ОБЛАСТЬ ТЕХНИКИ
Настоящее изобретение относится к толерогенным иммунотерапевтическим ДНК-вакцинам для уменьшения антигенспецифичной реактивности Т-клеток.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
Согласно традиционным подходам вакцинации очищенный белок/антиген инъецируется человеку/пациенту/животному для того, чтобы специфично стимулировать иммунные ответы на данный белок/антиген. Данный подход вакцинации имеет тенденцию к влиянию, главным образом, на продукцию антител, тогда как Т-клетки имеют тенденцию не подвергаться значимому влиянию, за исключением генерации памяти Т-клеток на антиген. Традиционные подходы вакцинации, таким образом, не считаются подходящими в связи с лечением и/или предупреждением заболеваний, возбуждаемых Т-клетками, таких как, например, диабет 1 типа (T1D), поскольку активация Т-клеток, особенно Т-клеток CD8+, считается этиологическим фактором данного заболевания. Экспериментальные подходы с толерогенными вакцинами на основе белков имели в качестве мишени, главным образом, В-клетки, продуцирующие антитела, а не релевантные для заболевания Т-клетки.
Вакцины на основе ДНК, в отличие от вакцин на основе белка, обычно представляют собой плазмиды, кодирующие конкретные антигены - данные плазмиды поглощаются клетками в организме хозяина («трансфицированными»). Данные трансфицированные клетки хозяина затем продуцируют антиген и перерабатывают антиген на маленькие фрагменты (эпитопы Т-клеток) для презентации иммунной системе, в частности, циркулирующим Т-клеткам. Поскольку Т-клетки могут выявлять только данные маленькие фрагменты антигена, а не целые белки, данный подход предпочтительно приводит к модификации Т-клеточных ответов, особенно в отношении Т-клеток CD8+ (или цитотоксических Т-клеток) - ключевых драйверов, например, T1D патологии. Таким образом, ДНК-вакцины, а не белковые вакцины, подходят для индуцирования Т-клеточных ответов. Тогда как в настоящее время не доступны ДНК-вакцины для человеческого применения, имеются три стимулирующие плазмидные ДНК-вакцины, лицензированные для ветеринарного применения, индуцирующие иммунитет на вирус инфекционной анемии у лошадей, вирус Западного Нила и некоторые раковые заболевания собак.
В отличие от стимулирующих ДНК-вакцин, толерогенные иммунотерапевтические ДНК-вакцины предназначены для подавления иммунореактивности в отношении антигена, а не для активации иммунных ответов против него. Данные вакцины не стимулируют иммунитет против закодированного антигена или не изменяют тип стимуляции (например, как делают антигенные десенсибилизирующие подходы вакцинации против аллергий), но вместо этого вызывают истощение и/или недостаток функции, и/или гибель Т-клеток, реагирующих на собственный антиген. Для того чтобы делать так, антиген должен презентироваться иммунной системе без костимулирующих или воспалительных эффектов, которые, в противном случае, примировали бы стимулирующие иммунные ответы. Данный подход презентирования антигена, подлежащего игнорированию иммунной системой или лишению иммуногенности, мог бы иметь ценность в лечении аутоиммунных заболеваний, так как таким образом осуществлялось бы нацеливание на специфический механизм заболевания, а не на системное подавление всего иммунного ответа. Толерогенная иммунотерапевтическая ДНК-вакцина, таким образом, представляет собой мягкий способ модулирования нежелательных иммунных ответов.
Конечной целью T1D-специфичной толерогенной иммунотерапевтической ДНК-вакцины является сохранение функции бета-клеток и эндгенной продукции инсулина. Это может происходить через предупреждение или задержку заболевания (что особенно ценно в когортах детей и молодых взрослых, где мониторинг затруднен, и «нормальность» жизни является главным стимулом для пациента) или продление «фазы медового месяца» минимального мониторинга и применения инсулина, что часто происходит в течение первых шести месяцев после постановки диагноза T1D.
В то время как известно то, что вакцины на основе ДНК являются безопасными, ни одна из (стимулирующих или толерогенных) ДНК-вакцин, которые были протестированы в клинических исследованиях, не имеет достаточной эффективности в качестве отдельного подхода для лечения, например, T1D. Толерогенные ДНК-вакцины, известные в данной области, показали малую эффективность и типично требуют высокоискусственных систем для индукции желательных эффектов. Таким образом, в данной области имеется потребность в толерогенных иммунотерапевтических ДНК-вакцинах со значительно увеличенной эффективностью, без нарушения профиля безопасности и предпочтительно также без потребности в неудобной схеме введения.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к мультицистронному вектору/плазмиде, который соэкспрессирует/кодирует антиген, удерживаемый в клетке, такой как инсулин, а также секретируемые иммуномодуляторы, такие как TGF-β (трансформирующий фактор роста-β), IL-10 (интерлейкин-10) и возможно IL-2. Настоящее изобретение, кроме того, относится к иммунотерапевтическим ДНК-вакцинам, содержащим такие плазмиды, а также к таким фармацевтическим препаратам и их наборам. Настоящее изобретение, наконец, относится к медицинскому применению таких продуктов, а также к способам получения таких плазмид.
Иммунотерапевтические вакцины на основе плазмид/ДНК, описанные в данном документе, имеют терапевтический потенциал в лечении аутоиммунных заболеваний, которые, главным образом, возбуждаются Т-клетками, таких как, например, диабет типа 1 (T1D).
В одном аспекте согласно настоящему изобретению предложена плазмида, которая кодирует:
i. инсулиновый антиген;
ii. TGF-β; и
iii. IL-10.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
Фиг. 1. Карта кольцевой плазмиды.
Фиг. 2. Карта мРНК и транслированного белка для продуктов вектора плазмиды из Фиг. 1.
Фиг. 3. Стабильность плазмиды к деформации сдвига на трех инъекционных пропусканиях через иглу G30 (30-го калибра).
Фиг. 4. Подтверждение фенотипа сохранения плазмиды посредством роста при 30°C (пассажи 1-50 с использованием 17-часовой инкубации и пассажи 51-100 с использованием 22-часовой инкубации).
ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Автор настоящего изобретения предложил в данном документе одиночный вектор, который управляет экспрессией многих секретируемых цитокинов, а также антигена, сохраняющегося в клетке, от одного промотора/мультицистронной мРНК.
Иммунотерапевтическая ДНК-вакцинация одиночным вектором, кодирующим все компоненты данной терапии в одной клетке, является весьма предпочтительной по сравнению с иммунотерапевтической вакцинацией смесью отдельных векторов/плазмид, каждый из которых управляет экспрессией одного компонента, так как случайная трансфекция клеток разными векторами не гарантирует экспрессии всех компонентов или даже любого конкретного соотношения компонентов из данной конкретной трансфицированной клетки.
Трансфекция одной мультицистронной плазмидой/вектором приводит к специфично сконструированному местному окружению/микроокружению вокруг трансфицированной клетки. Данным способом к антигенам могут быть добавлены комбинации иммуномодуляторов таким образом, что они потенцируют желательный иммунологический эффект одиночных Т-клеток без потребности в высоких системных дозах иммуномодулятора, которые в противном случае могли бы вызывать нежелательные явления и широкую иммунодепрессию.
Это местное ограничение продукции иммуномодулятора клетками-хозяевами, трансфицированными иммунотерапевтическими ДНК-вакцинами, обеспечивает безопасное применение высокоэффективных цитокиновых гормонов, которые являются синергичными в отношении модификации Т-клеточных ответов, но не могут дозироваться или достаточно часто для эффекта и/или титроваться с получением желательного ответа без неприемлемых нежелательных явлений.
Например, известно то, что и интерлейкин-10 (IL-10), и трансформирующий фактор роста-бета 1 (TGF-β1) способны индуцировать регуляторные Т-клетки (Treg) из наивных Т-клеток CD4+. Однако комбинация IL-10/TGF-β1 обеспечивает синергический эффект (в 15-20 раз более эффективный) в индуцировании Treg, чем один из двух цитокинов один (US 6083919 A), и данная комбинация, кроме того, приводит к иммунотолерантности в более широкой популяции клеток-мишеней, чем один из двух цитокинов один (Zeller JC, Panoskaltsis-Mortari A, Murphy WJ, et. al. 1999 J Immunol. 163(7):3684-91).
Кроме того, известно то, что интерлейкин-2 (IL-2) как размножает, так и стабилизирует Treg, но, с другой стороны, также может способствовать воспалительным ответам. Комбинация IL-2 и IL-10, однако, приводит к подавляющим Treg, а не к воспалительной стимуляции. Поскольку циркулирующие Т-клетки сталкиваются с клетками, которые трансфицированы описанной в данном документе иммунотерапевтической ДНК-вакциной, они временно подвергаются воздействию субоптимальных концентраций IL-10 и IL-2. Данные циркулирующие Т-клетки слегка отклоняются в направлении толерантности, и если они также являются реактивными в отношении соэкспрессируемого антигена (например, инсулина), они будут связываться с трансфицированной клеткой и, таким образом, получать более продолжительное воздействие иммуномодулятора, и, кроме того, они также будут получать другой сигнал, который программирует/переобучает их в отношении подавляющих эффектов. Данным способом те Т-клетки, которые отвечают на кодируемый антиген, селективно переобучаются до подавляющего фенотипа при их столкновении с трансфицированной клеткой.
Описанные в данном документе иммунотерапевтические вакцины на основе плазмид/векторов/ДНК, таким образом, сконструированы для индукции антигенспецифичных Treg, накапливающихся в сайтах аутоиммунитета, для ослабления заболевания (например, поджелудочной железы при T1D), а не для прямого влияния на заболевание через экспрессируемые цитокиновые гормоны.
Помимо антигена (инсулина в примере T1D), вектор/оперон/плазмида, описанные в данном документе, кодируют по меньшей мере два цитокина (например, TGF-β1 и IL-10), которые совместно синергетически подавляют антигенпрезентирующие клетки, а также функцию Т-клеток и управляют индукцией Treg. Данный эффект усиливается, если он также происходит в комбинации с эффективным воздействием антигена.
В одном воплощении TGF-β1 представляет собой конститутивно активную форму, которая не требует процессинга или воспалительного окружения для функции. В то время как Treg могут продуцироваться из наивных Т-клеток посредством воздействия антигена и TGF-β1, Treg, однако, являются «пластичными», означая то, что они могут дедифференцироваться и превращаться в эффекторные клетки Th17, и затем вызывать большую или меньшую аутоиммунную деструкцию. Комбинация IL-10 с TGF-β1, помимо того, что она является более мощным иммуномодулятором, подавляет среду, которая продуцировала бы патогенные клетки Th17, а не Treg.
В одном воплощении мультицистронный вектор, описанный в данном документе, также кодирует, помимо антигена, IL-2, TGF-β1 и IL-10. IL-2 увеличивает число Treg и стабилизирует их фенотип (предотвращает дедифференциацию Treg в эффекторные Т-клетки), и, таким образом, увеличивает их функциональную продолжительность жизни в воспаленных тканях-мишенях.
Три данных цитокина (TGF-β1, IL-10 и IL-2) в комбинации с антигеном, таким образом, имеют хорошо известные синергетические эффекты в отношении индуцирования толерантности посредством следующих механизмов: (i) значительно усиленная генерация антигенспецифичных подавляющих Treg, (ii) большая продолжительность жизни Treg и (iii) большая эффективность на индивидуальную клетку Treg в подавлении воспаления/аутореактивности. Однако требующиеся концентрации системно инфундируемого очищенного цитокина имели бы целый ряд серьезных или может быть даже летальных побочных эффектов, таких как: (i) смертельный фиброз из-за избытка TGF-β1, (ii) гриппоподобные симптомы, (iii) синдром утечки из капилляров из-за избытка IL-2, (iv) широкая иммунодепрессия, приводящая к хроническим инфекциям, (v) усиленное развитие опухоли, а также (vi) анемия из-за избытка IL-10.
Посредством соэкспрессии данных цитокинов от того же самого вектора/плазмиды и, следовательно, той же самой клеткой, презентирующей антиген иммунной системе, данный вектор достигает желательной местной среды для индукции толерантности без системного действия и соответствующих побочных эффектов, которые в противном случае возникали бы из-за введения очищенного цитокина в высокой дозе.
Инъекция «голой»/«обнаженной» плазмидной/векторной ДНК (вектора и одного буфера) приводит к очень низкому поглощению и показателю трансфекции - меньше, чем одна из примерно 100000 плазмидных молекул трансфицирует клетку - тогда как остальные деградируют и, таким образом, остаются без какого-либо биологического эффекта. Эта крайне низкая эффективность трансфекции обеспечивает механизм безопасности для распространения и ограничения трансфицированных клеток.
Введение системно активных количеств любого из данных цитокинов, либо посредством введения зрелых белков, либо посредством высокоэффективной трансдукции вирусного вектора, сделало бы затруднительным или даже невозможным титрование для получения безопасной и эффективной дозы. Ограничение общего воздействия до очень маленькой системной дозы, распределенной в некоторых из высокоэкспрессирующих микросред, приводит к имеющему значительные преимущества профилю безопасности и эффективности.
Комбинация антигена и данных трех цитокинов, описанных в данном документе, дает эффективную защиту от развития T1D и даже, по-видимому, способна стабильно обращать развитие заболевания. Из-за низкой эффективности трансфекции инъекции голой ДНК-плазмиды/вектора, очень мало клеток продуцируют данные рекомбинантные белки, и, таким образом, отсутствует выявляемое изменение уровней сывороточных цитокинов из цитокинов, кодируемых плазмидой/вектором - и, следовательно, отсутствует выявляемая иммуностимуляция или иммунодепрессия в отношении любых других антигенов, чем антиген, кодируемый плазмидой/вектором (препроинсулин). Это приводит к желательному профилю безопасности.
Обычно ДНК-вакцины плохо работают в связи с подкожной (п.к.) инъекцией, и, следовательно, типично вводятся с использованием внутримышечной инъекции (часто с электропорацией) или, в качестве альтернативы, с использованием внутрикожного безыгольного впрыскивания, требующего сложного прибора, а также существенного обслуживания и калибровки. Поскольку большинство проблем с побочными эффектами при использовании внутримышечной инъекции связаны с адъювантом (раздражение в месте инъекции), они, следовательно, не являются предметом беспокойства для описанного в данном документе формата иммунотерапевтической вакцины на основе голой ДНК. Кроме того, инъецируемые объемы обычно являются относительно малыми и, следовательно, не вызывают значительных мышечных растяжений и боли. В одном воплощении инъецируемые объемы составляют 1 мл или менее. В другом воплощении инъецируемые объемы составляют приблизительно 0,6 или 0,5 мл. Независимо от этого, предложенная в данном документе мультицитокиновая плазмида/вектор неожиданно, по-видимому, дает защиту от T1D даже при введении посредством п.к. пути, обеспечивая, посредством этого, многие потенциальные форматы дозировки для пациентов.
Помимо обеспечения местной синергии посредством кодирования всех трех или четырех транслированных продуктов одной плазмидой/вектором и от одного промотора, с предложением описанной в данном документе мультицистронной плазмиды дополнительно упрощаются регуляторная нагрузка и критерии выпуска лекарственного средства.
В отличие от этого, если каждый из белковых продуктов продуцируется от отдельной плазмиды, тогда синергетическая ценность соэкспрессии от той же самой трансфицированной клетки потенциально терялась или уменьшалась бы, так как трансфекция каждой плазмидой/вектором была бы независимым событием, вероятно нацеленным на разные клетки. При продукции трех-четырех рекомбинантных белков от двух, трех или четырех индивидуальных плазмид/векторов, любые синергетические эффекты в местном окружении трансфицированной клетки потенциально теряются; кроме того, таким образом, были бы необходимыми несколько индивидуальных клинических испытаний (одно для каждой плазмиды и каждой комбинации). Продуцирование всех белков от одной плазмиды/вектора и одной мРНК облегчает требования для анализа многих индивидуальных молекул и определения идеальных соотношений совместной упаковки, присущих формату множественной плазмиды/вектора.
В данном документе можно использовать любой формат вектора, подходящий для настоящего изобретения, такой как плазмиды (реплицирующиеся или пассивные), миникольца, линейные векторы (MiLV), вирусные векторы (как интегрирующие [например, лентивирусные], так и неинтегрирующие [например, аденовирусные], космиды, бактериальные искусственные хромосомы (ВАС), человеческие искусственные хромосомы (НАС) и т.д.
Кроме того, в данном документе можно использовать любой пермиссивный способ усиления трансфекции, например, электропорацию, сонопорацию (ультразвуковое усиление с усилением контраста микропузырьками или без него), агрегаты липид/полимер, гидродинамику (давление через большой инъекционный объем), биобаллистику / генную пушку (депонирование через кожу посредством сжатого газа) и т.д.
В одном воплощении в данном документе используется нереплицирующаяся эписомная плазмидная ДНК из-за: i) множества копий мРНК, образующихся в результате трансфекции одной плазмиды, и ii) увеличенных стабильности и функции плазмидных нуклеиновых кислот по сравнению с мРНК и другими форматами ДНК-векторов. Таким образом, в то время как экспрессионные системы и на основе мРНК, и ДНК могут обеспечивать внутриклеточную доставку и колокализацию, системы на основе плазмид обеспечивают больший контроль и стойкость дозировки.
В одном воплощении плазмиды/векторы кодируют четыре белка:
i) антиген,
ii) TGF бета 1 (TGF-β1),
iii) интерлейкин-10 и
iv) интерлейкин-2.
В одном воплощении антиген представляет собой нацеленный в эндосомы релевантный для T1D антиген, такой как инсулин или GAD65. Эндосомное нацеливание может осуществляться, например, посредством слияния Ii/CD74, слияния LIMPII/SCARB или слияния с трансферриновым рецептором.
В одном воплощении TGF-β1 представляет собой активированную форму.
Экспрессия четырех белков от одной плазмиды/вектора возможна, например, если желательные последовательности разделяются либо А) отдельными промоторами, Б) последовательностями IRES (внутренний сайт посадки рибосомы), которые рекрутируют новую рибосому для трансляции каждого сегмента, или В) вирусными последовательностями 2А (например, последовательности 2A FMDV или 2А TaV), которые транслируются и индуцируют приостановку/перескакивание рибосомы, что приводит к продукции отдельных полипептидов от одной открытой рамки считывания. Однако на практике каждая из этих стратегий является комплексной и сложной в осуществлении.
Экспрессия четырех независимых белков от одной плазмиды/вектора легче всего достигается посредством наличия отдельного промотора для каждого гена. Однако данный формат имеет значительные недостатки в том, что он А) приводит к очень большой, нестабильной и сложной в получении плазмиде из-за избыточной длины многих промоторов, Б) приводит к непредсказуемому поведению транслируемых белков по отношению друг к другу (они больше не продуцируются в фиксированных соотношениях друг с другом), В) каждый промотор может подвергаться независимому сайленсингу, приводя к селективной экспрессии некоторых генов, но не других, требующихся для полной эффективности, и Г) отсутствует простота регуляции. В отличие от этого, элементы IRES и последовательности 2А работают на уровнях мРНК и трансляции, и воспроизводимо соэкспрессируют фиксированные отношения каждого белка от одного промотора.
Каждый из четырех классов элементов IRES имеет разные требования к кофакторам для функционирования, а также разные требования к последовательности для трансляции работающего ниже гена. Например, IRES EMCV (вирус эндомиокардита) представляет собой IRES типа 1 из 630 пар оснований, который использует все эукариотические факторы инициации трансляции, тогда как IRES CrPv (вирус паралича сверчка) представляет собой IRES типа 4 из 200 пар оснований, который не имеет требующихся кофакторов, но использует нестандартный кодон инициации.
При использовании элементов IRES из других классов они мешают друг другу, таким образом, что каждый тип элемента IRES можно использовать только один раз в каждой плазмиде, и при использовании вместе разные типы элементов IRES ослабляют друг друга (уменьшают эффективность) способами, которые трудно прогнозировать.
Кроме того, осуществление перетасовки комбинаций ген/IRES приводит к непредсказуемым соотношениям транслированных продуктов, так как взаимодействия генов с элементами IRES не являются статичными, но зависимыми от контекста на фланкирующих нуклеотидных последовательностях. Кроме того, элементы IRES накладывают ограничения на первые несколько положений аминокислот в точке инициации или сразу после нее. Например, IRES CrPv требует того, чтобы первая аминокислота была аланином, а не стандартным метионином, и IRES EMCV не может переносить аминокислот P, W, C, R или K в пределах первых трех кодонов. В одном воплощении для приспособления к ограничениям N-концевых аминокислот, накладываемых IRES EMCV, ДНК-вакцина содержит трехаланиновое удлинение до N-конца гена IL-10.
Кроме того, каждый элемент IRES содержит значительное число пар оснований, варьирующее от 230 п.о. до более 700 п.о.; включение многих элементов IRES, таким образом, увеличивает размер и сложность плазмид/векторов в такой степени, что многие становятся нестабильными и сложными в промышленном производстве из-за спонтанных делеций и рекомбинаций. Кроме того, из-за высокой степени присутствия вторичной структуры, которую придают элементы IRES на транскрибированных мРНК, которые содержат их, они увеличивают вероятность активации рецепторов распознавания патогенов (Dabo S, Meurs EF. 2012 Viruses 4(11):2598-635.) в трансфицированной клетке, и продуцирование стимулирующих эффектов противодействует индукции толерантности, которая намечена.
Последовательности 2А, в отличие от элементов IRES, не взаимодействуют друг с другом и, следовательно, обеспечивают стабильную и согласованную эффективность. Однако они сами транслируются и, следовательно, влияют на сворачивание, функцию и стабильность конечных транслированных белковых продуктов. Все последовательности 2А приводят к значительному С-концевому слиянию (19-22 ак) на 5'-конце последовательностей, подлежащих отделению, и также начинают 3'-последовательность с пролина. Некоторые белки являются пермиссивными для данных модификаций, а некоторые - нет, приводя к практическим ограничениям применения последовательностей 2А. Например, продукт интерлейкин-10 является пермиссивным для 2А хвоста, но и интерлейкин-2, и TGF-β1 неправильно сворачивается и теряет функцию при экспрессии выше от метки 2А. Следовательно, в то время как возможно экспрессировать несколько независимых белков, разделенных последовательностями 2А, два из четырех белков, описанных в данном документе, не могут заканчиваться метками 2А, и, следовательно, должны использоваться другие стратегии.
Поскольку каждый тип аминокислотной последовательности 2А модифицирует рибосомальную функцию во время трансляции белка, он будет иметь разные эффективности в двух центральных свойствах семейства 2А, а именно: (i) разделении расположенных рядом генных продуктов и (ii) процессивности (повторной инициации) во второй генный продукт. Разные последовательности 2А имеют разные эффективности в генерации рибосомальной приостановки, которая разрушает пептидный остов (приводя к двум отдельным белкам), а также разные эффективности в повторной инициации пептидного синтеза второго генного продукта.
Способность последовательностей 2А разделять белковые продукты и повторно инициировать трансляцию белка зависит от аминокислотной последовательности 2А (Donnelly ML, Hughes LE, Luke G, et. al. 2001 J Gen Virol. 82(Pt 5):1027-41). Небольшие вариации в аминокислотных последовательностях 2А приводят к значительно отличающимся смесям разделенных и слитых фланкирующих генных продуктов, варьирующим от менее 5% (больше 95% слито) до полностью разделенных (0% слитых или 100% разделенных).
Кроме того, автор данного изобретения открыл в данном изобретении то, что смежные аминокислотные последовательности, кодирующие два фланкирующих белковых продукта, также влияют на эффективность повторной инициации и разделения последовательностей 2А, приводя к значительным отклонениям от описанных результатов. Эффективность повторной инициации, таким образом, варьирует в зависимости от типа использованной аминокислотной последовательности 2А, а также от окружения, предоставленного смежными аминокислотными последовательностями, и, таким образом, отношение генного продукта пре-2А и разделения белков будет определяться как использованной аминокислотной последовательностью 2А, так и ее контекстом.
В одном воплощении «2A FMDV» вставлена между последовательностью, кодирующей антиген, и последовательностью, кодирующей TGF-β1, описанными в данном документе, приводя к 100%-ному разделению, а также к отношению 1:1 данных белковых продуктов.
В другом воплощении «2A TaV» может быть вставлена между последовательностью, кодирующей IL-10, и последовательностью, кодирующей IL-2, описанными в данном документе, приводя к примерно 50% раздельных продуктов, а также к отношению данных белковых продуктов 10 к 6. Каждая трансфицированная клетка, таким образом, доставляет относительно низкую дозу интерлейкина-2, которая не способна стимулировать эффекторные Т-клетки, и более высокую дозу инерлейкина-10 для отклонения Т-клеток в направлении фенотипа Treg. Поскольку продукция слитого IL-10/IL-2 является неблагоприятной, были предприняты попытки сконструировать повышенную эффективность расщепления сегмента 2А TaV. Попытка того, чтобы сегменту 2А предшествовал «изолирующий сегмент», который представляет собой элемент, который удлиняет транслированную область выше 2А TaV для уменьшения влияния на элемент 2А расположенной выше последовательности, не улучшила разделение. В другой попытке решить проблему слияния добавляли расположенный выше разобщающий сегмент с последовательностью транслированного белка GSG; однако, данный подход привел только к добавочному улучшению эффективности расщепления.
Слияния цитокинов, как таковые, возникающие в результате разделения генов, кодирующих IL-10 и IL-2, посредством 2А TAV, по-видимому, являются иммуногенными.
В другом воплощении описанный в данном документе вектор/плазмида имеет сегмент «Р 2А». Разделение генов, кодирующих IL-10 и IL-2, посредством Р2А приводит к полному или почти полному разделению белковых продуктов, а также к по меньшей мере в два раза большему (или может быть даже вплоть до в четыре или пять раз большему) отношению IL-10 по сравнению с IL-2.
Для того чтобы устранить недостатки систем только IRES и только 2А, описанных выше, четыре описанные в данном документе последовательности кДНК (антиген, TGF-β1, IL-10, IL-2) организованы парами до и после одиночного IRES. Каждая пара дополнительно разделена последовательностью 2А, которая индуцирует перескакивание рибосомы и продукцию независимых белков от каждой последовательности в паре полипептидов. Поскольку TGF-β1 и IL-2 могут не находиться на N-концевой стороне слияния, один из них должен заканчиваться в центральном сайте IRES, а другой должен заканчивать транслируемую часть последовательности мРНК.
Хронология/последовательность экспрессируемых белков и элементов IRES/2A в данном документе, следовательно, может быть следующей: (i) антиген, (ii) 2A FMDV, (iii) TGF бета 1, (iv) IRES, (v) IL-10, (vi) P 2A и (vii) IL-2. Как следствие, все четыре белка могут независимо экспрессироваться от одного оперона/генного сегмента стабильным и предсказуемым способом. Поскольку каждый из данных белков экспрессируется от одной мРНК, отношения каждого продукта фиксируются - например, невозможно получать избыток IL-2 над IL-10.
Кроме применения комбинации элементов IRES и 2А для разделения закодированных генов, альтернативным решением в данном документе, могло бы быть применение двухнаправленного промотора для получения 2 мРНК - каждая из данных мРНК кодировала бы пару белков, а не все четыре в одной молекуле мРНК. Следовательно, можно сконструировать эквивалентные расположения с использованием пар экспрессионных кассет, подходящим образом организованных около двухнаправленного промотора млекопитающего и с использованием разделительных последовательностей 2А и/или элементов IRES. Данный подход, однако, ассоциирован с недостатками, прежде всего обусловленными большим размеров двухнаправленных промоторов, но также потенциальной повышенной регуляторной нагрузкой, имеющей отдельные элементы мРНК, включенные в один медицинский продукт. В предпочтительных воплощениях в данном документе, следовательно, используется один промотор и комбинация элеметов IRES и 2А, а не двухнаправленный промотор.
В теории некоторые последовательности 2А могли бы быть заменены внутриклеточными эндогенными протеазочувствительными последовательностями. Однако автор данного изобретения в данном документе открыл то, что такие протеазы ассоциированы со значительными недостатками (например, недостаток описанной функции, приводящий к секреции слитых белковых продуктов).
Для того чтобы антиген подвергался процессингу и презентировался иммунной системе в пределах местного окружения цитокиновых гормонов, кодируемых плазмидой, данный антиген должен удерживаться в пределах трансфицированной клетки. В случае диабета типа 1 продукция активного инсулина потенциально приводила бы к нежелательному снижению глюкозы в крови, если бы он секретировался или иным образом высвобождался из трансфицированной клетки.
Для того чтобы избежать секреции антигена, любые сигналы секреции могут быть удалены из последовательности, кодирующей антиген, например, из последовательности нуклеиновой кислоты, кодирующей препроинсулин, удаляется последовательность, кодирующая сигнал секреции, таким образом, был бы генерирован проинсулин, а не препроинсулин, обеспечивая, таким образом, накопление антигена внутри трансфицированной клетки. В то время как данный транслируемый антигенный продукт (например, инсулин) активно не секретировался бы, он мог бы высвобождаться во время лизиса из-за некроза, возникающего из-за атаки Т-клетками CD8+. Кроме того, сигнальная последовательность инсулина представляет собой область, для которой известно то, что она содержит эпитопы, релевантные для заболевания (потенциально индуцируя аутоиммунитет), и включение сигнальной последовательности, следовательно, обеспечивает индукцию более широкой толерантности и более высокую вероятность ослабления заболевания.
Кроме того, цитоплазматическое удерживание антигена обеспечивает лишь процессинг посредством протеасомы и презентацию через путь МНС (главный комплекс гистосовместимости) класса I, который выявляет внутриклеточные патогены посредством Т-клеток CD8+. Поскольку Т-клетки CD4+ являются важными соучастниками провоспалительных цитокинов, и большинство или даже все Treg, подавляющие аутоиммунитет, представляют собой CD4+, расширение презентации антигена для включения MHC класса II, который распознается Т-клетками CD4+, может быть полезным.
Процессинг MHC класса II и стимулирование Т-клеток CD4+ обычно не включают внутриклеточный антиген, так как доступ к данному пути осуществляется через эндоцитоз внеклеточного антигена. Обычно белковые продукты, продуцируемые в пределах трансфицированной клетки, презентируются только через внутриклеточный / протеасомный путь процессинга по умолчанию и MHC класса I, приводя к эффектам Т-клеток CD8+, но не к эффектам Т-клеток CD4+. Для того чтобы нацеливать и Т-клетки CD4+, и CD8+ для иммуномодуляции предпочтительное воплощение также включает факторы, приводящие к презентации MHC класса II.
В принципе, для индукции презентации MHC класса II антиген может быть слит с любым партнером, который направляет данное слияние в эндосомный компартмент, но имеются функциональные различия в активности и воздействии. Слияния с трансферриновым рецептором, также известным как белковый рецептор, транспортирующий железо, циклируют от плазматической мембраны/внеклеточного пространства до эндосомы и, следовательно, также могут экспонировать целому антигену другие иммунные клнтки, такие как В-клетки, макрофаги и т.д.. Слияния LimpII/SCARB нацелены непосредственно в эндосому, но предпочтительно в раннюю эндосому, и иногда приводят к чрезмерному процессингу и общему разрушению антигена. Слияния li (CD74), использующие такой же шапероновый сигнал, что и MHC класса II для локализации в поздней эндосоме, доставляют антиген и MHC класса II в такие же везикулы на той же самой стадии развития и максимизируют вероятность осуществления эффективной презентации антигена в контексте MHC класса II. Кроме того, даже с эндосомной сортировкой из слияний Ii последовательность секреции препроинсулина должна быть сделана неактивной, или антиген также секретировался бы и терялся бы перед процессингом.
Блокада секреции инсулинового антигена, в качестве альтернативы, осуществлялась в данном документе посредством мутирования двух аминокислот, требующихся для удаления сигнала секреции, посредством SRP (частица распознавания сигнала) на шероховатом эндоплазматическом ретикулуме. Мутации Ala (A) до Glu (E) полностью отменяли созревание и секрецию пре-проинсулина при поддержании требующейся эпитопной структуры антигена для индукции наилучшей толерантности.
В одном воплощении в данном изобретении используется плазмидная ДНК-вакцина. Данная плазмида выращивается/реплицируется, например, в E. coli и выделяется/очищается из среды и затем готовится в жидких препаратах, например, в воде, физиологическом растворе, жидких препаратах на основе PBS (фосфатно-солевой буферный раствор) или в виде лиофилизированного порошка для внутрикожного безыгольного впрыскивания, интраназального введения или ингаляции. В одном воплощении плазмиду в данном изобретении готовят в водном фармацевтическом препарате, возможно содержащем стабилизаторы. Для продукции плазмиды можно использовать любую подходящую микробную систему.
Стабилизаторы в препарате включают хелаторы, такие как EDTA (этилендиаминтетрауксусная кислота), EGTA (этиленгликольтетрауксусная кислота) или DPTA, но не ограничиваются ими, для захвата Mg++ и Fe+++, которые, в противном случае, могут участвовать в деградации ДНК, и/или цитрат, который защищает плазмиду от эффектов неспецифичной деградации. В одном воплощении плазмида, описанная в данном документе, может быть приготовлена в изотоничном PBS или, в качестве альтернативы, в TRIS плюс цитрат плюс EDTA. Такие плазмиды имеют преимущества стабильности, легкости в продукции, безопасности и удобства в применении.
В другом воплощении в связи с настоящим изобретением могли бы быть добавлены агенты для доставки, такие как вирус, липиды, липосомы, соупаковывающие препараты и т.д. Однако применение агентов для доставки в данном изобретении может иметь потенциальные проблемы с иммунитетом, интеграцией вируса и т.д.
Определения
Антиген: иммунотерапевтическая ДНК-вакцина в данном документе кодирует антиген. Антиген в данном документе может представлять собой любой тип белка, ассоциированного с иммуногенным заболеванием, или его фрагмент, который может распознаваться Т-клеточным компонентом иммунной системы. Например, в случае лечения или предупреждения диабета типа 1 можно использовать инсулиновый антиген. В одном примере инсулиновый антиген представляет собой иммунодоминантный пептид InsB 9-23. Для иммунотерапевтических ДНК-вакцин против рассеянного склероза в данном документе в качестве антигена можно использовать миелиновый основной белок (МВР), миелиновый олигодендроцитарный белок (MOG) и/или протеолипидный белковый антиген (PLP). Для применения в иммунотерапевтических ДНК-вакцинах в данном документе подходят последовательности, кодирующие аналогичный белковый антиген, для репрезентативных антигенов из алопеции, полимиозита/дерматомиозита, целиакии-спру и белковых аллергенов (например, белок арахиса ara h 2).
Нацеливание антиг ена: в одном воплощении антиген в данном документе нацелен в эндосомы. Антигены в данном документе включают цельный белок, предбелки с недостаточностью секреции или их функциональный или иммунодоминантный пептидный фрагмент.
Например, инсулиновый антиген в данном документе представляет собой антиген для применения в иммуномодулирующей терапии, а не агент, понижающий уровнь глюкозы. Он, следовательно, не должен быть полностью подвергнутым процессингу/созревшим или секретированным для убеждения в том, что он презентируется на молекулах МНС циркулирующим Т-клеткам. Иммунотерапевтическая ДНК-вакцина в данном документе, следовательно, не приводит к повышенным уровням инсулина в крови, но скорее к усиленной презентации антигенов иммунной системе, в частности, Т-клеткам.
Следовательно, инсулиновый антиген в данном документе может представлять собой фрагменты, кодирующие маленький иммунодоминантный пептид (например, пептид 9-23 цепи В инсулина, включая пептиды со сдвинутым регистром, демонстрирующие эквивалентные Т-клеточные эпитопы), цельный проинсулин, у которого отсутствует требующаяся последовательность секреции, но который является интактным в иных отношениях, или пре-проинсулиновые мутеины, которые содержат последовательность секреции, но модифицируются для предупреждения секреторной функции.
Примеры инсулиновых антигенов в данном документе включают:
Мышиный проинсулин (SEQ ID NO 1):
FVNQHLCGSHLVEALYLVCGERGFFYTPKTRREAEDLQVGQVELGGGPGAGSLQPLALEGSLQKRGIVEQCCTSICSLYQLENYCN
Человеческий проинсулин (SEQ ID NO 2):
FVNQHLCGSHLVEALYLVCGERGFFYTPKTRREAEDLQVGQVELGGGPGAGSLQPLALEGSLQKRGIVEQCCTSICSLYQLENYCN
Модифицированный мышиный пре-проинсулин, который не секретируется (замены по отношению к пре-проинсулину дикого типа показаны жирным шрифтом и подчеркиванием (SEQ ID NO 3)):
MALWMRLLPLLALLALWGPDPEQEFVNQHLCGSHLVEALYLVCGERGFFYTPKTRREAEDLQVGQVELGGGPGAGSLQPLALEGSLQKRGIVEQCCTSICSLYQLENYCN
Модифицированный человеческий пре-проинсулин, который не секретируется (замены по отношению к пре-проинсулину дикого типа показаны жирным шрифтом и подчеркиванием (SEQ ID NO 4)):
MALWMRLLPLLALLALWGPDPEQEFVNQHLCGSHLVEALYLVCGERGFFYTPKTRREAEDLQVGQVELGGGPGAGSLQPLALEGSLQKRGIVEQCCTSICSLYQLENYCN
Мышиный пре-проинсулин дикого типа (SEQ ID NO 5):
ALWMRLLPLLALLALWGPDPAQAFVNQHLCGSHLVEALYLVCGERGFFYTPKTRREAEDLQVGQVELGGGPGAGSLQPLALEGSLQKRGIVEQCCTSICSLYQLENYCN
Человеческий пре-проинсулин дикого типа (SEQ ID NO 6):
MALWMRLLPLLALLALWGPDPAQAFVNQHLCGSHLVEALYLVCGERGFFYTPKTRREAEDLQVGQVELGGGPGAGSLQPLALEGSLQKRGIVEQCCTSICSLYQLENYCN
Инсулиновый пептид «InsB 9-23», идентичный между мышиным и человеческим:
SHLVEALYLVCGERG (SEQ ID NO 7)
Модифицированный InsB 9-23 (замены по отношению к InsB 9-23 дикого типа показаны жирным шрифтом и подчеркиванием (SEQ ID NO 8) и (SEQ ID NO 27)):
SHLVEALYLVCGEEG и SHLVEALYLVCGGEG
Инсулиновые антигены, описанные в данном документе, таким образом, могут накапливаться в цитозоле трансфицированной клетки-хозяина и, таким образом, презентироваться посредством МНС класса I или высвобождаться при цитолизе.
Эндосомальное нацеливание, приводящее к презентации МНС класса II, может осуществляться в данном изобретении посредством слияния последовательности антигена с лидерными последовательностями, которые образуют трансмембранные сегменты с цитоплазматическими последовательностями «YXX∅», в которых Y представляет собой тирозин, Х представляет собой любую аминокислоту, и ∅ представляет собой объемную гидрофобную аминокислоту, такую как триптофан или изолейцин, «[DE]XXXL[LI]», где D и Е представляют собой аспарагиновую кислоту или глутаминовую кислоту, соответственно, тогда как L и I представляют собой лейцин и изолейцин, соответственно, или эндосомальные/лизосомальные сигналы сортировки «DXXLL», которые подчеркнуты в следующих типичных последовательностях. Домены белка, которые включают данные сигналы, следовательно, нацелены или циклируют в эндосому/лизосому, включают: трансферриновый рецептор, LimpII или CD74, также известный как инвариантная цепь, шаперон МНС II или li или любой аналогичный домен.
Примеры доменов эндосомального нацеливания в данном документе включают:
Мышиный CD74 / эндосомальный домен нацеливания инвариантной цепи (li) (SEQ ID NO 9):
MDDQRDLISNHEQLPILGNRPREPERCSRGALYTGVSVLVALLLAGQATTAYFLYQQQGRLDKLTITSQNLQLESLRMKLP
Человеческий CD74 / эндосомальный домен нацеливания инвариантной цепи (li) (SEQ ID NO 10):
MHRRRSRSCREDQKPVMDDQRDLISNNEQLPMLGRRPGAPESKCSRGALYTGFSILVTLLLAGQATTAYFLYQQQGRLDKLTITSQNLQLESLRMKLP, но не ограничиваются ими.
Диабет типа 1: диабет типа 1 (T1D) считается хроническим аутоиммунным заболеванием, где аутоагрессивные Т-клетки инфильтрируют островки Лангерганса в поджелудочной железе и играют важную роль посредством специфичного разрушения популяции инсулинпродуцирующих бета-клеток. Как только разрушается значительное число островковых клеток, пониженное количество инсулина или полное отсутствие инсулина будет приводить к инсулиновой недостаточности и гипергликемии у пациента. Пациенты с T1D, таким образом, не способны продуцировать достаточно инсулина, и регулярные инъекции данного гормона необходимы на протяжении всей жизни. У некоторых пациентов с диабетом типа 1 диагностируется «диабет типа 1,5», «латентный аутоиммунный диабет»/LADA, «двойной диабет» и т.д., которые представляют собой диабетические заболевания, имеющие симптомы и диабета типа 1, и диабета типа 2 - все диабетические заболевания, имеющие черты и диабета типа 1, и типа 2, таким образом, также содержатся в данном документе в термине «диабет типа 1».
Толерогенная ДНК-вакцина: основанные на ДНК иммунотерапевтические вакцины/векторы/плазмиды, описанные в данном документе, сконструированы для выключения или осуществления понижающей регуляции части иммунной системы, ответственной за разрушение нормальных здоровых «собственных» клеток, и, таким образом, предупреждают или уменьшают интенсивность аутоиммунитета на основе Т-клеток.
Подразумевается то, что термин «иммунотерапевтическая ДНК-вакцина» в том виде, в котором он используется в данном документе, означает соединение или композицию, содержащую молекулу ДНК, и которая вводится субъекту для того, чтобы снизить риск развития у указанного субъекта одного или более чем одного заболевания.
В некоторых воплощениях иммунотерапевтические вакцины на основе ДНК, описанные в данном документе, представляют собой плазмиды/векторы, кодирующие конкретные антигены. После вакцинации данные плазмиды поглощаются, другими словами, трансфицируются в антигенпрезентирующие клетки в организме хозяина. Данные «трансфицированные» клетки-хозяева затем продуцируют антиген и презентируют маленькие фрагменты данного антигена иммунной системе, в частности Т-клеткам. Данный подход приводит к модификации специфичных ответов Т-клеток на кодируемый антиген, а также к минимальной модификации иммунных ответов на другие (незакодированные или «иррелевантные») антигены. Только очень мало клеток-хозяев типично трансформируется плазмидой/вектором ДНК-вакцины, описанной в данном документе, означая то, что вероятно меньше, чем одна из ста тысяч, одна из пятиста тысяч или даже меньше, чем одна из миллиона молекул плазмиды/вектора, в конечном счете, поступает в клетку-хозяина. ДНК-вакцины, описанные в данном документе, таким образом, представляют собой очень мягкий и специфичный подход для модулирования иммунных ответов на антигены, такие как инсулин, у пациентов с T1D или пациентов, подверженных риску развития T1D.
Плазмида: плазмида представляет собой маленькую молекулу ДНК, которая чаще всего находится в бактериях в виде маленьких, кольцевых, двухцепочечных молкул ДНК. Искусственные плазмиды широко используются в качестве векторов в молекулярном клонировании, служа для управления репликацией последовательностей рекомбинантной ДНК в организмах-хозяевах. Плазмиды можно конструировать так, чтобы они подходили для применения в качестве иммунотерапевтических ДНК-вакцин. Плазмиды считаются репликонами - элементами ДНК, способными автономно реплицироваться в подходящем хозяине. Плазмиды могут передаваться от одной бактерии к другой бактерии, которая могла бы принадлежать к такому же или к другому виду бактерии, посредством трех главных механизмов: трансформация, трансдукция и конъюгация. Плазмиды ДНК-вакцины могут поглощаться клеткой-хозяином посредством пассивной трансформации - обычно с относительно низкой скоростью. Плазмиды, описанные в данном документе, эффективно реплицируются, но не управляют экспрессией белка в бактериях. Плазмиды, описанные в данном документе, кроме того, управляют экспрессией белка, но не репликацией плазмиды у человека и других млекопитающих, например, мышей. В одном воплощении в данном документе используется вектор pVAX1 (Invitrogen/LifeTechnologies) в качестве каркаса для вставки элементов, которые являются частью настоящего изобретения. Другие подходящие векторые каркасы в данном документе включают любой остов вектора, содержащий эукариотический промоторный элемент, прокариотический многокопийный репликатор и систему селекции для поддержания плазмиды.
Селективный ген и система селекции: в одном аспекте иммунотерапевтические ДНК-вакцины, описанные в данном документе, содержат селективный ген/селективный маркер для целей изготовления. Селективный маркер в данном документе представляет собой, например, ген, который придает устойчивость к клеточному токсину, например, антибиотику, такому как ампициллин, канамицин, хлорамфеникол, стрептомицин и т.д.
Другие типы подходящих систем селекции в данном документе включают, например, условно-летальные системы сайленсинга (например, системы типа CcdA/CcdB или ParD/ParE Hok/Sok) или последовательности, которые комплементируют геномный дефект в продукции штамма клеток и, таким образом, обеспечивают рост в противном случае нежизнеспособного хозяина (например, ауксотрофная комплементация dapD- или pyrF-, комплементация инициации трансляции infA- и т.д.).
Продукционные клетки, несущие вакцину на основе плазмиды/ДНК, которая включает селективный маркер, будут выживать при воздействии токсина/антибиотика/условия, тогда как клетки, которые не смогли поглотить последовательности плазмиды, погибнут. В одном воплощении ДНК-вакцины, как таковые, описанные в данном документе, содержат последовательность нуклеиновой кислоты, кодирующую селективный маркер, для того чтобы обеспечивать более высокий выход/чистоту и более эффективную продукцию/репликацию в продукционных клетках, таких как E. coli.
В то время как селекция с антибиотиком представляет собой обычную лабораторную стратегию, могут иметься преимущества, ассоциированные с системами селекции без антибиотиков, например, в связи с более эффективными регуляторными процессами. В то время как в данном изобретении также могут использоваться векторы, которые не содержат селективного механизма, такие как миникольца, синтетические линейные векторы и т.д., данные воплощения ассоциированы с определенными недостатками в продукции, в частности, из-за повышенных затрат на продукцию и контроль качества.
Примеры стратегий комплементации («спасения») известны в предшествующем уровне техники, однако, данные стратегии страдают от разных недостатков.
Системы метаболической комплементации, такие как системы dapD [биосинтез лизина] или pyrF [биосинтез уридина], часто приводят к «перекрестному кормлению» во время продукции E. coli в высокой плотности, где бактерия, содержащая плазмиду, будет продуцировать и секретировать избыток требующегося соединения и, посредством этого, «расслаблять» давление отбора в отношении соседних бактерий без плазмиды.
Другим примером подходящей системы селекции в данном изобретении являются плазмиды, кодирующие важные белки, как, например, infA, кодирующая IF1 / фактор инициации 1, который требуется для синтеза белка. В данной системе селекции перекрестное кормление не происходит, так как белок infA не секретируется. Однако не возможно дополнительно модифицировать плазмиду или размножить клетки, дефицитные по плазмиде, так как отсутствует способ экзогенной комплементации требующегося белка/infA (J Bacteriol. 1994 Jan;176(1):198-205 и J Biotechnol. 2004 Jul 1;111(1):17-30).
Для того чтобы обойти недостатки, ассоциированные с системой селекции infA, в данном документе была предложена альтернативная система селекции с температурочувствитель ным переключением трансляции (или «термосенсором») из гена белка инвазии prfA L. monocytogenes (Cell. 2002 Sep 6;110(5):551-61). Посредством размещения образующей шпильку части последовательности РНК «термосенсора» выше геномных копий infA E. coli посредством стандартной технологии генной инженерии его экспрессия становится регулируемой посредством контроля температуры ферментации, обеспечивая медленный рост клеток, не содержащих плазмиду, при 37°C и быструю гибель клеток при температурах меньше 30°C. Трансформация сконструированного термочувствительного продукционного штамма E. coli плазмидами, экспрессирующими infA дикого типа, таким образом, обеспечивает полные нормальные скорости роста при всех температурах, обеспечивая бесплазмидное размножение при 37°C, а также строгий отбор на плазмиду при 30°C. Кроме того, данная система не генерирует давления отбора для того, чтобы E. coli дикого типа сохранила плазмиду, и она, таким образом, теряется в пределах 8 часов в культуре, обеспечивая отсутствие сохранения в среде терапевтической плазмиды.
Нуклеотидная последовательность infA E. coli дикого типа (SEQ ID NO 11):
ATGGCCAAAGAAGACAATATTGAAATGCAAGGTACCGTTCTTGAAACGTTGCCTAATACCATGTTCCGCGTAGAGTTAGAAAACGGTCACGTGGTTACTGCACACATCTCCGGTAAAATGCGCAAAAACTACATCCGCATCCTGACGGGCGACAAAGTGACTGTTGAACTGACCCCGTACGACCTGAGCAAAGGCCGCATTGTCTTCCGTAGTCGCTGA
Последовательность белка IF1 E. coli дикого типа, образующегося в результате трансляции гена infA (исходный метионин/М не включен в слияние prfA - (SEQ ID NO 12)):
MAKEDNIEMQGTVLETLPNTMFRVELENGHVVTAHISGKMRKNYIRILTGDKVTVELTPYDLSKGRIVFRSR
Продукционные линии клеток E. coli, используемые в данном изобретении для продукции плазмид иммунотерапевтических ДНК-вакцин, могут, таким образом, содержать следующую нуклеотидную последовательность термочувствительного prfA:
Нуклеотидная последовательность prfA L. monocytogenes дикого типа («шпилька термосенсора») (последовательность Шайна Дальгарно подчеркнута, начало ATG выделено жирным шрифтом - (SEQ ID NO 13)):
TGTAAAAAACATCATTTAGCGTGACTTTCTTTCAACAGCTAACAATTGTTGTTACTGCCTAATGTTTTTAGGGTATTTTAAAAAAGGGCGATAAAAAACGATTGGGGGATGAGAAATGAACGCTCAA
Последовательность белка prfA L. monocytogenes дикого типа (слитая выше IF1 E. coli, возникающая в результате трансляции SEQ ID NO 13):
MNAQ
Репликатор («Ori»): репликатор, также именуемый точкой начала репликации, представляет собой конкретную последовательность в геноме, в которой инициируется репликация нити ДНК. В одном воплощении сайты репликатора в данном документе включают «Ori pUC», который обеспечивает репликацию в бактериальной продукционной линии клеток E. coli, но не в клетках-хозяевах млекопитающих, т.е. в клетках из организма вакцинированного субъекта/человека/пациента. Другие подходящие в данном документе бактериальные репликаторы включают R6K, pBR322, ColE1, pMB1, 15A, pSC101 и т.д., но не ограничиваются ими. В одном аспекте репликатор в данном документе представляет собой высококопийную версию, которая дает высокое отношение плазмида/биомасса для более эффективной продукции. В данном документе также могут использоваться векторы, которые не содержат репликатор, такие как миникольца, синтетические линейные векторы и т.д.
Промотор: промотор представляет собой область ДНК, которая инициирует транскрипцию конкретного гена. Промоторы находятся около сайтов начала транскрипции генов, на той же самой нити и выше на ДНК, ближе к 5'-концу смысловой нити. Для того чтобы происходила транскрипция РНК-полимераза должна прикрепляться к ДНК около гена. Промоторы содержат специфические последовательности ДНК, такие как элементы ответа, которые обеспечивают безопасный исходный сайт связывания для РНК-полимеразы и для транскрипционных факторов, которые рекрутируют РНК-полимеразу. Транскрипционные факторы имеют специфические активаторные или репрессорные последоваельности, которые присоединяются к специфичным промоторам и регулируют экспрессию генов. Промоторы, таким образом, представляют собой критически важные элементы, которые могут работать согласованно с другими регуляторными областями, такими как энхансеры, сайленсеры, пограничные элементы/изоляторы, для управления уровнем транскрипции данного гена. Классический промотор управляет продукцией одной матричной РНК (мРНК), тогда как двухнаправленные промоторы, описанные в данном документе, управляют продукцией двух мРНК, непосредственно прилегащих к промотору, как выше, так и ниже промотора.
В одном воплощении в данном документе используются эукариотические промоторы. Эукариотические промоторы не обязательно подчиняются правилу один ген/один промотор, как, например, некоторые вирусные промоторы, а также промоторы, которые демонстрируют широкую экспрессию (т.е. не имеют узкие специфичности в отношени типа клеток, как, например, экспрессия только в нейронах). Примеры промоторов, описанных в данном документе, которые способны управлять широкой транскрипцией молекул мРНК больших мультигенных семейств, включают: вирусный немедленный-ранний (IE) промотор CMV (цитомегаловирус) и промотор SV40; промоторы эндогенного EF1a, PGK1, Ubc и бета-актина; и синтетические промоторы, такие как гибридный промотор CAG. Существует много других подходящих промоторов млекопитающих, и еще больше конструируют посредством усилий синтетической биологии. Любой промотор, который приводит к желательным характеристикам экспрессии в человеческих клетках, может использоваться в описанных в данном документе плазмидах иммунотерапевтической ДНК-вакцины.
Энхансеры: энхансеры представляют собой элементы ДНК, которые увеличивают эффективность промоторов в продуцировании мРНК-транскриптов. Энхансеры в данном документе могут быть соответствующими (например, энхансер SV40/промотор CMV) или несоответствующими. В данном изобретении может использоваться любая подходящая комбинация энхансера/промотора для эукариотической функции.
Эукариотическое начало трансляции: эукариотическую последовательность начала трансляции обычно называют консенсусной последовательностью «Козака». Последовательность Козака на молекуле мРНК распознается рибосомой в качестве сайта начала трансляции, от которого кодируется белок. Для эукариотической рибосомы требуется данная последовательность или ее вариация для инициации трансляции белка. Последовательности Козака являются вырожденными или вариабельными и редко соответствуют консенсусным последовательностям. На самом деле, консенсусные последовательности Козака типично являются менее эффективными, чем варианты дикого типа, выделенные из мРНК млекопиающих. В то время как слабые последовательности Козака регулярно выделяют из природных мРНК, и они, вероятно, играют роль в контроле трансляции малочисленных белков, описанные в данном документе иммунотерапевтические ДНК-вакцины предпочтительно кодируют последовательность Козака средней или высокой эффективности. Примеры полезных последовательностей Козака в данном документе включают следующую нуклеотидную последовательность: gccRccATGG (SEQ ID NO 14), где основания, указанные строчными буквами, представляют собой самые обычные нуклеотиды, но они могут варьировать, тогда как нуклеотиды, указанные заглавными буквами, являются фиксированными (R представляет собой код неопределенности IUPAC (Международный союз теоретической и прикладной химии) для оснований А или G), и ATG показывает сайт начала трансляции - кодон метионина в положении +1.
Сигнал эндосомальной сортировки: эндосома представляет собой ограниченный мембраной компартмент внутри эукариотической клетки. Некоторые белки могут транспортироваться в эндосомы и деградировать там до пептидных фрагментов. Данные пептидные фрагменты могут связываться с молекулами МНС, присутствующими в эндосоме, с образованием комплексов МНС/пептид, которые могут затем транспортироваться к поверхности клетки для того, чтобы презентироваться циркулирующим Т-клеткам, в частности, Т-клекам CD4+. Сортировка белков в эндосомы опосредуется сигналами, присутствующими в цитозольных доменах белков. Эндосомальные сигналы обычно представляют собой короткие линейные аминокислотные последовательности. Описанные в данном документе антигены предпочтительно нацеливаются в эндосомы с использованием сигнала эндосомальной сортировки, такого как, например, сигналы эндосомальной/лизосомальной сортировки YXX∅, [DE]XXXL[LI] или DXXLL. Сигналы эндосомальной сортировки включают разные встречающиеся в природе или синтетические сигналы эндосомальной сортировки. Примеры в данном документе включают сигналы эндосомальной сортировки, присутствующие на Cd74/инвариантной цепи/li, LimpII/SCARB или трансферриновом рецепторе. Может быть использован любой домен эндосомального нацеливания, который является фармацевтически приемлемым и обеспечивает желательную функцию. Слияние таких доменов эндосомального нацеливания с антигенами направляет их в эндосомальный компартмент при трансляции для повышенной эффективности. Эндосомальная сортировка антигенов обеспечивает процессинг и презентацию иммунной системе в комплексах МНС класса II, помимо конститутивной презентации в комплексах МНС класса I, для более полной и надежной индукции толерантности и возможного размножения Treg (которое не может осуществляться посредством комплексов МНС класса I / антиген). В одном воплощении толерогенные ДНК-вакцины, описанные в данном документе, кодируют слияние антигена с CD74/инвариантной цепью/li для управления эндосомальным нацеливанием и презентацией антигена посредством МНС класса II.
Интроны: интроны представляют собой некодирующие последовательности в пределах мРНК. Известно то, что некоторые интроны значительно увеличивают трансляцию и функцию мРНК. Соответственно, в данном изобретении также может использоваться включение последовательностей интронов. Могут использоваться стандартные интроны, такие как интрон бета-глобина, или любой интрон, подчиняющийся правилам сплайсинга у млекопитающих, такой как MCM7. В одном воплощении описанные в данном документе векторы иммунотерапевтической ДНК-вакцины содержат последовательности, кодирующие один или более чем один интрон. В другом воплощении векторы иммунотерапевтической ДНК-вакцины, описанные в данном документе, не имеют последовательностей, кодирующих интроны.
Метка рибосомальной приостановки: в связи с настоящим изобретением может быть полезным включение одной или более чем одной последовательности метки рибосомальной приостановки между последовательностями, кодирующими белок, в описанном в данном документе векторе/плазмиде иммунотерапевтической ДНК-вакцины для того, чтобы разделять белковые продукты.
Примером является вирусная «метка 2А FMDV» (метка вируса ящура 2А). Транслированной аминокислотной последовательностью 2А FMDV является APVKQTLNFDLLKLAGDVESNPGP - (SEQ ID NO 15). Метка 2А FMDV способна приостанавливать и повторно инициировать рибосому. Отношение транслированного продукта до и после метки 2А FMDV является близким к 1:1, и образующиеся белковые продукты обычно полностью разделяются. Данные типы рибосомальных меток ранее использовали в связи с соэкспрессией двух разных доменов, например, тяжелой цепи и легкой цепи при продукции рекомбинантного антитела. Однако автор настоящего изобретения сделал неожиданное открытие того, что они являются полезными в связи с мультицистронными ДНК-вакцинами как для отделения фланкирующих продуктов, так и для контроля отношений экспрессированных белков из-за собственых эффективностей рибосомальной повторной инициации. Метки последовательности, которые благоприятствуют отношению транслированных продуктов 1:1, в данном изобретении предпочтительно вставляются между двумя последовательностями, кодирующими белки, которые должны предпочтительно продуцироваться в отношении 1:1 (или близком к нему), как, например, инсулиновый антиген и мощный цитокин, такой как, например, TGF-β.
Другим примером метки последовательности рибосомальной приостановки в данном документе является вирусная метка последовательноси «2A TaV» (2A вируса Thosea asigna - транслированная аминокислотная последовательность 2A TaV: RAEGRGSLLTCGDVEENPGP (SEQ ID NO 16). Сообщается то, что отношение транслированного продукта до/выше и после/ниже данной метки составляет 50:1 (или около того). Автор настоящего изобретения сделал неожиданное открытие того, что, в то время как данный тип метки может использоваться для контроля уровней экспрессии в случаях, где жизненно важным является то, что один транслированный продукт абсолютно преобладает над другим, отделение фланкирующих цитокиновых продуктов составляет меньше, чем 50% относительно последовательностей, раскрытых в литературе, и отношение экспрессии, таким образом, составляет примерно 10:6. В связи с настоящим изобретением метка последовательности рибосомальной приостановки типа 2А должна предпочтительно приводить к разным уровням экспрессии двух белков, кодируемых тем же самым вектором/плазмидой. Экспрессия маленьких количеств плейотропного цитокина (такого как IL-2) по отношению к противовоспалительному цитокину, такому как IL-10, является желательной в данном изобретении, а слитые продукты - нежелательными.
Другим примером аминокислотной последовательности метки рибосомальной приостановки в данном документе является вирусная последовательность «2A P» (2A свиного тешовируса-1, ATNFSLLKQAGDVEENPGP - (SEQ ID NO 17)). Последовательности 2А Р функционируют подходящим образом в данном изобретении при вставке между IL-10 и IL-2, приводя к почти полному разделению с отношением экспрессии больше 5:1 между IL-10 и IL-2.
В качестве альтернативы, в данном изобретении можно использовать протеазочувствительные последовательности, обеспечивающие эндогенное расщепление между полибелками, экспрессируемыми плазмидой. Для разделения белковых продуктов в данном изобретении можно использовать фуринчувствительную последовательность (распознающую мотивы RAKR) или карбоксипептидазочувствительную последовательность (распознающую мотивы RRRR, RKRR или RRKR). Однако автор настоящего изобретения сделал неожиданное открытие того, что ни фуриновые, ни карбоксипептидазные расщепляемые последовательности не приводят к разделенным продуктам в данном изобретении, таким образом, приводя к секреции нежелательных слитых белков IL-10/IL-2.
TGF-b/β/β1 (трансформирующий фактор роста бета/β1): TGF-β представляет собой секретируемый белок, который контролирует пролиферацию, клеточную дифференциацию и другие функции в большинстве клеток. TGF-β представляет собой очень мощный цитокин со значительными эффектами на судьбу и фенотип клетки способами, зависимыми от контекста, например, в зависимости от других одновременно получаемых цитокиновых сигналов. Эндогенный TGF-β продуцируется в латентной форме, ассоциированной с наружной поверхностью мембраны продуцирующей клетки, и требует активации (например, воспалительными макрофагами, экспрессирующими CD36 и протеиназу плазмин) для созревания и высвобождения активной формы. В одном воплощении TGF-β в данном документе представляет собой модифицированную форму, которая является конститутивно активной. Это достигается заменой цистеинов в положениях 223 и 225 аминокислотами, не способными образовать дисульфидные мостики. Например, для замены цистеинов в положениях 223 и 225 используются серин или валин. Это приводит к структуре активного пробелка, который высвобождается в местную микросреду.
Последовательность человеческого эндогенного TGF-β1 - SEQ ID NO 18:
MPPSGLRLLLLLLPLLWLLVLTPGRPAAGLSTCKTIDMELVKRKRIEAIRGQILSKLRLASPPSQGEVPPGPLPEAVLALYNSTRDRVAGESAEPEPEPEADYYAKEVTRVLMVETHNEIYDKFKQSTHSIYMFFNTSELREAVPEPVLLSRAELRLLRLKLKVEQHVELYQKYSNNSWRYLSNRLLAPSDSPEWLSFDVTGVVRQWLSRGGEIEGFRLSAHCSCDSRDNTLQVDINGFTTGRRGDLATIHGMNRPFLLLMATPLERAQHLQSSRHRRALDTNYCFSSTEKNCCVRQLYIDFRKDLGWKWIHEPKGYHANFCLGPCPYIWSLDTQYSKVLALYNQHNPGASAAPCCVPQALEPLPIVYYVGRKPKVEQLSNMIVRSCKCS.
Последовательность модифицированного человеческого TGF-β1, который является конститутивно активным и секретируется (замены по отношению к TGF-β1 дикого типа показаны жирным шрифтом и подчеркнуты) - SEQ ID NO 19:
MPPSGLRLLLLLLPLLWLLVLTPGRPAAGLSTCKTIDMELVKRKRIEAIRGQILSKLRLASPPSQGEVPPGPLPEAVLALYNSTRDRVAGESAEPEPEPEADYYAKEVTRVLMVETHNEIYDKFKQSTHSIYMFFNTSELREAVPEPVLLSRAELRLLRLKLKVEQHVELYQKYSNNSWRYLSNRLLAPSDSPEWLSFDVTGVVRQWLSRGGEIEGFRLSAHVSVDSRDNTLQVDINGFTTGRRGDLATIHGMNRPFLLLMATPLERAQHLQSSRHRRALDTNYCFSSTEKNCCVRQLYIDFRKDLGWKWIHEPKGYHANFCLGPCPYIWSLDTQYSKVLALYNQHNPGASAAPCCVPQALEPLPIVYYVGRKPKVEQLSNMIVRSCKCS.
Другой последовательностью модифицированного человеческого TGF-β1, которую можно использовать, является SEQ ID NO 25:
MPPSGLRLLLLLLPLLWLLVLTPGRPAAGLSTCKTIDMELVKRKRIEAIRGQILSKLRLASPPSQGEVPPGPLPEAVLALYNSTRDRVAGESAEPEPEPEADYYAKEVTRVLMVETHNEIYDKFKQSTHSIYMFFNTSELREAVPEPVLLSRAELRLLRLKLKVEQHVELYQKYSNNSWRYLSNRLLAPSDSPEWLSFDVTGVVRQWLSRGGEIEGFRLSAHSSSDSRDNTLQVDINGFTTGRRGDLATIHGMNRPFLLLMATPLERAQHLQSSRHRRALDTNYCFSSTEKNCCVRQLYIDFRKDLGWKWIHEPKGYHANFCLGPCPYIWSLDTQYSKVLALYNQHNPGASAAPCCVPQALEPLPIVYYVGRKPKVEQLSNMIVRSCKCS.
Последовательность терминатора: терминатор транскрипции представляет собой отрезок последовательности нуклеиновой кислоты, который маркирует конец гена во время транскрипции. Высвобождение транскрипционного комплекса высвобождает РНК-полимеразу и связанный транскрипционный аппарат для начала транскрипции новых мРНК. Кроме того, те же самые клеточные факторы добавляют нематричный «поли-А хвост», который значительно увеличивает время жизни и функциональность мРНК. Пример подходящего терминатора транскрипции включает в данном документе терминатор «bGH_PA»:
CGACTGTGCCTTCTAGTTGCCAGCCATCTGTTGTTTGCCCCTCCCCCGTGCCTTCCTTGACCCTGGAAGGTGCCACTCCCACTGTCCTTTCCTAATAAAATGAGGAAATTGCATCGCATTGTCTGAGTAGGTGTCATTCTATTCTGGGGGGTGGGGTGGGGCAGGACAGCAAGGGGGAGGATTGGGAAGACAATAGCAGGCATGCTGGGGATGCGGTGGGCTCTATGG (SEQ ID NO 20).
В данном изобретении может использоваться любая приемлемая последовательность терминатора. Вариации включают применение двух разных фланкирующих последовательностей терминатора в случае двухнапавленных промоторов, продуцирующих две противоположно ориентированные мРНК.
В одном воплощении плазмида по изобретению имеет последовательность, как изложено в SEQ ID NO 24.
Во втором воплощении плазмида по изобретению имеет последовательность SEQ ID NO 26: полная (неаннотированная) последовательность плазмиды:
GACTCTTCGCGATGTACGGGCCAGATATACGCGTTGACATTGATTATTGACTAGTTATTAATAGTAATCAATTACGGGGTCATTAGTTCATAGCCCATATATGGAGTTCCGCGTTACATAACTTACGGTAAATGGCCCGCCTGGCTGACCGCCCAACGACCCCCGCCCATTGACGTCAATAATGACGTATGTTCCCATAGTAACGCCAATAGGGACTTTCCATTGACGTCAATGGGTGGACTATTTACGGTAAACTGCCCACTTGGCAGTACATCAAGTGTATCATATGCCAAGTACGCCCCCTATTGACGTCAATGACGGTAAATGGCCCGCCTGGCATTATGCCCAGTACATGACCTTATGGGACTTTCCTACTTGGCAGTACATCTACGTATTAGTCATCGCTATTACCATGGTGATGCGGTTTTGGCAGTACATCAATGGGCGTGGATAGCGGTTTGACTCACGGGGATTTCCAAGTCTCCACCCCATTGACGTCAATGGGAGTTTGTTTTGGCACCAAAATCAACGGGACTTTCCAAAATGTCGTAACAACTCCGCCCCATTGACGCAAATGGGCGGTAGGCGTGTACGGTGGGAGGTCTATATAAGCAGAGCTCTCTGGCTAACTAGAGAACCCACTGCTTACTGGCTTATCGAAATTAATACGACTCACTATAGGGAGACCCAAGCTGGCTAGCGTTTAAACTTAAGCTTGGTACCGAGCTCGGATCCACTAGTCCAGTGTGGTGGAATTCTGCACTGCAGCTCGCATCTCTCCTTCACGCGCCCGCCGCCCTACCTGAGGCCGCCATCCACGCCGGTTGAGTCGCGTTCTGCCGCCTCCCGCCTGTGGTGCCTCCTGAACTGCGTCCGCCGTCTAGGTAAGTTTAAAGCTCAGGTCGAGACCGGGCCTTTGTCCGGCGCTCCCTTGGAGCCTACCTAGACTCAGCCGGCTCTCCACGCTTTGCCTGACCCTGCTTGCTCAACTCTAGGTAAGTTAATGAGACAGATAGAAACTGGTCTTGTAGAAACAGAGTAGTCGCCTGCTTTTCTGCCAGGTGCTGACTTCTCTCCCCTGGGCTTTTTTCTTTTTCTCAGGTTGAAAAGAAGAAGACGAAGAAGACGAAGAAGACAAACCGTCGTCGACTGCCATGCGCCGCTGATTAACGCCGCCACCATGGCCCACCGACGCAGATCCAGAAGCTGCCGTGAGGACCAGAAGCCCGTGATGGATGATCAGAGGGACCTTATCTCTAACAATGAACAACTGCCAATGCTCGGCAGACGGCCTGGGGCCCCGGAGAGCAAGTGCAGCAGAGGAGCCTTGTACACGGGGTTCTCCATTTTAGTGACTCTCCTTCTCGCCGGCCAAGCTACCACCGCCTACTTTCTGTACCAACAGCAAGGCAGACTAGACAAACTGACAATCACAAGCCAGAACCTTCAGCTGGAGTCTCTGCGGATGAAGCTGCCCGCTTTGTGGATGAGATTGCTTCCTCTACTTGCTCTCCTGGCGCTCTGGGGACCTGACCCCGAGCAAGAGTTTGTTAATCAGCACCTGTGTGGGAGTCATCTGGTGGAGGCACTCTATTTAGTGTGCGGAGAGAGGGGCTTCTTCTACACTCCAAAGACCAGACGGGAGGCCGAAGACCTTCAAGTGGGGCAAGTAGAACTGGGTGGCGGACCCGGTGCCGGGAGCCTTCAGCCGCTCGCCCTGGAGGGCTCTCTTCAGAAACGCGGCATCGTGGAGCAGTGTTGCACATCCATTTGCTCACTCTACCAGCTGGAGAACTACTGCAACGGAAGCGGAGTGAAGCAGACGTTGAATTTTGATTTGTTGAAGTTGGCGGGGGATGTGGAGAGCAATCCGGGGCCGATGCCCCCTAGTGGCCTCAGACTTTTGTTATTGTTATTACCGCTTTTATGGCTCTTGGTGCTGACACCGGGCCGTCCGGCTGCTGGCTTGTCGACTTGTAAGACAATTGATATGGAATTGGTGAAACGAAAACGGATTGAGGCCATCCGAGGACAGATTTTGAGCAAGCTGCGGCTTGCCTCGCCACCCTCGCAAGGGGAAGTCCCACCCGGACCTCTACCAGAAGCAGTCCTAGCGCTGTACAACAGTACAAGAGATAGAGTGGCCGGGGAATCCGCAGAACCAGAGCCTGAGCCTGAAGCCGATTATTATGCAAAGGAAGTGACTAGGGTCCTGATGGTCGAGACCCATAACGAAATCTACGACAAATTCAAACAAAGTACCCACTCTATCTACATGTTCTTCAACACCAGTGAGCTAAGAGAAGCCGTGCCCGAACCTGTGCTTCTTTCCCGCGCAGAACTCCGCCTCTTGAGACTCAAATTGAAAGTTGAACAACACGTAGAGCTTTACCAGAAATACTCTAATAATTCATGGCGATATCTTTCTAATCGTCTCCTCGCCCCATCTGACAGCCCTGAATGGCTCTCCTTCGACGTTACGGGAGTTGTGCGCCAGTGGCTCAGCAGAGGCGGAGAGATAGAGGGCTTTCGGCTGAGCGCACATAGCTCTAGCGACTCAAGGGACAACACATTGCAAGTGGATATTAACGGTTTTACAACTGGACGGAGAGGGGACCTGGCGACCATCCACGGCATGAATAGACCTTTCCTGCTGCTGATGGCTACTCCCCTGGAGAGGGCACAGCACTTACAGTCTTCCAGACACCGGCGCGCCCTGGATACAAACTACTGCTTCAGCTCCACCGAAAAGAACTGTTGCGTGCGGCAGCTGTACATTGACTTCAGAAAGGATCTGGGCTGGAAGTGGATTCATGAGCCCAAGGGGTATCATGCCAACTTCTGTCTTGGGCCATGCCCATACATCTGGTCACTGGATACCCAGTACTCCAAAGTTCTGGCCTTGTACAATCAACACAACCCTGGAGCTTCCGCCGCTCCTTGCTGTGTGCCCCAAGCCCTAGAGCCCCTGCCCATCGTTTATTATGTCGGACGCAAGCCCAAAGTAGAACAGCTATCAAATATGATCGTGAGAAGCTGCAAGTGTAGCTGATAAACGCGTCGAGCATGCATCTAGGGCGGCCAATTCCGCCCCTCTCCCCCCCACCCCTCTCCCTCCCCCCCCCCTAACGTTACTGGCCGAAGCCGCTTGGAATAAGGCCGGTGTGCGTTTGTCTATATGTTATTTTCCACCATATTGCCGTCTTTTGGCAATGTGAGGGCCCGGAAACCTGGCCCTGTCTTCTTGACGAGCATTCCTAGGGGTCTTTCCCCTCTCGCCAAAGGAATGCAAGGTCTGTTGAATGTCGTGAAGGAAGCAGTTCCTCTGGAAGCTTCTTGAAGACAAACAACGTCTGTAGCGACCCTTTGTAGACAGCGGAACCCCCCACCTGGCGATAGATGCCTCTGCGGCCAAAAGCCACGTGTATAAGATACACCTGCAAAGGCGGCACAACCCCAGTGCCACGTTGTGAGTTGGATAGTTGTGGAAAGAGTCAAATGGCTCTCCTCAAGCGTATTCAACAAGGGGCTGAAGGATGCCCAGAAGGTACCCCATTGTATGGGATCTGATCTGGGGCCTCGGTGCACATGCTTTACATGTGTTTAGTCGAGGTTAAAAAACGTCTAGGCCCCCCGAACCACGGGGACGTGGTTTTCCTTTGAAAAACACGATGATAATATGATGCACAGCTCAGCACTGCTCTGTTGCCTGGTCCTCCTGACTGGGGTGAGGGCCAGCCCAGGCCAGGGCACCCAGTCTGAGAACAGCTGCACCCACTTCCCAGGCAACCTGCCTAACATGCTTCGAGATCTCCGAGATGCCTTCAGCAGAGTGAAGACTTTCTTTCAAATGAAGGATCAGCTGGACAACTTGTTGTTAAAGGAGTCCTTGCTGGAGGACTTTAAGGGTTACCTGGGTTGCCAAGCCTTGTCTGAGATGATCCAGTTTTACCTGGAGGAGGTGATGCCCCAAGCTGAGAACCAAGACCCAGACATCAAGGCGCATGTGAACTCCCTGGGGGAGAACCTGAAGACCCTCAGGCTGAGGCTACGGCGCTGTCATCGATTTCTTCCCTGTGAAAACAAGAGCAAGGCCGTGGAGCAGGTGAAGAATGCCTTTAATAAGCTCCAAGAGAAAGGCATCTACAAAGCCATGAGTGAGTTTGACATCTTCATCAACTACATAGAAGCCTACATGACAATGAAGATACGAAACGGGAGCGGCGCTACTAACTTCAGCCTGCTGAAGCAGGCTGGAGACGTGGAGGAGAACCCTGGACCTATGTACAGAATGCAGCTGCTGAGCTGCATCGCCCTGAGCCTGGCCCTGGTGACCAACAGCGCACCCACGTCCTCTAGCACCAAGAAGACCCAGTTACAGTTGGAGCATCTACTTTTAGACCTGCAAATGATTTTGAACGGCATCAACAACTACAAGAATCCTAAACTTACTCGCATGCTTACCTTCAAATTTTACATGCCCAAGAAGGCCACCGAACTGAAGCACTTGCAATGTCTGGAGGAAGAACTCAAGCCGCTGGAGGAAGTTCTCAACCTCGCGCAGTCCAAGAATTTCCACCTCCGGCCAAGAGACCTGATCAGTAACATTAATGTGATAGTGCTGGAGCTGAAGGGAAGCGAGACTACATTTATGTGCGAGTACGCCGATGAAACCGCTACAATCGTCGAGTTCCTGAATAGATGGATCACATTTTGCCAGTCAATTATCTCTACTCTGACATGATAACTCGAGGTCTAGAGGGCCCGTTTAAACCCGCTGATCAGCCTCGACTGTGCCTTCTAGTTGCCAGCCATCTGTTGTTTGCCCCTCCCCCGTGCCTTCCTTGACCCTGGAAGGTGCCACTCCCACTGTCCTTTCCTAATAAAATGAGGAAATTGCATCGCATTGTCTGAGTAGGTGTCATTCTATTCTGGGGGGTGGGGTGGGGCAGGACAGCAAGGGGGAGGATTGGGAAGACAATAGCAGGCATGCTGGGGATGCGGTGGGCTCTATGGCTTCTACTGGGCGGTTTTATGGACAGCAAGCGAACCGGAATTGCCAGCTGGGGCGCCCTCTGGTAAGGTTGGGAAGCCCTGCAAAGTAAACTGGATGGCTTTCTCGCCGCCAAGGATCTGATGGCGCAGGGGATCAAGCTCTGATCAAGAGACAGGATGAGGATCGTTTCGCATGGCCAAAGAAGACAATATTGAAATGCAAGGTACCGTTCTTGAAACGTTGCCTAATACCATGTTCCGCGTAGAGTTAGAAAACGGTCACGTGGTTACTGCACACATCTCCGGTAAAATGCGCAAAAACTACATCCGCATCCTGACGGGCGACAAAGTGACTGTTGAACTGACCCCGTACGACCTGAGCAAAGGCCGCATTGTCTTCCGTAGTCGCTGATAAATTATTAACGCTTACAATTTCCTGATGCGGTATTTTCTCCTTACGCATCTGTGCGGTATTTCACACCGCATACAGGTGGCACTTTTCGGGGAAATGTGCGCGGAACCCCTATTTGTTTATTTTTCTAAATACATTCAAATATGTATCCGCTCATGAGACAATAACCCTGATAAATGCTTCAATAATAGCACGTGCTAAAACTTCATTTTTAATTTAAAAGGATCTAGGTGAAGATCCTTTTTGATAATCTCATGACCAAAATCCCTTAACGTGAGTTTTCGTTCCACTGAGCGTCAGACCCCGTAGAAAAGATCAAAGGATCTTCTTGAGATCCTTTTTTTCTGCGCGTAATCTGCTGCTTGCAAACAAAAAAACCACCGCTACCAGCGGTGGTTTGTTTGCCGGATCAAGAGCTACCAACTCTTTTTCCGAAGGTAACTGGCTTCAGCAGAGCGCAGATACCAAATACTGTTCTTCTAGTGTAGCCGTAGTTAGGCCACCACTTCAAGAACTCTGTAGCACCGCCTACATACCTCGCTCTGCTAATCCTGTTACCAGTGGCTGCTGCCAGTGGCGATAAGTCGTGTCTTACCGGGTTGGACTCAAGACGATAGTTACCGGATAAGGCGCAGCGGTCGGGCTGAACGGGGGGTTCGTGCACACAGCCCAGCTTGGAGCGAACGACCTACACCGAACTGAGATACCTACAGCGTGAGCTATGAGAAAGCGCCACGCTTCCCGAAGGGAGAAAGGCGGACAGGTATCCGGTAAGCGGCAGGGTCGGAACAGGAGAGCGCACGAGGGAGCTTCCAGGGGGAAACGCCTGGTATCTTTATAGTCCTGTCGGGTTTCGCCACCTCTGACTTGAGCGTCGATTTTTGTGATGCTCGTCAGGGGGGCGGAGCCTATGGAAAAACGCCAGCAACGCGGCCTTTTTACGGTTCCTGGGCTTTTGCTGGCCTTTTGCTCACATGTTCTT.
В третьем воплощении плазмида по изобретению имеет последовательность SEQ ID NO 28: полная (неаннотированная) последовательность плазмиды:
GACTCTTCGCGATGTACGGGCCAGATATACGCGTTGACATTGATTATTGACTAGTTATTAATAGTAATCAATTACGGGGTCATTAGTTCATAGCCCATATATGGAGTTCCGCGTTACATAACTTACGGTAAATGGCCCGCCTGGCTGACCGCCCAACGACCCCCGCCCATTGACGTCAATAATGACGTATGTTCCCATAGTAACGCCAATAGGGACTTTCCATTGACGTCAATGGGTGGACTATTTACGGTAAACTGCCCACTTGGCAGTACATCAAGTGTATCATATGCCAAGTACGCCCCCTATTGACGTCAATGACGGTAAATGGCCCGCCTGGCATTATGCCCAGTACATGACCTTATGGGACTTTCCTACTTGGCAGTACATCTACGTATTAGTCATCGCTATTACCATGGTGATGCGGTTTTGGCAGTACATCAATGGGCGTGGATAGCGGTTTGACTCACGGGGATTTCCAAGTCTCCACCCCATTGACGTCAATGGGAGTTTGTTTTGGCACCAAAATCAACGGGACTTTCCAAAATGTCGTAACAACTCCGCCCCATTGACGCAAATGGGCGGTAGGCGTGTACGGTGGGAGGTCTATATAAGCAGAGCTCTCTGGCTAACTAGAGAACCCACTGCTTACTGGCTTATCGAAATTAATACGACTCACTATAGGGAGACCCAAGCTGGCTAGCGTTTAAACTTAAGCTTGGTACCGAGCTCGGATCCACTAGTCCAGTGTGGTGGAATTCTGCAGCTCGCATCTCTCCTTCACGCGCCCGCCGCCCTACCTGAGGCCGCCATCCACGCCGGTTGAGTCGCGTTCTGCCGCCTCCCGCCTGTGGTGCCTCCTGAACTGCGTCCGCCGTCTAGGTAAGTTTAAAGCTCAGGTCGAGACCGGGCCTTTGTCCGGCGCTCCCTTGGAGCCTACCTAGACTCAGCCGGCTCTCCACGCTTTGCCTGACCCTGCTTGCTCAACTCTAGGTAAGTTAATGAGACAGATAGAAACTGGTCTTGTAGAAACAGAGTAGTCGCCTGCTTTTCTGCCAGGTGCTGACTTCTCTCCCCTGGGCTTTTTTCTTTTTCTCAGGTTGAAAAGAAGAAGACGAAGAAGACGAAGAAGACAAACCGTCGTCGACTGCCATGCGCCGCTGATTAACGCCGCCACCATGGCCCACCGACGCAGATCCAGAAGCTGCCGTGAGGACCAGAAGCCCGTGATGGATGATCAGAGGGACCTTATCTCTAACAATGAACAACTGCCAATGCTCGGCAGACGGCCTGGGGCCCCGGAGAGCAAGTGCAGCAGAGGAGCCTTGTACACGGGGTTCTCCATTTTAGTGACTCTCCTTCTCGCCGGCCAAGCTACCACCGCCTACTTTCTGTACCAACAGCAAGGCAGACTAGACAAACTGACAATCACAAGCCAGAACCTTCAGCTGGAGTCTCTGCGGATGAAGCTGCCCGCTTTGTGGATGAGATTGCTTCCTCTACTTGCTCTCCTGGCGCTCTGGGGACCTGACCCCGAGCAAGAGTTTGTTAATCAGCACCTGTGTGGGAGTCATCTGGTGGAGGCACTCTATTTAGTGTGCGGAGAGAGGGGCTTCTTCTACACTCCAAAGACCAGACGGGAGGCCGAAGACCTTCAAGTGGGGCAAGTAGAACTGGGTGGCGGACCCGGTGCCGGGAGCCTTCAGCCGCTCGCCCTGGAGGGCTCTCTTCAGAAACGCGGCATCGTGGAGCAGTGTTGCACATCCATTTGCTCACTCTACCAGCTGGAGAACTACTGCAACGGAAGCGGAGTGAAGCAGACGTTGAATTTTGATTTGTTGAAGTTGGCGGGGGATGTGGAGAGCAATCCGGGGCCGATGCCCCCTAGTGGCCTCAGACTTTTGTTATTGTTATTACCGCTTTTATGGCTCTTGGTGCTGACACCGGGCCGTCCGGCTGCTGGCTTGTCGACTTGTAAGACAATTGATATGGAATTGGTGAAACGAAAACGGATTGAGGCCATCCGAGGACAGATTTTGAGCAAGCTGCGGCTTGCCTCGCCACCCTCGCAAGGGGAAGTCCCACCCGGACCTCTACCAGAAGCAGTCCTAGCGCTGTACAACAGTACAAGAGATAGAGTGGCCGGGGAATCCGCAGAACCAGAGCCTGAGCCTGAAGCCGATTATTATGCAAAGGAAGTGACTAGGGTCCTGATGGTCGAGACCCATAACGAAATCTACGACAAATTCAAACAAAGTACCCACTCTATCTACATGTTCTTCAACACCAGTGAGCTAAGAGAAGCCGTGCCCGAACCTGTGCTTCTTTCCCGCGCAGAACTCCGCCTCTTGAGACTCAAATTGAAAGTTGAACAACACGTAGAGCTTTACCAGAAATACTCTAATAATTCATGGCGATATCTTTCTAATCGTCTCCTCGCCCCATCTGACAGCCCTGAATGGCTCTCCTTCGACGTTACGGGAGTTGTGCGCCAGTGGCTCAGCAGAGGCGGAGAGATAGAGGGCTTTCGGCTGAGCGCACATAGCTCTAGCGACTCAAGGGACAACACATTGCAAGTGGATATTAACGGTTTTACAACTGGACGGAGAGGGGACCTGGCGACCATCCACGGCATGAATAGACCTTTCCTGCTGCTGATGGCTACTCCCCTGGAGAGGGCACAGCACTTACAGTCTTCCAGACACCGGCGCGCCCTGGATACAAACTACTGCTTCAGCTCCACCGAAAAGAACTGTTGCGTGCGGCAGCTGTACATTGACTTCAGAAAGGATCTGGGCTGGAAGTGGATTCATGAGCCCAAGGGGTATCATGCCAACTTCTGTCTTGGGCCATGCCCATACATCTGGTCACTGGATACCCAGTACTCCAAAGTTCTGGCCTTGTACAATCAACACAACCCTGGAGCTTCCGCCGCTCCTTGCTGTGTGCCCCAAGCCCTAGAGCCCCTGCCCATCGTTTATTATGTCGGACGCAAGCCCAAAGTAGAACAGCTATCAAATATGATCGTGAGAAGCTGCAAGTGTAGCTGATAAACGCGTCGAGCATGCATCTAGGGCGGCCAATTCCGCCCCTCTCCCCCCCACCCCTCTCCCTCCCCCCCCCCTAACGTTACTGGCCGAAGCCGCTTGGAATAAGGCCGGTGTGCGTTTGTCTATATGTTATTTTCCACCATATTGCCGTCTTTTGGCAATGTGAGGGCCCGGAAACCTGGCCCTGTCTTCTTGACGAGCATTCCTAGGGGTCTTTCCCCTCTCGCCAAAGGAATGCAAGGTCTGTTGAATGTCGTGAAGGAAGCAGTTCCTCTGGAAGCTTCTTGAAGACAAACAACGTCTGTAGCGACCCTTTGTAGACAGCGGAACCCCCCACCTGGCGATAGATGCCTCTGCGGCCAAAAGCCACGTGTATAAGATACACCTGCAAAGGCGGCACAACCCCAGTGCCACGTTGTGAGTTGGATAGTTGTGGAAAGAGTCAAATGGCTCTCCTCAAGCGTATTCAACAAGGGGCTGAAGGATGCCCAGAAGGTACCCCATTGTATGGGATCTGATCTGGGGCCTCGGTGCACATGCTTTACATGTGTTTAGTCGAGGTTAAAAAACGTCTAGGCCCCCCGAACCACGGGGACGTGGTTTTCCTTTGAAAAACACGATGATAATATGATGCACAGCTCAGCACTGCTCTGTTGCCTGGTCCTCCTGACTGGGGTGAGGGCCAGCCCAGGCCAGGGCACCCAGTCTGAGAACAGCTGCACCCACTTCCCAGGCAACCTGCCTAACATGCTTCGAGATCTCCGAGATGCCTTCAGCAGAGTGAAGACTTTCTTTCAAATGAAGGATCAGCTGGACAACTTGTTGTTAAAGGAGTCCTTGCTGGAGGACTTTAAGGGTTACCTGGGTTGCCAAGCCTTGTCTGAGATGATCCAGTTTTACCTGGAGGAGGTGATGCCCCAAGCTGAGAACCAAGACCCAGACATCAAGGCGCATGTGAACTCCCTGGGGGAGAACCTGAAGACCCTCAGGCTGAGGCTACGGCGCTGTCATCGATTTCTTCCCTGTGAAAACAAGAGCAAGGCCGTGGAGCAGGTGAAGAATGCCTTTAATAAGCTCCAAGAGAAAGGCATCTACAAAGCCATGAGTGAGTTTGACATCTTCATCAACTACATAGAAGCCTACATGACAATGAAGATACGAAACGGGAGCGGCGCTACTAACTTCAGCCTGCTGAAGCAGGCTGGAGACGTGGAGGAGAACCCTGGACCTATGTACAGAATGCAGCTGCTGAGCTGCATCGCCCTGAGCCTGGCCCTGGTGACCAACAGCGCACCCACGTCCTCTAGCACCAAGAAGACCCAGTTACAGTTGGAGCATCTACTTTTAGACCTGCAAATGATTTTGAACGGCATCAACAACTACAAGAATCCTAAACTTACTCGCATGCTTACCTTCAAATTTTACATGCCCAAGAAGGCCACCGAACTGAAGCACTTGCAATGTCTGGAGGAAGAACTCAAGCCGCTGGAGGAAGTTCTCAACCTCGCGCAGTCCAAGAATTTCCACCTCCGGCCAAGAGACCTGATCAGTAACATTAATGTGATAGTGCTGGAGCTGAAGGGAAGCGAGACTACATTTATGTGCGAGTACGCCGATGAAACCGCTACAATCGTCGAGTTCCTGAATAGATGGATCACATTTTGCCAGTCAATTATCTCTACTCTGACATGATAACTCGAGTCTAGAGGGCCCGTTTAAACCCGCTGATCAGCCTCGACTGTGCCTTCTAGTTGCCAGCCATCTGTTGTTTGCCCCTCCCCCGTGCCTTCCTTGACCCTGGAAGGTGCCACTCCCACTGTCCTTTCCTAATAAAATGAGGAAATTGCATCGCATTGTCTGAGTAGGTGTCATTCTATTCTGGGGGGTGGGGTGGGGCAGGACAGCAAGGGGGAGGATTGGGAAGACAATAGCAGGCATGCTGGGGATGCGGTGGGCTCTATGGCTTCTACTGGGCGGTTTTATGGACAGCAAGCGAACCGGAATTGCCAGCTGGGGCGCCCTCTGGTAAGGTTGGGAAGCCCTGCAAAGTAAACTGGATGGCTTTCTCGCCGCCAAGGATCTGATGGCGCAGGGGATCAAGCTCTGATCAAGAGACAGGATGAGGATCGTTTCGCATGGCCAAAGAAGACAATATTGAAATGCAAGGTACCGTTCTTGAAACGTTGCCTAATACCATGTTCCGCGTAGAGTTAGAAAACGGTCACGTGGTTACTGCACACATCTCCGGTAAAATGCGCAAAAACTACATCCGCATCCTGACGGGCGACAAAGTGACTGTTGAACTGACCCCGTACGACCTGAGCAAAGGCCGCATTGTCTTCCGTAGTCGCTGATAAATTATTAACGCTTACAATTTCCTGATGCGGTATTTTCTCCTTACGCATCTGTGCGGTATTTCACACCGCATACAGGTGGCACTTTTCGGGGAAATGTGCGCGGAACCCCTATTTGTTTATTTTTCTAAATACATTCAAATATGTATCCGCTCATGAGACAATAACCCTGATAAATGCTTCAATAATAGCACGTGCTAAAACTTCATTTTTAATTTAAAAGGATCTAGGTGAAGATCCTTTTTGATAATCTCATGACCAAAATCCCTTAACGTGAGTTTTCGTTCCACTGAGCGTCAGACCCCGTAGAAAAGATCAAAGGATCTTCTTGAGATCCTTTTTTTCTGCGCGTAATCTGCTGCTTGCAAACAAAAAAACCACCGCTACCAGCGGTGGTTTGTTTGCCGGATCAAGAGCTACCAACTCTTTTTCCGAAGGTAACTGGCTTCAGCAGAGCGCAGATACCAAATACTGTTCTTCTAGTGTAGCCGTAGTTAGGCCACCACTTCAAGAACTCTGTAGCACCGCCTACATACCTCGCTCTGCTAATCCTGTTACCAGTGGCTGCTGCCAGTGGCGATAAGTCGTGTCTTACCGGGTTGGACTCAAGACGATAGTTACCGGATAAGGCGCAGCGGTCGGGCTGAACGGGGGGTTCGTGCACACAGCCCAGCTTGGAGCGAACGACCTACACCGAACTGAGATACCTACAGCGTGAGCTATGAGAAAGCGCCACGCTTCCCGAAGGGAGAAAGGCGGACAGGTATCCGGTAAGCGGCAGGGTCGGAACAGGAGAGCGCACGAGGGAGCTTCCAGGGGGAAACGCCTGGTATCTTTATAGTCCTGTCGGGTTTCGCCACCTCTGACTTGAGCGTCGATTTTTGTGATGCTCGTCAGGGGGGCGGAGCCTATGGAAAAACGCCAGCAACGCGGCCTTTTTACGGTTCCTGGGCTTTTGCTGGCCTTTTGCTCACATGTTCTT.
В четвертом воплощении плазмида по изобретению имеет последовательность SEQ ID NO 29: полная (неаннотированная) последовательность плазмиды:
GACTCTTCGCGATGTACGGGCCAGATATACGCGTTGACATTGATTATTGACTAGTTATTAATAGTAATCAATTACGGGGTCATTAGTTCATAGCCCATATATGGAGTTCCGCGTTACATAACTTACGGTAAATGGCCCGCCTGGCTGACCGCCCAACGACCCCCGCCCATTGACGTCAATAATGACGTATGTTCCCATAGTAACGCCAATAGGGACTTTCCATTGACGTCAATGGGTGGACTATTTACGGTAAACTGCCCACTTGGCAGTACATCAAGTGTATCATATGCCAAGTACGCCCCCTATTGACGTCAATGACGGTAAATGGCCCGCCTGGCATTATGCCCAGTACATGACCTTATGGGACTTTCCTACTTGGCAGTACATCTACGTATTAGTCATCGCTATTACCATGGTGATGCGGTTTTGGCAGTACATCAATGGGCGTGGATAGCGGTTTGACTCACGGGGATTTCCAAGTCTCCACCCCATTGACGTCAATGGGAGTTTGTTTTGGCACCAAAATCAACGGGACTTTCCAAAATGTCGTAACAACTCCGCCCCATTGACGCAAATGGGCGGTAGGCGTGTACGGTGGGAGGTCTATATAAGCAGAGCTCTCTGGCTAACTAGAGAACCCACTGCTTACTGGCTTATCGAAATTAATACGACTCACTATAGGGAGACCCAAGCTGGCTAGCGTTTAAACTTAAGCTTGGTACCGAGCTCGGATCCACTAGTCCAGTGTGGTGGAATTCTGCAGCTCGCATCTCTCCTTCACGCGCCCGCCGCCCTACCTGAGGCCGCCATCCACGCCGGTTGAGTCGCGTTCTGCCGCCTCCCGCCTGTGGTGCCTCCTGAACTGCGTCCGCCGTCTAGGTAAGTTTAAAGCTCAGGTCGAGACCGGGCCTTTGTCCGGCGCTCCCTTGGAGCCTACCTAGACTCAGCCGGCTCTCCACGCTTTGCCTGACCCTGCTTGCTCAACTCTAGGTAAGTTAATGAGACAGATAGAAACTGGTCTTGTAGAAACAGAGTAGTCGCCTGCTTTTCTGCCAGGTGCTGACTTCTCTCCCCTGGGCTTTTTTCTTTTTCTCAGGTTGAAAAGAAGAAGACGAAGAAGACGAAGAAGACAAACCGTCGTCGACTGCCATGCGCCGCTGATTAACGCCGCCACCATGGCCCACCGACGCAGATCCAGAAGCTGCCGTGAGGACCAGAAGCCCGTGATGGATGATCAGAGGGACCTTATCTCTAACAATGAACAACTGCCAATGCTCGGCAGACGGCCTGGGGCCCCGGAGAGCAAGTGCAGCAGAGGAGCCTTGTACACGGGGTTCTCCATTTTAGTGACTCTCCTTCTCGCCGGCCAAGCTACCACCGCCTACTTTCTGTACCAACAGCAAGGCAGACTAGACAAACTGACAATCACAAGCCAGAACCTTCAGCTGGAGTCTCTGCGGATGAAGCTGCCCGCTTTGTGGATGAGATTGCTTCCTCTACTTGCTCTCCTGGCGCTCTGGGGACCTGACCCCGAGCAAGAGTTTGTTAATCAGCACCTGTGTGGGAGTCATCTGGTGGAGGCACTCTATTTAGTGTGCGGAGAGAGGGGCTTCTTCTACACTCCAAAGACCAGACGGGAGGCCGAAGACCTTCAAGTGGGGCAAGTAGAACTGGGTGGCGGACCCGGTGCCGGGAGCCTTCAGCCGCTCGCCCTGGAGGGCTCTCTTCAGAAACGCGGCATCGTGGAGCAGTGTTGCACATCCATTTGCTCACTCTACCAGCTGGAGAACTACTGCAACGGAAGCGGAGTGAAGCAGACGTTGAATTTTGATTTGTTGAAGTTGGCGGGGGATGTGGAGAGCAATCCGGGGCCGATGCCCCCTAGTGGCCTCAGACTTTTGTTATTGTTATTACCGCTTTTATGGCTCTTGGTGCTGACACCGGGCCGTCCGGCTGCTGGCTTGTCGACTTGTAAGACAATTGATATGGAATTGGTGAAACGAAAACGGATTGAGGCCATCCGAGGACAGATTTTGAGCAAGCTGCGGCTTGCCTCGCCACCCTCGCAAGGGGAAGTCCCACCCGGACCTCTACCAGAAGCAGTCCTAGCGCTGTACAACAGTACAAGAGATAGAGTGGCCGGGGAATCCGCAGAACCAGAGCCTGAGCCTGAAGCCGATTATTATGCAAAGGAAGTGACTAGGGTCCTGATGGTCGAGACCCATAACGAAATCTACGACAAATTCAAACAAAGTACCCACTCTATCTACATGTTCTTCAACACCAGTGAGCTAAGAGAAGCCGTGCCCGAACCTGTGCTTCTTTCCCGCGCAGAACTCCGCCTCTTGAGACTCAAATTGAAAGTTGAACAACACGTAGAGCTTTACCAGAAATACTCTAATAATTCATGGCGATATCTTTCTAATCGTCTCCTCGCCCCATCTGACAGCCCTGAATGGCTCTCCTTCGACGTTACGGGAGTTGTGCGCCAGTGGCTCAGCAGAGGCGGAGAGATAGAGGGCTTTCGGCTGAGCGCACATAGCTCTAGCGACTCAAGGGACAACACATTGCAAGTGGATATTAACGGTTTTACAACTGGACGGAGAGGGGACCTGGCGACCATCCACGGCATGAATAGACCTTTCCTGCTGCTGATGGCTACTCCCCTGGAGAGGGCACAGCACTTACAGTCTTCCAGACACCGGCGCGCCCTGGATACAAACTACTGCTTCAGCTCCACCGAAAAGAACTGTTGCGTGCGGCAGCTGTACATTGACTTCAGAAAGGATCTGGGCTGGAAGTGGATTCATGAGCCCAAGGGGTATCATGCCAACTTCTGTCTTGGGCCATGCCCATACATCTGGTCACTGGATACCCAGTACTCCAAAGTTCTGGCCTTGTACAATCAACACAACCCTGGAGCTTCCGCCGCTCCTTGCTGTGTGCCCCAAGCCCTAGAGCCCCTGCCCATCGTTTATTATGTCGGACGCAAGCCCAAAGTAGAACAGCTATCAAATATGATCGTGAGAAGCTGCAAGTGTAGCTGATAAACGCGTCGAGCATGCATCTAGGGCGGCCAATTCCGCCCCTCTCCCCCCCACCCCTCTCCCTCCCCCCCCCCTAACGTTACTGGCCGAAGCCGCTTGGAATAAGGCCGGTGTGCGTTTGTCTATATGTTATTTTCCACCATATTGCCGTCTTTTGGCAATGTGAGGGCCCGGAAACCTGGCCCTGTCTTCTTGACGAGCATTCCTAGGGGTCTTTCCCCTCTCGCCAAAGGAATGCAAGGTCTGTTGAATGTCGTGAAGGAAGCAGTTCCTCTGGAAGCTTCTTGAAGACAAACAACGTCTGTAGCGACCCTTTGTAGACAGCGGAACCCCCCACCTGGCGATAGATGCCTCTGCGGCCAAAAGCCACGTGTATAAGATACACCTGCAAAGGCGGCACAACCCCAGTGCCACGTTGTGAGTTGGATAGTTGTGGAAAGAGTCAAATGGCTCTCCTCAAGCGTATTCAACAAGGGGCTGAAGGATGCCCAGAAGGTACCCCATTGTATGGGATCTGATCTGGGGCCTCGGTGCACATGCTTTACATGTGTTTAGTCGAGGTTAAAAAACGTCTAGGCCCCCCGAACCACGGGGACGTGGTTTTCCTTTGAAAAACACGATGATAATATGATGCACAGCTCAGCACTGCTCTGTTGCCTGGTCCTCCTGACTGGGGTGAGGGCCAGCCCAGGCCAGGGCACCCAGTCTGAGAACAGCTGCACCCACTTCCCAGGCAACCTGCCTAACATGCTTCGAGATCTCCGAGATGCCTTCAGCAGAGTGAAGACTTTCTTTCAAATGAAGGATCAGCTGGACAACTTGTTGTTAAAGGAGTCCTTGCTGGAGGACTTTAAGGGTTACCTGGGTTGCCAAGCCTTGTCTGAGATGATCCAGTTTTACCTGGAGGAGGTGATGCCCCAAGCTGAGAACCAAGACCCAGACATCAAGGCGCATGTGAACTCCCTGGGGGAGAACCTGAAGACCCTCAGGCTGAGGCTACGGCGCTGTCATCGATTTCTTCCCTGTGAAAACAAGAGCAAGGCCGTGGAGCAGGTGAAGAATGCCTTTAATAAGCTCCAAGAGAAAGGCATCTACAAAGCCATGAGTGAGTTTGACATCTTCATCAACTACATAGAAGCCTACATGACAATGAAGATACGAAACGGGAGCGGCGCTACTAACTTCAGCCTGCTGAAGCAGGCTGGAGACGTGGAGGAGAACCCTGGACCTATGTACAGAATGCAGCTGCTGAGCTGCATCGCCCTGAGCCTGGCCCTGGTGACCAACAGCGCACCCACGTCCTCTAGCACCAAGAAGACCCAGTTACAGTTGGAGCATCTACTTTTAGACCTGCAAATGATTTTGAACGGCATCAACAACTACAAGAATCCTAAACTTACTCGCATGCTTACCTTCAAATTTTACATGCCCAAGAAGGCCACCGAACTGAAGCACTTGCAATGTCTGGAGGAAGAACTCAAGCCGCTGGAGGAAGTTCTCAACCTCGCGCAGTCCAAGAATTTCCACCTCCGGCCAAGAGACCTGATCAGTAACATTAATGTGATAGTGCTGGAGCTGAAGGGAAGCGAGACTACATTTATGTGCGAGTACGCCGATGAAACCGCTACAATCGTCGAGTTCCTGAATAGATGGATCACATTTTGCCAGTCAATTATCTCTACTCTGACATGATAACTCGAGTCTAGAGGGCCCGTTTAAACCCGCTGATCAGCCTCGACTGTGCCTTCTAGTTGCCAGCCATCTGTTGTTTGCCCCTCCCCCGTGCCTTCCTTGACCCTGGAAGGTGCCACTCCCACTGTCCTTTCCTAATAAAATGAGGAAATTGCATCGCATTGTCTGAGTAGGTGTCATTCTATTCTGGGGGGTGGGGTGGGGCAGGACAGCAAGGGGGAGGATTGGGAAGACAATAGCAGGCATGCTGGGGATGCGGTGGGCTCTATGGCTTCTACTGGGCGGTTTTATGGACAGCAAGCGAACCGGAATTGCCAGCTGGGGCGCCCTCTGGTAAGGTTGGGAAGCCCTGCAAAGTAAACTGGATGGCTTTCTCGCCGCCAAGGATCTGATGGCGCAGGGGATCAAGCTCTGATCAAGAGACAGGATGAGGATCGTTTCGCATGGCCAAAGAAGACAATATTGAAATGCAAGGTACCGTTCTTGAAACGTTGCCTAATACCATGTTCCGCGTAGAGTTAGAAAACGGTCACGTGGTTACTGCACACATCTCCGGTAAAATGCGCAAAAACTACATCCGCATCCTGACGGGCGACAAAGTGACTGTTGAACTGACCCCGTACGACCTGAGCAAAGGCCGCATTGTCTTCCGTAGTCGCTGATAAATTATTAACGCTTACAATTTCCTGATGCGGTATTTTCTCCTTACGCATCTGTGCGGTATTTCACACCGCATACAGGTGGCACTTTTCGGGGAAATGTGCGCGGAACCCCTATTTGTTTATTTTTCTAAATACATTCAAATATGTATCCGCTCATGAGACAATAACCCTGATAAATGCTTCAATAATAGCACGTGCTAAAACTTCATTTTTAATTTAAAAGGATCTAGGTGAAGATCCTTTTTGATAATCTCATGACCAAAATCCCTTAACGTGAGTTTTCGTTCCACTGAGCGTCAGACCCCGTAGAAAAGATCAAAGGATCTTCTTGAGATCCTTTTTTTCTGCGCGTAATCTGCTGCTTGCAAACAAAAAAACCACCGCTACCAGCGGTGGTTTGTTTGCCGGATCAAGAGCTACCAACTCTTTTTCCGAAGGTAACTGGCTTCAGCAGAGCGCAGATACCAAATACTGTTCTTCTAGTGTAGCCGTAGTTAGGCCACCACTTCAAGAACTCTGTAGCACCGCCTACATACCTCGCTCTGCTAATCCTGTTACCAGTGGCTGCTGCCAGTGGCGATAAGTCGTGTCTTACCGGGTTGGACTCAAGACGATAGTTACCGGATAAGGCGCAGCGGTCGGGCTGAACGGGGGGTTCGTGCACACAGCCCAGCTTGGAGCGAACGACCTACACCGAACTGAGATACCTACAGCGTGAGCTATGAGAAAGCGCCACGCTTCCCGAAGGGAGAAAGGCGGACAGGTATCCGGTAAGCGGCAGGGTCGGAACAGGAGAGCGCACGAGGGAGCTTCCAGGGGGAAACGCCTGGTATCTTTATAGTCCTGTCGGGTTTCGCCACCTCTGACTTGAGCGTCGATTTTTGTGATGCTCGTCAGGGGGGCGGAGCCTATGGAAAAACGCCAGCAACGCGGCCTTTTTACGGTTCCTGGGCTTTTGCTGGCCTTTTGCTCACATGTTCTT
Термин «GLP-1 (глюкагонподобный пептид-1)/пептид GLP-1/пептид-агонист GLP-1R» в том виде, в котором он используется в данном документе, относится к описанным в данном документе молекулам/пептидам/белкам/вариантам/агонистам GLP-1, которые представляют собой молекулы, имеющие функцию агониста GLP-1R, означая то, что они представляют собой агонисты рецептора GLP-1. Данный класс лекарственных средств обычно используется для лечения диабета, в частности, диабета типа 2. Аминокислотная последовательность зрелого «человеческого GLP-1» представляет собой: HAEGTFTSDVSSYLEGQAAKEFIAWLVKGRG (SEQ ID NO: 21).
Термин «аналог GLP-1» в том виде, в котором он используется в данном документе, относится к пептиду или соединению, которое представляет собой вариант GLP-1 (SEQ ID NO: 15). Термины «аналог GLP-1» и «аналог» могут использоваться в данном документе взаимозаменяемо.
Аналоги GLP-1 могут быть описаны посредством ссылки на i) число аминокислотных остатков в человеческом GLP-1 (SEQ ID NO: 15), которое соответствует аминокислотному остатку, который модифицирован (т.е. соответствующее положение в GLP-1 (SEQ ID NO: 15)), и ii) реальную модификацию.
Термин производные GLP-1 относится к производным аналогов GLP-1. Термин «производное» в том виде, в котором он используется в данном документе в контексте аналога GLP-1, означает химически модифицированный аналог GLP-1, в котором один или более чем один заместитель был ковалентно присоединен к аналогу GLP-1. Термин «заместитель» в том виде, в котором он используется в данном документе, означает химическую группировку или группу/боковую группу, конъюгированную с белком/агонистом/аналогом GLP-1. Производное может содержать одну или более чем одну модификацию, выбранную из амидов, углеводов, алкильных групп, ацильных групп, сложных эфиров и тому подобного.
В некоторых воплощениях заместитель ковалентно присоединен через аминокислотный остаток в указанном полипептиде, например, в одном из положений аминокислот, выбранном из группы, состоящей из положения 22, 23, 27, 34, 35 и 36.
В некоторых воплощениях производное GLP-1 содержит заместитель, содержащий липофильную группировку. Термин «липофильная группировка» в том виде, в котором он используется в данном документе, означает алифатическую или циклическую углеводородную группировку с более чем 6 и менее чем 30 атомами углерода, где указанная углеводородная группировка может содержать дополнительные заместители.
Примеры агонистов GLP-1 включают (но не ограничиваются) экзенатидом, лираглутидом, ликсисентидом, албиглутидом, дулаглутидом, таспоглутидом и семаглутидом. Иммунотерапевтические ДНК-вакцины с использованием плазмид в данном документе можно исходно объединять с параллельной обработкой агонистом GLP-1R при лечении, например, пациентов с недавним началом T1D. Совместное введение GLP-1 может быть хроническим или временным и включает пероральные пути, помимо парентеральных путей.
Лираглутид: (SEQ ID NO 22):
Семаглутид (SEQ ID NO 23):
Фармацевтические композиции в данном документе предпочтительно представляют собой водные препараты, содержащие по меньшей мере 50% воды, более предпочтительно, по меньшей мере 60% воды, более предпочтительно, по меньшей мере 75% воды, более предпочтительно, по меньшей мере 90% воды, более предпочтительно, по меньшей мере 95% воды и наиболее предпочтительно по меньшей мере 99% воды. Фармацевтические композиции в данном документе, в качестве альтернативы, могут представлять собой сухие препараты, такие как лиофилизированные препараты, предназначенные для разведения, ингаляции, закапывания в нос, внутрикожного введения и т.д.
Фармацевтические препараты в данном документе предпочтительно вводятся без применения способов усиления трансформации, таких как электропорация. В одном воплощении фармацевтические препараты предназначены для парентерального введения, например, подкожного введения, внутрикожного введения, внутривенного введения, внутримышечного введения и т.д. В другом воплощении фармацевтические композиции в данном документе могут дополнительно вводиться местно, перорально, ректально или посредством ингаляции.
Фармацевтические композиции в данном документе предпочтительно приготовлены без добавления каких-либо конденсрующих агентов или других эксципиентов, которые могут индуцировать локальные реакции. Препараты в данном документе предпочтительно содержат ловушки свободных радикалов (например, 1% этанол) и/или хелаторы, такие как, например, ловушки двухвалентных катионов (например, EDTA [CAS #60-00-4], EGTA [CAS #67-42-5] или DPTA [CAS #67-43-6]), для того, чтобы увеличивать стабильность водной плазмидной ДНК. Фармацевтические композиции в данном документе, кроме того, могут находиться в форме физиологического раствора и/или буферного раствора, или содержат физиологический раствор и/или содержат буферный раствор (например, PBS - фосфатно-солевой буферный раствор, буфер TRIS или эквивалентные фармацевтически приемлемые буферы). Фармацевтические препараты в данном документе предпочтительно не содержат каких-либо адъювантов, а также других типичных ингредиентов вакцин, таких как, например, гидроксид алюминия, фенол, сорбит, силикон и т.д.
Введение: иммунотерапевтическая ДНК-вакцина в данном документе может вводиться пациенту с T1D или пациенту, подверженному риску развития T1D. Данная вакцина может вводиться, например, на ежесуточной основе, через сутки, дважды в неделю, один раз в неделю, дважды в месяц, один раз в месяц, каждый второй месяц, четыре раза в год или один раз в год - частота может корректироваться согласно общим или индивидуальным потребностям. Иммунотерапия в данном документе может быть хронической. Продолжительность терапии может составлять, например, один месяц, два месяца, три месяца, 6 месяцев, один год, два года, три года, пять лет, шесть лет, семь лет, восемь лет, девять лет или 10 лет.
Воплощения
Следующие воплощения иллюстрируют данное изобретение, и их не следует понимать каким-либо ограничивающим образом. Понятно, что все воплощения можно объединять любыми возможными способами.
1. Плазмида, которая кодирует:
i. антиген;
ii. TGF-β; и
iii. IL-10.
2. Плазмида по воплощению 1, в которой указанный антиген представляет собой инсулиновый антиген.
3. Плазмида, которая соэкспрессирует/кодирует (предпочтительно от одного оперона): (i) антиген, такой как, например, инсулиновый антиген; (ii) TGF-β/TGF-β1 (такой как конститутивно активная форма); и (iii) IL-10.
4. Плазмида по любому из воплощений 1-3, где указанный инсулиновый антиген выбран из группы, состоящей из: проинсулина, не способного к секреции пре-проинсулина, или их функционального, или иммунодоминантного пептидного фрагмента.
5. Плазмида по любому из воплощений 1-4, где указанный инсулиновый антиген выбран из группы, состоящей из: проинсулина, пре-проинсулина и их функционального, или иммунодоминантного пептидного фрагмента.
6. Плазмида по любому из воплощений 1-5, где указанный инсулиновый антиген представляет собой нацеленный в эндосомы инсулин.
7. Плазмида по любому из воплощений 1-6, где указанная плазмида экспрессирует инсулиновый антиген и TGF-β в соотношении примерно 1:1.
8. Плазмида по любому из воплощений 1-7, где указанная плазмида экспрессирует инсулиновый антиген и TGF-β в количестве по меньшей мере в 200 раз меньшем, чем IL-10.
9. Плазмида по любому из воплощений 1-8, где указанная плазмида экспрессирует инсулиновый антиген и TGF-β в количестве по меньшей мере в 2 раз меньшем, чем IL-10.
10. Плазмида по любому из воплощений 1-9, где указанная плазмида, кроме того, соэкспрессирует интерлейкин-2 (IL-2).
11. Плазмида по любому из воплощений 1-10, где указанная плазмида экспрессирует избыток IL-10 и IL-2 по сравнению с антигеном (например, инсулином) и TGF-β.
12. Плазмида по любому из воплощений 1-11, где указанная плазмида экспрессирует по меньшей мере примерно однократный, двухкратный, пятикратный или по меньшей мере примерно стократный избыток IL-10 и IL-2 по сравнению с TGF-β и инсулиновым антигеном (отношение IL-10 плюс IL-2 к инсулину плюс TGF-β может составлять по меньшей мере 1:1 или 2:1, или 5:1, или 100:1).
13. Плазмида по любому из воплощений 1-12, где указанная плазмида экспрессирует по меньшей мере примерно стократный, двухсоткратный, пятисоткратный или по меньшей мере примерно тысячекратный избыток IL-10 и IL-2 по сравнению с TGF-β и инсулиновым антигеном (отношение IL-10 плюс IL-2 к инсулину плюс TGF-β может составлять по меньшей мере 100:1 или 200:1, или 500:1, или 1000:1).
14. Плазмида по любому из воплощений 1-13, где указанная плазмида экспрессирует IL-10 и IL-2 в отношении примерно 1:1 - 100:1, таком как, например, 1:1 - 50:1, таком как, например, 1:1 - 25:1, таком как, например, 1:1 - 10:1, в качестве альтернативы - 1:1 - 3:1, в качестве альтернативы - 1:1 - 2:1. В качестве альтернативы, отношение между экспрессируемым IL-10 и экспрессируемым IL-2 может составлять примерно 1:1; 1:0,9; 1:0,8; 1:0,7; 1:0,6; 1:0,5; 1:0,4; 1:0,3; 1:0,2 или 1:0,1.
15. Плазмида по любому из воплощений 1-14, где указанная плазмида содержит: (i) элемент 2А FMDV, отделяющий последовательность, кодирующую инсулиновый антиген, и последовательность, кодирующую TGF-β, (ii) элемент IRES EMCV, отделяющий последовательность, кодирующую TGF-β, и последовательность, кодирующую IL-10, и (iii) элемент 2А, отделяющий последовательность, кодирующую IL-10, и последовательность, кодирующую IL-2.
16. Плазмида по любому из воплощений 1-15, где указанная плазмида содержит:
(i) элемент 2А (такой как элемент 2А FMDV или 2А Р), отделяющий последовательность, кодирующую инсулиновый антиген, и последовательность, кодирующую TGF-β,
(ii) элемент IRES EMCV (в качестве альтернативы, двухнаправленный промотор), отделяющий последовательность, кодирующую TGF-β, и последовательность, кодирующую IL-10 (предпочтительно три аминокислоты аланин кодируются немедленно N-терминально по отношению к гену IL-10), и
(iii) элемент 2А (такой как элемент 2А Р), отделяющий последовательность, кодирующую IL-10, и последовательность, кодирующую IL-2.
17. Плазмида по любому из воплощений 1-16, в которой последовательность, кодирующая TGF-β, кодирует конститутивно активный TGF-β, предпочтительно конститутивно активный человеческий TGF-β1.
18. Плазмида по любому из воплощений 1-17, где указанная плазмида содержит: (i) нацеленную в эндосомы кодирующую последовательность препроинсулина, (ii) элемент 2А FMDV, (iii) последовательность, кодирующую TGF-β, (iv) элемент IRES EMCV, (v) последовательность, кодирующую IL-10, (vi) элемент 2А Р, (vii) последовательность, кодирующую IL-2, (viii) элемент полиаденилирования / терминации, (ix) селективный ген, (х) репликатор, (xi) эукариотический промоторный элемент, (xii) эукариотическую последовательность начала трансляции, (xiii) последовательность эндосомальной сортировки и (xiv) возможно интрон.
19. Плазмида по любому из воплощений 1-18, где указанная плазмида содержит следующие элементы:
(i) промотор (такой как промотор IE CMV),
(ii) интрон (локализованный в пределах некодирующей лидерной последовательности) и
(iii) эукариотическая последовательность начала трансляции (такая как элемент Козака),
(iv) последовательность, кодирующая антиген, нацеленный в эндосомы (такая как последовательность, кодирующая нацеленный в эндосомы человеческий пре-проинсулин, дефектный в отношении секреции),
(v) элемент 2А FMDV, предпочтительно отделяющий последовательность, кодирующую антиген, и последовательность, кодирующую TGF-β,
(vi) последовательность, кодирующая TGF-β (как, например, последовательность, кодирующая конститутивно активный человеческий TGF-β, предпочтительно последовательность, кодирующая конститутивно активный человеческий TGF-β1),
(vii) элемент IRES EMCV (или, в качестве альтернативы, двухнаправленный эукариотический промотор), где указанный элемент IRES EMCV отделяет последовательность, кодирующую TGF-β, и последовательность, кодирующую IL-10,
(viii) последовательность, кодирующая IL-10 (как, например, последовательность, кодирующая человеческий IL-10 с N-концевым добавлением трех аминокислот аланинов),
(ix) элемент 2А, такой как элемент 2А Р, где указанный элемент 2А отделяет последовательность, кодирующую IL-10, и последовательность, кодирующую IL-2,
(х) последовательность, кодирующая IL-2 (как, например, последовательность, кодирующая человеческий IL-2),
(xi) терминирующий элемент (такой как терминирующий элемент bGH_PA),
(xii) селективный ген (такой как последовательность, кодирующая канамицин, или последовательность, кодирующая infA wt (дикого типа)),
(xiii) репликатор (такой как прокариотический репликатор, такой как, например, pUC ori).
20. Плазмида по воплощению 18, в которой элементы (i)-(xiii) организованы по порядку экспрессии.
21. Плазмида по любому из воплощений 1-20, в которой последовательность ДНК плазмиды является такой, как изложено в SEQ ID NO 24, или по существу как изложено в SEQ ID NO 24.
22. Плазмида по воплощению 21, в которой в описанной в данном документе SEQ ID NO 24 делается несколько минорных модификаций, приводящих, например, к одной, двум, трем или четырем аминокислотным заменам в одном или более чем одном антигене и/или цитокине.
23. Плазмида по любому из воплощений 1-20, в которой последовательность ДНК плазмиды является такой, как изложено в SEQ ID NO:26 или модификации SEQ ID NO:26, приводящей, например, к одной, двум, трем или четырем аминокислотным заменам в одном или более чем одном антигене и/или цитокине, или модификации SEQ ID NO:26, которая приводит к экспрессии таких же полипептидных последовательностей, как и от SEQ ID NO:26.
24. Плазмида по любому из воплощений 1-20, в которой последовательность ДНК плазмиды является такой, как изложено в SEQ ID NO:26 или модификации SEQ ID NO:26, имеющей меньше, чем 100 оснований, которые отличаются от SEQ ID NO:26.
25. Плазмида по любому из воплощений 1-20, в которой последовательность ДНК плазмиды является такой, как изложено в SEQ ID NO:28 или модификации SEQ ID NO:28, приводящей, например, к одной, двум, трем или четырем аминокислотным заменам в одном или более чем одном антигене и/или цитокине, или модификации SEQ ID NO:28, которая приводит к экспрессии таких же полипептидных последовательностей, как и от SEQ ID NO:28.
26. Плазмида по любому из воплощений 1-20, в которой последовательность ДНК плазмиды является такой, как изложено в SEQ ID NO:28 или модификации SEQ ID NO:28, имеющей меньше, чем 100 оснований, которые отличаются от SEQ ID NO:28.
27. Плазмида по любому из воплощений 1-20, в которой последовательность ДНК плазмиды является такой, как изложено в SEQ ID NO:29 или модификации SEQ ID NO:29, приводящей, например, к одной, двум, трем или четырем аминокислотным заменам в одном или более чем одном антигене и/или цитокине, или модификации SEQ ID NO:29, которая приводит к экспрессии таких же полипептидных последовательностей, как и от SEQ ID NO:29.
28. Плазмида по любому из воплощений 1-20, в которой последовательность ДНК плазмиды является такой, как изложено в SEQ ID NO:29 или модификации SEQ ID NO:29, имеющей меньше, чем 100 оснований, которые отличаются от SEQ ID NO:29.
29. Плазмида по любому из воплощений 1-20, где указанная плазмида содержит ген TGF-β, содержащий SEQ ID NO:25 или SEQ ID NO:25, имеющую меньше, чем 10 замен оснований.
30. Плазмида по любому из воплощений 1-29 для применения в отсрочке или предупреждении диабета типа I.
31. Плазмида по любому из воплощений 1-30 для внутримышечного, внутрикожного, интраназального или подкожного введения.
32. Плазмида по воплощению 31 для подкожного введения.
33. Плазмида по воплощению 31 для внутримышечной инъекции.
34. Плазмида по любому из воплощений 1-33 для применения в лечении медицинского состояния у субъекта, такого как диабет типа I, диабет типа I с ранним началом или повышенный риск развития диабета типа I (включая состояния типа диабета типа 1,5).
35. Иммунотерапевтическая ДНК-вакцина, содержащая плазмиду по любому из воплощений 1-34.
36. Иммунотерапевтическая ДНК-вакцина по воплощению 35 для применения в отсрочке или предупреждении диабета типа I.
37. Иммунотерапевтическая ДНК-вакцина по любому из воплощений 35-36 для внутримышечного, внутрикожного, интраназального или подкожного введения.
38. Иммунотерапевтическая ДНК-вакцина по воплощению 37 для подкожного введения.
39. Иммунотерапевтическая ДНК-вакцина по воплощению 37 для внутримышечного введения.
40. Иммунотерапевтическая ДНК-вакцина по любому из воплощений 35-39, используемая в ассоциации с или параллельно с другими типами медицинских лечений, таких как, например, терапия бета-клетками/бета-стволовыми клетками, пересадка бета-клеток/бета-стволовых клеток и т.д. для продления выживания и эффективности пересаженных клеток.
41. Фармацевтическая композиция, содержащая иммунотерапевтическую ДНК-вакцину по любому из воплощений 34-39 или плазмиду по любому из воплощений 1-34, где указанная фармацевтическая композиция содержит физиологический раствор и/или буфер, и/или хелатор.
42. Фармацевтическая композиция, содержащая иммунотерапевтическую ДНК-вакцину по любому из воплощений 35-40 или плазмиду по любому из воплощений 1-34, где указанная фармацевтическая композиция содержит физиологический раствор и/или буфер, и/или хелатор, и/или этанол.
43. Фармацевтическая композиция по любому из воплощений 41-42, в которой объем/процентное содержание по объему этанола составляет меньше, чем 5%, меньше, чем 4%, меньше, чем 3%, меньше, чем 2% или меньше, чем 1%.
44. Фармацевтическая композиция по любому из воплощений 41-43, где указанная композиция не содержит какого-либо вируса, липидного соупаковочного агента или конденсирующего агента.
45. Фармацевтическая композиция по любому из воплощений 41-44, где указанная композиция дополнительно содержит агонист GLP-1R.
46. Фармацевтическая композиция по любому из воплощений 41-44, где указанная композиция дополнительно содержит аналог GLP-1/агонист GLP-1R.
47. Фармацевтическая композиция по любому из воплощений 45-46, где указанный аналог GLP-1 или указанный агонист GLP-1R выбран из лираглутида, семаглутида или их смеси.
48. Набор, содержащий фармацевтическую композицию по любому из воплощений 41-47 и фармацевтическую композицию, содержащую аналог GLP-1/агонист GLP-1R (например, лираглутид и/или семаглутид).
49. Способ получения плазмиды по любому из воплощений 1-34, где указанный способ включает (i) инкубирование клетки-хозяина, такой как клетка-хозяин бактериального происхождения, такой как, например, E. coli), трансфицированной указанной плазмидой, при подходящих условиях и (ii) выделение/очистка указанной плазмиды.
50. Способ по воплощению 49, где указанная клетка-хозяин представляет собой термочувствительный штамм E. coli infA.
51. Способ отсрочки начала диабета типа 1 (T1D) или его симптомов у пациента, подвергающегося риску развития T1D, или у которого недавно диагностирован T1D, причем указанный способ включает введение иммунотерапевтической ДНК-вакцины, содержащей плазмиду по любому из воплощений 1-31, возможно в комбинации с аналогом GLP-1/агонистом GLP-1R.
52. Способ сохранения функции бета-клеток и/или эндогенной продукции инсулина у индивида, включающий введение иммунотерапевтической ДНК-вакцины, содержащей плазмиду по любому из воплощений 1-34, возможно в комбинации с аналогом GLP-1/агонистом GLP-1R.
53. Способ лечения диабетика, включающий введение вакцины, содержащей плазмиду по любому из воплощений 1-34, возможно в комбинации с аналогом GLP-1/агонистом GLP-1R (например, лираглутидом и/или семаглутидом).
54. Вакцина для предупреждения или отсрочки начала симптомов диабета типа-1 (T1D) у пациента, подвергающегося риску развития, или у которого недавно диагностирован T1D, причем указанная вакцина содержит плазмиду по любому из воплощений 1-34.
55. Способ уменьшения дозировки инсулина у индивида, имеющего диабет типа-1 (T1D), или человека, подвергающегося риску развития T1D, включающий введение иммунотерапевтической ДНК-вакцины, содержащей плазмиду по любому из воплощений 1-33, возможно в комбинации с аналогом GLP-1/агонистом GLP-1R (например, лираглутидом и/или семаглутидом).
ПРИМЕРЫ
Не страдающие ожирением диабетические мыши (мышиная модель NOD диабета типа 1): иммунная функция в аутоиммунитете основывается на сложной сети клеточных взаимодействий, которые не могут быть адекватно оценены in vitro.
Оценки подавления и/или лечения заболевания в данном документе проводятся в мышиной модели NOD, данная модель представляет собой полигенную модель со спонтанным началом, где у большинства мышей развиваются повышенные концентрации глюкозы в крови (BVG - значение глюкозы в крови), определенное из использования укола в хвостовую вену и ручного измерителя) в возрасте от 12 до 30 недель. Заболеваемость и прогрессирование заболевания являются непредсказуемыми с общей заболеваемостью, варьирующей от 60% до 95% в возрасте 30 недель (WoA), и прогрессированием от постановки диагноза (два последовательных показания BVG больше 250) до смерти (два последовательных показания BVG 600 или выше), варьирующим от 2 суток до 4 недель. Повтор повышенных BVG при последовательных показаниях необходим, так как мышам дают пищу и воду без ограничений, что приводит к умеренной вариабельности BGV помимо той, которая вызвана иммунопатологией.
Примером нуклеотидной последовательности плазмиды в данном документе является:
SEQ ID NO 24: полная (неаннотированная) последовательность плазмиды (6401 пара оснований)
GACTCTTCGCGATGTACGGGCCAGATATACGCGTTGACATTGATTATTGACTAGTTATTAATAGTAATCAATTACGGGGTCATTAGTTCATAGCCCATATATGGAGTTCCGCGTTACATAACTTACGGTAAATGGCCCGCCTGGCTGACCGCCCAACGACCCCCGCCCATTGACGTCAATAATGACGTATGTTCCCATAGTAACGCCAATAGGGACTTTCCATTGACGTCAATGGGTGGACTATTTACGGTAAACTGCCCACTTGGCAGTACATCAAGTGTATCATATGCCAAGTACGCCCCCTATTGACGTCAATGACGGTAAATGGCCCGCCTGGCATTATGCCCAGTACATGACCTTATGGGACTTTCCTACTTGGCAGTACATCTACGTATTAGTCATCGCTATTACCATGGTGATGCGGTTTTGGCAGTACATCAATGGGCGTGGATAGCGGTTTGACTCACGGGGATTTCCAAGTCTCCACCCCATTGACGTCAATGGGAGTTTGTTTTGGCACCAAAATCAACGGGACTTTCCAAAATGTCGTAACAACTCCGCCCCATTGACGCAAATGGGCGGTAGGCGTGTACGGTGGGAGGTCTATATAAGCAGAGCTCTCTGGCTAACTAGAGAACCCACTGCTTACTGGCTTATCGAAATTAATACGACTCACTATAGGGAGACCCAAGCTGGCTAGCGTTTAAACTTAAGCTTGGTACCGAGCTCGGATCCACTAGTCCAGTGTGGTGGAATTCTGCACTGCAGCTCGCATCTCTCCTTCACGCGCCCGCCGCCCTACCTGAGGCCGCCATCCACGCCGGTTGAGTCGCGTTCTGCCGCCTCCCGCCTGTGGTGCCTCCTGAACTGCGTCCGCCGTCTAGGTAAGTTTAAAGCTCAGGTCGAGACCGGGCCTTTGTCCGGCGCTCCCTTGGAGCCTACCTAGACTCAGCCGGCTCTCCACGCTTTGCCTGACCCTGCTTGCTCAACTCTAGGTAAGTTAATGAGACAGATAGAAACTGGTCTTGTAGAAACAGAGTAGTCGCCTGCTTTTCTGCCAGGTGCTGACTTCTCTCCCCTGGGCTTTTTTCTTTTTCTCAGGTTGAAAAGAAGAAGACGAAGAAGACGAAGAAGACAAACCGTCGTCGACTGCCATGCGCCGCTGATTAACGCCGCCACCATGGCCCACCGACGCAGATCCAGAAGCTGCCGTGAGGACCAGAAGCCCGTGATGGATGATCAGAGGGACCTTATCTCTAACAATGAACAACTGCCAATGCTCGGCAGACGGCCTGGGGCCCCGGAGAGCAAGTGCAGCAGAGGAGCCTTGTACACGGGGTTCTCCATTTTAGTGACTCTCCTTCTCGCCGGCCAAGCTACCACCGCCTACTTTCTGTACCAACAGCAAGGCAGACTAGACAAACTGACAATCACAAGCCAGAACCTTCAGCTGGAGTCTCTGCGGATGAAGCTGCCCGCTTTGTGGATGAGATTGCTTCCTCTACTTGCTCTCCTGGCGCTCTGGGGACCTGACCCCGAGCAAGAGTTTGTTAATCAGCACCTGTGTGGGAGTCATCTGGTGGAGGCACTCTATTTAGTGTGCGGAGAGAGGGGCTTCTTCTACACTCCAAAGACCAGACGGGAGGCCGAAGACCTTCAAGTGGGGCAAGTAGAACTGGGTGGCGGACCCGGTGCCGGGAGCCTTCAGCCGCTCGCCCTGGAGGGCTCTCTTCAGAAACGCGGCATCGTGGAGCAGTGTTGCACATCCATTTGCTCACTCTACCAGCTGGAGAACTACTGCAACGGAAGCGGAGTGAAGCAGACGTTGAATTTTGATTTGTTGAAGTTGGCGGGGGATGTGGAGAGCAATCCGGGGCCGATGCCCCCTAGTGGCCTCAGACTTTTGTTATTGTTATTACCGCTTTTATGGCTCTTGGTGCTGACACCGGGCCGTCCGGCTGCTGGCTTGTCGACTTGTAAGACAATTGATATGGAATTGGTGAAACGAAAACGGATTGAGGCCATCCGAGGACAGATTTTGAGCAAGCTGCGGCTTGCCTCGCCACCCTCGCAAGGGGAAGTCCCACCCGGACCTCTACCAGAAGCAGTCCTAGCGCTGTACAACAGTACAAGAGATAGAGTGGCCGGGGAATCCGCAGAACCAGAGCCTGAGCCTGAAGCCGATTATTATGCAAAGGAAGTGACTAGGGTCCTGATGGTCGAGACCCATAACGAAATCTACGACAAATTCAAACAAAGTACCCACTCTATCTACATGTTCTTCAACACCAGTGAGCTAAGAGAAGCCGTGCCCGAACCTGTGCTTCTTTCCCGCGCAGAACTCCGCCTCTTGAGACTCAAATTGAAAGTTGAACAACACGTAGAGCTTTACCAGAAATACTCTAATAATTCATGGCGATATCTTTCTAATCGTCTCCTCGCCCCATCTGACAGCCCTGAATGGCTCTCCTTCGACGTTACGGGAGTTGTGCGCCAGTGGCTCAGCAGAGGCGGAGAGATAGAGGGCTTTCGGCTGAGCGCACATGTATCTGTGGACTCAAGGGACAACACATTGCAAGTGGATATTAACGGTTTTACAACTGGACGGAGAGGGGACCTGGCGACCATCCACGGCATGAATAGACCTTTCCTGCTGCTGATGGCTACTCCCCTGGAGAGGGCACAGCACTTACAGTCTTCCAGACACCGGCGCGCCCTGGATACAAACTACTGCTTCAGCTCCACCGAAAAGAACTGTTGCGTGCGGCAGCTGTACATTGACTTCAGAAAGGATCTGGGCTGGAAGTGGATTCATGAGCCCAAGGGGTATCATGCCAACTTCTGTCTTGGGCCATGCCCATACATCTGGTCACTGGATACCCAGTACTCCAAAGTTCTGGCCTTGTACAATCAACACAACCCTGGAGCTTCCGCCGCTCCTTGCTGTGTGCCCCAAGCCCTAGAGCCCCTGCCCATCGTTTATTATGTCGGACGCAAGCCCAAAGTAGAACAGCTATCAAATATGATCGTGAGAAGCTGCAAGTGTAGCTGATAAACGCGTCGAGCATGCATCTAGGGCGGCCAATTCCGCCCCTCTCCCCCCCACCCCTCTCCCTCCCCCCCCCCTAACGTTACTGGCCGAAGCCGCTTGGAATAAGGCCGGTGTGCGTTTGTCTATATGTTATTTTCCACCATATTGCCGTCTTTTGGCAATGTGAGGGCCCGGAAACCTGGCCCTGTCTTCTTGACGAGCATTCCTAGGGGTCTTTCCCCTCTCGCCAAAGGAATGCAAGGTCTGTTGAATGTCGTGAAGGAAGCAGTTCCTCTGGAAGCTTCTTGAAGACAAACAACGTCTGTAGCGACCCTTTGTAGACAGCGGAACCCCCCACCTGGCGATAGATGCCTCTGCGGCCAAAAGCCACGTGTATAAGATACACCTGCAAAGGCGGCACAACCCCAGTGCCACGTTGTGAGTTGGATAGTTGTGGAAAGAGTCAAATGGCTCTCCTCAAGCGTATTCAACAAGGGGCTGAAGGATGCCCAGAAGGTACCCCATTGTATGGGATCTGATCTGGGGCCTCGGTGCACATGCTTTACATGTGTTTAGTCGAGGTTAAAAAACGTCTAGGCCCCCCGAACCACGGGGACGTGGTTTTCCTTTGAAAAACACGATGATAATATGGCTGCCGCTCATTCTAGTGCCCTTCTTTGCTGCCTGGTCCTGCTCACCGGGGTGCGAGCTAGCCCTGGACAAGGGACACAATCCGAAAACTCGTGCACCCACTTCCCGGGCAACCTCCCTAACATGCTGAGGGACCTCCGTGATGCCTTCAGTAGAGTGAAGACGTTCTTCCAAATGAAAGATCAGTTAGATAACCTGCTCCTGAAGGAGTCACTCTTAGAAGACTTCAAAGGATACCTCGGCTGCCAAGCACTTAGCGAGATGATTCAATTCTACTTAGAAGAAGTCATGCCTCAAGCTGAGAATCAAGACCCCGACATCAAAGCTCATGTGAATTCTTTGGGAGAAAATTTGAAGACTTTGCGGCTGCGGCTGCGGAGATGTCACCGCTTTCTGCCCTGTGAGAACAAATCAAAAGCGGTCGAGCAAGTTAAGAATGCCTTCAATAAGCTACAAGAGAAGGGCATCTACAAAGCAATGAGCGAGTTTGATATCTTTATCAATTACATTGAAGCCTACATGACAATGAAGATTAGGAATGCCGCGGGGAGCGGCGCTACTAACTTCAGCCTGCTGAAGCAGGCTGGAGACGTGGAGGAGAACCCTGGACCTATGTACAGAATGCAGCTGCTGAGCTGCATCGCCCTGAGCCTGGCCCTGGTGACCAACAGCGCACCCACGTCCTCTAGCACCAAGAAGACCCAGTTACAGTTGGAGCATCTACTTTTAGACCTGCAAATGATTTTGAACGGCATCAACAACTACAAGAATCCTAAACTTACTCGCATGCTTACCTTCAAATTTTACATGCCCAAGAAGGCCACCGAACTGAAGCACTTGCAATGTCTGGAGGAAGAACTCAAGCCGCTGGAGGAAGTTCTCAACCTCGCGCAGTCCAAGAATTTCCACCTCCGGCCAAGAGACCTGATCAGTAACATTAATGTGATAGTGCTGGAGCTGAAGGGAAGCGAGACTACATTTATGTGCGAGTACGCCGATGAAACCGCTACAATCGTCGAGTTCCTGAATAGATGGATCACATTTTGCCAGTCAATTATCTCTACTCTGACATGATAACTCGAGGTCTAGAGGGCCCGTTTAAACCCGCTGATCAGCCTCGACTGTGCCTTCTAGTTGCCAGCCATCTGTTGTTTGCCCCTCCCCCGTGCCTTCCTTGACCCTGGAAGGTGCCACTCCCACTGTCCTTTCCTAATAAAATGAGGAAATTGCATCGCATTGTCTGAGTAGGTGTCATTCTATTCTGGGGGGTGGGGTGGGGCAGGACAGCAAGGGGGAGGATTGGGAAGACAATAGCAGGCATGCTGGGGATGCGGTGGGCTCTATGGCTTCTACTGGGCGGTTTTATGGACAGCAAGCGAACCGGAATTGCCAGCTGGGGCGCCCTCTGGTAAGGTTGGGAAGCCCTGCAAAGTAAACTGGATGGCTTTCTCGCCGCCAAGGATCTGATGGCGCAGGGGATCAAGCTCTGATCAAGAGACAGGATGAGGATCGTTTCGCATGGCCAAAGAAGACAATATTGAAATGCAAGGTACCGTTCTTGAAACGTTGCCTAATACCATGTTCCGCGTAGAGTTAGAAAACGGTCACGTGGTTACTGCACACATCTCCGGTAAAATGCGCAAAAACTACATCCGCATCCTGACGGGCGACAAAGTGACTGTTGAACTGACCCCGTACGACCTGAGCAAAGGCCGCATTGTCTTCCGTAGTCGCTGATAAATTATTAACGCTTACAATTTCCTGATGCGGTATTTTCTCCTTACGCATCTGTGCGGTATTTCACACCGCATACAGGTGGCACTTTTCGGGGAAATGTGCGCGGAACCCCTATTTGTTTATTTTTCTAAATACATTCAAATATGTATCCGCTCATGAGACAATAACCCTGATAAATGCTTCAATAATAGCACGTGCTAAAACTTCATTTTTAATTTAAAAGGATCTAGGTGAAGATCCTTTTTGATAATCTCATGACCAAAATCCCTTAACGTGAGTTTTCGTTCCACTGAGCGTCAGACCCCGTAGAAAAGATCAAAGGATCTTCTTGAGATCCTTTTTTTCTGCGCGTAATCTGCTGCTTGCAAACAAAAAAACCACCGCTACCAGCGGTGGTTTGTTTGCCGGATCAAGAGCTACCAACTCTTTTTCCGAAGGTAACTGGCTTCAGCAGAGCGCAGATACCAAATACTGTTCTTCTAGTGTAGCCGTAGTTAGGCCACCACTTCAAGAACTCTGTAGCACCGCCTACATACCTCGCTCTGCTAATCCTGTTACCAGTGGCTGCTGCCAGTGGCGATAAGTCGTGTCTTACCGGGTTGGACTCAAGACGATAGTTACCGGATAAGGCGCAGCGGTCGGGCTGAACGGGGGGTTCGTGCACACAGCCCAGCTTGGAGCGAACGACCTACACCGAACTGAGATACCTACAGCGTGAGCTATGAGAAAGCGCCACGCTTCCCGAAGGGAGAAAGGCGGACAGGTATCCGGTAAGCGGCAGGGTCGGAACAGGAGAGCGCACGAGGGAGCTTCCAGGGGGAAACGCCTGGTATCTTTATAGTCCTGTCGGGTTTCGCCACCTCTGACTTGAGCGTCGATTTTTGTGATGCTCGTCAGGGGGGCGGAGCCTATGGAAAAACGCCAGCAACGCGGCCTTTTTACGGTTCCTGGGCTTTTGCTGGCCTTTTGCTCACATGTTCTT
Пример 1 - плазмиды, кодирующие антиген, по сравнению с плазмидами, кодирующими антиген плюс IL-10
В предыдущем уровне техники было предложено то, что обеднение иммуностимулирующими последовательностями CpG в остове плазмиды потребовалось бы для эффективного ДНК-иммунотерапевтического лечения T1D. Данный эксперимент, таким образом, был смоделирован после опубликованных ранее экспериментов (2008 J Immunol. 181(12):8298-307).
Мышам NOD давали восемь еженедельных доз плазмиды, начиная с недели 9 (возраст): давали либо пустой вектор (pVAX1, 50 мкг), либо pVAX1- проинсулиновый Ag (антиген) (не нацеленный в эндосомы, не препроинсулин), либо обедненный CpG pVAX1-проинсулиновый Ag, либо бицистронную конструкцию pVAX1-IL10-IRES-проинсулиновый Ag в эквимолярных отношениях.
Все введения были внутримышечными в левый квадрицепс под изофлурановой анестезией и содержали только плазмиду в PBS плюс EDTA. BGV оценивали у всех мышей на еженедельной основе, и заболеваемость диабетом типа 1 подсчитывали на основе двух показаний BGV больше 250 мг/дл. Мышей оценивали до возраста 30 недель или до достижения BGV 600, с последующим умерщвлением.
Результаты из данного эксперимента (таблица 1) демонстрируют то, что А) обеднение CpG не является ни необходимым, ни полезным для эффективности, Б) включение иммуномодулирующих цитокинов значимо увеличивает эффективность, и В) остов плазмиды (пустой вектор) является эквивалентным необработанным группам.
Таблица 1: заболеваемость T1D у мышей NOD в возрасте 30 недель
Пример 2 - экспрессируемые белковые продукты, образующиеся от плазмид, кодирующих антиген, IL-10, IL-2 и TGF-β
Создавали мультицистронные плазмиды для соэкспрессии TGF-β, IL-10 и, возможно, IL-2. Клетки Freestyle293 временно трансфицировали и культивировали в бессывороточных средах. Супернатанты отбирали и подвергали количественному измерению ELISA (твердофазный иммуноферментный анализ) через 72 часа.
Результаты в Таблице 2 ниже показывают то, что: А) от одного вектора достигается экспрессия многих независимых цитокинов, Б) продуцируются значимые количества каждого цитокина и в ожидаемых соотношениях, В) небольшие изменения последовательности значимо улучшают экспрессию IL-10 от первого поколения плазмиды IL10/проинсулин и Г) ни остов плазмиды (пустой вектор), ни эндосомальное нацеливание антигена (IIAg) не индуцирует продукцию или неправильную регуляцию цитокина.
Таблица 2: количественное измерение ELISA экспрессируемых белковых продуктов
Пример 3 - влияние TGF-β и IL-2 на подавление заболевания
Мультицистронные плазмиды оценивали в отношении предупреждения заболевания у мышей NOD как и в Примере 1, за исключением того, что дозирование продолжали один раз в неделю до умерщвления (начало диабета) или до недели 30. Одну мышь из каждой группы (исходно n равно 24) высылали для полной некропсии после 10 недель дозирования - включающей патологию на 10 стандартных перфузируемых тканях, полный анализ крови и клиническую химию. Кроме слабого разрушения мышцы и повторного роста, обусловленных механической травмой в месте инъекции, не было отклонений от животных, не подвергавшихся дозированию.
Результаты в Таблице 3, приведенной ниже, показывают то, что: А) добавление TGFb значительно увеличивает эффективность, Б) включение интерлейкина-2 может увеличивать эффективность и не индуцирует патологию, В) хроническое дозирование плазмид, экспрессирующих IL-10 и антиген, увеличивает эффективность предупреждения заболевания и Г) хроническое дозирование плазмид, экспрессирующих TGFβ, IL-10 и IL-2, увеличивает эффективность, не приводя к каким-либо сигналам, связанным с безопасностью.
Таблица 3: заболеваемость T1D у мышей NOD.
Пример 4. Оценка элементов IRES, интронов, а также подкожного введения
Мультицистронные плазмиды оценивали на предупреждение заболевания у мешей NOD так же, как и в Примере 3, за исключением того, что дозирование начиналось раньше (в неделю 5) для того, чтобы лучше имитировать хроническое педиатрическое введение. Помимо подтверждения содержащих интроны плазмид pVAX1-IIAg/TGFβ/IL10 и pVAX1-IIAg/TGFβ/IL10/IL2, проверяли другие контрольные группы. В частности, на предмет ожидаемых увеличений эффективности оценивали другой сегмент IRES (CrPV [из вируса паралича сверчка], в отличие от EMCV [из вируса энцефаломиокардита]), также как и делецию сегмента интрона для оценки его необходимости. Из-за очевидного отсутствия эффективности по сравнению с родительской плазмидой (pVAX1-IIAg/TGFβ/IL10/IL2) исследования групп CrPV и не содержащих интрон (н.и. - нет интрона) прекращали рано. Кроме того, когорта мышей, использованных в данном эксперименте, испытывала более быстрое прогрессирование заболевания, чем предыдущие когорты, со временем от постановки диагноза до умерщвления, составляющим в среднем 1,25 недели, а не 2,75 из предыдущих экспериментов. Наконец, добавляли группу с подкожным введением. Данной группе дозировали плазмиду для трех цитокинов (pVAX1-IIAg/TGFβ/IL10/IL2) с инъекцией один раз в неделю в п.к. (подкожное) пространство в загривок шеи без анестезии.
Результаты в Таблице 4 демонстрируют то, что: А) элементы IRES EMCV обеспечивают значительно лучшую эффективность, чем IRES CrPV, Б) включение интрона (в данной плазмиде расположен в пределах области эндосомального нацеливания CD74) значительно увеличивает эффективность, В) в то время как включение IL-2 дает минимальную пользу в условиях легкого заболевания, его присутствие значительно увеличивает эффективность и надежность лечения в условиях агрессивного заболевания, и Г) подкожное дозирование, которое является неэффективным при большинстве применений ДНК-вакцин, здесь демонстрирует умеренную эффективность и значительную задержку заболевания даже без оптимизации.
Таблица 4: заболеваемость T1D у мышей NOD
Пример 5. Сравнение имеющихся в продаже систем селекции без антибиотика с селекцией с антибиотиком
Был оценен альтернативный остов плазмиды с целью удаления устойчивости к канамицину для соответствия руководству Европейского агенства по оценке лекарственных средств. Такую же вставку (IIAg/TGFβ/IL10/IL2, включающую интрон) клонировали в «наноплазмидный» остов NTC9385R Nature Technology. Образующуюся плазмиду оценивали в мышах NOD так же, как и в Примере 3, за исключением того, что лечение начиналось в неделю 11 (позднее начало) и рано заканчивалось из-за неуспеха плазмиды на основе NTC9385R.
Результаты в Таблице 5 ниже показывают то, что: А) изменения в отношении системы селекции остова плазмиды неожиданно индуцируют значительные изменения эффективности плазмиды и Б) позднее начало в отношении лечения приводит к ранним превращениям. Данные из других, связанных экспериментов показывают то, что дозирование с использованием данных плазмид толерогенной ДНК-вакцины требует от двух до четырех недель для того, чтобы иметь эффективность, таким образом, что позднее начало в отношении лечения приводит к нескольким ранним случаям диабета перед тем, как лечение станет эффективным.
Таблица 5: заболеваемость T1D у мышей NOD
Пример 6. Эффективность подавления заболевания с использованием плазмид с антигеном и без него
Для определения роли закодированного антигена в функции плазмиды были проведены два эксперимента (Примеры 6 и 7). Альтернативную плазмиду оценивали с целью удаления области, кодирующей антиген (пре-проинсулин), при сохранении нацеливающего домена CD74 и всех трех секретируемых цитокинов. Образующуюся плазмиду оценивали в мышах NOD так же, как в Примере 3, за исключением того, что лечение начиналось в неделю 11 (позднее начало).
Данный эксперимент демонстрирует то, что для полной эффективности требуется антигенная часть, и что не просто продукция цитокинов управляет функцией плазмиды. Это один из двух критериев, необходимых для демонстрации антигенспецифичности лечения.
Таблица 6: заболеваемость T1D у мышей NOD
Пример 7. Влияние описанной в данном документе иммунотерапии антигеном на эффективность неродственных антигенных вакцин
Для определения роли закодированного антигена в функции плазмиды были проведены два эксперимента (Примеры 6 и 7). У мышей NOD либо проводили имитацию лечения с использованием инъекции PBS, либо лечили их плазмидой pVAX1-IIAg/TGFβ/IL10/IL2 так же, как и в Примере 3. После четырех доз (т.е. в возрасте 13 недель) каждую мышь в.б. (внутрибрюшинно) иммунизировали 50 мкг иррелевантного антигена (овальбумин курицы, OVA) в 100 мкл суспензии квасцов 1:1. Имитацию лечения или лечение плазмидами продолжали один раз в неделю до умерщвления через три недели (21 сутки) после иммунизации, причем в данное время отбирали сыворотку. Определяли антитела с переключением класса (общие IgG и IgG2a) против овальбуминового антигена посредством имеющихся в продаже наборов ELISA (твердофазный иммуноферментный анализ). Не наблюдали значимых различий между группами, которых лечили плазмидой, или с имитацией лечения по их общим уровням IgG против OVA, ни одна из групп не продуцировала IgG2a проив OVA.
Результаты в Таблице 7 ниже показывают то, что в то время как плазмида подавляет иммунные ответы, связанные с заболеванием-мишенью, она не подавляет иммунореактивность по отношению к неродственным антигенам (т.е. любым антигенам, не кодируемым данной плазмидой). Это является вторым из двух критериев, необходимых для демонстрации антигенспецифичности лечения. Поскольку лечение педиатрических пациентов будет включать сопутствующее введение стандартных детских вакцин, это является значительным преимуществом над системной / общей иммунодепрессией посредством таких агентов, как метотрексат или циклоспорин А.
Таблица 7: ответ на иррелевантный антиген у мышей NOD, которые получали иммунотерапевтическую ДНК-вакцинацию против T1D
Данные значения приводят к незначимому значению р 0,377 и доверительному интервалу от -1,99 до 4,87. Данные результаты показывают то, что лечение иммуномодулирующей плазмидой не влияет на иммунный ответ на другие антигены, не кодируемые плазмидой и, следовательно, не приводит к широкой или системной иммунодепрессии.
Пример 8. Индивидуальные белковые продукты, экспрессируемые от плазмиды
Элемент 2А TaV приводил в данном документе к неожиданным слитым продуктам IL-10 плюс IL-2 (данные не показаны), и, следовательно, были оценены другие стратегии разделения. Исходные методики разделения включали расположенные выше удлинения последовательности 2А TaV (приводящие к быстрой деградации и отсутствию секретированного IL-10) и также сайт расщепления карбоксипептидазой (который индуцировал гибель линий трансфицированных клеток). Дополнительными оцененными стратегиями разделения были GSG-2A TaV, фуриновый сайт расщепления, фуриновый сайт, с последующим 2А TaV, 2A P и 2А Е (вирус ринита лошади А).
Клетки Freestyle293 временно трансфицировали и культивировали в бессывороточной среде. И клеточные осадки, и супернатанты отбирали и подвергали полуколичественному многоцветному вестерн-блоттингу через 72 часа.
Результаты в Таблице 8, приведенной ниже, показывают то, что: А) неожиданно, сайты протеолитического расщепления между генами IL-10 и IL-2 не могут функционировать, Б) метки GSG (разобщающие последовательности) между IL-10 и IL-2 являются предпочтительными по отношению к удлиненным изолирующим последовательностям, В) 2А Р является предпочтительной по отношению либо к 2А TaV, либо к 2А Е и Г) последовательности 2А могут иметь значительные и неожиданные влияния на деградацию и секрецию экспрессируемых, расположенных выше белков, таких как IL-10.
Таблица 8: разделение экспрессированных белковых продуктов IL-10 и IL-2
Пример 9. Сравнение имеющейся в продаже системы селекции с термочувствительной системой селекции, предложенной в данном документе, а также сравнение между плазмидами, кодирующими IL-2, и плазмидами, не кодирующими IL-2 (подкожное введение)
Остовы плазмид создавали и оценивали с целью удаления устойчивости к канамицину для соответствия руководству Европейского агентсва по оценке лекарственных средств. Откорректированную вставку (IIAg/GSG-2A FMDV /TGFβ/ IRES EMCV/IL10/GSG-2A P/IL2, включающую интрон в расположенной выше некодирующей области) клонировали либо в модифицированный/минимально модифицированный вектор pVAX1, содержащий селективный маркер «RNA-OUT» от Nature Technology, либо эквивалентный минимально модифицированный вектор pVAX1, кодирующий infA дикого типа («pNN»), в качестве остовов. Кроме того, получали плазмиды либо содержащие дополнительный энхансерный элемент SV40, либо дефицитные по IL-2. Образующиеся плазмиды оценивали у мышей NOD также, как и в Примере 3, за исключением того, что введение было п.к. либо один раз в неделю, либо три раза в неделю (предпочтительно).
Результаты, показанные в Таблице 9 и 10 ниже, демонстрируют то, что: А) имеющаяся в продаже замена RNA-OUT на устойчивость к антибиотику канамицину в остове pVAX1 все еще неожиданно недостаточно эффективно работает, Б) комплементация infA системы селекции без антибиотика работает эквивалентно родительскому вектору pVAX1, В) интерлейкин-2 требуется для оптимальной эффективности, Г) добавление энхансерного элемента SV40 не улучшает эффективность и Д) откорректированная тройная цитокиновая вставка сохраняет полную функциональность.
Таблица 9: заболеваемость T1D у мышей NOD
30 недель
Таблица 10: заболеваемость T1D у мышей NOD
Пример 10. Проверка надежности эффекта толерантности после прекращения введения плазмиды
В предыдущем эксперименте (представленном в Таблице 9) группу IIAg/FMDV/TGFβ/IL10/2A P/IL2 не умерщвляли в возрасте 30 недель, но прекращали дозирование плазмиды. Значения глюкозы в крови отслеживали в течение дополнительных десяти (10) недель до общего возраста 40 недель для оценки того, индуцировала ли плазмида надежное состояние толерантности, или было ли необходимым для эффективности непрерывное дозирование.
Результаты, показанные в Таблице 11 ниже, указывают на то, что непрерывное дозирование требуется для надежности толерантности, так как стабильное состояние без заболевания до возраста 30 недель быстро ухудшалось после прерывания дозирования. Это указывает на полезный профиль безопасности, так как также ожидалось бы то, что любые вредные события, с которыми можно сталкиваться при дозировании плазмиды, прекращались бы с дозированием.
Таблица 11: заболеваемость T1D у мышей NOD после прекращения дозирования плазмиды
Пример 11. Проверка стабильности и надежности плазмиды при инъекции
Ключевым вопросом с введением плазмиды является деградация при введении. В случае инъекции силы сдвига, с которыми сталкиваются большие и вязкие молекулы плазмиды, проходящие через тонкую иглу под давлением, приводят к разрушению ковалентно замкнутой кольцевой структуры плазмиды, делая ее линейной и подверженной как пониженной способности к трансфекции, так и быстрой деструкции. Большинство плазмид имеют от 5 до 15% деградации до линейных форм при инъекции через иглы размеров, приемлемых для клинического применения, что приводит либо к пониженной эффективности, либо к необходимости больших исходных доз для компенсации потери. Несколько типов структур последовательности, которые могут приводить к разворачиванию плазмиды и чувствительности к деградации под напряжением сдвига, были преднамеренно минимизированы в раскрытых плазмидах с целью увеличения прочности и надежности при использовании протоколов инъекции. Для того чтобы оценить деградацию плазмиды под напряжением сдвига, которая может варьировать с изменением вязкости и, следовательно, концентрации, лидирующую плазмиду человека ресуспендировали в буфере Tris EDTA до концентраций 5, 7 и 9 мг/мл и три раза пропускали через иглу G30 (выталкивали, повторно втягивали в шприц, затем повторно выталкивали), и один (1) микрограмм образцов разделяли на агарозном геле относительно контрольных образцов, которые не проходили процесс инъекции.
Результаты, показанные на Фиг. 3, неожиданно показывают то, что плазмида заметно не деградирует посредством трех инъекционных пропусканий при любой протестированной концентрации или вязкости. Деградация плазмиды визуализировалась бы и как размазывание меньших полос (между главной суперспиральной полосой при 6 т.п.н. и меньшей полосой примесей процесса внизу геля или грубо при 600 п.н.). Такие мазки линеаризации / деградации не видны для любого образца, пропускаемого через процесс инъекции. Эта надежная физическая стабильность при дозировании является весьма желательной и большей, чем предполагаемая или описанная ранее в литературе.
Пример 12. Подтверждение сохранения плазмиды с использованием системы комплементации infA
Для того чтобы подтвердить то, что система селекции сохранения плазмиды на основе infA функционировала как желательно, бактерии, трансформированные плазмидой, выращивали на протяжении 100 пассажей (грубо 36 удвоений/поколений на пассаж, в течение всего 3600 поколений проверяли потенциальный дрейф или потерю плазмиды). Пассажи 1-100 получали по 11 в неделю - по 2 пассажа в будний день при 37°C и один в каждый выходной при 30°C. Все проводили в жидкой среде LB, не содержащей животных продуктов (сойтон Teknova), дополненной 15 микрограммами/мл наладиксовой кислоты (селекция на штамм на основе DH5a, не на присутствие плазмиды). Из каждого пассажа получали глицериновые маточные растворы и сохраняли, пока не было получено 100 пассажей для сопутствующего процессинга.
Соскобы глицериновых маточных растворов использовали для инокуляции 5 мл выращеных в течение ночи культур, которые перерабатывали посредством инструкций поставщика на наборах для минипрепаратов от Qiagen с использованием многократного вакуума (либо 16, либо 32 культуры на прогон, из- за ограничений размера геля). Не делали попыток получения показаний ОП600 (оптическая плотность при 600 нм) для нормирования введения клеток, и все препараты получали на основе стандартных объемов. Один микролитр каждого минипрепарата подвергали расщеплению PstI/Xhol для отделения остова (приблизительно 2,4 т.п.н.) от вставки (приблизительно 4 т.п.н.) без поправки на концентрацию плазмиды, получающуюся из каждого минипрепарата. Разделение на каждом геле проводили с фланкирующими лестницами Tridye 2-Log (NEB https://www.neb.com/products/n3200-2-log-dna-ladder-01-100-kb), первой полосой образца - нерасщепленной плазмидой, и визуализировали с использованием красителя SybrSafe. На изображениях гелей, несмотря на отсутствие контроля качества нуклеиновых кислот, все полосы продуктов расщепления демонстрируют и присутствие, и ожидаемую картину расщепления для плазмиды (видна на изображениях для пассажей 1-16, 17-48, 49-80 и 81-100).
В качестве дополнительного подтверждения глицериновые маточные растворы для пассажей 1-100 также вводили в качестве внутреннего стандарта на 50 сектор чашек с LB агаром, не содержащим антибиотик и животных продуктов, и инкубировали в течение ночи при 30°C. Не делали попытки контроля инокулята внутреннего стандарта. Как показано на Фиг. 4, все репрезентативные внутренние стандарты на основе глицериновых маточных растворов приводили к заметному росту и, таким образом, сохранению плазмиды.
Пример 13. Пригодность для увеличения масштаба с использованием системы комплементации infA
Для того чтобы подтвердить то, что система селекции сохранения плазмиды на основе infA функционировала как желательно в производственном масштабе, бактерии, трансформированные плазмидой, использовали в 50 л пилотном ферментере с периодической загрузкой, работающем со специфической стадией сдвига температуры, увеличивающей выход. Использовали минимальную среду с добавлением дрожжевого экстракта, уменьшая скорость удвоения до 0,88/час. Подпитку начинали в 17 ч 00 мин после инокуляции, и регуляцию растворенного кислорода на уровне 30% осуществляли последовательным увеличением каскадных параметров рО2 (перемешивание в 32 ч 15 мин, приложение давления в 40 ч 30 мин, затем ток воздуха в 45 ч 40 мин). Скорость увеличения биомассы ослабевала немедленно после сдвига до 42°C, как и ожидалось. Количество продуцированной плазмидной ДНК было оценено на уровне 1,03 плюс/минус 0,17 г/л с использованием методики экстракции плазмиды в малом масштабе, имитирующей немедленный выход после лизиса.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Ново Ноордиск А/С
<120> Толерогенная ДНК-вакцина
<130> 150042WO01
<150> US 62/415717
<151> 2016-11-01
<150> EP 17150037.4
<151> 2017-01-02
<150> EP 17169019.1
<151> 2017-05-02
<150> EP 17177289.0
<151> 2017-06-22
<150> EP 17198041.0
<151> 2017-10-24
<160> 29
<170> PatentIn version 3.5
<210> 1
<211> 86
<212> ПРТ
<213> mus musculus
<400> 1
Phe Val Asn Gln His Leu Cys Gly Ser His Leu Val Glu Ala Leu Tyr
1 5 10 15
Leu Val Cys Gly Glu Arg Gly Phe Phe Tyr Thr Pro Lys Thr Arg Arg
20 25 30
Glu Ala Glu Asp Leu Gln Val Gly Gln Val Glu Leu Gly Gly Gly Pro
35 40 45
Gly Ala Gly Ser Leu Gln Pro Leu Ala Leu Glu Gly Ser Leu Gln Lys
50 55 60
Arg Gly Ile Val Glu Gln Cys Cys Thr Ser Ile Cys Ser Leu Tyr Gln
65 70 75 80
Leu Glu Asn Tyr Cys Asn
85
<210> 2
<211> 86
<212> ПРТ
<213> homo sapiens
<400> 2
Phe Val Asn Gln His Leu Cys Gly Ser His Leu Val Glu Ala Leu Tyr
1 5 10 15
Leu Val Cys Gly Glu Arg Gly Phe Phe Tyr Thr Pro Lys Thr Arg Arg
20 25 30
Glu Ala Glu Asp Leu Gln Val Gly Gln Val Glu Leu Gly Gly Gly Pro
35 40 45
Gly Ala Gly Ser Leu Gln Pro Leu Ala Leu Glu Gly Ser Leu Gln Lys
50 55 60
Arg Gly Ile Val Glu Gln Cys Cys Thr Ser Ile Cys Ser Leu Tyr Gln
65 70 75 80
Leu Glu Asn Tyr Cys Asn
85
<210> 3
<211> 110
<212> ПРТ
<213> синтетическая
<400> 3
Met Ala Leu Trp Met Arg Leu Leu Pro Leu Leu Ala Leu Leu Ala Leu
1 5 10 15
Trp Gly Pro Asp Pro Glu Gln Glu Phe Val Asn Gln His Leu Cys Gly
20 25 30
Ser His Leu Val Glu Ala Leu Tyr Leu Val Cys Gly Glu Arg Gly Phe
35 40 45
Phe Tyr Thr Pro Lys Thr Arg Arg Glu Ala Glu Asp Leu Gln Val Gly
50 55 60
Gln Val Glu Leu Gly Gly Gly Pro Gly Ala Gly Ser Leu Gln Pro Leu
65 70 75 80
Ala Leu Glu Gly Ser Leu Gln Lys Arg Gly Ile Val Glu Gln Cys Cys
85 90 95
Thr Ser Ile Cys Ser Leu Tyr Gln Leu Glu Asn Tyr Cys Asn
100 105 110
<210> 4
<211> 110
<212> ПРТ
<213> синтетическая
<400> 4
Met Ala Leu Trp Met Arg Leu Leu Pro Leu Leu Ala Leu Leu Ala Leu
1 5 10 15
Trp Gly Pro Asp Pro Glu Gln Glu Phe Val Asn Gln His Leu Cys Gly
20 25 30
Ser His Leu Val Glu Ala Leu Tyr Leu Val Cys Gly Glu Arg Gly Phe
35 40 45
Phe Tyr Thr Pro Lys Thr Arg Arg Glu Ala Glu Asp Leu Gln Val Gly
50 55 60
Gln Val Glu Leu Gly Gly Gly Pro Gly Ala Gly Ser Leu Gln Pro Leu
65 70 75 80
Ala Leu Glu Gly Ser Leu Gln Lys Arg Gly Ile Val Glu Gln Cys Cys
85 90 95
Thr Ser Ile Cys Ser Leu Tyr Gln Leu Glu Asn Tyr Cys Asn
100 105 110
<210> 5
<211> 109
<212> ПРТ
<213> mus musculus
<400> 5
Ala Leu Trp Met Arg Leu Leu Pro Leu Leu Ala Leu Leu Ala Leu Trp
1 5 10 15
Gly Pro Asp Pro Ala Gln Ala Phe Val Asn Gln His Leu Cys Gly Ser
20 25 30
His Leu Val Glu Ala Leu Tyr Leu Val Cys Gly Glu Arg Gly Phe Phe
35 40 45
Tyr Thr Pro Lys Thr Arg Arg Glu Ala Glu Asp Leu Gln Val Gly Gln
50 55 60
Val Glu Leu Gly Gly Gly Pro Gly Ala Gly Ser Leu Gln Pro Leu Ala
65 70 75 80
Leu Glu Gly Ser Leu Gln Lys Arg Gly Ile Val Glu Gln Cys Cys Thr
85 90 95
Ser Ile Cys Ser Leu Tyr Gln Leu Glu Asn Tyr Cys Asn
100 105
<210> 6
<211> 110
<212> ПРТ
<213> homo sapiens
<400> 6
Met Ala Leu Trp Met Arg Leu Leu Pro Leu Leu Ala Leu Leu Ala Leu
1 5 10 15
Trp Gly Pro Asp Pro Ala Gln Ala Phe Val Asn Gln His Leu Cys Gly
20 25 30
Ser His Leu Val Glu Ala Leu Tyr Leu Val Cys Gly Glu Arg Gly Phe
35 40 45
Phe Tyr Thr Pro Lys Thr Arg Arg Glu Ala Glu Asp Leu Gln Val Gly
50 55 60
Gln Val Glu Leu Gly Gly Gly Pro Gly Ala Gly Ser Leu Gln Pro Leu
65 70 75 80
Ala Leu Glu Gly Ser Leu Gln Lys Arg Gly Ile Val Glu Gln Cys Cys
85 90 95
Thr Ser Ile Cys Ser Leu Tyr Gln Leu Glu Asn Tyr Cys Asn
100 105 110
<210> 7
<211> 15
<212> ПРТ
<213> homo sapiens
<400> 7
Ser His Leu Val Glu Ala Leu Tyr Leu Val Cys Gly Glu Arg Gly
1 5 10 15
<210> 8
<211> 15
<212> ПРТ
<213> синтетическая
<400> 8
Ser His Leu Val Glu Ala Leu Tyr Leu Val Cys Gly Glu Glu Gly
1 5 10 15
<210> 9
<211> 81
<212> ПРТ
<213> mus musculus
<400> 9
Met Asp Asp Gln Arg Asp Leu Ile Ser Asn His Glu Gln Leu Pro Ile
1 5 10 15
Leu Gly Asn Arg Pro Arg Glu Pro Glu Arg Cys Ser Arg Gly Ala Leu
20 25 30
Tyr Thr Gly Val Ser Val Leu Val Ala Leu Leu Leu Ala Gly Gln Ala
35 40 45
Thr Thr Ala Tyr Phe Leu Tyr Gln Gln Gln Gly Arg Leu Asp Lys Leu
50 55 60
Thr Ile Thr Ser Gln Asn Leu Gln Leu Glu Ser Leu Arg Met Lys Leu
65 70 75 80
Pro
<210> 10
<211> 98
<212> ПРТ
<213> homo sapiens
<400> 10
Met His Arg Arg Arg Ser Arg Ser Cys Arg Glu Asp Gln Lys Pro Val
1 5 10 15
Met Asp Asp Gln Arg Asp Leu Ile Ser Asn Asn Glu Gln Leu Pro Met
20 25 30
Leu Gly Arg Arg Pro Gly Ala Pro Glu Ser Lys Cys Ser Arg Gly Ala
35 40 45
Leu Tyr Thr Gly Phe Ser Ile Leu Val Thr Leu Leu Leu Ala Gly Gln
50 55 60
Ala Thr Thr Ala Tyr Phe Leu Tyr Gln Gln Gln Gly Arg Leu Asp Lys
65 70 75 80
Leu Thr Ile Thr Ser Gln Asn Leu Gln Leu Glu Ser Leu Arg Met Lys
85 90 95
Leu Pro
<210> 11
<211> 219
<212> ДНК
<213> E. coli
<400> 11
atggccaaag aagacaatat tgaaatgcaa ggtaccgttc ttgaaacgtt gcctaatacc 60
atgttccgcg tagagttaga aaacggtcac gtggttactg cacacatctc cggtaaaatg 120
cgcaaaaact acatccgcat cctgacgggc gacaaagtga ctgttgaact gaccccgtac 180
gacctgagca aaggccgcat tgtcttccgt agtcgctga 219
<210> 12
<211> 72
<212> ПРТ
<213> E. coli
<400> 12
Met Ala Lys Glu Asp Asn Ile Glu Met Gln Gly Thr Val Leu Glu Thr
1 5 10 15
Leu Pro Asn Thr Met Phe Arg Val Glu Leu Glu Asn Gly His Val Val
20 25 30
Thr Ala His Ile Ser Gly Lys Met Arg Lys Asn Tyr Ile Arg Ile Leu
35 40 45
Thr Gly Asp Lys Val Thr Val Glu Leu Thr Pro Tyr Asp Leu Ser Lys
50 55 60
Gly Arg Ile Val Phe Arg Ser Arg
65 70
<210> 13
<211> 127
<212> ДНК
<213> L. monocytogenes
<400> 13
tgtaaaaaac atcatttagc gtgactttct ttcaacagct aacaattgtt gttactgcct 60
aatgttttta gggtatttta aaaaagggcg ataaaaaacg attgggggat gagaaatgaa 120
cgctcaa 127
<210> 14
<211> 10
<212> ДНК
<213> синтетическая
<400> 14
gccrccatgg 10
<210> 15
<211> 24
<212> ПРТ
<213> синтетическая
<400> 15
Ala Pro Val Lys Gln Thr Leu Asn Phe Asp Leu Leu Lys Leu Ala Gly
1 5 10 15
Asp Val Glu Ser Asn Pro Gly Pro
20
<210> 16
<211> 20
<212> ПРТ
<213> Thosea asigna
<400> 16
Arg Ala Glu Gly Arg Gly Ser Leu Leu Thr Cys Gly Asp Val Glu Glu
1 5 10 15
Asn Pro Gly Pro
20
<210> 17
<211> 19
<212> ПРТ
<213> синтетическая
<400> 17
Ala Thr Asn Phe Ser Leu Leu Lys Gln Ala Gly Asp Val Glu Glu Asn
1 5 10 15
Pro Gly Pro
<210> 18
<211> 390
<212> ПРТ
<213> homo sapiens
<400> 18
Met Pro Pro Ser Gly Leu Arg Leu Leu Leu Leu Leu Leu Pro Leu Leu
1 5 10 15
Trp Leu Leu Val Leu Thr Pro Gly Arg Pro Ala Ala Gly Leu Ser Thr
20 25 30
Cys Lys Thr Ile Asp Met Glu Leu Val Lys Arg Lys Arg Ile Glu Ala
35 40 45
Ile Arg Gly Gln Ile Leu Ser Lys Leu Arg Leu Ala Ser Pro Pro Ser
50 55 60
Gln Gly Glu Val Pro Pro Gly Pro Leu Pro Glu Ala Val Leu Ala Leu
65 70 75 80
Tyr Asn Ser Thr Arg Asp Arg Val Ala Gly Glu Ser Ala Glu Pro Glu
85 90 95
Pro Glu Pro Glu Ala Asp Tyr Tyr Ala Lys Glu Val Thr Arg Val Leu
100 105 110
Met Val Glu Thr His Asn Glu Ile Tyr Asp Lys Phe Lys Gln Ser Thr
115 120 125
His Ser Ile Tyr Met Phe Phe Asn Thr Ser Glu Leu Arg Glu Ala Val
130 135 140
Pro Glu Pro Val Leu Leu Ser Arg Ala Glu Leu Arg Leu Leu Arg Leu
145 150 155 160
Lys Leu Lys Val Glu Gln His Val Glu Leu Tyr Gln Lys Tyr Ser Asn
165 170 175
Asn Ser Trp Arg Tyr Leu Ser Asn Arg Leu Leu Ala Pro Ser Asp Ser
180 185 190
Pro Glu Trp Leu Ser Phe Asp Val Thr Gly Val Val Arg Gln Trp Leu
195 200 205
Ser Arg Gly Gly Glu Ile Glu Gly Phe Arg Leu Ser Ala His Cys Ser
210 215 220
Cys Asp Ser Arg Asp Asn Thr Leu Gln Val Asp Ile Asn Gly Phe Thr
225 230 235 240
Thr Gly Arg Arg Gly Asp Leu Ala Thr Ile His Gly Met Asn Arg Pro
245 250 255
Phe Leu Leu Leu Met Ala Thr Pro Leu Glu Arg Ala Gln His Leu Gln
260 265 270
Ser Ser Arg His Arg Arg Ala Leu Asp Thr Asn Tyr Cys Phe Ser Ser
275 280 285
Thr Glu Lys Asn Cys Cys Val Arg Gln Leu Tyr Ile Asp Phe Arg Lys
290 295 300
Asp Leu Gly Trp Lys Trp Ile His Glu Pro Lys Gly Tyr His Ala Asn
305 310 315 320
Phe Cys Leu Gly Pro Cys Pro Tyr Ile Trp Ser Leu Asp Thr Gln Tyr
325 330 335
Ser Lys Val Leu Ala Leu Tyr Asn Gln His Asn Pro Gly Ala Ser Ala
340 345 350
Ala Pro Cys Cys Val Pro Gln Ala Leu Glu Pro Leu Pro Ile Val Tyr
355 360 365
Tyr Val Gly Arg Lys Pro Lys Val Glu Gln Leu Ser Asn Met Ile Val
370 375 380
Arg Ser Cys Lys Cys Ser
385 390
<210> 19
<211> 390
<212> ПРТ
<213> синтетическая
<400> 19
Met Pro Pro Ser Gly Leu Arg Leu Leu Leu Leu Leu Leu Pro Leu Leu
1 5 10 15
Trp Leu Leu Val Leu Thr Pro Gly Arg Pro Ala Ala Gly Leu Ser Thr
20 25 30
Cys Lys Thr Ile Asp Met Glu Leu Val Lys Arg Lys Arg Ile Glu Ala
35 40 45
Ile Arg Gly Gln Ile Leu Ser Lys Leu Arg Leu Ala Ser Pro Pro Ser
50 55 60
Gln Gly Glu Val Pro Pro Gly Pro Leu Pro Glu Ala Val Leu Ala Leu
65 70 75 80
Tyr Asn Ser Thr Arg Asp Arg Val Ala Gly Glu Ser Ala Glu Pro Glu
85 90 95
Pro Glu Pro Glu Ala Asp Tyr Tyr Ala Lys Glu Val Thr Arg Val Leu
100 105 110
Met Val Glu Thr His Asn Glu Ile Tyr Asp Lys Phe Lys Gln Ser Thr
115 120 125
His Ser Ile Tyr Met Phe Phe Asn Thr Ser Glu Leu Arg Glu Ala Val
130 135 140
Pro Glu Pro Val Leu Leu Ser Arg Ala Glu Leu Arg Leu Leu Arg Leu
145 150 155 160
Lys Leu Lys Val Glu Gln His Val Glu Leu Tyr Gln Lys Tyr Ser Asn
165 170 175
Asn Ser Trp Arg Tyr Leu Ser Asn Arg Leu Leu Ala Pro Ser Asp Ser
180 185 190
Pro Glu Trp Leu Ser Phe Asp Val Thr Gly Val Val Arg Gln Trp Leu
195 200 205
Ser Arg Gly Gly Glu Ile Glu Gly Phe Arg Leu Ser Ala His Val Ser
210 215 220
Val Asp Ser Arg Asp Asn Thr Leu Gln Val Asp Ile Asn Gly Phe Thr
225 230 235 240
Thr Gly Arg Arg Gly Asp Leu Ala Thr Ile His Gly Met Asn Arg Pro
245 250 255
Phe Leu Leu Leu Met Ala Thr Pro Leu Glu Arg Ala Gln His Leu Gln
260 265 270
Ser Ser Arg His Arg Arg Ala Leu Asp Thr Asn Tyr Cys Phe Ser Ser
275 280 285
Thr Glu Lys Asn Cys Cys Val Arg Gln Leu Tyr Ile Asp Phe Arg Lys
290 295 300
Asp Leu Gly Trp Lys Trp Ile His Glu Pro Lys Gly Tyr His Ala Asn
305 310 315 320
Phe Cys Leu Gly Pro Cys Pro Tyr Ile Trp Ser Leu Asp Thr Gln Tyr
325 330 335
Ser Lys Val Leu Ala Leu Tyr Asn Gln His Asn Pro Gly Ala Ser Ala
340 345 350
Ala Pro Cys Cys Val Pro Gln Ala Leu Glu Pro Leu Pro Ile Val Tyr
355 360 365
Tyr Val Gly Arg Lys Pro Lys Val Glu Gln Leu Ser Asn Met Ile Val
370 375 380
Arg Ser Cys Lys Cys Ser
385 390
<210> 20
<211> 228
<212> ДНК
<213> синтетическая
<400> 20
cgactgtgcc ttctagttgc cagccatctg ttgtttgccc ctcccccgtg ccttccttga 60
ccctggaagg tgccactccc actgtccttt cctaataaaa tgaggaaatt gcatcgcatt 120
gtctgagtag gtgtcattct attctggggg gtggggtggg gcaggacagc aagggggagg 180
attgggaaga caatagcagg catgctgggg atgcggtggg ctctatgg 228
<210> 21
<211> 31
<212> ПРТ
<213> homo sapiens
<400> 21
His Ala Glu Gly Thr Phe Thr Ser Asp Val Ser Ser Tyr Leu Glu Gly
1 5 10 15
Gln Ala Ala Lys Glu Phe Ile Ala Trp Leu Val Lys Gly Arg Gly
20 25 30
<210> 22
<211> 31
<212> ПРТ
<213> синтетическая
<400> 22
His Ala Glu Gly Thr Phe Thr Ser Asp Val Ser Ser Tyr Leu Glu Gly
1 5 10 15
Gln Ala Ala Lys Glu Phe Ile Ala Trp Leu Val Arg Gly Arg Gly
20 25 30
<210> 23
<211> 31
<212> ПРТ
<213> синтетическая
<220>
<221> MISC_FEATURE
<222> (2)..(2)
<223> 2-аминоизомасляная кислота
<400> 23
His Xaa Glu Gly Thr Phe Thr Ser Asp Val Ser Ser Tyr Leu Glu Gly
1 5 10 15
Gln Ala Ala Lys Glu Phe Ile Ala Trp Leu Val Arg Gly Arg Gly
20 25 30
<210> 24
<211> 6401
<212> ДНК
<213> синтетическая
<400> 24
gactcttcgc gatgtacggg ccagatatac gcgttgacat tgattattga ctagttatta 60
atagtaatca attacggggt cattagttca tagcccatat atggagttcc gcgttacata 120
acttacggta aatggcccgc ctggctgacc gcccaacgac ccccgcccat tgacgtcaat 180
aatgacgtat gttcccatag taacgccaat agggactttc cattgacgtc aatgggtgga 240
ctatttacgg taaactgccc acttggcagt acatcaagtg tatcatatgc caagtacgcc 300
ccctattgac gtcaatgacg gtaaatggcc cgcctggcat tatgcccagt acatgacctt 360
atgggacttt cctacttggc agtacatcta cgtattagtc atcgctatta ccatggtgat 420
gcggttttgg cagtacatca atgggcgtgg atagcggttt gactcacggg gatttccaag 480
tctccacccc attgacgtca atgggagttt gttttggcac caaaatcaac gggactttcc 540
aaaatgtcgt aacaactccg ccccattgac gcaaatgggc ggtaggcgtg tacggtggga 600
ggtctatata agcagagctc tctggctaac tagagaaccc actgcttact ggcttatcga 660
aattaatacg actcactata gggagaccca agctggctag cgtttaaact taagcttggt 720
accgagctcg gatccactag tccagtgtgg tggaattctg cactgcagct cgcatctctc 780
cttcacgcgc ccgccgccct acctgaggcc gccatccacg ccggttgagt cgcgttctgc 840
cgcctcccgc ctgtggtgcc tcctgaactg cgtccgccgt ctaggtaagt ttaaagctca 900
ggtcgagacc gggcctttgt ccggcgctcc cttggagcct acctagactc agccggctct 960
ccacgctttg cctgaccctg cttgctcaac tctaggtaag ttaatgagac agatagaaac 1020
tggtcttgta gaaacagagt agtcgcctgc ttttctgcca ggtgctgact tctctcccct 1080
gggctttttt ctttttctca ggttgaaaag aagaagacga agaagacgaa gaagacaaac 1140
cgtcgtcgac tgccatgcgc cgctgattaa cgccgccacc atggcccacc gacgcagatc 1200
cagaagctgc cgtgaggacc agaagcccgt gatggatgat cagagggacc ttatctctaa 1260
caatgaacaa ctgccaatgc tcggcagacg gcctggggcc ccggagagca agtgcagcag 1320
aggagccttg tacacggggt tctccatttt agtgactctc cttctcgccg gccaagctac 1380
caccgcctac tttctgtacc aacagcaagg cagactagac aaactgacaa tcacaagcca 1440
gaaccttcag ctggagtctc tgcggatgaa gctgcccgct ttgtggatga gattgcttcc 1500
tctacttgct ctcctggcgc tctggggacc tgaccccgag caagagtttg ttaatcagca 1560
cctgtgtggg agtcatctgg tggaggcact ctatttagtg tgcggagaga ggggcttctt 1620
ctacactcca aagaccagac gggaggccga agaccttcaa gtggggcaag tagaactggg 1680
tggcggaccc ggtgccggga gccttcagcc gctcgccctg gagggctctc ttcagaaacg 1740
cggcatcgtg gagcagtgtt gcacatccat ttgctcactc taccagctgg agaactactg 1800
caacggaagc ggagtgaagc agacgttgaa ttttgatttg ttgaagttgg cgggggatgt 1860
ggagagcaat ccggggccga tgccccctag tggcctcaga cttttgttat tgttattacc 1920
gcttttatgg ctcttggtgc tgacaccggg ccgtccggct gctggcttgt cgacttgtaa 1980
gacaattgat atggaattgg tgaaacgaaa acggattgag gccatccgag gacagatttt 2040
gagcaagctg cggcttgcct cgccaccctc gcaaggggaa gtcccacccg gacctctacc 2100
agaagcagtc ctagcgctgt acaacagtac aagagataga gtggccgggg aatccgcaga 2160
accagagcct gagcctgaag ccgattatta tgcaaaggaa gtgactaggg tcctgatggt 2220
cgagacccat aacgaaatct acgacaaatt caaacaaagt acccactcta tctacatgtt 2280
cttcaacacc agtgagctaa gagaagccgt gcccgaacct gtgcttcttt cccgcgcaga 2340
actccgcctc ttgagactca aattgaaagt tgaacaacac gtagagcttt accagaaata 2400
ctctaataat tcatggcgat atctttctaa tcgtctcctc gccccatctg acagccctga 2460
atggctctcc ttcgacgtta cgggagttgt gcgccagtgg ctcagcagag gcggagagat 2520
agagggcttt cggctgagcg cacatgtatc tgtggactca agggacaaca cattgcaagt 2580
ggatattaac ggttttacaa ctggacggag aggggacctg gcgaccatcc acggcatgaa 2640
tagacctttc ctgctgctga tggctactcc cctggagagg gcacagcact tacagtcttc 2700
cagacaccgg cgcgccctgg atacaaacta ctgcttcagc tccaccgaaa agaactgttg 2760
cgtgcggcag ctgtacattg acttcagaaa ggatctgggc tggaagtgga ttcatgagcc 2820
caaggggtat catgccaact tctgtcttgg gccatgccca tacatctggt cactggatac 2880
ccagtactcc aaagttctgg ccttgtacaa tcaacacaac cctggagctt ccgccgctcc 2940
ttgctgtgtg ccccaagccc tagagcccct gcccatcgtt tattatgtcg gacgcaagcc 3000
caaagtagaa cagctatcaa atatgatcgt gagaagctgc aagtgtagct gataaacgcg 3060
tcgagcatgc atctagggcg gccaattccg cccctctccc ccccacccct ctccctcccc 3120
cccccctaac gttactggcc gaagccgctt ggaataaggc cggtgtgcgt ttgtctatat 3180
gttattttcc accatattgc cgtcttttgg caatgtgagg gcccggaaac ctggccctgt 3240
cttcttgacg agcattccta ggggtctttc ccctctcgcc aaaggaatgc aaggtctgtt 3300
gaatgtcgtg aaggaagcag ttcctctgga agcttcttga agacaaacaa cgtctgtagc 3360
gaccctttgt agacagcgga accccccacc tggcgataga tgcctctgcg gccaaaagcc 3420
acgtgtataa gatacacctg caaaggcggc acaaccccag tgccacgttg tgagttggat 3480
agttgtggaa agagtcaaat ggctctcctc aagcgtattc aacaaggggc tgaaggatgc 3540
ccagaaggta ccccattgta tgggatctga tctggggcct cggtgcacat gctttacatg 3600
tgtttagtcg aggttaaaaa acgtctaggc cccccgaacc acggggacgt ggttttcctt 3660
tgaaaaacac gatgataata tggctgccgc tcattctagt gcccttcttt gctgcctggt 3720
cctgctcacc ggggtgcgag ctagccctgg acaagggaca caatccgaaa actcgtgcac 3780
ccacttcccg ggcaacctcc ctaacatgct gagggacctc cgtgatgcct tcagtagagt 3840
gaagacgttc ttccaaatga aagatcagtt agataacctg ctcctgaagg agtcactctt 3900
agaagacttc aaaggatacc tcggctgcca agcacttagc gagatgattc aattctactt 3960
agaagaagtc atgcctcaag ctgagaatca agaccccgac atcaaagctc atgtgaattc 4020
tttgggagaa aatttgaaga ctttgcggct gcggctgcgg agatgtcacc gctttctgcc 4080
ctgtgagaac aaatcaaaag cggtcgagca agttaagaat gccttcaata agctacaaga 4140
gaagggcatc tacaaagcaa tgagcgagtt tgatatcttt atcaattaca ttgaagccta 4200
catgacaatg aagattagga atgccgcggg gagcggcgct actaacttca gcctgctgaa 4260
gcaggctgga gacgtggagg agaaccctgg acctatgtac agaatgcagc tgctgagctg 4320
catcgccctg agcctggccc tggtgaccaa cagcgcaccc acgtcctcta gcaccaagaa 4380
gacccagtta cagttggagc atctactttt agacctgcaa atgattttga acggcatcaa 4440
caactacaag aatcctaaac ttactcgcat gcttaccttc aaattttaca tgcccaagaa 4500
ggccaccgaa ctgaagcact tgcaatgtct ggaggaagaa ctcaagccgc tggaggaagt 4560
tctcaacctc gcgcagtcca agaatttcca cctccggcca agagacctga tcagtaacat 4620
taatgtgata gtgctggagc tgaagggaag cgagactaca tttatgtgcg agtacgccga 4680
tgaaaccgct acaatcgtcg agttcctgaa tagatggatc acattttgcc agtcaattat 4740
ctctactctg acatgataac tcgaggtcta gagggcccgt ttaaacccgc tgatcagcct 4800
cgactgtgcc ttctagttgc cagccatctg ttgtttgccc ctcccccgtg ccttccttga 4860
ccctggaagg tgccactccc actgtccttt cctaataaaa tgaggaaatt gcatcgcatt 4920
gtctgagtag gtgtcattct attctggggg gtggggtggg gcaggacagc aagggggagg 4980
attgggaaga caatagcagg catgctgggg atgcggtggg ctctatggct tctactgggc 5040
ggttttatgg acagcaagcg aaccggaatt gccagctggg gcgccctctg gtaaggttgg 5100
gaagccctgc aaagtaaact ggatggcttt ctcgccgcca aggatctgat ggcgcagggg 5160
atcaagctct gatcaagaga caggatgagg atcgtttcgc atggccaaag aagacaatat 5220
tgaaatgcaa ggtaccgttc ttgaaacgtt gcctaatacc atgttccgcg tagagttaga 5280
aaacggtcac gtggttactg cacacatctc cggtaaaatg cgcaaaaact acatccgcat 5340
cctgacgggc gacaaagtga ctgttgaact gaccccgtac gacctgagca aaggccgcat 5400
tgtcttccgt agtcgctgat aaattattaa cgcttacaat ttcctgatgc ggtattttct 5460
ccttacgcat ctgtgcggta tttcacaccg catacaggtg gcacttttcg gggaaatgtg 5520
cgcggaaccc ctatttgttt atttttctaa atacattcaa atatgtatcc gctcatgaga 5580
caataaccct gataaatgct tcaataatag cacgtgctaa aacttcattt ttaatttaaa 5640
aggatctagg tgaagatcct ttttgataat ctcatgacca aaatccctta acgtgagttt 5700
tcgttccact gagcgtcaga ccccgtagaa aagatcaaag gatcttcttg agatcctttt 5760
tttctgcgcg taatctgctg cttgcaaaca aaaaaaccac cgctaccagc ggtggtttgt 5820
ttgccggatc aagagctacc aactcttttt ccgaaggtaa ctggcttcag cagagcgcag 5880
ataccaaata ctgttcttct agtgtagccg tagttaggcc accacttcaa gaactctgta 5940
gcaccgccta catacctcgc tctgctaatc ctgttaccag tggctgctgc cagtggcgat 6000
aagtcgtgtc ttaccgggtt ggactcaaga cgatagttac cggataaggc gcagcggtcg 6060
ggctgaacgg ggggttcgtg cacacagccc agcttggagc gaacgaccta caccgaactg 6120
agatacctac agcgtgagct atgagaaagc gccacgcttc ccgaagggag aaaggcggac 6180
aggtatccgg taagcggcag ggtcggaaca ggagagcgca cgagggagct tccaggggga 6240
aacgcctggt atctttatag tcctgtcggg tttcgccacc tctgacttga gcgtcgattt 6300
ttgtgatgct cgtcaggggg gcggagccta tggaaaaacg ccagcaacgc ggccttttta 6360
cggttcctgg gcttttgctg gccttttgct cacatgttct t 6401
<210> 25
<211> 390
<212> ПРТ
<213> синтетическая
<400> 25
Met Pro Pro Ser Gly Leu Arg Leu Leu Leu Leu Leu Leu Pro Leu Leu
1 5 10 15
Trp Leu Leu Val Leu Thr Pro Gly Arg Pro Ala Ala Gly Leu Ser Thr
20 25 30
Cys Lys Thr Ile Asp Met Glu Leu Val Lys Arg Lys Arg Ile Glu Ala
35 40 45
Ile Arg Gly Gln Ile Leu Ser Lys Leu Arg Leu Ala Ser Pro Pro Ser
50 55 60
Gln Gly Glu Val Pro Pro Gly Pro Leu Pro Glu Ala Val Leu Ala Leu
65 70 75 80
Tyr Asn Ser Thr Arg Asp Arg Val Ala Gly Glu Ser Ala Glu Pro Glu
85 90 95
Pro Glu Pro Glu Ala Asp Tyr Tyr Ala Lys Glu Val Thr Arg Val Leu
100 105 110
Met Val Glu Thr His Asn Glu Ile Tyr Asp Lys Phe Lys Gln Ser Thr
115 120 125
His Ser Ile Tyr Met Phe Phe Asn Thr Ser Glu Leu Arg Glu Ala Val
130 135 140
Pro Glu Pro Val Leu Leu Ser Arg Ala Glu Leu Arg Leu Leu Arg Leu
145 150 155 160
Lys Leu Lys Val Glu Gln His Val Glu Leu Tyr Gln Lys Tyr Ser Asn
165 170 175
Asn Ser Trp Arg Tyr Leu Ser Asn Arg Leu Leu Ala Pro Ser Asp Ser
180 185 190
Pro Glu Trp Leu Ser Phe Asp Val Thr Gly Val Val Arg Gln Trp Leu
195 200 205
Ser Arg Gly Gly Glu Ile Glu Gly Phe Arg Leu Ser Ala His Ser Ser
210 215 220
Ser Asp Ser Arg Asp Asn Thr Leu Gln Val Asp Ile Asn Gly Phe Thr
225 230 235 240
Thr Gly Arg Arg Gly Asp Leu Ala Thr Ile His Gly Met Asn Arg Pro
245 250 255
Phe Leu Leu Leu Met Ala Thr Pro Leu Glu Arg Ala Gln His Leu Gln
260 265 270
Ser Ser Arg His Arg Arg Ala Leu Asp Thr Asn Tyr Cys Phe Ser Ser
275 280 285
Thr Glu Lys Asn Cys Cys Val Arg Gln Leu Tyr Ile Asp Phe Arg Lys
290 295 300
Asp Leu Gly Trp Lys Trp Ile His Glu Pro Lys Gly Tyr His Ala Asn
305 310 315 320
Phe Cys Leu Gly Pro Cys Pro Tyr Ile Trp Ser Leu Asp Thr Gln Tyr
325 330 335
Ser Lys Val Leu Ala Leu Tyr Asn Gln His Asn Pro Gly Ala Ser Ala
340 345 350
Ala Pro Cys Cys Val Pro Gln Ala Leu Glu Pro Leu Pro Ile Val Tyr
355 360 365
Tyr Val Gly Arg Lys Pro Lys Val Glu Gln Leu Ser Asn Met Ile Val
370 375 380
Arg Ser Cys Lys Cys Ser
385 390
<210> 26
<211> 6389
<212> ДНК
<213> синтетическая
<400> 26
gactcttcgc gatgtacggg ccagatatac gcgttgacat tgattattga ctagttatta 60
atagtaatca attacggggt cattagttca tagcccatat atggagttcc gcgttacata 120
acttacggta aatggcccgc ctggctgacc gcccaacgac ccccgcccat tgacgtcaat 180
aatgacgtat gttcccatag taacgccaat agggactttc cattgacgtc aatgggtgga 240
ctatttacgg taaactgccc acttggcagt acatcaagtg tatcatatgc caagtacgcc 300
ccctattgac gtcaatgacg gtaaatggcc cgcctggcat tatgcccagt acatgacctt 360
atgggacttt cctacttggc agtacatcta cgtattagtc atcgctatta ccatggtgat 420
gcggttttgg cagtacatca atgggcgtgg atagcggttt gactcacggg gatttccaag 480
tctccacccc attgacgtca atgggagttt gttttggcac caaaatcaac gggactttcc 540
aaaatgtcgt aacaactccg ccccattgac gcaaatgggc ggtaggcgtg tacggtggga 600
ggtctatata agcagagctc tctggctaac tagagaaccc actgcttact ggcttatcga 660
aattaatacg actcactata gggagaccca agctggctag cgtttaaact taagcttggt 720
accgagctcg gatccactag tccagtgtgg tggaattctg cactgcagct cgcatctctc 780
cttcacgcgc ccgccgccct acctgaggcc gccatccacg ccggttgagt cgcgttctgc 840
cgcctcccgc ctgtggtgcc tcctgaactg cgtccgccgt ctaggtaagt ttaaagctca 900
ggtcgagacc gggcctttgt ccggcgctcc cttggagcct acctagactc agccggctct 960
ccacgctttg cctgaccctg cttgctcaac tctaggtaag ttaatgagac agatagaaac 1020
tggtcttgta gaaacagagt agtcgcctgc ttttctgcca ggtgctgact tctctcccct 1080
gggctttttt ctttttctca ggttgaaaag aagaagacga agaagacgaa gaagacaaac 1140
cgtcgtcgac tgccatgcgc cgctgattaa cgccgccacc atggcccacc gacgcagatc 1200
cagaagctgc cgtgaggacc agaagcccgt gatggatgat cagagggacc ttatctctaa 1260
caatgaacaa ctgccaatgc tcggcagacg gcctggggcc ccggagagca agtgcagcag 1320
aggagccttg tacacggggt tctccatttt agtgactctc cttctcgccg gccaagctac 1380
caccgcctac tttctgtacc aacagcaagg cagactagac aaactgacaa tcacaagcca 1440
gaaccttcag ctggagtctc tgcggatgaa gctgcccgct ttgtggatga gattgcttcc 1500
tctacttgct ctcctggcgc tctggggacc tgaccccgag caagagtttg ttaatcagca 1560
cctgtgtggg agtcatctgg tggaggcact ctatttagtg tgcggagaga ggggcttctt 1620
ctacactcca aagaccagac gggaggccga agaccttcaa gtggggcaag tagaactggg 1680
tggcggaccc ggtgccggga gccttcagcc gctcgccctg gagggctctc ttcagaaacg 1740
cggcatcgtg gagcagtgtt gcacatccat ttgctcactc taccagctgg agaactactg 1800
caacggaagc ggagtgaagc agacgttgaa ttttgatttg ttgaagttgg cgggggatgt 1860
ggagagcaat ccggggccga tgccccctag tggcctcaga cttttgttat tgttattacc 1920
gcttttatgg ctcttggtgc tgacaccggg ccgtccggct gctggcttgt cgacttgtaa 1980
gacaattgat atggaattgg tgaaacgaaa acggattgag gccatccgag gacagatttt 2040
gagcaagctg cggcttgcct cgccaccctc gcaaggggaa gtcccacccg gacctctacc 2100
agaagcagtc ctagcgctgt acaacagtac aagagataga gtggccgggg aatccgcaga 2160
accagagcct gagcctgaag ccgattatta tgcaaaggaa gtgactaggg tcctgatggt 2220
cgagacccat aacgaaatct acgacaaatt caaacaaagt acccactcta tctacatgtt 2280
cttcaacacc agtgagctaa gagaagccgt gcccgaacct gtgcttcttt cccgcgcaga 2340
actccgcctc ttgagactca aattgaaagt tgaacaacac gtagagcttt accagaaata 2400
ctctaataat tcatggcgat atctttctaa tcgtctcctc gccccatctg acagccctga 2460
atggctctcc ttcgacgtta cgggagttgt gcgccagtgg ctcagcagag gcggagagat 2520
agagggcttt cggctgagcg cacatagctc tagcgactca agggacaaca cattgcaagt 2580
ggatattaac ggttttacaa ctggacggag aggggacctg gcgaccatcc acggcatgaa 2640
tagacctttc ctgctgctga tggctactcc cctggagagg gcacagcact tacagtcttc 2700
cagacaccgg cgcgccctgg atacaaacta ctgcttcagc tccaccgaaa agaactgttg 2760
cgtgcggcag ctgtacattg acttcagaaa ggatctgggc tggaagtgga ttcatgagcc 2820
caaggggtat catgccaact tctgtcttgg gccatgccca tacatctggt cactggatac 2880
ccagtactcc aaagttctgg ccttgtacaa tcaacacaac cctggagctt ccgccgctcc 2940
ttgctgtgtg ccccaagccc tagagcccct gcccatcgtt tattatgtcg gacgcaagcc 3000
caaagtagaa cagctatcaa atatgatcgt gagaagctgc aagtgtagct gataaacgcg 3060
tcgagcatgc atctagggcg gccaattccg cccctctccc ccccacccct ctccctcccc 3120
cccccctaac gttactggcc gaagccgctt ggaataaggc cggtgtgcgt ttgtctatat 3180
gttattttcc accatattgc cgtcttttgg caatgtgagg gcccggaaac ctggccctgt 3240
cttcttgacg agcattccta ggggtctttc ccctctcgcc aaaggaatgc aaggtctgtt 3300
gaatgtcgtg aaggaagcag ttcctctgga agcttcttga agacaaacaa cgtctgtagc 3360
gaccctttgt agacagcgga accccccacc tggcgataga tgcctctgcg gccaaaagcc 3420
acgtgtataa gatacacctg caaaggcggc acaaccccag tgccacgttg tgagttggat 3480
agttgtggaa agagtcaaat ggctctcctc aagcgtattc aacaaggggc tgaaggatgc 3540
ccagaaggta ccccattgta tgggatctga tctggggcct cggtgcacat gctttacatg 3600
tgtttagtcg aggttaaaaa acgtctaggc cccccgaacc acggggacgt ggttttcctt 3660
tgaaaaacac gatgataata tgatgcacag ctcagcactg ctctgttgcc tggtcctcct 3720
gactggggtg agggccagcc caggccaggg cacccagtct gagaacagct gcacccactt 3780
cccaggcaac ctgcctaaca tgcttcgaga tctccgagat gccttcagca gagtgaagac 3840
tttctttcaa atgaaggatc agctggacaa cttgttgtta aaggagtcct tgctggagga 3900
ctttaagggt tacctgggtt gccaagcctt gtctgagatg atccagtttt acctggagga 3960
ggtgatgccc caagctgaga accaagaccc agacatcaag gcgcatgtga actccctggg 4020
ggagaacctg aagaccctca ggctgaggct acggcgctgt catcgatttc ttccctgtga 4080
aaacaagagc aaggccgtgg agcaggtgaa gaatgccttt aataagctcc aagagaaagg 4140
catctacaaa gccatgagtg agtttgacat cttcatcaac tacatagaag cctacatgac 4200
aatgaagata cgaaacggga gcggcgctac taacttcagc ctgctgaagc aggctggaga 4260
cgtggaggag aaccctggac ctatgtacag aatgcagctg ctgagctgca tcgccctgag 4320
cctggccctg gtgaccaaca gcgcacccac gtcctctagc accaagaaga cccagttaca 4380
gttggagcat ctacttttag acctgcaaat gattttgaac ggcatcaaca actacaagaa 4440
tcctaaactt actcgcatgc ttaccttcaa attttacatg cccaagaagg ccaccgaact 4500
gaagcacttg caatgtctgg aggaagaact caagccgctg gaggaagttc tcaacctcgc 4560
gcagtccaag aatttccacc tccggccaag agacctgatc agtaacatta atgtgatagt 4620
gctggagctg aagggaagcg agactacatt tatgtgcgag tacgccgatg aaaccgctac 4680
aatcgtcgag ttcctgaata gatggatcac attttgccag tcaattatct ctactctgac 4740
atgataactc gaggtctaga gggcccgttt aaacccgctg atcagcctcg actgtgcctt 4800
ctagttgcca gccatctgtt gtttgcccct cccccgtgcc ttccttgacc ctggaaggtg 4860
ccactcccac tgtcctttcc taataaaatg aggaaattgc atcgcattgt ctgagtaggt 4920
gtcattctat tctggggggt ggggtggggc aggacagcaa gggggaggat tgggaagaca 4980
atagcaggca tgctggggat gcggtgggct ctatggcttc tactgggcgg ttttatggac 5040
agcaagcgaa ccggaattgc cagctggggc gccctctggt aaggttggga agccctgcaa 5100
agtaaactgg atggctttct cgccgccaag gatctgatgg cgcaggggat caagctctga 5160
tcaagagaca ggatgaggat cgtttcgcat ggccaaagaa gacaatattg aaatgcaagg 5220
taccgttctt gaaacgttgc ctaataccat gttccgcgta gagttagaaa acggtcacgt 5280
ggttactgca cacatctccg gtaaaatgcg caaaaactac atccgcatcc tgacgggcga 5340
caaagtgact gttgaactga ccccgtacga cctgagcaaa ggccgcattg tcttccgtag 5400
tcgctgataa attattaacg cttacaattt cctgatgcgg tattttctcc ttacgcatct 5460
gtgcggtatt tcacaccgca tacaggtggc acttttcggg gaaatgtgcg cggaacccct 5520
atttgtttat ttttctaaat acattcaaat atgtatccgc tcatgagaca ataaccctga 5580
taaatgcttc aataatagca cgtgctaaaa cttcattttt aatttaaaag gatctaggtg 5640
aagatccttt ttgataatct catgaccaaa atcccttaac gtgagttttc gttccactga 5700
gcgtcagacc ccgtagaaaa gatcaaagga tcttcttgag atcctttttt tctgcgcgta 5760
atctgctgct tgcaaacaaa aaaaccaccg ctaccagcgg tggtttgttt gccggatcaa 5820
gagctaccaa ctctttttcc gaaggtaact ggcttcagca gagcgcagat accaaatact 5880
gttcttctag tgtagccgta gttaggccac cacttcaaga actctgtagc accgcctaca 5940
tacctcgctc tgctaatcct gttaccagtg gctgctgcca gtggcgataa gtcgtgtctt 6000
accgggttgg actcaagacg atagttaccg gataaggcgc agcggtcggg ctgaacgggg 6060
ggttcgtgca cacagcccag cttggagcga acgacctaca ccgaactgag atacctacag 6120
cgtgagctat gagaaagcgc cacgcttccc gaagggagaa aggcggacag gtatccggta 6180
agcggcaggg tcggaacagg agagcgcacg agggagcttc cagggggaaa cgcctggtat 6240
ctttatagtc ctgtcgggtt tcgccacctc tgacttgagc gtcgattttt gtgatgctcg 6300
tcaggggggc ggagcctatg gaaaaacgcc agcaacgcgg cctttttacg gttcctgggc 6360
ttttgctggc cttttgctca catgttctt 6389
<210> 27
<211> 15
<212> ПРТ
<213> синтетическая
<400> 27
Ser His Leu Val Glu Ala Leu Tyr Leu Val Cys Gly Gly Glu Gly
1 5 10 15
<210> 28
<211> 6383
<212> ДНК
<213> синтетическая
<400> 28
gactcttcgc gatgtacggg ccagatatac gcgttgacat tgattattga ctagttatta 60
atagtaatca attacggggt cattagttca tagcccatat atggagttcc gcgttacata 120
acttacggta aatggcccgc ctggctgacc gcccaacgac ccccgcccat tgacgtcaat 180
aatgacgtat gttcccatag taacgccaat agggactttc cattgacgtc aatgggtgga 240
ctatttacgg taaactgccc acttggcagt acatcaagtg tatcatatgc caagtacgcc 300
ccctattgac gtcaatgacg gtaaatggcc cgcctggcat tatgcccagt acatgacctt 360
atgggacttt cctacttggc agtacatcta cgtattagtc atcgctatta ccatggtgat 420
gcggttttgg cagtacatca atgggcgtgg atagcggttt gactcacggg gatttccaag 480
tctccacccc attgacgtca atgggagttt gttttggcac caaaatcaac gggactttcc 540
aaaatgtcgt aacaactccg ccccattgac gcaaatgggc ggtaggcgtg tacggtggga 600
ggtctatata agcagagctc tctggctaac tagagaaccc actgcttact ggcttatcga 660
aattaatacg actcactata gggagaccca agctggctag cgtttaaact taagcttggt 720
accgagctcg gatccactag tccagtgtgg tggaattctg cagctcgcat ctctccttca 780
cgcgcccgcc gccctacctg aggccgccat ccacgccggt tgagtcgcgt tctgccgcct 840
cccgcctgtg gtgcctcctg aactgcgtcc gccgtctagg taagtttaaa gctcaggtcg 900
agaccgggcc tttgtccggc gctcccttgg agcctaccta gactcagccg gctctccacg 960
ctttgcctga ccctgcttgc tcaactctag gtaagttaat gagacagata gaaactggtc 1020
ttgtagaaac agagtagtcg cctgcttttc tgccaggtgc tgacttctct cccctgggct 1080
tttttctttt tctcaggttg aaaagaagaa gacgaagaag acgaagaaga caaaccgtcg 1140
tcgactgcca tgcgccgctg attaacgccg ccaccatggc ccaccgacgc agatccagaa 1200
gctgccgtga ggaccagaag cccgtgatgg atgatcagag ggaccttatc tctaacaatg 1260
aacaactgcc aatgctcggc agacggcctg gggccccgga gagcaagtgc agcagaggag 1320
ccttgtacac ggggttctcc attttagtga ctctccttct cgccggccaa gctaccaccg 1380
cctactttct gtaccaacag caaggcagac tagacaaact gacaatcaca agccagaacc 1440
ttcagctgga gtctctgcgg atgaagctgc ccgctttgtg gatgagattg cttcctctac 1500
ttgctctcct ggcgctctgg ggacctgacc ccgagcaaga gtttgttaat cagcacctgt 1560
gtgggagtca tctggtggag gcactctatt tagtgtgcgg agagaggggc ttcttctaca 1620
ctccaaagac cagacgggag gccgaagacc ttcaagtggg gcaagtagaa ctgggtggcg 1680
gacccggtgc cgggagcctt cagccgctcg ccctggaggg ctctcttcag aaacgcggca 1740
tcgtggagca gtgttgcaca tccatttgct cactctacca gctggagaac tactgcaacg 1800
gaagcggagt gaagcagacg ttgaattttg atttgttgaa gttggcgggg gatgtggaga 1860
gcaatccggg gccgatgccc cctagtggcc tcagactttt gttattgtta ttaccgcttt 1920
tatggctctt ggtgctgaca ccgggccgtc cggctgctgg cttgtcgact tgtaagacaa 1980
ttgatatgga attggtgaaa cgaaaacgga ttgaggccat ccgaggacag attttgagca 2040
agctgcggct tgcctcgcca ccctcgcaag gggaagtccc acccggacct ctaccagaag 2100
cagtcctagc gctgtacaac agtacaagag atagagtggc cggggaatcc gcagaaccag 2160
agcctgagcc tgaagccgat tattatgcaa aggaagtgac tagggtcctg atggtcgaga 2220
cccataacga aatctacgac aaattcaaac aaagtaccca ctctatctac atgttcttca 2280
acaccagtga gctaagagaa gccgtgcccg aacctgtgct tctttcccgc gcagaactcc 2340
gcctcttgag actcaaattg aaagttgaac aacacgtaga gctttaccag aaatactcta 2400
ataattcatg gcgatatctt tctaatcgtc tcctcgcccc atctgacagc cctgaatggc 2460
tctccttcga cgttacggga gttgtgcgcc agtggctcag cagaggcgga gagatagagg 2520
gctttcggct gagcgcacat agctctagcg actcaaggga caacacattg caagtggata 2580
ttaacggttt tacaactgga cggagagggg acctggcgac catccacggc atgaatagac 2640
ctttcctgct gctgatggct actcccctgg agagggcaca gcacttacag tcttccagac 2700
accggcgcgc cctggataca aactactgct tcagctccac cgaaaagaac tgttgcgtgc 2760
ggcagctgta cattgacttc agaaaggatc tgggctggaa gtggattcat gagcccaagg 2820
ggtatcatgc caacttctgt cttgggccat gcccatacat ctggtcactg gatacccagt 2880
actccaaagt tctggccttg tacaatcaac acaaccctgg agcttccgcc gctccttgct 2940
gtgtgcccca agccctagag cccctgccca tcgtttatta tgtcggacgc aagcccaaag 3000
tagaacagct atcaaatatg atcgtgagaa gctgcaagtg tagctgataa acgcgtcgag 3060
catgcatcta gggcggccaa ttccgcccct ctccccccca cccctctccc tccccccccc 3120
ctaacgttac tggccgaagc cgcttggaat aaggccggtg tgcgtttgtc tatatgttat 3180
tttccaccat attgccgtct tttggcaatg tgagggcccg gaaacctggc cctgtcttct 3240
tgacgagcat tcctaggggt ctttcccctc tcgccaaagg aatgcaaggt ctgttgaatg 3300
tcgtgaagga agcagttcct ctggaagctt cttgaagaca aacaacgtct gtagcgaccc 3360
tttgtagaca gcggaacccc ccacctggcg atagatgcct ctgcggccaa aagccacgtg 3420
tataagatac acctgcaaag gcggcacaac cccagtgcca cgttgtgagt tggatagttg 3480
tggaaagagt caaatggctc tcctcaagcg tattcaacaa ggggctgaag gatgcccaga 3540
aggtacccca ttgtatggga tctgatctgg ggcctcggtg cacatgcttt acatgtgttt 3600
agtcgaggtt aaaaaacgtc taggcccccc gaaccacggg gacgtggttt tcctttgaaa 3660
aacacgatga taatatgatg cacagctcag cactgctctg ttgcctggtc ctcctgactg 3720
gggtgagggc cagcccaggc cagggcaccc agtctgagaa cagctgcacc cacttcccag 3780
gcaacctgcc taacatgctt cgagatctcc gagatgcctt cagcagagtg aagactttct 3840
ttcaaatgaa ggatcagctg gacaacttgt tgttaaagga gtccttgctg gaggacttta 3900
agggttacct gggttgccaa gccttgtctg agatgatcca gttttacctg gaggaggtga 3960
tgccccaagc tgagaaccaa gacccagaca tcaaggcgca tgtgaactcc ctgggggaga 4020
acctgaagac cctcaggctg aggctacggc gctgtcatcg atttcttccc tgtgaaaaca 4080
agagcaaggc cgtggagcag gtgaagaatg cctttaataa gctccaagag aaaggcatct 4140
acaaagccat gagtgagttt gacatcttca tcaactacat agaagcctac atgacaatga 4200
agatacgaaa cgggagcggc gctactaact tcagcctgct gaagcaggct ggagacgtgg 4260
aggagaaccc tggacctatg tacagaatgc agctgctgag ctgcatcgcc ctgagcctgg 4320
ccctggtgac caacagcgca cccacgtcct ctagcaccaa gaagacccag ttacagttgg 4380
agcatctact tttagacctg caaatgattt tgaacggcat caacaactac aagaatccta 4440
aacttactcg catgcttacc ttcaaatttt acatgcccaa gaaggccacc gaactgaagc 4500
acttgcaatg tctggaggaa gaactcaagc cgctggagga agttctcaac ctcgcgcagt 4560
ccaagaattt ccacctccgg ccaagagacc tgatcagtaa cattaatgtg atagtgctgg 4620
agctgaaggg aagcgagact acatttatgt gcgagtacgc cgatgaaacc gctacaatcg 4680
tcgagttcct gaatagatgg atcacatttt gccagtcaat tatctctact ctgacatgat 4740
aactcgagtc tagagggccc gtttaaaccc gctgatcagc ctcgactgtg ccttctagtt 4800
gccagccatc tgttgtttgc ccctcccccg tgccttcctt gaccctggaa ggtgccactc 4860
ccactgtcct ttcctaataa aatgaggaaa ttgcatcgca ttgtctgagt aggtgtcatt 4920
ctattctggg gggtggggtg gggcaggaca gcaaggggga ggattgggaa gacaatagca 4980
ggcatgctgg ggatgcggtg ggctctatgg cttctactgg gcggttttat ggacagcaag 5040
cgaaccggaa ttgccagctg gggcgccctc tggtaaggtt gggaagccct gcaaagtaaa 5100
ctggatggct ttctcgccgc caaggatctg atggcgcagg ggatcaagct ctgatcaaga 5160
gacaggatga ggatcgtttc gcatggccaa agaagacaat attgaaatgc aaggtaccgt 5220
tcttgaaacg ttgcctaata ccatgttccg cgtagagtta gaaaacggtc acgtggttac 5280
tgcacacatc tccggtaaaa tgcgcaaaaa ctacatccgc atcctgacgg gcgacaaagt 5340
gactgttgaa ctgaccccgt acgacctgag caaaggccgc attgtcttcc gtagtcgctg 5400
ataaattatt aacgcttaca atttcctgat gcggtatttt ctccttacgc atctgtgcgg 5460
tatttcacac cgcatacagg tggcactttt cggggaaatg tgcgcggaac ccctatttgt 5520
ttatttttct aaatacattc aaatatgtat ccgctcatga gacaataacc ctgataaatg 5580
cttcaataat agcacgtgct aaaacttcat ttttaattta aaaggatcta ggtgaagatc 5640
ctttttgata atctcatgac caaaatccct taacgtgagt tttcgttcca ctgagcgtca 5700
gaccccgtag aaaagatcaa aggatcttct tgagatcctt tttttctgcg cgtaatctgc 5760
tgcttgcaaa caaaaaaacc accgctacca gcggtggttt gtttgccgga tcaagagcta 5820
ccaactcttt ttccgaaggt aactggcttc agcagagcgc agataccaaa tactgttctt 5880
ctagtgtagc cgtagttagg ccaccacttc aagaactctg tagcaccgcc tacatacctc 5940
gctctgctaa tcctgttacc agtggctgct gccagtggcg ataagtcgtg tcttaccggg 6000
ttggactcaa gacgatagtt accggataag gcgcagcggt cgggctgaac ggggggttcg 6060
tgcacacagc ccagcttgga gcgaacgacc tacaccgaac tgagatacct acagcgtgag 6120
ctatgagaaa gcgccacgct tcccgaaggg agaaaggcgg acaggtatcc ggtaagcggc 6180
agggtcggaa caggagagcg cacgagggag cttccagggg gaaacgcctg gtatctttat 6240
agtcctgtcg ggtttcgcca cctctgactt gagcgtcgat ttttgtgatg ctcgtcaggg 6300
gggcggagcc tatggaaaaa cgccagcaac gcggcctttt tacggttcct gggcttttgc 6360
tggccttttg ctcacatgtt ctt 6383
<210> 29
<211> 6383
<212> ДНК
<213> синтетическая
<400> 29
gactcttcgc gatgtacggg ccagatatac gcgttgacat tgattattga ctagttatta 60
atagtaatca attacggggt cattagttca tagcccatat atggagttcc gcgttacata 120
acttacggta aatggcccgc ctggctgacc gcccaacgac ccccgcccat tgacgtcaat 180
aatgacgtat gttcccatag taacgccaat agggactttc cattgacgtc aatgggtgga 240
ctatttacgg taaactgccc acttggcagt acatcaagtg tatcatatgc caagtacgcc 300
ccctattgac gtcaatgacg gtaaatggcc cgcctggcat tatgcccagt acatgacctt 360
atgggacttt cctacttggc agtacatcta cgtattagtc atcgctatta ccatggtgat 420
gcggttttgg cagtacatca atgggcgtgg atagcggttt gactcacggg gatttccaag 480
tctccacccc attgacgtca atgggagttt gttttggcac caaaatcaac gggactttcc 540
aaaatgtcgt aacaactccg ccccattgac gcaaatgggc ggtaggcgtg tacggtggga 600
ggtctatata agcagagctc tctggctaac tagagaaccc actgcttact ggcttatcga 660
aattaatacg actcactata gggagaccca agctggctag cgtttaaact taagcttggt 720
accgagctcg gatccactag tccagtgtgg tggaattctg cagctcgcat ctctccttca 780
cgcgcccgcc gccctacctg aggccgccat ccacgccggt tgagtcgcgt tctgccgcct 840
cccgcctgtg gtgcctcctg aactgcgtcc gccgtctagg taagtttaaa gctcaggtcg 900
agaccgggcc tttgtccggc gctcccttgg agcctaccta gactcagccg gctctccacg 960
ctttgcctga ccctgcttgc tcaactctag gtaagttaat gagacagata gaaactggtc 1020
ttgtagaaac agagtagtcg cctgcttttc tgccaggtgc tgacttctct cccctgggct 1080
tttttctttt tctcaggttg aaaagaagaa gacgaagaag acgaagaaga caaaccgtcg 1140
tcgactgcca tgcgccgctg attaacgccg ccaccatggc ccaccgacgc agatccagaa 1200
gctgccgtga ggaccagaag cccgtgatgg atgatcagag ggaccttatc tctaacaatg 1260
aacaactgcc aatgctcggc agacggcctg gggccccgga gagcaagtgc agcagaggag 1320
ccttgtacac ggggttctcc attttagtga ctctccttct cgccggccaa gctaccaccg 1380
cctactttct gtaccaacag caaggcagac tagacaaact gacaatcaca agccagaacc 1440
ttcagctgga gtctctgcgg atgaagctgc ccgctttgtg gatgagattg cttcctctac 1500
ttgctctcct ggcgctctgg ggacctgacc ccgagcaaga gtttgttaat cagcacctgt 1560
gtgggagtca tctggtggag gcactctatt tagtgtgcgg agagaggggc ttcttctaca 1620
ctccaaagac cagacgggag gccgaagacc ttcaagtggg gcaagtagaa ctgggtggcg 1680
gacccggtgc cgggagcctt cagccgctcg ccctggaggg ctctcttcag aaacgcggca 1740
tcgtggagca gtgttgcaca tccatttgct cactctacca gctggagaac tactgcaacg 1800
gaagcggagt gaagcagacg ttgaattttg atttgttgaa gttggcgggg gatgtggaga 1860
gcaatccggg gccgatgccc cctagtggcc tcagactttt gttattgtta ttaccgcttt 1920
tatggctctt ggtgctgaca ccgggccgtc cggctgctgg cttgtcgact tgtaagacaa 1980
ttgatatgga attggtgaaa cgaaaacgga ttgaggccat ccgaggacag attttgagca 2040
agctgcggct tgcctcgcca ccctcgcaag gggaagtccc acccggacct ctaccagaag 2100
cagtcctagc gctgtacaac agtacaagag atagagtggc cggggaatcc gcagaaccag 2160
agcctgagcc tgaagccgat tattatgcaa aggaagtgac tagggtcctg atggtcgaga 2220
cccataacga aatctacgac aaattcaaac aaagtaccca ctctatctac atgttcttca 2280
acaccagtga gctaagagaa gccgtgcccg aacctgtgct tctttcccgc gcagaactcc 2340
gcctcttgag actcaaattg aaagttgaac aacacgtaga gctttaccag aaatactcta 2400
ataattcatg gcgatatctt tctaatcgtc tcctcgcccc atctgacagc cctgaatggc 2460
tctccttcga cgttacggga gttgtgcgcc agtggctcag cagaggcgga gagatagagg 2520
gctttcggct gagcgcacat agctctagcg actcaaggga caacacattg caagtggata 2580
ttaacggttt tacaactgga cggagagggg acctggcgac catccacggc atgaatagac 2640
ctttcctgct gctgatggct actcccctgg agagggcaca gcacttacag tcttccagac 2700
accggcgcgc cctggataca aactactgct tcagctccac cgaaaagaac tgttgcgtgc 2760
ggcagctgta cattgacttc agaaaggatc tgggctggaa gtggattcat gagcccaagg 2820
ggtatcatgc caacttctgt cttgggccat gcccatacat ctggtcactg gatacccagt 2880
actccaaagt tctggccttg tacaatcaac acaaccctgg agcttccgcc gctccttgct 2940
gtgtgcccca agccctagag cccctgccca tcgtttatta tgtcggacgc aagcccaaag 3000
tagaacagct atcaaatatg atcgtgagaa gctgcaagtg tagctgataa acgcgtcgag 3060
catgcatcta gggcggccaa ttccgcccct ctccccccca cccctctccc tccccccccc 3120
ctaacgttac tggccgaagc cgcttggaat aaggccggtg tgcgtttgtc tatatgttat 3180
tttccaccat attgccgtct tttggcaatg tgagggcccg gaaacctggc cctgtcttct 3240
tgacgagcat tcctaggggt ctttcccctc tcgccaaagg aatgcaaggt ctgttgaatg 3300
tcgtgaagga agcagttcct ctggaagctt cttgaagaca aacaacgtct gtagcgaccc 3360
tttgtagaca gcggaacccc ccacctggcg atagatgcct ctgcggccaa aagccacgtg 3420
tataagatac acctgcaaag gcggcacaac cccagtgcca cgttgtgagt tggatagttg 3480
tggaaagagt caaatggctc tcctcaagcg tattcaacaa ggggctgaag gatgcccaga 3540
aggtacccca ttgtatggga tctgatctgg ggcctcggtg cacatgcttt acatgtgttt 3600
agtcgaggtt aaaaaacgtc taggcccccc gaaccacggg gacgtggttt tcctttgaaa 3660
aacacgatga taatatgatg cacagctcag cactgctctg ttgcctggtc ctcctgactg 3720
gggtgagggc cagcccaggc cagggcaccc agtctgagaa cagctgcacc cacttcccag 3780
gcaacctgcc taacatgctt cgagatctcc gagatgcctt cagcagagtg aagactttct 3840
ttcaaatgaa ggatcagctg gacaacttgt tgttaaagga gtccttgctg gaggacttta 3900
agggttacct gggttgccaa gccttgtctg agatgatcca gttttacctg gaggaggtga 3960
tgccccaagc tgagaaccaa gacccagaca tcaaggcgca tgtgaactcc ctgggggaga 4020
acctgaagac cctcaggctg aggctacggc gctgtcatcg atttcttccc tgtgaaaaca 4080
agagcaaggc cgtggagcag gtgaagaatg cctttaataa gctccaagag aaaggcatct 4140
acaaagccat gagtgagttt gacatcttca tcaactacat agaagcctac atgacaatga 4200
agatacgaaa cgggagcggc gctactaact tcagcctgct gaagcaggct ggagacgtgg 4260
aggagaaccc tggacctatg tacagaatgc agctgctgag ctgcatcgcc ctgagcctgg 4320
ccctggtgac caacagcgca cccacgtcct ctagcaccaa gaagacccag ttacagttgg 4380
agcatctact tttagacctg caaatgattt tgaacggcat caacaactac aagaatccta 4440
aacttactcg catgcttacc ttcaaatttt acatgcccaa gaaggccacc gaactgaagc 4500
acttgcaatg tctggaggaa gaactcaagc cgctggagga agttctcaac ctcgcgcagt 4560
ccaagaattt ccacctccgg ccaagagacc tgatcagtaa cattaatgtg atagtgctgg 4620
agctgaaggg aagcgagact acatttatgt gcgagtacgc cgatgaaacc gctacaatcg 4680
tcgagttcct gaatagatgg atcacatttt gccagtcaat tatctctact ctgacatgat 4740
aactcgagtc tagagggccc gtttaaaccc gctgatcagc ctcgactgtg ccttctagtt 4800
gccagccatc tgttgtttgc ccctcccccg tgccttcctt gaccctggaa ggtgccactc 4860
ccactgtcct ttcctaataa aatgaggaaa ttgcatcgca ttgtctgagt aggtgtcatt 4920
ctattctggg gggtggggtg gggcaggaca gcaaggggga ggattgggaa gacaatagca 4980
ggcatgctgg ggatgcggtg ggctctatgg cttctactgg gcggttttat ggacagcaag 5040
cgaaccggaa ttgccagctg gggcgccctc tggtaaggtt gggaagccct gcaaagtaaa 5100
ctggatggct ttctcgccgc caaggatctg atggcgcagg ggatcaagct ctgatcaaga 5160
gacaggatga ggatcgtttc gcatggccaa agaagacaat attgaaatgc aaggtaccgt 5220
tcttgaaacg ttgcctaata ccatgttccg cgtagagtta gaaaacggtc acgtggttac 5280
tgcacacatc tccggtaaaa tgcgcaaaaa ctacatccgc atcctgacgg gcgacaaagt 5340
gactgttgaa ctgaccccgt acgacctgag caaaggccgc attgtcttcc gtagtcgctg 5400
ataaattatt aacgcttaca atttcctgat gcggtatttt ctccttacgc atctgtgcgg 5460
tatttcacac cgcatacagg tggcactttt cggggaaatg tgcgcggaac ccctatttgt 5520
ttatttttct aaatacattc aaatatgtat ccgctcatga gacaataacc ctgataaatg 5580
cttcaataat agcacgtgct aaaacttcat ttttaattta aaaggatcta ggtgaagatc 5640
ctttttgata atctcatgac caaaatccct taacgtgagt tttcgttcca ctgagcgtca 5700
gaccccgtag aaaagatcaa aggatcttct tgagatcctt tttttctgcg cgtaatctgc 5760
tgcttgcaaa caaaaaaacc accgctacca gcggtggttt gtttgccgga tcaagagcta 5820
ccaactcttt ttccgaaggt aactggcttc agcagagcgc agataccaaa tactgttctt 5880
ctagtgtagc cgtagttagg ccaccacttc aagaactctg tagcaccgcc tacatacctc 5940
gctctgctaa tcctgttacc agtggctgct gccagtggcg ataagtcgtg tcttaccggg 6000
ttggactcaa gacgatagtt accggataag gcgcagcggt cgggctgaac ggggggttcg 6060
tgcacacagc ccagcttgga gcgaacgacc tacaccgaac tgagatacct acagcgtgag 6120
ctatgagaaa gcgccacgct tcccgaaggg agaaaggcgg acaggtatcc ggtaagcggc 6180
agggtcggaa caggagagcg cacgagggag cttccagggg gaaacgcctg gtatctttat 6240
agtcctgtcg ggtttcgcca cctctgactt gagcgtcgat ttttgtgatg ctcgtcaggg 6300
gggcggagcc tatggaaaaa cgccagcaac gcggcctttt tacggttcct gggcttttgc 6360
tggccttttg ctcacatgtt ctt 6383
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| РЕКОМБИНАНТНЫЕ ВАКЦИНЫ ОТ FMDV И ИХ ПРИМЕНЕНИЕ | 2015 |
|
RU2745373C2 |
| РЕКОМБИНАНТНЫЕ ВЕКТОРЫ, ЭКСПРЕССИРУЮЩИЕ АНТИГЕНЫ ВИРУСА ПТИЧЬЕГО ГРИППА, И ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2761869C2 |
| ВАКЦИНЫ НА ОСНОВЕ ВИРУСОПОДОБНЫХ ЧАСТИЦ (VLP) СОБАЧЬЕГО ПАРВОВИРУСА (CPV) И ИХ ПРИМЕНЕНИЕ | 2016 |
|
RU2710854C1 |
| ХИМЕРНЫЙ БЕЛОК, СИНТЕТИЧЕСКАЯ ДНК, КОДИРУЮЩАЯ УКАЗАННЫЙ БЕЛОК, ЭКСПРЕССИОННЫЙ ВЕКТОР, ШТАММ-ПРОДУЦЕНТ СИНТЕТИЧЕСКОЙ ДНК И СПОСОБ ПОЛУЧЕНИЯ ПЛАЗМИДНОЙ ДНК | 2016 |
|
RU2736472C2 |
| Векторы для экспрессии простатоассоциированных антигенов | 2014 |
|
RU2650860C2 |
| ПРОТИВОРАКОВЫЕ ВАКЦИНЫ, НАПРАВЛЕННЫЕ НА MUC16, И ИХ ПРИМЕНЕНИЕ | 2018 |
|
RU2750689C1 |
| ПРОТИВОРАКОВЫЕ ВАКЦИНЫ, НАПРАВЛЕННЫЕ НА MUC16, И ИХ ПРИМЕНЕНИЕ | 2018 |
|
RU2777918C2 |
| БЕЛОК И ВАКЦИНА ПРОТИВ ИНФЕКЦИИ SARS-CoV-2 | 2020 |
|
RU2815060C1 |
| ПЕПТИДНЫЕ ВАКЦИНЫ ПРОТИВ ИНТЕРЛЕЙКИНА-31 | 2019 |
|
RU2786441C2 |
| СЛИТЫЙ БЕЛОК, СОДЕРЖАЩИЙ РЕЦЕПТОР TGF-БЕТА, И ЕГО ФАРМАЦЕВТИЧЕСКОЕ ПРИМЕНЕНИЕ | 2018 |
|
RU2776204C1 |
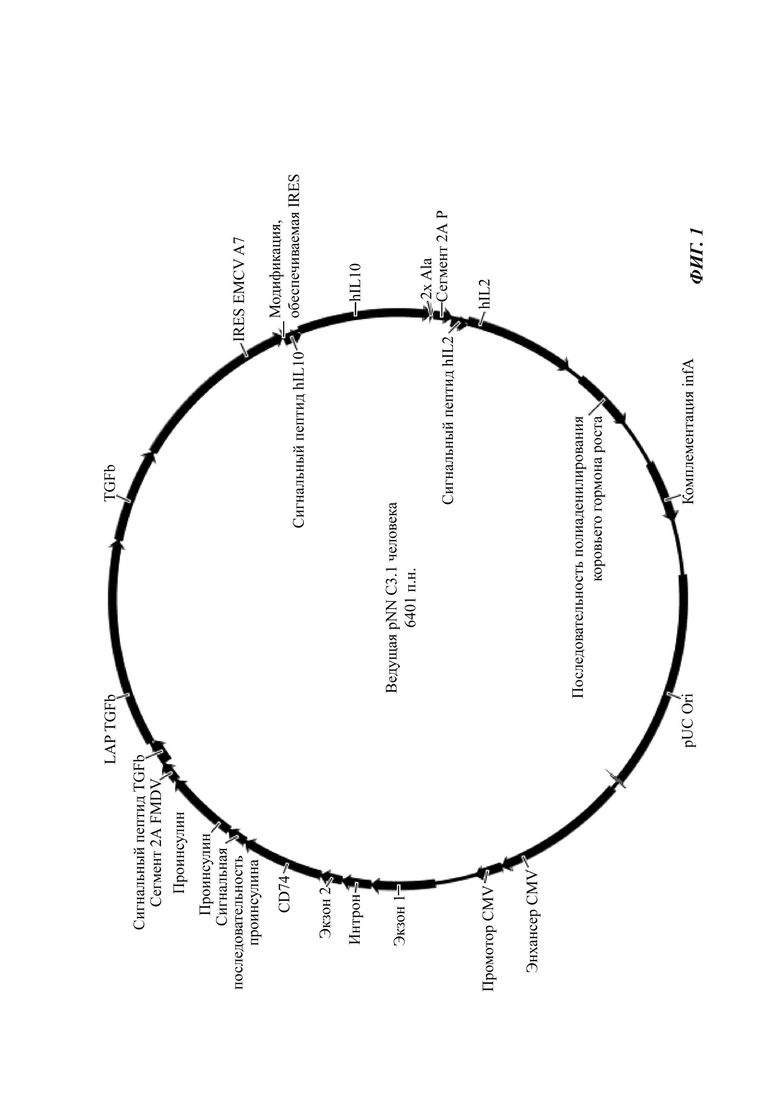
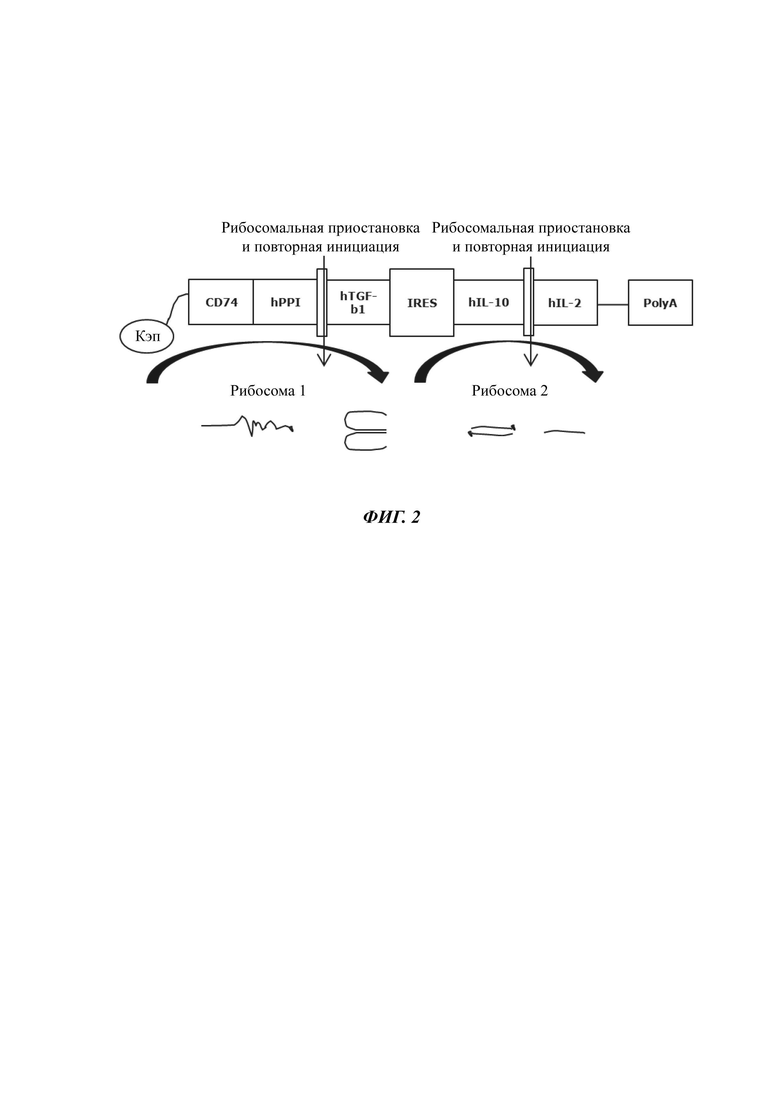
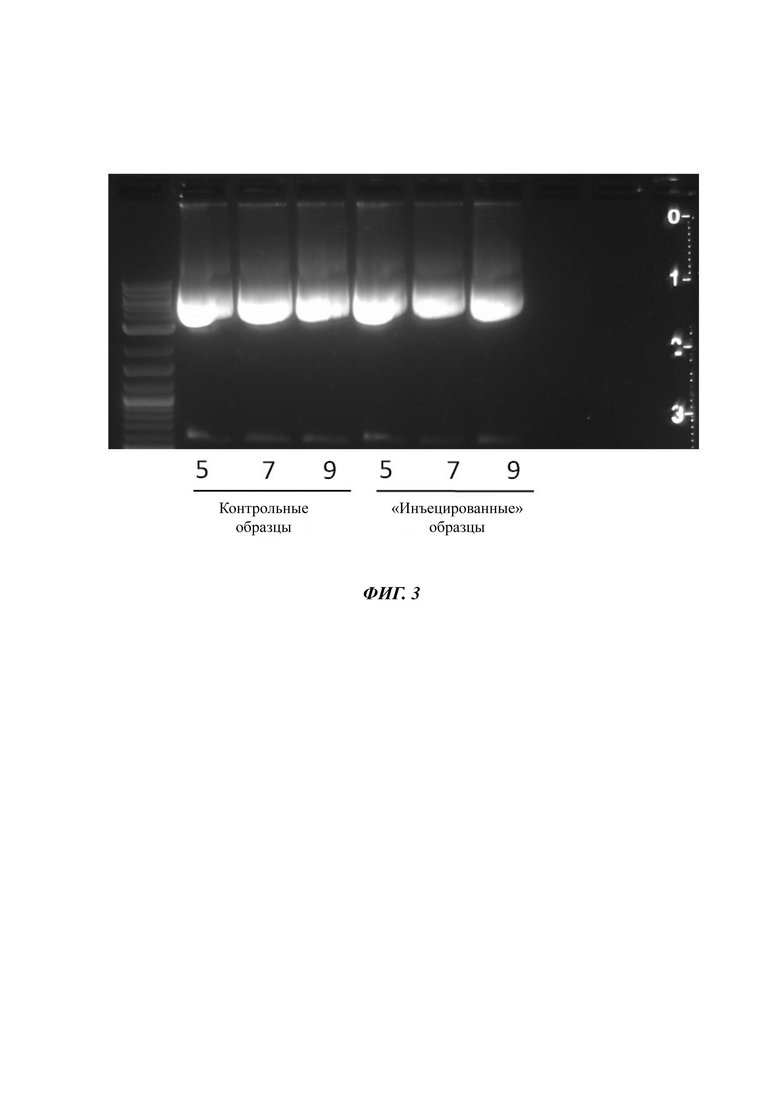

Изобретение относится к области биотехнологии, конкретно к плазмидам, кодирующим инсулиновые антигены и цитокины, и может быть использовано в медицине для лечения или предупреждения диабета типа I. Экспрессионная плазмида содержит i) нуклеотидную последовательность, кодирующую инсулиновый антиген, ii) нуклеотидную последовательность, кодирующую TGF-β, iii) нуклеотидную последовательность, кодирующую IL-10, и iv) нуклеотидную последовательность, кодирующую IL-2. При этом предложенная плазмида также содержит элемент 2А FMDV, отделяющий последовательность, кодирующую инсулиновый антиген, и последовательность, кодирующую TGF-β, элемент IRES EMCV, отделяющий последовательность, кодирующую TGF-β, и последовательность, кодирующую IL-10, и элемент 2А, отделяющий последовательность, кодирующую IL-10, и последовательность, кодирующую IL-2. Изобретение позволяет проводить придающую толерантность иммунизацию для эффективного предупреждения и/или отсрочки диабета типа I. 3 н. и 7 з.п. ф-лы, 4 ил., 11 табл., 13 пр.
1. Экспрессионная плазмида, которая содержит:
i) нуклеотидную последовательность, кодирующую инсулиновый антиген;
ii) нуклеотидную последовательность, кодирующую TGF-β;
iii) нуклеотидную последовательность, кодирующую IL-10; и
iv) нуклеотидную последовательность, кодирующую IL-2,
где указанная плазмида также содержит:
- элемент 2А FMDV, отделяющий последовательность, кодирующую инсулиновый антиген, и последовательность, кодирующую TGF-β,
- элемент IRES EMCV, отделяющий последовательность, кодирующую TGF-β, и последовательность, кодирующую IL-10, и
- элемент 2А, отделяющий последовательность, кодирующую IL-10, и последовательность, кодирующую IL-2.
2. Плазмида по п. 1, где указанный инсулиновый антиген выбран из группы, состоящей из проинсулина, пре-проинсулина и их функционального или иммунодоминантного пептидного фрагмента.
3. Плазмида по любому из пп. 1, 2, где указанный инсулиновый антиген представляет собой инсулин, нацеленный в эндосомы.
4. Плазмида по любому из пп. 1-3, где указанная плазмида экспрессирует инсулиновый антиген и TGF-β в по меньшей мере в 2 раза меньшем количестве, чем IL-10.
5. Плазмида по любому из пп. 1-4, где последовательность, кодирующая TGF-β, кодирует конститутивно активный TGF-β.
6. Иммунотерапевтическая ДНК-вакцина для лечения или предупреждения диабета типа I, содержащая эффективное количество плазмиды по любому из пп. 1-5.
7. Иммунотерапевтическая ДНК-вакцина по п. 6 для подкожного введения.
8. Иммунотерапевтическая ДНК-вакцина по п. 6 для внутримышечного введения.
9. Фармацевтическая композиция для лечения или предупреждения диабета типа I, содержащая эффективное количество иммунотерапевтической ДНК-вакцины по п. 6 или плазмиды по любому из пп. 1-5, где указанная фармацевтическая композиция содержит физиологический раствор и/или буфер, и/или хелатор.
10. Фармацевтическая композиция по п. 9, где указанный буфер не содержит какого- либо вируса, липидного сопакующего агента или конденсирующего агента.
| WO 2016057986 A1, 14.04.2016 | |||
| GOTTLIEB P | |||
| ET AL., Clinical optimization of antigen specific modulation of type 1 diabetes with the plasmid DNA platform, CLINICAL IMMUNOLOGY, 2013, v | |||
| Подъемник для выгрузки и нагрузки барж сплавными бревнами, дровами и т.п. | 1919 |
|
SU149A1 |
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| US 2002193330 A1, 19.12.2002 | |||
| TORRES-AGUILAR H | |||
| ET AL., IL-10/TGF-Î-Treated Dendritic Cells, Pulsed with Insulin, Specifically Reduce | |||

