Предпосылки изобретения
Зрелая красная клетка крови или эритроцит ответственна за транспорт кислорода в системе кровообращения позвоночных. Эритроциты содержат высокие концентрации гемоглобина, белка, который связывает кислород в легких при относительно высоком парциальном давлении кислорода (pO2), и доставляет кислород к частям организма с относительно низким pO2.
Зрелые эритроциты продуцируются плюрипотентными гематопоэтическими стволовыми клетками в процессе, называемом эритропоэзом. Постнатальный эритропоэз происходит главным образом в костном мозге и в красной пульпе селезенки. Координированное действие различных сигнальных путей регулирует баланс пролиферации, дифференциации, выживания и гибели клеток. В нормальных условиях эритроциты продуцируются со скоростью, которая поддерживает постоянную массу эритроцитов в организме, и продукция может повышаться или уменьшаться в ответ на различные стимулы, в том числе повышенное или пониженное напряжение кислорода или тканевую потребность. Процесс эритропоэза начинается с образования коммитированных клеток-предшественников линии дифференциации и проходит серию определенных типов клеток-предшественников. Конечные стадии эритропоэза происходят по мере того как ретикулоциты высвобождаются в кровоток и утрачивают их митохондрии и рибосомы, в то же время приобретая морфологию зрелого эритроцита. Повышенный уровень ретикулоцитов или повышенное соотношение ретикулоцитов:эритроцитов в крови указывает на повышенную скорость продукции эритроцитов.
Хорошо известно, что эритропоэтин (EPO) является наиболее значимым положительным регулятором постнатального эритропоэза у позвоночных. EPO регулирует компенсаторный эритропоэтический ответ на сниженное напряжение кислорода в тканях (гипоксию) и низкие уровни эритроцитов или низкие уровни гемоглобина. У людей повышенные уровни EPO способствуют образованию эритроцитов путем стимуляции образования эритроидных клеток-предшественников в костном мозге и селезенке. У мышей, EPO усиливает эритропоэз главным образом в селезенке.
Воздействия EPO опосредуются рецептором на клеточной поверхности, относящимся к суперсемейству цитокиновых рецепторов. Ген рецептора EPO человека кодирует трансмембранный белок из 483 аминокислот, тогда как считается, что активный рецептор EPO существует в виде мультимерного комплекса даже в отсутствие лиганда (см. патент США № 6319499). Клонированный полноразмерный рецептор EPO, экспрессированный в клетках млекопитающих, связывает EPO со сродством, аналогичным сродству нативного рецептора на эритроидных клетках-предшественниках. Связывание EPO с его рецептором вызывает конформационное изменение, приводящее к активации рецептора и биологическим эффектам, включая повышенную пролиферацию незрелых эритробластов, повышенную дифференциацию незрелых эритробластов и уменьшенный апоптоз эритроидных клеток-предшественников (Liboi et al., 1993, Proc Natl Acad Sci USA 90:11351-11355; Koury et al., 1990, Science 248:378-381).
Различные формы рекомбинантного EPO используются врачами для повышения уровней эритроцитов при разнообразных клинических ситуациях и, в частности, для лечения анемии. Анемия представляет собой хорошо изученное состояние, характеризуемое более низкими, чем в норме, уровнями гемоглобина или эритроцитов в крови. В некоторых случаях, анемия вызвана первичным расстройством продукции или стабильности эритроцитов. Чаще, анемия является вторичной в отношении заболеваний других систем (Weatherall & Provan (2000) Lancet 355, 1169-1175). Анемия может возникнуть в результате пониженной скорости продукции или повышенной скорости разрушения эритроцитов или потерей эритроцитов вследствие кровотечения. Анемия может возникнуть в результате различных расстройств, к которым относится, например, хроническая почечная недостаточность, химиотерапия, миелодиспластический синдром, ревматоидный артрит и трансплантация костного мозга.
Лечение EPO обычно приводит к повышению содержания гемоглобинов примерно на 1-3 г/дкл у здоровых людей в течение нескольких недель. При введении больным с анемией, эта схема лечения часто позволяет существенно повысить уровни гемоглобина и эритроцитов и приводит к улучшению качества жизни и продлевает жизнь больному. EPO невсегда эффективен, и многие индивиды устойчивы даже к его высоким дозам (Horl et al. (2000) Nephrol Dial Transplant 15, 43-50). Более чем у 50% пациентов, страдающих злокачественными заболеваниями, имеется неадекватная реакция на EPO, приблизительно у 10% пациентов с тяжелой стадией болезни почек наблюдается пониженная реакция на EPO (Glaspy et al. (1997) J Clin Oncol 15, 1218-1234; Demetri et al. (1998) J Clin Oncol 16, 3412-3425), и менее чем 10% пациентов с миелодиспластическим синдромом благоприятно реагируют на лечение EPO (Estey (2003) Curr Opin Hematol 10, 60-67). При наличии некоторых факторов, в том числе воспаление, дефицит железа и витаминов, нарушенный диализ, токсическое воздействие алюминия и гиперпаратиреоз, можно прогнозировать слабую реакцию на лечение. Молекулярные механизмы устойчивости к EPO пока неясны. Недавние доказательства свидетельствуют о том, что более высокие дозы EPO могут быть связаны с повышенным риском сердечно-сосудистых заболеваний, ростом опухоли и смертности пациентов некоторых популяций (Krapf et al., 2009, Clin J Am Soc Nephrol 4:470-480; Glaspy, 2009, Annu Rev Med 60:181-192). Поэтому было рекомендовано введение терапевтических соединений на основе EPO (средства, стимулирующие эритропоэтин, ESA) в самой низкой дозе, достаточной для того, чтобы исключить необходимость трансфузий эритроцитов (Jelkmann et al., 2008, Crit Rev Oncol. Hematol 67:39-61).
Таким образом, задачей настоящего описания являются альтернативные способы повышения уровней эритроцитов у пациентов, которые смогли бы обеспечить возможность применения сниженных доз активаторов рецепторов эритропоэтина.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Частично, описание демонстрирует, что ловушки GDF могут использоваться в комбинации (например, вводиться в одно и то же время или в различное время, но в целом таким образом, чтобы достичь перекрывающих фармакологических эффектов) с активаторами рецепторов EPO для повышения уровней эритроцитов (эритропоэза) или лечения анемии у пациентов. Частично, в описание показано, что ловушка GDF может вводиться в комбинации с активатором рецептора EPO для синергического повышения образования эритроцитов у пациента. Таким образом, эффект такого комбинированного лечения может быть значительно выше, чем сумма эффектов ловушки GDF и активатора рецептора EPO при введении их отдельно в соответствующих дозах. В определенных вариантах осуществления, такой синергизм может быть эффективным, поскольку он обеспечивает получение заданных уровней эритроцитов, при использовании более низких доз активатора рецептора EPO, таким образом, избегая возможных побочных эффектов или других проблем, связанных с более высокими уровнями активации рецептора EPO.
Активатор рецептора EPO может стимулировать эритропоэз прямым контактом и активацией рецептора EPO. В определенных вариантах осуществления, активатор рецептора EPO представляет собой соединение из класса соединений на основе последовательности длиной 165 аминокислотных остатков нативного EPO и в целом известных как агенты, стимулирующие эритропоэз (ESA), примерами которых являются эпоэтин альфа, эпоэтин бета, эпоэтин дельта и эпоэтин омега. В других вариантах осуществления, ESA включают синтетические белки EPO (SEP) и производные EPO с непептидными модификациями, которые придают желательные фармакокинетические свойства (более длинный период полувыведения из циркуляции), примерами которых являются дарбепоэтин альфа и метоксиполиэтиленгликоль-эпоэтин бета. В определенных вариантах осуществления, активатор рецептора EPO может представлять собой агонист рецептора EPO, который не содержит полипептидную основную цепь EPO, или в целом не классифицируется как ESA. К таким агонистам рецепторов EPO могут относиться, без ограничения, пептидные и непептидные миметики EPO, агонистические антитела, нацеливающие на рецептор EPO, слитые белки, содержащие домен миметика EPO и ограниченные агонисты рецептора эритропоэтина продолжительного действия (EREDLA).
В определенных вариантах осуществления, активатор рецептора EPO может стимулировать эритропоэз косвенно, без контакта с самим рецептором EPO, путем усиления продукции эндогенного EPO. Например, индуцируемые гипоксией транскрипционные факторы (HIF) представляют собой эндогенные стимуляторы генной экспрессии EPO, которые подавлены (дестабилизированы) в нормоксических условиях клеточными регуляторными механизмами. Отчасти, изобретение относится к повышению эритропоэза у пациента с помощью комбинированного лечения ловушкой GDF и косвенным активатором рецептора EPO со свойствами, стабилизирующими HIF (индуцируемый гипоксией фактор), таким как ингибитор пропил-гидроксилазы.
Варианты полипептидов ActRIIB, обладающие значительно сниженным сродством к активину (например, активину A и/или активину B) по сравнению с другими лигандами ActRIIB, такими как GDF11 и/или миостатин, называются ловушками GDF. Пока нет других указаний, варианты ActRIIB, описанные в настоящей заявке, представляют собой ловушки GDF. В частности, в описании изобретения показано, что ловушка GDF, которая представляет собой растворимую форму полипептида ActRIIB с кислым остатком в положении 79 последовательности SEQ ID NO:1, при введении in vivo, повышает содержание эритроцитов в крови. Поэтому, в определенных вариантах осуществления изобретение относится к способам применения ловушек GDF для повышения содержания эритроцитов и гемоглобина у пациентов и для лечения расстройств, связанных с низким содержанием эритроцитов и гемоглобина у таких пациентов. Как описано в заявке на патент США № 12/012652, который включен в настоящее описание в качестве ссылки, ловушки GDF могут использоваться для повышения мышечной массы и снижения жировой массы.
В определенных аспектах настоящее изобретение относится к ловушкам GDF, которые представляют собой варианты полипептидов ActRIIB, имеющие амино- и карбокси-концевые усечения и изменения в последовательностях. Необязательно, конструирование ловушек GDF по изобретению для предпочтительного антагонизма в отношении одного или нескольких лигандов рецепторов ActRIIB, таких как GDF8 (также называемого миостатином), GDF11, Nodal и BMP7 (также называемого OP-1). Примеры ловушек GDF включают набор вариантов, полученных из ActRIIB, которые обладают значительно пониженным сродством к активину. Такие варианты проявляют желаемые эффекты на эритроциты, в то же время, снижая воздействия на другие ткани. Примеры таких вариантов включают варианты с кислой аминокислотой (например, аспарагиновой кислотой, D, или глютаминовой кислотой, E) в положении, соответствующем положению 79 последовательности SEQ ID NO:1. В определенных вариантах осуществления полипептид-ловушка GDF содержит аминокислотную последовательность, которая содержит, состоит из или по существу состоит из аминокислотной последовательности SEQ ID NO:7, 26, 28, 29, 32, 37 или 38, полипептидов, которые по меньшей мере на 80%, 85%, 90%, 95%, 97%, 98% или 99% идентичны любому из указанных выше.
В определенных аспектах изобретение относится к фармацевтическим препаратам, содержащим ловушку GDF, которая связывается с лигандом ActRIIB, например, GDF8, GDF11, активином (например, активином B), BMP7 или Nodal, и фармацевтически приемлемый носитель. Необязательно, ловушка GDF связывается с лигандом ActRIIB со значением Kd, равным менее чем 10 мкм, менее чем 1 мкмоль, менее чем 100 наномоль, менее чем 10 наномоль или менее чем 1 наномоль. Необязательно, ловушка GDF ингибирует передачу сигнала ActRIIB, например, события внутриклеточной передачи сигналов, запускаемой лигандом ActRIIB. Ловушка GDF, которая может использоваться в таком препарате, может представлять собой любую ловушку GDF, описанную в настоящей заявке, в том числе, например, ловушки GDF с аминокислотной последовательностью, выбранной из SEQ ID NO:2, 3, 7, 11, 26, 28, 29, 32, 37, 38 или 40, или ловушки GDF с аминокислотной последовательностью, которая по меньшей мере на 80%, 85%, 90%, 95%, 97% или 99% идентична аминокислотной последовательности, выбранной из SEQ ID NO:2, 3, 7, 11, 26, 28, 29, 32, 37, 38 или 40, или ловушки GDF с аминокислотной последовательностью, которая по меньшей мере на 80%, 85%, 90%, 95%, 97% или 99% идентична аминокислотной последовательности, выбранной из SEQ ID NO:2, 3, 7, 11, 26, 28, 29, 32, 37, 38 или 40, где положение, соответствующее L79 в последовательности SEQ ID NO:1, представляет собой кислую аминокислоту. Предпочтительная ловушка GDF, которая может использоваться в таком препарате состоит из, или по существу состоит из аминокислотной последовательности SEQ ID NO:26. Ловушка GDF может содержать функциональный фрагмент природного полипептида ActRIIB, такой как фрагмент, содержащий по меньшей мере 10, 20 или 30 аминокислот последовательности, выбранной из SEQ ID NO:2, 3, 7, 11, 26, 28, 29, 32, 37, 38 или 40, или последовательности SEQ ID NO:2, лишенной C-концевых 1, 2, 3, 4, 5 или 10-15 аминокислот, и лишенной 1, 2, 3, 4 или 5 аминокислот на N-конце. Предпочтительный полипептид содержит усечение относительно SEQ ID NO: 2 или 40 из 2-5 аминокислот на N-конце и не более 3 аминокислот на C-конце. Ловушка GDF может содержать одно или более изменений в аминокислотной последовательности полипептида ActRIIB (например, в лиганд-связывающем домене) относительно природного полипептида ActRIIB. Изменение аминокислотной последовательности может, например, изменить гликозилирование полипептида при продукции в клетке млекопитающего, насекомого или другой эукариотической клетке, или изменить протеолитическое расщепление полипептида по сравнению с природным полипептидом ActRIIB.
Ловушка GDF может представлять собой слитый белок, который имеет, в качестве одного домена, полипептид ActRIIB (например, лиганд-связывающий домен ActRIIB с одним или более изменениями последовательности), и один или более дополнительных доменов, которые придают желательное свойство, такое как улучшенная фармакокинетика, более простая очистка, нацеливание на определенные ткани и т.д. Например, домен слитого белка может улучшить одну или более характеристик устойчивости in vivo, периода полувыведения in vivo, захвата/введения, локализации или распределения в тканях, образования белковых комплексов, мультимеризации слитого белка и/или очистки. Слитые белки ловушки GDF могут содержать домен Fc-фрагмента иммуноглобулина (дикого типа или мутантные) или сывороточный альбумин. В определенных вариантах осуществления, слитая ловушка GDF содержит относительно неструктурированный линкер, расположенный между доменом Fc-фрагмента и внеклеточным доменом ActRIIB. Неструктурированный линкер может соответствовать неструктурированной области длиной приблизительно 15 аминокислот на C-конце внеклеточного домена ActRIIB («хвосте»), или он может представлять собой искусственную последовательность длиной 3-5, 15, 20, 30, 50 или более аминокислот, которые относительно свободны от вторичной структуры. Линкер может быть обогащен остатками глицина или пролина и, поэтому, может содержать, например, повторяющиеся последовательности треонина/серина и глицинов (например, синглеты или повторы TG4 (SEQ ID NO:13) или SG4 (SEQ ID NO:14)) или серии трех глицинов. Слитый белок может содержать подпоследовательность очистки, такую как эпитопный таг, таг FLAG, последовательность полигистидина и слитый GST. В определенных вариантах осуществления, слитая ловушка GDF содержит лидерную последовательность. Лидерная последовательность может представлять собой лидерную последовательность ActRIIB или гетерологичную лидерную последовательность. В определенных вариантах осуществления, лидерная последовательность представляет собой лидерную последовательность тканевого активатора плазминогена (TPA). В одном из вариантов осуществления, слитый белок ловушки GDF содержит аминокислотную последовательность, представленную формулой A-B-C. Часть B представляет собой усеченный на N- и C-конце полипептид ActRIIB, состоящий из аминокислотной последовательности, соответствующей аминокислотам 25-131 последовательности SEQ ID NO: 2 или 40. Части A и C могут независимо представлять собой ноль, одну или более одной аминокислоты, а части и A, и C являются гетерологичными для B. Части A и/или C могут быть присоединены к части B через линкерную последовательность.
Необязательно, ловушка GDF содержит вариант полипептида ActRIIB, имеющий один или несколько модифицированных аминокислотных остатков, выбранных из: гликозилированной аминокислоты, ПЭГилированной аминокислоты, фарнезилированной аминокислоты, ацетилированной аминокислоты, биотинилированной аминокислоты, аминокислоты, конъюгированной с липидной частью, аминокислоты, конъюгированной с органическим агентом, используемом для получения производных. Фармацевтический препарат может также содержать одно или более дополнительных соединений, таких как соединение, которое используется для лечения расстройства, связанного с ActRIIB. Предпочтительно, фармацевтический препарат по существу лишен пирогенов. В целом, предпочтительно, чтобы ловушка GDF была экспрессирована в линии клеток млекопитающих, которая подходящим образом опосредует природное гликозилирование ловушки GDF с тем, чтобы уменьшить вероятность неблагоприятного иммунного ответа у пациента. Успешно использовались линии клеток человека и CHO, и ожидается, что могут использоваться другие привычные векторы экспрессии млекопитающих.
В определенных аспектах настоящее изобретение относится к упакованным фармацевтическим средствам, содержащим фармацевтический препарат, описанный в настоящей заявке, и вкладыш с описанием применения для повышения содержания эритроцитов у человека.
В определенных аспектах настоящее изобретение относится к ловушкам GDF, которые представляют собой растворимые полипептиды ActRIIB, содержащие измененный лиганд-связывающий (например, связывающий GDF8) домен. Ловушки GDF с измененными лиганд-связывающими доменами могут содержать, например, одну или несколько мутаций в таких аминокислотных остатках как E37, E39, R40, K55, R56, Y60, A64, K74, W78, L79, D80, F82 и F101 ActRIIB человека (нумерация указана относительно SEQ ID NO:1). Необязательно наличие измененного лиганд-связывающего домена может повысить селективность к лиганду, такому как GDF8/GDF11, по сравнению с лиганд-связывающим доменом дикого типа рецептора ActRIIB. Для иллюстрации, в настоящем описании показано, что такие мутации повышают селективность измененного лиганд-связывающего домена к GDF11 (и, следовательно, предположительно, GDF8) по сравнению с активином: K74Y, K74F, K74I, L79D, L79E и D80I. Следующие мутации обладают противоположным эффектом, повышая соотношение связывания активина по сравнению с GDF11: D54A, K55A, L79A и F82A. Общая активность связывания (GDF11 и активина) может быть повышена за счет введения «хвостовой» области или, предположительно, неструктурированной линкерной области, а также благодаря мутации K74A. Другие мутации, которые вызывают общее снижение сродства связывания лиганда, включают: R40A, E37A, R56A, W78A, D80K, D80R, D80A, D80G, D80F, D80M и D80N. Мутации могут комбинироваться для достижения желательных эффектов. Например, многие мутации, которые влияют на соотношение связывания GDF11:Активина, оказывают общий негативный эффект на связывание лигандов, и поэтому, они могут комбинироваться с мутациями, которые в целом увеличивают связывание лигандов для получения улучшенного связывающего белка с селективностью в отношении лиганда. В иллюстративном варианте осуществления, ловушка GDF представляет собой полипептид ActRIIB, содержащий мутацию L79D или L79E, возможно, в сочетании с дополнительными аминокислотными заменами, добавлениями или делециями.
Необязательно, ловушка GDF, содержащая измененный лиганд-связывающий домен, возможно, имеет отношение значения Kd для связывания активина к значению Kd для связывания GDF8, которое по меньшей мере в 2, 5, 10 или даже в 100 раз больше относительно отношения для лиганд-связывающего домена дикого типа. Необязательно, ловушка GDF, содержащая измененный лиганд-связывающий домен, возможно, имеет отношение значения IC50 для ингибирования активина к значению IC50 для ингибирования GDF8/GDF11, то есть, по меньшей мере в 2, 5, 10 или даже в 100 раз больше относительно лиганд-связывающего домена дикого типа ActRIIB. Необязательно, ловушка GDF, содержащая измененный лиганд-связывающий домен, ингибирует GDF8/GDF11 со значением IC50 по меньшей мере в 2, 5, 10, или даже в 100 раз больше, чем значение IC50 для ингибирования активина. Такие ловушки GDF могут представлять собой слитые белки, которые содержат домен Fc-фрагмента иммуноглобулина (или дикого типа, или мутантного). В некоторых случаях, растворимые ловушки GDF по изобретению представляют собой антагонисты (ингибиторы) GDF8 и/или GDF11.
Возможны и другие типы ловушек GDF, такие как нижеследующие. Слитый белок ловушки GDF, содержащий часть, полученную из последовательности ActRIIB SEQ ID NO: 1 или 39, и вторую полипептидную часть, где часть, полученная из ActRIIB, соответствует последовательности, начинающейся у любой из аминокислот 21-29 последовательности SEQ ID NO:1 или 39 (возможно, начинающейся у аминокислот 22-25 последовательности SEQ ID NO:1 или 39), и заканчивающейся у любой из аминокислот 109-134 последовательности SEQ ID NO:1 или 39, и где слитый белок ловушки GDF ингибирует передачу сигналов активином, миостатином и/или GDF11 в анализе на основе клеток. Описанный выше слитый белок ловушки GDF, где белок, полученный из ActRIIB, соответствует последовательности, начинающейся у любой из аминокислот 20-29 последовательности SEQ ID NO:1 или 39 (возможно, начинающейся у аминокислот 22-25 последовательности SEQ ID NO:1 или 39), и заканчивающейся у любой из аминокислот 109-133 последовательности SEQ ID NO:1 или 39. Описанный выше слитый белок ловушки GDF, где часть, полученная из ActRIIB, соответствует последовательности, начинающейся у любой из аминокислот 20-24 последовательности SEQ ID NO:1 или 39 (возможно, начинающейся у аминокислот 22-25 последовательности SEQ ID NO:1 или 39), заканчивающейся у любой из аминокислот 109-133 последовательности SEQ ID NO:1 или 39. Описанный выше слитый белок ловушки GDF, где часть, полученная из ActRIIB, соответствует последовательности, начинающейся у любой из аминокислот 21-24 последовательности SEQ ID NO:1 или 39, и заканчивающейся у любой из аминокислот 109-134 последовательности SEQ ID NO:1 или 39. Описанный выше слитый белок ловушки GDF, где часть, полученная из ActRIIB, соответствует последовательности, начинающейся у любой из аминокислот 20-24 последовательности SEQ ID NO: 1 или 39, и заканчивающейся у любой из аминокислот 118-133 последовательности SEQ ID NO: 1 или 39. Описанный выше слитый белок ловушки GDF, где часть, полученная из ActRIIB, соответствует последовательности, начинающейся у любой из аминокислот 21-24 последовательности SEQ ID NO:1 или 39, и заканчивающейся у любой из аминокислот 118-134 последовательности SEQ ID NO:1 или 39. Описанный выше слитый белок ловушки GDF, где часть, полученная из ActRIIB, соответствует последовательности, начинающейся у любой из аминокислот 20-24 последовательности SEQ ID NO:1 или 39, и заканчивающейся у любой из аминокислот 128-133 последовательности SEQ ID NO:1 или 39. Описанный выше слитый белок ловушки GDF, где часть, полученная из ActRIIB, соответствует последовательности, начинающейся у любой из аминокислот 20-24 последовательности SEQ ID NO:1 или 39, и заканчивающейся у любой из аминокислот 128-133 последовательности SEQ ID NO:1 или 39. Описанный выше слитый белок ловушки GDF, где часть, полученная из ActRIIB, соответствует последовательности, начинающейся у любой из аминокислот 21-29 последовательности SEQ ID NO:1 или 39, и заканчивающейся у любой из аминокислот 118-134 последовательности SEQ ID NO:1 или 39. Описанный выше слитый белок ловушки GDF, где часть, полученная из ActRIIB, соответствует последовательности, начинающейся у любой из аминокислот 20-29 последовательности SEQ ID NO:1 или 39, и заканчивающейся у любой из аминокислот 118-133 последовательности SEQ ID NO:1 или 39. Описанный выше слитый белок ловушки GDF, где часть, полученная из ActRIIB, соответствует последовательности, начинающейся у любой из аминокислот 21-29 последовательности SEQ ID NO: 1 или 39, и заканчивающейся у любой из аминокислот 128-134 последовательности SEQ ID NO:1 или 39. Описанный выше слитый белок ловушки GDF, где часть, полученная из ActRIIB, соответствует последовательности, начинающейся у любой из аминокислот 20-29 последовательности SEQ ID NO:1, и заканчивающейся у любой из аминокислот 128-133 последовательности SEQ ID NO:1 или 39. К удивлению, конструкции, начинающиеся у аминокислот 22-25 последовательности SEQ ID NO: 1 или 39, имеют уровни активности, большие чем белки, имеющие полный внеклеточный домен ActRIIB человека. В предпочтительном варианте осуществления слитый белок ловушки GDF содержит, по существу состоит из или состоит из аминокислотной последовательности, начинающейся в аминокислотном положении 25 последовательности SEQ ID NO:1 или 39, и заканчивающейся в аминокислотном положении 131 последовательности SEQ ID NO: 1 или 39. В других предпочтительных вариантах осуществления полипептид-ловушка GDF состоит из, или по существу состоит из аминокислотной последовательности SEQ ID NO: 7, 26, 28, 29, 32, 37 или 38. Любой из указанных выше слитых белков ловушки GDF могут быть получены в виде гомодимера. Любые из указанных выше слитых белков ловушки GDF могут иметь гетерологичную часть, которая содержит константную область из тяжелой цепи IgG, такую как Fc-домен. Любые из указанных выше слитых белков ловушки GDF могут содержать кислую аминокислоту в положении, соответствующем положению 79 последовательности SEQ ID NO:1, возможно, в комбинации с одним или более дополнительных аминокислотных замен, делеций или вставок относительно SEQ ID NO:1.
Возможны и другие белки ловушки GDF, такие как нижеприведенные. Белок ловушки GDF, содержащий аминокислотную последовательность, которая по меньшей мере на 80% идентична последовательности аминокислот 29-109 последовательности SEQ ID NO:1 или 39, где положение, соответствующее 64 последовательности SEQ ID NO:1 представляет собой R или K, и где белок ловушки GDF ингибирует передачу сигналов активином, миостатином и/или GDF11 в анализе на клеточной основе. Указанный выше белок ловушки GDF, где по меньшей мере одно изменение относительно последовательности SEQ ID NO: 1 или 39 расположено вне лиганд-связывающего кармана. Указанный выше белок ловушки GDF, где по меньшей мере одно изменение относительно последовательности SEQ ID NO:1 или 39 представляет собой консервативное изменение, расположенное внутри лиганд-связывающего кармана. Указанный выше белок ловушки GDF, где по меньшей мере одно изменение относительно последовательности SEQ ID NO:1 или 39 представляет собой изменение в одном или более положений, выбранных из группы, состоящей из K74, R40, Q53, K55, F82 и L79. Указанный выше белок ловушки GDF, где белок содержит по меньшей мере одну последовательность N-X-S/T в положении, отличном от эндогенной последовательности N-X-S/T ActRIIB, и в положении вне лиганд-связывающего кармана.
Возможны и другие ловушки GDF, такие как нижеприведенные. Белок ловушки GDF, содержащий аминокислотную последовательность, которая по меньшей мере на 80% идентична последовательности аминокислот 29-109 последовательности SEQ ID NO:1 или 39, и где белок содержит по меньшей мере одну последовательность N-X-S/T в положении, отличном от эндогенной последовательности N-X-S/T ActRIIB, и в положении, вне лиганд-связывающего кармана. Указанную выше ловушку GDF, где белок ловушки GDF содержит N в положении 24 последовательности SEQ ID NO: 1 или 39, и S или T в положении, соответствующем положению 26 последовательности SEQ ID NO:1 или 39, и где ловушка GDF ингибирует передачу сигналов активином, миостатином и/или GDF11 в анализе на клеточной основе. Указанная выше ловушка GDF, где белок ловушки GDF содержит R или K в положении, соответствующем положению 64 последовательности SEQ ID NO:1 или 39. Указанная выше ловушка GDF, где белок ActRIIB содержит D или E в положении, соответствующем положению 79 последовательности SEQ ID NO:1 или 39, и где ловушка GDF ингибирует передачу сигналов активином, миостатином и/или GDF11 в анализе на клеточной основе. Указанная выше ловушка GDF, где по меньшей мере одно изменение в отношении последовательности SEQ ID NO: 1 или 39 представляет собой консервативное изменение, расположенное внутри лиганд-связывающего кармана. Указанная выше ловушка GDF, где по меньшей мере одно изменение в отношении последовательности SEQ ID NO:1 или 39 представляет собой изменение в одном или более положениях, выбранных из группы, состоящей из K74, R40, Q53, K55, F82 и L79. Указанная выше ловушка GDF, где белок представляет собой слитый белок, кроме того, включающий гетерологичную часть. Любой из указанных выше слитых белков ловушки GDF может быть получен в виде гомодимера. Любой из указанных выше слитых белков ловушки GDF может иметь гетерологичную часть, которая содержит константную область из тяжелой цепи IgG, такой как Fc-домен.
В определенных аспектах, изобретение относится к нуклеиновым кислотам, кодирующим полипептид-ловушку GDF. Выделенный полинуклеотид может содержать кодирующую последовательность растворимого полипептида-ловушки GDF, такую как описано выше. Например, выделенная нуклеиновая кислота может содержать последовательность, кодирующую ловушку GDF, содержащую внеклеточный домен (например, лиганд-связывающий домен) полипептида ActRIIB, имеющего одну или более изменений в последовательности, и последовательность, которая бы кодировала часть или весь трансмембранный домен, и/или цитоплазматический домен полипептида ActRIIB, но стоп-кодон, расположенный внутри трансмембранного домена или цитоплазматического домена, или расположенный между внеклеточным доменом и трансмембранным доменом или цитоплазматическим доменом. Например, выделенный полинуклеотид, кодирующий ловушку GDF, может содержать полноразмерную полинуклеотидную последовательность ActRIIB, такую как SEQ ID NO:4, имеющую одно или более изменений, или частично усеченный вариант, причем указанный выделенный полинуклеотид, кроме того, содержит кодон окончания транскрипции по меньшей мере за 600 нуклеотидов до 3’-конца, или расположенный иначе таким образом, чтобы трансляция полинуклеотида привела к получению внеклеточного домена, возможно, слитого с усеченной частью полноразмерного ActRIIB. Описанные в настоящей заявке нуклеиновые кислоты могут быть функционально связаны с промотором для экспрессии, и настоящее изобретение относится к клеткам, трансформированным такими рекомбинантными полинуклеотидами. Предпочтительно, клетка представляет собой клетку млекопитающего, такую как клетка CHO.
В определенных аспектах, изобретение относится к способам получения полипептида-ловушки GDF. Такой способ может предусматривать экспрессирование любой из описанных в настоящей заявке нуклеиновых кислот (например, SEQ ID NO: 5, 25, 27, 30 или 31) в подходящей клетке, такой как клетка яичника китайского хомячка (CHO). Такой способ может включать: a) культивирование клетки в условиях, подходящих для экспрессии полипептида-ловушки GDF, где указанная клетка трансформируется экспрессирующей конструкцией ловушки GDF; и b) выделение экспрессированного таким образом полипептида-ловушки GDF. Полипептиды-ловушки GDF могут быть выделены в виде неочищенных, частично очищенных или высокоочищенных фракций с использованием любой из хорошо известных методик получения белка из клеточных культур.
В определенных аспектах, полипептид-ловушка GDF по настоящему изобретению может использоваться в способе стимуляции продукции эритроцитов или повышения содержания эритроцитов у индивида. В определенных вариантах осуществления, изобретение относится к способам лечения расстройства, связанного с низкими количествами эритроцитов или низкими уровнями гемоглобина (например, анемии), или для стимуляции продукции эритроцитов у пациентов, при необходимости. Способ может предусматривать введение индивиду эффективного количества полипептида-ловушки GDF. В определенных аспектах, изобретение относится к применению полипептидов-ловушек GDF для получения лекарственного средства для лечения расстройства или состояния по настоящему изобретению.
В определенных аспектах, изобретение относится к способам введения пациенту полипептида-ловушки GDF. Частично, в описании продемонстрировано, что полипептиды-ловушки GDF могут использоваться для повышения содержания эритроцитов и уровней гемоглобина. Полипептиды-ловушки GDF могут также использоваться для лечения или профилактики других видов терапевтического применения, таких как стимуляция мышечного роста. В определенных случаях, при введении полипептида-ловушки GDF для стимуляции роста мышц, может быть желателен контроль воздействий на эритроциты во время введения полипептида-ловушки GDF или определение или коррекция дозировки полипептида-ловушки GDF для снижения нежелательных воздействий на эритроциты. Например, повышение содержания эритроцитов, гемоглобина или уровней гематокрита может вызвать повышение артериального давления.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Материалы патента или заявки содержат по меньшей мере один исполненный в цвете чертеж. Экземпляры данного патента или патентной заявки с цветным чертежом (чертежами) может быть по запросу и оплате необходимой пошлины представлен в Патентное Ведомство.
На фиг. 1 показано выравнивание внеклеточных доменов ActRIIA человека (SEQ ID NO:15) и ActRIIB человека (SEQ ID NO:2) с остатками, которые установлены в настоящем описании на основании комбинированного анализа множественных кристаллических структур ActRIIB и ActRIIA для непосредственного контакта с лигандом (лиганд-связывающим карманом), указанным прямоугольными рамками.
На фиг. 2 показано выравнивание множества последовательностей белков ActRIIB различных позвоночных и ActRIIA человека (SEQ ID NO:16-23).
На фиг. 3 показана полная аминокислотная последовательность ActRIIB(L79D 20-134)-hFc (SEQ ID NO:11) ловушки GDF, включающая лидерную последовательность TPA (двойное подчеркивание), внеклеточный домен ActRIIB (остатки 20-134 в SEQ ID NO:1; подчеркнуты), и домен hFc. Аспартат, замещенный в положении 79 в нативной последовательности, подчеркнут двойной линией и выделен, как и глицин, выявленный секвенированием как N-концевой остаток в зрелом слитом белке.
На фиг. 4 показана нуклеотидная последовательность, кодирующая ActRIIB(L79D 20-134)-hFc. SEQ ID NO: 25 соответствует смысловой нити, а SEQ ID NO: 33 соответствует антисмысловой нити. Лидер TPA (нуклеотиды 1-66) подчеркнут двойной линией, а внеклеточный домен ActRIIB (нуклеотиды 76-420) подчеркнут одинарной линией.
На фиг. 5 показана полная аминокислотная последовательность усеченного ActRIIB(L79D 25-131)-hFc (SEQ ID NO:26) ловушки GDF, включающая лидер TPA (подчеркнут двойной линией), внеклеточный домен усеченного ActRIIB (остатки 25-131 в SEQ ID NO:1; подчеркнута), и домен hFc. Аспартат, замещенный в положении 79 нативной последовательности, подчеркнут двойной линией и выделен, как и глутамат, выявленный секвенированием как N-концевой остаток в зрелом слитом белке.
На фиг. 6 показана нуклеотидная последовательность, кодирующая ActRIIB(L79D 25-131)-hFc. SEQ ID NO:27 соответствует смысловой нити, а SEQ ID NO: 34 соответствует антисмысловой нити. Лидер TPA (нуклеотиды 1-66) подчеркнут двойной линией, а внеклеточный домен усеченного ActRIIB (нуклеотиды 76-396) подчеркнут одинарной линией. Также показана аминокислотная последовательность для внеклеточного домена ActRIIB (остатки 25-131 в SEQ ID NO:1).
На фиг. 7 показана аминокислотная последовательность усеченного ActRIIB(L79D 25-131)-hFc ловушки GDF без лидера (SEQ ID NO:28). Внеклеточный домен усеченного ActRIIB (остатки 25-131 в SEQ ID NO:1) подчеркнут. Аспартат, замещенный в положении 79 нативной последовательности, подчеркнут двойной линий, как и глутамат, выявленный секвенированием как N-концевой остаток в зрелом слитом белке.
На фиг. 8 показана аминокислотная последовательность усеченного ActRIIB(L79D 25-131) ловушки GDF без лидера, домена hFc и линкера (SEQ ID NO:29). Аспартат, замещенный в положении 79 нативной последовательности подчеркнут и выделен, как и глутамат, выявленный секвенированием как N-концевой остаток в зрелом слитом белке.
На фиг. 9 показана альтернативная нуклеотидная последовательность, кодирующая ActRIIB(L79D 25-131)-hFc. SEQ ID NO:30 соответствует смысловой нити, и SEQ ID NO:35 соответствует антисмысловой нити. Лидер TPA (нуклеотиды 1-66) подчеркнут двойной линией, внеклеточный домен усеченного ActRIIB (нуклеотиды 76-396) подчеркнут одинарной линией, и замещения в нуклеотидной последовательности дикого типа внеклеточного домена подчеркнуты двойной линией и выделены (сравнить с SEQ ID NO:27, фиг. 6). Показана также аминокислотная последовательность внеклеточного домена ActRIIB (остатки 25-131 в SEQ ID NO:1).
На фиг. 10 показаны нуклеотиды 76-396 (SEQ ID NO:31) альтернативной нуклеотидной последовательности, показанной на фиг. 9 (SEQ ID NO:30). Так же подчеркнуты и выделены подобные нуклеотидные замещения, указанные на фиг. 9. SEQ ID NO:31 кодирует только внеклеточный домен усеченного ActRIIB (соответствующий остаткам 25-131 в SEQ ID NO:1) с замещением L79D, например, ActRIIB(L79D 25-131).
На фиг. 11 показано воздействие ActRIIB(L79D 25-131)-hFc на концентрацию гемоглобина на модели анемии у мыши, вызванной химиотерапией. Данные представляют собой средние величины ±SEM (стандартная ошибка средней). **, P<0,01 в сравнении с паклитакселом в ту же точку времени. Данная ловушка GDF компенсировала анемию, вызванную лечением паклитакселом.
На фиг. 12 показано воздействие ActRIIB(L79D 25-131)-hFc на уровни эритроцитов (RBC) на модели хронической болезни почек у мыши, вызванной односторонней нефрэктомией (NEPHX). Данные представляют собой средние величины ±SEM. ***, P<0,001 в сравнении с исходным уровнем. Эта ловушка GDF устраняла вызванную нефрэктомией анемию, наблюдавшуюся у контрольных мышей.
На фиг. 13 показано воздействие ActRIIB(L79D 25-131)-hFc на уровни эритроцитов (RBC), гемоглобина (HGB) и гематокрита (HCT) модели хронической болезни почек у мыши, вызванной односторонней нефрэктомией (NEPHX). Данные представляют собой средние изменения от исходного уровня в течение 4 недель (± SEM). *, P<0,05; **, P<0,01; ***, P<0,001 в сравнении с контролем NEPHX. Эта ловушка GDF предотвращала связанное с нефрэктомией снижение параметров эритроцитов, увеличивая каждый на величину, аналогичную величине у мышей с интактными почками (ложно оперированных).
На фиг. 14 показано воздействие ActRIIB(L79D 25-131)-hFc на уровни эритроцитов (RBC) на модели анемии у крысы, вызванной острой кровопотерей. Удаление крови происходило в день -1, при введении в дни 0 и 3. Данные представляют собой средние величины ± SEM. **, P<0,01; ***, P<0,001 в сравнении с носителем в ту же точку времени. Эта ловушка GDF улучшала скорость и степень выздоровления при анемии, вызванной кровопотерей.
На фиг. 15 показано воздействие лечения ActRIIB(L79D 20-134)-hFc (серые столбцы) или ActRIIB(L79D 25-131)-hFc (черные столбы) на абсолютное изменение концентрации эритроцитов от исходного уровня у яванских макак. VEH = носитель. Данные представляют собой средние величины +SEM. n=4-8 на группу.
На фиг. 16 показано воздействие лечения ActRIIB(L79D 20-134)-hFc (серые столбцы) или ActRIIB(L79D 25-131)-hFc (черные столбы) на абсолютное изменение гематокрита от исходного уровня у яванских макак. VEH = носитель. Данные представляют собой средние величины +SEM. n=4-8 на группу.
На фиг. 17 показано воздействие лечения ActRIIB(L79D 20-134)-hFc (серые столбцы) или ActRIIB(L79D 25-131)-hFc (черные столбцы) на абсолютное изменение концентрации гемоглобина от исходного уровня у яванских макак. VEH = носитель. Данные представляют собой средние величины +SEM. n=4-8 на группу.
На фиг. 18 показано воздействие лечения ActRIIB(L79D 20-134)-hFc (серые столбцы) или ActRIIB(L79D 25-131)-hFc (черные столбцы) на абсолютное изменение концентрации ретикулоцитов в циркулирующей крови от исходного уровня у яванских макак. VEH = носитель. Данные представляют собой средние величины +SEM. n=4-8 на группу.
На фиг. 19 показано воздействие комбинированного лечения эритропоэтином (EPO) и ActRIIB(L79D 25-131)-hFc в течение 72 часов на гематокрит у мышей. Данные представляют собой средние величины ± SEM (n=4 на группу), и средние величины, которые значимо отличаются друг от друга (p<0,05, непарный t-критерий) обозначены другими буквами. Комбинированное лечение повышало гематокрит на 23%, по сравнению с носителем, синергическое увеличение больше чем сумма отдельных эффектов EPO и ActRIIB(L79D 25-131)-hFc.
На фиг. 20 показано воздействие комбинированного лечения EPO и ActRIIB(L79D 25-131)-hFc в течение 72 часов на величины концентрации гемоглобина у мышей. Данные представляют собой средние величины ± SEM (n=4 на группу), и средние величины, которые значимо отличаются друг от друга (p<0,05) обозначены другими буквами. Комбинированное лечение повышало величины концентрации гемоглобина на 23%, по сравнению с носителем, что также представляло собой синергический эффект.
На фиг. 21 показано воздействие комбинированного лечения EPO и ActRIIB(L79D 25-131)-hFc в течение 72 часов на величины концентрации эритроцитов у мышей. Данные представляют собой средние величины ± SEM (n=4 на группу), и средние величины, которые значимо отличаются друг от друга (p<0,05) обозначены другими буквами. Комбинированное лечение повышало величины концентрации эритроцитов на 20%, по сравнению с носителем, что также представляло собой синергический эффект.
На фиг. 22 показано воздействие комбинированного лечения EPO и ActRIIB(L79D 25-131)-hFc в течение 72 часов на количества эритропоэтических клеток-предшественников в селезенке мыши. Данные представляют собой средние величины ± SEM (n=4 на группу), и средние величины, которые значимо отличаются друг от друга (p<0,01) обозначены другими буквами. В то время, как один EPO резко увеличивал число базофильных эритробластов (BasoE) за счет созревания предшественников поздней стадии, комбинированное лечение повышало количество BasoE в меньшей, но все же значимой степени, в то же время, поддерживая не уменьшенное созревание предшественников поздней стадии.
ДЕТАЛЬНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
1. Обзор
EPO представляет собой гликопротеиновый гормон, участвующий в росте и созревании эритроидных клеток-предшественников в эритроциты. EPO продуцируется печенью во время внутриутробной жизни и почками у взрослых. Сниженная продукция EPO, которая обычно возникает у взрослых вследствие почечной недостаточности, приводит к анемии. EPO был получен способами генной инженерии на основании экспрессии и секреции белка из клетки-хозяина, трансфицированной геном EPO. Введение такого рекомбинантного EPO было эффективно при лечении анемии. Например, Eschbach et al. (1987, N Engl J Med 316:73) описывают применение EPO для коррекции анемии, вызванной хронической почечной недостаточностью.
Эффекты EPO опосредуются через его связывание с рецептором клеточной поверхности, относящимся к суперсемейству цитокиновых рецепторов, и обозначенным рецептором EPO, и его активацию. Рецепторы EPO человека и мыши были клонированы и экспрессированы (D’Andrea et al., 1989, Cell 57:277; Jones et al., 1990, Blood 76:31; Winkelman et al., 1990, Blood 76:24; WO 90/08822/патент США № 5278065). Ген рецептора EPO человека кодирует трансмембранный белок длиной 483 аминокислоты, содержащий внеклеточный домен длиной приблизительно 224 аминокислоты и проявляет идентичность аминокислотной последовательности приблизительно 82% с рецептором EPO мыши (см. патент США № 6319499). Клонированный полноразмерный рецептор EPO, экспрессированный в клетках млекопитающих (66-72 кДа) связывает EPO со сродством (KD=100-300 нМ), аналогичным сродству нативного рецептора на эритроидных клетках-предшественниках. Таким образом, считается, что такая форма содержит основную детерминанту связывания EPO и называется рецептором EPO. По аналогии с другими тесно связанными цитокиновыми рецепторами, считается, что рецептор EPO димеризируется после связывания агониста. Тем не менее, детальная структура рецептора EPO, который может представлять собой многомерный комплекс, и специфический механизм его активации полностью не выяснены (патент США № 6319499).
Активация рецептора EPO приводит к нескольким биологическим эффектам. Они включают повышенную пролиферацию незрелых эритробластов, увеличенную дифференциацию незрелых эритробластов и уменьшенный апоптоз у эритроидных клеток-предшественников (Liboi et al., 1993, Proc Natl Acad Sci USA 90:11351-11355; Koury et al., 1990, Science 248:378-381). По-видимому, пути передачи сигналов рецепторов EPO, опосредующие пролиферацию и дифференциацию, являются индивидуальными (Noguchi et al., 1988, Mol Cell Biol 8:2604; Patel et al., 1992, J Biol Chem 1992, 267:21300; Liboi et al., там же). Некоторые данные свидетельствуют о том, что для того, чтобы опосредовать сигнал дифференциации может потребоваться вспомогательный белок (Chiba et al., 1993, Nature 362:646; Chiba et al., 1993, Proc Natl Acad Sci USA 90:11593); однако существует ряд противоречивых данных относительно роли вспомогательных белков при дифференциации, поскольку конститутивно активируемая форма рецептора может стимулировать как пролиферацию, так и дифференциацию (Pharr et al., 1993, Proc Natl Acad Sci USA 90:938).
Активаторы рецепторов EPO включают средства, стимулирующие эритропоэз (ESA), а также соединения на основе EPO. Примером первого является димерный агонист на основе пептида, ковалентно связанный с полиэтиленгликолем (патентованное название Гематид), который продемонстрировал стимулирующие эритропоэз свойства у здоровых добровольцев и у пациентов и с хронической болезнью почек, и эндогенными анти-EPO антителами (Stead et al., 2006, Blood 108:1830-1834; Macdougall et al., 2009, N Engl J Med 361:1848-1855). Другие примеры включают ESA не на пептидной основе (Qureshi et al., 1999, Proc Natl Acad Sci USA 96:12156-12161).
Активаторы рецепторов EPO также включают соединения, которые косвенно стимулируют эритропоэз, не вступая в контакт с самим рецептором EPO, путем усиления продукции эндогенного EPO. Например, индуцируемые гипоксией транскрипционные факторы (HIF) представляют собой эндогенные стимуляторы генной экспрессии EPO, которые подавлены (дестабилизированы) в нормоксических условиях клеточными регуляторными механизмами. Поэтому, ингибиторы ферментов пролил-гидроксилазы HIF были исследованы для выявления индуцирующей EPO активности in vivo. Другие непрямые активаторы рецептора EPO включают ингибиторы фактора транскрипции GATA-2 (Nakano et al., 2004, Blood 104:4300-4307), который тонически ингибирует генную экспрессию EPO, и ингибиторы фосфатазы гематопоэтических клеток (HCP или SHP-1), которая функционирует в качестве отрицательного регулятора трансдукции сигналов рецепторов EPO (Klingmuller et al., 1995, Cell 80:729-738).
Суперсемейство трансформирующего фактора роста-бета (TGF-бета) содержит разнообразные факторы роста, которые разделяют общие элементы последовательности и структурные мотивы. Известно, что эти белки оказывают биологические эффекты на большое разнообразие типов клеток и у позвоночных, и беспозвоночных. Члены суперсемейства выполняют важные функции во время эмбрионального развития при формировании типа и спецификации ткани и могут влиять на разнообразные процессы дифференциации, включая адипогенез, миогенез, хондрогенез, кардиогенез, гематопоэз, нейрогенез и дифференциацию эпителиальных клеток. Семейство делится на две общие ветви: ветви BMP/GDF и TGF-бета/Активина/BMP10, члены которых оказывают разнообразные, часто дополняющие эффекты. Путем манипулирования активностью члена семейства TGF-бета, часто можно вызвать значительные физиологические изменения в организме. Например, пьемонская и бельгийская голубая породы коров несут мутацию утраты функции у гена GDF8 (также называемого миостатином), которая вызывает выраженное увеличение мышечной массы (Grobet et al., Nat Genet. 1997, 17(1):71-4). Кроме того, у людей неактивные аллели GDF8 связаны с увеличенной мышечной массой и, по сообщениям, исключительную силу (Schuelke et al., N Engl J Med 2004, 350:2682-8).
Сигналы TGF-β опосредуются гетеромерными комплексами рецепторов серин/треонин-киназы I типа и II типа, которые фосфорилируют и активируют находящиеся ниже по ходу транскрипции белки Smad после стимуляции лигандом (Massagué, 2000, Nat. Rev. Mol. Cell Biol. 1:169-178). Эти рецепторы I типа и II типа представляют собой трансмембранные белки, состоящие из лиганд-связывающего внеклеточного домена с обогащенной цистеином областью, трансмембранного домена и цитоплазматического домена с прогнозируемой серин/треониновой специфичностью. Рецепторы I типа существенны для передачи сигналов. Рецепторы II типа требуются для связывания лигандов и для экспрессии рецепторов I типа. Рецепторы активина I и II типа образуют устойчивый комплекс после связывания лиганда, приводя к фосфорилированию рецепторов I типа рецепторами II типа.
Два связанных рецептора II типа (ActRII), ActRIIA и ActRIIB, были идентифицированы как рецепторы II типа для активинов (Mathews and Vale, 1991, Cell 65:973-982; Attisano et al., 1992, Cell 68: 97-108). Кроме активинов, ActRIIA и ActRIIB могут биохимически взаимодействовать с несколькими другими белками семейства TGF-β, включая BMP7, Nodal, GDF8 и GDF11 (Yamashita et al., 1995, J. Cell Biol. 130:217-226; Lee and McPherron, 2001, Proc. Natl. Acad. Sci. 98:9306-9311; Yeo and Whitman, 2001, Mol. Cell 7: 949-957; Oh et al., 2002, Genes Dev. 16:2749-54). ALK4 представляет собой первичный рецептор I типа для активинов, в частности, для активина A, и ALK-7 также может служить в качестве рецептора для активинов, в частности, для активина B. В определенных вариантах осуществления, настоящее изобретение относится к антагонизму действия в отношении лиганда рецепторов ActRIIB (также называемого лигандом ActRIIB) полипептидом-ловушкой GDF по изобретению. Иллюстративные лиганды рецепторов ActRIIB включают некоторые члены семейства TGF-β, такие как активин,Nodal, GDF8, GDF11 и BMP7.
Активины представляют собой димерные полипептидные факторы роста, которые относятся к суперсемейству TGF-бета. Существуют три основные формы активина (A, B и AB), которые представляют собой гомо/гетеродимеры двух тесно связанных β субъединиц (соответственно βAβA, βBβB и βAβB). Геном человека также кодирует активин C и активин E, которые в первую очередь экспрессированы в печени, и также известны гетеродимерные формы, содержащие βC или βE. В суперсемействе TGF-бета, активины представляет собой уникальные и многофункциональные факторы, которые могут стимулировать продукцию гормонов в клетках яичников и плаценты, поддерживать выживание нейронных клеток, положительно или отрицательно влиять на ход клеточного цикла, в зависимости от типа клеток, и вызывать мезодермальную дифференциацию по меньшей мере у эмбрионов земноводных (DePaolo et al., 1991, Proc Soc Ep Biol Med. 198:500-512; Dyson et al., 1997, Curr Biol. 7:81-84; Woodruff, 1998, Biochem Pharmacol. 55:953-963). Кроме того, было обнаружено, что фактор эритроидной дифференциации (EDF), выделенный из стимулированных моноцитарных лейкозных клеток человека, идентичен активину A (Murata et al., 1988, PNAS, 85:2434). Предполагалось, что активин A стимулирует эритропоэз в костном мозге. В нескольких тканях, передаче сигналов активина противодействует его родственный гетеродимер, ингибин. Например, во время высвобождения фолликулостимулирующего гормона (ФСГ) из гипофиза, активин стимулирует секрецию и синтез ФСГ, в то время как ингибин предотвращает секрецию и синтез ФСГ. Другие белки, которые могут регулировать биологическую активность активина и/или связываться с активином, включают фоллистатин (FS), связанный с фоллистатином белок (FSRP) и α2-макроглобулин.
Белки Nodal выполняют функции при индукции и образовании мезодермы и эндодермы, а также последующей организации осевых структур, таких как сердце и желудок, при раннем эмбриогенезе. Было продемонстрировано, что дорзальная ткань в развивающемся эмбрионе позвоночного вносит вклад преимущественно в осевые структуры спинной струны и прехордальной пластины в то время как он вовлекает окружающие клетки в образование не осевых эмбриональных структур. Представляется, что Nodal передает сигналы через рецепторы и I типа, и II типа, и внутриклеточные эффекторы, известные как белки Smad. Недавние исследования подтверждают идею о том, что ActRIIA и ActRIIB служат в качестве рецепторов II типа для Nodal (Sakuma et al., Genes Cells. 2002, 7:401-12). Предполагается, что лиганды Nodal взаимодействуют с их кофакторами (например, крипто) для активации рецепторов активина I типа и II типа, которые фосфорилируют Smad2. Белки Nodal участвуют во многих явлениях, играющих решающую роль при раннем развитии эмбриона позвоночных, в том числе образование мезодермы, переднее структурирование и спецификацию левой-правой оси. Экспериментальные данные продемонстрировали, что передача Nodal сигналов активирует pAR3-Lux, репортерную люциферазу, который, как было показано, специфически реагирует на активин и TGF-бета. Однако Nodal неспособен индуцировать pTlx2-Lux, репортер, специфически реагирующий на костные морфогенетические белки. Недавно полученные данные обеспечивают прямое биохимическое доказательство, что нодальная передача сигналов опосредуется обоими Smad пути активина-TGF-бета, Smad2 и Smad3. Еще одни данные показали, что внеклеточный крипто белок требуется для нодальной передачи сигналов, делая ее отличной от передачи сигналов активина или TGF-бета.
Фактор-8 роста и дифференциации (GDF8) также известен как миостатин. GDF8 представляет собой негативный регулятор массы скелетных мышц. GDF8 высоко экспрессирован в развивающейся и взрослой скелетной мышце. Нулевая мутация GDF8 у трансгенных мышей характеризуется выраженной гипертрофией и гиперплазией скелетной мышцы (McPherron et al., Nature, 1997, 387:83-90). Аналогичные увеличения массы скелетных мышц очевидны при природных мутациях GDF8 у крупного рогатого скота (Ashmore et al., 1974, Growth, 38:501-507; Swatland and Kieffer, J. Anim. Sci., 1994, 38:752-757; McPherron and Lee, Proc. Natl. Acad. Sci. USA, 1997, 94:12457-12461; и Kambadur et al., Genome Res., 1997, 7:910-915) и, что поразительно, у людей (Schuelke et al., N Engl J Med 2004;350:2682-8). Исследования также показали, что мышечная атрофия, связанная с ВИЧ инфекцией у людей, сопровождается увеличением экспрессии белка GDF8 (Gonzalez-Cadavid et al., PNAS, 1998, 95:14938-43). Кроме того, GDF8 может модулировать продукцию специфичных для мышц ферментов (например, креатинкиназы) и модулировать клеточную пролиферацию миобластов (WO 00/43781). Пропептид GDF8 может не ковалентно связываться с димером зрелого домена GDF8, инактивируя его биологическую активность (Miyazono et al. (1988) J. Biol. Chem., 263: 6407-6415; Wakefield et al. (1988) J. Biol. Chem., 263; 7646-7654; и Brown et al. (1990) Growth Factors, 3: 35-43). Другие белки, которые связываются с GDF8 или структурно связанными белками и ингибируют их биологическую активность, включают фоллистатин и, потенциально, связанные с фоллистатином белки (Gamer et al. (1999) Dev. Biol., 208: 222-232).
Фактор-11 роста и дифференциации (GDF11), также известный как BMPII, представляет собой секретируемый белок (McPherron et al., 1999, Nat. Genet. 22: 260-264). GDF11 экспрессирован в хвостовой почке, почке конечности, верхнечелюстной и нижнечелюстной арках и ганглиях дорзальных корешков во время развития мыши (Nakashima et al., 1999, Mech. Dev. 80: 185-189). GDF11 играет уникальную роль в структурировании и мезодермальной, и нервной ткани (Gamer et al., 1999, Dev Biol., 208:222-32). Было показано, что GDF11 представляет собой негативный регулятор хондрогенеза и миогенеза в развивающейся конечности цыпленка (Gamer et al., 2001, Dev Biol. 229:407-20). Экспрессия GDF11 в мышце также свидетельствует о его роли в регуляции роста мышц, аналогично GDF8. Кроме того, экспрессия GDF11 в головном мозге предполагает, что GDF11 может также обладать видами активности, которые связаны с функцией нервной системы. Представляет интерес, что как было обнаружено, GDF11 ингибирует нейрогенез в обонятельном эпителии (Wu et al., 2003, Neuron. 37:197-207). Следовательно, GDF11 может применяться in vitro и in vivo при лечении таких заболеваний как заболевания мышц и нейродегенеративные заболевания (например, боковой амиотрофический склероз).
Хорошо известно, что костный морфогенетический белок (BMP7), также называемый остеогенным белком-1 (OP-1), индуцирует образование хряща и кости. Кроме того, BMP7 регулирует широкую серию физиологических процессов. Например, BMP7 может представлять собой остеоиндуктивный фактор, ответственный за феномен эпителиального остеогенеза. Обнаружено также, что BMP7 играет роль в регуляции кальция и костном гомеостазе. Подобно активину, BMP7 связывается с рецепторами II типа, ActRIIA и ActRIIB. Однако BMP7 и активин задействуют определенные рецепторы I типа в гетеромерные рецепторные комплексы. Основной наблюдаемый рецептор BMP7 I типа представлял собой ALK2, в то время как активин связывался исключительно с ALK4 (ActRIIB). BMP7 и активин вызывали отчетливые биологические реакции и активировали различные пути Smad (Macias-Silva et al., 1998, J Biol Chem. 273:25628-36).
Как продемонстрировано в настоящем описании, полипептид-ловушка GDF, который представляет собой вариантный полипептид ActRIIB (ActRIIB), более эффективен для повышения содержания эритроцитов in vivo, по сравнению с полипептидом растворимых ActRIIB дикого типа и оказывает благоприятные эффекты на разнообразных моделях анемии. Кроме того, известно, что применение полипептида-ловушки GDF в комбинации с активатором рецептора EPO вызывает существенное повышение образования эритроцитов. Следует отметить, что гематопоэз представляет собой сложный процесс, регулируемый разнообразными факторами, включая эритропоэтин, G-CSF и гомеостаз железа. Термин «повышение содержания эритроцитов» и «стимуляция образования эритроцитов» относятся к клинически наблюдаемым измеряемым показателям, таким как измеряемые величины гематокрита, количества эритроцитов и гемоглобина, и предназначены быть нейтральными в отношении механизма, посредством которого происходят такие изменения.
Кроме стимуляции уровней эритроцитов, полипептиды ловушки GDF могут использоваться для различных видов терапевтического использования, включая, например, усиление мышечного роста (см. Публикации PCT № WO 2006/012627 и WO 2008/097541, которые полностью включены в настоящее описание в качестве ссылки). В определенных случаях, при введении полипептида-ловушки GDF для увеличения мышц, может быть желательно уменьшение или минимизация воздействий на эритроциты. Путем мониторинга различных гематологических параметров у пациентов, получающих лечение полипептидом-ловушкой GDF или являющихся претендентами на такое лечение, соответствующая дозировка (включая количества вводимого полипептида и частоту введения) может быть определена на основании потребностей отдельного пациента, исходных гематологических параметров и цели лечения. Кроме того, ход лечения и его воздействия на один или более гематологических параметров с течением времени могут использоваться при наблюдении за пациентами, которым вводится полипептид-ловушка GDF, путем содействия уходу за пациентом, определения соответствующей поддерживающей дозы (и количества, и частоты) и т.д.
Термины, используемые в данном описании, в целом имеют их обычные значения в данной области, в пределах контекста настоящего изобретения и в определенном контексте, где используется каждый термин. Определенные термины обсуждаются ниже и в других местах в описании для предоставления дополнительного руководства практикующим специалистам при описании композиций и способов по изобретению, и объяснения способов их получения и применения. Объем или значение любого использования термина будет очевидно из контекста описания, в котором используется термин.
«Примерно» и «приблизительно» в целом обозначают приемлемую степень ошибки для измеряемого количества с учетом сущности или точности измерений. Обычно, иллюстративные степени ошибки находятся в пределах 20%, предпочтительно, в пределах 10%, более предпочтительно, в пределах 5% данной величины или диапазона величин.
Альтернативно, и, в частности, в биологических системах, термины «примерно» и «приблизительно» могут означать величины, которые находятся в пределах порядка величины, предпочтительно, в пределах 5-кратного, более предпочтительно, 2-кратной данной величины. Если нет других указаний, то числовые величины, приведенные в настоящем описании, являются приблизительными, означая, что если нет конкретного указания, то может подразумеваться термин «примерно» или «приблизительно».
Способы по изобретению могут включать стадии сравнения последовательностей друг с другом, включая сравнение последовательности дикого типа с одним или более мутантами (вариантами последовательности). Такие сравнения обычно включают выравнивание полимерных последовательностей, например, с использованием программ и/или алгоритмов выравнивания последовательностей, которые хорошо известны в данной области (например, BLAST, FASTA и MEGALIGN и многие другие). Опытный специалист в данной области может легко понять, что при таких выравниваниях, когда мутация содержит вставку или делецию остатка, выравнивание последовательностей введет «гэп» (обычно представленный тире или буквой «A») в полимерной последовательности, не содержащей вставленный или делетированный остаток.
Термин «гомологичный» во всех его грамматических формах и вариантах написания относится к связи между двумя белками, которые имеют «общее эволюционное происхождение», включая белки из суперсемейств в одном и том же виде организма, а также гомологичные белки от различных видов организма. Такие белки (и кодирующие их нуклеиновые кислоты) имеют гомологию последовательностей, отраженную в сходстве их последовательностей, или с точки зрения процентной идентичности, или присутствием определенных остатков или мотивов и сохраненных положений.
Термин «сходство последовательностей» во всех грамматических формах относится к степени идентичности или соответствия между последовательностями нуклеиновых кислот или аминокислот, которые могут разделять или не разделять общее эволюционное происхождение.
Однако при обычном использовании и в настоящем применении термин «гомологичная» при модификации таким наречием «высоко», может относиться к сходству последовательностей и может относиться или не относиться к общему эволюционному происхождению.
2. Полипептиды-ловушки GDF
В определенных аспектах, изобретение относится к полипептидам ловушки GDF, например, вариантными полипептидами растворимых ActRIIB, включая, например, фрагменты, функциональные варианты и модифицированные формы полипептидов ActRIIB. В определенных вариантах осуществления, полипептиды ловушки GDF имеют по меньшей мере одну сходную или такую же биологическую активность, как соответствующий полипептид ActRIIB дикого типа. Например, полипептид-ловушка GDF по изобретению может связываться с лигандом ActRIIB (например, активином A, активином AB, активином B, Nodal, GDF8, GDF11 или BMP7) и ингибировать его функцию. Полипептид-ловушка GDF, возможно, увеличивает уровни эритроцитов. Примеры полипептидов ловушки GDF содержат полипептиды предшественников ActRIIB человека (SEQ ID NO:1 или 39), имеющие одно или более изменений последовательности, и растворимые полипептиды ActRIIB человека (например, SEQ ID NO:2, 3, 7, 11, 26, 28, 29, 32, 37, 38, 40 и 41), имеющие одно или более изменений последовательностей. Ловушка GDF относится к полипептиду ActRIIB, имеющему пониженное сродство к активну по сравнению с другими лигандами ActRIIB, в том числе, например, с GDF11 и/или миостатином.
Используемый в настоящем описании термин «ActRIIB» относится к семейству белков рецептора активина IIb типа (ActRIIB) любого вида и вариантов, полученных из таких белков ActRIIB, путем мутагенеза или другой модификации. Под ссылкой на ActRIIB следует понимать ссылку на любую из идентифицированных в настоящее время форм. Члены семейства ActRIIB представляют собой в целом трансмембранные белки, состоящие из лиганд-связывающего внеклеточного домена с обогащенной цистеином областью, трансмембранный домен и цитоплазматический домен с прогнозируемой активностью серин/треонин-киназы. Аминокислотные последовательности растворимого внеклеточного домена ActRIIA человека (предоставленные для сравнения) и растворимого внеклеточного домена ActRIIB показаны на фиг. 1.
Термин «полипептид ActRIIB» включает полипептиды, содержащие любой природный полипептид члена семейства ActRIIB, а также любые их варианты (включая мутанты, фрагменты, слитые и пептидомиметические формы), которые сохраняют полезную активность. См., например, документ WO 2006/012627. Например, полипептиды ActRIIB содержат полипептиды, полученные из последовательности любого известного ActRIIB, имеющие последовательность, которая по меньшей мере на 80% идентична последовательности полипептида ActRIIB, и, возможно по меньшей мере идентична на 85%, 90%, 95%, 97%, 99% или более. Например, полипептид ActRIIB может связываться с белком ActRIIB и/или активином и ингибировать их функцию. Полипептид ActRIIB, который представляет собой ловушку GDF, может быть выбран по активности содействия образованию эритроцитов in vivo. Примеры полипептидов ActRIIB включают полипептид-предшественник ActRIIB человека (SEQ ID NO:1 и 39) и растворимые полипептиды ActRIIB человека (например, SEQ ID NO:2, 3, 7, 11, 26, 28, 29, 32, 37, 38, 40 и 41). Пока нет определенных иных обозначений, нумерация аминокислот для всех связанных с ActRIIB полипептидов, описанных в настоящей заявке, основана на нумерации для SEQ ID NO:1.
Белок-предшественник ActRIIB человека имеет следующую последовательность: MTAPWVALALLWGSLWPGSGRGEAETRECIYYNANWELERTNQSGLERCEGEQDKRLHCYASWRNSSGTIELVKKGCWLDDFNCYDRQECVATEENPQVYFCCCEGNFCNERFTHLPEAGGPEVTYEPPPTAPTLLTVLAYSLLPIGGLSLIVLLAFWMYRHRKPPYGHVDIHEDPGPPPPSPLVGLKPLQLLEIKARGRFGCVWKAQLMNDFVAVKIFPLQDKQSWQSEREIFSTPGMKHENLLQFIAAEKRGSNLEVELWLITAFHDKGSLTDYLKGNIITWNELCHVAETMSRGLSYLHEDVPWCRGEGHKPSIAHRDFKSKNVLLKSDLTAVLADFGLAVRFEPGKPPGDTHGQVGTRRYMAPEVLEGAINFQRDAFLRIDMYAMGLVLWELVSRCKAADGPVDEYMLPFEEEIGQHPSLEELQEVVVHKKMRPTIKDHWLKHPGLAQLCVTIEECWDHDAEARLSAGCVEERVSLIRRSVNGTTSDCLVSLVTSVTNVDLPPKESSI (SEQ ID NO:1)
Сигнальный пептид подчеркнут одной чертой; внеклеточный домен выделен жирным шрифтом и потенциальные сайты N-связанного гликозилирования заключены в рамки.
В литературе также имеется сообщение о форме с аланином в положении 64, имеющей представленную ниже последовательность: MTAPWVALALLWGSLWPGSGRGEAETRECIYYNANWELERTNQSGLERCEGEQDKRLHCYASWANSSGTIELVKKGCWLDDFNCYDRQECVATEENPQVYFCCCEGNFCNERFTHLPEAGGPEVTYEPPPTAPTLLTVLAYSLLPIGGLSLIVLLAFWMYRHRKPPYGHVDIHEDPGPPPPSPLVGLKPLQLLEIKARGRFGCVWKAQLMNDFVAVKIFPLQDKQSWQSEREIFSTPGMKHENLLQFIAAEKRGSNLEVELWLITAFHDKGSLTDYLKGNIITWNELCHVAETMSRGLSYLHEDVPWCRGEGHKPSIAHRDFKSKNVLLKSDLTAVLADFGLAVRFEPGKPPGDTHGQVGTRRYMAPEVLEGAINFQRDAFLRIDMYAMGLVLWELVSRCKAADGPVDEYMLPFEEEIGQHPSLEELQEVVVHKKMRPTIKDHWLKHPGLAQLCVTIEECWDHDAEARLSAGCVEERVSLIRRSVNGTTSDCLVSLVTSVTNVDLPPKESSI (SEQ ID NO:39)
Последовательность растворимого (внеклеточного), обработанного ActRIIB человека имеет следующий вид:
GRGEAETRECIYYNANWELERTNQSGLERCEGEQDKRLHCYASWRNSSGTIELVKKGCWLDDFNCYDRQECVATEENPQVYFCCCEGNFCNERFTHLPEAGGPEVTYEPPPTAPT (SEQ ID NO:2)
Последовательность альтернативной формы с A64 имеет следующий вид:
GRGEAETRECIYYNANWELERTNQSGLERCEGEQDKRLHCYASWANSSGTIELVKKGCWLDDFNCYDRQECVATEENPQVYFCCCEGNFCNERFTHLPEAGGPEVTYEPPPTAPT (SEQ ID NO:40)
В некоторых условиях, белок может быть получен с последовательностью «SGR…» на N-конце. C-концевой «хвост» внеклеточного домена подчеркнут. Последовательность с делетированным «хвостом» (последовательность Δ15) имеет следующую структуру:
GRGEAETRECIYYNANWELERTNQSGLERCEGEQDKRLHCYASWRNSSGTIELVKKGCWLDDFNCYDRQECVATEENPQVYFCCCEGNFCNERFTHLPEA (SEQ ID NO:3)
Альтернативная форма с A64 представляет собой, как показано ниже:
GRGEAETRECIYYNANWELERTNQSGLERCEGEQDKRLHCYASWANSSGTIELVKKGCWLDDFNCYDRQECVATEENPQVYFCCCEGNFCNERFTHLPEA (SEQ ID NO: 41)
В некоторых условиях, белок может быть получен с последовательностью «SGR…» на N-конце. Последовательность нуклеиновой кислоты, кодирующая белок-предшественник ActRIIB человека, имеет следующую структуру: (нуклеотиды 5-1543 Генного банка, регистрация NM_001106) (показанная последовательность предусматривает аланин в положении 64, и может быть модифицирована для введения аргинина вместо аланина)
ATGACGGCGCCCTGGGTGGCCCTCGCCCTCCTCTGGGGATCGCTGTGGCCCGGCTCTGGGCGTGGGGAGGCTGAGACACGGGAGTGCATCTACTACAACGCCAACTGGGAGCTGGAGCGCACCAACCAGAGCGGCCTGGAGCGCTGCGAAGGCGAGCAGGACAAGCGGCTGCACTGCTACGCCTCCTGGGCCAACAGCTCTGGCACCATCGAGCTCGTGAAGAAGGGCTGCTGGCTAGATGACTTCAACTGCTACGATAGGCAGGAGTGTGTGGCCACTGAGGAGAACCCCCAGGTGTACTTCTGCTGCTGTGAAGGCAACTTCTGCAACGAGCGCTTCACTCATTTGCCAGAGGCTGGGGGCCCGGAAGTCACGTACGAGCCACCCCCGACAGCCCCCACCCTGCTCACGGTGCTGGCCTACTCACTGCTGCCCATCGGGGGCCTTTCCCTCATCGTCCTGCTGGCCTTTTGGATGTACCGGCATCGCAAGCCCCCCTACGGTCATGTGGACATCCATGAGGACCCTGGGCCTCCACCACCATCCCCTCTGGTGGGCCTGAAGCCACTGCAGCTGCTGGAGATCAAGGCTCGGGGGCGCTTTGGCTGTGTCTGGAAGGCCCAGCTCATGAATGACTTTGTAGCTGTCAAGATCTTCCCACTCCAGGACAAGCAGTCGTGGCAGAGTGAACGGGAGATCTTCAGCACACCTGGCATGAAGCACGAGAACCTGCTACAGTTCATTGCTGCCGAGAAGCGAGGCTCCAACCTCGAAGTAGAGCTGTGGCTCATCACGGCCTTCCATGACAAGGGCTCCCTCACGGATTACCTCAAGGGGAACATCATCACATGGAACGAACTGTGTCATGTAGCAGAGACGATGTCACGAGGCCTCTCATACCTGCATGAGGATGTGCCCTGGTGCCGTGGCGAGGGCCACAAGCCGTCTATTGCCCACAGGGACTTTAAAAGTAAGAATGTATTGCTGAAGAGCGACCTCACAGCCGTGCTGGCTGACTTTGGCTTGGCTGTTCGATTTGAGCCAGGGAAACCTCCAGGGGACACCCACGGACAGGTAGGCACGAGACGGTACATGGCTCCTGAGGTGCTCGAGGGAGCCATCAACTTCCAGAGAGATGCCTTCCTGCGCATTGACATGTATGCCATGGGGTTGGTGCTGTGGGAGCTTGTGTCTCGCTGCAAGGCTGCAGACGGACCCGTGGATGAGTACATGCTGCCCTTTGAGGAAGAGATTGGCCAGCACCCTTCGTTGGAGGAGCTGCAGGAGGTGGTGGTGCACAAGAAGATGAGGCCCACCATTAAAGATCACTGGTTGAAACACCCGGGCCTGGCCCAGCTTTGTGTGACCATCGAGGAGTGCTGGGACCATGATGCAGAGGCTCGCTTGTCCGCGGGCTGTGTGGAGGAGCGGGTGTCCCTGATTCGGAGGTCGGTCAACGGCACTACCTCGGACTGTCTCGTTTCCCTGGTGACCTCTGTCACCAATGTGGACCTGCCCCCTAAAGAGTCAAGCATCTAA (SEQ ID NO:4)
Последовательность нуклеиновой кислоты, кодирующая растворимый полипептид (внеклеточного) ActRIIB человека, имеет следующую структуру: (показанная последовательность предусматривает аланин в положении 64, и может быть модифицирована для предоставления аргинина вместо аланина):
GGGCGTGGGGAGGCTGAGACACGGGAGTGCATCTACTACAACGCCAACTGGGAGCTGGAGCGCACCAACCAGAGCGGCCTGGAGCGCTGCGAAGGCGAGCAGGACAAGCGGCTGCACTGCTACGCCTCCTGGGCCAACAGCTCTGGCACCATCGAGCTCGTGAAGAAGGGCTGCTGGCTAGATGACTTCAACTGCTACGATAGGCAGGAGTGTGTGGCCACTGAGGAGAACCCCCAGGTGTACTTCTGCTGCTGTGAAGGCAACTTCTGCAACGAGCGCTTCACTCATTTGCCAGAGGCTGGGGGCCCGGAAGTCACGTACGAGCCACCCCCGACAGCCCCCACC (SEQ ID NO:5)
В определенном варианте осуществления, изобретение относится к полипептидам-ловушкам GDF, которые представляют собой вариантные формы растворимых полипептидов ActRIIB. Как описано в настоящей заявке, термин «растворимый полипептид ActRIIB» в целом относится к полипептидам, содержащим внеклеточный домен белка ActRIIB. Термин «растворимый полипептид ActRIIB», используемый в настоящем описании, включает любой природный внеклеточный домен белка ActRIIB, а также любой его вариант (включая мутанты, фрагменты и пептидомиметические формы), которые сохраняют полезную активность. Например, внеклеточный домен белка ActRIIB связывается с лигандом и является в целом растворимым. Примеры растворимых полипептидов ActRIIB включают растворимые полипептиды ActRIIB (например, SEQ ID NO:22, 3, 7, 11, 26, 28, 29, 32, 37, 38, 40 и 41). Другие примеры растворимых полипептидов ActRIIB содержат сигнальную последовательность в дополнение к внеклеточному домену белка ActRIIB, см. пример 1. Сигнальная последовательность может представлять собой нативную сигнальную последовательность ActRIIB, или сигнальную последовательность из другого белка, такую как сигнальная последовательность тканевого активатора плазминогена (TPA) или сигнальная последовательность мелиттина медоносной пчелы (HBM).
В описании идентифицированы функционально активные части и варианты ActRIIB. Заявители выяснили, что Fc-слитый белок, имеющий последовательность, описанную Hilden et al. (Blood. 1994 Apr 15;83(8):2163-70), который имеет Аланин в положении, соответствующем аминокислоте 64 последовательности SEQ ID NO:1 (A64), имеет относительно низкое сродство к активину и GDF-11. Напротив, такой же Fc-слитый белок с Аргинином в положении 64 (R64) имеет сродство к активину и GDF-11 в диапазоне от низкого наномолярного до высокого пикомолярного. Поэтому, в настоящем описании последовательность с R64 используется в качестве контрольной последовательности дикого типа для ActRIIB человека.
В статье Attisano et al. (Cell. 1992 Jan 10; 68(1):97-108) показано, что делеция узла пролина на C-конце внеклеточного домена ActRIIB снижает сродство рецептора к активину. ActRIIB-Fc слитый белок, содержащий аминокислоты 20-119 последовательности SEQ ID NO:1, «ActRIIB(20-119)-Fc», имеет пониженное связывание с GDF-11 и активином относительно ActRIIB(20-134)-Fc, который включает область узла пролина и полный юкстамембранный домен. Однако ActRIIB(20-129)-Fc белок сохраняет аналогичную, но несколько пониженную активность по сравнению с диким типом, даже если область узла пролина разорвана. Таким образом, предполагается, что все внеклеточные домены ActRIIB, которые останавливаются у аминокислоты 134, 133, 132, 131, 130 и 129, активны, но конструкции, останавливающиеся у 134 или 133, могут быть наиболее активны. Аналогичным образом, не предполагается, что мутации в любом из остатков 129-134 изменят сродство связывания лигандов крупными краями. В подтверждение этого, мутации P129 и P130 по существу не уменьшают связывание лигандов. Поэтому, ожидается, что полипептид-ловушка GDF, который представляет собой ActRIIB-Fc слитый белок, и может закончиться уже аминокислотой 109 (конечным цистеином), но, однако, образует окончание у аминокислот 109 и 119 или между ними, имеет пониженное связывание лигандов. Аминокислота 119 слабо сохранена и поэтому легко изменяется или усекается. Формы, заканчивающиеся у аминокислоты 128 или позднее, сохраняют активность связывания лигандов. Формы, заканчивающиеся у аминокислот 119 и 127 или между ними, имеют промежуточную способность связывания. Может быть желательно использование любой из указанных форм, в зависимости от клинических или экспериментальных условий.
Ожидается, что на N-конце ActRIIB, белок, начинающийся с аминокислоты 29 или ранее, сохранит лигнад-связывающую активность. Аминокислота 29 представляет начальный цистеин. Мутация аланина в аспарагин в положении 24 вводит N-связанную последовательность гликозилирования без существенного воздействия на связывание лигандов. Это подтверждает, что мутации в области между сигнальным пептидом расщепления и поперечно сшитой цистеином областью, соответствующей 20-29, хорошо переносимы. В частности, конструкции, начинающиеся в положении 20, 21, 22, 23 и 24, сохранят активность, и ожидается, что конструкции, начинающиеся в положениях 25, 26, 27, 28 и 29, также сохранят активность. Данные, представленные в разделе «Примеры», демонстрируют, что, к удивлению, конструкция, начинающаяся с аминокислоты 22, 23, 24 или 25, будет иметь наибольшую активность.
Взятая вместе, активная часть ActRIIB содержит аминокислоты 29-109 последовательности SEQ ID NO:1, и конструкции ловушки GDF могут, например, содержать часть ActRIIB, начинающуюся с остатка, соответствующего аминокислотам 20-29 последовательности SEQ ID NO:1 или 39, и заканчивающуюся в положении, соответствующем аминокислотам 109-134 последовательности SEQ ID NO:1 или 39. Другие примеры включают конструкции, которые начинаются в положении от 20-29 или 21-29 и заканчиваются в положении от 119-134, 119-133, 129-134, или 129-133 последовательности SEQ ID NO:1 или 39. Другие примеры включают конструкции, которые начинаются в положении от 20-24 (или 21-24, или 22-25) и заканчиваются в положении от 109-134 (или 109-133), 119-134 (или 119-133) или 129-134 (или 129-133) последовательности SEQ ID NO: 1 или 39. Предусматриваются также варианты в указанных диапазонах, в частности, те, которые по меньшей мере на 80%, 85%, 90%, 95% или 99% идентичны соответствующей части последовательности SEQ ID NO:1 или 39. В определенных вариантах осуществлении, полипептид-ловушка GDF содержит, по существу состоит из или состоит из полипептида, имеющего аминокислотную последовательность, которая по меньшей мере на 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотным остаткам 25-131 последовательности SEQ ID NO:1 или 39. В определенных вариантах осуществления, пептид-ловушка GDF содержит, по существу состоит из или состоит из полипептида, имеющего аминокислотную последовательность по меньшей мере на 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичную последовательности SEQ ID NO:7, 26, 28, 29, 32, 37 или 38. В предпочтительных вариантах осуществления, пептид-ловушка GDF состоит из или по существу состоит из аминокислотной последовательности SEQ ID NO:7, 26, 28, 29, 32, 37 или 38.
Описание включает результаты анализа композитных структур ActRIIB, показанных на фиг. 1, демонстрирующие, что лиганд-связывающий карман ограничивается остатками Y31, N33, N35, L38 по T41, E47, E50, Q53 по K55, L57, H58, Y60, S62, K74, W78 по N83, Y85, R87, A92 и E94 по F101. Ожидается, что в указанных положениях, будут допустимы консервативные мутации, хотя мутация K74A хорошо переносима, как и R40A, K55A, F82A и мутации в положении L79. R40 представляет собой K у шпорцевой лягушки, указывая на то, что основные аминокислоты в данном положении будут допустимы. Q53 представляет собой R в бычьем ActRIIB и K в ActRIIB шпорцевой лягушки, и поэтому аминокислоты, включающие R, K, Q, N и H, будут допустимы в данном положении. Таким образом, общая формула для белка-ловушки GDF представляет собой ту, которая включает аминокислоты 29-109 последовательности SEQ ID NO:1 или 39, но, возможно, начинающийся в положении в диапазоне от 20-24 или 22-25, и заканчивающийся в положении в диапазоне от 129-134, и включающий не более чем 1, 2, 5, 10 или 15 консервативных аминокислотных изменений в лиганд-связывающем кармане, и ноль, одно или более неконсервативных изменений в положениях 40, 53, 55, 74, 79 и/или 82 в лиганд-связывающем кармане. Такой белок может сохранить более чем 80%, 90%, 95% или 99% идентичности последовательности с последовательностью аминокислот 29-109 SEQ ID NO: 1 или 39. Сайты вне связывающего кармана, в которых вариабельность может быть особенно хорошо переносима, включают амино и карбокси концы внеклеточного домена (как отмечено выше) и положения 42-46 и 65-73. Изменение аспарагина на аланин в положении 65 (N65A) действительно улучшает связывание лиганда в среде A64, и, таким образом, ожидается, что оно не окажет вредного воздействия на связывание лиганда в среде R64. Такое изменение, вероятно, устраняет гликозилирование в N65 в среде A64, таким образом, демонстрируя, что значительное изменение в данной области, вероятно, является переносимым. В то время как изменение R64A плохо переносимо, R64K хорошо переносимо, и, таким образом, другой основный остаток, такой как H, может быть переносим в положении 64.
ActRIIB хорошо сохранен почти у всех позвоночных при полном сохранении крупных фрагментов секвенирования внеклеточного домена. Многие из лигандов, которые связываются с ActRIIB, также высоко сохранны. Соответственно, сравнения последовательностей ActRIIB организмов различных позвоночных обеспечивают возможность выявления остатков, которые могут быть изменены. Поэтому, активный вариантный полипептид ActRIIB человека, используемый в качестве ловушки GDF, может включать одну или более аминокислот в соответствующих положениях из последовательности ActRIIB другого позвоночного, или может содержать остаток, который аналогичен остатку в последовательности человека или другого позвоночного. Следующие примеры иллюстрируют этот подход к определению активного варианта ActRIIB. L46 представляет собой валин в ActRIIB шпорцевой лягушки, и, таким образом, данное положение может быть изменено, и возможно его изменение на другой гидрофобный остаток, такой как V, I или F, или неполярный остаток, такой как A. E52 представляет собой K у шпорцевой лягушки, указывая на то, что этот сайт может быть устойчив к широкому разнообразию изменений, включая полярные остатки, такие как E, D, K, R, H, S, T, P, G, Y и, вероятно, A. T93 представляет собой K у шпорцевой лягушки, указывая на то, что в данном положении переносится широкое структурное изменение, причем предпочтительны полярные остатки, такие как S, K, R, E, D, H, G, P, G и Y. F108 представляет собой Y у шпорцевой лягушки, и поэтому, Y или другая гидрофобная группа, такая как I, V или L должна быть переносима. E111 представляет собой K у шпорцевой лягушки, указывая на то, что заряженные остатки будут переносимы в данном положении, включая D, R, K и H, а также Q и N. R112 представляет собой K у шпорцевой лягушки, указывая на то, что основные остатки переносимы в данном положении, включая R и H. A в положении 119 относительно слабо сохранен и появляется в виде P у грызунов и V у шпорцевой лягушки, таким образом, по существу любая аминокислота должна быть переносимой в данном положении.
Описание демонстрирует, что добавление еще одного сайта для N-сцепленного гликозилирования (N-X-S/T) увеличивает период полувыведения из сыворотки слитого белка ActRIIB-Fc относительно формы ActRIIB(R64)-Fc. Путем введения аспарагина в положение 24 (конструкция A24N), создается последовательность NXT, которая обеспечивает более длительный период полувыведения. Другие последовательности NX(T/S) обнаруживаются в положениях 42-44 (NQS) и 65-67 (NSS), хотя последние могут не быть эффективно гликозилированы R в положении 64. Последовательности N-X-S/T могут быть в целом введены в положения вне лиганд-связывающего кармана, определенного на фиг. 1. Особенно подходящие сайты для введения не эндогенных последовательностей N-X-S/T содержат аминокислоты 20-29, 20-24, 22-25, 109-134, 120-134 или 129-134. Последовательности N-X-S/T могут также вводиться в линкер между последовательностью ActRIIB и Fc или другими компонентом слияния. Такой сайт может быть введен с минимальным усилием путем введения N в правильное положение относительно предсуществующего S или T, или путем введения S или T в положение, соответствующее предсуществующему N. Таким образом, желательные изменения, которые создали бы N-сцепленный сайт гликозилирования, представляют собой: A24N, R64N, S67N (возможно, в комбинации с изменением N65A), E106N, R112N, G120N, E123N, P129N, A132N, R112S и R112T. Любой S, который прогнозируется как гликозилированный, может быть изменен в T без создания иммуногенного сайта, ввиду защиты, обеспечиваемой гликозилированием. Аналогичным образом, любой T, который прогнозируется как гликозилированный, может быть изменен в S. Таким образом, предусматриваются изменения S67T и S44T. Аналогичным образом, в варианте A24N, может использоваться изменение S26T. Соответственно, ловушка GDF может представлять собой вариант ActRIIB, имеющий одну или более дополнительных, не эндогенных N-сцепленных гликолизированных консенсусных последовательностей.
Положение L79 ActRIIB может быть изменено для придания измененных свойств связывания активина-миостатина (GDF-11). L79A или L79P снижает связывание GDF-11 в большей степени, чем связывание активина. L79E или L79D сохраняет связывание GDF-11. Примечательно, что варианты L79E и L79D имеют значительно пониженное связывание активина. Эксперименты in vivo указывают на то, что эти неактивные рецепторы сохраняют значительную способность повышать содержание эритроцитов, но проявляют уменьшенные эффекты на другие ткани. Указанные данные демонстрируют желательность и осуществимость получения полипептидов с пониженными эффектами на активин. В иллюстративных вариантах осуществления, в способах, описанных в настоящей заявке, используется полипептид-ловушка GDF, который представляет собой вариантный полипептид ActRIIB, содержащий кислую аминокислоту (например, D или E) в положении, соответствующем положению 79 последовательности SEQ ID NO: 1 или 39, возможно, в комбинации с одним или более дополнительных аминокислотных замен, добавлений или делеций.
Описанные изменения могут комбинироваться различными путями. Кроме того, результаты описанной в настоящей заявке программы мутагенеза, указывают на то, что имеются положения аминокислот в ActRIIB, сохранение которых часто благоприятно. Они включают положение 64 (основная аминокислота), положение 80 (кислая или гидрофобная аминокислота), положение 78 (гидрофобная и, в частности, триптофан), положение 37 (кислая и, в частности, аспарагиновая и глутаминовая кислота), положение 56 (основная аминокислота), положение 60 (гидрофобная аминокислота, в частности, фенилаланин или тирозин). Таким образом, в каждом из вариантов, описанных в настоящей заявке, в описании представлен набор аминокислот, которые могут сохранены. Другие положения, сохранение которых может быть желательно, представляют собой следующие: положение 52 (кислая аминокислота), положение 55 (основная аминокислота), положение 81 (кислая аминокислота), 98 (полярная или заряженная, в частности, E, D, R или K).
В определенных вариантах осуществления, выделенные фрагменты полипептидов ActRIIB могут быть получены путем скрининга рекомбинантных полипептидов, полученных из соответствующего фрагмента нуклеиновой кислоты, кодирующей полипептид ActRIIB (например, SEQ ID NO:4 и 5). Кроме того, фрагменты могут быть химически синтезированы с использованием технологий, известных в данной области, таких как обычные химические способы твердофазного пептидного синтеза по методу Merrifield с использованием f-Moc (9-флуоренилметоксикарбонила) или t-Boc (трет-бутоксикарбонила). Фрагменты могут быть получены (рекомбинантными методами или химическим синтезом) и тестироваться для идентификации тех пептидильных фрагментов, которые могут функционировать, например, в качестве антагонистов (ингибиторов) или агонистов (активаторов) белка ActRIIB или лиганда ActRIIB.
В определенных вариантах осуществления, пептид-ловушка GDF представляет собой вариантный полипептид ActRIIB, имеющий аминокислотную последовательность, которая по меньшей мере на 75% идентична аминокислотной последовательности, выбранной из SEQ ID NO:2, 3, 7, 11, 26, 28, 29, 32, 37, 38, 40 или 41. В определенных случаях, ловушка GDF имеет аминокислотную последовательность по меньшей мере на 80%, 85%, 90%, 95%, 97%, 98%, 99% или 100% идентичную аминокислотной последовательности, выбранной из последовательности SEQ ID NO:2, 3, 7, 11, 26, 28, 29, 32, 37, 38, 40 или 41. В определенных вариантах осуществления, ловушка GDF содержит, по существу состоит из или состоит из аминокислотной последовательности по меньшей мере на 80%, 85%, 90%, 95%, 97%, 98%, 99% или 100% идентичной аминокислотной последовательности, выбранной из последовательности SEQ ID NO:2, 3, 7, 11, 26, 28, 29, 32, 37, 38, 40 или 41, где положение, соответствующее L79 последовательности SEQ ID NO:1, представляет собой кислую аминокислоту (например, аминокислотный остаток D или E).
В определенных вариантах осуществления, настоящее изобретение предусматривает получение функциональных вариантов путем модификации структуры полипептида-ловушки GDF для таких целей как повышение терапевтической эффективности, или устойчивости (например, при хранении ex vivo и устойчивости к протеолитическому разрушению in vivo). Полипептиды-ловушки GDF могут быть также получены заменами, делецией или добавлением аминокислот. Например, целесообразно ожидать, что изолированное замещение лейцина изолейцином или валином, аспартата глутамином, треонина серином или аналогичное замещение аминокислоты структурно связанной аминокислотой (например, консервативные мутации) не окажут существенного эффекта на биологическую активность полученной молекулы. Консервативные замены представляют собой те, которые имеют место в пределах семейства аминокислот, которые родственны своими боковыми цепями. То, приводит ли изменение аминокислотной последовательности полипептида-ловушки GDF к получению функционального варианта, можно легко определить оценкой способности полипептида-ловушки GDF вызывать реакцию в клетках относительно немодифицированного полипептида-ловушки GDF или полипептида ActRIIB дикого типа, или связываться с одним или более лигандами, такими как активин, GDF-11 или миостатин, по сравнению с немодифицированным полипептидом-ловушкой GDF или полипептидом ActRIIB дикого типа.
В определенных специфических вариантах осуществления, настоящее изобретение предусматривает получение мутаций во внеклеточном домене (также называемом лиганд-связывающим доменом) полипептида ActRIIB с тем, чтобы полипептид ActRIIB имел измененную лиганд-связывающую активность (например, сродство связывания или специфичность связывания). В определенных случаях, такие пептиды-ловушки GDF имеют измененное (повышенное или пониженное) сродство связывания к определенному лиганду. В других случаях, полипептиды-ловушки GDF имеют измененную специфичность связывания с лигандами ActRIIB.
Например, в описании представлены полипептиды-ловушки GDF, которые, предпочтительно, связываются с GDF8/GDF11 относительно активинов. В описании, кроме того, установлена необходимость таких полипептидов для снижения вне целевых эффектов, хотя такие селективные варианты могут быть менее желательны для лечения тяжелых заболеваний, где очень большие приросты уровней эритроцитов могут потребоваться для терапевтического эффекта, и где приемлем некоторый уровень вне целевого эффекта. Например, аминокислотные остатки белка ActRIIB, такие как E39, K55, Y60, K74, W78, D80 и F101, находятся в лиганд-связывающем кармане и опосредуют связывание с его лигандами, такими как активин и GDF8. Таким образом, настоящее изобретение относится к ловушке GDF, содержащей измененный лиганд-связывающий домен (например, GDF8-связывающий домен) рецептора ActRIIB, который содержит одну или более мутаций в указанных аминокислотных остатках. Возможно наличие у измененного лиганд-связывающего домена повышенной селективности к такому лиганду как GDF8, относительно лиганд-связывающего домена дикого типа рецептора ActRIIB. Эти мутации увеличивают селективность измененного лиганд-связывающего домена к GDF8 относительно активина. Возможно, измененный лиганд-связывающий домен имеет отношение значения Kd для связывания активина к значению Kd для связывания GDF8, величина которого по меньшей мере в 2, 5, 10 или даже 100 раз больше, чем для лиганд-связывающего домена дикого типа. Возможно, измененный лиганд-связывающий домен имеет отношение значения IC50 для ингибирования активина к значению IC50 для ингибирования GDF8, величина которого по меньшей мере в 2, 5, 10 или даже 100 больше чем для лиганд-связывающего домена дикого типа. Возможно, измененный лиганд-связывающий домен ингибирует GDF8 при значении IC50, величина которого по меньшей мере в 2, 5, 10 или даже 100 больше, чем значение IC50 для ингибирования активина.
В качестве специфического примера, положительно заряженный аминокислотный остаток Asp (D80) лиганд-связывающего домена ActRIIB может быть мутирован в другой аминокислотный остаток для получения полипептида-ловушки GDF, который, предпочтительно, связывается с GDF8, но не с активином. Предпочтительно, остаток D80 меняют на аминокислотный остаток, выбранный из группы, состоящей из незаряженного аминокислотного остатка, отрицательно заряженного аминокислотного остатка, и гидрофобного аминокислотного остатка. В качестве еще одного специфического примера, гидрофобный остаток, L79, может быть изменен в кислую аминокислоту аспарагиновую кислоту или глутаминовую кислоту для значительного снижения связывания активина, в то же время, при сохранении связывания GDF11. Как будет понятно специалистам в данной обалсти, большинство из описанных мутаций, вариантов или модификаций могут быть осуществлены на уровне нуклеиновых кислот или, в некоторых случаях, путем посттрансляционной модификации или химического синтеза. Такие технологии хорошо известны в данной области.
В определенных вариантах осуществления, настоящее изобретение предусматривает полипептиды-ловушки GDF, имеющие специфические мутации в ActRIIB с тем, чтобы изменить гликозилирование полипептида ActRIIB. Иллюстративные сайты гликозилирования в полипептидах-ловушках GDF показаны на фиг. 1 (например, подчеркнутые сайты NX(S/T)). Такие мутации могут быть выбраны так, чтобы ввести или устранить один или более сайтов гликозилирования, такие как O-сцепленные или N-сцепленные сайты гликозилирования. Сайты распознавания аспарагин-сцепленной гликолизации в целом содержат трипептидную последовательность, аспарагин-X-треонин (где «X» представляет собой любую аминокислоту), которая специфически распознается соответствующими ферментами гликозилирования. Изменение может быть также осуществлено добавлением или замещением одного или более остатков серина или треонина в последовательность полипептида ActRIIB (для O-сцепленных сайтов гликозилирования). Разнообразные аминокислотные замены или делеции в одном или обоих из положений первой и третьей аминокислоты сайта распознавания гликозилирования (и/или аминокислотной делеции во втором положении) приводит к отсутствию гликозилирования в модифицированной трипептидной последовательности. Другим средством увеличения числа углеводородных частей на полипептиде-ловушке GDF является химическое или ферментное соединение гликозидов с полипептидом-ловушкой GDF. В зависимости от используемого типа соединения, сахар(а) могут быть присоединены к (a) аргинину и гистидину; (b) свободным карбоксильным группам; (c) свободным сульфгидрильным группам, таким как сульфгидрильные группы цистеина; (d) свободным гидроксильным группам, таким как гидроксильные группы серина, треонина или гидроксипролина; (e) ароматическим остаткам, таким как ароматические остатки фенилаланина, тирозина или триптофана; или (f) амидная группа глутамина. Эти способы описаны в документе WO 87/05330 и в публикации Aplin and Wriston (1981) CRC Crit. Rev. Biochem., pp. 259-306, включенных в настоящее описание путем ссылки. Удаление одной или более углеводородных частей, присутствующих на полипептиде-ловушке GDF, может осуществляться химически и/или ферментативно. Химическое дегликозилирование может включать, например, воздействие на полипептид-ловушку GDF трифторметансульфоновой кислотой или эквивалентным соединением. Данная обработка приводит к расщеплению большинства или всех сахаров, за исключением связывающего сахара (N-ацетилглюкозамина или N-ацетилгалактозамина), в то же время, оставляя аминокислотную последовательность интактной. Химическое дегликозилирование, кроме того, описано Hakimuddin et al. (1987) Arch. Biochem. Biophys. 259:52 и Edge et al. (1981) Anal. Biochem. 118:131. Ферментное расщепление углеводородных частей на полипептиде-ловушке GDF может быть достигнуто использованием разнообразных эндо- и экзогликозидаз, как описано Thotakura et al. (1987) Meth. Enzymol. 138:350. Последовательность полипептида-ловушки GDF может при целесообразности корригироваться, в зависимости от требуемого типа системы экспрессии, поскольку все клетки млекопитающих, дрожжей, насекомых и растений могут внести отличающиеся типы гликозилирования, которое может осуществляться аминокислотной последовательностью пептида. В целом, полипептиды-ловушки GDF для использования у людей будут экспрессированы в линии клеток млекопитающих, которая обеспечивает должное гликозилирование, такой как линии клеток HEK293 или CHO, хотя ожидается, что могут также использоваться другие экспрессирующие линии клеток млекопитающих.
Данное описание, кроме того, предусматривает способ образования вариантов, в частности, наборов комбинаторных вариантов полипептида-ловушки GDF, включая, возможно, усеченные варианты; пулы комбинаторных мутаций, в частности, могут использоваться для идентификации последовательностей ловушки GDF. Целью скрининга таких комбинаторных библиотек может быть образование, например, вариантов полипептида-ловушки GDF, которые имеют измененные свойства, такие как измененную фармакокинетику, или измененное связывание лигандов. Ниже описаны разнообразные скрининговые анализы, и такие анализы могут использоваться для оценки вариантов. Например, может проводиться скрининг варианта полипептида-ловушки GDF для выявления его способности связываться с полипептидом ActRIIB, предотвращать связывание лиганда ActRIIB с полипептидом ActRIIB или препятствовать передаче сигналов, вызванной лигандом ActRIIB.
Активность полипептида-ловушки GDF или его вариантов может также тестироваться в анализе in vivo или на клетках. Например, может анализироваться воздействие варианта полипептида-ловушки GDF на экспрессию генов, участвующих в гематопоэзе. При необходимости, это может быть выполнено в присутствии одного или более рекомбинантных лигандных белков ActRIIB (например, активина), и клетки могут быть трансфицированы с тем, чтобы получить полипептид-ловушку GDF и/или его варианты, и, возможно, лиганд ActRIIB. Аналогичным образом, полипептид-ловушка GDF может вводиться мыши или другому животному, и один или более показателей крови, таких как количество эритроцитов, уровни гемоглобина, уровни гематокрита, запасы желаза или количество ретикулоцитов, могут оцениваться с использованием признанных способов.
Могут быть образованы варианты комбинаторного происхождения, которые имеют селективную активность в отношении контрольного полипептида-ловушки GDF. Такие вариантные белки при экспрессии из конструкций рекомбинантной РНК могут использоваться в протоколах генной терапии. Аналогичным образом, мутагенез может обеспечить получение вариантов, которые имеют внутриклеточные периоды полувыведения, резко отличающиеся от соответствующего немодифицированного полипептида-ловушки GDF. Например, измененному белку может быть придана или большая, или меньшая устойчивость к протеолитическому расщеплению или другим процессам, которые приводят к разрушению или иной инактивации немодифицированного полипептида-ловушки GDF. Такие варианты и гены, которые их кодируют, могут использоваться для изменения уровней полипептида-ловушки GDF путем модуляции периода полувыведения полипептидов-ловушек GDF. Например, короткий период полувыведения может вызвать более преходящие биологические эффекты, и, являясь частью индуцируемой системы экспрессии, может обеспечить возможность более жесткой регуляции уровней рекомбинантного полипептида-ловушки GDF внутри клетки. В слитом белке Fc, мутации могут быть внесены в линкер (если он присутствует) и/или часть Fc для изменения периода полувыведения белка.
В определенных вариантах осуществления, полипептиды-ловушки GDF по изобретению могут, кроме того, включать посттрансляционные модификации в дополнение к любым природным модификациям в полипептидах ActRIIB. Такие модификации включают, без ограничения, ацетилирование, карбоксилирование, гликозилирование, фосфорилирование, липидирование и ацилирование. В результате, полипептиды-ловушки GDF могут содержать не аминокислотные элементы, такие как полиэтиленгликоли, липиды, поли- или моносахариды и фосфаты. Воздействия таких не аминокислотных элементов на функциональность полипептида-ловушки GDF может тестироваться, как описано в настоящей заявке для других вариантов полипептидов-ловушек GDF. Когда полипептид-ловушка GDF продуцируется в клетках расщеплением появляющейся формой полипептида-ловушки GDF, то посттрансляционный процессинг может быть также важным для коррекции укладки и/или функции белка. Различные клетки (такие как CHO, HeLa, MDCK, 293, WI38, NIH-3T3 или HEK293) имеют специфическую клеточную механику и характерные механизмы для таких посттрансляционных видов активности и могут быть выбраны для обеспечения правильной модификации и процессинга полипептидов-ловушек GDF.
В определенных аспектах, полипептиды-ловушки GDF включают слитые белки, имеющие по меньшей мере часть полипептида ActRIIB и один или более слитых доменов. Хорошо известные примеры таких слитых доменов включают, без ограничения, полигистидин, Glu-Glu, глутатион-S-трансферазу (GST), тиоредоксин, белок A, белок G, константную область тяжелой цепи иммуноглобулина (например, Fc), мальтоза-связывающий белок (MBP) или сывороточный альбумин человека. Слитый домен может быть выбран так, чтобы придать желательное свойство. Например, некоторые слитые домены особенно полезны для выделения слитых белков аффинной хроматографией. С целью аффинной очистки, используются релевантные матрицы для аффинной хроматографии, такие как глютатион-, амилаза- и никель- или кобальт-конъюгированные смолы. Много таких матриц имеется в форме «наборов», таких как система очистки GST Pharmacia и система QIAexpressTM (Qiagen), используемые со слитыми партнерами (HIS6). В качестве другого примера, слитый домен может быть выбран с тем, чтобы содействовать выявлению полипептидов-ловушек GDF. Примеры таких доменов выявления включают различные флуоресцентные белки (например, GFP), а также «эпитопные метки», которые представляют собой обычно короткие пептидные последовательности, к которым имеется специфическое антитело. Хорошо известные эпитопные метки, для которых вполне доступны специфические моноклональные антитела, включают FLAG, гемагглютинин (HA) вируса гриппа и метки c-myc. В некоторых случаях, слитые домены имеют сайт протеазного расщепления, такой как для Фактора Xa или Тромбина, который обеспечивает возможность релевантной протеазе частично расщеплять слитые белки и, посредством этого, высвобождать из них рекомбинантные белки. Высвобожденные белки могут быть затем выделены из слитого домена последующим хроматографическим разделением. В определенных предпочтительных вариантах осуществления, полипептид-ловушка GDF слит с доменом, который стабилизирует полипептид-ловушку GDF in vivo («стабилизирующий» домен). Под «стабилизацией» подразумевается все, что увеличивает период полувыведения из сыворотки, независимо от того, происходит ли это вследствие уменьшенного разрушения, уменьшенного выведение или другого фармакокинетического эффекта. Известно, что слияния с частью Fc иммуноглобулина придают желательные фармакокинетические свойства широкому диапазону белков. Аналогичным образом, слияния с сывороточным альбумином человека могут придать желательные свойства. Другие типы слитых доменов, которые могут быть выбраны, включают мультимеризирующие (например, димеризирующие, тетрамеризирующие) домены и функциональные домены (которые придают дополнительную функцию, такую как дальнейшее повышение уровней эритроцитов).
В качестве специфического примера, настоящее изобретение относится к ловушке GDF, которая представляет собой слитый белок ActRIIB-Fc, который содержит внеклеточный (например, лиганд-связывающий) домен полипептида ActRIIB, слитого с Fc доменом. Последовательность иллюстративного Fc домена показана ниже (SEQ ID NO:6).
THTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVD(A)VSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCK(A)VSNKALPVPIEKTISKAKGQPREPQVYTLPPSREEMTKNQVSLT CLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGPFFLYSKLTVDKSRWQQGNVFSCSVMHEALHN(A)HYTQKSLSLSPGK*
Возможно, Fc домен имеет одну или более мутаций в таких остатках как Asp-265, лизин 322 и Asn-434. В определенных случаях, мутантный Fc домен, имеющий одну или более из указанных мутаций (например, мутацию Asp-265), имеет пониженную способность связывания с рецептором Fcγ относительно Fc домена дикого типа. В других случаях, мутантный Fc домен, имеющий одну или более из указанных мутаций (например, мутацию Asn-434), имеет увеличенную способность связывания с Fc-рецептором (FcRN), относящимся к I классу MHC (главного комплекса гистосовместимости), относительно Fc домена дикого типа.
Понятно, что различные элементы слитых белков могут быть расположены любым образом, который согласуется с желательной функциональностью. Например, полипептид-ловушка GDF может быть расположен на стороне C-конца от гетерологичного домена или, альтернативно, гетерологичный домен может быть расположен на стороне C-конца от полипептида-ловушки GDF. Домен полипептида-ловушки GDF и гетерологичный домен необязательно должны соседствовать в слитом белке, и дополнительные домены или аминокислотные последовательности могут быть включены на стороне C- или N-конца от любого домена или между доменами.
В определенных вариантах осуществления, слитый белок ловушки GDF содержит аминокислотную последовательность, представленную в формуле A-B-C. Часть B представляет собой усеченный на N- и C-конце полипептид ActRIIB, состоящий из аминокислотной последовательности, соответствующей аминокислотам 26-132 последовательности SEQ ID NO: 26. Части A и C могут представлять собой независимо ноль, одну или более чем одну аминокислоту, и обе части A и C, в случае их присутствия, являются гетерологичными в отношении B. Части A и/или C могут быть присоединены к части B через линкерную последовательность. Иллюстративные линкеры включают короткие полипептидные линкеры, такие как 2-10, 2-5, 2-4, 2-3 глициновых остатка, такие как, например, линкер Gly-Gly-Gly. Другие подходящие линкеры описаны ниже в настоящей заявке. В определенных вариантах осуществления, слитый белок ловушки GDF содержит аминокислотную последовательность, представленную в формуле A-B-C, где A представляет собой лидерную последовательность, B состоит из аминокислот 26-132 SEQ ID NO: 26, и C представляет собой часть полипептида, которая усиливает одну или более характеристик из устойчивости in vivo, периода полувыведения in vivo, захвата/введения, локализации или распределения в ткани, образования белковых комплексов и/или очистки. В определенных вариантах осуществления, слитый белок ловушки GDF содержит аминокислотную последовательность, представленную в формуле A-B-C, где A представляет собой лидерную последовательность TPA, B состоит из аминокислот 26-132 последовательности SEQ ID NO: 26, и C представляет собой домен Fc-фрагмента иммуноглобулина. Предпочтительный слитый белок ловушки GDF содержит аминокислотную последовательность, представленную в SEQ ID NO:26.
В определенных вариантах осуществления, полипептиды-ловушки GDF по настоящему изобретению содержат одну или более модификаций, которые способны стабилизировать полипептиды-ловушки GDF. Например, такие модификации увеличивают период полувыведения in vitro полипептидов-ловушек GDF, увеличивают период полувыведения из циркулирующей крови полипептидов-ловушек GDF или уменьшают протеолитическое разрушение полипептидов-ловушек GDF. Такие стабилизирующие модификации включают, без ограничения, слитые белки (включая, например, слитые белки, содержащие полипептид-ловушку GDF и домен-стабилизатор), модификации сайта гликозилирования (включая, например, добавление сайта гликолизирования в полипептид-ловушку GDF) и модификации углеводородной части (включая, например, удаление углеводородных частей из полипептида-ловушки GDF). В случае слитых белков, полипептид-ловушка GDF слит с доменом-стабилизатором, таким как молекула IgG (например, домен Fc-фрагмента). Используемый в настоящем описании термин «домен-стабилизатор» относится не только к слитому домену (например, Fc), как в случае слитых белков, но также включает небелковые модификации, такие как углеводородная часть или небелковый полимер, такой как полиэтиленгликоль.
В определенных вариантах осуществления, настоящее изобретение обеспечивает получение выделенных и/или очищенных форм полипептидов-ловушек GDF, которые изолированы от других белков или иным образом лишены их.
В определенных вариантах осуществления, полипептиды-ловушки GDF (немодифицированные или модифицированные) по изобретению могут быть получены разнообразными технологиями, известными в данной области. Например, такие полипептиды-ловушки GDF могут быть синтезированы с использованием стандартных технологий химии белков, таких как технологии, описанные в руководствах Bodansky, M. Principles of Peptide Synthesis, Springer Verlag, Berlin (1993) и Grant G. A. (ed.), Synthetic Peptides: A User's Guide, W. H. Freeman and Company, New York (1992). Кроме того, в продаже имеются автоматизированные пептидные синтезаторы (например, Advanced ChemTech Model 396; Milligen/Biosearch 9600). Альтернативно, полипептиды-ловушки GDF, их фрагменты или варианты могут быть получены рекомбинантными технологиями с использованием различных экспрессионных систем (например, E. coli, клеток яичников китайских хомячков (CHO), клеток COS, бакуловируса), как хорошо известно в данной области. В еще одном варианте осуществления, модифицированные или немодифицированные полипептиды-ловушки GDF могут быть получены расщеплением полученных рекомбинантной технологией полипептидов-ловушек GDF путем использования, например, протеазы, например, трипсина, термолизина, химотрипсина, пепсина или парного превращающего основную аминокислоту фермента (PACE). Компьютерный анализ (с использованием коммерчески доступного программного обеспечения, например, MacVector, Omega, PCGene, Molecular Simulation, Inc.) можно использовать для идентификации сайтов протеолитического расщепления. Альтернативно, такие полипептиды-ловушки GDF могут быть получены из полипептидов-ловушек полноразмерной GDF, полученных рекомбинантной технологией, такой как стандартные технологии, известные в данной области, например, химическим расщеплением (например, цианогенбромидом, гидроксиламином).
3. Нуклеиновые кислоты, кодирующие полипептиды-ловушки GDF
В определенных аспектах, изобретение относится к выделенным и/или рекомбинантным нуклеиновым кислотам, кодирующим любой из полипептидов-ловушек GDF, описанных в настоящей заявке. SEQ ID NO: 4 кодирует природный полипептид-предшественник ActRIIB, в то время как SEQ ID NO:5 кодирует растворимый полипептид ActRIIB, а SEQ ID NO:25, 27, 30 и 31 кодирует растворимые ловушки GDF. Нуклеиновые кислоты по изобретению могут быть однонитевыми или двухнитевыми. Такие нуклеиновые кислоты могут представлять собой молекулы ДНК или РНК. Эти нуклеиновые кислоты могут, например, использоваться в способах получения полипептидов-ловушек GDF или в качестве прямых терапевтических средств (например, при подходе в виде генной терапии).
В определенных аспектах, нуклеиновые кислоты по изобретению, кодирующие полипептиды-ловушки GDF, кроме того, как подразумевается, включают нуклеиновые кислоты, которые представляют собой варианты SEQ ID NO:5, 25, 27, 30 и 31. Вариантные нуклеотидные последовательности включают последовательности, которые отличаются замещениями, добавлениями или делециями одного или более нуклеотидов, такие как аллельные варианты, и поэтому включают кодирующие последовательности, которые отличаются от нуклеотидной последовательности, кодирующей последовательности, обозначенной в SEQ ID NO:5, 25, 27, 30 и 31.
В определенных вариантах осуществления, изобретение относится к последовательностям выделенных или рекомбинантных нуклеиновых кислот, которые по меньшей мере на 80%, 85%, 90%, 95%, 97%, 98%, 99% или 100% идентичны SEQ ID NO: 5, 25, 27, 30 или 31. Среднему специалисту в данной области понятно, что последовательности нуклеиновых кислот, комплементарные SEQ ID NO: 5, 25, 27, 30 или 31, и вариантам SEQ ID NO: 5, 25, 27, 30 или 31, также входят в объем настоящего изобретения. В других вариантах осуществления, последовательности нуклеиновых кислот по изобретению могут быть выделены, рекомбинированы и/или слиты с гетерологичной нуклеотидной последовательностью или в библиотеке ДНК.
В других вариантах осуществления, нуклеиновые кислоты по изобретению также включают нуклеотидные последовательности, которые гибридизируются в очень жестких условиях с нуклеотидной последовательностью, обозначенной в SEQ ID NO: 5, 25, 27, 30 или 31, комплементарной последовательностью SEQ ID NO: 5, 25, 27, 30 или 31, или их фрагментами. Как обсуждается выше, среднему специалисту в данной области будет понятно, что целесообразные жесткие условия, которые содействуют гибридизации ДНК, могут варьироваться. Например, можно выполнять гибридизацию при 6,0 x хлорида натрия/цитрата натрия (SSC) примерно при 45ºC, с последующим промыванием 2,0 x SSC при 50ºC. Например, концентрация соли на стадии промывания может быть выбрана из низкой жесткости примерно 2,0 x SSC при 50ºC до высокой жесткости 0,2 x SSC при 50ºC. Кроме того, температура на стадии промывания может увеличиваться от условий низкой жесткости при комнатной температуре, примерно 22ºC, до условий высокой жесткости примерно при 65ºC. И температура, и концентрация соли могут варьироваться, или температура или концентрация соли может удерживаться на постоянном уровне, в то время как другой параметр изменяется. В одном варианте осуществления, изобретение относится к нуклеиновым кислотам, которые гибридизируются в условиях низкой жесткости 6 x SSC при комнатной температуре с последующим промыванием при 2 x SSC и комнатной температуре.
Выделенные нуклеиновые кислоты, которые отличаются от нуклеиновых кислот, представленных SEQ ID NO: 5, 25, 27, 30 или 31, ввиду дегенерации генетического кода, также входят в объем изобретения. Например, ряд аминокислот обозначен более чем одним триплетом. Кодоны, которые определяют одну и ту же аминокислоту, или синонимы (например, CAU и CAC, являются синонимами для гистидина) могут привести к «молчащим» мутациям, которые не воздействуют на аминокислотную последовательность белка. В определенных вариантах осуществления, полипептид-ловушка GDF будет кодироваться альтернативной нуклеотидной последовательностью. Альтернативные нуклеотидные последовательности являются дегенеративными в отношении нативной последовательности нуклеиновой кислоты ловушки GDF, но все же кодируют тот же слитый белок. В определенных вариантах осуществления, ловушка GDF, имеющая SEQ ID NO:26, кодируется альтернативной последовательностью нуклеиновой кислоты, включающей SEQ ID NO:30. Однако ожидается, что полиморфизмы последовательности ДНК, которые ведут к изменениям аминокислотных последовательностей белков по изобретению, будут существовать среди клеток млекопитающих. Специалисту понятно, что указанные изменения в одном или более нуклеотидов (до примерно 3-5% нуклеотидов) нуклеиновых кислот, кодирующих конкретный белок, существуют среди индивидов данного вида вследствие природного аллельного изменения. Любое и все такие нуклеотидные изменения и полученные в результате полиморфизмы аминокислот входят в объем данного изобретения.
В определенных вариантах осуществления, рекомбинантные нуклеиновые кислоты по изобретению могут быть функционально связаны с одной или более регуляторных нуклеотидных последовательностей в экспрессионной конструкции. Регуляторные нуклеотидные последовательности в целом будут соответствовать клетке-хозяину, используемой для экспрессии. В данной области известны многочисленные типы соответствующих векторов экспрессии и подходящих регуляторных последовательностей для разнообразных клеток-хозяев. Обычно, указанные одна или более регуляторных нуклеотидных последовательностей включают, без ограничения, промоторные последовательности, лидерные или сигнальные последовательности, рибосомальные сайты связывания, сайта начала транскрипции и терминации транскрипции, сайты начала трансляции и терминации трансляции, последовательности энхансеров или активаторов. Конститутивные или индуцируемые промоторы, известные в данной области, предусмотрены изобретением. Промоторы могут представлять собой или природные промоторы, или гибридные промоторы, которые комбинируют элементы более чем одного промотора. Экспрессионная конструкция может присутствовать в клетке на эписоме, такой как плазмида, или экспрессионная конструкция может быть вставлена в хромосому. В предпочтительном варианте осуществления, вектор экспрессии содержит селекционируемый маркерный ген для обеспечения возможности отбора трансформированных клеток-хозяев. Селекционируемые маркерные гены хорошо известны в данной области и варьируются в зависимости от используемой клетки-хозяина.
В определенных аспектах изобретения, нуклеиновая кислота по изобретению предоставлена в векторе экспрессии, содержащем нуклеотидную последовательность, кодирующую полипептид-ловушку GDF, и функционально связанную по меньшей мере с одной регуляторной последовательностью. Регуляторные последовательности признаны в данной области и выбираются для направления экспрессии полипептида-ловушки GDF. Соответственно, термин «регуляторная последовательность» включает промоторы, энхансеры и другие элементы регуляции экспрессии. Иллюстративные регуляторные последовательности описаны в руководстве Goeddel; Gene Expression Technology: Methods in Enzymology, Academic Press, San Diego, CA (1990). Например, любая из широкого разнообразия последовательностей регуляции экспрессии, которые регулируют экспрессию последовательности ДНК при функциональной связи с ней, может использоваться в указанных векторах для экспрессии последовательностей ДНК, кодирующих полипептид-ловушку GDF. Такие полезные последовательности регуляции экспрессии включают, например, ранние или поздние промоторы SV40, промотор tet, немедленный ранний промотор аденовируса или цитомегаловируса, промоторы RSV (респираторного синцитиального вируса), систему lac оперона, систему trp, систему TAC или TRC, промотор T7, чья экспрессия направляется T7 РНК полиперазой, основные операторные и промоторные области фага лямбда, контрольные области белка fd покрытия, промотор для 3-фосфоглицерат-киназы или других гликолитических ферментов, промоторы кислой фосфатазы, например, Pho5, промоторы спаривания α-факторов дрожжей, многогранный промотор системы бакуловируса и другие последовательности, которые, как известно, регулируют экспрессию генов прокариотических или эукариотических клеток или их вирусов, и их различные комбинации. Следует понимать, что конструкция вектора экспрессии может зависеть от таких факторов как выбор подлежащей трансформации клетки-хозяина и/или типа белка, который желательно экспрессировать. Кроме того, следует также учитывать число копий вектора, способность регулировать число копий и экспрессию любого другого белка, кодируемого вектором, такого как антибиотические маркеры.
Рекомбинантная нуклеиновая кислота по изобретению может быть получена лигированием клонированного гена или его части в вектор, подходящий для экспрессии в любых из прокариотических клеток, эукариотических клеток (дрожжей, птиц, насекомых или млекопитающих) или в обоих. Носитель экспрессии для получения рекомбинантного полипептида-ловушки GDF включают плазмиды и другие векторы. Например, подходящие векторы включают плазмиды типов: pBR322-дериватизированные плазмиды, pEMBL-дериватизированные плазмиды, pEX-дериватизированные плазмиды, pBTac-дериватизированные плазмиды и pUC-дериватизированные плазмиды для экспрессии в прокариотических клетках, таких как E. coli.
Некоторые векторы экспрессии млекопитающих содержат обе прокариотические последовательности для содействия распространению вектора в бактериях, и одну или более эукариотических транскрипционных единиц, которые экспрессированы в эукариотических клетках. Векторы, полученные из pcDNAI/amp, pcDNAI/neo, pRc/CMV, pSV2gpt, pSV2neo, pSV2-dhfr, pTk2, pRSVneo, pMSG, pSVT7, pko-neo и pHyg, являются примерами векторов экспрессии млекопитающих, подходящих для трансфекции эукариотических клеток. Некоторые из указанных векторов модифицированы последовательностями из бактериальных плазмид, таких как pBR322, для содействия репликации и отбора устойчивости к лекарственным средствам и у прокариотических, и эукариотических клеток. Альтернативно, дериваты вирусов, таких как вирус бычьей папилломы (BPV-1), или вирус Эпштейна-Барра (pHEBo, pREP-derived и p205), могут использоваться для преходящей экспрессии белков в эукариотических клетках. Примеры других вирусных (включая ретровирусные) систем экспрессии можно найти ниже в описании систем доставки генной терапии. Различные способы, используемые при получении плазмид и при трнасформации организмов хозяев хорошо известны в данной области. Другие подходящие системы экспрессии и для прокариотических, и для эукариотических клеток, а также общие рекомбинантные процедуры можно найти в руководстве Molecular Cloning A Laboratory Manual, 2nd Ed., ed. by Sambrook, Fritsch and Maniatis (Cold Spring Harbor Laboratory Press, 1989) Главы 16 и 17. В некоторых случаях, может быть желательно экспрессировать рекомбинантные полипептиды путем использования бакуловирусной системы экспрессии. Примеры таких бакуловирусных систем экспрессии включают pVL-дериватизированные векторы (такие как pVL1392, pVL1393 и pVL941), pAcUW-дериватизированные векторы (такие как pAcUW1), и pBlueBac-дериватизированные векторы (такие как содержащий ß-gal pBlueBac III).
В предпочтительном варианте осуществления, вектор будет предназначен для получения полипептидов-ловушек GDF в клетках CHO, такой как вектор Pcmv-Script (Stratagene, La Jolla, Calif.), векторы pcDNA4 (Invitrogen, Carlsbad, Calif.) и векторы pCI-neo (Promega, Madison, Wisc.). Как будет очевидно, генные конструкции по изобретению могут использоваться для вызова экспрессии полипептидов-ловушек GDF по изобретению в клетках, размножаемых в культуре, например, для получения белков, включающих слитые белки или вариантные белки, для очистки.
Настоящее изобретение также относится к клетке-хозяину, трансфицированной рекомбинантным геном, включающим кодирующую последовательность (например, SEQ ID NO:4, 5, 25, 27, 30 или 31) для одного или более из полипептидов-ловушек GDF по изобретению. Клетка-хозяин может представлять собой любую прокариотическую или эукариотическую клетку. Например, полипептид-ловушка GDF по изобретению может быть экспрессирован в бактериальных клетках, таких как E. coli, клетки насекомых (например, с использованием бакуловирусной системы экспрессии), дрожжей или клеток млекопитающих. Другие подходящие клетки-хозяева известны специалистам в данной области.
Соответственно, настоящее изобретение, кроме того, относится к способам получения полипептидов-ловушек GDF по изобретению. Например, клетка-хозяин, трансфицированная вектором экспрессии, кодирующим полипептид-ловушку GDF, может культивироваться в соответствующих условиях для обеспечения возможности осуществления экспрессии полипептида-ловушки GDF. Полипептид-ловушка GDF может быть секретирован и выделен из смеси клеток и среды, содержащей полипептид-ловушку GDF. Альтернативно, полипептид-ловушка GDF может удерживаться цитоплазматически или во фракции мембраны, и клетки могут быть собраны, лизированы, и белок может быть выделен. Культура клеток включает клетки-хозяева, среды и другие побочные продукты. Подходящие среды для клеточной культуры хорошо известны в данной области. Полипептиды-ловушки GDF по изобретению могут быть выделены из среды клеточной культуры, клеток-хозяев или их обоих с использованием технологий, известных в области очистки белков, включая ионообменную хроматографию, гель-фильтрационную хроматографию, ультрафильтрацию, электрофорез и иммуноаффинную очистку антителами, специфическими к конкретным эпитопам полипептидов-ловушек GDF. В предпочтительном варианте осуществления, полипептид-ловушка GDF представляет собой слитый белок, содержащий домен, который облегчает его очистку.
В другом варианте осуществления, слитый ген, кодирующий лидерную последовательность очистки, такую как последовательность сайта расщепления поли-(His)/энтерокиназой на N-конце желательной части рекомбинантного полипептида-ловушки GDF, может обеспечить возможность очистки экспрессированного слитого белка аффинной хроматографией с использованием смолы металла Ni2+. Затем лидерная последовательность очистки может быть удалена обработкой энтерокиназой для получения очищенного полипептида-ловушки GDF (например, см. Hochuli et al., (1987) J. Chromatography 411:177; and Janknecht et al., PNAS USA 88:8972).
Технологии получения слитых генов хорошо известны. По существу, соединение различных фрагментов ДНК, кодирующих различные полипептидные последовательности, выполняется в соответствии с обычными методиками с использованием тупых и зигзагообразных концов для лигирования, расщепления рестрикционными ферментами для получения соответствующих концов, по мере необходимости, заполнения когезивных концов, обработки щелочной фосфатазы во избежание нежелательного соединения, и ферментативного лигирования. В другом варианте осуществления, слитый ген может быть синтезирован обычными технологиями, включающими автоматизированные синтезаторы ДНК. Альтернативно, может проводиться ПЦР-амплификация генных фрагментов с использованием якорных праймеров, которые образуют дополнительные выступы между двумя последовательными генными фрагментами, которые могут в последующем гибридизироваться для образования химерной генной последовательности (см., например, Current Protocols in Molecular Biology, eds. Ausubel et al., John Wiley & Sons: 1992).
4. Скрининговые анализы
В определенных аспектах, настоящее изобретение относится к применению полипептидов-ловушек GDF по изобретению (например, растворимых вариантных полипептидов ActRIIB) для идентификации соединений (агентов), которые являются агонистами или антагонистами полипептидов ActRIIB. Соединения, идентифицированные посредством скрининга, могут тестироваться для оценки их способности модулировать уровни эритроцитов, гемоглобина и/или ретикулоцитов in vivo или in vitro. Эти соединения могут тестироваться, например, на экспериментальных моделях у животных.
Существуют многочисленные подходы к скринингу терапевтических средств для увеличения уровней эритроцитов или гемоглобина путем нацеливание на передачу сигналов ActRIIB. В определенных вариантах осуществления, скрининг соединений с высокой пропускной способностью может проводиться для идентификации агентов, которые стимулируют опосредованные ActRIIB воздействия на выбранные линии клеток. В определенных вариантах осуществления, анализ проводится для скрининга и идентификации соединений, которые специфически ингибируют или снижают связывание полипептида ActRIIB с его партнером связывания, таким как лиганд ActRIIB (например, активином, Nodal, GDF8, GDF11 или BMP7). Альтернативно, анализ может использоваться для идентификации соединений, которые усиливают связывание полипептида ActRIIB с его партнером связывания, таким как лиганд ActRIIB. В еще одном варианте осуществления, соединения могут быть идентифицированы по их способности взаимодействовать с полипептидом ActRIIB.
В свете настоящего описания, указанным требованиям могут удовлетворять разнообразные форматы анализа, которые конкретно не описаны в настоящей заявке, но тем не менее понятны среднему специалисту в данной области. Как описано в настоящей заявке, тестируемые соединения (агенты) по изобретению могут быть созданы любым комбинаторным химическим способом. Альтернативно, соединения по изобретению могут представлять собой природные биомолекулы, синтезированные in vivo или in vitro. Соединения (агенты), подлежащие тестированию для выявления их способности действовать в качестве модуляторов тканевого роста, могут быть получены, например, бактериями, дрожжами, растениями или другими организмами (например, природными продуктами), получены химически (например, мелкие молекулы, включающие пептидомиметики), или получены рекомбинантной технологией. Тестируемые соединения, предусмотренные настоящим изобретением, включают не пептидильные органические молекулы, пептиды, полипептиды, пептидомиметики, сахара, гормоны и молекулы нуклеиновых кислот. В определенном варианте осуществления, тестируемый агент представляет собой небольшую органическую молекулу, имеющую молекулярную массу менее чем примерно 2000 Дальтон.
Тестируемые соединения по изобретению могут быть получены в виде отдельных, дискретных химических структурных единиц или получены в библиотеках большей сложности, таких как полученные комбинаторными химическими способами. Данные библиотеки могут включать, например, спирты, алкилгалиды, амины, амиды, сложные эфиры, альдегиды, простые эфиры и другие классы органических соединений. Представление тестируемых соединений в систему тестирования может осуществляться или в выделенной форме, или в виде смесей соединений, особенно на первоначальных стадиях скрининга. Возможна дериватизация соединений другими соединениями и наличие в них дериватизирующих групп, которые содействуют выделению соединений. Неограничивающие примеры дериватизирующих групп включают биотин, флуоресцеин, дигоксигенин, зеленый флуоресцентный белок, изотопы, полигистидин, магнитные гранулы, глутатион-S-трансферазу (GST), фотоактивируемые поперечно-сшивающие агенты или их комбинации.
Во многих программах скрининга лекарственных средств, в которых тестируются библиотеки соединений и природных экстрактов, желательны анализы с высокой пропускной способностью для максимизации числа обследованных соединений в данный период времени. Анализы, которые выполняются в бесклеточных системах, таких, которые могут быть получены очищенными или полуочищенными белками, часто предпочтительны в качестве «первичных» скринингов, потому что они могут быть получены для обеспечения возможности быстрой разработки и относительно легкого выявления изменения молекулярной мишени, которое опосредуется тестируемым соединением. Кроме того, эффектам клеточной токсичности или биодоступности тестируемого соединения можно в целом пренебречь в системе in vitro, причем анализ вместо этого сфокусирован в первую очередь на эффект лекарственного средства на молекулярную мишень, который может проявляться изменением сродства связывания между полипептидом ActRIIB и его партнером связывания (например, лигандом ActRIIB).
Просто для иллюстрации, в иллюстративном скрининговом анализе по настоящему изобретению, представляющее интерес соединение контактирует с выделенным и очищенным полипептидом ActRIIB, который обычно способен связываться с лигандом ActRIIB, в зависимости от цели анализа. Затем к смеси соединения и полипептида ActRIIB добавляется композиция, содержащая лиганд ActRIIB. Выявление и количественная оценка комплексов ActRIIB/лиганда ActRIIB обеспечивает средство для определения эффективности соединения при ингибировании (или потенцировании) образования комплекса между полипептидом ActRIIB и связывающим его белком. Эффективность соединения можно оценить построением кривых зависимости реакции от дозы по данным, полученным с использованием различных концентраций тестируемого соединения. Кроме того, можно также выполнять контрольный анализ для получения исходного уровня для сравнения. Например, в контрольном анализе выделенный и очищенный лиганд ActRIIB добавляется к композиции, содержащей полипептид ActRIIB, и образование комплекса ActRIIB/лиганда ActRIIB количественно определяется в отсутствие тестируемого соединения. Следует понимать, что в целом, порядок, в котором могут смешиваться реагенты, может варьироваться и могут смешиваться одновременно. Кроме того, вместо очищенных белков, клеточные экстракты и лизаты могут использоваться для предоставления подходящей бесклеточной аналитической системы.
Образование комплекса между полипептидом ActRIIB и связывающим его белком может быть выявлено разнообразными способами. Например, модуляция образования комплексов может быть количественно определена с использованием, например, детектируемо меченых белков, таких как меченый радиоактивной меткой (например, 32P, 35S, 14C или 3H), меченый флуоресцентной меткой (например, FITC), или меченый ферментной меткой полипептид ActRIIB или связывающий его белок, путем иммунного анализа или хроматографической детекцией.
В определенных вариантах осуществления, настоящее изобретение предусматривает применение анализов по поляризации флуоресценции и анализов резонансного переноса энергии флуоресценции (FRET) при прямом или косвенном измерении степени взаимодействия между полипептидом ActRIIB и связывающим его белком. Кроме того, другие типы детекции, такие как те, которые основаны на оптических волноводах (Публикация PCT WO 96/26432 и патент США № 5677196), поверхностном плазмонном резонансе (SPR), датчиках поверхностного заряда и поверхностных силовых датчиках, совместимы со многими вариантами осуществления изобретения.
Кроме того, настоящее изобретение предусматривает применение анализа взаимодействия ловушки, также известного как «двухгибридный анализ», для идентификации агентов, которые нарушают или потенцируют взаимодействие между полипептидом ActRIIB и его партнером связывания. См., например, патент США № 5283317; Zervos et al. (1993) Cell 72:223-232; Madura et al. (1993) J Biol Chem 268:12046-12054; Bartel et al. (1993) Biotechniques 14:920-924; и Iwabuchi et al. (1993) Oncogene 8:1693-1696). В определенном варианте осуществления, настоящее изобретение предусматривает использование обращенных двух гибридных систем для идентификации соединений (например, мелких молекул или пептидов), которые диссоциируют взаимодействие между полипептидом ActRIIB и связывающим его белком. См., например, Vidal and Legrain, (1999) Nucleic Acids Res 27:919-29; Vidal and Legrain, (1999) Trends Biotechnol 17:374-81; и патенты США №№ 5525490; 5955280; и 5965368.
В определенных вариантах осуществления, соединения по изобретению идентифицируются их способностью взаимодействовать с полипептидом ActRIIB. Взаимодействие между соединением и полипептидом ActRIIB может быть ковалентным или не ковалентным. Например, такое взаимодействие может быть идентифицировано на белковом уровне с использованием биохимических способов in vitro, включая фото-поперечную сшивку, связывание меченого радиоактивной меткой лиганда и аффинную хроматографию (Jakoby WB et al., 1974, Methods in Enzymology 46: 1). В определенных случаях, скрининг соединений может проводиться в анализе на основе механизма, таком как анализ для детекции соединений, которые связываются с полипептидом ActRIIB. Он может включать осуществление твердофазного или жидкофазного связывания. Альтернативно, ген, кодирующий полипептид ActRIIB, может быть трансфицирован репортерной системой (например, β-галактозидазой, люциферазой или зеленым флуоресцентным белком) в клетку и скрининг может быть проведен в сравнении с библиотекой, предпочтительно, скринингом с высокой пропускной способностью, или в сравнении с отдельными членами библиотеки. Может использоваться другой механизм на основе анализов связывания, например, анализы связывания, которые выявляют изменения свободной энергии. Анализы связывания могут выполняться при мишени, фиксированной к лунке, грануле или чипу, или захваченной иммобилизованным антителом или пептизированной капиллярным электрофорезом. Связанные соединения могут быть обычно выявлены с использованием калориметрии или флуоресценции или поверхностного плазмонного резонанса.
5. Примеры терапевтического применения
В определенных вариантах осуществления, полипептиды-ловушки GDF по настоящему изобретению могут использоваться для повышения содержания эритроцитов у млекопитающих, таких как грызуны и приматы, и, в частности, люди-пациенты. Кроме того, как показано в настоящем описании, полипептиды-ловушки GDF могут использоваться в комбинации с активаторами рецепторов EPO для достижения повышения содержания эритроцитов при более низких диапазонах доз. Это может быть благоприятно для снижения известных вне целевых эффектов и рисков, связанных с высокими дозами активаторов рецепторов EPO. В определенных вариантах осуществления, настоящее изобретение относится к способам лечения или профилактики анемии у индивида, при необходимости, путем введения индивиду терапевтически эффективного количества полипептида-ловушки GDF или комбинации (или сопутствующей терапии) полипептида-ловушки GDF и активатора рецепторов EPO. Эти способы могут использоваться для терапевтического и профилактического лечения млекопитающих и, в частности, людей.
Полипептиды-ловушки GDF могут использоваться в комбинации с активаторами рецепторов EPO для снижения требуемой дозы данных активаторов у пациентов, которые восприимчивы к побочным эффектам EPO. Первичными побочными эффектами ЕРО являются избыточное увеличение уровней гематокрита или гемоглобина и полицитемия. Повышенные уровни гематокрита могут привести к гипертонии (конкретнее, к усугублению гипертонии) и сосудистому тромбозу. Другие побочные эффекты EPO, о которых сообщалось, и некоторые из которых связаны с гипертонией, представляют собой головные боли, подобный гриппу синдром, обструкцию шунтов, инфаркты миокарда и церебральные судороги вследствие тромбоза, гипертензивную энцефалопатию и эритроцитарную аплазию (Singibarti, (1994) J. Clin Investig 72(suppl 6), S36-S43; Horl et al. (2000) Nephrol Dial Transplant 15(suppl 4), 51-56; Delanty et al. (1997) Neurology 49, 686-689; Bunn (2002) N Engl J Med 346(7), 522-523).
Быстрое воздействие описанных в настоящей заявке полипептидов-ловушек GDF на уровни эритроцитов указывают на то, что эти агенты действуют механизмом, отличным от EPO. Соответственно, эти антагонисты могут использоваться для увеличения уровней эритроцитов и гемоглобина у пациентов, которые не достаточно реагируют на EPO. Например, полипептид-ловушка GDF может быть благоприятна для пациента, у которого введение от обычной до увеличенной дозы EPO (>300 МЕ/кг/неделю) не приводит к увеличению уровня гемоглобина до целевого уровня. Пациенты с неадекватной реакцией на EPO обнаруживаются при всех типах анемии, но большие количества пациентов, не реагирующих на EPO, наблюдалось особенно часто у пациентов со злокачественными заболеваниями и пациентов с конечной стадией заболевания почек. Неадекватная реакция на EPO может быть или конститутивной (т.е., наблюдаемой после первого лечения EPO), или приобретенной (например, наблюдаемой после повторного лечения EPO).
Используемый в настоящем описании термин терапевтическое средство, которое «предотвращает» расстройство или состояние, относится к соединению, которое в статистическом образце снижает частоту возникновения расстройства или состояния у получавшей лечение выборки пациентов относительно не получавшей лечение контрольной выборки, или задерживает начало или уменьшает тяжесть одного или более симптомов расстройства или состояния относительно не получавшей лечение контрольной выборки. Используемый в настоящем описании термин «лечение» включает профилактику названного состояния или облегчение или устранение состояния после его окончательного установления. В любом случае, профилактика или лечение могут различаться в зависимости от диагноза, установленного врачом или другим медицинским работником, и предполагаемого результата введения терапевтического средства.
Как показано в настоящем описании, полипептиды-ловушки GDF, возможно, в комбинации с активатором рецепторов EPO, могут использоваться для повышения уровней эритроцитов, гемоглобина или ретикулоцитов у здоровых индивидов, и такие полипептиды-ловушки GDF могут применяться у выбранных популяций пациентов. Примеры соответствующих популяций пациентов включают пациентов с нежелательно низкими уровнями эритроцитов или гемоглобина, таких как пациенты, страдающие анемией, и пациенты с риском развития нежелательно низких уровней эритроцитов или гемоглобина, такие как пациенты, у которых предполагается проведение больших хирургических вмешательств или других процедур, которые могут привести к существенной кровопотере. В одном из вариантов осуществления, пациента с адекватными уровнями клеток крови лечат полипептидом-ловушкой GDF для увеличения уровней эритроцитов, и затем производят забор крови и ее хранение для использования позднее при трансфузиях.
Полипептид-ловушка GDF, возможно, в комбинации с активатором рецепторов EPO, описанной в настоящей заявке, может использоваться для увеличения уровня эритроцитов у пациентов, имеющих анемию. При наблюдении уровней гемоглобина у людей, уровень менее чем нормальный для соответствующей категории возраста и пола, может быть показателем анемии, хотя учитываются индивидуальные вариации. Например, уровень гемоглобина 12 г/дкл в целом считается нижним пределом нормального уровня у общей взрослой популяции. Потенциальные причины включают кровопотерю, алиментарную дистрофию, реакцию на медикаментозную терапию, различные проблемы с костным мозгом и многие заболевания. Конкретнее, анемию связывали с разнообразными расстройствами, которые включают, например, хроническую почечную недостаточность, миелодиспластический синдром, ревматоидный артрит, трансплантацию костного мозга. Анемия может быть также связана со следующими состояниями: солидные опухоли (например, рак молочных желез, рак легких, рак ободочной кишки); опухоли лимфоидной системы (например, хронический лимфолейкоз, неходжкинские и Ходжкинские лимфомы); опухоли гематопоэтической системы (например, лейкоз, миелодиспластический синдром, множественная миелома); лучевая терапия; химиотерапия (например, схемы, включающие платину); воспалительные и аутоиммунные заболевания, включая, без ограничения, ревматоидный артрит, другие воспалительные артриты, системную красную волчанку (SLE), острые или хронические заболевания кожи (например, псориаз), воспалительные кишечные заболевания (например, болезнь Крона и язвенный колит); острые или хронические заболевания почек или недостаточность, включая идиопатические или врожденные состояния; острые или хронически заболевания печени; острая или хроническая кровопотеря; ситуации, когда переливание эритроцитов невозможно вследствие наличия у пациентов алло- или аутоантител и/или по религиозным причинам (например, у некоторых свидетелей Иеговы), инфекции (например, малярию, остеомиелит); гемоглобинопатии, включая, например, серповидноклеточное заболевание, талассемии; применение наркотиков или наркомания, например, злоупотребление алкоголя; педиатрические пациенты с анемией в результате любой причины во избежание трансфузии; и пациенты пожилого возраста или пациенты с предшествующими сердечно-легочными заболеваниями, имеющие анемию, которые не могут получать трансфузии вследствие опасений циркуляторной перегрузки.
Полипептиды-ловушки GDF, возможно, в комбинации с активатором рецепторов EPO, были бы подходящими для лечения анемий гипопролиферативного костного мозга, которые обычно связаны с небольшим изменением морфологии эритроцитов (RBC). Гипопролиферативные анемии включают: 1) анемию при хронических заболеваниях, 2) анемию при заболеваниях почек, и 3) анемию, связанную с гипометаболическими состояниями. При каждом из указанных типов, уровни эндогенного эритропоэтина являются несоответственно низкими для наблюдаемой степени анемии. Другие гипопролиферативные анемии включают: 4) раннюю стадию железодефицитной анемии и 5) анемию, вызванную повреждением костного мозга. При указанных типах, уровни эндогенного эритропоэтина являются несоответственно повышенными для наблюдаемой степени анемии.
Самым распространенным типом является анемия при хронических заболеваниях, которая охватывает воспаление, инфекцию, повреждение ткани и такие состояния как злокачественные заболевания, и отличается и низкими уровнями эритропоэтина, и неадекватной реакцией на эритропоэтин в костном мозге (Adamson, 2008, Harrison’s Principles of Internal Medicine, 17th ed.; McGraw Hill, New York, pp 628-634). Многие факторы могут способствовать развитию анемии, связанной со злокачественными заболеваниями. Некоторые связаны с самим патологическим процессом и генерированием воспалительных цитокинов, таких как интерлейкин-1, интерферон-гамма и фактор некроза опухолей (Bron et al., 2001, Semin Oncol 28(Suppl 8):1-6). Среди своих эффектов, воспаление индуцирует ключевой регулирующий обмен железа пептид гепцидин, посредством этого ингибируя экспорт железа из макрофагов, и в целом ограничивая доступность железа для эритропоэза (Ganz, 2007, J Am Soc Nephrol 18:394-400). Кровопотеря различными путями может также способствовать развитию анемии, связанной со злокачественными заболеваниями. Частота анемии вследствие прогрессирования злокачественного заболевания варьируется в зависимости от типа злокачественного заболевания в диапазоне от 5% при раке предстательной железы до 90% множественной миеломе. Анемия, связанная со злокачественными заболеваниями, имеет глубокие последствия для пациентов, включая усталость и сниженное качество жизни, сниженную эффективность лечения и повышенную смертность.
Хроническое заболевание почек связано с гипопролиферативной анемией, которая варьируется по тяжести в зависимости от степени поражения почек. Такая анемия в первую очередь вызвана неадекватной продукцией эритропоэтина и сниженным выживанием эритроцитов. Хроническое заболевание почек обычно прогрессирует постепенно в течение периода нескольких лет или десятилетий в заболевание конечной стадии (Стадии-5), и при этом для выживания пациента требуется гемодиализ или трансплантация почки. Анемия часто развивается на ранней стадии этого процесса и усугубляется по мере прогрессирования заболевания. Клинические последствия анемии при болезни почек хорошо документированы и включают развитие гипертрофии левого желудочка, нарушенную когнитивную функцию, сниженное качество жизни и измененную иммунную функцию (Levin et al., 1999, Am J Kidney Dis 27:347-354; Nissenson, 1992, Am J Kidney Dis 20(Suppl 1):21-24; Revicki et al., 1995, Am J Kidney Dis 25:548-554; Gafter et al., 1994, Kidney Int 45:224-231). Как продемонстрировано заявителями на модели хронического заболевания почек у мыши (см. пример ниже), полипептид-ловушка GDF, возможно, в комбинации с активатором рецепторов EPO, может применяться для лечения анемии при заболеваниях почек.
Многие состояния, приводящие к гипометаболическому состоянию, могут вызвать гипопролиферативную анемию от легкой до умеренной степени тяжести. Среди таких состояний имеются состояния эндокринной недостаточности. Например, анемия может возникнуть при болезни Аддисона, гипотиреозе, гиперпаратиреозе или у самцов, которые кастрированы или получали лечение эстрогеном. Анемия от легкой до умеренной степени тяжести может также возникнуть при сниженном потреблении белка с рационом, состоянии, особенно распространенным у пожилых индивидов. Наконец, анемия может развиться у пациентов с хроническими заболеваниями почек, возникшими почти по любой причине (Adamson, 2008, Harrison’s Principles of Internal Medicine, 17th ed.; McGraw Hill, New York, pp 628-634).
Анемия, возникающая в результате острой кровопотери достаточного объема, такая как в результате травмы или послеродового кровотечения, известна как острая постгеморрагическая анемия. Острая кровопотеря первоначально вызывает гиповолемию без анемии, поскольку имеется пропорциональное истощение запасов RBC наряду с другими ингредиентами крови. Однако гиповолемия быстро запускает физиологические механизмы, которые сдвигают жидкость из внесосудистого в сосудистый компартмент, что приводит к гемодилюции и анемии. Если кровопотеря носит хронический характер, то она постепенно истощает запасы железа организма и, в конечном счете, приводит к дефициту железа. Как продемонстрировано заявителями на модели мыши (см. пример ниже), полипептид-ловушка GDF, возможно, в комбинации с активатором рецепторов EPO, может использоваться для ускорения восстановления от анемии после острой кровопотери.
Железодефицитная анемия представляет собой конечную стадию в постепенном прогрессировании увеличивающегося дефицита железа, которая включает отрицательный баланс железа и железодефицитный эритропоэз в качестве промежуточных стадий. Дефицит железа может возникнуть в результате увеличенной потребности в железе, уменьшенного потребления железа или увеличенной потери железа, что, например, имеет место при таких состояниях как беременность, неадекватный рацион, нарушение кишечного всасывания, острое или хроническое воспаление и острая или хроническая кровопотеря. При анемии данного типа от легкой до умеренной степени тяжести, костный мозг остается гипопролиферативным, и морфология эритроцитов в значительной степени нормальная; однако даже незначительная анемия может привести к появлению некоторого числа микроцитарных, гипохромных эритроцитов, и переход к тяжелой железодефицитной анемии сопровождается гиперпролиферацией костного мозга и все более распространенными микроцитарными и гипохромными эритроцитами (Adamson, 2008, Harrison’s Principles of Internal Medicine, 17th ed.; McGraw Hill, New York, pp 628-634). Соответствующая терапия железодефицитной анемии зависит от ее причины и тяжести при пероральных препаратах железа, парентеральных препаративных формах железа и трансфузии эритроцитов в качестве ее основных обычных вариантов. Полипептид-ловушка GDF, возможно, в комбинации с активатором рецепторов EPO, могут использоваться для лечения хронических железодефицитных анемий отдельно или в комбинации с обычными терапевтическими подходами, в частности, для лечения анемий многофакторного происхождения.
Гипопролиферативные анемии могут возникать в результате первичной дисфункции или недостаточности костного мозга, а не дисфункции, вызванной воспалением, инфекцией или прогрессированием злокачественных заболеваний. Показательным примером была бы миелосупрессия, вызванная противораковыми химиотерапевтическими лекарственными средствами или лучевой терапией злокачественных заболеваний. Широкий обзор клинических испытаний обнаружил, что легкая анемия может возникать у 100% пациентов после химиотерапии, тогда как более тяжелая анемия может возникать у числа таких пациентов, достигающего 80% (Groopman et al., 1999, J Natl Cancer Inst 91:1616-1634). Миелосупрессивные лекарственные средства включают: 1) алкилирующие агенты, такие как азотистые иприты (например, мелфалан) и нитрозомочевины (например, стрептозоцин); 2) антиметаболиты, такие как антагонисты фолиевой кислоты (например, метотрексат), аналоги пурина (например, тиогуанин) и аналоги пиримидина (например, гемцитабин); 3) цитотоксические антибиотики, такие как антрациклины (например, доксорубицин); 4) ингибиторы киназы (например, гефитиниб); 5) митотические ингибиторы, такие как таксаны (например, паклитаксел) и алкалоиды винки (например, винорелбин); 6) моноклональные антитела (например, ритуксимаб); и 7) ингибиторы топоизомеразы (например, топотекан и этопозид). Как продемонстрировано на модели анемии у мыши, вызванной химиотерапией (см. Пример ниже), полипептид-ловушка GDF, возможно, в комбинации с активатором рецепторов EPO, может использоваться для лечения анемии, вызванной химиотерапевтическими средствами и/или лучевой терапией.
Полипептид-ловушка GDF, возможно, в комбинации с активатором рецепторов EPO, был бы также целесообразен для лечения анемий, связанных с нарушением созревания эритроцитов, которые частично характеризуются имеющими уменьшенный размер (микроцитарными), имеющими избыточный размер (макроцитарными), деформированными или аномально окрашенными (гипохромными) эритроцитами.
Пациентов можно лечить с использованием схемы введения, предназначенной для восстановления у пациента целевого уровня гемоглобина, обычно от примерно 10 г/дкл до примерно 12,5 г/дкл, а обычно примерно 11,0 г/дкл (см. также Jacobs et al. (2000) Nephrol Dial Transplant 15, 15-19), хотя более низкие целевые уровни могут вызвать меньшее количество побочных эффектов на сердечно-сосудистую систему. Альтернативно, уровни гематокрита (процентной доли объема образца крови, занимаемой клетками) можно использовать в качестве показателя состояния эритроцитов. Уровни гематокрита для здоровых индивидов находятся от 41 до 51% для взрослых мужчин и от 35 до 45% для взрослых женщин. Целевые уровни гематокрита обычно составляют примерно 30-33%. Кроме того, уровни гемоглобина/гематокрита варьируются от индивида к индивиду. Таким образом, оптимально, целевой уровень гемоглобина/гематокрита может быть индивидуализирован для каждого пациента.
В определенных вариантах осуществления, настоящее изобретение относится к способам ведения пациента, который получал лечение или является претендентом на лечение пептидом-ловушкой GDF, путем измерения одного или более гематологических параметров у пациента. Гематологические параметры могут использоваться для оценки соответствующей дозировки для пациента, который является претендентом на лечение полипептидом-ловушкой GDF, для мониторинга гематологических параметров во время лечения полипептидом-ловушкой GDF для оценки, следует ли корригировать дозировку во время лечения полипептидом-ловушкой GDF и/или для оценки соответствующей поддерживающей дозы полипептида-ловушки GDF. Если один или более гематологических параметров находятся за пределами нормального уровня, то дозировку полипептида-ловушки GDF можно снизить, его введение можно отсрочить или закончить.
Гематологические параметры, которые можно измерить в соответствии со способами по настоящему изобретению, включают, например, уровни эритроцитов, артериальное давление, запасы железа и другие агенты, обнаруживаемые в биологических жидкостях, которые коррелируются с повышенными уровнями эритроцитов, с использованием признанных в данной области способов. Такие параметры можно определять с использованием образца крови, взятого у пациента. Увеличения уровней эритроцитов, уровней гемоглобина и/или уровней гематокрита вызывают повышение артериального давления.
В одном из вариантов осуществления, если один или более гематологических параметров находятся за пределами нормального диапазона или на высокой стороне нормального значения у пациента, который является претендентом на лечение полипептидом-ловушкой GDF, то начало введения полипептида-ловушки GDF может быть отсрочено до возвращения гематологических параметров к нормальному или приемлемому уровню или естественным путем, или посредством терапевтического вмешательства. Например, если у пациента-претендента имеется гипертония или прегипертоническое состояние, то пациента можно лечить средствами, снижающими артериальное давление, для снижения артериального давления пациента. Можно использовать любое средство, снижающее артериальное давление, целесообразное для состояния отдельного пациента, включая, например, диуретики, адренергические ингибиторы (включая альфа-блокаторы и бета-блокаторы), сосудорасширяющие средства, блокаторы кальциевых каналов, ингибиторы ангиотензин-превращающего фермента (ACE), или блокаторы рецепторов ангиотензина-II. Артериальную гипертонию можно альтернативно лечить с использованием диета и схемы физической нагрузки. Аналогичным образом, если у пациента-претендента на лечение имеются запасы железа, которые ниже нормальных, или на низкой стороне нормы, то пациента можно лечить соответствующей схемой рациона и/или добавок железа до тех пор, пока запасы железа пациента не вернутся к нормальному или приемлемому уровню. Для пациентов, имеющих уровни эритроцитов и/или уровни гемоглобина выше нормального, введение полипептида-ловушки GDF может быть отложено до тех пор, пока уровни вернутся к нормальному или приемлемому значению.
В определенных вариантах осуществления, если у пациента, который является претендентом на лечение полипептидом-ловушкой GDF, один или более гематологических параметров находятся за пределами нормального диапазона, или на высокой стороне нормы, то начало введение можно не откладывать. Однако количество дозировки или частота введения полипептида-ловушки GDF могут быть установлены на уровень, который снизил бы риск неприемлемого увеличения гематологических параметров, возникающего после введения полипептида-ловушки GDF. Альтернативно, для пациента может быть разработана терапевтическая схема, которая комбинирует полипептид-ловушку GDF с терапевтическим средством, которое корригирует нежелательный уровень гематологического параметра. Например, если у пациента имеется повышенное артериальное давление, то может быть разработана терапевтическая схема, включающая введение полипептида-ловушки GDF и средство, снижающее артериальное давление. Для пациента, имеющего запасы железа ниже желательного уровня, может быть разработана терапевтическая схема введения полипептида-ловушки GDF и пополнения запасов железа.
В одном из вариантов осуществления, исходные параметр(ы) для одного или более гематологических параметров могут быть установлены для пациента, который является претендентом на лечение полипептидом-ловушкой GDF, и соответствующая схема введения может устанавливаться для данного пациента на основании исходных величин(ы). Альтернативно, исходные параметры, установленные на основании медицинского анамнеза пациента, могут использоваться для сообщения пациенту соответствующей схемы введения полипептида-ловушки GDF. Например, если у здорового пациента имеется установленное значение исходного артериального давления, которое выше определенного нормального диапазона, то может не быть необходимости в доведении артериального давления пациента до диапазона, который считается нормальным для общей популяции, перед лечением полипептидом-ловушкой GDF. Исходные величины одного или более гематологических параметров пациента перед лечением полипептидом-ловушкой GDF могут также использоваться в качестве релевантных сравнительных величин для мониторинга любых изменений гематологических параметров во время лечения полипептидом-ловушкой GDF.
В определенных вариантах осуществления, один или более гематологических параметров измеряются у пациентов, которые получают лечение полипептидом-ловушкой GDF. Гематологические параметры могут использоваться для мониторинга пациента во время лечения и обеспечения возможности коррекции или прекращения введения полипептида-ловушки GDF или дополнительного введения другого терапевтического средства. Например, если введение полипептида-ловушки GDF приводит к повышению артериального давления, уровня эритроцитов или уровня гемоглобина, или к снижению запасов железа, то доза полипептида-ловушки GDF может быть снижена по количеству или частоте введения для уменьшения эффектов полипептида-ловушки GDF на один или более гематологических параметров. Если введение полипептида-ловушки GDF приводит к изменению одного или более гематологических параметров, что неблагоприятно для пациента, то введение полипептида-ловушки GDF может быть закончено или временно, до возврата гематологических параметров до приемлемого уровня, или постоянно. Аналогичным образом, если один или более гематологических параметров не доводятся до уровня в пределах приемлемого диапазона после снижения дозы или частоты введения полипептида-ловушки GDF, то введение может быть прекращено. В качестве альтернативы, или в дополнение к снижению или прекращению введения полипептида-ловушки GDF, пациенту может вводиться дополнительное терапевтическое средство, которое корригирует нежелательный уровень гематологических параметров, такое как, например, средство, снижающее артериальное давление, или добавка железа. Например, если у пациента, получающего лечение полипептидом-ловушкой GDF, имеется повышенное артериальное давление, то введение полипептида-ловушки GDF может продолжаться на том же уровне, и средство, снижающее артериальное давление, добавляется к схеме лечения, введение полипептида-ловушки GDF может быть снижено (например, по количеству и/или частоте), и средство, снижающее артериальное давление, добавляется к схеме лечения, или введение полипептида-ловушки GDF может быть прекращено, и пациент может получать лечение средством, снижающим артериальное давление.
В определенных вариантах осуществления, пациенты, получающие лечение полипептидом-ловушкой GDF, или пациенты-претенденты на лечение полипептидом-ловушкой GDF, представляют собой пациентов, нуждающихся в мышечном росте, таких как пациентов, страдающих нервно-мышечным расстройством или дегенеративным мышечным расстройством, или имеющих риск их развития. Например, пациенты или пациенты-претенденты могут страдать или иметь риск развития болезни Лу Герига (бокового амиотрофического склероза), синдрома раковой анорексии-кахексии, мышечной дистрофии, застойного обструктивного легочного заболевания (COPD) (и мышечной дистрофии, связанной с COPD), синдрома мышечной дистрофии, саркопении или кахексии. Мышечная дистрофия относится к группе дегенеративных мышечных заболеваний, характеризуемых постепенным ослаблением и деградацией скелетных мышц и иногда сердца и дыхательных мышц. Иллюстративные мышечные дистрофии, которые можно лечить схемой, включающей полипептиды-ловушки GDF по изобретению, включают: мышечную дистрофию Дюшенна (DMD), мышечную дистрофию Бекера (BMD), мышечную дистрофию Эмери-Дрейфесса (EDMD), мышечную дистрофию Лимба-Джирдла (LGMD), лице-лопаточно-плечевую мышечную дистрофию (FSH или FSHD) (также известную как мышечная дистрофия Ландузи-Дежерина), миотоническую дистрофию (MMD) (также известную как болезнь Стейнерта), глазоглоточную мышечную дистрофию (OPMD), дистальную мышечную дистрофию (DD), врожденную мышечную дистрофию (CMD).
6. Фармацевтические композиции
В определенных вариантах осуществления, соединения (например, полипептид-ловушка GDF) по настоящему изобретению составляются с фармацевтически приемлемым носителем. Например, полипептид-ловушка GDF может вводиться отдельно или в качестве компонента фармацевтической препаративной формы (терапевтической композиции). Соединения по изобретению могут составляться для введения любым подходящим путем для применения в медицине или ветеринарии.
В определенных вариантах осуществления, способ лечения по изобретению включает введение композиции системно или местно в виде имплантата или устройства. При введении, терапевтическая композиция для применения в данном изобретении является, конечно, беспирогенной, физиологически приемлемой форме. Терапевтически полезные средства, кроме полипептида-ловушки GDF, включение которых также возможно в композицию, как описано выше, могут вводиться одновременно или последовательно с объектами по изобретению (например, полипептидами-ловушками GDF) в способах по изобретению.
Обычно, соединения вводятся парентерально. Фармацевтические композиции, подходящие для парентерального введения, могут включать один или более полипептидов-ловушек GDF в комбинации с одним или более фармацевтически приемлемых стерильных изотонических водных или неводных растворов, дисперсий, суспензий или эмульсий, или стерильных порошков, влагосодержание которых может восстанавливаться стерильными инъецируемыми растворами или дисперсиями непосредственно перед применением, которые могут содержать антиоксиданты, буферы, бактериостатические агенты, растворенные вещества, которые придают препаративной форме изотоничность с кровью предполагаемого реципиента, или суспендирующие агенты или загустители. Примеры подходящих водных и неводных носителей, которые могут использоваться в фармацевтических композициях по изобретению, включают воду, этанол, полиолы (такие как глицерол, пропиленгликоль, полиэтиленгликоль и тому подобные) и их подходящие смеси, растительные масла, такие как оливковое масло, и инъецируемые органические сложные эфиры, такие как этилолеат. Должная текучесть может поддерживаться, например, путем использования покрывающих материалов, таких как лецитин, поддержанием требуемого размера частиц в случае дисперсий и путем использования поверхностно-активных веществ.
Кроме того, композиция может быть инкапсулирована или инъецирована в форме для доставки в целевой участок ткани (например, костный мозг). В определенных вариантах осуществления, композиции по настоящему изобретению могут включать матрицу, способную доставлять одно или более терапевтических соединений (например, полипептид-ловушку GDF) в целевой участок ткани (например, костный мозг), обеспечивающую структуру для развития ткани и, оптимально, способную к резорбции в организм. Например, матрица может обеспечить медленное высвобождение полипептидов-ловушек GDF. Такие матрицы могут быть образованы из материалов, используемых в настоящее время для других видов применения имплантируемых медицинских материалов.
Выбор матричного материала основывается на биосовместимости, биоразлагаемости, механических свойствах, косметическом внешнем виде и свойствах поверхности раздела. Конкретное применение композиции по изобретению определяет соответствующую препаративную форму. Потенциальные матрицы для композиций могут представлять собой биологически разлагаемый и химически определенный сульфат кальция, трикальцийфосфат, гидроксиапатит, полиуксусную кислоту и полиангидриды. Другие потенциальные материалы являются биологически разлагаемыми и биологически хорошо определенными, такими как костный или дермальный коллаген. Еще одни матрицы состоят из чистых белков или компонентов внеклеточной матрицы. Другие потенциальные матрицы являются не биологически разлагаемыми и химически определенными, такими как спекшийся гидроксиапатит, биокерамика, алюминаты или другие керамические материалы. Матрицы могут состоять из комбинаций любого из указанных выше типов материала, такого как полиуксусная кислота и гидроксиапатит или коллаген и трикальцийфосфат. Биокерамика может изменяться по композиции, такой как кальций-алюминат-фосфат, и переработке для изменения размера пор, размера частиц и биологической разлагаемости.
В определенных вариантах осуществления, в способах по изобретению, композиции могут вводиться перорально, например, в форме капсул, облаток, пилюль, таблеток, пастилок (с использованием ароматизированной основы, обычно сахарозы и акации или трагаканта), порошков, гранул или в виде раствора или суспензии в водной или неводной жидкости, или в виде эмульсии масла-в-воде или воды-в-масле, или в виде эликсира или сиропа, или в виде пастилок (с использованием инертного основания, такого как желатин и глицерин, или сахароза и акация) и/или в виде полосканий для ротовой полости и тому подобных, причем каждая содержит заданное количество средства в виде активного ингредиента. Средство может также вводиться в виде болюса, электуария или пасты.
В твердых лекарственных формах для перорального введения (капсулы, таблетки, пилюли, драже, порошки, гранулы и тому подобные), одно или более терапевтических соединений по настоящему изобретению могут смешиваться с одним или более фармацевтически приемлемых носителей, таких как цитрат натрия или дикальцийфосфат, и/или одним из следующих: (1) наполнителей или агентов, увеличивающих объем, таких как крахмалы, лактоза, сахароза, глюкоза, маннит и/или кремневая кислота; (2) связывающих агентов, таких как, например, карбоксиметилцеллюлоза, альгинаты, желатин, поливинилпирролидон, сахароза и/или акация; (3) увлажнителей, таких как глицерол; (4) разрыхлителей, таких как агар-агар, карбонат кальция, крахмал картофеля или тапиоки, альгиновая кислота, определенные силикаты и карбонат натрия; (5) агентов, задерживающих растворение, таких как парафин; (6) ускорителей абсорбции, таких как соединения четвертичного аммония; (7) смачивающих агентов, таких как, например, цетиловый спирт и глицерол моностеарат; (8) поглотителей, таких как каолин и бентонитовая глина; (9) смазывающих агентов, таких как тальк, стеарат кальция, стеарат магния, твердые полиэтиленгликоли, лаурилсульфат натрия и их смеси; и (10) красящих агентов. В случае капсул, таблеток и пилюль, фармацевтические композиции могут также содержать забуферивающие агенты. Твердые композиции аналогичного типа могут также использоваться в качестве наполнителей в мягких и твердых заполненных желатиновых капсулах с использованием таких наполнителей как лактоза или молочные сахара, а также полиэтиленгликоли с высокой молекулярной массой и тому подобные.
Жидкие лекарственные формы для перорального введения включают фармацевтически приемлемые эмульсии, микроэмульсии, растворы, суспензии, сиропы и эликсиры. В дополнение к активному ингредиенту, жидкие лекарственные формы могут содержать инертные разбавители, обычно используемые в данной области, такие как вода или другие растворители, солюбилизирующие агенты и эмульгаторы, такие как этиловый спирт, изопропиловый спирт, этилкарбонат, этилацетат, бензиловый спирт, бензилбензоат, пропиленгликоль, 1,3-бутиленгликоль, масла (в частности, масло хлопковых семян, арахисовое, кукурузное, масло зародышей пшеницы, оливковое, касторовое и кунжутное масла), глицерол, тетрагидрофуриловый спирт, полиэтиленгликоли и сложные эфиры жирных кислот сорбитана, и их смеси. Кроме инертных разбавителей, пероральные композиции могут также включать адъюванты, такие как смачивающие агенты, эмульгирующие и суспендирующие агенты, подслащивающие, ароматизирующие, красящие, одорирующие и консервирующие вещества.
Суспензии, в дополнение к активным соединениям, могут содержать суспендирующие агенты, такие как этоксилированные изостеариловые спирты, полиоксиэтиленсорбитан и сложные эфиры сорбитана, микрокристаллическую целлюлозу, метагидроксид алюминия, бентонит, агар-агар, и трагакант и их смеси.
Композиции по изобретению могут также содержать адъюванты, такие как консерванты, смачивающие агенты, эмульгирующие агенты и диспергирующие агенты. Предотвращение действия микроорганизмов может быть обеспечено включением различных антибактериальных и противогрибковых средств, например, парабена, хлорбутанола, фенолсорбиновой кислоты и тому подобных. Может быть также желательным включение в композиции изотонических агентов, таких как сахара, хлорид натрия и тому подобные. Кроме того, длительное всасывание инъецируемой фармацевтической формы может быть обеспечено включением агентов, которые задерживают всасывание, таких как моностеарат алюминия и желатин.
Понятно, что схема введения будет определяться лечащим врачом, учитывающим различные факторы, которые модифицируют действие обсуждаемых соединений по изобретению (т.е., полипептида-ловушки GDF). Различные факторы включают, без ограничения, количество эритроцитов, уровень гемоглобина у пациента или другие диагностические оценки, желательное целевое количество эритроцитов, возраст, пол и рацион питания пациента, тяжесть любого заболевания, которое может способствовать сниженному уровню эритроцитов, время введения и другие клинические факторы. Добавление других известных факторов роста к конечной композиции может также воздействовать на дозировку. Прогресс может контролироваться периодической оценкой уровней эритроцитов и гемоглобина, а также оценками уровней ретикулоцитов и другими показателями гематопоэтического процесса.
В определенных вариантах осуществления, настоящее изобретение также относится к генной терапии для получения полипептидов-ловушек GDF in vivo. Такая терапия достигла бы своего терапевтического эффекта введением последовательностей полипептидов-ловушек GDF в клетки или ткани, имеющие перечисленные выше расстройства. Доставка последовательностей полипептидов-ловушек GDF может быть достигнута с использованием рекомбинантного вектора экспрессии, такого как химерный вирус или коллоидная дисперсионная система. Предпочтительным для терапевтической доставки последовательностей полипептидов-ловушек GDF является использование нацеленных липосом.
Различные вирусные векторы, которые могут использоваться для генной терапии в соответствии с положениями настоящего изобретения, включают аденовирус, вирус герпеса, вирус коровьей оспы или вирус РНК, такой как ретровирус. Ретировирусный вектор может представлять собой производное ретровируса мыши или птицы. Примеры ретровирусных векторов, в которые может быть введен один инородный ген, включают без ограничения: вирус лейкоза Молони мыши (MoMuLV), вирус саркомы Харвея мыши (HaMuSV), вирус опухоли молочных желез мыши (MuMTV) и вирус саркомы Рауса (RSV). Ряд дополнительных ретровирусных векторов может включать множественные гены. Все указанные векторы могут переносить или включать ген для отбираемого маркера с тем, чтобы могли быть идентифицированы и получены трансдуцированные клетки. Ретровирусные векторы могут быть получены специфичными для целевого объекта путем прикрепления, например, сахара, гликолипида или белка. Предпочтительное нацеливание осуществляется использованием антитела. Специалистам в данной области будет понятно, что специфические полинуклеотидные последовательности могут быть вставлены в ретровирусный геном или прикреплены к вирусной оболочке для обеспечения возможности специфичной целевой доставки ретровирусного вектора, содержащего полипептид-мишень GDF.
Альтернативно, клетки тканевой культуры могут непосредственно трансфицироваться плазмидами, кодирующими ретровирусные структурные гены gag, pol и env, обычной трансфекцией фосфатом кальция. Затем эти клетки трансфицируются векторной плазмидой, содержащей представляющие интерес гены. Полученные в результаты клетки высвобождают ретровирусный вектор в культуральную среду.
Другая система нацеленной доставки полинуклеотидов-ловушек GDF представляет собой коллоидную дисперсионную систему. Коллоидные дисперсионные системы включают комплексы макромолекул, нанокапсулы, микросферы, гранулы и системы на основе липидов, включая эмульсии масла-в-воде, мицеллы, смешанные мицеллы и липосомы. Предпочтительная коллоидная система по настоящему изобретению представляет собой липосому. Липосомы представляют собой искусственные мембранные пузырьки, которые могут использоваться в качестве носителей доставки in vitro и in vivo. РНК, ДНК и интактные вирионы могут быть инкапсулированы в водную внутреннюю среду и доставлены в клетки в биологически активной форме (см., например, Fraley, et al., Trends Biochem. Sci., 6:77, 1981). Способы эффективного переноса генов с использованием липосомного носителя известны в данной области, см., например, Mannino, et al., Biotechniques, 6:682, 1988. Композиция липосомы представляет собой обычно комбинацию фосфолипидов, обычно в комбинации со стероидами, в частности, холестерином. Могут также использоваться другие фосфолипиды или другие липиды. Физические характеристики липосом зависят от pH, ионной силы и присутствия двухвалентных катионов.
Примеры липидов, используемых при получении липосом, включают фосфатидильные соединения, такие как фосфатидилглицерол, фосфатидилхолин, фосфатидилсерин, фосфатидилэтаноламин, сфинголипиды, цереброзиды и ганглиозиды. Иллюстративные фосфолипиды включают яичный фосфатидилхолин, дипальмитоилфлсфатидилхолин и дистеароилфосфатидилхолин. Нацеливание липосом также возможно на основании, например, органной специфичности, клеточной специфичности и специфичности к органеллам и известно в данной области.
ПРИМЕРЫ
Теперь, после общего описания изобретения, его можно будет легче понять путем ссылки на следующие примеры, которые предназначены просто для целей иллюстрации определенных вариантов осуществления и вариантов осуществления настоящего изобретения, и не предназначены для ограничения изобретения.
Пример 1. Образование ловушки GDF.
Заявители конструировали ловушку GDF следующим образом. Полипептид, имеющий модифицированный внеклеточный домен ActRIIB со значительно сниженным связыванием активина A относительно GDF11 и/или миостатина (как следствие замена лейцина на аспартат в положении 79 в SEQ ID NO:1) был слит с доменом Fc-фрагмента человека или мыши с минимальным линкером (три глициновых аминокислоты) между ними. Конструкции называются соответственно ActRIIB(L79D 20-134)-hFc и ActRIIB(L79D 20-134)-mFc. Альтернативные формы с глутаматом, а не аспартатом в положении 79 функционировали аналогичным образом (L79E). Альтернативные формы с аланином, а не валином в положении 226 в отношении SEQ ID NO: 7, показанной ниже, были также получены и функционировали эквивалентно во всех тестированных отношениях. Аспартат в положении 79 (относительно SEQ ID NO:1, или положении 60 относительно последовательности SEQ ID NO:7) выделен ниже серым фоном. Валин в положении 226 относительно последовательности SEQ ID NO:7 также выделен серым фоном ниже.
Ловушка GDF ActRIIB(L79D 20-134)-hFc, очищенная из линий клеток CHO (SEQ ID NO:7), показана ниже.
GRGEAETRECIYYNANWELERTNQSGLERCEGEQDKRLHCYASWRNSSGTIELVKKGCWDDDFNCYDRQECVATEENPQVYFCCCEGNFCNERFTHLPEAGGPEVTYEPPPTAPTGGGTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPVPIEKTISKAKGQPREPQVYTLPPSREEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK
Полученная из ActRIIB часть ловушки GDF имеет аминокислотную последовательность, представленную ниже (SEQ ID NO:32), и эта часть может использоваться в качестве мономера или в качестве не-Fc слитого белка в качестве мономера, димера или комплекса большего порядка.
GRGEAETRECIYYNANWELERTNQSGLERCEGEQDKRLHCYASWRNSSGTIELVKKGCWDDDFNCYDRQECVATEENPQVYFCCCEGNFCNERFTHLPEAGGPEVTYEPPPTAPT (SEQ ID NO:32)
Белок ловушки GDF был экспрессирован в линиях клеток CHO. Рассматривались три различных лидерных последовательности:
(i) Меллитин медоносных пчел (HBML): MKFLVNVALVFMVVYISYIYA (SEQ ID NO:8)
(ii) Активатор тканевого плазминогена (TPA): MDAMKRGLCCVLLLCGAVFVSP (SEQ ID NO:9)
(iii) Нативная: MTAPWVALALLWGSLCAGS (SEQ ID NO:10).
В выбранной форме используется лидер TPA и имеет следующую необработанную аминокислотную последовательность:
MDAMKRGLCCVLLLCGAVFVSPGASGRGEAETRECIYYNANWELERTNQSGLERCEGEQDKRLHCYASWRNSSGTIELVKKGCWDDDFNCYDRQECVATEENPQVYFCCCEGNFCNERFTHLPEAGGPEVTYEPPPTAPTGGGTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPVPIEKTISKAKGQPREPQVYTLPPSREEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK (SEQ ID NO: 11)
Этот полипептид кодируется следующей последовательностью нуклеиновой кислоты (SEQ ID NO:12):
A TGGATGCAAT GAAGAGAGGG CTCTGCTGTG TGCTGCTGCT GTGTGGAGCA GTCTTCGTTT CGCCCGGCGC CTCTGGGCGT GGGGAGGCTG AGACACGGGA GTGCATCTAC TACAACGCCA ACTGGGAGCT GGAGCGCACC AACCAGAGCG GCCTGGAGCG CTGCGAAGGC GAGCAGGACA AGCGGCTGCA CTGCTACGCC TCCTGGCGCA ACAGCTCTGG CACCATCGAG CTCGTGAAGA AGGGCTGCTG GGACGATGAC TTCAACTGCT ACGATAGGCA GGAGTGTGTG GCCACTGAGG AGAACCCCCA GGTGTACTTC TGCTGCTGTG AAGGCAACTT CTGCAACGAG CGCTTCACTC ATTTGCCAGA GGCTGGGGGC CCGGAAGTCA CGTACGAGCC ACCCCCGACA GCCCCCACCG GTGGTGGAAC TCACACATGC CCACCGTGCC CAGCACCTGA ACTCCTGGGG GGACCGTCAG TCTTCCTCTT CCCCCCAAAA CCCAAGGACA CCCTCATGAT CTCCCGGACC CCTGAGGTCA CATGCGTGGT GGTGGACGTG AGCCACGAAG ACCCTGAGGT CAAGTTCAAC TGGTACGTGG ACGGCGTGGA GGTGCATAAT GCCAAGACAA AGCCGCGGGA GGAGCAGTAC AACAGCACGT ACCGTGTGGT CAGCGTCCTC ACCGTCCTGC ACCAGGACTG GCTGAATGGC AAGGAGTACA AGTGCAAGGT CTCCAACAAA GCCCTCCCAG TCCCCATCGA GAAAACCATC TCCAAAGCCA AAGGGCAGCC CCGAGAACCA CAGGTGTACA CCCTGCCCCC ATCCCGGGAG GAGATGACCA AGAACCAGGT CAGCCTGACC TGCCTGGTCA AAGGCTTCTA TCCCAGCGAC ATCGCCGTGG AGTGGGAGAG CAATGGGCAG CCGGAGAACA ACTACAAGAC CACGCCTCCC GTGCTGGACT CCGACGGCTC CTTCTTCCTC TATAGCAAGC TCACCGTGGA CAAGAGCAGG TGGCAGCAGG GGAACGTCTT CTCATGCTCC GTGATGCATG AGGCTCTGCA CAACCACTAC ACGCAGAAGA GCCTCTCCCT GTCTCCGGGT AAATGA
Очистка может быть достигнута серией стадий колоночной хроматографии, включая, например, три или более из следующих, в любом порядке: хроматография белка A, хроматография на Q-сефарозе, хроматография на фенилсефарозе, хроматография с исключением по размеру и катионообменная хроматография. Очистка может завершаться вирусной фильтрацией и буферным обменом. В одном примере схемы очистки, среду клеточной культуры пропускают через колонку белка A, промывают в 150 мМ Tris/NaCl (pH 8,0), затем промывают 50 мМ Tris/NaCl (pH 8,0) и элюируют 0,1 M глицином, pH 3,0. Элюат с низким pH держат при комнатной температуре в течение 30 минут в качестве стадии вирусной очистки. Затем элюат нейтрализуют и пропускают через ионообменную колонку Q-сефарозы и промывают в 50 мМ Tris pH 8,0, 50 мМ NaCl, и элюируют в 50 мМ Tris pH 8,0, при концентрации NaCl от 150 мМ до 300 мМ. Затем элюат загружают в 50 мМ Tris pH 8,0, 1,1 M сульфата аммония и пропускают через колонку фенилсафарозы, промывают и элюируют в 50 мМ Tris pH 8,0 с сульфатом аммония от 150 до 300 мМ. Элюат диализируют и фильтруют для использования.
Дополнительные ловушки GDF (слитые белки ActRIIB-Fc, модифицированные с тем, чтобы уменьшить соотношение между связыванием активина A и миостатином или GDF11) описаны в документах PCT/US2008/001506 и WO 2006/012627, включенных в настоящее описание путем ссылки.
Пример 2. Биологический анализ передачи сигналов, опосредованной GDF-11 и активином.
Анализ репортерного гена A-204 использовали для оценки воздействий белков ActRIIB-Fc и ловушек GDF на передачу сигналов GDF-11 и активином A. Клеточная линия: рабдомиосаркома человека (происходящая из мышцы). Репортерный вектор: pGL3(CAGA)12 (Описанный в публикации Dennler et al, 1998, EMBO 17: 3091-3100). Мотив CAGA12 присутствует в генах, реагирующих на TGF-бета (ген PAI-1), и поэтому этот вектор в целом используется для факторов, передающих сигналы через Smad2 и 3.
День 1: Распределить клетки A-204 в 48-луночный планшет.
День 2: Клетки A-204 трансфицируют 10 мкг pGL3(CAGA)12 или pGL3(CAGA)12 (10 мкг)+ pRLCMV (1 мкг) и фугеном.
День 3: Добавить факторы (растворенные в среде + 0,1% BSA). Ингибиторы необходимо предварительно инкубировать с факторами в течение 1 ч перед добавлением к клеткам. Через 6 ч, клетки промывают PBS (солевым раствором с фосфатным буфером) и лизируют клетки.
За этим следует анализ люциферазы. В отсутствие каких-либо ингибиторов, активин A проявил 10-кратную стимуляцию экспрессии репортерного гена и ED50 ~2 нг/мл. GDF-11: 16-кратная стимуляция, ED50: ~1,5 нг/мл.
ActRIIB(20-134) представляет собой мощный ингибитор активности активина, GDF-8 и GDF-11 в данном анализе. В данном анализе также тестировали варианты.
Пример 3. Ингибирование N-концевых и C-концевых усечений GDF-11
Были получены варианты ActRIIB(20-134)-hFc с усечениями на N-конце и/или C-конце, и их тестировали на наличие активности в качестве ингибиторов GDF-11 и активина. Их активность показана ниже (по данным измерения в кондиционированных средах):
C-концевые усечения ActRIIB-hFc:
Как видно, усечения трех (заканчивающихся …PPT), шести (заканчивающихся …YEP) или более аминокислот на C-конце вызывает трехкратное или большее уменьшение активности молекулы. Усечение конечных 15 аминокислот части ActRIIB вызывает большую потерю активности (см. WO2006/012627).
Амино концевые усечения получали в теневом фенотипе белка ActRIIB(20-131)-hFc. Их активность показана ниже (по данным измерения в кондиционированных средах):
N-концевые усечения ActRIIB-hFc:
(GRG…)
(RGE…)
(GEA…)
(EAE…)
(AET…)
Соответственно, усечения двух, трех или четырех аминокислот с N-конца ведут к получению более активного белка, чем версии с внеклеточным полноразмерным доменом. Дополнительные эксперименты показывают, что усечение пяти аминокислот ActRIIB(25-131)-hFc имеют активность, эквивалентную не усеченной форме, и дополнительные делеции на N-конце продолжают нарушать активность белка. Поэтому, оптимальные конструкции будут иметь C-конец, заканчивающийся между аминокислотами 133-134 последовательности SEQ ID NO:1, и N-конец, начинающийся с аминокислот 22-24 последовательности SEQ ID NO:1. N-конец, соответствующий аминокислотам 21 или 25, обеспечит активность, которая аналогична конструкции ActRIIB(20-134)-hFc. Эти усечения могут также использоваться в контексте ловушек GDF, таких как вариант L79D или L79E.
Пример 4. Варианты ActRIIB-Fc, активность на клеточной основе.
Активность белков ActRIIB-Fc и ловушек GDF тестировали в анализе на клеточной основе, как описано выше. Результаты суммированы в представленной ниже таблице. Некоторые варианты тестировали в других конструкциях с C-концевым усечением. Как обсуждалось выше, усечения пяти или пятнадцати аминокислот вызывало снижение активности. Ловушки GDF (варианты L79D и L79E) показали существенную утрату связывания активина при сохранении ингибирования GDF-11 почти дикого типа.
Связывание растворимого ActRIIB-Fc с GDF11 и Активином A:
(приблизительно 10-8 M KI)
(приблизительно 10-8 M KI)
(приблизительно 10-6 M KI)
(приблизительно 10-6 M KI)
++ Умеренная активность (приблизительно 1×10-7 KI)
+++ Хорошая активность (дикого типа) (приблизительно 1×10-8 KI)
++++ Активность, большая чем дикого типа
Несколько вариантов оценивали в отношении периода полувыведения из сыворотки у крыс. ActRIIB(20-134)-Fc имеет период полувыведения из сыворотки приблизительно 70 часов. ActRIIB(A24N 20-134)-Fc имеет период полувыведения из сыворотки приблизительно 100-150 часов. Вариант A24N имеет величины активности в анализе на клеточной основе (см. выше) и анализах in vivo (см. ниже), которые эквивалентны молекуле дикого типа. В сочетании с более длинным периодом полувыведения, это означает, что с течением времени, вариант A24N даст больший эффект на единицу белка, чем молекула дикого типа. Вариант A24N и любой из других указанных выше тестированных вариантов могут комбинироваться с молекулами ловушки GDF, такими как варианты L79D или L79E.
Пример 5. Связывание GDF-11 и Активина A.
Связывание определенных белков ActRIIB-Fc и ловушек GDF с лигандами тестировали в анализе BiaCoreTM.
Варианты ActRIIB-Fc или белок дикого типа захватывали на систему с использованием анти-hFc антитела. Лиганды, вводимые инъекцией, протекали по захваченным рецепторным белкам. Результаты суммированы в представленных ниже таблицах.
Специфичность связывания лигандов вариантов IIB.
Эти данные подтверждают данные анализа на клеточной основе, демонстрируя, что вариант A24N сохраняет связывающую лиганды активность, которая аналогична активности молекулы ActRIIB(20-134)-hFc, и что молекула L79D или L79E сохраняет связывание миостатина и GDF11, но проявляет выражено сниженное (количественно не определяемое) связывание с Активином A.
Были получены и тестированы другие варианты, как сообщается в документе WO2006/012627 (полностью включенном в настоящее описание путем ссылки) см., например, стр. 59-60, где используются лиганды, соединенные с устройством, и протекание рецептора по соединенным лигандам. В особенности, K74Y, K74F, K74I (и, предположительно, другие гидрофобные замены в K74, например, K74L) и D80I, вызывают уменьшение отношения связывания Активина A к связыванию GDF11, относительно молекулы K74 дикого типа. Ниже в таблице представлены данные в отношении указанных вариантов:
Варианты растворимого ActRIIB-Fc, связывающиеся с GDF11 и Активином A (анализ BiaCore)
(+)
(+)
(+++)
(+++)
(++++)
+++++
+++++
-- < 1/5 связывания дикого типа
- ~ 1/2 связывания дикого типа
+ Дикий тип
++ увеличение связывания < чем в 2 раза
+++ увеличение связывания ~ в 5 раз
++++ увеличение связывания ~ в 10 раз
+++++ увеличение связывания ~ в 40 раз
Пример 6. ActRIIB-hFc стимулирует эритропоэз у не человекообразных приматов
ActRIIB(20-134)-hFc (IgG1) вводили один раз в неделю в течение 1 месяца самцам и самкам яванских макак подкожной инъекцией. 48 яванских макак (по 24 каждого пола) включали в одну из четырех групп лечения (6 животных/пол/группу), и им подкожно инъецировали или носитель, или ActRIIB-hFc в дозе 3, 10 или 30 мг/кг один раз в неделю в течение 4 недель (всего 5 доз). Оценивавшиеся параметры включали общую клиническую патологию (гематологию, клиническую химию, свертывание и анализ мочи). ActRIIB-hFc вызывал статистически значимое увеличение средних абсолютных величин ретикулоцитов к 15-му дню у получавших лечение животных. К 36-му дню, ActRIIB-hFc вызывал несколько гематологических изменений, включая повышенные средние абсолютные величины ретикулоцитов и величины ширины распределения эритроцитов и более низкие средние величины концентрации корпускулярного гемоглобина. Воздействию подверглись все группы лечения и представители обоих полов. Эти эффекты согласуются с положительным эффектом ActRIIB-hFc на высвобождение незрелых ретиколуцитов из костного мозга. Этот эффект устранялся после вымывания лекарственного препарата из организма получавших лечение животных (к 56-му дню исследования). Соответственно, заявители пришли к выводу, что ActRIIB-hFc стимулирует эритропоэз.
Пример 7. ActRIIB-mFc повышает аспекты эритропоэза у мышей путем стимуляции эритропоэтической активности селезенки
В данном исследовании анализировали эффекты введения in vivo ActRIIB(20-134)-mFc на частоту встречаемости гематопоэтических клеток-предшественников в костном мозге и селезенке. Одной группе мышей C57BL/6 инъецировали PBS в качестве контроля, а второй группе мышей вводили две дозы ActRIIB-mFc по 10 мг/кг, и животных обеих групп умерщвляли через 8 дней. Периферическую кровь использовали для выполнения полного анализа крови и бедренные кости и селезенки использовали для выполнения клоногенных анализов in vitro для оценки содержания лимфоидных, эритроидных и миелоидных клеток-предшественников в каждом органе. В короткой временной рамке данного исследования, не наблюдались значимые изменения уровней эритроцитов, гемоглобина или лейкоцитов у получавших лечение мышей. В бедренных костях не было различия чисел имеющих ядра клеток или содержания клеток-предшественников в контрольной группе и группах лечения. В селезенках, у группы, получавшей соединение, отмечено статистически значимое увеличение числа колоний зрелых эритроидных клеток-предшественников (CFU-E) на чашку, частоты встречаемости и общего числа клеток-предшественников на селезенку. Кроме того, наблюдалось и увеличение числа миелоидных клеток (CFU-GM), незрелых эритроидных клеток (BFU-E) и общего числа клеток-предшественников на селезенку.
Животные:
В данном исследовании использовали 16 самок мышей C57BL/6 в возрасте 6-8 недель. Восьми мышам в 1-й и 3-й день подкожно инъецировали тестируемое соединение ActRIIB-mFc в дозе 10 мг/кг, и восьми мышам в качестве контроля подкожно инъецировали солевой раствор с фосфатным буфером (PBS) в объеме 100 мкл на мышь. Всех мышей умерщвляли через 8 дней после первой инъекции в соответствии с релевантными Инструкциями по уходу за животными. Образцы периферической крови (PB) у отдельных животных брали пункцией сердца и использовали для полного анализа крови и определения лейкоцитарной формулы (CBC/Diff). У каждой мыши производили забор бедренных костей и селезенок.
Выполненные тесты:
CBC/Diff подсчет
PB у каждой мыши брали посредством сердечной пункции и помещали в соответствующие пробирки с микроконтейнерами. Образцы посылали в клиническую лабораторию для анализа на счетчике CellDyn 3500.
Клоногенные анализы
Клоногенные клетки-предшественники миелоидных, эритроидных и лимфоидных линий дифференциации оценивали с использованием описанных ниже in vitro систем сред на основе метилцеллюлозы.
Клетки-предшественники зрелых эритроидных клеток:
Клоногенные клетки-предшественники зрелых эритроидных (CFU-E) линий дифференциации культивировали в MethoCultTM 3334, среде на основе метилцеллюлозы, содержащей рекомбинантный человеческий (rh) эритропоэтин (3 ЕД/мл).
Лимофидные клетки-предшественники:
Клоногенные клетки-предшественники лимофидной (CFU-pre-B) линии дифференциации культивировали в MethoCult® 3630, среде на основе метилцеллюлозы, содержащей rh интерлейкин 7 (10 нг/мл).
Миелоидные и незрелые эритроидные клетки-предшественники:
Клоногенные клетки-предшественники гранулоцитарно-моноцитарных (CFU-GM), эритроидных (BFU-E) и мультипотентных (CFU-GEMM) линий дифференциации культивировали в MethoCultTM 3434, сред на основе метилцеллюлозы, содержащей рекомбинантный мышиный (rm) фактор стволовых клеток (50 нг/мл), rh интерлейкин 6 (10 нг/мл), rm интерлейкин 3 (10 нг/мл) и rh эритропоэтин (3 ЕД/мл).
Методы:
Бедренные кости мыши и селезенки обрабатывали с использованием стандартных последовательностей операций. Коротко, костный мозг получали промыванием полости бедренной кости, модифицированной по Искову средой Дульбекко, содержащей 2% фетальную телячью сыворотку (IMDM 2% FBS), используя иглу 21 калибра и шприц объемом 1 см3. Клетки селезенки получали продавливанием ткани селезенки через фильтр 70 мкМ и промыванием фильтра IMDM 2% FBS. Затем выполняли подсчет ядерных клеток в 3% ледяной уксусной кислоте в суспензиях одиночных клеток с использованием счетной камеры Нойбауэра с тем, чтобы можно было подсчитать общее количество клеток на орган. Для удаления загрязняющих эритроцитов, все клетки селезенки затем разбавляли трехкратным объемом литического буфера хлорида аммония и инкубировали на льду в течение 10 минут. Затем клетки промывали и ресуспендировали в IMDM 2% FBS и выполняли второй подсчет клеток для определения концентрации клеток после лизиса.
Получали запасы клеток и добавляли в каждый состав сред на основе метилцеллюлозы для получения оптимальных концентраций посева на чашки для каждой ткани в каждый состав среды. Клетки костного мозга высевали на чашки в концентрации 1×105 клеток на чашку в среду MethoCultTM 3334 для оценки зрелых эритроидных клеток-предшественников, 2×105 клеток на чашку в среду MethoCultTM 3630 для оценки лимфоидных клеток-предшественников и 3×104 клеток на чашку в среду MethoCultTM 3434 для оценки незрелых эритроидных и миелоидных клеток-предшественников. Клетки селезенки высевали на чашки в концентрации 4×105 клеток на чашку в среду MethoCultTM 3334 для оценки зрелых эритроидных клеток-предшественников, 4×105 клеток на чашку в среду MethoCultTM 3630 для оценки лимфоидных клеток-предшественников и 2×105 клеток на чашку в среду MethoCultTM 3434 для оценки незрелых эритроидных и миелоидных клеток-предшественников. Культуры, высеянные на чашки в трех повторениях, инкубировали при 37ºC, 5% CO2 до выполнения подсчета и оценки колоний подготовленным персоналом. Зрелые эритроидные клетки-предшественники культивировали в течение 2 дней, лимфоидные клетки-предшественники культивировали в течение 7 дней и зрелые эритроидные и миелоидные клетки-предшественники культивировали в течение 12 дней.
Анализ:
Среднюю величину +/- 1 стандартное отклонение рассчитывали для культур в трех повторениях культур клоногенных анализов и для контроля и групп лечения для всех наборов данных.
Частоту колониеобразующих клеток (CFC) в каждой ткани рассчитывали следующим образом:
Количество клеток, высеянных на чашку
Среднюю CFC рассчитывали на чашку
Общую CFC на бедренную кость или селезенку рассчитывали следующим образом:
Общая рассчитанная CFC×количество ядерных клеток на бедренную кость или селезенку (после лизиса эритроцитов)
Число культивированных ядерных клеток
Стандартные t-критерии использовали для оценки наличия различий среднего числа клеток или гематопоэтических клеток-предшественников между РBS контрольными мышами и мышами, получавшими лечение соединением. Вследствие возможной субъективности подсчета колоний, значимой считали величину p менее чем 0,01. Средние величины (+/- SD) для каждой группы показаны в таблицах ниже.
Гематологические параметры
(л/л)
CFC из бедренной кости и селезенки
Лечение мышей ActRIIB(20-134)-mFc, в короткой временной рамке настоящего исследования, не привел к значимому увеличению содержания эритроцитов или гемоглобина. Однако воздействие на содержание клеток-предшественников было заметным. В бедренных костях не было различия числа ядерных клеток или содержания клеток-предшественников между контролем и группами лечения. В селезенках, в группе, получавшей лечение соединением, наблюдалось статистически значимое увеличение числа ядерных клеток перед лизисом эритроцитов и числа колоний зрелых эритроидных клеток-предшественников (CFU-E) на чашку, частоты встречаемости и общего числа клеток-предшественников на селезенку. Кроме того, наблюдалось увеличение числа миелоидных (CFU-GM), незрелых эритроидных (BFU-E) и общего числа клеток-предшественников на селезенку. Соответственно, ожидается, что в течение более длительного времени, лечение ActRIIB(20-134)-mFc может привести к повышенному содержанию эритроцитов и гемоглобина.
Пример 8: Ловушка GDF увеличивает уровни эритроцитов in vivo
12-недельных мышей C57BL/6NTac включали в одну из двух групп лечения (N=10). Мышам вводили или носитель, или вариантный полипептид ActRIIB («Ловушку GDF») [ActRIIB(L79D 20-134)-hFc] путем подкожной инъекции (SC) в дозе 10 мг/кг дважды в неделю в течение 4 недель. В конце исследования, забор цельной крови производили пункцией сердца в пробирки, содержащие EDTA (этилендиаминтетрауксусную кислоту) и анализировали для определения распределения клеток с использованием гематологического анализатора HM2 (Abaxis, Inc).
Обозначение групп
Лечение ловушкой GDF не оказывало статистически значимый эффект на число лейкоцитов (WBC), по сравнению с контролями носителя. Количества эритроцитов (RBC) увеличились у получавших лечение групп относительно контролей (см. таблицу ниже). И содержание гемоглобина (HGB), и гематокрит (HCT) были также увеличены вследствие дополнительных эритроцитов. Средняя ширина эритроцитов (RDW) была больше у получавших лечение животных, указывая на увеличение пула незрелых эритроцитов. Поэтому, лечение ловушкой GDF ведет к увеличению количества эритроцитов без выявляемых эффектов на популяции лейкоцитов.
Гематологические результаты
Пример 9: Ловушка GDF превосходит ActRIIB-Fc для увеличения уровней эритроцитов In vivo .
19-недельных самцов мышей C57BL/6NTac методом случайной выборки включали в одну из трех групп. Мышам вводили носитель (10 мМ солевого раствора, забуференного Tris-буфером, TBS), ActRIIB(20-134)-mFc дикого типа или ловушку GDF ActRIIB(L79D 20-134)-hFc путем подкожной инъекции дважды в неделю в течение трех недель. Забор крови осуществляли щечным кровопусканием в исходном состоянии и после трех недель введения и анализировали для определения распределения клеток с использованием гематологического анализатора (HM2, Abaxis, Inc.)
Лечение ActRIIB-Fc или ловушкой GDF не оказывало значимого эффекта на количества лейкоцитов (WBC), по сравнению с контролями носителя. Все показатели: количество эритроцитов (RBC), уровни гематокрита (HCT) и гемоглобина были повышены у мышей, получавших лечение ловушкой GDF, по сравнению или с контролями, или с конструкцией дикого типа (см. таблицу ниже). Поэтому, в прямом сравнении, ловушка GDF содействует увеличению содержания эритроцитов в значимо большей степени, чем белок ActRIIB-Fc дикого типа. Действительно, в данном эксперименте белок ActRIIB-Fc дикого типа не вызывал статистически значимого увеличения количества эритроцитов, свидетельствуя о том, что для выявления данного эффекта потребовалось бы более длительное введение или более высокие дозы.
Гематологические результаты после трех недель введения
Пример 10. Получение ловушки GDF с усеченным внеклеточным доменом ActRIIB
Как описано в примере 1, ловушка GDF, называемая ActRIIB(L79D 20-134)-hFc, была получена N-концевым слиянием лидера TPA с внеклеточным доменом ActRIIB (остатки 20-134 в последовательности SEQ ID NO:1), содержащим замещение лейцина на аспартат (в остатке 79 в последовательности SEQ ID NO:1) и C-концевым слиянием домена Fc человека с минимальным линкером (тремя глициновыми остатками) (фиг. 3). Нуклеотидная последовательность, соответствующая данному слитому белку, показана на фиг. 4.
Ловушка GDF с внеклеточным доменом усеченного ActRIIB, называемая ActRIIB(L79D 25-131)-hFc, была получена N-концевым слиянием лидера TPA с усеченным внеклеточным доменом (остатки 25-131 в SEQ ID NO:1), содержащим замещение лейцина на аспартат (в остатке 79 в последовательности SEQ ID NO:1) и C-концевым слиянием домена Fc человека с минимальным линкером (тремя глициновыми остатками) (фиг. 5). Нуклеотидная последовательность, соответствующая данному слитому белку, показана на фиг. 6.
Пример 11. Селективное связывание лигандов ловушкой GDF с внеклеточным доменом ActRIIB с двойным усечением
Сродство ловушек GDF и других белков ActRIIB-hFc к нескольким лигандам оценивали in vitro прибором Biacore™. Результаты суммированы в таблице ниже. Величины Kd получали аффинной подгонкой в стационарном состоянии вследствие очень быстрой ассоциации и диссоциации комплекса, что препятствовало точному определению kon и koff.
Селективность вариантов ActRIIB-hFc в отношение лигандов:
(Kd e-11)
Ловушка GDF с усеченным внеклеточным доменом, ActRIIB(L79D 25-131)-hFc, имела равную или превосходящую селективность в отношении лигандов, проявляемую более длинным вариантом, ActRIIB(L79D 20-134)-hFc, с выраженной утратой связывания активина A и активина B и почти полным сохранением связывания GDF11, по сравнению с контрагентами ActRIIB-hFc, не имеющими замещение L79D. Следует отметить, что одно усечение (без замещения L79D) не изменяло селективность среди лигандов, проявившуюся в данном случае [сравнить ActRIIB(L79 25-131)-hFc с ActRIIB(L79 20-134)-hFc].
Пример 12. Получение ActRIIB(L79D 25-131)-hFc с альтернативными нуклеотидными последовательностями
Для получения ActRIIB(L79D 25-131)-hFc, внеклеточный домен ActRIIB человека с замещением аспартата в нативном положении 79 (SEQ ID NO:1) и с N-концевым и C-концевым усечениями (остатки 25-131 в SEQ ID NO:1) повергали N-концевому слиянию с лидерной последовательностью TPA вместо нативного лидера ActRIIB, и C-концевому слиянию с доменом Fc человека через минимальный линкер (три глициновых остатка) (Фиг. 5). Одна нуклеотидная последовательность, кодирующая этот слитый белок, показана на фиг. 6 (SEQ ID NO:27), а альтернативная нуклеотидная последовательность, кодирующая точно такой же слитый белок, показана на фиг. 9 (SEQ ID NO:30). Этот белок был экспрессирован и очищен с использованием методологии, описанной в примере 1.
Пример 13. Ловушка GDF с усеченным внеклеточным доменом ActRIIB увеличивает пролиферацию эритроидных предшественников у мышей
ActRIIB(L79D 25-131)-hFc оценивали для определения его эффекта на пролиферацию эритроидных предшественников. Самцов мышей C57BL/6 (в возрасте 8 недель) лечили введением ActRIIB(L79D 25-131)-hFc (10 мг/кг, подкожно; n=6) или носителя (TBS; n=6) в 1-й и 4-й день, затем их подвергали эвтаназии на 8-ой день для забора селезенок, большеберцовых костей, бедренных костей и крови. Клетки селезенки и костного мозга изолировали, разбавляли в модифицированной по Искову среде Дульбекко, содержащей 5% фетальную телячью сыворотку, суспендировали в специализированной среде на основе метилцеллюлозы и культивировали в течение или 2, или 12 дней для оценки уровней клоногенных предшественников, соответственно, на стадии колониеобразующих единиц эритроидного ряда (CFU-E) бурст-образующих единиц эритроидного ряда (BFU-E). Среда на основе метилцеллюлозы для определения BFU-E (MethoCult M3434, компания Stem Cell Technologies) включала рекомбинантный фактор стволовых клеток мыши, интерлейкин-3 и интерлейкин-6, которые не присутствовали в метилцеллюлозной среде для определения CFU-E (MethoCult M3334, Stem Cell Technologies), тогда как обе среды, среди других ингредиентов, содержали эритропоэтин. И для BFU-E, и для CFU-E, число колоний, полученных из каждого образца ткани, определяли в двух повторениях в культуральных планшетах, и статистический анализ результатов был основан на числе мышей на группу лечения.
В происходящих из селезенки культуры от мышей, получавших лечение ActRIIB(L79D 25-131)-hFc, имелось вдвое большее число колоний CFU-E, чем в соответствующих культурах от контрольных мышей (P<0,05), тогда как число колоний BFU-E значимо не отличалось при лечении in vivo. Число колоний CFU-E или BFU-E из культур костного мозга также значимо не отличались при лечении. Как ожидалось, увеличенные количества колоний CFU-E в происходящих из селезенки культурах сопровождались высоко значимыми (P < 0,001) изменениями уровня эритроцитов (увеличение на 11,6%), концентрации гемоглобина (увеличение на 12%) и уровня гематокрита (увеличение на 11,6%) при эвтаназии мышей, получавших лечение ActRIIB(L79D 25-131)-hFc, по сравнению с контролями. Эти результаты указывают на то, что введение in vivo ловушки GDF с усеченным внеклеточным доменом ActRIIB может стимулировать пролиферацию эритроидных клеток-предшественников в качестве части его общего эффекта на увеличение уровней эритроцитов.
Пример 14. Ловушка GDF с усеченным внеклеточным доменом Trap ActRIIB компенсирует вызванную химиотерапией анемию у мышей
Заявители исследовали эффект ActRIIB(L79D 25-131)-hFc на эритропоэтические параметры на модели анемии у мыши, вызванной химиотерапией на основе паклитаксела, который ингибирует клеточное деление блокированием полимеризации микротрубочек. Самцов мышей C57BL/6 (8-недельных) включали в одну из четырех групп лечения:
1) паклитаксел (25 мг/кг, внутрибрюшинно)
2) ActRIIB(L79D 25-131)-hFc (10 мг/кг, внутрибрюшинно)
3) паклитаксел + ActRIIB(L79D 25-131)-hFc
4) носитель (TBS).
Паклитаксел вводили в 0 день, тогда как ActRIIB(L79D 25-131)-hFc или носитель вводили в 0 и 3-й дни. Образцы крови брали для анализа CBC от отдельных когорт в 1-ый, 3-й и 5-й дни, и результаты по группам лечения 1-3 (указанным выше) были выражены в виде различия в процентах от группы носителя в данную точку времени. Истощение вследствие токсичности паклитаксела представляло собой проблему в когорте лечения одним паклитакселом на 3-й день (где n = 1); в других случаях, n = 3-5 на группу лечения на точку времени. По сравнению с носителем, один паклитаксел уменьшал концентрацию гемоглобина почти на 13% в 5-й день, тогда как добавление ActRIIB(L79D 25-131)-hFc предотвращало данное вызванное паклитакселом снижение (фиг. 11). Аналогичные эффекты наблюдали для уровней гематокрита и эритроцитов. В отсутствие паклитаксела, ActRIIB(L79D 25-131)-hFc увеличивал концентрацию гемоглобина на 10%, по сравнению с носителем на 3-й и 5-й дни (фиг. 11). Таким образом ловушка GDF с усеченным внеклеточным доменом ActRIIB может увеличить уровни эритроцитов в степени, достаточной для компенсации вызванной химиотерапией анемии.
Пример 15. Ловушка GDF с усеченным внеклеточным доменом ActRIIB устраняет анемию, вызванную нефрэктомией у мышей
Заявители исследовали эффект ActRIIB(L79D 25-131)-hFc на анемию на модели хронического заболевания почек у мыши, вызванного нефрэктомией. Самцов мышей C57BL/6 (11-недельных) подвергали или ложной операции, или односторонней нефрэктомии для снижения способности продукции эритропоэтина. Мышам давали возможность в течение недели восстановиться после операции и затем лечили введением дважды в неделю ActRIIB(L79D 25-131)-hFc (10 мг/кг, внутрибрюшинно; n=15 на состояние) или носителя (TBS; n=15 на состояние) в общем, в течение 4 недель. Образцы крови брали перед началом введения и после 4 недель лечения. В то время как у получавших лечение носителем мышей проявлялось значительное снижение количества эритроцитов в течение 4-недельного периода лечения, лечение ActRIIB(L79D 25-131)-hFc не только предотвращало снижение, но увеличивало уровни эритроцитов на 17% (P < 0,001) выше исходного уровня (фиг. 12), несмотря на пониженную почечную способность продукции эритропоэтина. У мышей после нефрэктомии, ActRIIB(L79D 25-131)-hFc также вызывал значимое увеличение от исходного уровня концентрации гемоглобина и уровня гематокрита и, что примечательно, стимулировал каждый из указанных эритропоэтических параметров приблизительно в такой же степени в условиях нефрэктомии, как и в условиях ложной операции (фиг. 13). Таким образом, ловушка GDF с усеченным внеклеточным доменом ActRIIB может увеличить уровни эритроцитов в степени, достаточной для устранения анемии на модели хронического заболевания почек.
Пример 16. Ловушка GDF с усеченным внеклеточным доменом ActRIIB улучшает восстановление после анемии, вызванной кровопотерей у крыс
Заявители исследовали эффект ActRIIB(L79D 25-131)-hFc на эритропоэтические параметры на модели анемии у крыс, вызванной острой кровопотерей (острой постгеморрагической анемии). Самцам крыс Sprague-Dawley (приблизительно 300 г) устанавливали в яремную вену хронический катетер от поставщика (Harlan). В -1 день, 20% всего объема крови удаляли у каждой крысы в течение 5-минутного периода через катетер под изофлюрановой анестезией. Объем удаленной крови был основан на величине общего объема крови, рассчитанной в соответствии со следующим соотношением, выведенным Lee и сотрудниками (J Nucl Med 25:72-76, 1985) для крыс с массой тела более чем 120 г:
Общий объем крови (мл) = 0,062 × массу тела (г) + 0,0012
Равный объем солевого раствора с фосфатным буфером замещали через катетер во время удаления крови. Крыс лечили ActRIIB(L79D 25-131)-hFc (10 мг/кг, подкожно; n=5) или носителем (TBS; n=5) в 0-й и 3-й день. Образцы крови для анализа CBC удаляли через катетер в -1-й (исходный уровень), 0-й, 2-ой, 4-ый и 6-ой дни.
Контрольные крысы реагировали на 20% кровопотерю падением почти на 15% уровней эритроцитов к 0-му дню. Эти уровни оставались значительно ниже, чем исходные, во 2-ой и 4-ый день, и полностью не восстановились к 6-му дню (фиг. 14). Хотя у крыс, получавших лечение ActRIIB(L79D 25-131)-hFc, проявилось почти идентичное падение уровней эритроцитов после 20% кровопотери, у этих крыс затем проявилось полное восстановление таких уровней ко 2-му дню с последующим дальнейшим подъемом на 4-ый и 6-ой дни, что представляет высоко значимое улучшение относительно контрольных уровней в соответствующие точки времени (фиг. 14). Аналогичные результаты были получены для концентрации гемоглобина. Эти данные демонстрируют, что ловушка GDF с усеченным внеклеточным доменом ActRIIB может вызывать более быстрое восстановление уровней эритроцитов при анемии, вызванной острым кровотечением.
Пример 17. Ловушка GDF с усеченным внеклеточным доменом ActRIIB увеличивает уровни эритроцитов у не человекообразных приматов
Две ловушки GDF, ActRIIB(L79D 20-134)-hFc и ActRIIB(L79D 25-131)-hFc, оценивали для выявления способности стимулировать продукцию эритроцитов у яванских макак. Макак лечили подкожными инъекциями ловушки GDF (10 мг/кг; n=4 самца/4 самки) или носителя (n=2 самца/2 самки) в 1-ый и 8-ой дни. Образцы крови брали в 1-ый день (исходный уровень перед лечением), 3, 8, 15, 29 и 44-ый дни и анализировали для определения уровней эритроцитов (фиг. 15), гематокрита (фиг. 16), уровней гемоглобина (фиг. 17) и уровней ретикулоцитов (фиг. 18). У макак, получавших носитель, проявлялись сниженные уровни эритроцитов, гематокрита и гемоглобина во все точки времени после лечения, что является ожидаемым эффектом повторного взятия образцов крови. Напротив, лечение ActRIIB(L79D 20-134)-hFc или ActRIIB(L79D 25-131)-hFc увеличивали указанные параметры к первой точке времени после лечения (3-й день) и поддерживали их на существенно повышенных уровнях в течение исследования (фиг. 15-17). Важно, что уровни ретикулоцитов у макак, получавших лечение ActRIIB(L79D 20-134)-hFc или ActRIIB(L79D 25-131)-hFc, были существенно повышены в 8-ой, 15-ый и 29-ый дни, по сравнению с носителем (фиг. 18). Данный результат демонстрирует, что лечение ловушкой GDF увеличивало продукцию предшественников эритроцитов, приводя к повышенным уровням эритроцитов.
Взятые вместе, эти данные демонстрируют, что усеченные ловушки GDF, а также полноразмерные варианты, могут использоваться в качестве селективных антагонистов GDF11 и потенциально связанных лигандов для увеличения образования эритроцитов in vivo.
Пример 18. Ловушка GDF, полученная из ActRIIB5
Другие авторы сообщали об альтернативной, растворимой форме ActRIIB (обозначенной как ActRIIB5), в которой экзон 4, включающий трансмембранный домен ActRIIB, был замещен другой C-концевой последовательностью (WO2007/053775).
Последовательность нативного ActRIIB5 человека без его лидера представлена ниже:
GRGEAETRECIYYNANWELERTNQSGLERCEGEQDKRLHCYASWRNSSGTIELVK
KGCWLDDFNCYDRQECVATEENPQVYFCCCEGNFCNERFTHLPEAGGPEGPWAST
TIPSGGPEATAAAGDQGSGALWLCLEGPAHE
(SEQ ID NO:36)
Замещение лейцина на аспартат или другие кислотные замещения могут выполняться в нативном положении 79 (подчеркнутые и выделенные серым фоном), как описано, для конструкции вариантного ActRIIB5(L79D), который имеет следующую последовательность:
GRGEAETRECIYYNANWELERTNQSGLERCEGEQDKRLHCYASWRNSSGTIELVK
KGCWDDDFNCYDRQECVATEENPQVYFCCCEGNFCNERFTHLPEAGGPEGPWAST
TIPSGGPEATAAAGDQGSGALWLCLEGPAHE
(SEQ ID NO:37)
Данный вариант может быть соединен с Fc линкером человека TGGG для получения слитого белка ActRIIB5(L79D)-hFc человека со следующей последовательностью:
GRGEAETRECIYYNANWELERTNQSGLERCEGEQDKRLHCYASWRNSSGTIELVK
KGCWDDDFNCYDRQECVATEENPQVYFCCCEGNFCNERFTHLPEAGGPEGPWAST
TIPSGGPEATAAAGDQGSGALWLCLEGPAHETGGGTHTCPPCPAPELLGGPSVFL
FPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYN
STYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLP
PSREEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLY
SKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK
(SEQ ID NO: 38)
Данная конструкция может быть экспрессирована в клетках CHO.
Пример 19. Эффекты у мышей комбинированного лечения EPO и ловушкой GDF с усеченным внеклеточным доменом ActRIIB
EPO вызывает образование эритроцитов путем повышения пролиферации эритроидных клеток-предшественников, тогда как ловушки GDF могут потенциально воздействовать на образование эритроцитов путями, которые дополняют или усиливают эффекты EPO. Поэтому, заявители исследовали эффект комбинированного лечения EPO и ActRIIB(L79D 25-131)-hFc на эритропоэтические параметры. Самцам мышей C57BL/6 (9-недельным) однократно внутрибрюшинно инъецировали один рекомбинантный EPO человека (эпоэтин альфа, 1800 единиц/кг), один ActRIIB(L79D 25-131)-hFc (10 мг/кг), и EPO, и ActRIIB(L79D 25-131)-hFc, или носитель (солевой раствор, забуференный Tris-буфером). Мышей подвергали эвтаназии через 72 часа после введения для забора крови, селезенок и бедренных костей.
Селезенки и бедренные кости обрабатывали для получения эритроидных клеток-предшественников для анализа проточной цитометрией. После удаления, селезенку измельчали в модифицированной по Искову среде Дульбекко, содержащей 5% фетальную телячью сыворотку, и механически разрушали проталкиванием через сетчатый фильтр с отверстиями размером 70 мкм поршнем от стерильного шприца емкостью 1 мл. Бедренные кости очищали от любой остаточной мышечной или соединительной ткани и концы торцевали для обеспечения возможности взятия костного мозга путем промывания остающегося тела бедренной кости модифицированной по Искову средой Дульбекко, содержащей 5% фетальную телячью сыворотку, через иглу 21 калибра, соединенную с 3-миллилитровым шприцем. Клеточные суспензии центрифугировали (2000 об/мин в течение 10 мин), и клеточный осадок после центрифугирования ресуспендировали в PBS, содержащем 5% фетальную телячью сыворотку. Клетки (106) из каждой ткани инкубировали с анти-мышиным IgG для блокировки неспецифического связывания, затем инкубировали с флуоресцентно мечеными антителами против клеточных поверхностных маркеров CD71 мыши (рецептор трансферрина) и Ter119 (антигена, связанного с гликофорином A клеточной поверхности), промывали и анализировали проточной цитометрией. Мертвые клетки в образцах исключали из анализа противоокрашиванием пропидием йодидом. Эритроидную дифференциацию в селезенке или костном мозге оценивали по степени мечения CD71, которая уменьшается с течением дифференциации, и мечения Ter119, которое увеличивается во время конечной эритроидной дифференциации, начиная со стадии проэритробластов (Socolovsky et al., 2001, Blood 98:3261-3273; Ying et al., 2006, Blood 108:123-133). Таким образом, проточную цитометрию использовали для определения числа проэритробластов (CD71highTer119low), базофильных эритробластов (CD71highTer119high), полихроматофильных + ортохроматофильных эритробластов (CD71medTer119high) и поздних ортохроматофильных эритробластов + ретикулоцитов (CD71lowTer119high), как описано.
Комбинированное лечение EPO и ActRIIB(L79D 25-131)-hFc привело к удивительно энергичному увеличению содержания эритроцитов. В 72-часовой временной рамке данного эксперимента, ни EPO, ни ActRIIB(L79D 25-131)-hFc отдельно значимо не увеличивали гематокрит, по сравнению с носителем, в то время как комбинированное лечение двумя средствами привело к увеличению гематокрита почти на 25%, что неожиданно являлось синергическим эффектом, т.е., большим, чем сумма их отдельных эффектов (фиг. 19). Синергия данного типа в целом считается доказательством того, что отдельные средства действуют посредством различных клеточных механизмов. Аналогичные результаты также наблюдались для концентраций гемоглобина (фиг. 20) и концентраций эритроцитов (фиг. 21), причем каждый из указанных параметров также синергически увеличивался комбинированным лечением.
Анализ уровней эритроидных клеток-предшественников выявил более сложный тип. У мышей, селезенка считается первичным органом, ответственным за индуцируемый («стрессовый») эритропоэз. Проточно-цитометрический анализ ткани селезенки через 72 ч выявил, что EPO значительно изменял профиль эритропоэтических клеток-предшественников, по сравнению с носителем, увеличивая число базофильных эритробластов более чем на 170% за счет поздних клеток-прешественников (поздних ортохроматофильных эритробластов + ретикулоцитов), которые уменьшились более чем на одну треть (фиг. 22). Важно, что комбинированное лечение значительно увеличивало содержание базофильных эритробластов, по сравнению с носителем, но в меньшей степени, чем один EPO, в то же время, поддерживая не уменьшенное созревание клеток-предшественников поздней стадии (фиг. 22). Таким образом, комбинированное лечение EPO и ActRIIB(L79D 25-131)-hFc увеличивало эритропоэз посредством сбалансированного усиления пролиферации и созревания клеток-предшественников. В отличие от селезенки, профиль клеток-предшественников в костном мозге после комбинированного лечения определяемо не отличался от профиля после применения одного EPO. Заявители прогнозируют по профилю селезеночных клеток-предшественников, что комбинированное лечение привело бы повышенным уровням ретикулоцитов и сопровождалось бы длительным повышением уровней зрелых эритроцитов, если бы эксперимент был продолжен свыше 72 ч.
Взятые вместе, указанные данные демонстрируют, что ловушка GDF с усеченным внеклеточным доменом ActRIIB может вводиться в комбинации с EPO для синергического увеличения образования эритроцитов in vivo. Действуя посредством дополнительного, но неопределенного механизма, ловушка GDF может смягчать сильный пролиферативный эффект одного активатора рецепторов EPO и все же обеспечивает возможность достижения целевых уровней эритроцитов более низкими дозами активатора рецепторов EPO, посредством этого избегая потенциальных побочных эффектов или других проблем, связанных с более высокими уровнями активации рецепторов EPO.
Включение путем ссылки
Все публикации и патенты, указанные в настоящем описании, полностью включены в него путем ссылки как если бы каждая отдельная публикация или патент был специально и отдельно указан как включенный путем ссылки.
Хотя были обсуждены определенные варианты осуществления предмета изобретения, приведенное выше описание является иллюстративным, а не ограничивающим. Многие изменения станут очевидными для специалистов в данной области после рассмотрения данного описания и представленной ниже формулы изобретения. Полный объем изобретения должен определяться ссылкой на формулу изобретения наряду с полным объемом ее эквивалентов и на описание наряду с такими изменениями.






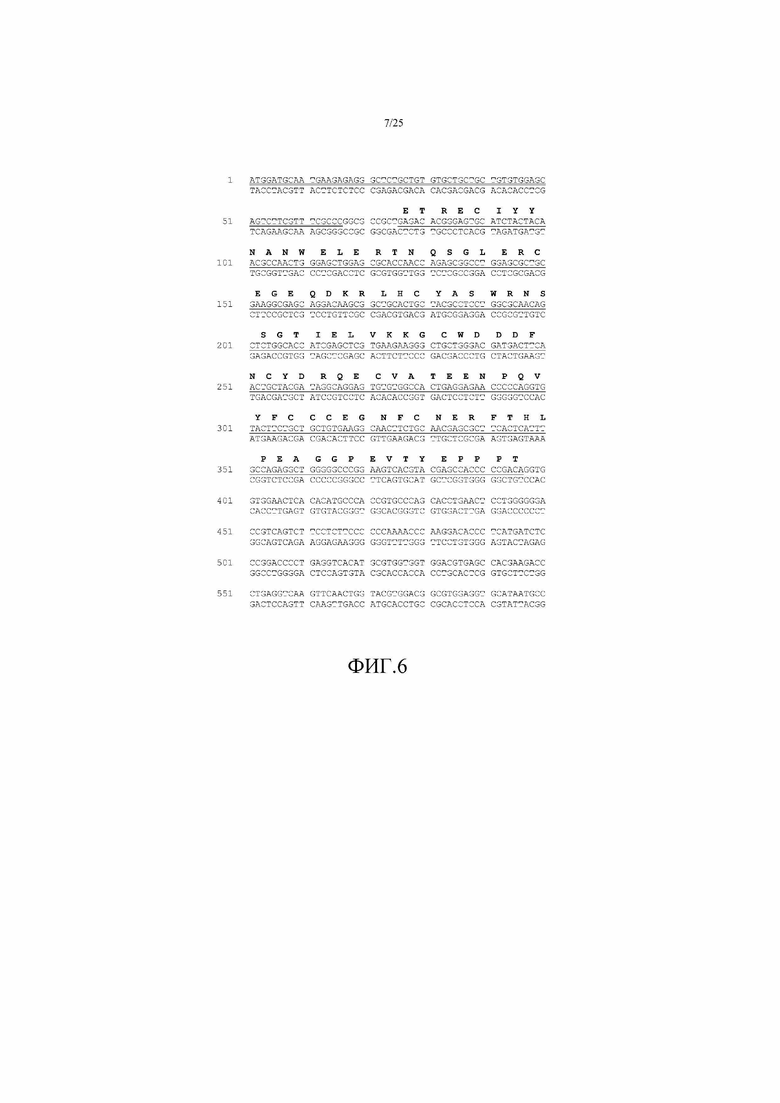







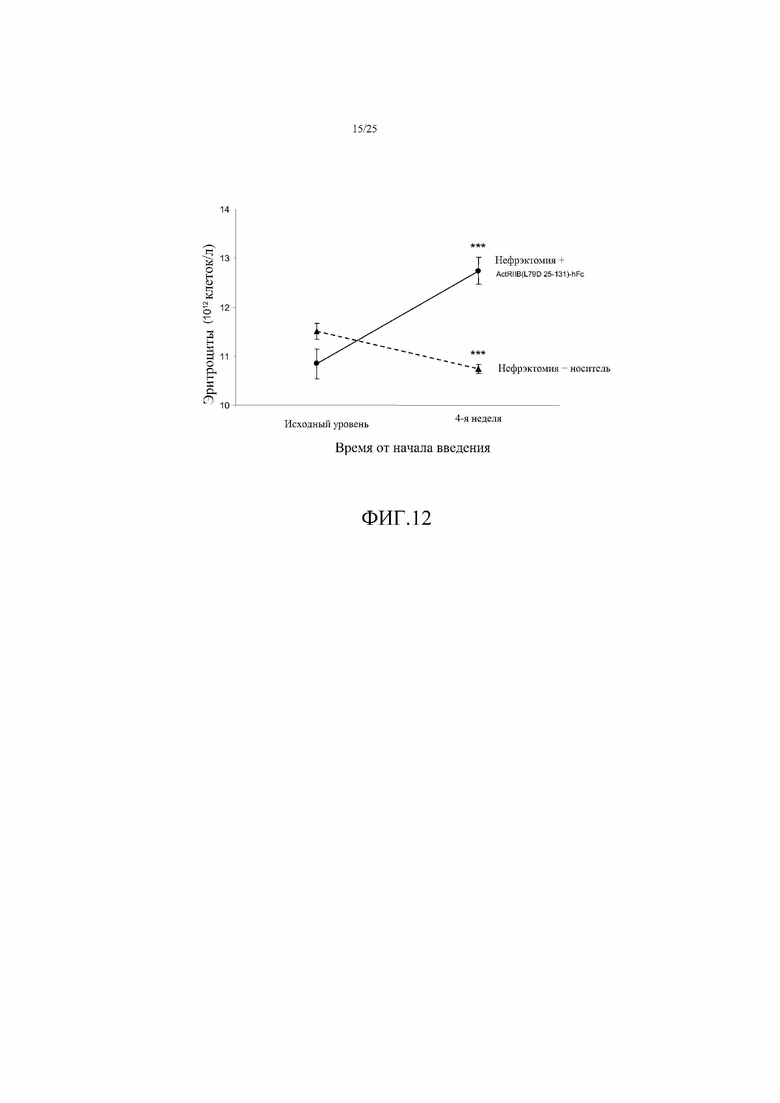
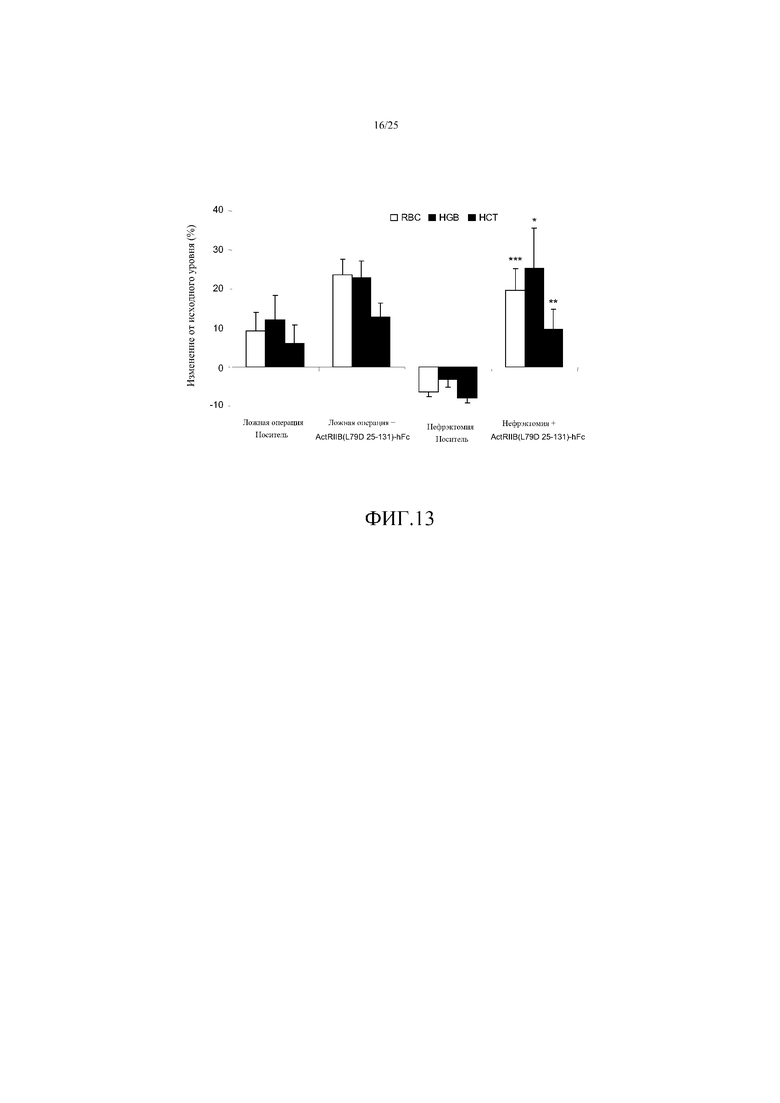
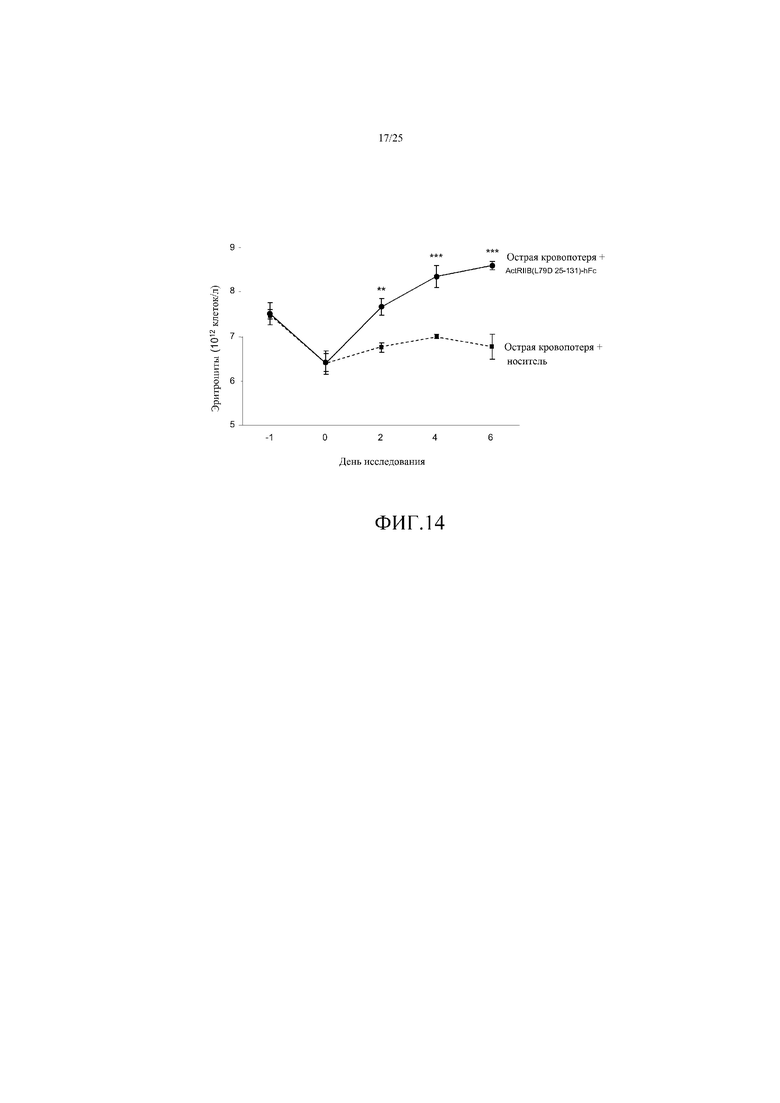



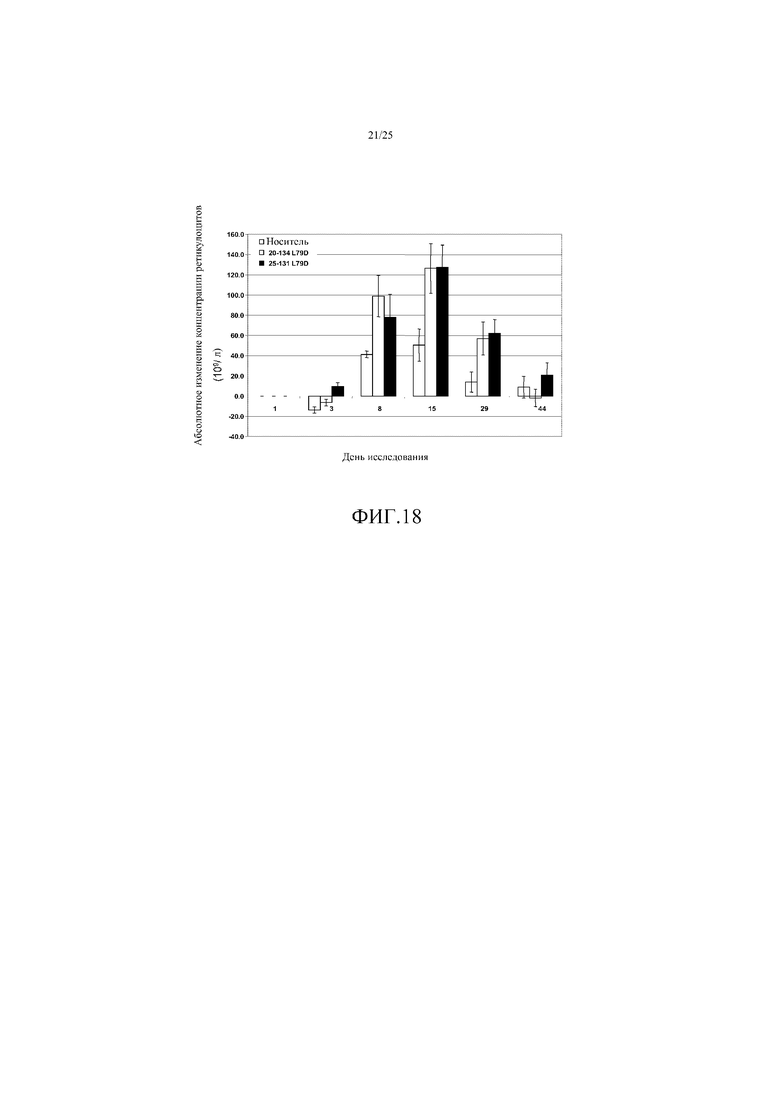
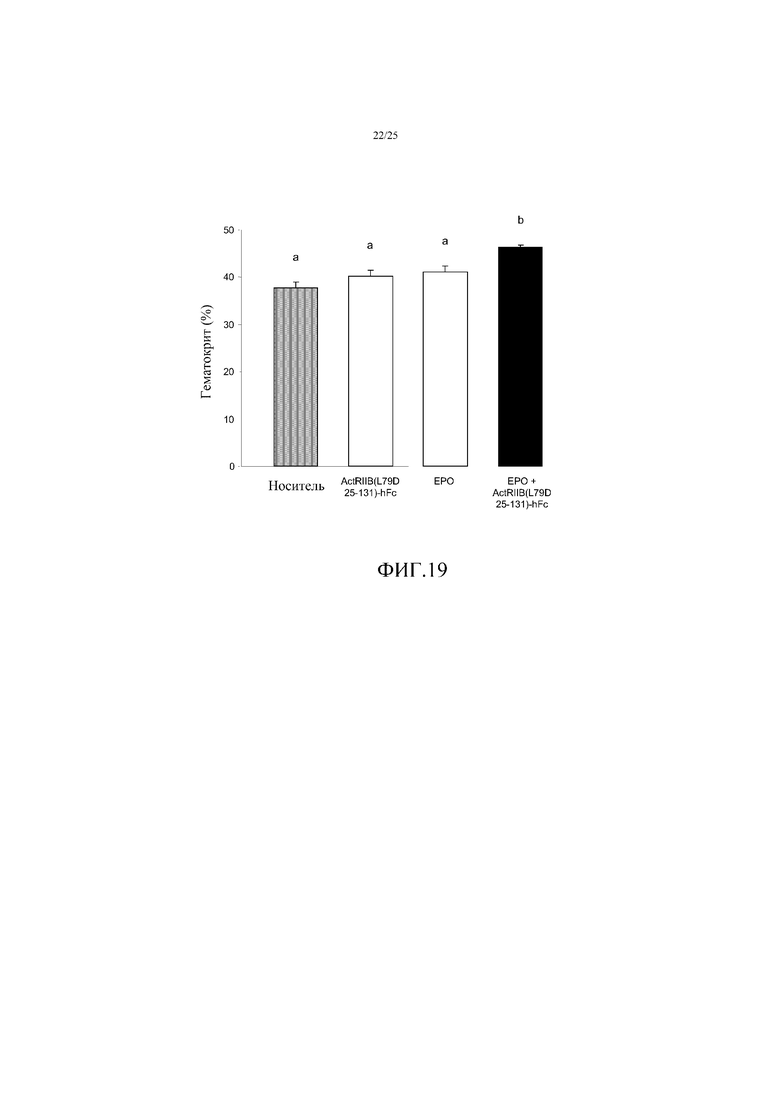
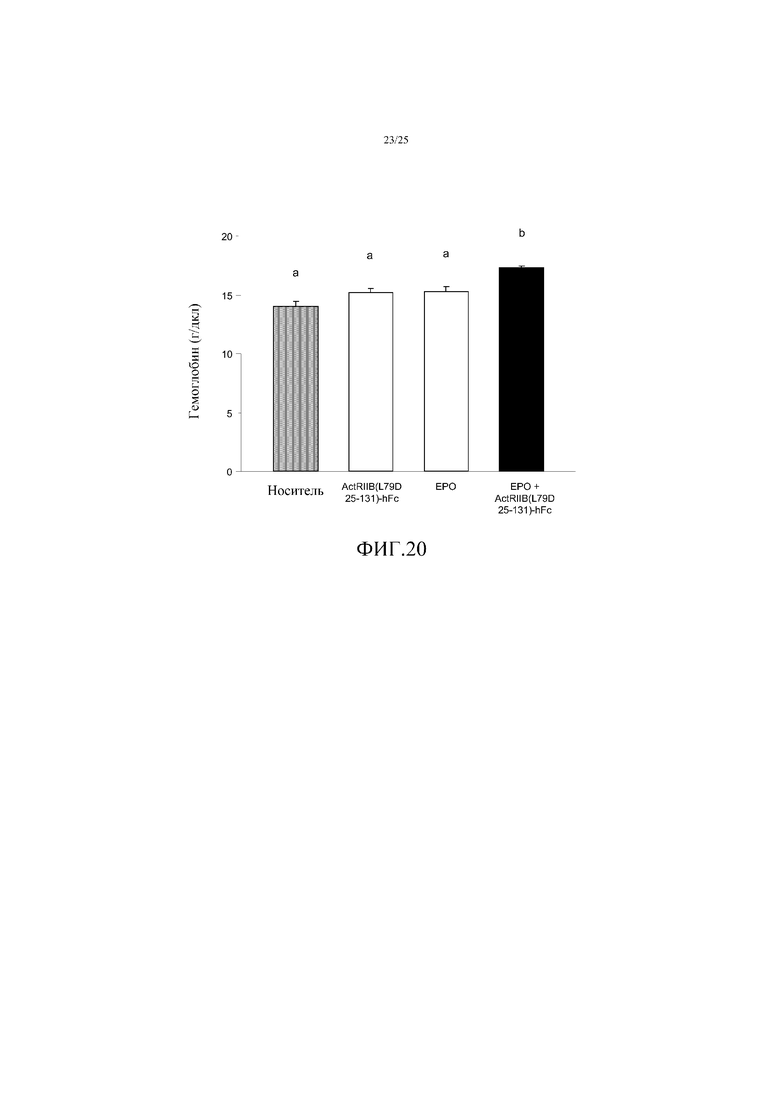

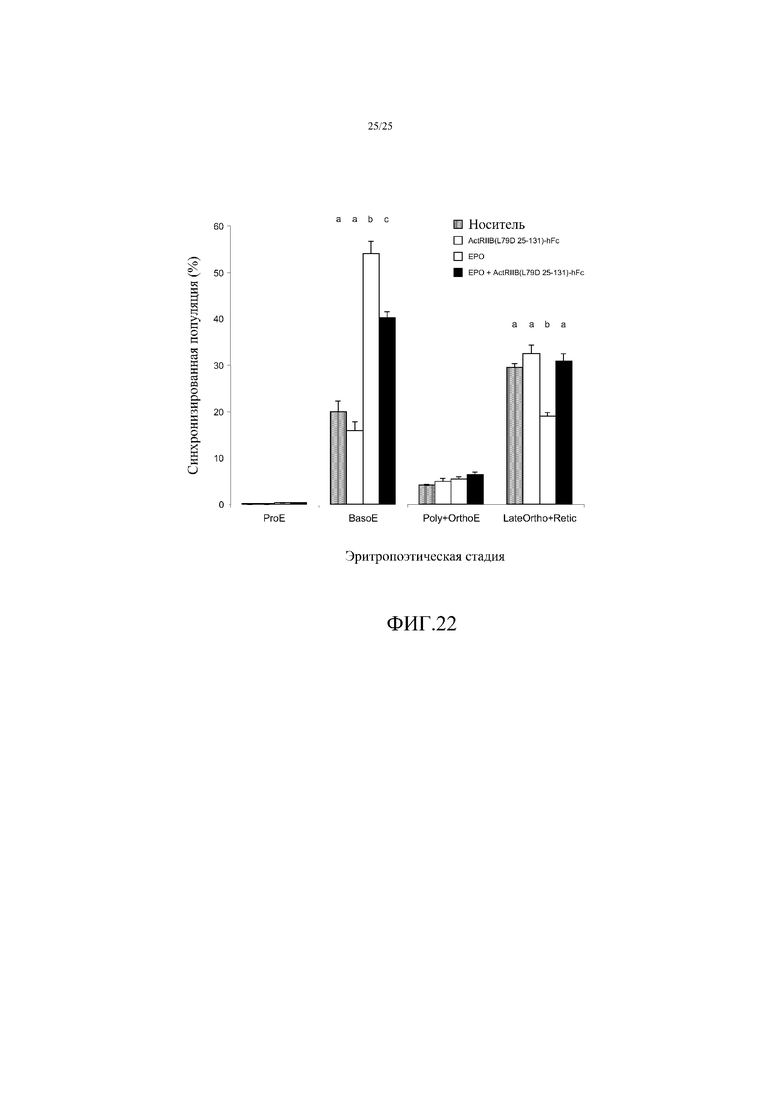
Изобретение относится к области биотехнологии, конкретно к получению выделенного полипептида ловушки GDF, содержащего аминокислотную последовательность SEQ ID NO:28, и может быть использовано в медицине. Полученный полипептид ловушки GDF может быть использован в составе фармацевтической композиции для лечения анемии. 7 н. и 7 з.п. ф-лы, 22 ил., 11 табл., 19 пр.
1. Выделенный полипептид ловушки GDF, содержащий аминокислотную последовательность SEQ ID NO:28.
2. Выделенный полипептид по п.1, который содержит аминокислотную последовательность, состоящую из SEQ ID NO:28.
3. Выделенный полипептид по п.1 или 2, который состоит из аминокислотной последовательности SEQ ID NO:28.
4. Выделенная нуклеиновая кислота, кодирующая полипептид по любому из пп.1-3.
5. Выделенная нуклеиновая кислота по п.4, которая содержит нуклеотидную последовательность SEQ ID NO:30 и нуклеотидную последовательность SEQ ID NO:27.
6. Генетическая конструкция нуклеиновой кислоты, кодирующая полипептид по любому из пп.1-3, содержащая нуклеиновую кислоту по любому из пп.4-5 и один или несколько регуляторов последовательностей, с которыми она функционально связана.
7. Вектор экспрессии, содержащий выделенную нуклеиновую кислоту по любому из пп.4-6.
8. Культивируемая клетка-хозяин для получения полипептида по любому из пп.1-3, содержащая вектор экспрессии по п.7.
9. Культивируемая клетка-хозяин по п.8, которая является клеткой млекопитающего.
10. Культивируемая клетка-хозяин по п.9, где клетка млекопитающего представляет собой клетку СНО.
11. Способ получения полипептида по любому из пп.1-3, включающий:
а) культивирование клетки-хозяина в условиях, подходящих для экспрессии полипептида, и
b) выделение экспрессированного полипептида,
где клетка-хозяин:
i) представляет собой клетку-хозяин по любому из пп.8-10,
ii) содержит вектор экспрессии, который содержит нуклеиновую кислоту, содержащую нуклеотидную последовательность SEQ ID NO:30, или
iii) содержит вектор экспрессии, который содержит нуклеиновую кислоту, содержащую нуклеотидную последовательность SEQ ID NO:27.
12. Способ по п.11, где клетка-хозяин является клеткой млекопитающего.
13. Способ по п.12, где клетка млекопитающего представляет собой клетку СНО.
14. Композиция для лечения анемии, содержащая эффективное количество полипептида по любому из пп.1-3 и фармацевтически приемлемый носитель.
| WO 2008076437, 13.11.2008 | |||
| HARRISON C.A | |||
| et al | |||
| Antagonists of activin signaling: mechanisms and potential biological applications, Trends in Endocrinology & Metabolism, 2005, V | |||
| Устройство для электрической сигнализации | 1918 |
|
SU16A1 |
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| COERVER K.A | |||
| et al | |||
| Activin signaling through activin receptor type II causes the cachexia-like symptoms in inhibin-deficient mice, Molecular | |||

