Изобретение относится к сельскому хозяйству, а именно к птицеводству, и может быть использовано при оценке качества суточных цыплят.
Целью изобретения является расширения области применения изобретения и повышение его точности.
Суточный молодняк птицы обычно оценивают по внешним признакам. Так, согласно ОСТ 10329-2003 «Суточный молодняк кур. Технические условия» суточный молодняк кур по внешним признакам должен соответствовать следующим требованиям: хорошие подвижность и устойчивость на ногах, активная реакция на звук при постукивании), хорошо выраженный рефлекс клевания; голова - широкая, пропорциональная; клюв - правильной формы, пигментированный; глаза - круглые, выпуклые, блестящие; корпус (на ощупь) - плотный; спина - ровная, умеренно длинная, широкая; грудная кость - киль длинный, упругий; живот (на ощупь) - мягкий, подобранный; плюсны - прямые, крепкие, пигментированные; крылья - плотно прижаты к туловищу; пух - полностью подсохший, равномерно распределенный по телу, гладкий, шелковистый; пупочное кольцо - плотно закрытое; клоака - чистая, розовая, влажная. Масса суточных мясных цыплят для промышленного стада должна быть не менее 32 г, для племенного - 34 г. При использовании требований указанного ОСТа все же трудно предсказать будущую продуктивность цыплят, так как многие изменения на биохимическом уровне не проявляются во внешних признаках, который можно оценить визуально.
Наши исследования убедительно показали, что антиоксидантная активность является интегральным показателем способности организма птиц противостоять стрессовым ситуациям, вызывающим патологические изменения в организме, вызванные избыточным образованием активных форм кислорода (Suraietal., 2019; Surai, 2018; 2006; 2002). Например, известно что в физиологических условиях в каждой клетке каждый день образуется около 20 миллиардов свободных радикалов, способных повреждать все типы биологических молекул, включая жиры, белки и нуклеиновые кислоты, что приводит к нарушению важнейших физиологических функций и биохимических процессов. У сельскохозяйственных животных и птиц это приводит к снижению продуктивности, иммунокомпетентности и их репродуктивных характеристик. При этом, в стресс-уловиях количество образовавшихся свободных радикалов может многократно возрастать (Surai et al., 2019; 2019а).
Для защиты от пагубного влияния свободных радикалов и токсических продуктов их метаболизма в процессе эволюции в организме животных, включая птиц, были развиты защитные механизмы, которые получили обобщенное название «антиоксидантаная система» организма (Surai, 2018; Suraietal., 2019; 2019а). Данная система состоит из множества различных антиоксидантов, часть из которых синтезируется в самом организме птиц (антиоксидантные ферменты, глутатион, тиоредоксин, аскорбиновая кислота и др.), другие - поступают с кормом (витамин Е, каротиноиды, селен и др.). Часть антиоксидантов, например витамины Е и С, способны непосредственно улавливать свободные радикалы и обрывать цепи переокисления. Другие антиоксиданты, например, карнитин и таурин, способствуют поддержанию структурной целостности митохондрий-главного места образования свободных радикалов. К тому же, целая группа антиоксидантных ферментов, включая супероксиддисмутазу (СОД), глутатионпероксидазу и каталазу, участвует в детоксикации свободных радикалов и превращении их в безвредные нетоксичные соединения. В целом, СОД является главным ферментом первой линии антиоксидантной защиты, определяющим эффективность всей антиоксидантной системы. При этом характерным признаком данного фермента является его участие в адаптационных изменениях в организме в ответ на стрессовую ситуацию (Lewandowskietal., 2018; Zouetal., 2017). Таким образом, активность данного фермента, участвующего в детоксикации свободных радикалов на самом первом этапе их образования, является определяющим фактором эффективности антиоксидантной защиты. При этом, данный фермент является индуцируемым ферментом, т.е. его активность может повышаться, в ответ на возникающий избыток образования свободных радикадлов. Это во многом происходит за счет активации ядерных факторов транскрипции, включая Nrf2 (Suraietal., 2019).
В последние годы было установлено, что в организме человека и животных существует ряд генов, которые отвечают за поддержание гомеостаза в стресс-условиях и являются ключевыми факторами адаптации к стрессу. Такие гены получили название ВИТАГЕНЫ (Surai and Fisinin, 2016с; 2016d; Surai, 2016; 2018; Suraietal., 2017; 2019). Данные гены ответственны за синтез защитных молекул, включая СОД, белки теплового шока, элементы тиоредоксиновой (тиоредоксин, тиоредоксинредуктаза и пероксиредоксин) и глутатионовой (глутатион, глутатионпероксидаза, глутатионредуктаза) систем и сиртуины. Согласно данному представлению активация упомянутых витагенов приводит к дополнительному синтезу защитных молекул, которые восстанавливают нарушенный баланс/гомеостаз в условиях стресса. Важным элементом данной концепции является представление о том, что на молекулярном уровне большинство стрессов, встречающихся в коммерческих условиях производства яиц и мяса птицы связаны с окислительным стрессом (Surai, 2002; 2006; 2015; 2015а; 2015b; 2015с; 2016; Surai and Fisinin, 2015; 2016а; 2016b). Таким образом, средовые и технологчисккие стрессы (Surai and Fisinin, 2016а), также как и кормовые и внутренние/биологические стрессы (Surai and Fisinin, 2016b) у птиц на клеточном уровне вызывают окислительный стресс в результате избыточного образования свободных радикалов и недостаточно эффективной антиоксидантной защиты. При этом оксилительный стресс рассматривается в качестве пускового механизма многих нарушений в организме птицы, включая иммуносупрессию, снижение продуктивных и воспроизводительных качеств. При этом наши исследования убедительно продемонстрировали, что витагены могут регулироваться различными кормовыми добавками поступающими с кормом или водой (Suraietal., 2017). Таким образом были разработаны теоретические подходы к новой системе предотвращения стрессов или же борьбы со стрессами в современном птицеводстве (Сурай и др., 2018).
Следует особо подчеркнуть, что промышленное птицеводство сопряжено с целым рядом стрессов, включая средовые стрессы (отклонение от оптимальной температуры, повышенное количество аммиака и др.), кормовые (микотоксины и окисленные жиры в кормах), технологические (посадка цыплят в корпус, прореживание, смена рационов, сортировка птицы, вакцинация и др.) и биологические/внутренние (бактериальные или вирусные инфекции, высокая скорость роста и др.) стрессы (Surai and Fisinin. 2016а; 2016b). В этом отношении стресс вылупления в инкубаторе является определяющим. Ведь цыпленок вылупляется в условиях повышенной температуры и влажности, после проклева скорлупы количество доступного кислорода существенно увеличивается, что является предрасполагающими факторами к развитию окислительного стресса (Surai, 1999, 2002). Нарушения антиоксидантного (редокс) баланса, вызванного окислительным стрессом в инкубатории (Karadasetal., 2011) отрицательно сказываются на дальнейшем росте и развитии цыплят.
Во всех вышеупомянутых случаях возникает потребность в усиленной защите организма от окислительного стресса, которая осуществляется путем активации витагенов (Surai and Fisinin, 2016с; 2016d; Suraietal., 2017) с последующим синтезом защитных молекул. Во многих случаях активация витагенов опосредована через повышение экспрессии ядерного фактора транскрипции Nrf2 (Suraietal., 2019). Если же антиоксидантная система не полностью справляется с защитой, то возникает окислительный стресс, нарушается редокс потенциал и сигнализация в клетках организма, происходит повреждение биологических молекул, что неизбежно влечет за собой снижение продуктивных и воспроизводительных качеств птицы. Таким образом, процесс инкубации яиц сопряжен с повышенным образованием свободных радикалов (Surai, 1999а; 2002, Surai et al., 2016; Surai, 2018). Любые нарушения в процессе инкубации отражаются на жизнеспособности цыплят, но обнаружить их визуально у суточных цыплят практически невозможно. Это касается, прежде всего, свежевылупившихся цыплят. При этом устойчивость цыплят к окислительному стрессу, определяемая эффективным ответом со стороны витагенов и антиоксидантной системы на избыточное образование свободных радикалов, является основой развития иммунитета, поддержания здоровья и будущей продуктивности (Surai, 2018; Surai and Fisinin, 2015). В этом отношении, активность фермента СОД может рассматриваться в качестве основного «предсказателя» устойчивости свежевылупившихся цыплят к окислительному стрессу. Наши исследования показали, что активность данного фермента в тканях развивающихся эмбрионов птиц изменяется в зависимости от сроков инкубации и имеет тканевую специфичность (Surai, 1999), имеет высокую экспрессию в мозге свежевылупившихся цыплят (Suraietal., 1999), по-разному экспрессируется в различных участках желудочно-кишечного тракта цыплят (Suraietal., 2018) и снижается в условиях токсикоза, вызванного избытком витамина А в рационе (Surai et al., 2000). В рамках вышеупомянутой новой концепции витагенов, т.е. генов ответственных за адаптацию к стрессовым ситуациям, активация одного из витагенов (в нашем случае СОД) в ответ на стрессовую ситуацию запускает целый каскад сигнальных событий, включая активацию факторов транскрипции (т.е. Nrf2, Surai et al., 2019), и приводит с одной стороны, к удалению избытка свободных радикалов, а с другой стороны- к дополнительному синтезу защитных молекул, помогающих справиться с удалением свободных радикалов и починкой поврежденных молекул, например, за счет белков теплового шока, являющихся важными витагенами. В конечном итоге, это приводит к улучшению адаптационной способности организма. Здесь роль СОД является критической.
Таким образом, принимая во внимание выше указанные особенности роста и развития цыплят, современное птицеводство нуждается в дополнительных методах оценки качества суточного молодняка, отражающего не только их жизнеспособность, но и адаптационную способность к преодолению окислительного стресса. Именно устойчивость цыплят к окислительному стрессу, возникающему в процессе их вывода и в раннем постнатальном развитии, определяет оптимальное развитие иммунной, пищеварительной и других систем организма (Surai and Fisinin, 2015), которые определяют такие важные показатели эффективности птицеводства, как среднесуточный привес и конверсия корма.
При этом известно, что все антиоксиданты работают сообща, эффективно взаимодействуя друг с другом. В последние годы появился термин «редокс-баланс» клетки или организма, который по своей сути является балансом между антиоксидантами и прооксидантами (Suraietal., 2019). При этом, роль СОД в поддержании антиоксидантной защиты и редокс-баланса в клетке/организме птиц трудно переоценить (Surai, 2016). Следует особо подчеркнуть роль данного фермента у свежевылупившихся цыплят (Surai, 1999), в условиях, когда антиоксидантная система организма еще полностью не сформирована и адаптационные возможности организма весьма ограничены (Surai, 1999 а; 2002).
Прототипом заявленного изобретения является «Способ оценки качества суточного молодняка птицы», включающий выборочный убой цыплят и определение биохимических параметров в биологическом материале, где в качестве биологического материала используют плазму крови, а в качестве биохимических параметров используют общую антиоксидантную активность (ОАА) и количество конечных продуктов перекисного окисления липидов (TBARS), при этом при величине отношения OAA/TBARS>800 цыплят относят к 1-й категории, при отношении OAA/TBARS 300-800 цыплят относят ко второй категории и при отношении OAA/TBARS<300 цыплят относят к третьей категории, непригодной для выращивания стадии оценки качества [Патент РФ 2699719, 09.09.2019].
Недостатком данного способа является ограниченность сферы его применения из-за отсутствия учета параметров, определяющих адаптационную устойчивость свежевылупившихся цыплят к окислительному стрессу. Это, прежде всего, распространяется на различные нарушения при хранении инкубационных яиц или их инкубации, которые отрицательно сказываются на жизнеспособности вылупившихся цыплят. Тем не менее, по нашим наблюдениям у таких цыплят изменения в отношении OAA/TBARS не всегда очевидны. Т.е. в большинстве случаев способ-прототип дает возможность сортировки цыплят по категориям, но в отдельных случаях, его возможности ограничены. Известно, что промышленное производство цыплят связано с множеством стрессовых ситуаций на разных этапах технологической цепочки, включая сбор инкубационных яиц, их хранение и транспортировку, процесс инкубации, вывод в инкубатории, транспортировку к месту выращивания и посадку в птичник для выращивания. При этом все упомянутые стрессы сопряжены с окислительным стрессом на молекулярном уровне (Surai and Fisinin, 2015; 2016а; 2016b). Таким образом, устойчивость к окислительному стрессу является важнейшим показателем качества суточных цыплят. Учет данного показателя может существенно расширить область применения данного изобретения и повысить его точность.
В частности, наши исследования показали, что именно устойчивость свежевылупившихся цыплят к окислительному стрессу, в качестве дополнительного фактора оценки качества цыплят позволяет дать более точный прогноз их будущей продуктивности, по сравнению со способом-прототипом. Это открывает новые возможности оценки различных нарушений технологии кормления родительского стада или же инкубации яиц (долговременное хранение инкубационных яиц перед закладкой, нарушение режимов инкубации, ранняя или поздняя выборка из инкубатора и др.) отрицательно сказывающихся на росте и развитии вылупившихся цыплят. При этом, использование соотношения активности СОД, главного фермента первой линии антиоксидантной защиты, регулируемого на уровне витагенов, к количеству конечных продуктов перекисного окисления липидов, как результат поддержания редокс-баланса, является обнаруженным нами новым показателем, позволяющим прогнозировать устойчивость свежевылупившихся цыплят к окислительному стрессу. Это, в свою очередь, дает возможность делать боле точный прогноз будущей продуктивности растущих цыплят и при необходимости применять специальные приемы коррекции антиоксидантной защиты путем активации витагенов. По нашим наблюдениям именно новый метод позволяет обнаружить нарушения в адаптационной способности цыплят, вызванные вышеупомянутыми факторами и предложить технологические решения, связанные с использованием витаген-активирующих добавок для коррекции указанных нарушений и предотвращения потери продуктивности и экономических потерь.
В разработанном нами новом способе поставленная цель достигается тем, что в способе оценки суточного молодняка птицы, включающем выборочный убой и определение биохимических параметров в биологическом материале, где в качестве биологического материала используют плазму крови, а в качестве биохимических параметров используют общую антиоксидантную активность (ОАА) и количество конечных продуктов перекисного окисления липидов (TBARS), с целью повышения его точности и расширения области применения изобретения за счет учета адаптационной способности и устойчивости цыплят к окислительному стрессу, дополнительно определяют активность витаген-регулируемых ферментов. При этом из витаген-регулируемых ферментов используют активность супероксиддисмутазы и используют соотношение активности супероксиддисмутазы к количеству конечных продуктов перекисного окисления липидов в качестве дополнительного параметра оценки качества цыплят.
Наиболее существенными отличительными от прототипа признаками предлагаемого технического решения являются:
1. В качестве дополнительного биохимического параметра используют активность витаген-регулируемых ферментов и в качестве такого фермента используют активность главного антиоксидантного фермента, регулируемого на уровне витагенов, супероксиддисмутазы.
2. Конечную оценку качества суточных цыплят проводят по отношению активности СОД к количеству конечных продуктов ПОЛ.
Это дает высокую точность определения за счет учета как положительных (антиоксидантная активность), так и отрицательных (перекисное окисление липидов) параметров, существенно влияющих на жизнеспособность молодняка. При этом соотношение активности СОД к количеству конечных продуктов перекисного окисления липидов позволяет дополнительно определить адаптационную способность и устойчивость цыплят к окислительному стрессу, являющемуся молекулярным механизмом большинства технологических, средовых, кормленческих и биологических стрессов в современном птицеводстве (Surai, 2018).
Выбор в качестве дополнительного биохимического параметра активности СОД определяется тем, что в наших исследованиях экспериментально доказано, что адаптация к окислительному стрессу происходит на уровне генов, которые получили название витагенов, которые кодируют синтез таких антиоксидантных молекул как СОД, гемоксигеназа, белки теплового шока, антиоксиданты тиоредоксинового и глутатионового циклов и сиртуины (Surai, 2016; Surai and Fisinin, 2016c; 2016d; Surai et al., 2017; Surai and Kochish, 2017; Suraietal., 2019). При этом в организме суточного цыпленка антиоксидантный статус зависит от баланса между антиоксидантами и прооксидантами и именно активность СОД, витаген-индуцируемого фермента первой линии антиоксидантной защиты в плазме крови наиболее точно отражает резистентность свежевылупившихся цыплят к окислительному стрессу, являющемуся молекулярным механизмом основных коммерческих стрессов в промышленном птицеводстве. При этом, как показали наши исследования, именно соотношение активности СОД (показателя антиоксидантной защиты на витагенном уровне) к количеству конечных продуктов ПОЛ (показателя, отражающего конечный результат антиоксидант-прооксидантного баланса в организме) позволяет эффективно прогнозировать устойчивость свежевылупившихся цыплят к окислительному стрессу и их адаптационную способность.
Таким образом, высокая активность СОД свидетельствует о высоком потенциале антиоксидатной системы и системы витагенов по обеспечению защиты от окислительного стресса у свежевылупившихся цыплят. С другой стороны, количество конечных продуктов ПОЛ в плазме крови отражает конечный результат действия антиоксидантной защиты и адаптации к стрессу. При этом увеличение количества TBARS происходит в стресс-условиях, когда антиоксидантная система не полностью справляется с окислительным стрессом, возникающим в силу различных стресс-факторов (Surai, 2018). Это, как правило, связано со снижением жизнеспособности цыплят, с ухудшением их роста и развития.
Выбор отношения активности СОД/TBARS в качестве дополнительного параметра оценки качества суточных цыплят базируется на результатах наших исследований по взаимосвязи активности СОД, количества TBARSb плазме крови цыплят и их жизнеспособностью при различных срессовых воздействиях. Именно данное соотношение наиболее полно отражает адаптационную способность и устойчивость организма суточных цыплят к окислительному стрессу и вместе с соотношением OAO/TBARS может эффективно прогнозировать их дальнейшую жизнеспособность, которая напрямую зависит от данного параметра.
В свете обнаруженных нами новых фактов регуляции антиоксидантной защиты на уровне активации витагенов и, учитывая особенности суточных цыплят, где антиоксидантная система еще незрелая и активность СОД играет решающую роль в поддержании его жизнеспособности и устойчивости к окислительному стрессу представляется наиболее обоснованным выбор данного биохимического показателя (СОД/TBARS) для улучшения оценки качества суточных цыплят, включая повышение его точности и расширение сферы применения способа.
Таким образом, выбор из биохимических параметров активности СОД в качестве дополнительного параметра оценки позволяет повысить точность и расширить сферу применения данного метода за счет учета важнейшего показателя жизнеспособности, каким является адаптационная способность и устойчивость к окислительному стрессу. Для определения обоих биохимических параметров (СОД и TBARS) существуют коммерческие наборы реактивов, что дает возможность стандартизировать данные определения.
Обоснованность данного выбора определяется тем, что активность СОД плазмы крови отражает эффективность важнейшего звена антиоксидантной системы, регулируемой на уровне витагенов. При этом высокая активность СОД является важнейшим элементом адаптации цыпленка к стрессовым условиям внешней среды (Surai, 2016; 2018). Учитывая тот факт, что производство яиц и мяса в промышленном птицеводстве сопряжено с целым рядом стрессовых ситуаций на всех этапах жизни и развития цыпленка, от момента его вылупления из яйца до убоя на мясо (Surai and Fisinin, 2016; 2016а), эффективность антиоксидантной защиты и активность витагенов рассматривается в качестве решающего фактора стрессустойчивости и жизнеспособности цыплят. Таким образом, с одной стороны, пониженная активность СОД плазмы крови свидетельствует о неадекватной антиоксидантной защите и потенциальных проблемах с устойчивостью к окислительному стрессу и жизнеспособностью при дальнейшем выращивании таких цыплят. При этом следует иметь ввиду, что соотношение СОД/TBARS учитывает как эффективность антиоксидантной защиты, так и результат этой защиты, т.е по сути дела отражает редокс-баланс в организме, который является важнейшим регулятором многих физиологических функций, определяет экспрессию важнейших генов, включая витагены, ответственные за адаптацию организма к стрессам (Surai and Fisinin, 2016; Surai, 2018; Surai et al., 2019). С другой стороны, высокая активность СОД плазмы крови свидетельствует о высокой адаптационной способности и устойчивости данных цыплят к окислительному стрессу и они способны преодолевать различные коммерческие стрессы с минимальными потерями. Это касается, в первую очередь, стрессов, связанных с выводом цыплят, их нахождением в инкубатории до отправки на посадку (иногда до 24-36 часов) и самой посадки на выращивание в корпусе. Физиология свежевылупившегося цыпленка устроена так, что в этот период многие из его систем (включая пищеварительную, иммунную, нервную) незрелые и активно развиваются (Surai and Fisinin, 2015). Именно оптимальный редокс баланс, обеспечивающий активацию важнейших ядерных факторов транскрипции, включая Nrf2, и поддерживаемый высокой активностью витагенов (СОД является одним из таких витагенов), позволяет обеспечить цыплятам высокую резистентность к окислительному стрессу, молекулярному механизму большинства коммерческих стрессов при выращивании цыплят и поддержать последующую высокую жизнеспособность.
Перекисное окисление липидов (ПОЛ) является результатом нарушения редокс-баланса (антиоксидант-прооксидантного баланса) в организме в целом и в индивидуальных тканях в частности (Conradetal., 2018). ПОЛ в мембранах приводит к нарушению их структуры и биологических свойств. В то же время, перекиси липидов оказывают токсическое действие на многие процессы в организме, вызывая воспаление и апоптоз (Gaschler and Stockwell, 2017). Таким образом, в результате ПОЛ происходит накопление конечных продуктов окисления липидов, включая различные перекиси, которые реагируя с тиобарбитуровой кислотой образуют цветной комплекс, который может быть определен фотометрически (TBARS). Следовательно, определяя TBARS в плазме крови можно следить за конечным результатом антиоксидантной защиты, и редокс баланса т.е. повышение TBARS свидетельствует об окислительном стрессе и возможном повреждении жизненно-важных органов и тканей в организме цыпленка (Surai, 2002). Кроме того, высокий уровень TBARS может свидетельствовать о том, что в процессе вывода цыплят были определенные нарушения, например повышенная температура или влажность, что сказывается на дальнейшем развитии цыплят, но визуально обнаружить эти изменения невозможно.
Предлагаемое соотношение СОД/TBARS является наиболее точным индикатором потенциальной адаптационной способности и устойчивости к окислительному стрессу свежевылупившихся цыплят и состояния антиоксидантной защиты в организме, и оно может служить дополнительным фактором предсказания здоровья и будущей продуктивности растущих цыплят-бройлеров, тем самым повышая точность и расширяя сферу применения данного способа.
Известно, что в первые дни жизни цыпленка происходит становление всех основных функций организма, у молодняка с низкой антиоксидантной активностью обнаруживается, как правило, различные нарушения в метаболизме, вызванные как неполноценностью инкубационных яиц, так и нарушениями в процессе инкубации (Suraietal., 1999; Surai and Fisinin, 2015; Suraietal., 2016).
Таким образом, предлагаемый способ оценки суточных цыплят основан на результатах изучения биологических основ развития цыпленка в первые дни жизни и на использовании впервые обнаруженного нами явления -адаптационного изменения активности витагенов и антиоксидантной активности плазмы крови и ее связи с основными функциями свежевылупившегося цыпленка.
Использование заявляемого способа позволит, с одной стороны, более точно прогнозировать физиологическое состояние молодняка сельскохозяйственной птицы и отбирать для выращивания наиболее полноценных цыплят, что в условиях промышленного птицеводства значительно повысит рентабельность отрасли за счет повышенных среднесуточных привесов, лучшей конверсии и сохранности цыплят-бройлеров. С другой стороны, ситуация с цыплятами, характеризующиеся пониженным показателем предлагаемого индекса и соответственно сниженной стрессоустойчивостью, может быть исправлена за счет скармливания или выпаивания специальных антиоксидантных витаген-регулирующих добавок/композиций (Сурай и др., 2017; Григорьева и др., 2017; Сурай и др., 2018).
Предлагаемый способ позволит более точно устанавливать причины плохого роста, развития, нарушения иммунитета и других отрицательных последствий стресса в условиях промышленного птицеводства.
Обоснованность предлагаемого критерия определяется тем, что активность СОД является критическим фактором антиоксидантной защиты, регулируемом на уровне витагенов и факторов транскрипции, и играет ключевую роль в адаптации к стрессу и стрессоустойчивости в целом, во многом определяющих процессы роста и развития молодняка в условиях окислительного стресса и увеличения уровня ПОЛ в их тканях.
Учитывая тот факт, что в первую неделю жизни цыплят концентрация основных антиоксидантов, например, витамина Е и каротиноидов, в их тканях существенно снижается (Surai et al., 1998), то высокая активность СОД и низкое количество TBARS является наиболее эффективным показателем адаптационной способности и устойчивости к окислительному стрессу в данный период отнтогенеза и, соответственно, наиболее точным прогноз-фактором для будущего роста, развития и сохранности растущего молодняка.
Таким образом, предлагаемый способ оценки качества суточного молодняка учитывает ряд обнаруженных нами принципиальных особенностей обмена веществ, включая механизмы антиоксидантной защиты на уровне активации витагенов в первые дни жизни цыплят и дает возможность объективно оценить качество суточных цыплят, отбираемого на выращивание.
Пример 1. (Пример осуществления способа)
Для оценки качества цыплят берут партию суточного молодняка в 200 голов. По комплексу экстерьерных признаков цыплята данной партии оценены как кондиционные. Методом случайного отбора от партии берут 10 голов цыплят для биохимического анализа, от которых отбирают кровь с гепарином и получают плазму путем центрифугирования. В плазме крови определяют общую антиоксидантную активность (ОАА) и концентрацию конечных продуктов ПОЛ (TBARS).
Определение общей антиоксидантной активности плазмы крови проводят с использованием метода основанного на подавлении антиоксидантами исследуемого образца образования окрашенных продуктов при окислении хромогенных субстратов.
При этом используется реакция, связанная с одноэлектронным окислением стабильного радикала 2,2'-азинобис(3-этилбензтиазолин-6-сульфониевой кислоты) (АБТС), в результате которой образуется стабильный катион-радикал АБТС+зелено-голубого цвета с максимумом поглощения λ=600 нм (Miller et al., 1993). Смысл исследования ОАА заключается в ингибировании окисления этого субстрата при инкубации с пероксидазой (метмиоглобин) и оксидазой (пероксид водорода) антиоксидантами биологической жидкости. Увеличение коэффициента абсорбции АБТС+ после фиксированного времени реакции тем ниже, чем больше антиоксидантов присутствует в образце. Большим преимуществом метода является, его высокая стандартизируемость и, соответственно, возможность сравнения данных, полученных исследователями в разных лабораториях (Rice-Evans, 2000). Набор для выполнения метода выпускается английской фирмой Randox под названием TAS (Total Antioxidant Status).
Таким образом, в наших исследованиях использовался коммерческий набор реактивов для определения общей антиоксидантной активности (ОАА) английской фирмы Randox, который широко используется в разных странах мира, как в медицинской практике, так и в научно-исследовательских работах.
В тест-набор входят следующие компоненты.
R1. Буфер -
Фосфатный буфер 80 mmol/1, рН 7.4
R2. Хромоген-
Метмиоглобин: 6.1 μmol/1
ABTS®: 610 μmol/1
R3. Субстрат-
Перекись водорода (в стабилизированной форме): 250 μmol/1
Стандарт для калибровки-
Тролокс: 6-гидрокси-2,5,7,8-тетраметилхроман-2-карбоксиловая кислота
Приготовление реагентов для исследования
R1. Буфер
Содержимое флакона готово к использованию
R2. Хромоген
Содержимое флакона с хромогеном R2 растворяется в10 мл буфера R1.
R3. Субстрат
1 мл субстрата R3 растворяется в 1.5 мл буфера R1.
Калибровочный стандарт
Содержимое флакона со стандартом растворяется в 1 мл дважды ионизированной воды
Процедура:
Длина волны - 600 nm
Кювета: 1 cm
Измерение: против воздуха
Измерение в контрольной пробе проводят следующим образом: в кювету добавляют
1 мл хромогена и 20 мкл плазмы крови и проводят измерение интенсивности поглощения на ФЭКе (или спектрофотометре) при 600 нм. Далее в кювету добавляют 0.2 мл субстрата и приводят второе измерение через 3 минуты после добавления субстрата.
Аналогичным образом проводят измерение со стандартным раствором Тролокса, добавляя в кювету вместо плазмы крови вышеупомянутый раствор калибровочного стандарта.
В холостой пробе проводят измерение при использовании вместо плазмы 20 мкл дистиллированной воды.
Расчет антиоксидантной активности проводят путем сравнения изменения поглощения в кювете с опытным образцом, по сравнению со стандартом и холостой пробой и активность выражают в микромолях на мл плазмы крови.
Определение продуктов перекисного окисления липидов, реагирующих стиобарбитуровой кислотой в плазме крови цыплят.
В процессе перекисного окисления липидов образуются смесь различных альдегидов и других продуктов. При этом малоновый диальдегид (МДА) используется в качестве основного показателя для оценки ПОЛ. Наиболее распространенный метод анализа продуктов ПОЛ основан на их реакции с тиобарбитуровой кислотой (ТБК). Тест с TBK(TBARS) очень чувствителен. С его помощью можно улавливать наномолярные концентрации МДА. Установлено, что между липидной пероксидацией и МДА существуют количественные взаимосвязи и продукты, образованные при проведении ТБК-теста (TBARS), свидетельствуют о присутствии и количестве липидных перекисей.
В наших исследованиях использовался метод определении количества конечных продуктов ПОЛ по реакции с тиобарбитуровой кислотой, описанный Ohkawa et al., (1979) и используемый нами во многочисленных исследованиях (Surai et al., 1996; Surai, 2000; Blount et al., 2004 и др.).
Принцип метода. Исследование интенсивности ПОЛ ведут, определяя один из продуктов перекисного окисления - МДА - с помощью ТБК (TBARS - тест). МДА и ТБК при высокой температуре и кислом значении рН образуют окрашенный триметиновый комплекс, содержащий одну молекулу МДА и две молекулы ТБК. Максимум поглощения комплекса приходится на 532 нм.
Оборудование. Водяная баня, центрифуга со скоростью 6000 об./мин, колориметр или спектрофотометр.
Реактивы и их приготовление.
1. 0.8%-ный раствор додецил сульфата натрия
2. 20%-ная кислота с рН 3.5 (рН доводят до нужной величины с помощью КОН)
3. 0.8%-ны раствор тиобарбитуровой кислоты (ТБА)
4. Бутанол
Ход определения. К 0,2 мл плазмы крови приливают 0.2 мл 0.8%-ного додецилсульфата натрия и 1.5 мл 20%-ной уксусной кислоты и 1.5 мл 0.8%-ного раствора ТБА. Пробы перемешиваются и помещаются в водяную баню при 95°С на 60 минут. Затем пробы охлаждают в холодной воде, добавляют 4 мл бутанола. Тщательно перемешивают и центрифугируют 10 мин при 3000 об./мин. Оптическую плотность верхней бутанольной фазы измеряют при длине волны 532 нм против холостой пробы, содержащей вместо сыворотки крови воду. Расчет содержания конечных продуктов ПОЛ, реагирующих с ТБК, производят по калибровочному графику, построенному с использованием 1,1,3,3-тетраметоксипропана (1,1,3,3-Tetramethoxypropane, Sigma). Полученные результаты выражают в микромолях на миллилитр (μмоль/мл) плазмы крови.
Для разделения суточных цыплят на категории согласно способу-прототипу используют соотношение ОАА (в микромолях на мл плазмы крови) к показателю TBARS (в наномолях на мл плазмы крови). При этом при данном соотношении >800 цыплят относят к первой категории, при соотношении 300-800 - ко второй категории и при соотношении ниже 300 цыплят относят к третей категории (брак) не пригодных для выращивания.
В предлагаемом способе в дополнение к определению вышеизложенных показателей (ОАА и TBARS) также определяют активность СОД в плазме крови.
В наших исследованиях использовался коммерческий набор реактивов для определения активности супероксиддисмутазы (RANSOD) английской фирмы Randox, который широко используется в разных странах мира, как в медицинской практике, так и в научно-исследовательских работах.
В тест-набор входят следующие компоненты.
Rla Смешанный субстрат - 5 х 20 мл
Rib Буфер 1 х 105 мл
R2 Ксантиноксидаза 3 х 10 мл
CAL - Стандарт 5 х 10 мд
Принцип метода
При исследовании активности СОД используется ксантин и фермент ксантиноксидаза для образования/генерирования супероксид-радикала, который реагирует с 2-(4-иодофенил)-3-(4-нитрофенол)-5-фенилтетразолий хлоридом (INT) с образованием окрашенного в красный цвет формазана. При этом активность СОД определяют, как степень ингибирования указанной реакции.
Состав реагентов
Rla Смешанный субстрат
Ксантин - 0.05 mmol/1
I.N.T. - 0.025 mmol
R1b Буфер
CAPS - 40 mmol/1, pH 10.2
EDTA - 0.94 mmol/1
R2 Ксантиноксидаза - 80 U/1
CAL Стандарт СОД
Процедура определения активности
50 микролитров плазмы крови смешивают с 1.7 мл смешанного субстрата (Rla) в 1 см кварцевой кювете. Поглощение при 505 нм измеряют с 30 секундным интервалом в течение 3 минут при 37°С после добавления в кювету 0.25 мл ксантин-оксидаза (80 Ед/литр) и перемешивания содержимого. Подавление поглощения/цвета образуемого формазана за счет СОД плазмы крови сравнивают со стандартом (CAL), который включен в набор реагентов для определения СОД. Активность фермента выражали в Ед/мл.
При соотношении СОД/TBARS на уровне 7 и выше цыплят относят к группе с повышенной адаптационной способностью, при показателях СОД/TBARS <7 цыплят относят к группе с пониженной адаптационной способностью, которая требует дополнительного внимания со стороны менеджмента предприятия.
Проведенные исследования показали, что при оценке качества цыплят, полученных из яиц, соответствующих стандартам практики современного мясного птицеводства со сроком хранения яиц перед инкубацией не превышающих 7 дней, оба способа дали сравнимые результаты и показали их действенность.
Пример 2. Для проверки сравнительной эффективности определения качества суточных цыплят по способу прототипа и по предлагаемому способу использовали цыплят, выведенных из яиц с разным сроком хранения перед закладкой. По нашим предварительным наблюдениям хранение яиц перед закладкой в течение более 7 дней связано со снижением выводимости и при хранении более 14 дней наблюдается снижение жизнеспособности вылупленных цыплят. Мы предположили, что одной из причин снижения жизнеспособности цыплят является снижение активности витагенов в результате длительного хранения яиц перед закладкой как результат окисления белка. Поэтому для сравнения мы использовали 2 группы цыплят по 200 голов, одна группа цыплят выведена из яиц, хранившихся перед инкубацией в течение 4 дней, другая группа- цыплята, выведенные из яиц, хранившихся в течение 18 дней перед инкубацией. Все отобранные в группы цыплята были визуально оценены как кондиционные. Далее проводили их оценку по способу прототипа, отбирая 10 цыплят и определяя соотношение OAA/TBARS, как описано выше и по предложенному способу, дополнительно определяя соотношение СОД/TBARS. Полученные результаты представлены в таблице 1.
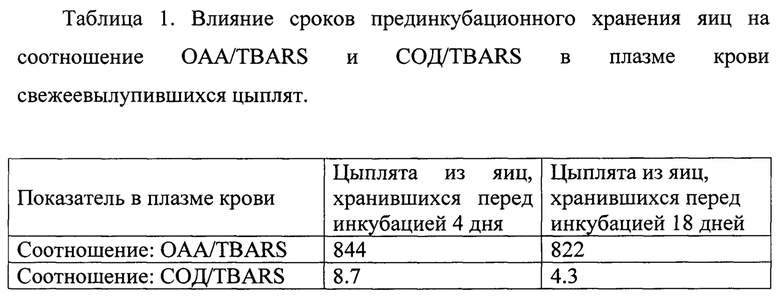
Как видно из приведенных данных, при оценке качества цыплят по способу прототипа цыплята обеих групп были отнесены к 1-й категории, и различия между группами по показателю OAA/TBARS были статистически не достоверными. Оценка цыплят по предлагаемому способу выявила высоко достоверную (Р<0.001) разницу по показателю СОД/TBARS, который различался между группами в 2 раза. Таким образом, результаты данных исследований продемонстрировали не способность способа-прототипа уловить разницу между цыплятами, выведенными из яиц с разным сроком хранения перед инкубацией и различающихся по их адаптационной способности при последующем выращивании. При этом, предлагаемый способ позволил уловить данную разницу, подтверждая повышение точности способа и расширение его возможности использования, включая оценку цыплят, выведенных из яиц длительного хранения.
Пример 3. (Сравнительные испытания предлагаемого способа и прототипа). Для повторного сравнения оценки цыплят по способу-прототипу и предлагаемому способу были отобраны по 200 цыплят с аналогичных групп, т.е. цыплят, вылупившихся из яиц, хранившихся 4 дня и 18 дней соответственно. Цыплята были оценены по способу-прототипу и по предлагаемому способу и далее были помещены в птичник для выращивания для подтверждения эффективности предлагаемого способа в оценке адаптационной способности цыплят. Полученные результаты представлены в таблице 2.
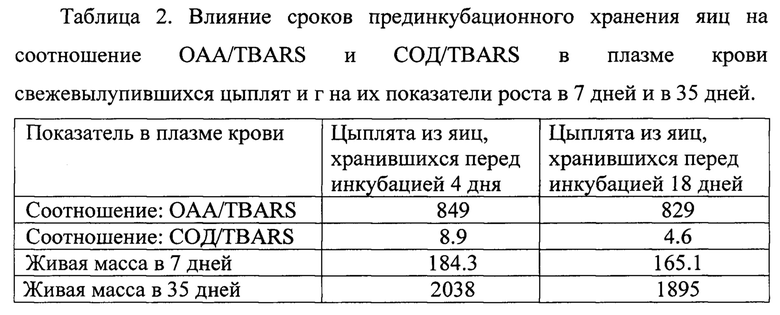
Полученные результаты подтвердили предположение о том, что оценка цыплят по способу прототипу не дает возможность определить адаптационную способность цыплят, которая была нарушена вследствие длительного хранения яиц перед инкубацией. В то же время, оценка качества цыплят с использованием предлагаемого способа позволяет выявить такие нарушения. Так, разница в живой массе цыплят в 7-дневном возрасте составила более 19 г, что при дальнейшем выращивании транслировалось в 143 г на каждом бройлере. Таким образом, не выявив вовремя нарушения в адаптивной способности цыплят, в результате длительного хранения яиц перед инкубацией, производитель мяса понесет существенные убытки. Полученные результаты подтвердили неприменимость способа-прототипа для оценки качества цыплят, полученных из яиц длительного хранения. При этом, предлагаемый способ позволил с высокой вероятностью разделить таких цыплят на две группы, что дает возможность работникам птицефабрики использовать систему дополнительной поддержки таких цыплят, включая выпойку витаген-регулируюших композиций (Сурай и др. 2018). Следует подчеркнуть, что в условиях промышленного птицеводства хранение яиц перед инкубацией более 7-10 дней встречается достаточно часто и оценка качества цыплят, выведенных из таких яиц достаточно затруднена. В этом отношении предлагаемый способ оценки качества суточных цыплят повышает точность оценки и расширяет возможности применения способа.
Пример 4. (Демонстрация возможности дополнительной поддержки цыплят, со сниженной адаптационно способностью). Исследования были проведены аналогично примеру 3, но при этом использовали 4 группы цыплят по 200 голов с целью определить возможность дополнительной поддержки цыплят, вылупившихся из яиц длительного хранения путем выпаивания витаген-регулирующего препарата. Для этого использовали выпойку препарата Меджик Антистресс Микс/PerforMax (Feed-Food. Ltd., Великобритания), содержащего витаген-активирующую смесь нутриентов, в течение 4 дней (возраст 3-6 дней) из расчета 1 г/л питьевой воды. Полученные результаты представлены в таблице 3. 
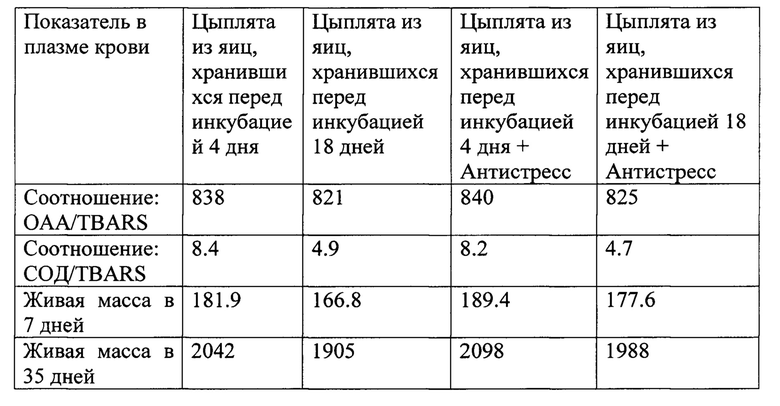
Как видно из приведенных данных, полученные данные подтвердили наше предположение о том, что путем использования витаген-регулирующей добавки можно частично снизить отрицательное действие стресса продолжительного хранения яиц на рост и развитие цыплят, вылупившихся из таких яиц. При этом использование заявляемого способа оценки качества суточных цыплят является важнейшим инструментом выявления различных нарушений и своевременного принятия решений по снижению их отрицательных последствий. Так, разница в живой массе цыплят в 7 дневном возрасте составила 15.1 г между первой и второй группами. При этом использование витаген-стимулирующей добавки сократило эту разницу до 4.3 г. То же самое наблюдалось при выращивании цыплят до 35-дневного возраста. Длительное хранение яиц перед инкубацией привело к потере 137 г живой массы при убое цыплят. При использовании витаген-регулирующей добавки данная разница сократилась до 54 г. Данные испытания показали, что при посадке на выращивание цыплят, со сниженной адаптационной способностью из-за длительного прединкубационного их хранения, есть возможность существенно улучшить их рост и развитие за счет активации витагенов, ответственных за адаптацию цыплят к стрессам. Таким образом, предлагаемый способ оценки качества суточных цыплят повышает точность способа и расширяет сферу использования данного способа за счет определения адаптационной способности цыплят, нарушенной вследствие различных стрессовых ситуаций. Наши предварительные данные свидетельствуют о том, что поддержка цыплят витаген-стимулирующей добавкой оказывает положительное действие и в других стрессовых ситуациях, когда адаптационная способность цыплят снижена, например при длительном (>24 часов) хранении цыплят в инкубатории перед их отправкой для посадки на выращивание в птичнике. Заключение
В примерах 1-4 показана осуществимость способа и его преимущества перед известными аналогами.
Список литературы
1. Григорьева М.А., Величко О.А., Шабалдин С.В., Фисинин В.И., Сурай П.Ф. (2017).
2. Регуляция активности витагенов как новая антистрессовая стратегия в птицеводстве: обоснование и производственный опыт. Сельскохозяйственная Биология 52,4: 716-730.
3. ОСТ 10329-2003 «Суточный молодняк кур. Технические условия».
4. Сурай П.Ф., Фисинин В.И., Шацких Е.В., Латыпова Е.Н. (2017). Современные методы борьбы со стрессами в птицеводстве и свиноводстве: концепция витагенов в действии. Сфера. Технологии, корма, ветеринария 5,2: 40-43.
5. Сурай П.Ф., Кочиш И.И., Фисинин В.И., Грозина А.А., Шацких Е.В. (2018). Молекулярные механизмы поддержания здоровья кишечника птицы: Роль микробиоты. Монография. Сельскохозяйственные технологии, 344 стр.
6. Blount, J.D., Houston, D.C., Surai, P.F., &,А.Р. (2004). Egg-laying capacity is limited by carotenoid pigment availability in wild gulls Larus fuscus. Proceedings of the Royal Society of London B: Biological Sciences, 271 (Suppl 3), S79-S81.
7. Conrad M, Kagan VE, Bayir H, Pagnussat GC, Head B, Traber MG, Stockwell BR. (2018).
8. Regulation of lipid peroxidation and ferroptosis in diverse species. Genes Dev. 32(9-10):602-619.
9. Gaschler MM, Stockwell BR. (2017). Lipid peroxidation in cell death. Biochem Biophys Res Commun. 482 (3):419-425.
10. Karadas F, Surai PF and sparks NH. (2011). Changes in broiler chick tissue concentrations of lipid-soluble antioxidants immediately post-hatch. Comp Biochem Physiol A Mol Integr Physiol. 160, 1:68-71.
11. Lewandowski L, Kepinska M, Milnerowicz H. (2019. The copper-zinc superoxide dismutase activity in selected diseases. Eur J Clin Invest. 49(1): el3036.
12. Miller, N.J., Rice-Evans., C, Davies, M.J., Gopinathan, V., & Milner, A. (1993). A novel method for measuring antioxidant capacity and its application to monitoring the antioxidant status in premature neonates. Clinical science, 84 (4), 407-412.
13. Ohkawa H, Ohishi N, Yagi K. (1979). Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal Biochem. 95 (2):351-8.
14. Rice-Evans CA. (2000). easurement of total antioxidant activity as a marker of antioxidant status in vivo: procedures and limitations. Free Radic Res. 2000 Nov; 33 Suppl:S59-66.
15. Ross 308 Parent Stock Nutrition Specification. 2016.
16. Surai P.F. (1999) Tissue-specific changes in the activities of antioxidant enzymes during the development of the chicken embryo. British Poultry Science, 40: 397-405.
17. Surai P. (1999a) Vitamin E in avian reproduction. Poultry and Avian Biology Reviews, 10: 1-60.
18. Surai P.F. (2000) Effect of the selenium and vitamin E content of the maternal diet on the antioxidant system of the yolk and the developing chick. British Poultry Science, 41: 235-243
19. Surai P.F. (2006). Selenium in Nutrition and health. Nottingham University Press, Nottingham, UK
20. Surai P.F. (2006). Natural antioxidants in avian nutrition and reproduction. Nottingham University Press, Nottingham, UK
21. Surai P.F. (2015). Antioxidant Action of Carnitine: Molecular Mechanisms and Practical Applications. EC Veterinary Science 2.1: 66-84.
22. Surai P.F. (2015a). Carnitine Enigma: From Antioxidant Action to Vitagene Regulation. Part 1. Absorption, Metabolism and Antioxidant Activities" J. Veter. Sci. Med. 3, 2: 14.
23. Surai P.F. (2015b). Carnitine Enigma: From Antioxidant Action to Vitagene Regulation. Part 2. Transcription Factors and Practical Applications J. Veter. Sci. Med. 3, 2: 17.
24. Surai P.F. (2015c). Silymarin as a Natural Antioxidant: An Overview of the Current Evidence and Perspectives. Antioxidants 4: 204-247.
25. Surai PF. (2016). Antioxidant Systems in Poultry Biology: Superoxide Dismutase. Journal of Animal Nutrition 1,1:8.
26. Surai P.F. (2018). Selenium in poultry nutrition and health. Wageningen Academic Publishers, Wageningen, The Netherlands
27. Surai P.F., FISININ V.I. (2015). Antioxidant-Prooxidant Balance in the Intestine: Applications in Chick Placement and Pig Weaning. J. Veter. Sci. Med. 3, 1: 1-16.
28. Surai P.F., Fisinin V.I. (2016a). Vitagenes in poultry production. Part 1. Technological and environmental stresses. World's Poultry Science Journal 72: 721-734.
29. Surai P.F., Fisinin V.I. (2016 b). Vitagenes in poultry production. Part 2. Nutritional and Internal stresses. World's Poultry Science Journal 72: 761-772.
30. Surai P.F., Fisinin V.I. (2016c). Vitagenes in poultry production. Part 3. Vitagene concept development. World's Poultry Science Journal 72: 793-804.
31. Surai, P.F. and Fisinin, V.I. (2016d). Antioxidant System regulation: From Vitamins to Vitagenes. In: Handbook of Cholesterol, edited by: Ronald Ross Watson and Fabien De Meester, Wageningen Academic Publishers, pp. 451-481.
32. Surai PF., Fisinin V.I., Karadas F. (2016). Antioxidant Systems in Chick Embryo Development. Part 1. Vitamin E, Carotenoids and Selenium. Animal Nutrition 2: 1-11.
33. Surai P., Ionov I., Kuchmistova E., Noble R., Speake B. (1998) The relationship between the levels of α-tocopherol and carotenoids in the maternal feed, yolk and neonatal tissues: Comparison between the chicken, turkey, duck and goose. Journal of the Science of Food and Agriculture, 76: 593-598.
34. Surai P.F. and Kochish I. (2017). Antioxidant systems and vitagenes in poultry biology: Heat Shock Proteins. In: Heat Shock Proteins in Veterinary. Edited by Alexzander A. A. Asea and Dr. Punit Kaur, Springer, Switzerland, pp. 123-177.
35. Surai P.F., Kochish I.I., Fisinin V.I., Kidd M.K. (2019). Antioxidant Defence Systems andOxidative Stress in Poultry Biology: An Update. Antioxidants (Basel).8, 7. pii: E235.
36. Surai P.F., Kochish I.I., Fisinin V.I., Juniper D.T. (2019a). Revisiting Oxidative Stress and the Use of Organic Selenium in Dairy Cow Nutrition. Animals (Basel) 9, 7, pii: E462.
37. Surai P.F., Kochish I.I., Fisinin V.I. (2017). Antioxidant systems in poultry biology: Nutritionalmodulation of vitagenes. European Journal of Poultry Science 81,1612-9199.
38. Surai P.F., Kochish I.I., Shapovalov S.O. (2018). Superoxide dismutase activity in chicken gut. Programme and Summaries. WPSA UK Branch Annual Meeting, Dublin, 2018, p.010.
39. Surai P.F., Kuklenko Т., Ionov I., Noble R., Sparks N. (2000) Effect of vitamin A on antioxidant system of the chick during early post-natal development. British Poultry Science, 41: 454-458.
40. Surai P.F., Speake B.K., Noble R.C. and Sparks N.H.C. (1999). Tissue-specific antioxidant profiles and susceptibility to lipid peroxidation of the newly hatched chick. Biology Trace ElementResearch, 68: 63-78.
41. Surai P., Noble R., Speake B. (1996). Tissue-specific differences in antioxidant distribution and susceptibility to lipid peroxidation during development of the chick embryo. Biochem. Biochys. Acta, 1304: 1-10.
42. Тота K, Bamelis F, De Ketelaere B, Bruggeman V, Moraes VM, Buyse J, Onagbesan O, Decuypere E. (2003). Effects of egg storage time on spread of hatch, chick quality, and chick juvenile growth. Poult Sci. 82 (5):736-41.
43. Zou X, Ratti BA, O'Brien JG, Lautenschlager SO, Gius DR, Bonini MG, Zhu Y. (2017) Manganese superoxide dismutase (SOD2): is there a center in the universe of mitochondrial redox signaling? J BioenergBiomembr. 2017 Aug;49(4):325-333.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ОЦЕНКИ СУТОЧНОГО МОЛОДНЯКА ПТИЦЫ | 2018 |
|
RU2699719C1 |
| Способ предупреждения и подавления интенсификации липопероксидации антиоксидантом глутатионом для улучшения критериев качества эмбрионов и перепелят в условиях промышленного производства | 2021 |
|
RU2772655C1 |
| СПОСОБ ОПТИМИЗАЦИИ ГОМЕОСТАЗА У ЭМБРИОНОВ И МОЛОДНЯКА КУР | 2019 |
|
RU2706563C1 |
| СПОСОБ ПОВЫШЕНИЯ И СИНХРОНИЗАЦИИ ВЫВОДА ЦЫПЛЯТ ПОСРЕДСТВОМ ПРОФИЛАКТИКИ ОКСИДАТИВНОГО СТРЕССА У ЭМБРИОНОВ КУР | 2014 |
|
RU2567898C2 |
| СПОСОБ РЕАНИМАЦИИ ЭМБРИОНОВ ПЕРЕПЕЛОВ И СТИМУЛЯЦИИ ИХ НЕСПЕЦИФИЧЕСКОЙ РЕЗИСТЕНТНОСТИ | 2020 |
|
RU2759674C1 |
| ПРЕМИКС ДЛЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ПТИЦ И СПОСОБ ЕГО СКАРМЛИВАНИЯ | 2006 |
|
RU2332022C1 |
| СПОСОБ ПРОФИЛАКТИКИ СТРЕСС-ИНДУЦИРОВАННЫХ НАРУШЕНИЙ КАК ЗАЛОГ ОПТИМИЗАЦИИ СТАНОВЛЕНИЯ МЕХАНИЗМОВ АДАПТАЦИИ У ЭМБРИОНОВ И МОЛОДНЯКА КУР | 2018 |
|
RU2700473C1 |
| Способ повышения качественных и количественных результатов промышленной инкубации яиц кур при трансовариальном введении инозитола | 2022 |
|
RU2815970C1 |
| СПОСОБ ОПТИМИЗАЦИИ ЭНЕРГЕТИЧЕСКОГО ОБМЕНА В ОРГАНИЗМЕ ЭМБРИОНОВ КУР ПРИ ИСКУССТВЕННОЙ ИНКУБАЦИИ | 2021 |
|
RU2777481C1 |
| СПОСОБ КОРРЕКЦИИ НЕГАТИВНЫХ ПОСЛЕДСТВИЙ ОКСИДАТИВНОГО СТРЕССА ЦЫПЛЯТ-БРОЙЛЕРОВ В УСЛОВИЯХ ПРОМЫШЛЕННОГО ПРОИЗВОДСТВА | 2024 |
|
RU2826272C1 |
Изобретение относится к области биотехнологии. Изобретение представляет собой способ оценки качества суточных цыплят, включающий выборочный убой и определение биохимических параметров в биологическом материале, где в качестве биологического материала используют плазму крови, а в качестве биохимических параметров используют общую антиоксидантную активность (ОАА) и количество конечных продуктов перекисного окисления липидов (TBARS), с целью повышения его точности и расширения области применения изобретения за счет учета адаптационной способности и устойчивости цыплят к окислительному стрессу, дополнительно определяют активность витаген-регулируемых ферментов. При этом из витаген-регулируемых ферментов используют активность супероксиддисмутазы и используют соотношение активности супероксиддисмутазы к количеству конечных продуктов перекисного окисления липидов в качестве дополнительного параметра оценки качества цыплят. 3 табл., 4 пр.
Способ оценки качества суточных цыплят, включающий выборочный убой и определение биохимических параметров в биологическом материале, где в качестве биологического материала используют плазму крови, а в качестве биохимических параметров используют общую антиоксидантную активность (ОАА) и количество конечных продуктов перекисного окисления липидов (TBARS), отличающийся тем, что дополнительно определяют активность витаген-регулируемых ферментов, при этом из витаген-регулируемых ферментов используют активность супероксиддисмутазы и используют соотношение активности супероксиддисмутазы к количеству конечных продуктов перекисного окисления, при этом при соотношении активности супероксиддисмутазы к количеству конечных продуктов перекисного окисления 7 и выше, цыплят относят к группе с повышенной адаптационной способностью, при показателях меньше 7, цыплят относят к группе с пониженной адаптационной способностью.
| СПОСОБ ОЦЕНКИ СУТОЧНОГО МОЛОДНЯКА ПТИЦЫ | 2018 |
|
RU2699719C1 |
| СУРАЙ П.Ф., Фисинин В.И., Шацких Е.В., Латыпова Е.Н | |||
| Автомобиль-сани, движущиеся на полозьях посредством устанавливающихся по высоте колес с шинами | 1924 |
|
SU2017A1 |
| Современные методы борьбы со стрессами в птицеводстве и свиноводстве: концепция витагенов в действии | |||
| Сфера | |||
| Технологии, корма, ветеринария 5,2: 40-43 | |||
| SURAI P.F | |||
| Металлический водоудерживающий щит висячей системы | 1922 |
|
SU1999A1 |

