Область техники
Изобретение относится к области биотехнологии, в частности предложена рекомбинантная плазмидная ДНК, кодирующая химерное антитело против фактора некроза опухоли-альфа человека.
Уровень техники
В литературе описано несколько работ по экспрессии антител в животных клетках и по созданию культур клеток-продуцентов антител на основе клеток грызунов и приматов, например на основе линии клеток СНО (клетки яичника китайского хомячка).
Известна плазмида pYR-GCEVH, содержащая тяжелую цепь антитела НСАb к раку кишечника человека, и плазмида pYR-GCEVL, содержащая легкую цепь антитела к раку кишечника человека [Xiong H., Ran Y., Xing J., Yang X., Li Y. and Chen Z. Expression vectors for human-mouse chimeric antibodies// Journal of Biochemistry and Molecular Biology. - 2005. - V.38(4). - P.414-419]. Плазмида pYR-GCEVL включает в себя ген устойчивости к неомицину/генетицину под управлением раннего промотора вируса SV40 с удаленным энхансером. Транскрипция гена легкой цепи находится под контролем CMV на 5' конце и терминатора бычьего гормона роста (BGHT), на 3' конце находится сайт полиаденилирования. Плазмида pYR-GCEVH включает в себя ген dhfr под управлением раннего промотора вируса SV40. Транскрипция гена легкой цепи находится под контролем CMV на 5' конце и терминатора бычьего гормона роста (BGHT), на 3' конце находится сайт полиаденилирования.
Культура клеток dhfr (-) СНО (АТСС, США), дефицитных по дигидрофолатредуктазе (ДГФР), была котрансфецирована с использованием равных количеств плазмид pYR-GCEVH, pYR-GCEVL. После проведения нескольких раундов селекции была получена культура, клоны которой имели продукцию химерных антител против рака кишечника человека от 30 до 100 и более мкг/мл кондиционированной среды [Xiong и др., 2005].
Известны плазмиды phCMV-VHRhD-g1C-neo и phCMV-VLRhD-KR-neo, кодирующие антитела против RhD антигена. Плазмида phCMV-VHRhD-g1C-neo содержит вариабельную и константную области тяжелой цепи под контролем главного раннего энхансера цитомегаловируса и промотор начала инициации транскрипции рекомбинантных тяжелых химерных цепей антитела, а также бактериальный ген устойчивости к неомицину под контролем раннего промотора SV40. Плазмида phCMV-VLRhDKR-neo содержит вариабельную и константную области легкой цепи под контролем раннего энхансера цитомегаловируса и промотор начала инициации транскрипции рекомбинантных легких химерных цепей антитела. В данной плазмиде ген устойчивости к неомицину под контролем раннего промотора SV40 был заменен геном dhfr из плазмиды pSV2-dhfr (Subramani S., Mulligan R., Berg P. Expression of the mouse dihydrofolatereductase complementary deoxyribonucleic acid in simian virus 40 vectors //Mol Cell Biol - 1981. - V.1(9). - P.854-864). В обе плазмиды были внесены секреторные последовательности перед вариабельными доменами цепей антител, отделяющими их от интронных последовательностей. После проведения трансфекции клеточной линии СНО, дефицитной по гену ДГФР, и нескольких раундов селекции клонов, полученных из трансформированной культуры CHO-mAb с использованием МТХ (до 1000 нМ) и селективного антибиотика G418 (600 мкг/мл), были получены относительно устойчивые клоны с продукцией до 110 мкг/мл химерных антител [Chusainow J., Yang Y. S, Yeo J.H.M., Toh P.C., Asvadi P., Wong N.S.C., Miranda G.S. Yap. A Study of Monoclonal Antibody-Producing СНО Cell Lines: What Makes a Stable High Producer // Biotechnology and Bioengineering. - 2009. - V.102(4). - P.1182-1196].
Известны плазмиды pOptiVEC-L и pcDNA3.3-L, содержащие в своем составе ген легкой цепи антитела против ФНО-альфа человека [Radko B.V., Boitchenko V.E., Nedospasov S.A., Korobko V.G. Characterization of the genes encoding variable light and heavy chains of the high-affinity monoclonal antibody against human tumor necrosis factor// Russ J Immunol. - 2002. - V.7(4). P. 371-4] под контролем промотора CMV, а также OptiVEC-H, pcDNA3.3-H, содержащие в своем составе ген легкой цепи антитела против ФНО-альфа человека под контролем промотора CMV. Плазмиды pcDNA3.3-L и pcDNA3.3-H содержат в своем составе также ген устойчивости к селективному антибиотику G418, а плазмиды pOptiVEC-L и pOptiVEC-H содержат в своем составе также миниген ДГФР мыши. После трансфекции экспрессирующими векторами pOptiVEC-L и pcDNA3.3-H клеток линии СНО DG44 и селекции в среде без нуклеотидов получен клон клеток, секретирующий химерные антитела против ФНО-альфа человека в среду культивирования с выходом 2 мкг на мл кондиционированной среды. Путем проведения нескольких раундов амплификации в присутствии возрастающих концентраций метотрексата и селективного антибиотика G418 была получена линия-продуцент, стабильно секретирующая химерные антитела против ФНО-альфа человека с выходом до 24,5 мкг/мл [Балабашин Д.С., Зайцева-Зотова Д.С., Топорова В.А., Панина А.А., Марквичева Е.А., Свирщевская Е.В., Алиев Т.К. Способы увеличения продукции рекомбинантных антител в клеточных линиях СНО DG44 // Современные проблемы науки и образования. - 2011. - №5, прототип].
Недостатком данного продуцента, в котором используются 2 различные плазмиды, имеющие в своем составе гены тяжелой и легкой цепей антитела является то, что интегрированные в них гены цепей антител против ФНО-альфа человека находятся в клетке под контролем одинаковых промоторов и терминаторов. Также различные генетические маркеры, позволяющие за счет селективного давления увеличивать продукцию cis-ориентированных генов в клетках-продуцентах, имеют различную активность, что может влиять на количество экспрессируемого белкового продукта каждой из плазмид. Данное обстоятельство может изменять соотношение легкой и тяжелой цепей и приводить к токсичности белкового продукта для клеток-продуцентов. При использовании метода селекции клеточной линии-продуцента с помощью метотрексата увеличение уровня экспрессии будет происходить только для той цепи антитела, которая находится в непосредственной близости от селективного маркера, гена ДГФР.
Раскрытие изобретения
Задачей, решаемой авторами настоящего изобретения, является создание технологии для ускорения и повышения экономичности процесса получения рекомбинантных химерных антител против ФНО-альфа человека.
Технический результат достигается за счет увеличения уровня биологического синтеза антител к ФНО-альфа клетками-продуцентами при использовании одного селективного агента (метотрексата) вместо двух.
Поставленная задача решается путем конструирования рекомбинантной плазмидной ДНК, названной авторами настоящего изобретения pBiPr-ABTNF, содержащей:
- фрагмент плазмиды pOptiVEC™-TOPO®, включающий промотор/энхансер ранних генов цитомегаловируса человека (CMV), внутренний сайт связывания рибосом (IRES) вируса энцефаломиокардита (EMCV), ген ДГФР, сигнал полиаденилирования тимидинкиназы вируса герпеса, сайт начала репликации в Е.coli из плазмиды pUC, ген β-лактамазы, а также следующие модификации - единичный сайт узнавания рестриктазы MluI вместо сайта узнавания рестриктазы SalI в положении 23, единичные сайты узнавания рестриктаз NheI и XhoI после промотора CMV для клонирования ДНК легкой цепи антитела;
- фрагмент ДНК, включающий фланкированную сайтами рестрикции NheI и XhoI ДНК легкой цепи химерного антитела к фактору некроза опухоли-альфа человека под контролем промотора/энхансера ранних генов цитомегаловируса человека (CMV);
- MluI/MluI - фрагмент ДНК, содержащий ДНК тяжелой цепи химерного антитела к фактору некроза опухоли-альфа человека под контролем гибридного промотора hEF1-HTLV и сигнал полиаденилирования.
Частным вариантом настоящего изобретения является упомянутая выше рекомбинантная плазмидная ДНК, характеризующаяся тем, что указанная ДНК тяжелой цепи химерного антитела к фактору некроза опухоли-альфа человека кодирует тяжелую цепь с аминокислотной последовательностью SEQ ID NO:1.
Частным вариантом настоящего изобретения является упомянутая выше рекомбинантная плазмидная ДНК, характеризующаяся тем, что указанная ДНК тяжелой цепи химерного антитела к фактору некроза опухоли-альфа человека имеет нуклеотидную последовательность SEQ ID NO:2.
Частным вариантом настоящего изобретения является упомянутая выше рекомбинантная плазмидная ДНК, характеризующаяся тем, что указанная ДНК легкой цепи химерного антитела к фактору некроза опухоли-альфа человека кодирует легкую цепь с аминокислотной последовательностью SEQ ID NO:3.
Частным вариантом настоящего изобретения является упомянутая выше рекомбинантная плазмидная ДНК, характеризующаяся тем, что указанная ДНК легкой цепи химерного антитела к фактору некроза опухоли-альфа человека имеет нуклеотидную последовательность SEQ ID NO:4.
Частным вариантом настоящего изобретения является упомянутая выше рекомбинантная плазмидная ДНК, характеризующаяся тем, что указанный промотор/энхансер ранних генов цитомегаловируса человека (CMV) имеет нуклеотидную последовательность SEQ ID NO:5.
Частным вариантом настоящего изобретения является упомянутая выше рекомбинантная плазмидная ДНК, характеризующаяся тем, что указанный гибридный промотор hEF1-HTLV имеет нуклеотидную последовательность SEQ ID NO:6.
Также настоящее изобретение представляет способ получения линии эукариотических клеток-продуцентов химерного антитела против фактора некроза опухоли-альфа человека путем трансфекции клеток упомянутой выше рекомбинантной плазмидной ДНК.
Частным вариантом настоящего изобретения является указанный способ получения, характеризующийся тем, что для получения линии эукариотических клеток-продуцентов используют линию клеток яичника китайского хомячка СНО.
Частным вариантом настоящего изобретения является указанный способ получения, характеризующийся тем, для получения линии эукариотических клеток-продуцентов используют линию клеток яичника китайского хомячка СНО.
Также настоящее изобретение представляет линию эукариотических клеток-продуцент химерного антитела против фактора некроза опухоли-альфа человека, полученную путем трансфекции упомянутой выше рекомбинантной плазмидной ДНК.
Частным вариантом настоящего изобретения является указанная линия клеток, представляющая собой линию клеток яичника китайского хомячка СНО, полученную путем трансфекции линии клеток СНО DG44 упомянутой выше рекомбинантной плазмидной ДНК.
Также настоящее изобретение представляет способ получения химерного антитела против фактора некроза опухоли-альфа человека, включающий:
- культивирование в питательной среде упомянутой выше линии клеток,
- выделение полученного целевого белка из культуральной жидкости.
В частном варианте воплощения настоящего изобретения указанная плазмида может представлять собой плазмиду, кодирующую полипептиды со свойствами тяжелой и легкой цепей химерного антитела к ФНО-альфа человека с молекулярной массой 2,83 Md (7,514 т.п.о.), и, например, состоять из:
фрагмента плазмиды pOptiVEC™-TOPO® длиной 4,402 т.п.о., включающего промотор/энхансер ранних генов цитомегаловируса человека (CMV), обеспечивающий высокий уровень экспрессии целевых белков в клетках млекопитающих, внутренний сайт связывания рибосом (IRES) вируса энцефаломиокардита (EMCV) для кэп-независимой трансляции гена ДГФР, ген ДГФР для ауксотрофной селекции трансфецированных клеток СНО DG44 и геномной амплификации стабильных клеточных линий с использованием метатрексата, сигнал полиаденилирования тимидинкиназы вируса герпеса для правильной терминации и процессинга рекомбинантных транскриптов, сайт начала репликации в Е.coli из плазмиды pUC, ген β-лактамазы, а также следующие модификации - единичный сайт узнавания рестриктазы MluI вместо сайта узнавания рестриктазы SalI в положении 23, единичные сайты узнавания рестриктаз NheI и XhoI после промотора CMV для клонирования кДНК легкой цепи антитела;
NheI/XhoI - фрагмента ДНК длиной 0,767 т.п.о., включающего фланкированную сайтами рестрикции NheI и XhoI кДНК легкой цепи химерного антитела к ФНО-альфа человека;
MluI/MluI - фрагмента ДНК длиной 2,325 т.п.о., включающего гибридный промотор hEF1-HTLV из плазмиды pMG, ген тяжелой цепи химерного антитела к ФНО-альфа человека и сигнал полиаденилирования гена фактора роста быка BGH polyA из плазмиды pBudCE4.1
Наличие в плазмиде pBiPr-ABTNF активного гена ДГФР, находящегося под контролем IRES, позволяет проводить селекцию и амплификацию чужеродных последовательностей, интегрированных в геном клетки СНО DG44 (dhfr-/- вариант линии клеток СНО-K1), в среде, содержащей метотрексат. Сочетание процессов трансфекции, селекции и амплификации позволяет получить линию клеток продуцентов CHO-S11, содержащую в своем геноме интегрированные копии генов химерного антитела против ФНО-альфа человека и стабильно секретирующую рекомбинантные антитела в культуральную жидкость. Наличие генов тяжелой и легкой цепей антитела в составе одной плазмиды позволяет осуществлять одновременную амплификацию генов легкой и тяжелой цепей рекомбинантного антитела, встроенных в геном клеток СНО, при воздействии метотрексатом.
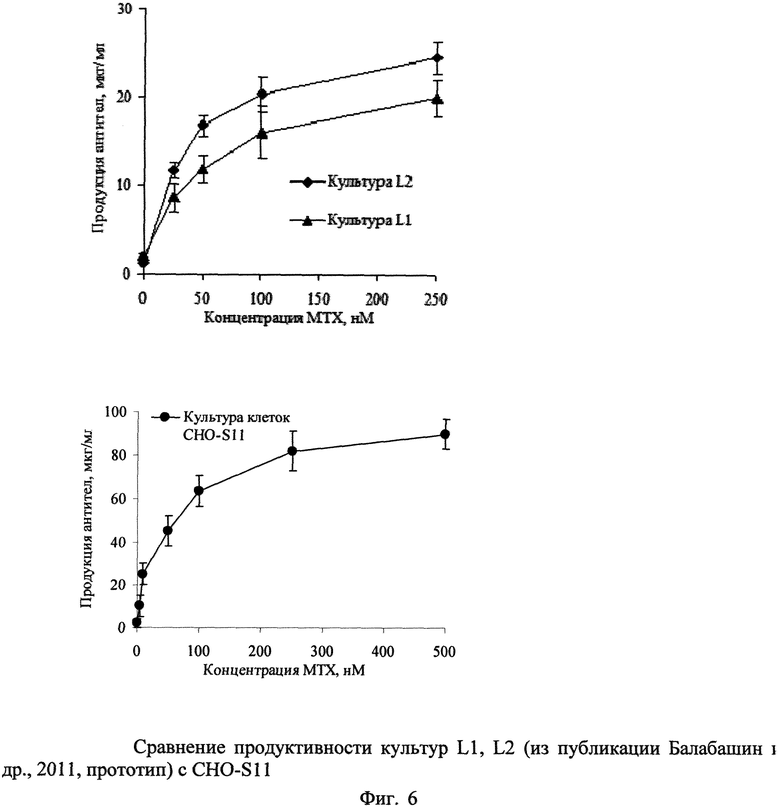
Полезность предлагаемого изобретения определяется тем, что культура, трансфецированная плазмидой pBiPr-ABTNF, значительно быстрее проходит процедуру селекции с использованием метотрексата в концентрации до 500 нМ в сравнении с культурой, трансфецированной двумя отдельными плазмидами, которой необходимы котрансфекция и дополнительный этап селекции с помощью добавления 500 мкг на мл питательной среды селективного антибиотика генетицина (G418). Линия-продуцент CHO-S11 отличается от линии клеток СНО DG44 по признакам, передаваемым последовательностями ДНК, использованными для трансфекции, т.е. является устойчивой к высоким дозам метотрексата (500 нМ) и синтезирует рекомбинантные антитела к ФНО-альфа человека с выходом до 90 мг/л культуральной среды.
Продукция антитела может быть осуществлена в эукариотических клетках. Примером эукариотической клетки, пригодной для продукции полноразмерного антитела, согласно настоящему изобретению являются, но не ограничиваются ими, клетки яичников Cricetulus griseus (СНО). Фраза "клетки яичников Cricetulus griseus" означает, что указанные эукариотические клетки классифицируют как клетки яичников С.griseus (CHO) в соответствии с классификацией, известной специалисту в данной области биотехнологии. Примерами клеток яичников Cricetulus griseus (CHO), применимых в рамках настоящего изобретения, являются, но не ограничиваются ими, клетки яичников Cricetulus griseus (CHO) клетки CHO DG44 (Invitrogen).
Способом согласно настоящему изобретению является способ получения полноразмерного гуманизированного антитела, включающий выращивание трансформированных клеток эукариот в питательной среде и выделение полученных антител из культуральной жидкости. В настоящем изобретении выращивание, накопление и очистка антител из культуральной жидкости может быть осуществлена методом, сходным с традиционными методами ферментации, когда некий белок производится с использованием трансформированных клеток.
Выращивание клеток эукариот осуществляют в атмосфере 5% CO2 в режиме культивирования с перемешиванием в синтетических средах, таких как среда OptiCHO, в течение 3-6 суток.
После выращивания твердые компоненты, такие как клетки, могут быть удалены из культуральной жидкости методом центрифугирования или фильтрации с использованием мембраны, а затем антитела могут быть выделены и очищены методом осаждения с солями, с использованием сульфата натрия или сульфата аммония, аффинной хроматографии, ионообменной хроматографии, и т.п.
Предложенные рекомбинантная плазмида pBiPr-ABTNF и способ получения линии культивируемых клеток CHO, основанный на использовании рекомбинантной плазмиды pBiPr-ABTNF, впервые получены авторами данного изобретения, в научной и патентной литературе не описаны, следовательно, можно сделать вывод о соответствии технического решения критериям изобретения "новизна" и "изобретательский уровень".
Краткое описание чертежей (Фигур)
Так, на Фиг.1 дана физическая карта рекомбинантной плазмиды pBiPr-ABTNF со следующими обозначениями: bla promoter - промотор гена β-лактамазы, BGH polyA - сигнал полиаденилирования бычьего гормона роста, chim H - ген тяжелой цепи химерного антитела к ФНО-альфа, signal - последовательность сигнального пептида, HTLV hEF1 - промотор эукариотического фактора элонгации трансляции 1 альфа, CMV promoter - промотор/энхансер ранних генов цитомегаловируса человека, CMV Forward priming site - сайт отжига праймера CMV Forward, chim L - ген легкой цепи химерного антитела к ФНО-альфа, EMCV IRES reverse primer binding site - сайт отжига праймера EMCV IRES reverse primer, EMCV IRES - внутренний сайт связывания рибосом (IRES) вируса энцефаломиокардита, DHFR - ген дигидрофолатредуктазы, TK polyA - сигнал полиаденилирования тимидинкиназы вируса герпеса, pUC origin - последовательность, отвечающая за репликацию плазмиды pUC, Ampicillin (bla) resistance gene - ген устойчивости к антибиотику ампициллину. MluI, SphI, XhoI, SacII, NheI, NdeI, XbaI, SalI, BglII, PvuI - сайты эндонуклеаз рестрикции.
На Фиг.2 дана структура олигонуклеотидного дуплекса Plink и использованных олигонуклеотидов Plink1 и Plink2 со следующими обозначениями: EcoRI, EcoRV, XhoI - сайты эндонуклеаз рестрикции.
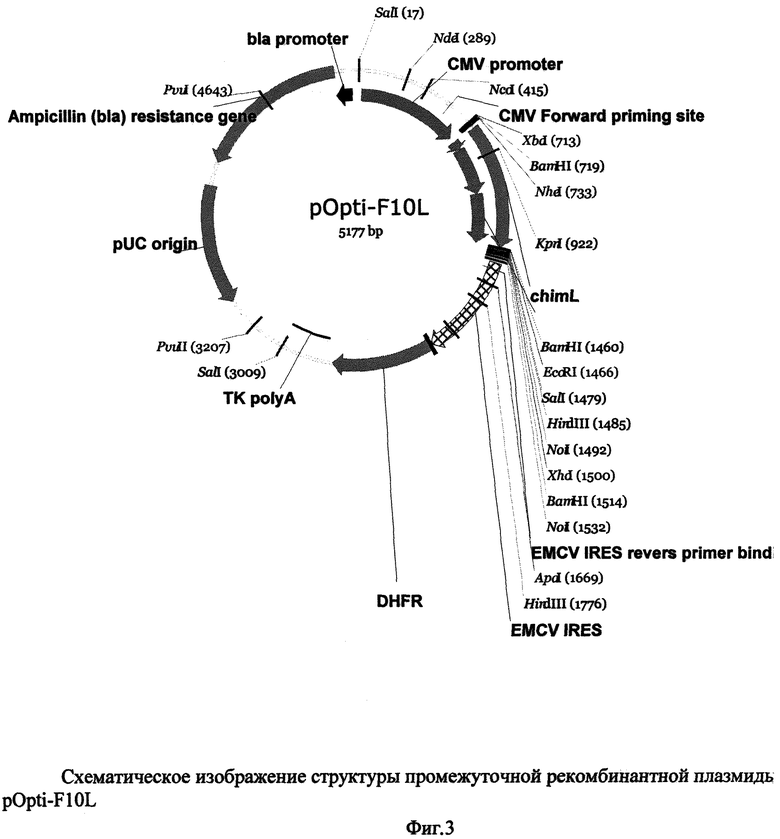
На Фиг.3 дано схематическое изображение структуры промежуточной рекомбинантной плазмиды pOpti-F10L со следующими обозначениями: bla promoter - промотор гена β-лактамазы, CMV promoter - промотор/энхансер ранних генов цитомегаловируса человека, CMV Forward priming site - сайт отжига праймера CMV Forward, chim L - ген легкой цепи химерного антитела к ФНО-альфа, EMCV IRES reverse primer binding site - сайт отжига праймера EMCV IRES reverse primer, EMCV IRES - внутренний сайт связывания рибосом (IRES) вируса энцефаломиокардита, DHFR - ген дигидрофолатредуктазы, ТК polyA - сигнал полиаденилирования тимидинкиназы вируса герпеса, pUC origin - последовательность, отвечающая за репликацию плазмиды pUC, Ampicillin (bla) resistance gene - ген устойчивости к антибиотику ампициллину. SalI, NdeI, NcoI, XbaI, BamHI, NheI, KpnI, EcoRI, SalI, HindIII, NotI, XhoI, ApaI, PvuII, PvuI - сайты эндонуклеаз рестрикции.
На Фиг.4 дана структура области встроенного сайта узнавания рестриктазы MluI со следующими обозначениями: SalI, MluI - сайты эндонуклеаз рестрикции.
На Фиг.5 дана структура ПЦР-фрагментов, содержащих гибридный промотор hEF1-HTLV, фланкированный сайтами узнавания рестриктаз KpnI и MluI на 5'-конце и NheI на 3'-конце (А), и сигнал полиаденилирования гена фактора роста быка BGH polyA, фланкированный сайтами узнавания рестриктаз NheI и XhoI на 5'-конце и MluI и PstI на 3'-конце (Б).
На Фиг.6 дано сравнение продуктивности культур L1, L2 (из публикации Балабашин и др., 2011, прототип) с CHO-S11.
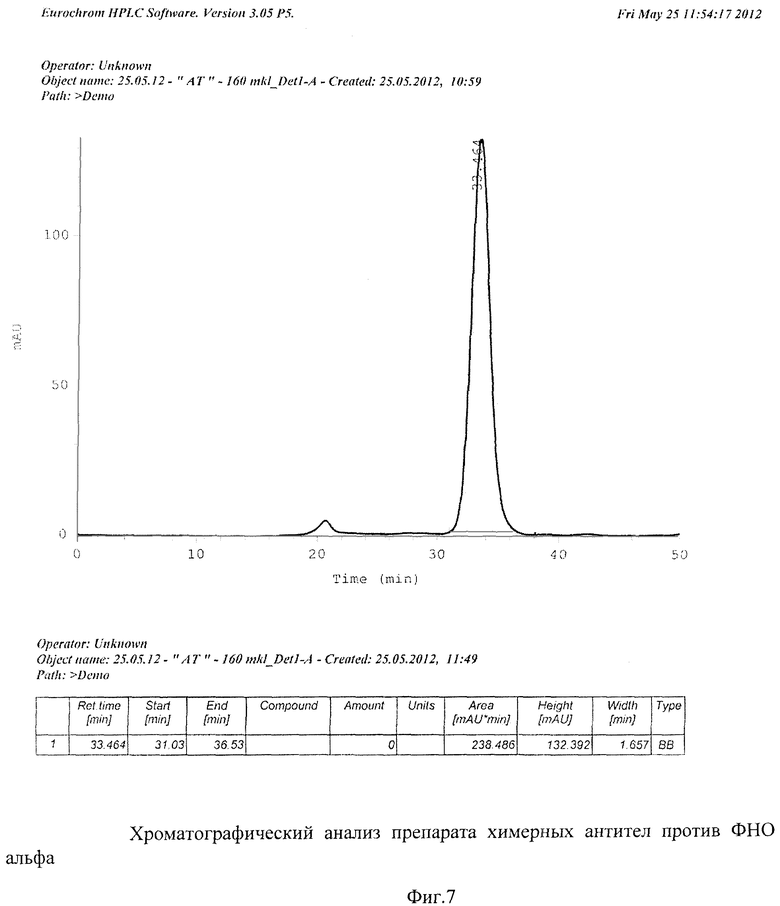
На Фиг.7 дан хроматографический анализ препарата рекомбинантных антител против ФНО-альфа.
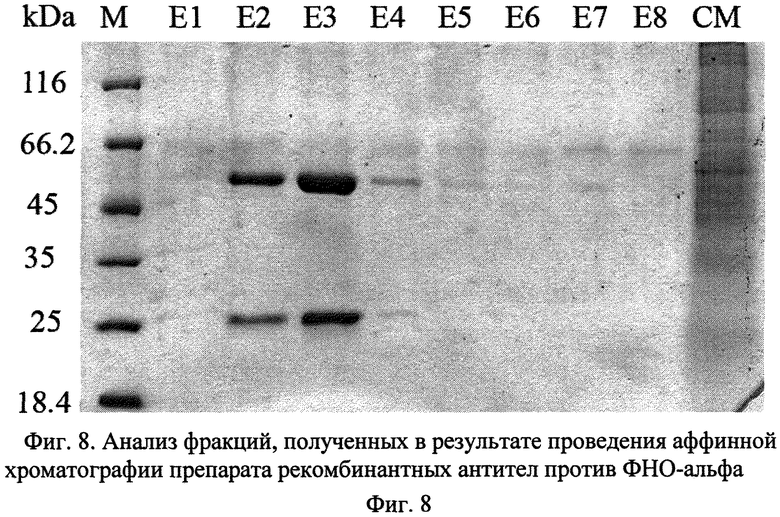
На Фиг.8 дана электрофореграмма фракций выделения химерных антител против ФНО-альфа человека со следующими обозначениями: М - белковые стандарты молекулярного веса; Е1-Е8 - фракции после элюции; CM - образец культуральной среды; kDa - единица молекулярного веса, килодальтон. Ниже следуют примеры осуществления предлагаемого изобретения.
Осуществление изобретения
Пример 1. Конструирование рекомбинантной плазмидной ДНК pBiPr-ABTNF
Пример 1а. Конструирование промежуточной рекомбинантной плазмидной ДНК pOpti-F10L-MluI
В вектор pOptiVEC™-TOPO® из набора pOptiVEC™-TOPO® ТА Cloning Kit (Invitrogen) с помощью синтетического олигонуклеотидного дуплекса Plink (Фиг.2) вводят сайты узнавания рестриктаз NheI и XhoI и после лигирования и трансформации клеток Escherichia coli отбирают клоны с правильной ориентацией дуплекса. Выделяют плазмидную ДНК из клонов и анализируют рестрикционным анализом. Отбирают плазмидные ДНК, содержащие нужный набор рестрикционных фрагментов. Определяют нуклеотидную последовательность отобранных ДНК и окончательно отбирают плазмидные ДНК, в которых нуклеотидная последовательность между промотором CMV и IRES (в сайте ТОРО) содержит последовательность дуплекса Plink в нужной ориентации.
Из экспрессирующей библиотеки кДНК, полученной на основе мРНК мышиной гибридомы F10, продуцирующей антитела к ФНО-альфа человека, были выделены и охарактеризованы клоны, содержащие гены вариабельных областей тяжелой и легкой цепей антитела к ФНО-альфа человека [Radko и др., 2002].
С помощью метода ПЦР на матрице хромосомной ДНК человека получают фрагменты, содержащие гены константных областей легкой (каппа) и тяжелой (IgG1) цепей антитела человека.
Кодирующую последовательность химерной легкой цепи получают методом SOE-ПЦР со специфическими олигонуклеотидными праймерами, с помощью которых на 5'-конец гена вводят сайт узнавания рестриктазы NheI, а на 3'-конец - сайт XhoI. После обработки ПЦР-фрагмента соответствующими рестриктазами кодирующую последовательность легкой цепи химерного антитела F10 клонируют в вектор pOptiVEC с введенными сайтами узнавания рестриктаз NheI и XhoI.
Отбирают плазмидные ДНК, содержащие нужный набор рестрикционных фрагментов. Определяют нуклеотидную последовательность отобранных клонов промежуточной рекомбинантной плазмидной ДНК pOpti-F10L секвенированием по двум цепям по методу Сэнгера. Структура полученной промежуточной плазмиды pOpti-F10L приведена в Фиг.3.
Сайт SalI в позиции 17 плазмиды pOpti-F10L заменяют на сайт MluI для введения гена тяжелой цепи антитела вместе с промотором и сайтом полиаденилирования. Для этого плазмиду pOpti-F10L обрабатывают рестриктазами PvuI и XbaI, продукты реакции разделяют электрофорезом в 1%-ном агарозном геле, фрагменты PvuI-XbaI 1,247 и 3,930 т.п.о. вырезают из геля и выделяют. Фрагмент PvuI-XbaI 3,930 т.п.о. оставляют в качестве вектора. Фрагмент PvuI-XbaI 1,247 т.п.о. обрабатывают рестриктазой SalI, и фрагменты PvuI-SalI 0,551 т.п.о. и SalI-XbaI 0,696 т.п.о. очищают электрофорезом и выделением из геля.
Праймер MluI-1 фосфорилируют и отжигают сам на себя. Фрагменты PvuI-XbaI 3,930 т.п.о., PvuI-SalI 0,551 т.п.о. и SalI-XbaI 0,696 т.п.о. соединяют с олигонуклеотидом MluI-1 при помощи лигазной реакции. Плазмидную ДНК отобранных клонов выделяют и отбирают по наличию сайта узнавания рестриктазы MluI. Окончательно структуру промежуточной рекомбинантной плазмидной ДНК р3б1 подтверждают определением нуклеотидной последовательности в области встроенного фрагмента, содержащего сайт узнавания рестриктазы MluI. Структура места замены показана в Фиг.4.
Пример 1б. Конструирование промежуточной плазмиды pSK-EF1-chimF10H-BGH
Гибридный промотор hEF1-HTLV, фланкированный сайтами узнавания рестриктаз KpnI и MluI на 5'-конце и NheI на 3'-конце, амплифицируют на матрице плазмиды pMG. Прохождение реакции и количество фрагмента контролируют по появлению в продуктах реакции при разделении их электрофорезом в 1,5%-ном агарозном геле фрагмента длиной 0,566 т.п.о. Структура полученного фрагмента показана в Фиг.5.
Полученный в результате ПЦР фрагмент длиной 0,566 т.п.о., содержащий гибридный промотор hEF1-HTLV, обрабатывают рестриктазами NheI и KpnI, из полученного гидролизата выделяют фрагмент ДНК длиной 0,550 т.п.о.
Сигнал полиаденилирования гена фактора роста быка BGH polyA, фланкированный сайтами узнавания рестриктаз NheI и XhoI на 5'-конце и MluI и PstI на 3'-конце, амплифицируют на матрице плазмиды pBudCE4.1. Прохождение реакции и количество фрагмента контролируют по появлению в продуктах реакции при разделении их электрофорезом в 1,5%-ном агарозном геле фрагмента длиной 0,255 т.п.о. Структура полученного фрагмента показана в Фиг.5.
Полученный в результате ПЦР фрагмент длиной 0,255 т.п.о., содержащий сигнал полиаденилирования гена фактора роста быка BGH polyA, обрабатывают рестриктазами NheI и PstI, из полученного гидролизата выделяют фрагмент ДНК длиной 0,247 т.п.о. после электрофоретического разделения в 1,5%-ном агарозном геле.
Плазмидную ДНК pBlueScriptSK+ (Stratagene) обрабатывают рестриктазами KpnI и PstI, из полученного гидролизата выделяют линеаризованную плазмидную ДНК длиной 2,910 т.п.о. после электрофоретического разделения в 0,8%-ном агарозном геле.
Векторную часть pBlueScriptSK+/KpnI-PstI 2,910 т.п.о., фрагмент KpnI-NheI длиной 0,550 т.п.о., содержащий гибридный промотор hEFI-HTLV, и фрагмент NheI-PstI длиной 0,247 т.п.о., содержащий сигнал полиаденилирования гена фактора роста быка BGH polyA, сшивают при помощи лигазной реакции и клонируют. Плазмидную ДНК отобранных клонов выделяют и отбирают рестриктным анализом по наличию двух сайтов узнавания рестриктазы MluI (наличие в продуктах реакции фрагмента длиной 0,785 т.п.о.). Окончательно структуру промежуточной рекомбинантной плазмидной ДНК pSK+/hEF1-HTLV-BGH подтверждают определением нуклеотидной последовательности в области встроенных фрагментов секвенированием по методу Сэнгера с олигонуклеотидными праймерами M13-20 и M13R (состав праймеров приведен в Таблице 1).
С помощью метода ПЦР на 5'- и 3'-конец гена вариабельной области тяжелой цепи антитела вводят сайты узнавания рестриктаз NheI и ApaI, соответственно. Аналогично на 5'- и 3'-конец гена константной области тяжелой цепи антитела человека вводят сайты узнавания рестриктаз ApaI и XhoI. После обработки соответствующими рестриктазами оба фрагмента совместно сшивают с фрагментом длиной 3,701 т.п.о., полученным из предобработанной рестриктазами NheI и XhoI плазмидой pSK+/hEF1-HTLV-BGH при помощи лигазной реакции и клонируют. Плазмидную ДНК полученных клонов выделяют и отбирают рестриктным анализом по наличию двух сайтов узнавания рестриктазы MluI (наличие в продуктах реакции фрагмента ДНК длиной 2,325 т.п.о. по сравнению с 0,785 т.п.о. в случае плазмиды pSK+/hEF1-HTLV-BGH), а также по наличию в продуктах гидролиза рестриктазами NheI и XhoI фрагмента длиной 1,546 т.п.о., содержащего ген тяжелой цепи химерного антитела F10 к ФНО-альфа человека. Окончательно структуру промежуточной рекомбинантной плазмидной ДНК pSK-EF1-chimF10H-BGH подтверждают определением нуклеотидной последовательности в области встроенных фрагментов секвенированием по методу Сэнгера с олигонуклеотидными праймерами M13-20 и M13R (состав праймеров приведен в Таблице 1).
Пример 1в. Конструирование рекомбинантной плазмиды pBiPr-ABTNF
Рекомбинантную плазмидную ДНК р361 обрабатывают рестриктазой MluI, из полученного гидролизата выделяют линеаризованную плазмидную ДНК длиной 5,189 т.п.о. после электрофоретического разделения в 0,8%-ном агарозном геле.
Промежуточную рекомбинантную плазмидную ДНК pSK-EF1-chimF10H-BGH обрабатывают рестриктазой MluI, из полученного гидролизата выделяют фрагмент ДНК длиной 2,325 т.п.о. после электрофоретического разделения в 1%-ном агарозном геле.
Векторную часть плазмидной ДНК р361/MluI длиной 5,189 т.п.о. и MluI/MluI - фрагмент ДНК длиной 2,325 т.п.о., включающий в себя гибридный промотор hEF1-HTLV из плазмиды pMG, ген тяжелой цепи химерного антитела F10 к ФНО-альфа человека и сигнал полиаденилирования гена фактора роста быка BGH polyA из плазмиды pBudCE4.1, сшивают при помощи лигазной реакции и клонируют. Плазмидную ДНК полученных клонов выделяют и отбирают рестриктным анализом по наличию двух сайтов узнавания рестриктазы MluI (наличие в продуктах реакции фрагмента ДНК длиной 2,325 т.п.о. по сравнению с 0,785 т.п.о. в случае плазмиды pSK+/hEF1-HTLV-BGH). Ориентацию встроенного фрагмента определяют с помощью метода ПЦР с олигонуклеотидными праймерами CMVrev1 и HTLV-R. Окончательно структуру рекомбинантной плазмидной ДНК pBiPr-ABTNF подтверждают определением нуклеотидной последовательности секвенированием по методу Сэнгера с олигонуклеотидными праймерами pOpti-4319-4343 и CMVrev1 (состав праймеров приведен в Таблице 1).
Пример 2. Получение линии СНО-продуцента химерных антител к ФНО-альфа человека с применением плазмиды pBiPr-ABTNF
Для получения стабильного продуцента рекомбинантных антител человека проводят трансфекцию клеток яичника китайского хомячка CHODG44 dhfr - плазмидой pBiPr-ABTNF. Клетки культивируют в среде CD DG44 (Invitrogen) с добавлением 200 мМ раствора L-Глутамина до конечной концентрации 8 мМ и содержащей 0,18% (v/v) Pluronic F-68 (Invitrogen). Во флаконы Эрленмейера объемом 125 мл засевают 30 мл клеточной суспензии (4,5 млн клеток) при постоянном помешивании на орбитальном шейкере с частотой 130 об/мин и через 20-24 ч проводят трансфекцию с использованием реагента Freestyle MAX (Invitrogen) [Freestyle MAX Reagent Protocol, http://tools.invitrogen.com/content/sfs/manuals/freestyle_max_reagent_man.pdf]. Плазмидную ДНК добавляют к клеткам в виде ДНК-липосомного преципитата. Преципитат готовят следующим образом: при комнатной температуре во флакон А вносят 18 мкг ДНК плазмиды в 1200 мкл среды OptiPRO SFM (Invitrogen), перемешивают, добавляют 15 мкл реагента Freestyle MAX (Invitrogen), инкубируют от 10 до 20 минут при комнатной температуре. После инкубации перемешивают пипетированием и по каплям вносят в культуральный флакон. Культуральный флакон инкубируют при температуре 37°С, 98% влажности, в атмосфере 8% СО2 и непрерывном перемешивании 130 об/мин.
Через 48 ч после трансфекции клетки отмывают и помещают в ростовую среду CD OptiCHO (Invitrogen) (без тимидина и гипоксантина) с добавлением 200 мМ раствора L-Глутамина до конечной концентрации 8 мМ и содержащей 0,18% (v/v) Pluronic F-68 на 48 ч. Клетки инкубируют в селективной среде в течение двух недель, смену среды и поддержание концентрации клеток в среде проводят каждые трое суток.
Через 14 дней после помещения в селективные условия в культуре остаются клетки, способные существовать без добавления в среду тимидина и гипоксантина. При достижении культурой времени удвоения популяции 24 часа производят полную замену среды на среду CD DG44, содержащей 8 мМ L-Глутамина, 0,18% Pluronic F-68, 50 нМ метотрексата.
Через 14 дней после помещения в среду, содержащую метотрексат, остаются только клетки, способные существовать в присутствии 50 нМ метотрексата. При достижении культурой времени удвоения популяции 24 часа производят полную замену среды на среду CD DG44, содержащую 8 мМ L-Глутамина, 0,18% (v/v) Pluronic F-68, 100 нМ метотрексата.
Данную процедуру увеличения концентрации метотрексата проводят до тех пор, пока не будет достигнута устойчивость клеток к концентрации метотрексата, равной 500 нМ (Фиг.6)
Таким образом, получают культуру CHO-S11, клетки которой стабильно продуцируют антитела к ФНО-альфа человека в культуральную жидкость с выходом до 90 мкг/мл среды при культивировании в условиях перемешивания со скоростью 130 об/мин.
Пример 3. Получение, выделение и очистка белкового продукта с помощью линии клеток СНО, продуцирующей химерные антитела к ФНО-альфа человека
Получение белкового продукта производили в колбах Эрленмейера различного объема без рассекателей, в ростовой среде CD OptiCHO (без тимидина и гипоксантина), с добавлением 200 мМ раствора L-Глутамина до конечной концентрации 8 мМ и 0,18% (v/v) Pluronic F-68, без добавления дополнительных количеств питательных веществ, при постоянном перемешивании с частотой 135 об/мин, при температуре 37°С, 98% влажности, в атмосфере 8% CO2. Культивирование для получения белкового продукта производили не менее 14 дней.
После окончания культивирования для получения продукта химерных антител к ФНО-альфа человека от культуры CHO-S11 полученную кондиционированную среду от культуры CHO-S11 центрифугируют при 4000 об/мин в течение 30 мин. Супернатант смешивают в соотношении 4:1 с Буфером для нанесения (50 мМ Трис-HCl, рН 7,0) и наносят на подготовленную Белок А-агарозную колонку (Gibco BRL, США). При элюции используют Буфер для элюции (100 мМ глицина, рН 3,0) и Нейтрализующий буфер (1 М Трис-HCl, рН 8,0). Колонку с 2 мл Белок А-агарозы промывают 3 раза по 4 мл Буфером для нанесения. Далее наносят смесь супернатанта кондиционированной среды и Буфера для нанесения. Образец наносят при комнатной температуре с помощью перистальтического насоса. После нанесения образца колонку промывают 3 раза по 20 мл Буфером для нанесения. Смывание антител с колонки производят с помощью 6 фракций по 2 мл Буфера для элюции. К образцу добавляют Нейтрализующий буфер в соотношении 1 часть буфера на 9 частей прошедшего через колонку Буфера для элюции.
Полученные образцы выделенных антител диализуют против ФСБ (фосфатно-солевой буфер, 137 мМ NaCl, 2,7 мМ KCl, 10 мМ Na2HPO4, 1,76 мМ KH2PO4, рН 7,4) и хранят при температуре +4°С.
Оценку гомогенности и степени очистки препарата проводят как с использованием электрофоретического метода, так и аналитической эксклюзионной ВЭЖХ.
Препараты рекомбинантных антител, полученных в ходе выделения с помощью аффинной хроматографии на Белок А-агарозе, анализируют на хроматографической ВЭЖХ системе Knauer (Германия), оснащенной колонкой Супердекс-200-10/300GL. Исследование проводят в протекающем буфере А (100 мМ Трис-HCl, 150 мМ NaCl, рН 7,5). Детекцию осуществляют при длине волны 230 нм УФ-детектором 2600 (UV detector 2600).
В Фиг.7 представлена хроматограмма рекомбинантных антител против ФНО-альфа. Элюция антител с колонки происходит в интервале 31-34 мин от момента старта проведения хроматографического анализа. Данные хроматографического анализа свидетельствуют о более чем 98%-ной чистоте полученного препарата рекомбинантных антител.
Электрофорез проводят в 10%-ном полиакриламидном геле. В качестве контроля используют неокрашенный белковый маркер молекулярных весов (Fermentas, Великобритания). Перед внесением к 15 мкл образца антител добавляют 5 мкл Буфера для нанесения с 2-меркаптоэтанолом (2-МЭ) (200 мМ Трис-HCl, рН 6,8, 400 мМ 2-МЭ, 4% натрия додецилсульфат, 0,01% бромофеноловый синий, 40%-ный глицерин). Образцы для внесения на гель инкубируют при температуре 95°С в течение 5 мин. Антитела наносят в Буфере для нанесения, как содержащем 2-МЭ, так и в отсутствие него.
Полученные образцы продукта химерных антител к ФНО-альфа человека характеризуются следующими показателями:
- гомогенность препарата не менее 98% (по данным гель-электрофореза в 10%-ном полиакриламидном геле с денситометрией) (Фиг.8);
- молекулярная масса - 148000 Да (по данным гель-электрофореза с белковыми маркерами молекулярных весов);
- молекулярная масса легкой цепи антитела 24000 Да, молекулярная масса тяжелой цепи антитела 50000 Да (по данным гель-электрофореза с белковыми маркерами молекулярных весов).
| название | год | авторы | номер документа |
|---|---|---|---|
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pBiPr-ABIgA2m1F16-ht ДЛЯ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО ИММУНОГЛОБУЛИНА А ИЗОТИПА IGA2m1 | 2016 |
|
RU2671477C2 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pBiPr-ABIgA1FI6-Intht ДЛЯ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО ИММУНОГЛОБУЛИНА А ИЗОТИПА IgA1 | 2016 |
|
RU2664184C2 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pBiPr-ABIgA1FI6-ht ДЛЯ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО ИММУНОГЛОБУЛИНА А ИЗОТИПА IGA1 | 2016 |
|
RU2656142C1 |
| СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО ИММУНОГЛОБУЛИНА IGA2m1-ИЗОТИПА В КЛЕТКАХ МЛЕКОПИТАЮЩИХ | 2022 |
|
RU2801178C1 |
| Способ получения димерной формы иммуноглобулина IgA1-изотипа в клетках млекопитающих | 2023 |
|
RU2822890C1 |
| Способ получения димерной формы мутантного иммуноглобулина IgA2m1-изотипа в клетках млекопитающих | 2023 |
|
RU2822889C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pOptiVEC-MBP-Fc, КОДИРУЮЩАЯ КОНСТАНТНЫЙ ФРАГМЕНТ ИММУНОГЛОБУЛИНА ЧЕЛОВЕКА, СЛИТНОГО С ФРАГМЕНТОМ ОСНОВНОГО БЕЛКА МИЕЛИНА, ЛИНИИ ЭУКАРИОТИЧЕСКИХ КЛЕТОК - ПРОДУЦЕНТОВ УКАЗАННОГО БЕЛКА И СПОСОБ ПОЛУЧЕНИЯ БЕЛКА MBP-Fc ДЛЯ ТЕРАПИИ РАССЕЯННОГО СКЛЕРОЗА | 2016 |
|
RU2679055C2 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pCLm4/hygro-14D5, КОДИРУЮЩАЯ ПОЛИПЕПТИД СО СВОЙСТВАМИ ЛЕГКОЙ ЦЕПИ ХИМЕРНОГО АНТИТЕЛА ПРОТИВ ВИРУСА КЛЕЩЕВОГО ЭНЦЕФАЛИТА, И РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pCHm2-14D5, КОДИРУЮЩАЯ ПОЛИПЕПТИД СО СВОЙСТВАМИ ТЯЖЕЛОЙ ЦЕПИ ХИМЕРНОГО АНТИТЕЛА ПРОТИВ ВИРУСА КЛЕЩЕВОГО ЭНЦЕФАЛИТА, ХИМЕРНОЕ АНТИТЕЛО, ОБЕСПЕЧИВАЮЩЕЕ ЭКСТРЕННУЮ ПРОФИЛАКТИКУ КЛЕЩЕВОГО ЭНЦЕФАЛИТА У МЫШЕЙ | 2013 |
|
RU2550252C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pСL1, КОДИРУЮЩАЯ ПОЛИПЕПТИД СО СВОЙСТВАМИ ЛЕГКОЙ ЦЕПИ АНТИТЕЛА ЧЕЛОВЕКА ПРОТИВ ВИРУСА ЭБОЛА, РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК рСН1, КОДИРУЮЩАЯ ПОЛИПЕПТИД СО СВОЙСТВАМИ ТЯЖЕЛОЙ ЦЕПИ УКАЗАННОГО АНТИТЕЛА, И ИХ ПРИМЕНЕНИЕ | 2004 |
|
RU2285043C2 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pcL37, КОДИРУЮЩАЯ ПОЛИПЕПТИД СО СВОЙСТВАМИ ЛЕГКОЙ ЦЕПИ АНТИТЕЛА ЧЕЛОВЕКА ПРОТИВ ВИРУСА ОСПОВАКЦИНЫ, РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pcH37, КОДИРУЮЩАЯ ПОЛИПЕПТИД СО СВОЙСТВАМИ ТЯЖЕЛОЙ ЦЕПИ УКАЗАННОГО АНТИТЕЛА, И ИХ ПРИМЕНЕНИЕ | 2006 |
|
RU2317330C2 |






Группа изобретений относится к области биотехнологии. Предложена рекомбинантная плазмидная ДНК, кодирующая химерное антитело против фактора некроза опухолей-альфа человека (ФНО-альфа), на основе плазмиды pOptiVECTM-TOPO®. Рассмотрена линия эукариотических клеток в качестве продуцента антитела против ФНО-альфа, способ ее получения путем трансфекции плазмидной ДНК по изобретению, а также способ получения химерного антитела против ФНО-альфа путем культивирования линии клеток по изобретению. Настоящее изобретение обеспечивает увеличение уровня синтеза антител против ФНО-альфа клетками-продуцентами. 4 н. и 8 з.п. ф-лы, 8 ил., 1 табл., 3 пр.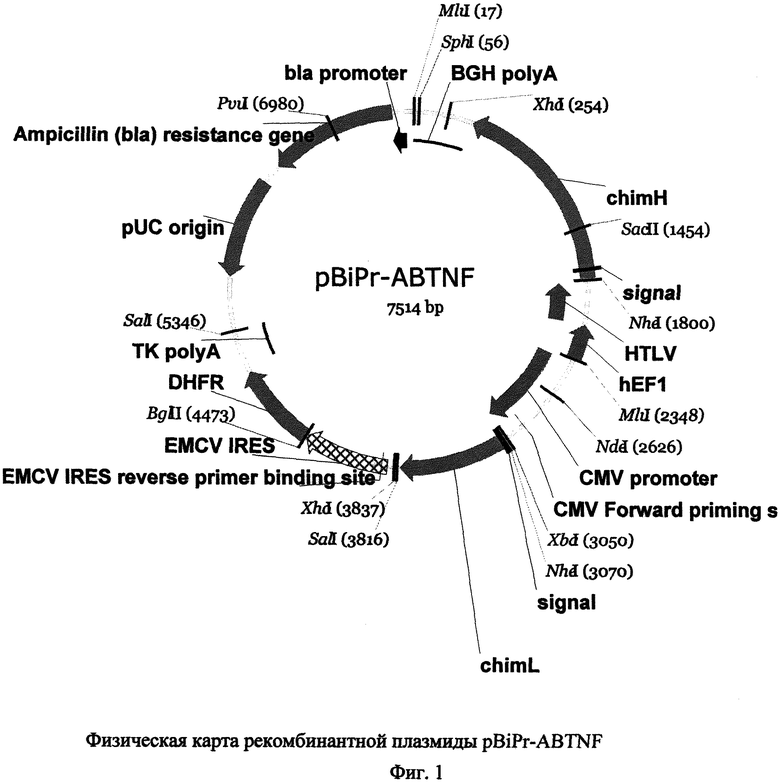
1. Рекомбинантная плазмидная ДНК, кодирующая химерное антитело против фактора некроза опухоли-альфа человека, содержащая:
- фрагмент плазмиды pOptiVEC™-TOPO®, включающий промотор/энхансер ранних генов цитомегаловируса человека (CMV), внутренний сайт связывания рибосом (IRES) вируса энцефаломиокардита (EMCV), ген ДГФР, сигнал полиаденилирования тимидинкиназы вируса герпеса, сайт начала репликации в Е.coli из плазмиды pUC, ген β-лактамазы, а также следующие модификации - единичный сайт узнавания рестриктазы MluI вместо сайта узнавания рестриктазы SalI в положении 23, единичные сайты узнавания рестриктаз NheI и XhoI после промотора CMV для клонирования ДНК легкой цепи антитела;
- фрагмент ДНК, включающий фланкированную сайтами рестрикции NheI и XhoI ДНК легкой цепи химерного антитела к фактору некроза опухоли-альфа человека под контролем промотора/энхансера ранних генов цитомегаловируса человека (CMV);
- MluI/MluI - фрагмент ДНК, содержащий ДНК тяжелой цепи химерного антитела к фактору некроза опухоли-альфа человека под контролем гибридного промотора hEF1-HTLV и сигнал полиаденилирования.
2. Рекомбинантная плазмидная ДНК по п.1, характеризующаяся тем, что указанная ДНК тяжелой цепи химерного антитела к фактору некроза опухоли-альфа человека кодирует тяжелую цепь с аминокислотной последовательностью SEQ ID NO:1.
3. Рекомбинантная плазмидная ДНК по п.1, характеризующаяся тем, что указанная ДНК тяжелой цепи химерного антитела к фактору некроза опухоли-альфа человека имеет нуклеотидную последовательность SEQ ID NO:2.
4. Рекомбинантная плазмидная ДНК по п.1, характеризующаяся тем, что указанная ДНК легкой цепи химерного антитела к фактору некроза опухоли-альфа человека кодирует легкую цепь с аминокислотной последовательностью SEQ ID NO:3.
5. Рекомбинантная плазмидная ДНК по п.1, характеризующаяся тем, что указанная ДНК легкой цепи химерного антитела к фактору некроза опухоли-альфа человека имеет нуклеотидную последовательность SEQ ID NO:4.
6. Рекомбинантная плазмидная ДНК по п.1, характеризующаяся тем, что указанный промотор/энхансер ранних генов цитомегаловируса человека (CMV) имеет нуклеотидную последовательность SEQ ID NO:5.
7. Рекомбинантная плазмидная ДНК по п.1, характеризующаяся тем, что указанный гибридный промотор hEF1-HTLV имеет нуклеотидную последовательность SEQ ID NO:6.
8. Способ получения линии эукариотических клеток-продуцентов химерного антитела против фактора некроза опухоли-альфа человека путем трансфекции клеток рекомбинантной плазмидной ДНК по любому из пп.1-6.
9. Способ получения по п.8, характеризующийся тем, что для получения линии эукариотических клеток-продуцентов используют линию клеток яичника китайского хомячка СНО.
10. Линия эукариотических клеток-продуцент химерного антитела против фактора некроза опухоли-альфа человека, полученная путем трансфекции рекомбинантной плазмидной ДНК по любому из пп.1-7.
11. Линия клеток по п.10, представляющая собой линию клеток яичника китайского хомячка СНО, полученную путем трансфекции линии клеток СНО DG44 рекомбинантной плазмидной ДНК по любому из пп.1-7.
12. Способ получения химерного антитела против фактора некроза опухоли-альфа человека, включающий:
- культивирование в питательной среде линии клеток по любому из пп.10-11,
- выделение полученного целевого белка из культуральной жидкости.
| БАЛАБАШИН Д.С | |||
| и др | |||
| Приспособление для плетения проволочного каркаса для железобетонных пустотелых камней | 1920 |
|
SU44A1 |

