УРОВЕНЬ ТЕХНИКИ
Витамины представляют собой важные микронутриенты, необходимые для поддержания роста и размножения клеток. Клетки млекопитающих не могут их синтезировать, и поэтому млекопитающие должны получать витамины из своего рациона. Напротив, бактерии, грибы и растения синтезируют витамины. Основная функция витаминов заключается в том, чтобы действовать как кофакторы или коферменты в различных ферментативных реакциях, таких как цикл Кребса, гликолиз, синтез аминокислот и биосинтез ацетил-КоА.
Недостаточность витаминов напрямую связана с многочисленными заболеваниями. Например, острая недостаточность витамина В1 у людей приводит к болезни, называемой бери-бери, что, в свою очередь, может привести к неврологическим и сердечно-сосудистым нарушениям со смертельным исходом. Более того, у мышей, у которых отсутствуют гены, участвующие в процессах поглощения витаминов, проявляются тяжелые симптомы. Например, нокаут митохондриального транспортера витамина B1 Slc25a19 вызывает гибель эмбриона, пороки развития ЦНС и анемию (Lindhurst и др., 2006). У мышей, у которых отсутствует транспортер витаминов Н и В5 (пантотенат), наблюдают замедление роста, пониженную плотность костной ткани, уменьшенную длину кости и летальность через 10 недель (Ghosal и др., 2012). Недостаточность цитоплазматической или митохондриальной активности, которая может быть связана с метаболизмом витаминов, может также изменять функции клеток или организма. Например, нокаут генов пантотенатной киназы мыши (PANK) приводит к нарушению митохондриального и клеточного дыхания, а также к недостаточности коэнзима A (Brunetti и др., 2012; Garcia и др., 2012).
Клетки яичника китайского хомячка (СНО) широко применяют в промышленных процессах для получения рекомбинантных терапевтических белков. Жизнеспособность клеток СНО и других эукариотических клеток, применяемых в промышленных процессах (NS0, клетки почки детеныша хомяка (BHK) и клетки почки эмбриона человека 293 (HEK-293)), зависит от поглощения витаминов. Аналогичным образом, жизнеспособность первичных клеток, таких как клетки человека, применяемые для генной терапии или терапии на основе клеток и для регенеративной медицины, также зависит от поглощения витаминов.
Оптимизацию сред для культивирования для клеток и клеточных линий часто выполняют для получения более высокого выхода рекомбинантных белков. Недавние исследования, определяющие изменения в центральном метаболизме, которые сопровождают рост и образование моноклональных антител, выявили регуляторную связь между клеточным метаболизмом, метаболитами среды и ростом клеток (Dean и др., 2013). Например, исследование было направлено на установление контроля за циклом деления клеток посредством снижения определенных питательных веществ или посредством непосредственного контроля за регуляторами клеточного цикла, поскольку чрезмерный рост и деление клеток отрицательно влияют на выработку белка (см. Du и др., 2014 и ссылки в данном документе). Однако эти вмешательства часто сопровождаются нежелательным воздействием на качество и/или на посттрансляционный процессинг указанного рекомбинантного белка (Nam и др., 2008; Sajan и др., 2010; Sampathkumar и др., 2006; Trummer и др., 2006).
Другие усилия по улучшению отбора трансформированных клеточных линий были сосредоточены на разработке новых молекулярных маркеров, которые не требуют какой-либо устойчивости к токсическому соединению-антибиотику. Например, повышенную экспрессию компонентов путей биосинтеза нуклеотидов или аминокислот, таких как дегидрофолатредуктаза или глутаминсинтаза, применяли для метаболического отбора экспрессирующих белок рекомбинантных клеток путем включения их кодирующих последовательностей в векторы экспрессии (Cacciatore и др., 2010, Birch и Racher 2006, WO 2009/080759, публикация патента СШКА 20100330572, которая полностью включена в настоящее описание посредством ссылки, как и все ссылки, приведенные в настоящем документе). Например, указанную кодирующую последовательность транспортера фолата применяли для отбора клеток с повышенной экспрессией трансгена (Rothem и др., 2005). Хотя данный подход позволял получить повышенную экспрессию представляющих фармакологический интерес белков, несколько исследований сообщали о нестабильных уровнях экспрессии, например, когда его применяли для амплификации количества копий трансгена (Schlatter и др., 2005; Chusainow и др., 2009).
Клеточный метаболизм и рост клеток млекопитающих могут напрямую зависеть от доступности витаминов. Таким образом, в области техники существует необходимость в модуляции метаболизма и/или роста культивируемых клеток путем установления контроля за поглощением витаминов, экспрессией генов метаболизма витаминов, и/или концентрацией конкретных витаминов в среде для культивирования в целом с целью улучшения экспрессии терапевтических белков. Также существует необходимость в альтернативных способах отбора клеток. Настоящее изобретение направлено на решение одной или нескольких из этих задач, а также других задач в данной области техники.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
На Фиг. 1 представлен обзор переноса витаминов в клетки и органеллы млекопитающих.
На Фиг. 2 А-В представлен рост СНО-М в обедненной по витаминам среде по сравнению с необедненной средой. Клетки в количестве до 50000 клеток/мл высевали в 500 мкл среды для культивирования B-CDmin, дополненной или недополненной витаминами В1, В5 или Н (см. Таблицу 2), или в полную среду (SFM). Клетки культивировали в 24-луночных планшетах в течение указанного времени без встряхивания. Плотность (А) и жизнеспособность клеток (анализ исключения трипанового синего, панель (В)) измеряли после 3, 6 и 10 дней культивирования.
На Фиг. 3 представлен рост СНО-М в обедненных по витаминам средах. Клетки (5000 клеток/мл) переносили в 150 мкл среды с различными концентрациями витамина В1, В5 или Н (от 0 до 1X, см. Таблицу 2). Применяли 96-луночные планшеты и измеряли рост после 9 дней культивирования посредством измерения оптической плотности (OD) при 600 нм. Заштрихованные (с правым наклоном) пунктирные темные и заштрихованные (с левым наклоном) столбцы указывают на модуляцию концентраций B1, В5 и Н соответственно.
На Фиг. 4 представлено влияние обеднения по витамину В5 на рост и жизнеспособность клона клеток СНО-М, секретирующего антитела, в культуре с периодической подпиткой. Секретирующий трастузумаб клон клеток СНО-М выращивали на полной среде (черные квадраты) или на среде, обедненной по витаминам (серые треугольники, 5000:1 об.: об., смесь B-CDmin и полной CD-среды), обе дополненные 6 мМ глутамина. Дозы той же среды для культивирования добавляли на 3, 6, 7, 8, 9, 10 и 13 день. Культуры анализировали на плотность жизнеспособных клеток (VCD, сплошные линии) и жизнеспособность клеток (% жизнеспособных клеток, пунктирные линии).
На Фиг. 5 представлено влияние обеднения по витамину В5 на титр иммуноглобулина клона клеток СНО-М в культурах с периодической подпиткой. Секретирующий трастузумаб клон клеток СНО-М, выращенный на полной среде (черные квадраты) или на среде, обедненной по витамину В5, см. Фиг. 4, анализировали на титр антитела, секретируемого в среде для культивирования, при помощи анализа ИФА с применением метода «двойного сэндвича».
На Фиг. 6 представлены уровни мРНК SLC5A6 в стабильных линиях СНО-М. Поликлональные популяции, трансфицированные указанным количеством вектора экспрессии Slc5a6, были отобраны по их устойчивости к пуромицину, а уровни мРНК Slc5a6 определяли при помощи ОТ-кПЦР. Накопление транскрипта SLC5A6 нормировали по уровню накопления мРНК GAPDH, и оно представлено относительно эндогенного уровня мРНК SLC5A6 нетрансформированных клеток, применяемых в качестве контроля, который был установлен на 1. 0 нг указывает на клетки, трансфицированные исключительно при помощи GFP и векторов экспрессии с устойчивостью к пуромицину, тогда как С обозначает нетрансформированные контрольные клетки.
На Фиг. 7 представлено воздействие SLC5A6 на рост СНО-М в ограниченных по витаминам условиях. Клетки в количестве 20000 клеток/мл высевали в 500 мкл среды В-CDmin, дополненной указанными количествами витамина В5 и Н и B1 (1X). Применяли 24-луночные планшеты и измеряли рост после 6 дней культивирования посредством измерения плотности жизнеспособных клеток. Звездочки обозначают достоверную разницу (р<0,05) между трансфицированными и нетрансфицированными клетками при одних и тех же условиях роста и среде для культивирования.
На Фиг. 8 А-В представлены результаты отбора трансфицированных клеток СНО-М с применением обедненной по В5 среды и транспортера SLC5A6. Трансфекцию осуществляли с применением увеличивающегося количества вектора Slc5a6 (0, 50, 250 и 1000 нг) совместно с GFP и пуромициновыми плазмидами. После отбора стабильной поликлональной популяции в обедненной по В5 среде (10-3Х), все положительно экспрессирующие GFP клетки (серые столбцы: все клетки GFP +) и клетки-экспрессоры с высоким уровнем экспрессии GFP (черные столбцы, клетки-экспрессоры с высокой экспрессией GFP) количественно оценивали с применением способа сортировки клеток с возбуждением флуоресценции (FACS) (А). Среднее значение флуоресценции для одних и тех же клеток также количественно оценивали при помощи FACS (В). С обозначает ^трансформированные клетки СНО-М, применяемые в качестве контроля. Количество клеток, выживших в процессе отбора, было слишком низким для количественной оценки при трансфекции ДНК-носителя (0 нг) или 50 нг указанного вектора экспрессии Slc5a6.
На Фиг. 9 А-С представлены графики FACS, на которых представлено обогащение всех GFP +- клеток и клеток с высокой экспрессией GFP из стабильных популяций поликлональных клеток, совместно трансфицированных с применением SLC5A6, GFP и плазмид экспрессии с устойчивостью к пуромицину (puro). Осуществляли первичный отбор трансфицированных клеток при помощи пуромицина с последующим вторичным отбором путем культивирования в средах, содержащих либо избыточную (В5 10Х/Н 10-4Х), либо лимитирующую (В5 10-3Х/Н 10-4х) концентрацию витамина В5, или их культивировали с применением неселективной среды для культивирования (В5 1Х/Н 10-4Х) в качестве контроля. (А): FACS-профили GFP-флуоресценции трансфицированных СНО-М после культивирования указанных клеток в течение 7 дней в средах, содержащих различную концентрацию витамина В5 (10-3Х, 1X или 10Х), как указано. Окно 1 представляет собой все GFP-экспрессирующие клетки, в то время как Окно 2 ограничено клетками, экспрессирующими GFP на самом высоком уровне. Обогащение GFP +- флуоресцентных клеток (В) и геометрическое среднее для GFP-флуоресценции указанных клеток (С) представлены для пулов поликлональных клеток, совместно трансфицированных с применением различных количеств векторов экспрессии Slc5a6 (например, 0 нг, 100 нг и 250 нг, как указано), и с применением GFP и векторов с устойчивостью к пуромицину.
На Фиг. 10 представлен рабочий процесс экспериментального отбора клеток. Клетки СНО-М совместно трансфицировали с применением указанного SLC5A6 и плазмиды легкой цепи IgG и конструкции с устойчивостью к пуромицину и тяжелой цепи IgG, после чего указанную культуру расщепляли и отбирали либо в присутствии пуромицина (условие А), либо в среде для культивирования, обедненной по витаминам (минимальная среда, условие В), или применяли двойной отбор (AD и BD). За этим следовали анализы секреции иммуноглобулина полученных пулов поликлональных клеток. После отбора часть указанных клеток переносили в неселективную среду для культивирования для пассажа в течение 10-недельного исследования стабильности экспрессии (А+, В+) или в биореактор с периодической загрузкой (AD+ и BD+).
На Фиг. 11 представлены результаты секреции иммуноглобулина популяциями клеток, отобранных с применением пуромицина или обеднения по витаминам, или с применением обоих вариантов. Общее количество пулов поликлональных клеток отбирали путем обеспечения роста в полной среде, содержащей 5 мкг/мл пуромицина (А+), в минимальной среде (В1+) или путем двойного отбора в минимальной среде и в присутствии 5 мкг/мл пуромицина (B1D+ и B2D+), как представлено на Фиг. 10. Анализировали две независимые популяции клеток для режима отбора В, называемого В1 и В2, что приводит к получению дважды отобранных популяции B1D+ и B2D+ соответственно. Две независимые популяции анализировали для режима отбора BD+. Отобранные пулы выращивали на полной среде и подпитку добавляли на 3 день и на день от 6 до 10. Образцы анализировали на титр секретируемого антитела при помощи анализа ИФА с применением метода «двойного сэндвича».
На Фиг. 12 представлены результаты секреции иммуноглобулина популяцией клеток, отобранных с применением пуромицина или обеднения по витаминам. (А) Клетки анализировали на 39 день культивирования после режима отбора, представленного на Фиг. 10. Клеточную секрецию обнаруживали при помощи комплекса флуоресцентных антител, который связывает секретируемое терапевтическое антитело, которое проявляется на клеточной поверхности и секретируется. Черные столбцы и левая шкала указывают на процент клеток, которые секретируют указанное антитело в поликлональных популяциях. Средняя интенсивность флуоресценции клеточной поверхности, указывающая на уровень секреции отдельных клеток, представлена в виде белых столбцов произвольных единицах измерения (AU) на правой шкале. Анализировали две независимые популяции для режима отбора В+(В1+, В2+). (В). Популяции клеток панели А анализировали на определение уровней мРНК тяжелой цепи (Hc) и легкой цепи (Lc) IgG или транспортера витамина В5 (SLC5A6).
На Фиг. 13 представлена стабильность секреции иммуноглобулина популяций, отобранных с применением пуромицина или обеднения по витаминам. Пулы поликлональных клеток из Фиг. 8, отобранные с применением пуромицина (А+) или обеднения по витамину В5 (В+), поддерживали в полной среде и осуществляли их пассаж два раза в неделю для проведения исследований на стабильность экспрессии. Удельную производительность указанных популяций клеток, выраженную в пикограммах секретируемого антитела на клетку и в день (pcd), измеряли после каждого пассажа в течение 10 недель.
На Фиг. 14 представлены результаты исследования образования иммуноглобулина в культурах популяций клеток с периодической подпиткой, отобранных с применением пуромицина или обеднения по витаминам. Отобранные пулы выращивали на полной среде в культурах с периодической подпиткой, и подпитку добавляли на 3 день и на день от 6 до 10. Образцы анализировали на определение плотности жизнеспособных клеток (VCD) и на жизнеспособность (% жизнеспособных клеток, пунктирные линии) (А), и на определение титра секретируемого антитела при помощи анализа ИФА с применением метода «двойного сэндвича». (В)
На Фиг. 15 представлены кодирующие последовательности (CDS) различных СНО-М-генов витаминов.
На Фиг. 16 представлены аминокислотные последовательности указанных на Фиг. 15 СНО-М-генов витаминов.
На Фиг. 17 представлен натрий-зависимый мультивитаминный транспортер (SMVT) SLC5A6 при компьютерном моделировании прогноза для трансмембранных областей (определен через веб-сайт Центра анализа биологических последовательностей, Технический университет Дании, март 2015 г.).
На Фиг. 18 представлен протокол отбора клеток с высокой экспрессией при помощи осуществления совместной трансфекции вектора экспрессии для транспортера витамина SLC5a6 (справа) и культивирования клеток в селективной среде для культивирования, обедненной по витаминам. Клетки СНО совместно трансфицировали при помощи GFP или векторов экспрессии легкой и тяжелой цепи IgG и плазмиды с устойчивостью к пуромицину либо в отсутствии (условие А), либо в присутствии (условия В и С) вектора экспрессии SLC5a6. Указанные культуры затем отбирали либо в присутствии пуромицина (условия А и В), либо путем культивирования в обедненной по витаминам среде для культивирования, содержащей лимитирующие (В5 10-3Х/Н 10-4Х) концентрации витаминов (условие С). Следует обратить внимание на то, что перечеркнутый круг указывает на то, что клетки, которые не трансфицировали с применением вектора экспрессии SLC5a6, не выживали при отборе в среде для культивирования с недостатком витаминов. После отбора клетки культивировали в неселективной среде для культивирования до проведения анализа при помощи FACS или исследований секреции иммуноглобулина полученных пулов поликлональных клеток (Фиг. 19-20), или во время образования и анализа моноклональных популяций (Фиг. 20-21).
На Фиг. 19 представлено обогащение клеток, экспрессирующих репортерный белок GFP, трансфицированных согласно протоколу, представленному на Фиг. 18. Анализ с применением цитофлуорометрии для определения уровня флуоресценции GFP (А) показал высокий уровень поликлональных популяций после отбора, основанного на витаминной депривации (кружок С). Обогащение GFP-положительных флуоресцентных клеток (В) и геометрическое среднее GFP-флуоресценции указанных клеток (С) представлены для пулов поликлональных клеток.
На Фиг. 20 представлено обогащение клеток, экспрессирующих терапевтический иммуноглобулин (нежели, чем GFP) на высоких уровнях в поликлональных популяциях после отбора на основе обеднения витаминами, согласно протоколов, представленных на Фиг. 18. Уровни образования клеток, отобранных путем витаминной депривации (обозначенные С), выше уровня поликлональных клеток (панель А) и для 10 случайно отобранных клонов клеток, полученных путем предельных разведений (панель В) (см. также легенду Фиг. 18).
На Фиг. 21 представлен высокий уровень секреции IgG, полученный при помощи окрашивания клеточной поверхности для одного из IgG-продуцирующих клонов (Клон С_а), полученных путем отбора с применением витаминов (А), стабильности образования для двух таких клонов (Клоны С_а и С_b) (В), а также высокая плотность жизнеспособных клеток и уровни образования двух указанных клонов в условиях культуры с периодической подпиткой (С и D) по сравнению с ранее полученным суперпродуцентным рефересным клоном, выращенным параллельно (BS03).
На Фиг. 22 представлен отбор (при помощи антибиотика или путем культивирования в среде, обедненной по витаминам («метаболический»)) поликлональных популяций, экспрессирующих различные терапевтические белки, одно легко-экспрессирующееся антитело (А и В) и один трудно-экспрессирующийся белок (интерферон-бета, панель С). Это показывает универсальность системы отбора для осуществления отбора клеток, продуцирующих целевые терапевтические белки на улучшенных уровнях по сравнению с традиционным отбором антибиотиков.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к эукариотической системе экспрессии, содержащей:
- по меньшей мере один первый полинуклеотид, кодирующий по меньшей мере один белок метаболизма витаминов под контролем по меньшей мере одной первой регуляторной последовательности и
- под контролем по меньшей мере одной второй регуляторной последовательности, по меньшей мере одного сайта расщепления эндонуклеазой рестрикции и/или по меньшей мере одного второго полинуклеотида, кодирующего по меньшей мере один целевой продукт (продукт, представляющий интерес).
По меньшей мере один белок метаболизма витаминов может представлять собой белок транспорта витаминов. По меньшей мере один второй полинуклеотид может быть встроен в указанный по меньшей мере один сайт расщепления эндонуклеазой рестрикции. Указанный белок транспорта витаминов может транспортировать растворимый витамин, такой как витамин B1l, В5 и/или Н. Указанным транспортным белком для витаминов может являться THTR-1 (тиаминовый транспортер-1), THTR-2 (тиаминовый транспортер-2), ТРС (тиаминпирофосфатный носитель), ТРК (тиаминпирофосфокиназа) и/или, в частности, SMVT (натрий-зависимый мультивитаминный транспортер). Вектор экспрессии может содержать указанную систему экспрессии. В частности, отдельный вектор может содержать указанный по меньшей мере один первый и указанный по меньшей мере один второй полинуклеотид.
Указанная первая и/или вторая регуляторная последовательность может представлять собой промоторы, энхансеры, области контроля локуса (LCR), участки прикрепления к ядерному матриксу (MAR), участки прикрепления к ядерному скаффолду (SAR), инсуляторные элементы и/или ДНК, ассоциированные с ядерным матриксом.
Настоящее изобретение также относится к набору, содержащему в одном контейнере эукариотическую систему экспрессии, раскрытую в настоящем описании (в частности, на один или более векторов), а во втором контейнере - инструкции по применению указанной системы. Указанный набор может дополнительно содержать среду для культивирования клеток, предпочтительно содержащую лимитирующую и/или насыщающую концентрацию по меньшей мере одного витамина, такого как витамин B1, В5 и/или Н.
Настоящее изобретение также относится к рекомбинантной эукариотической клетке, содержащей описанную в настоящем документе систему экспрессии и/или содержащей повышающую или понижающую мутацию в белке метаболизма витаминов и полинуклеотид (второй полинуклеотид), кодирующий целевой продукт, или регуляторную последовательность, регулирующую экспрессию полинуклеотида, кодирующего указанный белок метаболизма витаминов, при этом указанный белок метаболизма витаминов является необязательным для указанной клетки. Указанной клеткой может являться клетка яичника китайского хомячка (СНО). По меньшей мере один первый полинуклеотид может быть мутирован/содержать повышающую или понижающую мутацию. Указанный белок метаболизма витаминов может влиять на метаболизм витамина и/или связывать данный витамин внутри клетки. Указанный белок метаболизма витаминов может представлять собой пантотенатную 1, 2 и/или 3 и/или тиаминпирофосфаткиназу, такую как TPK1 (тиаминпирофосфаткиназа 1).
Участвующий в метаболизме витаминов белок может представлять собой селектируемый маркер для указанной рекомбинантной эукариотической клетки, и указанная рекомбинантная эукариотическая клетка может продуцировать и, предпочтительным образом, секретировать целевой продукт.
Настоящее изобретение также относится к среде для культивирования эукариотической клетки, содержащей описанные в настоящем документе рекомбинантные эукариотические клетки, предпочтительно поликлональные, предпочтительно экспрессирующие (i) белок транспорта витаминов в качестве селектируемого маркера и (ii) целевой белок. Указанная среда может быть средой с лимитирующей концентрацией В5 или средой с насыщающей концентрацией В5, но с лимитирующей средой Н.
Настоящее изобретение также относится к способу культивирования и необязательно отбору рекомбинантных эукариотических клеток, включающему:
получение раскрытой в настоящем документе системы экспрессии,
получение эукариотических клеток, при этом жизнеспособность, рост и/или деление указанных эукариотических клеток зависит от поглощения витамина,
введение указанной системы экспрессии в указанные эукариотические клетки для образования указанных рекомбинантных эукариотических клеток, экспрессирующих указанный белок метаболизма витаминов и указанный целевой белок,
культивирование указанных эукариотических клеток в среде для культивирования клеток, например, среде с лимитирующей концентрацией В5, или среде с насыщающей концентрацией В5, но среде с лимитирующей концентрацией или среде с нелимитирующей концентрацией Н, или среде с насыщающей концентрацией Н, но среде с лимитирующей или среде с нелимитирующей концентрацией В5, и,
необязательно, отбор при помощи указанного белка метаболизма витаминов, который предпочтительным образом экспрессируется на поверхности указанных рекомбинантных эукариотических клеток, причем указанные рекомбинантные эукариотические клетки стабильно экспрессируют целевой продукт.
Среда для отбора, раскрытая в настоящем описании, может являться средой с лимитирующей концентрацией В5 или с насыщающей концентрацией В5, но средой с лимитирующей или нелимитирующей концентрацией Н.
Настоящее изобретение также относится к применению участвующего в синтезе витаминов белка и его кодирующей ДНК последовательности в качестве селективного маркера для отбора рекомбинантных эукариотических клеток, стабильно экспрессирующих целевой продукт, при этом жизнеспособность, рост и/или деление указанной клетки может зависеть от поглощения витамина.
Настоящее изобретение также относится к среде для культивирования, содержащей по меньшей мере один витамин:
- в концентрации менее, чем 10 нМ и/или
- в концентрации 20 мкМ или более, при этом указанный по меньшей мере один витамин является незаменимым витамином.
По меньшей мере один витамин может представлять собой витамин B1, В5 и/или Н. Среда для культивирования может содержать одну или более рекомбинантных эукариотических клеток, экспрессирующих, предпочтительно секретирующих, целевой белок. Указанный целевой белок может представлять собой терапевтический белок. Рост и/или деление клеток можно остановить, и целевой белок можно получить на максимальном уровне остановки (MAL в [г/л]), который превышает максимальный уровень (ML в [г/л]) белка, экспрессия которого осуществляется указанными клетками, при выращивании на среде, предпочтительно на стандартной среде, в которой рост не останавливают, при этом величина MAL более, чем в 1,5 раза больше значения ML, более, чем в 2 раза больше значения ML или даже более, чем в 2,5 или 3 раза больше значения ML.
Настоящее изобретение также относится к способу получения целевого белка, включающему:
(a) трансформацию эукариотических клеток при помощи системы экспрессии, раскрытой в настоящем описании, с получением рекомбинантных эукариотических клеток;
(b) культивирование указанных рекомбинантных эукариотических клеток в среде для культивирования, в которой жизнеспособность и/или рост или деление рекомбинантных эукариотических клеток зависит от активности одного или более белков метаболизма витаминов;
(c) отбор рекомбинантных эукариотических клеток, экспрессирующих указанный один или более белков метаболизма витаминов, при этом указанный белок метаболизма витаминов является селектируемым маркером для получения отобранных рекомбинантных эукариотических клеток предпочтительно тогда, когда указанные рекомбинантные эукариотические клетки являются частью популяции моноклональных клеток (происходящих от одной клетки); а также
(d) очистку указанного целевого белка, полученного из указанных отобранных рекомбинантных эукариотических клеток или из их среды для культивирования, содержащей указанные выбранные рекомбинантные эукариотические клетки.
Участвующий в метаболизме витаминов белок может представлять собой белок транспорта витаминов, предпочтительным образом осуществляющий перенос витаминов В5, В1 и/или Н, и указанная среда для культивирования может быть лимитирующей и/или насыщенной по одному из более указанных витаминов. Указанный белок транспорта витаминов может представлять собой SMVT, а среда для культивирования может являться средой с лимитирующей концентрацией В5 или средой с насыщающей концентрацией В5, но средой с лимитирующей концентрацией Н.
Настоящее изобретение также относится к клеткам, способам, системам и векторам экспрессии, раскрытым в настоящем описании, при этом указанный SMVT-белок кодируется геном Slc5a6 или его производным и/или при этом указанные эукариотические клетки являются частью популяции моноклональных клеток.
Настоящее изобретение также в целом больше относится к оценке того, можно ли применять строгие требования к витаминам для эукариотических клеток как инструменту для отбора трансформированных клеток, в частности, трансформированных клеток, которые стабильно экспрессируют высокие уровни целевого гена при его совместной экспрессии с геном, участвующим в поглощении витаминов. Настоящее изобретение также в целом больше относится к оценке того, можно ли применять среды для культивирования, обедненные или обогащенные витаминами, для дальнейшего улучшения образования белка такими клетками.
Один конкретный вариант реализации настоящего изобретения относится к снижению доступности витамина В5 на поздней фазе образования рекомбинантного белка для замедления деления клеток и, таким образом, увеличению уровня терапевтических белков, образуемых в биореакторе.
Еще один конкретный вариант реализации настоящего изобретения также относится к клонированию и экспрессии мультивитаминного транспортера Slc5a6 (SMVT), участвующего в поглощении как витамина В5, так и Н в клетке, в частности, в клетках СНО-М. Настоящее изобретение также относится к клеткам, осуществляющим гиперэкспрессию данного витаминного транспортера, для обеспечения более быстрого роста и более высокой жизнеспособности в средах с лимитирующими концентрациями витамина В5 по сравнению с нетрансформированными клетками. Настоящее изобретение также относится к совместной экспрессии SLC5A6 в качестве селективного маркера для получения клеточных линий, обладающих более высокими уровнями образования рекомбинантного белка. Кроме того, настоящее изобретение относится к гиперэкспрессии SLC5A6 в клетках для обеспечения более лучшей жизнеспособности клеток даже в необедненной среде для культивирования, что, предпочтительным образом, способствует еще более благоприятным уровням экспрессии терапевтических белков.
ОБСУЖДЕНИЕ РАЗЛИЧНЫХ И ПРЕДПОЧТИТЕЛЬНЫХ ВАРИАНТОВ РЕАЛИЗАЦИИ
Определения
Система экспрессии эукариот согласно настоящему изобретению содержит элементы, которые позволяют осуществлять экспрессию целевого гена в эукариотических клетках, таких как клетках СНО, предпочтительно, клетках СНО K1, предпочтительно, клетках СНО-М. В целом, указанная эукариотическая система экспрессии содержит, по меньшей мере, один вектор экспрессии. Однако эукариотическая система экспрессии может также являться частью генома эукариотической клетки. Система/вектор экспрессии содержит регуляторные последовательности, такие как промоторы, энхансеры, области контроля локуса (LCR), участки прикрепления к ядерному матриксу (MAR), участки прикрепления к ядерному скаффолду (SAR), инсуляторные элементы и/или ассоциированные с ядерным матриксом ДНК, которые приводят к эффективной транскрипции трансгена, интегрированного в указанную систему экспрессии. Эти регуляторные последовательности, как и любые другие последовательности, упоминаемые в настоящем описании, часто являются гетерологичными (т.е. чужеродными по отношению к используемой клетке-хозяину, например, полученными от вида клетки, отличного от используемой клетки-хозяина) или, будучи гомологичными (т.е. эндогенными по отношению к используемой клетке-хозяину), присутствуют в разной геномной области(-тях), в отличие от других присущих клеткам последовательностей (здесь и далее упоминаемые как «гетеролокальные»). Вектор экспрессии может также содержать точку начала репликации.
Первый полинуклеотид, кодирующий по меньшей мере один белок метаболизма витаминов и второй полинуклеотид, кодирующий по меньшей мере один целевой продукт согласно настоящему изобретению, добавляют к эукариотической клетке для создания рекомбинантной эукариотической клетки. Гены или белки, присущие эукариотической клетке, не добавляют в указанную клетку, однако они существуют в клетке независимо от любого вида трансформации. Однако, как это будет понятно специалисту в данной области, первый и второй полинуклеотиды могут являться копиями присущего клетке гена, такими как гетеролокальные копии указанного гена. Во многих случаях предпочтительно, чтобы некоторые или все кодирующие последовательности ДНК (CDS) гена дикого типа образовывали полинуклеотиды согласно настоящему изобретению, включая первый полинуклеотид, кодирующий по меньшей мере один белок метаболизма витаминов.
В контексте настоящего описания термин «плазмида» и «вектор» используются взаимозаменяемо, поскольку плазмида является наиболее часто применяемой формой вектора. Тем не менее, указанное изобретение предназначено для включения таких других форм векторов экспрессии, включая, но не ограничиваясь ими, как вирусные векторы (например, репликации дефектных ретровирусов, аденовирусы и аденоассоциированные вирусы) или транспортируемые векторы, которые выполняют эквивалентные функции. В настоящем документе трансформация относится к введению векторной ДНК в любую клетку, независимо от применяемых средств или типа вектора.
«Целевой ген» или «трансген», который в настоящем описании также упоминается как «полинуклеотид, кодирующий целевой продукт», кодирует, например, «целевой белок» (структурный или регуляторный белок). Указанный целевой белок часто является терапевтическим белком. В контексте настоящего описания термин «белок» относится в целом к пептидам и полипептидам, содержащим более, чем приблизительно десять аминокислот. Указанные белки могут являться «гомологичными» по отношению к хозяину (т.е. эндогенными в отношении используемой клетки-хозяина) или «гетерологичными» (т.е. чужеродными в отношении используемой клетки-хозяина), такими как белок человека, продуцируемый дрожжами. Указанный белок можно получить в виде нерастворимого агрегата или в виде растворимого белка в периплазматическом пространстве или цитоплазме клетки или во внеклеточной среде. Примеры указанных терапевтических белков включают гормоны, такие как гормон роста или эритропоэтин (ЕРО), факторы роста, такие как эпидермальный фактор роста, обезболивающие вещества, такие как энкефалин, ферменты, такие как химотрипсин, рецепторы или антитела (например, моноклональный иммуноглобулин Трастузумаб (IgG)). Гены, которые в целом применяют в качестве маркера для визуализации, например, зеленый флуоресцентный белок, также являются подходящими трансгенами. Трансген также может кодировать, например, регуляторную РНК, такую как киРНК (siRNA). Гетеролокальный полинуклеотид может образовывать гомологичный белок или РНК. Во многих случаях предпочтительно, чтобы некоторые или все кодирующие последовательности ДНК (CDS) гена дикого типа образовывали полинуклеотиды согласно настоящему изобретению, включая второй полинуклеотид, кодирующий по меньшей мере один представляющий интерес продукт.
Эукариотические клетки, применяемые в контексте настоящего изобретения, включают, но не ограничиваются ими, вышеупомянутые клетки СНО-М (доступные от SELEXIS SA) и другие клетки, которые являются подходящими для производства белка в промышленном масштабе. Эти клетки хорошо известны специалистам и они происходят, например, от Cricetulus griseus, Cercopithecus aethiops, Homo sapiens, Mesocricetus auratus, Mus musculus и Chlorocebus. Соответствующие клеточные линии известны как СНО-клетки (яичник китайского хомяка), COS-клетки (клеточная линия, полученная из почек обезьян (африканская зеленая обезьяна), Vero-клетки (эпителиальные клетки почек, выделенные из африканской зеленой обезьяны), Hela-клетки (линию получили из клеток рака шейки матки, взятых от Генриетты Лакс (Henrietta Lacks)), BHK-клетки (клетки почек детеныша хомяка, HEK-клетки (почка эмбриона человека), NSO-клетки (линия клеток миеломы мышей), С127-клетки (линия неозлокачественных клеток мыши), PerC6.RTM-клетки (клеточная линия человека, Cracell), САР-клетки ( Amniocyte Production) и Sp-2/0-клетки (клетки миеломы мыши). Эукариотические клетки, применяемые в контексте настоящего изобретения, могут также, например, представлять собой первичные клетки человека, включая гемопоэтические стволовые клетки, такие как клетки, полученные из клеток костного мозга или стволовых клеток, таких как эмбриональные стволовые клетки (ES), индуцированные полипотентные стволовые клеток (iPS) или дифференцированные клетки, полученные из ES- или iPS-клеток.
Amniocyte Production) и Sp-2/0-клетки (клетки миеломы мыши). Эукариотические клетки, применяемые в контексте настоящего изобретения, могут также, например, представлять собой первичные клетки человека, включая гемопоэтические стволовые клетки, такие как клетки, полученные из клеток костного мозга или стволовых клеток, таких как эмбриональные стволовые клетки (ES), индуцированные полипотентные стволовые клеток (iPS) или дифференцированные клетки, полученные из ES- или iPS-клеток.
Белок метаболизма витаминов согласно настоящему изобретению представляет собой белок, который либо снижает, либо увеличивает доступность витамина или его применение в клетке.
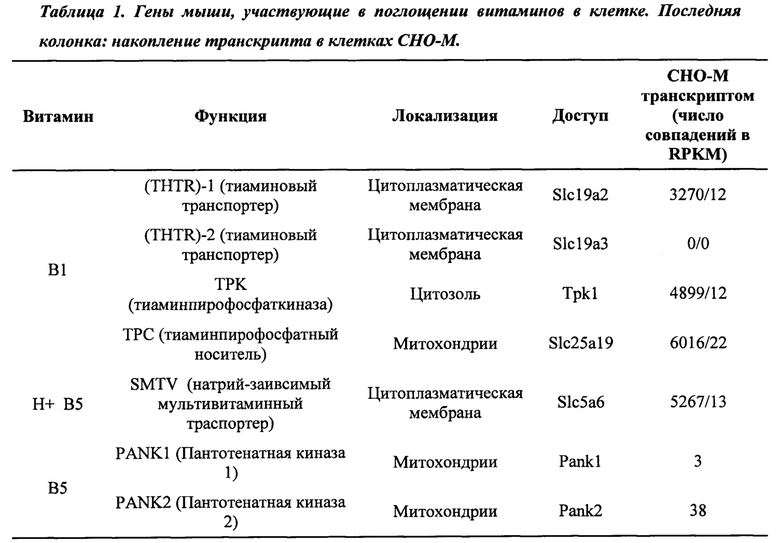
Одним предпочтительным белком метаболизма витаминов является белок транспорта витаминов, который в целом является белком, связанным с мембраной, и переносит витамины, доступные в среде для культивирования, в клетку. В Таблице 1 приведены примеры этих белков под заголовком «Функция». Как можно увидеть из данной таблицы, два цитоплазматических и один митохондриальный транспортер охарактеризованы для витамина B1 (SLC19A2 [SEQ ID №: 24], SLC19A3 [SEQ ID №: 25] и SLC25A19 [SEQ ID №: 27]), тогда как один цитоплазматический транспортер охарактеризован как для витамина В5, так и для витамина Н, называемый натрий-мультивитаминный транспортер SLC5A6 [SEQ ID №: 21].
Другие примеры белков метаболизма витаминов включают пантотенатные киназы 1, 2 или 3, кодируемые геном PANK1 [SEQ ID №:. 22], PANK2 [SEQ ID №: 23] и PANK3 [SEQ ID №: 35, 36], и TPK1 (тиаминпирофосфат-киназу 1), кодируемую геном TPK1 [SEQ ID №: 26]. Пантотенатные киназы являются ключевым регулятивными ферментами в биосинтезе коэнзима А (КоА); гомодимерный белок TPK1 катализирует преобразование тиамина в тиаминпирофосфат. Специалисту в данной области техники будет понятно, что другие белки метаболизма витаминов также являются частью настоящего изобретения.
Для клеток, растущих в полной среде для культивирования, будут доступны все витамины в стандартных концентрациях. Стандартные концентрации упоминаются в настоящем описании как 1X. Стандартные концентрации для витаминов B1, В5 и Н (1X) были установлены на уровне 7,5 мкМ, 2,5 мкМ и 0,5 мкМ соответственно. Определили, что у В5 диапазон значений концентрации, лимитирующий рост клеток СНО, составляет от 10-4Х до 10-3Х (0,25-2,5 нм), тогда как концентрации 10-2Х и выше способствуют нормальному росту культуры. Определили, что значения лимитирующих концентраций В1 для клеток СНО составляли от 10-5Х (15 пМ) до 10-4Х (150 пМ), тогда как для Н они были ниже, чем 10-5Х (5 пМ).
В среде, содержащей лимитирующую концентрацию (лимитирующая среда или обедненная среда) указанного витамина, концентрация составляет менее, чем 1X, например, 10-1Х, 10-2Х, 10-3Х, 10-4Х, 10-5Х, относительно указанной стандартной концентрации соответствующего витамина, присутствующего в полной среде (1X). Концентрация витамина считается насыщающей, если указанная концентрация превышает концентрацию в стандартной референсной среде (в настоящем описании также упоминаемая как «насыщенная среда») (например, в полной среде обнаруживают количество 2Х, 3Х, 4Х, 5Х или 10Х).
Среды для культивирования клеток, содержащие лимитирующую и/или насыщающую концентрацию витамина, являются частью настоящего изобретения. Например, среда может быть обеднена по отношению к одному витамину, но насыщена по отношению к другому витамину.
В лимитирующей среде рост и/или деление указанных клеток можно остановить, и целевой белок можно получить на максимальном уровне остановки («MAL» в [г/л]). Величина MAL может превышать максимальный уровень («ML» в [г/л]) белка, который экспрессируют клетки того же типа при выращивании на среде, такой как стандартная среда, в которой их рост не останавливают. В некоторых вариантах реализации настоящего изобретения величина MAL более, чем 1,5 раза большая ML, более, чем в 2 большая ML или даже более, чем в 2,5 или 3 раза больше величины ML. Например, хотя ML целевого белка, такого как антитело, которое экспрессируется рекомбинантными клетками, такими как рекомбинантные клетки СНО в стандартной среде, составляет приблизительно 1 г/л IgG, MAL целевого белка, такого как антитело, которое экспрессируется рекомбинантными клетками, такими как рекомбинантные клетки СНО в стандартной среде, составляет около 3 г/л IgG или более.
Участвующий в метаболизме витаминов белок, включая белок транспорта витаминов, может представлять собой полноразмерный белок дикого типа или может являться мутированным, в том числе посредством точечных мутаций, замен, вставок, добавок и/или терминальных или внутренних делеций или инверсий. Хотя участвующий в метаболизме белок может по отношению к определенной последовательности содержать мутацию, которая обладает (i) активностью, соответствующей белку дикого типа (нейтральная мутация), белок метаболизма витаминов/белок транспорта витаминов в контексте настоящего изобретения упоминается как мутированный, когда мутация вызывает (ii) измененную активность/стабильность по сравнению с белком дикого типа, которая включает повышенную активность («повышающая мутация») (например, более, чем 10%, более, чем 20%, более, чем 30%, более, чем 40%, более, чем 50%, более, чем 60%, более, чем 70%, более, чем 80%, более, чем 90% или более, чем 100%) или пониженную активность/стабильность («понижающая мутация») (например, менее, чем 10%, менее, чем 20%, менее, чем 30%, менее, чем 40%, менее, чем 50%, менее, чем 60%, менее, чем 70%, менее, чем 80%, менее, чем 90%, менее, чем или на 100%). Независимо от того, является ли конкретная мутация повышающей или понижающей мутацией, это можно легко оценить путем проведения стандартных исследований, доступных в данной области техники. Мутированный белок метаболизма витаминов является результатом мутации в по меньшей мере первом полинуклеотиде, кодирующем белок метаболизма витаминов. Аналогичным образом, мутация в последовательности, регулирующей экспрессию указанного первого полипептида, называется повышающей мутацией, когда уровень экспрессии указанного полипептида, кодируемого полинуклеотидом, больше или более стабилен (например, на 10%, 20%, 30%, 40%, 50% или более), чем в том случае, когда последовательность, регулирующая экспрессию указанного первого полипептида, не содержит указанную мутацию. Мутация в последовательности, регулирующей экспрессию указанного первого полипептида, называется понижающей мутацией, когда уровень экспрессии полипептида, кодируемого полинуклеотидом, меньше уровня первого полинуклеотида или менее стабилен (например, на 10%, 20%, 30%, 40%, 50% или менее), чем в том случае, когда последовательность, регулирующая экспрессию указанного первого полипептида, не содержит указанную мутацию. Повышающие мутации в последовательностях, регулирующих экспрессию первого полипептида, также могут соответствовать добавлению MAR, SAR, LCR и/или инсуляторного элемента в дополнение к последовательностям энхансера и промотора для повышения уровня экспрессии или стабильности белка, кодируемого указанным полинуклеотидом.
Желаемые модификации или мутации в полипептиде можно осуществить с применением любых методов, известных в данной области техники. Методы рекомбинантной ДНК для введения таких изменений в белковую последовательность хорошо известны в данной области техники. В определенных вариантах реализации указанные модификации производят путем сайт-направленного мутагенеза полинуклеотида, кодирующего белок или последовательность, регулирующую (регуляторные последовательности, определенные выше) его экспрессию. Другие методы введения мутаций обсуждаются в Molecular Cloning: The Laboratory Manual, 2nd Ed. под редакцией Sambrook, Fritsch и Maniatis (Cold Spring Harbor Laboratory Press: 1989); труде «Methods in Enzymnology» (Academic Press, Inc., Нью-Йорк); Ausubel и др. Current Protocols in Molecular Biology (John Wiley & Sons, Inc., Нью-Йорк, 1999); каждый из которых включен в настоящее описание посредством ссылки.
Хорошо известны, в частности, понижающие мутации в промоторах и других регуляторных последовательностях, присущих клетке. Указанная мутация снижает сродство факторов транскрипции к участку промотора, понижая скорости транскрипции. Однако мутации в участках промотора также могут быть нейтральными или вызывать повышающие мутации.
Полинуклеотиды и белки, обладающие более, чем 50%, 60%, 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% идентичностью последовательности к последовательностями полинуклеотидов и белков, раскрытых в настоящем документе, в частности, те, которые раскрыты на Фиг. 15 и 16, также являются частью настоящего изобретения либо по отдельности, либо как часть любой системы (например, векторов и клеток), способа и набора, раскрытых в настоящем документе. На Фиг. 15, в частности, представлена CDS (кодирующая последовательность ДНК) соответствующего гена, эрго (ergo), части гена ДНК или РНК, состоящего из экзонов, которая кодирует соответствующую последовательность белка/аминокислотную последовательность (см. Фиг. 16). Полинуклеотиды согласно настоящему изобретению могут отличаться от любой последовательности дикого типа по меньшей мере одним, двумя, тремя, четырьмя, пятью, шестью, семью, восемью, девятью или более нуклеотидами. Во многих случаях предпочтительными являются полинуклеотиды, образованные CDS соответствующего гена или кДНК.
Термин «идентичность последовательности» относится к измерению идентичности последовательностей нуклеотидов или аминокислотных последовательностей. В целом, указанные последовательности выравнивают таким образом, что достигается совпадение высшего порядка. Термин «идентичность» сам по себе имеет признанное значение в данной области техники и ее можно рассчитать с применением опубликованных методов, (см., например: Computational Molecular Biology, Lesk, A. M., ed., Oxford University Press, Нью-Йорк, 1988; Biocomputing: Informatics and Genome Projects, Smith, D. W., ed., Academic Press, Нью-Йорк, 1993; Computer Analysis of Sequence Data, Part I, Griffin, A. M. и Griffin, H. G., eds., Humana Press, New Jersey, 1994; Sequence Analysis in Molecular Biology, von Heinje, G., Academic Press, 1987, и Sequence Analysis Primer, Gribskov, M. и Devereux, J., eds., M Stockton Press, Нью-Йорк, 1991). Хотя и существует ряд способов измерения идентичности между двумя полинуклеотидными или полипептидными последовательностями, термин «идентичность» хорошо известен квалифицированным специалистам (Carillo, Н. & Lipton, D., SIAM. J. Applied. Math. 48: 1073 (1988)).
Если ли какая-либо конкретная молекула нуклеиновой кислоты является по меньшей мере на 50%, 60%, 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% идентичной, например, последовательности нуклеиновой кислоты SMTV [SEQ ID №: 21] или ее части, то это можно определить традиционным образом с применением известных компьютерных программ, таких как программное обеспечение DNAsis (Hitachi Software, Сан Бруно, штат Калифорния) для первоначального выравнивания последовательностей с последующим применением программного обеспечения ESEE версии 3.0 (последовательности ДНК/белков) (cabot@trog.mbb.sfu.ca) для множественных выравниваний последовательностей.
Если аминокислотная последовательность является по меньшей мере на 50%, 60%, 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% идентичной, например, последовательности SEQ ID №: 28 или ее части, то это можно определить традиционным образом с применением известных компьютерных программ, таких как программа BESTFIT (Wisconsin Sequence Analysis Package, Версия 8 для Unix, Genetics Computer Group, University Research Park, 575 Science Drive, Мэдисон, штат Висконсин, 53711). BESTFIT использует алгоритм локальной гомологии по Smith и Waterman, Advances in Applied Mathematics 2: 482-489 (1981) для нахождения лучшего сегмента гомологии между двумя последовательностями.
При применении DNAsis, ESEE, BESTFIT или любой другой программы выравнивания последовательностей для определения того, является ли конкретная последовательность, например, на 95% идентичной референсной последовательности согласно настоящему изобретению, параметры устанавливают таким образом, что процент идентичности вычисляют в сравнении с полной длиной референсной последовательности нуклеиновой кислоты или аминокислотной последовательности, и при этом допускаются пробелы в гомологии величиной не более 5% от общего числа нуклеотидов в референсной последовательности.
Рекомбинантная эукариотическая клетка согласно настоящему изобретению представляет собой эукариотическую клетку, содержащую трансген, определенный выше.
Незаменимым витамином согласно настоящему изобретению является витамин, который незаменим для роста, деления и/или жизнеспособности клеток.
Системы экспрессии в целом содержат селектируемый маркерный ген, который способствует отбору эукариотических клеток (клеток-хозяев), трансформированных при помощи векторов, содержащих полинуклеотид, кодирующий целевой белок. Селектируемый маркер (или «селектируемый маркерный белок»), который экспрессируется геном, часто основывается на устойчивости к антибиотикам. Например, селекционную кассету экспрессии, обеспечивающую устойчивость к пуромицину, можно применять для идентификации клеток путем добавления пуромицина, успешную трансформацию которых осуществляли при помощи указанной кассеты. Тем не менее, также возможен отбор без какой-либо устойчивости к антибиотикам. Примерами селектируемых маркеров такого типа являются дигидрофолатредуктаза (DHFR) и глутаминсинтетаза (GS). Отбор происходит, например, в условиях отсутствия метаболитов, например, глицина, гипоксантина и тимидина для DHFR и глутамина для GS. Отбор выживших клеток включает одну или более копий трансформированной плазмиды в геноме клетки. В контексте настоящего изобретения белок метаболизма витаминов/белок транспорта витаминов может служить в качестве селектируемого маркера либо по отдельности, либо в комбинации с другими селектируемыми маркерами. Таким образом, в своей простейшей форме в среде с недостатком одного витамина рекомбинантные эукариотические клетки, экспрессирующие соответствующий белок транспорта витаминов в качестве селектируемого маркера, могут расти лучше, чем клетки, неэкспрессирующие соответствующий белок транспорта витаминов. Однако, как обсуждается в настоящем документе, даже в стандартной среде белки транспорта витаминов обеспечивают преимущество роста и, следовательно, их можно применять в качестве селектируемого маркера. Системы экспрессии согласно настоящему изобретению могут содержать в качестве селектируемых маркеров белок(-и) метаболизма витаминов / белок(-и) транспорта витаминов в дополнение к селектируемым маркерным генам на основе, например, устойчивости к антибиотикам.
Аналогичным образом, мутация в последовательности, регулирующей экспрессию указанного первого полипептида, называется повышающей мутацией, когда уровень экспрессии указанного полипептида, кодируемого полинуклеотидом, больше или более стабилен (например, на 10%, 20%, 30%, 40%, 50% или более), чем в том случае, когда последовательность, регулирующая экспрессию указанного первого полипептида, не содержит указанную мутацию. Мутация в последовательности, регулирующей экспрессию указанного первого полипептида, называется понижающей мутацией, когда уровень экспрессии полипептида, кодируемого полинуклеотидом, меньше уровня первого полинуклеотида или менее стабилен (например, на 10%, 20%, 30%, 40%, 50% или менее), чем в том случае, когда последовательность, регулирующая экспрессию указанного первого полипептида, не содержит указанную мутацию.
Желаемые модификации или мутации в полипептиде можно осуществить с применением любых методов, известных в данной области техники. Методы рекомбинантной ДНК для введения таких изменений в белковую последовательность хорошо известны в данной области техники. В определенных вариантах реализации указанные модификации производят путем сайт-направленного мутагенеза полинуклеотида, кодирующего белок или последовательность, регулирующую (регуляторные последовательности, определенные выше) его экспрессию. Другие методы введения мутаций обсуждаются в Molecular Cloning: The Laboratory Manual, 2nd Ed. под редакцией Sambrook, Fritsch и Maniatis (Cold Spring Harbor Laboratory Press: 1989); трактате «Methods in Enzymnology» (Academic Press, Inc., Нью-Йорк); Ausubel и др. Current Protocols in Molecular Biology (John Wiley & Sons, Inc., Нью-Йорк, 1999); каждый из которых включен в настоящее описание посредством ссылки.
Хорошо известны, в частности, понижающие мутации в промоторах и других регуляторных последовательностях, присущих клетке. Указанная мутация снижает сродство факторов транскрипции к участку промотора, понижая скорости транскрипции. Мутации в участках промотора могут быть нейтральными, вызывать понижающие или повышающие мутации. Аналогичным образом, мутации в, например, гене белка метаболизма витаминов, такого как белка транспорта витаминов, могут быть нейтральными, быть понижающими или повышающими мутациями.
1 - Воздействие переноса присутствующего в лимитирующей концентрации витамина на рост клеток СНО и экспрессию рекомбинантного белка
Первым этапом использования витаминов культивируемыми клетками млекопитающих является их поглощение клетками из среды для культивирования. Витамин В1 (тиамин), В5 (пантотенат) и Н (В8 или биотин) являются растворимыми витаминами, которые транспортируются в цитоплазму, а затем в митохондрии, где они действуют как участвующие в метаболизме кофакторы (Фиг. 1). Два цитоплазматических и один митохондриальный транспортер охарактеризован для витамина B1 (SLC19A2, SLC19A3 и SLC25A19), тогда как один цитоплазматический транспортер охарактеризован как для витамина В5, так и для витамина Н, называемый натрий-мультивитаминный транспортер SLC5A6 (Таблица 1).
1.1. Определение лимитирующих рост концентраций витамина
Чтобы оценить воздействие лимитирующей концентрации витамина на рост клеток, среду для культивирования клеток, специфически образом обедненную по витаминам В1, В5 и Н, называемую B-CDmin, получили из коммерчески доступной среды для роста (среда для роста BalanCD СНО, IRVINE SCIENTIFIC INC). Клетки СНО-М, высеянные на среду В-CDmin, как и ожидалось, не могли поддерживать деление клеток (Фиг. 2А). В динамике размер клеток понижался, и указанные клетки начинали терять жизнеспособность после 6 дней инкубации в среде, не содержащей витаминов (Фиг. 2В). Затем среду B-CDmin дополняли известными количествами витаминов, устанавливая значения стандартных концентраций для B1, В5 и Н (1X) при 7,5 мкМ, 2,5 мкМ и 0,5 мкМ соответственно, обнаруживаемых в обычно применяемых полных средах (Таблица 2). В среде для культивирования с недостатком только витамина В5 клетки не делились, а их жизнеспособность снижалась через 6 дней, как и в среде B-CDmin (Фиг. 2А и 2В). Когда среду обедняли либо по витамину В1, либо по витамину Н, клетки могли делиться на 3-6 день соответственно, хотя рост культуры в целом уменьшался в обедненной по витамину Н среде по сравнению с указанной полной средой. В связи с этим авторы настоящего изобретения пришли к выводу о том, что В5 может быть наиболее лимитирующим для роста клеток в краткосрочной перспективе, поскольку он должен постоянно присутствовать в среде для культивирования для поддержания деления клеток.

Обедненную среду B-CDmin дополняли меньшей концентрацией каждого витамина по отдельности, чтобы определить диапазон концентраций, лимитирующих рост СНО-М. Витамин В5 был незаменим для роста СНО-М, с диапазоном лимитирующих рост концентраций приблизительно от 10-4Х до 10-3Х (0,25-2,5 нМ), тогда как 10-2Х и более высокие концентрации способствовали нормальному росту культуры (Фиг. 3 и данные не представлены). Лимитирующие концентрации витамина В1 наблюдали в диапазоне от 10-5Х (15 пМ) до 10-4Х (150 пМ), тогда как для витамина Н они были ниже, чем 10-5Х (5 пМ). Интересно отметить то, что в присутствии витамина Н при низкой концентрации (10-5Х), плотность клеток была несколько выше, чем наблюдаемая плотность в полной среде. Поскольку витамины В5 и Н применяли в качестве одно и того же транспортера для проникновения в клетку, и в связи с тем, что витамин В5 является наиболее лимитирующим для роста клеток, снижение концентрации витамина Н ниже уровня насыщения могло увеличить доступность транспортера для витамина В5, что может способствовать достижению витамином В5 более высоких внутриклеточных уровней по сравнению с клетками, выращенными в полной среде.
1.2. Воздействие лимитирующей рост концентрации витамина В5 на экспрессию и модификации белка
Затем оценивали, можно ли остановление роста, наблюдаемое при обеднении В5, применять для прекращения или замедления деления клеток в условиях образования белка с тем, чтобы, возможно, увеличить образование белка, с применением культуры с периодической подпиткой, поддерживаемой в биореакторах со спин-трубкой. Клон клеток, полученный из СНО-М, экспрессирующий терапевтический белок, показал увеличение количества клеток до 8 дня при выращивании в полной среде, после чего жизнеспособность клеток и количество жизнеспособных клеток снижалось, как это обычно наблюдают в этих условиях культивирования (Фиг. 4). Однако в условиях с лимитирующей концентрацией витамина число клеток, которое перестало увеличиваться с 7 дня, и высокая жизнеспособность клеток сохранялись до 14 дня, что свидетельствуе о том, что указанные клетки использовали доступность лимитирующего рост витамина из среды и эндогенный клеточный В5-пул для ограниченного числа делений клеток прежде чем он становится лимитирующим рост фактором.
Титр антитела, секретируемого в супернатанте клеточной культуры, увеличивался до уровня не более, чем 3 г/л до 9 дня в культуре полной среды, после чего он уменьшался (Фиг. 5), что и ожидали, исходя из уменьшенной жизнеспособности клеток, отмеченной ранее (Фиг. 4). Тем не менее, накопление указанного антитела продолжалось до 15 дня культурования, выполненного с применением обедненной по витаминам среды, в которой оно достигло уровней больших, чем 6 г/л (Фиг. 5). В целом авторы настоящего исследования пришли к выводу о том, что витаминную депривацию можно применять для остановки роста клеток в биореакторе с тем, чтобы продлить продолжительность жизнеспособности клеток и секреции антитела, тем самым обеспечивая очень высокие титры терапевтического антитела. Этот подход можно в целом применять для улучшения образования рекомбинантного белка.
1.3. Воздействие увеличивающегося переноса витамина В5 на рост клеток
Основываясь на выводах о том, что рост клеток можно ингибировать либо посредством отсутствия витамина В5, либо при помощи высоких концентраций витамина Н, которые могут конкурировать с В5 за их общий транспортер, или высоких концентраций витамина В5, которые могут конкурировать с Н за их общий транспортер, было высказано предположение о том, что гиперэкспрессия общего транспортера Slc5a6 может обеспечить клетки преимуществом роста и/или может привести к более высокому уровню жизнеспособности клеток. Таким образом, авторы настоящего изобретения клонировали кДНК СНО-М, кодирующую мультивитаминный транспортер Slc5a6, и другие транспортеры витамина В1, указанные в Таблице 1, и осуществляли их вставку под контролем сильного промотора GAPDH и элемента эпигенетического активатора MAR 1-68, расположенного рядом с селекционной кассетой экспрессии, обеспечивающей устойчивость к пуромицину. Клетки СНО-М совместно трансформировали при помощи данной Slc5a6-конструкции с вектором экспрессии GFP и с селективной по отношению к пуромицину плазмидой, после чего получали стабильные поликлональные популяции в результате отбора устойчивых к пуромицину клеток. В популяциях клеток СНО-М, трансформированных при помощи увеличивающегося количества вектора экспрессии, наблюдали накопление транскрипта Сlс5а на уровне не более, чем в 100 раз превышающим при сравнении уровень эндогенной экспрессии (Фиг. 6).
Популяции клеток, гиперэкспрессирующих SLC5A6, затем выращивали без проведения отбора с применением пуромицина в среде B-CDmin, дополненной различными концентрациями витаминов В5 и Н. Как и ранее, деление клеток в условиях отсутствия витамина В5 после 6 дней культивирования было почти остановлено, независимо от уровня гиперэкспрессии транспортера или присутствия витамина Н (Фиг. 7). Однако клетки, трансформированные при помощи плазмиды экспрессии транспортера, достигали достоверно более высоких уровней плотности в условиях лимитирующих концентраций В5 (10-3Х) и при низкой концентрации витамина Н (10-4Х). Наибольший рост наблюдали у клеток, совместно трансформированных при помощи 100 нг вектора экспрессии транспортера (Фиг. 7), что свидетельствует о том, что был достигнут оптимальный уровень экспрессии транспортера.
Интересно отметить, что когда витамин В5 добавляли в 10Х избытке в присутствии низкого количества витамина Н (10Х В5, 10-4Х Н), рост нетрансформированных клеток сильно ингибировался относительно культуры этих клеток в полной среде (1X В5, 10-4Х Н). Однако клетки, эспрессирующие самый высокий уровень транспортера, росли достоверно больше, чем те, которые экспрессировали указанный транспортер на более низких уровнях в присутствии избытка витамина В5 (10Х В5, 10-4Х Н). Это также подтверждает то, что наличие конкуренции между двумя витаминами за их общий транспортер при насыщающей концентрации витамина В5 может ингибировать поглощение низких количеств витамина Н в среде для культивирования, тем самым ограничивая рост до тех пор, пока не осуществляется гиперэкспрессия транспортера. В целом было сделано заключение о том, что гиперэкспрессия транспортера SLC5A6 может обеспечить преимущество роста в присутствии либо лимитирующих концентраций витамина В5, либо, наоборот, в присутствии насыщающих концентраций витамина В5, но с лимитирующими количествами витамина Н. Было высказано предположение о том, что, таким образом, это можно применять для распознавания клеток, которые экспрессируют повышенные количества транспортера относительно тех клеток, которые экспрессируют его на более низких уровнях.
2 Применение экспрессии транспортера SLC5A6 (SMVT) в качестве селективного маркера для трансформированных клеток
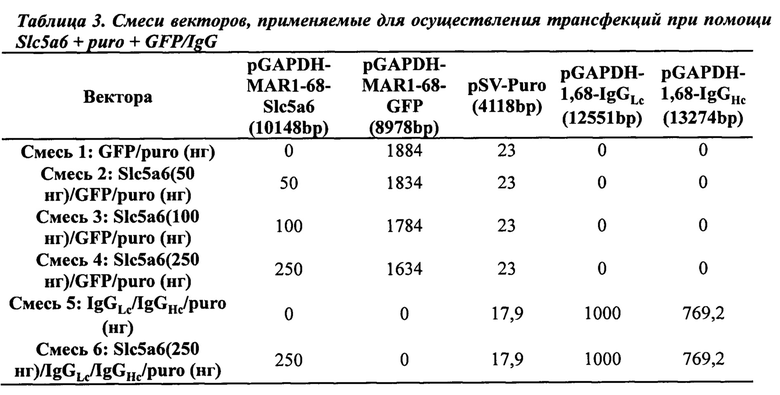
Экспрессию при совместно трансформированном векторе GFP количественно оценивали для определения того, может ли совместная трансформация транспортера Slc5a6 увеличивать общие уровни экспрессии трансгена. Клетки, содержащие интегрированные в их геном плазмиды и стабильно экспрессирующие трансгены, отбирали либо путем культивирования в среде с лимитирующей концентрацией витамина В5, либо в присутствии пуромицина. Процент GFP-экспрессирующих флуоресцентных клеток, а также уровни интенсивности клеточной флуоресценции, сначала оценивали после отбора путем витаминной депривации по В5. После отбора в присутствии лимитирующих количеств витамина В5 (10-3Х) наибольшую долю как GFP-положительных клеток, так и средних уровней флуоресценции получили при совместном трансформировании клеток 250 нг плазмиды экспрессии SLC5A6 (Фиг. 8). Трансформация с применением более высоких уровней плазмиды (1000 нг) вектора Slc5a6 приводила к получению сходного количества GFP-положительных клеток и несколько более низкой флуоресценции, тогда как более низкое количество плазмиды (50 нг) не позволяло получить достаточного количества клеток для проведения количественной оценки. Это свидетельствует о том, что совместную трансформацию этого гена транспортера витамина можно применять в качестве селектируемого маркера для стабильной трансформации путем осуществления совместной трансформации небольшого количества плазмиды SLC5A6 с большим количеством конструкции, экспрессирующей целевой белок (Таблица 3). Небольшое количество плазмиды SLC5A6 обычно составляет 1000 нг, 250 нг, 100 нг или менее (см., например, Фиг. 6, 8 и 9). Как представлено в Таблице 3, более высокие количества конструкции, экспрессирующей целевой белок, могут варьироваться от более, чем двукратного до более, чем 15-кратного увеличения от количества вектора экспрессии белка метаболизма витаминов, включая более, чем 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13 или 14-кратные увеличения. Менее благоприятные результаты получали, когда проводили аналогичные эксперименты с транспортными кДНК для витамина В1 (данные не представлены), что было ожидаемого исходя из того, что витамин В5 является более лимитирующим, чем витамин В1.
Когда клетки отбирали с применением пуромицина в среде, содержащей нелимитирующую концентрацию витамина В5, GFP-флуоресцентные клетки, как и ожидалось, получали независимо от экспрессии Slc5a6. Тем не менее, клетки с наиболее высоким уровнем флуоресценции часто получали при совместной трансформации 250 нг вектора экспрессии Slc5a6 (данные не представлены). Это свидетельствует о том, что транспортер витамина может обеспечить клетки, которые осуществляют его экспрессию на более высоких уровнях даже в нелимитирующей среде для культивирования, избирательным преимуществом. Когда после отбора с применением пуромицина следовало дальнейшее культивирование в среде с лимитирующей концентрацией витамина В5, наблюдали чрезмерно высокие уровни экспрессии у большинства клеток, экспрессирующих транспортер SLC5a6 (Фиг. 9А). Количественная оценка общего процентиля GFP-экспрессирующих клеток (Окно 1 на Фиг. 9А) или клеток с высокой экспрессией (Окно 2 на Фиг. 9А) показала, что более 80% клеток экспрессировали GFP на очень высоких уровнях после трансформации 100 или 250 нг плазмиды экспрессии Slc5a6 либо при осуществлении отбора клеток в обедненной по витамину В5 среде, либо в условиях избытка витамина В5 (Фиг. 9В). Уровни экспрессии GFP также увеличивались более чем в два раза, когда отбор витамина В5 осуществляли после отбора с применением пуромицина по сравнению с осуществлением отбора только с применением пуромицина (при сравнении 0 нг Slc5a6 и 1X В5, 10-4Х Н с 100 или 250 нг SLC5A6 и 10Х В5; 10-4Х Н или 10-3Х В5, 10-4Х Н, Фиг. 9С). В целом, это свидетельствует о том, что Slc5a6 и опосредованный витаминами отбор можно также применять в сочетании с отбором при помощи антибиотиков, чтобы предпочтительным образом отбирать клетки, которые опосредуют самые высокие уровни экспрессии трансгена.
Этот подход применяли для экспрессии трансгена, кодирующего терапевтический рекомбинантный белок, а именно моноклональный иммуноглобулин Трастузумаб (IgG). Клетки совместно трансформировали плазмидой, кодирующей как Slc5a6, так и легкую цепь иммуноглобулина, а также другим вектором, экспрессирующим маркер устойчивости к пуромицину и тяжелую цепь иммуноглобулина. Затем клетки отбирали при различных режимах витаминной депривации по В5 или обработки пуромицином (Фиг. 10), а секретируемый IgG Трастузумаб определяли путем окрашивания поверхности клеток с применением флуоресцентного антитела против IgG.
В первую очередь оценивали, какие из условий отбора позволяют получить популяции поликлональных клеток, демонстрирующих самые высокие уровни секреции IgG в супернатантах культур с периодической подпиткой. Клетки, отобранные при помощи пуромицина, позволяли получить только самые низкие уровни секретируемого IgG (условие А+, Фиг. 11). Клетки, отобранные путем витаминной депривации по В5 (В1+) или витаминной депривации с последующим добавлением пуромицина в минимальную среду (B1D+ и B2D+), позволяли получить сравнительно высокие уровни IgG. Клетки, отобранные при помощи пуромицина с последующей витаминной депривацией по В5, позволяли получить титры IgG с промежуточным значением. Учитывая то, что проведение отбора при помощи пуромицина в дополнение к витаминной депривации не позволяло получить достоверное увеличение по сравнению с отбором, при котором обеспечивают депривацию витамина по В5 (Фиг. 11, сравнение B1D+ с В1+); причем далее уделяли внимание проведению анализа клеток, отобранных только путем витаминной депривации с применением клеток, отобранных при помощи пуромицина в качестве контролей.
Наибольшая доля IgG-экспрессирующих клеток в диапазоне от 80 до 90% и наиболее высокие уровни флуоресценции клеточной поверхности наблюдали для пулов поликлональных клеток, отобранных путем витаминной депривации (Фиг. 12А, условие В+). Высокие и вместе с тем сбалансированные уровни мРНК тяжелой и легкой цепи IgG получали при отборе с применением витамина В5, а уровни мРНК транспортера Slc5a6, экспрессию которого осуществляли для целей отбора, оказались довольно низкими по сравнению с уровнями IgG (Фиг. 12В). Обнаружили, что скорость секреции IgG была приблизительно в 3 раза выше для поликлональных популяций, отобранных путем витаминной депривации, по сравнению с отбором с применением антибиотиков, и обнаружили, что экспрессия иммуноглобулина стабильна при продленной культуре в неселективной полной среде, даже когда его секреция была на самых высоких уровнях (Фиг. 13). Когда эти популяции поликлональных клеток оценивали в культурах с периодической подпиткой с применением полной среды для культивирования, титры, превышающие 8 г/л, получали для популяций, отобранных путем витаминной депривации, тогда как титр, полученный при отборе с применением пуромицина, составлял 2 г/л (Фиг. 14). Таким образом, отбор путем витаминной депривации и основанный на гиперэкспрессии SLC5a6 отбор поликлональных популяций позволили получить исключительно высокие титры белка в диапазоне накопления IgG, который получают только случайным образом после утомительной и трудоемкой сортировки и отбора наиболее продуктивных моноклональных популяций.
Пример процесса отбора клеток представлен на Фиг. 18. СНО-клетки совместно трансфицировали в присутствии (условие А) или отсутствии (условия В и С) вектора экспрессии SLC5a6 совместно с GFP или плазмидами легкой или тяжелой цепи IgG или с устойчивостью к пуромицину, после чего отбор указанной культуры проводили либо в присутствии пуромицина (условия А и В), либо в среде для культивирования с недостатком витаминов, содержащей лимитирующие (В5 10-3Х/Н 10-4Х) концентрации витаминов (условие С). Перечекнутый круг указывает на то, что клетки, которые не трансфицировали при помощи вектора экспрессии SLC5a6, не выживали при отборе в среде для культивирования с недостатком витаминов. Можно увидеть, что применяемая здесь плазмида GFP также содержит последовательность MAR. После отбора клетки культивировали в неселективной среде для культивирования до проведения анализа с применением FACS или анализа на определение уровней секреции иммуноглобулина полученных пулов поликлональных клеток (Фиг. 19-20) или во время образования и анализа моклональных популяций (Фиг. 20-21).
Экспрессирующие GFP популяции поликлональных клеток, полученных в указанном процессе, изображенном на Фиг. 18, культивировали в течение 9 дней в неселективной среде и анализировали (Фиг. 19). Анализ с применением цитофлуорометрии для определения флуоресценции GFP обеспечивал профили флуоресценции FACS, на которых представлено обогащение всех GFP + клеток и клеток с высокой экспрессией GFP, полученных из стабильных популяций поликлональных клеток, совместно трансфицированных плазмидами экспрессии Slc5a6, GFP и с устойчивостью к пуромицину (рurо), и отобранных путем культивирования с пуромицином (условия А и В, см. Фиг. 18) или в среде для культивирования с недостатком витаминов (условие С). Долю клеток и среднюю флуоресценцию всех GFP-положительных клеток и высоко флуоресцентных клеток определяли из клеток, ограниченных таким образом, как представлено на Фиг. 19В, и геометрическое среднее GFP-флуоресценции указанных клеток представлено для пулов поликлональных клеток (Фиг. 19С). Можно увидеть, что отбор с применением витамина В5 клеток, трансфицированных при помощи вектора экспрессии SLC5a6, обеспечивал достоверное обогащение GFP-флуоресцентных клеток среди клеток с высокой экспрессией GFP (Фиг. 19В) и достоверно увеличивал геометрическое среднее GFP-флуоресценции.
На Фиг. 20 представлена удельная производительность иммуноглобулина популяций клеток, отобранных с применением пуромицина или путем витаминной депривации. На Фиг. 20А получили общий пул поликлональных клеток, экспрессирующих терапевтический IgG, как показано для условий А, В и С Фиг. 18, и проанализировали удельную производительность IgG. Удельная производительность представлена в пикограммах секретируемого антитела на клетку и в день (данные являются результатом трех независимых биологических экспериментов. Две звездочки: Р<0,05, односторонний Т-тест с равными дисперсиями). На Фиг. 20В представлены результаты, полученные с применением десяти клонов, которые случайным образом выделили путем предельных разведений популяций клеток, полученных при условиях отбора В и С, и определяли удельную производительность IgG. В этом же случае удельная производительность культур клеток при условии С была достоверно выше, чем удельная производительность культур клеток при условии В.
Также анализировали отобранные клеточные клоны. В частности, проанализировали два клона (С_а и С_b), полученные путем предельного разведения пула поликлональных клеток, экспрессирующих SLC5A6 и терапевтический IgG, и отобранные путем витаминной депривации (условие С на Фиг. 18 и 20). Секретируемый IgG, проявляемый на поверхности клетки, метили путем инкубации с люминесцентным антителом, направленным против IgG, и клетки анализировали при помощи цитофлуорометрии, как представлено на Фиг. 21А. Для сравнения представлены профили флуоресценции исходного пула поликлональных клеток С и производного клона С_а. На Фиг. 21В представлена стабильность экспрессии иммуноглобулина двух моноклональных популяций, отобранных путем витаминной депривации. Клоны С_а и С_b поддерживали в полной неселективной среде и осуществляли их пассаж два раза в неделю в течение 30 дней. Удельную производительность популяций клеток, экспрессирующих (в пикограммах) секретируемое антитело на клетку и в день (PCD), анализировали в указанные моменты времени. Можно увидеть, что клоны показали высокую стабильность (уровни PCD снижаются не более, чем на 50%, не более, чем на 40%, не более, чем на 30% не более, чем на 20% или даже не более, чем на 10% или на 5% от исходного уровня при поддержании в полной неселективной среде и пассажах два раза в неделю в течение 30 дней). На Фиг. 21С и D представлены результаты исследований образования иммуноглобулинов культурами клонов С_а и С_b с периодической подпиткой. Указанные клоны выращивали в полной среде для культивирования с периодической подпиткой, и подпитку добавляли на 3, 6, 8 и 10 день. Образцы анализировали на плотность жизнеспособных клеток (Фиг. 21С) и на титр секретируемого антитела при помощи анализа ИФА с применением метода «двойного сэндвича» (Фиг. 21D). В качестве эталона применяли клон BS03 с высокой экспрессией IgG и нетрансфицированные родительские клетки СНО-М. Можно увидеть, что оба клона хорошо представлены относительно клона BS03 с высокой экспрессией IgG.
На Фиг. 22 представлен отбор популяций клеток, продуцирующих различные рекомбинантные белки на высоких уровнях при помощи совместной трансфекции SLC5a6 и отбора путем витаминной депривации. На Фиг. 22А представлены титры, полученные из культур популяций поликлональных клеток с периодической подпиткой, экспрессирующих легко экспрессирующийся IgG, а именно Герцептин, после проведения отбора либо при помощи пуромицина («антибиотик»), либо путем культивирования в обедненной по витаминам среде («метаболический»). На Фиг. 22 В представлены результаты определения процента клеток, экспрессирующих Герцептин, а также средние уровни секреции при помощи изображения колонии. Титры, полученные из популяций поликлональных клеток, продуцирующих трудно экспрессирующийся белок, а именно интерферон-бета, отобранные путем добавления антибиотиков или витаминной депривации, представлены на Фиг. 22С. Можно увидеть, что в особенности титры, полученные из популяций поликлональных клеток, продуцирующих трудно экспрессирующийся белок, здесь интерферон-бета, отобранных путем витаминной депривации, в 3-5 раз превышали те, которые отбирали путем добавления антибиотиков.
Специалисту в данной области будет понятно, что можно осуществить гиперэкспрессию других генов метаболизма белков для аналогичных целей, как представлено, например, на Фиг. 1 и в Таблице 1. Результаты, полученные с применением витамина В5 и Slc5a6, приведены здесь в качестве примеров, тогда как применение транспортеров для других витаминов (например, В1) или применение других генов метаболизма витамина В5 (например, PANK, см. Таблицу 1), также возможно и входит в объем настоящего изобретения.
Аналогичным образом, клетки-хозяева могут быть сконструированы для осуществления экспрессии транспортера и других генов на более низких уровнях для образования клеточных линий с еще более сильными селекционными свойствами. Наконец, применение сред для культивирования клеток с недостатком витаминов B1, В5 или Н или их комбинаций, применяемых в данном исследовании, представляет собой общий подход, который можно применять для увеличения уровней продуцирования клетками, независимо от того, были ли они сконструированы для осуществления гиперэкспрессии одного или более генов метаболизма витаминов, как представлено на Фиг. 14, но и также при применении клеток, модификацию которых не осуществить при уровнях экспрессии генов витаминов, представленных на Фиг. 5. Для специалиста в данной области техники и в пределах объемах настоящего изобретения также будет очевидно, что этот подход можно применять для получения высоких уровней терапевтического белка in vitro с применением культивируемых линий клеток, таких как клетки СНО-М, например, в биореакторе, но и также in vivo с применением первичных клеток, таких как клетки человека, для препаратов на основе генов или клеток, а также для регенеративной медицины.
Вышеприведенное показывает, что популяции поликлональных или моноклональных клеток, продуцирующих рекомбинантные белки на гомогенных и очень высоких уровнях, можно получить с применением кодирующих последовательностей, экспрессирующих белки метаболизма витаминов в качестве селективных маркеров. Было показано, что витаминную депривацию во время режима работы биореактора с периодической загрузкой можно применять для улучшения жизнеспособности клонов клеток и их производительности с точки зрения титра секретируемых рекомбинантных терапевтических белков. Интересно то, что эти эффекты получали путем снижения уровней, например, витаминов В5 или Н, но и также тогда, когда уровни одного из указанных витаминов повышали выше насыщающих уровней. Этот более поздний эффект отметили тогда, когда повышение концентрации В5 выше обычных уровней способствовало отбору клеток, которые экспрессируют высокие уровни селективного гена SLC5a6 при выращивании в присутствии низких количеств витамина Н. Таким образом, оптимальный режим отбора также можно разработать путем повышения концентрации витамина или путем изменения относительных уровней двух витаминов, которые используют один и тот же мембранный транспортер. Таким образом, описанный здесь подход имеет большое значение для отбора и идентификации клонов клеток, которые продуцируют целевой белок до более высоких и стабильных уровней, и, таким образом, применяется меньше усилий и сокращается время скрининга, а также для повышения уровней образования белка и жизнеспособности клеток независимо от происхождения клетки или разработки генов витаминов.
Материалы и Способы
Последовательности генов витаминов и векторные конструкции ДНК
Геномные последовательности и последовательности кДНК витаминов определяли после выравнивания гомологичных генов у мышей SCL5A6, SLC19A2, SLC19A3, TPK1, SLC25A19 с применением программного обеспечения NCBI BLAST. Последовательность транскрипции и накопление соответствующих генов определяли с применением базы данных экспрессии генов SELEXIS СНО-М. CDS (кодирующие последовательности ДНК) и последовательности белков перечислены на Фиг. 15 и Фиг. 16 соответственно.
Библиотеку кДНК СНО-М (SURE СНО-М Cell Line™ (SELEXIS Inc., Сан-Франциско, США)) амплифицировали при помощи обратной транскрипции, полученной из 1 мкг полной РНК, выделенной из 106 клеток СНО-М (набор РНК NucleoSpin™, Macherey-Nagel) с применением Superscript Reverse Transcription Enzyme II и случайных праймеров (Goscript Reverse Transcription System, PROMEGA).
Кодирующие витамин последовательности (CDS) клонировали в вектор pGAPDH-MAR 1-68-GFP, вырезая ген зеленого флуоресцентного белка (GFP) и заменяя его на CDS витамина. Векторы были конструировали следующим образом: CDS амплифицировали из библиотеки кДНК СНО-М при помощи ПЦР (ДНК-полимеразы с высокой достоверностью PHUSION, Finnzymes, THERMO FISHER SCIENTIFIC) от ATG до Stop с применением праймеров, несущих сайт рестрикции HinIII/XbaI для SCL5A6, HinIIIll/FseI для SLC19A2, NcoI/XbaI для SLC19A3, HinIII/XbaI для TPK1, HinIII/XbaI для SLC25A19 (Таблица 4). Затем кДНК-продукты и векторы pGAPDH дважды расщепляли при помощи соответствующих рестриктаз. Наконец, кДНК лигировали в вектор pGAPDH-MAR 1-68, где GFP-последовательность вырезали после расщепления с применением тех же рестриктаз.

Вектор pGAPDH-MAR 1-68-GFP был описан ранее (Girod и др., 2007; Hart и Laemmli, 1998; Grandjean и др., 2011). GFP-белок экспрессировали с применением кассеты экспрессии эукариот, состоящей из энхансера цитомегаловируса человека (CMV) и промотора глицеральдегидов 3-фосфатдегидрогеназы (GAPDH), расположенной выше по каскаду относительно кодирующей последовательности, за которой следует сигнал полиаденилирования вируса обезьян 40 (SV40), терминатор гастрина человека и энхансер SV40 (Le Fourn и др., 2013).
Вектор pSV-puro содержит ген устойчивости к пуромицину (puro) под контролем промотора SV40, происходящего из pRc/RSV-плазмиды (INVITROGEN/LIFE TECHNOLOGIES).
Векторы экспрессии иммуноглобулинов 1-68 filled-IgG1-Lc и 1-68-filled-IgG1-Hc представляли собой те вектора, которые были описаны ранее.
Исследования культуры клеток, стабильной трансформации и стабильных поликлональных линий
Суспензию клеток яичника китайского хомячка (СНО-М) поддерживали в суспензионной культуре в среде без содержания сыворотки SFM4CHO-M Hyclone (SFM, ThermoScientific™), дополненной L-глутамином (РАА, Австрия) и НТ-добавкой (GIBCO, INVITROGEN LIFE SCIENCES), при 37°С, 5% CO2 в увлажненном воздухе. Другой средой для клеток, применяемой для данных экспериментов, являются BalanCD СНО-М Growth А (B-CDfull, Irvine Scientific) и Deficient BalanCD CHO-M Growth A (B-CDmin; Irvine Scientific), дополненные витамином B1 (тиамина гидрохлорид; SIGMA ALDRICH), витамином B5 (кальций DL-пантотенат, TCI) и витамином Н (биотин, SIGMA ALDRICH).
Клетки СНО-М трансформировали при помощи векторов экспрессии расщепляемого PvuI SLC5A6, GFP, пуромицина, IgG1-Hc или IgG1-Lc (см. смеси векторов в Таблице 3) путем электропорации согласно рекомендациям производителя (NEONDEVICES, INVITROGEN).
GFP- и IgG1-продуцирующие линии поликлональных клеток, экспрессирующих Slc5a6 и GFP или IgG, отбирали для дальнейших экспериментов следующим образом: за один день до осуществления трансформации клетки выращивали при концентрации 300000 клеток/мл в селективных по В5 средах, которые состояли из среды B-CDmin, дополненной 7,5 мкМ В1 (1X), 250 нМ В5 (10-3Х) и 5 мкМ Н 10-4Х). После трансформации клетки непосредственно инкубировали в 24-луночном планшете с селективной по В5 средой в течение 24 часов, затем переносили в несколько лунок в зависимости от экспериментов. Для отбора при помощи пуромицина клетки высевали в SFM-среду, дополненную 10 мг/мл пуромицина в течение 2 недель, затем переносили в лунку с SFM-средой в течение 5 дней, затем в 50 мл центрифужные пробирки (spin tubes) с SFM-средой.
Для отбора при помощи витамина В5 клетки высевали в селективную по В5 среду в течение 7-9 дней, затем переносили в неселективные среды SFM, как при отборе при помощи пуромицина.
Для двойного отбора клеток при помощи пуромицина и затем витамина В5 стабильные линии поликлональных клеток сначала отбирали при помощи пуромицина, затем клетки высевали в концентрации 20000 клеток/мл в 24-луночный планшет в селективную по В5 среду в течение 7 дней (применяли среду B-CDfull в качестве отрицательного контроля), затем переносили в лунки с полной SFM-средой в течение 7 дней, а затем высевали в центрифужную пробирку с SFM-средой.
Процент флуоресцентных клеток и интенсивность флуоресценции GFP-положительных клеток определяли при помощи FACS-анализа с применением проточного цитометра CyAn ADP (BECKMAN COULTER). Концентрации иммуноглобулина в супернатантах культуры клеток измеряли при помощи анализа ИФА с применением метода «двойного сэндвича». Накопление транскрипта Slc5a6, GFP, IgG1Lc и IgG1Hc подтверждали при помощи ОТ-количественных ПЦР-исследований перед проведением анализов. Поверхностное окрашивание, титр IgG и предельное разведение осуществляли согласно Le Fourn и др. (2014).
Количественный ПЦР-анализ
Для проведения количественного ПЦР-анализа (кПЦР) общую РНК экстрагировали из 106 клеток и обратно транскрибировали в кДНК. Накопление транскриптов количественно оценивали при помощи кПЦР с применением полимеразного набора SYBR Green-Taq от Eurogentec Inc и аппарата ПЦР ABI Prism 7700 PCR (Applied Biosystems) и с применением праймеров Slc5a6-qRT-F и Slc5a6-qRT-R, перечисленных в Таблице 4. Уровни транскрипции нормировали по уровням конститутивного гена GAPDH.
Статистический анализ
Результаты выражены как средние значения ± стандартная ошибка среднего (SEM). Статистический анализ проводили с применением двухстороннего t-теста Стьюдента. Звездочки на панелях Фигур относятся к статистическим вероятностям. Статистические значения вероятности менее 0,05 считались достоверными.
Понятно то, что системы (векторы/клетки и т.д.), способы и наборы согласно настоящему изобретению могут быть включены в виде множества вариантов реализации, только некоторые из которых раскрыты в настоящем документе. Специалисту будет понятно, что существуют другие варианты реализации и они не отклоняются от сущности изобретения. Таким образом, описанные варианты реализации являются иллюстративными и не должны истолковываться как ограничивающие.
Список литературы
Birch, J. R. и Racher, A.J. (2006) Antibody production. Adv. Drug Deliv. Rev., 58, 671-85.
Cacciatore, J.J., Chasin, L.А. и Leonard, E.F. (2010). Gene amplification and vector engineering to achieve rapid and high-level therapeutic protein production using the Dhfr-based CHO cell selection system. Biotechnol. Adv., 28, 673-81.
Brunetti D, Dusi S, Morbin M, Uggetti A, Moda F,  I, Giordano C,
I, Giordano C,  G, Cozzi A, Levi S, Hayflick S, Tiranti V. (2012). Pantothenate kinase-associated neurodegeneration: altered mitochondria membrane potential and defective respiration in Pank2 knock-out mouse model. Hum. Mol. Genet. 21:5294-305.
G, Cozzi A, Levi S, Hayflick S, Tiranti V. (2012). Pantothenate kinase-associated neurodegeneration: altered mitochondria membrane potential and defective respiration in Pank2 knock-out mouse model. Hum. Mol. Genet. 21:5294-305.
Chusainow, J., Yang, Y.S., Yeo, J.H., Toh, P.С.и др., (2009). A study of monoclonal antibody-producing CHO cell lines: what makes a stable high producer? Biotechnol. Bioeng. 102, 1182-1196.
Dean, J. и P. Reddy (2013). «Metabolic analysis of antibody producing CHO cells in fed-batch production». Biotechnology and bioengineering 110(6): 1735-1747.
Du Z, Treiber D, McCarter JD, Fomina-Yadlin D, Saleem RA, McCoy RE, Zhang Y, Tharmalingam T, Leith M, Follstad BD, Dell B, Grisim B, Zupke C, Heath C, Morris AE, Reddy P. (2015). Use of a small molecule cell cycle inhibitor to control cell growth and improve specific productivity and product quality of recombinant proteins in CHO cell cultures. Biotechnol. Bioeng. 112: 141-155.
Garcia M., Leonardi R, Zhang YM, Rehg JE, Jackowski S. (2012). Germline deletion of pantothenate kinases 1 and 2 reveals the key roles for CoA in postnatal metabolism. PLoS One. 7:e40871.
P.A. Girod, D.Q. Nguyen, D. Calabrese, S. Puttini, M. Grandjean, D. Martinet, A. Regamey, D. Saugy, J.S. Beckmann, P. Bucher, N. Mermod (2007). Genome-wide prediction of matrix attachment regions that increase gene expression in mammalian cells. Nat. Methods, 4 (2007), pp. 747-753.
Ghosal, A., N. Lambrecht, и др. (2013). «Conditional knockout of the Slc5a6 gene in mouse intestine impairs biotin absorptions American journal of physiology. Gastrointestinal and liver physiology 304(1): G64-71.
M. Grandjean, P.A. Girod, D. Calabrese, K. Kostyrko, M. Wicht, F. Yerly, C. Mazza, J.S. Beckmann, D. Martinet, N. Mermod (2011). High-level transgene expression by homologous recombination-mediated gene transfer. Nucleic Acids Res., 39, p. e104.
C.M. Hart, U.K. Laemmli (1998). Facilitation of chromatin dynamics by SARs. Curr. Opin. Genet. Dev., 8, pp. 519-525.
Jostock, T. (2011) Expression of Antibody in Mammalian Cells Cell Engineering, 7,1-24.
V. Le Fourn, P.A. Girod, M. Buceta, A. Regamey, N. Mermod (2014). CHO cell engineering to prevent polypeptide aggregation and improve therapeutic protein secretion. Metab. Eng., 21, pp. 91-102.
Nam JH, Zhang F, Ermonval M, Linhardt RJ, Sharfstein ST. (2008). The effects of culture conditions on the glycosylation of secreted human placental alkaline phosphatase produced in Chinese hamster ovary cells. Biotechnol. Bioeng. 100:1178-1192.
Rothem, L., B. Berman, и др. (2005). «The reduced folate carrier gene is a novel selectable marker for recombinant protein overexpression». Molecular pharmacology 68(3): 616-624.
Sajan E, Matanguihan R, Heidemann R, Abu-Absi S, Asuncion W, Yamasaki G, Wu X, Chen J, Murphy JE, Zhang C. (2010). The effect of bioreactor pH and temperature on protein glycosylation in perfusion cultures of mammalian cells. ESACT Proc. 4:785-788.
Sampathkumar SG, Jones MB, Meledeo MA, Campbell CT, Choi SS, Hida K, Gomutputra P, Sheh A, Gilmartin T, Head SR., и др. (2006). Targeting glycosylation pathways and the cell cycle: Sugar-dependent activity of butyrate-carbohydrate cancer prodrugs. Chem. Biol. 13:1265-1275.
Schlatter, S., Stansfield, S.H., Dinnis, D.M., Racher, A.J., и др., (2005) On the optimal ratio of heavy to light chain genes for efficient recombinant antibody production by CHO cells. Biotechnol. Prog. 21, 122-133.
Trummer E, Fauland K, Seidinger S, Schriebl K, Lattenmayer C, Kunert R, Vorauer-Uhl K, Weik R, Borth N, Katinger H, и др. (2006). Process parameter shifting: Part II. Biphasic cultivation-A tool for enhancing the volumetric productivity of batch processes using Epo-Fc expressing CHO cells. Biotechnol. Bioeng. 94:1045-1052.
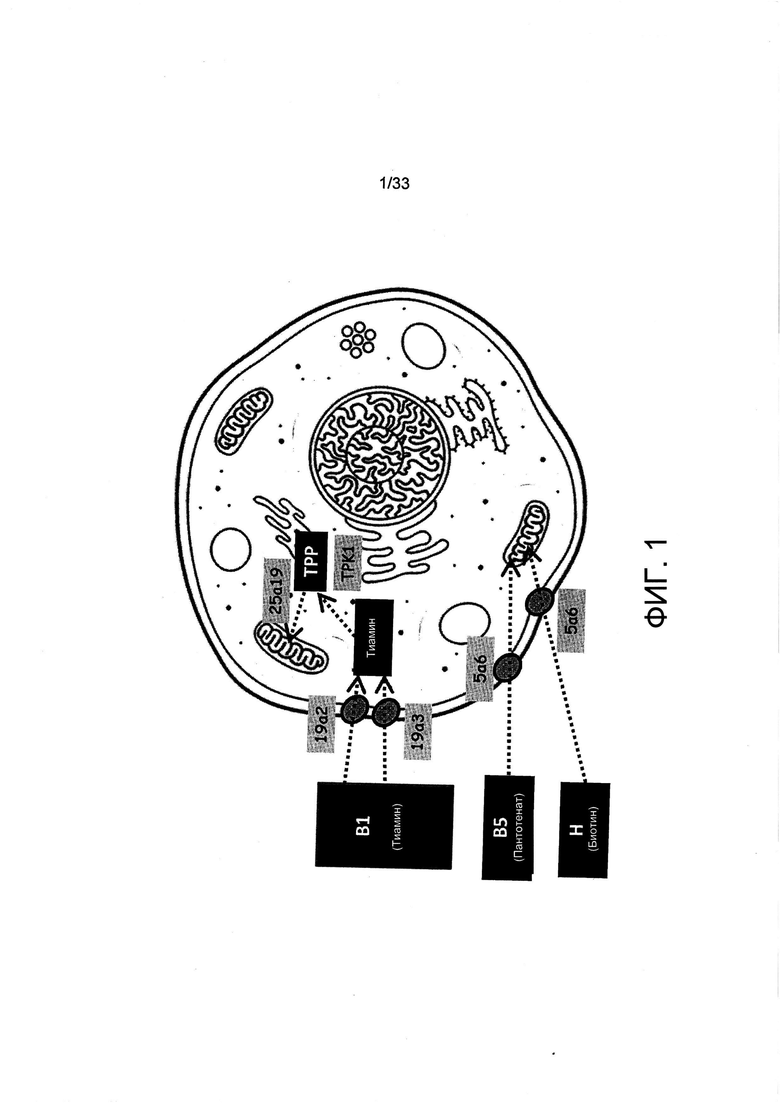
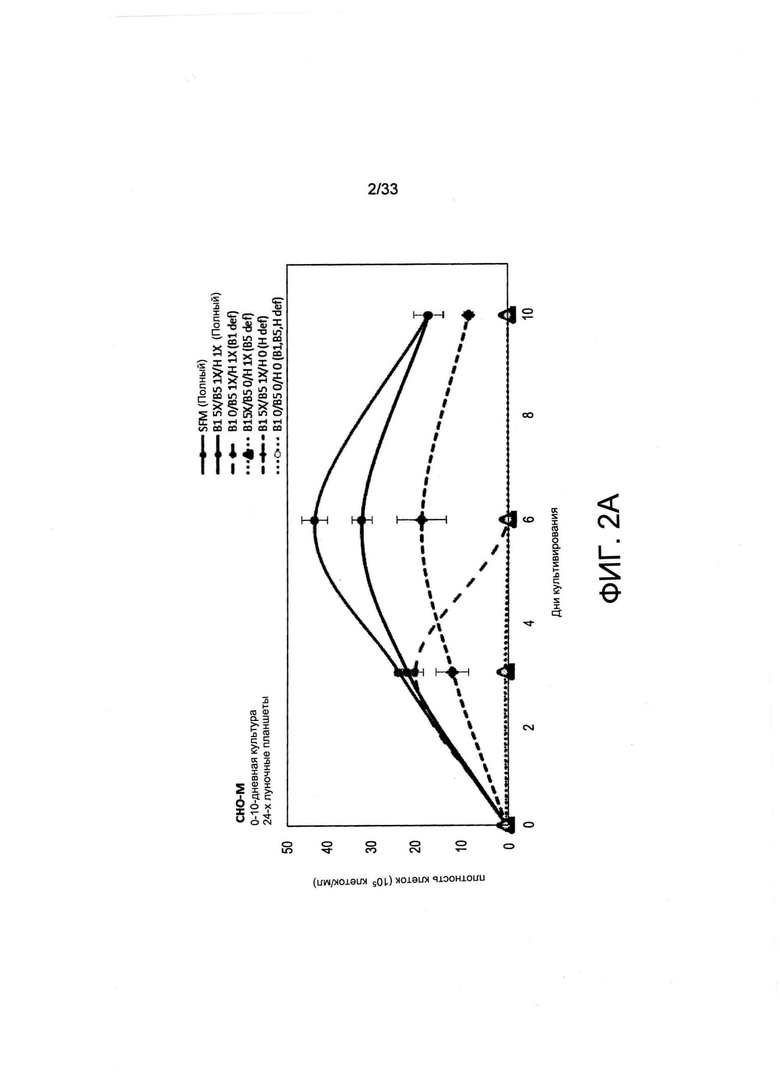
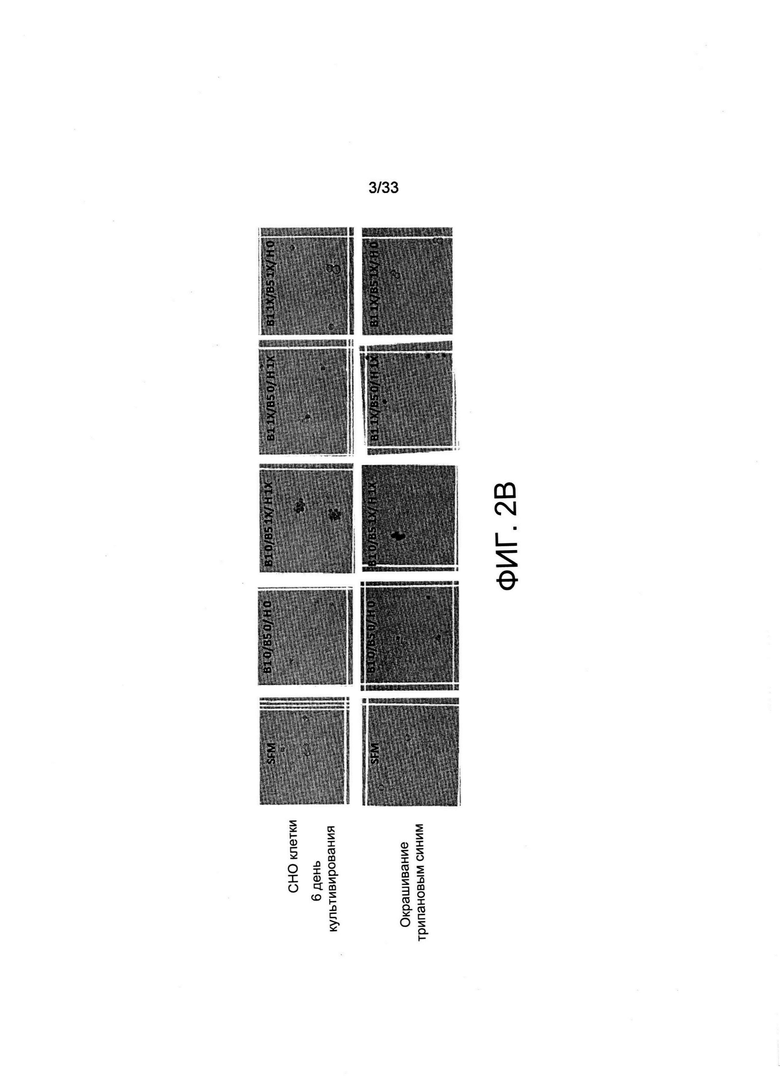
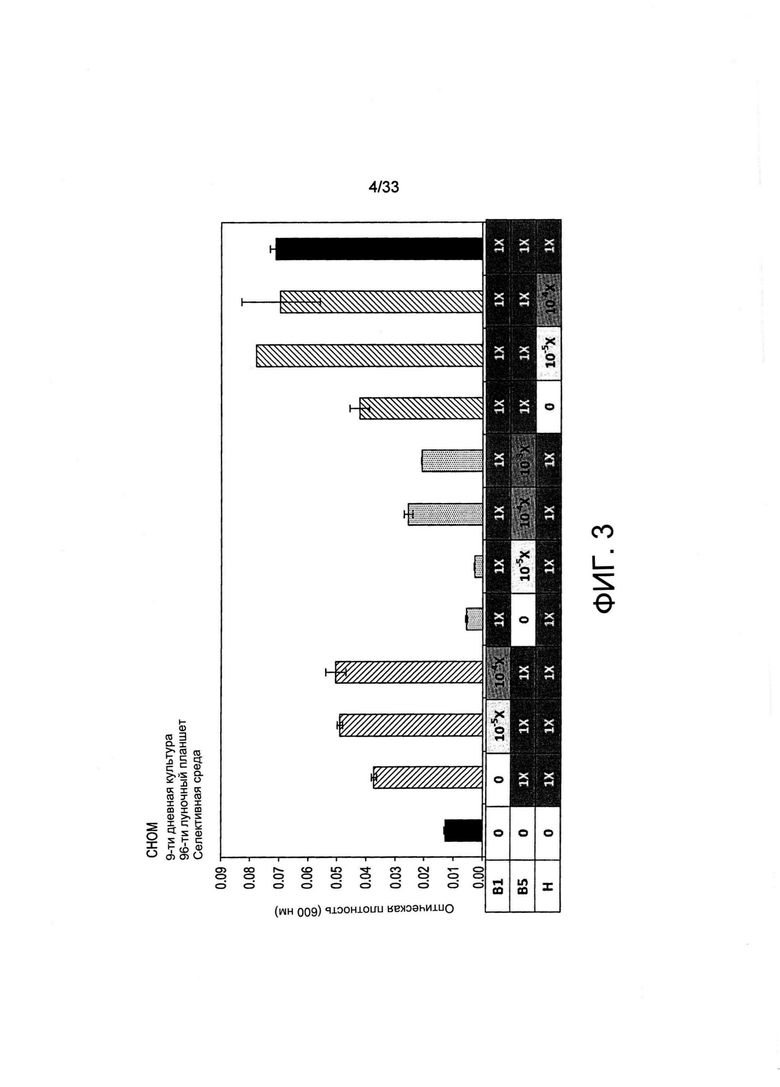
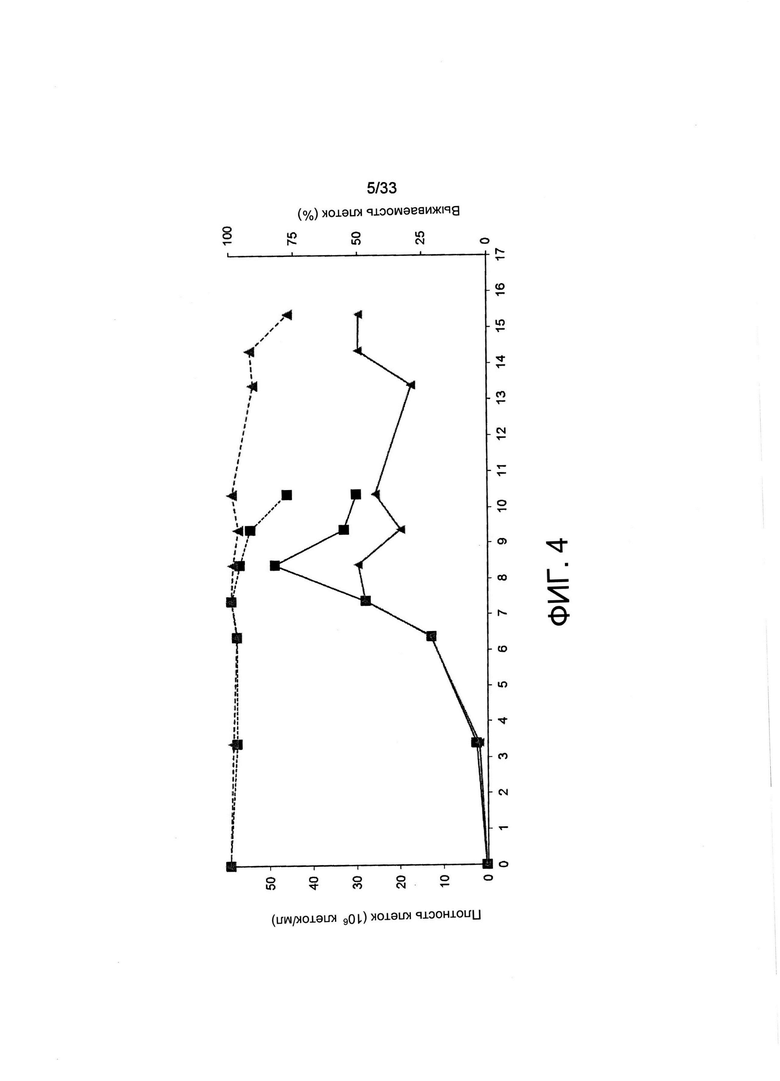
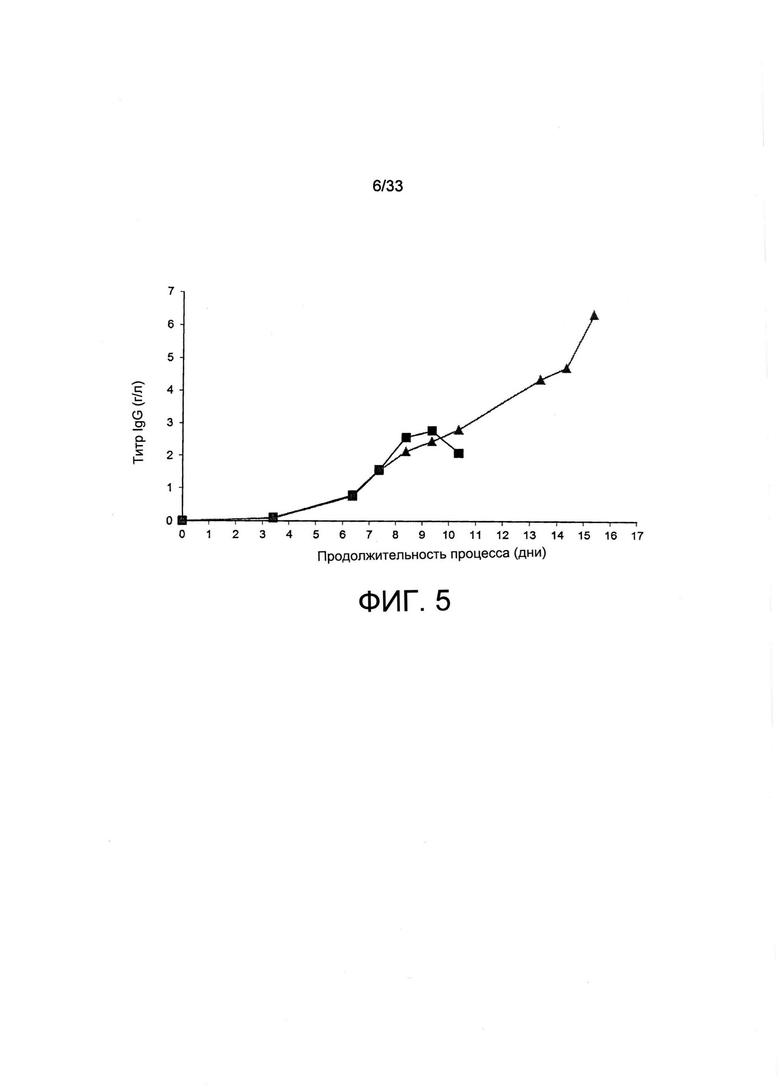
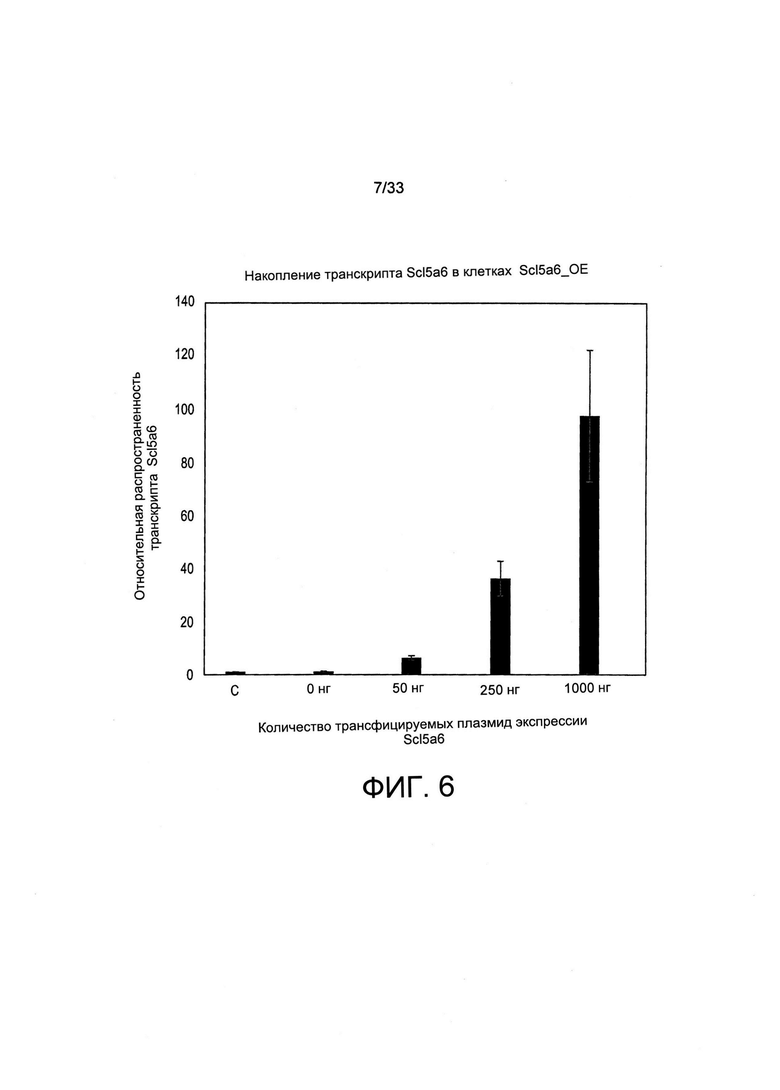
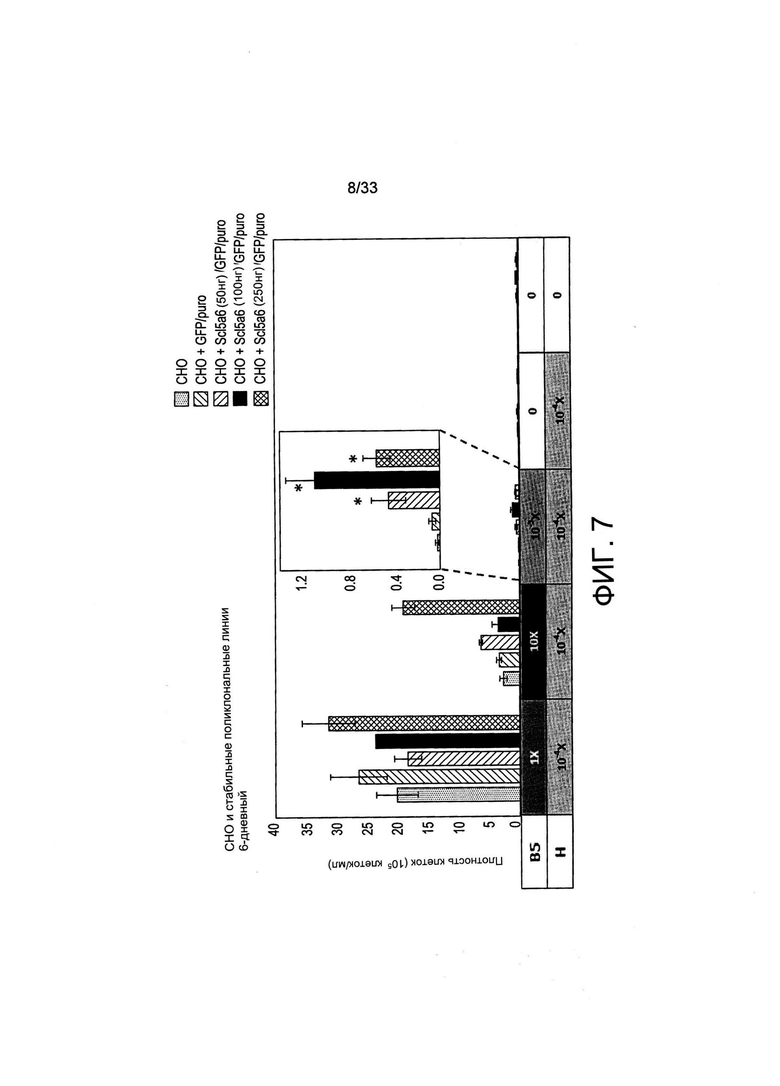
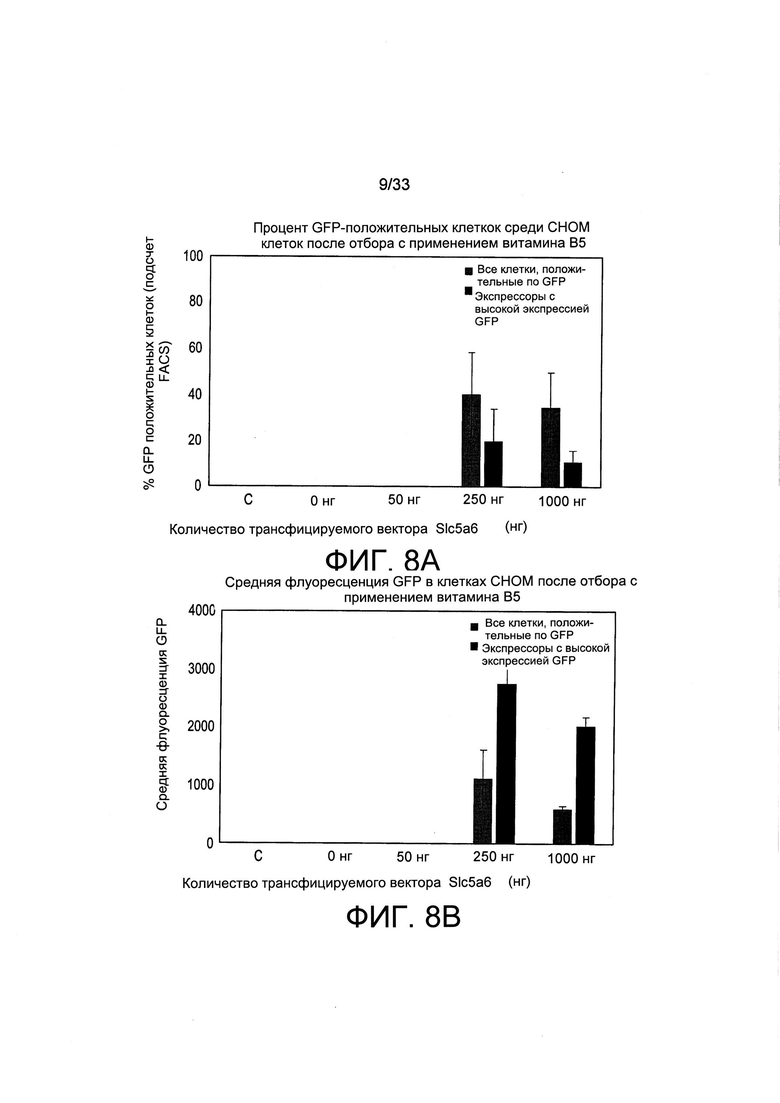
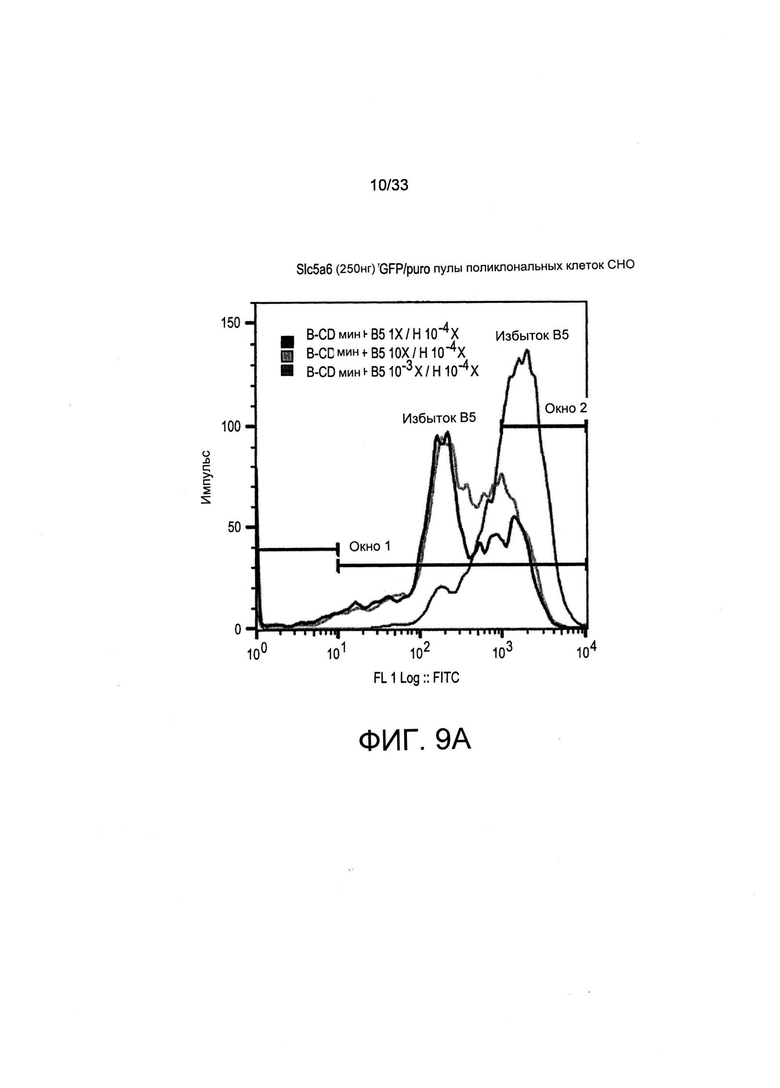
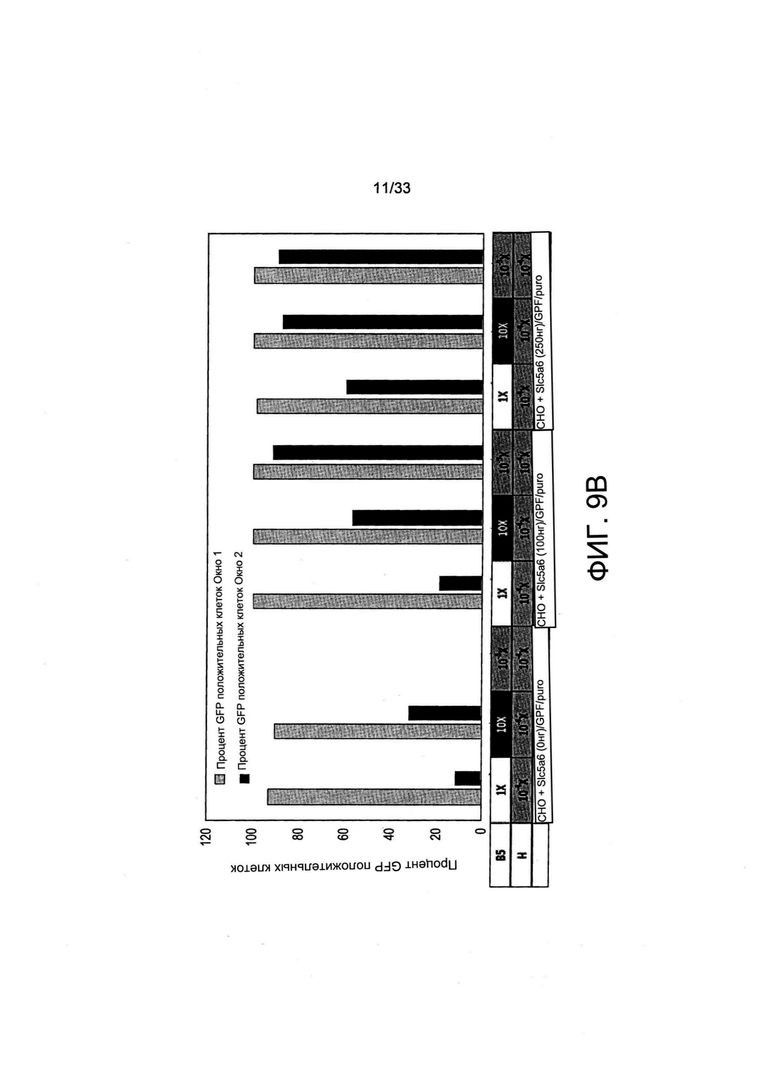

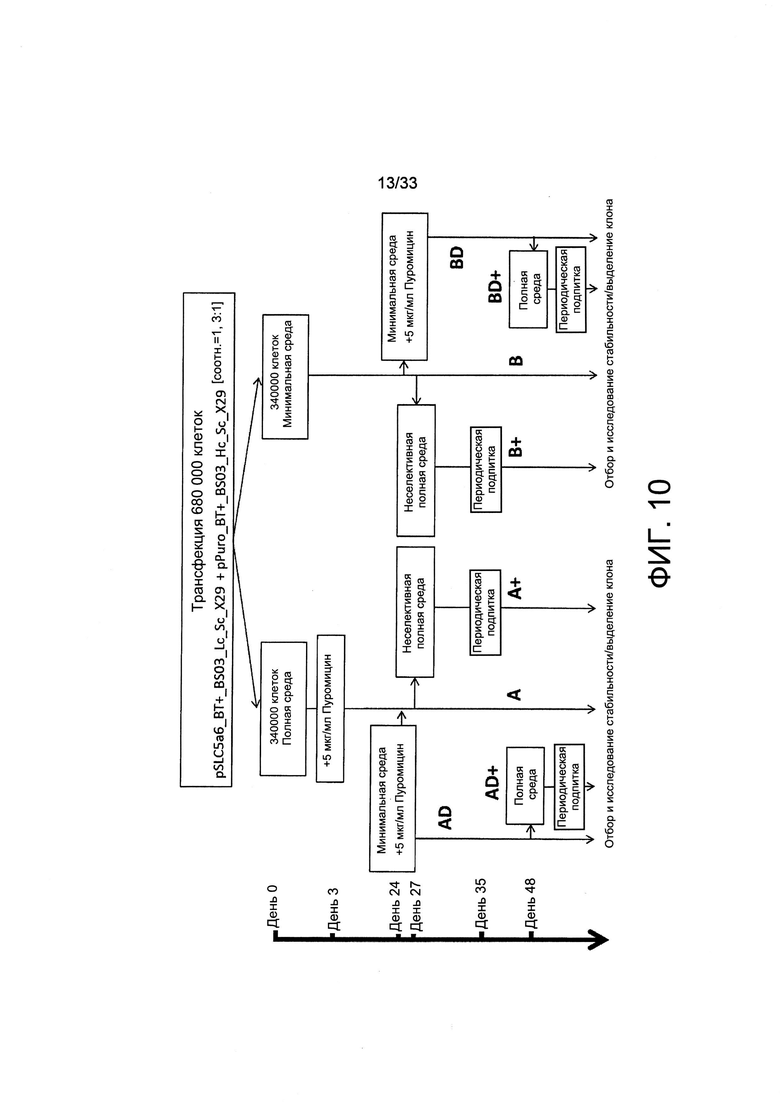
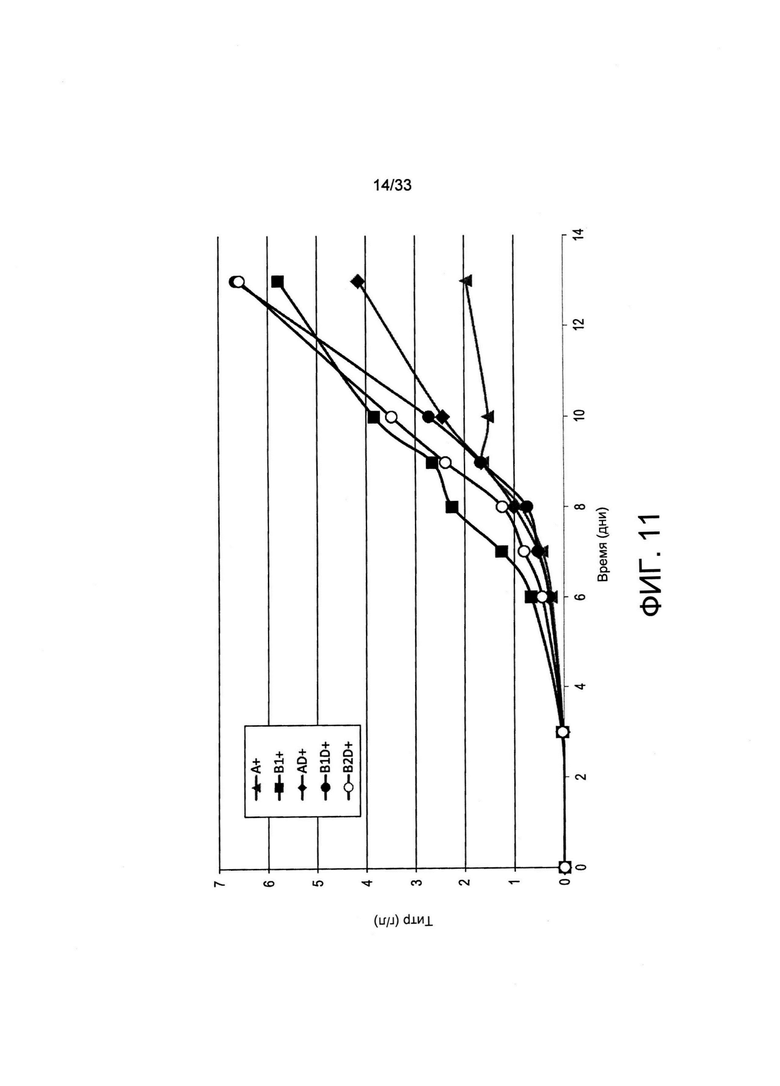
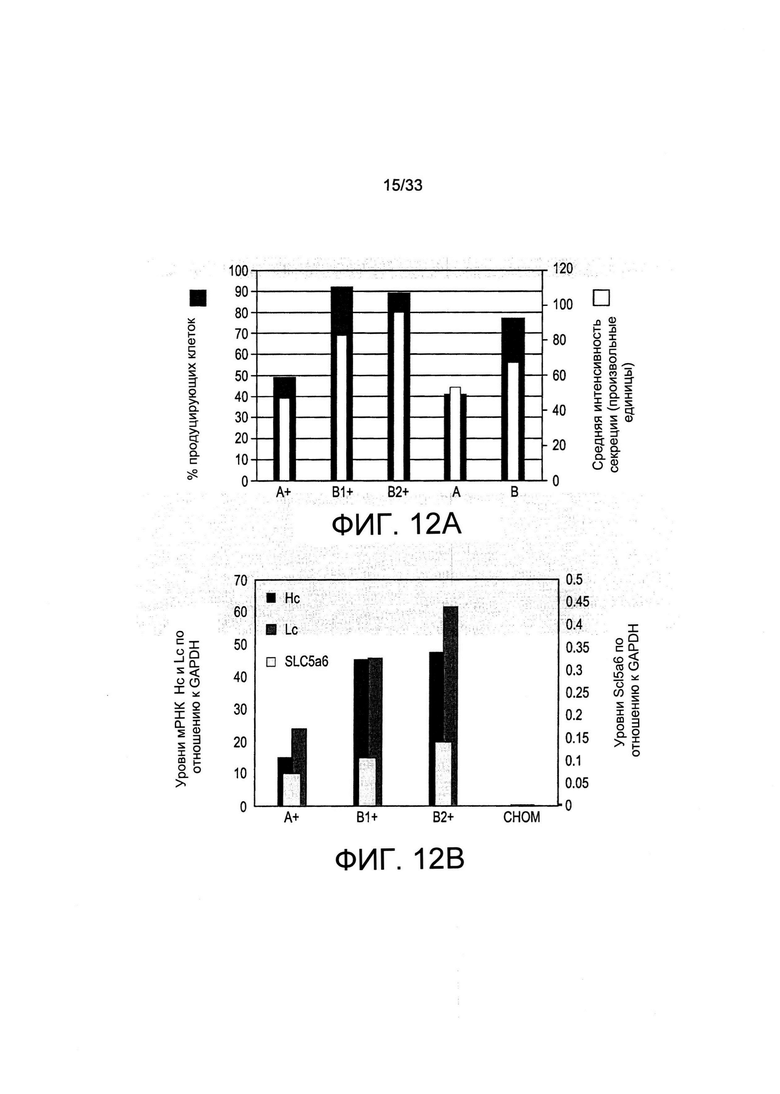
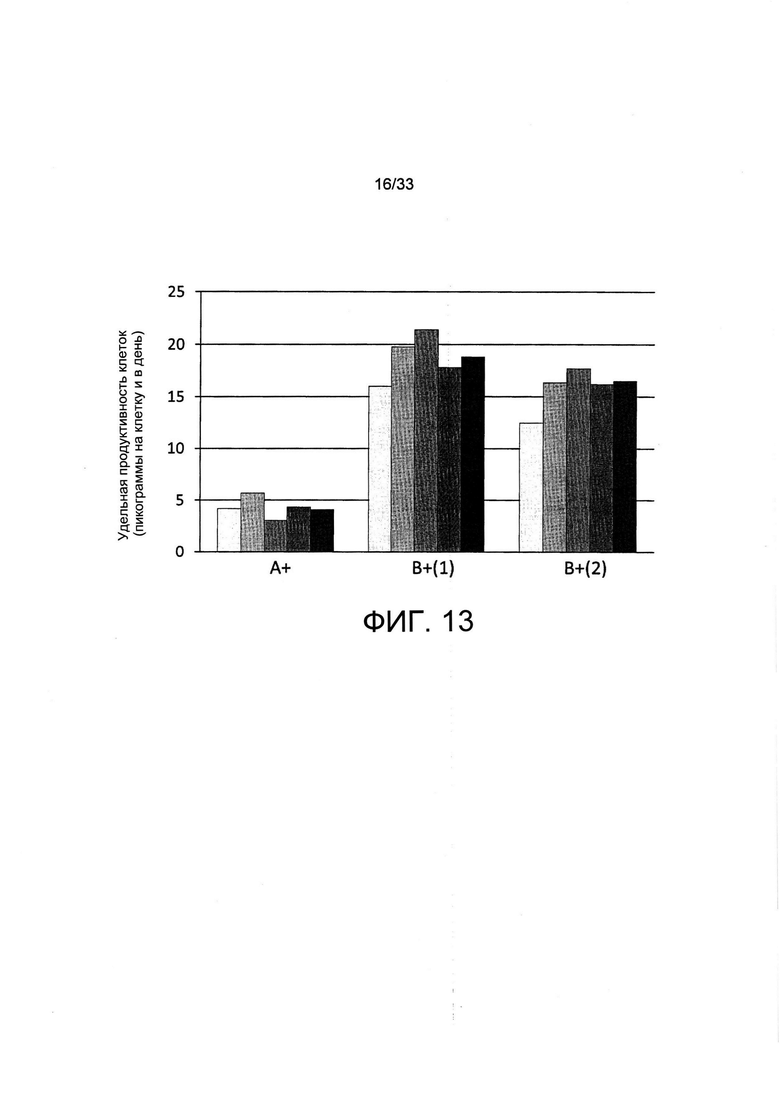
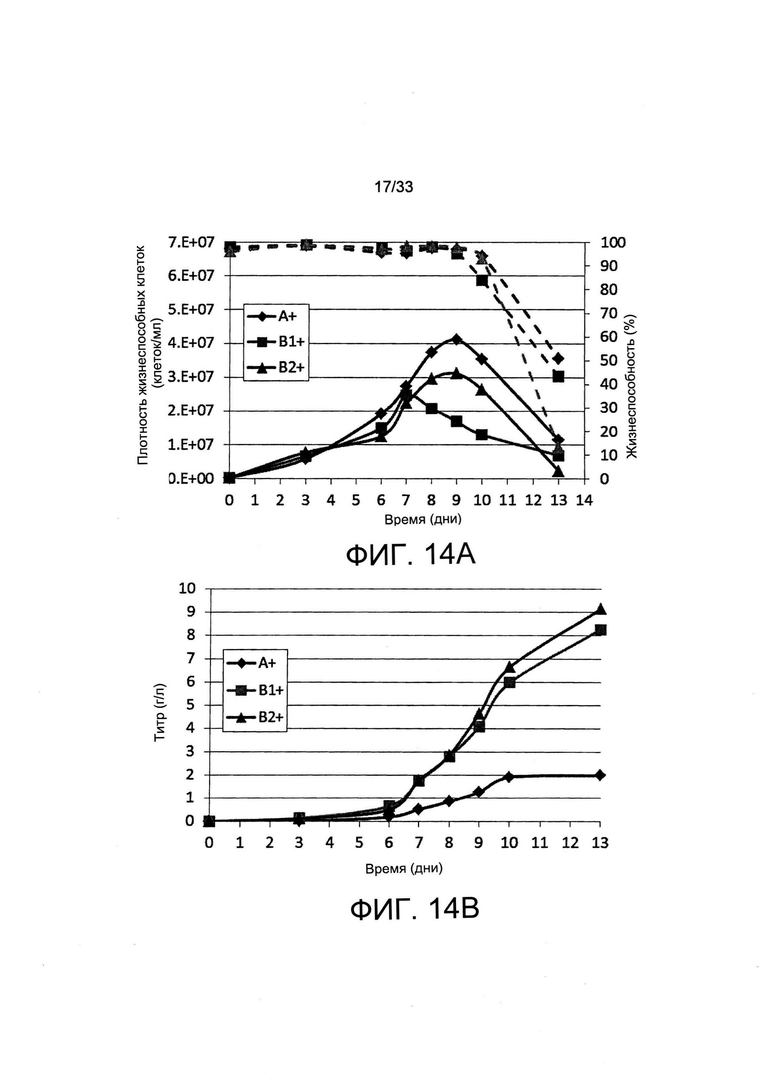









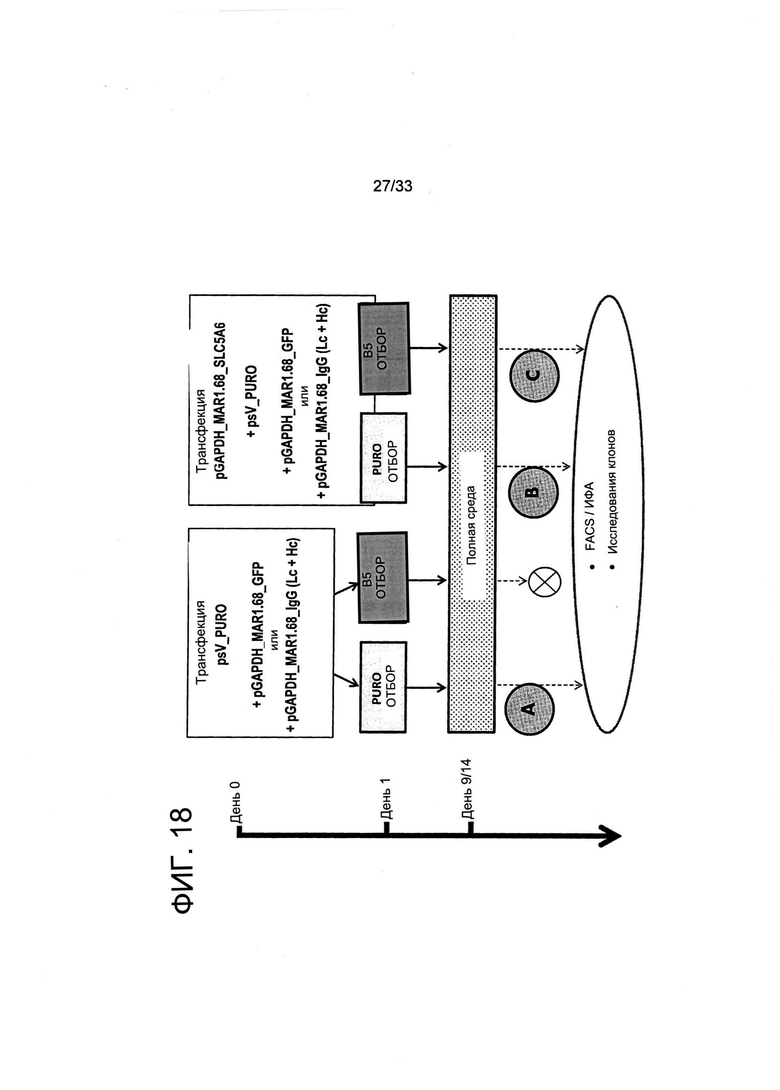
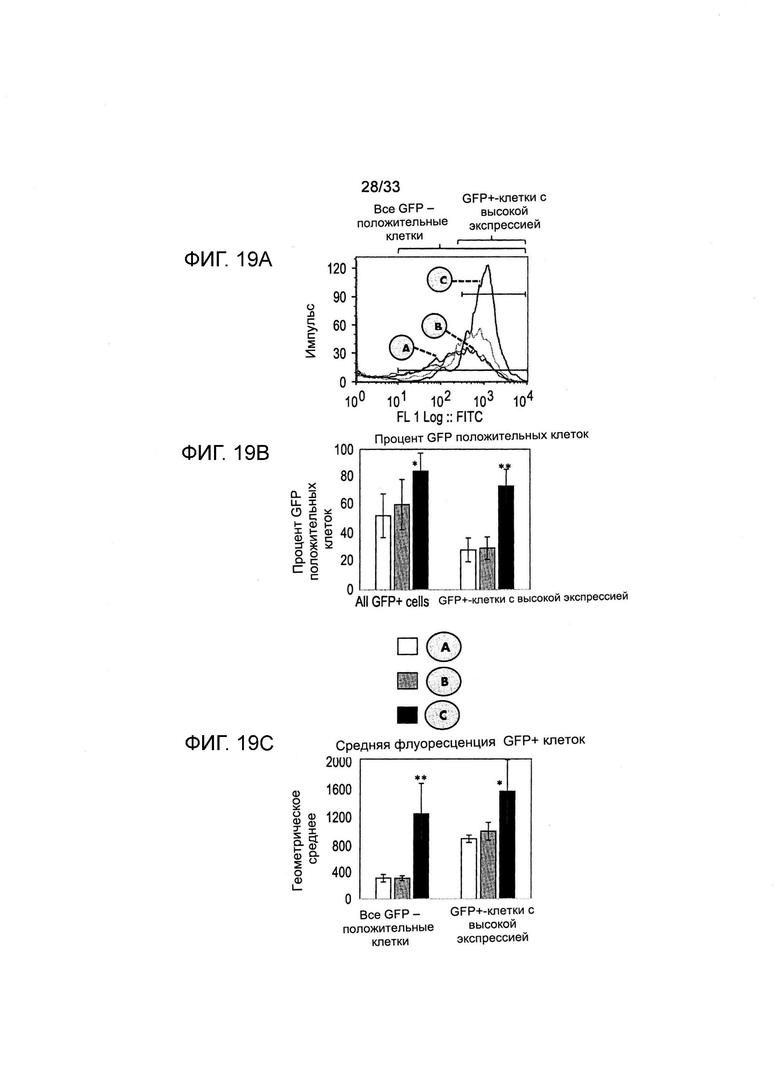
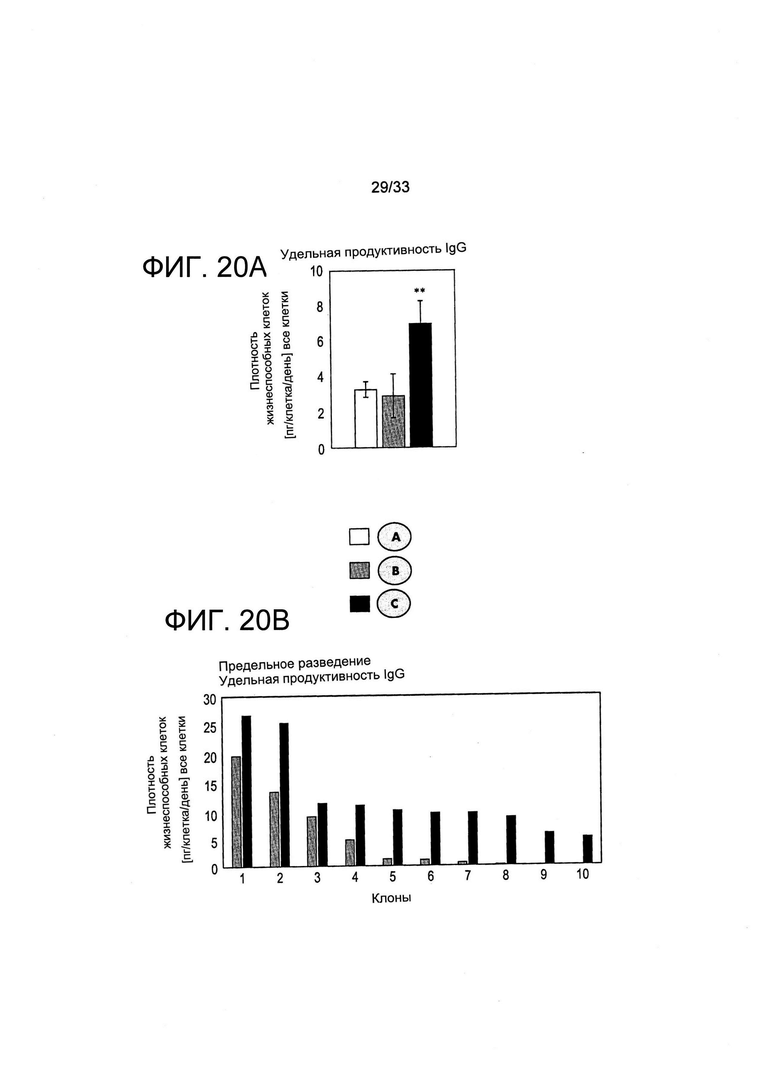
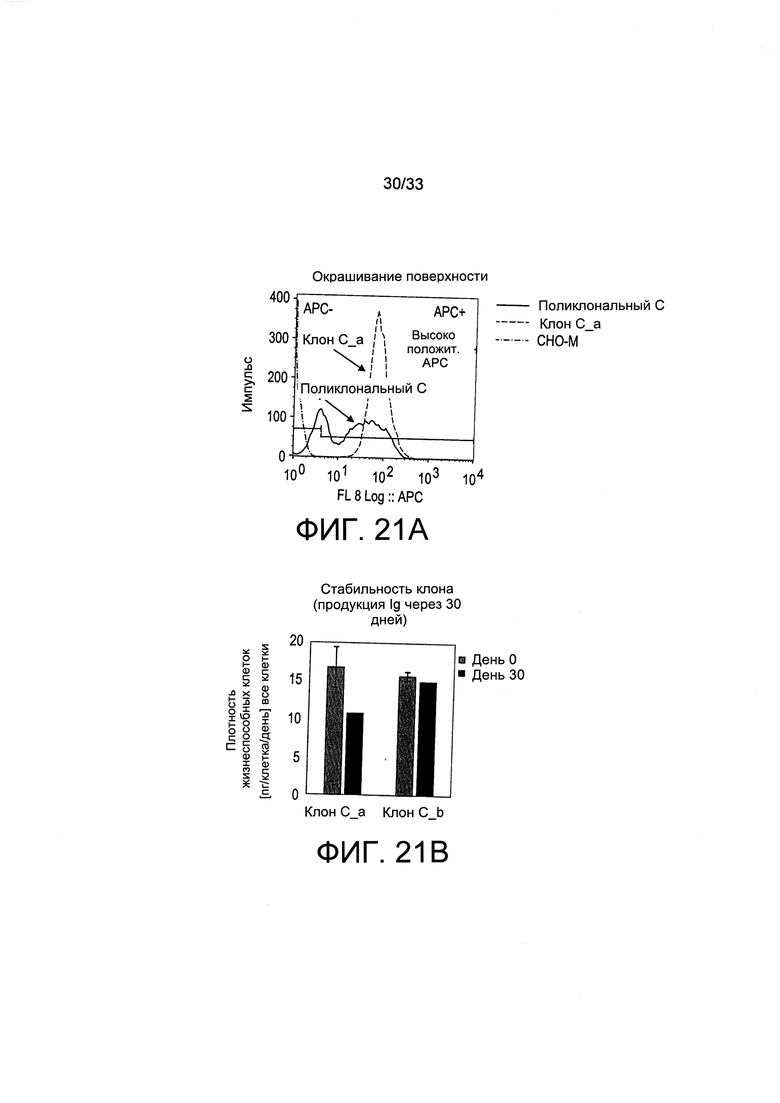
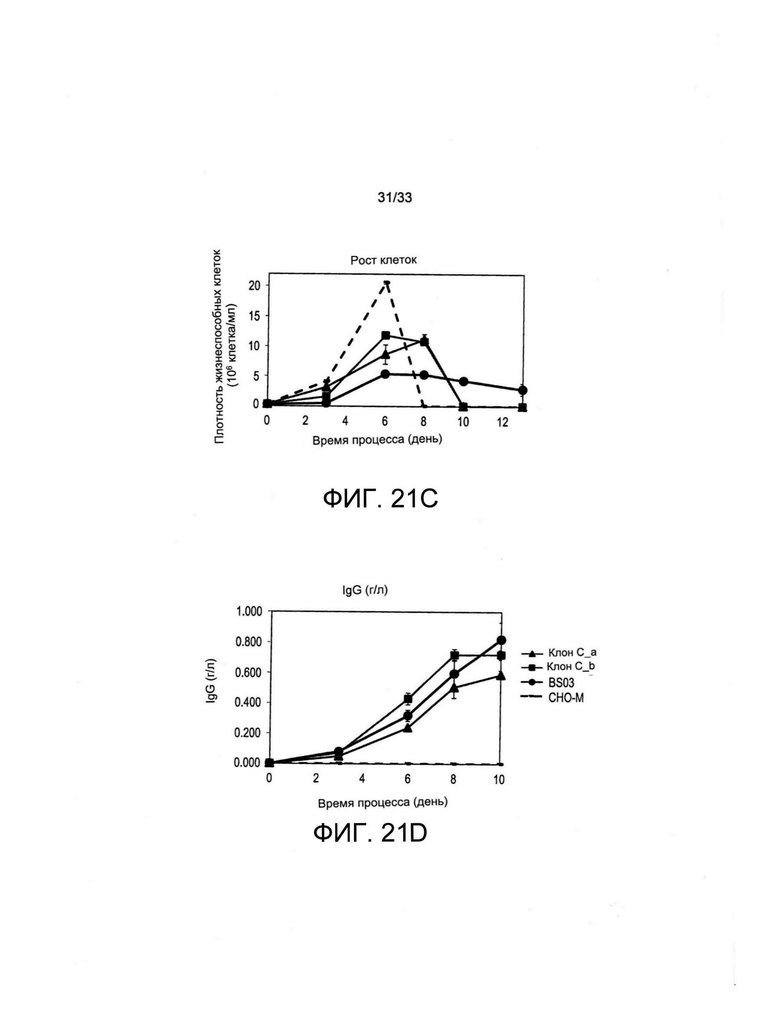
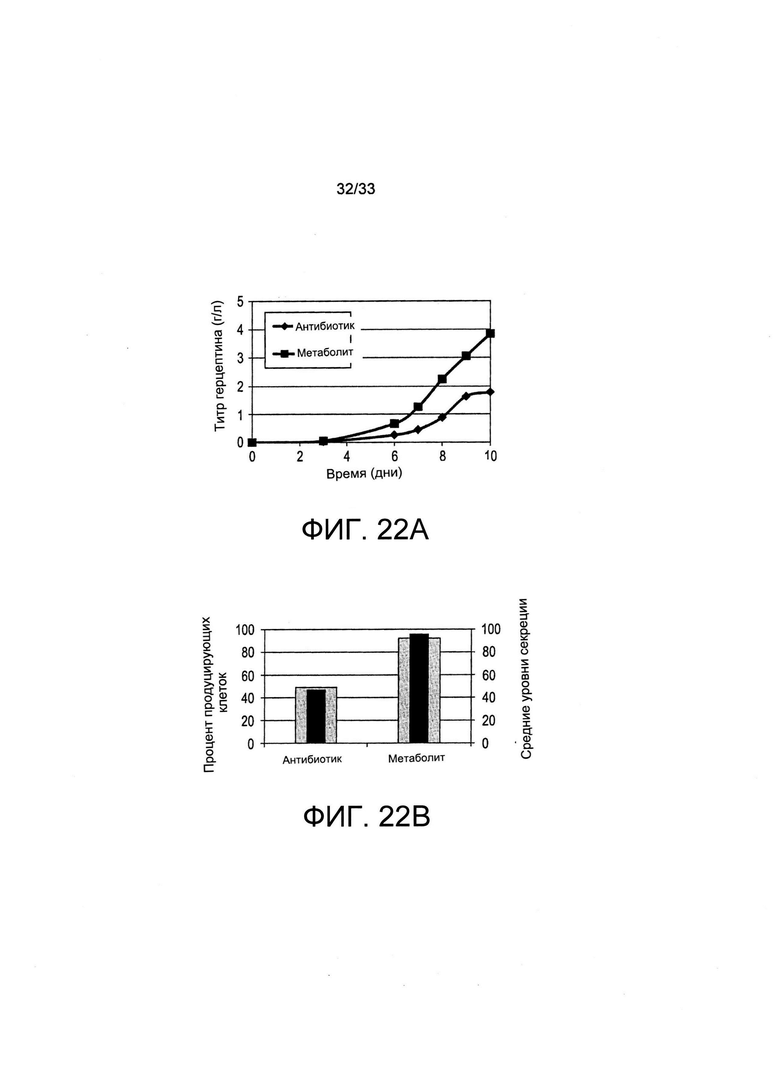
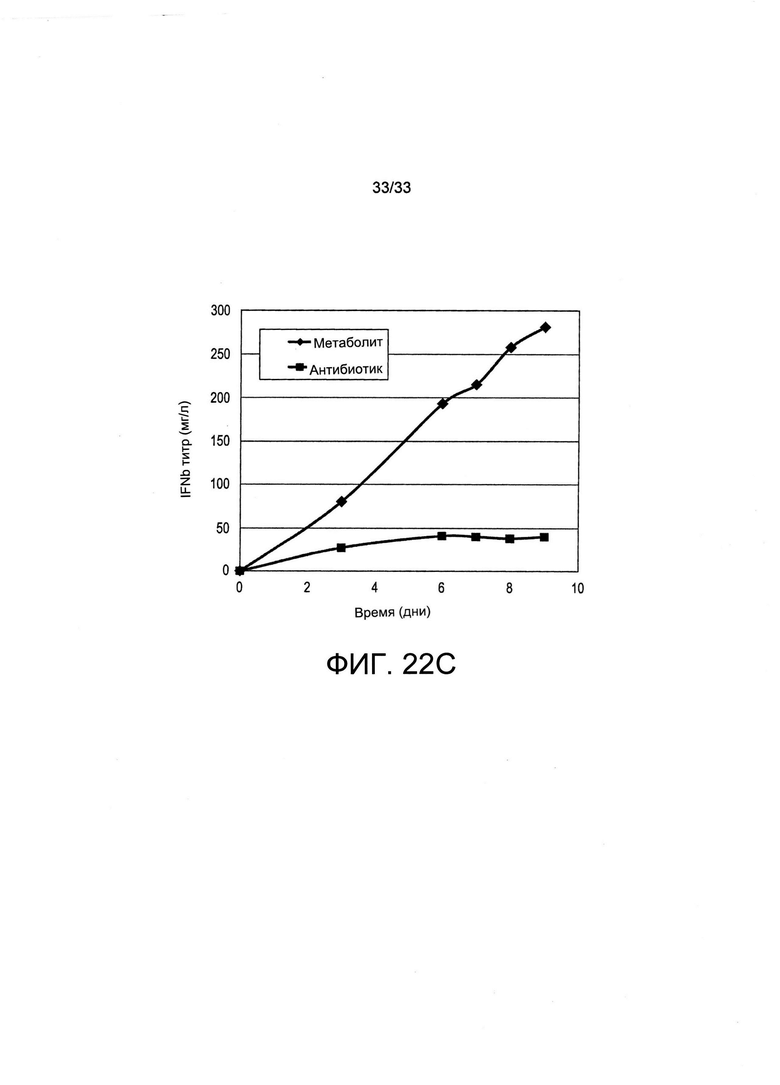
Изобретение относится к биотехнологии и представляет собой эукариотические системы экспрессии и способы отбора линий клеток млекопитающих, которые продуцируют целевые белки, такие как терапевтические белки. Указанные системы и способы способствуют простому и быстрому отбору клеток, опосредующих высокие уровни продукции рекомбинантного белка. Указанные системы и способы снижают усилия и время, необходимые для обеспечения доступности нового терапевтического белка для пациентов, а также увеличивают производительность клеток в биореакторе. 8 н. и 24 з.п. ф-лы, 22 ил., 4 табл.
1. Система для оптимизации экспрессии целевого белка в эукариотических клетках, содержащая:
- по меньшей мере один первый полинуклеотид, кодирующий по меньшей мере один белок метаболизма витаминов под контролем по меньшей мере одной первой регуляторной последовательности, и
- под контролем по меньшей мере одной второй регуляторной последовательности, по меньшей мере один сайт расщепления эндонуклеазой рестрикции и/или по меньшей мере один второй полинуклеотид, кодирующий по меньшей мере один целевой белок.
2. Система по п. 1, отличающаяся тем, что указанный по меньшей мере один белок метаболизма витаминов представляет собой белок транспорта витаминов.
3. Система по п. 1, отличающаяся тем, что указанный по меньшей мере один второй полинуклеотид встроен в указанный по меньшей мере один сайт расщепления эндонуклеазой рестрикции.
4. Система по п. 2 или 3, отличающаяся тем, что указанный белок транспорта витаминов переносит растворимый витамин, такой как витамин B1, B5 и/или H.
5. Система по любому из пп. 2-4, отличающаяся тем, что указанный белок транспорта витаминов представляет собой THTR-1, THTR-2, TPK, TPC и/или SMVT (натрий-зависимый мультивитаминный транспортер).
6. Система по любому из пп. 2-5, отличающаяся тем, что указанный белок транспорта витаминов представляет собой SMVT.
7. Система по любому из предшествующих пунктов, отличающаяся тем, что указанная система представляет собой по меньшей мере один вектор экспрессии.
8. Система по п. 7, отличающаяся тем, что отдельный вектор содержит указанный по меньшей мере один первый и указанный по меньшей мере один второй полинуклеотид.
9. Система по любому из предшествующих пунктов, отличающаяся тем, что указанная первая и/или указанная вторая регуляторная последовательность представляют собой промоторы, энхансеры, области контроля локуса (LCR), участки прикрепления к ядерному матриксу (MAR), участки прикрепления к ядерному скаффолду (SAR), инсуляторные элементы и/или ДНК, ассоциированные с ядерным матриксом.
10. Набор для трансфекции эукариотической клетки, содержащий в одном контейнере указанную систему по любому из предшествующих пунктов и содержащий во втором контейнере инструкции по применению указанной системы.
11. Набор по п. 10, дополнительно содержащий среду для культивирования клеток, предпочтительно содержащую лимитирующую и/или насыщающую концентрацию по меньшей мере одного витамина.
12. Набор по п. 11, отличающийся тем, что указанная среда для культивирования клеток содержит лимитирующую и/или насыщающую концентрацию витамина B1, B5 и/или H.
13. Рекомбинантная эукариотическая клетка для экспрессии целевого белка, содержащая систему по любому из пп. 1-9 и/или
содержащая повышающую или понижающую мутацию белка метаболизма витаминов; при этом указанная рекомбинантная эукариотическая клетка содержит полинуклеотид, кодирующий целевой белок, и при этом указанный белок метаболизма витаминов является необязательно присущим указанной клетке.
14. Рекомбинантная эукариотическая клетка по п. 13, отличающаяся тем, что указанная клетка представляет собой клетку яичника китайского хомячка (CHO).
15. Рекомбинантная эукариотическая клетка по п. 13 или 14, отличающаяся тем, что указанный по меньшей мере один первый полинуклеотид или последовательность, регулирующая экспрессию указанного по меньшей мере одного первого полинуклеотида, содержит повышающую или понижающую мутацию.
16. Рекомбинантная эукариотическая клетка по п. 13 или 14, отличающаяся тем, что указанный белок метаболизма витаминов нарушает метаболизм витаминов и/или связывает витамин внутри клетки.
17. Рекомбинантная эукариотическая клетка по п. 16, отличающаяся тем, что указанный белок метаболизма витаминов представляет собой пантотенатную киназу 1, 2 и/или 3 и/или тиаминпирофосфаткиназу, такую как TPK1 (тиаминпирофосфаткиназа 1).
18. Рекомбинантная эукариотическая клетка по любому из пп. 13-17, содержащая указанную систему, причем указанный белок метаболизма витаминов является селектируемым маркером для указанной рекомбинантной эукариотической клетки и указанная рекомбинантная эукариотическая клетка продуцирует и предпочтительно секретирует указанный целевой белок.
19. Среда для культивирования эукариотических клеток, содержащая указанные рекомбинантные эукариотические клетки по любому из пп. 13-18, экспрессирующие (i) белок транспорта витаминов в качестве селектируемого маркера и (ii) указанный целевой белок.
20. Среда для культивирования клеток по п. 19, отличающаяся тем, что указанная среда представляет собой среду с лимитирующей концентрацией B5 или среду с насыщающей концентрацией B5, но с лимитирующей концентрацией или нелимитирующей концентрацией H; или среду с насыщающей концентрацией H, но с лимитирующей или нелимитирующей концентрацией B5, предпочтительно среду с лимитирующей концентрацией B5 или среду с насыщающей концентрацией B5, но лимитирующей концентрацией H.
21. Способ культивирования и, необязательно, отбора рекомбинантных эукариотических клеток, включающий:
- обеспечение системы для оптимизации экспрессии по любому из пп. 1-9,
- обеспечение эукариотических клеток, причем жизнеспособность, рост и/или деление указанных эукариотических клеток зависит от поглощения витаминов,
- введение указанной системы в указанные эукариотические клетки для получения указанных рекомбинантных эукариотических клеток, экспрессирующих указанный белок метаболизма витаминов,
- культивирование указанных эукариотических клеток в среде для культивирования клеток и
- необязательно отбор посредством указанного белка метаболизма витаминов, который предпочтительным образом экспрессируется на поверхности указанных рекомбинантных эукариотических клеток.
22. Способ по п. 21, отличающийся тем, что указанные рекомбинантные эукариотические клетки стабильно экспрессируют по меньшей мере один целевой белок, кодируемый по меньшей мере одним вторым полинуклеотидом.
23. Способ по п. 21, отличающийся тем, что указанная среда представляет собой среду с лимитирующей концентрацией B5 или среду с насыщающей концентрацией B5, но с лимитирующей концентрацией H.
24. Применение белка метаболизма витаминов в качестве селективного маркера для отбора рекомбинантной эукариотической клетки, стабильно экспрессирующей целевой белок, при этом жизнеспособность, рост и/или деление указанной клетки зависит от поглощения витаминов.
25. Среда для культивирования для отбора рекомбинантной эукариотической клетки, стабильно экспрессирующей целевой белок, путем применения белка метаболизма витаминов в качестве селективного маркера, содержащая по меньшей мере один витамин:
- в концентрации менее чем 10 нМ и/или
- в концентрации 20 мкМ или более, при этом указанный по меньшей мере один витамин представляет собой незаменимый витамин.
26. Среда для культивирования по п. 25, отличающаяся тем, что указанный по меньшей мере один витамин представляет собой витамин B1, B5 и/или H.
27. Среда для культивирования по п. 25 или 26, отличающаяся тем, что указанная среда для культивирования содержит по меньшей мере одну рекомбинантную эукариотическую клетку, экспрессирующую, предпочтительно секретирующую, целевой белок.
28. Среда для культивирования по п. 27, отличающаяся тем, что указанный целевой белок представляет собой терапевтический белок.
29. Среда для культивирования по п. 27 или 28 или способ обеспечения указанной среды для культивирования, отличающиеся тем, что рост и/или деление указанных клеток останавливают и образование указанного целевого белка происходит на максимальном уровне остановки (MAL в [г/Л]), который превышает максимальный уровень (ML в [г/л]) белка, экспрессируемого указанными клетками, при выращивании на среде, предпочтительно на стандартной среде, в которой рост не останавливают, при этом значение MAL более чем в 1,5 больше значения ML, более чем в 2 раза больше значения ML или даже более чем в 2,5 или 3 раза больше значения ML.
30. Способ получения целевого белка, включающий:
(а) трансформацию эукариотических клеток системой для оптимизации экспрессии по любому из пп. 1-9, при этом указанная система содержит по меньшей мере один полинуклеотид, кодирующий по меньшей мере один целевой белок, для получения рекомбинантных эукариотических клеток;
(b) культивирование указанных рекомбинантных эукариотических клеток в среде для культивирования, в которой жизнеспособность, рост и/или деление указанных рекомбинантных эукариотических клеток зависят от активности по меньшей мере одного белка метаболизма витаминов;
(с) отбор рекомбинантных эукариотических клеток, экспрессирующих указанный по меньшей мере один белок метаболизма витаминов, при этом указанный белок метаболизма витаминов является селектируемым маркером для получения отобранных рекомбинантных эукариотических клеток, и
(d) очистку указанного целевого белка, полученного из указанных отобранных рекомбинантных эукариотических клеток или из их среды для культивирования, содержащей указанные отобранные рекомбинантные эукариотические клетки.
31. Способ по п. 30, отличающийся тем, что указанный белок метаболизма витаминов представляет собой белок транспорта витаминов, предпочтительно переносящий витамины B5, B1 и/или H, и указанная среда для культивирования содержит лимитирующую концентрацию и/или насыщающую концентрацию по меньшей мере одного из указанных витаминов.
32. Способ по п. 31, отличающийся тем, что указанный белок транспорта витаминов представляет собой SMVT и указанная среда для культивирования представляет собой среду с лимитирующей концентрацией B5 или среду с насыщающей концентрацией B5, но среду с лимитирующей концентрацией H.
| WO 2010097240, 02.09.2010 | |||
| WO 2009080759, 02.07.2009 | |||
| ГЕНЫ CORYNEBACTERIUM GLUTAMICUM, КОДИРУЮЩИЕ БЕЛКИ, УЧАСТВУЮЩИЕ В МЕТАБОЛИЗМЕ УГЛЕРОДА И ПРОДУЦИРОВАНИИ ЭНЕРГИИ | 2005 |
|
RU2321634C2 |

