Область изобретения
Настоящее изобретение относится к области молекулярной биологии растений, главным образом, к области скрещивания трансгенных культур в исследовании сельскохозяйственной биотехнологии. В частности, настоящее изобретение относится к трансгенному объекту - сое DBN8002, устойчивому к насекомым и толерантному к гербициду глюфосинат, и последовательностям нуклеиновой кислоты для выявления того, содержит ли биологический образец конкретный трансгенный объект - сою DBN8002, и способу его выявления.
Предшествующий уровень техники
Соя (Glycine max) представляет собой одну из пяти главных сельскохозяйственных культур в мире. Для улучшения агрономических признаков и ее качества к сое применяют биотехнологию. Толерантность к гербицидам, особенно толерантность к гербициду глифосат, является важным агрономическим признаком при производстве сои. Например, существуют успешные объекты сои, такие как GTS40-3-2 и MON89788, которые в различных местах выращивались на больших площадях посадки сои, как например, в США. Еще одним важным агрономическим признаком является устойчивость к насекомым, главным образом устойчивость к насекомым Lepidoptera. Например, существуют успешные объекты сои, такие как MON87701, которые в различных местах выращивались на больших площадях посадки сои, как например, в Бразилии. Стоит упомянуть, что белки Vip имеют механизм действия, отличный от белков Cry, поскольку они представляют собой инсектицидные белки на протяжении вегетационного периода и могут быть использованы в качестве средства для эффективного управления насекомыми, устойчивыми к белкам Cry. Устойчивость к Lepidoptera сое может придавать экспрессия генов устойчивости к Lepidoptera в растениях сои посредством способа трансгенеза. Кроме того, гербицид глюфосинат имеет механизм действия, отличный от гербицида глифосат, поскольку гербицид глюфосинат представляет собой неселективный гербицид контактного типа и может использоваться как средство эффективного борьбы с сорняками, толерантными к глифосату. Толерантность к гербициду глюфосинат сое может придавать экспрессия генов толерантности к гербициду глюфосинат (например, PAT) в растениях сои посредством способа трансгенеза.
Очень важно конструировать экспрессионный вектор, содержащий функциональные чужеродные гены (гены Vip3Aa и PAT), подходящие для трансформации сельскохозяйственных культур сои и для получения соответствующих коммерческих трансгенных объектов сои. На сегодняшний момент все еще отсутствует успешный случай применения белков Vip для осуществления борьбы с насекомыми у растений сои, в то время как толерантность к гербицидам в качестве важного агрономического признака при производстве сои является практически незаменимой. Таким образом, хороший коммерчески подходящий объект трансформации сои требует всестороннего рассмотрения факторов, таких как дизайн вектора для генов Vip3Aa и PAT в растениях сои, взаимодействия между двумя экспрессионными кассетами, эффективность устойчивости к насекомым, эффективность толерантности к гербицидам и эффекты, оказываемые на урожайность и другие физиологические показатели растений, таким образом, чтобы гены Vip3Aa и PAT могли экспрессироватья в соответствующих количествах в сое и выполнять их соответствующие функции, не влияя на урожайность и другие физиологические показатели объекта-сои.
Известно, что на экспрессию экзогенных генов в растениях влияют их положения на хромосомах, возможно из-за структуры хроматина (например, гетерохроматина), или близости элементов регуляции транскрипции (например, энхансера) к сайту интеграции. По данной причине, часто необходимо осуществлять скрининг большого количества объектов для идентификации объекта, который может быть коммерциализирован (а именно, объекта, в котором возможно экспрессируется вводимый целевой ген). Например, у растений и других организмов наблюдали, что может существовать большое различие среди объектов в отношении уровня экспрессии вводимого гена; и могут также существовать различия в отношении пространственных и временных профилей экспрессии, например, различия в относительной экспрессии трансгена в разных растительных тканях, которые отражаются в возможном противоречии между фактическим профилем экспрессии и ожидаемым профилем экспрессии в зависимости от элементов, регулирующих транскрипцию, в вводимой генетической конструкции. По данной причине, обычно существует необходимость в получении сотен-тысяч разных объектов и скрининге данных объектов в отношении одного единственного объекта, который обладает ожидаемым уровнем и профилем экспрессии трансгена в коммерческих целях. Объект, который имеет ожидаемые уровни или профили экспрессии трансгена, является полезным для интрогрессии трансгена в другой генетический фон в результате полового ауткроссинга с использованием традиционных способов скрещивания. Потомство, получаемое посредством такого скрещивания, сохраняет характеристики экспрессии трансгена исходного трансформанта. Данную стратегию используют для обеспечения надежной экспрессии гена в целом ряде сортов, которые хорошо адаптированы к местным условиям выращивания.
Было бы преимущественным иметь способность выявлять наличие конкретного объекта для определения того, содержит ли потомство полового скрещивания целевой ген. Кроме того, способ выявления конкретного объекта был бы полезным для соблюдения родственных правил, таких как правила, требующие предпродажного одобрения и маркирующие продукты питания, происходящие из рекомбинантных сельскохозяйственных растений. Возможно выявлять наличие трансгена любыми хорошо известными способами выявления полинуклеотида, такими как полимеразная цепная реакция (ПЦР) или ДНК-гибридизация с использованием полинуклеотидных зондов. Данные способы выявления обычно сфокусированы на часто используемых генетических элементах, таких как промоторы, терминаторы, маркерные гены. В результате, такие способы могут не быть полезными для проведения различия между разными объектами, особенно объектами, полученными с использованием одной и той же ДНК-конструкции, если не известна последовательность хромосомной ДНК, прилегающая к вставленной трансгенной ДНК («фланкирующая ДНК»). Таким образом, пара праймеров, охватывающая соединение вставленного трансгена и фланкирующей ДНК, обычно используется для идентификации конкретного трансгенного объекта посредством ПЦР, в частности, первый праймер, содержащийся во вставленной последовательности, и второй праймер, содержащийся во вставленной последовательности.
Краткое изложение сущности изобретения
Целью настоящего изобретения является предложение последовательностей нуклеиновой кислоты для выявления растения сои DBN8002 и способов его выявления. Трансгенный объект - соя DBN8002 обладает хорошей устойчивостью к насекомым, а также хорошей толерантностью к гербициду глюфосинат. Способы выявления могут точно и быстро идентифицировать то, содержит ли биологический образец молекулу ДНК трансгенного объекта - сои DBN8002.
Для достижения указанной выше цели, согласно настоящему изобретению предложена последовательность нуклеиновой кислоты, содержащая по меньшей мере 11 последовательных нуклеотидов в положениях 1-642 SEQ ID NO: 3 или комплементарной ей последовательности, и по меньшей мере 11 последовательных нуклеотидов в положениях 643-1524 SEQ ID NO: 3 или комплементарной ей последовательности; и/или по меньшей мере 11 последовательных нуклеотидов в положениях 1-347 SEQ ID NO: 4 или комплементарной ей последовательности, и по меньшей мере 11 последовательных нуклеотидов в положениях 348-656 SEQ ID NO: 4, или комплементарной ей последовательности.
Предпочтительно, последовательность нуклеиновой кислоты содержит 22-25 последовательных нуклеотидов в положениях 1-642 SEQ ID NO: 3 или комплементарной ей последовательности и 22-25 последовательных нуклеотидов в положениях 643-1524 SEQ ID NO: 3 или комплементарной ей последовательности; и/или 22-25 последовательных нуклеотидов в положениях 1-347 SEQ ID NO: 4 или комплементарной ей последовательности и 22-25 последовательных нуклеотидов в положениях 348-656 SEQ ID NO: 4 или комплементарной ей последовательности.
Предпочтительно, последовательность нуклеиновой кислоты содержит SEQ ID NO: 1 или комплементарную ей последовательность, и/или SEQ ID NO: 2 или комплементарную ей последовательность.
Указанная SEQ ID NO: 1 или комплементарная ей последовательность представляет собой последовательность из 22 нуклеотидов, которая расположена около соединения со вставкой на 5'-конце вставленной последовательности в трансгенном объекте - сое DBN8002. Указанная SEQ ID NO: 1 или комплементарная ей последовательность охватывает последовательность геномной ДНК, фланкирующую участок вставки сои, и ДНК-последовательность на 5'-конце вставленной последовательности. Таким образом, включение SEQ ID NO: 1 или комплементарной ей последовательности могло бы отождествляться с наличием трансгенного объекта - сои DBN8002. Указанная SEQ ID NO: 2 или комплементарная ей последовательность представляет собой последовательность из 22 нуклеотидов, которая расположена около соединения со вставкой на 3'-конце вставленной последовательности в трансгенном объекте - сое DBN8002. Указанная SEQ ID NO: 2 или комплементарная ей последовательность охватывает ДНК-последовательность на 3'-конце вставленной последовательности и геномную ДНК-последовательность, фланкирующую участок вставки сои. Таким образом, включение SEQ ID NO: 2 или комплементарной ей последовательности могло бы отождествляться с наличием трансгенного объекта - сои DBN8002.
Предпочтительно, последовательность нуклеиновой кислоты содержит SEQ ID NO: 3 или комплементарную ей последовательность и/или SEQ ID NO: 4 или комплементарную ей последовательность.
Последовательность нуклеиновой кислоты по настоящему изобретению может представлять собой последовательность, которая содержит по меньшей мере 11 или более последовательных полинуклеотидов в любом участке вставляемой последовательности Т-ДНК в SEQ ID NO: 3 или комплементарной ей последовательности (первая последовательность нуклеиновой кислоты) или по меньшей мере 11 или более последовательных полинуклеотидов в любом участке 5'-фланкирующей области геномной ДНК сои в SEQ ID NO: 3 или комплементарной ей последовательности (вторая последовательность нуклеиновой кислоты). Кроме того, последовательность нуклеиновой кислоты может представлять собой последовательность, гомологичную или комплементарную участку SEQ ID NO: 3, содержащей всю SEQ ID NO: 1. При совместном использовании первая последовательность нуклеиновой кислоты и вторая последовательность нуклеиновой кислоты может действовать в качестве пары ДНК-праймеров в способе амплификации ДНК, при котором получается продукт амплификации. Если продукт амплификации, получаемый посредством использования указанной пары ДНК-праймеров в способе амплификации ДНК, содержит SEQ ID NO: 1, может быть диагностировано наличие трансгенного объекта - сои DBN8002 или его потомства. SEQ ID NO: 3 или комплементарная ей последовательность представляет собой последовательность из 1524 нуклеотидов, которая расположена вокруг соединения со вставкой на 5'-конце последовательности Т-ДНК, вставляемой в трансгенный объект сои DBN8002. SEQ ID NO: 3 или комплементарная ей последовательность состоит из 642 нуклеотидов из геномной 5'-фланкирующей последовательности сои (нуклеотиды 1-642 SEQ ID NO: 3), 384 нуклеотидов из ДНК-последовательности конструкции pDBN4006 (нуклеотиды 643-1026 SEQ ID NO: 3) и 498 нуклеотидов из последовательности точки начала транскрипции prAtAct2 (нуклеотиды 1027-1524 SEQ ID NO: 3). Таким образом, включение SEQ ID NO: 3 или комплементарной ей последовательности могло бы отождествляться с наличием трансгенного объекта - сои DBN8002.
Последовательность нуклеиновой кислоты может представлять собой последовательность из по меньшей мере 11 или более последовательных полинуклеотидов в любом участке вставляемой последовательности Т-ДНК в SEQ ID NO: 4 или комплементарной ей последовательности (третья последовательность нуклеиновой кислоты), или по меньшей мере 11 или более последовательных полинуклеотидов в любом участке 3'-фланкирующей области геномной ДНК сои в SEQ ID NO: 4 или комплементарной ей последовательности (четвертая последовательность нуклеиновой кислоты). Кроме того, последовательность нуклеиновой кислоты может представлять собой последовательность, гомологичную или комплементарную участку SEQ ID NO: 4, содержащей всю SEQ ID NO: 2. При использовании совместно, третья последовательность нуклеиновой кислоты и четвертая последовательность нуклеиновой кислоты могут действовать в качестве пары ДНК-праймеров в способе амплификации ДНК, посредством которого образуется продукт амплификации. Если продукт амплификации, образованный посредством использования указанной пары ДНК-праймеров в способе амплификации ДНК, содержит SEQ ID NO: 2, может быть диагностировано наличие трансгенного объекта - сои DBN8002 или его потомства. SEQ ID NO: 4 или комплементарная ей последовательность представляет собой последовательность из 656 нуклеотидов, которая расположена вокруг соединения со вставкой на 3'-конце вставляемой последовательности Т-ДНК в трансгенном объекте - сое DBN8002. SEQ ID NO: 4 или комплементарная ей последовательность состоит из 145 нуклеотидов из ДНК-последовательности терминатора транскрипции t35S (нуклеотиды 1-145 SEQ ID NO: 4), 202 нуклеотидов из ДНК-последовательности конструкции pDBN4006 (нуклеотиды 146-347 SEQ ID NO: 4) и 309 нуклеотидов из геномной 3'-фланкирующей последовательности сои (нуклеотиды 348-656 SEQ ID NO: 4). Таким образом, включение SEQ ID NO: 4 или комплементарной ей последовательности могло бы отождествляться с наличием трансгенного объекта сои DBN8002.
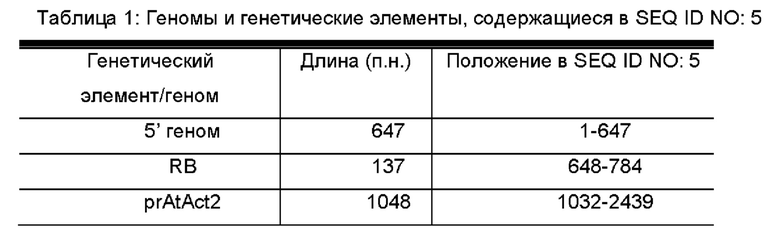
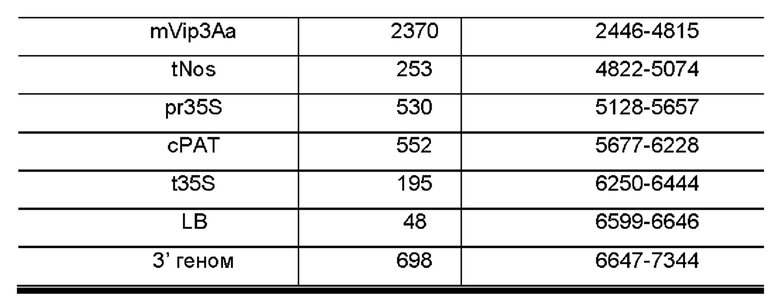
Кроме того, последовательность нуклеиновой кислоты содержит SEQ ID NO: 5 или комплементарную ей последовательность.
Указанная SEQ ID NO: 5 или комплементарная ей последовательность представляет собой последовательность из 7344 нуклеотидов, которая характеризует трансгенный объект - сою DBN8002. Конкретные геномы и генетические элементы, содержащиеся в SEQ ID NO: 5, показаны в Таблице 1. Включение SEQ ID NO: 5 или комплементарной ей последовательности могло бы отождествляться с наличием трансгенного объекта - сои DBN8002.
Как хорошо известно специалистам в данной области, первая, вторая, третья и четвертая последовательности нуклеиновых кислот могут состоять не только из ДНК, но также могут содержать РНК, смесь ДНК и РНК или комбинацию ДНК, РНК и других нуклеотидов или их аналогов, которые не работают в качестве матриц для одной или более полимераз. Кроме того, зонды или праймеры по настоящему изобретению должны иметь длину по меньшей мере примерно 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21 или 22 последовательных нуклеотидов и могут быть отобраны из нуклеотидов, как изложено в SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 3, SEQ ID NO: 4 и SEQ ID NO: 5. При выборе из нуклеотидов, как изложено в SEQ ID NO: 3, SEQ ID NO: 4 и SEQ ID NO: 5, зонды и праймеры могут иметь длину по меньшей мере от примерно 21 до примерно 50 или более последовательных нуклеотидов.
Последовательности нуклеиновой кислоты или их комплементарные последовательности могут быть использованы в способах амплификации ДНК для получения ампликонов, которые используются для выявления наличия трансгенного объекта - сои DBN8002 или его потомства в биологическом образце. Последовательности нуклеиновых кислот или их комплементарные последовательности могут быть использованы в способах выявления нуклеотидов для выявления наличия трансгенного объекта - сои DBN8002 или его потомства в биологическом образце.
Для достижения указанной выше цели согласно настоящему изобретению также предложен способ выявления наличия ДНК, соответствующей трансгенному объекту - сое DBN8002 в образце, включающий:
- приведение образца, подлежащего выявлению, в контакт с по меньшей мере двумя праймерами для амплификации целевого продукта амплификации в реакции амплификации нуклеиновой кислоты;
- проведение реакции амплификации нуклеиновой кислоты; и
- выявление наличия целевого продукта амплификации;
где целевой продукт амплификации содержит последовательность нуклеиновой кислоты.
Предпочтительно, целевой продукт амплификации содержит SEQ ID NO: 1 или комплементарную ей последовательность, SEQ ID NO: 2 или комплементарную ей последовательность, SEQ ID NO: 6 или комплементарную ей последовательность и/или SEQ ID NO: 7 или комплементарную ей последовательность.
В частности, праймеры включают первый праймер и второй праймер, где первый праймер выбран из SEQ ID NO: 1, SEQ ID NO: 8 и SEQ ID NO: 10; и второй праймер выбран из SEQ ID NO: 2, SEQ ID NO: 9 и SEQ ID NO: 11.
Для достижения указанной выше цели согласно настоящему изобретению также предложен способ выявления наличия ДНК, соответствующей трансгенному объекту-сое DBN8002 в образце, включающий:
- приведение образца, подлежащего выявлению, в контакт с зондом, где зонд содержит последовательность нуклеиновой кислоты;
- гибридизацию образца, подлежащего выявлению, с зондом в жестких условиях гибридизации; и
- выявление гибридизации образца, подлежащего выявлению, с зондом.
Жесткие условия могут представлять собой гибридизацию при 65°С в растворе 6 x SSC (цитрат натрия) и 0,5% SDS (от англ. sodium dodecyl sulfate - додецилсульфат натрия) с последующей промывкой на мембране в растворе 2 x SSC и 0,1% SDS и растворе 1 x SSC и 0,1% SDS (каждым по одному разу).
Предпочтительно, зонд содержит SEQ ID NO: 1 или комплементарную ей последовательность, SEQ ID NO: 2 или комплементарную ей последовательность, SEQ ID NO: 6 или комплементарную ей последовательность и/или SEQ ID NO: 7 или комплементарную ей последовательность.
Возможно, по меньшей мере один зонд мечен по меньшей мере одним флуорофором.
Для достижения указанной выше цели согласно настоящему изобретению также предложен способ выявления наличия ДНК, соответствующей трансгенному объекту - сое DBN8802 в образце, включающий:
- приведение образца, подлежащего выявлению, в контакт с маркерной молекулой нуклеиновой кислоты, где маркерная молекула нуклеиновой кислоты содержит последовательность нуклеиновой кислоты;
- гибридизацию образца, подлежащего выявлению, с маркерной молекулой нуклеиновой кислоты в жестких условиях гибридизации;
- выявление гибридизации образца, подлежащего выявлению, с маркерной молекулой нуклеиновой кислоты, и дополнительно проведение анализа скрещивания с помощью маркера для определения того, связана ли генетически устойчивость к насекомым и/или толерантность к гербицидам с маркерной молекулой нуклеиновой кислоты.
Предпочтительно, маркерная молекула нуклеиновой кислоты содержит по меньшей мере одну последовательность, выбранную из группы, состоящей из SEQ ID NO: 1 или комплементарной ей последовательности, SEQ ID NO: 2 или комплементарной ей последовательности и/или SEQ ID NO: 6-11 или комплементарных им последовательностей.
Для достижения указанной выше цели, согласно настоящему изобретению также предложен набор для выявления ДНК, содержащий по меньшей мере одну молекулу ДНК, где молекула ДНК содержит последовательность нуклеиновой кислоты, и молекула ДНК может действовать в качестве ДНК-праймера или зонда, специфичного в отношении трансгенного объекта - сои DBN8002 или его потомства.
Предпочтительно, молекула ДНК содержит SEQ ID NO: 1 или комплементарную ей последовательность, SEQ ID NO: 2 или комплементарную ей последовательность, SEQ ID NO: 6 или комплементарную ей последовательность и/или SEQ ID NO: 7 или комплементарную ей последовательность.
Для достижения указанной выше цели согласно настоящему изобретению также предложена растительная клетка, содержащая последовательность нуклеиновой кислоты, кодирующую белок устойчивости к насекомым Vip3Aa, последовательность нуклеиновой кислоты, кодирующую белок толерантности к гербициду глюфосинат PAT, и последовательность нуклеиновой кислоты специфичной области, где последовательность нуклеиновой кислоты данной специфичной области содержит последовательность, как изложено в SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 6 и/или SEQ ID NO: 7.
Предпочтительно, растительная клетка содержит последовательность нуклеиновой кислоты, кодирующую белок устойчивости к насекомым Vip3Aa, последовательность нуклеиновой кислоты, кодирующую белок толерантности к гербициду глюфосинат PAT, и последовательность нуклеиновой кислоты специфичной области, где последовательность нуклеиновой кислоты специфичной области содержит последовательность, как изложено в SEQ ID NO: 3 и/или SEQ ID NO: 4.
Предпочтительно, растительная клетка содержит последовательно SEQ ID NO: 1, последовательность нуклеиновой кислоты в положениях 1032-6444 SEQ ID NO: 5 и SEQ ID NO: 2 или содержит последовательность, как изложено в SEQ ID NO: 5.
Для достижения упомянутой выше цели согласно настоящему изобретению также предложен способ защиты растения сои от нашествия насекомых, включающий предоставление по меньшей мере одной трансгенной растительной клетки сои в рацион целевого насекомого, где трансгенная растительная клетка сои содержит в своем геноме последовательность, как изложено в SEQ ID NO: 1, и/или SEQ ID NO: 2; и попадание внутрь трансгенной растительной клетки сои ингибирует дальнейшее питание целевого насекомого данным трансгенным растением - соей.
Предпочтительно, трансгенная растительная клетка сои сдержит в своем геноме последовательность, как изложено в SEQ ID NO: 3 и/или SEQ ID NO: 4.
Предпочтительно, трансгенная растительная клетка сои последовательно содержит в своем геноме SEQ ID NO: 1, последовательность нуклеиновой кислоты в положениях 1032-6444 SEQ ID NO: 5 и SEQ ID NO: 2 или содержит SEQ ID NO: 5.
Для достижения указанной выше цели согласно настоящему изобретению также предложен способ защиты растения сои от повреждения, обусловленного гербицидом, или борьбы с сорняками в поле, в котором посажено растение сои, включающий внесение эффективного количества гербицида глюфосинат в поле, в котором посажено по меньшей мере одного трансгенное растение сои, где трансгенное растение сои содержит в своем геноме последовательность, как изложено в SEQ ID NO: 1 и/или SEQ ID NO: 2, и трансгенное растение сои обладает толерантностью к гербициду глюфосинат.
Предпочтительно, трансгенное растение сои содержит в своем геноме последовательность, как изложено в SEQ ID NO: 3 и/или SEQ ID NO: 4.
Предпочтительно, трансгенное растение сои последовательно содержит в своем геноме SEQ ID NO: 1, последовательность нуклеиновой кислоты в положениях 1032-6444 SEQ ID NO: 5 и SEQ ID NO: 2, или содержит последовательность, как изложено в SEQ ID NO: 5.
Для достижения указанной выше цели согласно настоящему изобретению также предложен способ скрещивания растения сои, устойчивого к насекомым и/или толерантного к гербициду глюфосинат, включающий:
- посадку по меньшей мере одного семени сои, где данное семя сои содержит в своем геноме последовательность нуклеиновой кислоты, кодирующую белок устойчивости к насекомым Vip3Aa, и/или последовательность нуклеиновой кислоты, кодирующую белок толерантности к гербициду глюфосинат PAT, и последовательность нуклеиновой кислоты специфичной области, или семя сои содержит в своем геноме последовательность нуклеиновой кислоты, как изложено в SEQ ID NO: 5;
- выращивание семени сои в растение сои; и
- заражение растения сои целевым насекомым и/или опрыскивание растения сои эффективным количеством гербицида глюфосинат и затем осуществление сбора растения с уменьшенным повреждением растения, по сравнению с другими растениями, которые не содержат последовательность нуклеиновой кислоты специфичной области;
где последовательность нуклеиновой кислоты специфичной области представляет собой последовательность, как изложено в SEQ ID NO: 1 и/или SEQ ID NO: 2; предпочтительно, последовательность нуклеиновой кислоты специфичной области представляет собой последовательность, как изложено в SEQ ID NO: 3 и/или SEQ ID NO: 4.
Для достижения упомянутой выше цели согласно настоящему изобретению также предложен способ получения растения сои, устойчивого к насекомым и/или толерантного к гербициду глюфосинат, включающий:
- введение последовательности нуклеиновой кислоты, кодирующей белок устойчивости к насекомым Vip3Aa, и/или последовательности нуклеиновой кислоты, кодирующей белок толерантности к гербициду глюфосинат PAT, и последовательности нуклеиновой кислоты специфичной области, содержащейся в геноме первого растения сои, или последовательности нуклеиновой кислоты, как изложено в SEQ ID NO: 5, содержащейся в геноме первого растения сои, во второе растение сои, получая таким образом множество растений потомства; и
- отбор растений потомства, содержащих последовательность нуклеиновой кислоты специфичной области, которые также устойчивы к насекомым и/или толерантны к гербициду глюфосинат;
которые также являются устойчивыми к насекомым и/или толерантными к гербициду глюфосинат;
где последовательность нуклеиновой кислоты специфичной области представляет собой последовательность, как изложено в SEQ ID NO: 1 и/или SEQ ID NO: 2; предпочтительно, последовательность нуклеиновой кислоты специфичной области представляет собой последовательность, как изложено в SEQ ID NO: 3 и/или SEQ ID NO: 4.
Предпочтительно, способ включает:
- половое скрещивание трансгенного объекта - сои DBN8002 с растением сои, которое не обладает признаком устойчивости к насекомым и/или толерантности к гербициду глюфосинат, с получением, таким образом, множества растений потомства;
- отбор растений потомства, содержащих последовательность нуклеиновой кислоты специфичной области;
- заражение растений потомства целевым насекомым и/или обработка растений потомства глюфосинатом; и
- отбор растений потомства, которые являются устойчивыми к насекомым и/или толерантными к гербициду глюфосинат.
Для достижения указанной выше цели, согласно настоящему изобретению также предложен сельскохозяйственный продукт или товар, полученный из трансгенного объекта - сои DBN8002, где сельскохозяйственный продукт или товар представляет собой лецитин, жирные кислоты, глицерин, стерины, соевые хлопья, соевую муку, соевые белки или их концентраты, соевые масла, белковые волокна сои, продукты свертывания соевого молока или соевый творог.
В последовательностях нуклеиновой кислоты для выявления растений сои и способах их выявления согласно настоящему изобретению предложены следующие определения и способы для лучшего определения настоящего изобретения и для руководства для специалистов в данной области в реализации настоящего изобретения. Если не указано иное, термины нужно понимать в соответствии с общепринятым применением специалистами в данной области.
Термин «соя» относится к Glycine max и содержит все сорта растений, которые могут быть сопряжены с соей, включая дикорастущие виды сои.
Термин «содержащий», «включают» или «содержат» означает «включающий, но не ограниченный…».
Термин «растение» включает целое растение, растительные клетки, органы растений, протопласты растений, культуры тканей из растительных клеток, из которых растения могут быть регенерированы, растительные куллусы, скопления растительных клеток и растительные клетки, которые являются интактными в растениях или частях растения, где данные части растения могут представлять собой, например, зародыши, пыльцу, семязачатки, семена, листья, цветки, ветки, плоды, стебли, корни, корневые кончики или пыльники. Следует понимать, что в пределах объема настоящего изобретения, части трансгенных растений включают растительные клетки, протопласты, ткани, каллусы, зародыши, цветки, стебли, пложы, листья и корни, но не ограничиваются ими. Упомянутые выше части растений происходят из трансгенных растений или их потомства, которые предварительно трансформированы молекулыми ДНК по настоящему изобретению, и, таким образом, по меньшей мере частично состоят из трансгенных клеток.
Термин «ген» относится к фрагменту нуклеиновой кислоты, который экспрессирует конкретный белок, включающему регуляторные последовательности, предшествующие (5' некодирующие последовательности) и следующие за (3' некодирующие последовательности) кодирующей последовательностью. «Нативный ген» относится к гену, который обнаружен в природе со своими собственными регуляторными последовательностями. «Химерный ген» относится к любому гену, который не является нативным геном, содержащему регуляторные и кодирующие последовательности, которые не обнаружены в природе. «Эндогенный ген» относится к нативному гену в своем природном положении в геноме организма. «Экзогенный ген» представляет собой чужеродный ген, в настоящее время присутствующий в геноме организма, который не содержит его в природе, и также относится к гену, вводимому в рецепторную клетку способом трансгенеза. Экзогенные гены могут включать нативные гены или химерные гены, вставленные в организм, не являющийся нативным. «Трансген» представляет собой ген, который был введен в геном способом трансформации. «Участок вставки» или «целевой участок» относится к участку в растительном геноме, в который вставлена рекомбинантная ДНК.
«Фланкирующая ДНК» может содержать или геном, находящийся в природе в организме, таком как растение, или экзогенную (гетерологичную) ДНК, введенную способом трансформации, например, фрагмент, ассоциированный с объектом трансформации. Таким образом, фланкирующая ДНК может включать комбинацию нативной ДНК и экзогенной ДНК. В том виде, в котором он используется в данном документе, термин «фланкирующая ДНК», также называемая «фланкирующей областью», «фланкирующей последовательностью», «фланкирующей геномной последовательностью» или «фланкирующей геномной ДНК», относится к последовательности из по меньшей мере 3, 5, 10, 11, 15, 20, 50, 100, 200, 300, 400, 1000, 1500, 2000, 2500 или 5000 пар оснований или больше, которая расположена или непосредственно выше или ниже исходно вставленной экзогенной молекулы ДНК и прилегает к ней. Когда данная фланкирующая область расположена ниже, она может также называться «3' фланкирующей областью» или «фланкирующей областью левой границы» и тому подобное. Когда данная фланкирующая область расположена выше, она может также называться «5' фланкирующей областью» или «фланкирующей областью правой границы» и тому подобное.
Способы трансформации, приводящие к случайной интеграции экзогенной ДНК, будут приводить к получению трансформантов, содержащих разные фланкирующие области, которые характерны для каждого трансформанта. Когда рекомбинантную ДНК вводят в растение посредством традиционного скрещивания, ее фланкирующие области обычно не будут изменены. Трансформанты также будут содержать уникальные соединения вставки гетерологичной ДНК и фрагмента геномной ДНК или двух фрагментов геномной ДНК или двух фрагментов гетерологичной ДНК. «Соединение» представляет собой точку, в которой связаны два конкретных фрагмента ДНК. Например, соединение существует в положении, в котором ДНК-вставка связана с фланкирующей ДНК. Точка соединения также существует в трансформированном организме, в котором два фрагмента ДНК связаны вместе до некоторой степени, которая модифицирована по сравнению со степенью, обнаруженной в нативном организме. «Область соединения» или «последовательность соединения» относится к ДНК, которая содержит точку соединения.
Согласно настоящему изобретению предложен трансгенный объект - соя, называемый DBN8002, и его потомство. Трансгенный объект - сою DBN8002 также называют растением сои DBN8002, включая растения и семена трансгенного объекта - сои DBN8002 вместе с растительными клетками или их возобновляемыми частями, где данные части растения трансгенного объекта - сои DBN8002 включают клетки, пыльцу, семязачатки, цветки, почки, корни, стебли, листья, стручки и продукты, происходящие из растения сои DBN8002, такие как соевый жмых, порошки и масла, конкретно лецитин, жирные кислоты, глицерин, стерины, пищевые масла, обезжиренные соевые хлопья, обезжиренная и пропеченная соевая мука, продукты свертывания соевого молока, соевый творог, концентраты соевых белков, выделенные соевые белки, гидролизованные растительные белки, организованные соевые белки и волокна соевых белков, но не ограничиваются ими.
Трансгенный объект - соя DBN8002 по настоящему изобретению содержит ДНК-конструкцию, которая при экспрессии в растительных клетках придает трансгенному объекту - сое DBN8002 устойчивость к насекомым и толерантность к гербициду глюфосинат. ДНК-конструкция содержит две экспрессионные кассеты, расположенные последовательно. Первая экспрессионная кассета содержит подходящий промотор для экспрессии в растении и подходящую сигнальную последовательность полиаденилирования, где промотор функционально связан с последовательностью нуклеиновой кислоты белка Vip3Aa, который главным образом устойчив к насекомым Lepidoptera. Вторая экспрессионная кассета содержит подходящий промотор для экспрессии в растении и подходящую сигнальную последовательность полиаденилирования, где промотор функционально связан с геном, кодирующим фосфинотрицин-N-ацетилтрансферазу (PAT - от англ. phosphinothricin N-acetyltransferase), и последовательность нуклеиновой кислоты белка PAT устойчива к гербициду глюфосинат. Кроме того, промотор может представлять собой подходящий промотор, выделенный из растений, включая конститутивный, индуцибельный и/или тканеспецифичный промоторы. Подходящий промотор включает, но не ограничивается промотором 35S вируса мозаики цветной капусты (CaMV - от англ. Cauliflower mosaic virus), промотором 35S вируса мозаики норичника (FMV - от англ. figwort mosaic virus), промотором убиквитина, промотором актина, промотором нопалинсинтетазы Agrobacteríum tumefaciens (NOS - от англ. nopaline synthetase), промотором октопинсинтетазы (OCS - от англ. octopine synthetase), промотором вируса желтой курчавости листьев Cestrum, промотором пататина, промотором рибулозо-1,5-бисфосфаткарбоксилазы/оксигеназы (RuBisCO), промотором глутатион-S-трансферазы (GST - от англ. glutathione S-transferase), промотором Е9, промотором GOS, промотором alcA/alcR, промотором Agrobacteríum rhizogenes RolD и промотором Suc2 Arabidopsís thalíana. Сигнальная последовательность полиаденилирования может представлять собой подходящую сигнальную последовательность полиаденилирования, которая работает у растений. Подходящая сигнальная последовательность полиаденилирования включает, но не ограничивается сигнальной последовательностью полиаденилирования, происходящей из гена нопалинсинтетазы (NOS) Agrobacteríum tumefaciens, сигнальной последовательностью полиаденилирования, происходящей из терминатора 35S вируса мозаики цветной капусты (CaMV), сигнальной последовательностью полиаденилирования, происходящей из гена ингибитора протеазы II (PIN II), и сигнальной последовательностью полиаденилирования, происходящей из гена α-тубулина.
Кроме того, экспрессионная кассета может дополнительно содержать другие генетические элементы, включая, но, не ограничиваясь энхансером и сигнальным пептидом/транзитным пептидом. Энхансер, усиливающий уровень экспрессии гена, включает, но не ограничивается активатором трансляции вируса гравировки табака (TEV - от англ. Tobacco Etch Virus), энхансером CaMV35S и энхансером FMV35S. Сигнальный пептид/транзитный пептид могут направлять белок Vip3Aa и/или белок PAT для осуществления транспорта к внеклеточному пространству или в конкретную внутриклеточную органеллу или компартмент. Например, последовательность, кодирующую транзитный пептид хлоропласта используют для нацеливания на хлоропласт, или последовательности удерживания «KDEL» используют для нацеливания на эндоплазматический ретикулум.
Указанный ген mVip3Aa может быть выделен из Bacillus thuringiensis (сокр. Bt), и нуклеотидную последовательность гена mVip3Aa можно менять посредством оптимизации кодонов или другими способами, таким образом, чтобы улучшать стабильность и доступность транскрипта в трансформированных клетках.
Термин «Lepidoptera», включающий как мотыльков, так и бабочек, представляет собой самый большой отряд насекомых-вредителей в сельском и лесном хозяйстве. Он включает, например, Agrotis ypsiion (Rottemberg), Helicoverpa armígera (Hubner, Prodenia litura, Athetis lepigone и Conogethes punctiferalis.
Ген фосфинотрицин-N-ацетилтрансферазы (PAT) может представлять собой фермент, выделенный из штамма Streptomyces viridochromogenes, который катализирует превращение L-фосфинотрицина в его неактивную форму посредством ацетилирования таким образом, что придает растению толерантность к гербициду глюфосинат. Фосфинотрицин (РТС, 2-амино-4-метилфосфинил-масляная кислота) представляет собой ингибитор глутаминсинтетазы. РТС представляет собой структурную единицу антибиотика 2-амино-4-метилфосфинилаланилаланина. Данный трипептид (РТТ) является активным в отношении грамположительных и грамотрицательных бактерий и гриба Botrytis cinérea. Ген фосфинотрицин-N-ацетилтрансферазы (PAT) может также служить селективным маркерным геном.
«Глюфосинат» (также известный как фосфинотрицин) относится к аммоний-2-амино-4-[гидрокси(метил)фосфинил]бутирату. Обработки «гербицидом глюфосинат» относятся к обработкам любой композицией гербицида, содержащей глюфосинат. Выбор норм внесения для композиции глюфосината находится в пределах компетентности обычного агротехника таким образом, чтобы реализовать биологически эффективное количество. Обработка поля, содержащего растительный материал от трансгенного объекта - сои DBN8002, любой композицией гербицида, содержащей глюфосинат, будет осуществлять борьбу с ростом сорняков в поле и не будет влиять на рост или урожай растительного материала, происходящего из трансгенного объекта - сои DBN8002.
ДНК-конструкцию вводят в растение способами трансформации, включая, но, не ограничиваясь трансформацией, опосредованной Agrobacteríum, баллистической трансформацией и подходом к трансформации «путь пыльцевой трубки».
Трансформация, опосредованная агробактерией, является широко используемым способом трансформации растений. Экзогенную ДНК, подлежащую введению в растение, клонируют в вектор между консенсусными последовательностями левой и правой границ, а именно, область Т-ДНК. Вектором трансформируют клетки Agrobacteríum, которые впоследствии используют для инфицирования растительных тканей. Область Т-ДНК в векторе, содержащую экзогенную ДНК, вставляют в растительный геном.
Баллистическая трансформация представляет собой бомбардировку растительных клеток вектором, содержащим экзогенную ДНК (биолистическая трансформация, опосредованная частицами).
Трансформация «путь пыльцевой трубки» представляет собой способ переноса экзогенной ДНК в зародышевый мешок путем, опосредованным нуцеллусом, с использованием природной пыльцевой трубки (также известен как передающая ткань пыльцевой трубки), образованной после опыления растений.
После трансформации необходимо регенерировать трансгенное растение из трансформированной растительной ткани и выбрать потомство с экзогенной ДНК, используя подходящий маркер.
ДНК-конструкция представляет собой сборку молекул ДНК, связанных вместе, которая обеспечивает одну или более экспрессионных кассет. ДНК-конструкция предпочтительно представляет собой плазмиду, которая может самореплицироваться в бактериальной клетке и содержит разные сайты распознавания эндонуклеазой рестрикции, которые полезны для введения молекул ДНК, которые обеспечивают функциональные генетические элементы, а именно, промоторы, интроны, лидерные последовательности, кодирующие последовательности, 3'-области терминации и другие последовательности. Экспрессионные кассеты, содержащиеся в ДНК-конструкции, содержат необходимые генетические элементы, которые необходимы для транскрипции матричной РНК и могут быть сконструированы для экспрессии в прокариотических клетках или эукариотических клетках. Наиболее предпочтительно, экспрессионные кассеты по настоящему изобретению сконструированы для экспрессии в растительных клетках.
Трансгенный «объект» получают в результате трансформации растительных клеток гетерологичной ДНК-конструкцией, включая стадии осуществления вставки экспрессионной кассеты нуклеиновой кислоты, содержащей по меньшей мере один целевой ген, в геном растения способом трансгенеза для создания популяции растений, регенерации данной популяции растений и отбора конкретного растения, имеющего характеристики вставки в конкретное положение генома. Термин «объект» относится к исходному трансформанту и его потомству, которые включают гетерологичную ДНК. Термин «объект» также относится к потомству, полученному в результате полового скрещивания исходного трансформанта и отдельно взятого объекта других сортов, которое включает гетерологичную ДНК. Даже после повторного обратного скрещивания с родителем при обратном скрещивании, вставленная ДНК и фланкирующая геномная ДНК из родителя исходного трансформанта все еще находятся в том же положении на хромосоме в потомстве, полученном в результате скрещивания. Термин «объект» также относится к ДНК-последовательности из исходного трансформанта, содержащей вставленную ДНК и фланкирующие геномные последовательности, непосредственно прилегающие к вставленной ДНК, где последовательность ДНК, как будут ожидать, переносится в потомство, которое получают в результате полового скрещивания родительской линии, которая включает вставленную ДНК (например, исходный трансформант и его потомство, полученное в результате самоопыления), и родительской линии, которая не содержит вставленной ДНК, и получает вставленную ДНК, включающую целевой ген.
Термин «рекомбинация», в том виде, в котором он используется в данном документе, относится к форме ДНК и/или белка и/или организма, которая обычно не будет обнаружена в природе, и в таком качестве создается в результате вмешательства человека. Такое вмешательство человека может приводить к получению рекомбинантной молекулы ДНК и/или рекомбинантного растения. «Рекомбинантная молекула ДНК» создана посредством искусственной комбинации двух иначе разделенных сегментов последовательности, например, посредством химического синтеза или посредством манипулирования выделенными сегментами нуклеиновой кислоты с использованием технологии генной инженерии. Технология работы с нуклеиновыми кислотами хорошо известна в данной области.
Термин «трансген» включает любую клетку, клеточную линию, каллус, ткань, часть растения или растение, генотип которого изменен вследствие наличия гетерологичной нуклеиновой кислоты. «Трансген» включает данные трансгенные организмы, исходно измененные таким образом, а также потомство, полученное из исходного трансгенного организма в результате полового скрещивания или бесполого размножения. Термин «трансген», в том виде, в котором он используется в данном документе, не охватывает (хромосомного или внехромосомного) изменения генома традиционными способами скрещивания растений или в результате происходящих в природе событий, таких как случайное перекрестное опыление, нерекомбинантное вирусное инфицирование, нерекомбинантная бактериальная трансформация, нерекомбинантная транспозиция или спонтанная мутация.
Термин «гетерологичный», в том виде, в котором он используется в данном документе, относится к первой молекуле, которая в нормальных условиях не обнаруживается в сочетании со второй молекулой в природе. Например, молекула может происходить из первого вида и быть вставлена в геном второго вида. Молекула, таким образом, будет гетерологична в отношении хозяина и искусственно введена в геном клетки-хозяина.
Трансгенный объект - соя DBN8002, обладающий устойчивостью к насекомому Lepidoptera и толерантностью к гербициду глюфосинат, можно скрещивать посредством следующих стадий: во-первых, половое скрещивание первого родительского растения сои, состоящего из растения сои, культивируемого из трансгенного объекта - сои DBN8002 и его потомства, со вторым родительским растением сои, у которого отсутствует устойчивость к насекомому Lepidoptera и толерантность к гербициду глюфосинат, с получением, таким образом, множества растений первого поколения, где трансгенный объект - соя DBN8002 и его потомство получены в результате трансформации с использованием экспрессионной кассеты по настоящему изобретению, которая несет устойчивость к насекомым Lepidoptera и толерантность к гербициду глюфосинат; и затем отбор растения потомства, которое устойчиво к нашествию насекомых Lepidoptera и/или устойчиво к гербициду глюфосинат, с осуществлением скрещивания, таким образом, растения сои, которое устойчиво к нашествию насекомых Lepidoptera и устойчиво к гербициду глюфосинат. Данные стадии могут дополнительно включать обратное скрещивание растения потомства, устойчивого к насекомому Lepidoptera и/или устойчивого к глюфосинату, со вторым родительским растением сои или третьем родительским растением сои, затем осуществление скрининга потомства посредством заражения насекомыми Lepidoptera, применения гербицида глюфосинат или идентификации молекулярных маркеров (например, молекула ДНК, содержащая соединения, идентифицируемые от 5'- и 3'-концов вставляемой последовательности в трансгенном объекте - сое DBN8002), ассоциированных с признаком, с получением, таким образом, растения сои, устойчивого к насекомому Lepidoptera и толерантного к гербициду глюфосинат.
Также следует понимать, что два разных трансгенных растения могут быть также скрещены с получением потомства, которое содержит два независимых и отдельно добавляемых экзогенных гена. Самоопыление соответствующего потомства может приводить к получению растений потомства, которые гомозиготны в отношении обоих добавляемых экзогенных генов. Обратное скрещивание с родительским растением и ауткроссинг с нетрансгенным растением, как описано ранее, также рассматриваются, как и вегетативное размножение.
Термин «зонд» означает выделенную молекулу нуклеиновой кислоты, которая присоединена к общепринятой выявляемой метке или репортерной молекуле, например, радиоактивному изотопу, лиганду, хемилюминесцентному агенту или ферменту. Такой зонд комплементарен цепи целевой нуклеиновой кислоты, в настоящем изобретении, цепи ДНК из генома трансгенного объекта - сои DBN8002, независимо от того, происходит ли геномная ДНК из трансгенного объекта сои DBN8002 или семени или из растения или семени или его экстракта, происходящего из трансгенного объекта - сои DBN8002. Зонды по настоящему изобретению включают не только дезоксирибонуклеиновые или рибонуклеиновые кислоты, а также полиамиды и другие вещества зонда, которые специфично связываются с целевой ДНК-последовательностью и могут быть использованы для выявления наличия той целевой ДНК-последовательности.
Термин «праймер» представляет собой фрагмент выделенной молекулы нуклеиновой кислоты, который отжигается на комплементарной цепи целевой ДНК в результате гибридизации нуклеиновой кислоты с образованием гибрида между праймером и цепью целевой ДНК, затем удлиняется вдоль цепи целевой ДНК под действием полимеразы (например, ДНК-полимераза). Пары праймеров по настоящему изобретению относятся к их применению в амплификации целевой последовательности нуклеиновой кислоты, например, посредством полимеразной цепной реакции (ПЦР) или других общепринятых способов амплификации нуклеиновой кислоты.
Зонды и праймеры обычно представляют собой 11 полинуклеотидов или более в длину, предпочтительно 18 полинуклеотидов или более, более предпочтительно 24 полинуклеотида или более, наиболее предпочтительно 30 полинуклеотидов или более. Такие зонды и праймеры специфично гибридизуются с целевой последовательностью в условиях гибридизации высокой жесткости. Предпочтительно, зонды и праймеры по настоящему изобретению обладают полной идентичностью ДНК-последовательностей с последовательными нуклеиновыми кислотами в целевой последовательности, хотя зонд, который отличается от целевой последовательности ДНК и сохраняет способность к гибридизации с целевой последовательностью ДНК в условиях высокой жесткости, может быть сконструирован традиционными способами.
Праймеры и зонды на основе фланкирующей геномной ДНК и вставленной последовательности по настоящему изобретению могут быть определены традиционными способами, например, посредством выделения соответствующей молекулы ДНК из растительного материала, происходящего из трансгенного объекта сои DBN8002, и определения последовательности нуклеиновой кислоты молекулы ДНК. Молекула ДНК содержит трансгенную вставленную последовательность и геномные фланкирующие последовательности сои, и фрагмент молекулы ДНК может быть использован в качестве праймера или зонда.
Зонд и праймер на основе нуклеиновой кислоты по настоящему изобретению гибридизуются с целевой последовательностью ДНК в жестких условиях. Любой общепринятый способ гибридизации или амплификации нуклеиновой кислоты может быть использован для идентификации наличия ДНК из трансгенного объекта - сои DBN8002 в образце. Молекулы нуклеиновых кислот или их фрагменты могут специфично гибридизоваться с другими молекулами нуклеиновых кислот в определенных обстоятельствах. В том виде, в котором они используются в данном документе, две молекулы нуклеиновой кислоты могут специфично гибридизоваться друг с другом, если данные две молекулы могут образовывать структуру антипараллельной, двухцепочечной нуклеиновой кислоты. Говорят, что молекула нуклеиновой кислоты представляет собой «комплементарную последовательность» другой молекулы нуклеиновой кислоты, если они демонстрируют полную комплементарность. В контексте данного документа, говорят, что две молекулы нуклеиновых кислот демонстрируют «полную комплементарность», если каждый нуклеотид молекулы нуклеиновой кислоты комплементарен соответствующему нуклеотиду другой молекулы нуклеиновой кислоты. Говорят, что две молекулы нуклеиновой кислоты являются «минимально комплементарными», если они могут гибридизоваться друг с другом с достаточной стабильностью, таким образом, что они могут отжигаться и связываться друг с другом в по меньшей мере традиционных условиях «низкой жесткости». Аналогично, говорят, что две молекулы нуклеиновой кислоты обладают «комплементарностью», если они могут гибридизоваться одна с другой с достаточной стабильностью, таким образом, что они могут отжигаться и связываться друг с другом в общепринятых условиях «высокой жесткости». Отступления от полной комплементарности позволительны, при условии, что такие отступления не полностью предотвращают образование двумя молекулами двухцепочечной структуры. Для того, чтобы служить праймером или зондом, молекула нуклеиновой кислоты только должна обладать достаточной комплементарностью в последовательности, таким образом, чтобы она могла образовывать стабильную двухцепочечную структуру при конкретных используемых концентрациях растворителя и соли.
В контексте данного документа по существу гомологичная последовательность представляет собой молекулу нуклеиновой кислоты, которая будет специфично гибридизоваться с комплементарной цепью другой равноценной молекулой нуклеиновой кислоты в условиях высокой жесткости. Соответствующие жесткие условия, которые стимулируют гибридизацию ДНК, известны специалистам в данной области, например, обработка 6,0 x хлоридом натрия/цитратом натрия (SSC) при примерно 45°С с последующей промывкой 2,0 x SSC при 50°С. Например, концентрация солей на стадии промывки может составлять от примерно 2,0 x SSC при 50°С в условиях низкой жесткости до примерно 0,2 x SSC при 50°С в условиях высокой жесткости. Кроме того, температуру на стадии промывки можно повышать с комнатной температуры (примерно 22°С) в условиях низкой жесткости до примерно 65°С в условиях высокой жесткости. Как температуру, так и концентрацию солей можно менять, или одну из них можно поддерживать постоянной, в то время как другая переменная меняется. Предпочтительно, молекула нуклеиновой кислоты по настоящему изобретению будет специфично гибридизоваться с одной или более молекулами нуклеиновых кислот SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 3, SEQ ID NO: 4, SEQ ID NO: 5, SEQ ID NO: 6 и SEQ ID NO: 7, или с их комплементарными последовательностями или любым фрагментом указанных выше последовательностей в условиях умеренной жесткости, например, при примерно 2,0 x SSC и примерно 65°С. Более предпочтительно, молекула нуклеиновой кислоты по настоящему изобретению будет специфично гибридизоваться с одной или более молекулами нуклеиновой кислоты SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 3, SEQ ID NO: 4, SEQ ID NO: 5, SEQ ID NO: 6 и SEQ ID NO: 7 или их комплементарными последовательностями или любым фрагментом указанных выше последовательностей в условиях высокой жесткости. Предпочтительная маркерная молекула нуклеиновой кислоты по настоящему изобретению содержит SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 6 и SEQ ID NO: 7 или их комплементарные последовательности или любой фрагмент указанных выше последовательностей. Еще одна предпочтительная маркерная молекула нуклеиновой кислоты по настоящему изобретению обладает 80% - 100% или 90% - 100% идентичностью последовательностей с SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 6 и SEQ ID NO: 7 или их комплементарными последовательностями или любым фрагментом указанных выше последовательностей. SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 6 и SEQ ID NO: 7 могут быть использованы в качестве маркеров в способах скрещивания растений для идентификации потомства генетического скрещивания. Гибридизацию зонда с целевой молекулой ДНК можно выявлять любым способом, хорошо известным специалистам в данной области, включая флуоресцентные метки, радиоактивные метки, метки на основе антител и хемилюминесцентные метки, но, не ограничиваясь ими.
Рассматривая амплификацию целевой последовательности нуклеиновой кислоты с использованием конкретных праймеров для амплификации (например, ПЦР), «жесткие условия» представляют собой условия, которые позволяют праймеру гибридизоваться только с целевой последовательностью нуклеиновой кислоты в тепловой реакции амплификации ДНК, где праймер имеет последовательность дикого типа, соответствующую целевой последовательности нуклеиновой кислоты (или комплементарной ей последовательности), и таким образом мог бы связываться с ней, предпочтительно, с получением уникального продукта амплификации, а именно, ампликона.
Термин «специфично связывающийся с (целевой последовательностью)» указывает на то, что в жестких условиях гибридизации зонд или праймер гибридизуется только с целевой последовательностью в образце, содержащем целевую последовательность.
В контексте данного документа термин «ампликон» относится к продукту амплификации нуклеиновой кислоты целевой последовательности нуклеиновой кислоты, используемой в качестве матрицы нуклеиновой кислоты. Например, для определения того, получено ли растение сои в результате полового скрещивания трансгенного объекта - сои DBN8002 по настоящему изобретению, или содержит ли образец сои, собранный с поля, трансгенный объект - сою DBN8002, или содержит ли экстракт сои (такой как крупа, мука или масло) трансгенный объект - сою DBN8002, ДНК, выделяемую из образца растительной такни сои, или экстракт можно подвергать способу амплификации нуклеиновой кислоты с использованием пары праймеров с получением ампликона, который позволяет диагностировать наличие ДНК трансгенного объекта - сои DBN8002. Пара праймеров включает первый праймер, происходящий из фланкирующих последовательностей в растительном геноме, прилегающих к участку вставки вставленной экзогенной ДНК, и второй праймер, происходящий из вставленной экзогенной ДНК. Ампликон имеет длину и последовательность, которая также позволяет диагностировать трансгенный объект - сою DBN8002. Ампликон может иметь длину, равную сумме объединенной длины пары праймров плюс одной пары нуклеотидных оснований или предпочтительно плюс примерно 50 пар нуклеотидных оснований или более предпочтительно плюс примерно 250 пар нуклеотидных оснований или наиболее предпочтительно плюс примерно 450 пар нуклеотидных оснований или больше.
В качестве альтернативы, пара праймеров может происходить из фланкирующих геномных последовательностей на обеих сторонах вставленной ДНК, таким образом, чтобы получать ампликон, который включает полную вставленную нуклеотидную последовательность. Одна из пар праймеров, происходящая из последовательности растительного генома, может быть расположена на расстоянии от вставленной ДНК-последовательности, и данное расстояние может колебаться в пределах от одной пары нуклеотидных оснований до примерно двадцати тысяч пар нуклеотидных оснований. В том виде, в котором он используется в данном документе, термин «ампликон» конкретно исключает димеры прймеров, образованные в тепловой реакции амплификации ДНК.
Реакция амплификации нуклеиновой кислоты может быть выполнена любым из разных способов амплификации нуклеиновой кислоты, известных в данной области, включая полимеразную цепную реакцию (ПЦР). Множество способов амплификации известно специалистам в данной области. Способы ПЦР-амплификации разработаны для амплификации вплоть до 22 т.п.н. геномной ДНК и вплоть до 42 т.п.н. ДНК бактериофага. Данные способы, а также другие способы амплификации ДНК, известные в данной области, могут быть использованы в настоящем изобретении. Вставленная экзогенная последовательность ДНК и фланкирующие последовательности ДНК из трансгенного объекта-сои DBN8002 можно амплифицировать на геноме трансгенного объекта - сои DBN8002 с использованием предложенных последовательностей праймеров. После амплификации ПЦР-ампликон или клонированную ДНК секвенируют посредством способов стандартного секвенирования.
Наборы для выявления ДНК, которые основаны на способах амплификации ДНК, содержат молекулы ДНК, используемые в качестве праймеров, которые гибридизуются специфично с целевой ДНК и амплифицируют диагностический ампликон в соответствующих условиях реакции. Набор может обеспечивать способ выявления на основе агарозного геля или много известных способов в данной области для выявления диагностического ампликона. Согласно настоящему изобретению предложен набор, который содержит ДНК-праймеры, которые гомологичны или комплементарны любой части генома сои в SEQ ID NO: 3 или SEQ ID NO: 4, и любой части трансгенной вставленной области в SEQ ID NO: 5. Пара праймеров, которую конкретно идентифицируют как полезную в способе амплификации ДНК, содержит SEQ ID NO: 8 и SEQ ID NO: 9, которые амплифицируют диагностический ампликон, гомологичный участку 5'-области трансгена/генома трансгенного объекта сои DBN8002, где ампликон содержит SEQ ID NO: 1. Другие молекулы ДНК, полезные в качестве ДНК-праймеров, могут быть выбраны из SEQ ID NO: 5.
Ампликон, полученный данными способами, можно выявлять посредством множества методик. Один из таких способов представляет собой генетический анализ Bit, в котором конструируют олигонуклеотидную цепь ДНК, которая охватывает вставленную последовательность ДНК и соседние фланкирующие последовательности геномной ДНК. Олигонуклеотидная цепь иммобилизована в лунках микролуночного планшета. После ПЦР-амплификации целевой области, (используя два праймера соответственно для вставленной последовательности и соседней фланкирующей геномной последовательности), одноцепочечный ПЦР-продукт можно гибридизовать с иммобилизованным олигонуклеотидом, и он может служить в качестве матрицы для реакции достройки по одному основанию с использованием ДНК-полимеразы и специфично меченых ddNTP (англ. dideoxynucleotide triphosphate - дидезоксинуклеотид трифосфат) для следующего ожидаемого основания. Результат может быть получен флуоресцентным способом или способом на основе ELISA (от англ. enzyme-linked immunosorbent assay -твердофазный иммуноферментный анализ). Сигнал указывает на наличие вставленной/фланкирующей геномной последовательности, которая демонстрирует, что амплификация, гибридизация и достройка по одному основанию являются успешными.
Еще одним способом является методика пиросеквенирования. В данном способе конструируют олигонуклеотидную цепь, которая охватывает участок соединения вставленной последовательности ДНК и прилегающей геномной ДНК. Олигонуклеотидная цепь гибридизуется с одноцепочечным ПЦР-продуктом из целевой области (с использованием двух праймеров, соответственно, для вставленной последовательности и прилегающей фланкирующей геномной последовательности) и инкубировали вместе с ДНК-полимеразой, АТФ, сульфурилазой, люциферазой, апиразой, адинозин-5'-фосфосульфатом и люциферином. dNTP (от англ. deoxynucleoside triphosphate - дезоксинуклеозидтрифосфат) добавляют отдельно и измеряют полученный световой сигнал. Световой сигнал указывает на наличие вставленной/фланкирующей последовательности, что демонстрирует то, что амплификация, гибридизация и достройка по одному или многим основаниям являются успешными.
Явление поляризации флуоресценции, как описано Chen et al. (Genome Res., 1999, 9: 492-498), также является способом, который может быть использован для выявления ампликона по настоящему изобретению. Для использования данного способа должна быть сконструирована олигонуклеотидная цепь, которая охватывает участок соединения вставленной последовательности ДНК и прилегающей геномной ДНК. Олигонуклеотидную цепь гибридизуют с одноцепочечным ПЦР-продуктом из целевой области (используя два праймера соответственно для вставленной последовательности и прилегающей фланкирующей геномной последовательности) и инкубируют вместе с ДНК-полимеразой и флуоресцентно меченым ddNTP. Достройка по одному основанию приводит к включению ddNTP. Такое включение может быть измерено как изменение в поляризации с использованием флуориметра. Изменение в поляризации указывает на наличие вставленной/фланкирующей последовательности, которое демонстрирует, что амплификация, гибридизация и достройка по одному основанию являются успешными.
Taqman описан, как способ выявления и количественного анализа наличия ДНК-последовательности и полностью описан в инструкциях, предоставленных изготовителем. В настоящее время он кратко проиллюстрирован, как изложено ниже. Сконструирован олигонуклеотидный зонд на основе FRET (от англ. fluorescence resonance energy transfer - резонансный перенос энергии флуоресценции), который охватывает участок соединения вставленной последовательности ДНК и прилегающей фланкирующей геномной последовательности. FRET-зонд и ПЦР-праймеры (с использованием двух праймеров соответственно для вставленной последовательности и прилегающей фланкирующей геномной последовательности) подвергают циклам реакции в присутствии термостабильной полимеразы и dNTP. Гибридизация FRET-зонда приводит к расщеплению между флуоресцентной группировкой и группировкой гашения на FRET-зонде и высвобождению флуоресцентной группировки. Образование флуоресцентного сигнала указывает на наличие вставленной/фланкирующей последовательности, которое демонстрирует, что амплификация и гибридизация являются успешными.
Основываясь на принципе гибридизации, подходящие методики выявления растительных материалов из трансгенного объекта - сои DBN8002 также включают саузерн-блоттинг, нозерн-блоттинг и гибридизацию in situ. В частности, подходящие технологии включают инкубирование зонда с образцом, их промывку для удаления несвязанного зонда и выявление того, гибридизован ли зонд. Способ выявления зависит от типа метки, присоединенной к зонду, например, радиоактивно меченый зонд может быть выявлен посредством экспонирования и проявления рентгеновской пленки, или ферментативно меченый зонд может быть выявлен в результате превращения субстрата с эффектом изменения окраски.
Применение молекулярных маркеров в выявлении последовательностей описано Tyangi et al. (Nature. Biotech., 1996, 14: 303-308), которое кратко описано следующим образом. Конструируют FRET-олигонуклеотидный зонд, который охватывает участок соединения вставленной ДНК-последовательности и прилегающей геномной фланкирующей последовательности. Из-за уникальной структуры FRET-зонда он содержит вторичную структуру, которая сохраняет флуоресцентную группировку и группировку гашения в непосредственной близости. FRET-зонд и ПЦР-праймеры (с использованием двух праймеров соответственно для вставленной последовательности и фланкирующей геномной последовательности) подвергают прохождению циклов реакции в присутствии термостабильной полимеразы и dNTP. После успешной ПЦР-амплификации, гибридизация FRET-зонда с целевой последовательностью приводит к потере вторичной структуры зонда, таким образом, что флуоресцентная группировка и группировка гашения пространственно разделяются, и генерируется сигнал флуоресценции. Генерирование сигнала флуоресценции указывает на наличие вставленной/фланкирующей последовательности, которое демонстрирует, что амплификация и гибридизация являются успешными.
Согласно другим описанным способам, таким как микрогидродинамика, предложены способы и устройства для выделения и амплификации ДНК-образцов. Оптические красители используют для выявления и определения конкретных молекул ДНК. Устройства на основе нанотрубки, которые содержат электронный сенсор для выявления молекул ДНК или наногранулы для связывания конкретных молекул ДНК, и которые, таким образом, могут быть выявлены, полезны для выявления молекул ДНК по настоящему изобретению.
Наборы для выявления ДНК могут быть разработаны с использованием композиций, описанных в данном документе, и способов, описанных или известных в области выявления ДНК. Наборы полезны для идентификации того, содержит ли образец ДНК трансгенного объекта - сои DBN8002, и могут применяться для скрещивания растений сои, содержащих ДНК трансгенного объекта сои DBN8002. Наборы могут содержать ДНК праймеры или зонды, которые гомологичны или комплементарны по меньшей мере участку SEQ ID NO: 1, 2, 3, 4 или 5, или содержат другие ДНК праймеры или зонды, гомологичные или комплементарные ДНК, содержащейся в трансгенных генетических элементах ДНК, и данные ДНК-последовательности могут быть использованы в реакциях амплификации ДНК или в качестве зондов в способах гибридизации ДНК. Участок соединения ДНК-структуры трансгенной вставленной последовательности и геномной последовательности сои, содержащийся в геноме сои и проиллюстрированный на Фиг. 1 и в Таблице 1, содержит следующее: фланкирующая геномная область сои DBN8002, прилегающая к 5'-концу трансгенной вставленной последовательности; участок вставленной последовательности от области правой границы (RB) Agrobacteríum; первая экспрессионная кассета, состоящая из промотора ACTIN2 арабидопсиса (prAtAct2), функционально связанного с геном mVip3Aa устойчивости к насекомым Bacillus thuringiensis, дополнительно функционально связанным с терминатором нопалинсинтетазы (tNos); вторая экспрессионная кассета, состоящая из промотора вируса мозаики цветной капусты (pr35S), функционально связанного с геном фосфинотрицин-N-ацетилтрансферазы, устойчивой к глюфосинату (сРАТ) Streptomyces, дополнительно функционально связанным с терминатором вируса мозаики цветной капусты 35S (t35S); фрагмент вставленной последовательности от области левой границы (LB) Agrobacteríum; и фланкирующая геномная область растения сои DBN8002 на 3'-конце трансгенной вставленной последовательности (SEQ ID NO: 5). В способах амплификации ДНК молекулы ДНК, полезные в качестве праймеров, могут представлять собой любой фрагмент трансгенной вставленной последовательности, происходящей из трансгенного объекта - сои DBN8002, или любой фрагмент, происходящий из фланкирующей последовательности геномной ДНК сои в трансгенном объекте - сое DBN8002.
Трансгенный объект - соя DBN8002 может быть использован в сочетании с другими трансгенными сортами сои, например, трансгенными сортами сои, толерантными к гербициду (такому как глифосат и дикамба), или трансгенными сортами сои, несущими другие гены устойчивости к насекомым. Разные сочетания этих разных трансгенных объектов, при использовании для скрещивания с трансгенным объектом - соей DBN8002 по настоящему изобретению, могут обеспечивать улучшенные гибридные трансгенные сорта сои, устойчивые к разным насекомым и толерантные к разным гербицидам. Данные сорта демонстрируют лучшие показатели, такие как повышенная урожайность, по сравнению с нетрансгенными сортами и сортами, трансгенными по одному признаку.
Трансгенный объект - соя DBN8002 по настоящему изобретению устойчив к повреждению в результате поглощения, обусловленного насекомыми-вредителями Lepidoptera, и устойчив к фитотоксичности сельскохозяйственных гербицидов, содержащих глюфосинат. Растение сои, трансгенное по двум признакам, экспрессирует белок Vip3Aa Bacillus thuringiensis, обеспечивающий устойчивость к повреждению в результате поглощения, обусловленного насекомыми-вредителями Lepidoptera (такими как Cianis bilineata), и экспрессирует белок - глюфосинат-устойчивую фосфинотрицин-N-ацетил-трансферазу (PAT) Streptomyces, придающий растению устойчивость к глюфосинату. Соя, трансгенная по двум признакам, имеет следующие преимущества: 1) избегают экономических потерь, обусловленных насекомыми-вредителями Lepidoptera (такими как Cianis bilineata и Prodenia litura), где Cianis bilineata и Prodenia litura являются основными насекомыми-вредителями в областях посадки сои; 2) внесение сельскохозяйственных гербицидов, содержащих глюфосинат, придает сое способность к контролю над сорняками широкого спектра; и 3) продуктивность сои не снижается. Кроме того, трансгены, кодирующие признаки устойчивости к насекомым и устойчивости к глюфосинату, связаны с одним и тем же сегментом ДНК, и существуют в одном локусе генома трансгенного объекта - сои DBN8002, которые обеспечивают усиленную эффективность скрещивания и позволяют отследить трансгенные вставленные фрагменты в размножающейся популяции и ее потомстве посредством использования молекулярных маркеров. Между тем, в способах выявления по настоящему изобретению SEQ ID NO: 1 или комплементарная ей последовательность, SEQ ID NO: 2 или комплементарная ей последовательность, SEQ ID NO: 6 или комплементарная ей последовательность или SEQ ID NO: 7 или комплементарная ей последовательность могут быть использованы в качестве ДНК-праймеров или зондов для получения продуктов амплификации, позволяющих диагностировать трансгенный объект- сою DBN8002 или его потомство, и могут быстро, точно и стабильно идентифицировать наличие растительных материалов из трансгенного объекта - сои DBN8002.
Краткое описание последовательностей
SEQ ID NO: 1 последовательность длиной 22 нуклеотида, которая расположена рядом с соединением со вставкой на 5'-конце вставленной последовательности в трансгенном объекте - сое DBN8002, где нуклеотиды 1-11 и нуклеотиды 12-22 расположены, соответственно, по обе стороны от участка вставки генома сои;
SEQ ID NO: 2 последовательность длиной 22 нуклеотида, которая расположена рядом с соединением со вставкой на 3'-конце вставленной последовательности в трансгенном объекте - сое DBN8002, где нуклеотиды 1-11 и нуклеотиды 12-22 расположены, соответственно, по обе стороны от участка вставки генома сои;
SEQ ID NO: 3 последовательность из 1524 нуклеотидов, которая расположена рядом с соединением со вставкой на 5'-конце вставленной последовательности в трансгенном объекте - сое DBN8002;
SEQ ID NO: 4 последовательность из 656 нуклеотидов, которая расположена рядом с соединением со вставкой на 3'-конце вставленной последовательности в трансгенном объекте - сое DBN8002;
SEQ ID NO: 5 полная последовательность Т-ДНК, 5'- и 3'-концевые геномные фланкирующие последовательности сои;
SEQ ID NO: 6 последовательность, расположенная в SEQ ID NO: 3, охватывающая ДНК последовательность конструкции pDBN4006 и последовательность точки начала транскрипции prAtAct2;
SEQ ID NO: 7 последовательность, расположенная в SEQ ID NO: 4, охватывающая последовательность терминатора транскрипции t35S и ДНК-последовательность конструкции pDBN4006;
SEQ ID NO: 8 первый праймер для амплификации SEQ ID NO: 3;
SEQ ID NO: 9 второй праймер для амплификации SEQ ID NO: 3;
SEQ ID NO: 10 первый праймер для амплификации SEQ ID NO: 4;
SEQ ID NO: 11 второй праймер для амплификации SEQ ID NO: 4;
SEQ ID NO: 12 праймер с 5'-фланкирующей геномной последовательности;
SEQ ID NO: 13 праймер с Т-ДНК, который соединяется с SEQ ID NO: 12;
SEQ ID NO: 14 праймер с 3'-фланкирующей геномной последовательности, который может быть использован в паре с SEQ ID NO: 12, для выявления того, является ли трансген гомозиготным или гетерозиготным;
SEQ ID NO: 15 праймер с Т-ДНК, который соединяется с SEQ ID NO: 14;
SEQ ID NO: 16 первый праймер для выявления гена mVip3Aa в Taqman;
SEQ ID NO: 17 второй праймер для выявления гена mVip3Aa BTaqman;
SEQ ID NO: 18 зонд для выявления гена mVip3Aa в Taqman;
SEQ ID NO: 19 первый праймер для выявления гена РАТв Taqman;
SEQ ID NO: 20 второй праймер для выявления гена РАТв Taqman;
SEQ ID NO: 21 зонд для выявления гена PAT в Taqman;
SEQ ID NO: 22 первый праймер для эндогенного гена лектина сои;
SEQ ID NO: 23 второй праймер для эндогенного гена лектина сои;
SEQ ID NO: 24 зонд для гена mVip3Aa в анализе на основе саузерн-блоттинга;
SEQ ID NO: 25 зонд для гена РАТ в анализе на основе саузерн-блоттинга;
SEQ ID NO: 26 праймер с Т-ДНК, который имеет такое же направление, как SEQ ID NO: 13;
SEQ ID NO: 27 праймер с Т-ДНК, который имеет направление, противоположное SEQ ID NO: 13, и используется для получения фланкирующей последовательности;
SEQ ID NO: 28 праймер с Т-ДНК, которые имеет направление, противоположное SEQ ID NO: 13, и используется для получения фланкирующей последовательности;
SEQ ID NO: 29 праймер с Т-ДНК, который имеет такое же направление, как SEQ ID NO: 15;
SEQ ID NO: 30 праймер с Т-ДНК, который имеет направление, противоположное SEQ ID NO: 15, и используется для получения фланкирующей последовательности;
SEQ ID NO: 31 праймер с Т-ДНК, который имеет направление, противоположное SEQ ID NO: 15, и используется для получения фланкирующей последовательности.
Технические решения по настоящему изобретению будут дополнительно подробно описаны ниже со ссылкой на прилагаемые графические материалы и примеры.
Краткое описание графических материалов
Фиг. 1 представляет собой структурную схему участков соединения трансгенных вставленных последовательностей и генома сои, и схему относительных положений последовательностей нуклеиновых кислот для выявления растения сои DBN8002 (в отношении схемы относительных положений, пожалуйста, см. Wm82.a2 RefGen) в последовательностях нуклеиновой кислоты для выявления растений сои DBN8002 и способов его выявления согласно настоящему изобретению.
Фиг. 2 представляет собой структурную схему рекомбинантного экспрессионного вектора pDBN4006 в последовательностях нуклеиновой кислоты для выявления растения сои DBN8002 и способов его выявления согласно настоящему изобретению;
На Фиг. 3 показан эффект биоанализа трансгенного объекта - сои DBN8002 против Helicoverpa armigera (Hubner) в последовательностях нуклеиновой кислоты для выявления растения сои DB8002 и способов его выявления согласно настоящему изобретению;
На Фиг. 4 показан эффект биоанализа трансгенного объекта - сои DBN8002 против Spodoptera litura, в последовательностях нуклеиновой кислоты для выявления растения сои DB8002 и способов его выявления согласно настоящему изобретению;
На Фиг. 5 показан эффект биоанализа трансгенного объекта - сои DBN8002 против Spodoptera exigua в последовательностях нуклеиновой кислоты для выявления растения сои DB8002 и способов его выявления согласно настоящему изобретению;
На Фиг. 6 показан эффект биоанализа трансгенного объекта - сои DBN8002 против Cianis bilineata в последовательностях нуклеиновой кислоты для выявления растения сои DB8002 и способов его выявления согласно настоящему изобретению.
На Фиг. 7 показан эффект на месте использования трансгенного объекта - сои DBN8002, инокулированного Helicoverpa armigera (Hubner), в последовательностях нуклеиновой кислоты для выявления растения сои DB8002 и способов его выявления согласно настоящему изобретению;
На Фиг. 8 показано действие на месте использования трансгенного объекта - сои DBN8002 в последовательностях нуклеиновой кислоты для выявления растения сои DB8002 и способов его выявления согласно настоящему изобретению в условиях встречающихся в природе Spodoptera exigua;
На Фиг. 9 показан эффект на месте использования трансгенного объекта - сои DBN8002 в последовательностях нуклеиновой кислоты для выявления растения сои DB8002 и способов его выявления согласно настоящему изобретению в условиях встречающихся в природе Spodoptera litura;
На Фиг. 10 показан эффект биоанализа трансгенного объекта - сои DBN8002 против Spodoptera frugiperda в последовательностях нуклеиновой кислоты для выявления растения сои DB8002 и способов его выявления согласно настоящему изобретению.
Конкретные воплощения изобретения
Технические решения последовательностей нуклеиновой кислоты для выявления растения сои DBN8002 и способов его выявления согласно настоящему изобретению будут дополнительно проиллюстрированы ниже со ссылкой на конкретные примеры.
Пример 1: Клонирование и трансформация
1.1 Клонирующий вектор
Рекомбинантный экспрессионный вектор pDBN4006 (как показано на Фиг. 2) конструировали посредством использования стандартных методик клонирования генов. Вектор pDBN4006 содержит две трансгенные экспрессионные кассеты, расположенные последовательно: первая экспрессионная кассета, состоящая из промотора ACTIN2 Arabidopsis (prAtAct2), функционально связанного с геном устойчивости к насекомым mVip3Aa Bacillus thuringiensis (CN103509808 В), дополнительно функционально связанным с терминатором нопалинсинтетазы (tNos); вторая экспрессионная кассета, состоящая из промотора вируса мозаики цветной капусты (pr35S), функционально связанного с геном устойчивой к глюфосинату фосфинотрицин-N-ацетил-трансферазы (сРАТ) Streptomyces, дополнительно функционально связанным с терминатором транскрипции вируса мозаики цветной капусты 35S (pr35S).
Вектором pDBN4006 трансформировали Agrobacterium LBA4404 (Invitrogen, Chicago, США; кат. №:18313-015) посредством способа на основе жидкого азота, и 4-[гидрокси(метил)фосфинил]-DL-гомоаланин использовали в качестве селективного маркера для осуществления скрининга трансформированных клеток.
1.2 Трансформация растений
Трансформацию проводили традиционным способом трансформации, опосредованным Agrobacterium, включающим: совместную культивацию тканей семядольного узла сои, культивируемых в стерильных условиях, с Agrobacterium, как описано в Примере 1.1, для переноса Т-ДНК в рекомбинантном экспрессионном векторе pDBN4006 в хромосомы сои, таким образом, чтобы получить трансгенный объект - сою DBN8002.
Кратко, трансформация сои, опосредованная Agrobacterium, включает: проращивание зрелых семян сои в среде для прорастания сои (3,1 г/л соли В5, витамина В5, 20 г/л сахарозы и 8 г/л агара; рН 5,6); осуществление инокуляции семян на среде для прорастания; и культивирование их в следующих условиях: температура 25±1°С и фотопериод (свет/темнота) 16 ч/8 ч через 4-6 суток после прорастания, сбор свежих зеленых стерильных проростков сои с набухшим семядольным узлом; отрезание гипокотелей приблизительно 3-4 мм ниже семядольного узла; получение продольных отрезков через семядоли и удаление апикальных почек, латеральных почек и зародышевых корней; поранение на уровне семядольного узла посредством задней части лезвия скальпеля и приведение тканей семядольного узла с порезами в контакт с суспензией Agrobacterium; где Agrobacterium может переносить нуклеотидную последовательность гена mVip3Aa и нуклеотидную последовательность гена PAT в ткани семядольного узла с порезами (стадия 1: стадия инфицирования). На данной стадии ткани семядольного узла предпочтительно погружали в суспензию Agrobacterium (OD660=0,5-0,8, среда для инфицирования (2,15 г/л соли MS, витамина В5, 20 г/л сахарозы, 10 г/л глюкозы, 40 мг/л ацетосирингона (AS - от англ. acetosyringone), 4 г/л 2-морфолиноэтансульфоновой кислоты (MES - от англ. 2-morpholine ethanesulfonic acid) и 2 мг/л зеатина (ZT); рН 5,3) для начала инфицирования. Ткани семядольного узла совместно культивировали с Agrobacterium на протяжении периода времени (3 суток) (стадия 2: стадия совместного культивирования). Предпочтительно, ткани семядольного узла культивировали в твердой среде (содержащей 4,3 г/л соли MS, витамина В5, 20 г/л сахарозы, 10 г/л глюкозы, 4 г/л MES, 2 мг/л ZT и 8 г/л агара; рН 5,6) после стадии инфицирования. После стадии совместного культивирования может существовать возможная стадия «выделения». На данной «стадии выделения» среда для выделения (3,1 г/л соли В5, витамина В5, 1 г/л MES, 30 г/л сахарозы, 2 мг/л ZT, 8 г/л агара, 150 мг/л цефалоспорина, 100 мг/л глутаминовой кислоты и 100 мг/л аспарагиновой кислоты; рН 5,6) содержит по меньшей мере один антибиотик (150-250 мг/л цефалоспорина), который, как известно, ингибирует рост Agrobacterium, одновременно не содержит никакого селективного агента для растительных трансформантов (стадия 3: стадия выделения). Предпочтительно, блоки ткани, регенерируемые из семядольного узла, культивировали в твердой среде, содержащей антибиотик, но не содержащей селективный агент для устранения Agrobacterium и обеспечения стадии выделения инфицированных клеток. Затем, блоки тканей, регенерированные из семядольного узла, культивировали на среде, содержащей селективный агент (4-[(гидрокси(метил)фосфинил)]-DL-гомоаланин), и отбирали растущие трансформированные каллусы (стадия 4: стадия селекции). Предпочтительно, блоки тканей, регенерированные из семядольного узла, культивировали в твердой селективной среде (3,1 г/л соли В5, витамина В5, 1 г/л MES, 30 г/л сахарозы, 1 мг/л 6-бензиладенина (6-ВАР - от англ. benzyladenine), 8 г/л агара, 150 мг/л цефалоспорина, 100 мг/л глутаминовой кислоты, 100 мг/л аспарагиновой кислоты и 10 мг/л 4-(гидрокси(метил)фосфинил)-DL-гомоаланина, рН 5,6), содержащей селективный агент, который приводил к непрерывному росту трансформированных клеток. Затем, трансформированные клетки регенерировали в растения (стадия 5: стадия регенерации). Предпочтительно, блоки тканей, регенерированные из семядольного узла, растущего на среде, содержащей селективный агент, культивировали в твердой среде (среда для дифференцировки В5 и среда для укоренения В5) для регенерации растений.
Устойчивые ткани, полученные в результате скрининга, переносили на среду для дифференцировки В5 (3,1 г/л соли В5, витамина В5, 1 г/л MES, 30 г/л сахарозы, 1 мг/л ZT, 8 г/л агара, 150 мг/л цефалоспорина, 50 мг/л глутаминовой кислоты, 50 мг/л аспарагиновой кислоты, 1 мг/л гиббереллина, 1 мг/л ауксина и 5 мг/л 4-(гидрокси(метил)фосфинил)-DL-гомоаланина; рН 5,6) и культивировали для дифференцировки при 25°С. Дифференцированные проростки переносили на среду для укоренения В5 (3,1 г/л соли В5, витамина В5, 1 г/л MES, 30 г/л сахарозы, 8 г/л агара, 150 мг/л цефалоспорина и 1 мг/л индолил-3-масляной кислоты (IBA - от англ. indole-3-butyric acid)) и культивировали при 25°С. Когда проростки достигали примерно 10 см в высоту, их перемещали в теплицу и культивировали до плодоношения. В теплице их культивировали при 26°С в течение 16 часов и затем при 20°С в течение 8 часов/сутки.
1.3 Идентификация и осуществление скрининга трансгенных объектов
В общей сложности получали 288 независимых трансгенных Т0 растений. Для осуществления отбора в результате скрининга трансгенного объекта с оптимальными параметрами, 288 независимых трансгенных Т0 растений, указанных выше, перемещали в теплицу для трансплантации, культивации и размножения для обеспечения трансгенных одиночных T1 растений.
Поскольку процесс генетической трансформации сои с использованием зрелых семян сои и глюфосината в качестве селективного агента имеет тенденцию к созданию ложно позитивных трансгенных объектов, поколение T1 опрыскивали глюфосинатом для идентификации позитивных трансгенных объектов, и получали в общей сложности 154 позитивных трансгенных одиночных растений. Посредством анализа TaqMan™ у 154 указанных выше трансгенных растений сои выявляли наличие однокопийных генов mVip3Aa и PAT и отсутствие каркасных последовательностей вектора; и получали в общей сложности 90 трансгенных одиночных растений. Посредством анализа участков трангенных вставок посредством скрининга получали всего 24 трансгенных одиночных растений, в которых данные последовательности с обеих сторон Т-ДНК были интактными, Т-ДНК не была вставлена в важные гены генома сои, и вставка гена не приводила к какой-либо большой открытой рамке считывания (ORF - от англ. open reading frame). Посредством оценки и сравнения устойчивости в отношении основных целевых насекомых (таких как Helicoverpa armigera (Hubner), Prodenia litura и Spodoptera exigua), посредством скрининга получали в общей сложности 21 трансгенное одиночное растение с хорошей устойчивостью к насекомым. Поскольку генетическая трансформация и вставка гена может влиять на агрономические признаки растений сои (такие как мощность проростков, период размножения, высота или полегание растений), указанное выше 21 трансгенное одиночное растение Т2-поколения высаживали в поле для идентификации эффективности агрономического признака трансгенных одиночных растений Т2 на разных фазах (фаза прорастания - фаза полного цветения, исходная фаза образования зерна - фаза спелости). Посредством самоопыления и обратного скрещивания, осуществляют скрининг в отношении того, могут ли агрономические признаки, молекулярная биология, устойчивость к целевым насекомым и толерантность трансгенных растений сои к гербициду глюфосинат стабильно наследоваться в условиях разных поколений, разных географических условий окружающей среды и/или разных фоновых веществ, и трансгенный объект - сою DBN8002 отбирали в качестве прекрасного объекта, который имеет одну копию трансгенов (см. Пример 2), обладает хорошей устойчивостью к насекомым, толерантностью к гербициду глюфосинат и эффективностью агрономических признаков (см. Примеры 6 и 7).
Пример 2: Выявление трансгенного объекта - сои DBN8002 посредством TaqMan
Используя набор для выделения растительной ДНК (набор DNeasy Plant Maxi, Qiagen), геномную ДНК выделяли из листа (примерно 100 мг) трагсгенного объекта - сои DBN8002 в качестве образца, и число копий генов mVip3Aa и PAT выявляли методом флуоресцентной количественной ПЦР с использованием зонда Taqman. Между тем, растение сои дикого типа в качестве контроля подвергали выявлению и анализу в соответствии с указанными выше способами. Эксперименты проводили в трехкратной повторности, и рассчитывали среднее значение.
Конкретный способ выглядит следующим образом:
Стадия 1: 100 мг листа трансгенного объекта - сои DBN8002 растирали до гомогената в ступке с жидким азотом, и каждый образец приготовляли в трехкратной повторности;
Стадия 2: геномные ДНК указанных выше образцов выделяли с использованием набора для выделения ДНК растений (набор DNeasy Plant Maxi, Qiagen, за подробными способами, пожалуйста, см. инструкции по применению продукта);
Стадия 3: концентрации геномных ДНК указанных выше образцов измеряли, используя ультрамикроспектрофотометр (NanoDrop 2000, Thermo Scientific);
Стадия 4: концентрации геномных ДНК указанных выше образцов доводили до одного и того же значения концентрации, находящегося в интервале от 80 до 100 нг/мкл;
Стадия 5: число копий образцов идентифицировали способом флуоресцентной количественной ПЦР с использованием зонда Taqman, где идентифицированный образец с известным числом копий использовали в качестве стандарта, и образец из растения сои дикого типа использовали в качестве контроля. Каждый образец тестировали в трехкратной повторности, и рассчитывали среднее значение. Последовательности праймеров и зондов для флуоресцентной количественной ПЦР выглядят следующим образом:
Следующие праймеры и зонд используют для выявления последовательности гена mVip3Aa:
Праймер 1: cgaatacagaaccctgtcggc, как изложено в SEQ ID NO: 16 в перечне последовательностей;
Праймер 2: cgtgaggaaggtctcagaaatgac, как изложено в SEQ ID NO: 17 в перечне последовательностей;
Зонд 1: cgacgatggcgtgtatatgcctcttgg, как изложено в SEQ ID NO: 18 в перечне последовательностей;
Следующие праймеры и зонд используют для выявления последовательности гена PAT.
Праймер 3: gagggtgttgtggctggtattg, как изложено в SEQ ID NO: 19 в перечне последовательностей;
Праймер 4: tctcaactgtccaatcgtaagcg, как изложено в SEQ ID NO: 20 в перечне последовательностей;
Зонд 2: cttacgctgggccctggaaggctag, как изложено в SEQ ID NO: 21 в перечне последовательностей.
Система ПЦР-реакции содержит:
где 50 × смесь праймеров/зонда содержит 45 мкл каждого праймера в концентрации 1 мМ, 50 мкл зонда в концентрации 100 мкМ и 860 мкл 1 × ТЕ буфера (10 мМ Tris-HCl, 1 мМ ЭДТА (Этилендиаминтетрауксусная кислота); рН 8,0), и хранили в янтарной пробирке при 4°С.
Условия ПЦР-реакции включают следующее:

Данные анализировали, используя программное обеспечение быстрой системы флуоресцентной количественной ПЦР в реальном времени (Applied Biosystems 7900НТ Fast Real-Time PCR System SDS v2.3, Applied Biosystems). Результаты демонстрируют, что полученный трансгенный объект - соя DBN8002 является однокопийным.
Пример 3: Анализ участка вставки трансгенного объекта - сои DBN8002
3.1 Выделение геномной ДНК
ДНК выделяли общепринятым способом на основе СТАВ (от англ. cetyl trimethyl ammonium bromide - цетилтриметиламмония бромид): после того, как 2 г молодого листа трансгенного объекта - сои DBN8002 растирали до порошка в жидком азоте, добавляли 0,5 мл СТАВ буфера для выделения ДНК (20 г/л СТАВ, 1,4 M NaCl, 100 мМ Tris-HCl и 20 мМ ЭДТА (эдетовая кислота) с подведением рН до рН 8,0 с помощью NaOH), предварительно нагретого при 65°С, затем их тщательно перемешивали и проводили выделение при 65°С в течение 90 мин; 0,5 объема фенола и 0,5 объема хлороформа добавляли и смешивали посредством переворачивания; смесь центрифугировали при 12000 об./мин. (оборот/минута) в течение 10 мин; супернатант отбирали пипеткой, и добавляли 2 объема абсолютного этанола; центрифужную пробирку мягко встряхивали и затем оставляли стоять при 4°С в течение 30 мин; центрифужную пробирку снова центрифугировали при 12000 об./мин в течение 10 мин таким образом, что ДНК была собрана на дне пробирки; супернатант отбрасывали, и осадок промывали 1 мл 70%-ого этанола (массовая концентрация); смесь центрифугировали при 12000 об./мин. в течение 5 мин; осадок сушили в вакууме или просушивали в сверхчистом боксе; и полученный осадок ДНК растворяли в соответствующем количестве буфера ТЕ и хранили при -20°С.
3.2 Анализ фланкирующих последовательностей ДНК
Измеряли концентрацию указанного выше образца выделенной ДНК. Концентрация образца, подлежащего тестированию, составляет 80-100 нг/мкл. Геномную ДНК расщепляли, используя рестриктазы EcoR I (анализ 5'-конца) и EcoR V (анализ 3'-конца), соответственно. В каждую систему ферментативного расщепления добавляли 26,5 мкл геномной ДНК, 0,5 мкл указанных выше рестриктаз и 3 мкл буфера для ферментативного расщепления (все используемые рестриктазы представляют собой ферменты с подобранными к ним буферами, или воспроизведенные буферы из компании NEB (в настоящее время известные как NEBCutSmart), и подвергали расщеплению в течение 1 часа. После расщепления к системе ферментативного расщепления добавляли 70 мкл абсолютного этанола, хранили на ледяной бане в течение 30 мин и центрифугировали при 12000 об./мин. в течение 7 мин. После отбрасывания супернатанта осадок просушивали, затем к нему добавляли 8,5 мкл дважды дистиллированной воды, 1 мкл 10 × буфера для Т4 ДНК лигазы (ΝΕΒ Т4 DNA Ligase Reaction Buffer; для того, чтобы узнать его конкретный состав, пожалуйста, зайдите на веб-сайты NEB или см. https://www.neb.com/products/restriction-endonucleases или https://www.neb.com/products/b0202-t4-dna-ligase-reaction-buffer) и 0,5 мкл Т4-ДНК лигазы и затем лигировали в течение ночи при 4°С. 5'- и 3'-концевые геномные ДНК выделяли посредством ПЦР-амплификации, используя целый ряд вложенных праймеров. Конкретно, комбинация праймеров для выделения 5'-концевой геномной ДНК включает SEQ ID NO: 13 и SEQ ID NO: 26 в качестве первых праймеров, SEQ ID NO: 27 и SEQ ID NO: 28 в качестве вторых праймеров и SEQ ID NO: 13 в качестве праймера для секвенирования. Комбинация праймеров для выделения 3'-концевой геномной ДНК содержит SEQ ID NO: 15 и SEQ ID NO: 29 в качестве первых праймеров, SEQ ID NO: 30 и SEQ ID NO: 31 в качестве вторых праймеров и SEQ ID NO: 15 в качестве праймера для секвенирования. Условия ПЦР-реакции показаны в Таблице 3.
Продукт амплификации, полученный в результате указанной выше ПЦР-амплификации, подвергали электрофорезу в агарозном геле с массовой долей 2,0% для выделения ПЦР-продукта амплификации; впоследствии, целевой фрагмент выделяли из агарозного матрикса с использованием набора для выделения из геля (Набор для выделения из геля QIAquick, кат. №_28704, Qiagen Inc., Valencia, СА). Затем, очищенный ПЦР-продукт амплификации секвенировали (например, с использованием ABI PrismTM 377, РЕ Biosystems, Foster City, СА) и анализировали (например, используя программное обеспечение для анализа последовательностей DNASTAR, DNASTAR Inc., Madison, WI).
5'- и 3'-фланкирующие последовательности и последовательности соединения подтверждали, используя стандартные способы на основе ПЦР. 5'-фланкирующие последовательности и последовательности соединения подтверждали, используя SEQ ID NO: 8 или SEQ ID NO: 12 в комбинации с SEQ ID NO: 9, SEQ ID NO: 13 или SEQ ID NO: 26. 3'-фланкирующие последовательности и последовательности соединения подтверждали, используя SEQ ID NO: 11 или SEQ ID NO: 14 в комбинации с SEQ ID NO: 10, SEQ ID NO: 15 или SEQ ID NO: 29. Системы ПЦР-реакции и условия амплификации показаны в Таблицах 2 и 3. Специалистам в данной области будет понятно, что другие последовательности праймеров могли бы быть также использованы для подтверждения фланкирующих последовательностей и последовательностей соединения.
ДНК-секвенирование ПЦР-продуктов амплификации предоставляет ДНК, которую можно было бы использовать для конструирования других молекул ДНК, которые можно было бы использовать в качестве праймеров и зондов для идентификации растений или семян сои, происходящих из трансгенного объекта - сои DBN8002.
Обнаружили, что вставленная последовательность трансгенного объекта-сои DBN8002 фланкирована с правой границы (5'-фланкирующая последовательность) геномной последовательностью сои, показанной в нуклеотидах 1-647 SEQ ID NO: 5, и фланкирована на левой границе (3'-фланкирующая последовательность) геномной последовательностью сои, показанной в нуклеотидах 6647-7344 SEQ ID NO: 5. 5'-последовательность соединения изложена в SEQ ID NO: 1 и 3'-последовательность соединения изложена в SEQ ID NO: 2.
3.3. ПЦР-анализ зиготности
Последовательность соединения представляет собой молекулу относительно короткого полинуклеотида, которая представляет собой новую ДНК-последовательность и позволяет диагностировать ДНК трансгенного объекта - сои DBN8002 при выявлении в выявлении и анализе полинуклеотидов. Последовательности соединения в SEQ ID NO: 1 и SEQ ID NO: 2, соответственно, представляют собой полинуклеотиды из 11 нуклеотидов с обеих сторон участка вставки трансгенного фрагмента и геномной ДНК сои в трансгенном объекте - сое DBN8002. Более длинные или более короткие полинуклеотидные последовательности соединения могут быть выбраны из SEQ ID NO: 3 или SEQ ID NO: 4. Последовательности соединения (SEQ ID NO: 1, используемая как 5'-область соединения, и SEQ ID NO: 2, используемая как 3'-область соединения) полезны в способе выявления ДНК в качестве молекул ДНК-зондов или ДНК-праймеров. Последовательности соединения SEQ ID NO: 6 и SEQ ID NO: 7 также были новыми последовательностями ДНК трансгенного объекта - сои DBN8002, и могли бы также использоваться для выявления наличия трансгенного объекта - сои DBN8002 в качестве молекул ДНК-зондов или ДНК-праймеров. SEQ ID NO: 6 (нуклеотиды 911-1129 SEQ ID NO: 3) охватывает ДНК-последовательность и последовательность точки начала транскрипции prAtAct2 в конструкции pDBN4006, и SEQ ID NO: 7 (нуклеотиды 1-243 SEQ ID NO: 4) охватывает последовательность терминации транскрипции t35S и ДНК-последовательность в конструкции pDBN4006.
Кроме того, ампликон получали посредством использования по меньшей мере одного праймера, происходящего из SEQ ID NO: 3 или SEQ ID NO: 4, и с помощью указанного праймера, при использовании в способе на основе ПЦР, образовывался ампликон, позволяющий диагностировать трансгенный объект - сою DBN8002.
Конкретно, продукт ПЦР-амплификации образовывался с 5'-конца трансгенной вставленной последовательности, содержащей участок геномной ДНК на 5'-конце, фланкирующей вставленную последовательность Т-ДНК в геном растительного материала, происходящего из трансгенного объекта - сои DBN8002. ПЦР-продукт амплификации содержит SEQ ID NO: 3. Для ПЦР-амплификации конструировали праймер 5 (SEQ ID NO: 8), который может гибридизоваться с последовательностью геномной ДНК на 5'-конце, фланкирующей трансгенную вставленную последовательность, и праймер 6 (SEQ ID NO: 9), который использовался в паре с праймером 5 и был расположен в последовательности точки начала транскрипции prAtAct2 вставленной последовательности Т-ДНК.
Продукт ПЦР-амплификации образовывался с 3'-конца трансгенной вставленной последовательности, содержащей участок геномной ДНК, фланкирующей 3'-конец вставленной последовательности Т-ДНК в геноме растительного материала, происходящего из трансгенного объекта - сои DBN8002. ПЦР-продукт содержал SEQ ID NO: 4. Для ПЦР-амплификации конструировали праймер 7 (SEQ ID NO: 10), который был расположен в последовательности терминации транскрипции t35S вставленной последовательности Т-ДНК, и праймер 8 (SEQ ID NO: 11), который использовался в паре с праймером 7 и может гибридизоваться с последовательностью геномной ДНК, фланкирующей 3'-конец трансгенной вставленной последовательности.
Условия амплификации ДНК, описанные в Таблицах 2 и 3, могли быть использованы в упомянутых выше ПЦР-анализах зиготности для получения ампликона, позволяющего диагностировать трансгенный объект - сою DBN8002. Выявление ампликонов могло быть проведено посредством использования термоциклера, такого как Stratagene Robocycler, MJ Engine, Perkin-Elmer 9700 или Eppendorf Mastercycler Gradien, или с помощью способов и приборов, известных специалистам в данной области.
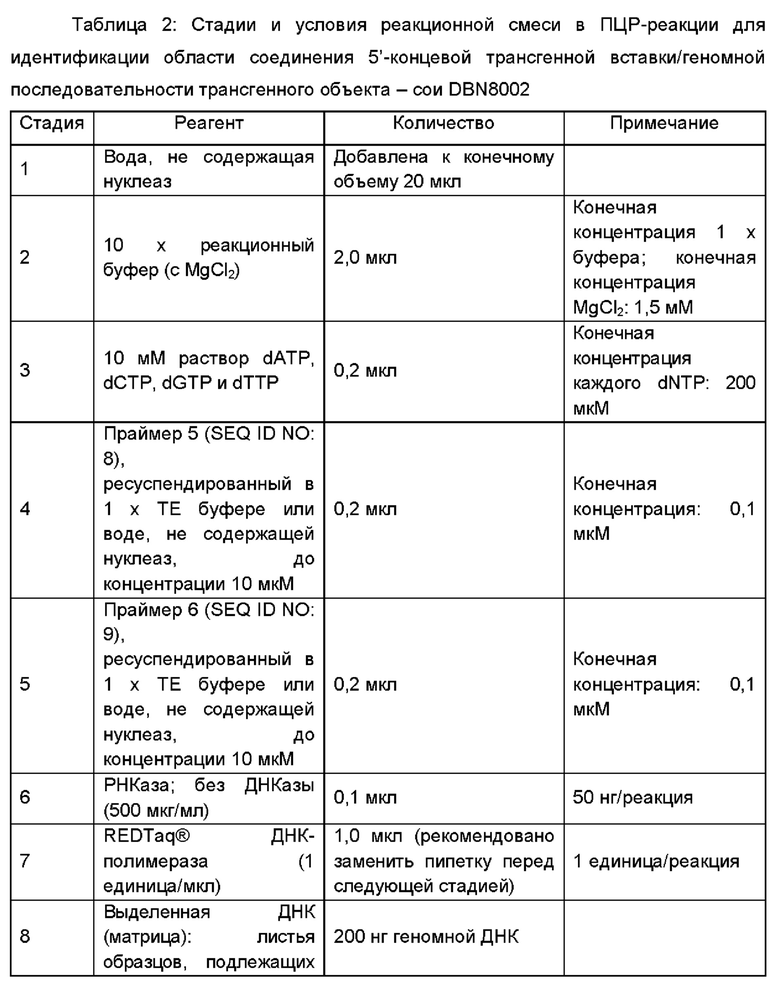
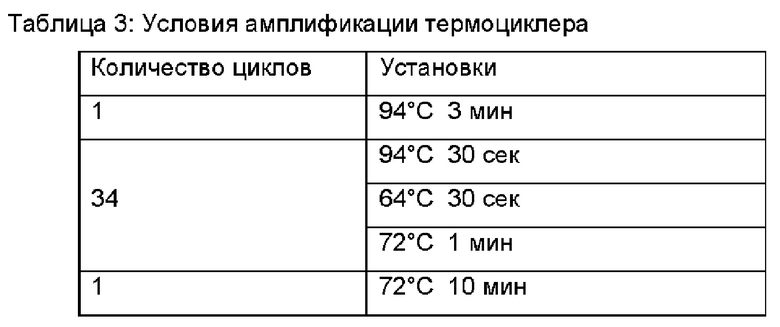
Реакционные растворы аккуратно смешивали, и при отсутствии «горячей крышки» в термоциклере, сверху каждого реакционного раствора добавляли 1-2 капли минерального масла. Реакцию ПЦР проводили в термоциклере, таком как Stratagene Robocycler (Stratagene, La Jolla, СА), MJ Engine (MJ R-Biorad, Hercules, CA), Perkin-Elmer 9700 (Perkin Elmer, Бостон, MA) или Eppendorf Mastercycler Gradient (Eppendorf, Гамбург, Германия) с использованием параметров циклов, показанных в Таблице 3. Термоциклер MJ Engine или Eppendorf Mastercycler Gradient следует запускать в расчетном режиме. Скорость изменения температуры термоциклера Perkin-Elmer 9700 должна быть установлена на максимальном значении при запуске.
Результаты экспериментов показывают следующее: при использовании в ПЦР-реакции геномной ДНК трансгенного объекта - сои DBN8002, праймеры 5 и 6 (SEQ ID NO: 8 и 9) приводят к получению фрагмента продукта амплификации, размером 1524 п.н., но когда данные праймеры использовали в ПЦР-реакциях с геномной ДНК нетрансформированной сои и геномной ДНК сои, не являющейся DBN8002, фрагмент не амплифицировался; при использовании в ПЦР-реакции геномной ДНК трансгенного объекта - сои DBN8002, праймеры 7 и 8 (SEQ ID NO: 10 и 11) приводят к получению фрагмента продукта амплификации, размером 656 п.н., но когда данные праймеры использовали в ПЦР-реакциях с геномной ДНК нетрансформированной сои и геномной ДНК сои, не являющейся DBN8002, фрагмент не амплифицировался.
ПЦР-анализы в отношении зиготности также можно было использовать для идентификации того, является ли материал, полученный из трансгенного объекта - сои DBN8002, гомозиготным или гетерозиготным. Праймер 9 (SEQ ID NO: 12), праймер 10 (SEQ ID NO: 13) и праймер 11 (SEQ ID NO: 14) использовали в реакции амплификации для получения ампликона, позволяющего диагностировать трансгенный объект - сою DBN8002. Условия амплификации ДНК, описанные в Таблицах 4 и 5, могли быть использованы в указанных выше анализах зиготности с получением ампликона, позволяющего диагностировать трансгенный объект - сою DBN8002.
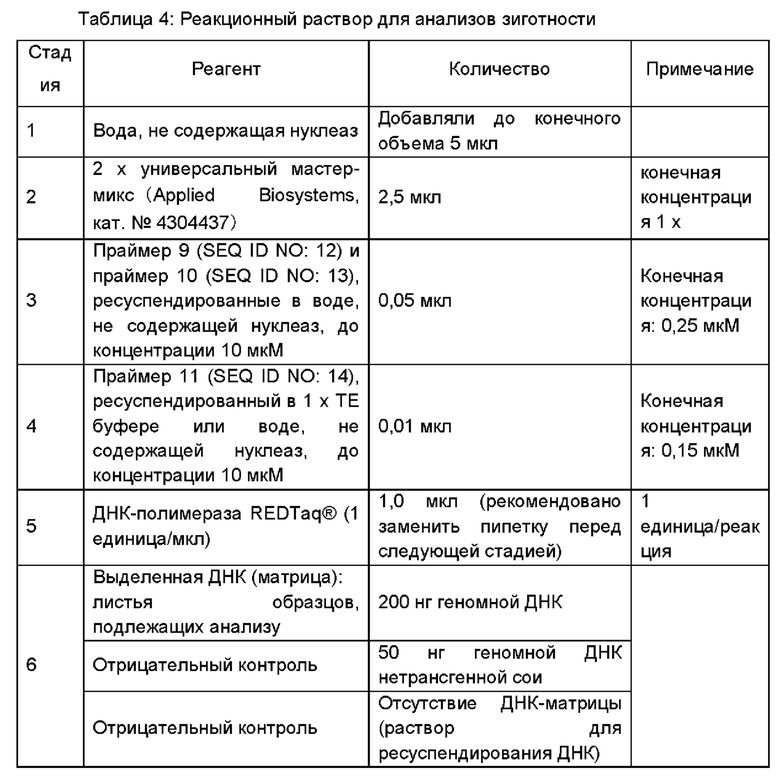
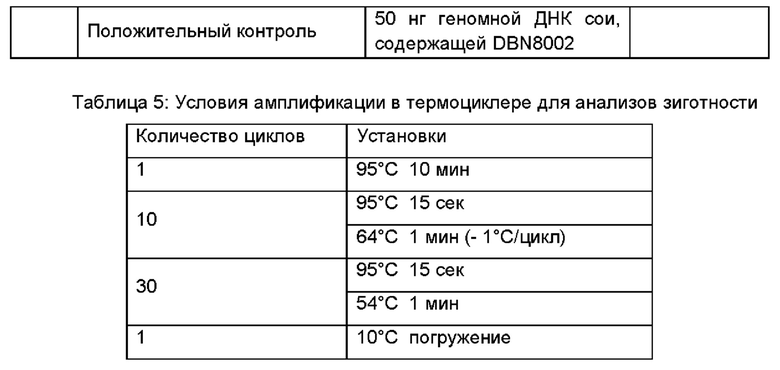
ПЦР-реакцию проводили в термоциклере, таком как Stratagene Robocycler (Stratagene, La Jolla, СА), MJ Engine (MJ R-Biorad, Hercules, CA), Perkin-Elmer 9700 (Perkin Elmer, Бостон, MA) или Eppendorf Mastercycler Gradient (Eppendorf, Гамбург, Германия) с использованием параметров циклов, показанных в Таблице 5. Термоциклер MJ Engine или Eppendorf Mastercycler Gradient следует запускать в расчетном режиме. Скорость изменения температуры термоциклера Perkin-Elmer 9700 следует установить на уровне максимального значения при запуске.
В реакции амплификации биологический образец, содержащий ДНК-матрицу, содержит ДНК, позволяющую диагностировать наличие трансгенного объекта - сои DBN8002. В качестве альтернативы, реакция амплификации будет приводить к получению двух разных ДНК-ампликонов, образованных из биологического образца, содержащего ДНК, происходящую из генома сои, и ДНК, происходящая из генома сои, является гетерозиготной в отношении соответствующей аллели вставленной ДНК, присутствующей в трансгенном объекте - сое DBN8002. Данные два разных ампликона будут соответствовать первому ампликону (SEQ ID NO: 12 и SEQ ID NO: 14), происходящему из локуса генома сои дикого типа, и второй ампликон (SEQ ID NO: 12 и SEQ ID NO: 13), позволяющий диагностировать наличие ДНК трансгенного объекта - сои DBN8002. Образец ДНК сои, который образует только один единственный ампликон, соответствующий второму ампликону, как описано в отношении гетерозиготного генома, мог бы свидетельствовать о наличии трансгенного объекта - сои DBN8002 в данном образце, и данный образец получали из семян сои, гомозиготных в отношении соответствующей аллели ДНК-вставки, присутствующей в трансгенном растении сои DBN8002.
Следует отметить, что пары праймеров трансгенного объекта - сои DBN8002 используют для получения ампликона, позволяющего диагностировать геномную ДНК трансгенного объекта - сои DBN8002. Данные пары праймеров включают, но не ограничиваются праймерами 5 и 6 (SEQ ID NO: 8 и SEQ ID NO: 9) и праймерами 7 и 8 (SEQ ID NO: 10 и SEQ ID NO: 11), и их используют в способе амплификации ДНК. Кроме того, праймеры контроля 12 и 13 (SEQ ID NO: 22 и 23) для амплификации эндогенного гена сои включают в качестве внутреннего стандарта для условий реакции. Анализ образца выделения ДНК трансгенного объекта - сои DBN8002 должен включать экстракт ДНК ткани из трансгенного объекта - сои DBN8002 в качестве положительного контроля, экстракт ДНК из растения сои, которое не является трансгенным объектом - соей DBN8002 в качестве отрицательного контроля, и отрицательный контроль без экстракта ДНК сои в качестве матрицы. Помимо данных пар праймеров можно использовать любую пару праймеров из SEQ ID NO: 3 или комплементарной ей последовательности или SEQ ID NO: 4 или комплементарной ей последовательности. Когда данные праймеры используют для реакции амплификации ДНК, они, соответственно, образуют ампликоны, содержащие SEQ ID NO: 1 или SEQ ID NO: 2 и позволяют диагностировать ткани, происходящие из трансгенного объекта - сои DBN8002. Условия амплификации ДНК, как описано в Таблицах 2-5, могут быть использованы в получении ампликона, позволяющего диагностировать трансгенный объект - сою DBN8002, посредством использования подходящих пар праймеров. Экстракт ДНК растений или семян сои, который приводит к образованию ампликона, позволяющего диагностировать трансгенный объект - сою DBN8002 в способе амплификации ДНК, и экстракт ДНК растений или семян сои, который, как предполагают, содержит трансгенный объект - сою DBN8002, или продукт, полученный из трансгенного объекта-сои DBN8002, можно использовать в качестве матрицы для амплификации для определения наличия трансгенного объекта - сои DBN8002.
Пример 4: Выявление трансгенного объекта - сои DBN8002 посредством саузерн-блоттинга
4.1 Выделение ДНК для применения в саузерн-блоттинге
Приблизительно от 5 до 10 г ткани листа растирали в жидком азоте с использованием ступки и пестика. От 4 до 5 г растертой ткани листа ресуспендировали в 20 мл лизирующего СТАВ-буфера (100 мМ Tris-HCl, рН 8,0, 20 мМ ЭДТА рН 8,0, 1,4 M NaCl, 0,2% (об./об.) β-меркаптоэтанола, 2% (масс./об.) СТАВ), и инкубировали при температуре 65°С в течение 60 минут. Во время инкубации образец однородно перемешивали посредством переворачивания один раз каждые 10 минут. После инкубации добавляли равный объем смеси фенол/хлороформ/изоамиловый спирт (25:24:1) и аккуратно перемешивали посредством переворачивания для экстракции, и затем центрифугировали при скорости вращения 4000 об./мин. в течение 20 минут. Водную фазу собирали и повторно экстрагировали один раз равным объемом смеси хлороформ/изоамиловый спирт (24:1). Водную фазу повторно собирали и добавляли с равным объемом изопропанола. Смесь перемешивали однородно, оставляли стоять при температуре -20°С в течение 1 часа для осаждения ДНК и затем центрифугировали при скорости вращения 4000 об./мин. в течение 5 минут с получением осадка ДНК, который впоследствии ресуспендировали в 1 мл ТЕ буфера (10 мМ Tris-HCl, 1 мМ ЭДТА, рН 8,0). Для деградации какой-либо возможной РНК, ДНК инкубировали при температуре 37°С в течение 30 минут с 40 мкл 10 мг/мл РНКазы А, центрифугировали при скорости вращения 4000 об./мин. в течение 5 минут и затем центрифугировали при скорости вращения 12000 об./мин. в течение 10 минут для осаждения ДНК в присутствии 0,1 объема 3 M ацетата натрия (рН 5,2) и 2 объемов абсолютного этанола. После отбрасывания супернатанта осадок промывали 1 мл 70% (об./об.) этанола и сушили при комнатной температуре и затем ДНК повторно растворяли в 1 мл ТЕ буфера.
4.2 Расщепление рестриктазами
Концентрацию геномной ДНК в упомянутом выше образце измеряли посредством использования спектрофотометра в сверхмикрообъемах (NanoDrop 2000, Thermo Scientific).
В 100 мкл реакционной системы 5 мкг ДНК расщепляли каждый раз. Геномную ДНК расщепляли соответственно рестриктазами Mfe I и Neo I, используя фрагмент последовательностей генов mVip3Aa и PAT в Т-ДНК в качестве зондов. Для каждого фермента продукты расщепления инкубировали в течение ночи при соответствующей температуре. Образец крутили на центрифужном вакуумном концентраторе (speed Vacuum, Thermo Scientific) для уменьшения объема до 20 мкл.
4.3 Электрофорез в геле
Загрузочный краситель бромфенолового синего добавляли к каждому образцу из Примера 4.2, и каждый образец загружали на 0,7%-ный агарозный гель, содержащий этидия бромид, и проводили разделение посредством электрофореза для выделения в ТАЕ электрофорезном буфере (40 мМ Tris-уксусная кислота, 2 мМ ЭДТА, рН 8,5). Электрофорез в геле запускали в течение ночи при напряжении 20 В.
После электрофореза гель обрабатывали 0,25 M HCl в течение 10 минут для осуществления депуринизации ДНК и затем обрабатывали денатурирующим раствором (1,5 M NaCl, 0,5 M NaOH) и нейтрализующим раствором (1,5 M NaCl, 0,5 M Tris-HCl, рН 7,2) в течение 30 минут, соответственно. 5 x SSC (3 M NaCl, 0,3 M цитрат натрия, рН 7,0) наливали в фарфоровую чашку, накрывали куском стекла и затем последовательно помещали мостик из смоченной фильтровальной бумаги, гель, положительно заряженную нейлоновую мембрану (Roche, кат. №11417240001), три кусочка фильтровальной бумаги, бумажную башню и тяжелый объект. После мембранного переноса при комнатной температуре в течение ночи нейлоновую мембрану дважды промывали деионизированной водой, и ДНК иммобилизировали на мембране посредством ультрафиолетового сшивающего средства (UVP, UV Crosslinker CL-1000).
4.4 Гибридизация
Подходящую ДНК-последовательность амплифицировали посредством ПЦР для получения зонда. ДНК-зонды представляли собой SEQ ID NO: 24 или SEQ ID NO: 25, или последовательности, частично гомологичные или комплементарные указанным выше последовательностям. DIG-мечение зондов, саузерн-блоттинг, промывку мембраны и другие операции проводили посредством использования стартового набора для мечения и выявления ДНК II (Roche, кат. №11585614910). В отношении конкретных способов, пожалуйста, см. их инструкции по применению продукта. Наконец, рентгеновскую пленку (Roche, кат №11666916001) использовали для выявления положения, в котором связан зонд.
Каждый саузерн-блоттинг включал два контрольных образца: (1) ДНК из негативного (нетрансформированного) сегреганта, который использовали для идентификации любых эндогенных последовательностей сои, которые могли гибридизоваться с элемент-специфичным зондом; и (2) ДНК из негативного сегреганта, в который была введена Hind Ill-расщепленная плазмида pDBN4006, в количестве, эквивалентном одиночному числу копий в зависимости от длины зонда, который использовали в качестве положительного контроля для того, чтобы продемонстрировать чувствительность эксперимента, когда в геноме сои выявляли одну копию гена.
Данные по гибридизации предоставляют подтверждающее доказательство для поддержки ПЦР-анализа TaqMan™, а именно, растение сои DBN8002 содержит гены mVip3Aa и PAT в одной копии. Посредством использования зонда для гена mVip3Aa, ферментативное расщепление Mfe I и Neo I, соответственно, приводит к образованию одиночных полос, размером примерно 5,5 т.п.н. и 11 т.п.н.; и посредством использования зонда для гена PAT, ферментативное расщепление Mfe I и Nco I, соответственно, приводит к образованию одиночных полос, размером примерно 2,5 т.п.н. и 10 т.п.н. Это указывает на то, что гены mVip3Aa и PAT соответственно находились в растении сои DBN8002 в одной копии. Кроме того, полоса гибридизации не была получена с использованием зонда каркаса. Это указывало на то, что каркасная последовательность вектора pDBN4006 не была введена в геном растения сои DBN8002 во время трансформации.
Пример 5: Выявление экспрессии белка в трансгенном объекте - сое DBN8002 посредством ELISA
Диапазоны экспрессии белков Vip3Aa и PAT в трансгенном объекте - сое DBN8002 можно выявить посредством ELISA.
2 мг лиофилизированного листа трансгенного объекта сои DBN8002 взвешивали и брали в качестве образца. Лист измельчали в жидком азоте и затем к нему добавляли 1 мл экстракционного буфера (8 г/л NaCl, 0,27 г/л KH2PO4, 1,42 г/л Na2HPO4, 0,2 г/л KCl и 5,5 мл/л Tween-20; рН 7,4). Смесь перемешивали однородно, давали отстояться при температуре 4°С в течение 30 минут и затем центрифугировали при скорости вращения 12000 об./мин. в течение 10 минут. Супернатант отбирали посредством пипетки и разводили указанным выше экстракционным буфером до подходящего разведения. 80 мкл разведенного супернатанта собирали для выявления посредством ELISA.
Количества белка (белки Vip3Aa и PAT) в образце в расчете на сухую массу листа измеряли и анализировали посредством наборов для выявления на основе ELISA (твердофазный иммуноферментный анализ) (компания ENVIRLOGIX, набор для Vip3Aa (АР085) и набор для PAT (АР014)). За подробностями способов, пожалуйста, см. их инструкции по применению продукта. Между тем, лист растения сои дикого типа (нетрансгенного, NGM) использовали в качестве контрольного образца. Выявление и анализ проводили в соответствии с указанным выше способом, с 6 повторностями на растение.
Результаты эксперимента по содержаниям белка (белки Vip3Aa и PAT) трансгенного объекта - сои DBN8002 представляли собой такие, как показано в Таблице 6. Средние уровни экспрессии белков Vip3Aa в трансгенном объекте - сое DBN8002 и листьях растения сои дикого типа в расчете на сухую массу листа (мкг/г) составляли 14,49 и 0, соответственно; и средние уровни экспрессии белков PAT в трансгенном объекте - сое DBN8002 и листьях растений сои дикого типа в растете на сухую массу листа (мкг/г) составляли 227,29 и 0, соответственно.

Пример 6: Выявление устойчивости объекта к насекомым 6.1
Биологический анализ растения сои DBN8002 в Китае
Два вида растений, трансгенный объект - сою DBN8002 и растение сои дикого типа (нетрансгенное, NGM), анализировали, используя Helicoverpa armigera (CBW), Spodoptera litura (TCW), Spodoptera exigua (BAW) и Cianis bilineata (BHM) посредством следующих биологических анализов:
Получали вторые сверху листья в фазу V3 из двух типов растений, трансгенного объекта - сои DBN8002 и растения сои дикого типа (нетрансгенного, NGM), соответственно. Их промывали стерильной водой и отсасывали воду досуха с помощью марли. Затем, с удалением жилок, листья сои нарезали на формы, размером примерно 2,5 см × 3 см. 1-3 кусочка листа (число листьев определили в зависимости от потребления листьев насекомым) помещали на фильтровальную бумагу, увлажненную дистиллированной водой, на дно круглых пластмассовых чашек Петри. 10 только что выведенных личинок посредством искусственного кормления помещали в каждую чашку. Затем, чашки Петри накрывали крышками и хранили в течение 3 суток в условиях, включающих температуру 26-28°С, относительную влажность 70%-80% и фотопериод (свет/темнота) 16:8, до получения статистического результата. Три индикатора, скорость развития личинки, смертность анализируемых насекомых и долю повреждения листа, статистически анализировали с получением суммарного балла признака устойчивости (общий показатель: 300 баллов). Суммарный балл признака устойчивости = 100 × смертность + [100 × смертность + 90 × (число только что выведенных насекомых/общее количество инокулированных личинок) + 60 × (число только что выведенных насекомых отрицательного контроля/общее количество инокулированных личинок) + 10 × (число насекомых отрицательного контроля/общее количество инокулированных личинок)] + 100 × (1 - доля повреждения листа), где общее количество инокулированных личинок относится к общему числу инокулированных личинок, а именно, 10 личинок в каждой чашке; скорость развития личинки отражена в формуле суммарного балла признака устойчивости; и доля повреждения листа относится к доле площади питания насекомого в общей площади листа. В случае каждого насекомого, тестировали пять растений из трансгенного объекта - сои DBN8002 и растения сои дикого типа (нетрансгенного, NGM), по 6 повторностей на растение. Результаты показаны в Таблицах 7-8 и на Фиг. 3-6, соответственно.
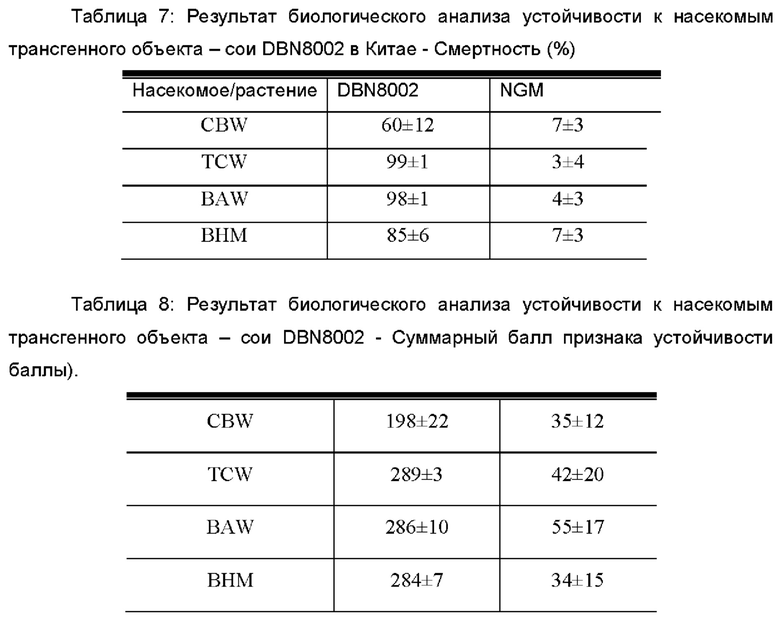
Результаты показывают, что трансгенный объект - соя DBN8002 демонстрирует значимо более высокую смертность анализируемых насекомых и суммарный балл признака устойчивости, чем NGM, что указывает на то, что трансгенный объект - соя DBN8002 обладает хорошей устойчивостью в отношении Helicoverpa armigera, Spodoptera litura, Spodoptera exigua и Cianis bilineata.
6.2 Испытание в полевых условиях трансгенного объекта - сои DBN8002 в Китае
Трансгенный объект - сою DBN8002 и растение сои дикого типа (нетрансгенное, NGM) сажали в поле. Использовали рандомизированную схему блоков с 3 повторностями. Площадь участка составляла 30 м (5 м × 6 м), расстояние между рядами - 60 см и расстояние между растениями - 10 см. Растения традиционно культивировали и регулировали, избегая распыления инсектицидов на протяжении всего периода выращивания.
(1) Helicoverpa armigera
Растения сои подвергали лишь природному заражению на площади с серьезной распространенностью в природе Helicoverpa armigera (условия для возникновения повреждения насекомыми в природе: наиболее сильное повреждение происходит с июня по июль, и оптимальная температура для роста насекомых-вредителей находится в интервале от 20 до 30°С). Когда растения сои дорастали до фазы V3 (трехлистной) за ними наблюдали для исследования листьев NGM, которыми питались личинки Helicoverpa armigera. Когда второй и третий верхние листья NGM не употреблялись, степень повреждения (степень повреждения = суммарная площадь повреждения листа в каждом единичном растении/общая площадь листьев растения × 100%) Helicoverpa armigera на растениях сои исследовали поочередно. Результат по устойчивости трансгенного объекта - сои DBN8002 в отношении Helicoverpa armigera показан в Таблице 9.

Результаты показывают, что в условиях распространенности в природе Helicoverpa armigera, по сравнению с NGM, трансгенный объект - соя DBN8002 демонстрирует значимо сниженную степень повреждения Helicoverpa armigera, что указывает на то, что трансгенный объект - соя DBN8002 обладает хорошей устойчивостью к Helicoverpa armigera. Эффект на месте использования трансгенного объекта - сои DBN8002 в условиях распространенности в природе Helicoverpa armigera показан на Фиг. 7.
(2) Spodoptera exigua
Растения сои подвергали лишь природному заражению на площади с серьезной распространенностью в природе Spodoptera exigua (условия для возникновения повреждения насекомыми в природе: наиболее сильное повреждение происходит с июня по июль, и оптимальная температура для роста насекомых-вредителей находится в интервале от 20 до 30°С). Когда растения сои дорастали до фазы V3, за ними наблюдали для исследования листьев NGM, которыми питались личинки Spodoptera exigua. Когда второй и третий верхние листья NGM не употреблялись, степень повреждения (степень повреждения = суммарная площадь повреждения листа в каждом единичном растении/общая площадь листа растений × 100%) Spodoptera exigua на растениях сои исследовали поочередно. Результат по устойчивости трансгенного объекта - сои DBN8002 к Spodoptera exigua показан в Таблице 10.

Результаты показывают, что в условиях распространенности в природе Spodoptera exigua, по сравнению с NGM, трансгенный объект - соя DBN8002 демонстрирует значимо уменьшенную степень повреждения Spodoptera exigua, что, таким образом, указывает на то, что трансгенный объект - соя DBN8002 обладает хорошей устойчивостью к Spodoptera exigua. Эффект на месте использования трансгенного объекта - сои DBN8002 в условиях распространенности в природе Spodoptera exigua показан на Фиг. 8.
(3) Spodoptera litura
Искусственную инокуляцию два раза проводили в фазу V3 растений сои. 100 растений вблизи центрального района в каждом участке отбирали и подвергали искусственной инокуляции. Второй сверху лист каждого растения сои подвергали инокуляции примерно 10 только что вылупившимися личинками посредством искусственного кормления. Спустя 3 суток после инокуляции, вторую инокуляцию проводили такой же плотностью личинок, как и в первую инокуляцию. Спустя 5-21 суток после инокуляции, поочередно исследовали площади листьев, съеденные личинками растения. Исследование обычно проводили через 14 суток после инокуляции. Если степень повреждения (степень повреждения = суммарная площадь повреждения листа в каждом единичном растении/общая площадь листа растений × 100%) листьев в NGM достигала 15%, ее считали общезначимой. Если степень повреждения не достигала 15%, исследование могло быть надлежащим образом отложено; однако, если степень повреждения все еще не достигала 15%, спустя 21 сутки, данную инокуляцию считали недостоверной. Рассчитывали среднее значение степени повреждения Spodoptera litura на листьях сои на протяжении фазы V3 растений сои в каждом участке. Результат устойчивости трансгенного объекта - сои DBN8002 в отношении Spodoptera litura показан в Таблице 11.

Результаты показывают, что в условиях искусственной инокуляции трансгенный объект - соя DBN8002 демонстрирует значимо меньшую степень повреждения, по сравнению с NGM, что, таким образом, указывает на то, что трансгенный объект - соя DBN8002 обладает хорошей устойчивостью к Spodoptera litura. Эффект на месте использования трансгенного объекта - сои DBN8002, инокулированного Spodoptera litura, показан на Фиг. 9.
6.3. Биологический анализ растения сои DBN8002 в Аргентине
Два вида растений, трансгенный объект - сою DBN8002 и растение сои дикого типа (нетрансгенное, NGM), анализировали, используя Chrysodeixis includens (SBL), Rachiplusia пи, (SFL), Spodoptera frugiperda (FAW) и Spodoptera cosmioides (BLAW) посредством следующих биологических анализов:
Получали вторые верхние листья в фазу V3 из двух типов растений, трансгенного объекта - сои DBN8002 и растения сои дикого типа (нетрансгенного, NGM), соответственно. Их промывали стерильной водой и отсасывали воду досуха с помощью марли. Затем, с удалением жилок, листья сои нарезали на круглые формы, диаметром примерно 1 см. 1-3 кусочка (число листьев определили в зависимости от потребления листьев насекомым) круглого листа помещали на фильтровальную бумагу, увлажненную дистиллированной водой, в лунках планшетов для биологического анализа (как показано на Фиг. 10). Одну только что выведенную личинку помещали в каждую лунку. Затем, планшеты накрывали крышками и хранили в течение 5 суток в условиях, включающих температуру 26-28°С, относительную влажность 70%-80% и фотопериод (свет/темнота) 16:8, до получения статистического результата. Статистически анализировали смертность анализируемых насекомых и степень повреждения листьев (степень повреждения листа относится к доле площади листа, съеденной насекомыми, в суммарной площади листа). В случае каждого насекомого, тестировали 6 растений с эквивалентной силой роста из трансгенного объекта - сои DBN8002 и растения сои дикого типа (нетрансгенного, NGM), по 32 лунки для биологического анализа на растение. Результаты показаны в Таблице 12 и на Фиг. 10, (Spodoptera frugiperda).
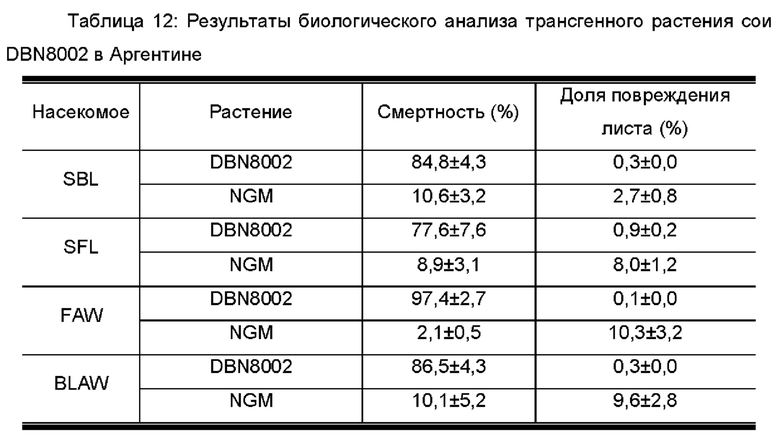
Результаты показывают, что трансгенный объект - соя DBN8002 демонстрирует значимо более высокую смертность анализируемых насекомых упомянутых выше видов насекомых, по сравнению с NGM, что указывает на то, что трансгенный объект - соя DBN8002 обладает хорошей устойчивостью к Chrysodeixis includens (SBL), Rachiplusia nu (SFL), Spodoptera frugiperda (FAW) и Spodoptera cosmioides (BLAW) (типичные насекомые-вредители, обнаруженные на сое в Южной Америке).
6.4 Анализ в полевых условиях трансгенного объекта - сои DBN8002 в Аргентине
Два вида растений, трансгенный объект - сою DBN8002 и растение сои дикого типа (нетрансгенное, NGM), сажали в поле; и проводили биологические анализы в поле in vivo, используя Chrysodeixis includens (SBL), Rachiplusia nu (SFL), Anticarsia gemmatalis (VBC) и Spodoptera frugiperda (FAW) в соответствии со следующим способом.
Большие клетки для биологического анализа (тип ячейки сита) получали в поле. Каждую клетку для биологического анализа использовали для анализа только одного насекомого. Отдельные клетки для биологического анализа не приводили во взаимодействие, и их дополнительно физически разделяли посредством искусственно посаженной кукурузы и сорняков, растущих в природе в поле. Трансгенный объект - сою DBN8002 и растение сои дикого типа (нетрансгенное, NGM) случайным образом сажали в каждую отдельную клетку для биологического анализа, с 3 повторностями на тип растения, и каждая повторность высаживалась в один ряд (длина ряда: 3 м, 30 растений на ряд, расстояние между рядами: 50 см). Растения традиционно культивировали и осуществляли контроль, избегая распыления инсектицидов на протяжении всего периода выращивания. Когда растения дорастали до примерно фазы V5 (пятилистная), подходящее количество взрослых насекомых упомянутых выше видов отсаживали в клетку. Спустя 10 суток, исследовали долю повреждения листьев (которая относится к доле площади листа, которой питались насекомые, в суммарной площади листа). Результаты показаны в Таблице 13.
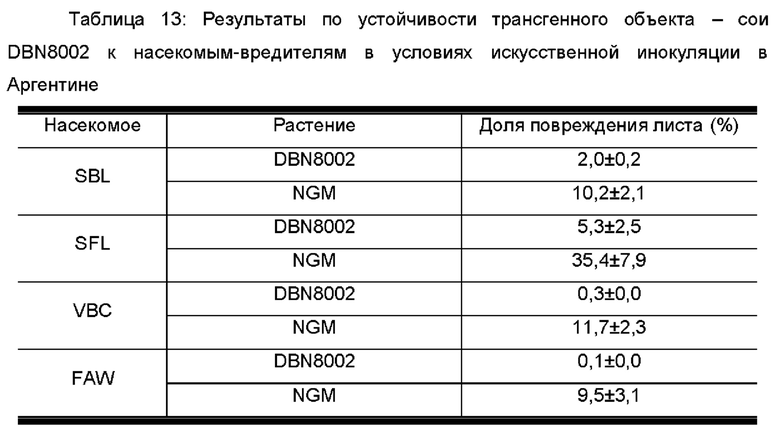
Результаты показывают, что в условиях искусственной инокуляции трансгенный объект - соя DBN8002 демонстрирует меньшую степень повреждения листа, по сравнению с NGM, что указывает на то, что трансгенный объект - соя DBN8002 обладает хорошей устойчивостью к Chrysodeixis includens (SBL), Rachiplusia nu (SFL), Anticarsia gemmatalis (VBC) и Spodoptera frugiperda (FAW) (типичные насекомые-вредители, обнаруженные на сое в Южной Америке).
Пример 7: Выявление толерантности объекта к гербициду
Гербицид Basta (в котором активным ингредиентом является 18%-ный водный раствор глюфосината) выбирали для нанесения распылением в эксперименте. Использовали рандомизированную схему блоков с тремя повторностями. Площадь участка составляла 15 м2 (5 м × 3 м), расстояние между рядами - 60 см и расстояние между растениями - 25 см. Растения традиционно культивировали и осуществляли регуляцию, и среди данных участков устанавливали интервал в один метр. Трансгенный объект - сою DBN8002 подвергали следующим двум обработкам: (1) отсутствие нанесения распылением, а распыление равного объема чистой воды при распылении гербицида во время обработки (2); и (2) нанесение распылением гербицида Basta в фазу V2-V3 листа (двух- или трехлистная) в дозе 800 г а.и./га (а.и./га означает «активный ингредиент на гектар»). Следует отметить, что гербицид глюфосинат (такой как Basta) представляет собой гербицид контактного типа; если с гербицидом обращаются несоответствующим образом в поле, например, локальная аккумуляция слишком большого количества раствора гербицида, может встречаться симптом фитотоксичности; это не означает, что устойчивость трансгенного объекта - сои DBN8002 нарушена; и гербициды глюфосинат в разных концентрациях и композициях также применимы к следующему заключению, если они превращаются в указанное выше эквивалентное количество активного ингредиента - глюфосината.
Исследование симптомов фитотоксичности соответственно проводили через 1 неделю и 2 недели после применения, и урожайность участков определяли в момент сбора урожая. Градация симптомов фитотоксичности показана в Таблице 14. Толерантность объекта трансформации к гербициду оценивали посредством использования степени повреждения от гербицида в качестве индикатора. В частности, степень повреждения от гербицида (%)=Σ (число растений одного и того же уровня повреждения × номер уровня) / (общее число растений × самый высокий уровень), где степень повреждения от гербицида относится к степени повреждения от глюфосината, которую определяли в соответствии с результатом исследования фитотоксичности через две недели после обработки глюфосинатом; и уровень устойчивости сои к гербициду оценивали на основе степени повреждения от гербицида (глюфосинат). Урожайность сои каждого участка получали посредством взвешивания общего урожая (по массе) зерен сои 3 рядов в середине каждого участка. Разницу в урожайности у разных обработок оценивали как процент урожайности. Процент урожайности (%) = урожайность с нанесением распылением/урожайность без нанесения распылением. Результаты по устойчивости трансгенного объекта - сои DBN8002 к гербициду и урожайности сои показаны в Таблице 15.
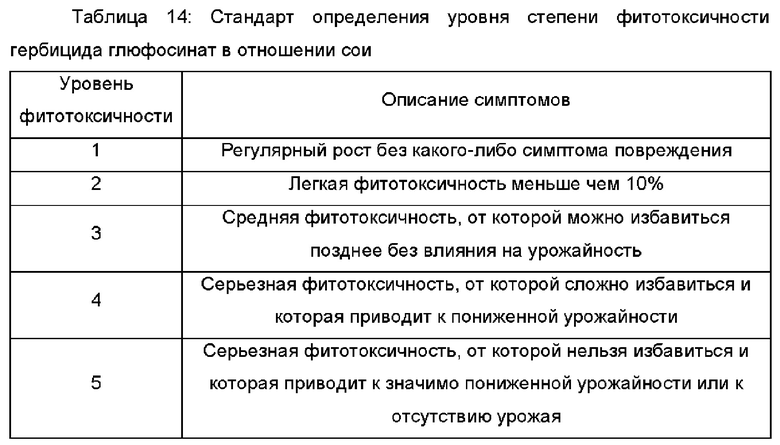

Результаты показывают, что, в отношении степени повреждения гербицидом глюфосинат: степень повреждения трансгенного объекта - сои DBN8002, которого обрабатывали гербицидом глюфосинат (800 г а.и./га), равняется 0. Таким образом, трансгенный объект - соя DBN8002 обладает хорошей толерантностью к гербициду глюфосинат.
В отношении урожайности, отсутствует значимое различие в урожайности трансгенного объекта - сои DBN8002 между обработкой без нанесения распылением и обработкой с нанесением распылением 800 г а.и./га глюфосината. Таким образом, это дополнительно показывает, что трансгенный объект - соя DBN8002 обладает хорошей толерантностью к гербициду глюфосинат, и урожайность не снижается.
Пример 8
Сельскохозяйственные продукты или товары могут быть получены из трансгенного объекта - сои DBN8002. Если в сельскохозяйственных продуктах или товарах выявляется достаточная экспрессия, ожидают, что они содержат нуклеотидные последовательности, которые могут позволить диагностировать наличие материала из трансгенного объекта - сои DBN8002 в сельскохозяйственных продуктах или товарах. Сельскохозяйственные продукты или товары включают, но не ограничиваются соевым жмыхом, порошками и маслами, конкретно лецитином, жирными кислотами, глицерином, стеринами, пищевыми маслами, обезжиренными соевыми хлопьями, обезжиренной и пропеченной соевой мукой, продуктами свертывания соевого молока, соевым творогом, концентратами соевых белков, выделенными соевыми белками, гидролизованными растительными белками, организованными соевыми белками, волокнами соевых белков и любыми другими пищевыми продуктами, предназначенными для потребления в качестве источника пищи животным, и т.д. Способы и/или наборы для выявления нуклеиновой кислоты на основе зондов или пар праймеров могут быть разработаны для выявления нуклеотидных последовательностей из трансгенного объекта - сои DBN8002 в биологическом образце, таких как нуклеотидные последовательности, показанные в SEQ ID NO: 1 или SEQ ID NO: 2, где последовательность зонда или последовательность праймера выбрана из последовательностей, как показано в SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 3, SEQ ID NO: 4 и SEQ ID NO: 5, или их фрагментов, таким образом, чтобы диагностировать наличие трансгенного объекта - сои DBN8002.
В заключение, трансгенный объект - соя DBN8002 по настоящему изобретению обладает хорошей устойчивостью к насекомым Lepidoptera и высокой толерантностью к гербициду глюфосинат без снижения урожайности, и способы выявления по настоящему изобретению могут точно и быстро идентифицировать то, содержит ли биологический образец молекулы ДНК трансгенного объекта - сои DBN8002.
Семена, соответствующие трансгенному объекту - сое DBN8002, депонировали в соответствии с Будапештским договором в центре общей микробиологии китайского комитета по управлению коллекциями культур микроорганизмов (сокращенно CGMCC, адрес: No.3, No.1 Precinct, Beichen West Road, Chaoyang District. Beijing, Institute of Microbiology, Academy of Sciences of China, zip code 100101) 19 февраля 2019 года, классификация и номенклатура которых представляют собой сою (Glycine max), и статус депонирования представляет собой «жизнеспособный», и номер доступа представляет собой CGMCC №17299. Депозит буде депонирован в депозитарии на протяжении 30 лет.
Наконец, следует отметить, что приведенные выше Примеры используют только лишь для иллюстрации технических решений настоящего изобретения, а не ограничения настоящего изобретения. Несмотря на то, что настоящее изобретение подробно описано со ссылкой на предпочтительные Примеры, специалисты в данной области должны понимать, что технические решения настоящего изобретения могли бы быть модифицированы или заменены эквивалентно без отступления от сущности и объема технических решений настоящего изобретения.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> BEIJING DABEINONG BIOTECHNOLOGY CO., LTD.
<120> ПОСЛЕДОВАТЕЛЬНОСТЬ НУКЛЕИНОВОЙ КИСЛОТЫ ДЛЯ ВЫЯВЛЕНИЯ РАСТЕНИЯ
СОИ DBN8002 И СПОСОБ ЕЕ ВЫЯВЛЕНИЯ
<130> FP1190646P
<160> 31
<170> SIPOSequenceListing 1.0
<210> 1
<211> 22
<212> ДНК
<213> Искусственная последовательность из 22 нуклеотидов в длину,
которая расположена около соединения со вставкой на 5’-конце
вставленной последовательности в DBN8002
<400> 1
aaattaatta ctcaaacact ga
22
<210> 2
<211> 22
<212> ДНК
<213> Искусственная последовательность из 22 нуклеотидов в длину,
которая расположена около соединения со вставкой на 3’-конце
вставленной последовательности в DBN8002
<400> 2
ttacaccaca ataattatgt gt
22
<210> 3
<211> 1524
<212> ДНК
<213> Искусственная последовательность из 1524 нуклеотидов в длину,
которая расположена около соединения со вставкой на 5’-конце
вставленной последовательности в DBN8002
<400> 3
tccaaaccaa accgcatgtt aatttattgt ttggttcgaa tgtaattttg agaagaaaaa
60
aaattgctca aaccgcaccg cgaacaccct aatggtattg aagtgttttt ttttataaaa
120
aaattaacaa aaggaaaata atgataaatt taactttttt ttataaatgc acttcacttt
180
ttaatttgga gaaattaaat gagaaaattg tataaaagtt gaagtgcata attatgtatt
240
ttttaatttt ttttttatct tcttttcttc tatgaatact taataagaaa cttgttcaaa
300
cataatgtat ataattttat gcatagaata acataattag agataaaagg aaattaattc
360
acaacagcta accaaagtcg tgttataatt cattaaccac ctaaatctat taaggtcgac
420
ctagtgatat aaaatatttg agtagtataa ggtctttggt tggattatcg gtaccaccat
480
tatatacaca taaaatcact aaccatacat gttggttaag taggttatga agtcttagat
540
aatgattgta aatcaaatga ctaattattt gagatctcat tttatgttag agtaagaatc
600
aatttttgtt taaaaaaaat atttagatac caaattaatt actcaaacac tgatagttta
660
aactgaaggc gggaaacgac aatctgatca tgagcggaga attaagggag tcacgttatg
720
acccccgccg atgacgcggg acaagccgtt ttacgtttgg aactgacaga accgcaacgc
780
tgcaggaatt ggccgcaggt ggatttgtat taaactaatg actaattagt ggcactagcc
840
tcaccgactt cgcagacgag gccgctaagt cgcagctacg ctctcaacgg cactgactag
900
gtagtttaaa cgtgcactta attaaggtac cgggaattta aatcccggga ggtctcgcag
960
acctagctag ttagaatccc gagacctaag tgactagggt cacgtgaccc tagtcactta
1020
aagcttgtcg acaaaattta gaacgaactt aattatgatc tcaaatacat tgatacatat
1080
ctcatctaga tctaggttat cattatgtaa gaaagttttg acgaatatgg cacgacaaaa
1140
tggctagact cgatgtaatt ggtatctcaa ctcaacatta tacttatacc aaacattagt
1200
tagacaaaat ttaaacaact attttttatg tatgcaagag tcagcatatg tataattgat
1260
tcagaatcgt tttgacgagt tcggatgtag tagtagccat tatttaatgt acatactaat
1320
cgtgaatagt gaatatgatg aaacattgta tcttattgta taaatatcca taaacacatc
1380
atgaaagaca ctttctttca cggtctgaat taattatgat acaattctaa tagaaaacga
1440
attaaattac gttgaattgt atgaaatcta attgaacaag ccaaccacga cgacgactaa
1500
cgttgcctgg attgactcgg ttta
1524
<210> 4
<211> 656
<212> ДНК
<213> Искусственная последовательность из 656 нуклеотидов в длину,
которая расположена около соединения со вставкой на 3’-конце
вставленной последовательности в DBN8002
<400> 4
gtgagtagtt cccagataag ggaattaggg ttcttatagg gtttcgctca tgtgttgagc
60
atataagaaa cccttagtat gtatttgtat ttgtaaaata cttctatcaa taaaatttct
120
aattcctaaa accaaaatcc agtggcctgc agggaattct taattaagtg cacgcggccg
180
cctacttagt caagagcctc gcacgcgact gtcacgcggc caggatcgcc tcgtgagcct
240
cgcaatctgt acctagttta gctagttagg acgttaacag ggacgcgcct ggccgtatcc
300
gcaatgtgtt attaagttgt ctaagcgtca atttgtttac accacaataa ttatgtgtaa
360
cgcttgtttt gctatatatg ggacacttga ggaatgacat tattagttac gtaacttgca
420
ttaattaagt attcatgtgt ttgcataaaa aaatagtttt ctcatttcca tattagattt
480
tggaaaaaaa aaattcataa tcaaacttat acttttgttt atcctaaagc aaacacttcg
540
agagggttga attgtttctc atatgtgaag ttcattcaac cacgaatgtt tttttcgcaa
600
tatttacaag gtatgtctaa ttgatccgag taaaataagt tcaacagtct ctcttg
656
<210> 5
<211> 7344
<212> ДНК
<213> Искусственная последовательность -полная последовательность
Т-ДНК, 5’- и 3’-концевые геномные фланкирующие последовательности сои
<400> 5
aaaaatccaa accaaaccgc atgttaattt attgtttggt tcgaatgtaa ttttgagaag
60
aaaaaaaatt gctcaaaccg caccgcgaac accctaatgg tattgaagtg ttttttttta
120
taaaaaaatt aacaaaagga aaataatgat aaatttaact tttttttata aatgcacttc
180
actttttaat ttggagaaat taaatgagaa aattgtataa aagttgaagt gcataattat
240
gtatttttta attttttttt tatcttcttt tcttctatga atacttaata agaaacttgt
300
tcaaacataa tgtatataat tttatgcata gaataacata attagagata aaaggaaatt
360
aattcacaac agctaaccaa agtcgtgtta taattcatta accacctaaa tctattaagg
420
tcgacctagt gatataaaat atttgagtag tataaggtct ttggttggat tatcggtacc
480
accattatat acacataaaa tcactaacca tacatgttgg ttaagtaggt tatgaagtct
540
tagataatga ttgtaaatca aatgactaat tatttgagat ctcattttat gttagagtaa
600
gaatcaattt ttgtttaaaa aaaatattta gataccaaat taattactca aacactgata
660
gtttaaactg aaggcgggaa acgacaatct gatcatgagc ggagaattaa gggagtcacg
720
ttatgacccc cgccgatgac gcgggacaag ccgttttacg tttggaactg acagaaccgc
780
aacgctgcag gaattggccg caggtggatt tgtattaaac taatgactaa ttagtggcac
840
tagcctcacc gacttcgcag acgaggccgc taagtcgcag ctacgctctc aacggcactg
900
actaggtagt ttaaacgtgc acttaattaa ggtaccggga atttaaatcc cgggaggtct
960
cgcagaccta gctagttaga atcccgagac ctaagtgact agggtcacgt gaccctagtc
1020
acttaaagct tgtcgacaaa atttagaacg aacttaatta tgatctcaaa tacattgata
1080
catatctcat ctagatctag gttatcatta tgtaagaaag ttttgacgaa tatggcacga
1140
caaaatggct agactcgatg taattggtat ctcaactcaa cattatactt ataccaaaca
1200
ttagttagac aaaatttaaa caactatttt ttatgtatgc aagagtcagc atatgtataa
1260
ttgattcaga atcgttttga cgagttcgga tgtagtagta gccattattt aatgtacata
1320
ctaatcgtga atagtgaata tgatgaaaca ttgtatctta ttgtataaat atccataaac
1380
acatcatgaa agacactttc tttcacggtc tgaattaatt atgatacaat tctaatagaa
1440
aacgaattaa attacgttga attgtatgaa atctaattga acaagccaac cacgacgacg
1500
actaacgttg cctggattga ctcggtttaa gttaaccact aaaaaaacgg agctgtcatg
1560
taacacgcgg atcgagcagg tcacagtcat gaagccatca aagcaaaaga actaatccaa
1620
gggctgagat gattaattag tttaaaaatt agttaacacg agggaaaagg ctgtctgaca
1680
gccaggtcac gttatcttta cctgtggtcg aaatgattcg tgtctgtcga ttttaattat
1740
ttttttgaaa ggccgaaaat aaagttgtaa gagataaacc cgcctatata aattcatata
1800
ttttcctctc cgctttgaat tgtctcgttg tcctcctcac tttcatcagc cgttttgaat
1860
ctccggcgac ttgacagaga agaacaagga agaagactaa gagagaaagt aagagataat
1920
ccaggagatt cattctccgt tttgaatctt cctcaatctc atcttcttcc gctctttctt
1980
tccaaggtaa taggaacttt ctggatctac tttatttgct ggatctcgat cttgttttct
2040
caatttcctt gagatctgga attcgtttaa tttggatctg tgaacctcca ctaaatcttt
2100
tggttttact agaatcgatc taagttgacc gatcagttag ctcgattata gctaccagaa
2160
tttggcttga ccttgatgga gagatccatg ttcatgttac ctgggaaatg atttgtatat
2220
gtgaattgaa atctgaactg ttgaagttag attgaatctg aacactgtca atgttagatt
2280
gaatctgaac actgtttaag gttagatgaa gtttgtgtat agattcttcg aaactttagg
2340
atttgtagtg tcgtacgttg aacagaaagc tatttctgat tcaatcaggg tttatttgac
2400
tgtattgaac tctttttgtg tgtttgcagc tcataaaaag agctcatgaa caagaacaac
2460
accaagctct ccacacgggc acttccctcc tttattgact actttaatgg catctatggg
2520
tttgctacgg ggatcaagga cattatgaac atgatcttca agacagacac tggcggggat
2580
cttacgctcg acgagattct taagaatcag caactcctga acgatatctc tggcaagctg
2640
gacggcgtga atgggtcact taacgacctc atcgctcagg ggaatctcaa cacagaactg
2700
tctaaggaga tcctcaagat tgcaaatgag cagaaccaag ttcttaatga tgtgaacaat
2760
aagctcgacg ccatcaacac aatgcttcgc gtgtacctcc caaagattac tagcatgctc
2820
tcggacgtca tgaagcagaa ctacgcgctg tcccttcaaa ttgagtatct gagcaagcag
2880
cttcaagaaa tctcggacaa gctggatatc attaatgtga acgtcctcat caacagcacc
2940
ctgacggaga ttacaccggc gtaccagagg atcaagtatg tgaatgagaa gttcgaggaa
3000
ctcacttttg ctacagaaac ttccagcaag gtcaagaagg atggctcacc agccgacatc
3060
ctggatgagc ttacagaact cactgagctg gcgaagtccg tgaccaagaa tgacgtcgat
3120
ggcttcgagt tttacctgaa cacgttccac gacgttatgg tgggcaacaa tctttttggg
3180
cggagcgctc tcaagactgc atcggaactg atcaccaagg agaacgttaa gacgagcggc
3240
tcggaggtcg ggaatgttta caacttcctt atcgtcctca ccgcactcca ggcccaagcg
3300
tttctcacgc tgaccacctg ccgcaagctc ctcggcctcg cagacatcga ttacacctcc
3360
atcatgaacg agcacctgaa caaggagaag gaggagttcc gcgtgaatat ccttccgaca
3420
ctctcgaaca ctttttctaa tccaaactac gctaaggtca agggctccga cgaagatgca
3480
aagatgatcg ttgaggccaa gcctggccat gcgctcatcg ggttcgagat ttctaacgac
3540
tcaattaccg tgctgaaggt ctacgaggcg aagctcaagc agaattatca agtggacaag
3600
gattctctgt cagaggttat ctacggcgac atggataagc tgctttgccc tgatcagtcc
3660
gagcaaatct actatacgaa caatattgtc ttccccaacg aatacgtgat caccaagatt
3720
gactttacga agaagatgaa gacactccgg tacgaggtga cggctaactt ctatgattcg
3780
tctacgggcg agatcgacct caacaagaag aaggtcgaat catccgaggc cgaatacaga
3840
accctgtcgg cgaacgacga tggcgtgtat atgcctcttg gggtcatttc tgagaccttc
3900
ctcacgccca tcaatggctt tgggctccag gcagatgaga actcccgcct gatcaccctt
3960
acgtgcaaga gctacctcag ggagctgctg cttgccaccg acctctctaa caaggaaacg
4020
aagctgatcg ttccgccatc aggcttcatc tccaatattg tggagaacgg gtcaattgag
4080
gaagataatc tggaaccgtg gaaggctaac aataagaacg catacgttga ccacacaggc
4140
ggggtgaatg gcactaaggc gctctatgtg cataaggatg gtggcatctc ccagttcatt
4200
ggcgacaagc tgaagccgaa gacagaatac gtgattcaat atactgtgaa gggcaagcca
4260
agcatccacc tcaaggatga gaacacaggg tacatccatt acgaagatac taacaacaac
4320
ctggaggact accagacaat caataagagg ttcacaactg gcactgacct gaagggggtc
4380
tatcttattc tcaagtccca gaatggcgat gaggcctggg gcgacaactt catcattctc
4440
gaaatctccc ctagcgagaa gctcctgagc cccgagctga ttaacaccaa taactggaca
4500
tccactggca gcacgaatat ctcggggaac accctgacgc tttaccaggg cgggagaggc
4560
attctgaagc agaacctcca actggattcg ttctctacct acagagtcta tttttcagtt
4620
tccggcgacg cgaatgtgcg catcaggaac tcgcgggaag tcctcttcga gaagagatac
4680
atgtctggcg ctaaggatgt gtcagaaatg ttcaccacga agtttgagaa ggacaacttt
4740
tatatcgaac tgtcccaagg gaataacctc tacggcggcc ccattgttca tttttacgac
4800
gtgagcatca agtgaggcgc cgatcgttca aacatttggc aataaagttt cttaagattg
4860
aatcctgttg ccggtcttgc gatgattatc atataatttc tgttgaatta cgttaagcat
4920
gtaataatta acatgtaatg catgacgtta tttatgagat gggtttttat gattagagtc
4980
ccgcaattat acatttaata cgcgatagaa aacaaaatat agcgcgcaaa ctaggataaa
5040
ttatcgcgcg cggtgtcatc tatgttacta gatcggcgcg ccagtaagtg actagagtca
5100
cgtgacccta gtcacttaaa tcctaggcca tggagtcaaa gattcaaata gaggacctaa
5160
cagaactcgc cgtaaagact ggcgaacagt tcatacagag tctcttacga ctcaatgaca
5220
agaagaaaat cttcgtcaac atggtggagc acgacacgct tgtctactcc aaaaatatca
5280
aagatacagt ctcagaagac caaagggcaa ttgagacttt tcaacaaagg gtaatatccg
5340
gaaacctcct cggattccat tgcccagcta tctgtcactt tattgtgaag atagtggaaa
5400
aggaaggtgg ctcctacaaa tgccatcatt gcgataaagg aaaggccatc gttgaagatg
5460
cctctgccga cagtggtccc aaagatggac ccccacccac gaggagcatc gtggaaaaag
5520
aagacgttcc aaccacgtct tcaaagcaag tggattgatg tgatatctcc actgacgtaa
5580
gggatgacgc acaatcccac tatccttcgc aagacccttc ctctatataa ggaagttcat
5640
ttcatttgga gaggacaggg tacccgggga tccaccatgt ctccggagag gagaccagtt
5700
gagattaggc cagctacagc agctgatatg gccgcggttt gtgatatcgt taaccattac
5760
attgagacgt ctacagtgaa ctttaggaca gagccacaaa caccacaaga gtggattgat
5820
gatctagaga ggttgcaaga tagataccct tggttggttg ctgaggttga gggtgttgtg
5880
gctggtattg cttacgctgg gccctggaag gctaggaacg cttacgattg gacagttgag
5940
agtactgttt acgtgtcaca taggcatcaa aggttgggcc taggatccac attgtacaca
6000
catttgctta agtctatgga ggcgcaaggt tttaagtctg tggttgctgt tataggcctt
6060
ccaaacgatc catctgttag gttgcatgag gctttgggat acacagcccg gggtacattg
6120
cgcgcagctg gatacaagca tggtggatgg catgatgttg gtttttggca aagggatttt
6180
gagttgccag ctcctccaag gccagttagg ccagttaccc agatctgagt cgacctgcag
6240
gcatgcccgc tgaaatcacc agtctctctc tacaaatcta tctctctcta taataatgtg
6300
tgagtagttc ccagataagg gaattagggt tcttataggg tttcgctcat gtgttgagca
6360
tataagaaac ccttagtatg tatttgtatt tgtaaaatac ttctatcaat aaaatttcta
6420
attcctaaaa ccaaaatcca gtggcctgca gggaattctt aattaagtgc acgcggccgc
6480
ctacttagtc aagagcctcg cacgcgactg tcacgcggcc aggatcgcct cgtgagcctc
6540
gcaatctgta cctagtttag ctagttagga cgttaacagg gacgcgcctg gccgtatccg
6600
caatgtgtta ttaagttgtc taagcgtcaa tttgtttaca ccacaataat tatgtgtaac
6660
gcttgttttg ctatatatgg gacacttgag gaatgacatt attagttacg taacttgcat
6720
taattaagta ttcatgtgtt tgcataaaaa aatagttttc tcatttccat attagatttt
6780
ggaaaaaaaa aattcataat caaacttata cttttgttta tcctaaagca aacacttcga
6840
gagggttgaa ttgtttctca tatgtgaagt tcattcaacc acgaatgttt ttttcgcaat
6900
atttacaagg tatgtctaat tgatccgagt aaaataagtt caacagtctc tcttgaataa
6960
tcaatagaaa tttttaaaca attatcttat tttcaattca attattaatg ttatgactat
7020
tacatttaat attttatctt taatttctct ttcatatcat ttatgtataa aattaaaact
7080
ttataattat taattaaaag ttaataatta aaactatact aaaatgcata aatatatata
7140
ataaaattat aaaatattaa aatcttaata attatttttt caatcattat attatttatt
7200
cacgttagga aaaatatata tgctgtaatt ttaattttta tattttttat ctataacttt
7260
taattaaaat ttatactttt atctataatt tttaattaaa atataatgaa aatgttggaa
7320
aagtaaagta ttatacttag gagt
7344
<210> 6
<211> 219
<212> ДНК
<213> Искусственая последовательность -последовательность,
расположенная в SEQ ID NO: 3, охватывающая ДНК последовательность
конструкции pDBN4006 и последовательность точки начала транскрипции
prAtAct2
<400> 6
cgtgcactta attaaggtac cgggaattta aatcccggga ggtctcgcag acctagctag
60
ttagaatccc gagacctaag tgactagggt cacgtgaccc tagtcactta aagcttgtcg
120
acaaaattta gaacgaactt aattatgatc tcaaatacat tgatacatat ctcatctaga
180
tctaggttat cattatgtaa gaaagttttg acgaatatg
219
<210> 7
<211> 243
<212> ДНК
<213> Искусственая последовательность- последовательность,
расположенная в SEQ ID NO: 4, охватывающая последовательность
терминатора транскрипции t35S и ДНК-последовательность конструкции
pDBN4006;
<400> 7
gtgagtagtt cccagataag ggaattaggg ttcttatagg gtttcgctca tgtgttgagc
60
atataagaaa cccttagtat gtatttgtat ttgtaaaata cttctatcaa taaaatttct
120
aattcctaaa accaaaatcc agtggcctgc agggaattct taattaagtg cacgcggccg
180
cctacttagt caagagcctc gcacgcgact gtcacgcggc caggatcgcc tcgtgagcct
240
cgc
243
<210> 8
<211> 20
<212> ДНК
<213> Искусственая последовательность - первый праймер для
амплификации SEQ ID NO: 3
<400> 8
tccaaaccaa accgcatgtt
20
<210> 9
<211> 19
<212> ДНК
<213> Искусственая последовательность - второй праймер для
амплификации SEQ ID NO: 3
<400> 9
taaaccgagt caatccagg
19
<210> 10
<211> 20
<212> ДНК
<213> Искусственая последовательность -первый праймер для
амплификации SEQ ID NO: 4
<400> 10
gtgagtagtt cccagataag
20
<210> 11
<211> 20
<212> ДНК
<213> Искусственая последовательность -второй праймер для
амплификации SEQ ID NO: 4
<400> 11
caagagagac tgttgaactt
20
<210> 12
<211> 26
<212> ДНК
<213> Искусственая последовательность - праймер с 5’-фланкирующей
геномной последовательности
<400> 12
gagatctcat tttatgttag agtaag
26
<210> 13
<211> 24
<212> ДНК
<213> Искусственая последовательность-праймер с T-ДНК, который
соединяется с SEQ ID NO: 12
<400> 13
ctacctagtc agtgccgttg agag
24
<210> 14
<211> 21
<212> ДНК
<213> Искусственая последовательность - 14 праймер с 3’-фланкирующей
геномной последовательности, который может быть использован в паре с
SEQ ID NO: 12, для выявления того, является ли трансген гомозиготным
или гетерозиготным
<400> 14
agaaacaatt caaccctctc g
21
<210> 15
<211> 21
<212> ДНК
<213> Искусственая последовательность - праймер с T-ДНК, который
соединяется с SEQ ID NO: 14
<400> 15
cctaaaacca aaatccagtg g
21
<210> 16
<211> 21
<212> ДНК
<213> Искусственая последовательность - первый праймер для выявления
гена mVip3Aa в Taqman
<400> 16
cgaatacaga accctgtcgg c
21
<210> 17
<211> 24
<212> ДНК
<213> Искусственая последовательность - второй праймер для выявления
гена mVip3Aa в Taqman
<400> 17
cgtgaggaag gtctcagaaa tgac
24
<210> 18
<211> 27
<212> ДНК
<213> Искусственая последовательностье-зонд для выявления гена
mVip3Aa в Taqman
<400> 18
cgacgatggc gtgtatatgc ctcttgg
27
<210> 19
<211> 22
<212> ДНК
<213> Искусственая последовательность-первый праймер для выявления
гена PAT в Taqman
<400> 19
gagggtgttg tggctggtat tg
22
<210> 20
<211> 23
<212> ДНК
<213> Искусственая последовательность-второй праймер для выявления
гена PAT в Taqman
<400> 20
tctcaactgt ccaatcgtaa gcg
23
<210> 21
<211> 25
<212> ДНК
<213> Искусственая последовательность-зонд для выявления гена PAT в
Taqman
<400> 21
cttacgctgg gccctggaag gctag
25
<210> 22
<211> 25
<212> ДНК
<213> Искусственая последовательность-первый праймер для эндогенного
гена лектина сои
<400> 22
tgccgaagca accaaacatg atcct
25
<210> 23
<211> 25
<212> ДНК
<213> Искусственая последовательность-второй праймер для эндогенного
гена лектина сои;
<400> 23
tgatggatct gatagsattg acgtt
25
<210> 24
<211> 346
<212> ДНК
<213> Искусственая последовательность - зонд для гена mVip3Aa в
анализе на основе саузерн-блоттинга
<400> 24
acagaactca ctgagctggc gaagtccgtg accaagaatg acgtcgatgg cttcgagttt
60
tacctgaaca cgttccacga cgttatggtg ggcaacaatc tttttgggcg gagcgctctc
120
aagactgcat cggaactgat caccaaggag aacgttaaga cgagcggctc ggaggtcggg
180
aatgtttaca acttccttat cgtcctcacc gcactccagg cccaagcgtt tctcacgctg
240
accacctgcc gcaagctcct cggcctcgca gacatcgatt acacctccat catgaacgag
300
cacctgaaca aggagaagga ggagttccgc gtgaatatcc ttccga
346
<210> 25
<211> 310
<212> ДНК
<213> Искусственая последовательность -зонд для гена PAT в анализе
на основе саузерн-блоттинга
<400> 25
cagacttaaa accttgcgcc tccatagact taagcaaatg tgtgtacaat gtggatccta
60
ggcccaacct ttgatgccta tgtgacacgt aaacagtact ctcaactgtc caatcgtaag
120
cgttcctagc cttccagggc ccagcgtaag caataccagc cacaacaccc tcaacctcag
180
caaccaacca agggtatcta tcttgcaacc tctctagatc atcaatccac tcttgtggtg
240
tttgtggctc tgtcctaaag ttcactgtag acgtctcaat gtaatggtta acgatatcac
300
aaaccgcggc
310
<210> 26
<211> 20
<212> ДНК
<213> Искусственая последовательность - праймер с T-ДНК, который
имеет такое же направление, как SEQ ID NO: 13
<400> 26
cgtgacccta gtcacttagg
20
<210> 27
<211> 23
<212> ДНК
<213> Искусственая последовательность - праймер c T-ДНК, который
имеет направление, противоположное SEQ ID NO: 13, и используется для
получения фланкирующей последовательности
<400> 27
ctgtggtcga aatgattcgt gtc
23
<210> 28
<211> 19
<212> ДНК
<213> Искусственая последовательность -праймер c T-ДНК, который
имеет направление, противоположное SEQ ID NO: 13, и используется для
получения фланкирующей последовательности
<400> 28
cgctctttct ttccaaggt
19
<210> 29
<211> 20
<212> ДНК
<213> Искусственая последовательность -праймер с T-ДНК, который
имеет такое же направление, как SEQ ID NO: 15
<400> 29
ttaggccagt tacccagatc
20
<210> 30
<211> 22
<212> ДНК
<213> Искусственая последовательность -праймер с T-ДНК, который
имеет направление, противоположное SEQ ID NO: 15, и используется для
получения фланкирующей последовательности
<400> 30
aaccttgcgc ctccatagac tt
22
<210> 31
<211> 20
<212> ДНК
<213> Искусственая последовательность -праймер с T-ДНК, который
имеет направление, противоположное SEQ ID NO: 15, и используется для
получения фланкирующей последовательности
<400> 31
ccagccacaa caccctcaac
20
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ПОСЛЕДОВАТЕЛЬНОСТЬ НУКЛЕИНОВОЙ КИСЛОТЫ ДЛЯ ВЫЯВЛЕНИЯ РАСТЕНИЯ КУКУРУЗЫ DBN9501 И СПОСОБ ЕГО ВЫЯВЛЕНИЯ | 2020 |
|
RU2815214C2 |
| Нуклеиновокислотная последовательность для обнаружения наличия трансгенного трансформанта сои DBN9004 в биологическом образце, набор, содержащий такую последовательность, и способ такого обнаружения | 2017 |
|
RU2743397C2 |
| РАСТЕНИЕ КУКУРУЗЫ DBN9858, УСТОЙЧИВОЕ К ГЕРБИЦИДУ, И НУКЛЕОТИДНАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ И СПОСОБ ДЛЯ ЕЕ ВЫЯВЛЕНИЯ | 2016 |
|
RU2712730C2 |
| РАСТЕНИЕ МАИСА DBN9936 И СПОСОБ ПРИМЕНЕНИЯ В ДЕТЕКТИРОВАНИИ ЕГО ПОСЛЕДОВАТЕЛЬНОСТИ НУКЛЕИНОВОЙ КИСЛОТЫ | 2016 |
|
RU2707527C2 |
| СПОСОБ ОБНАРУЖЕНИЯ ТРАНСФОРМАНТА СОИ рDAB9582.814.19.1 | 2012 |
|
RU2622079C2 |
| ПАКЕТИРОВАННОЕ СОБЫТИЕ 8264.44.06.1 С УСТОЙЧИВОСТЬЮ К ГЕРБИЦИДАМ, СВЯЗАННЫЕ ТРАНСГЕННЫЕ ЛИНИИ СОИ И ИХ ДЕТЕКТИРОВАНИЕ | 2011 |
|
RU2608650C2 |
| ОБЪЕКТ СОИ 9582.814.19.1, ПРИДАЮЩИЙ УСТОЙЧИВОСТЬ К НАСЕКОМЫМ И УСТОЙЧИВОСТЬ К ГЕРБИЦИДАМ | 2012 |
|
RU2628099C2 |
| РАСТЕНИЯ СОИ, УСТОЙЧИВОЙ К ГЕРБИЦИДАМ, И СПОСОБЫ ИХ ИДЕНТИФИКАЦИИ | 2010 |
|
RU2639520C2 |
| ТРАНСГЕННЫЙ ОБЪЕКТ СОИ MON 87708 И СПОСОБЫ ЕГО ПРИМЕНЕНИЯ | 2010 |
|
RU2624025C2 |
| ДЕТЕКТИРОВАНИЕ AAD-12-СОБЫТИЯ 416 У СОИ | 2010 |
|
RU2573898C2 |
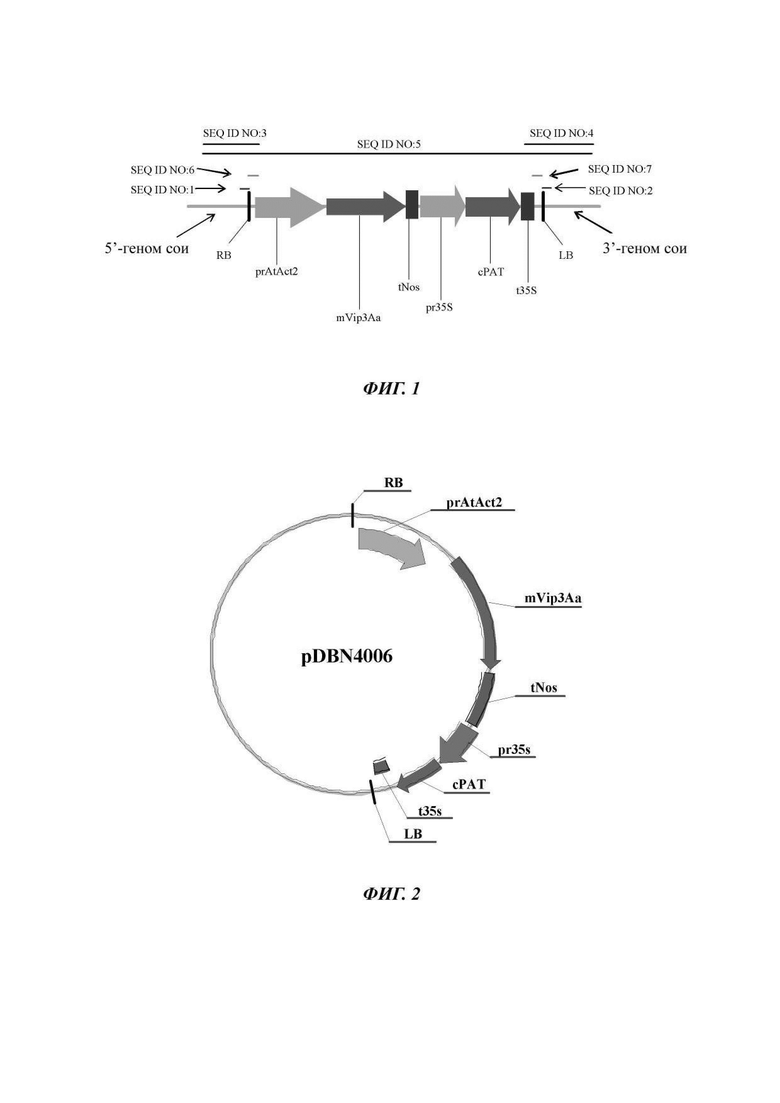
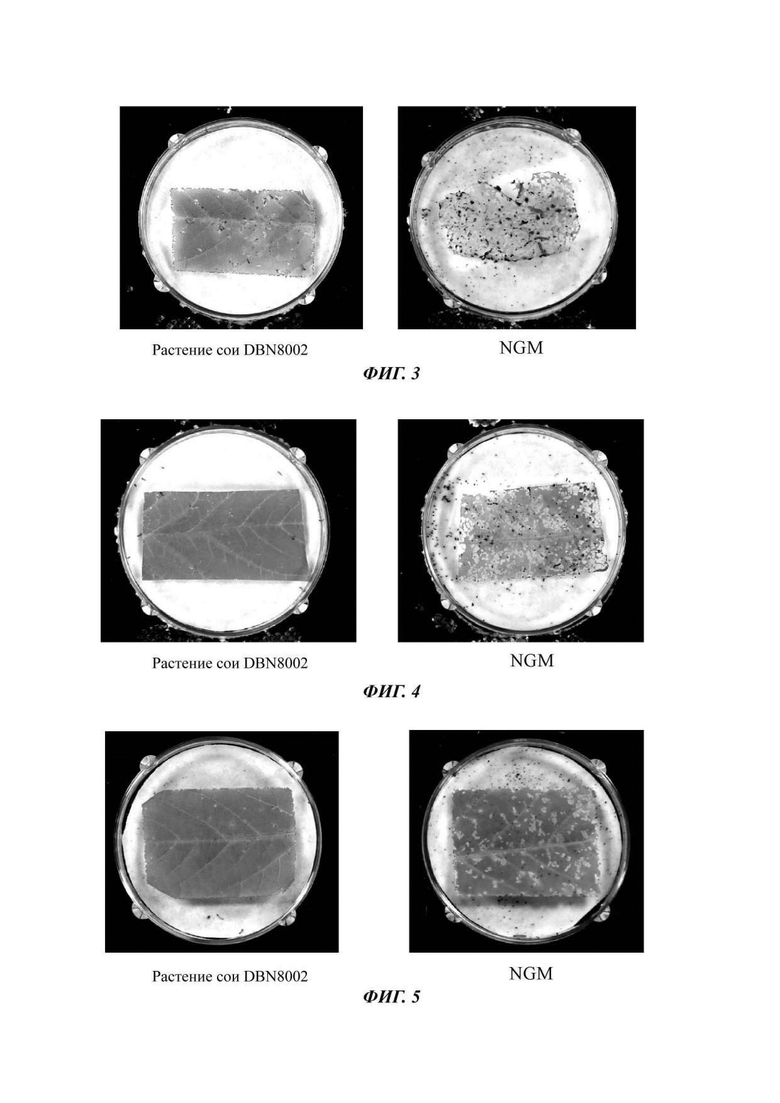
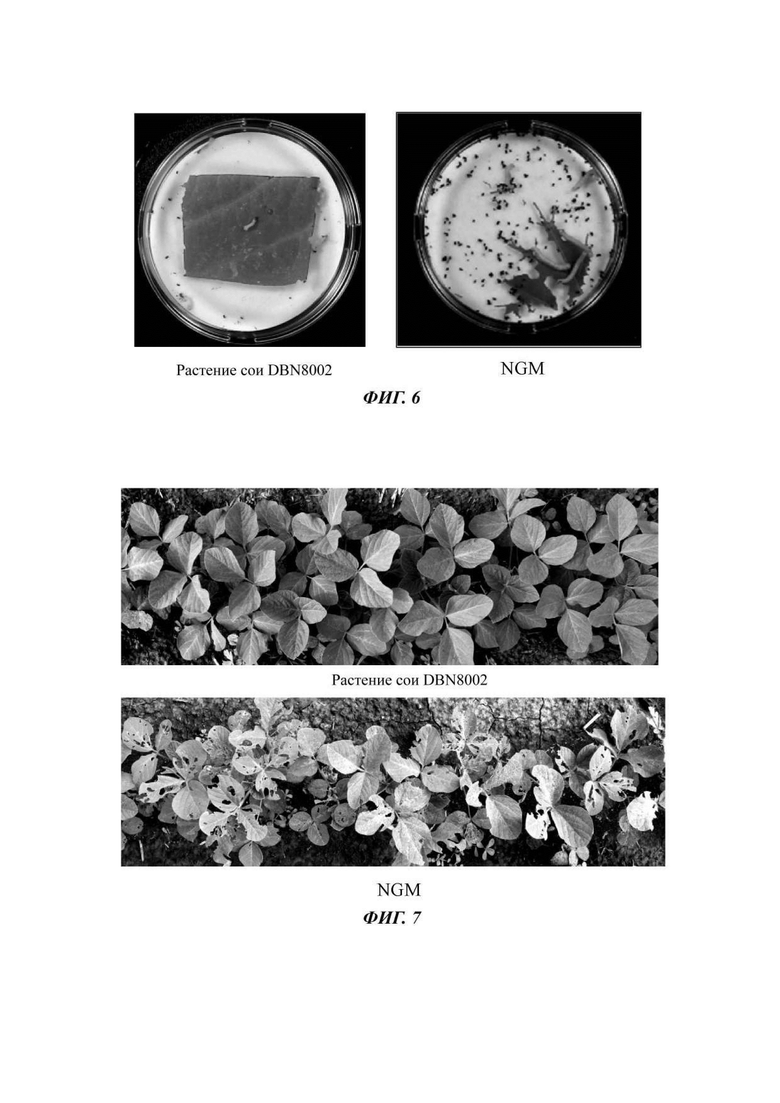


Изобретение относится к области биохимии, в частности к молекуле нуклеиновой кислоты для выявления наличия ДНК трансгенного объекта. Раскрыт набор для выявления ДНК с помощью указанной молекулы ДНК; сельскохозяйственный продукт; товар. Также раскрыт способ выявления наличия ДНК, соответствующей трансгенному объекту – сое; способ защиты растения сои от нашествия целевых насекомых и способ борьбы с сорняками в поле с помощью указанной молекулы нуклеиновой кислоты; способ получения растения сои с помощью указанной молекулы нуклеиновой кислоты. Изобретение позволяет получать растения сои, обладающие устойчивостью к насекомым Lepidoptera и толерантностью к гербициду глюфосинат. 12 н. и 14 з.п. ф-лы, 10 ил., 15 табл., 8 пр.
1. Молекула нуклеиновой кислоты для выявления наличия ДНК трансгенного объекта - сои в образце, отличающаяся тем, что последовательность нуклеиновой кислоты молекулы нуклеиновой кислоты содержит SEQ ID NO: 1 или комплементарную ей последовательность и/или SEQ ID NO: 2 или комплементарную ей последовательность, и где трансгенный объект - соя содержит SEQ ID NO: 1, последовательность нуклеиновой кислоты нуклеотидов 1032-6444 SEQ ID NO: 5 и SEQ ID NO: 2 последовательно или SEQ ID NO: 5.
2. Молекула нуклеиновой кислоты по п. 1, отличающаяся тем, что последовательность нуклеиновой кислоты молекулы нуклеиновой кислоты содержит SEQ ID NO: 3 или комплементарную ей последовательность и/или SEQ ID NO: 4 или комплементарную ей последовательность.
3. Молекула нуклеиновой кислоты по п. 2, отличающаяся тем, что указанная последовательность нуклеиновой кислоты молекулы нуклеиновой кислоты содержит SEQ ID NO: 5 или комплементарную ей последовательность.
4. Способ выявления наличия ДНК, соответствующей трансгенному объекту - сое, в образце, отличающийся тем, что он включает:
- приведение образца, подлежащего выявлению, в контакт с по меньшей мере двумя праймерами для амплификации целевого продукта амплификации в реакции амплификации нуклеиновой кислоты, где указанные два праймера содержат SEQ ID NO: 1 и комплементарную последовательность SEQ ID NO: 2, SEQ ID NO: 8 и SEQ ID NO: 9 или SEQ ID NO: 10 и SEQ ID NO: 11;
- проведение реакции амплификации нуклеиновой кислоты; и
- выявление наличия целевого продукта амплификации;
где целевой продукт амплификации содержит молекулу нуклеиновой кислоты по любому из пп. 1-3 и где трансгенный объект - соя содержит SEQ ID NO: 1, последовательность нуклеиновой кислоты нуклеотидов 1032-6444 SEQ ID NO: 5 и SEQ ID NO: 2 последовательно,или SEQ ID NO: 5.
5. Способ выявления наличия ДНК, соответствующей трансгенному объекту - сое, в образце по п. 4, отличающийся тем, что целевой продукт амплификации дополнительно содержит SEQ ID NO: 6 или комплементарную ей последовательность и/или SEQ ID NO: 7 или комплементарную ей последовательность.
6. Способ выявления наличия ДНК, соответствующей трансгенному объекту - сое, в образце, отличающийся тем, что он включает:
- приведение образца, подлежащего выявлению, в контакт с зондом, где зонд содержит молекулу нуклеиновой кислоты по п. 1 или 2;
- подвергание образца, подлежащего выявлению, и зонда жестким условиям гибридизации; и
- выявление гибридизации образца, подлежащего выявлению, с зондом,
где трансгенный объект - соя содержит SEQ ID NO: 1, последовательность нуклеиновой кислоты нуклеотидов 1032-6444 SEQ ID NO: 5 и SEQ ID NO: 2 последовательно или SEQ ID NO: 5.
7. Способ выявления наличия ДНК, соответствующей трансгенному объекту - сое, в образце по п. 6, отличающийся тем, что зонд дополнительно содержит SEQ ID NO: 6 или комплементарную ей последовательность и/или SEQ ID NO: 7 или комплементарную ей последовательность.
8. Способ выявления наличия ДНК, соответствующей трансгенному объекту - сое, в образце по п. 6 или 7, отличающийся тем, что по меньшей мере один зонд мечен по меньшей мере одним флуорофором.
9. Способ выявления наличия ДНК, соответствующей трансгенному объекту - сое, в образце, отличающийся тем, что он включает:
- приведение образца, подлежащего выявлению, в контакт с маркерной молекулой нуклеиновой кислоты, где маркерная молекула нуклеиновой кислоты содержит молекулу нуклеиновой кислоты по п. 1 или 2;
- подвергание образца, подлежащего выявлению, и маркерной молекулы нуклеиновой кислоты жестким условиям гибридизации;
- выявление гибридизации образца, подлежащего выявлению, с маркерной молекулой нуклеиновой кислоты и дальнейшее проведение анализа скрещивания с помощью маркера для определения того, связана ли генетически устойчивость к насекомым и/или толерантность к гербициду с маркерной молекулой нуклеиновой кислоты;
где трансгенный объект - соя содержит SEQ ID NO: 1, последовательность нуклеиновой кислоты нуклеотидов 1032-6444 SEQ ID NO: 5 и SEQ ID NO: 2 последовательно или SEQ ID NO: 5.
10. Способ выявления наличия ДНК, соответствующей трансгенному объекту - сое, в образце по п. 9, отличающийся тем, что маркерная молекула нуклеиновой кислоты дополнительно содержит по меньшей мере одну последовательность, выбранную из группы, состоящей из SEQ ID NO: 6-11 или комплементарных им последовательностей.
11. Набор для выявления ДНК для выявления наличия ДНК трансгенного объекта - сои в образце, отличающийся тем, что содержит пару праймеров ДНК, специфичных для трансгенного объекта - сои или его потомства, где указанные ДНК праймеры содержат молекулу нуклеиновой кислоты по п. 1 или 2 и где трансгенный объект - соя содержит SEQ ID NO: 1, последовательность нуклеиновой кислоты нуклеотидов 1032-6444 SEQ ID NO: 5 и SEQ ID NO: 2 последовательно или SEQ ID NO: 5.
12. Набор для выявления ДНК по п. 11, отличающийся тем, что ДНК праймеры дополнительно содержат SEQ ID NO: 6 или комплементарную ей последовательность и/или SEQ ID NO: 7 или комплементарную ей последовательность.
13. Набор для выявления ДНК для выявления наличия ДНК трансгенного объекта - сои в образце, отличающийся тем, что содержит зонд и реагенты для гибридизации, где зонд является специфичным для трансгенного объекта - сои или его потомства и содержит молекулу нуклеиновой кислоты по п. 1 или 2 и где трансгенный объект - соя содержит SEQ ID NO: 1, последовательность нуклеиновой кислоты нуклеотидов 1032-6444 SEQ ID NO: 5 и SEQ ID NO: 2 последовательно или SEQ ID NO: 5.
14. Способ защиты растения сои от нашествия целевых насекомых, ингибированных белком Vip3Aa, отличающийся тем, что включает предоставление по меньшей мере одной трансгенной растительной клетки сои в рацион целевого насекомого, где трансгенная растительная клетка сои содержит в своем геноме последовательность, как изложено в SEQ ID NO: 1 и/или SEQ ID NO: 2; при этом попадание внутрь трансгенной растительной клетки сои ингибирует дальнейшее питание целевого насекомого трансгенным растением сои.
15. Способ защиты растения сои от нашествия целевых насекомых, ингибированных белком Vip3Aa, по п. 14, отличающийся тем, что трансгенная растительная клетка сои сдержит в своем геноме последовательность, как изложено в SEQ ID NO: 3 и/или SEQ ID NO: 4.
16. Способ защиты растения сои от нашествия целевых насекомых, ингибированных белком Vip3Aa, по п. 15, отличающийся тем, что трансгенная растительная клетка сои последовательно содержит в своем геноме SEQ ID NO: 1, последовательность нуклеиновой кислоты в положениях 1032-6444 SEQ ID NO: 5 и SEQ ID NO: 2 или содержит последовательность, как изложено в SEQ ID NO: 5.
17. Способ защиты растения сои от повреждения, обусловленного гербицидом, отличающийся тем, что включает внесение эффективного количества гербицида глюфосинат в поле, в котором посажено по меньшей мере одно трансгенное растение сои, где трансгенное растение сои содержит в своем геноме последовательность, как изложено в SEQ ID NO: 1 и/или SEQ ID NO: 2, и трансгенное растение сои обладает толерантностью к гербициду глюфосинат.
18. Способ защиты растения сои от повреждения, обусловленного гербицидом, по п.17, отличающийся тем, что трансгенное растение сои содержит в своем геноме последовательность, как изложено в SEQ ID NO: 3 и/или SEQ ID NO: 4.
19. Способ защиты растения сои от повреждения, обусловленного гербицидом, по п.18, отличающийся тем, что трансгенное растение сои последовательно содержит в своем геноме SEQ ID NO: 1, последовательность нуклеиновой кислоты в положениях 1032-6444 SEQ ID NO: 5 и SEQ ID NO: 2 или содержит последовательность, как изложено в SEQ ID NO: 5.
20. Способ борьбы с сорняками в поле, в котором посажено растение сои, отличающийся тем, что включает внесение эффективного количества гербицида глюфосинат в поле, в котором посажено по меньшей мере одно трансгенное растение сои, где трансгенное растение сои содержит в своем геноме последовательность, как изложено в SEQ ID NO: 1 и/или SEQ ID NO: 2, и трансгенное растение сои обладает толерантностью к гербициду глюфосинат.
21. Способ борьбы с сорняками в поле, в котором посажено растение сои, по п. 20, отличающийся тем, что трансгенное растение сои содержит в своем геноме последовательность, как изложено в SEQ ID NO: 3 и/или SEQ ID NO: 4.
22. Способ борьбы с сорняками в поле, в котором посажено растение сои, по п. 21, отличающийся тем, что трансгенное растение сои последовательно содержит в своем геноме SEQ ID NO: 1, последовательность нуклеиновой кислоты в положениях 1032-6444 SEQ ID NO: 5 и SEQ ID NO: 2 или содержит последовательность, как изложено в SEQ ID NO: 5.
23. Способ получения растения сои, устойчивого к целевым насекомым и толерантного к гербициду глюфосинат, где целевые насекомые ингибированы белком Vip3Aa, при этом способ включает:
- введение последовательности нуклеиновой кислоты, кодирующей белок устойчивости к насекомым Vip3Aa, и/или последовательности нуклеиновой кислоты, кодирующей белок толерантности к гербициду глюфосинат PAT, и последовательности нуклеиновой кислоты специфичной области в геноме первого растения сои или последовательности нуклеиновой кислоты, как изложено в SEQ ID NO: 5, в геноме первого растения сои во второе растение сои, получая таким образом множество растений потомства; и
- отбор растений потомства, содержащих последовательность нуклеиновой кислоты специфичной области, которые также устойчивы к насекомым и толерантны к гербициду глюфосинат;
где последовательность нуклеиновой кислоты специфичной области представляет собой последовательность, как изложено в SEQ ID NO: 1 и/или SEQ ID NO: 2.
24. Способ получения растения сои, устойчивого к целевым насекомым и толерантного к гербициду глюфосинат, по п. 23, отличающийся тем, что последовательность нуклеиновой кислоты специфичной области представляет собой последовательность, как изложено в SEQ ID NO: 3 и/или SEQ ID NO: 4.
25. Сельскохозяйственный продукт, полученный из трансгенного объекта - сои, отличающийся тем, что трансгенный объект - соя находится в указанном сельскохозяйственном продукте и указанный сельскохозяйственный продукт представляет собой лецитин, жирные кислоты, глицерин, стерины, соевые хлопья, соевую муку, соевые белки или их концентраты, соевые масла, волокна соевых белков, продукты свертывания соевого молока или соевый творог, и где трансгенный объект - соя содержит SEQ ID NO: 1, последовательность нуклеиновой кислоты нуклеотидов 1032-6444 SEQ ID NO: 5 и SEQ ID NO: 2 последовательно или SEQ ID NO: 5.
26. Товар, полученный из трансгенного объекта - сои, отличающийся тем, что трансгенный объект - соя находится в указанном товаре и указанный товар представляет собой лецитин, жирные кислоты, глицерин, стерины, соевые хлопья, соевую муку, соевые белки или их концентраты, соевые масла, волокна соевых белков, продукты свертывания соевого молока или соевый творог, и где трансгенный объект - соя содержит SEQ ID NO: 1, последовательность нуклеиновой кислоты нуклеотидов 1032-6444 SEQ ID NO: 5 и SEQ ID NO: 2 последовательно или SEQ ID NO: 5.
| WO 2017215328 A1, 21.12.2017 | |||
| WO 2014201235 A2, 18.12.2014 | |||
| ЛИНИИ ТРАНСГЕННОЙ СОИ, ГЕНЕТИЧЕСКОЕ СОБЫТИЕ 8264.42.32.1, УСТОЙЧИВОЕ К ГЕРБИЦИДАМ С ПАКЕТИРОВАННЫМИ ГЕНАМИ НА ЕГО ОСНОВЕ, И ИХ ДЕТЕКТИРОВАНИЕ | 2012 |
|
RU2636021C2 |

