Область изобретения
В целом изобретение относится к области неинвазивного пренатального скрининга и диагностики. В изобретении предложен способ выявления наличия или отсутствия анеуплоидий, в частности, трисомии хромосомы 21, 18 и 13, в эмбрионе по образцу крови, взятому у матери на ранней стадии беременности.
Уровень техники
На сегодняшний день пренатальное исследование является неотъемлемым компонентом акушерской практики. Основной целью пренатального исследования является скрининг в отношении эмбриональных анеуплоидий, таких как трисомия хромосомы 21 (Т21, синдром Дауна), трисомия 18 (Т18, синдром Эдвардса) и трисомия 13 (Т13, синдром Патау). Хотя большинство эмбрионов с анеуплоидией приводят к прерыванию беременности во время развития эмбриона, Т21 характеризуется наивысшим уровнем выживаемости, следовательно, пренатальное выявления Т21 считается наиболее важным генетическим исследованием в целях пренатального скрининга или диагностики. Доступны надежные инвазивные пренатальные исследования, однако, из-за связанного с ними риска в настоящее время их проводят только в случаях беременности с высокой степенью риска. Разработка надежного способа неинвазивного пренатального исследования (НИПТ) эмбриональных анеуплоидий, в частности, трисомий, на данный момент является наиболее важной задачей наблюдения беременности.
С точки зрения медицины интерес также могут представлять другие трисомии, например, трисомия хромосомы 8, 9, 16 или 22. Также может быть важным пренатальное исследование эмбриональных моносомий, например, моносомии Х-хромосомы (синдром Тернера).
Обнаружение циркулирующей внеклеточной эмбриональной ДНК (вкэДНК) в материнской крови (Lo et al., 1997) создало возможность разработки неинвазивных процессов, в которых используются эмбриональные нуклеиновые кислоты из образца материнской периферической крови, для выявления эмбриональных хромосомных аномалий. ВкэДНК составляет приблизительно менее чем 10% общей циркулирующей внеклеточной ДНК (вкДНК) в плазме материнской крови, при этом недавно было обнаружено, что в материнской крови присутствует полный эмбриональный геном в форме вкэДНК, который, таким образом, является перспективным материалом для НИПТ.
В нескольких документах (Chiu et al., 2008, Fan et al., 2008, Chiu et al, 2011, Sehnert et al., 2011, Lau et al., 2102 и Bianchi et al., 2012, EP 2183693, EP 2366031, US 8296076) раскрыты способы, в которых используется секвенирование ДНК из материнской крови для получения информации по содержанию абберантных хромосом в эмбрионе. Во всех вышеуказанных способах используется анализ общей вкДНК в плазме материнской крови без необходимости выделения эмбрион-специфической ДНК, вкэДНК. Эти способы основаны на выявлении дополнительной копии хромосомы, чтобы иметь возможность различать нормальные случаи и случаи трисомии. В случае эмбриона с трисомией число копий трисомической хромосомы в материнской крови немного повышено по сравнению с другими хромосомами. Аналогичный подход можно применять для выявления моносомий, при этом число копий моносомической хромосомы понижено. Появление массового параллельного секвенирования ДНК (МПС) позволило проводить секвенирование больших количеств молекул ДНК и, таким образом, с недавних пор для неинвазивного выявления эмбриональной анеуплоидии по материнской крови применяется секвенирование ДНК следующего поколения (ССП).
В общем случае выявление эмбриональной трисомии методом ССП проводят с помощью следующего процесса. Сначала проводят секвенирование короткого участка на одном конце каждой молекулы ДНК из плазмы материнской крови, а полученные считываемые фрагменты последовательностей картируют против стандартного человеческого генома, чтобы определить хромосомное происхождение каждой последовательности. Далее количество меток (т.е. картированных считываемых фрагментов) из представляющей интерес хромосомы (например, хромосомы 21) сравнивают с какой-либо стандартной хромосомой (т.е. с количеством меток другой отдельной хромосомы или выбранной группы хромосом), обычно посредством соотношения таких величин, и проводят сравнение полученного в результате соотношения для исследуемого образца и эуплоидных контрольных образцов.
В ЕР 2183693 (Lo, Y.M., D. et al., права переданы Chinese University of Hong Kong, HK) раскрыт способ, в котором соответствующие количества клинически релевантной хромосомы и фоновой хромосомы определяют по результатам МПС и рассчитывают процентное содержание последовательностей, картированных на хромосоме 21. Было обнаружено, что такое процентное содержание существенно выше в образце от беременной женщины, несущей эмбрион с трисомией 21, по сравнению с женщиной с нормальным эмбрионом. В этом способе z-показатель не имел весомого значения.
В US 8296076 (Fan, Ch. Н.М. and Quake, S.R., права переданы The Board of Trustees of the Leland Stanford Junior University, US) раскрыт способ диагностики эмбриональной анеуплоидии на основании секвенирования. Подсчитывая количество меток, картированных на заданном окне в каждой хромосоме, можно выявить излишек или недостаток содержания любой хромосомы в ДНК из плазмы материнской крови, вызванный наличием анеуплоидного эмбриона.
Способы из Chiu et al., 2008, Chiu et al., 2011, Sehnert et al., 2011, Lau et al., 2012 основаны на секвенировании полного генома с помощью МПС и определении z-показателя. Хотя z-показатель является общепринятым в качестве стандартного параметра, используемого для выявления анеуплоидных образцов, существуют различия в его определении. В Chiu et al., 2011 раскрыт подход с применением меток, картированных на всех хромосомах, используемых в качестве стандарта для определения z-показателя, в Lau et al., 2012 раскрыт подход с применением меток, картированных на какой-либо конкретной хромосоме, например, 14, в качестве стандарта для Т21, а в Sehnert et al., 2011 в качестве оптимального внутреннего стандарта для хромосомы 21 была выбрана хромосома 9.
В общем случае существует три основных этапа определения анеуплоидии эмбриона по образцу материнской крови, плазмы или сыворотки: 1) получение образца ДНК и библиотеки ДНК, 2) секвенирование и 3) анализ данных по последовательности. В последние несколько лет был достигнут значительный прогресс в секвенировании, однако на первом и третьем этапе остается простор для улучшения, которое может привести к снижению затрат и оказать большое влияние на качество исследования.
Существуют различные подходы для улучшения первого этапа, как описано, например, в ЕР 2366031 (Rava, R.P. et al., права переданы Verinata Health, Inc., US). В этом документе раскрыт способ пренатального скрининга и диагностики эмбриональной хромосомной анеуплоидии на основе ССП, включающий новый протокол для получения библиотек секвенирования из материнского образца. Новый подход для получения библиотек секвенирования включает последовательные этапы репарации концов, присоединения dA и лигирования адаптеров в указанных нуклеиновых кислотах, а указанные последовательные этапы не включают очистку продуктов репарации концов перед этапом присоединения dA и не включают очистку продуктов присоединения dA перед этапом лигирования адаптеров. Способ позволяет определить вариацию числа копий (CNV) любой представляющей интерес последовательности. В этом способе z-показатель не имел весомого значения.
Другой вариант улучшения раскрыт в WO 2011/051283 (Benz, М. et al., права переданы Lifecodexx, AG, DE). Раскрытый в ней способ неинвазивной диагностики хромосомной анеуплоидии улучшен путем обогащения и количественной оценки выбранных последовательностей вкДНК в образце материнской крови.
Несмотря на существование нескольких способов неинвазивного выявления эмбриональной анеуплоидии, до сих пор существует потребность в улучшенном способе или альтернативном способе, который был бы по меньшей мере таким же чувствительным и специфическим, как и существующие на сегодняшний день способы.
Описание изобретения
В настоящем изобретении предложен альтернативный и надежный способ, который применим по меньшей мере к практической реализации неинвазивного пренатального скрининга анеуплоидий, таких как трисомии или моносомии, предпочтительно, трисомии хромосомы 13, 18 или 21. Настоящий способ улучшен по сравнению со способами известного уровня техники, в особенности в части обработки данных секвенирования. Благодаря новому и изобретательскому подходу этот способ обеспечивает альтернативный метод определения z-показателя, который обеспечивает лучшее разграничение между эуплоидными и анеуплоидными образцами, т.е. улучшает надежность исследования. Кроме того, данный способ требует относительно небольшое количество данных секвенирования и, следовательно, такой способ является относительно недорогим и его могут позволить даже небольшие учреждения здравоохранения.
В нижеследующем описании будут разъяснены основные признаки способа согласно настоящему изобретению; однако это не означает, что изобретение должно включать все описанные в данном документе признаки и аспекты. Объем правовой охраны определяется патентной формулой изобретения, прилагаемой к данному описанию. Настоящее изобретение станет полностью понятно для специалиста после изучения нижеследующего описания вместе с примерами, в которых некоторые конкретные признаки и аспекты будут разъяснены более подробно, и прилагающимися фигурами.
Используемые в данном документе технические и научные термины имеют то же значение, которое обычно подразумевается специалистами в области техники медицины, молекулярной биологии, генетики, биоинформатики и пренатальной диагностики, если не указано иное.
Краткое изложение способа в соответствии с настоящим изобретением
Настоящее изобретение относится к способу определения анеуплоидии эмбриона по образцу материнской крови, содержащему смесь молекул нуклеиновых кислот эмбриона и матери, при этом смесь молекул нуклеиновых кислот эмбриона и матери представляет собой молекулы внеклеточной ДНК, а указанный способ включает четыре основных этапа: 1) получение и обработку образцов материнской крови, 2) получение образца ДНК и библиотеки ДНК, 3) секвенирование и 4) обработку данных по последовательностям для получения величины, которая имеет прогнозную значимость. Существенной частью способа является получение и обработка проверочных данных, т.е. обработка группы эуплоидных образцов, которые обрабатывают тем же способом, что и исследуемые образцы. Результат, полученный при обработке проверочных образцов, служит в качестве контрольных данных на этапе 4), упомянутом выше. Термин образец крови употребляется в широком значении и включает как саму кровь, так и образец плазмы или сыворотки.
Способ в соответствии с настоящим изобретением можно применять в отношении анеуплоидии любого типа, например, моносомии или трисомии, например, анеуплоидии хромосом 8, 9, 13, 16, 18, 21, 22 и X, в частности, трисомии хромосом 13, 18 и 21.
Основное улучшение и преимущество настоящего способа относится к этапу обработки и анализа данных по последовательностям. Определение значения (z-показателя), которое является определяющим для данного способа (сравнение этого конкретного значения с заданным пороговым значением позволяет определить наличие анеуплоидии, такой как трисомия или моносомия), проводят на основании модели полиномиального распределения меток на хромосомах или бинах человеческого генома. Как продемонстрировано в примерах, значения z-показателя для трисомических образцов, определенные способом согласно настоящему изобретению, существенно выше, чем значения, определенные любым из способов известного уровня техники (для образцов Т21 мы получили медианную разницу значения z-показателя -1,895, -1,595, -2,485 при сравнении Chiu et al., 2008, Sehnert et al., 2011, Lau et al., 2012, соответственно, с нашим способом с дифференциальным р-значением <0,05 для всего известного уровня техники 
Отправной точкой настоящего способа является обработка материнского образца, который представляет собой периферическую кровь, следовательно, способ является неинвазивным. Периферическая кровь содержит внеклеточную ДНК (вкДНК), которая представляет собой смесь материнской и эмбриональной ДНК (вкэДНК). Значение имеет вкэДНК; следовательно, эмбриональную ДНК можно обогатить (путем выбора более короткого фрагмента как in silico, так и физически). Далее образец ДНК (все еще содержащий как материнскую, так и эмбриональную ДНК, т.е. смешанный образец) подвергают массовому параллельному секвенированию, проводимому с применением подхода ССП, чтобы получить огромное количество коротких считываемых фрагментов последовательностей. Эти считываемые фрагменты служат метками, т.е. они соответствуют определенной геномной области хромосомы. В целях данного способа полный человеческий геном (24 хромосомы вместе, представляющие ДНК с более чем 3×109 п.о.) делят на неперекрывающиеся 1 м.п.о. бины (т.е. приблиз. 3×103 бинов), а настоящий способ учитывает как метки, картированные на хромосомы, так и эти бины. Затем, просто говоря, подсчитывая число меток, картированных на конкретном представляющем интерес бине (например, бине, связанном с потенциально триплоидной хромосомой, такой как хромосома 21), и сравнивая это число с числом меток, картированных на какой-либо стандартной хромосоме (или группе стандартных хромосом или их частей, т.е. конкретной группе стандартных бинов), можно выявить отклонения в представляющей интерес хромосоме или ее частях.
Существенно частью способа является получение и обработка проверочных образцов, т.е. обработка группы эуплоидных образцов, которые обрабатывают тем же путем, что и исследуемые образцы. Группа проверочных данных служит в качестве основания для выбора стандартных хромосом или их частей. Выбор правильного стандарта, а также в значительной степени случайной проверочной группы, является наиболее важным, как следует из наших собственных экспериментов, а также из документов известного уровня техники, перечисленных выше. Следовательно, наш подход включает новый и улучшенный способ отбора внутреннего стандарта, в частности, хромосом внутреннего стандарта и бинов внутреннего стандарта.
Кроме того, настоящий способ включает определение значения z-показателя на основании нового подхода, который до настоящего времени не применялся. Вкратце, z-показатель рассчитывают по значениям среднего и стандартного отклонения, определяемым на основании полиномиального распределения меток на бинах. И наконец, сравнение z-показателя с заданным граничным значением (например, 3) свидетельствует о наличии или отсутствии анеуплоидии.
Настоящий полиномиальный подход является, в принципе, альтернативным способом определения z-показателя; при этом он основан на данных, значительно более общих, чем способы известного уровня техники. В частности, первая часть улучшения настоящего способа состоит в более общем выборе стандартной группы хромосом. Мы не только рассматриваем в качестве внутреннего стандарта любую комбинацию хромосом (исключая комбинации с хромосомами 13, 18 и 21 вследствие потенциальной трисомии), но также делаем возможным наличие только частей хромосом, т.е. бинов (с разрешением 1 м.п.о.). Таким образом, мы можем лучше охарактеризовать эуплоидную популяцию и, следовательно, более легко обнаружить отклонения, т.е. анеуплоидные образцы. Следовательно, первое преимущество нашего подхода состоит в том, что внутренним стандартом может быть любая группа хромосом или частей хромосом с разрешением в 1 м.п.о. бинов (вместо одной конкретной хромосомы для всех аутосом, как в способах известного уровня техники). Другим преимуществом настоящего способа является то, что наш полиномиальный подход основан на математически обоснованном и комплексном моделировании распределения меток на бинах и соответствующего соотношения анеуплоидной, например, трисомической хромосомы и внутренних стандартных бинов. Полиномиальный подход приводит к получению намного большего z-показателя для трисомических образцов по сравнению со способами известного уровня техники, и в то же время не приводит к значительному изменению z-показателей эуплоидных образцов. Таким образом, повышается надежность исследования. Кроме того, настоящий способ позволяет оценить эмбриональную фракцию вместе с погрешностью этой оценки. Хотя оценка эмбриональной фракции была ранее представлена в Rava et al., 2014, Hudecova et al., 2014, и Yu et al., 2014, до этого времени не было представлено погрешности этой оценки. Кроме того, настоящий способ позволяет оценить минимальное число картированных меток, при котором ожидается, что анеуплоидный, например, трисомический, образец с определенной эмбриональной фракцией продемонстрирует z-показатель, соответствующий по меньшей мере некоторому заданному значению (например, пороговому значению). Эта информация может оказаться полезной при разрешении случаев с низкой эмбриональной фракцией и приближенным к пороговому значением z-показателя.
Существует практическая необходимость в автоматизации методов пренатальных исследований или диагностики. Способ согласно настоящему изобретению может быть в значительной мере автоматизирован. По меньшей мере биоинформатическую часть способа (т.е. обработку данных секвенирования и все последующие определения и расчеты) можно осуществлять с помощью любой подходящей компьютерной системы, такой как, например, ПК, оборудованный процессором, периферическими устройствами ввода/вывода (например, портами, интерфейсами), памятью (например, системной памятью, жестким диском), клавиатурой, монитором, мышкой и т.д. Предпочтительно компьютерная система может иметь возможность обмена данными с системой секвенирования, обеспечивая данные по последовательностям, предпочтительно в форме множества считываемых фрагментов последовательностей (через проводную или беспроводную сеть, блютуз, интернет, облако и т.д.). Это означает, что компьютерная система выполнена с возможностью получать данные по последовательностям от системы секвенирования. Подходящие компьютерные системы, а также средства для соединения с системой секвенирования хорошо известны специалистам в данной области техники.
По меньшей мере часть способа, в частности, биоинформатическая часть способа, может быть реализована в виде программного кода, т.е. множества инструкций (компьютерной программы), предназначенных для выполнения процессором компьютерной системы. Код может находиться на машиночитаемом носителе для хранения или передачи, таком как, например, RAM, ROM, жесткий диск, SDS, CD, DVD, флэш-память и т.д. Кроме того, код может быть передан через любую подходящую проводную, оптическую или беспроводную сеть, например, через интернет. Например, пользователь (потребитель) может загрузить целую компьютерную программу через интернет.
Следовательно, настоящее изобретение также относится к компьютерному программному продукту, включающему машиночитаемый носитель, содержащий множество инструкций для управления компьютерной системой для осуществления по меньшей мере части способа в соответствии с изобретением, предпочтительно части, начинающейся с этапа получения информации по последовательностям после этапа случайного секвенирования, осуществляемого автоматической системой секвенирования. Указанная выше компьютерная программа предпочтительно может быть установлена на компьютере системы секвенирования.
Предмет настоящего изобретения определен в пп. 1-13 прилагаемой формулы изобретения.
Признаки и преимущества настоящего изобретения станут понятны специалисту в данной области техники после изучения нижеследующего подробного описания, примеров и фигур.
Подробное описание способа
СПОСОБ
Способ представляет собой способ определения z-показателя на основании модели полиномиального распределения картированных меток на бинах. Следует отметить, что в этом разделе мы в качестве примера пишем в основном о трисомии, в частности, хромосомы 21, однако, способ можно применять в отношении любой анеуплоидии, моносомии или другой трисомии, в частности, трисомии хромосомы 13 и 18, аналогичным образом.
Предварительным условием обработки исследуемого образца является выбор и обработка соответствующей проверочной группы (т.е. группы проверочных образцов) с целью выбора соответствующего внутреннего стандарта для оценки исследуемых образцов. В общем случае проверочная группа должна включать эуплоидные женские образцы, а также эуплоидные мужские образцы, причем образцы обязательно должны обрабатываться тем же путем, что и исследуемые образцы.
ПРОВЕРОЧНЫЕ ОБРАЗЦЫ
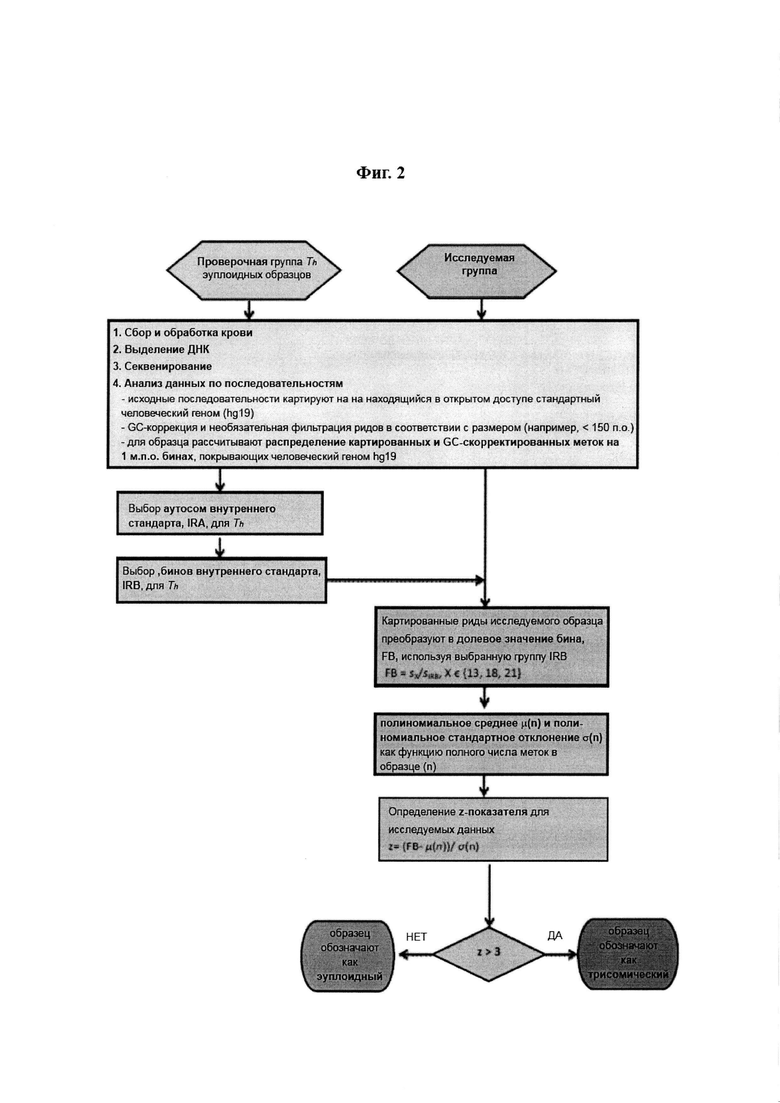
Выбирают эуплоидные мужские и женские образцы, чтобы сформировать проверочную группу, обозначаемую Th. Для каждого образца из проверочной группы проводят следующие этапы:
1. Сбор и обработка образцов крови,
2. Выделение ДНК,
3. Секвенирование,
4. Для каждого образца из проверочной группы применяют следующие этапы обработки данных по последовательностям:
(a) Исходные считываемые фрагменты картируют на находящийся в открытом доступе стандартный человеческий геном (hg19, Genome Reference Consortium Human Build 37 (GRCh37), Feb. 2009, доступ сборки GenBank: GCA_000001405.1, доступ сборки RefSeq: GCF_000001405.13), используя алгоритм Bowtie2 (Langmead and Salzberg, 2012).
(b) Оставляют только считываемые фрагменты, которые можно картировать только на одной геномной локации не более чем с одним несовпадением.
(c) Коррекцию GC проводят в соответствии с Liao et al., 2014 (в нашем случае - без внутрисерийной нормировки).
(d) Необязательно: Фильтруют считываемые фрагменты в соответствии с их размером (например, <150 п.о.) (Minarik et al., 2015).
(e) Стандартный геном hg19 разбивают на 1 м.п.о. последовательные и неперекрывающиеся бины с картированными ДНК-метками, распределенными на бинах в соответствии с их расположением.
Фильтрацию по размеру на этапе 4d можно использовать, чтобы искусственно увеличить эмбриональную фракцию и, как следствие, также z-показатель трисомических образцов. Однако обычно это происходит за счет снижения общего количества меток в образце, так как метки, превышающие принятый порог, отбраковываются (подробную информацию см. в Minarik et al., 2015).
Кроме того, бины внутреннего стандарта, обозначаемые IRB, должны быть выбраны до определения значения z-показателя исследуемого образца, что осуществляется посредством выбора аутосом внутреннего стандарта, обозначаемых IRA, с применением проверочной группы Th и генетического алгоритма. Следовательно, способ включает этапы:
5. Выбор аутосом внутреннего стандарта, IRA, с применением проверочной группы Th.
(a) Любая комбинация аутосом (за исключением комбинаций с аутосомами 13, 18 и 21 вследствие потенциальной анеуплоидии) считается потенциальным внутренним стандартом (всего 524 287 кандидатных комбинаций).
(b) Для каждой потенциальной комбинации внутреннего стандарта рассчитывают коэффициент вариации (CV) (дополнительно см. Расчет коэффициента вариации кандидатной комбинации аутосом внутреннего стандарта, ниже).
(c) Кандидатные комбинации аутосом внутреннего стандарта располагают в соответствии с их значением CV в порядке возрастания.
(d) Комбинации внутреннего стандарта, чьи значения CV меньше или равны кратному 1,1 значению наименьшей величины CV, выбирают в качестве IRA. Предпочтительно выбирают по меньшей мере сто (т.е. первую сотню) этих комбинаций (они представляют собой наилучшие аутосомные комбинации внутреннего стандарта в терминах значений CV).
6. Выбор бинов внутреннего стандарта, IRB, с помощью генетического алгоритма с применением проверочной группы Th (дополнительно см. Выбор бинов внутреннего стандарта, ниже).
Расчет коэффициента вариации кандидатной комбинации аутосом внутреннего стандарта
Пусть М=(mi,j)kx22 является проверочной матрицей, связанной с проверочной группой Th, где k - размер Th, и mi,j соответствует числу меток, картированных (и GC-скорректированных) на j-ой аутосоме для i-го образца Th, a ir={0, 1}22 является кандидатной комбинацией внутреннего стандарта (ir[i]=1 в случае выбора для внутреннего стандарта i-ой аутосомы, i∈{1, 2, …, 22}, в ином случае ir[i]=0). Также пусть Mi обозначает i-ый ряд матрицы М. Нормированное значение i-го образца (т.е. i-ый ряд матрицы М) определяют как mi,x/Mi⋅ir, где x - анеуплоидная хромосома, которую мы хотим исследовать. Мы обозначаем нормированную величину образца i как υi. Пусть Vir=(υ1, υ2, …, υk). Тогда значение CV кандидатной комбинации внутреннего стандарта ir определяется как
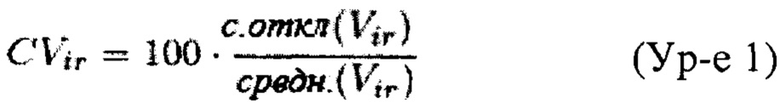
Выбор бинов внутреннего стандарта (генетический алгоритм)
Пусть М=(mi,j)kxt является матрицей, связанной с проверочной группой Th, где k - размер Th, t - число 1 м.п.о. бинов, которые покрывают геном hg19, а mi,j соответствует числу меток, картированных на j-ом бине для i-го образца Th. Также пусть Mi обозначает i-ый ряд матрицы М. Кандидатная комбинация бинов внутреннего стандарта (IRB) представляет собой любую комбинацию колонок матрицы М за исключением комбинаций, которые содержат бины, связанные с хромосомами 13, 18 и 21 (эти бины не должны присутствовать в выбранных IRB вследствие потенциальной трисомии; другие трисомии являются очень редкими). Пусть ir={0, 1}t такой кандидатный IRB (ir[j]=1 в случае выбора для внутреннего стандарта j-го бина, j∈{1, 2, …, t}, в ином случае ir[j]=0). Кроме того, пусть бинарный вектор υ21 обозначает бины, связанные с хромосомой 21, аналогично вектору ir (так же и для хромосом 13 и 18 или других анеуплоидий). Для каждого кандидатного IRB мы рассчитываем долевое значение бина FBi для каждого образца в Th из i-го ряда матрицы М как

где ⋅ представляет скалярное произведение. Пусть Vir=(FB1, FB2, …, FBk) соответствует долевым значениям бинов для всех образцов в Th. Мы приписываем каждому кандидатному IRB коэффициент размаха Стьюдента (SRC), который подается в виде

Значение SRC описывает разброс значений FB проверочной группы. Помимо значения SRC мы также приписываем каждому кандидатному IRB значение CV, которое рассчитывается аналогично Расчету коэффициента вариации кандидатной комбинации аутосом внутреннего стандарта. Мы хотим найти такой IRB, который имеет наименьшие возможные значения SRC и CV. Однако группа всех IRB-кандидатов включает приблизительно 23000 членов и, следовательно, для них всех невозможно рассчитать значения SRC и CV. Следовательно, чтобы выбрать хороший IRB, мы используем генетический алгоритм (генетические алгоритмы хорошо известны специалисту в данной области техники, а мы используем программу DEAP, представленную в Fortin et al., 2012). В частности, исходная популяция IRB-кандидатов, применяемых в генетическом алгоритме, состоит из двух групп. Первая группа представляет собой случайную выборку бинов из всех аутосом, за исключением бинов из аутосом 13, 18 и 21 вследствие потенциальной трисомии, в то время как вторая группа базируется на заранее определенных аутосомах внутреннего стандарта (IRA). Таким образом, в популяции кандидатных IRB уже находятся хорошие представители (то есть, из группы IRA), а генетический алгоритм может только усилить предыдущие комбинации из выборки IRA, например, путем выбора только некоторых бинов комбинаций IRA.
Возможно, генетический алгоритм не выберет лучший IRB в терминах значений SRC и CV, но полученный в результате IRB будет (часто в значительной мере) лучше, чем IRA. Это связано с тем, что IRB делает возможным наличие только частей хромосом в стандарте в противоположность целым хромосомам в случае IRA.
Нам бы хотелось подчеркнуть, что выбранный IRB зависит от выбранной представляющей интерес хромосомы (например, хромосомы 21), образцов в проверочной группе, а также в некоторой степени от природы генетических алгоритмов. Таким образом, многоразовое проведение этой части процесса наиболее вероятно приведет к получению разных IRB. По этой причине советуется повторять этот процесс много раз (число раз ограничено только временем, которое мы отводим на эту часть процесса) и выбрать наилучший имеющийся IRB.
ИССЛЕДУЕМЫЙ ОБРАЗЕЦ
Исследуемый образец обрабатывают точно таким же путем, что и проверочные образцы на этапах от 1 до 4, как подробно описано выше. Кроме того, используя бины внутреннего стандарта, IRB, определение которых приведено выше, проводят следующие этапы:
5. Соответствующие метки исследуемого образца преобразуют в долевое значение бина (FB), используя выбранную группу IRB (см. Выбор бинов внутреннего стандарта)

где s21 обозначает число меток исследуемого образца, картированных на бинах хромосомы 21 (или другой представляющей интерес хромосомы), a sIRB обозначает число меток исследуемого образца, картированных на бинах IRB.
6. Рассчитывают полиномиальное среднее μ(n) и стандартное отклонение σ(n) для исследуемого образца (дополнительно см. Расчет полиномиального среднего и стандартного отклонения)
7. Оценивают исследуемый образец (дополнительно см. Оценка исследуемого образца)
Расчет полиномиального среднего и стандартного отклонения
Сначала следует отметить, что под полиномиальным средним и стандартным отклонением мы не подразумеваем среднее и стандартное отклонение полиномиального распределения. Скорее это среднее и стандартное отклонение новой случайной переменной, полученной из полиномиального распределения, определение которой мы приводим в разделе Математическая модель далее в этом описании.
Пусть IRB является группой выбранных бинов внутреннего стандарта. Пусть Th является группой проверочных эуплоидных образцов размером k. Пусть М=(mi,j)kxt является матрицей, связанной с проверочной группой Th, где k - размер Th, t - число 1 м.п.о. бинов, которые покрывают человеческий геном hg19, а mi,j соответствует числу меток, картированных на j-ом бине для i-го образца Th. Пусть mi, i∈{1, 2, …, k}, обозначает сумму i-го ряда матрицы М. Пусть Р=(pi,j)kxt является матрицей, причем pi,j=mi,j/mi, i∈{1, 2, …, k}, j∈{1, 2, …, t}, где mi,j является элементом из матрицы М. Кроме того, пусть pj, j∈{1, 2, …, m}, обозначает среднее значение j-ой колонки матрицы Р и пусть p=(р1, …, pt). Мы интерпретируем значение pj, j∈{1, 2, …, m}, как возможность того, что фрагмент ДНК будет картирован на j-ом бине. Следует отметить, что этот вектор дает проверочная группа Th и он является единственным значением, которое необходимо проверять в нашем способе. Мы наблюдали, что эти значения меняются при разной лабораторной обработке образцов, следовательно, проверочные образцы Th и исследуемые образцы необходимо готовить одинаковым образом. Пусть PX, X∈{13, 18, 21} в зависимости от трисомии, которую необходимо исследовать, a PIRB является суммой вероятностей для бинов, связанных с X-ой хромосомой, и IRB, соответственно. Мы определяем полиномиальное среднее μ(n) и полиномиальное стандартное отклонение σ(n) как
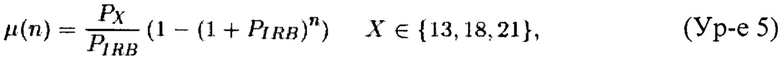

где
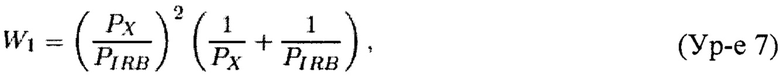
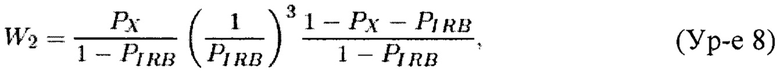
и n - общее количество картированных меток (теперь включая метки из бинов, связанных с хромосомами 13, 18 и 21) образца, который мы хотим исследовать. Следует отметить, что исследуемые образцы с разным n характеризуются разным полиномиальным средним и стандартным отклонением. Таким образом, исследование персонализировано для каждого образца.
Оценка данных исследования
Пусть FB является долевым значением бина в образце, который мы хотим исследовать (см. Выбор бинов внутреннего стандарта). Пусть n является общим числом картированных ДНК-меток (включая метки из бинов, связанных с хромосомами 13, 18 и 21) для этого образца. Учитывая случай трисомии, который мы хотим исследовать, мы рассчитываем соответствующее полиномиальное среднее μ(n) и полиномиальное стандартное отклонение σ(n) (см. выше Расчет полиномиального среднего и стандартного отклонения). После этого мы рассчитываем z-показатель для исследуемого образца

и если z превышает заданную величину, согласно настоящему способу образец помечается как трисомический (обычно для z>3).
Эмбриональная фракция
Кроме того, приблизительная эмбриональная фракция ƒ исследуемого образца подается в виде

где Р21 и PIRB взяты из Расчета полиномиального среднего и стандартного отклонения (аналогично для других трисомий), a FB является долевым значением бина из Выбора бинов внутреннего стандарта.
Погрешность оценки эмбриональной фракции
Пусть p=(р1, …, pt) взято из Расчета полиномиального среднего и стандартного отклонения. Пусть Q(f)=(q_1(f), q_2(f), q_3(f), …, q_t(f)), где

где S_21 - сумма p_j, связанных с хромосомой 21. i принадлежит диапазону от 1 до t, a f является переменной, которая может принадлежать диапазону от 0 до 1. Для других трисомий уравнение имеет такой же вид. Для моносомии уравнение меняется на

Мы преобразуем вектор Q(f) в вектор Q'(f)=(Q_X, Q_IRB, 1 - Q_X - Q_IRB), где Q_X - сумма значений q_i(f), связанных с хромосомой 21 или другой анеуплоидной хромосомой, a Q_IRB - сумма значений q_i(f), связанных с бинами IRB. Следует отметить, что до тех пор, пока мы не определим значение f, векторы Q(f) и Q'(f) являются символическими. Используя значения Q_X и Q_IRB вместо Р_Х и P_IRB, мы определяем значения μ(n, ƒ) и σ(n, f) аналогично значениям μ(n) и σ(n) из Расчета полиномиального среднего и стандартного отклонения. Следует отметить, что мы пишем μ(n, ƒ) вместо μ(n), так как мы еще не установили значение f.
Пусть FB является долевым значением бина в образце, с которым мы на данный момент работаем (см. Выбор бинов внутреннего стандарта). Решая уравнение

для f (напомним, что n является общим числом меток исследуемого образца), мы получаем нижнюю границу для эмбриональной фракции исследуемого образца. Решая уравнение

для f, мы получаем верхнюю границу для эмбриональной фракции исследуемого образца.
Природа этих границ следующая. Так как секвенирование является случайным процессом в том смысле, что выбор секвенируемых фрагментов происходит случайно, может оказаться, что два эксперимента по секвенированию одного образца приведут к получению разного числа секвенируемых фрагментов из, скажем, хромосомы 21. По этой причине z-показатель образца может варьироваться в разных экспериментах по секвенированию (с учетом одинакового выбора IRB), и то же самое касается полученной эмбриональной фракции. Эти границы устанавливаются таким образом, чтобы 95% экспериментов по секвенированию одного образца приводили к получению эмбриональной фракции в этих границах. Таким образом, для ранее рассчитанной эмбриональной фракции f, эти границы дают нам вероятный предел погрешности для этого значения.
Минимальное количество меток для надежного прогноза
Для эуплоидных образцов ожидаемое долевое значение бина (FB) составляет приблизительно р'1/p'2 (по Теореме 1). В качестве примера, для образца Т21 с эмбриональной фракцией ƒ это значение FB приблизительно равно (1+ƒ/2)p'1/p'2, так как в трисомических образцах параметр р'1 увеличен на количество ƒ/2р'1 из третьей копии эмбриональной хромосомы 21, в то время как другие хромосомы не меняются (при условии, что в образце не присутствуют другие трисомии). Также вводится нормирующий фактор 1/(1+ƒ/2 р'1), так как, повышая вероятностный параметр р'1, мы нарушаем уравнение р'1+р'2+р'3=1.
Однако этот фактор исключается из соотношения р'1/р'2. Таким образом, по Теоремам 1 и 2 (см. далее) ожидаемый z-показатель трисомического образца с эмбриональной фракцией ƒ составляет (примите во внимание, что мы пренебрегли членом W2/n2 из Теоремы 2 в целях упрощения)
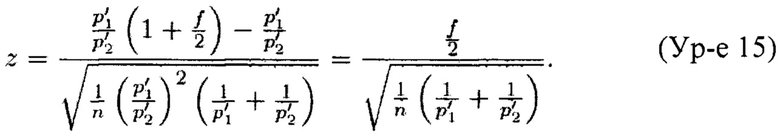
Мы хотим, чтобы z составляло по меньшей мере некоторое k. Кроме того, z-показатели эуплоидных образцов распределены по обе стороны вокруг 0 (0 является ожидаемым z-показателем), и то же самое относится к трисомическим образцам (конечно, такие z-показатели будут распределены вокруг другого среднего значения). Следовательно, для трисомического образца существует возможность, что его долевое значение бина FB не будет равным (1+ƒ/2) р'1/p'2; оно может быть как выше, так и ниже. Таким образом, мы хотим получить
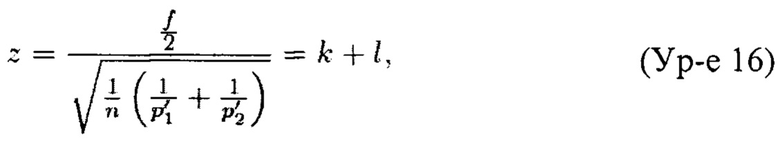
где k - z-показатель, которого мы бы хотели достичь, а 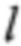 корректирует возможность получения показателя для образца s ниже ожидаемой (как в случае эуплоидного образца, который имеет z-показатель ниже 0). Так как z-показатели эуплоидных образцов подлежат стандартному нормальному распределению (это предположение было положительным образом подтверждено статическими исследованиями), 95% z-показателей этих образцов должны попадать в интервал [-2; 2], поэтому установление
корректирует возможность получения показателя для образца s ниже ожидаемой (как в случае эуплоидного образца, который имеет z-показатель ниже 0). Так как z-показатели эуплоидных образцов подлежат стандартному нормальному распределению (это предположение было положительным образом подтверждено статическими исследованиями), 95% z-показателей этих образцов должны попадать в интервал [-2; 2], поэтому установление 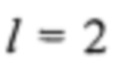 кажется обоснованным. Решая последнее уравнение для n, мы получаем требуемое общее количество соответствующих фрагментов ДНК для ожидаемого z-показателя, составляющего по меньшей мере k, как
кажется обоснованным. Решая последнее уравнение для n, мы получаем требуемое общее количество соответствующих фрагментов ДНК для ожидаемого z-показателя, составляющего по меньшей мере k, как
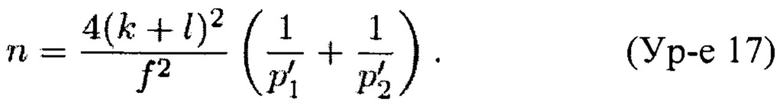
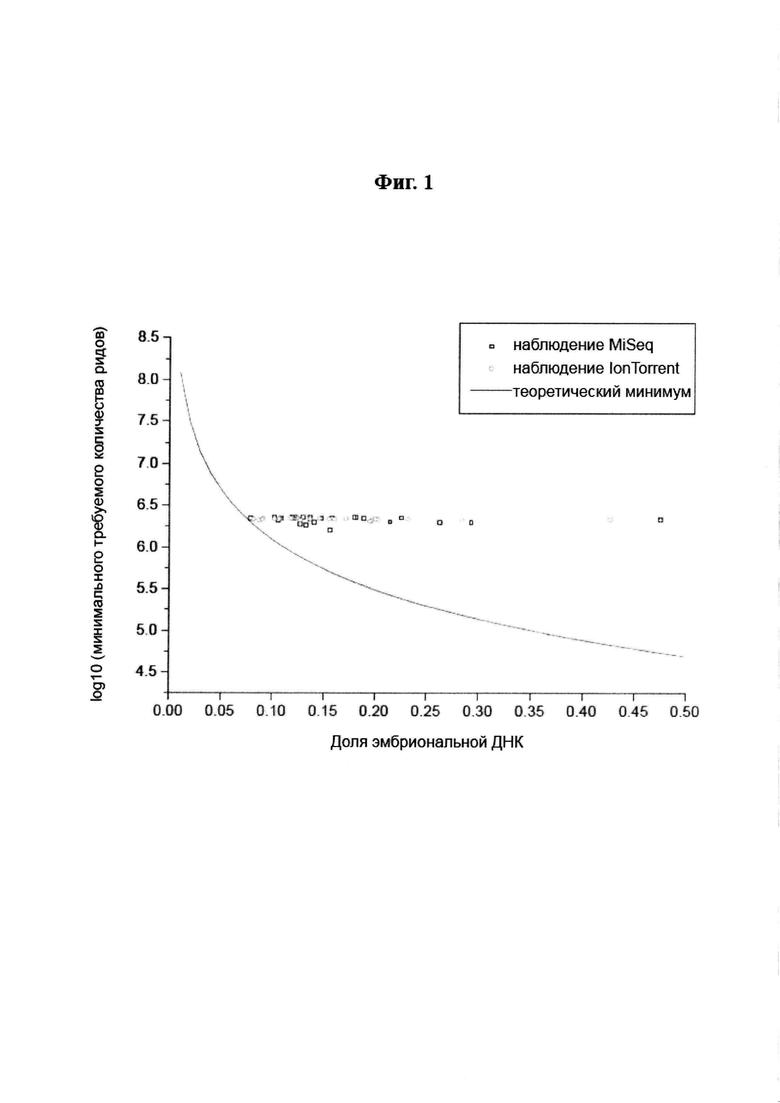
С k=4 и мы получаем минимальное количество меток около 1,24 миллиона для эмбриональной фракции ƒ=0,1. Более общий график приведен на Фиг. 1.
РАЗРАБОТКА ПОЛИНОМИАЛЬНОЙ МОДЕЛИ
БИОЛОГИЧЕСКАЯ МОТИВАЦИЯ
Согласно наблюдениям относительное представление генома (FGR) конкретной хромосомы зависит (помимо прочего) от ее размера (Chiu et al., 2008; Ehrich et al., 2011). Из этого можно заключить, в математическом смысле, что в случае секвенирования образца плазмы материнской крови и последующего картирования и подсчета этих последовательностей для каждой хромосомы результатом этого (случайного) процесса является вектор из 24 чисел, по одному для каждой хромосомы, и мы можем рассматривать этот вектор как случайный вектор, который получен из полиномиального распределения с параметрами (p1, p2, …, p24; n), где pi, i∈{1, 2, …, 24}, является вероятностью того, что конкретная секвенированная ДНК-метка будет картирована на i-ой хромосоме, а n - число всех картированных меток. Следовательно, мы формулируем следующее наблюдение.
Наблюдение: Данные секвенирования (т.е. картированные вкДНК-метки) полиномиально распределены на хромосомах, при этом вероятностные параметры сильно коррелируют с длиной хромосом.
Переход от хромосом к 1 м.п.о. бинам является несложным, только в этом случае число параметров pi равно не 24, а числу бинов, которые покрывают человеческий геном hg19 (мы обозначаем это число как t).
МАТЕМАТИЧЕСКАЯ МОДЕЛЬ
Пусть X является случайным вектором из полиномиального распределения. Мы обозначаем новую случайную переменную Y как соотношение 2-х или более элементов полиномиального случайного вектора X. Например, для X=(х1, x2, x3, …, x20) Y может быть определен как Y=(x1+x2)/(x2+x3+x10). Чтобы смоделировать ситуацию, описанную в Биологической мотивации, мы предполагаем, что случайный вектор X=(x1, x2, …, xt) соответствует числу меток, картированных на 1 м.п.о. бинах, покрывающих человеческой геном hg19. В случае выявления трисомии числитель Y представляет собой число меток, картированных на хромосоме 21 (или 13 или 18), а знаменатель Y представляет собой сумму меток, картированных на бинах IRB. Однако это определение Y является без нужды обобщенным и сложным. Это связано с тем, что чтобы получить значение Y для конкретного образца, в действительности не важно, например, на какой конкретном бине IRB картирована какая-либо ДНК-метка; нам только нужно различать три класса меток: 1) картированные на бине, связанном с анеуплоидной хромосомой, 2) картированные на одном из бинов внутреннего стандарта и 3) все остальные. Следовательно, мы можем заменить полиномиальный случайный вектор X новым полиномиальным случайным вектором X'=(x'1, x'2, x'3), соответствующим трем указанным классам, со следующими новыми вероятностными параметрами, подаваемыми в виде



где PX и PIRB были определены выше в Расчете полиномиального среднего и стандартного отклонения. В этом смысле теперь мы можем упростить определение Y как Y=x'1/x'2. И наконец, чтобы избежать проблем с нулем в числителе Y, мы изменяем определение Y следующим образом

Так как значения, приписываемые случайному вектору X'=(x'1, x'2, x'3) имеют порядок 104 и более, +1 в знаменателе Y оказывает пренебрежимо малое влияние. В заявке данной модели нам необходимо знать значения E[Y] и Var[Y].
Теорема 1. Пусть X'=(x'1, x'2, x'3) является случайным вектором из полиномиального распределения с параметрами (p'1, p'2, p'3; n). Ожидаемое значение случайной переменной Y, задаваемой Ур-ем 21, составляет
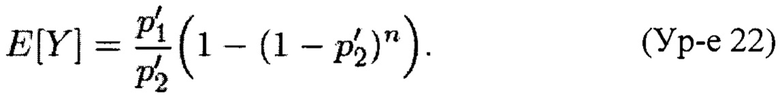
Теорема 2. Пусть X'=(x'1, x'2, x'3) является случайным вектором из полиномиального распределения с параметрами (p'1, p'2, p'3; n). Для n≥105 и 1>p'1, p'2≥0,01 вариация случайной переменной Y, задаваемой Ур-ем 21, может быть аппроксимирована как

где
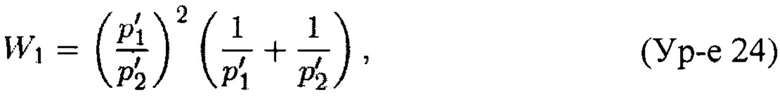
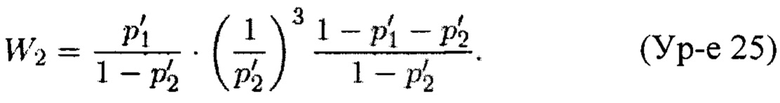
Погрешность этого приближения составляет менее 0,01% от истинной величины.
Заданный исследуемый образец, характеризуемый своим долевым значением бина FB, нормируют, используя значения E[Y] и (Var[Y])-1/2. В применяемых на данный момент способах определения z-показателя (указанных выше и описанных ниже), если нормированное значение для образца превышало 3, его считали трисомическим. Это пороговое значение получено из предположения, что нормированные значения распределены в соответствии со стандартным нормальным распределением. Также мы исследовали это предположение положительным образом, что означает, что случайная переменная z, задаваемая Ур-ем 9, подлежит стандартному нормальному распределению. По этой причине установление порогового значения 3, которое на данный момент является стандартным, подходит также и для нашего способа.
Краткое описание графических материалов
Фиг. 1: Оценка необходимой глубины секвенирования циркулирующей внеклеточной ДНК в плазме материнской крови для определения эмбриональной анеуплоидии 21-й аутосомы как функции доли эмбриональной ДНК. Теоретический минимум основан на Ур-ии 17 и значениях k=3 (минимальный z-показатель для того, чтобы назвать образец трисомическим), 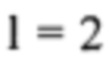 (задаваемый дисперсией эуплоидных z-показателей вокруг нуля), а р'1=0,0128, р'2=0,128 (задаваемые проверочными данными MiSeq). Для IonTorrent кривая теоретического минимума почти идентична и не отображена на графике. Нанесенные на график точки соответствуют наблюдаемым образцам Т21 на MiSeq (квадраты) и IonTorrent (круги).
(задаваемый дисперсией эуплоидных z-показателей вокруг нуля), а р'1=0,0128, р'2=0,128 (задаваемые проверочными данными MiSeq). Для IonTorrent кривая теоретического минимума почти идентична и не отображена на графике. Нанесенные на график точки соответствуют наблюдаемым образцам Т21 на MiSeq (квадраты) и IonTorrent (круги).
Фиг. 2: Схема основных этапов способа неинвазивного пренатального выявления эмбриональной хромосомной анеуплоидии по образцам материнской крови в соответствии с настоящим изобретением.
Примеры
Пример 1
Методики секвенирования, применяемые при реализации изобретения
Анализ последовательностей
Массовое параллельное секвенирование необходимо для применения способа в соответствии с изобретением. Способ был специально разработан и апробирован для небольших настольных систем секвенирования следующего поколения, чтобы сделать возможным уменьшение начальных затрат на лабораторную установку сервиса НИПТ. Способ был апробирован на системе MiSeq (Illumina, Inc., San Diego, CA, USA) и Ion Torrent PGM (Life Technologies Corp., San Francisco, CA, USA).
Коммерчески доступные устройства секвенирования вместе с соответствующими протоколами и реагентами, рекомендуемыми поставщиком, применяли в качестве иллюстративного примера, однако специалисту в данной области техники известно большое количество различных способов секвенирования и их вариаций, которые также можно применять для практической реализации настоящего изобретения.
Применяли набор для секвенирования Illumina, версия 3, и набор реагентов MiSeq v3 (MS-102-3003).
В наших экспериментах в среднем получали 5,829±1,026 миллионов и 3,098±0,951 миллионов исходных считываемых фрагментов после анализа секвенирования на IonTorrent PGM и MiSeq, соответственно. Так как применяли протоколы с 200 п.о. одиночными концами и 2×100 п.о. спаренными концами на Ion Torrent PGM и Miseq, мы смогли определить распределение размеров фрагментов нативной ДНК. Средние длины считываемых фрагментов в 179±7 п.о. и 172±7 п.о. были записаны на IonTorrent PGM и MiSeq, соответственно. Чтобы получить сопоставимые условия для сравнения двух исследуемых настольных платформ ССП на этапе апробации, полученные исходные количества считываемых фрагментов нормировали и случайным образом выбирали максимум 3 миллиона считываемых фрагментов для всех образцов для последующих этапов. После качественной фильтрации считываемых фрагментов, картирования считываемых фрагментов, отбора уникально картированных считываемых фрагментов, исключения меток, которые были картированы на области с покрытием, более чем в 10 раз большим, чем среднее покрытие, и GC-коррекции среднее количество так называемых конечных меток снижалось от 3 миллионов на образец до 2,160±0,065 миллионов и 2,099±0,221 миллионов меток для IonTorrent PGM и MiSeq, соответственно (подробную информацию см. в Minarik et al., 2015).
Пример 2
Процедура получения образцов для секвенатора MiSeq (Illumina Inc., San Diego, CA, USA))
Полный способ получения данных по последовательностям описан ниже в виде простого перечня основных процедур, так как применялись в основном стандартные рутинные лабораторные процедуры. Также протоколы для конкретных этапов хорошо известны специалистам в области молекулярной биологии, пренатальных исследований и биоинформатики, и такому специалисту известны возможные модификации процедур. Также многие специальные протоколы предоставляются производителем наборов или платформы для секвенирования. Более подробное описание лабораторных процедур можно найти в Minarik et al. 2015)
1. СБОР ОБРАЗЦОВ
Способ был апробирован на образцах, полученных на 12-й неделе беременности. 2×10 мл образцы крови собирали в K3-ЭДТУ пробирки и обрабатывали во время следующего этапа в течение 48 ч.
2. ПЛАЗМОСЕПАРАЦИЯ
Проводили двухэтапную плазмосепарацию, чтобы получить оптимальные результаты исследования. Дополнительные подробности процедуры можно найти в Minarik et al. 2015.
3. ВЫДЕЛЕНИЕ ДНК
Применяли набор Qiagen Blood Mini Kit. Образцы плазмы обрабатывали незамедлительно или хранили и обрабатывали в течение 1 месяца после процедуры плазмосепарации, что не оказывало видимого влияния на результаты исследований. Дополнительные подробности процедуры можно найти в Minarik et al. 2015.
4. ПОЛУЧЕНИЕ БИБЛИОТЕКИ
Процедура включает следующие этапы:
a. ИЗМЕРЕНИЕ КОНЦЕНТРАЦИИ ВЫДЕЛЕННОЙ ДНК
b. РЕПАРАЦИЯ КОНЦОВ
c. ОТБОР ПО РАЗМЕРУ
d. А-ПРИСОЕДИНЕНИЕ, ЛИГИРОВАНИЕ АДАПТОРА
e. ПЦР-АМПЛИФИКАЦИЯ
f. КОЛИЧЕСТВЕННАЯ ОЦЕНКА БИБЛИОТЕКИ
5. АНАЛИЗ СЕКВЕНИРОВАНИЯ
Способ был апробирован со следующими настройками анализа секвенирования:
Тип анализа: только FastQ
Тип библиотеки: Truseq LT
Секвенирование спаренных концов: Да
Длина секвенирования: 2×100
Приблизительное время секвенирования: 24h
Файлы FastQ загружены на веб-сервис через веб-браузер Firefox. Сгенерированные отчеты можно впоследствии загрузить, используя тот же самый веб-сервис.
Процедура с платформой Ion Torrent
При обработке образцов для платформы Ion Torrent (Life Technologies Corp., San Francisco, CA, USA) этапы 1 и 2 обязательно идентичны, остальные сходны или разные, что является отличительной чертой этой конкретной платформы секвенирования. Так как протоколы доступны от производителя или опубликованы в другом месте, специалисту известно, как получить данные по последовательностям, используя эту платформу.
Пример 3
Апробация нового способа с двумя платформами секвенирования и сравнение с другими способами определения z-показателя
На данный момент существует несколько способов выявления трисомии на основании определения z-показателя. Мы решили сравнить способ в соответствии с настоящим изобретением, обозначаемый MUL, с тремя наиболее общепринятыми уже применяемыми способами. Мы назвали эти способы по авторам статей, в которых они описаны:
1. CHIU (способ описан в Chiu et al., 200%),
2. LAU (способ описан в Lau et al., 2011), и
3. SEH (способ описан в Sehnert et al., 2011)
Способ CHIU, кратко резюмируя, основан на относительном представлении генома (FGR) хромосомы 21, которое представляет собой соотношение r21 числа меток, картированных (после GC-коррекции) на хромосоме 21, и общего числа картированных меток (после GC-коррекции и с исключением меток, картированных на половых хромосомах). Использовали среднее и стандартное отклонение r21 контрольных образцов, чтобы нормировать значение r21 образца с неизвестным кариотипом
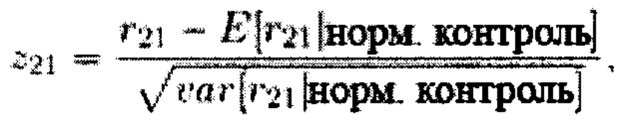
и если нормированное значение z21 превышает 3, образец классифицируют как трисомический (это связано с тем, что Chiu et al., 2008 предположили, что нормированные значения подлежат стандартному нормальному распределению).
Sehnert et al., 2011 и Lau et al., 2012 предложили другой алгоритм, в котором рассчитывают соотношение qk числа меток, картированных на хромосоме 21, и числа меток, картированных на какой-либо другой хромосоме (так называемой хромосоме внутреннего стандарта). В то время как Sehnert et al., 2011 выбрали хромосому 9 в качестве оптимального внутреннего стандарта для хромосомы 21, Lau et al., 2012 выбрали хромосому 14. Как и в случае со способом CHIU, использовали среднее и стандартное отклонение q21 контрольных образцов, чтобы нормировать значение q21 образца с неизвестным кариотипом
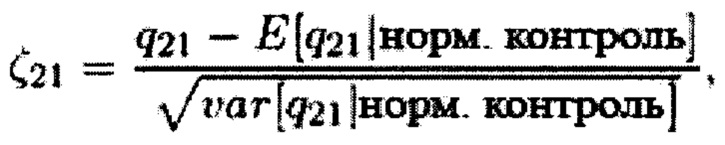
и если нормированное значение ζ21 превышает 3, образец классифицируют как трисомический (это снова было предположено Sehnert et al., 2011 и Lau et al., 2012, что нормированные значения ζ подлежат стандартному нормальному распределению).
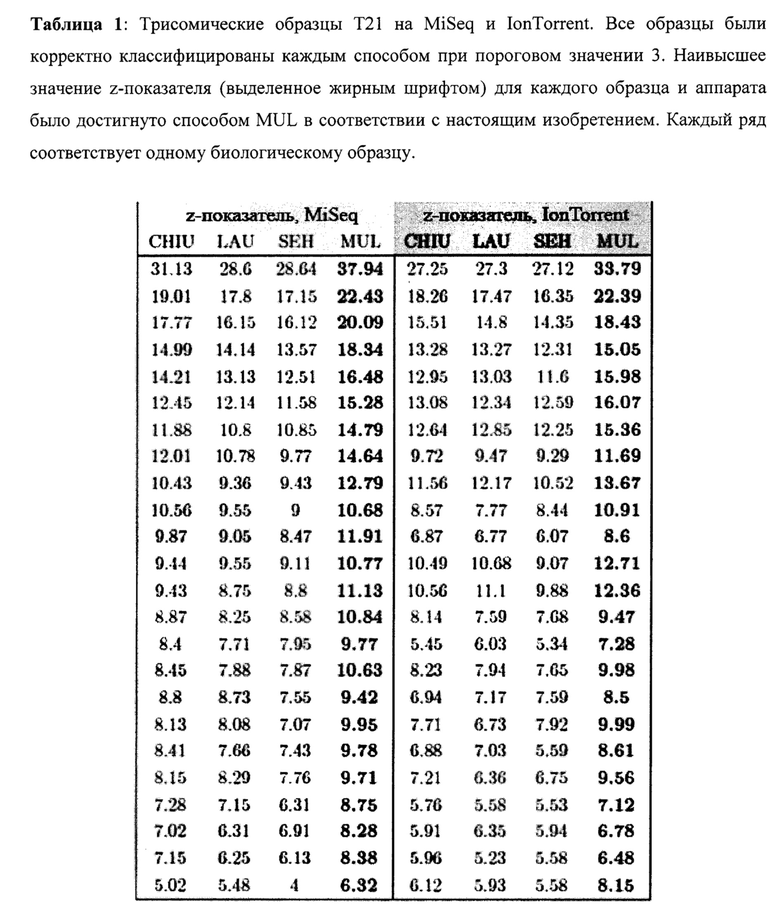
Для всех способов определения z-показателя, включая способ MUL в соответствии с настоящим изобретением (схематически изображенный на Фиг. 2), входные данные по количеству меток были одинаковыми (см. выше СПОСОБ, ПРОВЕРОЧНЫЕ ОБРАЗЦЫ, без необязательного этапа отбора по размеру). Проверочная группа состояла из 60 эуплоидных образцов, одинаковых для всех способов. Из оставшегося 71 исследуемого образца было 24 случая Т21 и 47 эуплоидных случаев. Кроме того, бины внутреннего контроля IRB для нашего способа MUL были найдены, как описано выше (см. СПОСОБ, ПРОВЕРОЧНЫЕ ОБРАЗЦЫ). В Таблице 1 отображены z-показатели образцов Т21 из нашей исследуемой группы Т21 (наивысшие значения выделены жирным). В любом из обсуждаемых способов не наблюдалось ложно-отрицательных результатов с пороговым значением 3.
Наблюдаемая разница в z-показателях между MiSeq и IonTorrent может быть связана с разницей между обработкой биологических образцов, погрешностями приборов или другими неизвестными факторами.
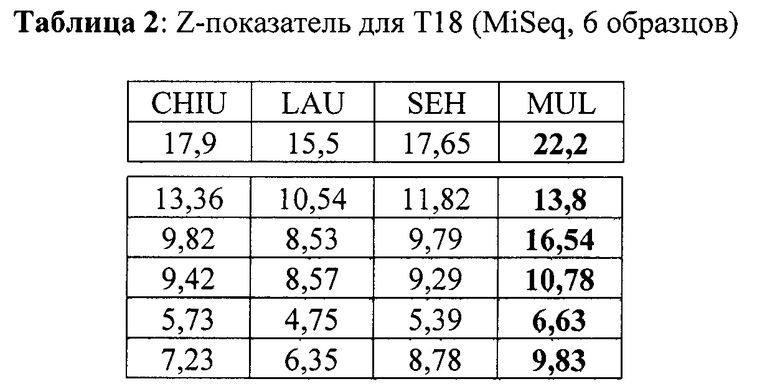
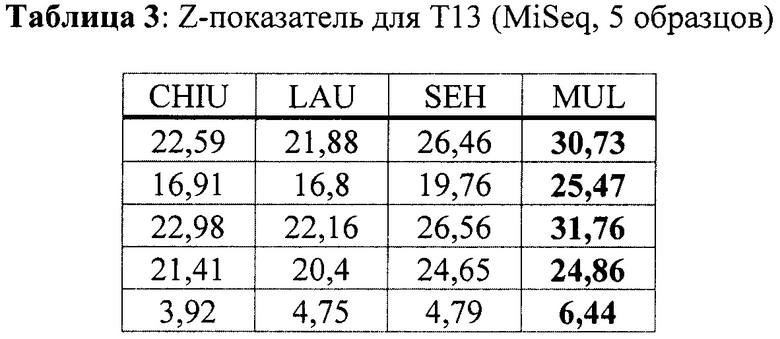
Кроме того, в Таблице 2 продемонстрированы результаты для 6 образцов с трисомией хромосомы 18, а в Таблице 3 продемонстрированы результаты для 5 образцов с трисомией хромосомы 13. Данные по последовательностям для образцов Т18 и Т13 были получены с помощью платформы MiSeq.
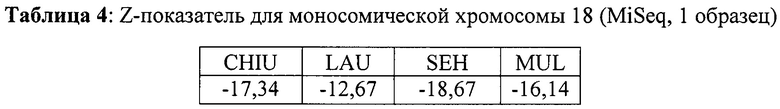
Кроме того, анализировали один образец с моносомией хромосомы 18. Результаты продемонстрированы в Таблице 4. Значения z-показателей отрицательны в случае моносомии, так как в этом случае недостает одной копии хромосомы 18. В результате количество меток, картированных на хромосоме 18, меньше, чем в нормальных образцах с двумя копиями хромосомы 18, что и демонстрирует значение z-показателя.
Естественно, по одному эксперименту нельзя сделать далекоидущих выводов, однако, пример продемонстрировал, что нашим способом можно выявлять моносомию, по меньшей мере также хорошо, как способами известного уровня техники.
Как продемонстрировано в Таблицах 1-4, значения z-показателей трисомических образцов, определенные способом согласно настоящему изобретению, значительно выше, чем значения, определенные любым из способов известного уровня техники. Например, для трисомических образцов Т21 мы получили медианную разницу значений z-показателей - 1,895, -1,595, -2,485 при сравнении CHIU, SEH и LAU, соответственно, с нашим способом с дифференциальным р-значением <0,05 для всего известного уровня техники в соответствии с критерием множественного сравнения Данна), в то время как значения для нормальных (не трисомических) образцов являются в значительной степени идентичными (для хромосомы 21 мы получили медианную разницу с нашим способом -0,15, -0,05, -0,21 для CHIU, SEH и LAU, соответственно), что означает, что отличие z-показателей трисомических образцов от порогового значения увеличено в случае настоящего изобретения по сравнению со способами известного уровня техники, в то время как отличие эуплоидных образцов от этого порогового значения преимущественно сохраняется.
ССЫЛКИ
Непатентная литература
Bianchi, D.W., L.D. Platt, J.D. Goldberg, A.Z. Abuhamad, A.J. Sehnert, and R.P. Rava, Genome-wide fetal aneuploidy detection by maternal plasma DNA sequencing. Obstet Gynecol, 2012. 119(5): p. 890-901.
Chiu, R.W., K.A. Chan, Y. Gao, V.Y. Lau, W. Zheng, T.Y. Leung, С.H. Foo, B. Xie, N.B. Tsui, F.M. Lun, et al., Noninvasive prenatal diagnosis of fetal chromosomal aneuploidy by massively parallel genomic sequencing of dna in maternal plasma. Proceedings of the National Academy of Sciences, 105(51): 20458-20463, 2008.
Chiu, R.W., R. Akolekar, Y.W. Zheng, T.Y. Leung, H. Sun, K.C. Chan, F.M. Lun, A.T. Go, E.T. Lau, W.W. To, W.C. Leung, R.Y. Tang, S.K. Au-Yeung, H. Lam, Y.Y. Kung, X. Zhang, J.M. van Vugt, R. Minekawa, M.H. Tang, J. Wang, C.B. Oudejans, Т.K. Lau, K.H. Nicolaides, and Y.M. Lo, Non-invasive prenatal assessment of trisomy 21 by multiplexed maternal plasma DNA sequencing: large scale validity study. BMJ, 2011. 342: p. c7401.
Ehrich, M., C. Deciu, T. Zwiefelhofer, J.A. Tynan, L. Cagasan, R. Tim, V. Lu, R. McCullough, E. McCarthy, A.O. Nygren, J. Dean, L. Tang, D. Hutchison, T. Lu, H. Wang, V. Angkachatchai, P. Oeth, C.R. Cantor, A. Bombard, and D. van den Boom, Noninvasive detection of fetal trisomy 21 by sequencing of DNA in maternal blood: a study in a clinical setting. Am J Obstet Gynecol, 2011. 204(3): p. 205 e1-11.
Fan, H.C., Y.J. Blumenfeld, U. Chitkara, L. Hudgins, and S.R. Quake, Noninvasive diagnosis of fetal aneuploidy by shotgun sequencing DNA from maternal blood. Proc Natl Acad Sci USA, 2008. 105(42): p. 16266-71.
Fortin, F.-A., et al. "DEAP: Evolutionary algorithms made easy." The Journal of Machine Learning Research 13.1 (2012): 2171-2175.
Hudecova, I., D. Sahota, M. Heung, Y. Jin, W.S. Lee, T.Y. Leung, Y.M.D. Lo, and R.W. Chiu., Maternal plasma fetal dna fractions in pregnancies with low and high risks for fetal chromosomal aneuploidies. PloS one, 9(2): e88484, 2014.
Langmead, В., Salzberg, S., Fast gapped-read alignment with Bowtie 2. Nature Methods. 2012, 9: 357-359
Lau, Т.K., F. Chen, X. Pan, R.K. Pooh, F. Jiang, Y. Li, H. Jiang, X. Li, S. Chen, and X. Zhang. Noninvasive prenatal diagnosis of common fetal chromosomal aneuploidies by maternal plasma DNA sequencing. Journal of Maternal-Fetal and Neonatal Medicine, 25(8): 1370 {1374, 2012.
Liao, С, A.H. Yin, C.F. Peng, F. Fu, J.X. Yang, R. Li, Y.Y. Chen, D.H. Luo, Y.L. Zhang, Y.M. Ou, J. Li, J. Wu, M.Q. Mai, R. Hou, F. Wu, H. Luo, D.Z. Li, H.L. Liu, X.Z. Zhang, and K. Zhang, Noninvasive prenatal diagnosis of common aneuploidies by semiconductor sequencing. Proc Natl Acad Sci USA, 2014. 111(20): p. 7415-20.
Lo, Y.M., N. Corbetta, P.F. Chamberlain, V. Rai, I.L. Sargent, C.W. Redman, and J.S. Wainscoat, Presence of fetal DNA in maternal plasma and serum. Lancet, 1997. 350(9076): p. 485-7.
Rava, R.P., A. Srinivasan, A.J. Sehnert, and D.W. Bianchi. Circulating fetal cell-free DNA fractions differ in autosomal aneuploidies and monosomy X. Clinical chemistry, 60(1): 243-250, 2014.
Minarik, G., Repiska, G., Hyblova, M., Nagyova, E., Soltys, K., Budis, J. et al. Utilization of Benchtop Next Generation Sequencing Platforms Ion Torrent PGM and MiSeq in Noninvasive Prenatal Testing for Chromosome 21 Trisomy and Testing of Impact of In Silico and Physical Size Selection on Its Analytical Performance. PLoS One. 2015; 10(12): e0144811. doi: 10.1371/journal.pone.0144811.
Sehnert, A.J., B. Rhees, D. Comstock, E. de Feo, G. Heilek, J. Burke, and R.P. Rava, Optimal detection of fetal chromosomal abnormalities by massively parallel DNA sequencing of cell-free fetal DNA from maternal blood. Clin Chem, 2011. 57(7): p. 1042-9.
Yu, S.C. et al., Size-based molecular diagnostics using plasma DNA for noninvasive prenatal testing. Proceedings of the National Academy of Sciences 111.23 (2014): 8583-8588.
Патентные документы
EP 2183693
EP 2366031
US 8296076
WO 2011/051283
| название | год | авторы | номер документа |
|---|---|---|---|
| НЕИНВАЗИВНОЕ ОБНАРУЖЕНИЕ ГЕНЕТИЧЕСКОЙ АНОМАЛИИ ПЛОДА | 2011 |
|
RU2589681C2 |
| НЕИНВАЗИВНЫЙ ДИАГНОСТИЧЕСКИЙ ТЕСТ ДНК ДЛЯ ОБНАРУЖЕНИЯ АНЕУПЛОИДИИ | 2012 |
|
RU2638456C2 |
| Способ неинвазивного пренатального скрининга анеуплоидий плода | 2019 |
|
RU2712175C1 |
| СПОСОБ ОПРЕДЕЛЕНИЯ ИСТОЧНИКА АНЕУПЛОИДНЫХ КЛЕТОК ПО КРОВИ БЕРЕМЕННОЙ ЖЕНЩИНЫ | 2016 |
|
RU2674700C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ВЫСОКОМУЛЬТИПЛЕКСНОЙ ПЦР | 2012 |
|
RU2650790C2 |
| СПОСОБ НЕИНВАЗИВНОЙ ПРЕНАТАЛЬНОЙ ДИАГНОСТИКИ АНЕУПЛОИДИЙ ПЛОДА | 2015 |
|
RU2627673C2 |
| СПОСОБ НЕИНВАЗИВНОЙ ПРЕНАТАЛЬНОЙ ДИАГНОСТИКИ АНЕУПЛОИДИЙ ПЛОДА | 2014 |
|
RU2583830C2 |
| СПОСОБ ОПРЕДЕЛЕНИЯ АНЕУПЛОИДИИ ПЛОДА В ОБРАЗЦЕ КРОВИ БЕРЕМЕННОЙ ЖЕНЩИНЫ | 2021 |
|
RU2777072C1 |
| СПОСОБ НЕИНВАЗИВНОЙ ДИАГНОСТИКИ АНЕУПЛОИДИЙ ПЛОДА МЕТОДОМ СЕКВЕНИРОВАНИЯ | 2014 |
|
RU2543155C1 |
| СПОСОБ И УСТРОЙСТВО ДЛЯ ОПРЕДЕЛЕНИЯ ФРАКЦИИ ВНЕКЛЕТОЧНЫХ НУКЛЕИНОВЫХ КИСЛОТ В БИОЛОГИЧЕСКОМ ОБРАЗЦЕ И ИХ ПРИМЕНЕНИЕ | 2015 |
|
RU2699728C2 |
Изобретение относится к биотехнологии. Предложен способ скрининга анеуплоидий. Способ включает четыре основных этапа: 1) получение и обработку образцов материнской крови, 2) получение образца ДНК и библиотеки ДНК, 3) секвенирование и 4) обработку данных по последовательностям для получения величины, которая имеет прогнозную значимость. Существенной частью способа является получение и обработка проверочных данных, т.е. обработка группы эуплоидных образцов, которые обрабатывают тем же способом, что и исследуемые образцы. Результат, полученный при обработке проверочных образцов, служит в качестве контрольных данных на этапе 4). В настоящем изобретении предложен альтернативный и надежный способ, который применим по меньшей мере к практической реализации неинвазивного пренатального скрининга анеуплоидий, таких как трисомии или моносомии, предпочтительно трисомии хромосомы 13, 18 или 21. Благодаря новому и изобретательскому подходу этот способ обеспечивает альтернативный метод определения z-показателя, который обеспечивает лучшее разграничение между эуплоидными и анеуплоидными образцами, т.е. улучшает надежность исследования. Кроме того, данный способ требует относительно небольшое количество данных секвенирования и, следовательно, такой способ является относительно недорогим и его могут позволить даже небольшие учреждения здравоохранения. 9 з.п. ф-лы, 4 табл., 3 пр., 2 ил.
1. Способ определения эмбриональной анеуплоидии представляющей интерес хромосомы по образцу материнской крови, содержащему смесь молекул внеклеточной ДНК эмбрионального и материнского происхождения, включающий этапы, в которых:
a) выделяют внеклеточную ДНК из материнской крови;
b) проводят случайное секвенирование по меньшей мере части множества молекул внеклеточной ДНК, содержащихся в крови, при этом указанная часть содержит ДНК, соответствующую по меньшей мере одной представляющей интерес хромосоме и множеству предположительно эуплоидных хромосом, с получением, таким образом, множества считываемых последовательностей;
c) картируют считываемые последовательности на стандартный человеческий геном с получением, таким образом, множества картированных меток;
d) разбивают указанный стандартный человеческий геном на 1 м.п.о. последовательные и неперекрывающиеся бины;
e) определяют распределение меток на указанных бинах и целых хромосомах в соответствии с их расположением на карте;
f) определяют долевое значение бина FB в виде соотношения между числом меток тестируемого образца, картированных на бинах представляющей интерес хромосомы в образце, и числом меток, картированных на бинах внутреннего стандарта, IRB;
g) определяют полиномиальное среднее µ(n) и полиномиальное стандартное отклонение σ(n) образца, причем μ(n) и σ(n) вычисляются для рандомной величины, определенной как отношение между количеством меток, картированных на представляющую интерес хромосому в образце, и суммой меток, картированных на бинах внутреннего стандарта IRB, при этом распределение вероятностей картирования меток на представляющую интерес хромосому и на бины внутреннего стандарта IRB является полиномиальным, и параметры указанного полиномиального распределения определяются из обучающего набора образцов эуплоидной крови;
h) определяют значение z-показателя в виде разницы между долевым значением бина FB и полиномиальным средним μ(n), деленной на полиномиальное стандартное отклонение σ(n); и
i) сравнивают значение z-показателя по меньшей мере с одним пороговым значением для того, чтобы определить наличие анеуплоидии для представляющей интерес хромосомы;
при этом в качестве предварительного условия для проведения этапов f)-i) проверочную группу эуплоидных образцов крови подвергают в значительной степени сходным процедурам, состоящим из этапов a)-e) выше, а затем проводят следующие этапы:
f') выбирают аутосомы внутреннего стандарта IRA из кандидатных аутосом внутреннего стандарта на основании их коэффициента вариации CV, при этом исключены потенциально анеуплоидные хромосомы;
g') выбирают с помощью генетического алгоритма бины внутреннего стандарта IRB из двух кандидатных групп, причем первая кандидатная группа представляет собой группу из случайно выбранных бинов, при этом исключены бины, соответствующие потенциально анеуплоидным хромосомам, а вторая кандидатная группа представляет собой группу бинов, соответствующих лучшим аутосомам внутреннего стандарта IRA, определенным на предыдущем этапе f'.
2. Способ по п. 1, отличающийся тем, что представляющая интерес хромосома является хромосомой 8, 9, 13, 16, 18, 21, 22 и X.
3. Способ по п. 1 или 2, отличающийся тем, что анеуплоидия является трисомией хромосомы 13, хромосомы 18 или хромосомы 21.
4. Способ по любому из пп. 1-3, отличающийся тем, что проверочная группа эуплоидных образцов крови содержит эуплоидные мужские и эуплоидные женские образцы.
5. Способ по любому из пп. 1-4, дополнительно включающий этап фильтрации, в котором ограничивают размер считываемых последовательностей, приемлемый для дальнейшей обработки.
6. Способ по любому из пп. 1-5, отличающийся тем, что при выборе аутосом внутреннего стандарта IRA для каждой потенциальной комбинации внутреннего стандарта рассчитывают коэффициент вариации CV, кандидатные комбинации внутреннего стандарта аутосом располагают в соответствии с их значением CV в порядке возрастания, а затем те комбинации внутреннего стандарта, значение CV которых меньше или равно кратному 1,1 значению наименьшей величины CV, выбирают в качестве IRA, при этом для дальнейшей обработки в качестве IRA выбирают предпочтительно первую сотню этих комбинаций.
7. Способ по любому из пп. 1-6, отличающийся тем, что при выборе бинов внутреннего стандарта IRB с помощью генетического алгоритма первую кандидатную группу выбирают из кандидатной группы на основании их значений FB, используя коэффициент размаха Стьюдента, и в то же время на основании их значений CV.
8. Способ по любому из пп. 1-7, дополнительно включающий этап, в котором определяют эмбриональную фракцию.
9. Способ по п. 8, дополнительно включающий этап, в котором определяют погрешность оценки эмбриональной фракции.
10. Способ по любому из пп. 1-9, дополнительно включающий этап, в котором определяют минимальное количество меток для надежного прогнозирования анеуплоидии.
| CHIU ROSSA W | |||
| Noninvasive prenatal diagnosis of fetal chromosomal aneuploidy by massively parallel genomic sequencing of DNA in maternal plasma | |||
| Прибор для равномерного смешения зерна и одновременного отбирания нескольких одинаковых по объему проб | 1921 |
|
SU23A1 |
| LAU TZE KIN | |||
| Noninvasive prenatal diagnosis of common fetal chromosomal aneuploidies by maternal plasma DNA sequencing | |||
| The Journal of Maternal-Fetal & | |||

