Область техники
Изобретение относится к биотехнологии, в частности к композиции для бессывороточных сред для увеличения продуктивности клеток млекопитающих при их культивировании, и может быть использовано при суспензионном культивировании клеток млекопитающих, а также для получения рекомбинантных антител IgG1 при экспрессии в клетках млекопитающих.
Уровень техники
В настоящее время разработано и успешно применяется несколько композиций и добавок для бессывороточных сред для культивирования широкого спектра эукариотических клеток, и, в частности, клеток линии СНО и ее производных. Однако необходимо отметить, что различные клеточные линии, полученные из исходной линии СНО, имеют специфический метаболизм и особенности использования питательных элементов. Известно, что при высокой плотности клеток (культуры) продукция рекомбинантных антител может снижаться из-за истощения среды культивирования клетками и вследствие этого рост культуры замедляется и останавливается, а культура погибает. С гибелью культуры останавливается специфическая продукция. Однако для целей промышленного культивирования фармацевтической индустрии требуется оптимизация уровня экспрессии иммуноглобулинов в СНО клетках. Некоторые авторы также сообщают об увеличении продукции терапевтических белков в клетках СНО и улучшении роста клеток СНО при внесении в бессывороточную среду культивирования добавок, представляющих собой гидролизаты белков растительного происхождения, в частности, - гидролизата белка риса (G. Bare*, Н. Charlier, L. De Nus, F. Verhoeye, Y-J. Schneider, S. Agathos and P. Thonart (2001) Effects of a Rice Protein Hydrolysate on Growth of CHO Cells and Production of Human Interferon-γ in a Serum-Free Medium. In: Lindner-Olsson E., Chatzissavidou N.,  E. (eds) Animal Cell Technology: From Target to Market. ESACT Proceedings, vol 1. Springer, Dordrecht). Подбор условий внесения гидролизата белка риса в среду культивирования позволил авторам разработать стратегию добавления гидролизата растительного происхождения к бессывороточной среде культивирования (Mols J, Peeters-Joris С, Agathos SN, Schneider YJ (2004) Origin of rice protein hydrolysates added to protein-free media alters secretion and extracellular proteolysis of recombinant interferon-gamma as well as CHO-320 cell growth. Biotechnol Lett. Jul; 26(13):1043-6).
E. (eds) Animal Cell Technology: From Target to Market. ESACT Proceedings, vol 1. Springer, Dordrecht). Подбор условий внесения гидролизата белка риса в среду культивирования позволил авторам разработать стратегию добавления гидролизата растительного происхождения к бессывороточной среде культивирования (Mols J, Peeters-Joris С, Agathos SN, Schneider YJ (2004) Origin of rice protein hydrolysates added to protein-free media alters secretion and extracellular proteolysis of recombinant interferon-gamma as well as CHO-320 cell growth. Biotechnol Lett. Jul; 26(13):1043-6).
По литературным данным гидролизаты дрожжей, как пример гидролизатов неживотного происхождения, способствуют значительному приросту продуктивности клеток линии СНО в бессывороточной среде культивирования (Mathilde Mosser, Isabelle Chevalot, Eric Olmos, Fabrice Blanchard, Romain Kapel, Eric Oriol, Ivan Marc and Annie (March 2013) Combination of yeast hydrolysates to improve CHO cell growth and IgG production Cytotechnology. Aug; 65(4): 629-641). Ряд других исследований подтверждают способность гидролизатов неживотного происхождения увеличивать клеточную плотность в процессе культивирования и специфическую продуктивность линий клеток на основе клеток СНО (Gupta AJ, Gruppen Н, Maes D, Boots JW, Wierenga PA. (2013) Factors causing compositional changes in soy protein hydrolysates and effects on cell culture functionality. J Agric Food Chem. Nov 13; 61(45):10613-25. doi: 10.1021/jf403051z. Epub 2013 Oct 29. Richardson Jl, Shah B, Bondarenko PV, Bhebe P, Zhang Z, Nicklaus M, Kombe MC. (2015) Metabolomics analysis of soy hydrolysates for the identification of productivity markers of mammalian cells for manufacturing therapeutic proteins. Biotechnol Prog. 2015 Mar-Apr; 31(2):522-31. doi: 10.1002/btpr.2050. Epub 2015 Jan 27.)
Известен процесс культивирования клеток животных, продуцирующих сложные белки, в котором один пептон растительного происхождения или комбинация пептонов растительного происхождения подается в клеточную культуру, а также способ снижения токсического эффекта избытка аминокислот во время процесса культивирования клеток животных, продуцирующих сложные белки /СА 2610315, C12N 5/0018, ⋅2006-12-07/.
Известен способ, описывающий культивирование клеток-продуцентов в бессывороточной среде с применением растительных гидролизатов /JP 2014110794 А C12N 9/6437⋅2014-06-19/. Описывает способ крупномасштабного получения полипептида в клетках эукариот, содержащихся в бессывороточной культуральной жидкости, причем указанный способ включает I фазу размножения указанных клеток, где клетки размножаются в первой жидкости для культивирования клеток, и II фазу продуцирования указанных клеток, где клетки присутствуют во второй жидкости для культивирования клеток, где каждая из жидкостей для культивирования клеток содержит гидролизат растительного белка и где соотношение между концентрацией гидролизата растительного белка (С1) в первой жидкости для культивирования клеток, а концентрация гидролизата растительного белка (С2) во второй жидкости для культивирования клеток составляет, по меньшей мере, 1,5:1 (C1:С2).
Известен способ предсказания уровня продукции целевого белка при использовании гидролизатов растительного сырья и дрожжевого экстракта /US 2016327484 A1, G01N 21/6486, 2016-11-10/. Способ прогнозирования производительности характеристики гидролизата растительного или дрожжевого происхождения заключается в том, что растительный или дрожжевой гидролизат образца измеряют с применением двумерной флуоресцентной спектроскопии в виде порошка. Указанный способ содержит следующие этапы представления модели, основанные на заранее определенном значении производственного параметра. Для этой цели применяют учебный комплект, состоящий из заранее определенного производственного интересующего параметра (например, показателя объемной продуктивности, титра вируса или числа клеток) и используются данные флуоресцентной спектроскопии. Данные спектроскопической флуоресценции коррелируют со значениями производственного параметра, представляющего интерес для получения калибровочной модели или модели параметров применения многомерного анализа данных.
В перечисленных способах используется соевый или дрожжевой экстракт, методика получения которых не показана в полном объеме, уже достаточно полно описана в литературе или показатели ростовых характеристик, продуктивности и жизнеспособности недостаточно высоки в сравнении с полученным гидролизатом белка подсолнечника.
Известные питательные среды и ростовые добавки, предлагаемые для культивирования клеток СНО фирмами-производителями «Lonza», «Invitrogen», являются затратными с точки зрения финансов и времени доставки, так как изготавливаются за рубежом и импортируется в Российскую Федерацию. Например, среда CD OptiCHO («Invitrogen», США) предназначена для культивирования клеток линии СНО DG44, уже имеющих устойчивую продукцию рекомбинантного белка. Однако для масштабного биотехнологического производства требуется большое количество среды и добавок, из-за чего возрастает стоимость препаратов на основе антител.
Известна среда IMDM (https://www.sigmaaldrich.com/content/dam/sigma-aldrich/docs/Sigma/Formulation/i2510for.pdf) с определенным химическим составом, используемая для культивирования ряда клеточных культур при дополнении IMDM 10% фетальной сыворотки коров. Известна композиция ITS (Insulin-Transferrin-Selenium Supplement), включающая в себя в определенной концентрации белки инсулин, трансферрин и низкомолекулярный компонент селенит натрия, являющаяся стократным концентратом (https://assets.thermofisher.com/TFS-Assets/LSG/manuals/2672.pdf). Известно сочетание, позволяющее обеспечить рост клеток на среде IMDM при дополнении последней однократной концентрацией добавки ITS. Известен также ряд коммерческих добавок в бессывороточную среду культивирования клеток линии СНО, включающих гидролизаты растительного происхождения. Данные добавки являются комплексными. Состав таких композиций является коммерческой тайной и не раскрывается в публикациях.
Раскрытие изобретения
Техническая проблема, решаемая заявляемым изобретением, заключается в разработке композиций бессывороточной среды культивирования на основе IMDM, что дает возможность использовать их для эффективной экспрессии стабильных клеточных линий продуцентов как IgG1, так и других рекомбинантных белков. Особенно это актуально для клеточных линий, продуцирующих IgG1, которые по причине сложностей в сборке полноценных молекул показывают более низкую продукцию в сравнении с продуцентами других рекомбинантных белков. Данная особенность проявляется в связи со сложностью синтеза длинных белковых последовательностей легкой и тяжелой цепей антитела, входящих в состав полноразмерного антитела, и сборки полноценного антитела из отдельных цепей полноразмерных молекул антитела. Позволяет заменить дорогостоящие питательные среды и добавки для культивирования клеточных линий продуцентов рекомбинантных белков, таких как IgG1.
Техническим результатом, достигаемым настоящим изобретением, является повышение специфической продуктивности при культивировании клеточных линий-продуцентов антител на основе СНО клеток в бессывороточных средах на основе IMDM.
Техническим результатом, достигаемым при решении заявленной технической проблемы, является повышение уровня продукции линией-продуцентом целевого белка культивирования в среду культивирования при культивировании на бессывороточных средах, что дает возможность использовать данную среду культивирования для эффективной экспрессии целевого белка культивирования в среду культивирования. Под термином «гидролизат» предполагается смесь компонентов (пептидов и аминокислот), полученных в результате проведения реакции гидролиза белковых порошков растений.
Для решения заявленной технической проблемы и обозначенного технических результатов предлагается композиция для повышения специфической продуктивности клеточных линий-продуцентов антител на основе СНО клеток, содержащая IMDM и L-глутамин, Pluronic F-68, инсулин, трансферрин, селенит натрия, и гидролизата белков подсолнечника, при следующем соотношении исходных компонентов на 1 литр:
L-глутамина - 1,168 г,
Pluronic F-68 - 1 г,
инсулин - 0,01 г,
трансферрин - 0,0055 г,
селенит натрия - 0,0000067 г,
гидролизат белка подсолнечника 0,07 г,
среда IMDM - остальное до одного литра.
При этом компоненты, входящие в состав гидролизата белка подсолнечника, имеют молекулярную массу не более 5 кДа.
Предлагается также способ получения указанной композиции, включающий этап приготовления белков-растворов, экстрагирование белков, гидролиз, центрифугирование, при этом на этапе приготовления белков-растворов смешивают исходные компоненты, приготовление растворов белков-субстратов осуществляют в буфере, экстрагированный белок отделяют центрифугированием с последующим качественным и количественным анализом и подвергают гидролизу с папаином, останавливают реакцию с последующим количественным анализом, выпавший осадок удаляют, а экстрагированный белок фильтруют на мембране для отсечения молекул с молекулярной массой выше 5 кДа.
Для получения композиции используют четырехкратный раствор исходных компонентов, состоящий из 0,004 М ЭДТА⋅Na2 (рН 6,2); 2 М NaCl; 0,05 М Cys⋅HCl; 4Х кратный 0,004 М ЭДТА.
Буферные растворы гидролизата белка подсолнечника используют с исходной концентрацией 0,7 мг/мл.
Приготовление растворов белков-субстратов осуществляют в буфере 0.001 М ЭДТА⋅Na2 при рН 6,2, в концентрации 10-25 мг/мл, экстракцией в течение 8 часов при комнатной температуре на качалке (150 об/мин).
Центрифугирование осуществляют при 12 тыс. об/мин в течение 40 мин.
Для качественного анализа определяют содержание белка по методу Брэдфорд с использованием реактива Bio-Rad, контроль экстрагированных растительных белков осуществляют электрофорезом в 12% ПААГ. Для что качественного контроля отбирают аликвоту белкового раствора, содержащую 30-50 мкг белка, 1/20 часть белкового раствора оставляют для контроля.
Папаин предварительно растворяют в буфере 0.001 М ЭДТА, 0.005 М Cys, 0.06 мМ β-меркаптоэтанол, выдерживают в течение 30 мин перед добавлением в растворы белков-субстратов.
Протеолитическую реакцию останавливают кипячением на водяной бане в течение 10 мин с последующим охлаждением реакционной смеси при +7 С.
Качественный контроль гидролизованных папаином растительных белков осуществляют электрофорезом в 12% ПААГ (восстанавливающие условия) с отбором пробы из реакционной смеси в тех же объемах как после их экстракции из порошка.
Для удаления осадка осуществляют доведение рН реакционной смеси до 7,2 и центрифугирование 12 тыс. об/мин, 40 мин.
Полученную смесь пропускают через ячейку с мембраной для отсечения компонентов с молекулярной массой выше 5 кДа.
Количественно определяют полученный раствор методом Бредфорд, а качественный контроль осуществляют электрофорезом.
Дополнительно для повышения плотности культуры клеток, повышения продолжительности их времени жизни в культуре и продуктивности осуществляют интервальное внесение раствор гидролизата белка подсолнечника. Для интервального внесения используют исходный раствор гидролизата белка подсолнечника в объеме изначального внесения, пересчитанного на объем культивирования.
Изобретение включает разработанные многокомпонентные добавки к базовым бессывороточным средам IMDM (среда Искова IMDM - модификация среды Дульбекко) или DMEM, которые включают в себя раствор гидролизата белка подсолнечника. Данное изобретение увеличивает продуктивность суспензионной культуры, повышает плотность культуры, увеличивает продолжительности времени культивирования и повышает уровень экспрессии рекомбинантных антител класса IgG1.
Данное решение было найдено при использовании композиций к базовой бессывороточной среде. В качестве базовой бессывороточной среды использовали среду IMDM производства «Gibco», США; «БиолоТ», РФ и «ПанЭко», РФ.
Заявляемая композиция для предотвращения агрегирования, повышения плотности и однородности культуры, повышению продуктивности включает растворы гидролизата белка подсолнечника с исходной концентрацией 0,7 мг/мл.
Наилучший результат достигается при использовании композиции, содержащей гидролизат белка подсолнечника, при этом раствор гидролизата белка подсолнечника содержится в исходной концентрации 0,7 мг/мл.
Впервые было установлено, что использование предлагаемой многокомпонентной добавки к базовой бессывороточной среде позволяет повысить плотность, время культивирования и продуктивность клеточных линий-продуцентов рекомбинантных антител клеток при их суспензионном культивировании.
Краткое описание чертежей
Изобретение иллюстрируют следующие графические материалы:
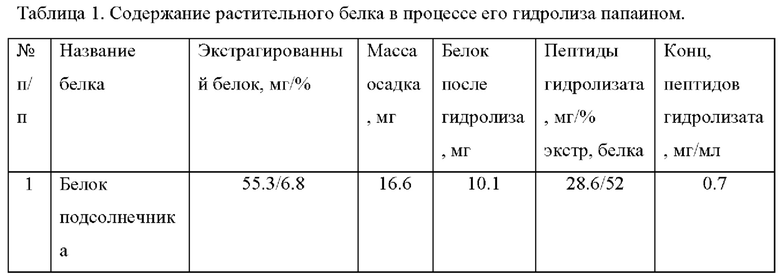
Фиг. 1. Гистограмма зависимости концентрации и жизнеспособности линии клеток СНО DG44, продуцирующей рекомбинантные моноклональные антитела IgG1 - изотипа, во времени при добавлении гидролизата белка подсолнечника в среду культивирования IMDM с добавлением однократного ITS (инсулина, трансферрина и селенита натрия). Каждая из сред была дополнена 8 Мм L-глутамина и 0,1% Pluronic F-68.
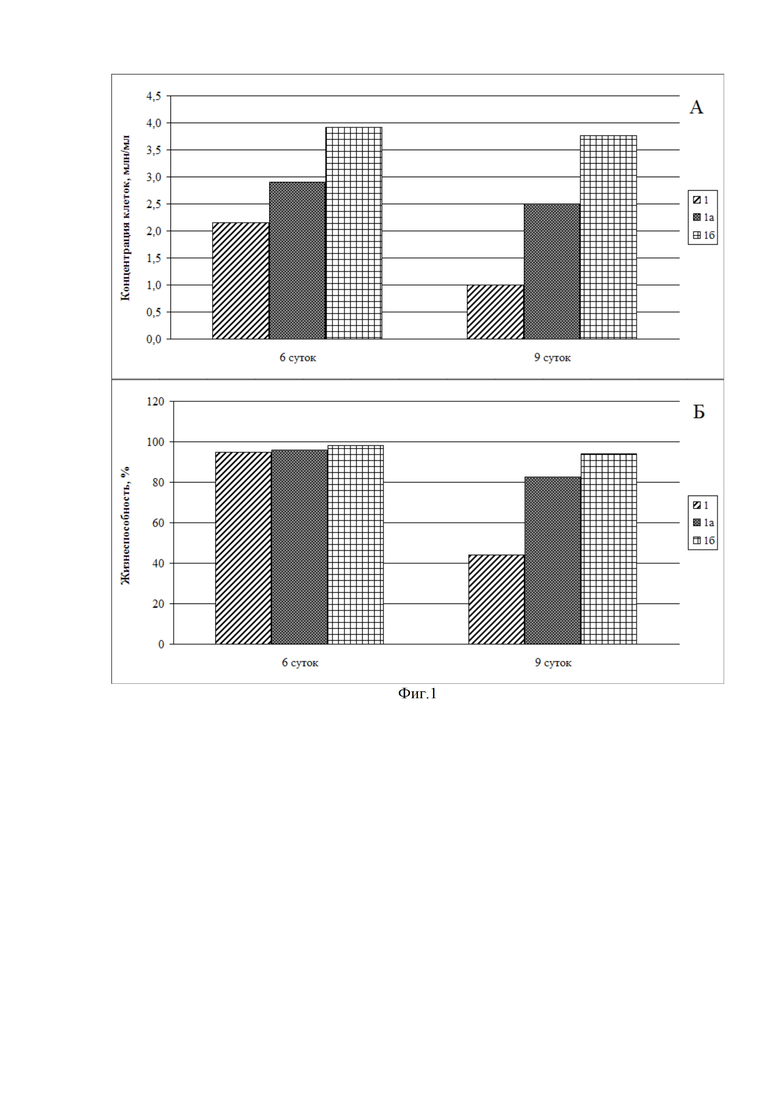
Фиг. 2. Гистограмма зависимости продуктивности рекомбинантных моноклональных антител IgG1-изотипа стабильной клеточной линией-продуцентом СНО DG44 во времени при добавлении гидролизата белка подсолнечника в среду культивирования на основе IMDM. Каждая из сред была дополнена 8 Мм L-глутамина и 0,1% Pluronic F-68.
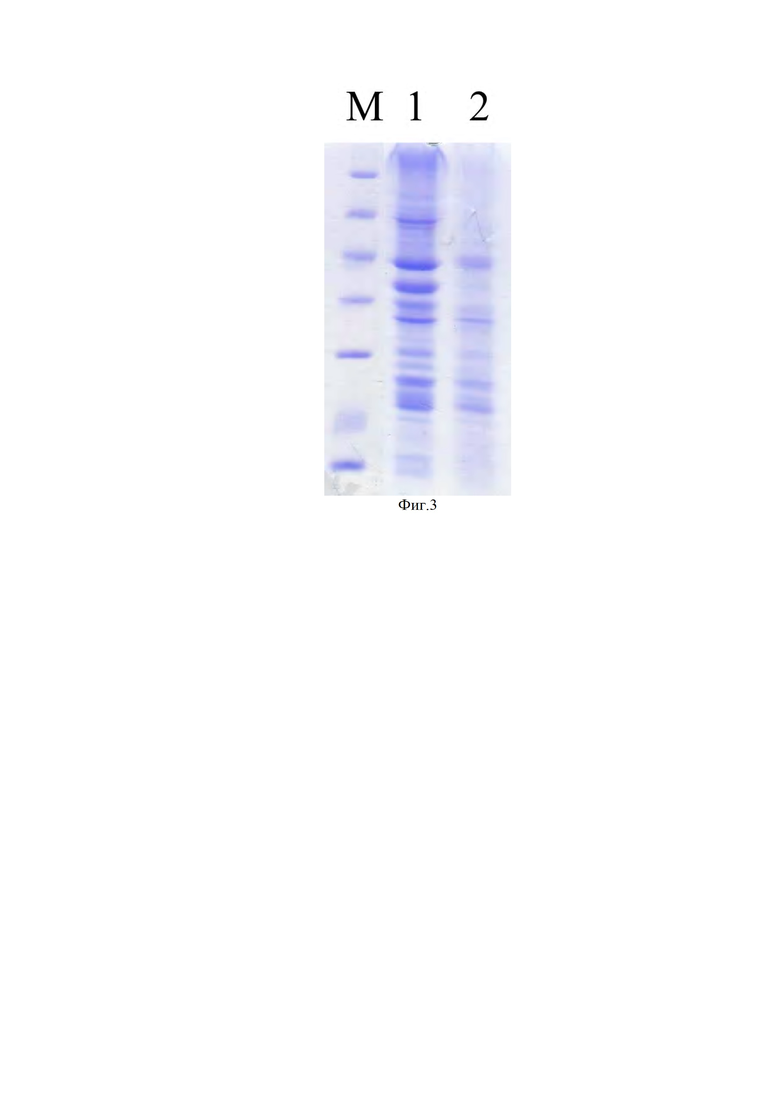
Фиг. 3. Электрофореграмма в 12% ПААГ, в восстанавливающих условиях, водного экстракта белка подсолнечника до (1) и после (2) гидролиза папаином.
Осуществление изобретения
Изобретение иллюстрируется примерами конкретного выполнения, которые более подробно описывают возможность осуществления заявляемого изобретения с достижением заявляемого технического результата.
Методика получения композиции из гидролизата белка подсолнечника, предназначенной для повышения специфической продуктивности клеточных линий-продуцентов антител на основе СНО клеток, следующая.
1. Растворы 4Х кратный 0,004 М ЭДТА⋅Na2 (рН 6,2); 2 М NaCl; 0,05 М Cys⋅HCl; 4Х кратный 0,004 М ЭДТА.
2. Приготовление растворов белков-субстратов (белковых порошков растений) проводили в буфере 0.001 М ЭДТА⋅Na2 (рН 6,2), в концентрации 10-25 мг/мл, экстракцией в течение ночи при комнатной температуре на качалке (150 об/мин).
3. Экстрагированный белок из белковых порошков растений отделяли центрифугированием при 12 тыс. об/мин в течение 40 мин. Далее использовали для гидролиза надосадочную жидкость, где определяли содержание белка по методу Брэдфорд (реактив Bio-Rad). Оценивали содержание экстрагированного белка в порошке. Качественный контроль экстрагированных растительных белков проводили электрофорезом в 12% ПААГ (восстанавливающие условия), отбирая аликвоту белкового раствора, содержащую 30-50 мкг белка. 1/20 часть белкового раствора оставляли для контроля, которую использовали на всех стадиях гидролиза без добавления папаина.
4. Папаин предварительно растворяли в буфере (0.001 М ЭДТА, 0.005 М Cys, 0.06 мМ β-меркаптоэтанол), выдерживали 30 мин (активация папаина) перед добавлением в растворы белков-субстратов.
5. Реакционная смесь (для каждого вида растительного белка в отдельности) содержала белок-субстрат и папаин в соотношении белок : папаин = 1:20 (по количеству белка) в буфере 0,001 М ЭДТА⋅Na2 (рН 6.2) с 0,005 М Cys. Смесь выдерживали в течение 20 ч при 39 С в качалке (150 об/мин). Протеолитическую реакцию останавливали кипячением на водяной бане в течение 10 мин, затем охлаждали реакционную смесь при +7 С.
6. Качественный контроль гидролизованных папаином растительных белков проводили электрофорезом в 12% ПААГ (восстанавливающие условия), отбирая пробы из реакционной смеси в тех же объемах как после их экстракции из порошка.
7. Доводили рН реакционной смеси до 7,2, выпавший осадок удаляли центрифугированием (12 тыс. об/мин, 40 мин). Определяли содержание белка по методу Бредфорд (Bio-Rad) в надосадочной жидкости и оценивали процент гидролиза белка по сравнению с соответствующим контролем без папаина, пропускали ее через ячейку с мембраной для отсечения молекул с молекулярной массой выше 5 кДа.
Количество полученных пептидов и аминокислот в гидролизате (молекулярной массой менее 5 кДа) определяли по количеству гидролизованного белка.
АНАЛИЗ ПОЛУЧЕННЫХ РЕЗУЛЬТАТОВ
В предварительных результатах по гидролизу папаином растворенного порошка подсолнечника в буфере мы установили, что основная часть белка, подвергающегося гидролизу папаином, находится в водной фазе. Более того, твердая фаза порошка частично сорбировала папаин и выводила его из реакции гидролиза.
После экстракции белка из порошка проводили его количественную (Таблица 1) и качественную характеристику (Фиг. 3). Далее после их гидролиза папаином, останавливали реакцию кипячением, доводили рН до 7,2, чтобы в дальнейшем использовать в средах культивирования клеток, выпавший осадок удаляли центрифугированием и дальше не использовали. Отбирали пробы для качественной характеристики гидролизованного белка из супернатанта (надосадочная жидкость). Затем, надосадочную жидкость пропускали через ячейку с мембраной для отсечения молекул с молекулярной массой выше 5 кДа. Для верного подсчета гидролизованного белка (пептидов) осадок, полученный после гидролиза, подсушивали при +65°С, взвешивали и вычитали из количества первоначально экстрагированного белка. Белок, полученный при концентрировании на мембране выше 5 кДа (белок после гидролиза, таблица), также вычитали из количества первоначально экстрагированного белка. Белок, который составил разницу в результате вычитания осадка и белка с молекулярной массой более 5 кДа, мы отнесли гидролизованному белку на низкомолекулярные пептиды и аминокислоты, входящие в гидролизат, который использовали в качестве добавок неживотного происхождения в исследовании их влияния на пролиферативную активность клеток-продуцентов.
8. По результатам качественного контроля (Фиг. 3) мы отметили, что экстрагированные белки подсолнечника равномерно гидролизуются на низкомолекулярные пептиды и аминокислоты: полностью исчезают только белки с молекулярной массой. 66 кДа, 40 и 35 кДа.
В соответствии с указанной выше методикой получают композиции на основе гидролизата белков подсолнечника, имеющую следующее компонентов:
Водный раствор L-глутамина (200 мМ) - 40 мл;
Водный раствор Pluronic F-68 (10%) - 10 мл;
Комплексная белковая добавка ITS - 10 мл;
Буферный раствор гидролизата белков подсолнечника (0,7 мг/мл) - 100 мл (состав подтвержден разницей между образцами, прошедшими процедуру гидролиза и не пошедшими таковой, а также фильтрованием через полупроницаемую мембрану с порами, не пропускающими частицы крупнее 5 кДа);
Среды IMDM до одного литра (850 мл).
Пример 1. Повышение концентрации и жизнеспособности клеток стабильной клеточной линии СНО DG44, продуцирующей рекомбинантные моноклональные антитела IgG1-изотипа, при добавлении в бессывороточную среду культивирования композиции на основе гидролизата белка подсолнечника, содержащая IMDM и L-глутамин, ITS, Pluronic F-68 и гидролизата белков подсолнечника, при следующем соотношении компонентов на 1 литр:
L-глутамина - 1,168 г,
Pluronic F-68 - 1 г,
инсулин - 0,01 г,
трансферрин - 0,0055 г,
селенита натрия - 0,0000067 г,
гидролизат белка подсолнечника 0,07 г
среда IMDM - остальное до одного литра.
В опыте используют суспензионную стабильную клеточную линию - продуцент рекомбинантных антител IgG1-изотипа, на основе клеточной линии СНО DG44 (dhfr-/-) (клетки яичников китайского хомячка). Клеточные культуры выращивают на бессывороточной базовой среде IMDM ("БиолоТ", РФ). Для определения влияния на продуктивность, плотность и жизнеспособность клеточных культур культивирования, эксперимент проводят в формате культур в 6-луночных планшетах в стандартных условиях выращивания при 37°С и 8% CO2 и 96% влажности при непрерывном перемешивании со скоростью 130 об/мин.
Для сравнения эффекта влияния на рост клеточных культур-продуцентов антител IgG1-изотипа в качестве дополнительного источника питательных веществ, используют гидролизат белка подсолнечника и добавку инсулина-трансферрин-селенита натрия (ITS) фирмы «Gibco». Стандартная добавка ITS обычно применяется для повышения плотности культуры и стимулирования роста, однако является недешевым реагентом, что повышает затраты на производство терапевтических иммуноглобулинов.
Для изучения влияния на рост СНО клеток, в среду для культивирования в IMDM добавляют гидролизат белка подсолнечника: в исходной концентрации 0,7 мг/мл. Раствор гидролизата может быть добавлен более одного раза в пропорции 1:10 от объема культуры, в которую он добавляется.
В качестве контроля используют культуру клеток, культивируемую в среде с добавлением инсулина-трансферрина-селенита (ITS). Увеличение плотности клеточной культуры происходит на 5-й день культивирования, причем увеличение роста находится в зависимости от количества разовых подкормок культуры. На Фиг. 1А показано увеличение плотности культуры клеток при добавлении гидролизата подсолнечника в среду IMDM с ITS через 6 суток по сравнению с контрольным образцом. Плотность культуры линии клеток СНО DG44 (dhfr-/-), продуцирующей рекомбинантные моноклональные антитела IgG1-изотипа, увеличивалась в 1,3 раза, после однократного добавления, через 24 ч после инокуляции культуры, в среду IMDM с ITS раствора гидролизата белка подсолнечника в исходной концентрации 0,7 мг/мл, в пропорции 1:10 от объема культуры. Плотность культуры линии клеток СНО DG44 (dhfr-/-), продуцирующей рекомбинантные моноклональные антитела IgG1-изотипа, увеличивалась в 1,8 раза, после троекратного добавления каждые 48 ч после первого добавления раствора, в базовую среду IMDM с ITS раствора гидролизата белка подсолнечника в исходной концентрации 0,7 мг/мл, в пропорции 1:10 от объема культуры.
Применение в качестве ростовой добавки раствора гидролизата белка подсолнечника показывает также его пролонгированное действие на поддержание плотности клеточной культуры и времени роста в течение 9 дней в среде IMDM с ITS. При отсутствии гидролизата белка подсолнечника в среде IMDM с ITS культивирования на 9-10-й день культивирования происходит существенное уменьшение плотности клеток и практическая гибель клеточной культуры (Фиг. 1А).
На Фиг. 1Б продемонстрировано влияние гидролизата белка подсолнечника в исходной концентрации 0,7 мг/мл на жизнеспособность линии клеток СНО DG44 (dhfr-/-), продуцирующей рекомбинантные моноклональные антитела IgG1-изотипа, через 6 и 9 суток. Измерение концентрации клеток и их жизнеспособности проводят с использованием красителя 0,4% раствора в воде трипанового синего в камере Горяева. Во всех образцах этих клеток через 9 суток культивирования на среде IMDM с ITS наблюдали высокую степень жизнеспособности: свыше 82,8% и 94% для однократного и троекратного добавления раствора гидролизата белка подсолнечника в среду культивирования, соответственно (Фиг. 1Б).
Пример 2. Влияние композиции раствора гидролизата белка подсолнечника на увеличение продуктивности стабильной клеточной линии СНО DG44 (dhfr-/-), продуцирующей рекомбинантные моноклональные антитела формы IgG1-изотипа
В опыте используют суспензионную стабильную клеточную линию - продуцент рекомбинантных антител IgG1-изотипа, на основе клеточной линии СНО DG44 (dhfr-/-) (клетки яичников китайского хомячка). Клеточные культуры выращивают на бессывороточной базовой среде IMDM ("ПанЭко", РФ). Для определения влияния на продуктивность, плотность и жизнеспособность клеточных культур культивирования, эксперимент проводят в формате статических культур в 6-луночных планшетах в стандартных условиях выращивания при 37 С и 8% CO2 и 96% влажности при непрерывном перемешивании на орбитальном шейкере с частотой 130 об/мин и диаметром орбиты равным 20 мм.
Для определения влияния на продукцию антител IgG1 в бессывороточную базовую среду IMDM фирмы "ПанЭко" добавляют раствор гидролизата белка подсолнечника в исходной концентрации 0,7 мг/мл в количестве в пропорции 10% от исходного объема среды культивирования. В качестве контрольного образца используют культуру клеток, выращенную в среде IMDM с ITS без добавок.
Продуктивность определяют методом ИФА на 6-й, 9-й и 14-й дни культивирования. На Фиг. 2 показано влияние композиции на увеличение продуктивности по сравнению с контрольным образцом, а также соотношение концентраций рекомбинантного антитела изотипа IgG1 при однократном и троекратном внесении гидролизата белка подсолнечника. Внесение гидролизата в случае однократного внесения производили через 24 часа после высевания культуры клеток в питательную среду, а в случае троекратного внесения - через 24, 72, 120 часов (1, 3, 5 дни культивирования). В контрольный образец вносился буфер, использованный для приготовления гидролизата, рН=7,4, в том же объеме, что и для внесения гидролизата троекратно. Буфер вносился троекратно. При использовании гидролизата белка подсолнечника увеличение концентрации антител по сравнению с контролем составило 1,87 раза для однократного внесения гидролизата и 4,63 раза для его троекратного внесения.
Пример 3. Методика получения гидролизатов неживотного происхождения
Использованные реактивы:
- белок подсолнечника (Biotechnologies, РФ);
- этилендиаминтетрауксусная кислота;
- гидроксид натрия;
- цистеин-HCl;
- хлорид натрия;
- β-меркаптоэтанол;
- мембрана 5 кДа (Millipore, США).
Для оценки уровня экспрессии рекомбинантных белков и степени их очистки необходимы:
- персульфат аммония;
- N,N,N,N'-тетраметилетилендиамин;
- акриламид, бис-акриламид;
- додецилсульфат натрия;
- Кумасси синий R250;
- уксусная кислота;
- этиловый спирт;
- предокрашенный белковый маркер MW 19.0-118.0 кДа (Thermo Scientific, США).
Для очистки гидролизатов от высокомолекулярных примесей необходимы:
- Система ультрафильтрации Amicon 50 с мембраной
- рН-метр (Hanna Instruments, Германия);
- прибор для элетрофореза mini Protein (Bio-Rad, США)
- система высокой очистки воды (Milli-Q, Millipore);
- весы лабораторные;
- рН-метр;
- мешалки магнитные для приготовления буферных растворов;
- центрифуги «микро» и «мини» с возможностью охлаждения до 4°С для концентрирования образцов и центрифугирования растворов (AG Eppendorf Centrifuge 5810 R, Германия).
Таким образом, заявленная композиция на основе гидролизата белков подсолнечника, содержащая IMDM, ITS, L-глутамина и раствор гидролизата, при добавлении бессывороточную среду культивирования позволяет повысить концентрации и жизнеспособности клеток стабильной клеточной линии СНО, увеличить продуктивность стабильной клеточной линии СНО.
| название | год | авторы | номер документа |
|---|---|---|---|
| КОМПОЗИЦИЯ НА ОСНОВЕ ГИДРОЛИЗАТА БЕЛКА ГОРОХА ДЛЯ ПОВЫШЕНИЯ СПЕЦИФИЧЕСКОЙ ПРОДУКТИВНОСТИ КЛЕТОЧНЫХ ЛИНИЙ ПРОДУЦЕНТОВ И СПОСОБ ЕЁ ПОЛУЧЕНИЯ | 2019 |
|
RU2741088C1 |
| Композиция для предотвращения агрегирования и повышения однородности культуры, увеличения продуктивности клеточных линий-продуцентов рекомбинантных белков | 2018 |
|
RU2731988C2 |
| Композиция на основе янтарной кислоты и аспарагина для повышения продуктивности клеточных линий-продуцентов рекомбинантных белков | 2018 |
|
RU2728377C2 |
| СПОСОБ ПОЛУЧЕНИЯ СТАБИЛЬНОЙ КЛЕТОЧНОЙ ЛИНИИ-ПРОДУЦЕНТА РЕКОМБИНАНТНЫХ БЕЛКОВ | 2020 |
|
RU2768735C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pBiPr-ABIgA1FI6-ht ДЛЯ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО ИММУНОГЛОБУЛИНА А ИЗОТИПА IGA1 | 2016 |
|
RU2656142C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pBiPr-ABIgA2m1F16-ht ДЛЯ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО ИММУНОГЛОБУЛИНА А ИЗОТИПА IGA2m1 | 2016 |
|
RU2671477C2 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pBiPr-ABIgA1FI6-Intht ДЛЯ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО ИММУНОГЛОБУЛИНА А ИЗОТИПА IgA1 | 2016 |
|
RU2664184C2 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК, КОДИРУЮЩАЯ ХИМЕРНОЕ АНТИТЕЛО ПРОТИВ ФАКТОРА НЕКРОЗА ОПУХОЛИ-АЛЬФА ЧЕЛОВЕКА, ЛИНИЯ ЭУКАРИОТИЧЕСКИХ КЛЕТОК-ПРОДУЦЕНТ ХИМЕРНОГО АНТИТЕЛА И СПОСОБ ПОЛУЧЕНИЯ ХИМЕРНОГО АНТИТЕЛА | 2013 |
|
RU2555533C9 |
| СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО ИММУНОГЛОБУЛИНА IGA2m1-ИЗОТИПА В КЛЕТКАХ МЛЕКОПИТАЮЩИХ | 2022 |
|
RU2801178C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pOptiVEC-MBP-Fc, КОДИРУЮЩАЯ КОНСТАНТНЫЙ ФРАГМЕНТ ИММУНОГЛОБУЛИНА ЧЕЛОВЕКА, СЛИТНОГО С ФРАГМЕНТОМ ОСНОВНОГО БЕЛКА МИЕЛИНА, ЛИНИИ ЭУКАРИОТИЧЕСКИХ КЛЕТОК - ПРОДУЦЕНТОВ УКАЗАННОГО БЕЛКА И СПОСОБ ПОЛУЧЕНИЯ БЕЛКА MBP-Fc ДЛЯ ТЕРАПИИ РАССЕЯННОГО СКЛЕРОЗА | 2016 |
|
RU2679055C2 |
Группа изобретений относится к биотехнологии. Композиция для повышения специфической продуктивности клеточных линий-продуцентов антител на основе СНО клеток содержит следующие исходные компоненты на 1 л: L-глутамин в количестве 1,168 г, Pluronic F-68 в количестве 1 г, инсулин в количестве 0,01 г, трансферрин в количестве 0,0055 г, селенит натрия в количестве 0,0000067 г, гидролизат белка подсолнечника в количестве 0,07 г и среду IMDM - остальное до 1 л. Способ получения композиции включает следующие этапы. Готовят белки путем смешивания исходных компонентов в виде их растворов в буфере с исходной концентрацией 0,7 мг/мл. Экстрагируют белки и центрифугируют. Осуществляют качественный и количественный анализ и подвергают гидролизу с папаином. Останавливают реакцию с последующим количественным анализом. Выпавший осадок удаляют, а раствор экстрагированного белка фильтруют на мембране для отсечения компонентов гидролизата с молекулярной массой выше 5 кДа. Группа изобретений обеспечивает повышение специфической продуктивности при культивировании СНО клеток в бессывороточных средах на основе IMDM. 2 н. и 13 з.п. ф-лы, 1 табл., 3 пр., 4 ил.
1. Композиция для повышения специфической продуктивности клеточных линий - продуцентов антител на основе СНО клеток, содержащая IMDM и L-глутамин, Pluronic F-68, инсулин, трансферрин, селенит натрия и гидролизата белков подсолнечника, при следующем соотношении исходных компонентов на 1 л:
L-глутамина - 1,168 г,
Pluronic F-68 - 1 г,
инсулин - 0,01 г,
трансферрин - 0,0055 г,
селенита натрия - 0,0000067 г,
гидролизат белка подсолнечника 0,07 г
среда IMDM - остальное до одного литра.
2. Композиция по п. 1, характеризующаяся тем, что компоненты, входящие в состав гидролизата белка подсолнечника, имеют молекулярную массу не более 5 кДа.
3. Способ получения композиции по п. 1, включающий этап приготовления белков-растворов, экстрагирование белков, гидролиз, центрифугирование, при этом на этапе приготовления белков-растворов смешивают исходные компоненты в виде их растворов, приготовление растворов белков-субстратов осуществляют в буфере с исходной концентрацией 0,7 мг/мл, экстрагированный белок отделяют центрифугированием с последующим качественным и количественным анализом и подвергают гидролизу с папаином, останавливают реакцию с последующим количественным анализом, выпавший осадок удаляют, а раствор экстрагированного белка фильтруют на мембране для отсечения компонентов гидролизата с молекулярной массой выше 5 кДа.
4. Способ по п. 3, отличающийся тем, что используют четырехкратный раствор исходных компонентов, состоящий из 0,004 М ЭДТА⋅Na2 (рН 6,2); 2 М NaCl; 0,05 М Cys⋅HCl; 4Х кратный 0,004 М ЭДТА.
5. Способ по п. 3, отличающийся тем, что приготовление растворов белков-субстратов осуществляют в буфере 0,001 М ЭДТА⋅Na2 при рН 6,2, в концентрации 10-25 мг/мл, экстракцией в течение 8 ч при комнатной температуре на качалке (150 об/мин).
6. Способ по п. 3, отличающийся тем, что центрифугирование осуществляют при 12 тыс. об/мин в течение 40 мин.
7. Способ по п. 3, отличающийся тем, что для качественного анализа определяют содержание белка по методу Брэдфорд с использованием реактива Bio-Rad.
8. Способ по п. 3, отличающийся тем, что качественный контроль экстрагированных растительных белков осуществляют электрофорезом в 12% ПААГ.
9. Способ по п. 9, отличающийся тем, что для качественного контроля отбирают аликвоту белкового раствора, содержащую 30-50 мкг белка, а 1/20 часть белкового раствора оставляют для контроля.
10. Способ по п. 3, отличающийся тем, что папаин предварительно растворяют в буфере 0,001 М ЭДТА, 0,005 М Cys, 0,06 мМ β-меркаптоэтанол, выдерживают в течение 30 мин перед добавлением в растворы белков-субстратов.
11. Способ по п. 3, отличающийся тем, что протеолитическую реакцию останавливают кипячением на водяной бане в течение 10 мин с последующим охлаждением реакционной смеси при +7°С.
12. Способ по п. 3, отличающийся тем, что качественный контроль гидролизованных папаином растительных белков осуществляют электрофорезом в 12% ПААГ (восстанавливающие условия) с отбором пробы из реакционной смеси в тех же объемах, как после их экстракции из порошка.
13. Способ по п. 3, отличающийся тем, что для удаления осадка осуществляют доведение рН реакционной смеси до 7,2 и центрифугирование при 12 тыс. об/мин в течение 40 мин.
14. Способ по п. 3, отличающийся тем, что полученную смесь пропускают через ячейку с мембраной для отсечения пептидов с молекулярной массой выше 5 кДа.
15. Способ по п. 3, отличающийся тем, что для повышения плотности культуры клеток, повышения продолжительности их времени жизни в культуре и продуктивности дополнительно осуществляют интервальное внесение раствор гидролизата белка подсолнечника в объеме изначального внесения гидролизата, пересчитанного в пропорции на объем культивирования.
| БАЛАБАШИН Д.С., Анализ базовых сред для культивирования линий клеток и получения рекомбинантных белков, Сборник материалов международных научно-практических конференций, ИП Коротких Алиса Анатольевна, Москва, 2018, с.125-130 | |||
| АЛИЕВ Т.К | |||
| и др., Влияние различных компонентов среды культивирования на продукцию рекомбинантных антител в СНО клетках в |

