ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Данное изобретение относится к области пептидных вакцин и из применений в клинических и научных процедурах, включая диагностические процедуры. Пептидные вакцины по данному изобретению могут найти применение для иммунизации и/или защиты млекопитающего, такого как кошка, собака, лошадь или человек, против заболевания, опосредуемого IL-31.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
Согласно определению рабочей группы Американского колледжа ветеринарной дерматологии, атопический дерматит представляет собой «воспалительное сопровождающееся зудом аллергическое заболевание кожи с характерными клиническими признаками и генетической предрасположенностью» (Olivry, et al. Veterinary Immunology and Immunopathology 2001; 81: 143-146). Рабочая группа также признала, что у собак заболевание ассоциировалось с аллерген-специфичным IgE (Olivry, et al. 2001, см. выше; Marsella & Olivry Clinics in Dermatology 2003; 21: 122-133). Сильный прурит наряду с вторичной алопецией и эритемой являются наиболее заметными и тревожащими симптомами для владельцев домашних животных.
Возможные факторы, вовлеченные в аллергический дерматит, многочисленны и плохо изучены. Атопический дерматит могут инициировать присутствующие в пище компоненты (Picco, et al. Vet Dermatol. 2008; 19: 150-155), а также аллергены из окружающей среды, такие как блохи, пылевые клещи, амброзия, растительные экстракты и т.д. Важную роль также играют генетические факторы. И хотя никаких подтверждений предрасположенности в отношении породы нет, полагают, что имеется некоторое влияние наследственности в повышении предрасположенности к атопическому дерматиту (Sousa & Marsella Veterinary Immunology and Immunopathology 2001; 81: 153-157; Schwartzman, et al. Clin. Exp. Immunol. 1971; 9: 549-569).
Согласно оценкам, распространенность атопического дерматита от общей популяции собак составляет 10% (Marsella & Olivry 2003 см. выше; Scott, et al. Canadian Veterinary Journal 2002; 43: 601-603; Hillier Veterinary Immunology and Immunopathology 2001; 81: 147-151). В целом, примерно 4,5 миллиона собак подвержены воздействию этого хронического и продолжающегося в течение всей жизни состояния. По всей видимости, уровень заболеваемости увеличивается. Полагали, что есть предрасположенность в отношении породы и пола, однако в зависимости от географического региона могут быть сильные различия (Hillier, 2001, см. выше; Picco, et al. 2008, см. выше).
У кошек аллергический дерматит представляет собой воспалительное сопровождающееся зудом заболевание кожи, которое, как считается, вызвано аномальной реакцией иммунной системы на вещества, которые не вызывают реакции у здоровых кошек. Наиболее характерным признаком аллергического дерматита у кошек является хронический рецидивирующий зуд. Типичные клинические проявления аллергического дерматита у кошек включают самоиндуцированную алопецию, милиарный дерматит, комплекс эозинофильной гранулемы (включая бляшки, гранулемы и вялотекущую язву) и сфокусированный зуд головы и шеи, характеризующийся экскориациями, эрозиями и/или язвами. Предрасположенность в отношении пород и пола не была продемонстрирована, а молодые кошки кажутся более склонными к заболеванию (Hobi et al. Vet Dermatol 2011 22: 406-413; Ravens et al. Vet Dermatol 2014; 25: 95-102; Buckely In Practice 2017; 39: 242-254).
Современные методы лечения кошек с диагнозом аллергический дерматит зависят от тяжести клинических признаков, продолжительности заболевания и предпочтений хозяина и включают аллерген-специфическую иммунотерапию и противозудные препараты, такие как глюкокортикоиды и циклоспорины (Buckley, см. выше). Иммунотерапия эффективна для некоторых пациентов, но требует частых инъекций, и клиническое улучшение может не наблюдаться в течение 6-9 месяцев (Buckley, см. выше). Иммунодепрессанты, такие как глюкокортикоиды и циклоспорины, как правило, эффективны, однако их длительное применение часто приводит к нежелательным побочным эффектам.
Атопический дерматит у лошадей считается потенциальной причиной зуда. В развитии атопического дерматита лошадей все больше признается роль аллергенов окружающей среды. Заболевание может быть сезонным или несезонным, в зависимости от аллергена(ов). О предрасположенности в отношении возраста, пород и пола не сообщалось. В предварительной работе в Школе ветеринарной медицины Калифорнийского университета в Дэвисе (SVM-UCD) средний возраст начала заболевания составлял 6,5 лет, чистокровные были самой распространенной породой, составлявшей 25% лошадей, а самцы (обычно мерины) преобладали над кобылами почти вдвое; однако эти данные относились только к 24 лошадям и еще не сравнивались с популяцией клиники в целом. Зуд, часто затрагивающий морду, задние ноги или туловище, является наиболее частым клиническим признаком атопического дерматита лошадей. Могут присутствовать алопеция, эритема, крапивница и папулы. Крапивница может быть довольно тяжелой, но не сопровождаться зудом. У лошади может быть семейная предрасположенность к крапивному атопическому дерматиту. У лошадей может быть вторичная пиодермия, которая проявляется избыточным шелушением, мелкими эпидермальными воротничками или папулами, покрытыми коркой («милиарный дерматит»). Диагностика атопического дерматита основана на клинических признаках и исключении других диагнозов, особенно гиперчувствительности к насекомым (мокрецам) (White Clin Tech Equine Pract 2005; 4: 311-313; Fadok Vet Clin Equine 2013; 29 541-550). В настоящее время проводят как симптоматическое лечение атопического дерматита у лошадей, подавляя воспаление и зуд, вызванные аллергической реакцией, так и этиотропное (т.е. выявляя аллергены, ответственные за заболевание, и создавая аллергенспецифичные вакцины). Симптоматический подход обычно необходим в краткосрочной перспективе, для комфортного самочувствия пациента и сведения к минимуму самотравмирования. Данный подход основан на использовании комбинации местной и системной терапии, включая антигистаминные препараты, незаменимые жирные кислоты, пентоксифиллин и глюкокортикоиды. Первичный подход к контролю аллергии на факторы окружающей среды включает идентификацию аллергенов, вызывающих реакцию гиперчувствительности. Дерматологи считают, что аллерген-специфическая иммунотерапия может помочь лошадям с атопией. Однако, как правило, у большинства лошадей улучшение наблюдается только после первых 6 месяцев иммунотерапии (Marsella Vet Clin Equine 2013; 29: 551-557). Кроме того, длительное применение иммунодепрессантов у лошадей может привести к нежелательным побочным эффектам.
Было показано, что интерлейкин-31 (IL-31), цитокин, продуцируемый Т-хелперными клетками 2 типа, вызывает зуд у людей, мышей и собак (Bieber N Engl J Med 2008; 358: 1483-1494; Dillon et al. Nat Immunol 2004; 5:752-60; US Patent No. 8,790,651 to Bammert et al.; Gonzalez et al. Vet Dermatl. 2013; 24(1): 48-53). IL-31 связывается с корецептором, состоящим из рецептора IL-31 A (IL-31 RA) и рецептора онкостатина М (OSMR) (Dillon et al. 2004, см. выше, и Bilsborough et al. J Allergy Clin Immunol. 2006 117(2):418-25). Активация рецептора приводит к фосфорилированию STAT через рецептор(ы) JAK. Экспрессия корецептора обнаружена в макрофагах, кератиноцитах и в ганглиях задних корешков.
Недавно было обнаружено, что IL-31 играет роль при дерматите, кожном зуде, аллергии и гиперчувствительности дыхательных путей. Cytopoint®, моноклональное антитело против собачьего IL-31, производимое Zoetis Inc., Parsippany, NJ, снижает зуд и кожные поражения у собак с атопическим дерматитом (Gonzalez et al. 2013, см. выше, Michels et al. Vet Dermatol. 2016; Dec; 27(6): 478-e129). Было бы желательно предоставить альтернативные подходы к профилактике и лечению IL-31-опосредованных заболеваний у млекопитающих. Было бы особенно желательно предоставить вакцины для уменьшения кожного зуда и повреждений кожи у собак, кошек, лошадей и людей с атопическим дерматитом.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
В одном воплощении настоящее изобретение относится к вакцинной композиции для иммунизации и/или защиты млекопитающего от расстройства, опосредованного IL-31, где композиция включает: комбинацию полипептида-носителя и по меньшей мере одного мимотопа, выбранного из мимотопа IL-31 кошки, мимотопа IL-31 собаки, мимотопа IL-31 лошади или мимотопа IL-31 человека, и адъюванта.
В одном воплощении вакцинной композиции мимотоп IL-31 собаки представляет собой и/или содержит в качестве своей части аминокислотную последовательность SVPADTFECKSF (SEQ ID NO: 186), SVPADTFERKSF (SEQ ID NO: 187), NSSAILPYFRAIRPLSDKNIIDKIIEQLDKLKF (SEQ ID NO: 192), APTHQLPPSDVRKIILELQPLSRG (SEQ ID NO: 196), TGVPES (SEQ ID NO: 200) или их варианты, которые сохраняют связывание с анти-1L-31 агентом.
В другом воплощении вакцинной композиции мимотоп IL-31 кошки представляет собой и/или содержит в качестве своей части аминокислотную последовательность SMPADNFERKNF (SEQ ID NO: 188), NG SAILPYFRAIRPLSDKNTIDKIIEQLDKLKF (SEQ ID NO: 193), APAHRLQPSDIRKIILELRPM SKG (SEQ ID NO: 197), IGLPES (SEQ ID NO: 201) или их варианты, которые сохраняют связывание с анти-1L-31 агентом.
В другом воплощении вакцинной композиции мимотоп IL-31 лошади представляет собой и/или содержит в качестве своей части аминокислотную последовательность SMPTDNFERKRF (SEQ ID NO: 189), NS SAILPYFKAISPSLNNDKSLYIIEQLDKLNF (SEQ ID NO: 194), GPIYQLQPKEIQAIIVELQNLS KK (SEQ ID NO: 198), KGVQKF (SEQ ID NO: 202) или их варианты, которые сохраняют связывание с анти-1L-31 агентом.
В одном воплощении вакцинной композиции мимотоп IL-31 человека представляет собой и/или содержит в качестве своей части аминокислотную последовательность SVPTDTHECKRF (SEQ ID NO: 190), SVPTDTHERKRF (SEQ ID NO: 191), HSPAIRAYLKTIRQLDNKSVIDEIIEHLDKLIF (SEQ ID NO: 195), LPVRLLRPSDDVQKIVEELQSLSKM (SEQ ID NO: 199), KGVLVS (SEQ ID NO: 203) или их варианты, которые сохраняют связывание с анти-1L-31 агентом.
В одном воплощении мимотоп, содержащийся в композиции вакцины, связывается с антителом против IL-31 или его антигенсвязывающей частью, которая специфически связывается с областью белка IL-31 млекопитающих, задействованной во взаимодействии белка IL-31 со своим ко-рецептором. В одном воплощении на связывание указанного антитела с указанной областью влияют мутации в области, связывающей эпитоп 15Н05, выбранные из группы, состоящей из:
а) области между аминокислотными остатками 124 и 135 последовательности IL-31 кошки, представленной SEQ ID NO: 157 (кошачий IL-31 дикого типа);
б) области между аминокислотными остатками 124 и 135 последовательности IL-31 собаки, представленной SEQ ID NO: 155 (собачий IL-31); и
в) области между аминокислотными остатками 118 и 129 последовательности IL-31 лошади, представленной SEQ ID NO: 165 (лошадиный IL-31).
В конкретном воплощении мимотоп связывается с антителом против IL-31 или его антигенсвязывающей частью, содержащей по меньшей мере одну из следующих комбинаций последовательностей участков, определяющих комплементарность (CDR):
1) антитело 15Н05: CDR1 вариабельной области тяжелой цепи (VH) с последовательностью SYTIH (SEQ ID NO: 1), VH-CDR2 с последовательностью NINPTSGYTENNQRFKD (SEQ ID NO: 2), VH-CDR3 с последовательностью WGFKYDGEWSFDV (SEQ ID NO: 3),CDR1 вариабельной области легкой цепи (VL) с последовательностью RASQGISIWLS (SEQ ID NO: 4), VL-CDR2 с последовательностью KASNLHI (SEQ ID NO: 5) и VL-CDR3 с последовательностью LQSQTYPLT (SEQ ID NO: 6);
2) антитело ZIL1: CDR1 вариабельной области тяжелой цепи (VH) с последовательностью SYGMS (SEQ ID NO: 13), VH-CDR2 с последовательностью HINSGGSSTYYADAVKG (SEQ ID NO: 14), VH-CDR3 с последовательностью VYTTLAAFWTDNFDY (SEQ ID NO: 15), вариабельной области легкой цепи (VL) последовательности SGSTNNIGIL A AT (SEQ ID NO: 16), VL-CDR2 с последовательностью SDGNRPS (SEQ ID NO: 17) и VL-CDR3 с последовательностью QSFDTTLDAYV (SEQ ID NO: 18);
3) антитело ZIL8: VH-CDR1 с последовательностью DYAMS (SEQ ID NO: 19), VH-CDR2 с последовательностью GIDSVGSGTSYADAVKG (SEQ ID NO: 20), VH-CDR3 с последовательностью GFPGSFEH (SEQ ID NO: 21), VL-CDR1 с последовательностью TGSSSNIGSGYVG (SEQ ID NO: 22), VL-CDR2 с последовательностью YNSDRPS (SEQ ID NO: 23), VL-CDR3 с последовательностью SVYDRTFNAV (SEQ ID NO: 24);
4) антитело ZIL9: VH-CDR1 с последовательностью SYDMT (SEQ ID NO: 25), VH-CDR2 с последовательностью DVNSGGTGTAYAVAVKG (SEQ ID NO: 26), VH-CDR3 с последовательностью LGVRDGLSV (SEQ ID NO: 27), VL-CDR1 с последовательностью SGESLNEYYTQ (SEQ ID NO: 28), VL-CDR2 с последовательностью RDTERPS (SEQ ID NO: 29), VL-CDR3 с последовательностью ESAVDTGTLV (SEQ ID NO: 30);
5) антитело ZIL11: VH-CDR1 с последовательностью TYVMN (SEQ ID NO: 31), VH-CDR2 с последовательностью SINGGGSSPTYADAVRG (SEQ ID NO: 32), VH-CDR3 с последовательностью SMVGPFDY (SEQ ID NO: 33), VL-CDR1 с последовательностью SGESLSNYYAQ (SEQ ID NO: 34), VL-CDR2 с последовательностью KDTERPS (SEQ ID NO: 35), VL-CDR3 с последовательностью ESAVSSDTIV (SEQ ID NO: 36);
6) антитело ZIL69: VH-CDR1 с последовательностью SYAMK (SEQ ID NO: 37), VH-CDR2 с последовательностью TINNDGTRTGYADAVRG (SEQ ID NO: 38), VH-CDR3 с последовательностью GNAESGCTGDHCPPY (SEQ ID NO: 39), VL-CDR1 с последовательностью SGESLNKYYAQ (SEQ ID NO: 40), VL-CDR2 с последовательностью KDTERPS (SEQ ID NO: 41), VL-CDR3 с последовательностью ESAVSSETNV (SEQ ID NO: 42); 7) антитело ZIL94: VH-CDR1 с последовательностью TYFMS (SEQ ID NO: 43), VH-CDR2 с последовательностью LISSDGSGTYYADAVKG (SEQ ID NO: 44), VH-CDR3 с последовательностью FWRAFND (SEQ ID NO: 45), VL-CDR1 с последовательностью GLNSGSVSTSNYPG (SEQ ID NO: 46), VL-CDR2 с последовательностью DTGSRPS (SEQ ID NO: 47), VL-CDR3 с последовательностью SLYTDSDILV (SEQ ID NO: 48);
8) антитело ZIL154: VH-CDR1 с последовательностью DRGMS (SEQ ID NO: 49), VH-CDR2 с последовательностью YIRYDGSRTDYADAVEG (SEQ ID NO: 50), VH-CDR3 с последовательностью WDGSSFDY (SEQ ID NO: 51), VL-CDR1 с последовательностью KASQSLLHSDGNTYLD (SEQ ID NO: 52), VL-CDR2 с последовательностью KVSNRDP (SEQ ID NO: 53), VL-CDR3 с последовательностью MQAIHFPLT (SEQ ID NO: 54);
9) антитело ZIL159: VH-CDR1 с последовательностью SYVMT (SEQ ID NO: 55), VH-CDR2 с последовательностью GFNSEGSRTAYADAVKG (SEQ ID NO: 56), VH-CDR3 с последовательностью GDIVATGTSY (SEQ ID NO: 57), VL-CDR1 последовательности SGETLNRF YTQ (SEQ ID NO: 58), VL-CDR2 с последовательностью KDTERPS (SEQ ID NO: 59), VL-CDR3 с последовательностью KSAVSIDVGV (SEQ ID NO: 60);
10) антитело ZIL171: VH-CDR1 с последовательностью TYVMN (SEQ ID NO: 61), VH-CDR2 с последовательностью SINGGGSSPTYADAVRG (SEQ ID NO: 62), VH-CDR3 с последовательностью SMVGPFDY (SEQ ID NO: 63), VL-CDR1 с последовательностью SGKSLSYYYAQ (SEQ ID NO: 64), VL-CDR2 с последовательностью KDTERPS (SEQ ID NO: 65), VL-CDR3 с последовательностью ESAVSSDTIV (SEQ ID NO: 66); или
11) вариант (1) - (10), отличающийся от соответствующего родительского антитела 15Н05, ZIL1, ZIL8, ZIL9, ZIL11, ZIL69, ZIL94, ZIL154, ZIL159 или ZIL171 добавлением, делецией и/или заменой одного или более чем одного аминокислотного остатка по меньшей мере в одном из CDR1, CDR2 или CDR3 VH или VL.
В некоторых воплощениях мимотоп, используемый в вакцинных композициях по настоящему изобретению, связывается с антителом против IL-31 или его антигенсвязывающей частью, которая связывается с кошачьим IL-31, при этом антитело включает цепь VL, содержащую изменения каркасной области 2 (FW2), выбранные из следующего: аспарагин вместо лизина в положении 42, изолейцин вместо валина в положении 43, валин вместо лейцина в положении 46, аспарагин вместо лизина в положении 49 и их комбинации, где положения относятся к нумерации SEQ ID NO: 127 (FEL_15H05_VL1).
В одном воплощении вакцинных композиций, описанных выше, мимотоп представляет собой мимотоп с ограниченной конформационной свободой. В конкретном воплощении мимотоп с ограниченной конформационной свободой представляет собой химически связанный циклический пептид.
В некоторых воплощениях описанных выше вакцинных композиций мимотоп химически конъюгирован с полипептидом-носителем. В других воплощениях полипепид-носитель и мимотоп являются частью рекомбинантного слитого белка.
В одном воплощении вакцинных композиций, описанных выше, полипептид-носитель, который комбинирован с мимотопом, включает бактериальный анатоксин или его производное, гемоцианин фиссуреллы (KLH) или вирусоподобную частицу. В одном воплощении мимотоп комбинирован с бактериальным анатоксином или производным, выбранным из столбнячного анатоксина, дифтерийного анатоксина, столбнячного анатоксина, белкового комплекса наружной мембраны N. meningitidis группы В, экзотоксина Pseudomonas или нетоксичного мутанта дифтерийного токсина (CRM197). В другом воплощении мимотоп комбинирован с вирусоподобной частицей, выбранной из HBsAg, HBcAg, бактериофага Qbeta E.coli, вируса Norwalk, вируса чумы собак (CDV) или гемагглютинина (НА) вируса гриппа. В конкретном воплощении мимотоп комбинирован с полипептидом-носителем, который содержит или состоит из CRM197.
В одном воплощении адъювант, содержащийся в описанных выше вакцинных композициях по настоящему изобретению, выбран из адъюванта масло в воде, адъюванта полимер и вода, адъюванта вода в масле, адъюванта гидроксида алюминия, адъюванта витамин Е и их комбинаций.
В одном воплощении адъювант представляет собой состав, содержащий сапонин, стерол, четвертичное аммониевое соединение и полимер. В конкретном воплощении сапонин представляет собой Quil А или его очищенную фракцию, стерол представляет собой холестерин, четвертичное аммониевое соединение представляет собой бромид диметилдиоктадециламмония (DDA), и полимер представляет собой полиакриловую кислоту.
В другом воплощении адъювант включает комбинацию одного или нескольких выделенных иммуностимулирующих олигонуклеотидов, стерола и сапонина. В конкретном воплощении один или несколько выделенных иммуностимулирующих олигонуклеотидов содержат CpG, стерол представляет собой холестерин, а сапонин представляет собой Quil А или его очищенную фракцию.
В данном изобретении также предложен способ защиты млекопитающего от заболевания, опосредованного IL-31. Такой способ включает введение млекопитающему вакцинной композиции согласно настоящему изобретению. В одном воплощении млекопитающее, которому вводят вакцину согласно настоящему изобретению, выбрано из собаки, кошки, лошади или человека. В конкретном воплощении вакцинная композиция включает мимотоп пептида IL-31, который вводят млекопитающему в дозе от приблизительно 10 мкг до приблизительно 100 мкг или соответствующей дозе, чтобы вызвать эквивалентный иммунный ответ. В одном воплощении вакцинная композиция включает мимотоп IL-31, который вводят млекопитающему, такому как кошка, в дозе приблизительно 10 мкг.
В одном воплощении заболевание, опосредованное IL-31, представляет собой состояние зуда или аллергическое состояние. В некоторых воплощениях состояние зуда или аллергическое состояние представляет собой состояние, сопровождающееся зудом, выбранное из атопического дерматита, экземы, псориаза, склеродермии и зуда. В других воплощениях зуд или аллергическое состояние представляет собой аллергическое состояние, выбранное из аллергического дерматита, летней экземы, крапивницы, эмфиземы легких, воспалительного заболевания дыхательных путей, рецидивирующей обструкции дыхательных путей, гиперчувствительности дыхательных путей, хронической обструктивной болезни легких и воспалительных процессов, вызванных аутоиммунитетом. В других воплощениях расстройство, опосредованное IL-31, представляет собой прогрессирование опухоли. В некоторых воплощениях расстройство, опосредованное IL-31, представляет собой эозинофильную болезнь или мастоцитомы.
В настоящем документе также предложен способ качественного или количественного определения антитела против IL-31 в образце. Такой способ включает инкубирование образца, содержащего антитело против IL-31, по меньшей мере, с одним мимотопом, выбранным из мимотопа кошачьего IL-31, мимотопа собачьего IL-31, мимотопа лошадиного IL-31 и мимотопа человеческого IL-31 и качественное или количественное определение в образце анти-IL-31 агента.
В одном из воплощений мимотоп собачьего IL-31, используемый в способе качественного или количественного определения антитела против IL-31 в образце, представляет собой и/или содержит в качестве своей части аминокислотную последовательность SVPADTFECKSF (SEQ ID NO: 186), SVPADTFERKSF (SEQ ID NO: 187), NSSAILPYFRAIRPLSDKNIIDKIIEQLDKLKF (SEQ ID NO: 192), APTHQLPPSDVRKIILELQPLSRG (SEQ ID NO: 196), TGVPES (SEQ ID NO: 200) или их варианты, которые сохраняют связывание с анти-IL-31 агентом.
В другом воплощении мимотоп кошачьего IL-31, используемый в таком способе, представляет собой и/или содержит в качестве своей части аминокислотную последовательность SMPADNFERKNF (SEQ ID NO: 188), NGSAILPYFRAIRPLSDKNTIDKIIEQLDKLKF (SEQ ID NO: 193), APAHRLQPSDIRKIILELRPM SKG (SEQ ID NO: 197), IGLPES (SEQ ID NO: 201) или их варианты, которые сохраняют связывание анти-IL-31 агентом.
В следующем воплощении мимотоп лошадиного IL-31, используемый в таком способе, представляет собой и/или содержит в качестве своей части аминокислотную последовательность SMPTDNFERKRF (SEQ ID NO: 189), NSSAILPYFKAISPSLNNDKSLYIIEQLDKLNF (SEQ ID NO: 194), GPIYQLQPKEIQAIIVELQNLS KK (SEQ ID NO: 198), KGVQKF (SEQ ID NO: 202) или их варианты, которые сохраняют связывание с анти-IL-31 агентом.
Кроме того, мимотоп человеческого IL-31, используемый в таком способе, представляет собой и/или содержит в качестве своей части аминокислотную последовательность SVPTDTHECKRF (SEQ ID NO: 190), SVPTDTHERKRF (SEQ ID NO: 191), HSPAIRAYLKTIRQLDNKSVIDEIIEHLDKLIF (SEQ ID NO: 195), LPVRLLRPSDDVQKIVEELQSLSKM (SEQ ID NO: 199), KGVLVS (SEQ ID NO: 203) или их варианты, которые сохраняют связывание анти-IL-31 агентом.
В одном из вариантов вышеописанного способа диагностики мимотоп представляет собой реагент для захвата, связанный с твердой поверхностью. В одном воплощении образец добавляют к мимотопу, представляющему собой реагент для захвата, и затем добавляют вторичные детектирующие реагенты для определения количества антитела в образце.
В данном изобретении также предложен способ определения количества IL-31 в образце от млекопитающего. Такой способ включает инкубацию образца от млекопитающего, содержащего IL-31, с комплексом меченое антитело против IL-31: Мимотоп IL-31, связанным с твердой поверхностью, где мимотоп в комплексе выбран из группы, состоящей из мимотопа кошачьего IL- 31, мимотопа собачьего IL-31, мимотопа лошадиного IL-31 и мимотопа человеческого IL-31; и определение уровня IL-31 в образце, где меченое антитело против IL-31 в комплексе имеет более низкую аффинность к мимотопу в комплексе, по сравнению с его аффинностью к IL-31 в образце. В одном варианте этого способа этап определения включает измерение сигнала, исходящего от меченого антитела, которое высвобождается с твердой поверхности, когда IL-31 в образце связывается с меченым антителом против IL-31 комплекса, при этом уровень IL-31 в образце обратно пропорционален сигналу.
В одном воплощении мимотоп собачьего IL-31, используемый в способе определения количества IL-31 в образце, представляет собой и/или содержит в качестве своей части аминокислотную последовательность SVPADTFECKSF (SEQ ID NO: 186), SVPADTFERKSF (SEQ ID NO: 187), NSSAILPYFRAIRPLSDKNIIDKIIEQLDKLKF (SEQ ID NO: 192), APTHQLPPSDVRKIILELQPLSRG (SEQ ID NO: 196), TGVPES (SEQ ID NO: 200) или их варианты, которые сохраняют связывание с анти-IL-31 агентом.
В другом воплощении мимотоп кошачьего IL-31, используемый в таком способе, представляет собой и/или содержит в качестве своей части аминокислотную последовательность SMPADNFERKNF (SEQ ID NO: 188), NGSAILPYFRAIRPLSDKNTIDKIIEQLDKLKF (SEQ ID NO: 193), APAHRLQPSDIRKIILELRPM SKG (SEQ ID NO: 197), IGLPES (SEQ ID NO: 201) или их варианты, которые сохраняют связывание с анти-IL-31 агентом.
В еще одном воплощении мимотоп лошадиного IL-31, используемый в таком способе, представляет собой и/или содержит в качестве своей части аминокислотную последовательность SMPTDNFERKRF (SEQ ID NO: 189), NSSAILPYFKAISPSLNNDKSLYIIEQLDKLNF (SEQ ID NO: 194), GPIYQLQPKEIQAIIVELQNLS KK (SEQ ID NO: 198), KGVQKF (SEQ ID NO: 202) или их варианты, которые сохраняют связывание с анти-IL-31 агентом.
В следующем воплощении мимотоп человеческого IL-31, используемый в таком способе, представляет собой и/или содержит в качестве своей части аминокислотную последовательность SVPTDTHECKRF (SEQ ID NO: 190), SVPTDTHERKRF (SEQ ID NO: 191), HSPAIRAYLKTIRQLDNKSVIDEIIEHLDKLIF (SEQ ID NO: 195), LPVRLLRPSDDVQKIVEELQSLSKM (SEQ ID NO: 199), KGVLVS (SEQ ID NO: 203) или их варианты, которые сохраняют связывание с анти-IL-31 агентом.
В некоторых воплощениях описанных выше способов диагностики по изобретению мимотоп связывается с антителом к IL-31 или его антигенсвязывающей частью, которая специфически связывается с областью белка IL-31 млекопитающих, задействованной во взаимодействии белка IL-31 со своим ко-рецептором. В одном воплощении способов диагностики по изобретению на связывание указанного антитела с указанной областью влияют мутации в области эпитопа связывания 15Н05, выбранные из группы, состоящей из:
а) области между аминокислотными остатками 124 и 135 последовательности IL-31 кошки, представленной SEQ ID NO: 157 (кошачий IL-31 дикого типа);
б) области между аминокислотными остатками 124 и 135 последовательности IL-31 собаки, представленной SEQ ID NO: 155 (собачий IL-31); и
в) области между аминокислотными остатками 118 и 129 последовательности IL-31 лошади, представленной SEQ ID NO: 165 (лошадиный IL-31).
В одном конкретном воплощении любых способов диагностики по данному изобретению мимотоп связывается с антителом против IL-31 или его антигенсвязывающей частью, содержащей по меньшей мере одну из следующих комбинаций последовательностей участков, определяющих комплементарность (CDR):
1) антитело 15Н05: CDR1 вариабельной области тяжелой цепи (VH) с последовательностью SYTIH (SEQ ID NO: 1), VH-CDR2 с последовательностью NINPTSGYTENNQRFKD (SEQ ID NO: 2), VH-CDR3 с последовательностью WGFKYDGEWSFDV (SEQ ID NO: 3),CDR1 вариабельной области легкой цепи (VL) с последовательностью RASQGISIWLS (SEQ ID NO: 4), VL-CDR2 с последовательностью KASNLHI (SEQ ID NO: 5) и VL-CDR3 с последовательностью LQSQTYPLT (SEQ ID NO: 6);
2) антитело ZIL1: CDR1 вариабельной области тяжелой цепи (VH) с последовательностью SYGMS (SEQ ID NO: 13), VH-CDR2 с последовательностью HINSGGSSTYYADAVKG (SEQ ID NO: 14), VH-CDR3 с последовательностью VYTTLAAFWTDNFDY (SEQ ID NO: 15), вариабельной области легкой цепи (VL) с последовательностью SGSTNNIGILAAT (SEQ ID NO: 16), VL-CDR2 с последовательностью SDGNRPS (SEQ ID NO: 17) и VL-CDR3 с последовательностью QSFDTTLDAYV (SEQ ID NO: 18);
3) антитело ZIL8: VH-CDR1 с последовательностью DYAMS (SEQ ID NO: 19), VH-CDR2 с последовательностью GIDSVGSGTSYADAVKG (SEQ ID NO: 20), VH-CDR3 с последовательностью GFPGSFEH (SEQ ID NO: 21), VL-CDR1 с последовательностью TGSSSNIGSGYVG (SEQ ID NO: 22), VL-CDR2 с последовательностью YNSDRPS (SEQ ID NO: 23), VL-CDR3 с последовательностью SVYDRTFNAV (SEQ ID NO: 24);
4) антитело ZIL9: VH-CDR1 с последовательностью SYDMT (SEQ ID NO: 25), VH-CDR2 с последовательностью DVNSGGTGTAYAVAVKG (SEQ ID NO: 26), VH-CDR3 с последовательностью LGVRDGLSV (SEQ ID NO: 27), VL-CDR1 с последовательностью SGESLNEYYTQ (SEQ ID NO: 28), VL-CDR2 с последовательностью RDTERPS (SEQ ID NO: 29), VL-CDR3 с последовательностью ESAVDTGTLV (SEQ ID NO: 30);
5) антитело ZIL11: VH-CDR1 с последовательностью TYVMN (SEQ ID NO: 31), VH-CDR2 с последовательностью SINGGGSSPTYADAVRG (SEQ ID NO: 32), VH-CDR3 с последовательностью SMVGPFDY (SEQ ID NO: 33), VL-CDR1 с последовательностью SGESLSNYYAQ (SEQ ID NO: 34), VL-CDR2 с последовательностью KDTERPS (SEQ ID NO: 35), VL-CDR3 с последовательностью ESAVSSDTIV (SEQ ID NO: 36);
6) антитело ZIL69: VH-CDR1 с последовательностью SYAMK (SEQ ID NO: 37), VH-CDR2 с последовательностью TINNDGTRTGYADAVRG (SEQ ID NO: 38), VH-CDR3 с последовательностью GNAESGCTGDHCPPY (SEQ ID NO: 39), VL-CDR1 с последовательностью SGESLNKYYAQ (SEQ ID NO: 40), VL-CDR2 с последовательностью KDTERPS (SEQ ID NO: 41), VL-CDR3 с последовательностью ESAVSSETNV (SEQ ID NO: 42);
7) антитело ZIL94: VH-CDR1 с последовательностью TYFMS (SEQ ID NO: 43), VH-CDR2 с последовательностью LISSDGSGTY YADA VKG (SEQ ID NO: 44), VH-CDR3 с последовательностью FWRAFND (SEQ ID NO: 45), VL-CDR1 с последовательностью GLNSGSVSTSNYPG (SEQ ID NO: 46), VL-CDR2 с последовательностью DTGSRPS (SEQ ID NO: 47), VL-CDR3 с последовательностью SLYTDSDILV (SEQ ID NO: 48);
8) антитело ZIL154: VH-CDR1 с последовательностью DRGMS (SEQ ID NO: 49), VH-CDR2 с последовательностью YIRYDGSRTDYADAVEG (SEQ ID NO: 50), VH-CDR3 с последовательностью WDGSSFDY (SEQ ID NO: 51), VL-CDR1 с последовательностью KASQSLLHSDGNTYLD (SEQ ID NO: 52), VL-CDR2 с последовательностью KVSNRDP (SEQ ID NO: 53), VL-CDR3 с последовательностью MQAIHFPLT (SEQ ID NO: 54);
9) антитело ZIL159: VH-CDR1 с последовательностью SYVMT (SEQ ID NO: 55), VH-CDR2 с последовательностью GINSEGSRTAYADAVKG (SEQ ID NO: 56), VH-CDR3 с последовательностью GDIVATGTSY (SEQ ID NO: 57), VL-CDR1 с последовательностью SGETLNRFYTQ (SEQ ID NO: 58), VL-CDR2 с последовательностью KDTERPS (SEQ ID NO: 59), VL-CDR3 с последовательностью KSAVSIDVGV (SEQ ID NO: 60);
10) антитело ZIL171: VH-CDR1 с последовательностью TYVMN (SEQ ID NO: 61), VH-CDR2 с последовательностью SINGGGSSPTYADAVRG (SEQ ID NO: 62), VH-CDR3 с последовательностью SMVGPFDY (SEQ ID NO: 63), VL-CDR1 с последовательностью SGKSLSYYYAQ (SEQ ID NO: 64), VL-CDR2 последовательности KDTERPS (SEQ ID NO: 65), VL-CDR3 с последовательностью ESAVSSDTIV (SEQ ID NO: 66); или
11) вариант (1) (10), отличающийся от соответствующего родительского антитела 15Н05, ZIL1, ZIL8, ZIL9, ZIL11, ZIL69, ZIL94, ZIL154, ZIL159 или ZIL171 добавлением, делецией и/или заменой одного или более чем одного аминокислотного остатка по меньшей мере в одном из CDR1, CDR2 или CDR3 VH или VL.
В некоторых воплощениях мимотоп, используемый в способах диагностики по настоящему изобретению, связывается с антителом против IL-31 или его антигенсвязывающей частью, которая связывается с кошачьим IL-31, при этом антитело включает цепь VL, содержащую изменения каркасной области 2 (FW2), выбранные из следующего: аспарагин вместо лизина в положении 42, изолейцин вместо валина в положении 43, валин вместо лейцина в положении 46, аспарагин вместо лизина в положении 49 и их комбинации, где положения относятся к нумерации SEQ ID NO: 127 (FEL_15H05_VL1).
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
Фиг. 1 представляет собой выравнивание, показывающее сохранение аминокислотной последовательности между IL-31 от разных биологических видов. В частности, представлено сравнение SEQ ID NO: 155 (IL-31 собаки), SEQ ID NO: 157 (IL-31 кошки), SEQ ID NO: 165 (IL-31 лошади) и SEQ ID NO: 181 (IL-31 человека). Также указан процент идентичности аминокислотных последовательностей собачьего, кошачьего, лошадиного и человеческого IL-31.
На Фиг. 2 подробно показана аффинность, с которой антитела-кандидаты с CDR мышиного происхождения связывают кошачий и собачий IL-31, при использовании поверхностного плазмонного резонанса (SPR) в системе Biacore (Biacore Life Sciences (GE Healthcare), Уппсала, Швеция).
Фиг. 3 представляет собой таблицу, показывающую эффективность (IC50 (мкг/мл)) антител-кандидатов с CDR мышиного происхождения, по данным анализов на клетках собак и кошек. В частности, оценивали способность антител-кандидатов ингибировать опосредованное IL-31 фосфорилирование STAT в макрофагоподобных клетках DH-82 собаки или FCWF4 кошки.
На Фиг. 4 показаны результаты, полученные для связывания моноклональных антител-кандидатов с CDR собачьего происхождения с различными белками при использовании методов непрямого ELISA и Biacore. Для непрямого ELISA оценивали связывание (ELISA OD) с кошачьим IL-31 дикого типа и мутантным кошачьим IL-31 15Н05, который имел мутации в области эпитопа, распознаваемого моноклональным антителом 15Н05. Для подтверждения связывания проводили анализ biacore с использованием белков IL-31 собаки, кошки, лошади, человека, мутантных белков 15Н05 и 11Е12 кошачьего IL-31 в качестве поверхностей и единственной исследуемой концентрации антитела. Мутант 11Е12 кошачьего IL-31 имел мутации в области эпитопа, распознаваемого моноклональным антителом 11Е12.
На Фиг. 5 - Фиг. 5А показано выравнивание последовательности VL мышиного антитела 11Е12 (SEQ ID NO: 73), сравнивающее ранее раскрытые канинизированные последовательности 11Е12, обозначенные как Can_11E12_VL_cUn_1 (SEQ ID NO: 182) и CAN_llE12_VL_cUn_FW2 (SEQ ID NO: 184) с фелинизированными версиями, обозначенными как FEL_11Е12_VH1 (SEQ ID NO: 111) и FEL_11E12_VL1_FW2 (SEQ ID NO: 117). Под выравниванием на Фиг.5А отмечены точки, показывающие положения соответствующих изменений в Fel_11E12_VL1, которые были необходимы для восстановления аффинности этого антитела к белку IL-31. На Фиг. 5В показано выравнивание последовательности VL мышиного антитела 15Н05, обозначенной в данном документе как MU_15Н05_VL (SEQ ID NO: 69), с последовательностями VL фелинизированного 15Н05, обозначенными в данном документе как FE1_15H05_VL1 (SEQ ID NO: 127) и FE1_15H05_VL_FW2 (SEQ ID NO: 135). Точки под выравниванием на рисунке 5 В указывают на необходимые изменения в VL фелинизированного 15Н05 (Fe1_15Н05_VL1), которые требовались не только для восстановления, но и для улучшения его аффинности к собачьему и кошачьему IL-31 по сравнению с мышиными и химерными формами этого антитела.
На Фиг.6 - Фиг.6А показано выравнивание кошачьего IL-31 дикого типа (SEQ ID NO: 157) с мутантами 15Н05 (SEQ ID NO: 163) и 11Е12 (SEQ ID NO: 161) с выделением положений, в которых происходят замены на аланин. На Фиг.6 В показана построенная по гомологии модель кошачьего IL-31, на которой выделены положения двух аминокислот, участвующих в связывании антител 11Е12 (сайт 1) и 15Н05 (сайт 2). Фиг.6С представляет собой график, показывающий результаты, полученные для связывания моноклональных антител 11Е12 и 15Н05 с кошачьим IL-31 дикого типа и с мутантными белками IL-31 15Н05 (SEQ ID NO: 163) и 11Е12 (SEQ ID NO: 161), когда в качестве иммобилизованных антигенов использовали белок дикого типа и указанные мутанты.
Фиг. 7 представляет собой графики, показывающие оценки конкурентного связывания mAb 15Н05 и 11Е12 с использованием Biacore. На Фиг. 7А показаны данные конкурентного связывания для мышиных антител 15Н05 и 11Е12 с собачьим IL-31. На Фиг. 7В показаны данные конкурентного связывания антител 15Н05 и 11Е12 с иммобилизованным на поверхности кошачьим IL-31.
Фиг. 8 представляет собой график, показывающий результаты, полученные для связывания отдельных субъединиц рецепторов OSMR и IL-31Ra с кошачьим IL-31 дикого типа и с мутантными белками IL-31 15Н05 (SEQ ID NO: 163) и 11Е12 (SEQ ID NO: 161), когда в качестве иммобилизованных антигенов использовали белок дикого типа и указанные мутанты.
Фиг. 9 представляет собой график, демонстрирующий предварительные данные по эффективности химеры мышиного:кошачьего 11Е12, химеры мышиного:кошачьего 15Н05 и фелинизированного 11Е12 (кошачий 11Е12 1.1) на модели индуцированного IL-31 зуда у кошек.
Фиг. 10 представляет собой график, демонстрирующий оценку эффективности in vivo фелинизированного антитела 15Н05 против IL-31, обозначенного ZTS-361, на модели с провокацией зуда у кошек. На Фиг. 10А показано исходное поведение до провокации зуда в группе Т01, получавшей плацебо, и группе Т02, получавшей антитело ZTS-361, начиная со дня - 7 до дня 28, где день 0 был днем введения антитела группе Т02. На Фиг. 10В показана эффективность антитела ZTS-361, демонстрирующего значительное снижение зуда, наблюдавшееся на 7 день (р менее 0,0001), 21 день (р менее 0,0027) и 28 день (р менее 0,0238) после индукции IL-31 по сравнению с контролем, получавшим плацебо.
Фиг. 11 - Фиг. 11А представляет собой график, демонстрирующий уровни IL-31 в плазме у принадлежащих клиентам животных среди собак с атопическим и аллергическим дерматитом по сравнению с нормальными лабораторными животными. Фиг. 11В представляет собой график, демонстрирующий результаты недавнего исследования для определения уровней IL-31 в сыворотке у кошек с предполагаемым диагнозом аллергический дерматит (АД) из нескольких географических областей США. Фиг. 11С представляет собой график, демонстрирующий фармакокинетический профиль собачьего IL-31 у собак после подкожного введения собачьего IL-31 в дозе 1,75 мкг/кг.
На Фиг. 12 представлена таблица, показывающая результаты сканирующего мутагенеза собачьего IL-31 с полной заменой аминокислот, указанных на Фиг. 12. Каждое изображенное положение в полноразмерном собачьем белке IL-31 (SEQ ID NO: 155) отдельно заменяли одной из других возможных 19 аминокислот и оценивали связывание антитела 15Н05 с помощью непрямого ELISA. Для сравнения показаны соответствующие области кошачьего (SEQ ID NO: 157), лошадиного (SEQ ID NO: 165) и человеческого IL-31 (SEQ ID NO: 181).
Фиг. 13 - Фиг. 13A представляет собой таблицу, показывающую последовательности и химические линкеры различных пептидов с ограниченной конформационной свободой. Пептид ZTS-561 содержит аминокислотную последовательность N-TEISVPADTFERKSFILT-C, которая соответствует положениям 121-138 последовательности SEQ ID NO: 155 с заменой аргинина (R) на цистеин (С) в положении номер 132. Пептид ZTS-562 содержит аминокислотную последовательность N-EISVPADTFERKSF-C, которая соответствует положениям 122-135 последовательности SEQ ID NO: 155 с заменой аргинина (R) на цистеин (С) в положении номер 132. Пептид ZTS-563 содержит аминокислотную последовательность N-AKVSMPADNFERKNFILT-C, которая соответствует положениям 121-138 последовательности SEQ ID NO: 157 с заменой треонина (Т) на аланин (А) в положении номер 138. Пептид ZTS-564 содержит аминокислотную последовательность N-TEISVPADTFERKSFILT-C, которая соответствует положениям 121-138 последовательности SEQ ID NO: 155. Каждый из пептидов ZTS-561, ZTS-562, ZTS-563 и ZTS-564 также включает N и С концевые цистеины, как изображено, для облегчения конъюгации с использованием свободных тиоловых групп. На Фиг. 13В показаны результаты оценки аффинности каждого из пептидов ZTS-561, ZTS-562, ZTS-563 и ZTS-564, которые независимо конъюгировали с полипептидом-носителем (CRM-197). Для оценки аффинности каждый пептид независимо иммобилизовали на поверхности чипа biacore и определяли KD для фелинизированного mAb против IL-31 15Н05 (ZTS-927).
На Фиг. 14 изображена схема исследования для определения иммуногенности, проведенного для оценки способности CRM-197-конъюгированных мимотопов IL-31 вызывать эпитоп-специфический иммунный ответ, направленный на соответствующую область белка IL-31, где связываются антитело 15Н05 и другие описанные в данном документе антитела к IL-31.
На Фиг. 15 приведены графики, показывающие сывороточные титры, полученные после вакцинации собак собачьими и кошачьими мимотопами IL-31 15Н05 и кошачьим полноразмерным белком IL-31, представленные по группам воздействия, показывающие ответ в каждый день, когда бралась сыворотка. На Фиг. 15А показаны средние титры антител собак на полноразмерный кошачий белок IL-31 (SEQ ID NO: 159). На Фиг. 15В показаны средние титры антител собак на полноразмерный кошачий мутант IL-31 15Н05 (SEQ ID NO: 163). На Фиг. 15С показаны средние титры антител собак на полноразмерный собачий IL-31 (SEQ ID NO: 155). На Фиг. 15D показаны средние титры антител собак на полноразмерный лошадиный IL-31 (SEQ ID NO: 165). На Фиг. 15Е показаны средние титры антител собак на полноразмерный человеческий IL-31 (SEQ ID NO: 181).
На Фиг. 16 - Фиг. 16А изображена схема исследования для определения иммуногенности, проведенного для оценки способности CRM-197-конъюгированного полноразмерного собачьего белка IL-31 или мимотопов вызывать иммунный ответ у лабораторных собак породы бигль. Каждый из описанных в данном документе мимотопов разработан, чтобы вызывать эпитоп-специфический иммунный ответ, направленный на соответствующую область белка IL-31, где связываются антитело 15Н05 и другие описанные в данном документе антитела к IL-31. Последовательности и химические линкеры пептидов различных мимотопов показаны как группы Т02-Т04. Пептид ZTS-420 содержит аминокислотную последовательность N-TEISVPADTFERKSFILT-C, которая соответствует положениям 121-138 последовательности SEQ ID NO: 155 с заменой аргинина (R) на цистеин (С) в положении номер 132. Пептид ZTS-421 содержит аминокислотную последовательность N-TNISVPTDTHECKRFILT-C, которая соответствует положениям 122-139 последовательности SEQ ID NO: 181. Пептид ZTS-766 содержит аминокислотную последовательность N-NSSAILPYFRAIRPLSDKNIIDKIIEQLDKLKF-C, которая соответствует положениям 83-115 последовательности SEQ ID NO: 155. Каждый из пептидов ZTS-420, ZTS-421 и ZTS-766 также включает N и С концевые цистеины, как изображено, для облегчения конъюгации с использованием свободных тиоловых групп. ZTS-766 также содержит дополнительную спейсерную последовательность из трех аминокислот (GSG) рядом с N концевым цистеином. На Фиг. 16В показаны гомологичные последовательности, позволяющие сравнить мимотоп спирали ВС собачьего IL-31 (ZTS-766) с соответствующей последовательностью кошачьего, лошадиного и человеческого IL-31 и включает номер референтной последовательности и аминокислотное положение для каждого из них.
На Фиг. 17 приведены графики, показывающие сывороточные титры, полученные после вакцинации собак мимотопами собачьего и человеческого IL-31 15Н05, мимотопом спирали ВС собачьего IL-31 и кошачьим полноразмерным белком IL-31, представленные по группам воздействия, показывающие ответ в каждый день, когда проводили забор сыворотки. Собакам осуществляли введение в дни 0, 28 и 56, указанные стрелками. На фигуре 17А представлены средние титры собачьих антител к полноразмерному собачьему белку IL-31 (SEQ ID NO: 155). На Фиг. 17В показаны средние титры собачьих антител к полноразмерному человеческому IL-31 (SEQ ID NO: 181) в дни 0, 42 и 84 только в группе Т03. У собак в группе Т03 (мимотоп человеческого 15Н05) отсутствовала CRAR на собачий IL-31 (данные не приведены).
На Фиг. 18 - Фиг. 18А изображена схема исследования для определения иммуногенности, проведенного для оценки способности CRM-197-конъюгированного полноразмерного кошачьего белка IL-31 или мимотопов вызывать иммунный ответ у лабораторных кошек. Все группы воздействия получали смесь адъювантов, включающую гликолипидный адъювант Bay R1005 (N- (2-дезокси-2-L-лейциламино-β-D-глюкопиранозил)-N-окгадецилдодеканоиламидгидроацетат), а также олигонуклеотиды CpG.
Каждый из описанных в данном документе мимотопов разработан, чтобы вызывать эпитоп-специфический иммунный ответ, направленный на соответствующую область белка IL-31, где связываются антитело 15Н05 и другие описанные в данном документе антитела к IL-31. Последовательности и химические линкеры пептидов различных мимотопов показаны как группы Т02-Т05. Пептид ZTS-563 содержит аминокислотную последовательность N-AKVSMPADNFERKNFILT-C, которая соответствует положениям 121-138 последовательности SEQ ID NO: 157 с заменой треонина (Т) на аланин (А) в положении номер 138. Пептид ZTS-418 содержит аминокислотную последовательность N-TEVSMPTDNFERKRFILT-C, которая соответствует положениям 115-132 последовательности SEQ ID NO: 165. Пептид ZTS-423 содержит аминокислотную последовательность N-NGSAILPYFRAIRPLSDKNTIDKIIEQLDKLKF-C, которая соответствует положениям 83-115 последовательности SEQ ID NO: 157. Пептид ZTS-422 содержит аминокислотную последовательность N-AKVSMPADNFERKNFILT-C, которая соответствует положениям 121-138 последовательности SEQ ID NO: 157 с заменой треонина (Т) на аланин (А) в положении номер 138. Каждый из пептидов ZTS-563, ZTS-418, ZTS-423 и ZTS-422 также включает N и С концевые цистеины, как изображено, для облегчения конъюгации с использованием свободных тиоловых групп.ZTS-422 также содержит дополнительный линкер аминогексановой кислоты (Ahx) между двумя N-концевыми цистеинами. ZTS-423 также содержит дополнительную спейсерную последовательность из трех аминокислот (GSG) рядом с N концевым цистеином. На Фиг. 18В показаны средние титры кошачьих антител к полноразмерному кошачьему IL-31 (SEQ ID NO: 157) во всех группах воздействия, кроме Т03. У кошек в группе Т03 (мимотоп лошадиного 15Н05) отсутствовала CRAR на кошачий IL-31 (данные не приведены).
На Фиг. 19А представлена аминокислотная последовательность минимального эпитопа, связанного с антителом М14 против собачьего IL-31 согласно WO 2018/156367 (Kindred Biosciences, Inc.). Приведено сравнение последовательностей от разных биологических видов, указаны их идентификационные номера и относительные положения аминокислот. На Фигуре 19В показана эта минимальная аминокислотная последовательность собачьего IL-31, выделенная черным прямоугольником. На данной Фиг. также показано выравнивание последовательностей в области, окружающей белок, и указаны относительные положения соответствующих аминокислот в референтной последовательности.
На Фиг. 20 показан фрагмент белка IL-31 с петлей, образованной сближением спирали А с находящейся за ней последовательностью случайной спирали, которая имеет схожие с петлей 15Н05 положение и структуру. Показано сравнение аминокислотных последовательностей различных биологических видов и указаны идентификационные номера последовательностей и положения аминокислот.
На Фиг. 21А показаны аминокислотные последовательности трех пептидных мимотопов лошадиного IL-31, представляющие различные ключевые области эпитопа на белке. Мимотоп 15Н05 содержит аминокислотную последовательность N-TEVSMPTDNFERKRFILT-C, которая соответствует положениям 115-132 последовательности SEQ ID NO: 165. Мимотоп спирали ВС содержит аминокислотную последовательность N-NSSAILPYFKAISPSLNNDKSLYIIEQLDKLNF-C, которая соответствует положениям 77-109 последовательности SEQ ID NO: 165. Мимотоп спирали А содержит аминокислотную последовательность N-GPIYQLQPKEIQAIIVELQNLSKK-C, которая соответствует положениям 20-43 последовательности SEQ ID NO: 165. Мимотоп 15Н05 также включает N и С концевые цистеины, как изображено, для облегчения конъюгации с использованием свободных тиоловых групп. Все три мимотопа также содержат дополнительную спейсерную последовательность из трех аминокислот (GSG) рядом с N концевой биотиновой группой, выделенной в последовательностях жирным шрифтом и подчеркиванием. Показаны соответствующие положения каждого аминокислотного остатка в SEQ ID NO: 165. На Фиг. 21В показаны результаты анализа связывания с применением биослойной интерферометрии. Указанные мимотопы иммобилизовали на стрептавидиновые биосенсоры и использовали для исследования различных разведений сыворотки мышей. Использовали сыворотку мышей, вакцинированных белком IL-31 лошади (SEQ ID NO: 165) или контрольную сыворотку мышей, вакцинированных посторонним белком.
Определения
Перед подробным описанием настоящего изобретения будут приведены определения нескольких терминов, используемых в контексте данного изобретения. В дополнение к этим терминам в описании при необходимости приводятся определения других терминов. Если не указано иное, термины уровня техники, используемые в этом описании, будут иметь общепринятое в данной области значение.
Как использовано в описании и формуле изобретения, формы единственного числа включают ссылки на множественное число, если контекст ясно не предусматривает иное. Например, ссылка на «антитело» включает множество таких антител. В качестве другого примера можно привести упоминание «мимотопа», «мимотопа IL-31» и т.п., включающих множество таких мимотопов.
Подразумевается, что использованный в данном описании термин «содержащий» означает, что композиции и способы включают перечисленные элементы, но не исключая другие элементы.
Используемый в данном документе термин «вакцинная композиция» включает по меньшей мере один антиген или иммуноген в фармацевтически приемлемом носителе, пригодном для индукции иммунного ответа у хозяина. Вакцинные композиции можно вводить в дозах и способами, хорошо известными специалистам в области медицины или ветеринарии, принимая во внимание такие факторы, как возраст, пол, вес, биологический вид и состояние млекопитающего-реципиента, а также путь введения. Путь введения может быть чрескожным, через слизистую оболочку (например, пероральным, назальным, анальным, вагинальным) или парентеральным путем (внутрикожным, трансдермальным, внутримышечным, подкожным, внутривенным или внутрибрюшинным). Вакцинные композиции можно вводить отдельно или можно вводить совместно или последовательно с другими видами лечения или терапиями. Лекарственные формы могут включать суспензии, сиропы или эликсиры, а также препараты для парентерального, подкожного, внутрикожного, внутримышечного или внутривенного введения (например, введения путем инъекции), такие как стерильные суспензии или эмульсии. Вакцинные композиции можно вводить в виде спрея, или смешивать с пищей и/или водой, или доставлять в смеси с подходящим носителем, разбавителем или эксципиентом, таким как стерильная вода, физиологический раствор, глюкоза или тому подобное. Композиции могут содержать вспомогательные вещества, такие как смачивающие или эмульгирующие вещества, вещества, буферизующие вещества для поддержания рН, адъюванты, желирующие или повышающие вязкость добавки, консерванты, корригенты, красители и тому подобное, в зависимости от пути введения и желаемого препарата. Для приготовления подходящих препаратов без излишних экспериментов можно обращаться к стандартным фармацевтическим текстам, таким как «Remington's Pharmaceutical Sciences»(1990).
Термин «иммунный ответ», используемый в данном документе, относится к ответу, вызываемому у животного или человека. Иммунный ответ может относиться к клеточному иммунитету (CMI), гуморальному иммунитету или может включать и то, и другое. Настоящее изобретение также предполагает ответ, ограниченный частью иммунной системы. Как правило, «иммунный ответ» включает один или несколько из следующих эффектов: выработка или активация антител, В-клеток, хелперных Т-клеток, супрессорных Т-клеток и/или цитотоксических Т-клеток и/или гамма-дельта Т-клеток, специфически направленных на антиген или антигены, включенные в представляющую интерес композицию или вакцину, без ограничения. Предпочтительно, хозяин проявляет либо терапевтический, либо защитный иммунный ответ, так что усиливается устойчивость к заболеванию или нарушению и/или уменьшается клиническая тяжесть заболевания. Такая защита будет демонстрироваться либо уменьшением, либо отсутствием симптомов, обычно проявляемых пораженным хозяином, более быстрым временем восстановления и/или пониженным титром антигена (например, IL-31) у пораженного хозяина.
Термин «защитный», используемый в данном документе, означает обеспечение терапевтического иммунного ответа млекопитающего-хозяина, так что у хозяина-млекопитающего усиливается устойчивость к заболеванию или нарушению и/или уменьшается клиническая тяжесть заболевания.
Используемый в данном документе термин «иммуногенность» означает способность вызывать иммунный ответ против антигена или антигенов у млекопитающего-хозяина. Этот иммунный ответ составляет основу защитного иммунитета, вызываемого вакциной против конкретного антигена.
В данном контексте термины «иммунизировать», «иммунизация» и им подобные означают процесс, посредством которого млекопитающее приобретает иммунитет или становится устойчивым к заболеванию, обычно путем введения вакцины. Вакцины стимулируют собственную иммунную систему млекопитающего, чтобы защитить его от последующего заболевания.
Используемый в данном документе термин «адъювант» означает композицию, состоящую из одного или нескольких веществ, которые усиливают иммунный ответ на антиген(ы). Механизм действия адъюванта полностью не известен. Считается, что некоторые адъюванты усиливают иммунный ответ за счет медленного высвобождения антигена, в то время как другие адъюванты сами по себе обладают сильной иммуногенностью и, как полагают, действуют синергетически.
Эпитоп, как использовано в данном описании, относится к антигенной детерминанте, распознаваемой CDR антитела. Другими словами, эпитоп относится к тому участку любой молекулы, который способно распознавать и с которым может связываться антитело. Если не указано иное, термин «эпитоп», использованный в данном описании, относится к области IL-31, с которой анти-IL-31 агент вступает в реакцию.
«Антиген» представляет собой молекулу или участок молекулы, с которыми может связываться антитело, которые помимо этого способно распознавать и с которыми может связываться антитело (соответствующая область связывания антитела может быть названа паратопом). В общем случае, эпитопы состоят из химически активных поверхностных группировок молекул, например боковых цепей аминокислот или Сахаров, и имеют определенные пространственные структурные характеристики, а также определенные характеристики в отношении заряда. Эпитопы представляют собой антигенные детерминанты белка, распознаваемые иммунной системой. Компонентами иммунной системы, распознающей эпитопы, являются антитела, Т-клетки и В-клетки. Эпитопы Т-клеток экспонируются на поверхности антигенпрезентирующих клеток (АРС) и обычно имеют длину 8-11 (МНС класс I) или более 15 (МНС класс II) аминокислот. Распознавание экспонируемого комплекса МНС-пептид Т-клетками имеет решающее значение для их активации. Эти механизмы позволяют адекватно распознавать собственные и «чужие» белки, такие как бактериальные и вирусные. Независимые аминокислотные остатки, которые не обязательно являются смежными, способствуют взаимодействию со связывающей бороздой АРС и последующему распознаванию Т-клеточным рецептором (Janeway, Travers, Walport, Immunobiology: The Immune System in Health and Disease. 5-е издание New York: Garland Science; 2001). Эпитопы, которые распознаются растворимыми антителами и рецепторами В-клеток, ассоциированными с клеточной поверхностью, сильно различаются по длине и степени непрерывности (Sivalingam and Shepherd, Immunol. 2012 Jul;51(3-4):304-309 9). И вновь, даже линейные эпитопы или эпитопы, обнаруженные в непрерывном участке белковой последовательности, часто будут иметь несмежные аминокислоты, которые представляют ключевые точки контакта с паратопами антител или В-клеточным рецептором. Эпитопы, распознаваемые антителами и В-клетками, могут быть конформационными и содержать аминокислоты, составляющие общую область контакта на белке в трехмерном пространстве, и зависят от третичных и четвертичных структурных особенностей белка. Эти остатки часто находятся в пространственно различных областях первичной аминокислотной последовательности.
Используемый в данном документе термин «мимотоп» означает линейный пептид или пептид с ограниченной конформационной свободой, который имитирует эпитоп антигена. Мимотоп может иметь первичную аминокислотную последовательность, способную вызывать эффекторный ответ Т-клеток, и/или трехмерную структуру, необходимую для связывания В-клеток, что приводит к созреванию приобретенного иммунного ответа у животного. Антитело к данному эпитопу антигена распознает мимотоп, который имитирует этот эпитоп. В качестве альтернативы, мимотоп IL-31 может обозначаться в данном документе мимотопом пептида IL-31. В некоторых воплощениях мимотоп (линейный или с ограниченной конформационной свободой) для применения в композициях и/или способах по настоящему изобретению представляет собой и/или включает в качестве своей части пептид, длина которого составляет от приблизительно 5 аминокислотных остатков до приблизительно 40 аминокислотных остатков.
Термин «специфически», применительно к связыванию с антителом, относится к высокоавидному и/или высокоаффинному связыванию антитела с конкретным антигеном, т.е. полипептидом или эпитопом. Во многих воплощениях конкретным антигеном является антиген (или фрагмент либо субфракция антигена), используемый для иммунизации животного-хозяина, из которого были выделены антитело-продуцирующие клетки. Специфическое связывание антитела с антигеном является более сильным, чем связывание того же антитела с другими антигенами. Антитела, специфически связывающиеся с полипептидом, могут обладать способностью связывать другие полипептиды на низком, однако же детектируемом уровне (например 10% или меньше от связывания, показанного в отношении представляющего интерес полипептида). Такое слабое связывание или фоновое связывание легко отличить от специфического связывания антитела с исследуемым полипептидом, например посредством использования подходящих контролей. В общем случае, специфические антитела связываются с антигеном с аффинностью связывания с KD 10-7 М или менее, например, 10-8 М или менее (например, 10-9 М или менее, 10-10 или менее, 10-11 или менее, 10-12 или менее, или 10-13 и т.д.).
Использованный в данном описании термин «антитело» относится к интактному иммуноглобулину, имеющему две легкие и две тяжелые цепи. Таким образом, единичное выделенное антитело или фрагмент может представлять собой поликлональное антитело, моноклональное антитело, синтетическое антитело, рекомбинантное антитело, химерное антитело, гетерохимерное антитело, канинизированное антитело, фелинизированное антитело, полностью собачье антитело, полностью кошачье антитело, полностью лошадиное антитело или полностью человеческое антитело. Термин «антитело» предпочтительно относится к моноклональным антителам и их фрагментам (например, включая, антигенсвязывающие части антитела, без ограничения) и их иммунологическим связывающим эквивалентам, которые могут связываться с белком IL-31 и его фрагментами или модифицированными фрагментами. Такие фрагменты и модифицированные фрагменты IL-31 могут включать мимотопы пептида IL-31, используемые в различных воплощениях этого изобретения. Например, антитело к данному эпитопу на IL-31 будет распознавать мимотоп пептида IL-31, который имитирует этот эпитоп. Термин «антитело» используется для обозначения как гомогенной молекулярной субстанции, так и смеси, как например, сывороточного продукта, состоящего из множества различных молекулярных субстанций.
«Нативные антитела» и «нативные иммуноглобулины» обычно представляют собой гетеротетрамерные гликопротеины с молекулярной массой примерно 150000 дальтон, составленные из двух идентичных легких (L) цепей и двух идентичных тяжелых (Н) цепей. Каждая легкая цепь связана с тяжелой цепью одной ковалентной дисульфидной связью, тогда как число дисульфидных связей у тяжелых цепей разных изотипов иммуноглобулинов варьирует. Каждая тяжелая и легкая цепь также имеет правильно расположенные внутрицепочечные дисульфидные мостики. Каждая тяжелая цепь имеет на одном конце вариабельный домен (VH), за которым следует несколько константных доменов. Каждая легкая цепь имеет вариабельный домен на одном конце (VL) и константный домен на своем другом конце; константный домен легкой цепи выравнен относительно первого константного домена тяжелой цепи, а вариабельный домен легкой цепи выравнен относительно вариабельного домена тяжелой цепи. Считается, что определенные аминокислотные остатки образуют поверхность контакта между вариабельными доменами легкой и тяжелой цепей.
Термин «фрагмент антитела» относится к структуре меньшей, чем интактное антитело, включая, без ограничения, выделенную одиночную цепь антитела, конструкцию Fv, конструкцию Fab, конструкцию Fc, последовательность вариабельной области легкой цепи или области, определяющей комплементарность (CDR) и т.д. Например, фрагмент антитела может включать антигенсвязывающую часть антитела.
Термин «вариабельная» область включает каркас и CDR (известные еще как гипервариабельные области) и относится к тому факту, что некоторые участки вариабельных доменов значительно отличаются в последовательности среди антител и используются при связывании каждого конкретного антитела со своим конкретным антигеном и определяют специфичность этого антитела. Однако вариабельность не распределена равномерно по всем вариабельным доменам антител. Она сконцентрирована в трех называемых гипервариабельными областями сегментах вариабельных доменов как легкой цепи, так и тяжелой цепи. Более высококонсервативные участки вариабельных доменов называют каркасной областью (FR). Каждый из вариабельных доменов нативных тяжелых и легких цепей содержит многочисленные FR, в большинстве случаев принимающие β-складчатую конфигурацию, соединенные тремя гипервариабельными областями, которые образуют петли, соединяющие и в некоторых случаях образующие часть α-складчатой структуры. Гипервариабельные области в каждой цепи удерживаются вместе в непосредственной близости друг от друга посредством FR и вместе с гипервариабельными областями из другой цепи вносят вклад в образование антигенсвязывающего сайта антител (см. Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, Md. (1991), pages 647-669). Константные домены непосредственно не вовлечены в связывание антитела с антигеном, однако демонстрируют различные эффекторные функции, как например участие антитела в антителозависимой клеточной токсичности.
Термин «гипервариабельная область» при использовании в данном документе относится к аминокислотным остаткам антитела, которые отвечают за связывание антигена. Гипервариабельная область содержит аминокислотные остатки из «определяющей комплементарность области» или «CDR» (Kabat, et al. (1991), см. выше) и/или эти остатки из «гипервариабельной петли» (Chothia and Lesk J. Mol. Biol. 196:901-917 (1987). «Каркасные остатки», или остатки «FR» (от англ. Framework), представляют собой такие остатки вариабельных доменов, которые отличаются от остатков гипервариабельных областей, определенных выше.
Расщепление антител папаином дает два идентичных антигенсвязывающих фрагмента, называемых «Fab»-фрагментами, каждый с одним антигенсвязывающим сайтом, и остаточный «Fc-фрагмент, название которого отражает его способность легко кристаллизоваться. Обработка пепсином дает фрагмент F(ab')2, который имеет два сайта связывания антигена и все еще способен к образованию перекрестных сшивок между антигенами.
«Fv» - это минимальный фрагмент антитела, который содержит полный сайт распознавания и связывания антигена. Эта область состоит из димера одной тяжелой цепи и одного вариабельного домена легкой цепи, находящихся в тесной нековалентной связи. Именно в этой конфигурации три гипервариабельные области каждого вариабельного домена взаимодействуют для определения антигенсвязывающего сайта на поверхности димера VH-VL. В совокупности антигенсвязывающую специфичность антителу придают шесть гипервариабельных областей. Однако, даже один вариабельный домен (или половина Fv, содержащая только три гипервариабельных области, специфичных к антигену) обладает способностью распознавать и связывать антиген, хотя и с более низкой аффинностью, чем целый сайт связывания.
Fab-фрагмент также содержит константный домен легкой цепи и первый константный домен (СН1) тяжелой цепи. Fab'-фрагменты отличаются от Fab-фрагментов тем, что у них добавлено несколько остатков на карбоксильном конце CH1-домена тяжелой цепи, включая один или более чем один цистеин из шарнирной области антитела. Fab'-SH в данном описании представляет собой обозначение для Fab', в котором остаток(ки) цистеина константных доменов несет(ут) свободную тиоловую группу. F(ab')2-фрагменты антител изначально получают в виде пар Fab'-фрагментов, которые содержат между собой шарнирные цистеины. Кроме этого известны другие методы химического сочетания фрагментов антител.
«Легкие цепи» антител (иммуноглобулинов) любых видов позвоночных на основании аминокислотных последовательностей их константных доменов могут быть отнесены к одному из двух четко различающихся типов, называемых каппа (κ) и лямбда (λ).
В зависимости от аминокислотной последовательности константного домена их тяжелых цепей иммуноглобулины можно отнести к разным классам. В настоящее время существует пять основных классов иммуноглобулинов: IgA, IgD, IgE, IgG и IgM, и некоторые из них могут быть далее разделены на подклассы (изотипы), например, IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2 (обозначения для мыши и человека). Константные домены тяжелой цепи, которые соответствуют различным классам иммуноглобулинов, называются альфа, дельта, эпсилон, гамма и мю, соответственно. Структуры субъединиц и трехмерные конфигурации различных классов иммуноглобулинов хорошо известны для многих видов. Распространенность индивидуальных изотипов и функциональные активности, ассоциированные с этими константными доменами, являются видоспецифичными и должны быть определены экспериментально.
«Моноклональное антитело», как оно определено в данном описании, представляет собой антитело, продуцируемое клетками одного клона (конкретно, гибридомными клетками одного клона), и поэтому относится к одному чистому гомогенному типу антитела. Все моноклональные антитела, полученные из одного и того же клона, идентичны и имеют одинаковую антигенную специфичность. Термин «моноклональный» относится к клеткам одного клона, одной клетке и потомству этой клетки.
«Полностью собачье антитело», согласно определению в данном документе, представляет собой моноклональное антитело, продуцируемое клоном клеток (обычно, клеточной линией СНО) и, следовательно, один чистый гомогенный тип антитела. Антитела из единичных В-клеток иммунизированных млекопитающих, таких как собаки, создают в виде рекомбинантных белков IgG после идентификации последовательностей их вариабельных доменов. Пересадка этих вариабельных доменов на константные домены собачьего происхождения (константные домены тяжелой цепи и легкой цепи каппа или лямбда) приводит к получению рекомбинантных полностью собачьих антител. Все полностью собачьи моноклональные антитела, полученные из одного и того же клона, идентичны и имеют одинаковую антигенную специфичность. Термин «моноклональный» относится к клеткам одного клона, одной клетке и потомству этой клетки.
«Полностью кошачье антитело», согласно определению в данном документе, представляет собой моноклональное антитело, продуцируемое клоном клеток (обычно клеточной линией СНО) и, следовательно, один чистый гомогенный тип антитела. Антитела из единичных В-клеток иммунизированных млекопитающих, таких как собаки, создают в виде рекомбинантных белков IgG после идентификации последовательностей их вариабельных доменов. Пересадка этих вариабельных доменов на константные домены кошачьего происхождения (константные домены тяжелой цепи и легкой цепи каппа или лямбда) приводит к образованию рекомбинантных полностью кошачьих антител. Все полностью кошачьи моноклональные антитела, полученные из одного и того же клона, идентичны и имеют одинаковую антигенную специфичность. Термин «моноклональный» относится к клеткам одного клона, одной клетке и потомству этой клетки.
«Полностью лошадиное антитело», согласно определению в данном документе, представляет собой моноклональное антитело, продуцируемое клоном клеток (обычно клеточной линией СНО) и, следовательно, один чистый гомогенный тип антитела. Антитела из единичных В-клеток иммунизированных млекопитающих, таких как собаки, создают в виде рекомбинантных белков IgG после идентификации последовательностей их вариабельных доменов. Пересадка этих вариабельных доменов на константные домены лошади (константные домены тяжелой цепи и легкой цепи каппа или лямбда) приводит к образованию рекомбинантных полностью конских антител. Все полностью лошадиные моноклональные антитела, полученные из одного и того же клона, идентичны и имеют одинаковую антигенную специфичность. Термин «моноклональный» относится к клеткам одного клона, одной клетке и потомству этой клетки.
«Полностью человеческое антитело», согласно определению в данном документе, представляет собой моноклональное антитело, продуцируемое клоном клеток (обычно клеточной линией СНО) и, следовательно, один чистый гомогенный тип антитела. Антитела из единичных В-клеток иммунизированных млекопитающих, таких как собаки, создают в виде рекомбинантных белков IgG после идентификации последовательностей их вариабельных доменов. Пересадка этих вариабельных доменов на человеческие константные домены (константные домены тяжелой цепи и легкой цепи каппа или лямбда) приводит к образованию рекомбинантных полностью человеческих антител. Все полностью человеческие моноклональные антитела, полученные из одного и того же клона, идентичны и имеют одинаковую антигенную специфичность. Термин «моноклональный» относится к клеткам одного клона, одной клетке и потомству этой клетки.
Моноклональные антитела в данном документе, в частности, включают «химерные» антитела (иммуноглобулины), в которых часть тяжелой и/или легкой цепи идентична или гомологична соответствующим последовательностям в антителах, происходящих от определенного биологического вида, в то время как остальная часть цепи (цепей) идентична или гомологична соответствующим последовательностям в антителах, происходящих от другого биологического вида, а также фрагментам таких антител, если они проявляют желаемую биологическую активность. Обычно химерные антитела представляют собой антитела, гены легкой и тяжелой цепей которых сконструированы, как правило, с помощью генной инженерии, из генов вариабельной и константной областей антитела, принадлежащих к разным биологическим видам. Например, гены вариабельных сегментов моноклонального антитела мыши могут быть присоединены к генам константных сегментов антитела собаки. В одном воплощении химерного мышиного:собачьего IgG сайт связывания антигена происходит от мыши, в то время как Fc-часть является собачьей.
«Канинизированные» формы антител, не являющихся собачьими (например, мышиные), представляют собой антитела, полученные с помощью генной инженерии, которые содержат минимальную последовательность, происходящую из иммуноглобулина, не являющегося собачьим. Канинизированные антитела представляют собой последовательности иммуноглобулина собаки (реципиентное антитело), в которых остатки гипервариабельной области реципиента заменены остатками гипервариабельной области биологического вида, не являющегося собакой (донорское антитело), такого как мышь, имеющими желаемую специфичность, аффинность и связывающую способность. В некоторых случаях остатки каркасной области (FR) последовательностей собачьего иммуноглобулина заменяют соответствующими остатками, не являющимися собачьими. Кроме того, канинизированные антитела могут включать остатки, которых нет в реципиентном антителе или в донорском антителе. Такие модификации осуществляют для дальнейшего улучшения характеристик антител. В общем, канинизированное антитело будет включать по существу все, из по меньшей мере, одного, а обычно из двух вариабельных доменов, в которых все или по существу все гипервариабельные области соответствуют таковым из последовательности иммуноглобулина, отличного от собачьего, и все или по существу все FR относятся к последовательности собачьего иммуноглобулина. Канинизированное антитело возможно также будет содержать всю константную область иммуноглобулина (Fc) или по меньшей мере ее участок, обычно таковую из последовательности собачьего иммуноглобулина. В одном из воплощений модификации антител с целью увеличения их сходства с естественными антителами, или канинизации мышиного IgG, мышиные CDR прививают на каркасные участки собачьего происхождения.
«Фелинизированные» формы антител, не являющихся кошачьими (например, мышиные), представляют собой антитела, полученные с помощью генной инженерии, которые содержат минимальную последовательность, происходящую из иммуноглобулина, не являющегося кошачьим. Фелинизированные антитела представляют собой последовательности иммуноглобулина кошки (реципиентное антитело), в которых остатки гипервариабельной области реципиента заменены остатками гипервариабельной области биологического вида, не являющегося кошкой (донорское антитело), такого как мышь, обладающими желаемой специфичностью, аффинностью и связывающей способностью. В некоторых случаях остатки каркасной области (FR) последовательностей кошачьих иммуноглобулинов заменяют соответствующими остатками, не являющимися кошачьими. Кроме того, фелинизированные антитела могут включать остатки, которых нет в реципиентном антителе или в донорском антителе. Такие модификации осуществляют для дальнейшего улучшения характеристик антител. В общем, фелинизированное антитело будет включать по существу все, из по меньшей мере, одного, а обычно из двух вариабельных доменов, в которых все или по существу все гипервариабельные области соответствуют таковым из последовательности иммуноглобулина, отличного от кошачьего, и все или по существу все FR относятся к последовательности кошачьего иммуноглобулина. Фелинизированное антитело возможно также будет содержать всю константную область иммуноглобулина (Fc) или по меньшей мере ее участок, обычно таковую из последовательности кошачьего иммуноглобулина.
«Эквинизированные» формы антител, не являющихся конскими (например, мышиных), представляют собой антитела, полученные с помощью генной инженерии, которые содержат минимальную последовательность, происходящую из иммуноглобулина, не являющегося лошадиным. Эквинизированные антитела представляют собой последовательности иммуноглобулина лошади (реципиентное антитело), в которых остатки гипервариабельной области реципиента заменены остатками гипервариабельной области биологического вида, не являющегося лошадью (донорское антитело), такого как мышь, обладающими желаемой специфичностью, аффинностью и связывающей способностью. В некоторых случаях остатки каркасной области (FR) последовательностей иммуноглобулина лошади заменяют соответствующими остатками, не являющимися лошадиными. Кроме того, эквинизированные антитела могут включать остатки, которых нет в реципиентном антителе или в донорском антителе. Такие модификации осуществляют для дальнейшего улучшения характеристик антител. В общем, эквинизированное антитело будет включать по существу все, из по меньшей мере одного, а обычно из двух вариабельных доменов, в которых все или по существу все гипервариабельные области соответствуют таковым из последовательности иммуноглобулина, отличного от лошадиного, и все или по существу все FR относятся к последовательности иммуноглобулина лошади. Эквинизированное антитело, возможно также будет содержать всю константную область иммуноглобулина (Fc) или по меньшей мере ее участок, обычно таковую из последовательности лошадиного иммуноглобулина.
«Гуманизированные» формы антител, не являющихся человеческими (например, мышиных), представляют собой антитела, полученные с помощью генной инженерии, которые содержат минимальную последовательность, происходящую из иммуноглобулина, не являющегося человеческим. Гуманизированные антитела представляют собой последовательности иммуноглобулина человека (реципиентное антитело), в которых остатки гипервариабельной области реципиента заменены остатками гипервариабельной области из биологического вида, не являющегося человеком (донорское антитело), такого как мышь, обладающими желаемой специфичностью, аффинностью и связывающей способностью. В некоторых случаях остатки каркасной области (FR) последовательностей иммуноглобулина человека заменяют соответствующими остатками, не являющимися человеческими. Кроме того, гуманизированные антитела могут включать остатки, которых нет в реципиентном антителе или в донорском антителе. Такие модификации осуществляют для дальнейшего улучшения характеристик антител. В общем, гуманизированное антитело будет включать по существу все, из по меньшей мере, одного, а обычно из двух вариабельных доменов, в которых все или по существу все гипервариабельные области соответствуют таковым из последовательности иммуноглобулина, отличного от человеческого, и все или по существу все FR относятся к последовательности человеческого иммуноглобулина. Гуманизированное антитело, возможно также будет содержать всю константную область иммуноглобулина (Fc) или по меньшей мере ее участок, обычно таковую из последовательности человеческого иммуноглобулина.
«Полностью собачьи» антитела - это антитела, созданные с помощью генной инженерии, которые не содержат последовательности, полученной из иммуноглобулина, не являющегося собачьим. Полностью собачьи антитела представляют собой последовательности собачьего иммуноглобулина (реципиентное антитело), в которых остатки гипервариабельной области происходят из встречающегося в природе собачьего антитела (донорское антитело), имеющего желаемую специфичность, аффинность и связывающую способность. В некоторых случаях остатки каркасной области (FR) последовательностей собачьего иммуноглобулина заменяют соответствующими остатками, не являющимися собачьими. Кроме того, полностью собачьи антитела могут включать остатки, которые отсутствуют в реципиентном антителе или в донорском антителе, например, включая, изменения в CDR для модификации аффинности, но не ограничиваясь ими. Такие модификации осуществляют для дальнейшего улучшения характеристик антител. В общем, полностью собачье антитело будет включать по существу все из по меньшей мере одного, а обычно из двух вариабельных доменов, в которых все или по существу все гипервариабельные области соответствуют таковым из последовательности собачьего иммуноглобулина, и все или по существу все FR относятся к последовательности собачьего иммуноглобулина. Полностью собачье антитело, также будет содержать всю константную область иммуноглобулина (Fc) или по меньшей мере ее участок, обычно таковую из последовательности собачьего иммуноглобулина.
«Полностью кошачьи» антитела - это антитела, созданные с помощью генной инженерии, которые не содержат последовательности, полученной из иммуноглобулина, не являющегося кошачьим. Полностью кошачьи антитела представляют собой последовательности кошачьего иммуноглобулина (реципиентное антитело), в которых остатки гипервариабельной области происходят из встречающегося в природе кошачьего антитела (донорское антитело), имеющего желаемую специфичность, аффинность и связывающую способность. В некоторых случаях остатки каркасной области (FR) последовательностей кошачьих иммуноглобулинов заменяют соответствующими остатками, не являющимися кошачьими. Кроме того, полностью кошачьи антитела могут включать остатки, которые не отсутствуют в реципиентном антителе или в донорском антителе, например, включая изменения в CDR для модификации аффинности, но не ограничиваясь ими. Такие модификации осуществляют для дальнейшего улучшения характеристик антител. В общем, полностью кошачье антитело будет включать по существу все из по меньшей мере одного, а обычно двух вариабельных доменов, в которых все или по существу все гипервариабельные области соответствуют таковым из последовательности иммуноглобулина кошки, и все или по существу все FR относятся к последовательности иммуноглобулина кошки. Полностью кошачье антитело, возможно, также будет содержать всю константную область иммуноглобулина (Fc) или по меньшей мере ее участок, обычно таковую из последовательности кошачьего иммуноглобулина.
«Полностью лошадиные» антитела - это антитела, созданные с помощью генной инженерии, которые не содержат последовательности, полученной из иммуноглобулина, не являющегося лошадиным. Полностью лошадиные антитела представляют собой последовательности иммуноглобулина лошади (реципиентное антитело), в которых остатки гипервариабельной области происходят из встречающегося в природе лошадиного антитела (донорское антитело), имеющего желаемую специфичность, аффинность и связывающую способность. В некоторых случаях остатки каркасной области (FR) последовательностей иммуноглобулина лошади заменяют соответствующими остатками, не являющимися лошадиными. Кроме того, полностью лошадиные антитела могут включать остатки, которые отсутствуют в реципиентном антителе или в донорском антителе, например, включая изменения в CDR для модификации аффинности, но не ограничиваясь ими. Такие модификации осуществляют для дальнейшего улучшения характеристик антител. В общем, полностью лошадиное антитело будет включать по существу все из по меньшей мере одного, а обычно из двух вариабельных доменов, в которых все или по существу все гипервариабельные области соответствуют таковым из последовательности иммуноглобулина лошади, и все или по существу все FR относятся к последовательности иммуноглобулина лошади. Полностью лошадиное антитело, возможно, также будет содержать всю константную область иммуноглобулина (Fc) или по меньшей мере ее участок, обычно таковую из последовательности лошадиного иммуноглобулина.
«Полностью человеческие» антитела - это антитела, созданные с помощью генной инженерии, которые не содержат последовательности, полученной из иммуноглобулина, не являющегося человеческим. Полностью человеческие антитела представляют собой последовательности иммуноглобулина человека (реципиентное антитело), в которых остатки гипервариабельной области происходят из встречающегося в природе человеческого антитела (донорское антитело), имеющего желаемую специфичность, аффинность и связывающую способность. В некоторых случаях остатки каркасной области (FR) последовательностей иммуноглобулина человека заменяют соответствующими остатками, не являющимися человеческими. Кроме того, полностью человеческие антитела могут включать остатки, которые отсутствуют в реципиентном антителе или в донорском антителе, например, включая изменения в CDR для модификации аффинности, но не ограничиваясь ими. Такие модификации осуществляют для дальнейшего улучшения характеристик антител. В общем, полностью человеческое антитело будет включать по существу все из, по меньшей мере, одного, а обычно из двух вариабельных доменов, в которых все или по существу все гипервариабельные области соответствуют таковым из последовательности иммуноглобулина человека, и все или по существу все FR относятся к последовательности человеческого иммуноглобулина. Полностью человеческое антитело, также будет содержать всю константную область иммуноглобулина (Fc) или по меньшей мере ее участок, обычно таковую из последовательности человеческого иммуноглобулина.
Термин «гетерохимерное», согласно определению в данном документе, относится к антителу, в котором одна из цепей антитела (тяжелая или легкая) является, например, канинизированной, фелинизированной, эквинизированной или гуманизированной, в то время как другая является химерной. В одном воплощении фелинизированная вариабельная область тяжелой цепи (где все CDR являются мышиными, а все FR являются кошачьими) спарена с химерной вариабельной областью легкой цепи (где все CDR являются мышиными и все FR являются мышиными). В этом воплощении как вариабельные области тяжелой цепи, так и вариабельные области легкой цепи слиты с константной областью кошачьего происхождения.
Используемый в данном документе термин «вариант» относится к последовательности пептида, полипептида или нуклеиновой кислоты, кодирующей пептид или полипептид, которая имеет одну или несколько консервативных аминокислотных вариаций или других незначительных модификаций, так что соответствующий пептид или полипептид имеет по существу эквивалентную функцию, при сравнении с пептидом или полипептидом дикого типа. Обычно вариантны пептидных мимотопов для применения в настоящем изобретении будут иметь по меньшей мере 30% идентичности с родительским мимотопом, более предпочтительно по меньшей мере 50%, более предпочтительно по меньшей мере 75%, более предпочтительно по меньшей мере 80%, более предпочтительно по меньшей мере 85% более предпочтительно по меньшей мере 90% и наиболее предпочтительно по меньшей мере 95% идентичности последовательности с родительским мимотопом.
«Вариант» антитела против IL-31 в настоящем документе относится к молекуле, которая отличается по аминокислотной последовательности от аминокислотной последовательности «родительского» антитела против IL-31 вследствие добавления, делеции и/или замены одного или более чем одного аминокислотного остатка в последовательности родительского антитела и сохраняет по меньшей мере одну желаемую активность родительского антитела против IL-31. Желаемые активности могут включать способность специфически связываться с антигеном, способность снижать, ингибировать или нейтрализовать активность IL-31 у животного и способность ингибировать опосредованный IL-31 сигнальный путь pSTAT в исследованиях на клетках. В одном воплощении вариант содержит одну или несколько аминокислотных замен в одной или нескольких гипервариабельных и/или каркасных областях родительского антитела. Например, вариант может содержать по меньшей мере одну, например, от приблизительно одной до приблизительно десяти и предпочтительно от приблизительно двух до приблизительно пяти замен в одной или нескольких гипервариабельных и/или каркасных областях родительского антитела. Обычно вариант будет иметь аминокислотную последовательность, имеющую идентичность аминокислотной последовательности с последовательностями вариабельных доменов тяжелой или легкой цепи родительского антитела по меньшей мере 50%, более предпочтительно, по меньшей мере 65%, более предпочтительно, по меньшей мере 75%, более предпочтительно, по меньшей мере 80%, более предпочтительно, по меньшей мере 85%, более предпочтительно, по меньшей мере 90% и наиболее предпочтительно, идентичность последовательностей по меньшей мере 95%. Идентичность или гомологию по отношению к этой последовательности в данном документе определяют как процент аминокислотных остатков в последовательности-кандидате, которые идентичны остаткам родительского антитела, после выравнивания последовательностей и введения пробелов, если необходимо, для достижения максимального процента идентичности последовательностей. Ни одно из N-концевых, С-концевых или внутренних удлинений, делеций или вставок в последовательность антитела не следует считать влияющими на идентичность или гомологию последовательности. Вариант сохраняет способность связывать IL-31 и предпочтительно имеет желаемую активность, которая превосходит таковую родительского антитела. Например, вариант может иметь более сильную аффинность связывания, повышенную способность снижать, ингибировать или нейтрализовать активность IL-31 у животных и/или повышенную способность ингибировать опосредованный IL-31 сигнальный путь pSTAT в исследованиях на клетках.
«Вариант» нуклеиновой кислоты в данном документе относится к молекуле, которая отличается последовательностью от «родительской» нуклеиновой кислоты. Расхождения в полинуклеотидных последовательностях могут быть результатом мутационных изменений, таких как делеций, замены или добавления одного или нескольких нуклеотидов. Каждое из этих изменений в заданной последовательности может происходить по отдельности или в комбинации, один или несколько раз.
«Родительское» антитело в данном документе представляет собой антитело, которое кодируется аминокислотной последовательностью, используемой для получения варианта. В одном воплощении родительское антитело имеет собачью каркасную область и константную(ые) область(и) собачьего антитела, если таковые присутствуют. Например, родительское антитело может быть канинизированным или собачьим антителом. В качестве другого примера, родительское антитело может быть фелинизированным или кошачьим антителом. В качестве еще одного примера, родительское антитело может быть эквинизированным или лошадиным антителом. В другом примере родительское антитело может быть гуманизированным или человеческим антителом. В еще одном примере родительское антитело представляет собой мышиное моноклональное антитело.
Термины «антигенсвязывающая область», «антигенсвязывающая часть» и т.п., используемые в описании и формуле изобретения, относятся к той части молекулы антитела, которая содержит аминокислотные остатки, которые взаимодействуют с антигеном и придают антителу его специфичность и сродство к антигену. Связывающая область антитела включает «каркасные» аминокислотные остатки, необходимые для поддержания правильной конформации антигенсвязывающих остатков. Антигенсвязывающая часть антитела согласно настоящему изобретению может альтернативно обозначаться в данном описании IL-31-специфическим пептидом или полипептидом или, например, анти-IL-31 пептидом или полипептидом.
Термин «выделенный» означает, что вещество (например, антитело или нуклеиновая кислота) отделен от компонентов его естественного окружения и/или выделен. Загрязняющие компоненты его естественной окружающей среды представляют собой вещества, которые могут мешать диагностическому или терапевтическому использованию вещества, и могут включать ферменты, гормоны и другие белковые или небелковые растворенные вещества. Что касается нуклеиновой кислоты, выделенная нуклеиновая кислота может включать ту, которая отделена от 5'-3'-последовательностей, с которыми она обычно связана в хромосоме. В предпочтительных воплощениях вещество будет очищено до более чем 95% по массе вещества и, наиболее предпочтительно, до более чем 99% по массе. Выделенное вещество включает вещество in situ в рекомбинантных клетках, поскольку по меньшей мере один компонент естественной среды вещества будет отсутствовать. Однако обычно выделенное вещество получают с применением по меньшей мере одной стадии очистки.
Слово «метка», когда оно используется в данном документе, относится к детектируемому соединению или композиции, которая напрямую или опосредовано конъюгирована, например, с антителом, нуклеиновой кислотой или мимотопом. Метка может быть обнаружена сама по себе (например, радио изотопные метки или флуоресцентные метки) или, в случае ферментативной метки, может катализировать химическое изменение субстратного соединения или композиции, которые поддаются обнаружению.
Термины «нуклеиновая кислота», «полинуклеотид», «молекула нуклеиновой кислоты» и т.п.могут использоваться в данном документе взаимозаменяемо и относиться к ряду нуклеотидных оснований (также называемых «нуклеотидами») в ДНК и РНК. Нуклеиновая кислота может содержать дезоксирибонуклеотиды, рибонуклеотиды и/или их аналоги. Термин «нуклеиновая кислота» включает, например, одноцепочечные и двухцепочечные молекулы. Нуклеиновая кислота может представлять собой, например, ген или фрагмент гена, экзоны, интроны, молекулу ДНК (например, кДНК), молекулу РНК (например, мРНК), рекомбинантные нуклеиновые кислоты, плазмиды и другие векторы, праймеры и зонды. Термин включает как 5'-3' (смысловые), так и 3'-5' (антисмысловые) полинуклеотиды.
«Субъект» или «пациент» относится к млекопитающему, нуждающемуся в лечении, на которое могут воздействовать молекулы изобретения. Млекопитающие, которых можно лечить в соответствии с изобретением, включают позвоночных, среди которых особенно предпочтительными примерами являются млекопитающие, такие как собаки, кошки, лошади и люди.
«Терапевтически эффективное количество» (или «эффективное количество») относится к количеству активного ингредиента, например, вещества по изобретению, достаточному для достижения полезных или желаемых результатов при введении субъекту или пациенту. Эффективное количество можно вводить за одно или несколько введений, применений или дозировок. Терапевтически эффективное количество композиции согласно изобретению может легко определить специалист в данной области. В контексте этого изобретения «терапевтически эффективное количество» представляет собой количество, которое вызывает объективно измеряемое изменение одного или нескольких параметров, связанных с лечением опосредованного IL-31 расстройства, такого как сопровождающееся зудом состояние, аллергическое состояние или прогрессирование опухоли, включая клиническое улучшение симптомов. Несомненно, терапевтически эффективное количество будет варьироваться в зависимости от конкретного субъекта и состояния, которое лечат, веса и возраста субъекта, тяжести патологического состояния, конкретного выбранного соединения, схемы введения, времени введения, способа введения и тому подобного, все из перечисленного может легко определить специалист в данной области.
Используемый в данном документе термин «терапевтический» охватывает полный спектр лечения заболевания или нарушения. «Терапевтический» агент по настоящему изобретению может действовать профилактическим или превентивным способом, включая те, которые включают процедуры, предназначенные для воздействия на животных, у которых может быть выявлена подверженность риску (фармакогенетика); или способом, который носит улучшающий или лечебный характер; или может замедлять скорость или степень прогрессирования по меньшей мере одного симптома заболевания или расстройства, лечение которого осуществляют.
«Лечение», «лечить» и т.п. относятся как к терапевтическому воздействию, так и к профилактическим или превентивным мерам. Животные, нуждающиеся в лечении, включают тех, которые уже имеют заболевание, а также тех, у которых заболевание необходимо предотвратить. Термины «лечение» заболевания или расстройства или «лечить» заболевание или расстройство включают предотвращение или защиту от заболевания или расстройства (то есть предотвращение развития клинических симптомов); ингибирование заболевания или расстройства (т.е. остановку или подавление развития клинических симптомов) и/или облегчение заболевания или расстройства (т.е. обратное развитие клинических симптомов). Очевидно, что не всегда возможно провести различие между «предотвращением» и «подавлением» заболевания или нарушения, поскольку результирующее событие или события могут быть неизвестными или латентными. Соответственно, под «профилактикой» следует понимать тип «лечения», который включает как «предотвращение», так и «подавление». Таким образом, термин «лечение» включает в себя «профилактику».
Термин «аллергическое состояние» определяется в данном документе как нарушение или заболевание, вызванное взаимодействием между иммунной системой и веществом, чужеродным для организма. Это чужеродное вещество называется «аллергеном». Общие аллергены включают аэроаллергены, такие как пыльца, пыль, плесень, белки пылевых клещей, слюна, попадающая при укусах насекомых и т.д. Примеры аллергических состояний включают следующие: аллергический дерматит, летняя экзема, крапивница, эмфизема легких, воспалительное заболевание дыхательных путей, рецидивирующая обструкция дыхательных путей, гиперчувствительность дыхательных путей, хроническая обструктивная болезнь легких и воспалительные процессы, возникающие вследствие аутоиммунных заболеваний, таких как синдром раздраженного кишечника (СРК), но не ограничиваются ими.
Термин «сопровождающееся зудом состояние» определяется в данном документе как заболевание или нарушение, характеризующееся сильным зудом, вызывающим желание потереть или почесать кожу для получения облегчения. Примеры сопровождающихся зудом состояний включают следующие: атопический дерматит, аллергический дерматит, экзема, псориаз, склеродермия и зуд, но не ограничиваются ими.
В контексте настоящего описания термины «клетка», «линия клеток» и «культура клеток» могут использоваться взаимозаменяемо. Все эти термины также включают их потомство, то есть любые возможные последующие поколения. Понятно, что все потомство не может быть идентичным из-за преднамеренных или случайных мутаций. В контексте экспрессии гетерологичной последовательности нуклеиновой кислоты «клетка-хозяин» относится к прокариотической или эукариотической клетке (например, бактериальным клеткам, клеткам дрожжей, клеткам млекопитающих и клеткам насекомых), находящимся in vitro или in vivo. Например, клетки-хозяева могут находиться у трансгенного животного. Клетка-хозяин может использоваться в качестве реципиента для векторов и может включать любой трансформируемый организм, который способен реплицировать вектор и/или экспрессировать гетерологичную нуклеиновую кислоту, кодируемую вектором.
«Композиция» предназначена для обозначения комбинации активного вещества и другого соединения или композиции, которая может быть инертной (например, метка) или активной, такой как адъювант.
В контексте настоящего описания термины «фармацевтически приемлемый носитель» и «фармацевтически приемлемая несущая среда» являются взаимозаменяемыми и относятся к жидкой несущей среде, содержащей антигены вакцины, которую можно вводить хозяину без побочных эффектов. Фармацевтически приемлемые носители, подходящие для использования в изобретении, хорошо известны специалистам в данной области. Такие носители включают воду, физиологический раствор, забуференный физиологический раствор, фосфатный буфер, спиртовые/водные растворы, эмульсии или суспензии, без ограничения. Модно добавлять другие стандартно используемые разбавители, адъюванты и эксципиенты, в соответствии со стандартной практикой. Такие носители могут включать этанол, полиолы и их подходящие смеси, растительные масла и органические сложные эфиры для инъекций. Также могут использоваться буферы и агенты, регулирующие рН. Буферы включают соли, полученные из органической кислоты или основания, без ограничения. Типичные буферы включают соли органических кислот, такие как соли лимонной кислоты, например цитраты, аскорбиновой кислотой, глюконовой кислоты, гистидин-HC1, угольной кислоты, винной кислоты, янтарной кислоты, уксусной кислоты или фталевой кислоты, Трис, гидрохлорид триметиламина или фосфатные буферы, без ограничения. Носители для парентерального введения могут включать раствор хлорида натрия, раствор Рингера с декстрозой, декстрозу, трегалозу, сахарозу и хлорид натрия, Рингер-лактат или жирные масла. Носители для внутривенного введения могут включать вещества для восполнения жидкости и питательных веществ, вещества для восполнения электролитов, такие как средства на основе раствора Рингера с декстрозой и т.п. Консерванты и другие добавки, например, такие как противомикробные средства, антиоксиданты, хелатирующие агенты (например, EDTA), инертные газы и тому подобное, также могут входить в фармацевтические носители. Настоящее изобретение не ограничивается выбором носителя. Приготовление этих фармацевтически приемлемых композиций из описанных выше компонентов, имеющих соответствующие рН, изотоничность, стабильность и другие обычные характеристики, находится в компетенции специалистов в данной области. См., например, такие документы, как Remington: The Science and Practice of Pharmacy, 20-e издание, Lippincott Williams & Wilkins, publ., 2000 и The Handbook of Pharmaceutical Excipients, 4-е издание, дополненное, под ред. R. С.Rowe et al, APhA Publications, 2003.
Термин «консервативная аминокислотная замена» указывает на любую аминокислотную замену данного аминокислотного остатка, где замещающий остаток настолько химически подобен данному остатку, что не приводит к существенному снижению функции полипептида (например, ферментативной активности). Консервативные аминокислотные замены широко известны в данной области, и их примеры описаны, например, в патентах US 6790639, 6774107, 6194167 или 5350576. В предпочтительном воплощении консервативной аминокислотной заменой будет любая замена, которая встречается в одной из следующих шести групп
• 1. Небольшие алифатические, по существу, неполярные остатки: Ala, Gly, Pro, Ser и Thr;
• 2. Крупные алифатические неполярные остатки: Ile, Leu и Val; Met;
• 3. Полярные отрицательно заряженные остатки и их амиды: Asp и Glu;
• 4. Амиды полярных отрицательно заряженных остатков: Asn и Gln; His;
• 5. Полярные положительно заряженные остатки: Arg и Lys; His; и
• 6. Крупные ароматические остатки: Trp и Tyr; Phe.
В предпочтительном воплощении консервативная аминокислотная замена будет представлять собой любую замену из следующих, которые приведены в виде пар нативных остатков (консервативных замен): Ala (Ser); Arg (Lys); Asn (Gin; His); Asp (Glu); Gln (Asn); Glu (Asp); Gly (Pro); His (Asn; Gln); Ile (Leu; Val); Leu (Ile; Val); Lys (Arg; Gln; Glu); Met (Leu; Ile); Phe (Met; Leu; Tyr); Ser (Thr); Thr (Ser); Trp (Tyr); Tyr (Trp; Phe) и Val (Ile; Leu).
Подобно тому, как полипептид может содержать консервативную аминокислотную замену(ы), полинуклеотид может содержать консервативную(ые) замену(ы) кодона. Замена кодона считается консервативной, если при экспрессии она дает консервативную аминокислотную замену, как описано выше, в полинуклеотид ах согласно настоящему изобретению также может применяться замещение вырожденного кодона, которое не приводит к замене аминокислот. Таким образом, например, полинуклеотид, кодирующий выбранный полипептид, который может применяться в воплощении настоящего изобретения, может быть мутирован путем замены вырожденного кодона, чтобы приблизиться к частоте использования кодона, демонстрируемой экспрессирующей клеткой-хозяином, трансформируемой им, или для улучшения экспрессии другим способом.
СВЕДЕНИЯ, ПОДТВЕРЖДАЮЩИЕ ВОЗМОЖНОСТЬ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
Следует понимать, что изобретение не ограничено конкретными способами, протоколами и реагентами и т.п., описанными в данном документе, и поэтому могут варьировать. Также следует понимать, что используемая в данном документе терминология служит исключительно для описания конкретных воплощений и не предназначена для ограничения объема изобретения, который определяется исключительно формулой изобретения.
Если не указано иное, все научные и технические термины, используемые в данном документе применительно к композициям вакцины и антителам, описанным в данном документе, имеют обычное значение, известное специалистам в области техники. Кроме того, если из контекста не следует иное, термины в единственном числе включают и термины во множественном числе, а термины во множественном числе включают термины в единственном числе. Обычно используемые термины и способы культивирования клеток и тканей, молекулярной биологии, а также химии белков и олиго- или полинуклеотидов и гибридизации, описанные в данном документе, хорошо известны и широко используются в данной области.
Стандартные методы используются для рекомбинантной ДНК, синтеза олигонуклеотидов, культивирования тканей и трансфекции (например, электропорация, липофекция). Ферментативные реакции и способы очистки осуществляются в соответствии со спецификациями производителя или в соответствии с общепринятой практикой в данной области, или как описано в данном документе. Вышеупомянутые методы и процедуры обычно выполняются в соответствии со стандартными способами, хорошо известными в данной области техники и описанными в различных общих и более конкретных документах, которые цитируются и обсуждаются в настоящем описании, см., например, Sambrook et al. MOLECULAR CLONING: LAB. MANUAL (3rd ed., Cold Spring Harbor Lab. Press, Cold Spring Harbor, N.Y., 2001) and Ausubel et al. Current Protocols in Molecular Biology (New York: Greene Publishing Association/Wiley Interscience), 1993. В данном изобретении термины и лабораторные процедуры и методы, относящиеся к аналитической химии, химии органического синтеза, медицинской и фармацевтической химии, описанные в данном документе, хорошо известны и широко применяются в данной области техники. Стандартные методы используются для химического синтеза, химического анализа, составления и доставки фармацевтических препаратов, а также лечения пациентов.
За исключением рабочих примеров или там, где не указано иное, следует понимать, что все числа, выражающие количества ингредиентов или реакционные условия, использованные в данном описании, во всех случаях скорректированы с использованием термина «примерно».
Все патенты и другие обнаруженные публикации явным образом включены в данное описание посредством ссылки с целью изложения и раскрытия, например, методологий, описанных в таких публикациях, которые могут быть использованы применительно к настоящему изобретению. Эти публикации приведены исключительно ввиду их раскрытия до даты подачи настоящей заявки.
Композиции
В данном изобретении предложены мимотопы (пептиды) IL-31 и их варианты, а также их применения в клинических и научных методиках, включая диагностические методики. В контексте настоящего описания такой мимотоп IL-31 представляет собой линейный пептид или пептид с ограниченной конформационной свободой, который имитирует эпитоп антигена. Анти-IL-31 антитело к данному эпитопу антигена IL-31 будет распознавать мимотоп IL-31, который имитирует этот эпитоп.
Мимотопы (пептиды) IL-31 используются в вакцинных композициях согласно настоящему изобретению. Такие вакцинные композиции могут применяться для защиты млекопитающего от расстройства, опосредованного IL-31, такого как сопровождающееся зудом или аллергическое состояние. В некоторых воплощениях сопровождающееся зудом или аллергическое состояние, опосредованное IL-31, представляет собой сопровождающееся зудом состояние, выбранное из атопического дерматита, экземы, псориаза, склеродермии и зуда. В других воплощениях сопровождающееся зудом или аллергическое состояние, опосредованное IL-31, представляет собой аллергическое состояние, выбранное из аллергического дерматита, летней экземы, крапивницы, эмфиземы легких, воспалительного заболевания дыхательных путей, рецидивирующей обструкции дыхательных путей, гиперчувствительности дыхательных путей, хронической обструктивной болезни легких и воспалительных процессов, вызванных аутоиммунитетом. В других воплощениях расстройство, опосредованное IL-31, представляет собой прогрессирование опухоли. В некоторых воплощениях расстройство, опосредованное IL-31, представляет собой эозинофильную болезнь или мастоцитомы.
В одном воплощении вакцинная композиция по настоящему изобретению включает комбинацию полипептида-носителя и по меньшей мере одного мимотопа, выбранного из мимотопа кошачьего IL-31, мимотопа собачьего IL-31, мимотопа лошадиного IL-31 и мимотопа человеческого IL-31 и адъюванта. В некоторых воплощениях вакцинные композиции по настоящему изобретению могут включать более одного мимотопа IL-31 данного биологического вида или даже комбинацию мимотопов IL-31 от разных биологических видов. В некоторых воплощениях мимотоп (линейный или с ограниченной конформационной свободой) для использования в композициях и/или способах по настоящему изобретению представляет собой и/или включает в качестве части пептид, длина которого составляет от приблизительно 5 аминокислотных остатков до приблизительно 40 аминокислотных остатков.
В одном воплощении по меньшей мере один мимотоп, используемый в композициях и способах настоящего изобретения, выбран из мимотопа IL-31 15Н05, мимотопа области спирали ВС IL-31, мимотопа области спирали A IL-31, мимотопа области петли АВ IL- 31 или любой их комбинации.
В одном воплощении по меньшей мере один мимотоп для использования в композициях и способах настоящего изобретения вызывает выработку антител, которые нейтрализуют биоактивность IL-31.
В другом воплощении вакцинные композиции по настоящему изобретению способны вызывать направленный иммунный ответ для выработки у млекопитающего антител, направленных против по меньшей мере одного нейтрализующего эпитопа на IL-31, но не против эпитопов на IL-31, не являющихся нейтрализующими.
В одном воплощении вакцинная композиция включает мимотоп собачьего IL-31, который представляет собой и/или содержит в качестве своей части аминокислотную последовательность SVPADTFECKSF (SEQ ID NO: 186), SVPADTFERKSF (SEQ ID NO: 187), NSSAILPYFRAIRPLSDKNIIDKIIEQLDKLKF (SEQ ID NO: 192), APTHQLPPSDVRKIILELQPLSRG (SEQ ID NO: 196), TGVPES (SEQ ID NO: 200) или их варианты, которые сохраняют связывание с анти-IL-31 агентом.
В другом воплощении вакцинная композиция включает мимотоп кошачьего IL-31, который представляет собой и/или содержит в качестве своей части аминокислотную последовательность SMPADNFERKNF (SEQ ID NO: 188), NGSAILPYFRAIRPLSDKNTIDKIIEQLDKLKF (SEQ ID NO: 193), APAHRLQPSDIRKIILELRPM SKG (SEQ ID NO: 197), IGLPES (SEQ ID NO: 201) или их варианты, которые сохраняют связывание с анти-IL-31 агентом.
В еще одном воплощении вакцинная композиция включает мимотоп лошадиного IL-31, который представляет собой и/или содержит в качестве своей части аминокислотную последовательность SMPTDNFERKRF (SEQ ID NO: 189), NSSAILPYFKAISPSLNNDKSLYIIEQLDKLNF (SEQ ID NO: 194), GPIYQLQPKEIQAIIVELQNLS KK (SEQ ID NO: 198), KGVQKF (SEQ ID NO: 202) или их варианты, которые сохраняют связывание с анти-IL-31 агентом.
В следующем воплощении вакцинная композиция включает мимотоп человеческого IL-31, который представляет собой и/или содержит в качестве своей части аминокислотную последовательность SVPTDTHECKRF (SEQ ID NO: 190), SVPTDTHERKRF (SEQ ID NO: 191), HSPAIRAYLKTIRQLDNKSVIDEIIEHLDKLIF (SEQ ID NO: 195), LPVRLLRPSDDVQKIVEELQSLSKM (SEQ ID NO: 199), KGVLVS (SEQ ID NO: 203) или их варианты, которые сохраняют связывание с анти-IL-31 агентом.
В одном воплощении мимотоп, используемый в вакцинных композициях по данному изобретению, представляет собой мимотоп с ограниченной конформационной свободой. В одном воплощении такой мимотоп с ограниченной конформационной свободой представляет собой химически связанный циклический пептид.
Линейные мимотопы IL-31 могут быть синтезированы химическим путем или получены рекомбинантными способами. Мимотопы IL-31 с ограниченной конформационной свободой, такие как химически связанный циклический пептид, могут быть синтезированы химическим путем или могут быть получены с использованием комбинации химического синтеза и рекомбинантной технологии.
В некоторых воплощениях мимотоп IL-31, используемый в композиции вакцины, химически конъюгирован с полипептидом-носителем. В других воплощениях полипептид-носитель и мимотоп являются частью рекомбинантного слитого белка.
Полипептид-носитель, который комбинирован с мимотопом IL-31, может представлять собой или может включать в качестве своей части бактериальный токсоид или его производное, гемоцианин фиссуреллы (KLH) или вирусоподобную частицу. В качестве неограничивающих примеров бактериальный анатоксин или производное может представлять собой столбнячный анатоксин, дифтерийный анатоксин, столбнячный анатоксин, комплекс белков внешней мембраны из N. meningitidis группы В, экзотоксин Pseudomonas или нетоксичный мутант дифтерийного токсина (CRM197). В качестве других неограничивающих примеров вирусоподобная частица может быть HBsAg, HBcAg, бактериофагом Qbeta E.coli, вирусом Norwalk, вирусом чумы собак (CDV) или гемагглютинином (НА) вируса гриппа. В одном предпочтительном воплощении мимотоп IL-31 находится в комбинации с полипептидом-носителем, который включает или состоит из CRM197.
Композиции вакцины согласно настоящему изобретению включают по меньшей мере один адъювант или адъювантный состав, как будет более подробно описано ниже.
Вакцины по настоящему изобретению могут быть составлены в соответствии с принятым соглашением, чтобы включать фармацевтически приемлемые носители для животных, включая людей (если применимо), такие как стандартные буферы, стабилизаторы, разбавители, консерванты и/или растворители, а также могут быть составлены для обеспечения пролонгированного высвобождения. Разбавители включают воду, физиологический раствор, декстрозу, этанол, глицерин и тому подобное. Добавки для изотоничности включают, среди прочего, хлорид натрия, декстрозу, маннит, сорбит и лактозу. Стабилизаторы, среди прочего, включают альбумин. Другие подходящие несущие среды и добавки для вакцин, включая те, которые особенно полезны при составлении модифицированных живых вакцин, известны или будут очевидны специалистам в данной области техники. См., например, Remington's Pharmaceutical Science, 18-е изд., 1990, Mack Publishing, чье содержание включено в данное описание путем ссылки.
Вакцины по настоящему изобретению могут дополнительно включать один или более чем один дополнительный иммуномодулирующий компонент, например, такой как адъювант или цитокин. Типы подходящих адъювантов для применения в композициях по настоящему изобретению включают следующее: адъювант типа масло в воде, адъювант полимер и вода, адъювант вода в масле, адъювант гидроксид алюминия, адъювант витамин Е и их комбинации. Некоторые конкретные примеры адъювантов включают полный адъювант Фрейнда, неполный адъювант Фрейнда, Corynebacterium parvum, бациллу Кальмета-Герена, гель гидроксид алюминия, глюкан, сульфат декстрана, оксид железа, альгинат натрия, бакто-адъювант, некоторые синтетические полимеры, такие как полиаминокислоты и сополимеры аминокислот, блок-сополимер (CytRx, Атланта, Джорджия), QS-21 (Cambridge Biotech Inc., Cambridge Mass.), SAF-M (Chiron, Emeryville CA), Адъювант AMPHIGEN®, сапонин, Quil А или другую фракцию сапонинов, монофосфориллипид А и адъювант авридин липид-амин (N,N-диоктадецил-N',N'-бис(2-гидроксиэтил) -пропандиамин), «РЕГРЕССИИ» (Vetrepharm, Athens, GA), парафиновое масло, систему адъювантов RIBI (Ribi Inc., Hamilton, Mont.), мурамилдипептид и тому подобные, без ограничения.
Неограничивающие примеры эмульсий масло-в-воде, которые могут применяться в вакцине по изобретению, включают модифицированные составы SEAM62 и SEAM 1/2. Модифицированный SEAM62 представляет собой эмульсию масло-в-воде, содержащую 5% (об./об.) сквалена (Sigma), 1% (об./об.) детергента SPAN® 85 (ICI Surfactants), 0,7% (об./об.) детергента TWEEN® 80 (ICI Surfactants), 2,5% (об./об.) этанола, 200 мкг/мл Quil А, 100 мкг/мл холестерина и 0,5% (об./об.) лецитина. Модифицированный SEAM 1/2 представляет собой эмульсию масло-в-воде, содержащую 5% (об./об.) сквалена, 1% (об./об.) детергента SPAN® 85, 0,7% (об./об.) детергента Tween 80, 2,5% (об./об.) этанола, 100 мкг/мл Quil А и 50 мкг/мл холестерина.
Другим примером адъюванта, который может применяться в композиции по изобретению, является масло SP. Используемый в описании и формуле изобретения термин «масло SP» обозначает масляную эмульсию, содержащую блок-сополимер полиоксиэтилена и полиоксипропилена, сквалан, полиоксиэтиленсорбитанмоноолеат и забуференный солевой раствор. Блок-сополимеры полиоксиэтилена и полиоксипропилена представляют собой поверхностно-активные вещества, которые способствуют суспендированию твердых и жидких компонентов. Эти поверхностно-активные вещества доступны для приобретения в виде полимеров под торговым названием Pluronic®. Предпочтительным поверхностно-активным веществом является полоксамер 401, который доступен для приобретения под торговым названием Pluronic® L-121. Обычно масляная эмульсия SP представляет собой иммуностимулирующую адъювантную смесь, которая будет включать от приблизительно 1 до 3% об./об. блок-сополимера, от приблизительно 2 до 6% об./об. сквалана, более конкретно от приблизительно 3 до 6% сквалана и приблизительно 0,1-0,5% об./об. полиоксиэтиленсорбитанмоноолеата, а остальная часть представляет собой забуференный солевой раствор.
«Иммуномодуляторы», которые могут быть включены в вакцину, включают, например, иммуностимулирующие олигонуклеотиды, один или несколько интерлейкинов, интерферонов или других известных цитокинов. В одном воплощении адъювант может быть производным циклодекстрина или полианионным полимером, таким как те, что описаны в патентах US 6165995 и 6610310, соответственно.
В одном воплощении адъювант представляет собой состав, содержащий сапонин, стерол, четвертичное аммониевое соединение и полимер. В конкретном воплощении сапонин представляет собой Quil А или его очищенную фракцию, стерол представляет собой холестерин, четвертичное аммониевое соединение представляет собой бромид диметилдиоктадециламмония (DDA), а полимер представляет собой полиакриловую кислоту.
В другом воплощении адъювант включает комбинацию одного или нескольких выделенных иммуностимулирующих олигонуклеотидов, стерола и сапонина. В конкретном воплощении один или несколько выделенных иммуностимулирующих олигонуклеотидов содержат CpG, стерол представляет собой холестерин, а сапонин представляет собой Quil А или его очищенную фракцию. В данном контексте адъювант ZA-01, упомянутый в разделе примеров, включает Quil А (сапонин), холестерин, CpG и разбавитель.
В другом воплощении полезный адъювант для использования в композициях по настоящему изобретению включает CpG-содержащие иммуностимулирующие олигонуклеотиды. CpG-содержащие олигонуклеотиды описаны, например, в патенте US 8580280. В одном конкретном воплощении адъювант для использования в настоящем изобретении представляет собой смесь, включающую по меньшей мере один гликолипидный адъювант и CpG-содержащие олигонуклеотиды. Конкретным примером пригодного адъюванта является смесь, которая включает гликолипидный адъювант Bay R1005 (N-(2-дезокси-2-L-лейциламино-β-D-глюкопиранозил)-N-октадецилдодеканоиламидгидроацетат), а также олигонуклеотиды CpG.
В одном варианте адъювант или смесь адъювантов добавляют в количестве от примерно 100 мкг до примерно 10 мг на дозу. В другом воплощении адъювант/смесь адъювантов добавляют в количестве от примерно 200 мкг до примерно 5 мг на дозу. В еще одном воплощении адъювант/смесь адъювантов добавляют в количестве от примерно 300 мкг до примерно 1 мг/дозу.
С появлением методов молекулярной биологии и рекомбинантной технологии стало возможным получать вышеупомянутые пептиды и полипептиды рекомбинантными способами и, таким образом, генерировать последовательности генов, которые кодируют конкретные аминокислотные последовательности, обнаруженные в структуре пептида или полипептида. В одном воплощении пептид представляет собой мимотоп IL-31 или, по меньшей мере, является частью мимотопа IL-31. В другом воплощении полипептид представляет собой полипептид-носитель, который присутствует в комбинации с мимотопом IL-31. В еще одном воплощении полипептид представляет собой антитело, такое как антитело, с которым связывается мимотоп IL-31, используемый в композиции вакцины или диагностических способах по настоящему изобретению. Такие антитела можно получить либо путем клонирования последовательностей генов, кодирующих полипептидные цепи указанных антител, либо путем прямого синтеза указанных полипептидных цепей со сборкой синтезированных цепей с образованием активных тетрамерных (H2L2) структур со сродством к конкретным эпитопам и антигенным детерминантам. Это позволило получить готовые антитела с последовательностями, характерными для нейтрализующих антител из разных видов и источников.
Независимо от источника антител или того, как они конструируются рекомбинантно, или как они синтезируются, in vitro или in vivo, с использованием трансгенных животных, массовых клеточных культур лабораторного или промышленного масштаба, с использованием трансгенных растений или путем прямого химического синтеза, без использования живых организмов на какой-либо стадии процесса, все антитела имеют сходную общую трехмерную структуру. Эта структура часто обозначается как H2L2 и относится к тому факту, что антитела обычно содержат две легких аминокислотных цепи (L) и две тяжелых аминокислотных цепи (Н). Обе цепи имеют области, способные взаимодействовать со структурно комплементарной антигенной мишенью. Области, взаимодействующие с мишенью, называются «вариабельными» или «V»-областями и характеризуются различиями в аминокислотной последовательности с антителами иной антигенной специфичности. Вариабельные области Н или L цепей содержат аминокислотные последовательности, способные специфически связываться с антигенными мишенями.
«Антигенсвязывающая область» или «антигенсвязывающая часть» антитела относится к той части молекулы антитела, которая содержит аминокислотные остатки, которые взаимодействуют с антигеном и придают антителу его специфичность и сродство к антигену. Связывающая область антитела включает «каркасные» аминокислотные остатки, необходимые для поддержания правильной конформации антигенсвязывающих остатков. Антигенсвязывающая часть антитела, упомянутая в описании и формуле изобретения, может обозначаться в данном документе, например, как IL-31-специфический пептид или полипептид или как анти-IL-31 пептид или полипептид.
В вариабельных областях Н- или L-цепей, которые обеспечивают антигенсвязывающие области, есть более маленькие последовательности, названные «гипервариабельными» из-за их крайней изменчивости у антител с разной специфичностью. Такие гипервариабельные области также называют «областями, определяющими комплементарность» или «CDR» областями. Эти участки CDR определяют основную специфичность антитела к конкретной структуре антигенной детерминанты.
CDR представляют собой несмежные участки аминокислот в вариабельных областях, но было обнаружено, что положения этих критических аминокислотных последовательностей аналогичны в аминокислотных последовательностях вариабельных областей тяжелой и легкой цепи, независимо от вида. Каждая вариабельная область тяжелой и легкой цепи всех антител имеет три участка CDR, каждый из которых не является смежным с другими.
У всех видов млекопитающих пептиды антител содержат константные (т.е. высококонсервативные) и вариабельные области, а внутри последних есть CDR и так называемые «каркасные области», состоящие из аминокислотных последовательностей в составе вариабельной области тяжелой или легкой цепь, но за пределами CDR.
Что касается антигенной детерминаты, распознаваемой CDR областями антитела, ее также называют «эпитопом». «Другими словами, эпитоп относится к той части любой молекулы, которая способна распознаваться и связываться антителом (соответствующая связывающая область антитела может называться паратопом).
«Антиген» представляет собой молекулу или часть молекулы, способную связываться с антителом, которое дополнительно способно индуцировать у животного продуцирование антитела, способного связываться с эпитопом этого антигена. Антиген может иметь один или более чем один эпитоп. Вышеупомянутая специфическая реакция указывает на то, что антиген будет реагировать высокоселективным образом со своим соответствующим антителом, а не с множеством других антител, которые могут быть вызваны другими антигенами.
Подразумевается, что упомянутые в данном документе антитела включают как интактные молекулы иммуноглобулина, так и их части, фрагменты, пептиды и их производные, такие как, например, Fab, Fab', F(ab')2, Fv, Fse, области CDR, паратопы или любую часть (например, полипептид) или пептидную последовательность антитела, способную связывать антиген или эпитоп. Говорят, что антитело «способно связывать» молекулу, если оно способно специфически реагировать с молекулой, тем самым связывая молекулу с антителом.
Упомянутые в данном документе антитела также включают химерные антитела, гетерохимерные антитела, канинизированные антитела, фелинизированные антитела, эквинизированные антитела, гуманизированные антитела, полностью собачьи антитела, полностью кошачьи антитела, полностью лошадиные антитела, полностью человеческие антитела, а также их фрагменты, части, области, пептиды или производные, полученные любым известным способом, таким как ферментативное расщепление, синтез пептидов или рекомбинантные методы, без ограничения. Такие упомянутые в данном документе антитела способны специфически связываться по меньшей мере с одним из собачьего IL-31, кошачьего IL-31, лошадиного IL-31 или человеческого IL-31. Фрагменты или части антител могут не иметь Fc-фрагмента интактного антитела, быстрее выводиться из кровотока и могут меньше неспецифически связываться с тканью по сравнению с интактным антителом. Примеры фрагментов антител могут быть получены из интактных антител с использованием способов, хорошо известных в данной области техники, например, протеолитическим расщеплением такими ферментами, как папаин (для получения Fab-фрагментов) или пепсин (для получения F(ab')2 фрагментов). См., например, Wahl et al., 24 J. Nucl. Med. 316-25 (1983). Части антител могут быть получены любым из вышеуказанных способов или могут быть получены путем экспрессии части рекомбинантной молекулы. Например, область(и) CDR рекомбинантного антитела можно выделить и субклонировать в соответствующий вектор экспрессии. Например, область(и) CDR рекомбинантного антитела можно выделить и субклонировать в соответствующий вектор экспрессии. См., например, патент US №6680053.
Нуклеотидные и аминокислотные последовательности клонов 15Н05, ZIL1, ZIL8, ZIL9, ZIL11, ZIL69, ZIL94, ZIL154, ZIL159 и ZIL171
В некоторых воплощениях данного изобретения предложены мимотопы IL-31, которые связываются с новыми моноклональными антителами, которые связываются по меньшей мере с одним из собачьего IL-31, кошачьго IL-31 или лошадиного IL-31. Такие моноклональные антитела можно использовать в диагностических способах по настоящему изобретению вместе с мимотопом IL-31. В одном воплощении моноклональное антитело, упомянутое в описании и формуле изобретения, связывается с собачьим IL-31, кошачьим IL-31 или лошадиным IL-31 и предотвращает его связывание и активацию его корецепторного комплекса, содержащего рецептор A IL-31 (IL-31 Ra) и онкостатин-М-специфический рецептор (OsmR или IL-31Rb). Примеры таких моноклональных антител обозначены в данном документе как «15Н05», «ZIL1», «ZIL8», «ZIL9», «ZIL11», «ZIL69», «ZIL94», «ZIL154», «ZIL159» и «ZIL171», в соответствии с номерами, присвоенными их клонам. В данном документе «15Н05», «ZIL1», «ZIL8», «ZIL9», «ZIL11», «ZIL69», «ZIL94», «ZIL154», «ZIL159» и «ZIL171» также относятся к части моноклонального антитела, паратопу или CDR, которые специфически связываются с эпитопом IL-31, обозначенным как 15Н05, ZIL1, ZIL8, ZIL9, ZIL11, ZIL69, ZIL94, ZIL154, ZIL159 и ZIL171 благодаря его способности связывать антитела 15Н05, ZIL1, ZIL8, ZIL9, ZIL11, ZIL69, ZIL94, ZIL154, ZIL159 и ZIL171, соответственно. Несколько рекомбинантных, химерных, гетерохимерных, канинизированных, фелинизированных, эквинизированных, полностью собачьих, полностью кошачьих и/или полностью лошадиных форм 15Н05, ZIL1, ZIL8, ZIL9, ZIL11, ZIL69, ZIL94, ZIL154, ZIL159 и ZIL171, описанных в данном документе, могут обозначаться таким же названием.
В одном воплощении вакцинная композиция по данному изобретению включает мимотоп, который связывается с антителом против IL-31 или его антигенсвязывающей частью, которая специфически связывается с областью белка IL-31 млекопитающих, задействованной во взаимодействии белка IL-31 со своим ко-рецептором. В одном воплощении на связывание указанного антитела с указанной областью влияют мутации в области эпитопа, связывающейся с 15Н05, выбранной из: а) области между аминокислотными остатками 124 и 135 последовательности кошачьего IL-31, представленной SEQ ID NO: 157 (кошачий_IL-31_дикий тип); б) области между аминокислотными остатками 124 и 135 последовательности собачьего IL-31, представленной SEQ ID NO: 155 (собачий_IL-31) и в) области между аминокислотными остатками 118 и 129 последовательности лошадиного IL-31, представленной SEQ ID NO: 165 (лошадиный_IL-31). В одном воплощении мимотоп для применения в композициях по данному изобретению связывается с антителом против IL-31 или его антигенсвязывающей частью, которая специфически связывается с вышеупомянутой областью эпитопа, распознаваемой 15Н05.
В одном конкретном воплощении композиции вакцины по данному изобретению мимотоп связывается с антителом против IL-31 или его антигенсвязывающей частью, включающей по меньшей мере одну из следующих комбинаций последовательностей областей, определяющих комплементарность (CDR):
1) антитело 15Н05: CDR1 вариабельной области тяжелой цепи (VH) с последовательностью SYTIH (SEQ ID NO: 1), VH-CDR2 с последовательностью NINPTSGYTENNQRFKD (SEQ ID NO: 2), VH-CDR3 с последовательностью WGFKYDGEWSFDV (SEQ ID NO: 3), CDR1 вариабельной области легкой цепи (VL) с последовательностью RASQGISIWLS (SEQ ID NO: 4), VL-CDR2 с последовательностью KASNLHI (SEQ ID NO: 5) и VL-CDR3 с последовательностью LQSQTYPLT (SEQ ID NO: 6);
2) антитело ZIL1: CDR1 вариабельной области тяжелой цепи (VH) с последовательностью SYGMS (SEQ ID NO: 13), VH-CDR2 с последовательностью HINSGGSSTYYADAVKG (SEQ ID NO: 14), VH-CDR3 с последовательностью VYTTLAAFWTDNFDY (SEQ ID NO: 15), вариабельной области легкой цепи (VL) с последовательностью SGSTNNIGILAAT (SEQ ID NO: 16), VL-CDR2 с последовательностью SDGNRPS (SEQ ID NO: 17) и VL-CDR3 с последовательностью QSFDTTLDAYV (SEQ ID NO: 18);
3) антитело ZIL8: VH-CDR1 с последовательностью DYAMS (SEQ ID NO: 19), VH-CDR2 с последовательностью GIDSVGS GTS YADA VKG (SEQ ID NO: 20), VH-CDR3 с последовательностью GFPGSFEH (SEQ ID NO: 21), VL-CDR1 с последовательностью TGSSSNIGSGYVG (SEQ ID NO: 22), VL-CDR2 с последовательностью YNSDRPS (SEQ ID NO: 23), VL-CDR3 с последовательностью SVYDRTFNAV (SEQ ID NO: 24);
4) антитело ZIL9: VH-CDR1 с последовательностью SYDMT (SEQ ID NO: 25), VH-CDR2 с последовательностью DVNSGGTGTAYAVAVKG (SEQ ID NO: 26), VH-CDR3 с последовательностью LGVRDGLSV (SEQ ID NO: 27), VL-CDR1 с последовательностью SGESLNEYYTQ (SEQ ID NO: 28), VL-CDR2 с последовательностью RDTERPS (SEQ ID NO: 29), VL-CDR3 с последовательностью ESAVDTGTLV (SEQ ID NO: 30);
5) антитело ZIL11: VH-CDR1 с последовательностью TYVMN (SEQ ID NO: 31), VH-CDR2 с последовательностью SINGGGSSPTYADAVRG (SEQ ID NO: 32), VH-CDR3 с последовательностью SMVGPFDY (SEQ ID NO: 33), VL-CDR1 с последовательностью SGESLSNYYAQ (SEQ ID NO: 34), VL-CDR2 с последовательностью KDTERPS (SEQ ID NO: 35), VL-CDR3 с последовательностью ESAVSSDTIV (SEQ ID NO: 36);
6) антитело ZIL69: VH-CDR1 с последовательностью SYAMK (SEQ ID NO: 37), VH-CDR2 с последовательностью TINNDGTRTGYADAVRG (SEQ ID NO: 38), VH-CDR3 с последовательностью GNAESGCTGDHCPPY (SEQ ID NO: 39), VL-CDR1 с последовательностью SGESLNKYYAQ (SEQ ID NO: 40), VL-CDR2 с последовательностью KDTERPS (SEQ ID NO: 41), VL-CDR3 с последовательностью ESAVSSETNV (SEQ ID NO: 42);
7) антитело ZIL94: VH-CDR1 с последовательностью TYFMS (SEQ ID NO: 43), VH-CDR2 с последовательностью LIS SDGSGTY YADA VKG (SEQ ID NO: 44), VH-CDR3 с последовательностью FWRAFND (SEQ ID NO: 45), VL-CDR1 с последовательностью GLNSGSVSTSNYPG (SEQ ID NO: 46), VL-CDR2 с последовательностью DTGSRPS (SEQ ID NO: 47), VL-CDR3 с последовательностью SLYTDSDILV (SEQ ID NO: 48);
8) антитело ZIL154: VH-CDR1 с последовательностью DRGMS (SEQ ID NO: 49), VH-CDR2 с последовательностью YIRYDGSRTDYADAVEG (SEQ ID NO: 50), VH-CDR3 с последовательностью WDGSSFDY (SEQ ID NO: 51), VL-CDR1 с последовательностью KASQSLLHSDGNTYLD (SEQ ID NO: 52), VL-CDR2 с последовательностью KVSNRDP (SEQ ID NO: 53), VL-CDR3 с последовательностью MQAIHFPLT (SEQ ID NO: 54);
9) антитело ZIL159: VH-CDR1 с последовательностью SYVMT (SEQ ID NO: 55), VH-CDR2 с последовательностью GINSEGSRTAYADAVKG (SEQ ID NO: 56), VH-CDR3 с последовательностью GDIVATGTSY (SEQ ID NO: 57), VL-CDR1 с последовательностью SGETLNRFYTQ (SEQ ID NO: 58), VL-CDR2 с последовательностью KDTERPS (SEQ ID NO: 59), VL-CDR3 с последовательностью KSAVSIDVGV (SEQ ID NO: 60);
10) антитело ZIL171: VH-CDR1 с последовательностью TYVMN (SEQ ID NO: 61), VH-CDR2 с последовательностью SINGGGSSPTYADAVRG (SEQ ID NO: 62), VH-CDR3 с последовательностью SMVGPFDY (SEQ ID NO: 63), VL-CDR1 с последовательностью SGKSLSYYYAQ (SEQ ID NO: 64), VL-CDR2 с последовательностью KDTERPS (SEQ ID NO: 65), VL-CDR3 с последовательностью ESAVSSDTIV (SEQ ID NO: 66); или
11) вариант (1)-(10), отличающийся от соответствующего родительского антитела 15Н05, ZIL1, ZIL8, ZIL9, ZIL11, ZIL69, ZIL94, ZIL154, ZIL159 или ZIL171 добавлением, делецией и/или заменой одного или более чем одного аминокислотного остатка по меньшей мере в одном из CDR1, CDR2 или CDR3 VH или VL.
В одном воплощении мимотоп IL-31, используемый в композиции вакцины, связывается с антителом, которое специфически связывается с кошачьим IL-31, где антитело связывается с областью между аминокислотными остатками 125 и 134 последовательности IL-31 кошки, представленной SEQ ID NO: 157 (кошачий_IL-31_дикого типа). В некоторых воплощениях такое антитело включает цепь VL, содержащую изменения каркасной области 2 (FW2), выбранные из следующих: аспарагин вместо лизина в положении 42, изолейцин вместо валина в положении 43, валин вместо лейцина в положении 46, аспарагин вместо лизина в положении 49 и их комбинаций, где положения относятся к нумерации SEQ ID NO: 127 (FEL_15Н05_VL1).
В некоторых воплощениях мимотоп связывается с антителом, характеризующимся тем, что:
1) антитело ZIL1 включает по меньшей мере одно из следующего:
a) вариабельную область легкой цепи, содержащей CAN-ZIL1_VL:
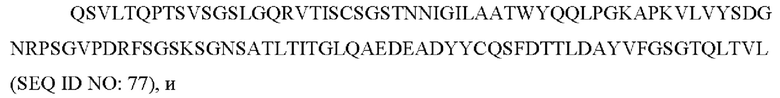
b) вариабельную область тяжелой цепи, содержащей CAN-ZIL1_VH:

2) антитело ZIL8 включает по меньшей мере одно из следующего:
a) вариабельную область легкой цепи, содержащей CAN-ZIL8_VL:
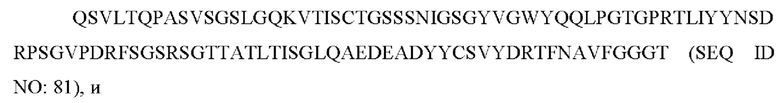
b) вариабельную область тяжелой цепи, содержащей CAN-ZIL8_VH:
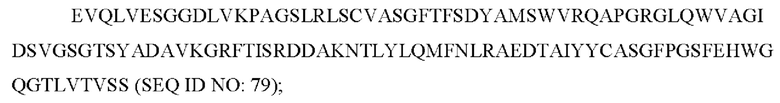
3) антитело ZIL9 включает по меньшей мере одно из следующего:
а) вариабельную область легкой цепи, содержащей CAN-ZIL9_VL:
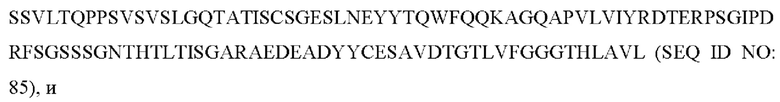
б) вариабельную область тяжелой цепи, содержащей CAN-ZIL9_VH:
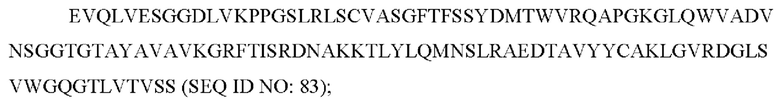
4) антитело ZIL11 включает по меньшей мере одно из следующего:
а) вариабельную область легкой цепи, содержащей CAN-ZIL11_VL:
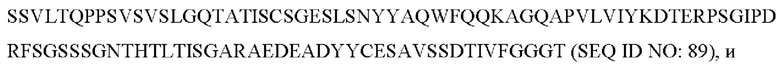
б) вариабельную область тяжелой цепи, содержащей CAN-ZIL11_VH:

5) антитело ZIL69 включает по меньшей мере одно из следующего:
а) вариабельную область легкой цепи, содержащей CAN-ZIL169_VL:
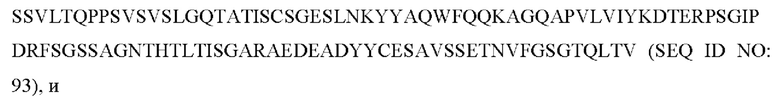
б) вариабельную область тяжелой цепи, содержащей CAN-ZIL69_VH:
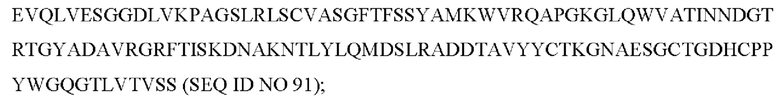
6) антитело ZIL94 включает по меньшей мере одно из следующего:
а) вариабельную область легкой цепи, содержащей CAN-ZIL94_VL:
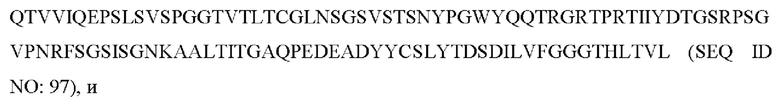
б) вариабельную область тяжелой цепи, содержащей CAN-ZIL94_VH:
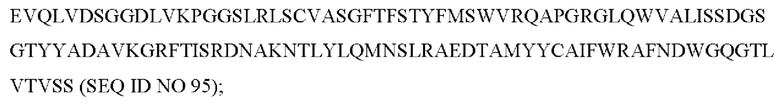
7) антитело ZIL154 включает по меньшей мере одно из следующего:
а) вариабельную область легкой цепи, содержащей CAN-ZIL154_VL:
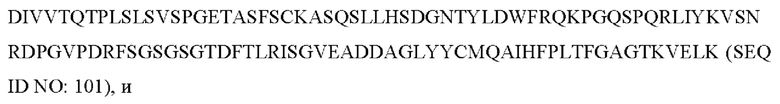
б) вариабельную область тяжелой цепи, содержащей CAN-ZIL154_VH:
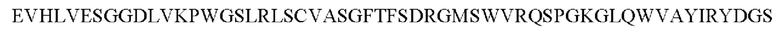
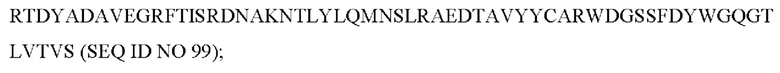
8) антитело ZIL159 включает по меньшей мере одно из следующего:
а) вариабельную область легкой цепи, содержащей CAN-ZIL159_VL:
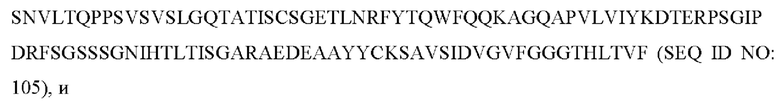
б) вариабельную область тяжелой цепи, содержащей CAN-ZIL159_VH:
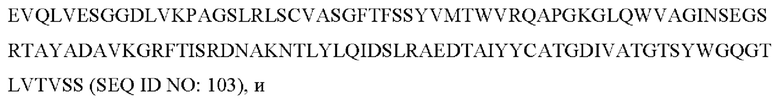
9) антитело ZIL171 включает по меньшей мере одно из следующего:
а) вариабельную область легкой цепи, содержащей CAN-ZIL171_VL:
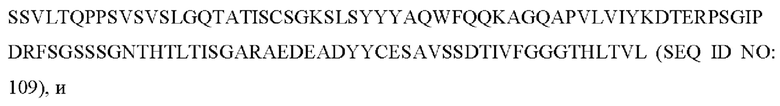
б) вариабельную область тяжелой цепи, содержащей CAN-ZIL171_VH:
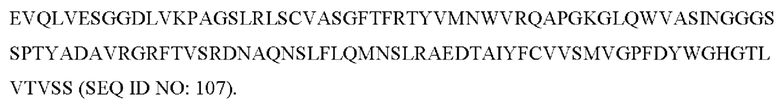
Клетка-хозяин может быть использована для получения антитела, описанного выше. Такие антитела можно использовать в диагностических процедурах, описанных как часть этого изобретения, хотя диагностические процедуры не ограничиваются этими конкретными антителами.
Нуклеотидные последовательности, кодирующие вариабельные области легкой и тяжелой цепей антитела против IL-31, можно использовать для получения антител против IL-31, описанных в данном документе. Такие нуклеотидные последовательности включают любую нуклеотидную последовательность, которая кодирует аминокислотную последовательность антител 15Н05, ZIL1, ZIL8, ZIL9, ZIL11, ZIL69, ZIL94, ZIL154, ZIL159 или ZIL171 или IL-31-специфических полипептидов или их пептидов, но не ограничиваются ими. Кроме того, в одном воплощении нуклеотидные последовательности, кодирующие мимотопы IL-31, можно использовать для рекомбинантного получения мимотопов отдельно или как части слитого белка вместе с полипептидом-носителем. Альтернативно или в качестве дополнения мимотопы можно синтезировать химическим путем.
В некоторых воплощениях выделенная нуклеиновая кислота может быть использована для получения полезного антитела (такого как антитело, используемое в одном из диагностических методов, описанных в настоящем документе), где последовательность нуклеиновой кислоты кодирует по меньшей мере одну из следующих комбинаций последовательностей определяющих комплементарность участков (CDR) вариабельной области тяжелой цепи:
1) 15Н05: CDR1 вариабельной области тяжелой цепи (VH) с последовательностью SYTIH (SEQ ID NO: 1), VH-CDR2 с последовательностью NINPTSGYTENNQRFKD (SEQ ID NO: 2) и VH-CDR3 с последовательностью WGFKYDGEWSFDV (SEQ ID NO: 3);
2) ZIL1: VH-CDR1 с последовательностью SYGMS (SEQ ID NO: 13), VH-CDR2 с последовательностью HINSGGSSTYYADAVKG (SEQ ID NO: 14) и VH-CDR3 с последовательностью VYTTLAAFWTDNFDY (SEQ ID NO: 15);
3) ZIL8: VH-CDR1 с последовательностью DYAMS (SEQ ID NO: 19), VH-CDR2 с последовательностью GIDSVGSGTSYADAVKG (SEQ ID NO: 20) и VH-CDR3 с последовательностью GFPGSFEH (SEQ ID NO: 21);
4) ZIL9: VH-CDR1 с последовательностью SYDMT (SEQ ID NO: 25), VH-CDR2 с последовательностью DVNSGGTGTAYAVAVKG (SEQ ID NO: 26) и VH-CDR3 с последовательностью LGVRDGLSV (SEQ ID NO: 27);
5) ZIL11: VH-CDR1 с последовательностью TYVMN (SEQ ID NO: 31), VH-CDR2 с последовательностью SINGGGSSPTYADAVRG (SEQ ID NO: 32) и VH-CDR3 с последовательностью SMVGPFDY (SEQ ID NO: 33);
6) ZIL69: VH-CDR1 с последовательностью SYAMK (SEQ ID NO: 37), VH-CDR2 с последовательностью TINNDGTRTGYADAVRG (SEQ ID NO: 38) и VH-CDR3 с последовательностью GNAESGCTGDHCPPY (SEQ ID NO: 39);
7) ZIL94: VH-CDR1 с последовательностью TYFMS (SEQ ID NO: 43), VH-CDR2 с последовательностью LISSDGSGTYYADAVKG (SEQ ID NO: 44) и VH-CDR3 с последовательностью FWRAFND (SEQ ID NO: 45)
8) ZIL154: VH-CDR1 с последовательностью DRGMS (SEQ ID NO: 49), VH-CDR2 с последовательностью YIRYDGSRTDYADAVEG (SEQ ID NO: 50) и VH-CDR3 с последовательностью WDGSSFDY (SEQ ID NO: 51);
9) ZIL159: VH-CDR1 с последовательностью SYVMT (SEQ ID NO: 55), VH-CDR2 с последовательностью GINSEGSRTAYADAVKG (SEQ ID NO: 56) и VH-CDR3 с последовательностью GDIVATGTSY (SEQ ID NO: 57);
10) ZIL171: VH-CDR1 с последовательностью TYVMN (SEQ ID NO: 61), VH-CDR2 с последовательностью SINGGGSSPTYADAVRG (SEQ ID NO: 62) и VH-CDR3 с последовательностью SMVGPFDY (SEQ ID NO: 63), или
11) вариант (1)-(10), отличающийся от CDR соответствующего родительского антитела 15Н05, ZIL1, ZIL8, ZIL9, ZIL11, ZIL69, ZIL94, ZIL154, ZIL159 или ZIL171 добавлением, делецией и/или заменой одного или более чем одного аминокислотного остатка по меньшей мере в одном из CDR1, CDR2 или CDR3 VH.
В другом воплощении выделенная нуклеиновая кислота содержит последовательность нуклеиновой кислоты, кодирующей по меньшей мере одну из следующих комбинаций последовательностей определяющих комплементарность участков (CDR) вариабельной области легкой цепи:
1) 15Н05: CDR1 вариабельной области легкой цепи (VL) с последовательностью RASQGISIWLS (SEQ ID NO: 4), VL-CDR2 с последовательностью KASNLHI (SEQ ID NO: 5) и VL-CDR3 с последовательностью LQSQTYPLT (SEQ ID NO: 6);
2) ZIL1: VL-CDR1 с последовательностью SGSTNNIGILAAT (SEQ ID NO: 16), VL-CDR2 с последовательностью SDGNRPS (SEQ ID NO: 17 и VL-CDR3 с последовательностью QSFDTTLDAYV (SEQ ID NO: 18);
3) ZIL8: VL-CDR1 с последовательностью TGSSSNIGSGYVG (SEQ ID NO: 22), VL-CDR2 с последовательностью YNSDRPS (SEQ ID NO: 23) и VL-CDR3 с последовательностью SVYDRTFNAV (SEQ ID NO: 24);
4) ZIL9: VL-CDR1 с последовательностью SGESLNEYYTQ (SEQ ID NO: 28), VL-CDR2 с последовательностью RDTERPS (SEQ ID NO: 29) и VL-CDR3 с последовательностью ESAVDTGTLV (SEQ ID NO: 30);
5) ZIL11: VL-CDR1 с последовательностью SGESLSNYYAQ (SEQ ID NO: 34), VL-CDR2 с последовательностью KDTERPS (SEQ ID NO: 35) и VL-CDR3 с последовательностью ESAVSSDTIV (SEQ ID NO: 36);
6) ZIL69: VL-CDR1 с последовательностью SGESLNKYYAQ (SEQ ID NO: 40), VL-CDR2 с последовательностью KDTERPS (SEQ ID NO: 41) и VL-CDR3 с последовательностью ESAVSSETNV (SEQ ID NO: 42);
7) ZIL94: VL-CDR1 с последовательностью GLNSGSVSTSNYPG (SEQ ID NO: 46), VL-CDR2 с последовательностью DTGSRPS (SEQ ID NO: 47) и VL-CDR3 с последовательностью SLYTDSDILV (SEQ ID NO: 48);
8) ZIL154: VL-CDR1 с последовательностью KASQSLLHSDGNTYLD (SEQ ID NO: 52), VL-CDR2 с последовательностью KVSNRDP (SEQ ID NO: 53) и VL-CDR3 с последовательностью MQAIHFPLT (SEQ ID NO: 54);
9) ZIL159: VL-CDR1 с последовательностью SGETLNRFYTQ (SEQ ID NO: 58), VL-CDR2 с последовательностью KDTERPS (SEQ ID NO: 59) и VL-CDR3 с последовательностью KSAVSIDVGV (SEQ ID NO: 60);
10) ZIL171: VL-CDR1 с последовательностью SGKSLSYYYAQ (SEQ ID NO: 64), VL-CDR2 с последовательностью KDTERPS (SEQ ID NO: 65) и VL-CDR3 с последовательностью ESAVSSDTIV (SEQ ID NO: 66) или
11) вариант (1)-(10), отличающийся от CDR соответствующего родительского антитела 15Н05, ZIL1, ZIL8, ZIL9, ZIL11, ZIL69, ZIL94, ZIL154, ZIL159 или ZIL171 добавлением, делецией и/или заменой одного или более чем одного аминокислотного остатка по меньшей мере в одном из CDR1, CDR2 или CDR3 VL.
В еще одном воплощении выделенная нуклеиновая кислота, используемая при производстве описанного в данном документе антитела, содержит последовательность нуклеиновой кислоты, кодирующую вышеописанные последовательности определяющих комплементарность участков (CDR) вариабельной области легкой цепи, а также описанные выше последовательности CDR вариабельной области тяжелой цепи соответствующего родительского антитела 15Н05, ZIL1, ZIL8, ZIL9, ZIL11, ZIL69, ZIL94, ZIL154, ZIL159 или ZIL171 или их вариантов.
Вектор, включающий по меньшей мере одну из вышеописанных нуклеиновых кислот, может быть использован при производстве этих антител. Как будет описано более подробно ниже, последовательность нуклеиновой кислоты, кодирующая по меньшей мере одну из описанных выше комбинаций последовательностей определяющих комплементарность участков (CDR) вариабельной области тяжелой цепи, может содержаться в том же векторе вместе с последовательностью нуклеиновой кислоты, кодирующей по меньшей мере одну из описанных выше комбинаций последовательностей CDR вариабельной области легкой цепи. В альтернативном варианте последовательность нуклеиновой кислоты, кодирующая по меньшей мере одну из описанных выше комбинаций последовательностей CDR вариабельной области тяжелой цепи и последовательность нуклеиновой кислоты, кодирующая по меньшей мере одну из описанных выше комбинаций последовательностей CDR вариабельной области тяжелой цепи, могут содержаться в разных векторах.
По причине вырожденности генетического кода для кодирования конкретной аминокислоты может использоваться более чем один кодон. Используя генетический код, можно идентифицировать одну или несколько различных нуклеотидных последовательностей, каждая из которых способна кодировать аминокислоту. Вероятность того, что конкретный олигонуклеотид на самом деле будет представлять собой фактическую последовательность, кодирующую XXX, можно оценить, учитывая аномальные отношения спаривания оснований и частоту, с которой конкретный кодон фактически используется (для кодирования конкретной аминокислоты) в эукариотических или прокариотических клетках, экспрессирующих антитело против IL-31 или его часть, специфичную для IL-31. Такие «правила использования кодонов» изложены Lathe, et al., 183 J. Molec. Biol. 1-12 (1985). Используя «правила использования кодонов» Lathe, можно идентифицировать одну нуклеотидную последовательность или набор нуклеотидных последовательностей, которые содержат теоретическую «наиболее вероятную» нуклеотидную последовательность, способную кодировать анти-IL-31 последовательности. Также предполагается, что кодирующие области антитела также могут быть получены путем изменения существующих генов антител с использованием стандартных молекулярно-биологических методов, которые приводят к вариантам (агонистам) антител и пептидов, описанных в данном документе. Такие варианты включают делеции, добавления и замены в аминокислотной последовательности антител против IL-31 или IL-31-специфических полипептидов или пептидов (таких как части или фрагменты антител), без ограничения. Кроме того, варианты пептидных мимотопов, описанных в данном документе, могут быть получены путем изменения нуклеотидной последовательности, кодирующей родительский пептидный мимотоп.
Например, один класс замен представляет собой консервативные аминокислотные замены. Такие замены представляют собой замены, при которых заданная аминокислота в антителе против IL-31, IL-31-специфичном полипептиде или пептиде заменяется другой аминокислотой со схожими характеристиками. Аналогично, мимотопы IL-31, которые связываются с таким антителом или его антигенсвязывающей частью, могут включать консервативные замены или другие типы аминокислотных замен. Как правило, консервативные замены представляют собой замены друг на друга алифатических аминокислот Ala, Val, Leu и Ile; взаимные замены гидроксильных остатков Ser и Thr, замену отрицательно заряженных остатков Asp и Glu, замену между амидными остатками Asn и Gin, замену положительно заряженных остатков Lys и Arg, замены между ароматическими остатками Phe, Tyr и тому подобное. Указания относительно того, какие аминокислотные изменения, скорее всего, будут фенотипически незаметными, можно найти в Bowie et al., 247 Science 1306-10 (1990).
Вариантные или агонистические антитела к IL-31 или IL-31-специфические полипептиды или пептиды могут быть полностью функциональными или могут быть лишены одной или более чем одной активности. Аналогично, вариантные или агонистические мимотопы IL-31 могут быть полностью функциональными или могут быть лишены одной или более чем одной активности. Полностью функциональные варианты обычно содержат только консервативные вариации или вариации остатков или областей, не являющихся ключевыми. Функциональные варианты также могут содержать замену аналогичных аминокислот, которая не приводит к изменению или приводит к незначительному изменению функции. В альтернативном варианте такие замены могут в некоторой степени положительно или отрицательно повлиять на функцию. Нефункциональные варианты обычно содержат одну или несколько неконсервативных аминокислотных замен, делеций, вставок, инверсий, или усечения, или замены, вставки, инверсии или делеций в ключевом остатке или ключевой области.
Аминокислоты, которые необходимы для функционирования, могут быть идентифицированы способами, известными в данной области, такими как сайт-специфический мутагенез или аланин-сканирующий мутагенез. Cunningham et al., 244 Science 1081-85 (1989). Последняя процедура вводит одиночные мутации каждого остатка в молекуле с заменой на аланин. Затем у полученных мутантных молекул исследуют биологическую активность, такую как связывание эпитопа или ADCC активность in vitro. Сайты, которые имеют ключевое значение для связывания лиганд-рецептор, также можно определить с помощью структурного анализа, такого как кристаллография, ядерный магнитный резонанс или фотоаффинное мечение. Smith et al., 224 J. Mol. Biol. 899-904 (1992); de Vos et al., 255 Science 306-12 (1992).
Кроме того, полипептиды часто содержат аминокислоты, отличные от двадцати «встречающихся в природе» аминокислот. Более того, многие аминокислоты, включая концевые аминокислоты, могут быть модифицированы в ходе естественных процессов, таких как процессинг и другие посттрансляционные модификации, или способами химической модификации, хорошо известными в данной области техники. Известные модификации включают ацетилирование, ацилирование, АДФ-рибозилирование, амидирование, ковалентное присоединение флавина, ковалентное присоединение гема, ковалентное присоединение нуклеотида или производного нуклеотида, ковалентное присоединение липида или производного липида, ковалентное присоединение фосфотидилинозитола, образование поперечных сшивок, циклизацию, образование дисульфидных связей, деметилирование, образование ковалентных сшивок, образование цистина, образование пироглутамата, формилирование, гамма-карбоксилирование, гликозилирование, образование GPI-якоря (гликозилфосфатидилинозитольного якоря), гидроксилирование, иодирование, метилирование, миристиолирование (ацетилирование остатком миристиновой кислоты), окисление, протеолиз, фосфорилирование, пренилирование, рацемизацию, селеноилирование, сульфатирование, опосредованное транспортной РНК добавление аминокислот к белкам, такое как аргинилирование и убиквитинирование, но не ограничиваются ими.
Такие модификации хорошо известны специалистам в данной области техники и подробно описаны в научной литературе. Некоторые особенно распространенные модификации, например, гликозилирование, присоединение липидов, сульфатирование, гамма-карбоксилирование остатков глутаминовой кислоты, гидроксилирование и АДФ-рибозилирование, описаны в большинстве базовых текстов, таких как Proteins-Structure and Molecular Properties (2-е изд., ТЕ Creighton, WH Freeman & Co., NY, 1993). На эту тему имеется множество подробных обзоров, таких как Wold, Posttranslational Covalent Modification of proteins, 1-12 (Johnson, ed., Academic Press, NY, 1983); Seifter et al. 182 Meth. Enzymol. 626-46 (1990); and Rattan et al. 663 Ann. NY Acad. Sci. 48-62 (1992).
Соответственно, IL-31-специфические антитела, полипептиды и пептиды, описанные в данном документе, а также мимотопы пептида IL-31, описанные в настоящем документе, также охватывают производные или аналоги, в которых замещенный аминокислотный остаток не кодируется генетическим кодом.
Аналогично, добавления и замены в аминокислотной последовательности, а также описанные вариации и модификации могут быть в равной степени применимы к аминокислотной последовательности антигена IL-31 и/или его эпитопа или пептидов, и, таким образом, охватываются настоящим изобретением.
Производные антител и мимотопов
В объем данного изобретения входят производные антител и мимотопов. «Производное» антитела или мимотопа содержит дополнительные химические группировки, обычно не являющиеся частью белка или пептида. Ковалентные модификации белка или пептида входят в объем данного изобретения. Такие модификации могут быть введены в молекулу в результате взаимодействия целевых аминокислотных остатков антитела или мимотопа с органическим дериватизирующим агентом, который способен реагировать с выбранными боковыми цепями или концевыми остатками. Например, дериватизация бифункциональными агентами, хорошо известными в данной области, может применяться для образования поперечных сшивок между антителом или фрагментом или мимотопом и нерастворимой в воде матрицей-носителем или другими макромолекулярными носителями.
Производные также включают радиоактивно меченные моноклональные антитела или мимотопы. Например, с радиоактивным йодом (125I, 131I), углеродом (14С), серой (35S), индием (111In), тритием (3Н) и т.п.; конъюгаты моноклональных антител с биотином или авидином, с ферментами, такими как пероксидаза хрена, щелочная фосфатаза, бефа-D-галактозидаза, глюкозооксидаза, глюкоамилаза, ангидраза карбоновых кислот, ацетилхолинэстераза, лизоцим, малатдегидрогеназа или глюкозо-6-фосфатдегидрогеназа; а также конъюгаты моноклональных антител с биолюминесцентными агентами (такими как люцифераза), хемилюминесцентными агентами (такими как сложные эфиры акридина) или флуоресцентными агентами (такими как фикобилипротеины). Аналогично, в некоторых воплощениях мимотопы могут быть мечеными.
Другое производное бифункционального антитела представляет собой биспецифическое антитело, полученное путем объединения частей двух отдельных антител, распознающих две разные антигенные группы. Это может быть достигнуто с помощью образования поперечных сшивок или рекомбинантных методов.
Кроме того, к антителу или его части или к мимотопам IL-31, описанным в настоящем документе, могут быть добавлены группировки для увеличения времени полужизни in vivo (например, путем увеличения времени выведения из кровотока). Такие методы включают, например, добавление группировок ПЭГ (также обозначаемое ПЭГилированием) и хорошо известны в данной области техники. См. заявку на патент US №20030031671.
Рекомбинантная экспрессия антител, мимотопов и полипептидов-носителей
В некоторых воплощениях нуклеиновые кислоты, кодирующие рассматриваемое моноклональное антитело или слитый белок, содержащий как мимотоп, так и полипептид-носитель, вводят непосредственно в клетку-хозяина и инкубируют клетку в условиях, достаточных для индукции экспрессии кодируемого антитела или слитого белка. После того, как рассматриваемые нуклеиновые кислоты были введены в клетку, клетку обычно инкубируют, обычно при 37°С, иногда в условиях отбора, в течение приблизительно 1-24 часов, чтобы обеспечить экспрессию антитела или слитого белка, несущего пептидный мимотоп и полипептид-носитель. В одном воплощении антитело или слитый белок секретируются в надосадочную жидкость среды, в которой выращивают клетки.
Традиционно моноклональные антитела продуцировали в виде нативных молекул в линиях мышиных гибридом. В дополнение к этой технологии настоящее изобретение обеспечивает экспрессию рекомбинантной ДНК моноклональных антител. Это позволяет производить канинизированные, фелинизированные, эквинизированные, гуманизированные, полностью собачьи, полностью кошачьи, полностью лошадиные и полностью человеческие антитела, а также спектр производных антител и слитых белков в выбранных биологических видах-хозяевах.
Последовательность нуклеиновой кислоты, кодирующая по меньшей мере одно антитело против IL-31, его часть или его специфический к IL-31 полипептид, или последовательность нуклеиновой кислоты, кодирующая в качестве своей части по меньшей мере один мимотоп пептида IL-31, который связывается с таким антителом или его частью, могут быть рекомбинированы с ДНК вектора в соответствии с общепринятыми методами, включая тупые концы или липкие концы для лигирования, расщепление рестрикционным ферментом для получения подходящих концов, заполнение липких концов при необходимости, обработку щелочной фосфатазой, чтобы избежать нежелательного соединения, и лигирование при помощи соответствующих лигаз. Техника таких манипуляций изложена, например, Maniatis et al., MOLECULAR CLONING, LAB. MANUAL, (Cold Spring Harbor Lab. Press, NY, 1982 and 1989) и Ausubel et al. 1993, см. выше, и может использоваться для конструирования последовательностей нуклеиновых кислот, которые кодируют молекулу моноклонального антитела или его антигенсвязывающую область или мимотоп пептида IL-31.
Молекула нуклеиновой кислоты, такая как ДНК, считается «способной к экспрессии» полипептида, если она содержит нуклеотидные последовательности, которые содержат информацию для регулирования транскрипции и трансляции, и такие последовательности «функционально связаны» с нуклеотидными последовательностями, кодирующими полипептид. Функциональная связь представляет собой связь, в которой регуляторные последовательности ДНК и последовательность ДНК, которая должна быть экспрессирована, связаны таким образом, чтобы обеспечить экспрессию генов в виде пептидов против IL-31 или частей антител, или в виде слитых белков, несущих мимотопы IL-31 в выделяемых количествах. Точная природа регуляторных областей, необходимых для экспрессии генов, может варьироваться от организма к организму, как хорошо известно в области техники. См., например, Sambrook et al., 2001, выше, Ausubel et al., 1993, выше.
Настоящее изобретение, соответственно, охватывает экспрессию антитела против IL-31 или IL-31-специфического полипептида или пептида, или слитого белка, включающего мимотоп IL-31, в прокариотических или эукариотических клетках. Подходящие хозяева включают бактериальных или эукариотических хозяев, включая бактерии, дрожжи, насекомые, грибы, клетки птиц и млекопитающих in vivo или in situ, или клетки-хозяева, происходящие из млекопитающих, насекомых, птиц или дрожжей. Клетка или ткань млекопитающего могут происходить из организма человека, примата, хомяка, кролика, мыши, коровы, свиньи, овцы, лошади, козы, собаки или кошки, но могут быть использованы клетки любых других млекопитающих.
В одном воплощении введенная нуклеотидная последовательность будет включена в плазмидный или вирусный вектор, способный к автономной репликации в реципиенте-хозяине. Для этой цели можно использовать множество векторов. См., например, Ausubel et al., 1993, выше. Факторы, важные при выборе конкретной плазмиды или вирусного вектора, включают: легкость, с которой клетки-реципиенты, содержащие вектор, могут быть распознаны и отобраны из тех клеток-реципиентов, которые не содержат вектор; количество копий вектора, которые желательны в конкретном хозяине; и желательна ли возможность переноса вектора между клетками-хозяевами разных биологических видов.
Примеры прокариотических векторов, известных в данной области техники, включают плазмиды, такие как плазмиды, способные к репликации в Е. coli (например, такие как pBR322, ColE1, pSC101, pACYC 184, .pi. VX). Такие плазмиды описаны, например, Maniatis et al., 1989, выше; Ausubel et al, 1993, выше. Плазмиды Bacillus включают pC194, pC221, pT127 и др. Такие плазмиды раскрыты Gryczan в THE MOLEC. BIO. OF THE BACILLI 307-329 (Academic Press, NY, 1982). Подходящее Плазмиды Streptomyces включают pIJ101 (Kendall et al., 169 J. Bacteriol. 4177-83 (1987)) и бактериофаги Streptomyces, такие как .phi.C31 (Chater et al., in SIXTH INT'L SYMPOSIUM ON ACTINOMYCETALES BIO. 45-54 (Akademiai Kaido, Budapest, Hungary 1986). Обзор плазмид Pseudomonas представлен John et al., 8 Rev. Infect. Dis. 693-704 (1986); Izaki, 33 Jpn. J. Bacteriol. 729-42 (1978) и Ausubel et al., 1993, см. выше.
В качестве альтернативы, элементы экспрессии генов, применяющиеся для экспрессии кДНК, кодирующей антитела или пептиды против IL-31, или слитые белки, описанные в настоящем документе, включают (а) вирусные промоторы транскрипции и их энхансерные элементы, такие как ранний промотор SV40 (Okayama et al., 3 Mol. Cell. Biol. 280 (1983)), LTR вируса саркомы Рауса (Gorman et al., 79 Proc. Natl. Acad. Sci., USA 6777 (1982)) и LTR вируса мышиного лейкоза Молони (Grosschedl et al., 41 Cell 885 (1985)); (б) участки сплайсинга и сайты полиаденилирования, такие как те, которые происходят из поздно транскрибируемой области SV40 (Okayama et al., МСВ, 3: 280 (1983) и (в) сайты полиаденилирования, такие как в SV40 (Okayama et al., 1983, выше), но не ограничиваются ими.
Гены кДНК иммуноглобулина могут быть экспрессированы, как описано Weidle et al., 51 (1) Gene 21-29 (1987), с использованием в качестве элементов, регулирующих экспрессию, раннего промотора SV40 и его энхансера, энхансеров промотора Н-цепи иммуноглобулина мыши, элементов, регулирующих сплайсинг мРНК поздно транскрибируемой области SV40, вставочной последовательности S-глобина кролика, сайтов полиаденилирования S-глобина и иммуноглобулина кролика и элементов полиаденилирования SV40.
Для генов иммуноглобулинов, состоящих из части кДНК, части геномной ДНК (Whittle et al., 1 Protein Engin. 499-505 (1987)), промотором транскрипции может быть цитомегаловирус человека, энхансеры промотора могут быть энхансеры цитомегаловируса и иммуноглобулина мыши/человека, а области, регулирующие сплайсинг мРНК, и сайт полиаденилирования могут быть хромосомными последовательностями нативного иммуноглобулина.
В одном воплощении для экспрессии генов кДНК в клетках грызунов промотор транскрипции представляет собой последовательность вирусных LTR, энхансеры промотора транскрипции представляют собой либо энхансер тяжелой цепи иммуноглобулина мыши, либо энхансер вирусных LTR, область сплайсинга содержит интрон превышающий 31 п.н., и области полиаденилирования и терминации транскрипции происходят из нативной хромосомной последовательности, соответствующей синтезируемой цепи иммуноглобулина. В других воплощениях последовательности кДНК, кодирующие другие белки, комбинируют с перечисленными выше элементами экспрессии для достижения экспрессии белков в клетках млекопитающих.
Каждый слитый ген может быть собран или встроен в вектор экспрессии. Клетки-реципиенты, способные экспрессировать продукт гена химерной цепи иммуноглобулина, затем трансфицируют по отдельности пептидом против IL-31 или геном, кодирующим химерную Н или химерную L цепь, или совместно трансфицируют геном химерной Н цепи и химерной L цепи. Трансфицированные клетки-реципиенты культивируют в условиях, которые позволяют экспрессироваться встроенным генам, и экспрессированные цепи иммуноглобулина или интактные антитела или фрагменты выделяют из культуры.
В одном воплощении слитые гены, кодирующие анти-IL-31 пептид или химерные Н- и L-цепи, или их части, собраны в отдельные векторы экспрессии, которые затем используются для совместной трансфекции клетки-реципиента. В альтернативном варианте слитые гены, кодирующие химерные Н- и L-цепи, могут быть собраны в одном и том же векторе экспрессии.
Для трансфекции векторов экспрессии и получения химерного антитела линия клеток-реципиентов может быть клеткой миеломы. Клетки миеломы могут синтезировать, собирать и секретировать иммуноглобулины, кодируемые трансфицированными генами иммуноглобулинов, и обладают механизмом гликозилирования иммуноглобулина. Клетки миеломы можно выращивать в культуре или в брюшной полости мыши, где секретируемый иммуноглобулин может быть получен из асцитной жидкости. Другие подходящие клетки-реципиенты включают лимфоидные клетки, такие как В-лимфоциты, имеющие или не имеющие человеческое происхождение, клетки гибридомы, имеющие или не имеющие человеческое происхождение, или клетки межвидовых гетерогибридом.
Вектор экспрессии, несущий нуклеотидную последовательность, кодирующую химерные, канинизированные, фелинизированные, эквинизированные, гуманизированные, полностью собачьи, полностью кошачьи, полностью лошадиные или полностью человеческие последовательности конструкции антител против IL-31 или IL-31-специфический полипептид или пептид (например, антигенсвязывающую часть антител, описанных в данном документе), или вектор экспрессии, несущий нуклеотидную последовательность, кодирующую слитый белок, описанный в настоящем документе, могут быть введены в подходящую клетку-хозяина любым из множества подходящих способов, включая такие биохимические средства, как трансформация, трансфекция, конъюгация, слияние протопластов, осаждение фосфатом кальция, с применением поликатионов, таких как диэтиламиноэтил (DEAE) декстран, и такими механическими средствами, как электропорация, прямая микроинъекция и бомбардировка микрочастицами. Johnston et al., 240 Science 1538-1541 (1988).
Дрожжи могут обеспечить существенные преимущества перед бактериями при производстве Н- и L-цепей иммуноглобулина. Дрожжи осуществляют посттрансляционные модификации пептидов, включая гликозилирование. В настоящее время существует ряд стратегий рекомбинантной ДНК, в которых используются последовательности сильных промоторов и плазмиды с большим числом копий, которые можно использовать для получения желаемых белков в дрожжах. Дрожжи распознают лидерные последовательности клонированных продуктов генов млекопитающих и секретируют пептиды, несущие лидерные последовательности (т.е. препептиды). Hitzman et al., 11th Int'l Conference on Yeast, Genetics & Molec. Biol. (Montpelier, France, 1982).
Системы экспрессии генов дрожжей можно рутинно оценивать на предмет уровней продукции, секреции и стабильности анти-IL-31 пептидов, антител и собранных мышиных и химерных, гетерохимерных, канинизированных, фелинизированных, эквинизированных, гуманизированных, полностью собачьих, полностью кошачьих, полностью лошадиных или полностью человеческих антител, их фрагментов и областей. Может быть использована любая из серии систем экспрессии дрожжевых генов, включающих промоторные и терминирующие элементы активно экспрессируемых генов, кодирующих гликолитические ферменты, вырабатываемые в больших количествах при выращивании дрожжей в среде, богатой глюкозой. Известные гликолитические гены также могут обеспечивать очень эффективные сигналы, контролирующие транскрипцию. Например, могут быть использованы сигналы промотора и терминатора гена фосфоглицераткиназы (PGK). Для оценки оптимальных экспрессионных плазмид для экспрессии клонированных кДНК иммуноглобулинов в дрожжах можно использовать ряд подходов. См. Vol. II DNA Cloning, 45-66, (Glover, ed.,) IRL Press, Oxford, UK 1985).
В качестве хозяев для продукции молекул антител или пептидов или слитых белков, описанных в данном изобретении, можно также использовать штаммы бактерий. С такими хозяевами-бактериями используют плазмидные векторы, содержащие репликон и контрольные последовательности, происходящие от видов, совместимых с клеткой-хозяином. Вектор несет сайт репликации, а также специфические гены, которые способны обеспечивать фенотипический отбор трансформированных клеток. Для оценки экспрессионных плазмид для получения мышиных, химерных, гетерохимерных, канинизированных, фелинизированных, эквинизированных, гуманизированных, полностью собачьих, полностью кошачьих, полностью лошадиных или полностью человеческих антител, фрагментов и областей или цепей антител, кодируемых клонированными кДНК иммуноглобулинов в бактериях, можно использовать ряд подходов (см. Glover, 1985 см. выше; Ausubel, 1993 см. выше; Sambrook, 2001 см. выше; Colligan et al., eds. Current Protocols in Immunology, John Wiley & Sons, NY, NY (1994-2001); Colligan et al., eds. Current Protocols in Protein Science, John Wiley & Sons, NY, NY (1997-2001).
Клетки млекопитающих-хозяев можно выращивать in vitro или in vivo. Клетки млекопитающих обеспечивают посттрансляционные модификации молекул белка иммуноглобулина, включая удаление лидерного пептида, сворачивание и сборку Н- и L-цепей, гликозилирование молекул антител и секрецию функционального белка антитела.
Клетки млекопитающих, которые могут применяться в качестве хозяев для продукции белков антител, в дополнение к клеткам лимфоидного происхождения, описанным выше, включают клетки фибр областного происхождения, такие как клетки Vero (АТСС CRL 81) или СНО-K1 (АТСС CRL 61).
Для экспрессии клонированных генов Н- и L-цепей анти-IL-31 пептидов в клетках млекопитающих доступно множество систем векторов (см. Glover, 1985, выше). Для получения полных антител H2L2 можно использовать разные подходы. Можно совместно экспрессировать Н- и L-цепи в одних и тех же клетках для достижения внутриклеточной ассоциации и связывания Н- и L-цепей в полные тетрамерные антитела H2L2 и/или анти-IL-31 пептиды. Совместная экспрессия может происходить при использовании одной и той же или разных плазмид в одном хозяине. Гены для Н- и L-цепей и/или анти-IL-31 пептидов могут быть помещены в одну и ту же плазмиду, которой затем трансфицируют клетки, тем самым напрямую отбирая клетки, экспрессирующие обе цепи. В альтернативном варианте клетки можно сначала трансфицировать плазмидой, кодирующей одну цепь, например L-цепь, после чего трансфицировать полученную клеточную линию плазмидой Н-цепи, содержащей второй селектируемый маркер. Клеточные линии, продуцирующие анти-IL-31 пептиды и/или молекулы H2L2 любым путем, могут быть трансфицированы плазмидами, кодирующими дополнительные копии пептидов, Н, L или Н+L цепей в сочетании с дополнительными селектируемыми маркерами для создания клеточных линий с улучшенными свойствами, такими как более высокая продукция собранных молекул антител H2L2 или повышенная стабильность трансфицированных клеточных линий.
Для длительного получения рекомбинантных антител с высоким выходом может использоваться стабильная экспрессия. Например, могут быть сконструированы клеточные линии, которые стабильно экспрессируют молекулу антитела. Вместо использования векторов экспрессии, которые содержат точки начала репликации вирусов, клетки-хозяев а можно трансформировать экспрессионными кассетами, кодирующими иммуноглобулин и селектируемый маркер. После введения чужеродной ДНК сконструированным клеткам можно дать возможность расти в течение 1-2 дней в обогащенной среде, а затем перевести на селективную среду. Селектируемый маркер в рекомбинантной плазмиде придает устойчивость при отборе и позволяет клеткам стабильно интегрировать плазмиду в хромосому и расти с образованием колоний, которые, в свою очередь, можно клонировать и размножать в клеточных линиях. Такие сконструированные клеточные линии могут быть особенно эффективны при скрининге и оценке соединений/компонентов, которые прямо или косвенно взаимодействуют с молекулой антитела.
После получения антитела его можно очистить любым известным в данной области способом очистки молекулы иммуноглобулина, например, хроматографией (например, ионообменной, аффинной, особенно на основе аффинности к специфическому антигену после хроматографии с белком А, и гель-фильтрационной колоночной хроматографией), центрифугированием, методом на основе дифференциальной растворимости или любым другим стандартным способом очистки белков. Во многих воплощениях антитела секретируются из клетки в культуральную среду и их собирают из культуральной среды.
Практическое применение
Вакцинные композиции по настоящему изобретению можно использовать, например, для лечения и/или защиты от IL-31-опосредованных заболеваний, таких как сопровождающиеся зудом и/или аллергические состояния у млекопитающих, таких как собаки, кошки, лошади и люди. Фармацевтические композиции по настоящему изобретению применимы для парентерального введения, например, подкожно, внутримышечно или внутривенно. В данном документе описаны другие подходящие способы введения.
Вакцины по настоящему изобретению можно вводить либо в виде самостоятельных терапевтических агентов, либо в комбинации с другими терапевтическими агентами. Их можно вводить отдельно, но обычно их вводят с фармацевтическим носителем, выбранным на основе выбранного пути введения и стандартной фармацевтической практики.
Введение описанных в данном документе вакцинных композиций может осуществляться любыми подходящими способами, включая парентеральную инъекцию (например, внутрибрюшинную, подкожную или внутримышечную инъекцию), перорально или путем местного введения вакцин на поверхность дыхательных путей. Местное введение на поверхность дыхательных путей может осуществляться интраназальным введением (например, с помощью пипетки, тампона или ингалятора). Местное введение вакцин на поверхность дыхательных путей также может осуществляться путем ингаляционного введения, например, путем создания респирабельных частиц фармацевтического состава (включая твердые и жидкие частицы), содержащих вакцины в виде аэрозольной суспензии, и затем вдыхания субъектом респирабельных частиц. Способы и устройства для введения вдыхаемых частиц фармацевтических составов хорошо известны, и можно использовать любой общепринятый метод. Пероральное введение может быть, например, в форме жидкого или твердого препарата для приема внутрь.
В некоторых предпочтительных воплощениях вакцины вводят путем парентеральной инъекции. Для парентерального введения вакцины могут быть приготовлены в виде раствора, суспензии, эмульсии или лиофилизированного порошка в сочетании с фармацевтически приемлемым парентеральным носителем. Например, носитель может быть раствором комбинации мимотопа и полипептида-носителя (например, конъюгата мимотопа) или их коктейлем, растворенным в приемлемом носителе, таком как водный носитель, такими несущими средами являются вода, физиологический раствор, раствор Рингера, раствор декстрозы, трегалозы или сахарозы или 5% сывороточный альбумин, 0,4% физиологический раствор, 0,3% глицин и тому подобное. Также можно использовать липосомы и неводные носители, такие как жирные масла. Эти растворы стерильны и обычно не содержат твердых частиц. Эти композиции можно стерилизовать общепринятыми, хорошо известными методами стерилизации. Композиции могут содержать фармацевтически приемлемые вспомогательные вещества, необходимые для приближения к физиологическим условиям, такие как регуляторы рН и буферные агенты, вещества для регулирования тоничности и тому подобные, например ацетат натрия, хлорид натрия, хлорид калия, хлорид кальция, лактат натрия и т.д. Также, согласно описанию в данном документе, вакцинные композиции по настоящему изобретению включают адъювант или адъювантный состав. Концентрация конъюгата мимотопа в этих вакцинных композициях может широко варьироваться, например от менее чем примерно 0,5%, обычно от по меньшей мере, приблизительно 1% до 15% или 20% по массе, и будет выбираться в первую очередь на основе объемов жидкости, вязкости и т.д. в соответствии с выбранным конкретным способом введения. Носитель или лиофилизированный порошок могут содержать добавки, которые поддерживают изотоничность (например, хлорид натрия, маннит) и химическую стабильность (например, буферы и консерванты). Состав стерилизуют с помощью общепринятых методов.
Фактические способы получения композиций для парентерального введения будут известны или очевидны специалистам в данной области техники и описаны более подробно, например, в REMINGTON'S PHARMA. SCI. (15th ed., Mack Pub. Co., Easton, Pa., 1980).
Вакцины по данному изобретению можно лиофилизировать для хранения и восстанавливать в подходящем носителе перед использованием. Можно применять любые подходящие способы лиофилизации и восстановления. Специалисты в данной области поймут, что лиофилизация и восстановление могут привести к различной степени потери активности и что их используемые уровни, возможно, придется скорректировать для компенсации потери.
Композиции, содержащие настоящие мимотопы IL-31 (например, конъюгаты мимотопа IL-31) или их смесь, можно вводить для предотвращения рецидивов и/или терапии существующего заболевания. Подходящие фармацевтические носители описаны в последнем издании REMINGTON'S PHARMACEUTICAL SCIENCES, стандартном справочном тексте в данной области техники.
При терапевтическом применении композиции вводят субъекту, уже страдающему заболеванием, в количестве, достаточном для излечения или, по меньшей мере, частичной остановки или облегчения заболевания и его осложнений. Количество, достаточное для достижения этого, определяется как «терапевтически эффективная доза» или «терапевтически эффективное количество». Эффективные количества для такого применения будут зависеть от тяжести заболевания и общего состояния собственной иммунной системы субъекта. Терапевтически эффективное количество вакцинной композиции согласно изобретению может легко определить специалист в данной области техники.
Вводимая доза, конечно, будет варьироваться в зависимости от известных факторов, таких как фармакодинамические характеристики конкретного агента, а также способа и пути его введения; возраста, состояния здоровья и массы реципиента; характера и степени симптомов, вида сопутствующего лечения, частоты лечения и желаемого эффекта.
В качестве неограничивающего примера лечение связанных с IL-31 патологий у собак, кошек, лошадей или людей может осуществляться в виде введения вакцин по настоящему изобретению раз в две недели или раз в месяц в диапазоне доз, описанном выше.
Однократное или многократное введение вакцинных композиций можно проводить в соответствии с уровнями доз и схемой, выбранными лечащим врачом или ветеринаром. В любом случае фармацевтические составы должны обеспечивать количество вакцинных композиций по настоящему изобретению, достаточное для эффективного лечения субъекта.
Диагностическое применение
В настоящем изобретении также предложены мимотопы IL-31 и антитела против IL-31 для использования в диагностических методах определения IL-31 или антител к IL-31 в образцах от млекопитающих, включая образцы от млекопитающих, у которых известно о наличии или подозревается наличие сопровождающегося зудом и/или аллергического состояния.
Например, в настоящем изобретении предложен способ качественного или количественного определения антитела против IL-31 в образце. Такой способ включает инкубирование образца, содержащего антитело против IL-31, по меньшей мере, с одним мимотопом, таким как мимотоп IL-31 кошки, мимотоп IL-31 собаки, мимотоп IL-31 лошади и мимотоп IL-31 человека, и качественное и/или количественное определение анти-IL-31 агента в образце.
В одном воплощении мимотоп собачьего IL-31, используемый в способе качественного и/или количественного определения IL-31 в образце, представляет собой и/или содержит в качестве своей части аминокислотную последовательность SVPADTFECKSF (SEQ ID NO: 186), SVPADTFERKSF (SEQ ID NO: 187), NSSAILPYFRAIRPLSDKNIIDKIIEQLDKLKF (SEQ ID NO: 192), APTHQLPPSDVRKIILELQPLSRG (SEQ ID NO: 196), TGVPES (SEQ ID NO: 200) или их варианты, которые сохраняют связывание с анти-IL-31 агентом.
В другом воплощении мимотоп кошачьего IL-31, используемый в таком способе, представляет собой и/или содержит как часть аминокислотную последовательность SMPADNFERKNF (SEQ ID NO: 188), NGSAILPYFRAIRPLSDKNTIDKIIEQLDKLKF (SEQ ID NO: 193), APAHRLQPSDIRKIILELRPMSKG (SEQ ID NO: 197), IGLPES (SEQ ID NO: 201) или их варианты, которые сохраняют связывание анти-IL-31 агентом.
В следующем воплощении мимотоп лошадиного IL-31, используемый в таком способе, представляет собой и/или включает в качестве своей части аминокислотную последовательность SMPTDNFERKRF (SEQ ID NO: 189), NSSAILPYFKAISPSLNNDKSLYIIEQLDKLNF (SEQ ID NO: 194), GPIYQLQPKEIQAIIVELQNLSKK (SEQ ID NO: 198), KGVQKF (SEQ ID NO: 202) или их варианты, которые сохраняют связывание с анти-IL-31 агентом.
Кроме того, мимотоп человеческого IL-31, используемый в таком способе, представляет собой и/или содержит как часть аминокислотную последовательность SVPTDTHECKRF (SEQ ID NO: 190), SVPTDTHERKRF (SEQ ID NO: 191), HSPAIRAYLKTIRQLDNKSVIDEIIEHLDKLIF (SEQ ID NO: 195), LPVRLLRPSDDVQKIVEELQSLSKM (SEQ ID NO: 199), KGVLVS (SEQ ID NO: 203) или их варианты, которые сохраняют связывание анти-IL-31 агентом.
В одном из вариантов вышеописанного способа диагностики мимотоп представляет собой реагент для захвата, связанный с твердой поверхностью. В одном воплощении образец добавляют к мимотопу, представляющему собой реагент для захвата, и затем добавляют вторичные детектирующие реагенты для определения количества антитела в образце.
В данном изобретении также предложен способ определения количества IL-31 в образце от млекопитающего. Такой способ будет полезен для определения IL-31 от многих биологических видов. Такой способ включает инкубацию образца от млекопитающего, содержащего IL-31, с комплексом меченое антитело против IL-31: Мимотоп IL-31, связанным с твердой поверхностью, где мимотоп в комплексе выбран из группы, состоящей из мимотопа кошачьего IL- 31, мимотопа собачьего IL-31, мимотопа лошадиного IL-31 и мимотопа человеческого IL-31; и определение уровня IL-31 в образце, где меченое антитело против IL-31 в комплексе имеет более низкую аффинность к мимотопу в комплексе, по сравнению с его аффинностью к IL-31 в образце. В одном варианте этого способа стадия определения включает измерение сигнала, исходящего от меченого антитела, которое высвобождается с твердой поверхности, когда IL-31 в образце связывается с меченым антителом против IL-31 комплекса, при этом уровень IL-31 в образце обратно пропорционален сигналу.
В одном воплощении мимотоп собачьего IL-31, используемый в способе определения количества IL-31 в образце, представляет собой и/или содержит в качестве своей части аминокислотную последовательность SVPADTFECKSF (SEQ ID NO: 186), SVPADTFERKSF (SEQ ID NO: 187), NSSAILPYFRAIRPLSDKNIIDKIIEQLDKLKF (SEQ ID NO: 192), APTHQLPPSDVRKIILELQPLSRG (SEQ ID NO: 196), TGVPES (SEQ ID NO: 200) или их варианты, которые сохраняют связывание с анти-IL-31 агентом.
В другом воплощении мимотоп кошачьего IL-31, используемый в таком способе, представляет собой и/или содержит как часть аминокислотную последовательность SMPADNFERKNF (SEQ ID NO: 188), NGSAILPYFRAIRPLSDKNTIDKIIEQLDKLKF (SEQ ID NO: 193), APAHRLQPSDIRKIILELRPMSKG (SEQ ID NO: 197), IGLPES (SEQ ID NO: 201) или их варианты, которые сохраняют связывание анти-IL-31 агентом.
В еще одном воплощении мимотоп IL-31 лошади, используемый в таком способе, представляет собой и/или содержит в качестве своей части аминокислотную последовательность SMPTDNFERKRF (SEQ ID NO: 189), NSSAILPYFKAISPSLNNDKSLYIIEQLDKLNF (SEQ ID NO: 194), GPIYQLQPKEIQAIIVELQNLSKK (SEQ ID NO: 198), KGVQKF (SEQ ID NO: 202) или их варианты, которые сохраняют связывание с анти-IL-31 агентом.
В следующем воплощении мимотоп IL-31 человека, используемый в таком способе, представляет собой и/или содержит в качестве своей части аминокислотную последовательность SVPTDTHECKRF (SEQ ID NO: 190), SVPTDTHERKRF (SEQ ID NO: 191), HSPAIRAYLKTIRQLDNKSVIDEIIEHLDKLIF (SEQ ID NO: 195), LPVRLLRPSDDVQKIVEELQSLSKM (SEQ ID NO: 199), KGVLVS (SEQ ID NO: 203) или их варианты, которые сохраняют связывание анти-IL-31 агентом.
В некоторых воплощениях любого из описанных выше способов диагностики по изобретению мимотоп связывается с антителом против IL-31 или его антигенсвязывающей частью, которая специфически связывается с областью белка IL-31 млекопитающих, задействованной во взаимодействии белка IL-31 со своим ко-рецептором. В одном воплощении способов диагностики по изобретению на связывание указанного антитела с указанной областью влияют мутации в области эпитопа, связывающейся с 15Н05, выбранные из группы, состоящей из:
а) области между аминокислотными остатками 124 и 135 последовательности IL-31 кошки, представленной SEQ ID NO: 157 (кошачий IL-31 дикого типа);
б) области между аминокислотными остатками 124 и 135 последовательности IL-31 собаки, представленной SEQ ID NO: 155 (собачий IL-31); и
в) области между аминокислотными остатками 118 и 129 последовательности IL-31 лошади, представленной SEQ ID NO: 165 (лошадиный IL-31).
В одном воплощении способов диагностики по данному изобретению мимотоп связывается с антителом против IL-31 или его антигенсвязывающей частью, которая специфически связывается с вышеупомянутой областью эпитопа 15Н05. В одном конкретном воплощении любых способов диагностики данного изобретения мимотоп связывается с антителом против IL-31 или его антигенсвязывающей частью, содержащей по меньшей мере одну из следующих комбинаций последовательностей участков, определяющих комплементарность (CDR):
1) антитело 15Н05: CDR1 вариабельной области тяжелой цепи (VH) с последовательностью SYTIH (SEQ ID NO: 1), VH-CDR2 с последовательностью NINPTSGYTENNQRFKD (SEQ ID NO: 2), VH-CDR3 с последовательностью WGFKYDGEWSFDV (SEQ ID NO: 3), CDR1 вариабельной области легкой цепи (VL) с последовательностью RASQGISIWLS (SEQ ID NO: 4), VL-CDR2 с последовательностью KASNLHI (SEQ ID NO: 5) и VL-CDR3 с последовательностью LQSQTYPLT (SEQ ID NO: 6);
2) антитело ZIL1: CDR1 вариабельной области тяжелой цепи (VH) с последовательностью SYGMS (SEQ ID NO: 13), VH-CDR2 с последовательностью HINSGGSSTYYADAVKG (SEQ ID NO: 14), VH-CDR3 с последовательностью VYTTLAAFWTDNFDY (SEQ ID NO: 15), вариабельной области легкой цепи (VL) с последовательностью SGSTNNIGILAAT (SEQ ID NO: 16), VL-CDR2 с последовательностью SDGNRPS (SEQ ID NO: 17) и VL-CDR3 с последовательностью QSFDTTLDAYV (SEQ ID NO: 18);
3) антитело ZIL8: VH-CDR1 с последовательностью DYAMS (SEQ ID NO: 19), VH-CDR2 с последовательностью GIDSVGSGTSYADAVKG (SEQ ID NO: 20), VH-CDR3 с последовательностью GFPGSFEH (SEQ ID NO: 21), VL-CDR1 с последовательностью TGSSSNIGSGYVG (SEQ ID NO: 22), VL-CDR2 с последовательностью YNSDRPS (SEQ ID NO: 23), VL-CDR3 с последовательностью SVYDRTFNAV (SEQ ID NO: 24);
4) антитело ZIL9: VH-CDR1 с последовательностью SYDMT (SEQ ID NO: 25), VH-CDR2 с последовательностью DVNSGGTGTAYAVAVKG (SEQ ID NO: 26), VH-CDR3 с последовательностью LGVRDGLSV (SEQ ID NO: 27), VL-CDR1 с последовательностью SGESLNEYYTQ (SEQ ID NO: 28), VL-CDR2 с последовательностью RDTERPS (SEQ ID NO: 29), VL-CDR3 с последовательностью ESAVDTGTLV (SEQ ID NO: 30);
5) антитело ZIL11: VH-CDR1 с последовательностью TYVMN (SEQ ID NO: 31), VH-CDR2 с последовательностью SINGGGSSPTYADAVRG (SEQ ID NO: 32), VH-CDR3 с последовательностью SMVGPFDY (SEQ ID NO: 33), VL-CDR1 с последовательностью SGESLSNYYAQ (SEQ ID NO: 34), VL-CDR2 с последовательностью KDTERPS (SEQ ID NO: 35), VL-CDR3 с последовательностью ESAVSSDTIV (SEQ ID NO: 36);
6) антитело ZIL69: VH-CDR1 с последовательностью SYAMK (SEQ ID NO: 37), VH-CDR2 с последовательностью TINNDGTRTGYADAVRG (SEQ ID NO: 38), VH-CDR3 с последовательностью GNAESGCTGDHCPPY (SEQ ID NO: 39), VL-CDR1 с последовательностью SGESLNKYYAQ (SEQ ID NO: 40), VL-CDR2 с последовательностью KDTERPS (SEQ ID NO: 41), VL-CDR3 с последовательностью ESAVSSETNV (SEQ ID NO: 42);
7) антитело ZIL94: VH-CDR1 с последовательностью TYFMS (SEQ ID NO: 43), VH-CDR2 с последовательностью LISSDGSGTYYADAVKG (SEQ ID NO: 44), VH-CDR3 с последовательностью FWRAFND (SEQ ID NO: 45), VL-CDR1 с последовательностью GLNSGSVSTSNYPG (SEQ ID NO: 46), VL-CDR2 с последовательностью DTGSRPS (SEQ ID NO: 47), VL-CDR3 с последовательностью SLYTDSDILV (SEQ ID NO: 48);
8) антитело ZIL154: VH-CDR1 с последовательностью DRGMS (SEQ ID NO: 49), VH-CDR2 с последовательностью YIRYDGSRTDYADAVEG (SEQ ID NO: 50), VH-CDR3 с последовательностью WDGSSFDY (SEQ ID NO: 51), VL-CDR1 с последовательностью KASQSLLHSDGNTYLD (SEQ ID NO: 52), VL-CDR2 с последовательностью KVSNRDP (SEQ ID NO: 53), VL-CDR3 с последовательностью MQAIHFPLT (SEQ ID NO: 54);
9) антитело ZIL159: VH-CDR1 с последовательностью SYVMT (SEQ ID NO: 55), VH-CDR2 с последовательностью GINSEGSRTAYADAVKG (SEQ ID NO: 56), VH-CDR3 с последовательностью GDIVATGTSY (SEQ ID NO: 57), VL-CDR1 с последовательностью SGETLNRFYTQ (SEQ ID NO: 58), VL-CDR2 с последовательностью KDTERPS (SEQ ID NO: 59), VL-CDR3 с последовательностью KSAVSIDVGV (SEQ ID NO: 60);
10) антитело ZIL171: VH-CDR1 с последовательностью TYVMN (SEQ ID NO: 61), VH-CDR2 с последовательностью SINGGGSSPTYADAVRG (SEQ ID NO: 62), VH-CDR3 с последовательностью SMVGPFDY (SEQ ID NO: 63), VL-CDR1 с последовательностью SGKSLSYYYAQ (SEQ ID NO: 64), VL-CDR2 с последовательностью KDTERPS (SEQ ID NO: 65), VL-CDR3 с последовательностью ESAVSSDTIV (SEQ ID NO: 66); или
11) вариант (1)-(10), отличающийся от соответствующего родительского антитела 15Н05, ZIL1, ZIL8, ZIL9, ZIL11, ZIL69, ZIL94, ZIL154, ZIL159 или ZIL171 добавлением, делецией и/или заменой одного или более чем одного аминокислотного остатка по меньшей мере в одном из CDR1, CDR2 или CDR3 VH или VL.
В некоторых воплощениях мимотоп, используемый в способах диагностики по настоящему изобретению, связывается с антителом против IL-31 или его антигенсвязывающей частью, которая связывается с кошачьим IL-31, при этом антитело включает цепь VL, содержащую изменения каркасной области 2 (FW2), выбранные из следующего: аспарагин вместо лизина в положении 42, изолейцин вместо валина в положении 43, валин вместо лейцина в положении 46, аспарагин вместо лизина в положении 49 и их комбинации, где положения относятся к нумерации SEQ ID NO: 127 (FEL_15Н05_VL1).
Антитела, полипептиды и/или пептиды против IL-31 по настоящему изобретению и пептидные мимотопы IL-31 применимы для иммуноанализов, которые определяют или количественно определяют IL-31 или антитела против IL-31 в образце. Иммуноанализ на IL-31 обычно включает инкубацию клинического или биологического образца в присутствии детектируемого меченного высокоаффинного (высокоавидного) анти-IL-31 антитела, полипептида или пептида по настоящему изобретению, способного селективно связываться с IL-31, и детектирование меченого полипептида, пептида или антитела, которые связались в образце. В предпочтительном воплощении мимотоп IL-31 связывается с твердой поверхностью и используется для захвата меченого антитела против IL-31, так что комплекс меченое антитело против IL-31: мимотоп IL-31 становится привязанным к твердой поверхности. Меченое антитело к IL-31 в составе комплекса имеет более низкую аффинность к мимотопу в составе комплекса, по сравнению с его аффинностью к IL-31 в образце. Таким образом, уровень IL-31 в образце можно определить путем измерения сигнала, исходящего от меченого антитела, которое высвобождается с твердой поверхности, когда IL-31 в образце связывается с меченым антителом к IL-31 комплекса антитело к IL-31: мимотоп IL-31. В этом случае уровень IL-31 в образце обратно пропорционален сигналу. В области техники хорошо известны различные методы клинических исследований. См. например, IMMUNOASSAYS FOR THE 80'S (Voller et al., eds., Univ. Park, 1981). Такие образцы включают биоптаты ткани, образцы крови, сыворотки и фекалий или жидкости, забранные у животных и исследованные методом ELISA, как описано ниже.
В некоторых воплощениях связывание антигена с антителом выявляется без использования твердой подложки. Например, связывание антигена с антителом можно определять в жидкофазном анализе.
В других воплощениях пептидный мимотоп IL-31 или антитело, полипептид или пептид против IL-31 можно, например, фиксировать на нитроцеллюлозе или другой твердой подложке, способной иммобилизовать клетки, клеточные частицы или растворимые белки. Затем подложку можно промыть подходящими буферами с последующей обработкой детектируемым меченным специфическим к IL-31 полипептидом, пептидом или антителом. Затем твердофазную подложку можно промыть буфером второй раз для удаления несвязанного полипептида, пептида или антитела. Затем количество связанной метки на твердой подложке можно определять с помощью известных этапов способа.
«Твердофазная подложка» или «носитель» относится к любой подложке, способной связывать полипептид, пептид, антиген или антитело. Хорошо известные подложки или носители включают стекло, полистирол, полипропилен, полиэтилен, поливинилиденфторид (PVDF), декстран, нейлон, амилазы, природные и модифицированные целлюлозы, полиакриламиды, агарозы и магнетит. По своей природе носитель может быть либо растворимым до некоторой степени, либо нерастворимым для целей настоящего изобретения. Материал подложки может иметь практически любую возможную структурную конфигурацию при условии, что связанная молекула способна связываться с мимотопом пептида IL-31, IL-31 или антителом против IL-31. Предполагается, что мимотоп IL-31, связанный с подложкой, может сам быть конъюгирован с полипептидом-носителем, если это желательно. Таким образом, конфигурация подложки может быть сферической, как у бусины, или цилиндрической, как на внутренней поверхности пробирки, или на внешней поверхности стержня. В качестве альтернативы поверхность может быть плоской, такой как лист, культуральная чашка, тест-полоска и т.д. Например, подложки могут включать бусины из полистирола. Специалисты в данной области техники знают много других подходящих носителей для связывания антитела, полипептида, пептида или антигена или могут подобрать их с помощью обычных экспериментов.
На этапах хорошо известного метода можно определить связывающую активность данной партии мимотопа или анти-IL-31 полипептида, пептида и/или антитела. Специалисты в данной области могут определить рабочие и оптимальные условия анализа с помощью обычных экспериментов.
Детектируемое мечение IL-31-специфического полипептида, пептида и/или антитела, а также мечение мимотопа пептида IL-31 (или его конъюгата) можно осуществлять несколькими различными способами, включая связывание с ферментом для использования в иммуноферментном анализе (EIA) или твердофазном иммуноферментном анализе (ELISA). Связанный фермент вступает в реакцию с экспонированным субстратом с образованием химической группировки, которая может быть обнаружена, например, спектрофотометрическими, флуориметрическими или визуальными средствами. Ферменты, которые можно использовать для детектируемого мечения IL-31-специфических антител или мимотопов, описанных в настоящем документе, включают малатдегидрогеназу, стафилококковую нуклеазу, дельта-5-стероид-изомеразу, алкогольдегидрогеназу дрожжей, альфа-глицерофосфатдегидрогеназу, триозофосфатизомеразу, пероксидазу хрена, щелочную фосфатазу, аспарагиназу, глюкозооксидазу, бета-галактозидазу, рибонуклеазу, уреазу, каталазу, глюкозо-6-фосфатдегидрогеназу, глюкоамилазу и ацететилхолинэстеразу, но не ограничиваются ими.
Путем радиоактивного мечения IL-31-специфичных антител или мимотопов, можно обнаружить IL-31 с помощью радиоиммуноанализа (РИА). См. Work et al., LAB. TECHNIQUES & BIOCHEM. IN MOLEC. Bio. (No. Holland Pub. Co., NY, 1978). Радиоактивный изотоп может быть обнаружен такими средствами, как использование гамма-счетчика или сцинтилляционного счетчика или авторадиографии. Изотопы, которые особенно эффективны для целей настоящего изобретения, включают 3H, 125I, 131I, 35S, 14С и 125I.
Также можно пометить IL-31-специфические антитела или мимотопы флуоресцентным соединением. Когда флуоресцентно меченое антитело подвергается воздействию света соответствующей длины волны, его присутствие может быть обнаружено по флуоресценции. К наиболее часто используемым флуоресцентным метящим соединениям относятся изотиоцианат флуоресцеина, родамин, фикоэритрин, фикоцианин, аллофикоцианин, о-фтальдегид и флуорескамин.
Специфические к IL-31 антитела или мимотопы также могут быть помечены детектируемой меткой с использованием металлов, излучающих флуоресценцию, таких как 125Eu или другие из ряда лантаноидов. Эти металлы могут быть присоединены к IL-31-специфическим антителам или мимотопам с использованием таких хелатирующих металлы групп, как диэтилентриаминпентауксусная кислота (DTPA) или этилендиаминтетрауксусная кислота (EDTA).
Специфические к IL-31 антитела также могут быть помечены детектируемой меткой путем связывания с хемилюминесцентным соединением. Присутствие хемилюминесцентно меченого антитела или мимотопа затем определяют путем обнаружения люминесценции, возникающей в ходе химической реакции. Примерами полезных хемилюминесцентных метящих соединений являются люминол, изолюминол, тероматический сложный эфир акридиния, имидазол, соли акридиния и сложный эфир щавелевой кислоты.
Аналогичным образом, биолюминесцентное соединение можно использовать для мечения мимотопа или IL-31-специфического антитела, его части, фрагмента, полипептида или его производного. Биолюминесценция - это тип хемилюминесценции, обнаруживаемый в биологических системах, в которых белок, обладающий каталитической активностью, увеличивает эффективность хемилюминесцентной реакции. Присутствие биолюминесцентного белка определяют путем обнаружения люминесценции. Важными биолюминесцентными соединениями для целей маркировки являются люциферин, люцифераза и экворин.
Обнаружение мимотопа, IL-31-специфического антитела, части, фрагмента, полипептида или производного можно осуществлять с помощью сцинтилляционного счетчика, например, если обнаруживаемая метка представляет собой источник гамма-излучения, или с помощью флуориметра, например, если метка представляет собой флуоресцентное вещество. В случае ферментной метки обнаружение может быть выполнено колориметрическими методами, в которых используется субстрат для фермента. Детекцию также можно осуществлять путем визуального сравнения интенсивности ферментативной реакции с субстратом и аналогичным образом приготовленными стандартами.
Для целей настоящего изобретения IL-31, который определяют с помощью вышеуказанных исследований, может присутствовать в биологическом образце. Можно использовать любой образец, содержащий IL-31. Например, образец представляет собой биологическую жидкость, например, такую как кровь, сыворотка, лимфа, моча, кал, воспалительный экссудат, спинномозговая жидкость, амниотическая жидкость, экстракт или гомогенат ткани и тому подобное. Изобретение не ограничивается исследованиями с использованием только этих образцов, однако рядовой специалист в данной области техники ознакомившись с данным описанием может установить подходящие условия, которые позволяют использовать другие образцы.
Определение in situ можно осуществлять путем взятия гистологического образца у животного-субъекта и добавления меченого антитела (отдельно или в комплексе с мимотопом IL-31, описанным в настоящем документе) к такому образцу. Также предполагается, что антитело в составе комплекса может включать только часть антитела. Антитело (или его часть) может быть представлено в виде меченого антитела (или его части), нанесенного или наслоенного на биологический образец. С помощью такой процедуры можно определить не только присутствие IL-31, но и распределение IL-31 в исследуемой ткани. Используя настоящее изобретение, рядовые специалисты легко поймут, что любой из широкого разнообразия гистологических методов (таких как процедуры окрашивания) можно модифицировать для такого определения in situ.
Мимотоп, антитело, фрагмент или производное по настоящему изобретению можно адаптировать для применения в иммунометрическом анализе, также известном как «двухсайтовый» или «сэндвич-анализ». В типичном иммунометрическом анализе некоторое количество немеченого антитела (или фрагмента антитела) связывают с твердой подложкой, которая нерастворима в тестируемой жидкости, и добавляют определенное количество детектируемого меченного растворимого антитела, что позволяет обнаружить и/или определить количество тройного комплекса, образованного между связанным с твердой фазой антителом, антигеном и меченым антителом.
Антитела или комплексы антитело: мимотоп можно использовать для количественного или качественного определения IL-31 в образце или для обнаружения присутствия клеток, экспрессирующих IL-31. Это может быть достигнуто с помощью иммунофлуоресцентных методов с использованием флуоресцентно меченного антитела или комплекса антитело: мимотоп (см. ниже) в сочетании с флуоресцентной микроскопией, проточной цитометрией или флуорометрическим детектированием. Для диагностических целей антитела могут быть как мечеными, так и немечеными. Немеченые антитела можно использовать в комбинации с другими мечеными антителами (вторыми антителами), которые вступают в реакцию с антителом, такими как антитела, специфичные для константных областей иммуноглобулина собак или кошек. В качестве альтернативы, антитела могут быть помечены напрямую. Можно использовать самые разные метки, такие как радионуклиды, флюоресцирующие агенты, ферменты, субстраты ферментов, кофакторы ферментов, ингибиторы ферментов, лиганды (особенно гаптены) и т.д. Доступны многочисленные типы иммуноанализов, такие как те, что обсуждались ранее, и они хорошо известны специалистам в данной области.
В одном воплощении диагностический метод для определения IL-31 представляет собой иммунохроматографический анализ. Он также известен как иммунохроматографический анализ, быстрая иммуно-миграция (RIM™) или тест-полоска. Иммунохроматографический анализ - это, по сути, иммуноанализ, адаптированный для функционирования вдоль одной оси, чтобы соответствовать формату тест-полоски. На основе нескольких вариантов этой технологии разработаны коммерческие продукты, но все они работают по одному и тому же основному принципу. Типичная тест-полоска состоит из следующих компонентов: (1) площадка для образца - абсорбирующая площадка, на которую наносят исследуемый образец; (2) площадка с конъюгатом или реагентом - она содержит антитела, специфичные к целевому аналиту, конъюгированные с окрашенными частицами (обычно частицами коллоидного золота или латексными микросферами); (3) реакционная мембрана - обычно гидрофобная мембрана из нитроцеллюлозы или ацетата целлюлозы, на которой иммобилизованы антитела против целевого анализируемого вещества в виде линии, пересекающей мембрану, образуя зону захвата или тестовую линию (также может присутствовать зона контроля, содержащая антитела, специфичные к конъюгированным антителам); и (4) впитывающая зона или резервуар для отходов - дополнительная абсорбирующая площадка, предназначенная для продвижения образца по реакционной мембране за счет капиллярных сил и его сбора. Компоненты полоски обычно прикреплены к инертному материалу основы и могут быть представлены в формате простой полоски или имеющей пластиковый корпус с отверстием для пробы и окошком реакции, где видны зона захвата и зона контроля. В микробиологических исследованиях используют два основных типа иммунохроматографического анализа: сэндвич-тесты с двумя антителами и конкурентные тесты. В сэндвич-тестах с двумя антителами образец мигрирует от площадки для образца через площадку с конъюгатом, где любой присутствующий целевой аналит связывается с конъюгатом. Затем образец продолжает мигрировать через мембрану, пока не достигнет зоны захвата, где комплекс мишень/конъюгат будет связываться с иммобилизованными антителами с появлением на мембране видимой линии. Затем образец перемещается дальше по полоске, пока не достигнет зоны контроля, где связывается избыток конъюгата и на мембране появляется вторая видимая линия. Эта контрольная линия указывает на то, что образец прошел через мембрану, как и предполагалось. Две четкие линии на мембране - положительный результат. Одна линия в зоне контроля - отрицательный результат. Конкурентные анализы отличаются от сэндвич-тестов с двумя антителами тем, что площадка для конъюгата содержит антитела, которые уже связаны с целевым аналитом или с его аналогом. Если целевой аналит присутствует в образце, он, следовательно, не будет связываться с конъюгатом и останется немеченым. По мере того как образец перемещается вдоль мембраны и достигает зоны захвата, избыток немеченого аналита связывается с иммобилизованными антителами и блокирует захват конъюгата, так что видимые линии не появляются. Несвязанный конъюгат затем свяжется с антителами в зоне контроля с появлением видимой контрольной линии. Единственная контрольная линия на мембране - положительный результат. Две видимые линии в зоне захвата и контроля - отрицательный результат. Однако, если избыток немеченого целевого аналита отсутствует, в зоне захвата может появиться слабая линия, указывающая на сомнительный результат. Существует ряд разновидностей технологии иммунохроматографического анализа. Зона захвата на мембране может содержать не антитела, а иммобилизованные антигены или ферменты - в зависимости от целевого аналита. Для создания мультиплексного теста можно использовать несколько зон захвата. Например, были разработаны коммерческие тест-полоски, способные определять Шига токсины ST1 и ST2 энтерогеморрагических штаммов Е. coli отдельно в одном образце.
Важно отметить, что описанные в данном документе мимотопы и антитела могут применяться при диагностике зуда и/или аллергии у собак, кошек или лошадей. Более конкретно, антитело в составе комплекса антитело:мимотоп может связываться с IL-31 в образце и помогать обнаруживать избыточную экспрессию IL-31 у млекопитающих, включая домашних животных. Таким образом, описанные в данном документе антитела, которые можно использовать в сочетании с мимотопом, могут служить важным инструментом иммуногистохимии. В одном воплощении в данном документе предложена схема анализа, посредством которой мимотоп (пептид) IL-31 используют для захвата антитела по настоящему изобретению, меченого для определения в анализе. Это захваченное антитело будет иметь более низкую аффинность к присоединенному мимотопу, по сравнению с аффинностью к нативному IL-31, циркулирующему у биологического вида-хозяина. В этом воплощении жидкость, полученную от биологического вида-хозяина, инкубируют с комплексом меченое антитело:мимотоп, связанным с твердой поверхностью. IL-31, присутствующий в исследуемой жидкости, полученной от биологического вида-хозяина, будет иметь более высокое сродство к антителу, таким образом меченое антитело будет высвобождаться с твердой поверхности и может быть удалено на этапах промывки. Таким образом, уровень IL-31 в тестовой жидкости может быть обратно пропорционален сигналу, генерируемому на поверхности, с которой связан мимотоп. Предполагается, что такой анализ может применяться в качестве диагностического теста в исследованиях или в клинике для измерения IL-31.
Описанные в данном документе антитела и мимотопы можно использовать в чипах, очень удобных для измерения профиля экспрессии генов.
Наборы
В объем настоящего изобретения также включены наборы для практического применения рассматриваемых терапевтических методов и диагностических методов. В одном воплощении набор согласно настоящему изобретению, по меньшей мере, включает вакцинную композицию согласно настоящему изобретению. В одном воплощении вакцина по настоящему изобретению может быть представлена в контейнере, обычно в лиофилизированной форме. В другом воплощении набор согласно настоящему изобретению может включать компоненты, необходимые для осуществления диагностических методов согласно настоящему изобретению. Например, набор по настоящему изобретению может включать в качестве одного из своих компонентов мимотоп IL-31, согласно описанию в данном документе, такой как мимотоп кошачьего IL-31, мимотоп собачьего IL-31, мимотоп лошадиного IL-31 или мимотоп человеческого IL-31. Такой мимотоп может уже быть связан с твердой поверхностью. Набор согласно настоящему изобретению может также включать антитела. Антитела, которые могут быть конъюгированы с меткой или токсином или неконъюгированы, обычно включаются в наборы с буферами, такими как трис, фосфат, карбонат и т.д., стабилизаторами, биоцидами, инертными белками, например, сывороточным альбумином и тому подобным. Обычно эти материалы будут присутствовать в количестве менее 5% мас. в расчете на количество активного антитела и обычно присутствуют в общем количестве по меньшей мере примерно 0,001% мас. снова в расчете на концентрацию антител. Часто бывает желательно включить инертный наполнитель или эксципиент для разбавления активных ингредиентов, где эксципиент может присутствовать в количестве от приблизительно 1% до 99% по массе от общей композиции. Если в анализе используется второе антитело, способное связываться с первичным антителом, оно обычно находится в отдельном флаконе. Второе антитело обычно конъюгировано с меткой и составляется с препаратами антител, описанными выше, аналогичным образом. Наборы обычно также включают набор инструкций по применению.
В одном воплощении набор по настоящему изобретению представляет собой набор тест-полосок (набор для иммунохроматографического анализа), который можно использовать для определения в образце IL-31, такого как белок IL-31 собаки, кошки, лошади или человека. Такая тест-полоска обычно включает площадку для образца, на которую наносят исследуемый образец; площадку с конъюгатом или реагентом, содержащую антитело или антитело:мимотоп, специфичные к собачьему, кошачьему, лошадиному или человеческому IL-31, где антитело или комплекс антитело:мимотоп конъюгированы с окрашенными частицами (обычно частицами коллоидного золота); реакционная мембрана, на которой антитела к IL-31 или комплекс антитело:мимотоп иммобилизованы в виде линии, пересекающей мембрану, образуя зону захвата или тестовую линию (также может присутствовать зона контроля, содержащая антитела или комплекс антитело:мимотоп, к которым специфичны конъюгированные антитела); и еще одну абсорбирующую площадку, предназначенную для продвижения образца через реакционную мембрану за счет капиллярных сил и его сбора. Набор тест-полосок обычно также включает инструкции по применению.
Далее изобретение будет описано ниже на примерах, не ограничивающих объем изобретения. В приведенном ниже разделе Примеры и в графических материалах любые данные, представленные для антител, содержащих в своем обозначении «11Е12», приведены для сравнения с антителами по настоящему изобретению.
Примеры
1. Пример 1.
1.1. Получение собачьего интерлейкина 31 (cIL-31) в клетках яичника китайского хомячка (СНО)
Консервативность аминокислотной последовательности белка интерлейкин 31 различается среди гомологичных видов (Фиг. 1), но считается, что он имеет общую структурную архитектуру с другими членами семейства цитокинов типа I (Boulay et al. 2003, Immunity. Aug; 19(2):159-632003; Dillon et al. 2004 Nat Immunol. Jul; 5(7):752-60). Топология пучков типа «вверх-вниз» важна для способа распознавания рецептора, общего для этих цитокинов (Dillon et al. см. выше, Cornelissen et al. 2012 Eur J Cell Biol. Jun-Jul; 91(6-7):552-66). Из-за различий в последовательностях белка IL-31 у разных биологических видов невозможно предсказать, будут ли антитела, выработанные против одного вида, перекрестно реагировать с другими, учитывая различную частоту встречаемости аминокислот в эпитопах и локальный аминокислотный состав. Как следствие, для данной работы рассматривали множество форм белка IL-31, представляющих множество видов и систем экспрессии. Собачий белок IL-31 (cIL-31) получали для использования в качестве иммуногена и реагента для проверки аффинности и эффективности вариантов антител. Рекомбинантный cIL-31 получали в клетках СНО с использованием системы CHROMOS АСЕ (искусственная экспрессия хромосом) (Chromos Molecular Systems, Inc., Burnaby, British Columbia) с образованием секретируемого собачьего белка IL-31, имеющего последовательность (SEQ ID NO: 155; Canine_IL-31), соответствующей нуклеотидной последовательностью для которой является (SEQ ID NO: 156; Canine_IL-31). Кондиционированную среду из 400 мл клеточной культуры (линия клеток СНО) получали и диализовали против 10 объемов буфера QA (20 мМ Трис рН 8,0, 20 мМ NaCl) в течение 4,5 часов. Диализованную среду фильтровали через фильтр с диаметром пор 0,2 мкм и загружали со скоростью 1 мл/мин на колонку SOURCE™ Q (GE Healthcare, Uppsala, Sweden), предварительно уравновешенную буфером QA. Белок элюировали с использованием многоступенчатого линейного градиента. Большая часть cIL-31 оставалась в проточной фракции (FT), небольшое количество cIL-31 элюировалось в начале градиента. Подлинность белка ранее была подтверждена вестерн-блоттингом и масс-спектрометрическим (МС) анализом триптического гидролизата. Белок во фракции FT концентрировали в 4-5 раз и диализовали в течение ночи против забуференного фосфатом физиологического раствора (PBS) при 4°С. После диализа в PBS проверяли стабильность белка. Преципитации не наблюдалось, и через несколько дней при 4°С протеолиза не наблюдалось. В результате дегликозилирования с использованием N-гликозидазы F получали белок, мигрирующий при электрофорезе в полиакриламидном геле в присутствии додецилсульфата натрия (SDS-PAGE) в виде единственной полосы, соответствующей приблизительно 15 кДа. Концентрацию белка определяли с помощью бицинхонинового анализа (анализ ВСА) с бычьим сывороточным альбумином (BSA) в качестве стандарта (ThermoFisher Scientific, Inc., Rockford, IL). Белковый раствор делили на аликвоты, быстро замораживали (жидкий N2) и хранили при -80°С.
1.2. Транзиторная экспрессия кошачьего интерлейкина 31 (fIL-31) дикого типа и мутантного в клетках СНО
Чтобы помочь в идентификации антител со способностью связывать подходящие эпитопы, белки кошачьего IL-31 дикого типа и мутантные экспрессировали в системе экспрессии млекопитающих для продукции, очистки и исследования аффинности в анализах на клетках. Сайт связывания антитела 11Е12 на IL-31 был описан ранее (патент US 8790651, Bammert, et al.). В данном документе приведена характеристика нового сайта связывания на IL-31, распознаваемого антителом 15Н05. Диким типом обозначают полноразмерный кошачий белок IL-31 без изменений нативных аминокислотных остатков. Мутантные белки обозначали в соответствии с названиями антител к ним (11Е12 и 15Н05), указывающим на мутации аминокислот белка IL-31, которые (будучи измененными) влияют на связывание с каждым соответствующим антителом. Идентификация соответствующих мутаций, необходимых для кошачьего белка IL-31 15Н05, описана ниже в разделе 1.10. Задача состояла в том, чтобы изменить аминокислоты в эпитопе IL-31 и наблюдать потерю связывания с каждым соответствующим антителом. Затем в ходе скрининга можно провести сравнение, чтобы посмотреть, связываются ли новые антитела-кандидаты с белком дикого типа, но не с мутантом. Затем можно отсортировать новые варианты антител по их связыванию с тем же эпитопом, что и антитело 11Е12 или 15Н05, или с похожим эпитопом.
Экспрессионные конструкции оптимизировали по частоте использования кодонов и синтезировали для экспрессии в клетках яичников китайского хомячка (СНО). Синтезированные гены клонировали в pD2529 (вектор ATUM) для использования в транзиторной экспрессии. Кошачий белок IL-31 дикого типа представлен (SEQ ID NO: 157; Feline_IL-31_wildtype), ему соответствует нуклеотидная последовательность (SEQ ID NO: 158; Feline_IL-31_wildtype). Мутантный белок кошачьего IL-31 11Е12 представлен (SEQ ID NO: 161; Feline_IL-31_11Е12_mutant), соответствующей нуклеотидной последовательностью для него является (SEQ ID NO: 162; Feline_IL-31_11E12_mutant). Мутантный белок кошачьего IL-31 15Н05 представлен (SEQ ID NO: 163; Feline_IL-31_15H05_mutant), соответствующей нуклеотидной последовательностью для него является (SEQ ID NO: 164; Feline_IL-31_15Н05_mutant). Рекомбинантные кошачьи белки IL-31 экспрессировали в клетках ExpiCHO-S™ (ThermoFisher Scientific, Inc., Rockford, IL), следуя протоколу Max Titer от производителя для транзиторной экспрессии в клетках СНО. Через двенадцать дней после трансфекции клетки центрифугировали и фильтровали для захвата секретируемого белка из кондиционированной среды. Для каждой конструкции (дикого типа и мутантов) 120 мл кондиционированной среды (из культуры клеток СНО, профильтрованной через фильр с размером пор 0,2 мкм) доводили до 30 мСм/см добавлением NaCl, 5 мМ имидазола и рН до 7,4. Каждый образец среды объединяли с 5 мл смолы HisPur Cobalt (ThermoFisher Scientific, Inc., Rockford, IL), уравновешенной 5 мМ имидазола, 20 мМ фосфата натрия, 300 мМ NaCl, рН 7,4. Каждый образец и смолу оставляли перемешиваться при 4°С в течение ночи. Смолы собирали (и отделяли от несвязанной фракции) путем пропускания через колонки BioRad Econcolumn (Bio-Rad, Hercules, CA). Смолы промывали 5×5 мл буфера (как указано выше), а затем элюировали 5×5 мл 500 мМ имидазола в том же буфере. Фракции оценивали с помощью SDS-PAGE. Концентрацию белка измеряли с помощью анализа ВСА согласно стандартным методам.
1.3. Получение кошачьего интерлейкина 31 (fIL-31) в Е. coli
Рекомбинантный кошачий белок IL-31 получали в экспрессирующем хозяине Е. coli для использования в качестве реагента в анализе и в экспериментах in vivo, чтобы спровоцировать у кошек ответную реакцию в виде зуда. Ген, представляющий кошачий IL-31, синтезировали для оптимальной экспрессии в Е. coli. Создавали экспрессирующие конструкции с полноразмерным геном кошачьего IL-31, содержащим N-концевую гексагистидиновую метку для детекции и очистки. Этот белок кошачьего IL-31 представлен (SEQ ID NO: 159; Feline_IL-31_E_coli), соответствующей нуклеотидной последовательностью для которой является (SEQ ID NO: 160; Feline_IL-31_Е_coli). Плазмиды с подтвержденной последовательностью использовали для трансформации Е. coli BL21 DE3 (Invitrogen Corp., Carlsbad, СА) и последующей экспрессии белка.
Клеточную массу Е. coli (262,3 г) лизировали следующим образом. Клеточную массу ресуспендировали в 500 мл 50 мМ Трис рН 8, фильтровали через фильтр из нержавеющей сетки для удаления частиц, а затем лизировали путем двукратного пропускания через микрофлюидизатор при 8,963 Мпа (1300 фунтов на квадратный дюйм). Лизат (объемом приблизительно 1200 мл) делили на четыре флакона, центрифугировали при 12000 g в течение 20 минут при 10°С. Супернатант декантировали и отбрасывали. Каждый осадок промывали путем растворения в 300 мл 5 мМ EDTA, 0,5% Triton Х-100, рН 9,0, а затем центрифугировали при 12000 g, в течение 50 минут при 10°С. Супернатант декантировали и отбрасывали. Промытый осадок хранили при -20°С до сворачивания и выделения.
Перед выделением один из осадков промывали водой для удаления остаточного детергента, а затем центрифугировали при 10000 g, в течение 20 минут при 4°С. Снова декантировали супернатант. Наконец, промытый осадок солюбилизировали в 60 мл 50 мМ фосфата натрия, 300 мМ NaCl, 6 М гуанидин-HCl, 5 мМ имидазола, рН 7,4. Осадое оставляли перемешиваться при комнатной температуре в течение приблизительно 25 минут перед повторным центрифугированием при 10000 g, в течение 20 минут при 4°С. На этот раз супернатант декантировали и оставляли для дальнейшей обработки. Осадок (ресуспендированный в воде до исходного объема) оставляли только для SDS-PAGE. Для повышения чистоты перед сворачиванием выполняли первичную аффинную хроматографию с иммобилизованными ионами металла (IMAC). В этом случае 15 мл Ni-NTA Superflow (Qiagen Inc, Germantown, MD, кат. номер 30450, предварительно уравновешенную тем же буфером) добавляли к осветленному супернатанту и оставляли перемешиваться при комнатной температуре в течение приблизительно 90 минут. Несвязанную фракцию декантировали и оставляли для SDS-PAGE. Смолу IMAC промывали 5 мМ имидазолом, 50 мМ фосфатом натрия, 300 мМ NaCl, 6 М гуанидин-HCl, рН 7,4 (таким же, как буфер для солюбилизации). Смолу элюировали (сначала 7,5 мл, а затем несколько раз по 15 мл, элюирование белка отслеживали с помощью анализа по Брэдфорду) 200 мМ имидазола, 50 мМ фосфата натрия, 300 мМ NaCl, 6 М гуанидин-HCl, рН 7,4. Элюированные фракции, содержащие белок (по Брэдфорду), объединяли (125 мл) для дальнейшей обработки.
Сворачивание белка IL-31 осуществляли следующим образом. IL-31 восстанавливали добавлением дитиотрейтола до конечной концентрации 10 мМ и оставляли перемешиваться при комнатной температуре в течение 2 часов. Затем разбавленный образец разбавляли по каплям в 2500 мл (20Х объем) PBS + 1 М NaCl при быстром перемешивании. Теоретическая концентрация мочевины в этот момент должна была составлять примерно 0,4 М. Остаток мочевины медленно удаляли путем диализа в PBS, осуществляя 3 замены (каждый раз в объеме 4 л) при 4°С в течение ночи. После диализа образец фильтровали через фильтр с размером пор 0,2 мкм для удаления любого несвернутого/осажденного белка.
Далее образец очищали во втором раунде IMAC, на этот раз с элюированием линейным градиентом. К образцу добавляли 15 мл смолы Ni-NTA Superflow и оставляли для связывания партиями путем перемешивания (с магнитной мешалкой) в течение ночи при 4°С. И вновь, несвязанную фракцию декантировали и оставляли. Смолу Ni-NTA Superflow помещали в колонку XK16 (GE Healthcare Lifesciences, Marlborough, MA) и подключали к системе хроматографии марки АКТА (GE Healthcare Lifesciences, Marlborough, MA). Затем колонку промывали 50 мМ Трис, 300 мМ NaCl, рН 8,2 и элюировали с помощью 150 мл линейного градиента от 0 до 500 мМ имидазола в промывочном буфере. Фракции анализировали с помощью SDS-PAGE. Фракции, имеющие IL-31 достаточной чистоты, объединяли и снова заменяли буфер путем диализа в PBS, осуществляя 3 замены (каждый раз по 2 л) при 4°С в течение ночи. Наконец, свернутый и очищенный образец собирали после диализа, стерилизовали фильтрованием, измеряли концентрацию, делили на аликвоты, мгновенно замораживали в бане с сухим льдом/изопропанолом и хранили при -80°С.
1.4. Способ определения аффинности антител против IL-31 к IL-31 с помощью поверхностного плазмонного резонанса
Аффинность, с которой mAb-кандидаты связывают собачий и кошачий IL-31, определяли с использованием поверхностного плазмонного резонанса (SPR) в системе Biacore (Biocore Life Sciences (GE Healthcare), Uppsala, Sweden). Чтобы избежать различий в аффинности, связанных с различной подготовкой поверхности, которая может возникнуть при иммобилизации антител на поверхностях, использовали стратегию, при которой IL-31 непосредственно конъюгировали с поверхностью. Иммобилизацию осуществляли путем присоединения через аминогруппы 5 мкг/мл IL-31 с использованием N-гидроксисукцинимида (NHS)/1-этил-3-(3-диметиламинопропил)карбодиимида (EDC). Чипы блокировали этаноламином и оценивали аффинность, с которой все mAb-кандидаты связывались с иммобилизованным IL-31. Для аппроксимации всех кривых использовали модель 1:1. Константы аффинности (KD) менее 1×10-11 М (1Е-11 М) меньше нижнего предела количественного определения прибора. Результаты измерений аффинности описаны в данном документе.
1.5. Способ оценки эффективности антител против IL-31 по ингибированию передачи сигналов PSTAT3, индуцированной собачьим и кошачьим IL-3L в макрофагальных клетках собак и кошек
Чтобы идентифицировать кандидатов с ингибирующей активностью, оценивали способность антител влиять на фосфорилирование STAT3, опосредованное IL-31, в анализе на клетках собак или кошек. Фосфорилирование STAT3 определяли в макрофагоподобных клетках DH-82 собак (АТСС® CRL-10389™) или FCWF4 кошек (АТСС CRL-2787). Клетки DH82 и FCWF4 примировали собачьим интерфероном гамма (R&D Systems, Minneapolis, MN) в концентрации 10 нг/мл в течение 24 часов или кошачьим интерфероном гамма (R&D Systems, Minneapolis, MN) при 125 нг/мл в течение 96 часов, соответственно, для увеличения экспрессии рецепторов. Оба типа клеток содержали в отсутствии сыворотки в течение 2 часов перед воздействием IL-31 и mAb. Используя два независимых метода, оценивали способность всех mAb-кандидатов ингибировать фосфорилирование STAT3, индуцированное 1 мкг/мл собачьего или 0,2 мкг/мл кошачьего IL-31. Также проводили исследования для демонстрации перекрестной реактивности цитокинов собак и кошек и перекрестной функциональной способности антител ингибировать передачу сигналов у обоих биологических видов. Чтобы обеспечить образование комплекса, до стимуляции клеток проводили совместную инкубацию mAb и цитокина IL-31 в течение одного часа. Стимуляцию клеток IL-31 осуществляли в течение пяти минут. Фосфорилирование STAT3 измеряли с использованием технологии AlphaLISA SureFire ULTRA™ (Perkin Elmer, Waltham, MA). В случае, когда концентрация и чистота антител неизвестны, проводили качественное определение способности супернатантов гибридомы ингибировать фосфорилирование STAT3 после 1-часовой совместной инкубации с 1 мг/мл собачьего или 0,2 мг/мл кошачьего IL-31. Эффективность индивидуальных моноклональных антител, определяемая их способностью ингибировать опосредованное IL-31 фосфорилирование STAT3 в этих исследованиях, считали ключевым критерием отбора для дальнейшего совершенствования отобранных антител. Термин «эффективность» относится к значению IC50, рассчитанному на основе этих исследований, и представляет собой концентрацию антитела, при которой передача сигналов, индуцированная IL-31, снижается до половины от максимального значения. Повышенная эффективность, описанная в данном документе, коррелирует с более низким значением IC50.
1.6. Нахождение мышиных и собачьих моноклональных антител, распознающих собачий и кошачий интерлейкин 31
Мышей и собак иммунизировали рекомбинантным собачьим IL-31 (SEQ ID No. 155) с целью нахождения антител. Титры сывороточных антител от иммунизированных животных определяли с использованием твердофазного иммуноферментного анализа (ELISA). Собачий или кошачий IL-31 (50 нг/лунку) иммобилизовали на полистироловых планшетах для микротитрования и использовали в качестве антигена для захвата. Сыворотку иммунизированных животных разводили в забуференном фосфатом физиологическом растворе с 0,05% твин-20 (PBST). Присутствие антител против IL-31 определяли с помощью подходящего вторичного антитела, меченного HRP. После добавления хромогенного субстрата (SureBlue Reserve ТМВ, 1-компонентного субстрата перокидазы для микропланшетов, KPL, Inc., Gaithersburg, MD) и десятиминутной инкубации при комнатной температуре (RT) реакцию останавливали добавлением 100 мкл 0,1 N HCl. Определяли оптическую плотность (OD) в каждой лунке при длине волны 450 нм. Отбирали антитела по их способности связывать собачий и кошачий IL-31 с использованием ELISA. В некоторых случаях во время отбора проводили дополнительную характеризацию с использованием ELISA с мутантной формой кошачьего белка IL-31 в качестве антигена для захвата. Выбирали клетки, продуцирующие антитела с желаемыми свойствами связывания и ингибирования, для анализа последовательности РНК-транскриптов, представляющих вариабельные области тяжелых (VH) и легких (VL) цепей IgG.
В случае мышиных антител для слияния использовали донорские спленоциты от одной мыши CF-1, давшей ответную реакцию, а супернатанты гибридомы подвергали скринингу на антитела, связывающиеся с собачьим или кошачьим белком IL-31, методом ELISA. Это позволило идентифицировать одно мышиное антитело Mu-15Н05, имеющее аффинность к обоим видам IL-31 в суб-наномолярном диапазоне (Фиг. 2А). Мышиное анти-IL-31 антитело 15Н05 дополнительно субклонировали для получения гибридомы, продуцирующей гомогенное антитело, и для секвенирования вариабельных областей тяжелых и легких цепей. Последовательности вариабельных областей мышиного антитела к IL-31, определенные для антитела 15Н05, представлены ниже: вариабельная область тяжелой цепи 15Н05 (SEQ ID NO: 67; MU-15H05-VH), соответствующей нуклеотидной последовательностью для которой является (SEQ ID NO: 68; MU-15H05-VH), вариабельная область легкой цепи 15Н05 (SEQ ID NO: 69; MU-15H05-VL), соответствующей нуклеотидной последовательностью для которой является (SEQ ID NO: 70; MU-15H05-VL).
Помимо мышиного антитела 15Н05, дополнительное внимание было уделено мышиному антителу 11Е12, которое ранее было описано в патенте US 8790651, выданном Bammert, et al. В настоящем документе приведены данные, показывающие способность антитела 11Е12 связывать как собачьи, так и кошачьи белки IL-31 с высокой аффинностью. Способность 11Е12 связывать кошачий IL-31 сделала это антитело подходящим кандидатом для фелинизации и потенциального терапевтического применения у кошек. Последовательности вариабельных областей мышиного антитела к IL-31, определенные ранее для антитела 11Е12, представлены ниже: вариабельная область тяжелой цепи 11Е12 (SEQ ID NO: 71; MU-11E12-VH), соответствующей нуклеотидной последовательностью для которой является (SEQ ID NO: 72; MU-11E12-VH), вариабельная область легкой цепи 11Е12 (SEQ ID NO: 73; MU-11E12-VL), соответствующей нуклеотидной последовательностью для которой является (SEQ ID NO: 74; MU-11E12-VL).
Собак, имеющих повышенные титры антител к IL-31 после вакцинации, отбирали для анализа популяций В-клеток, продуцирующих антитела с желаемыми фенотипами. В-клетки для дальнейшего анализа получали из РВМС, костного мозга, селезенки или лимфатических узлов. Одиночные В-клетки разделяли по отдельным лункам и анализировали на наличие секретируемых IgG, способных связываться с собачьим IL-31 дикого типа и мутантными формами 11Е12 и 15Н05 (AbCellera, Vancouver, ВС), используя методы, описанные в US 2012/0009671 А1, US 2016/0252495 A1, US 9188593, WO 2015/176162 А9 и WO 2016/123692 A1.
Эта стратегия скрининга основана на известных областях белка IL-31, которые имеют решающее значение для связывания и передачи сигнала через его корецепторный комплекс. Отбор этих мутантных белков для скрининга описан в разделе 1.2 настоящей заявки. После RT-PCR осуществляли секвенирование вариабельных доменов тяжелой и легкой цепей IgG из отдельных В-клеток-кандидатов. Этот скрининг привел к идентификации девяти собачьих антител, выбранных для дальнейших исследований. Последовательности вариабельных областей этих собачьих антител против IL-31 являются следующими: вариабельная область тяжелой цепи ZIL1 (SEQ ID NO: 75; CAN-ZIL1_VH), соответствующей нуклеотидной последовательностью для которой является (SEQ ID NO: 76; CAN-ZIL1_VH), вариабельная область легкой цепи ZIL1 (SEQ ID NO: 77; CAN-ZIL1_VL), соответствующей нуклеотидной последовательностью для которой является (SEQ ID NO: 78; CAN-ZIL1_VL); вариабельная область тяжелой цепи ZIL8 (SEQ ID NO: 79; CAN-ZIL8_VH), соответствующей нуклеотидной последовательностью для которой является (SEQ ID NO: 80; CAN-ZIL8_VH), вариабельная область легкой цепи ZIL8 (SEQ ID NO: 81; CAN-ZIL8_VL), соответствующей нуклеотидной последовательностью для которой является (SEQ ID NO: 82; CAN-ZIL8_VL); вариабельная область тяжелой цепи ZIL9 (SEQ ID NO: 83; CAN-ZIL9_VH), соответствующей нуклеотидной последовательностью для которой является (SEQ ID NO: 84; CAN-ZIL9_VH), вариабельная область легкой цепи ZIL9 (SEQ ID NO: 85; CAN-ZIL9_VL), соответствующей нуклеотидной последовательностью для которой является (SEQ ID NO: 86; CAN-ZIL9_VL); вариабельная область тяжелой цепи ZIL11 (SEQ ID NO: 87; CAN-ZIL11_VH), соответствующей нуклеотидной последовательностью для которой является (SEQ ID NO: 88; CAN-ZIL11_VH), вариабельная область легкой цепи ZIL11 (SEQ ID NO: 89; CAN-ZIL11_VL), соответствующей нуклеотидной последовательностью для которой является (SEQ ID NO: 90; CAN-ZIL11_VL); вариабельная область тяжелой цепи ZIL69 (SEQ ID NO: 91; CAN-ZIL69_VH), соответствующей нуклеотидной последовательностью для которой является (SEQ ID NO: 92; CAN-ZIL69_VH), вариабельная область легкой цепи ZIL69 (SEQ ID NO: 93; CAN-ZIL69_VL), соответствующей нуклеотидной последовательностью для которой является (SEQ ID NO: 94; CAN-ZIL69_VL); вариабельная область тяжелой цепи ZIL94 (SEQ ID NO: 95; CAN-ZIL94_VH), соответствующей нуклеотидной последовательностью для которой является (SEQ ID NO: 96; CAN-ZIL94_VH), вариабельная область легкой цепи ZIL94 (SEQ ID NO: 97; CAN-ZIL94_VL), соответствующей нуклеотидной последовательностью для которой является (SEQ ID NO: 98; CAN-ZIL94_VL); вариабельная область тяжелой цепи ZIL154 (SEQ ID NO:99; CAN-ZIL154_VH), соответствующей нуклеотидной последовательностью для которой является (SEQ ID NO: 100; CAN-ZIL154_VH), вариабельная область легкой цепи ZIL154 (SEQ ID NO: 101; CAN-ZIL154_VL), соответствующей нуклеотидной последовательностью для которой является (SEQ ID NO: 102; CAN-ZIL154_VL); вариабельная область тяжелой цепи ZIL159 (SEQ ID NO: 103; CAN-ZIL159_VH), соответствующей нуклеотидной последовательностью для которой является (SEQ ID NO: 104; CAN-ZIL159_VH), вариабельная область легкой цепи ZIL159 (SEQ ID NO: 105; CAN-ZIL159_VL), соответствующей нуклеотидной последовательностью для которой является (SEQ ID NO: 106; CAN-ZIL159_VL); вариабельная область тяжелой цепи ZIL171 (SEQ ID NO: 107; CAN-ZIL171_VH), соответствующей нуклеотидной последовательностью для которой является (SEQ ID NO: 108; CAN-ZIL171_VH), вариабельная область легкой цепи ZIL171 (SEQ ID NO: 109; CAN-ZIL171_VL), соответствующей нуклеотидной последовательностью для которой является (SEQ ID NO: 110; CAN-ZIL171_VL).
Вышеупомянутые девять моноклональных антител, которые отбирали для дальнейшей характеризации, могут упоминаться в другом месте описания, в графических материалах или в формуле изобретения как ZIL1, ZIL8, ZIL8, ZIL11, ZIL69, ZIL94, ZIL154, ZIL159 и ZIL171.
1.7. Конструирование рекомбинантных химерных и полностью собачьих антител
Вариабельные домены антител отвечают за связывание антигена. Ожидается, что пересадка полного вариабельного домена в соответствующую константную область окажет незначительное влияние или не окажет никакого влияния на способность антитела связывать иммуноген IL-31. Для одновременного подтверждения того, что была идентифицирована правильная последовательность вариабельных областей тяжелой и легкой цепей и для получения гомогенного материала, сконструировали экспрессирующие векторы для получения рекомбинантных химерных или полностью собачьих антител в экспрессирующих системах млекопитающих. Описанные в данном документе химерные антитела состоят из вариабельной последовательности (как CDR, так и каркасной области) антитела от биологического вида-хозяина, пересаженной к соответствующим константным областям тяжелой и легкой цепи молекулы IgG кошки или собаки (например: вариабельная область мышиного происхождения:константная область собачьего происхождения обозначаются как мышиное:собачье химерное антитело). Полностью собачьи антитела, описанные в данном документе, состоят из последовательности вариабельной области (как CDR, так и каркасной области) антитела от биологического вида-хозяина (собаки), привитых к соответствующим константным областям тяжелой и легкой цепи молекулы собачьего IgG. Синтетические последовательности ДНК были сконструированы для последовательностей вариабельной области тяжелой цепи (VH) и вариабельной области легкой цепи (VL) выбранных антител. Эти последовательности содержат уникальные сайты рестрикции эндонуклеазами, консенсусную последовательность Козака и N-концевую лидерную последовательность для секреции, облегчающую экспрессию и секрецию рекомбинантного антитела из линии клеток млекопитающих.
Для мышиных:кошачьих химерных антител каждую соответствующую вариабельную область клонировали в плазмиду для экспрессии в клетках млекопитающих, содержащую константные области либо тяжелой цепи IgG кошки (SEQ ID NO: 173; Feline_НС_AlleleA_1), соответствующей нуклеотидной последовательностью, для которой является (SEQ ID NO: 174; Feline_HC_AlleleA_1), либо легкой цепи (SEQ ID NO: 175; Feline_LC_Kappa_G_minus), соответствующей нуклеотидной последовательностью для которой является (SEQ ID NO: 176; Feline_LC_G_K_minus). Для мышиных:собачьих химерных антител или полностью собачьих антител каждую соответствующую вариабельную область клонировали в плазмиду для экспрессии в клетках млекопитающих, содержащую константные области либо тяжелой цепи IgG собаки (SEQ ID NO: 177; Canine_HC_65_1), соответствующей нуклеотидной последовательностью, для которой является (SEQ ID NO: 178; Canine_НС_65_1), либо легкой цепи (SEQ ID NO: 179; Canine_LC_Kappa), соответствующей нуклеотидной последовательностью для которой является (SEQ ID NO: 180; Canine_LC_Kappa). Плазмидами, кодирующими каждую тяжелую и легкую цепь, под контролем промотора CMV, совместно трансфицировали клетки HEK 293 с использованием стандартных методов. После шести дней экспрессии химерные mAb очищали из 50 мл надосадочной жидкости транзиторно трансфицированных клеток HEK293FS с использованием смолы MabSelect Sure protein A (GE Flealthcare, Uppsala, Sweden) в соответствии со стандартными методами очистки белков Элюированные фракции объединяли, концентрировали до приблизительно 500 мкл, используя центрифужное устройство Nanosep Omega с номинальным отсечением по молекулярной массе 10000 (Pall Corp., Port Washington, NY), диализовали в течение ночи при 4°С в 1×PBS, рН 7,2 и хранили при 4°С для дальнейшего использования. Аффинность и эффективность отобранных рекомбинантных антител в анализах на клетках описаны ниже.
На Фиг. 2 подробно показана аффинность антител с CDR мышиного происхождения, исследованная с использованием biacore. На Фиг. 2а показана аффинность мышиных анти IL-31 антител 11Е12 и 15Н05 и соответствующая аффинность кошачьих и собачьих химерных форм к поверхности с кошачьим и собачьим IL-31. Эти наблюдения подтверждают правильность последовательности обоих мышиных антител и указывают на то, что преобразование в химерную форму приводит к получению антител с эквивалентной или более высокой аффинностью по сравнению с родительским мышиным антителом, за исключением мышиной:кошачьей химеры 15Н05, которая потеряла некоторое сродство к IL-31 обоих видов в результате преобразования в химерную форму.
Также исследовали активность полностью мышиных и химерных форм антител 11Е12 и 15Н05 в отношении клеток собак и кошек, как описано в разделе 1.5. На Фиг. 3 показаны результаты этих анализов. Активность мышиных антител 11Е12 и 15Н05 в отношении клеток собак и кошек исследовали с использованием как собачьего, так и кошачьего IL-31 для стимуляции передачи сигналов. Эффективность обоих мышиных антител в отношении как собачьих, так и кошачьих клеток при использовании кошачьего цитокина была сопоставима, за исключением 15Н05 против IL-31 кошки в кошачьих клетках FCWF4, который демонстрировал небольшое увеличение IC50. 15Н05 мыши было способно блокировать передачу сигналов собачьего IL-31 как в кошачьих, так и в собачьих клетках, при этом эффективность в отношении собачьих клеток была немного выше. Эти результаты показывают, что соответствующие эпитопы, распознаваемые этими антителами, существуют как на собачьем, так и на кошачьем IL-31, и связывание этих антител способно нейтрализовать рецептор-опосредованную клеточную передачу сигналов в соответствующих клеточных линиях обоих биологических видов.
На Фиг. 3 также показана эффективность отобранных химер в анализах на обоих типах клеток. Преобразование мышиных антител в кошачьи и собачьи химеры оказывало минимальное влияние на эффективность против кошачьего IL-31 в анализе на клетках кошек (диапазон IC50 1,15-3,45 мкг/мл). Сходные результаты наблюдались, когда эти химеры тестировали в отношении передачи сигнала кошачьего IL-31 на собачьей клеточной линии DH82, при этом для химерного мышиного:собачьего антитела 15Н05 наблюдалось небольшое увеличение активности (IC50=0,71 мкг/мл). В целом наблюдалось увеличение значений IC50 в отношении собачьего IL-31 как в клетках собак, так и в клетках кошек. Мышиная:кошачья химера 15Н05 была немного менее эффективной в анализе этого формата по сравнению с мышиной:собачьей формой (IC50 28,61 против 12,49 мкг/мл). Преобразование в собачьи и кошачьи химерные формы привело к минимальным изменениям эффективности, что согласовывалось с данными для мышиных антител.
Описанные выше антитела, выработанные одиночными В-клетками иммунизированных собак, сконструировали в виде рекомбинантных белков IgG после идентификации последовательностей их вариабельных доменов. Пересадка этих вариабельных доменов на тяжелую цепь Fc собачьего происхождения (изотип 65_1) привела к созданию рекомбинантных полностью собачьих антител. Представляло интерес идентифицировать дополнительные собачьи антитела, связывающие кошачий IL-31 дикого типа и имеющие пониженное связывание с мутантным кошачьим IL-31 15Н05 (то есть направленными против эпитопа 15Н05). Такие антитела, полученные из этого альтернативного источника (собачьи вместо мышиных), обеспечивают дополнительные паратопы (часть антитела, которая распознает белок IL-31, включает CDR), распознающие эпитоп 15Н05, и таким образом увеличивают выбор антител с различными физическими свойствами.
На Фиг. 4 показаны результаты связывания этих рекомбинантных собачьих антител с различными белками, полученные с использованием методов ELISA и Biacore. Связывание антител с кошачьим IL-31 дикого типа и мутантным белком 15Н05 оценивали методом непрямого ELISA. Все девять собачьих моноклональных антител (ZIL1, ZIL8, ZIL8, ZIL11, ZIL69, ZIL94, ZIL154, ZIL159 и ZIL171) были способны связываться с кошачьим IL-31 дикого типа, и на связывание влияли мутации в области эпитопа, распознаваемой mAb 15Н05, что подтверждало правильный связывающий фенотип, установленный во время первоначального скрининга, использованного для их нахождения. Для сравнения, антитело 11Е12 связывалось с кошачьим IL-31 дикого типа, и на его связывание не влияли мутации в области эпитопа, распознаваемой 15Н05, о чем свидетельствуют данные на Фиг. 4. Для подтверждения связывания проводили анализ biacore с использованием белков IL-31 собак, кошек, лошадей, человека, мутантного кошачьего белка IL-31 и мутантного кошачьего белка 11Е12, иммобилизованными на поверхности, и единственной исследуемой концентрации антитела. Все протестированные антитела связывались с кошачьим IL-31 дикого типа, что согласовывалось с данными, полученными с помощью ELISA. В соответствии с данными, описанными выше в этом разделе, мышиные антитела 11Е12 и 15Н05 связывались с иммобилизованным на поверхности IL-31 собак и кошек. Еще три антитела демонстрировали такое свойство двойного связывания: ZIL69 (частичное связывание с собачьим), ZIL94 и ZIL159. Из этой группы из девяти полностью собачьих антител только ZIL1 и ZIL9 перекрестно реагировали с лошадиным IL-31.
Следует отметить, что антитело 15Н05 было единственным из всех проанализированных в данном документе антител, которое связывалось с собачьим, кошачьим и лошадиным IL-31, что указывает на некоторую степень консервативности эпитопа у трех биологических видов. Напротив, ни одно из описанных в данном документе антител не связывалось с человеческим IL-31. Для проверки данных ELISA, демонстрирующих различное связывание антител с кошачьим IL-31 дикого типа и с двумя белками с мутациями в эпитопах, распознаваемых 15Н05 (мутант 15Н05) или 11Е12 (мутант 11Е12), проводили дополнительные эксперименты biacore. Как и ожидалось, контрольное мышиное антитело 11Е12 связывалось с мутантом IL-31 15Н05 и не связывалось с мутантом IL-31 11Е12 из-за мутаций в эпитопе. Аналогично, мышиное 15Н05 не связывалось с мутантом 15Н05 и сохраняло связывание с мутантом IL-31 11Е12, что служило дополнительным различием отдельных связывающих эпитопов, распознаваемые этими двумя антителами. Мутация 15Н05 влияла на все полностью собачьи антитела, за исключением ZIL94, ZIL154 и ZIL171 (частичное влияние), что согласовывалось с результатами ELISA. Различия в результатах можно объяснить различиями в двух методиках анализа. Кроме того, было показано, что мутация 11Е12 влияет на связывание трех антител: ZIL1 (частичное влияние), ZIL8 и ZIL159. Эти результаты указывают на то, что на эпитоп, распознаваемый этими антителами, влияют изменения в обеих областях белка IL-31. В совокупности, эти результаты подтверждают характеристику девяти антител, происходящих из В-клеток собак, связывающихся с областью белка IL-31 кошки, которая распознается антителом 15Н05.
1.8. Фелинизация мышиных антител 11Е12 и 15Н05 и оптимизация аффинностей связывания
Образование антител к лекарствам (ADA) может быть связано с потерей эффективности любого биотерапевтического белка, включая моноклональные антитела. Тщательное изучение литературы показало, что модификация моноклональных антител с целью увеличения их сходства с естественными антителами того или иного биологического вида может снизить склонность mAb к иммуногенности, хотя можно найти примеры иммуногенных полностью человеческих mAb и неиммуногенных химерных mAb. Чтобы помочь снизить риски, связанные с образованием ADA к моноклональным антителам против IL-31, предложенным в настоящем документе, использовали стратегию фелинизации. Эта стратегия фелинизации основана на нахождении наиболее подходящей последовательности антител зародышевой линии кошек для пересадки CDR. После глубокого изучения всех доступных последовательностей зародышевой линии кошек как для вариабельной области тяжелой, так и легкой цепи, отобрали кандидатные зародышевые линии на основе их гомологии с мышиными mAb, и CDR из мышиных mAb-предшественников использовали для замены нативных кошачьих CDR. Задача состояла в том, чтобы сохранить высокую аффинность и активность в отношении клеток с использованием каркасных областей кошачьих антител, чтобы минимизировать возможную иммуногенность in vivo. Фелинизированные mAb экспрессировали и характеризовали по их аффинности к кошачьему IL-31 и их эффективности в исследованиях на клетках. В случае, если фелинизированное антитело теряет способность связывать IL-31, проводили системный анализ, чтобы идентифицировать: 1) цепь, ответственную за потерю функции, 2) каркасную область, ответственную за потерю функции, и 3) аминокислоту (аминокислоты), ответственную (ые) за потерю функции.
Создали синтетические нуклеотидные конструкции, представляющие фелинизированные вариабельные области тяжелой и легкой цепей mAb 11Е12 и 15Н05. После субклонирования каждой вариабельной области цепи в плазмиды, содержащие соответствующую тяжелую цепь кошачьего происхождения или константную область каппа, плазмидами котрансфицировали клетки HEK 293 для экспрессии антител. Первоначальные попытки фелинизации антитела 11Е12 были сосредоточены на использовании одной каркасной области VH кошачьего происхождения (SEQ ID NO: 111; FEL_11E12_VH1), соответствующей нуклеотидной последовательностью которой является (SEQ ID NO: 112; FEL_11E12_VH1), независимо спаренной с каркасными областями VL (SEQ ID NO: 113; FEL_11E12_VL1), соответствующей нуклеотидной последовательностью которой является (SEQ ID NO: 114; FEL_11E12_VL1), и (SEQ ID NO: 115; FEL_11E12_VL2), соответствующей нуклеотидной последовательностью которой является (SEQ ID NO: 116; FEL_11E12_VL2), с образованием Feline 11E12 1.1 и Feline 11E12 1.2, соответственно. Данная попытка модификации моноклональных антител с целью увеличения их сходства с естественными кошачьими антителами привела к потере аффинности mAb Feline 11Е12 1.1 как к кошачьим, так и к собачьим белкам IL-31 и к полной потере связывания mAb Feline 11Е12 1.2 по сравнению с мышиной формой антитела (Фиг. 2b). Эффективность этих антител, модифицированных с целью увеличения их сходства с естественными кошачьими антителами, проверяли в исследованиях на собачьих клетках DH82 и кошачьих клетках FCWF4 с использованием цитокина IL-31 кошки. Эффективность фелинизированного 11Е12 1.1 в отношении кошачьего IL-31 в исследовании на кошачьих FCWF была приблизительно в два раза ниже по сравнению с мышиной версией антитела. У этого антитела наблюдалась полная потеря эффективности в анализе на клетках, что согласовывалось с потерей аффинности фелинизированного 11Е12 1.2 (Фиг. 3). В попытке восстановить потерю аффинности при фелинизации, предприняли стратегию, аналогичную той, которую использовали в предыдущем эксперименте по канинизации ортолога mAb 11Е12 (патент US 8790651, Bammert, et al.). Для создания антитела Feline 11Е12 VL1 FW2 произвели замену фелинизированной каркасной области 2 (FW2) антитела Feline 11Е12 VL1 мышиной FW2 из (SEQ ID NO: 73; Mu_11Е12_VL), соответствующей нуклеотидной последовательностью которой является (SEQ ID NO: 74; Mu_11E12_VL). Кроме того, произвели одиночную замену в положении 46 кошачьей VL (K46Q) для получения (SEQ ID NO: 119; FEL_11E12_VL1_K46Q), соответствующей нуклеотидной последовательностью которой является (SEQ ID NO: 120; FEL_11Е12_VL1_K46Q). Спаривание вышеуказанных VL с Fel_11Е12_VH1 привело к получению Feline 11Е12 1.1 FW2 и Feline 11Е12 1.1 K46Q, соответственно. Изменение FW2 привело к восстановлению аффинности Feline 11Е12 1.1 FW2 к кошачьему белку IL-31 и KD, эквивалентной таковым мышиной и химерной форм (Фиг. 2А и 2 В). Однако, эти изменения негативно сказались на аффинности Feline 11Е12 1.1 FW2 к собачьему белку IL-31, что указывает на четкое различие в природе способности антитела 11Е12 связывать этот эпитоп на кошачьем и собачьем цитокинах. Единственная аминокислотная замена в Feline 11Е12 1.1 K46Q не смогла повлиять на аффинность этого антитела. Увеличение аффинности антитела 11Е12 1.1 FW2 к кошачьему белку IL-31 приводило к увеличению эффективности в отношении кошачьего цитокина в исследовании на собачьих клетках DH82 (Фиг. 3).
Усилия по фелинизации мышиного антитела 15Н05 были сосредоточены на комбинировании трех кошачьих каркасных областей VH с тремя кошачьих каркасными областями VL, с образованием в общей сложности 9 фелинизированных mAb. FEL_15Н05_VH1 (SEQ ID NO: 121; FEL_15H05_VH1), соответствующей нуклеотидной последовательностью которой является (SEQ ID NO: 122; FEL_15H05_VH1), комбинировали с (SEQ ID NO: 127; FEL_15H05_VL1), соответствующей нуклеотидной последовательностью которой является (SEQ ID NO: 128; FEL_15H05_VL1), (SEQ ID NO: 129; FEL_15H05_VL2), соответствующей нуклеотидной последовательностью которой является (SEQ ID NO: 130; FEL_15Н05_VL2), и (SEQ ID NO: 131; FEL_15H05_VL3), соответствующей нуклеотидной последовательностью которой является (SEQ ID NO: 132; FEL_15H05_VL3), с образованием Feline 15Н05 1.1, Feline 15Н05 1.2 и Feline 15Н05 1.3, соответственно. FEL_15Н05_VH2 (SEQ ID NO: 123; FEL_15H05_VH2), соответствующей нуклеотидной последовательностью которой является (SEQ ID NO: 124; FEL_15H05_VH2), комбинировали с (SEQ ID NO: 127; FEL_15H05_VL1), соответствующей нуклеотидной последовательностью которой является (SEQ ID NO: 128; FEL_15H05_VL1), (SEQ ID NO: 129; FEL_15H05_VL2), соответствующей нуклеотидной последовательностью которой является (SEQ ID NO: 130; FEL_15H05_VL2), и (SEQ ID NO: 131; FEL_15H05_VL3), соответствующей нуклеотидной последовательностью которой является (SEQ ID NO: 132; FEL_15H05_VL3), с образованием Feline 15Н05 2.1, Feline 15H05 2.2 и Feline 15H05 2.3, соответственно. FEL_15H05_VH3 (SEQ ID NO: 125; FEL_15H05_VH3), соответствующей нуклеотидной последовательностью которой является (SEQ ID NO: 126; FEL_15H05_VH3), коибинировали с (SEQ ID NO: 127; FEL_15H05_VL1), соответствующей нуклеотидной последовательностью которой является (SEQ ID NO: 128; FEL_15H05_VL1), (SEQ ID NO: 129; FEL_l5H05_VL2), соответствующей нуклеотидной последовательностью которой является (SEQ ID NO: 130; FEL_15H05_VL2), и (SEQ ID NO: 131; FEL_15H05_VL3), соответствующей нуклеотидной последовательностью которой является (SEQ ID NO: 132; FEL_15H05_VL3), с образованием Feline 15Н05 3.1, Feline 15H05 3.2 и Feline 15H05 3.3, соответственно. Аналогично результатам с антителом 11Е12, первая попытка фелинизации антитела 15Н05 привела к потере аффинности к кошачьему белку IL-31 по сравнению с мышиным 15Н05 и нейтральному эффекту по сравнению с химерным мышиным:кошачьим антителом 15Н05 (Фиг. 2А и 2С). Аналогично результатам связывания фелинизированного антитела 11Е12 с собачьим IL-31, некоторые комбинации каркасных областей VH и VL кошачьего 15Н05 оказывали нейтральное или положительное влияние на аффинность к собачьему IL-31 (см. Фиг. 2С Feline 15Н05 1.1, 2.2 и 3.2).
В попытке восстановить аффинность фелинизированного антитела 15Н05 каждую фелинизированную VH 15Н05 объединяли в пары с мышиной VL 15Н05 с образованием гетерохимерных антител. FEL_15H05_VH1 (SEQ ID NO: 121; FEL_15Н05_VH1), соответствующей нуклеотидной последовательностью которой является (SEQ ID NO: 122; FEL_15Н05_VH1), комбинировали с MU_15Н05_VL (SEQ ID NO: 69; MU_15H05_VL), соответствующей нуклеотидной последовательностью которой является (SEQ ID NO: 70; MU_15H05_VL), с образованием Feline 15H05 VH1 mouse VL. FEL_15H05_VH2 (SEQ ID NO: 123; FEL_15H05_VH2), соответствующей нуклеотидной последовательностью которой является (SEQ ID NO: 124; FEL_15H05_VH2), комбинировали с MU_15H05_VL (SEQ ID NO: 69; MU_15H05_VL), соответствующей нуклеотидной последовательностью которой является (SEQ ID NO: 70; MU_15H05_VL), с образованием Feline 15Н05 VH2 mouse VL. FEL_15H05_VH3 (SEQ ID NO: 125; FEL_15H05_VH3), соответствующей нуклеотидной последовательностью которой является (SEQ ID NO: 126; FEL_15H05_VH3), комбинировали с MU_15H05_VL (SEQ ID NO: 69; MU_15H05_VL), соответствующей нуклеотидной последовательностью которой является (SEQ ID NO: 70; MU_15H05_VL), с образованием Feline 15Н05 VH3 mouse VL. Анализировали аффинность этих гетерохимерных антител с фелинизированными VH и мышиными VL к собачьему и кошачьему IL-31. Объединение в пары фелинизированных VH1 и VH3 15Н05 с мышиными VL 15Н05 восстанавливало аффинность к кошачьему IL-31 до эквивалентной или более высокой, чем у мышиной и химерной форм. Эта тенденция к повышению аффинности наблюдалась также по отношению к собачьему белку IL-31 (Фиг. 2А и 2С).
Для дальнейшего анализа положений в каркасных областях 15Н05, ответственных за потерю аффинности, одну фелинизированную VH из 15Н05 (FEL_15H05_VH1) объединяли в пары с мышиной VL из 15Н05 с отдельными заменами в каркасной области. FEL_15H05_VH1 (SEQ ID NO: 122; FEL_15H05_VH1), соответствующей нуклеотидной последовательностью которой является (SEQ ID NO: 123; FEL_15Н05_VH1), комбинировали независимо с FEL_15Н05_VL1_FW1 (SEQ ID NO: 133; FEL_15H05_VL1_FW1), соответствующей нуклеотидной последовательностью которой является (SEQ ID NO: 134; FEL_15H05_VL1_FW1), FEL_15H05_VL1_FW2 (SEQ ID NO: 135; FEL_15H05_VL1_FW2) соответствующей нуклеотидной последовательностью которой является (SEQ ID NO: 136; FEL_15H05_VL1_FW2), и FEL_15H05_VL1_FW3 (SEQ ID NO: 137; FEL_15H05_VL1_FW3), соответствующей нуклеотидной последовательностью которой является (SEQ ID NO: 138; FEL_15H05_VL1_FW3), с образованием Feline 15H05 1.1 FW1, Feline 15H05 1.1 FW2 и Feline 15H05 1.1 FW3, соответственно. Замена мышиной FW1 15Н05 на Feline 15Н05 1.1 негативно сказывалась на аффинности как к кошачьему, так и к собачьему IL-31, однако, когда мышиную FW2 или FW3 заменяли на Feline 15Н05 1.1, достигалась отличная аффинность к собачьему и кошачьему IL-31, при этом FW2 демонстрировала превосходящие свойства в отношении обоих биологических видов (Фиг. 2С). Для определения степени изменения аффинности с помощью этого подхода осуществляли дополнительные попарные замены каркасных областей. FEL_15H05_VH1 (SEQ ID NO: 121; FEL_15H05_VH1), соответствующей нуклеотидной последовательностью которой является (SEQ ID NO: 122; FEL_15H05_VH1), комбинировали независимо с FEL_15H05_VL1_FW1_2 (SEQ ID NO: 139; FEL_15H05_VL1_FW1_FW2), соответствующей нуклеотидной последовательностью которой является (SEQ ID NO: 140; FEL_15H05_VL1_FW1_FW2), FEL_15H05_VL1_FW2_3 (SEQ ID NO: 143; FEL_15H05_VL1_FW2_FW3), соответствующей нуклеотидной последовательностью которой является (SEQ ID NO: 144; FEL_15H05_VL1_FW2_FW3), и FEL_15H05_VL1_FW1_3 (SEQ ID NO: 141; FEL_15H05_VL1_FW1_FW3), соответствующей нуклеотидной последовательностью которой является (SEQ ID NO: 142; FEL_15H05_VL1_FW1_FW3), с образованием Feline 15H05 1.1 FW1_2, Feline 15H05 1.1 FW2_3 и Feline 15H05 1.1 FW1_3, соответственно. Интересно, что замена одной мышиной FW1 негативно сказывалась на аффинности, тогда как результатом комбинации FW1 с FW2 или FW3 была хорошая аффинность как к кошачьему, так и к собачьему IL-31 (Фиг. 2С).
Наконец, была предпринята попытка минимизировать количество обратных мутаций в кошачьих каркасных областях, начиная с наиболее перспективных комбинаций фелинизированных последовательностей VH и VL. Для этого FEL_15Н05_VH1 (SEQ ID NO: 121; FEL_15Н05_VH1), соответствующей нуклеотидной последовательностью которой является (SEQ ID NO: 122; FEL_15Н05_VH1), комбинировали независимо с FEL_15H05_VL1_FW2_K42N (SEQ ID NO: 145; FEL_15H05_VL1_FW2_K42N), соответствующей нуклеотидной последовательностью которой является (SEQ ID NO: 146; FEL_15H05_VL1_FW2_K42N), FEL_15H05_VL1_FW2_V43I (SEQ ID NO: 147; FEL_15H05_VL1_FW2_V43I), соответствующей нуклеотидной последовательностью которой является (SEQ ID NO: 148; FEL_15H05_VL1_FW2_V43I), FEL_15H05_VL1_FW2_L46V (SEQ ID NO: 149; FEL_15Н05_VL1_FW2_L46V), соответствующей нуклеотидной последовательностью которой является (SEQ ID NO: 150; FEL_15Н05_VL1_FW2_L46V), FEL_15H05_VL1_FW2_Y49N (SEQ ID NO: 151; FEL_15H05_VL1_FW2_Y49N), соответствующей нуклеотидной последовательностью которой является (SEQ ID NO: 152; FEL_15H05_VL1_FW2_Y49N) и FEL_15H05_VL1_FW2_K42N_V43I (SEQ ID NO: 153; FEL_15H05_VL1_FW2_K42N_V43I), соответствующей нуклеотидной последовательностью которой является (SEQ ID NO: 154; FEL_15H05_VL1_FW2_K42N_V43I), с образованием Feline 15H05 1.1 K42N, Feline 15H05 1.1 V43I, Feline 15H05 1.1 L46V, Feline 15H05 1.1 Y49N и Feline 15H05 1.1 K42N_V43I, соответственно. В то время как замена всей мышиной каркасной области FW2 на таковую Felinized 15Н05 VL1 привела к получению антитела с прекрасной аффинностью к собачьему и кошачьему IL-31 (Фиг. 2С, Feline 15Н05 1.1 FW2), отдельные обратные мутации аминокислотных остатков FW2 имели нейтральный или негативный эффект, указывающий на то, что все 4 замены необходимы для поддержания оптимальной третичной структуры для позиционирования CDR на эпитопе IL-31. Благодаря повышенной аффинности фелинизированного 15Н05 1.1 FW2 к кошачьему и собачьему IL-31 это антитело отобрали для дальнейшей работы.
На Фиг. 5А показано выравнивание последовательности VL мышиного антитела 11Е12, где ранее указанная канинизированная последовательность 11Е12 сравнивается с фелинизированными версиями. Под выравниванием точками отмечены положения соответствующих изменений в Fel 11Е12 VL1, которые были необходимы для восстановления аффинности этого антитела к белку IL-31. Аналогичным образом на Фиг. 5В показаны необходимые изменения фелинизированной VL 15Н05 (Fel_15H05_VL1), которые требовались не только для восстановления, но и для улучшения его аффинности к собачьему и кошачьему IL-31 по сравнению с мышиной и химерными формами этого антитела.
1.9. Получение клеточных линий, экспрессирующих фелинизированные антитела против IL-31 из плазмид, кодирующих глутаминсинтетазу (GS)
В качестве кандидата для создания стабильных клеточных линий, которые будут производить гомогенные антитела для дальнейшей характеристики выбрали фелинизированное 15Н05 1.1 FW2. Гены, кодирующие фелинизированные тяжелые и легкие цепи для продукции клеточных линий, клонировали в плазмиды GS рЕЕ 6.4 и рЕЕ 12.4, соответственно (Lonza, Базель, Швейцария). Полученные плазмиды расщепляли в соответствии с протоколом производителя и лигировали вместе с образованием единой плазмиды для экспрессии в клетках млекопитающих. Для ZTS-927 тяжелую цепь (SEQ ID NO: 121; FEL_15H05_VH1), соответствующей нуклеотидной последовательностью для которой является (SEQ ID NO: 122; FEL_15Н05_VH1), комбинировали с константой областью тяжелой цепи IgG кошки (SEQ ID NO: 171; Feline_НС_AlleleA_wt), соответствующей нуклеотидной последовательностью для которой является (SEQ ID NO: 172; Feline_HC_AlleleA_wt). Для ZTS-927 тяжелую цепь (SEQ ID NO: 135; FEL-15H05-VL1_FW2), соответствующей нуклеотидной последовательностью для которой является (SEQ ID NO: 136; FEL-15H05-VL1_FW2), комбинировали с константой областью легкой цепи IgG кошки (SEQ ID NO: 175; Feline LC Kappa G minus), соответствующей нуклеотидной последовательностью для которой является (SEQ ID NO: 176; Feline_LC_Kappa_G_minus). Для ZTS-927 тяжелую цепь (SEQ ID NO: 121; FEL_15H05_VH1), соответствующей нуклеотидной последовательностью для которой является (SEQ ID NO: 122; FEL_15Н05_VH1), комбинировали с константой областью тяжелой цепи IgG кошки (SEQ ID NO: 173; Feline_НС_AlleleA_1), соответствующей нуклеотидной последовательностью для которой является (SEQ ID NO: 174; Feline_HC_AlleleA_1). Для ZTS-927 тяжелую цепь (SEQ ID NO: 135; FEL-15H05-VL1_FW2), соответствующей нуклеотидной последовательностью для которой является (SEQ ID NO: 136; FEL-15H05-VL1_FW2), комбинировали с константой областью легкой цепи IgG кошки (SEQ ID NO: 175; Feline_LC_Kappa_G_minus), соответствующей нуклеотидной последовательностью для которой является (SEQ ID NO: 176; Feline_LC_Kappa_G_minus).
Чтобы продемонстрировать транзиторную продукцию антитела, каждую плазмиду использовали для трансфекции клеток HEK 293, и осуществляли экспрессию в культурах различного масштаба. Белок выделяли из кондиционированной среды HEK с использованием аффинной хроматографии с белком А в соответствии со стандартными методами очистки белка. Среду наносили на хроматографическую смолу и элюировали путем изменения рН. Перед использованием доводили рН элюированного белка, диализовали и стерилизовали фильтрованием. ZTS-361 впоследствии использовали для оценки на модели кошачьего зуда для оценки эффективности in vivo. Исследовали аффинность и эффективность антител, полученных из одной плазмиды GS, ZTS-927 и ZTS-361. На Фиг. 2D показаны результаты оценки аффинности этих антител с использованием biacore. Аффинность ZTS-927 и ZTS-361 к кошачьему IL-31 в высокой степени согласуется с аффинностью мышиной и химерной форм родительского мышиного mAb 15Н05. Определяли эффективность этих двух антител против собачьего и кошачьего IL-31 тестах с использованием как собачьих, так и кошачьих клеток (Фиг. 3). В соответствии с предыдущими результатами, значения IC50 были пропорционально выше при использовании собачьей формы IL-31 с обоими типами клеток. Значения IC50 для ZTS-927 и ZTS-361 в отношении кошачьего IL-31 также в высокой степени согласовались со значениями, полученными для химерной и мышиной форм антитела, что указывало на то, что конечная фелинизированная версия mAb 15Н05, продуцированная из одной плазмиды GS, была пригодна для создания клеточной линии.
Для создания стабильной клеточной линии, продуцирующей антитела-кандидаты, плазмиду GS линеаризовали перед трансфекцией рестрикционным ферментом Pvul, который разрезает один сайт в остове плазмиды. Клетки GS-CHOK1SV (клон 144Е12) трансфицировали линеаризованной плазмидной ДНК посредством электропорации. После трансфекции клетки помещали в 48-луночные планшеты (48WP) для создания стабильных пулов. Когда пулы в 48WP достигали конфлюентности по меньшей мере 50%, анализировали экспрессию IgG в 100 мкл супернатанта с использованием биосенсоров ForteBio Octet и protein A (Pall ForteBio, Fremont, CA). Клоны с наилучшей экспрессией помещали в 6-луночные планшеты (6 WP), а затем во встряхиваемые колбы (SF) объемом 125 мл. После того, как клетки адаптировались к культивированию в суспензии в колбах объемом 125 мл, 2 флакона каждого пула клеточной линии помещали в банк для хранения в жидком азоте. Поскольку продуцирующие клеточные линии должны происходить от одного клона, 3 пула с наилучшей экспрессией субклонировали методом предельного разведения в 96-луночных культуральных планшетах. Чтобы подтвердить происхождение от одного клона и избежать второго раунда предельного разведения, осуществляли визуализацию 96-луночных планшетов с помощью Molecular Devices Clone-Select Imager (CSI) (Molecular Devices LLC, San Jose, CA), который фиксирует изображения отдельных клеток и их последующий рост. Клоны отбирали на основе изображений CSI, роста и продуктивности в 96WP.
Чтобы оценить рост и продуктивность клеточной культуры, пулы с максимальной экспрессией дополнительно оценивали в 14 культуре с подпиткой в 125 мл SF. Клетки высевали в среду представляющую собой среду CD СНО компании Life Technologies с добавлением 4 аминокислот в виде добавки собственного производства CDF v6.2 и 10% глюкозы. После 14-дневной культуры с подпиткой пулы центрифугировали и тАВ, продуцируемые CD СНО, выделяли фильтрованием супернатанта через мембрану из полиэфирсульфона (PES) с размером пор 0,20 мкм с последующей очисткой.
Типичная очистка включает загрузку двух литров кондиционированной среды (из культуры клеток СНО, профильтрованной через фильтр с диаметром пор 0,2 мкм) в колонку с MabSelect объемом 235 мл (GE healthcare, каталожный номер 17-5199-02). Колонка была предварительно уравновешена PBS. Время удерживания загружаемого образца составляло более 2,5 минут. После загрузки колонку снова промывали PBS, а затем 25 мМ ацетатом натрия при нейтральных значениях рН. Колонку элюировали 25 мМ уксусной кислотой, рН 3,6, а затем десорбировали 250 мМ уксусной кислоты, 250 мМ хлорида натрия, рН приблизительно 2,2. Фракции (50 мл) собирали во время стадий элюирования и десорбирования. На всех стадиях определяли УФ-поглощение при А280. Пиковые фракции объединяли, рН доводили до примерно 5,5 добавлением 20 мМ ацетата натрия, а затем диализовали путем трех замен буфера. Диализат собирали, стерилизовали фильтрованием и хранили при 4°С.
1.10. Идентификация эпитопа на IL-31, распознаваемого антителом 15Н05
Изучение эпитопа IL-31, который распознается антителом, имеет решающее значение для понимания механизма, с помощью которого оно нейтрализует связывание цитокина с корецептором IL-31 Ra: OSMR. Кроме того, изучение эпитопа позволяет оптимизировать аффинность связывания антител и создание миметиков пептидных эпитопов (мимотопов), которые могут иметь широкое применение в качестве аналитических реагентов для захвата и в качестве субъединичных вакцин для индукции соответствующего целенаправленного иммунного ответа (но не ограничивается этим). Многоступенчатый процесс с использованием технологии CLIPS (химическое присоединение пептидов к каркасам) (Timmerman et al. J Mol Recognit. 2007; 20(5): 283-299) использовали для идентификации и оптимизации пептида, способного связываться с паратопом mAb 15Н05 (Pepscan, Lelystad Netherlands). Аффинность mAb 15H05 как к собачьим, так и к кошачьим белкам IL-31 является высокой (Фиг. 2, MU-15H05), поэтому первичная последовательность IL-31 обоих биологических видов сочли подходящей для решения этих задач. Для идентификации пептидов, способных связывать mAb 15Н05, создали пептидную библиотеку, репрезентирующую собачий белок IL-31, и использовали в непрямом ELISA. После идентификации пептидов, первичные аминокислотные последовательности которых представляют область связывания mAb 15Н05 на IL-31, провели целенаправленный анализ с полной заменой аминокислот в пептидах, представляющих сегмент IL-31, заменяя каждую из 12 аминокислот в этой связывающейся с mAb 15Н05 области 19-ю другими возможными аминокислотными остатками. Этот анализ был необходим для идентификации ключевых аминокислотных остатков IL-31, участвующих в связывании mAb 15Н05, а также показал, где замены в первичной последовательности собачьего происхождения приводили к усилению связывания антител.
Аминокислоты собачьего белка IL-31, которые распознаются антителом 11Е12, были описаны ранее (патент US 8790651, Bammert, et al.). В этом документе описан мутационный анализ собачьего белка IL-31, показывающий положения на собачьем белке IL-31, в которых замены на аланин влияют на связывание mAb 11Е12. На основании анализа с полной заменой, описанного выше для mAb 15Н05, и имеющихся сведений о связывающем эпитопе 11Е12, создали мутантные формы кошачьего белка IL-31 путем замены аланином двух ключевых остатков в эпитопе, распознаваемом каждым антителом (мутанты описано в разделе 1.2 выше). Мутации каждого эпитопа обозначили по названию антитела, которое распознает сайт мутации (мутант 11Е12 и 15Н05 в отличие от нативной последовательности белка дикого типа).
На Фиг. 6А показано выравнивание кошачьего IL-31 дикого типа (SEQ ID NO: 157) с мутантами 15Н05 (SEQ ID NO: 163) и 11Е12 (SEQ ID NO: 161), выделены положения, в которых произведены замены на аланин. IL-31 принадлежит к семейству цитокинов IL-6 с четырьмя антипараллельными пучками спиралей, имеющими архитектуру типа «вверх и вниз» (база данных CATH, Dawson et al., 2017 Nucleic Acids Res. 2017 Jan 4; 45 (Database issue): D289-D295). На основе гомологиии со структурой 1Р9М человеческого IL-6 создали модель (Boulanger et al. 2003 Science. Jun 27; 300(5628):2101-4) с использованием программного обеспечения МОЕ (Chemical Computing Group, Montreal, QC, Canada). На Фиг. 6B показана построенная на основе гомологии модель кошачьего IL-31, на которой выделены положения аминокислот, участвующих в связывании антител 11Е12 (сайт 1) и 15Н05 (сайт 2). Сайты связывания для каждого антитела, по-видимому, расположены в отдельных положениях белка IL-31.
Чтобы определить влияние способности mAb 11Е12 и 15Н05 связывать эти мутантные формы кошачьего IL-31, провели непрямой ELISA с использованием мутантов, непосредственно нанесенных на планшет для иммуноанализа. На Фиг. 6С показаны результаты этого ELISA, демонстрирующие, что mAb 11Е12 и 15Н05 способны связываться с кошачьим IL-31 дикого типа в анализе такого формата. Когда для захвата использовали мутант 11Е12, связывание mAb 11Е12 сильно ослаблялось, а связывание mAb 15Н05 частично ослаблялось. Предыдущий анализ эпитопа 11Е12 на собачьем IL-31 (описанный в патенте US 8790651, выданный Bammert et al.) показал, что при мутации на аланин на связывание mAb влияют 4 аминокислотных остатка, поэтому в данном случае мутация 2 остатков может быть недостаточной для полного устранения высокоаффинного связывания mAb 11Е12 в таком формате ELISA. Незначительное ослабление связывания mAb 15Н05 с мутантом 11Е12, вероятно, связано с трансляционными эффектами мутаций из-за движения двух передних спиралей, воздействующих на сайт связывания 15Н05. Мутации, разработанные для воздействия на связывание mAb 15Н05 (мутант 15Н05), показывают полную потерю способности mAb 1505 связывать этот мутант IL-31 в ELISA. В отличие от мутанта 11Е12, изменения в случайной спирали, распознаваемой mAb 15Н05 (мутант 15Н05), не влияли на связывание mAb 11Е12, что дополнительно подтверждало различие между двумя эпитопами (Фиг. 6С).
1.11. Оценка mAb 15Н05 и 11Е12 методом конкурентного связывания с использованием biacore
Чтобы дополнительно охарактеризовать эпитопы IL-31, связанные с mAb 15Н05 и 11Е12, проводили эксперименты с блокированием с использованием biacore, иммобилизуя на поверхности чипа белок IL-31 и затем последовательно пропуская антитела. На Фиг. 7 показано относительное связывание каждого антитела с IL-31 после захвата 11Е12 или 15Н05. Столбцы, обозначенные HBS-EP (буфер для анализа), указывают на максимальный сигнал, полученный при связывании каждого антитела с иммобилизованным на поверхности IL-31 в отсутствие конкуренции. На Фиг. 7А показаны результаты конкурентного связывания мышиных антител 15Н05 и 11Е12 с собачьим IL-31. Эти результаты ясно показывают, что антитела 15Н05 и 11Е12 способны связываться с собачьим IL-31 в присутствии друг друга, что указывает на то, что они распознают различные эпитопы на белке. Сенсограммы, приведенные на Фиг. 7А, показывают, что оба антитела очень медленно диссоциируют с этой новообразованной поверхностью чипа biacore, поэтому при добавлении того же антитела дополнительные сайты связывания не могут быть заняты (данные не показаны).
На Фиг. 7 В показаны результаты конкурентного связывания антител 15Н05 и 11Е12 с иммобилизованным на поверхности кошачьим IL-31, вновь демонстрирующие, что распознаваемые ими эпитопы не перекрываются. Связывание дополнительных антител в присутствии того же самого антитела является результатом увеличения скорости диссоциации из-за более низкого качества используемой поверхности. Наблюдалось увеличение скорости диссоциации, см. значения KD на новообразованных поверхностях с иммобилизованным кошачьим IL-31, приведенные на Фиг. 2.
Эти результаты дополнительно подтверждают данные картирования эпитопа в разделе 1.10, указывающие на то, что CDR, содержащиеся в антителе MU-15H05, распознают иной эпитоп нежели MU-11E12. Эпитоп, распознаваемый антителом 15Н05, отличается от распознаваемого антителом 11Е12, описанным в патенте US 8790651 (Bammert, et al.), и является новой мишенью на белке IL-31 для нейтрализации активности этого цитокина у многих биологических видов. Эти данные подчеркивают различное пространственное расположение сайтов связывания, описанное построенной на основе гомологии моделью кошачьего IL-31 (Фиг. 6 В), и подтверждают гипотезу о том, что эта сторона цитокина является ключевой для взаимодействия с рецепторным комплексом IL-31 Ra:OSMR.
1.12. Синтез и характеристика растворимого кошачьего корецептора IL-31 (IL-31 RA и OSMR)
Было показано, что гетеромерный рецепторный комплекс человеческого IL-31, состоящий из субъединиц IL31Ra и OSMR, необходим для опосредованной IL-31 межклеточной активации пути JAK-STAT и вовлечен в атопические заболевания кожи (Dillon et al. 2004 Nat Immunol. Jul; 5(7):752-60, Dreuw et al. 2004 J Biol Chem. 279:36112-36120; and Diveu et al. 2004 Eur Cytokine Netw. 15:291-302). Позднее было описано, что субъединица IL-31 Ra человека опосредует первоначальное связывание, которое происходит, когда IL-31 находится в контакте с рецепторами клеточной поверхности, и это событие является необходимым условием для привлечения OSMR с последующим образованием высокоаффинного корецепторного комплекса (Le Saux et al., 2010 J Biol Chem. Jan 29;285(5):3470-7). В данном документе авторы изобретения приводят доказательства того, что кошачий белок IL-31 способен независимо связываться как с OSMR, так и с IL-31 Ra. Эти данные являются новыми и имеют важное значение для понимания того, как белок IL-31 взаимодействует с корецептором IL-31Ra:OSMR, а также для биологической роли IL-31, поскольку он независимо взаимодействует с отдельными субъединицами.
Чтобы понять, как IL-31 связывается со своим корецептором, и охарактеризовать ингибирующие свойства идентифицированных антител, синтезировали две формы рецептора. Обе отдельные субъединицы рецептора IL-31, IL-31Ra (SEQ ID NO: 169; Feline_IL31Ra_HIgG1_Fc_X1_Fn3), соответствующей нуклеотидной последовательностью которой является (SEQ ID NO: 170; Feline_IL31Ra_HIgG1_Fc_X1_Fn3) и субъединица OSMR (SEQ ID NO: 167; Feline_OSMR_hIgG1_Fc), соответствующей нуклеотидной последовательностью которой является (SEQ ID NO: 168; Feline_OSMR_hIgG1_Fc), были сконструированы в слитой с Fc IgG1 человека форме. По гомологии с человеческими гомологами, идентифицировали связывающий цитокины домен, фибронектин-подобный домен III типа и Ig-подобный домен. Для оценки индивидуальных субъединиц рецептора внеклеточные домены OSMR и IL-31Ra (с его ожидаемым N-концевым проксимальным фибронектин-подобным доменом III типа) были созданы в слитой с Fc IgG1 человека форме, оба с использованием их естественных сигнальных пептидов. Все синтетические кассеты клонировали в pcDNA3.1, экспрессировали в системе ExpiCHO и очищали, как описано выше.
Чтобы проанализировать способность этих форм рецепторов связывать белок IL-31 дикого типа и мутантные белки IL-31, выполняли непрямой ELISA путем нанесения 100 мкл каждого соответствующего белка на планшет immulon 2НВ (1 мкг/мл) в карбонатном/бикарбонатном буфере (sigma С3041-100САР) и инкубировали при 4°С в течение ночи. Затем планшеты для ELISA блокировали 5% блокирующим буфером NFDM в PBST в течение 1 часа при комнатной температуре с последующим связыванием каждой рецепторной конструкции в нескольких концентрациях при комнатной температуре в течение 1 часа. После промывания PBST присутствие связанного рецептора (слитого с Fc) определяли путем инкубации с мышиным антителом против человеческого IgG1 (Lifetech А10684, разведение 1:500) в течение 1 часа при комнатной температуре. Лунки снова промывали PBST и проявляли субстратом для микролунок KPL sureblue 3,3',5,5'-тетра-метилбензидин (ТМВ). На Фиг. 8 показаны результаты этого непрямого ELISA с использованием для захвата кошачьего белка IL-31 дикого типа и мутантных форм. Эти данные демонстрируют способность кошачьего IL-31 дикого типа независимо связываться с субъединицами рецептора IL-31Ra и OSMR. Эти данные контрастируют с предыдущими сообщениями, показывающими, что белок IL-31 первоначально связывается с субъединицей IL-31Ra и далее рекрутирует в этот сайт OSMR. Поскольку биологическая роль IL-31 еще не определена, очень важно понять динамику связывания рецептора и возможные последствия ослабления его функции при таких заболеваниях, как атопический дерматит. По этой причине эти данные также принимали во внимание при характеристике связывания антител с эпитопами, способными нарушать способность IL-31 распознавать IL-31Ra и OSMR.
В разделе 1.2 описано ослабленное связывание антител 11Е12 и 15Н05 с мутантами, у которых ключевые аминокислоты в их сайтах связывания преобразованы в аланин (мутанты 11Е12 и 15Н05, соответственно). Поэтому представляло большой интерес понять влияние этих мутаций на способность связываться с отдельными субъединицами рецептора IL-31Ra и OSMR. На Фиг. 8 показано, что мутация в сайте связывания 11Е12 или 15Н05 полностью нарушает способность к связыванию IL-31 Ra и OSMR, что указывает на то, что оба антитела связывают эпитопы, которые необходимы для взаимодействия IL-31 с обеими субъединицами рецептора. Отсутствие связывания также может быть следствием изменений в конформации IL-31 в результате мутации, однако эти мутанты все еще способны связываться с антителом, что предполагает, что это не так. Этот ключевой факт подтверждает способность обоих антител 11Е12 и 15Н05 (и их производных) распознавать эпитопы на IL-31, которые нейтрализуют способность цитокинов передавать сигнал через его корецептор и дополнительно блокировать связывание цитокина с любым клеточным рецептором в ходе этого процесса. Эти данные подтверждают, что найдены антитела, которые способны удалять IL-31 из кровотока и делать его неспособным связываться с находящимися на поверхности клетки или растворимыми формами рецепторов.
1.13. Оценка химерных антител на модели с провокацией зуда in vivo с помощью кошачьего IL-31
Способность антитела эффективно нейтрализовать свою мишень можно оценивать in vitro путем исследования связывания с соответствующим эпитопом на белке-мишени с соответствующей аффинностью и эффективностью в анализах на клетках, которые можно экстраполировать на эффективность in vivo. Выше описаны шаги, предпринятые для характеристики двух серий антител, полученных из мышиных mAb-предшественников 11Е12 и 15Н05. В разделе 1.7 описано создание химерных мышиных:кошачьих форм mAb 11Е12 и 15Н05, аффинность которых к собачьему и кошачьему IL-31 сопоставима с исходным мышиным моноклональным антителом (Фиг. 2А). Химерные мышиные:кошачьи формы 11Е12 и 15Н05 также имели сопоставимые значения IC50, свидетельствующие об ингибировании индуцированной кошачьим IL-31 передачи сигналов pSTAT3 в макрофагальных клетках собак и кошек (Фиг. 3). Во время процесса фелинизации, описанного в разделе 1.8, мышиный mAb 11Е12 преобразовали в фелинизированную версию (Feline 11Е12 1.1), приведшую к потере аффинности к собачьему и кошачьему IL-31 (Фиг. 3) и потере эффективности против передачи сигналов кошачьим IL-31 в клетках собак и кошек (Фиг. 3). Перед оптимизацией фелинизированных антител 11Е12 и 15Н05, описанной в разделе 1.8, было интересно понять способность этих предварительно фелинизированных и химерных форм нейтрализовать вызывающую зуд активность кошачьего IL-31 в модели на кошках. Представлял интерес фармакодинамический эффект этих различных антител на нейтрализацию зуда и понимание какой-либо корреляции с аффинностью, эффективностью в анализах на клетках или распознаванием эпитопа, которые могут повлиять на эффективность. В будущем, дальнейшую оптимизацию можно осуществлять, опираясь на диапазон эффективности в исследованиях на клетках in vitro, коррелирующая с эффективностью in vivo в модели с провокацией зуда.
Чтобы протестировать предварительную эффективность мышиной:кошачьей химеры 11Е12, мышиной:кошачьей химеры 15Н05 и фелинизированного 11Е12 (Feline 11Е12 1.1), разработали модель индуцированного IL-31 зуда у кошек. После внутривенного введения кошачьего IL-31 в дозе 0,5 мкг/кг (SEQ ID NO: 159; Feline_IL-31_Е_coli), соответствующей нуклеотидной последовательностью для которого является (SEQ ID NO: 160; Feline_IL-31_Е_coli), у кошек транзиторно наблюдалось поведение, сопряженное с зудом, включающее облизывание, жевание, царапание и потрясывание головой или телом, без ограничения. Трение о клетку не считалось активностью, сопряженной с зудом. Наблюдения за зудом проводились обученным исследователем за 30 минут до введения белка IL-31 и в течение 1 часа после. В этом исследовании первоначальную провокацию кошачьим IL-31 проводили за 1 месяц до введения антитела. В день 0 антитело в дозе 0,5 мг/кг объединяли с 0,5 мкг/кг кошачьего IL-31 при комнатной температуре за 60 минут до введения каждому животному предварительно связанного комплекса. В качестве контроля включали контроль «без mAb». При такой дозе mAb антитело находится в молярном избытке к цитокину. Сопряженную с зудом активность контролировали, как описано, на 0, 7 и 21 дни. Результаты на Фиг. 9 показывают значительное улучшение (р менее 0,05) оценки зуда при использовании химерного мышиного:кошачьего mAb 15Н05 на дни 0, 7 и 21 по сравнению с плацебо, служившего контролем. Хотя мышиное:кошачье химерное антитело 11Е12 изначально продемонстрировало тенденцию к эффективности в день 0, достоверного уменьшения зуд в какой-либо момент времени по сравнению с плацебо не достигалось. Кошачье 11Е12 1.1 не уменьшало зуд в день 0 и не демонстрировало тенденции к эффективности по сравнению с плацебо, поэтому дальнейшие провокации IL-31 на 7 и 21 дни не выполняли.
В совокупности эти результаты демонстрируют четкое различие между активностями этих антител при отсутствии эффективности кошачьего 11Е12 1.1 в предотвращении сопряженного с зудом поведения кошек, вызванного IL-31. Потеря аффинности и эффективности кошачьего 11Е12 1.1, вероятно, привела к недостаточной эффективности in vivo. При сравнении результатов эффективности мышиной:кошачьей химерной формы 11Е12 и мышиной:кошачьей химерной формы 15Н05 различие было менее выраженным. Химерные формы обоих mAb имели значения KD, сопоставимые с их мышиным предшественником, при этом аффинность мышиной:кошачьей формы 11Е12 в отношении как кошачьего, так и собачьего IL-31 была несколько выше (Фиг. 2А). Тем не менее, эта более высокая аффинность не приводила непосредственно к увеличению эффективности, поскольку IC50 в отношении индуцированной кошачьим IL-31 передачи сигнала pSTAT3 в клетках FCWF4 кошек у химерной мышиной:кошачьей формы 15Н05 была приблизительно в два раза выше, чем у химерой мышиной: кошачьей формы 11Е12 (Фиг. 3). Эти данные позволяют предположить, что способ, которым CDR антитела 15Н05 распознают кошачий IL-31, позволяет лучше нейтрализовать передачу цитокинами сигнала через свой корецептор, что, в свою очередь, делает его более эффективным в блокировании зуда у кошек. Различия в IC50, наблюдаемые в этих анализах на клетках, представляются перспективными для прогнозирования эффективности in vivo и для выявления незначительных различий в узнавании эпитопа антителами как одной, так и разных серий.
1.14. Оценка эффективности фелинизированного антитела 15Н05 против IL-31 на модели с провокацией зуда у кошек in vivo
На основании положительных результатов, полученных в отношении эффективности химерной мышиной: кошачьей формы 15Н05, описанных выше, проводили дополнительные эксперименты по увеличению аффинности и эффективности фелинизированного 15Н05 (описанного выше в разделе 1.8). Систематическая замена каркасных участков вариабельной области легкой цепи в кошачьем антителе 15Н05 1.1 позволила найти Feline 15Н05 1.1 FW2, имеющее повышенную аффинность как к кошачьему, так и к собачьему IL-31 по сравнению с мышиным 15Н05 (Фиг. 2). Объединение тяжелой и легкой цепей Feline 15Н05 1.1 FW2 в единую плазмиду привело к образованию антител ZTS-927 и ZTS-361, продуцированных в системах экспрессии HEK и СНО. Аффинность и эффективность обоих антител, полученных в результате экспрессии из одной плазмиды, также описаны на Фиг. 2 и 3, соответственно.
Эффективность полностью фелинизированного mAb ZTS-361 против кошачьего IL-31 оценивали по его способности нейтрализовать индуцированный IL-31 зуд на модели у кошек in vivo. На Фиг. 10А показано исходное поведение до провокации зуда в группе Т01, получавшей плацебо, и группе Т02, получавшей антитело ZTS-361, начиная с дня -7 до дня 28, где день 0 был днем введения антитела группе Т02. Как показано на этом графике, сопряженное с зудом поведение, которое оценивали в группах Т01 и Т02 до провокации IL-31, варьировалось незнчительно, при этом количество сопряженных с зудом явлений в течение 30-минутного периода наблюдения составляло от 0 до 10. Отличием этого исследования от предварительной модели с провокацией зуда у кошек, описанной выше в разделе 1.13, было то, что в день 0 кошкам вводили 4 мг/кг ZTS-361 подкожно без кошачьего IL-31, образующего предварительно связанный комплекс. Это является более точной оценкой эффективности, поскольку антитело ZTS-361 будет находиться в циркуляции в течение семи дней до первой провокации IL-31, требующей, чтобы количество антитела, воздействующего на организм, было достаточным для связывания и нейтрализации циркулирующего IL-31.
В данном исследовании сопряженное с зудом поведение оценивали на 7, 21 и 28 дни в течение 1 часа после провокации 0,5 мкг/кг белка IL-31 внутривенно. На Фиг. 10 В показана эффективность антитела ZTS-361, демонстрирующего значительное снижение зуда, наблюдавшееся на 7 день (р менее 0,0001), 21 день (р менее 0,0027) и 28 день (р менее 0,0238) после провокации IL-31 по сравнению с контролем, получавшим плацебо. Результаты, полученные в этой модели с провокацией, подтверждают предыдущие наблюдения, демонстрирующие эффективность химерной мышиной:кошачьей формы 15Н05, и подтверждают выявленную в тестах на клетках эффективность и важность эпитопа на кошачьем IL-31, распознаваемого CDR 15Н05. Эти данные также подтверждают способность антитела ZTS-361 нейтрализовать зуд, вызванный кошачьим IL-31 in vivo, и позволяют предположить, что это антитело может служить терапевтическим средством при лечении опосредованных IL-31 заболеваний у кошек, включая атопический дерматит.
Последние данные по изучению уровней IL-31 в плазме у животных, принадлежащих клиентам, показывают повышенное количество цитокинов в циркуляции у собак с атопическим и аллергическим дерматитом по сравнению с нормальными лабораторными собаками породы бигль (Фиг. 11А). Недавно было проведено исследование для определения сывороточных уровней IL-31 у кошек с предположительным диагнозом аллергического дерматита (AD) в нескольких различных географических регионах США. На Фиг. 11В показаны результаты этой оценки, показывающие, что как и у собак с атопическим и аллергическим дерматитом, средний уровень циркулирующего IL-31 у 73 обследованных кошек с этим предполагаемым диагнозом, составлял 8799 фг/мл в отличие от 205 фг/мл у 17 контрольных кошек сопоставимого возраста. Чтобы понять уровни собачьего IL-31 в предыдущем исследовании с построением модели, анализировали фармакокинетический профиль собачьего IL-31 у собак после подкожного введения дозы 1,75 мкг/кг. На Фиг. 11С показаны пиковые уровни в плазме в течение первого часа, достигающие максимума приблизительно 30 нг/мл и сохраняющиеся на уровне приблизительно 400 пг/мл через три часа. На основании этих результатов разумно полагать, что при внутривенном введении 0,5 мкг/кг кошачьего IL-31, использованном в этой модели на кошках, его количество в циркуляции будет намного превышать количество, наблюдаемое при естественном течении болезни у собак и кошек.
2. Пример 2. Характеристика и использование мимотопов IL-31 в вакцинах и диагностике
2.1. Аминокислотные остатки собачьего IL-31, задействованные в связывании с антителом 15Н05
Как описано в разделе 1.10 данной заявки, выполняли сканирующий мутагенез собачьего белка IL-31 с полной заменой аминокислот, указанных на Фиг. 12. Каждое положение в данной области IL-31 по-отдельности заменяли в полноразмерном белке одной из других возможных 19 аминокислот и оценивали связывание антитела 15Н05 с помощью непрямого ELISA. При тех заменах, которые не влияли на связывание, сигнал в ELISA был эквивалентным сигналу с белком IL-31 (или более высоким), в то время как при заменах, влияющих на связывание антитела, сигнал в анализе был более низким (или отсутствовал). Как подробно показано на Фиг. 12, некоторые положения собачьего IL-31 были толерантными к определенным заменам указанных аминокислот (SEQ ID NO: 155; положения 124, 125, 129 и 132-135), а другие - нет (SEQ ID NO: 155; положения 126, 127, 128, 130 и 131). Для сравнения показана соответствующая область кошачьего (SEQ ID NO: 157), лошадиного (SEQ ID NO: 165) и человеческого IL-31 (SEQ ID NO: 181). Результаты мутаций собачьего IL-31 можно экстраполировать на другие биологические виды для создания гомологичных пептидов. Это точное картирование положений в области эпитопа на собачьем IL-31 позволило сконструировать как линейные, так и пептиды с ограниченной конформационной свободой, имитирующие сайт связывания на белке IL-31, распознаваемый антителом 15Н05. Эти результаты в сочетании с моделью кошачьего IL-31, описанной в разделе 1.10 (Фиг. 6В), показывают, что эпитоп, распознаваемый mAb 15Н05, представляет собой участок последовательно расположенных аминокислот, которые сближены в случайной спирали, образующей четвертый спиральный домен цитокина. Экспонирование этого эпитопа позволяет связываться mAb 15Н05 как с линейными, так и с пептидами с ограниченной конформационной свободой, при этом с большей аффинностью с формой с ограниченной конформационной свободой. Результаты мутаций, описанные выше (раздел 1.12), дополнительно подчеркивают важность расположения этого эпитопа (и эпитопа 11Е12, ранее описанного в патенте US 8790651, Bammert et al.) на поверхности белка IL-31 для связывания с корецепторным комплексом.
2.2. Характеристика пептидных миметиков. представляющих эпитоп на IL-31, распознаваемый антителом 15Н05
Как упоминалось ранее, конструирование миметиков пептидных эпитопов (мимотопов) может найти широкое применение в качестве аналитических реагентов для захвата и в качестве субъединичных вакцин для индукции соответствующего целенаправленного иммунного ответа для выработки у животных антител, направленных на нейтрализацию эпитопа на IL-31. Для достижения этой цели сконструировали собачьи и кошачьи пептиды и охарактеризовали их аффинность к антителу ZTS-927 и их способность блокировать способность антитела ZTS-361 подавлять опосредованную IL-31 передачу сигналов от рецептора на клетках FCFW4 кошек. Конструирование антител ZTS-927 и ZTS-361 (оба содержат CDR мышиного антитела 15Н05) описано выше в разделе 1.9. Пептид ZTS-561 содержит аминокислотную последовательность N-TEISVPADTFERKSFILT-C, которая соответствует положениям 121-138 последовательности SEQ ID NO: 155 с заменой аргинина (R) на цистеин (С) в положении номер 132. Пептид ZTS-561 также включает N и С концевые цистеины для облегчения конъюгации с использованием свободных тиоловых групп. Пептид ZTS-562 содержит аминокислотную последовательность N-EISVPADTFERKSF-C, которая соответствует положениям 122-135 последовательности SEQ ID NO: 155 с заменой аргинина (R) на цистеин (С) в положении номер 132. Пептид ZTS-562 также включает N и С концевые цистеины для облегчения конъюгации с использованием свободных тиоловых групп. Пептид ZTS-563 содержит аминокислотную последовательность N- AKVSMPADNFERKNFILT-C, которая соответствует положениям 121-138 последовательности SEQ ID NO: 157 с заменой треонина (Т) на аланин (А) в положении номер 138. Пептид ZTS-563 также включает N и С концевые цистеины для облегчения конъюгации с использованием свободных тиоловых групп. Пептид ZTS-564 содержит аминокислотную последовательность N-TEISVPADTFERKSFILT-C, которая соответствует положениям 121-138 последовательности SEQ ID NO: 155. Пептид ZTS-564 также включает N и С концевые цистеины для облегчения конъюгации с использованием свободных тиоловых групп. Многоступенчатый процесс с использованием технологии CLIPS (Timmerman et al. J Mol Recognit. 2007; 20(5): 283-299) использовали для идентификации и оптимизации этих четырех пептидов, способных связываться с паратопом mAb 15Н05 (Pepscan, Lelystad Netherlands). С целью создания иммуногенов эти четыре пептида (изображенные на Фиг. 13А) независимо конъюгировали с белком-носителем, который представляет собой неактивную мутантную (нетоксичную) форму дифтерийного токсина (CRM 197), с использованием стандартной химии перекрестного сшивания. Для оценки аффинности каждый пептид независимо иммобилизовали на поверхности чипа biacore и определяли KD для фелинизированного mAb 15Н05 против IL-31 (ZTS-927) (Фиг. 13В). Все четыре пептида связывали ZTS-927 с аффинностью в наномолярном диапазоне, что указывает на то, что они являются близким отображением сайта связывания на полноразмерном IL-31. Для оценки эффективности каждого пептида в анализе с использованием FCWF-4 (кошачьи макрофагоподобные клетки) конъюгированные или неконъюгированные пептиды в различных концентрациях инкубировали при 37°С в течение 1 часа с 0,2 мкМ (6,5 мкг/мл) mAb ZTS-361 и затем добавляли кошачий IL-31. Рассчитывали значения IC50, откладывая эффект в процентах (ось у), определяемый как способность пептида связывать и блокировать ингибирование mAb ZTS-361 опосредованного белком IL-31 фосфорилирования STAT3 в кошачьих макрофагах FCWF-4, в зависимости от возрастающих концентраций пептида (ось х). Пептид ZTS-564 имел пониженную растворимость в растворе, что, вероятно, приводило к неэффективной конъюгации, низкой плотности эпитопа и низкой эффективности. Пептид ZTS-561 имел низкую эффективность в конъюгированной форме, но сохранял хорошую эффективность в неконъюгированной форме (IC60 примерно 1,7 мкг/мл). ZTS-562 и ZTS-563 демонстрировали отличную эффективность, будучи неконъюгированными, значения IC50 составляли 1,046 и 1,742 мкг/мл, соответственно. После конъюгации их эффективность снизилась приблизительно в 3 раза, при этом значения IC50 для ZTS-562 и ZTS-563 составили 3,024 мкг/мл и 3,384 мкг/мл, соответственно (Фиг. 13В). Способность этих пептидов блокировать высокоаффинное связывание mAb ZTS-361 с белком IL-31 была весьма многообещающей и послужила дополнительным доказательством возможности их применения в качестве миметиков соответствующего эпитопа на IL-31 (далее обозначаемых мимотопами IL-31 15Н05). Эти мимотопы IL-31 15Н05 дополнительно исследовали на предмет возможности их применения в качестве иммуногенов для индукции иммунного ответа против IL-31.
2.3. Дизайн исследования для определения сывороточных титров к IL-31 после иммунизации собак породы бигль мимотопами собачьего и кошачьего IL-31 15Н05 и полноразмерным кошачьим белком IL-31
Исследование для определения иммуногенности проводили для оценки способности конъюгированных с CRM-197 мимотопов IL-31 15Н05 вызывать эпитоп-специфический иммунный ответ, направленный на соответствующую область белка IL-31, с которой связывается антитело 15Н05 и нейтрализует способность цитокинов активировать корецептор IL-31Ra:OSMR. Схема исследования приведена на Фиг. 14.
Чистокровным самцам собак породы бигль подкожно вводили конъюгированные мимотопы IL-31 15Н05 с адъювантом ZA-01. На диаграмме ниже показана схема эксперимента в каждой группе. Включали контрольные группы, получавшие только CRM-197 с адъювантом ZA-01 (Т01) и конъюгированный с CRM-197 кошачий IL-31 (SEQ ID NO: 159; Feline_IL-31_E_coli), соответствующей нуклеотидной последовательностью для которого является (SEQ ID NO: 160; Feline_IL-31_E_coli). Каждый мимотоп с адъювантом или контроль вводили подкожно в в дозе 10 мкг в дни 0, 28 и 56 (0,5 мл раствора 20 мкг/мл). Кровь на сыворотку брали в день 0 (перед введением), 7, 12, 28 (перед введением), 35, 42, 49, 56 (перед введением), 63, 70, 77 и 84. Кроме того, в 35 и 84 дни у каждого животного забирали примерно 40 мл крови в пробирки с гепаринатом лития и выделяли мононуклеарные клетки периферической крови (РВМС) с использованием стандартного метода. РВМС подвергали криоконсервации до дальнейших исследований антиген-специфических В-клеток.
2.4. Сывороточные титры, полученные после вакцинации собак собачьими и кошачьими мимотопами IL-31 15Н05 и кошачьим полноразмерным белком IL-31
У каждого животного в каждый день исследования оценивали сывороточные титры, указанные выше в разделе 2.3. Титры определяли с помощью непрямого ELISA, где в каждом соответствующем исследовании для захвата использовали полноразмерный белок IL-31. В каждой исследуемой группе сыворотку анализировали на связывание с кошачьим белком IL-31, кошачьим мутантным белком 15Н05, а также с собачьим, лошадиным и человеческим белками IL-31. Задача состояла в том, чтобы понять иммунный ответ, вызываемый кошачьим белком IL-31 (SEQ ID NO: 159; Feline_IL-31_E_coli) или мимотопами пептида 15Н05, против IL-31 нескольких биологических видов, аминокислотные последовательности которых имеют ряд идентичностей (Фиг. 1А). В группе воздействия 2 (полноразмерный кошачий IL-31 с CRM) представлен адаптивный иммунный ответ на множественные эпитопы, охватывающие всю последовательность белка. Использование полноразмерного белка в качестве иммуногена будет приводить к образованию антител, которые являются как нейтрализующими, так и не нейтрализующими по отношению к биологической активности IL-31. В предыдущей работе на мышах, описывающей идентификацию антител, нейтрализующих передачу сигналов IL-31, показано, что процент этих антител невелик и, следовательно, большая часть поликлонального ответа на полноразмерный белок будет не нейтрализующего типа (US8790651, Bammert, и другие.). Что касается вакцины, то образование не нейтрализующих антител к IL-31 может иметь неблагоприятные последствия для безопасности и эффективности. Ненейтрализующие антитела могут вызывать увеличение количества биоактивного IL-31 в кровотоке в результате связывания в комплексы цитокинов и антител. Эти комплексы могут обеспечить существование в кровотоке мономерных или агрегированных форм IL-31, позволяя рецептор-связывающей части IL-31 взаимодействовать с корецептором IL-31:OSMR. Усиление передачи сигналов pSTAT в результате увеличения количества IL-31 в кровотоке усугубит зуд при таком заболевании, как атопический дерматит (Gonzales et al. Vet Dermatol. 2013 Feb; 24(1):48-53. ell-2).
На Фиг. 15 A-E приведены графики, на которых показаны сывороточные титры в ответ на каждый белок IL-31, представленные по группам воздействия, показывающие ответ в каждый день, когда проводили забор сыворотки. Титры сыворотки в ответ на IL-31 от разных биологических видов исследовали, чтобы понять выраженность перекрестной реакции антител (CRAR), которая может иметь место. Максимальное разведение, испытанное для каждого образца сыворотки, составляло 1: 50000, поэтому, когда титр превышал это значение, его обозначали как 50000. Для ясности ниже будет приведено описание графических материалов, где показан ответ на каждый белок IL-31, используемый в твердофазном иммуноферментном анализе с захватом, в отдельных группах воздействия.
Учитывая высокий процент идентичности между гомологами, ожидалось, что у собак, вакцинированных полноразмерным кошачьим IL-31 (Т02), полученная поликлональная сыворотка будет связывать белок от нескольких биологических видов. На Фиг. 15А показаны титры собачьих антител, которые связывались с кошачьим IL-31. Анализ группы Т02 показывает, что после введения третьей дозы формировался умеренный и устойчивый гуморальный ответ к полноразмерному кошачьему белку IL-31, который сохранялся до завершения исследования на 84 день. При изучении ответа в группе Т02 на мутантный кошачий белок 15Н05 (SEQ ID NO: 163; Feline_IL-31_15H05_mutant) наблюдался аналогичный профиль, при этом титры на 63, 70 и 77 день были еще выше (Фиг. 15В). На Фиг. 15С показаны титры в ответ на собачий IL-31. При изучении титров в группе Т02 можно оценить выраженность CRAR на собачий белок IL-31 у вакцинированных собак. Перед введением третьей дозы кошачьего IL-31 с CRM, ответ не наблюдался. После введения 3 дозы в 56-й день наблюдалась транзиторная CRAR с 63 до 77 дней, возвращавшаяся практически к исходному уровню к 84-му дню. Величина ответа на собачий белок была аналогична ответу на кошачий, однако его продолжительность была короче. Интересно, что CRAR на лошадиный и человеческий IL-31 была пренебрежимо малой или незначительной, соответственно (Фиг. 15D и Е, 63 и 84 дни для человеческого). Таким образом, иммунный ответ собак на кошачий IL-31 с CRM был наиболее устойчивым и длительным по сравнению с ответом на собственно кошачий белок IL-31. Аминокислотные последовательности кошачьего и собачьего IL-31 имеют идентичность 76%, что, по-видимому, является достаточно высоким уровнем для развития CRAR на собачий белок. Лошадиный и человеческий IL-31 идентичны кошачьему на 57 и 49%, соответственно, что вызывает лишь незначительную CRAR в случае человеческого белка.
Мимотоп IL-31 15Н05 ZTS-561 представляет собой сайт связывания на собачьем IL-31, распознаваемый антителом 15Н05. Гуморальный ответ у собак, вакцинированных ZTS-561 с CRM, обозначен как Т03 на Фиг. 15 А-Е. Задача в данном случае состояла в том, чтобы оценить гуморальный ответ на этот специфический участок на IL-31, который, как известно, участвует во взаимодействии цитокинов с его рецептором. Фокусированный иммунный ответ на конкретный эпитоп означает, что антитела, направленные на этот участок белка, будут нейтрализовать его биологическуюй активность. ZTS-561 CRM представляет собой 20-мер с ограниченной конформационной свободой, представляющий часть собачьего белка IL-31, распознаваемую антителами с CDR идентичными мышиному антителу 15Н05 (SEQ ID NO: 67; MU_15Н05_VH), соответствующей нуклеотидной последовательностью для которого является (SEQ ID NO: 68; MU_15H05_VH), спаренный с VL (SEQ ID NO: 69; MU_15H05_VL), соответствующей нуклеотидной последовательностью для которой является (SEQ ID NO: 70; MU_15H05_VL). Мимотоп ZTS-561 не вызывал выраженный ответ на кошачий IL-31 на протяжении всего исследования (Фиг. 15А), включая мутантный кошачий IL-31 15Н05 (Фиг. 15 В). Напротив, ZTS-561 с CRM вызывал у собак очень сильный иммунный ответ на собачий белок IL-31, начиная с 35 дня после второй инъекции, сохранявшийся до завершения исследования на 84 день (Фиг. 15С). CRAR, вызванная ZTS-561, на лошадиный IL-31, была пренебрежимо малой, а на человеческий наблюдался лишь незначительный ответ на 56 день исследования (Фиг. 15D и Е). Интересно отметить, что даже несмотря на то, что кошачий и собачий белки имеют высокую степень идентичности в этой области белка (Фиг. 1), после вакцинации собак собачьим мимотопом 15Н05, видоспецифический иммунный ответ был направлен на белок IL-31 собаки.
ZTS-561 CRM представляет собой усеченную 16-мерную версию ZTS-561 с ограниченной конформационной свободой, также представляющую часть собачьего белка IL-31, распознаваемую антителами с CDR идентичными мышиному антителу 15Н05 (SEQ ID NO: 67; MU_15Н05_VH), соответствующей нуклеотидной последовательностью для которой является (SEQ ID NO: 68; MU_15H05_VH), спаренную с VL (SEQ ID NO: 69; MU_15H05_VL), соответствующей нуклеотидной последовательностью для которой является (SEQ ID NO: 70; MU_15Н05_VL). Результаты ответа собак на ZTS-562 представлены как группа Т04 на Фиг. 15 А-Е. Интересно, что CRAR, вызванная этой более короткой версией, была более выраженной, приводя к умеренным титрам против кошачьего белка, начиная с 35 по 70 дни (Фиг. 15А). Некоторая реакция также наблюдалась на мутантный белок IL-31 15Н05 между 35 и 63 днями (Фиг. 15В). Ответ против собачьего IL-31, вызванный этим мимотопом, был выдающимся, начинаясь с 35-го дня после второй дозы и сохраняясь до завершения исследования на 84-й день. ZTS-562 CRM не вызывал CRAR на лошадиный и человеческий белки, что согласовывалось с другими результатами с собачьим пептидом ZTS-561 CRM.
ZTS-563 CRM представляет собой 18-мер с ограниченной конформационной свободой и является единственным мимотопом, представляющим часть кошачьего белка IL-31, распознаваемую антителами с CDR идентичными мышиному антителу 15Н05 (SEQ ID NO: 67; MU_15H05_VH), соответствующей нуклеотидной последовательностью для которой является (SEQ ID NO: 68; MU_15H05_VH), спаренную с VL (SEQ ID NO: 69; MU_15H05_VL), соответствующей нуклеотидной последовательностью для которой является (SEQ ID NO: 70; MU_15H05_VL). Результаты ответа собак на ZTS-563 представлены как группа Т05 на Фиг. 15 А-Е. Кошачий мимотоп (ZTS-563) вызвал у собак ответ, специфический к кошачьему IL-31, что согласовывалось с предыдущими данными, демонстрирующими видоспецифический ответ на собачий мимотоп. На Фиг. 15А показан титр антител против кошачьего IL-31 в ответ на вакцинацию ZTS-563, достигающий более 1:50000 на 35-й день и сохраняющийся на этом уровне до 77-го дня, незначительно снижаясь на 84-й день. Сравнивая группу воздействия Т05 (ZTS-563) на Фиг. 15А и 15В, можно четко увидеть разницу между титрами в ответ на кошачий IL-31 и мутантный кошачий IL-31 15H05. Снижение титров на мутантный белок с течением времени (по сравнению с белком дикого типа) указывает на то, что значительная часть иммунного ответа направлена на очень специфическую часть белка, представленную мимотопом ZTS-563. Примечательно, что ответ против собачьего IL-31 был от умеренного до низкого, что дополнительно подтверждало видоспецифичность ответа, генерируемого иммунной системой собак, на незначительные различия в аминокислотных последовательностях двух биологических видов. ZTS-563 является единственным мимотопом, вакцинация которым вызывала у собак CRAR на лошадиный белок IL-31. Эти результаты показывают, что незначительные изменения в последовательности мимотопа могут обуславливать видовую специфичность, а также могут вызывать межвидовой иммунный ответ. Понимание этих свойств полезно для разработки вакцины, направленной против IL-31, с использованием этой технологии для применения у одного или многих биологических видов.
И, наконец, ZTS-564 CRM представляет собой 18-мер с ограниченной конформационной свободой, идентичный ZTS-561, в котором использовали альтернативный линкер mT2b (Фиг. 15A). ZTS-564 CRM представляет собой часть собачьего белка IL-31, распознаваемую антителами с CDR идентичными мышиному антителу 15Н05 (SEQ ID NO: 67; MU_15H05_VH), соответствующей нуклеотидной последовательностью для которой является (SEQ ID NO: 68; MU_15H05_VH), спаренную с VL (SEQ ID NO: 69; MU_15Н05_VL), соответствующей нуклеотидной последовательностью для которой является (SEQ ID NO: 70; MU_15Н05_VL). Результаты ответа собак на ZTS-564 представлены как группа Т06 на Фиг. 15 А-Е. Этот мимотоп практически не вызывал у собак ответа на кошачий IL-31 (Фиг. 15А и 15В), что согласовывалось с другими наблюдениями. Ответ против собачьего IL-31, вызванный ZTS-564, был очень устойчивым. На Фиг. 15С показаны все группы воздействия в этом исследовании, Т06 (ZTS-564) - единственная группа, в которой развивался иммунный ответ против собачьего IL-31 после однократной дозы. Титры антител против собачьего IL-31, полученные после второй и третьей доз, соответствовали максимальному ответу в ходе исследования (более 1:50000), начиная с 35 дня и до завершения исследования на 84-й день. CRAR на лошадиный IL-31 не наблюдалась, однако этот мимотоп вызывал единственный стойкий ответ на человеческий IL-31, наблюдавшийся среди групп воздействия. Примечательно, что такие незначительные различия в химических характеристиках линкера и, возможно, более строгие конформационные ограничения, накладываемые линкером mT2b, обеспечивают более целенаправленный ответ против IL-31, потенциально снижая потребность в более частом введении.
Данные этого исследования показывают, что пептидные миметики, представляющие сайт связывания нейтрализующего антитела 15Н05 против IL-31, способны вызывать у животного иммунный ответ, и этот иммунный ответ направлен против эпитопа, распознаваемого CDR антитела 15Н05. Из этих данных следует, что дальнейшая характеристика этой антисыворотки с использованием рекомбинантных корецепторов IL-31 может применяться для определения нейтрализующей фракции IL-31, образующейся в ходе этого поликлонального ответа. Эти результаты также позволяют предположить, что данный подход может быть полезен при применении в качестве вакцины против расстройств, опосредованных IL-31, таких как атопический дерматит.
2.5. Идентификация нейтрализующих IL-31 антител, образованных отдельными В-клетками, выделенными из плазматических клеток собак породы бигль, иммунизированных мимотопами IL-31 15Н05
Как описано выше, кровь собирали на 35 и 84 дни исследования с целью выделения РВМС. Благодаря устойчивости ответа, направленного против эпитопа, распознаваемого 15Н05 (Фиг. 15А и 15В), для дальнейшей оценки антитело-положительных В-клеток использовали РВМС от одной собаки Т05, вакцинированной ZTS-563 с CRM (кошачий мимотоп). Проводили скрининг активированных В-клеток памяти с использованием антитела против Fc собачьего IgG, связанного с микрогранулами для нахождения клеток, секретирующих антитела. Одновременно оценивали способность секретируемых IgG связывать кошачий IL-31 дикого типа и связываться с мутантным кошачьим IL-31 15Н05. По результатам такого первичного скрининга из этой популяции клеток РВМС отобрали 7 вариантов. Из этих 7 вариантов 3 не связывались с мутантом 15Н05, что указывало на то, что в результате иммунизации мимотопом IL-31 15Н05 ZTS-563 эти В-клетки вырабатывают антитела, наиболее точно узнающие эпитоп 15Н05 (данные не показаны). После секвенирования вариабельной тяжелой и легкой цепей этих 7 вариантов конструировали рекомбинантные полностью собачьи версии, экспрессировали в клетках HEK и очищали, как описано выше. При повторном скрининге этих 7 рекомбинантных собачьих IgG остался единственный вариант (ZIL1), который сохранил связывание с кошачьим белком IL-31 (Фиг. 4). Кроме того, связывание ZIL1 с мутантом IL-31 15Н05 снижено по результатам методов ELISA и Biacore, что указывает на то, что это антитело связывается с той же областью эпитопа, что и антитело 15Н05. Дополнительные варианты, полученные непосредственно из В-клеток собак, описанные в разделе 1.6 и на Фиг. 4 (ZIL8 - ZIL171), получали от собак, иммунизированных полноразмерным кошачьим IL-31, а также из других источников тканей, описанных выше в данном документе. Только антитело ZIL1 происходило из РВМС после вакцинации пептидным мимотопом.
Важным аспектом этих экспериментов является способность идентифицировать В-клетки, секретирующие антитела, специфические к эпитопу 15Н05, в кровотоке собаки после иммунизации пептидным мимотопом из белка IL-31. Эти результаты обосновывают применение пептида, имитирующего область эпитопа на IL-31, которая, как известно, имеет отношение к способу ингибирующего действия антитела 15Н05. Поскольку антитела с CDR, происходящими из mAb 15Н05, способны предотвращать опосредованный IL-31 зуд in vivo, эти результаты дополнительно подтверждают концепцию иммунизации таким миметиком, разработанным для выработки направленного на эпитоп иммунного ответа в качестве вакцины для предотвращения IL-31-опосредованных заболеваний, таких как атопический дерматит.
2.6. Применение мимотопа IL-31 15Н05 в качестве реагента для захвата для определения подлинности и эффективности антитела 15Н05 и его производных
Мимотоп IL-31 15Н05 представляет собой сайт связывания на белке IL-31, который распознается CDR мышиного mAb-предшественника 15Н05. Как упоминалось ранее, желательно иметь такой реагент для использования в формате ELISA (или другого анализа) для мониторинга процесса продуцирования антител на протяжении всего производства. Предполагается, что такие описанные мимотопы будут полезны, когда производят партию продуцируемого антитела против IL-31 и пептидный мимотоп может использоваться в качественном и количественном анализе полученного антитела.
2.7. Применение мимотопа IL-31 15Н05 в качестве диагностического средства для измерения уровней антител или определения уровней IL-31 у вида-хозяина
Кроме того, в данном документе предлагается использование описанного пептидного мимотопа в качестве реагента в анализе для измерения количества циркулирующего антитела в организме хозяина после лечения терапевтическим средством опосредованного IL-31 расстройства, такого как атопический дерматит. Биологическую жидкость животного добавляют непосредственно к мимотопу, который связан с твердой поверхностью, а затем добавляют соответствующие вторичные детектирующие реагенты для количественного определения антитела. Кроме того, предложена схема анализа, в котором мимотоп используют для захвата антитела, меченого для определения в анализе. Это захваченное антитело будет иметь более низкую аффинность к присоединенному мимотопу, по сравнению с аффинностью к нативному IL-31, циркулирующему у биологического вида-хозяина. В этом воплощении жидкость, полученную от биологического вида-хозяина, инкубируют с комплексом меченое антитело:мимотоп, связанным с твердой поверхностью. IL-31, присутствующий в исследуемой жидкости, полученной от биологического вида-хозяина, будет иметь более высокое сродство к антителу, таким образом, меченое антитело будет высвобождаться с твердой поверхности и может быть удалено на этапах промывки. Таким образом, уровень IL-31 в тестовой жидкости может быть обратно пропорционален сигналу, генерируемому на поверхности, с которой связан мимотоп. Предполагается, что такой анализ может применяться в качестве диагностического теста в исследованиях или в клинике для измерения IL-31.
2.8. Сывороточные титры против IL-31 после иммунизации собак породы бигль мимотопами IL-31 и полноразмерным собачьим белком IL-31
Второе серологическое исследование проводили с использованием чистокровных стерилизованных самцов собак породы бигль как и в схеме исследования, описанной в разделе 2.3, однако в этом исследовании сравнивали разные мимотопы Чистокровным самцам собак породы бигль подкожно вводили конъюгированные мимотопы IL-31 с адъювантом ZA-01. Включали контрольную группу, получавшую только конъюгированный с CRM-197 собачий IL-31 (SEQ ID NO: 155; Canine IL-31), соответствующей нуклеотидной последовательностью для которого является (SEQ ID NO: 156; Canine IL-31) в ZA-01 (T01). Каждый мимотоп с адъювантом или контроль вводили подкожно в в дозе 10 мкг в дни 0, 28 и 56 (0,5 мл раствора 20 мкг/мл). Кровь на сыворотку брали в день -1,0 (перед введением), 14, 28 (перед введением), 42, 56 (перед введением), 70 и 84. Кроме того, в -1, 35 и 63 дни у каждого животного забирали примерно 40 мл крови в пробирки с гепаринатом лития и обрабатывали для выделения РВМС с использованием стандартного метода. РВМС подвергали криоконсервации до дальнейших исследований антиген-специфических В-клеток.
В группе воздействия 1 (полноразмерный собачий IL-31 с CRM) представлен адаптивный иммунный ответ на множественные эпитопы, охватывающие всю последовательность белка, замысел был таким же, как и при использовании полноразмерного кошачьего белка, описанном в разделе 2.4. На Фиг. 16А представлена таблица, в которой указаны исследуемые группы воздействия. В дополнение к полноразмерному белку, описанному для Т01, использовали два мимотопа, представляющих эпитоп 15Н05 у собаки (Т02, ZTS-420) и человека (Т03, ZTS-421). ZTS-420 похож на ZTS-561, описанный ранее в серологическом исследовании на кошках, однако конформационная свобода ZTS-420 ограничена дисульфидной связью, образованной цистеинами на N- и С-концах пептида, в отличие от ZTS-561, конформационная свобода которого ограничена линкером mT2a. ZTS-421 представляет собой гомологичный участок человеческого белка IL-31 (SEQ ID NO: 181; HumanlL-31),с конформационная свобода которого ограничена связыванием N- и С-концов с линкером mT2b, как показно на Фиг. 13А. Ссылки на ключевые аминокислотные последовательности, участвующие в связывании антитела 15Н05, можно найти на Фиг. 12. Четвертая группа была включена для изучения иммуногенного потенциала ключевой области связывания антител на собачьем IL-31, описанном ранее, с использованием двух антител, которые, как известно, нейтрализуют опосредованную IL-31 передачу сигналов pSTAT и опосредованный IL-31 зуд у собак (US8790651, Bammert, et al.). Эта область на кошачьем IL-31 выделена в модели на основе гомологии, показанной на Фиг. 6 В. Принятая модель IL-31 представляет собой цитокин с четырьмя антипараллельными спиральными доменами, образующими топологию типа «вверх-вниз». В дальнейшем описание структуры белка IL-31 будет приведено для этих четырех спиралей на собачьем IL-31 (SEQ ID NO: 155; Canine IL-31), соответствующей нуклеотидной последовательностью для которого является (SEQ ID NO: 156; Canine_IL-31), с учетом соответствующих положений на гомологичных белках IL-31 других биологических видов (Фиг. 1). Спираль А состоит из последовательности примерно от 33 до 59 аминокислоты, спираль В состоит из последовательности примерно от 83 до 98 аминокислоты, спираль С состоит из последовательности примерно от 101 до 114 аминокислоты, а спираль D состоит из последовательности примерно от 129 до 156 аминокислоты. Приблизительно между 97 и 101 аминокислотами находится область петли. Петля, следующая за спиралью А, расположена примерно от 57 до 62 аминокислоты, а петля, предшествующая спирали D, находится примерно от аминокислоты 126 до 129. Любая промежуточная последовательность, в которой отсутствует предсказанная вторичная структура, будет обозначаться случайной спиралью. Группа воздействия 4 получала ZTS-766, представляющий собой мимотоп, представляющий спирали В и С собачьего IL-31, включавший N-концевой остаток цистеина для облегчения связывания с белком-носителем CRM-197. Выравнивание пептидных последовательностей этой области белка IL-31 показано на Фиг. 16В, где сравниваются собачий, кошачий, лошадиный и человеческий белки с указанием соответствующих идентификационных номеров последовательности и положений аминокислот.
Титры определяли с помощью непрямого ELISA, где в каждом соответствующем исследовании для захвата использовали полноразмерный белок IL-31. В каждой исследуемой группе сыворотку анализировали на связывание с собачьим и человеческим белками IL-31 (Фиг. 17А и 17В, соответственно). У собак, вакцинированных CRM-197, конъюгированным с полноразмерным собачьим белком IL-31 (Т01), наблюдалось умеренное увеличение титра на 42-й день после введения второй дозы на 28-й день. В этой группе наблюдалось снижение титра в ходе исследования даже после введения третьей дозы на 56-й день. Учитывая общий ответ на все эпитопы белка IL-31 (как нейтрализующий, так и не нейтрализующий) в сочетании с низкими титрами, маловероятно, что полноразмерный белок IL-31 является перспективным кандидатом для разработки вакцины. Группа 2 в этом исследовании получала ZTS-420, представляющий собой собачий мимотоп 15Н05, похожий на ZTS-561, описанный в разделе 2.4, однако конформационная свобода ZTS-420 ограничена дисульфидной связью между цистеинами, добавленными на N- и С-конце пептида, в отличие от ZTS-561, конформационная свобода которго ограничена линкером mT2a. Этот мимотоп не смог вызвать устойчивый иммунный ответ в отличие от формы с конформационной свободой, ограниченной mT2a (см. Фиг. 17А и 15С). Наблюдалось умеренное увеличение титра на 84-й день после введения второй дозы на 56-й день. Возможно, циклизация дисульфидной связью некорректна или видоизменяется во время конъюгации с CRM-197, что приводит к неоптимальной презентации иммуногена клеткам иммунной системы собак. Группа 4, получавшая ZTS-766, представляющий спирали В и С собачьего IL-31, давала наиболее устойчивый ответ, проявляющийся после введения второй дозы на 28 день и нарастающий до 84 дня до завершения исследования. Учитывая способность антител, распознающих эту ранее описанную последовательность, нейтрализовать IL-31, этот мимотоп представляет собой перспективный кандидат для вакцины, предотвращающей заболевания, опосредованные IL-31. Группа воздействия 3 получала ZTS-421, представляющий собой эпитоп 15Н05, в котором использовали последовательность человеческого IL-31 в этой области мимотопа. Интересно, что ни у одной из собак, вакцинированных этим мимотопом, не развивался ответ против собачьего белка IL-31 (данные не показаны), однако после введения второй и третьей доз наблюдался иммунный ответ на белок IL-31 человека (Фиг. 17 В). Учитывая сходство последовательностей между ключевой областью эпитопа мимотопа 15Н05 (Фиг. 12), такая специфичность собачьего ответа против человеческого IL-31 является превосходной.
2.9. Сывороточные титры против IL-31 после иммунизации лабораторных кошек кошачьими и лошадиными мимотопами IL-31 и полноразмерным кошачьим белком IL-31
Второе серологическое исследование проводили с использованием лабораторных кошек, как и в схеме исследования, описанной в разделе 2.3, однако в этом исследовании сравнивали кошачий и лошадиный мимотопы. Лабораторным кошкам подкожно вводили конъюгированные мимотопы IL-31 со смесью, включающей гликолипидный адъювант Bay R1005 (N-(2-дезокси-2-L-лейциламино-β-D-глюкопиранозил)-N-октадецилдодеканоиламидгидроацетат), а также олигонуклеотиды CpG. Включали контрольную группу, получавшую конъюгированный с CRM-197 кошачий IL-31 (SEQ ID NO: 157; Feline_IL-31_wildtype), соответствующей нуклеотидной последовательностью для которого является (SEQ ID NO: 158; Feline_IL-31_wildtype) в смеси адъювантов, включающей гликолипидный адъювант Bay R1005 (N-(2-дезокси-2-L-лейциламино-β-D-глюкопиранозил)-N-октадецилдодеканоиламидгидроацетат), а также олигонуклеотиды CpG. (Т01). Каждый мимотоп с адъювантом или контроль вводили подкожно в в дозе 10 мкг в дни 0, 28 и 56 (0,5 мл раствора 20 мкг/мл). Кровь на сыворотку брали в день -14, 0 (перед введением), 28 (перед введением), 42, 56 (перед введением), 70 и 84. Кроме того, в 35 и 63 дни у каждого животного забирали примерно 40 мл крови в пробирки с гепаринатом лития и обрабатывали для выделения РВМС с использованием стандартного метода. РВМС подвергали криоконсервации до дальнейших исследований антиген-специфических В клеток. На Фиг. 18А представлены исследуемые группы воздействия. ZTS-563 (Т02) описан в данном документе в разделе 2.4 как иммуноген, который использовался в предыдущем серологическом исследовании на собаках. ZTS-563 представляет собой мимотоп 15Н05, конформационная свобода которого ограничена mT2a, конъюгированный с CRM-197. Т03 (ZTS-418) представляет собой мимотоп 15Н05 с лошадиной последовательностью (для сравнения см. гомологичную собачью версию ZTS-420 на Фиг. 16А). Группа воздействия 4 получала ZTS-423, пептидный мимотоп, представляющий спираль ВС, описанную в разделе 2.8, с последовательностью кошачьего IL-31. Группа воздействия 5 получала ZTS-422, кошачий мимотоп 15Н05 с линкером mT2b из аминогексановой кислоты (Ahx). На Фиг. 18В показаны результаты сывороточных титров антител в группах воздействия Т01, Т02, Т04 и Т05 (в Т03 отсутствовала CRAR на кошачий IL-31, данные не показаны) в -14, 42 и 84 дни на полноразмерный кошачий IL-31, полученные с применением непрямого ELISA. И вновь конъюгированная форма полноразмерного белка IL-31 (Т01) показала самый слабый гуморальный ответ у кошек, при котором титры на полноразмерный IL-31 никогда не превышали 1: 20000 на протяжении всего исследования. Ответ в группе Т02 (ZTS-563) был умеренным на протяжении всего исследования, демонстрируя дозозависимое увеличение до 84 дня, что указывает на то, что кошачий эпитоп 15Н05, презентированный таким образом, может быть подходящей вакциной. Средние титры у кошек в ответ на полноразмерный кошачий белок IL-31 после трех доз ZTS-423 (Т04) дозозависимо повышаются до более чем 1:100000 после введения второй и третьей доз, что указывает на выдающийся иммунный ответ на очень значимую область эпитопа. ZTS-422 (Т05), представляющий мимотоп 15Н05 с линкером AhX mT2b, также показывает устойчивый иммунный ответ у кошек с титрами, превышающими 1:100 000, после введения второй и третьей дозы. Эпитоп 15Н05 в этой форме, несомненно, является надлежащим отображением этой области белка IL-31 и представляет собой перспективный мимотоп для вакцины, нейтрализующей активность IL-31 in vivo.
2.10. Аспекты последовательности и структуры, которые необходимо учитывать при создании мимотопов. предназначенных для применения в качестве вакцин
В данном документе описаны несколько пептидных отображений эпитопов белка IL-31 (мимотопов) с уникальными последовательностями, соответствующими аминокислотам биологического вида, от которого они происходят. Конечная задача при создании вакцины - воспроизведение эпитопов, благодаря чему иммунная система распознает их и вырабатывает устойчивый и специфический ответ. Создание вакцины облегчается добавлением белков-носителей, таких как CRM-197, и добавлением в состав адъювантов, но не ограничивается этим. В данном документе описаны примеры эпитопов, которые были идентифицированы на основе свойств антител, с которыми они связываются. Ключевые эпитопы - это такие области белка IL-31, которые при связывании с антителом не способны в дальнейшем взаимодействовать с рецепторным комплексом IL-31RA: OSMR и, следовательно, не способны вызывать ответную сигнальную реакцию pSTAT в культуре клеток или in vivo. Таким образом, блокада передачи сигналов, опосредованной рецептором IL-31, представляет собой подход к предотвращению и/или лечению расстройств, опосредованных IL-31, таких как атопический дерматит.
Картирование сайтов связывания антител на белках с использованием мутационных методов является эффективным способом идентификации ключевых остатков, участвующих в распознавании антигена антителом, как было ранее описано для IL-31 в US8790651, Bammert, et al. Основываясь на этих данных, удалось создать спирали ВС собачьих и кошачьих мимотопов, которые описаны в данном документе как эффективные иммуногены, вызывающие устойчивые ответы против IL-31 у собак и кошек. Другой способ с использованием слитого белка GST собачьего IL-31 был недавно описан для картирования антитела М14 против собачьего IL-31 (WO 2018/156367 (Kindred Biosciences, Inc). Эти авторы стремились определить минимальную последовательность эпитопа, распознаваемую антителом М14, состоящую из аминокислот PSDX1X2KI (SEQ ID NO 155, аминокислоты 34-40), где X представляет собой любую аминокислоту. Описание этой последовательности в сравнении с гомологичными видами IL-31 можно найти на Фиг. 19А. Дальнейшее описание, включая фланкирующую последовательность, окружающую вышеуказанное, приведено на Фиг. 19В. После идентификации минимального связывающего фрагмента, указанного на Фиг. 19В (аминокислоты 34-42, выделенные серым прямоугольником), авторы произвели замены на аланин в каждом положении, используя данный пептидный фрагмент, слитый с GST. На основании этих данных был описан указанный выше минимальный связывающий фрагмент М14. Фундаментальный недостаток этого подхода заключается в том, что свойства описанного связывающего фрагмента определяются его структурой в рамках слитого с GST белка. Не привязываясь к единой теории, предполагают, что аминокислотная последовательность, распознаваемая антителом М14, является частью упорядоченного альфа-спирального домена, описываемого в данном документе как спираль А. В пептидах и белках существуют альфа-спирали, которые поддерживаются водородными связями между кислородом карбонильной группы и азотом аминогруппы остова (правило Кори-Полинга, химический словарь, 2008), но не ограничиваются таковыми. Считается, что минимальный связывающий фрагмент, описанный для антитела M14, не является адекватным описанием эпитопа, поскольку нет подтверждений его связывающих свойств в отсутствие каркаса GST. Более того, в составе последовательности, окружающей и включающий указанный эпитоп M14, содержится большое количество неполярных аминокислот (I, L, V, Р, G, А, М) (Фиг. 19В). Физические свойства этих аминокислот будут приводить к нерастворимости пептида в воде и неупорядоченной вторичной структуре, в отсутствие встроенных между ними полярных или заряженных аминокислот. Таким образом, в данном документе предполагается, что свойства минимального связывающего фрагмента антитела М14, описанного в WO 2018/156367 (Kindred Biosciences, Inc.) зависят от слияния с GST, а не присущи самому пептиду.
В настоящем документе описано несколько пептидных отображений эпитопов IL-31, их свойства присущи самим пептидам, существующим независимо, а не в составе слитого белка. Это проиллюстрировано в разделе 2.2 (Фиг. 13В), где показаны связывающие и ингибирующие свойства мимотопов класса 15Н05 как в конъюгированной, так и в неконъюгированной форме. Помимо характеристик вторичной структуры пептидов, еще одним ключевым аспектом при создании вакцин, необходимым для соответствующей презентации на поверхности Т-клеток, является первичная аминокислотная последовательность. Соответствующие аминокислотные последовательности в сочетании с белком-носителем, имеющим В- и Т-клеточные эпитопы, будут вызывать иммунный ответ, направленный на ключевые участки белка IL-31. Множество областей IL-31 описано в данном документе как подходящие для выработки направленного иммунного ответа у собак и кошек. Успех мимотопов вакцины зависит от факторов, описанных в данном документе, и в конечном итоге определяется эффективностью ответа in vivo. Тем не менее, на основании изучения нескольких участков эпитопов на IL-31, описанных в данном документе, предполагается, что могут существовать другие такие эпитопы, которые могут оказаться подходящими мимотопами вакцины. Антитело 15Н05 распознает петлю, предшествующую спирали D, показанную на Фиг. 6 В как сайт 2. Предполагают, что и другие петли белка, возможно, представляют эпитопы, доступные для антител. Например, петля, образованная сближением спирали А с находящейся за ней последовательностью случайной спирали, имеет схожие с петлей 15Н05 положение и структуру. Эта петля АВ описана на Фиг. 20, где приведено сравнение первичных аминокислотных последовательностей нескольких биологических видов. Не желая ограничиваться этим примером как единственным, полагают, что другие подобные области белка могут иметь общие иммуногенные характеристики.
2.11. Сывороточные титры к мимотопам лошадиного IL-31 после вакцинации мышей полноразмерным лошадиным белком IL-31
Мышей иммунизировали полноразмерным лошадиным IL-31 (SEQ ID NO: 165; Equine_IL-31), соответствующей нуклеотидной последовательностью которого является (SEQ ID NO: 166; Canine_IL-31), конъюгированным с CRM-197, аналогично способу, описанному в разделе 1.6 настоящего описания. Пептиды, конъюгированные с биотином, представляющие три описанных в данном документе области эпитопа, сконструировали для исследования связывания методом биослойной интерферометрии (Octet, ForteBio). Описание этих пептидов приведено на Фиг. 21А. Каждый пептид содержит N-концевой биотин со спейсерной последовательностью из трех аминокислот (GSG), подписанной на рисунке жирным шрифтом с подчеркиванием. На рисунке также указан номер соответствующего положения в аминокислотной последовательности SEQ ID NO 165. Мимотоп 15Н05 включает два концевых остатка цистеина (также выделены жирным шрифтом и подчеркнуты) для облегчения циклизации по дисульфидной связи. На Фиг. 21В показаны результаты биослойной интерферометрии, где пептиды, описанные на Фиг. 21А, иммобилизовали на поверхности сенсора, покрытой стрептавидином, а затем использовали для исследования нескольких разведений мышиных антител против лошадиного IL-31 или контрольной мышиной сыворотки. Контрольную мышиную сыворотку получали от мыши, вакцинированной посторонним белком. Ответ, описываемый в данном документе как амплитуда сигнала после 120 секунд связывания с антисывороткой, представлен на оси у Фиг. По этим данным можно оценить иммунный ответ, развивающийся в результате презентации эпитопов полноразмерного лошадиного белка IL-31. Кроме того, способность этих мимотопов IL-31 распознаваться в ходе таких иммунных ответов можно оценить по связыванию. Эти данные показывают, что все три описанных пептида-мимотопа (15Н05, ВС-спираль и А-спираль) распознаются как надлежащие иммуногены в результате процессинга и презентации лошадиного белка IL-31 in vivo. При связывании контрольной сыворотки с каждым мимотопом наблюдался минимальный сигнал, за исключением спирали А, которая давала некоторый сигнал, зависящий от разведения. Подобно тому, как презентация описанных в данном документе мимотопов вызывает иммунные ответы на полноразмерный белок, в данном эксперименте подтвердили развитие иммунного ответа на мимотопы при иммунизации белком, произведя реципрокную проверку этих эпитопов.
--->
SEQUENCE LISTING
<110> Zoetis Services LLC
Bammert, Gary F.
Dunham, Steven A.
<120> PEPTIDE VACCINES AGAINST INTERLEUKIN-31
<130> ZP000229A
<150> US 62/643,921
<151> 2018-03-16
<160> 203
<170> PatentIn version 3.5
<210> 1
<211> 5
<212> PRT
<213> Mus musculus
<400> 1
Ser Tyr Thr Ile His
1 5
<210> 2
<211> 17
<212> PRT
<213> Mus musculus
<400> 2
Asn Ile Asn Pro Thr Ser Gly Tyr Thr Glu Asn Asn Gln Arg Phe Lys
1 5 10 15
Asp
<210> 3
<211> 13
<212> PRT
<213> Mus musculus
<400> 3
Trp Gly Phe Lys Tyr Asp Gly Glu Trp Ser Phe Asp Val
1 5 10
<210> 4
<211> 11
<212> PRT
<213> Mus musculus
<400> 4
Arg Ala Ser Gln Gly Ile Ser Ile Trp Leu Ser
1 5 10
<210> 5
<211> 7
<212> PRT
<213> Mus musculus
<400> 5
Lys Ala Ser Asn Leu His Ile
1 5
<210> 6
<211> 9
<212> PRT
<213> Mus musculus
<400> 6
Leu Gln Ser Gln Thr Tyr Pro Leu Thr
1 5
<210> 7
<211> 5
<212> PRT
<213> Mus musculus
<400> 7
Tyr Tyr Asp Ile Asn
1 5
<210> 8
<211> 17
<212> PRT
<213> Mus musculus
<400> 8
Trp Ile Phe Pro Gly Asp Gly Gly Thr Lys Tyr Asn Glu Thr Phe Lys
1 5 10 15
Gly
<210> 9
<211> 14
<212> PRT
<213> Mus musculus
<400> 9
Ala Arg Gly Gly Thr Ser Val Ile Arg Asp Ala Met Asp Tyr
1 5 10
<210> 10
<211> 15
<212> PRT
<213> Mus musculus
<400> 10
Arg Ala Ser Glu Ser Val Asp Asn Tyr Gly Ile Ser Phe Met His
1 5 10 15
<210> 11
<211> 7
<212> PRT
<213> Mus musculus
<400> 11
Arg Ala Ser Asn Leu Glu Ser
1 5
<210> 12
<211> 9
<212> PRT
<213> Mus musculus
<400> 12
Gln Gln Ser Asn Lys Asp Pro Leu Thr
1 5
<210> 13
<211> 5
<212> PRT
<213> Canis familiaris
<400> 13
Ser Tyr Gly Met Ser
1 5
<210> 14
<211> 17
<212> PRT
<213> Canis familiaris
<400> 14
His Ile Asn Ser Gly Gly Ser Ser Thr Tyr Tyr Ala Asp Ala Val Lys
1 5 10 15
Gly
<210> 15
<211> 15
<212> PRT
<213> Canis familiaris
<400> 15
Val Tyr Thr Thr Leu Ala Ala Phe Trp Thr Asp Asn Phe Asp Tyr
1 5 10 15
<210> 16
<211> 13
<212> PRT
<213> Canis familiaris
<400> 16
Ser Gly Ser Thr Asn Asn Ile Gly Ile Leu Ala Ala Thr
1 5 10
<210> 17
<211> 7
<212> PRT
<213> Canis familiaris
<400> 17
Ser Asp Gly Asn Arg Pro Ser
1 5
<210> 18
<211> 11
<212> PRT
<213> Canis familiaris
<400> 18
Gln Ser Phe Asp Thr Thr Leu Asp Ala Tyr Val
1 5 10
<210> 19
<211> 5
<212> PRT
<213> Canis familiaris
<400> 19
Asp Tyr Ala Met Ser
1 5
<210> 20
<211> 17
<212> PRT
<213> Canis familiaris
<400> 20
Gly Ile Asp Ser Val Gly Ser Gly Thr Ser Tyr Ala Asp Ala Val Lys
1 5 10 15
Gly
<210> 21
<211> 8
<212> PRT
<213> Canis familiaris
<400> 21
Gly Phe Pro Gly Ser Phe Glu His
1 5
<210> 22
<211> 13
<212> PRT
<213> Canis familiaris
<400> 22
Thr Gly Ser Ser Ser Asn Ile Gly Ser Gly Tyr Val Gly
1 5 10
<210> 23
<211> 7
<212> PRT
<213> Canis familiaris
<400> 23
Tyr Asn Ser Asp Arg Pro Ser
1 5
<210> 24
<211> 10
<212> PRT
<213> Canis familiaris
<400> 24
Ser Val Tyr Asp Arg Thr Phe Asn Ala Val
1 5 10
<210> 25
<211> 5
<212> PRT
<213> Canis familiaris
<400> 25
Ser Tyr Asp Met Thr
1 5
<210> 26
<211> 17
<212> PRT
<213> Canis familiaris
<400> 26
Asp Val Asn Ser Gly Gly Thr Gly Thr Ala Tyr Ala Val Ala Val Lys
1 5 10 15
Gly
<210> 27
<211> 9
<212> PRT
<213> Canis familiaris
<400> 27
Leu Gly Val Arg Asp Gly Leu Ser Val
1 5
<210> 28
<211> 11
<212> PRT
<213> Canis familiaris
<400> 28
Ser Gly Glu Ser Leu Asn Glu Tyr Tyr Thr Gln
1 5 10
<210> 29
<211> 7
<212> PRT
<213> Canis familiaris
<400> 29
Arg Asp Thr Glu Arg Pro Ser
1 5
<210> 30
<211> 10
<212> PRT
<213> Canis familiaris
<400> 30
Glu Ser Ala Val Asp Thr Gly Thr Leu Val
1 5 10
<210> 31
<211> 5
<212> PRT
<213> Canis familiaris
<400> 31
Thr Tyr Val Met Asn
1 5
<210> 32
<211> 17
<212> PRT
<213> Canis familiaris
<400> 32
Ser Ile Asn Gly Gly Gly Ser Ser Pro Thr Tyr Ala Asp Ala Val Arg
1 5 10 15
Gly
<210> 33
<211> 8
<212> PRT
<213> Canis familiaris
<400> 33
Ser Met Val Gly Pro Phe Asp Tyr
1 5
<210> 34
<211> 11
<212> PRT
<213> Canis familiaris
<400> 34
Ser Gly Glu Ser Leu Ser Asn Tyr Tyr Ala Gln
1 5 10
<210> 35
<211> 7
<212> PRT
<213> Canis familiaris
<400> 35
Lys Asp Thr Glu Arg Pro Ser
1 5
<210> 36
<211> 10
<212> PRT
<213> Canis familiaris
<400> 36
Glu Ser Ala Val Ser Ser Asp Thr Ile Val
1 5 10
<210> 37
<211> 5
<212> PRT
<213> Canis familiaris
<400> 37
Ser Tyr Ala Met Lys
1 5
<210> 38
<211> 17
<212> PRT
<213> Canis familiaris
<400> 38
Thr Ile Asn Asn Asp Gly Thr Arg Thr Gly Tyr Ala Asp Ala Val Arg
1 5 10 15
Gly
<210> 39
<211> 15
<212> PRT
<213> Canis familiaris
<400> 39
Gly Asn Ala Glu Ser Gly Cys Thr Gly Asp His Cys Pro Pro Tyr
1 5 10 15
<210> 40
<211> 11
<212> PRT
<213> Canis familiaris
<400> 40
Ser Gly Glu Ser Leu Asn Lys Tyr Tyr Ala Gln
1 5 10
<210> 41
<211> 7
<212> PRT
<213> Canis familiaris
<400> 41
Lys Asp Thr Glu Arg Pro Ser
1 5
<210> 42
<211> 10
<212> PRT
<213> Canis familiaris
<400> 42
Glu Ser Ala Val Ser Ser Glu Thr Asn Val
1 5 10
<210> 43
<211> 5
<212> PRT
<213> Canis familiaris
<400> 43
Thr Tyr Phe Met Ser
1 5
<210> 44
<211> 17
<212> PRT
<213> Canis familiaris
<400> 44
Leu Ile Ser Ser Asp Gly Ser Gly Thr Tyr Tyr Ala Asp Ala Val Lys
1 5 10 15
Gly
<210> 45
<211> 7
<212> PRT
<213> Canis familiaris
<400> 45
Phe Trp Arg Ala Phe Asn Asp
1 5
<210> 46
<211> 14
<212> PRT
<213> Canis familiaris
<400> 46
Gly Leu Asn Ser Gly Ser Val Ser Thr Ser Asn Tyr Pro Gly
1 5 10
<210> 47
<211> 7
<212> PRT
<213> Canis familiaris
<400> 47
Asp Thr Gly Ser Arg Pro Ser
1 5
<210> 48
<211> 10
<212> PRT
<213> Canis familiaris
<400> 48
Ser Leu Tyr Thr Asp Ser Asp Ile Leu Val
1 5 10
<210> 49
<211> 5
<212> PRT
<213> Canis familiaris
<400> 49
Asp Arg Gly Met Ser
1 5
<210> 50
<211> 17
<212> PRT
<213> Canis familiaris
<400> 50
Tyr Ile Arg Tyr Asp Gly Ser Arg Thr Asp Tyr Ala Asp Ala Val Glu
1 5 10 15
Gly
<210> 51
<211> 8
<212> PRT
<213> Canis familiaris
<400> 51
Trp Asp Gly Ser Ser Phe Asp Tyr
1 5
<210> 52
<211> 16
<212> PRT
<213> Canis familiaris
<400> 52
Lys Ala Ser Gln Ser Leu Leu His Ser Asp Gly Asn Thr Tyr Leu Asp
1 5 10 15
<210> 53
<211> 7
<212> PRT
<213> Canis familiaris
<400> 53
Lys Val Ser Asn Arg Asp Pro
1 5
<210> 54
<211> 9
<212> PRT
<213> Canis familiaris
<400> 54
Met Gln Ala Ile His Phe Pro Leu Thr
1 5
<210> 55
<211> 5
<212> PRT
<213> Canis familiaris
<400> 55
Ser Tyr Val Met Thr
1 5
<210> 56
<211> 17
<212> PRT
<213> Canis familiaris
<400> 56
Gly Ile Asn Ser Glu Gly Ser Arg Thr Ala Tyr Ala Asp Ala Val Lys
1 5 10 15
Gly
<210> 57
<211> 10
<212> PRT
<213> Canis familiaris
<400> 57
Gly Asp Ile Val Ala Thr Gly Thr Ser Tyr
1 5 10
<210> 58
<211> 11
<212> PRT
<213> Canis familiaris
<400> 58
Ser Gly Glu Thr Leu Asn Arg Phe Tyr Thr Gln
1 5 10
<210> 59
<211> 7
<212> PRT
<213> Canis familiaris
<400> 59
Lys Asp Thr Glu Arg Pro Ser
1 5
<210> 60
<211> 10
<212> PRT
<213> Canis familiaris
<400> 60
Lys Ser Ala Val Ser Ile Asp Val Gly Val
1 5 10
<210> 61
<211> 5
<212> PRT
<213> Canis familiaris
<400> 61
Thr Tyr Val Met Asn
1 5
<210> 62
<211> 17
<212> PRT
<213> Canis familiaris
<400> 62
Ser Ile Asn Gly Gly Gly Ser Ser Pro Thr Tyr Ala Asp Ala Val Arg
1 5 10 15
Gly
<210> 63
<211> 8
<212> PRT
<213> Canis familiaris
<400> 63
Ser Met Val Gly Pro Phe Asp Tyr
1 5
<210> 64
<211> 11
<212> PRT
<213> Canis familiaris
<400> 64
Ser Gly Lys Ser Leu Ser Tyr Tyr Tyr Ala Gln
1 5 10
<210> 65
<211> 7
<212> PRT
<213> Canis familiaris
<400> 65
Lys Asp Thr Glu Arg Pro Ser
1 5
<210> 66
<211> 10
<212> PRT
<213> Canis familiaris
<400> 66
Glu Ser Ala Val Ser Ser Asp Thr Ile Val
1 5 10
<210> 67
<211> 122
<212> PRT
<213> Mus musculus
<400> 67
Gln Val Gln Leu Gln Gln Ser Ala Ala Glu Leu Ala Arg Pro Gly Ala
1 5 10 15
Ser Val Lys Met Ser Cys Lys Thr Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Thr Ile His Trp Ile Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Asn Ile Asn Pro Thr Ser Gly Tyr Thr Glu Asn Asn Gln Arg Phe
50 55 60
Lys Asp Lys Thr Thr Leu Thr Val Asp Arg Ser Ser Asn Thr Ala Tyr
65 70 75 80
Leu Gln Leu His Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Phe Cys
85 90 95
Ala Arg Trp Gly Phe Lys Tyr Asp Gly Glu Trp Ser Phe Asp Val Trp
100 105 110
Gly Ala Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 68
<211> 366
<212> DNA
<213> Mus musculus
<400> 68
caggtccagc tgcagcagtc tgcagctgaa ctggcaagac ctggggcctc agtgaagatg 60
tcctgcaaga cttctggcta cacatttact tcctacacga tacactggat aaaacagagg 120
cctggacagg gtctggaatg gattggaaac attaatccca ccagtggata cactgagaac 180
aatcagaggt tcaaggacaa gaccacattg actgtagaca gatcctccaa cacagcctat 240
ttgcaactgc acagcctgac atctgaggac tctgcggtct atttctgtgc aagatggggc 300
tttaaatatg acggagaatg gtccttcgat gtctggggcg cagggaccac ggtcaccgtc 360
tcctca 366
<210> 69
<211> 107
<212> PRT
<213> Mus musculus
<400> 69
Asp Ile Gln Met Asn Gln Ser Pro Ser Ser Leu Ser Ala Ser Leu Gly
1 5 10 15
Asp Thr Ile Thr Val Thr Cys Arg Ala Ser Gln Gly Ile Ser Ile Trp
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Asn Ile Pro Lys Val Leu Ile
35 40 45
Asn Lys Ala Ser Asn Leu His Ile Gly Val Pro Pro Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr His Phe Thr Leu Thr Ile Thr Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys Leu Gln Ser Gln Thr Tyr Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Asn
100 105
<210> 70
<211> 321
<212> DNA
<213> Mus musculus
<400> 70
gacatccaaa tgaaccagtc tccatccagt ctgtctgcat ccctcggaga cacaatcacc 60
gtcacttgcc gtgccagtca gggcatcagt atttggttaa gctggtacca gcagaaacca 120
ggaaatattc ctaaagtatt gatcaataag gcttccaact tgcacatagg agtcccacca 180
aggtttagtg gcagtggatc tggaacacat ttcacattaa ctatcaccag cctacagcct 240
gaagacattg ccacttacta ctgtctacag agtcaaactt atcctctcac gttcggaggg 300
gggaccaagc tggaaataaa c 321
<210> 71
<211> 121
<212> PRT
<213> Mus musculus
<400> 71
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Leu Ser Cys Lys Ala Ser Gly Tyr Thr Phe Lys Tyr Tyr
20 25 30
Asp Ile Asn Trp Val Arg Gln Arg Pro Glu Gln Gly Leu Glu Trp Ile
35 40 45
Gly Trp Ile Phe Pro Gly Asp Gly Gly Thr Lys Tyr Asn Glu Thr Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Thr Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Arg Leu Thr Ser Glu Asp Ser Ala Val Tyr Phe Cys
85 90 95
Ala Arg Gly Gly Thr Ser Val Ile Arg Asp Ala Met Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Ser Val Thr Val Ser Ser
115 120
<210> 72
<211> 363
<212> DNA
<213> Mus musculus
<400> 72
caggttcagc tgcagcagtc tggagctgaa ctggtaaagc ctggggcttc agtgaagttg 60
tcctgcaagg cttctggcta caccttcaaa tactatgata taaactgggt gaggcagagg 120
cctgaacagg gacttgagtg gattggatgg atttttcctg gagatggtgg tactaagtac 180
aatgagacgt tcaagggcaa ggccacactg actacagaca aatcctccag cacagcctac 240
atgcagctca gcaggctgac atctgaggac tctgctgtct atttctgtgc aagagggggg 300
acttcggtga taagggatgc tatggactac tggggtcaag gaacctcagt caccgtctcc 360
tca 363
<210> 73
<211> 111
<212> PRT
<213> Mus musculus
<400> 73
Asp Ile Val Leu Thr Gln Ser Pro Ala Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Gln Arg Ala Thr Ile Ser Cys Arg Ala Ser Glu Ser Val Asp Asn Tyr
20 25 30
Gly Ile Ser Phe Met His Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Arg Ala Ser Asn Leu Glu Ser Gly Ile Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Arg Thr Asp Phe Thr Leu Thr Ile Asn
65 70 75 80
Pro Val Glu Thr Asp Asp Val Ala Thr Tyr Tyr Cys Gln Gln Ser Asn
85 90 95
Lys Asp Pro Leu Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys
100 105 110
<210> 74
<211> 333
<212> DNA
<213> Mus musculus
<400> 74
gacattgtgc tgacccaatc tccagcttct ttggctgtgt ctctagggca gagggccacc 60
atctcctgca gagccagcga aagtgttgat aattatggca ttagttttat gcactggtac 120
cagcagaaac caggacagcc acccaaactc ctcatctatc gtgcatccaa cctagaatct 180
gggatccctg ccaggttcag tggcagtggg tctaggacag acttcaccct caccattaat 240
cctgtggaga ctgatgatgt tgcaacctat tactgtcagc aaagtaataa ggatccgctc 300
acgttcggtg ctgggaccaa gctggagctg aaa 333
<210> 75
<211> 124
<212> PRT
<213> Canis familiaris
<400> 75
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Val Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Gln Trp Val
35 40 45
Ala His Ile Asn Ser Gly Gly Ser Ser Thr Tyr Tyr Ala Asp Ala Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Val Glu Val Tyr Thr Thr Leu Ala Ala Phe Trp Thr Asp Asn Phe Asp
100 105 110
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 76
<211> 372
<212> DNA
<213> Canis familiaris
<400> 76
gaggtgcagc tggtggagtc tgggggagac ctggtgaagc ctggggggtc cctgagactc 60
tcctgtgtgg cttctggatt caccttcagt agttatggca tgagctgggt ccgccaggct 120
ccagggaagg ggctgcagtg ggtcgcacac attaacagtg gtggaagtag cacatactac 180
gcagacgctg tgaagggacg attcaccatc tccagagaca acgccaagaa cacgctctat 240
ctgcagatga acagcctgag agctgaggac acggccgtct attactgtgt ggaggtttac 300
actacgttag ctgcattctg gacagacaat tttgactact ggggccaggg aaccctggtc 360
accgtctcct ca 372
<210> 77
<211> 110
<212> PRT
<213> Canis familiaris
<400> 77
Gln Ser Val Leu Thr Gln Pro Thr Ser Val Ser Gly Ser Leu Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Thr Asn Asn Ile Gly Ile Leu
20 25 30
Ala Ala Thr Trp Tyr Gln Gln Leu Pro Gly Lys Ala Pro Lys Val Leu
35 40 45
Val Tyr Ser Asp Gly Asn Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Asn Ser Ala Thr Leu Thr Ile Thr Gly Leu Gln
65 70 75 80
Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Gln Ser Phe Asp Thr Thr Leu
85 90 95
Asp Ala Tyr Val Phe Gly Ser Gly Thr Gln Leu Thr Val Leu
100 105 110
<210> 78
<211> 330
<212> DNA
<213> Canis familiaris
<400> 78
cagtctgtgc tgactcagcc gacctcagtg tcggggtccc ttggccagag ggtcaccatc 60
tcctgctctg gaagcacgaa caacatcggt attcttgctg cgacctggta ccaacaactc 120
ccaggaaagg cccctaaagt cctcgtgtac agtgatggga atcgaccgtc aggggtccct 180
gaccggtttt ccggctccaa gtctggcaac tcagccaccc tgaccatcac tgggcttcag 240
gctgaggacg aggctgatta ttactgccag tcctttgata ccacgcttga tgcttacgtg 300
ttcggctcag gaacccaact gaccgtcctt 330
<210> 79
<211> 117
<212> PRT
<213> Canis familiaris
<400> 79
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Ala Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Val Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Arg Gly Leu Gln Trp Val
35 40 45
Ala Gly Ile Asp Ser Val Gly Ser Gly Thr Ser Tyr Ala Asp Ala Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Phe Asn Leu Arg Ala Glu Asp Thr Ala Ile Tyr Tyr Cys
85 90 95
Ala Ser Gly Phe Pro Gly Ser Phe Glu His Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 80
<211> 351
<212> DNA
<213> Canis familiaris
<400> 80
gaggtgcagc tggtggagtc tgggggagac ctggtgaagc ctgcagggtc cctgagactg 60
tcctgtgtgg cctctggatt caccttcagt gactatgcca tgagctgggt ccgccaggct 120
cctgggaggg gactgcagtg ggtcgcaggt attgacagtg ttggaagtgg cacaagctac 180
gcagacgctg tgaagggccg attcacaatc tccagagacg acgccaagaa cacactgtat 240
ctgcagatgt tcaacctgag agccgaggac acggccatat attactgtgc gagcgggttc 300
cctgggtcct ttgagcactg gggccagggc accctggtca ccgtctcctc a 351
<210> 81
<211> 104
<212> PRT
<213> Canis familiaris
<400> 81
Gln Ser Val Leu Thr Gln Pro Ala Ser Val Ser Gly Ser Leu Gly Gln
1 5 10 15
Lys Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Ile Gly Ser Gly
20 25 30
Tyr Val Gly Trp Tyr Gln Gln Leu Pro Gly Thr Gly Pro Arg Thr Leu
35 40 45
Ile Tyr Tyr Asn Ser Asp Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Arg Ser Gly Thr Thr Ala Thr Leu Thr Ile Ser Gly Leu Gln
65 70 75 80
Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Val Tyr Asp Arg Thr Phe
85 90 95
Asn Ala Val Phe Gly Gly Gly Thr
100
<210> 82
<211> 327
<212> DNA
<213> Canis familiaris
<400> 82
cagtctgtac tgactcagcc ggcctcagtg tctgggtccc tgggccagaa ggtcaccatc 60
tcctgcactg gaagtagttc caacattggt agtggttatg tgggctggta ccagcagctc 120
ccaggaacag gccccagaac cctcatctat tataacagtg accgaccttc gggggtcccc 180
gatcgattct ctggctccag gtcaggcacc acagcaaccc tgaccatctc tggactccag 240
gctgaggacg aggctgatta ttactgctca gtatatgaca ggactttcaa tgctgtgttc 300
ggcggaggca cccacctgac cgtcctc 327
<210> 83
<211> 118
<212> PRT
<213> Canis familiaris
<400> 83
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Pro Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Val Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Thr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Gln Trp Val
35 40 45
Ala Asp Val Asn Ser Gly Gly Thr Gly Thr Ala Tyr Ala Val Ala Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Lys Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Leu Gly Val Arg Asp Gly Leu Ser Val Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser
115
<210> 84
<211> 354
<212> DNA
<213> Canis familiaris
<400> 84
gaggtgcagc tggtggagtc tgggggagac ctggtgaagc ctccagggtc cctgagactg 60
tcctgtgtgg cctctggatt caccttcagc agttatgaca tgacctgggt ccgccaggct 120
cctgggaagg gactgcagtg ggtcgcagat gttaacagtg gtggaactgg cacggcctac 180
gcagtcgctg tgaagggccg attcaccatc tccagagaca acgccaagaa aacactctat 240
ttacagatga acagcctgag agccgaagac acggccgttt attattgtgc gaaactaggt 300
gtgagagatg gtctttctgt ctggggccag ggcaccctgg tcaccgtctc ctcg 354
<210> 85
<211> 107
<212> PRT
<213> Canis familiaris
<400> 85
Ser Ser Val Leu Thr Gln Pro Pro Ser Val Ser Val Ser Leu Gly Gln
1 5 10 15
Thr Ala Thr Ile Ser Cys Ser Gly Glu Ser Leu Asn Glu Tyr Tyr Thr
20 25 30
Gln Trp Phe Gln Gln Lys Ala Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Arg Asp Thr Glu Arg Pro Ser Gly Ile Pro Asp Arg Phe Ser Gly Ser
50 55 60
Ser Ser Gly Asn Thr His Thr Leu Thr Ile Ser Gly Ala Arg Ala Glu
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Glu Ser Ala Val Asp Thr Gly Thr Leu
85 90 95
Val Phe Gly Gly Gly Thr His Leu Ala Val Leu
100 105
<210> 86
<211> 321
<212> DNA
<213> Canis familiaris
<400> 86
tccagtgtgc tgactcagcc tccctcggta tcagtgtctc tgggacagac agcaaccatc 60
tcctgctctg gagagagtct gaatgaatat tatacacaat ggttccagca gaaggcaggc 120
caagcccctg tcttggtcat atatagggac actgagcggc cctctgggat ccctgaccga 180
ttctctggct ccagttcagg gaacacacac accctaacca tcagcggggc tcgggccgag 240
gacgaggctg actattactg cgagtcagcg gtcgacactg gaacccttgt ctttggcgga 300
ggcacccacc tggccgtcct c 321
<210> 87
<211> 117
<212> PRT
<213> Canis familiaris
<400> 87
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Ala Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Val Ala Ser Gly Phe Thr Phe Arg Thr Tyr
20 25 30
Val Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Gln Trp Val
35 40 45
Ala Ser Ile Asn Gly Gly Gly Ser Ser Pro Thr Tyr Ala Asp Ala Val
50 55 60
Arg Gly Arg Phe Thr Val Ser Arg Asp Asn Ala Gln Asn Ser Leu Phe
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Phe Cys
85 90 95
Val Val Ser Met Val Gly Pro Phe Asp Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 88
<211> 351
<212> DNA
<213> Canis familiaris
<400> 88
gaggtgcagc tggtggagtc tgggggagac ctggtgaagc ctgcagggtc cctgagactg 60
tcctgtgtgg cctctggatt caccttcagg acctatgtca tgaactgggt ccgccaggct 120
cctgggaagg ggctgcaatg ggtcgcaagt attaacggtg gtggaagtag cccaacctac 180
gcagacgctg tgaggggccg attcaccgtc tccagggaca acgcccagaa ctcactgttt 240
ctgcagatga acagcctgag agccgaggac acagccgtgt atttttgtgt cgtgtcgatg 300
gttgggccct tcgactactg gggccaaggg accctggtca ccgtgtcctc a 351
<210> 89
<211> 102
<212> PRT
<213> Canis familiaris
<400> 89
Ser Ser Val Leu Thr Gln Pro Pro Ser Val Ser Val Ser Leu Gly Gln
1 5 10 15
Thr Ala Thr Ile Ser Cys Ser Gly Glu Ser Leu Ser Asn Tyr Tyr Ala
20 25 30
Gln Trp Phe Gln Gln Lys Ala Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Lys Asp Thr Glu Arg Pro Ser Gly Ile Pro Asp Arg Phe Ser Gly Ser
50 55 60
Ser Ser Gly Asn Thr His Thr Leu Thr Ile Ser Gly Ala Arg Ala Glu
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Glu Ser Ala Val Ser Ser Asp Thr Ile
85 90 95
Val Phe Gly Gly Gly Thr
100
<210> 90
<211> 321
<212> DNA
<213> Canis familiaris
<400> 90
tccagtgtgc tgactcagcc tccctcggta tcagtgtctc tggggcagac agcaaccatc 60
tcctgctctg gagagagtct gagtaactat tatgcacaat ggttccagca gaaggcaggc 120
caagcccctg tgttggtcat atataaggac actgagcggc cctctgggat ccctgaccga 180
ttctctggct ccagttcagg gaacacacac accctgacca tcagcggggc tcgggccgag 240
gacgaggctg actattactg tgagtcagca gtcagttctg atactattgt gttcggcgga 300
ggcacccacc tgaccgtcct c 321
<210> 91
<211> 124
<212> PRT
<213> Canis familiaris
<400> 91
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Ala Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Val Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Lys Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Gln Trp Val
35 40 45
Ala Thr Ile Asn Asn Asp Gly Thr Arg Thr Gly Tyr Ala Asp Ala Val
50 55 60
Arg Gly Arg Phe Thr Ile Ser Lys Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asp Ser Leu Arg Ala Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Lys Gly Asn Ala Glu Ser Gly Cys Thr Gly Asp His Cys Pro Pro
100 105 110
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 92
<211> 372
<212> DNA
<213> Canis familiaris
<400> 92
gaggtgcagc tggtggagtc tgggggagac ctggtgaagc ctgcggggtc cctgagactg 60
tcctgtgtgg cctctggatt caccttcagt agttatgcca tgaaatgggt ccgccaggct 120
cctgggaagg ggctgcagtg ggtcgcgact attaacaatg atggaaccag aacaggctac 180
gcagacgctg tgaggggccg attcaccatc tccaaagaca acgccaaaaa cacactgtat 240
ctgcagatgg acagcctgag agccgacgac acggccgtct attactgtac aaagggcaat 300
gccgaatccg gctgtactgg tgatcactgt cctccctact ggggccaggg aaccctggtc 360
accgtctcct ca 372
<210> 93
<211> 107
<212> PRT
<213> Canis familiaris
<400> 93
Ser Ser Val Leu Thr Gln Pro Pro Ser Val Ser Val Ser Leu Gly Gln
1 5 10 15
Thr Ala Thr Ile Ser Cys Ser Gly Glu Ser Leu Asn Lys Tyr Tyr Ala
20 25 30
Gln Trp Phe Gln Gln Lys Ala Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Lys Asp Thr Glu Arg Pro Ser Gly Ile Pro Asp Arg Phe Ser Gly Ser
50 55 60
Ser Ala Gly Asn Thr His Thr Leu Thr Ile Ser Gly Ala Arg Ala Glu
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Glu Ser Ala Val Ser Ser Glu Thr Asn
85 90 95
Val Phe Gly Ser Gly Thr Gln Leu Thr Val Leu
100 105
<210> 94
<211> 321
<212> DNA
<213> Canis familiaris
<400> 94
tccagtgtgc tgactcagcc tccctcggta tcagtgtctc tgggacagac agcaaccatc 60
tcctgctctg gagagagtct gaataaatat tatgcacaat ggttccaaca gaaggcaggc 120
caagcccctg tgttggtcat atataaggac actgagcggc cctctgggat ccctgaccga 180
ttctccggct ccagtgcagg caacacacac accctgacca tcagcggggc tcgggccgag 240
gacgaggctg actattactg cgagtcagca gtcagttctg aaactaacgt gttcggctca 300
ggaacccaac tgaccgtcct t 321
<210> 95
<211> 116
<212> PRT
<213> Canis familiaris
<400> 95
Glu Val Gln Leu Val Asp Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Val Ala Ser Gly Phe Thr Phe Ser Thr Tyr
20 25 30
Phe Met Ser Trp Val Arg Gln Ala Pro Gly Arg Gly Leu Gln Trp Val
35 40 45
Ala Leu Ile Ser Ser Asp Gly Ser Gly Thr Tyr Tyr Ala Asp Ala Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Ile Phe Trp Arg Ala Phe Asn Asp Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser
115
<210> 96
<211> 348
<212> DNA
<213> Canis familiaris
<400> 96
gaggtacaac tggtggactc tgggggagac ctggtgaagc ctggggggtc cctgagactc 60
tcctgtgtgg cctctggatt caccttcagt acctacttca tgtcctgggt ccgccaggct 120
ccagggaggg ggcttcagtg ggtcgcactt attagcagtg atggaagtgg cacatactac 180
gcagacgctg tgaagggccg attcaccatc tccagagaca atgccaagaa cacgctgtat 240
ctgcagatga acagcctgag agccgaggac acggctatgt attactgtgc gatattctgg 300
cgggccttta acgactgggg ccagggcacc ctggtcaccg tctcctca 348
<210> 97
<211> 110
<212> PRT
<213> Canis familiaris
<400> 97
Gln Thr Val Val Ile Gln Glu Pro Ser Leu Ser Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys Gly Leu Asn Ser Gly Ser Val Ser Thr Ser
20 25 30
Asn Tyr Pro Gly Trp Tyr Gln Gln Thr Arg Gly Arg Thr Pro Arg Thr
35 40 45
Ile Ile Tyr Asp Thr Gly Ser Arg Pro Ser Gly Val Pro Asn Arg Phe
50 55 60
Ser Gly Ser Ile Ser Gly Asn Lys Ala Ala Leu Thr Ile Thr Gly Ala
65 70 75 80
Gln Pro Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Leu Tyr Thr Asp Ser
85 90 95
Asp Ile Leu Val Phe Gly Gly Gly Thr His Leu Thr Val Leu
100 105 110
<210> 98
<211> 330
<212> DNA
<213> Canis familiaris
<400> 98
cagactgtgg taatccagga gccatcactc tcagtgtctc caggagggac agtcacactc 60
acatgtggcc tcaactctgg gtcagtctcc acaagtaatt accctggctg gtaccagcag 120
acccgaggcc ggactcctcg cacgattatc tacgacacag gcagtcgccc ctctggggtc 180
cctaatcgct tctccggatc catctctgga aacaaagccg ccctcaccat cacaggagcc 240
cagcccgagg atgaggctga ctattactgt tccttatata cggatagtga cattcttgtt 300
ttcggcggag gcacccacct gaccgtcctc 330
<210> 99
<211> 117
<212> PRT
<213> Canis familiaris
<400> 99
Glu Val His Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Trp Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Val Ala Ser Gly Phe Thr Phe Ser Asp Arg
20 25 30
Gly Met Ser Trp Val Arg Gln Ser Pro Gly Lys Gly Leu Gln Trp Val
35 40 45
Ala Tyr Ile Arg Tyr Asp Gly Ser Arg Thr Asp Tyr Ala Asp Ala Val
50 55 60
Glu Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Trp Asp Gly Ser Ser Phe Asp Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 100
<211> 351
<212> DNA
<213> Canis familiaris
<400> 100
gaggtgcatt tggtggagtc tgggggagac ctggtgaagc cttgggggtc cttgagactg 60
tcctgtgtgg cctctggatt cacctttagt gatcgtggca tgagctgggt ccgtcagtct 120
ccagggaagg ggctgcagtg ggtcgcatat attaggtatg atgggagtag gacagactac 180
gcagacgctg tggagggccg attcaccatc tccagagaca acgccaagaa cacgctctac 240
ctgcagatga acagcctgag agccgaggac acggccgtgt attactgtgc gagatgggac 300
ggtagttctt ttgactattg gggccagggc accctggtca ccgtctcctc a 351
<210> 101
<211> 112
<212> PRT
<213> Canis familiaris
<400> 101
Asp Ile Val Val Thr Gln Thr Pro Leu Ser Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Thr Ala Ser Phe Ser Cys Lys Ala Ser Gln Ser Leu Leu His Ser
20 25 30
Asp Gly Asn Thr Tyr Leu Asp Trp Phe Arg Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Arg Leu Ile Tyr Lys Val Ser Asn Arg Asp Pro Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Arg Ile
65 70 75 80
Ser Gly Val Glu Ala Asp Asp Ala Gly Leu Tyr Tyr Cys Met Gln Ala
85 90 95
Ile His Phe Pro Leu Thr Phe Gly Ala Gly Thr Lys Val Glu Leu Lys
100 105 110
<210> 102
<211> 336
<212> DNA
<213> Canis familiaris
<400> 102
gatattgtcg tgacacagac cccgctgtcc ctgtccgtca gccctggaga gactgcctcc 60
ttctcctgca aggccagtca gagcctcctg cacagtgatg gaaacacgta tttggattgg 120
ttccgacaga agccaggcca gtctccacag cgtttgatct acaaggtctc caacagagac 180
cctggggtcc cagacaggtt cagtggcagc gggtcaggga cagatttcac cctgagaatc 240
agcggagtgg aggctgacga tgctggactt tattactgca tgcaagcaat acactttcct 300
ctgacgttcg gagcaggaac caaggtggag ctcaaa 336
<210> 103
<211> 119
<212> PRT
<213> Canis familiaris
<400> 103
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Ala Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Val Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Val Met Thr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Gln Trp Val
35 40 45
Ala Gly Ile Asn Ser Glu Gly Ser Arg Thr Ala Tyr Ala Asp Ala Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Ile Asp Ser Leu Arg Ala Glu Asp Thr Ala Ile Tyr Tyr Cys
85 90 95
Ala Thr Gly Asp Ile Val Ala Thr Gly Thr Ser Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 104
<211> 357
<212> DNA
<213> Canis familiaris
<400> 104
gaggtgcagc tggtggagtc tgggggagac ctggtgaagc ctgcagggtc cctgagactg 60
tcctgtgtgg cctctggatt caccttcagt agttatgtca tgacctgggt ccgccaggct 120
cctgggaagg gactgcagtg ggtcgcaggc attaatagtg aggggagtag gacagcctac 180
gcagacgctg tgaagggccg attcaccatc tccagagaca acgccaagaa tacactttat 240
ctacaaatag acagcctgag agccgaggac acggccatat attactgtgc gacaggcgat 300
atagtagcga ctggtacttc gtattggggc cagggcaccc tggtcaccgt ctcctca 357
<210> 105
<211> 107
<212> PRT
<213> Canis familiaris
<400> 105
Ser Asn Val Leu Thr Gln Pro Pro Ser Val Ser Val Ser Leu Gly Gln
1 5 10 15
Thr Ala Thr Ile Ser Cys Ser Gly Glu Thr Leu Asn Arg Phe Tyr Thr
20 25 30
Gln Trp Phe Gln Gln Lys Ala Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Lys Asp Thr Glu Arg Pro Ser Gly Ile Pro Asp Arg Phe Ser Gly Ser
50 55 60
Ser Ser Gly Asn Ile His Thr Leu Thr Ile Ser Gly Ala Arg Ala Glu
65 70 75 80
Asp Glu Ala Ala Tyr Tyr Cys Lys Ser Ala Val Ser Ile Asp Val Gly
85 90 95
Val Phe Gly Gly Gly Thr His Leu Thr Val Phe
100 105
<210> 106
<211> 321
<212> DNA
<213> Canis familiaris
<400> 106
tccaatgtac tgactcagcc tccctcggta tcagtgtctc tgggacagac agcaaccatc 60
tcctgctctg gagagactct gaatagattt tatacacaat ggttccagca gaaggcaggc 120
caagcccctg tgttggtcat atataaggac actgagcggc cctctgggat ccctgaccga 180
ttctccggct ccagttcagg gaacatacac accctgacca tcagcggggc tcgggccgag 240
gacgaggctg cctattactg caagtcagca gtcagtattg atgttggtgt gttcggcgga 300
ggcacccacc tgaccgtctt c 321
<210> 107
<211> 117
<212> PRT
<213> Canis familiaris
<400> 107
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Ala Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Val Ala Ser Gly Phe Thr Phe Arg Thr Tyr
20 25 30
Val Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Gln Trp Val
35 40 45
Ala Ser Ile Asn Gly Gly Gly Ser Ser Pro Thr Tyr Ala Asp Ala Val
50 55 60
Arg Gly Arg Phe Thr Val Ser Arg Asp Asn Ala Gln Asn Ser Leu Phe
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Ile Tyr Phe Cys
85 90 95
Val Val Ser Met Val Gly Pro Phe Asp Tyr Trp Gly His Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 108
<211> 351
<212> DNA
<213> Canis familiaris
<400> 108
gaggtgcagc tggtggagtc tgggggagac ctggtgaagc ctgcagggtc cctgagactg 60
tcctgtgtgg cctctggatt caccttcagg acctatgtca tgaactgggt ccgccaggct 120
cctgggaagg ggctgcaatg ggtcgcaagt attaacggtg gtggaagtag cccaacctac 180
gcagacgctg tgaggggccg attcaccgtc tccagggaca acgcccagaa ctcactgttt 240
ctgcagatga acagcctgag agccgaggac acagccatat atttttgtgt cgtgtcgatg 300
gttgggccct tcgactactg gggccatggg accctggtca ccgtgtcctc a 351
<210> 109
<211> 107
<212> PRT
<213> Canis familiaris
<400> 109
Ser Ser Val Leu Thr Gln Pro Pro Ser Val Ser Val Ser Leu Gly Gln
1 5 10 15
Thr Ala Thr Ile Ser Cys Ser Gly Lys Ser Leu Ser Tyr Tyr Tyr Ala
20 25 30
Gln Trp Phe Gln Gln Lys Ala Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Lys Asp Thr Glu Arg Pro Ser Gly Ile Pro Asp Arg Phe Ser Gly Ser
50 55 60
Ser Ser Gly Asn Thr His Thr Leu Thr Ile Ser Gly Ala Arg Ala Glu
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Glu Ser Ala Val Ser Ser Asp Thr Ile
85 90 95
Val Phe Gly Gly Gly Thr His Leu Thr Val Leu
100 105
<210> 110
<211> 321
<212> DNA
<213> Canis familiaris
<400> 110
tccagtgtgc tgactcagcc tccctcggta tcagtgtctc tggggcagac agcaaccatc 60
tcctgctctg gaaagagtct gagttactat tatgcacaat ggttccagca gaaggcaggc 120
caagcccctg tgttggtcat atataaggac actgagcggc cctctgggat ccctgaccga 180
ttctctggct ccagttcagg gaacacacac accctgacca tcagcggggc tcgggccgag 240
gacgaggctg actattactg tgagtcagca gtcagttctg atactattgt gttcggcgga 300
ggcacccacc tgaccgtcct c 321
<210> 111
<211> 121
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность вариабельной области тяжелой цепи
фелинизированного моноклонального антитела от Mus musculus и
Felis catus
<400> 111
Gln Val Leu Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Ile Phe Cys Lys Ala Ser Gly Tyr Ser Phe Thr Tyr Tyr
20 25 30
Asp Ile Asn Trp Leu Arg Gln Ala Pro Glu Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Phe Pro Gly Asp Gly Gly Thr Lys Tyr Asn Glu Thr Phe
50 55 60
Lys Gly Arg Leu Thr Leu Thr Ala Asp Thr Ser Thr Asn Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Ala Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Gly Gly Thr Ser Val Ile Arg Asp Ala Met Asp Tyr Trp Gly
100 105 110
Gln Gly Ala Leu Val Thr Val Ser Ser
115 120
<210> 112
<211> 363
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность, кодирующая последовательность
вариабельной области тяжелой цепи фелинизированного моноклонального
антитела от Mus musculus и Felis catus
<400> 112
caggtgctgc tggtccagtc aggagcagag gtaaaaaagc ccggggcgag tgtcaagatt 60
ttctgtaagg cctccggata ctcttttacg tattacgata ttaactggct tcgccaggcc 120
cctgagcagg ggctcgaatg gatgggttgg atattccccg gagatggggg aaccaagtac 180
aacgaaacct tcaaggggag gctgaccctg actgcagata ccagcacgaa cacagtgtat 240
atggagttgt cctcactgcg atctgctgat actgccatgt actactgcgc tcgcggcggc 300
acttcagtta tcagggatgc catggactat tgggggcagg gcgcactcgt cactgtctcg 360
agc 363
<210> 113
<211> 106
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность вариабельной области легкой цепи
фелинизированного моноклонального антитела от Mus musculus и
Felis catus
<400> 113
Glu Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Pro Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Ser Val Asp Asn Tyr
20 25 30
Gly Ile Ser Phe Met His Trp Tyr Gln Gln Lys Pro Gly Lys Val Pro
35 40 45
Lys Leu Leu Ile Tyr Arg Ala Ser Asn Leu Glu Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Glu Pro Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Ser Asn
85 90 95
Lys Asp Pro Leu Thr Phe Gly Gln Gly Thr
100 105
<210> 114
<211> 333
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность, кодирующая последовательность
вариабельной области легкой цепи фелинизированного моноклонального
антитела от Mus musculus и Felis catus
<400> 114
gaaatccaga tgacacaatc tcccagctcc ctcagcgcat ctcctggcga cagggtaacc 60
atcacctgcc gcgccagcga gtcagtagac aactatggca tatccttcat gcactggtat 120
caacaaaagc ccgggaaagt ccccaaactg ttgatttaca gagcaagcaa tctcgagtca 180
ggagtcccat ctcgcttctc tggttccggt tccggaaccg acttcactct gacaatttct 240
tctctggagc ccgaggatgc cgctacatat tactgtcagc aaagcaataa agatccactg 300
accttcggac agggtaccaa gctggagatc aaa 333
<210> 115
<211> 106
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность вариабельной области легкой цепи
фелинизированного моноклонального антитела от Mus musculus и
Felis catus
<400> 115
Glu Val Val Leu Thr Gln Ser Ser Ala Phe Leu Ser Arg Thr Leu Lys
1 5 10 15
Glu Lys Ala Thr Ile Thr Cys Arg Ala Ser Glu Ser Val Asp Asn Tyr
20 25 30
Gly Ile Ser Phe Met His Trp Tyr Gln Gln Lys Pro Asn Gln Ala Pro
35 40 45
Lys Leu Leu Val Lys Arg Ala Ser Asn Leu Glu Ser Gly Ala Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Pro Glu Pro Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Ser Asn
85 90 95
Lys Asp Pro Leu Thr Phe Gly Gln Gly Thr
100 105
<210> 116
<211> 318
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность, кодирующая вариабельную область
легкой цепи фелинизированного моноклонального антитела от
Mus musculus и Felis catus
<400> 116
gaggtggtgc tgactcagag tagcgcgttt ctgtctcgga ccctgaaaga gaaagctacc 60
atcacgtgca gggcaagcga gagcgtggac aactatggta tcagcttcat gcattggtat 120
cagcagaaac ctaatcaggc gcctaagctg ctcgtgaaaa gagcctccaa ccttgagagc 180
ggcgcaccat caaggttttc aggaagtggc agcgggacag acttcaccct tacaatctct 240
agtccagagc cggaggacgc agctacctac tattgccagc aatccaataa agacccgttg 300
acattcggcc aaggtacc 318
<210> 117
<211> 106
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность вариабельной области легкой цепи
фелинизированного моноклонального антитела от Mus musculus и
Felis catus
<400> 117
Glu Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Pro Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Ser Val Asp Asn Tyr
20 25 30
Gly Ile Ser Phe Met His Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Arg Ala Ser Asn Leu Glu Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Glu Pro Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Ser Asn
85 90 95
Lys Asp Pro Leu Thr Phe Gly Gln Gly Thr
100 105
<210> 118
<211> 318
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность, кодирующая вариабельную область
легкой цепи фелинизированного моноклонального антитела от
Mus musculus и Felis catus
<400> 118
gagatccaga tgacccagtc tccatcctca ctgagtgcta gcccggggga tcgagtgact 60
ataacatgtc gggccagtga atcagtggac aactatggaa tcagttttat gcactggtat 120
cagcagaagc ccggccagcc accgaagctg ttgatttatc gcgcaagcaa tctggagtca 180
ggagtgccct ctagattttc tgggagcggt tctggcacag atttcacact cacaatatca 240
tccttggaac cggaagacgc agccacatac tattgccagc agagtaacaa ggaccctttg 300
acttttggcc agggtacc 318
<210> 119
<211> 106
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность вариабельной области легкой цепи
фелинизированного моноклонального антитела от Mus musculus и
Felis catus
<400> 119
Glu Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Pro Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Ser Val Asp Asn Tyr
20 25 30
Gly Ile Ser Phe Met His Trp Tyr Gln Gln Lys Pro Gly Gln Val Pro
35 40 45
Lys Leu Leu Ile Tyr Arg Ala Ser Asn Leu Glu Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Glu Pro Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Ser Asn
85 90 95
Lys Asp Pro Leu Thr Phe Gly Gln Gly Thr
100 105
<210> 120
<211> 318
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность,кодирующая вариабельную область
легкой цепи фелинизированного моноклонального антитела от
Mus musculus и Felis catus
<400> 120
gagatccaga tgacccagtc tccatcctca ctgagtgcta gcccggggga tcgagtgact 60
ataacatgtc gggccagtga atcagtggac aactatggaa tcagttttat gcactggtat 120
cagcagaagc ccggccaggt cccgaagctg ttgatttatc gcgcaagcaa tctggagtca 180
ggagtgccct ctagattttc tgggagcggt tctggcacag atttcacact cacaatatca 240
tccttggaac cggaagacgc agccacatac tattgccagc agagtaacaa ggaccctttg 300
acttttggcc agggtacc 318
<210> 121
<211> 122
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность вариабельной области тяжелой цепи
фелинизированного моноклонального антитела от Mus musculus и
Felis catus
<400> 121
Gln Val Leu Leu Val Gln Ser Gly Ala Glu Val Arg Thr Pro Gly Ala
1 5 10 15
Ser Val Lys Ile Phe Cys Lys Ala Ser Gly Tyr Ser Phe Thr Ser Tyr
20 25 30
Thr Ile His Trp Leu Arg Gln Ala Pro Ala Gln Gly Leu Glu Trp Met
35 40 45
Gly Asn Ile Asn Pro Thr Ser Gly Tyr Thr Glu Asn Asn Gln Arg Phe
50 55 60
Lys Asp Arg Leu Thr Leu Thr Ala Asp Thr Ser Thr Asn Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Ala Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Trp Gly Phe Lys Tyr Asp Gly Glu Trp Ser Phe Asp Val Trp
100 105 110
Gly Ala Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 122
<211> 366
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность, кодирующая вариабельную область
тяжелой цепи фелинизированного моноклонального антитела от Mus musculus и
Felis catus
<400> 122
caagtgcttc tggtgcaaag cggggcggaa gttaggaccc caggagcctc agtaaaaatt 60
ttttgtaagg catccggcta cagtttcacc agctacacta ttcactggct gaggcaggcc 120
ccggcccaag ggctggagtg gatgggaaat atcaatccca cgtctggcta tacagagaat 180
aaccaaaggt ttaaggatag gctgactctg acagctgaca catcaaccaa tacggcatac 240
atggagctct cctctctccg gagtgccgac accgccatgt actactgtgc tcggtggggg 300
tttaaatacg atggcgagtg gagcttcgac gtgtggggcg cgggcacaac cgtgaccgtc 360
tcgagc 366
<210> 123
<211> 122
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность вариабельной области тяжелой цепи от Mus musculus и
Felis catus
<400> 123
Gln Val Leu Leu Val Gln Ser Gly Ala Glu Val Arg Thr Pro Gly Ala
1 5 10 15
Ser Val Lys Ile Phe Cys Lys Ala Ser Gly Tyr Gly Phe Thr Ser Tyr
20 25 30
Thr Ile His Trp Val Arg Gln Ser Pro Ala Gln Gly Leu Glu Trp Met
35 40 45
Gly Asn Ile Asn Pro Thr Ser Gly Tyr Thr Glu Asn Asn Gln Arg Phe
50 55 60
Lys Asp Arg Leu Thr Leu Thr Ala Asp Thr Ser Thr Asn Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Ala Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Trp Gly Phe Lys Tyr Asp Gly Glu Trp Ser Phe Asp Val Trp
100 105 110
Gly Ala Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 124
<211> 366
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность, кодирующая последовательность
вариабельной области тяжелой цепи фелинизированного моноклонального
антитела от Mus musculus и Felis catus
<400> 124
caggtgctgc tcgtgcagag cggagccgaa gtgaggacac ccggtgcgag tgtaaaaatt 60
ttttgcaagg caagcggcta cgggtttaca tcctatacca tccactgggt gaggcagtcc 120
ccagcgcagg gacttgaatg gatgggaaat attaatccaa caagcgggta tactgaaaac 180
aaccaaagat ttaaggacag actgacactc accgcagata catctacaaa tacagcctac 240
atggagttgt cttccctgcg gagtgccgac acggctatgt actactgtgc tcggtggggg 300
tttaagtatg atggcgaatg gtccttcgac gtctggggag ctggaaccac cgtgaccgtc 360
tcgagc 366
<210> 125
<211> 122
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность вариабельной области тяжелой цепи от Mus musculus и
Felis catus
<400> 125
Gln Val Leu Leu Val Gln Ser Gly Ala Glu Val Arg Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Ile Phe Cys Lys Ala Ser Gly Tyr Ser Phe Thr Ser Tyr
20 25 30
Thr Ile His Trp Leu Arg Gln Ala Pro Glu Gln Gly Leu Glu Trp Met
35 40 45
Gly Asn Ile Asn Pro Thr Ser Gly Tyr Thr Glu Asn Asn Gln Arg Phe
50 55 60
Lys Asp Arg Leu Thr Leu Thr Ala Asp Thr Ser Thr Asn Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Ala Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Trp Gly Phe Lys Tyr Asp Gly Glu Trp Ser Phe Asp Val Trp
100 105 110
Gly Ala Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 126
<211> 366
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность, кодирующая последовательность
вариабельной области тяжелой цепи фелинизированного моноклонального
антитела от Mus musculus и Felis catus
<400> 126
caggtcctct tggttcaaag cggagccgaa gtccgaaaac cgggtgcctc agtgaaaatc 60
ttctgtaagg cctccggcta tagtttcacg agttacacaa tccactggct gcgacaggca 120
ccagagcagg gactggagtg gatgggaaat ataaatccga cgtctgggta cacagaaaac 180
aaccagagat tcaaggatag attgacactg accgcggata ctagtacaaa tacggcttac 240
atggaactgt cctcactccg gtcagccgac accgccatgt attactgtgc tcgctggggg 300
ttcaagtatg atggagagtg gagcttcgac gtatggggag ccggaaccac tgtgaccgtc 360
tcgagc 366
<210> 127
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность вариабельной области легкой цепи фелинизированного
моноклонального антитела от Mus musculus и Felis catus
<400> 127
Glu Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Pro Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser Ile Trp
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Lys Val Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Asn Leu His Ile Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Ala Ala Thr Tyr Tyr Cys Leu Gln Ser Gln Thr Tyr Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 128
<211> 321
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность, кодирующая последовательность
вариабельной области легкой цепи фелинизированного моноклонального
антитела от Mus musculus и Felis catus
<400> 128
gaaattcaga tgactcaatc accttcatct ttgagtgcat cacccggaga tagagttaca 60
atcacctgca gggcgagtca agggatctcc atatggttgt catggtatca gcagaaaccc 120
ggaaaggtcc cgaaactctt gatctacaag gcctctaacc tgcacattgg cgtgccaagc 180
cgattcagcg ggagcggaag tggcaccgac tttactctga cgatcagttc actggagccc 240
gaggacgctg caacatacta ttgtctgcaa tctcagacct accctctcac ctttggagga 300
ggtaccaagc tggagatcaa a 321
<210> 129
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность вариабельной области легкой цепи фелинизированного
моноклонального антитела от Mus musculus и Felis catus
<400> 129
Glu Ile Thr Met Thr Gln Ser Pro Gly Ser Leu Ala Gly Ser Pro Gly
1 5 10 15
Gln Gln Val Thr Met Asn Cys Arg Ala Ser Gln Gly Ile Ser Ile Trp
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Gln His Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Asn Leu His Ile Gly Val Pro Asp Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Asn Leu Gln Ala
65 70 75 80
Glu Asp Val Ala Ser Tyr Tyr Cys Leu Gln Ser Gln Thr Tyr Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 130
<211> 321
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность, кодирующая последовательность
вариабельной области легкой цепи фелинизированного моноклонального
антитела от Mus musculus и Felis catus
<400> 130
gaaattacca tgacacaaag ccccggctcc ctggccggct cccccggaca gcaagtgacc 60
atgaattgtc gggccagcca gggaatttct atatggctct cttggtatca gcaaaaaccc 120
ggacagcacc ctaaacttct gatctacaaa gcaagtaact tgcacatcgg cgtccctgat 180
cgattcagtg gctcaggttc cggtacagat tttactctta ccatcagcaa tctgcaggct 240
gaggatgtgg caagctatta ctgtctccaa agtcagactt accctctgac atttgggggc 300
ggtaccaagc tggagatcaa a 321
<210> 131
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность вариабельной области легкой цепи фелинизированного
моноклонального антитела от Mus musculus и Felis catus
<400> 131
Glu Val Val Leu Thr Gln Ser Ser Ala Phe Leu Ser Arg Thr Leu Lys
1 5 10 15
Glu Lys Ala Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser Ile Trp
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Asn Gln Ala Pro Lys Leu Leu Val
35 40 45
Lys Lys Ala Ser Asn Leu His Ile Gly Ala Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Pro Glu Pro
65 70 75 80
Glu Asp Ala Ala Thr Tyr Tyr Cys Leu Gln Ser Gln Thr Tyr Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Gly Asp Gln
100 105
<210> 132
<211> 322
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность, кодирующая последовательность
вариабельной области легкой цепи фелинизированного моноклонального
антитела от Mus musculus и Felis catus
<400> 132
gaggtagtgc tgactcagtc ctccgccttc ttgtcaagaa ctctcaaaga gaaagcaaca 60
atcacttgtc gggcgtctca agggatatca atttggctga gctggtatca gcagaaacca 120
aatcaagcgc cgaaactgct ggtgaagaag gcctccaatc tccacattgg cgcacccagc 180
aggttttccg gcagtggctc tggcacagat ttcactctga ccatcagctc acccgagccc 240
gaagacgccg ctacatacta ttgcttgcaa tcccagacat accccctgac ttttggggga 300
ggtaccaagc tgggagatca aa 322
<210> 133
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность вариабельной области легкой цепи фелинизированного
моноклонального антитела от Mus musculus и Felis catus
<400> 133
Asp Ile Gln Met Asn Gln Ser Pro Ser Ser Leu Ser Ala Ser Leu Gly
1 5 10 15
Asp Thr Ile Thr Val Thr Cys Arg Ala Ser Gln Gly Ile Ser Ile Trp
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Lys Val Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Asn Leu His Ile Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Ala Ala Thr Tyr Tyr Cys Leu Gln Ser Gln Thr Tyr Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 134
<211> 321
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность, кодирующая последовательность
вариабельной области легкой цепи фелинизированного моноклонального
антитела от Mus musculus и Felis catus
<400> 134
gacattcaga tgaatcagtc tcctagctca ctgtcagcca gccttggaga caccattaca 60
gtcacttgca gggcgagtca agggatctcc atatggttgt catggtatca gcagaaaccc 120
ggaaaggtcc cgaaactctt gatctacaag gcctctaacc tgcacattgg cgtgccaagc 180
cgattcagcg ggagcggaag tggcaccgac tttactctga cgatcagttc actggagccc 240
gaggacgctg caacatacta ttgtctgcaa tctcagacct accctctcac ctttggagga 300
ggtaccaagc tggagatcaa a 321
<210> 135
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность вариабельной области легкой цепи фелинизированного
моноклонального антитела от Mus musculus и Felis catus
<400> 135
Glu Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Pro Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser Ile Trp
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Asn Ile Pro Lys Val Leu Ile
35 40 45
Asn Lys Ala Ser Asn Leu His Ile Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Ala Ala Thr Tyr Tyr Cys Leu Gln Ser Gln Thr Tyr Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 136
<211> 321
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность, кодирующая последовательность
вариабельной области легкой цепи фелинизированного моноклонального
антитела от Mus musculus и Felis catus
<400> 136
gaaattcaga tgactcaatc accttcatct ttgagtgcat cacccggaga tagagttaca 60
atcacctgca gggcgagtca agggatctcc atatggttgt catggtatca gcagaaaccg 120
ggcaatatcc caaaggtgct gattaacaag gcctctaacc tgcacattgg cgtgccaagc 180
cgattcagcg ggagcggaag tggcaccgac tttactctga cgatcagttc actggagccc 240
gaggacgctg caacatacta ttgtctgcaa tctcagacct accctctcac ctttggagga 300
ggtaccaagc tggagatcaa a 321
<210> 137
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность вариабельной области легкой цепи фелинизированного
моноклонального антитела от Mus musculus и Felis catus
<400> 137
Glu Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Pro Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser Ile Trp
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Lys Val Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Asn Leu His Ile Gly Val Pro Pro Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr His Phe Thr Leu Thr Ile Thr Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys Leu Gln Ser Gln Thr Tyr Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 138
<211> 321
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность, кодирующая последовательность
вариабельной области легкой цепи фелинизированного моноклонального
антитела от Mus musculus и Felis catus
<400> 138
gaaattcaga tgactcaatc accttcatct ttgagtgcat cacccggaga tagagttaca 60
atcacctgca gggcgagtca agggatctcc atatggttgt catggtatca gcagaaaccc 120
ggaaaggtcc cgaaactctt gatctacaag gcctctaacc tgcacattgg ggtcccccca 180
aggttcagcg gatctggatc cgggacccac tttactctga ccataacaag cctgcagcct 240
gaagacattg ctacctatta ctgcctgcaa tctcagacct accctctcac ctttggagga 300
ggtaccaagc tggagatcaa a 321
<210> 139
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность вариабельной области легкой цепи фелинизированного
моноклонального антитела от Mus musculus и Felis catus
<400> 139
Asp Ile Gln Met Asn Gln Ser Pro Ser Ser Leu Ser Ala Ser Leu Gly
1 5 10 15
Asp Thr Ile Thr Val Thr Cys Arg Ala Ser Gln Gly Ile Ser Ile Trp
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Asn Ile Pro Lys Val Leu Ile
35 40 45
Asn Lys Ala Ser Asn Leu His Ile Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Ala Ala Thr Tyr Tyr Cys Leu Gln Ser Gln Thr Tyr Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 140
<211> 321
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность, кодирующая последовательность
вариабельной области легкой цепи фелинизированного моноклонального
антитела от Mus musculus и Felis catus
<400> 140
gacattcaga tgaatcagtc tcctagctca ctgtcagcca gccttggaga caccattaca 60
gtcacttgca gggcgagtca agggatctcc atatggttgt catggtatca gcagaaaccg 120
ggcaatatcc caaaggtgct gattaacaag gcctctaacc tgcacattgg cgtgccaagc 180
cgattcagcg ggagcggaag tggcaccgac tttactctga cgatcagttc actggagccc 240
gaggacgctg caacatacta ttgtctgcaa tctcagacct accctctcac ctttggagga 300
ggtaccaagc tggagatcaa a 321
<210> 141
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность вариабельной области легкой цепи фелинизированного
моноклонального антитела от Mus musculus и Felis catus
<400> 141
Asp Ile Gln Met Asn Gln Ser Pro Ser Ser Leu Ser Ala Ser Leu Gly
1 5 10 15
Asp Thr Ile Thr Val Thr Cys Arg Ala Ser Gln Gly Ile Ser Ile Trp
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Lys Val Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Asn Leu His Ile Gly Val Pro Pro Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr His Phe Thr Leu Thr Ile Thr Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys Leu Gln Ser Gln Thr Tyr Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 142
<211> 321
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность, кодирующая последовательность
вариабельной области легкой цепи фелинизированного моноклонального
антитела от Mus musculus и Felis catus
<400> 142
gacattcaga tgaatcagtc tcctagctca ctgtcagcca gccttggaga caccattaca 60
gtcacttgca gggcgagtca agggatctcc atatggttgt catggtatca gcagaaaccc 120
ggaaaggtcc cgaaactctt gatctacaag gcctctaacc tgcacattgg ggtcccccca 180
aggttcagcg gatctggatc cgggacccac tttactctga ccataacaag cctgcagcct 240
gaagacattg ctacctatta ctgcctgcaa tctcagacct accctctcac ctttggagga 300
ggtaccaagc tggagatcaa a 321
<210> 143
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность вариабельной области легкой цепи фелинизированного
моноклонального антитела от Mus musculus и Felis catus
<400> 143
Glu Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Pro Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser Ile Trp
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Asn Ile Pro Lys Val Leu Ile
35 40 45
Asn Lys Ala Ser Asn Leu His Ile Gly Val Pro Pro Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr His Phe Thr Leu Thr Ile Thr Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys Leu Gln Ser Gln Thr Tyr Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 144
<211> 321
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность, кодирующая последовательность
вариабельной области легкой цепи фелинизированного моноклонального
антитела от Mus musculus и Felis catus
<400> 144
gaaattcaga tgactcaatc accttcatct ttgagtgcat cacccggaga tagagttaca 60
atcacctgca gggcgagtca agggatctcc atatggttgt catggtatca gcagaaaccg 120
ggcaatatcc caaaggtgct gattaacaag gcctctaacc tgcacattgg ggtcccccca 180
aggttcagcg gatctggatc cgggacccac tttactctga ccataacaag cctgcagcct 240
gaagacattg ctacctatta ctgcctgcaa tctcagacct accctctcac ctttggagga 300
ggtaccaagc tggagatcaa a 321
<210> 145
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность вариабельной области легкой цепи фелинизированного
моноклонального антитела от Mus musculus и Felis catus
<400> 145
Glu Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Pro Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser Ile Trp
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Asn Val Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Asn Leu His Ile Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Ala Ala Thr Tyr Tyr Cys Leu Gln Ser Gln Thr Tyr Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 146
<211> 321
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность, кодирующая последовательность
вариабельной области легкой цепи фелинизированного моноклонального
антитела от Mus musculus и Felis catus
<400> 146
gaaatccaga tgacacagtc ccccagtagc ctttccgctt caccgggcga tagagtcact 60
attacgtgca gggcctccca gggtatttct atctggctga gctggtatca gcagaagccc 120
ggtaatgtgc caaagctctt gatctacaag gcatctaacc ttcatatcgg agtgccctca 180
agatttagtg ggtcaggcag cggaaccgat ttcacattga ccattagttc tctggaacca 240
gaggacgctg ccacttacta ctgcctgcag tcccaaacat accctttgac ttttgggggg 300
ggtaccaagc tggagatcaa a 321
<210> 147
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность вариабельной области легкой цепи фелинизированного
моноклонального антитела от Mus musculus и Felis catus
<400> 147
Glu Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Pro Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser Ile Trp
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Lys Ile Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Asn Leu His Ile Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Ala Ala Thr Tyr Tyr Cys Leu Gln Ser Gln Thr Tyr Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 148
<211> 321
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность, кодирующая последовательность
вариабельной области легкой цепи фелинизированного моноклонального
антитела от Mus musculus и Felis catus
<400> 148
gagattcaga tgacccagag cccatcaagc ctctccgctt cccccggaga ccgggtgacc 60
atcacatgca gagcttcaca gggaatctca atctggctca gctggtatca gcagaagcca 120
ggcaagattc cgaagttgct tatctataag gccagtaacc tgcatatcgg agttccatca 180
agattcagtg gtagcggaag tgggacagat ttcactctca ccatcagctc cctcgaacca 240
gaggacgctg caacttacta ctgcctgcag tcccagacat atccacttac tttcggcggg 300
ggtaccaagc tggagatcaa a 321
<210> 149
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность вариабельной области легкой цепи фелинизированного
моноклонального антитела от Mus musculus и Felis catus
<400> 149
Glu Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Pro Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser Ile Trp
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Lys Val Pro Lys Val Leu Ile
35 40 45
Tyr Lys Ala Ser Asn Leu His Ile Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Ala Ala Thr Tyr Tyr Cys Leu Gln Ser Gln Thr Tyr Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 150
<211> 322
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность, кодирующая последовательность
вариабельной области легкой цепи фелинизированного моноклонального
антитела от Mus musculus и Felis catus
<400> 150
gagattcaga tgactcagag cccatctagt ctctctgcat ctcccggaga cagagttacg 60
atcacctgca gggctagcca agggatatca atttggctgt cctggtatca gcaaaaacct 120
ggcaaagtgc caaaggtctt gatttacaaa gcatccaatt tgcacatcgg cgtccctagt 180
cgcttttccg ggtctggtag cggcaccgac ttcaccctca ccataagctc actcgagccg 240
gaagatgccg ctacttacta ttgcctgcag tctcagactt accccctgac tttcggcgga 300
ggtaccaagc tggagatcaa ac 322
<210> 151
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность вариабельной области легкой цепи фелинизированного
моноклонального антитела от Mus musculus и Felis catus
<400> 151
Glu Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Pro Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser Ile Trp
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Lys Val Pro Lys Leu Leu Ile
35 40 45
Asn Lys Ala Ser Asn Leu His Ile Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Ala Ala Thr Tyr Tyr Cys Leu Gln Ser Gln Thr Tyr Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 152
<211> 321
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность, кодирующая последовательность
вариабельной области легкой цепи фелинизированного моноклонального
антитела от Mus musculus и Felis catus
<400> 152
gagatccaga tgacgcagag ccctagcagc ctctctgcat ccccaggaga cagagtaaca 60
attacctgtc gcgccagcca gggaatatct atatggctgt catggtatca acagaaaccg 120
ggaaaggttc caaagctctt gatcaataag gctagcaatc tgcatattgg agtgccctcc 180
cgcttctctg gtagcggaag tggcacagat ttcaccctga ccattagtag tctggagcct 240
gaggatgcgg ccacctacta ctgcctccag tcccaaacct atcccctgac cttcggagga 300
ggtaccaagc tggagatcaa a 321
<210> 153
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность вариабельной области легкой цепи фелинизированного
моноклонального антитела от Mus musculus и Felis catus
<400> 153
Glu Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Pro Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser Ile Trp
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Asn Ile Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Asn Leu His Ile Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Ala Ala Thr Tyr Tyr Cys Leu Gln Ser Gln Thr Tyr Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 154
<211> 321
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность, кодирующая последовательность
вариабельной области легкой цепи фелинизированного моноклонального
антитела от Mus musculus и Felis catus
<400> 154
gaaattcaga tgactcagag tcctagcagc ctgtccgcaa gcccaggtga ccgagtcacc 60
ataacctgca gggccagtca ggggatctcc atatggctct cttggtatca acagaaaccc 120
ggcaatatcc ctaagctcct gatttataaa gcgtcaaatc tgcatatcgg ggtgccatca 180
agattctctg ggtccggctc aggaaccgac tttaccctga ccatttcttc tctcgaaccc 240
gaggatgccg ccacctatta ttgccttcaa agccagacat acccattgac cttcggcggc 300
ggtaccaagc tggagatcaa a 321
<210> 155
<211> 159
<212> PRT
<213> Canis familiaris
<400> 155
Met Leu Ser His Thr Gly Pro Ser Arg Phe Ala Leu Phe Leu Leu Cys
1 5 10 15
Ser Met Glu Thr Leu Leu Ser Ser His Met Ala Pro Thr His Gln Leu
20 25 30
Pro Pro Ser Asp Val Arg Lys Ile Ile Leu Glu Leu Gln Pro Leu Ser
35 40 45
Arg Gly Leu Leu Glu Asp Tyr Gln Lys Lys Glu Thr Gly Val Pro Glu
50 55 60
Ser Asn Arg Thr Leu Leu Leu Cys Leu Thr Ser Asp Ser Gln Pro Pro
65 70 75 80
Arg Leu Asn Ser Ser Ala Ile Leu Pro Tyr Phe Arg Ala Ile Arg Pro
85 90 95
Leu Ser Asp Lys Asn Ile Ile Asp Lys Ile Ile Glu Gln Leu Asp Lys
100 105 110
Leu Lys Phe Gln His Glu Pro Glu Thr Glu Ile Ser Val Pro Ala Asp
115 120 125
Thr Phe Glu Cys Lys Ser Phe Ile Leu Thr Ile Leu Gln Gln Phe Ser
130 135 140
Ala Cys Leu Glu Ser Val Phe Lys Ser Leu Asn Ser Gly Pro Gln
145 150 155
<210> 156
<211> 477
<212> DNA
<213> Canis familiaris
<400> 156
atgctctccc acacaggacc atccaggttt gccctgttcc tgctctgctc tatggaaacc 60
ttgctgtcct cccatatggc acccacccat cagctaccac caagtgatgt acgaaaaatc 120
atcttggaat tacagccctt gtcgagggga cttttggaag actatcagaa gaaagagaca 180
ggggtgccag aatccaaccg taccttgctg ctgtgtctca cctctgattc ccaaccacca 240
cgcctcaaca gctcagccat cttgccttat ttcagggcaa tcagaccatt atcagataag 300
aacattattg ataaaatcat agaacagctt gacaaactca aatttcaaca tgaaccagaa 360
acagaaattt ctgtgcctgc agatactttt gaatgtaaaa gcttcatctt gacgatttta 420
cagcagttct cggcgtgcct ggaaagtgtg tttaagtcac taaactctgg acctcag 477
<210> 157
<211> 165
<212> PRT
<213> Artificial Sequence
<220>
<223> Аминокислотная последовательность, представляющая кошачий IL-31
дикого типа с C-концевой гистидиновой меткой
<400> 157
Met Leu Ser His Ala Gly Pro Ala Arg Phe Ala Leu Phe Leu Leu Cys
1 5 10 15
Cys Met Glu Thr Leu Leu Pro Ser His Met Ala Pro Ala His Arg Leu
20 25 30
Gln Pro Ser Asp Ile Arg Lys Ile Ile Leu Glu Leu Arg Pro Met Ser
35 40 45
Lys Gly Leu Leu Gln Asp Tyr Leu Lys Lys Glu Ile Gly Leu Pro Glu
50 55 60
Ser Asn His Ser Ser Leu Pro Cys Leu Ser Ser Asp Ser Gln Leu Pro
65 70 75 80
His Ile Asn Gly Ser Ala Ile Leu Pro Tyr Phe Arg Ala Ile Arg Pro
85 90 95
Leu Ser Asp Lys Asn Thr Ile Asp Lys Ile Ile Glu Gln Leu Asp Lys
100 105 110
Leu Lys Phe Gln Arg Glu Pro Glu Ala Lys Val Ser Met Pro Ala Asp
115 120 125
Asn Phe Glu Arg Lys Asn Phe Ile Leu Ala Val Leu Gln Gln Phe Ser
130 135 140
Ala Cys Leu Glu His Val Leu Gln Ser Leu Asn Ser Gly Pro Gln His
145 150 155 160
His His His His His
165
<210> 158
<211> 495
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность, представляющая ген кошачьего IL-31
дикого типа с кодируемой C-концевой гистидиновой меткой
<400> 158
atgctttcac acgctggacc agcccgattc gccctcttcc tcctctgctg tatggagact 60
ctgttgccgt cccacatggc cccggcacat aggctgcagc cgtctgacat ccggaagatc 120
attctcgaac ttcgccccat gtcgaagggg ttgctgcaag actacctgaa gaaggagatc 180
ggcctgcccg aaagcaacca ctcctcgctg ccttgcctgt caagcgattc ccagctgccc 240
cacattaacg gttccgccat cctcccgtac ttccgggcca tcagaccact gtcggacaag 300
aacaccatcg acaagatcat tgaacagctg gacaagctga agtttcagcg cgagcctgaa 360
gccaaagtgt ccatgcccgc cgataacttc gagcggaaga atttcattct cgcggtgctg 420
cagcagttct ccgcgtgcct ggagcacgtc ctgcaatccc tgaacagcgg acctcagcac 480
caccatcacc accat 495
<210> 159
<211> 148
<212> PRT
<213> Artificial Sequence
<220>
<223> Аминокислотная последовательность, представляющая кошачий белок IL-31
с N-концевой гистидиновой меткой
<400> 159
Met Arg Gly Ser His His His His His His Gly Ser Ser His Met Ala
1 5 10 15
Pro Ala His Arg Leu Gln Pro Ser Asp Ile Arg Lys Ile Ile Leu Glu
20 25 30
Leu Arg Pro Met Ser Lys Gly Leu Leu Gln Asp Tyr Leu Lys Lys Glu
35 40 45
Ile Gly Leu Pro Glu Ser Asn His Ser Ser Leu Pro Cys Leu Ser Ser
50 55 60
Asp Ser Gln Leu Pro His Ile Asn Gly Ser Ala Ile Leu Pro Tyr Phe
65 70 75 80
Arg Ala Ile Arg Pro Leu Ser Asp Lys Asn Thr Ile Asp Lys Ile Ile
85 90 95
Glu Gln Leu Asp Lys Leu Lys Phe Gln Arg Glu Pro Glu Ala Lys Val
100 105 110
Ser Met Pro Ala Asp Asn Phe Glu Arg Lys Asn Phe Ile Leu Ala Val
115 120 125
Leu Gln Gln Phe Ser Ala Cys Leu Glu His Val Leu Gln Ser Leu Asn
130 135 140
Ser Gly Pro Gln
145
<210> 160
<211> 444
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность, представляющая ген кошачьего IL-31
с кодируемой N-концевой гистидиновой меткой
<400> 160
atgagaggat cccatcacca tcaccaccac ggctcatctc atatggcccc cgcacatcgc 60
ctgcagccga gtgacattcg taaaattatc ttggagctgc gcccgatgtc caagggctta 120
ctgcaggatt atctgaagaa agagatcggg ctgcctgaaa gcaaccatag tagcctgccg 180
tgtttatcgt ctgatagcca gttaccacac atcaatggct ctgcgatttt gccctacttt 240
cgcgccatcc gtccgctgtc cgataaaaat accatcgaca aaattatcga acaactggat 300
aaattgaagt ttcagcgcga gcctgaagcg aaagtttcga tgccagcmga taacttcgaa 360
cgcaaaaact ttattttagc ggtgttgcag cagttttctg cctgtctgga acacgtgctc 420
cagtcactca atagtgggcc acaa 444
<210> 161
<211> 165
<212> PRT
<213> Artificial Sequence
<220>
<223> Амнокислотная последовательность, представляющая мутантный кошачий
белок IL-31 11E12 с C-концевой гистидиновой меткой
<400> 161
Met Leu Ser His Ala Gly Pro Ala Arg Phe Ala Leu Phe Leu Leu Cys
1 5 10 15
Cys Met Glu Thr Leu Leu Pro Ser His Met Ala Pro Ala His Arg Leu
20 25 30
Gln Pro Ser Asp Ile Arg Lys Ile Ile Leu Glu Leu Arg Pro Met Ser
35 40 45
Lys Gly Leu Leu Gln Asp Tyr Leu Lys Lys Glu Ile Gly Leu Pro Glu
50 55 60
Ser Asn His Ser Ser Leu Pro Cys Leu Ser Ser Asp Ser Gln Leu Pro
65 70 75 80
His Ile Asn Gly Ser Ala Ile Leu Pro Tyr Phe Arg Ala Ile Arg Pro
85 90 95
Leu Ser Asp Lys Asn Thr Ile Ala Lys Ile Ala Glu Gln Leu Asp Lys
100 105 110
Leu Lys Phe Gln Arg Glu Pro Glu Ala Lys Val Ser Met Pro Ala Asp
115 120 125
Asn Phe Glu Arg Lys Asn Phe Ile Leu Ala Val Leu Gln Gln Phe Ser
130 135 140
Ala Cys Leu Glu His Val Leu Gln Ser Leu Asn Ser Gly Pro Gln His
145 150 155 160
His His His His His
165
<210> 162
<211> 495
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность, представляющая ген мутантного
кошачьего IL-31 11E12 с кодируемой C-концевой гистидиновой меткой
<400> 162
atgctctctc acgccggtcc tgcccggttc gcactgttcc tcctctgttg catggagact 60
ctgcttccct cccacatggc accggcccat agactgcagc cgtccgacat cagaaagatc 120
atccttgaat tgcgccctat gagcaagggg ctgctgcagg attacctgaa aaaggagatc 180
ggcctgccgg aatcgaacca cagctcactg ccatgcctgt cctccgactc gcaactgccc 240
cacatcaatg gatccgccat tctgccgtac ttccgcgcta ttcggcctct ctccgacaag 300
aacaccatcg ccaagattgc cgagcagctg gataagctga agttccagag ggagccagaa 360
gccaaggtgt ccatgcccgc tgacaacttc gagcggaaga actttatcct cgcggtgctg 420
cagcagttct cagcgtgcct cgaacacgtc ttgcaaagcc tgaactcggg accccagcac 480
caccaccatc atcac 495
<210> 163
<211> 165
<212> PRT
<213> Artificial Sequence
<220>
<223> Аминокислотная последовательность, представляющая мутантный кошачий
белок IL-31 15H05 с C-концевой гистидиновой меткой
<400> 163
Met Leu Ser His Ala Gly Pro Ala Arg Phe Ala Leu Phe Leu Leu Cys
1 5 10 15
Cys Met Glu Thr Leu Leu Pro Ser His Met Ala Pro Ala His Arg Leu
20 25 30
Gln Pro Ser Asp Ile Arg Lys Ile Ile Leu Glu Leu Arg Pro Met Ser
35 40 45
Lys Gly Leu Leu Gln Asp Tyr Leu Lys Lys Glu Ile Gly Leu Pro Glu
50 55 60
Ser Asn His Ser Ser Leu Pro Cys Leu Ser Ser Asp Ser Gln Leu Pro
65 70 75 80
His Ile Asn Gly Ser Ala Ile Leu Pro Tyr Phe Arg Ala Ile Arg Pro
85 90 95
Leu Ser Asp Lys Asn Thr Ile Asp Lys Ile Ile Glu Gln Leu Asp Lys
100 105 110
Leu Lys Phe Gln Arg Glu Pro Glu Ala Lys Val Ser Met Ala Ala Ala
115 120 125
Asn Phe Glu Arg Lys Asn Phe Ile Leu Ala Val Leu Gln Gln Phe Ser
130 135 140
Ala Cys Leu Glu His Val Leu Gln Ser Leu Asn Ser Gly Pro Gln His
145 150 155 160
His His His His His
165
<210> 164
<211> 495
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательностьс, представляющая ген мутантного
Кошачьего IL-31 15H05 с кодируемой C-концевой гистидиновой меткой
<400> 164
atgctctctc acgccggtcc tgcccggttc gcactgttcc tcctctgttg catggagact 60
ctgcttccct cccacatggc accggcccat agactgcagc cgtccgacat cagaaagatc 120
atccttgaat tgcgccctat gagcaagggg ctgctgcagg attacctgaa aaaggagatc 180
ggcctgccgg aatcgaacca cagctcactg ccatgcctgt cctccgactc gcaactgccc 240
cacatcaatg gatccgccat tctgccgtac ttccgcgcta ttcggcctct ctccgacaag 300
aacaccatcg acaagattat tgagcagctg gataagctga agttccagag ggagccagaa 360
gccaaggtgt ccatggccgc tgccaacttc gagcggaaga actttatcct cgcggtgctg 420
cagcagttct cagcgtgcct cgaacacgtc ttgcaaagcc tgaactcggg accccagcac 480
caccaccatc atcac 495
<210> 165
<211> 152
<212> PRT
<213> Artificial Sequence
<220>
<223> Аминокислотная последовательность лошадиного белка IL-31 с
C-концевой гистидиновой меткой
<400> 165
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Val His Ser Gly Pro Ile Tyr Gln Leu Gln Pro Lys Glu Ile Gln Ala
20 25 30
Ile Ile Val Glu Leu Gln Asn Leu Ser Lys Lys Leu Leu Asp Asp Tyr
35 40 45
Leu Asn Lys Glu Lys Gly Val Gln Lys Phe Asp Ser Asp Leu Pro Ser
50 55 60
Cys Phe Thr Ser Asp Ser Gln Ala Pro Gly Asn Ile Asn Ser Ser Ala
65 70 75 80
Ile Leu Pro Tyr Phe Lys Ala Ile Ser Pro Ser Leu Asn Asn Asp Lys
85 90 95
Ser Leu Tyr Ile Ile Glu Gln Leu Asp Lys Leu Asn Phe Gln Asn Ala
100 105 110
Pro Glu Thr Glu Val Ser Met Pro Thr Asp Asn Phe Glu Arg Lys Arg
115 120 125
Phe Ile Leu Thr Ile Leu Arg Trp Phe Ser Asn Cys Leu Glu His Arg
130 135 140
Ala Gln His His His His His His
145 150
<210> 166
<211> 456
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность гена лошадиного IL-31 с кодируемой
C-концевой гистидиновой меткой
<400> 166
atgggctggt cctgcatcat tctgtttctg gtggccacag ccaccggcgt gcactctgga 60
cctatctatc agctgcagcc caaagagatc caggccatca tcgtggaact gcagaacctg 120
agcaagaagc tgctggacga ctacctgaac aaagaaaagg gcgtgcagaa gttcgacagc 180
gacctgccta gctgcttcac cagcgattct caggcccctg gcaacatcaa cagcagcgcc 240
atcctgcctt acttcaaggc catctctccc agcctgaaca acgacaagag cctgtacatc 300
atcgagcagc tggacaagct gaacttccag aacgcccctg aaaccgaggt gtccatgcct 360
accgacaact tcgagcggaa gcggttcatc ctgaccatcc tgcggtggtt cagcaactgc 420
ctggaacaca gagcccagca ccaccaccat caccat 456
<210> 167
<211> 652
<212> PRT
<213> Artificial Sequence
<220>
<223> Аминокислотная последовательность внеклеточного домена кошачьего OSMR,
слитого с Fc человеческого IgG1
<400> 167
Met Ala Leu Phe Ser Ala Phe Gln Thr Thr Phe Leu Leu Ala Leu Leu
1 5 10 15
Ser Leu Lys Thr Tyr Gln Ser Glu Val Leu Ser Glu Pro Leu Ser Leu
20 25 30
Ala Pro Glu Ser Leu Glu Val Ser Ile Asp Ser Ala Arg Gln Cys Leu
35 40 45
His Leu Lys Trp Ser Val His Asn Leu Ala Tyr His Gln Glu Leu Lys
50 55 60
Met Val Phe Gln Ile Glu Ile Ser Arg Ile Lys Thr Ser Asn Val Ile
65 70 75 80
Trp Val Glu Asn Tyr Ser Thr Thr Val Lys Arg Asn Gln Val Leu Arg
85 90 95
Trp Ser Trp Glu Ser Lys Leu Pro Leu Glu Cys Ala Lys His Ser Val
100 105 110
Arg Met Arg Gly Ala Val Asp Asp Ala Gln Val Pro Glu Leu Arg Phe
115 120 125
Trp Ser Asn Trp Thr Ser Trp Glu Glu Val Asp Val Gln Ser Ser Leu
130 135 140
Gly His Asp Pro Leu Phe Val Phe Pro Lys Asp Lys Leu Val Glu Glu
145 150 155 160
Gly Ser Asn Val Thr Ile Cys Tyr Val Ser Arg Ser His Gln Asn Asn
165 170 175
Ile Ser Cys Tyr Leu Glu Gly Val Arg Met His Gly Glu Gln Leu Asp
180 185 190
Pro Asn Val Cys Val Phe His Leu Lys Asn Val Pro Phe Ile Arg Glu
195 200 205
Thr Gly Thr Asn Ile Tyr Cys Lys Ala Asp Gln Gly Asp Val Ile Lys
210 215 220
Gly Ile Val Leu Phe Val Ser Lys Val Phe Glu Glu Pro Lys Asp Phe
225 230 235 240
Ser Cys Glu Thr Arg Asp Leu Lys Thr Leu Asn Cys Thr Trp Ala Pro
245 250 255
Gly Ser Asp Ala Gly Leu Leu Thr Gln Leu Ser Gln Ser Tyr Thr Leu
260 265 270
Phe Glu Ser Phe Ser Gly Lys Lys Thr Leu Cys Lys His Lys Ser Trp
275 280 285
Cys Asn Trp Gln Val Ser Pro Asp Ser Gln Glu Met Tyr Asn Phe Thr
290 295 300
Leu Thr Ala Glu Asn Tyr Leu Arg Lys Arg Ser Val His Leu Leu Phe
305 310 315 320
Asn Leu Thr His Arg Val His Pro Met Ala Pro Phe Asn Val Phe Val
325 330 335
Lys Asn Val Ser Ala Thr Asn Ala Thr Met Thr Trp Lys Val His Ser
340 345 350
Ile Gly Asn Tyr Ser Thr Leu Leu Cys Gln Ile Glu Leu Asp Gly Glu
355 360 365
Gly Lys Val Ile Gln Lys Gln Asn Val Ser Val Lys Val Asn Gly Lys
370 375 380
His Leu Met Lys Lys Leu Glu Pro Ser Thr Glu Tyr Ala Ala Gln Val
385 390 395 400
Arg Cys Ala Asn Ala Asn His Phe Trp Lys Trp Ser Glu Trp Thr Arg
405 410 415
Arg Asn Phe Thr Thr Ala Glu Ala Ala Asp Lys Thr His Thr Cys Pro
420 425 430
Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe
435 440 445
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
450 455 460
Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe
465 470 475 480
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
485 490 495
Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr
500 505 510
Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
515 520 525
Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala
530 535 540
Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg
545 550 555 560
Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly
565 570 575
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro
580 585 590
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser
595 600 605
Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln
610 615 620
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
625 630 635 640
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
645 650
<210> 168
<211> 2076
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность, кодирующая внеклеточный домен
кошачьего OSMR, слитый с Fc человеческого IgG1
<400> 168
atggccctgt tcagcgcctt ccagaccacc ttcctgctgg ccctgctgag cctgaaaacc 60
taccagagcg aggtgctgag cgagcccctg tctctggccc ctgagagcct ggaagtgtcc 120
atcgacagcg ccagacagtg cctgcacctg aagtggagcg tgcacaacct ggcctaccac 180
caggaactga agatggtgtt ccagatcgag atcagccgga tcaagaccag caacgtgatc 240
tgggtggaaa actacagcac caccgtgaag cggaaccagg tgctgcggtg gtcctgggag 300
tctaagctgc ctctggaatg cgccaagcac agcgtgcgga tgagaggcgc cgtggatgat 360
gcccaggtgc ccgagctgag attctggtcc aactggacct cctgggaaga ggtggacgtg 420
cagtctagcc tgggccacga ccccctgttc gtgttcccca aggacaagct ggtggaagag 480
ggctccaacg tgaccatctg ctacgtgtcc agaagccacc agaacaacat cagctgctac 540
ctggaaggcg tgcgcatgca cggcgagcag ctggacccta acgtgtgcgt gttccacctg 600
aagaacgtgc ccttcatcag agagacaggc accaacatct actgcaaggc cgaccagggc 660
gacgtgatca agggcatcgt gctgtttgtg tccaaggtgt tcgaggaacc caaggacttc 720
agctgcgaga cacgggatct gaaaaccctg aactgtacct gggcccctgg ctccgatgcc 780
ggactgctga ctcagctgtc ccagagctac accctgttcg agagcttcag cggcaaaaag 840
accctgtgca agcacaagag ctggtgcaac tggcaagtgt cccccgatag ccaggaaatg 900
tacaacttca ccctgaccgc cgagaactac ctgcggaaga gatccgtgca tctgctgttc 960
aacctgaccc acagagtgca ccccatggcc cccttcaacg tgttcgtgaa gaatgtgtcc 1020
gccaccaacg ccaccatgac atggaaggtg cacagcatcg gcaactactc caccctgctg 1080
tgtcagatcg agctggacgg cgagggcaaa gtgatccaga aacagaacgt gtcagtgaaa 1140
gtgaacggca agcacctgat gaagaagctg gaacccagca ccgagtacgc cgcccaggtg 1200
cgctgtgcca acgccaacca cttctggaag tggagtgaat ggacccggcg gaacttcacc 1260
acagccgaag ccgccgctga gaacgaggtg tccacaccta tgcaggccct gaccaccaac 1320
aaggacgacg acaacatcct gttccgggac tccgccaatg ccaccagcct gcctgtgcag 1380
gatagcagct ctgtgctgcc cgccaagccc gagaacatct cctgcgtgtt ctactacgag 1440
gaaaacttca cttgcacctg gtcccccgag aaagaggcca gctacacctg gtacaaagtg 1500
aagagaacct acagctacgg ctacaagagc gacatctgcc ccagcgacaa cagcaccaga 1560
ggcaaccaca ccttctgcag ctttctgccc cccaccatca ccaaccccga caactacacc 1620
atccaggtgg aagcccagaa cgccgacggc atcatcaagt ccgacatcac ccactggtcc 1680
ctggacgcca tcacaaagat cgagcccccc gagatcttct ccgtgaagcc tgtgctgggc 1740
gtgaagagga tggtgcagat caagtggatc cggcccgtgc tggccccagt gtctagcacc 1800
ctgaagtaca ccctgcggtt caagaccgtg aacagcgcct actggatgga agtgaatttc 1860
accaaagagg acatcgaccg ggacgagaca tacaatctga ccggactgca ggccttcaca 1920
gagtacgtgc tggctctgag atgcgccacc aaagaatcca tgttttggag cggctggtcc 1980
caggaaaaga tgggcaccac cgaagagggt aagcctatcc ctaaccctct cctcggtctc 2040
gattctacgc gtaccggtca tcatcaccat caccat 2076
<210> 169
<211> 474
<212> PRT
<213> Artificial Sequence
<220>
<223> Аминокислотная последовательность кошачьего IL-31-Ra, слитого с Fc
человеческого IgG1
<400> 169
Met Met Trp Pro Gln Val Trp Gly Leu Glu Ile Gln Phe Ser Pro Gln
1 5 10 15
Pro Ala Cys Ile Asp Leu Gly Met Met Trp Ala His Ala Leu Trp Thr
20 25 30
Leu Leu Leu Leu Cys Lys Phe Ser Leu Ala Val Leu Pro Ala Lys Pro
35 40 45
Glu Asn Ile Ser Cys Val Phe Tyr Tyr Glu Glu Asn Phe Thr Cys Thr
50 55 60
Trp Ser Pro Glu Lys Glu Ala Ser Tyr Thr Trp Tyr Lys Val Lys Arg
65 70 75 80
Thr Tyr Ser Tyr Gly Tyr Lys Ser Asp Ile Cys Pro Ser Asp Asn Ser
85 90 95
Thr Arg Gly Asn His Thr Phe Cys Ser Phe Leu Pro Pro Thr Ile Thr
100 105 110
Asn Pro Asp Asn Tyr Thr Ile Gln Val Glu Ala Gln Asn Ala Asp Gly
115 120 125
Ile Ile Lys Ser Asp Ile Thr His Trp Ser Leu Asp Ala Ile Thr Lys
130 135 140
Ile Glu Pro Pro Glu Ile Phe Ser Val Lys Pro Val Leu Gly Val Lys
145 150 155 160
Arg Met Val Gln Ile Lys Trp Ile Arg Pro Val Leu Ala Pro Val Ser
165 170 175
Ser Thr Leu Lys Tyr Thr Leu Arg Phe Lys Thr Val Asn Ser Ala Tyr
180 185 190
Trp Met Glu Val Asn Phe Thr Lys Glu Asp Ile Asp Arg Asp Glu Thr
195 200 205
Tyr Asn Leu Thr Gly Leu Gln Ala Phe Thr Glu Tyr Val Leu Ala Leu
210 215 220
Arg Cys Ala Thr Lys Glu Ser Met Phe Trp Ser Gly Trp Ser Gln Glu
225 230 235 240
Lys Met Gly Thr Thr Glu Glu Asp Lys Thr His Thr Cys Pro Pro Cys
245 250 255
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
260 265 270
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
275 280 285
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
290 295 300
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
305 310 315 320
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
325 330 335
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
340 345 350
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
355 360 365
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
370 375 380
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
385 390 395 400
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
405 410 415
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
420 425 430
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
435 440 445
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
450 455 460
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
465 470
<210> 170
<211> 1422
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность, кодирующая кошачий IL-31 Ra, слитый
с Fc человеческого IgG1
<400> 170
atgatgtggc cacaagtgtg gggcctggag atccagttca gcccccagcc tgcctgcatc 60
gatctgggca tgatgtgggc tcacgctctg tggaccctgc tgctgctgtg caagttttcc 120
ctggccgtgc tgcccgctaa gcctgagaac atcagctgcg tgttctacta tgaggagaac 180
ttcacctgta catggtcccc cgagaaggag gctagctata cctggtacaa ggtgaagaga 240
acatacagct atggctacaa gtctgatatc tgccccagcg acaactctac ccgcggcaat 300
cacacattct gttcttttct gccccctacc atcacaaacc ctgataatta taccatccag 360
gtggaggccc agaacgctga tggcatcatc aagtctgaca tcacccattg gtccctggac 420
gccatcacaa agatcgagcc acccgagatt ttctccgtga agcccgtgct gggcgtgaag 480
aggatggtgc agatcaagtg gatcaggcct gtgctggctc cagtgtccag caccctgaag 540
tatacactga gattcaagac cgtgaactcc gcttactgga tggaggtgaa cttcaccaag 600
gaggacatcg atagggacga gacctataat ctgacaggcc tgcaggcctt caccgagtac 660
gtgctggccc tgaggtgcgc tacaaaggag tccatgtttt ggtccggctg gagccaggag 720
aagatgggca ccacagagga ggataagacc cacacatgcc ctccatgtcc agctccagag 780
ctgctgggag gaccaagcgt gttcctgttt ccacctaagc ctaaggacac cctgatgatc 840
tctcgcaccc ctgaggtgac atgcgtggtg gtggacgtgt cccacgagga cccagaggtg 900
aagtttaact ggtatgtgga tggcgtggag gtgcataatg ccaagaccaa gcctagagag 960
gagcagtata acagcaccta ccgcgtggtg tctgtgctga cagtgctgca tcaggactgg 1020
ctgaacggca aggagtacaa gtgcaaggtg agcaataagg ccctgcctgc tccaatcgag 1080
aagaccatct ctaaggctaa gggacagcca agggagccac aggtgtatac actgccaccc 1140
agccgggagg agatgaccaa gaaccaggtg tctctgacat gtctggtgaa gggcttctac 1200
ccatctgata tcgctgtgga gtgggagtcc aatggccagc ccgagaacaa ttataagacc 1260
acacctccag tgctggattc tgacggctcc ttctttctgt actccaagct gaccgtggac 1320
aagagcaggt ggcagcaggg caacgtgttt tcttgctccg tgatgcatga ggctctgcac 1380
aatcattaca cacagaagag cctgtctctg tccccaggca ag 1422
<210> 171
<211> 335
<212> PRT
<213> Felis catus
<400> 171
Ala Ser Thr Thr Ala Pro Ser Val Phe Pro Leu Ala Pro Ser Cys Gly
1 5 10 15
Thr Thr Ser Gly Ala Thr Val Ala Leu Ala Cys Leu Val Leu Gly Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ala Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Met Val Thr Val Pro Ser Ser Arg Trp Leu Ser Asp Thr
65 70 75 80
Phe Thr Cys Asn Val Ala His Pro Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Thr Val Arg Lys Thr Asp His Pro Pro Gly Pro Lys Pro Cys Asp Cys
100 105 110
Pro Lys Cys Pro Pro Pro Glu Met Leu Gly Gly Pro Ser Ile Phe Ile
115 120 125
Phe Pro Pro Lys Pro Lys Asp Thr Leu Ser Ile Ser Arg Thr Pro Glu
130 135 140
Val Thr Cys Leu Val Val Asp Leu Gly Pro Asp Asp Ser Asp Val Gln
145 150 155 160
Ile Thr Trp Phe Val Asp Asn Thr Gln Val Tyr Thr Ala Lys Thr Ser
165 170 175
Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu
180 185 190
Pro Ile Leu His Gln Asp Trp Leu Lys Gly Lys Glu Phe Lys Cys Lys
195 200 205
Val Asn Ser Lys Ser Leu Pro Ser Pro Ile Glu Arg Thr Ile Ser Lys
210 215 220
Ala Lys Gly Gln Pro His Glu Pro Gln Val Tyr Val Leu Pro Pro Ala
225 230 235 240
Gln Glu Glu Leu Ser Arg Asn Lys Val Ser Val Thr Cys Leu Ile Lys
245 250 255
Ser Phe His Pro Pro Asp Ile Ala Val Glu Trp Glu Ile Thr Gly Gln
260 265 270
Pro Glu Pro Glu Asn Asn Tyr Arg Thr Thr Pro Pro Gln Leu Asp Ser
275 280 285
Asp Gly Thr Tyr Phe Val Tyr Ser Lys Leu Ser Val Asp Arg Ser His
290 295 300
Trp Gln Arg Gly Asn Thr Tyr Thr Cys Ser Val Ser His Glu Ala Leu
305 310 315 320
His Ser His His Thr Gln Lys Ser Leu Thr Gln Ser Pro Gly Lys
325 330 335
<210> 172
<211> 1005
<212> DNA
<213> Felis catus
<400> 172
gcctccacca cggccccatc ggtgttccca ctggccccca gctgcgggac cacatctggc 60
gccaccgtgg ccctggcctg cctggtgtta ggctacttcc ctgagccggt gaccgtgtcc 120
tggaactccg gcgccctgac cagcggtgtg cacaccttcc cggccgtcct gcaggcctcg 180
gggctgtact ctctcagcag catggtgaca gtgccctcca gcaggtggct cagtgacacc 240
ttcacctgca acgtggccca cccgcccagc aacaccaagg tggacaagac cgtgcgcaaa 300
acagaccacc caccgggacc caaaccctgc gactgtccca aatgcccacc ccctgagatg 360
cttggaggac cgtccatctt catcttcccc ccaaaaccca aggacaccct ctcgatttcc 420
cggacgcccg aggtcacatg cttggtggtg gacttgggcc cagatgactc cgatgtccag 480
atcacatggt ttgtggataa cacccaggtg tacacagcca agacgagtcc gcgtgaggag 540
cagttcaaca gcacctaccg tgtggtcagt gtcctcccca tcctacacca ggactggctc 600
aaggggaagg agttcaagtg caaggtcaac agcaaatccc tcccctcccc catcgagagg 660
accatctcca aggccaaagg acagccccac gagccccagg tgtacgtcct gcctccagcc 720
caggaggagc tcagcaggaa caaagtcagt gtgacctgcc tgatcaaatc cttccacccg 780
cctgacattg ccgtcgagtg ggagatcacc ggacagccgg agccagagaa caactaccgg 840
acgaccccgc cccagctgga cagcgacggg acctacttcg tgtacagcaa gctctcggtg 900
gacaggtccc actggcagag gggaaacacc tacacctgct cggtgtcaca cgaagctctg 960
cacagccacc acacacagaa atccctcacc cagtctccgg gtaaa 1005
<210> 173
<211> 335
<212> PRT
<213> Artificial Sequence
<220>
<223> Аминокислотная последовательность тяжелой цепи кошачьего происхождения,
генномодифицированной для изменения эффекторной функции антитела
<400> 173
Ala Ser Thr Thr Ala Pro Ser Val Phe Pro Leu Ala Pro Ser Cys Gly
1 5 10 15
Thr Thr Ser Gly Ala Thr Val Ala Leu Ala Cys Leu Val Leu Gly Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ala Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Met Val Thr Val Pro Ser Ser Arg Trp Leu Ser Asp Thr
65 70 75 80
Phe Thr Cys Asn Val Ala His Pro Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Thr Val Arg Lys Thr Asp His Pro Pro Gly Pro Lys Pro Cys Asp Cys
100 105 110
Pro Lys Cys Pro Pro Pro Glu Ala Ala Gly Ala Pro Ser Ile Phe Ile
115 120 125
Phe Pro Pro Lys Pro Lys Asp Thr Leu Ser Ile Ser Arg Thr Pro Glu
130 135 140
Val Thr Cys Leu Val Val Asp Leu Gly Pro Asp Asp Ser Asp Val Gln
145 150 155 160
Ile Thr Trp Phe Val Asp Asn Thr Gln Val Tyr Thr Ala Lys Thr Ser
165 170 175
Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu
180 185 190
Pro Ile Leu His Gln Asp Trp Leu Lys Gly Lys Glu Phe Lys Cys Lys
195 200 205
Val Asn Ser Lys Ser Leu Pro Ser Pro Ile Glu Arg Thr Ile Ser Lys
210 215 220
Ala Lys Gly Gln Pro His Glu Pro Gln Val Tyr Val Leu Pro Pro Ala
225 230 235 240
Gln Glu Glu Leu Ser Arg Asn Lys Val Ser Val Thr Cys Leu Ile Lys
245 250 255
Ser Phe His Pro Pro Asp Ile Ala Val Glu Trp Glu Ile Thr Gly Gln
260 265 270
Pro Glu Pro Glu Asn Asn Tyr Arg Thr Thr Pro Pro Gln Leu Asp Ser
275 280 285
Asp Gly Thr Tyr Phe Val Tyr Ser Lys Leu Ser Val Asp Arg Ser His
290 295 300
Trp Gln Arg Gly Asn Thr Tyr Thr Cys Ser Val Ser His Glu Ala Leu
305 310 315 320
His Ser His His Thr Gln Lys Ser Leu Thr Gln Ser Pro Gly Lys
325 330 335
<210> 174
<211> 1005
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность, кодирующая тяжелую цепь кошачьего
происхождения, генномодифицированную для изменения эффекторной
функции антитела
<400> 174
gcctccacca cggccccatc ggtgttccca ctggccccca gctgcgggac cacatctggc 60
gccaccgtgg ccctggcctg cctggtgtta ggctacttcc ctgagccggt gaccgtgtcc 120
tggaactccg gcgccctgac cagcggtgtg cacaccttcc cggccgtcct gcaggcctcg 180
gggctgtact ctctcagcag catggtgaca gtgccctcca gcaggtggct cagtgacacc 240
ttcacctgca acgtggccca cccgcccagc aacaccaagg tggacaagac cgtgcgcaaa 300
acagaccacc caccgggacc caaaccctgc gactgtccca aatgcccacc ccctgaggcg 360
gctggagcac cgtccatctt catcttcccc ccaaaaccca aggacaccct ctcgatttcc 420
cggacgcccg aggtcacatg cttggtggtg gacttgggcc cagatgactc cgatgtccag 480
atcacatggt ttgtggataa cacccaggtg tacacagcca agacgagtcc gcgtgaggag 540
cagttcaaca gcacctaccg tgtggtcagt gtcctcccca tcctacacca ggactggctc 600
aaggggaagg agttcaagtg caaggtcaac agcaaatccc tcccctcccc catcgagagg 660
accatctcca aggccaaagg acagccccac gagccccagg tgtacgtcct gcctccagcc 720
caggaggagc tcagcaggaa caaagtcagt gtgacctgcc tgatcaaatc cttccacccg 780
cctgacattg ccgtcgagtg ggagatcacc ggacagccgg agccagagaa caactaccgg 840
acgaccccgc cccagctgga cagcgacggg acctacttcg tgtacagcaa gctctcggtg 900
gacaggtccc actggcagag gggaaacacc tacacctgct cggtgtcaca cgaagctctg 960
cacagccacc acacacagaa atccctcacc cagtctccgg gtaaa 1005
<210> 175
<211> 110
<212> PRT
<213> Artificial Sequence
<220>
<223> Аминокислотная последовательность легкой каппа-цепи кошачьего
происхождения, генномодифицированной для нокаута гликозилирования
(G-) в положении 103
<400> 175
Arg Ser Asp Ala Gln Pro Ser Val Phe Leu Phe Gln Pro Ser Leu Asp
1 5 10 15
Glu Leu His Thr Gly Ser Ala Ser Ile Val Cys Ile Leu Asn Asp Phe
20 25 30
Tyr Pro Lys Glu Val Asn Val Lys Trp Lys Val Asp Gly Val Val Gln
35 40 45
Asn Lys Gly Ile Gln Glu Ser Thr Thr Glu Gln Asn Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Met Ser Ser Thr Glu Tyr Gln
65 70 75 80
Ser His Glu Lys Phe Ser Cys Glu Val Thr His Lys Ser Leu Ala Ser
85 90 95
Thr Leu Val Lys Ser Phe Gln Arg Ser Glu Cys Gln Arg Glu
100 105 110
<210> 176
<211> 330
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность, кодирующая легкую каппа-цепь
кошачьего происхождения, генномодифицированную для нокаута
гликозилирования (G-)
<400> 176
cggagtgatg ctcagccatc tgtctttctc ttccaaccat ctctggacga gttacataca 60
ggaagtgcct ctatcgtgtg catattgaat gacttctacc ccaaagaggt caatgtcaag 120
tggaaagtgg atggcgtagt ccaaaacaaa ggcatccagg agagcaccac agagcagaac 180
agcaaggaca gcacctacag cctcagcagc accctgacga tgtccagtac ggagtaccaa 240
agtcatgaaa agttctcctg cgaggtcact cacaagagcc tggcctccac cctcgtcaag 300
agcttccaga ggagcgagtg tcagagagag 330
<210> 177
<211> 335
<212> PRT
<213> Canis familiaris
<400> 177
Ala Ser Thr Thr Ala Pro Ser Val Phe Pro Leu Ala Pro Ser Cys Gly
1 5 10 15
Ser Thr Ser Gly Ser Thr Val Ala Leu Ala Cys Leu Val Ser Gly Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ser Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ser Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Met Val Thr Val Pro Ser Ser Arg Trp Pro Ser Glu Thr
65 70 75 80
Phe Thr Cys Asn Val Ala His Pro Ala Ser Lys Thr Lys Val Asp Lys
85 90 95
Pro Val Pro Lys Arg Glu Asn Gly Arg Val Pro Arg Pro Pro Asp Cys
100 105 110
Pro Lys Cys Pro Ala Pro Glu Met Leu Gly Gly Pro Ser Val Phe Ile
115 120 125
Phe Pro Pro Lys Pro Lys Asp Thr Leu Leu Ile Ala Arg Thr Pro Glu
130 135 140
Val Thr Cys Val Val Val Asp Leu Asp Pro Glu Asp Pro Glu Val Gln
145 150 155 160
Ile Ser Trp Phe Val Asp Gly Lys Gln Met Gln Thr Ala Lys Thr Gln
165 170 175
Pro Arg Glu Glu Gln Phe Asn Gly Thr Tyr Arg Val Val Ser Val Leu
180 185 190
Pro Ile Gly His Gln Asp Trp Leu Lys Gly Lys Gln Phe Thr Cys Lys
195 200 205
Val Asn Asn Lys Ala Leu Pro Ser Pro Ile Glu Arg Thr Ile Ser Lys
210 215 220
Ala Arg Gly Gln Ala His Gln Pro Ser Val Tyr Val Leu Pro Pro Ser
225 230 235 240
Arg Glu Glu Leu Ser Lys Asn Thr Val Ser Leu Thr Cys Leu Ile Lys
245 250 255
Asp Phe Phe Pro Pro Asp Ile Asp Val Glu Trp Gln Ser Asn Gly Gln
260 265 270
Gln Glu Pro Glu Ser Lys Tyr Arg Thr Thr Pro Pro Gln Leu Asp Glu
275 280 285
Asp Gly Ser Tyr Phe Leu Tyr Ser Lys Leu Ser Val Asp Lys Ser Arg
290 295 300
Trp Gln Arg Gly Asp Thr Phe Ile Cys Ala Val Met His Glu Ala Leu
305 310 315 320
His Asn His Tyr Thr Gln Glu Ser Leu Ser His Ser Pro Gly Lys
325 330 335
<210> 178
<211> 1005
<212> DNA
<213> Canis familiaris
<400> 178
gcctcaacaa ctgctcctag cgtgtttccc ctggccccta gctgcggaag tacctcaggc 60
agcacagtgg ccctggcttg tctggtgtct ggatatttcc ctgagccagt gaccgtgagt 120
tggaacagcg gctctctgac ctccggggtg cacacatttc catctgtgct gcagtctagt 180
ggcctgtact ccctgtcaag catggtgact gtgccttcct ctaggtggcc atcagaaact 240
ttcacctgca acgtggccca tcccgccagc aagaccaaag tggacaagcc cgtgcctaaa 300
agggagaatg gaagggtgcc aagaccacct gattgcccta agtgtccagc tccagaagcg 360
gcgggagcac caagcgtgtt catctttcca cccaagccca aagacacact gctgattgct 420
agaactcccg aggtgacctg cgtggtggtg gacctggatc cagaggaccc cgaagtgcag 480
atctcctggt tcgtggatgg gaagcagatg cagacagcca aaactcagcc tcgggaggaa 540
cagtttaacg gaacctatag agtggtgtct gtgctgccaa ttggacacca ggactggctg 600
aagggcaaac agtttacatg caaggtgaac aacaaggccc tgcctagtcc aatcgagagg 660
actatttcaa aagctagggg acaggctcat cagccttccg tgtatgtgct gcctccatcc 720
cgggaggaac tgtctaagaa cacagtgagt ctgacttgtc tgatcaaaga tttctttccc 780
cctgacattg atgtggagtg gcagagcaat gggcagcagg agccagaatc caagtacaga 840
accacaccac cccagctgga cgaagatggc tcctatttcc tgtacagtaa gctgtcagtg 900
gacaaatcta ggtggcagcg cggggatacc tttatctgcg ccgtgatgca cgaggctctg 960
cacaatcatt acacacaaga aagtctgtca catagccccg gcaag 1005
<210> 179
<211> 106
<212> PRT
<213> Canis familiaris
<400> 179
Arg Asn Asp Ala Gln Pro Ala Val Tyr Leu Phe Gln Pro Ser Pro Asp
1 5 10 15
Gln Leu His Thr Gly Ser Ala Ser Val Val Cys Leu Leu Asn Ser Phe
20 25 30
Tyr Pro Lys Asp Ile Asn Val Lys Trp Lys Val Asp Gly Val Ile Gln
35 40 45
Asp Thr Gly Ile Gln Glu Ser Val Thr Glu Gln Asp Lys Asp Ser Thr
50 55 60
Tyr Ser Leu Ser Ser Thr Leu Thr Met Ser Ser Thr Glu Tyr Leu Ser
65 70 75 80
His Glu Leu Tyr Ser Cys Glu Ile Thr His Lys Ser Leu Pro Ser Thr
85 90 95
Leu Ile Lys Ser Phe Gln Arg Ser Glu Cys
100 105
<210> 180
<211> 318
<212> DNA
<213> Canis familiaris
<400> 180
aggaacgacg cccagcctgc tgtgtatctg tttcagccct cccctgatca gctgcacact 60
ggctctgcta gtgtggtgtg tctgctgaac agcttctacc caaaggatat caatgtgaag 120
tggaaagtgg acggcgtgat ccaggatact gggattcagg agtccgtgac cgaacaggac 180
aaagattcaa catatagcct gagctccact ctgaccatgt ctagtaccga gtacctgagc 240
cacgaactgt attcctgcga gatcactcat aagtccctgc cctctaccct gatcaagagc 300
ttccagagat cagagtgt 318
<210> 181
<211> 164
<212> PRT
<213> Homo sapiens
<400> 181
Met Ala Ser His Ser Gly Pro Ser Thr Ser Val Leu Phe Leu Phe Cys
1 5 10 15
Cys Leu Gly Gly Trp Leu Ala Ser His Thr Leu Pro Val Arg Leu Leu
20 25 30
Arg Pro Ser Asp Asp Val Gln Lys Ile Val Glu Glu Leu Gln Ser Leu
35 40 45
Ser Lys Met Leu Leu Lys Asp Val Glu Glu Glu Lys Gly Val Leu Val
50 55 60
Ser Gln Asn Tyr Thr Leu Pro Cys Leu Ser Pro Asp Ala Gln Pro Pro
65 70 75 80
Asn Asn Ile His Ser Pro Ala Ile Arg Ala Tyr Leu Lys Thr Ile Arg
85 90 95
Gln Leu Asp Asn Lys Ser Val Ile Asp Glu Ile Ile Glu His Leu Asp
100 105 110
Lys Leu Ile Phe Gln Asp Ala Pro Glu Thr Asn Ile Ser Val Pro Thr
115 120 125
Asp Thr His Glu Cys Lys Arg Phe Ile Leu Thr Ile Ser Gln Gln Phe
130 135 140
Ser Glu Cys Met Asp Leu Ala Leu Lys Ser Leu Thr Ser Gly Ala Gln
145 150 155 160
Gln Ala Thr Thr
<210> 182
<211> 111
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность вариабельной области легкой цепи канинизированного
моноклонального антитела от Mus musculus и Canis
<400> 182
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ala Ser Glu Ser Val Asp Asn Tyr
20 25 30
Gly Ile Ser Phe Met His Trp Phe Gln Gln Lys Pro Gly Gln Ser Pro
35 40 45
Gln Arg Leu Ile Tyr Arg Ala Ser Asn Leu Glu Ser Gly Val Pro Asp
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Arg Ile Ser
65 70 75 80
Arg Val Glu Ala Asp Asp Ala Gly Val Tyr Tyr Cys Gln Gln Ser Asn
85 90 95
Lys Asp Pro Leu Thr Phe Gly Ala Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 183
<211> 333
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность, кодирующая вариабельную область
легкой цепи канинизированного моноклонального антитела от
Mus musculus и Canis
<400> 183
gacatcgtga tgacccagac ccccctgagc ctgagcgtgt cccctggcga gcctgccagc 60
atcagctgca gagccagcga gagcgtggac aactacggca tcagcttcat gcactggttc 120
cagcagaagc ccggccagag cccccagcgg ctgatctaca gagccagcaa cctggaaagc 180
ggcgtgcccg atcggtttag cggctctggc agcggcaccg acttcaccct gcggatctct 240
cgggtggaag ccgatgacgc cggagtgtac tactgccagc agagcaacaa ggaccccctg 300
acctttggcg ccggtaccaa gctggagatc aag 333
<210> 184
<211> 111
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность вариабельной области легкой цепи канинизированного
моноклонального антитела от Mus musculus и Canis
<400> 184
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ala Ser Glu Ser Val Asp Asn Tyr
20 25 30
Gly Ile Ser Phe Met His Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Arg Ala Ser Asn Leu Glu Ser Gly Val Pro Asp
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Arg Ile Ser
65 70 75 80
Arg Val Glu Ala Asp Asp Ala Gly Val Tyr Tyr Cys Gln Gln Ser Asn
85 90 95
Lys Asp Pro Leu Thr Phe Gly Ala Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 185
<211> 333
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность, кодирующая вариабельную область легкой
цепи канинизированного моноклонального антитела от Mus musculus и Canis
<400> 185
gatatagtga tgacacaaac tcctctcagt ctttccgtat caccgggaga accggcttcc 60
atttcctgtc gggcctcaga gtctgtggac aactacggga tatccttcat gcactggtat 120
cagcagaaac ccggccagcc ccctaaactc cttatttaca gggccagtaa tctggaaagc 180
ggtgtgcccg atcgatttag cggttccggg agcggcacag atttcaccct gcgaatctct 240
agagttgaag cggatgatgc aggagtatat tactgccagc aatccaataa ggatcccctt 300
acattcggcg cgggtaccaa gctggagatc aag 333
<210> 186
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность мимотопа собачьего IL-31 15H05
<400> 186
Ser Val Pro Ala Asp Thr Phe Glu Cys Lys Ser Phe
1 5 10
<210> 187
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность мимотопа собачьего IL-31 15H05
<400> 187
Ser Val Pro Ala Asp Thr Phe Glu Arg Lys Ser Phe
1 5 10
<210> 188
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность мимотопа кошачьего IL-31 15H05
<400> 188
Ser Met Pro Ala Asp Asn Phe Glu Arg Lys Asn Phe
1 5 10
<210> 189
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность мимотопа лошадиного IL-31 15H05
<400> 189
Ser Met Pro Thr Asp Asn Phe Glu Arg Lys Arg Phe
1 5 10
<210> 190
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность мимотопа человеческого IL-31 15H05
<400> 190
Ser Val Pro Thr Asp Thr His Glu Cys Lys Arg Phe
1 5 10
<210> 191
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность мимотопа человеческого IL-31 15H05
<400> 191
Ser Val Pro Thr Asp Thr His Glu Arg Lys Arg Phe
1 5 10
<210> 192
<211> 33
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность мимотопа области спирали BC собачьего IL-31
<400> 192
Asn Ser Ser Ala Ile Leu Pro Tyr Phe Arg Ala Ile Arg Pro Leu Ser
1 5 10 15
Asp Lys Asn Ile Ile Asp Lys Ile Ile Glu Gln Leu Asp Lys Leu Lys
20 25 30
Phe
<210> 193
<211> 33
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность мимотопа области спирали BC кошачьего IL-31
<400> 193
Asn Gly Ser Ala Ile Leu Pro Tyr Phe Arg Ala Ile Arg Pro Leu Ser
1 5 10 15
Asp Lys Asn Thr Ile Asp Lys Ile Ile Glu Gln Leu Asp Lys Leu Lys
20 25 30
Phe
<210> 194
<211> 33
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность мимотопа области спирали BC лошадиного IL-31
<400> 194
Asn Ser Ser Ala Ile Leu Pro Tyr Phe Lys Ala Ile Ser Pro Ser Leu
1 5 10 15
Asn Asn Asp Lys Ser Leu Tyr Ile Ile Glu Gln Leu Asp Lys Leu Asn
20 25 30
Phe
<210> 195
<211> 33
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность мимотопа области спирали BC человеческого IL-31
<400> 195
His Ser Pro Ala Ile Arg Ala Tyr Leu Lys Thr Ile Arg Gln Leu Asp
1 5 10 15
Asn Lys Ser Val Ile Asp Glu Ile Ile Glu His Leu Asp Lys Leu Ile
20 25 30
Phe
<210> 196
<211> 24
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность мимотопа области спирали A собачьего IL-31
<400> 196
Ala Pro Thr His Gln Leu Pro Pro Ser Asp Val Arg Lys Ile Ile Leu
1 5 10 15
Glu Leu Gln Pro Leu Ser Arg Gly
20
<210> 197
<211> 24
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность мимотопа области спирали A кошачьего IL-31
<400> 197
Ala Pro Ala His Arg Leu Gln Pro Ser Asp Ile Arg Lys Ile Ile Leu
1 5 10 15
Glu Leu Arg Pro Met Ser Lys Gly
20
<210> 198
<211> 24
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность мимотопа области спирали A лошадиного IL-31
<400> 198
Gly Pro Ile Tyr Gln Leu Gln Pro Lys Glu Ile Gln Ala Ile Ile Val
1 5 10 15
Glu Leu Gln Asn Leu Ser Lys Lys
20
<210> 199
<211> 25
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность мимотопа области спирали A человеческого IL-31
<400> 199
Leu Pro Val Arg Leu Leu Arg Pro Ser Asp Asp Val Gln Lys Ile Val
1 5 10 15
Glu Glu Leu Gln Ser Leu Ser Lys Met
20 25
<210> 200
<211> 6
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность мимотопа области спирали AB собачьего IL-31
<400> 200
Thr Gly Val Pro Glu Ser
1 5
<210> 201
<211> 6
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность мимотопа области спирали AB кошачьего IL-31
<400> 201
Ile Gly Leu Pro Glu Ser
1 5
<210> 202
<211> 6
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность мимотопа области спирали AB лошадиного IL-31
<400> 202
Lys Gly Val Gln Lys Phe
1 5
<210> 203
<211> 6
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность мимотопа области спирали AB человеческого IL-31
<400> 203
Lys Gly Val Leu Val Ser
1 5
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА К ИНТЕРЛЕЙКИНУ-31 ДЛЯ ВЕТЕРИНАРНОГО ПРИМЕНЕНИЯ | 2019 |
|
RU2795411C2 |
| АНТИ-IL31 АНТИТЕЛА ДЛЯ ПРИМЕНЕНИЯ В ВЕТЕРИНАРИИ | 2020 |
|
RU2835543C1 |
| АНТИ-IL31 АНТИТЕЛА ДЛЯ ПРИМЕНЕНИЯ В ВЕТЕРИНАРИИ | 2018 |
|
RU2795485C2 |
| АНТИТЕЛА К РЕЦЕПТОРУ IL4 ДЛЯ ПРИМЕНЕНИЯ В ВЕТЕРИНАРИИ | 2019 |
|
RU2837141C2 |
| АНТИТЕЛА К СОБАЧЬЕМУ АЛЬФА-РЕЦЕПТОРУ ИНТЕРЛЕЙКИНА-4 | 2020 |
|
RU2840044C1 |
| АНТИТЕЛА К СОБАЧЬЕМУ АЛЬФА-РЕЦЕПТОРУ ИНТЕРЛЕЙКИНА-4 | 2020 |
|
RU2838943C1 |
| АНТИТЕЛА К ПАРВОВИРУСУ ДЛЯ ПРИМЕНЕНИЯ В ВЕТЕРИНАРИИ | 2020 |
|
RU2830441C2 |
| МОЛЕКУЛА РЕЦЕПТОРА IL4/IL13 ДЛЯ ВЕТЕРИНАРНОГО ПРИМЕНЕНИЯ | 2018 |
|
RU2795591C2 |
| ВАРИАНТЫ FC-ОБЛАСТИ С ИЗМЕНЕННЫМ СВЯЗЫВАНИЕМ С НЕОНАТАЛЬНЫМ FC-РЕЦЕПТОРОМ (FCRN) ДЛЯ ПРИМЕНЕНИЯ В ВЕТЕРИНАРИИ | 2019 |
|
RU2830231C2 |
| ИММУНОЦИТОКИНЫ И ИХ ПРИМЕНЕНИЕ | 2021 |
|
RU2818371C1 |
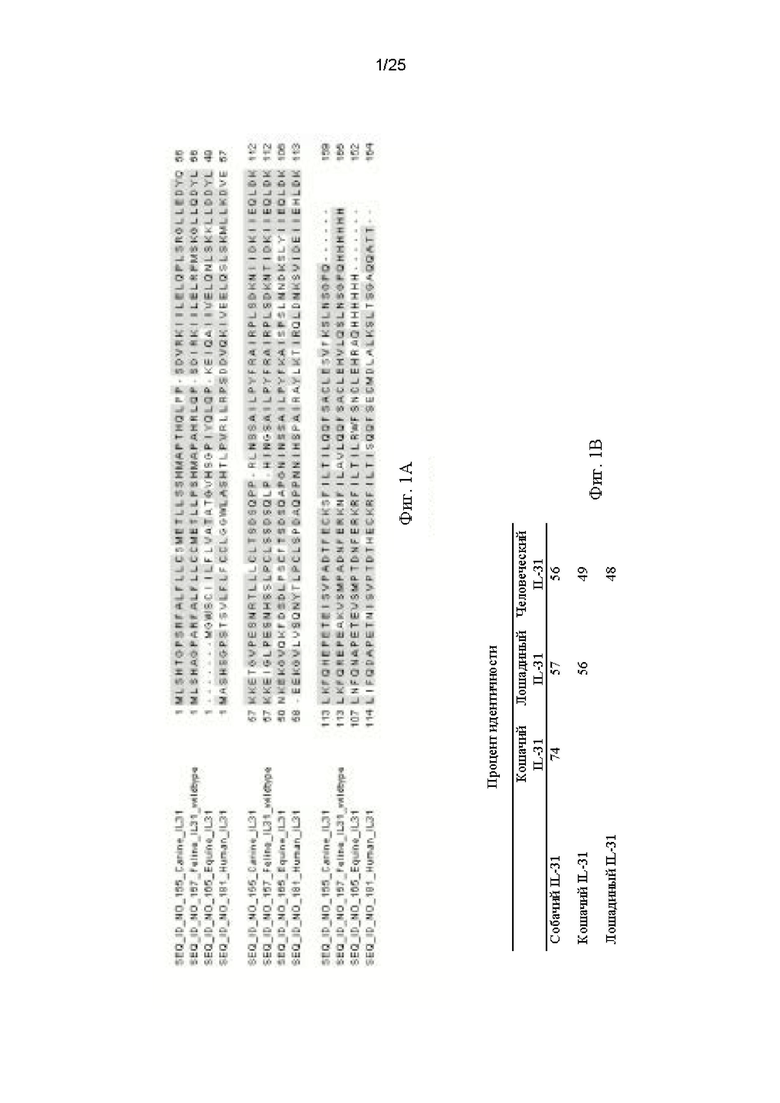

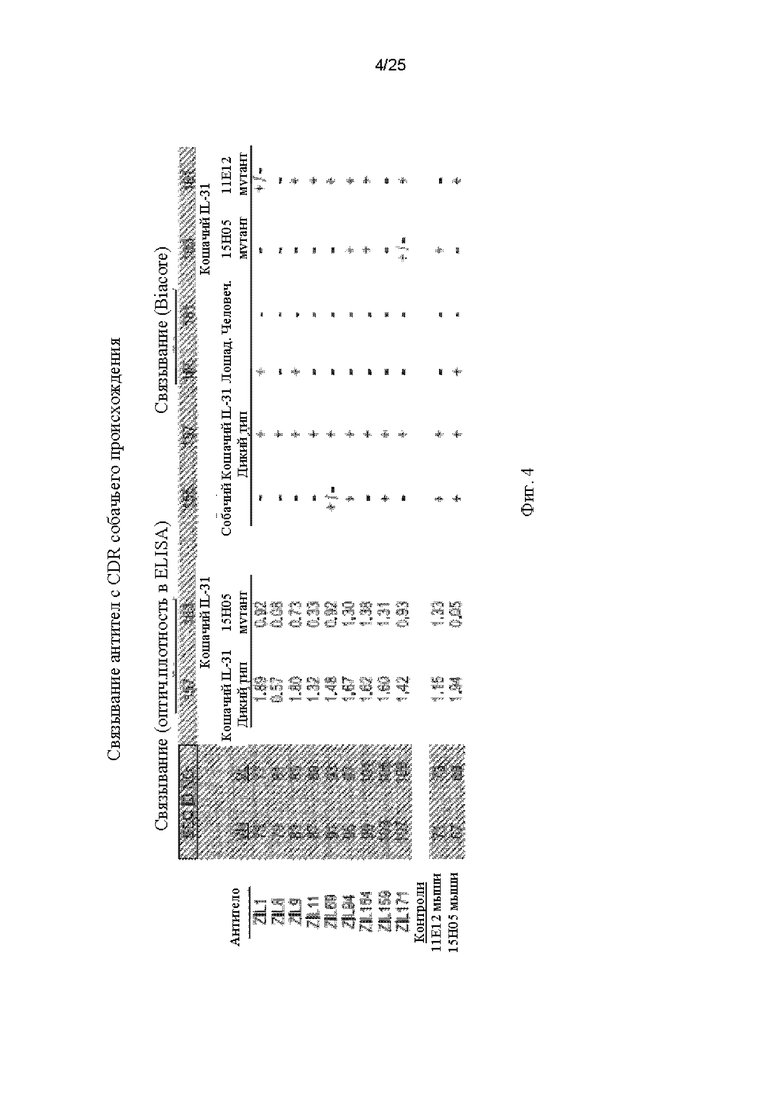

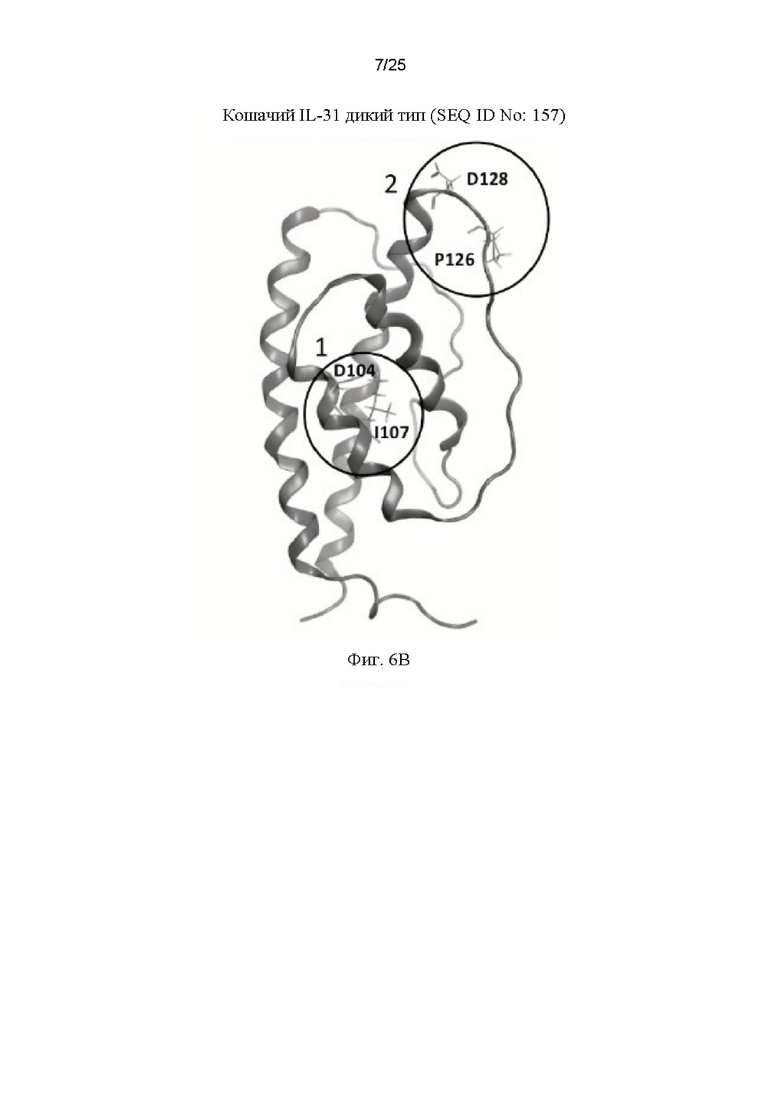
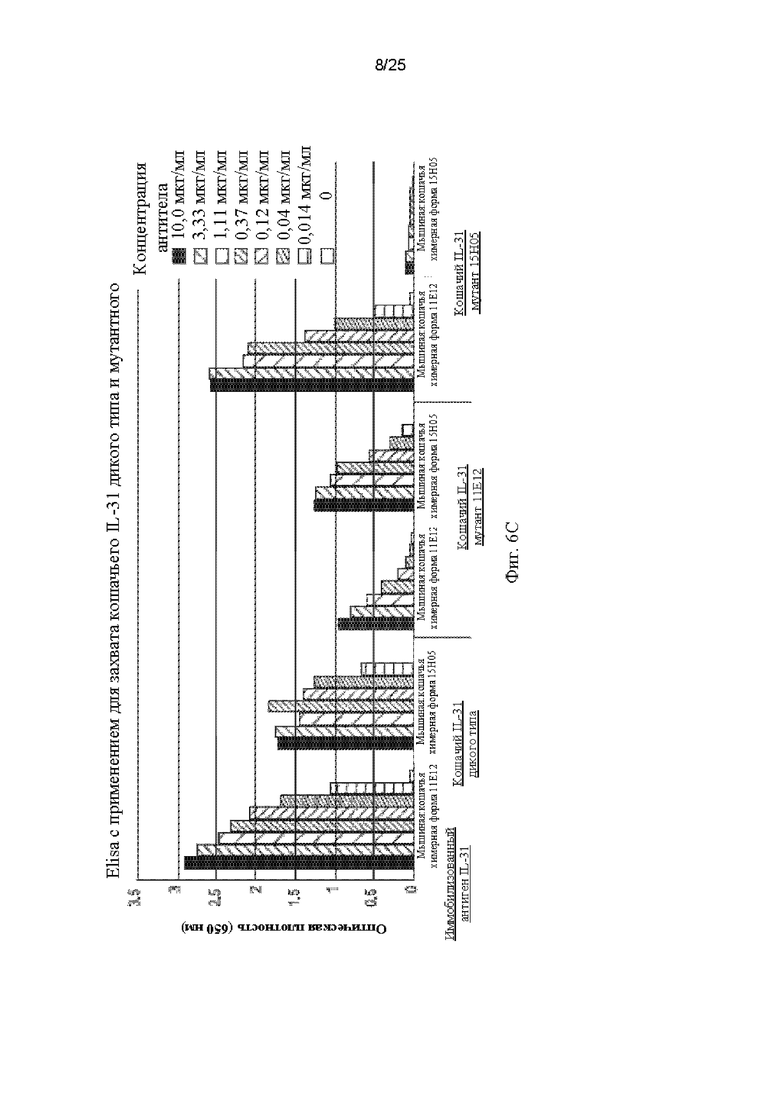
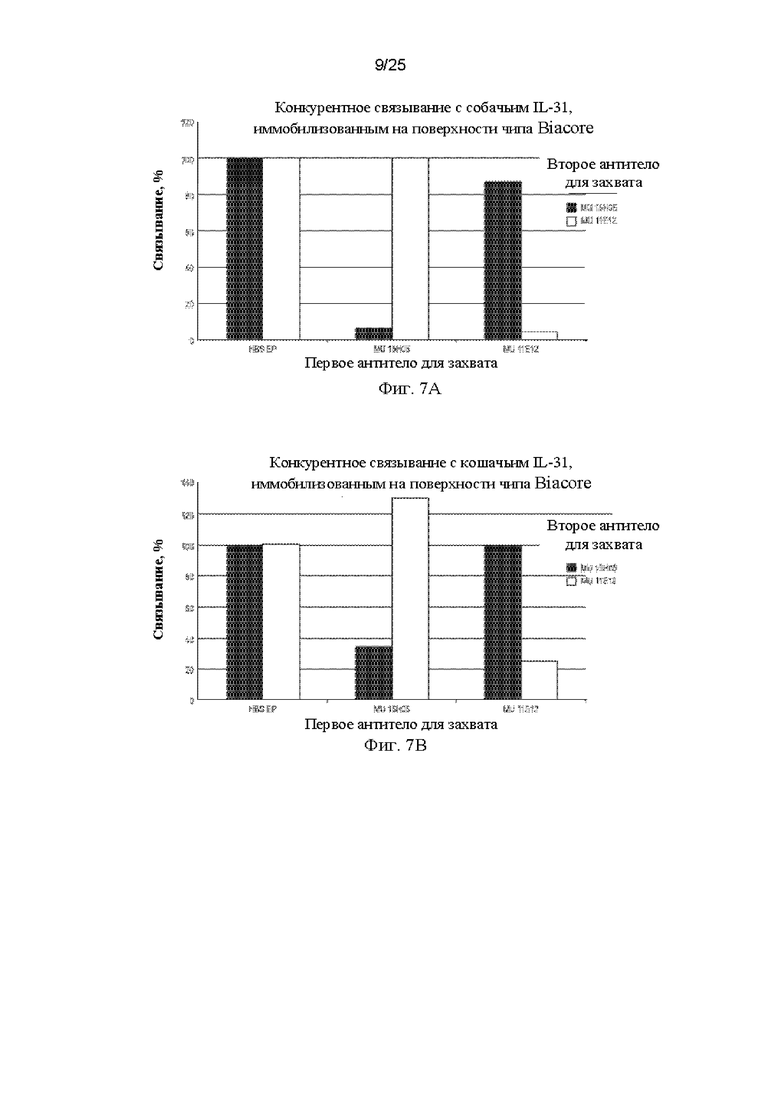
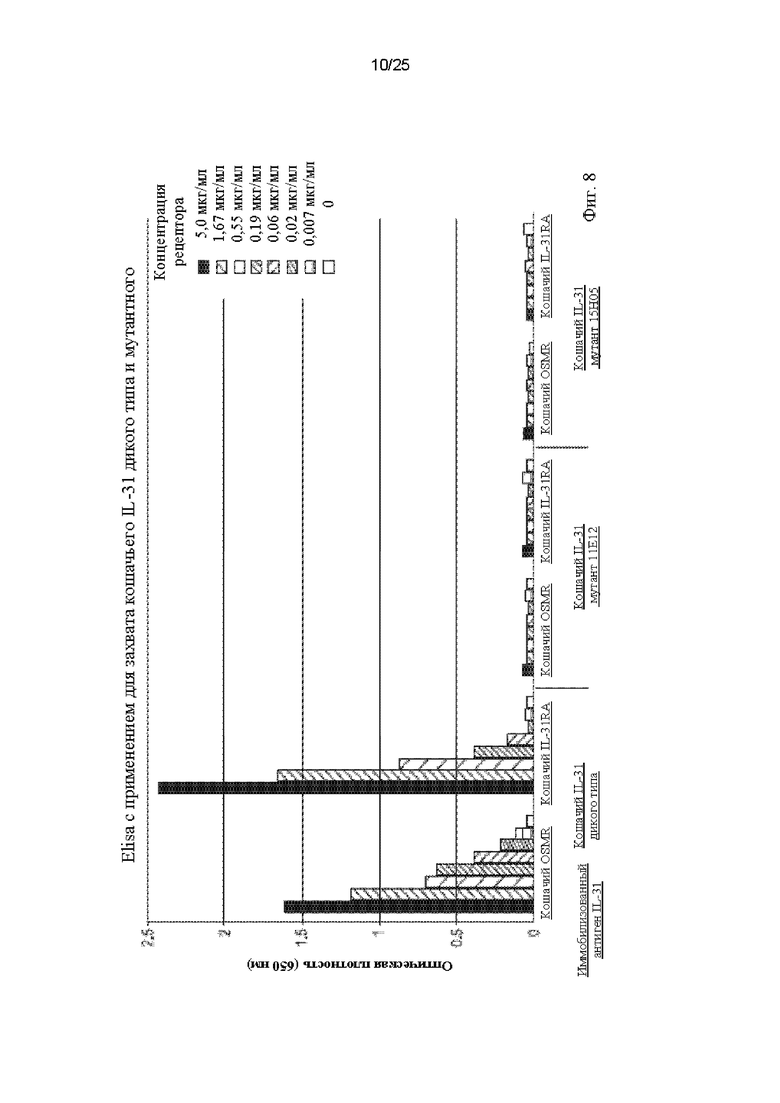
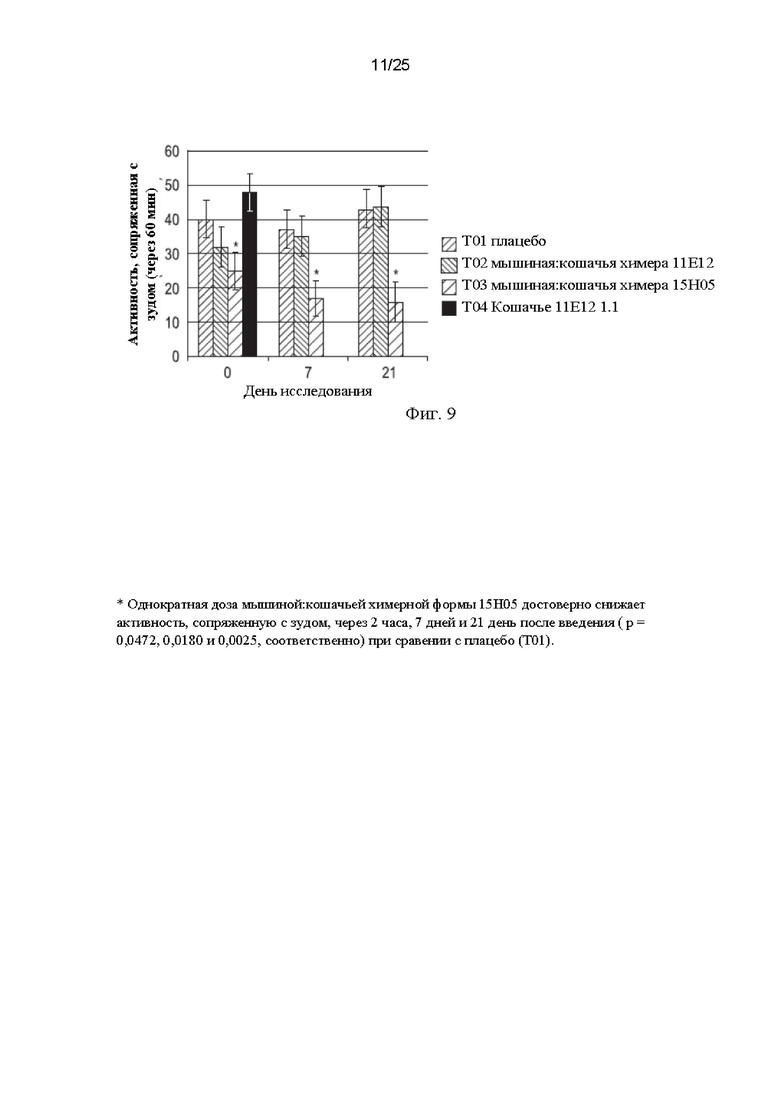
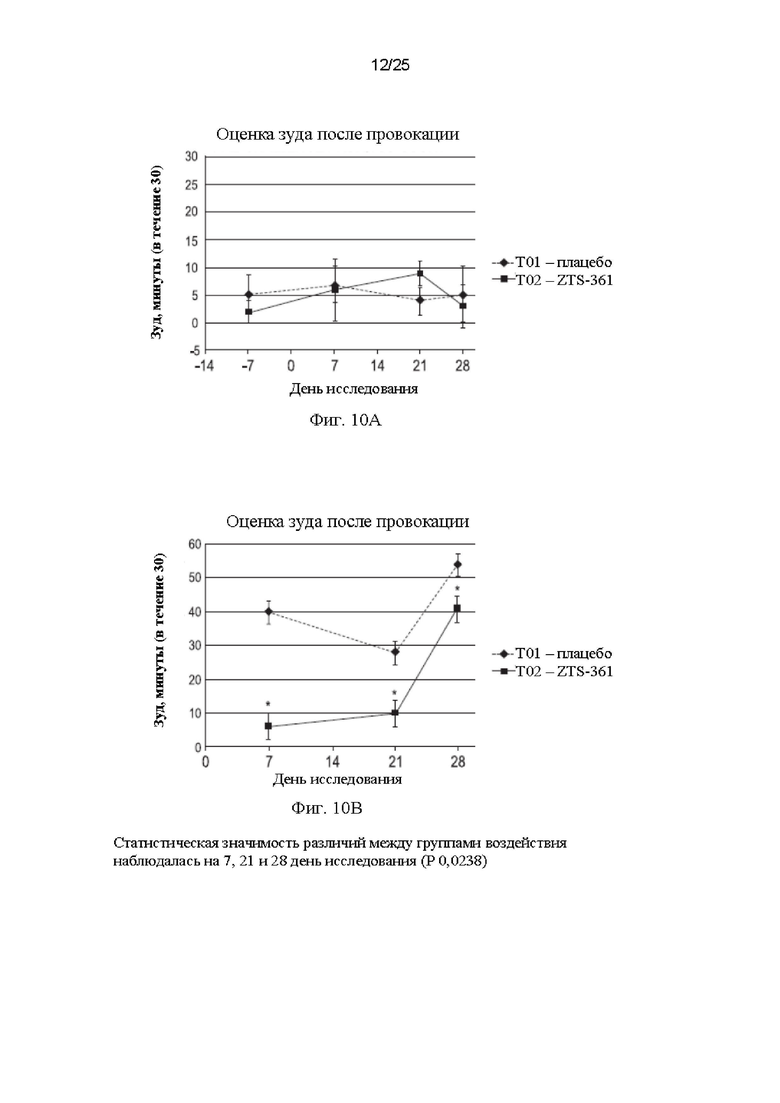
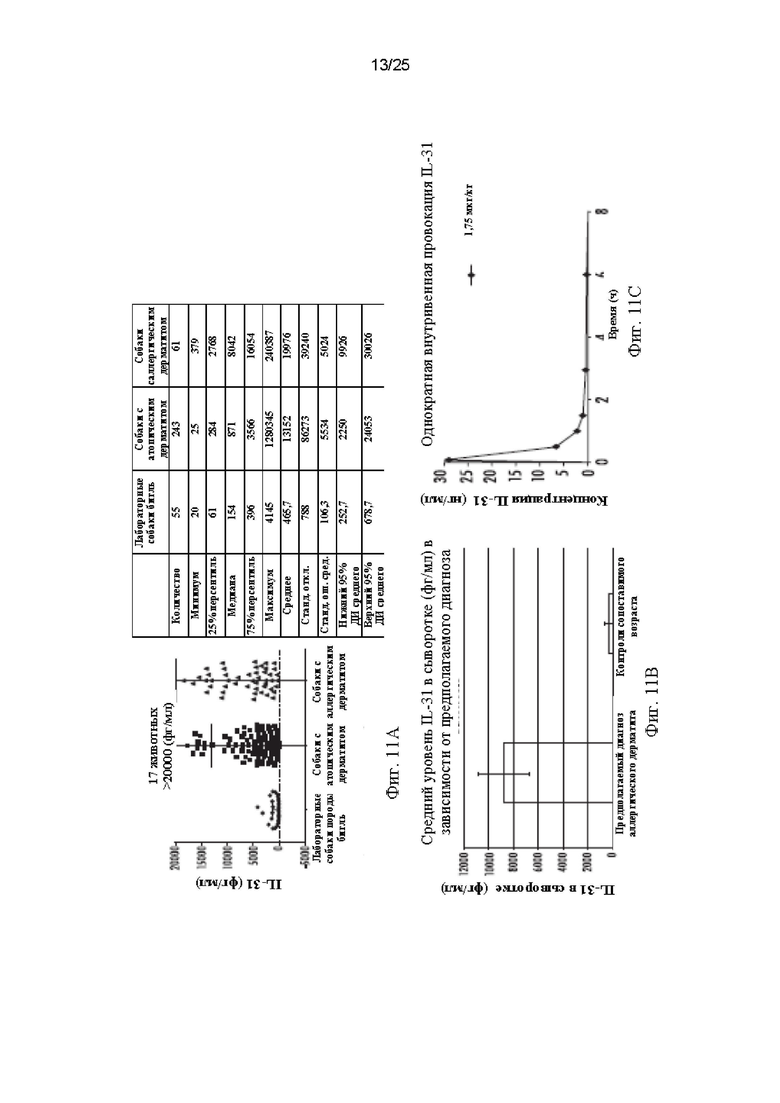
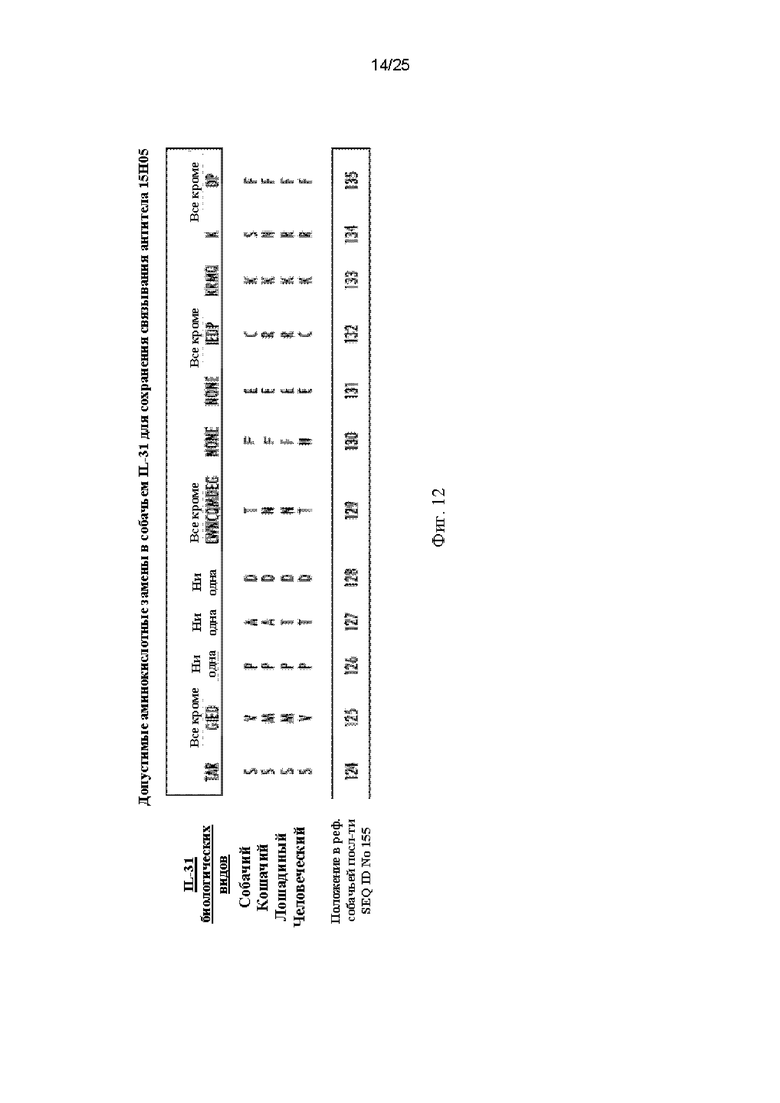
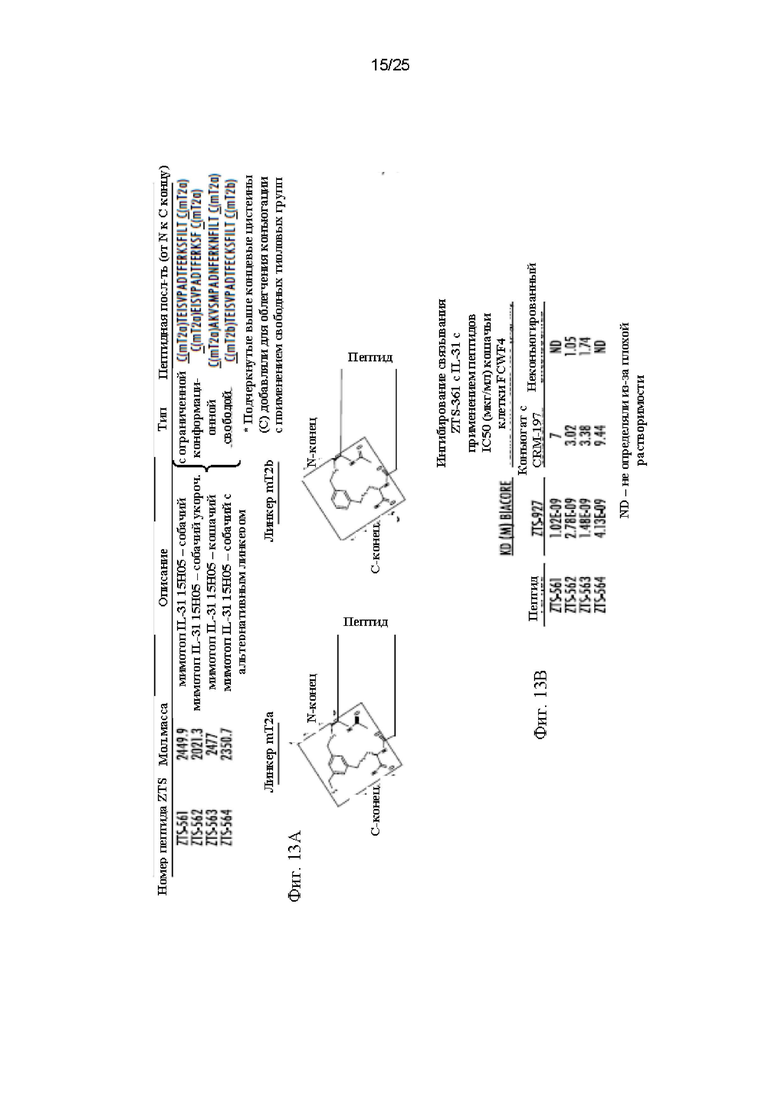
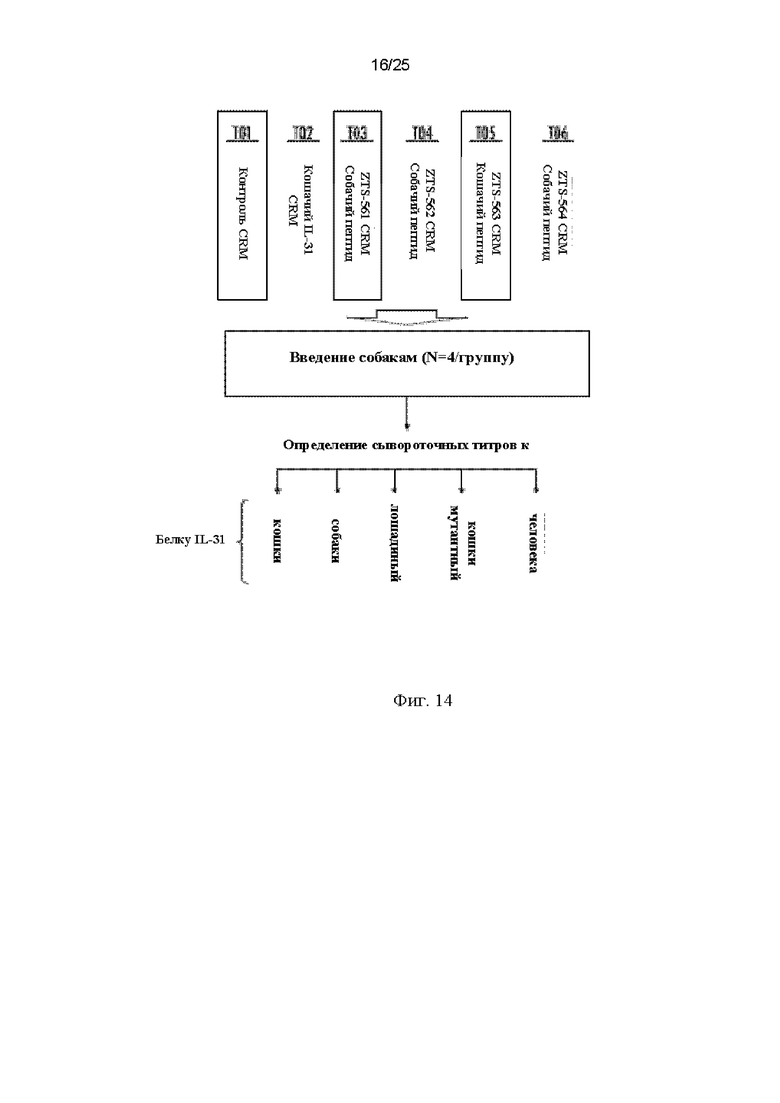

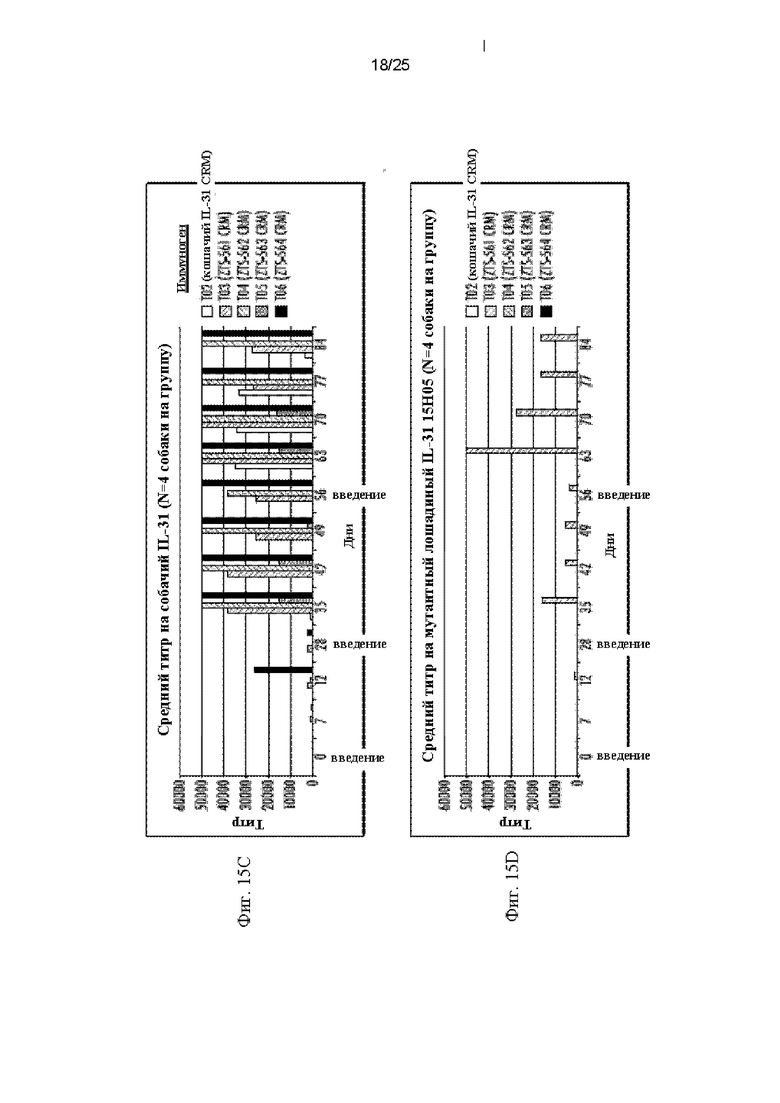
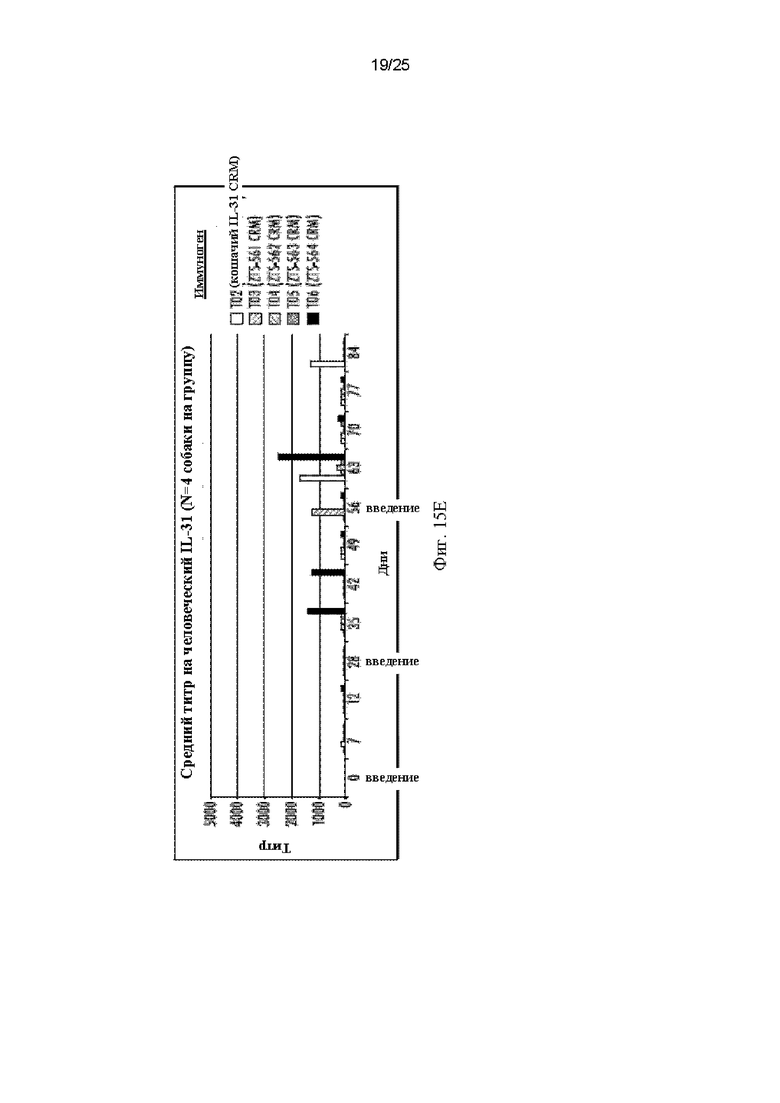

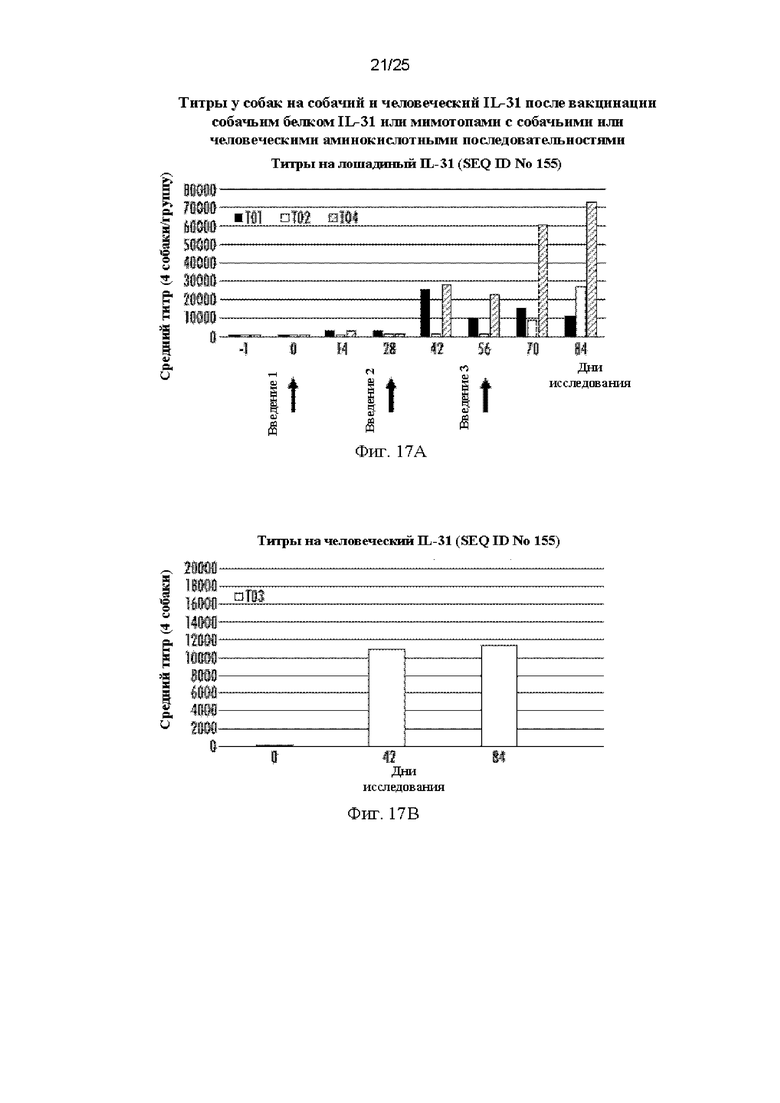

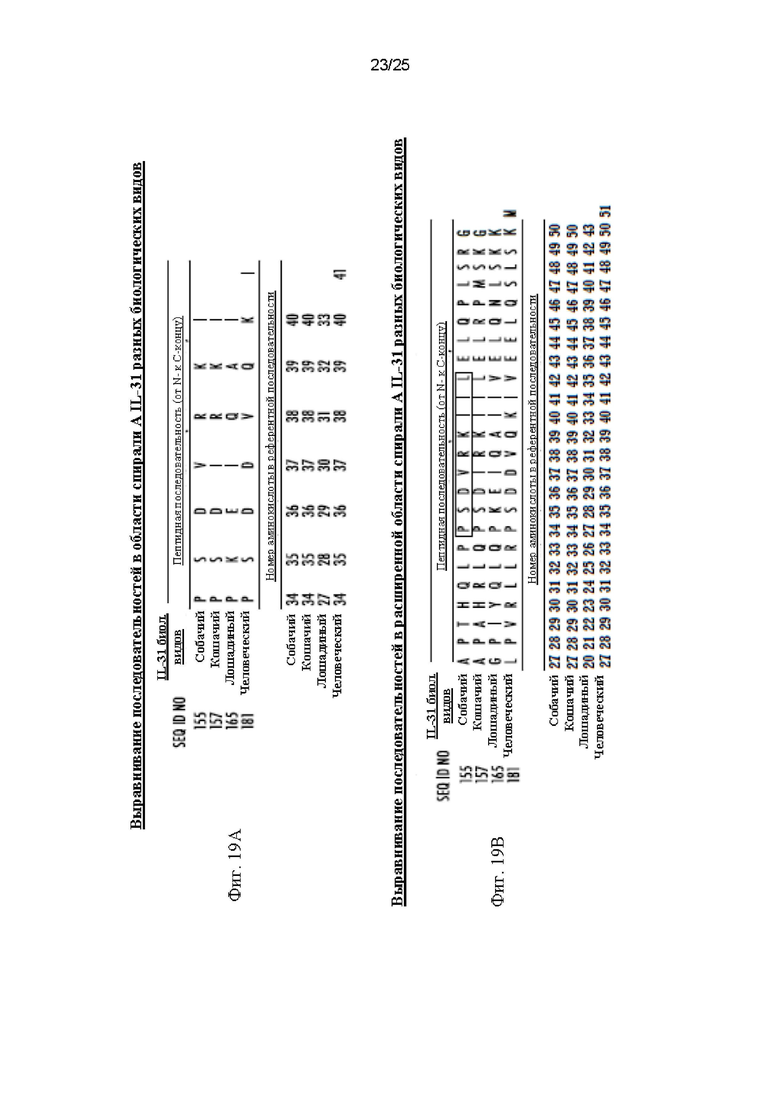


Изобретение относится к ветеринарии. Предложена вакцинная композиция для иммунизации и/или защиты млекопитающего - собаки, кошки или лошади - от опосредованного IL-31 заболевания, где указанная композиция содержит комбинацию полипептида-носителя и по меньшей мере одного мимотопа пептида IL-31, и адъювант. Мимотоп пептида IL-31 имеет длину от приблизительно 5 до приблизительно 40 аминокислот и выбран из группы, состоящей из мимотопа собачьего IL-31, мимотопа кошачьего IL-31, мимотопа лошадиного IL-31 и мимотопа человеческого IL-31. Представлен способ защиты млекопитающего от заболевания, опосредованного IL-31. Способ включает введение млекопитающему представленной вакцинной композиции. Изобретение позволяет получить вакцину для уменьшения кожного зуда и повреждений кожи у собак, кошек, лошадей с атопическим дерматитом. 2 н. и 18 з.п. ф-лы, 39 ил., 2 пр.
1. Вакцинная композиция для иммунизации и/или защиты млекопитающего - собаки, кошки или лошади - от опосредованного IL-31 заболевания, где указанная композиция содержит:
комбинацию полипептида-носителя и по меньшей мере одного мимотопа пептида IL-31, и
адъювант, где мимотоп пептида IL-31 имеет длину от приблизительно 5 до приблизительно 40 аминокислот и выбран из группы, состоящей из:
1) мимотопа собачьего IL-31, который представляет собой или содержит в качестве своей части аминокислотные последовательности SVPADTFECKSF (SEQ ID NO: 186), SVPADTFERKSF (SEQ ID NO: 187), NSSAILPYFRAIRPLSDKNIIDKIIEQLDKLKF (SEQ ID NO: 192), APTHQLPPSDVRKIILELQPLSRG (SEQ ID NO: 196), TGVPES (SEQ ID NO: 200) или их варианты, которые сохраняют связывание с анти-IL-31 агентом;
2) мимотопа кошачьего IL-31, который представляет собой или содержит в качестве своей части аминокислотные последовательности SMPADNFERKNF (SEQ ID NO: 188), NGSAILPYFRAIRPLSDKNTIDKIIEQLDKLKF (SEQ ID NO: 193), APAHRLQPSDIRKIILELRPMSKG (SEQ ID NO: 197), IGLPES (SEQ ID NO: 201) или их варианты, которые сохраняют связывание с анти-IL-31 агентом;
3) мимотопа лошадиного IL-31, который представляет собой или содержит в качестве своей части аминокислотнын последовательности SMPTDNFERKRF (SEQ ID NO: 189), NSSAILPYFKAISPSLNNDKSLYIIEQLDKLNF (SEQ ID NO: 194), GPIYQLQPKEIQAIIVELQNLS KK (SEQ ID NO: 198), KGVQKF (SEQ ID NO: 202) или их варианты, которые сохраняют связывание с анти-IL-31 агентом; и
4) мимотопа человеческого IL-31, который представляет собой или содержит в качестве своей части аминокислотные последовательности SVPTDTHECKRF (SEQ ID NO: 190), SVPTDTHERKRF (SEQ ID NO: 191), HSPAIRAYLKTIRQLDNKSVIDEIIEHLDKLIF (SEQ ID NO: 195), LPVRLLRPSDDVQKIVEELQSLSKM (SEQ ID NO: 199), KGVLVS (SEQ ID NO: 203) или их варианты, которые сохраняют связывание с анти-IL-31 агентом.
2. Вакцинная композиция по п. 1, где мимотоп связывается с антителом против IL-31 или его антигенсвязывающей частью, которое(ая) специфически связывается с областью белка IL-31 млекопитающих, вовлеченнной во взаимодействие белка IL-31 со своим ко-рецептором.
3. Вакцинная композиция по п. 2, где на связывание указанного антитела с указанной областью влияют мутации в области эпитопа связывания с 15Н05, выбранные из группы, состоящей из:
а) области между аминокислотными остатками 124 и 135 последовательности IL-31 кошки, представленной SEQ ID NO: 157 (кошачий IL-31 дикого типа);
б) области между аминокислотными остатками 124 и 135 последовательности IL-31 собаки, представленной SEQ ID NO: 155 (собачий IL-31); и
в) области между аминокислотными остатками 118 и 129 последовательности IL-31 лошади, представленной SEQ ID NO: 165 (лошадиный IL-31).
4. Вакцинная композиция по п. 2, где мимотоп связывается с антителом против IL-31 или его антигенсвязывающей частью, содержащей по меньшей мере одну из следующих комбинаций последовательностей участков, определяющих комплементарность (CDR):
1) антитело 15Н05: CDR1 вариабельной области тяжелой цепи (VH) с последовательностью SYTIH (SEQ ID NO: 1), VH-CDR2 с последовательностью NINPTSGYTENNQRFKD (SEQ ID NO: 2), VH-CDR3 с последовательностью WGFKYDGEWSFDV (SEQ ID NO: 3), CDR1 вариабельной области легкой цепи (VL) с последовательностью RASQGISIWLS (SEQ ID NO: 4), VL-CDR2 с последовательностью KASNLHI (SEQ ID NO: 5) и VL-CDR3 с последовательностью LQSQTYPLT (SEQ ID NO: 6);
2) антитело ZIL1: CDR1 вариабельной области тяжелой цепи (VH) с последовательностью SYGMS (SEQ ID NO: 13), VH-CDR2 с последовательностью HINSGGSSTYYADAVKG (SEQ ID NO: 14), VH-CDR3 с последовательностью VYTTLAAFWTDNFDY (SEQ ID NO: 15), CDR1 вариабельной области легкой цепи (VL) с последовательностью SGSTNNIGILAAT (SEQ ID NO: 16), VL-CDR2 с последовательностью SDGNRPS (SEQ ID NO: 17) и VL-CDR3 с последовательностью QSFDTTLDAYV (SEQ ID NO: 18);
3) антитело ZIL8: VH-CDR1 с последовательностью DYAMS (SEQ ID NO: 19), VH-CDR2 с последовательностью GIDSVGSGTSYADAVKG (SEQ ID NO: 20), VH-CDR3 с последовательностью GFPGSFEH (SEQ ID NO: 21), VL-CDR1 с последовательностью TGSSSNIGSGYVG (SEQ ID NO: 22), VL-CDR2 с последовательностью YNSDRPS (SEQ ID NO: 23), VL-CDR3 с последовательностью SVYDRTFNAV (SEQ ID NO: 24);
4) антитело ZIL9: VH-CDR1 с последовательностью SYDMT (SEQ ID NO: 25), VH-CDR2 с последовательностью DVNSGGTGTAYAVAVKG (SEQ ID NO: 26), VH-CDR3 с последовательностью LGVRDGLSV (SEQ ID NO: 27), VL-CDR1 с последовательностью SGESLNEYYTQ (SEQ ID NO: 28), VL-CDR2 с последовательностью RDTERPS (SEQ ID NO: 29), VL-CDR3 с последовательностью ESAVDTGTLV (SEQ ID NO: 30);
5) антитело ZIL11: VH-CDR1 с последовательностью TYVMN (SEQ ID NO: 31), VH-CDR2 с последовательностью SINGGGSSPTYADAVRG (SEQ ID NO: 32), VH-CDR3 с последовательностью SMVGPFDY (SEQ ID NO: 33), VL-CDR1 с последовательностью SGESLSNYYAQ (SEQ ID NO: 34), VL-CDR2 с последовательностью KDTERPS (SEQ ID NO: 35), VL-CDR3 с последовательностью ESAVSSDTIV (SEQ ID NO: 36);
6) антитело ZIL69: VH-CDR1 с последовательностью SYAMK (SEQ ID NO: 37), VH-CDR2 с последовательностью TINNDGTRTGYADAVRG (SEQ ID NO: 38), VH-CDR3 с последовательностью GNAESGCTGDHCPPY (SEQ ID NO: 39), VL-CDR1 с последовательностью SGESLNKYYAQ (SEQ ID NO: 40), VL-CDR2 с последовательностью KDTERPS (SEQ ID NO: 41), VL-CDR3 с последовательностью ESAVSSETNV (SEQ ID NO: 42);
7) антитело ZIL94: VH-CDR1 с последовательностью TYFMS (SEQ ID NO: 43), VH-CDR2 с последовательностью LISSDGSGTYYADAVKG (SEQ ID NO: 44), VH-CDR3 с последовательностью FWRAFND (SEQ ID NO: 45), VL-CDR1 с последовательностью GLNSGSVSTSNYPG (SEQ ID NO: 46), VL-CDR2 с последовательностью DTGSRPS (SEQ ID NO: 47), VL-CDR3 с последовательностью SLYTDSDILV (SEQ ID NO: 48);
8) антитело ZIL154: VH-CDR1 с последовательностью DRGMS (SEQ ID NO: 49), VH-CDR2 с последовательностью YIRYDGSRTDYADAVEG (SEQ ID NO: 50), VH-CDR3 с последовательностью WDGSSFDY (SEQ ID NO: 51), VL-CDR1 с последовательностью KASQSLLHSDGNTYLD (SEQ ID NO: 52), VL-CDR2 с последовательностью KVSNRDP (SEQ ID NO: 53), VL-CDR3 с последовательностью MQAIHFPLT (SEQ ID NO: 54);
9) антитело ZIL159: VH-CDR1 с последовательностью SYVMT (SEQ ID NO: 55), VH-CDR2 с последовательностью GINSEGSRTAYADAVKG (SEQ ID NO: 56), VH-CDR3 с последовательностью GDIVATGTSY (SEQ ID NO: 57), VL-CDR1 с последовательностью S GETLNRFYTQ (SEQ ID NO: 58), VL-CDR2 с последовательностью KDTERPS (SEQ ID NO: 59), VL-CDR3 с последовательностью KSAVSIDVGV (SEQ ID NO: 60);
10) антитело ZIL171: VH-CDR1 с последовательностью TYVMN (SEQ ID NO: 61), VH-CDR2 с последовательностью SINGGGSSPTYADAVRG (SEQ ID NO: 62), VH-CDR3 с последовательностью SMVGPFDY (SEQ ID NO: 63), VL-CDR1 с последовательностью SGKSLSYYYAQ (SEQ ID NO: 64), VL-CDR2 с последовательностью KDTERPS (SEQ ID NO: 65), VL-CDR3 с последовательностью ESAVSSDTIV (SEQ ID NO: 66); или
11) вариант 1)-10), отличающийся от соответствующего родительского антитела 15Н05, ZIL1, ZIL8, ZIL9, ZIL11, ZIL69, ZIL94, ZIL154, ZIL159 или ZIL171 добавлением, делецией и/или заменой одного или более чем одного аминокислотного остатка по меньшей мере в одном из CDR1, CDR2 или CDR3 VH или VL.
5. Вакцинная композиция по любому из пп. 1-4, где мимотоп представляет собой мимотоп с ограниченной конформационной свободой.
6. Вакцинная композиция по п. 5, где мимотоп с ограниченной конформационной свободой представляет собой химически связанный циклический пептид.
7. Вакцинная композиция по любому из пп. 1-6, где мимотоп химически конъюгирован с полипептидом-носителем.
8. Вакцинная композиция по любому из пп. 1-6, где полипептид-носитель и мимотоп являются частью рекомбинантного слитого белка.
9. Вакцинная композиция по любому из пп. 1-8, где полипептид-носитель содержит бактериальный анатоксин или его производное, гемоцианин фиссуреллы (KLH) или вирусоподобную частицу.
10. Вакцинная композиция по п. 9, где бактериальный анатоксин или производное представляет собой столбнячный анатоксин, дифтерийный анатоксин, столбнячный анатоксин, комплекс белков внешней мембраны N. meningitidis группы В, экзотоксин Pseudomonas или нетоксичный мутант дифтерийного токсина (CRM197).
11. Вакцинная композиция по п. 9, где вирусоподобная частица представляет собой HBsAg, HBcAg, бактериофаг Qbeta E. coli, вирус Norwalk, вирус чумы собак (CDV) или гемагглютинин (НА) вируса гриппа.
12. Вакцинная композиция по п. 10, где полипептид-носитель содержит или представляет собой CRM197.
13. Вакцинная композиция по любому из пп. 1-12, где адъювант выбран из группы, состоящей из адъюванта масло в воде, адъюванта полимер и вода, адъюванта вода в масле, адъюванта гидроксид алюминия, адъюванта витамин Е и их комбинаций.
14. Вакцинная композиция по любому из пп. 1-13, где адъювант представляет собой состав, содержащий сапонин, стерол, четвертичное аммониевое соединение и полимер.
15. Вакцинная композиция по п. 14, где сапонин представляет собой Quil А или его очищенную фракцию, стерол представляет собой холестерин, четвертичное аммониевое соединение представляет собой бромид диметилдиоктадециламмония (DDA) и полимер представляет собой полиакриловую кислоту.
16. Вакцинная композиция по любому из пп. 1-13, где адъювант содержит комбинацию одного или нескольких выделенных иммуностимулирующих олигонуклеотидов, стерола и сапонина.
17. Вакцинная композиция по п. 16, где один или несколько выделенных иммуностимулирующих олигонуклеотидов содержат CpG, стерол представляет собой холестерин, а сапонин представляет собой Quil А или его очищенную фракцию.
18. Способ защиты млекопитающего от заболевания, опосредованного IL-31, включающий введение млекопитающему вакцинной композиции по любому из пп. 1-17.
19. Способ по п. 18, где млекопитающее выбрано из группы, состоящей из собаки, кошки и лошади.
20. Способ по п. 18, где мимотоп IL-31, содержащийся в вакцинной композиции, вводят млекопитающему в дозе от приблизительно 10 мкг до приблизительно 100 мкг.
| Автомобиль-сани, движущиеся на полозьях посредством устанавливающихся по высоте колес с шинами | 1924 |
|
SU2017A1 |
| Автомобиль-сани, движущиеся на полозьях посредством устанавливающихся по высоте колес с шинами | 1924 |
|
SU2017A1 |
| Многоступенчатая активно-реактивная турбина | 1924 |
|
SU2013A1 |
| Пломбировальные щипцы | 1923 |
|
SU2006A1 |
| СЕРГЕЕВ О.В., и др, Синтетические пептидные вакцины, Вопросы вирусологии, том 61, N1, 2016, с.5-8, DOI: 10.18821/0507-4088-2016-61-1-5-8. | |||

