Область техники изобретения
Настоящее изобретение относится к нуклеотидной последовательности для обнаружения толерантного к гербицидам растения сои DBN9004 и к способу его обнаружения, в частности, к соевому растению DBN9004, толерантному к глифосату и глюфосинату, и способу обнаружения того, содержится ли молекула ДНК конкретного трансгенного объекта сои DBN9004 в биологическом образце.
Уровень техники
N-фосфонометилглицин, также известный как глифосат, представляет собой неселективный гербицид длительного системного действия с широким спектром активности. Глифосат является конкурентным ингибитором фосфоенолпировиноградной кислоты (phosphoenolpyruvic acid (PEP)), субстрата 5-енолпирувилшикимат-3-фосфатсинтазы (5-enolpyruvylshikimate-3-phosphate synthase (EPSPS)) при синтезе, и может ингибировать катализируемое EPSPS превращение двух субстратов, а именно, PEP и 3-фосфошикимата, в 5-енолпирувилшикимат-3-фосфокиксимат, блокируя путь биосинтеза шикимовой кислоты, предшественника ароматических аминокислот в пути биосинтеза, тем самым вызывая гибель растений и бактерий путем воздействия на биосинтез белка.
Толерантность к глифосату может быть достигнута путем экспрессии модифицированного EPSPS. Модифицированный EPSPS имеет более низкое сродство к глифосату и поэтому в присутствии глифосата EPSPS сохраняет свою каталитическую активность, обеспечивая толерантость к глифосату.
Соя (Glycine max) является одной из пяти мировых основных сельскохозяйственных культур. Толерантность к гербицидам, особенно, толерантность к глифосатному гербициду, является важной агрономически полезным свойством при производстве сои. Толерантность сои к глифосатному гербициду может быть достигнута с помощью способов трансгенной экспрессии толерантных к гербициду глифосату генов (EPSPS, CP4) в соевых растениях, таких как соевый объект GTS40-3-2 и соевый объект MON89788.
В последние годы широкое распространение толерантных к глифосату сельскохозяйственных систем и все более широкое применение глифосата привели к широкому распространению устойчивых к глифосату сорняков. В районах, в которых фермеры сталкиваются с устойчивыми к глифосату сорняками или сорняками, которые превращаются в труднее уничтожаемые виды сорняков, фермеры могут компенсировать слабое действие глифосата путем смешивания или чередования с другими гербицидами, способными уничтожать не поддающиеся действию глифосата сорняки.
Глюфосинат является несистемным и неселективным гербицидом, относящимся к фосфинотрициновым гербицидам. Главным образом, он используется для обработки почвы после появления всходов с целью уничтожения однолетних или многолетних широколиственных сорняков, и уничтожение сорняков осуществляется за счет необратимого ингибирования L-фосфономицином (активный ингредиент в глюфосинате) глютаминсинтазы (фермент, необходимый для детоксикации аммиака в растениях). В отличие от глифосата, который может убивать корни растений, глюфосинат может сначала убивать листья и попадать в ксилему растений транспирацией, и по скорости действия находится в ряду между паракватом и глифосатом.
L-фосфинотрицин превращается в его неактивную форму посредством ацетилирования, катализируемого фосфинотрицин-N-ацетилтрансферазой (phosphinothricin N-acetyltransferase (PAT)), выделенной из Streptomyces. Гены, экспрессирующие оптимизированные для растений формы PAT, использовались в соевых бобах, чтобы придать соевым бобам, таким как объект сои A5547-127, толерантность по отношению к глюфосинатному гербициду. Таким образом, использование глюфосинатных гербицидов в сочетании с глюфосинат-толерантными растениями может служить неселективным средством для эффективного контроля над устойчивыми к глифосату сорняками.
В будущем, в связи с популяризацией устойчивых к насекомым трансгенных соевых бобов и их крупномасштабными посевами, у небольшого числа выживших насекомых/вредителей может вырабатываться устойчивость после нескольких поколений размножения. Совместная посадка гербицид-толерантных трансгенных соевых бобов в виде обладающих устойчивостью к насекомым трансгенных соевых бобов совместно с не обладающими устойчивостью к насекомым трансгенных соевых бобов в определенном соотношении может задерживать выработку устойчивости к насекомым/вредителям.
Известно, что на экспрессию чужеродных генов в растениях влияет их локализация в геноме растения, вероятно, вследствие структуры хроматина (например, гетерохроматина) или близости регулирующих транскрипцию элементов (например, энхансеров) к сайту интеграции. По указанной причине часто необходимо исследовать большое число объектов, чтобы обнаружить объекты, которые могут быть коммерциализированы (т. е. объекты, в которых введенные целевые гены могут быть оптимально экспрессированы). Например, установлено, что растения и другие организмы могут характеризоваться широким диапазоном изменения уровней экспрессии введенного гена в разных объектах; кроме того, может иметь место различие в пространственных или временных паттернах экспрессии, например, различие в относительной экспрессии трансгенов в разных тканях растения, причем такое различие проявляется в том, что фактическая картина экспрессии может быть несовместимой с паттерном экспрессии, ожидаемым исходя из регулирующих транскрипцию элементов, присутствующих в конструкциях введенного гена. Поэтому обычно необходимо получать сотни разных объектов, которые исследуют с целью обнаружения единственного объекта, характеризующегося требуемыми уровнями экспрессии трансгена и паттернами экспрессии, подходящими для промышленных целей. Объекты с ожидаемыми уровнями экспрессии трансгена и паттернами экспрессии трансгена могут быть использованы для введения трансгенов в другие генетические среды с помощью половой гетеротипической гибридизации с использованием стандартных способов скрещивания. Получаемое таким гибридизационным подходом потомство сохраняет характеристики экспрессии трансгена первичных объектов. Применение данной стратегии может обеспечить надежную экспрессию генов у многих сортов, и данные сорта могут быть хорошо адаптированы к местным условиям произрастания.
Был бы полезен способ, позволяющий обнаружить наличие конкретного объекта и определить возможность сохранения в потомстве гибрида представляющих интерес трансгенов. Кроме того, способы обнаружения конкретного объекта должны помогать удовлетворять соответствующим требованиям, например, официальной апробации до поступления на рынок и маркировки продуктов питания, получаемых из рекомбинантных сельскохозяйственных культур. Присутствие трансгенов можно обнаружить любым известным способом обнаружения полинуклеотидов, например, полимеразных цепных реакций (ПЦР) или гибридизации ДНК с использованием полинуклеотидного зонда. Данные способы обнаружения обычно основаны на часто используемых генетических элементах, таких как промоторы, терминаторы и гены-маркеры. Таким образом, если не известна последовательность хромосомной ДНК («фланкирующей ДНК»), расположенная рядом со вставочной трансгенной ДНК, такие способы, описанные выше, не смогут различать разные объекты, в частности, такие объекты, которые были получены при использовании одной и той же конструкции ДНК. Поэтому пара праймеров, которые перекрывают сайт коньюгации вставленного трансгена и фланкирующей ДНК, в частности, первый праймер, содержащий фланкирующую последовательность, и второй праймер, содержащий вставленную последовательность, часто используется для идентификации конкретных трансгенных объектов с помощью ПЦР в режиме реального времени.
Сущность изобретения
Задача настоящего изобретения состоит в том, чтобы обеспечить нуклеотидную последовательность для обнаружения толерантного к гербицидам растения сои DBN9004 и способ такого обнаружения, причем данный трансгенный объект сои DBN9004 имеет лучшую толерантность к глифосатному гербициду и глюфосинатному гербициду, и способ обнаружения может точно и быстро определить, содержится ли молекула ДНК конкретного трансгенного объекта сои DBN9004 в биологическом образце.
Для достижения вышеупомянутой цели настоящее изобретение обеспечивает молекулу нуклеиновой кислоты, имеющую нижеприведенную нуклеотидную последовательность, причем данная нуклеотидная последовательность содержит по меньшей мере 11 последовательных нуклеотидов в SEQ ID NO: 3 или комплементарной ей последовательности, и/или по меньшей мере 11 последовательных нуклеотидов в SEQ ID NO: 4 или комплементарной ей последовательности.
Предпочтительно нуклеотидная последовательность содержит SEQ ID NO: 1 или комплементарную ей последовательность и/или SEQ ID NO: 2 или комплементарную ей последовательность.
Кроме того, нуклеотидная последовательность содержит SEQ ID NO: 3 или комплементарную ей последовательность и/или SEQ ID NO: 4 или комплементарную ей последовательность.
Кроме того, нуклеотидная последовательность содержит SEQ ID NO: 5 или комплементарную ей последовательность.
SEQ ID NO: 1 или комплементарная ей последовательность представляет собой последовательность длиной 22 нуклеотида, расположенную на 5'-конце вставочной последовательности вблизи сайта конъюгации вставки в трансгенном объекте сои DBN9004, и SEQ ID NO: 1 или комплементарная ей последовательность перекрывает фланкирующую последовательность геномной ДНК соевого вставочного сайта и последовательность ДНК на 5'-конце вставочной последовательности; таким образом, включение SEQ ID NO: 1 или комплементарной ей последовательности может быть идентифицировано как присутствие трансгенного объекта сои DBN9004. SEQ ID NO: 2 или комплементарная ей последовательность представляет собой последовательность длиной 22 нуклеотида, расположенную на 3'-конце вставочной последовательности вблизи сайта конъюгации вставки в трансгенном объекте сои DBN9004, и SEQ ID NO: 2 или комплементарная ей последовательность перекрывает последовательность ДНК на 3'-конце введенной последовательности и фланкирующую последовательность геномной ДНК соевого вставочного сайта; таким образом, включение SEQ ID NO: 2 или комплементарной ей последовательности может быть идентифицировано как присутствие трансгенного объекта сои DBN9004.
В настоящем изобретении нуклеотидная последовательность может представлять собой по меньшей мере 11 или более последовательных полинуклеотидов (первая нуклеотидная последовательность) любой части трансгенной вставочной последовательности в SEQ ID NO: 3 или комплементарной ей последовательности, или по меньшей мере 11 или более последовательных полинуклеотидов (вторая нуклеотидная последовательность) любой части 5'-фланкирующей области геномной ДНК сои в SEQ ID NO: 3 или комплементарной ей последовательности. Нуклеотидная последовательность может дополнительно быть частью, гомологичной или комплементарной последовательности SEQ ID NO: 3, содержащей интактную SEQ ID NO: 1. Когда первая нуклеотидная последовательность и вторая нуклеотидная последовательность используются вместе, данные нуклеотидные последовательности в качестве пары ДНК-праймеров могут быть использованы в способе амплификации ДНК для получения продукта амплификации. Присутствие трансгенного объекта сои DBN9004 или его потомства может быть установлено, когда продукт амплификации, полученный в способе амплификации ДНК с использованием пары ДНК-праймеров, представляет собой продукт амплификации, содержащий SEQ ID NO: 1. Специалисту в данной области техники хорошо известно, что первая и вторая последовательности нуклеиновых кислот не обязательно состоят исключительно из ДНК, но могут также содержать РНК, смесь ДНК и РНК или сочетание ДНК, РНК или других нуклеотидов или их аналогов, которые не являются шаблонами одной или нескольких полимераз. Кроме того, зонды или праймеры, описанные в настоящем изобретении, должны иметь длину, составляющую по меньшей мере около 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21 или 22 последовательных нуклеотидов, которые могут быть выбраны из нуклеотидов, показанных как последовательности SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 3, SEQ ID NO: 4 и SEQ ID NO: 5. Когда зонды и праймеры выбраны из нуклеотидов, показанных как последовательности SEQ ID NO: 3, SEQ ID NO: 4 и SEQ ID NO: 5, то данные зонды и праймеры могут быть последовательными нуклеотидами, составляющими по меньшей мере от около 21 до около 50 или более нуклеотидов в длину. SEQ ID NO: 3 или комплементарная ей последовательность представляет собой последовательность длиной 1207 нуклеотидов, расположенную на 5'-конце вставочной последовательности вблизи сайта конъюгации вставки в трансгенном объекте сои DBN9004, и SEQ ID NO: 3 или комплементарная ей последовательность состоит из фланкирующей последовательности геномной ДНК сои длиной 740 нуклеотидов (нуклеотиды 1-740 из SEQ ID NO: 3), 74 нуклеотидов в последовательности ДНК конструкции pDBN4003 (нуклеотиды 741-814 из SEQ ID NO: 3), терминаторной последовательности t35S длиной 195 нуклеотидов (нуклеотиды 815-1009 из SEQ ID NO: 3), векторной спейсерной последовательности длиной 21 нуклеотидов (нуклеотиды 1010-1030 из SEQ ID NO: 3) и 5'-концевой последовательности ДНК фосфатинотрицин-N-ацетилтрансферазы cPAT длиной 177 нуклеотидов (нуклеотиды 1031-1207 из SEQ ID NO: 3) (нуклеотиды 815-1207 из SEQ ID NO: 3); таким образом, включение SEQ ID NO: 3 или комплементарной ей последовательности может быть идентифицировано как присутствие трансгенного объекта сои DBN9004.
Нуклеотидная последовательность может представлять собой по меньшей мере 11 или более последовательных полинуклеотидов (третья нуклеотидная последовательность) любой части трансгенной вставочной последовательности в SEQ ID NO: 4 или комплементарной ей последовательности или по меньшей мере 11 или более последовательных полинуклеотидов (четвертая нуклеотидная последовательность) любой части 3'-фланкирующей области геномной ДНК сои в SEQ ID NO: 4 или комплементарной ей последовательности. нуклеотидная последовательность может дополнительно быть частью, гомологичной или комплементарной SEQ ID NO: 4, содержащей интактную SEQ ID NO: 2. Когда третья нуклеотидная последовательность и четвертая нуклеотидная последовательность используются вместе, данные нуклеотидные последовательности в качестве пары ДНК-праймеров могут быть использованы в способе амплификации ДНК для получения продукта амплификации. Присутствие трансгенного объекта сои DBN9004 или его потомства может быть диагностировано, когда продукт амплификации, полученный в способе амплификации ДНК с использованием пары ДНК-праймеров, представляет собой продукт амплификации, содержащий SEQ ID NO: 2. SEQ ID NO: 4 или комплементарная ей последовательность представляет собой последовательность с длиной 631 нуклеотидов, расположенную на 3'-конце вставочной последовательности вблизи сайта конъюгации вставки в трансгенном объекте сои DBN9004, и SEQ ID NO: 4 или комплементарная ей последовательность состоит из последовательности промотора prGm17gTsf1 длиной 226 нуклеотидов (нуклеотиды 1-226 из SEQ ID NO: 4), 61 нуклеотид в ДНК-последовательности конструкции pDBN4003 (нуклеотиды 227-287 из SEQ ID NO: 4), конъюгирующей последовательности из 6 нуклеотидов (нуклеотиды 288-293 из SEQ ID NO: 4) и фланкирующей последовательности геномной ДНК сои длиной 338 нуклеотидов (нуклеотиды 294-631 из SEQ ID NO: 4); таким образом, включение SEQ ID NO: 4 или комплементарной ей последовательности может быть идентифицировано как присутствие трансгенного объекта сои DBN9004.
SEQ ID NO: 5 или комплементарная ей последовательность представляет собой последовательность из 7059 нуклеотидов в длину, которая характеризует трансгенный объект сои DBN9004, геномы и генетические элементы, конкретно содержащиеся в ней, являются такими, как показано в таблице 1. Включение SEQ ID NO: 5 или комплементарной ей последовательности может быть идентифицировано как присутствие трансгенного объекта сои DBN9004.
Таблица 1. Геномы и генетические элементы, содержащиеся в SEQ ID NO: 5
Нуклеотидные последовательности или комплементарные им последовательности могут быть использованы в способе амплификации ДНК для получения ампликонов, причем обнаружение ампликонов диагностирует присутствие трансгенного объекта сои DBN9004 или его потомства в биологическом образце; и последовательности нуклеиновых кислот или комплементарные им последовательности могут быть использованы в способах обнаружения нуклеотидов для обнаружения присутствия трансгенного объекта сои DBN9004 или его потомства в биологическом образце.
Для достижения вышеупомянутой цели настоящее изобретение также обеспечивает способ обнаружения присутствия ДНК трансгенного объекта сои DBN9004 в образце, включающий:
приведение исследуемого образца в контакт по меньшей мере с двумя типами праймеров для амплификации целевого продукта амплификации в реакции амплификации нуклеиновой кислоты;
осуществление реакции амплификации нуклеиновой кислоты; а также
обнаружение присутствия целевого продукта амплификации;
целевой продукт амплификации содержит по меньшей мере 11 последовательных нуклеотидов в SEQ ID NO: 3 или комплементарной ей последовательности и/или по меньшей мере 11 последовательных нуклеотидов в SEQ ID NO: 4 или комплементарной ей последовательности.
Кроме того, целевой продукт амплификации содержит последовательные нуклеотиды в положениях 1-11 или положениях 12-22 в SEQ ID NO: 1 или комплементарной ей последовательности и/или последовательные нуклеотиды в положениях 1-11 или положениях 12-22 в SEQ ID NO: 2 или комплементарной ей последовательности.
Кроме того, целевой продукт амплификации содержит по меньшей мере одно, выбранное из: SEQ ID NO: 1 или комплементарной ей последовательности, SEQ ID NO: 2 или комплементарной ей последовательности, SEQ ID NO: 6 или комплементарной ей последовательности и SEQ ID NO: 7 или комплементарной ей последовательности.
В вышеупомянутых технических решениях по меньшей мере один из праймеров содержит нуклеотидную последовательность или ее фрагмент или последовательность, комплементарную ей или данному фрагменту.
В частности, праймеры содержат первый праймер и второй праймер, причем первый праймер выбран из SEQ ID NO: 8 и SEQ ID NO: 10; и второй праймер выбран из SEQ ID NO: 9 и SEQ ID NO: 11.
Для достижения вышеупомянутой цели настоящее изобретение также обеспечивает способ обнаружения присутствия ДНК трансгенного объекта сои DBN9004 в образце, включающий:
приведение исследуемого образца в контакт с зондом, причем данный зонд содержит по меньшей мере 11 последовательных нуклеотидов в SEQ ID NO: 3 или комплементарной ей последовательности и/или по меньшей мере 11 последовательных нуклеотидов в SEQ ID NO: 4 или комплементарной ей последовательности;
гибридизацию исследуемого образца с зондом в строгих условиях гибридизации; а также
детектирование состояния гибридизации между образцом, подлежащим детектированию, и зондом.
Строгие условия могут представлять собой гибридизацию в растворе 6,0-кратного объема SSC (цитрат натрия) и 0,5% SDS (лаурил сульфат натрия) при 65°C, затем однократную промывку мембраны 2,0-кратным объемом SSC, 0,1% SDS, и 1,0-кратным объемом SSC, 0,1% SDS, соответственно.
Кроме того, зонд содержит последовательные нуклеотиды в положениях 1-11 или положениях 12-22 в SEQ ID NO: 1 или комплементарной ей последовательности и/или последовательные нуклеотиды в положениях 1-11 или положениях 12-22 в SEQ ID NO: 2 или комплементарной ей последовательности.
Кроме того, зонд содержит, по меньшей мере, одно, выбранное из: SEQ ID NO: 1 или комплементарной ей последовательности, SEQ ID NO: 2 или комплементарной ей последовательности, SEQ ID NO: 6 или комплементарной ей последовательности и SEQ ID NO: 7 или комплементарной ей последовательности.
Необязательно, по меньшей мере один из зондов помечен по меньшей мере одним флуорофором.
Для достижения вышеупомянутой цели настоящее изобретение также обеспечивает способ обнаружения присутствия ДНК трансгенного объекта сои DBN9004 в образце, включающий:
приведение исследуемого образца в контакт с помощью молекулы маркерной нуклеиновой кислоты, причем данная маркерная молекула нуклеиновой кислоты содержит по меньшей мере 11 последовательных нуклеотидов в SEQ ID NO: 3 или комплементарной ей последовательности и/или по меньшей мере 11 последовательных нуклеотидов в SEQ ID NO: 4 или комплементарной ей последовательности;
гибридизацию исследуемого образца с молекулой маркерной нуклеиновой кислоты в строгих условиях гибридизации; а также
детектирование состояния гибридизации исследуемого образца с помощью молекулы маркерной нуклеиновой кислоты и дополнительное определение генетической связи между толерантностью к глифосату и/или толерантностью к глюфосинату и молекулой маркерной нуклеиновой кислоты генетическим способом с помощью анализа полового скрещивания с использованием маркера.
Кроме того, маркерная молекула нуклеиновой кислоты содержит последовательные нуклеотиды в положениях 1-11 или положениях 12-22 в SEQ ID NO: 1 или комплементарной ей последовательности и/или последовательные нуклеотиды в положениях 1-11 или положениях 12-22 в SEQ ID NO: 2 или комплементарной ей последовательности.
Кроме того, маркерная молекула нуклеиновой кислоты содержит по меньшей мере одно, выбранное из: SEQ ID NO: 1 или комплементарной ей последовательности, SEQ ID NO: 2 или комплементарной ей последовательности, SEQ ID NO: 6 или комплементарной ей последовательности и SEQ ID NO: 7 или комплементарной ей последовательности.
Для достижения вышеупомянутой цели настоящее изобретение также обеспечивает набор для обнаружения ДНК, содержащий по меньшей мере одну молекулу ДНК, причем данная молекула ДНК содержит по меньшей мере 11 последовательных нуклеотидов гомологичной последовательности SEQ ID NO: 3 или комплементарной ей последовательности, и/или по меньшей мере 11 последовательных нуклеотидов гомологичной последовательности SEQ ID NO: 4 или комплементарной ей последовательности, и может быть использована в качестве ДНК-праймера или зонда, специфичного для трансгенного объекта сои DBN9004 или его потомства.
Кроме того, молекула ДНК содержит последовательные нуклеотиды в положениях 1-11 или положениях 12-22 в SEQ ID NO: 1 или комплементарной ей последовательности и/или последовательные нуклеотиды в положениях 1-11 или положениях 12-22 в SEQ ID NO: 2 или комплементарной ей последовательности.
Кроме того, молекула ДНК содержит по меньшей мере одно, выбранное из: гомологичной последовательности SEQ ID NO: 1 или комплементарной ей последовательности, гомологичной последовательности SEQ ID NO: 2 или комплементарной ей последовательности, гомологичной последовательности SEQ ID NO: 6 или комплементарной ей последовательности и гомологичной последовательности ID NO: 7 или комплементарной ей последовательности.
Для достижения вышеупомянутой цели настоящее изобретение также относится к клетке или части растения, содержащей нуклеотидную последовательность, кодирующую толерантный к глифосату белок EPSPS, нуклеотидную последовательность, кодирующую толерантный к глюфосинату белок РАТ, и нуклеотидную последовательность определенной области. Нуклеотидная последовательность определенной области содержит, по меньшей мере, одно, выбранное из: последовательностей, представленных SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 6 и SEQ ID NO: 7.
Для достижения вышеупомянутой цели настоящее изобретение также обеспечивает способ получения растения сои, толерантного к глифосатному гербициду и/или глюфосинатному гербициду, включающий введение в геном растения сои нуклеотидной последовательности, кодирующей толерантный к глифосату белок EPSPS, и/или нуклеотидной последовательности, кодирующей толерантный к глюфосинату белок РАТ, и нуклеотидной последовательности определенной области, выбранной из по меньшей мере одной нуклеотидной последовательности из последовательностей, представленных SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 3, SEQ ID NO: 4, SEQ ID NO: 5, SEQ ID NO: 6 и SEQ ID NO: 7.
В частности, способ получения растения сои, толерантного к глифосатному гербициду и/или глюфосинатному гербициду, включает:
Половую гибридизацию первого родительского растения сои трансгенного объекта сои DBN9004, которое является толерантным к глифосатному гербициду и/или глюфосинатному гербициду, со вторым родительским соевым растением, в котором отсутствует толерантность к глифосатному гербициду и/или глюфосинатному гербициду, с производством большого числа потомства данных растений;
обработку потомства данных растений глифосатным гербицидом и/или глюфосинатным гербицидом; а также
отбор потомства данных растений, которое является толерантным к глифосату и/или глюфосинату.
Для достижения вышеупомянутой цели настоящее изобретение также относится к способу культивирования растения сои, толерантного к глифосатному гербициду и/или глюфосинатному гербициду, содержащему:
посадку по меньшей мере одного семени сои, причем геном семени сои содержит нуклеотидную последовательность, кодирующую толерантный к глифосату белок EPSPS, и/или нуклеотидную последовательность, кодирующую толерантный к глюфосинату белок РАТ, и нуклеотидную последовательность определенной области;
выращивание семени сои в растение сои; а также
опрыскивание растения сои эффективной дозой глифосатного гербицида и/или глюфосинатного гербицида и сбор растений, характеризующихся меньшим повреждением растений по сравнению с другими растениями, не имеющими нуклеотидной последовательности определенной области;
нуклеотидная последовательность определенной области выбрана из по меньшей мере одной нуклеотидной последовательности из последовательностей, представленных SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 3, SEQ ID NO: 4, SEQ ID NO: 5, SEQ ID NO: 6 и SEQ ID NO: 7.
Для достижения вышеупомянутой цели настоящее изобретение также обеспечивает способ защиты растения от повреждений, вызванных гербицидом, включая внесение гербицида, содержащего эффективную дозу глифосата и/или глюфосината, в поле, предназначенное для посадки по меньшей мере одного трансгенного растения сои, причем трансгенное растение сои содержит в своем геноме по меньшей мере одну нуклеотидную последовательность, выбранную из последовательностей, представленных SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 3, SEQ ID NO: 4, SEQ ID NO: 5, SEQ ID NO: 6 и SEQ ID NO: 7, и трансгенное растение сои обладает толерантостью к глифосатному гербициду и/или глюфосинатному гербициду.
Для достижения вышеупомянутой цели настоящее изобретение также обеспечивает способ борьбы с полевыми сорняками, включающий внесение гербицида, содержащего эффективную дозу глифосата и/или глюфосината, на поле, предназначенное для посадки по меньшей мере одного трансгенного растения сои, причем трансгенное растение сои содержит в своем геноме по меньшей мере одну нуклеотидная последовательность, выбранную из последовательностей, представленных SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 3, SEQ ID NO: 4, SEQ ID NO: 5, SEQ ID NO: 6 и SEQ ID NO: 7, и трансгенное растение сои обладает толерантностью к глифосатному гербициду и/или глюфосинатному гербициду.
Для достижения вышеупомянутой цели настоящее изобретение также обеспечивает способ борьбы с устойчивыми к глифосату сорняками в поле для толерантного к глифосату растения, включающий внесение гербицида, содержащего эффективную дозу глифосата и/или глюфосината, на поле, предназначенное для посадки по меньшей мере одного толерантного к глифосату трансгенного растения сои, причем толерантное к глифосату трансгенное растение сои содержит в своем геноме по меньшей мере одну нуклеинововокислотная последовательность, выбранную из последовательностей, представленных SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 3, SEQ ID NO: 4, SEQ ID NO: 5, SEQ ID NO: 6 и SEQ ID NO: 7, и толерантное к глифосату трансгенное растение сои обладает в то же время толерантностью к глюфосинатному гербициду.
Для достижения вышеупомянутой цели настоящее изобретение также обеспечивает способ задержки выработки устойчивости к насекомым, включающий выращивание по меньшей мере одного трансгенного растения сои, обладающего толерантностью к глифосату и/или глюфосинату, в поле, предназначенное для посадки растения сои, устойчивого к воздействию насекомых, причем трансгенное растение сои, имеющее толерантность к глифосату и/или глюфосинату, содержит в своем геноме по меньшей мере одну нуклеотидную последовательность, выбранную из последовательностей SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 3, SEQ ID NO: 4, SEQ ID NO: 5, SEQ ID NO: 6 и SEQ ID NO: 7.
Для достижения вышеупомянутой цели настоящее изобретение также относится к сельскохозяйственному продукту или товару, содержащему полинуклеотид SEQ ID NO: 1 или SEQ ID NO: 2, причем сельскохозяйственным продуктом или товаром является лецитин, жирная кислота, глицерин, стерол, соевые хлопья, соевая мука, соевый белок или его концентрат, соевое масло, соевое белковое волокно, соевый молочный сгусток или соевый творог.
Относительно нуклеотидной последовательности для обнаружения толерантного к гербициду растения сои DBN9004 и способа такого обнаружения согласно настоящему изобретению, следующие определения и способы могут способствовать пониманию настоящеего изобретения и помочь специалисту в данной области техники осуществить настоящее изобретение на практике, и если не указано иное, термины в настоящем изобретении имеют значения, известные специалисту в данной области техники.
Термин «соя» означает вид Glycine max и включает все сорта данного вида, которые могут быть получены путем селекции растения сои, включая виды дикой сои.
Термин «содержать» или «включать» относится к «включая, но не ограничиваясь».
Термин «растение» включает в себя цельное растение, растительную клетку, растительный орган, растительный протопласт, культуру ткани растительных клеток, из которой может быть регенерировано растение, каллус растения, растительную колонию и интактную растительную клетку в растении или части растения, такую как эмбрион, пыльца, яйцеклетка, семя, лист, цветок, ветка, плод, стебель, корень, корневой наконечник и пыльник. Следует понимать, что часть трансгенного растения в рамках настоящего изобретения включает, но не ограничивается ими, растительную клетку, протопласт, ткань, каллус, эмбрион и цветок, стебель, плод, лист и корень, а вышеупомянутые части растений получены из трансгенных растений, которые предварительно трансформируются молекулой ДНК по настоящему изобретению и, таким образом, по меньшей мере частично состоят из трансгенных клеток или их потомков.
Термин «ген» относится к фрагменту нуклеиновой кислоты, который экспрессирует конкретный белок, включая регуляторную последовательность (5'-некодирующая последовательность) перед кодирующей последовательностью и регуляторную последовательность (3'-некодирующая последовательность) после кодирующей последовательности, Термин «природный ген» относится к гену, который, как обнаружено, имеет свои собственные природные регуляторные последовательности. Термин «химерный ген» относится к любому гену, который не является природным геном, который содержит не встречающиеся в природе регуляторные и кодирующие последовательности. Термин «эндогенный ген» относится к природному гену, который находится в своем естественном положении в геноме организма. Термин «экзогенный ген» представляет собой чужеродный ген, который теперь присутствует в геноме организма, но первоначально не присутствует, а также относится к гену, который вводится в рецепторную клетку через трансгенные стадии. Экзогенный ген может содержать природный ген или химерный ген, введенный в неприродный организм. Термин «трансген» представляет собой ген, который был введен в геном посредством процедуры трансформации. Сайт, в котором рекомбинантная ДНК была вставлена в растительный геном, может упоминаться как «инсерционный сайт», «сайт вставки» или «целевой сайт».
Термин «фланкирующая ДНК» может относиться к геному, который существует в природе, например, в организме растения, или экзогенной (гетерологичной) ДНК, введенной в процессе трансформации, такой как фрагмент, связанный с объектом трансформации. Таким образом, фланкирующая ДНК может включать комбинацию природных и экзогенных ДНК. В настоящем изобретении «фланкирующая область» или «фланкирующая последовательность» или «геномная пограничная область» или «геномная пограничная последовательность» относится к последовательности, имеющей по меньшей мере 3, 5, 10, 11, 15, 20, 50, 100, 200, 300, 400, 1000, 1500, 2000, 2500 или 5000 пар оснований в длину, или к более длинной последовательности, которая располагается непосредственно перед или после (в направлении от места инициации транскрипции) первоначально экзогенно вставленной молекулы ДНК и рядом с первоначально экзогенно вставленной молекулой ДНК. Когда фланкирующая область располагается после места инициации транскрипции в направлении 5'-3', ее также можно назвать «левым пограничным флангом» или «3'-флангом» или «3'-геномной пограничной областью», или «3'-геномной пограничной последовательностью» и тому подобное. Когда фланкирующая область располагается перед сайтом инициации транскрипции в направлении 5'-3', ее также можно назвать «границей правого фланга» или «5'-флангом» или «5'-геномной пограничной областью» или «5''-геномной пограничной последовательностью», и тому подобное.
Процедура трансформации, вызывающая случайную интеграцию экзогенной ДНК, приведет к объектам, содержащим разные фланкирующие области, которые конкретно содержатся в каждом объекте. Когда рекомбинантную ДНК вводят в растение общепринятой гибридизацией, ее фланкирующая область обычно не изменяется. Объекты также будут содержать уникальные сайты конъюгации между гетерогенной вставочной ДНК и сегментом геномной ДНК, или между двумя сегментами геномных ДНК, или между двумя сегментами гетерогенных ДНК. «Сайт конъюгации (conjugate site)» представляет собой сайт, в котором связаны два специфических фрагмента ДНК. Например, сайт конъюгации существует в том месте, где ДНК-вставка связана с фланкирующей ДНК. Сайты конъюгации также присутствуют в трансформированных организмах, в которых два фрагмента ДНК связаны между собой таким образом, как найдено в модифицированных природных организмах. «Конъюгированная ДНК» или «область конъюгации» относится к ДНК, содержащей сайт конъюгации.
Настоящее изобретение относится к трансгенному объекту сои, названному DBN9004, и его потомству, трансгенный объект сои DBN9004 представляет собой растение сои DBN9004, которое включает в себя растения и семена трансгенного объекта сои DBN9004 и его растительные клетки или регенерируемые части. Растительные части трансгенного объекта сои DBN9004 включают, но не ограничиваются ими, клетки, пыльцу, яйцеклетки, цветы, почки, корни, стебли, листья, стручки и продукты, полученные из растения сои DBN9004, такие как соевые лепешки, порошки и масла, в частности, лецитин, жирные кислоты, глицерин, стерины, пищевые масла, обезжиренные соевые хлопья, обезжиренные и обожженные соевые муки, соевые молочные сгустки, соевый творог, концентраты соевых белков, изолированные соевые белки, гидролизованные растительные белки, организованные соевые белки и соевые белковые волокна.
Трансгенный объект сои DBN9004 по настоящему изобретению включает конструкцию ДНК, причем трансгенный объект сои DBN9004 приобретает толерантность к глифосатному гербициду и глюфосинатному гербициду, когда конструкция ДНК экспрессируется в растительных клетках. Конструкция ДНК содержит две тандемные кассеты экспрессии, причем первая кассета экспрессии содержит подходящий промотор и подходящую сигнальную последовательность полиаденилирования для экспрессии в растении, промотор может быть функционально связан с геном, кодирующим 5-енолпирусилшикиммет-3-фосфатсинтазу (EPSPS), и EPSPS является толерантной к глифосатному гербициду. Вторая кассета экспрессии содержит подходящий промотор и подходящую сигнальную последовательность полиаденилирования для экспрессии в растении, промотор может быть функционально связан с геном, кодирующим фосфинотрицин-N-ацетилтрансферазу (PAT), и нуклеотидная последовательность белка PAT является толерантной к глюфосинатному гербициду. Кроме того, промотор может быть подходящим промотором, выделенным из растения, включая конститутивные, индуцируемые и/или тканеспецифические промоторы, и подходящий промотор включает, но не ограничивается ими, промотор 35S мозаичного вируса цветной капусты (cauliflower mosaic virus (CaMV)), промотор 35S вируса мозаики смоковницы (figwort mosaic virus (FMV)), промотор Tsf1, промотор убиквитина, промотор актина, промотор нопалинсинтазы (nopaline synthase (NOS)) Agrobacterium tumefaciens, промотор октопинсинтазы (octopine synthase (OCS)), промотор вируса желтой курчавости листьев цеструма Cestrum, промотор пататина, промотор рибулозо-1,5-бисфосфат-карбоксилазы/оксигеназы (ribulose-1,5-bisphosphate carboxylase/oxygenase (RuBisCO)), промотор глутатион-S-трансферазы (glutathione S-transferase (GST)), промотор E9, промотор GOS, промотор alcA/alcR, промотор RolD Agrobacterium rhizogenes и промотор Suc2 Arabidopsis. Сигнальная последовательность полиаденилирования может быть подходящей сигнальной последовательностью полиаденилирования, которая функционирует в растениях, и подходящая сигнальная последовательность полиаденилирования включает, но не ограничивается ими, сигнальную последовательность полиаденилирования, полученную из гена нопалинсинтазы (NOS) Agrobacterium tumefaciens, сигнальные последовательности полиаденилирования, полученные из терминатора 35S мозаичного вируса цветной капусты (CaMV) и полученные из терминатора E9 гранулозы-1,5-бисфосфаткарбоксилазы/оксигеназы, сигнальную последовательность полиаденилирования, полученную из гена ингибитора протеазы II (PIN II) и сигнальную последовательность полиаденилирования, полученную из гена α-тубулина.
Кроме того, кассета экспрессии может также включать другие генетические элементы, включая, но не ограничиваясь ими, энхансеры и сигнальные пептиды/транзитные пептиды. Энхансер может увеличивать уровень экспрессии гена, и энхансер включает, но не ограничивается ими, фактор активации трансляции вируса гравировки табака (tobacco etch virus (TEV)), энхансер CaMV35S и энхансер FMV35S. Сигнальный пептид/транзитный пептид может направлять белок EPSPS и/или белок PAT для переноса во внеклеточную или внутриклеточную специфическую органеллу или компартмент, например, нацеливание на хлоропласт осуществляется с использованием последовательности, кодирующей транзитный пептид хлоропласта, или нацеливание на эндоплазматический ретикулум осуществляется с использованием удерживающей последовательности «KDEL».
Ген 5-енолпирувильшикимат-3-фосфатсинтазы (EPSPS) может быть выделен из штамма CP4 Agrobacterium tumefaciens, а полинуклеотиды, кодирующие EPSPS, могут быть изменены путем оптимизации кодонов или другими способами для достижения цели повышения стабильности и доступности транскриптов в трансформированных клетках. Ген 5-енолпирувильшикимат-3-фосфатсинтазы (EPSPS) может также использоваться в качестве селективного гена-продуцента.
«Глифосат» относится к N-фосфонометилглицину и его солям. Обработка «глифосатным гербицидом» относится к проведению обработки с использованием любого гербицидного препарата, содержащего глифосат. Выбор нормы расхода определенного препарата глифосата для достижения эффективной биологической дозы не будет превышать навыков обычного агрономического специалиста. Обработка поля, включающего растительные материалы, полученные из толерантного к гербицидам растения сои DBN9004 с использованием любого гербицидного препарата, содержащего глифосат, будет контролировать рост сорняков в поле и не повлияет на рост или выход растительных материалов, полученных из толерантного к гербицидам растения сои DBN9004.
Ген фосфатинотрицин-N-ацетилтрансферазы (PAT), выделенный из Streptomyces viridochromogenes, катализирует превращение L-фосфинотрицина в его неактивную форму путем ацетилирования, что обеспечивает толерантность растений к глюфосинатному гербициду. Фосфинотрицин (PTC, 2-амино-4-метилфосфономасляная кислота) является ингибитором глутаминсинтетазы. PTC является структурной единицей антибиотика 2-амино-4-метилфосфоно-аланил-аланина, данный трипептид (PTT) обладает активностью против грамположительных и грамотрицательных бактерий и грибов Botrytis cinerea. Ген фосфинотрицин-N-ацетилтрансферазы (PAT) может также служить в качестве селективного маркерного гена.
«Глюфосинат» (также известный как фосфинотрицин) относится к 2-амино-4-[гидрокси(метил)фосфонил]бутирату аммония. Обработка «глюфосинатным гербицидом» относится к проведению обработки с использованием любого препарата гербицида, содержащего глюфосинат. Выбор нормы расхода определенного препарата глюфосината для достижения эффективной биологической дозы не будет превышать навыков обычного агрономического специалиста. Обработка поля, включающего растительные материалы, полученные из толерантного к гербицидам растения сои DBN9004 с использованием любого препарата, содержащего глюфосинат, будет контролировать рост сорняков в поле и не повлияет на рост или выход растительных материалов, полученных из толерантного к гербицидам растения сои DBN9004.
Конструкцию ДНК вводят в растение с использованием способа трансформации, включающего, но не ограничиваясь этим, Agrobacterium-опосредованную трансформацию, биолистическую трансформацию с помощью «генного ружья» и трансформацию с использованием пыльцевых трубок.
Способ Agrobacterium-опосредованной трансформации является распространенным способом трансформации растений. Экзогенная ДНК, вводимая в растение, клонируется между левой и правой пограничными консенсусными последовательностями вектора, т. е. в области Т-ДНК. Клетка Agrobacterium трансформируется вектором, а затем используется для заражения растительной ткани, и область Т-ДНК вектора, содержащая экзогенную ДНК, вводится в геном растения.
Способ трансформации с помощью «генной пушки» представляет собой способ использования носителя, содержащего экзогенную ДНК, для бомбардировки растительной клетки (particle-mediated bio-projectile transformation).
Способ трансформации с использованием пыльцевых трубок является способом переноса экзогенной ДНК в эмбриональный мешок через путь нуцеллуса с использованием пути естественных пыльцевых трубок (также называемого проводником пыльцевой трубкой), образованных опылением растений.
После трансформации трансгенные растения должны быть регенерированы из трансформированной растительной ткани, и потомки с экзогенной ДНК отобраны с использованием соответствующих маркеров.
Конструкция ДНК представляет собой объединение молекул ДНК, которые соединены друг с другом, и данное объединение обеспечивает одну или более экспрессионных кассет. Конструкция ДНК предпочтительно представляет собой плазмиду, способную к самовоспроизводству внутри бактериальной клетки и содержащую различные сайты рестрикционных ферментов, и сайты рестрикционных ферментов, содержащиеся в них, используются для введения молекул ДНК, обеспечивающих функциональные генные элементы, то есть промотор, интрон, лидерную последовательность, кодирующую последовательность, 3'-терминаторную область и другие последовательности. Кассета экспрессии, содержащаяся в конструкции ДНК, содержит генные элементы, которые необходимы для обеспечения транскрипции информационной РНК, и экспрессионная кассета может быть сконструирована так, чтобы экспрессироваться в прокариотических клетках или эукариотических клетках. Кассеты экспрессии по настоящему изобретению сконструированы так, чтобы наиболее предпочтительно экспрессироваться в растительных клетках.
Трансгенный «объект» получают путем трансформации растительной клетки конструкцией гетерологичной ДНК, то есть включая создание популяции растений путем инсерции кассеты экспрессии нуклеиновой кислоты, содержащей представляющий интерес ген, в геном растения трансгенными способами, регенерацию популяции растений и отбор конкретного растения, характеризующегося наличием инсерции в определенном положении в геноме. Термин «объект» включает исходный объект гетерологичной ДНК и потомков данного объекта. Термин «объект» также относится к потомству, полученному половым скрещиванием объекта и растения другого сорта, содержащему гетерологичную ДНК, даже если после повторного обратного полового скрещивания с родительской формой, с которой гибрид скрещивается вновь, введенная ДНК и фланкирующая геномная ДНК из трансформированных родительских форм также присутствует в потомстве гибрида в том же положении в хромосоме. Термин «объект» также относится к последовательности ДНК из первичного объекта, данная последовательность ДНК содержит введенную ДНК и фланкирующую геномную последовательность рядом с введенной ДНК, которые должны быть переданы потомству в результате полового скрещивания одной родительской формы, включающей введенную ДНК (например, первичный объект и его потомство, полученное в результате самоопыления), с родительской формой, не содержащей введенную ДНК, и данное потомство получает введенную ДНК, содержащую представляющий интерес ген.
«Рекомбинант» в настоящем изобретении относится к формам ДНК и/или белка и/или организма, которые обычно не встречаются в природе и, таким образом, продуцируется искусственным вмешательством. Такое искусственное вмешательство может приводить к образованию рекомбинантных молекул ДНК и/или рекомбинантных растений. «Молекула рекомбинантной ДНК» получается путем искусственного объединения двух сегментов последовательности, которые изолированы в других случаях, например, сегментов нуклеиновой кислоты, химически синтезированных или выделенных способами генно-инженерных манипуляций. Способы осуществления манипуляций с нуклеиновой кислотой хорошо известны.
Термин «трансген» включает любую клетку, клеточную линию, каллус, ткань, растительную часть или растение, их генотипы будут варьироваться в зависимости от присутствия гетерологичной нуклеиновой кислоты, и трансген содержит трансгенное тело, которое первоначально изменено как таковое, и потомство индивидуально получено из первоначальных трансгенных тел половым скрещиванием или бесполым размножением. В настоящем изобретении термин "трансген" не включает изменения геномов (хромосомные или внехромосомные), вызванные стандартными способами селекции растений или встречающимися в природе событиями, и встречающимися в природе событиями являются, например, рандомизированное перекрестное оплодотворение, нерекомбинантная вирусная инфекция, нерекомбинантная бактериальная трансформация, нерекомбинантная транспозиция или спонтанная мутация.
В настоящем изобретении «гетерологичный» относится к первой молекуле, про которую известно, что в природных условиях она, как правило, не объединена со второй молекулой. Например, молекула может происходить из первого сорта и вводиться в геном второго сорта. Таким образом, данная молекула является гетерологичной по отношению к хозяину и искусственно вводится в геном клетки-хозяина.
Культивирование трансгенного объекта сои DBN9004, который является толерантным к глифосатному гербициду и глюфосинатному гербициду, достигается с помощью следующих стадий: во-первых, половая гибридизация первого родительского растения сои со вторым родительским соевым растением для получения разнообразного первого поколения гибридов растений, причем первое родительское растение сои состоит из растений сои, которые культивируются из трансгенного объекта сои DBN9004 и его потомства, которое получают путем трансформации с помощью экспрессионной кассеты по настоящему изобретению, несущей толерантность к глифосатному гербициду и глюфосинатному гербициду, и второе родительское растение сои не имеет толерантности к глифосатному гербициду и/или глюфосинатному гербициду; затем отбирают растения потомства, которые являются толерантными к применению глифосатного гербицида и/или глюфосинатного гербицида, тем самым культивируя растение сои, толерантное к глифосатному гербициду и глюфосинатным гербицидам. Данные стадии могут дополнительно включать в себя обратное скрещивание растения-потомка, имеющего толерантность к применению глифосатного гербицида и/или глюфосинатного гербицида, со вторым родительским соевым растением или третьим родительским соевым растением, затем отбор потомства путем применения глифосатного гербицида и глюфоситатного гербицида или путем идентификации молекулярных маркеров (таких как молекула ДНК, содержащая сайт конъюгации, идентифицированное на 5'-конце и 3'-конце вставочной последовательности в трансгенном объекте сои DBN9004), связанных с свойствами, тем самым получая растение сои, которое является толерантным к глифосатному гербициду и глюфосинатному гербициду.
Следует также понимать, что два разных трансгенных растения также могут быть скрещены для получения потомков, содержащих два независимых, отдельно добавленных экзогенных гена. Растения-потомки, которые являются гомозиготами по двум добавленным экзогенным генам, могут быть получены из самоопыления соответствующего потомства. Также можно проводить обратное скрещивание родительских растений и ауткроссинг с нетрансгенными растениями, как описано выше, и бесполое размножение также представляет собой то же самое.
Культуры трансгенных соевых бобов со встроенным геном Bt-белка могут убивать, например, Lepidoptera насекомых/вредителей, но имеются также несколько выживших насекомых/вредителей, которые после нескольких поколений размножения могут производить устойчивых насекомых/вредителей, устойчивых к Bt-белку. Чтобы решить проблемы устойчивости насекомых/вредителей, Агентство по охране окружающей среды США (US Environmental Protection Agency) дает указания относительно применения трансгенных соевых культур, то есть необходимо обеспечить определенную часть соевых растений-защитников (shelter) (которые могут представлять собой не устойчивые к насекомым трансгенные соевые растения (которые представлять собой не устойчивые к вредителям трансгенные соевые растения (например, толерантные к гербицидам трансгенные соевые растения или не устойчивые к представляющим интерес вредителям трансгенные соевые растения или нетрансгенные соевые растения)). Когда большинство насекомых/вредителей погибает на соответствующих устойчивых к насекомым трансгенных соевых растениях, часть насекомых/вредителей выживает на соевых растениях-защитниках, что гарантирует, что число насекомых/вредителей, не обладающих устойчивостью, преобладает. Таким образом, даже если сохранилось небольшое количество устойчивых насекомых/вредителей, устойчивые гены значительно уменьшаются в количестве после спаривания устойчивых насекомых/вредителей с преобладающим количеством устойчивых насекомых/вредителей.
Термин «зонд» означает выделенную молекулу нуклеиновой кислоты, к которой присоединена стандартная детектируемая метка или репортерная молекула, например, радиоизотоп, лиганд, хемилюминесцентное вещество или фермент. Такой зонд является комплементарным цепи нуклеиновой кислоты-мишени. В настоящем изобретении зонд является комплементарным цепи ДНК из генома трансгенного объекта сои DBN9004, независимо от того, получена ли геномная ДНК из трансгенного объекта сои DBN9004 или его семени, или получена из растений или семян или экстрактов трансгенного объекта сои DBN9004. Зонд по настоящему изобретению включает не только дезоксирибонуклеиновую кислоту или рибонуклеиновую кислоту, но также полиамиды и другие вещества зонда, которые специфически связываются с последовательностью ДНК-мишени и могут быть использованы для обнаружения присутствия представляющей интерес последовательности ДНК-мишени.
Термин «праймер» означает выделенную молекулу нуклеиновой кислоты, которая связывается с цепью комплементарной ДНК-мишени путем гибридизации и отжига нуклеиновой кислоты, образует гибрид между праймером и цепью ДНК-мишени и затем удлиняется по цепи ДНК-мишени под действием полимеразы (например, ДНК-полимеразы). Пары праймеров по настоящему изобретению относится к их применению в амплификации последовательностей нуклеиновых кислот-мишеней, например, при помощи полимеразной цепной реакции (ПЦР) или других стандартных способов амплификации нуклеиновых кислот.
Длины зонда и праймера обычно составляют 11 полинуклеотидов или более, предпочтительно 18 полинуклеотидов или более, более предпочтительно 24 полинуклеотида или более и наиболее предпочтительно 30 полинуклеотидов или более. Такие зонд и праймер специфически гибридизуют с последовательностью-мишенью в очень строгих условиях гибридизации. Хотя стандартными способами могут быть созданы зонды, которые отличаются от последовательностей ДНК-мишеней и сохраняют способность гибридизировать с последовательностями ДНК-мишеней, зонды и праймеры по настоящему изобретению предпочтительно обладают полным сходством ДНК-последовательности с последовательными нуклеиновыми кислотами нуклеиновых кислот-мишеней.
Праймеры и зонды фланкирующих последовательностей геномной ДНК и вставочных последовательностей по настоящему изобретению могут быть определены стандартными способами, например, путем выделения соответствующей молекулы ДНК из растительных материалов, полученных из трансгенного объекта сои DBN9004, и определения нуклеотидной последовательности молекулы ДНК. Молекула ДНК включает трансгенную вставочную последовательность и фланкирующую последовательность генома сои, а фрагмент молекулы ДНК можно использовать в качестве праймера или зонда.
Нуклеинокислотные зонды и праймеры по настоящему изобретению гибридизуются с последовательностью ДНК-мишени в строгих условиях. Любой традиционный способ гибридизации или амплификации нуклеиновой кислоты может быть использован для идентификации присутствия ДНК, полученной из трансгенного объекта сои DBN9004 в образце. Молекула нуклеиновой кислоты или ее фрагмент способна специфически гибридизоваться с другими молекулами нуклеиновой кислоты при определенных обстоятельствах. Как используется в настоящем изобретении, если две молекулы нуклеиновых кислот способны образовывать антипараллельную двухцепочечную структуру нуклеиновой кислоты, то можно считать, что данные две молекулы нуклеиновой кислоты могут специфически гибридизироваться друг с другом. Если две молекулы нуклеиновой кислоты являются полностью комплементарными, то одна молекула нуклеиновой кислоты из данных двух молекул называется "комплементом" другой молекулы нуклеиновой кислоты. Как используется в настоящем изобретении, если каждый нуклеотид одной молекулы нуклеиновой кислоты является комплементарным соответствующему нуклеотиду другой молекулы нуклеиновой кислоты, то данные две молекулы нуклеиновой кислоты проявляют «полную комплементарность». Если две молекулы нуклеиновой кислоты способны гибридизироваться друг с другом с достаточной степенью устойчивости для связывания и сохранения гибрида по меньшей мере в стандартных условиях «пониженной строгости», то данные две молекулы нуклеиновой кислоты считаются «минимально комплементарными». Аналогичным образом, если две молекулы нуклеиновой кислоты способны гибридизироваться друг с другом с достаточной степенью устойчивости для связывания и сохранения гибрида по меньшей мере в стандартных условиях «высокой строгости», то данные две молекулы нуклеиновой кислоты называются «комплементарными». Отклонение от полной комплементарности допустимо, если такое отклонение полностью не предотвращает способность двух молекул образовывать двухцепочечную структуру. Молекула нуклеиновой кислоты, которая может быть использована в качестве праймера или зонда, должна иметь достаточно комплементарную последовательность, способную образовывать устойчивую двухцепочечную структуру при определенных концентрациях растворителя и соли.
Как используется в настоящем изобретении, по существу гомологичная последовательность представляет собой сегмент молекулы нуклеиновой кислоты, причем данная молекула нуклеиновой кислоты может специфически гибридизироваться с комплементарной цепью другого сегмента соответствующей молекулы нуклеиновой кислоты в строгих условиях гибридизации. Подходящими строгими условиями, которые способствуют гибридизации ДНК, являются, например, обработка 6,0-кратным объемом хлорида натрия/цитрата натрия (sodium chloride/sodium citrate (SSC)) при температуре около 45°C с последующей промывкой 2,0-кратным объемом SSC при 50°C, данные условия хорошо известны специалисту в данной области техники. Например, концентрация соли на стадии промывки может быть выбрана в диапазоне от условий пониженной строгости, включающих использование около 2,0-кратного объема SSC при 50°C, до строгих условий, включающих использование около 0,2-кратного объема SSC при 50°C. Кроме того, температура на стадии промывки может быть повышена от комнатной температуры, около 22°C, создаваемой в условиях пониженной строгости, около 65°C, применяемой в очень строгих условиях. Могут быть изменены как температура, так и концентрация соли, либо один из параметров, температура или концентрация соли, может оставаться постоянным при изменении другого параметра. Предпочтительно, молекула нуклеиновой кислоты по настоящему изобретению может специфически гибридизоваться в умеренно строгих условиях, например, при около 2,0-кратного объема SSC и около 65°C, с одной или более молекулами нуклеиновой кислоты последовательностей SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 3,SEQ ID NO: 4, SEQ ID NO: 5, SEQ ID NO: 6 и SEQ ID NO: 7, или комплементарной им последовательности, или любого из фрагментов вышеупомянутых последовательностей. Более предпочтительно, молекула нуклеиновой кислоты по настоящему изобретению может специфически гибридизоваться в очень строгих условиях с одной или более молекулами нуклеиновой кислоты последовательностей SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 3, SEQ ID NO: 4, SEQ ID NO: 5, SEQ ID NO: 6 и SEQ ID NO: 7, или комплементарной им последовательности, или любого из фрагментов вышеупомянутых последовательностей. В настоящем изобретении предпочтительная маркерная молекула нуклеиновой кислоты имеет SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 6 или SEQ ID NO: 7 или комплементарную им последовательность или представляет собой любой фрагмент вышеупомянутых последовательностей. Другая предпочтительная маркерная молекула нуклеиновой кислоты по настоящему изобретению имеет от 80 до 100% или от 90% до 100% идентичности последовательности с последовательностями SEQ ID NO: 1, SEQ ID NO:2, SEQ ID NO: 6 или SEQ ID NO: 7, или с комплементарной им последовательностью, или с любым фрагментом вышеупомянутых последовательностей. SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 6 и SEQ ID NO: 7 могут использоваться в качестве маркеров в процессе селекции растений для идентификации потомков генетических гибридов. Гибридизацию зонда с молекулой ДНК-мишени можно обнаружить любым способом, хорошо известным специалисту в данной области, включая, но не ограничиваясь ими, флуоресцентные метки, радиоактивные метки, метки на основе антител и хемилюминесцентные метки.
В случае амплификации (например, с помощью ПЦР) последовательности-мишени нуклеиновой кислоты с использованием конкретной пары праймеров для амплификации, «строгие условия» представляют собой условия, в которых праймеры могут гибридизоваться только с последовательностью-мишенью нуклеиновой кислоты в реакции термической амплификации ДНК, с которой праймер, имеющий соответствующую последовательность дикого типа (или комплементарную ей последовательность), соответствующую последовательности-мишени нуклеиновой кислоты, способен связываться и предпочтительно продуцировать уникальный продукт амплификации, то есть ампликон.
Термин «специфичное связывание с (целевой последовательностью)» означает, что зонд или праймер гибридизирует в строгих условиях гибридизации только с последовательностью-мишенью в образце, включающем такую последовательность-мишень.
Как используется в настоящем изобретении, «амплифицированная ДНК» или «ампликон», означает продукт амплификации нуклеиновой кислоты последовательности-мишени нуклеиновой кислоты, которая является частью матрицы нуклеиновой кислоты. Например, для определения того, получено ли растение сои в результате полового скрещивания трансгенного объекта сои DBN9004 по настоящему изобретению, или что образец сои, собранный с поля, содержит трансгенный объект сои DBN9004, или что соевый экстракт, такой как соевая мука, соевый порошок или соевые масла, содержит трансгенный объект сои DBN9004, ДНК, экстрагированная из образца ткани растения сои или ее экстракта, может быть подвергнута амплификации нуклеиновой кислоты с использованием пары праймеров для продуцирования ампликонов, позволяющих диагностировать наличие ДНК трансгенного объекта сои DBN9004. Пара праймеров содержит один праймер, полученный из фланкирующей последовательности, локализованной в геноме растения рядом с инсерционным сайтом вставленной гетерологичной ДНК, и второй праймер, полученный из вставленной экзогенной ДНК. Трансгенный объект сои DBN9004 также можно диагностировать на основании длины и последовательности ампликона. Длина ампликона может быть равна общей длине пар праймеров плюс одна пара нуклеотидных оснований, предпочтительно плюс около 50 пар нуклеотидных оснований, более предпочтительно плюс около 250 пар нуклеотидных оснований, наиболее предпочтительно плюс около 450 пар нуклеотидных оснований или более.
Необязательно, пары праймеров могут быть получены из фланкирующих геномных последовательностей с обеих сторон вставленной ДНК с образованием ампликона, содержащего всю нуклеотидную последовательность вставки. Один член пары пары праймеров, полученный из геномной последовательности растения, может быть расположен на расстоянии от вставленной последовательности ДНК, и данное расстояние может варьироваться от одной пары нуклеотидных оснований до около 20000 пар нуклеотидных оснований. Использование термина «ампликон» специально исключает димеры праймеров, образованные при выполнении термической реакции амплификации ДНК.
Нуклеиновая кислота может быть амплифицирована любым из способов амплификации нуклеиновой кислоты, известных в данной области техники, включая полимеразную цепную реакцию (ПЦР). Специалистам в данной области техники хорошо известны различные способы амплификации нуклеиновых кислот. Был разработан способ амплификации ПЦР для амплификации до 22 т.п.н.о. геномной ДНК и до 42 т.п.н.о. ДНК бактериофага. В настоящем изобретении могут быть использованы данные способы и другие способы амплификации ДНК, известные в данной области техники. Геном трансгенного объекта сои DBN9004 может быть амплифицирован с использованием вставочной экзогенной последовательности ДНК и фланкирующей ДНК-последовательности, полученной из трансгенного объекта сои DBN9004, с предоставленными праймерными последовательностями, и после амплификации может быть выполнено стандартное секвенирование ДНК ампликона ПЦР или клонированной ДНК.
Наборы для обнаружения ДНК, основанные на способах амплификации ДНК, содержат молекулы ДНК-праймеров, которые специфически гибридизуются с ДНК-мишенью в соответствующих реакционных условиях и амплифицируют диагностические ампликоны. Набор может обеспечить способ обнаружения на основе агарозного геля или ряд способов, известных в данной области техники для обнаружения диагностических ампликонов. Набор, содержащий ДНК-праймер, гомологичный или комплементарный любой части соевой геномной области, приведенной в последовательности SEQ ID NO: 3 или SEQ ID NO: 4, и гомологичный или комплементарный любой части трансгенной инсерционной области, приведенной в последовательности SEQ ID NO: 5, обеспечивается настоящим изобретением. В частности, SEQ ID NO: 8 и SEQ ID NO: 9 идентифицированы как пара праймеров, которые применяются в способе амплификации ДНК и амплифицируют диагностический ампликон, гомологичный части 5’-трансгенной/геномной области трансгенного объекта сои DBN9004, причем данный ампликон содержит SEQ ID NO: 1. Другие молекулы ДНК, используемые в качестве ДНК-праймеров, могут быть выбраны из SEQ ID NO: 5.
Ампликоны, полученные данными способами, могут быть обнаружены различными способами. Одним из таких способов является генетический двоичный анализ (Genetic Bit Analysis), с использованием олигонуклеотида ДНК, перекрывающего вставленную последовательность ДНК и смежную фланкирующую геномную последовательность ДНК. Данный олигонуклеотид иммобилизуют в лунках микропланшета, и после ПЦР-амплификации представляющей интерес области (с использованием одного праймера во вставленной последовательности и одного праймера в смежной фланкирующей геномной последовательности) одноцепочечный продукт ПЦР может быть гибридизован иммобилизованным олигонуклеотидом и использован в качестве матрицы для выполнения на одно основание с использованием ДНК-полимеразы и меченых ddNTP, специфичных к предполагаемому следующему основанию. Считывание может быть произведено при помощи флуоресцентного анализа или ELISA. Сигнал указывает на наличие вставочной/фланкирующей последовательности вследствие успешной амплификации, гибридизации и удлинения на одно основание.
Другим способом является техника пиросеквенирования. При выполнении данного способа создают олигонуклеотид, который перекрывает сайт конъюгации смежной геномной ДНК и вставленной ДНК. Олигонуклеотидную цепь гибридизуют с одноцепочечным ПЦР-продуктом из представляющей интерес области (с использованием одного праймера во вставочной последовательности и одного праймера в соседней фланкирующей геномной последовательности), затем инкубируют с ДНК-полимеразой, АТФ, сульфилазой, люциферазой, апиразой, аденозин-5’-фосфосульфатом и люциферином. Соответственно, добавляют dNTP, в результате чего возникают оптические сигналы, которые измеряют. Световой сигнал свидетельствует о наличии вставочной/фланкирующей последовательности вследствие успешной амплификации, гибридизации и удлинения на одно или несколько оснований.
Явление поляризации флуоресценции, описанное Chen et al. (Genome Res. 9: 492-498, 1999) является еще одним способом, который может быть использован для обнаружения ампликонов по настоящему изобретению. В соответствии с данным способом необходимо создать олигонуклеотидную цепь, которая перекрывает сайт конъюгации вставленной последовательности ДНК и расположенной рядом геномной ДНК. Олигонуклеотидную цепь гибридизуют с одноцепочечным ПЦР-продуктом из представляющей интерес области (с использованием одного праймера во вставочной последовательности и одного праймера в соседней фланкирующей геномной последовательности), затем инкубируют с ДНК-полимеразой и меченным флуоресцентным красителем ddNTP. Удлинение на одно основание вызывает включение ddNTP. Такое включение можно измерить в виде изменения поляризации при помощи флуорометра. Изменение поляризации представляет наличие вставочной/фланкирующей последовательности вследствие успешной амплификации, гибридизации и удлинения на одно основание.
Taqman является способом обнаружения и количественного определения наличия последовательности ДНК и подробно описан в инструкциях производителя. В кратком изложении, представленном ниже, способ включает создание олигонуклеотидного зонда FRET, который перекрывает сайт конъюгации вставочной последовательности ДНК и фланкирующей геномной последовательности. Зонд FRET и праймеры для ПЦР области (с использованием одного праймера во вставочной последовательности и одного праймера в соседней фланкирующей геномной последовательности) подвергают циклической обработке в присутствии термостабильных полимераз и dNTP. Гибридизация зонда FRET приводит к расщеплению флуоресцентного фрагмента и гасящего фрагмента зонда FRET и высвобождению флуоресцентной части. Появление флуоресцентного сигнала свидетельствует о наличии вставочной/фланкирующей последовательности вследствие успешной амплификации и гибридизации.
Подходящие способы для обнаружения растительных материалов, полученных из толерантного к гербицидам трансгенного объекта сои DBN9004, могут также включать гибридизацию с Саузерн-блотингом, гибридизацию с Нозерн-блоттингом и гибридизацию in situ на основе принципа гибридизации. В частности, подходящий способ включает инкубацию зонда и образца, промывку для удаления несвязанного зонда и обнаружение гибридизации зонда. Описанный выше способ обнаружения зависит от типа маркера, присоединенного к зонду, например, радиоактивно меченный зонд может быть обнаружен при экспонировании и проявлении рентгеновской пленки, или меченый ферментом зонд может быть обнаружен путем достижения изменения цвета через преобразование субстратов.
Tyangi et al. (Nat. Biotech, 14: 303-308, 1996) предложили применение молекулярных маркеров для обнаружения последовательности. Как кратко изложено ниже, создан олигонуклеотидный зонд FRET, который перекрывает сайт конъюгации вставочной последовательности ДНК и фланкирующей геномной последовательности. Уникальная структура зонда FRET обеспечивает вторичную структуру, содержащую флуоресцентную и гасящую части в непосредственной близости друг от друга. Зонд FRET и праймеры для ПЦР области (с использованием одного праймера во вставочной последовательности и одного праймера в соседней фланкирующей геномной последовательности) подвергают циклической обработке в присутствии термостабильных полимераз и dNTP. После успешной амплификации способом ПЦР гибридизация зонда FRET с последовательностью-мишенью вызывает разворачивание вторичной структуры зонда и пространственное разделение флуоресцентной и гасящей частей, приводящее к возникновению флуоресцентного сигнала. Появление флуоресцентного сигнала свидетельствует о наличии вставочной/фланкирующей последовательности вследствие успешной амплификации и гибридизации.
Другие описанные способы, такие как microfluidics, обеспечивают способы и устройства для выделения и амплификации образца ДНК. Люминесцентные красители используются для обнаружения и определения конкретных молекул ДНК. Основанное на использовании нанотрубок устройство, содержащее электронный датчик для обнаружения молекулы ДНК или наношариков (nanobeads), к которым присоединена конкретная молекула ДНК, которая может быть таким образом обнаружена, применимо для обнаружения молекулы ДНК по настоящему изобретению.
Набор для обнаружения ДНК может быть разработан с использованием композиций, описанных в настоящем изобретении, и способов, описанных или известных в области обнаружения ДНК. Набор облегчает идентификацию присутствия ДНК трансгенного объекта сои DBN9004 в образце и может также использоваться для культивирования растений сои, содержащих ДНК трансгенного объекта сои DBN9004. Набор может содержать ДНК-праймеры или зонды, гомологичные или комплементарные, по меньшей мере, части последовательностей SEQ ID NO: 1, 2, 3, 4 или 5 или содержащие другие ДНК-праймеры или зонды, гомологичные или комплементарные ДНК, содержащейся в трансгенных генетических элементах ДНК, и данные последовательности ДНК могут быть использованы для реакций амплификации ДНК или в качестве зондов в способах гибридизации ДНК. Структура ДНК, содержащаяся в геноме сои, сайта конъюгации вставочной трансгенной последовательности и гена сои, описанная на фигурах 1 и в таблице 1, включает: фланкирующую геномную область растения сои DBN9004, расположенную на 5'-конце трансгенной вставочной последовательности; часть вставочной последовательности из левой пограничной области (left boundary (LB)) Agrobacterium; первую кассету экспрессии, состоящую из промотора 35S (pr35S) вируса мозаики цветной капусты, содержащего тандемный повтор энхансерной области, который может быть функционально связан с толерантной к глюфосинату фосфинотрицин-N-ацетилтрансферазой (cPAT) из Streptomyces и затем функционально связан с терминатором 35S (t35S) вируса мозаики цветной капусты; вторую кассету экспрессии, состоящую из соевого гена Tsf1 (кодирующего фактор элонгации EF-1α) (prGm17gTsf1), который может быть функционально связан с кодирующей последовательностью (spAtCTP2) транзитного пептида хлоропласта из Arabidopsis для EPSPS, затем функционально связан с глифосат-толерантной 5-енол-пирувильшиким-3-фосфатсинтазой (cEPSPS) штамма Agrobacterium CP4, и затем функционально связан с 3'-нетранслируемой последовательностью (tPse9), полученной из рибулоза-1,5-бисфосфат-карбоксилазы гороха; часть вставочной последовательности из правой пограничной области (right boundary (RB)) Agrobacterium и фланкирующую геномную область растения сои DBN9004, расположенную на 3'-конце трансгенной вставочной последовательности (SEQ ID NO: 5). В способе амплификации ДНК молекулой ДНК в качестве праймера может быть любая часть, полученная из трансгенной вставочной последовательности в соевом растении DBN9004, или может быть любая часть, полученная из области ДНК фланкирующего соевого генома трансгенного объекта сои DBN9004.
Трансгенный объект сои DBN9004 может быть объединен с другими трансгенными сортами сои, например, соей с толерантностью к гербицидам (например, 2,4-D, дикамба и т.д.), или с трансгенными сортами сои, несущими другие гены, устойчивые к насекомым (например, Cry1Ac, Cry2Ab и т. д.). Все упомянутые разные трансгенные объекты можно скрещивать в различных сочетаниях вместе с трансгенным объектом сои DBN9004 по настоящему изобретению, чтобы обеспечить улучшенный гибридный трансгенный сорт сои, устойчивый к различным вредителям и толерантный к различным гербицидам. Данные сорта могут обладать улучшенными характерными особенностями, такими как повышенная урожайность по сравнению с нетрансгенным сортом и трансгенным сортом с одним полезным свойством.
В настоящем изобретении обеспечивается нуклеотидная последовательность для обнаружения толерантного к гербицидам растения сои DBN9004 и способа его обнаружения. Трансгенный объект сои DBN9004 является толерантным к фитотоксическим эффектам сельскохозяйственных гербицидов, содержащих глифосат и/или глюфосинат. Обладающее двумя дополнительными свойствами растение сои экспрессирует устойчивый к глифосату белок 5-енолпирувилшиктим-3-фосфатсинтазу (EPSPS) из штамма CP4 Agrobacterium, который обеспечивает толерантность растения к глифосату, и экспрессирует устойчивый к глюфосинату белок фосфиномицин-N-ацетилтрансферазу (PAT) из Streptomyces, который обеспечивает устойчивость растений к глюфосинату. Обладающее двумя дополнительными свойствами растение сои имеет следующие преимущества: 1) оно обеспечивает контроль широкого спектра сорняков при применении к соевым культурам глифосатсодержащих сельскохозяйственных гербицидов; 2) применение глюфосинатных гербицидов в сочетании со свойствами толерантности к глюфосинату (смешанных или чередующихся с глифосатными гербицидами) может служить неселективным средством для эффективной борьбы с устойчивыми к глифосату сорняками; 3) совместная посадка гербицид-устойчивых трансгенных соевых бобов в виде не устойчивых к насекомым трансгенных соевых бобов совместно с устойчивыми к насекомым трансгенными соевыми бобами в определенном соотношении может задержать индукцию толерантности у насекомых/вредителей; и 4) отсутствует падение урожайности сои. К тому же, гены, кодирующие свойства толерантности к глифосату и толерантности к глюфосинату, конъюгированы в одном и том же участке ДНК и присутствуют в одном локусе генома сои DBN9004, что обеспечивает повышенную эффективность при скрещивании и позволяет использовать молекулярные маркеры для отслеживания трансгенных вставленных фрагментов в разводимой популяции и ее потомстве. Между тем, SEQ ID NO: 1 или комплементарная ей последовательность, SEQ ID NO: 2 или комплементарная ей последовательность, SEQ ID NO: 6 или комплементарная ей последовательность или SEQ ID NO: 7 или комплементарная ей последовательность могут быть использованы в качестве ДНК-праймеров или зондов в способе обнаружения по настоящему изобретению для получения продукта амплификации, который диагностируется как трансгенный объект сои DBN9004 или его потомство, и для быстрого, точного и стабильного определения присутствия растительных материалов, полученных из трансгенного объекта сои DBN9004.
Краткое описание последовательности
SEQ ID NO: 1 Последовательность с длиной 22 нуклеотидов на 5'-конце вставочной последовательности вблизи сайта конъюгации вставки в трансгенном объекте сои DBN9004, причем нуклеотиды в положениях 1-11 и нуклеотиды в положениях 12-22 расположены по обе стороны от вставочного сайта в геноме сои, соответственно;
SEQ ID NO: 2 Последовательность длиной 22 нуклеотида на 3'-конце вставочной последовательности вблизи вставочного сайта конъюгации трансгенного объекта сои DBN9004, причем нуклеотиды в положениях 1-11 и нуклеотиды в положениях 12-22 расположены по обе стороны от вставочного сайта в геноме сои, соответственно;
SEQ ID NO: 3 Последовательность с длиной 1207 нуклеотидов на 5'-конце вставочной последовательности вблизи вставочного сайта конъюгации трансгенного объекта сои DBN9004;
SEQ ID NO: 4 Последовательность с длиной 631 нуклеотидов на 3'-конце вставочной последовательности вблизи вставочного сайта конъюгациитрансгенного объекта сои DBN9004;
SEQ ID NO: 5 Полная последовательность Т-ДНК, 5'- и 3'-фланкирующие геномные последовательности сои;
SEQ ID NO: 6 Последовательность, которая находится в SEQ ID NO: 3 и перекрывает нуклеотидную последовательность в последовательности ДНК конструкции pDBN4003 и последовательность терминации транскрипции t35S;
SEQ ID NO: 7 Последовательность, которая находится в SEQ ID NO: 4 и перекрывает последовательность промотора prGm17gTsf1 и нуклеотидную последовательность в последовательности ДНК конструкции pDBN4003;
SEQ ID NO: 8 Первый праймер, амплифицирующий SEQ ID NO: 3;
SEQ ID NO: 9 Второй праймер, амплифицирующий SEQ ID NO: 3;
SEQ ID NO: 10 Первый праймер, амплифицирующий SEQ ID NO: 4;
SEQ ID NO: 11 Второй праймер, амплифицирующий SEQ ID NO: 4;
SEQ ID NO: 12 Праймер на 5'-фланкирующей геномной последовательности;
SEQ ID NO: 13 Праймер, расположенный на Т-ДНК и спаренный с SEQ ID NO: 12;
SEQ ID NO: 14 Праймер на 3'-фланкирующей геномной последовательности, который спарен с SEQ ID NO: 12 для определения того, является ли трансген гомозиготой или гетерозиготой;
SEQ ID NO: 15 Праймер, расположенный на Т-ДНК и спаренный с SEQ ID NO: 14;
SEQ ID NO: 16 Taqman праймер 1 для обнаружения EPSPS;
SEQ ID NO: 17 Taqman primer 2 для обнаружения EPSPS;
SEQ ID NO: 18 Taqman зонд 1 для обнаружения EPSPS;
SEQ ID NO: 19 Taqman праймер 3 для обнаружения PAT;
SEQ ID NO: 20 Taqman праймер 4 для обнаружения PAT;
SEQ ID NO: 21 Taqman зонд 2 для обнаружения PAT;
SEQ ID NO: 22 Первый праймер эндогенного соевого гена убиквитина;
SEQ ID NO: 23 Второй праймер эндогенного соевого гена убиквитина;
SEQ ID NO: 24 Зонд для EPSPS при обнаружении способом Саузерн-блот-гибридизации;
SEQ ID NO: 25 Зонд для PAT при обнаружении способом Саузерн-блот-гибридизации;
SEQ ID NO: 26 Праймер, расположенный на Т-ДНК в одном и том же направлении с SEQ ID NO: 13;
SEQ ID NO: 27 Праймер, расположенный на Т-ДНК в противоположном направлении к SEQ ID NO: 13 и используемый для получения фланкирующей последовательности;
SEQ ID NO: 28 Праймер, расположенный на Т-ДНК в противоположном направлении к SEQ ID NO: 13 и используемый для получения фланкирующей последовательности;
SEQ ID NO: 29 Праймер, расположенный на Т-ДНК в одном и том же направлении с SEQ ID NO: 15;
SEQ ID NO: 30 Праймер, расположенный на Т-ДНК в противоположном направлении к SEQ ID NO: 15 и используемый для получения фланкирующей последовательности;
SEQ ID NO: 31 Праймер, расположенный на Т-ДНК в противоположном направлении к SEQ ID NO: 15 и используемый для получения фланкирующей последовательности.
Техническое решение настоящего изобретения далее подробно описано на чертежах и в примерах ниже.
Описание чертежей
Фигура 1 представляет собой структурную схему сайта конъюгации трансгенной вставочной последовательности и генома сои в соответствии с нуклеотидной последовательностью для обнаружения толерантного к гербицидам растения сои DBN9004 и способа такого обнаружения по настоящему изобретению;
Фигура 2 представляет собой структурную схему рекомбинантного экспрессирующего вектора pDBN4003 в соответствии с нуклеотидной последовательностью для обнаружения толерантного к гербицидам растения сои DBN9004 и способа такого обнаружения по настоящему изобретению.
Конкретные варианты осуществления
Технические решения нуклеинововокислотной последовательности для обнаружения толерантного к гербицидам растения сои DBN9004 и способа такого обнаружения в соответствии с настоящим изобретением будут дополнительно проиллюстрированы ниже в конкретных вариантах осуществления.
Пример 1. Клонирование и трансформация
1.1. Клонирование вектора
Рекомбинантный экспрессирующий вектор pDBN4003 был создан с использованием стандартных способов клонирования генов (как показано на фигуре 2). Вектор pDBN4003 содержит две тандемные трансгенные экспрессионные кассеты, причем первая кассета экспрессии состоит из гена Tsf1 сои (кодирующего фактор элонгации EF-1α) (prGm17gTsf1), который может быть функционально связан с кодирующей последовательностью (spAtCTP2) транзитного пептида хлоропласта из Arabidopsis для EPSPS, затем функционально связан с глифосат-толерантной 5-енол-пирувильшиким-3-фосфатсинтазой (cEPSPS) штамма Agrobacterium CP4, и затем функционально связан с 3’-нетранслируемой последовательностью (tPse9), полученной из рибулоза-1,5-бисфосфат-карбоксилазы гороха; и вторая кассета экспрессии состоит из промотора 35S вируса мозаики цветной капусты (pr35S), содержащего тандемный повтор энхансерной области, который может быть функционально связан с толерантной к глюфосинату фосфинотрицин-N-ацетилтрансферазой (cPAT) из Streptomyces и затем функционально связан с терминатором 35S (t35S) вируса мозаики цветной капусты.
Agrobacterium LBA4404 (Invitrogen, Чикаго, США, кат. №:18313-015) трансформировали вектором pDBN4003 способом с жидким азотом и трансформированные клетки подвергали скринингу с использованием 5-енол-пирувильшикимат-3-фосфатсинтазы (EPSPS) в качестве селективного маркера.
1.2. Трансформация растений
Трансформацию проводили обычным способом инфекции с помощью Agrobacterium, и стерильную культивированную ткань соевого семядольного узла выращивали совместно с Agrobacterium, описанной в примере 1.1, для введения Т-ДНК в созданном рекомбинантном экспрессирующем векторе pDBN4003 в хромосому сои для получения трансгенного объекта сои DBN9004.
В отношении Agrobacterium-опосредованной трансформации сои, в кратком изложении, зрелые семена сои проращивали в культуральной среде для прорастания сои (3,1 г/л соли B5, витамин B5, 20 г/л сахарозы и 8 г/л агара, рН 5,6), и семена инокулировали в культуральную среду прорастания и культивировали в следующих условиях: при температуре 25±1°C; и фотопериод (свет/темнота) 16 ч/8 ч. По прошествии от 4 до 6 дней прорастания стерильные саженцы сои, разбухшие на ярко-зеленых семядольных узлах, вырезали, гипокотилярные оси обрезали на 3-4 миллиметра ниже семядольных узлов, семядоли разрезали продольно и удаляли апикальные почки, боковые бутоны и семенные корни. Повреждение наносилось в семядольном узле с использованием спинки скальпеля, ткани поврежденных семядольных узлов подвергали контакту с суспензией Agrobacterium, причем Agrobacterium может переносить нуклеотидную последовательность гена EPSPS и нуклеотидную последовательность гена PAT в ткани поврежденных семядольных узлов (стадия 1: стадия инфицирования). На данной стадии ткани семядольных узлов предпочтительно погружали в суспензию Agrobacterium (OD660=0,5-0,8, культуральная среда для инфицирования (2,15 г/л соли MS, витамин B5, 20 г/л сахарозы, 10 г/л глюкозы, 40 мг/л ацетозиригона (AS), 4 г/л 2-морфолиновой этансульфоновой кислоты (MES) и 2 мг/л зеатина (ZT), pH 5,3)), чтобы инициировать инфицирование. Ткани семядольных узлов культивировали совместно с Agrobacterium в течение определенного периода времени (3 дня) (стадия 2: стадия совместного культивирования). Предпочтительно ткани семядольных узлов культивировали в твердой культуральной среде (4,3 г/л соли МС, витамин В5, 20 г/л сахарозы, 10 г/л глюкозы, 4 г/л 2-морфолинэтансульфоновой кислоты (MES), 2 мг/л зеатина и 8 г/л агара, рН 5,6) после стадии инфицирования. После данной стадии совместного культивирования проводили дополнительную стадию «восстановления». На стадии «восстановления» может быть по меньшей мере один антибиотик (от 150 до 250 мг/л цефалоспорина), о котором известно, что он ингибирует рост Agrobacterium в культуральной среде для восстановления (3,1 г/л соли B5, витамин B5, 1 г/л 2-морфолинэтансульфоновой кислоты (MES), 30 г/л сахарозы, 2 мг/л зеатина (ZT), 8 г/л агара, 150 мг/л цефалоспорина, 100 мг/л глутаминовой кислоты и 100 мг/л аспарагиновой кислоты, рН 5,6), без добавления селективного агента для растительного объекта (стадия 3: стадия восстановления). Предпочтительно, тканевые блоки, регенерированные из семядольных узлов, культивировали в твердой культуральной среде с антибиотиком, но без селективного агента, чтобы удалить Agrobacterium и обеспечить стадию восстановления инфицированных клеток. Затем тканевые блоки, регенерированные из семядольных узлов, культивировали в культуральной среде, содержащей селективный агент (глифосат), и отбирали растущие трансформированные каллусы (стадия 4: стадия отбора). Предпочтительно, тканевые блоки, регенерированные из семядольных узлов, культивировали в скринирующей твердой культуральной среде (3,1 г/л соли B5, витамин B5, 1 г/л 2-морфолинэтансульфоновой кислоты (MES), 30 г/л сахарозы, 1 мг/л 6-бензиладенина (6-BAP), 8 г/л агара, 150 мг/л цефалоспорина, 100 мг/л глутаминовой кислоты, 100 мг/л аспарагиновой кислоты и 10 мг/л соли глифосата с изопропиламином, pH 5,6) с помощью селективного агента, что приводит к непрерывному росту трансформированных клеток. Затем растения регенерировали из трансформированных клеток (стадия 5: стадия регенерации). Предпочтительно, тканевые блоки, регенерированные из семядольных узлов, выращенных в культуральной среде, содержащей селективный агент, культивировали в твердой культуральной среде (культуральная среда дифференцировки B5 и культуральная среда B5) для регенерации растений.
Отобранные устойчивые ткани переносили на культуральную среду дифференцировки B5 (3,1 г/л соли B5, витамин B5, 1 г/л 2-морфолинэтансульфоновой кислоты (MES), 30 г/л сахарозы, 1 мг/л зеатина (ZT), 8 г/л агара, 150 мг/л цефалоспорина, 50 мг/л глутаминовой кислоты, 50 мг/л аспарагиновой кислоты, 1 мг/л гиббереллина, 1 мг/л ауксина и 10 мг/л соли глифосата с изопропиламином, рН 5,6) и культивировали при 25°C для дифференцировки. Дифференцированные сеянцы переносили на культуральную среду B5 (3,1 г/л соли B5, витамин B5, 1 г/л 2-морфолина этансульфоновой кислоты (MES), 30 г/л сахарозы, 8 г/л агара, 150 мг/л цефалоспорина и 1 мг/л индол-3-масляной кислоты (IBA)), культивировали в культуральной среде для развития корней до достижения высоты около 10 см при 25°C и переносили в теплицу для культивирования до плодоношения. В теплице растения культивировали при 26°C в течение 16 часов, а затем культивировали при 20°C в течение 8 часов каждый день.
1.3. Идентификация и скрининг трансгенных объектов
Было получено 288 независимых трансгенных растений T0.
Присутствие или отсутствие генов EPSPS и PAT в регенерированных трансгенных соевых растениях анализировали с помощью анализа TaqMan™ (см. Пример 2) и характеризовали число копий в толерантных к глифосату и глутозинату штаммах. При скрининге был отобран, как очень эффективный, объект DBN9004 с однокопийным трансгеном, хорошей устойчивостью к глифосатному гербициду, устойчивостью к глюфозинатному гербициду и агрономически свойствами (см. Пример 6).
Пример 2. Обнаружение трансгенного объекта сои DBN9004 с помощью анализа TaqMan.
Около 100 мг листьев отбирали в качестве образца от трансгенного объекта сои DBN9004 и экстрагировали для получения геномной ДНК с использованием набора для экстракции ДНК растений (DNeasy Plant Maxi Kit, Qiagen). Числа копий гена EPSPS и гена PAT устанавливали способом количественной ПЦР с флуоресцентным зондом Taqman. В то же время, в качестве контролей использовали растения сои дикого типа, которые детектировали и анализировали в соответствии с вышеупомянутым способом. Эксперименты проводили с тройными повторами, результаты которых усредняли.
Конкретные процедуры были следующими:
Стадия 11. 100 мг листьев отбирали от трансгенного объекта сои DBN9004 и измельчали в гомогенат в ступке с жидким азотом, и для каждого образца выполнялись тройные повторы;
Стадия 12. Геномные ДНК вышеупомянутых образцов экстрагировали с использованием набора DNeasy Plant Mini Kit фирмы Qiagen, и описание конкретного способа может содержаться в руководстве по его продукту;
Стадия 13. Концентрации геномных ДНК вышеупомянутых образцов детектировали с использованием прибора NanoDrop 2000 (Thermo Scientific);
Стадия 14. Концентрации геномных ДНК вышеупомянутых образцов доводили до согласованного значения концентрации, которое составляло от 80 до 100 нг/мкл;
Стадия 15. Числа копий у образцов идентифицировали с использованием способа количественной ПЦР с флуоресцентным зондом Taqman, где образцы, для которых числа копий были идентифицированы и известны, были взяты в качестве стандартов, образцы растений сои дикого типа были взяты в качестве контроля и тройные повторы были взяты для каждого образца и усреднены; последовательности флуоресцентных праймеров для количественной ПЦР и флуоресцентного зонда были следующими:
для определения последовательности гена EPSPS использовали следующие праймеры и зонд:
праймер 1: TTGGTGCTAACCTTACCGTTGAG, показанный как SEQ ID NO: 16 в списке последовательностей;
праймер 2: GCTTACCACGACCTTCAAGACG, показанный как SEQ ID NO: 17 в списке последовательностей;
зонд 1: CTGATGCTGACGGTGTGCGTACCATC, показанный как SEQ ID NO: 18 в списке последовательностей;
для определения последовательности гена PAT использовали следующие праймеры и зонд:
праймер 3: CAGTTGAGATTAGGCCAGCTACAG, показанный как SEQ ID NO: 19 в перечне последовательностей;
праймер 4: TTCACTGTAGACGTCTCAATGTAATGG, показанный как SEQ ID NO: 20 в перечне последовательностей;
зонд 2: CAGCTGATATGGCCGCGGTTTGTG, показанный как SEQ ID NO: 21 в списке последовательностей;
Реакционная система ПЦР:
50 × смесь праймеров/зондов содержит 45 мкл каждого праймера в концентрации 1 мМ, 50 мкл зонда в концентрации 100 мкМ и 860 мкл буфера L1 × TE и хранится при 4°C в amber пробирке.
Условия реакции ПЦР:

Данные анализировали с использованием программного обеспечения SDS 2.3 (Applied Biosystems), и был получен однокопийный трансгенный объект сои DBN9004.
Пример 3. Обнаружение трансгенного объекта сои DBN9004
3.1. Экстракция геномной ДНК
Экстракцию ДНК проводили в соответствии со стандартно используемым способом CTAB (cetyltrimethylammonium bromide (цетилтриметиламмонийбромид; ЦТАБ)): взятие 2 г молодых листьев от трансгенного объекта сои DBN9004 и измельчение в порошок в жидком азоте с последующим добавлением 0,5 мл CTAB-буфера для экстракции ДНК (20 г/л CTAB, 1,4 М NaCl, 100 мМ Tris-HCl, 20 мМ ЭДТА (этилендиаминтетрауксусная кислота), с рН, доведенным до 8,0 с помощью NaOH), предварительно нагретого при температуре 65°C, и тщательное смешивание и затем экстрагирование при температуре 65°C в течение 90 минут; добавление 0,5-кратного объема фенола и 0,5-кратного объема хлороформа, затем переворачивание для равномерного смешивания; центрифугирование при 12000 об/мин (обороты в минуту) в течение 10 минут; пипетирование супернатанта, 2-кратный объем безводного этанола, осторожное встряхивание центрифужной пробирки и выдерживание при температуре 4°C в течение 30 минут; повторное центрифугирование со уровенью вращения 12000 об/мин в течение 10 минут; сбор ДНК в нижней части пробирки; отбрасывание супернатанта и промывание осадка 1 мл этанола с массовой концентрацией 70%; центрифугирование со уровенью вращения 12000 об/мин в течение 5 минут; сушка под вакуумом или высушивание на суперчистом столе; и растворение осадка ДНК в соответствующем количестве ТЕ-буфера (10 мМ Трис-HCl, 1 мМ ЭДТА, рН 8,0) и хранение при температурном режиме -20°C.
3.2. Анализ фланкирующей последовательности ДНК
Концентрацию вышеупомянутого экстрагированного образца ДНК измеряли таким образом, чтобы концентрация исследуемого образца составляла от 80 до 100 нг/мкл. Геномную ДНК расщепляли выбранными рестрикционными ферментами PsiI, DraI и TaqI (для 5'-концевого анализа) и AflII, TaqI и PsiI (для 3'-концевого анализа), соответственно. 26,5 мкл геномной ДНК, 0,5 мкл вышеупомянутого рестрикционного фермента и 3 мкл буфера для ферментативного расщепления добавляли в каждую систему ферментативного расщепления и расщепляли в течение 1 часа. После завершения ферментативного расщепления 70 мл абсолютного этанола добавляли в систему расщепления фермента, затем помещали на ледяную баню в течение 30 минут, центрифугировали со уровенью вращения 12000 об/мин в течение 7 минут, супернатант отбрасывали, сушили, после чего добавляли 8,5 мкл дважды дистиллированной воды (dd H2O), 1 мкл 10X T4-DNA-лигазного буфера (NEB T4 DNA Ligase Reaction Buffer, и его конкретную формулу можно найти на веб-сайте NEB или https://www.neb.com/продукты/restriction-endonucleases и https://www.neb.com/products/b0202-t4-dna-ligase-reaction-buffer), и 0,5 мкл T4-DNA-лигазы для лигирования при температуре 4°C в течение ночи. Амплификацию посредством ПЦР проводили с помощью ряда «вложенных праймеров» для выделения 5'- и 3'-трансгенных/геномных ДНК. Конкретно, сочетание праймеров для выделения 5'-трансгенной/геномной ДНК содержит SEQ ID NO: 13 и SEQ ID NO: 26 в качестве первого праймера, SEQ ID NO: 27 и SEQ ID NO: 28 в качестве второго праймера и SEQ ID NO: 13 в качестве праймера для секвенирования. Конкретно, сочетание праймеров для выделения 3'-трансгенной/геномной ДНК содержит SEQ ID NO: 15 и SEQ ID NO: 29 в качестве первого праймера, SEQ ID NO: 30 и SEQ ID NO: 31 в качестве второго праймера и SEQ ID NO: 15 в качестве праймера для секвенирования, и условия реакции ПЦР были таковы, как показано в таблице 3.
Полученные ампликоны подвергали электрофорезу на 2,0% агарозном геле для отделения продукта реакции ПЦР, затем целевые фрагменты выделяли из агарозного матрикса с использованием набора для извлечения из геля (набор для экстракции геля QIAquick Gel Extraction Kit, №28704, Qiagen Inc., Valencia, CA). Затем очищенный продукт ПЦР секвенировали (например, ABI PrismTM 377, PE Biosystems, Foster City, CA) и анализировали (например, программное обеспечение для анализа последовательностей DNASTAR, DNASTAR Inc., Madison, WI).
5'- и 3'-фланкирующие последовательности и контактные последовательности идентифицировали с использованием стандартного способа ПЦР. 5'-фланкирующую последовательность и контактную последовательность можно идентифицировать с использованием SEQ ID NO: 8 или SEQ ID NO: 12, в сочетании с SEQ ID NO: 9, SEQ ID NO: 13 или SEQ ID NO: 26. 3'-фланкирующая последовательность и контактная последовательность могут быть идентифицированы с использованием SEQ ID NO: 11 или SEQ ID NO: 14, в сочетании с SEQ ID NO: 10, SEQ ID NO: 15 или SEQ ID NO: 29. Реакционная система ПЦР и условия амплификации были таковы, как приведено в таблице 2 и таблице 3. Специалисту в данной области техники будет понятно, что для идентификации фланкирующих последовательностей и контактных последовательностей могут также использоваться другие праймерные последовательности.
Секвенирование ДНК продукта ПЦР обеспечивает ДНК, которая может быть использована для создания других молекул ДНК, которые могут быть использованы в качестве праймеров и зондов для идентификации растений или семян сои, полученных из трансгенного объекта сои DBN9004.
Было обнаружено, что позиции 1-1268 нуклеотидов в SEQ ID NO: 5, как было показано, представляют собой правый пограничный фланг (5'-фланкирующая последовательность) вставочной последовательности трансгенного объекта сои DBN9004 в геномной последовательности сои, и положения 6068-7059 нуклеотидов SEQ ID NO: 5, как было показано, представляют собой левый пограничный фланг (3'-фланкирующая последовательность) вставочной последовательности трансгенного объекта сои DBN9004 в геномной последовательности сои. 5'-конъюгированная последовательность была представлена в SEQ ID NO: 1, и 3'-конъюгированная последовательность была представлена в SEQ ID NO: 2.
3.3. ПЦР-анализ конъюгации
Конъюгированная последовательность представляет собой относительно короткую полинуклеотидную молекулу, которая представляет собой новую последовательность ДНК, которая является диагностической для ДНК трансгенного объекта сои DBN9004, когда конъюгированная последовательность обнаруживается в анализе полинуклеиновой кислоты. Конъюгированные последовательности в SEQ ID NO: 1 и SEQ ID NO: 2 представляют собой 11 полинуклеотидов с каждой стороны вставочного сайта трансгенных фрагментов и геномной ДНК сои в трансгенном объекте сои DBN9004. Более длинная или более короткая полинуклеотидная конъюгированная последовательность может быть выбрана из SEQ ID NO: 3 или SEQ ID NO: 4. Конъюгированные последовательности (5'-связывающая область SEQ ID NO: 1 и 3'-связывающая область SEQ ID NO: 2) применимы в качестве ДНК-зондов или молекул ДНК-праймеров в способах обнаружения ДНК. Конъюгированные последовательности SEQ ID NO: 6 и SEQ ID NO: 7 также являются новыми последовательностями ДНК в трансгенном объекте сои DBN9004 и могут также использоваться в качестве ДНК-зондов или молекул ДНК-праймеров для обнаружения присутствия ДНК трансгенного объекта сои DBN9004. SEQ ID NO: 6 (позиции 741-872 нуклеотидов в SEQ ID NO: 3) перекрывает последовательность ДНК конструкции pDBN4003 и терминаторную последовательность t35S, а SEQ ID NO: 7 (позиции 170-287 нуклеотидов в SEQ ID NO: 4) перекрывает последовательность промотора prGm17gTsf1 и последовательность ДНК конструкции pDBN4003.
Кроме того, ампликон генерируют, используя по меньшей мере один праймер из SEQ ID NO: 3 или SEQ ID NO: 4, и праймер при использовании в способе ПЦР используется для генерирования диагностического ампликона трансгенного объекта сои DBN9004.
В частности, продукт ПЦР образуется с 5'-конца трансгенной вставочной последовательности и представляет собой часть геномной ДНК, которая фланкирует 5'-конец введенной последовательности Т-ДНК в геноме, содержащем растительные материалы, полученные из трансгенного объекта сои DBN9004. Данный продукт ПЦР содержит SEQ ID NO: 3. Для осуществления амплификации посредством ПЦР созданы праймер 5 (SEQ ID NO: 8), гибридизованный с последовательностью геномной ДНК, фланкирующей 5'-конец трансгенной вставочной последовательности, и праймер 6 (SEQ ID NO: 9), который спарен с праймером 5 и расположен в трансгенной последовательности терминации транскрипции t35S.
Продукт ПЦР образуется с 3'-конца трансгенной вставочной последовательности и содержит часть геномной ДНК, которая фланкирует 3'-конец введенной последовательности Т-ДНК в геноме, содержащем растительные материалы, полученные из трансгенного объекта сои DBN9004. Данный продукт ПЦР содержит SEQ ID NO: 4. Для осуществления амплификации посредством ПЦР созданы праймер 8 (SEQ ID NO: 11), гибридизованный с последовательностью геномной ДНК, фланкирующей 3'-конец трансгенной вставочной последовательности, и праймер 7 (SEQ ID NO: 10), который спарен с праймером 8 и расположен на 3’-конце вставки, в промоторной последовательности prGm17gTsf1.
Условия амплификации ДНК, описанные в таблицах 2 и 3, могут быть использованы для вышеупомянутой конъюгации с помощью ПЦР, для получения диагностического ампликона трансгенного объекта сои DBN9004. Обнаружение ампликонов может быть выполнено с использованием Stratagene Robocycler, MJ Engine, Perkin-Elmer 9700 или Eppendorf Mastercycler Gradien thermocycler и т.д., или способами и устройствами, известными специалисту в данной области техники.
Таблица 2. Стадии ПЦР и условия реакционной смеси для идентификации 5’-трансгенной вставочной/геномной конъюгированной области трансгенного объекта сои DBN9004
Таблица 3. Условия термоциклера Perkin-Elmer 9700
Материалы осторожно смешивали, если на термоциклере не было изоляционного колпачка, поверх каждого реакционного раствора можно было добавить 1-2 капли минерального масла. ПЦР проводили с использованием вышеупомянутых циклических параметров (таблица 3) в термоциклере Stratagene Robocycler (Stratagene, La Jolla, CA), MJ Engine (MJ R-Biorad, Hercules, CA), Perkin-Elmer 9700 (Perkin Elmer, Boston, MA) или Eppendorf Mastercycler Gradient (Eppendorf, Hamburg, Germany). Термоциклер MJ Engine или Eppendorf Mastercycler Gradient должен запускаться в расчетном режиме. При работе термоциклера Perkin-Elmer 9700 должна быть установлена максимальная уровень подъема температуры.
Экспериментальные результаты показали, что: когда в реакции ПЦР геномной ДНК трансгенного объекта сои DBN9004 были использованы праймеры 5 и 6 (SEQ ID NO: 8 и 9), будет получен продукт амплификации, представляющий собой фрагмент длиной 1207 п.н.о.; когда данные праймеры были использованы в реакциях ПЦР нетрансформированной геномной ДНК сои и геномной ДНК не из DBN9004, ни один фрагмент не будет амплифицироваться; Когда в реакции ПЦР геномной ДНК трансгенного объекта сои DBN9004 были использованы праймеры 7 и 8 (SEQ ID NO: 10 и 11), будет получен продукт амплификации, представляющий собой фрагмент длиной 631 п.н.о.; когда данные праймеры были использованы в реакциях ПЦР нетрансформированной геномной ДНК сои и геномной ДНК не из DBN9004, ни один фрагмент не будет амплифицироваться.
ПЦР-анализ конъюгации также может быть использован для определения того, является ли материал, полученный из трансгенного объекта сои DBN9004, гомозиготой или гетерозиготой. Праймер 9 (SEQ ID NO: 12), праймер 10 (SEQ ID NO: 13) и праймер 11 (SEQ ID NO: 14) использовали для реакции амплификации для получения диагностического ампликона трансгенного объекта сои DBN9004. Условия амплификации ДНК, описанные в таблицах 4 и 5, могут быть использованы для вышеупомянутого анализа конъюгации для получения диагностического ампликона трансгенного объекта сои DBN9004.
Таблица 4. Реакционный раствор для анализа конъюгации
листья анализируемого образца
Таблица 5. Условия термоциклера Perkin-Elmer 9700 для анализа конъюгации
ПЦР проводили с использованием вышеупомянутых циклических параметров (таблица 3) в термоциклере Stratagene Robocycler (Stratagene, La Jolla, CA), MJ Engine (MJ R-Biorad, Hercules, CA), Perkin-Elmer 9700 (Perkin Elmer, Boston, MA) или Eppendorf Mastercycler Gradient (Eppendorf, Hamburg, Germany). Термоциклер MJ Engine или Eppendorf Mastercycler Gradient должен запускаться в расчетном режиме. При работе термоциклера Perkin-Elmer 9700 должна быть установлена максимальная уровень подъема температуры.
В реакции амплификации биологический образец, содержащий матричную ДНК, содержит ДНК для диагностики присутствия трансгенного объекта сои DBN9004 в образце; или реакция будет производить два разных ДНК-ампликона из биологического образца, содержащего ДНК, полученную из генома сои, ДНК, полученная из генома сои, является гетерозиготной по отношению к аллелю, соответствующему вставочной ДНК, присутствующей в трансгенном объекте сои DBN9004. Два таких разных ампликона будут соответствовать первому ампликону, полученному из локуса гена генома дикого типа, и второму ампликону, который диагностирует присутствие ДНК трансгенного объекта сои DBN9004. Получают только образец ДНК сои, соответствующий единственному ампликону второго ампликона, описанного для гетерозиготного генома, таким образом, присутствие в образце трансгенного объекта сои DBN9004 может быть диагностировано и определено, и образец производится семенами сои, которые являются гомозиготными относительно аллеля, соответствующего вставочной ДНК, присутствующей в трансгенном соевом растении DBN9004.
Следует отметить, что пары праймеров трансгенного объекта сои DBN9004 были использованы для генерации ампликонов, которые являются диагностическими для геномной ДНК трансгенного объекта сои DBN9004. Данные пары праймеров включают, но не ограничиваются ими, праймеры 5 и 6 (SEQ ID NO: 8 и 9) и праймеры 7 и 8 (SEQ ID NO: 10 и 11) для использования в способе амплификации ДНК, описанном выше. Кроме того, одна, служащая контролем праймерная пара праймеров 12 и 13 (SEQ ID NO: 22 и 23), используемая для амплификации эндогенного гена сои, была включена в качестве внутреннего стандарта для условий проведения реакции. Анализ образца экстрагированной ДНК трансгенного объекта сои DBN9004 должен включать в себя положительный контроль экстракта ДНК ткани трансгенного объекта сои DBN9004, отрицательный контроль экстракта ДНК, полученный из нетрансгенного объекта сои DBN9004, и отрицательный контроль, не содержащий экстракт матричной ДНК сои. В дополнение к данным парам праймеров также можно использовать любую из праймерных пар из SEQ ID NO: 3 или SEQ ID NO: 4 или комплементарных им последовательностей, и когда они используются в реакциях амплификации ДНК, будут продуцироваться ампликоны, содержащие SEQ ID NO: 1 или SEQ ID NO: 2, которые являются диагностическими по отношению к тканям, полученным из трансгенного объекта растения сои DBN9004, соответственно. Условия амплификации ДНК, описанные в таблицах от 2 до 5, могут быть использованы для получения диагностических ампликонов трансгенного объекта сои DBN9004 с подходящими праймерными парами. При тестировании способом амплификации ДНК, экстракт, содержащий ДНК растения сои или семян сои, которая продуцирует ампликон, диагностический для трансгенного объекта сои DBN9004 и, следовательно, предполагается, что она содержит трансгенный объект сои DBN9004, или продукт, полученный из трансгенного объекта сои DBN9004, может использоваться в качестве матрицы для амплификации, чтобы определить наличие трансгенного объекта сои DBN9004.
Пример 4. Обнаружение трансгенного объекта сои DBN9004 способом саузерн-блот-гибридизации
4.1. Экстракция ДНК для саузерн-блот-гибридизации
Анализ способом саузерн-блот проводили с использованием трансформированных гомозиготных объектов генерации T4, T5 и T6. Ткань растения измельчали в жидком азоте с использованием ступки и пестика. От 4 до 5 г измельченной растительной ткани ресуспендировали в 20 мл буфера CTAB для лизиса (100 мМ Трис, рН 8,0, 20 мМ ЭДТА, рН 8,0, 1,4 М NaCl, 0,2% об./об. β-меркаптоэтанола, 2% мас./об. поливинил-пирролидона) и инкубировали при температуре 65°C в течение 60 минут. В течение инкубационного периода образец переворачивали для равномерного смешивания один раз каждые 10 минут. После инкубации добавляли равный объем хлороформ/изоамиловый спирт (24: 1) и осторожно смешивали переворачиванием, затем центрифугировали со уровенью вращения 4000 об/мин в течение 20 минут. Водную фазу собирали и выдерживали при температуре -20°C в течение 1 часа после добавления равного объема изопропанола, чтобы осадить ДНК, затем центрифугировали со уровенью вращения 4000 об/мин в течение 5 минут, чтобы получить осадок ДНК, который затем ресуспендировали в 1 мл ТЕ-буфера. Для деградации любой присутствующей РНК ДНК инкубировали при температуре 37°C в течение 30 минут с 5 мкл РНКазы А с концентрацией 30 мг/мл, затем центрифугировали со уровенью вращения 4000 об/мин в течение 5 минут и затем центрифугировали со уровенью вращения 14000 об/мин в течение 10 минут для осаждения ДНК в присутствии 0,1 объема 3 М ацетата натрия и 2 объемов безводного этанола. После удаления надосадочной жидкости осадок промывали 1 мл 70% (об./об.) этанола, высушивали и затем повторно растворяли в 1 мл ТЕ-буфера. Концентрации геномных ДНК вышеупомянутых образцов были определены с использованием ультрамикроспектрофотометра (NanoDrop 2000, Thermo Scientific).
4.2. Расщепление рестрикционными ферментами
В 100 мкл реакционной системы каждый раз расщепляли 5 мкг ДНК. Геномную ДНК расщепляли рестрикционными ферментами EcoRI и EcoRV, соответственно, с частичными последовательностями EPSPS и PAT на Т-ДНК в качестве зондов. Для каждого фермента продукты расщепления инкубировали в течение ночи при соответствующей температуре. Образец вращали с помощью центробежного вакуумного концентратора (скоростной вакуум, Thermo Scientific) для уменьшения объема до 20 мкл.
4.3. Гель-электрофорез
Загрузочный краситель бромфенол синий добавляли к каждому образцу из примера 4.2, и каждый образец загружали в 0,7% агарозный гель, содержащий бромид этидия, затем разделяли электрофорезом в буфере для электрофореза TAE (40 мМ Трис-уксусная кислота, 2 мМ ЭДТА, рН 8,0), и гель-электрофорез проводили в течение ночи при 20 вольтах.
Гель обрабатывали денатурирующим раствором (1,5 М NaCl, 0,5 М NaOH) и нейтрализующим раствором (1,5 М NaCl, 0,5 М Трис, рН 7,2) в течение 30 мин, соответственно. 5-кратный раствор SSC выливали в фарфоровое блюдо, поверх клали стеклянную пластину, а затем последовательно помещали мокрый мостик из фильтровальной бумаги, гель, положительно заряженную нейлоновую мембрану (Roche, Cat. No. 11417240001), трехслойную фильтровальную бумагу, бумажные тампоны и тяжелый предмет. После проведения транс-блоттинга в течение ночи при комнатной температуре нейлоновую мембрану промывали в 2-кратном растворе SSC в течение 10 секунд и ДНК иммобилизировали на мембране с помощью УФ-сшивания.
4.4. Гибридизация
Соответствующую последовательность ДНК амплифицировали с помощью ПЦР для подготовки зонда. Зонды ДНК представляют собой SEQ ID NO: 24 и SEQ ID NO: 25 или являются гомологичными или комплементарными вышеупомянутым последовательностям. DIG-маркировку зондов, саузерн-блот-гибридизацию, обнаружение сигнала и другие операции выполняли с использованием высокоэффективного набора для маркировки дигоксином и набора для обнаружения ДНК II (DIG High Prime DNA labelling and Detection Starter Kit II kit, Roche, кат. №1585614910), и конкретные способы описаны в спецификациях продукта.
Каждый саузерн-блот содержит два контрольных образца: (1) ДНК из отрицательного (нетрансформированного) сегреганта, который используется для идентификации любой эндогенной последовательности сои, которая может гибридизоваться с зондом, специфичным для элемента; (2) расщепленную Hind III плазмиду pDBN4003, эквивалентную одной копии на основе длины зонда, которая используется в качестве положительного контроля гибридизации и используется для иллюстрации чувствительности эксперимента.
Данные по гибридизации предоставили убедительные доказательства для поддержки результатов анализа ПЦР способом TaqMan™, т. е. растение сои DBN9004 содержит одиночные копии генов EPSPS и PAT. Одиночные полосы размером около 11 т.о. и 13 т.о. были получены путем расщепления EcoRI и EcoRV с использованием EPSPS-зонда, соответственно; одиночные полосы размером около 8 т.о. и 3,2 т.о. были получены путем расщепления EcoRI и EcoRV с использованием PAT-зонда, соответственно. Это означало, что каждая копия EPSPS и PAT присутствует в соевом трансформированном объекте DBN9004.
Пример 5. Обнаружение трансгенного объекта сои, названного белком, с помощью ELISA
Диапазон экспрессии белков EPSPS и PAT в трансгенном объекте сои DBN9004 может быть выявлен с помощью ELISA.
В качестве образца брали 2 мг свежих листьев трансгенного объекта сои DBN9004 и измельчали в жидком азоте с последующим добавлением 1 мл экстракционного буфера (8 г/л NaCl, 0,27 г/л KH2PO4, 1,42 г/L Na2HPO4, 0,2 г/л KCl и 5,5 мл/л Tween-20, pH 7,4), затем центрифугировали со уровенью вращения 4000 об/мин в течение 10 минут, разбавляя супернатант в 40 раз экстракционным буфером и отобрав 80 мкл разбавленного супернатанта для обнаружения ELISA.
Соотношения количеств белков (белка EPSPS и белка PAT) в образце по массе свежих листьев измеряли и анализировали с помощью наборов ELISA (твердофазный иммуноферментный анализ) (ENVIRLOGIX Company, набор EPSPS и набор PAT). Конкретные способы даны в спецификациях продуктов. В то же время листья растений сои дикого типа (нетрансгенные, NGM; НГМ) использовались в качестве контролей, а также детектировались и анализировались в соответствии с вышеупомянутым способом, и для каждого растения устанавливались шестикратные повторы.
Экспериментальные результаты по содержанию белков (белка EPSPS и белка PAT) трансгенного объекта сои DBN9004 приведены в таблице 6. Соотношения средних значений экспрессии EPSPS-белков в свежих листьях трансгенного объекта сои DBN9004 и соевых растений дикого типа по массе свежих листьев (мкг/г) были определены как 230,9 и 0, соответственно; и соотношения средних величин экспрессии PAT-белков в свежих листьях трансгенного объекта сои DBN9004 и соевых растений дикого типа по массе свежих листьев (мкг/г) составляли соответственно 200,1 и 0.
Таблица 6. Средние результаты измерения величин экспрессии (мкг/г) белков трансгенного объекта сои DBN9004
Пример 6. Обнаружение толерантности к гербицидам трансгенного объекта сои DBN9004
В данном эксперименте для опрыскивания использовали гербицид Roundup (водный агент, содержащий соль 41% глифосата-изопропиламмония) и гербицид Basta (с активным ингредиентом 18% глюфосината). Использовали случайную блочную схему с 3 повторами. Площадь участка составляет 18 м2 (5 м × 3,6 м), с шагом в 60 см, расстояние между рядами 8 см, обычное культивирование и уход, а также широкая зона изоляции размером 1 м между участками. Следующие три типа обработки были проведены на трансгенном объекте сои DBN9004: 1) гербицид не распыляли, а сорняки искусственно уничтожали; 2) гербицид Roundup распыляли в дозе 1680 г э.к./га. (э.к./га. обозначает «эквивалент кислоты активного ингредиента на гектар») во время стадии развития листьев V3, затем гербицид Roundup снова распыляли во время стадии R2 (стадия с полным цветением) в той же дозе; 3) гербицид Basta распыляли в дозе 800 г а.и./га. (а.и./га. обозначает «активный ингредиент на гектар») во время стадии развития листьев V3, затем гербицид Basta снова распыляли во время стадии V6 в той же дозе; 4) гербицид Basta распыляли в дозе 800 г/га во время стадии развития листьев V3, затем гербицид Roundup распыляли в дозе 1680 г а.е./га во время стадии R2. Параллельный контрольный эксперимент проводили с соевыми растениями дикого типа (нетрансгенными, NGM). Следует отметить, что способы перерасчета глифосатных гербицидов разного содержания и форм дозировки в форму эквивалента глифосатной кислоты и перерасчета глюфосинатных растворов разных концентраций в эквивалент активного ингредиента глюфосината, описанные выше, использовались и при получении нижеприведенных выводов.
Симптомы фитотоксичности были исследованы через 1 неделю и 2 недели после применения, а урожайность сои на участке был измерена при уборке. Оценка симптомов фитотоксичности была такой, как показано в таблице 7. Уровень повреждения гербицидами использовали в качестве показателя для оценки толерантности к гербицидам трансформированного объекта. В частности, уровень повреждения гербицидами (%) = Σ (число поврежденных растений одного и того же уровня × номер уровня)/(общее число растений × самый высокий уровень); где уровень повреждения гербицидами включает в себя уровень повреждения глифосатом и уровень повреждения глюфосинатом, и уровень повреждения гербицидами определяли на основании результатов исследования фитотоксичности через 2 недели после обработки глифосатом или глюфосинатом. Урожайность сои на каждом участке представляет собой общую урожайность (массу) соевых зерен, взвешенных в трех средних рядах каждого участка. Разница в урожайности между различными обработками измеряется в виде процента урожайности, где процент урожайности (%) = урожайность с опрыскиванием/урожайность без опрыскивания. Результаты толерантности к гербицидам трансгенного объекта сои DBN9004 и урожайности сои были такими, как показано в таблице 8.
Таблица 7. Стандарты классификации степени фитотоксичности у сои, вызванной гербицидами
Таблица 8. Результаты толерантности к гербицидам трансгенного объекта сои DBN9004 и результаты теста на урожайность
Результаты показали, что в отношении уровня повреждения гербицидами (глифосатом и глюфосинатом): 1) уровень повреждения трансгенного объекта сои DBN9004 составлял по существу 0 при обработке глифосатным гербицидом (1680 г а.е./га); 2) уровень повреждения трансгенного объекта сои DBN9004 также составлял, по существу, 0 при обработке глюфосинатным гербицидом (800 г а.и./га); 3) уровень повреждения трансгенного объекта сои DBN9004 также был равен 0 при обработке глюфосинатным гербицидом (800 г а.и./га) и глифосатным гербицидом (1680 г а.е./га); таким образом, трансгенный объект сои DBN9004 обладает хорошей толерантностью к гербицидам (глифосат и глюфосинат).
Что касается урожайности: урожайность трансгенного объекта сои DBN9004 слегка возросла при трех обработках опрыскиванием глифосатным гербицидом (1680 г а.е./га), глюфосинатным гербицидом (800 г а.и./га) и глюфосинатным гербицидом (800 г а.и./га) + глифосатным гербицидом (1680 г а.е./га) по сравнению с обработкой без опрыскивания, тем самым, дополнительно указывая на то, что трансгенный объект сои DBN9004 обладает хорошей толерантностью к гербицидам (глифосат и глюфосинат).
Пример 7.
Например, сельскохозяйственные продукты или товары могут быть произведены из трансгенного объекта сои DBN9004. Если в сельскохозяйственных продуктах или товарах обнаружена достаточная степень экспрессии, можно ожидать, что сельскохозяйственные продукты или товары содержат нуклеотидную последовательность, способную диагностироваться на присутствие материала трансгенного объекта сои DBN9004 в сельскохозяйственных продуктах или товарах. Продукты или товары включают, но не ограничиваются ими, продукты, полученные из растения сои DBN9004, такие как соевые лепешки, порошки и масла, в частности, лецитины, жирные кислоты, глицерин, стерины, пищевые масла, обезжиренные соевые хлопья, обезжиренная и запеченная соевая мука, соевые молочные сгустки, соевый творог, концентраты соевых белков, изолированные соевые белки, гидролизованные растительные белки, организованные соевые белки, соевые белковые волокна и т.д. Может быть разработан способ обнаружения нуклеиновой кислоты и/или набор на основе зонда или праймерной пары для обнаружения нуклеотидной последовательности трансгенного объекта сои DBN9004, как показано, например, в SEQ ID NO: 1 или SEQ ID NO: 2, в биологическом образце, причем последовательность зонда или последовательность праймера выбрана из последовательностей, представленных SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 3, SEQ ID NO: 4 и SEQ ID NO: 5, или их фрагмента или комплементарной им последовательности, для диагностики присутствия трансгенного объекта сои DBN9004.
В заключение, трансгенный объект сои DBN9004 по настоящему изобретению обладает большей толерантностью к глифосатному гербициду и глюфосинатному гербициду и не влияет на урожайность, и способ обнаружения может точно и быстро определить, содержится ли молекула ДНК трансгенного объекта сои DBN9004 в биологическом образце.
Семена, соответствующие трансгенному объекту сои DBN9004, были депонированы в Центре общей микробиологической культуры Китая (China General Microbiological Culture Collection Center, сокращенно CGMCC, адрес: Institute of Microbiology, Chinese Academy of Sciences, No. 3, No. 1 Courtyard, West Beichen Road, Chaoyang District, Beijing, Postcode: 100101) 27 ноября 2015 года, причем по классификации и номенклатуре они являются соей (Glycine max), с номером доступа CGMCC №11171. Коллекции будут храниться в хранилище в течение 30 лет.
Наконец, следует отметить, что приведенные выше варианты осуществления используются только в целях иллюстрации, а не для ограничения технического решения настоящего изобретения; и хотя настоящее изобретение было подробно описано со ссылкой на предпочтительные варианты осуществления, специалист в данной области техники должно быть очевидно, что модификации или эквивалентные замены для технического решения настоящего изобретения могут быть сделаны без отхода от сущности и объема технического решения настоящего изобретения.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> БЭЙДЖИНГ ДАБЕЙНОНГ ТЕКНОЛОДЖИ ГРУП КО., ЛТД.
БЭЙДЖИНГ ДАБЕЙНОНГ БИОТЕКНОЛОДЖИ КО., ЛТД.
<120> Нуклеиновокислотная последовательность для обнаружения наличия трансгенного трансформанта сои DBN9004 в биологическом образце, набор, содержащий такую последовательность, и способ такого обнаружения
<130> FP1170131P
<160> 31
<170> PatentIn version 3.5
<210> 1
<211> 22
<212> DNA
<213> Artificial Sequence
<220>
<223> A sequence with a length of 22 nucleotides located at the 5ЎЇ terminus of the inserted sequence near the insertion conjugation site in the transgenic soybean event DBN9004
<400> 1
tcacagagtt gtaaacaaat tg 22
<210> 2
<211> 22
<212> DNA
<213> Artificial Sequence
<220>
<223> A sequence with a length of 22 nucleotides located at the 3ЎЇ terminus of the inserted sequence near the insertion conjugation site in the transgenic soybean event DBN9004
<400> 2
tttgaagaca tatttttgtg ga 22
<210> 3
<211> 1207
<212> DNA
<213> Artificial Sequence
<220>
<223> A sequence with a length of 1207 nucleotides located at the 5ЎЇ terminus of the inserted sequence near the insertion conjugation site in the transgenic soybean event DBN9004
<400> 3
cgatagatta cgaataatag cgaaagatct gaaagaaatt tttcatgttc ttttatgaat 60
atgagtttga ctcttcttaa agttatataa aatttcgttg gttggacatt tccttttatg 120
gattaattac gtaattgtga cgacatgaaa ctgcagttta aactcactat aatttcttca 180
ataaatcatt actacacctg acatagtatt tactattttt aaaagtgttt aattttggat 240
tattattagt gtaacgaatt gaattaatta ttttcaaaaa catcactttt ttataaataa 300
aatcctaagg aataatttat tttatgggta gataaaattg attttaagta attgaatgaa 360
atactcatag aaaacattga ataaaattgg aaactatgtc aaaattaatc tcaattatta 420
attcaaatgg gaatacatgc tttttattta ttaaaattac gagaattaaa tttctaacaa 480
aaataaacag gatttgaatt tgtttatcaa acatacaatt acatttttga gtttgatctt 540
gtttatttta aagttgagta aaactatttt tgatataatt aattttaaac tcttggacct 600
gattaaaata tatatatata ttaaaatatg tattcgttat taaaaataca atacaaagaa 660
aatagacggg acttgatgaa agaagagcgt tgcatttacg ctccataaac gtgtgctttc 720
attacggggt cacagagttg taaacaaatt gacgcttaga caacttaata acacattgcg 780
gatacggcca tgctggccgc ccatcatggg cgcgccactg gattttggtt ttaggaatta 840
gaaattttat tgatagaagt attttacaaa tacaaataca tactaagggt ttcttatatg 900
ctcaacacat gagcgaaacc ctataagaac cctaattccc ttatctggga actactcaca 960
cattattata gagagagata gatttgtaga gagagactgg tgatttcagc gggcatgcct 1020
gcaggtcgac tcagatctgg gtaactggcc taactggcct tggaggagct ggcaactcaa 1080
aatccctttg ccaaaaacca acatcatgcc atccaccatg cttgtatcca gctgcgcgca 1140
atgtaccccg ggctgtgtat cccaaagcct catgcaacct aacagatgga tcgtttggaa 1200
ggcctat 1207
<210> 4
<211> 631
<212> DNA
<213> Artificial Sequence
<220>
<223> A sequence with a length of 631 nucleotides located at the 3ЎЇ terminus of the inserted sequence near the insertion conjugation site in the transgenic soybean event DBN9004
<400> 4
gaagaagctg cgctaaaacc ctagccgcaa gagaagaaga gtgaacaaag gtgtgaagag 60
aagtaatgcg acgagggtta gagggtttat ttatagctac aactcttttc ttctgtttac 120
gaaattaccc ttactaattt accctagttc cgtttgtatc cttctctgct ttcgtagcga 180
caattgaaac gctcgctaaa gcgcgtgctt ctcacgtctc gtagttaagc ttgataacgc 240
ggccgctcta gggcgcgccg ggtcccgttt aaactatcag tgtttgaaga catatttttg 300
tggatctagg ttcatcacct acatcttttt ctttctattt taatttaaat atttgtttaa 360
ttctcattaa ttttctttcg gtcatttctt tccacataat tacccattaa atttgagtaa 420
ctacagctct aaaaatcttt tttttaaaat gataattatt acaacaacaa caacaacaac 480
aacgccttat cccactaggt ggggtcgact acatggatca acttccgtca taatgttcta 540
tcaagtacca tacttctatc caaatcatta agttcgaggt ccttcttgat aacctctctt 600
ataatctttt tgggtcttcc tctgcctcga a 631
<210> 5
<211> 7059
<212> DNA
<213> Artificial Sequence
<220>
<223> Entire T-DNA sequence, 5ЎЇ and 3ЎЇ flanking soybean genome sequences
<400> 5
atgtttccct ttttcccgac ccacttctcc ttctatccct tcctatttat aacacctaat 60
tatctatttt attaaagttt tataaataat tcatttattt aataataata attttttaaa 120
atataattga tagtatttta ttttattttt taattattta tcaatatatt ctttaaattt 180
agagtttttt ttttctaaaa ctgattaata caaaaaacat tataaattaa ttcttctaaa 240
aaattaaata ttattttaaa cttagtattc ttataaattg gaatagagta ttatactact 300
cactgttctg tacctttcaa ctttcaattt ggctccctcc actgcttaaa ttctactctc 360
gtcttccatt actggcgtcc ttaattatgc ctttttctta atttcttcat atctcaactc 420
attcataggc ttttgcttat ttattattac gcaaactcgc ctaacatgta ttttctatta 480
aattaatcga tcttgtttta ctaggacaaa aacatatttg ttagatgtcg atagattacg 540
aataatagcg aaagatctga aagaaatttt tcatgttctt ttatgaatat gagtttgact 600
cttcttaaag ttatataaaa tttcgttggt tggacatttc cttttatgga ttaattacgt 660
aattgtgacg acatgaaact gcagtttaaa ctcactataa tttcttcaat aaatcattac 720
tacacctgac atagtattta ctatttttaa aagtgtttaa ttttggatta ttattagtgt 780
aacgaattga attaattatt ttcaaaaaca tcactttttt ataaataaaa tcctaaggaa 840
taatttattt tatgggtaga taaaattgat tttaagtaat tgaatgaaat actcatagaa 900
aacattgaat aaaattggaa actatgtcaa aattaatctc aattattaat tcaaatggga 960
atacatgctt tttatttatt aaaattacga gaattaaatt tctaacaaaa ataaacagga 1020
tttgaatttg tttatcaaac atacaattac atttttgagt ttgatcttgt ttattttaaa 1080
gttgagtaaa actatttttg atataattaa ttttaaactc ttggacctga ttaaaatata 1140
tatatatatt aaaatatgta ttcgttatta aaaatacaat acaaagaaaa tagacgggac 1200
ttgatgaaag aagagcgttg catttacgct ccataaacgt gtgctttcat tacggggtca 1260
cagagttgta aacaaattga cgcttagaca acttaataac acattgcgga tacggccatg 1320
ctggccgccc atcatgggcg cgccactgga ttttggtttt aggaattaga aattttattg 1380
atagaagtat tttacaaata caaatacata ctaagggttt cttatatgct caacacatga 1440
gcgaaaccct ataagaaccc taattccctt atctgggaac tactcacaca ttattataga 1500
gagagataga tttgtagaga gagactggtg atttcagcgg gcatgcctgc aggtcgactc 1560
agatctgggt aactggccta actggccttg gaggagctgg caactcaaaa tccctttgcc 1620
aaaaaccaac atcatgccat ccaccatgct tgtatccagc tgcgcgcaat gtaccccggg 1680
ctgtgtatcc caaagcctca tgcaacctaa cagatggatc gtttggaagg cctataacag 1740
caaccacaga cttaaaacct tgcgcctcca tagacttaag caaatgtgtg tacaatgtgg 1800
atcctaggcc caacctttga tgcctatgtg acacgtaaac agtactctca actgtccaat 1860
cgtaagcgtt cctagccttc cagggcccag cgtaagcaat accagccaca acaccctcaa 1920
cctcagcaac caaccaaggg tatctatctt gcaacctctc tagatcatca atccactctt 1980
gtggtgtttg tggctctgtc ctaaagttca ctgtagacgt ctcaatgtaa tggttaacga 2040
tatcacaaac cgcggccata tcagctgctg tagctggcct aatctcaact ggtctcctct 2100
ccggagacat ggtggatccc cgggtaccct gtcctctcca aatgaaatga acttccttat 2160
atagaggaag ggtcttgcga aggatagtgg gattgtgcgt catcccttac gtcagtggag 2220
atatcacatc aatccacttg ctttgaagac gtggttggaa cgtcttcttt ttccacgatg 2280
ctcctcgtgg gtgggggtcc atctttggga ccactgtcgg cagaggcatc ttcaacgatg 2340
gcctttcctt tatcgcaatg atggcatttg taggagccac cttccttttc cactatcttc 2400
acaataaagt gacagatagc tgggcaatgg aatccgagga ggtttccgga tattaccctt 2460
tgttgaaaag tctcaattgc cctttggtct tctgagactg tatctttgat atttttggag 2520
tagacaagcg tgtcgtgctc caccatgttg acgaagattt tcttcttgtc attgagtcgt 2580
aagagactct gtatgaactg ttcgccagtc tttacggcga gttctgttag gtcctctatt 2640
tgaatctttg actccatggc cactgcaggt cacgatcgat gcggccgctt cgagtggctg 2700
caggtcgatt gatgcatgtt gtcaatcaat tggcaagtca taaaatgcat taaaaaatat 2760
tttcatactc aactacaaat ccatgagtat aactataatt ataaagcaat gattagaatc 2820
tgacaaggat tctggaaaat tacataaagg aaagttcata aatgtctaaa acacaagagg 2880
acatacttgt attcagtaac atttgcagct tttctaggtc tgaaaatata tttgttgcct 2940
agtgaataag cataatggta caactacaag tgttttactc ctcatattaa cttcggtcat 3000
tagaggccac gatttgacac atttttactc aaaacaaaat gtttgcatat ctcttataat 3060
ttcaaattca acacacaaca aataagagaa aaaacaaata atattaattt gagaatgaac 3120
aaaaggacca tatcattcat taactcttct ccatccattt ccatttcaca gttcgatagc 3180
gaaaaccgaa taaaaaacac agtaaattac aagcacaaca aatggtacaa gaaaaacagt 3240
tttcccaatg ccataatact caaactcagt aggattctgg tgtgtgcgca atgaaactga 3300
tgcattgaac ttgacgaacg ttgtcgaaac cgatgatacg aacgaaagct ctagctagag 3360
gatccggtac cgagctcgaa ttcttgagct catcaagcag ccttagtgtc ggagagttcg 3420
atcttagctc caagaccagc catcaaatcc atgaactctg ggaagctagt agcgatcata 3480
gtagcatcat caacagtaac agggttttca gaaacgagac ccataacgag gaagctcata 3540
gcgatacggt gatcgaggtg ggtagcgaca gctgctccag aagcgttacc gagaccctta 3600
ccgtcaggac gaccacgcac gacgagagaa gtctcacctt catcgcaatc aacaccgttg 3660
agcttgagac cgtttgcgac agcagaaaga cggtcgcttt ccttaacacg gagttcttcc 3720
aaaccgttca taacggtagc accttcagcg aatgcagctg caacagcgag aattggatac 3780
tcgtcgatca tagaaggagc acggtcttct ggaacagtaa cacccttcaa agtagaagaa 3840
cgaacacgca agtcagccac gtcttctcca ccagcaagac gtgggttgat cacttcgatg 3900
tcggcaccca tttcctgcag agtcaagatg agaccagtac gggttgggtt catcaaaacg 3960
ttaaggatgg tgacgtcgga acctggaaca agcaaggcag caaccaatgg gaaagcagta 4020
gaggatggat cacctggaac atcaatcact tgaccggtga gcttaccacg accttcaaga 4080
cggatggtac gcacaccgtc agcatcagtc tcaacggtaa ggttagcacc aaaaccttga 4140
agcatctttt cagtgtggtc acgagtcatg attggctcga taacagtggt gatacctggg 4200
gtgttgagac cagcaagcag aacagcggac ttcacttgag cggaagccat aggtaccctg 4260
taggtgattg gcgttggagt ctttggtcca cgcaaggtaa ctggaagacg atcaccgtct 4320
tcagacttca cctgcacacc catttcgcga agtgggttca acacacgacc cattggacgc 4380
ttagtgagag aagcgtcacc aatgaaagtg ctatcgaaat cgtaaacacc aacaagaccc 4440
atagtcaaac ggcaaccagt tgcagcgtta ccgaaatcga gaggagcctc aggagcaagg 4500
agtccaccgt taccaacacc atcaatgatc caagtatcac cttccttacg gattctggca 4560
cccatagctt gcatagcctt accagtgttg ataacatctt caccttccaa aagaccggtg 4620
atacgagttt caccgctagc gagacctcca aacatgaagg acctgtggga gatagacttg 4680
tcacctggaa tacggacggt tccagaaaga ccagaggact tacgagcagt tgctggacgg 4740
ctgcttgcac cgtgaagcat gcacgccgtg gaaacagaag acatgacctt aagaggacga 4800
agctcagagc caattaacgt catcccactc ttcttcaatc cccacgacga cgaaatcgga 4860
taagctcgtg gatgctgctg cgtcttcaga gaaaccgata agggagattt gcgttgactg 4920
gatttcgaga gattggagat aagagatggg ttctgcacac cattgcagat tctgctaact 4980
tgcgccatgg cttccttaaa tctgcaaaaa tccagaaaaa aaaaaagttg tcaaaaccat 5040
aacacatcaa cacaccatat aaagtaaaaa taaaagttta acaacattaa gacctttgtc 5100
attacaaaac ctttcataag cataaaaaga ttaagatcta aatacccacc atacaacaag 5160
cccaaaaata ggtatgctca atcgtaacgc atcaaactca gtatgattcc aacagataaa 5220
aaaatagaca taacaaaatt tatggcactc ccatacataa gatcgagata aaacaagggc 5280
atgtttagca cactgtgacg tatatagatc tattcatcag taaccaaaaa acttgagctt 5340
gaaccgatta aaagtaacag ataatagatc tattcatcag taaccaaaaa actttagctt 5400
gaatcgatta aagtaacagt taatagatct attcatcagt aaccaaaaac tttcttgaac 5460
caaaaactta tataagatct ggatccaaag atttgaatat aacaagcaaa taaaagcctt 5520
accaaaaaaa ctttaaatca aaagtaataa aaaaaacact tttgaaaaca ccatagcacc 5580
gttattaaca actcgaacga aaagaagaag aataagcaca tactacacaa acaaaattga 5640
tctctgaaac atggataaaa ttgaagacaa cgacgacaga ttcagaaaaa acatattgca 5700
agcaacataa accaatagaa taaacacgaa tagaaggaat aaaatccata gaacggtggt 5760
agataacgaa cctagaagaa gctgcgctaa aaccctagcc gcaagagaag aagagtgaac 5820
aaaggtgtga agagaagtaa tgcgacgagg gttagagggt ttatttatag ctacaactct 5880
tttcttctgt ttacgaaatt acccttacta atttacccta gttccgtttg tatccttctc 5940
tgctttcgta gcgacaattg aaacgctcgc taaagcgcgt gcttctcacg tctcgtagtt 6000
aagcttgata acgcggccgc tctagggcgc gccgggtccc gtttaaacta tcagtgtttg 6060
aagacatatt tttgtggatc taggttcatc acctacatct ttttctttct attttaattt 6120
aaatatttgt ttaattctca ttaattttct ttcggtcatt tctttccaca taattaccca 6180
ttaaatttga gtaactacag ctctaaaaat ctttttttta aaatgataat tattacaaca 6240
acaacaacaa caacaacgcc ttatcccact aggtggggtc gactacatgg atcaacttcc 6300
gtcataatgt tctatcaagt accatacttc tatccaaatc attaagttcg aggtccttct 6360
tgataacctc tcttataatc tttttgggtc ttcctctgcc tcgaattgtt tgctttctct 6420
ccatctggtc tactctcctc actacagagt ctaccggttt tctctctaca tgcccaaacc 6480
acctaagtct attttccacc atcttatcta caataggcgc tactccaacc ctctctctaa 6540
tagctccgtt tctaatttta tcctgtcgag tcttaccaca catccaccgc aacatcctca 6600
tctccactac acctacttta gtctcatgtt ggctcttgac cgcccaacat tctgttccgt 6660
acaaaatcgc cagtcttacc gcagtcccat aaaactttcc ctttagcttg atcggtacct 6720
ttgcatcaca taacaccccc gatgcttttc tccatttcat caatcctgct tgaatgcgat 6780
gattcacatc cccttcaatt tctccatcat cctgtattac agacccaaga tatttaaacc 6840
gtgtgacttg agggataata tggtctccta ttttcacctc tgagttagat accctcctac 6900
ttttgttgaa cttacattcc atatactccg atttgcttct gcttaggcga aagccatgtg 6960
tttctagagc tcgtctccaa gtttccaacc tctcattcaa ctcctccctc gactctccaa 7020
ggaggactat gtcatctaca aaaagcatgc atctcggcg 7059
<210> 6
<211> 132
<212> DNA
<213> Artificial Sequence
<220>
<223> A sequence which is located within SEQ ID NO: 3 and spans the nucleotide sequence in the pDBN4003 construct DNA sequence and t35S transcription termination sequence
<400> 6
taaacaaatt gacgcttaga caacttaata acacattgcg gatacggcca tgctggccgc 60
ccatcatggg cgcgccactg gattttggtt ttaggaatta gaaattttat tgatagaagt 120
attttacaaa ta 132
<210> 7
<211> 118
<212> DNA
<213> Artificial Sequence
<220>
<223> A sequence which is located within SEQ ID NO: 4 and spans the prGm17gTsf1 promoter sequence and the nucleotide sequence in the pDBN4003 construct DNA sequence
<400> 7
tttcgtagcg acaattgaaa cgctcgctaa agcgcgtgct tctcacgtct cgtagttaag 60
cttgataacg cggccgctct agggcgcgcc gggtcccgtt taaactatca gtgtttga 118
<210> 8
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> A first primer amplifying SEQ ID NO: 3
<400> 8
cgatagatta cgaataatag 20
<210> 9
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> A second primer amplifying SEQ ID NO: 3
<400> 9
ataggccttc caaacgatcc 20
<210> 10
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> A first primer amplifying SEQ ID NO: 4
<400> 10
gaagaagctg cgctaaaacc 20
<210> 11
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> A second primer amplifying SEQ ID NO: 4
<400> 11
ttcgaggcag aggaagaccc 20
<210> 12
<211> 25
<212> DNA
<213> Artificial Sequence
<220>
<223> A primer on 5ЎЇ flanking genome sequence
<400> 12
acaatacaaa gaaaatagac gggac 25
<210> 13
<211> 25
<212> DNA
<213> Artificial Sequence
<220>
<223> A primer located on T-DNA and paired with SEQ ID NO: 12
<400> 13
gagcatataa gaaaccctta gtatg 25
<210> 14
<211> 21
<212> DNA
<213> Artificial Sequence
<220>
<223> A primer on 3ЎЇ flanking genome sequence, which is paired with SEQ ID NO: 12 to detect whether the transgene is a homozygote or a heterozygote
<400> 14
gggcatgtag agagaaaacc g 21
<210> 15
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> A primer located on T-DNA and paired with SEQ ID NO: 14
<400> 15
attgaaacgc tcgctaaagc 20
<210> 16
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<223> Taqman primer 1 for detecting EPSPS
<400> 16
ttggtgctaa ccttaccgtt gag 23
<210> 17
<211> 22
<212> DNA
<213> Artificial Sequence
<220>
<223> Taqman primer 2 for detecting EPSPS
<400> 17
gcttaccacg accttcaaga cg 22
<210> 18
<211> 26
<212> DNA
<213> Artificial Sequence
<220>
<223> Taqman probe 1 for detecting EPSPS
<400> 18
ctgatgctga cggtgtgcgt accatc 26
<210> 19
<211> 24
<212> DNA
<213> Artificial Sequence
<220>
<223> Taqman primer 3 for detecting PAT
<400> 19
cagttgagat taggccagct acag 24
<210> 20
<211> 27
<212> DNA
<213> Artificial Sequence
<220>
<223> Taqman primer 4 for detecting PAT
<400> 20
ttcactgtag acgtctcaat gtaatgg 27
<210> 21
<211> 24
<212> DNA
<213> Artificial Sequence
<220>
<223> Taqman probe 2 for detecting PAT
<400> 21
cagctgatat ggccgcggtt tgtg 24
<210> 22
<211> 25
<212> DNA
<213> Artificial Sequence
<220>
<223> A first primer of soybean endogenous gene of ubiquitin
<400> 22
agcatttgaa ctacattctc cttga 25
<210> 23
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<223> A second primer of soybean endogenous gene of ubiquitin
<400> 23
tgcagctaca caaatctaaa tcc 23
<210> 24
<211> 506
<212> DNA
<213> Artificial Sequence
<220>
<223> A probe of EPSPS in Southern blot hybridization detection
<400> 24
ccacgacctt caagacggat ggtacgcaca ccgtcagcat cagtctcaac ggtaaggtta 60
gcaccaaaac cttgaagcat cttttcagtg tggtcacgag tcatgattgg ctcgataaca 120
gtggtgatac ctggggtgtt gagaccagca agcagaacag cggacttcac ttgagcggaa 180
gccataggta ccctgtaggt gattggcgtt ggagtctttg gtccacgcaa ggtaactgga 240
agacgatcac cgtcttcaga cttcacctgc acacccattt cgcgaagtgg gttcaacaca 300
cgacccattg gacgcttagt gagagaagcg tcaccaatga aagtgctatc gaaatcgtaa 360
acaccaacaa gacccatagt caaacggcaa ccagttgcag cgttaccgaa atcgagagga 420
gcctcaggag caaggagtcc accgttacca acaccatcaa tgatccaagt atcaccttcc 480
ttacggattc tggcacccat agcttg 506
<210> 25
<211> 310
<212> DNA
<213> Artificial Sequence
<220>
<223> A probe of PAT in Southern blot hybridization detection
<400> 25
cagacttaaa accttgcgcc tccatagact taagcaaatg tgtgtacaat gtggatccta 60
ggcccaacct ttgatgccta tgtgacacgt aaacagtact ctcaactgtc caatcgtaag 120
cgttcctagc cttccagggc ccagcgtaag caataccagc cacaacaccc tcaacctcag 180
caaccaacca agggtatcta tcttgcaacc tctctagatc atcaatccac tcttgtggtg 240
tttgtggctc tgtcctaaag ttcactgtag acgtctcaat gtaatggtta acgatatcac 300
aaaccgcggc 310
<210> 26
<211> 25
<212> DNA
<213> Artificial Sequence
<220>
<223> A primer located on T-DNA in the same direction with SEQ ID NO: 13
<400> 26
gggttcttat agggtttcgc tcatg 25
<210> 27
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<223> A primer located on T-DNA in the opposite direction to SEQ ID NO: 13 and used to obtain a flanking sequence
<400> 27
gcctcatgca acctaacaga tgg 23
<210> 28
<211> 21
<212> DNA
<213> Artificial Sequence
<220>
<223> A primer located on T-DNA in the opposite direction to SEQ ID NO: 13 and used to obtain a flanking sequence
<400> 28
ccaacatcat gccatccacc a 21
<210> 29
<211> 27
<212> DNA
<213> Artificial Sequence
<220>
<223> A primer located on T-DNA in the same direction with SEQ ID NO: 15
<400> 29
cctagttccg tttgtatcct tctctgc 27
<210> 30
<211> 29
<212> DNA
<213> Artificial Sequence
<220>
<223> A primer located on T-DNA in the opposite direction to SEQ ID NO: 15 and used to obtain a flanking sequence
<400> 30
gtttgtgtag tatgtgctta ttcttcttc 29
<210> 31
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<223> A primer located on T-DNA in the opposite direction to SEQ ID NO: 15 and used to obtain a flanking sequence
<400> 31
ctgaatctgt cgtcgttgtc ttc 23
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| РАСТЕНИЕ МАИСА DBN9936 И СПОСОБ ПРИМЕНЕНИЯ В ДЕТЕКТИРОВАНИИ ЕГО ПОСЛЕДОВАТЕЛЬНОСТИ НУКЛЕИНОВОЙ КИСЛОТЫ | 2016 |
|
RU2707527C2 |
| ПОСЛЕДОВАТЕЛЬНОСТЬ НУКЛЕИНОВОЙ КИСЛОТЫ ДЛЯ ВЫЯВЛЕНИЯ РАСТЕНИЯ СОИ DBN8002 И СПОСОБ ЕЕ ВЫЯВЛЕНИЯ | 2019 |
|
RU2818368C2 |
| РАСТЕНИЕ КУКУРУЗЫ DBN9858, УСТОЙЧИВОЕ К ГЕРБИЦИДУ, И НУКЛЕОТИДНАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ И СПОСОБ ДЛЯ ЕЕ ВЫЯВЛЕНИЯ | 2016 |
|
RU2712730C2 |
| ПАКЕТИРОВАННОЕ СОБЫТИЕ 8264.44.06.1 С УСТОЙЧИВОСТЬЮ К ГЕРБИЦИДАМ, СВЯЗАННЫЕ ТРАНСГЕННЫЕ ЛИНИИ СОИ И ИХ ДЕТЕКТИРОВАНИЕ | 2011 |
|
RU2608650C2 |
| ПОСЛЕДОВАТЕЛЬНОСТЬ НУКЛЕИНОВОЙ КИСЛОТЫ ДЛЯ ВЫЯВЛЕНИЯ РАСТЕНИЯ КУКУРУЗЫ DBN9501 И СПОСОБ ЕГО ВЫЯВЛЕНИЯ | 2020 |
|
RU2815214C2 |
| ОБЪЕКТ КУКУРУЗЫ MON 87411 | 2013 |
|
RU2648155C2 |
| ГЕНЕТИЧЕСКАЯ МОДИФИКАЦИЯ MON89788 СОИ И СПОСОБЫ ЕЕ ОПРЕДЕЛЕНИЯ | 2006 |
|
RU2411720C2 |
| ЛИНИИ ТРАНСГЕННОЙ СОИ, ГЕНЕТИЧЕСКОЕ СОБЫТИЕ 8264.42.32.1, УСТОЙЧИВОЕ К ГЕРБИЦИДАМ С ПАКЕТИРОВАННЫМИ ГЕНАМИ НА ЕГО ОСНОВЕ, И ИХ ДЕТЕКТИРОВАНИЕ | 2012 |
|
RU2636021C2 |
| РАСТЕНИЯ СОИ, УСТОЙЧИВОЙ К ГЕРБИЦИДАМ, И СПОСОБЫ ИХ ИДЕНТИФИКАЦИИ | 2010 |
|
RU2639520C2 |
| ЭЛИТНОЕ СОБЫТИЕ ЕЕ-GM3 И СПОСОБЫ И НАБОРЫ ДЛЯ ИДЕНТИФИКАЦИИ ТАКОГО СОБЫТИЯ В БИОЛОГИЧЕСКИХ ОБРАЗЦАХ | 2010 |
|
RU2639530C2 |
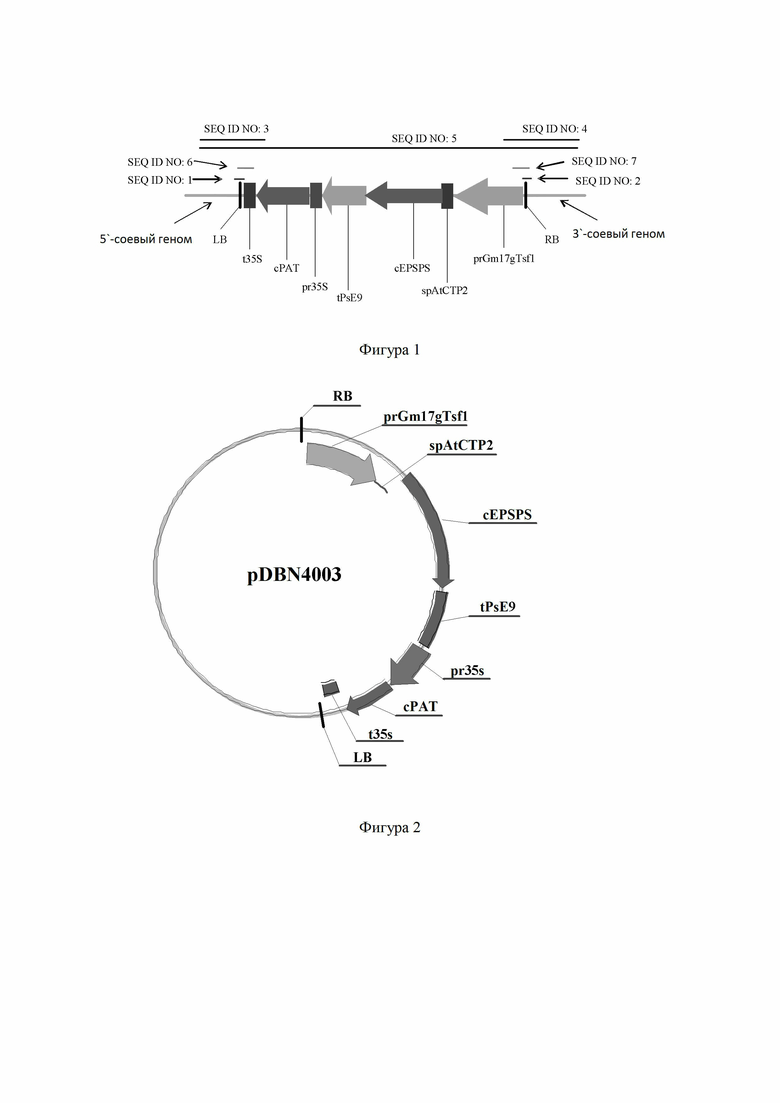
Изобретение относится к области биохимии, в частности к молекуле нуклеиновой кислоты для обнаружения присутствия ДНК трансгенного объекта сои. Также раскрыты способы обнаружения присутствия ДНК трансгенного объекта сои в образце, получение растения сои, культивирования растения сои, защиты растения от повреждения, борьбы с сорняками, с помощью указанной молекулы нуклеиновой кислоты. Раскрыты набор для обнаружения присутствия ДНК трансгенного объекта сои в образце, растительная клетка, часть растения, сельскохозяйственный продукт или товар, содержащие указанную молекулу нуклеиновой кислоты. Изобретение позволяет эффективно получать растения, толерантные к глифосату и/или глюфосинату. 14 н. и 10 з.п. ф-лы, 8 табл.,7 пр., 2 ил.
1. Молекула нуклеиновой кислоты для обнаружения присутствия ДНК трансгенного объекта сои в образце, причем нуклеотидная последовательность молекулы нуклеиновой кислоты содержит SEQ ID NO: 1 или комплементарную ей последовательность и/или SEQ ID NO: 2 или комплементарную ей последовательность, причем трансгенный объект сои представляет собой растение сои, содержащее конструкцию ДНК, имеющую последовательность, как показано в SEQ ID NO: 5.
2. Молекула нуклеиновой кислоты по п. 1, в которой нуклеотидная последовательность содержит SEQ ID NO: 3 или комплементарную ей последовательность и/или SEQ ID NO: 4 или комплементарную ей последовательность.
3. Молекула нуклеиновой кислоты по п. 2, в которой нуклеотидная последовательность содержит SEQ ID NO: 5 или комплементарную ей последовательность.
4. Способ обнаружения присутствия ДНК трансгенного объекта сои в образце, причем данный способ включает: приведение исследуемого образца в контакт по меньшей мере с двумя типами праймеров для амплификации целевого продукта амплификации в реакции амплификации нуклеиновой кислоты;
осуществление реакции амплификации нуклеиновой кислоты; и
обнаружение присутствия целевого продукта амплификации;
причем целевой продукт амплификации содержит SEQ ID NO: 1 или комплементарную ей последовательность и/или SEQ ID NO: 2 или комплементарную ей последовательность, и трансгенный объект сои представляет собой растение сои, содержащее конструкцию ДНК, имеющее последовательность, как показано в SEQ ID NO: 5.
5. Способ обнаружения присутствия ДНК трансгенного объекта сои в образце по п. 4, в котором целевой продукт амплификации дополнительно содержит по меньшей мере одно, выбранное из: SEQ ID NO: 6 или комплементарной ей последовательности и SEQ ID NO: 7 или комплементарной ей последовательности.
6. Способ обнаружения присутствия ДНК трансгенного объекта сои в образце по п. 4 или 5, в котором по меньшей мере один из праймеров содержит нуклеотидную последовательность по любому из пп. 1-3 или ее фрагмент или комплементарную ей последовательность.
7. Способ обнаружения присутствия ДНК трансгенного объекта сои в образце по п. 6, в котором праймеры содержат первый праймер, выбранный из SEQ ID NO: 8 и SEQ ID NO: 10, и второй праймер, выбранный из SEQ ID NO: 9 и SEQ ID NO: 11.
8. Способ обнаружения присутствия ДНК трансгенного объекта сои в образце, причем данный способ включает:
приведение исследуемого образца в контакт с зондом, причем данный зонд содержит SEQ ID NO: 1 или комплементарную ей последовательность и/или SEQ ID NO: 2 или комплементарную ей последовательность;
гибридизацию исследуемого образца с зондом в строгих условиях гибридизации; и
обнаружение состояния гибридизации между исследуемым образцом и зондом;
причем трансгенный объект сои представляет собой растение сои, содержащее конструкцию ДНК, имеющее последовательность, как показано в SEQ ID NO: 5.
9. Способ обнаружения присутствия ДНК трансгенного объекта сои в образце по п. 8, в котором данный зонд дополнительно содержит по меньшей мере одно, выбранное из: SEQ ID NO: 6 или комплементарной ей последовательности, и SEQ ID NO: 7 или комплементарной ей последовательности.
10. Способ обнаружения присутствия ДНК трансгенного объекта сои в образце по п. 8 или 9, в котором по меньшей мере один из зондов помечен по меньшей мере одним флуорофором.
11. Способ обнаружения присутствия ДНК трансгенного объекта сои в образце, причем данный способ включает:
приведение исследуемого образца в контакт с молекулой маркерной нуклеиновой кислоты, причем маркерная молекула нуклеиновой кислоты содержит SEQ ID NO: 1 или комплементарную ей последовательность и/или SEQ ID NO: 2 или комплементарную ей последовательность;
гибридизацию исследуемого образца с маркерной молекулой нуклеиновой кислоты в строгих условиях гибридизации; и
детектирование состояния гибридизации исследуемого образца с маркерной молекулой нуклеиновой кислоты и дальнейшее определение генной связи между толерантностью к глифосату и/или толерантностью к глюфосинату и маркерной молекулой нуклеиновой кислоты генетическим путем с помощью анализа скрещивания с использованием маркера;
причем трансгенный объект сои представляет собой растение сои, содержащее конструкцию ДНК, имеющее последовательность, как показано в SEQ ID NO: 5.
12. Способ обнаружения присутствия ДНК трансгенного объекта сои в образце по п. 11, в котором маркерная молекула нуклеиновой кислоты дополнительно содержит по меньшей мере одно, выбранное из: SEQ ID NO: 6 или комплементарной ей последовательности и SEQ ID NO: 7 или комплементарной ей последовательности.
13. Набор для обнаружения присутствия ДНК трансгенного объекта сои в образце, содержащий по меньшей мере одну молекулу ДНК, причем молекула ДНК содержит последовательность, гомологичную SEQ ID NO: 1 или комплементарной ей последовательности, и последовательность, гомологичную SEQ ID NO: 2 или комплементарной ей последовательности, и она может служить в качестве ДНК-праймера или зонда, специфичного для трансгенного объекта сои или его потомства; и причем трансгенный объект сои представляет собой растение сои, содержащее конструкцию ДНК, имеющее последовательность, как показано в SEQ ID NO: 5.
14. Набор по п. 13, в котором молекула ДНК дополнительно содержит по меньшей мере одно, выбранное из: последовательности, гомологичной SEQ ID NO: 6 или комплементарной ей последовательности, и последовательности, гомологичной SEQ ID NO: 7 или комплементарной ей последовательности.
15. Растительная клетка для обеспечения толерантной к глифосатному и глюфосинатному гербициду растительной клетки, причем она содержит нуклеотидную последовательность, кодирующую толерантный к глифосату белок EPSPS, нуклеотидную последовательность, кодирующую толерантный к глюфосинату белок РАТ, и нуклеотидную последовательность определенной области, причем нуклеотидная последовательность определенной области содержит по меньшей мере одно, выбранное из последовательностей, представленных в SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 6 и SEQ ID NO: 7.
16. Часть растения для обеспечения толерантной к глифосатному и глюфосинатному гербициду части растения, причем она содержит нуклеотидную последовательность, кодирующую толерантный к глифосату белок EPSPS, нуклеотидную последовательность, кодирующую толерантный к глюфосинату белок РАТ, и нуклеотидную последовательность определенной области, причем нуклеотидная последовательность определенной области содержит по меньшей мере одно, выбранное из последовательностей, представленных в SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 6 и SEQ ID NO: 7.
17. Способ получения растения сои, толерантного к глифосатному гербициду и/или глюфосинатному гербициду, причем данный способ включает введение в геном растения сои нуклеотидной последовательности, кодирующей толерантный к глифосату белок EPSPS, и/или нуклеотидной последовательности, кодирующей толерантный к глюфосинату белок РАТ, и нуклеотидной последовательности определенной области, выбранной по меньшей мере из одной нуклеотидной последовательности последовательностей, представленных в SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 3, SEQ ID NO: 4, SEQ ID NO: 6 и SEQ ID NO: 7; предпочтительно, данный способ включает введение в геном растения сои нуклеотидной последовательности, представленной в SEQ ID NO: 5.
18. Способ получения растения сои, толерантного к глифосатному гербициду и/или глюфосинатному гербициду, по п. 17, причем данный способ дополнительно включает: половую гибридизацию первого родительского растения сои трансгенного объекта сои, которое является толерантным к глифосатному гербициду и/или глюфосинатному гербициду, со вторым родительским растением сои, которое не обладает толерантностью к глифосату и/или глюфосинату, для получения большого числа потомков-растений, причем трансгенный объект сои представляет собой растение сои, содержащее конструкцию ДНК, имеющее последовательность, как показано в SEQ ID NO: 5; обработку потомков-растений глифосатным гербицидом и/или глюфосинатным гербицидом; и отбор потомков-растений, которые являются толерантными к глифосату и/или глюфосинату.
19. Способ культивирования растения сои, толерантного к глифосатному гербициду и/или глюфосинатному гербициду, причем данный способ включает: посадку по меньшей мере одного семени сои, причем геном семени сои содержит нуклеотидную последовательность, кодирующую толерантный к глифосату белок EPSPS, и/или нуклеотидную последовательность, кодирующую толерантный к глюфосинату белок РАТ, и нуклеотидную последовательность определенной области;
выращивание семени сои в растение сои; и
опрыскивание растения сои эффективной дозой глифосатного гербицида и/или глюфосинатного гербицида и сбор урожая растения, менее поврежденного по сравнению с другими растениями без нуклеотидной последовательности определенной области; нуклеотидная последовательность определенной области выбрана по меньшей мере из одной нуклеотидной последовательности из последовательностей, представленных в SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 3, SEQ ID NO: 4, SEQ ID NO: 6 и SEQ ID NO: 7;
предпочтительно, способ включает:
посадку по меньшей мере одного семени сои, причем геном семени сои содержит нуклеотидную последовательность, представленную в SEQ ID NO: 5;
выращивание семени сои в растение сои; а также
опрыскивание растения сои эффективным количеством глифосатного гербицида и сбор урожая урожая растения, менее поврежденного по сравнению с другими растениями без SEQ ID NO: 5.
20. Способ защиты растения от повреждения, вызванного гербицидом и/или борьбой с сорняками в поле, причем данный способ включает нанесение гербицида, содержащего эффективную дозу глифосата и/или глюфосината, на поле, предназначенное для посадки по меньшей мере одного трансгенного растения сои, где трансгенное растение сои содержит в своем геноме по меньшей мере одну нуклеотидную последовательность, выбранную из последовательностей, представленных в SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 3, SEQ ID NO: 4, SEQ ID NO: 5, SEQ ID NO: 6 и SEQ ID NO: 7, и трансгенное растение сои обладает толерантностью к глифосатному гербициду и/или глюфосинатному гербициду.
21. Способ борьбы с устойчивыми к глифосату сорняками в поле для толерантного к глифосату растения, причем данный способ включает внесение гербицида, содержащего эффективную дозу глюфосината, на поле, предназначенное для посадки по меньшей мере одного толерантного к глифосату трансгенного растения сои, где толерантное к глифосату трансгенное растение сои содержит в своем геноме по меньшей мере одну нуклеотидную последовательность, выбранную из последовательностей, представленных в SEQ ID NO:1, SEQ ID NO: 2, SEQ ID NO: 3, SEQ ID NO: 4, SEQ ID NO: 5, SEQ ID NO: 6 и SEQ ID NO: 7, и толерантное к глифосату трансгенное растение сои в то же время обладает толерантностью к глюфосинатному гербициду.
22. Способ задержки выработки устойчивости к насекомым, причем данный способ включает выращивание по меньшей мере одного трансгенного растения сои, обладающего толерантностью к глифосату и/или глюфосинату, в поле, предназначенном для посадки растения сои, устойчивого к воздействию насекомых, где трансгенное растение сои, имеющее толерантность к глифосату и/или глюфосинату, содержит в своем геноме по меньшей мере одну нуклеотидную последовательность, выбранную из последовательностей, представленных в SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 3, SEQ ID NO: 4, SEQ ID NO: 5, SEQ ID NO: 6 и SEQ ID NO: 7.
23. Сельскохозяйственный продукт, содержащий полинуклеотид SEQ ID NO: 1 или SEQ ID NO: 2, причем данным сельскохозяйственным продуктом является лецитин, жирная кислота, глицерин, стерол, хлопья сои, соевая мука, соевый белок или его концентрат, соевое масло, соевое белковое волокно, соевый молочный сгусток или соевый творог.
24. Сельскохозяйственный товар, содержащий полинуклеотид SEQ ID NO: 1 или SEQ ID NO: 2, причем данным сельскохозяйственным товаром является лецитин, жирная кислота, глицерин, стерол, хлопья сои, соевая мука, соевый белок или его концентрат, соевое масло, соевое белковое волокно, соевый молочный сгусток или соевый творог.
| УСТОЙЧИВЫЕ К ГЕРБИЦИДАМ РАСТЕНИЯ ПОДСОЛНЕЧНИКА | 2009 |
|
RU2551781C2 |
| Лапка к швейным машинам с приспособлением для вдевания нити в ушко иглы | 1929 |
|
SU19029A1 |
| KR 101354996 B1, 03.02.2014 | |||
| DE 0069332974 D1, 18.06.2003. | |||

