ОБЛАСТЬ ТЕХНИКИ
Настоящее изобретение направлено на обеспечение конструкций нуклеиновых кислот и белков, которые вовлечены или воздействуют на метаболические пути, опосредующие или влияющие на метаболизм клетки, например, на перемещение через мембрану эндоплазматического ретикулума (ЭР) и/или секрецию через цитоплазматическую мембрану, а также на способы влияния на клеточный метаболизм. Настоящее изобретение также направлено на получение и применение рекомбинантных клеток млекопитающих, в которых, например, модифицированы перемещение/секреция большого разнообразия гетерологичных белков (продуктов экспрессии трансгенов). Указанные способы, конструкции нуклеиновых кислот, как правило, разработаны таким образом, чтобы улучшить экспрессию трансгена.
УРОВЕНЬ ТЕХНИКИ
Получение терапевтических белков биотехнологическими способами, а также генная и клеточная терапия зависят от успешной экспрессии трансгенов, внедренных в эукариотическую клетку. Для успешной экспрессии трансгена, как правило, требуется встраивание трансгена в хромосому хозяина, при этом указанная экспрессия ограничена, среди прочего, количеством встроенных копий трансгена и эпигенетическими эффектами, которые могут вызывать низкий уровень или нестабильность транскрипции и/или высокую клональную изменчивость. Недостаточный или пониженный транспорт продукта экспрессии трансгена из клетки также часто ограничивает получение терапевтических белков, а также генную и клеточную терапию.
Публикации и другие материалы, включая патенты, заявки на патент и номера доступа, используемые в данной заявке для иллюстрации настоящего изобретения и, в частности, для предоставления дополнительных подробностей касательно осуществления настоящего изобретения, полностью включены в данную заявку посредством ссылки.
Для повышения и стабилизации экспрессии трансгена в клетках млекопитающих все чаще применяют эпигенетические регуляторы, для того чтобы защитить трансгены от отрицательных эффектов положения (Bell и Felsenfeld, 1999). Данные эпигенетические регуляторы включают граничные или инсуляторные элементы, регуляторные области локусов (LCR), стабилизирующие и антирепрессорные (STAR) элементы, универсальные элементы, открывающие хроматин (UCOE), и упомянутые выше участки прикрепления к матриксу (MAR). Все эти эпигенетические регуляторы применялись для получения рекомбинантного белка в линиях клеток млекопитающих (Zahn-Zabal и др., 2001; Kim и др., 2004) и в способах генной терапии (Agarwal и др., 1998; Allen и др., 1996; Castilla и др., 1998).
Процессинг и транспорт из клетки продукта экспрессии трансгена часто сопряжен с различными затруднениями. Клетка, которая снабжена лишь средствами для процессинга и транспорта ее нативных белков, может быстро испытать перегрузку от транспорта некоторых типов продуктов экспрессии трансгенов, особенно когда они продуцируются на аномально высоких уровнях, что часто желательно, что приводит к агрегации продукта внутри клетки и/или, например, препятствует правильному фолдингу функционального белкового продукта.
Для преодоления затруднений, связанных с транспортом и процессингом, использовались различные подходы. Например, сконструированы клетки СНО с улучшенными секреторными свойствами путем экспрессии белков семейства SM Munc18c или Sly1, которые действуют как регуляторы транспорта мембранных везикул и, следовательно, экзоцитоза секретируемых белков (публикация заявки на патент США 20090247609). Х-бокс-связывающий белок 1 (Xbp1) - фактор транскрипции, который регулирует дифференцировку секреторных клеток и поддержание и экспансию ЭР, или различные протеиндисульфидизомеразы (PDI) применяли для снижения стресса ЭР и повышения секреции белка (Mohan и др. 2007). Другие попытки повышения секреции белка включали экспрессию шаперонов ERp57, кальнексина, кальретикулина и BiP1 в клетках СНО (Chung и др., 2004). Было показано, что экспрессия белка, индуцированного Холодовым шоком, в частности, индуцированного холодом РНК-связывающего белка (CIRP), повышает выход рекомбинантного гамма-интерферона. Также были предприняты попытки сверхэкспрессировать белки секреторных комплексов. Тем не менее, например, Lakkaraju и др. (2008) сообщили, что экспрессия экзогенного SRP14 в человеческих клетках дикого типа (например, в клетках, которые не были сконструированы для экспрессии низких уровней SRP14) не улучшала эффективность секреции секретируемого белка щелочной фосфатазы.
Таким образом, существует потребность в эффективной, надежной экспрессии трансгена, например, при получении рекомбинантного белка и для генной терапии. Также существует потребность в успешном транспорте продукта экспрессии трансгена за пределы клетки.
На удовлетворение данной и других потребностей, существующих в данной области техники, направлены варианты реализации настоящего изобретения.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение в одном варианте реализации направлено на рекомбинантную молекулу нуклеиновой кислоты, содержащую:
(a) 5'- и 3'-транспозон-специфичный инвертированный концевой повтор (ITR),
(b) по меньшей мере одну последовательность нуклеиновой кислоты, кодирующую белок экспрессии/процессинга трансгена (ТЕР), или функциональную РНК ТЕР, расположенную между 5'- и 3'-ITR, которая находится под контролем промотора, и
(c) возможно по меньшей мере один трансген, также расположенный между 5'- и 3'-ITR, который находится под контролем трансгенного промотора, при этом указанная молекула нуклеиновой кислоты необязательно является частью вектора.
Указанная рекомбинантная молекула нуклеиновой кислоты может содержать по меньшей мере один эпигенетический регуляторный элемент, в частности, по меньшей мере один элемент MAR (участок прикрепления к матриксу).
Элемент MAR может быть расположен между 5'- и 3'-ITR. Трансген, такой как ген устойчивости к антибиотику или ген, кодирующий иммуноглобулин, возможно под контролем дополнительного промотора, может быть расположен между 5'- и 3'-ITR, например, между 5'-ITR и MAR.
Белок ТЕР или функциональная РНК ТЕР может представлять собой белок или функциональную РНК, которая непосредственно или опосредованно вовлечена во встраивание последовательностей нуклеиновых кислот в геном, в процессинг или трансляцию продукта трансгенной РНК или участвует в перемещении в ЭР, секреции, процессинге, фолдинге, транспорте ЭР-Гольджи-плазматическая мембрана, гликозилировании и/или другой посттрансляционной модификации белков, таких как продукты экспрессии трансгенов.
Белок ТЕР может представлять собой белок пути секреции белка, белок путей рекомбинации или репарации ДНК, белок процессинга белка или метаболический белок, включая шапероны, такие как BiP, или комбинацию перечисленных белков.
Белок ТЕР может представлять собой один или более из следующих белков пути секреции белка: SRP14 человека (hSRP14), hSec61α1, hSec61β, hSec61γ, hSRP54, hSRP9, hSRPRα, hSRPβ и hCANX.
Белок ТЕР также может соответствовать одной или более из следующих последовательностей аминокислот белков пути секреции белка: hSRP14 с последовательностью SEQ ID NO: 13, hSec61α1 с последовательностью SEQ ID NO: 15, hSec61β с последовательностью SEQ ID NO: 17, hSec61γ с последовательностью SEQ ID NO: 19, hSRP54 с последовательностью SEQ ID NO: 21, hSRP9 с последовательностью SEQ ID NO: 23, hSRPRα с последовательностью SEQ ID NO: 25, hSRPβ с последовательностью SEQ ID NO: 27 и hCANX с последовательностью SEQ ID NO: 29 и/или может соответствовать последовательностям аминокислот, идентичным более чем на 80%, 90%, 95% или 98% указанным последовательностям.
Белок ТЕР может представлять собой один или более из следующих белков процессинга белка или метаболических белков: hUCP4, hCMPSAT, ST6Gal1 крысы (rST6Gal1), hCOMSC, Т-синтазу человека, hP4HA1, hP4HB, hGILZ, hCyPB, hNRF2, hHK1, hPDI, hPIN1, hSEPW1, hCALR, hDDOST, hHSP40, hATP5A1, hSERCA2, hPDIA4, hHSC70/HSPA8, hHYOU1, hCMP-SAS, беклин-1 человека, hERdj3, CHO-AGE, hWip1, hRTP4, hREEP2, hDPM1 и hDRiP78.
Белок ТЕР также может соответствовать одной или более из следующих последовательностей аминокислот белков процессинга белка или метаболических белков: hUCP4 с последовательностью SEQ ID NO: 31, hCMPSAT с последовательностью SEQ ID NO: 33, rST6Gal1 с последовательностью SEQ ID NO: 35, hCOMSC с последовательностью SEQ ID NO: 37, Т-синтазы человека с последовательностью SEQ ID NO: 39, hP4HA1 с последовательностью SEQ ID NO: 41, hP4HB с последовательностью SEQ ID NO: 43, hGILZ с последовательностью SEQ ID NO: 45, hCyPB с последовательностью SEQ ID NO: 47, hNRF2 с последовательностью SEQ ID NO: 49, hHK1 с последовательностью SEQ ID NO: 51, hPDI с последовательностью SEQ ID NO: 53, hPIN1 с последовательностью SEQ ID NO: 55, hSEPW1 с последовательностью SEQ ID NO: 57, hCALR с последовательностью SEQ ID NO: 59, hDDOST с последовательностью SEQ ID NO: 62, hHSP40 с последовательностью SEQ ID NO: 64, hATP5A1 с последовательностью SEQ ID NO: 66, hSERCA2 с последовательностью SEQ ID NO: 68, hPDIA4 с последовательностью SEQ ID NO: 70, hHSC70 /HSPA8 с последовательностью SEQ ID NO: 72, hHYOU1 с последовательностью SEQ ID NO: 74, hCMP-SAS с последовательностью SEQ ID NO: 76, беклина-1 человека с последовательностью SEQ ID NO: 78, hERdj3 с последовательностью SEQ ID NO: 80, CHO-AGE с последовательностью SEQ ID NO: 82, hWip1 с последовательностью SEQ ID NO: 84, hRTP4 с последовательностью SEQ ID NO: 86, hREEP2 с последовательностью SEQ ID NO: 88, hDPM1 с последовательностью SEQ ID NO: 90 и hDRiP78 с последовательностью SEQ ID NO: 92 и/или может соответствовать последовательностям аминокислот, идентичным более чем на 80%, 90%, 95% или 98% указанным последовательностям.
Белок ТЕР может представлять собой шаперон, в частности, белок BiP, а конкретнее, модифицированное производное белка BiP дрозофилы (DroBiP), последовательность которого идентична на 80%, 90%, 95% или 100% последовательности SEQ ID NO: 60.
Элемент MAR можно выбрать из последовательностей SEQ ID NO: 1 (MAR 1-68), 2 (MAR 1_6), 3 (MAR X_S29), 4 (MAR S4), 5 (MAR лизоцима цыпленка), или он предпочтительно представляет собой сконструированный, в частности, перестроенный аналог, и/или его последовательность по меньшей мере на 80%, 90%, 95%, 98%, 99% или 100% идентична любой из последовательностей SEQ ID NO: с 1 по 5 или любой из последовательностей SEQ ID NO: с 6 по 10.
Функциональная РНК ТЕР внутри указанной клетки может включать/состоять из последовательности нуклеиновых кислот, кодирующей функциональную РНК, предпочтительно микроРНК или короткую шпилечную РНК (кшРНК), которая препятствует экспрессии по меньшей мере одного белка пути рекомбинации или репарации ДНК, такого как, но не ограничиваясь перечисленными: Rad51, Rad51B, Rad51C, Rad51D, Xrcc2, Xrcc3, Rad52, Rad54, Brca1, Brca2, циклин D1, Ercc, MDC1, Bard1, лигаза 1, Mre11 и/или 53BP1.
Функциональная РНК ТЕР также может препятствовать экспрессии генов, последовательность которых по меньшей мере на 80%, 90%, 95%, 98% или 100% идентична Rad51 с последовательностью SEQ ID NO: 93, Rad51B с последовательностью SEQ ID NO: 94, Rad51C с последовательностью SEQ ID NO: 95, Rad51D с последовательностью SEQ ID NO: 96, Xrcc2 с последовательностью SEQ ID NO: 99, Xrcc3 с последовательностью SEQ ID NO: 100, Rad52 с последовательностью SEQ ID NO: 97, Rad54 с последовательностью SEQ ID NO: 98, Brca1 с последовательностью SEQ ID NO: 101, Brca2 с последовательностью SEQ ID NO: 102, циклину D1 с последовательностью SEQ ID NO: 103, Ercc1 с последовательностью SEQ ID NO: 104, MDC1 с последовательностью SEQ ID NO: 105, Bard1 с последовательностью SEQ ID NO: 106, лигазе 1 с последовательностью SEQ ID NO: 107, Mre11 с последовательностью SEQ ID NO: 108 и/или 53BP1 с последовательностью SEQ ID NO: 109.
Длина рекомбинантной молекулы нуклеиновой кислоты может составлять по меньшей мере 5000, 6000, 7000, 8000, 90000 или 10000 п.о.
5'- и 3'-ITR могут представлять собой 5'- и 3'-ITR транспозона Спящая Красавица или предпочтительно транспозона PiggyBac.
После первой трансфекции клетки млекопитающего одной из указанных рекомбинантных молекул нуклеиновых кислот и второй последующей трансфекции дополнительной рекомбинантной молекулой нуклеиновой кислоты, содержащей трансген, встраивание и/или экспрессия трансгена могут быть повышены в указанной клетке по сравнению с клеткой, не подвергнутой указанной первой трансфекции.
Последовательность, кодирующая ТЕР, или функциональная РНК ТЕР, упомянутые в данной заявке, могут представлять собой часть вектора, включая вектор экспрессии. Указанный вектор может содержать единичный элемент MAR, два или более элементов MAR, при этом указанный элемент(ы) может быть расположен между 5'- и 3'-ITR.
Например, указанный вектор может содержать два элемента MAR. Первый элемент MAR может быть расположен в 3'-5' направлении от ТЕР или функциональной РНК ТЕР и второй элемент MAR может быть расположен в 5'-3' направлении от ТЕР или функциональной РНК ТЕР, при этом указанный первый элемент MAR может включать элемент MAR 1_6 и/или элемент, последовательность которого по меньшей мере на 80%, 90%, 95%, 98%, 99% или 100% идентична последовательности SEQ ID NO: 2, в частности, перестроенные MAR на основе MAR 1-6, а конкретнее, элементы, последовательности которых по меньшей мере на 80%, 90%, 95%, 98%, 99% или 100% идентичны последовательности SEQ ID NO: 8 (MAR 1_6R2), и второй элемент MAR может включать элемент MAR 1-68 и/или элемент, последовательность которого по меньшей мере на 80%, 90%, 95%, 98%, 99% или 100% идентична последовательности SEQ ID NO: 1.
Вектор также может содержать единичный элемент MAR. Указанный единичный элемент MAR может быть расположен в 5'-3' направлении от ТЕР или функциональной РНК ТЕР, при этом указанный единичный элемент MAR может представлять собой MAR 1-68, или элемент MAR Х-29, и/или элемент, последовательность которого по меньшей мере на 80%, 90%, 95%, 98%, 99% или 100% идентична последовательности SEQ ID NO: 1 или 3, в частности, перестроенный MAR на основе MAR 1-68 или MAR Х-29, в частности, элемент, последовательность которого по меньшей мере на 80%, 90%, 95%, 98%, 99% или 100% идентична последовательности SEQ ID NO: 6, 7 или 10 (MAR 1_68R, 1_68R2 или X_29R3) или 9, и может предпочтительно представлять собой элемент MAR Х-29 и/или элемент, последовательность которого по меньшей мере на 80%, 90%, 95%, 98%, 99% или 100% идентична последовательности SEQ ID NO: 3.
ТЕР или функциональная РНК ТЕР может находиться под контролем промотора EF1-альфа, и за ним возможно следует поли(А) сигнал гормона роста быка (BGH).
Вектор может содержать промотор(ы) и/или энхансер(ы) или продукты их слияния, такие как промотор(ы) и/или энхансер(ы) GAPDH, SV40p, CMV, EF1-альфа клеток СНО, Actb СНО и/или Hspa5 СНО, или сконструированные продукты их слияния, такие как CGAPDH.
Промоторы, которые являются частью вектора, могут представлять собой GAPDH с последовательностью SEQ ID NO: 111, SV40p с последовательностью SEQ ID NO: 114, CMVp с последовательностью SEQ ID NO: 113, EF1-альфа СНО с последовательностью SEQ ID NO: 112, Actb СНО с последовательностью SEQ ID NO: 115, Hspa5 СНО с последовательностью SEQ ID NO: 116, и/или продукты их слияния, такие как CGAPDH с последовательностью SEQ ID NO: 11, или их последовательности нуклеиновых кислот могут быть идентичны указанным последовательностям более чем на 80%, 90%, 95% или 98%.
Настоящее изобретение также направлено на способ экспрессии ТЕР или функциональной РНК ТЕР, включающий:
обеспечение рекомбинантной клетки млекопитающего, содержащей трансген, и вектор представляет собой вектор экспрессии, который экспрессирует ТЕР или функциональную РНК ТЕР, при этом ТЕР или функциональная РНК ТЕР, экспрессируемая с указанного вектора, возможно повышает экспрессию трансгена в указанной клетке млекопитающего на по меньшей мере 10%, по меньшей мере 20%, по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60% или по меньшей мере 70%.
Указанный вектор может содержать единичный элемент MAR Х-29 и/или последовательность нуклеиновой кислоты, которая по меньшей мере на 80%, 90%, 95%, 98%, 99% или 100% идентична последовательности SEQ ID NO: 3, и при этом через более чем 4, 5, 6, 7, 8, 9, 10, 11, 12, 13 или 14 недель культивирования ТЕР или функциональная РНК ТЕР, экспрессируемая с указанного вектора, может повышать экспрессию интересующего гена на по меньшей мере 10%, по меньшей мере 20%, по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60% или по меньшей мере 70%.
Настоящее изобретение также направлено на рекомбинантную клетку млекопитающего, содержащую не более чем 20, 15, 10 или 5 рекомбинантных молекул нуклеиновых кислот, предпочтительно встроенных в геном указанной клетки в виде единичных копий.
Выше отмечено, что белок ТЕР может представлять собой один или более из следующих белков пути секреции белка: hSRP14, hSec61α1, hSec61β, hSec61γ, hSRP54, hSRP9, hSRPRα, hSRPβ и hCANX.
Белок ТЕР также может соответствовать одной или более из следующих последовательностей аминокислот белков пути секреции белка: hSRP14 с последовательностью SEQ ID NO: 13, hSec61α1 с последовательностью SEQ ID NO: 15, hSec61β с последовательностью SEQ ID NO: 17, hSec61γ с последовательностью SEQ ID NO: 19, hSRP54 с последовательностью SEQ ID NO: 21, hSRP9 с последовательностью SEQ ID NO: 23, hSRPRα с последовательностью SEQ ID NO: 25, hSRPβ с последовательностью SEQ ID NO: 27 и hCANX с последовательностью SEQ ID NO: 29, и/или может соответствовать последовательностям аминокислот, идентичным более чем на 80%, 90%, 95% или 98% указанным последовательностям.
Белок ТЕР может представлять собой один или более из следующих белков процессинга белка или метаболических белков: hUCP4, hCMPSAT, rST6Gal1, hCOMSC, T-синтазу человека, hP4HA1, hP4HB, hGILZ, hCyPB, hNRF2, hHK1, hPDI, hPIN1, hSEPW1, hCALR, hDDOST, hHSP40, hATP5A1, hSERCA2, hPDIA4, hHSC70/HSPA8, hHYOU1, hCMP-SAS, беклин-1 человека, hERdj3, CHO-AGE, hWip1, hRTP4, hREEP2, hDPM1 и hDRiP78.
Белок ТЕР также может соответствовать одной или более из следующих последовательностей аминокислот белков процессинга белка или метаболических белков: hUCP4 с последовательностью SEQ ID NO: 31, hCMPSAT с последовательностью SEQ ID NO: 33, rST6Gal1 с последовательностью SEQ ID NO: 35, hCOMSC с последовательностью SEQ ID NO: 37, Т-синтазы человека с последовательностью SEQ ID NO: 39, hP4HA1 с последовательностью SEQ ID NO: 41, hP4HB с последовательностью SEQ ID NO: 43, hGILZ с последовательностью SEQ ID NO: 45, hCyPB с последовательностью SEQ ID NO: 47, hNRF2 с последовательностью SEQ ID NO: 49, hHK1 с последовательностью SEQ ID NO: 51, hPDI с последовательностью SEQ ID NO: 53, hPIN1 с последовательностью SEQ ID NO: 55, hSEPWI с последовательностью SEQ ID NO: 57, hCALR с последовательностью SEQ ID NO: 59, hDDOST с последовательностью SEQ ID NO: 62, hHSP40 с последовательностью SEQ ID NO: 64, ИАТР5А1 с последовательностью SEQ ID NO: 66, hSERCA2 с последовательностью SEQ ID NO: 68, hPDIA4 с последовательностью SEQ ID NO: 70, hHSC70/HSPA8 с последовательностью SEQ ID NO: 72, hHYOU1 с последовательностью SEQ ID NO: 74, hCMP-SAS с последовательностью SEQ ID NO: 76, беклина-1 человека с последовательностью SEQ ID NO: 78, hERdj3 с последовательностью SEQ ID NO: 80, CHO-AGE с последовательностью SEQ ID NO: 82, hWip1 с последовательностью SEQ ID NO: 84, hRTP4 с последовательностью SEQ ID NO: 86, hREEP2 с последовательностью SEQ ID NO: 88, hDPM1 с последовательностью SEQ ID NO: 90 и hDRiP78 с последовательностью SEQ ID NO: 92, и/или может соответствовать последовательностям аминокислот, идентичным более чем на 80%, 90%, 95% или 98% указанным последовательностям.
Белок ТЕР может представлять собой шаперон, в частности, белок BiP, а конкретнее, сконструированное производное белка BiP дрозофилы (DroBiP), последовательность которого на 80%, 90%, 95% или 100% идентична последовательности SEQ ID NO: 60.
Функциональная РНК ТЕР внутри рекомбинантной клетки млекопитающего может включать/состоять из последовательности нуклеиновой кислоты (последовательностей нуклеиновых кислот), кодирующей функциональную РНК, предпочтительно микроРНК или кшРНК, которая препятствует экспрессии по меньшей мере одного белка рекомбинации, предпочтительно гена гомологичной рекомбинации, такого как, но не ограничиваясь перечисленными: Rad51, Rad51B, Rad51C, Rad51D, Xrcc2, Xrcc3, Rad52, Rad54, Brca1, Brca2, циклин D1, Ercc1, MDC1, Bard1, лигаза 1, Mre11 и/или 53BP1. Последовательность нуклеиновых кислот может быть по меньшей мере на 80%, 90%, 95%, 98% или 100% идентична Rad51 с последовательностью SEQ ID NO: 93, Rad51B с последовательностью SEQ ID NO: 94, Rad51C с последовательностью SEQ ID NO: 95, Rad51D с последовательностью SEQ ID NO: 96, Xrcc2 с последовательностью SEQ ID NO: 99, Xrcc3 с последовательностью SEQ ID NO: 100, Rad52 с последовательностью SEQ ID NO: 97, Rad54 с последовательностью SEQ ID NO: 98, Brca1 с последовательностью SEQ ID NO: 101, Brca2 с последовательностью SEQ ID NO: 102, циклину D1 с последовательностью SEQ ID NO: 103, Ercc1 с последовательностью SEQ ID NO: 104, MDC1 с последовательностью SEQ ID NO: 105, Bard1 с последовательностью SEQ ID NO: 106, лигазе 1 с последовательностью SEQ ID NO: 107, Mre11 с последовательностью SEQ ID NO: 108 и/или 53BP1 с последовательностью SEQ ID NO: 109.
Рекомбинантная клетка млекопитающего может представлять собой первичную стволовую клетку, клетку хомячка, например, клетку яичника китайского хомячка (СНО), или клетку человека, например, клетку НЕK293.
Настоящее изобретение также направлено на рекомбинантную клетку млекопитающего, содержащую:
а.) по меньшей мере одну функциональную РНК ТЕР и/или по меньшей мере одну рекомбинантную последовательность нуклеиновой кислоты, кодирующую белок ТЕР или кодирующую функциональную РНК ТЕР,
и
b.) рекомбинантную молекулу нуклеиновой кислоты, содержащую:
(i) по меньшей мере один интересующий трансген, и
(ii) возможно, элемент MAR.
Выше отмечено, что белок ТЕР может представлять собой один или более из следующих белков пути секреции белка: hSRP14, hSec61α1, hSec61β, hSec61γ, hSRP54, hSRP9, hSRPRα, hSRPβ и hCANX.
Белок ТЕР также может соответствовать одной или более из следующих последовательностей аминокислот белков пути секреции белка: hSRP14 с последовательностью SEQ ID NO: 13, hSec61α1 с последовательностью SEQ ID NO: 15, hSec61β с последовательностью SEQ ID NO: 17, hSec61γ с последовательностью SEQ ID NO: 19, hSRP54 с последовательностью SEQ ID NO: 21, hSRP9 с последовательностью SEQ ID NO: 23, hSRPRα с последовательностью SEQ ID NO: 25, hSRPβ с последовательностью SEQ ID NO: 27, и hCANX с последовательностью SEQ ID NO: 29 и/или может соответствовать последовательностям аминокислот, идентичным более чем на 80%, 90%, 95% или 98% указанным последовательностям.
Белок ТЕР может представлять собой один или более из следующих белков процессинга белка или метаболических белков: hUCP4, hCMPSAT, rST6Gal1, hCOMSC, T-синтазу человека, hP4HA1, hP4HB, hGILZ, hCyPB, hNRF2, hHK1, hPDI, hPIN1, hSEPW1, hCALR, hDDOST, hHSP40, hATP5A1, hSERCA2, hPDIA4, hHSC70 /HSPA8, hHYOU1, hCMP-SAS, беклин-1 человека, hERdj3, CHO-AGE, hWip1, hRTP4, hREEP2, hDPM1 и hDRiP78.
Белок ТЕР также может соответствовать одной или более из следующих последовательностей аминокислот белков процессинга белка или метаболических белков: hUCP4 с последовательностью SEQ ID NO: 31, hCMPSAT с последовательностью SEQ ID NO: 33, rST6Gal1 с последовательностью SEQ ID NO: 35, hCOMSC с последовательностью SEQ ID NO: 37, Т-синтазы человека с последовательностью SEQ ID NO: 39, hP4HA1 с последовательностью SEQ ID NO: 41, hP4HB с последовательностью SEQ ID NO: 43, hGILZ с последовательностью SEQ ID NO: 45, hCyPB с последовательностью SEQ ID NO: 47, hNRF2 с последовательностью SEQ ID NO: 49, hHK1 с последовательностью SEQ ID NO: 51, hPDI с последовательностью SEQ ID NO: 53, hPIN1 с последовательностью SEQ ID NO: 55, hSEPW1 с последовательностью SEQ ID NO: 57, hCALR с последовательностью SEQ ID NO: 59, hDDOST с последовательностью SEQ ID NO: 62, hHSP40 с последовательностью SEQ ID NO: 64, ИАТР5А1 с последовательностью SEQ ID NO: 66, hSERCA2 с последовательностью SEQ ID NO: 68, hPDIA4 с последовательностью SEQ ID NO: 70, hHSC70/HSPA8 с последовательностью SEQ ID NO: 72, hHYOU1 с последовательностью SEQ ID NO: 74, hCMP-SAS с последовательностью SEQ ID NO: 76, беклина-1 человека с последовательностью SEQ ID NO: 78, hERdj3 с последовательностью SEQ ID NO: 80, CHO-AGE с последовательностью SEQ ID NO: 82, hWip1 с последовательностью SEQ ID NO: 84, hRTP4 с последовательностью SEQ ID NO: 86, hREEP2 с последовательностью SEQ ID NO: 88, hDPM1 с последовательностью SEQ ID NO: 90 и hDRiP78 с последовательностью SEQ ID NO: 92 и/или может соответствовать последовательностям аминокислот, идентичным более чем на 80%, 90%, 95% или 98% указанным последовательностям.
Белок ТЕР может представлять собой шаперон, в частности, белок BiP, а конкретнее, синтетическое производное белка BiP дрозофилы (DroBiP), последовательность которого на 80%, 90%, 95% или 100% идентична последовательности SEQ ID NO: 60.
Функциональная РНК может представлять собой временно трансфицированную миРНК или кшРНК, которая транскрибируется с указанной по меньшей мере одной изолированной последовательности нуклеиновой кислоты, при этом указанная миРНК или указанная процессированная кшРНК представляет собой антисмысловую РНК длиной 20, 21, 22, 23, 24 или 25 пар оснований, которая полностью комплементарна 20, 21, 22, 23, 24 или 25 последовательным нуклеотидам мРНК по меньшей мере одного целевого гена, который является частью рекомбинантного пути НГСК (негомологичного соединения концов), ГР (гомологичной рекомбинации), ОМСК (опосредованного микрогомологией соединения концов) или представляет собой белок репарации ДНК, такой как MDC1 (медиатор контрольных точек повреждения ДНК 1).
Указанный по меньшей мере один целевой ген, который может быть частью:
- репарации ДНК и НГСК, представляет собой
53ВР1 (белок 1, связывающий опухолевый супрессор р53),
- ГР, представляет собой
Rad51 (белок репарации ДНК RAD51), Rad51B (гомолог 2 белка репарации ДНК RAD51), Rad51C (гомолог 3 белка репарации ДНК RAD51),
Rad51D (гомолог 4 белка репарации ДНК RAD51), Rad52 (белок репарации ДНК RAD52), Rad54 (белок репарации и рекомбинации ДНК RAD54),
Xrcc2 (репарация после облучения рентгеновскими лучами, исправляющая (комплементирующая) нарушенную репарацию в клетках китайского хомячка 2),
Xrcc3 (репарация после облучения рентгеновскими лучами, комплементирующая нарушенную репарацию в клетках китайского хомячка 3),
Brca1 (рак молочной железы 1, раннее начало),
Brca2 (рак молочной железы 2, раннее начало),
Bard1 (ассоциированный с BRCA1 RING-домен 1),
- ОМСК, представляет собой
Ercc1 (эксцизионная репарация, кросс-комплементирующая нарушенную репарацию у грызунов, группа комплементации 1),
Mre11 (мейотическая рекомбинация 11)
лигазу 1 (ДНК-лигаза 1),
и/или
представляет собой белок репарации ДНК MDC1.
Целевые гены могут представлять собой нуклеиновые кислоты, последовательность которых по меньшей мере на 80%, 90%, 95%, 98% или 100% идентична Rad51 с последовательностью SEQ ID NO: 93, Rad51B с последовательностью SEQ ID NO: 94, Rad51C с последовательностью SEQ ID NO: 95, Rad51D с последовательностью SEQ ID NO: 96, Xrcc2 с последовательностью SEQ ID NO: 99, Xrcc3 с последовательностью SEQ ID NO: 100, Rad52 с последовательностью SEQ ID NO: 97, Rad54 с последовательностью SEQ ID NO: 98, Brca1 с последовательностью SEQ ID NO: 101, Brca2 с последовательностью SEQ ID NO: 102, циклину D1 с последовательностью SEQ ID NO: 103, Ercd с последовательностью SEQ ID NO: 104, MDC1 с последовательностью SEQ ID NO: 105, Bard1 с последовательностью SEQ ID NO: 106, лигазе 1 с последовательностью SEQ ID NO: 107, Mre11 с последовательностью SEQ ID NO: 108 и/или 53BP1 с последовательностью SEQ ID NO: 109.
С указанного по меньшей мере одного трансгена можно экспрессировать терапевтический белок, такой как иммуноглобулин, гормон, такой как эритропоэтин, или фактор роста, и при этом необязательно встраивание и/или экспрессия трансгена повышена в рекомбинантной клетке млекопитающего по сравнению с клеткой, не содержащей указанную рекомбинантную молекулу(ы) нуклеиновой кислоты.
Рекомбинантная клетка млекопитающего может содержать по меньшей мере две функциональные РНК ТЕР, при этом одна или обе РНК ТЕР представляют собой временно трансфицированные миРНК, или экспрессируются с указанной изолированной последовательности(ей) нуклеиновой кислоты, кодирующей функциональную РНК ТЕР.
Рекомбинантная клетка млекопитающего может содержать элемент MAR.
Настоящее изобретение также направлено на способ трансфекции клеток млекопитающих, в частности, клеток хомячка, включающий:
трансфекцию, возможно при первой трансфекции, указанных клеток млекопитающих
(i) по меньшей мере одной из указанных рекомбинантных молекул нуклеиновых кислот по любому из пп. 1-13 и/или
(ii) по меньшей мере одной изолированной функциональной РНК ТЕР и по меньшей мере одним трансгеном, который необязательно является частью рекомбинантной молекулы нуклеиновой кислоты, которой возможно трансфицируют при второй, последующей трансфекции, возможно вместе с изолированной нуклеиновой кислотой или мРНК, экспрессирующей транспозазу, которая узнает указанные 5'- и 3'-ITR.
Рекомбинантную клетку млекопитающего можно трансфицировать более чем одной молекулой нуклеиновой кислоты, включая по меньшей мере две, по меньшей мере три или по меньшей мере четыре из указанных рекомбинантных молекул нуклеиновых кислот, кодирующих один, два или три из следующих белков: hSRP14, hSec61α1, hSec61β, hSec61γ, hSRP54, hSRP9, hSRPRα, hSRPβ и hCANX.
Ее также можно трансфицировать hSRP14 с последовательностью SEQ ID NO: 13, hSec61α1 с последовательностью SEQ ID NO: 15, hSec61β с последовательностью SEQ ID NO: 17, hSec61γ с последовательностью SEQ ID NO: 19, hSRP54 с последовательностью SEQ ID NO: 21, hSRP9 с последовательностью SEQ ID NO: 23, hSRPRα с последовательностью SEQ ID NO: 25, hSRPβ с последовательностью SEQ ID NO: 27, и hCANX с последовательностью SEQ ID NO: 29, и/или последовательностями аминокислот, идентичными более чем на 80%, 90%, 95% или 98% указанным последовательностям.
Любые из рекомбинантных молекул нуклеиновых кислот могут представлять собой часть вектора, при этом можно осуществить котрансфекцию указанными векторами.
Котрансфекция векторами, кодирующими несколько белков ТЕР, предпочтительно котрансфекция рекомбинантными молекулами нуклеиновых кислот, кодирующими белки SRP14, SRP9 и SRP54, может повышать встраивание и/или экспрессию трансгена в указанной клетке по сравнению с клеткой, не подвергнутой такой котрансфекции. Котрансфекция векторами, содержащими последовательности нуклеиновых кислот, которые по меньшей мере на 80%, 90%, 95%, 98% или 100% идентичны последовательностям SEQ ID NO: 12, SEQ ID NO: 22, SEQ ID NO: 20, также входит в объем настоящего изобретения.
Можно получить множество указанных клеток млекопитающих, которые стабильно экспрессируют указанный белок ТЕР или функциональную РНК ТЕР, чтобы получить рекомбинантные клетки млекопитающих, и при этом указанное количество рекомбинантных клеток млекопитающих может не зависеть от наличия указанного элемента MAR. Указанные клетки млекопитающих можно трансфицировать два и возможно три раза.
Предпочтительно, по меньшей мере 30%, 40% или 45% указанных клеток млекопитающих могут стать рекомбинантными клетками млекопитающих и экспрессировать указанный трансген.
Клетку млекопитающего можно трансфицировать указанной по меньшей мере одной изолированной функциональной РНК ТЕР и вектором, содержащим указанный по меньшей мере один трансген и необязательно 3'-ITR и 5'-ITR, фланкирующие указанный по меньшей мере один трансген, возможно вместе с изолированной нуклеиновой кислотой или мРНК, экспрессирующей транспозазу, которая узнает указанные 5'- и 3'-ITR.
Трансген может представлять собой терапевтический белок, такой как иммуноглобулин, гормон, цитокин или фактор роста.
Рекомбинантная молекула нуклеиновой кислоты может необязательно содержать маркер отбора, и при этом указанный по меньшей мере один трансген экспрессируется
(а) без отбора по указанному маркеру, или
(b) с отбором по указанному маркеру, например, путем добавления селекционного агента в культуральную среду, и
(c) в отсутствие транспозазы, или
(d) в присутствии транспозазы.
Функциональная РНК ТЕР может кодироваться рекомбинантными последовательностями нуклеиновых кислот, кодирующими кшРНК или микроРНК, или может включать/состоять из миРНК, которая препятствует экспрессии по меньшей мере одного гена гомологичной рекомбинации, такого как, но не ограничиваясь перечисленными: Rad51, Rad51B, Rad51C, Rad51D, Xrcc2, Xrcc3, Rad52, Rad54, Brca1, Brca2, циклин D1, Ercc1, MDC1, Bard1, лигаза 1, Mre11 и/или 53BP1.
Функциональная РНК ТЕР также может препятствовать экспрессии генов, последовательность которых по меньшей мере на 80%, 90%, 95%, 98% или 100% идентична Rad51 с последовательностью SEQ ID NO: 93, Rad51B с последовательностью SEQ ID NO: 94, Rad51C с последовательностью SEQ ID NO: 95, Rad51D с последовательностью SEQ ID NO: 96, Xrcc2 с последовательностью SEQ ID NO: 99, Xrcc3 с последовательностью SEQ ID NO: 100, Rad52 с последовательностью SEQ ID NO: 97, Rad54 с последовательностью SEQ ID NO: 98, Brca1 с последовательностью SEQ ID NO: 101, Brca2 с последовательностью SEQ ID NO: 102, циклину D1 с последовательностью SEQ ID NO: 103, Ercc1 с последовательностью SEQ ID NO: 104, MDC1 с последовательностью SEQ ID NO: 105, Bard1 с последовательностью SEQ ID NO: 106, лигазе 1 с последовательностью SEQ ID NO: 107, Mre11 с последовательностью SEQ ID NO: 108 и/или 53BP1 с последовательностью SEQ ID NO: 109.
Встраивание и/или экспрессию трансгена можно повысить в такой клетке по сравнению с клеткой, не трансфицированной указанными изолированными молекулами нуклеиновых кислот и/или по меньшей мере одной из указанных изолированных функциональных РНК ТЕР.
Настоящее изобретение также направлено на набор, в одном контейнере содержащий по меньшей мере один вектор, содержащий любую из указанных рекомбинантных молекул нуклеиновых кислот согласно пп. 1-13, и во втором необязательном контейнере содержащий вектор, кодирующий подходящую транспозазу, и в дополнительном контейнере содержащий инструкцию по применению указанного вектора или векторов.
Предложен указанный выше набор, в котором более чем один вектор предусмотрен в одном или более контейнерах и в котором белки ТЕР представляют собой по меньшей мере два из следующих белков: шаперон, SRP14, SRP9, SRP54, SR или транслокон.
Предложен указанный выше набор, в котором функциональная(ые) РНК ТЕР внутри указанного вектора(ов) включает(ют)/состоит(ят) из последовательностей нуклеиновых кислот, кодирующих микроРНК, миРНК или кшРНК, которые препятствуют экспрессии по меньшей мере одного гена гомологичной рекомбинации, такого как, но не ограничиваясь перечисленными: Rad51, Rad51B, Rad51C, Rad51D, Xrcc2, Xrcc3, Rad52, Rad54, Brca1, Brca2, циклин D1, Ercc1, MDC1, Bard1, лигаза 1, Mre11 и/или 53BP1, и предпочтительно в дополнительном контейнере указанный набор содержит одну или более миРНК, которые препятствуют экспрессии по меньшей мере одного другого гена гомологичной рекомбинации, такого как, но не ограничиваясь перечисленными: Rad51, Rad51B, Rad51C, Rad51D, Xrcc2, Xrcc3, Rad52, Rad54, Brca1, Brca2, циклин D1, Ercc, MDC1, Bard1, лигаза 1, Mre11 и/или 53BP1.
Ген гомологичной рекомбинации может соответствовать нуклеиновым кислотам, последовательность которых по меньшей мере на 80%, 90%, 95%, 98% или 100% идентична Rad51 с последовательностью SEQ ID NO: 93, Rad51B с последовательностью SEQ ID NO: 94, Rad51C с последовательностью SEQ ID NO: 95, Rad51D с последовательностью SEQ ID NO: 96, Xrcc2 с последовательностью SEQ ID NO: 99, Xrcc3 с последовательностью SEQ ID NO: 100, Rad52 с последовательностью SEQ ID NO: 97, Rad54 с последовательностью SEQ ID NO: 98, Brca1 с последовательностью SEQ ID NO: 101, Brca2 с последовательностью SEQ ID NO: 102, циклину D1 с последовательностью SEQ ID NO: 103, Ercc1 с последовательностью SEQ ID NO: 104, MDC1 с последовательностью SEQ ID NO: 105, Bard1 с последовательностью SEQ ID NO: 106, лигазе 1 с последовательностью SEQ ID NO: 107, Mre11 с последовательностью SEQ ID NO: 108 и/или 53BP1 с последовательностью SEQ ID NO: 109.
Настоящее изобретение также направлено на применение рекомбинантных нуклеиновых кислот, описанных в данной заявке, и/или рекомбинантных клеток млекопитающих, описанных в данной заявке, предпочтительно для повышения встраивания и/или экспрессии трансгена.
Настоящее изобретение также направлено на вектор экспрессии, содержащий:
(a) трансген, который фланкирован в 3'-5' направлении промотором и в 5'-3' направлении сигналом полиаденилирования,
(b) единичный элемент MAR в 5'-3' направлении от сигнала полиаденилирования, или
(c) первый элемент MAR в 3'-5' направлении от интересующего трансгена и второй элемент MAR в 5'-3' направлении от места встраивания указанного трансгена.
Единичный или первый и второй элементы MAR можно выбрать из элементов MAR 1_68, 1_6, 1_6R2, 1_68R, 1_68R2, X_29R3 или X_29 или элементов, последовательности которых по меньшей мере на 80%, 90%, 95%, 98%, 99% или 100% идентичны последовательностям SEQ ID NO: 1, 2, 3, 6, 7, 8, 9 или 10.
Единичный или первый и второй элемент(ы) MAR можно выбрать из перестроенных элементов MAR 1_6R2, 1_68R, 1_68R2 или X_29R3 или элементов, последовательности которых по меньшей мере на 80%, 90%, 95%, 98%, 99% или 100% идентичны последовательностям SEQ ID NO: 6, 7, 8, или 10, при этом необязательно указанный элемент(ы) MAR повышает экспрессию интересующего трансгена на по меньшей мере 10%, по меньшей мере 20%, по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60% или по меньшей мере 70% по сравнению с их неперестроенными аналогами.
Промотор может представлять собой промотор EF1-альфа и сигнал полиаденилирования представляет собой поли(А) сигнал BGH.
Указанный вектор может содержать промотор(ы) и/или энхансер(ы) или продукты их слияния, такие как GAPDH, CGAPD, CSV40p, CMVp, СНО Ef1-альфа, Actb СНО или HspaS СНО.
Промоторы могут представлять собой промоторы GAPDH с последовательностью SEQ ID NO: 111, CGAPDH с последовательностью SEQ ID NO: 11, SV40p с последовательностью SEQ ID NO: 114, CMVp с последовательностью SEQ ID NO: 113, EF1-альфа СНО с последовательностью SEQ ID NO:112, Actb СНО с последовательностью SEQ ID NO: 115 и/или HspaS СНО с последовательностью SEQ ID NO: 116 и последовательности нуклеиновых кислот, идентичные более чем на 80%, 90%, 95% или 98% указанным последовательностям.
Промотор может представлять собой промотор GAPDH и включать энхансер CMV.
Первый и/или второй MAR, энхансер, промотор, интересующий трансген и сигнал полиаденилирования могут быть расположены между 5'- и 3'-ITR.
В некотором варианте реализации указанный вектор экспрессии может содержать:
(a) единичный элемент MAR в 5'-3' направлении от сигнала полиаденилирования, при этом указанный единичный элемент MAR предпочтительно представляет собой элемент MAR 1-68 или MARX-29 и/или элемент, последовательность которого по меньшей мере на 80%, 90%, 95%, 98%, 99% или 100% идентична последовательности SEQ ID NO: 1 или 3, в частности, перестроенный MAR на основе MAR 1-68 или MAR Х-29, в частности, элемент, последовательность которого по меньшей мере на 80%, 90%, 95%, 98%, 99% или 100% идентична последовательностям SEQ ID NO: 6, 7 или 10 (элементы MAR 1_68R, 1_68R2 или X_29R3) или SEQ ID NO: 9, и предпочтительно представляет собой элемент MAR Х-29, и/или его последовательность по меньшей мере на 80%, 90%, 95%, 98%, 99% или 100% идентична последовательности SEQ ID NO: 3, или
(b) первый элемент MAR в 3'-5' направлении от интересующего трансгена и второй элемент MAR в 5'-3' направлении от указанного интересующего трансгена, при этом указанный первый элемент MAR предпочтительно включает элемент 1_6, и/или его последовательность по меньшей мере на 80%, 90%, 95%, 98%, 99% или 100% идентична последовательности SEQ ID NO: 2, в частности, идентична перестроенным элементам MAR на основе MAR 1-6, в частности, элементам, последовательности которых по меньшей мере на 80%, 90%, 95%, 98%, 99% или 100% идентичны последовательности SEQ ID NO: 8 (MAR 1_6R2), и второй элемент MAR, который предпочтительно включает элемент MAR 1-68, и/или его последовательность по меньшей мере на 80%, 90%, 95%, 98%, 99% или 100% идентична последовательности SEQ ID NO: 1.
Вектор экспрессии может содержать единичный элемент MAR, и указанный единичный элемент MAR может быть расположен в 5'-3' направлении от сайта полиаденилирования и представляет собой MAR 1-68 или MAR Х-29, и/или его последовательность по меньшей мере на 80%, 90%, 95%, 98%, 99% или 100% идентична последовательности SEQ ID NO: 1 или 3, в частности, идентична перестроенным элементам MAR на основе MAR 1-68 или MAR Х-29, в частности, элементам, последовательность которых по меньшей мере на 80%, 90%, 95%, 98%, 99% или 100% идентична последовательностям SEQ ID NO: 6, 7 или 10 (элементы MAR 1_68R, 1_68R2 или X_29R3) или 9, и может предпочтительно представлять собой полученный из MAR Х-29 элемент, и/или его последовательность по меньшей мере на 80%, 90%, 95%, 98%, 99% или 100% идентична SEQ ID NO: 3.
Указанный первый элемент MAR может находиться в 3'-5' направлении от интересующего трансгена и второй элемент MAR может находиться в 5'-3' направлении от указанного интересующего трансгена, при этом указанный первый элемент MAR может включать элемент MAR 1_6, и/или его последовательность по меньшей мере на 80%, 90%, 95%, 98%, 99% или 100% идентична последовательности SEQ ID NO 2, и указанный второй элемент MAR может включать элемент MAR 1_68, и/или его последовательность может быть по меньшей мере на 80%, 90%, 95%, 98%, 99% или 100% идентична SEQ ID NO: 1.
Настоящее изобретение также направлено на способ экспрессии трансгена, включающий:
обеспечение рекомбинантной клетки млекопитающего, содержащей один из упомянутых выше векторов, содержащих указанный трансген и экспрессирующих указанный трансген, при этом указанный элемент(ы) MAR может повышать экспрессию указанного трансгена предпочтительно на по меньшей мере 10%, по меньшей мере 20%, по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60% или по меньшей мере 70%. Указанный вектор может содержать единичный элемент MAR Х_29 и/или нуклеиновую кислоту, последовательность которой по меньшей мере на 80%, 90%, 95%, 98%, 99% или 100% идентична последовательности SEQ ID NO: 3, и при этом через более чем 4, 5, 6, 7, 8, 9, 10, 11, 12, 13 или 14 недель культивирования указанный элемент MAR может повышать экспрессию интересующего трансгена на по меньшей мере 10%, по меньшей мере 20%, по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60% или по меньшей мере 70%.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Фиг. 1. Конструирование вектора, несущего транспозон.
Для того, чтобы проверить, может ли влиять добавление элемента MAR к транспозону РВ (PiggyBac) на эффективность транспозиции и экспрессию трансгена, и чтобы оценить, оказывало ли расположение MAR в конструкции какое-либо влияние на данные эффекты, разработали серию донорных конструкций транспозонов, содержащих ген зеленого флуоресцентного белка (GFP) и ген устойчивости к пуромицину (Puro), в которые встроили MAR 1_68 или контрольную нейтральную спейсерную последовательность ДНК в различные положения в указанной плазмиде. Исходную плазмиду, несущую транспозон, Puro-GFP без вставки использовали в качестве контроля транспозиции, чтобы отличить вклад увеличенного размера транспозона от влияния добавления MAR или спейсерной последовательности.
Фиг. 2. Векторы, несущие транспозоны: эффективность транспозиции.
Эффективность транспозиции различных конструкций, несущих транспозоны, измеряли путем оценки (А) процента клеток, экспрессирующих GFP, после трансфекции и трех недель культивирования без селекции антибиотиком и (В) путем подсчета устойчивых к пуромицину колоний.
Фиг. 3. Векторы, несущие транспозоны: уровень экспрессии.
Анализ уровня экспрессии, обеспеченного различными векторами, несущими транспозоны, трансфицированными совместно (+РВ) или без (-РВ) плазмиды, экспрессирующей транспозазу, путем измерения уровней флуоресценции GFP в клетках СНО после 3 недель культивирования без селекции (А) или при селекции (В) по устойчивости к пуромицину после трансфекции, с учетом флуоресценции только GFP-положительных клеток.
Фиг. 4. Влияние MAR и транспозазы на встраивание трансгена в геном.
Количество встроенных копий GFP-трансгена определяли, применяя количественную ПЦР, и значения нормировали по отношению к клеточному гену В2М, применяя геномную ДНК, выделенную из не подвергнутых селекции клеток СНО (А), или устойчивых к пуромицину клеток (В), полученных как описано в подписях к фигурам 1-3. Значения представляют средние значения ± стандартная ошибка среднего (n=3). *Р<0,05.
Фиг. 5. Экспрессия трансгена относительно трансгена.
Оценка внутреннего экспрессионного потенциала векторов независимо от их способности встраиваться в геном, без селекции (А) и при селекции (В) пуромицином.
Фиг. 6. Влияние экспрессии секреторных белков с векторов на основе транспозонов и плазмидных векторов на экспрессию рекомбинантного белка (трансгена). Векторы на основе транспозонов или обычные плазмидные векторы сконструировали для экспрессии секреторных белков SRP9, SRP14, SRP54, альфа и бета субъединиц (SR) рецептора SRP или транслокона. Векторы на основе транспозонов котрансфицировали с вектором экспрессии транспозазы PiggyBac (правая панель), тогда как не основанные на транспозонах плазмидные векторы трансфицировали отдельно (левая панель) в клон клетки, экспрессирующей антитело инфликсимаб, как описано в данном тексте. После трех недель культивирования при селекции (левая панель) или без селекции (правая панель) анализировали уровни секретируемого антитела инфликсимаба в культуральных супернатантах. Можно увидеть, что удельная продуктивность, которая представляет собой относительную экспрессию в клетке, содержащей последовательность, кодирующую белок экспрессии/процессинга трансгена (ТЕР), или функциональную РНК ТЕР, повышалась при применении вектора, несущего транспозон, от 0,25-1,5 до 1-2,5, соответственно, по сравнению с исходной клеткой без ТЕР.
Фиг. 7. Экспрессия рекомбинантного белка в суспензиях подвергнутых электро по рации клеток СНО-М.
(A) Клетки СНО-М, которые подвергали электропорации один или два раза несущим MAR Х-29 вектором, экспрессирующим GFP и транспозон, в присутствии (+РВ) или отсутствие (-РВ) транспозазы piggyBac. Показан процент стабильно экспрессирующих GFP клеток после 3 недель культивирования, осуществленного в отсутствие селекции.
(B) Сренее значение флуоресценции GFP в GFP-положительных клетках.
(C) кДНК, кодирующие легкие и тяжелые цепи антитела бевацизумаба (Beva), адалимумаба (Adal) и ритуксимаба (Ritu), встраивали вместо GFP в плазмиды, несущие транспозон, содержащие MAR Х29. Клетки СНО-М подвергали электропорации три раза с интервалами по 12 дней конструкциями, несущими транспозоны, содержащими легкую и тяжелую цепь, вместе с вектором экспрессии, несущим транспозазу piggyBac. Показаны уровни иммуноглобулина, секретируемого в культуральные супернатанты совокупностью поликлональных клеток, которых растили без селекции (незакрашенные столбики). В качестве альтернативы, не подвергнутые селекции поликлональные популяции клеток подвергали сортировке путем пэннинга (просеивания) клеток, на поверхностях которых были выставлены иммуноглобулины, применяя магнитные микрогранулы. Показаны уровни иммуноглобулинов, секретируемых не подвергнутыми сортировке популяциями (закрашенные столбики).
(D) Популяции трансфицированных клеток подвергали сортировке для обнаружения экспрессирующих иммуноглобулины колоний, применяя устройство, собирающее колонии, и по два клона, экспрессирующих каждый из трех указанных иммуноглобулинов, растили в культурах с подпиткой в биореакторах в виде центрифужных пробирок. Показаны уровни секретируемых иммуноглобулинов, и их определяли также, как и для панели (С).
Фиг.8. Гетерологичная экспрессия SRP14 улучшает секрецию трастузумаба и восстанавливает секрецию инфликсимаба.
Клоны HP и LP СНО-К1, экспрессирующие иммуноглобулины трастузумаб (А) или инфликсимаб (В) на наиболее высоких из полученных уровней, повторно стабильно трансфицировали вектором экспрессии SRP14, и выделяли моноклональные популяции. Оценивали рост и продуктивность клеток в условиях периодической культуры для полученных субклонов, обозначенных от А до Е. Концентрацию клеток (клетки/мл) и титр IgG (мкг/мл) наносили на график для каждого дня отбора образцов на протяжении 7 дней культивирования. (С) Распределение удельной продуктивности субклонов трастузумаба (HP) и инфликсимаба (LP) после трансфекции вектором экспрессии SRP14 (дорожки S) по сравнению с таковой для исходных клонов HP и LP (-). (D) Определяли относительные уровни мРНК SRP14 для 5 отдельных субклонов А-Е SRP14-LP и исходного контрольного клона LP, и наносили их на график в зависимости от удельной продуктивности IgG для 4 полученных культур. Средние значения и стандартные отклонения уровней мРНК и удельной продуктивности выражали в виде кратности превышения соответствующих значений для контрольного клона LP.
Фиг. 9. Гетерологичная экспрессия SRP14 опосредует высокий выход с трудом экспрессируемого иммуноглобулина в процессе получения.
Трансфицированные вектором, несущим SRP14, субклон HP В, экспрессирующий трастузумаб (А), и субклон LP Е, экспрессирующий инфликсимаб (В), проанализированные на фиг. 8, культивировали в вентилируемых встряхиваемых колбах объемом 125 мл в культурах с подпиткой с рабочим объемом 25 мл и определяли концентрацию жизнеспособных клеток и титр IgG в течение 11-дневного периода.
Фиг. 10. Экспрессия SRP14 нарушает агрегацию легких цепей в клонах клеток СНО.
(А) Собранные путем центрифугирования супернатанты и осадки пермеабилизованных Triton Х100 (Тx100) клеток анализировали с помощью ЭФ в ПААГ/ДСН, как изображено на панелях с обозначением растворимых в Тх-100 и нерастворимых в Тх-100 агрегатов, соответственно, для субклона Е SRP14-LP, полученного из LP, и субклона В SRP14-HP, полученного из HP (дорожки S), или для субклонов СНО, экспрессирующих контрольный белок GFP (дорожки G). Стрелками показаны недостаточно процессированная свободная легкая цепь (LC) и агрегировавшая (агр.) LC. (В) Осуществляли анализ с вытеснением для различных промежуточных вариантов агрегации LC, НС и IgG, полученных в клоне Е SRP14-LP и LP-контрольном клоне Е, результаты которого показаны на данной фигуре.
Фиг. 11. Влияние совместной экспрессии субъединиц SRP, SR и транслокона на секрецию иммуноглобулина.
(А) Клон LP Е, экспрессирующий инфликсимаб, повторно трансфицировали различными комбинациями векторов экспрессии на основе транспозонов, несущих SRP, SR и транслокон. Удельную продуктивность полученных популяций клеток затем оценивали при периодическом культивировании и представили в виде % от значений в пикограммах на клетку в день (pcd) для LP-контрольных клеток. На диаграммах размаха представлены средняя, верхняя и нижняя квартили нормированных удельных продуктивностей, определенных в день 3 для независимых серий культивирования. (В) Субклон клетки Е, экспрессирующий SRP14 и продуцирующий инфликсимаб, повторно трансфицировали различными комбинациями векторов экспрессии на основе транспозонов, несущих SR и транслокон. Удельная продуктивность популяций клеток представлена аналогично панели А.
Фиг. 12. Модель восстановления секреции инфликсимаба экспрессирующими SRP14 клонами
Модель фолдинга и секреции IgG клонами с низкой продуцирующей способностью до (А) и после (В) сверхэкспрессии субъединиц SRP/транслокона. Полученные результаты свидетельствуют о том, что вновь синтезированная LC, продуцируемая клонами с низкой продуцирующей способностью, недостаточно процесс и руется и имеет неправильный фолдинг. Недостаточный процессинг сигнального пептида LC инфликсимаба может приводить к насыщению аппарата котрансляционной транслокации ЭР (панель А, номер 1). Ее агрегация в ЭР внутри не способных к сборке IgG агрегировавших форм LC (панель А, номер 2) вызывает ЭР-стресс и вызывает образование подобной аутофагосоме структуры (панель А, номер 3). Сверхэкспрессия SRP14 и других компонентов белков SRP/транслокона полностью восстанавливала процессинг и секрецию инфликсимаба IgG (панель В). Активность SRP14, останавливающая элонгацию, возможно задерживает перемещение LC в ЭР в процессе трансляции ее мРНК (панель В, номер 1). Это, в свою очередь, будет способствовать правильному процессингу LC и ее правильному взаимодействию с осуществляющими фолдинг шаперонами в ЭР (панель В, номер 2). Поддержание вновь синтезированной LC в состоянии, позволяющем сборку IgG, таким образом, восстанавливает высокий выход секреции полностью собранных антител (панель В, номер 3).
Фиг. 13. Влияние подавления ГР и НГСК молекулами миРНК на экспрессию.
Кратность различий в проценте GFP-положительных клеток (по сравнению с клетками, трансфицированными контрольной плазмидой GFP, представленными на данной фигуре как 1,0), представляющем частоты событий рекомбинации в необработанных клетках (ложных), клетках, обработанных миРНК отрицательного контроля (миОтр), миРНК против факторов НГСК (миРНК против Ku70, Ku80 и каталитической субъединицы ДНК-зависимой протеинкиназы (миКu70+80+ДНК-ПКкс)) и миРНК против ГР (MnRad51). На дорожках GFP показан положительный контроль экспрессирующих GFP клеток. На дорожках, обозначенных «нерасщепленные ГР» и «нерасщепленные НГСК», показаны клетки отрицательного контроля, т.е. клетки, трансфицированные кольцевыми репортерными плазмидами ГР и НГСК. На дорожках, обозначенных «ГР I-Scel» и «НГСК I-Scel», представлены клетки, трансфицированные расщепленными Seel репортерными плазмидами, которые восстанавливают экспрессию GFP после репарации расщепления ДНК путем гомологичной рекомбинации или негомологичного соединения концов, соответственно. На данной фигуре показана эффективность ингибирования ГР или НГСК молекулами миРНК, о чем свидетельствует процент GFP-положительных клеток, который нормировали на процент dsRed-положительных клеток и выразили в виде кратности изменения относительно процента контрольных экспрессирующих GFP клеток, который приравняли к 1. Среднее значение для 3 экспериментов, усы показывают стандартную ошибку среднего. Статистическую значимость определяли с помощью непарного критерия Стьюдента; уровень значимости р<0,05 (*) и р<0,01 (**).
Фиг. 14. Влияние элементов MAR на подавление НГСК молекулами миРНК.
Показана кратность повышения экспрессии и встраивания GFP в клетки СНО, обработанные молекулами миРНК, направленными против факторов НГСК, и повторно трансфицированные плазмидами GFP или MAR-GFP. Средняя флуоресценция GFP, число копий и флуоресценция на копию GFP показаны в виде кратности превышения результата, полученного для необработанных клеток (обозначенных как «ложные»), трансфицированных плазмидой GFP. А) Результаты проточной цитометрии, В) анализ числа копий GFP в геноме с помощью количественной ПЦР, С) средняя флуоресценция каждого встроенного гена GFP (рассчитанная для каждого эксперимента как отношение экспрессии к числу копий). Среднее значение для 3 или более экспериментов; статистическую значимость определяли с помощью непарного критерия Стьюдента. Звездочками указаны значимые различия между обработанным миРНК образцом и соответствующим необработанным контролем; уровни значимости: р<0,05 (*), р<0,01 (**); усы показывают стандартную ошибку среднего.
Фиг. 15. Влияние элементов MAR на подавление ГР молекулами миРНК.
Показана кратность повышения экспрессии и встраивания GFP в клетки СНО, обработанные молекулами миРНК, направленными против факторов ГР, и повторно трансфицированные плазмидами GFP или MAR-GFP. Средняя флуоресценция GFP, число копий и флуоресценция на копию GFP показаны в виде кратности превышения результата, полученного для необработанных клеток (обозначенных как «ложные»), трансфицированных плазмидой GFP. А) Результаты проточной цитометрии, В) анализ числа копий GFP в геноме с помощью количественной ПЦР, С) средняя флуоресценция каждого встроенного гена GFP (рассчитанная для каждого эксперимента как отношение экспрессии к числу копий). Среднее значение для 3 или более экспериментов; статистическую значимость определяли с помощью непарного критерия Стьюдента. Звездочками указаны значимые различия между обработанным миРНК образцом и соответствующим необработанным контролем; уровни значимости: р<0,05 (*), р<0,01 (**); усы показывают стандартную ошибку среднего.
Фиг. 16. Влияние элементов MAR на подавление ОМСК молекулами миРНК.
Показана экспрессия и встраивание GFP в клетки СНО, обработанные молекулами миРНК, направленными против факторов ОМСК (и некоторых факторов ГР), и повторно трансфицированные плазмидами GFP (А) или MAR-GFP (В). Средняя флуоресценция GFP, число копий и флуоресценция на копию GFP показаны в виде кратности превышения результата, полученного для необработанных клеток (обозначенных как «ложные»), трансфицированных плазмидой GFP. На фигурах показаны результаты проточной цитометрии. Показано среднее значение для количества экспериментов, указанных внизу под графиком. Клетки, трансфицированные миMDC1, экспрессировали GFP даже без MAR на уровне, в 11,8 раз превышающем таковой для клеток, не трансфицированных миMDC. Особенно хороших результатов также можно было добиться для некоторых плазмид, которые не содержали MAR, а именно, для миВаrd1 и миLigl.
Фиг. 17. Влияние опосредованного миРНК подавления белка ГР.
На фигуре показано, что можно добиться более высокой экспрессии GFP и иммуноглобулина клетками СНО-М, стабильно экспрессирующими кшРНК, направленную против Rad51. Клетки СНО-М трансфицировали вектором экспрессии, полученным на основе транспозона PiggyBac, экспрессирующим кшРНК, направленную против Rad51, и полученную поликлональную популяцию клеток, а также клоны клеток, полученные из нее, повторно трансфицировали плазмидой, экспрессирующей GFP, наряду с исходными клетками СНО-М. Флуоресценцию GFP исходных СНО-М, популяции клеток, экспрессирующих Rad51-кшРНК, и полученных клонов оценивали через 10 дней после отбора на основании стабильной экспрессии гена GFP и гена устойчивости к пуромицину. Профили флуоресценции двух наиболее флуоресцирующих клонов показаны рядом с таковыми для объединенной популяции клеток и исходных клеток (А), а также процент клеток в секторах М1, М2 и М3 через 10 дней после отбора на основании устойчивости к пуромицину (В), который изображен на панели А в виде горизонтальных полос, отмеченных как 1, 2 и 3. Количественное соотношение клеток М3 с высоким уровнем экспрессии отслеживали в течение дополнительных 68 дней культивирования без селекции, чтобы показать, что можно получить более стабильную экспрессию на более высоком уровне в клонах клеток, экспрессирующих кшРНК, по сравнению с исходными клетками СНО-М (С). В качестве альтернативы, экспрессионными плазмидами, кодирующими легкие и тяжелые цепи антитела инфликсимаба, трансфицировали типичные клоны, и оценивали удельную продуктивность секретируемого иммуноглобулина после селекции в течение дополнительных трех недель культивирования без антибиотика.
Фиг. 18. Влияние различных рекомбинантных элементов MAR человека, расположенных в 3'-5' направлении, на процентиль клеток с высоким и очень высоким уровнем продукции (% М3/М2), которое оценивали с помощью анализа методом сортировки клеток с возбуждением флуоресценции (FACS) по флуоресценции GFP для двух конструкций MAR.
(А) Указанные элементы MAR представляли собой перестроенные производные MAR Х-29 (X_29R2 (SEQ ID NO: 9), X_29R3 (SEQ ID NO: 10)), MAR 1-42 (1_42R2Bis, 1_42R3), MAR 1-6 (1_6R2 (SEQ ID NO: 8), 1_6R3) или MAR 1-68 (1_68R2 (SEQ ID NO: 7)), о чем свидетельствуют названия указанных конструкций. (В) Типичные профили FACS, полученные для лучших расположенных в 3'-5' направлении элементов MAR (MAR 1_68R (SEQ ID NO: 6)).
Фиг. 19. Стабильность экспрессии для двух векторов MAR.
Поликлональные популяции, сконструированные из векторов, содержащих производные MAR 1_68R2, 1_6R2 и X_29R3, исследовали в течение 5 недель культивирования без селекции, и еженедельно оценивали флуоресценцию GFP на протяжении данного периода. Оценивали процентиль субпопуляции М3: элемент MAR 1_6R2, расположенный в 3'-5' направлении, и неперестроенный MAR 1-68, расположенный в 5'-3' направлении, представляли собой лучшую исследованную комбинацию для вектора с двумя элементами MAR. Также показаны субпопуляции М1 и М2.
Фиг. 20. Векторы экспрессии, содержащие единичный генетический элемент.
Исследовали MAR 1_68 и Х_29 и использовали их в комбинации с репликатором LmnB2. Элементы MAR расположили в 5'-3' направлении от кассеты экспрессии трансгена, и оценивали в анализе трансфекции трансгеном на протяжении двух месяцев. Поликлональную популяцию стабильно трансфицированных клеток подвергали отбору по устойчивости к антибиотику в течение двух недель и исследовали флуоресценцию GFP с помощью анализа методом сортировки клеток с возбуждением флуоресценции (FACS) в течение семи недель. Соотношение клеток М3 с высоким уровнем продукции показано на графике (А), тогда как на графике (В) показаны типичные профили FACS.
Фиг. 21. Векторы экспрессии, содержащие единичный генетический элемент Х-29.
Анализ стабильности вектора Х_29: показали, что вектор экспрессии, содержащий единичный Х_29 в 5'-3' направлении от кассеты экспрессии, оказался стабильным и позволял получить очень высокую процентиль субпопуляций М2 и М3 даже через 14 недель культивирования (27 пассажей).
Фиг. 22. Сравнительный анализ стабильно трансфицированных популяций СНО после 24 недель селекции антибиотиком.
Вектор с единичным MAR Х_29, расположенным в 5'-3' направлении от кассеты экспрессии (Puro_CGAPD_GFP_гастрин_X29), повышает появление клеток, экспрессирующих высокий уровень GFP, а также стабильность экспрессии с течением времени по сравнению с вектором с двумя элементами MAR: с 1_6R2 в качестве MAR, расположенного в 3'-5' направлении, и 1_68 в качестве MAR, расположенного в 5'-3' направлении (Puro_1_6R2_CGAPD_GFP_гастрин_1_68).
ПОДРОБНОЕ ОПИСАНИЕ РАЗЛИЧНЫХ И ПРЕДПОЧТИТЕЛЬНЫХ ВАРИАНТОВ РЕАЛИЗАЦИИ НАСТОЯЩЕГО ИЗОБРЕТЕНИЯ
Термин трансген, используемый в контексте настоящего изобретения, представляет собой изолированную последовательность дезоксирибонуклеотидов (ДНК), кодирующую данный зрелый белок (также называемую в данном тексте ДНК, кодирующей белок), предшественника белка или функциональную РНК, которая не кодирует белок (некодирующую РНК). Трансген выделяют и встраивают в клетку для получения продукта трансгена. Некоторые предпочтительные трансгены согласно настоящему изобретению представляют собой трансгены, кодирующие иммуноглобулины (Ig), Fc-слитые белки и другие белки, в частности, белки с терапевтической активностью ("биотерапевтические средства"). Например, некоторые иммуноглобулины, такие как инфликсимаб (ремикейд) или другие секретируемые белки, такие как фактор коагуляции VIII, достаточно трудно экспрессировать вследствие клеточных затруднений, преимущественно неохарактеризованных. С помощью рекомбинантных молекул нуклеиновых кислот, векторов и способов согласно настоящему изобретению данные затруднения можно обнаружить и/или раскрыть. Это, как правило, позволяет повысить количество терапевтических белков, которые можно получить, и/или их качество, например, их процессинг и гомогенность посттрансляционных модификаций, таких как гликозилирование.
В данном тексте термин трансген не должен, в контексте ДНК, кодирующей белок, включать нетранскрибируемые, фланкирующие участки, такие как сигналы инициации транскрипции РНК, сайты полиаденилирования, промоторы или энхансеры. Другие предпочтительные трансгены включают последовательности ДНК, кодирующие функциональные РНК. Таким образом, термин трансген в контексте настоящего изобретения относится к последовательности ДНК, которую вводят в клетку, такую как эукариотическая клетка-хозяин, путем трансфекции (которая включает, в контексте настоящего изобретения, также трансдукцию, т.е., внедрение посредством вирусных векторов), и которая кодирует интересующий продукт, также называемый в данном тексте "продуктом экспрессии трансгена", например, "гетерологичными белками". Указанный трансген может быть функционально присоединен к последовательности, которая кодирует сигнальный пептид, который, в свою очередь, опосредует и/или способствует перемещению и/или секреции через эндоплазматический ретикулум и/или цитоплазматическую мембрану и удаляется до или в процессе секреции.
Малые интерферирующие РНК (миРНК) представляют собой двунитевые молекулы РНК, как правило, длиной 20-25 пар оснований, которые играют роль в РНК-интерференции (РНКи) путем интерференции с экспрессией определенных генов с комплементарной последовательностью нуклеотидов. миРНК можно непосредственно ввести в клетки или можно экспрессировать в клетке посредством вектора. Изолированная миРНК ТЕР, упоминаемая в данном тексте, представляет собой такую миРНК длиной 20-25 пар оснований, которую обычно вводят непосредственно в клетку, т.е., без экспрессии посредством нуклеиновой кислоты, которую внедрили в клетку.
Малая/короткая шпилечная РНК (кшРНК) представляет собой последовательность РНК, которая образует плотный шпилечный изгиб, который можно применять для замалчивания экспрессии целевого гена посредством РНКи. Экспрессию кшРНК в клетках обычно осуществляют путем доставки плазмид или вирусных векторов, таких как ретровирусные векторы. Для того, чтобы получить молекулы кшРНК, последовательность миРНК обычно модифицируют, чтобы ввести короткую петлю между двумя нитями миРНК. Нуклеиновую кислоту, кодирующую кшРНК, затем доставляют посредством вектора в клетку, и она транскрибируется в виде короткой шпилечной РНК (кшРНК), которую, возможно, дайсер обычным способом процессирует в функциональную миРНК.
Ми/кшРНК способна зависимым от последовательности способом снижать экспрессию целевого гена. КшРНК может гибридизоваться с участком транскрипта мРНК, кодирующего продукт целевого гена, тем самым ингибируя экспрессию целевого гена посредством РНК-интерференции. Бифункциональные кшРНК имеют больше одной мишени, например, кодирующую область, а также некоторые нетранслируемые области мРНК. Встраивание в геном клетки способствует длительному или конститутивному замалчиванию гена, которое может распространяться на дочерние клетки.
МикроРНК (миРНК) представляет собой малую молекулу РНК, например, длиной от 20 до 24 нуклеотидов, в частности, 22 нуклеотида, которая участвует в транскрипционной и посттранскрипционной регуляции экспрессии генов посредством спаривания с комплементарными последовательностями внутри мРНК. Замалчивание гена может происходить либо путем ингибирования транскрипции трансгена, либо путем деградации мРНК, либо путем предотвращения трансляции мРНК. МикроРНК можно экспрессировать путем доставки плазмид или вирусных векторов, таких как ретровирусные векторы. В качестве альтернативы, можно синтезировать молекулы РНК, ингибирующие или имитирующие микроРНК, и непосредственно трансфицировать ими клетки.
"Последовательность, кодирующая белок экспрессии/процессинга трансгена (ТЕР) или функциональную РНК ТЕР" позволяет экспрессировать или экспрессировать на повышенном уровне данный белок ТЕР после его переноса в клетку, тогда как последовательность, кодирующая некодирующие функциональные молекулы РНК, ингибирует экспрессию клеточных белков, соответственно. Белки ТЕР могут быть идентичны или аналогичны клеточным белкам, или они могут представлять собой белки из отличной клетки или вида. Клеточные белки, экспрессия которых, например, ингибируется функциональными молекулами РНК, представляют собой конститутивные белки клетки, в которую ввели функциональные молекулы РНК. Белок ТЕР также может дополнять экспрессию другого клеточного белка и, в результате, предпочтительно, повышает экспрессию трансгена. Указанные белки могут участвовать в рекомбинации; в процессах трансляции мРНК; в перемещении в ЭР, секреции, процессинге или фолдинге полипептидов, в транспорте ЭР-Гольджи-плазматическая мембрана, в гликозилировании и/или другой посттрансляционной модификации. Функциональные молекулы РНК включают, например, миРНК, кшРНК, микроРНК, сплайсированную в форме лассо РНК, малую временную смысловую РНК (мвРНК), антисмысловую РНК (аРНК), РНК рибозима и другие РНК, в частности, такие, которые могут подавлять экспрессию целевого гена. В особо предпочтительном варианте реализации данные белки вовлечены в "путь секреции белка" или в "пути рекомбинации", но также включают некоторые белки процессинга белка или метаболические белки, описанные ниже.
Функциональные РНК ТЕР можно не только экспрессировать с последовательности нуклеиновой кислоты, описанной выше, но их можно непосредственно ввести в клетку. Это, в частности, справедливо для изолированных миРНК ТЕР.
Термин "изолированная молекула нуклеиновой кислоты" в контексте настоящего изобретения эквивалентен термину "рекомбинантная молекула нуклеиновой кислоты", т.е. молекула нуклеиновой кислоты, которая не существует в данной форме в природе, но которую сконструировали из частей, которые существуют в природе.
Последовательность нуклеиновой кислоты, такая как ДНК или РНК, комплементарна другой ДНК или РНК, если нуклеотиды, например, из двух нитей однонитевых ДНК или двух нитей однонитевых РНК, могут образовывать стабильные водородные связи, например, водородную связь между гуанином (G) и цитозином (С). В клетке комплементарное спаривание оснований позволяет, например, клеткам копировать информацию от одного поколения к другому. При РНК-интерференции (РНКи) комплементарное спаривание оснований позволяет замалчивание или полный нокаут некоторых целевых генов. По существу, последовательность миРНК, кшРНК или микроРНК специфично снижает или нарушает экспрессию целевого гена путем выравнивания одинарной нити РНК (например, антисмысловой нити в миРНК) с РНК, в частности, мРНК клетки-хозяина. Степень комплементарности между двумя нитями нуклеиновой кислоты может изменяться от полной комплементарности (каждый нуклеотид напротив противоположного) до частичной комплементарности (50%, 60%, 70%, 80%, 90% или 95%). Степень комплементарности определяет стабильность комплекса и, таким образом, то, насколько успешно ген можно, например, нокаутировать. Таким образом, полная или по меньшей мере 95% комплементарность является предпочтительной.
Активность молекул миРНК при РНКи в значительной мере зависит от их способности связываться с индуцируемым РНК комплексом сайленсинга (RISC). За связыванием дуплекса миРНК с RISC следует разворачивание и расщепление смысловой нити эндонуклеазами. Оставшийся комплекс антисмысловая нить-RISC затем может связываться с целевыми мРНК для инициации замалчивания транскрипции.
В контексте настоящего изобретения трансгены, определение которых приведено выше, экспрессируют, как правило, белки, которые желательно получить в больших количествах, например, для применения в фармацевтике, тогда как последовательности, кодирующие белки ТЕР/функциональные РНК ТЕР, или сами функциональные молекулы РНК, разработаны таким образом, чтобы помогать экспрессии таких трансгенов либо непосредственно, либо опосредованно. "Типичный перечень белков ТЕР, экспрессированных с применением векторов, несущих транспозоны", приведен в таблице А. Для специалиста в данной области должно быть очевидно, что огромное количество таких белков было описано в данной области, и в таблице А представлены как идентификационные номера последовательностей NCBI для соответствующих белков, так и последовательности нуклеиновых кислот, кодирующие их. В последней колонке приведены идентификаторы последовательностей для некоторых из данных последовательностей. Для специалиста в данной области должно быть очевидно, что варианты белков, а также последовательности, идентичные им более чем на 80%, 90%, 95% или 98%, входят в объем настоящего изобретения.
"Типичный перечень кшРНК, экспрессируемых с применением, например, определенных векторов, несущих транспозоны piggybac", приведен в таблице В. Для специалиста в данной области должно быть очевидно, что такие кшРНК можно легко сконструировать после того, как выбран целевой ген. Например, любой из известных генов пути рекомбинации представляет собой готовый целевой ген. Тем не менее, другие гены, такие как гены, кодирующие белки, представленные в таблице А, могут представлять собой готовые мишени для миРНК, полученных из таких кшРНК. Таблица С представляет собой перечень примеров миРНК (смысловая нить) и примеров кшРНК, полученных из соответствующих миРНК. Антисмысловую нить миРНК в конечном счете применяют, чтобы блокировать и/или вызвать деградацию клеточной мРНК. Это, как правило, приводит к снижению уровней белка, кодируемого указанной мРНК.
Идентичность означает степень родства между двумя последовательностями нуклеотидов, которую определяют по идентичности пар между двумя нитями таких последовательностей, например, целой и полной последовательности. Идентичность можно легко рассчитать. Несмотря на то, что существует множество способов измерения идентичности между двумя последовательностями нуклеотидов, термин "идентичность" хорошо известен квалифицированным специалистам (Computational Molecular Biology, Lesk, A.M., ред., Oxford University Press, Нью-Йорк, 1988; Biocomputing: Informatics and Genome Projects, Smith, D. W., ред., Academic Press, Нью-Йорк, 1993; Computer Analysis of Sequence Data, часть I, Griffin, A. M., и Griffin, H. G., ред., Human Press, Нью-Джерси, 1994; Sequence Analysis in Molecular Biology, von Heinje, G., Academic Press, 1987; и Sequence Analysis Primer, Gribskov, M. и Devereux, J., ред., M Stockton Press, Нью-Йорк, 1991). Способы, широко используемые для определения идентичности между двумя последовательностями, включают, но не ограничены способами, описанными в Guide to Huge Computers, Martin J. Bishop, ред., Academic Press, Сан-Диего, 1994, и Carillo, H., и Lipman, D., SIAM J Applied Math. 48: 1073 (1988). Разработаны предпочтительные способы определения идентичности, чтобы получить наибольшие совпадения между двумя исследуемыми последовательностями. Такие способы закодированы в компьютерных программах. Предпочтительные способы на основе компьютерных программ для определения идентичности между двумя последовательностями включают, но не ограничены пакетом программ GCG (Genetics Computer Group, Мэдисон, Висконсин) (Devereux, J., и др., Nucleic Acids Research 12(1). 387 (1984)), BLASTP, BLASTN, FASTA (Altschul и др. (1990); Altschul и др. (1997)). Хорошо известный алгоритм Smith Waterman также можно применять для определения идентичности.
В качестве примера, нуклеиновая кислота, включающая последовательность нуклеотидов, "идентичную" по меньшей мере, например, на 95% эталонной последовательности нуклеотидов, означает, что последовательность нуклеотидов нуклеиновой кислоты идентична эталонной последовательности за исключением того, что указанная последовательность нуклеотидов может содержать до пяти точечных мутаций на каждые 100 нуклеотидов эталонной последовательности нуклеотидов. Другими словами, чтобы получить последовательность нуклеотидов, по меньшей мере на 95% идентичную эталонной последовательности нуклеотидов, до 5% нуклеотидов в указанной эталонной последовательности можно удалить или заменить на другой нуклеотид, или множество нуклеотидов, составляющее до 5% от всех нуклеотидов в эталонной последовательности, можно вставить в эталонную последовательность. Данные мутации эталонной последовательности могут встречаться в 5'- или 3'-концевых положениях эталонной последовательности нуклеотидов или в любом месте между этими концевыми положениями, либо разбросанные отдельно среди нуклеотидов в эталонной последовательности, либо расположенные в одной или более непрерывных группах внутри эталонной последовательности. Идентичности последовательностей более чем на приблизительно 60%, приблизительно 70%, приблизительно 75%, приблизительно 85% или приблизительно 90% для любой последовательности, описанной в данной заявке (например, путем указания SEQ ID и/или номера доступа), также входят в объем настоящего изобретения.
Последовательность нуклеиновой кислоты, по существу идентичная другой последовательности нуклеиновой кислоты, относится к последовательности, содержащей точечные мутации, делеции или вставки в последовательности, которые не оказывают или оказывают несущественное влияние на соответствующий описанный способ и часто представлены одной, двумя, тремя или четырьмя мутациями на 100 п.о.
Настоящее изобретение относится к вариантам как полинукпеотида, так и полипептида. "Вариант" относится к полинуклеотиду или полипептиду, отличающемуся от описанного полинукпеотида или полипептида, но сохранившему его существенные свойства. Как правило, варианты в целом очень сходны и на многих участках идентичны полинуклеотиду или полипептиду согласно настоящему изобретению. Указанные варианты могут включать изменения в кодирующих областях, некодирующих областях или в обоих указанных типах областей. Особенно предпочтительны полинуклеотидные варианты, включающие изменения, которые вызывают молчащие замены, вставки или делеции, но не изменяют свойства или активности кодированного полипептида. Нуклеотидные варианты, полученные путем молчащих замен, вследствие вырожденности генетического кода, являются предпочтительными. Более того, варианты, в которых 5-10, 1-5 или 1-2 аминокислоты, описанные в данной заявке, заменены, удалены или вставлены в любой комбинации, также предпочтительны. В объем настоящего изобретения также входят аллельные варианты указанных полинуклеотидов. Аллельный вариант обозначает любую из двух или более альтернативных форм гена, занимающих один и тот же хромосомный локус. Аллельная вариация в природе возникает в результате мутации и может приводить к полиморфизму внутри популяции. Мутации в гене могут быть молчащими (без изменений в кодируемом полипептиде) или могут кодировать полипептиды, последовательности аминокислот которых изменены. Аллельный вариант полипептида представляет собой полипептид, кодируемый аллельным вариантом гена. Варианты любых из молекул нуклеиновых кислот, описанных в данной заявке, входят в объем настоящего изобретения.
Последовательность промотора или сам промотор представляет собой последовательность нуклеиновой кислоты, которая узнается клеткой-хозяином для экспрессии определенной последовательности нуклеиновой кислоты. Последовательность промотора включает последовательности, контролирующие транскрипцию, которые регулируют экспрессию указанного полинукпеотида. Промотор может представлять собой любую последовательность нуклеиновой кислоты, которая проявляет транскрипционную активность в выбранной клетке хозяина, включая мутантные, укороченные и гибридные промоторы, и его можно получить из генов, кодирующих внеклеточные или внутриклеточные полипептиды, либо гомологичные, либо гетерологичные для клетки хозяина. Промоторы согласно настоящему изобретению включают индуцируемые и неиндуцируемые промоторы. Последовательность нуклеиновой кислоты находится под контролем промотора, когда промотор выполняет свою функцию на указанной нуклеиновой кислоте.
CGAPDH (также называемый в данном тексте C_GAPDH) представляет собой слитые энхансер и промотор, которые включают промотор GAPDH человека и энхансер немедленно-раннего гена CMV человека. В одном варианте реализации для его получения промотор GAPDH человека и его 5'-нетранслируемую область (НТО) амплифицировали путем ПЦР из геномной ДНК клеток НЕК293 человека. Полученный продукт помещали в 5'-3' направлении от энхансера немедленно-раннего гена CMV человека. См. пример последовательности в SEQ ID NO: 11. Последовательности, которые на по меньшей мере 80%, 90%, 95%, 98%, 99% или 100% идентичны последовательности SEQ ID NO: 11, также входят в объем настоящего изобретения. Другой желательный промотор(ы) и/или энхансер(ы) или продукты их слияния представляют собой, но не ограничены перечисленными: энхансер немедленно-раннего гена CMV человека, промотор GAPDH человека, промотор EFI-альфа человека, промотор CMV, промотор SV40, промотор Actb СНО или промотор HspaS СНО. Данные элементы хорошо известны в данной области, и примеры последовательностей перечислены в SEQ ID NO: 110-116. Для специалиста в данной области должно быть очевидно, что их варианты также входят в объем настоящего изобретения, также, как и элемент, последовательность которого по меньшей мере на 80%, 90%, 95%, 98%, 99% или 100% идентична любой из последовательностей SEQ ID NO: 110-116.
"Транспозон" представляет собой мобильный генетический элемент, который эффективно перемещается между векторами и хромосомами с помощью механизма "вырезать и вставить" или "копировать и вставить". В процессе транспозиции транспозаза (например, транспозаза РВ в системе транспозона PiggyBac) узнает транспозон-специфичные последовательности инвертированных концевых повторов (ITR), расположенные на обоих концах транспозона (существуют 5'- и а 3'-ITR в любой системе транспозона), перемещает содержимое из исходных сайтов и встраивает их в хромосомные сайты, такие как хромосомные сайты ТТАА. Мощная активность системы транспозона PiggyBac позволяет легко перемещать интересующие гены, расположенные между двумя ITR, в целевые геномы. Система транспозона PiggyBac описана, например, в 2010/0154070, которая полностью включена в данную заявку посредством ссылки.
Элементы MAR (конструкции MAR, последовательности MAR, S/MAR или просто MAR) принадлежат к более широкой группе эпигенетических регуляторных элементов, которые также включают граничные или инсуляторные элементы, такие как cHS4, регуляторные области локусов (LCR), стабилизирующие и антирепрессорные (STAR) элементы, универсальные элементы, открывающие хроматин (UCOE), или модификаторы гистонов, такие как гистоновая дезацетилаза (HDAC).
Элементы MAR могут быть определены на основании идентифицированного MAR, на котором они первично основаны: конструкция MAR S4, соответственно, представляет собой элементы MAR, большинство нуклеотидов которых (50% и более, предпочтительно 60%, 70% или 80%) основаны на MAR S4. Некоторые простые мотивы последовательности, такие как мотивы с высоким содержанием А и Т, часто обнаруживают внутри элементов MAR. Другие широко распространенные мотивы представляют собой мотивы А-бокс, Т-бокс, мотивы раскручивания спирали ДНК, сайты связывания SATB1 (Н-бокс, А/Т/С25) и консенсусные сайты топоизомеразы II для позвоночных или дрозофилы.
MAR, как правило, описывают как последовательности в ДНК эукариотических хромосом, к которым присоединяется ядерный матрикс. Свойства MAR лишь отчасти определяются их первичной структурой. Например, известно, что типичная первичная структура, обнаруженная в элементах MAR, таких как АТ-богатые области, приводит к образованию третичных структур, а именно, некоторых изгибов, которые определяют функцию MAR. Таким образом, MAR часто определяют не только по их первичной структуре, но также по их вторичной, третичной структуре, например, степени их изогнутости и/или физическим свойствам, таким как температура плавления.
Обогащенная АТ/ТА-динуклеотидами область изгиба ДНК (здесь и далее называемая "АТ-богатой областью"), обнаруживаемая в большинстве элементов MAR, представляет собой область изгиба ДНК, содержащую большое количество А и Т, в частности, в виде динуклеотидов AT и ТА. В предпочтительном варианте реализации она содержит по меньшей мере 10% динуклеотида ТА, и/или по меньшей мере 12% динуклеотида AT на участке из 100 непрерывных пар оснований, предпочтительно, по меньшей мере 33% динуклеотида ТА, и/или по меньшей мере 33% динуклеотида AT на участке из 100 непрерывных пар оснований (или на соответствующем более коротком участке при меньшей длине АТ-богатой области), и при этом имеет изогнутую вторичную структуру. Тем не менее, "АТ-богатые области" могут быть настолько короткими, как приблизительно 30 нуклеотидов или менее, но предпочтительно их длина составляет приблизительно 50 нуклеотидов, приблизительно 75 нуклеотидов, приблизительно 100 нуклеотидов, приблизительно 150, приблизительно 200, приблизительно 250, приблизительно 300, приблизительно 350 или приблизительно 400 нуклеотидов или более.
Некоторые сайты связывания также часто имеют относительно высокое содержание А и Т, такие как сайты связывания SATB1 (Н-бокс, А/Т/С25) и консенсусные сайты топоизомеразы II для позвоночных (RNYNNCNNGYNGKTNYNY) или дрозофилы (GTNWAYATTNATNNR). Тем не менее, область сайта связывания (модуль), в частности, область TFBS, которая содержит кластер сайтов связывания, можно легко отличить от областей, обогащенных динуклеотидами AT и ТА ("АТ-богатых областей"), от элементов MAR, богатых содержанием А и Т, путем сравнения паттерна кривизны указанных областей. Например, средняя степень кривизны MAR 1_68 человека может превышать приблизительно 3,8 или приблизительно 4,0, тогда как средняя степень кривизны участка TFBS может быть ниже приблизительно 3,5 или приблизительно 3,3. Области идентифицированного MAR также можно подтвердить с помощью альтернативных способов, таких как, но не ограничиваясь определением относительных температур плавления, описанных в других местах в данной заявке. Тем не менее, такие значения видоспецифичны и, следовательно, могут изменяться от вида к виду и могут, например, быть ниже. Таким образом, градусы кривизны соответствующих областей, богатых динуклеотидами AT и ТА, могут быть ниже, например, от приблизительно 3,2 до приблизительно 3,4, или от приблизительно 3,4 до приблизительно 3,6, или от приблизительно 3,6 до приблизительно 3,8, и градусы кривизны областей TFBS могут быть пропорционально ниже, например, ниже приблизительно 2,7, ниже приблизительно 2,9, ниже приблизительно 3,1, ниже приблизительно 3,3. Квалифицированный специалист выберет, соответственно, меньшие размеры окна в программе SMAR Scan II.
Элемент MAR, конструкция MAR, последовательность MAR, S/MAR или просто MAR согласно настоящему изобретению представляет собой последовательность нуклеотидов, одно или более (например, два, три или четыре) свойств которой, таких как описанные выше, аналогичны таковым у встречающихся в природе "SAR" или "MAR". Предпочтительно у такого элемента MAR, конструкции MAR, последовательности MAR, S/MAR или просто MAR по меньшей мере одно свойство способствует экспрессии белка любого гена, находящегося под влиянием указанного MAR. Элемент MAR, как правило, также обладает свойством изолированной и/или очищенной нуклеиновой кислоты, предпочтительно проявляющей активность MAR, в частности, проявляющей активность модуляции транскрипции, предпочтительно активность усиления транскрипции, но также проявляющей, например, активность стабилизации экспрессии и/или другие активности.
Термины элемент MAR, конструкция MAR, последовательность MAR, S/MAR или просто MAR также включают, в некоторых вариантах реализации, улучшенные конструкции MAR, которые обладают свойствами, которые обеспечивают улучшение по сравнению со встречающимся в природе и/или идентифицированным MAR, на котором может быть основана конструкция MAR согласно настоящему изобретению. Такие свойства включают, но не ограничены перечисленными: уменьшенную длину по сравнению с полноразмерным встречающимся в природе и/или обнаруженным MAR, повышенную экспрессию гена/транскрипции, повышенную стабильность экспрессии, тканеспецифичность, способность к индуцированию или комбинацию перечисленных свойств. Соответственно, элемент MAR, который подвергли улучшению, например, содержит менее чем приблизительно 90%, предпочтительно менее чем приблизительно 80%, еще более предпочтительно менее чем приблизительно 70%, менее чем приблизительно 60%, или менее чем приблизительно 50% от количества нуклеотидов идентифицированной последовательности MAR. Элемент MAR может повышать экспрессию гена и/или транскрипцию трансгена после трансформации подходящей клетки указанной конструкцией.
Элемент MAR предпочтительно встраивают в 3'-5' направлении от промоторной области, с которой функционально связан или может быть функционально связан интересующий ген. Тем не менее, в некоторых вариантах реализации предпочтительно, чтобы элемент MAR был расположен как в 3'-5' направлении, так и в 5'-3' направлении, или только в 5'-3' направлении от интересующего гена/последовательности нуклеиновой кислоты. Другие разнообразные цис- и/или транс-расположения MAR также входят в объем настоящего изобретения.
Термин «синтетический», когда его применяют в контексте элемента MAR, относится к MAR, разработка которого включала больше изменений, чем простая перестановка, дупликация и/или делеция последовательностей/областей или части областей идентифицированных MAR или MAR, основанных на них. В частности, синтетические MAR/элементы MAR как правило включают одну или более, предпочтительно одну область идентифицированного MAR, которая, тем не менее, в некотором варианте реализации могла быть синтезирована или модифицирована, а также специально разработанные, хорошо охарактеризованные элементы, такие как единичный TFBS или серия TFBS, которые, в предпочтительном варианте реализации, получают синтетическим путем. Данные разработанные элементы во многих вариантах реализации относительно коротки, в частности, их длина, как правило, составляет не больше чем приблизительно 300 п.о., предпочтительно не больше чем приблизительно 100, приблизительно 50, приблизительно 40, приблизительно 30, приблизительно 20 или приблизительно 10 п.о. Данные элементы в некоторых вариантах реализации могут быть мультимеризованы. Такие синтетические элементы MAR также входят в объем настоящего изобретения, и должно быть очевидно, что, как правило, в настоящем описании следует понимать, что все, что указано как применимое к "элементу MAR", в равной степени применимо к синтетическому элементу MAR.
Функциональные фрагменты последовательностей нуклеотидов обнаруженных элементов MAR также включены при условии, что они сохраняют функции элемента MAR, описанные выше.
Некоторые предпочтительные идентифицированные элементы MAR включают, но не ограничены перечисленными: MAR 1_68, MAR Х_29, MAR 1_6, MAR S4, MAR S46, включая все их перестановки, описанные в WO 2005040377 и публикации патента США 20070178469, которые особенно включены в настоящую заявку посредством ссылки для описания последовательностей данных и других элементов MAR. MAR лизоцима цыпленка также представляет собой предпочтительный вариант реализации (см., патент США номер 7129062, который также особенно включен в данную заявку для описания элементов MAR).
Если указано, что вектор содержит единичный MAR, это означает, что внутри указанного вектора присутствует один MAR и нет других MAR либо такого же, либо отличного типа или структуры.
В некоторых вариантах реализации настоящего изобретения присутствует несколько MAR, которые могут иметь одинаковый или отличный тип или структуру, все из которых могут быть расположены в 5'-3' направлении от интересующего гена. Это называется кластером единичных MAR.
Если указано, что что-либо, например, количество клеток, стабильно экспрессирующих полипептид, "не зависит" от присутствия, например, последовательности, то указанная последовательность не оказывает влияния (например, на количество клеток, стабильно экспрессирующих полипептид) до любой статистически значимой степени.
Трансген или последовательность, кодирующая белок экспрессии/процессинга трансгена или функциональную РНК согласно настоящему изобретению, часто является частью вектора.
Вектор согласно настоящему изобретению представляет собой молекулу нуклеиновой кислоты, способную переносить другую нуклеиновую кислоту, такую как трансген, которую нужно экспрессировать с помощью данного вектора, с которым она была связана, как правило, в который она была встроена. Например, плазмида представляет собой тип вектора, ретровирус или лентивирус представляет собой другой тип вектора. В предпочтительном варианте реализации настоящего изобретения вектор линеаризуют перед трансфекцией. Вектор экспрессии содержит регуляторные элементы или находится под контролем таких регуляторных элементов, которые разработаны таким образом, чтобы способствовать транскрипции и/или экспрессии последовательности нуклеиновой кислоты, которую несет указанный вектор экспрессии. Регуляторные элементы включают энхансеры и/или промоторы, но также различные другие элементы, описанные в данной заявке (см. также раздел "Разработка вектора").
Последовательность вектора представляет собой последовательность ДНК или РНК указанного вектора, за исключением любых "других" нуклеиновых кислот, таких как трансгены, а также генетических элементов, таких как элементы MAR.
Эукариотическую клетку, включая клетку млекопитающего, такую как рекомбинантная клетка млекопитающего/эукариотическая клетка-хозяин, согласно настоящему изобретению можно поддерживать в условиях культуры клеток. Лишь некоторые из примеров данного типа клеток представляют собой эукариотические клетки-хозяева, не относящиеся к приматам, такие как клетки яичника китайского хомячка (СНО) и клетки почки детеныша хомячка (BHK, АТСС CCL 10). Эукариотические клетки-хозяева примата включают, например, клетки карциномы шейки матки человека (HELA, АТСС CCL 2) и линию клеток почки обезьяны CV1, трансформированную SV40 (COS-7, АТСС CRL-1587). Рекомбинантная эукариотическая клетка-хозяин или рекомбинантная клетка млекопитающего означает клетку, которую модифицировали, например, путем трансфекции, например, трансгенной последовательностью, и/или путем мутации. Эукариотические клетки-хозяева или рекомбинантные клетки млекопитающих способны осуществлять посттранскрипционные модификации белков, экспрессируемых указанными клетками. В некоторых вариантах реализации настоящего изобретения клеточный аналог в эукариотической (например, не относящейся к примату) клетке-хозяине полностью функционален, т.е., не был, например, инактивирован путем мутации. Предпочтительно, чтобы трансгенная последовательность (например, примата) экспрессировалась дополнительно к ее клеточному аналогу (например, не относящемуся к примату).
Трансфекция согласно настоящему изобретению представляет собой внедрение нуклеиновой кислоты в реципиентную эукариотическую клетку, например, не ограничиваясь перечисленными, путем электропорации, липофекции, как правило с помощью невирусного вектора (опосредованная вектором трансфекция) или с помощью химических способов, включая применение поликатионных липидов. Трансфекция, не опосредованная вектором, включает, например, непосредственное внедрение изолированных миРНК ТЕР в клетку. Во временно трансфицированной клетке, например, миРНК сохраняется лишь временно. В контексте настоящего изобретения можно осуществить первую трансфекцию по меньшей мере одной молекулой нуклеиновой кислоты с последовательностью, кодирующей белок экспрессии/процессинга трансгена (ТЕР) или функциональную РНК ТЕР, или, в качестве альтернативы, непосредственно функциональной РНК ТЕР (например, миРНК), и вторую последующую трансфекцию нуклеиновой кислотой, кодирующей трансген. Как первую, так и вторую трансфекцию можно повторить. Например, миРНК, внедренная в процессе первой трансфекции, действует, в частности, ингибирует, рекомбинационный белок (белок, который участвует в событиях рекомбинации в трансфицированной клетке). После этого в процессе второй последующей трансфекции вводят трансген.
Транскрипция означает синтез РНК по матрице ДНК. "Транскрипционно активный" относится, например, к трансгену, который транскрибируется. Трансляция представляет собой процесс, с помощью которого по РНК синтезируется белок.
Улучшение секреции измеряют по сравнению со значением, полученным для контрольной клетки, которая не содержит соответствующую трансгенную последовательность. Любое статистически значимое улучшение по сравнению со значением для контроля называют стимуляцией.
Маркер отбора представляет собой нуклеиновую кислоту, которая содержит ген, продукт которого придает устойчивость к селекционному агенту - антибиотику (например, хлорамфениколу, ампициллину, гентамицину, стрептомицину, тетрациклину, канамицину, неомицину, пуромицину) или способность расти на селективных средах (например, с DHFR (дигидрофолатредуктазой)).
Класс белков, известных как шапероны, определяют как белки, которые связываются и стабилизируют в противном случае нестабильный конформационный изомер другого белка и, путем контролируемого связывания и высвобождения, способствуют его правильной судьбе in vivo, будь то фолдинг, сборка олигомера, транспорт в конкретный клеточный компартмент или утилизация путем деградации. BiP (также известный как GRP78, белок, связывающий тяжелую цепь Ig, и Kar2p в дрожжах) представляет собой распространенный шаперон массой приблизительно 70 кДа из семейства hsp 70, находящийся в эндоплазматическом ретикулуме (ER), который, среди прочих функций, способствует транспорту в секреторной системе и способствует фолдингу белков. Протеиндисульфидизомераза (PDI) представляет собой белок-шаперон, находящийся в ЭР, который участвует в катализе образования дисульфидной связи при посттрансляционном процессинге белков.
КЛЕТОЧНАЯ МЕТАБОЛИЧЕСКАЯ ИНЖЕНЕРИЯ
В клеточной метаболической инженерии, например, модифицируют процессы, свойственные экспрессирующей клетке. Например, некоторые белки секреторного пути, например, экспрессируют сверх меры. В качестве альтернативы, изменяют события рекомбинации путем влияния на пути рекомбинации.
ПУТЬ СЕКРЕЦИИ БЕЛКА
Секреция белков представляет собой процесс, общий для организмов из всех трех царств. Для такого сложного пути секреции в первую очередь требуется перемещение белка из цитозоля через цитоплазматическую мембрану клетки. Для достижения белком конечного назначения необходимо несколько этапов и различных факторов. В клетках млекопитающих такой путь секреции включает два основных макромолекулярных ансамбля: сигналузнающую частицу (SRP) и секреторный комплекс (Sec-комплекс или транслокон). SRP состоит из шести белков с массами 9, 14, 19, 54, 68 и 72 кДа и 7S РНК, а транслокон представляет собой тороидальную частицу, состоящую из Sec61αβγ, Sec62 и Sec63. Номера доступа (в скобках), представленные для человеческого варианта некоторых из данных белков, описаны далее: hSRP14 (номер доступа Х73459.1); hSRP9 (NM_001130440); hSRP54 (NM_003136); hSRPRα (NM_003139); hSRPRβ (NM_021203); hSEC61α1 (NM_013336); hSEC61 β (L25085.1); hSEC61 γ (AK311845,1).
Первый этап секреции белка зависит от сигнальных пептидов, которые содержат определенную пептидную последовательность на аминоконце полипептида, которая опосредует перемещение образующегося белка через мембрану и в просвет эндоплазматического ретикулума (ЭР). На данном этапе сигнальный пептид, который выходит из лидирующей транслирующей рибосомы, взаимодействует с субъединицей частицы SRP, которая узнает сигнальный пептид, а именно, с SRP54. Связывание SRP с сигнальным пептидом блокирует дальнейшую элонгацию образующегося полипептида, что приводит к остановке трансляции. Белки SRP-9 и -14 необходимы для остановки элонгации (Walter и Blobel 1981). На втором этапе комплекс рибосома-образующийся полипептид-SRP стыкуется с мембраной ЭР посредством взаимодействия SRP54 с рецептором SRP (SR) (Gilmore, Blobel и др. 1982; Pool, Stumm и др. 2002). SR представляет собой гетеродимерный комплекс, содержащий два белка, SRα и SRβ, которые проявляют ГТФазную активность (Gilmore, Walter и др. 1982). Взаимодействие SR с SRP54 зависит от связывания ГТФ (Connolly, Rapiejko и др. 1991). SR координирует высвобождение SRP из комплекса рибосома-образующийся полипептид и ассоциацию сайта выхода из рибосомы с комплексом Sec61 (транслоконом). Растущий образующийся полипептид проникает в ЭР через канал транслокона, и трансляция возобновляется с обычной скоростью. Рибосома остается связанной на цитоплазматической стороне транслокона до тех пор, пока трансляция не завершится. Дополнительно к рибосомам, транслоконы тесно связаны с рибофорином на цитоплазматической стороне и с шаперонами, такими как кальретикулин и кальнексин, и протеиндисульфидизомеразами (PDI) и олигосахарид-трансферазой на стороне просвета ЭР. После внедрения растущего образующегося полипептида в просвет ЭР сигнальный пептид отщепляется от пре-белка ферментом, называемым сигнальной пептидазой, что приводит к высвобождению зрелого белка в ЭР. После посттрансляционной модификации, правильному фолдингу и мультимеризации белки покидают ЭР и мигрируют в аппарат Гольджи, а затем в секреторные везикулы. Слияние секреторных везикул с плазматической мембраной приводит к высвобождению содержимого везикулы во внеклеточное пространство.
Удивительно, что секретируемые белки в ходе эволюции обрели определенные сигнальные последовательности, которые хорошо подходят для их перемещения через мембрану клетки. Различные последовательности, обнаруженные как отдельные сигнальные пептиды, могут взаимодействовать уникальными путями с секреторным аппаратом. Сигнальные последовательности преимущественно гидрофобны по своей природе, и это свойство может способствовать получению из образующегося пептида секреторного белка. Дополнительно к гидрофобному участку аминокислот последовательности большинства сигналов секреции млекопитающих обладают множеством общих свойств. Различные сигнальные пептиды отличаются по эффективности, с которой они направляют секрецию гетерологичных белков, но были обнаружены несколько сигнальных пептидов секреции (т.е. сигнальных последовательностей из интерлейкина, иммуноглобулина, рецептора гистосовместимости и т.д.), которые можно применять, чтобы направлять секрецию гетерологичных рекомбинантных белков. Несмотря на сходства, данные последовательности не оптимальны для стимуляции эффективной секреции некоторых белков, которые трудно поддаются экспрессии, вследствие того, что нативный сигнальный пептид не может правильно функционировать вне нативного контекста, или из-за различий, связанных с клеткой-хозяином или процессом секреции. Выбор подходящей сигнальной последовательности для эффективной секреции гетерологичного белка может дополнительно осложняться взаимодействием последовательностей внутри отщепленного сигнального пептида с другими участками зрелого белка (Johansson, Nilsson и др. 1993).
ПУТИ РЕКОМБИНАЦИИ
Пути рекомбинации, также известные как пути рекомбинации ДНК, представляют собой клеточные пути, которые приводят к репарации повреждения ДНК, например, к соединению концов молекулы ДНК после двухнитевых разрывов хромосомы и к обмену или слиянию последовательностей ДНК между хромосомными и нехромосомными молекулами ДНК, как, например, при кроссинговере хромосом при мейозе или перестройке генов иммуноглобулинов в лимфоцитарных клетках. Существует три основных пути рекомбинации: путь гомологичной рекомбинации (ГР), путь негомологичного соединения концов (НГСК), путь опосредованного микрогомологией соединения концов (ОМСК) и альтернативного соединения концов (Альт-СК).
МЕХАНИЗМЫ ГОМОЛОГИЧНОЙ РЕКОМБИНАЦИИ (ГР), НЕГОМОЛОГИЧНОГО СОЕДИНЕНИЯ КОНЦОВ (НГСК) И ОПОСРЕДОВАННОГО МИКРОГОМОЛОГИЕЙ СОЕДИНЕНИЯ КОНЦОВ (ОМСК)
Трансгены используют аппарат рекомбинации для встраивания в двухнитевой разрыв в геноме хозяина,
Двухнитевые разрывы (ДНР) представляют собой биологически наиболее разрушительный тип геномного повреждения, потенциально приводящий к гибели клетки или большому разнообразию генетических перестроек. Точная репарация необходима для успешного поддержания и передачи генетической информации.
Существует два основных механизма репарации ДНР: негомологичное соединение концов (НГСК) и гомологичная рекомбинация (ГР). Третий механизм, называемый опосредованным микрогомологией соединением концов (ОМСК), часто начинает действовать, когда два основных механизма репарации ДНР претерпевают неудачу. Гомологичная рекомбинация представляет собой процесс генетического обмена между последовательностями ДНК, которые гомологичны друг другу, и действует преимущественно в течение фаз S/G2 клеточного цикла, тогда как НГСК просто соединяет два разорванных конца ДНК, обычно при отсутствии гомологии последовательностей, и действует на всех фазах клеточного цикла, но имеет особое значение во время G0 - G1 и ранней S фазы митотических клеток (Wong и Capecchi, 1985; Delacote и Lopez, 2008). У позвоночных ГР, НГСК и ОМСК различным образом участвуют в репарации ДНР, в зависимости от природы ДНР и фазы клеточного цикла (Takata и др., 1998).
НГСК: основные механизмы
По существу, молекулярный механизм процесса НГСК кажется простым: 1) набор ферментов захватывает разорванную молекулу ДНК, 2) образуется молекулярный мостик, который сближает два конца ДНК, и 3) разорванные молекулы заново лигируются. Аппарат НГСК в клетках млекопитающих включает два белковых комплекса для осуществления таких реакций: гетеродимер Ku80/Ku70, связанный с ДНК-ПКкс (каталитической субъединицей ДНК-зависимой протеинкиназы), и ДНК-лигаза IV с ее кофактором XRCC4 (комплементирующий ген 4 репарации рентгеновских повреждений у китайского хомячка) и множеством белковых факторов, таких как Artemis (Артемида) и XLF (XRCC4-подобный фактор; или Cernunnos (Цернун)) (Delacote и др., 2002). НГСК часто считают допускающая ошибки репарацией ДНР, так как она просто соединяет два разорванных конца ДНК, обычно при отсутствии гомологии последовательностей, и порождает небольшие вставки и делеции (Moore и Haber, 1996; Wilson и др., 1999). НГСК обеспечивает механизм репарации ДНР на всем протяжении клеточного цикла, но особое значение имеет в течение G0 - G1 и ранней S-фазы митотических клеток (Takata и др., 1998; Delacote и Lopez, 2008). Репарацию ДНР посредством НГСК наблюдают в организмах начиная с бактерий и заканчивая млекопитающими, что указывает на ее консервативный характер в процессе эволюции.
После образования ДНР ключевым этапом в пути репарации НГСК является физическое сближение разорванных концов ДНК. НГСК запускается при взаимодействии гетеродимерного белкового комплекса Ku70/80 с обоими концами разорванной молекулы ДНК, чтобы захватить, сблизить концы и создать каркас для сборки других ключевых факторов НГСК. Связанный с ДНК гетеродимерный комплекс Ku привлекает в место ДНР ДНК-ПКкс, белок массой 460 кДа, принадлежащий к семейству PIKK (фосфоинозитид-3-киназа-подобному семейству протеинкиназ) (Gottlieb и Jackson, 1993), и активирует его серин/треонинкиназную функцию (Yaneva и др., 1997). Две молекулы ДНК-ПКкс с двух концов ДНР взаимодействуют, таким образом образуя молекулярный мостик между обоими разорванными концами ДНК и ингибируя их деградацию (DeFazio и др., 2002). Затем концы ДНК можно непосредственно лигировать, хотя большинство концов, возникших в результате ДНР, должны быть надлежащим образом процессированы перед лигированием (Nikjoo и др., 1998). В зависимости от природы разрыва может потребоваться действие различных комбинаций ферментов процессинга для получения совместимых выступов путем заполнения брешей, удаления поврежденной ДНК или вторичных структур, окружающих разрыв. Данный этап в процессе НГСК считают ответственным за случайную потерю нуклеотидов, связанную с репарацией НГСК. Один ключевой фермент процессинга концов при НГСК млекопитающих представляет собой Artemis, член суперсемейства ферментов металло-β-лактамаз, ген которого оказался мутированным у большинства пациентов с радиочувствительным тяжелым комбинированным иммунодефицитом (SCID) (Moshous и др., 2001). Artemis обладает как 5'→3' экзонуклеазной активностью, так и ДНК-ПКкс-зависимой эндонуклеазной активностью по отношению к ДНК, содержащей дн-он переходы (переходы от двух нитей к одной), и шпилькам ДНК (Ма и др., 2002). Его активность также регулируется протеинкиназой ATM. Таким образом, похоже, что Artemis, вероятно, участвует во множестве реакций на повреждение ДНК. Тем не менее, похоже, что Artemis участвует в репарации лишь подгруппы повреждений ДНК, так как не наблюдали значительного дефекта репарации ДНР в клетках, лишенных Artemis (Wang и др., 2005, Darroudi и др., 2007).
Бреши в ДНК должны быть заполнены, чтобы репарация стала возможной. Добавление нуклеотидов в ДНР могут осуществлять только полимеразы μ и λ (Lee и др., 2004; Сарр и др., 2007). Посредством взаимодействия с XRCC4 полинукпеотидкиназа (PNK) также привлекается к концам ДНК, чтобы позволить полимеризацию, а также лигирование ДНК (Koch и др., 2004). Наконец, НГСК завершается путем лигирования концов ДНК, этапа, осуществляемого комплексом, содержащим XRCC4, ДНК-лигазу IV и XLF (Grawunder и др., 1997). Другие лигазы могут частично заменять ДНК-лигазу IV, так как НГСК может происходить в отсутствие XRCC4 и лигазы IV (Yan и др., 2008). Более того, исследования показали, что XRCC4 и лигаза IV не играют роли вне НГСК, тогда как, напротив, KU принимает участие в других процессах, таких как транскрипция, апоптоз и реакции на микроокружение (Monferran и др., 2004; Müller и др., 2005; Downs и Jackson, 2004).
НГСК можно ослабить или прекратить различными способами, многие из которых непосредственно затрагивают упомянутые выше белки (например, гетеродимер Ku80/Ku70, ДНК-ПКкс, но, в особенности, ДНК-лигазу IV, XRCC4, Artemis и XLF (XRCC4-подобный фактор; или Cernunnos), PIKK (фосфоинозитид-3-киназа-подобное семейство протеинкиназ).
ГР: основные механизмы
Гомологичная рекомбинация (ГР) представляет собой очень точный механизм репарации. Гомологичная хроматида служит в качестве матрицы для репарации разорванной нити. ГР происходит в фазах S и G2 клеточного цикла, когда сестринские хроматиды становятся доступными. Классическая ГР главным образом характеризуется тремя этапами: 1) удаление 5'-нуклеотидов с разорванных концов, 2) внедрение нити и обмен с гомологичным дуплексом ДНК и 3) расхождение промежуточных продуктов (интермедиатов) рекомбинации. Репарация ДНР может завершаться различными путями, в зависимости от способности осуществлять внедрение нити, которые включают путь зависимого от синтеза ДНК отжига нитей (ЗСОН), классическую репарацию двухнитевого разрыва (РДНР) (Szostak и др., 1983), вызванную разрывом репликацию (ВРР) и, в качестве альтернативы, путь однонитевого отжига (ОНО). Все механизмы ГР взаимосвязаны и включают много общих ферментативных этапов.
Первый этап из всех реакций ГР соответствует удалению 5'-конца разорванной нити ДНК нуклеазами с помощью комплекса MRN (MRE11, RAD50, NBN (ранее NBS1, синдром разрывов Неймеген 1)) и CtIP (CtBP-взаимодействующий белок) (Sun и др., 1991; White и Haber, 1990). Полученная в результате этого совокупность последовательностей с 3'-одноцепочечными ДНР может искать гомологичную последовательность. Внедрение гомологичного дуплекса осуществляют с помощью нуклеофиламента, состоящего из 3'-онДНК, покрытой белком рекомбиназой RAD51 (Benson и др., 1994). Для сборки RAD51-филамента требуется привлечение ре пли кати в но го белка A (RPA), гетеротримерного онДНК-связывающего белка, вовлеченного в метаболические процессы ДНК, связанные с онДНК, у эукариотов (Wold, 1997, Song и Sung, 2000). Затем RAD51 взаимодействует с RAD52, который обладает кольцеобразной структурой (Shen и др., 1996), чтобы вытеснить молекулы RPA и способствовать загрузке RAD51 (Song и sung, 2000). Rad52 важен для процессов рекомбинации у дрожжей (Symington, 2002). Тем не менее, у позвоночных, похоже, скорее BRCA2 (белок чувствительности к раку молочной железы 2 типа), чем RAD52 играет важную роль во внедрении и обмене нитей (Davies и Pellegrini, 2007; Esashi и др., 2007). Взаимодействие RAD51/RAD52 стабилизируется путем связывания с RAD54. RAD54 также играет роль в созревании интермедиатов рекомбинации после образования D-петли (Bugreev и др., 2007). С другой стороны, BRCA1 (рак молочной железы 1) взаимодействует с BARD1 (ассоциированный с BRCA1 RING-домен 1) и ВАСН1 (гомолог 1 ВТВ и CNC), чтобы осуществить лигазную и геликазную активность репарации ДНР, соответственно (Greenberg и др., 2006). BRCA1 также взаимодействует с CtIP CDK-зависимым образом и подвергается убиквитинированию в ответ на повреждение ДНК (Limbo и др., 2007). Следовательно, BRCA1, CtIP и комплекс MRN играют роль в активации ГР-опосредованной репарации ДНК в S и G2 фазах клеточного цикла.
Внедрение нуклеофиламента приводит к образованию гетеродуплекса, называемого петлей замещения (D-петлей), и включает замещение одной нити дуплекса внедряющейся нитью и ее спаривание с другой нитью дуплекса. Затем несколько путей ГР могут завершить репарацию, используя гомологичную последовательность в качестве матрицы для замещения последовательности, окружающей ДНР. В зависимости от используемого механизма взаимные обмены (кроссинговеры) между гомологичной матрицей и разорванной молекулой ДНК могут быть или не быть связаны с репарацией ГР. Кроссинговеры могут приводить к значительным генетическим последствиям, таким как геномные перестройки или утрата гетерозиготности.
В гомологичной рекомбинации также участвуют пять паралогов Rad51: Xrcc2, Xrcc3, Rad51B, Rad51C, Rad51D (Suwaki и др., 2011). Паралоги Rad51 образуют два типа комплексов: один, названный BCDX2, содержит Rad51B, Rad51C, Rad51D и Xrcc2; другой содержит Rad51C и Xrcc3 (СХ3) (Masson и др., 2001). Предположили, что первый комплекс участвует в образовании и/или стабилизации комплекса Rad51-ДНК (Masson и др., 2001). Похоже, что роль второго комплекса состоит в миграции ветви и расщеплении структуры Холлидея (Liu и др., 2007).
Ранее сообщалось, что увеличение ГР по сравнению с НГСК (см. публикацию патента США 20120231449, которая полностью включена в данную заявку посредством ссылки) можно применять, чтобы усилить и/или способствовать экспрессии трансгена.
Настоящее изобретение направлено на уменьшение или выключение ГР. ГР можно уменьшить или выключить различными способами, многие из которых непосредственно затрагивают упомянутые выше белки (например, белки комплекса MRN (MRE11, RAD50, NBN (ранее NBS1, синдром разрывов Неймеген 1)) и CtIP (CtBP-взаимодействующий белок), RAD51, белок репликации A (RPA), Rad52, BRCA2 (белок чувствительности к раку молочной железы 2 типа), RAD54, BRCA1 (рак молочной железы 1), взаимодействующий с BARD1 (ассоциированный с BRCA1 RING-домен 1), ВАСН1 (гомолог 1 ВТВ и CNC)). Настоящее изобретение сосредоточено на получении молекул РНК, таких как миРНК, для достижения данной цели.
Опосредованное микрогомологией соединение концов (ОМСК)
Когда другие пути рекомбинации не справляются или не активны, ДНР могут подвергаться репарации с помощью другого допускающего ошибки механизма репарации, называемого опосредованным микрогомологией соединением концов (ОМСК). Данный путь все еще требуется полностью охарактеризовать, и иногда его также называют альтернативным соединением концов (альт-СК), хотя не ясно, действительно ли в основе данных двух процессов лежит один и тот же механизм. Наиболее отличительное свойство данного пути, которое отличается от НГСК, состоит в использовании микрогомологии по 5-25 п.о. в процессе выравнивания разорванных нитей ДНК (McVey и Lee, 2008).
ОМСК может происходить на любой стадии клеточного цикла и не зависит от основных факторов НГСК и ГР, т.е. генов Ku70, лигазы IV и Rad52 (Boboila и др., 2010; Yu и McVey, 2010; Lee и Lee, 2007; Ma и др., 2003). Вместо того, чтобы использовать для инициации ОМСК собственный набор белков, наиболее важные применяемые для инициации белки являются компонентами комплекса MRN (MRX в дрожжах), содержащего Mre11, Rad50 и Nbs1 (Xrs2 в дрожжах), также участвующего в первых этапах ГР (Ма и др., 2003). Помимо комплекса MRN многие другие факторы были предложены в качестве участников в ОМСК, например, СТВР-взаимодействующий белок (CtIP; Yun и Hiom, 2009), поли(АДФ-рибоза)-полимераза 1 (PARP1), комплекс лигаза III/Xrcc1, лигаза I (Audebert и др., 2004), ДНК-полимераза θ (Yu и McVey, 2010) и комплекс ERCC1/XPF (Ma и др., 2003). Тем не менее, еще многие другие белки принимают участие в данном процессе.
Предположили, что в отсутствие других белков, связывающися с концом ДНК (таких как Ku или Rad51), ДНР узнаются PARP1, который затем инициирует их репарацию посредством ОМСК (McVey и Lee, 2008). Процесс репарации, аналогично ГР, начинается с удаления нуклеотидов с 5'- к 3'-концу, которое открывает короткие участки гомологии на каждой стороне от разрыва. Данный этап процессинга осуществляется комплексом MRN и регулируется CtIP (Mladenov и IIiakis, 2011). Комплементарные участки (присутствующие во фрагментах 3'-онДНК) спариваются и некомплементарные фрагменты (флэпы) удаляются (Yu и McVey, 2010), вероятно, комплексом ERCC1/XPF. Бреши (если такие есть) затем заполняются полимеразой (например, ДНК-полимеразой θ или δ (Yu и McVey, 2010; Lee и Lee, 2007)) и разрывы соединяются лигазой I или комплексом лигаза III/Xrcc1.
В отсутствие участков непосредственной микрогомологии на концах ДНК, что наиболее часто встречается, более удаленный фрагмент подвергнутой репарации молекулы можно скопировать, применяя точную ДНК-полимеразу (например, полимеразу 6). Данный дуплицированный участок затем участвует в выравнивании концов ДНК, что приводит к образованию вставки в месте соединения. Данный более сложный вариант опосредованной микрогомологией репарации назвали синтез-зависимой ОМСК (СЗ-ОМСК) (Yu и McVey, 2010).
Хотя ОМСК считали альтернативным путем рекомбинационной репарации, было показано, что он очень эффективен в процессе рекомбинации переключения классов IgH в В-лимфоцитах (Boboila и др., 2010), позволяя предложить, что он может играть большую роль, чем просто быть запасным механизмом. Также возможно, что некоторые ДНР, например, несовместимые выступы или тупые концы (которые плохо поддаются репарации посредством НГСК и/или ГР), могут более эффективно репарироваться посредством ОМСК (Zhang и Paull, 2005).
В ТАБЛИЦЕ D перечислены некоторые из ключевых генов в каждом из трех путей, которые, следовательно, также являются ключевыми мишенями для влияния на каждый из трех путей (см. также публикацию патента США 20120231449, которая полностью включена в данную заявку посредством ссылки). В указанную таблицу также включены белки, участвующие в репарации ДНК, такие как MDC1 и MHS2. MDC1 необходим для активации в ответ на повреждение ДНК контрольной точки в S-фазе и контрольной точки фазы G2/M клеточного цикла. Тем не менее, MDC1 также функционирует в опосредованной Rad51 гомологичной рекомбинации путем удерживания Rad51 в хроматине.
"Нокдаун" в контексте настоящего изобретения означает, что экспрессия целевого гена снижается, например, на 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90% или более. Полное подавление означает, что больше нет детектируемой экспрессии целевого гена. В таблице D также показаны результаты, полученные для некоторых мишеней для нокдауна. Для специалиста в данной области должно быть очевидно, что существуют вариации в последовательностях нуклеиновых кислот указанных мишеней, следовательно, варианты генов, в частности, варианты, последовательности которых идентичны на 80%, 90% или 95% последовательностям мишеней, входят в объем настоящего изобретения.
ПРОЦЕССИНГ БЕЛКА И МЕТАБОЛИЧЕСКИЕ БЕЛКИ
Данная категория белков, которые можно применять для клеточной метаболической инженерии, не принадлежит ни к пути секреции белка, ни к пути рекомбинации, но иным образом влияет на процессы, свойственные экспрессирующей клетке.
Белки процессинга белка или метаболические белки часто представляют собой ферменты, такие как шапероны (см. выше определения шаперонов), протеин-изомеразы, добавляющие сахар ферменты (например, сиалил- или гликозилтра нефе разы) или фосфатазы, или белки, контролирующие уровень энергии клетки или функции митохондрий.
В ТАБЛИЦЕ А приведен перечень белков, которые экспрессировали (экспр.) и/или экспрессию которых "подавляли" (нокдаун, НД), под подзаголовком "Процессинг белка и метаболические белки".
РАЗРАБОТКА ВЕКТОРА
Среди невирусных векторов транспозоны особенно привлекательны благодаря их способности встраивать единичные копии последовательностей ДНК с высокой частотой в многочисленные локусы внутри генома хозяина. Сообщалось, что, в отличие от вирусных векторов, некоторые транспозоны преимущественно не встраиваются близко к клеточным генам, и они, таким образом, с меньшей вероятностью приводят к повреждающим мутациям. Более того, транспозоны легко получить и легко с ними обращаться, включая, как правило, донорную плазмиду, несущую транспозон, которая содержит перемещаемую ДНК, фланкированную последовательностями инвертированных повторов, и экспрессирующую транспозазу вспомогательную плазмиду или мРНК. Разработали несколько систем транспозонов для перемещения ДНК в различных линиях клеток, не препятствуя эндогенным копиям транспозона. Например, транспозон PiggyBac (РВ), изначально выделенный из металловидки серой, эффективно переносит целевую ДНК в различные клетки млекопитающих.
Эпигенетические регуляторные элементы можно применять, чтобы защитить перемещаемую ДНК от нежелательных эпигенетических эффектов, когда ее помещают рядом с трансгеном в плазмидные векторы. Например, предложили элементы, называемые участками прикрепления к матриксу (MAR), чтобы повысить встраивание в геном и транскрипцию перемещаемой ДНК, при этом предотвращая замалчивание гетерохроматина, примером которых является эффективный MAR 1-68 человека. Они также могут действовать как инсуляторы и, тем самым, предотвращать активацию соседних клеточных генов. Элементы MAR, таким образом, применяют, чтобы опосредовать высокий уровень продолжительной экспрессии в контексте плазмидных или вирусных векторов.
В данной заявке показано, что при правильной разработке вектора благоприятные свойства эпигенетических регуляторов, в частности, элементов MAR, можно комбинировать с таковыми у векторов на основе транспозонов.
Транспозоны и векторы на основе транспозонов согласно настоящему изобретению можно применять в клеточной метаболической инженерии, например, для экспрессии секреторных белков из различных секреторных путей, описанных в данной заявке. Они также особенно полезны, когда требуется несколько раундов внедрения перемещаемой ДНК. Это подтвердили, когда тестировали множество белков секреторного пути клетки, когда трансфекция множеством векторов и/или множество следующих друг за другом циклов трансфекции могут исчерпать доступный антибиотик или помешать другим способам селекции. Возможность быстро экспрессировать терапевтические белки без необходимости селекции антибиотиком также представляет особый интерес, например, когда нужно экспрессировать множество кандидатных терапевтических белков с целью скрининга, поскольку можно получить значительные количества белков из не подвергнутых селекции популяций клеток через 2-3 недели после трансфекции. В частности, содержащие MAR векторы, несущие транспозоны, таким образом, представляют собой перспективное дополнение к доступному на сегодняшний день арсеналу векторов экспрессии.
Выбранные экспериментальные подходы позволили (в противоположность подходам, в которых полагаются на анализы на основе антибиотиков) провести различие между эффектами, основанными на (1) числе копий перемещаемой ДНК, и эффектами, основанными на (2) уровни экспрессии перемещаемой ДНК.
MAR 1-68 был особенно эффективен, когда он был расположен посередине между ITR транспозона PiggyBac, так как он не снижал эффективность транспозиции. MAR Х-29 также хорошо работал на краях транспозона, не снижая эффективность транспозиции или экспрессии.
Примечательно, что степень MAR-опосредованной активации перемещенных генов была меньше, по сравнению с таковой для самопроизвольного встраивания плазмиды. Более того, уровень экспрессии, нормированный, например, на копии трансгена, был выше при применении транспозонов, чем полученный в результате самопроизвольного встраивания плазмид в отсутствие транспозазы. Данный эффект наблюдали независимо от размера конструкций, присутствия MAR или силы промотора. Этого будут ожидать, так как транспозиция часто может происходить в геномных локусах, относительно позволяющих экспрессию, например, благодаря тому, что структуры хроматина могут быть более доступны как для транспозазы, так и для факторов транскрипции. В этом отношении, предшествующие исследования позволили предположить, что транспозоны преимущественно могут встраиваться внутрь интронов генов, в промоторы или в геномные локусы с более низкой способностью к замалчиванию, хотя данное утверждение остается темой для обсуждения. В качестве альтернативы, совместное встраивание множества копий плазмиды в один и тот же геномный локус, которое происходит в результате событий самопроизвольного встраивания, может приводить к образованию гетерохроматина и к замалчиванию повторяемых последовательностей, которому будет препятствовать MAR, тогда как встраивание единственной копии транспозона может в меньшей степени способствовать такому опосредованному хроматином замалчиванию. Кроме того, встраивание транспозонов в множество независимых геномных локусов повышает вероятность того, что по меньшей мере одна копия встроилась в благоприятное геномное окружение и экспрессируется, тогда как обнаружили, что встраивание плазмиды происходит преимущественно только в один геномный локус.
Наиболее высокие уровни экспрессии перемещаемой ДНК, например, трансген/TER, получили для содержащего MAR транспозона, когда он был связан с сильным промотором. Неожиданно было обнаружено, что высокие уровни экспрессии можно получить при перемещении незначительного числа копий ДНК, например, не более чем 20, 15, 10 или 5. Если, тем не менее, удается добиться высокой продуктивности, меньшее количество встроенных, например, перемещаемых копий ДНК является преимуществом, так как благодаря этому снижается вероятность возникновения точечных мутаций в одном или в подгруппе указанных трансгенов, возникающих в результате спонтанных событий мутагенеза. Кроме того, опосредованные транспозазой события встраивания менее мутагенны, чем механизмы репарации и рекомбинации ДНК, вовлеченные в самопроизвольное встраивание плазмиды, которые могут приводить к встраиванию неполных или перестроенных копий трансгена.
Высокая эффективность встраивания в геном с помощью транспозона piggyBac (РВ) также полезна, когда количество целевых клеток ограничено, например, для невирусного переноса терапевтических генов в первичные стволовые клетки для получения клональных популяций, например, для основанных на клеточной системе методов лечения или регенеративной медицины. В данном контексте, физиологические уровни экспрессии с нескольких перемещенных копий ДНК и частое возникновение событий транспозиции, таким образом, избавляющие от необходимости селекции антибиотиком, являются преимуществом, поскольку использование генов устойчивости к антибиотикам и/или ненадежной, например, экспрессии трансгена может вызывать сомнения относительно безопасности.
Влияние включения MAR на эффективность транспозиции
Так как устойчивость к антибиотику не обязательно отражает эффективную экспрессию трансгена, в качестве индикатора использовали зеленый флуоресцентный белок (GFP), экспрессируемый с производного сильного клеточного промотора GAPDH. Для того, чтобы проверить, может ли добавление элемента MAR в транспозон РВ влиять на эффективность транспозиции и экспрессию трансгена, и чтобы оценить, оказывает ли расположение MAR в конструкции какое-либо влияние на данные эффекты, разработали серию донорных конструкций, несущих транспозоны, содержащих GFP и ген устойчивости к пуромицину (Puro), в которые встроили MAR 1-68 или контрольную нейтральную спейсерную последовательность ДНК в различные положения в указанной плазмиде (фигура 1). Исходную несущую транспозон плазмиду Puro-GFP без вставки использовали в качестве контроля транспозиции, чтобы отличить вклад увеличенного размера транспозона от влияния добавления MAR или спейсерной последовательности.
В присутствии транспозазы наблюдали наиболее высокий уровень экспрессии GFP в не подвергнутых отбору клетках, когда MAR был расположен посередине, но не тогда, когда MAR помещали в 5'-3' направлении от последовательности, кодирующей GFP, и не тогда, когда его вставляли вне перемещенной последовательности, как и ожидалось (фигура 3А). При селекции пуромицином MAR-опосредованная активация уменьшалась, как с транспозазой, так и без нее, тогда как средние значения экспрессии GFP были повышены на один порядок величины (фигура 3В). Полученные результаты подтвердили, что отбор пуромицином позволял получить лишь небольшое число клеток, в которых уровни экспрессии наиболее высоки, что предполагали выше по подсчету событий транспозиции. Дополнительно показано, что векторы на основе транспозонов, содержащие MAR, расположенный по центру, позволяли получить уровни экспрессии, аналогичные таковым для такой же плазмиды, трансфицированной без транспозаз.
Влияние включения MAR на число копий встроенного транспозона
Более высокие уровни флуоресценции GFP могут возникать в результате повышенной транскрипции трансгенов и/или встраивания большего числа копий трансгена. Это оценивали путем определения количества встроенных в геном копий трансгена, внедренных с помощью различных типов векторов. Общую геномную ДНК выделяли из объединенных популяций клеток, либо после цитофлуори метрической сортировки флуоресцирующих клеток из не подвергнутых селекции популяций, либо после отбора по устойчивости к пуромицину. Число копий трансгена определяли с помощью анализа методом количественной полимеразной цепной реакции (qPCR) последовательности, кодирующей GFP, по сравнению с клеточным геном β2-микроглобулина (В2М). В отсутствие отбора антибиотиком среднее количество встроенных трансгенов, либо транспозазой, либо клеточными ферментами рекомбинации, было сходным, около 1-6 копий на геном, и на него значительно не влияли MAR или контрольная последовательность (фиг. 4А). Тем не менее, наименьшее число копий получали, когда MAR вставляли на границе транспозона, что поддерживает наше более раннее заключение, что он уменьшает транспозицию в данном положении. После селекции пуромицином клеток с высоким уровнем экспрессии количество перемещенных трансгенов оказалось в аналогичном диапазоне 2-7 копий (фиг. 4В). Тем не менее, количество копий трансгенов, встроенных в отсутствие транспозазы, как правило, было значительно выше и находилось в диапазоне от 6 до 14 копий. Это можно легко объяснить тем фактом, что самопроизвольное встраивание обычно приводит к встраиванию конкатемеров нескольких копий плазмиды в один геномный локус (результаты не представлены), и что большее число копий трансгенов должно приводить к более высоким уровням экспрессии, если клетки, подвергнутые влиянию замалчивания, были удалены путем отбора антибиотиком. Наряду с предшествующим заключением, что отбор антибиотиком позволяет получить преимущественно клетки с высоким уровнем экспрессии, это также свидетельствует о том, что самопроизвольное встраивание плазмиды приводит к большей вариабельности числа копий трансгена, чем при применении векторов на основе транспозонов.
Уровень экспрессии GFP затем нормировали на число копий гена, чтобы оценить внутренний экспрессионный потенциал указанных векторов независимо от их способности встраиваться в геном. В целом, получали более низкий уровень экспрессии на копию трансгена для не подвергнутых селекции клеток или для подвергнутых селекции антибиотиком клеток, трансфицированных без транспозазы или с MAR в центральном положении, что указывает на то, что на экспрессию трансгена влияет как включение эпигенетического регуляторного элемента, так и способ встраивания трансгена (фиг. 5). Уровень экспрессии на копию гена, как правило, повышался при применении транспозазы, когда проводили оценку различных векторов и комбинаций элементов, и данный эффект наблюдали как при отборе антибиотиком, так и без него. Наиболее высокие уровни экспрессии на копию трансгена получали после отбора антибиотиком клеток, в которые был внедрен вектор, несущий транспозон, содержащий элемент MAR в центральном положении, в присутствии транспозазы. Включение MAR непосредственно в 5'-3' направлении от последовательности, кодирующей GFP, значительно не повышало экспрессию трансгена, что было отмечено ранее для абсолютных уровней экспрессии.
Наконец, оценивали, может ли благоприятное влияние MAR 1-68 на экспрессию быть ограничено использованием сильного промотора GAPDH человека, применяемого в данном случае, или оно также будет иметь место с другими промоторами. Таким образом, мы заменили промотор GAPDH человека, запускающий экспрессию GFP, на более слабый ранний промотор вируса обезьян 40 (SV40). Применение более слабого промотора позволяло получить сравнимое количество GFP-положительных клеток и встроенных трансгенов, указывая на то, что экспрессия трансгена не изменяет эффективность транспозиции (не представленные результаты, фиг. 2А и фиг. 4В). Тем не менее, абсолютные уровни экспрессии были ниже при применении промотора SV40 (не представленные результаты по сранению с фиг. 3В). Кроме того, уровень экспрессии, нормированный на число копий транспозона, снижался в 4,6 раз при применении промотора SV40 в отсутствие MAR и в 3,1 раза в присутствии MAR 1-68 (результаты не представлены). Это свидетельствует о том, что MAR мог частично, но не полностью, предотвращать снижение экспрессии, возникшее в результате применения более слабого промотора, даже в присутствии транспозазы. В целом, можно показать, что нескольких встроенных копий достаточно для получения высокого уровня экспрессии трансгена с помощью транспозонов, и что наиболее высокий уровень экспрессии на трансген получали, когда, в данном контексте, MAR-68 помещали в 3'-5' направлении от сильного промотора.
Клетки СНО-М подвергали электропорации один или два раза одним вектором на основе транспозонов, содержащим трансген MAR Х_29. Эффективность транспозиции была наиболее высокой после электропорации (стабильная экспрессия наблюдалась в 30% - 45% клеток). Тем не менее, уровни экспрессии трансгена были аналогичны таковым при химической трансфекции, при которой наблюдали меньшее количество положительных клеток и, следовательно, более низкую эффективность транспозиции. На фиг. 7 показаны результаты для легких и тяжелых цепей терапевтических иммуноглобулинов, встроенных в 3'-5' направлении от MAR Х_29, и были получены титры в диапазоне от 1 до 8 мкг/мл. Указанные уровни дополнительно повышались до 23-55 мкг/мл после сортировки экспрессирующих клеток (фиг. 7С).
Экспрессию трансгенов также можно существенно повысить, часто независимо от применения транспозонов, путем разработки определенного вектора, в частности, с применением определенного элемента(ов) MAR в определенных положениях относительно трансгена и, предпочтительно, комбинации данного элемента(ов) MAR с промоторами, энхансерами или продуктами их слияния.
Соответствующий вектор может содержать элементы MAR, которые фланкируют кассету экспрессии трансгена. Например, указанный вектор может содержать, например, элементы MAR (один или более), расположенные в 3'-5' направлении, и элементы MAR (один или более), расположенные в 5'-3' направлении, например, один MAR, расположенный в 3'-5' направлении от кассеты экспрессии трансгена, и один MAR, расположенный в 5'-3' направлении от нее (фиг. 18А, фиг. 18В, фиг. 19). Указанный вектор может содержать встроенный ген устойчивости к пуромицину под контролем промотора SV40. Трансген может находиться под контролем промотора GAPDH человека, слитого с энхансером немедленно-ранних генов цитомегаловируса (CMV) человека (в частности, слитый промотор CGAPD, обсуждаемый выше).
Наиболее высокий процентиль клеток с высоким и очень высоким уровнем продукции (% М3/М2), который оценивали по флуоресценции GFP с помощью анализа FACS и наименьшей вариабельности, можно было получить при применении 1_6R2, 1_68R2 и X_29R3 в качестве MAR, расположенных в 3'-5' направлении (свыше 80%, 80% и свыше 80%). Таким образом, процентили клеток с высоким и очень высоким уровнем продукции (% М3/М2), составляющие более чем 70%, более чем 75% или более чем 80%, входят в объем настоящего изобретения. Для специалиста в данной области должно быть очевидно, что допустимы некоторые отклонения от определенной последовательности данных элементов MAR. Соответственно, векторы, содержащие последовательности нуклеиновых кислот, более чем на 80%, 85%, 90%, 95% идентичные последовательностям SEQ ID NO: 6, 7, 8, 9 и 10, входят в объем настоящего изобретения (фиг. 18А, фиг. 18В).
Затухание экспрессии в биореакторе и/или в отсутствие селекционного давления часто ограничивает получение интересующего белка. Векторы, содержащие производные 1_68R2, 1_6R2 и X_29R3 MAR в качестве MAR, расположенных в 3'-5' направлении, исследовали на протяжении периода культивирования 5 недель в отсутствие селекции, и флуоресценцию GFP оценивали еженедельно в течение данного периода. При рассмотрении процентиля субпопуляции М3 обнаружили, что элемент 1_6R2 в качестве MAR, расположенного в 3'-5' направлении, и неперестроенный MAR 1-68 в качестве MAR, расположенного в 5'-3' направлении, оказались наилучшей из исследованных комбинаций в векторах с по меньшей мере одним элементом MAR, расположенным в 3'-5' направлении, и одним элементом MAR, расположенным в 5'-3' направлении (значительно выше 80% после более 2, 3, 4 недель) (см. фиг. 19).
Разработанный аналогичным образом вектор также может содержать, например, только элементы MAR (один или более), расположенные в 5'-3' направлении, например, один MAR, расположенный в 5'-3' направлении от кассеты экспрессии трансгенов (фиг. 20А, 20В, 21 и 22), и не содержать MAR, расположенного в 3'-5' направлении. Указанный вектор также, в данном случае, может содержать встроенный ген устойчивости к пуромицину под контролем промотора SV40. Трансген может находиться под контролем промотора GAPDH человека, слитого с энхансером немедленно-ранних генов цитомегаловируса (CMV) человека. См., например, SEQ ID NO: 11 и другие последовательности, идентичные более чем на 80%, 85%, 90% или 95% указанной последовательности. В такой комбинации с единичным MAR, где в качестве MAR выступал Х_29, добились превосходных результатов. Процентиль клеток с высоким уровнем экспрессии GFP (определенная, как описано выше), а также стабильность экспрессии с течением времени (определенная, как описано выше) были лучше, чем, например, таковые для векторов с высоким уровнем экспрессии, в которых элементы MAR фланкировали кассету экспрессии трансгена, а именно, для вектора, содержащего MAR 1_6R2, расположенный в 3'-5' направлении, и неперестроенный MAR 1-68, расположенный в 5'-3' направлении (см. фиг. 22). Данное открытие противопоставляется общепринятым предположениям, что элементы MAR наиболее эффективны, когда они фланкируют трансген (см. патент США 5731178). Стабильность экспрессии означает, что интересующая ДНК, например, трансген, экспрессируется популяцией клеток даже после некоторого периода времени, например, после более чем 4, 5, 6, 7, 8, 9, 10, 11, 12, 13 или 14 недель, на уровне, сравнимом (не более чем на 20%, 10 или 5% меньшем) и, особенно, даже более высоком, чем в течение двух недель от начала экспрессии. Часто стабильная экспрессия связана с высокой процентилью (например, более чем 80%) субпопуляций клеток с высоким уровнем экспрессии.
КЛЕТОЧНАЯ МЕТАБОЛИЧЕСКАЯ ИНЖЕНЕРИЯ: СЕКРЕТОРНЫЕ БЕЛКИ
Секреция гетерологичных белков, таких как IgG, снижается при неправильном процессинге полипептида и низкой продукции IgG в культивируемых клетках, таких как клетки СНО.
Наблюдали, что вызывалась экспрессия индуцированных стрессом шаперонов, таких как BiP, и что шаперон правильно локализовался в ЭР и был способен взаимодействовать с цепями предшественника IgG. Впрочем, IgG, содержащие определенные вариабельные последовательности, такие, которые обнаруживаются в инфликсимабе, тем не менее, неправильно процессировались и собирались, что приводило к низкой секреции. Следовательно, активация реакции несвернутых белков (UPR) в данных клетках оставалась неэффективной для восстановления существенного уровня иммуноглобулина.
Было показано, что SRP14 вовлечен в молекулярный этап секреторного пути, который является лимитирующим в клетках СНО, экспрессирующих сверх меры экзогенный белок. Примечательно, что данный лимитирующий этап также наблюдается для легко экспрессируемого трастузумаба, что легко приводит к получению клонов с высоким уровнем экспрессии. Данное заключение следовало за открытием, что экспрессия SRP14 человека легко восстанавливала экспрессию LP клонов, но она также повышала секрецию легко экспрессируемого IgG. Обнаружили, что экспрессия SRP14 повышает процессинг и доступность предшественников LC и НС и позволяет получить сравнимые уровни секреции обоих типов IgG. В целом, продемонстрировали, что SRP14 может быть, как правило, лимитирующим, когда секретируемые белки, такие как IgG, чрезмерно экспрессируются в клетках СНО.
Сильный эффект, полученный при экспрессии SRP14 в клетках СНО, наблюдаемый в данном исследовании, был неожиданным и позволил предположить, что SRP14 вызывает длительную задержку элонгации LC в клонах, с трудом продуцирующих IgG (фиг. 12В, пункт 1).
Предшествующее исследование показало, что сигнальный пептид, который выходит из транслирующей рибосомы, первый взаимодействует с субъединицей SRP54 частицы SRP, тогда как ассоциация с SRP9 и SRP14 может блокировать дальнейшую элонгацию образующегося полипептида, что приводит к остановке трансляции (Walter и Blobel 1981). На втором этапе комплекс рибосома-образующийся полипептид-SRP стыкуется с мембраной ЭР посредством взаимодействия с рецептором SR (Gilmore и др., 1982; Walter и др., 1982). SR затем может координировать высвобождение SRP из комплекса рибосома-образующийся полипептид и ассоциацию сайта выхода из рибосомы с каналом транслокона, через который растущий образующийся полипептид проникает в ЭР (Lakkaraju и др., 2008). Затем сопряженное с трансляцией перемещение может возобновиться, что приводит к удалению сигнального пептида и к синтезу правильно процессированных и секретируемых полипептидов.
Для правильного процессинга с трудом поддающихся экспрессии IgG может потребоваться необычно длинная трансляционная пауза, если кинетика стыковки с ЭР может быть медленнее для конкретных комбинаций последовательностей вариабельного домена IgG и сигнального пептида, вследствие неблагоприятных структур образующегося пептида. Таким образом, считали, что, в свою очередь, модуляция кинетики остановки трансляции путем экспрессии экзогенного компонента SRP14 человека улучшает правильную стыковку с ЭР и перемещение пре-LC в ЭР и, таким образом, восстанавливает эффективный процессинг сигнального пептида (фиг. 12В, пункт 2). Соответственно, снижение уровней SRP14 в клетках человека приводит к отсутствию задержки элонгации трансляции в полисомах, что может привести к чрезмерному увеличению длины образующихся полипептидов сверх критического значения, после которого SRP больше не сможет правильно нацелить секретируемый белок на ЭР.
Таким образом, аффинность SRP14 и возможно также SRP54 в клетках СНО к сигнальным последовательностям гетерологичного белка инфликсимаба человека будет снижена, что приведет к неправильной стыковке с ЭР и/или к элонгации образующегося пептида до того, как произойдет правильная стыковка. Это будет исправлено путем сверхэкспрессии SRP14 человека, увеличения периода времени, в течение которого остановленный комплекс рибосома-SRP может искать правильно организованный сайт стыковки на ЭР, несмотря на 'молекулярный затор' из сверхэкспрессированных белков IgG, происходящий на входе в ЭР.
Соответственно, сверхэкспрессия SR и транслокона, которые могут повышать вместительность ЭР в отношении транслокации, также приводила к улучшению секреции, даже в отсутствие сверхэкспрессии SRP14 человека. Наконец, продемонстрировали, что метаболическая инженерия секреторного пути посредством коэкспрессии комбинаций SRP человека, транслокона и субъединиц SR, приводит к дополнительному улучшению способности клетки секретировать белок, позволяя получить даже более высокие уровни секреции. В целом, пришли к заключению, что белки SRP, их рецепторы и транслокон могут быть, как правило, лимитирующими, когда секретируемые белки, такие как иммуноглобулины человека, чрезмерно экспрессируются в клетках СНО.
Немногое известно об относительном содержании SRP и компонентов мембраны ЭР по сравнению с секретируемыми белками и рибосомами в различных типах клеток, но в клетках, экспрессирующих большие количества рекомбинантного белка, потенциально могут возникать нарушения транслокации. Например, SR и/или транслокон могут стать лимитирующими, когда секретируемые белки экспрессируется на аномально высоких уровнях, или SRP14 может встречаться в клетках СНО на субстехиометрических уровнях по сравнению с другими субъединицами SRP. Соответственно, SRP9 и SRP14 присутствуют в 20-кратном избытке по сравнению с другими белками SRP в клетках приматов, но не в клетках мышей, и сверхэкспрессия SRP14 человека в нормальных клетках человека не повышает эффективность секреции щелочной фосфатазы. Более того, SRP14 человека крупнее, чем SRP14 грызуна, так как он содержит аланин-богатый хвост на С-конце, который не обнаружен в SRP14 грызуна. Таким образом, включение более крупного SRP14 человека в SRP СНО может приводить к образованию функциональной химеры SRP с более высокой активностью, с доминирующе-положительным эффектом.
Открытие того, что экспрессия цитозольных компонентов SRP, таких как SRP14, приводит к эффективному процессингу и секреции сверхэкспрессированных белков в клетках СНО, указывает на затруднение, которое можно использовать для увеличения выхода рекомбинантного белка. Данное затруднение ограничивает экспрессию различных и неродственных IgG и, возможно, также множества других моноклональных антител и производных, которые составляют несомненно наиболее распространенный класс рекомбинантных терапевтических белков.
Анализ интермедиатов секреции и возможных стрессовых реакций клетки с последующим систематическим поиском вышележащих лимитирующих действующих факторов, которые вызывают такую стрессовую реакцию, а затем, в конечном счете, разработкой метаболизма секреции клеток СНО привел к лучшему пониманию метаболических ограничений в данных клетках и того, как их обходить.
Гетерологичная экспрессия SRP14 восстанавливает секрецию и процессинг LC
Клоны IgG HP (с высоким уровнем продукции) и LP (с низким уровнем продукции) котрансфицировали с вектором, кодирующим компонент SRP14 SRP, и с плазмидой устойчивости к неомицину. Отдельные клетки из устойчивой к неомицину популяции клеток отделяли с помощью метода серийных разведений, а затем исследовали их рост и секрецию иммуноглобулина в партии чашек для культивирования при взбалтывании. Экспрессирующие SRP14 полученные из клонов LP субклоны секретировали значительно более высокие количества антитела, чем исходные клоны во всей культуре, и позволяли получить титры иммуноглобулина, аналогичные таковым для субклонов HP, экспрессирующих SRP14 (фиг. 8А и 8В, левые панели).
Экспрессия SRP14 не влияла на жизнеспособность клеток, но, похоже, она замедляла и продлевала рост культуры HP-клеток до аналогичных плотностей клеток (фиг. 8А и 8В, правые панели). Собрали культуральные супернатанты различных субклонов и анализировали в них концентрацию антитела. На фиг. 8С показано, что экспрессия SRP14 повышала секрецию из LP-клеток, что приводило к 7-кратному увеличению удельной продуктивности IgG. Более того, экзогенная экспрессия SRP14 также улучшала секрецию IgG из НР-субклонов, что приводило к 30% увеличению удельной продуктивности. Примечательно, что отдельные субклоны, экспрессирующие SRP14, секретировали трудно- и легкоэкспрессируемые IgG на по существу идентичных средних уровнях, при этом срединные значения удельных продуктивностей превышали 30 пикограмм на клетку в день (pcd). Такие очень высокие уровни секреции IgG поддерживались в течение более чем 6 месяцев культивирования, указывая на то, что это является постоянным свойством клеток, экспрессирующих SRP14.
Для того, чтобы дополнительно исследовать взаимосвязь между экспрессией SRP14 и продуктивностью в отношении IgG, анализировали уровни мРНК SRP14 в 5 отдельных экспрессирующих SRP14 LP-субклонах с помощью сравнительной количественной ПЦР. На фиг. 8D показано, что субклоны сверхэкспрессировали SRP14 на уровнях, находящихся в диапазоне от 50 до почти 200-кратных по сравнению с таковыми для эндогенных мРНК SRP14 в клетках СНО. Это сопровождалось улучшением секреции IgG в 4-6 раз по сравнению с LP контрольным клоном клетки. Примечательно, что наибольшую удельную продуктивность получили у субклона с промежуточным уровнем повышения экспрессии SRP14, приблизительно в 100 раз относительно уровня эндогенной мРНК SRP14 в клетках СНО (SRP14-LP субклон Е, результаты не представлены). Это подразумевает взаимозависимость уровня сверхэкспрессии SRP14 и удельной продуктивности IgG вплоть до порогового уровня SRP14, соответствующего 100-кратному повышению относительно эндогенного уровня экспрессии. Это позволило предположить, что другие компоненты секреторного пути, в свою очередь, могут стать лимитирующими при очень высоких уровнях SRP14, и что для оптимальной экспрессии IgG может быть необходима сбалансированная экспрессия компонентов данного пути.
Для того, чтобы проверить, может ли повышенная удельная продуктивность, полученная в процессе оценки клональной линии клеток, применяться в производственном процессе, лучшие экспрессирующие HP и LP субклоны SRP14 исследовали в чашках для культивирования при взбалтывании в условиях периодической культуры с подпиткой (т.е. LP субклон Е и HP субклон В, см. фиг. 8). Экспрессирующий SRP14 LP субклон позволял получить большие количества жизнеспособных клеток и титры иммуноглобулина, сходные с экспрессирующим SRP14 HP субклоном, при этом максимальная концентрация клеток составляла 8×106 клеток/мл и максимальный титр был выше 2 г на литр по окончании производственного цикла (фиг. 9А и В).
Затем исследовали вклад сверхэкспрессии SRP14 в синтез иммуноглобулина для двух данных субклонов. Результаты показали, что экспрессия SRP14 человека в субклоне, полученном из LP, позволяла получить нормально процессированную и зрелую LC, способную к фолдингу и сборке IgG (фиг. 10А, LP, дорожка S по сравнению с дорожкой -). Миграция свободной НС не изменилась, указывая на то, что экспрессия SRP14 действовала, в частности, на неправильно процессированную LC с трудом поддающегося экспрессии белка. Удивительно, что экспрессия SRP14 полностью нарушала накопление агрегировавшей LC в нерастворимой в Triton X фракции (фиг. 9А, нижняя панель). Экспрессия контрольного белка GFP не улучшала растворимость белка, а также не восстанавливала правильный процессинг LC (фиг. 10А, дорожка G для LP клеток). Экспрессия SRP14 не влияла на паттерн миграции НС и LC, полученный для HP субклона, и наблюдали небольшое влияние на количество свободных цепей и полностью собранного IgG по сравнению с контролями (фиг. 10А, дорожка G для HP клеток).
Осуществляли анализы с вытеснением на основе циклогексимида, чтобы исследовать кинетику фолдинга и сборки IgG, а также судьбу агрегатов IgG в экспрессирующем SRP14 субклоне, продуцирующем инфликсимаб. В противоположность исходным LP клеткам, содержащим агрегировавшие LC, неспособные участвовать в сборке IgG, экспрессирующий SRP14 LP субклон более не накапливал нерастворимые в Тх-100 LC (фиг. 9В, нижняя панель). Тем не менее, свободная LC оставалась в небольших количествах по сравнению со свободной НС, что также было отмечено для HP клеток, указывая на то, что она быстро включалась в состав HC-LC димеров и зрелого IgG и что она, возможно, ограничивала сборку IgG (фиг. 9А и 9В). В совокупности данные результаты позволяют предположить, что SRP14 может играть важную роль в процессинге LC в LP клетках, и что дополнительная экспрессия белка SRP может улучшить секрецию с трудом поддающихся экспрессии и легко поддающихся экспрессии IgG до аналогичных уровней.
Инженерия перемещения в ЭР улучшает секрецию рекомбинантного IgG
В связи с тем, что сверхэкспрессия гетерологичного SRP14 повышала секрецию IgG до данного порога, сделали вывод, что другие компоненты секреторного пути, которые взаимодействуют непосредственно или опосредованно с SRP14, могут стать лимитирующими в SRP14-LP субклонах. Следовательно, мы исследовали, может ли сверхэкспрессия других компонентов секреторного пути также улучшать экспрессию IgG, либо отдельно, либо в комбинации с SRP14. Данные компоненты включали белки SRP9 и SRP54 человека, которые входят в состав комплекса SRP вместе с SRP14, субъединицы рецептора SRP (SR) и субъединицы транслокона Sec61α, β и γ.
В первой серии экспериментов наилучший LP клон, а именно LP клон Е на фиг. 1, трансфицировали векторами экспрессии, кодирующими белки SRP или белки транслокона, отдельно или в комбинации. Затем оценивали продукцию IgG в полученных поликлональных популяциях LP клеток при периодическом культивировании. Экспрессия компонентов SRP или белков транслокона повышала секрецию иммуноглобулина данными повторно трансфицированными поликлональными популяциями LP клеток (фиг. 11А и не представленные результаты). По сравнению с экспрессией SRP14 отдельно сверхэкспрессия комбинаций белков SRP или транслокона улучшала удельную продуктивность трансфицированных поликлональных популяций LP клеток на дополнительные 20% - 40% (фиг. 11А, сравнение срединных значений). Полученные результаты явно свидетельствуют о том, что определенные комбинации более эффективно восстанавливали секрецию инфликсимаба, чем экспрессия SRP14 отдельно, такие как комбинации, состоящие из экспрессии трех полипептидов SRP и его рецептора (SR), или из совместной экспрессии SR и транслокона (фиг. 11А).
Также оценивали, можно ли дополнительно оптимизировать экспрессирующий SRP14 LP субклон Е путем экспрессии комбинаций SR и/или транслокона. По сравнению с 30 pcd для SRP14-LP субклона Е, поликлональные популяции клеток, подвергнутые отбору после трансфекции белками SR и транслоконом, позволяли получить удельные продуктивности выше 60 pcd с трудом поддающегося экспрессии иммуноглобулина (фиг. 10В). Пришли к заключению, что векторы, экспрессирующие белки SR и транслокон, также можно применять для получения клонов с повышенными удельными продуктивностями по сравнению с SRP14-LP клоном E, и что можно успешно применять подход, основанный на сериях последовательных циклов трансфекции и селекции.
КЛЕТОЧНАЯ МЕТАБОЛИЧЕСКАЯ ИНЖЕНЕРИЯ: подавление пути рекомбинации и экспрессии ТЕР и функциональной РНК ТЕР
Для того, чтобы повлиять на рекомбинацию, можно подвергнуть замалчиванию важные гены рекомбинации ДНК. Мишенями для нокдауна являются гены, известные из литературы как важные для определенных путей рекомбинационной репарации у млекопитающих (таблица D). Поскольку большинство из данных генов необходимы для выживаемости и развития клетки, постоянное их замалчивание может привести к снижению жизнеспособности клетки. Следовательно, предпочтительно временное замалчивание данных целевых генов с применением РНК-интерференции (РНКи).
В клетках, обработанных смесью миРНК против факторов, вовлеченных в первые этапы НГСК, т.е. Ku 70, Ku 80 и ДНК-ПКкс (один дуплекс миРНК на белок), частота событий НГСК была значительно ниже, чем в необработанных клетках. Взамен, в данных клетках ГР была более эффективной, что приводило к уменьшению отношения НГСК к ГР (от 2,7:1 до 1,3:1). Напротив, в клетках, обработанных миРНК, направленной против незаменимого фактора ГР - Rad51, ГР-зависимое восстановление уровня GFP почти полностью прекращалось, что повышало отношение НГСК : ГР до 5:1. Похоже, что данные результаты свидетельствуют о том, что репортерный анализ ГР и НГСК обладает достаточной чувствительностью, чтобы его можно было применять в экстрахромосомной форме. Они также дополнительно подвергают сомнению надежность используемых ранее мутантных линий клеток СНО (а именно, клеток 51D1, изначально опубликованных как ГР-отрицательные клетки, но, по-видимому, способных осуществлять ГР согласно данному анализу, результаты не представлены).
Повышение экспрессии и встраивания GFP в присутствии MAR. В параллельных экспериментах клетки СНО, обработанные различными миРНК, направленными против ГР или против НГСК, трансфицировали плазмидами, содержащими GFP или MAR-GFP. После двух недель отбора антибиотиком анализировали экспрессию и встраивание GFP в данных клетках с помощью FACS и количественной ПЦР, соответственно. При всех исследованных условиях добавление MAR приводило к 4-5-кратному повышению экспрессии GFP по сравнению с клетками, трансфицированными плазмидой без MAR (фиг. 13, фиг. 14А), что согласовалось с предшествующими сообщениями (Grandjean и др., 2011). Это повышение сопровождалось приблизительно 3-кратным увеличением числа копий встроенного GFP в клетках, трансфицированных плазмидой MAR-GFP, по сравнению с клетками, которые получили вектор без MAR (фиг. 13, фиг. 14В). Также наблюдалось 2-кратное увеличение средней флуоресценции GFP на копию гена (фиг. 13, фиг. 14С), возможно, вследствие более благоприятной локализации сайта встраивания (например, на участке, богатом эухроматином). Следовательно, можно предположить, что элементы MAR обладают способностью направлять гены в геномные локусы, позволяющие экспрессию гена. Комбинация миРНК против ГР с MAR (миРНК: RAD51, Rad51C и Brca1) приводила к изменению среднего уровня экспрессии GFP больше, чем в 11 раз, тогда как одна миРНК против ГР (миРНК: Rad51) приводила к изменению меньше, чем в 9 раз. Другие комбинации, такие как комбинация миРНК против ОМСК с MAR, также приводили к улучшенной экспрессии по сравнению с одной миРНК (результаты не представлены). Соответственно, комбинация различных миРНК (2, 3, 4, 5, 6, 8, 9, 10) к мишеням в том же или отличных метаболических путях, в частности, в пути рекомбинации, входит в объем настоящего изобретения.
Отсутствие влияния подавления гена НГСК на экспрессию и встраивание трансгена.
Обработка миРНК против белков НГСК, похоже, значительно не влияла на стабильную экспрессию трансгена. Также не наблюдалось существенного изменения числа копий GFP в геноме по сравнению с необработанными клетками (за исключением клеток, обработанных миРНК против 53ВР1, но только в отсутствие MAR, что, возможно, указывает на эффект, не свойственный рекомбинации посредством пути НГСК).
Повышение экспрессии и встраивания трансгена в отсутствие факторов ГР. В противоположность подавлению факторов НГСК, присутствие миРНК против белков ГР часто приводило к существенному повышению уровней стабильной экспрессии GFP по сравнению с необработанными клетками (за исключением Brca2, для которого наблюдалось существенное снижение уровня, но снова лишь в отсутствие MAR) (фиг. 15А). Как и в случае с замалчиванием генов НГСК, присутствие MAR приводило к повышению уровней стабильной экспрессии и встраивания GFP, а также к повышению уровней экспрессии на копию гена. На этот раз, тем не менее, замалчивание факторов ГР увеличивало данный эффект в 5-7 раз (наиболее ярким было подавление Rad51, приводящее к уровням экспрессии GFP, большим в 7,4 раза) (фиг. 15А), что может свидетельствовать о том, что белки ГР противодействуют положительному влиянию элемента MAR на экспрессию трансгена. В отсутствие MAR повышение экспрессии GFP коррелировало с увеличенным числом копий GFP в геноме (результаты не представлены). Неожиданно, этого не наблюдали в присутствии MAR, что указывает на то, что подавление белка ГР не влияло на опосредованное MAR увеличение числа копий (за исключением миРНК Rad51D). Похоже, это позволяет предположить, что отсутствие функционального пути репарации ГР не увеличивает количество событий рекомбинации, вызванных MAR. Вместо этого, оно стимулирует встраивание MAR и трансгена в более благоприятный локус, позволяя ему более эффективно экспрессироваться. Эта точка зрения также поддерживается повышенной экспрессией отдельных копий GFP в присутствии MAR в клетках, обработанных миРНК против ГР (фиг. 14С). Другая возможность состоит в том, что количество копий плазмиды, встроенных в геном, уже находится на максимальном уровне в контрольных клетках (при количестве плазмидной ДНК, используемом здесь), и его невозможно дополнительно увеличить даже в условиях, более благоприятных для MAR.
В совокупности, данные результаты позволяют предположить, что процесс опосредованного MAR встраивания трансгена преимущественно опосредован путем, противоположным гомологичной рекомбинации, хотя, вероятно, не путем НГСК, поскольку подавление его компонентов не влияло на встраивание или экспрессию. Можно предположить, что данный альтернативный путь менее активен в присутствии функционального пути ГР, но становится более важным, если ГР не работает.
ЭКСПРЕССИЯ кшРНК ПРОТИВ ГР ДЛЯ ПОВЫШЕНИЯ ЭКСПРЕССИИ ТЕРАПЕВТИЧЕСКИХ БЕЛКОВ
Три миРНК, нацеленные на Rad51, превратили в последовательности кшРНК, которые могут образовывать шпилечные структуры, и последовательность, кодирующую кшРНК, встраивали в вектор, несущий транспозон piggyBac, под контролем слитых энхансера GAPDH и промотора CMV, за которыми следовал MAR Х-29, но без гена селекции антибиотиком. Адаптированные для суспензии клетки СНО-М трансфицировали три раза донорной плазмидой, несущей транспозон, и вектором экспрессии транспозазы, после чего произвольно собирали 30 отдельных клонов клеток, применяя устройство ClonePix. Исходные клетки, а также экспрессирующую кшРНК популяцию клеток и клоны, повторно трансфицировали плазмидой, экспрессирующей GFP (а именно, конструкцией Puro-GFP-MAR Х_29), а затем осуществляли селекцию пуромицином политональных популяций экспрессирующих GFP клеток. Сравнение профилей флуоресценции GFP свидетельствовало о том, что получили более высокое соотношение клеток со средним - высоким уровнем флуоресценции (популяция клеток М2) или очень высоким уровнем флуоресценции (популяция клеток М3) из популяции клеток, трансфицированных вектором, несущим кшРНК, по сравнению с исходными клетками СНО-М (фиг. 17А).
Несколько клонов, экспрессирующих кшРНК, опосредовали очень высокие уровни GFP, при этом более 80% устойчивых к антибиотику клеток находились в субпопуляции М3 с высоким уровнем флуоресценции через 10 дней после трансфекции, например, клон 16 и клон 26 (фиг. 17А и В). Высокие уровни флуоресценции GFP поддерживались в данных двух клонах после 35 дней дополнительного культивирования без селекции (фиг. 17С). Напротив, клон 17 не экспрессировал очень высокие уровни GFP в день 10, и экспрессия GFP, похоже, была нестабильной (фиг. 17В и С). Промежуточные уровни экспрессии и промежуточную стабильность получили для клона 8 и клона 22. Данные клоны также трансфицировали экспрессионными плазмидами, кодирующими легкую и тяжелую цепь с трудом поддающегося экспрессии терапевтического антитела инфликсимаба. По-прежнему, клоны 16 и 26 продуцировали наиболее высокие уровни антитела, за ними следовали клоны 8 и 22, тогда как клон 17 экспрессировал количества иммуноглобулина, которые были близки к 0,5-1 pcd, полученным для исходных клеток СНО-М (фиг. 17D и не представленные результаты). В клонах клеток, демонстрирующих наиболее значительно сниженные уровни мРНК Rad51, обнаруживали высокие уровни экспрессии как GFP, так и иммуноглобулина инфликсимаба, указывая на то, что повышенная экспрессия трансгена происходила в результате пониженной экспрессии рекомбинационного белка (результаты не представлены).
Таким образом, можно добиться более высоких уровней и более стабильной продукции секретируемых терапевтических белков, таких как инфликсимаб, клетками, экспрессирующими кшРНК, нацеленную на Rad51, или клетками, временно трансфицированными миРНК, такими как миРНК, нацеленная на Rad51.
МАТЕРИАЛЫ И МЕТОДЫ
Плазмиды и векторы ДНК
Вектор экспрессии, несущий транспозазу РВ, pCS2+U5V5PBU3, содержит последовательность, кодирующую транспозазу РВ, окруженную 5'- и 3'-нетранслируемыми концевыми участками (НТО) гена β-глобина Xenopus laevis. Данную плазмиду сконструировали, как описано далее: фрагмент 3'-НТО размером 317 п.о. из pBSSK/SB10 (любезно предоставленный доктором S. Ivies) встраивали в pCS2+U5 (Invitrogen/life Technologies, Пейсли, Великобритания) с получением pCS2+U5U3. Последовательность, кодирующую транспозазу РВ (2067 п.о., номер доступа в GenBank: EF587698), синтезировали в ATG:biosynthetic (Мерцхаузен, Германия) и клонировали в каркас pCS2+U5U3 между двумя НТО. Контрольный вектор РВ соответствует немодифицированной плазмиде pCS2+U5 (фигура 1, левая панель).
Векторы, несущие различные транспозоны, используемые в данном исследовании, получали путем вставки 3'- и 5'-инвертированных концевых повторов (ITR) РВ размером 235 п.о. и 310 п.о., соответственно, синтезированных в ATG:biosynthetic (Мерцхаузен, Германия), в плазмиду pBluescript SK (pBSK ITR3'-ITR5', фигура 1, правая панель). Затем вставляли ген устойчивости к пуромицину (PuroR) под контролем промотора SV40 из плазмиды pRc/RSV (Invitrogen/Life Technologies) между двумя указанными ITR. Элементы MAR 1-68 и MAR Х-29, гены устойчивости к пуромицину и GFP, применяемые в данном исследовании, были такие, как описано ранее. Векторы экспрессии иммуноглобулинов и последовательности, кодирующие SRP9, SRP14, SRP54, SRPR-альфа, SRPR-бета, SEC61A1, SEC61B и SEC61G, были такие, как описано в Le Fourn и др. (Metab, Eng., Epub, 1 февраля 2013 г.)
GFP, иммуноглобулин или секреторные белки экспрессировали, применяя эукариотическую кассету экспрессии, состоящую из энхансера CMV человека и промотора GAPDH человека, расположенных в 3'-5' направлении от кодирующей последовательности, за которыми следовал сигнал полиаденилирования SV40, терминатор гастрина человека и энхансер SV40 (см. Le Fourn и др., 2013). Кассеты экспрессии и/или элементы MAR встраивали между последовательностями ITR или в каркас бактериального вектора, как показано на фигуре 1 и в описаниях к фигуре, применяя стандартные способы клонирования.
Культивирование клеток и анализ трансфекции
Линию клеток DG44 СНО культивировали в DMEM: F12 (Gibco), дополненной гипоксантином/тимидином (НТ, Gibco) и 10% фетальной бычьей сывороткой (FBS, Gibco). Трансфекцию осуществляли, применяя PEI (JetPRIME, Polyplus Transfection), следуя инструкциям производителя. Клетки трансфицировали различными количествами плазмидной ДНК, содержащей транспозазу РВ (в диапазоне от 0 до 1500 нг), для экспериментов с титрованием или котрансфицировали оптимальным соотношением 300 нг плазмиды, экспрессирующей транспозазу РВ, и 300 нг донорной плазмиды, несущей транспозон. Через два дня после трансфекции клетки переносили в несколько чашек Петри в зависимости от эксперимента. Для анализа не подвергнутых селекции трансфицированных клеток СНО, клетки пересевали без селекции антибиотиком в течение 3 недель и процент флуоресцентных клеток и интенсивность флуоресценции GFP-положительных клеток определяли с помощью анализа FACS, применяя проточный цитометр CyAn ADP (Beckman Coulter). Для анализа числа копий гена в не подвергнутых селекции клетках стабильные GFP-положительные клетки СНО подвергали сортировке, применяя FACSAriall. Для количественного анализа устойчивых к антибиотику колоний, 50000 трансфицированных клеток высевали в чашки диаметром 100 мм и подвергали селекции 5 мкг/мл пуромицина в течение 2 недель. Затем устойчивые колонии фиксировали и окрашивали 0,7% метиленовым синим в 70% EtOH в течение 10 мин, и подсчитывали колонии диаметром >0,5 мм. Для исследования экспрессии GFP клетки подвергали селекции в течение двух недель перед анализом FACS флуоресценции GFP, описанным выше.
Клетки СНО-М поддерживали в суспензионной культуре в бессывороточной среде SFM4CHO Hyclone (ThermoScientific), дополненной L-глутамином (РАА, Австрия) и добавкой НТ (Gibco, Invitrogen life sciences), при 37°C, 5% CO2 в увлажненном воздухе. Несущие транспозон донорные плазмиды переносили в данные клетки путем электропорации согласно рекомендациям производителя (Neon devices, Invitrogen). Проводили количественный анализ секреции иммуноглобулина периодическими культурами, как описано ранее (см. Le Fourn и др., 2013). Вкратце, популяции клеток, экспрессирующих иммуноглобулины, оценивали при периодическом культивировании в минибиореакционных пробирках объемом 50 мл (ТРР, Швейцария) при 37°С во влажной камере при 5% CO2 в течение 7 дней. Концентрации иммуноглобулина в культуральных супернатантах измеряли с помощью «сэндвич»-иммуноферментного анализа (ИФА).
Анализы числа копий генов методом количественной ПЦР (qPCR)
Общую ДНК выделяли из популяций стабильных клеток СНО после анализов транспозиции, применяя набор DNeasy Tissue (Qiagen, Хильден, Германия), следуя протоколу производителя. Число копий встроенных в геном трансгенов оценивали, применяя 6 нг геномной ДНК, с помощью количественной ПЦР, применяя набор с SYBR Green-Taq полимеразой от Eurogentec Inc и устройство для проведения ПЦР ABI Prism 7700 (Applied Biosystems). Праймеры GFP-прямой: ACATTATGCCGGACAAAGCC и GFP-обратный: TTGTTTGGTAATGATCAGCAAGTTG использовали для определения количества гена GFP, тогда как праймеры В2М-прямой: ACCACTCTGAAGGAGCCCA и В2М-обратный: GGAAGCTCTATCTGTGTCAA использовали для амплификации гена бета-2 микроглобулина. Для ампликона, полученного с помощью праймеров В2М, обнаружили одно совпадение на гаплоидный геном СНО после выравнивания с нашей сборкой генома СНО, применяя программное обеспечение NCBI BLAST. Так как СНО являются почти диплоидными клетками, приблизительно подсчитали, что В2М присутствует в 2 копиях на геном. Соотношения числа копий целевого гена GFP рассчитывали по сравнению с таковым для эталонного гена В2М, как описано ранее.
Сортировка и анализ экспрессирующих иммуноглобулин клеток
Для сортировки магнитным способом экспрессирующих IgG клеток, трансфицированные клетки СНО-М высевали при концентрации клеток 3×105 клеток в мл в среде SFM4CHO (Thermo Scientific), дополненной 8 мМ L-глутамином и добавкой 1 × НТ (оба от Gibco), называемой полной средой. После 4 дней в культуре 2×106 клеток промывали, ресуспендировали в ФБР и инкубировали с биотинилированным IgG человека (KPL216-1006) в конечной концентрации 3 мкг/мл вместе с 30 мкл заранее промытых покрытых стрептавидином гранул MyOne Т1 Dynabeads (Invitrogen), на вращающемся колесе в течение 30 минут при комнатной температуре. Смесь клеток и гранул затем помещали на магнит, чтобы отделить помеченные клетки от не помеченных клеток. Гранулы промывали 4 раза фосфатно-солевым буферным раствором (ФБР). После последней промывки ФБР гранулы и клетки ресуспендировали в 500 мкл заранее нагретой полной среды, переносили в 24-луночный планшет и инкубировали при 37°С при 5% CO2. После 24 ч отделенные методом магнитной сортировки поликлональные клетки отделяли от гранул и продолжали инкубацию до тех пор, пока клетки не достигли достаточной плотности для их размножения в 50 мл центрифужных биореакторах ТРР (TECHNO PLASTIC PRODUCTS AG, Швейцария).
В качестве альтернативы, выделяли два клона из не подвергнутых сортировке и не подвергнутых селекции популяций, экспрессирующих каждый из трех IgG, применяя устройство clonePix. Вкратце, для иммобилизации отдельных клеток применяли полутвердые среды, и колонии, секретирующие большие количества IgG, собирали через десять дней после заливки. Данные линии клеток пересевали каждые 3-4 дня в центрифужные биореакторы при плотности 3×105 клеток/мл в содержащую пептон ростовую среду (SFM4CHO, дополненную 8 мМ глутамином, от Hyclone) и инкубировали во влажной камере при 37°С и 5% СО2 при вращении на 180 об/мин.
При оценке поликлональных популяций клеток титры IgG определяли для клеток, которые высевали при концентрации 1×105 клеток в мл и растили в течение 6 дней в 5 мл полной среды в 50 мл центрифужных биореакторах. В качестве альтернативы, культуры клональных популяций во встряхиваемых колбах инокулировали при плотности 3×105 клеток/мл в среде SFM4CHO, чтобы начать производственный процесс периодического культивирования с подпиткой. Производство периодических культур с подпиткой осуществляли в культуре объемом 25 мл во встряхиваемых колбах объемом 125 мл или в культуре объемом 5 мл в культуральных пробирках ТРР объемом 50 мл во влажных камерах при 37°С и 5% CO2 при взбалтывании при 150 об/мин (встряхиваемые колбы и центрифужные пробирки объемом 125 мл). Производство осуществляли в течение десяти дней с подпиткой 16% от исходного объема культуры концентрированной подпитки с определенным химическим составом (Hyclone, Cell Boost 5, 52 г/л) в дни ноль, три и шесть - восемь. В процессе культивирования не применяли подпитку глутамином и глюкозой. Жизнеспособность и концентрацию жизнеспособных клеток (VCD) в культуре измеряли ежедневно, применяя устройство GUAVA (Millipore). Двойной анализ «сэндвич»-ИФА применяли, чтобы определить концентрации МАТ, секретируемых в культуральные среды.
Плазмиды и относительный количественный анализ ПЦР
Клонирующие векторы, применяемые в данном исследовании, представляли собой векторы для экспрессии в млекопитающем SLXplasmid_082 от Selexis. Последовательность люциферазы контрольного вектора pGL3 (Promega) заменяли на эукариотическую кассету экспрессии, состоящую из энхансера CMV человека и промотора GAPDH человека, расположенную в 3'-5' направлении от последовательности, кодирующей EGFP, за которой следовал сигнал полиаденилирования SV40, терминатор гастрина человека и энхансер SV40. Два полученных из MAR генетических элемента человека фланкировали указанную кассету экспрессии, и ген устойчивости к пуромицину экспрессировался под контролем промотора SV40, при этом SLXplasmid_082 отличается по типу элемента MAR, расположенного в 3'-5' направлении от указанной кассеты экспрессии (hMAR 1-68 и hMAR X-29; Girod и др., 2007).
кДНК тяжелых и легких цепей трастузумаба и инфликсимаба клонировали в вектор экспрессии, чтобы заменить EGFP. Вектор, несущий кассеты экспрессии обеих тяжелой и легкой цепей каждого IgG, получали путем объединения кассет экспрессии тяжелой и легкой цепей на одном плазмидном векторе. Последовательности сигнальных пептидов из всех тяжелых и легких цепей, а также константные области легких цепей идентичны. Константные области тяжелых цепей отличаются в нескольких положениях аминокислот (варианты DEL по сравнению с ЕЕМ).
Праймеры для амплификации методом ПЦР и номера доступа в GenBank кДНК SRP9, SRP14, SRP54, SRPR-альфа, SRPR-бета, SEC61A1 и SEC61B перечислены в других местах в данном тексте. Продукты ПЦР, кодирующие секреторные белки, клонировали в вектор, чтобы заменить последовательность EGFP. Когда несколько секреторных белков экспрессировали совместно, инвертированные концевые последовательности транспозона piggyBac встраивали в векторы, чтобы заключить в них кассету экспрессии, и полученные в результате этого векторы котрансфицировали с вектором экспрессии, несущим транспозазу piggyBac, чтобы улучшить встраивание трансгена и избавиться от необходимости отбора антибиотиком.
Обычный вектор экспрессии, несущий транспозазу РВ, который представлял собой pCS2+U5V5PBU3, содержащий последовательность, кодирующую транспозазу РВ, окруженную 5'- и 3'-нетранслируемыми концевыми областями (НТО) гена β-глобина Xenopus laevis, применяли в смежных экспериментах. Данную плазмиду конструировали, как описано далее: фрагмент 3'-НТО размером 317 п.о. из pBSSK/SB10 встраивали в pCS2+U5 (Invitrogen/Life Technologies, Пейсли, Великобритания) с получением pCS2+U5U3. Последовательность, кодирующую транспозазу РВ (2067 п.о., номер доступа в GenBank: EF587698), синтезировали в ATG:biosynthetic (Мерцхаузен, Германия) и клонировали в каркас pCS2+U5U3 между двумя НТО. Контрольный вектор РВ соответствовал немодифицированной плазмиде pCS2+U5.
Векторы, несущие различные транспозоны, получали путем вставки 3'- и 5'-инвертированных концевых повторов РВ (ITR) размером 235 п.о. и 310 п.о., соответственно, синтезированных в ATG:biosynthetic (Мерцхаузен, Германия), в плазмиду pBluescript SK (pBSK ITR3'-ITR5'). Ген фосфотрансферазы неомицина (NeoR) под контролем промотора SV40 из плазмиды pRc/RSV (Invitrogen/Life Technologies) затем вставляли между двумя указанными ITR. Элементы MAR 1-68 и MAR Х-29, гены устойчивости к пуромицину и GFP, применяемые в данном исследовании, были такие, как описано ранее (Girod и др. 2007; Grandjean и др. 2011; Hart и Laemmli 1998). Векторы экспрессии иммуноглобулинов и последовательности, кодирующие SRP9, SRP14, SRP54, SRPR-альфа, SRPR-бета, SEC61A1 и SEC61B, описаны в данной заявке. Секреторные белки экспрессировали, применяя эукариотическую кассету экспрессии, состоящую из энхансера CMV человека и промотора GAPDH человека, расположенных в 3'-5' направлении от кодирующей последовательности, за которыми следовал сигнал полиаденилирования SV40, терминатор гастрина человека и энхансер SV40. Кассеты экспрессии и/или элементы MAR встраивали между последовательностями ITR или в каркас бактериального вектора, применяя стандартные способы клонирования.
Системы транспозона PiggyBac, включая подходящие 3'- и 5'-ITR, а также транспозазу, например, доступны для приобретения у SYSTEM BIOSCIENCE.
Для относительного анализа методом количественной ПЦР общую РНК выделяли из 1×105 клеток и транскрибировали с помощью обратной транскриптазы в кДНК, применяя набор FastLane Cell cDNA (Qiagen), следуя инструкциям производителя. Осуществляли количественный анализ экспрессии SRP14 и GAPDH с помощью количественной ПЦР, применяя Rotor Gene Q (Qiagen) и LightCycler®480 SYBR Green I Master (Roche), с применением праймеров, перечисленых в приложенной таблице S1. Уровни информационной РНК SRP14 нормировали на таковые для GAPDH, применяя программное обеспечение Rotor-Gene Q Series (Qiagen).
Культивирование клеток, стабильная трансфекция и субклонирование линий клеток СНО
Суспензионные клетки яичника китайского хомячка (СНО-K1) поддерживали в бессывороточной среде SFM4CHO Hyclone (ThermoScientific), дополненной L-глутамином (РАА, Австрия) и добавкой НТ (Gibco, Invitrogen life sciences), при 37°C, 5% CO2 в увлажненном воздухе. Клетки СНО-K1 трансфицировали векторами экспрессии тяжелых и легких цепей трастузумаба или инфликсимаба, несущими ген устойчивости к пуромицину, путем электропорации согласно рекомендациям производителя (Neon devices, Invitrogen). Через два дня клетки переносили в планшеты Т75 в среду, содержащую 10 мкг/мл пуромицина, и указанные клетки дополнительно культивировали в условиях селекции в течение двух недель. Затем получали стабильные отдельные клоны клеток, экспрессирующих IgG трастузумаб и инфликсимаб, с помощью метода серийных разведений, растили и анализировали эффективность их роста и уровни продукции IgG. Продуцирующие IgG трастузумаб и инфликсимаб клоны клеток, экспрессирующих наиболее высокие уровни IgG, отбирали для дополнительных биохимических экспериментов. Некоторые из данных клонов затем котрансфицировали путем электропорации вектором, экспрессирующим SRP14, и плазмидой, несущей ген устойчивости к неомицину. Клетки затем культивировали в среде, содержащей 300 мкг/мл G418, в течение двух недель, как описано выше. Стабильные клоны выделяли с помощью ограниченного разведения и подтверждали экспрессию SRP14 с помощью анализов методом количественной ПЦР перед выращиванием культуры для биохимического анализа.
Периодическое культивирование и периодическое культивирование с подпиткой
Эффективность роста и продукции для отдельных клонов, экспрессирующих трастузумаб и инфликсимаб, оценивали при периодическом культивировании в минибиореакторе объемом 50 мл (ТРР, Швейцария) при 37°С во влажной камере при 5% CO2 в течение 7 дней. В день 3, день 4 и день 7 культивирования клеток определяли концентрацию и жизнеспособность клеток, применяя систему для проточной цитометрии Guava EasyCyte (Millipore). Титр IgG в культуральных супернатантах измеряли с помощью «сэндвич»-ИФА. Плотность клеток (Cv*мл-1) и значения титра IgG (мкг*мл-1) наносили на график в указанный день отбора образцов в процессе эксперимента. Удельную продуктивность IgG клонов, экспрессирующих трастузумаб и инфликсимаб, определяли как падение концентрации IgG в зависимости от суммарного количества жизнеспособных клеток (IVCD), рассчитанное в день 3 - день 7 (фаза продукции), и выражали в виде пг на клетку в день (pcd).
Для периодических культур с подпиткой клетки высевали при плотности 0,3×106 клеток/мл во встряхиваемые колбы объемом 125 мл в 25 мл бессывороточной среды SFM4CHO Hyclone. Культуры поддерживали при 37°С и 5% СО2 при встряхивании. Культуры подпитывали ежедневно коммерчески доступной подпиткой Hyclone (ThermoScientific). Плотности клеток и продукцию IgG оценивали ежедневно.
Анализ экспрессии и агрегации белков
Растворимые цитоплазматические белки экстрагировали путем пермеабилизации клеток 1% Triton Х-100 в буфере ФБР в присутствии коктейля ингибиторов протеаз (Roche, inc). После инкубации в течение 30 мин на льду клетки центрифугировали 10 мин при 14000 об/мин. Супернатант назвали фракцией "растворимых цитозольных и ЭР белков". Осадок растворяли путем обработки ультразвуком в буфере Лэммли с мочевиной (62,5 мМ Трис, 2% ДСН, 8 М мочевина, 5% глицерин, краситель бромфеноловый синий), что позволяло получить нерастворимую фракцию агрегировавших и везикулярных белков. Растворимую и нерастворимую фракции затем разбавляли буфером Лэммли с добавлением или без 2-меркаптоэтанола и кипятили 8 мин при 95°С. Образцы в восстанавливающем и невосстанавливающем буферах разделяли на 10% или градиентном 4-10% акриламидных гелях с помощью электрофореза в полиакриламидном геле с добавлением додецилсульфата натрия (ЭФ в ПААГ/ДСН), соответственно.
Белки затем переносили на нитроцеллюлозную мембрану. После блокирования в 5% молоке, разбавленном в TBS-Tween (20 мМ Трис, 0,5 М NaCl, 0,1% Tween 20), мембраны анализировали для обнаружения различных белков, применяя следующие первичные антитела: антитело осла против IgG (H+L) человека, конъюгированное с пероксидазой хрена (JK immunoresearch, №709 035 149, 1:5000), поликлональные антитела кролика против BiP человека (Cell signalling, BiP, С50 В12, 1:2000), моноклональное антитело мыши против CHOP человека (Cell signalling, CHOP, L63F7, 1:500), поликлональное антитело козы против GAPDH человека. После инкубации в течение ночи при 4°С каждую мембрану обрабатывали конъюгированными с пероксидазой хрена HRP IgG против антител кролика или IgG против антител мыши (Cell signalling, 1:20000). Белки, специфично узнаваемые каждым антителом, детектировали, применяя реагенты для улучшенной хемилюминесценции (ECL) и экспонирование с пленкой для ECL (Amersham Biosciences).
Эксперименты с вытеснением белков на основе циклогексимида
Эксперименты с вытеснением на основе циклогексимида осуществляли на клонах СНО-K1 с высоким (HP) и низким (LP) уровнем продукции IgG. Равные количества клеток высевали на 6-луночные планшеты в полной культуральной среде, дополненной 100 мкМ циклогексимида (Sigma). В различные моменты времени клетки собирали и лизировали в ФБР, 1% Triton Х-100. Растворимые и нерастворимые в Тх фракции затем разделяли на 4-10% акриламидном геле с помощью ПААГ/ДСН в невосстановительных условиях и подвергали иммуноблоттингу с антителом к IgG человека.
Анализы дифференциального фракционирования детергентом
Фракционирование цитозольных и мембранных белков осуществляли путем дифференциального экстрагирования детергентом из осадка клеток. Клетки сначала промывали в 1 мл ФБР, и плазматические мембраны HP и LP клеток пермеабилизовали в буфере КНМ (110 мМ KAc, 20 мМ HEPES, 2 мМ MgCl2, рН 7,2), содержащем 0,01% дигитонин (Sigma), в течение 10 мин в присутствие или без 1% Triton Х-100. Полупермеабилизованные клетки однократно промывали в буфере КНМ и добавляли трипсин до конечной концентрации 50 мкг/мл на 10 мин при комнатной температуре, чтобы расщепить растворимые белки. Расщепление трипсином останавливали путем добавления 1 мМ фенилметилсульфонилфторида (PMSF) и 4 мМ 4-(2-аминоэтил)-бензосульфонилфторида (AEBSF). Клетки собирали путем центрифугирования и растворимые белки экстрагировали в присутствии Triton Х-100 и ингибитора протеаз, как описано а разделе 2.4. Восстанавливающий буфер Лэммли, содержащий 2-меркаптоэтанол, добавляли к осадку и фракциям супернатанта, которые затем подвергали ЭФ в 8% ПААГ/ДСН. Осуществляли иммуноблоттинг, чтобы детектировать белки IgG и BiP.
Перекрестное сшивание белков и анализ методом вестерн-блоттинга
Клетки, продуцирующие инфликсимаб на низком уровне (LP), однократно промывали в ФБР и инкубировали с добавлением или без 1 мМ кросслинкера дитиобис(сукцинимидилпропионата) (DSP) (ThermoScientific) в течение 30 мин на льду. Перекрестное сшивание гасили путем инкубации с добавлением 50 мМ Трис-HCl (рН 7,4) в течение 10 мин перед экстрагированием белка в буфере ФБР, содержащем 1% Triton Х-100. После центрифугирования в течение 10 мин при 14000 об/мин в микроцентрифуге нерастворимую в Triton Х-100 фракцию или цельный экстракт белка анализировали с помощью ЭФ в ПААГ/ДСН в восстановительных условиях, подвергали иммуноблоттингу и окрашивали антителами против BiP и LC. Равные количества белков из нерастворимых в Тх фракций анализировали параллельно.
Должно быть очевидно, что способы и композиции согласно настоящему изобретению могут быть включены в виде различных вариантов реализации, лишь незначительное количество которых описано в данной заявке. Для специалиста должно быть очевидно, что существуют другие варианты реализации, которые не выходят за рамки сущности настоящего изобретения. Таким образом, описанные варианты реализации являются иллюстративными и не должны истолковываться как ограничительные.

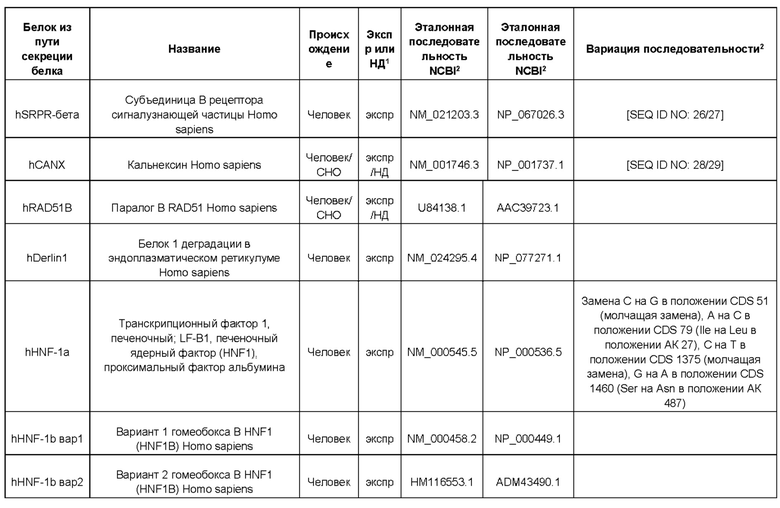
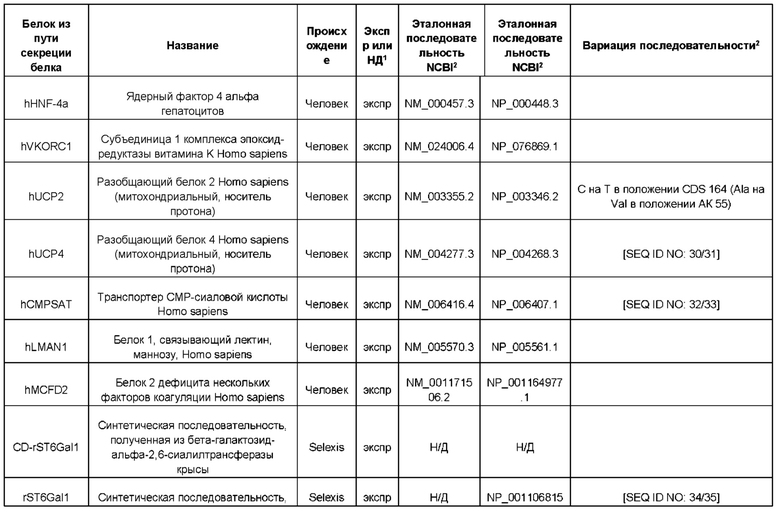
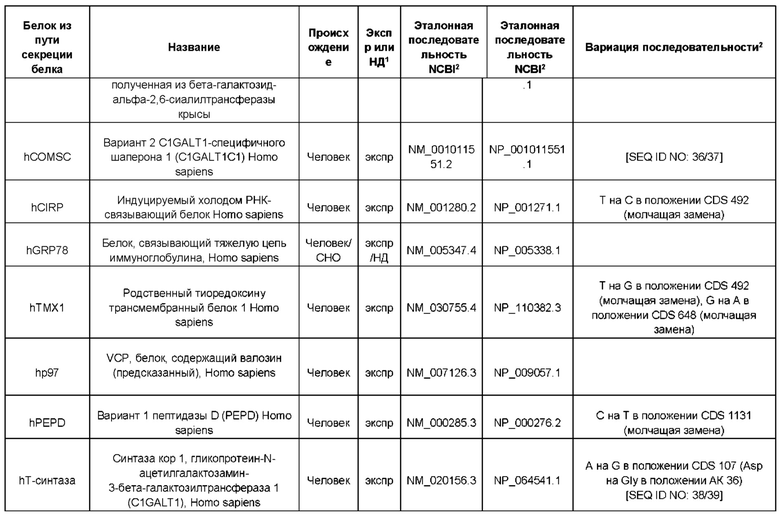
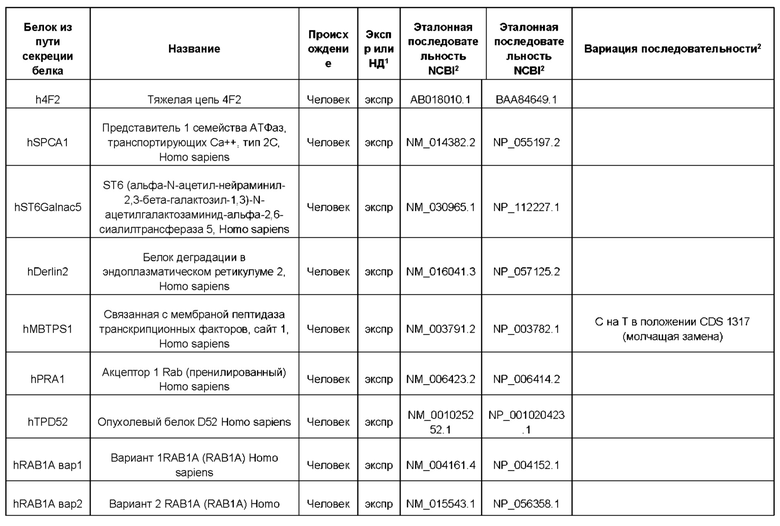
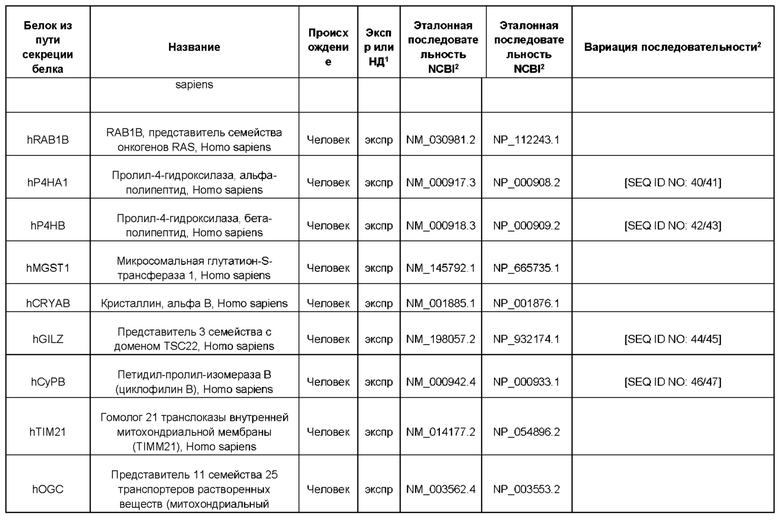
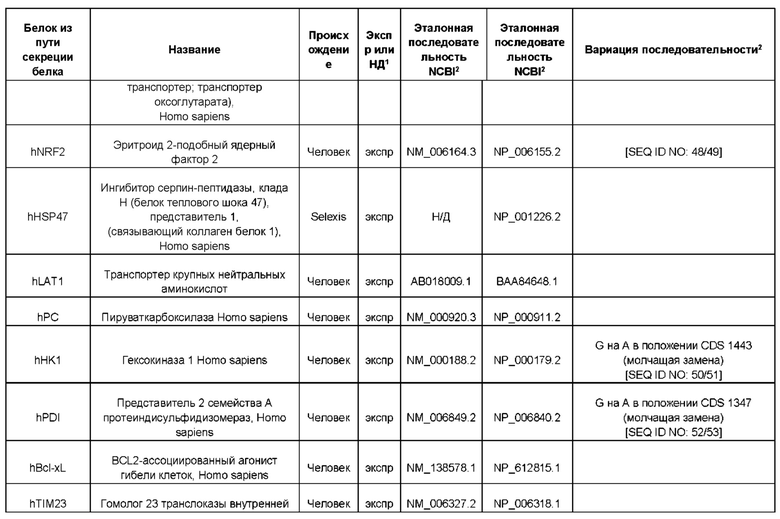
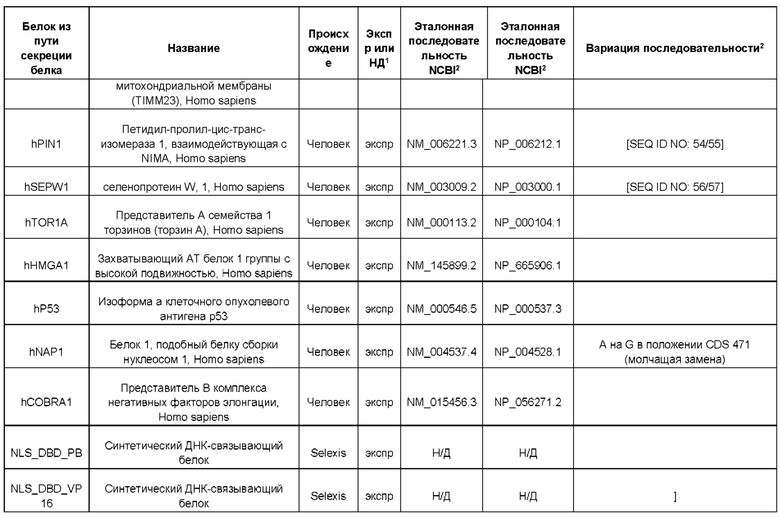
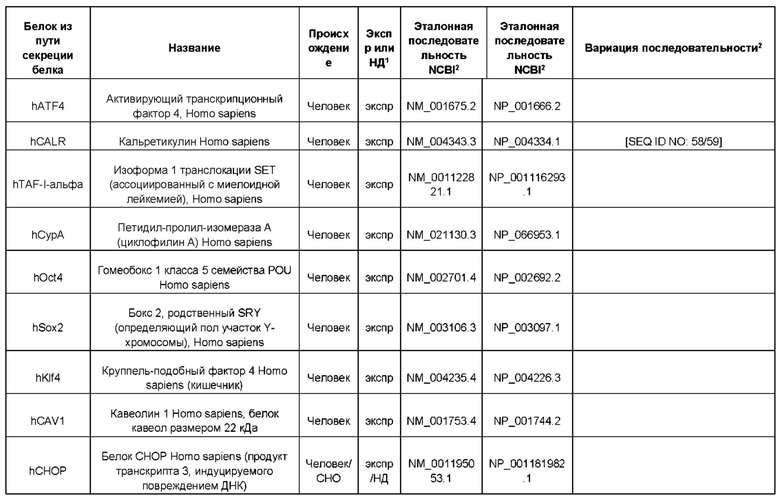
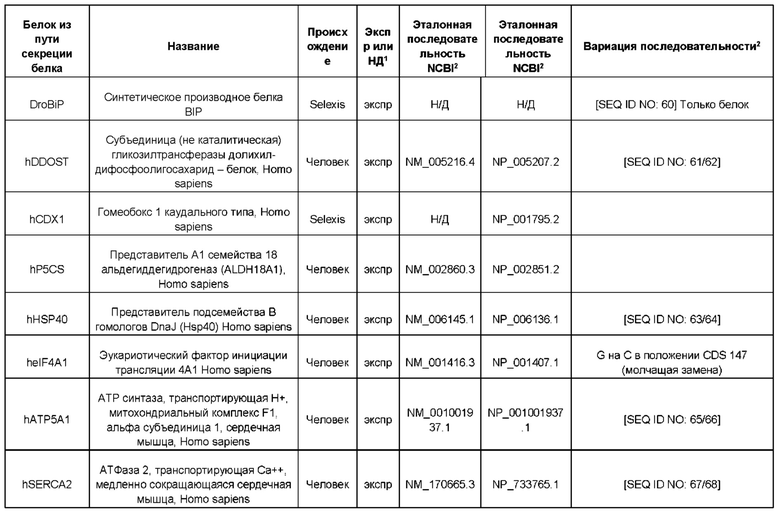
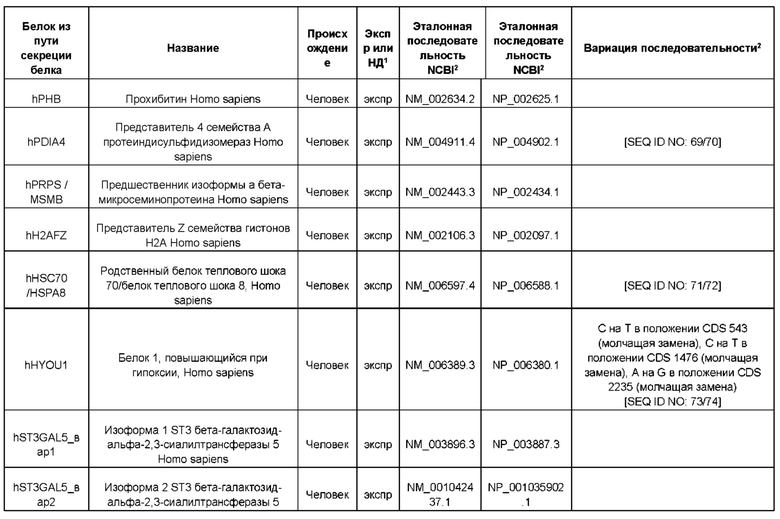
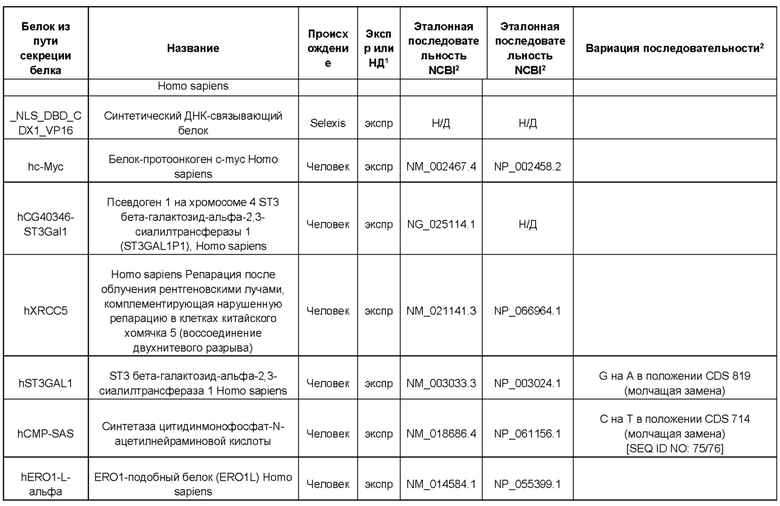
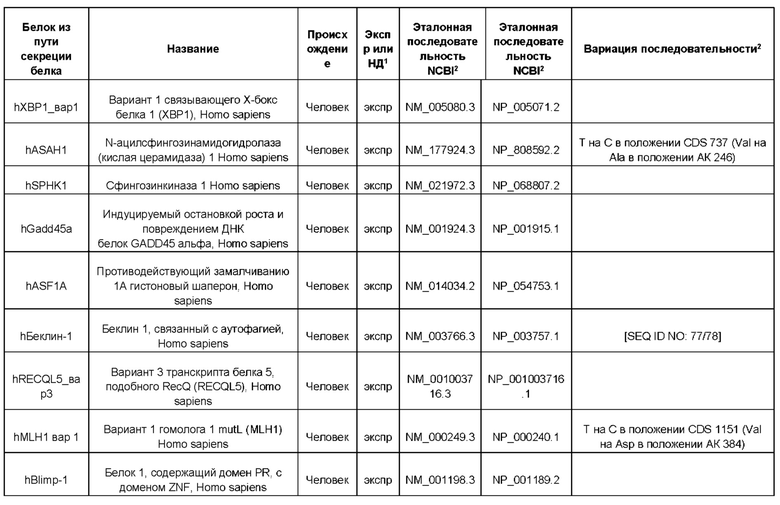
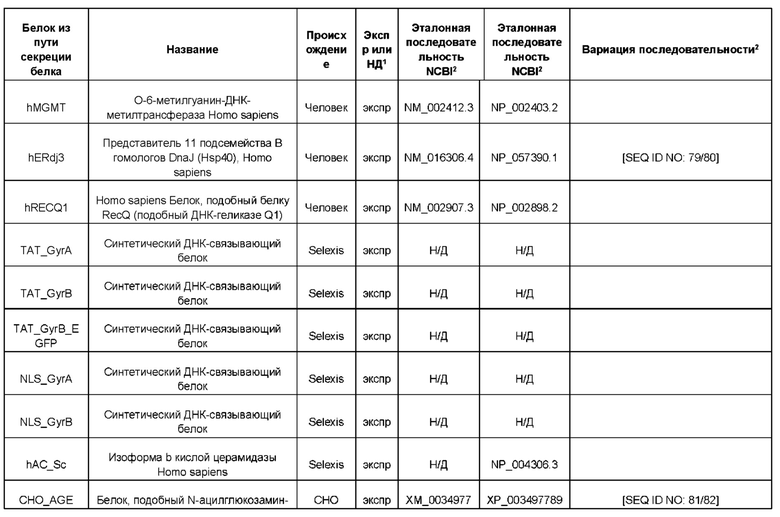
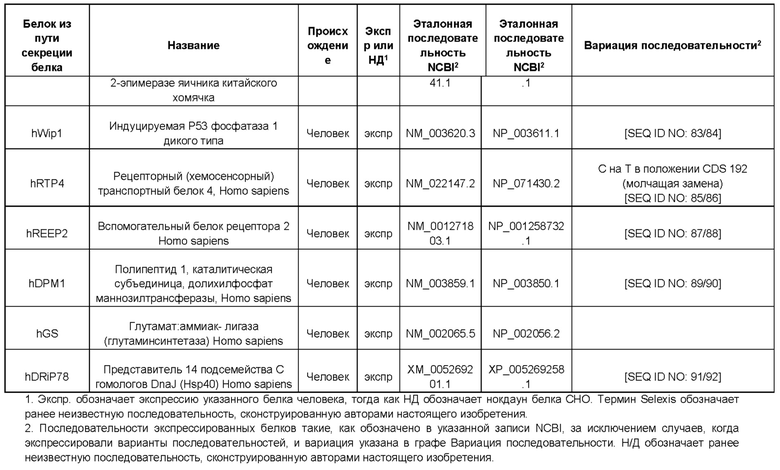



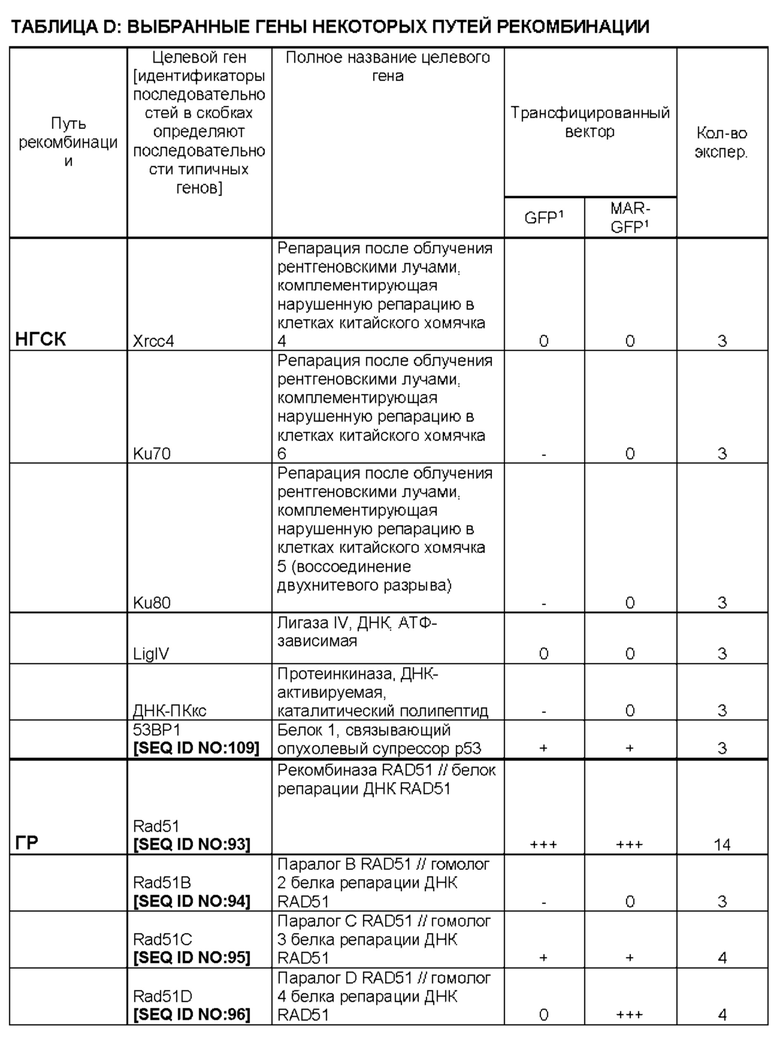



| название | год | авторы | номер документа |
|---|---|---|---|
| УЛУЧШЕННЫЕ ЭКСПРЕССИЯ И ПРОЦЕССИНГ ТРАНСГЕНА | 2014 |
|
RU2711505C2 |
| УЛУЧШЕННЫЕ ЭУКАРИОТИЧЕСКИЕ КЛЕТКИ ДЛЯ ПОЛУЧЕНИЯ БЕЛКА И СПОСОБЫ ИХ ПОЛУЧЕНИЯ | 2016 |
|
RU2752529C2 |
| ЭКСПРЕССИОННАЯ СИСТЕМА ДЛЯ ПОВЫШЕНИЯ УРОВНЯ ЭКСПРЕССИИ ГЕНА, МОЛЕКУЛА НУКЛЕИНОВОЙ КИСЛОТЫ, КЛЕТКА, ТРАНСГЕННОЕ ЖИВОТНОЕ И НАБОР | 2007 |
|
RU2469089C2 |
| СОЗДАНИЕ ГАПЛОИДНЫХ РАСТЕНИЙ И УСОВЕРШЕНСТВОВАНИЕ СЕЛЕКЦИИ РАСТЕНИЙ | 2010 |
|
RU2571927C2 |
| МОДИФИКАЦИЯ B-КЛЕТОК | 2018 |
|
RU2783116C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ЛЕЧЕНИЯ БОЛЕЗНИ ХАНТИНГТОНА | 2015 |
|
RU2691102C2 |
| ПРИМЕНЕНИЕ ВИТАМИНОВ И ГЕНОВ И БЕЛКОВ МЕТАБОЛИЗМА ВИТАМИНОВ ДЛЯ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО БЕЛКА В КЛЕТКАХ МЛЕКОПИТАЮЩИХ | 2016 |
|
RU2736705C2 |
| SYNP161, ПРОМОТОР ДЛЯ СПЕЦИФИЧЕСКОЙ ЭКСПРЕССИИ ГЕНОВ В ПАЛОЧКОВЫХ ФОТОРЕЦЕПТОРАХ | 2016 |
|
RU2758211C2 |
| ПОЛУЧЕНИЕ ВИРУСНОГО ВЕКТОРА | 2014 |
|
RU2658487C2 |
| Нуклеиновая кислота, имеющая промоторную активность, и ее применение | 2022 |
|
RU2818112C2 |












































































































































































































































































































































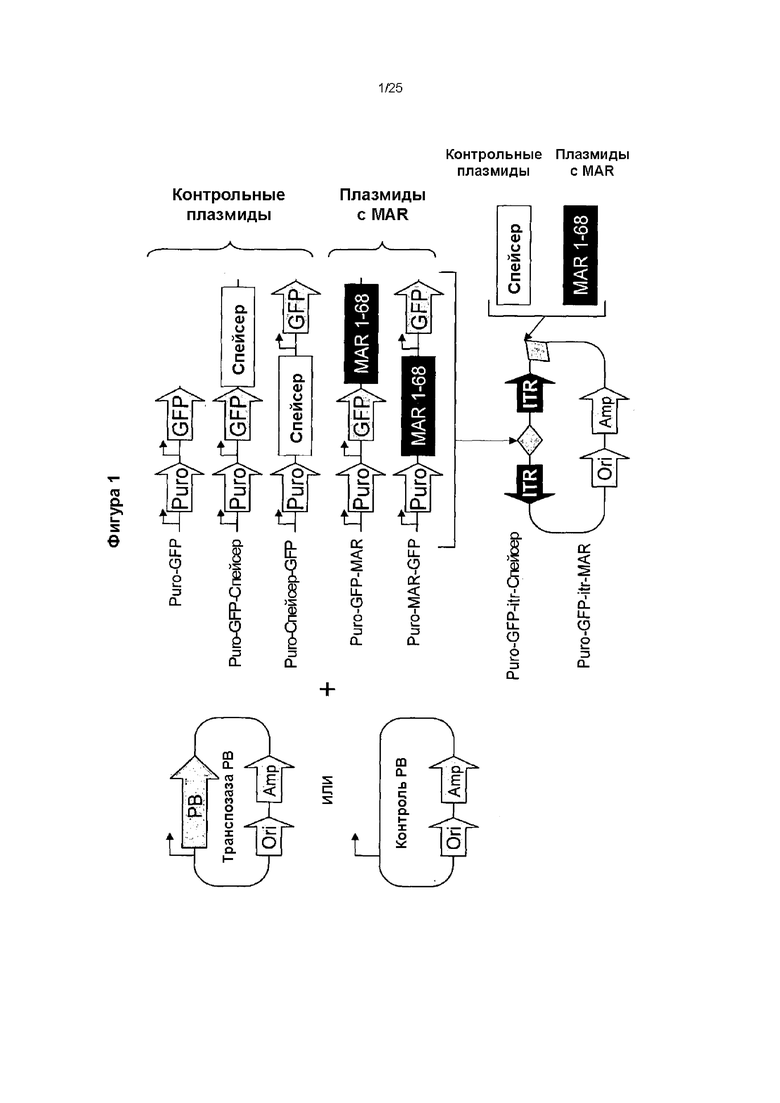
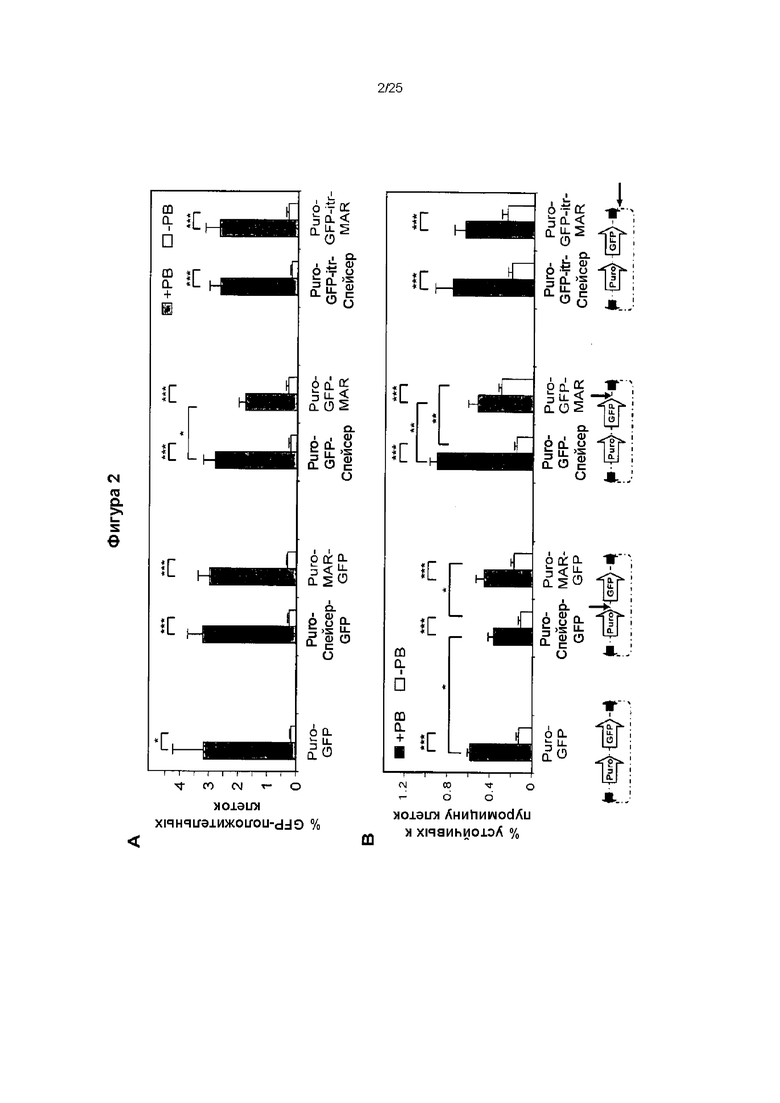


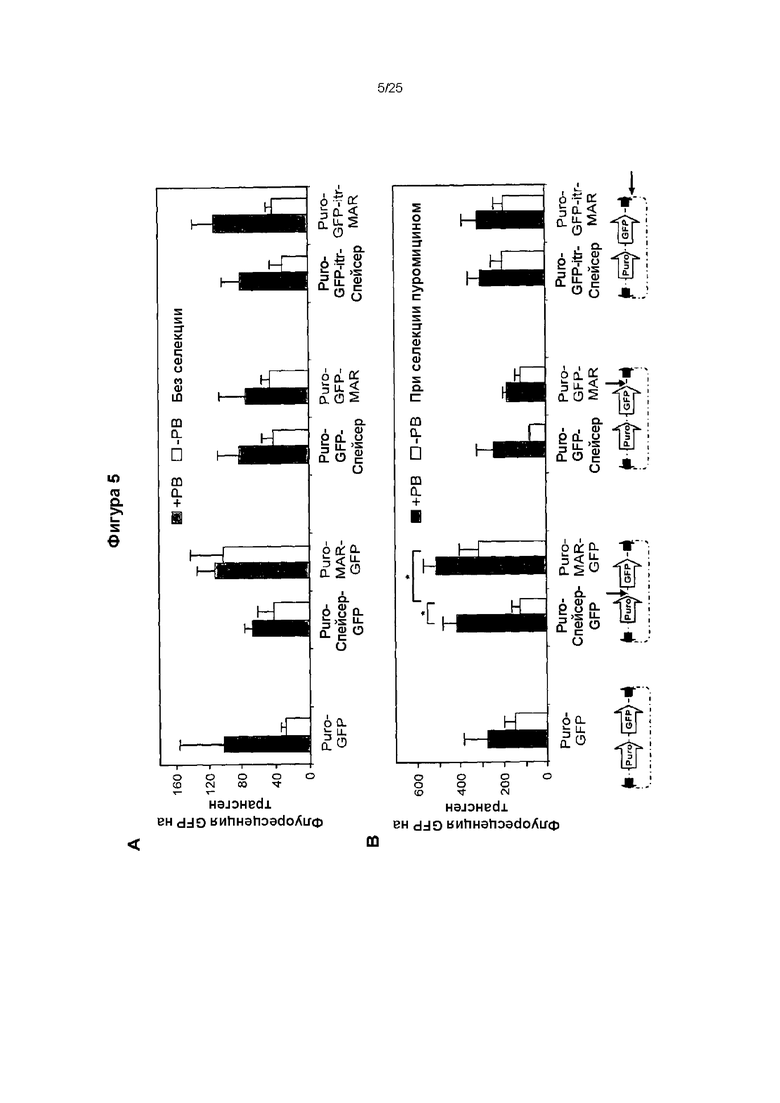
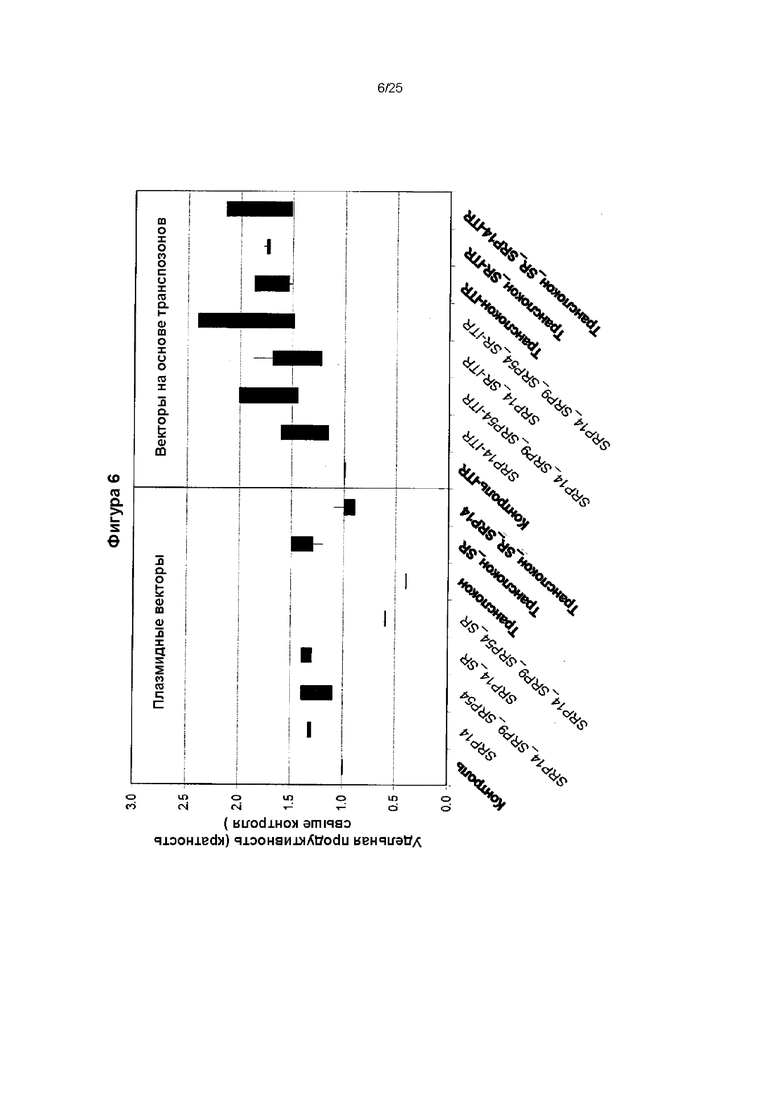
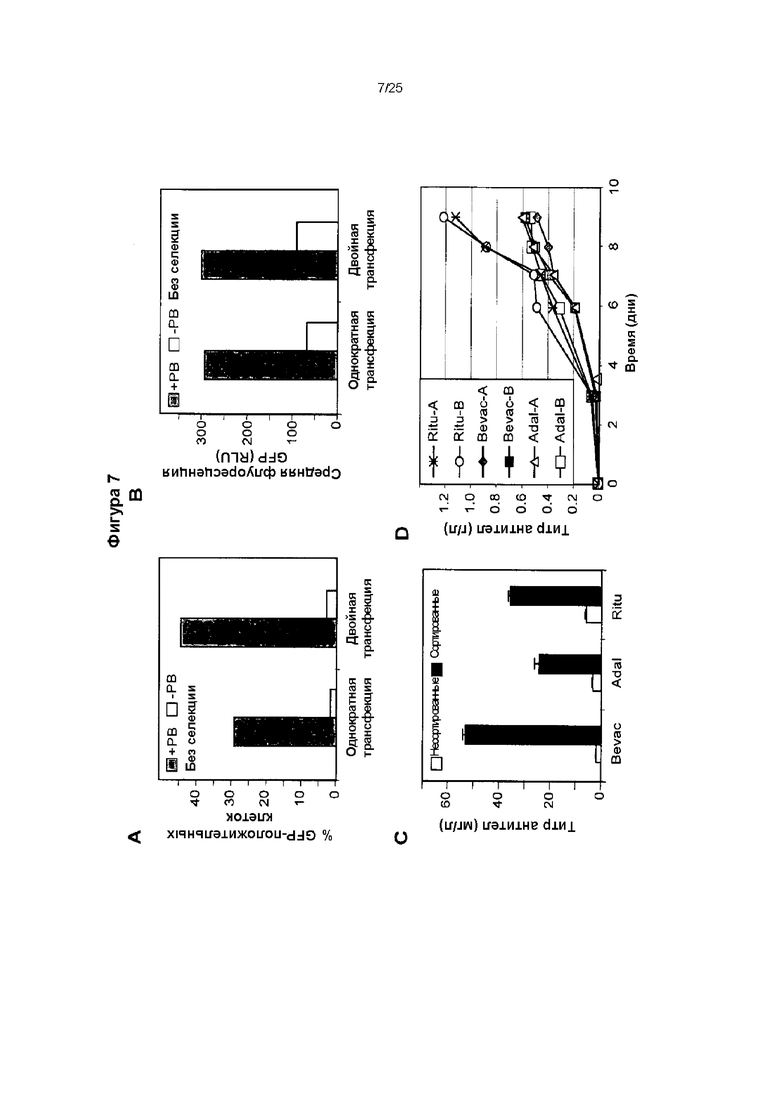

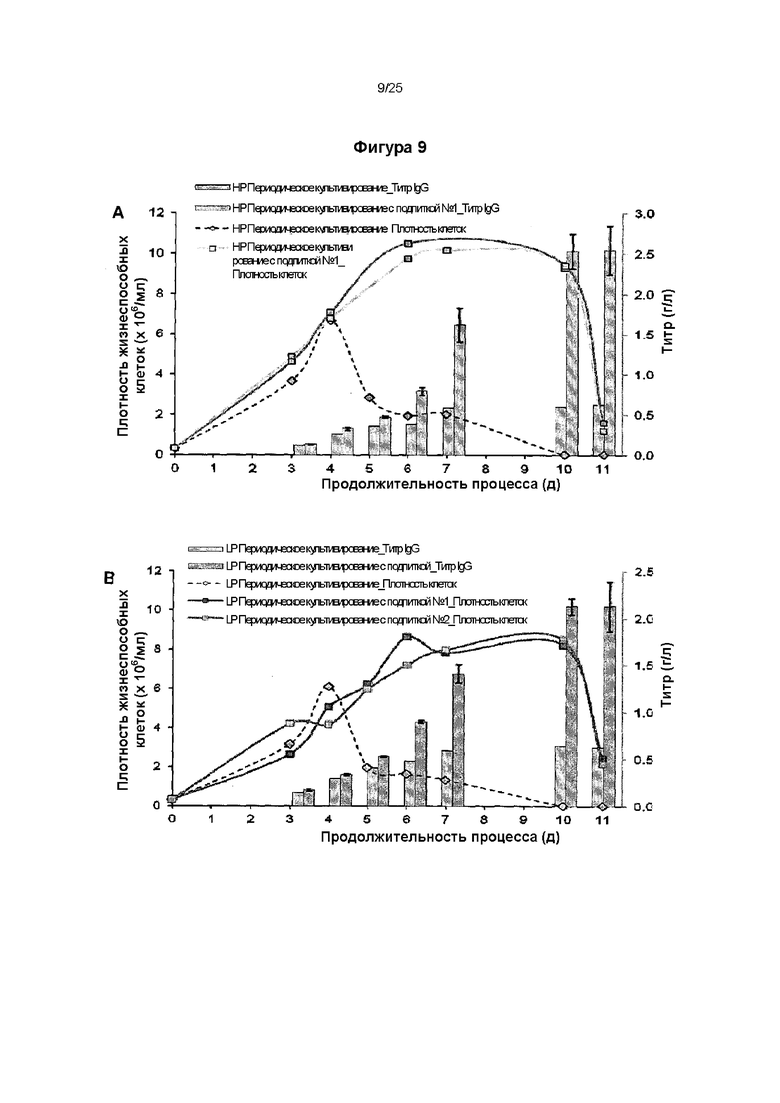

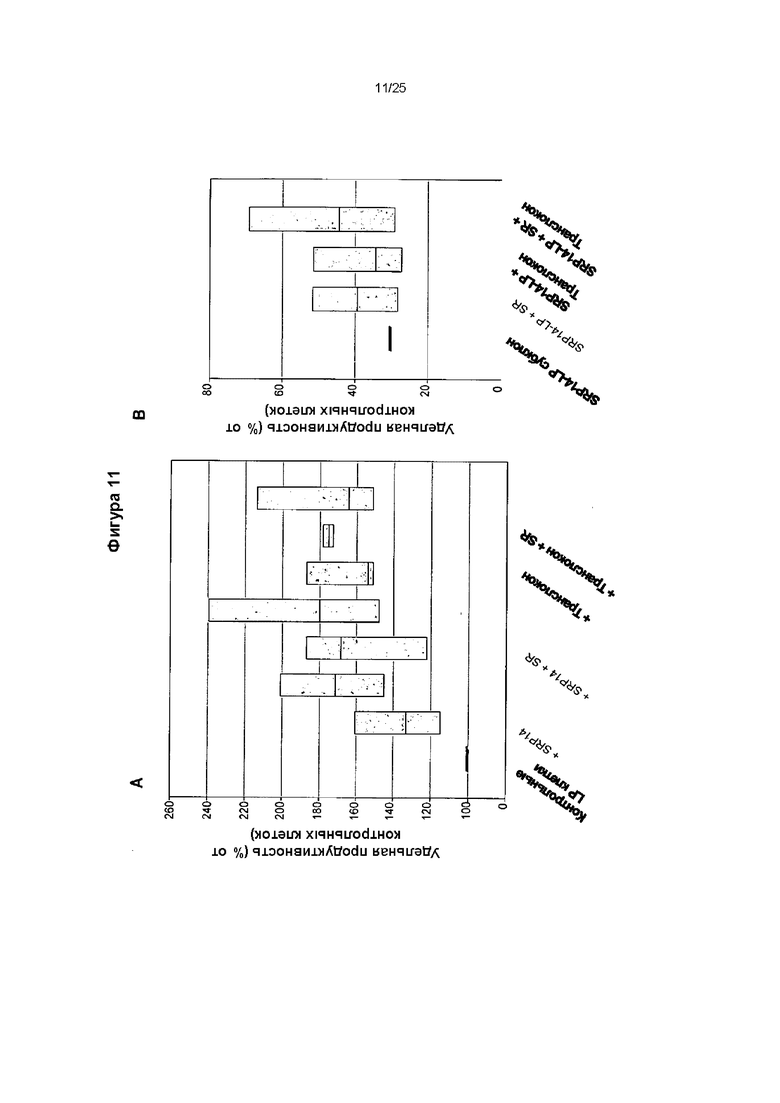
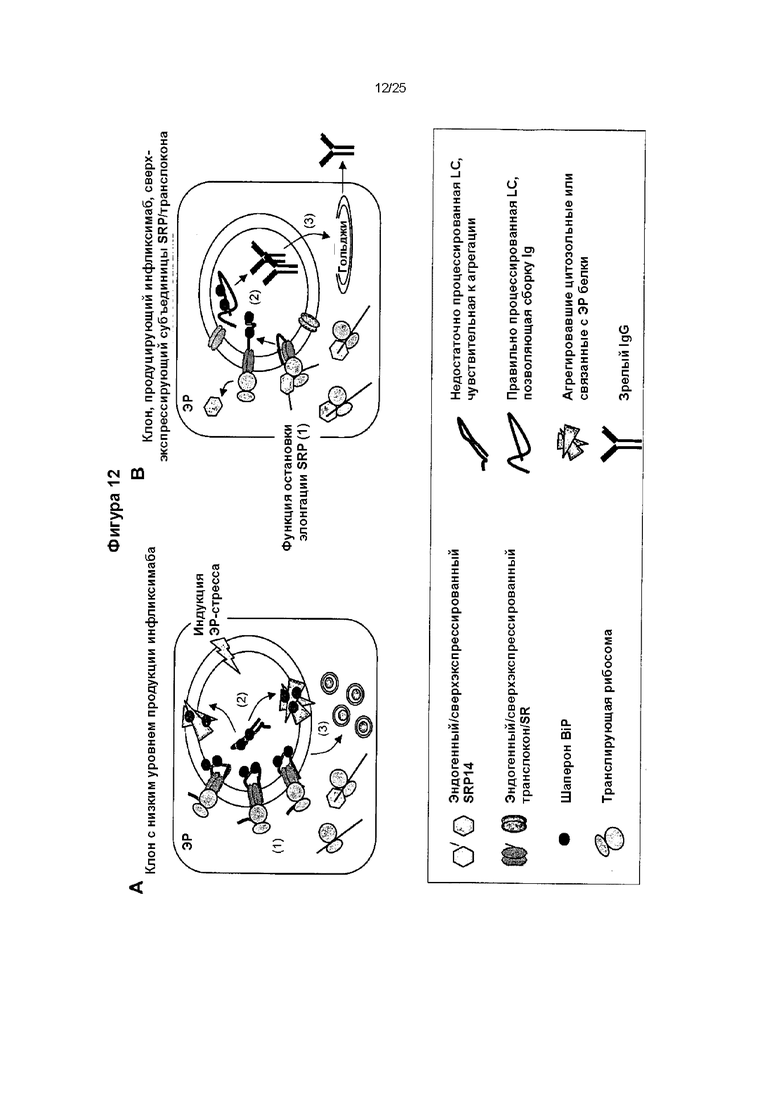
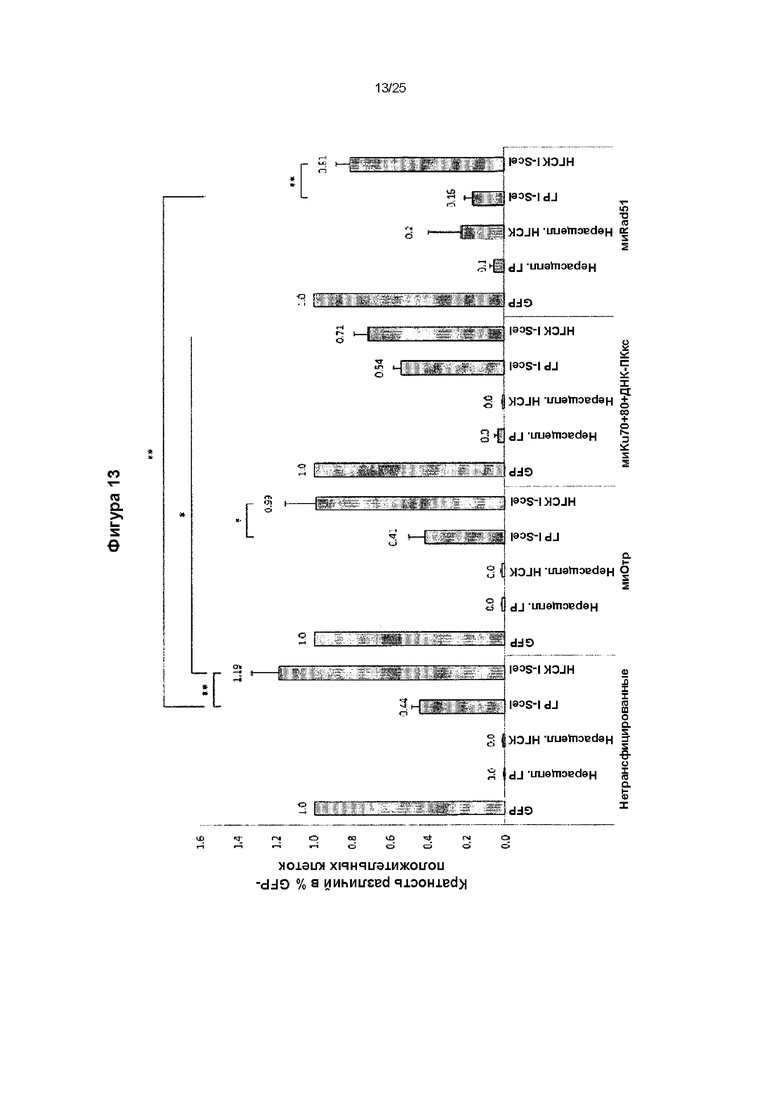
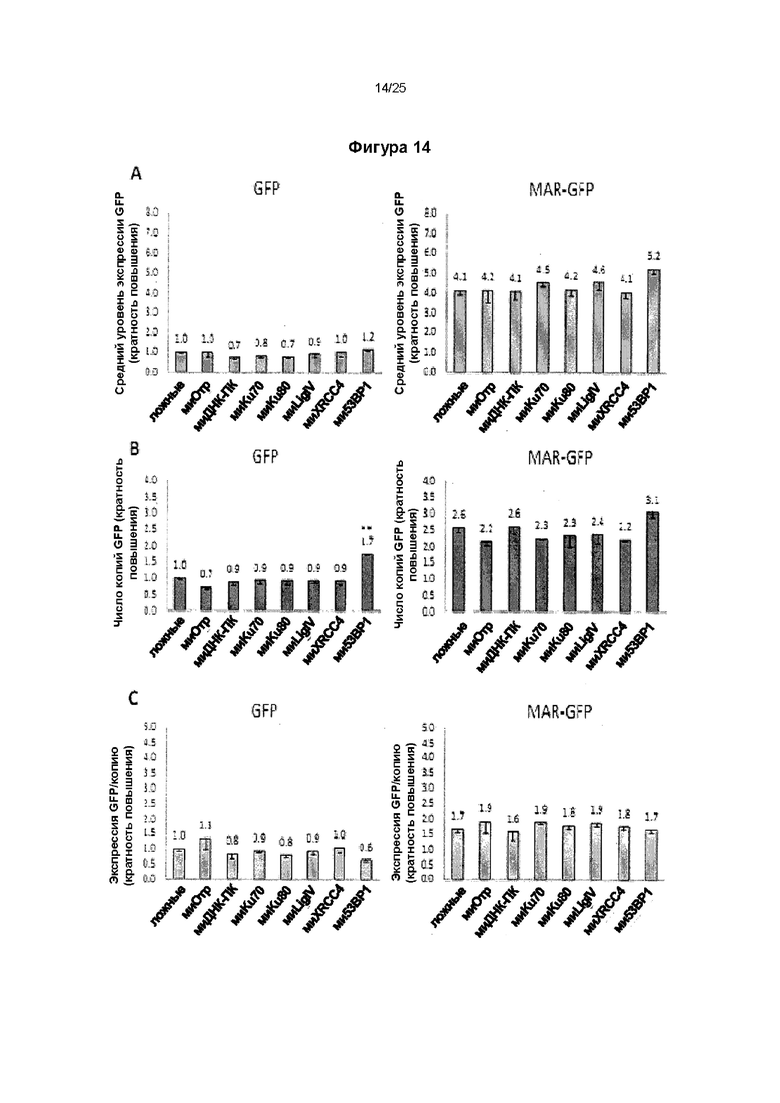



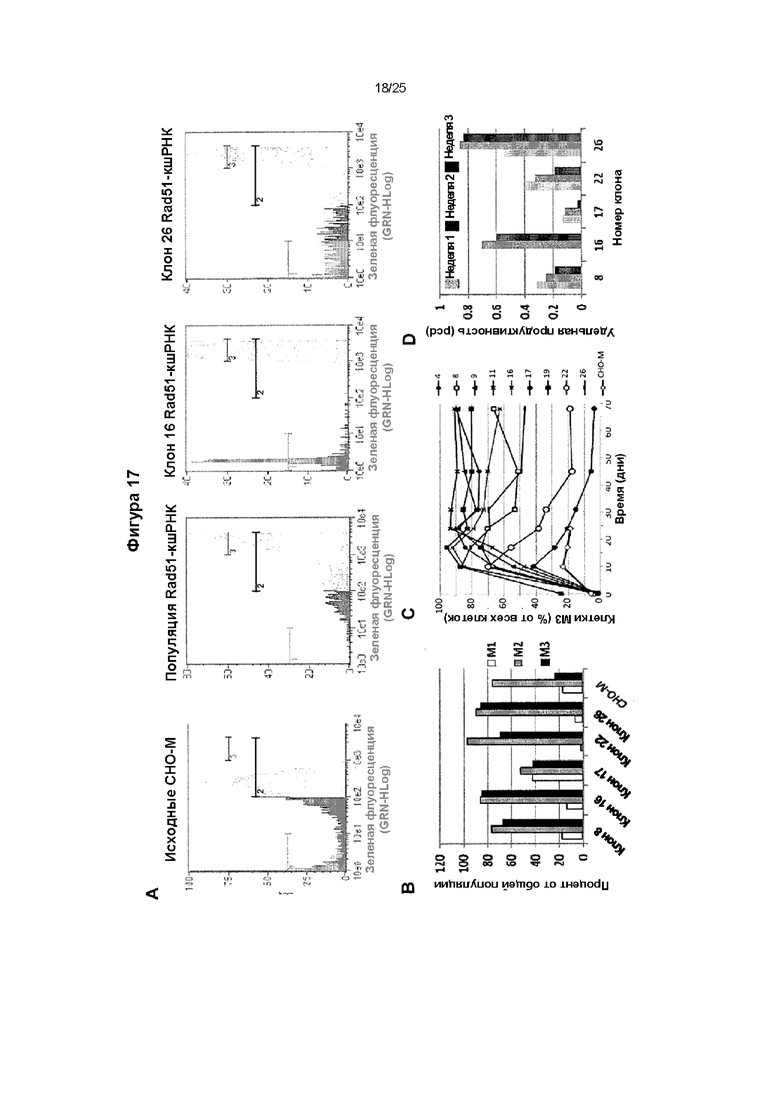
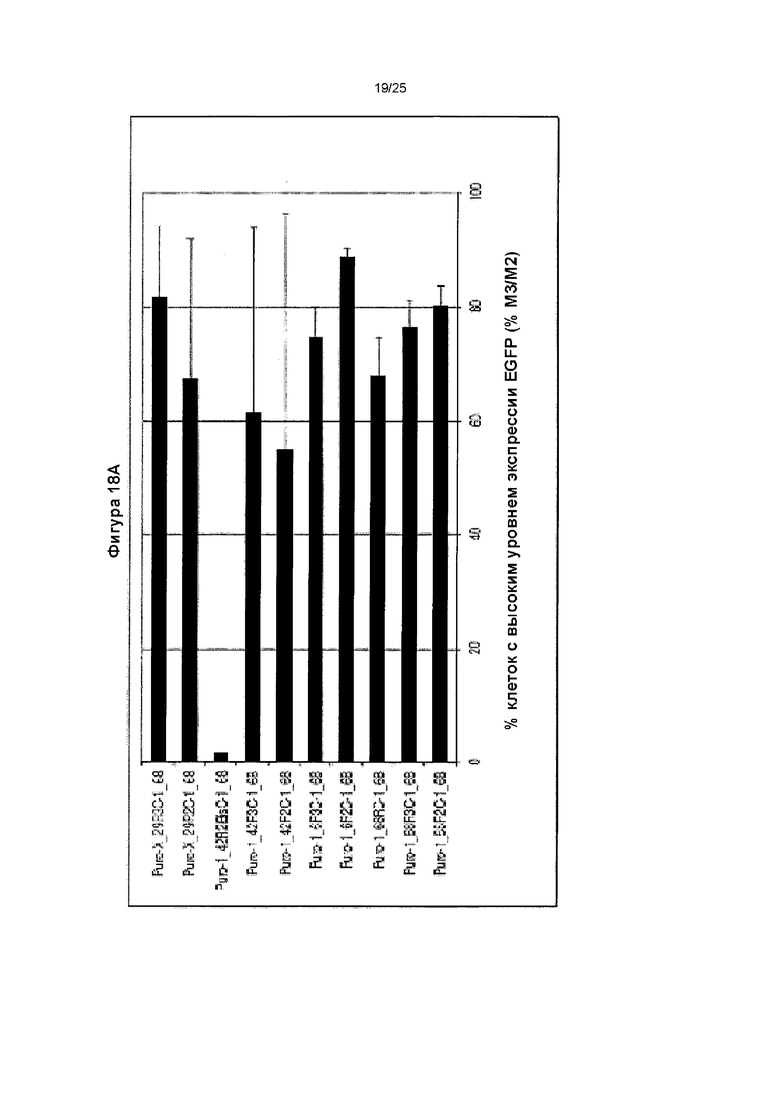

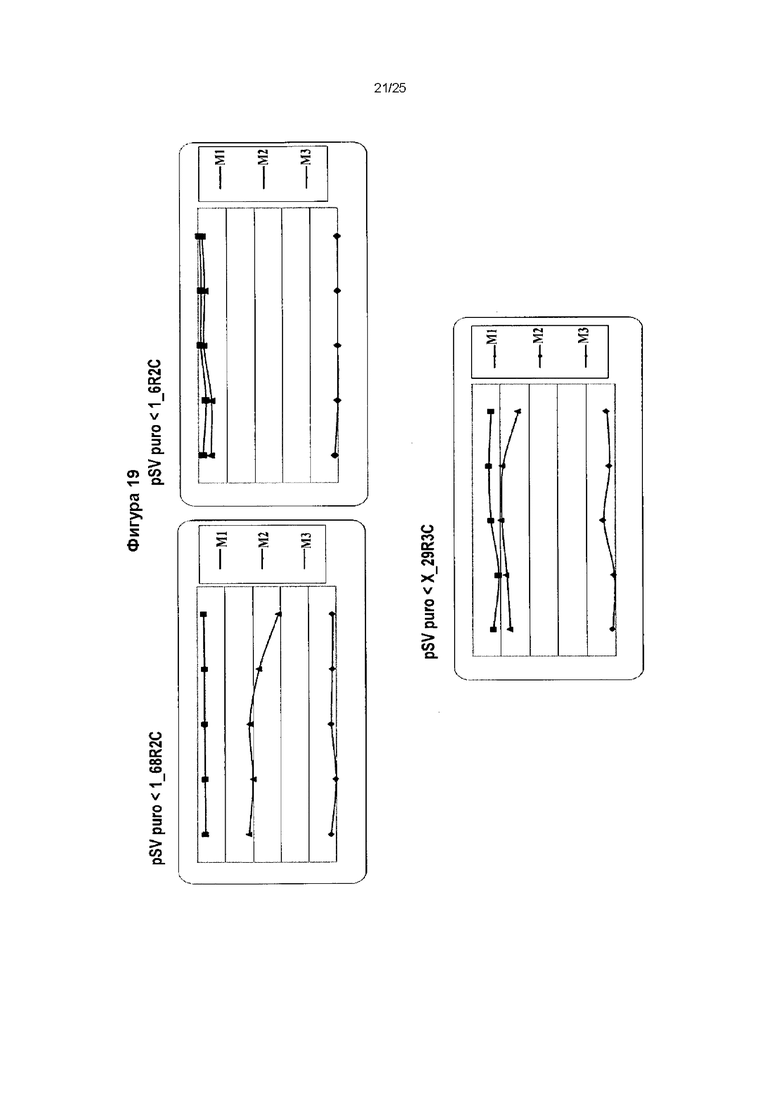
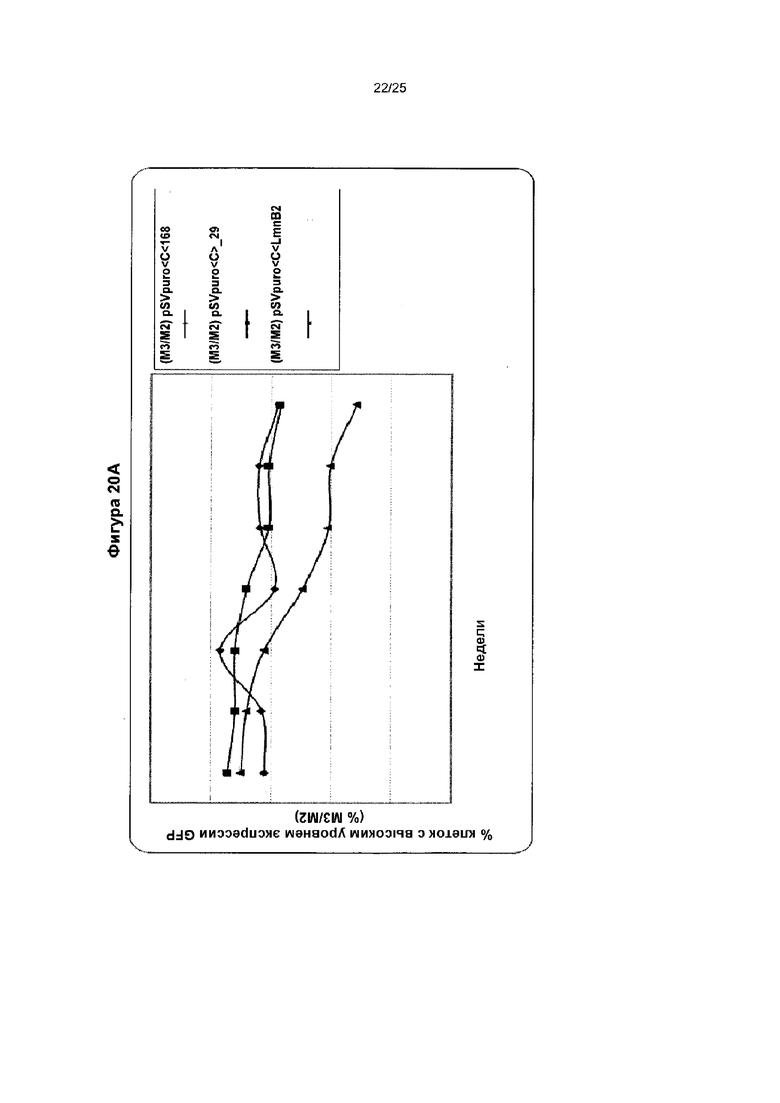
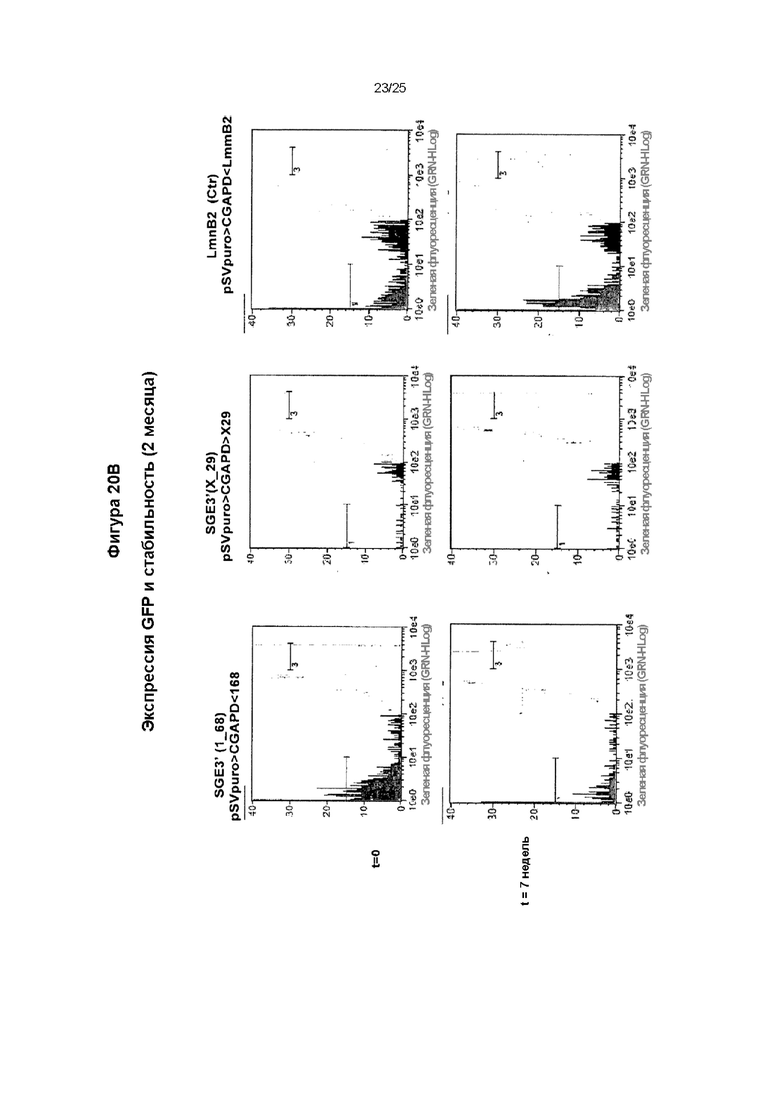


Изобретение относится к области биотехнологии, конкретно к рекомбинантной молекуле нуклеиновой кислоты для увеличения эффективности продукции трансгена, экспрессируемого в клетке млекопитающего. Рекомбинантная молекула нуклеиновой кислоты согласно настоящему изобретению сконструирована таким образом, что включает 5’-3’-транспозон-специфичный концевой повтор ITR, нуклеиновую кислоту, кодирующую белок трансгена TEP или РНК TEP, а также дополнена эпигенетическим регуляторным элементом. Кроме того, описаны рекомбинантный экспрессионный вектор, клетка-хозяин, способ экспрессии кодируемого ТЕР, способ трансфекции клетки и набор для осуществления трансфекции клетки. В сравнении с известными аналогами изобретение демонстрирует преимущества в отношении величины и качества экспрессии, включая высокую стабильность экспрессии трансгена и, при необходимости, транспорт экспрессированного продукта наружу из клетки. 9 н. и 11 з.п. ф-лы, 4 табл., 22 ил.
1. Рекомбинантная молекула нуклеиновой кислоты для увеличения эффективности продукции трансгена, экспрессируемого в клетке млекопитающего, содержащая:
5’- и 3’-транспозон-специфичные инвертированные концевые повторы (ITR);
по меньшей мере одну последовательность нуклеиновой кислоты, кодирующую белок экспрессии/процессинга трансгена (TEP), расположенную между указанными 5’- и 3’-ITR, и которая находится под контролем промотора; и
по меньшей мере один участок прикрепления к матриксу (MAR), который расположен между 5’- и 3’-ITR; при этом:
указанный белок экспрессии/процессинга трансгена (TEP) представляет собой hSERCA2, hCALR, hSPCA1, hNRF2, hP4HA1, hP4HB, hCOSMC или hPDI;
указанный участок прикрепления к матриксу (MAR) выбран из SEQ ID NOs: 1 (MAR 1-68), 2 (MAR 1_6), 3 (MAR X_S29), 4 (MAR S4), 5 (MAR лизоцима курицы), или представляет собой сконструированный, в частности, перестроенный аналог, последовательность которого по меньшей мере на 80%, 90%, 95%, 98%, 99% или 100% идентична любой из последовательностей SEQ ID NO: с 1 по 5.
2. Рекомбинантная молекула нуклеиновой кислоты по п. 1, которая дополнительно содержит по меньшей мере один трансген, расположенный между указанными 5’- и 3’-ITR, который находится под контролем промотора трансгена.
3. Вектор экспрессии, содержащий рекомбинантную молекулу нуклеиновой кислоты по любому из пп. 1, 2.
4. Вектор по п. 3, отличающийся тем, что указанный вектор содержит единичный элемент MAR или два или более элементов MAR, при этом указанный элемент(ы) расположен между указанными 5’- и 3’-ITR.
5. Вектор по п. 3, отличающийся тем, что указанный вектор содержит два элемента MAR, при этом первый элемент MAR расположен в 3’-5’ направлении от TEP, и второй элемент MAR расположен в 5’-3’ направлении от TEP, при этом указанный первый элемент MAR включает элемент MAR 1_6 или элемент, последовательность которого по меньшей мере на 80%, 90%, 95%, 98%, 99% или 100% идентична последовательности SEQ ID NO: 2, в частности, перестроенные MAR на основе MAR 1-6, более конкретно - элементы, последовательности которых по меньшей мере на 80%, 90%, 95%, 98%, 99% или 100% идентичны SEQ ID NO: 8 (MAR 1_6R2), и второй элемент MAR включает элемент MAR 1-68 или элемент, последовательность которого по меньшей мере на 80%, 90%, 95%, 98%, 99% или 100% идентична последовательности SEQ ID NO: 1.
6. Вектор по п. 3, отличающийся тем, что указанный вектор содержит единичный элемент MAR, при этом указанный единичный элемент MAR расположен в 5’-3’ направлении от TEP, и при этом единичный элемент MAR представляет собой MAR 1-68, или элемент MAR X-29, или элемент, последовательность которого по меньшей мере на 80%, 90%, 95%, 98%, 99% или 100% идентична последовательности SEQ ID NO: 1 или 3, в частности, перестроенный MAR на основе MAR 1-68 или MAR X-29, в частности, элемент, последовательность которого по меньшей мере на 80%, 90%, 95%, 98%, 99% или 100% идентична последовательности SEQ ID NO: 6, 7 или 10 (элементы MAR 1_68R, 1_68R2 или X_29R3) или SEQ ID NO: 9, и предпочтительно представляет собой элемент MAR X-29 или элемент, последовательность которого по меньшей мере на 80%, 90%, 95%, 98%, 99% или 100% идентична последовательности SEQ ID NO: 3.
7. Способ экспрессии трансгена и белка экспрессии/процессинга трансгена (TEP), включающий:
обеспечение рекомбинантной клетки млекопитающего, содержащей трансген, и введение вектора по любому из пп. 3-6 в указанную рекомбинантную клетку млекопитающего с обеспечением экспрессии указанного трансгена и TEP в указанной клетке, причём экспрессия TEP с помощью указанного вектора может повышать экспрессию трансгена в указанной клетке млекопитающего на по меньшей мере 10%, по меньшей мере 20%, по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60% или по меньшей мере 70%.
8. Способ по п. 7, отличающийся тем, что указанный вектор содержит единичный элемент MAR X-29 или последовательность нуклеиновой кислоты, которая по меньшей мере на 80%, 90%, 95%, 98%, 99% или 100% идентична последовательности SEQ ID NO: 3, и при этом через более чем 4, 5, 6, 7, 8, 9, 10, 11, 12, 13 или 14 недель культивирования TEP, экспрессируемый с указанного вектора, повышает экспрессию представляющего интерес гена на по меньшей мере 10%, по меньшей мере 20%, по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60% или по меньшей мере 70%.
9. Рекомбинантная клетка млекопитающего для экспрессии трансгена, содержащая не более чем 20, 15, 10 или 5 рекомбинантных молекул нуклеиновых кислот по п. 1 или 2.
10. Рекомбинантная клетка по п. 9, отличающаяся тем, что указанные рекомбинантные молекулы предпочтительно встроены в геном указанной клетки в виде единичных копий.
11. Рекомбинантная клетка млекопитающего для экспрессии трансгена, содержащая:
а) рекомбинантную молекулу нуклеиновой кислоты по п. 1, где указанная молекула нуклеиновой кислоты встроена в геном указанной клетки млекопитающего; и
б) рекомбинантную молекулу нуклеиновой кислоты, содержащую:
(i) по меньшей мере один представляющий интерес трансген; и
(ii) элемент MAR, где указанный элемент MAR выбран из SEQ ID NOs: 1 (MAR 1-68), 2 (MAR 1_6), 3 (MAR X_S29), 4 (MAR S4), 5 (MAR лизоцима курицы), или представляет собой сконструированный, в частности, перестроенный аналог, последовательность которого по меньшей мере на 80%, 90%, 95%, 98%, 99% или 100% идентична любой из последовательностей SEQ ID NO: с 1 по 5.
12. Способ трансфекции клетки млекопитающего in vitro, включающий:
трансфекцию указанной клетки млекопитающего по меньшей мере одной, по меньшей мере двумя, по меньшей мере тремя или по меньшей мере четырьмя рекомбинантными молекулами нуклеиновой кислоты по любому из пп. 1 и 2.
13. Способ по п. 12, отличающийся тем, что трансфекцию по меньшей мере одной указанной рекомбинантной молекулой нуклеиновой кислоты осуществляют в качестве первой трансфекции, и при этом в качестве второй последующей трансфекции осуществляют трансфекцию нуклеиновой кислотой, кодирующей трансген.
14. Способ по п. 12 или 13, отличающийся тем, что указанная клетка млекопитающего представляет собой клетку хомячка.
15. Набор для осуществления трансфекции клетки, который в одном контейнере содержит по меньшей мере один вектор, содержащий рекомбинантную молекулу нуклеиновой кислоты по любому из пп. 1 и 2, и в дополнительном контейнере содержит инструкцию по применению указанного вектора или векторов.
16. Набор по п. 15, отличающийся тем, что указанный набор дополнительно к указанным двум контейнерам содержит в ещё одном дополнительном контейнере вектор, кодирующий подходящую транспозазу.
17. Применение рекомбинантной нуклеиновой кислоты по любому из пп. 1 или 2 и/или рекомбинантной клетки млекопитающего по любому из пп. 9-11 для повышения встраивания трансгена в геном клетки хозяина и/или экспрессии трансгена in vitro.
18. Рекомбинантная клетка млекопитающего для экспрессии трансгена, которая принадлежит к клеточной линии и содержит:
А) по меньшей мере одну рекомбинантную молекулу нуклеиновой кислоты, кодирующую белок экспрессии/процессинга трансгена (TEP), где указанный TEP представляет собой hSERCA2, hSPCA1 или hNRF2; и
B) рекомбинантную молекулу нуклеиновой кислоты, содержащую:
(i) по меньшей мере один представляющий интерес трансген; и
(ii) элемент MAR.
19. Рекомбинантная клетка млекопитающего по п. 18, где указанный по меньшей мере один трансген экспрессирует терапевтический белок, выбранный из иммуноглобулина, гормона, такого как эритропоэтин, или фактора роста, и где, необязательно, в указанной рекомбинантной клетке млекопитающего интеграция и/или экспрессия трансгена повышена по сравнению с клеткой, не содержащей указанную рекомбинантную(ые) молекулу(ы) нуклеиновой кислоты.
20. Рекомбинантная клетка млекопитающего по любому из пп. 18, 19, отличающаяся тем, что указанная клетка представляет собой первичную стволовую клетку, клетку хомячка, например клетку СНО, или клетку человека, такую как клетка НЕК293.
| US 2004235011 A1, 25.11.2004 | |||
| EP 0743365 A2, 20.11.1996 | |||
| WO 2011033375 A2, 24.03.2011 | |||
| WO 2005010046 A1, 03.02.2005 | |||
| MATASCI M | |||
| et al | |||
| "CHO cell lines generated by PiggyBac transposition", BMC PROCEEDINGS, 2011, v | |||
| Кипятильник для воды | 1921 |
|
SU5A1 |
| Способ очистки нефти и нефтяных продуктов и уничтожения их флюоресценции | 1921 |
|
SU31A1 |
| АДЕНОВИРУСНЫЙ/АЛЬФА-ВИРУСНЫЙ ГИБРИДНЫЙ ВЕКТОР ДЛЯ ЭФФЕКТИВНОГО ВВЕДЕНИЯ И ЭКСПРЕССИИ ТЕРАПЕВТИЧЕСКИХ ГЕНОВ В ОПУХОЛЕВЫЕ КЛЕТКИ | 2005 |
|
RU2394104C2 |
