Изобретение относится к медицинской вирусологии и может быть использовано в здравоохранении для определения репродуктивной активности вирусов гриппа в составе тривалентной и квадривалентной живой гриппозной вакцины (ЖГВ).
Сезонные ЖГВ включают три штамма вируса гриппа - A/H1N1, A/H3N2 и В, которые содержат гены гемагглютинина (НА) и нейраминидазы (NA) от эпидемических вирусов и 6 генов внутренних и неструктурных белков от холодоадаптированных доноров аттенуации: А/Ленинград/134/17/57(H2N2) для вирусов гриппа типа А [Александрова Г.И. Применение метода генетической рекомбинации для получения вакцинных штаммов вируса
гриппа // Вопр. Вирусол. - 1977. - №4. - С. 387-395.] и В/СССР/60/69 для вирусов гриппа типа В [Alexandrova G.I., Maassab H.F., Kendal А.Р. et al Laboratory properties of cold-adapted influenza В live vaccine strains developed in the US and USSR, and their B/Ann Arbor/1/86 cold-adapted reassortant vaccine candidates // Vaccine. - 1990. - Vol. 8. - p. 61-64].
Начиная с 1988 г. в мире наблюдается циркуляция двух генетически и антигенно различных линий вирусов гриппа В, филогенетически представленных штаммами В/Виктория/1/87 (В/Виктория) и В/Ямагата/16/88 (В/Ямагата). Вирусы обеих антигенных линий циркулировали повсеместно до 1990 года, после чего выявлялись преимущественно вирусы группы В/Ямагата.
Начиная с 1997 г. вирусы группы В/Виктория вновь появились в циркуляции [Luo С., Morishita Т., Satou K., et al. / Evolutionary pattern of influenza В viruses based on the HA and NS genes during 1940 to 1999: origin of the NS genes after 1997 // Arch. Virol. 1999. - 144(10). - P. 1881-91.]. Поскольку в тривалентную гриппозную вакцину включен только один штамм вируса гриппа В, неэффективность вакцинации против гриппа В в последние годы привела к значительному увеличению случаев, госпитализаций и смертей (до 970000 случаев в год, с 8200 госпитализациями и 485 смертельными случаями в США) [Reed С, Meltzer MI, Finelli L, Fiore A. Public health impact of including two lineages of influenza В in a quadrivalent seasonal influenza vaccine. Vaccine 2012; 30: 1993-1998.].
В связи с этим Всемирной Организацией Здравоохранения (ВОЗ) было рекомендовано применение квадривалентных гриппозных вакцин. В отделе вирусологии ФГБНУ «ИЭМ» проводятся исследования квадривалентной живой гриппозной вакцины, включающей два вируса гриппа В - В/Виктория и В/Ямагата [Desheva Y.A., Smolonogina Т.А., Doroshenko Е.М., Rudenko L.G. VoprVirusol, 61(1): 16-20, (2016)].
Включение дополнительных вакцинных штаммов в состав ЖГВ требует разработки методики для определения их репродуктивной активности в
составе поливалентных вакцинных препаратов. Согласно требованиям Государственного стандарта качества лекарственного продукта Министерства Здравоохранения Российской Федерации препарат тривалентной ЖГВ должен обладать инфекционной активностью не менее 106,9 ЭИД50/0,5 мл (50% эмбриональная инфекционная доза) для штаммов вируса гриппа типа А и не менее 106,4 ЭИД50/0,5 мл для штамма вируса гриппа типа В.
При производстве тривалентной ЖГВ определение репродуктивной активности вакцинных вирусов гриппа проводят с лиофилизированными полуфабрикатами моновакцин на развивающихся куриных эмбрионах 10-12-дневного возраста. Монопрепараты объединяются в тривалентный препарат, вследствие чего может изменяться их инфекционная активность. Определение инфекционных титров вирусов в составе тривалентной или квадривалентной вакцины является трудоемким процессом.
Для определения инфекционной активности каждого из штаммов вируса гриппа в составе поливалентных ЖГВ требуется использование поликлональных антисывороток или моноклональных антител для блокирования инфекционности других вирусов [Romanova J.R., Ermachenko Т.А., Alexandrova G.I., & Tannock G.A. Interference between cold-adapted (ca) influenza A and В vaccine reassortants or between ca reassortants and wild-type strains in eggs and mice //Vaccine. - 1994. - T. 12. - №. 1. - C. 23-27].
Метод получения моноклональных антител, разработанный в 1975 г. Kohler и Milstein [Kohler G, Milstein С. Derivation of specific antibodyproducing tissue culture and tumor lines by cell fusion. Eur J Immunol 1976; 6, 511-9.], позволил определять инфекционную активность каждого штамма вируса гриппа в составе тривалентных ЖГВ реакцией непрямой иммунофлуоресценции (РНИФ) с использованием специфических моноклональных антител к белку нуклеопротеина (NP).
Однако эти методики являются продолжительными и затратными, поскольку получение моноклональных антител является длительным
процессом, и могут быть недостаточно точными, если антитела не являются высокоспецифичными. Поэтому развитие быстрого и точного метода определения титров вирусов гриппа в составе тривалентной вакцины, основанного на молекулярно-клеточных технологиях, необходимо для своевременного и эффективного производства вакцины.
Метод ПЦР в реальном времени (ПЦР-РВ) широко применяется для оценки экспрессии генов возбудителей инфекций и определения вирусной нагрузки при различных вирусных инфекциях. В настоящее время ПЦР-РВ используется для идентификации вирусов гриппа в текущем надзоре за эпидемической ситуацией.
Описана методика дифференцирования вирусов гриппа В по двум антигенным линиям, В/Виктория и В/Ямагата с помощью ПЦР-РВ [Biere В, Bauer В, Schweiger В. Differentiation of influenza В virus lineages Yamagata and Victoria by real-time PCR. J Clin Microbiol 2010; 48: 1425-1427.] для применения в текущем эпидемическом надзоре.
Метод титрования моновалентных ЖГВ на основе технологии гидролизируемых зондов (Taqman) описан в статье Yang Zang, et al. [Development of one-step real-time PCR assay for titrating trivalent live attenuated influenza vaccines. Yang Zang et al. Human Vaccines & Immunotherapeutics 10:12, 3642-3648; December 2014].
Три моновалентных вакцинных штамма (A/H1N1, A/H3N2 и В), рекомендованные ВОЗ, с известным титром ЭИД50 использовались для проведения анализа методом обратно-транскриптазной полимеразно-цепной реакции в реальном времени (ОТ-ПЦР-РВ). Репродукцию вирусов определяли серийными разведениями путем заражения клеток MDCK, инфекционные титры определяли методом ОТ-ПЦР-РВ и сравнивали с методом ЭИД50, для подтверждения эффективности метода.
Результаты показали, что анализ ОТ-ПЦР-РВ, выполненный с набором специфических праймеров и зонда для каждого из вирусов, был сопоставим с традиционным методом ЭИД50 и может быть принят в качестве
альтернативы ЭИД50 для титрования моновалентных штаммов ЖГВ. Тем не менее не было проведено изучение репродукции вирусов в составе тривалентной вакцины. К тому же эта методика не предусматривает различий вирусов гриппа В антигенных линий В/Виктория и В/Ямагата в составе квадривалентной ЖГВ, так как для изучения репродукции единственного компонента В в составе тривалентной вакцины использовали праймеры и зонд к гену нуклеопротеина (NP), который является единым у реассортантов как линии Виктория, так и линии Ямагата, поскольку приобретен от донора аттенуации В/СССР/60/69.
Задачей, на решение которой направлено заявляемое изобретение, является создание путем определения инфекционных титров вакцинных вирусов методом ОТ-ПЦР-РВ с применением технологии гидролизируемых олигонуклеотидных зондов (Taqman).
Поставленная задача решалась путем разработки праймеров и зондов, специфичных к гемагглютинину вирусов гриппа А и В, подбора оптимальных условий проведения реакции, разработке стандартов для количественного анализа.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Способ оценки репродуктивной активности вирусов гриппа в составе тривалентной и квадривалентной живой гриппозной вакцины заключается в том, что предварительно проводят сокультивирование вакцинных штаммов в клеточной культуре либо осуществляют иммунизацию поливалентной живой гриппозной вакциной in vivo, после чего проводят одношаговую обратно-транскриптазную полимеразную цепную реакцию в реальном времени с использованием праймеров и зондов, характеризующихся нуклеотидными последовательностями SEQ ID No: 1 - SEQ ID No: 25, гомологичными к вирусам гриппа А, В/Виктория и В/Ямагата, при подобранных оптимальных условиях проведения реакции с разработанными стандартами для относительного количественного анализа методом разницы пороговых циклов.
Состав исследуемых вирусов представлен в таблице 1.

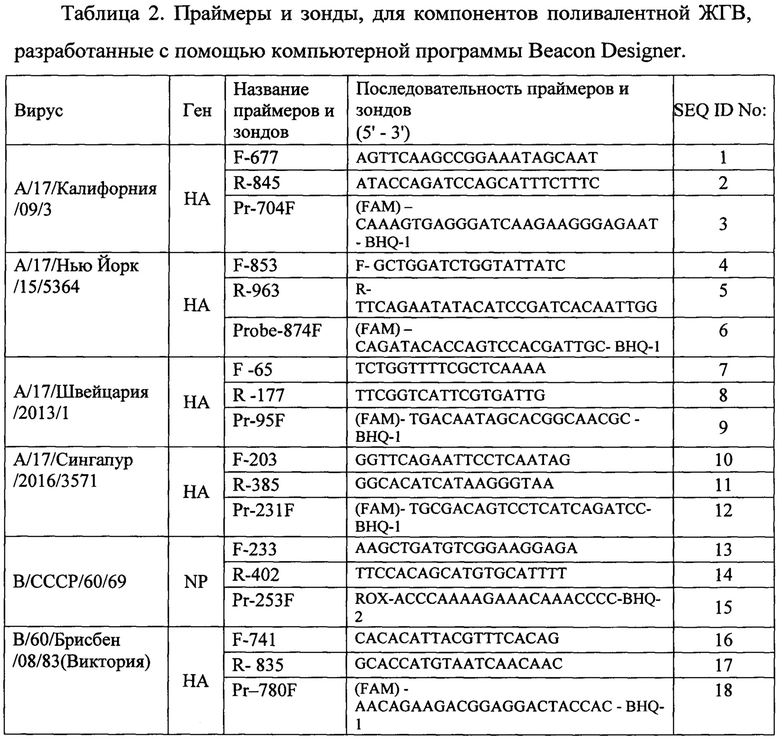
Состав олигонуклеотидных праймеров и зондов представлен в таблице 2.

Вирусы гриппа культивировали в 10-дневных развивающихся куриных эмбрионах (РКЭ) при 33°С 48 часов для вирусов гриппа А и 72 часа для вирусов гриппа В. Инфекционную активность вирусов определяли методом ЭИД50 и в клеточной культуре MDCK. 50% эмбриональную инфекционную дозу (ЭИД50) рассчитывали по методу Рида и Менча [Reed L.J. Simple method of estimating fifty percent endpoints/ L.J. Reed, H.A. Muench // American Journal of Hygiene. - 1938. - P. 493-497]. Титры вируса, определенные методом ОТ-ПЦР-РВ в клетках MDCK, выражали в log10 разведений исходного вирусного материала.
Далее проводили построение стандартных кривых и осуществляли оценку эффективности проведения реакции. В качестве стандартных образцов для ОТ-ПЦР-РВ-анализа тривалентной ЖГВ использовали РНК, выделенную из различных моновалентных вакцинных штаммов (A/H1N1, A/H3N2, В) с известными титрами ЭИД50, указанными в паспорте тривалентной вакцины. Штаммы тривалентной ЖГВ, используемые для одношагового анализа ОТ-ПЦР-РВ, включали вакцинные вирусы А/17/Калифорния/09/38 (H1N1), А/17/Швейцария/2013/1 (H3N2) и В/60/Пхукет/2013/26 (Ямагата), подготовленные на основе рекомендованных ВОЗ эпидемических вирусов для использования в эпидемическом сезоне 2015-2016 гг.
Инфекционные титры, указанные в паспорте тривалентной вакцины составили 7,1 lg ЭИД50/0,5 мл для A/H1N1, 8,4 lg ЭИД50/0,5 мл для A/H3N2 и 7,9 lg ЭИД50/0,5 мл для В. Выделение вирусной РНК производилось с
использованием набора реактивов QIAamp Viral RNA Mini Kit (QIAGEN) из серийных разведений РНК, выделенной из вируссодержащей аллантоисной жидкости с соответствующим указанному в паспорте на вакцину титром ЭИД50/0,5 мл. Полученные образцы РНК хранились при температуре -20°С и были готовы к дальнейшему использованию. ОТ-ПЦР-РВ выполняли в дупликатах с помощью наборов Superscript® III One-Step RT-PCR System (Thermo Fisher Scientific, Уолтем, США) согласно инструкции производителя.
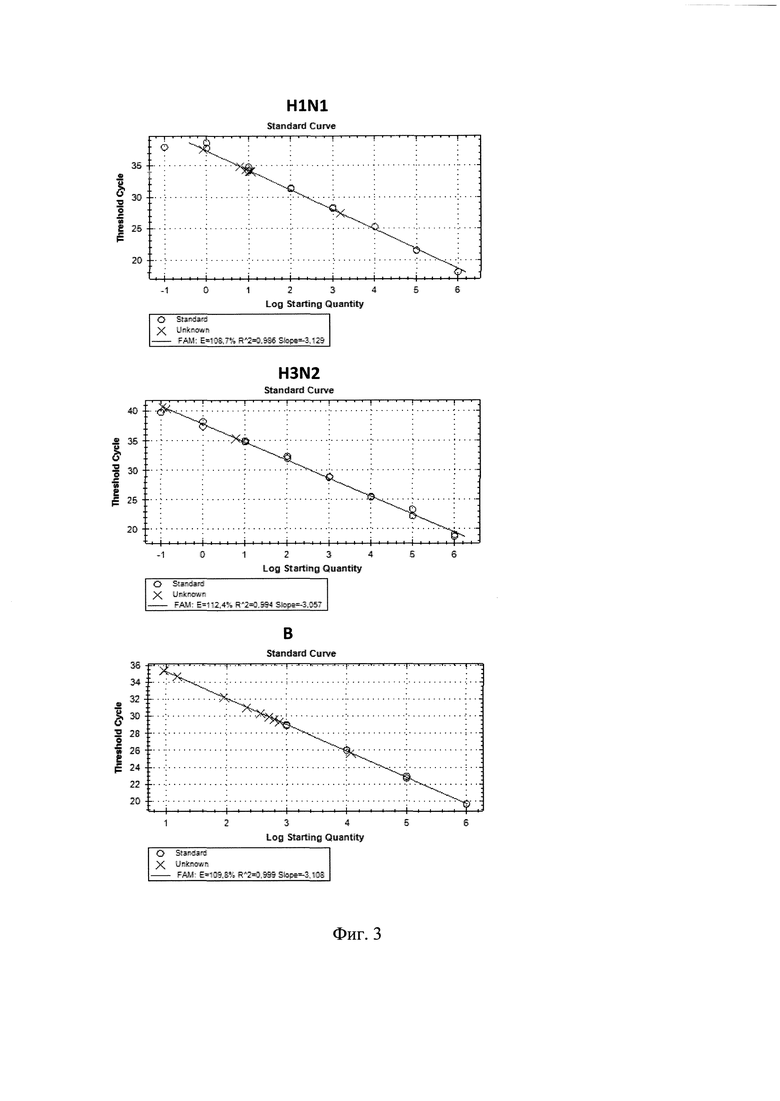
На фиг. 1 приведены калибровочные графики с результатами измерения количества копий в образцах РНК вакцинных вирусов гриппа А/17/Калифорния/09/38 (H1N1), А/17/Швейцария/2013/1 (H3N2) и В/60/Пхукет/2013/26 (Ямагата).
Показано, что все три флуоресцентных сигнала были положительными при выявлении к-ДНК вирусов A/H1N1, A/H3N2 и В. Эти результаты показали, что праймеры и зонды, используемые в этом тесте, были специфичными. Для точного определения эффективности реакции ПЦР выполняли не меньше пяти серийных 10-кратных разведений вирусной РНК. При построении стандартной кривой коэффициент корреляции (R2), построенный путем нанесения значений Ct на 10-кратные серийные разведения образца вирусной РНК, содержащие 102-108 копий, показал значения в пределах 0,95-0,99; эффективность (Е) прохождения реакций для каждой мишени составила от 92 до 105%, что можно увидеть на фиг.(1). Данные результаты подтвердили высокую специфичность и эффективность анализа.
Пример 1. Оценка репродуктивной активности тривалентной вакцины в клетках MDCK
Инфекционную активность в клетках MDCK определяли методом ОТ-ПЦР-РВ с РНК, выделенной из культуральных надосадков MDCK. Для этого в 96-луночные панели для клеточных культур с монослоем MDCK вносили падающие серийные разведения вакцинных вирусов гриппа А и В с
известным титром ЭИД50/0,5 мл (по 3 лунки на разведение) или серий тривалентной ЖГВ (Микроген). Надосадки отбирали через 24 часа после инкубации при 33°С в атмосфере С02. Выделение вирусной РНК производилось с использованием набора реактивов QIAamp Viral RNA Mini Kit (QIAGEN) по инструкции производителя.
Для определения титров вируса методом иммуноферментного анализа (ИФА) оставшийся клеточный монослой фиксировали 80% раствором ацетона в фосфатном буфере, затем обрабатывали мышиными моноклональными антителами, специфичными к гемагглютинину вирусов гриппа подтипов H1, Н3 и В, приведенными в Таблице 3.

Моноклональные антитела использовали в концентрации 0.1 мкг/мл. После окрашивания выполняли ИФА с козьими анти-IgG антителами, меченными пероксидазой хрена и ТМВ-субстратом, реакцию останавливали с помощью 1Н H2SO4. Экспрессию гемагглютинина вирусов на поверхности клеток определяли на планшетном спектрофотометре при длине волны 450 нм. Пороговым значением для определения наличия вируса было среднее значение оптической плотности (ОП) для трех негативных лунок, не содержащих вируса плюс три среднеквадратических отклонения (СКО). 50% культуральную инфекционную дозу (КИД50) рассчитывали по методу Рида и Менча [Reed L.J. Simple method of estimating fifty percent endpoints / L.J. Reed, H. A. Muench // American Journal of Hygiene. - 1938. - P. 493-497].
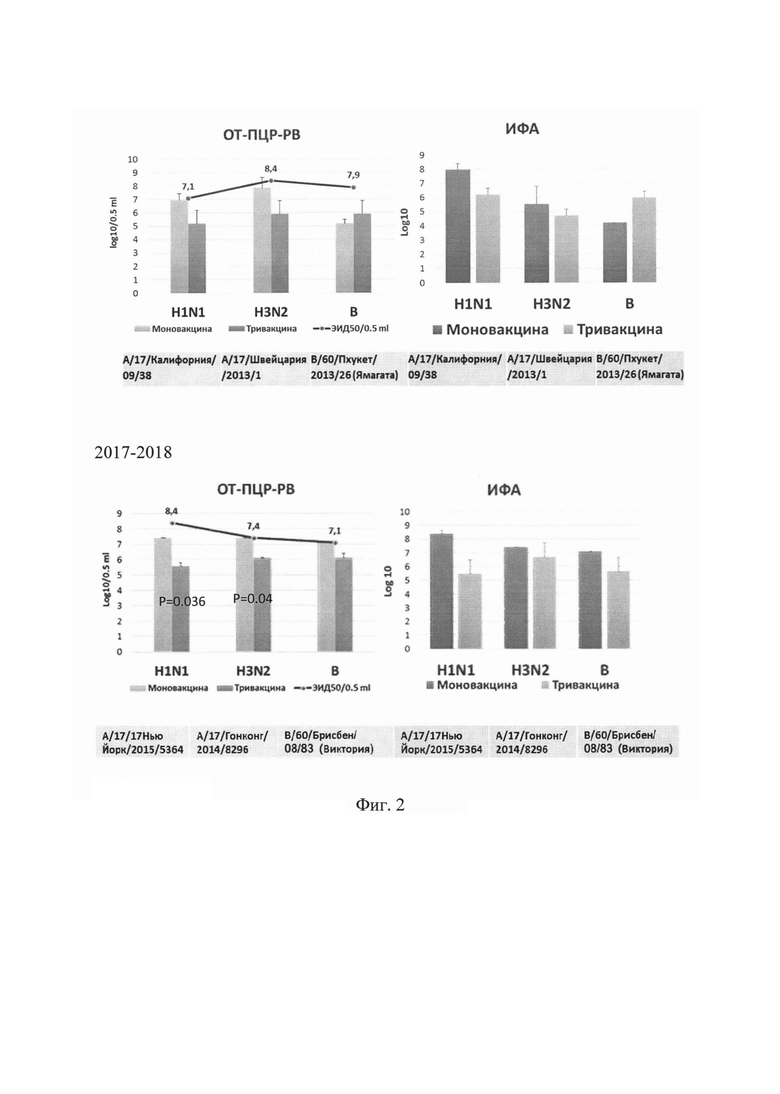
На фиг. 2 приведены результаты определения инфекционной активности вакцинных вирусов, входящих в состав ЖГВ, для эпидемических сезонов
2015-2016 и 2017-2018 гг. в культуре клеток MDCK методом ОТ-ПЦР-РВ и ИФА в сравнении с инфекционными титрами монопрепаратов (ЭИД50/0,5 мл), предоставленными НПО «Микроген». Представлены усредненные данные трех и более независимых экспериментов.
Показано снижение репродукции в культуре клеток MDCK штаммов вирусов A/H1N1 (А/17/Калифорния/09/38 и A/17/Нью Йорк/15/5364) в составе тривалентной вакцины по сравнению с соответствующими монопрепаратами в среднем в 100 раз, однако различия не были статистически значимыми.
Репродукция штаммов вирусов гриппа A/H3N2 (А/17/Швейцария/2013/1 и А/17/Гонконг/2014/8296) в составе тривалентной вакцины снижалась в 10-30 раз по сравнению с монопрепаратами.
Репродукция вакцинного штамма вируса гриппа В антигенной линии Ямагата (В/60/Пхукет/2013/26) для эпидемического сезона 2015-2016 гг. была даже несколько выше в сравнении с моновакциной, а репродукция вакцинного штамма вируса гриппа В антигенной линии Виктория (В/60/Брисбен/08/83) для 2017-2018 гг. в составе тривалентной вакцины понижалась незначительно.
Таким образом, показано, что наибольшему угнетению в составе тривалентной вакцины подвергался вирус А/17/Калифорния/09/38(/H1N1)pdm09. Эти данные могут объяснять сниженную иммуногенность компонента A/H1N1pdm, входящего в состав тривалентной вакцины, как показано в ряде зарубежных исследований американской ЖГВ компании "Medimmune" на основе донора аттенуации А/Энн Ap6op/6/60(H2N2), тем не менее, снижение иммуногенности не влияло на эффективность ЖГВ [Chen Z. Generation of live attenuated novel influenza virus A/California/7/09 (H1N1) vaccines with high yield in embryonated chicken eggs / Z. Chen, W. Wang, H. Zhou // J Virol. - 2010. - №84. - P. 44-51].
Частично снижение репродукции вирусов гриппа А может объясняться влиянием вирусов гриппа В, которые, как было показано ранее, способны до
некоторой степени подавлять репродукцию вирусов гриппа А в гетерогенных смесях [Daniel R. Functional Analysis of PA Binding by Influenza A Virus PB1: Effects on Polymerase Activity and Viral Infectivity / R. Daniel, Perez and Ruben O. Donis // Journal of Virology. - 2001. - №84. - P. 45-52].
Продемонстрированы различия в репродуктивной активности в клеточной культуре MDCK вирусов гриппа антигенных В/Виктория и В/Ямагата. Так, вирус В/60/Пхукет/2013/26 (линия Ямагата) репродуцировался в составе тривалентной вакцины не хуже, если не лучше, чем в виде моновакцины. В отношении вируса В/60/Брисбен/08/83 (Виктория) показано незначительное снижение репродуктивной активности в составе тривалентной вакцины по сравнению с монопрепаратом.
Пример 2. Оценка репродукции тривалентной ЖГВ на мышах
Мыши линии СВА были иммунизированы тривалентной ЖГВ состава 2017-2018 г., содержащей вакцинные вирусы A/17/Нью Йорк/15/5364 (H1N1), А/17/Гонконг/2014/8296(H3N2) и В/60/Брисбен/08/83(Виктория). На 3-й день после интраназальной иммунизации были получены образцы тканей легких и носовых ходов. Выделение вирусной РНК проводили, как указано выше после гомогенизации образцов в лизирующем буфере. ОТ-ПЦР-РВ проводили, как указано выше. В этом исследовании использовали праймеры и зонды, входящие в панель для типирования вируса гриппа методом ОТ-ПЦР-РВ (каталожный № FluRUO-01, FR-198) полученные из сотрудничающего центра ВОЗ по эпиднадзору, эпидемиологии и контролю гриппа (Центры по контролю и профилактике заболеваний, Атланта, Джорджия, США). Для построения стандартной кривой использовали серийные разведения вирусных РНК, выделенных из вакцинных вирусов с известным инфекционным титром ЭИД50/0,5 мл, установленным в куриных эмбрионах. Для построения стандартных кривых были выполнены 10-кратные разведения образцов (РНК, выделенная из вирусного материала с известным инфекционным титром ЭИД50), содержащих 102-106 копий.
На фиг. 3 приведен пример использования стандартной кривой для определения числа копий мРНК ( - стандарт,
- стандарт, 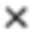 - образец).
- образец).
Кривая построена как зависимость логарифма исходного числа копий транскрипта стандартного вирусного материала относительно соответствующих им значений Ct (порогового цикла).
Если эффективность амплификации приближается к 100%, то удвоение матрицы происходит в каждом цикле амплификации. Поэтому относительную количественную оценку содержания вирусов в органах можно определить умножением концентрации стандарта на уровень изменения экспрессии гена гемагглютинина [Ермилова Е.В. и др. Количественный анализ экспрессии генов. - 2010] по формуле:
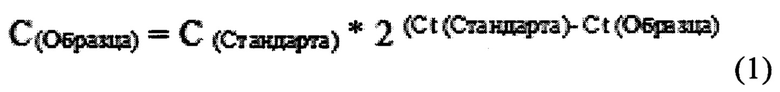
где С(Образца) - концентрация образца, С(Стандарта) - концентрация стандарта, Ct - пороговый цикл.
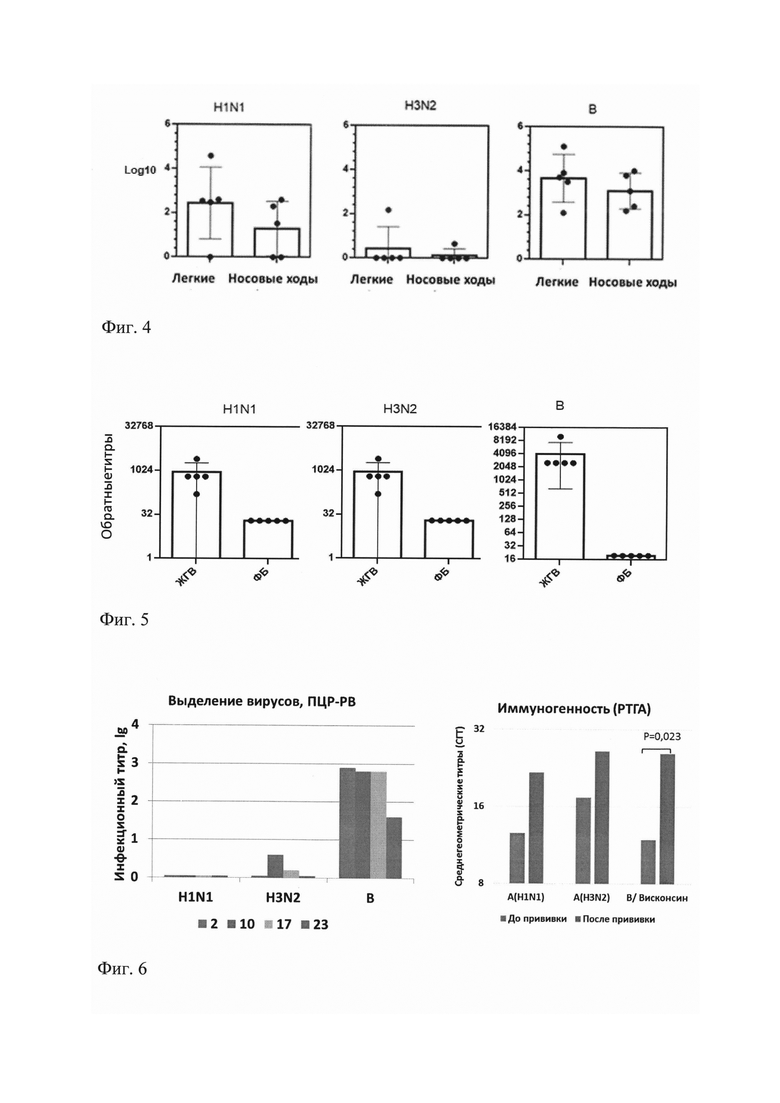
На фиг. 4 представлены титры вирусов гриппа A/17/Нью Йорк/15/5364 (H1N1), А/17/Гонконг/2014/8296 (H3N2), В/60/Брисбен/08/83 (Виктория) в дыхательных путях иммунизированных мышей через 48 часов после интраназальной иммунизации. Представлены индивидуальные титры вирусов, выраженные в logio, средние значения и среднеквадратические отклонения.
Показано, что вирус A/H3N2 практически не репродуцировался в легких и носовых ходах мышей, что соответствует данным о низкой инфекционности эпидемических вирусов вирусов A/H3N2 для мышей [Baz М. et al. Synergistic рА and НА mutations confer mouse adaptation of a contemporary A/H3N2 influenza virus //Scientific reports. - 2019. - T. 9. - №. 1. - C. 1-14].
Тем не менее, это обстоятельство не оказывало влияния на снижение иммуногенности вируса A/H3N2 по сравнению с другими компонентами вакцины.
На фиг. 5 показаны результаты выявления вирус-специфических IgG в сыворотках крови, полученных через 21 день после иммунизации мышей тривалентной ЖГВ. Представлены индивидуальные титры IgG, специфичных к вирусам А/17/Нью-Йорк/15/5364 (H1N1), А/17/Гонконг/2014/8296 (H3N2) и В/60/Брисбен/08/83 (Виктория), а также среднегеометрические титры и среднеквадратическое отклонение.
Показано, что титры вирус-специфических IgG к вирусу A/H3N2 были не ниже, чем к вирусу A/H1N1, который репродуцировался в дыхательных путях мышей намного лучше вируса A/H3N2. Таким образом, низкий уровень репродукции вируса в дыхательных путях мышей не был препятствием для развития иммунного ответа на вакцинацию. Наиболее высокий уровень репродукции вируса гриппа В в легких и носовых ходах мышей соответствовал высоким титрам сывороточных антител к этому вирусу.
Таким образов, метод ОТ-ПЦР-РВ позволяет определять титры вакцинных вирусов в составе тривалентной вакцины при иммунизации лабораторных животных.
Пример 3. Оценка репродукции вирусов А и В в носовых ходах привитых пациентов после иммунизации тривалентной ЖГВ
Была изучена репродукция вакцинных вирусов при введении лицам, привитым сезонной тривалентной ЖГВ. Для этого использовали архивные материалы - носовые смывы, полученные в октябре 2012 г. при исследовании прививочных свойств коммерческой тривалентной ЖГВ. В исследовании приняли участие здоровые лица мужского и женского пола в возрасте от 18 до 56 лет с неизвестным анамнезом в отношении заболевания гриппом и противогриппозной вакцинации. На каждого привитого было получено информированное согласие. У привитых в поствакцинальный период (3-и сутки после вакцинации) проводили взятие клинических материалов (мазки из полости носа) для последующих вирусологических исследований. Носовые смывы хранили при -70°С. Выделение РНК и постановку ОТ-ПЦР-
РВ проводили как указано выше. В качестве стандарта использовали РНК, выделенную из соответствующего вакцинного вируса с известным титром ЭИД50/0,5 мл. Иммуногенность оценивали в реакции торможения гемагглютинации (РТГА) с сыворотками крови, полученными на 21 день после иммунизации. РТГА проводили с тестовыми антигенами, входившими в состав вакцины (см. табл. 1).
На фиг. 6 представлена относительная количественная оценка выделения вируса у лиц, привитых тривалентной вакциной состава 2012-2013 г. на 3-й день после иммунизации (изоляты №2, 10, 17, 23) и результаты изучения иммуногенности.
В состав вакцины входили вакцинные штаммы А/Калифорния/7/2009(H1N1)pdm09, А/Виктория/361/2011(H3N2) и В/Висконсин/1/2010 (Ямагата) с титрами ЭИД50/0,5 мл 9,1, 9,7 и 8,8 соответственно. Вирус A/H1N1 не был выделен ни у одного из привитых, вирус A/H3N2 - у двух, а вирус В - у всех обследованных. Выделение вируса у привитых наблюдалось в намного меньших титрах, по сравнению с титрами вируса, указанными в паспорте на тривакцину.
Дополнительно было проанализировано формирование сывороточных антител к вакцинным вирусам после иммунизации. Оказалось, что ни у одного из обследованных не отмечено прироста поствакцинальных антител к вирусу A/H1N1, у трех из четырех (75%) отмечено 2-4-кратное повышение антител к вирусу А/H3N2, у двух из четырех (50%) наблюдался двукратный прирост антител к вирусу В (Р≤0,05). Эти результаты соответствовали результатам выделения вирусов по данным ОТ-ПЦР-РВ.
Пример 4. Оценка репродукции вирусов В/Виктория и В/Ямагата в клеточной культуре МДСК
Сочетанное заражение вирусами гриппа В/60/Флорида/04/18 (Ямагата), В/60/Малайзия/04/898 (Виктория), В/60/Висконсин/2010/125 (Ямагата) и В/60/Брисбен/08/83 (Виктория) проводили на сформировавшемся монослое MDCK в 24-х луночных планшетах для клеточных культур. Вирусы вносили
с множественностью инфекции (МОИ) 0,1-0,01/1 мл. Надосадки отбирали через 24 часа после заражения. РНК выделяли из надосадочной жидкости. Для построения стандартных кривых были сделаны 10-кратные разведения образцов (РНК, выделенная из вирусного материала с известным инфекционным титром ЭИД50), содержащих 102-106 копий.
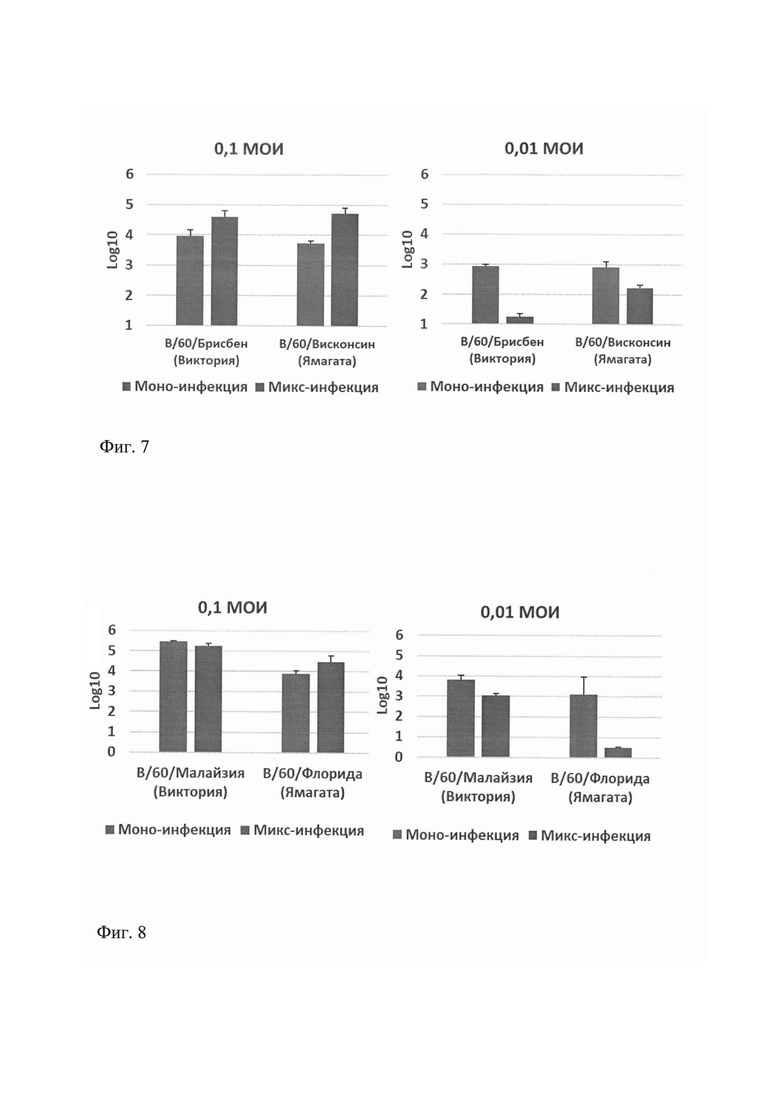
Показано, что при МОИ 0,1 вирусы гриппа В линий В/Виктория и В/Ямагата репродуцировались в составе смешанного препарата не хуже, если не лучше, чем в виде монопрепарата. При снижении заражающей дозы вирусов до 0,01 МОИ репродукция как В/Виктория, так и В/Ямагата в составе смеси снижалась. Таким образом, при создании квадривалентной ЖГВ объединение двух вакцинных вирусов гриппа В с титрами не ниже, чем указанные в стандарте на фармпрепарат, не приводит к снижению репродуктивной активности (фиг. 7 и фиг. 8).
Заключение
Метод ОТ-ПЦР-РВ с использованием гидролизируемых олигонуклеотидных флуоресцентных зондов (Taqman) является чувствительным специфичным для изучения репродукции вакцинных вирусов в культуре клеток MDCK в составе тривалентной или квадривалентной вакцины. Применение данного метода дает возможность изучения взаимного влияния компонентов тривалентной вакцины даже при сниженном уровне репродукции вакцинных вирусов в клетках млекопитающих по сравнению с репродукцией в КЭ.
Метод позволяет проводить оценку репродукции вакцинных вирусов гриппа А и В в составе тривалентной вакцины в пробах, полученных из дыхательных путей лабораторных животных и пациентов после иммунизации тривалентной ЖГВ.
Метод позволяет изучать взаимное влияние вирусов гриппа В антигенных линий Виктория и Ямагата для включения в состав квадривалентной вакцины.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> ФГБНУ "ИЭМ"
<120> Способ оценки репродуктивной активности вирусов гриппа в составе тривалентной и квадривалентной живой гриппозной вакцины
<140>
<141>
<150>
<151>
<160> 25
<170>
<210> 1
<211> 22
<212> Нуклеотидная последовательность
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидная последовательность гемагглютинина вируса гриппа
A/17/Калифорния/2009/38(H1N1)pdm, прямой -677
<400> 1
agttcaagcc ggaaatagca at 22
<210> 2
<211> 23
<212> Нуклеотидная последовательность
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидная последовательность гемагглютинина вируса гриппа
A/17/Калифорния/2009/38(H1N1) pdm, обратный-845
<400> 2
ataccagatc cagcatttct ttc 23
<210> 3
<211> 23
<212> Нуклеотидная последовательность
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидная последовательность гемагглютинина вируса гриппа A/17/Калифорния/2009/38(H1N1) pdm, проба, прямой 704
<400> 3
caaagtgagg gatcaagaag ttc 23
<210> 4
<211> 18
<212> Нуклеотидная последовательность
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидная последовательность гемагглютинина вируса гриппа
А/17/Нью Йорк/15/5364 (H1N1) pdm09, прямой - 853
<400> 4
gctggatctg gtattatc 18
<210> 5
<211> 28
<212> Нуклеотидная последовательность
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидная последовательность гемагглютинина вируса гриппа
А/17/Нью Йорк/15/5364 (H1N1) pdm09, обратный - 963
<400> 5
ttcagaatat acatccgatc acaattgg 28
<210> 6
<211> 23
<212> Нуклеотидная последовательность
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидная последовательность гемагглютинина специфичная к вирусу гриппа А/17/Нью Йорк/15/5364 (H1N1) pdm09 Проба 874
<400> 6
cagatacacc agtccacgat tgc 23
<210> 7
<211> 18
<212> Нуклеотидная последовательность
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидная последовательность гемагглютинина специфичная к вирусу гриппа A/17/Швейцария/2013/1 (H3N2) прямой 65
<400> 7
tctggttttc gctcaaaa 18
<210> 8
<211> 18
<212> Нуклеотидная последовательность
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидная последовательность гемагглютинина специфичная к вирусу гриппа A/17/Швейцария/2013/1 (H3N2) обратный 177
<400> 8
ttcggtcatt cgtgattg 18
<210> 9
<211> 20
<212> Нуклеотидная последовательность
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидная последовательность гемагглютинина специфичная к вирусу гриппа A/17/Швейцария/2013/1 (H3N2) Проба 95
<400> 9
tgacaatagc acggcaacgc 20
<210> 10
<211> 20
<212> Нуклеотидная последовательность
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидная последовательность гемагглютинина вируса гриппа A/17/Сингапур/2016/ 3571 (H3N2), прямой - 203
<400> 10
ggttcagaat tcctcaatag 20
<210> 11
<211> 18
<212> Нуклеотидная последовательность
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидная последовательность гемагглютинина вируса гриппа A/17/Сингапур/2016/ 3571 (H3N2), обратный - 385
<400> 11
ggcacatcat aagggtaa 18
<210> 12
<211> 22
<212> Нуклеотидная последовательность
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидная последовательность гемагглютинина вируса гриппа A/17/Сингапур/2016/ 3571 (H3N2), проба, прямой - 231
<400> 12
tgcgacagtc ctcatcagat cc 22
<210> 13
<211> 20
<212> Нуклеотидная последовательность
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидная последовательность белка нуклепротеина (NP) вируса гриппа В/СССР/60/69, прямой - 233
<400> 13
aagctgatgt cggaaggaga 20
<210> 14
<211> 20
<212> Нуклеотидная последовательность
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидная последовательность белка нуклепротеина (NP) вируса гриппа В/СССР/60/69, обратный - 402
<400> 14
ttccacagca tgtgcatttt 20
<210> 15
<211> 20
<212> Нуклеотидная последовательность
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидная последовательность белка нуклепротеина (NP) вируса гриппа В/СССР/60/69, проба, прямой - 253
<400> 15
acccaaaaga aacaaacccc 20
<210> 16
<211> 19
<212> Нуклеотидная последовательность
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидная последовательность гемагглютинина вируса гриппа В/60/Брисбен/08/83 (Виктория), прямой - 741
<400> 16
cacacattac gtttcacag 19
<210> 17
<211> 19
<212> Нуклеотидная последовательность
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидная последовательность гемагглютинина вируса гриппа В/60/Брисбен/08/83 (Виктория), обратный-835
<400> 17
gcaccatgta atcaacaac 19
<210> 18
<211> 23
<212> Нуклеотидная последовательность
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидная последовательность гемагглютинина вируса гриппа В/60/Брисбен/08/83 (Виктория), проба, прямой – 780
<400> 18
aacagaagac ggaggactac cac 23
<210> 19
<211> 25
<212> Нуклеотидная последовательность
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидная последовательность гемагглютинина вируса гриппа В/60/Висконсин/2010/125 (Ямагата), прямой - 705
<400> 19
ccacacatta tgtttctcag attgg 25
<210> 20
<211> 21
<212> Нуклеотидная последовательность
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидная последовательность гемагглютинина вируса гриппа В/60/Висконсин/2010/125 (Ямагата), обратный- 864
<400> 20
tgaggcaaca aaacacctct t 21
<210> 21
<211> 20
<212> Нуклеотидная последовательность
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидная последовательность гемагглютинина вируса гриппа В/60/Висконсин/2010/125 (Ямагата), проба, прямой - 794
<400> 21
gatgcaaaaa cctgggaaaa 20
<210> 22
<211> 24
<212> Нуклеотидная последовательность
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидная последовательность гемагглютинина вирусов гриппа В Виктория и Ямагата, прямой- 432
<400> 22
accctacara mttggaacyt cagg 24
<210> 23
<211> 19
<212> Нуклеотидная последовательность
<220>
<223> Олигонуклеотидная последовательность гемагглютинина вирусов гриппа В Виктория и Ямагата, обратный - 479
<400> 23
acagcccaag ccattgttg 19
<210> 24
<211> 24
<212> Нуклеотидная последовательность
<213> Искусственная последовательность
<220>
<223> <213> Искусственная последовательность
<400> 24
aaatcccgtt tccattggta atgt 24
<210> 25
<211> 19
<212> Нуклеотидная последовательность
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидная последовательность гемагглютинина вируса гриппа B/Ямагата, прямой- 439
<400> 25
aatccgmtyt tactggtag 19
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ШТАММ ВИРУСА ГРИППА ДЛЯ ПРОИЗВОДСТВА ЖИВОЙ ГРИППОЗНОЙ ИНТРАНАЗАЛЬНОЙ ВАКЦИНЫ ДЛЯ ВЗРОСЛЫХ И ДЕТЕЙ | 2011 |
|
RU2464311C1 |
| РЕАССОРТАНТНЫЙ ШТАММ ВИРУСА ГРИППА RN 1/09-SWINE A(H7N1) ДЛЯ ОПРЕДЕЛЕНИЯ АНТИТЕЛ К НЕЙРАМИНИДАЗЕ ПРИ ГРИППОЗНОЙ ИНФЕКЦИИ И ВАКЦИНАЦИИ | 2010 |
|
RU2428476C1 |
| ШТАММ ВИРУСА ГРИППА А/Гонконг/1/68/162/35 (H3N2)-УНИВЕРСАЛЬНЫЙ ДОНОР ВНУТРЕННИХ ГЕНОВ ДЛЯ РЕАССОРТАНТОВ И РЕАССОРТАНТНЫЕ ШТАММЫ А/СПБ/ГК/09 (H1N1) И А/НК/Astana/6:2/2010 (H5N1), ПОЛУЧЕННЫЕ НА ЕГО ОСНОВЕ | 2011 |
|
RU2511431C2 |
| ШТАММ ВИРУСА ГРИППА ДЛЯ ПРОИЗВОДСТВА ЖИВОЙ И ИНАКТИВИРОВАННОЙ ГРИППОЗНОЙ ВАКЦИНЫ | 2011 |
|
RU2464309C1 |
| ШТАММ ВИРУСА ГРИППА ГКВ 2389 ДЛЯ ПОЛУЧЕНИЯ ЖИВОЙ ИНТРАНАЗАЛЬНОЙ И ИНАКТИВИРОВАННОЙ ГРИППОЗНОЙ ВАКЦИНЫ | 2006 |
|
RU2318871C1 |
| Рекомбинантный вакцинный штамм для живой интраназальной вакцины, обеспечивающей сочетанную профилактику гриппозной и коронавирусной инфекций | 2022 |
|
RU2782531C1 |
| ШТАММ ВИРУСА ГРИППА А/17/ВИСКОНСИН/05/84(Н3N2) ДЛЯ ПРОИЗВОДСТВА ЖИВОЙ ГРИППОЗНОЙ ИНТРАНАЗАЛЬНОЙ ВАКЦИНЫ ДЛЯ ВЗРОСЛЫХ И ДЕТЕЙ | 2007 |
|
RU2340671C1 |
| РЕАССОРТАНТНЫЙ ШТАММ ВИРУСА ГРИППА А/17/НОВАЯ КАЛЕДОНИЯ/99/76(H1N1) ДЛЯ ПРОИЗВОДСТВА ЖИВОЙ ГРИППОЗНОЙ ИНТРАНАЗАЛЬНОЙ ВАКЦИНЫ | 2011 |
|
RU2464310C1 |
| РЕАССОРТАНТНЫЙ ШТАММ ВИРУСА ГРИППА RN9/13-HUMAN A(H6N9) ДЛЯ ОПРЕДЕЛЕНИЯ АНТИТЕЛ К НЕЙРАМИНИДАЗЕ ПРИ ГРИППОЗНОЙ ИНФЕКЦИИ И ВАКЦИНАЦИИ | 2014 |
|
RU2587629C1 |
| ШТАММ ВИРУСА ГРИППА А/17/ВИКТОРИЯ/2011/89 (H3N2) ДЛЯ ПРОИЗВОДСТВА ЖИВОЙ ГРИППОЗНОЙ ИНТРАНАЗАЛЬНОЙ ВАКЦИНЫ ДЛЯ ВЗРОСЛЫХ И ДЛЯ ДЕТЕЙ | 2012 |
|
RU2532844C2 |

Изобретение относится к области биотехнологии. Разработан способ оценки репродуктивной активности вирусов гриппа в составе тривалентной и квадривалентной живой гриппозной вакцины, основанный на одношаговой ОТ-ПЦР-РВ с использованием гидролизируемых олигонуклеотидных флуоресцентных зондов. ОТ-ПЦР-РВ с использованием разработанных специфичных праймеров и олигонуклеотидных зондов является чувствительной и специфичной для изучения репродукции вакцинных вирусов в культуре клеток MDCK в составе тривалентной или квадривалентной вакцины. Применение ОТ-ПЦР-РВ дает возможность изучения взаимного влияния компонентов поливалентной ЖГВ даже при сниженном уровне репродукции вакцинных вирусов в клетках млекопитающих по сравнению с репродукцией в КЭ. Разработанный способ позволяет проводить оценку репродукции вакцинных вирусов гриппа А и В в составе тривалентной вакцины в пробах, полученных из дыхательных путей лабораторных животных и пациентов после иммунизации тривалентной ЖГВ. Способ позволяет оценивать взаимное влияние вирусов гриппа В антигенных линий Виктория и Ямагата для включения их в состав поливалентных вакцин. 3 табл., 8 ил.
Способ оценки репродуктивной активности вирусов гриппа А и В, входящих в состав поливалентных живых гриппозных вакцин, заключающийся в том, что предварительно проводят сокультивирование вакцинных штаммов в клеточной культуре либо осуществляют иммунизацию поливалентной живой гриппозной вакциной in vivo, после чего проводят одношаговую обратно-транскриптазную полимеразную цепную реакцию в реальном времени с использованием праймеров и зондов, характеризующихся нуклеотидными последовательностями SEQ ID No: 1 - SEQ ID No: 25, гомологичными к вирусам гриппа А, В/Виктория и В/Ямагата, при подобранных оптимальных условиях проведения реакции с разработанными стандартами для относительного количественного анализа методом разницы пороговых циклов.
| Kohler G., et al., Derivation of specific antibodyproducing tissue culture and tumor lines by cell fusion, Eur J Immunol 1976; 6, 511-9 | |||
| Romanova J.R., et al., Interference between cold-adapted (ca) influenza A and В vaccine reassortants or between ca reassortants and wild-t strains in eggs and mice, Vaccine | |||
| Прибор для охлаждения жидкостей в зимнее время | 1921 |
|
SU1994A1 |
| - T | |||
| Способ гальванического снятия позолоты с серебряных изделий без заметного изменения их формы | 1923 |
|
SU12A1 |
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| - C | |||
| Прибор для равномерного смешения зерна и одновременного отбирания нескольких одинаковых по объему проб | 1921 |
|
SU23A1 |

