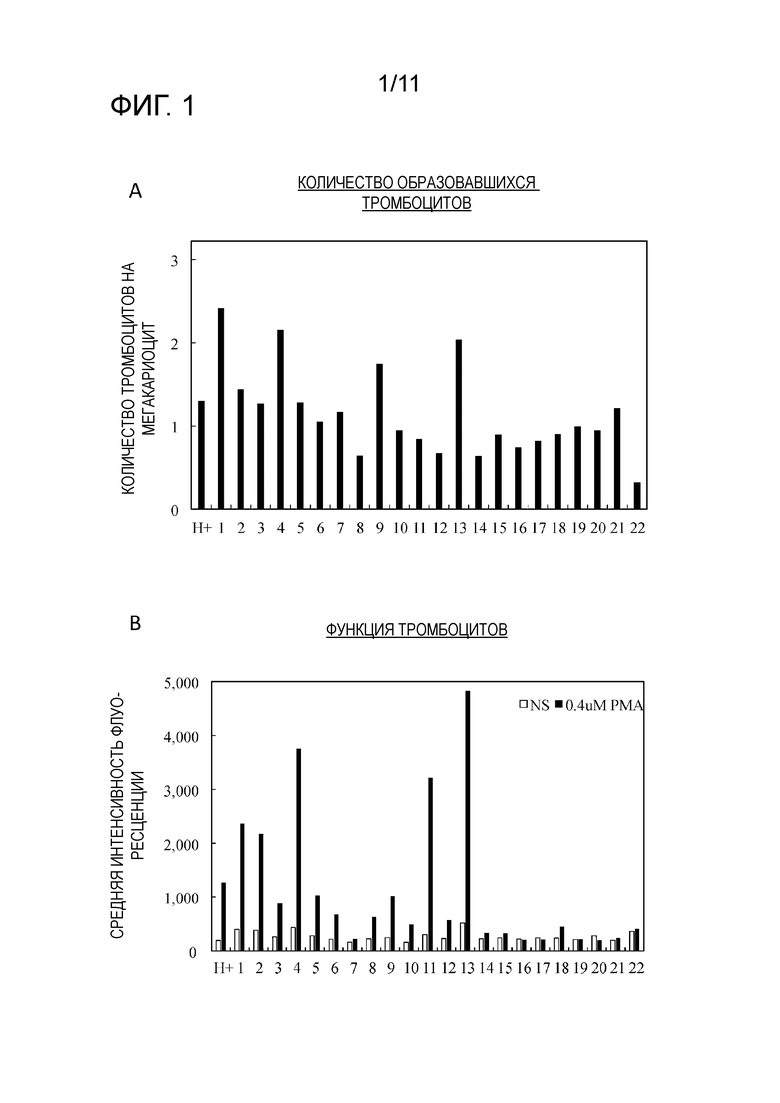
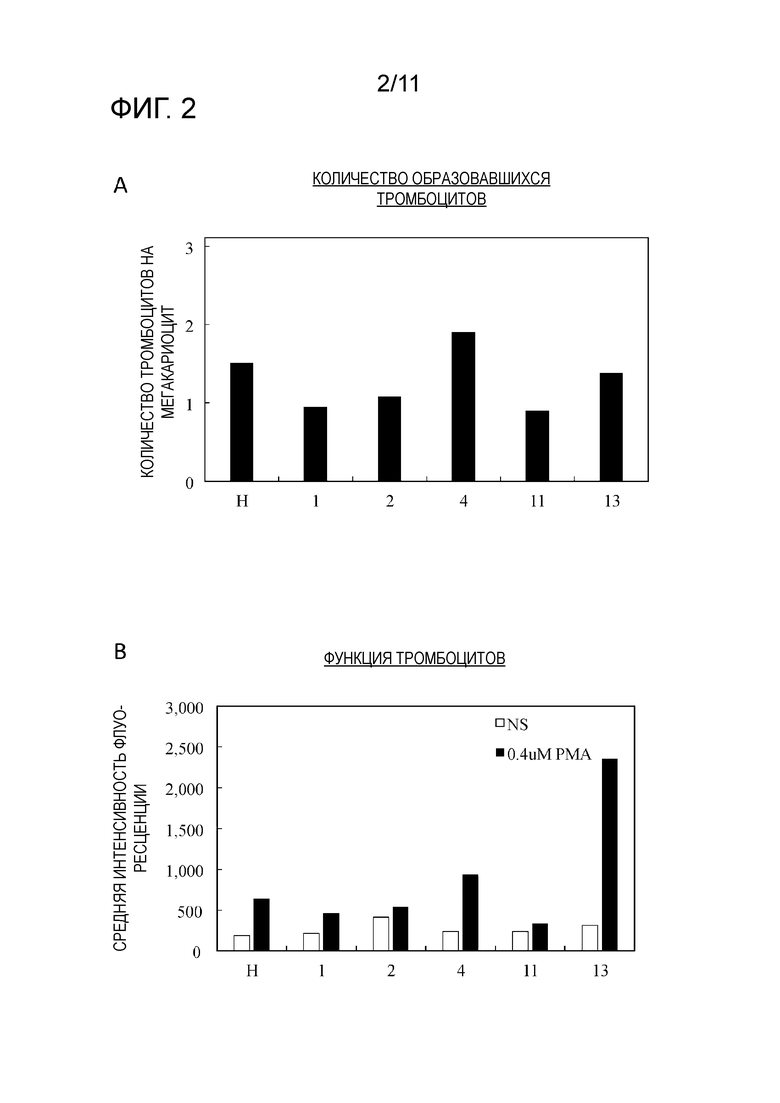
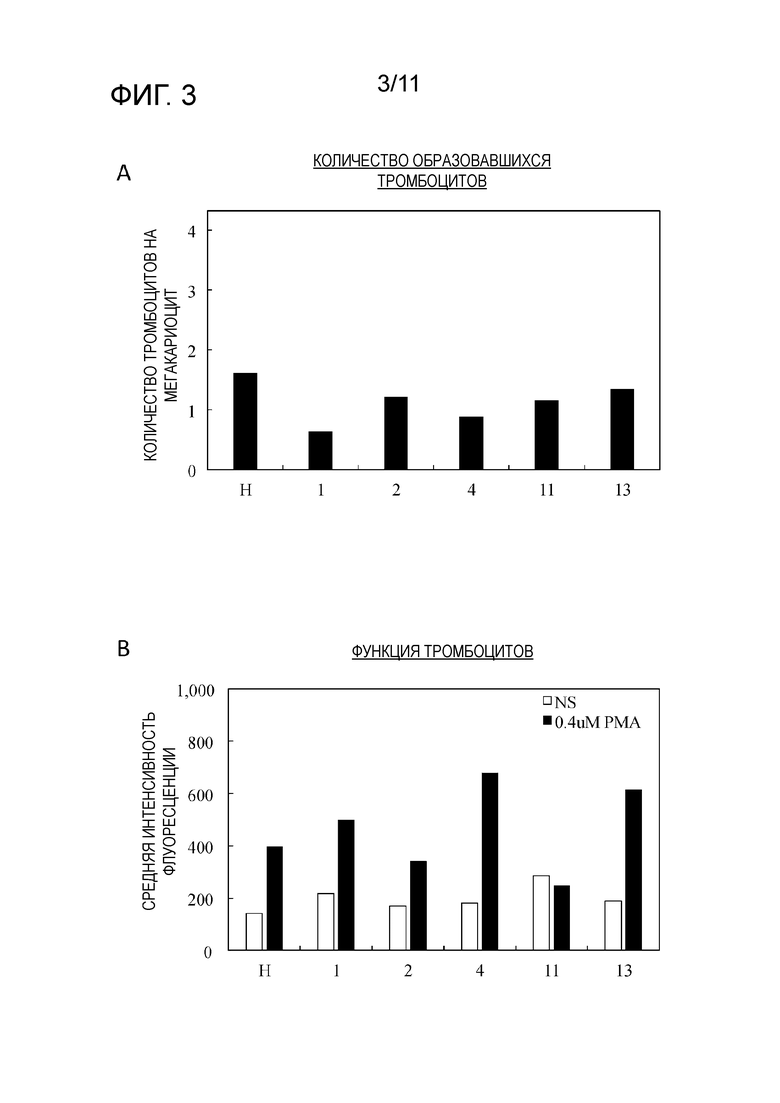
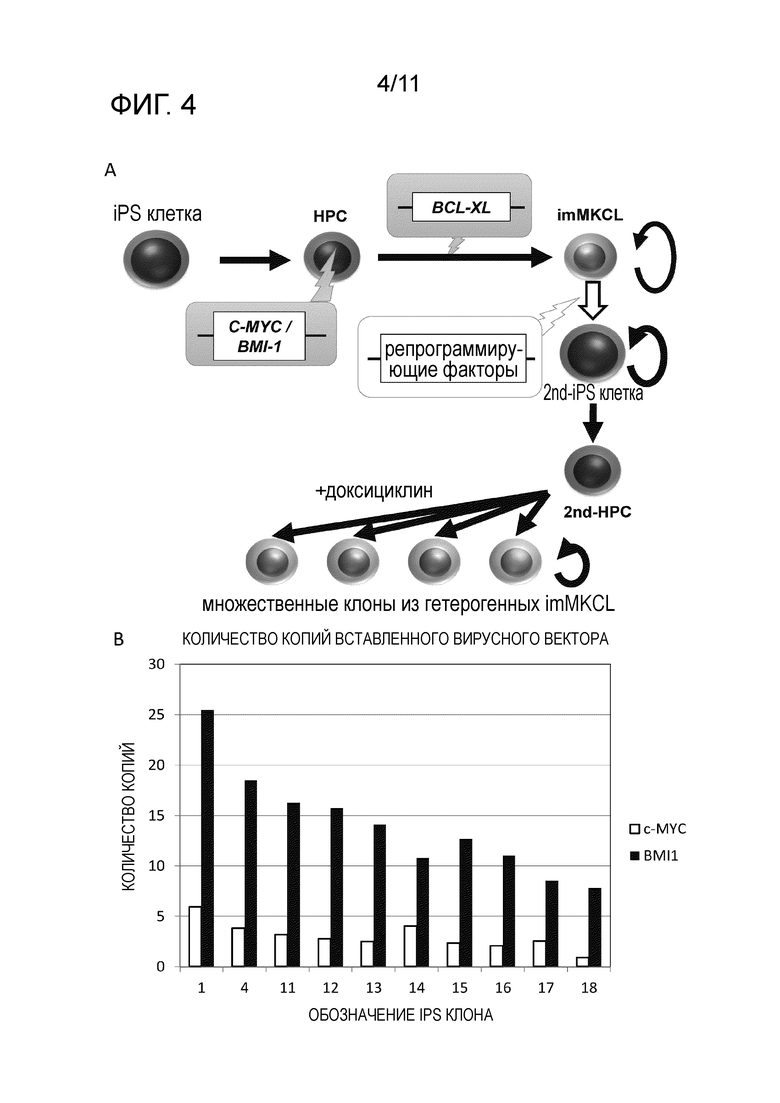
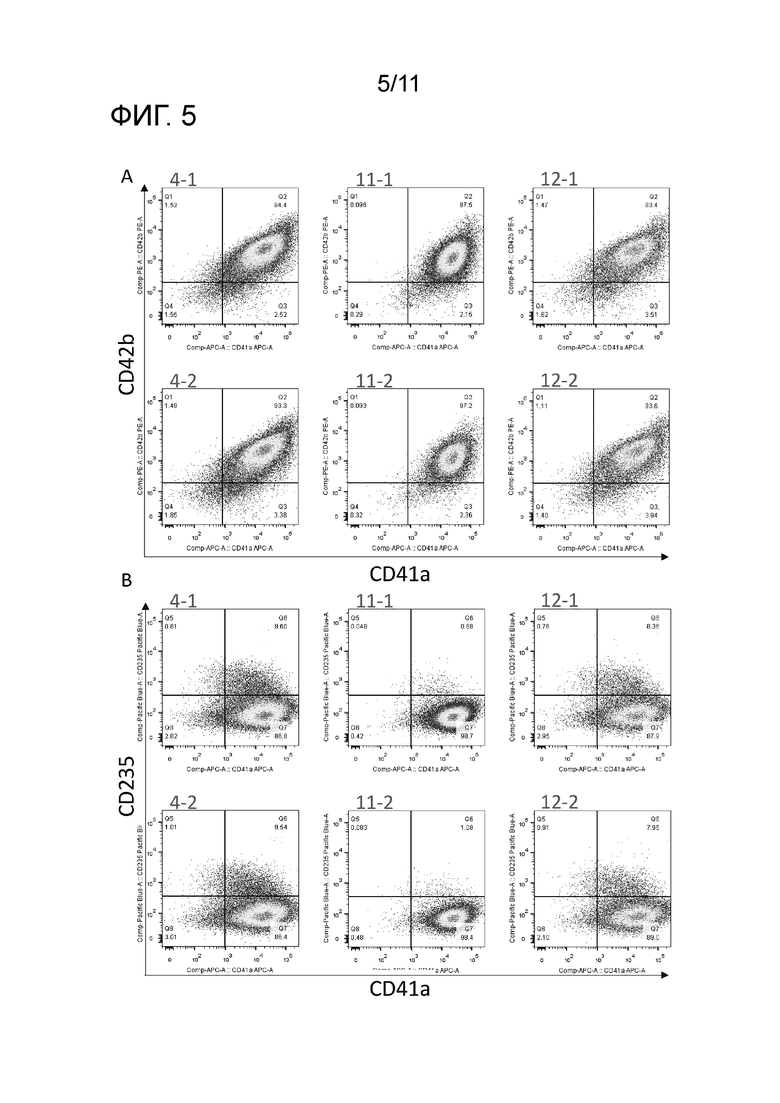
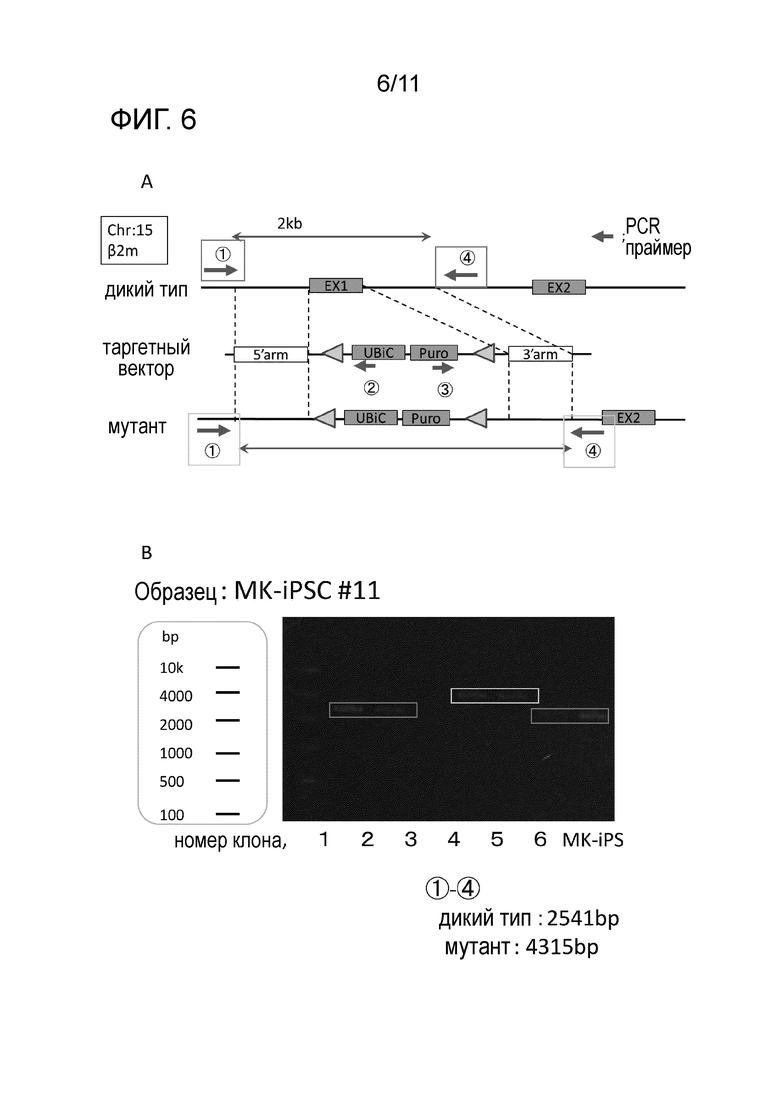
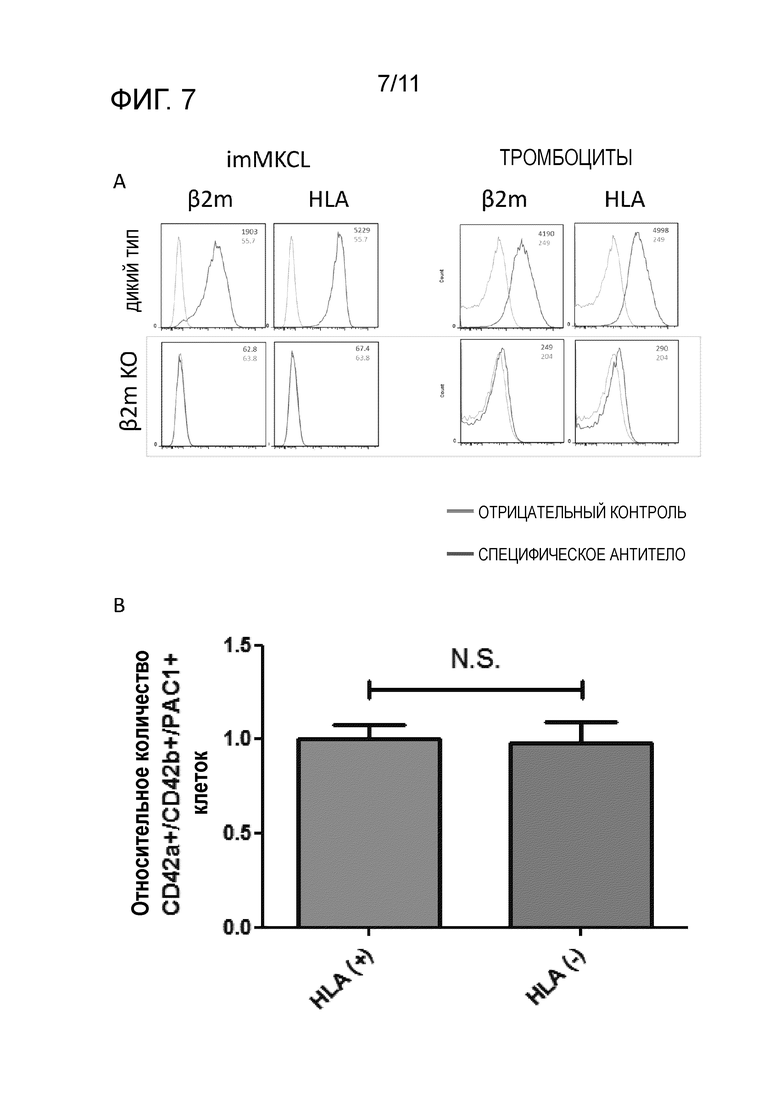
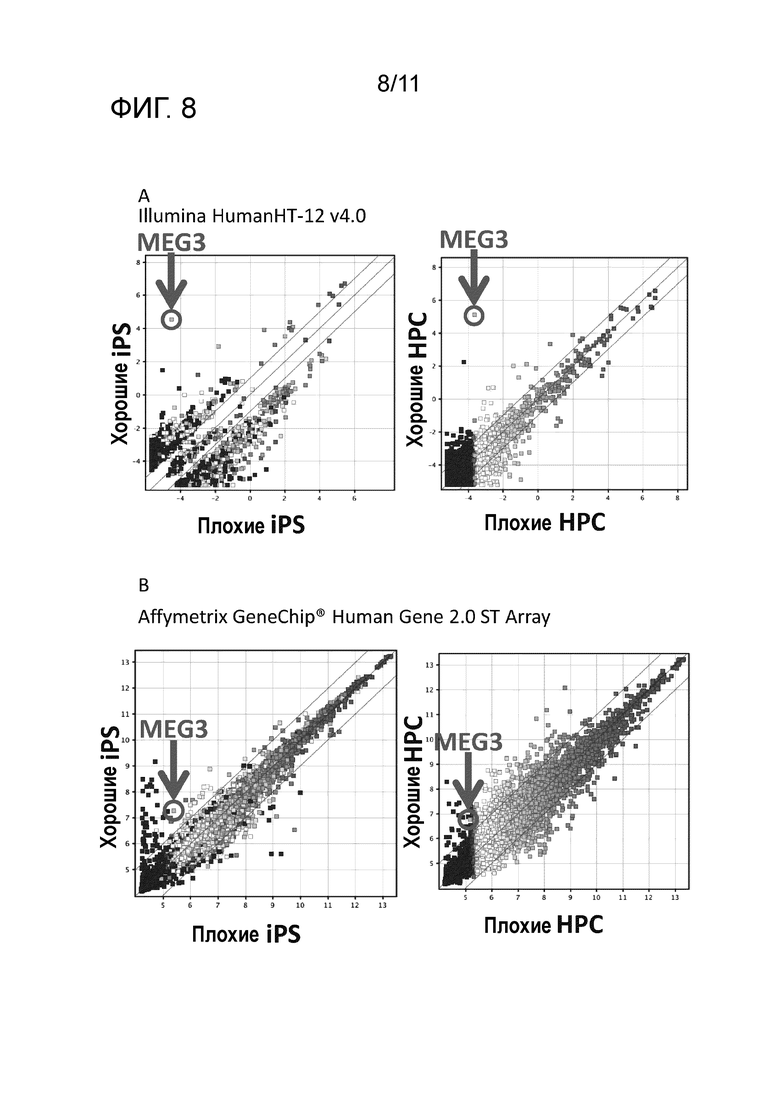
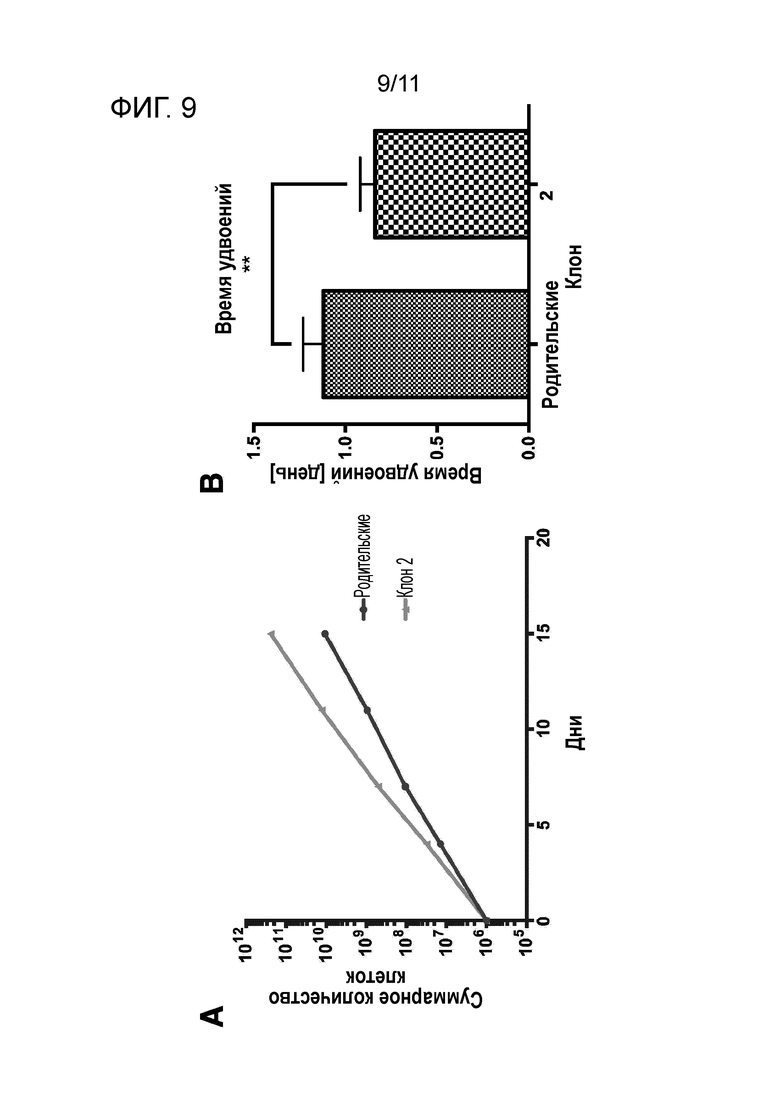
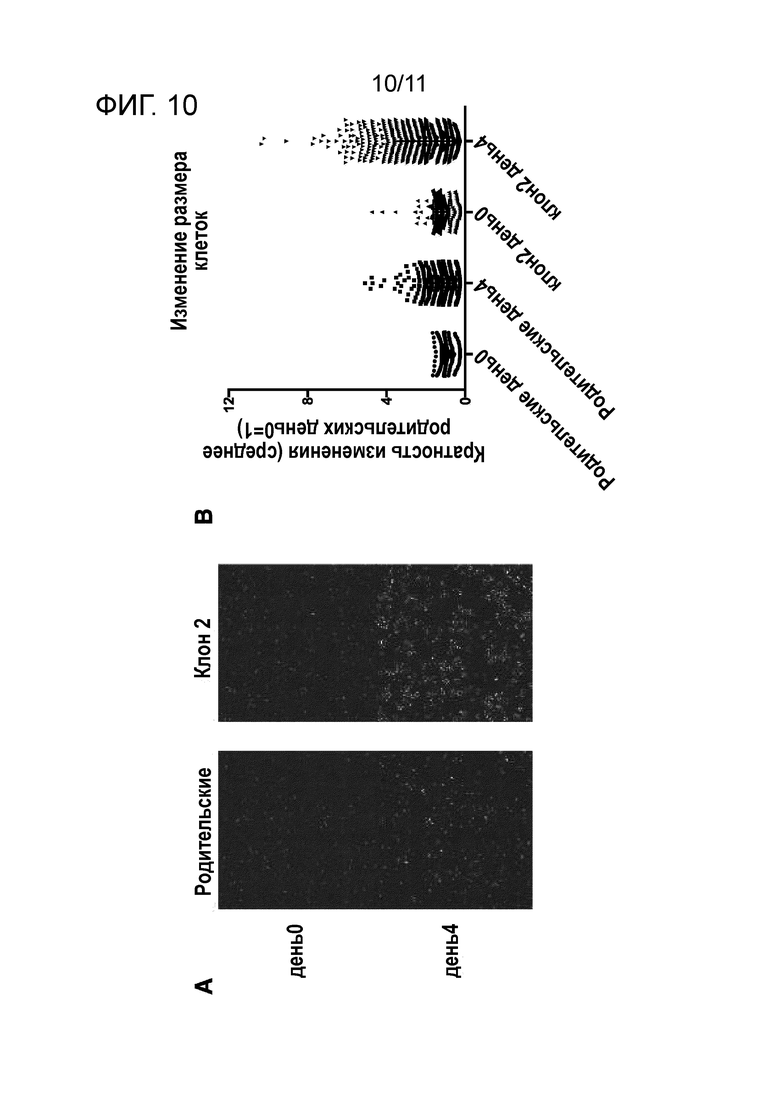
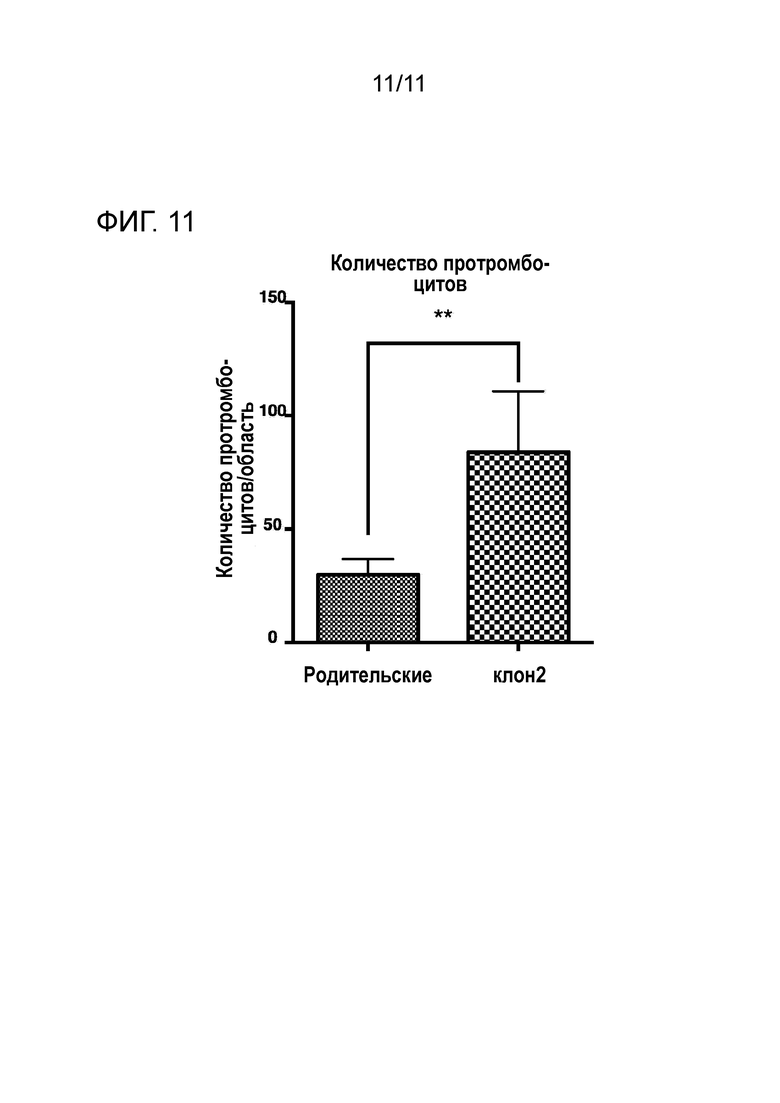
Изобретение относится к области биохимии, в частности к способу получения клонов вторичных стволовых клеток из клеток-предшественников мегакариоцитов, предусматривающему дедифференцировку клеток-предшественников мегакариоцитов для образования колонии стволовых клеток, а также выделения из нее вторичных стволовых клеток. Также раскрыт способ получения клеток-предшественников мегакариоцитов, предусматривающий индукцию дифференцировки выделенных клонов вторичных стволовых клеток в клетки-предшественники мегакариоцитов. Изобретение также относится к способу получения тромбоцитов, предусматривающему обеспечение созревания индуцированных к дифференцировке клеток-предшественников мегакариоцитов в мегакариоциты и высвобождения тромбоцитов. Изобретение позволяет эффективно получать тромбоциты из клеток-предшественников мегакариоцитов. 3 н. и 7 з.п. ф-лы, 11 ил., 1 табл., 3 пр.
1. Способ получения клонов вторичных стволовых клеток из клеток-предшественников мегакариоцитов, который содержит этапы:
(i) дедифференцировки клеток-предшественников мегакариоцитов для образования колонии стволовых клеток, где клетки-предшественники мегакариоцитов дифференцируют из стволовых клеток, которые экспрессируют по меньшей мере один экзогенный ген, выбранный из группы, состоящей из онкогенов, генов поликомб и генов, подавляющих апоптоз; и
(ii) выделения вторичных стволовых клеток, несущих по меньшей мере один экзогенный ген, встроенный в их хромосому, из колонии стволовых клеток, образованной на этапе (i).
2. Способ по п. 1, дополнительно включающий сравнение эффективности индуцирования дифференцировки изолированных клонов вторичных стволовых клеток в клетки-предшественники мегакариоцитов с эффективностью индуцирования дифференцировки стволовых клеток перед клонированием.
3. Способ по п. 1, где онкогенами являются гены семейства MYC, генами поликомб являются Bmi1 и генами, подавляющими апоптоз, являются гены BCL-XL.
4. Способ в соответствии с п.1, где изолируют вторичные стволовые клетки, экспрессирующие MEG3.
5. Способ в соответствии с п.1, где экзогенный ген функционально связан с отвечающим на препарат промотором.
6. Способ в соответствии с п.1, где дедифференцировку на этапе (i) проводят посредством введения репрограммирующего фактора, выбранного из группы, состоящей из OCT3/4, SOX2 и KLF4.
7. Способ получения клеток-предшественников мегакариоцитов, который содержит этап:
(i) дедифференцировки клеток-предшественников мегакариоцитов для образования колонии стволовых клеток, где клетки-предшественники мегакариоцитов дифференцируют из стволовых клеток, которые экспрессируют по меньшей мере один экзогенный ген, выбранный из группы, состоящей из онкогенов, генов поликомб и генов, подавляющих апоптоз; и
(ii) выделения вторичных стволовых клеток, несущих по меньшей мере один экзогенный ген, встроенный в их хромосому, из колонии стволовых клеток, образованной на этапе (i);
(iii) индукции дифференцировки выделенных клонов вторичных стволовых клеток в клетки-предшественники мегакариоцитов.
8. Способ получения тромбоцитов, который содержит этапы:
(i)дедифференцировки клеток-предшественников мегакариоцитов для образования колонии стволовых клеток, где клетки-предшественники мегакариоцитов дифференцируют из стволовых клеток, которые экспрессируют по меньшей мере один экзогенный ген, выбранный из группы, состоящей из онкогенов, генов поликомб и генов, подавляющих апоптоз;
(ii) выделения вторичных стволовых клеток, несущих по меньшей мере один экзогенный ген, встроенный в их хромосому, из колонии стволовых клеток, образованной на этапе (i);
(iii) индукции дифференцировки выделенных клонов вторичных стволовых клеток в клетки-предшественники мегакариоцитов; и
(iv) обеспечение созревания индуцированных к дифференцировке клеток-предшественников мегакариоцитов в мегакариоциты и высвобождения тромбоцитов.
9. Способ по п. 8, дополнительно включающий инициирование клонов стволовых клеток быть дефицитными по HLA.
10. Способ по п. 8, где стволовые клетки, которые экспрессируют по меньшей мере один экзогенный ген, являются гематопоэтическими клетками-предшественниками.
| NAOYA TAKAYAMA et al., Transient activation of c-MYC expression is critical for efficient platelet generation from human induced pluripotent stem cells, J | |||
| Exp | |||
| Med., 2010, Vol | |||
| Станок для изготовления из дерева круглых палочек | 1915 |
|
SU207A1 |
| Насос | 1917 |
|
SU13A1 |
| ТЕЛЕГРАФНЫЙ КЛАВИАТУРНЫЙ ПЕРЕДАТЧИК ЗНАКОВ МОРЗЕ | 1925 |
|
SU2817A1 |
| US 2012009158 A1, 12.01.2012 | |||
| US 2012034192 A1, 09.02.2012 | |||
| WO 2014123242 A1, 14.08.2014 | |||
| Механическая форсунка | 1928 |
|
SU14166A1 |

