Настоящее изобретение относится к молекулам ДНК, которые могут транскрибироваться в мРНК, несущую новые UTR-последовательности, которые обеспечивают содержащим их молекулам РНК преимущества, объединяющие их очень малую длину и в то же время способность обеспечивать высокую эффективность трансляции. Кроме того, настоящее изобретение относится к векторам, содержащим указанную молекулу ДНК, и к клеткам-хозяевам, содержащим указанный вектор. Кроме того, настоящее изобретение относится к соответствующим молекулам РНК, которые содержат указанные UTR. Кроме того, настоящее изобретение относится к фармацевтической композиции, содержащей описанную молекулу РНК и необязательно фармацевтически приемлемый носитель, а также к применению описанных UTR для трансляции кодирующей области молекулы РНК в полипептид или белок, кодируемый указанной кодирующей областью.
В последние годы повысилась значимость матричной РНК (мРНК) в качестве нового лекарственного средства. В отличие от генных терапий на основе ДНК, для мРНК не требуется транспортирование в ядро, она непосредственно транслируется в белок в цитоплазме (J Control Release, 150, 2011, сс.238-247 и Eur J Pharm Biopharm, 71, 2009, cc. 484-489). Это делает мРНК безопасной, позволяя избегать потенциального инсерционного мутагенеза, риск возникновения которого является маловероятным, но существует при применении медикаментозных средств для генной терапии на основе ДНК. Поэтому терапии на основе мРНК рассматриваются в качестве перспективных альтернатив генной и белковой замещающих терапий для широкого спектра медицинских показаний (J Control Release, 150, 2011, сс.238-247; Eur J Pharm Biopharm, 71, 2009, cc. 484-489; Nat Biotech, 29, 2011, cc. 154-157 и Nat Rev Genet, 12, 2011, cc. 861-874). Однако требуется преодолеть сильную иммуногенность, а также ограниченную стабильность канонической мРНК для достижения в дальнейшем возможности ее клинического применения. В этом плане стабильность мРНК и прежде всего скорость трансляции мРНК является имеющим решающее значение параметром при рассмотрении возможности медицинских применений, поскольку это определяет, например, дозирование и интервалы между введением доз лекарственных средств на основе мРНК.
Несколько стратегий доказали свою ценность как в отношении повышения стабильности, так и в отношении снижения иммуногенного ответа, запускаемого введением мРНК в клетки или организмы. Среди них следует упомянуть включение химически модифицированных нуклеотидов (Current Opinion in Drug Discovery and Development, 10, 2007, c. 523). Kormann с соавторами продемонстрировали, что замена всего лишь 25% остатков уридина и цитидина на 2-тиоуредин и 5-метилцитидин является достаточной для повышения стабильности мРНК, а также для уменьшения активации врожденного иммунитета, запускаемого вводимой извне мРНК in vitro (WO 2012/0195936; WO 2007024708 A2).
Кроме того, описано, что нетранслируемые области (UTR) в мРНК играют существенную роль как в регуляции стабильности мРНК, так и в трансляции мРНК. Известно, что UTR влияют на инициацию, элонгацию и терминацию трансляции, а также на стабилизацию и внутриклеточную локализацию мРНК, посредством взаимодействия с РНК-связывающими белками (Briefings in Bioinformatics, 1, 2000, сс.236-249 и Cold Spring Harbor Monograph Archive, 48, 2007, cc. 87-128). UTR в зависимости от специфических мотивов, присутствующих в ней, может либо повышать, либо понижать кругооборот мРНК (Cell. Mol. Life Sci., 69, 2012, cc. 3613-3634; Nucleic Acids Research, 33, 2005, D141-D146; Science, 309, 2005, cc. 1514-1518 и Current Protein & Peptide Science, 13, 2012, cc. 294-304). Опубликованы современные данные о времени полужизни мРНК и сответствующих UTR-последовательностей (Nucleic Acids Research, 39, 2011, cc. 556-566 и Nucleic acids research, 37, e115).
UTR представляют собой участки молекулы мРНК, расположенные в обратном направлении относительно стартового кодона и в прямом направлении относительно стоп-кодона мРНК, т.е. последовательности, которые не транслируются. Эти области транскрибируются с кодирующей областью и, следовательно, являются экзонами, поскольку присутствуют в зрелой мРНК. UTR, расположенную в обратном направлении относительно стартового кодона мРНК, называют 5' UTR, и после транскрипции она несут среди прочего последовательности, которые соответствуют (оставшимся 3'-) частям промотора, а также так называемой последовательности Козак.
Консенсусная последовательность Козак, консенсус Козак или последовательность Козак представляет собой последовательность, которая, как известно, встречается в мРНК эукариотических организмов и имеет консенсус (gcc)gccRccAUGG. Консенсусная последовательность Козак играет основную роль в инициации процесса трансляции. Последовательность названа в честь впервые описавшей ее Мэрилин Козак. Эта последовательность в молекуле мРНК распознается рибосомой в сайте инициации трансляции, из который белок кодируется указанной молекулой мРНК. Эта последовательность или ее возможная вариация требуется для рибосомы для инициации трансляции. Последовательность на основе обобщения данных из широкого разнообразия источников, проанализированных Козак (примерно в целом 699), обозначают следующим образом (gcc)gccRccAUGG, где строчными буквами обозначены наиболее часто встречающееся основание в положении, в котором или редко изменяется, если вообще изменяется, символ «R», который обозначает пурин (аденин или гуанин), всегда присутствует в указанном положении (причем, по данным основание тем не менее может варьироваться; «AUGG» является постоянной Козак аденин встречается чаще); а роль последовательности в скобках ((gcc)) не установлена.
Консенсусная последовательность Козак исходно была обозначена как ACCAUGG на основе анализа воздействия точечных мутаций вокруг инициирующего кодона (AUG, где А в этом контексте соответствует положению +1) на трансляцию гена препроинсулина. Более подробное изучение мутагенеза мРНК 699 позвоночных животных привело к получению данных о том, что в консенсусной последовательности GCCGCCACCAUGG, в которой А, расположенный в обратном направлении относительно стартового кодона AUG в положении -3, может также представлять собой G (Nucleic Acids Res., 15 (20), 1987, сс. 8125-8148). Исследования трансляции препроинсулина и альфа-глобина в эукариотических клетках продемонстрировали, что важнейшим для инициации эффективной трансляции является пурин (как правило, А) в положении -3, а, если пурин удалить, то важнейшим является G в положении +4 (J. Cell Biol., 108, 1989, сс. 2292-41). Количество белка, синтезированного с молекулы мРНК сильно зависит последовательности элемента Козак: стартовый кодон AUG, кодирующий N-концевой метионин белка, является наиболее важным. Для строгого консенсуса нуклеотиды в положениях +4 (G) и -3 (А или G) оба должны соответствовать консенсусу. Приемлемая консенсусная последовательность имеет только один из этих двух сайтов, а слабая консенсусная последовательность не удовлетворяет ни одному из требований к положениям +4 или -3. Два остатка цитидина в положениях -1 и -2 не являются в значительной степени консервативными (Cell, 44 (2), 1986, cc. 283-292), в то время как G в положении -6 является важным для инициации трансляции (Br. J. Haematol., 124(2), 2004, cc. 224-231).
Хотя из существующего уровня техники уже известны средства и методы повышения стабильности мРНК, снижения иммуногенного ответа, запускаемого мРНК, введенной в клетки или организмы, и повышения эффективности экспрессии (т.е. эффективности транскрипции и/или трансляции), все еще существует необходимость в усовершенствованиях, прежде всего касательно дополнительных средств повышения эффективности экспрессии (т.е. эффективности транскрипции и/или трансляции), поскольку эффективность экспрессии является важнейшим параметром для предполагаемых медицинских применений, поскольку она определяет, например, дозирование и интервалы между введением доз лекарственных средств на основе мРНК и, в конце концов, определяет биодоступность конечного продукта, т.е. кодируемого пептида или белка. В то же время существует постоянная необходимость в дополнительном снижении стоимости производства лекарственных средств на основе мРНК, повышении выхода полученных молекул мРНК и увеличении доступного пространства в полученной молекуле мРНК для фактического трансгена, т.е. для кодирующей области, которая кодирует требуемый полипептид.
Настоящая заявка направлена на решение указанной задачи, что описано в вариантах осуществления изобретения, представленных в формуле изобретения.
В частности, при создании настоящего изобретения неожиданно было установлено, что существует возможность уменьшения размера UTR-последовательности до последовательности «минимальной UTR», снижая тем самым стоимость получения лекарственных средств на основе мРНК, повышая выход полученных молекул мРНК и увеличивая доступное пространство в полученной молекуле мРНК для фактического трансгена, т.е. для кодирующей области, которая кодирует требуемый полипептид. Кроме того, одновременно при создании изобретения неожиданно было установлено, что указанная минимальная UTR-последовательность сохраняет или даже повышает уровень экспрессии по сравнению с каноническими UTR-последовательностями, при этом установлено, что модификации в указанной минимальной UTR-последовательности даже повышают уровень экспрессии молекулы мРНК.
Эти данные привели к получению вариантов, охарактеризованных в формуле изобретения, в частности к получению молекул ДНК, которые обеспечивали получение молекул РНК, несущих указанную последовательность «минимальной UTR», а также к получению соответствующих молекул РНК.
В первом объекте изобретения соответствующие молекулы описаны на уровне ДНК, в то время как ниже во втором объекте изобретения соответствующие молекулы описаны на уровне РНК.
Таким образом, первым объектом настоящего изобретения является молекула ДНК, которая может транскрибироваться в мРНК, где указанная молекула ДНК содержит одну цепь со следующими элементами:
(а) кодирующая область, включая стартовый кодон на ее 5'-конце, которая кодирует полипептид; и
(б) расположенная непосредственно в обратном направлении относительно указанной кодирующей последовательности последовательность, выбранная из группы, которая состоит из:
(б1) R1-CGCCACC (SEQ ID NO: 1);
или последовательности, где в указанной последовательности С в положении 6 SEQ ID NO: 1 заменен на А и С в положении 7 SEQ ID NO: 1 заменен на G; и/или А в положении 5 SEQ ID NO: 1 заменен на G; и
(б2) R1-CNGCCACC (SEQ ID NO: 2), в которой нуклеотид N в положении SEQ ID NO: 2 представляет собой нуклеотид, выбранный из группы, которая состоит из Т, G, С или А;
или последовательности, где в указанной последовательности С в положении 7 SEQ ID NO: 2 заменен на А и С в положении 8 SEQ ID NO: 2 заменен на G; и/или А в положении 6 SEQ ID NO: 2 заменен на G,
где R1 обозначает промотор, который распознается ДНК-зависимой РНК-полимеразой;
или содержит комплементарную цепь.
Последовательность ДНК называют «смысловой», если последовательность является такой же, что и у копии матричной РНК, которая транслируется в белок. Последовательность противоположной комплементарной цепи называют «антисмысловой» последовательностью. Молекула ДНК, предлагаемая в настоящем изобретении, указанная выше в пунктах (а) и (б), представлена со ссылкой на смысловую цепь, при этом специалист в данной области легко может определить соответствующую комплементарную антисмысловую цепь с использованием правил спариваний оснований.
Молекула ДНК, предлагаемая в настоящем изобретении, представляет собой молекулу ДНК, которая может транскрибироваться в молекулу мРНК. Транскрипция представляет собой первую стадию генной экспрессии, при которой конкретный сегмент молекулы ДНК копируется в молекулу мРНК с помощью фермента РНК-полимеразы. В процессе транскрипции последовательность ДНК считывается РНК-полимеразой, которая продуцирует комплементарную антипараллельную цепь РНК, называемую первичным транскриптом.
В качестве матрицы для транскрипции служит только одна из двух цепей ДНК. Антисмысловая цепь ДНК в процессе транскрипции считывается ДНК-зависимой РНК-полимеразой в направлении от 3'-конца к 5'-концу (3'→5'). Комплементарная РНК создается в противоположном направлении, т.е. направлении 5'→3', соответствуя последовательности смысловой цепи, за исключением замены тимина на урацил. Это направление обусловлено тем, что РНК-полимераза может добавлять нуклеотиды только к 3'-концу растущей цепи мРНК. Не являющуюся матрицей смысловую цепь ДНК называют кодирующей цепью, поскольку она имеет такую же последовательность, что и вновь созданный РНК-транскрипт (за исключением замены тимина на урацил). Она представляет собой цепь, которую используют по соглашению и в контексте настоящего изобретения при презентации ДНК-последовательности.
Молекула ДНК, предлагаемая в настоящем изобретении, может быть двухцепочечной или одноцепочечной или частично двухцепочечной и частично одноцепочечной.
Молекула ДНК, предлагаемая в настоящем изобретении, содержит два основных модуля (которые обозначают также как «элементы»), т.е. (а) кодирующую область, которая кодирует полипептид и которая включает стартовый кодон на ее 5'-конце, и (б) расположенную непосредственно в обратном направлении относительно указанной кодирующей области последовательность, указанную выше в подпунктах (б1) или (б2). Указанная молекула ДНК при транскрипции приводит к получению мРНК с чрезвычайно короткой UTR-последовательностью, которая обеспечивает описанные выше преимущества.
Кроме того, молекула ДНК, предлагаемая в настоящем изобретении, предпочтительно содержит последовательность, которая при транскрипции в мРНК, приводит к получению UTR, расположенной в прямом направлении относительно кодирующей области. Таким образом, молекула ДНК, предлагаемая в настоящем изобретении, предпочтительно несет кодирующую область, а также последовательности, которые после транскрипции приводят к получению (5' и 3') нетранслируемых областей (UTR) в созданной молекуле мРНК.
Понятие «кодирующая область, включающая стартовый кодон на ее 5'-конце» при применении в контексте настоящего изобретения относится к последовательности ДНК, которая состоит из кодонов, которые транскрибируются в молекулу мРНК с помощью ДНК-зависимой РНК-полимеразы, при этом соответствующая молекула мРНК может расшифровываться и транслироваться в белки с помощью рибосомы в соответствии с информацией, представленной в виде «генетического кода». Кодирующие области, как правило, начинаются стартовым кодоном на их 5'-конце и заканчиваются стоп-кодоном. Как правило, стартовый кодон представляет собой триплет ATG (соответствующий триплету AUG на уровне РНК), а стоп-кодон представляет собой ТАА, TAG или TGA (соответствующий UAA, UAG или UGA на уровне РНК). Помимо способности кодировать белок, части кодирующих областей могут служить в качестве регуляторных последовательностей в пре-мРНК, в качестве энхансеров сплайсинга экзонов или сайленсеров сплайсинга экзонов. Кодирующую область гена, которая кодирует полипептид или белок, применяемую согласно настоящему изобретению, называют также кодирующей последовательностью или CDS (сокращение от обозначения «кодирующая последовательность ДНК»), и она представляет собой часть генной ДНК или РНК, состоящую из экзонов, которая кодирует полипептид или белок. Кодирующая область в мРНК фланкирована 5'-нетранслируемой областью (5' UTR) и 3'-нетранслируемой областью (3' UTR), которые также являются частью экзонов. Кроме того, молекулы мРНК могут содержать также так называемый 5'-кэп и поли-А-хвост. 5'-кэп, 5' UTR, 3' UTR и поли-А-хвост представляют собой области молекулы мРНК, которые не транслируются в белок.
Понятие «нетранслируемая область» или «UTR» при применении в контексте настоящего изобретения относится к участкам мРНК, расположенным в обратном направлении относительно стартового кодона и в прямом направлении относительно стоп-кодона, которые не транслируются и, поэтому их обозначают как пять-штрих нетранслируемая область (5' UTR) и три-штрих нетранслируемая область (3' UTR) соответственно. Эти области транскрибируются с кодирующей области и, таким образом, являются экзонными, поскольку присутствуют в зрелой мРНК.
В контексте настоящего изобретения 3'-нетранслируемая область (3 'UTR) означает участок матричной РНК (мРНК), расположенный непосредственно за кодоном терминации трансляции. 3' UTR может содержать регуляторные области в 3'-нетранслируемой области, которые, как известно, влияют на полиаденилирование и стабильность мРНК. Многие 3' UTR содержат также богатые AU элементы (ARE). Кроме того, 3' UTR может предпочтительно содержать последовательность AAUAAA, которая контролирует добавление нескольких сотен остатков аденина, которые называют поли(А)-хвостом, на конец мРНК-транскрипта.
5'-нетранслируемая область (5' UTR) (которую обозначают также как лидерная последовательность или лидер РНК) представляет собой область мРНК, расположенную непосредственно в обратном направлении относительно стартового кодона. 5' UTR начинается с сайта инициации транскрипции и заканчивается за один нуклеотид (nt) до стартового кодона (как правило, AUG в мРНК) кодирующей области. У эукариот длина 5' UTR, как правило, составляет от 100 до нескольких тысяч нуклеотидов, однако иногда у эукариот встречаются более короткие UTR.
Согласно настоящему изобретению последовательность между промотором и кодирующей областью (которая указана выше в подпунктах (б1) или (б2)) является чрезвычайно короткой и приводит к образованию после транскрипции молекулы мРНК с очень короткой «минимальной» UTR-последовательностью.
Один модуль молекулы ДНК, т.е. «кодирующая область, включающая стартовый кодон на ее 5'-конце, которая кодирует полипептид» (модуль (а)), не ограничен конкретным модулем и может представлять собой любую требуемую кодирующую область, которая должна экспрессироваться в данной клетке. Так, указанный модуль может представлять собой кодирующую область, которая кодирует требуемый полипептид, т.е. требуемый конечный продукт. Настоящее изобретение не имеет ограничений касательно «кодирующей области, включающей стартовый кодон на ее 5'-конце, которая кодирует полипептид», поскольку природа кодирующей области зависит от требуемого продукта, который должен продуцироваться в клетке. Указанная кодирующая область может также представлять собой нуклеотидную последовательность, которая отличается от известной встречающейся в естественных условиях последовательности и содержит мутации (т.е. точечные мутации, инсерционную мутацию, делеции и их комбинации). Кроме того, указанная кодирующая область может частично или полностью представлять собой последовательность с оптимизированными кодонами, происходящую из встречающейся в естественных условиях последовательности, которую можно применять в качестве модуля (а). Оптимизация кодонов представляет собой методику, предназначенную для повышения до максимума экспрессии белка путем повышения эффективности трансляции мРНК, полученной из представляющего интерес гена. Известно, что во встречающихся в естественных условиях генах доступные кодоны используются не случайным образом, а отдается определенное предпочтение конкретным кодонам для одной и той же аминокислоты. Таким образом, вырожденность генетического кода - т.е. ситуация, когда одна аминокислота может кодироваться несколькими кодонами - приводит к трансформации нуклеотидной последовательности представляющего интерес гена в набор предпочтительных кодонов одного и того же или другого вида.
Как отмечалось выше, модуль (а) не ограничен конкретным модулем и может представлять собой любую требуемую кодирующую область, которая должна экспрессироваться в данной клетке. Таким образом, в контексте настоящего изобретения под «кодирующей областью» следует понимать любую молекулу полидезоксирибонуклеотида, которая при интродукции в клетку может транскрибироваться в молекулу мРНК, которая может транслироваться в полипептид/белок или его фрагмент. В контексте настоящего описания понятия «полипептид» и «белок» относятся к любому типу аминокислотной последовательности, т.е. цепи из двух или большего количества аминокислот, каждая из которых сцеплена через пептидные связи, понятия включают также пептиды и слитые белки.
В предпочтительном варианте осуществления изобретения «кодирующая область, включающая стартовый кодон на ее 5'-конце, которая кодирует полипептид» содержит дезоксирибонуклеотидную последовательность, которая кодирует полипептид/белок или его фрагмент, функция которого в клетке или в окружении клетки является необходимой или благоприятной, например, белок, отсутствие которого или дефектная форма которого «запускает» заболевание или расстройство, поставка которого может изменять или предупреждать заболевание или расстройство, или белок, который может усиливать процесс, благоприятный для организма, в клетке или ее окружении. Кодирующая область может содержать последовательность для полного белка или его функционального варианта. Кроме того, дезоксирибонуклеотидная последовательность кодирующей области может кодировать белок, действующий в качестве фактора, индуктора, регулятора, стимулятора или фермента, или его функциональный фрагмент, при этом указанный белок представляет собой белок, функция которого необходима для лечения нарушения, в частности, метаболического нарушения, или для инициации процессов in vivo, таких как формирование новых кровеносных сосудов, тканей и т.д. В контексте настоящего описания под функциональным вариантом подразумевается фрагмент, который в клетке может принимать на себя функцию белка, функция которого необходима для клетки или отсутствие или дефектная форма которого являются патогенными.
В предпочтительном варианте осуществления изобретения «кодирующая область, включающая стартовый кодон на ее 5'-конце, которая кодирует полипептид» кодирует терапевтически или фармацевтически активный полипептид или белок, обладающий терапевтическим или профилактическим действием. Саму молекулу ДНК, предлагаемую в настоящем изобретении, которая может транскрибироваться в мРНК, которая содержит указанную «кодирующую область, включающую стартовый кодон на ее 5'-конце, которая кодирует полипептид», можно использовать в терапии на основе нуклеиновых кислот в аналогичных вариантах применения. В этом контексте согласно изобретению транскрипция и трансляция молекулы ДНК, предлагаемой в настоящем изобретении, в мРНК и затем в полипептид или белок может требоваться для компенсации или дополнения экспрессии эндогенного гена, в частности, в случаях, когда эндогенный ген является дефектным или молчащим, что приводит к отсутствию продукта генной экспрессии, к недостаточному количеству или к дефектному или дисфунциональному продукту, как, например, в случае многих метаболических и наследственных заболеваний, таких как, среди прочего, муковисцидоз, гемофилия или мышечная дистрофия. Транскрипция и трансляция молекулы ДНК, предлагаемой в настоящем изобретении, в мРНК и затем в полипептид или белок может требоваться также для получения продукта экспрессии, который взаимодействует или создает помехи эндогенного клеточному процессу, такому как регуляция генной экспрессии, трансдукция сигналов и другие клеточные процессы. Транскрипция и трансляция молекулы ДНК, предлагаемой в настоящем изобретении, в мРНК и затем в полипептид или белок может требоваться также для вызывания иммунного ответа в контексте организма, в котором присутствуют или могут присутствовать трансфектированные или трансдуцированные клетки. Примерами являются генетическая модификация антигенпрезентирующих клеток, таких как дендритные клетки, для презентации ими антигена для целей вакцинации. Другим примером является транскрипция и трансляция молекулы ДНК, предлагаемой в настоящем изобретении, в мРНК и затем в полипептид или белок, в которой указанная кодирующая область кодирует цитокины. Это может, например, требоваться при опухолях для генерации опухольспецифического иммунного ответа. Кроме того, транскрипция и трансляция молекулы ДНК, предлагаемой в настоящем изобретении, в мРНК и затем в полипептид или белок, может требоваться для создания in vivo или ex vivo кратковременно генетически модифицированных клеток для клеточных терапий, таких как модифицированные Т-клетки или клетки-предшественники, или стволовые клетки, или другие клетки для регенеративной медицины.
В других предпочтительных вариантах осуществления изобретения «кодирующая область, включающая стартовый кодон на ее 5'-конце, которая кодирует полипептид» может кодироваться белок, принимающий участие в процессах роста и ангиогенеза, который необходим, например, для контролируемой регенерации и который может образовываться специфически в результате интродукции молекулы РНК, предлагаемой в изобретении. Это может, например, являться полезным в процессах роста или для лечения дефектов кости, дефектов ткани и в контексте имплантации и трансплантации.
Как уже упоминалось, молекулу ДНК и, в частности, соответствующим образом транскрибируемую молекулу РНК, предлагаемую в настоящем изобретении, которая содержит «кодирующую область, включающую стартовый кодон на ее 5'-конце, которая кодирует полипептид», можно применять соответственно в любом случае, когда полипептид или белок, который должен в естественных условиях присутствовать в организме, но он отсутствует или присутствует в дефектной форме или в слишком малом количестве из-за дефектов гена или заболеваний, должен пополняться в организме. Белки и кодирующие их гены, дефицит или дефект которых связан с заболеванием, известны. Согласно настоящему изобретению можно применять интактную версию кодирующей области, которая кодирует интактный полипептид или белок.
Известны многочисленные генетические нарушения, вызываемые мутацией одного гена, и они являются кандидатами для использования в отношении их терапевтических подходов на основе мРНК. Нарушения, вызываемые мутацией в одном гене, типа муковисцидоза, гемофилии и многих других, могут быть доминантными или рецессивными касательно вероятности того, что определенный признак может появляться у потомства. В то время как доминантный аллель определяет проявление фенотипа у индивидуумов, которые имеют только одну копию аллеля, в случае рецессивного аллеля индивидуум должен иметь две копии, по одной от каждого родителя, для проявления признака. В противоположность этому, полигенные нарушения вызываются двумя или большим количеством генов, и проявление соответствующего заболевания часто является плавным и связано с факторами окружающей среды. Примерами полигенных нарушений являются гипертензия, повышенный уровень холестерина, рак, нейродегенеративные нарушения, психические расстройства и др. В таких случаях терапевтическая мРНК, характерная для одного или нескольких указанных генов, также может оказывать благоприятное воздействие на таких пациентов. Кроме того, генетическое нарушение не обязательно должно определяться генами пациента, а может также вызываться новыми мутациями. И в этих случаях также терапевтическая мРНК, характерная для правильной генной последовательности, может оказывать благоприятное воздействие на пациентов.
В онлайн-каталоге представлено 22993 названия человеческих генов и генетических нарушений в сочетании с соответствующими генами и дано описание их фенотипов, доступных на веб-странице ONIM (Online Mendelian Inheritance in Man) (http://onim.org); последовательности каждого из них доступны из базы данных Uniprot database (http://www.uniprot.org). В качестве примеров (но не ограничиваясь только ими) ниже в таблице 1 представлен перечень некоторых врожденных заболеваний и соответствующего(их) гена(ов). Из-за высокой степени влияния на клеточные пути передачи сигналов мутации в некоторых генах вызывают несколько патогенных симптомов, из которых в таблице 1 указан только один характерный симптом.
В некоторых вариантах осуществления настоящего изобретения терапевтический белок выбирают из клеточных белков, перечисленных в таблице 1. Так, молекула ДНК, предлагаемая в изобретении, может кодировать терапевтический клеточный белок, где кодируемый терапевтический белок представляет собой один из перечисленных в таблице 1 или его гомолог.
В другом варианте осуществления настоящего изобретения терапевтический белок выбирают из секретируемых белков, перечисленных в таблице 1. Так, молекула ДНК, предлагаемая в изобретении, может кодировать терапевтический слитый белок, где кодируемый терапевтический белок или его гомолог представляет один из перечисленных в таблице 1, а второй белок представляет собой сигнальный пептид, который обеспечивает секрецию терапевтического белка. Сигнальный пептид представляет собой короткую, как правило, состоящую из 5-30 аминокислот аминокислотную последовательность, которая присутствует на N-конце указанного терапевтического белка и направляет слитый белок в секреторный путь клетки через определенные органеллы (т.е. эндоплазматический ретикулум, аппарат Гольджи или эндосомы). Таким образом, указанный слитый белок секретируется из клетки или из клеточной органеллы или встраивается в клеточную мембрану (например, многократно пронизывающие мембрану трансмембранные белки) клеточного компартмента или присутствует на клеточной поверхности.
Так, в предпочтительных вариантах осуществления настоящего изобретения «кодирующая область, включающая стартовый кодон на ее 5'-конце, которая кодирует полипептид» (модуль (а)), может кодировать следующие гены (но не ограничиваясь только ими) которые вызывают, предопределяют или защищают от заболеваний. Примеры указанных нарушений, которые можно лечить (или предупреждать), включают (но не ограничиваясь только ими) те, для которых указанный опосредующий их полипептид, белок или пептид выбран из группы соединений, состоящей из указанных ниже в таблице 1.
В некоторых вариантах осуществления изобретения «кодирующая область, включающая стартовый кодон на ее 5'-конце, которая кодирует полипептид» может транскрибироваться и транслироваться в частичный или полноразмерный белок, обладающий клеточной активностью на уровне, равном или превышающем уровень нативного белка. В некоторых вариантах осуществления изобретения «кодирующая область, включающая стартовый кодон на ее 5'-конце, которая кодирует полипептид» кодирует терапевтически или фармацевтически активный полипептид, белок или пептид, обладающий терапевтическим или профилактическим действием, где указанный полипептид, белок или пептид выбирают из группы, состоящей из указанных ниже в таблице 1. «Кодирующую область, включающую стартовый кодон на ее 5'-конце, которая кодирует полипептид» можно применять для экспрессии частичного или полноразмерного белка, обладающего клеточной активностью на уровне, равном или превышающем уровень нативного белка. Это может обеспечивать лечение заболеваний, при которых может быть показано введение молекулы РНК.


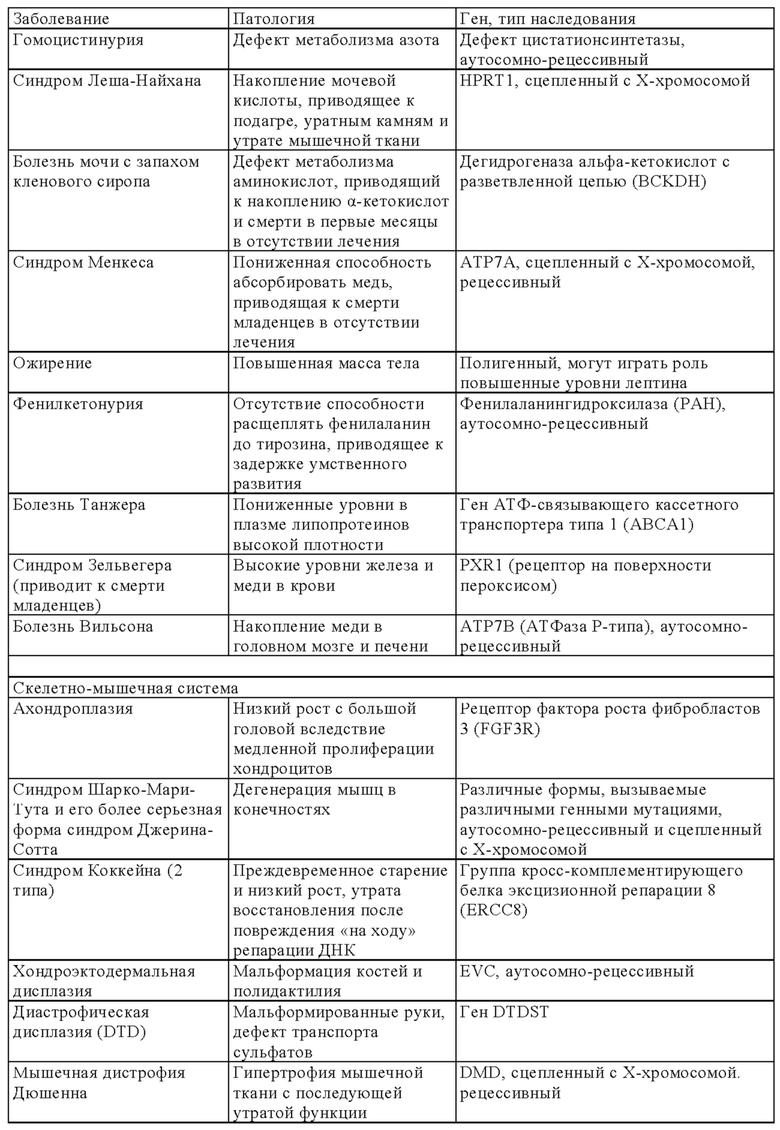

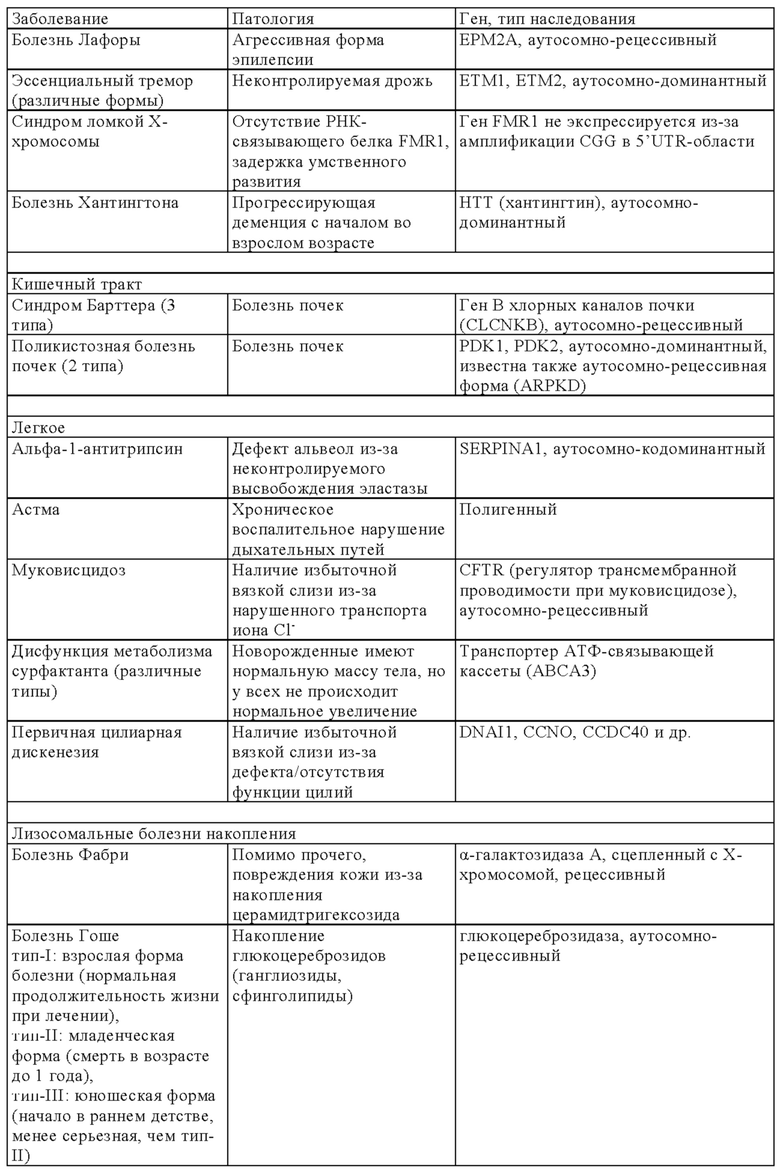
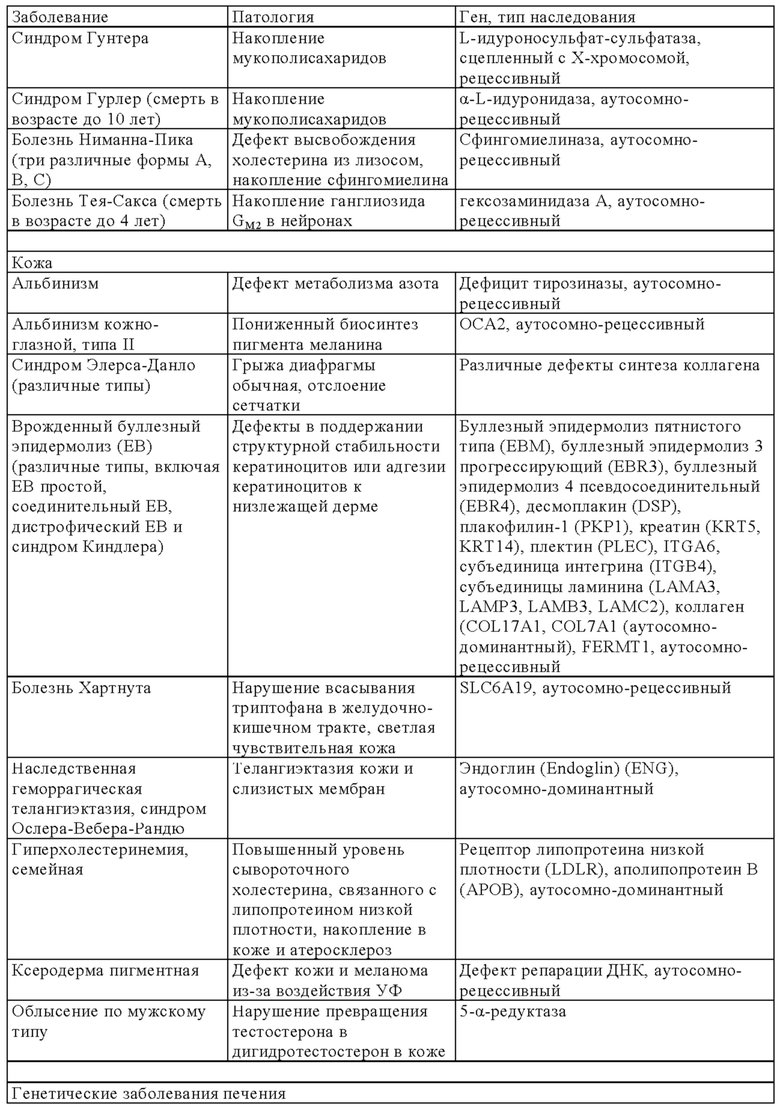
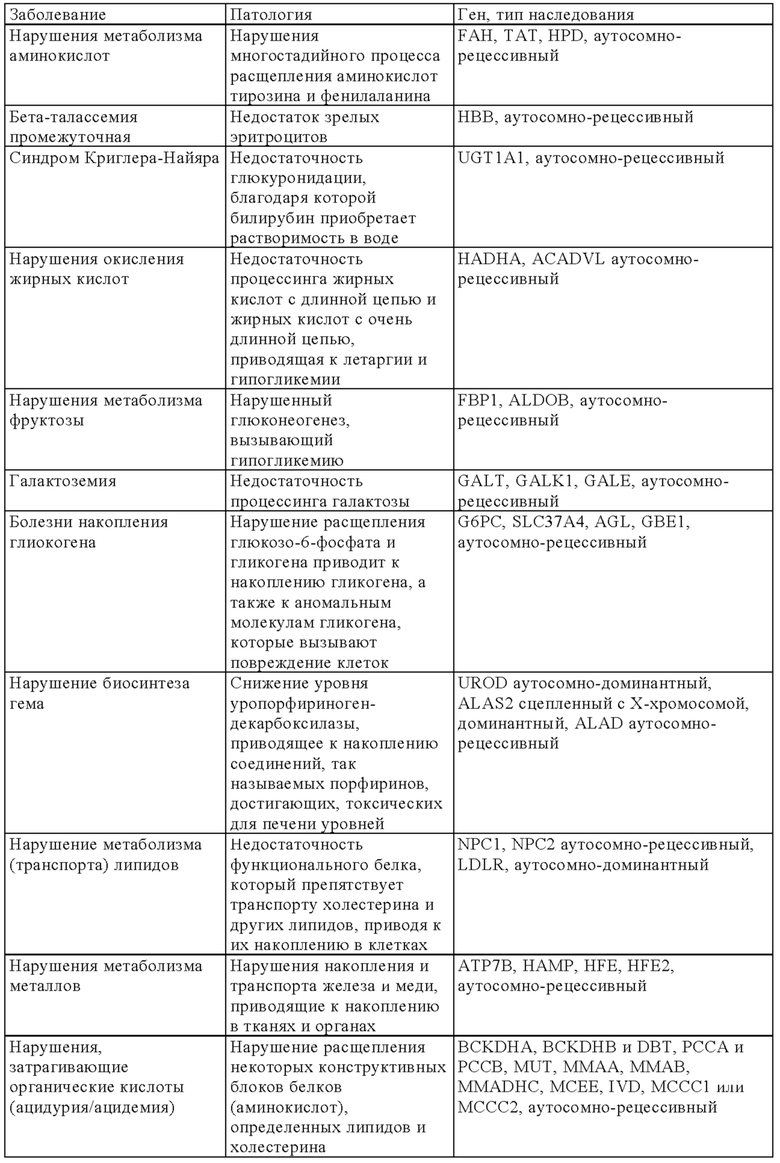
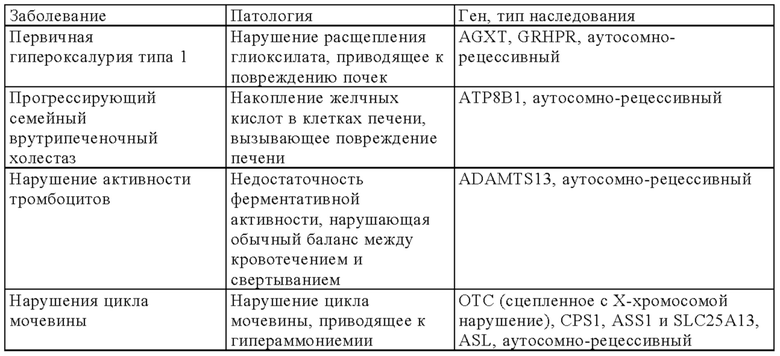
Выше в таблице 1 приведены примеры генов, дефект которых приводит к заболеванию, которое можно лечить с помощью молекулы РНК, транскрибируемой с молекулы ДНК, предлагаемой в настоящем изобретении, где молекула ДНК (и соответствующая транскрибируемая молекула РНК) содержит «кодирующую область, включающую стартовый кодон на ее 5'-конце, которая кодирует полипептид», которая кодирует интактную версию белка или его функциональный фрагмент описанного выше дефектного гена. В наиболее предпочтительных вариантах осуществления изобретения наследственные заболевания, которые можно отметить, включают, например, те, которые поражают легкие, такие как дефицит SPB (белок сурфактанта В), дефицит АВСА3, муковисцидоз и дефицит α1-антитрипсина, или те, которые влияют на белки плазмы (например, врожденный гемохроматоз (дефицит гепсицида), тромботическая тромбоцитопеническая пурпура (ТРР, дефицит ADAMTS 13) и вызывают дефекты свертывания (например, гемофилия а и b) и дефекты системы комплемента (например, дефицит белка С), иммунные дефекты, такие, например, как SCID (вызываемый мутациями в различных генах, таких как: RAG1, RAG2, JAK3, IL7R, CD45, CD3δ, CD3ε) или дефицитом или отсутствием аденозиндезаминазы, например (ADA-SCID), септический грануломатоз (например, вызываемый мутациями гена gp-91-phox, гена p47-phox, гена р67-phox или гена р33-phox) и болезни накопления типа болезни Гоше, болезни Фабри, болезни Краббе, MPS I, MPS II (синдром Гунтера), MPS VI, болезнь накопления гликогена типа II или мукополисахаридоз.
Другие нарушения, для которых можно применять предлагаемую в настоящем изобретении «кодирующую область, включающую стартовый кодон на ее 5'-конце, которая кодирует полипептид», включают такие нарушения, как SMN1-связанная спинальная мышечная атрофия (SMA); амиотрофический боковой склероз (ALS); GALT-связанная галактоземия; муковисцидов (CF); SLC3A1-связанные нарушения, включая цистинурию; COL4A5-связанные нарушения, включая синдром Альпорта; дефицит галактоцереброзидазы; сцепленные с Х-хромосомой адренолейкодистрофия и адреномиелоневропатия; атаксия Фридрейха; болезнь Пелицеуса-Мерцбахера; связанный с TSC1 и TSC2 туберозный склероз; синдром Санфилиппо В (MPS IIIB); CTNS-связанный цистиноз; FMR1-связанные нарушения, которые включают синдром ломкой X-хромосомы, ассоциированный с ломкой Х-хромосомой синдром тремора/атаксии и связанный с ломкой Х-хромосомой синдром преждевременного истощения яичников; синдром Прадера-Вилли; наследственная геморрагическая телангиэктазия (AT); болезнь Ниманна-Пика типа С1; связанные с нейрональным цероидным липофусцинозом заболевания, включая ювенильный нейрональный цероидный липофусциноз (JNCL), ювенильная форма болезни Баттена, болезнь Сантавуори-Халтиа, болезнь Янского-Билыповского и дефициты РТТ-1 и ТРР1; связанная с EIF2B1, EIF2B2, EIF2B3, EIF2B4 и EIF2B5 атаксия у детей с гипомиелинизацией центральной нервной системы/разрушением белого вещества; связанная с CACNA1A и CACNB4 эпизодическая атаксия типа 2; связанные с МЕСР2 нарушения, включающие классический синдром Ретта, МЕСР2-связанная серьезная неонатальная энцефалопатия и синдром РРМ-Х; CDKL5-связанный атипичный синдром Ретта; болезнь Кеннеди (SBMA); связанная с Notch-3 церебральная аутосомно-доминантная артериопатия с субкортикальным инфарктом и лейкоэнцефалопатией (CADASIL); связанные с захватом SCN1A и SCN1B нарушения; связанные с полимеразой G нарушения, которые включают синдром Альперса-Хуттенлохера, связанная POLG сенсорная атаксическая нейропатия, дизартрия и офтальмопарез, и аутосомно-доминантные и рецессивные прогрессирующие экстернальные офтальмоплегии с делециями митохондриальной ДНК; Х-сцепленная гиперплазия надпочечников; X-сцепленная агаммаглобулинемия; болезнь Фабри; и болезнь Вильсона.
При всех указанных заболеваниях белок, например, фермент, является дефектным, и его можно улучшать путем обработки РНК, транскрибируемой с молекулы ДНК, предлагаемой в настоящем изобретении, что делает доступным белок, кодируемый дефектным геном. Транскрипт-замещающие терапии/фермент-замещающие терапии не влияют на лежащий в основе генетический дефект, а повышают концентрацию фермента, дефицит которого имеется в организме пациента. Например, при болезни Помпе транскрипт-замещающая терапия/фермент-замещающая терапия обеспечивает пополнение дефицита лизосомного фермента кислой альфа-глюкозидазы (GAA).
Таким образом, примерами белков, которые могут кодироваться «кодирующей областью, включающей стартовый кодон на ее 5'-конце, которая кодирует полипептид» модуля (а), согласно изобретению являются (но не ограничиваясь только ими) эритропоэтин (ЕРО), гормон роста (соматотропин, hGH), регулятор трансмембранной проводимости при муковисцидозе (CFTR), факторы роста, такие как GM-SCF, G-CSF, MPS, белок С, гепцидин, АВСА3 и белок сурфактанта В. Другими примерами заболеваний, которые можно лечить с помощью РНК, предлагаемой в изобретении, являются гемофилия А/В, болезнь Фабри, CGD, ADAMTS13, болезнь Гурлер, опосредуемая Х-хромосомой А-γ-глобулинемия, связанный с аденозиндеаминазой иммунодефицит и респираторный дистресс-синдром у новорожденных, который связан с SP-B. Наиболее предпочтительно «кодирующая область, включающая стартовый кодон на ее 5'-конце, которая кодирует полипептид», молекулы ДНК, предлагаемой в изобретении, содержит последовательность белка сурфактанта В (SP-B) или эритропоэтина. Другие примеры белков, которые могут кодироваться «кодирующей областью, включающей стартовый кодон на ее 5'-конце, которая кодирует полипептид», молекулы ДНК, предлагаемой в изобретении, являются факторы роста, такие как человеческий гормон роста hGH, ВМР-2 или факторы ангиогенеза.
Альтернативно этому, нуклеиновые кислоты могут кодировать полноразмерные антитела или имеющие меньший размер антитела (например, и тяжелую, и легкую цепи) для придания иммунитета индивидууму. В другом варианте осуществления изобретения «кодирующая область, включающая стартовый кодон на ее 5'-конце, которая кодирует полипептид», может кодировать функциональное моноклональное или поликлональное антитело, которое можно применять для таргетинга и/или инактивации биологической мишени (например, стимуляторного цитокина, такого как фактор некроза опухоли). Аналогично этому, «кодирующая область, включающая стартовый кодон на ее 5'-конце, которая кодирует полипептид», может кодировать, например, функциональные антитела против нефротического фактора, которые можно применять для лечения мембранопролиферативного гломерулонефрита типа II или острого гемолитического уремического синдрома, или альтернативно этому, может кодировать антитела к сосудистому эндотелиальному фактору роста (VEGF), которые можно применять для лечения опосредуемых VEGF заболеваний, таких как рак.
Модуль (а), т.е. «кодирующая область, включающая стартовый кодон на ее 5'-конце, которая кодирует полипептид», может представлять собой кодирующую область, которая кодирует полипептид или белок, который можно применять в технологиях редактирования генома. Редактирование генома представляет собой тип генетической инженерии, при котором ДНК встраивают, изымают путем делеции или заменяют в геноме организма с использованием нуклеаз. Эти нуклеазы создают сайтспецифические двухцепочечные разрывы (DSB) в требуемых локусах в геноме. Индуцированный двухцепочечный разрыв репарируется с помощью негомологичного соединения концов или гомологичной рекомбинации, что приводит к целевым мутациям в геноме, тем самым «редактируя» геном. В данной области известны многочисленные системы редактирования, в которых используются различные полипептиды или белки, например, система CRISPR-Cas, мегануклеазы, нуклеазы с цинковыми пальцами (цинковопальцевые нуклеазы) (ZFN) и нуклеазы на основе эффекторов, напоминающих факторы транскрипции (TALEN). Методы генетической инженерии обобщены в Trends in Biotechnology, 31 (7), 2013, cc. 397-405.
Таким образом, в предпочтительном варианте осуществления изобретения «кодирующая область, включающая стартовый кодон на ее 5'-конце, которая кодирует полипептид» содержит дезоксирибонуклеотидную последовательность, которая кодирует полипептид или белок из семейства белков Cas (CRISPR-ассоциированный белок), предпочтительно Cas9 (CRISPR-ассоциированный белок 9). Белки из семейства Cas, предпочтительно Cas9, можно применять в основанных на CRISPR/Cas9 методах и/или в технологиях редактирования генома на основе CRISPR/Cas9. Основанные на CRISPR-Cas системы для редактирования, регуляции и таргетинга генома обобщены в Nat. Biotechnol., 32(4), 2014, сс. 347-355.
В другом предпочтительном варианте осуществления изобретения «кодирующая область, включающая стартовый кодон на ее 5'-конце, которая кодирует полипептид», содержит дезоксирибонуклеотидную последовательность, которая кодирует мегануклеазу. Мегануклеазы представляют собой эндодезоксирибонуклеазы, которые в отличие от «канонических» эндодезоксирибонуклеаз распознают крупный сайт узнавания (например, двухцепочечную последовательность ДНК, состоящую из 12-40 пар оснований). При этом соответствующие сайты встречаются в любом конкретном геноме лишь несколько раз, предпочтительно только один раз. Таким образом, мегануклеазы рассматриваются в качестве наиболее специфических встречающихся в естественных условиях рестриктаз, и поэтому они являются наиболее приемлемыми инструментами для технологий редактирования генома.
В другом предпочтительном варианте осуществления изобретения «кодирующая область, включающая стартовый кодон на ее 5'-конце, которая кодирует полипептид», содержит дезоксирибонуклеотидную последовательность, которая кодирует цинковопальцевую нуклеазу (ZFN). ZFN представляют собой искусственные рестриктазы, созданные путем слияния ДНК-связывающего домена цинковых пальцев с ДНК-расщепляющим доменом. Домены цинковых пальцев можно создавать для таргетинга специфических требуемых последовательностей ДНК и поэтому указанные цинковопальцевые нуклеазы могут таргетировать уникальные последовательности в сложных геномах. С учетом преимущества механизма репарации эндогенной ДНК ZFN можно применять для точного изменения генома высших организмов, и поэтому они являются приемлемыми инструментами для технологий редактирования генома.
В другом предпочтительном варианте осуществления изобретения «кодирующая область, включающая стартовый кодон на ее 5'-конце, которая кодирует полипептид», содержит дезоксирибонуклеотидную последовательность, которая кодирует нуклеазу на основе эффекторов, напоминающих факторы транскрипции (TALEN). TALEN представляют собой рестриктазы, которые создают для разрезания специфических последовательностей ДНК. TALEN представляют собой слитые белки, в который ДНК-связывающий домен напоминающего фактор транскрипции (TAL)-эффектора слит с ДНК-расщепляющим доменом нуклеазы. Эффекторы, напоминающие факторы транскрипции (TALE), можно создавать для связывания практически с любой требуемой последовательностью ДНК. Таким образом, путем объединения с нуклеазой можно разрезать ДНК в специфических требуемых локусах.
Молекула ДНК, предлагаемая в настоящем изобретении, содержит в качестве второго модуля (б) последовательность, локализованную непосредственно в обратном направлении относительно кодирующей последовательности.
Более конкретно, молекула ДНК, предлагаемая в настоящем изобретении, содержит модуль (б) непосредственно в обратном направлении относительно указанной кодирующей последовательности, где указанный модуль (б) представляет собой последовательность, выбранную из группы, которая состоит из:
(б1) R1-CGCCACC (SEQ ID NO: 1);
или последовательности, где в указанной последовательности С в положении 6 SEQ ID NO: 1 заменен на А и С в положении 7 SEQ ID NO: 1 заменен на G; и/или А в положении 5 SEQ ID NO: 1 заменен на G; и
(б2) R1-CNGCCACC (SEQ ID NO: 2), в которой нуклеотид N в положении 2 SEQ ID NO: 2 представляет собой нуклеотид, выбранный из группы, которая состоит из Т, G, С или А;
или последовательности, где в указанной последовательности С в положении 7 SEQ ID NO: 2 заменен на А и С в положении 8 SEQ ID NO:2 заменен на G; и/или А в положении 6 SEQ ID NO: 2 заменен на G,
где R1 обозначает промотор, который распознается ДНК-зависимой РНК-полимеразой.
Последовательности, указанные выше в пункте (б) в настоящем описании, не ограничены конкретной(ыми) указанной(ыми) выше последовательностью(ями), но могут также относиться к последовательности(ям), которая(ые) содержит(ат) дополнительный(ые) нуклеотид(ы), где дополнительный(ые) нуклеотид(ы) можно добавлять на 5'-конец R1 в описанной(ых) выше последовательности(ях). Дополнительный(ые) нуклеотид(ы) содержит полинуклеотидные цепи, содержащие вплоть до 0 (отсутствие изменений), 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 нуклеотидов вплоть до 20 нуклеотидов. Более предпочтительно 11, 12, 13, 14, 15, 16, 18 или 19 нуклеотидов добавляют на 5'-конец. Еще более предпочтительно добавляют на 5'-конец вплоть до 30 нуклеотидов.
Поскольку добавление нуклеотидов в обратном направлении относительно промотора R1 не должно изменять указанные выше функциональные свойства UTR, предлагаемой(ых) в изобретении, то дополнительные нуклеотиды могут также иметь длину вплоть до 40, 50, 60, 70, 80, 90 или даже 100 нуклеотидов или даже еще больше, вплоть до 200, 300, 400 или 500 нуклеотидов.
Как отмечалось выше, двухцепочечная молекула ДНК содержит две антипараллельные цепи, из которых одну цепь называют «смысловой» цепью, если она имеет последовательность, такую же, что и копия матричной РНК, которая транслируется в белок. Последовательность противоположной комплементарной цепи называют «антисмысловой» последовательностью. Таким образом, понятие «молекула ДНК, предлагаемая в настоящем изобретении» относится не только к вышеописанной молекуле ДНК, которая соответствует мРНК, содержащей одну цепь с описанными выше элементами (а) и (б), но также и к молекуле ДНК, содержащей комплементарную цепь, т.е. антисмысловую цепь, которая может транскрибироваться в мРНК. Указанную комплементарную цепь молекулы ДНК, предлагаемой в настоящем изобретении, определяют со ссылкой на антисмысловую цепь, которую легко можно определять на основе правил спаривания оснований.
Молекула ДНК, предлагаемая в настоящем изобретении, содержит также в модуле (б) промотор R1, который распознается ДНК-зависимой РНК-полимеразой. Предпочтительно указанный промотор R1 непосредственно связан с остальной последовательностью, указанной в подпункте (б1) или (б2), т.е. без присутствии каких-либо промежуточных нуклеотидов.
На природу промотора R1, который распознается ДНК-зависимой РНК-полимеразой, не накладываются конкретные ограничения. Любой промотор (и его варианты) можно применять, если соответствующая ДНК-зависимая РНК-полимераза может распознавать соответствующую последовательность. В данной области известны многочисленные РНК-полимеразы (которые называют также ДНК-зависимыми РНК-полимеразами и часто сокращенно обозначают как RNAP или RNApol). Эти ферменты обладают способностью продуцировать первичный РНК-транскрипт. Как описано выше, ДНК-зависимые РНК-полимеразы обладают способностью синтезировать цепи РНК с использованием ДНК в качестве матриц в процессе, называемом транскрипцией. ДНК-зависимая РНК-полимераза инициирует транскрипцию на специфических последовательностях ДНК, известных как промоторы. Затем она продуцирует цепь РНК, комплементарную цепи матричной ДНК. Процесс добавления нуклеотидов к цепи РНК известен как элонгация. Таким образом, в контексте настоящего изобретения понятие «распознавание (узнавание)» предпочтительно означает (но не ограничиваясь указанным), что ДНК-зависимая РНК-полимераза обладает способностью специфически обнаруживать/связывать соответствующую ей промоторную последовательность R1. Указанное понятие относится также к способности ДНК-зависимой РНК-полимеразы инициировать транскрипцию и затем продуцировать молекулу РНК в процессе элонгации.
Специалист в данной области с помощью известных методов может определять, обладает ли ДНК-зависимая РНК-полимераза способностью распознавать соответствующий промотор. Кроме того, с помощью методов, хорошо известных в данной области, предназначенных для оценки взаимодействий белка/ДНК, можно идентифицировать соответствующую (неизвестную) промоторную последовательность R1 для данной ДНК-зависимой РНК-полимеразы и наоборот.
Так, способность ДНК-зависимой РНК-полимеразы распознавать/связывать ее промотор R1 и предпочтительно способность инициировать транскрипцию можно определять методами, известными в данной области, например, описанными в Journal of Biological Chemistry, 268(26), 1993, cc. 19299-19304, a данные об открытых многочисленных ДНК-зависимых РНК-полимеразах обобщены в Journal of Biological Chemistry, 280(52), 2005, cc. 42477-42485).
В предпочтительном варианте осуществления изобретения промотор R1, который распознается ДНК-зависимой РНК-полимеразой, представляет собой промотор бактериофага.
Только в качестве примера, следует упомянуть, что в данной области известно, что ДНК-зависимая РНК-полимераза Т7 распознает последовательность TAATACGACTCACTATAGGGAGA (SEQ ID NO: 3), ДНК-зависимая РНК-полимераза Т3 распознает последовательность AATTAACCCTCACTAAAGGGAGA (SEQ ID NO: 4), ДНК-зависимая РНК-полимераза SP6 распознает последовательность ATTTAGGTGACACTATAGAAG (SEQ ID NO: 5) и ДНК-зависимая РНК-полимераза K11 распознает последовательность AATTAGGGCACACTATAGGGA (SEQ ID NO: 6). Однако эти примеры даны только с целью иллюстрации, поскольку настоящее изобретение не ограничено указанными промоторами и соответствующими ДНК-зависимыми РНК-полимеразами. Фактически можно применять любой промотор (и его варианты), если ДНК-зависимая РНК-полимераза, предпочтительно ДНК-зависимая РНК-полимераза бактериофага, может распознавать соответствующую последовательность.
В предпочтительном варианте осуществления изобретения R1 выбирают из группы, которая состоит из:
(I) TAATACGACTCACTATAGGGAGA (SEQ ID NO: 3) или последовательности, имеющей 1-6 замен по сравнению с SEQ ID NO: 3, и которая распознается ДНК-зависимой РНК-полимеразой Т7;
(II) AATTAACCCTCACTAAAGGGAGA (SEQ ID NO: 4) или последовательности, имеющей 1-6 замен по сравнению с SEQ ID NO: 4, и которая распознается ДНК-зависимой РНК-полимеразой Т3;
(III) ATTTAGGTGACACTATAGAAG (SEQ ID NO: 5) или последовательности, имеющей 1-6 замен по сравнению с SEQ ID NO: 5, и которая распознается ДНК-зависимой РНК-полимеразой SP6; и
(IV) AATTAGGGCACACTATAGGGA (SEQ ID NO: 6) или последовательности, имеющей 1-6 замен по сравнению с SEQ ID NO: 6, и которая распознается ДНК-зависимой РНК-полимеразой K11.
В другом предпочтительном варианте осуществления изобретения последовательность может представлять собой последовательность, имеющую 1-3, 4 или 5 замен, если соответствующая последовательность все еще может распознаваться ДНК-зависимой РНК-полимеразой Т7, Т3, SP6 и K11 соответственно. В более предпочтительном варианте осуществления изобретения последовательность может представлять собой последовательность, имеющую 1-2 замены, если соответствующая последовательность все еще может распознаваться ДНК-зависимой РНК-полимеразой Т7, Т3, SP6 и K11 соответственно. Более предпочтительно последовательность может представлять собой последовательность, имеющую 1 замену, если соответствующая последовательность все еще может распознаваться ДНК-зависимой РНК-полимеразой Т7, Т3, SP6 и K11 соответственно.
В других вариантах осуществления изобретения промоторные последовательности R1, которые распознаются ДНК-зависимой РНК-полимеразой, не ограничены конкретно какой-либо из последовательностей SEQ ID NO: 3, SEQ ID NO: 4, SEQ ID NO: 5 или SEQ ID NO: 6 или последовательностями, имеющими 1-6 замен по сравнению с ними, а также могут представлять собой последовательности, имеющие 1-7, 8, 9, 10, 11 или 12 замен, если соответствующая последовательность все еще может распознаваться ДНК-зависимой РНК-полимеразой Т7, Т3, SP6 и K11 соответственно.
В предпочтительном варианте осуществления изобретения из вышеуказанной(ых) замены(н) в последовательностях
TAATACGACTCACTATAGGGAGA (SEQ ID NO: 3),
AATTAACCCTCACTAAAGGGAGA (SEQ ID NO: 4),
ATTTAGGTGACACTATAGAAG (SEQ ID NO: 5) или
AATTAGGGCACACTATAGGGA (SEQ ID NO: 6) замены в 5 нуклеотидах «САСТА» в положениях 11-12 в вышеуказанных последовательностях SEQ ID NO: 3-6 исключены, поскольку указанные 5 нуклеотидов являются консервативными для четырех последовательностей.
В другом предпочтительном варианте осуществления изобретения из вышеуказанной(ых) замены(н) в последовательностях SEQ ID NO: 3-6 замена нуклеотида «Т» в положении 4 в вышеуказанных последовательностях SEQ ID NO: 3-6 исключена, поскольку указанный нуклеотид является консервативным для четырех последовательностей.
В другом предпочтительном варианте осуществления изобретения из вышеуказанной(ых) замены(н) в последовательностях SEQ ID NO: 3-6 замена нуклеотида «А» в положении 5 в вышеуказанных последовательностях SEQ ID NO: 3-6 исключена, поскольку указанный нуклеотид является консервативным для четырех последовательностей.
В другом предпочтительном варианте осуществления изобретения из вышеуказанной(ых) замены(н) в последовательностях SEQ ID NO: 3-6 замена нуклеотида «G» в положении 18 в вышеуказанных последовательностях SEQ ID NO: 3-6 исключена, поскольку указанный нуклеотид является консервативным для четырех последовательностей.
Способность ДНК-зависимой РНК-полимеразы Т7, Т3, SP6 и K11 распознавать/связывать ее промотор R1 можно определять методами, известными в данной области, которые описаны выше.
В более предпочтительном варианте осуществления изобретения молекула ДНК, предлагаемая в настоящем изобретении, представляет собой молекулу ДНК, которая содержит модуль (б1) непосредственно в обратном направлении относительно указанной кодирующей последовательности, где в указанном модуле (б1) нуклеотид N в положении 2 SEQ ID NO: 2 представляет собой нуклеотид, выбранный из группы, которая состоит из Т, G или С, и нуклеотид N не представляет собой А.
В еще более предпочтительном варианте осуществления изобретения указанный нуклеотид N в положении 2 SEQ ID NO: 2 представляет собой Т.
В предпочтительном варианте осуществления изобретения молекула ДНК, предлагаемая в настоящем изобретении, представляет собой молекулу ДНК, в которой нуклеотид, расположенный непосредственно в прямом направлении относительно стартового кодона, представляет собой нуклеотид G. В другом предпочтительном варианте осуществления изобретения молекула ДНК, предлагаемая в настоящем изобретении, представляет собой молекулу ДНК, в которой нуклеотид, расположенный непосредственно в прямом направлении относительно стартового кодона, представляет собой нуклеотид, выбранный из группы, которая состоит из А, Т и С.
В еще более предпочтительном варианте осуществления изобретения молекула ДНК, предлагаемая в настоящем изобретении, представляет собой молекулу ДНК, которая содержит модуль (б1), указанный выше, где указанный модуль (б1) представляет собой последовательность, в которой С в положении 6 SEQ ID NO: 1 заменен на А и С в положении 7 SEQ ID NO: 1 заменен на G; и/или А в положении 5 SEQ ID NO: 1 заменен на G и в которой нуклеотид, расположенный непосредственно в прямом направлении относительно стартового кодона, представляет собой нуклеотид, выбранный из группы, которая состоит из А, Т и С.
В другом еще более предпочтительном варианте осуществления изобретения молекула ДНК, предлагаемая в настоящем изобретении, представляет собой молекулу ДНК, которая содержит модуль (б2), указанный выше, где указанный модуль (б2) представляет собой последовательность, в которой С в положении 7 SEQ ID NO: 2 заменен на А и С в положении 8 SEQ ID NO: 2 заменен на G; и/или А в положении 6 SEQ ID NO: 2 заменен на G и в которой нуклеотид, расположенный непосредственно в прямом направлении относительно стартового кодона, представляет собой нуклеотид, выбранный из группы, которая состоит из А, Т и С.
В молекулярной биологии и генетике понятия «в обратном направлении» и «прямом направлении» оба характеризуют относительное положение в молекуле ДНК. В контексте настоящего изобретения в обратном направлении означает по направлению к 5'-концу смысловой цепи молекулы ДНК, а в прямом направлении означает по направлению к 3'-концу молекулы.
Таким образом, в настоящем изобретении последовательность, указанная выше в пункте (б), локализована непосредственно в обратном направлении относительно кодирующей области, указанной в пункте (а), более конкретно, непосредственно в обратном направлении относительно стартового кодона кодирующей области. Так, «непосредственно в обратном направлении» в данном контексте означает, что нет никакого(их) другого(их) нуклеотида(ов) между последовательностью, указанной в пункте (б), и кодирующей последовательностью, которая начинается со стартового кодона. Так, кодирующая область, которая начинается со стартового кодона, непосредственно примыкает к последовательности, указанной выше в пункте (б).
Молекулы ДНК, предлагаемые в настоящем изобретении, можно создавать/синтезировать рекомбинантно (например, с помощью системы in vivo или in vitro) или с помощью синтеза (например, с помощью ПЦР-реакции или химической реакции) с использованием методов, хорошо известных специалисту в данной области.
Молекула ДНК, предлагаемая в настоящем изобретении, предпочтительно представляет собой рекомбинантную молекулу нуклеиновой кислоты, т.е. она состоит из элементов, которые не встречаются в естественных условиях в такой комбинации. Молекула нуклеиновой кислоты, предлагаемая в изобретении, может быть синтетической или полусинтетической.
Молекула ДНК может присутствовать в форме слитых ДНК-последовательностей модулей (а) и (б) (указанных выше с пунктах (а) и (б) соответственно) т.е. (слитой) молекулы ДНК, образованной путем объединения по меньшей мере двух нуклеотидных последовательностей, содержащих указанные модули. Как правило, что будет более подробно описано ниже, это можно осуществлять путем клонирования кДНК в экспрессионном векторе, в котором обеспечивается транскрипция в молекулу РНК. Таким образом, молекула ДНК, предлагаемая в настоящем изобретении, может представлять собой слитую последовательность ДНК, т.е. химерную молекулу, образованную путем соединения двух или большего количества полинуклеотидов через фосфатную группу из одного нуклеотида, связанную с 3'-углеродом другого нуклеотида, с формированием фосфодиэфирной связи между соответствующими концами одного модуля и концом другой молекулы. Таким путем молекулы ДНК, содержащие указанные по меньшей мере два модуля, соединяют вместе в форме молекулы ДНК. После клонирования в рамке считывания указанная рекомбинантная молекула ДНК может далее транскрибироваться в соответствующую нуклеотидную последовательность РНК, кодирующую указанную молекулу белка, полипептида или фермента.
Молекулу ДНК, предлагаемую в настоящем изобретении, можно интродуцировать в вектор, предпочтительно экспрессионный вектор, стандартными методами молекулярной биологии (см., например, Sambrook и др., Molecular Cloning, A laboratory manual, 2-ое изд., 1989). Под понятием «вектор», такой как «экспрессионный вектор» или «клонирующий вектор», в контексте настоящего изобретения подразумевается кольцевая двухцепочечная единица ДНК, которая реплицируется внутри клетки вне зависимости от хромосомной ДНК и которую применяют в качестве носителя для переноса генетического материала в клетку, где она может реплицироваться и/или экспрессироваться (т.е. транскрибироваться в РНК и транслироваться в аминокислотную последовательность). Вектор, содержащий чужеродную ДНК, обозначают как рекомбинантная ДНК. Обычно сам вектор представляет собой последовательность ДНК, которая, как правило, состоит из вставки (например, молекулы нуклеиновой кислоты/молекулы ДНК, предлагаемой в настоящем изобретении) и более крупной последовательности, которая служит в качестве «каркаса» вектора. Плазмиды в контексте настоящего изобретения наиболее часто применяют в бактериях и их используют в исследованиях рекомбинантной ДНК для переноса генов между клетками, они представляют собой субпопуляцию «векторов», когда это понятие употребляется в контексте настоящего изобретения.
Специалисту в данной области должно быть очевидно, что дополнительные регуляторные последовательности можно добавлять в молекулу ДНК, предлагаемую в изобретении. Например, можно применять энхансеры транскрипции и/или последовательности, обеспечивающие индукцию экспрессии. Приемлемая индуцибельная система представляет собой, например, регулируемую тетрациклином генную экспрессию, например, описанную у Gossen и Bujard, Proc. Natl. Acad. Sci. USA 89, 1992, cc. 5547-5551) и Gossen, Trends Biotech. 12, 1994, cc. 58-62, или индуцируемую дексаметазоном систему генной экспрессии, описанную, например, у Crook, EMBO J. 8, 1989, сс. 513-519.
Настоящее изобретение относится также к вектору, предпочтительно экспрессионному вектору, который содержит молекулу ДНК, предлагаемую в настоящем изобретении.
Вектор, предлагаемый в настоящем изобретении, может представлять собой, например, плазмиду, космиду, вирус, бактериофаг или другой вектор, например, общепринятый для применения в генной инженерии, и может содержать дополнительные гены, такие как маркерные гены, обеспечивающие отбор указанного вектора в приемлемой клетке-хозяине и в приемлемых условиях.
Молекула ДНК, предлагаемая в настоящем изобретении, предпочтительно содержит также поли-А-сигнал, обеспечивающий терминацию транскрипции и стабилизацию транскрипта путем добавления поли-А-хвоста.
Молекулы ДНК и векторы, предлагаемые в изобретении, можно создавать для непосредственной интродукции или интродукции посредством липосом, вирусных векторов (например, аденовирусных, ретровирусных), электропорации, баллистических (например, с помощью генной пушки) или других систем доставки в клетку. Кроме того, можно применять бакуловирусную систему в качестве эукариотической экспрессионной системы для молекул нуклеиновых кислот, предлагаемых в изобретении.
Настоящее изобретение относится также к клетке-хозяину, которая содержит вектор, предлагаемый в настоящем изобретении. Таким образом, настоящее изобретение относится к хозяину, трансфектированному или трансформированному вектором, предлагаемым в изобретении, или хозяину, не представляющему собой человека, который несет вектор, предлагаемый в настоящем изобретении, т.е. к клетке-хозяину или хозяину, генетически модифицированной/модифицированному молекулой ДНК, предлагаемой в изобретении, или вектором, который содержит указанную молекулу ДНК. Понятие «генетически модифицирована/модифицирован» означает, что клетка-хозяин или хозяин содержит в дополнение к ее встречающемуся в естественных условиях геному молекулу ДНК или вектор, предлагаемую/предлагаемый в изобретении, интродуцированную/интродуцированный в клетку или хозяина или в один из ее/его предшественников/родителей. Молекула ДНК или вектор может присутствовать в генетически модифицированной клетке-хозяине или генетически модифицированном хозяине либо в виде независимой молекулы вне генома, предпочтительно в виде молекулы, обладающей способностью к репликации, либо может быть стабильно интегрирована/интегрирован в геном клетки-хозяина или хозяина. Трансформацию клетку-хозяина вектором, предлагаемым в изобретении, можно осуществлять стандартными методами, например, описанными у Sambrook и Russell, Molecular Cloning: A Laboratory Manual, изд-во CSH Press, Cold Spring Harbor, NY, USA, 2001; Methods in Yeast Genetics, A Laboratory Course Manual, изд-во Cold Spring Harbor Laboratory Press, 1990. Клетку-хозяина культивируют в питательных средах, удовлетворяющих требованиям конкретной применяемой клетки-хозяина, в частности, касательно значения рН, температуры, концентрации солей, аэрации, антибиотиков, витаминов, микроэлементов и т.д.
Клетка-хозяин, предлагаемая в настоящем изобретении, может представлять собой любую прокариотическую или эукариотическую клетку. Приемлемыми прокариотическими клетками являются клетки, которые обычно применяют для клонирования, типа Е. coli или Bacillus subtilis. Кроме того, к эукариотическим клеткам относятся, например, клетки грибов или животных. Примерами приемлемых грибных клеток являются клетки дрожжей, предпочтительно рода Saccharomyces и наиболее предпочтительно вида Saccharomyces cerevisiae. Приемлемыми клетками животных являются, например, клетки насекомых, клетки позвоночных животных, предпочтительно клетки млекопитающих, такие, например, как НЕК293, NSO, CHO,COS-7, MDCK, U2-OSHela, NIH3T3, MOLT-4, Jurkat, PC-12, PC-3, IMR, NT2N, Sk-n-sh, CaSki, С33А. Другие приемлемые клеточные линии, известные в данной области, представляют собой линии клеток, которые можно получать из депозитариев клеточных линий, типа, например, Немецкая коллекция микроорганизмов и клеточных культур (Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH (DSMZ)) или Американская коллекция типовых культур (American Type Culture Collection (АТСС)). Согласно настоящему изобретению также считается, что первичные клетки/клеточные культуры могут функционировать в качестве клеток-хозяев. Указанные клетки получают прежде всего из насекомых (типа насекомых вида p. Drosophila или p. Blatta) или млекопитающих (типа человека, свиней, мышей или крыс).Указанные клетки-хозяева могут содержать также клетки из и/или имеющие происхождение из клеточных линий типа линий клеток нейробластомы. Указанные выше первичные клетки хорошо известны в данной области и включают среди прочего первичные астроциты, (смешанные) культуры клеток спинного мозга или гиппокампа.
Настоящее изобретение относится также к композиции, которая содержит молекулу ДНК, предлагаемую в настоящем изобретении, вектор, предлагаемый в настоящем изобретении, или клетку-хозяина, предлагаемую в настоящем изобретении.
Вторым объектом настоящего изобретения является молекула РНК, которая содержит
(а) кодирующую область, включающую стартовый кодон на ее 5'-конце, которая кодирует полипептид; и
(б) расположенную непосредственно в обратном направлении относительно указанной кодирующей последовательности UTR, выбранную из группы, которая содержит:
(б1) UTR-последовательности
R2-CGCCACC (SEQ ID NO: 1),
или последовательности, где в указанной UTR-последовательности С в положении 6 SEQ ID NO: 1 заменен на А и С в положении 7 SEQ ID NO: 1 заменен на G; и/или А в положении 5 SEQ ID NO: 1 заменен на G; и
(б2) UTR-последовательности
R2-CNGCCACC (SEQ ID NO: 2), в которой нуклеотид N в положении 2 SEQ ID NO: 2 представляет собой нуклеотид, выбранный из группы, которая состоит из Т, G, С или А; или последовательности, где в указанной последовательности С в положении 7 SEQ ID NO: 2 заменен на А и С в положении 8 SEQ ID NO:2 заменен на G; и/или А в положении 6 SEQ ID NO: 2 заменен на G,
где R2 обозначает последовательность РНК, соответствующую части промоторной области, начинающейся с нуклеотида, с которого ДНК-зависимая РНК-полимераза инициирует синтез РНК.
Молекула рибонуклеиновой кислоты (РНК), применяемая в контексте настоящего изобретения, означает полимерную молекулу, которая собрана в виде цепи нуклеотидов, обозначенных как G, A, U и С. Каждый нуклеотид в РНК содержит сахар рибозу, атомы углерода которой пронумерованы от 1' до 5'. Азотистое основание, присоединенное к 1'-положению, как правило, представляет собой аденин (А), цитозин (С), гуанин (G) или урацил (U). В полимерной молекуле РНК фосфатная группа присоединена к 3'-положению одного из остатков рибозы и 5'-положению следующего остатка. Таким образом, нуклеотиды в полимерной молекуле РНК ковалентно связаны друг с другом, при этом фосфатная группа одного нуклеотида связывается с 3'-углеродом следующего нуклеотида, образуя тем самым фосфодиэфирную связь. Таким образом, цепь РНК имеет 5'-конец и 3'-конец, обозначение относится к атомам углерода в рибозном кольце. Принято, что понятия «в обратном направлении» и «в прямом направлении» относятся к 5'→3' направлению транскрипции РНК. Предпочтительно молекула РНК представляет собой молекулу матричной РНК (мРНК). мРНК относится к большому семейству молекул РНК, которые переносят генетическую информацию с ДНК на рибозим, где они определяют аминокислотную последовательность белковых продуктов генной экспрессии. В результате транскрипции первичного транскрипта мРНК (обозначена пре-мРНК) с помощью РНК-полимеразы, процессинга, зрелая мРНК транслируется в полимер аминокислот: белок, как это обобщено в центральной догме молекулярной биологии. Аналогично ДНК, генетическая информация мРНК закодирована в последовательности нуклеотидов, которые организованы в виде кодонов, состоящих из трех оснований каждый. Каждый кодон кодирует конкретную аминокислоту за исключением стоп-кодонов, которые завершают синтез белка.
Молекула РНК, предлагаемая в настоящем изобретении, содержат два основных модуля, указанных выше в пунктах (а) и (б). Кроме того, молекула РНК, предлагаемая в настоящем изобретении, предпочтительно содержит UTR на ее 3'-конце. Таким образом, молекула РНК, предлагаемая в настоящем изобретении, напоминает с точки зрения ее структуры «нормальную (каноническую)» молекулу мРНК, которая встречается в природе, несущую кодирующую область, а также (5'- и 3'-) нетранслируемые области (UTR) и необязательно поли-А-хвост.
Понятие «кодирующая область, включающая стартовый кодон на ее 5'-конце», применяемое согласно настоящему изобретению, относится к последовательности, которая состоит из кодонов, которые расшифровываются и транслируются в белок с помощью рибосомы в соответствии с информацией, предоставляемой генетическим кодом. Кодирующие области обычно начинаются со стартового кодона на их 5'-конце и оканчиваются стоп-кодоном. Как правило, стартовый кодон представляет собой триплет AUG, а стоп-кодон представляет собой UAA, UAG или UGA. Помимо того, что они кодируют белок, части кодирующих областей могут служить в качестве регуляторных последовательностей в пре-мРНК, таких как энхансеры сплайсинга экзонов или сайленсеры сплайсинга экзонов. Кодирующую область гена, которая кодирует полипептид или белок, применяемую согласно настоящему изобретению, обозначают также как кодирующая последовательность или CDS (из кодирующей последовательности ДНК), и она представляет собой часть генной ДНК или РНК, состоящую из экзонов, которые кодируют полипептид или белок. Кодирующая область в мРНК фланкирована 5'-нетранслируемой областью (5' UTR) и 3'-нетранслируемой областью (3' UTR), которые также являются частью экзонов. Кроме того, молекулы мРНК могут содержать также так называемый 5'-кэп и поли-А-хвост. 5'-кэп, 5' UTR, 3' UTR и поли-А-хвост представляют собой области молекулы мРНК, которые не транслируются в белок.
Понятие «нетранслируемая область» или «UTR» при применении в контексте настоящего изобретения относится к участкам мРНК, расположенным в обратном направлении относительно стартового кодона и в прямом направлении относительно стоп-кодона, которые не транслируются и, поэтому, их обозначают как пять-штрих нетранслируемая область (5'UTR) и три-штрих нетранслируемая область (3'UTR) соответственно. Эти области транскрибируются с кодирующей области и, таким образом, являются экзонными, поскольку присутствуют в зрелой мРНК.
В контексте настоящего изобретения 3'-нетранслируемая область (3'UTR) означает участок матричной РНК (мРНК), расположенный непосредственно за кодоном терминации трансляции. 3'UTR может содержать регуляторные области в 3'-нетранслируемой области, которые, как известно, влияют на полиаденилирование и стабильность мРНК. Многие 3'UTR содержат также богатые AU элементы (ARE). Кроме того, 3'UTR может предпочтительно содержать последовательность AAUAAA, которая контролирует добавление нескольких сотен остатков аденина, которые называют поли(А)-хвостом, на конец мРНК-транскрипта.
В контексте настоящего изобретения 5'-нетранслируемая область (5'UTR) (которую обозначают также как лидерная последовательность или лидер РНК) представляет собой область мРНК, расположенную непосредственно в обратном направлении относительно стартового кодона. 5'UTR начинается с сайта инициации транскрипции и заканчивается за один нуклеотид (nt) до стартового кодона (как правило, AUG) кодирующей области. У эукариот длина 5'UTR, как правило, составляет от 100 до нескольких тысяч нуклеотидов, однако иногда у эукариот встречаются более короткие UTR.
Согласно настоящему изобретению 5'UTR является чрезвычайно короткой, поскольку объектом настоящего изобретения является создание минимальной UTR-последовательности.
Молекула РНК, предлагаемая в настоящем изобретении, может содержать также поли-А-хвост. Поли-А-хвост представляет собой длинную последовательность адениновых нуклеотидов (часто несколько сотен), добавленную к 3'-концу пре-мРНК с помощью процесса, называемого полиаденилированием. Указанный хвост усиливает экспорт из ядра и трансляцию и защищает мРНК от расщепления. Полиаденилирование представляет собой добавление поли(А)-хвоста к матричной РНК. Поли(А)-хвост состоит из множества аденозинфосфатов; другими словами, представляет собой участок РНК, состоящий только из адениновых оснований. У эукариот полиаденилирование является частью процесса, приводящего в получению зрелой матричной РНК (мРНК) для трансляции.
Один модуль молекулы РНК, т.е. «кодирующая область, включающая стартовый кодон на ее 5'-конце, которая кодирует полипептид» (модуль (а)), не ограничен конкретным модулем и может представлять собой любую требуемую кодирующую область, которая должна экспрессироваться в данной клетке. В предпочтительных вариантах осуществления изобретения понятие «кодирующая область, включающая стартовый кодон на ее 5'-конце, которая кодирует полипептид» (модуль (а)), применимо также mutatis mutandis (с необходимыми изменениями) к молекуле РНК, предлагаемой в настоящем изобретении, которая описана выше в контексте молекулы ДНК, предлагаемой в настоящем изобретении.
Молекула РНК, предлагаемая в настоящем изобретении, содержит модуль (б) непосредственно в обратном направлении относительно указанной кодирующей последовательности, где указанный модуль (б) представляет собой UTR, выбранную из группы, которая состоит из:
(б1) UTR-последовательности
R2-CGCCACC (SEQ ID NO: 1),
или последовательности, где в указанной последовательности UTR С в положении 6 SEQ ID NO: 1 заменен на А и С в положении 7 SEQ ID NO: 1 заменен на G; и/или А в положении 5 SEQ ID NO: 1 заменен на G; и
(б2) UTR-последовательности
R2-CNGCCACC (SEQ ID NO: 2), в которой нуклеотид N в положении 2 SEQ ID NO: 2 представляет собой нуклеотид, выбранный из группы, которая состоит из Т, G, С или А; или последовательности, где в указанной последовательности С в положении 7 SEQ ID NO: 2 заменен на А и С в положении 8 SEQ ID NO: 2 заменен на G; и/или А в положении 6 SEQ ID NO: 2 заменен на G,
где R2 обозначает последовательность РНК, соответствующую части промоторной области, начинающейся с нуклеотида, с которого ДНК-зависимая РНК-полимераза инициирует синтез РНК.
Природа R2 не ограничена конкретно. Можно применять любую последовательность РНК, соответствующую части промоторной области, начинающейся с нуклеотида, с которого ДНК-зависимая РНК-полимераза инициирует синтез РНК. Специалист в данной области может легко определить положение, соответствующее частям промоторной области, начинающейся с нуклеотида, с которого ДНК-зависимая РНК-полимераза инициирует синтез РНК. Указанная РНК-последовательность R2 представляет собой последовательность промотора, которая соответствует транскрибируемой части промотора, т.е. фактически присутствует в транскрипте после транскрипции.
В предпочтительном варианте осуществления изобретения промотор R2 представляет собой последовательность РНК, соответствующую части промоторной области, начинающейся с нуклеотида, с которого происходящая из бактериофага ДНК-зависимая РНК-полимераза инициирует синтез РНК.
В предпочтительном варианте осуществления изобретения промотор R2 представляет собой последовательность РНК, соответствующую части промоторной области, начинающейся с нуклеотида, с которого ДНК-зависимая РНК-полимераза Т7, ДНК-зависимая РНК-полимераза Т3, ДНК-зависимая РНК-полимераза SP6 или ДНК-зависимая РНК-полимераза K11 инициирует синтез РНК.
Для иллюстрации указанного в качестве примеров (но не ограничиваясь только ими) R2 имеет последовательность, подчеркнутую в приведенных ниже промоторных последовательностях TAATACGACTCACTATAGGGAGA (SEQ ID NO: 3; т.е. промотор, распознаваемый ДНК-зависимой РНК-полимеразой Т7), AATTAACCCTCACTAAAGGGAGA (SEQ ID NO: 4; т.е. промотор, распознаваемый ДНК-зависимой РНК-полимеразой Т3), ATTTAGGTGACACTATAGAAG (SEQ ID NO: 5; т.е. промотор, распознаваемый ДНК-зависимой РНК-полимеразой SP6) и AATTAGGGCАСАСТАТАGGGA (SEQ ID NO: 6; т.е. промотор, распознаваемый ДНК-зависимой РНК-полимеразой K11). Подчеркнутые последовательности соответствуют части указанного промотора, с которой ДНК-зависимая РНК-полимераза инициирует синтез РНК и, таким образом, которая фактически присутствует в молекуле РНК (т.е. в транскрипте) после транскрипции.
UTR-последовательность(и), имеющая(ие) любую из вышеуказанных замен по сравнению с UTR-последовательностью R2-CGCCACC (SEQ ID NO: 1) или по сравнению с UTR-последовательностью R2-CNGCCACC (SEQ ID NO: 2), может приводить к созданию молекулы РНК, имеющей такую же или сходную, предпочтительно более высокую, эффективность трансляции по сравнению с молекулой РНК, которая содержит UTR-последовательность R2-CGCCACC (SEQ ID NO: 1), и с молекулой РНК, которая содержит UTR-последовательность R2-CNGCCACC (SEQ ID NO: 2) соответственно. Эффективность трансляции данной молекулы РНК, содержащей UTR, указанную в настоящем описании, специалист в данной области может определять с помощью методов, которые известны в данной области и описаны ниже.
Эффективность трансляции представляет собой скорость трансляции мРНК в полипептиды или белки внутри клеток. Эффективность трансляции данной мРНК измеряют как количество белков или полипептидов, транслированных с мРНК в единицу времени. Трансляция представляет собой процесс, в котором клеточные рибосомы создают белки и является хорошо известным специалисту в данной области. В целом, при трансляции матричная РНК (мРНК), которая получена в результате транскрипции с ДНК, расшифровывается рибосомой с образованием конкретной аминокислотной цепи или полипептида, или белка.
Таким образом, эффективность трансляции данной молекулы РНК, которая несет модифицированную последовательность UTR с любой из указанных выше замен, предпочтительно является такой же или более высокой по сравнению с эффективностью трансляции данной РНК, но которая не несет UTR R2-CGCCACC (SEQ ID NO: 1) или R2-CNGCCACC (SEQ ID NO: 2) соответственно, указанную выше в настоящем описании. Следовательно, количество белков или полипептидов, которые кодируются кодирующей областью молекулы РНК, несущей модифицированную последовательность UTR с любой из указанных выше замен, которые транслируются с РНК в единицу времени, является по меньшей мере таким же или предпочтительно более высоким, чем количество белков или полипептидов, которые кодируются кодирующей областью молекулы РНК, несущей UTR R2-CGCCACC (SEQ ID NO: 1) или R2-CNGCCACC (SEQ ID NO: 2) соответственно, указанные выше в настоящем описании, которые транслируются с РНК в единицу времени.
Эффективность трансляции в контексте настоящего изобретения предпочтительно представляет собой количество мРНК, транслируемой в белок внутри клетки в определенный момент времени, относительно количества мРНК, кодирующей соответствующий белок в указанной клетке в этот же момент времени. Таким образом, эффективность трансляции представляет собой отношение количества мРНК, транслируемой в белок в клетке в определенный момент времени, и количества мРНК, кодирующей соответствующий белок. Оба параметра, т.е. количество мРНК, транслируемой в белок в клетке, и количество мРНК, кодирующей соответствующий белок, можно определять методами, известными в данной области. В качестве примеров (но не ограничиваясь только ими) количество мРНК, транслируемой в белок в клетке, можно определять, например, с помощью проточной цитометрии (FC), а количество мРНК, кодирующей соответствующий белок, можно измерять, например, с помощью qПЦР.
UTR, указанная(ые) выше в настоящем описании в пункте (б), не ограничена(ы) конкретно указанными выше конкретными последовательностями, но могут включать также UTR-последовательность(и), которая(ые) содержит(ат) последовательность, имеющую добавление(я) нуклеотида(ов) по сравнению с указанными последовательностями, где дополнительный(ые) нуклеотид(ы) можно добавлять на 5'-конец вышеописанной(ых) UTR. Дополнительный(ые) нуклеотид(ы) содержит(ат) полинуклеотидные цепи, содержащие вплоть до 0 (нет изменений), 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 нуклеотидов, предпочтительно вплоть до 20 нуклеотидов. Более предпочтительно добавляют 11, 12, 13, 14, 15, 16, 18 или 19 нуклеотидов на 5'-конец. Еще более предпочтительно вплоть до 30 нуклеотидов добавляют на 5'-конец.
В свете разумного предположения о том, что добавление нуклеотидов, вероятно, не изменяет указанные выше функциональные свойства соответствующей(их) UTR, добавленные нуклеотиды могут иметь также длину вплоть до 40, 50, 60, 70, 80, 90 или даже 100 нуклеотидов или даже более, вплоть до 200, 300, 400 или 500 нуклеотидов, если указанные последовательности имеют сходную способность (в понятиях описанной выше эффективности трансляции) с UTR, указанными выше в настоящем описании в пункте (б).
В предпочтительном варианте осуществления изобретения UTR, указанная выше в настоящем описании в подпункте (б1), имеет максимальную длину 11, 12 или 13 нуклеотидов. Предпочтительно UTR, указанная выше в настоящем описании в подпункте (б1), имеет максимальную длину 13 нуклеотидов, если R2 обозначает GGGAGA (SEQ ID NO: 7) или GGGAGA (SEQ ID NO: 8).
Предпочтительно UTR, указанная выше в настоящем описании в подпункте (б1), имеет максимальную длину 11 нуклеотидов, если R2 обозначает GAAG (SEQ ID NO: 9) или GGGA (SEQ ID NO: 10).
В другом предпочтительном варианте осуществления изобретения UTR, указанная выше в настоящем описании в подпункте (б2), имеет максимальную длину 12, 13 или 14 нуклеотидов. Предпочтительно UTR, указанная выше в настоящем описании в подпункте (б2), имеет максимальную длину 14 нуклеотидов, если R2 обозначает GGGAGA (SEQ ID NO: 7) или GGGAGA (SEQ ID NO: 8).
Предпочтительно UTR, указанная выше в настоящем описании в подпункте (б2), имеет максимальную длину 12 нуклеотидов, если R2 обозначает GAAG (SEQ ID NO: 9) или GGGA (SEQ ID NO: 10).
Молекулы РНК, предлагаемые в настоящем изобретении, содержащие описанную(ые) выше UTR, можно создавать/синтезировать рекомбинантно (например, с помощью системы in vivo или in vitro) или с помощью синтеза с использованием методов, хорошо известных специалисту в данной области.
Для транскрипции РНК in vitro, как правило, требуется линейная ДНК-матрица, содержащая двухцепочечную промоторную область, с которой ДНК-зависимая РНК-полимераза связывается и инициирует синтез РНК, в то время как кодирующая область может быть двухцепочечной или одноцепочечной. В том случае, когда линейная ДНК-матрица содержит одноцепочечную кодирующую область, антимысловая цепь (т.е. цепь, которая считывается ДНК-зависимой полимеразой) кодирующей области представляет собой часть матрицы. Общепринятые ДНК-зависимые РНК-полимеразы представляют собой полимеразу Т7, полимеразу Т3, полимеразу SP6 и полимеразу K11. Полноразмерные последовательности соответствующих им промоторов представлены в SEQ ID NO: 3-6.
Матрицы для транскрипции, предназначенные для транскрипции in vitro, включают, например кДНК-матрицы, синтезированные из РНК-предшественника, матрицы, созданные с помощью ПЦР, химически синтезированные олигонуклеотиды и плазмидные конструкции. Многие широко применяемые плазмидные клонирующие векторы несут промоторы для полимеразы фагов, локализованные на каждой стороне сайта множественного клонирования, что обеспечивает транскрипцию любой цепи нуклеотидной последовательности, встроенной в сайт множественного клонирования. Широко применяемые клонирующие векторы включают, например, векторы pCRII фирмы Invitrogen, pGEM фирмы Promega и pBluescript фирмы Stratagene. Представители семейства векторов pTRIPLEscript фирмы Ambion содержат все три промотора для фаговых полимераз в тандеме (на одной и той же стороне сайта множественного клонирования), что позволяет применять любую из трех полимераз SP6, Т7 или Т3.
Молекулы РНК, предлагаемые в настоящем изобретении, можно получать рекомбинантно, например, с использованием систем in vivo, с помощью методов, известных специалистам в данной области.
Альтернативно этому, молекулы РНК, предлагаемые в настоящем изобретении, можно получать в системе in vitro, используя, например, систему транскрипции in vitro. Системы транскрипции in vitro являются общеизвестными и в них, как правило, требуется очищенная линейная ДНК-матрица, содержащая последовательность ДНК, «кодирующую» молекулу РНК, где указанная последовательность ДНК находится под контролем соответствующего промотора. Кроме того, для системы транскрипции in vitro также, как правило, требуются рибонуклеозидтрифосфаты, буферная система, которая включает ДТТ и ионы магния, и соответствующая РНК-полимераза, которая обладает ферментативной активностью для транскрипции in vitro последовательности ДНК в соответствующую молекулу РНК, предлагаемую в настоящем изобретении.
Кроме того, молекулы РНК можно синтезировать химически, например, путем общепринятого химического синтеза с использованием автоматического синтезатора нуклеотидных последовательностей, применяя твердофазную подложку и стандартные методики, или путем химического синтеза соответствующих ДНК-последовательностей и последующей их транскрипции in vitro или in vivo.
В соответствии с вышеизложенным, в настоящем изобретении предложены молекулы РНК/молекулы полирибонуклеиновых кислот, предпочтительно молекулы модифицированных полирибонуклеиновых кислот, где один модуль указанной молекулы РНК, т.е. «кодирующая область, включающая стартовый кодон на ее 5'-конце» (модуль (а)), кодирует полипептид. Понятия «нуклеиновая кислота» и «полинуклеотид» применяют взаимозаменяемо, и они включают любое соединение и/или субстанцию, которые содержат полимер из нуклеотидов. Понятие нуклеотид включает дезоксинуклеотиды и рибонуклеотиды. Понятия рибонуклеиновая кислота и полирибонуклеотид применяют взаимозаменяемо, и они в некоторых вариантах осуществления изобретения включают любое соединение и/или субстанцию, которые содержат полимер из нуклеотидов, в котором более 50% нуклеотидов представляют собой рибонуклеотиды. В некоторых вариантах осуществления изобретения полирибонуклеотиды содержат полимер из нуклеотидов, в котором более 60%, 70%, 75%, 80%, 90%, более 95%, более 99% или 100% нуклеотидов представляют собой рибонуклеотиды. Полирибонуклеотиды, в которых один или несколько нуклеотидов представляют собой модифицированные нуклеотиды, можно обозначать как модифицированные полирибонуклеотиды. Однако понятие полирибонуклеотиды может включать модифицированные полирибонуклеотиды.
Последовательности молекул РНК/полирибонуклеотидов можно получать, например, из любой приемлемой нуклеиновой кислоты, которая содержит генетическую информацию представляющего интерес гена. Примеры нуклеиновых кислот включают геномную ДНК, РНК или кДНК из любой бактериальной клетки или клетки архей, содержащей представляющий(ие) интерес ген(ы). Полинуклеотиды можно получать из нуклеиновых кислот, несущих мутантные гены и полиморфизмы. Молекула РНК/полирибонуклеотид, предлагаемая/предлагаемый в настоящем изобретении, содержит последовательность, которая не ограничена конкретно указанной последовательностью и может содержать в качестве модуля (а) любую требуемую кодирующую область, которая экспрессируется в данной клетке. В предпочтительном варианте осуществления изобретения указанная последовательность может представлять собой кодирующую область для требуемого полипептида/белка, указанного выше. Предпочтительно в свете вышесказанного молекула РНК /полирибонуклеотид дополнительно содержит нетранслируемую последовательность, расположенную в обратном направлении (5') относительно стартового кодона модуля (а), нетранслируемую последовательность, расположенную в прямом направлении (3') относительно стоп-кодона модуля (а) или как нетранслируемую последовательность, расположенную в обратном направлении (5') относительно стартового кодона модуля (а), так и нетранслируемую последовательность, расположенную в прямом направлении (3') относительно стоп-кодона модуля (а). В предпочтительном варианте осуществления изобретения молекула РНК/полирибонуклеотид предлагаемая/предлагаемый в настоящем изобретении, может представлять собой модифицированную молекулу РНК/модифицированный полирибонуклеотид.
Помимо четырех классических рибонуклеотидов, а именно, аденозина, гуанозина, цитидина и уридина, существуют многочисленные аналоги каждого из указанных нуклеотидных оснований. Иногда в настоящем описании и в литературе указанные аналоги или молекулы РНК/полирибонуклеотиды, которые включают один или несколько из указанных аналогов, обозначают как модифицированные (например, модифицированные нуклеотиды или модифицированные рибонуклеотиды). Некоторые аналоги отличаются от описанных выше канонических нуклеотидных оснований, однако они еще могут существовать в природе. Другие аналоги не встречаются в естественных условиях. Под объем изобретения подпадает любой тип аналога.
В некоторых вариантах осуществления изобретения молекулы РНК/полирибонуклеотиды, предлагаемые в настоящем изобретении, содержат нуклеотидные аналоги (например, полирибонуклеотид содержит модифицированный полирибонуклеотид). Примеры нуклеотидных аналогов представлены ниже (например, аналоги U; аналоги С; аналоги А; аналоги G). Кроме того, в некоторых вариантах осуществления изобретения молекула РНК/полирибонуклеотид или другая нуклеиновая кислота, предложенная в настоящем описании, может содержать также (в дополнительном или альтернативном варианте) модификации в фосфодиэфирном каркасе или в связи между нуклеотидными основаниями. Примеры нуклеиновых кислот, которые могут образовывать часть или всю молекулу РНК/часть или весь полирибонуклеотид, предложенную/предложенный в настоящем описании, включают (но не ограничиваясь только ими) рибонуклеиновые кислоты (РНК), дезоксирибонуклеиновые кислоты (ДНК), треозо-нуклеиновые кислоты (ТНК), нуклеиновые кислоты гликоля (ГНК), пептидные нуклеиновые кислоты (РНК), «закрытые» нуклеиновые кислоты (ЗНК, включая ЗНК, имеющие бета-D-рибо-конфигурацию, альфа-ЗНК, имеющие альфа-L-рибо-конфигурацию (диастереомер ЗНК), 2'-амино-ЗНК, имеющие 2'-амино-функционализацию и 2'-амино-альфа-ЗНК, имеющие 2'-амино-функционализацию) или их гибриды.
В некоторых вариантах осуществления изобретения модификация может затрагивать один или несколько нуклеозид(ов) или каркас нуклеиновой кислоты/молекулы полинуклеотида. В некоторых вариантах осуществления изобретения модификация может затрагивать как нуклеозид, так и связь в каркасе. В некоторых вариантах осуществления изобретения модификацию можно создавать в полинуклеотиде in vitro. В некоторых вариантах осуществления изобретения модифицированный рибонуклеотид/нуклеотид можно синтезировать также после транскрипции путем ковалентной модификации классических/встречающихся в естественных условиях рибонуклеотидов/нуклеотидов.
Молекула РНК/полирибонуклеотид, предлагаемая/предлагаемый в настоящем изобретении, может представлять собой модифицированную молекулу РНК/модифицированный полирибонуклеотид и в некоторых вариантах осуществления изобретения может содержать аналоги пуринов и/или аналоги пиримидинов. В некоторых вариантах осуществления изобретения модифицированная молекула РНК/модифицированный полирибонуклеотид, предлагаемая/предлагаемый в настоящем изобретении, содержат пиримидиновый аналог, такой как аналог уридина и/или аналог цитидина. В некоторых вариантах осуществления изобретения модифицированная молекула РНК/модифицированный полирибонуклеотид, предлагаемая/предлагаемый в настоящем изобретении, содержат аналог уридина и аналог цитидина. В некоторых вариантах осуществления изобретения модифицированная молекула РНК/модифицированный полирибонуклеотид, предлагаемая/предлагаемый в настоящем изобретении, не содержат аналоги аденозина и/или аналоги гуанозина. В некоторых вариантах осуществления изобретения молекула РНК/полирибонуклеотид, предлагаемая/предлагаемый в настоящем изобретении, содержат один тип аналога уридина и один тип аналога цитидина (например, один тип аналога, но не одну молекула аналога, при этом процентное содержание одного аналога может представлять собой любое из нескольких указанных в настоящем описании процентных содержаний). В других вариантах осуществления изобретения молекула РНК/полирибонуклеотид содержит более одного типа аналога уридина и/или цитидина и необязательно, если они присутствуют, один или несколько аналогов аденозина и/или гуанозина (или ни одного из них, или оба).
В некоторых случаях модифицированный уридин (например, аналог уридина) выбирают из 2-тиоуридина, 5'-метилуридина, псевдоуридина, 5-йодуридина (I5U), 4-тиоуридина (S4U), 5-бромуридина (Br5U), 2'-метил-2'-дезоксиуридина (U2'm), 2'-амино-2'-дезоксиуридина (U2'NH2), 2'-азидо-2'-дезоксиуридина (U2'N3) и 2'-фтор-2'-дезоксиуридина (U2'F). В некоторых случаях модифицированный цитидин (например, аналог цитидина) выбирают из 5-метилцитидина, 3-метилцитидина, 2-тиоцитидина, 2'-метил-2'-дезоксицитидина (C2'm), 2'-амино-2'-дезоксицитидина (C2'NH2), 2'-фтор-2'-дезоксицитидина (C2'F), 5-йодцитидина (I5C), 5-бромцитидина (Br5C) и 2'-азидо-2'-дезоксицитидина (C2'N3). Следует отметить, что вышеуказанная ссылка на аналоги относится также к аналогам в их 5'-трифосфатной форме. В некоторых вариантах осуществления изобретения аналог цитидина представляет собой 5-йодцитидин и аналог уридина представляет собой 5-йодуридин.
В некоторых вариантах осуществления изобретения молекула РНК/полирибонуклеотид представляет собой модифицированную молекулу РНК/модифицированный полирибонуклеотид. В некоторых случаях модифицированная молекула РНК/модифицированный полирибонуклеотид по меньшей мере на 25% является более стабильной/стабильным по сравнению с немодифицированной молекулой РНК/немодифицированным полирибонуклеотидом. В некоторых случаях модифицированная молекула РНК/модифицированный полирибонуклеотид может быть по меньшей мере на 30% более стабильной/стабильным, по меньшей мере на 35% более стабильной/стабильным, по меньшей мере на 40% более стабильной/стабильным, по меньшей мере на 45% более стабильной/стабильным, по меньшей мере на 50% более стабильной/стабильным, по меньшей мере на 55% более стабильной/стабильным, по меньшей мере на 60% более стабильной/стабильным, по меньшей мере на 65% более стабильной/стабильным, по меньшей мере на 70% более стабильной/стабильным, по меньшей мере на 75% более стабильной/стабильным, по меньшей мере на 80% более стабильной/стабильным, по меньшей мере на 85% более стабильной/стабильным, по меньшей мере на 90% более стабильной/стабильным или по меньшей мере на 95% более стабильной/стабильным по сравнению с немодифицированной молекулой РНК/немодифицированным полирибонуклеотидом. В некоторых вариантах осуществления изобретения стабильность оценивают in vivo. В некоторых вариантах осуществления изобретения стабильность оценивают in vitro. В некоторых вариантах осуществления изобретения стабильность оценивают количественно, измеряя время полужизни полирибонуклеотида.
Молекула РНК/полирибонуклеотид, предлагаемая/предлагаемый в настоящем изобретении, может иметь нуклеотиды, которые были модифицированы в одинаковой форме или также смесь различных модифицированных нуклеотидов. Модифицированные нуклеотиды могут иметь модификации, которые встречаются или не встречаются в естественных условиях в матричной РНК. Можно применять смесь различных модифицированных нуклеотидов. Например, один или несколько модифицированных нуклеотидов в молекуле РНК/полирибонуклеотиде могут иметь встречающиеся в естественных условиях модификации, в то время как другая часть может иметь модификации, которые не встречаются в естественных условиях в мРНК. Кроме того, некоторые модифицированные нуклеотиды могут иметь модификацию оснований, а другие модифицированные нуклеотиды иметь модификацию сахара. Также возможен случай, когда все модификации представляют собой модификации оснований или все модификации представляют собой модификации сахара или любую их приемлемую смесь. В некоторых случаях стабильность модифицированной молекулы РНК/модифицированного полирибонуклеотида можно избирательно оптимизировать путем изменения природы модифицированных оснований в модифицированном полирибонуклеотиде.
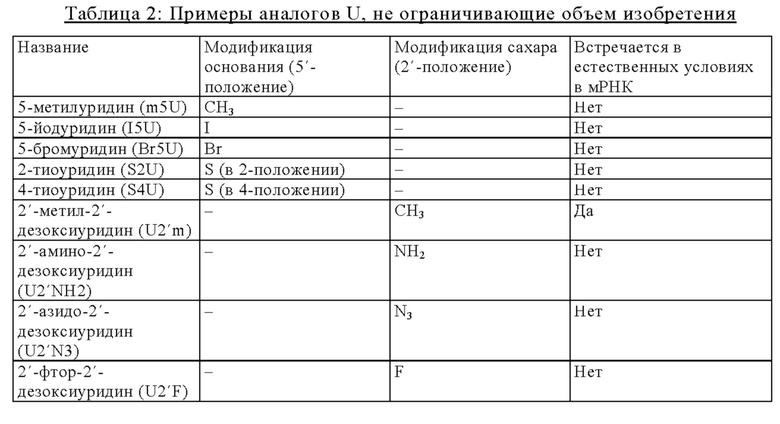

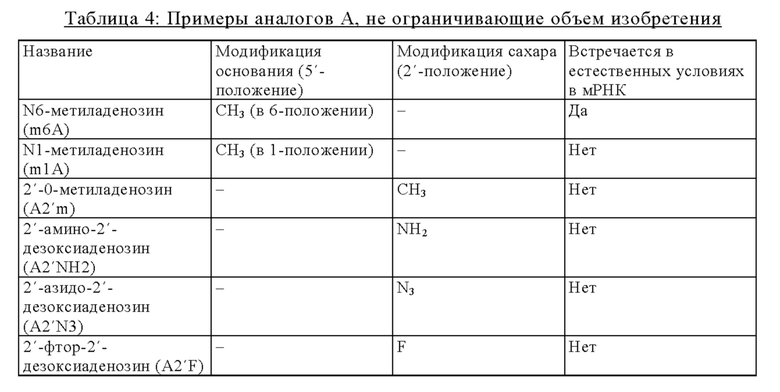
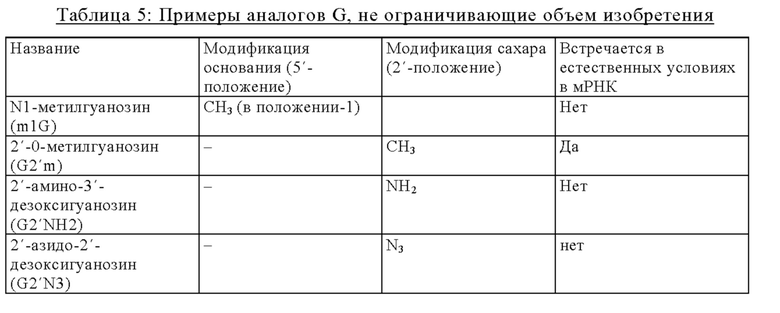

В некоторых вариантах осуществления изобретения аналог (например, модифицированный нуклеотид) можно выбирать из группы, содержащей пиридин-4-он-рибонуклеозид, 5-йодуридин, 5-йодцитидин, 5-азауридин, 2'-амино-2'-дезоксицитидин, 2'-фтор-2'-дезоксицитидин, 2-тио-5-азауридин, 2-тиоуридин, 4-тиопсевдоуридин, 2-тиопсевдоуридин, 5-гидроксиуридин, 3-метилуридин, 5-карбоксиметилуридин, 1-карбоксиметилпсевдоуридин, 5-пропинилуридин, 1-пропинилпсевдоуридин, 5-тауринометилуридин, 1-тауринометилпсевдоуридин, 5-тауринометил-2-тиоуридин, 1-тауринометил-4-тиоуридин, 5-метилуридин, 1-метилпсевдоуридин, 4-тио-1-метилпсевдоуридин, 2-тио-1-метилпсевдоуридин, 1-метил-1-деазапсевдоуридин, 2-тио-1-метил-1-деазапсевдоуридин, дигидроуридин, дигидропсевдоуридин, 2-тиодигидроуридин, 2-тиодигидропсевдоуридин, 2-метоксиуридин, 2-метокси-4-тиоуридин, 4-метоксипсевдоуридин, 4-метокси-2-тиопсевдоуридин, 5-азацитидин, псевдоизоцитидин, 3-метил-цитидин, N4-ацетилцитидин, 5-формилцитидин, 5-метилцитидин, N4-метилцитидин, 5-гидроксиметилцитидин, 1-метил-псевдоизоцитидин, пирролоцитидин, пирролопсевдоизоцитидин, 2-тиоцитидин, 2-тио-5-метилцитидин, 4-тиопсевдоизоцитидин, 4-тио-1-метилпсевдоизоцитидин, 4-тио-1-метил-1-деазапсевдоизоцитидин, 1-метил-1-деазапсевдоизоцитидин, зебуларин, 5-азазебуларин, 5-метилзебуларин, 5-аза-2-тиозебуларин, 2-тиозебуларин, 2-метоксицитидин, 2-метокси-5-метилцитидин, 4-метоксипсевдоизоцитидин, 4-метокси-1-метилпсевдоизоцитидин, 2-аминопурин, 2,6-диаминопурин, 7-деазааденин, 7-деаза-8-азааденин, 7-деаза-2-аминопурин, 7-деаза-8-аза-2-аминопурин, 7-деаза-2,6-диаминопурин, 7-деаза-8-аза-2,6-диаминопурин, 1-метиладенозин, N6-метиладенозин, N6-изопентениладенозин, N6-(цис-гидроксиизопентенил)аденозин, 2-метилтио-N6-(цис-гидроксиизопентенил)аденозин, N6-глицинилкарбамоиладенозин, N6-треонилкарбамоиладенозин, 2-метилтио-N6-треонилкарбамоиладенозин, N6,N6-диметиладенозин, 7-метиладенин, 2-метилтиоаденин, 2-метокси-аденин, инозин, 1-метилинозин, виозин, вибутозин, 7-деазагуанозин, 7-деаза-8-азагуанозин, 6-тиогуанозин, 6-тио-7-деазагуанозин, 6-тио-7-деаза-8-азагуанозин, 7-метилгуанозин, 6-тио-7-метилгуанозин, 7-метилинозин, 6-метоксигуанозин, 1-метилгуанозин, N2-метилгуанозин, N2,N2-диметилгуанозин, 8-оксогуанозин, 7-метил-8-оксогуанозин, 1-метил-6-тиогуанозин, N2-метил-6-тиогуанозин и N2,N2-диметил-6-тиогуанозин.
В некоторых вариантах осуществления изобретения модифицированная молекула РНК/модифицированный полирибонуклеотид, предлагаемая/предлагаемый в настоящем изобретении, не включает псевдоуридин. В некоторых варианта осуществления изобретения модифицированная молекула РНК/модифицированный полирибонуклеотид, предлагаемая/ предлагаемый в настоящем изобретении, не включает 5-метилцитидин. В некоторых вариантах осуществления изобретения модифицированная молекула РНК/модифицированный полирибонуклеотид, предлагаемая/ предлагаемый в настоящем изобретении, не включает 5-метилуридин. В некоторых вариантах осуществления изобретения модифицированная молекула РНК/модифицированный полирибонуклеотид, предлагаемая/ предлагаемый в настоящем изобретении, содержит аналоги U и аналоги С, где указанные аналоги U все могут представлять собой одинаковый аналог или могут представлять собой различные аналоги (например, более, чем один тип аналога), и указанные аналоги С все могут представлять собой одинаковый аналог или могут представлять собой различные аналоги (например, более чем один тип аналога). В некоторых вариантах осуществления изобретения модифицированная молекула РНК/модифицированный полирибонуклеотид, предлагаемая/предлагаемый в настоящем изобретении, не включает аналоги аденозина и аналоги гуанозина.
Как описано подробно в настоящем описании, когда молекула РНК/полирибонуклеотид содержит модифицированный полирибонуклеотид, то аналоги могут присутствовать в определенной пропорции относительно нуклеотидов в соединении (например, определенный процент конкретного нуклеотидного основания может приходиться на долю аналога, указанного в настоящем описании).
Молекула РНК/полирибонуклеотид, которая/который содержит по меньшей мере один модифицированный нуклеотид, представляет собой модифицированную молекулу РНК/модифицированный полирибонуклеотид. В некоторых вариантах осуществления изобретения по меньшей мере примерно 5% модифицированной молекулы РНК/модифицированного полирибонуклеотида включает модифицированный или не встречающийся в естественных условиях (например, аналоги или модифицированные) аденозин, цитидин, гуанозин или уридин, например, аналоги нуклеотидов, представленные в настоящем описании. В некоторых случаях по меньшей мере 5%, 10%, 15%, 20%, 25%, 30%, 40%, 45%, 50% модифицированной молекулы РНК/модифицированного полирибонуклеотида включает модифицированный или не встречающийся в естественных условиях (например, аналоги или модифицированные) аденозин, цитидин, гуанозин или уридин. В некоторых случаях максимум примерно 50%, 45%, 40%, 35%, 30%, 25%, 20%, 15%, 10%, 5% модифицированной молекулы РНК/модифицированного полирибонуклеотида включает модифицированный или не встречающийся в естественных условиях аденозин, цитидин, гуанозин или уридин.
В предпочтительном варианте осуществления изобретения молекула РНК, предлагаемая в настоящем изобретении, содержит комбинацию модифицированных и немодифицированных нуклеотидов. Предпочтительно молекула РНК, предлагаемая в настоящем изобретении, содержит комбинацию модифицированных и немодифицированных нуклеотидов, описанную в WO 2011/012316. Указанные молекулы РНК также известны и поступают в продажу под названием «SNIM®-RNA». Опубликованы данные о том, что молекула РНК, описанная в WO 2011/012316, обладает повышенной стабильностью и пониженной иммуногенностью. В предпочтительном варианте осуществления изобретения в такой модифицированной молекуле РНК модифицированы от 5 до 50% цитидиновых нуклеотидов и от 5 до 50% уридиновых нуклеотидов. Содержащие аденозин и гуанозин нуклеотиды могут быть немодифицироваными. Аденозиновые и гуанозиновые нуклеотиды могут быть немодифицированными или частично модифицированными, предпочтительно они присутствуют в немодифицированной форме. Предпочтительно от 10 до 35% цитидиновых и уридиновых нуклеотидов являются модифицированными, и наиболее предпочтительно содержание модифицированных цитидиновых нуклеотидов находится в пределах от 7,5 до 25% и содержание модифицированных уридиновых нуклеотидов находится в пределах от 7,5 до 25%. Было установлено, что фактически наличие относительно небольшого содержания, например, лишь 10%, модифицированных и цитидиновых, и уридиновых нуклеотидов, позволяет достигать требуемых свойств. Наиболее предпочтительно, когда модифицированные цитидиновые нуклеотиды представляют собой 5-метилцитидиновые остатки, а модифицированные уридиновые нуклеотиды представляют собой 2-тиоуридиновые остатки. Наиболее предпочтительно содержание модифицированных цитидиновых нуклеотидов и содержание модифицированных уридиновых нуклеотидов составляет 25% соответственно.
В некоторых других вариантах осуществления изобретения в указанной модифицированной молекуле РНК/молекуле полирибонуклеотида от 5 до 50% цитидинов представляют собой аналоги С и от 5 до 50% уридинов представляют собой аналоги U. В некоторых вариантах осуществления изобретения в указанной модифицированной молекуле полирибонуклеотида от 5 до 40% цитидинов представляют собой аналоги С и от 5 до 40% уридинов представляют собой аналоги U. В некоторых вариантах осуществления изобретения в указанной модифицированной молекуле РНК/молекуле полирибонуклеотида от 5 до 30% цитидинов представляют собой аналоги С и от 5 до 30% уридинов представляют собой аналоги U. В некоторых вариантах осуществления изобретения в указанной модифицированной молекуле РНК/молекуле полирибонуклеотида от 10 до 30% цитидинов представляют собой аналоги С и от 10 до 30% уридинов представляют собой аналоги U. В некоторых вариантах осуществления изобретения в указанной модифицированной молекуле полирибонуклеотида от 5 до 20% цитидинов представляют собой аналоги С и от 5 до 20% уридинов представляют собой аналоги U. В некоторых вариантах осуществления изобретения в указанной модифицированной молекуле РНК/молекуле полирибонуклеотида от 5 до 10% цитидиновых нуклеотидов и от 5 до 10% уридиновых нуклеотидов являются модифицированными. В некоторых вариантах осуществления изобретения в указанной модифицированной молекуле РНК/молекуле полирибонуклеотида 25% цитидиновых нуклеотидов и 25% уридиновых нуклеотидов являются модифицированными. В некоторых вариантах осуществления изобретения содержащие аденозин и гуанозин нуклеотиды могут быть немодифицированными. В некоторых вариантах осуществления изобретения аденозиновые и гуанозиновые нуклеотиды могут быть немодифицрованными или частично модифицированными, но предпочтительно они присутствуют в немодифицированной форме.
Как указано выше, в некоторых вариантах осуществления изобретения аналоги U относятся к одному типу аналога U. В некоторых вариантах осуществления изобретения аналоги U относятся к двум или большему количеству типов аналогов U. В некоторых вариантах осуществления изобретения аналоги С относятся к одному типу аналога С. В некоторых вариантах осуществления изобретения аналоги С относятся к двум или большему количеству типов аналогов С.
В некоторых вариантах осуществления изобретения процентное содержание цитидинов в молекуле РНК/полирибонуклеотиде, которые представляют собой аналоги цитидина, не такое же как процентное содержание уридинов в молекуле РНК/полирибонуклеотиде, которые представляют собой аналоги уридина. В некоторых вариантах осуществления изобретения процентное содержание аналогов цитидина ниже, чем процентное содержание аналогов уридина. Как отмечалось выше, аналоги аденозина и гуанозина могут присутствовать или отсутствовать, но в некоторых вариантах осуществления изобретения аналоги аденозина и аналоги гуанозина отсутствуют. В некоторых вариантах осуществления изобретения полирибонуклеотиды, представленные в описании, содержат менее 15%, менее 10%, менее 5% или менее 2% аналогов аденозина, аналогов гуанозина или их обоих.
В некоторых вариантах осуществления изобретения молекула РНК/полирибонуклеотид, предлагаемая/предлагаемый в настоящем изобретении, содержит аналоги цитидина и аналоги уридина, и от 5 до 20% цитидинов представляют собой аналоги цитидина и от 25 до 45% уридинов представляют собой аналоги уридина. Другими словами, молекула РНК/полирибонуклеотид содержит модифицированные и немодифицированные цитидины и модифицированные и немодифицированные уридины, и от 5 до 20% цитидинов приходится на долю аналогов цитидина, а от 25 до 45% уридинов приходится на долю аналогов уридина. В других вариантах осуществления изобретения молекула РНК/полирибонуклеотид содержит от 5 до 10% аналогов цитидина и от 30 до 40% аналогов уридина, например, 7-9% аналогов цитидина, например, примерно 7, 7,5 или 8% и, например, 32-38% аналогов уридина, например, примерно 33, 34, 35, 36%.
В некоторых вариантах осуществления изобретения можно применять любые аналоги уридина и аналоги цитидина, указанные в настоящем описании, необязательно за исключением псевдоуридина. В некоторых вариантах осуществления изобретения аналог цитидина содержит или состоит из (например, в случае, когда состоит, то применяют один тип аналога) 5-йодцитидина, а аналог уридина содержит или состоит из (например, в случае, когда состоит, то применяют один тип аналога) 5-йодуридина.
В некоторых вариантах осуществления изобретения в любом из вышеупомянутых случаев процент аналогов данного нуклеотида относится к исходному проценту (например, к проценту аналогов в исходной реакции, такой как исходная реакция транскрипции in vitro). В некоторых вариантах осуществления изобретения в любом из вышеупомянутых случаев процент аналогов данного нуклеотида относится к выходу (например, к проценту в синтезированном или транскрибированном соединении).
Молекулы РНК/молекулы полирибонуклеотидов, предлагаемые в настоящем изобретении, можно получать рекомбинантно в системах in vivo с помощью методов, известных специалистам в данной области, которые более подробно дополнительно описаны ниже.
Альтернативно этому, модифицированные молекулы полирибонуклеотидов, предлагаемых в настоящем изобретении, можно получать в системе in vitro, используя, например, систему транскрипции in vitro, которая более подробно дополнительно описана ниже. Для системы транскрипции in vitro для получения молекул РНК/полирибонуклеотидов требуется исходная смесь модифицированных и немодифицированных нуклеозидтрифосфатов для получения модифицированных молекул РНК/полирибонуклеотидов с требуемыми согласно настоящему изобретению свойствами. В некоторых вариантах осуществления изобретения от 5 до 50% цитидинов представляют собой аналоги цитидина в указанной исходной смеси и от 5 до 50% уридинов представляют собой аналоги уридина в указанной исходной смеси. В некоторых вариантах осуществления изобретения от 5 до 40% цитидинов представляют собой аналоги цитидина в указанной исходной смеси и от 5 до 40% уридинов представляют собой аналоги уридина в указанной исходной смеси. В некоторых вариантах осуществления изобретения от 5 до 30% цитидинов представляют собой аналоги цитидина в указанной исходной смеси и от 5 до 30% уридинов представляют собой аналоги уридина в указанной исходной смеси. В некоторых вариантах осуществления изобретения от 5 до 30% цитидинов представляют собой аналоги цитидина в указанной исходной смеси и от 10 до 30% уридинов представляют собой аналоги уридина в указанной исходной смеси. В некоторых вариантах осуществления изобретения от 5 до 20% цитидинов представляют собой аналоги цитидина в указанной исходной смеси и от 5 до 20% уридинов представляют собой аналоги уридина в указанной исходной смеси. В некоторых вариантах осуществления изобретения от 5 до 10% цитидинов представляют собой аналоги цитидина в указанной исходной смеси и от 5 до 10% уридинов представляют собой аналоги уридина в указанной исходной смеси. В некоторых вариантах осуществления изобретения 25% цитидинов представляют собой аналоги цитидина в указанной исходной смеси и 25% уридинов представляют собой аналоги уридина в указанной исходной смеси. В некоторых вариантах осуществления изобретения исходная смесь не содержит аналоги аденозина и/или гуанозина. В других вариантах осуществления изобретения необязательно исходная смесь содержит один или несколько аналогов аденозина и/или гуанозина (или не один из них, или оба).
В некоторых вариантах осуществления изобретения процент цитидинов в исходной смеси, представляющих собой аналоги цитидина, не равен проценту уридинов в исходной смеси, представляющих собой аналоги уридина. В некоторых вариантах осуществления изобретения процент аналогов цитидина в исходной смеси ниже, чем процент аналогов уридина в исходной смеси. Как указано выше, аналоги аденозина и гуанозина могут присутствовать или отсутствовать в исходной смеси, но в некоторых вариантах осуществления изобретения аналоги аденозина и аналоги гуанозина отсутствуют в исходной смеси.
В некоторых вариантах осуществления изобретения исходная смесь нуклеотидов для применения с системе транскрипции in vitro, в которой получают молекулу РНК/полирибонуклеотид, предлагаемую/предлагаемый в настоящем изобретении, содержит аналоги цитидина и аналоги уридина, и от 5 до 20% цитидинов в исходной смеси представляют собой аналоги цитидина и от 25 до 45% уридинов в исходной смеси представляют собой аналоги уридина. Другими словами, исходная смесь содержит модифицированные и немодифицированные цитидины и модифицированные и немодифицированные уридины и от 5 до 20% цитидинов в исходной смеси приходится на долю аналогов цитидина, а от 25 до 45% уридинов в исходной смеси приходится на долю аналогов уридина. В других вариантах осуществления изобретения исходная смесь содержит от 5 до 10% аналогов цитидина и от 30 до 40% аналогов уридина, например, 7-9% аналогов цитидина, например, 7, 7,5 или 8%, и, например, 32-38% аналогов уридина, например, 33, 34, 35, 36%.
В некоторых вариантах осуществления изобретения можно применять любой из аналогов уридина и аналогов цитидина, указанных в настоящем описании, необязательно за исключением псевдоуридина. В некоторых вариантах осуществления изобретения аналог цитидина содержит или состоит из 5-йодцитидина (например, в случае, когда применяют один тип аналога С), а аналог уридина содержит или состоит из 5-йодуридина (например, в случае, когда применяют один тип аналога U).
Примеры аналогов представлены в таблице выше. Должно быть очевидно, что для модифицированных полирибонуклеотидов, кодирующих требуемый полипептид (модуль (а)), аналоги и уровень модификации касаются, если не указано иное, полного полирибонуклеотида, который кодирует требуемый полипептид (модуль (а)), включая 5'- и 3'-нетранслируемые области (например, уровень модификации определяется исходными соотношениями аналогов в реакции транскрипции in vitro, при этом, аналоги могут быть включены в положения, которые транскрибируются).
Кроме того, модифицированные молекулы РНК/молекулы полирибонуклеотидов можно химически синтезировать, например, с помощью общепринятого химического синтеза с использованием автоматического синтезатора нуклеотидных последовательностей с применением твердофазной подложки и стандартных методик или с помощью химического синтеза соответствующих последовательностей ДНК и последующей их транскрипции in vitro или in vivo.
В молекулярной биологии и генетике понятия «в обратном направлении» и «в прямом направлении» оба относятся к относительному положению в молекуле РНК. В контексте настоящего изобретения в обратном направлении обозначает в направлении 5'-конца молекулы РНК, а в прямом направлении обозначает в направлении 3'-конца молекулы.
Таким образом, согласно настоящему изобретению UTR, указанная в пункте (б), выше, локализована непосредственно в обратном направлении относительно кодирующей области, указанной в пункте (а), более конкретно непосредственно в обратном направлении относительно стартового кодона кодирующей области. Так, «непосредственно в обратном направлении» в данном контексте означает, что нет никакого(их) другого(их) нуклеотида(ов) между последовательностью, указанной в пункте (б), и кодирующей последовательностью, которая начинается со стартового кодона. Таким образом, кодирующая область, которая начинается со стартового кодона, непосредственно примыкает к указанной последовательности UTR.
Молекула РНК может присутствовать в форме слитых РНК-последовательностей модулей (а) и (б) (которые указаны выше в пунктах (а) и (б) соответственно), т.е. (слитая) молекула РНК, которая образуется в результате экспрессии гибридного гена, создается путем объединения по меньшей мере двух нуклеотидных последовательностей, кодирующих указанные модули. Как правило, что будет более подробно объяснено ниже, это можно осуществлять путем клонирования кДНК в экспрессионном векторе, который обеспечивает транскрипцию в молекулу РНК. Таким образом, молекулу ДНК, кодирующую молекулу РНК, предлагаемую в настоящем изобретении, можно сливать с последовательностью ДНК, т.е. получать химерную молекулу, образованную путем соединения двух или большего количества полинуклеотидов через фосфатную группу одного нуклеотида, связанную с 3'-углеродом другого нуклеотида, с образованием фосфодиэфирной связи между соответствующими концами одного модуля и концом другой молекулы. Таким путем указанные выше молекулы ДНК, кодирующие указанные по меньшей мере два модуля, объединяют в форме молекулы ДНК. Указанную рекомбинантную молекулу ДНК затем транскрибируют в соответствующую нуклеотидную последовательность РНК.
В одном из предпочтительных вариантов осуществления изобретения R2 выбирают из группы, состоящей из:
(I) GGGAGA (SEQ ID NO: 7);
(II) GGGAGA (SEQ ID NO: 8);
(III) GAAG (SEQ ID NO: 9); и
(IV) GGGA (SEQ ID NO: 10).
В предпочтительном варианте осуществления изобретения молекула РНК, содержащая последовательность R2-CNGCCACC (SEQ ID NO: 2), представляет собой молекулу РНК, в которой нуклеотид N в положении 2 SEQ ID NO: 2 представляет собой нуклеотид, выбранный из группы, состоящей из U, G или С, и в которой нуклеотид N не представляет собой А.
В другом предпочтительном варианте осуществления изобретения указанный нуклеотид N в положении 2 SEQ ID NO: 2 представляет собой U.
В предпочтительном варианте осуществления изобретения молекула РНК, предлагаемая в настоящем изобретении, представляет собой молекулу РНК, в которой нуклеотид, расположенный непосредственно в прямом направлении относительно стартового кодона, не представляет собой нуклеотид G. В другом предпочтительном варианте осуществления изобретения молекула РНК, предлагаемая в настоящем изобретении, представляет собой молекулу РНК, в которой нуклеотид, расположенный непосредственно в прямом направлении относительно стартового кодона, представляет собой нуклеотид, выбранный из группы, состоящей из A, U и С.
В еще более предпочтительном варианте осуществления изобретения молекула РНК, предлагаемая в настоящем изобретении, представляет собой молекулу РНК, которая содержит модуль (б1), указанный выше, где указанный модуль (б1) представляет собой последовательность, в которой С в положении 6 SEQ ID NO: 1 заменен на А и С в положении 7 SEQ ID NO: 1 заменен на G; и/или А в положении 5 SEQ ID NO: 1 заменен на G, и в которой нуклеотид, расположенный непосредственно в прямом направлении относительно стартового кодона, представляет собой нуклеотид, выбранный из группы, состоящей из A, U и С.
В другом еще более предпочтительном варианте осуществления изобретения молекула РНК, предлагаемая в настоящем изобретении, представляет собой молекулу РНК, которая содержит модуль (б2), указанный выше, где указанный модуль (б2) представляет собой последовательность, в которой С в положении 7 SEQ ID NO: 2 заменен на А и С в положении 8 SEQ ID NO: 2 заменен на G; и/или А в положении 6 SEQ ID NO: 2 заменен на G и в которой нуклеотид, расположенный непосредственно в прямом направлении относительно стартового кодона, представляет собой нуклеотид, выбранный из группы, состоящей из A, U и С.
Как отмечалось выше, консенсусная последовательность Козак (gcc)gccRccAUGG может, среди прочего, быть вариантной касательно нуклеотида в положении -3 (т.е. на расстоянии 3 нуклеотидов в обратном направлении относительно стартового кодона AUG), ее обозначают как «R», если в этом положении находится пурин (т.е. аденин или гуанин). В описанных выше UTR нуклеотид, соответствующий этому положению, представляет собой «А». Однако настоящее изобретение относится также к молекулам РНК, содержащим соответствующую UTR, в которой в этом положении находится «G».
Таким образом, в предпочтительном варианте осуществления изобретения молекула РНК, предлагаемая в настоящем изобретении, содержит UTR, указанную выше в подпункте (б1), где в UTR-последовательности, указанной в (б1), А в положении 5 SEQ ID NO: 1 заменен на G; или в UTR-последовательности, указанной в (б2), А в положении 6 SEQ ID NO: 2 заменен нa G.
Как отмечалось выше, молекула РНК, предлагаемая в настоящем изобретении, может содержать также поли-А-хвост. В контексте настоящего описания понятие «поли-А-хвост» относится к последовательности адениновых нуклеотидов, локализованной на 3'-конце РНК. Поли-А-хвост, как правило, добавляется к 3'-концу РНК с помощью процесса, называемого полиаденилированием. Таким образом, настоящее изобретение относится к любой из вышеописанных РНК, где молекула РНК содержит поли-А-хвост на 3'-конце.
Длина поли-А-хвоста не ограничена конкретной величиной. Так, в предпочтительных вариантах осуществления изобретения молекула РНК, предлагаемая в настоящем изобретении, содержит поли-А-хвост на 3'-конце, где поли-А-хвост состоит по меньшей мере из 50, 60, 70, 80, 90, 100 или 110 нуклеотидов. В более предпочтительном варианте осуществления изобретения молекула РНК, предлагаемая в настоящем изобретении, содержит поли-А-хвост на 3'-конце, где поли-А-хвост состоит по меньшей мере из 120 нуклеотидов. В других предпочтительных вариантах осуществления изобретения молекула РНК, предлагаемая в настоящем изобретении, содержит поли-А-хвост на 3'-конце, где поли-А-хвост состоит по меньшей мере из 150, 200, 250, 300, 350, 400, 500, 600, 700, 800, 900 или 1000 нуклеотидов.
В случае, когда молекулу РНК, предлагаемую в настоящем изобретении, получают методом транскрипции in vitro, который дополнительно представлен ниже в настоящем описании, то поли-А-хвост локализован на 3'-конце РНК, примыкающем к UTR, на 3'-конце молекулы РНК, при этом плазмиду, которая несет молекулу РНК, предлагаемую в настоящем изобретении, линеаризуют перед транскрипцией in vitro в прямом направлении относительной поли-А-хвоста для того, чтобы гарантировать, что транскрибируемая in vitro молекула РНК будет содержать указанный поли-А-хвост.
Как указано выше, молекула РНК, предлагаемая в настоящем изобретении, может присутствовать в форме слитых последовательностей РНК в виде модулей (а) и (б), т.е. (слитой) молекулы РНК, полученной в результате транскрипции гибридного гена, созданного путем объединения по меньшей мере двух нуклеотидных последовательностей, кодирующих указанные модули. Как правило, это осуществляют путем клонирования кДНК в экспрессионном векторе, который обеспечивает транскрипцию полной молекулы РНК. Известны различные методы создания слитых конструкций, включая синтез, гибридизацию и/или амплификацию нуклеиновых кислот с получением синтетической двухцепочечной молекулы нуклеиновой кислоты, которая «кодирует» молекулу РНК, предлагаемую в настоящем изобретении. Указанная двухцепочечная молекула нуклеиновой кислоты (т.е. молекула ДНК) несет на одной цепи (т.е. на кодирующей или смысловой цепи) последовательность ДНК, соответствующую молекуле РНК, предлагаемой в настоящем изобретении, и, следовательно, «кодирует» молекулу РНК, предлагаемую в настоящем изобретении. Другими словами, указанная двухцепочечная молекула нуклеиновой кислоты/ДНК содержит на одной цепи генетическую информацию, которая соответствует транскрибируемой молекуле РНК, предлагаемой в настоящем изобретении, которая указана выше в настоящем описании. Понятие «кодировать» или «кодирующая» в контексте настоящего изобретения применяют не только в его общепринятом значении, т.е. касательно ДНК гена, который кодирует белок (и, следовательно, генетическая информация которого может транслироваться в полипептид или аминокислотную последовательность белка). Точнее в понятиях, применяемых в настоящем изобретении, в конструкции, в которой индивидуальные последовательности ДНК, кодирующие модули (а) и (б), «слиты» или связаны в одну (химерную) молекулу ДНК, конструкция содержит также компонент (т.е. модуль (б)), который не транслируется в белок. При этом, последовательность ДНК, соответствующая модулю (б), дает информацию, т.е. «кодирует» структуру 5' UTR, и поэтому понятие «кодирует» в настоящем изобретении относится также к генетической информации о UTR, которые могут экспрессироваться, т.е. транскрибироваться, если, например, присутствуют в двухцепочечной молекуле нуклеиновой кислоты. Таким образом, понятие «кодирует» в контексте настоящего изобретения, хотя его, как привило, применяют только касательно кодирования/экспрессии белка, следует понимать таким образом, что молекула нуклеиновой кислоты может транскрибироваться в соответствующую молекулу РНК, которая несет части, кодирующие белок или полипептид (т.е. модуль (а)), и части «кодирующие» UTR (т.е. модуль (б)), при этом последние представляют собой конечный продукт при экспрессии, поскольку UTR не транслируются в белки или полипептиды. Указанную двухцепочечную нуклеиновую кислоту можно встраивать в экспрессионные векторы стандартными методами молекулярной биологии (см., например, Sambrook и др., Molecular Cloning, A laboratory manual, 2-ое изд., 1989). Под понятием «вектор», таким как «экспрессионный вектор» или «клонирующий вектор», в контексте настоящего изобретения подразумевается кольцевая двухцепочечная единица ДНК, которая реплицируется в клетке вне зависимости от хромосомной ДНК и которую применяют в качестве носителя для переноса генетического материала в клетку, в которой она может реплицироваться и/или экспрессироваться (т.е. транскрибироваться в РНК и транслироваться в аминокислотную последовательность). Вектор, содержащий чужеродную ДНК, обозначают как рекомбинантная ДНК. Вектор сам, как правило, представляет собой ДНК-последовательность, которая обычно состоит из вставки (т.е. модуля (б), который не транслируется в белок, и модуля (а), т.е. кодирующей области) и более крупной последовательности, которая служит в качестве «каркаса» вектора. Плазмиды в контексте настоящего изобретения наиболее часто присутствуют в бактериях и их применяют в исследованиях рекомбинантной ДНК для переноса генов между клетками, и они рассматриваются как субпопуляция «векторов» в контексте настоящего изобретения.
Таким образом, настоящее изобретение относится также к молекуле нуклеиновой кислоты, которая кодирует молекулу РНК, предлагаемую в настоящем изобретении.
Нуклеиновая кислота представляет собой, например, ДНК, которая кодирует два основных модуля (т.е. модуль (а) и модуль (б)) молекулы РНК, предлагаемой в настоящем изобретении. Вышеуказанная молекула нуклеиновой кислоты, предлагаемая в настоящем изобретении, предпочтительно представляет собой рекомбинантную молекулу нуклеиновой кислоты. Молекула нуклеиновой кислоты, предлагаемая в изобретении, может быть синтетической или полусинтетической.
Специалисту в данной области должно быть очевидно, что дополнительные регуляторные последовательности можно добавлять к молекуле нуклеиновой кислоты, предлагаемой в изобретении, которая кодирует молекулу РНК. Например, можно применять энхансеры транскрипции и/или последовательности, которые обеспечивают индуцибельную экспрессию. Приемлемой системой индуцибельной экспрессии является например, регулируемая геном тетрациклина экспрессии, описанная, например, у Gossen и Bujard, Proc. Natl. Acad. Sci. USA 89, 1992, cc. 5547-5551) и у Gossen, Trends Biotech. 12, 1994, cc. 58-62, или индуцируемая геном дексаметазона система, описанная, например, у Crook, EMBO J. 8, 1989, сс. 513-519.
Настоящее изобретение относится также к вектору, предпочтительно, экспрессионному вектору, который содержит молекулу нуклеиновой кислоты, предлагаемую в настоящем изобретении.
Касательно векторов, содержащих молекулу нуклеиновой кислоты, которая кодирует молекулу РНК, предлагаемую в настоящем изобретении, то к ним применимы с необходимыми изменениями условия, описанные выше в контексте векторов, которые содержат молекулу ДНК, предлагаемую в настоящем изобретении.
Настоящее изобретение относится также к клетке-хозяину, которая содержит вектор, предлагаемый в настоящем изобретении. Таким образом, настоящее изобретение относится к хозяину, трансфектированному или трансформированному вектором, предлагаемым в изобретении, или хозяину, не представляющему собой человека, который несет вектор, предлагаемый в настоящем изобретении, т.е. к клетке-хозяину или хозяину, генетически модифицированной/модифицированному молекулой нуклеиновой кислоты, предлагаемой в изобретении, или вектором, который содержит указанную молекулу нуклеиновой кислоты.
Касательно клетки-хозяина, которая содержит вектор, содержащий молекулу нуклеиновой кислоты, кодирующую молекулу РНК, предлагаемую в настоящем изобретении, то к ней применимы с необходимыми изменениями такие же условия, которые описаны выше в контексте клеток-хозяев, которые содержат векторы, содержащие описанную выше молекулу ДНК, предлагаемую в настоящем изобретении.
Настоящее изобретение относится также к способам получения молекулы РНК, предлагаемой в настоящем изобретении, путем культивирования клетки-хозяина, несущей экспрессионный вектор, который кодирует индивидуальные модули, предлагаемые в настоящем изобретении, или полную молекулу РНК, предлагаемую в изобретении, в культуральной среде и выделения молекулы РНК из клетки-хозяина или культуральной среды. Настоящее изобретение может относиться также к способу получения молекулы РНК, предлагаемой в настоящем изобретении, включающему культивирование клетки-хозяина, предлагаемой в настоящем изобретении, и необязательно выделение молекулы РНК из культуры.
Методы выделения и/или последующей очистки молекулы РНК, предлагаемой в настоящем изобретении, известны специалисту в данной области.
Настоящее изобретение относится также к способам получения молекулы РНК, предлагаемой в настоящем изобретении, с использованием реакции in vitro с помощью методов, известных специалисту в данной области. Более конкретно, молекулу РНК, предлагаемую в настоящем изобретении, можно получать in vitro с использованием системы транскрипции in vitro. Системы транскрипции in vitro хорошо известны и, как правило, для них требуется очищенная линейная ДНК-матрица, содержащая последовательность ДНК, которая «кодирует» модуль (б) и модуль (а), описанные выше, в который указанная последовательность ДНК находится под контролем соответствующего промотора. Кроме того, для системы транскрипции in vitro также обычно требуются рибонуклеотидтрифосфаты, буферная систем, которая включает ДТТ и ионы магния, и соответствующая РНК-полимераза, которая обладает ферментативной активностью для обеспечения транскрипции in vitro последовательности ДНК в молекулу РНК, предлагаемую в настоящем изобретении.
Методы, которые обычно применяют для получения молекул РНК с использованием транскрипции in vitro, хорошо известны специалисту в данной области и описаны, например, в Methods Mol. Biol. 703, 2011, сс. 29-41.
Как указано выше, в случае, когда молекулу РНК, предлагаемую в настоящем изобретении, получают с помощью метода транскрипции in vitro, который дополнительно будет описан ниже, то вышеуказанный поли-А-хвост может представлять собой часть молекулы РНК, предлагаемой в настоящем изобретении (и он необязательно исходно локализован в клонирующем векторе), и он локализован на 3'-конце РНК, например, примыкает к UTR на 3'-конце молекулы РНК. В случае, когда молекулу РНК, предлагаемую в настоящем изобретении, получают с помощью метода транскрипции in vitro, то плазмиду, которая несет молекулу РНК, предлагаемую в настоящем изобретении, линеаризуют перед транскрипцией in vitro в прямом направлении относительно поли-А-хвоста для гарантии того, что транскрибируемая in vitro молекула РНК будет содержать указанный поли-А-хвост.
Альтернативно этому, молекулу РНК, предлагаемую в настоящем изобретении, можно также синтезировать химически, например, путем общепринятого химического синтеза с использованием автоматического синтезатора нуклеотидных последовательностей, применяя твердофазную подложку и стандартные методики.
Настоящее изобретение относится также к способам получения с использованием реакции in vitro молекулы РНК, предлагаемой в настоящем изобретении, с помощью методов, известных специалисту в данной области, и изложенных выше, и выделения молекулы РНК из реакционной смеси.
Методы выделения и/или последующей очистки молекулы РНК, предлагаемой в настоящем изобретении, известны специалисту в данной области.
Молекулу РНК, предлагаемую в настоящем изобретении, можно легко применять в системах трансляции in vitro, известных в данной области, для эффективной экспрессии любого требуемого полипептида или белка, кодируемого кодирующей областью модуля (а).
Системы трансляции in vitro известны в данной области и их можно непосредственно использовать в сочетании с молекулой РНК, предлагаемой в настоящем изобретении. Альтернативно этому, указанные системы трансляции in vitro можно объединять с описанными выше системами транскрипции in vitro. Соответствующие бесклеточные системы для in vitro транскрипции и/или in vitro трансляции известны и доступны. Эти бесклеточные системы для синтеза белков (который обозначают также как in vitro синтез белков или сокращенно CFPS) обеспечивают экспрессию/получение полипептида или белка с использованием биологического механизма без применения живых клеток. В этих системах окружение системы синтеза белка in vitro не ограничено клеточной стенкой или условиями гомеостаза, необходимыми для поддержания жизнеспособности клеток, и обеспечивает непосредственный доступ и контроль условий трансляции, которые обладают преимуществами для нескольких вариантов применения, включая оптимизацию производства белка, оптимизацию комплексообразования белка, изучение синтеза белка, включение не встречающихся в естественных условиях аминокислот, широкомасштабный скрининг и синтетическую биологию. Обычные компоненты бесклеточной реакции включают клеточный экстракт, источник энергии, источник поступления аминокислот, кофакторы, такие как магний, и ДНК или РНК, кодирующую требуемый полипептид или белок. Клеточный экстракт можно получать путем лизиса представляющих интерес клеток и центрифугирования для отделения клеточных стенок, геномной ДНК и другого клеточного дебриса. Оставшийся продукт представляет собой необходимые для клетки структуры, включая рибосомы, аминоацил-тРНК-синтетазы, факторы инициации и элонгации трансляции, нуклеазы и т.д. В бесклеточной системе для синтеза полипептидов или белков, начинающегося с ДНК (т.е. в системе, которая включает стадию in vitro транскрипции и in vitro трансляции), как правило, применяют два типа ДНК, т.е. либо плазмиды, либо линейные матрицы для экспрессии (LET). В бесклеточной системе для синтеза полипептидов или белков, начинающегося с РНК (т.е. в системе, которая включает только стадию in vitro трансляции), РНК можно применять непосредственно. Для указанных бесклеточных реакций in vitro требуется источник энергии, который, как правило, представляет собой отдельную смесь, которая содержит необходимый источник энергии, наряду с источником поступления аминокислот, которые добавляют к экстракту в реакционную смесь. Общепринятыми источниками энергии являются фосфоенолпируват, ацетилфосфат и креатинфосфат. Общепринятыми клеточными экстрактами, которые обычно используют, являются экстракты, полученные из Escherichia coli (ЕСЕ), кроличьих ретикулоцитов (RRL), ростков пшеницы (WGE) и клеток насекомых (ICE). Все указанные экстракты поступают в продажу.
Таким образом, настоящее изобретение относится также к применению молекулы РНК, предлагаемой в настоящем изобретении, для in vitro трансляции требуемого полипептида или белка, кодируемого кодирующей областью, которая содержится в указанной молекуле РНК.
В предпочтительных вариантах осуществления изобретения, касающихся применения молекулы РНК, предлагаемой в настоящем изобретении, применение является таким же с необходимыми изменениями, которое описано выше в контексте молекулы РНК, указанной выше.
Молекулы РНК, указанные выше, наиболее предпочтительно применять при медицинских показаниях и для лечения определенного заболевания и, прежде всего, в терапиях на основе РНК. Таким образом, настоящее изобретение относится также к фармацевтической композиции, которая содержит молекулу РНК, предлагаемую в настоящем изобретении, молекулу нуклеиновой кислоты, предлагаемую в настоящем изобретении, вектор, предлагаемый в настоящем изобретении, или клетку-хозяина, предлагаемую в настоящем изобретении, и необязательно фармацевтически приемлемый носитель.
Понятие «лечение» и подобные понятия применяют в контексте настоящего описания, как правило, в качестве средства для достижения требуемого фармакологического и/или физиологического действия. Таким образом, лечение согласно настоящем изобретению может относиться к лечению (острых) состояний определенного заболевания, но может относиться также к профилактическому лечению в понятиях полного или частичного предупреждения заболевания или его симптома. Предпочтительно понятие «лечение» следует рассматривать как терапевтическое в понятиях полного или частичного излечения заболевания и/или побочного действия, и/или симптомов, связанных с заболеванием. «Острое» в этом контексте означает, что у индивидуума проявляются симптомы заболевания. Другими словами, подлежащий лечению индивидуум остро нуждается в лечении, и понятие «лечение острого состояния (неотложное лечение)» в контексте настоящего изобретения относится к мерам, предпринимаемым для неотложного лечения заболевания после начала заболевания или рецидива заболевания. Лечение может также быть профилактическим или превентивным лечением, т.е. относиться к мерам предупреждения заболевания, например, для предупреждения инфекции и/или начала заболевания.
Фармацевтическую композицию, предлагаемую в настоящем изобретении, можно применять с использованием широкого диапазона классов форм введения, известных специалисту в данной области. Введение может быть системным, местным, оральным, с помощью аэрозоля, с использованием таблеток, игольной инъекции, ингаляторов, кремов, пен, гелей, лосьонов, мазей (но не ограничиваясь только ими).
Как указано, настоящее изобретение относится к фармацевтической композиции, содержащей в эффективном количестве молекулу РНК (или молекулу нуклеиновой кислоты, вектор или клетку-хозяина), предлагаемую в настоящем изобретении, которая описана выше, и по меньшей мере один фармацевтически приемлемый эксципиент или носитель.
Эксципиент или носитель представляет собой неактивную субстанцию, включенную в композицию наряду с действующим веществом, т.е. молекулой РНК (или молекулой нуклеиновой кислоты, вектором или клеткой-хозяином), предлагаемой в настоящем изобретении, с целью увеличения объема композиций, которые содержат эффективные действующие вещества. Эксципиенты часто обозначают как «наполнители», «присадки» или «разбавители». Увеличение объема обеспечивает удобство и точность распределения лекарственной субстанции при приготовлении лекарственной формы. Они могут служить также различным повышающим терапевтическое действие целям, таким как облегчение абсорбции или растворимости лекарственного средства, или другим фармакокинетическим требованиям. Эксципиенты могут также оказывать благоприятное действие на процесс производства, облегчать обращение с действующим веществом, например, повышать текучесть порошка или обеспечивать снижение прилипания, в дополнение к повышению стабильности in vitro, например, в результате предупреждения денатурации в течение ожидаемого срока хранения. Выбор соответствующих эксципиентов зависит также от пути введения и лекарственной формы, а также от действующего вещества и других факторов.
Таким образом, фармацевтическая композиция, содержащая в эффективном количестве молекулу РНК (или молекулу нуклеиновой кислоты, вектор или клетку-хозяина), предлагаемую в настоящем изобретении, может быть твердой, жидкой или газообразной и может среди прочего иметь форму порошка(ов), таблетки(ок), раствора(ов) или аэрозоля(ей). Предпочтительно, чтобы указанная фармацевтическая композиция необязательно содержала фармацевтически приемлемый носитель и/или разбавитель.
Примеры приемлемых фармацевтических носителей, эксципиентов и/или разбавителей хорошо известны в данной области и включают забуференные фосфатом физиологические растворы, воду, эмульсии, такие как эмульсии типа масло/вода, различные типы смачивающих агентов, стерильные растворы и т.д. Композиции, содержащие указанные носители, можно приготавливать хорошо известными общепринятыми методами. Указанные фармацевтические композиции можно вводить индивидууму в приемлемой дозе, т.е. в «эффективном количестве», которое легко может определять специалист в данной области известными в данной области методами. Режим дозирования должен определять лечащий врач, и он зависит от клинических факторов. Как хорошо известно в области медицины, дозы для любого пациента зависят от многих факторов, включая размер пациента или индивидуума, площадь поверхности тела, возраст, конкретное вводимое соединение, пол, время и путь введения, общее состояние здоровья и другие лекарственные средства, которые применяют одновременно.
Таким образом, предпочтительно молекулу РНК (или молекулу нуклеиновой кислоты, вектор или клетку-хозяина), предлагаемую в настоящем изобретении, включают в эффективном количестве. Понятие «эффективное количество» относится к количеству, достаточному для индукции заметного терапевтического ответа у индивидуума, которому вводят терапевтическую композицию. Согласно вышеизложенному содержание молекулы РНК (или молекулы нуклеиновой кислоты, вектора или клетки-хозяина), предлагаемой в настоящем изобретении, в фармацевтической композиции не ограничено конкретно, если его достаточно для указанного выше лечения, но предпочтительно составляет 0,0000001-10% в пересчете на массу всей композиции. Кроме того, молекулу РНК (или молекулу нуклеиновой кислоты, вектор или клетку-хозяина), указанную в настоящем описании, предпочтительно применяют в носителе. Как правило, в носителе применяют соответствующее количество фармацевтически приемлемой соли для придания композиции изотоничности. Примерами носителей являются (но не ограничиваясь только ими) физиологический раствор, раствор Рингера и раствор декстрозы. Предпочтительно приемлемые эксципиенты, носители или стабилизаторы являются нетоксичными в применяемых дозах и концентрациях, и они включают буферы, такие как цитратный, фосфатный, и другие органические кислоты; солеобразующие противоионы, например, натрий и калий; низкомолекулярные (>10 аминокислотных остатков) полипептиды; белки, например сывороточный альбумин или желатин; гидрофильные полимеры, например, поливинилпирролидон; аминокислоты, такие как гистидин, глутамин, лизин, аспарагин, аргинин или глицин; углеводы, включая глюкозу, маннозу или декстрины; моносахариды, дисахариды; другие сахара, например, сахароза, маннит, трегалоза или сорбит; хелатирующие агенты, например, ЭДТА; неионогенные поверхностно-активные вещества, например, Твин, плюроники или полиэтиленгликоль; антиоксиданты, включая метионин, аскорбиновую кислоту и токоферол; и/или консерванты, например, октадецилдиметилбензиламмонийхлорид; гексаметонийхлорид; бензалконийхлорид, бензетонийхлорид; фенол, бутиловый или бензиловый спирт; алкилпарабены, например, метил- или пропилпарабен; катехол; резокринол; циклогексанол; 3-пентанол и м-крезол). Приемлемые носители и их формы описаны более подробно в  Pharmaceutical Sciences, 17-ое изд., изд-во Mack Publishing Со, 1985.
Pharmaceutical Sciences, 17-ое изд., изд-во Mack Publishing Со, 1985.
Можно осуществлять мониторинг успеха терапевтической обработки путем периодической оценки. Молекула РНК (или молекула нуклеиновой кислоты, вектор или клетка-хозяин), предлагаемая в настоящем изобретении, или фармацевтическая композиция, предлагаемая в изобретении, может иметь форму стерильных водных или неводных растворов, суспензий и эмульсий, а также кремов и суппозиториев. Примерами неводных растворителей являются пропиленгликоль, полиэтиленгликоль, растительные масла, такие как оливковое масло, и органические сложные эфиры, такие как этилолеат. Водные носители включают воду, спиртовые/водные растворы, эмульсии или суспензии, включая соляной раствор и забуференные среды. Могут присутствовать также консерванты и другие добавки, такие, например, как противомикробные средства, антиоксиданты, хелатирующие агенты и инертные газы и т.п. Кроме того, фармацевтическая композиция, предлагаемая в изобретении, может содержать также другие агенты в зависимости от предполагаемого применения фармацевтической композиции. Указанные агенты могут представлять собой, например, полиоксиэтиленсорбитанмонолаурат, поступающий в продажу под торговым знаком Твин, пропиленгликоль, ЭДТА, цитрат, сахарозу, а также другие агенты, пригодные для требуемого применения фармацевтической композиции, которые хорошо известны специалисту в данной области.
Согласно настоящему изобретению понятие «фармацевтическая композиция» относится к композиции, предназначенной для введения пациенту, предпочтительно человеку.
Фармацевтическую композицию, предлагаемую в настоящем изобретении, можно применять в терапиях на основе РНК. Как указано выше, молекула РНК, предлагаемая в настоящем изобретении, которая содержит «кодирующую область, кодирующую полипептид», можно применять в терапиях на основе РНК, в которых «кодирующая область, кодирующая полипептид», кодирует терапевтически или фармацевтически активный полипептид или белок, обладающий терапевтическим или профилактическим действием. Таким образом, в предпочтительных вариантах осуществления изобретения фармацевтическую композицию, предлагаемую в настоящем изобретении, можно применять в терапиях на основе РНК для лечения или предупреждения заболевания, указанного выше в таблице 1. Таким образом, терапии на основе РНК, предлагаемые в настоящем изобретении, можно применять для лечения или предупреждения заболевания, указанного выше в таблице 1.
Таким образом, фармацевтическую композицию, предлагаемую в настоящем изобретении, можно применять в терапиях на основе РНК, в случаях, когда дефекты генов, указанные выше в таблице 1, приводят к заболеванию, которое можно лечить или предупреждать с помощью транскрипт-заместительной терапии/фермент-заместительной терапии с использованием молекулы РНК, предлагаемой в настоящем изобретении, где молекула РНК содержит «кодирующую полипептид область», которая кодирует интактную версию белка или его функциональный фрагмент, которые компенсируют указанный дефектный ген. В наиболее предпочтительных вариантах осуществления изобретения фармацевтическую композицию, предлагаемую в настоящем изобретении, можно применять в терапиях на основе РНК для лечения или предупреждения лизосомных болезней типа болезни Гоше, болезни Фабри, MPS I, MPS II (синдром Гунтера), MPS VI и болезней накопления гликогена, таких, например, как болезнь накопления гликогена типа I (болезнь фон-Гирке), типа II (болезнь Помпе), типа III (болезнь Кори), типа IV (болезнь Андерсена), типа V (болезнь Мак-Ардли), типа VI (болезнь Герса), типа VII (болезнь Таури), типа VIII, типа IX, типа X, типа XI (синдром Фанкони-Бикеля), типа XI или типа 0. Благоприятное действие транскрипт-замещающих терапий/фермент-замещающих терапий не влияет на лежащий в основе генетический дефект, но повышают концентрацию фермента, дефицит которого имеет место у пациента. Так, например, при болезни Помпе транскрипт-замещающая терапия/фермент-замещающая терапия обеспечивает пополнение дефицита лизосомного фермента кислой альфа-глюкозидазы (GAA).
В других предпочтительных вариантах осуществления изобретения фармацевтическую композицию, предлагаемую в настоящем изобретении, можно применять в терапиях на основе РНК, предлагаемых в настоящем изобретении, в которых «кодирующая область, которая кодирует полипептид», кодирует терапевтически или фармацевтически активный полипептид, белок или пептид, обладающий терапевтическим или профилактическим действием, где указанный полипептид, белок или пептид выбран из группы молекул, которые кодируются генами, указанными в таблице 1.
В других предпочтительных вариантах осуществления изобретения терапии на основе РНК, предлагаемые в настоящем изобретении, можно применять для лечения рака, сердечно-сосудистых заболеваний, вирусной инфекции, иммунной дисфункции, аутоиммунного заболевания, неврологического нарушения, наследственных метаболических нарушений или генетического нарушения, или любого заболевания, при котором продуцируемый в клетке белок или белковый фрагмент может оказывать благоприятное действие на пациента. Примеры рака включают рак головы и шеи, рак молочной железы, рак почки, рак мочевого пузыря, рак легкого, рак предстательной железы, рак кости, рак головного мозга, рак шейки матки, анальный рак, рак ободочной кишки, колоректальный рак, рак аппендикса, рак глаза, рак желудка, лейкоз, лимфому, рак печени, рак кожи, рак яичника, рак пениса, рак поджелудочной железы, рак яичек, рак щитовидной железы, вагинальный рак, рак вульвы, рак эндометрия, рак сердца и саркому.
Примеры сердечно-сосудистых заболеваний включают атеросклероз, коронарную болезнь сердца, легочную болезнь сердца и кардиомиопатию.
Примеры иммунных дисфункций и аутоиммунных заболеваний включают (но не ограничиваясь только ими) ревматические болезни, рассеянный склероз и астму.
Примеры вирусных инфекций включают (но, не ограничиваясь только ими) инфекции, вызываемые вирусом иммунодефицита человека, вирусом герпеса простого, вирусом папилломы человека, а также вирусом гепатита В и С.
Примеры неврологических нарушений включают (но не ограничиваясь только ими) болезнь Паркинсона, рассеянный склероз и деменции.
Примеры наследственных метаболических нарушений включают (но не ограничиваясь только ими) болезнь Гоше и фенилкетонурию.
Изобретение относится также к способу осуществления основанной на РНК терапии. Так, настоящее изобретение относится к способу лечения заболевания, такого как рак, сердечно-сосудистое заболевание, вирусная инфекция, иммунная дисфункция, аутоиммунное заболевание, неврологическое нарушение, наследственные метаболические нарушения или генетическое нарушение, с помощью терапии на основе РНК. В предпочтительных вариантах осуществления изобретения способ лечения можно применять с необходимыми изменениями аналогично описанному выше в контексте молекулы РНК или фармацевтической композиции, предназначенной для применения в описанной выше терапии на основе РНК.
В предпочтительном варианте осуществления изобретения в качестве субъекта можно рассматривать млекопитающее, такое как собака, кошка, свинья, корова, лошадь, грызун, например, крыса, мышь и морская свинка, или примат, например, горилла, шимпанзе, и человек. В наиболее предпочтительном варианте осуществления изобретения субъект представляет собой человека.
Как отмечено выше, молекулы РНК, описанные выше, являются наиболее ценными для медицинских целей и для лечения определенного заболевания и, в частности, для терапий на основе РНК. Таким образом, настоящее изобретение относится также к фармацевтической композиции, которая содержит молекулу РНК, молекулу нуклеиновой кислоты, вектор или клетку-хозяина, предлагаемые в настоящем изобретении, и необязательно фармацевтически приемлемый носитель.
Кроме того, в терапиях на основе РНК часто требуется «выключение» («молчание») действия молекулы РНК на некоторой стадии.
Это можно осуществлять, например, с применением механизма РНКi (РНК-интерференция), с использованием цепи нуклеиновой кислоты, комплементарной UTR-последовательности, предлагаемой в настоящем изобретении. Фактически малый размер минимальных UTR, предлагаемых в настоящем изобретении, делает этот подход выполнимым, поскольку указанные UTR не образуют вторичных или третичных структур и не присутствуют в здоровых клетках. Таким образом, комплементарную цепь указанной UTR-последовательности можно с успехом применять в медицинских целях после лечения вышеуказанных заболеваний или после применения вышеуказанных терапий на основе РНК с использованием фармацевтической композиции, предлагаемой в настоящем изобретении, вызывая «выключение» терапевтических молекул РНК, предлагаемых в настоящем изобретении.
Таким образом, РНКi-подход также предусматривается в контексте настоящего изобретения для применения в препарате фармацевтической композиции для «выключения» действия терапевтических молекул РНК, предлагаемых в настоящем изобретении.
Понятие «РНК-интерференция» или «ингибирование РНК» (РНКi/iРНК) относится к применению двухцепочечной РНК для таргетинга расщепления специфических РНК, приводя тем самым к «выключению» их экспрессии. Предпочтительные ингибирующие молекулы РНК можно выбирать из группы, состоящей из двухцепочечной РНК (dsРНК), РНКi, siРНК, shРНК и stРНК. dsРНК, соответствующую последовательности гена, синтезируют in vitro и интродуцируют в клетку. dsРНК можно также интродуцировать в клетку в форме вектора, экспрессирующего последовательность гена-мишени в смысловой или антисмысловой ориентации, например, в форме мРНК-шпильки. Смысловые и антисмысловые последовательности можно также экспрессировать в различных векторах, при этом индивидуальные антисмысловые и смысловые молекулы образуют двухцепочечную РНК после их экспрессии. В данной области известно, что в некоторых случаях экспрессия последовательности в смысловой ориентации или даже промоторной последовательности достаточно для начала формирования dsРНК и затем siРНК благодаря внутренним механизмам амплификации в клетке. Таким образом, все средства и методы, которые приводят к снижению активности полипептида или белка, кодируемого кодирующей областью, можно применять согласно настоящему изобретению, например, смысловые конструкции, антисмысловые конструкции, конструкции в виде шпилек, смысловые и антисмысловые молекулы и их комбинации, можно применять для создания/интродукции указанных siРНК. dsРНК входит во встречающийся в естественных условиях, но пока не окончательно понятный процесс, включающий высоко консервативную нуклеазу Dicer, которая расщепляет молекулы-предшественники dsРНК с образованием коротких интерферирующих РНК (siРНК). Создание и получение siРНК, а также метод ингибирования экспрессии гена-мишени описаны, среди прочено в WO 02/055693, Wei, Dev. Biol. 15, 2000, сс. 239-255; La Count, Biochem. Paras. 111, 2000, cc. 67-76; Baker, Curr. Biol. 10, 2000, cc. 1071-1074; Svoboda, Development 127, 2000, cc. 4147-4156 или у Marie, Curr. Biol. 10, 2000, cc. 289-292. Затем указанные siРНК образуют последовательность-специфическую часть РНК-индуцируемого комплекса выключения гена (RISC), мнококомпонентую нуклеазу, которая расщепляет гомологи матричных РНК для запуска их выключения). У Elbashir, EMBO J. 20, 2001, сс. 6877-6888 продемонстрировано, что дуплексы состоящих из 21 нуклеотида РНК можно применять в клеточной культуре для препятствия экспрессии гена в клетках млекопитающих. Уже известно, что РНКi очень эффективно опосредуется siРНК в клетках млекопитающих, но создание стабильных клеточных линий или не относящихся к людям трансгенных животных оказалось ограниченным. Однако для стабильной экспрессии можно применять новые поколения векторов, например, РНК, образующие короткие шпильки (shРНК). Стабильная экспрессия siРНК в клетках млекопитающих продемонстрирована, среди прочего, у Brummelkamp Science 296, 2002, сс. 550-553. Кроме того, у Paul, Nat. Biotechnol. 20, 2002, сс. 505-508 продемонстрирована эффективная экспрессия малой интерферирующей РНК в клетках человека. РНК-интерференция в результате экспрессии коротких интерферирующих РНК и образующих шпильки РНК в клетках млекопитающих продемонстрирована также у Yu, PNAS 99, 2002, сс. 6047-6052. Подход на основе shРНК для «выключения» гена хорошо известен в данной области и может включать применение stРНК (малые временные РНК); см., среди прочего, у Paddison, Genes Dev, 16, 2002, сс. 948-958. Эти подходы могут основываться на применении векторов, например, в качестве иллюстрации можно применять вектор pSUPER или векторы РНК polIII, описанные, среди прочего, у Yu, 2002, loc. cit.; Miyagishi, 2002, loc. cit. или Brummelkamp, 2002, loc. cit. Предусматривается, что регуляторные последовательности, предлагаемые в настоящем изобретении, применяют путем, аналогичным пути применения систем на основе векторов pSUPER или РНК polIII.
Методы выведения и конструирования siРНК известны в данной области и описаны у Elbashir, Methods 26, 2002, сс. 199-213, на веб-сайтах в Интернете имеющихся на рынке продавцов siРНК, например, фирмы Qiagen GmbH (https://www1.qiagen.com/GeneGlobe/Default.aspx); фирмы Dharmacon (www.dharmacon.com); фирмы Xeragon Inc. (http://www.dharmacon.com/Default.aspx) и фирмы Ambion (www.ambion.com), или на веб-сайте исследовательской группы Tom Tuschl (http://www.rockefeller.edu/labheads/tuschl/sirna.html). Кроме того, доступны программы онлайн для выведения siРНК на основе данной последовательности мРНК (например, http://www.ambion.com/techlib/misc/siRNA_finder.html or http://katahdin.cshl.org:9331/RNAi/html/rnai.html). Остатки уридина во втором нуклеотиде (2-nt) выступающего 3'-конца можно заменять на 2'-дезокситимидин без утраты активности, что существенно снижает стоимость синтеза РНК и может также повышать устойчивость дуплексов siРНК при применении на клетках млекопитающих (Elbashir, 2001, loc. cit). siРНК можно синтезировать также ферментативно, используя РНК-полимеразу Т7 или другие РНК-полимеразы (Donze, Nucleic Acids Res 30, 2002: e46). Дуплексы коротких РНК, которые могут обусловливать эффективную РНК-интерференцию (esiРНК), можно получать также путем гидролиза с помощью RNазы III Escherichia coli (Yang, PNAS 99, 2002, cc. 9942-9947). Кроме того, разработаны экспрессионные векторы для экспрессии двухцепочечных siРНК, соединенных с помощью петель из образующих малую шпильку РНК, в эукариотических клетках (см., например, (Brummelkamp, Science 296, 2002, cc. 550-553). Все указанные конструкции можно создавать с помощью указанных выше программ. Кроме того, для предсказания последовательности siРНК можно применять поступающие в продажу инструменты для прогнозирования последовательностей, включенные в программы для анализа последовательности, или отдельно поступающие в продажу, например, инструмент для конструирования siРНК (siRNA Design Tool), доступный на сайте www.oligoEngine.com (Сиэтл, шт. Вашингтон).
Таким образом, специфические интерферирующие РНК можно применять согласно настоящему изобретению в качестве антагонистов/сайленсеров экспрессии и/или функции полипептида или белка, кодируемого кодирующей областью молекулы РНК, предлагаемой в настоящем изобретении. Указанные siРНК образуются комплементарной/антисмысловой и смысловой цепью, при этом, антисмысловая/смысловая цепь предпочтительно содержит по меньшей мере 10, более предпочтительно по меньшей мере 12, более предпочтительно по меньшей мере 14, более предпочтительно по меньшей мере 16, более предпочтительно по меньшей мере 18, более предпочтительно по меньшей мере 19, 20, 21 или 22 нуклеотида. В еще более предпочтительном варианте осуществления изобретения антисмысловая/смысловая цепь предпочтительно содержит 25 или более нуклеотидов.
Как указано выше, методы получения siРНК, которые можно применять согласно настоящему изобретению, хорошо известны в данной области. С учетом представленной в настоящем описании идеи специалист в данной области легко может не только получать указанные siРНК, но также и оценивать способность siРНК обладать антагонизирующим/ингибирующим/вызывающим «молчание» действием в отношении полипептида или белка, кодируемого кодирующей областью молекулы РНК, предлагаемой в настоящем изобретении. В контексте настоящего описания подразумевается, что описанные выше siРНК приводят к расщеплению молекулы РНК, предлагаемой в настоящем изобретении, которая несет кодирующую область, кодирующую полипептид или белок и модуль UTR, и таким образом к снижению уровня полипептида/белка, который кодируется кодирующей областью молекулы РНК, предлагаемой в настоящем изобретении.
Таким образом, настоящее изобретение относится к молекуле РНК, которая является комплементарной UTR, предлагаемой в настоящем изобретении, которая указана выше в настоящем описании.
В предпочтительном варианте осуществления изобретения указанная молекула РНК комплементарна UTR, предлагаемой в настоящем изобретении, содержит последовательность CAUGGUGGCGUCUCCC (SEQ ID NO: 11) или последовательность с 1-4 заменами по сравнению с SEQ ID NO: 11.
В другом предпочтительном варианте осуществления изобретения указанная молекула РНК, комплементарная UTR, предлагаемой в настоящем изобретении, содержит последовательность CAUGGUGGCNGUCUCCC (SEQ ID NO: 12), в которой нуклеотид N в положении 10 SEQ ID NO: 12 представляет собой нуклеотид, выбранный из группы, которая состоит из U, G, С или А, или последовательность с 1-4 заменами по сравнению с SEQ ID NO: 12 и которая обладает антагонизирующим /ингибирующим/вызывающим «молчание» действием в отношении полипептида или белка, кодируемого кодирующей областью молекулы РНК, предлагаемой в настоящем изобретении.
В предпочтительном варианте осуществления изобретения указанная молекула РНК, комплементарная UTR, предлагаемой в настоящем изобретении, содержит последовательность CAUCUUGGCGUCUCCC (SEQ ID NO: 13) или последовательность с 1-4 заменами по сравнению с SEQ ID NO: 13.
В другом предпочтительном варианте осуществления изобретения указанная молекула РНК, комплементарная UTR, предлагаемой в настоящем изобретении, содержит последовательность CAUCUUGGCNGUCUCCC (SEQ ID NO: 14), в которой нуклеотид N в положении 10 SEQ ID NO: 14 представляет собой нуклеотид, выбранный из группы, которая состоит из U, G, С или А, при этом более предпочтительным является А, или последовательность с 1-4 заменами по сравнению с SEQ ID NO: 14 и которая обладает антагонизирующим/ингибирующим/вызывающим «молчание» действием в отношении полипептида или белка, кодируемого кодирующей областью молекулы РНК, предлагаемой в настоящем изобретении.
В предпочтительном варианте осуществления изобретения указанная молекула РНК, комплементарная UTR, предлагаемой в настоящем изобретении, содержит последовательность CAUGGCGGCGUCUCCC (SEQ ID NO: 15) или последовательность с 1-4 заменами по сравнению с SEQ ID NO: 15.
В другом предпочтительном варианте осуществления изобретения указанная молекула РНК, комплементарная UTR, предлагаемой в настоящем изобретении, содержит последовательность CAUGGCGGCNGUCUCCC (SEQ ID NO: 16), в которой нуклеотид N в положении 10 SEQ ID NO: 16 представляет собой нуклеотид, выбранный из группы, которая состоит из U, G, С или А, или последовательность с 1-4 заменами по сравнению с SEQ ID NO: 16 и которая обладает антагонизирующим /ингибирующим/вызывающим «молчание» действием в отношении полипептида или белка, кодируемого кодирующей областью молекулы РНК, предлагаемой в настоящем изобретении.
В предпочтительном варианте осуществления изобретения указанная молекула РНК, комплементарная UTR, предлагаемой в настоящем изобретении, содержит последовательность CAUCUCGGCGUCUCCC (SEQ ID NO: 17) или последовательность с 1-4 заменами по сравнению с SEQ ID NO: 17.
В другом предпочтительном варианте осуществления изобретения указанная молекула РНК, комплементарная UTR, предлагаемой в настоящем изобретении, содержит последовательность CAUCUCGGCNGUCUCCC (SEQ ID NO: 18), в которой нуклеотид N в положении 10 SEQ ID NO: 18 представляет собой нуклеотид, выбранный из группы, которая состоит из U, G, С или А, при этом А является более предпочтительным, или последовательность с 1-4 заменами по сравнению с SEQ ID NO: 18 и которая обладает антагонизирующим /ингибирующим/вызывающим «молчание» действием в отношении полипептида или белка, кодируемого кодирующей областью молекулы РНК, предлагаемой в настоящем изобретении.
В другом предпочтительном варианте осуществления изобретения указанная молекула РНК, комплементарная UTR, предлагаемой в настоящем изобретении, содержит последовательность CAUGGUGGCGUCCC (SEQ ID NO: 19) или последовательность с 1-4 заменами по сравнению с SEQ ID NO: 19.
В другом предпочтительном варианте осуществления изобретения указанная молекула РНК, комплементарная UTR, предлагаемой в настоящем изобретении, содержит последовательность CAUGGUGGCNGUCCC (SEQ ID NO: 20), в которой нуклеотид N в положении 10 SEQ ID NO: 20 представляет собой нуклеотид, выбранный из группы, которая состоит из U, G, С или А, или последовательность с 1-4 заменами по сравнению с SEQ ID NO: 20 и которая обладает антагонизирующим/ингибирующим/вызывающим «молчание» действием в отношении полипептида или белка, кодируемого кодирующей областью молекулы РНК, предлагаемой в настоящем изобретении.
В предпочтительном варианте осуществления изобретения указанная молекула РНК, комплементарная UTR, предлагаемой в настоящем изобретении, содержит последовательность CAUCUUGGCGUCCC (SEQ ID NO: 21) или последовательность с 1-4 заменами по сравнению с SEQ ID NO: 21.
В другом предпочтительном варианте осуществления изобретения указанная молекула РНК, комплементарная UTR, предлагаемой в настоящем изобретении, содержит последовательность CAUCUUGGCNGUCCC (SEQ ID NO: 22), в которой нуклеотид N в положении 10 SEQ ID NO: 22 представляет собой нуклеотид, выбранный из группы, которая состоит из U, G, С или А, при этом А является более предпочтительным, или последовательность с 1-4 заменами по сравнению с SEQ ID NO: 22 и которая обладает антагонизирующим/ингибирующим/вызывающим «молчание» действием в отношении полипептида или белка, кодируемого кодирующей областью молекулы РНК, предлагаемой в настоящем изобретении.
В предпочтительном варианте осуществления изобретения указанная молекула РНК, комплементарная UTR, предлагаемой в настоящем изобретении, содержит последовательность CAUGGCGGCGUCCC (SEQ ID NO: 23) или последовательность с 1-4 заменами по сравнению с SEQ ID NO: 23.
В другом предпочтительном варианте осуществления изобретения указанная молекула РНК, комплементарная UTR, предлагаемой в настоящем изобретении, содержит последовательность CAUGGCGGCNGUCCC (SEQ ID NO: 24), в которой нуклеотид N в положении 10 SEQ ID NO: 24 представляет собой нуклеотид, выбранный из группы, которая состоит из U, G, С или А, или последовательность с 1-4 заменами по сравнению с SEQ ID NO: 24 и которая обладает антагонизирующим/ингибирующим/вызывающим «молчание» действием в отношении полипептида или белка, кодируемого кодирующей областью молекулы РНК, предлагаемой в настоящем изобретении.
В предпочтительном варианте осуществления изобретения указанная молекула РНК, комплементарная UTR, предлагаемой в настоящем изобретении, содержит последовательность CAUCUCGGCGUCCC (SEQ ID NO: 25) или последовательность с 1-4 заменами по сравнению с SEQ ID NO: 25.
В другом предпочтительном варианте осуществления изобретения указанная молекула РНК, комплементарная UTR, предлагаемой в настоящем изобретении, содержит последовательность CAUCUCGGCNGUCCC (SEQ ID NO: 26), в которой нуклеотид N в положении 10 SEQ ID NO: 26 представляет собой нуклеотид, выбранный из группы, которая состоит из U, G, С или А, при этом А является более предпочтительным, или последовательность с 1-4 заменами по сравнению с SEQ ID NO: 26 и которая обладае антагонизирующим/ингибирующим/вызывающим «молчание» действием в отношении полипептида или белка, кодируемого кодирующей областью молекулы РНК, предлагаемой в настоящем изобретении.
В другом предпочтительном варианте осуществления изобретения указанная молекула РНК, комплементарная UTR, предлагаемой в настоящем изобретении, содержит последовательность CAUGGUGGCGCUUC (SEQ ID NO: 27) или последовательность с 1-4 заменами по сравнению с SEQ ID NO: 27.
В другом предпочтительном варианте осуществления изобретения указанная молекула РНК, комплементарная UTR, предлагаемой в настоящем изобретении, содержит последовательность CAUGGUGGCNGCUUC (SEQ ID NO: 28), в которой нуклеотид N в положении 10 SEQ ID NO: 28 представляет собой нуклеотид, выбранный из группы, которая состоит из U, G, С или А, или последовательность с 1-4 заменами по сравнению с SEQ ID NO: 28 и которая обладает антагонизирующим/ингибирующим/вызывающим «молчание» действием в отношении полипептида или белка, кодируемого кодирующей областью молекулы РНК, предлагаемой в настоящем изобретении.
В предпочтительном варианте осуществления изобретения указанная молекула РНК, комплементарная UTR, предлагаемой в настоящем изобретении, содержит последовательность CAUCUUGGCGCUUC (SEQ ID NO: 29) или последовательность с 1-4 заменами по сравнению с SEQ ID NO: 29.
В другом предпочтительном варианте осуществления изобретения указанная молекула РНК, комплементарная UTR, предлагаемой в настоящем изобретении, содержит последовательность CAUCUUGGCNGCUUC (SEQ ID NO: 30), в которой нуклеотид N в положении 10 SEQ ID NO: 30 представляет собой нуклеотид, выбранный из группы, которая состоит из U, G, С или А, при этом А является более предпочтительным, или последовательность с 1-4 заменами по сравнению с SEQ ID NO: 30 и которая обладает антагонизирующим/ингибирующим/вызывающим «молчание» действием в отношении полипептида или белка, кодируемого кодирующей областью молекулы РНК, предлагаемой в настоящем изобретении.
В предпочтительном варианте осуществления изобретения указанная молекула РНК, комплементарная UTR, предлагаемой в настоящем изобретении, содержит последовательность CAUGGCGGCGCUUC (SEQ ID NO: 31) или последовательность с 1-4 заменами по сравнению с SEQ ID NO: 31.
В другом предпочтительном варианте осуществления изобретения указанная молекула РНК, комплементарная UTR, предлагаемой в настоящем изобретении, содержит последовательность CAUGGCGGCNGCUUC (SEQ ID NO: 32), в которой нуклеотид N в положении 10 SEQ ID NO: 32 представляет собой нуклеотид, выбранный из группы, которая состоит из U, G, С или А, или последовательность с 1-4 заменами по сравнению с SEQ ID NO: 32 и которая обладает антагонизирующим/ингибирующим/вызывающим «молчание» действием в отношении полипептида или белка, кодируемого кодирующей областью молекулы РНК, предлагаемой в настоящем изобретении.
В предпочтительном варианте осуществления изобретения указанная молекула РНК, комплементарная UTR, предлагаемой в настоящем изобретении, содержит последовательность CAUCUCGGCGCUUC (SEQ ID NO: 33) или последовательность с 1-4 заменами по сравнению с SEQ ID NO: 33.
В другом предпочтительном варианте осуществления изобретения указанная молекула РНК, комплементарная UTR, предлагаемой в настоящем изобретении, содержит последовательность CAUCUCGGCNGCUUC (SEQ ID NO: 34), в которой нуклеотид N в положении 10 SEQ ID NO: 34 представляет собой нуклеотид, выбранный из группы, которая состоит из U, G, С или А, при этом А является более предпочтительным, или последовательность с 1-4 заменами по сравнению с SEQ ID NO: 34 и которая обладает антагонизирующим/ингибирующим/вызывающим «молчание» действием в отношении полипептида или белка, кодируемого кодирующей областью молекулы РНК, предлагаемой в настоящем изобретении.
Другой предпочтительный вариант осуществления настоящего изобретения относится к молекуле РНК, выбранной из группы, которая состоит из SEQ ID NO: 11-34, которая несет дополнительный(ые) нуклеотид(ы) на 5'-конце, простирающемся за триплет, комплементарный стартовому кодону, и которая комплементарна последовательностям требуемого полипептида или белка, кодируемого кодирующей областью молекулы РНК, предлагаемой в настоящем изобретении. Предпочтительно комплементарные последовательности, которые содержат указанные выше последовательности, комплементарные UTR-последовательностям, предлагаемым в настоящем изобретении (т.е. молекула РНК, выбранная из группы, которая состоит из SEQ ID NO: 11-34), предпочтительно содержат по меньшей мере 15, более предпочтительно по меньшей мере 16, более предпочтительно по меньшей мере 17, более предпочтительно по меньшей мере 18, более предпочтительно по меньшей мере 19, более предпочтительно по меньшей мере 20, 21, 22, 23 или 24 нуклеотида. В еще более предпочтительном варианте осуществления изобретения эти последовательности содержат 25, 30, 35, 40 или большее количество нуклеотидов. Удлинение на 5'-конце может оказаться желательным для повышения специфичности комплементарной последовательности, что предупреждает нежелательные побочные действия.
Другой предпочтительный вариант осуществления настоящего изобретения относится не только к вышеуказанным молекулам РНК, но также к молекуле РНК, выбранной из группы, которая состоит из SEQ ID NO: 11-34, которая содержит вплоть до 5%, 10%, 20% или 30% ошибочных спариваний (мисмэтчи) относительно молекул РНК, описанных выше. Кроме того, молекулы РНК можно химически модифицировать согласно представленным выше в настоящем описании методами.
Настоящее изобретение относится также к набору, содержащему молекулу ДНК, предлагаемую в настоящем изобретении, молекулу РНК, предлагаемую в настоящем изобретении, молекулу нуклеиновой кислоты, предлагаемую в настоящем изобретении, вектор, предлагаемый в настоящем изобретении, или клетку-хозяина, предлагаемую в настоящем изобретении. Касательно предпочтительных вариантов осуществления изобретения, то к ним относится с необходимыми изменениями, то, что было указано выше в контексте молекулы ДНК, молекулы РНК, молекулы нуклеиновой кислоты, вектора или клетки-хозяина, предлагаемых в настоящем изобретении. Целесообразно, когда набор, предлагаемый в настоящем изобретении, дополнительно содержит необязательно (а) буфер(ы), растворы для хранения и/или другие реагенты или материалы, требуемые для осуществления указанных выше и ниже вариантов применения и способов. Кроме того, компоненты набора, предлагаемого в изобретении, могут быть упакованы индивидуально в пузырьки или банки или находиться в комбинации в контейнерах или в упаковках в виде микроконтейнера. Набор, предлагаемый в настоящем изобретении, целесообразно применять, среди прочего, для осуществления способов, предлагаемых в изобретении, или для получения молекулы РНК, предлагаемой в изобретении, и можно использовать в различных областях применения, указанных в настоящем описании, например, в вариантах применения, которые указаны выше и ниже. Другой компонент, который можно включать в набор, представляет собой инструкции, предназначенные для человека, который применяет указанный набор. Приготовление наборов осуществляют предпочтительно с использованием стандартных процедур, которые известны специалисту в данной области.
Настоящее изобретение относится также к применению UTR, указанной выше в настоящем описании, для трансляции кодирующей области молекулы РНК в полипептид или белок, кодируемый указанной кодирующей областью.
Более предпочтительной вариант осуществления настоящего изобретения относится также к применению UTR, указанной выше в настоящем описании, для повышения эффективности трансляции кодирующей области молекулы РНК в полипептид или белок, кодируемый указанной кодирующей областью.
Касательно предпочтительных вариантов осуществления изобретения, то к ним относится с необходимыми изменениями, то, что было указано выше в контексте молекулы РНК, предлагаемой в настоящем изобретении.
Предпочтительные варианты осуществления настоящего изобретения относятся к следующим вариантам, которые охарактеризованы ниже в пунктах 1-20:
1. Молекула ДНК, которая может транскрибироваться в мРНК, содержащая одну цепь со следующими элементами:
(а) кодирующая область, включая стартовый кодон на ее 5'-конце, которая кодирует полипептид; и
(б) расположенная непосредственно в обратном направлении относительно указанной кодирующей последовательности последовательность, выбранная из группы, которая состоит из:
(б1) R1-CGCCACC (SEQ ID NO: 1);
или последовательности, где в указанной последовательности С в положении 6 SEQ ID NO: 1 заменен на А и С в положении 7 SEQ ID NO: 1 заменен на G; и/или А в положении 5 SEQ ID NO: 1 заменен на G; и
(б2) R1-CNGCCACC (SEQ ID NO: 2), в которой нуклеотид N в положении 2 SEQ ID NO: 2 представляет собой нуклеотид, выбранный из группы, которая состоит из Т, G, С или А;
или последовательности, где в указанной последовательности С в положении 7 SEQ ID NO: 2 заменен на А и С в положении 8 SEQ ID NO: 2 заменен на G; и/или А в положении 6 SEQ ID NO: 2 заменен на G,
где R1 обозначает промотор, который распознается ДНК-зависимой РНК-полимеразой;
или содержит комплементарную цепь.
2. Молекула ДНК по п. 1, в которой промотор, распознаваемый ДНК-зависимой РНК-полимеразой, выбирают из группы, которая состоит из:
(I) TAATACGACTCACTATAGGGAGA (SEQ ID NO: 3) или последовательности, которая содержит 1-6 замен по сравнению с SEQ ID NO: 3 и которая распознается ДНК-зависимой РНК-полимеразой Т7;
(II) AATTAACCCTCACTAAAGGGAGA (SEQ ID NO: 4) или последовательности, которая содержит 1-6 замен по сравнению с SEQ ID NO: 4 и которая распознается ДНК-зависимой РНК-полимеразой Т3;
(III) ATTTAGGTGACACTATAGAAG (SEQ ID NO: 5) или последовательности, которая содержит 1-6 замен по сравнению с SEQ ID NO: 5 и которая распознается ДНК-зависимой РНК-полимеразой SP6; и
(IV) AATTAGGGCACACTATAGGGA (SEQ ID NO: 6) или последовательности, которая содержит 1-6 замен по сравнению с SEQ ID NO: 6 и которая распознается ДНК-зависимой РНК-полимеразой K11.
3. Молекула ДНК по п. 1 или п. 2, в которой нуклеотид N в положении 2 SEQ ID NO: 2 представляет собой нуклеотид, выбранный из группы, которая состоит из Т, G или С, и в которой нуклеотид N не представляет собой А.
4. Молекула ДНК по п. 3, в которой указанный нуклеотид N в положении 2 SEQ ID NO: 2 представляет собой Т.
5. Вектор, содержащий молекулу ДНК по п. 4.
6. Клетка-хозяин, содержащая вектор по п. 5.
7. Композиция, содержащая:
молекулу ДНК по одному из п.п. 1-4, вектор по п. 5 или клетку-хозяина по п. 6.
8. Молекула РНК, которая содержит
(а) кодирующую область, включающую стартовый кодон на ее 5'-конце, которая кодирует полипептид; и
(б) расположенную непосредственно в обратном направлении относительно указанной кодирующей последовательности UTR, выбранную из группы, которая содержит:
(б1) UTR-последовательности
R2-CGCCACC (SEQ ID NO: 1),
или последовательности, где в указанной последовательности UTR С в положении 6 SEQ ID NO: 1 заменен на А и С в положении 7 SEQ ID NO: 1 заменен на G; и/или А в положении 5 SEQ ID NO: 1 заменен на G; и
(б2) UTR-последовательности
R2-CNGCCACC (SEQ ID NO: 2), в которой нуклеотид N в положении 2 SEQ ID NO: 2 представляет собой нуклеотид, выбранный из группы, которая состоит из Т, G, С или А; или последовательности, где в указанной последовательности С в положении 7 SEQ ID NO: 2 заменен на А и С в положении 8 SEQ ID NO: 2 заменен на G; и/или А в положении 6 SEQ ID NO: 2 заменен на G,
где R2 обозначает последовательность РНК, соответствующую части промоторной области, начинающейся с нуклеотида, с которого ДНК-зависимая РНК-полимераза инициирует синтез РНК.
9. Молекула РНК по п. 8, в которой R2 выбирают из группы, состоящей из:
(I) GGGAGA (SEQ ID NO: 7);
(II) GGGAGA (SEQ ID NO: 8);
(III) GAAG (SEQ ID NO: 9); и
(IV) GGGA (SEQ ID NO: 10).
10. Молекула РНК по п. 8 или п. 9, в которой нуклеотид N в положении 2 SEQ ID NO: 2 представляет собой нуклеотид, выбранный из группы, которая состоит из U, G или С, и в которой нуклеотид N не представляет собой А.
11. Молекула РНК по п. 10, в которой указанный нуклеотид N в положении 2 SEQ ID NO: 2 представляет собой U.
12. Молекула РНК по одному из п.п. 8-11, где молекула РНК содержит поли-А-хвост на 3'-конце.
13. Молекула РНК по одному из п.п. 8-12, в которой поли-А-хвост состоит по меньшей мере из 120 нуклеотидов.
14. Молекула нуклеиновой кислоты, кодирующая молекулу РНК по одному из п.п. 8-13.
15. Вектор, содержащий молекулу нуклеиновой кислоты по п. 14.
16. Клетка-хозяин, содержащая вектор по п. 15.
17. Фармацевтическая композиция, содержащая молекулу РНК по одному из п.п. 8-13, молекулу нуклеиновой кислоты по п. 14, вектор по п. 15 или клетку-хозяина по п. 16 и необязательно фармацевтически приемлемый носитель.
18. Фармацевтическая композиция по п. 17, предназначенная для применения в терапиях на основе РНК.
19. Набор, содержащий молекулу ДНК по одному из п.п. 1-4, молекулу РНК по одному из п.п. 8-13, молекулу нуклеиновой кислоты по п. 14, вектор по п. 5 или п. 15 или клетку-хозяина по п. 6 или п. 16.
20. Применение UTR по п. 8(б) для трансляции кодирующей области молекулы РНК в полипептид или белок, кодируемый указанной кодирующей областью.
На чертежах показано:
на фиг. 1 - последовательности, несущие последовательность «минимальной UTR», наряду с названием соответствующих люциферазных репортерных конструкций, применяемых в настоящем изобретении. Последовательности несут части промотора Т7 и элемент Козак, за которым следует стартовый кодон ATG. Первые 10 оснований, включающие последовательность TATA, и последующие 6 оснований (GGGAGA) представляют собой последовательности, полученные из промотора Т7, а остальные основания, расположенные в обратном направлении относительно стартового кодона ATG, относятся к элементу Козак (GCCACC). «Sp30» обозначает произвольную последовательность, состоящую из 30 нуклеотидов. Подчеркнутая последовательность в последовательности №9 представляет собой последовательность 5' UTR из человеческого альфа-глобина («hAg»), состоящую из 30 нуклеотидов.
Последовательности 1-9 на фиг. 1 соответствуют SEQ ID NO: 37-45 соответственно;
на фиг. 2А и Б - данные, демонстрирующие, что дополнительный «С» в «минимальной UTR» имеет важное значение (последовательность №1 и №2 на фиг. 1). Линию клеток человеческого альвеолярного эпителия (А549) и линию клеток человеческой печеноночноклеточной карциномы (HepG2) высевали с плотностью 20000 клеток/лунку и 40000 клеток/лунку соответственно в 96-луночный планшет. Через 24 ч после посева клетки трансфектировали различными, кодирующими люциферазу конструкциями SNIM-РНК (последовательность №1 и №2 на фиг. 1), используя липофектамин 2000. Экспрессию люциферазы измеряли через 24 ч после трансфекции. Величины представлены в виде среднего значения ± SD по 3 повторностям и в виде графика зависимости от применяемой для трансфекции дозы, и данные проанализированы с помощью программы GraphPad Prism. При изучении КАК на клетках А549, так и на клетках HepG2, делеция С приводила к снижению уровня экспрессии. В результате этого указанный дополнительный С включали при создании всех дополнительных конструкций;
на фиг. 3 - данные о воздействии индивидуальных указанных нуклеотидов и демонстрирующие роль расстояния между дополнительным «С» и элементом Козак при анализе на трансфектированных клетках А549. Клетки трансфектировали и анализ люциферазы осуществляли согласно процедуре, описанной в разделе «Материалы и методы». Поскольку результаты, полученные при использовании более высоких доз, выпадали из линейного диапазона, представлены данные только для доз, не превышающих 62,5 нг/лунку. 5'UTR из человеческого альфа-глобина применяли в качестве положительного контроля. Эксперименты по трансфекции осуществляли с использованием молекул SNIM-РНК, несущих последовательности №№3-8 (см. фиг. 1) соответственно. Линию клеток человеческого альвеолярного эпителия (А549) высевали с плотностью 20000 клеток/лунку в 96-луночный планшет. Через 24 ч после посева клетки трансфектировали различными кодирующими люциферазу конструкциями SNIM-РНК (последовательности №№3-8 на фиг. 1), используя липофектамин 2000. Экспрессию люциферазы измеряли через 24 ч после трансфекции. Строили графики зависимости от применяемой для трансфекции дозы, и данные анализировали с помощью программы GraphPad Prism. Величины представлены в виде среднего значения ± SD по 3 повторностям.
При изучении на линии клеток альвеолярного эпителия (А549) установлено, что встраивание дополнительного «А» между С и элементом Козак (последовательность №3 на фиг. 1) приводило к существенному снижению уровня экспрессии (фиг. 3). Встраивание одного «Т» между С и элементом Козак (последовательность №4 на фиг. 1) приводило к получению уровней экспрессии, сопоставимых с уровнями, полученными при применении 5'UTR человеческого альфа-глобина, который применяли в качестве положительного контроля;
на фиг. 4 - данные о воздействии индивидуальных указанных нуклеотидов и демонстрирующие роль расстояния между дополнительным «С» и элементом Козак при анализе на трансфектированных клетках HepG2. Клетки трансфектировали и анализ люциферазы осуществляли согласно процедуре, описанной в разделе «Материалы и методы». Поскольку результаты, полученные при использовании более высоких доз, выпадали из линейного диапазона, представлены данные только для доз, не превышающих 62,5 нг/лунку. Эксперименты по трансфекции осуществляли с использованием последовательностей №№3-8, представленных на фиг. 1. Линию клеток человеческой печеноночноклеточной карциномы (HepG2) высевали с плотностью 40000 клеток /лунку в 96-луночный планшет. Через 24 ч после посева клетки трансфектировали различными кодирующими люциферазу конструкциями SNIM-РНК (последовательности №№3-8 на фиг. 1), используя липофектамин 2000. Экспрессию люциферазы измеряли через 24 ч после трансфекции. Строили графики зависимости от применяемой для трансфекции дозы, и данные анализировали с помощью программы GraphPad Prism. Величины представлены в виде среднего значения ± SD по 3 повторностям. Для обеих линий клеток (клетки А549 (фиг. 3) и HepG2 (фиг. 4) установлено, что встраивание дополнительного «А» между С и элементом Козак (последовательность №3 на фиг. 1) приводило к существенному снижению экспрессии (фиг. 3 и 4). Для обеих линий клеток установлено, что встраивание одного «Т» между С и элементом Козак (последовательность №4 на фиг. 1) приводило к получению уровней экспрессии, сопоставимых с уровнями, полученными при применении 5'UTR человеческого альфа-глобина, который применяли в качестве положительного контроля. При изучение на клетках HepG2 последовательность №1 (фиг. 1) также обладала такой же эффективностью;
на фиг. 5 - данные о воздействии TISU-(инициатор трансляции короткой 5'UTR) элемента на экспрессию люциферазы в клетках А549. Осуществляли детальное изучение зависимости от дозы и подгонку кривых для выбора кодирующих люциферазу конструкций. На основе представленных выше на фиг. 2-4 результатов вносили TISU-элемент в комбинацию с последовательностью №4 (фиг. 1), которая содержала два требуемых отличительных признака: (С между промотором Т7 и элементом Козак и дополнительный T между С и элементом Козак с получением последовательности №9, представленной на фиг. 1).
Линию клеток человеческого альвеолярного эпителия (А549) (фиг. 5А и Б) и линию клеток человеческой печеноночноклеточной карциномы (HepG2) (фиг. 5В и Г) высевали с плотностью 20000 клеток/лунку и 40000 клеток/лунку соответственно в 96-луночный планшет. Через 24 ч после посева клетки трансфектировали различными кодирующими люциферазу конструкциями SNIM-РНК, используя липофектамин 2000. Экспрессию люциферазы измеряли через 24 и 48 ч после трансфекции (фиг. 5Д). Строили графики зависимости от применяемой для трансфекции дозы, и данные анализировали с помощью программы GraphPad Prism. Представлены данные, полученные при трансфекции клеток А459 (А, Б) и HepG2 (В, Г) различными кодирующими люциферазу мРНК. Люциферазную активность измеряли через 24 ч (А, В) и 48 ч (Б, Г) после трансфекции. Величины представлены в виде среднего значения ± SD по 3 повторностям.
Для обеих линий клеток и для обоих моментов времени оценки установлена существенно более высокая экспрессия при использовании конструкции люциферазы, содержащей TISU-элемент (фиг. 5А-Г);
на фиг. 6 - данные о воздействии TISU-элемента на экспрессию люциферазы в клетках А549 (фиг. 6А) и HepG2 (фиг. 6Б). Линию клеток человеческого альвеолярного эпителия (А549) и линию клеток человеческой печеноночноклеточной карциномы (HepG2) высевали с плотностью 20000 клеток/лунку и 40000 клеток/лунку соответственно в 96-луночный планшет. Через 24 ч после посева клетки трансфектировали различными кодирующими люциферазу конструкциями SNIM-РНК, используя липофектамин 2000 (на Х-оси представлено количество в нг SNIM-РНК на лунку 96-луночного планшета). Экспрессию люциферазы измеряли через 24 ч после трансфекции. Строили графики зависимости от применяемой для трансфекции дозы, и данные анализировали с помощью программы GraphPad Prism. Представлены данные, полученные при трансфекции клеток А459 (А) и HepG2 (Б) различными кодирующими люциферазу мРНК;
на фиг. 7 - результаты экспериментов in vivo на мышах, полученные с использованием различных кодирующих люциферазу конструкций мРНК. Конструкции, кодирующие люциферазу, которые представлены на фиг. 7 (соответствующую последовательность UTR-элемента см. на фиг. 1), тестировали in vivo на мышах Balb/c (самки возрастом 6-8 недель). В этом варианте эксперимента тестировали также эффективность дополнительного UTR-элемента, для которого продемонстрирована способность повышать экспрессию трансгена (публикация международной заявки на патент WO 2012/170930 А1). Конструкцию люциферазы, содержащую указанный UTR-элемент, обозначали как Luc2-SUSA. 20 мкг соответствующей SNIM-РНК включали в комплекс с LF-44 и инъецировали внутривенно мышам Balb/c. Визуализацию in vivo осуществляли через 6 ч после инъекции, используя систему для визуализации IVIS, и строили график зависимости для величин, выраженных в фотонах/с/см2/ср. Результаты, полученные при визуализации целого животного, представлены на фиг. 7А, а результаты, полученные при визуализации всего органа, представлены на фиг. 7Б (печень), 7 В (легкое), 7Е (селезенка) соответственно.
Органы, изъятые из животных, замораживали в жидком азоте и гомогенизировали. Клетки лизировали, используя буфер для лизиса Трис-HCl, и измеряли активность люциферазы. Результаты представлены на фиг. 7Д (печень), 7Е (легкое), 7Ж (селезенка) соответственно.
Установлено, что инсерция TISU-элемента приводила к повышению экспрессии по сравнению с ранее описанными 5'- и 3'-UTR (публикация международной заявки на патент WO 2012/170930 А1). Добавление одного T между С и элементом Козак (последовательность №4, представленная на фиг. 1) приводила к получению уровней экспрессии, сопоставимыми с уровнями, полученными для UTR человеческого альфа-глобина (последовательность №8, представленная на фиг. 1). Добавление TISU-элемента в последовательность №4 (фиг. 1) дополнительно повышало экспрессию (последовательность №9, представленная на фиг. 1). При создании изобретения неожиданно было установлено, что воздействие UTR человеческого альфа-глобина не являлось специфическим для последовательности. 30 случайных нуклеотидных последовательностей поддерживали уровень экспрессии, сходный с уровнем, поддерживаемым 5'UTR человеческого альфа-глобина.
На основе результатов in vitro на клеточных линиях и экспериментов in vivo на мышах последовательности №№1, 4, 7 и 9 (фиг. 1) предложены в качестве перспективных последовательностей-кандидатов, несущих «минимальные UTR», для терапии на основе транскриптов. Указанные минимальные UTR-последовательности не оказывали отрицательных воздействий на выход РНК в процессе транскрипции in vitro, и образовавшаяся мРНК существенно более эффективно транслировалась по сравнению с мРНК, содержащими принятые в данной области UTR;
на фиг. 8 - данные о количестве лейкоцитов (WBC) (фиг. 8А), эритроцитов (RBC) (фиг. 8Б), тромбоцитов (фиг. 8В), уровне гемоглобина (фиг. 8Г) и о гематокрите (фиг. 8Д) у мышей, обработанных различными кодирующими люциферазу конструкциями мРНК. Эксперимент осуществляли в целом согласно методу, описанному на фиг. 7, а параметры крови анализировали, используя автоматический гематологический анализатор фирмы Sysmex KX-21N™ (штат Иллинойс, США);
на фиг. 9 - результаты экспериментов по оценке экспрессии с использованием содержащей TISU-элемент мРНК, кодирующей человеческий ЕРО, в сравнении с экспрессией при использовании мРНК, кодирующей человеческий ЕРО, которая содержит 5'- и 3'-UTR (описанные в публикации международной заявки на патент WO 2012/170930 А1: фиг. 1 и 2) (SUSA-UTR), для которых известна способность поддерживать очень высокий уровень экспрессии ЕРО.
Линию клеток человеческого альвеолярного эпителия (А549) и линию клеток человеческой печеноночноклеточной карциномы (HepG2) высевали с плотностью 20000 клеток/лунку и 40000 клеток/лунку соответственно в 96-луночный планшет. Через 24 ч после посева клетки трансфектировали, используя 250 нг различных кодирующих ЕРО конструкций SNIM-РНК с применением липофектамина 2000. Количество ЕРО определяли через 24 ч после трансфекции с помощью ELISA (ELISA-набор для количественного определения человеческого эритропоэтина (Human Erythropoietin Quantikine IVD Kit) фирмы R&D Systems (шт. Миннесота, США)) и данные анализировали с помощью GraphPad Prism. Величины представлены в виде среднего значения ± SD по 3 повторностям;
на фиг. 10 - результаты экспериментов по оценке экспрессии человеческой ОТС (орнитинтранскарбамилаза). Для человеческой ОТС сравнивали экспрессию при использовании содержащей TISU-элемент кодирующей hOTC мРНК и экспрессию кодирующей hOTC мРНК, содержащей 5' UTR человеческого альфа-глобина, для которой известен наиболее высокий уровень экспрессии по сравнению со всеми другими известными к настоящему времени комбинациями.
Линию клеток человеческой печеноночноклеточной карциномы (HepG2) высевали в 96-луночный планшет и через 24 ч после посева клетки трансфектировали различными кодирующими hOTC конструкциями SNIM-РНК, используя липофектамин 2000. Через 24 ч после трансфекции клетки лизировали и количество ОТС определяли с помощью Вестерн-блоттинга.
Содержащие как hAg, так и TISU-элемент кодирующие hOTC SNIM-РНК обеспечивали сходный уровень экспрессии hOTC (фиг. 10А). Ген винкулина применяли в качестве гена «домашнего хозяйства» и интенсивность полосы оценивали количественно и применяли в качестве внутреннего стандарта для расчета количества (фиг. 10Б);
на фиг. 11 - предсказанные вторичные структуры произвольного состоящего из 30 нуклеотидов спейсера, присутствующего в последовательности №7 (слева), и 5'UTR человеческого альфа-глобина, присутствующего в последовательности №8 (справа).
Другие объекты и преимущества изобретения будут описаны ниже в примерах, которые даны только с целью иллюстрации и никоим образом не направлены на ограничение изобретения. Каждая публикация, каждый патент, каждая заявка на патент или другой документ, процитированные в настоящем описании, полностью включены в него в качестве ссылки.
Примеры
I. Материалы и методы
Плазмидные векторы
Соответствующие последовательности 5' UTR в сочетании с последовательностями люциферазы с оптимизированными кодонами синтезировали на фирме GeneScriptG (шт. Нью-Джерси, США) и клонировали в pUC57-Kan (фирма GeneScript). В случае ЕРО (последовательность с оптимизированными кодонами человеческого эритропоэтина) и ОТС (последовательность с оптимизированными кодонами человеческой орнитинтранскармилазы) кодирующую последовательность гена люциферазы заменяли на кодирующую последовательность гена ЕРО (SEQ ID NO: 35) и ОТС (SEQ ID No: 36) соответственно. UTR-последовательности, применяемые в конструкциях в сочетании с указанной соответствующей репортерной люциферазной конструкцией, представлены на фиг. 1.
Получение мРНК
Для создания in vitro транскрибируемой мРНК (IVT мРНК) плазмиды линеаризовывали путем расщепления BstBI и очищали путем экстракции хлороформом и осаждения этанолом. Очищенные линейные плазмиды применяли в качестве матрицы для in vitro транскрипции с использованием системы для крупномасштабного производства РНК RiboMax (Large Scale RNA production System-T7) (фирма Promega, Германия). Добавляли в реакционную смесь анти-инвертированный аналог кэп-структуры (ARCA) для создания 5'-кэппированной мРНК, и мРНК полиаденилировали (фирма Thermo Scientific) с получением 3'-поли-А-хвоста.
Дополнительно для получения SNIM-мРНК добавляли химически модифицированные нуклеотиды, такие как метил-ЦТФ и тио-УТФ (фирма Jena Bioscience, Германия) до конечной концентрации АТФ : ЦТФ : УТФ : метил-ЦТФ : тио-УТФ : ГТФ 7,57 мМ : 5,68 мМ : 5,68 мМ : 1,89 мМ : 1,89 мМ : 1,21 мМ. Полную смесь IVT инкубировали при 37°С в течение 2 ч с последующим расщеплением ДНКазой I в течение 20 мин при 37°С. РНК осаждали ацетатом аммония (конечная концентрация 2,5 М) и промывали 70% EtOH. Стадию промывки осуществляли дважды. И, наконец, дебрис РНК ресуспендировали в свободной от РНКазы воде. Все мРНК верифицировали на 1% агарозных гелях. Транскрибированные РНК являлись химически модифицированными, поскольку примерно 25% остатков уридина представляли собой 2-тиоуридин (s2U) и примерно 25% остатков цитидина представляли собой 5-метилцитидин (m5С). Последовательности UTR представлены на фиг. 1.
In vitro трансфекция
Линию клеток человеческого альвеолярного эпителия (А549) и линию клеток человеческой печеноночноклеточной карциномы (HepG2) высевали с плотностью 20000 клеток/лунку и 40000 клеток/лунку соответственно в 96-луночный планшет. Через 24 ч после посева клетки трансфектировали различными кодирующими люциферазу конструкциями SNIM-РНК, применяя поступающий в продажу реагент для трансфекции Lipofectamine™ 2000 в соотношении 2,5 мкл Lipofectamine™ 2000 на 1 мкг мРНК (на Х-оси на фиг. 2-6 представлено количество в нг SNIM-РНК на лунку 96-луночного планшета). Комплекс получали следующим образом: Lipofectamine™ 2000 и мРНК растворяли по отдельности в среде для трансфекции OptiMEM, в каждом случае до достижения общего объема 45 мкл. Указанные смеси инкубировали при комнатной температуре в течение 5 мин. Раствор Lipofectamine™ 2000 смешивали с раствором мРНК, после чего инкубировали в течение еще 20 мин при комнатной температуре. Клетки инкубировали в общем объеме среды для трансфекции 90 мкл при 37°С (уровень СО2 5%) в течение 1 ч. Затем среду для трансфекции удаляли и клетки промывали ЗФР. После чего клетки повторно инкубировали в среде Лейбовица L-15, содержащей 10% FBS.
Культура клеток
Линию клеток человеческого альвеолярной аденокарциномы (А549, АТСС CCL-185) выращивали в среде Хэма F12K, дополненной 10% FBS. Линию клеток человеческой печеноночноклеточной карциномы (HepG2, АТСС НВ-8065) культивировали в среде DMEM, дополненной 10% фетальной бычьей сыворотки. Все линии клеток выращивали в увлажненной атмосфере с уровнем СО2 5%.
Измерение биолюминесценции
Люцифераза светлячка (FFL) представляет собой общепринятый белок, который эндогенно не присутствует у млекопитающих и который легко можно обнаруживать с помощью визуализации люминесценции. Люцифераза катализирует реакцию взаимодействия люциферина и кислорода, которая приводит к испусканию биолюминесценции.
Линию клеток человеческого альвеолярного эпителия (А549) и линию клеток человеческой печеноночноклеточной карциномы (HepG2) высевали с плотностью 20000 клеток/лунку и 40000 клеток/лунку соответственно в 96-луночный планшет. Через 24 ч после посева клетки трансфектировали различными кодирующими люциферазу конструкциями SNIM-РНК, применяя Липофектамин 2000 (на Х-оси представлено количество в нг SNIM-РНК на лунку 96-луночного планшета). Биолюминесценцию измеряли через 24 ч после трансфекции. Строили графики зависимости от применяемой для трансфекции дозы, и данные анализировали с помощью программы GraphPad Prism.
Для количественного определения экспрессии люциферазы в гомогенизированных тканевых лизатах органы изымали из животных, замораживали в жидком азоте, гомогенизировали и клетки лизировали с помощью буфера для лизиса (25 мМ Трис-HCl, рН 7,5 с 0,1% Тритон-Х100).
Животные
Самок мышей линии BALB/c возрастом 6-8 недель получали от фирмы Janvier, Route Des  SecsBP5, F-53940 Le Genest St. Иль, Франция, и содержали в свободных от специфических патогенов условиях. Мышам давали акклиматизироваться к окружающей среде вивария в течение по меньшей мере 7 дней до начала экспериментов. Все процедуры с животными были одобрены и контролировались местным комитетом по этике и их осуществляли в соответствии с нормативами немецкого закона о защите жизни животных.
SecsBP5, F-53940 Le Genest St. Иль, Франция, и содержали в свободных от специфических патогенов условиях. Мышам давали акклиматизироваться к окружающей среде вивария в течение по меньшей мере 7 дней до начала экспериментов. Все процедуры с животными были одобрены и контролировались местным комитетом по этике и их осуществляли в соответствии с нормативами немецкого закона о защите жизни животных.
Композиции на основе липидоидов
Приготавливали композиции липидоидов с мРНК следующим образом: С12-(2-3-2), DOPE, Chol и DSPE-PEG2k (массовое соотношение 3,6:0,18:0,76:1) растворяли в этаноле и быстро инъецировали в забуференный цитратом раствор (10 мМ лимонная кислота, 150 мМ NaCl, рН=4,5), содержащий химически модифицированную мРНК, которая кодирует люциферазу светлячка, при массовом отношении липид/мРНК 10,5 с получением конечной концентрации в этаноле 20% и подвергали диализу в противотоке воды. Образовавшиеся комплексы липидоид/мРНК представляли собой положительно заряженные наночастицы (92,6±0,7 нм; 21,0±0,2 мВ) и их инъецировали внутривенно в хвостовую вену обездвиженных мышей. Во втором эксперименте комплексы липидоид/мРНК вводили в ЗФР перед внутривенной инъекцией, что приводило к получению практически незаряженных наночастиц (91,5±0,6 нм; -0.7±0,2 мВ).
Измерение активности Luc у мышей с помощью визуализации люминисценции in vivo
Через 24 ч после обработки мышей анестезировали путем внутрибрюшинной инъекции медетомидина (11,5 мкг/кг массы тела (BW)), мидазолама (115 мкг/кг BW) и фентанила (1,15 мкг/кг BW). D-люциферин в качестве субстрата (3 мг/100 мкл ЗФР на мышь) вносили путем внутривенной инъекции. Биолюминесценцию измеряли через 10 мин с помощью системы визуализации IVIS 100 (фирма Xenogen, Аламида, США) и следующих задаваемых параметров камеры: Bin(HS), поле зрения 10, f1 f-стоп, биннинг для высокого разрешения и продолжительность экспозиции 5 мин. Оцифровку и анализ сигнала осуществляли с использованием версии 2.50 программы Living Image (фирма Xenogen, Аламида, США).
Анализ методом Вестерн-блоттинга белка ОТС
Замороженные планшеты подвергали оттаиванию и осуществляли лизис клеток непосредственно в планшете. Белки лизировали с помощью буфера для лизиса (25 мМ Трис, 0,1% Тритон-Х100, фирма Sigma-Aldrich, Германия), дополненного ингибитором протеазы (cOmplete, свободный от ЭДТА, фирма Roche Diagnostics, Германия) и ДНКазой (раствор ДНКазы I (2500 ед./мл), (фирма Thermo Fisher, США). После лизиса образцы смешивали с буфером для образца NuPage® LDS и восстановителем для образца (фирма Thermo Fisher, США) и выдерживали в течение 10 мин при 70°С. Гель-электрофорез осуществляли, используя 15 мкл лизата на NuPAGE 10% Бис-Трис миди-гели с помощью системы ХСеll4 SureLock™ миди, Bio-Rad Criterion™ (фирма Thermo Fisher, США). Белки переносили с помощью системы для переноса TransBlot® Turbo™ (фирма Biorad, Германия) в течение 30 мин. После переноса мембраны блокировали с помощью чистого желатина в течение 30 мин перед инкубацией мембраны в течение ночи при 4°C с первичным антителом, разведенным в чистом желатине в соотношении 1:2000 (поликлональное антитело к ОТС (Center), АР6928с-АВ, фирма Biocat, Германия). После трех стадий промывки чистым желатином добавляли конъюгированное с пероксидазой из хрена вторичное антитело (козье антитело против кроличьего IgG-HRP, sc-2004, фирма Santa Cruz Biotechnology, США), разведенное в соотношении 1:10000 в чистом желатине, в течение 1 ч при КТ. Мембрану промывали вновь три раза чистым желатином, после чего сигналы визуализировали с помощью набора хемилюминисцентных субстратов (Luminata Crescendo Western HRP substrate, фирма Merck Millipore, Германия) и визуализировали с использованием системы ChemiDoc™ MP (фирма Biorad, Германия).
Материалы
FBS, среду Лейбовица L-15 (фирма Gibco), Lipofectamine™ 2000 и OptiMEM (фирма Gibco) покупали у фирмы Invitrogen, Германия. Стерильный ЗФР приготавливали в лаборатории заявителей. Среду Хэма F-12K, DMEM и трипсин-ЭДТА покупали у фирмы с.с.pro GmbH, Германия.
II. Результаты
II.а Эксперименты с клеточными культурами
На фиг. 2А и Б продемонстрировано, что дополнительный «С» между промотором Т7 и элементом Козак имеет важное значение. Делеция указанного основания приводила к снижению экспрессии в обоих сравниваемых типах клеток. Для обеих конструкций (последовательность №1 и №2 на фиг. 1) для удобства сравнения полный диапазон доз и линейный диапазон (исключенные значения: дозы, превышающие 62,5 нг/лунку исключали из анализа) представлены по отдельности. В обеих линиях клеток А549 и HepG2 делеция С приводила к снижению экспрессии. По этой причине указанный дополнительный С включали при создании всех дополнительных конструкций.
Основываясь на результатах, полученных с использованием клеток А549 и HepG2, дополнительные эксперименты проводили с использованием конструкции, содержащей дополнительный «С» (последовательность №1: T7Luc2).
Фиг. 3 и фиг. 4
Последовательность №1 применяли в качестве матрицы и в эту последовательность включали либо один нуклеотид (А, Т, G или С: последовательности №№3-6 на фиг. 1 соответственно), либо произвольную последовательность длиной 30 нуклеотидов и которая не имела какой-либо прогнозируемой вторичной структуры (последовательность №7), либо 5' UTR из человеческого альфа-глобина (последовательность №8) между исследуемым «С» и элементом Козак.
Клетки трансфектировали и анализ люциферазы осуществляли согласно описанному в разделе «Материалы и методы». Поскольку более высокие дозы выпадали из линейного диапазона, представлены данные о дозовой зависимости только для дозы вплоть до 62,5 нг/лунку. 5'UTR из человеческого альфа-глобина применяли в качестве положительного контроля.
Обобщая полученные выше результаты, на фиг. 1-4 продемонстрировано, что дополнительный «С» между промотором Т7 и элементом Козак имеет важное значение для достижения высокого уровня экспрессии белка при применении минимализированной 5'UTR. Делеция нуклеотида приводила к снижению экспрессии. Встраивание дополнительного «А» между дополнительным «С» и элементом Козак оказывало отрицательное действие на экспрессию. Когда пиримидиновое основание, наиболее предпочтительно «Т», добавляли в указанное положение, то получали уровни, сопоставимые с уровнями, полученными при применении 5'UTR из hAg.
Затем осуществляли дополнительные эксперименты с целью:
- объяснения воздействия TISU-элемента при его объединении с обладающей наиболее высокой эффективностью последовательностью (последовательность №9), и
- определения того, является ли действие 5' UTR из hAg специфическим для последовательности действием, и является ли важным расстояние между 5'-кэпом и стартовым кодоном.
На фиг. 5 продемонстрировано воздействие TISU-элемента на экспрессию люциферазы в клетках А549. «TISU-элемент» включает «AG» вместо «СС» в последовательности №9, что продемонстрировано на фиг. 1, по сравнению с последовательностью №4, представленной на фиг. 1. В клетках А549 (фиг. 5А и Б), а также в клетках HepG2 (фиг. 5 В и Г) обнаружен существенно более высокий уровень экспрессии люциферазы при применении конструкции, кодирующей люциферазу, которая содержит TISU-элемент в сочетании с «С» из последовательности №1 и дополнительный «Т» между указанным «С» и элементом Козак, при оценке через 24 ч (А, В) и 48 ч (Б, Г) после трансфекции.
На фиг. 6 продемонстрированы результаты такого же эксперимента, результаты которого представлены на фиг. 5, но с добавлением 5'UTR, содержащей произвольную состоящую из 30 нуклеотидов последовательность, что позволяло осуществлять одновременное сравнение UTR человеческого альфа-глобина (последовательность №8 на фиг. 1) и произвольной последовательности такой же длины (последовательность №7 на фиг. 1). Экспрессию люциферазы измеряли в клетках HepG2 (фиг. 6А) и клетках А549 (фиг. 6Б) через 24 ч после трансфекции указанными SNIM-РНК.
На фиг. 9 представлены результаты экспериментов по оценке экспрессии с использованием содержащей TISU-элемент кодирующей hEPO мРНК в сравнении с мРНК, кодирующей hEPO, которая содержит 5'- и 3'-UTR (описанные в публикации международной заявки на патент WO 2012/170930 А1: фиг. 1 и 2) (SUSA-UTR),) которую применяли в качестве стандарта, после трансфекции клеток А549 и HepG2 соответствующей SNIM-РНК. Количества ЕРО определяли через 24 ч после трансфекции с помощью ELISA. Величины представлены в виде среднего значения ± SD по 3 повторностям.
При изучении на человеческих клетках А549 установлено, что включение TISU-элемента приводило к более высокому уровню экспрессии по сравнению с уровнем, достигаемым при включении 5'- и 3' UTR (фиг. 9А). Сопоставимые уровни экспрессии обнаружены при изучении на клетках HepG2 (фиг. 9Б). Этот результат является неожиданным, поскольку включение 5'- и 3' UTR SUSA делает РНК примерно на 200 нуклеотидов более длинными, чем при применении UTR, предлагаемой в настоящем изобретении.
На фиг. 10 представлены результаты экспериментов по оценке экспрессии человеческой ОТС. Для сравнения экспрессию при применении содержащей TISU-элемент кодирующей hOTC мРНК сопоставляли с экспрессией при применении кодирующей hOTC мРНК, содержащей 5' UTR человеческого альфа-глобина, для которой установлен наиболее высокий уровень экспрессии по сравнению со всеми известными к настоящему времени другими комбинациями.
Клетки HepG2 трансфектировали различными кодирующими hOTC конструкциями SNIM-РНК, лизировали через 24 ч и количество ОТС определяли с помощью Вестерн-блоттинга.
Содержащие как hAg, так TISU-элемент кодирующие hOTC SNIM-РНК обеспечивали сходный уровень экспрессии hOTC (фиг. 10А). Винкулин применяли в качестве гена «домашнего хозяйства» и интенсивность соответствующей ему полосы сравнивали с использованием денситометрии (фиг. 10Б).
II.б IV-применение конструкций Luc2 на мышах
Результаты представлены на фиг. 7 и фиг. 8.
Следующие конструкции применяли для IV-обработки мышей:
Luc2 (+8+А);
Luc2 (+8+Т);
Luc2 (+8+Т)+TISU;
Luc2-hAg;
Luc2-Sp30;
Luc2-SUSA UTR.
Получали комплекс, содержащий 20 мкг соответствующей SNIM-РНК и LF-44, и инъецировали IV мышам Balb/c. В качестве дополнительного контроля получали также последовательность Luc2, фланкированную энхансером человеческого CMV на 5'-конце (Luc2-SUSA) и 3'UTR человеческого гормона роста на 3'-конце. Данные о последовательностях, применяемых в качестве UTR в этой конструкции, получали из патента фирмы Shire (WO 2012/170930 A1: последовательность ID 1, фиг. 1.)
Визуализацию in vivo осуществляли через 6 ч после инъекции, применяя систему визуализации IVIS, и строили график зависимости для величин, выраженных в фотонах/с/см2/ср. Результаты, полученные при визуализации целого животного, представлены на фиг. 7А, а результаты, полученные при визуализации всего органа, представлены на фиг. 7Б (печень), 7 В (легкое), 7Е (селезенка) соответственно.
Органы, изъятые из животных, замораживали в жидком азоте, гомогенизировали, лизировали и измеряли активность люциферазы. Результаты представлены на фиг. 7Д (печень), 7Е (легкое), 7Ж (селезенка) соответственно.
Параметры крови животных анализировали, используя автоматический гематологический анализатор фирмы Sysmex KX-21N™: количество лейкоцитов (WBC) (фиг. 8А), эритроцитов (RBC) (фиг. 8Б), тромбоцитов (фиг. 8 В), уровень гемоглобина (фиг. 8Г) и гематокрит (фиг. 8Д) у мышей, обработанных различными кодирующими люциферазу конструкциями мРНК, существенно не отличались.
На фиг. 11 представлены предсказанные вторичные структуры произвольного состоящего из 30 нуклеотидов спейсера, присутствующего в последовательности №7 (слева), и 5'UTR человеческого альфа-глобина, присутствующего в последовательности №8 (справа). Хотя вторичные структуры обеих последовательностей даже не являются сходными, они приводят к сходным уровнями экспрессии (фиг. 6А и 6Б), которые в обоих случаях являются одинаково низкими по сравнению с T7Luc2(+8+T)-TISU.




















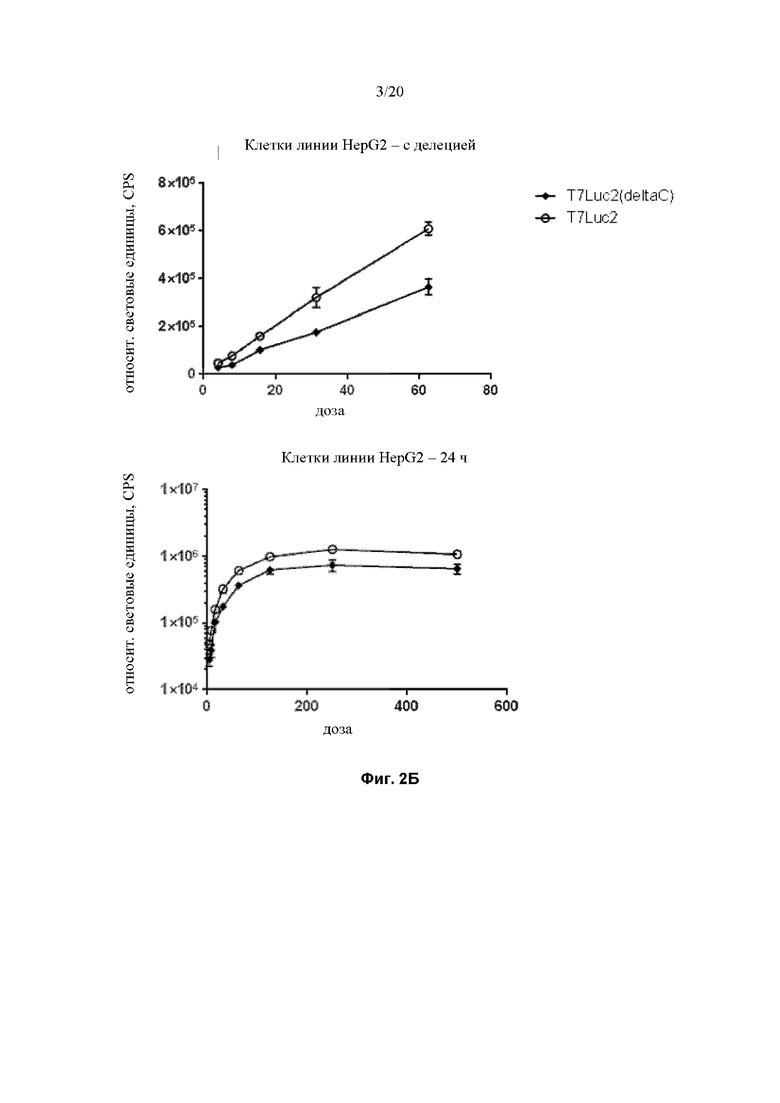
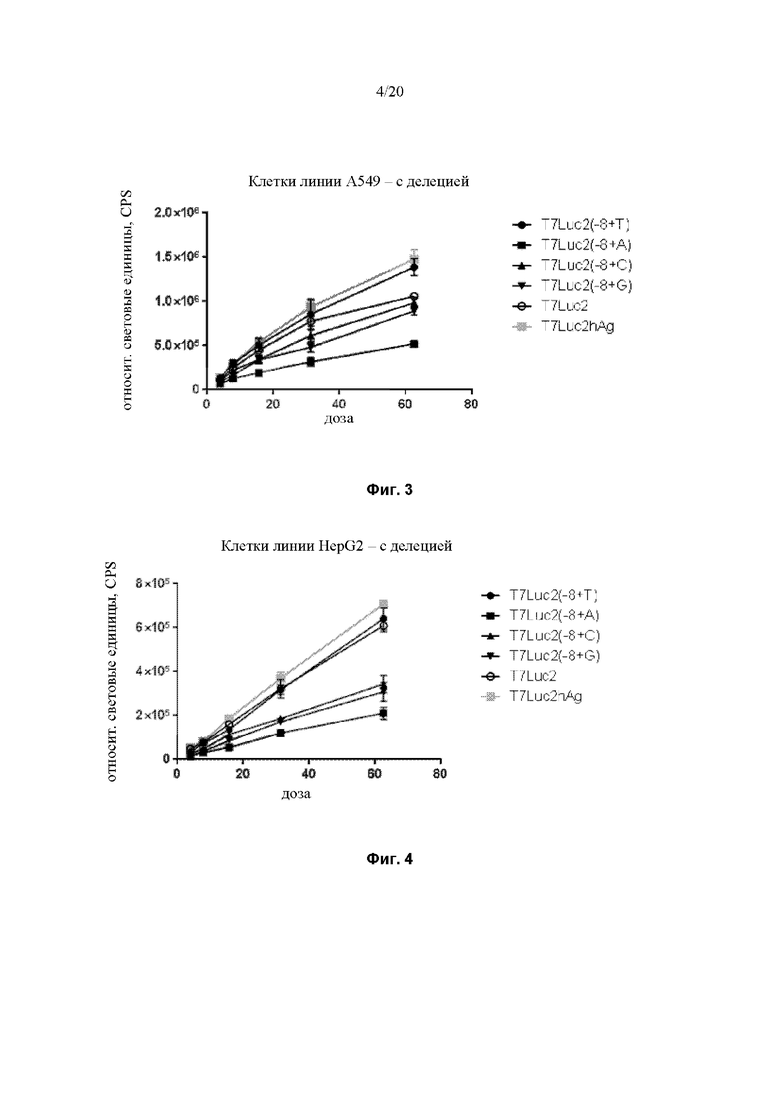


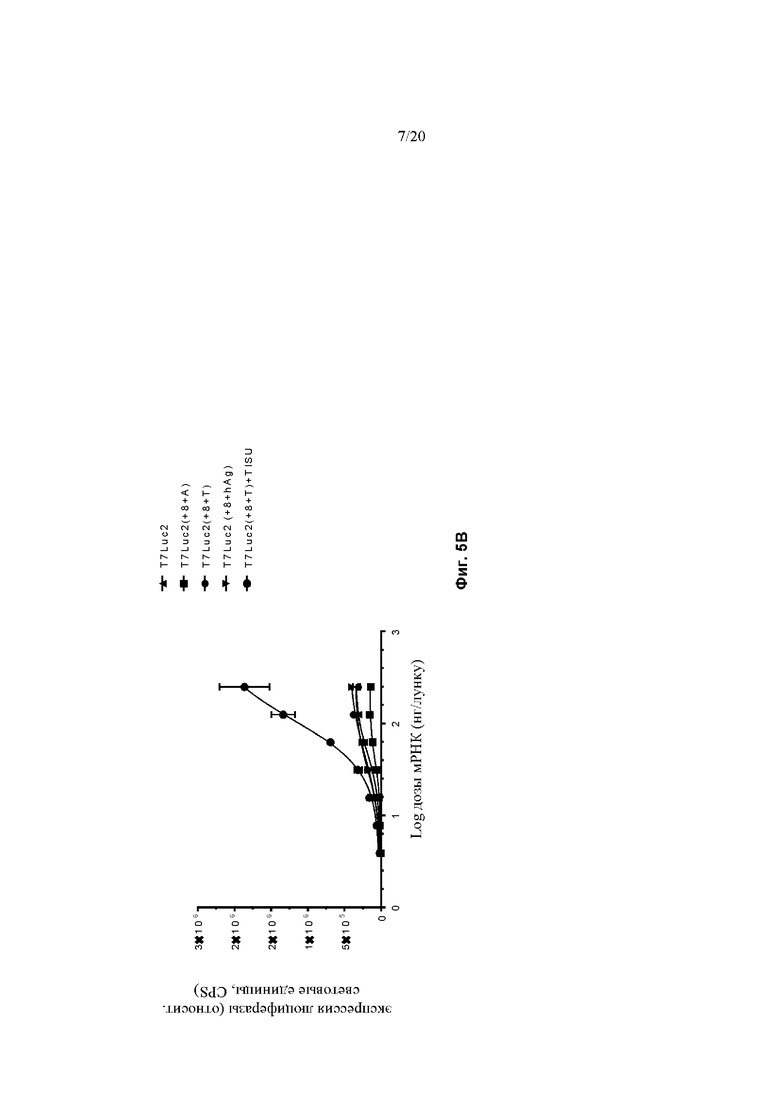
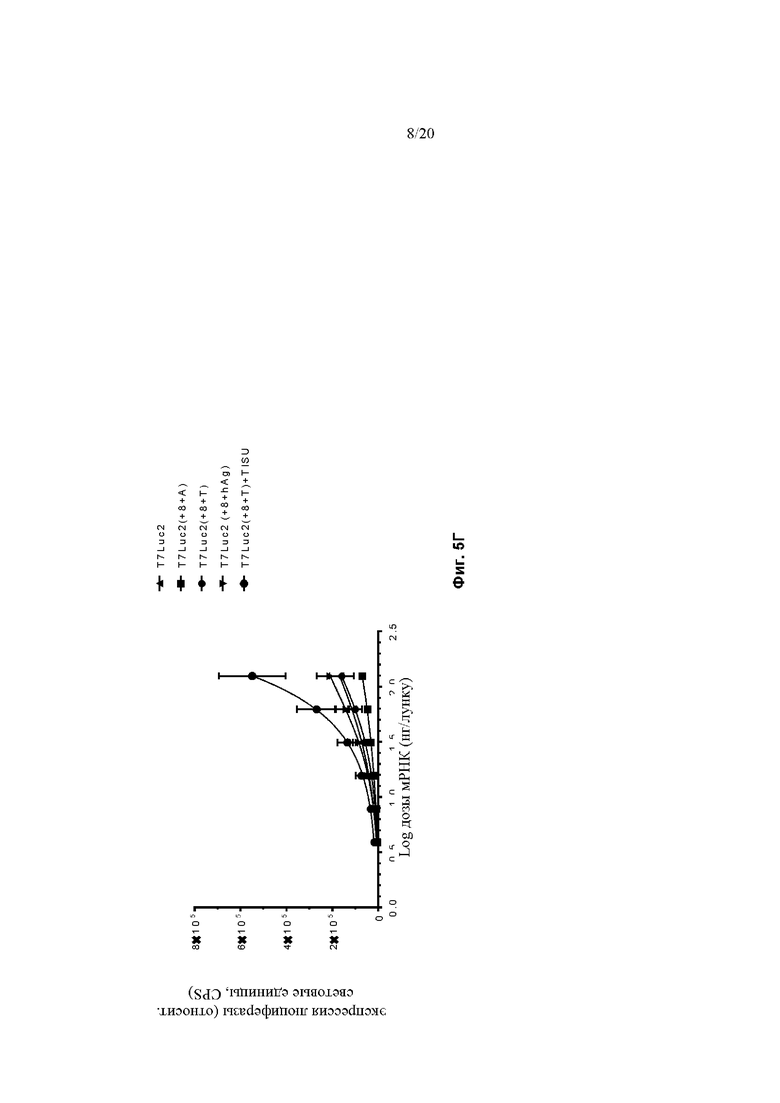



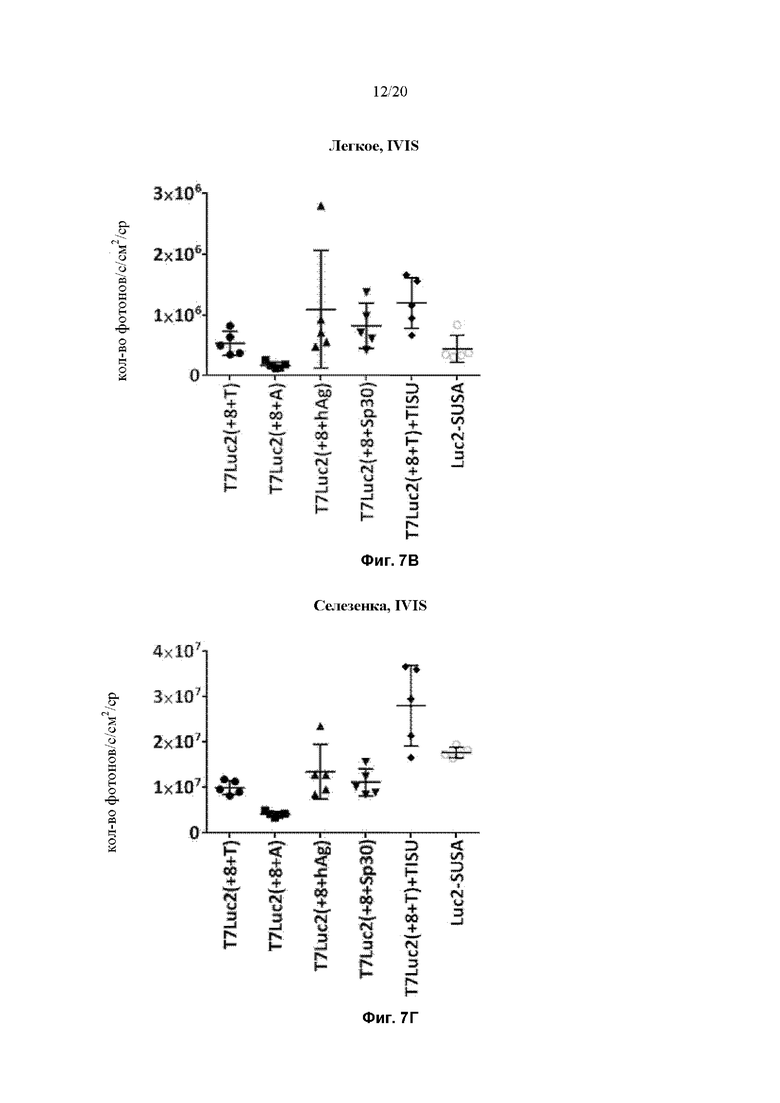
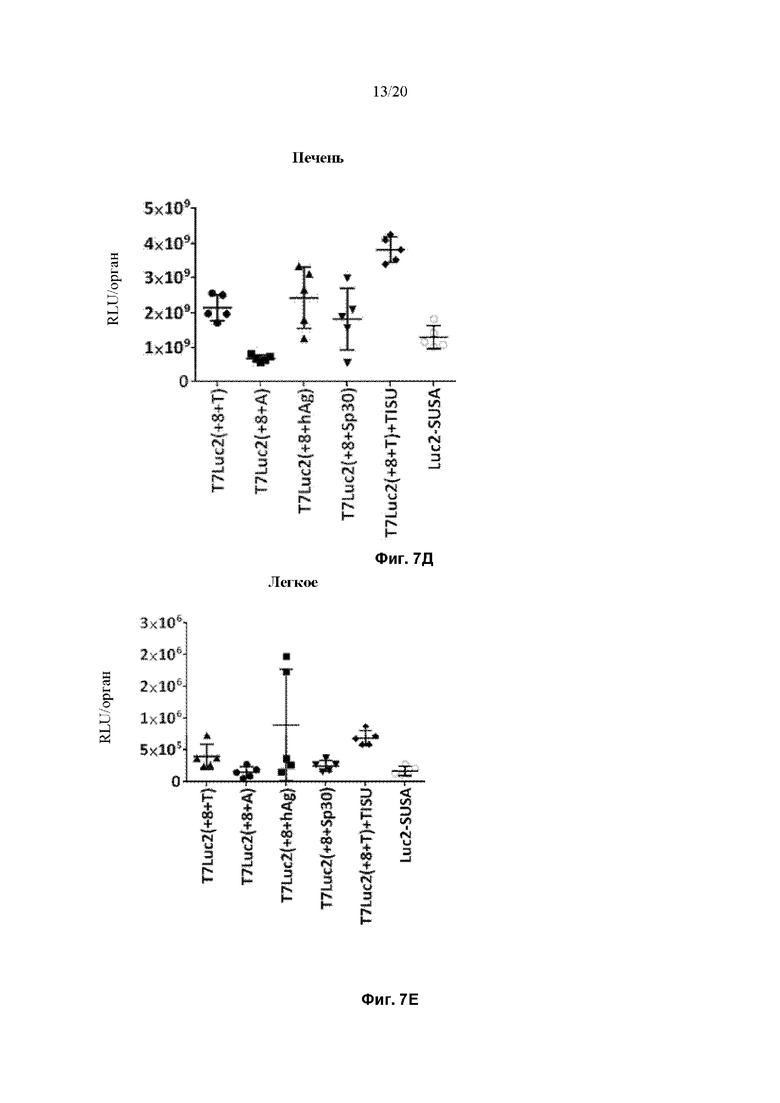
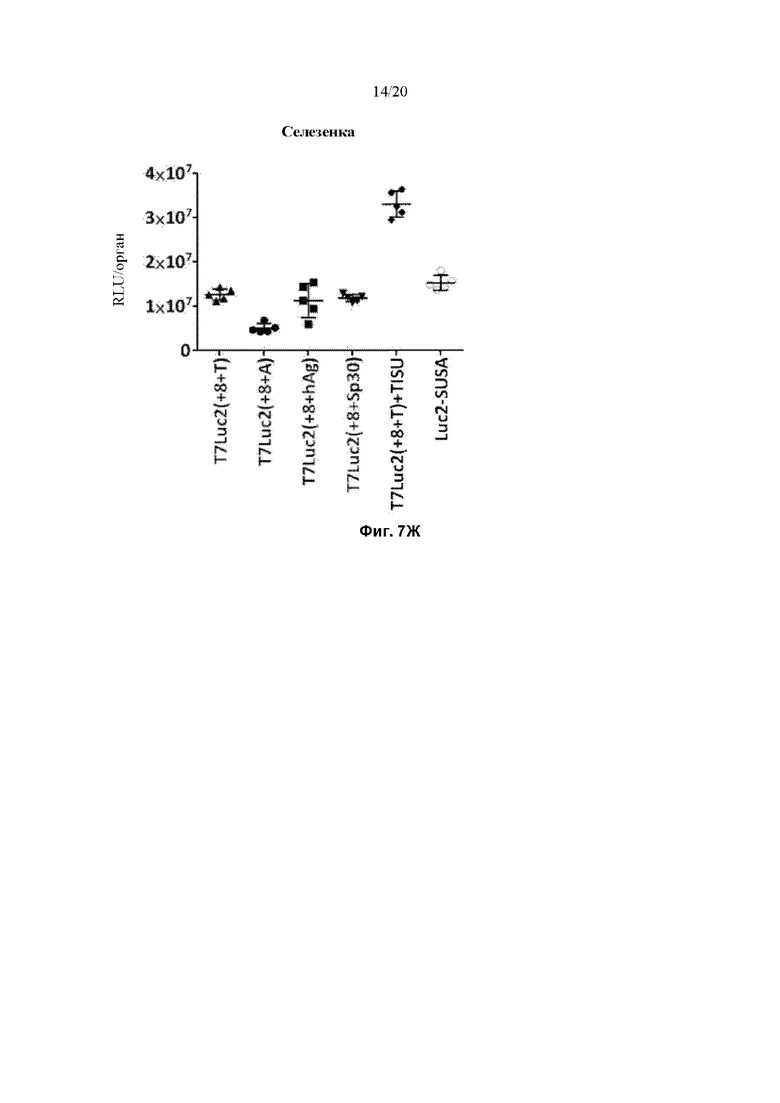
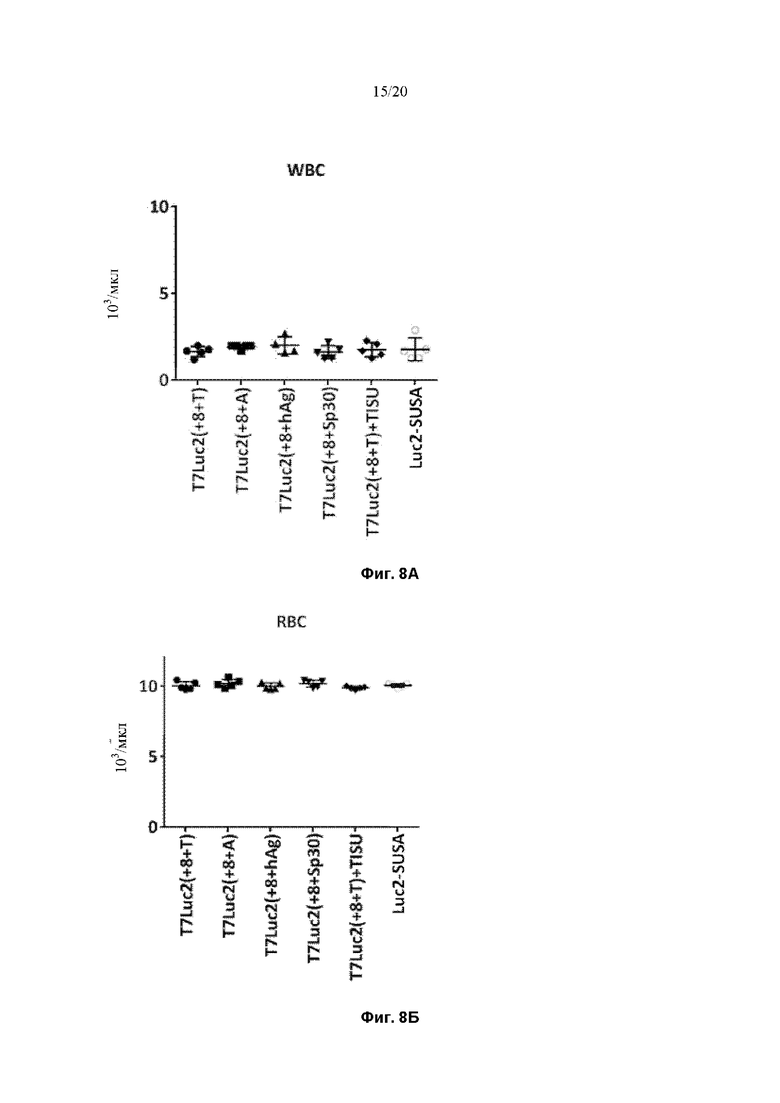
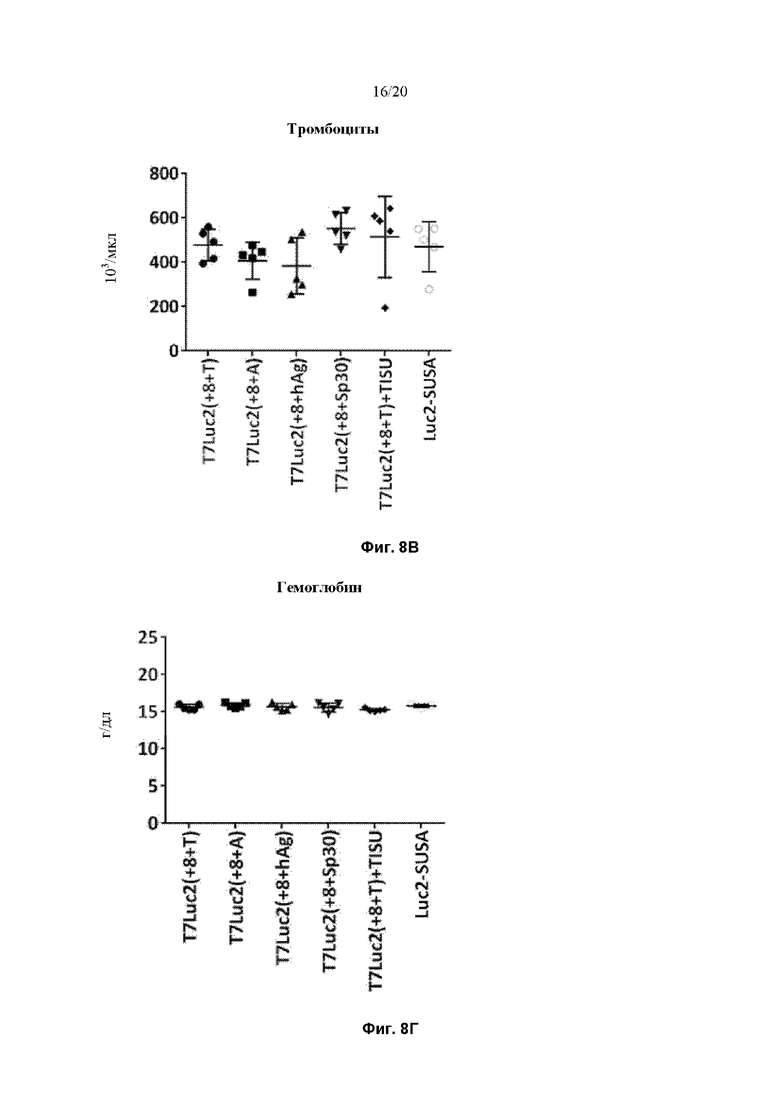

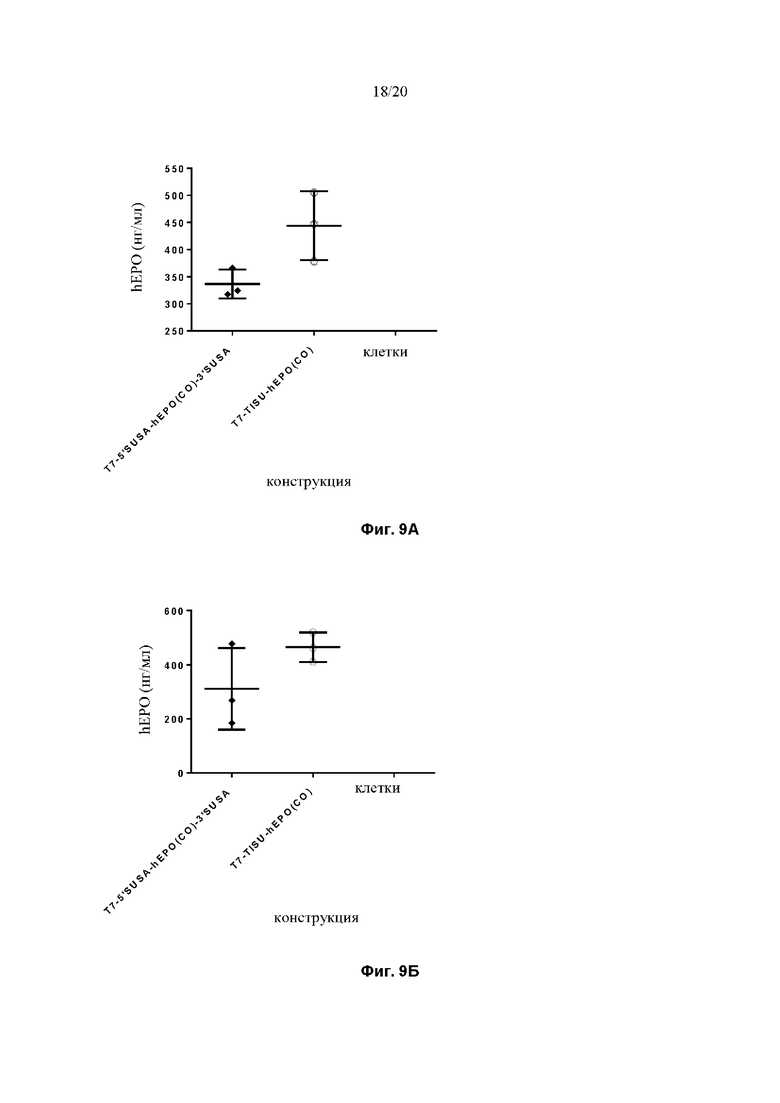
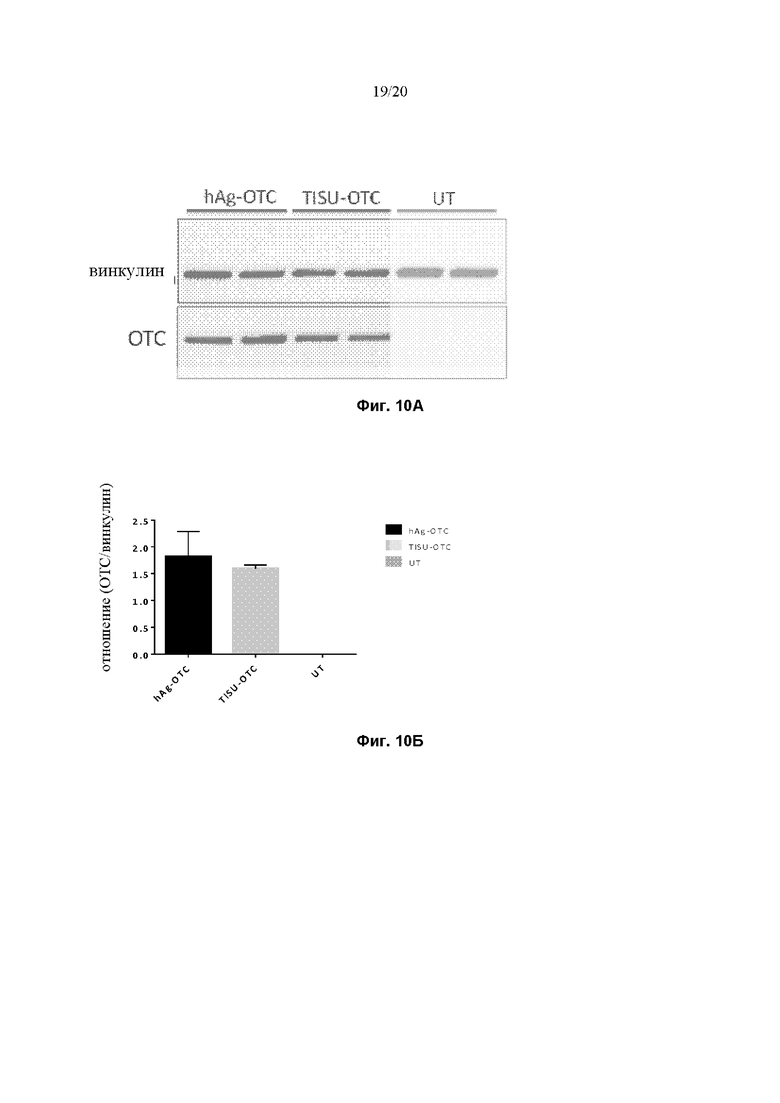
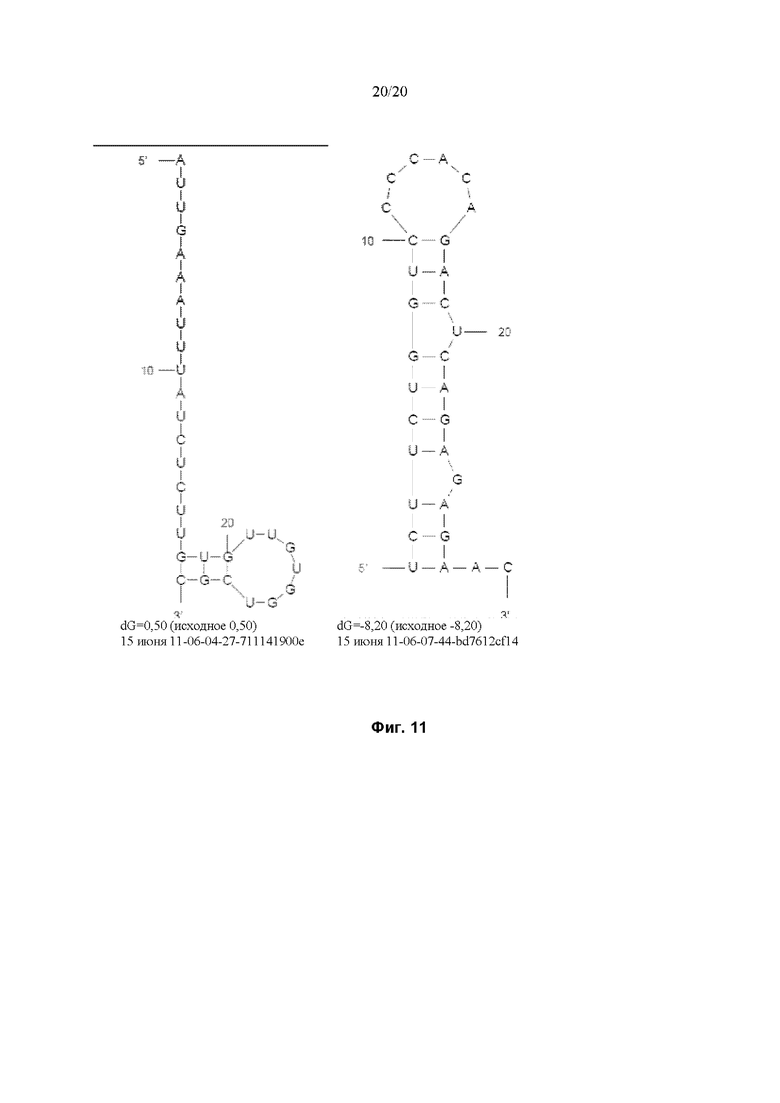
Изобретение относится к области биохимии, в частности к молекуле ДНК. Также раскрыты вектор экспрессии, содержащий указанную ДНК, клетка-хозяин, содержащая указанный вектор, композиция и набор, содержащие указанную молекулу ДНК. Раскрыто применение указанной молекулы ДНК. Изобретение позволяет эффективно увеличивать экспрессию терапевтических генов. 11 н. и 5 з.п. ф-лы, 1 табл., 28 ил., 1 пр.
1. Молекула ДНК, применяемая в генной терапии с целью увеличения экспрессии терапевтических генов, содержащая одну цепь со следующими элементами:
(a) кодирующая область, включая стартовый кодон на ее 5’-конце, которая кодирует полипептид, белок или пептид, обладающий профилактическим или терапевтическим действием;
(б) расположенная непосредственно в обратном направлении относительно указанной кодирующей последовательности последовательность, выбранная из группы, которая состоит из:
(б1) R1-CGCCACC (SEQ ID NO: 1);
или последовательности, где в указанной последовательности C в положении 6 SEQ ID NO: 1 заменен на A и C в положении 7 SEQ ID NO: 1 заменен на G; и/или A в положении 5 SEQ ID NO: 1 заменен на G; и
(б2) R1-CNGCCACC (SEQ ID NO: 2), в которой нуклеотид N в положении 2 SEQ ID NO: 2 представляет собой нуклеотид, выбранный из группы, которая состоит из T, G, C или A; или последовательности, где в указанной последовательности C в положении 7 SEQ ID NO: 2 заменен на A и C в положении 8 SEQ ID NO: 2 заменен на G; и/или A в положении 6 SEQ ID NO: 2 заменен на G,
где R1 обозначает промотор, который распознается ДНК-зависимой РНК-полимеразой;
или содержит комплементарную цепь, в которой промотор, распознаваемый ДНК-зависимой РНК-полимеразой, выбирают из группы, которая состоит из:
(I) TAATACGACTCACTATAGGGAGA (SEQ ID NO: 3), которая
распознается ДНК-зависимой РНК-полимеразой T7;
(II) AATTAACCCTCACTAAAGGGAGA (SEQ ID NO: 4), которая
распознается ДНК-зависимой РНК-полимеразой T3;
(III) ATTTAGGTGACACTATAGAAG (SEQ ID NO: 5), которая
распознается ДНК-зависимой РНК-полимеразой SP6; и
(IV) AATTAGGGCACACTATAGGGA (SEQ ID NO: 6, которая
распознается ДНК-зависимой РНК-полимеразой K11.
2. Молекула ДНК по п. 1, в которой нуклеотид N в положении 2 SEQ ID NO: 2 представляет собой нуклеотид, выбранный из группы, которая состоит из T, G или C, и в которой нуклеотид N не представляет собой A.
3. Молекула ДНК по п. 2, в которой указанный нуклеотид N в положении 2 SEQ ID NO: 2 представляет собой T.
4. Вектор экспрессии, содержащий молекулу ДНК по любому из пп. 1-3.
5. Клетка-хозяин, применяемая в генной терапии с целью увеличения экспрессии терапевтических генов, содержащая вектор по п. 4.
6. Композиция для генной терапии с целью увеличения экспрессии терапевтических генов, содержащая эффективное количество молекулы ДНК по любому из пп. 1-3, вектора по п. 4 или клетки-хозяина по п. 5.
7. Молекула РНК, применяемая в генной терапии с целью увеличения экспрессии терапевтических генов, которая содержит:
(a) кодирующую область, включающую стартовый кодон на ее 5’-конце, которая кодирует полипептид, белок или пептид, обладающий профилактическим или терапевтическим действием, и
(б) расположенную непосредственно в обратном направлении относительно указанной кодирующей последовательности UTR, выбранную из
группы, которая содержит:
(б1) UTR-последовательности
R2-CGCCACC (SEQ ID NO: 1),
или последовательности, где в указанной последовательности UTR C в положении 6 SEQ ID NO: 1 заменен на A и C в положении 7 SEQ ID NO: 1 заменен на G; и/или A в положении 5 SEQ ID NO: 1 заменен на G; и
(б2) UTR-последовательности
R2-CNGCCACC (SEQ ID NO: 2), в которой нуклеотид N в положении 2 SEQ ID NO: 2 представляет собой нуклеотид, выбранный из группы, которая состоит из U, G, C или A; или последовательности, где в указанной последовательности C в положении 7 SEQ ID NO: 2 заменен на A и C в положении 8 SEQ ID NO: 2 заменен на G; и/или A в положении 6 SEQ ID NO: 2 заменен на G,
где R2 обозначает последовательность РНК, соответствующую части промоторной области, начинающейся с нуклеотида, с которого ДНК-зависимая РНК-полимераза инициирует синтез РНК, в которой R2 выбирают из группы, состоящей из:
(I) GGGAGA (SEQ ID NO: 7);
(II) GGGAGA (SEQ ID NO: 8);
(III) GAAG (SEQ ID NO: 9); и
(IV) GGGA (SEQ ID NO: 10); и
где молекула РНК содержит поли-A-хвост на 3’-конце.
8. Молекула РНК по п. 7, в которой нуклеотид N в положении 2 SEQ ID NO: 2 представляет собой нуклеотид, выбранный из группы, которая состоит из U, G или C, и в которой нуклеотид N не представляет собой A.
9. Молекула РНК по п. 8, в которой указанный нуклеотид N в положении 2 SEQ ID NO: 2 представляет собой U.
10. Молекула РНК по любому из пп. 7-9, в которой поли-A-хвост состоит по меньшей мере из 120 нуклеотидов.
11. Молекула нуклеиновой кислоты, кодирующая молекулу РНК по любому из пп. 7-10.
12. Вектор экспрессии, содержащий молекулу нуклеиновой кислоты по п. 11.
13. Клетка-хозяин, применяемая в генной терапии с целью увеличения экспрессии терапевтических генов, содержащая вектор по п. 12.
14. Фармацевтическая композиция, предназначенная для генной терапии на основе РНК с целью увеличения экспрессии терапевтических генов, содержащая эффективное количество молекулы РНК по любому из пп. 7-10, молекулы нуклеиновой кислоты по п. 11, вектора по п. 12 или клетки-хозяина по п. 13 и фармацевтически приемлемый носитель.
15. Набор, предназначенный для генной терапии на основе РНК с целью увеличения экспрессии терапевтических генов, содержащий молекулу ДНК по любому из пп. 1-3, молекулу РНК по любому из пп. 7-10, молекулу нуклеиновой кислоты по п. 11, вектор по п. 4 или 12 или клетку-хозяина по п. 5 или 13 и инструкцию.
16. Применение UTR в составе молекулы РНК по п. 7(б) для трансляции кодирующей области молекулы РНК в полипептид, белок или пептид, обладающий профилактическим или терапевтическим действием, кодируемый указанной кодирующей областью.
| CN 104032031 A, 10.09.2014 | |||
| US 20110244519 A1, 06.10.2011 | |||
| ГЕНЫ CORYNEBACTERIUM GLUTAMICUM, КОДИРУЮЩИЕ БЕЛКИ, УЧАСТВУЮЩИЕ В МЕТАБОЛИЗМЕ УГЛЕРОДА И ПРОДУЦИРОВАНИИ ЭНЕРГИИ | 2005 |
|
RU2321634C2 |

