ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
[0001] Терапевтические средства на основе антител открывают значительные перспективы в лечении ряда заболеваний. Разрабатывается множество форматов, в том числе моноклональные, мышиные, химерные, гуманизированные, человеческие, полноразмерные, Fab, пэгилированные, содержащие радиоизотопную метку, конъюгированные с лекарственным средством, полиспецифические антитела и т.д. (см., например, Reichert, J.M., 2012, mAbs 4:3, 413-415; Nixon, A.E. et al., 2014, mAbs 6:1, 73-85; включенные в данный документ посредством ссылки). Из более чем 40 терапевтических средств на основе антител, которые прошли регистрацию для маркетинга в Соединенных Штатах Америки или Европе, все были получены с применением технологий, которые основаны на сборке генов традиционных антител, полученных от человека и/или отличных от человека источников (например, мыши), с помощью in vitro (например, фагового дисплея) или in vivo (например, генетически модифицированных животных) систем. Тем не менее, разработка особенно эффективных средств на основе антител, которые связывают мишени заболеваний, с трудом поддающихся лечению, остается сложной задачей.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[0002] В данном документе раскрыто признание того, что требуется конструирование животных, отличных от человека, чтобы обеспечить улучшенные in vivo системы для идентификации и разработки новых терапевтических средств на основе антител и, в некоторых вариантах осуществления, средств, представляющих собой антитела (например, моноклональные антитела и/или их фрагменты), которые можно применять для лечения множества заболеваний, характеризующихся наличием мишеней заболеваний, которые с трудом поддаются лечению. Кроме того, в данном документе раскрыто признание того, что требуются животные, отличные от человека, имеющие сконструированные кластер/участок разнообразия тяжелой цепи (DH) в пределах вариабельного участка тяжелой цепи иммуноглобулина (например, гетерологичного вариабельного участка тяжелой цепи иммуноглобулина), в частности, сконструированный кластер DH (или участок DH), содержащий нуклеотидные кодирующие последовательности, не встречающиеся в природе, в пределах вариабельного участка тяжелой цепи иммуноглобулина, и/или иным образом экспрессирующие, содержащие или продуцирующие антитела, содержащие определяющие комплементарность участки (CDR), которые характеризуются разнообразием, которое управляет связыванием с определенными антигенами, например, для применения в идентификации и разработке терапевтических средств на основе антител, которые могут нацеливаться, например, на трансмембранные или цитоплазматические полипептиды. В некоторых вариантах осуществления животные, отличные от человека, раскрываемые в данном документе, представляют собой in vivo системы для разработки антител и/или терапевтических средств на основе антител для введения людям.
[0003] В некоторых вариантах осуществления представлено животное, отличное от человека, геном которого, например геном зародышевой линии, содержит вариабельный участок тяжелой цепи иммуноглобулина, который включает сконструированный участок DH, где сконструированный участок DH включает одну или более нуклеотидных последовательностей, каждая из которых кодирует представляющий интерес полипептид, не являющийся иммуноглобулином, или его часть.
[0004] В другом аспекте животные, отличные от человека, геном которых, например геном зародышевой линии, модифицирован таким образом, что содержит вариабельный участок тяжелой цепи иммуноглобулина, который включает сконструированный участок DH, где сконструированный участок DH включает одну или более нуклеотидных последовательностей, каждая из которых кодирует представляющий интерес полипептид, не являющийся иммуноглобулином, или его часть, при этом он может быть дополнительно модифицирован так, чтобы экспрессировать единую перегруппированную легкую цепь, например, общую легкую цепь (ULC).
[0005] Генная последовательность единого перегруппированного вариабельного участка легкой цепи, функционально связанная с константным участком легкой цепи, также называемые как общая или универсальная легкая цепь (ULC), может кодироваться локусом легкой цепи, содержащим генную последовательность единого перегруппированного участка VL:JL. В некоторых вариантах осуществления локус легкой цепи содержит генную последовательность единого перегруппированного участка VL:JL, при этом последовательность участка VL представляет собой генную последовательность Vκ. В некоторых аспектах последовательность Vκ выбрана из Vκ1-39 или Vκ3-20. В некоторых аспектах последовательность JL представляет собой генную последовательность Jκ, например, последовательность Jκ1, последовательность Jκ2, последовательность Jκ3, последовательность Jκ4 или последовательность Jκ5 и т.д. В некоторых вариантах осуществления локус легкой цепи содержит последовательность единого перегруппированного участка Vκ:Jκ, выбранную из группы, состоящей из Vκ1-39Jκ5 и Vκ3-20Jκ1. В одном варианте осуществления локус легкой цепи содержит последовательность единого перегруппированного участка Vκ:Jκ, представляющую собой Vκ1-39Jκ5. В другом варианте осуществления локус легкой цепи содержит последовательность единого перегруппированного участка Vκ:Jκ, представляющую собой Vκ3-20Jκ1. В некоторых вариантах осуществления генная последовательность единого перегруппированного вариабельного участка функционально связана с геном константного участка легкой цепи, отличной от человеческой, например, эндогенным геном константного участка легкой цепи, отличной от человеческой. В другом варианте осуществления генная последовательность единого перегруппированного вариабельного участка функционально связанна с геном константного участка легкой цепи человека. В некоторых аспектах генная последовательность единого перегруппированного вариабельного участка представляет собой последовательность участка V:J человека, встроенную в эндогенный локус легкой цепи иммуноглобулина, вследствие чего полученное животное, отличное от человека, не содержит функциональных неперегруппированных генных сегментов V и/или J в одном или более локусах легкой цепи.
[0006] В некоторых вариантах осуществления представлены выделенные клетка или ткань, отличные от человеческих, геном которых содержит вариабельный участок тяжелой цепи иммуноглобулина, который включает сконструированный участок DH, где сконструированный участок DH включает одну или более нуклеотидных последовательностей, каждая из которых кодирует представляющий интерес полипептид, не являющийся иммуноглобулином, или его часть, и необязательно общую или универсальную легкую цепь. В некоторых вариантах осуществления клетка происходит из лимфоидной или миелоидной линии. В некоторых вариантах осуществления клетка представляет собой лимфоцит. В некоторых вариантах осуществления клетка выбрана из B-клетки, дендритной клетки, макрофага, моноцита и T-клетки. В некоторых вариантах осуществления ткань выбрана из ткани жировой клетчатки, мочевого пузыря, головного мозга, молочной железы, костного мозга, глаза, сердца, кишечника, почки, печени, легкого, лимфатического узла, мышцы, поджелудочной железы, плазмы крови, сыворотки крови, кожи, селезенки, желудка, вилочковой железы, яичка, яйцеклетки и их комбинации.
[0007] В некоторых вариантах осуществления представлена иммортализированная клетка, полученная из выделенной клетки, отличной от человеческой, описанной в данном документе.
[0008] В некоторых вариантах осуществления представлена эмбриональная стволовая (ES) клетка, отличная от человеческой, геном которой содержит вариабельный участок тяжелой цепи иммуноглобулина, который включает сконструированный участок DH, где сконструированный участок DH включает одну или более нуклеотидных последовательностей, каждая из которых кодирует представляющий интерес полипептид, не являющийся иммуноглобулином, или его часть, и необязательно общую легкую цепь. В некоторых вариантах осуществления эмбриональная стволовая клетка, отличная от человеческой, представляет собой эмбриональную стволовую клетку грызуна. В некоторых определенных вариантах осуществления эмбриональная стволовая клетка грызуна представляет собой эмбриональную стволовую клетку мыши и получена от мыши линии 129, линии C57BL или их гибрида. В некоторых определенных вариантах осуществления эмбриональная стволовая клетка грызуна представляет собой эмбриональную стволовую клетку мыши, и мышь является гибридом линий 129 и C57BL.
[0009] В некоторых вариантах осуществления представлена зародышевая клетка, отличная от человеческой, геном которой содержит вариабельный участок тяжелой цепи иммуноглобулина, который включает сконструированный участок DH, где сконструированный участок DH включает одну или более нуклеотидных последовательностей, каждая из которых кодирует представляющий интерес полипептид, не являющийся иммуноглобулином, или его часть, и необязательно общую легкую цепь. В некоторых вариантах осуществления зародышевая клетка, отличная от человеческой, представляет зародышевую клетку грызуна. В некоторых определенных вариантах осуществления эмбриональная клетка грызуна представляет собой эмбриональную клетку мыши и получена от мыши линии 129, линии C57BL или их гибрида. В некоторых определенных вариантах осуществления эмбриональная клетка грызуна представляет собой эмбриональную клетку мыши, и мышь является гибридом линий 129 и C57BL.
[0010] В некоторых вариантах осуществления представлено применение эмбриональной стволовой клетки или зародышевой клетки, отличных от человеческих, описываемых в данном документе для получения животного, отличного от человека. В некоторых определенных вариантах осуществления эмбриональная стволовая клетка или зародышевая клетка, отличные от человеческих, представляют собой мышиную эмбриональную стволовую клетку или зародышевую клетку мыши, и их применяют для получения мыши, содержащей вариабельный участок тяжелой цепи иммуноглобулина, который включает сконструированный участок DH, и необязательно общую легкую цепь, описываемую в данном документе. В некоторых определенных вариантах осуществления эмбриональная стволовая клетка или зародышевая клетка, отличные от человеческих, представляют собой эмбриональную стволовую клетку или зародышевую клетку крысы, и их применяют для получения крысы, содержащей вариабельный участок тяжелой цепи иммуноглобулина, который включает сконструированный участок DH, и необязательно общую легкую цепь, описываемую в данном документе.
[0011] В некоторых вариантах осуществления предусмотрен эмбрион, отличный от человеческого, содержащий эмбриональную стволовую клетку, отличную от человеческой, описанную в данном документе, созданный из нее, полученный из нее или образованный из нее. В некоторых определенных вариантах осуществления эмбрион, отличный от человеческого, представляет собой эмбрион грызуна; в некоторых вариантах осуществления эмбрион мыши; в некоторых вариантах осуществления эмбрион крысы.
[0012] В некоторых вариантах осуществления представлено применение эмбриона, отличного от человеческого, описанного в данном документе, для получения животного, отличного от человека. В некоторых определенных вариантах осуществления эмбрион, отличный от человеческого, представляют собой эмбрион мыши, и его применяют для получения мыши, содержащей вариабельный участок тяжелой цепи иммуноглобулина, который включает сконструированный участок DH, и необязательно общую легкую цепь, описываемую в данном документе. В некоторых определенных вариантах осуществления эмбрион, отличный от человеческого, представляют собой эмбрион крысы, и его применяют для получения крысы, содержащей вариабельный участок тяжелой цепи иммуноглобулина, который включает сконструированный участок DH, и необязательно общую легкую цепь, описываемую в данном документе.
[0013] В некоторых вариантах осуществления представлен набор, содержащий выделенную клетку или ткань, отличные от человеческих, описываемые в данном документе, иммортализированную клетку, описываемую в данном документе, эмбриональную стволовую клетку, отличную от человеческой, описываемую в данном документе, эмбрион, отличный от человеческого, описываемый в данном документе, или животное, отличное от человека, описываемое в данном документе.
[0014] В некоторых вариантах осуществления представлен набор, описываемый в данном документе, для применения в производстве и/или разработке лекарственного средства (например, антитела или его антиген-связывающего фрагмента) для терапии или диагностики.
[0015] В некоторых вариантах осуществления представлен набор, описываемый в данном документе, для применения в производстве и/или разработке лекарственного средства (например, антитела или его антиген-связывающего фрагмента) для лечения, профилактики или улучшения заболевания, нарушения или состояния.
[0016] В некоторых вариантах осуществления представлены трансген, конструкция нуклеиновой кислоты, ДНК-конструкция или нацеливающий вектор, описываемые в данном документе. В некоторых вариантах осуществления предусмотрен трансген, конструкция нуклеиновой кислоты, ДНК-конструкция или нацеливающий вектор, содержащие сконструированный участок DH, описываемый в данном документе. В некоторых определенных вариантах осуществления трансген, конструкция нуклеиновой кислоты, ДНК-конструкция или нацеливающий вектор содержат фрагмент ДНК, который включает одну или более нуклеотидных кодирующих последовательностей, описываемых в данном документе. В некоторых определенных вариантах осуществления трансген, конструкция нуклеиновой кислоты, ДНК-конструкция или нацеливающий вектор содержат сконструированный участок DH, который содержит одну или более нуклеотидных кодирующих последовательностей, выбранных из таблицы 3 или таблицы 4, при этом каждая одна или более нуклеотидных кодирующих последовательностей фланкированы последовательностью сигнала рекомбинации, выбранной из представленных на фигуре 2. В некоторых определенных вариантах осуществления трансген, конструкция нуклеиновой кислоты, ДНК-конструкция или нацеливающий вектор дополнительно содержат один или более селективных маркеров. В некоторых определенных вариантах осуществления трансген, конструкция нуклеиновой кислоты, ДНК-конструкция или нацеливающий вектор дополнительно содержат один или более сайтов для сайт-специфической рекомбинации (например, lox, Frt или их комбинации). В некоторых определенных вариантах осуществления трансген, конструкция нуклеиновой кислоты, ДНК-конструкция или нацеливающий вектор изображены на любой из фигур 3A, 3B, 4A, 4B, 7A, 7B, 8A и 8B.
[0017] В некоторых вариантах осуществления представлено применение трансгена, конструкции нуклеиновой кислоты, ДНК-конструкции или нацеливающего вектора, описываемых в данном документе, для получения эмбриональной стволовой клетки, отличной от человеческой, клетки, отличной от человеческой, эмбриона, отличного от человеческого, и/или животного, отличного от человека.
[0018] В некоторых вариантах осуществления представляющий интерес полипептид, не являющийся иммуноглобулином, представляет собой хемокиновый рецептор. В некоторых вариантах осуществления хемокиновый рецептор выбран из группы, состоящей из CC-хемокинового рецептора (илиβ-хемокинового рецептора), CXC-хемокинового рецептора, CX3C-хемокинового рецептора и XC-хемокинового рецептора. В некоторых вариантах осуществления хемокиновый рецептор представляет собой атипический хемокиновый рецептор (ACKR). В некоторых вариантах осуществления ACKR выбран из группы, состоящей из ACKR1, ACKR2, ACKR3 и ACKR4. В некоторых определенных вариантах осуществления ACKR представляет собой ACKR2 или D6 хемокиновый рецептор-ловушку.
[0019] В некоторых вариантах осуществления представляющий интерес полипептид, не являющийся иммуноглобулином, представляет собой токсин. В некоторых вариантах осуществления токсин представляет собой токсин, который обнаружен в яде тарантула, паука, скорпиона или морской анемоны.
[0020] В некоторых вариантах осуществления представляющий интерес полипептид, не являющийся иммуноглобулином, представляет собой конотоксин или токсин тарантула. В некоторых вариантах осуществления конотоксин выбран из группы, состоящей из α-конотоксина, δ-конотоксина, κ-конотоксина, μ-конотоксина, ω-конотоксина и их комбинаций. В некоторых определенных вариантах осуществления конотоксин представляет собой μ-конотоксин. В некоторых определенных вариантах осуществления токсин тарантула представляет собой ProTxI, ProTxII, хувентоксин-IV (HWTX-IV) или их комбинации.
[0021] В некоторых вариантах осуществления сконструированный участок DH включает 5, 10, 15, 20, 25 или больше нуклеотидных последовательностей, каждая из которых кодирует внеклеточную часть D6 хемокинового рецептора-ловушки, или каждая из которых кодирует часть конотоксина (например, μ-конотоксина), или каждая из которых кодирует часть токсина тарантула (например, ProTxI, ProTxII и т.д.) или их комбинации.
[0022] В некоторых вариантах осуществления сконструированный участок DH включает 25 нуклеотидных последовательностей, каждая из которых кодирует внеклеточную часть D6 хемокинового рецептора-ловушки, или включает 26 нуклеотидных последовательностей, каждая из которых кодирует часть конотоксина (например, μ-конотоксина) и/или токсина тарантула (например, ProTxI, ProTxII и т.д.).
[0023] В некоторых вариантах осуществления внеклеточная часть D6 хемокинового рецептора-ловушки выбрана из группы, состоящей из N-концевого участка, внеклеточной петли и их комбинаций.
[0024] В некоторых вариантах осуществления часть конотоксина, описываемая в данном документе, включает последовательность, которая содержит одну или более дисульфидных связей. В некоторых вариантах осуществления часть конотоксина, описываемая в данном документе, включает последовательность, в которой отсутствует одна или более дисульфидных связей по сравнению с последовательностью конотоксина, которая встречается в природе (например, эталонной или исходной последовательностью конотоксина). В некоторых вариантах осуществления часть конотоксина, описываемая в данном документе, включает последовательность, которая характеризуется определенным количеством и/или паттерном дисульфидных связей, которые являются одинаковыми или отличающимися по сравнению с последовательностью конотоксина, которая встречается в природе (например, эталонной или исходной последовательностью конотоксина).
[0025] В некоторых вариантах осуществления часть токсина тарантула, описываемая в данном документе, включает последовательность, которая содержит мотив цистеинового узла, встречающийся в последовательности токсина тарантула, обнаруженной в природе. В некоторых вариантах осуществления часть токсина тарантула, описываемая в данном документе, включает последовательность, которая представляет собой или содержит пептид(ы) цистеинового узла. В некоторых вариантах осуществления часть токсина тарантула, описываемая в данном документе, включает последовательность, в которой отсутствует одна или более дисульфидных связей по сравнению с последовательностью токсина тарантула, которая встречается в природе (например, эталонной или исходной последовательностью токсина тарантула). В некоторых вариантах осуществления часть токсина тарантула, описываемая в данном документе, включает последовательность, которая характеризуется определенным количеством и/или паттерном дисульфидных связей, которые являются одинаковыми или отличающимися по сравнению с последовательностью токсина тарантула, которая встречается в природе (например, эталонной или исходной последовательностью токсина тарантула).
[0026] В некоторых вариантах осуществления сконструированный участок DH включает 25 или 26 нуклеотидных последовательностей, которые по меньшей мере на 50%, 60%, 70%, 80%, 90%, 95% или по меньшей мере 98% идентичны нуклеотидной последовательности, которая находится в таблице 3 или таблице 4, и/или кодируют аминокислотную последовательность, которая представлена в таблице 3 или таблице 4. В некоторых вариантах осуществления сконструированный участок DH включает 25 или 26 нуклеотидных последовательностей, каждая из которых кодирует последовательность, которая практически идентична или идентична аминокислотной последовательности, которая находится в таблице 3 или таблице 4, или которая обладает той же самой функцией, что и аминокислотная последовательность, которая представлена в таблице 3 или 4.
[0027] В некоторых вариантах осуществления одна или более нуклеотидных последовательностей содержат одну или более нуклеотидных замен, которые увеличивают частоту возникновения соматической гипермутации в одной или более нуклеотидных последовательностях.
[0028] В некоторых вариантах осуществления сконструированный участок DH дополнительно включает первую и вторую последовательности сигнала рекомбинации, фланкирующие каждую из одной или более нуклеотидных последовательностей. В некоторых определенных вариантах осуществления первая последовательность сигнала рекомбинации содержит последовательность, которая по меньшей мере на 50%, 60%, 70%, 80%, 90%, 95% или по меньшей мере 98% идентична первой последовательности сигнала рекомбинации, которая представлена на фигуре 2. В некоторых определенных вариантах осуществления первая последовательность сигнала рекомбинации содержит последовательность, которая практически идентична или идентична первой последовательности сигнала рекомбинации, которая представлена на фигуре 2. В некоторых определенных вариантах осуществления вторая последовательность сигнала рекомбинации содержит последовательность, которая по меньшей мере на 50%, 60%, 70%, 80%, 90%, 95% или по меньшей мере 98% идентична второй последовательности сигнала рекомбинации, которая представлена на фигуре 2. В некоторых определенных вариантах осуществления вторая последовательность сигнала рекомбинации содержит последовательность, которая практически идентична или идентична второй последовательности сигнала рекомбинации, которая представлена на фигуре 2. В некоторых вариантах осуществления первая и вторая последовательности сигнала рекомбинации выбраны из представленных на фигуре 2.
[0029] В некоторых вариантах осуществления в геноме представленного животного, отличного от человека, отсутствует один или более эндогенных генных сегментов DH дикого типа. В некоторых определенных вариантах осуществления в геноме представленного животного, отличного от человека, отсутствуют все или практически все эндогенные генные сегменты DH дикого типа. В некоторых вариантах осуществления в геноме представленного животного, отличного от человека, отсутствуют одна или болееэндогенных последовательностей сигнала рекомбинации дикого типа.
[0030] В некоторых вариантах осуществления сконструированный участок DH содержит один или более генных сегментов DH человека дикого типа. В некоторых определенных вариантах осуществления сконструированный участок DH содержит генный сегмент DH6-25 человека. В некоторых определенных вариантах осуществления в сконструированном участке DH отсутствует генный сегмент DH6-25 человека.
[0031] В некоторых вариантах осуществления вариабельный участок тяжелой цепи иммуноглобулина функционально связан с константным участком тяжелой цепи иммуноглобулина.
[0032] В некоторых вариантах осуществления константный участок тяжелой цепи иммуноглобулина представляет собой эндогенный константный участок тяжелой цепи иммуноглобулина.
[0033] В некоторых вариантах осуществления вариабельный участок тяжелой цепи иммуноглобулина представляет собой неперегруппированный вариабельный участок тяжелой цепи иммуноглобулина человека, например, содержащий по меньшей мере один неперегруппированный генный сегмент VH человека (h) и/или по меньшей мере один неперегруппированный генный сегмент JH человека (h), фланкирующие сконструированный (e) участок DH. В некоторых вариантах осуществления вариабельный участок тяжелой цепи иммуноглобулина содержит несколько неперегруппированных генных сегментов VH человека (h) и/или несколько из одного неперегруппированного генного сегмента JH человека (h), фланкирующие сконструированный (e) участок DH. В некоторых вариантах осуществления неперегруппированный вариабельный участок тяжелой цепи иммуноглобулина человека функционально связан с константным участком тяжелой цепи иммуноглобулина, например, константным участком тяжелой цепи иммуноглобулина, отличного от человеческого, например, в эндогенном локусе тяжелой цепи, отличной от человеческой.
[0034] В некоторых вариантах осуществления вариабельный участок тяжелой цепи иммуноглобулина человека содержит перегруппированный вариабельный участок тяжелой цепи иммуноглобулина человека, где перегруппированная тяжелая цепь иммуноглобулина человека содержит по меньшей мере один неперегруппированный генный сегмент VH человека (h) и/или по меньшей мере один неперегруппированный генный сегмент JH человека (h), рекомбинированные со сконструированным (e) участком DH с образованием генной последовательности перегруппированного участка (h)VH/(e)DH/(h)JH, которая может быть функционально связана с константным участком тяжелой цепи. В некоторых вариантах осуществления такая рекомбинация происходит в B-клетке во время развития B-клетки.
[0035] Соответственно, описанное в данном документе животное, отличное от человека, может содержать:
(i) зародышевую клетку, содержащую неперегруппированный вариабельный участок тяжелой цепи человека, содержащий:
(a) по меньшей мере один или несколько неперегруппированных генных сегментов VH человека (h),
(b) по меньшей мере один или несколько неперегруппированных генных сегментов JH человека (h), и
(c) сконструированный (e) участок DH, фланкированный (a) и (b), где (a), (b) и (c) рекомбинируют с образованием последовательности перегруппированного вариабельного участка тяжелой цепи человека hVH/eDH/hJH; и
(ii) соматическую клетку, например, B-клетку, содержащую генную последовательность перегруппированного участка hVH/eDH/hJH человека, где генная последовательность перегруппированного участка hVH/eDH/hJH содержит последовательность, кодирующую CDR3, содержащую одну или более нуклеотидных последовательностей, которые кодируют представляющий интерес полипептид, не являющийся иммуноглобулином, или его часть, или его вариант, подвергнутый соматической гипермутации. В некоторых вариантах осуществления человеческий неперегруппированный или перегруппированный вариабельный участок тяжелой цепи человека функционально связан с константным участком тяжелой цепи, который может представлять собой константный участок тяжелой цепи, отличной от человеческой, например, в эндогенном локусе тяжелой цепи, отличной от человеческой. В некоторых вариантах осуществления вариабельный участок тяжелой цепи иммуноглобулина представляет собой вариабельный участок тяжелой цепи иммуноглобулина человека. В некоторых вариантах осуществления в В-клетке дополнительно экспрессируется генная последовательность перегруппированного участка hVH/eDH/hJH, которая функционально связана с константным участком тяжелой цепи, в виде полипептида, подобного тяжелой цепи иммуноглобулина, содержащего CDR3, содержащего представляющий интерес полипептид, не являющийся иммуноглобулином, его часть или его вариант, подвергнутый соматической гипермутации.
[0036] В некоторых вариантах осуществления описанное в данном документе животное, отличное от человека, дополнительно содержит локус гуманизированной легкой цепи. В некоторых вариантах осуществления раскрываемое в данном документе животное, отличное от человека, содержит
(i) в зародышевой клетке:
(a) локус тяжелой цепи иммуноглобулина, содержащий неперегруппированный вариабельный участок тяжелой цепи иммуноглобулина и константный участок тяжелой цепи иммуноглобулина, где неперегруппированный вариабельный участок тяжелой цепи иммуноглобулина содержит по меньшей мере один неперегруппированный генный сегмент VH (который может представлять собой неперегруппированный генный сегмент VH человека, например, hVH), сконструированный участок DH (который может включать генный сегмент DH человека и/или сконструированные генные сегменты DH, например, hDH) и по меньшей мере один неперегруппированный генный сегмент JH, необязательно сегмент человека, где генный сегмент(ы) VH, сконструированный участок DH и генный сегмент(ы) JH функционально связаны, вследствие чего они могут рекомбинировать, например, в В-клетке во время развития В-клетки, с образованием перегруппированной генной последовательности вариабельного участка тяжелой цепи иммуноглобулина hVH/eDH/hJH в функциональной связи с константным участок тяжелой цепи иммуноглобулина, и
(b) локус легкой цепи иммуноглобулина, содержащий генные сегменты VL и/или JL человека, и который может кодировать общую легкую цепь (человека); и
(ii) в соматической клетке, например B-клетке:
(a) генную последовательность перегруппированного вариабельного участка тяжелой цепи иммуноглобулина hVH/eDH/hJH в функциональной связи с константным участком тяжелой цепи иммуноглобулина, где сконструированный участок DH содержит последовательность, кодирующую представляющий интерес полипептид, не являющийся иммуноглобулином, или его часть, и где перегруппированная генная последовательность hVH/eDH/hJH содержит CDR3-кодирующую последовательность, содержащую одну или более нуклеотидных последовательностей, которые кодируют представляющий интерес полипептид, не являющийся иммуноглобулином, или его часть, или его вариант, подвергнутый соматической гипермутации, и
(b) локус гуманизированной и/или общей легкой цепи иммуноглобулина или его вариант, подвергнутый соматической гипермутации. В некоторых вариантах осуществления человеческий неперегруппированный или перегруппированный вариабельный участок тяжелой цепи человека функционально связан с константным участком тяжелой цепи, который может представлять собой константный участок тяжелой цепи, отличной от человеческой, например, в эндогенном локусе тяжелой цепи, отличной от человеческой. В некоторых вариантах осуществления вариабельный участок тяжелой цепи иммуноглобулина представляет собой вариабельный участок тяжелой цепи иммуноглобулина человека. В некоторых вариантах осуществления в В-клетке дополнительно экспрессируется генная последовательность перегруппированного участка hVH/eDH/hJH, функционально связанная с константным участком тяжелой цепи, в виде полипептида, подобного тяжелой цепи иммуноглобулина, содержащего CDR3, содержащий представляющий интерес полипептид, не являющийся иммуноглобулином, его часть или его вариант, подвергнутый соматической гипермутации, а также человеческую (гуманизированную) и/или общую легкую цепь в виде тетрамерного антигенсвязывающего белка, подобного иммуноглобулину, где тетрамер содержит димер из полипептидов, подобных тяжелой цепи иммуноглобулина, каждый из которых ковалентно связан с человеческой (гуманизированной) и/или общей легкой цепью.
[0037] В некоторых вариантах осуществления представлен способ получения животного, отличного от человека, геном которого содержит вариабельный участок тяжелой цепи иммуноглобулина, который включает сконструированный участок DH, при этом способ предусматривает (a) осуществление вставки фрагмента ДНК в эмбриональную стволовую клетку, отличную от человеческой, причем указанный фрагмент ДНК содержит одну или более нуклеотидных последовательностей, каждая из которых кодирует представляющий интерес полипептид, не являющийся иммуноглобулином, или его часть; (b) получение эмбриональной стволовой клетки, отличной от человеческой, образованной на (a); и (c) создание организма, отличного от человека, с применением эмбриональной стволовой клетки из (b).
[0038] В одном варианте осуществления способ получения животного, отличного от человека, раскрываемого в данном документе, предусматривает (a) получение первого животного, отличного от человека, геном которого содержит вариабельный участок тяжелой цепи иммуноглобулина, который включает сконструированный участок DH, раскрываемый в данном документе, и (b) скрещивание первого животного, отличного от человека, из (a) с вторым животным, отличным от человека, которое в одном аспекте может представлять собой другую линию первого животного, отличного от человека, где второе животное, отличное от человека, экспрессирует универсальную легкую цепь, и где результатом скрещивания является потомство, которое продуцирует, например, содержит генетически сконструированную тяжелую цепь, содержащую аминокислотную последовательность белка (или ее часть), не являющегося иммуноглобулином, например, в CDR3, и генетически сконструированную перегруппированную легкую цепь (единую перегруппированную легкую цепь; ULC).
[0039] В некоторых вариантах осуществления фрагмент ДНК включает 5, 10, 15, 20, 25 или больше нуклеотидных последовательностей, каждая из которых кодирует внеклеточную часть D6 хемокинового рецептора-ловушки, или каждая из которых кодирует часть конотоксина (например, μ-конотоксина), токсина тарантула или их комбинации. В некоторых определенных вариантах осуществления фрагмент ДНК включает 25 нуклеотидных последовательностей, каждая из которых кодирует внеклеточную часть D6 хемокинового рецептора-ловушки, или включает 26 нуклеотидных последовательностей, каждая из которых кодирует часть конотоксина (например, μ-конотоксина) и/или токсина тарантула (например, ProTxI, ProTxII и т.д.). В некоторых определенных вариантах осуществления фрагмент ДНК дополнительно содержит первую и вторую последовательности сигнала рекомбинации, фланкирующие каждую из 25 или 26 нуклеотидных последовательностей.
[0040] В некоторых вариантах осуществления представлен способ получения животного, отличного от человека, геном которого содержит вариабельный участок тяжелой цепи иммуноглобулина, который включает сконструированный участок DH, при этом способ предусматривает модификацию генома животного, отличного от человека, таким образом, что он содержит вариабельный участок тяжелой цепи иммуноглобулина, который включает сконструированный участок DH, причем сконструированный участок DH содержит одну или более нуклеотидных последовательностей, каждая из которых кодирует представляющий интерес полипептид, не являющийся иммуноглобулином, или его часть, с получением тем самым животного, отличного от человека.
[0041] В некоторых вариантах осуществления геном животного, отличного от человека, модифицируют таким образом, что он включает 5, 10, 15, 20, 25 или больше нуклеотидных последовательностей, каждая из которых кодирует внеклеточную часть D6 хемокинового рецептора-ловушки, или каждая из которых кодирует часть конотоксина (например, μ-конотоксина), токсина тарантула или их комбинации. В некоторых определенных вариантах осуществления геном животного, отличного от человека, модифицируют таким образом, что он включает 25 нуклеотидных последовательностей, каждая из которых кодирует внеклеточную часть D6 хемокинового рецептора-ловушки, или модифицируют таким образом, что он включает 26 нуклеотидных последовательностей, каждая из которых кодирует часть конотоксина (например, μ-конотоксина) и/или токсина тарантула (например, ProTxI, ProTxII и т.д.). В некоторых определенных вариантах осуществления геном животного, отличного от человека, модифицируют таким образом, что он дополнительно включает первую и вторую последовательности сигнала рекомбинации, фланкирующие каждую из 25 или 26 нуклеотидных последовательностей.
[0042] В некоторых вариантах осуществления представлен способ получения антитела в животном, отличного от человека, при этом способ предусматривает стадии: (a) иммунизации животного, отличного от человека, антигеном, причем животное, отличное от человека, имеет геном, содержащий вариабельный участок тяжелой цепи иммуноглобулина, который включает сконструированный участок DH, где сконструированный участок DH включает одну или более нуклеотидных последовательностей, каждая из которых кодирует представляющий интерес полипептид, не являющийся иммуноглобулином, или его часть; (b) содержания животного, отличного от человека, в условиях, подходящих для того, чтобы у животного, отличного от человека, вырабатывался иммунный ответ на антиген; и (c) выделения из животного, отличного от человека, или клетки, отличной от человеческой, антитела, которое связывает антиген. В некоторых определенных вариантах осуществления клетка, отличная от человеческой, представляет собой B-клетку. В некоторых определенных вариантах осуществления клетка, отличная от человеческой, представляет собой гибридому.
[0043] В некоторых вариантах осуществления представлено животное, отличное от человека, геном зародышевой клетки которого содержит (a) вариабельный участок тяжелой цепи иммуноглобулина человека, который содержит один или более неперегруппированных генных сегментов VH человека, сконструированный участок DH и один или более неперегруппированных генных сегментов JH человека, при этом сконструированный участок DH включает (i) одну или более нуклеотидных последовательностей, каждая из которых кодирует внеклеточную часть атипичного хемокинового рецептора (ACKR); и (ii) первую и вторую последовательности сигнала рекомбинации, фланкирующие каждую из одной или более нуклеотидных последовательностей из (i), так что один или более неперегруппированных генных сегментов VH человека, сконструированный участок DH и один или более неперегруппированных генных сегментов JH человека рекомбинируют, например, в B-клетке, вследствие чего животное, отличное от человека, содержит геном В-клетки, содержащий вариабельный участок тяжелой цепи иммуноглобулина человека, который содержит генную последовательность перегруппированного участка hVH/eDH/hJH; где вариабельный участок тяжелой цепи иммуноглобулина человека функционально связан с одним или более эндогенными генами константного участка тяжелой цепи иммуноглобулина, так что животное, отличное от человека, характеризуется тем, что при его иммунизации антигеном генная последовательность перегруппированного участка hVH/eDH/hJH, функционально связанная с геном константного участка тяжелой цепи иммуноглобулина, кодирует антитело, содержащее вариабельные домены тяжелой цепи человека, кодируемые одним из генных сегментов VH человека (или его частью), сконструированным участком DH (или его частью) и одним из генных сегментов JH человека (или его частью), функционально связанных с константными доменами тяжелой цепи животного, отличного от человека, кодируемыми одним или более эндогенными генами константного участка иммуноглобулина, и где антитело проявляет специфическое связывание с антигеном. В некоторых вариантах осуществления зародышевая клетка и, например, B-клетка животного, отличного от человека, дополнительно содержит локус легкой цепи иммуноглобулина, кодирующий общую легкую цепь, вследствие чего антитело дополнительно содержит общую легкую цепь.
[0044] В некоторых вариантах осуществления представлено животное, отличное от человека, геном зародышевой клетки которого содержит вариабельный участок тяжелой цепи иммуноглобулина человека, который содержит один или более неперегруппированных генных сегментов VH человека (h), сконструированный (e) участок DH и один или более генных сегментов JH человека (h), при этом сконструированный участок DH включает (i) одну или более нуклеотидных последовательностей, каждая из которых кодирует часть токсина (например, μ-конотоксина, токсина тарантула или их комбинаций); и (ii) первую и вторую последовательности сигнала рекомбинации, фланкирующие каждую из одной или более нуклеотидных последовательностей из (i), чтобы один или более неперегруппированных генных сегментов VH человека, сконструированный участок DH и один или более неперегруппированных генных сегментов JH человека рекомбинировали, например, в B-клетке, вследствие чего животное, отличное от человека, содержит геном В-клетки, содержащий вариабельный участок тяжелой цепи иммуноглобулина человека, который содержит генную последовательность перегруппированного участка hVH/eDH/hJH; где вариабельный участок тяжелой цепи иммуноглобулина человека функционально связан с одним или более эндогенными генами константного участка тяжелой цепи иммуноглобулина таким образом, что животное, отличное от человека, характеризуется тем, что при его иммунизации антигеном генная последовательность перегруппированного участка hVH/eDH/hJH, функционально связанная с геном константного участка тяжелой цепи иммуноглобулина, кодирует антитело, содержащее вариабельный домен тяжелой цепи человека, кодируемый одним из генных сегментов VH человека (или его частью), сконструированным участком DH (или его частью) и одним из генных сегментов JH человека (или его частью), функционально связанными с одним или более константными доменами тяжелой цепи, кодируемыми одним или более эндогенными генами константного участка иммуноглобулина, и где антитела проявляют специфическое связывание с антигеном. В некоторых вариантах осуществления зародышевая клетка и, например, B-клетка животного, отличного от человека, дополнительно содержит локус легкой цепи иммуноглобулина, кодирующий общую легкую цепь, вследствие чего антитело дополнительно содержит общую легкую цепь.
[0045] В некоторых вариантах осуществления представлено животное, отличное от человека, дополнительно содержащее вставку из одного или более генных сегментов VL человека и одного или более генных сегментов JL человека в эндогенном локусе легкой цепи. В некоторых вариантах осуществления сегменты VL и JL человека представляют собой генные сегменты Vκ и Jκ, и они вставлены в эндогенный локус легкой κ-цепи. В некоторых вариантах осуществления генные сегменты Vκ и Jκ человека функционально связаны с геном Cκ грызуна (например, геном Cκ мыши или крысы). В некоторых вариантах осуществления сегменты VL и JL человека представляют собой генные сегменты Vλ и Jλ, и они вставлены в эндогенный локус легкой λ-цепи. В некоторых вариантах осуществления генные сегменты Vλ и Jλ человека функционально связаны с геном Cλ грызуна (например, геном Cλ мыши или крысы). В некоторых вариантах осуществления генная последовательность единого перегруппированного вариабельного участка легкой цепи человека функционально связана с эндогенным геном константного участка легкой цепи, отличной от человеческой. В некоторых вариантах осуществления генная последовательность единого перегруппированного участка Vκ/Jκ человека функционально связана с эндогенным геном Cκ (например, геном Cκ мыши или крысы). В некоторых вариантах осуществления генная последовательность единого перегруппированного участка Vλ/Jλ функционально связана с эндогенным геном Cλ.
[0046] В некоторых вариантах осуществления представленное животное, отличное от человека, является гомозиготным, гетерозиготным или гемизиготным по сконструированному участку DH, описываемому в данном документе. В некоторых вариантах осуществления представленное животное, отличное от человека, является трансгенным по сконструированному участку DH, описываемому в данном документе.
[0047]Также в данном документе раскрыты клетки, например, B-клетки или гибридомы, полученные из них путем слияния с клеткой миеломы, при этом каждая содержит последовательность перегруппированного участка (h)VH/eDH/(h)JH, которая может быть функционально связана с константным участком тяжелой цепи человека или отличной от человеческой, содержащим один или более генов константного участка тяжелой цепи. Такая клетка, например, В-клетка, может быть выделена из животного, отличного от человека, например, грызуна (например, крысы, мыши и т.д.), описываемого в данном документе.
[0048] Также в данном документе описаны нуклеотидные последовательности, содержащий последовательность перегруппированного вариабельного участка (h)VH/eDH/(h)JH, которая может быть функционально связана с константным участком тяжелой цепи человека или отличной от человеческой, содержащим один или более генов константного участка тяжелой цепи. Такие нуклеотидные последовательности могут быть выделены из животного, отличного от человека, например, грызуна (например, крысы, мыши и т.д.), или клетки, отличной от человеческой, описываемых в данном документе.
[0049] В некоторых вариантах осуществления представлено применение животного, отличного от человека, описываемого в данном документе, для производства и/или разработки лекарственного средства или вакцины для применения в медицине, как, например, для применения в качестве лекарственного препарата.
[0050] В некоторых вариантах осуществления представлено применение животного, отличного от человека, описываемого в данном документе, при производстве лекарственного препарата для лечения заболевания, нарушения или состояния.
[0051] В некоторых вариантах осуществления представлено применение животного, отличного от человека, описываемого в данном документе, при производстве и/или разработке антитела, которое связывает хемокин или потенциал-зависимый натриевый (NaV) канал.
[0052] В некоторых вариантах осуществления представлено применение животного, отличного от человека, описываемого в данном документе, при производстве лекарственного препарата для лечения или выявления заболевания, характеризующегося экспрессией или функцией хемокина или потенциал-зависимого натриевого (NaV) канала (например, нарушенной экспрессией или функцией).
[0053] В некоторых вариантах осуществления представлено животное, отличное от человека, описываемое в данном документе, для применения при производстве и/или разработке лекарственного средства для терапии или диагностики.
[0054] В некоторых вариантах осуществления представлено животное, отличное от человека, описываемое в данном документе, для применения при производстве лекарственного препарата для лечения, предупреждения или уменьшения тяжести заболевания, нарушения или состояния.
[0055] В некоторых вариантах осуществления представлено животное, отличное от человека, описываемое в данном документе, для применения при производстве и/или разработке антитела, которое связывает хемокин или потенциал-зависимый натриевый (NaV) канал.
[0056] В некоторых вариантах осуществления заболевание, нарушение или состояние представляет собой воспалительное заболевание, нарушение или состояние. В некоторых вариантах осуществления заболевание, нарушение или состояние характеризуется экспрессией или функцией хемокинов (например, нарушенной экспрессией или функцией хемокинов).
[0057] В некоторых вариантах осуществления заболевание, нарушение или состояние представляет собой заболевание, нарушение или состояние в виде боли. В некоторых вариантах осуществления заболевание, нарушение или состояние характеризуется экспрессией или функцией ионных каналов (например, нарушенной экспрессией или функцией NaV-каналов).
[0058] Во многих вариантах осуществления животное, отличное от человека, представленное в данном документе, представляет собой грызуна; в некоторых вариантах осуществления мышь; в некоторых вариантах осуществления крысу.
[0059] Используемые в данной заявке термины «приблизительно» и «примерно» используются в качестве эквивалентов. Предполагается, что все числа, используемые в данной заявке вместе с термином «приблизительно/примерно» или без него, охватывают любые нормальные отклонения, понятные специалисту в соответствующей области.
[0060] Другие признаки, объекты и преимущества животных, отличных от человека, клеток, нуклеиновых кислот и композиций, раскрытых в данном документе, станут очевидны из подробного описания определенных вариантов осуществления, которое следует ниже. Однако следует понимать, что подробное описание, хотя и указывает на определенные варианты осуществления, приведено только в целях иллюстрации, а не ограничения.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
[0061] Включенные в данный документ графические материалы, которые состоят из следующих фигур, приведены только в иллюстративных целях, а не для ограничения.
[0062] На фигурах 1A–1D показана иллюстративная оптимизация выбранных кодирующих последовательностей D6 хемокинового рецептора-ловушки для введения «горячих точек» соматической гипермутации. Фигура 1A: оптимизированный Nterm домен из D6 хемокинового рецептора-ловушки с местоположениями природных (заштрихованная стрелка) и искусственных (стрелка с диагональной штриховкой) «горячих точек» RGYW, индуцированных активацией цитидиндезаминазы (AID); фигура 1B: оптимизированный домен EC1 из D6 хемокинового рецептора-ловушки с местоположениями природных (заштрихованная стрелка) и искусственных (стрелка с диагональной штриховкой) «горячих точек» RGYW, индуцированных активацией цитидиндезаминазы (AID); фигура 1C: оптимизированный домен EC2 из D6 хемокинового рецептора-ловушки с местоположениями природных (заштрихованная стрелка) и искусственных (стрелка с диагональной штриховкой) «горячих точек» RGYW AID; фигура 1D: оптимизированный домен EC3 из D6 хемокинового рецептора-ловушки с местоположениями естественных (заштрихованная прерывистая линия) и искусственных (линия с диагональной штриховкой) «горячих точек» RGYW AID.
[0063] На фигуре 2 показана таблица иллюстративных оптимизированных 5’- и 3’-последовательностей сигнала рекомбинации (RSS), разработанных для каждой нуклеотидной кодирующей последовательности, чтобы обеспечить эффективную частоту рекомбинации и равное использование нуклеотидных кодирующих последовательностей во время рекомбинации V(D)J. 5'-RSS консенсусной последовательности глобальной RSS (SEQ ID NO:51); 3'-RSS консенсусной последовательности глобальной RSS (SEQ ID NO:52); 5'-RSS консенсусной последовательности DH человека (SEQ ID NO:53); 3'-RSS консенсусной последовательности DH человека (SEQ ID NO:54); 5'-RSS консенсусной последовательности DH мыши (SEQ ID NO:55); 3'-RSS консенсусной последовательности DH мыши (SEQ ID NO:56);5'-RSS оптимизированной RSS (SEQ ID NO:57); 3'-RSS оптимизированной RSS (SEQ ID NO:58); 5'-RSS 1-1 opt (SEQ ID NO:59); 3'-RSS 1-1 opt (SEQ ID NO:60); 5'-RSS 1-7 opt (SEQ ID NO:61); 3'-RSS 1-7 opt (SEQ ID NO:62); 5'-RSS 1-14 ORF opt (SEQ ID NO:63); 3’-RSS 1-14 ORF opt (SEQ ID NO:64); 5'-RSS 1-20 opt (SEQ ID NO:65); 3'-RSS 1-20 opt (SEQ ID NO:66); 5'-RSS 1-26 opt (SEQ ID NO:67); 3'-RSS 1-26 opt (SEQ ID NO:68); 5'-RSS 2-2*02 opt (SEQ ID NO:69); 3'-RSS 2-2*02 opt (SEQ ID NO:70); 5’-RSS 2-8*01 opt (SEQ ID NO:71); 3'-RSS 2-8*01 opt (SEQ ID NO:72); 5'-RSS 2-15 opt (SEQ ID NO:73); 3'-RSS 2-15 opt (SEQ ID NO:74); 5'-RSS 2-21*02 opt (SEQ ID NO:75); 3’-RSS 2-21*02 opt (SEQ ID NO:76); 5'-RSS 3-3*01 opt (SEQ ID NO:77); 3'-RSS 3-3*01 opt (SEQ ID NO:78); 5'-RSS 3-9 opt (SEQ ID NO:79); 3'-RSS 3-9 opt (SEQ ID NO:80); 5'-RSS 3-10*01 opt (SEQ ID NO:81); 3’-RSS 3-10*01 opt (SEQ ID NO:82); 5'-RSS 3-16*02 opt (SEQ ID NO:83); 3’-RSS 3-16*02 opt (SEQ ID NO:84); 5'-RSS 3-22 opt (SEQ ID NO:85); 3'-RSS 3-22 opt (SEQ ID NO:86); 5'-RSS 4-4 opt (SEQ ID NO:87); 3'-RSS 4-4 opt (SEQ ID NO:88); 5'-RSS 4-11 ORF opt (SEQ ID NO:89); 3'-RSS 4-11 ORF opt (SEQ ID NO:90); 5'-RSS 4-17 opt (SEQ ID NO:91); 3’-RSS 4-17 opt (SEQ ID NO:92); 5'-RSS 4-23 ORF opt (SEQ ID NO:93); 3'-RSS 4-23 ORF opt (SEQ ID NO:94); 5'-RSS 5-5 opt (SEQ ID NO:95); 3'-RSS 5-5 opt (SEQ ID NO:96); 5'-RSS 5-12 opt (SEQ ID NO:97); 3'-RSS 5-12 opt (SEQ ID NO:98); 5'-RSS 5-18 opt (SEQ ID NO:99); 3’-RSS 5-18 opt (SEQ ID NO:100); 5' RSS 5-24 ORF opt (SEQ ID NO:101); 3'-RSS 5-24 ORF opt (SEQ ID NO:102); 5'-RSS 6-6 opt (SEQ ID NO:103); 3'-RSS 6-6 opt (SEQ ID NO:104); 5'-RSS 6-13 opt (SEQ ID NO:105); 3’-RSS 6-13 opt (SEQ ID NO:106); 5'-RSS 6-19 opt (SEQ ID NO:107); 3'-RSS 6-19 opt (SEQ ID NO:108); 5'-RSS 6-25 (не оптимизирована) (SEQ ID NO:109); 3'-RSS 6-25 (не оптимизирована) (SEQ ID NO:110); 5'-RSS 7-27 (не оптимизирована) (SEQ ID NO:111); 3'-RSS 7-27 (не оптимизирована) (SEQ ID NO:112). В случае консенсусной последовательности глобальной RSS и 7-27 жирным шрифтом и курсивом показано совпадение с консенсусной последовательностью глобальной RSS на основе RSS грызуна и человека из генных сегментов V, D и J иммуноглобулина и T-клеточного рецептора; в случае консенсусной последовательности DH мыши жирным шрифтом показано совпадение с консенсусной последовательностью DH иммуноглобулина мыши; в случае консенсусной последовательности DH человека, оптимизированной RSS и всех остальных RSS (например, 1-1 opt, 1-7 opt, 1-20 opt, и т.д.) жирным шрифтом показано совпадение с консенсусной последовательностью DH иммуноглобулина человека.
[0064] На фигурах 3A–3B показана иллюстрация, без соблюдения масштаба, иллюстративной стратегии для разработки нацеливающего вектора для интеграции в эмбриональные стволовые (ES) клетки грызуна для получения грызуна, геном которого содержит вариабельный участок тяжелой цепи иммуноглобулина, который включает сконструированный кластер разнообразия (т.е. участок DH), при этом кластер разнообразия включает одну или более нуклеотидных последовательностей, каждая из которых кодирует часть полипептида, не являющегося иммуноглобулином (например, внеклеточную часть D6 хемокинового рецептора-ловушки). Фигура 3A: указаны четыре исходные стадии: (1) de novo синтез кодирующих последовательностей D6, (2) расщепление и лигирование с помощью AgeI/EcoRI кассеты для отбора (например, по устойчивости к неомицину) и фрагмента ДНК D6, (3) расщепление с помощью SnaBI фрагментов ДНК D6 и расщепление с помощью NotI/AscI вектора BAC (pBacE3.6), и (4) одностадийная изотермальная сборка расщепленных фрагментов ДНК с получением непрерывного сконструированного кластера разнообразия с кодирующими последовательностями D6 хемокинового рецептора-ловушки вместо стандартных сегментов DH; фигура 3B: дополнительная стадия для получения нацеливающего вектора для интеграции в геном ES-клеток грызуна: (5) расщепление и лигирование с помощью PI-SceI/I-CeuI 25 синтетических кодирующих последовательностей D6 хемокинового рецептора-ловушки в клон BAC для присоединения к 5'- и 3'-плечам гомологии, содержащим ДНК VH и ДНК JH иммуноглобулина человека соответственно. Различные сайты для распознавания рестриктазами указаны для каждого из изображенных фрагментов ДНК. lp: loxP-сайт; neo: кассета для отбора по устойчивости к неомицину под управлением убиквитинового промотора; cm: кассета для отбора по устойчивости к хлорамфениколу; frt: целевая последовательность для распознавания флиппазой; hyg: кассета для отбора по устойчивости к гигромицину; Ei: интронный энхансер тяжелой цепи мыши; IgM: ген константного участка иммуноглобулина M мыши; lox: последовательность loxP-сайта в векторе pBACe3.6.
[0065] На фигурах 4A–4B показана иллюстрация, без соблюдения масштаба, альтернативной иллюстративной стратегии сборки кодирующих последовательностей D6 хемокинового рецептора-ловушки путем последовательного лигирования для разработки нацеливающего вектора для интеграции в эмбриональные стволовые (ES) клетки грызуна для получения грызуна, геном которого содержит вариабельный участок тяжелой цепи иммуноглобулина, который включает сконструированный кластер разнообразия (т.е. сконструированный участок DH), при этом кластер разнообразия включает одну или более нуклеотидных последовательностей, каждая из которых кодирует внеклеточную часть D6 хемокинового рецептора-ловушки. Фигура 4A: указаны четыре исходные стадии: (1) de novo синтеза кодирующих последовательностей D6, (2) расщепление и лигирование с помощью AgeI/EcoRI кассеты для отбора (например, по устойчивости к неомицину) и фрагмента ДНК D6, (3) расщепление и лигирование с помощью NotI/AscI фрагмента ДНК D6 в вектор BAC (pBacE3.6), и (4) расщепление с помощью PacI/Nsi и лигирование фрагментов D6 в остов вектора BAC; фигура 4B: две дополнительные стадии для получения нацеливающего вектора для интеграции в геном ES-клеток грызуна: (5) расщепление и лигирование с помощью PI-SceI/I-CeuI дополнительного фрагмента ДНК D6 в остов вектора BAC, и (6) расщепление и лигирование с помощью NsiI/I-CeuI фрагмента ДНК D6 с получением 25 синтетических кодирующих последовательностей D6 хемокинового рецептора-ловушки в остове вектора BAC. Различные сайты для распознавания рестриктазами указаны для каждого из изображенных фрагментов ДНК; lp: последовательность loxP-сайта; neo: кассета для отбора по устойчивости к неомицину под управлением убиквитинового промотора; cm: кассета для отбора по устойчивости к хлорамфениколу.
[0066] На фигуре 5 показана иллюстративная стратегия скрининга с использованием генетического материала из колоний, устойчивых к лекарственному средству, после электропорации, которых подвергали скринингу с помощью TAQMAN™ и кариотипированию. Названия и примерное местоположение, без соблюдения масштаба (линии, окруженные овалом), различных наборов праймеров/зондов (см. таблицу 7) указаны ниже различных показанных аллелей (без соблюдения масштаба). hyg: кассета для отбора по устойчивости к гигромицину под управлением убиквитинового промотора; neo: кассета для отбора по устойчивости к неомицину под управлением убиквитинового промотора; L: последовательность loxP-сайта; Frt: целевая последовательность для распознавания флиппазой.
[0067] На фигурах 6A–6L показана иллюстративная оптимизация выбранных кодирующих последовательностей μ-конотоксина и токсина тарантула для включения «горячих точек» соматической гипермутации. Фигура 6A: оптимизированная KIIIA fl μ-конотоксина с местоположениями искусственных (линия с диагональной штриховкой) «горячих точек» RGYW, индуцируемых активацией цитидиндезаминазы (AID); фигура 6B: оптимизированная KIIIA mini (вверху) и KIIIA midi (внизу) μ-конотоксина с местоположениями (линия с диагональной штриховкой) «горячих точек» RGYW, индуцируемых активацией цитидиндезаминазы (AID); фигура 6C: оптимизированная PIIIA fl μ-конотоксина с местоположениями искусственных (линия с диагональной штриховкой) «горячих точек» RGYW AID; фигура 6D: оптимизированная PIIIA mini (вверху) и PIIIA midi (внизу) μ-конотоксина с местоположениями искусственных (линия с диагональной штриховкой) «горячих точек» RGYW AID; фигура 6E: оптимизированная SMIIIA fl μ-конотоксина с местоположениями искусственных (линия с диагональной штриховкой) «горячих точек» RGYW AID; фигура 6F: оптимизированная SmIIIA mini (вверху) и SmIIIA midi (внизу) μ-конотоксина с местоположениями искусственных (линия с диагональной штриховкой) «горячих точек» RGYW AID; фигура 6G: оптимизированная ProTxII токсина тарантула с местоположениями искусственных (линия с диагональной штриховкой) «горячих точек» RGYW AID; фигура 6H: оптимизированная ProTxII C1SC4S (вверху), ProTxII C2SC5S (посередине) и ProTxII C3SC6S (внизу) токсина тарантула с местоположениями искусственных (линия с диагональной штриховкой) «горячих точек» RGYW AID; фигура 6I: оптимизированная SSRW-петля SmIIIA (слева), SSKW-петля SmIIIA (посередине) и RSRQ-петля PIIIA (справа) μ-конотоксина с местоположениями искусственных (линия с диагональной штриховкой) «горячих точек» RGYW AID; фигура 6J: оптимизированная KIIIA или SmIIIA mini/midi μ-конотоксина в местоположениях сегмента DH3 с местоположениями искусственных (линия с диагональной штриховкой) «горячих точек» RGYW AID; фигура 6K: оптимизированная SmIIIA или PIIIA mini/midi μ-конотоксина в местоположениях сегмента DH3 или DH1 с местоположениями искусственных (линия с диагональной штриховкой) «горячих точек» RGYW AID; фигура 6L: оптимизированные SSRW- или RSRQ-петли μ-конотоксина в местоположениях сегмента DH2 с местоположениями искусственных (линия с диагональной штриховкой) «горячих точек» RGYW AID.
[0068] На фигурах 7A–7B показана иллюстрация, без соблюдения масштаба, иллюстративной стратегии разработки нацеливающего вектора для интеграции в эмбриональные стволовые (ES) клетки грызуна для получения грызуна, геном которого содержит вариабельный участок тяжелой цепи иммуноглобулина, который включает сконструированный кластер разнообразия (т.е. сконструированный участок DH), при этом кластер разнообразия включает одну или более нуклеотидных последовательностей, каждая из которых кодирует часть полипептида, не являющегося иммуноглобулином (например, часть μ-конотоксина и/или токсина тарантула). Фигура 7A: указаны четыре исходные стадии: (1) de novo синтез кодирующих последовательностей токсина, (2) расщепление и лигирование с помощью AgeI/EcoRI кассеты для отбора (например, по устойчивости к неомицину) и фрагмента ДНК токсина (TX-DH1166), (3) расщепление с помощью SnaBI фрагментов ДНК токсина и расщепление с помощью NotI/AscI вектора BAC (pBacE3.6), и (4) одностадийная изотермальная сборка расщепленных фрагментов ДНК с получением сконструированного кластера разнообразия, содержащего непрерывные кодирующие последовательности токсина вместо одного или более, и необязательно, всех функциональных генных сегментов DH; фигура 7B: дополнительная стадия для получения нацеливающего вектора для интеграции в геном ES-клеток грызуна: (5) расщепление и лигирование с помощью PI-SceI/I-CeuI 26 синтетическихкодирующих последовательностей токсина в клон BAC для присоединения к 5'- и 3'-плечам гомологии, содержащим ДНК VH и JH иммуноглобулина человека соответственно. Различные сайты для распознавания рестриктазами указаны для каждого из изображенных фрагментов ДНК; lp: loxP-сайт; neo: кассета для отбора по устойчивости к неомицину под управлением убиквитинового промотора; cm: кассета для отбора по устойчивости к хлорамфениколу; frt: целевая последовательность для распознавания флиппазой; hyg: кассета для отбора по устойчивости к гигромицину; Ei: интронный энхансер тяжелой цепи мыши; IgM: ген константного участка иммуноглобулина M мыши; lox: loxP-сайт в векторе pBACe3.6.
[0069] На фигурах 8A–8B показана иллюстрация, без соблюдения масштаба, альтернативной иллюстративной стратегии для сборки кодирующих последовательностей токсина (например, μ-конотоксина и токсина тарантула) путем последовательного лигирования для разработки нацеливающего вектора для интеграции в эмбриональные стволовые (ES) клетки грызуна для получения грызуна, геном которого содержит вариабельный участок тяжелой цепи иммуноглобулина, который включает сконструированный кластер разнообразия (т.е. участок DH), при этом кластер разнообразия включает одну или более нуклеотидных последовательностей, каждая из которых кодирует часть пептидного токсина (например, μ-конотоксина и токсина тарантула ProTxII). Фигура 8A: указаны четыре исходные стадии: (1) de novo синтез кодирующих последовательностей токсина, (2) расщепление и лигирование с помощью AgeI/EcoRI кассеты для отбора (например, по устойчивости к неомицину) и фрагмента ДНК токсина (TX-DH1166), (3) расщепление и лигирование с помощью NotI/AscI фрагмента ДНК токсина в вектор BAC (pBacE3.6), и (4) расщепление и лигирование с помощью PacI/Nsi фрагментов ДНК токсина в остов вектора BAC; фигура 8B: две дополнительные стадии получения нацеливающего вектора для интеграции в геном ES-клеток грызуна: (5) расщепление и лигирование с помощью PI-SceI/I-CeuI дополнительного фрагмента ДНК токсина в остов вектора BAC, и (6) расщепление и лигирование с помощью NsiI/I-CeuI конечного фрагмента ДНК токсина с получением 26 синтетических кодирующих последовательностей токсина в остове вектора BAC. Различные сайты для распознавания рестриктазами указаны для каждого из изображенных фрагментов ДНК. lp: loxP-сайт; neo: кассета для отбора по устойчивости к неомицину под управлением убиквитинового промотора; cm: кассета для отбора по устойчивости к хлорамфениколу.
[0070] На фигурах 9A–9D показаны репрезентативные контурные графики лимфоцитов в селезенке, собранной у мышей VELOCIMMUNE® (VI) и мышей, гомозиготных по сконструированному участку DH, содержащему кодирующие последовательности токсина (6579ho/1293ho, «TX-DH ho»; линия грызунов с геномом, содержащим гомозиготный локус тяжелой цепи иммуноглобулина, содержащий несколько сегментов VH человека, сконструированных сегментов DH, включающих кодирующие последовательности токсина, вместо стандартных сегментов DH и сегментов JH, функционально связанных с константным участком тяжелой цепи иммуноглобулина грызуна, включающим энхансеры и регуляторные участки тяжелой цепи грызуна, и содержащий вставленную нуклеотидную последовательность, кодирующую один или более генов Adam6 мыши [например, патенты №№8642835 и 8697940]; и гомозиготный локус легкой κ-цепи иммуноглобулина, содержащий генные сегменты Vκ и Jκ человека, функционально связанные с геном участка Cκ грызуна, включающим энхансеры легкой κ-цепи грызуна), и они окрашены для выявления экспрессии различных клеточных маркеров на клеточной поверхности. Фигура 9A: репрезентативный контурный график лимфоцитов из селезенки, гейтированных по синглетам, демонстрирующим экспрессию CD19 (ось y) и CD3 (ось x). Фигура 9B: репрезентативный контурный график лимфоцитов из селезенки, гейтированных по CD19+ синглетам, демонстрирующим экспрессию иммуноглобулина D (IgD, ось y) и иммуноглобулина M (IgM, ось x); зрелые (CD19+ IgD+ IgMint) и переходные (CD19+ IgDint IgM) B-клетки указаны на каждом точечном графике. Фигура 9C: репрезентативный контурный график лимфоцитов из селезенки, гейтированных по CD19+ синглетам, демонстрирующим экспрессию легкой цепи Igλ (ось y) или Igκ (ось x). Фигура 9D: показан репрезентативный контурный график созревания В-клеток, демонстрирующих лимфоциты из синглетов селезенки, гейтированных по CD19+, и показывающих экспрессию [слева направо] CD93 (ось y) и B220 (ось х), IgM (ось y) и CD23 (ось х); CD21/35 (ось y) и IgM (ось х), B220 (ось y) и CD23 (ось х), а также IgD (ось y) и IgM (ось х). Верхний ряд: мыши VELOCIMMUNE®; нижний ряд: мыши TX-DH ho (6579ho/1293ho). Специфические популяции В-клеток указаны на каждом точечном графике: незрелые (CD19+ CD93+ B220+), зрелые (CD19+ CD93- B220+), T1 (CD19+ CD93+ B220+ IgM+ CD23-), T2 (CD19+ CD93+ B220+ IgM+ CD23+), T3 (CD19+ CD93+ B220+ IgMint CD23+), MZ (CD19+ CD93- B220+ CD21/35+ IgM+ CD23-), предшественники MZ (CD19+ CD93- B220+ CD21/35+ IgM+ CD23+), Fol I (CD19+ CD93- B220+ CD21/35int IgMint IgD+) и Fol II (CD19+ CD93- B220+ CD21/35int IgM+ IgD+).
[0071] На фигурах 10A–10D показаны репрезентативные контурные графики лимфоцитов в костном мозге, собранном у мышей VELOCIMMUNE® (VI) и мышей, гомозиготных по сконструированному участку DH, содержащему кодирующие последовательности токсина (6579ho/1293ho, «TX-DH ho»; линия грызунов с геномом, содержащим гомозиготный локус тяжелой цепи иммуноглобулина, содержащий несколько сегментов VH человека, сконструированных сегментов DH, включающих кодирующие последовательности токсина, вместо стандартных сегментов DH и сегментов JH, функционально связанных с константным участком тяжелой цепи иммуноглобулина грызуна, включающим энхансеры и регуляторные участки тяжелой цепи грызуна, и содержащим вставленную нуклеотидную последовательность, кодирующую один или более генов Adam6 мыши [например, патенты №№ 8642835 и 8697940]; и гомозиготный локус легкой κ-цепи иммуноглобулина, содержащий генные сегменты Vκ и Jκ человека, функционально связанные с геном участком Cκ грызуна, включающим энхансеры легкой κ-цепи грызуна), и они окрашены для выявления экспрессии различных клеточных маркеров на клеточной поверхности. Фигура 10A: репрезентативный контурный график лимфоцитов из костного мозга, гейтированных по синглетам, демонстрирующим экспрессию CD19 (ось y) и CD3 (ось x). Фигура 10B: репрезентативный контурный график лимфоцитов из костного мозга, гейтированных по CD19+ IgM- /lowIgD–, демонстрирующих экспрессию c-kit (ось y) и CD43 (ось х); пре- (c-kit- CD43-) и про-B (c-kit+ CD43+)-клетки указаны на каждом точечном графике. Фигура 10C: репрезентативный контурный график лимфоцитов из костного мозга, гейтированных по синглетам, демонстрирующим экспрессию IgM (ось y) и B220 (ось х); незрелые (IgMint to +B220int) и зрелые (IgMint to +B220+) B-клетки указаны на каждом точечном графике. Фигура 10D: репрезентативный контурный график лимфоцитов из костного мозга, гейтированных по CD19+IgMint to +B220int (верхний ряд) и CD19+IgMint/+ B220+ (нижний ряд), демонстрирующих экспрессию легкой цепи Igλ (ось y) или Igκ (ось х).
[0072] На фигурах 11A–11D показаны репрезентативные контурные графики лимфоцитов в селезенке, собранной у мышей VELOCIMMUNE® (VI) и мышей, гетерозиготных по сконструированному участку DH, содержащему кодирующие последовательности D6 (6590het, «D6-DH het»; линия грызунов с геномом, содержащим гетерозиготный локус тяжелой цепи иммуноглобулина, содержащий несколько сегментов VH человека, сконструированных сегментов DH, включающих кодирующие последовательности D6 хемокинового рецептора-ловушки, вместо стандартных сегментов DH и сегментов JH, функционально связанных с константным участком тяжелой цепи иммуноглобулина грызуна, включающим энхансеры и регуляторные участки тяжелой цепи грызуна, и содержащим вставленную нуклеотидную последовательность, кодирующую один или более генов Adam6 мыши [например, патенты №№ 8642835 и 8697940]; и они окрашены для выявления экспрессии различных клеточных маркеров на клеточной поверхности. Фигура 11A: репрезентативный контурный график лимфоцитов из селезенки, гейтированных по синглетам, демонстрирующим экспрессию CD19 (ось y) и CD3 (ось x). Фигура 11B: репрезентативный контурный график лимфоцитов из селезенки, гейтированных по CD19+ синглетам, демонстрирующим экспрессию IgD (ось y) и IgM (ось x); зрелые (CD19+ IgD+ IgMint) и переходные (CD19+ IgDint IgM+) B-клетки указаны на каждом точечном графике. Фигура 11C: репрезентативный контурный график лимфоцитов из селезенки, гейтированных по CD19+ синглетам, демонстрирующим экспрессию легкой цепи Igλ (ось y) или Igκ (ось x). Фигура 11D: показан репрезентативный контурный график созревания В-клеток, демонстрирующий лимфоциты из селезенки, гейтированные по CD19+ синглетам, и показывающих экспрессию [слева направо] CD93 (ось y) и B220 (ось х), IgM (ось y) и CD23 (ось х); CD21/35 (ось y) и IgM (ось х), B220 (ось y) и CD23 (ось х), а также IgD (ось y), и IgM (ось х). Верхний ряд: мыши VELOCIMMUNE®; нижний ряд: мыши D6-DH het (6590het). Специфические популяции В-клеток указаны на каждом точечном графике: незрелые (CD19+ CD93+ B220+), зрелые (CD19+ CD93- B220+), T1 (CD19+ CD93+ B220+ IgM+ CD23-), T2 (CD19+ CD93+ B220+ IgM+ CD23+), T3 (CD19+ CD93+ B220+ IgMint CD23+), MZ (CD19+ CD93- B220+ CD21/35+ IgM+ CD23-), предшественники MZ (CD19+ CD93- B220+ CD21/35+ IgM+ CD23+), Fol I (CD19+ CD93- B220+ CD21/35int IgMint IgD+) и Fol II (CD19+ CD93- B220+ CD21/35int IgM+ IgD+).
[0073] На фигурах 12A – 12D показаны репрезентативные контурные графики лимфоцитов в костном мозге, собранном у мышей VELOCIMMUNE® (VI) и мышей, гетерозиготных по сконструированному участку DH, содержащему кодирующие последовательности D6 (6590het, «D6-DH het»; линия грызунов с геномом, содержащим гетерозиготный локус тяжелой цепи иммуноглобулина, содержащий несколько сегментов VH человека, сконструированных сегментов DH, включающих кодирующие последовательности D6 хемокинового рецептора-ловушки, вместо стандартных сегментов DH и сегментов JH, функционально связанных с константным участком тяжелой цепи иммуноглобулина грызуна, включающим энхансеры и регуляторные участки тяжелой цепи грызуна, и содержащим вставленную нуклеотидную последовательность, кодирующую один или более генов Adam6 мыши [например, патенты №№ 8642835 и 8697940]; и они окрашены для выявления экспрессии различных клеточных маркеров на клеточной поверхности. Фигура 12A: репрезентативный контурный график лимфоцитов из костного мозга, гейтированных по синглетам, демонстрирующим экспрессию CD19 (ось y) и CD3 (ось x). Фигура 12B: репрезентативный контурный график лимфоцитов из костного мозга, гейтированных по CD19+IgM- to lowIgD–, демонстрирующих экспрессию c-kit (ось y) и CD43 (ось х); пре- (c-kit- CD43-) и про-B-(c-kit+CD43+) клетки указаны на каждом точечном графике. Фигура 12C: репрезентативный контурный график лимфоцитов из костного мозга, гейтированных по CD19+, демонстрирующих экспрессию IgM (ось y) и B220 (ось х); незрелые (IgMint to +B220int) и зрелые (IgMint to +B220+), пре- и про-B-клетки (IgM- to low B220int) указаны на каждом точечном графике. Фигура 12D: репрезентативный контурный график лимфоцитов из костного мозга, гейтированных по CD19+IgMint to + B220int (верхний ряд) и CD19+IgM int to +B220+ (нижний ряд), демонстрирующих экспрессию легкой цепи Igλ (ось y) или Igκ (ось х).
[0074] На фигурах 13A–13D показаны репрезентативные контурные графики лимфоцитов в селезенке, собранной у мышей VELOCIMMUNE® (VI) и мышей, гомозиготных по сконструированному участку DH, содержащему кодирующие последовательности D6 (6590ho/1293ho, «D6-DH ho»; линия грызунов с геномом, содержащим гетерозиготный локус тяжелой цепи иммуноглобулина, содержащий несколько сегментов VH человека, сконструированных сегментов DH, включающих кодирующие последовательности D6 хемокинового рецептора-ловушки, вместо стандартных сегментов DH и сегментов JH, функционально связанных с константным участком тяжелой цепи иммуноглобулина грызуна, включающим энхансеры и регуляторные участки тяжелой цепи грызуна, и содержащим вставленную нуклеотидную последовательность, кодирующую один или более генов Adam6 мыши [например, патенты №№ 8642835 и 8697940]; и гомозиготный локус легкой κ-цепи иммуноглобулина, содержащий генные сегменты Vκ и Jκ человека, функционально связанные с геном участком Cκ грызуна, включающим энхансеры легкой κ-цепи грызуна), и они окрашены для выявления экспрессии различных клеточных маркеров на клеточной поверхности. Фигура 13A: репрезентативный контурный график лимфоцитов из селезенки, гейтированных по синглетам, демонстрирующим экспрессию CD19 (ось y) и CD3 (ось x). Фигура 13B: репрезентативный контурный график лимфоцитов из селезенки, гейтированных по CD19+, демонстрирующих экспрессию IgD (ось y) и IgM (ось x); зрелые (CD19+ IgD+ IgMint) и переходные (CD19+ IgDint IgM+) B-клетки указаны на каждом точечном графике. Фигура 13C: репрезентативный контурный график лимфоцитов из селезенки, гейтированных по CD19+, демонстрирующих экспрессию легкой цепи Igλ (ось y) или Igκ (ось x). Фигура 13D: показан репрезентативный контурный график созревания В-клеток, демонстрирующий лимфоциты из селезенки, гейтированные по CD19+ синглетам, и показывающих экспрессию [слева направо] CD93 (ось y) и B220 (ось х), IgM (ось y) и CD23 (ось х); CD21/35 (ось y) и IgM (ось х), B220 (ось y) и CD23 (ось х), а также IgD (ось y), и IgM (ось х). Верхний ряд: мыши VELOCIMMUNE®; нижний ряд: мыши D6-DH ho (6590ho). Специфические популяции В-клеток указаны на каждом точечном графике: незрелые (CD19+ CD93+ B220+), зрелые (CD19+ CD93- B220+), T1 (CD19+ CD93+ B220+ IgM+ CD23-), T2 (CD19+ CD93+ B220+ IgM+ CD23+), T3 (CD19+ CD93+ B220+ IgMint CD23+), MZ (CD19+ CD93- B220+ CD21/35+ IgM+ CD23-), предшественники MZ (CD19+ CD93- B220+ CD21/35+ IgM+ CD23+), Fol I (CD19+ CD93- B220+ CD21/35int IgMint IgD+) и Fol II (CD19+ CD93- B220+ CD21/35int IgM+ IgD+).
[0075] На фигурах 14A–14D показаны репрезентативные контурные графики лимфоцитов в костном мозге, собранном у мышей VELOCIMMUNE® (VI) и мышей, гомозиготных по сконструированному участку DH, содержащему кодирующие последовательности D6 (6590ho/1293ho, «D6-DH ho»; линия грызунов с геномом, содержащим гетерозиготный локус тяжелой цепи иммуноглобулина, содержащий несколько сегментов VH человека, сконструированных сегментов DH, включающих кодирующие последовательности D6 хемокинового рецептора-ловушки, вместо стандартных сегментов DH и сегментов JH, функционально связанных с константным участком тяжелой цепи иммуноглобулина грызуна, включающим энхансеры и регуляторные участки тяжелой цепи грызуна, и содержащим вставленную нуклеотидную последовательность, кодирующую один или более генов Adam6 мыши [например, патенты №№ 8642835 и 8697940]; и гомозиготный локус легкой κ-цепи иммуноглобулина, содержащий генные сегменты Vκ и Jκ человека, функционально связанные с генным участком Cκ грызуна, включающим энхансеры легкой κ-цепи грызуна), и они окрашены для выявления экспрессии различных клеточных маркеров на клеточной поверхности. Фигура 14A: репрезентативный контурный график лимфоцитов из костного мозга, гейтированных по синглетам, демонстрирующим экспрессию CD19 (ось y) и CD3 (ось x). Фигура 14B: репрезентативный контурный график лимфоцитов из костного мозга, гейтированных по CD19+IgM- tolowIgD–, демонстрирующих экспрессию c-kit (ось y) и CD43 (ось х); пре- (c-kit- CD43-) и про-B-(c-kit+CD43+) клетки указаны на каждом точечном графике. Фигура 14C: репрезентативный контурный график лимфоцитов из костного мозга, гейтированных по CD19+, демонстрирующих экспрессию IgM (ось y) и B220 (ось х); незрелые (IgMint to +B220int) и зрелые (IgMint to +B220+), пре- и про-B-клетки (IgM- to low B220int) указаны на каждом точечном графике. Фигура 14D: репрезентативный контурный график лимфоцитов из костного мозга, гейтированных по CD19+IgMint to + B220int (верхний ряд) и CD19+IgM int to +B220+ (нижний ряд), демонстрирующих экспрессию легкой цепи Igλ (ось y) или Igκ (ось х).
[0076] На фигуре 15 показано репрезентативное использование кодирующих последовательностей токсина в сконструированном участке DH в амплифицированной РНК из селезенки и костного мозга (все семейства VH объединены, не отражает количественное использование VH) от трех мышей 6579ho/1293ho («TX-DH ho», выше). На оси y указано название каждой кодирующей последовательности токсина в пределах сконструированного участка DH. На оси х указана частота (процентное содержание последовательностей) каждой кодирующей последовательности токсина среди проанализированных ридов последовательностей.
[0077] На фигуре 16 показано репрезентативное процентное использование генных сегментов VH человека в амплифицированной РНК из селезенки и костного мозга (все семейства VH объединены, не отражает количественное использование VH) от трех мышей 6579ho/1293ho («TX-DH ho», выше). На оси х указано название каждого генного сегмента VH человека в пределах вариабельного участка гуманизированной тяжелой цепи.
[0078] На фигуре 17 показано репрезентативное процентное использование генных сегментов JH человека в амплифицированной РНК из селезенки и костного мозга (все семейства VH объединены, не отражает количественное использование JH) от трех мышей 6579ho/1293ho («TX-DH ho», выше). На оси х указано название каждого генного сегмента JH человека в пределах вариабельного участка гуманизированной тяжелой цепи.
[0079] На фигуре 18 показана репрезентативная частота использования выбранных кодирующих последовательностей D6 в сконструированном участке DH в амплифицированной РНК из селезенки и костного мозга (все семейства VH объединены, не отражает количественное использование VH) от трех мышей 6590het («D6-DH het», выше). На оси y указано название выбранных кодирующих последовательностей D6 в пределах сконструированного участка DH. На оси х показана частота (процентное содержание последовательностей) кодирующих последовательностей D6 среди проанализированных ридов последовательностей. BM: костный мозг.
[0080] На фигуре 19 показано репрезентативное процентное использование генных сегментов VH человека в амплифицированной РНК из селезенки и костного мозга (все семейства VH объединены, не отражает количественное использование VH) от трех мышей 6590het («D6-DH het», выше). На оси х указано название каждого генного сегмента VH человека в пределах вариабельного участка гуманизированной тяжелой цепи. BM: костный мозг.
[0081] На фигуре 20 показано репрезентативное процентное использование генных сегментов JH человека в амплифицированной РНК из селезенки и костного мозга (все семейства VH объединены, не отражает количественное использование JH) от трех мышей 6590het («D6-DH het», выше). На оси х указано название каждого генного сегмента JH человека в пределах вариабельного участка гуманизированной тяжелой цепи. BM: костный мозг.
[0082] На фигуре 21 показан титр, превышающий фоновый уровень (ось y), у контрольных животных и животных 6579HO/1634 после иммунизации с использованием сконструированной растворимой формой белка клеточной поверхности.
ОПРЕДЕЛЕНИЯ
[0083] При ознакомлении с настоящим изобретением специалисты в данной области будут осознавать различные модификации, которые могут быть эквивалентны таким описанным вариантам осуществления или иным образом находится в пределах объема настоящего изобретения. В целом, используемая в данном документе терминология соответствует значениям, понятным в данной области, если явно не указано иное. Точные определения некоторых терминов приведены в данном документе ниже; значения этих и других терминов в конкретных случаях по всему данного описания будут понятны специалистам в данной области из контекста. Дополнительные определения для следующих терминов и других терминов изложены по всему описанию. Ссылки, приведенные в данном описании, или соответствующие их части включены в данный документ посредством ссылки.
[0084] «Введение» обозначает введение композиции в субъекта или живую систему (например, в клетку, орган, ткань, организм или их соответствующий компонент или набор компонентов). Специалисты в данной области поймут, что путь введения может варьировать, например, в зависимости от субъекта или системы, которым будет вводиться композиция, природы композиции, цели введения и т. д. Например, в определенных вариантах осуществления введение субъекту-животному (например, человеку или грызуну) может быть бронхиальным (в том числе посредством бронхиальной инстилляции), трансбуккальным, энтеральным, внутрикожным, внутриартериальным, интрадермальным, внутрижелудочным, интрамедуллярным, внутримышечным, интраназальным, внутрибрюшинным, интратекальным, внутривенным, интравентрикулярным, чресслизистым, назальным, пероральным, ректальным, подкожным, подъязычным, местным, трахеальным (в том числе посредством интратрахеальной инстилляции), трансдермальным, вагинальным и/или витреальным. В некоторых вариантах осуществления введение может подразумевать прерывистое введение доз. В некоторых вариантах осуществления введение может включать непрерывное введение доз (например, перфузию) в течение по меньшей мере выбранного периода времени.
[0085] Термин «антитело» включает молекулы типичных иммуноглобулинов, содержащие четыре полипептидные цепи, две тяжелые (H) цепи (каждая из которых может содержать аминокислотную последовательность, кодируемую сконструированным кластером DH) и две легкие (L) цепи (каждая из которых может представлять собой общую легкую цепь), соединенных дисульфидными связями. Данный термин также включает иммуноглобулин, который является активным по отношению к антигену или его фрагменту. Подходящие антитела включают без ограничения человеческие антитела, приматизированные антитела, химерные антитела, моноклональное антитела, моноспецифические антитела, поликлональные антитела, полиспецифические антитела, неспецифические антитела, биспецифические антитела, мультиспецифические антитела, гуманизированные антитела, синтетические антитела, рекомбинантные антитела, гибридные антитела, подвергнутые мутации антитела, привитые конъюгированные антитела (т.е. антитела, конъюгированные или слитые с другими белками, радиоактивными метками, цитоксинами) и полученные in vitro антитела. Специалист в данной области с легкостью определит общие изотипы антител, например, антител, имеющих константный участок тяжелой цепи, выбранных из группы, состоящей из IgG, IgA, IgM, IgD и IgE и любых их подклассов (например, IgG1, IgG2, IgG3 и IgG4).
[0086] «Примерно», используемое в данном документе в отношении одного или более представляющих интерес значений обозначает значение, которое аналогично указанному эталонному значению. В некоторых вариантах осуществления термин «примерно» или «приблизительно» относится к диапазону значений, находящихся в пределах 25%, 20%, 19%, 18%, 17%, 16%, 15%, 14%, 13%, 12%, 11%, 10%, 9%, 8%, 7%, 6%, 5%, 4%, 3%, 2%, 1% или меньше в любую сторону (большую или меньшую) от указанного эталонного значения, если иное не указано или иное не очевидно из контекста (за исключением случаев, в которых такое число будет превышать 100% от возможного значения).
[0087] «Биологически активный» обозначает характеристику любого средства, обладающего активностью в биологической системе in vitro или in vivo (например, в организме). Например, средство, которое в случае присутствия в организме обладает биологическим эффектом в пределах этого организма, считается биологически активным. В конкретных вариантах осуществления, в которых белок или полипептид являются биологически активными, часть этого белка или полипептида, обладающая по меньшей мере одним общим с данным белком или полипептидом видом биологической активности, обычно называется «биологически активной» частью.
[0088] «Сравнимый» обозначает два или более средства, объекта, ситуации, наборы условий и т.д., которые могут не быть идентичны друг другу, но которые являются в достаточной степени похожими для обеспечения возможности их сравнения, чтобы на основании наблюдаемых различий или сходств можно было сделать разумные выводы. Средним специалистам в данной области из контекста будет понятно, какая степень идентичности требуется в любых заданных обстоятельствах для двух или более таких средств, объектов, ситуаций, наборов условий и т.д., чтобы они считались сравнимыми.
[0089] Фраза «определяющий комплементарность участок» или термин «CDR» включают аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты генов иммуноглобулина организма, которая в норме (т.e. у животного дикого типа) находится между двух каркасных участков в вариабельном участке легкой или тяжелой цепи молекулы иммуноглобулина (например, антитела или T-клеточного рецептора). CDR может кодироваться, например, последовательностью зародышевой линии, или последовательностью перегруппированного или неперегруппированного участка, и, например, наивной или зрелой B-клеткой или T-клеткой. CDR может быть подвергшимся соматической мутации (например, отличаться от последовательности, кодируемой в зародышевой линии животного), гуманизированным и/или модифицированным с помощью аминокислотных замен, добавлений или делеций. В некоторых случаях (например, в случае CDR3) CDR могут кодироваться двумя или более последовательностями (например, последовательностями зародышевой линии), которые не являются смежными (например, в случае последовательности неперегруппированной нуклеиновой кислоты), но они являются смежными в последовательности нуклеиновой кислоты в B-клетке, например, как результат сплайсинга или соединения последовательностей (например, V-D-J-рекомбинация с образованием CDR3 тяжелой цепи).
[0090] «Консервативный» в отношении консервативной аминокислотной замены обозначает замену аминокислотного остатка другим аминокислотным остатком, имеющим R-группу боковой цепи с аналогичными химическими свойствами (например, зарядом или гидрофобностью). В целом, консервативная аминокислотная замена практически не будет изменять представляющие интерес функциональные свойства белка, например, способность рецептора связываться с лигандом. Примеры групп аминокислот, которые имеют боковые цепи с аналогичными химическими свойствами, включают: алифатические боковые цепи, такие как у глицина, аланина, валина, лейцина и изолейцина; алифатические боковые цепи с гидроксильными группами, такие как у серина и треонина; амид-содержащие боковые цепи, такие как у аспарагина и глутамина; ароматические боковые цепи, такие как у фенилаланина, тирозина и триптофана; основные боковые цепи, такие как у лизина, аргинина и гистидина; кислотные боковые цепи, такие как у аспарагиновой кислоты и глутаминовой кислоты; и серосодержащие боковые цепи, такие как у цистеина и метионина. Группы консервативных аминокислотных замен включают, например, валин/лейцин/изолейцин, фенилаланин/тирозин, лизин/аргинин, аланин/валин, глутамат/аспартат и аспарагин/глутамин. В некоторых вариантах осуществления консервативная аминокислотная замена может представлять собой замену любого нативного остатка в белке аланином, применяемую, например, в аланин-сканирующем мутагенезе. В некоторых вариантах осуществления проводят консервативную замену, которая имеет положительное значение в матрице логарифмической функции правдоподобия PAM250, раскрытой в Gonnet, G.H. et al., 1992, Science 256:1443-1445, включенной в данный документ посредством ссылки. В некоторых вариантах осуществления замена представляет собой умеренно консервативную замену, где замена имеет неотрицательное значение в матрице логарифмической функции правдоподобия PAM250.
[0091] «Контроль» обозначает понятное в данной области значение «контроля», подразумевающее стандарт, с которым сравнивают результаты. Как правило, контроли используют для повышения целостности в экспериментах путем разделения переменных для того, чтобы сделать вывод об этих переменных. В некоторых вариантах осуществления контроль представляет собой реакцию или анализ, проводимые одновременно с тестовыми реакцией или анализом для обеспечения объекта сравнения. Термин «контроль» может относиться к «контрольному животному». «Контрольное животное» может иметь модификацию, описанную в данном документе, модификацию, отличную от описанной в данном документе, или не иметь модификации (т.е. в случае животного дикого типа). В одном эксперименте используют «тест» (т.е. тестируемую переменную). Во втором эксперименте, «контрольном», тестируемую переменную не используют. В некоторых вариантах осуществления контроль представляет собой исторический контроль (т.е. тест или анализ, проведенные ранее, или количество или результат, известные заранее). В некоторых вариантах осуществления контроль представляет собой или включает напечатанную или иным образом сохраненную запись. Контроль может представлять собой положительный контроль или отрицательный контроль.
[0092] «Разрушение» обозначает результат события гомологичной рекомбинации с молекулой ДНК (например, с эндогенной гомологичной последовательностью, такой как ген или локус гена). В некоторых вариантах осуществления разрушение может приводить к вставке, делеции, замене, замещению, миссенс-мутации или сдвигу рамки считывания в последовательности(ях) ДНК или любой их комбинации или может быть представлено такими. Вставки могут включать вставку целых генов, фрагментов генов, например, экзонов, которые по происхождению могут быть отличны от эндогенной последовательности (например, являются гетерологичной последовательностью), или кодирующих последовательностей, полученных или выделенных из конкретного представляющего интерес гена. В некоторых вариантах осуществления разрушение может приводить к повышению экспрессии и/или активности гена или продукта гена (например, белка, кодируемого геном). В некоторых вариантах осуществления разрушение может приводить к снижению экспрессии и/или активности гена или продукта гена. В некоторых вариантах осуществления разрушение может приводить к изменению последовательности гена или кодируемого продукта гена (например, кодируемого белка). В некоторых вариантах осуществления разрушение может приводить к усечению или фрагментации гена или кодируемого продукта гена (например, кодируемого белка). В некоторых вариантах осуществления разрушение может приводить к удлинению гена или кодируемого продукта гена. В некоторых таких вариантах осуществления за счет разрушения может обеспечиваться сборка слитого белка. В некоторых вариантах осуществления разрушение может приводить к изменению уровня экспрессии, но не активности гена или продукта гена. В некоторых вариантах осуществления разрушение может приводить к изменению активности, но не уровня экспрессии гена или продукта гена. В некоторых вариантах осуществления разрушение может не оказывать значительного влияния на уровень экспрессии гена или продукта гена. В некоторых вариантах осуществления разрушение может не оказывать значительного влияния на активность гена или продукта гена. В некоторых вариантах осуществления разрушение может не оказывать значительного влияния ни на уровень экспрессии, ни на активность гена или продукта гена.
[0093] «Определение», «измерение», «установление», «оценка», «анализ» и «проведение анализа» используются взаимозаменяемо для обозначения любой формы измерения, и они включают определение присутствия или отсутствия элемента. Эти термины включают как количественные, а также и/или качественные определения. Анализ может быть относительным или абсолютным. «Анализ на наличие» может представлять собой определение количества чего-либо присутствующего и/или определение того, присутствует или отсутствует ли оно.
[0094] «Эндогенный локус» или «эндогенный ген» обозначают генетический локус, находящийся в исходном или эталонном организме до введения изменения, разрушения, делеции, вставки, модификации, замещения или замены, описываемых в данном документе. В некоторых вариантах осуществления эндогенный локус содержит последовательность, частично или полностью, встречающуюся в природе. В некоторых вариантах осуществления эндогенный локус представляет собой локус дикого типа. В некоторых вариантах осуществления эталонный организм представляет собой организм дикого типа. В некоторых вариантах осуществления эталонный организм представляет собой организм, полученный с помощью генной инженерии. В некоторых вариантах осуществления эталонный организм представляет собой организм, разводимый в лаборатории (организм дикого типа или полученный с помощью генной инженерии).
[0095] «Эндогенный промотор» обозначает промотор, который в естественных условиях, например в организме дикого типа, связан с эндогенным геном.
[0096] «Сконструированный» обозначает, в целом, аспект проведения манипуляций человеком. Например, в некоторых вариантах осуществления полинуклеотид может считаться «сконструированным» в том случае, если две или более последовательности, которые не соединены друг с другом в этом порядке в природе, подвергнуты манипуляции человеком для непосредственного соединения друг с другом в сконструированном полинуклеотиде. В некоторых таких конкретных вариантах осуществления сконструированный полинуклеотид может содержать регуляторную последовательность, которая встречается в природе в функциональной связи с первой кодирующей последовательностью, но не в функциональной ассоциации со второй кодирующей последовательностью, которая связана человеком таким образом, что она функционально связана со второй кодирующей последовательностью. В качестве альтернативы или дополнительно, в некоторых вариантах осуществления первая и вторая последовательности нуклеиновых кислот, каждая из которых кодирует полипептидные элементы или домены, которые в природе не связаны друг с другом, могут быть связаны друг с другом в единый сконструированный полинуклеотид. Аналогично, в некоторых вариантах осуществления клетка или организм могут считаться «сконструированными» в том случае, если они были подвергнуты манипуляции таким образом, чтобы их генетическая информация была изменена (например, был введен новый генетический материал, который ранее отсутствовал, или ранее присутствовавший генетический материал был изменен или удален). Как понимается специалистами в данной области и является общепринятой практикой, потомство сконструированного полинуклеотида или клетки, как правило, по-прежнему называют «сконструированным», даже если фактическую манипуляцию выполняли на предыдущем объекте. Кроме того, как будет понятно специалистам в данной области, доступны различные методологии, посредством которых можно осуществлять «конструирование», описываемое в данном документе. Например, в некоторых вариантах осуществления «конструирование» может предусматривать отбор или разработку (например, последовательностей нуклеиновых кислот, полипептидных последовательностей, клеток, тканей и/или организмов) путем использования компьютерных систем, запрограммированных для проведения анализа или сравнения или, в иных случаях, для анализа, рекомендации и/или отбора последовательностей, изменений и т. д. В качестве альтернативы или дополнительно, в некоторых вариантах осуществления «конструирование» может включать применение методологий химического синтеза in vitro и/или методик рекомбинантных нуклеиновых кислот, таких как, например, амплификация нуклеиновой кислоты (например, посредством полимеразной цепной реакции), гибридизация, мутирование, трансформация, трансфекция и т.д., и/или любой из множества методологий контролируемого размножения. Как будет понятно специалистам в данной области, множество таких общепринятых методик (например, для рекомбинантной ДНК, синтеза олигонуклеотидов и культивирования и трансформации тканей (например, электропорация, липофекция и т.д.)) хорошо известно в данной области и описано в различных общих и более специфических литературных источниках, которые цитируются и/или обсуждаются во всем настоящем описании. См., например, Sambrook et al., Molecular Cloning: A Laboratory Manual (2d ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y., 1989).
[0097] «Ген» обозначает последовательность ДНК в хромосоме, которая кодирует продукт (например, РНК-продукт и/или полипептидный продукт). В некоторых вариантах осуществления ген содержит кодирующую последовательность (т. е. последовательность, которая кодирует конкретный продукт). В некоторых вариантах осуществления ген содержит некодирующую последовательность. В некоторых конкретных вариантах осуществления ген может содержать как кодирующую (например, экзонную), так и некодирующую (например, интронную) последовательности. В некоторых вариантах осуществления ген может содержать одну или более регуляторных последовательностей (например, промоторы, энхансеры и т.д.) и/или интронных последовательностей, которые, например, могут контролировать или влиять на один или более аспектов экспрессии гена (например, специфичную для типа клеток экспрессию, индуцируемую экспрессию и т.д.). Для ясности отметим, что используемый в настоящей заявке термин «ген» обычно обозначает часть нуклеиновой кислоты, которая кодирует полипептид; при этом данный термин может необязательно охватывать регуляторные последовательности, как будет понятно из контекста средним специалистам в данной области. Это определение не подразумевает исключение применения термина «ген» к единицам экспрессии, не кодирующим белок, но скорее поясняет то, что в большинстве случаев термин, используемый в данном документе, относится к нуклеиновой кислоте, кодирующей полипептид.
[0098] Фраза «генный сегмент» или «сегмент» включает ссылку на генный сегмент V (легкой или тяжелой цепи), или D, или J (легкой или тяжелой цепи) иммуноглобулина, что включает последовательности неперегруппированных участков в локусе иммуноглобулина (например, у человека и мыши), которые могут принимать участие в перегруппировке (опосредованной, например, эндогенными рекомбиназами) с образованием последовательности перегруппированного участка V/J (легкая цепь) или V/D/J (тяжелая цепь). Если не указано иное, то сегменты V, D и J содержат последовательности сигнала рекомбинации (RSS), которые обеспечивают возможность V/J-рекомбинации или V/D/J-рекомбинации в соответствии с правилом 12/23. Если не указано иное, то сегменты дополнительно содержат последовательности, с которыми они связаны в природе, или их функциональные эквиваленты (например, в случае сегментов V, это промотор(ы) и лидерная последовательность(и)).
[0099] Термин «зародышевая линия» в отношении последовательности нуклеиновой кислоты иммуноглобулина подразумевает последовательность нуклеиновой кислоты, которая может переходить в потомство, например, в геном зародышевой линии, который может находиться в зародышевой клетке.
[00109] Фраза «тяжелая цепь» или «тяжелая цепь иммуноглобулина» подразумевает последовательность тяжелой цепи иммуноглобулина, в том числе последовательность константного участка тяжелой цепи иммуноглобулина, из любого организма. Вариабельные домены тяжелой цепи включают три определяющих комплементарность участка (CDR) тяжелой цепи и четыре участка FR, если не указано иное. Фрагменты тяжелых цепей включают CDR, CDR и FR, а также их комбинации. В типичной тяжелой цепи после вариабельного домена (от N-конца к C-концу) расположен домен CH1, шарнир, домен CH2, домен CH3 и домен CH4 (в контексте IgM или IgE). Функциональный фрагмент тяжелой цепи подразумевает фрагмент, который способен специфически распознавать эпитоп (например, распознавать эпитоп с KD в микромолярном, наномолярном или пикомолярном диапазонах), который способен экспрессироваться и секретироваться клеткой и который содержит по меньшей мере один CDR. Вариабельный домен тяжелой цепи кодируется генной последовательностью вариабельного участка, который, как правило, содержит сегменты VH, DH и JH, полученные из репертуара сегментов VH, DH и JH, присутствующих в зародышевой линии. Последовательности, местоположения и номенклатуру для сегментов V, D и J тяжелой цепи у различных организмов можно просмотреть на веб-странице Международной информационной службы по иммуногенетике (IMGT) – www.imgt.org.
[00101] Фраза «легкая цепь» подразумевает последовательность легкой цепи иммуноглобулина из любого организма и, если не указано иное, предусматривает легкие каппа- и лямбда-цепи человека и VpreB, а также суррогатные легкие цепи. Вариабельные домены легкой цепи, как правило, включают три определяющих комплементарность участка (CDR) легкой цепи и четыре каркасных (FR) участка, если не указано иное. В целом, полноразмерная легкая цепь включает, от амино-конца к карбоксильному концу, вариабельный домен, который включает FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4, и константный домен легкой цепи. Вариабельный домен легкой цепи кодируется генной последовательностью вариабельного участка тяжелой цепи, который, как правило, содержит генные сегменты VL и JL, полученные из репертуара генных сегментов VL и JL, присутствующих в зародышевой линии. Последовательности, местоположения и номенклатуру для сегментов V и J легкой цепи у различных организмов можно просмотреть на веб-странице Международной информационной службы по иммуногенетике (IMGT) – www.imgt.org. Легкие цепи включают, например, цепи, которые не связывают селективно или первый, или второй эпитоп, селективно связанные эпитоп-связывающим белком, в котором они находятся. Легкие цепи также включают цепи, которые связывают и распознают или помогают тяжелой цепи связываться и распознавать один или более эпитопов, селективно связанных эпитоп-связывающим белком, в котором они находятся. Фраза легкая цепь подразумевает «общую легкую цепь», также называемую как «универсальная легкая цепь» (ULC).
[00102] Общие или универсальные легкие цепи (ULC) включают цепи, которые получены из локуса легкой цепи иммуноглобулина, содержащего последовательность, кодирующую единый перегруппированный вариабельный участок легкой цепи иммуноглобулина, функционально связанную с константным участком легкой цепи, при этом в результате экспрессии локуса легкой цепи иммуноглобулина продуцируется только легкая цепь, полученная из единого перегруппированного вариабельного участка легкой цепи иммуноглобулина, функционально связанного с константным участком легкой цепи, независимо от включения других последовательностей нуклеиновой кислоты, например, других генных сегментов легкой цепи, в локус легкой цепи иммуноглобулина. Универсальные легкие цепи включают ген Vκ1-39Jκ человека (например, ген Vκ1-39Jκ5) или ген Vκ3-20Jκ человека (например, ген Vκ3-20Jκ1), и включают их варианты, подвергшиеся соматической мутации (например, созреванию аффинности).
[00103] «Гетерологичный» обозначает средство или объект из другого источника. Например, при использовании в отношении полипептида, гена или продукта гена, присутствующих в конкретной клетке или организме, данный термин поясняет, что соответствующий полипептид или его фрагмент, ген или его фрагмент, или продукт гена или его фрагмент: 1) был сконструирован человеком; 2) был введен в клетку или организм (или их предшественника) человеком (например, с помощью генной инженерии), и/или 3) в естественных условиях не продуцируется соответствующей клеткой или организмом (например, соответствующим типом клетки или типом организма) или не присутствует в них. Используемый в данном документе термин «гетерологичный» также включает полипептид или его фрагмент, ген или его фрагмент, или продукт гена или его фрагмент, которые обычно присутствуют в конкретных нативной клетке или организме, но были модифицированы, например, за счет мутации или помещения под контроль не связанных в естественных условиях и, в некоторых вариантах осуществления, не являющихся эндогенными регуляторных элементов (например, промотора).
[00104] «Клетка-хозяин» обозначает клетку, в которую были введены гетерологичные (например, экзогенные) нуклеиновая кислота или белок. При ознакомлении с данным изобретением специалистам в данной области будет понятно, что такие термины обозначают не только конкретную рассматриваемую клетку, но также используются для обозначения потомства такой клетки. Поскольку в последующих поколениях вследствие влияния мутаций или окружающей среды могут происходить определенные модификации, такое потомство в действительности может не быть идентичным исходной клетке, но по-прежнему включается в объем термина «клетка-хозяин», используемого в данном документе. В некоторых вариантах осуществления клетка-хозяин представляет собой или предусматривает прокариотическую или эукариотическую клетку. Как правило, клетка-хозяин представляет собой любую клетку, подходящую для приема и/или продуцирования гетерологичных нуклеиновой кислоты или белка, независимо от того, к какому царству живой природы относится клетка. Иллюстративные клетки включают клетки прокариот и эукариот (одноклеточных или многоклеточных), бактериальные клетки (например, штаммы Escherichia coli, Bacillus spp., Streptomyces spp. и т.д.), клетки микобактерий, клетки грибов, клетки дрожжей (например, Saccharomyces cerevisiae, Schizosaccharomyces pombe, Pichia partoris, Pichia methanolica и т.д.), клетки растений, клетки насекомых (например, SF-9, SF-21, клетки насекомых, зараженные бакуловирусами, Trichoplusiani и т.д.), клетки животных, отличных от человека, клетки человека или продукты слияния клеток, такие как, например, гибридомы или квадромы. В некоторых вариантах осуществления клетка представляет собой клетку человека, обезьяны, человекообразной обезьяны, хомячка, крысы или мыши. В некоторых вариантах осуществления клетка является эукариотической и выбрана из следующих клеток: CHO (например, CHO K1, DXB-11 CHO, Veggie-CHO), COS (например, COS-7), клетки сетчатки, Vero, CV1, клетки почки (например, HEK293, 293 EBNA, MSR 293, MDCK, HaK, BHK), HeLa, HepG2, WI38, MRC 5, Colo205, HB 8065, HL-60 (например, BHK21), Jurkat, Daudi, A431 (эпидермальные), CV-1, U937, 3T3, L-клетки, клетки C127, SP2/0, NS-0, MMT 060562, клетки Сертоли, клетки BRL 3A, клетки HT1080, клетки миеломы, опухолевой клетки и линии клеток, происходящей из вышеупомянутых клеток. В некоторых вариантах осуществления клетка содержит один или более вирусных генов, например, клетка сетчатки, которая экспрессирует вирусный ген (например, клетка PER.C6®). В некоторых вариантах осуществления клетка-хозяин представляет собой или предусматривает выделенную клетку. В некоторых вариантах осуществления клетка-хозяин представляет собой часть ткани. В некоторых вариантах осуществления клетка-хозяин представляет собой часть организма.
[00105] «Идентичность», используемая применительно к сравнению последовательностей, обозначает идентичность, определяемую с помощью ряда различных алгоритмов, известных из уровня техники, которые можно применять для измерения идентичности нуклеотидных и/или аминокислотных последовательностей. В некоторых вариантах осуществления значения идентичности, описываемые в данном документе, определяют с применением выравнивания ClustalW v. 1.83 (медленного), в котором используются штраф за открытие гэпа 10,0, штраф за продление гэпа 0,1, а также с применением матрицы сходства Gonnet (MACVECTOR™ 10.0.2, MacVector Inc., 2008).
[00106] «In vitro» обозначает события, которые происходят в искусственной окружающей среде, например, в пробирке или реакционном сосуде, в культуре клеток и т.д., а не в пределах многоклеточного организма.
[00107] «In vivo» обозначает события, которые происходят в пределах многоклеточного организма, такого как человек и животное, отличное от человека. В контексте систем на основе клетки данный термин можно использовать для обозначения событий, которые происходят в пределах живой клетки (в противоположность, например, системам in vitro).
[00108] «Выделенный» обозначает вещество и/или объект, которые были (1) отделены по меньшей мере от некоторых компонентов, с которыми они были связаны при первоначальном продуцировании (будь то в природе и/или в экспериментальных условиях), и/или (2) сконструированы, продуцированы, получены и/или произведены человеком. Выделенные вещества и/или объекты могут быть отделены от приблизительно 10%, приблизительно 20%, приблизительно 30%, приблизительно 40%, приблизительно 50%, приблизительно 60%, приблизительно 70%, приблизительно 80%, приблизительно 90%, приблизительно 91%, приблизительно 92%, приблизительно 93%, приблизительно 94%, приблизительно 95%, приблизительно 96%, приблизительно 97%, приблизительно 98%, приблизительно 99% или более чем приблизительно 99% других компонентов, с которыми они были изначально связаны. В некоторых вариантах осуществления выделенные средства являются чистыми на приблизительно 80%, на приблизительно 85%, на приблизительно 90%, на приблизительно 91%, на приблизительно 92%, на приблизительно 93%, на приблизительно 94%, на приблизительно 95%, на приблизительно 96%, на приблизительно 97%, на приблизительно 98%, на приблизительно 99% или более чем на приблизительно 99%. Как используется в данном документе, вещество является «чистым», если оно практически не содержит других компонентов. В некоторых вариантах осуществления, как будет понятно специалистам в данной области, вещество может по-прежнему считаться «выделенным» или даже «чистым» после объединения с определенными другими компонентами, такими как, например, один или более носителей или наполнителей (например, буфер, растворитель, вода и т.д.); в таких вариантах осуществления процент выделения или чистоты вещества рассчитывают без учета таких носителей или наполнителей. В виде лишь одного примера, в некоторых вариантах осуществления биологический полимер, такой как полипептид или полинуклеотид, встречающийся в природе, считается «выделенным», если: a) он ввиду своего происхождения или источника получения не связан с некоторыми или всеми компонентами, сопровождающими его в его нативном состоянии в природе; b) он практически не содержит других полипептидов или нуклеиновых кислот того же вида от вида, который продуцирует их в природе; или c) он экспрессируется или иным образом связан с компонентами в клетке или другой системе экспрессии, не принадлежащей к виду, который продуцирует его в природе. Таким образом, например, в некоторых вариантах осуществления полипептид, синтезируемый химическим путем или синтезируемый в клеточной системе, отличной от той, которая продуцирует его в природе, считается «выделенным» полипептидом. В качестве альтернативы или дополнительно, в некоторых вариантах осуществления полипептид, который был подвергнут одной или более методикам очистки, может считаться «выделенным» полипептидом в той степени, в которой он был отделен от других компонентов: a) с которыми он связан в природе и/или b) с которыми он был связан при первоначальном продуцировании.
[00109] «Животное, отличное от человека» обозначает любой позвоночное живое существо, которое не является человеком. В некоторых вариантах осуществления животное, отличное от человека, представляет собой круглоротое, костную рыбу, хрящевую рыбу (например, акулу или ската), земноводное, пресмыкающееся, млекопитающее и птицу. В некоторых вариантах осуществления млекопитающее, отличное от человека, представляет собой примата, козу, овцу, свинью, собаку, корову или грызуна. В некоторых вариантах осуществления животное, отличное от человека, представляет собой грызуна, такого как крыса или мышь.
[00110] «Нуклеиновая кислота», в своем самом широком смысле, обозначает любое соединение и/или вещество, включенному или которое может быть включено в состав олигонуклеотидной цепи. В некоторых вариантах осуществления «нуклеиновая кислота» представляет собой соединение и/или вещество, включенное или которое может быть включено в состав олигонуклеотидной цепи с помощью фосфодиэфирной связи. Как будет очевидно из контекста, в некоторых вариантах осуществления «нуклеиновая кислота» обозначает отдельные остатки нуклеиновой кислоты (например, нуклеотиды и/или нуклеозиды); в некоторых вариантах осуществления «нуклеиновая кислота» обозначает олигонуклеотидную цепь, содержащую отдельные остатки нуклеиновой кислоты. В некоторых вариантах осуществления «нуклеиновая кислота» представляет собой или содержит РНК; в некоторых вариантах осуществления «нуклеиновая кислота» представляет собой или содержит ДНК. В некоторых вариантах осуществления «нуклеиновая кислота» представляет собой один или более остатков природной нуклеиновой кислоты, содержит их или состоит из них. В некоторых вариантах осуществления «нуклеиновая кислота» представляет собой один или более аналогов нуклеиновой кислоты, содержит их или состоит из них. В некоторых вариантах осуществления аналог нуклеиновой кислоты отличается от «нуклеиновой кислоты» тем, что в нем не используется фосфодиэфирный остов. Например, в некоторых вариантах осуществления «нуклеиновая кислота» представляет собой одну или более «пептидных нуклеиновых кислот», которые известны в данной области и имеют в остове пептидные связи вместо фосфодиэфирных связей, содержит их или состоит из них. В качестве альтернативы или дополнительно, в некоторых вариантах осуществления «нуклеиновая кислота» имеет одну или более фосфоротиоатных и/или 5'-N-фосфорамидитных связей вместо фосфодиэфирных связей. В некоторых вариантах осуществления «нуклеиновая кислота» представляет собой один или более природных нуклеозидов (например, аденозин, тимидин, гуанозин, цитидин, уридин, дезоксиаденозин, дезокситимидин, дезоксигуанозин и дезоксицитидин), содержит их или состоит из них. В некоторых вариантах осуществления «нуклеиновая кислота» представляет собой один или более аналогов нуклеозидов (например, 2-аминоаденозин, 2-тиотимидин, инозин, пирролопиримидин, 3-метиладенозин, 5-метилцитидин, C5-пропинилцитидин, C5-пропинилуридин, 2-аминоаденозин, C5-бромуридин, C5-фторуридин, C5-йодуридин, C5-пропинилуридин, C5-пропинилцитидин, C5-метилцитидин, 2-аминоаденозин, 7-деазааденозин, 7-деазагуанозин, 8-оксоаденозин, 8-оксогуанозин, O(6)-метилгуанин, 2-тиоцитидин, метилированные основания, интеркалированные основания и их комбинации), содержит их или состоит из них. В некоторых вариантах осуществления «нуклеиновая кислота» содержит один или более модифицированных сахарных остатков (например, 2’-фторрибозу, рибозу, 2’-дезоксирибозу, арабинозу и гексозу) по сравнению с сахарными остатками в природных нуклеиновых кислотах. В некоторых вариантах осуществления «нуклеиновая кислота» имеет нуклеотидную последовательность, которая кодирует функциональный продукт гена, такой как РНК или белок. В некоторых вариантах осуществления «нуклеиновая кислота» имеет нуклеотидную последовательность, которая кодирует фрагмент полипептида (например, пептид). В некоторых вариантах осуществления «нуклеиновая кислота» предусматривает один или более интронов. В некоторых вариантах осуществления «нуклеиновая кислота» предусматривает один или более экзонов. В некоторых вариантах осуществления «нуклеиновая кислота» предусматривает одну или более кодирующих последовательностей. В некоторых вариантах осуществления «нуклеиновую кислоту» получают посредством одного или более из выделения из природного источника, ферментативного синтеза путем полимеризации на основе комплементарной матрицы (in vivo или in vitro), воспроизводства в рекомбинантной клетке или системе и химического синтеза. В некоторых вариантах осуществления длина «нуклеиновой кислоты» составляет по меньшей мере 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25, 30, 35, 40, 45, 50, 55, 60, 65, 70, 75, 80, 85, 90, 95, 100, 110, 120, 130, 140, 150, 160, 170, 180, 190, 20, 225, 250, 275, 300, 325, 350, 375, 400, 425, 450, 475, 500, 600, 700, 800, 900, 1000, 1500, 2000, 2500, 3000, 3500, 4000, 4500, 5000 или более остатков. В некоторых вариантах осуществления «нуклеиновая кислота» является однонитевой; в некоторых вариантах осуществления «нуклеиновая кислота» является двухнитевой. В некоторых вариантах осуществления «нуклеиновая кислота» имеет нуклеотидную последовательность, содержащую по меньшей мере один элемент, который кодирует полипептид или его фрагмент или является комплементарной последовательностью для последовательности, которая кодирует полипептид или его фрагмент. В некоторых вариантах осуществления «нуклеиновая кислота» обладает ферментативной активностью.
[00111] «Функционально связанный» обозначает контактное положение, при котором описанные компоненты находятся во взаимосвязи, позволяющей им функционировать надлежащим им образом. Последовательность контроля, «функционально связанная» с кодирующей последовательностью, лигирована таким образом, что экспрессия кодирующей последовательности достигается в условиях, подходящих для функционирования последовательностей контроля. «Функционально связанные» последовательности включают как последовательности контроля экспрессию, смежные с представляющим интерес геном, так и последовательности контроля экспрессии, действующие в транс-положении или на некотором расстоянии c осуществлением контроля представляющего интерес гена. Используемый в данном документе термин «последовательность контроля экспрессии» обозначает полинуклеотидные последовательности, необходимые для воздействия на экспрессию и процессинга кодирующих последовательностей, с которыми они лигированы. «Последовательности контроля экспрессии» включают: соответствующие последовательности инициации и терминации транскрипции, промоторные и энхансерные последовательности; сигналы для эффективного процессинга РНК, такие как сигналы для сплайсинга и полиаденилирования; последовательности, стабилизирующие цитоплазматическую мРНК; последовательности, повышающие эффективность трансляции (т.е. консенсусная последовательность Козак); последовательности, повышающие стабильность белка; и, если это необходимо, последовательности, усиливающие секрецию белка. Природа таких последовательностей контроля отличается в зависимости от организма-хозяина. Например, у прокариот такие последовательности контроля обычно включают в себя промотор, сайт связывания рибосом и последовательность терминации транскрипции, тогда как у эукариот такие последовательности контроля, как правило, включают промоторы и последовательность терминации транскрипции. Подразумевается, что термин «последовательности контроля» включает компоненты, присутствие которых является необходимым для экспрессии и процессинга, а также может включать дополнительные компоненты, присутствие которых является благоприятным, например, лидерные последовательности и последовательности партнера по слиянию.
[00112] «Физиологические условия» включает понятное в данной области значение, обозначающее условия, при которых клетки или организмы живут и/или размножаются. В некоторых вариантах осуществления данный термин обозначает условия во внешней или внутренней среде организма, которые могут встречаться в природе в организме или клеточной системе. В некоторых вариантах осуществления физиологические условия представляют собой условия, присутствующие в пределах организма человека или животного, отличного от человека, в частности, те условия, присутствующие в месте и/или в пределах области хирургического вмешательства. Физиологические условия, как правило, включают, например, температурный диапазон, составляющий 20–40°C, атмосферное давление, составляющее 1, pH, составляющий 6–8, концентрацию глюкозы, составляющую 1–20 мМ, концентрацию кислорода при уровнях, равным атмосферным, и силу тяжести, которая встречается на земном шаре. В некоторых вариантах осуществления с условиями в лаборатории осуществляют манипуляции и/или их поддерживают при физиологических условиях. В некоторых вариантах осуществления физиологические условия встречаются в организме (например, животном, отличном от человека).
[00113] «Полипептид» обозначает любую полимерную цепь из аминокислот. В некоторых вариантах осуществления полипептид имеет аминокислотную последовательность, встречающуюся в природе. В некоторых вариантах осуществления полипептид имеет аминокислотную последовательность, не встречающуюся в природе. В некоторых вариантах осуществления полипептид имеет аминокислотную последовательность, которая содержит части, встречающиеся в природе отдельно друг от друга (т.е. из двух или более различных организмов, например, часть, принадлежащую человеку, и часть, отличную от человеческой). В некоторых вариантах осуществления полипептид имеет аминокислотную последовательность, которая является сконструированной в том смысле, что она разработана и/или получена в результате действий человека. В некоторых вариантах осуществления полипептид может содержать или состоять из нескольких фрагментов, каждый из которых встречается в том же исходном полипептиде в другом пространственном порядке относительно друг друга, чем порядок, встречающийся в представляющем интерес полипептиде (например, фрагменты, которые напрямую связаны в исходном полипептиде, могут быть пространственно разделены в представляющем интерес полипептиде или наоборот, и/или фрагменты могут присутствовать в представляющем интерес полипептиде в другом порядке, чем порядок в исходном), так что представляющий интерес полипептид является производным своего исходного полипептида.
[00114] «Рекомбинантный» обозначает полипептиды, которые разработаны, сконструированы, получены, экспрессированы, созданы или выделены с помощью рекомбинантных способов, такие как полипептиды, экспрессируемые с использованием рекомбинантного вектора экспрессии, вводимого путем трансфекции в клетку-хозяина, полипептиды, выделяемые из комбинаторной библиотеки рекомбинантных полипептидов человека (Hoogenboom H. R., 1997 TIB Tech. 15:62-70; Hoogenboom H., and Chames P., 2000, Immunology Today 21:371-378; Azzazy H., and Highsmith W. E., 2002, Clin. Biochem. 35:425-445; Gavilondo J. V., and Larrick J. W., 2002, BioTechniques 29:128-145), антитела, выделяемые из животного (например, мыши), трансгенного по генам иммуноглобулинов человека (см., например, Taylor, L. D., et al., 1992, Nucl. Acids Res. 20:6287-6295; Little M. et al., 2000, Immunology Today 21:364-370; Kellermann S. A. and Green L. L., 2002, Current Opinion in Biotechnology 13:593-597; Murphy, A.J., et al., 2014, Proc. Natl. Acad. Sci. U. S. A. 111(14):5153-5158), или полипептиды, которые получены, экспрессированы, созданы или выделены любыми другими способами, предусматривающими сплайсинг выбранных элементов последовательности друг с другом. В некоторых вариантах осуществления один или более из таких выбранных элементов последовательности встречается в природе. В некоторых вариантах осуществления один или более из таких выбранных элементов последовательности разработан in silico. В некоторых вариантах осуществления один или более из таких выбранных элементов последовательности получены в результате мутагенеза (например, in vivo или in vitro) известного элемента последовательности, например, из природного или синтетического источника. Например, в некоторых вариантах осуществления рекомбинантный полипептид содержит последовательности, встречающиеся в геноме (или полипептиде) представляющего интерес организма-источника (например, человека, мыши и т.д.). В некоторых вариантах осуществления рекомбинантный полипептид содержит последовательности, которые встречаются в природе отдельно друг от друга (т.е. из двух или более разных организмов, например, часть, принадлежащая человеку, и часть, отличная от человеческой) в двух разных организмах (например, человека и организме, отличном от человека). В некоторых вариантах осуществления рекомбинантный полипептид имеет аминокислотную последовательность, полученную в результате мутагенеза (например,in vitro или in vivo, например, в животном, отличном от человека), так что аминокислотные последовательности рекомбинантных полипептидов представляют собой последовательности, которые хоть и происходят из полипептидных последовательностей и являются родственными им, но в естественных условиях могут не существовать in vivo в пределах генома животного, отличного от человека.
[00115] Подразумевается, что «эталон» описывает стандартные или контрольные средство, животное, группу, индивидуума, популяцию, образец, последовательность или значение, с которыми сравнивают представляющие интерес средство, животное, группу, индивидуума, популяцию, образец, последовательность или значение. В некоторых вариантах осуществления эталонные средство, животное, группу, индивидуума, популяцию, образец, последовательность или значение тестируют и/или определяют практически одновременно с тестированием или определением представляющих интерес средства, животного, группы, индивидуума, популяции, образца, последовательности или значения. В некоторых вариантах осуществления эталонные средство, животное, группа, индивидуум, популяция, образец, последовательность или значение представляют собой исторический эталон, необязательно внесенный на материальный носитель. В некоторых вариантах осуществления эталон может обозначать контроль. Используемый в данном документе «эталон» может обозначать «эталонное животное». «Эталонное животное» может иметь модификацию, описываемую в данном документе, модификацию, которая отличается от описываемой в данном документе, или не иметь модификации (т.е. в случае животного дикого типа). Как будет понятно специалистам в данной области, эталонные средство, животное, группу, индивидуум, популяцию, образец, последовательность или значение обычно определяют или характеризуют в условиях, сравнимых с условиями, используемыми для определения или получения характеристик представляющих интерес средства, животного (например, млекопитающего), группы, индивидуума, популяции, образца, последовательности или значения.
[00116] Иммуноглобулины принимают участие в клеточном механизме, называемом соматической гипермутацией, в результате которого образуются варианты антитела с созревшей аффинностью, характеризующиеся высокой аффинностью в отношении своей мишени. Хотя соматическая гипермутация главным образом происходит в пределах CDR вариабельных участков антител, мутации преимущественно нацеливаются на определенные мотивы последовательности, которые называются «горячими точками», например, «горячие точки» RGYW, индуцируемые активацией цитидиндезаминазы (AID) (см., например, Li, Z. et al., 2004, Genes Dev. 18:1-11; Teng, G. and F.N. Papavasiliou, 2007, Annu. Rev. Genet. 41:107-20; включенные в данный документ посредством ссылки). Представляющие интерес полипептиды, не являющиеся иммуноглобулинами, или их части, раскрытые в данном документе, применимые для получения сконструированного участка DH, могут содержать одну или более природных и/или искусственных «горячих точек». Фраза «подвергшаяся соматической мутации» включает ссылку на последовательность нуклеиновой кислоты из В-клетки, которая перенесла переключение класса, где последовательность нуклеиновой кислоты вариабельного участка иммуноглобулина (например, нуклеотидная последовательность, кодирующая вариабельный домен тяжелой цепи или включающая последовательность CDR или FR тяжелой цепи) в В-клетке, подвергшейся переключению класса, не является идентичной последовательности нуклеиновой кислоты в В-клетке до переключения класса, как, например, имеется различие в последовательности нуклеиновой кислоты, кодирующей CDR или каркасный участок, у В-клетки, которая не перенесла переключение класса, и В-клетки, которая перенесла переключение класса. «Подвергшиеся соматической мутации» включает ссылку на последовательности нуклеиновой кислоты из B-клеток с созревшей аффинностью, которые не являются идентичными соответствующим последовательностям вариабельного участка тяжелой цепи иммуноглобулина в B-клетках, которые не являются клетками с созревшей аффинностью (т.е. последовательностям в геноме клеток зародышевой линии). Фраза «подвергшиеся соматической мутации» также включает ссылку на последовательность нуклеиновой кислоты вариабельного участка иммуноглобулина из В-клетки после воздействия на В-клетку представляющего интерес эпитопа, где последовательность нуклеиновой кислоты отличается от соответствующей последовательности нуклеиновой кислоты до воздействия на В-клетку представляющего интерес эпитопа. Фраза «подвергшиеся соматической мутации» относится к последовательностям связывающих белков, которые были получены в животном, например, мыши, имеющей последовательности нуклеиновой кислоты вариабельного участка иммуноглобулина человека, в ответ на заражение иммуногеном, и которые являются результатом процессов отбора, врожденно действующих в таком животном.
[00117] «Практически» обозначает качественное состояние проявления полной или практически полной меры или степени выраженности представляющих интерес характеристики или свойства. Рядовому специалисту в области биологии будет понятно, что биологические и химические явления редко, если это вообще происходит, продолжаются до окончания и/или протекают до завершения или достигают или скрывают абсолютный результат. Следовательно, термин «практически» используется в данном документе для охвата потенциального отсутствия завершенности, присущего многим биологическим и химическим явлениям.
[00118] «Практически гомологичный» обозначает сравнение аминокислотных последовательностей или последовательностей нуклеиновой кислоты. Как будет понятно специалистам в данной области, две последовательности обычно считаются «практически гомологичными», если они содержат гомологичные остатки в соответствующих положениях. Гомологичные остатки могут представлять собой идентичные остатки. В качестве альтернативы гомологичные остатки могут представлять собой неидентичные остатки с соответствующими сходными структурными и/или функциональными характеристиками. Например, как хорошо известно рядовым специалистам в данной области, определенные аминокислоты обычно классифицируют как «гидрофобные» или «гидрофильные» аминокислоты и/или как имеющие «полярные» или «неполярные» боковые цепи. Замена одной аминокислотой другой того же типа часто может считаться «гомологичной» заменой. Типичное распределение аминокислот по категориям обобщено ниже:
[00119] Из уровня техники хорошо известно, что аминокислотные последовательности или последовательности нуклеиновой кислоты можно сравнивать с помощью любого из множества алгоритмов, в том числе алгоритмов, доступных в коммерческих компьютерных программах, таких как BLASTN для нуклеотидных последовательностей и BLASTP, BLAST с гэпами и PSI-BLAST для аминокислотных последовательностей. Такие иллюстративные программы описаны в Altschul et al., 1990, J. Mol. Biol., 215(3): 403-410; Altschul, S. F. et al., 1997, Methods in Enzymology; Altschul, S. F. et al., 1997, Nucleic Acids Res., 25:3389-3402; Baxevanis, A.D., and B. F. F. Ouellette (eds.) Bioinformatics: A Practical Guide to the Analysis of Genes and Proteins, Wiley, 1998; и Misener et al. (eds.) Bioinformatics Methods and Protocols (Methods in Molecular Biology, Vol. 132), Humana Press, 1998. В дополнение к идентификации гомологичных последовательностей упомянутые выше программы, как правило, дают представление о степени гомологии. В некоторых вариантах осуществления две последовательности считаются практически гомологичными, если по меньшей мере 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более их соответствующих остатков являются гомологичными на протяжении рассматриваемого фрагмента из остатков. В некоторых вариантах осуществления рассматриваемый фрагмент представляет собой полную последовательность. В некоторых вариантах осуществления рассматриваемый фрагмент содержит по меньшей мере 9, 10, 11, 12, 13, 14, 15, 16, 17 или более остатков. В некоторых вариантах осуществления рассматриваемый фрагмент предусматривает смежные остатки вдоль полной последовательности. В некоторых вариантах осуществления рассматриваемый фрагмент предусматривает остатки, расположенные с промежутками вдоль полной последовательности, например, расположенные с промежутками остатки, сводимые вместе за счет складчатой конформации полипептида или его части. В некоторых вариантах осуществления рассматриваемый фрагмент содержит по меньшей мере 10, 15, 20, 25, 30, 35, 40, 45, 50 или более остатков.
[00120] «Практически идентичный» обозначает сравнение аминокислотных последовательностей или последовательностей нуклеиновых кислот. Как будет понятно рядовым специалистам в данной области, две последовательности обычно считаются «практически идентичными», если они содержат идентичные остатки в соответствующих положениях. Из уровня техники хорошо известно, что аминокислотные последовательности или последовательности нуклеиновой кислоты можно сравнивать с помощью любого из множества алгоритмов, в том числе алгоритмов, доступных в коммерческих компьютерных программах, таких как BLASTN для нуклеотидных последовательностей и BLASTP, BLAST с гэпами и PSI-BLAST для аминокислотных последовательностей. Такие иллюстративные программы описаны в Altschul et al., 1990, J. Mol. Biol., 215(3): 403-410; Altschul, S. F. et al., 1997, Methods in Enzymology; Altschul, S. F. et al., 1997, Nucleic Acids Res., 25:3389-3402; Baxevanis, A.D., and B. F. F. Ouellette (eds.) Bioinformatics: A Practical Guide to the Analysis of Genes and Proteins, Wiley, 1998; и Misener et al. (eds.) Bioinformatics Methods and Protocols (Methods in Molecular Biology, Vol. 132), Humana Press, 1998. В дополнение к идентификации идентичных последовательностей упомянутые выше программы, как правило, дают представление о степени идентичности. В некоторых вариантах осуществления две последовательности считаются практически идентичными, если по меньшей мере 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более их соответствующих остатков являются идентичными на протяжении рассматриваемого фрагмента из остатков. В некоторых вариантах осуществления рассматриваемый фрагмент представляет собой полную последовательность. В некоторых вариантах осуществления рассматриваемый фрагмент содержит по меньшей мере 10, 15, 20, 25, 30, 35, 40, 45, 50 или более остатков.
[00121] «Трансформация» обозначает любой способ, посредством которого экзогенную ДНК вводят в клетку-хозяина. Трансформация может происходить в природных или искусственных условиях с помощью различных способов, хорошо известных из уровня техники. Трансформация может быть обусловлена любым известным способом вставки чужеродных последовательностей нуклеиновой кислоты в прокариотическую или эукариотическую клетку-хозяина. В некоторых вариантах осуществления конкретную методику трансформации выбирают, исходя из клетки-хозяина, подлежащей трансформации, и она может включать без ограничения инфицирование вирусом, электропорацию, слияние, липофекцию. В некоторых вариантах осуществления «трансформированная» клетка является стабильно трансформированной в том смысле, что встроенная ДНК способна к репликации либо в виде автономно реплицирующейся плазмиды, либо в виде части хромосомы хозяина. В некоторых вариантах осуществления трансформированная клетка временно экспрессирует введенную нуклеиновую кислоту на протяжении ограниченных периодов времени.
[00122] «Нацеливающий вектор» или «нацеливающая конструкция» обозначает полинуклеотидную молекулу, содержащую нацеливающий участок. Нацеливающий участок содержит последовательность, которая идентична или практически идентична последовательности в целевых клетке, ткани или животном, и обеспечивает интеграцию нацеливающей конструкции в положение в пределах генома клетки, ткани или животного посредством гомологичной рекомбинации. Также предусмотрены нацеливающие участки, которые нацеливаются с помощью сайтов для распознавания сайт-специфической рекомбиназой (например, сайтов loxP или Frt). В некоторых вариантах осуществления нацеливающая конструкция, описанная в данном документе, дополнительно содержит последовательность нуклеиновой кислоты или ген, представляющие особенный интерес, селектируемый маркер, контрольные и/или регуляторные последовательности, а также другие последовательности нуклеиновой кислоты, обеспечивающие возможность рекомбинации, опосредованной добавлением экзогенных белков, которые способствуют или содействуют рекомбинации с вовлечением таких последовательностей. В некоторых вариантах осуществления нацеливающая конструкция дополнительно содержит представляющий интерес полный или неполный ген, где представляющий интерес ген является гетерологичным геном, который кодирует полный или неполный полипептид, имеющий функцию, сходную с таковой у белка, кодируемого эндогенной последовательностью. В некоторых вариантах осуществления нацеливающая конструкция дополнительно содержит представляющий интерес гуманизированный полный или неполный ген, где представляющий интерес гуманизированный ген кодирует полный или неполный полипептид, имеющий функцию, сходную с таковой у полипептида, кодируемого эндогенной последовательностью. В некоторых вариантах осуществления нацеливающая конструкция (или нацеливающий вектор) могут содержать последовательность нуклеиновой кислоты, в отношении которой проведена манипуляция человеком. Например, в некоторых вариантах осуществления нацеливающая конструкция (или нацеливающий вектор) могут быть сконструированы таким образом, что они содержат разработанный или рекомбинантный полинуклеотид, который содержит две или более последовательности, которые не связаны друг с другом в таком порядке в природе, пока не подвергаются манипуляции человеком для связывания напрямую друг с другом в сконструированном или рекомбинантном полинуклеотиде.
[00123] «Трансген» или «трансгенная конструкция» обозначают последовательность нуклеиновой кислоты (кодирующей, например, представляющий интерес полный или неполный полипептид), которую человек ввел в клетку с помощью таких способов, которые описаны в данном документе. Трансген может быть частично или полностью гетерологичным, т. е. чужеродным, для трансгенного животного или клетки, в которых он введен. Трансген может содержать одну или более последовательностей регуляции транскрипции и любую другую нуклеиновую кислоту, такую как интроны или промоторы, которые могут быть необходимы для экспрессии выбранной последовательности нуклеиновой кислоты.
[00124] «Трансгенное животное», «трансгенное животное, отличное от человека» или «Tg+» могут использоваться взаимозаменяемо и обозначать любое не встречающееся в природе животное, отличное от человека, в котором одна или более клеток животного, отличного от человека, содержат гетерологичную нуклеиновую кислоту и/или ген, кодирующий представляющий интерес полный или неполный полипептид. В некоторых вариантах осуществления гетерологичную нуклеиновую кислоту и/или ген вводят в клетку напрямую или опосредованно, путем введения в клетку-предшественника, путем преднамеренной генетической манипуляции, например, с помощью микроинъекции или путем инфицирования рекомбинантным вирусом. Термин «генетическая манипуляция» не включает классические методики скрещивания, а скорее относится к введению молекулы(молекул) рекомбинантной ДНК. Данная молекула может быть интегрирована в хромосому или может представлять собой внехромосомно реплицирующуюся ДНК. Термин «Tg+» включает животных, которые являются гетерозиготными или гомозиготными по гетерологичным нуклеиновой кислоте и/или гену, и/или животных, которые имеют одну или множество копий гетерологичных нуклеиновой кислоты и/или гена.
[00125] «Вариант» обозначает объект, демонстрирующий значительную структурную идентичность с эталонным объектом, но структурно отличающийся от эталонного объекта по присутствию или уровню одного или более химических фрагментов по сравнению с эталонным объектом. Во многих вариантах осуществления «вариант» также отличается от своего эталонного объекта функционально. Как правило, следует ли считать конкретный объект «вариантом» эталонного объекта, зависит от степени его структурной идентичности с эталонным объектом. Как будет понятно специалистам в данной области, любой биологический или химический эталонный объект имеет определенные характерные структурные элементы. «Вариант» по определению представляет собой отдельный химический объект с общими одним или более такими характерными структурными элементами. В виде лишь нескольких примеров, малая молекула может иметь характерный сердцевинный структурный элемент (например, макроциклическую сердцевину) и/или один или более характерных «подвешенных» фрагментов, так что вариант малой молекулы имеет общие сердцевинный структурный элемент и характерные «подвешенные» фрагменты, но отличается другими «подвешенными» фрагментами и/или типами связей (одинарные по сравнению с двойными, E по сравнению с Z и т. д.), присутствующими в пределах сердцевины, полипептид может иметь характерный элемент последовательности, состоящий из нескольких аминокислот, имеющих определенные положения по отношению друг к другу в линейном или трехмерном пространстве и/или способствующих осуществлению конкретной биологической функции, нуклеиновая кислота может иметь характерный элемент последовательности, состоящий из нескольких нуклеотидных остатков, имеющих определенные положения по отношению друг к другу в линейном или трехмерном пространстве. Например, «вариант полипептида» может отличаться от эталонного полипептида вследствие одного или более различий в аминокислотной последовательности и/или одного или более различий в химических фрагментах (например, углеводах, липидах и т. д.), прикрепленных ковалентной связью к полипептидному остову. В некоторых вариантах осуществления «вариант полипептида» демонстрирует общую идентичность последовательности с эталонным полипептидом, составляющую по меньшей мере 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97% или 99%. В качестве альтернативы или в дополнение в некоторых вариантах осуществления «вариант полипептида» не имеет по меньшей мере одного характерного элемента последовательности, общего с эталонным полипептидом. В некоторых вариантах осуществления эталонный полипептид обладает одним или более видами биологической активности. В некоторых вариантах осуществления «вариант полипептида» обладает одним или более видами биологической активности, общими с эталонным полипептидом. В некоторых вариантах осуществления у «варианта полипептида» отсутствует один или более видов биологической активности эталонного полипептида. В некоторых вариантах осуществления «вариант полипептида» демонстрирует пониженный уровень одного или более видов биологической активности по сравнению с эталонным полипептидом. Во многих вариантах осуществления представляющий интерес полипептид считается «вариантом» исходного или эталонного полипептида, если представляющий интерес полипептид имеет аминокислотную последовательность, идентичную последовательности у исходной молекулы, за исключением небольшого количества изменений в конкретных положениях последовательности. Как правило, в варианте заменены менее 20%, 15%, 10%, 9%, 8%, 7%, 6%, 5%, 4%, 3%, 2% остатков по сравнению с исходной молекулой. В некоторых вариантах осуществления «вариант» имеет 10, 9, 8, 7, 6, 5, 4, 3, 2 или 1 замену остатков по сравнению с исходной молекулой. Зачастую «вариант» имеет очень небольшое количество (например, менее 5, 4, 3, 2 или 1) замен функциональных остатков (т.е. остатков, принимающих участие в конкретном виде биологической активности). Кроме того, «вариант» обычно имеет не более 5, 4, 3, 2 или 1 добавления или делеции и зачастую не имеет добавлений или делеций по сравнению с исходной молекулой. Более того, любые добавления или делеции, как правило, охватывают менее приблизительно 25, приблизительно 20, приблизительно 19, приблизительно 18, приблизительно 17, приблизительно 16, приблизительно 15, приблизительно 14, приблизительно 13, приблизительно 10, приблизительно 9, приблизительно 8, приблизительно 7, приблизительно 6 и зачастую охватывают менее приблизительно 5, приблизительно 4, приблизительно 3 или приблизительно 2 остатков. В некоторых вариантах осуществления исходный или эталонный полипептид представляет собой полипептид, встречающийся в природе. Как будет понятно рядовым специалистам в данной области, несколько вариантов конкретного представляющего интерес полипептида обычно можно встретить в природе, особенно если представляющий интерес полипептид представляет собой полипептид возбудителя инфекции.
[00126] «Вектор» обозначает молекулу нуклеиновой кислоты, способную транспортировать другую нуклеиновую кислоту, с которой она связана. В некоторых вариантах осуществления векторы способны к внехромосомной репликации и/или экспрессии нуклеиновых кислот, с которыми они соединены в клетке-хозяине, такой как эукариотическая и/или прокариотическая клетка. Векторы, способные управлять экспрессией функционально связанных генов, называются в данном документе «векторами экспрессии».
[00127] «Дикий тип» включает его понятное в данной области значение, которое обозначает объект, обладающий структурой и/или активностью, встречающимися природе в «нормальном» (в противоположность мутантному, болезненному, измененному и т.д.) состоянии или окружении. Рядовым специалистам в данной области будет понятно, что гены и полипептиды дикого типа часто существуют в нескольких различных формах (например, аллелях).
ПОДРОБНОЕ ОПИСАНИЕ КОНКРЕТНЫХ ВАРИАНТОВ ОСУЩЕСТВЛЕНИЯ
[00128] В данном документе, среди прочих объектов, раскрыты трансгенные животные, отличные от человека, имеющие гетерологичный генетический материал, кодирующий одну или более частей (функциональных фрагментов, связывающих частей и т.д.) представляющего интерес полипептида, при этом гетерологичный генетический материал встроен в кластер разнообразия (т.е. участок DH) вариабельного участка тяжелой цепи иммуноглобулина, так что гетерологичный генетический материал функционально связан с вариабельным (VH) и соединяющими (JH) сегментами тяжелой цепи. Также предполагается, что такие животные, отличные от человека, проявляют способность вырабатывать антитела к мишеням заболеваний, которые с трудом поддаются лечению. Также предполагается, что у таким животных, отличные от человека, имеется популяция антител, характеризующихся вариабельными участками тяжелой цепи с повышенным разнообразием CDR3 по сравнению с популяцией антител с разнообразием CDR3 вариабельного участка тяжелой цепи иммуноглобулина, полученным за счет стандартных генных сегментов DH иммуноглобулина (или генных сегментов DH иммуноглобулина, которые встречаются в природе). Следовательно, описанные в данном документе животные, отличные от человека, могут быть применимы для разработки терапевтических средств на основе антител, которые связывают конкретные антигены, в частности, антигены, ассоциированные с низкой и/или со слабой иммуногенностью. В частности, в данном документе раскрыто введение иллюстративных нуклеотидных кодирующих последовательностей, каждая из которых кодирует часть представляющего интерес полипептида (например, внеклеточную часть атипичного хемокинового рецептора, часть конотоксина или часть токсина тарантула), в участок DH вариабельного участка тяжелой цепи иммуноглобулина, что приводит к экспрессии антител, имеющих вариабельные участки тяжелой цепи и, в частности, участки CDR3, полученные в результате V(D)J-рекомбинации при участии встроенной нуклеотидной кодирующей последовательности. В некоторых вариантах осуществления встроенными нуклеотидными кодирующими последовательностями представляющего интерес полипептида (например, атипичного хемокинового рецептора (ACKR), конотоксина, токсина тарантула или их комбинаций) заменены все или практически все стандартные сегменты DH (т.е. сегменты DH дикого типа) в пределах кластера разнообразия тяжелой цепи иммуноглобулина (т.е. участка DH), описываемого в данном документе. В некоторых вариантах осуществления встроенными нуклеотидными кодирующими последовательностями представляющего интерес полипептида (например, атипичного хемокинового рецептора (ACKR), конотоксина, токсина тарантула или их комбинаций) частично заменены один или более стандартных сегментов DH в пределах кластера разнообразия тяжелой цепи иммуноглобулина (т.е. участка DH), описываемого в данном документе. В некоторых вариантах осуществления нуклеотидные кодирующие последовательности представляющего интерес полипептида (например, атипичного хемокинового рецептора (ACKR), конотоксина, токсина тарантула или их комбинаций) встроены в один или более стандартных сегментов DH в пределах кластера разнообразия тяжелой цепи иммуноглобулина (т.е. участка DH), описываемого в данном документе, так что указанные нуклеотидные кодирующие последовательности фланкированы последовательностями, которые являются обычными или встречаются в природе в одном или более стандартных сегментах DH или связаны с ними. В некоторых вариантах осуществления один или более стандартных сегментов DH остаются интактными в пределах кластера разнообразия тяжелой цепи иммуноглобулина, описываемого в данном документе. В некоторых вариантах осуществления один или более стандартных сегментов DH подвергнуты делеции, удалены или иным образом приведены в нефункциональное состояние в кластере разнообразия тяжелой цепи иммуноглобулина, описываемом в данном документе. В некоторых определенных вариантах осуществления в кластере разнообразия тяжелой цепи иммуноглобулина, описываемом в данном документе, отсутствуют все или практически все стандартные сегменты DH. В некоторых определенных вариантах осуществления кластер разнообразия тяжелой цепи иммуноглобулина, описываемый в данном документе, содержит синтетические сегменты DH, созданные или полученные с использованием нуклеотидных кодирующих последовательностей, описанных в данном документе. Такие трансгенные животные, отличные от человека, обеспечивают in vivo систему для идентификации и разработки антител и/или терапевтических средств на основе антител, которые связывают мишени заболеваний, которая превосходит нацеливающие способности общепринятых технологий поиска новых лекарственных средств. Кроме того, такие трансгенные животные, отличные от человека, обеспечивают применимую систему на основе животной модели для разработки антител и/или терапевтических средств на основе антител, фокусирующихся или сконструированных для нарушения белок-белковых взаимодействий, которые являются ключевыми при различных заболеваниях и/или патологических состояниях, поражающих человека.
[00129] В некоторых вариантах осуществления описанные в данном документе животные, отличные от человека, содержат вариабельный участок тяжелой цепи иммуноглобулина, содержащий сконструированный кластер разнообразия (т.е. сконструированный участок DH), характеризующийся присутствием одной или более нуклеотидных кодирующих последовательностей, соответствующих части(ям) представляющего интерес полипептида, такого как, например, внеклеточный домен ACKR (например, D6 хемокиновый рецептор-ловушка), токсин (например, блокатор ионных каналов, такой как, например, конотоксин, токсин паукообразных, токсин тарантула, токсин морской анемоны или токсин скорпиона), сопряженный с G-белком рецептор, длинные CDR тяжелой цепи выбранных антител (например, нейтрализующих антител, которые связывают вирусы, в том числе, например, HIV, HCV, HPV, вирус гриппа и т.д.), агонисты рецептора глюкагон-подобного пептида-1 (например, эксенатид, лираглутид, ликсисенатид, альбиглутид, дулаглутид, таспоглутид и т.д.), генные сегменты разнообразия тяжелой цепи (DH) от видов, отличных от человека (например, птицы, курицы, коровы, кролика, свиньи и т.д.) и т.д. В таких вариантах осуществления антитела, содержащие CDR3, полученные в результате рекомбинации при участии таких нуклеотидных кодирующих последовательностей, можно охарактеризовать как обладающие повышенным разнообразием для управления связыванием с конкретными антигенами (например, с трансмембранными полипептидами). В некоторых вариантах осуществления антитела, продуцируемые описанными в данном документе животными, отличными от человека, имеют последовательность вариабельного участка тяжелой цепи иммуноглобулина, которая содержит участок CDR3, соответствующий пептиду, кодируемому одной или более нуклеотидными кодирующими последовательностями. В некоторых вариантах осуществления описанные в данном документе животные, отличные от человека, содержат вариабельный (VH) и соединяющий (JH) генные сегменты тяжелой цепи, функционально связанные с одной или более нуклеотидными кодирующими последовательностями, так что V(D)J-рекомбинация происходит между указанными VH, JH и одной или более нуклеотидными кодирующими последовательностями с образованием вариабельного участка тяжелой цепи, который связывает представляющий интерес антиген. В некоторых вариантах осуществления описанные в данном документе животные, отличные от человека, содержат несколько генных сегментов VH и JH, функционально связанных с 5, 10, 15, 20, 25 или больше (например, с несколькими) нуклеотидных кодирующих последовательностей в вариабельном участке тяжелой цепи иммуноглобулина в геноме животного, отличного от человека. Во многих вариантах осуществления сегменты VH и JH представляют собой генные сегменты VH человека и JH человека. В некоторых вариантах осуществления описанные в данном документе животные, отличные от человека, дополнительно содержат человеческий или гуманизированный локус легкой цепи иммуноглобулина (например, κи/или λ), так что животные, отличные от человека, продуцируют антитела, содержащие вариабельные участки человека (т. е. тяжелой и легкой цепи) и константные участки, отличные от человеческих. В некоторых определенных вариантах осуществления указанный локус легкой цепи иммуноглобулина человека или гуманизированной легкой цепи содержит генные сегменты VL и JL человека, функционально связанные с константным участком легкой цепи грызуна (например, Cκ или Cλ грызуна). В некоторых вариантах осуществления описанные в данном документе животные, отличные от человека, дополнительно содержат локус легкой цепи иммуноглобулина, описываемый в публикациях заявок на патенты США под №№ 2011-0195454 A1, 2012-0021409 A1, 2012-0192300 A1, 2013-0045492 A1, 2013-0185821 A1, 2013-0198880 A1, 2013-0302836 A1, 2015-0059009 A1; публикациях международных патентных заявок под №№ WO 2011/097603, WO 2012/148873, WO 2013/134263, WO 2013/184761, WO 2014/160179, WO 2014/160202; все из которых включены в данный документ посредством ссылки).
[00130] Различные аспекты композиций и способов подробно описаны в следующих разделах. Использование разделов не подразумевает ограничения любым вариантом осуществления. Каждый раздел можно применять относительно любого конкретно описанного варианта осуществления. В настоящей заявке использование «или» означает «и/или», если не указано иное.
V(D)J-рекомбинация
[00131] Серия событий рекомбинации при участии нескольких генетических компонентов служит для сборки иммуноглобулинов из упорядоченного массива генных сегментов (например, V, D и J). Известно, что данная сборка генных сегментов является неточной и, таким образом, разнообразие иммуноглобулинов достигается вследствие как комбинации разных генных сегментов, так и образования уникальных соединений за счет неточного соединения. Кроме того, разнообразие создается за счет процесса, известного как соматическая гипермутация, при котором последовательность вариабельного участка иммуноглобулинов изменяется с повышением аффинности и специфичности в отношении антигена. Иммуноглобулин представляет полипептид Y-образной формы, состоящий из двух идентичных легких и тяжелых цепей, каждая из которых имеет два структурных компонента: один вариабельный домен и один константный домен. Это именно вариабельные домены тяжелой и легкой цепей образованы за счет сборки генных сегментов, тогда как константные домены сливаются с вариабельными доменами за счет РНК-сплайсинга. Хотя механизм сборки (или связывания) генных сегментов в тяжелой и легкой цепях является схожим, в случае легких цепей требуется только одно событие соединения (т.е. V с J), тогда как в случае тяжелых цепей требуется два (т.е. D с J и V с DJ).
[00132] Сборка генных сегментов вариабельных участков для тяжелой и легкой цепей управляется консервативными последовательностями некодирущей ДНК, фланкирующими каждый генный сегмент, которые называются последовательностями сигнала рекомбинации (RSS), что обеспечивает возможность перестроек ДНК в точных местоположениях относительно кодирующих последовательностей V, D и J (см., например, Ramsden, D.A. et al., 1994, Nuc. Acids Res. 22(10):1785-96). Каждая RSS состоит из консервативного блока из семи нуклеотидов (гептамера), который является смежным с кодирующей последовательностью (например, сегментом V), после которой расположен консервативный спейсер (либо из 12, либо из 23 п.о.), и второго консервативного блока из девяти нуклеотидов (нонамера). Хотя допустима существенное отклонение последовательностей у разных индивидуумов, длина данных последовательностей, как правило, не варьирует. Рекомбинация между генными сегментами иммуноглобулинов следует правилу, которое обычно обозначается как правило 12/23, согласно которому генные сегменты, фланкированные RSS со спейсером длиной 12 п.о., как правило, соединяются с генным сегментом, фланкированным спейсером длиной 23 п.о. (см., например, Hiom, K. and M. Gellert, 1998, Mol. Cell. 1(7):1011-9). Сообщалось, что последовательность RSS оказывает влияние на эффективность и/или частоту рекомбинации с конкретным генным сегментом (см., например, Ramsden, D.A and G.E. Wu, 1991, Proc. Natl. Acad. Sci. U.S.A. 88:10721-5; Boubnov, N.V. et al., 1995, Nuc. Acids Res. 23:1060-7; Ezekiel, U.R. et al., 1995, Immunity 2:381-9; Sadofsky, M. et al., 1995, Genes Dev. 9:2193-9; Cuomo, C.A. et al., 1996, Mol. Cell Biol. 16:5683-90; Ramsden, D.A. et al., 1996, EMBO J 15:3197-3206). В действительности, во многих сообщениях отмечается чрезвычайно смещенное и вариабельное использование генных сегментов, в частности, сегментов DH, у разных индивидуумов.
[00133] В некоторых вариантах осуществления описанные в данном документе животные, отличные от человека, содержат одну или более RSS, фланкирующих кодирующие последовательности, которые являются оптимизированными для рекомбинации с генным сегментом V и/или J. В различных вариантах осуществления кодирующие последовательности встроены в локус тяжелой цепи иммуноглобулина вместо (или в пределах) стандартных генных сегментов DH (или генных сегментов DH, которые встречаются в природе). Оптимизации RSS можно достичь с использованием стандартных методик, известных из уровня техники, таких как, например, сайт-направленный мутагенез известных последовательностей RSS или in silico создание синтетические последовательностей RSS с последующим синтезом de novo. В качестве одного примера, RSS, которая ассоциирована с низкой или слабой эффективностью и/или частотой рекомбинации, можно оптимизировать путем сравнения RSS, которая ассоциирована с высокой или оптимальной эффективностью и/или частотой рекомбинации. В некоторых вариантах осуществления эффективность и/или частоту рекомбинации можно определить по частоте использования генных сегментов в популяции последовательностей антител (например, от индивидуума или группы индивидуумов; см., например, Arnaout, R. et al., 2011, PLoS One 6(8):e22365; Glanville, J. et al., 2011, Proc. Natl. Acad. Sci. U.S.A. 108(50):20066-71). Таким образом, в некоторых вариантах осуществления описанные в данном документе животные, отличные от человека, могут содержать одну или более оптимизированных RSS, фланкирующих кодирующие последовательности (или генные сегменты), так что рекомбинация кодирующих последовательностей (или генных сегментов) происходит с равной и приблизительно равной частотой. Иллюстративные оптимизированные RSS изложены на фигуре 2.
[00134] Сборка генных сегментов с образованием вариабельных участков тяжелой и легкой цепей приводит к образованию антигенсвязывающих участков (или сайтов) иммуноглобулинов. Такие антигенсвязывающие участки характеризуются, помимо всего прочего, присутствием гипервариабельных участков, которые обычно обозначаются как участки, определяющие комплементарность (CDR). В тяжелой и легкой цепях находится по три CDR (т. е. в общей сложности шесть CDR), причем оба CDR1 и CDR2 полностью кодируются генным сегментом V. Однако CDR3 кодируется последовательностью, образующейся в результате соединения сегментов V и J в случае легких цепей и сегментов V, D и J в случае тяжелых цепей. Таким образом, дополнительный генный сегмент, используемый в ходе рекомбинации для получения кодирующей последовательности вариабельного участка тяжелой цепи, существенно повышает разнообразие антигенсвязывающих сайтов тяжелых цепей.
Хемокины и хемокиновые рецепторы
[00135] Иммунный и воспалительный ответы представляют собой сложные биологические процессы, в которые вовлечены несколько типов иммунных клеток и молекулярных компонентов. Сообщалось, что миграция лейкоцитов является важным фактором при инициации, поддержании и завершении иммунного и воспалительного ответов, некоторые из которых достигаются за счет действия хемокинов и их рецепторов. В действительности, сообщалось о некоторых хемокинах и соответствующих рецепторах. Хемокиновые рецепторы имеют структуру, характеризующуюся семиспиральным трансмембранным доменом, который соединяется с G-белком для сигнальной трансдукции во внутриклеточный компартмент, и они подразделяются на множество разных семейств, соответствующих подмножествам хемокинов, с которыми они связываются: CC-хемокиновые рецепторы (β-хемокиновые рецепторы), CXC-хемокиновые рецепторы, CX3C-хемокиновые рецепторы и XC-хемокиновые рецепторы. В дополнение к данным стандартным хемокиновым рецепторам сообщалось о других хемокиновых рецепторах (под названием атипичные хемокиновые рецепторы или ACKR), которые имеют схожую структуру, но при этом у них отсутствует способность инициировать передачу сигнала в ответ на связывание лиганда (см., например, Bonecchi, R. et al., 2010, Curr. Top. Microbiol. Immunol. 341:15-36; Nibbs, R.J.B. and G.J. Graham, 2013, Nature Rev. 13:815-29).Иммунный и воспалительный ответы представляют собой сложные биологические процессы, в которые вовлечены несколько типов иммунных клеток и молекулярных компонентов. Сообщалось, что миграция лейкоцитов является важным фактором при инициации, поддержании и завершении иммунного и воспалительного ответов, некоторые из которых достигаются за счет действия хемокинов и их рецепторов. В действительности, сообщалось о некоторых хемокинах и соответствующих рецепторах. Хемокиновые рецепторы имеют структуру, характеризующуюся семиспиральным трансмембранным доменом, который соединяется с G-белком для сигнальной трансдукции во внутриклеточный компартмент, и они подразделяются на множество разных семейств, соответствующих подмножествам хемокинов, с которыми они связываются: CC-хемокиновые рецепторы (β-хемокиновые рецепторы), CXC-хемокиновые рецепторы, CX3C-хемокиновые рецепторы и XC-хемокиновые рецепторы. В дополнение к данным стандартным хемокиновым рецепторам сообщалось о других хемокиновых рецепторах (под названием атипичные хемокиновые рецепторы или ACKR), которые имеют схожую структуру, но при этом у них отсутствует способность инициировать передачу сигнала в ответ на связывание лиганда (см., например, Bonecchi, R. et al., 2010, Curr. Top. Microbiol. Immunol. 341:15-36; Nibbs, R.J.B. and G.J. Graham, 2013, Nature Rev. 13:815-29).
[00136] Среди множества ACKR ACKR2 (также известный как CCBP2 и D6 хемокиновый рецептор-ловушка) представляет собой семиспиральный трансмембранный белковый рецептор, аналогичный другим G-белок-сопряженным рецепторам, и кодируемый геном CCBP2 (Nibbs, R.J.B. et al., 1997, J. Biol. Chem. 272(51):32078-83). В отличие от других хемокиновых рецепторов ACKR2 содержит мотив DKYLEIV во второй внутриклеточной петле вместо канонического мотива DRYLAIV, и он не способен к передаче сигнала при участии Gαi-белков. ACKR2 экспрессируется на поверхности лимфатических эндотелиальных клеток кожи, кишечника и легких, а также на B-клетках и дендритных клетках. ACKR представляет собой промискуитетный рецептор для многих провоспалительных β-хемокинов (CC-хемокинов), но не связывает конститутивные CCL-хемокины. Было высказано предположение, что ACKR2 является фагоцитарным рецептором, который осуществляет интернализацию CC-хемокинов и нацеливает их на лизосомальную деградацию, ограничивая тем самым воспалительные ответы посредством элиминации CC-хемокинов (Jamieson, T. et al., 2005, Nature Immunol. 6(4):403-11; Bonecchi, R. et al., выше; Hansell, C.A.H. et al., 2011, Immunol. Cell Biol. 89(2):197-206; Nibbs, R.J.B. and G.J. Graham, выше).
Пептидные токсины
[00137] Токсины представляют собой встречающиеся в природе вещества, выявленные в растениях и животных, которые могут быть ядовитыми для людей. Например, улитки конусы продуцируют нейротоксические пептиды, называемые конотоксинами, содержащиеся в их яде, которые модулируют активность различных рецепторов у человека. В частности, было показано, что некоторые конотоксины модулируют ионные каналы (например, NaV-каналы). Как правило, конотоксины представляют собой пептиды длиной 10–30 аминокислот, которые включают одну или более дисульфидных связей, и их характеризуют на основании мишени, на которую они воздействуют: α-конотоксины (ацетилхолиновые рецепторы), δ-конотоксины (потенциал-зависимые натриевые каналы), κ-конотоксины (калиевые каналы), μ-конотоксины (потенциал-зависимые натриевые каналы) и ω-конотоксины (потенциал-зависимые кальциевые каналы). Известно, что конотоксины являются высокополиморфными у разных видов улиток и, вследствие этого, кодирующие их гены не являются консервативными (см., например, Terlau, H. and B.M. Olivera, 2004, Physiol. Rev. 84(1):41-68; Biggs, J.S. et al., 2010, Mol. Phylogenet. Evol. 56(1):1-12; Olivera, B.M. et al., 2012, Ann. N.Y. Acad. Sci. 1267(1):61-70; Wong, E.S. and K. Belov, 2012, Gene 496(1):1-7).
[00138] Сообщалось, что среди множества конотоксинов μ-конотоксины имеют два типа цистеиновых паттернов и оказывают действие на потенциал-зависимые натриевые каналы в мышечной ткани (Cruz, L.J. et al., 1985, J. Biol. Chem. 260(16):9280-8; Zeikus, R.D. et al., 1985, J. Biol. Chem. 260(16):9280-8; McIntosh, J.M. and R.M. Jones, 2001, Toxicon. 39(10):1447-51; Nielsen, K.J. et al., 2002, J. Biol. Chem. 277(30):27247-55; Floresca, C.Z., 2003, Toxicol. Appl. Pharmacol. 190(2):95-101; Priest, B.T. et al., 2007, Toxicon. 49(2):194-201; Schmalhofer, W.A. et al., 2008, Mol. Pharmacol. 74(5):1476-84; Ekberg, J. et al., 2008, Int. J. Biochem. Cell Biol. 40(11):2363-8). В действительности, μ-конотоксины были объектом исследования в отношении их потенциального фармакологического применения (см., например, Olivera, B.M. and R.W. Teichert, 2007, Mol. Interv. 7(5):251-60; Stevens, M. et al., 2012, J. Biol. Chem. 287(37):31382-92). Хотя было проведено несколько исследований для выявления механизма действия разных μ-конотоксинов, многое в нем остается неизвестным.
[00139] Другие примеры токсинов включают яд морских анемон, скорпионов, пауков и тарантулов, которые, как сообщалось, действуют на ионные каналы путем ингибирования активации и блокирования нейронной передачи. В действительности, были идентифицированы несколько токсинов скорпионов и была определена их структура (см., например, Rochat, H. and J. Gregoire, 1983, Toxicon. 21(1):153-62; Zhou, X.H. et al., 1989, Biochem. J. 257(2):509-17; Granier, C. et al., 1990, FEBS Lett. 261(2):423-6). Кроме того, с использованием токсинов из яда скорпионов были разработаны библиотеки на основе нейротоксинов для калиевых каналов (см., например, Takacs, Z. et al., 2009, Proc. Natl. Acad. Sci. U.S.A. 106(52):22211-6). В качестве еще одного примера сообщалось, что пептиды из яда тарантула специфически действуют на потенциал-зависимые натриевые каналы, такие как NaV1.5 и NaV1.7 (см., например, Priest, B.T. et al., 2007, Toxicon. 49(2):194-201; Xiao, Y. et al., 2010, Mol. Pharmacol. 78(6):1124-34). В данном документе описано обнаружение того, что особенно применимый набор нуклеотидных кодирующих последовательностей для создания сконструированного участка DH представляет собой или содержит нуклеотидные кодирующие последовательности из токсина тарантула и последовательности μ-конотоксина (см., например, таблицу 4). В данном документе раскрыто применение кодирующих последовательностей токсина, который воздействует на потенциал-зависимые натриевые каналы (например, NaV1.7), для создания сконструированного участка DH. Способы, описанные в данном документе, можно использовать для применения любого набора кодирующих последовательностей, полученных из любого требуемого пептидного токсина(ов) или комбинации пептидных токсинов (или фрагментов последовательностей пептидного токсина) из множества (т.е. двух, трех, четырех, пяти и т.д.) пептидных токсинов, если это необходимо.
Представленные in vivo системы
[00140] В данном документе раскрыто признание того, что конкретные антигены ассоциированы с низкой и/или слабой иммуногенностью и, следовательно, являются неудовлетворительными мишенями для терапевтических средств на основе антител. В действительности многие мишени заболеваний (например, трансмембранные белки) были охарактеризованы как с трудом поддающиеся лечению или не поддающиеся медикаментозной терапии. Таким образом, в данном документе раскрыто создание in vivo системы для разработки антител и терапевтических средств на основе антител, которые устраняют недостатки, связанные с общепринятыми технологиями поиска лекарственных средств. Настоящее изобретение конкретно демонстрирует конструкцию в трансгенном грызуне, геном которого содержит вариабельный участок тяжелой цепи иммуноглобулина, который включает сконструированный участок DH, при этом сконструированный участок DH включает одну или более гетерологичных нуклеотидных кодирующих последовательностей, каждая из которых кодирует часть (например, внеклеточную часть, связывающую часть, функциональный фрагмент и т. д.) гетерологичного полипептида или представляющего интерес пептида. Способы, описанные в данном документе, можно адаптировать для использования набора гетерологичных нуклеотидных кодирующих последовательностей, каждая из которых кодирует часть любого представляющего интерес полипептида или пептида (например, трансмембранного белка, токсина, G-белок сопряженного рецептора и т.д.) для создания сконструированного участка DH. После интеграции в вариабельный участок тяжелой цепи иммуноглобулина (т.е. помещения в функциональную связь с генными сегментами V и J и/или одним или более константными участками) сконструированный участок DH обеспечивается возможность рекомбинации генных сегментов (т.е. V и J) с одной или более гетерологичными нуклеотидными кодирующими последовательностями с образованием антител, характеризующихся тяжелыми цепями с добавленным разнообразием (т.е. разнообразием CDR3) для управления связыванием с конкретными антигенами.
[00141] В данном документе раскрыто признание того, что особенно применимые представляющие интерес гетерологичные полипептиды, из которых сконструирован набор нуклеотидных кодирующих последовательностей (или комбинаций нуклеотидных кодирующих последовательностей) для разработки сконструированного участка DH, описываемого в данном документе, включают хемокиновые рецепторы, конотоксины, токсины тарантулов и/или их комбинации.
[00142] В некоторых вариантах осуществления хемокиновые рецепторы включают CC-хемокиновые рецепторы, CXC-хемокиновые рецепторы, CX3C-хемокиновые рецепторы, XC-хемокиновые рецепторы и их комбинации.
[00143] В некоторых вариантах осуществления CC-хемокиновые рецепторы (также известные как β-хемокиновые рецепторы) включают CCR1, CCR2, CCR3, CCR4, CCR5, CCR6, CCR7, CCR8, CCR9, CCR10 и CCR11.
[00144] В некоторых вариантах осуществления CXC-хемокиновые рецепторы включают CXCR1, CXCR2, CXCR3, CXCR4, CXCR5, CXCR6 и CXCR7.
[00145] В некоторых вариантах осуществления CX3C-хемокиновые рецепторы включают CX3CR1.
[00146] В некоторых вариантах осуществления XC-хемокиновые рецепторы включают XCR1.
[00147] В некоторых вариантах осуществления конотоксины включают α-конотоксины, δ-конотоксины, κ-конотоксины, μ-конотоксины, ω-конотоксины и их комбинации.
[00148] В некоторых вариантах осуществления конотоксины представляют собой или предусматривают μ-конотоксины.
[00149] В некоторых вариантах осуществления токсины тарантула включают ProTxI, ProTxII, хувентоксин-IV (HWTX-IV) и их комбинации.
[00150] Не связывая себя какой-либо конкретной теорией, следует отметить, что представленные в данном документе данные демонстрируют, что в некоторых вариантах осуществления грызуны, геном которых содержит локус вариабельного участка тяжелой цепи иммуноглобулина, который включает сконструированный участок DH, характеризующийся включением одной или более гетерологичных нуклеотидных кодирующих последовательностей, полученных из внеклеточной части гетерологичного атипичного хемокинового рецептора (например, ACKR2, также известного как D6 хемокиновый рецептор-ловушка), эффективно образуют локус вариабельного участка тяжелой цепи иммуноглобулина, с помощью которого продуцируются антитела, характеризующиеся CDR3 с добавленным разнообразием для связывания лигандов гетерологичного атипичного хемокинового рецептора (например, гетерологичного D6 хемокинового рецептора-ловушки). Также следует отметить, что данные, представленные в данном документе, демонстрируют, что в некоторых вариантах осуществления грызуны, геном которых содержит локус вариабельного участка тяжелой цепи иммуноглобулина, который включает сконструированный участок DH, характеризующийся включением одной или более гетерологичных нуклеотидных кодирующих последовательностей, полученных из части одного или более токсинов (например, μ-конотоксина и/или ProTxII), эффективно образуют локус вариабельного участка тяжелой цепи иммуноглобулина, с помощью которого продуцируются антитела, характеризующиеся CDR3 с добавленным разнообразием для связывания гетерологичного потенциал-зависимого натриевого канала (например, гетерологичного NaV-канала).
[00151] В частности, настоящее изобретение, помимо прочего, конкретно демонстрирует иллюстративные нуклеотидные кодирующие последовательности атипичного хемокинового рецептора (ACKR) человека, такого как, например, ACKR2 (т.е. D6 хемокиновый рецептор-ловушка), которые особенно применимы для интеграции в участок DH локуса тяжелой цепи иммуноглобулина для получения антител, характеризующихся CDR3 с разнообразием, возникающим вследствие рекомбинации нуклеотидных кодирующих последовательностей с сегментами VH и JH, и которые блокируют несколько воспалительных цитокинов (например, CCL2, CCL3, CCL4, CCL5, CCL7, CCL8, CCL11, CCL12, CCL13, CCL14, CCL17, CCL22 и CCL3L1). Также в настоящем изобретении продемонстрированы иллюстративные нуклеотидные кодирующие последовательности из конотоксина (например, μ-конотоксина) и токсина тарантула (например, ProTxII), которые особенно применимы для интеграции в участок DH локуса тяжелой цепи иммуноглобулина для получения антител, характеризующихся CDR3 с разнообразием, возникающим вследствие рекомбинации нуклеотидных кодирующих последовательностей с сегментами VH и JH, и которые блокируют и/или ингибируют активацию и/или функционирование потенциал-зависимого натриевого канала(ов) (например, NaV1.7). Таким образом, по меньшей мере в некоторых вариантах осуществления настоящее изобретение охватывает разработку in vivo системы для получения антител и/или терапевтических средств на основе антител для мишеней заболеваний, которые с трудом поддаются лечению.
[00152] Иллюстративные ACKR человека (вместе с ассоциированными с ними лигандами) изложены в таблице 1 (см., например, Nibbs, R.J.B. and G.J. Graham, 2013, Nature Reviews 13:815-29). Иллюстративные токсины (вместе с ассоциированными с ними мишенями) изложены в таблице 2 (см., например, Terlau, H. and B.M. Olivera, 2004, Physiol. Rev. 84(1):41-68).
Кодирующие последовательности ACKR2 (D6 хемокинового рецептора-ловушки)
[00153] Иллюстративные нуклеотидные кодирующие последовательности (ДНК и аминокислотная последовательность (AA)) ACKR2 человека (D6 хемокинового рецептора-ловушки) для создания сконструированного участка DH, описываемого в данном документе, изложены в таблице 3. В наборе нуклеотидных кодирующих последовательностей ACKR2 человека, изложенных в таблице 3, охарактеризованы четыре внеклеточных домена, четыре внеклеточных домена с заменами Cys на Ser для удаления дисульфидных связей, четыре Cys-кроссовера (Nterm-EC3, EC3-Nterm, EC1-EC2, EC2-EC1), четыре Cys-кроссовера с заменами Cys на Ser для удаления дисульфидных связей, два слияния с петлями (с сохраненными Cys; Nterm+EC3, EC1+EC2) и семь частичных доменов.
[00154] Иллюстративные фрагменты ДНК, содержащие нуклеотидные кодирующие последовательности ACKR2 человека (D6 хемокинового рецептора-ловушки) для создания сконструированного участка DH, приведены ниже.
[00155] D6-DH1166 (SEQ ID NO:131) включает кодирующие последовательности D6, встроенные в положениях, соответствующих DH1–1 — DH6–6:
TACGTAGCCGTTTCGATCCTCCCGAATTGACTAGTGGGTAGGCCTGGCGGCCGCTGCCATTTCATTACCTCTTTCTCCGCACCCGACATAGATACCGGTGGATTCGAATTCTCCCCGTTGAAGCTGACCTGCCCAGAGGGGCCTGGGCCCACCCCACACACCGGGGCGGAATGTGTACAGGCCCCGGTCTCTGTGGGTGTTCCGCTAACTGGGGCTCCCAGTGCTCACCCCACAACTAAAGCGAGCCCCAGCCTCCAGAGCCCCCGAAGGAGATGCCGCCCACAAGCCCAGCCCCCATCCAGGAGGCCCCAGAGCTCAGGGCGCCGGGGCGGATTTTGTACAGCCCCGAGTCACTGTGCGGAAGGATGCTGTGGTTAGCTTTGGCAAAGTTTTCCTGCCACCACAGTGAGAAAAACTGTGTCAAAAACCGTCTCCTGGCCCCTGCTGGAGGCCGCGCCAGAGAGGGGAGCAGCCGCCCCGAACCTAGGTCCTGCTCAGCTCACACGACCCCCAGCACCCAGAGCACAACGGAGTCCCCATTGAATGGTGAGGACGGGGACCAGGGCTCCAGGGGGTCATGGAAGGGGCTGGACCCCATCCTACTGCTATGGTCCCAGTGCTCCTGGCCAGAACTGACCCTACCACCGACAAGAGTCCCTCAGGGAAACGGGGGTCACTGGCACCTCCCAGCATCAACCCCAGGCAGCACAGGCATAAACCCCACATCCAGAGCCGACTCCAGGAGCAGAGACACCCCAGTACCCTGGGGGACACCGACCCTGATGACTCCCCACTGGAATCCACCCCAGAGTCCACCAGGACCAAAGACCCCGCCCCTGTCTCTGTCCCTCACTCAGGACCTGCTGCGGGGCGGGCCATGAGACCAGACTCGGGCTTAGGGAACACCACTGTGGCCCCAACCTCGACCAGGCCACAGGCCCTTCCTTCCTGCCCTGCGGCAGCACAGACTTTGGGGTCTGTGCAGAGAGGAATCACAGAGGCCCCAGGCTGAGGTGGTGGGGGTGGAAGACCCCCAGGAGGTGGCCCACTTCCCTTCCTCCCAGCTGGAACCCACCATGACCTTCTTAAGATAGGGGTGTCATCCGAGGCAGGTCCTCCATGGAGCTCCCTTCAGGCTCCTCCCCGGTCCTCACTAGGCCTCAGTCCCGGCTGCGGGAATGCAGCCACCACAGGCACACCAGGCAGCCCAGACCCAGCCAGCCTGCAGTGCCCAAGCCCACATTCTGGAGCAGAGCAGGCTGTGTCTGGGAGAGTCTGGGCTCCCCACCGCCCCCCCGCACACCCCACCCACCCCTGTCCAGGCCCTATGCAGGAGGGTCAGAGCCCCCCATGGGGTATGGACTTAGGGTCTCACTCACGTGGCTCCCCTCCTGGGTGAAGGGGTCTCATGCCCAGATCCCCACAGCAGAGCTGGTCAAAGGTGGAGGCAGTGGCCCCAGGGCCACCCTGACCTGGACCCTCAGGCTCCTCTAGCCCTGGCTGCCCTGCTGTCCCTGGGAGGCCTGGACTCCACCAGACCACAGGTCCAGGGCACCGCCCATAGGTGCTGCCCACACTCAGTTCACAGGAAGAAGATAAGCTCCAGACCCCCAAGACTGGGACCTGCCTTCCTGCCACCGCTTGTAGCTCCAGACCTCCGTGCCTCCCCCGACCACTTACACACGGGCCAGGGAGCTGTTCCACAAAGATCAACCCCAAACCGGGACCGCCTGGCACTCGGGCCGCTGCCACTTCCCTCTCCATTTGTTCCCAGCACCTCTGTGCTCCCTCCCTCCTCCCTCCTTCAGGGGAACAGCCTGTGCAGCCCCTCCCTGCACCCCACACCCTGGGGAGGCCCAACCCTGCCTCCAGCCCTTTCTCCCCCGCTGCTCTTCCTGCCCATCCAGACAACCCTGGGGTCCCATCCCTGCAGCCTACACCCTGGTCTCCACCCAGACCCCTGTCTCTCCCTCCAGACACCCCTCCCAGGCCAACCCTGCACATGCAGGCCCTCCCCTTTTCTGCTGCCAGAGCCTCAGTTTCTACCCTCTGTGCCTACCCCCTGCCTCCTCCTGCCCACAACTCGAGCTCTTCCTCTCCTGGGGCCCCTGAGCCATGGCACTGACCGTGCACTCCCACCCCCACACTGCCCATGCCCTCACCTTCCTCCTGGACACTCTGACCCCGCTCCCCTCTTGGACCCAGCCCTGGTATTTCCAGGACAAAGGCTCACCCAAGTCTTCCCCATGCAGGCCCTTGCCCTCACTGCCCGGTTACACGGCAGCCTCCTGTGCACAGAAGCAGGGAGCTCAGCCCTTCCACAGGCAGAAGGCACTGAAAGAAATCGGCCTCCAGCACCCTGATGCACGTCCGCCTGTGTCTCTCACTGCCCGCACCTGCAGGGAGGCTCGGCACTCCCTGTAAAGACGAGGGATCCAGGCAGCAACATCATGGGAGAATGCAGGGCTCCCAGACAGCCCAGCCCTCTCGCAGGCCTCTCCTGGGAAGAGACCTGCAGCCACCACTGAACAGCCACGGAGCCCGCTGGATAGTAACTGAGTCAGTGACCGACCTGGAGGGCAGGGGAGCAGTGAACCGGAGCCCAGACCATAGGGACAGAGACCAGCCGCTGACATCCCGAGCCCCTCACTGGCGGCCCCAGAACACCGCGTGGAAACAGAACAGACCCACATTCCCACCTGGAACAGGGCAGACACTGCTGAGCCCCCAGCACCAGCCCTGAGAAACACCAGGCAACGGCATCAGAGGGGGCTCCTGAGAAAGAAAGGAGGGGAGGTCTCCTTCACCAGCAAGTACTTCCCTTGACCAAAAACAGGGTCCACGCAACTCCCCCAGGACAAAGGAGGAGCCCCCTGTACAGCACTGGGCTCAGAGTCCTCTCCCACACACCCTGAGTTTCAGACAAAAACCCCCTGGAAATCATAGTATCAGCAGGAGAACTAGCCAGAGACAGCAAGAGGGGACTCAGTGACTCCCGCGGGGACAGGAGGATTTTGTGGGGGCTCGTGTCACTGTGCTGCATACCCTGCTGGACCTGCAAGTATTCGGCAACTGTGAGGTTAGCCAGCATCTAGACTATGCCCACAGTGACACAGCCCCATTCAAAAACCCCTGCTGTAAACGCTTCCACTTCTGGAGCTGAGGGGCTGGGGGGAGCGTCTGGGAAGTAGGGCCTAGGGGTGGCCATCAATGCCCAAAACGCACCAGACTCCCCCCCAGACATCACCCCACTGGCCAGTGAGCAGAGTAAACAGAAAATGAGAAGCAGCTGGGAAGCTTGCACAGGCCCCAAGGAAAGAGCTTTGGCGGGTGTGCAAGAGGGGATGCGGGCAGAGCCTGAGCAGGGCCTTTTGCTGTTTCTGCTTTCCTGTGCAGATAGTTCCATAAACTGGTGTTCAAGATCGATGGCTGGGAGTGAGCCCAGGAGGACAGTGTGGGAAGGGCACAGGGAAGGAGAAGCAGCCGCTATCCTACACTGTCATCTTTCAAGAGTTTGCCCTGTGCCCACAATGCTGCATCATGGGATGCTTAACAGCTGATGTAGACACAGCTAAAGAGAGAATCAGTGAAATGGATTTGCAGCACAGATCTGAATAAATTCTCCAGAATGTGGAGCCACACAGAAGCAAGCACAAGGAAAGTGCCTGATGCAAGGGCAAAGTACAGTGTGTACCTTCAGGCTGGGCACAGACACTCTGAAAAGCCTTGGCAGGAACTCCCTGCAACAAAGCAGAGCCCTGCAGGCAATGCCAGCTCCAGAGCCCTCCCTGAGAGCCTCATGGGCAAAGATGTGCACAACAGGTGTTTCTCATAGCCCCAAACTGAGAATGAAGCAAACAGCCATCTGAAGGAAAACAGGCAAATAAACGATGGCAGGTTCATGAAATGCAAACCCAGACAGCCAGAAGGACAACAGTGAGGGTTACAGGTGACTCTGTGGTTGAGTTCATGACAATGCTGAGTAATTGGAGTAACAAAGGAAAGTCCAAAAAATACTTTCAATGTGATTTCTTCTAAATAAAATTTACAGCCGGCAAAATGAACTATCTTCTTAAGGGATAAACTTTCCACTAGGAAAACTATAAGGAAAATCAAGAAAAGGATGATCACATAAACACAGTGGTCGTTACTTCTACTGGGGAAGGAAGAGGGTATGAACTGAGACACACAGGGTTGGCAAGTCTCCTAACAAGAACAGAACAAATACATTACAGTACCTTGAAAACAGCAGTTAAAATTCTAAATTGCAAGAAGAGGAAAATGCACACAGCTGTGTTTAGAAAATTCTCAGTCCAGCACTGTTCATAATAGCAAAGACATTAACCCAGGTTGGATAAATAAACGATGACACAGGCAATTGCACAATGATACAGACATACATTCAGTATATGAGACATTGATGATGTATCCCCAAAGAAATGACTTTAAAGAGAAAAGGCCTGATATGTGGTGGCACTCACCTCCCTGGGCATCCCCGGACAGGCTGCAGGCACACTGTGTGGCAGGGCAGGCTGGTACCTGCTGGCAGCTCCTGGGGCCTGATGTGGAGCAGGCACAGAGCCGTATCCCCCCGAGGACATATACCCCCAAGGACGGCACAGTTGGTACATTCCGGAGACAAGCAACTCAGCCACACTCCCAGGCCAGAGCCCGAGAGGGACGCCCATGCACAGGGAGGCAGAGCCCAGCTCCTCCACAGCCAGCAGCACCCGTGCAGGGGCCGCCATCTGGCAGGCACAGAGCATGGGCTGGGAGGAGGGGCAGGGACACCAGGCAGGGTTGGCACCAACTGAAAATTACAGAAGTCTCATACATCTACCTCAGCCTTGCCTGACCTGGGCCTCACCTGACCTGGACCTCACCTGGCCTGGACCTCACCTGGCCTAGACCTCACCTCTGGGCTTCACCTGAGCTCGGCCTCACCTGACTTGGACCTTGCCTGTCCTGAGCTCACATGATCTGGGCCTCACCTGACCTGGGTTTCACCTGACCTGGGCTTCACCTGACCTGGGCCTCATCTGACCTGGGCCTCACTGGCCTGGACCTCACCTGGCCTGGGCTTCACCTGGCCTCAGGCCTCATCTGCACCTGCTCCAGGTCTTGCTGGAACCTCAGTAGCACTGAGGCTGCAGGGGCTCATCCAGGGTTGCAGAATGACTCTAGAACCTCCCACATCTCAGCTTTCTGGGTGGAGGCACCTGGTGGCCCAGGGAATATAAAAAGCCTGAATGATGCCTGCGTGATTTGGGGGCAATTTATAAACCCAAAAGGACATGGCCATGCAGCGGGTAGGGACAATACAGACAGATATCAGCCTGAAATGGAGCCTCAGGGCACAGGTGGGCACGGACACTGTCCACCTAAGCCAGGGGCAGACCCGAGTGTCCCCGCAGTAGACCTGAGAGCGCTGGGCCCACAGCCTCCCCTCGGTGCCCTGCTACCTCCTCAGGTCAGCCCTGGACATCCCGGGTTTCCCCAGGCCTGGCGGTAGGATTTTGTTGAGGTCTGTGTCACTGTGCATGGCAGCTACTGCCAGCCCGCAGCCACTGGCTACTGAGGATGCCGATTCTGAGAATAGCAGCTTCTACTACTATGACTACCTGGATGAAGTAGCTTTCATGCTCTGCCGGAAGGATGCTGTGGTTAGCTTTGGCAAAGTTTTCCTGCCACCACAGTGTCACAGAGTCCATCAAAAACCCATCCCTGGGAACCTTCTGCCACAGCCCTCCCTGTGGGGCACCGCCGCGTGCCATGTTAGGATTTTGACTGAGGACACAGCACCATGGGTATGGTGGCTACCGCAGCAGTGCAGCCCGTGACCCAAACACACAGGGCAGCAGGCACAACAGACAAGCCCACAAGTGACCACCCTGAGCTCCTGCCTGCCAGCCCTGGAGACCATGAAACAGATGGCCAGGATTATCCCATAGGTCAGCCAGACCTCAGTCCAACAGGTCTGCATCGCTGCTGCCCTCCAATACCAGTCCGGATGGGGACAGGGCTGGCCCACATTACCATTTGCTGCCATCCGGCCAACAGTCCCAGAAGCCCCTCCCTCAAGGCTGGGCCACATGTGTGGACCCTGAGAGCCCCCCATGTCTGAGTAGGGGCACCAGGAAGGTGGGGCTGGCCCTGTGCACTGTCCCTGCCCCTGTGGTCCCTGGCCTGCCTGGCCCTGACACCTGGGCCTCTCCTGGGTCATTTCCAAGACAGAAGACATTCCCAGGACAGCTGGAGCTGGGAGTCCATCATCCTGCCTGGCCGTCCTGAGTCCTGCGCCTTTCCAAACCTCACCCGGGAAGCCAACAGAGGAATCACCTCCCACAGGCAGAGACAAAGACCTTCCAGAAATCTCTGTCTCTCTCCCCAGTGGGCACCCTCTTCCAGGGCAGTCCTCAGTGATATCACAGTGGGAACCCACATCTGGATCGGGACTGCCCCCAGAACACAAGATGGCCCACAGGGACAGCCCCACAGCCCAGCCCTTCCCAGACCCCTAAAAGGCGTCCCACCCCCTGCATCTGCCCCAGGGCTCAAACTCCAGGAGGACTGACTCCTGCACACCCTCCTGCCAGACATCACCTCAGCCCCTCCTGGAAGGGACAGGAGCGCGCAAGGGTGAGTCAGACCCTCCTGCCCTCGATGGCAGGCGGAGAAGATTCAGAAAGGTCTGAGATCCCCAGGACGCAGCACCACTGTCAATGGGGGCCCCAGACGCCTGGACCAGGGCCTGCGTGGGAAAGGCCTCTGGGCACACTCAGGGGGATTTTGTGAAGGGTCCTCCCACTGTGCATGGCAGCTACTGCCAGCCCGCAGCCACTGGCTACTGAGGATGCCGATTCTGAGAATAGCAGCTTCTACTACTATGACTACCTGGATGAAGTAGCTTTCATGCTCTGCCGGAAGGATGCTGTGGTTAGCTTTGGCAAAGTTTTCCTGCCACTGCATACCCTGCTGGACCTGCAAGTATTCGGCAACTGTGAGGTTAGCCAGCATCTAGACTATGCCCACAGTGATGAACCCAGCATCAAAAACCGACCGGACTCCCAAGGTTTATGCACACTTCTCCGCTCAGAGCTCTCCAGGATCAGAAGAGCCGGGCCCAAGGGTTTCTGCCCAGACCCTCGGCCTCTAGGGACATCTTGGCCATGACAGCCCATGGGCTGGTGCCCCACACATCGTCTGCCTTCAAACAAGGGCTTCAGAGGGCTCTGAGGTGACCTCACTGATGACCACAGGTGCCCTGGCCCCTTCCCCACCAGCTGCACCAGACCCCGTCATGACAGATGCCCCGATTCCAACAGCCAATTCCTGGGGCCAGGAATCGCTGTAGACACCAGCCTCCTTCCAACACCTCCTGCCAATTGCCTGGATTCCCATCCCGGTTGGAATCAAGAGGACAGCATCCCCCAGGCTCCCAACAGGCAGGACTCCCACACCCTCCTCTGAGAGGCCGCTGTGTTCCGTAGGGCCAGGCTGCAGACAGTCCCCCTCACCTGCCACTAGACAAATGCCTGCTGTAGATGTCCCCACCTGGAAAATACCACTCATGGAGCCCCCAGCCCCAGGTACAGCTGTAGAGAGAGTCTCTGAGGCCCCTAAGAAGTAGCCATGCCCAGTTCTGCCGGGACCCTCGGCCAGGCTGACAGGAGTGGACGCTGGAGCTGGGCCCATACTGGGCCACATAGGAGCTCACCAGTGAGGGCAGGAGAGCACATGCCGGGGAGCACCCAGCCTCCTGCTGACCAGAGGCCCGTCCCAGAGCCCAGGAGGCTGCAGAGGCCTCTCCAGGGGGACACTGTGCATGTCTGGTCCCTGAGCAGCCCCCCACGTCCCCAGTCCTGGGGGCCCCTGGCACAGCTGTCTGGACCCTCTCTATTCCCTGGGAAGCTCCTCCTGACAGCCCCGCCTCCAGTTCCAGGTGTGGATTTTGTCAGGGGGTGTCACACTGTGCAGCTTCTTGTGCAAGCACAGTGGTGCTGCCCATATCAAAAACCAGGCCAAGTAGACAGGCCCCTGCTGTGCAGCCCCAGGCCTCCAGCTCACCTGCTTCTCCTGGGGCTCTCAAGGCTGCTGTTTTCTGCACTCTCCCCTCTGTGGGGAGGGTTCCCTCAGTGGGAGATCTGTTCTCAACATCCCACGGCCTCATTCCTGCAAGGAAGGCCAATGGATGGGCAACCTCACATGCCGCGGCTAAGATAGGGTGGGCAGCCTGGCGGGGACAGGACATCCTGCTGGGGTATCTGTCACTGTGCCTAGTGGGGCACTGGCTCCCAAACAACGCAGTCCTTGCCAAAATCCCCACGGCCTCCCCCGCTAGGGGCTGGCCTGATCTCCTGCAGTCCTAGGAGGCTGCTGACCTCCAGAATGGCTCCGTCCCCAGTTCCAGGGCGAGAGCAGATCCCAGGCCGGCTGCAGACTGGGAGGCCACCCCCTCCTTCCCAGGGTTCACTGCAGGTGACCAGGGCAGGAAATGGCCTGAACACAGGGATAACCGGGCCATCCCCCAACAGAGTCCACCCCCTCCTGCTCTGTACCCCGCACCCCCCAGGCCAGCCCATGACATCCGACAACCCCACACCAGAGTCACTGCCCGGTGCTGCCCTAGGGAGGACCCCTCAGCCCCCACCCTGTCTAGAGGACTGGGGAGGACAGGACACGCCCTCTCCTTATGGTTCCCCCACCTGGCTCTGGCTGGGACCCTTGGGGTGTGGACAGAAAGGACGCTTGCCTGATTGGCCCCCAGGAGCCCAGAACTTCTCTCCAGGGACCCCAGCCCGAGCACCCCCTTACCCAGGACCCAGCCCTGCCCCTCCTCCCCTCTGCTCTCCTCTCATCACCCCATGGGAATCCAGAATCCCCAGGAAGCCATCAGGAAGGGCTGAGGGAGGAAGTGGGGCCACTGCACCACCAGGCAGGAGGCTCTGTCTTTGTGAACCCAGGGAGGTGCCAGCCTCCTAGAGGGTATGGTCCACCCTGCCTATGGCTCCCACAGTGGCAGGCTGCAGGGAAGGACCAGGGACGGTGTGGGGGAGGGCTCAGGGCCCCGCGGGTGCTCCATCTTGGATGAGCCTATCTCTCTCACCCACGGACTCGCCCACCTCCTCTTCACCCTGGCCACACGTCGTCCACACCATCCTAAGTCCCACCTACACCAGAGCCGGCACAGCCAGTGCAGACAGAGGCTGGGGTGCAGGGGGGCCGACTGGGCAGCTTCGGGGAGGGAGGAATGGAGGAAGGGGAGTTCAGTGAAGAGGCCCCCCTCCCCTGGGTCCAGGATCCTCCTCTGGGACCCCCGGATCCCATCCCCTCCAGGCTCTGGGAGGAGAAGCAGGATGGGAGAATCTGTGCGGGACCCTCTCACAGTGGAATACCTCCACAGCGGCTCAGGCCAGATACAAAAGCCCCTCAGTGAGCCCTCCACTGCAGTGCTGGGCCTGGGGGCAGCCGCTCCCACACAGGATGAACCCAGCACCCCGAGGATGTCCTGCCAGGGGGAGCTCAGAGCCATGAAGGAGCAGGATATGGGACCCCCGATACAGGCACAGACCTCAGCTCCATTCAGGACTGCCACGTCCTGCCCTGGGAGGAACCCCTTTCTCTAGTCCCTGCAGGCCAGGAGGCAGCTGACTCCTGACTTGGACGCCTATTCCAGACACCAGACAGAGGGGCAGGCCCCCCAGAACCAGGGATGAGGACGCCCCGTCAAGGCCAGAAAAGACCAAGTTGCGCTGAGCCCAGCAAGGGAAGGTCCCCAAACAAACCAGGAGGATTTTGTAGGTGTCTGTGTCACTGTGCAAACCCATGAAAACCCCAAGGGAGTTTGGAACTGCCATGCCGATTTCGGCGGGCATGGCACCATTTGGAAGCTCTTCCTCCGGTTCCAGCAGAACCTGCTACCACAGTGACACTCGCCAGGTCAAAAACCCCATCCCAAGTCAGCGGAATGCAGAGAGAGCAGGGAGGACATGTTTAGGATCTGAGGCCGCACCTGACACCCAGGCCAGCAGACGTCTCCTGTCCACGGCACCCTGCCATGTCCTGCATTTCTGGAAGAACAAGGGCAGGCTGAAGGGGGTCCAGGACCAGGAGATGGGTCCGCTCTACCCAGAGAAGGAGCCAGGCAGGACACAAGCCCCCACGCGTGGGCTCGTAGTTTGACGTGCGTGAAGTGTGGGTAAGAAAGTACGTA
[00156] D6-D17613 (SEQ ID NO:132) включает кодирующие последовательности D6, встроенные в положениях, соответствующих DH1-7 — DH6-13:
GCGGCCGCTGCCATTTCATTACCTCTTTCTCCGCACCCGACATAGATTACGTAACGCGTGGGCTCGTAGTTTGACGTGCGTGAAGTGTGGGTAAGAAAGTCCCCATTGAGGCTGACCTGCCCAGAGGGTCCTGGGCCCACCCAACACACCGGGGCGGAATGTGTGCAGGCCTCGGTCTCTGTGGGTGTTCCGCTAGCTGGGGCTCACAGTGCTCACCCCACACCTAAAACGAGCCACAGCCTCCGGAGCCCCTGAAGGAGACCCCGCCCACAAGCCCAGCCCCCACCCAGGAGGCCCCAGAGCACAGGGCGCCCCGTCGGATTTTGTACAGCCCCGAGTCACTGTGCAGCTTCTTGCACAGTGAGAAAAGCTTCGTCAAAAACCGTCTCCTGGCCACAGTCGGAGGCCCCGCCAGAGAGGGGAGCAGCCACCCCAAACCCATGTTCTGCCGGCTCCCATGACCCCGTGCACCTGGAGCCCCACGGTGTCCCCACTGGATGGGAGGACAAGGGCCGGGGGCTCCGGCGGGTCGGGGCAGGGGCTTGATGGCTTCCTTCTGCCGTGGCCCCATTGCCCCTGGCTGGAGTTGACCCTTCTGACAAGTGTCCTCAGAGAGTCAGGGATCAGTGGCACCTCCCAACATCAACCCCACGCAGCCCAGGCACAAACCCCACATCCAGGGCCAACTCCAGGAACAGAGACACCCCAATACCCTGGGGGACCCCGACCCTGATGACTCCCGTCCCATCTCTGTCCCTCACTTGGGGCCTGCTGCGGGGCGAGCACTTGGGAGCAAACTCAGGCTTAGGGGACACCACTGTGGGCCTGACCTCGAGCAGGCCACAGACCCTTCCCTCCTGCCCTGGTGCAGCACAGACTTTGGGGTCTGGGCAGGGAGGAACTTCTGGCAGGTCACCAAGCACAGAGCCCCCAGGCTGAGGTGGCCCCAGGGGGAACCCCAGCAGGTGGCCCACTACCCTTCCTCCCAGCTGGACCCCATGTCTTCCCCAAGATAGGGGTGCCATCCAAGGCAGGTCCTCCATGGAGCCCCCTTCAGGCTCCTCTCCAGACCCCACTGGGCCTCAGTCCCCACTCTAGGAATGCAGCCACCACGGGCACACCAGGCAGCCCAGGCCCAGCCACCCTGCAGTGCCCAAGCCCACACCCTGGAGGAGAGCAGGGTGCGTCTGGGAGGGGCTGGGCTCCCCACCCCCACCCCCACCTGCACACCCCACCCACCCTTGCCCGGGCCCCCTGCAGGAGGGTCAGAGCCCCCATGGGATATGGACTTAGGGTCTCACTCACGCACCTCCCCTCCTGGGAGAAGGGGTCTCATGCCCAGATCCCCCCAGCAGCGCTGGTCACAGGTAGAGGCAGTGGCCCCAGGGCCACCCTGACCTGGCCCCTCAGGCTCCTCTAGCCCTGGCTGCCCTGCTGTCCCTGGGAGGCCTGGGCTCCACCAGACCACAGGTCTAGGGCACCGCCCACACTGGGGCCGCCCACACACAGCTCACAGGAAGAAGATAAGCTCCAGACCCCCAGGCCCGGGACCTGCCTTGCTGCTACGACTTCCTGCCCCAGACCTCGTTGCCCTCCCCCGTCCACTTACACACAGGCCAGGAAGCTGTTCCCACACAGACCAACCCCAGACGGGGACCACCTGGCACTCAGGTCACTGCCATTTCCTTCTCCATTCACTTCCAATGCCTCTGTGCTTCCTCCCTCCTCCTTCCTTCGGGGGAGCACCCTGTGCAGCTCCTCCCTGCAGTCCACACCCTGGGGAGACCCGACCCTGCAGCCCACACCCTGGGGAGACCTGACCCTCCTCCAGCCCTTTCTCCCCCGCTGCTCTTGCCACCCACCAAGACAGCCCTGGGGTCCTGTCCCTACAGCCCCCACCCAGTTCTCTACCTAGACCCGTCTTCCTCCCTCTAAACACCTCTCCCAGGCCAACCCTACACCTGCAGGCCCTCCCCTCCACTGCCAAAGACCCTCAGTTTCTCCTGCCTGTGCCCACCCCCGTGCTCCTCCTGCCCACAGCTCGAGCTCTTCCTCTCCTAGGGCCCCTGAGGGATGGCATTGACCGTGCCCTCGCACCCACACACTGCCCATGCCCTCACATTCCTCCTGGCCACTCCAGCCCCACTCCCCTCTCAGGCCTGGCTCTGGTATTTCTGGGACAAAGCCTTACCCAAGTCTTTCCCATGCAGGCCTGGGCCCTTACCCTCACTGCCCGGTTACAGGGCAGCCTCCTGTGCACAGAAGCAGGGAGCTCAGCCCTTCCACAGGCAGAAGGCACTGAAAGAAATCGGCCTCCAGCGCCTTGACACACGTCTGCCTGTGTCTCTCACTGCCCGCACCTGCAGGGAGGCTCGGCACTCCCTCTAAAGACGAGGGATCCAGGCAGCAGCATCACAGGAGAATGCAGGGCTACCAGACATCCCAGTCCTCTCACAGGCCTCTCCTGGGAAGAGACCTGAAGACGCCCAGTCAACGGAGTCTAACACCAAACCTCCCTGGAGGCCGATGGGTAGTAACGGAGTCATTGCCAGACCTGGAGGCAGGGGAGCAGTGAGCCCGAGCCCACACCATAGGGCCAGAGGACAGCCACTGACATCCCAAGCCACTCACTGGTGGTCCCACAACACCCCATGGAAAGAGGACAGACCCACAGTCCCACCTGGACCAGGGCAGAGACTGCTGAGACCCAGCACCAGAACCAACCAAGAAACACCAGGCAACAGCATCAGAGGGGGCTCTGGCAGAACAGAGGAGGGGAGGTCTCCTTCACCAGCAGGCGCTTCCCTTGACCGAAGACAGGATCCATGCAACTCCCCCAGGACAAAGGAGGAGCCCCTTGTTCAGCACTGGGCTCAGAGTCCTCTCCAAGACACCCAGAGTTTCAGACAAAAACCCCCTGGAATGCACAGTCTCAGCAGGAGAGCCAGCCAGAGCCAGCAAGATGGGGCTCAGTGACACCCGCAGGGACAGGAGGATTTTGTGGGGGCTCGTGTCACTGTGCTGCATACCCTGCTGGACCTGCAAGTATTCGGCAACAGTGAGGTTAGCCAGCATCTAGACTATGCCCACAGTGACACAGCCCCATTCAAAAACCCCTACTGCAAACGCATTCCACTTCTGGGGCTGAGGGGCTGGGGGAGCGTCTGGGAAATAGGGCTCAGGGGTGTCCATCAATGCCCAAAACGCACCAGACTCCCCTCCATACATCACACCCACCAGCCAGCGAGCAGAGTAAACAGAAAATGAGAAGCAAGCTGGGGAAGCTTGCACAGGCCCCAAGGAAAGAGCTTTGGCGGGTGTGTAAGAGGGGATGCGGGCAGAGCCTGAGCAGGGCCTTTTGCTGTTTCTGCTTTCCTGTGCAGAGAGTTCCATAAACTGGTGTTCGAGATCAATGGCTGGGAGTGAGCCCAGGAGGACAGCGTGGGAAGAGCACAGGGAAGGAGGAGCAGCCGCTATCCTACACTGTCATCTTTCGAAAGTTTGCCTTGTGCCCACACTGCTGCATCATGGGATGCTTAACAGCTGATGTAGACACAGCTAAAGAGAGAATCAGTGAGATGGATTTGCAGCACAGATCTGAATAAATTCTCCAGAATGTGGAGCAGCACAGAAGCAAGCACACAGAAAGTGCCTGATGCAAGGACAAAGTTCAGTGGGCACCTTCAGGCATTGCTGCTGGGCACAGACACTCTGAAAAGCCCTGGCAGGAACTCCCTGTGACAAAGCAGAACCCTCAGGCAATGCCAGCCCCAGAGCCCTCCCTGAGAGCCTCATGGGCAAAGATGTGCACAACAGGTGTTTCTCATAGCCCCAAACTGAGAGCAAAGCAAACGTCCATCTGAAGGAGAACAGGCAAATAAACGATGGCAGGTTCATGAAATGCAAACCCAGACAGCCACAAGCACAAAAGTACAGGGTTATAAGCGACTCTGGTTGAGTTCATGACAATGCTGAGTAATTGGAGTAACAAAGTAAACTCCAAAAAATACTTTCAATGTGATTTCTTCTAAATAAAATTTACACCCTGCAAAATGAACTGTCTTCTTAAGGGATACATTTCCCAGTTAGAAAACCATAAAGAAAACCAAGAAAAGGATGATCACATAAACACAGTGGTGGTTACTTCTGCTGGGGAAGGAAGAGGGTATGAACTGAGATACACAGGGTGGGCAAGTCTCCTAACAAGAACAGAACGAATACATTACAGTACCTTGAAAACAGCAGTTAAACTTCTAAATTGCAAGAAGAGGAAAATGCACACAGTTGTGTTTAGAAAATTCTCAGTCCAGCACTGTTCATAATAGCAAAGACATTAACCCAGGTCGGATAAATAAGCGATGACACAGGCAATTGCACAATGATACAGACATATATTTAGTATATGAGACATCGATGATGTATCCCCAAATAAACGACTTTAAAGAGATAAAGGGCTGATGTGTGGTGGCATTCACCTCCCTGGGATCCCCGGACAGGTTGCAGGCTCACTGTGCAGCAGGGCAGGCGGGTACCTGCTGGCAGTTCCTGGGGCCTGATGTGGAGCAAGCGCAGGGCCATATATCCCGGAGGACGGCACAGTCAGTGAATTCCAGAGAGAAGCAACTCAGCCACACTCCCCAGGCAGAGCCCGAGAGGGACGCCCACGCACAGGGAGGCAGAGCCCAGCACCTCCGCAGCCAGCACCACCTGCGCACGGGCCACCACCTTGCAGGCACAGAGTGGGTGCTGAGAGGAGGGGCAGGGACACCAGGCAGGGTGAGCACCCAGAGAAAACTGCAGACGCCTCACACATCCACCTCAGCCTCCCCTGACCTGGACCTCACTGGCCTGGGCCTCACTTAACCTGGGCTTCACCTGACCTTGGCCTCACCTGACTTGGACCTCGCCTGTCCCAAGCTTTACCTGACCTGGGCCTCAACTCACCTGAACGTCTCCTGACCTGGGTTTAACCTGTCCTGGAACTCACCTGGCCTTGGCTTCCCCTGACCTGGACCTCATCTGGCCTGGGCTTCACCTGGCCTGGGCCTCACCTGACCTGGACCTCATCTGGCCTGGACCTCACCTGGCCTGGACTTCACCTGGCCTGGGCTTCACCTGACCTGGACCTCACCTGGCCTCGGGCCTCACCTGCACCTGCTCCAGGTCTTGCTGGAGCCTGAGTAGCACTGAGGGTGCAGAAGCTCATCCAGGGTTGGGGAATGACTCTAGAAGTCTCCCACATCTGACCTTTCTGGGTGGAGGCAGCTGGTGGCCCTGGGAATATAAAAATCTCCAGAATGATGACTCTGTGATTTGTGGGCAACTTATGAACCCGAAAGGACATGGCCATGGGGTGGGTAGGGACATAGGGACAGATGCCAGCCTGAGGTGGAGCCTCAGGACACAGGTGGGCACGGACACTATCCACATAAGCGAGGGATAGACCCGAGTGTCCCCACAGCAGACCTGAGAGCGCTGGGCCCACAGCCTCCCCTCAGAGCCCTGCTGCCTCCTCCGGTCAGCCCTGGACATCCCAGGTTTCCCCAGGCCTGGCGGTAGGATTTTGTTGAGGTCTGTGTCACTGTGCATGGCAGCTACTGCCAGCCCGCAGCCACTGGCTACTGAGGATGCCGATTCTGAGAATAGCAGCTTCTACTACTATGACTACCTGGATGAAGTAGCTTTCATGCTCCACAGTGTCACAGAGTCCATCAAAAACCCATGCCTGGAAGCTTCCCGCCACAGCCCTCCCCATGGGGCCCTGCTGCCTCCTCAGGTCAGCCCCGGACATCCCGGGTTTCCCCAGGCTGGGCGGTAGGATTTTGTTGAGGTCTGTGTCACTGTGCCAAACCCATGAAAACCCCAAGGGAGTTTGGAACAGCCATGCCGATTTCGGCGGGCATGGCACCATTTGGAAGCTCTTCCTCCGGTTCCAGCAGAACCTGCTACCCACAGTGTCACAGAGTCCATCAAAAACCCATCCCTGGGAGCCTCCCGCCACAGCCCTCCCTGCAGGGGACCGGTACGTGCCATGTTAGGATTTTGATCGAGGAGACAGCACCATGGGTATGGTGGCTACCACAGCAGTGCAGCCTGTGACCCAAACCCGCAGGGCAGCAGGCACGATGGACAGGCCCGTGACTGACCACGCTGGGCTCCAGCCTGCCAGCCCTGGAGATCATGAAACAGATGGCCAAGGTCACCCTACAGGTCATCCAGATCTGGCTCCGAGGGGTCTGCATCGCTGCTGCCCTCCCAACGCCAGTCCAAATGGGACAGGGACGGCCTCACAGCACCATCTGCTGCCATCAGGCCAGCGATCCCAGAAGCCCCTCCCTCAAGGCTGGGCACATGTGTGGACACTGAGAGCCCTCATATCTGAGTAGGGGCACCAGGAGGGAGGGGCTGGCCCTGTGCACTGTCCCTGCCCCTGTGGTCCCTGGCCTGCCTGGCCCTGACACCTGAGCCTCTCCTGGGTCATTTCCAAGACAGAAGACATTCCTGGGGACAGCCGGAGCTGGGCGTCGCTCATCCTGCCCGGCCGTCCTGAGTCCTGCTCATTTCCAGACCTCACCGGGGAAGCCAACAGAGGACTCGCCTCCCACATTCAGAGACAAAGAACCTTCCAGAAATCCCTGCCTCTCTCCCCAGTGGACACCCTCTTCCAGGACAGTCCTCAGTGGCATCACAGCGGCCTGAGATCCCCAGGACGCAGCACCGCTGTCAATAGGGGCCCCAAATGCCTGGACCAGGGCCTGCGTGGGAAAGGCCTCTGGCCACACTCGGGGATTTTGTGAAGGGCCCTCCCACTGTGCCAGCTTCTTGAGCCATGCCGATTTCGGCGGGCATGGCACCATTTGGAAGCTCTTCCTCCGGTTCCAGCAGAACCTGCTACCACAGTGATGAACCCAGTGTCAAAAACCGGCTGGAAACCCAGGGGCTGTGTGCACGCCTCAGCTTGGAGCTCTCCAGGAGCACAAGAGCCGGGCCCAAGGATTTGTGCCCAGACCCTCAGCCTCTAGGGACACCTGGGTCATCTCAGCCTGGGCTGGTGCCCTGCACACCATCTTCCTCCAAATAGGGGCTTCAGAGGGCTCTGAGGTGACCTCACTCATGACCACAGGTGACCTGGCCCTTCCCTGCCAGCTATACCAGACCCTGTCTTGACAGATGCCCCGATTCCAACAGCCAATTCCTGGGACCCTGAATAGCTGTAGACACCAGCCTCATTCCAGTACCTCCTGCCAATTGCCTGGATTCCCATCCTGGCTGGAATCAAGAAGGCAGCATCCGCCAGGCTCCCAACAGGCAGGACTCCCGCACACCCTCCTCTGAGAGGCCGCTGTGTTCCGCAGGGCCAGGCCCTGGACAGTTCCCCTCACCTGCCACTAGAGAAACACCTGCCATTGTCGTCCCCACCTGGAAAAGACCACTCGTGGAGCCCCCAGCCCCAGGTACAGCTGTAGAGACAGTCCTCGAGGCCCCTAAGAAGGAGCCATGCCCAGTTCTGCCGGGACCCTCGGCCAGGCCGACAGGAGTGGACGCTGGAGCTGGGCCCACACTGGGCCACATAGGAGCTCACCAGTGAGGGCAGGAGAGCACATGCCGGGGAGCACCCAGCCTCCTGCTGACCAGAGGCCCGTCCCAGAGCCCAGGAGGCTGCAGAGGCCTCTCCAGGGAGACACTGTGCATGTCTGGTACCTAAGCAGCCCCCCACGTCCCCAGTCCTGGGGGCCCCTGGCTCAGCTGTCTGGGCCCTCCCTGCTCCCTGGGAAGCTCCTCCTGACAGCCCCGCCTCCAGTTCCAGGTGTGGATTTTGTCAGGCGATGTCACACTGTGCAGCTTCTTGAGCAAGCACAGTGGTGCCGCCCATATCAAAAACCAGGCCAAGTAGACAGGCCCCTGCTGCGCAGCCCCAGGCATCCACTTCACCTGCTTCTCCTGGGGCTCTCAAGGCTGCTGTCTGTCCTCTGGCCCTCTGTGGGGAGGGTTCCCTCAGTGGGAGGTCTGTGCTCCAGGGCAGGGATGATTGAGATAGAAATCAAAGGCTGGCAGGGAAAGGCAGCTTCCCGCCCTGAGAGGTGCAGGCAGCACCACGGAGCCACGGAGTCACAGAGCCACGGAGCCCCCATTGTGGGCATTTGAGAGTGCTGTGCCCCCGGCAGGCCCAGCCCTGATGGGGAAGCCTGTCCCATCCCACAGCCCGGGTCCCACGGGCAGCGGGCACAGAAGCTGCCAGGTTGTCCTCTATGATCCTCATCCCTCCAGCAGCATCCCCTCCACAGTGGGGAAACTGAGGCTTGGAGCACCACCCGGCCCCCTGGAAATGAGGCTGTGAGCCCAGACAGTGGGCCCAGAGCACTGTGAGTACCCCGGCAGTACCTGGCTGCAGGGATCAGCCAGAGATGCCAAACCCTGAGTGACCAGCCTACAGGAGGATCCGGCCCCACCCAGGCCACTCGATTAATGCTCAACCCCCTGCCCTGGAGACCTCTTCCAGTACCACCAGCAGCTCAGCTTCTCAGGGCCTCATCCCTGCAAGGAAGGTCAAGGGCTGGGCCTGCCAGAAACACAGCACCCTCCCTAGCCCTGGCTAAGACAGGGTGGGCAGACGGCTGTGGACGGGACATATTGCTGGGGCATTTCTCACTGTCACTTCTGGGTGGTAGCTCTGACAAAAACGCAGACCCTGCCAAAATCCCCACTGCCTCCCGCTAGGGGCTGGCCTGGAATCCTGCTGTCCTAGGAGGCTGCTGACCTCCAGGATGGCTCCGTCCCCAGTTCCAGGGCGAGAGCAGATCCCAGGCAGGCTGTAGGCTGGGAGGCCACCCCTGCCCTTGCCGGGGTTGAATGCAGGTGCCCAAGGCAGGAAATGGCATGAGCACAGGGATGACCGGGACATGCCCCACCAGAGTGCGCCCCTTCCTGCTCTGCACCCTGCACCCCCCAGGCCAGCCCACGACGTCCAACAACTGGGCCTGGGTGGCAGCCCCACCCAGACAGGACAGACCCAGCACCCTGAGGAGGTCCTGCCAGGGGGAGCTAAGAGCCATGAAGGAGCAAGATATGGGGCCCCCGATACAGGCACAGATGTCAGCTCCATCCAGGACCACCCAGCCCACACCCTGAGAGGAACGTCTGTCTCCAGCCTCTGCAGGTCGGGAGGCAGCTGACCCCTGACTTGGACCCCTATTCCAGACACCAGACAGAGGCGCAGGCCCCCCAGAACCAGGGTTGAGGGACGCCCCGTCAAAGCCAGACAAAACCAAGGGGTGTTGAGCCCAGCAAGGGAAGGCCCCCAAACAGACCAGGAGGATTTTGTAGGTGTCTGTGTCACTGTGCATGGCAGCTACTGCCAGCCCGCAGCCACTGGCTACTGAGGATGCCGATTCTGAGAATAGCAGCTTCTACTACTATGACTACCTGGATGAAGTAGCTTTCATGCTCAGCCGGAAGGATGCTGTGGTTAGCTTTGGCAAAGTTTTCCTGCCACCCACAGTGACACTCACCCAGTCAAAAACCCCATTCCAAGTCAGCGGAAGCAGAGAGAGCAGGGAGGACACGTTTAGGATCTGAGACTGCACCTGACACCCAGGCCAGCAGACGTCTCCCCTCCAGGGCACCCCACCCTGTCCTGCATTTCTGCAAGATCAGGGGCGGCCTGAGGGGGGGTCTAGGGTGAGGAGATGGGTCCCCTGTACACCAAGGAGGAGTTAGGCAGGTCCCGAGCACTCTTAATTAAACGACGCCTCGAATGGAACTACTACAACGAATGGTTGCTCTACGTAATGCATTCGCTACCTTAGGACCGTTATAGTTAGGCGCGCC
[00157] D6-DH114619 (SEQ ID NO:133) включает кодирующую последовательность D6, встроенную в положениях, соответствующих DH1–14 — DH6–19:
TACGTATTAATTAAACGACGCCTCGAATGGAACTACTACAACGAATGGTTGCTCTCCCCATTGAGGCTGACCTGCCCAGAGAGTCCTGGGCCCACCCCACACACCGGGGCGGAATGTGTGCAGGCCTCGGTCTCTGTGGGTGTTCCGCTAGCTGGGGCTCACAGTGCTCACCCCACACCTAAAATGAGCCACAGCCTCCGGAGCCCCCGCAGGAGACCCCGCCCACAAGCCCAGCCCCCACCCAGGAGGCCCCAGAGCTCAGGGCGCCCCGTCGGATTTTGTACAGCCCCGAGTCACTGTGCAAACCCATGAAAACCCCAAGGGAGTTTGGAACCACAGTGAGAATAGCTACGTCAAAAACCGTCCAGTGGCCACTGCCGGAGGCCCCGCCAGAGAGGGCAGCAGCCACTCTGATCCCATGTCCTGCCGGCTCCCATGACCCCCAGCACGCGGAGCCCCACAGTGTCCCCACTGGATGGGAGGACAAGAGCTGGGGATTCCGGCGGGTCGGGGCAGGGGCTTGATCGCATCCTTCTGCCGTGGCTCCAGTGCCCCTGGCTGGAGTTGACCCTTCTGACAAGTGTCCTCAGAGAGACAGGCATCACCGGCGCCTCCCAACATCAACCCCAGGCAGCACAGGCACAAACCCCACATCCAGAGCCAACTCCAGGAGCAGAGACACCCCAATACCCTGGGGGACCCCGACCCTGATGACTTCCCACTGGAATTCGCCGTAGAGTCCACCAGGACCAAAGACCCTGCCTCTGCCTCTGTCCCTCACTCAGGACCTGCTGCCGGGCGAGGCCTTGGGAGCAGACTTGGGCTTAGGGGACACCAGTGTGACCCCGACCTTGACCAGGACGCAGACCTTTCCTTCCTTTCCTGGGGCAGCACAGACTTTGGGGTCTGGGCCAGGAGGAACTTCTGGCAGGTCGCCAAGCACAGAGGCCACAGGCTGAGGTGGCCCTGGAAAGACCTCCAGGAGGTGGCCACTCCCCTTCCTCCCAGCTGGACCCCATGTCCTCCCCAAGATAAGGGTGCCATCCAAGGCAGGTGCTCCTTGGAGCCCCATTCAGACTCCTCCCTGGACCCCACTGGGCCTCAGTCCCAGCTCTGGGGATGAAGCCACCACAAGCACACCAGGCAGCCCAGGCCCAGCCACCCTGCAGTGCCCAAGCACACACTCTGGAGCAGAGCAGGGTGCCTCTGGGAGGGGCTGAGCTCCCCACCCCACCCCCACCTGCACACCCCACCCACCCCTGCCCAGCGGCTCTGCAGGAGGGTCAGAGCCCCACATGGGGTATGGACTTAGGGTCTCACTCACGTGGCTCCCATCATGAGTGAAGGGGCCTCAAGCCCAGGTTCCCACAGCAGCGCCTGTCGCAAGTGGAGGCAGAGGCCCGAGGGCCACCCTGACCTGGTCCCTGAGGTTCCTGCAGCCCAGGCTGCCCTGCTGTCCCTGGGAGGCCTGGGCTCCACCAGACCACAGGTCCAGGGCACCGGGTGCAGGAGCCACCCACACACAGCTCACAGGAAGAAGATAAGCTCCAGACCCCCAGGGCCAGAACCTGCCTTCCTGCTACTGCTTCCTGCCCCAGACCTGGGCGCCCTCCCCCGTCCACTTACACACAGGCCAGGAAGCTGTTCCCACACAGAACAACCCCAAACCAGGACCGCCTGGCACTCAGGTGGCTGCCATTTCCTTCTCCATTTGCTCCCAGCGCCTCTGTCCTCCCTGGTTCCTCCTTCGGGGGAACAGCCTGTGCAGCCAGTCCCTGCAGCCCACACCCTGGGGAGACCCAACCCTGCCTGGGGCCCTTCCAACCCTGCTGCTCTTACTGCCCACCCAGAAAACTCTGGGGTCCTGTCCCTGCAGTCCCTACCCTGGTCTCCACCCAGACCCCTGTGTATCACTCCAGACACCCCTCCCAGGCAAACCCTGCACCTGCAGGCCCTGTCCTCTTCTGTCGCTAGAGCCTCAGTTTCTCCCCCCTGTGCCCACACCCTACCTCCTCCTGCCCACAACTCTAACTCTTCTTCTCCTGGAGCCCCTGAGCCATGGCATTGACCCTGCCCTCCCACCACCCACAGCCCATGCCCTCACCTTCCTCCTGGCCACTCCGACCCCGCCCCCTCTCAGGCCAAGCCCTGGTATTTCCAGGACAAAGGCTCACCCAAGTCTTTCCCAGGCAGGCCTGGGCTCTTGCCCTCACTTCCCGGTTACACGGGAGCCTCCTGTGCACAGAAGCAGGGAGCTCAGCCCTTCCACAGGCAGAAGGCACTGAAAGAAATCGGCCTCCAGCACCTTGACACACGTCCGCCCGTGTCTCTCACTGCCCGCACCTGCAGGGAGGCTCCGCACTCCCTCTAAAGACAAGGGATCCAGGCAGCAGCATCACGGGAGAATGCAGGGCTCCCAGACATCCCAGTCCTCTCACAGGCCTCTCCTGGGAAGAGACCTGCAGCCACCACCAAACAGCCACAGAGGCTGCTGGATAGTAACTGAGTCAATGACCGACCTGGAGGGCAGGGGAGCAGTGAGCCGGAGCCCATACCATAGGGACAGAGACCAGCCGCTGACATCCCGAGCTCCTCAATGGTGGCCCCATAACACACCTAGGAAACATAACACACCCACAGCCCCACCTGGAACAGGGCAGAGACTGCTGAGCCCCCAGCACCAGCCCCAAGAAACACCAGGCAACAGTATCAGAGGGGGCTCCCGAGAAAGAGAGGAGGGGAGATCTCCTTCACCATCAAATGCTTCCCTTGACCAAAAACAGGGTCCACGCAACTCCCCCAGGACAAAGGAGGAGCCCCCTATACAGCACTGGGCTCAGAGTCCTCTCTGAGACACCCTGAGTTTCAGACAACAACCCGCTGGAATGCACAGTCTCAGCAGGAGAACAGACCAAAGCCAGCAAAAGGGACCTCGGTGACACCAGTAGGGACAGGAGGATTTTGTGGGGGCTCGTGTCACTGTGCAAACCCATGAAAACCCCAAGGGAGTTTGGAACTGCAAGCACAGTGACACAGACCCATTCAAAAACCCCTACTGCAAACACACCCACTCCTGGGGCTGAGGGGCTGGGGGAGCGTCTGGGAAGTAGGGTCCAGGGGTGTCTATCAATGTCCAAAATGCACCAGACTCCCCGCCAAACACCACCCCACCAGCCAGCGAGCAGGGTAAACAGAAAATGAGAGGCTCTGGGAAGCTTGCACAGGCCCCAAGGAAAGAGCTTTGGCGGGTGTGCAAGAGGGGATGCAGGCAGAGCCTGAGCAGGGCCTTTTGCTGTTTCTGCTTTCCTGTGCAGAGAGTTCCATAAACTGGTGTTCAAGATCAGTGGCTGGGAATGAGCCCAGGAGGGCAGTCTGTGGGAAGAGCACAGGGAAGGAGGAGCAGCCGCTATCCTACACTGTCATCTTTCAAAAGTTTGCCTTGTGACCACACTATTGCATCATGGGATGCTTAAGAGCTGATGTAGACACAGCTAAAGAGAGAATCAGTGAGATGAATTTGCAGCATAGATCTGAATAAACTCTCCAGAATGTGGAGCAGTACAGAAGCAAACACACAGAAAGTGCCTGATGCAAGGACAAAGTTCAGTGGGCACCTTCAGGCATTGCTGCTGGGCACAGACACTCTGAAAAGCCTTGGCAGGATCTCCCTGCGACAAAGCAGAACCCTCAGGCAATGCCAGCCCCAGAGCCCTCCCTGAGAGCGTCATGGGGAAAGATGTGCAGAACAGCTGATTATCATAGACTCAAACTGAGAACAGAGCAAACGTCCATCTGAAGAACAGTCAAATAAGCAATGGTAGGTTCATGCAATGCAAACCCAGACAGCCAGGGGACAACAGTAGAGGGCTACAGGCGGCTTTGCGGTTGAGTTCATGACAATGCTGAGTAATTGGAGTAACAGAGGAAAGCCCAAAAAATACTTTTAATGTGATTTCTTCTAAATAAAATTTACACCAGGCAAAATGAACTGTCTTCTTAAGGGATAAACTTTCCCCTGGAAAAACTACAAGGAAAATTAAGAAAACGATGATCACATAAACACAGTTGTGGTTACTTCTACTGGGGAAGGAAGAGGGTATGAGCTGAGACACACAGAGTCGGCAAGTCTCCAAGCAAGCACAGAACGAATACATTACAGTACCTTGAATACAGCAGTTAAACTTCTAAATCGCAAGAACAGGAAAATGCACACAGCTGTGTTTAGAAAATTCTCAGTCCAGCACTATTCATAATAGCAAAGACATTAACCCAGGTTGGATAAATAAATGATGACACAGGCAATTGCACAATGATACAGACATACATTTAGTACATGAGACATCGATGATGTATCCCCAAAGAAATGACTTTAAAGAGAAAAGGCCTGATGTGTGGTGGCACTCACCTCCCTGGGATCCCCGGACAGGTTGCAGGCACACTGTGTGGCAGGGCAGGCTGGTACATGCTGGCAGCTCCTGGGGCCTGATGTGGAGCAAGCGCAGGGCTGTATACCCCCAAGGATGGCACAGTCAGTGAATTCCAGAGAGAAGCAGCTCAGCCACACTGCCCAGGCAGAGCCCGAGAGGGACGCCCACGTACAGGGAGGCAGAGCCCAGCTCCTCCACAGCCACCACCACCTGTGCACGGGCCACCACCTTGCAGGCACAGAGTGGGTGCTGAGAGGAGGGGCAGGGACACCAGGCAGGGTGAGCACCCAGAGAAAACTGCAGAAGCCTCACACATCCACCTCAGCCTCCCCTGACCTGGACCTCACCTGGTCTGGACCTCACCTGGCCTGGGCCTCACCTGACCTGGACCTCACCTGGCCTGGGCTTCACCTGACCTGGACCTCACCTGGCCTCCGGCCTCACCTGCACCTGCTCCAGGTCTTGCTGGAACCTGAGTAGCACTGAGGCTGCAGAAGCTCATCCAGGGTTGGGGAATGACTCTGGAACTCTCCCACATCTGACCTTTCTGGGTGGAGGCATCTGGTGGCCCTGGGAATATAAAAAGCCCCAGAATGGTGCCTGCGTGATTTGGGGGCAATTTATGAACCCGAAAGGACATGGCCATGGGGTGGGTAGGGACATAGGGACAGATGCCAGCCTGAGGTGGAGCCTCAGGACACAGTTGGACGCGGACACTATCCACATAAGCGAGGGACAGACCCGAGTGTTCCTGCAGTAGACCTGAGAGCGCTGGGCCCACAGCCTCCCCTCGGTGCCCTGCTGCCTCCTCAGGTCAGCCCTGGACATCCCGGGTTTCCCCAGGCCAGATGGTAGGATTTTGTTGAGGTCTGTGTCACTGTGCATGGCAGCTACTGCCAGCCCGCAGCCACTGGCTACTGAGGATGCCGATTCTGAGAATAGCAGCTTCTACTACTATGACTACCTGGATGAAGTAGCTTTCATGCTCTGCGAGGTTAGCCAGCATCTAGACTATGCCCACAGTGTCACACGGTCCATCAAAAACCCATGCCACAGCCCTCCCCGCAGGGGACCGCCGCGTGCCATGTTACGATTTTGATCGAGGACACAGCGCCATGGGTATGGTGGCTACCACAGCAGTGCAGCCCATGACCCAAACACACAGGGCAGCAGGCACAATGGACAGGCCTGTGAGTGACCATGCTGGGCTCCAGCCCGCCAGCCCCGGAGACCATGAAACAGATGGCCAAGGTCACCCCACAGTTCAGCCAGACATGGCTCCGTGGGGTCTGCATCGCTGCTGCCCTCTAACACCAGCCCAGATGGGGACAAGGCCAACCCCACATTACCATCTCCTGCTGTCCACCCAGTGGTCCCAGAAGCCCCTCCCTCATGGCTGAGCCACATGTGTGAACCCTGAGAGCACCCCATGTCAGAGTAGGGGCAGCAGAAGGGCGGGGCTGGCCCTGTGCACTGTCCCTGCACCCATGGTCCCTCGCCTGCCTGGCCCTGACACCTGAGCCTCTTCTGAGTCATTTCTAAGATAGAAGACATTCCCGGGGACAGCCGGAGCTGGGCGTCGCTCATCCCGCCCGGCCGTCCTGAGTCCTGCTTGTTTCCAGACCTCACCAGGGAAGCCAACAGAGGACTCACCTCACACAGTCAGAGACAAAGAACCTTCCAGAAATCCCTGTCTCACTCCCCAGTGGGCACCTTCTTCCAGGACATTCCTCGGTCGCATCACAGCAGGCACCCACATCTGGATCAGGACGGCCCCCAGAACACAAGATGGCCCATGGGGACAGCCCCACAACCCAGGCCTTCCCAGACCCCTAAAAGGCGTCCCACCCCCTGCACCTGCCCCAGGGCTAAAAATCCAGGAGGCTTGACTCCCGCATACCCTCCAGCCAGACATCACCTCAGCCCCCTCCTGGAGGGGACAGGAGCCCGGGAGGGTGAGTCAGACCCACCTGCCCTCGATGGCAGGCGGGGAAGATTCAGAAAGGCCTGAGATCCCCAGGACGCAGCACCACTGTCAATGGGGGCCCCAGACGCCTGGACCAGGGCCTGCGTGGGAAAGGCCGCTGGGCACACTCAGGGGGATTTTGTGAAGGCCCCTCCCACTGTGCAGCTTCTTGTGCAAGCAAACCCATGAAAACCCCAAGGGAGTTTGGAACTGCCATGCCGATTTCGGCGGGCATGGCACCATTTGGAAGCTCTTCCTCCGGTTCCAGCAGAACCTGCTACCACAGTGATGAAACTAGCATCAAAAACCGGCCGGACACCCAGGGACCATGCACACTTCTCAGCTTGGAGCTCTCCAGGACCAGAAGAGTCAGGTCTGAGGGTTTGTAGCCAGACCCTCGGCCTCTAGGGACACCCTGGCCATCACAGCGGATGGGCTGGTGCCCCACATGCCATCTGCTCCAAACAGGGGCTTCAGAGGGCTCTGAGGTGACTTCACTCATGACCACAGGTGCCCTGGCCCCTTCCCCGCCAGCTACACCGAACCCTGTCCCAACAGCTGCCCCAGTTCCAACAGCCAATTCCTGGGGCCCAGAATTGCTGTAGACACCAGCCTCGTTCCAGCACCTCCTGCCAATTGCCTGGATTCACATCCTGGCTGGAATCAAGAGGGCAGCATCCGCCAGGCTCCCAACAGGCAGGACTCCCGCACACCCTCCTCTGAGAGGCCGCTGTGTTCCGCAGGGCCAGGCCCTGGACAGTTCCCCTCACCTGCCACTAGAGAAACACCTGCCATTGTCGTCCCCACCTGGAAAAGACCACTCGTGGAGCCCCCAGCCCCAGGTACAGCTGTAGAGAGACTCCCCGAGGGATCTAAGAAGGAGCCATGCGCAGTTCTGCCGGGACCCTCGGCCAGGCCGACAGGAGTGGACACTGGAGCTGGGCCCACACTGGGCCACATAGGAGCTCACCAGTGAGGGCAGGAGAGCACATGCCGGGGAGCACCCAGCCTCCTGCTGACCAGAGGCCCGTCCCAGAGCCCAGGAGGCTGCAGAGGCCTCTCCAGGGGGACACTGTGCATGTCTGGTCCCTGAGCAGCCCCCCACGTCCCCAGTCCTGGGGGCCCCTGGCACAGCTGTCTGGACCCTCCCTGTTCCCTGGGAAGCTCCTCCTGACAGCCCCGCCTCCAGTTCCAGGTGTGGATTTTGTCAGGGGGTGTCACACTGTGCTGCATACCCTGCTGGACCTGCAAGTATTCGGCAACTGTCGGAAGGATGCTGTGGTTAGCTTTGGCAAAGTTTTCCTGCCACCACAGTGGTGCTGCCCATATCAAAAACCAGGCCAAGTAGACAGGCCCCTGCTGTGCAGCCCCAGGCCTCCACTTCACCTGCTTCTCCTGGGGCTCTCAAGGTCACTGTTGTCTGTACTCTGCCCTCTGTGGGGAGGGTTCCCTCAGTGGGAGGTCTGTTCTCAACATCCCAGGGCCTCATGTCTGCACGGAAGGCCAATGGATGGGCAACCTCACATGCCGCGGCTAAGATAGGGTGGGCAGCCTGGCGGGGGACAGTACATACTGCTGGGGTGTCTGTCACTGTGCCTAGTGGGGCACTGGCTCCCAAACAACGCAGTCCTCGCCAAAATCCCCACAGCCTCCCCTGCTAGGGGCTGGCCTGATCTCCTGCAGTCCTAGGAGGCTGCTGACCTCCAGAATGTCTCCGTCCCCAGTTCCAGGGCGAGAGCAGATCCCAGGCCGGCTGCAGACTGGGAGGCCACCCCCTCCTTCCCAGGGTTCACTGGAGGTGACCAAGGTAGGAAATGGCCTTAACACAGGGATGACTGCGCCATCCCCCAACAGAGTCAGCCCCCTCCTGCTCTGTACCCCGCACCCCCCAGGCCAGTCCACGAAAACCAGGGCCCCACATCAGAGTCACTGCCTGGCCCGGCCCTGGGGCGGACCCCTCAGCCCCCACCCTGTCTAGAGGACTTGGGGGGACAGGACACAGGCCCTCTCCTTATGGTTCCCCCACCTGCCTCCGGCCGGGACCCTTGGGGTGTGGACAGAAAGGACACCTGCCTAATTGGCCCCCAGGAACCCAGAACTTCTCTCCAGGGACCCCAGCCCGAGCACCCCCTTACCCAGGACCCAGCCCTGCCCCTCCTCCCCTCTGCTCTCCTCTCATCACCCCATGGGAATCCGGTATCCCCAGGAAGCCATCAGGAAGGGCTGAAGGAGGAAGCGGGGCCGTGCACCACCGGGCAGGAGGCTCCGTCTTCGTGAACCCAGGGAAGTGCCAGCCTCCTAGAGGGTATGGTCCACCCTGCCTGGGGCTCCCACCGTGGCAGGCTGCGGGGAAGGACCAGGGACGGTGTGGGGGAGGGCTCAGGGCCCTGCGGGTGCTCCTCCATCTTCGGTGAGCCTCCCCCTTCACCCACCGTCCCGCCCACCTCCTCTCCACCCTGGCTGCACGTCTTCCACACCATCCTGAGTCCTACCTACACCAGAGCCAGCAAAGCCAGTGCAGACAAAGGCTGGGGTGCAGGGGGGCTGCCAGGGCAGCTTCGGGGAGGGAAGGATGGAGGGAGGGGAGGTCAGTGAAGAGGCCCCCTTCCCCTGGGTCCAGGATCCTCCTCTGGGACCCCCGGATCCCATCCCCTCCTGGCTCTGGGAGGAGAAGCAGGATGGGAGAATCTGTGCGGGACCCTCTCACAGTGGAATATCCCCACAGCGGCTCAGGCCAGACCCAAAAGCCCCTCAGTGAGCCCTCCACTGCAGTCCTGGGCCTGGGTAGCAGCCCCTCCCACAGAGGACAGACCCAGCACCCCGAAGAAGTCCTGCCAGGGGGAGCTCAGAGCCATGAAAGAGCAGGATATGGGGTCCCCGATACAGGCACAGACCTCAGCTCCATCCAGGCCCACCGGGACCCACCATGGGAGGAACACCTGTCTCCGGGTTGTGAGGTAGCTGGCCTCTGTCTCGGACCCCACTCCAGACACCAGACAGAGGGGCAGGCCCCCCAAAACCAGGGTTGAGGGATGATCCGTCAAGGCAGACAAGACCAAGGGGCACTGACCCCAGCAAGGGAAGGCTCCCAAACAGACGAGGAGGATTTTGTAGCTGTCTGTATCACTGTGCAGCTTCTTGTGCCATGCCGATTTCGGCGGGCATGGCACCATTTGGAAGCTCTTCCTCCGGTTCCAGCAGAACCTGCTACCACAGTGACACTCGCCAGGTCAAAAACCCCGTCCCAAGTCAGCGGAAGCAGAGAGAGCAGGGAGGACACGTTTAGGATCTGAGGCCGCACCTGACACCCAGGGCAGCAGACGTCTCCCCTCCAGGGCACCCTCCACCGTCCTGCGTTTCTTCAAGAATAGGGGCGGCCTGAGGGGGTCCAGGGCCAGGCGATAGGTCCCCTCTACCCCAAGGAGGAGCCAGGCAGGACCCGAGCACCGATGCATCTAACGCAGTCATGTAATGCTGGGTGACAGTCAGTTCGCCTACGTA
[00158] D6-DH120126 (SEQ ID NO:134) включает кодирующую последовательность D6, встроенную в положениях, соответствующих DH1-20 — DH1-26:
TACGTAATGCATCTAACGCAGTCATGTAATGCTGGGTGACAGTCAGTTCGCCTCCCCATTGAGGCTGACCTGCCCAGACGGGCCTGGGCCCACCCCACACACCGGGGCGGAATGTGTGCAGGCCCCAGTCTCTGTGGGTGTTCCGCTAGCTGGGGCCCCCAGTGCTCACCCCACACCTAAAGCGAGCCCCAGCCTCCAGAGCCCCCTAAGCATTCCCCGCCCAGCAGCCCAGCCCCTGCCCCCACCCAGGAGGCCCCAGAGCTCAGGGCGCCTGGTCGGATTTTGTACAGCCCCGAGTCACTGTGCATGCCGATTTCGGCGGGCATGGCACCATTTGGAAGCTCTTCCTCCGGTTCCAGCAGAACCTGCTACCACAGTGAGAAAAACTGTGTCAAAAACCGACTCCTGGCAGCAGTCGGAGGCCCCGCCAGAGAGGGGAGCAGCCGGCCTGAACCCATGTCCTGCCGGTTCCCATGACCCCCAGCACCCAGAGCCCCACGGTGTCCCCGTTGGATAATGAGGACAAGGGCTGGGGGCTCCGGTGGTTTGCGGCAGGGACTTGATCACATCCTTCTGCTGTGGCCCCATTGCCTCTGGCTGGAGTTGACCCTTCTGACAAGTGTCCTCAGAAAGACAGGGATCACCGGCACCTCCCAATATCAACCCCAGGCAGCACAGACACAAACCCCACATCCAGAGCCAACTCCAGGAGCAGAGACACCCCAACACTCTGGGGGACCCCAACCGTGATAACTCCCCACTGGAATCCGCCCCAGAGTCTACCAGGACCAAAGGCCCTGCCCTGTCTCTGTCCCTCACTCAGGGCCTCCTGCAGGGCGAGCGCTTGGGAGCAGACTCGGTCTTAGGGGACACCACTGTGGGCCCCAACTTTGATGAGGCCACTGACCCTTCCTTCCTTTCCTGGGGCAGCACAGACTTTGGGGTCTGGGCAGGGAAGAACTACTGGCTGGTGGCCAATCACAGAGCCCCCAGGCCGAGGTGGCCCCAAGAAGGCCCTCAGGAGGTGGCCACTCCACTTCCTCCCAGCTGGACCCCAGGTCCTCCCCAAGATAGGGGTGCCATCCAAGGCAGGTCCTCCATGGAGCCCCCTTCAGACTCCTCCCGGGACCCCACTGGACCTCAGTCCCTGCTCTGGGAATGCAGCCACCACAAGCACACCAGGAAGCCCAGGCCCAGCCACCCTGCAGTGGGCAAGCCCACACTCTGGAGCAGAGCAGGGTGCGTCTGGGAGGGGCTAACCTCCCCACCCCCCACCCCCCATCTGCACACAGCCACCTACCACTGCCCAGACCCTCTGCAGGAGGGCCAAGCCACCATGGGGTATGGACTTAGGGTCTCACTCACGTGCCTCCCCTCCTGGGAGAAGGGGCCTCATGCCCAGATCCCTGCAGCACTAGACACAGCTGGAGGCAGTGGCCCCAGGGCCACCCTGACCTGGCATCTAAGGCTGCTCCAGCCCAGACAGCACTGCCGTTCCTGGGAAGCCTGGGCTCCACCAGACCACAGGTCCAGGGCACAGCCCACAGGAGCCACCCACACACAGCTCACAGGAAGAAGATAAGCTCCAGACCCCAGGGCGGGACCTGCCTTCCTGCCACCACTTACACACAGGCCAGGGAGCTGTTCCCACACAGATCAACCCCAAACCGGGACTGCCTGGCACTAGGGTCACTGCCATTTCCCTCTCCATTCCCTCCCAGTGCCTCTGTGCTCCCTCCTTCTGGGGAACACCCTGTGCAGCCCCTCCCTGCAGCCCACACGCTGGGGAGACCCCACCCTGCCTCGGGCCTTTTCTACCTGCTGCACTTGCCGCCCACCCAAACAACCCTGGGTACGTGACCCTGCAGTCCTCACCCTGATCTGCAACCAGACCCCTGTCCCTCCCTCTAAACACCCCTCCCAGGCCAACTCTGCACCTGCAGGCCCTCCGCTCTTCTGCCACAAGAGCCTCAGGTTTTCCTACCTGTGCCCACCCCCTAACCCCTCCTGCCCACAACTTGAGTTCTTCCTCTCCTGGAGCCCTTGAGCCATGGCACTGACCCTACACTCCCACCCACACACTGCCCATGCCATCACCTTCCTCCTGGACACTCTGACCCCGCTCCCCTCCCTCTCAGACCCGGCCCTGGTATTTCCAGGACAAAGGCTCACCCAAGTCTTCCCCATGCAGGCCCTTGCCCTCACTGCCTGGTTACACGGGAGCCTCCTGTGCGCAGAAGCAGGGAGCTCAGCTCTTCCACAGGCAGAAGGCACTGAAAGAAATCAGCCTCCAGTGCCTTGACACACGTCCGCCTGTGTCTCTCACTGCCTGCACCTGCAGGGAGGCTCCGCACTCCCTCTAAAGATGAGGGATCCAGGCAGCAACATCACGGGAGAATGCAGGGCTCCCAGACAGCCCAGCCCTCTCGCAGGCCTCTCCTGGGAAGAGACCTGCAGCCACCACTGAACAGCCACGGAGGTCGCTGGATAGTAACCGAGTCAGTGACCGACCTGGAGGGCAGGGGAGCAGTGAACCGGAGCCCATACCATAGGGACAGAGACCAGCCGCTAACATCCCGAGCCCCTCACTGGCGGCCCCAGAACACCCCGTGGAAAGAGAACAGACCCACAGTCCCACCTGGAACAGGGCAGACACTGCTGAGCCCCCAGCACCAGCCCCAAGAAACACTAGGCAACAGCATCAGAGGGGGCTCCTGAGAAAGAGAGGAGGGGAGGTCTCCTTCACCATCAAATGCTTCCCTTGACCAAAAACAGGGTCCACGCAACTCCCCCAGGACAAAGGAGGAGCCCCCTGTACAGCACTGGGCTCAGAGTCCTCTCTGAGACAGGCTCAGTTTCAGACAACAACCCGCTGGAATGCACAGTCTCAGCAGGAGAGCCAGGCCAGAGCCAGCAAGAGGAGACTCGGTGACACCAGTCTCCTGTAGGGACAGGAGGATTTTGTGGGGGTTCGTGTCACTGTGCAAACCCATGAAAACCCCAAGGGAGTTTGGAACAGCAAGCACAGTGACACAACCCCATTCAAAAACCCCTACTGCAAACGCACCCACTCCTGGGACTGAGGGGCTGGGGGAGCGTCTGGGAAGTATGGCCTAGGGGTGTCCATCAATGCCCAAAATGCACCAGACTCTCCCCAAGACATCACCCCACCAGCCAGTGAGCAGAGTAAACAGAAAATGAGAAGCAGCTGGGAAGCTTGCACAGGCCCCAAGGAAAGAGCTTTGGCAGGTGTGCAAGAGGGGATGTGGGCAGAGCCTCAGCAGGGCCTTTTGCTGTTTCTGCTTTCCTGTGCAGAGAGTTCCATAAACTGGTATTCAAGATCAATGGCTGGGAGTGAGCCCAGGAGGACAGTGTGGGAAGAGCACAGGGAAGGAGGAGCAGCCGCTATCCTACACTGTCATCTTTTGAAAGTTTGCCCTGTGCCCACAATGCTGCATCATGGGATGCTTAACAGCTGATGTAGACACAGCTAAAGAGAGAATCAGTGAAATGGATTTGCAGCACAGATCTGAATAAATCCTCCAGAATGTGGAGCAGCACAGAAGCAAGCACACAGAAAGTGCCTGATGCCAAGGCAAAGTTCAGTGGGCACCTTCAGGCATTGCTGCTGGGCACAGACACTCTGAAAAGCACTGGCAGGAACTGCCTGTGACAAAGCAGAACCCTCAGGCAATGCCAGCCCTAGAGCCCTTCCTGAGAACCTCATGGGCAAAGATGTGCAGAACAGCTGTTTGTCATAGCCCCAAACTATGGGGCTGGACAAAGCAAACGTCCATCTGAAGGAGAACAGACAAATAAACGATGGCAGGTTCATGAAATGCAAACTAGGACAGCCAGAGGACAACAGTAGAGAGCTACAGGCGGCTTTGCGGTTGAGTTCATGACAATGCTGAGTAATTGGAGTAACAGAGGAAAGCCCAAAAAATACTTTTAATGTGATTTCTTCTAAATAAAATTTACACCCGGCAAAATGAACTATCTTCTTAAGGGATAAACTTTCCCCTGGAAAAACTATAAGGAAAATCAAGAAAACGATGATCACATAAACACAGTGGTGGTTACTTCTACTGGGGAAGGAAGAGGGTATGAGCTGAGACACACAGAGTCGGCAAGTCTCCTAACAAGAACAGAACAAATACATTACAGTACCTTGAAAACAGCAGTTAAACTTCTAAATCGCAAGAAGAGGAAAATGCACACACCTGTGTTTAGAAAATTCTCAGTCCAGCACTGTTCATAATAGCAAAGACATTAACCCAGGTTGGATAAATAAGCGATGACACAGGCAATTGCACAATGATACAGACATACATTCAGTATATGAGACATCGATGATGTATCCCCAAAGAAATGACTTTAAAGAGAAAAGGCCTGATGTGTGGTGGCAATCACCTCCCTGGGCATCCCCGGACAGGCTGCAGGCTCACTGTGTGGCAGGGCAGGCAGGCACCTGCTGGCAGCTCCTGGGGCCTGATGTGGAGCAGGCACAGAGCTGTATATCCCCAAGGAAGGTACAGTCAGTGCATTCCAGAGAGAAGCAACTCAGCCACACTCCCTGGCCAGAACCCAAGATGCACACCCATGCACAGGGAGGCAGAGCCCAGCACCTCCGCAGCCACCACCACCTGCGCACGGGCCACCACCTTGCAGGCACAGAGTGGGTGCTGAGAGGAGGGGCAGGGACACCAGGCAGGGTGAGCACCCAGAGAAAACTGCAGAAGCCTCACACATCCCTCACCTGGCCTGGGCTTCACCTGACCTGGACCTCACCTGGCCTCGGGCCTCACCTGCACCTGCTCCAGGTCTTGCTGGAGCCTGAGTAGCACTGAGGCTGTAGGGACTCATCCAGGGTTGGGGAATGACTCTGCAACTCTCCCACATCTGACCTTTCTGGGTGGAGGCACCTGGTGGCCCAGGGAATATAAAAAGCCCCAGAATGATGCCTGTGTGATTTGGGGGCAATTTATGAACCCGAAAGGACATGGCCATGGGGTGGGTAGGGACAGTAGGGACAGATGTCAGCCTGAGGTGAAGCCTCAGGACACAGGTGGGCATGGACAGTGTCCACCTAAGCGAGGGACAGACCCGAGTGTCCCTGCAGTAGACCTGAGAGCGCTGGGCCCACAGCCTCCCCTCGGGGCCCTGCTGCCTCCTCAGGTCAGCCCTGGACATCCCGGGTTTCCCCAGGCCTGGCGGTAGGATTTTGTTGAGGTCTGTGTCACTGTGCATGGCAGCTACTGCCAGCCCGCAGCCACTGGCTACTGAGGATGCCGATTCTGAGAATAGCAGCTTCTACTACTATGACTACCTGGATGAAGTAGCTTTCATGCTCAGCGAGGTTAGCCAGCATCTAGACTATGCCCACAGTGTCACAGAGTCCATCAAAAACCCATGCCTGGGAGCCTCCCACCACAGCCCTCCCTGCGGGGGACCGCTGCATGCCGTGTTAGGATTTTGATCGAGGACACGGCGCCATGGGTATGGTGGCTACCACAGCAGTGCAGCCCATGACCCAAACACACGGGGCAGCAGAAACAATGGACAGGCCCACAAGTGACCATGATGGGCTCCAGCCCACCAGCCCCAGAGACCATGAAACAGATGGCCAAGGTCACCCTACAGGTCATCCAGATCTGGCTCCAAGGGGTCTGCATCGCTGCTGCCCTCCCAACGCCAAACCAGATGGAGACAGGGCCGGCCCCATAGCACCATCTGCTGCCGTCCACCCAGCAGTCCCGGAAGCCCCTCCCTGAACGCTGGGCCACGTGTGTGAACCCTGCGAGCCCCCCATGTCAGAGTAGGGGCAGCAGGAGGGCGGGGCTGGCCCTGTGCACTGTCACTGCCCCTGTGGTCCCTGGCCTGCCTGGCCCTGACACCTGAGCCTCTCCTGGGTCATTTCCAAGACATTCCCAGGGACAGCCGGAGCTGGGAGTCGCTCATCCTGCCTGGCTGTCCTGAGTCCTGCTCATTTCCAGACCTCACCAGGGAAGCCAACAGAGGACTCACCTCACACAGTCAGAGACAACGAACCTTCCAGAAATCCCTGTTTCTCTCCCCAGTGAGAGAAACCCTCTTCCAGGGTTTCTCTTCTCTCCCACCCTCTTCCAGGACAGTCCTCAGCAGCATCACAGCGGGAACGCACATCTGGATCAGGACGGCCCCCAGAACACGCGATGGCCCATGGGGACAGCCCAGCCCTTCCCAGACCCCTAAAAGGTATCCCCACCTTGCACCTGCCCCAGGGCTCAAACTCCAGGAGGCCTGACTCCTGCACACCCTCCTGCCAGATATCACCTCAGCCCCCTCCTGGAGGGGACAGGAGCCCGGGAGGGTGAGTCAGACCCACCTGCCCTCAATGGCAGGCGGGGAAGATTCAGAAAGGCCTGAGATCCCCAGGACGCAGCACCACTGTCAATGGGGGCCCCAGACGCCTGGACCAGGGCCTGTGTGGGAAAGGCCTCTGGCCACACTCAGGGGGATTTTGTGAAGGGCCCTCCCACTGTGGAGGTTAGCCAGCATCTAGACTATGCCCACAGTGATGAAACCAGCATCAAAAACCGACCGGACTCGCAGGGTTTATGCACACTTCTCGGCTCGGAGCTCTCCAGGAGCACAAGAGCCAGGCCCGAGGGTTTGTGCCCAGACCCTCGGCCTCTAGGGACACCCGGGCCATCTTAGCCGATGGGCTGATGCCCTGCACACCGTGTGCTGCCAAACAGGGGCTTCAGAGGGCTCTGAGGTGACTTCACTCATGACCACAGGTGCCCTGGTCCCTTCACTGCCAGCTGCACCAGACCCTGTTCCGAGAGATGCCCCAGTTCCAAAAGCCAATTCCTGGGGCCGGGAATTACTGTAGACACCAGCCTCATTCCAGTACCTCCTGCCAATTGCCTGGATTCCCATCCTGGCTGGAATCAAGAGGGCAGCATCCGCCAGGCTCCCAACAGGCAGGACTCCCACACACCCTCCTCTGAGAGGCCGCTGTGTTCCGCAGGGCCAGGCCGCAGACAGTTCCCCTCACCTGCCCATGTAGAAACACCTGCCATTGTCGTCCCCACCTGGCAAAGACCACTTGTGGAGCCCCCAGCCCCAGGTACAGCTGTAGAGAGAGTCCTCGAGGCCCCTAAGAAGGAGCCATGCCCAGTTCTGCCGGGACCCTCGGCCAGGCCGACAGGAGTGGACGCTGGAGCTGGGCCCACACTGGGCCACATAGGAGCTCACCAGTGAGGGCAGGAGAGCACATGCCGGGGAGCACCCAGCCTCCTGCTGACCAGAGACCCGTCCCAGAGCCCAGGAGGCTGCAGAGGCCTCTCCAGGGGGACACAGTGCATGTCTGGTCCCTGAGCAGCCCCCAGGCTCTCTAGCACTGGGGGCCCCTGGCACAGCTGTCTGGACCCTCCCTGTTCCCTGGGAAGCTCCTCCTGACAGCCCCGCCTCCAGTTCCAGGTGTGGATTTTGTCAGGGGGTGCCACACTGTGCTGCATACCCTGCTGGACCTGCAAGTATTCGGCAACAGTCGGAAGGATGCTGTGGTTAGCTTTGGCAAAGTTTTCCTGCCACCACAGTGGTGCCGCCCATATCAAAAACCAGGCCAAGTAGACAGACCCCTGCCACGCAGCCCCAGGCCTCCAGCTCACCTGCTTCTCCTGGGGCTCTCAAGGCTGCTGTCTGCCCTCTGGCCCTCTGTGGGGAGGGTTCCCTCAGTGGGAGGTCTGTGCTCCAGGGCAGGGATGACTGAGATAGAAATCAAAGGCTGGCAGGGAAAGGCAGCTTCCCGCCCTGAGAGGTGCAGGCAGCACCACAGAGCCATGGAGTCACAGAGCCACGGAGCCCCCAGTGTGGGCGTGTGAGGGTGCTGGGCTCCCGGCAGGCCCAGCCCTGATGGGGAAGCCTGCCCCGTCCCACAGCCCAGGTCCCCAGGGGCAGCAGGCACAGAAGCTGCCAAGCTGTGCTCTACGATCCTCATCCCTCCAGCAGCATCCACTCCACAGTGGGGAAACTGAGCCTTGGAGAACCACCCAGCCCCCTGGAAACAAGGCGGGGAGCCCAGACAGTGGGCCCAGAGCACTGTGTGTATCCTGGCACTAGGTGCAGGGACCACCCGGAGATCCCCATCACTGAGTGGCCAGCCTGCAGAAGGACCCAACCCCAACCAGGCCGCTTGATTAAGCTCCATCCCCCTGTCCTGGGAACCTCTTCCCAGCGCCACCAACAGCTCGGCTTCCCAGGCCCTCATCCCTCCAAGGAAGGCCAAAGGCTGGGCCTGCCAGGGGCACAGTACCCTCCCTTGCCCTGGCTAAGACAGGGTGGGCAGACGGCTGCAGATAGGACATATTGCTGGGGCATCTTGCTCTGTGACTACTGGGTACTGGCTCTCAACGCAGACCCTACCAAAATCCCCACTGCCTCCCCTGCTAGGGGCTGGCCTGGTCTCCTCCTGCTGTCCTAGGAGGCTGCTGACCTCCAGGATGGCTTCTGTCCCCAGTTCTAGGGCCAGAGCAGATCCCAGGCAGGCTGTAGGCTGGGAGGCCACCCCTGTCCTTGCCGAGGTTCAGTGCAGGCACCCAGGACAGGAAATGGCCTGAACACAGGGATGACTGTGCCATGCCCTACCTAAGTCCGCCCCTTTCTACTCTGCAACCCCCACTCCCCAGGTCAGCCCATGACGACCAACAACCCAACACCAGAGTCACTGCCTGGCCCTGCCCTGGGGAGGACCCCTCAGCCCCCACCCTGTCTAGAGGAGTTGGGGGGACAGGACACAGGCTCTCTCCTTATGGTTCCCCCACCTGGCTCCTGCCGGGACCCTTGGGGTGTGGACAGAAAGGACGCCTGCCTAATTGGCCCCCAGGAACCCAGAACTTCTCTCCAGGGACCCCAGCCCGAGCACCCCCTTACCCAGGACCCAGCCCTGCCCCTCCTCCCCTCTGCTCTCCTCTCATCACTCCATGGGAATCCAGAATCCCCAGGAAGCCATCAGGAAGGGCTGAAGGAGGAAGCGGGGCCGCTGCACCACCGGGCAGGAGGCTCCGTCTTCGTGAACCCAGGGAAGTGCCAGCCTCCTAGAGGGTATGGTCCACCCTGCCTGGGGCTCCCACCGTGGCAGGCTGCGGGGAAGGACCAGGGACGGTGTGGGGGAGGGCTCAGGGCCCTGCAGGTGCTCCATCTTGGATGAGCCCATCCCTCTCACCCACCGACCCGCCCACCTCCTCTCCACCCTGGCCACACGTCGTCCACACCATCCTGAGTCCCACCTACACCAGAGCCAGCAGAGCCAGTGCAGACAGAGGCTGGGGTGCAGGGGGGCCGCCAGGGCAGCTTTGGGGAGGGAGGAATGGAGGAAGGGGAGGTCAGTGAAGAGGCCCCCCTCCCCTGGGTCTAGGATCCACCTTTGGGACCCCCGGATCCCATCCCCTCCAGGCTCTGGGAGGAGAAGCAGGATGGGAGATTCTGTGCAGGACCCTCTCACAGTGGAATACCTCCACAGCGGCTCAGGCCAGATACAAAAGCCCCTCAGTGAGCCCTCCACTGCAGTGCAGGGCCTGGGGGCAGCCCCTCCCACAGAGGACAGACCCAGCACCCCGAAGAAGTCCTGCCAGGGGGAGCTCAGAGCCATGAAGGAGCAAGATATGGGGACCCCAATACTGGCACAGACCTCAGCTCCATCCAGGCCCACCAGGACCCACCATGGGTGGAACACCTGTCTCCGGCCCCTGCTGGCTGTGAGGCAGCTGGCCTCTGTCTCGGACCCCCATTCCAGACACCAGACAGAGGGACAGGCCCCCCAGAACCAGTGTTGAGGGACACCCCTGTCCAGGGCAGCCAAGTCCAAGAGGCGCGCTGAGCCCAGCAAGGGAAGGCCCCCAAACAAACCAGGAGGTTTCTGAAGCTGTCTGTGTCACAGTCGGGCATAGCCACGGCTACCACAATGACACTGGGCAGGACAGAAACCCCATCCCAAGTCAGCCGAAGGCAGAGAGAGCAGGCAGGACACATTTAGGATCTGAGGCCACACCTGACACTCAAGCCAACAGATGTCTCCCCTCCAGGGCGCCCTGCCCTGTTCAGTGTTCCTGAGAAAACAGGGGCAGCCTGAGGGGATCCAGGGCCAGGAGATGGGTCCCCTCTACCCCGAGGAGGAGCCAGGCGGGAATCCCAGCCCCCTCCCCATTGAGGCCATCCTGCCCAGAGGGGCCCGGACCCACCCCACACACCCAGGCAGAATGTGTGCAGGCCTCAGGCTCTGTGGGTGCCGCTAGCTGGGGCTGCCAGTCCTCACCCCACACCTAAGGTGAGCCACAGCCGCCAGAGCCTCCACAGGAGACCCCACCCAGCAGCCCAGCCCCTACCCAGGAGGCCCCAGAGCTCAGGGCGCCTGGGTGGATTTTGTACAGCCCCGAGTCACTGTGCTGCATACCCTGCTGGACCTGCAAGTATTCGGCAACCACAGTGAGAAAAGCTATGTCAAAAACCGTCTCCCGGCCACTGCTGGAGGCCCAGCCAGAGAAGGGACCAGCCGCCCGAACATACGACCTTCCCAGACCTCATGACCCCCAGCACTTGGAGCTCCACAGTGTCCCCATTGGATGGTGAGGATGGGGGCCGGGGCCATCTGCACCTCCCAACATCACCCCCAGGCAGCACAGGCACAAACCCCAAATCCAGAGCCGACACCAGGAACACAGACACCCCAATACCCTGGGGGACCCTGGCCCTGGTGACTTCCCACTGGGATCCACCCCCGTGTCCACCTGGATCAAAGACCCCACCGCTGTCTCTGTCCCTCACTCAGGGCCTGCTGAGGGGCGGGTGCTTTGGAGCAGACTCAGGTTTAGGGGCCACCATTGTGGGGCCCAACCTCGACCAGGACACAGATTTTTCTTTCCTGCCCTGGGGCAACACAGACTTTGGGGTCTGTGCAGGGAGGACCTTCTGGAAAGTCACCAAGCACAGAGCCCTGACTGAGGTGGTCTCAGGAAGACCCCCAGGAGGGGGCTTGTGCCCCTTCCTCTCATGTGGACCCCATGCCCCCCAAGATAGGGGCATCATGCAGGGCAGGTCCTCCATGCAGCCACCACTAGGCAACTCCCTGGCGCCGGTCCCCACTGCGCCTCCATCCCGGCTCTGGGGATGCAGCCACCATGGCCACACCAGGCAGCCCGGGTCCAGCAACCCTGCAGTGCCCAAGCCCTTGGCAGGATTCCCAGAGGCTGGAGCCCACCCCTCCTCATCCCCCCACACCTGCACACACACACCTACCCCCTGCCCAGTCCCCCTCCAGGAGGGTTGGAGCCGCCCATAGGGTGGGGGCTCCAGGTCTCACTCACTCGCTTCCCTTCCTGGGCAAAGGAGCCTCGTGCCCCGGTCCCCCCTGACGGCGCTGGGCACAGGTGTGGGTACTGGGCCCCAGGGCTCCTCCAGCCCCAGCTGCCCTGCTCTCCCTGGGAGGCCTGGGCACCACCAGACCACCAGTCCAGGGCACAGCCCCAGGGAGCCGCCCACTGCCAGCTCACAGGAAGAAGATAAGCTTCAGACCCTCAGGGCCGGGAGCTGCCTTCCTGCCACCCCTTCCTGCCCCAGACCTCCATGCCCTCCCCCAACCACTTACACACAAGCCAGGGAGCTGTTTCCACACAGTTCAACCCCAAACCAGGACGGCCTGGCACTCGGGTCACTGCCATTTCTGTCTGCATTCGCTCCCAGCGCCCCTGTGTTCCCTCCCTCCTCCCTCCTTCCTTTCTTCCTGCATTGGGTTCATGCCGCAGAGTGCCAGGTGCAGGTCAGCCCTGAGCTTGGGGTCACCTCCTCACTGAAGGCAGCCTCAGGGTGCCCAGGGGCAGGCAGGGTGGGGGTGAGGCTTCCAGCTCCAACCGCTTCGCTACCTTAGGACCGTTATAGTTAGGCGCGCCGTCGACCAATTCTCATGTTTGACAGCTTATCATCGAATTTCTACGTA
Кодирующие последовательности токсина
[00159] Иллюстративные нуклеотидные кодирующие последовательности (ДНК и аминокислотная последовательность (AA)) токсинов (например, μ-конотоксина и токсина тарантула ProTxII) для создания сконструированного участка DH, описываемого в данном документе, изложены в таблице 4.
[00160] Иллюстративные фрагменты ДНК, содержащие нуклеотидные кодирующие последовательности токсина для создания сконструированного участка DH, приведены ниже.
[00161] TX-DH1166 (SEQ ID NO:232) включает кодирующие последовательности токсина, которые встроены в положениях, соответствующих DH1–1 — DH6–6:
TACGTAGCCGTTTCGATCCTCCCGAATTGACTAGTGGGTAGGCCTGGCGGCCGCTGCCATTTCATTACCTCTTTCTCCGCACCCGACATAGATACCGGTGGATTCGAATTCTCCCCGTTGAAGCTGACCTGCCCAGAGGGGCCTGGGCCCACCCCACACACCGGGGCGGAATGTGTACAGGCCCCGGTCTCTGTGGGTGTTCCGCTAACTGGGGCTCCCAGTGCTCACCCCACAACTAAAGCGAGCCCCAGCCTCCAGAGCCCCCGAAGGAGATGCCGCCCACAAGCCCAGCCCCCATCCAGGAGGCCCCAGAGCTCAGGGCGCCGGGGCGGATTTTGTACAGCCCCGAGTCACTGTGGAGAGAAGCTGCAATGGCAGACGCGGCTGCAGCAGCAGATGGAGCCGCGATCATAGCAGGTGCTGCCACAGTGAGAAAAACTGTGTCAAAAACCGTCTCCTGGCCCCTGCTGGAGGCCGCGCCAGAGAGGGGAGCAGCCGCCCCGAACCTAGGTCCTGCTCAGCTCACACGACCCCCAGCACCCAGAGCACAACGGAGTCCCCATTGAATGGTGAGGACGGGGACCAGGGCTCCAGGGGGTCATGGAAGGGGCTGGACCCCATCCTACTGCTATGGTCCCAGTGCTCCTGGCCAGAACTGACCCTACCACCGACAAGAGTCCCTCAGGGAAACGGGGGTCACTGGCACCTCCCAGCATCAACCCCAGGCAGCACAGGCATAAACCCCACATCCAGAGCCGACTCCAGGAGCAGAGACACCCCAGTACCCTGGGGGACACCGACCCTGATGACTCCCCACTGGAATCCACCCCAGAGTCCACCAGGACCAAAGACCCCGCCCCTGTCTCTGTCCCTCACTCAGGACCTGCTGCGGGGCGGGCCATGAGACCAGACTCGGGCTTAGGGAACACCACTGTGGCCCCAACCTCGACCAGGCCACAGGCCCTTCCTTCCTGCCCTGCGGCAGCACAGACTTTGGGGTCTGTGCAGAGAGGAATCACAGAGGCCCCAGGCTGAGGTGGTGGGGGTGGAAGACCCCCAGGAGGTGGCCCACTTCCCTTCCTCCCAGCTGGAACCCACCATGACCTTCTTAAGATAGGGGTGTCATCCGAGGCAGGTCCTCCATGGAGCTCCCTTCAGGCTCCTCCCCGGTCCTCACTAGGCCTCAGTCCCGGCTGCGGGAATGCAGCCACCACAGGCACACCAGGCAGCCCAGACCCAGCCAGCCTGCAGTGCCCAAGCCCACATTCTGGAGCAGAGCAGGCTGTGTCTGGGAGAGTCTGGGCTCCCCACCGCCCCCCCGCACACCCCACCCACCCCTGTCCAGGCCCTATGCAGGAGGGTCAGAGCCCCCCATGGGGTATGGACTTAGGGTCTCACTCACGTGGCTCCCCTCCTGGGTGAAGGGGTCTCATGCCCAGATCCCCACAGCAGAGCTGGTCAAAGGTGGAGGCAGTGGCCCCAGGGCCACCCTGACCTGGACCCTCAGGCTCCTCTAGCCCTGGCTGCCCTGCTGTCCCTGGGAGGCCTGGACTCCACCAGACCACAGGTCCAGGGCACCGCCCATAGGTGCTGCCCACACTCAGTTCACAGGAAGAAGATAAGCTCCAGACCCCCAAGACTGGGACCTGCCTTCCTGCCACCGCTTGTAGCTCCAGACCTCCGTGCCTCCCCCGACCACTTACACACGGGCCAGGGAGCTGTTCCACAAAGATCAACCCCAAACCGGGACCGCCTGGCACTCGGGCCGCTGCCACTTCCCTCTCCATTTGTTCCCAGCACCTCTGTGCTCCCTCCCTCCTCCCTCCTTCAGGGGAACAGCCTGTGCAGCCCCTCCCTGCACCCCACACCCTGGGGAGGCCCAACCCTGCCTCCAGCCCTTTCTCCCCCGCTGCTCTTCCTGCCCATCCAGACAACCCTGGGGTCCCATCCCTGCAGCCTACACCCTGGTCTCCACCCAGACCCCTGTCTCTCCCTCCAGACACCCCTCCCAGGCCAACCCTGCACATGCAGGCCCTCCCCTTTTCTGCTGCCAGAGCCTCAGTTTCTACCCTCTGTGCCTACCCCCTGCCTCCTCCTGCCCACAACTCGAGCTCTTCCTCTCCTGGGGCCCCTGAGCCATGGCACTGACCGTGCACTCCCACCCCCACACTGCCCATGCCCTCACCTTCCTCCTGGACACTCTGACCCCGCTCCCCTCTTGGACCCAGCCCTGGTATTTCCAGGACAAAGGCTCACCCAAGTCTTCCCCATGCAGGCCCTTGCCCTCACTGCCCGGTTACACGGCAGCCTCCTGTGCACAGAAGCAGGGAGCTCAGCCCTTCCACAGGCAGAAGGCACTGAAAGAAATCGGCCTCCAGCACCCTGATGCACGTCCGCCTGTGTCTCTCACTGCCCGCACCTGCAGGGAGGCTCGGCACTCCCTGTAAAGACGAGGGATCCAGGCAGCAACATCATGGGAGAATGCAGGGCTCCCAGACAGCCCAGCCCTCTCGCAGGCCTCTCCTGGGAAGAGACCTGCAGCCACCACTGAACAGCCACGGAGCCCGCTGGATAGTAACTGAGTCAGTGACCGACCTGGAGGGCAGGGGAGCAGTGAACCGGAGCCCAGACCATAGGGACAGAGACCAGCCGCTGACATCCCGAGCCCCTCACTGGCGGCCCCAGAACACCGCGTGGAAACAGAACAGACCCACATTCCCACCTGGAACAGGGCAGACACTGCTGAGCCCCCAGCACCAGCCCTGAGAAACACCAGGCAACGGCATCAGAGGGGGCTCCTGAGAAAGAAAGGAGGGGAGGTCTCCTTCACCAGCAAGTACTTCCCTTGACCAAAAACAGGGTCCACGCAACTCCCCCAGGACAAAGGAGGAGCCCCCTGTACAGCACTGGGCTCAGAGTCCTCTCCCACACACCCTGAGTTTCAGACAAAAACCCCCTGGAAATCATAGTATCAGCAGGAGAACTAGCCAGAGACAGCAAGAGGGGACTCAGTGACTCCCGCGGGGACAGGAGGATTTTGTGGGGGCTCGTGTCACTGTGAGGATATTGTAGCAGCAGATGGTGCTATACCCACAGTGACACAGCCCCATTCAAAAACCCCTGCTGTAAACGCTTCCACTTCTGGAGCTGAGGGGCTGGGGGGAGCGTCTGGGAAGTAGGGCCTAGGGGTGGCCATCAATGCCCAAAACGCACCAGACTCCCCCCCAGACATCACCCCACTGGCCAGTGAGCAGAGTAAACAGAAAATGAGAAGCAGCTGGGAAGCTTGCACAGGCCCCAAGGAAAGAGCTTTGGCGGGTGTGCAAGAGGGGATGCGGGCAGAGCCTGAGCAGGGCCTTTTGCTGTTTCTGCTTTCCTGTGCAGATAGTTCCATAAACTGGTGTTCAAGATCGATGGCTGGGAGTGAGCCCAGGAGGACAGTGTGGGAAGGGCACAGGGAAGGAGAAGCAGCCGCTATCCTACACTGTCATCTTTCAAGAGTTTGCCCTGTGCCCACAATGCTGCATCATGGGATGCTTAACAGCTGATGTAGACACAGCTAAAGAGAGAATCAGTGAAATGGATTTGCAGCACAGATCTGAATAAATTCTCCAGAATGTGGAGCCACACAGAAGCAAGCACAAGGAAAGTGCCTGATGCAAGGGCAAAGTACAGTGTGTACCTTCAGGCTGGGCACAGACACTCTGAAAAGCCTTGGCAGGAACTCCCTGCAACAAAGCAGAGCCCTGCAGGCAATGCCAGCTCCAGAGCCCTCCCTGAGAGCCTCATGGGCAAAGATGTGCACAACAGGTGTTTCTCATAGCCCCAAACTGAGAATGAAGCAAACAGCCATCTGAAGGAAAACAGGCAAATAAACGATGGCAGGTTCATGAAATGCAAACCCAGACAGCCAGAAGGACAACAGTGAGGGTTACAGGTGACTCTGTGGTTGAGTTCATGACAATGCTGAGTAATTGGAGTAACAAAGGAAAGTCCAAAAAATACTTTCAATGTGATTTCTTCTAAATAAAATTTACAGCCGGCAAAATGAACTATCTTCTTAAGGGATAAACTTTCCACTAGGAAAACTATAAGGAAAATCAAGAAAAGGATGATCACATAAACACAGTGGTCGTTACTTCTACTGGGGAAGGAAGAGGGTATGAACTGAGACACACAGGGTTGGCAAGTCTCCTAACAAGAACAGAACAAATACATTACAGTACCTTGAAAACAGCAGTTAAAATTCTAAATTGCAAGAAGAGGAAAATGCACACAGCTGTGTTTAGAAAATTCTCAGTCCAGCACTGTTCATAATAGCAAAGACATTAACCCAGGTTGGATAAATAAACGATGACACAGGCAATTGCACAATGATACAGACATACATTCAGTATATGAGACATTGATGATGTATCCCCAAAGAAATGACTTTAAAGAGAAAAGGCCTGATATGTGGTGGCACTCACCTCCCTGGGCATCCCCGGACAGGCTGCAGGCACACTGTGTGGCAGGGCAGGCTGGTACCTGCTGGCAGCTCCTGGGGCCTGATGTGGAGCAGGCACAGAGCCGTATCCCCCCGAGGACATATACCCCCAAGGACGGCACAGTTGGTACATTCCGGAGACAAGCAACTCAGCCACACTCCCAGGCCAGAGCCCGAGAGGGACGCCCATGCACAGGGAGGCAGAGCCCAGCTCCTCCACAGCCAGCAGCACCCGTGCAGGGGCCGCCATCTGGCAGGCACAGAGCATGGGCTGGGAGGAGGGGCAGGGACACCAGGCAGGGTTGGCACCAACTGAAAATTACAGAAGTCTCATACATCTACCTCAGCCTTGCCTGACCTGGGCCTCACCTGACCTGGACCTCACCTGGCCTGGACCTCACCTGGCCTAGACCTCACCTCTGGGCTTCACCTGAGCTCGGCCTCACCTGACTTGGACCTTGCCTGTCCTGAGCTCACATGATCTGGGCCTCACCTGACCTGGGTTTCACCTGACCTGGGCTTCACCTGACCTGGGCCTCATCTGACCTGGGCCTCACTGGCCTGGACCTCACCTGGCCTGGGCTTCACCTGGCCTCAGGCCTCATCTGCACCTGCTCCAGGTCTTGCTGGAACCTCAGTAGCACTGAGGCTGCAGGGGCTCATCCAGGGTTGCAGAATGACTCTAGAACCTCCCACATCTCAGCTTTCTGGGTGGAGGCACCTGGTGGCCCAGGGAATATAAAAAGCCTGAATGATGCCTGCGTGATTTGGGGGCAATTTATAAACCCAAAAGGACATGGCCATGCAGCGGGTAGGGACAATACAGACAGATATCAGCCTGAAATGGAGCCTCAGGGCACAGGTGGGCACGGACACTGTCCACCTAAGCCAGGGGCAGACCCGAGTGTCCCCGCAGTAGACCTGAGAGCGCTGGGCCCACAGCCTCCCCTCGGTGCCCTGCTACCTCCTCAGGTCAGCCCTGGACATCCCGGGTTTCCCCAGGCCTGGCGGTAGGATTTTGTTGAGGTCTGTGTCACTGTGGTATTACGATTGCAACTGCAGCAGATGGCGCGACCATAGCAGGTGCTGCTATTATACCCACAGTGTCACAGAGTCCATCAAAAACCCATCCCTGGGAACCTTCTGCCACAGCCCTCCCTGTGGGGCACCGCCGCGTGCCATGTTAGGATTTTGACTGAGGACACAGCACCATGGGTATGGTGGCTACCGCAGCAGTGCAGCCCGTGACCCAAACACACAGGGCAGCAGGCACAACAGACAAGCCCACAAGTGACCACCCTGAGCTCCTGCCTGCCAGCCCTGGAGACCATGAAACAGATGGCCAGGATTATCCCATAGGTCAGCCAGACCTCAGTCCAACAGGTCTGCATCGCTGCTGCCCTCCAATACCAGTCCGGATGGGGACAGGGCTGGCCCACATTACCATTTGCTGCCATCCGGCCAACAGTCCCAGAAGCCCCTCCCTCAAGGCTGGGCCACATGTGTGGACCCTGAGAGCCCCCCATGTCTGAGTAGGGGCACCAGGAAGGTGGGGCTGGCCCTGTGCACTGTCCCTGCCCCTGTGGTCCCTGGCCTGCCTGGCCCTGACACCTGGGCCTCTCCTGGGTCATTTCCAAGACAGAAGACATTCCCAGGACAGCTGGAGCTGGGAGTCCATCATCCTGCCTGGCCGTCCTGAGTCCTGCGCCTTTCCAAACCTCACCCGGGAAGCCAACAGAGGAATCACCTCCCACAGGCAGAGACAAAGACCTTCCAGAAATCTCTGTCTCTCTCCCCAGTGGGCACCCTCTTCCAGGGCAGTCCTCAGTGATATCACAGTGGGAACCCACATCTGGATCGGGACTGCCCCCAGAACACAAGATGGCCCACAGGGACAGCCCCACAGCCCAGCCCTTCCCAGACCCCTAAAAGGCGTCCCACCCCCTGCATCTGCCCCAGGGCTCAAACTCCAGGAGGACTGACTCCTGCACACCCTCCTGCCAGACATCACCTCAGCCCCTCCTGGAAGGGACAGGAGCGCGCAAGGGTGAGTCAGACCCTCCTGCCCTCGATGGCAGGCGGAGAAGATTCAGAAAGGTCTGAGATCCCCAGGACGCAGCACCACTGTCAATGGGGGCCCCAGACGCCTGGACCAGGGCCTGCGTGGGAAAGGCCTCTGGGCACACTCAGGGGGATTTTGTGAAGGGTCCTCCCACTGTGGAGAGGCTTAGCTGTGGCTTCCCTAAGAGCTGCCGCAGCAGGCAAAGCAAGCCTCACAGATGCTGCCACAGTGATGAACCCAGCATCAAAAACCGACCGGACTCCCAAGGTTTATGCACACTTCTCCGCTCAGAGCTCTCCAGGATCAGAAGAGCCGGGCCCAAGGGTTTCTGCCCAGACCCTCGGCCTCTAGGGACATCTTGGCCATGACAGCCCATGGGCTGGTGCCCCACACATCGTCTGCCTTCAAACAAGGGCTTCAGAGGGCTCTGAGGTGACCTCACTGATGACCACAGGTGCCCTGGCCCCTTCCCCACCAGCTGCACCAGACCCCGTCATGACAGATGCCCCGATTCCAACAGCCAATTCCTGGGGCCAGGAATCGCTGTAGACACCAGCCTCCTTCCAACACCTCCTGCCAATTGCCTGGATTCCCATCCCGGTTGGAATCAAGAGGACAGCATCCCCCAGGCTCCCAACAGGCAGGACTCCCACACCCTCCTCTGAGAGGCCGCTGTGTTCCGTAGGGCCAGGCTGCAGACAGTCCCCCTCACCTGCCACTAGACAAATGCCTGCTGTAGATGTCCCCACCTGGAAAATACCACTCATGGAGCCCCCAGCCCCAGGTACAGCTGTAGAGAGAGTCTCTGAGGCCCCTAAGAAGTAGCCATGCCCAGTTCTGCCGGGACCCTCGGCCAGGCTGACAGGAGTGGACGCTGGAGCTGGGCCCATACTGGGCCACATAGGAGCTCACCAGTGAGGGCAGGAGAGCACATGCCGGGGAGCACCCAGCCTCCTGCTGACCAGAGGCCCGTCCCAGAGCCCAGGAGGCTGCAGAGGCCTCTCCAGGGGGACACTGTGCATGTCTGGTCCCTGAGCAGCCCCCCACGTCCCCAGTCCTGGGGGCCCCTGGCACAGCTGTCTGGACCCTCTCTATTCCCTGGGAAGCTCCTCCTGACAGCCCCGCCTCCAGTTCCAGGTGTGGATTTTGTCAGGGGGTGTCACACTGTGTACAGCCAGAAGTGGATGTGGACTTGCGATAGTGAGAGGAAGTGCAGTGAGGGTATGGTATGCCGGCTGTGGTGTAAGAAGAAGCTCTGGCACAGTGGTGCTGCCCATATCAAAAACCAGGCCAAGTAGACAGGCCCCTGCTGTGCAGCCCCAGGCCTCCAGCTCACCTGCTTCTCCTGGGGCTCTCAAGGCTGCTGTTTTCTGCACTCTCCCCTCTGTGGGGAGGGTTCCCTCAGTGGGAGATCTGTTCTCAACATCCCACGGCCTCATTCCTGCAAGGAAGGCCAATGGATGGGCAACCTCACATGCCGCGGCTAAGATAGGGTGGGCAGCCTGGCGGGGACAGGACATCCTGCTGGGGTATCTGTCACTGTGCCTAGTGGGGCACTGGCTCCCAAACAACGCAGTCCTTGCCAAAATCCCCACGGCCTCCCCCGCTAGGGGCTGGCCTGATCTCCTGCAGTCCTAGGAGGCTGCTGACCTCCAGAATGGCTCCGTCCCCAGTTCCAGGGCGAGAGCAGATCCCAGGCCGGCTGCAGACTGGGAGGCCACCCCCTCCTTCCCAGGGTTCACTGCAGGTGACCAGGGCAGGAAATGGCCTGAACACAGGGATAACCGGGCCATCCCCCAACAGAGTCCACCCCCTCCTGCTCTGTACCCCGCACCCCCCAGGCCAGCCCATGACATCCGACAACCCCACACCAGAGTCACTGCCCGGTGCTGCCCTAGGGAGGACCCCTCAGCCCCCACCCTGTCTAGAGGACTGGGGAGGACAGGACACGCCCTCTCCTTATGGTTCCCCCACCTGGCTCTGGCTGGGACCCTTGGGGTGTGGACAGAAAGGACGCTTGCCTGATTGGCCCCCAGGAGCCCAGAACTTCTCTCCAGGGACCCCAGCCCGAGCACCCCCTTACCCAGGACCCAGCCCTGCCCCTCCTCCCCTCTGCTCTCCTCTCATCACCCCATGGGAATCCAGAATCCCCAGGAAGCCATCAGGAAGGGCTGAGGGAGGAAGTGGGGCCACTGCACCACCAGGCAGGAGGCTCTGTCTTTGTGAACCCAGGGAGGTGCCAGCCTCCTAGAGGGTATGGTCCACCCTGCCTATGGCTCCCACAGTGGCAGGCTGCAGGGAAGGACCAGGGACGGTGTGGGGGAGGGCTCAGGGCCCCGCGGGTGCTCCATCTTGGATGAGCCTATCTCTCTCACCCACGGACTCGCCCACCTCCTCTTCACCCTGGCCACACGTCGTCCACACCATCCTAAGTCCCACCTACACCAGAGCCGGCACAGCCAGTGCAGACAGAGGCTGGGGTGCAGGGGGGCCGACTGGGCAGCTTCGGGGAGGGAGGAATGGAGGAAGGGGAGTTCAGTGAAGAGGCCCCCCTCCCCTGGGTCCAGGATCCTCCTCTGGGACCCCCGGATCCCATCCCCTCCAGGCTCTGGGAGGAGAAGCAGGATGGGAGAATCTGTGCGGGACCCTCTCACAGTGGAATACCTCCACAGCGGCTCAGGCCAGATACAAAAGCCCCTCAGTGAGCCCTCCACTGCAGTGCTGGGCCTGGGGGCAGCCGCTCCCACACAGGATGAACCCAGCACCCCGAGGATGTCCTGCCAGGGGGAGCTCAGAGCCATGAAGGAGCAGGATATGGGACCCCCGATACAGGCACAGACCTCAGCTCCATTCAGGACTGCCACGTCCTGCCCTGGGAGGAACCCCTTTCTCTAGTCCCTGCAGGCCAGGAGGCAGCTGACTCCTGACTTGGACGCCTATTCCAGACACCAGACAGAGGGGCAGGCCCCCCAGAACCAGGGATGAGGACGCCCCGTCAAGGCCAGAAAAGACCAAGTTGCGCTGAGCCCAGCAAGGGAAGGTCCCCAAACAAACCAGGAGGATTTTGTAGGTGTCTGTGTCACTGTGAGCTGCAACTGCAGCAGCAAATGGAGCCGCGACCATAGCAGGTGCTGCCACAGTGACACTCGCCAGGTCAAAAACCCCATCCCAAGTCAGCGGAATGCAGAGAGAGCAGGGAGGACATGTTTAGGATCTGAGGCCGCACCTGACACCCAGGCCAGCAGACGTCTCCTGTCCACGGCACCCTGCCATGTCCTGCATTTCTGGAAGAACAAGGGCAGGCTGAAGGGGGTCCAGGACCAGGAGATGGGTCCGCTCTACCCAGAGAAGGAGCCAGGCAGGACACAAGCCCCCACGCGTGGGCTCGTAGTTTGACGTGCGTGAAGTGTGGGTAAGAAAGTACGTA
[00162] TX-DH17613 (SEQ ID NO:233) включает кодирующие последовательности токсина, которые встроены в положениях, соответствующих DH1-7 — DH6-13:
GCGGCCGCTGCCATTTCATTACCTCTTTCTCCGCACCCGACATAGATTACGTAACGCGTGGGCTCGTAGTTTGACGTGCGTGAAGTGTGGGTAAGAAAGTCCCCATTGAGGCTGACCTGCCCAGAGGGTCCTGGGCCCACCCAACACACCGGGGCGGAATGTGTGCAGGCCTCGGTCTCTGTGGGTGTTCCGCTAGCTGGGGCTCACAGTGCTCACCCCACACCTAAAACGAGCCACAGCCTCCGGAGCCCCTGAAGGAGACCCCGCCCACAAGCCCAGCCCCCACCCAGGAGGCCCCAGAGCACAGGGCGCCCCGTCGGATTTTGTACAGCCCCGAGTCACTGTGGAGAGATGCAATGGCAGACGCGGCTGCAGCAGATGGCGCGATCATAGCAGGTGCTGCCACAGTGAGAAAAGCTTCGTCAAAAACCGTCTCCTGGCCACAGTCGGAGGCCCCGCCAGAGAGGGGAGCAGCCACCCCAAACCCATGTTCTGCCGGCTCCCATGACCCCGTGCACCTGGAGCCCCACGGTGTCCCCACTGGATGGGAGGACAAGGGCCGGGGGCTCCGGCGGGTCGGGGCAGGGGCTTGATGGCTTCCTTCTGCCGTGGCCCCATTGCCCCTGGCTGGAGTTGACCCTTCTGACAAGTGTCCTCAGAGAGTCAGGGATCAGTGGCACCTCCCAACATCAACCCCACGCAGCCCAGGCACAAACCCCACATCCAGGGCCAACTCCAGGAACAGAGACACCCCAATACCCTGGGGGACCCCGACCCTGATGACTCCCGTCCCATCTCTGTCCCTCACTTGGGGCCTGCTGCGGGGCGAGCACTTGGGAGCAAACTCAGGCTTAGGGGACACCACTGTGGGCCTGACCTCGAGCAGGCCACAGACCCTTCCCTCCTGCCCTGGTGCAGCACAGACTTTGGGGTCTGGGCAGGGAGGAACTTCTGGCAGGTCACCAAGCACAGAGCCCCCAGGCTGAGGTGGCCCCAGGGGGAACCCCAGCAGGTGGCCCACTACCCTTCCTCCCAGCTGGACCCCATGTCTTCCCCAAGATAGGGGTGCCATCCAAGGCAGGTCCTCCATGGAGCCCCCTTCAGGCTCCTCTCCAGACCCCACTGGGCCTCAGTCCCCACTCTAGGAATGCAGCCACCACGGGCACACCAGGCAGCCCAGGCCCAGCCACCCTGCAGTGCCCAAGCCCACACCCTGGAGGAGAGCAGGGTGCGTCTGGGAGGGGCTGGGCTCCCCACCCCCACCCCCACCTGCACACCCCACCCACCCTTGCCCGGGCCCCCTGCAGGAGGGTCAGAGCCCCCATGGGATATGGACTTAGGGTCTCACTCACGCACCTCCCCTCCTGGGAGAAGGGGTCTCATGCCCAGATCCCCCCAGCAGCGCTGGTCACAGGTAGAGGCAGTGGCCCCAGGGCCACCCTGACCTGGCCCCTCAGGCTCCTCTAGCCCTGGCTGCCCTGCTGTCCCTGGGAGGCCTGGGCTCCACCAGACCACAGGTCTAGGGCACCGCCCACACTGGGGCCGCCCACACACAGCTCACAGGAAGAAGATAAGCTCCAGACCCCCAGGCCCGGGACCTGCCTTGCTGCTACGACTTCCTGCCCCAGACCTCGTTGCCCTCCCCCGTCCACTTACACACAGGCCAGGAAGCTGTTCCCACACAGACCAACCCCAGACGGGGACCACCTGGCACTCAGGTCACTGCCATTTCCTTCTCCATTCACTTCCAATGCCTCTGTGCTTCCTCCCTCCTCCTTCCTTCGGGGGAGCACCCTGTGCAGCTCCTCCCTGCAGTCCACACCCTGGGGAGACCCGACCCTGCAGCCCACACCCTGGGGAGACCTGACCCTCCTCCAGCCCTTTCTCCCCCGCTGCTCTTGCCACCCACCAAGACAGCCCTGGGGTCCTGTCCCTACAGCCCCCACCCAGTTCTCTACCTAGACCCGTCTTCCTCCCTCTAAACACCTCTCCCAGGCCAACCCTACACCTGCAGGCCCTCCCCTCCACTGCCAAAGACCCTCAGTTTCTCCTGCCTGTGCCCACCCCCGTGCTCCTCCTGCCCACAGCTCGAGCTCTTCCTCTCCTAGGGCCCCTGAGGGATGGCATTGACCGTGCCCTCGCACCCACACACTGCCCATGCCCTCACATTCCTCCTGGCCACTCCAGCCCCACTCCCCTCTCAGGCCTGGCTCTGGTATTTCTGGGACAAAGCCTTACCCAAGTCTTTCCCATGCAGGCCTGGGCCCTTACCCTCACTGCCCGGTTACAGGGCAGCCTCCTGTGCACAGAAGCAGGGAGCTCAGCCCTTCCACAGGCAGAAGGCACTGAAAGAAATCGGCCTCCAGCGCCTTGACACACGTCTGCCTGTGTCTCTCACTGCCCGCACCTGCAGGGAGGCTCGGCACTCCCTCTAAAGACGAGGGATCCAGGCAGCAGCATCACAGGAGAATGCAGGGCTACCAGACATCCCAGTCCTCTCACAGGCCTCTCCTGGGAAGAGACCTGAAGACGCCCAGTCAACGGAGTCTAACACCAAACCTCCCTGGAGGCCGATGGGTAGTAACGGAGTCATTGCCAGACCTGGAGGCAGGGGAGCAGTGAGCCCGAGCCCACACCATAGGGCCAGAGGACAGCCACTGACATCCCAAGCCACTCACTGGTGGTCCCACAACACCCCATGGAAAGAGGACAGACCCACAGTCCCACCTGGACCAGGGCAGAGACTGCTGAGACCCAGCACCAGAACCAACCAAGAAACACCAGGCAACAGCATCAGAGGGGGCTCTGGCAGAACAGAGGAGGGGAGGTCTCCTTCACCAGCAGGCGCTTCCCTTGACCGAAGACAGGATCCATGCAACTCCCCCAGGACAAAGGAGGAGCCCCTTGTTCAGCACTGGGCTCAGAGTCCTCTCCAAGACACCCAGAGTTTCAGACAAAAACCCCCTGGAATGCACAGTCTCAGCAGGAGAGCCAGCCAGAGCCAGCAAGATGGGGCTCAGTGACACCCGCAGGGACAGGAGGATTTTGTGGGGGCTCGTGTCACTGTGAGGATATTGTACTAATCGGAGCAGGCAGGGTGTATGCTATACCCACAGTGACACAGCCCCATTCAAAAACCCCTACTGCAAACGCATTCCACTTCTGGGGCTGAGGGGCTGGGGGAGCGTCTGGGAAATAGGGCTCAGGGGTGTCCATCAATGCCCAAAACGCACCAGACTCCCCTCCATACATCACACCCACCAGCCAGCGAGCAGAGTAAACAGAAAATGAGAAGCAAGCTGGGGAAGCTTGCACAGGCCCCAAGGAAAGAGCTTTGGCGGGTGTGTAAGAGGGGATGCGGGCAGAGCCTGAGCAGGGCCTTTTGCTGTTTCTGCTTTCCTGTGCAGAGAGTTCCATAAACTGGTGTTCGAGATCAATGGCTGGGAGTGAGCCCAGGAGGACAGCGTGGGAAGAGCACAGGGAAGGAGGAGCAGCCGCTATCCTACACTGTCATCTTTCGAAAGTTTGCCTTGTGCCCACACTGCTGCATCATGGGATGCTTAACAGCTGATGTAGACACAGCTAAAGAGAGAATCAGTGAGATGGATTTGCAGCACAGATCTGAATAAATTCTCCAGAATGTGGAGCAGCACAGAAGCAAGCACACAGAAAGTGCCTGATGCAAGGACAAAGTTCAGTGGGCACCTTCAGGCATTGCTGCTGGGCACAGACACTCTGAAAAGCCCTGGCAGGAACTCCCTGTGACAAAGCAGAACCCTCAGGCAATGCCAGCCCCAGAGCCCTCCCTGAGAGCCTCATGGGCAAAGATGTGCACAACAGGTGTTTCTCATAGCCCCAAACTGAGAGCAAAGCAAACGTCCATCTGAAGGAGAACAGGCAAATAAACGATGGCAGGTTCATGAAATGCAAACCCAGACAGCCACAAGCACAAAAGTACAGGGTTATAAGCGACTCTGGTTGAGTTCATGACAATGCTGAGTAATTGGAGTAACAAAGTAAACTCCAAAAAATACTTTCAATGTGATTTCTTCTAAATAAAATTTACACCCTGCAAAATGAACTGTCTTCTTAAGGGATACATTTCCCAGTTAGAAAACCATAAAGAAAACCAAGAAAAGGATGATCACATAAACACAGTGGTGGTTACTTCTGCTGGGGAAGGAAGAGGGTATGAACTGAGATACACAGGGTGGGCAAGTCTCCTAACAAGAACAGAACGAATACATTACAGTACCTTGAAAACAGCAGTTAAACTTCTAAATTGCAAGAAGAGGAAAATGCACACAGTTGTGTTTAGAAAATTCTCAGTCCAGCACTGTTCATAATAGCAAAGACATTAACCCAGGTCGGATAAATAAGCGATGACACAGGCAATTGCACAATGATACAGACATATATTTAGTATATGAGACATCGATGATGTATCCCCAAATAAACGACTTTAAAGAGATAAAGGGCTGATGTGTGGTGGCATTCACCTCCCTGGGATCCCCGGACAGGTTGCAGGCTCACTGTGCAGCAGGGCAGGCGGGTACCTGCTGGCAGTTCCTGGGGCCTGATGTGGAGCAAGCGCAGGGCCATATATCCCGGAGGACGGCACAGTCAGTGAATTCCAGAGAGAAGCAACTCAGCCACACTCCCCAGGCAGAGCCCGAGAGGGACGCCCACGCACAGGGAGGCAGAGCCCAGCACCTCCGCAGCCAGCACCACCTGCGCACGGGCCACCACCTTGCAGGCACAGAGTGGGTGCTGAGAGGAGGGGCAGGGACACCAGGCAGGGTGAGCACCCAGAGAAAACTGCAGACGCCTCACACATCCACCTCAGCCTCCCCTGACCTGGACCTCACTGGCCTGGGCCTCACTTAACCTGGGCTTCACCTGACCTTGGCCTCACCTGACTTGGACCTCGCCTGTCCCAAGCTTTACCTGACCTGGGCCTCAACTCACCTGAACGTCTCCTGACCTGGGTTTAACCTGTCCTGGAACTCACCTGGCCTTGGCTTCCCCTGACCTGGACCTCATCTGGCCTGGGCTTCACCTGGCCTGGGCCTCACCTGACCTGGACCTCATCTGGCCTGGACCTCACCTGGCCTGGACTTCACCTGGCCTGGGCTTCACCTGACCTGGACCTCACCTGGCCTCGGGCCTCACCTGCACCTGCTCCAGGTCTTGCTGGAGCCTGAGTAGCACTGAGGGTGCAGAAGCTCATCCAGGGTTGGGGAATGACTCTAGAAGTCTCCCACATCTGACCTTTCTGGGTGGAGGCAGCTGGTGGCCCTGGGAATATAAAAATCTCCAGAATGATGACTCTGTGATTTGTGGGCAACTTATGAACCCGAAAGGACATGGCCATGGGGTGGGTAGGGACATAGGGACAGATGCCAGCCTGAGGTGGAGCCTCAGGACACAGGTGGGCACGGACACTATCCACATAAGCGAGGGATAGACCCGAGTGTCCCCACAGCAGACCTGAGAGCGCTGGGCCCACAGCCTCCCCTCAGAGCCCTGCTGCCTCCTCCGGTCAGCCCTGGACATCCCAGGTTTCCCCAGGCCTGGCGGTAGGATTTTGTTGAGGTCTGTGTCACTGTGGTATTACGATTGCAACTGCAGCAGATGGGCTCGCGACCATAGCAGGTGCTGCTATTATAACCACAGTGTCACAGAGTCCATCAAAAACCCATGCCTGGAAGCTTCCCGCCACAGCCCTCCCCATGGGGCCCTGCTGCCTCCTCAGGTCAGCCCCGGACATCCCGGGTTTCCCCAGGCTGGGCGGTAGGATTTTGTTGAGGTCTGTGTCACTGTGGTATTACTATGAGAGATGCAATGGCAGACGCGGCTGCAGCAGATGGCGCGATCATAGCAGGTGCTGCTATTATAACCACAGTGTCACAGAGTCCATCAAAAACCCATCCCTGGGAGCCTCCCGCCACAGCCCTCCCTGCAGGGGACCGGTACGTGCCATGTTAGGATTTTGATCGAGGAGACAGCACCATGGGTATGGTGGCTACCACAGCAGTGCAGCCTGTGACCCAAACCCGCAGGGCAGCAGGCACGATGGACAGGCCCGTGACTGACCACGCTGGGCTCCAGCCTGCCAGCCCTGGAGATCATGAAACAGATGGCCAAGGTCACCCTACAGGTCATCCAGATCTGGCTCCGAGGGGTCTGCATCGCTGCTGCCCTCCCAACGCCAGTCCAAATGGGACAGGGACGGCCTCACAGCACCATCTGCTGCCATCAGGCCAGCGATCCCAGAAGCCCCTCCCTCAAGGCTGGGCACATGTGTGGACACTGAGAGCCCTCATATCTGAGTAGGGGCACCAGGAGGGAGGGGCTGGCCCTGTGCACTGTCCCTGCCCCTGTGGTCCCTGGCCTGCCTGGCCCTGACACCTGAGCCTCTCCTGGGTCATTTCCAAGACAGAAGACATTCCTGGGGACAGCCGGAGCTGGGCGTCGCTCATCCTGCCCGGCCGTCCTGAGTCCTGCTCATTTCCAGACCTCACCGGGGAAGCCAACAGAGGACTCGCCTCCCACATTCAGAGACAAAGAACCTTCCAGAAATCCCTGCCTCTCTCCCCAGTGGACACCCTCTTCCAGGACAGTCCTCAGTGGCATCACAGCGGCCTGAGATCCCCAGGACGCAGCACCGCTGTCAATAGGGGCCCCAAATGCCTGGACCAGGGCCTGCGTGGGAAAGGCCTCTGGCCACACTCGGGGATTTTGTGAAGGGCCCTCCCACTGTGGAGAGGCTTTGTGGCTTCCCTAAGAGCTGCAGCAGGCAAAAGCCTCACAGATGCTGCCACAGTGATGAACCCAGTGTCAAAAACCGGCTGGAAACCCAGGGGCTGTGTGCACGCCTCAGCTTGGAGCTCTCCAGGAGCACAAGAGCCGGGCCCAAGGATTTGTGCCCAGACCCTCAGCCTCTAGGGACACCTGGGTCATCTCAGCCTGGGCTGGTGCCCTGCACACCATCTTCCTCCAAATAGGGGCTTCAGAGGGCTCTGAGGTGACCTCACTCATGACCACAGGTGACCTGGCCCTTCCCTGCCAGCTATACCAGACCCTGTCTTGACAGATGCCCCGATTCCAACAGCCAATTCCTGGGACCCTGAATAGCTGTAGACACCAGCCTCATTCCAGTACCTCCTGCCAATTGCCTGGATTCCCATCCTGGCTGGAATCAAGAAGGCAGCATCCGCCAGGCTCCCAACAGGCAGGACTCCCGCACACCCTCCTCTGAGAGGCCGCTGTGTTCCGCAGGGCCAGGCCCTGGACAGTTCCCCTCACCTGCCACTAGAGAAACACCTGCCATTGTCGTCCCCACCTGGAAAAGACCACTCGTGGAGCCCCCAGCCCCAGGTACAGCTGTAGAGACAGTCCTCGAGGCCCCTAAGAAGGAGCCATGCCCAGTTCTGCCGGGACCCTCGGCCAGGCCGACAGGAGTGGACGCTGGAGCTGGGCCCACACTGGGCCACATAGGAGCTCACCAGTGAGGGCAGGAGAGCACATGCCGGGGAGCACCCAGCCTCCTGCTGACCAGAGGCCCGTCCCAGAGCCCAGGAGGCTGCAGAGGCCTCTCCAGGGAGACACTGTGCATGTCTGGTACCTAAGCAGCCCCCCACGTCCCCAGTCCTGGGGGCCCCTGGCTCAGCTGTCTGGGCCCTCCCTGCTCCCTGGGAAGCTCCTCCTGACAGCCCCGCCTCCAGTTCCAGGTGTGGATTTTGTCAGGCGATGTCACACTGTGTACTGCCAGAAGTGGATGTGGACTAGCGATAGTGAGAGGAAGTGCTGTGAGGGTATGGTAAGCCGGCTGTGGTGTAAGAAGAAGCTCTGGCACAGTGGTGCCGCCCATATCAAAAACCAGGCCAAGTAGACAGGCCCCTGCTGCGCAGCCCCAGGCATCCACTTCACCTGCTTCTCCTGGGGCTCTCAAGGCTGCTGTCTGTCCTCTGGCCCTCTGTGGGGAGGGTTCCCTCAGTGGGAGGTCTGTGCTCCAGGGCAGGGATGATTGAGATAGAAATCAAAGGCTGGCAGGGAAAGGCAGCTTCCCGCCCTGAGAGGTGCAGGCAGCACCACGGAGCCACGGAGTCACAGAGCCACGGAGCCCCCATTGTGGGCATTTGAGAGTGCTGTGCCCCCGGCAGGCCCAGCCCTGATGGGGAAGCCTGTCCCATCCCACAGCCCGGGTCCCACGGGCAGCGGGCACAGAAGCTGCCAGGTTGTCCTCTATGATCCTCATCCCTCCAGCAGCATCCCCTCCACAGTGGGGAAACTGAGGCTTGGAGCACCACCCGGCCCCCTGGAAATGAGGCTGTGAGCCCAGACAGTGGGCCCAGAGCACTGTGAGTACCCCGGCAGTACCTGGCTGCAGGGATCAGCCAGAGATGCCAAACCCTGAGTGACCAGCCTACAGGAGGATCCGGCCCCACCCAGGCCACTCGATTAATGCTCAACCCCCTGCCCTGGAGACCTCTTCCAGTACCACCAGCAGCTCAGCTTCTCAGGGCCTCATCCCTGCAAGGAAGGTCAAGGGCTGGGCCTGCCAGAAACACAGCACCCTCCCTAGCCCTGGCTAAGACAGGGTGGGCAGACGGCTGTGGACGGGACATATTGCTGGGGCATTTCTCACTGTCACTTCTGGGTGGTAGCTCTGACAAAAACGCAGACCCTGCCAAAATCCCCACTGCCTCCCGCTAGGGGCTGGCCTGGAATCCTGCTGTCCTAGGAGGCTGCTGACCTCCAGGATGGCTCCGTCCCCAGTTCCAGGGCGAGAGCAGATCCCAGGCAGGCTGTAGGCTGGGAGGCCACCCCTGCCCTTGCCGGGGTTGAATGCAGGTGCCCAAGGCAGGAAATGGCATGAGCACAGGGATGACCGGGACATGCCCCACCAGAGTGCGCCCCTTCCTGCTCTGCACCCTGCACCCCCCAGGCCAGCCCACGACGTCCAACAACTGGGCCTGGGTGGCAGCCCCACCCAGACAGGACAGACCCAGCACCCTGAGGAGGTCCTGCCAGGGGGAGCTAAGAGCCATGAAGGAGCAAGATATGGGGCCCCCGATACAGGCACAGATGTCAGCTCCATCCAGGACCACCCAGCCCACACCCTGAGAGGAACGTCTGTCTCCAGCCTCTGCAGGTCGGGAGGCAGCTGACCCCTGACTTGGACCCCTATTCCAGACACCAGACAGAGGCGCAGGCCCCCCAGAACCAGGGTTGAGGGACGCCCCGTCAAAGCCAGACAAAACCAAGGGGTGTTGAGCCCAGCAAGGGAAGGCCCCCAAACAGACCAGGAGGATTTTGTAGGTGTCTGTGTCACTGTGTGCAACTGCAGCAGATGGCGCGACCATAGCAGGTGCTGCCACAGTGACACTCACCCAGTCAAAAACCCCATTCCAAGTCAGCGGAAGCAGAGAGAGCAGGGAGGACACGTTTAGGATCTGAGACTGCACCTGACACCCAGGCCAGCAGACGTCTCCCCTCCAGGGCACCCCACCCTGTCCTGCATTTCTGCAAGATCAGGGGCGGCCTGAGGGGGGGTCTAGGGTGAGGAGATGGGTCCCCTGTACACCAAGGAGGAGTTAGGCAGGTCCCGAGCACTCTTAATTAAACGACGCCTCGAATGGAACTACTACAACGAATGGTTGCTCTACGTAATGCATTCGCTACCTTAGGACCGTTATAGTTAGGCGCGCC
[00163] TX-DH114619 (SEQ ID NO:234) включает кодирующую последовательность токсина, которая встроена в положениях, соответствующих DH1–14 — DH6–19:
TACGTATTAATTAAACGACGCCTCGAATGGAACTACTACAACGAATGGTTGCTCTCCCCATTGAGGCTGACCTGCCCAGAGAGTCCTGGGCCCACCCCACACACCGGGGCGGAATGTGTGCAGGCCTCGGTCTCTGTGGGTGTTCCGCTAGCTGGGGCTCACAGTGCTCACCCCACACCTAAAATGAGCCACAGCCTCCGGAGCCCCCGCAGGAGACCCCGCCCACAAGCCCAGCCCCCACCCAGGAGGCCCCAGAGCTCAGGGCGCCCCGTCGGATTTTGTACAGCCCCGAGTCACTGTGGAGAGATGCTGCAATGGCAGACGCGGCTGCAGCAGCAGATGGTGCCGCGATCATAGCAGGTGCTGCCACAGTGAGAATAGCTACGTCAAAAACCGTCCAGTGGCCACTGCCGGAGGCCCCGCCAGAGAGGGCAGCAGCCACTCTGATCCCATGTCCTGCCGGCTCCCATGACCCCCAGCACGCGGAGCCCCACAGTGTCCCCACTGGATGGGAGGACAAGAGCTGGGGATTCCGGCGGGTCGGGGCAGGGGCTTGATCGCATCCTTCTGCCGTGGCTCCAGTGCCCCTGGCTGGAGTTGACCCTTCTGACAAGTGTCCTCAGAGAGACAGGCATCACCGGCGCCTCCCAACATCAACCCCAGGCAGCACAGGCACAAACCCCACATCCAGAGCCAACTCCAGGAGCAGAGACACCCCAATACCCTGGGGGACCCCGACCCTGATGACTTCCCACTGGAATTCGCCGTAGAGTCCACCAGGACCAAAGACCCTGCCTCTGCCTCTGTCCCTCACTCAGGACCTGCTGCCGGGCGAGGCCTTGGGAGCAGACTTGGGCTTAGGGGACACCAGTGTGACCCCGACCTTGACCAGGACGCAGACCTTTCCTTCCTTTCCTGGGGCAGCACAGACTTTGGGGTCTGGGCCAGGAGGAACTTCTGGCAGGTCGCCAAGCACAGAGGCCACAGGCTGAGGTGGCCCTGGAAAGACCTCCAGGAGGTGGCCACTCCCCTTCCTCCCAGCTGGACCCCATGTCCTCCCCAAGATAAGGGTGCCATCCAAGGCAGGTGCTCCTTGGAGCCCCATTCAGACTCCTCCCTGGACCCCACTGGGCCTCAGTCCCAGCTCTGGGGATGAAGCCACCACAAGCACACCAGGCAGCCCAGGCCCAGCCACCCTGCAGTGCCCAAGCACACACTCTGGAGCAGAGCAGGGTGCCTCTGGGAGGGGCTGAGCTCCCCACCCCACCCCCACCTGCACACCCCACCCACCCCTGCCCAGCGGCTCTGCAGGAGGGTCAGAGCCCCACATGGGGTATGGACTTAGGGTCTCACTCACGTGGCTCCCATCATGAGTGAAGGGGCCTCAAGCCCAGGTTCCCACAGCAGCGCCTGTCGCAAGTGGAGGCAGAGGCCCGAGGGCCACCCTGACCTGGTCCCTGAGGTTCCTGCAGCCCAGGCTGCCCTGCTGTCCCTGGGAGGCCTGGGCTCCACCAGACCACAGGTCCAGGGCACCGGGTGCAGGAGCCACCCACACACAGCTCACAGGAAGAAGATAAGCTCCAGACCCCCAGGGCCAGAACCTGCCTTCCTGCTACTGCTTCCTGCCCCAGACCTGGGCGCCCTCCCCCGTCCACTTACACACAGGCCAGGAAGCTGTTCCCACACAGAACAACCCCAAACCAGGACCGCCTGGCACTCAGGTGGCTGCCATTTCCTTCTCCATTTGCTCCCAGCGCCTCTGTCCTCCCTGGTTCCTCCTTCGGGGGAACAGCCTGTGCAGCCAGTCCCTGCAGCCCACACCCTGGGGAGACCCAACCCTGCCTGGGGCCCTTCCAACCCTGCTGCTCTTACTGCCCACCCAGAAAACTCTGGGGTCCTGTCCCTGCAGTCCCTACCCTGGTCTCCACCCAGACCCCTGTGTATCACTCCAGACACCCCTCCCAGGCAAACCCTGCACCTGCAGGCCCTGTCCTCTTCTGTCGCTAGAGCCTCAGTTTCTCCCCCCTGTGCCCACACCCTACCTCCTCCTGCCCACAACTCTAACTCTTCTTCTCCTGGAGCCCCTGAGCCATGGCATTGACCCTGCCCTCCCACCACCCACAGCCCATGCCCTCACCTTCCTCCTGGCCACTCCGACCCCGCCCCCTCTCAGGCCAAGCCCTGGTATTTCCAGGACAAAGGCTCACCCAAGTCTTTCCCAGGCAGGCCTGGGCTCTTGCCCTCACTTCCCGGTTACACGGGAGCCTCCTGTGCACAGAAGCAGGGAGCTCAGCCCTTCCACAGGCAGAAGGCACTGAAAGAAATCGGCCTCCAGCACCTTGACACACGTCCGCCCGTGTCTCTCACTGCCCGCACCTGCAGGGAGGCTCCGCACTCCCTCTAAAGACAAGGGATCCAGGCAGCAGCATCACGGGAGAATGCAGGGCTCCCAGACATCCCAGTCCTCTCACAGGCCTCTCCTGGGAAGAGACCTGCAGCCACCACCAAACAGCCACAGAGGCTGCTGGATAGTAACTGAGTCAATGACCGACCTGGAGGGCAGGGGAGCAGTGAGCCGGAGCCCATACCATAGGGACAGAGACCAGCCGCTGACATCCCGAGCTCCTCAATGGTGGCCCCATAACACACCTAGGAAACATAACACACCCACAGCCCCACCTGGAACAGGGCAGAGACTGCTGAGCCCCCAGCACCAGCCCCAAGAAACACCAGGCAACAGTATCAGAGGGGGCTCCCGAGAAAGAGAGGAGGGGAGATCTCCTTCACCATCAAATGCTTCCCTTGACCAAAAACAGGGTCCACGCAACTCCCCCAGGACAAAGGAGGAGCCCCCTATACAGCACTGGGCTCAGAGTCCTCTCTGAGACACCCTGAGTTTCAGACAACAACCCGCTGGAATGCACAGTCTCAGCAGGAGAACAGACCAAAGCCAGCAAAAGGGACCTCGGTGACACCAGTAGGGACAGGAGGATTTTGTGGGGGCTCGTGTCACTGTGAGGATATTGTAGTGGTAGCAGCAGATGGGGTAGCTGCTACTCCCACAGTGACACAGACCCATTCAAAAACCCCTACTGCAAACACACCCACTCCTGGGGCTGAGGGGCTGGGGGAGCGTCTGGGAAGTAGGGTCCAGGGGTGTCTATCAATGTCCAAAATGCACCAGACTCCCCGCCAAACACCACCCCACCAGCCAGCGAGCAGGGTAAACAGAAAATGAGAGGCTCTGGGAAGCTTGCACAGGCCCCAAGGAAAGAGCTTTGGCGGGTGTGCAAGAGGGGATGCAGGCAGAGCCTGAGCAGGGCCTTTTGCTGTTTCTGCTTTCCTGTGCAGAGAGTTCCATAAACTGGTGTTCAAGATCAGTGGCTGGGAATGAGCCCAGGAGGGCAGTCTGTGGGAAGAGCACAGGGAAGGAGGAGCAGCCGCTATCCTACACTGTCATCTTTCAAAAGTTTGCCTTGTGACCACACTATTGCATCATGGGATGCTTAAGAGCTGATGTAGACACAGCTAAAGAGAGAATCAGTGAGATGAATTTGCAGCATAGATCTGAATAAACTCTCCAGAATGTGGAGCAGTACAGAAGCAAACACACAGAAAGTGCCTGATGCAAGGACAAAGTTCAGTGGGCACCTTCAGGCATTGCTGCTGGGCACAGACACTCTGAAAAGCCTTGGCAGGATCTCCCTGCGACAAAGCAGAACCCTCAGGCAATGCCAGCCCCAGAGCCCTCCCTGAGAGCGTCATGGGGAAAGATGTGCAGAACAGCTGATTATCATAGACTCAAACTGAGAACAGAGCAAACGTCCATCTGAAGAACAGTCAAATAAGCAATGGTAGGTTCATGCAATGCAAACCCAGACAGCCAGGGGACAACAGTAGAGGGCTACAGGCGGCTTTGCGGTTGAGTTCATGACAATGCTGAGTAATTGGAGTAACAGAGGAAAGCCCAAAAAATACTTTTAATGTGATTTCTTCTAAATAAAATTTACACCAGGCAAAATGAACTGTCTTCTTAAGGGATAAACTTTCCCCTGGAAAAACTACAAGGAAAATTAAGAAAACGATGATCACATAAACACAGTTGTGGTTACTTCTACTGGGGAAGGAAGAGGGTATGAGCTGAGACACACAGAGTCGGCAAGTCTCCAAGCAAGCACAGAACGAATACATTACAGTACCTTGAATACAGCAGTTAAACTTCTAAATCGCAAGAACAGGAAAATGCACACAGCTGTGTTTAGAAAATTCTCAGTCCAGCACTATTCATAATAGCAAAGACATTAACCCAGGTTGGATAAATAAATGATGACACAGGCAATTGCACAATGATACAGACATACATTTAGTACATGAGACATCGATGATGTATCCCCAAAGAAATGACTTTAAAGAGAAAAGGCCTGATGTGTGGTGGCACTCACCTCCCTGGGATCCCCGGACAGGTTGCAGGCACACTGTGTGGCAGGGCAGGCTGGTACATGCTGGCAGCTCCTGGGGCCTGATGTGGAGCAAGCGCAGGGCTGTATACCCCCAAGGATGGCACAGTCAGTGAATTCCAGAGAGAAGCAGCTCAGCCACACTGCCCAGGCAGAGCCCGAGAGGGACGCCCACGTACAGGGAGGCAGAGCCCAGCTCCTCCACAGCCACCACCACCTGTGCACGGGCCACCACCTTGCAGGCACAGAGTGGGTGCTGAGAGGAGGGGCAGGGACACCAGGCAGGGTGAGCACCCAGAGAAAACTGCAGAAGCCTCACACATCCACCTCAGCCTCCCCTGACCTGGACCTCACCTGGTCTGGACCTCACCTGGCCTGGGCCTCACCTGACCTGGACCTCACCTGGCCTGGGCTTCACCTGACCTGGACCTCACCTGGCCTCCGGCCTCACCTGCACCTGCTCCAGGTCTTGCTGGAACCTGAGTAGCACTGAGGCTGCAGAAGCTCATCCAGGGTTGGGGAATGACTCTGGAACTCTCCCACATCTGACCTTTCTGGGTGGAGGCATCTGGTGGCCCTGGGAATATAAAAAGCCCCAGAATGGTGCCTGCGTGATTTGGGGGCAATTTATGAACCCGAAAGGACATGGCCATGGGGTGGGTAGGGACATAGGGACAGATGCCAGCCTGAGGTGGAGCCTCAGGACACAGTTGGACGCGGACACTATCCACATAAGCGAGGGACAGACCCGAGTGTTCCTGCAGTAGACCTGAGAGCGCTGGGCCCACAGCCTCCCCTCGGTGCCCTGCTGCCTCCTCAGGTCAGCCCTGGACATCCCGGGTTTCCCCAGGCCAGATGGTAGGATTTTGTTGAGGTCTGTGTCACTGTGGTATTATGATTACGAGAGAGCTTGCAATGGCAGACGCGGCTGCAGCAGATGGGCTCGCGATCATAGCAGGTGCTGCTATCGTTATACCCACAGTGTCACACGGTCCATCAAAAACCCATGCCACAGCCCTCCCCGCAGGGGACCGCCGCGTGCCATGTTACGATTTTGATCGAGGACACAGCGCCATGGGTATGGTGGCTACCACAGCAGTGCAGCCCATGACCCAAACACACAGGGCAGCAGGCACAATGGACAGGCCTGTGAGTGACCATGCTGGGCTCCAGCCCGCCAGCCCCGGAGACCATGAAACAGATGGCCAAGGTCACCCCACAGTTCAGCCAGACATGGCTCCGTGGGGTCTGCATCGCTGCTGCCCTCTAACACCAGCCCAGATGGGGACAAGGCCAACCCCACATTACCATCTCCTGCTGTCCACCCAGTGGTCCCAGAAGCCCCTCCCTCATGGCTGAGCCACATGTGTGAACCCTGAGAGCACCCCATGTCAGAGTAGGGGCAGCAGAAGGGCGGGGCTGGCCCTGTGCACTGTCCCTGCACCCATGGTCCCTCGCCTGCCTGGCCCTGACACCTGAGCCTCTTCTGAGTCATTTCTAAGATAGAAGACATTCCCGGGGACAGCCGGAGCTGGGCGTCGCTCATCCCGCCCGGCCGTCCTGAGTCCTGCTTGTTTCCAGACCTCACCAGGGAAGCCAACAGAGGACTCACCTCACACAGTCAGAGACAAAGAACCTTCCAGAAATCCCTGTCTCACTCCCCAGTGGGCACCTTCTTCCAGGACATTCCTCGGTCGCATCACAGCAGGCACCCACATCTGGATCAGGACGGCCCCCAGAACACAAGATGGCCCATGGGGACAGCCCCACAACCCAGGCCTTCCCAGACCCCTAAAAGGCGTCCCACCCCCTGCACCTGCCCCAGGGCTAAAAATCCAGGAGGCTTGACTCCCGCATACCCTCCAGCCAGACATCACCTCAGCCCCCTCCTGGAGGGGACAGGAGCCCGGGAGGGTGAGTCAGACCCACCTGCCCTCGATGGCAGGCGGGGAAGATTCAGAAAGGCCTGAGATCCCCAGGACGCAGCACCACTGTCAATGGGGGCCCCAGACGCCTGGACCAGGGCCTGCGTGGGAAAGGCCGCTGGGCACACTCAGGGGGATTTTGTGAAGGCCCCTCCCACTGTGGAGAGGCTTGCTTGTGGCTTCCCTAAGAGCTGCAGCAGGCAAGCTAAGCCTCACAGATGCTGCCACAGTGATGAAACTAGCATCAAAAACCGGCCGGACACCCAGGGACCATGCACACTTCTCAGCTTGGAGCTCTCCAGGACCAGAAGAGTCAGGTCTGAGGGTTTGTAGCCAGACCCTCGGCCTCTAGGGACACCCTGGCCATCACAGCGGATGGGCTGGTGCCCCACATGCCATCTGCTCCAAACAGGGGCTTCAGAGGGCTCTGAGGTGACTTCACTCATGACCACAGGTGCCCTGGCCCCTTCCCCGCCAGCTACACCGAACCCTGTCCCAACAGCTGCCCCAGTTCCAACAGCCAATTCCTGGGGCCCAGAATTGCTGTAGACACCAGCCTCGTTCCAGCACCTCCTGCCAATTGCCTGGATTCACATCCTGGCTGGAATCAAGAGGGCAGCATCCGCCAGGCTCCCAACAGGCAGGACTCCCGCACACCCTCCTCTGAGAGGCCGCTGTGTTCCGCAGGGCCAGGCCCTGGACAGTTCCCCTCACCTGCCACTAGAGAAACACCTGCCATTGTCGTCCCCACCTGGAAAAGACCACTCGTGGAGCCCCCAGCCCCAGGTACAGCTGTAGAGAGACTCCCCGAGGGATCTAAGAAGGAGCCATGCGCAGTTCTGCCGGGACCCTCGGCCAGGCCGACAGGAGTGGACACTGGAGCTGGGCCCACACTGGGCCACATAGGAGCTCACCAGTGAGGGCAGGAGAGCACATGCCGGGGAGCACCCAGCCTCCTGCTGACCAGAGGCCCGTCCCAGAGCCCAGGAGGCTGCAGAGGCCTCTCCAGGGGGACACTGTGCATGTCTGGTCCCTGAGCAGCCCCCCACGTCCCCAGTCCTGGGGGCCCCTGGCACAGCTGTCTGGACCCTCCCTGTTCCCTGGGAAGCTCCTCCTGACAGCCCCGCCTCCAGTTCCAGGTGTGGATTTTGTCAGGGGGTGTCACACTGTGTACTGCCAGAAGTGGATGTGGACTTGCGATAGTGAGAGGAAGAGCTGTGAGGGTATGGTATGCCGGCTGTGGAGTAAGAAGAAGCTCTGGCACAGTGGTGCTGCCCATATCAAAAACCAGGCCAAGTAGACAGGCCCCTGCTGTGCAGCCCCAGGCCTCCACTTCACCTGCTTCTCCTGGGGCTCTCAAGGTCACTGTTGTCTGTACTCTGCCCTCTGTGGGGAGGGTTCCCTCAGTGGGAGGTCTGTTCTCAACATCCCAGGGCCTCATGTCTGCACGGAAGGCCAATGGATGGGCAACCTCACATGCCGCGGCTAAGATAGGGTGGGCAGCCTGGCGGGGGACAGTACATACTGCTGGGGTGTCTGTCACTGTGCCTAGTGGGGCACTGGCTCCCAAACAACGCAGTCCTCGCCAAAATCCCCACAGCCTCCCCTGCTAGGGGCTGGCCTGATCTCCTGCAGTCCTAGGAGGCTGCTGACCTCCAGAATGTCTCCGTCCCCAGTTCCAGGGCGAGAGCAGATCCCAGGCCGGCTGCAGACTGGGAGGCCACCCCCTCCTTCCCAGGGTTCACTGGAGGTGACCAAGGTAGGAAATGGCCTTAACACAGGGATGACTGCGCCATCCCCCAACAGAGTCAGCCCCCTCCTGCTCTGTACCCCGCACCCCCCAGGCCAGTCCACGAAAACCAGGGCCCCACATCAGAGTCACTGCCTGGCCCGGCCCTGGGGCGGACCCCTCAGCCCCCACCCTGTCTAGAGGACTTGGGGGGACAGGACACAGGCCCTCTCCTTATGGTTCCCCCACCTGCCTCCGGCCGGGACCCTTGGGGTGTGGACAGAAAGGACACCTGCCTAATTGGCCCCCAGGAACCCAGAACTTCTCTCCAGGGACCCCAGCCCGAGCACCCCCTTACCCAGGACCCAGCCCTGCCCCTCCTCCCCTCTGCTCTCCTCTCATCACCCCATGGGAATCCGGTATCCCCAGGAAGCCATCAGGAAGGGCTGAAGGAGGAAGCGGGGCCGTGCACCACCGGGCAGGAGGCTCCGTCTTCGTGAACCCAGGGAAGTGCCAGCCTCCTAGAGGGTATGGTCCACCCTGCCTGGGGCTCCCACCGTGGCAGGCTGCGGGGAAGGACCAGGGACGGTGTGGGGGAGGGCTCAGGGCCCTGCGGGTGCTCCTCCATCTTCGGTGAGCCTCCCCCTTCACCCACCGTCCCGCCCACCTCCTCTCCACCCTGGCTGCACGTCTTCCACACCATCCTGAGTCCTACCTACACCAGAGCCAGCAAAGCCAGTGCAGACAAAGGCTGGGGTGCAGGGGGGCTGCCAGGGCAGCTTCGGGGAGGGAAGGATGGAGGGAGGGGAGGTCAGTGAAGAGGCCCCCTTCCCCTGGGTCCAGGATCCTCCTCTGGGACCCCCGGATCCCATCCCCTCCTGGCTCTGGGAGGAGAAGCAGGATGGGAGAATCTGTGCGGGACCCTCTCACAGTGGAATATCCCCACAGCGGCTCAGGCCAGACCCAAAAGCCCCTCAGTGAGCCCTCCACTGCAGTCCTGGGCCTGGGTAGCAGCCCCTCCCACAGAGGACAGACCCAGCACCCCGAAGAAGTCCTGCCAGGGGGAGCTCAGAGCCATGAAAGAGCAGGATATGGGGTCCCCGATACAGGCACAGACCTCAGCTCCATCCAGGCCCACCGGGACCCACCATGGGAGGAACACCTGTCTCCGGGTTGTGAGGTAGCTGGCCTCTGTCTCGGACCCCACTCCAGACACCAGACAGAGGGGCAGGCCCCCCAAAACCAGGGTTGAGGGATGATCCGTCAAGGCAGACAAGACCAAGGGGCACTGACCCCAGCAAGGGAAGGCTCCCAAACAGACGAGGAGGATTTTGTAGCTGTCTGTATCACTGTGTGCAACTGCAGCAGATGGGCTCGCGACCATAGCAGGTGCTGCCACAGTGACACTCGCCAGGTCAAAAACCCCGTCCCAAGTCAGCGGAAGCAGAGAGAGCAGGGAGGACACGTTTAGGATCTGAGGCCGCACCTGACACCCAGGGCAGCAGACGTCTCCCCTCCAGGGCACCCTCCACCGTCCTGCGTTTCTTCAAGAATAGGGGCGGCCTGAGGGGGTCCAGGGCCAGGCGATAGGTCCCCTCTACCCCAAGGAGGAGCCAGGCAGGACCCGAGCACCGATGCATCTAACGCAGTCATGTAATGCTGGGTGACAGTCAGTTCGCCTACGTA
[00164] TX-DH120126 (SEQ ID NO:235) включает кодирующую последовательность токсина, которая встроена в положениях, соответствующих DH1-20 — DH1-26:
TACGTAATGCATCTAACGCAGTCATGTAATGCTGGGTGACAGTCAGTTCGCCTCCCCATTGAGGCTGACCTGCCCAGACGGGCCTGGGCCCACCCCACACACCGGGGCGGAATGTGTGCAGGCCCCAGTCTCTGTGGGTGTTCCGCTAGCTGGGGCCCCCAGTGCTCACCCCACACCTAAAGCGAGCCCCAGCCTCCAGAGCCCCCTAAGCATTCCCCGCCCAGCAGCCCAGCCCCTGCCCCCACCCAGGAGGCCCCAGAGCTCAGGGCGCCTGGTCGGATTTTGTACAGCCCCGAGTCACTGTGGAGAGAGCTTGCAATGGCAGACGCGGCTGCAGCAGATGGGCTCGCGATCATAGCAGGTGCTGCCACAGTGAGAAAAACTGTGTCAAAAACCGACTCCTGGCAGCAGTCGGAGGCCCCGCCAGAGAGGGGAGCAGCCGGCCTGAACCCATGTCCTGCCGGTTCCCATGACCCCCAGCACCCAGAGCCCCACGGTGTCCCCGTTGGATAATGAGGACAAGGGCTGGGGGCTCCGGTGGTTTGCGGCAGGGACTTGATCACATCCTTCTGCTGTGGCCCCATTGCCTCTGGCTGGAGTTGACCCTTCTGACAAGTGTCCTCAGAAAGACAGGGATCACCGGCACCTCCCAATATCAACCCCAGGCAGCACAGACACAAACCCCACATCCAGAGCCAACTCCAGGAGCAGAGACACCCCAACACTCTGGGGGACCCCAACCGTGATAACTCCCCACTGGAATCCGCCCCAGAGTCTACCAGGACCAAAGGCCCTGCCCTGTCTCTGTCCCTCACTCAGGGCCTCCTGCAGGGCGAGCGCTTGGGAGCAGACTCGGTCTTAGGGGACACCACTGTGGGCCCCAACTTTGATGAGGCCACTGACCCTTCCTTCCTTTCCTGGGGCAGCACAGACTTTGGGGTCTGGGCAGGGAAGAACTACTGGCTGGTGGCCAATCACAGAGCCCCCAGGCCGAGGTGGCCCCAAGAAGGCCCTCAGGAGGTGGCCACTCCACTTCCTCCCAGCTGGACCCCAGGTCCTCCCCAAGATAGGGGTGCCATCCAAGGCAGGTCCTCCATGGAGCCCCCTTCAGACTCCTCCCGGGACCCCACTGGACCTCAGTCCCTGCTCTGGGAATGCAGCCACCACAAGCACACCAGGAAGCCCAGGCCCAGCCACCCTGCAGTGGGCAAGCCCACACTCTGGAGCAGAGCAGGGTGCGTCTGGGAGGGGCTAACCTCCCCACCCCCCACCCCCCATCTGCACACAGCCACCTACCACTGCCCAGACCCTCTGCAGGAGGGCCAAGCCACCATGGGGTATGGACTTAGGGTCTCACTCACGTGCCTCCCCTCCTGGGAGAAGGGGCCTCATGCCCAGATCCCTGCAGCACTAGACACAGCTGGAGGCAGTGGCCCCAGGGCCACCCTGACCTGGCATCTAAGGCTGCTCCAGCCCAGACAGCACTGCCGTTCCTGGGAAGCCTGGGCTCCACCAGACCACAGGTCCAGGGCACAGCCCACAGGAGCCACCCACACACAGCTCACAGGAAGAAGATAAGCTCCAGACCCCAGGGCGGGACCTGCCTTCCTGCCACCACTTACACACAGGCCAGGGAGCTGTTCCCACACAGATCAACCCCAAACCGGGACTGCCTGGCACTAGGGTCACTGCCATTTCCCTCTCCATTCCCTCCCAGTGCCTCTGTGCTCCCTCCTTCTGGGGAACACCCTGTGCAGCCCCTCCCTGCAGCCCACACGCTGGGGAGACCCCACCCTGCCTCGGGCCTTTTCTACCTGCTGCACTTGCCGCCCACCCAAACAACCCTGGGTACGTGACCCTGCAGTCCTCACCCTGATCTGCAACCAGACCCCTGTCCCTCCCTCTAAACACCCCTCCCAGGCCAACTCTGCACCTGCAGGCCCTCCGCTCTTCTGCCACAAGAGCCTCAGGTTTTCCTACCTGTGCCCACCCCCTAACCCCTCCTGCCCACAACTTGAGTTCTTCCTCTCCTGGAGCCCTTGAGCCATGGCACTGACCCTACACTCCCACCCACACACTGCCCATGCCATCACCTTCCTCCTGGACACTCTGACCCCGCTCCCCTCCCTCTCAGACCCGGCCCTGGTATTTCCAGGACAAAGGCTCACCCAAGTCTTCCCCATGCAGGCCCTTGCCCTCACTGCCTGGTTACACGGGAGCCTCCTGTGCGCAGAAGCAGGGAGCTCAGCTCTTCCACAGGCAGAAGGCACTGAAAGAAATCAGCCTCCAGTGCCTTGACACACGTCCGCCTGTGTCTCTCACTGCCTGCACCTGCAGGGAGGCTCCGCACTCCCTCTAAAGATGAGGGATCCAGGCAGCAACATCACGGGAGAATGCAGGGCTCCCAGACAGCCCAGCCCTCTCGCAGGCCTCTCCTGGGAAGAGACCTGCAGCCACCACTGAACAGCCACGGAGGTCGCTGGATAGTAACCGAGTCAGTGACCGACCTGGAGGGCAGGGGAGCAGTGAACCGGAGCCCATACCATAGGGACAGAGACCAGCCGCTAACATCCCGAGCCCCTCACTGGCGGCCCCAGAACACCCCGTGGAAAGAGAACAGACCCACAGTCCCACCTGGAACAGGGCAGACACTGCTGAGCCCCCAGCACCAGCCCCAAGAAACACTAGGCAACAGCATCAGAGGGGGCTCCTGAGAAAGAGAGGAGGGGAGGTCTCCTTCACCATCAAATGCTTCCCTTGACCAAAAACAGGGTCCACGCAACTCCCCCAGGACAAAGGAGGAGCCCCCTGTACAGCACTGGGCTCAGAGTCCTCTCTGAGACAGGCTCAGTTTCAGACAACAACCCGCTGGAATGCACAGTCTCAGCAGGAGAGCCAGGCCAGAGCCAGCAAGAGGAGACTCGGTGACACCAGTCTCCTGTAGGGACAGGAGGATTTTGTGGGGGTTCGTGTCACTGTGAGCATATTGTCGGAGCAGGCAGTGCTATTCCCACAGTGACACAACCCCATTCAAAAACCCCTACTGCAAACGCACCCACTCCTGGGACTGAGGGGCTGGGGGAGCGTCTGGGAAGTATGGCCTAGGGGTGTCCATCAATGCCCAAAATGCACCAGACTCTCCCCAAGACATCACCCCACCAGCCAGTGAGCAGAGTAAACAGAAAATGAGAAGCAGCTGGGAAGCTTGCACAGGCCCCAAGGAAAGAGCTTTGGCAGGTGTGCAAGAGGGGATGTGGGCAGAGCCTCAGCAGGGCCTTTTGCTGTTTCTGCTTTCCTGTGCAGAGAGTTCCATAAACTGGTATTCAAGATCAATGGCTGGGAGTGAGCCCAGGAGGACAGTGTGGGAAGAGCACAGGGAAGGAGGAGCAGCCGCTATCCTACACTGTCATCTTTTGAAAGTTTGCCCTGTGCCCACAATGCTGCATCATGGGATGCTTAACAGCTGATGTAGACACAGCTAAAGAGAGAATCAGTGAAATGGATTTGCAGCACAGATCTGAATAAATCCTCCAGAATGTGGAGCAGCACAGAAGCAAGCACACAGAAAGTGCCTGATGCCAAGGCAAAGTTCAGTGGGCACCTTCAGGCATTGCTGCTGGGCACAGACACTCTGAAAAGCACTGGCAGGAACTGCCTGTGACAAAGCAGAACCCTCAGGCAATGCCAGCCCTAGAGCCCTTCCTGAGAACCTCATGGGCAAAGATGTGCAGAACAGCTGTTTGTCATAGCCCCAAACTATGGGGCTGGACAAAGCAAACGTCCATCTGAAGGAGAACAGACAAATAAACGATGGCAGGTTCATGAAATGCAAACTAGGACAGCCAGAGGACAACAGTAGAGAGCTACAGGCGGCTTTGCGGTTGAGTTCATGACAATGCTGAGTAATTGGAGTAACAGAGGAAAGCCCAAAAAATACTTTTAATGTGATTTCTTCTAAATAAAATTTACACCCGGCAAAATGAACTATCTTCTTAAGGGATAAACTTTCCCCTGGAAAAACTATAAGGAAAATCAAGAAAACGATGATCACATAAACACAGTGGTGGTTACTTCTACTGGGGAAGGAAGAGGGTATGAGCTGAGACACACAGAGTCGGCAAGTCTCCTAACAAGAACAGAACAAATACATTACAGTACCTTGAAAACAGCAGTTAAACTTCTAAATCGCAAGAAGAGGAAAATGCACACACCTGTGTTTAGAAAATTCTCAGTCCAGCACTGTTCATAATAGCAAAGACATTAACCCAGGTTGGATAAATAAGCGATGACACAGGCAATTGCACAATGATACAGACATACATTCAGTATATGAGACATCGATGATGTATCCCCAAAGAAATGACTTTAAAGAGAAAAGGCCTGATGTGTGGTGGCAATCACCTCCCTGGGCATCCCCGGACAGGCTGCAGGCTCACTGTGTGGCAGGGCAGGCAGGCACCTGCTGGCAGCTCCTGGGGCCTGATGTGGAGCAGGCACAGAGCTGTATATCCCCAAGGAAGGTACAGTCAGTGCATTCCAGAGAGAAGCAACTCAGCCACACTCCCTGGCCAGAACCCAAGATGCACACCCATGCACAGGGAGGCAGAGCCCAGCACCTCCGCAGCCACCACCACCTGCGCACGGGCCACCACCTTGCAGGCACAGAGTGGGTGCTGAGAGGAGGGGCAGGGACACCAGGCAGGGTGAGCACCCAGAGAAAACTGCAGAAGCCTCACACATCCCTCACCTGGCCTGGGCTTCACCTGACCTGGACCTCACCTGGCCTCGGGCCTCACCTGCACCTGCTCCAGGTCTTGCTGGAGCCTGAGTAGCACTGAGGCTGTAGGGACTCATCCAGGGTTGGGGAATGACTCTGCAACTCTCCCACATCTGACCTTTCTGGGTGGAGGCACCTGGTGGCCCAGGGAATATAAAAAGCCCCAGAATGATGCCTGTGTGATTTGGGGGCAATTTATGAACCCGAAAGGACATGGCCATGGGGTGGGTAGGGACAGTAGGGACAGATGTCAGCCTGAGGTGAAGCCTCAGGACACAGGTGGGCATGGACAGTGTCCACCTAAGCGAGGGACAGACCCGAGTGTCCCTGCAGTAGACCTGAGAGCGCTGGGCCCACAGCCTCCCCTCGGGGCCCTGCTGCCTCCTCAGGTCAGCCCTGGACATCCCGGGTTTCCCCAGGCCTGGCGGTAGGATTTTGTTGAGGTCTGTGTCACTGTGGTATTACTATGAGAGGCTTGCTTGTGGCTTCCCTAAGAGCTGCAGCAGGCAAGCTAAGCCTCACAGATGCTGCTATTACTACCACAGTGTCACAGAGTCCATCAAAAACCCATGCCTGGGAGCCTCCCACCACAGCCCTCCCTGCGGGGGACCGCTGCATGCCGTGTTAGGATTTTGATCGAGGACACGGCGCCATGGGTATGGTGGCTACCACAGCAGTGCAGCCCATGACCCAAACACACGGGGCAGCAGAAACAATGGACAGGCCCACAAGTGACCATGATGGGCTCCAGCCCACCAGCCCCAGAGACCATGAAACAGATGGCCAAGGTCACCCTACAGGTCATCCAGATCTGGCTCCAAGGGGTCTGCATCGCTGCTGCCCTCCCAACGCCAAACCAGATGGAGACAGGGCCGGCCCCATAGCACCATCTGCTGCCGTCCACCCAGCAGTCCCGGAAGCCCCTCCCTGAACGCTGGGCCACGTGTGTGAACCCTGCGAGCCCCCCATGTCAGAGTAGGGGCAGCAGGAGGGCGGGGCTGGCCCTGTGCACTGTCACTGCCCCTGTGGTCCCTGGCCTGCCTGGCCCTGACACCTGAGCCTCTCCTGGGTCATTTCCAAGACATTCCCAGGGACAGCCGGAGCTGGGAGTCGCTCATCCTGCCTGGCTGTCCTGAGTCCTGCTCATTTCCAGACCTCACCAGGGAAGCCAACAGAGGACTCACCTCACACAGTCAGAGACAACGAACCTTCCAGAAATCCCTGTTTCTCTCCCCAGTGAGAGAAACCCTCTTCCAGGGTTTCTCTTCTCTCCCACCCTCTTCCAGGACAGTCCTCAGCAGCATCACAGCGGGAACGCACATCTGGATCAGGACGGCCCCCAGAACACGCGATGGCCCATGGGGACAGCCCAGCCCTTCCCAGACCCCTAAAAGGTATCCCCACCTTGCACCTGCCCCAGGGCTCAAACTCCAGGAGGCCTGACTCCTGCACACCCTCCTGCCAGATATCACCTCAGCCCCCTCCTGGAGGGGACAGGAGCCCGGGAGGGTGAGTCAGACCCACCTGCCCTCAATGGCAGGCGGGGAAGATTCAGAAAGGCCTGAGATCCCCAGGACGCAGCACCACTGTCAATGGGGGCCCCAGACGCCTGGACCAGGGCCTGTGTGGGAAAGGCCTCTGGCCACACTCAGGGGGATTTTGTGAAGGGCCCTCCCACTGTGGAGAGGCTTTGCTGTGGCTTCCCTAAGAGCTGCCGCAGCAGGCAATGCAAGCCTCACAGATGCTGCCACAGTGATGAAACCAGCATCAAAAACCGACCGGACTCGCAGGGTTTATGCACACTTCTCGGCTCGGAGCTCTCCAGGAGCACAAGAGCCAGGCCCGAGGGTTTGTGCCCAGACCCTCGGCCTCTAGGGACACCCGGGCCATCTTAGCCGATGGGCTGATGCCCTGCACACCGTGTGCTGCCAAACAGGGGCTTCAGAGGGCTCTGAGGTGACTTCACTCATGACCACAGGTGCCCTGGTCCCTTCACTGCCAGCTGCACCAGACCCTGTTCCGAGAGATGCCCCAGTTCCAAAAGCCAATTCCTGGGGCCGGGAATTACTGTAGACACCAGCCTCATTCCAGTACCTCCTGCCAATTGCCTGGATTCCCATCCTGGCTGGAATCAAGAGGGCAGCATCCGCCAGGCTCCCAACAGGCAGGACTCCCACACACCCTCCTCTGAGAGGCCGCTGTGTTCCGCAGGGCCAGGCCGCAGACAGTTCCCCTCACCTGCCCATGTAGAAACACCTGCCATTGTCGTCCCCACCTGGCAAAGACCACTTGTGGAGCCCCCAGCCCCAGGTACAGCTGTAGAGAGAGTCCTCGAGGCCCCTAAGAAGGAGCCATGCCCAGTTCTGCCGGGACCCTCGGCCAGGCCGACAGGAGTGGACGCTGGAGCTGGGCCCACACTGGGCCACATAGGAGCTCACCAGTGAGGGCAGGAGAGCACATGCCGGGGAGCACCCAGCCTCCTGCTGACCAGAGACCCGTCCCAGAGCCCAGGAGGCTGCAGAGGCCTCTCCAGGGGGACACAGTGCATGTCTGGTCCCTGAGCAGCCCCCAGGCTCTCTAGCACTGGGGGCCCCTGGCACAGCTGTCTGGACCCTCCCTGTTCCCTGGGAAGCTCCTCCTGACAGCCCCGCCTCCAGTTCCAGGTGTGGATTTTGTCAGGGGGTGCCACACTGTGTACTGCCAGAAGTGGATGTGGACTTGCGATAGTGAGAGGAAGTGCTGTGAGGGTATGGTATGCCGGCTGTGGTGTAAGAAGAAGCTCTGGCACAGTGGTGCCGCCCATATCAAAAACCAGGCCAAGTAGACAGACCCCTGCCACGCAGCCCCAGGCCTCCAGCTCACCTGCTTCTCCTGGGGCTCTCAAGGCTGCTGTCTGCCCTCTGGCCCTCTGTGGGGAGGGTTCCCTCAGTGGGAGGTCTGTGCTCCAGGGCAGGGATGACTGAGATAGAAATCAAAGGCTGGCAGGGAAAGGCAGCTTCCCGCCCTGAGAGGTGCAGGCAGCACCACAGAGCCATGGAGTCACAGAGCCACGGAGCCCCCAGTGTGGGCGTGTGAGGGTGCTGGGCTCCCGGCAGGCCCAGCCCTGATGGGGAAGCCTGCCCCGTCCCACAGCCCAGGTCCCCAGGGGCAGCAGGCACAGAAGCTGCCAAGCTGTGCTCTACGATCCTCATCCCTCCAGCAGCATCCACTCCACAGTGGGGAAACTGAGCCTTGGAGAACCACCCAGCCCCCTGGAAACAAGGCGGGGAGCCCAGACAGTGGGCCCAGAGCACTGTGTGTATCCTGGCACTAGGTGCAGGGACCACCCGGAGATCCCCATCACTGAGTGGCCAGCCTGCAGAAGGACCCAACCCCAACCAGGCCGCTTGATTAAGCTCCATCCCCCTGTCCTGGGAACCTCTTCCCAGCGCCACCAACAGCTCGGCTTCCCAGGCCCTCATCCCTCCAAGGAAGGCCAAAGGCTGGGCCTGCCAGGGGCACAGTACCCTCCCTTGCCCTGGCTAAGACAGGGTGGGCAGACGGCTGCAGATAGGACATATTGCTGGGGCATCTTGCTCTGTGACTACTGGGTACTGGCTCTCAACGCAGACCCTACCAAAATCCCCACTGCCTCCCCTGCTAGGGGCTGGCCTGGTCTCCTCCTGCTGTCCTAGGAGGCTGCTGACCTCCAGGATGGCTTCTGTCCCCAGTTCTAGGGCCAGAGCAGATCCCAGGCAGGCTGTAGGCTGGGAGGCCACCCCTGTCCTTGCCGAGGTTCAGTGCAGGCACCCAGGACAGGAAATGGCCTGAACACAGGGATGACTGTGCCATGCCCTACCTAAGTCCGCCCCTTTCTACTCTGCAACCCCCACTCCCCAGGTCAGCCCATGACGACCAACAACCCAACACCAGAGTCACTGCCTGGCCCTGCCCTGGGGAGGACCCCTCAGCCCCCACCCTGTCTAGAGGAGTTGGGGGGACAGGACACAGGCTCTCTCCTTATGGTTCCCCCACCTGGCTCCTGCCGGGACCCTTGGGGTGTGGACAGAAAGGACGCCTGCCTAATTGGCCCCCAGGAACCCAGAACTTCTCTCCAGGGACCCCAGCCCGAGCACCCCCTTACCCAGGACCCAGCCCTGCCCCTCCTCCCCTCTGCTCTCCTCTCATCACTCCATGGGAATCCAGAATCCCCAGGAAGCCATCAGGAAGGGCTGAAGGAGGAAGCGGGGCCGCTGCACCACCGGGCAGGAGGCTCCGTCTTCGTGAACCCAGGGAAGTGCCAGCCTCCTAGAGGGTATGGTCCACCCTGCCTGGGGCTCCCACCGTGGCAGGCTGCGGGGAAGGACCAGGGACGGTGTGGGGGAGGGCTCAGGGCCCTGCAGGTGCTCCATCTTGGATGAGCCCATCCCTCTCACCCACCGACCCGCCCACCTCCTCTCCACCCTGGCCACACGTCGTCCACACCATCCTGAGTCCCACCTACACCAGAGCCAGCAGAGCCAGTGCAGACAGAGGCTGGGGTGCAGGGGGGCCGCCAGGGCAGCTTTGGGGAGGGAGGAATGGAGGAAGGGGAGGTCAGTGAAGAGGCCCCCCTCCCCTGGGTCTAGGATCCACCTTTGGGACCCCCGGATCCCATCCCCTCCAGGCTCTGGGAGGAGAAGCAGGATGGGAGATTCTGTGCAGGACCCTCTCACAGTGGAATACCTCCACAGCGGCTCAGGCCAGATACAAAAGCCCCTCAGTGAGCCCTCCACTGCAGTGCAGGGCCTGGGGGCAGCCCCTCCCACAGAGGACAGACCCAGCACCCCGAAGAAGTCCTGCCAGGGGGAGCTCAGAGCCATGAAGGAGCAAGATATGGGGACCCCAATACTGGCACAGACCTCAGCTCCATCCAGGCCCACCAGGACCCACCATGGGTGGAACACCTGTCTCCGGCCCCTGCTGGCTGTGAGGCAGCTGGCCTCTGTCTCGGACCCCCATTCCAGACACCAGACAGAGGGACAGGCCCCCCAGAACCAGTGTTGAGGGACACCCCTGTCCAGGGCAGCCAAGTCCAAGAGGCGCGCTGAGCCCAGCAAGGGAAGGCCCCCAAACAAACCAGGAGGTTTCTGAAGCTGTCTGTGTCACAGTCTGCTGCAACTGCAGCAGCAAATGGTGCCGCGACCATAGCAGGTGCTGCCACAATGACACTGGGCAGGACAGAAACCCCATCCCAAGTCAGCCGAAGGCAGAGAGAGCAGGCAGGACACATTTAGGATCTGAGGCCACACCTGACACTCAAGCCAACAGATGTCTCCCCTCCAGGGCGCCCTGCCCTGTTCAGTGTTCCTGAGAAAACAGGGGCAGCCTGAGGGGATCCAGGGCCAGGAGATGGGTCCCCTCTACCCCGAGGAGGAGCCAGGCGGGAATCCCAGCCCCCTCCCCATTGAGGCCATCCTGCCCAGAGGGGCCCGGACCCACCCCACACACCCAGGCAGAATGTGTGCAGGCCTCAGGCTCTGTGGGTGCCGCTAGCTGGGGCTGCCAGTCCTCACCCCACACCTAAGGTGAGCCACAGCCGCCAGAGCCTCCACAGGAGACCCCACCCAGCAGCCCAGCCCCTACCCAGGAGGCCCCAGAGCTCAGGGCGCCTGGGTGGATTTTGTACAGCCCCGAGTCACTGTGGGTATAGTGGGGAGAGGCTTTGTGGCTTCCCTAAGAGCTGCAGCAGGCAAAAGCCTCACAGATGCTGCAGCTACTACCACAGTGAGAAAAGCTATGTCAAAAACCGTCTCCCGGCCACTGCTGGAGGCCCAGCCAGAGAAGGGACCAGCCGCCCGAACATACGACCTTCCCAGACCTCATGACCCCCAGCACTTGGAGCTCCACAGTGTCCCCATTGGATGGTGAGGATGGGGGCCGGGGCCATCTGCACCTCCCAACATCACCCCCAGGCAGCACAGGCACAAACCCCAAATCCAGAGCCGACACCAGGAACACAGACACCCCAATACCCTGGGGGACCCTGGCCCTGGTGACTTCCCACTGGGATCCACCCCCGTGTCCACCTGGATCAAAGACCCCACCGCTGTCTCTGTCCCTCACTCAGGGCCTGCTGAGGGGCGGGTGCTTTGGAGCAGACTCAGGTTTAGGGGCCACCATTGTGGGGCCCAACCTCGACCAGGACACAGATTTTTCTTTCCTGCCCTGGGGCAACACAGACTTTGGGGTCTGTGCAGGGAGGACCTTCTGGAAAGTCACCAAGCACAGAGCCCTGACTGAGGTGGTCTCAGGAAGACCCCCAGGAGGGGGCTTGTGCCCCTTCCTCTCATGTGGACCCCATGCCCCCCAAGATAGGGGCATCATGCAGGGCAGGTCCTCCATGCAGCCACCACTAGGCAACTCCCTGGCGCCGGTCCCCACTGCGCCTCCATCCCGGCTCTGGGGATGCAGCCACCATGGCCACACCAGGCAGCCCGGGTCCAGCAACCCTGCAGTGCCCAAGCCCTTGGCAGGATTCCCAGAGGCTGGAGCCCACCCCTCCTCATCCCCCCACACCTGCACACACACACCTACCCCCTGCCCAGTCCCCCTCCAGGAGGGTTGGAGCCGCCCATAGGGTGGGGGCTCCAGGTCTCACTCACTCGCTTCCCTTCCTGGGCAAAGGAGCCTCGTGCCCCGGTCCCCCCTGACGGCGCTGGGCACAGGTGTGGGTACTGGGCCCCAGGGCTCCTCCAGCCCCAGCTGCCCTGCTCTCCCTGGGAGGCCTGGGCACCACCAGACCACCAGTCCAGGGCACAGCCCCAGGGAGCCGCCCACTGCCAGCTCACAGGAAGAAGATAAGCTTCAGACCCTCAGGGCCGGGAGCTGCCTTCCTGCCACCCCTTCCTGCCCCAGACCTCCATGCCCTCCCCCAACCACTTACACACAAGCCAGGGAGCTGTTTCCACACAGTTCAACCCCAAACCAGGACGGCCTGGCACTCGGGTCACTGCCATTTCTGTCTGCATTCGCTCCCAGCGCCCCTGTGTTCCCTCCCTCCTCCCTCCTTCCTTTCTTCCTGCATTGGGTTCATGCCGCAGAGTGCCAGGTGCAGGTCAGCCCTGAGCTTGGGGTCACCTCCTCACTGAAGGCAGCCTCAGGGTGCCCAGGGGCAGGCAGGGTGGGGGTGAGGCTTCCAGCTCCAACCGCTTCGCTACCTTAGGACCGTTATAGTTAGGCGCGCCGTCGACCAATTCTCATGTTTGACAGCTTATCATCGAATTTCTACGTA
ДНК-конструкции
[00165] Как правило, полинуклеотидную молекулу, содержащую одну или более нуклеотидных кодирующих последовательностей, каждая из которых кодирует представляющий интерес полипептид, не являющийся иммуноглобулином, или его часть (например, внеклеточную часть полипептида ACKR2 или часть пептидного токсина), встраивают в вектор, предпочтительно ДНК-вектор, с целью репликации полинуклеотидной молекулы в подходящей клетке-хозяине.
[00166] Вследствие их размера одну или более нуклеотидных кодирующих последовательностей можно клонировать напрямую из источников кДНК, доступных от коммерческих поставщиков, или конструировать in silico на основании опубликованных последовательностей, доступных в GenBank. В качестве альтернативы библиотеки бактериальной искусственной хромосомы (BAC) могут обеспечивать гетерологичные нуклеотидные кодирующие последовательности из представляющих интерес генов (например, гетерологичного гена ACKR2 или последовательности, кодирующей токсин). Библиотеки BAC содержат вставки со средним размером 100–150 т.о., и они способны нести вставки размером до 300 т.о. (Shizuya, et al., 1992, Proc. Natl. Acad. Sci., USA 89:8794-8797; Swiatek, et al., 1993, Genes and Development 7:2071-2084; Kim, et al., 1996, Genomics 34 213-218; включенные в данный документ посредством ссылки). Например, геномные библиотеки BAC для человека и мыши были сконструированы и являются коммерчески доступными (например, Invitrogen, Карлсбад, Калифорния). Геномные библиотеки BAC также могут служить источником гетерологичных кодирующих последовательностей, а также участков контроля транскрипции.
[00167] В качестве альтернативы гетерологичные нуклеотидные кодирующие последовательности можно выделить, клонировать и/или перенести из дрожжевых искусственных хромосом (YAC). Полный гетерологичный ген или локус можно клонировать и помещать в одну или более YAC. Если используют несколько YAC, и при этом они содержат перекрывающиеся гомологичные участки, их можно рекомбинировать внутри штаммов дрожжей-хозяев с получением единой конструкции, представляющей полный локус. Плечи YAC можно дополнительно модифицировать с помощью кассет для отбора млекопитающих путем их модифицирования, чтобы содействовать введению конструкций в эмбриональные стволовые клетки или эмбрионы с помощью способов, известных из уровня техники и/или описанных в данном документе.
[00168] Как описано выше, иллюстративные последовательности ДНК и аминокислотные последовательности для применения в создании сконструированного участка DH локуса тяжелой цепи иммуноглобулина представлены в таблицах 3 и 4 соответственно. Другие гетерологичные нуклеотидные кодирующие последовательности также можно найти в базе данных GenBank или в других базах данных последовательностей, известных из уровня техники. Например, последовательности мРНК и аминокислотные последовательности ACKR2 человека можно найти в GenBank под номерами доступа NM_001296.4 и NP_001287.2 соответственно, и они включены в данный документ посредством ссылки. Также, например, последовательности ДНК и аминокислотные последовательности α-конотоксина можно найти в GenBank под номерами доступа JX177132.1 и AFR68318.1 соответственно; δ-конотоксина в GenBank под номерами доступа KR013220.1 и AKD43185.1 соответственно; κ-конотоксина в GenBank под номерами доступа DQ311073.1 и ABD33865.1 соответственно; μ-конотоксина в GenBank под номерами доступа AY207469.1 и AAO48588.1 соответственно и/или ω-конотоксина в GenBank под номерами доступа M84612.1 и AAA81590.1 соответственно; все из которых включены в данный документ посредством ссылки. Кроме того, например, последовательности токсина тарантула Grammostola spatulata можно найти в GenBank под номерами доступа 1TYK_A и 1LUP_A, для SGTX-I можно найти в GenBank под номером доступа 1LA4_A, для хувентоксина-IV (HWTX-IV) можно найти в GenBank под номером доступа P83303.2, для Protoxin-I (ProTxI) в GenBank под номером доступа 2M9L_A, для Protoxin-2 (ProTxII) в GenBank под номером доступа P83476.1; все из которых включены в данный документ посредством ссылки.
[00169] В некоторых вариантах осуществления ДНК-конструкции, содержащие одну или более нуклеотидных кодирующих последовательностей, описываемых в данном документе, содержат последовательности ДНК ACKR2 человека, кодирующие внеклеточную часть полипептида ACKR2 человека, которые функционально связаны с последовательностями сигнала рекомбинации (RSS, т.е. фланкированы 5'-RSS и 3'-RSS) для рекомбинации с генными сегментами иммуноглобулина (например, VH и JH) в трансгенном животном, отличном от человека. В некоторых вариантах осуществления ДНК-конструкции, содержащие одну или более нуклеотидных кодирующих последовательностей, описываемых в данном документе, содержат последовательности ДНК токсина, кодирующие часть пептидного μ-конотоксина и/или токсина тарантула, функционально связанную с последовательностями сигнала рекомбинации (RSS, т.е. фланкированы 5'-RSS и 3'-RSS) для рекомбинации с генными сегментами иммуноглобулинов (например, VH и JH) в трансгенном животном, отличном от человека. Последовательности сигнала рекомбинации могут быть идентичны или практически идентичны последовательностям сигнала рекомбинации, встречающимися в природе (например, геномным), или они могут быть сконструированы человеком (например, оптимизированы). В некоторых вариантах осуществления RSS являются геномными по происхождению, и они включают последовательность или последовательности, которые встречаются в локусе тяжелой цепи иммуноглобулинов, встречающемся в природе (например, локусе тяжелой цепи иммуноглобулина человека или грызуна). Например, ДНК-конструкция может включать последовательности сигнала рекомбинации, расположенные в 5'-фланкирующем и/или 3'-фланкирующем участках нуклеотидной кодирующей последовательности, кодирующей представляющий интерес полипептид, не являющийся иммуноглобулином, или его часть (например, внеклеточную часть гетерологичного полипептида ACKR2 или часть пептидного токсина), которые функционально связаны с нуклеотидной кодирующей последовательностью таким образом, что возможна рекомбинация нуклеотидной кодирующей последовательности с генным сегментом VH и/или JH. В некоторых вариантах осуществления последовательности сигнала рекомбинации содержат последовательность, которая естественным образом ассоциирована со стандартным генным сегментом DH (т.е. RSS, встречающейся в природе). В некоторых вариантах осуществления последовательности сигнала рекомбинации содержат последовательность, которая естественным образом не ассоциирована со стандартным генным сегментом DH. В некоторых вариантах осуществления последовательности сигнала рекомбинации содержат последовательность, которая оптимизирована для рекомбинации с генными сегментами VH и JH. В некоторых вариантах осуществления последовательности сигнала рекомбинации, функционально связанные с одной или более нуклеотидными кодирующими последовательностями, каждая из которых кодирует представляющий интерес полипептид, не являющийся иммуноглобулином, или его часть (например, внеклеточную часть полипептида ACKR2 или часть пептидного токсина), обеспечивают рекомбинацию на уровне, в большей или меньшей степени аналогичном уровню рекомбинации у животного, от которых получена последовательность. Если для оптимизации рекомбинации одной или более нуклеотидных кодирующих последовательностей применяют дополнительные фланкирующие последовательности, то такие последовательности можно клонировать с применением существующих последовательностей в качестве зондов. Дополнительные последовательности, необходимые для максимального увеличения уровня рекомбинации и/или экспрессии вариабельного участка тяжелой цепи, содержащего нуклеотидную кодирующую последовательность полипептида, не являющегося иммуноглобулином (например, ACKR2 или токсина), можно получать из геномных последовательностей или других источников в зависимости от требуемого результата.
[00170] В различных вариантах осуществления каждая из одной или более нуклеотидных кодирующих последовательностей, описываемых в данном документе, фланкирована на 5' и/или 3' с помощью оптимизированных RSS. Иллюстративные оптимизированные RSS, которые можно использовать, приведены на фигуре 2 и описаны в примере 1.
[00171] В различных вариантах осуществления каждая из одной или более нуклеотидных кодирующих последовательностей, описываемых в данном документе, фланкирована на 5' с помощью оптимизированной RSS, имеющей последовательность, которая по меньшей мере на 50% (например, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или больше) идентична 5'-RSS, которая показана на фигуре 2.
[00172] В различных вариантах осуществления каждая из одной или более нуклеотидных кодирующих последовательностей, описываемых в данном документе, фланкирована на 5' с помощью RSS, имеющей последовательность, которая практически идентична или идентична 5’-RSS, которая показана на фигуре 2.
[00173] В различных вариантах осуществления каждая из одной или более нуклеотидных кодирующих последовательностей фланкирована на 3' с помощью RSS, имеющей последовательность, которая по меньшей мере на 50% (например, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или больше) идентична 3’-RSS, которая показана на фигуре 2.
[00174] В различных вариантах осуществления каждая из одной или более нуклеотидных кодирующих последовательностей, описываемых в данном документе, фланкирована на 3' с помощью RSS, имеющей последовательность, которая практически идентична или идентична 5'-RSS, которая показана на фигуре 2.
[00175] В различных вариантах осуществления каждая из одной или более нуклеотидных кодирующих последовательностей, описываемых в данном документе, фланкирована на 5' и 3' с помощью оптимизированных RSS, при этом каждая из 5'- и 3'-RSS имеют последовательность, которая по меньшей мере на 50% (например, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или больше) идентична 5'- и 3'-RSS, показанным на фигуре 2.
[00176] В различных вариантах осуществления каждая из одной или более нуклеотидных кодирующих последовательностей, описываемых в данном документе, фланкирована на 5' и 3' с помощью оптимизированных RSS, при этом каждая из 5'- и 3'-RSS имеет последовательность, которая практически идентична или идентична 5'- и 3'-RSS, показанным на фигуре 2.
[00177] В различных вариантах осуществления каждая из одной или более нуклеотидных кодирующих последовательностей, описываемых в данном документе, фланкирована на 5' и/или 3' с помощью RSS, которые выбраны из показанных на фигуре 2.
[00178] ДНК-конструкции можно получить при помощи способов, известных из уровня техники. Например, ДНК-конструкцию можно получить как часть большей плазмиды. Такой препарат позволяет эффективно клонировать и отбирать правильные конструкции, как известно из уровня техники. ДНК-фрагменты, содержащие одну или более нуклеотидных кодирующих последовательностей, описываемых в данном документе, можно расположить между подходящими сайтами рестрикции на плазмиде, так что их можно легко выделить из остальных последовательностей плазмиды для включения в требуемое животное.
[00179] Различные способы, используемые в получении плазмид и трансформации организмов-хозяев, известны из уровня техники. Другие подходящие систем экспрессии, подходящие как для прокариотических, так и для эукариотических клеток, а также общие процедуры рекомбинации см. в Molecular Cloning: A Laboratory Manual, 2nd Ed., ed. by Sambrook, J. et al., Cold Spring Harbor Laboratory Press: 1989.
Получение животных, отличных от человека, имеющих сконструированный участок DH
[00180] Представлены животные, отличные от человека, в которых экспрессируются антитела, характеризующиеся разнообразием CDR3 тяжелой цепи для управления связыванием с конкретными антигенами, что является результатом интеграции одной или более нуклеотидных кодирующих последовательностей, при этом каждая из одной или более нуклеотидных кодирующих последовательностей кодирует представляющий интерес полипептид, не являющийся иммуноглобулином, или его часть (например, внеклеточную часть атипичного хемокинового рецептора, такого как, например, ACKR2, или часть пептидного токсина), в вариабельный участок тяжелой цепи иммуноглобулина в геноме животного, отличного от человека. Подходящие примеры, описанные в данном документе, включают грызунов, в частности мышей.
[00181] В некоторых вариантах осуществления одна или более гетерологичных кодирующих последовательностей содержат генетический материал от гетерологичного вида (например, человека, пауков, скорпионов, улиток, тарантулов, морских анемон и т.д.), где каждая гетерологичная нуклеотидная кодирующая последовательность кодирует представляющий интерес полипептид, не являющийся иммуноглобулином, или его часть (например, внеклеточную часть полипептида ACKR или часть пептидного токсина), которая содержит кодируемую часть генетического материала от гетерологичного вида. В некоторых вариантах осуществления гетерологичные нуклеотидные кодирующие последовательности, описанные в данном документе, содержат нуклеотидные кодирующие последовательности от гетерологичного вида, которые кодируют представляющий интерес полипептид, не являющийся иммуноглобулином, или его часть (например, внеклеточную часть гетерологичного полипептида ACKR или часть пептидного токсина), при этом часть представляющего интерес полипептида, не являющегося иммуноглобулином, находится в тяжелой цепи иммуноглобулина, в частности, в CDR3 тяжелой цепи, то есть экспрессируется В-клеткой животного, отличного от человека, описываемого в данном документе. Также представлены животные, отличные от человека, эмбрионы, клетки и нацеливающие конструкции для получения животных, отличных от человека, эмбрионов, отличных от человеческих, и клеток, содержащих указанные гетерологичные кодирующие последовательности.
[00182] В различных вариантах осуществления одна или более гетерологичных нуклеотидных кодирующих последовательностей встроены в участок DH вариабельного участка тяжелой цепи иммуноглобулина в пределах генома животного, отличного от человека. В некоторых вариантах осуществления участок DH (или его часть) вариабельного участка тяжелой цепи иммуноглобулина не подвергнут делеции (т.е. является интактным). В некоторых вариантах осуществления участок DH (или его часть) вариабельного участка тяжелой цепи иммуноглобулина изменен, разрушен, подвергнут делеции или заменен одной или более гетерологичными нуклеотидными кодирующими последовательностями (например, одной или более гетерологичными нуклеотидными кодирующими последовательностями ACKR2 или одной или более гетерологичными нуклеотидными последовательностями, кодирующим токсин). В некоторых вариантах осуществления весь или практически весь участок DH заменен одной или более гетерологичными нуклеотидными кодирующими последовательностями; в некоторых определенных вариантах осуществления один или более стандартных генных сегментов DH не подвергали делеции или замене на участок DH вариабельного участка тяжелой цепи иммуноглобулина. В некоторых вариантах осуществления участок DH, описываемый в данном документе, представляет собой синтетический участок DH, при этом синтетический участок DH содержит одну или более гетерологичных нуклеотидных кодирующих последовательностей, описываемых в данном документе. В некоторых вариантах осуществления участок DH представляет собой участок DH человека. В некоторых вариантах осуществления участок DH представляет собой участок DH мыши. В некоторых вариантах осуществления сконструированный участок DH (или его часть), описываемый в данном документе, встроен в вариабельный участок тяжелой цепи иммуноглобулина, так что указанный сконструированный участок DH (или его часть) функционально связан с одним или более генными сегментами VH и/или одним или более генный сегментами JH. В некоторых вариантах осуществления одна или более гетерологичных нуклеотидных кодирующих последовательностей встроены в одну из двух копий вариабельного участка тяжелой цепи иммуноглобулина, что дает в результате животное, отличное от человека, которое является гетерозиготным по одной или более гетерологичным нуклеотидным кодирующим последовательностям (т. е. сконструированному участку DH). В некоторых вариантах осуществления представлено животное, отличное от человека, которое является гомозиготным по одной или более гетерологичным нуклеотидным кодирующим последовательностям (т.е. сконструированному участку DH). В некоторых вариантах осуществления представлено животное, отличное от человека, которое является гетерозиготным по одной или более гетерологичным нуклеотидным кодирующим последовательностям (т.е. сконструированному участку DH).
[00183] В некоторых вариантах осуществления описанное в данном документе животное, отличное от человека, содержит вариабельный участок тяжелой цепи иммуноглобулина человека, который включает участок DH, который содержит одну или более гетерологичных нуклеотидныхкодирующих последовательностей в пределах его генома (например, интегрированных случайным образом). Таким образом, такие животные, отличные от человека, могут быть описаны как имеющие трансген тяжелой цепи иммуноглобулина человека, содержащий сконструированный участок DH. Сконструированный участок DH можно выявить с применением разнообразных способов, в том числе, например, ПЦР, вестерн-блоттинга, Саузерн-блоттинга, определения полиморфизма длины рестрикционных фрагментов (RFLP) или анализа приобретения или потери аллеля. В некоторых вариантах осуществления описанное в данном документе животное, отличное от человека, является гетерозиготным по сконструированному участку DH, описываемому в данном документе. В некоторых вариантах осуществления описанное в данном документе животное, отличное от человека, является гомозиготным по сконструированному участку DH, описываемому в данном документе. В некоторых вариантах осуществления описанное в данном документе животное, отличное от человека, является гемизиготным по сконструированному участку DH, описываемому в данном документе. В некоторых вариантах осуществления описанное в данном документе животное, отличное от человека, содержит одну или более копий сконструированного участка DH, описываемого в данном документе.
[00184] В некоторых вариантах осуществления одна или более гетерологичных нуклеотидных кодирующих последовательностей ACKR, раскрытых в данном документе, являются гетерологичными нуклеотидными кодирующими последовательностями ACKR2. В некоторых вариантах осуществления одна или более гетерологичных нуклеотидных кодирующих последовательностей ACKR2 являются человеческими.
[00185] В некоторых вариантах осуществления одна или более гетерологичных нуклеотидных кодирующих последовательностей токсина, описанные в данном документе, являются гетерологичными нуклеотидными кодирующими последовательностями μ-конотоксина, гетерологичными нуклеотидными кодирующими последовательностями токсина тарантула и/или их комбинациями.
[00186] В различных вариантах осуществления одна или более гетерологичных нуклеотидных кодирующих последовательностей, описанных в данном документе, включают одну или более нуклеотидных кодирующих последовательностей, каждая из которых имеет последовательность, которая по меньшей мере на 50% (например, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или больше) идентична одной или более нуклеотидным кодирующим последовательностям, приведенным в таблице 3 или таблице 4.
[00187] В различных вариантах осуществления одна или более гетерологичных нуклеотидных кодирующих последовательностей, описанных в данном документе, включают одну или более нуклеотидных кодирующих последовательностей, каждая из которых содержит последовательность, которая практически идентична или идентична одной или более нуклеотидным кодирующим последовательностям, приведенным в таблице 3 или таблице 4.
[00188] В различных вариантах осуществления одна или более гетерологичных нуклеотидных кодирующих последовательностей, описанных в данном документе, выбраны из приведенных в таблице 3 и/или таблице 4.
[00189] В различных вариантах осуществления сконструированный участок DH, описанный в данном документе, содержит одну или более гетерологичных нуклеотидных кодирующих последовательностей, каждая из которых имеет последовательность, которая по меньшей мере на 50% (например, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или больше) идентична одной или более нуклеотидным кодирующим последовательностям, которые приведены в таблице 3 или таблице 4.
[00190] В различных вариантах осуществления сконструированный участок DH, описанный в данном документе, содержит одну или более гетерологичных нуклеотидных кодирующих последовательностей, каждая из которых имеет последовательность, которая практически идентична или идентична одной или более нуклеотидным кодирующим последовательностям, приведенным в таблице 3 или таблице 4.
[00191] В различных вариантах осуществления сконструированный участок DH, описанный в данном документе, содержит 5, 10, 15, 20 или 25 гетерологичных нуклеотидных кодирующих последовательностей ACKR2, каждая из которых имеет последовательность, которая идентична 5, 10, 15, 20 или 25 гетерологичным нуклеотидным кодирующим последовательностям ACKR2, приведенным в таблице 3.
[00192] В различных вариантах осуществления сконструированный участок DH, описанный в данном документе, содержит 5, 10, 15, 20, 25 или 26 гетерологичных нуклеотидных кодирующих последовательностей токсина, каждая из которых имеет последовательность, которая идентична 5, 10, 15, 20, 25 или 26 гетерологичным нуклеотидным кодирующим последовательностям токсина, приведенным в таблице 4.
[00193] В различных вариантах осуществления сконструированный участок DH, описанный в данном документе, содержит одну или более гетерологичных нуклеотидных кодирующих последовательностей, каждая из которых фланкирована 5'-последовательностью сигнала рекомбинации (5'-RSS) и 3'-последовательностью сигнала рекомбинации (3’-RSS), при этом каждая из 5-'RSS и 3'-RSS имеет последовательность, которая по меньшей мере на 50% (например, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или больше) идентична 5’-RSS и 3’-RSS, представленным на фигуре 2.
[00194] В различных вариантах осуществления сконструированный участок DH, описанный в данном документе, содержит одну или более гетерологичных нуклеотидных кодирующих последовательностей, каждая из которых фланкирована 5'-последовательностью сигнала рекомбинации (5'-RSS) и 3'-последовательностью сигнала рекомбинации (3'-RSS), при этом каждая из 5'-RSS и 3'-RSS имеет последовательность, которая практически идентична или идентична 5'-RSS и 3'-RSS, представленным на фигуре 2.
[00195] В различных вариантах осуществления сконструированный участок DH, описанный в данном документе, содержит одну или более гетерологичных нуклеотидных кодирующих последовательностей, каждая из которых фланкирована 5'-последовательностью сигнала рекомбинации (5'-RSS) и 3'-последовательностью сигнала рекомбинации (3'-RSS), при этом 5'-RSS и 3'-RSS выбраны из представленных на фигуре 2.
[00196] В различных вариантах осуществления сконструированный участок DH, описанный в данном документе, содержит
(a) SEQ ID NO:39, фланкированную на 5' с помощью SEQ ID NO:59, и фланкированную на 3' с помощью SEQ ID NO:60;
(b) SEQ ID NO:7, фланкированную на 5' с помощью SEQ ID NO:69, и фланкированную на 3' с помощью SEQ ID NO:70;
(c) SEQ ID NO:1, фланкированную на 5' с помощью SEQ ID NO:77, и фланкированную на 3' с помощью SEQ ID NO:78;
(d) SEQ ID NO:33, фланкированную на 5' с помощью SEQ ID NO:87, и фланкированную на 3' с помощью SEQ ID NO:88;
(e) SEQ ID NO:3, фланкированную на 5' с помощью SEQ ID NO:95, и фланкированную на 3' с помощью SEQ ID NO:96, и
(f) SEQ ID NO:5, фланкированную на 5' с помощью SEQ ID NO:103, и фланкированную на 3' с помощью SEQ ID NO:104.
[00197] В различных вариантах осуществления сконструированный участок DH, описанный в данном документе, содержит
(a) SEQ ID NO:41, фланкированную на 5' с помощью SEQ ID NO:61, и фланкированную на 3' с помощью SEQ ID NO:62;
(b) SEQ ID NO:15, фланкированную на 5' с помощью SEQ ID NO:71, и фланкированную на 3' с помощью SEQ ID NO:72;
(c) SEQ ID NO:37, фланкированную на 5' с помощью SEQ ID NO:79, и фланкированную на 3' с помощью SEQ ID NO:80;
(d) SEQ ID NO:13, фланкированную на 5' с помощью SEQ ID NO:81, и фланкированную на 3' с помощью SEQ ID NO:82;
(e) SEQ ID NO:29, фланкированную на 5' с помощью SEQ ID NO:89, и фланкированную на 3' с помощью SEQ ID NO:90;
(f) SEQ ID NO:11, фланкированную на 5' с помощью SEQ ID NO:97, и фланкированную на 3' с помощью SEQ ID NO:98, и
(g) SEQ ID NO:9, фланкированную на 5' с помощью SEQ ID NO:105, и фланкированную на 3' с помощью SEQ ID NO:106.
[00198] В различных вариантах осуществления сконструированный участок DH, описанный в данном документе, содержит
(a) SEQ ID NO:43, фланкированную на 5' с помощью SEQ ID NO:63, и фланкированную на 3' с помощью SEQ ID NO:64;
(b) SEQ ID NO:23, фланкированную на 5' с помощью SEQ ID NO:73, и фланкированную на 3' с помощью SEQ ID NO:74;
(c) SEQ ID NO:17, фланкированную на 5' с помощью SEQ ID NO:83, и фланкированную на 3' с помощью SEQ ID NO:84;
(d) SEQ ID NO:35, фланкированную на 5' с помощью SEQ ID NO:91, и фланкированную на 3' с помощью SEQ ID NO:92;
(e) SEQ ID NO:19, фланкированную на 5' с помощью SEQ ID NO:99, и фланкированную на 3' с помощью SEQ ID NO:100, и
(f) SEQ ID NO:21, фланкированную на 5' с помощью SEQ ID NO:107, и фланкированную на 3' с помощью SEQ ID NO:108.
[00199] В различных вариантах осуществления сконструированный участок DH, описанный в данном документе, содержит
(a) SEQ ID NO:45, фланкированную на 5' с помощью SEQ ID NO:65, и фланкированную на 3' с помощью SEQ ID NO:66;
(b) SEQ ID NO:31, фланкированную на 5' с помощью SEQ ID NO:75, и фланкированную на 3' с помощью SEQ ID NO:76;
(c) SEQ ID NO:25, фланкированную на 5' с помощью SEQ ID NO:85, и фланкированную на 3' с помощью SEQ ID NO:86;
(d) SEQ ID NO:49, фланкированную на 5' с помощью SEQ ID NO:93, и фланкированную на 3' с помощью SEQ ID NO:94;
(e) SEQ ID NO:27, фланкированную на 5' с помощью SEQ ID NO:101, и фланкированную на 3’ с помощью SEQ ID NO:102, и
(f) SEQ ID NO:47, фланкированную на 5' с помощью SEQ ID NO:67, и фланкированную на 3' с помощью SEQ ID NO:68.
[00200] В различных вариантах осуществления сконструированный участок DH, описанный в данном документе, содержит один или более фрагментов ДНК, каждый из которых имеет последовательность, которая по меньшей мере на 50% (например, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или больше) идентична фрагменту ДНК, выбранному из группы, состоящей из SEQ ID NO:131,SEQ ID NO:132,SEQ ID NO:133,SEQ ID NO:134 и их комбинаций.
[00201] В различных вариантах осуществления сконструированный участок DH, описанный в данном документе, содержит один или более фрагментов ДНК, каждый из которых имеет последовательность, которая практически идентична или идентична фрагменту ДНК, выбранному из группы, состоящей из SEQ ID NO:131, SEQ ID NO:132, SEQ ID NO:133,SEQ ID NO:134 и их комбинаций.
[00202] В различных вариантах осуществления сконструированный участок DH, описанный в данном документе, содержит любую из SEQ ID NO:131,SEQ ID NO:132, SEQ ID NO:133 и SEQ ID NO:134.
[00203] В различных вариантах осуществления сконструированный участок DH, описанный в данном документе, содержит SEQ ID NO:131,SEQ ID NO:132, SEQ ID NO:133 и SEQ ID NO:134. В некоторых определенных вариантах осуществления сконструированный участок DH, описанный в данном документе, содержит в направлении от 5' до 3': SEQ ID NO:131,SEQ ID NO:132, SEQ ID NO:133 и SEQ ID NO:134.
[00204] В различных вариантах осуществления сконструированный участок DH, описанный в данном документе, содержит
(a) SEQ ID NO:180, фланкированную на 5' с помощью SEQ ID NO:59, и фланкированную на 3' с помощью SEQ ID NO:60;
(b) SEQ ID NO:182, фланкированную на 5' с помощью SEQ ID NO:69, и фланкированную на 3' с помощью SEQ ID NO:70;
(c) SEQ ID NO:184, фланкированную на 5' с помощью SEQ ID NO:77, и фланкированную на 3' с помощью SEQ ID NO:78;
(d) SEQ ID NO:186, фланкированную на 5' с помощью SEQ ID NO:87, и фланкированную на 3' с помощью SEQ ID NO:88;
(e) SEQ ID NO:188, фланкированную на 5' с помощью SEQ ID NO:95, и фланкированную на 3' с помощью SEQ ID NO:96, и
(f) SEQ ID NO:190, фланкированную на 5' с помощью SEQ ID NO:103, и фланкированную на 3' с помощью SEQ ID NO:104.
[00205] В различных вариантах осуществления сконструированный участок DH, описанный в данном документе, содержит
(a) SEQ ID NO:192, фланкированную на 5' с помощью SEQ ID NO:61, и фланкированную на 3' с помощью SEQ ID NO:62;
(b) SEQ ID NO:194, фланкированную на 5' с помощью SEQ ID NO:71, и фланкированную на 3' с помощью SEQ ID NO:72;
(c) SEQ ID NO:196, фланкированную на 5' с помощью SEQ ID NO:79, и фланкированную на 3' с помощью SEQ ID NO:80;
(d) SEQ ID NO:198, фланкированную на 5' с помощью SEQ ID NO:81, и фланкированную на 3' с помощью SEQ ID NO:82;
(e) SEQ ID NO:200, фланкированную на 5' с помощью SEQ ID NO:89, и фланкированную на 3' с помощью SEQ ID NO:90;
(f) SEQ ID NO:202, фланкированную на 5' с помощью SEQ ID NO:97, и фланкированную на 3' с помощью SEQ ID NO:98, и
(g) SEQ ID NO:204, фланкированную на 5' с помощью SEQ ID NO:105, и фланкированную на 3' с помощью SEQ ID NO:106.
[00206] В различных вариантах осуществления сконструированный участок DH, описанный в данном документе, содержит
(a) SEQ ID NO:206, фланкированную на 5' с помощью SEQ ID NO:63, и фланкированную на 3' с помощью SEQ ID NO:64;
(b) SEQ ID NO:208, фланкированную на 5' с помощью SEQ ID NO:73, и фланкированную на 3' с помощью SEQ ID NO:74;
(c) SEQ ID NO:210, фланкированную на 5' с помощью SEQ ID NO:83, и фланкированную на 3' с помощью SEQ ID NO:84;
(d) SEQ ID NO:212, фланкированную на 5' с помощью SEQ ID NO:91, и фланкированную на 3' с помощью SEQ ID NO:92;
(e) SEQ ID NO:214, фланкированную на 5' с помощью SEQ ID NO:99, и фланкированную на 3' с помощью SEQ ID NO:100, и
(f) SEQ ID NO:216, фланкированную на 5' с помощью SEQ ID NO:107, и фланкированную на 3' с помощью SEQ ID NO:108.
[00207] В различных вариантах осуществления сконструированный участок DH, описанный в данном документе, содержит
(a) SEQ ID NO:218, фланкированную на 5' с помощью SEQ ID NO:65, и фланкированную на 3’ с помощью SEQ ID NO:66;
(b) SEQ ID NO:220, фланкированную на 5' с помощью SEQ ID NO:75, и фланкированную на 3' с помощью SEQ ID NO:76;
(c) SEQ ID NO:222, фланкированную на 5' с помощью SEQ ID NO:85, и фланкированную на 3' с помощью SEQ ID NO:86;
(d) SEQ ID NO:224, фланкированную на 5' с помощью SEQ ID NO:93, и фланкированную на 3' с помощью SEQ ID NO:94;
(e) SEQ ID NO:226, фланкированную на 5' с помощью SEQ ID NO:101, и фланкированную на 3' с помощью SEQ ID NO:102;
(f) SEQ ID NO:228, фланкированную на 5' с помощью SEQ ID NO:109, и фланкированную на 3' с помощью SEQ ID NO:110, и
g) SEQ ID NO:230, фланкированную с 5' с помощью SEQ ID NO:67, и фланкированную с 3' с помощью SEQ ID NO:68.
[00208] В различных вариантах осуществления сконструированный участок DH, описанный в данном документе, содержит один или более фрагментов ДНК, каждый из которых имеет последовательность, которая по меньшей мере на 50% (например, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или больше) идентична фрагменту ДНК, выбранному из группы, состоящей из SEQ ID NO:232,SEQ ID NO:233,SEQ ID NO:234,SEQ ID NO:235 и их комбинаций.
[00209] В различных вариантах осуществления сконструированный участок DH, описанный в данном документе, содержит один или более фрагментов ДНК, каждый из которых имеет последовательность, которая практически идентична или идентична фрагменту ДНК, выбранному из группы, состоящей из SEQ ID NO:232,SEQ ID NO:233,SEQ ID NO:234, SEQ ID NO:235 и их комбинаций.
[00210] В различных вариантах осуществления сконструированный участок DH, описанный в данном документе, содержит любую из SEQ ID NO:232,SEQ ID NO:233,SEQ ID NO:234 и SEQ ID NO:235.
[00211] В различных вариантах осуществления сконструированный участок DH, описанный в данном документе, содержит SEQ ID NO:232,SEQ ID NO:233, SEQ ID NO:234 и SEQ ID NO:235. В некоторых определенных вариантах осуществления сконструированный участок DH описанный в данном документе, содержит в направлении от 5' до 3': SEQ ID NO:232,SEQ ID NO:233,SEQ ID NO:234 и SEQ ID NO:235.
[00212] В различных вариантах осуществления антитело, продуцируемое описанным в данном документе животным, отличным от человека, содержит вариабельный участок тяжелой цепи, который включает CDR3, имеющий аминокислотную последовательность, которая является или содержит аминокислотную последовательность, которая по меньшей мере на 50% (например, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или больше) идентична аминокислотной последовательности, которая представлена в таблице 3 или таблице 4.
[00213] В различных вариантах осуществления антитело, продуцируемое описанным в данном документе животным, отличным от человека, содержит вариабельный участок тяжелой цепи, который включает CDR3, имеющий аминокислотную последовательность, которая является или содержит аминокислотную последовательность, которая практически идентична или идентична аминокислотной последовательности, которая представлена в таблице 3 или таблице 4.
[00214] В различных вариантах осуществления антитело, продуцируемое описанным в данном документе животным, отличным от человека, содержит вариабельный участок тяжелой цепи, который включает CDR3, имеющий аминокислотную последовательность, которая представляет собой вариант (т.е. ее подвергшийся соматической мутации вариант) аминокислотной последовательности, который является или содержит аминокислотную последовательность, которая представлена в таблице 3 или таблице 4.
[00215] Представлены композиции и способы получения животных, отличных от человека, геном которых содержит вариабельный участок тяжелой цепи иммуноглобулина, который включает сконструированный участок DH, где сконструированный участок DH включает одну или более нуклеотидных кодирующих последовательностей, каждая из которых кодирует часть представляющего интерес полипептида, в том числе нуклеотидные кодирующие последовательности специфических полиморфных форм представляющего интерес полипептида, аллельных вариантов представляющего интерес полипептида (например, отличия в одну аминокислоту) или изоформ представляющего интерес полипептида, которые получены вследствие альтернативно сплайсинга, в том числе композиции и способы получения животных, отличных от человека, которые экспрессируют антитела, содержащие вариабельный участок тяжелой цепи, который включает CDR3, имеющий аминокислотную последовательность, кодируемую такими нуклеотидными кодирующими последовательностями из локуса тяжелой цепи иммуноглобулина, который содержит генные сегменты VH и JH человека, функционально связанные с одним или более генами константного участка тяжелой цепи, отличной от человеческой. В некоторых вариантах осуществления также представлены композиции и способы получения животных, отличных от человека, которые экспрессируют такие антитела под контролем эндогенного энхансера(ов) и/или эндогенной регуляторной последовательности(ей). В некоторых вариантах осуществления также представлены композиции и способы получения животных, отличных от человека, которые экспрессируют такие антитела под контролем гетерологичного энхансера(ов) и/или гетерологичной регуляторной последовательности(ей). Способы включают осуществление вставки одной или более нуклеотидных кодирующих последовательностей, каждая из которых кодирует часть представляющего интерес полипептида, или одного или более фрагментов ДНК, каждый из которых содержит несколько нуклеотидных кодирующих последовательностей, каждая из которых кодирует часть представляющего интерес полипептида, в геном животного, отличного от человека, так что экспрессируется антитело, которое включает указанную часть представляющего интерес полипептида.
[00216] В некоторых вариантах осуществления способы включают осуществление вставки ДНК размером приблизительно 10050 п.о., которая включает шесть нуклеотидных кодирующих последовательностей, соответствующих внеклеточным частям гетерологичного полипептида ACKR2, ДНК размером приблизительно 9768 п.о., которая включает семь нуклеотидных кодирующих последовательностей, соответствующих внеклеточным частям гетерологичного полипептида ACKR2, ДНК размером приблизительно 9788 п.о., которая включает шесть нуклеотидных кодирующих последовательностей, соответствующих внеклеточным частям гетерологичного полипептида ACKR2, и/или ДНК размером приблизительно 11906 п.о., которая включает шесть нуклеотидных кодирующих последовательностей, соответствующих внеклеточным частям гетерологичного полипептида ACKR2. Вместе такая ДНК включает 25 кодирующих последовательностей, соответствующих внеклеточным частям гетерологичного полипептида ACKR2. В некоторых вариантах осуществления способы включают осуществление вставки ДНК, которая дополнительно содержит последовательности сигнала рекомбинации, которые фланкируют каждую из нуклеотидных кодирующих последовательностей, соответствующих внеклеточным частям гетерологичного полипептида ACKR2. Генетический материал, который включает нуклеотидные кодирующие последовательности, соответствующие внеклеточным частям гетерологичного полипептида ACKR2, и фланкирующие последовательности сигнала рекомбинации, описанные выше, может быть вставлен в геном животного, отличного от человека, за счет чего создается животное, отличное от человека, имеющее сконструированный участок DH, который содержит нуклеотидные кодирующие последовательности, соответствующие внеклеточным частям гетерологичного полипептида ACKR2, и последовательности сигнала рекомбинации, необходимые для обеспечения возможности рекомбинации со смежными генными сегментами VH и JH.
[00217] В некоторых вариантах осуществления способы включают осуществление вставки ДНК размером приблизительно 9812 п.о., которая включает шесть нуклеотидных кодирующих последовательностей, соответствующих частям одного или более токсинов (например, μ-конотоксина и/или токсина тарантула), ДНК размером приблизительно 9512 п.о, которая включает семь нуклеотидных кодирующих последовательностей, соответствующих частям одного или более токсинов (например, μ-конотоксина и/или токсина тарантула), ДНК размером приблизительно 9691 п.о., которая включает шесть нуклеотидных кодирующих последовательностей, соответствующих частям одного или более токсинов (например, μ-конотоксина и/или токсина тарантула), и/или ДНК размером приблизительно 11896 п.о., которая включает семь нуклеотидных кодирующих последовательностей, соответствующих частям одного или более токсинов (например, μ-конотоксина и/или токсина тарантула). Вместе такая ДНК включает 26 кодирующих последовательностей, соответствующих частям одного или более пептидных токсинов. В некоторых вариантах осуществления способы включают осуществление вставки ДНК, которая дополнительно содержит последовательности сигнала рекомбинации, которые фланкируют каждую из нуклеотидных кодирующих последовательностей, соответствующих частям одного или более пептидных токсинов. Генетический материал, который включает нуклеотидные кодирующие последовательности, соответствующие частям одного или более пептидных токсинов, и фланкирующие последовательности сигнала рекомбинации, описанные выше, может быть вставлен в геном животного, отличного от человека, за счет чего создается животное, отличное от человека, имеющее сконструированный участок DH, который содержит нуклеотидные кодирующие последовательности, соответствующие частям одного или более пептидных токсинов, и последовательности сигнала рекомбинации, необходимые для обеспечения возможности рекомбинации со смежными генными сегментами VH и JH.
[00218] При необходимости нуклеотидные кодирующие последовательности, соответствующие частям гетерологичного представляющего интерес полипептида, могут быть модифицированы для включения кодонов, которые оптимизированы для экспрессии в животном, отличном от человека (например, см. патенты США №№ 5670356 и 5874304). Кодон-оптимизированные последовательности представляют собой синтетические последовательности, и они предпочтительно кодируют идентичный полипептид (или биологически активный фрагмент полноразмерного полипептида, обладающий практически такой же активностью, что и полноразмерный полипептид), кодируемый исходным полинуклеотидом, не являющимся кодон-оптимизированным. В некоторых вариантах осуществления нуклеотидные кодирующие последовательности, соответствующие частям гетерологичного представляющего интерес полипептида, могут включать измененную последовательность, чтобы оптимизировать частоту использования кодонов для конкретного типа клетки (например, клетки грызуна). Например, кодоны нуклеотидных кодирующих последовательностей, соответствующих частям гетерологичного представляющего интерес полипептида, подлежащих вставке в геном животного, отличного от человека (например, грызуна), можно оптимизировать для экспрессии в клетке животного, отличного от человека. Такая последовательность может быть описана как кодон-оптимизированная последовательность.
[00219] Вставку нуклеотидных кодирующих последовательностей, соответствующих частям гетерологичного представляющего интерес полипептида, в участок DH осуществляют таким образом, что указанные нуклеотидные кодирующие последовательности, функционально связанные с генными сегментами VH и JH (например, несколькими генными сегментами VH и JH), приводят к относительно минимальной модификации генома, результатом чего является экспрессия антител, содержащих тяжелые цепи, которые характеризуются CDR3, имеющими аминокислотные последовательности, соответствующие частям гетерологичного представляющего интерес полипептида в животном, отличном от человека.
[00220] Способы получения трансгенных животных, отличных от человека, в том числе с помощью нокаутов и нокинов, хорошо известны в данной области (см., например, Gene Targeting: A Practical Approach, Joyner, ed., Oxford University Press, Inc. (2000)). Например, получение трансгенных грызунов необязательно может подразумевать разрушение генетических локусов одного или более эндогенных генов (или генных сегментов) грызуна и введение одного или более гетерологичных генов (или генных сегментов, или нуклеотидных кодирующих последовательностей) в геном грызуна, в некоторых вариантах осуществления в то же местоположение, что и эндогенный ген (или генные сегменты) грызуна. В некоторых вариантах осуществления нуклеотидные кодирующие последовательности, соответствующие частям гетерологичного представляющего интерес полипептида, вводят в участок DH из вставленного случайным образом локуса тяжелой цепи иммуноглобулина локус в геноме грызуна. В некоторых вариантах осуществления нуклеотидные кодирующие последовательности, соответствующие частям гетерологичного представляющего интерес полипептида, вводят в участок DH эндогенного локуса тяжелой цепи иммуноглобулина в геноме грызуна; в некоторых определенных вариантах осуществления эндогенный локус тяжелой цепи иммуноглобулина изменен, модифицирован или сконструирован таким образом, что он содержит генные сегменты человека (например, V и/или J), функционально связанные с одним или более генами константного участка (например, человека или мыши).
[00221] Представлена схематическая иллюстрация (не в масштабе) иллюстративной стратегии конструирования нацеливающего вектора для интеграции в эмбриональные стволовые (ES) клетки грызуна для получения грызуна, геном которого содержит вариабельный участок тяжелой цепи иммуноглобулина, который включает сконструированный кластер разнообразия (т.е. участок DH), при этом кластер разнообразия включает одну или более нуклеотидных кодирующих последовательностей, каждая из которых кодирует часть гетерологичного полипептида ACKR2 (например, внеклеточную часть D6 хемокинового рецептора-ловушки), как представлено на фигурах 3A и 3B, или каждая из которых кодирует часть пептидного токсина (например, часть μ-конотоксина и/или пептидного токсина тарантула), как представлено на фигурах 7A и 7B. Как показано на иллюстрации, фрагменты ДНК, каждый из которых содержит различное количество нуклеотидных кодирующих последовательностей, каждая из которых кодирует внеклеточную часть гетерологичного полипептида ACKR2 (фигуры 3A–3B) или каждая из которых кодирует часть пептидного токсина (фигуры 7A–7B), собраны вместе с применением изотермической сборки вместе с кассетой для отбора (например, по устойчивости к неомицину), фланкированной сайтами распознавания для сайт-специфической рекомбинации (например, loxP). Альтернативная стратегия, в которой используют последовательное лигирование фрагментов ДНК, проиллюстрирована на фигурах 4A и 4B (для нуклеотидных кодирующих последовательностей ACKR2) и 8A и 8B (для нуклеотидных кодирующих последовательностей токсина) соответственно. Фрагменты ДНК включают последовательности сигнала рекомбинации, которые фланкируют (на 5' и с 3') каждую из нуклеотидных кодирующих последовательностей, чтобы обеспечить рекомбинацию с генными сегментами VH и JH, после их интеграции в локус тяжелой цепи иммуноглобулина. Последовательности сигнала рекомбинации могут быть получены из геномных источников или, в качестве альтернативы, они могут быть оптимизированы, как описано в данном документе, для обеспечения эффективной рекомбинации нуклеотидных кодирующих последовательностей с функционально связанными генными сегментами VH и JH. Нуклеотидные кодирующие последовательности сами по себе могут быть оптимизированными, вследствие чего соматическая гипермутация во время созревания аффинности последовательностей антител происходит на уровне, примерно на уровне или выше уровня соматической гипермутации, наблюдаемой у животных дикого типа, отличных от человека, которые содержат локус тяжелой цепи иммуноглобулина, включающий стандартные генные сегменты DH, или животного, отличного от человека, которое является трансгенным по локусу тяжелой (и/или легкой цепи) иммуноглобулина человека, который включает стандартные генные сегменты DH человека. После сборки собранные фрагменты ДНК (т.е. сконструированный участок DH) лигируют в вектор BAC, который содержит геномную ДНК вариабельного участка тяжелой цепи (например, человека), для создания нацеливающего вектора для интеграции в локус тяжелой цепи иммуноглобулина. Лигирование осуществляют таким образом, что сконструированный участок DH фланкирован на 5' с помощью геномной ДНК VH и на 3' с помощью геномной ДНК JH и геномной ДНК константного участка тяжелой цепи (например, интронным энхансером и геном константного участка IgM), отличной от человеческой (например, грызуна). Окончательный нацеливающий вектор для встраивания в геном клетки, отличной от человеческой (например, эмбриональной стволовой клетки грызуна), содержит геномную ДНК VH (например, содержащую один или более генных сегментов VH), сконструированный участок DH, на 3' геномную ДНК JH и геномную ДНК константного участка тяжелой цепи, отличную от человеческой (например, грызуна), все из которых функционально связаны для обеспечения рекомбинации между генными сегментами VH, нуклеотидной кодирующей последовательностью в пределах сконструированного участка DH и генным сегментом JH после интеграции в геном животного, отличного от человека.
[00222] Нацеливающий вектор вводят в эмбриональные стволовые клетки грызуна (например, мыши) таким образом, что последовательность, содержащаяся в нацеливающем векторе (т.е. сконструированный участок DH), приводит к способности клетки, отличной от человеческой, или животного, отличного от человека (например, мыши), экспрессировать антитела, которые содержат CDR3, имеющие аминокислоты, кодируемые встроенными нуклеотидными кодирующими последовательностями.
[00223] Как описано в данном документе, получают трансгенного грызуна, у которого сконструированный участок DH был введен в эндогенный локус тяжелой цепи иммуноглобулина генома грызуна (например, эндогенный локус тяжелой цепи иммуноглобулина, сконструированный таким образом, что он содержит генные сегменты вариабельного участка человека). Иммуноглобулины экспрессируются на мышиных клетках, при этом иммуноглобулины имеют CDR3, содержащие аминокислоты, кодируемые нуклеотидными кодирующими последовательностями из сконструированного участка DH. Иммуноглобулины дополнительно содержат константные участки мыши и, следовательно, обеспечивают необходимые эффекторные функции для разных иммунных клеток в иммунной системе грызуна. Если эндогенный локус тяжелой цепи иммуноглобулина в геноме грызуна не подвергается нацеливанию под действием нацеливающего вектора, то сконструированный участок DH предпочтительно встраивается в локус тяжелой цепи иммуноглобулина в местоположение, отличное от местоположения эндогенного локуса тяжелой цепи иммуноглобулина мыши (например, встраивается случайным образом). В таких случаях эндогенный локус тяжелой цепи иммуноглобулина мыши можно подвергнуть делеции или иным образом сделать нефункциональным (например, путем нацеленной делеции, вставки, инверсии и т.д.), вследствие чего в антителах, продуцируемых животным, отличным от человека, используются генные сегменты и последовательности из локуса тяжелой цепи иммуноглобулина, содержащего встроенный сконструированный участок DH.
[00224] В некоторых вариантах осуществления геном описанного в данном документе животного, отличного от человека, дополнительно содержит один или более генов тяжелой и/или легкой цепей иммуноглобулина человека (см., например, патент США №8502018; патент США № 8642835; патент США № 8697940; патент США №8791323; и заявку на патент США № 2013/0096287 A1; включенные в данный документ посредством ссылки). В качестве альтернативы сконструированный участок DH можно вводить в эмбриональную стволовую клетку другой модифицированной линии, такой как, например, линия VELOCIMMUNE® (см., например, патент США №8502018 или патент США № 8642835; включенные в данный документ посредством ссылки). В некоторых вариантах осуществления сконструированный участок DH можно вводить в эмбриональную стволовую клетку модифицированной линии, описываемой в патентах США №№8697940 и 8642835; включенных в данный документ посредством ссылки.
[00225] В некоторых вариантах осуществления геном описанного в данном документе животного, отличного от человека, дополнительно содержит (например, за счет проведения кроссбридинга или стратегий множественного нацеливания) один или более генов или локусов легкой цепи иммуноглобулина человека (например, трансгенных, эндогенных и т.д.), описываемых в публикациях заявок на патенты США №№2011-0195454 A1, 2012-0021409 A1, 2012-0192300 A1, 2013-0045492 A1, 2013-0185821 A1, 2013-0198880 A1, 2013-0302836 A1, 2015-0059009 A1; публикациях международных патентных заявок №№ WO 2011/097603, WO 2012/148873, WO 2013/134263, WO 2013/184761, WO 2014/160179, WO 2014/160202; все из которых включены в данный документ посредством ссылки.
[00226] Было описано генетическое конструирование единой перегруппированной легкой цепи, например, легкой цепи, содержащей перегруппированный вариабельный участок легкой цепи. Например, получение универсальной легкой цепи (ULC) мыши, содержащей генную последовательность единого перегруппированного вариабельного участка VL:JL, и получение антиген-специфических антител в таких мышах, например, описано в заявках на патент США №№13/022759, 13/093156, 13/412936, 13/488628, 13/798310 и 13/948818 (публикации №№2011/0195454, 2012/0021409, 2012/0192300, 2013/0045492, US 20130185821 и US 20130302836 соответственно), каждая из которых включена в данный документ посредством ссылки во всей своей полноте. Сконструированная общая легкая цепь мыши, описанная в публикации заявки на патент США №№2011/0195454, 2012/0021409, 2012/0192300 и 2013/0045492, содержит последовательность нуклеиновой кислоты, кодирующую ограниченный репертуар вариантов легкой цепи, например, общую или универсальную легкую цепь «ULC», которая содержит не более двух генных сегментов VL, или последовательность единого перегруппированного вариабельного участка легкой цепи иммуноглобулина человека. Для обеспечения такого ограниченного репертуара мышь была сконструирована таким образом, чтобы сделать нефункциональной или практически нефункциональной ее способность создавать или перегруппировывать нативный вариабельный домен легкой цепи мыши. В одном аспекте это обеспечивается, например, за счет осуществления делеции генных сегментов вариабельного участка легкой цепи мыши. Как описано ранее, затем эндогенный локус мыши может быть модифицирован с помощью выбранных подходящих экзогенных генных сегментов вариабельного участка легкой цепи, предпочтительно генных сегментов вариабельного участка легкой цепи человека, функционально связанных с эндогенным константным доменом легкой цепи мыши таким образом, что такие экзогенные генные сегменты вариабельного участка могут объединяться с эндогенным геном константного участка легкой цепи мыши и они образуют перегруппированный ген обратно химерной легкой цепи (вариабельный участок человека, константный участок мыши). В различных вариантах осуществления вариабельный участок легкой цепи способен подвергаться соматической мутации. В различных вариантах осуществления для максимального увеличения способности вариабельного участка легкой цепи приобретать соматические мутации у мыши сохраняют соответствующий энхансер(ы). В одном аспекте при модификации локуса легкой κ-цепи мыши для замены эндогенных генных сегментов легкой κ-цепи мыши на генные сегменты легкой κ-цепи человека интронный энхансер κ-цепи мыши и 3'-энхансер мыши оставляют функциональными или неразрушенными.
[00227] Таким образом, представлена генетически сконструированная мышь, в которой экспрессируется ограниченный репертуар обратно химерных (вариабельный участок человека, константный участок мыши) легких цепей, ассоциированных с разнообразием обратно химерных (вариабельный участок человека, константный участок мыши) тяжелых цепей, содержащих аминокислоты, кодируемые генетически сконструированным участком DH (или его частью), описываемыми в данном документе. В различных вариантах осуществления эндогенные генные сегменты легкой κ-цепи мыши подвергнуты делеции и заменены единым (или двумя) перегруппированным участком легкой цепи человека, функционально связанным с эндогенным геном Cκ мыши. В некоторых вариантах осуществления для максимального увеличения уровня соматической гипермутации перегруппированного участка легкой цепи человека сохраняют интронный энхансер κ-цепи мыши и 3'-энхансер мыши. В различных вариантах осуществления мышь также содержит нефункциональный локус легкой κ-цепи, или он подвергнут делеции, или содержит делецию, которая делает локус не способным к образованию легкой κ-цепи.
[00228] Таким образом, в одном варианте осуществления в данном документе представлено животное, отличное от человека (например, грызун, например, мышь или крыса), которое содержит в своем геноме, например, в своей зародышевой линии, ограниченный репертуар предпочтительных вариабельных участков легкой цепи человека или единый перегруппированный вариабельный участок легкой цепи человека из ограниченного репертуара предпочтительных генных сегментов вариабельного участка легкой цепи человека, где животное, отличное от человека, также содержит в своем геноме, например, в своей зародышевой линии, вариабельный участок тяжелой цепи иммуноглобулина, который включает сконструированный участок DH, где сконструированный участок DH включает одну или более нуклеотидных последовательностей, каждая из которых кодирует представляющий интерес полипептид, не являющийся иммуноглобулином, или его часть.
[00229] Представлены генетически сконструированные животные, в которых экспрессируется ограниченный репертуар вариабельных доменов легкой цепи человека или единый вариабельный домен легкой цепи человека из ограниченного репертуара генных последовательностей вариабельного участка легкой цепи человека. В одном варианте осуществления последовательность единого перегруппированного участка V/J легкой цепи человека выбрана из Vκ1-39Jκ и Vκ3-20Jκ, например, Vκ1-39Jκ5 и Vκ3-20Jκ1. В некоторых вариантах осуществления животное, отличное от человека, раскрываемое в данном документе, содержит модифицированный локус легкой цепи, содержащий замену всех эндогенных функциональных генных сегментов VL и всех эндогенных функциональных генных сегментов JL на последовательность единого перегруппированного участка V/J легкой цепи, где последовательность единого перегруппированного участка V/J легкой цепи функционально связана с эндогенным геном константного участка легкой цепи. В некоторых вариантах осуществления модифицированный локус легкой цепи расположен в геноме зародышевой линии животного, отличного от человека. В одном варианте осуществления животное, отличное от человека, содержит в своем геноме зародышевой линии генную последовательность единого перегруппированного вариабельного участка легкой цепи, функционально связанную с генной последовательностью константного участка легкой цепи, где генная последовательность единого перегруппированного вариабельного участка легкой цепи содержит генные сегменты VL зародышевой линии человека и JL зародышевой линии человека, например, Vκ1-39 зародышевой линии человека и Jκ5 зародышевой линии человека или Vκ3-20 и Jκ1 зародышевой линии человека. В некоторых вариантах осуществления животное, отличное от человека, раскрываемое в данном документе, содержит B-клетку, например, В-клетку, которая не подверглась переключению класса, содержащую в своем геноме последовательность единого перегруппированного участка V/J легкой цепи, функционально связанную с эндогенным геном константного участка легкой цепи, где единый перегруппированный участок V/J легкой цепи не содержит соматические мутации по сравнению с последовательностью единого перегруппированного участка V/J легкой цепи, функционально связанной с эндогенным геном константного участка легкой цепи, обнаруженном в геноме зародышевого линии животного, отличного от человека. В других вариантах осуществления животное, отличное от человека, раскрываемое в данном документе, содержит B-клетку, например, В-клетку, которая подверглась переключению класса, содержащую в своем геноме последовательность единого перегруппированного участка V/J легкой цепи, функционально связанную с эндогенным геном константного участка легкой цепи, где единый перегруппированный участок V/J легкой цепи содержит соматические мутации по сравнению с последовательностью единого перегруппированного участка V/J легкой цепи, функционально связанную с эндогенным геном константного участка легкой цепи, обнаруженном в геноме зародышевого линии животного, отличного от человека.
[00230] Соответственно, представлено генетически модифицированное животное, отличное от человека, а также способы и композиции для получения данного животного, где генетическая модификация предусматривает сконструированный участок DH и локус единой перегруппированной легкой цепи, и где в данном животном дополнительно экспрессируется генетически сконструированная единая перегруппированная легкая цепь, например, сконструированная общая легкая цепь (ULC), которая может быть ассоциирована с тяжелой цепью, содержащей аминокислоты, кодируемые сконструированным участком DH в тканях или клетках животного, отличного от человека.
[00231] Трансгенное животное-основателя, отличное от человека, можно идентифицировать на основе присутствия в его геноме сконструированного участка DH и/или экспрессии антител, содержащих аминокислоты, кодируемые нуклеотидными кодирующими последовательностями, в тканях или клетках животного, отличного от человека. Затем трансгенное животное-основателя, отличное от человека, можно использовать для разведения дополнительных животных, отличных от человека, несущих сконструированный участок DH, получая тем самым серию животных, отличных от человека, каждое из которых несет одну или более копий сконструированного участка DH. Более того, трансгенных животных, отличных от человека, несущих сконструированный участок DH, при необходимости можно дополнительно скрещивать с другими трансгенными животными, отличными от человека, несущими другие трансгены (например, гены иммуноглобулина человека).
[00232] Можно также получать трансгенных животных, отличных от человека, содержащих выбранные системы, которые позволяют регулировать экспрессию трансгена или управлять ею. Иллюстративные системы включают систему рекомбиназ Cre/loxP из бактериофага P1 (см., например, Lakso, M. et al., 1992, Proc. Natl. Acad. Sci. U.S.A. 89:6232-6236) и систему рекомбиназ FLP/Frt из S. cerevisiae (O'Gorman, S. et al, 1991, Science 251:1351-1355). Таких животных можно получить путем создания «дважды» трансгенных животных, например путем спаривания двух трансгенных животных, одного с трансгеном, содержащим выбранную модификацию (например, сконструированный участок DH), и другого с трансгеном, кодирующим рекомбиназу (например, рекомбиназу Cre).
[00233] Животных, отличных от человека, описанных в данном документе, можно получить, как описано выше, или с использованием способов, известных из уровня техники, чтобы они содержали дополнительные человеческие или гуманизированные гены, зачастую в зависимости от предполагаемого применения животного, отличного от человека. При необходимости, генетический материал таких дополнительных человеческих или гуманизированных генов можно вводить посредством дальнейшего изменения генома клеток (например, эмбриональных стволовых клеток), имеющих генетические модификации, описываемые выше, или с помощью методик скрещивания, известных из уровня техники, с другими генетически модифицированными линиями. В некоторых вариантах осуществления описанных в данном документе животных, отличных от человека, получают таким образом, что они дополнительно содержат трансгенные гены тяжелой и легкой цепи иммуноглобулина человека (см., например, Murphy, A.J. et al., (2014) Proc. Natl. Acad. Sci. U.S.A. 111(14):5153-5158; патент США № 8502018; патент США № 8642835; патент США № 8697940; патент США № 8791323 и публикацию заявки на патент США № 2013/0096287 A1; включенные в данный документ посредством ссылки). В некоторых вариантах осуществления описанных в данном документе животных, отличных от человека, получают таким образом, что они дополнительно содержат один или более генов или локусов легкой цепи иммуноглобулина человека (например, трансгенных, эндогенных), описываемых в публикациях заявок на патент США №№ 2011-0195454 A1, 2012-0021409 A1, 2012-0192300 A1, 2013-0045492 A1, 2013-0185821 A1, 2013-0198880 A1, 2013-0302836 A1, 2015-0059009 A1; публикациях международных патентных заявок №№ WO 2011/097603, WO 2012/148873, WO 2013/134263, WO 2013/184761, WO 2014/160179, WO 2014/160202; все из которых включены в данный документ посредством ссылки.
[00234] В некоторых вариантах осуществления описанных в данном документе животных, отличных от человека, можно получить путем введения нацеливающего вектора, описываемого в данном документе, в клетку из модифицированной линии. В виде лишь одного примера, нацеливающий вектор, описываемый выше, может быть введен в мышь VELOCIMMUNE®. Мыши VELOCIMMUNE® экспрессируют антитела, которые имеют полностью человеческие вариабельные участки и константные участки мыши. В некоторых вариантах осуществления описанных в данном документе животных, отличных от человека, получают таким образом, что они дополнительно содержат гены иммуноглобулина человека (гены вариабельного и/или константного участка). В некоторых вариантах осуществления описанные в данном документе животные, отличные от человека, содержат сконструированный участок DH, описываемый в данном документе, и генетический материал от гетерологичного вида (например, человека), где генетический материал кодирует, полностью или частично, один или более вариабельных участков тяжелой и/или легкой цепей человека.
[00235] Например, описанные в данном документе животные, отличные от человека, содержащие сконструированный участок DH, могут дополнительно содержать (например, за счет кроссбридинга или стратегий множественного нацеливания) одну или более модификаций, которые описаны в Murphy, A.J. et al., (2014) Proc. Natl. Acad. Sci. U.S.A. 111(14):5153-5158; патенте США № 8502018; патенте США № 8642835; патенте США № 8697940; патенте США № 8791323; публикациях заявок на патент США №№ 2011-0195454 A1, 2012-0021409 A1, 2012-0192300 A1, 2013-0045492 A1, 2013-0096287 A1, 2013-0185821 A1, 2013-0198880 A1, 2013-0302836 A1, 2015-0059009 A1; публикациях международных патентных заявок №№ WO 2011/097603, WO 2012/148873, WO 2013/134263, WO 2013/184761, WO 2014/160179 или WO 2014/160202; при этом все данные заявки включены в данный документ посредством ссылки во всей своей полноте. В некоторых вариантах осуществления грызуна, содержащего сконструированный участок DH, описываемый в данном документе, скрещивают с грызуном, содержащим локус гуманизированного вариабельного участка тяжелой и/или легкой цепи иммуноглобулина (см., например, патент США № 8502018 или патент США № 8642835; включенные в данный документ посредством ссылки). В некоторых вариантах осуществления грызуна, содержащего сконструированный участок DH, описываемый в данном документе, скрещивают с грызуном, содержащим гены или локусы гуманизированной легкой цепи иммуноглобулина, как описано в публикациях заявок на патент США №№ 2011-0195454 A1, 2012-0021409 A1, 2012-0192300 A1, 2013-0045492 A1, 2013-0185821 A1, 2013-0198880 A1, 2013-0302836 A1, 2015-0059009 A1; публикациях международных патентных заявок №№ WO 2011/097603, WO 2012/148873, WO 2013/134263, WO 2013/184761, WO 2014/160179, WO 2014/160202; все из которых включены в данный документ посредством ссылки.
[00236] Хотя в данном документе подробно обсуждаются варианты осуществления, в которых сконструированный участок DH используется в мыши (т.е. мышь со сконструированными участком DH, функционально связанным с генными сегментами VH и JH человека, все из которых функционально связаны с одним или более генами константного участка тяжелой цепи мыши), также представлены другие животные, отличные от человека, которые содержат сконструированный участок DH. В некоторых вариантах осуществления такие животные, отличные от человека, содержат сконструированный участок DH, функционально связанный с эндогенными генными сегментами VH и JH. В некоторых вариантах осуществления такие животные, отличные от человека, содержат сконструированный участок DH, функционально связанный с гуманизированными генными сегментами VH и JH. Такие животные, отличные от человека, включают любых животных, которых можно генетически модифицировать для экспрессии антител, имеющих CDR3, которые включают аминокислоты, кодируемые нуклеотидными кодирующими последовательностями, соответствующими части представляющего интерес полипептида, раскрываемого в данном документе, в том числе, например, млекопитающих, например, мышь, крысу, кролика, свинью, представителя бычьих (например, корову, быка, буйвола), оленя, овцу, козу, курицу, кошку, собаку, хорька, примата (например, игрунку, макака-резуса) и т.д. Например, у тех животных, отличных от человека, у которых сложно получить подходящие генетически модифицируемые ES-клетки, для получения животного, отличного от человека, содержащего генетическую модификацию используют другие способы. Такие способы включают, например, модификацию генома клеток, отличных от ES-клеток (например, фибробласта или индуцированной плюрипотентной клетки), и использование переноса ядер соматических клеток (SCNT) для переноса генетически модифицированного генома в подходящую клетку, например, в энуклеированный ооцит, и вынашивание модифицированной клетки (например, модифицированного ооцита) животным, отличным от человека, в подходящих условиях для образования эмбриона.
[00237] Способы модификации генома животного, отличного от человека (например, свиньи, коровы, грызуна, курицы и т.д.), включают, например, использование нуклеазы с «цинковыми пальцами» (ZFN), эффекторной нуклеазы, подобной активатору транскрипции (TALEN), или Cas-белка (т.e. системы CRISPR/Cas) для модификации генома таким образом, чтобы он содержал сконструированный участок DH, описываемый в данном документе. Руководство по способам модификации генома зародышевой линии животного, отличного от человека, можно найти, например, в публикациях заявок на патент США №№ 2015-0376628 A1, US 2016-0145646 A1 и US 2016-0177339 A1; включенных в данный документ посредством ссылки.
[00238] В некоторых вариантах осуществления описанное в данном документе животное, отличное от человека, представляет собой млекопитающее. В некоторых вариантах осуществления описанное в данном документе животное, отличное от человека, представляет собой мелкое млекопитающее, например, из надсемейства Dipodoidea или Muroidea. В некоторых вариантах осуществления генетически модифицированное животное, описанное в данном документе, представляет собой грызуна. В некоторых вариантах осуществления грызун, описанный в данном документе, выбран из мыши, крысы и хомяка. В некоторых вариантах осуществления грызун, описанный в данном документе, выбран из надсемейства Muroidea. В некоторых вариантах осуществления генетически модифицированное животное, описанное в данном документе, относится к семейству, выбранному из Calomyscidae (например, мышевидные хомячки), Cricetidae (например, хомяки, крысы и мыши Нового света, полевки), Muridae (настоящие мыши и крысы, песчанки, иглистые мыши, косматые хомяки), Nesomyidae (лазающие мыши, скальные мыши, белохвостые крысы, малагасийские крысы и мыши), Platacanthomyidae (например, колючие соневидные хомяки) и Spalacidae (например, слепыши, бамбуковые крысы и цокоры). В некоторых определенных вариантах осуществления генетически модифицированный грызун, описанный в данном документе, выбран из настоящей мыши или крысы (семейство Muridae), песчанки, иглистой мыши и косматого хомяка. В некоторых определенных вариантах осуществления генетически модифицированная мышь, описанная в данном документе, является представителем семейства Muridae. В некоторых вариантах осуществления описанное в данном документе животное, отличное от человека, представляет собой грызуна. В некоторых определенных вариантах осуществления грызун, описанный в данном документе, выбран из мыши и крысы. В некоторых вариантах осуществления описанное в данном документе животное, отличное от человека, представляет собой мышь.
[00239] В некоторых вариантах осуществления описанное в данном документе животное, отличное от человека, представляет собой грызуна, который представляет собой мышь линии C57BL, выбранной из C57BL/A, C57BL/An, C57BL/GrFa, C57BL/KaLwN, C57BL/6, C57BL/6J, C57BL/6ByJ, C57BL/6NJ, C57BL/10, C57BL/10ScSn, C57BL/10Cr и C57BL/Ola. В некоторых определенных вариантах осуществления мышь, описанная в данном документе, принадлежит к линии 129, выбранной из группы, состоящей из линии 129P1, 129P2, 129P3, 129X1, 129S1 (например, 129S1/SV, 129S1/SvIm), 129S2, 129S4, 129S5, 129S9/SvEvH, 129/SvJae, 129S6 (129/SvEvTac), 129S7, 129S8, 129T1, 129T2 (см., например, Festing et al., 1999, Mammalian Genome 10:836; Auerbach, W. et al., 2000, Biotechniques 29(5):1024-1028, 1030, 1032). В некоторых определенных вариантах осуществления генетически модифицированная мышь, описанная в данном документе, представляет собой гибрид вышеупомянутой линии 129 и вышеупомянутой линии C57BL/6. В некоторых определенных вариантах осуществления мышь, описанная в данном документе, представляет собой гибрида вышеупомянутых линий 129 или гибрида вышеупомянутых линий BL/6. В некоторых определенных вариантах осуществления линия 129 в гибриде, описываемом в данном документе, представляет собой линию 129S6 (129/SvEvTac). В некоторых вариантах осуществления мышь, описанная в данном документе, представляет собой линию BALB, например линию BALB/c. В некоторых вариантах осуществления мышь, описанная в данном документе, представляет собой гибрида линии BALB и другой вышеупомянутой линии.
[00240] В некоторых вариантах осуществления описанное в данном документе животное, отличное от человека, представляет собой крысу. В некоторых определенных вариантах осуществления крыса, описанная в данном документе, выбрана из крысы линии Wistar, линии LEA, линии Sprague Dawley, линии Fischer, F344, F6 и Dark Agouti. В некоторых определенных вариантах осуществления линия крыс, описанная в данном документе, представляет собой гибрида двух или более линий, выбранных из группы, включающей Wistar, LEA, Sprague Dawley, Fischer, F344, F6 и Dark Agouti.
Способы, в которых используются животные, отличные от человека, имеющие сконструированный кластер разнообразия
[00241] Было разработано несколько in vitro и in vivo методик для получения терапевтических средств на основе антител. В частности, характерной особенностью in vivo методик является получение трансгенных животных (т.е. грызунов), содержащих гены иммуноглобулина человека, либо случайным образом встроенные в геном животного (например, см. патент США № 5569825), либо точно помещенные в эндогенный локус иммуноглобулина в функциональной связи с эндогенными константными участками иммуноглобулина животного (например, см. патенты США №№ 8502018; 8642835; 8697940 и 8791323). Оба подхода были эффективны в получении перспективных кандидатных терапевтических антител для применения у человека. Кроме того, оба подхода имеют преимущество над in vitro подходами в том, что кандидатные антитела выбирают из репертуаров антител, полученных in vivo, что включает отбор на аффинность и специфичность к антигену в пределах внутренней среды иммунной системы хозяина. При таком способе антитела связываются с естественным образом презентируемым антигеном (в пределах релевантных биологических эпитопов и поверхностей), а не с искусственными средами или при предсказаниях in silico , которые могут сопутствовать in vitro методикам. Несмотря на то, что с помощью in vivo технологий получают репертуары функциональных антител, создание антител к сложным (например, к трансмембранным полипептидам) или цитоплазматическим антигенам остается затруднительным. Кроме того, получение антител к полипептидам, которые характеризуются высокой степенью идентичности последовательности у разных видов (например, человека и мыши), остается проблемой вследствие иммунной толерантности.
[00242] Таким образом, среди прочих объектов в данном документе описано признание того, что конструкция на основе in vivo системы, характеризующейся продуцированием антител, имеющих добавленное разнообразие в CDR, в частности, CDR3, полученных из нестандартных последовательностей (т.е. нестандартных генных сегментов DH или генных сегментов DH, которые встречаются в природе) может быть создана с использованием одной или более нуклеотидных кодирующих последовательностей (т.е. синтетических последовательностей), каждая из которых представляет собой часть представляющего интерес полипептида. Такое добавленное разнообразие может управлять связыванием с конкретными антигенами (например, трансмембранными полипептидами). Например, описанные в данном документе антитела, которые блокируют один или более воспалительных цитокинов (например, β-хемокинов), можно получать в животных, отличных от человека, сконструированных таким образом, что они содержат локус тяжелой цепи иммуноглобулина, который содержит сконструированный участок DH, при этом сконструированный участок DH включает одну или более нуклеотидных кодирующая последовательностей, каждая из которых кодирует внеклеточную часть цитокинового рецептора (например, β-хемокинового рецептора). Также в данном документе описаны антитела, которые блокируют или ингибируют активность и/или функцию ионного канала (например, NaV канала), которые можно получать в животных, отличных от человека, сконструированных таким образом, что они содержат локус тяжелой цепи иммуноглобулина, который содержит сконструированный участок DH, при этом сконструированный участок DH включает одну или более нуклеотидных кодирующих последовательностей, каждая из которых кодирует часть токсина (например, μ-конотоксина и/или токсина тарантула). После рекомбинации генного сегмента VH, нуклеотидной кодирующей последовательности и генного сегмента JH образуется последовательность, кодирующая вариабельный участок тяжелой цепи, которая содержит участок CDR3, имеющий последовательность, частично кодируемую нуклеотидной кодирующей последовательностью, и, тем самым, за счет этого происходит связывание с конкретным антигеном, ассоциированным с нуклеотидной кодирующей последовательностью.
[00243] Описанных в данном документе животных, отличных от человека, можно использовать для получения человеческого антитела, при этом человеческое антитело содержит вариабельные домены, полученные из одной или более последовательностей нуклеиновой кислоты вариабельного участка, кодируемых генетическим материалом из клетки животного, отличного от человека, описываемого в данном документе. Например, описанное в данном документе животное, отличное от человека, иммунизируют β-хемокином (или полипептидом NaV канала, полным или частичным) в условиях и в течение времени, достаточного для того, чтобы у животного, отличного от человека, развился иммунный ответ на указанный β-хемокин (или полипептид NaV канала, полный или частичный). Антитела выделяют из животного, отличного от человека (или одной или более клеток, например, одной или более B-клеток), и их характеризуют с использованием различных анализов, посредством которых измеряют, например, аффинность, специфичность, проводят картирование эпитопов, определяют способность блокирования взаимодействия лиганд-рецептор, ингибирование активации рецепторов и т. д. В различных вариантах осуществления антитела, продуцируемые описанными в данном документе животными, отличными от человека, содержат один или более вариабельных доменов человека, которые получены из одной или более нуклеотидных последовательностей вариабельного участка человека, выделенных из животного, отличного от человека. В некоторых вариантах осуществления в животных, отличных от человека, описанных в данном документе, может быть индуцирована выработка антител к лекарственным средствам (например, антиидиотипического антитела).
[00244] Описанные в данном документе животные, отличные от человека, представляют улучшенную in vivo систему и источник биологических материалов (например, клеток) для продуцирования человеческих антител, которые применимы во множестве анализов. В различных вариантах осуществления описанных в данном документе животных, отличных от человека, используют для разработки терапевтических средств, которые нацеливаются на один или более цитокинов, и/или модулируют активность цитокинов, и/или модулируют взаимодействия цитокинов с другими партнерами по связыванию (например, цитокиновым рецептором). В различных вариантах осуществления описанных в данном документе животных, отличных от человека, используют для разработки терапевтических средств, которые нацеливаются на один или более ионных каналов, и/или модулируют активность ионных каналов, и/или модулируют взаимодействия ионных каналов с другими партнерами по связыванию. В различных вариантах осуществления описанных в данном документе животных, отличных от человека, используют для проведения идентификации, скрининга и/или разработки кандидатных терапевтических средств (например, антител, миРНК и т.д.), которые связывают один или более полипептидов цитокинов человека или ионных каналов (например, калиевого канала, кальциевого канала или натриевого канала). В различных вариантах осуществления описанных в данном документе животных, отличных от человека, используют для проведения скрининга и разработки кандидатных терапевтических средств (например, антител, миРНК и т.д.), которые блокируют активность одного или более полипептидов β-хемокина человека или который блокируют активность одного или более потенциал-зависимых натриевых (NaV) каналов человека. В различных вариантах осуществления описанных в данном документе животных, отличных от человека, используют для определения профиля связывания антагонистов и/или агонистов одного или более полипептидов β-хемокина человека или одного или более каналов NaV человека. В некоторых вариантах осуществления описанных в данном документе животных, отличных от человека, используют для определения эпитопа или эпитопов для одного или более кандидатных терапевтических антител, которые связывают один или более полипептидов β-хемокина человека или которые связывают один или более каналов NaV человека.
[00245] В различных вариантах осуществления описанных в данном документе животных, отличных от человека, используют для определения фармакокинетических профилей антител к β-хемокину или к NaV. В различных вариантах осуществления одно или более описанных в данном документе животных, отличных от человека, и одно или более контрольных или эталонных животных, отличных от человека, подвергают воздействию одного или более кандидатных терапевтических антител к β-хемокину или к NaV в различных дозах (например, 0,1 мг/кг, 0,2 мг/кг, 0,3 мг/кг, 0,4 мг/кг, 0,5 мг/кг, 1 мг/кг, 2 мг/кг, 3 мг/кг, 4 мг/кг, 5 мг/кг, 7,5 мг/кг, 10 мг/кг, 15 мг/кг, 20 мг/кг, 25 мг/кг, 30 мг/кг, 40 мг/кг или 50 мг/кг или больше). Кандидатные терапевтические антитела можно дозировать посредством любого необходимого пути введения, в том числе парентерального и непарентерального пути введения. Парентеральные пути предусматривают, например, внутривенный, внутриартериальный, интрапортальный, внутримышечный, подкожный, внутрибрюшинный, интраспинальный, интратекальный, интрацеребровентрикулярный, интракраниальный, интраплевральный или другие пути введения. Непарентеральные пути предусматривают, например, пероральный, интраназальный, трансдермальный, ингаляционный, ректальный, трансбуккальный, вагинальный, внутриглазной. Введение также можно осуществлять посредством непрерывной инфузии, местного применения, замедленного высвобождения из имплантатов (гелей, мембран и т.п.) и/или внутривенной инъекции. У животных, отличных от человека (гуманизированных и контрольных), отбирают кровь в различные моменты времени (например, 0 ч, 6 ч, 1 день, 2 дня, 3 дня, 4 дня, 5 дней, 6 дней, 7 дней, 8 дней, 9 дней, 10 дней, 11 дней или до 30 или более дней). Можно проводить различные анализы для определения фармакокинетических профилей вводимых кандидатных терапевтических антител с применением образцов, полученных от описываемых в данном документе животных, отличных от человека, в том числе без ограничения уровня общего IgG, ответа на терапевтические антитела, агглютинации и т.д.
[00246] В различных вариантах осуществления описанных в данном документе животных, отличных от человека, используют для измерения терапевтического эффекта блокирования или модулирования активности β-хемокина (или передачи сигнала с помощью β-хемокина, или взаимодействий, опосредованных β-хемокином) и эффекта в отношении генной экспрессии в результате клеточных изменений или плотности β-хемокиновых рецепторов в клетках описанных в данном документе животных, отличных от человека. В различных вариантах осуществления описанное в данном документе животное, отличное от человека, или клетки, выделенные из него, подвергают воздействию кандидатного терапевтического средства, которое связывает полипептид β-хемокина человека (или часть полипептида β-хемокина человека), и по истечении дополнительного периода времени проводят анализ в отношении эффектов на β-хемокин-зависимые процессы (или взаимодействия), например, взаимодействия лиганд-рецептор или передачу сигнала с помощью β-хемокина.
[00247] В различных вариантах осуществления описанных в данном документе животных, отличных от человека, используют для измерения терапевтического эффекта блокирования или модуляции активности NaV канала (или передачи сигнала с помощью NaV канала, или взаимодействий, опосредованных NaV, или потенциалов действия NaV канала) и эффекта в отношении генной экспрессии в результате клеточных изменений или плотности NaV каналов в клетках описываемых в данном документе животных, отличных от человека. В различных вариантах осуществления описанное в данном документе животное, отличное от человека, или клетки, выделенные из него, подвергают воздействию кандидатного терапевтического средства, которое связывает NaV канал человека (или часть NaV канала человека), и по истечении дополнительного периода времени проводят анализ в отношении эффектов на NaV канал-зависимые процессы (или взаимодействия), например, взаимодействия лиганд-рецептор или потенциалы действия NaV канала.
[00248] В различных вариантах осуществления описанные в данном документе животные, отличные от человека, могут быть использованы для получения человеческого антитела, которое оказывает действие на потенциал-зависимый натриевый канал (например, NaV1.7). Такое человеческое антитело можно тестировать и/или разрабатывать с помощью животного, отличного от человека, имеющего модификацию, отличную от описанной в данном документе, такую как, например, модификацию, описанную в патентах США №№ 8871996 и 8486647, включенную в данный документ посредством ссылки.
[00249] В описанных в данном документе животных, отличных от человека, экспрессируются антитела, которые связывают один или более β-хемокинов (или NaV каналов), поэтому можно получать клетки, линии клеток и культуры клеток, служащие в качестве источника антител к β-хемокину (или к NaV) для применения в анализах связывания и функциональных анализах, например, для анализа связывания или функции антагониста или агониста β-хемокина человека (или NaV канала), в частности, если антагонист или агонист является специфическим в отношении последовательности β-хемокина человека (последовательности NaV канала человека) или эпитопа, или, в качестве альтернативы, специфическим в отношении последовательности β-хемокина человека (последовательности NaV канала человека) или эпитопа, который осуществляет функцию во взаимодействии лиганд-рецептор (связывание). В различных вариантах осуществления эпитопы β-хемокина (или эпитопы NaV каналов), связанные с кандидатными терапевтическими антителами или миРНК, можно определять с применением клеток, выделенных от описанных в данном документе животных, отличных от человека.
[00250] Клетки описанных в данном документе животных, отличных от человека, можно выделять и использовать по ситуации или их можно поддерживать в культуре в течение многих поколений. В различных вариантах осуществления клетки описанного в данном документе животного, отличного от человека, подвергают иммортализации (например, посредством использования вируса) и поддерживают в культуре в течение неопределенного периода времени (например, в последовательных культурах).
[00251] Описанные в данном документе животные, отличные от человека, обеспечивают in vivo систему для получения вариантов антитела, которое связывает один или более полипептидов β-хемокина (или NaV каналов). Такие варианты включают антитела, обладающие требуемой функциональностью, специфичностью, низкой перекрестной реактивностью с общим эпитопом, который встречается у двух или более полипептидов β-хемокина (или NaV каналов). В некоторых вариантах осуществления описанных в данном документе животных, отличных от человека, используют для создания панелей антител для получения серии вариантов антител, которые подвергают скринингу в отношении необходимой или улучшенной функциональности.
[00252] Описанные в данном документе животные, отличные от человека, обеспечивают in vivo систему для создания библиотек антител к βхемокинам (или антител к NaV). Такие библиотеки обеспечивают источник последовательностей вариабельного участка тяжелой и легкой цепей, которые могут быть привиты на разные Fc-участки, исходя из необходимой эффекторной функции, и/или использоваться в качестве источника для созревания аффинности у последовательности вариабельного участка, с использованием методик, известных из уровня техники (например, сайт-направленный мутагенез, ПЦР с внесением ошибок и т.д.).
[00253] Описанные в данном документе животные, отличные от человека, обеспечивают in vivo систему для анализа и тестирования лекарственного средства или вакцины. В различных вариантах осуществления кандидатное лекарственное средство или вакцину можно доставлять одному или более описанным в данном документе животным, отличным от человека, с последующим наблюдением за животными, отличными от человека, для определения одного или более иммунных ответов на лекарственное средство или вакцину, профиля безопасности лекарственного средства или вакцины или эффекта в отношении заболевания или состояния и/или одного или более симптомов заболевания или состояния. Иллюстративные способы, используемые для определения профиля безопасности, включают измерения токсичности, оптимальной концентрации дозы, ответа с участием антитела (т.е. антитела к лекарственному средству), эффективности лекарственного средства или вакцины и возможных факторов риска. Такие лекарственные средства или вакцины можно улучшать и/или разрабатывать в таких животных, отличных от человека.
[00254] Эффективность вакцины можно определять с помощью ряда способов. Вкратце, описанных в данном документе животных, отличных от человека, вакцинируют с использованием способов, известных из уровня техники, а затем осуществляют контрольное заражение вакциной или вводят вакцину в уже инфицированное животное, отличное от человека. Ответ животного(ых), отличного от человека, на вакцину можно измерять с помощью мониторинга и/или проведения одного или более анализов в отношении животного(ых), отличного от человека (или клеток, выделенных из него), для определения эффективности вакцины. Ответ животного(ых), отличного от человека, на вакцину затем сравнивают с контрольными животными, используя один или более показателей, известных из уровня техники и/или описанных в данном документе.
[00255] Эффективность вакцины можно дополнительно определять с помощью анализов нейтрализации вируса. Вкратце, описанных в данном документе животных, отличных от человека, иммунизируют и сыворотку крови собирают в разные дни после иммунизации. Серийные разведения сыворотки крови предварительно инкубируют с вирусом, в течение этого времени антитела в сыворотке крови, специфические к вирусу, свяжутся с ним. Затем смесь вирус/сыворотка крови добавляют к пермиссивным клеткам для определения инфекционности с помощью анализа бляшкообразования или анализа микронейтрализации. Если антитела в сыворотке крови нейтрализуют вирус, то наблюдается меньшее количество бляшек или более низкий уровень относительных люциферазных единиц по сравнению с контрольной группой.
[00256] Описанные в данном документе животные, отличные от человека, обеспечивают улучшенную in vivo систему для разработки и определения характеристик терапевтических средств на основе антител для применения в лечении рака и/или воспалительных заболеваний. Воспаление в течение длительного времени ассоциировали с раком (см., например, обзор в Grivennikov, S.I. et al., 2010, Cell 140:883-99; Rakoff-Nahoum, S., 2006, Yale J. Biol. Med. 79:123-30). В действительности, окружение развивающейся опухоли d в некоторой степени характеризуется инфильтрацией разных медиаторов воспаления. Также хроническое воспаление может приводить к более высокой вероятности развития рака. Таким образом, в некоторых вариантах осуществления описанное в данном документе животное, отличное от человека, обеспечивает in vivo систему для разработки и/или идентификации противораковых и/или противовоспалительных терапевтических средств. В различных вариантах осуществления описанным в данном документе животным, отличным от человека, или контрольным животным, отличным от человека (например, имеющим генетическую модификацию, отличную от описанной в данном документе, или не имеющим генетической модификации, т.е. дикого типа), может быть имплантирована опухоль (или опухолевые клетки) с последующим введением одного или более кандидатных терапевтических средств. В некоторых вариантах осуществления кандидатные терапевтические средства могут предусматривать полиспецифическое антитело (например, биспецифическое антитело) или смесь антител. В некоторых вариантах осуществления кандидатные терапевтические средства предусматривают комбинированную терапию, такую как, например, введение доз двух или более моноспецифических антител последовательно или одновременно. Перед введением одного или более кандидатных терапевтических средств опухоли может быть предоставлено достаточное время для развития в одном или более местоположениях в пределах организма животного, отличного от человека. Пролиферацию, рост, выживаемость и т.д. опухолевых клеток можно измерять как перед введением, так и после введения кандидатного терапевтического средства(средств). При необходимости у животного, отличного от человека, также можно измерять цитотоксичность кандидатных терапевтических средств.
[00257] Описанные в данном документе животные, отличные от человека, обеспечивают улучшенную in vivo систему для разработки и определения характеристик терапевтических средств на основе антител для применения в лечении боли (например, нейропатической) или для применения в качестве болеутоляющего средства. В некоторых вариантах осуществления описанное в данном документе животное, отличное от человека, обеспечивает in vivo систему для разработки и/или идентификации болеутоляющих терапевтических средств. В различных вариантах осуществления описанные в данном документе животные, отличные от человека, или контрольные животные, отличные от человека (например, имеющие генетическую модификацию, отличную от описанной в данном документе [такую как, например, модификация, описываемая в патентах США №№ 8871996 и 8486647; включенных в данный документ посредством ссылки], или не имеющие генетической модификации, т.е. дикого типа) могут быть подвергнуты болевой стимуляции (например, инцизионной, химически индуцированной и т.д.; см., например, Recognition and Alleviation of Pain in Laboratory Animals. Washington D.C.: National Academies Press, 2009. Print) с последующим введением одного или более кандидатных терапевтических средств. В некоторых вариантах осуществления кандидатные терапевтические средства могут предусматривать полиспецифическое антитело (например, биспецифическое антитело) или смесь антител. В некоторых вариантах осуществления кандидатные терапевтические средства предусматривают комбинированную терапию, такую как, например, введение доз двух или более моноспецифических антител последовательно или одновременно. Перед введением одного или более кандидатных терапевтических средств может быть предоставлен достаточный период времени. Разные измерения и/или тесты, которые главным образом ассоциированы с оценкой обезболивания и/или эффективностью болеутоляющего терапевтического средства, могут регистрироваться и перед введением, и после введения кандидатного терапевтического средства(средств).
Наборы
[00258] Также в данном документе описаны упаковка или набор, содержащие один или более контейнеров, заполненных по меньшей мере одним животным, отличным от человека, клеткой, отличной от человеческой, фрагментом ДНК и/или нацеливающим вектором, описываемыми в данном документе. Наборы можно применять в любом применимом способе (например, в способе исследования). Необязательно, с таким контейнером(ами) может быть ассоциировано уведомление в форме, предписанной правительственным учреждением, регулирующим производство, применение или продажу фармацевтических препаратов или биологических продуктов, при этом уведомление отражает (а) одобрение органом по производству, применению или продаже для введения человеку, (b) указания для применения или и то, и другое, или договор, который регулирует передачу материалов и/или биологических продуктов (например, описываемых в данном документе животного, отличного от человека, или клетки, отличной от человеческой) между двумя или более организациями.
[00259] Другие признаки описанных вариантов осуществления станут очевидными в ходе следующих описаний иллюстративных вариантов осуществления, которые приведены в иллюстративных целях и не предназначены для их ограничения.
ПРИМЕРЫ
[00260] Следующие примеры приведены для того, чтобы обеспечить рядовых специалистов в данной области описанием того, как получать и применять способы и композиции по настоящему изобретению, и они не предназначены для ограничения объема того, что авторы настоящего изобретения рассматривают в качестве своего изобретения. Если не указано иное, температура указана в градусах Цельсия, а давление является атмосферным или близким к нему.
Пример 1. Разработка сконструированного кластера разнообразия, который включает нуклеотидные кодирующие последовательности D6 хемокинового рецептора-ловушки в пределах вариабельного участка тяжелой цепи иммуноглобулина
[00261] В данном примере показаны иллюстративные способы разработки нацеливающего вектора для вставки в геном животного, отличного от человека, такого как грызун (например, мышь). В частности, способы, описанные в данном примере, демонстрируют получение нацеливающего вектора для вставки в геном эмбриональных стволовых (ES) клеток грызуна (например, мыши), чтобы получить грызуна, геном которого содержит вариабельный участок тяжелой цепи иммуноглобулина, который включает сконструированный участок разнообразия (DH) тяжелой цепи, при этом сконструированный участок DH включает одну или более нуклеотидных последовательностей, каждая из которых кодирует полипептид, не являющийся иммуноглобулином, или его часть. В данном примере при разработке нацеливающего вектора для интеграции в вариабельный участок тяжелой цепи иммуноглобулина использовали кодирующие последовательности внеклеточного домена атипичного хемокинового рецептора (ACKR), например, D6 хемокинового рецептора-ловушки. Как описано ниже, кодирующие последовательности помещали в функциональную связь с вариабельными сегментами тяжелой цепи (VH) и соединяющими сегментами тяжелой цепи (JH) вместо стандартных генных сегментов DH, вследствие чего после VDJ-рекомбинации экспрессируются антитела, имеющие CDR3 VH, полученные из кодирующих последовательностей.
[00262] Нацеливающие векторы, содержащие кодирующие последовательности из внеклеточных доменов D6 хемокинового рецептора-ловушки для вставки в вариабельный участок тяжелой цепи иммуноглобулина, создавали с использованием технологии VELOCIGENE® (см., например, патент США №6586251 и Valenzuela et al., 2003, Nature Biotech. 21(6):652-659; включенные в данный документ посредством ссылки) и методик молекулярной биологии, известных из уровня техники. Способы, описанные в данном примере, можно применять для использования любого набора кодирующих последовательностей, полученных из любого требуемого полипептида, или комбинации кодирующих последовательностей (или фрагментов кодирующей последовательности), если это необходимо. Альтернативная и неограничивающая иллюстративная стратегия разработки нацеливающего вектора с использованием кодирующих последовательностей из внеклеточных доменов D6 хемокинового рецептора-ловушки представлена на фигурах 3 и 4.
[00263] Вкратце, серию из четырех фрагментов ДНК (D6-DH1116, D6-DH17613, D6-DH114619 и D6DH120126), каждый из которых содержал несколько кодирующих последовательностей D6, получали с помощью de novo синтеза ДНК (таблица 5; Blue Heron Biotech, Ботелл, Вашингтон). Различные сайты для рестриктаз и/или перекрывающиеся участки (~40 п.о., которые включали множество сайтов для рестриктаз) включали на концах каждого фрагмента ДНК для обеспечения возможности последующего тандемного клонирования (см. ниже). Единый сегмент DH человека (т.е. DH6–25) оставался интактным в участке DH, сконструированном таким образом, чтобы он содержал нуклеотидные кодирующие последовательности, соответствующие частям внеклеточного домена D6 хемокинового рецептора-ловушки. В некоторых вариантах осуществления генные сегменты иммуноглобулина (например, сегменты DH) могут быть ассоциированы с субоптимальными или дефектными последовательностями сигнала рекомбинации (например, последовательностями гептамера и/или нонамера), вследствие чего использование таких сегментов DH значительно меньше, чем использование сегментов DH, ассоциированных с последовательностями сигнала рекомбинации, характеризующимися как последовательности дикого типа, нормальные и/или не являющиеся дефектными. Таким образом, такие сегменты DH (например, DH6–25, последовательности RSS показаны на фиг. 2) могут быть оставлены интактными или их подвергают делеции при конструировании участка DH таким образом, чтобы он содержал нуклеотидные кодирующие последовательности, каждая из которых кодирует представляющий интерес полипептид, не являющийся иммуноглобулином, или его часть (например, D6 хемокиновый рецептор-ловушку).
[00264] Иммуноглобулины принимают участие в клеточном механизме, называемом соматической гипермутацией, в результате которого образуются варианты антитела с созревшей аффинностью, характеризующиеся высокой аффинностью в отношении своей мишени. Хотя соматическая гипермутация главным образом происходит в пределах CDR вариабельных участков антител, мутации преимущественно нацеливаются на определенные мотивы последовательности, которые называются «горячими точками», например, «горячие точки» RGYW, индуцируемые активацией цитидиндезаминазы (AID) (см., например, Li, Z. et al., 2004, Genes Dev. 18:1-11; Teng, G. and F.N. Papavasiliou, 2007, Annu. Rev. Genet. 41:107-20; включенные в данный документ посредством ссылки). Последовательность нуклеиновой кислоты в каждой кодирующей последовательности D6 по своей природе содержала последовательности таких «горячих точек», однако искусственные «горячие точки» вводили в выбранные кодирующие последовательности D6 для оптимизации потенциала соматической гипермутации во время клонального отбора В-клеточных рецепторов. Искусственные «горячие точки» для SHM вводили в кодирующие последовательности D6 in silico перед de novo синтезом. На фигуре 1 изложен иллюстративный анализ природных и искусственных «горячих точек», используемых в описанных в данном документе кодирующих последовательностях D6.
[00265] V(D)J-рекомбинацией управляют фланкирующие последовательности ДНК, называемые последовательностями сигнала рекомбинации (RSS), что обеспечивает возможность перестроек ДНК в точных местоположениях относительно кодирующих последовательностей V, D и J. Каждая RSS состоит из консервативного блока из семи нуклеотидов (гептамера), который является смежным с кодирующей последовательностью, после которой расположен неконсервативный спейсер (либо из 12, либо из 23 п.о.), и второго консервативного блока из девяти нуклеотидов (нонамера). Каждую из кодирующих последовательностей D6 разрабатывали с оптимизированными RSS для обеспечения большей частоты рекомбинации и равного использования. Вкратце, все гептамеры и нонамеры в последовательностях RSS DH заменяли на консенсусные последовательности (фигура 2). Последовательности RSS из нефункциональных сегментов DH, называемых «только ORF», подвергали репарации для обеспечения использования кодирующих последовательностей D6. Как описано в данном документе, «только ORF» представляют собой сегменты DH, которые включают кодирующую последовательность, которая может транслироваться в виде по меньшей мере одной открытой рамки считывания, но которые имеют по меньшей мере одну нефункциональную последовательность RSS (не распознаваемую рекомбиназой RAG). Авторы настоящего изобретения пришли к выводу, что оптимизация гептамеров и нонамеров не гарантировала того, что все кодирующие последовательности D6, будут использоваться с равной частотой, что обусловлено, по меньшей мере частично, непрогнозированными воздействиями спейсеров, кодирующих концевых последовательностей, фланкирующих межгенные последовательности и т.д. Как описано выше для «горячих точек» соматической гипермутации, оптимизированные RSS разрабатывали in silico перед de novo синтезом фрагментов ДНК D6.
[00266] Фрагменты ДНК D6 получали для тандемной сборки вместе, используя сайты для рестриктаз и перекрывающиеся участки, разработанные в каждом фрагменте. Фрагмент D6-DH1166 и плазмиду, несущую кассету для отбора по устойчивости к неомицину, подготавливали к лигированию с помощью рестрикционного расщепления с применением AgeI и EcoRI (фигура 3A, вверху). Расщепленную с помощью AgeI/EcoRI плазмиду и фрагмент D6-DH1166 лигировали вместе с использованием ДНК-лигазы согласно инструкциям производителя. Продукты лигирования и остальные фрагменты ДНК D6 расщепляли с помощью SnaBI. Вектор BAC (pBACE3.6) отдельно расщепляли с помощью NotI и AscI для вставки фрагментов ДНК D6 (фигура 3A, посередине). Продукт лигирования D6-DH1166/кассета для отбора по устойчивости к неомицину и остальные фрагменты ДНК D6 подвергали сборке, используя одностадийную изотермальную сборку, как описано ранее (фигура 3A, внизу; Barnes, W.M., 1994, Proc. Natl. Acad. Sci. 91(6):2216-20; Gibson, D.G. et al., 2009, Nat. Methods 6(5):343-5; Gibson, D.G. et al., 2010, Nat. Methods 7(11):901-3). Подвергнутый сборке фрагмент ДНК D6 (фигура 3B, вверху) затем расщепляли с помощью PI-SceI и I-CeuI и лигировали вместе с клоном BAC, содержащим 5'- и 3'-плечи гомологии, содержащие ~50 т.о. геномной ДНК вариабельного участка человека и кластер JH человека соответственно, посредством совместимых концов (фигура 3B, посередине). Окончательный нацеливающий вектор содержал, в направлении от 5' r 3', 5'-плечо гомологии размером 50 т.о., содержащее ДНК вариабельного участка человека, кассету для отбора по устойчивости к неомицину, фрагмент ДНК размером ~41 т.о, содержащий 25 внеклеточных кодирующих последовательностей D6 хемокинового рецептора-ловушки, кластер JH человека, интронный энхансер тяжелой цепи мыши (Ei), ген константного участка IgM мыши и lox-сайт (фигура 3B, внизу).
[00267] В качестве альтернативы фрагменты ДНК D6, описанные выше, могут быть собраны с использованием молекулярных методик, которые отличаются от описанных выше, однако известны из уровня техники. Например, поскольку фрагменты ДНК D6 расположены тандемно, при необходимости может быть получено множество разных структур (например, путем изменения порядка кодирующих последовательностей) за счет использования различных сайтов рестрикции/перекрывающихся участков, встроенных в концы фрагментов. Иллюстративный альтернативный способ сборки фрагментов ДНК D6, описанных выше, путем последовательного лигирования изложен на фигурах 4A и 4B.
[00268] Нацеливающий вектор D6-DH, описанный выше, вводили в мышиные эмбриональные стволовые (ES) клетки для получения модицифированных ES-клеток, содержащих локус тяжелой цепи иммуноглобулина, содержащий сконструированный кластер разнообразия, например, сконструированный участок DH.
SEQ ID NO:131
D6 EC3 (7)
D6 Nterm (1)
D6 Nterm+EC3 (33)
D6 EC1 (3)
D6 EC2 (5)
DH2-2
DH3-3
DH4-4
DH5-5
DH6-6
SEQ ID NO:132
D6 EC3-S (15)
D6 Nterm-N (37)
D6 EC2-S (13)
D6 EC1-EC2-S (29)
D6 EC1-S (11)
D6 Nterm-S (9)
DH2-8
DH3-9
DH3-10
DH4-11
DH5-12
DH6-13
SEQ ID NO:133
D6 EC2-EC1 (23)
D6 Nterm-EC3 (17)
D6 EC1+EC2 (35)
D6 EC3-Nterm (19)
D6 EC1-EC2 (21)
DH2-15
DH3-16
DH4-17
DH5-18
DH6-19
SEQ ID NO:134
D6 EC2-EC1-S (31)
D6 Nterm-EC3-S (25)
D6 EC3-C (49)
D6 EC3-Nterm-S (27)
D6 EC3-N (47)
DH2-21
DH3-22
DH4-23
DH5-24
DH1-26
Пример 2. Создание грызунов, имеющих сконструированный кластер разнообразия, который включает нуклеотидные кодирующие последовательности D6 хемокинового рецептора-ловушки в пределах вариабельного участка тяжелой цепи иммуноглобулина
[00269] В данном примере продемонстрировано получение животных, отличных от человека (например, грызунов), геном которых содержит вариабельный участок тяжелой цепи иммуноглобулина, который включает сконструированный участок разнообразия тяжелой цепи (DH), где сконструированный участок DH включает одну или более нуклеотидных последовательностей, каждая из которых кодирует часть полипептида, не являющегося иммуноглобулином, в частности, внеклеточную часть атипичного хемокинового рецептора (ACKR), такого как, например, D6 хемокиновый рецептор-ловушка.
[00270] Правильную сборку нацеливающего вектора D6-DH, описанного в примере 1, и подвергнутую нацеливанию вставку фрагментов ДНК D6 в кластер разнообразия ДНК BAC подтверждали путем секвенирования и проведения полимеразной цепной реакции на всем протяжении конструкции нацеливающего вектора, используя праймеры, изложенные в таблице 6. Подвергнутую нацеливанию ДНК BAC, подтвержденную полимеразной цепной реакцией, затем вводили в эмбриональные стволовые (ES) клетки мыши, представляющие собой гибрид F1 (129S6SvEvTac/C57BL6NTac), посредством электропорации с последующим культивированием в среде для отбора. ES-клетки, применяемые для электропорации, имели геном, который включал генные сегменты VH и JH человека, функционально связанные с константными участками тяжелой цепи иммуноглобулинов грызуна (например, IgM), в котором полностью или частично отсутствовали участок DH человека (т.е. он был подвергнут делеции), и он содержал вставленную последовательность, кодирующую один или более генов Adam6 мыши (см., например, патенты США №№ 8642835 и 8697940). Колонии, устойчивые к лекарственному средству, отбирали через 10 дней после электропорации и подвергали скринингу с использованием TAQMAN™ и кариотипированию в отношении корректного нацеливания, как описано ранее (Valenzuela et al., выше; Frendewey, D. et al., 2010, Methods Enzymol. 476:295-307), с использованием наборов праймер/зонд, которые выявляли правильность интеграции кодирующих последовательностей D6 в кластер разнообразия вариабельного участка иммуноглобулина (таблица 7 [F: прямой праймер, P: зонд, R: обратный праймер] и фигура 5).
[00271] Применяли способ VELOCIMOUSE® (DeChiara, T.M. et al., 2010, Methods Enzymol. 476:285-294; Dechiara, T.M., 2009, Methods Mol. Biol. 530:311-324; Poueymirou et al., 2007, Nat. Biotechnol. 25:91-99), при котором подвергнутые нацеливанию ES-клетки вводили в эмбрионы мышей Swiss Webster на стадии 8 бластомеров, не прошедших компактизацию, для получения здоровых мышей поколения F0, происходящих полностью из ES-клеток, гетерозиготных по сконструированному участку DH, и в которых экспрессируются антитела, содержащие вариабельные участки тяжелой цепи, которые включают участки CDR3, полученные в результате рекомбинации кодирующих последовательностей D6. Гетерозиготного самца поколения F0 скрещивали с самками C57Bl6/NTac для получения гетерозигот F1, которых скрещивали между собой с получением гомозигот поколения F2 и мышей дикого типа для фенотипических анализов.
[00272] Кассету для отбора по устойчивости к лекарственному средству необязательно можно удалить с помощью последующего добавления рекомбиназы (например, путем обработки с помощью Cre) или путем скрещивания с линией мышей Cre-deleter (см., например, публикацию международной патентной заявки № WO 2009/114400), чтобы удалить любую выбранную кассету, фланкированную lox, которая введена с помощью нацеливающей конструкции, которая не удаляется, например, на стадии ES-клеток или в эмбрионе. Необязательно кассета для отбора сохраняется в мышах.
[00273] В целом, в данном примере проиллюстрировано создание грызуна (например, мыши), геном которого содержит вариабельный участок тяжелой цепи иммуноглобулина, который включает сконструированный участок разнообразия тяжелой цепи (DH), при этом сконструированный участок DH характеризуется включением одной или более нуклеотидных последовательностей, каждая из которых кодирует внеклеточную часть атипичного хемокинового рецептора (ACKR), такого как, например, D6 хемокиновый рецептор-ловушка. Описанная в данном документе стратегия вставки кодирующих последовательностей D6 на место стандартных сегментов DH обеспечивает возможность конструирования грызуна, в котором экспрессируются несколько антител, каждое из которых содержит CDR3 тяжелой цепи, созданный за счет рекомбинации кодирующей последовательности D6. Благодаря присутствию сегментов VH и JH человека, грызуны, описанные в данном документе, обеспечивают in vivo систему для получения терапевтических средств на основе человеческих антител, которые характеризуются разнообразием, которое связывает один или более CCL, а в некоторых вариантах осуществления обеспечивают противовоспалительные лекарственные средства на основе антител для применения у человека.
Пример 3. Разработка сконструированного кластера разнообразия, который включает нуклеотидные кодирующие последовательности одного или более пептидных токсинов в пределах вариабельного участка тяжелой цепи иммуноглобулина
[00274] В данном примере показаны иллюстративные способы разработки нацеливающего вектора для вставки в геном животного, отличного от человека, такого как грызун (например, мышь). В частности, в способах, описанных в данном примере, продемонстрировано получение нацеливающего вектора для вставки в геном эмбриональных стволовых (ES) клеток грызуна (например, мыши) для получения грызуна, геном которого содержит вариабельный участок тяжелой цепи иммуноглобулина, который включает сконструированный участок разнообразия (DH) тяжелой цепи, при этом сконструированный участок DH включает одну или более нуклеотидных последовательностей, каждая из которых кодирует часть полипептида, не являющегося иммуноглобулином. В данном примере кодирующие последовательности двух токсинов (например, конотоксина, такого как, например, μ-конотоксин, и токсина тарантула, такого как, например, ProTxII) использовали в разработке нацеливающего вектора для интеграции в вариабельный участок тяжелой цепи иммуноглобулина. Как описано ниже, кодирующие последовательности помещали в функциональную связь с вариабельными сегментами тяжелой цепи (VH) и соединяющими сегментами тяжелой цепи (JH) вместо стандартных генных сегментов DH, вследствие чего после VDJ-рекомбинации экспрессируются антитела, имеющие CDR3, полученные из кодирующих последовательностей.
[00275] Нацеливающие векторы, содержащие кодирующие последовательности из μ-конотоксина и токсина тарантула (например, ProTxII) для вставки в вариабельный участок тяжелой цепи иммуноглобулина, создавали, как описано выше в примере 1. При необходимости, способы, описанные в данном документе, можно применять для использования любого набора кодирующих последовательностей, полученных из любого требуемого конотоксина (например, α-конотоксина, δ-конотоксина, κ-конотоксина, μ-конотоксина и/или ω-конотоксина), токсина тарантула (например, ProTxI, ProTxII, хувентоксина-IV [HWTX-IV] и т.д.), или комбинации кодирующих последовательностей (или фрагментов кодирующих последовательностей). Иллюстративная стратегия для разработки нацеливающего вектора с использованием кодирующих последовательностей из μ-конотоксина и токсина тарантула изложена на фигурах 7A и 7B.
[00276] Вкратце, серию из четырех фрагментов ДНК, каждый из которых содержал несколько кодирующих последовательностей μ-конотоксина (μCTX) и токсина тарантула (ProTxII), получали с помощью de novo синтеза ДНК (таблица 8; Blue Heron Biotech, Ботелл, Вашингтон). Как описано в примере 1, различные сайты для рестриктаз и/или перекрывающиеся участки включали на концах каждого фрагмента ДНК для обеспечения возможности последующего тандемного клонирования. В общей сложности 26 кодирующих последовательностей токсина использовали в разработке сконструированного участка DH, который включал замену полных кодирующих последовательностей DH и вставку в кодирующие последовательности DH (т. е. лишь частичную замену DH). Вставки из кодирующих последовательностей токсина осуществляли в положениях DH 1–26, 2–2, 2–8, 2–15, 2–21, 3–3, 3–9, 3–10, 3–16 и 3–22 с помощью соответствующих кодирующих последовательностей токсина, тогда как замену последовательности DH на кодирующие последовательности токсина осуществляли в положениях DH 1–1, 1–7, 1–14, 1–20, 4–4, 4–11, 4–17, 4–23, 5–5, 5–12, 5-18, 5-24, 6-6, 6-13, 6-19 и 6-25 (см. таблицу 8). В случае вставок последовательности токсина вставляли во вторую открытую рамку считывания (ORF2) и фланкировали консенсусными последовательностями DH (например, см. фигуру 2). Сохранение нативных последовательностей, фланкирующих DH, обеспечивает природную гибкую линкерную последовательность и/или применение дисульфидной связи в сегментах DH2 для циклизации кодируемого пептидного токсина. В отличие от сконструированного участка DH, разработанного с использованием кодирующих последовательностей D6, описанных в примере 1, DH6–25 замещали полноразмерным μCTX KIIIA (μCTX-KIIIA fl; SEQ ID NO:228, см. таблицу 8) в случае конструирования участка DH, который включает кодирующие последовательности токсина.
[00277] Как описано в примере 1, последовательность нуклеиновой кислоты в каждой кодирующей последовательности токсина по своей природе содержала последовательности таких «горячих точек», однако искусственные «горячие точки» вводили в выбранные кодирующие последовательности токсина in silico перед de novo синтезом для оптимизации потенциала соматической гипермутации во время клонального отбора B-клеточных рецепторов (см. фигуры 6A–6L). Кроме того, каждую из кодирующих последовательностей токсина разрабатывали с оптимизированными RSS in silico перед de novo синтезом для обеспечения большей частоты рекомбинации и равного использования (фигура 2). Сборку фрагментов ДНК токсина получали, как описано в примере 1 (см. фигуры 7A и 7B).
[00278] В качестве альтернативы фрагменты ДНК токсина, описанные выше, могут быть собраны с использованием молекулярных методик, которые отличаются от описанных выше, однако известны из уровня техники. Например, поскольку фрагменты ДНК токсина расположены тандемно, при необходимости может быть получено множество разных структур (например, путем изменения порядка кодирующих последовательностей) за счет использования различных сайтов рестрикции/перекрывающихся участков, встроенных в концы фрагментов. Иллюстративный альтернативный способ сборки фрагментов ДНК токсина, описанных выше, путем последовательного лигирования изложен на фигурах 8A и 8B.
[00279] Нацеливающий вектор TX-DH, описанный выше, вводили в мышиные эмбриональные стволовые (ES) клетки для получения модицифированных ES-клеток, содержащих локус тяжелой цепи иммуноглобулина, содержащий сконструированный кластер разнообразия.
SEQ ID NO:232
μCTX-CSSRWC (182)
μCTX-KIIIA mini (184)
μCTX-PIIIA C1SC4S (186)
ProTxII C1SC4S (188)
μCTX-KIIIA C1SC4S (190)
DH2-2
DH3-3
DH4-4
DH5-5
DH6-6
SEQ ID NO:233
μвставка CTX-RSRQ (194)
μCTX-KIIIA midi (196)
μCTX-SmIIIA mini (198)
μCTX-PIIIA mini (200)
ProTxII C2SC5S (202)
μCTX-KIIIA mini (204)
DH2-8
DH3-9
DH3-10
DH4-11
DH5-12
DH6-13
SEQ ID NO:234
μвставка CTX-SSRW (208)
μCTX-SmIIIA midi (210)
μCTX-PIIIA midi (212)
ProTxII C3SC6S (214)
μCTX-KIIIA midi (216)
DH2-15
DH3-16
DH4-17
DH5-18
DH6-19
SEQ ID NO:235
μCTX-CRSRQC (220)
μCTX-PIIIA midi (222)
μCTX-PIIIA fl (224)
ProTxII fl (226)
μCTX-KIIIA fl (228)
μCTX-PIIIA mini (230)
DH2-21
DH3-22
DH4-23
DH5-24
DH6-25
DH1-26
Пример 4. Создание грызунов, имеющих сконструированный кластер разнообразия, который включает нуклеотидные кодирующие последовательности одного или более пептидных токсинов в пределах вариабельного участка тяжелой цепи иммуноглобулина
[00280] В данном примере продемонстрировано получение животных, отличных от человека (например, грызунов), геном которых содержит вариабельный участок тяжелой цепи иммуноглобулина, который включает сконструированный участок разнообразия тяжелой цепи (DH), где сконструированный участок DH включает одну или более нуклеотидных последовательностей, каждая из которых кодирует часть полипептида, не являющегося иммуноглобулином, в частности, токсина, такого как, например, конотоксин (например, μ-конотоксин), токсин тарантула (например, ProTxII) или их комбинации.
[00281] Правильную сборку нацеливающего вектора TX-DH, описанного в примере 3, и подвергнутую нацеливанию вставку фрагментов ДНК токсина в кластер разнообразия ДНК BAC подтверждали путем секвенирования и проведения полимеразной цепной реакции на всем протяжении конструкции нацеливающего вектора, используя праймеры, изложенные в таблице 6. Подвергнутую нацеливанию ДНК BAC, подтвержденную полимеразной цепной реакцией, затем вводили в эмбриональные стволовые (ES) клетки мыши, представляющие собой гибрид F1 (129S6SvEvTac/C57BL6NTac), посредством электропорации с последующим культивированием в среде для отбора. ES-клетки, применяемые для электропорации, имели геном, который включал генные сегменты VH и JH человека, функционально связанные с константными участками тяжелой цепи иммуноглобулинов грызуна (например, IgM), в котором отсутствовали участок DH (т.е. был подвергнут делеции), и он содержал вставленную последовательность, кодирующую один или более генов Adam6 мыши (см., например, патенты США №№ 8642835 и 8697940). Колонии, устойчивые к лекарственному средству, отбирали через 10 дней после электропорации и подвергали скринингу с использованием TAQMAN™ и кариотипированию в отношении корректного нацеливания, как описано ранее (Valenzuela et al., выше; Frendewey, D. et al., 2010, Methods Enzymol. 476:295-307), с использованием наборов праймер/зонд, которые выявляли правильность интеграции кодирующих последовательностей токсина в кластер разнообразия вариабельного участка иммуноглобулина (таблица 7 [F: прямой праймер, P: зонд, R: обратный праймер] и фигура 5).
[00282] Применяли способ VELOCIMOUSE® (DeChiara, T.M. et al., 2010, Methods Enzymol. 476:285-294; Dechiara, T.M., 2009, Methods Mol. Biol. 530:311-324; Poueymirou et al., 2007, Nat. Biotechnol. 25:91-99), при котором подвергнутые нацеливанию ES-клетки вводили в эмбрионы мышей Swiss Webster на стадии 8 бластомеров, не прошедших компактизацию, для получения здоровых мышей поколения F0, происходящих полностью из ES-клеток, гетерозиготных по сконструированному участку DH, и в которых экспрессируются антитела, содержащие вариабельные участки тяжелой цепи, которые включают участки CDR3, полученные в результате рекомбинации кодирующих последовательностей токсина. Гетерозиготного самца поколения F0 скрещивали с самками C57Bl6/NTac для получения гетерозигот F1, которых скрещивали между собой с получением гомозигот поколения F2 и мышей дикого типа для фенотипических анализов.
[00283] Кассету для отбора по устойчивости к лекарственному средству необязательно можно удалить с помощью последующего добавления рекомбиназы (например, путем обработки с помощью Cre) или путем скрещивания с линией мышей Cre-deleter (см., например, публикацию международной патентной заявки № WO 2009/114400), чтобы удалить любую выбранную кассету, фланкированную lox, которая введена с помощью нацеливающей конструкции, которая не удаляется, например, на стадии ES-клеток или в эмбрионе. Необязательно кассета для отбора сохраняется в мышах.
[00284] В целом, в данном примере проиллюстрировано создание грызуна (например, мыши), геном которого содержит вариабельный участок тяжелой цепи иммуноглобулина, который включает сконструированный участок разнообразия тяжелой цепи (DH), при этом сконструированный участок DH характеризуется включением одной или более нуклеотидных последовательностей, каждая из которых кодирует часть пептидного токсина, такого как, например, μ-конотоксин, ProTxII или их комбинации. Описанная в данном документе стратегия вставки кодирующих последовательностей токсина на место стандартных сегментов DH обеспечивает возможность конструирования грызуна, в котором экспрессируются антитела, сконфигурированные таким образом, что они содержат CDR3 тяжелой цепи, которые созданы за счет рекомбинации кодирующих последовательностей токсина. Благодаря присутствию сегментов VH и JH человека, грызуны, описанные в данном документе, обеспечивают in vivo систему для получения терапевтических средств на основе человеческих антител, которые характеризуются разнообразием, которое управляет связыванием с ионными каналами (например, потенциал-зависимыми натриевыми каналами, такими как NaV1.7), а в некоторых вариантах осуществления обеспечивает болеутоляющие лекарственные средства на основе антител для применения у человека.
Пример 5. Фенотипическая оценка грызунов, имеющих сконструированный кластер разнообразия
[00285] В данном примере продемонстрировано определение характеристик разных популяций иммунных клеток в грызунах (например, мышах), геном которых содержит вариабельный участок тяжелой цепи иммуноглобулина, который включает сконструированный участок разнообразия тяжелой цепи (DH), где сконструированный участок DH включает одну или более нуклеотидных последовательностей, каждая из которых кодирует часть полипептида, не являющегося иммуноглобулином. В данном примере грызуны, гомозиготные по сконструированному участку DH, который включает кодирующие последовательности токсина (например, μ-конотоксинов и токсина тарантула), описываемые в примере 3, анализировали с помощью проточной цитометрии для идентификации иммунных клеток в собранных селезенке и костном мозге с помощью нескольких флуоресцентно-меченых антител. В данном примере также продемонстрирована идентификация иммунных клеток в собранной селезенке и костном мозге мышей, гетерозиготных по сконструированному участку DH, который включает кодирующие последовательности внеклеточной части ACKR2 (также известного как D6 хемокиновый рецептор-ловушка), описываемой в примере 1. Как описано ниже, у мышей, содержащих сконструированный участок DH, описанный в данном документе, продемонстрированы аналогичные уровни B-клеток в компартментах селезенки и костного мозга по сравнению с контрольными сконструированными мышами. Важно то, что в данном примере также продемонстрировано, что у мышей, содержащих сконструированный участок DH, описываемый в данном документе, продемонстрировано развитие и созревание В-клеток, не имеющих каких-либо дефектов или недостаточности в B-клеточном иммунном ответе, и они продуцируют антитела, содержащие участки CDR3, полученные в результате перестройки вставленных кодирующих последовательностей и смежных генных сегментов иммуноглобулина (т. е. VH и JH человека).
[00286] Вкратце, селезенки и бедренные кости собирали у мышей VELOCIMMUNE® (n=3, 26% C57BL/6; 22% 129S6/SvEvTac; 50% Balb/cAnNTac; см., например, патенты США №№ 8502018 и 8642835) и мышей 6579ho/1293ho («TX-DH ho», n=3; 38% C57BL/6NTac; 36% 129S6/SvEvTac; 25% Balb/cAnNTac; мыши, гомозиготные по локусу тяжелой цепи иммуноглобулина, содержащему несколько сегментов VH человека, сконструированных сегментов DH, включающих кодирующие последовательности токсин, вместо стандартных сегментов DH и сегментов JH, функционально связанных с константным участком тяжелой цепи иммуноглобулина грызуна, включающего энхансеры и регуляторные участки тяжелой цепи грызуна, и содержащему вставленную нуклеотидную последовательность, кодирующую один или более генов Adam6 мыши [например, патенты США №№ 8642835 и 8697940]; и гомозиготные по локусу легкой κ-цепи иммуноглобулина, содержащему генные сегменты Vκ и Jκ человека, функционально связанные с геном участка Cκ грызуна, включающего энхансеры легкой κ-цепи грызуна). Костный мозг собирали из бедренных костей с помощью центрифугирования. Эритроциты из селезенки и препараты костного мозга лизировали с помощью буфера для лизиса ACK (Gibco) с последующей промывкой с помощью 1xPBS с 2% FBS. Выделенные клетки (1x106) инкубировали с выбранными смесями антител в течение 30 мин. при +4°C: окрашивание 1: антитело крысы к CD43 мыши, конъюгированное с FITC (Biolegend 121206, клон 1B11), антитело крысы к с-kit мыши, конъюгированное с PE (Biolegend 105808, клон 2B8), антитело крысы к IgM мыши, конъюгированное с PeCy (eBiosciences 25-5790-82, клон II/41), антитело крысы к IgD мыши, конъюгированное с PerCP-Cy5.5 (Biolegend 405710, клон 11-26c.2a), антитело крысы к CD3 мыши, конъюгированное с PB (Biolegend 100214, клон 17-A2), антитело крысы к B220 мыши, конъюгированное с APC (eBiosciences 17-0452-82, клон RA3-6B2), и антитело крысы к CD19 мыши, конъюгированное с APC-H7 (BD 560143 клон 1D3); окрашивание 2: антитело крысы к легкой каппа-цепи мыши, конъюгированное с FITC (BD 550003, клон 187.1), антитело крысы к легкой лямбда-цепи мыши, конъюгированное с РЕ (Biolegend 407308, клон RML-42), антитело крысы к IgM мыши, конъюгированное с PeCy (eBiosciences 25-5790-82, клон II/41), антитело крысы к IgD мыши, конъюгированное с PerCP-Cy5.5 (Biolegend 405710, клон 11-26c.2a), антитело крысы к CD3 мыши, конъюгированное с PB (Biolegend 100214, клон 17-A2), антитело крысы к B220 мыши, конъюгированное с APC (eBiosciences 17-0452-82, клон RA3-6B2), и антитело крысы к CD19 мыши, конъюгированное с APC-H7 (BD 560143 клон 1D3); окрашивание 3: антитело крысы к CD23 мыши, конъюгированное с FITC (Biolegend 101606, клон B3B4), антитело крысы к CD93 мыши (Biolegend 136504, клон AA4.1), антитело крысы к IgM мыши, конъюгированное с PeCy (eBiosciences 25-5790-82, клон II/41), антитело крысы к IgD мыши, конъюгированное с PerCP-Cy5.5 (Biolegend 405710, клон 11-26c.2a), антитело крысы к CD19 мыши, конъюгированное с V450 (Biolegend 560375, клон 1D3), антитело крысы к CD21/35 мыши, конъюгированное с APC (BD 558658, клон 7G6), и антитело крысы к B220 мыши, конъюгированное с APC-eFl780 (eBioscences 47-0452-82, клон RA3-6B2). После окрашивания клетки промывали и фиксировали в 2% формальдегиде. Получение данных осуществляли на проточном цитометре BD LSRFORTESSA™ и анализировали с помощью программного обеспечения FLOWJO™. Репрезентативные результаты изложены на фигурах 9A–9D и фигурах 10A–10D.
[00287] В аналогичном эксперименте селезенки и бедренные кости собирали у мышей VELOCIMMUNE® (n=3, 26% C57BL/6; 23% 129SvEvTac; 51% Balb/cAnNTac; см., например, патенты США №№ 8502018 и 8642835) и мышей 6590het («D6-DH het», n=3; 75% C57BL/6; 25% 129SvEvTac; мышей, гетерозиготных по локусу тяжелой цепи иммуноглобулина, содержащему несколько сегментов VH человека, сконструированных сегментов DH, включающих кодирующие последовательности D6 хемокинового рецептора-ловушки, вместо стандартных сегментов DH и сегментов JH, функционально связанных с константным участком тяжелой цепи иммуноглобулина грызуна, включающего энхансеры и регуляторные участки тяжелой цепи грызуна, и содержащему вставленную нуклеотидную последовательность, кодирующую один или более генов Adam6 мыши [например, патенты США №№ 8642835 и 8697940]; и анализировали с помощью проточной цитометрии (как описано выше). Репрезентативные результаты изложены на фигурах 11A–11D и фигурах 12A–12D.
[00288] В другом аналогичном эксперименте селезенки и бедренные кости собирали у мышей VELOCIMMUNE® (n=3, 38% C57BL/6; 36% 129SvEvTac; 25% Balb/cAnNTac; см., например, патенты США №№ 8502018 и 8642835) и мышей 6590ho/1293ho («D6-DH ho», n=3; 38% C57BL/6; 36% 129SvEvTac; 25% Balb/cAnNTac; мыши, гомозиготные по локусу тяжелой цепи иммуноглобулина, содержащему несколько сегментов VH человека, сконструированных сегментов DH, включающих кодирующие последовательности D6 хемокинового рецептора-ловушки, вместо одного или более стандартных сегментов DH и сегментов JH, функционально связанных с константным участком тяжелой цепи иммуноглобулина грызуна, включающего энхансеры и регуляторные участки тяжелой цепи грызуна, и содержащему вставленную нуклеотидную последовательность, кодирующую один или более генов Adam6 мыши [например, патенты США №№ 8642835 и 8697940]; и гомозиготные по локусу легкой κ-цепи иммуноглобулина, содержащему генные сегменты Vκ и Jκ человека, функционально связанные с геном участка Cκ грызуна, включающего энхансеры легкой κ-цепи грызуна) и анализировали с помощью проточной цитометрии (как описано выше). Репрезентативные результаты изложены на фигурах 13A – 13D и фигурах 14A–14D.
[00289] Результаты показали, что у мышей TX-DH ho, продемонстрированы аналогичное процентное содержание B-клеток в компартментах селезенки и костного мозга по сравнению с контрольными сконструированными мышами. У некоторых мышей TX-DH ho наблюдали более высокое процентное содержание Igλ-положительных клеток (например, см. фигуры 9C и 10D). В основном созревание В-клеток в селезенке демонстрировало нормальное протекание у мышей TX-DH ho по сравнению с контрольными сконструированными мышами.
[00290] У мышей D6-DH het также продемонстрировано аналогичное процентное содержание В-клеток в компартментах как селезенки, так и костного мозга (фигуры 11A – 12D). У мышей D6-DH het также проявлялись аналогичные соотношения легких цепей по сравнению с контрольными мышами (фигуры 11C и 12D). У мыши D6-DH het также продемонстрировано в основном нормальное развитие и созревание B-клеток.
Пример 6. Использование генов человека и перестройка кодирующих последовательностей в сконструированных участках DH
[00291] В данном примере продемонстрировано определение использования генных сегментов V(D)J человека у грызунов, содержащих сконструированный участок DH, описываемый в данном документе, с применением анализа репертуара антител с помощью секвенирования нового поколения (NGS). В частности, секвенирование с применением RT-PCR проводили с РНК, выделенной из подвергнутой сортировке селезенки и не подвергнутого сортировке костного мозга от мышей, гомозиготных по сконструированному участку DH, содержащему кодирующие последовательности токсина, и мышей, гетерозиготных по сконструированному участку DH, содержащему кодирующие последовательности ACKR2. Как описано ниже, все сконструированные сегменты DH наблюдали в проанализированных последовательностях от мышей, несущих сконструированный участок DH, содержащий кодирующие последовательности токсина, вместо стандартных сегментов DH. Также ниже описано, что большинство (т. е. 16 из 25) сконструированных сегментов DH наблюдали в проанализированных последовательностях от мышей, несущих сконструированный участок DH, содержащей кодирующие последовательности внеклеточной части ACKR2 (D6 хемокинового рецептора-ловушки).
[00292] Вкратце, селезенку и костный мозг собирали у мышей 6579ho/1293ho («TX-DH ho», n=3; выше), и B-клетки подвергали положительному обогащению из общих спленоцитов с помощью магнитной сортировки клеток с помощью магнитных гранул с антителом к CD19 мыши и колонок MACS® (Miltenyi Biotech). Общую РНК выделяли из обогащенных B-клеток в подвергнутых сортировке спленоцитах и не подвергнутом сортировке общем костном мозге с помощью набора для выделения РНК RNeasy Plus (Qiagen) в соответствии с инструкциями производителя. Затем из общей РНК синтезировали кДНК тяжелой цепи иммуноглобулина (μ-цепь, IgM) с помощью обратной транскриптазы SuperScript III (Thermo Fisher Scientific) и праймера константного участка IgM мыши (таблица 9, IgM RT) в соответствии с инструкциями производителя. кДНК IgM амплифицировали с применением стратегии мультиплексной ПЦР, использующей смесь праймеров, специфичных к семейству VH, (таблица 9, VH multi-1, 2, 3, 4, 5 и 6), разработанных для отжига с лидерными последовательностями VH, и праймер (таблица 9, IgM PCR1), сконструированный для отжига с константным участком IgM. Продукты ПЦР, варьирующие в размере ~450–800 п.о., выделяли с использованием набора Pippin Prep (SAGE Science), а затем проводили вторую ПЦР с применением праймеров PCR2 (таблица 9, PCR2-F и PCR2-R, где «XXXXXX» представляет собой 6-нуклеотидный индекс для распознавания библиотек секвенирования и для обеспечения одновременного секвенирования множества библиотек), для присоединения адапторов и индексов секвенирования. Продукты ПЦР, варьирующие в размере ~490°п.о. – 800 п.о. выделяли, очищали и количественно подсчитывали с помощью qPCR с использованием набора для количественного определения библиотек KAPA (KAPA Biosystems) перед загрузкой в секвенатор MiSeq (Illumina) для секвенирования с помощью наборов реагентов MiSeq v3 (2x300 циклов). В таблице 9 изложены последовательности выбранных праймеров, используемых для разработки библиотеки репертуара.
[00293] Для проведения биоинформационного анализа исходные последовательности Illumina подвергали демультиплексированию и фильтровали на основе качества, длины и совпадения с соответствующим праймером гена константного участка. Перекрывающиеся риды парных концов сливали и анализировали с использованием собственного, разработанного в лаборатории процесса обработки данных. В данном процессе обработки данных использовали локально установленную IgBLAST (NCBI, v2.2.25+) для выравнивания последовательностей перегруппированных легких цепей с базой данных генных сегментов VH и JH зародышевой линии человека и отмечали продуктивные и непродуктивные связи, а также наличие стоп-кодонов. Последовательности CDR3 и предполагаемые нематричные нуклеотиды экстрагировали с использованием границ, определенных в системе Международной информационной службы по иммуногенетике (IMGT). Репрезентативные результаты изложены в таблице 10 (общие совпадения с кодирующими последовательностями токсина среди ридов последовательностей с использованием каждую смесь праймеров, специфичных для семейства VH) и на фигурах 15–17.
[00294] Данные результаты демонстрировали, что большинство проанализированных последовательностей содержало последовательности токсина в пределах участка CDR3 в последовательностях участка VH. В частности, приблизительно 87% последовательностей, соответствующих продуктивным перестройкам у мышей, несущих сконструированный участок DH, содержащий множество кодирующих последовательностей токсина, вместо стандартных сегментов DH, содержали аминокислотные последовательности токсина в пределах участков CDR3. Помимо этого, у данных мышей выявляли с различными частотами все 26 кодирующих последовательностей токсина, расположенных в месторасположении стандартных сегментов DH. Наиболее часто используемыми выявленными кодирующими последовательностями токсина были короткая петля из четырех (4)-аминокислот μCTX-SmIIIa (положение DH, DH2–15), вставка μCTX-SmIIIA midi (положение DH, DH3–16) и μCTX-PIIIA midi (положение DH, DH3–22).
[00295] В аналогичном эксперименте использование сегмента VH человека, сконструированного сегмента DH и сегмента JH человека анализировали у мышей 6590het («D6-DH het», n=3; выше) с применением анализа репертуара антител с помощью секвенирования нового поколения (NGS), описываемого выше. Секвенирование RT-PCR проводили с РНК, выделенной из подвергнутой сортировке селезенки и не подвергнутого сортировке костного мозга мышей c использованием стратегии мультиплексной ПЦР, описываемой выше. Репрезентативные результаты изложены в таблице 11 (количество аминокислотных (AA) последовательностей в рамке считывания среди ридов нуклеотидной последовательности (NT) в РНК, амплифицированной из костного мозга и селезенки), в таблице 12 (процент выбранных кодирующих последовательностей D6 в амплифицированной РНК из селезенки и костного мозга [объединены все семейства VH, не отражает количественное использование VH]) и на фигурах 18–20.
[00296] Результаты демонстрировали, что длина большинства кодирующих последовательностей D6, используемых в РНК, амплифицированной из костного мозга и селезенки, составляли от 30–40 нуклеотидов. В частности, самую длинную (например, Nterm+EC3 (216 нуклеотидов)) и самую короткую (например, EC1-N (9 нуклеотидов)) кодирующие последовательности D6 изначально не обнаруживали в последовательностях, проанализированных от мышей D6-DH het. Авторы настоящего изобретения наблюдали определенные ограничения при оценке использования кодирующих последовательностей D6 из сконструированного участка DH. Например, авторы настоящего изобретения отметили, что последовательность IgM, содержащая полноразмерные (или более длинные, чем полноразмерные) длинные кодирующие последовательности D6 (например, D6 Nterm+EC3, положение DH, DH4-4; SEQ ID NO:33), по-видимому, исключались из последовательностей вследствие стадии отбора по размеру в геле (см. выше), а более короткие кодирующие последовательности D6 (например, D6 EC1, положение DH, DH5-5; SEQ ID NO:3), по-видимому, исключались вследствие стадии отбора посредством «тримминга». В случае более коротких кодирующих последовательностей D6, однако, просмотр последовательностей показал, что данные кодирующие последовательности D6, в действительности, присутствовали среди последовательностей, полученных из амплифицированных РНК. Кроме того, авторы настоящего изобретения наблюдали неправильное выравнивание последовательностей при сравнении с базами данных из-за сходства между выбранными сегментами JH (например, JH1) и кодирующими последовательностями D6. Такое сходство последовательностей приводило к идентификации более коротких последовательностей DH и возможности добавления неоднозначных аннотаций в определенных базах данных. В результате этого для некоторых последовательностей потребовался отдельный анализ с использованием выравниваний, проведенных вручную. Среди всех проанализированных последовательностей от мышей D6-DH het, приблизительно 50–60% аминокислотных последовательностей находились внутри рамки считывания, и в них использовалась кодирующая последовательность D6 в пределах участков CDR3 амплифицированных тяжелых цепей. Кроме того, 16 из 25 сконструированных сегментов DH с D6 выявляли в последовательностях, амплифицированных из РНК (таблица 12), в то же время в проанализированных последовательностях не выявляли D6 Nterm (DH3-3), D6 Nterm+EC3 (DH4-4), D6 EC1 (DH5-5), D6 EC1-N (DH1-7), D6 EC1-S (DH5-12), D6 Nterm-S (DH6-13), D6 Nterm-EC3 (DH3-16), D6 EC1+EC2 (DH4-17) и D6 EC3-C (DH4-23). Как описано выше, использование некоторых таких кодирующих последовательностей D6, в частности, более длинных кодирующих последовательностей D6, не могло подтверждаться в проанализированном наборе последовательностей. В целом, значительных отличий в использовании кодирующих последовательностей D6 между образцами, полученными из костного мозга и селезенки, выявлено не было. Также, по-видимому, в проанализированных последовательностях отсутствовало смещение, обусловленное негативным отбором против перестройки кодирующих последовательностей D6.
[00297] В целом, в данном примере продемонстрировано, что у мышей, несущих сконструированный локус тяжелой цепи иммуноглобулина, который характеризуется присутствием сконструированного участка DH, содержащего кодирующие последовательности полипептида, не являющегося иммуноглобулином (например, D6 хемокинового рецептора-ловушки или одного или более токсинов), вместо стандартных сегментов DH, может происходить перестройка сконструированных кодирующих последовательностей со смежными сегментами VH и JH человека с образованием функциональных тяжелых цепей. Кроме того, у представленных сконструированных мышей продемонстрирован надежный репертуар антител, в которых используются множество кодирующих последовательностей, сконструированных в синтетических участках DH в связи с некоторыми семействами сегментов VH и JH человека.
TCACCATGGACTGSACCTGGA (SEQ ID NO:237)
CCATGGACACACTTTGYTCCAC (SEQ ID NO:238)
TCACCATGGAGTTTGGGCTGAGC (SEQ ID NO:239)
AGAACATGAAACAYCTGTGGTTCTT (SEQ ID NO:240)
ATGGGGTCAACCGCCATCCT (SEQ ID NO:241)
ACAATGTCTGTCTCCTTCCTCAT (SEQ ID NO:242)
GGGAAGACATTTGGGAAGGAC (SEQ ID NO:243)
GTGACTGGAGTTCAGACGTGTGCTCTTCCGATCT (SEQ ID NO:244)
ACACTCTTTCCCTACACGACGCTCTTCCGATCT (SEQ ID NO:245)
Пример 7. Продуцирование антител в грызунах, содержащих сконструированные участки DH
[00298] В данном примере продемонстрировано продуцирование антител в грызуне, геном которого содержит вариабельный участок тяжелой цепи иммуноглобулина, который включает сконструированный участок DH, описываемый в данном документе. При необходимости, для иммунизации грызунов, содержащих сконструированный участок DH, описываемый в данном документе, можно применять способы, описанные в данном примере, и/или способы иммунизации, хорошо известные из уровня техники, с использованием полипептидов или их фрагментов (например, пептидов, полученных из требуемого эпитопа), или комбинации полипептидов или их фрагментов.
[00299] Вкратце, когортам мышей, содержащих сконструированный участок DH, описываемый в данном документе, вводили представляющий интерес антиген с использованием способов иммунизации, известных из уровня техники. Гуморальный иммунный ответ контролировали с использованием иммунологического анализа ELISA (т.e. определения титра сыворотки).
[00300] Получение общей легкой цепи мыши (также называемой универсальной легкой цепью или ULC мыши), содержащей генную последовательность единого перестроенного вариабельного участка V:J (например, общая легкая цепь мыши Vκ1-39Jκ5 или Vκ3-20Jκ1) и получение антиген-специфических антител в таких мышах описано, например, в заявках на патент США №№ 13/022759, 13/093156, 13/412936, 13/488628, 13/798310 и 13/948818 (публикации №№ 2011/0195454, 2012/0021409, 2012/0192300, 2013/0045492, US 20130185821 и US 20130302836 соответственно), каждая из которых включена в данный документ посредством ссылки во всей своей полноте. Более конкретно, получали мышей, у которых генетически сконструированная легкая каппа-цепь Vκ1-39Jκ5 (1633 HO или 1634 HO) или генетически сконструированная легкая каппа-цепь Vκ3-20Jκ1 (1635 HO или 1636 HO) экспрессируется в их зародышевой линии.
[00301] Мышей VELOCIMMUNE®, содержащих единый перегруппированный участок легкой цепи зародышевой линии человека (ULC Vκ1-39Jκ5; 1633 или 1634) или (ULC Vκ3-20Jκ1; 1635 или 1636), скрещивали с мышами, несущими модифицированный константный участок IgG. В частности, таких мышей с ULC скрещивали с мышами, имеющими геном, содержащий гомозиготный локус тяжелой цепи иммуноглобулина, содержащий несколько сегментов VH человека, сконструированных сегментов DH, включающих кодирующие последовательности токсина, вместо стандартных сегментов DH и сегментов JH, функционально связанных с константным участком тяжелой цепи иммуноглобулина мыши, включающим энхансеры и регуляторные участки тяжелой цепи мыши, и содержащий встроенную нуклеотидную последовательность, кодирующую один или более генов Adam6 мыши [например, патенты США №№ 8642835 и 8697940], с получением мышей-потомков, гетерозиготных или гомозиготных по локусу тяжелой цепи, содержащему кодирующую последовательность токсина, и гетерозиготных или гомозиготных по универсальной легкой цепи. Как показано на фигуре 21, мыши, гомозиготные по сконструированному участку DH, содержащему кодирующие последовательности токсина, и гомозиготные по универсальной легкой цепи (6579HO/1634HO), имели гуморальный иммунный ответ после иммунизации растворимым белком по сравнению с контрольным животным (1460/1634).
Пример 8. Выделение клеток, экспрессирующих антитела, продуцируемые в грызунах, содержащих сконструированные участки DH , и/или нуклеиновых кислот, кодирующих такие антитела
[00302] При достижении требуемого иммунного ответа собирали спленоциты (и/или другую лимфатическую ткань) и сливали их с клетками миеломы мыши для сохранения их жизнеспособности и образования иммортализированных линий клеток гибридомы. Проводили скрининг (например, с помощью анализа ELISA) и отбор линий клеток гибридомы для идентификации линий клеток гибридомы, которые продуцировали антиген-специфические антитела. При необходимости, гибридомы можно дополнительно характеризовать в отношении относительной аффинности связывания и изотипу. С использованием данной методики получали несколько антиген-специфических химерных антител (т. e. антител, имеющих вариабельные домены человека и константные домены грызуна).
[00303] ДНК, кодирующую вариабельные участки тяжелой цепи и легких цепей, можно выделять и связать с требуемыми изотипами (константными участками) тяжелой цепи и легкой цепи для получения полностью человеческих антител. Такой белок антитела может продуцироваться в клетке, такой как клетка CHO. Полностью человеческие антитела затем характеризуют в отношении относительной аффинности связывания и/или активности нейтрализации представляющего интерес антигена.
[00304] ДНК, кодирующую антиген-специфические химерные антитела или вариабельные домены легкой и тяжелой цепей, можно выделять напрямую из антиген-специфических лимфоцитов. Изначально выделяли химерные антитела с высокой аффинностью, имеющие вариабельный участок человека и константный участок грызуна, и их характеризовали и подвергали отбору в отношение требуемых характеристик, включая аффинность, селективность, эпитоп и т.д. Константные участки грызуна заменяли на требуемые константные участки человека для получения полностью человеческих антител. Хотя выбранный константный участок может варьировать в зависимости от конкретного применения, у вариабельной области сохраняются характеристики высокой аффинности связывания с антигеном или специфичности к мишеням. Также антиген-специфические антитела выделяли напрямую из антиген-положительных B-клеток (из иммунизированных мышей) без слияния с клетками миеломы, как описано в патенте США №7582298, который специально включен в данный документ посредством ссылки во всей своей полноте. С помощью данного способа получали несколько полностью человеческих антиген-специфических антител (т.e. антител, обладающих вариабельными доменами человека и константными доменами человека).
ЭКВИВАЛЕНТЫ
[00305] Имея описанные таким образом некоторые аспекты по меньшей мере одного варианта осуществления настоящего изобретения, специалисты в данной области будут понимать, что различные изменения, модификации и улучшения без труда придут на ум специалистам в данной области. Подразумевается, что такие изменения, модификации и улучшения являются частью настоящего изобретения, и подразумевается, что они находятся в пределах сущности и объема настоящего изобретения. Соответственно, вышеприведенные описание и графические материалы представлены исключительно в качестве примера, и настоящее изобретение подробно описано в нижеследующей формуле изобретения.
[00306] Порядковые термины, такие как «первый», «второй», «третий» и т.д., используемые в формуле изобретения для модификации элемента формулы изобретения, сами по себе не означают какой-либо приоритет, предшествование или порядок расположения одного элемента формулы изобретения перед другим или временной порядок, в котором осуществляют действия способа, но используются исключительно в качестве отметок для разграничения одного элемента формулы изобретения, имеющего определенное название, от другого элемента, имеющего такое же название (за исключением используемого порядкового термина), для разграничения элементов формулы изобретения.
[00307] Формы единственного числа в описании и формуле изобретения, если четко не указано противоположное, следует понимать как охватывающие определяемые объекты в форме множественного числа. Признаки, раскрытые в пунктах формулы изобретения или частях описания, у которых между одним или более членами группы содержится «или», считаются отвечающими требованиям, если один, более одного или все члены группы присутствуют в указанном продукте или способе, используются в них или иным образом имеют отношение к ним, если не указано противоположное или иное не очевидно из контекста. Настоящее изобретение охватывает варианты осуществления, в которых только один член группы присутствует в указанном продукте или способе, используется в них или иным образом имеет отношение к ним. Настоящее изобретение также охватывает варианты осуществления, в которых более одного или все члены группы присутствуют в указанном продукте или способе, используются в них или иным образом имеют отношение к ним. Кроме того, следует понимать, что настоящее изобретение охватывает все варианты, комбинации и перестановки, в которых одно или более ограничений, элементов, условий, описательных терминов и т.д. из одного или более перечисленных пунктов формулы изобретения входят в другой пункт формулы изобретения, зависимый от того же основного пункта формулы изобретения (или, в зависимости от того, что применимо, любого другого пункта формулы изобретения), если не указано иное или если среднему специалисту в данной области не будет очевидным возникновение противоречия или несоответствия. В тех случаях, когда элементы представлены в виде перечней (например, в виде группы Маркуша или в аналогичном формате), следует понимать, что также раскрыта каждая подгруппа элементов, и любой элемент(ы) можно исключить из группы. В целом, следует понимать, что в тех случаях, когда настоящее изобретение или аспекты настоящего изобретения упоминаются как содержащие конкретные элементы, признаки и т.д., определенные варианты осуществления настоящего изобретения или аспекты настоящего изобретения содержат или фактически содержат такие элементы, признаки и т.д. С целью упрощения такие варианты осуществления не были в каждом случае специально прямо изложены в данном документе. Также следует понимать, что любой вариант осуществления или аспект настоящего изобретения может быть явным образом исключен из формулы изобретения независимо от упоминания конкретного исключения в настоящем описании.
[00308] Специалистам в данной области будут понятны типичные стандартные отклонения или ошибки, присущие значениям, получаемым в анализах или других процессах, описанных в данном документе.
[00309] Публикации, веб-сайты и другие справочные материалы, упоминаемые в данном документе для описания предпосылок настоящего изобретения и для предоставления дополнительных подробностей, относящихся к его практическому осуществлению, настоящим включены в данный документ посредством ссылки.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Ридженерон Фармасьютикалс Инк.
Макдональд, Линн
Макрайтер, Джон
Мерфи, Эндрю
<120> ГРЫЗУНЫ, ИМЕЮЩИЕ СКОНСТРУИРОВАННЫЙ УЧАСТОК РАЗНООБРАЗИЯ ТЯЖЕЛОЙ ЦЕПИ
<130> 017283.1066/10150WO01
<140> TBD
<141> 2017-01-12
<150> US 62/278127
<151> 2016-01-13
<160> 245
<170> PatentIn версия 3.5
<210> 1
<211> 150
<212> ДНК
<213> Homo sapiens
<400> 1
atggcagcta ctgccagccc gcagccactg gctactgagg atgccgattc tgagaatagc 60
agcttctact actatgacta cctggatgaa gtagctttca tgctctgccg gaaggatgct 120
gtggttagct ttggcaaagt tttcctgcca 150
<210> 2
<211> 50
<212> БЕЛОК
<213> Homo sapiens
<400> 2
Met Ala Ala Thr Ala Ser Pro Gln Pro Leu Ala Thr Glu Asp Ala Asp
1 5 10 15
Ser Glu Asn Ser Ser Phe Tyr Tyr Tyr Asp Tyr Leu Asp Glu Val Ala
20 25 30
Phe Met Leu Cys Arg Lys Asp Ala Val Val Ser Phe Gly Lys Val Phe
35 40 45
Leu Pro
50
<210> 3
<211> 15
<212> ДНК
<213> Homo sapiens
<400> 3
agcttcttgt gcaag 15
<210> 4
<211> 5
<212> БЕЛОК
<213> Homo sapiens
<400> 4
Ser Phe Leu Cys Lys
1 5
<210> 5
<211> 102
<212> ДНК
<213> Homo sapiens
<400> 5
caaacccatg aaaaccccaa gggagtttgg aactgccatg ccgatttcgg cgggcatggc 60
accatttgga agctcttcct ccggttccag cagaacctgc ta 102
<210> 6
<211> 34
<212> БЕЛОК
<213> Homo sapiens
<400> 6
Gln Thr His Glu Asn Pro Lys Gly Val Trp Asn Cys His Ala Asp Phe
1 5 10 15
Gly Gly His Gly Thr Ile Trp Lys Leu Phe Leu Arg Phe Gln Gln Asn
20 25 30
Leu Leu
<210> 7
<211> 66
<212> ДНК
<213> Homo sapiens
<400> 7
ctgcataccc tgctggacct gcaagtattc ggcaactgtg aggttagcca gcatctagac 60
tatgcc 66
<210> 8
<211> 22
<212> БЕЛОК
<213> Homo sapiens
<400> 8
Leu His Thr Leu Leu Asp Leu Gln Val Phe Gly Asn Cys Glu Val Ser
1 5 10 15
Gln His Leu Asp Tyr Ala
20
<210> 9
<211> 150
<212> ДНК
<213> Homo sapiens
<400> 9
atggcagcta ctgccagccc gcagccactg gctactgagg atgccgattc tgagaatagc 60
agcttctact actatgacta cctggatgaa gtagctttca tgctcagccg gaaggatgct 120
gtggttagct ttggcaaagt tttcctgcca 150
<210> 10
<211> 50
<212> БЕЛОК
<213> Homo sapiens
<400> 10
Met Ala Ala Thr Ala Ser Pro Gln Pro Leu Ala Thr Glu Asp Ala Asp
1 5 10 15
Ser Glu Asn Ser Ser Phe Tyr Tyr Tyr Asp Tyr Leu Asp Glu Val Ala
20 25 30
Phe Met Leu Ser Arg Lys Asp Ala Val Val Ser Phe Gly Lys Val Phe
35 40 45
Leu Pro
50
<210> 11
<211> 15
<212> ДНК
<213> Homo sapiens
<400> 11
agcttcttga gcaag 15
<210> 12
<211> 5
<212> БЕЛОК
<213> Homo sapiens
<400> 12
Ser Phe Leu Ser Lys
1 5
<210> 13
<211> 102
<212> ДНК
<213> Homo sapiens
<400> 13
caaacccatg aaaaccccaa gggagtttgg aacagccatg ccgatttcgg cgggcatggc 60
accatttgga agctcttcct ccggttccag cagaacctgc ta 102
<210> 14
<211> 34
<212> БЕЛОК
<213> Homo sapiens
<400> 14
Gln Thr His Glu Asn Pro Lys Gly Val Trp Asn Ser His Ala Asp Phe
1 5 10 15
Gly Gly His Gly Thr Ile Trp Lys Leu Phe Leu Arg Phe Gln Gln Asn
20 25 30
Leu Leu
<210> 15
<211> 66
<212> ДНК
<213> Homo sapiens
<400> 15
ctgcataccc tgctggacct gcaagtattc ggcaacagtg aggttagcca gcatctagac 60
tatgcc 66
<210> 16
<211> 22
<212> БЕЛОК
<213> Homo sapiens
<400> 16
Leu His Thr Leu Leu Asp Leu Gln Val Phe Gly Asn Ser Glu Val Ser
1 5 10 15
Gln His Leu Asp Tyr Ala
20
<210> 17
<211> 135
<212> ДНК
<213> Homo sapiens
<400> 17
atggcagcta ctgccagccc gcagccactg gctactgagg atgccgattc tgagaatagc 60
agcttctact actatgacta cctggatgaa gtagctttca tgctctgcga ggttagccag 120
catctagact atgcc 135
<210> 18
<211> 45
<212> БЕЛОК
<213> Homo sapiens
<400> 18
Met Ala Ala Thr Ala Ser Pro Gln Pro Leu Ala Thr Glu Asp Ala Asp
1 5 10 15
Ser Glu Asn Ser Ser Phe Tyr Tyr Tyr Asp Tyr Leu Asp Glu Val Ala
20 25 30
Phe Met Leu Cys Glu Val Ser Gln His Leu Asp Tyr Ala
35 40 45
<210> 19
<211> 81
<212> ДНК
<213> Homo sapiens
<400> 19
ctgcataccc tgctggacct gcaagtattc ggcaactgtc ggaaggatgc tgtggttagc 60
tttggcaaag ttttcctgcc a 81
<210> 20
<211> 27
<212> БЕЛОК
<213> Homo sapiens
<400> 20
Leu His Thr Leu Leu Asp Leu Gln Val Phe Gly Asn Cys Arg Lys Asp
1 5 10 15
Ala Val Val Ser Phe Gly Lys Val Phe Leu Pro
20 25
<210> 21
<211> 78
<212> ДНК
<213> Homo sapiens
<400> 21
agcttcttgt gccatgccga tttcggcggg catggcacca tttggaagct cttcctccgg 60
ttccagcaga acctgcta 78
<210> 22
<211> 26
<212> БЕЛОК
<213> Homo sapiens
<400> 22
Ser Phe Leu Cys His Ala Asp Phe Gly Gly His Gly Thr Ile Trp Lys
1 5 10 15
Leu Phe Leu Arg Phe Gln Gln Asn Leu Leu
20 25
<210> 23
<211> 39
<212> ДНК
<213> Homo sapiens
<400> 23
caaacccatg aaaaccccaa gggagtttgg aactgcaag 39
<210> 24
<211> 13
<212> БЕЛОК
<213> Homo sapiens
<400> 24
Gln Thr His Glu Asn Pro Lys Gly Val Trp Asn Cys Lys
1 5 10
<210> 25
<211> 135
<212> ДНК
<213> Homo sapiens
<400> 25
atggcagcta ctgccagccc gcagccactg gctactgagg atgccgattc tgagaatagc 60
agcttctact actatgacta cctggatgaa gtagctttca tgctcagcga ggttagccag 120
catctagact atgcc 135
<210> 26
<211> 45
<212> БЕЛОК
<213> Homo sapiens
<400> 26
Met Ala Ala Thr Ala Ser Pro Gln Pro Leu Ala Thr Glu Asp Ala Asp
1 5 10 15
Ser Glu Asn Ser Ser Phe Tyr Tyr Tyr Asp Tyr Leu Asp Glu Val Ala
20 25 30
Phe Met Leu Ser Glu Val Ser Gln His Leu Asp Tyr Ala
35 40 45
<210> 27
<211> 81
<212> ДНК
<213> Homo sapiens
<400> 27
ctgcataccc tgctggacct gcaagtattc ggcaacagtc ggaaggatgc tgtggttagc 60
tttggcaaag ttttcctgcc a 81
<210> 28
<211> 27
<212> БЕЛОК
<213> Homo sapiens
<400> 28
Leu His Thr Leu Leu Asp Leu Gln Val Phe Gly Asn Ser Arg Lys Asp
1 5 10 15
Ala Val Val Ser Phe Gly Lys Val Phe Leu Pro
20 25
<210> 29
<211> 78
<212> ДНК
<213> Homo sapiens
<400> 29
agcttcttga gccatgccga tttcggcggg catggcacca tttggaagct cttcctccgg 60
ttccagcaga acctgcta 78
<210> 30
<211> 26
<212> БЕЛОК
<213> Homo sapiens
<400> 30
Ser Phe Leu Ser His Ala Asp Phe Gly Gly His Gly Thr Ile Trp Lys
1 5 10 15
Leu Phe Leu Arg Phe Gln Gln Asn Leu Leu
20 25
<210> 31
<211> 39
<212> ДНК
<213> Homo sapiens
<400> 31
caaacccatg aaaaccccaa gggagtttgg aacagcaag 39
<210> 32
<211> 13
<212> БЕЛОК
<213> Homo sapiens
<400> 32
Gln Thr His Glu Asn Pro Lys Gly Val Trp Asn Ser Lys
1 5 10
<210> 33
<211> 216
<212> ДНК
<213> Homo sapiens
<400> 33
atggcagcta ctgccagccc gcagccactg gctactgagg atgccgattc tgagaatagc 60
agcttctact actatgacta cctggatgaa gtagctttca tgctctgccg gaaggatgct 120
gtggttagct ttggcaaagt tttcctgcca ctgcataccc tgctggacct gcaagtattc 180
ggcaactgtg aggttagcca gcatctagac tatgcc 216
<210> 34
<211> 72
<212> БЕЛОК
<213> Homo sapiens
<400> 34
Met Ala Ala Thr Ala Ser Pro Gln Pro Leu Ala Thr Glu Asp Ala Asp
1 5 10 15
Ser Glu Asn Ser Ser Phe Tyr Tyr Tyr Asp Tyr Leu Asp Glu Val Ala
20 25 30
Phe Met Leu Cys Arg Lys Asp Ala Val Val Ser Phe Gly Lys Val Phe
35 40 45
Leu Pro Leu His Thr Leu Leu Asp Leu Gln Val Phe Gly Asn Cys Glu
50 55 60
Val Ser Gln His Leu Asp Tyr Ala
65 70
<210> 35
<211> 117
<212> ДНК
<213> Homo sapiens
<400> 35
agcttcttgt gcaagcaaac ccatgaaaac cccaagggag tttggaactg ccatgccgat 60
ttcggcgggc atggcaccat ttggaagctc ttcctccggt tccagcagaa cctgcta 117
<210> 36
<211> 39
<212> БЕЛОК
<213> Homo sapiens
<400> 36
Ser Phe Leu Cys Lys Gln Thr His Glu Asn Pro Lys Gly Val Trp Asn
1 5 10 15
Cys His Ala Asp Phe Gly Gly His Gly Thr Ile Trp Lys Leu Phe Leu
20 25 30
Arg Phe Gln Gln Asn Leu Leu
35
<210> 37
<211> 105
<212> ДНК
<213> Homo sapiens
<400> 37
atggcagcta ctgccagccc gcagccactg gctactgagg atgccgattc tgagaatagc 60
agcttctact actatgacta cctggatgaa gtagctttca tgctc 105
<210> 38
<211> 35
<212> БЕЛОК
<213> Homo sapiens
<400> 38
Met Ala Ala Thr Ala Ser Pro Gln Pro Leu Ala Thr Glu Asp Ala Asp
1 5 10 15
Ser Glu Asn Ser Ser Phe Tyr Tyr Tyr Asp Tyr Leu Asp Glu Val Ala
20 25 30
Phe Met Leu
35
<210> 39
<211> 42
<212> ДНК
<213> Homo sapiens
<400> 39
cggaaggatg ctgtggttag ctttggcaaa gttttcctgc ca 42
<210> 40
<211> 14
<212> БЕЛОК
<213> Homo sapiens
<400> 40
Arg Lys Asp Ala Val Val Ser Phe Gly Lys Val Phe Leu Pro
1 5 10
<210> 41
<211> 9
<212> ДНК
<213> Homo sapiens
<400> 41
agcttcttg 9
<210> 42
<211> 3
<212> БЕЛОК
<213> Homo sapiens
<400> 42
Ser Phe Leu
1
<210> 43
<211> 33
<212> ДНК
<213> Homo sapiens
<400> 43
caaacccatg aaaaccccaa gggagtttgg aac 33
<210> 44
<211> 11
<212> БЕЛОК
<213> Homo sapiens
<400> 44
Gln Thr His Glu Asn Pro Lys Gly Val Trp Asn
1 5 10
<210> 45
<211> 66
<212> ДНК
<213> Homo sapiens
<400> 45
catgccgatt tcggcgggca tggcaccatt tggaagctct tcctccggtt ccagcagaac 60
ctgcta 66
<210> 46
<211> 22
<212> БЕЛОК
<213> Homo sapiens
<400> 46
His Ala Asp Phe Gly Gly His Gly Thr Ile Trp Lys Leu Phe Leu Arg
1 5 10 15
Phe Gln Gln Asn Leu Leu
20
<210> 47
<211> 36
<212> ДНК
<213> Homo sapiens
<400> 47
ctgcataccc tgctggacct gcaagtattc ggcaac 36
<210> 48
<211> 12
<212> БЕЛОК
<213> Homo sapiens
<400> 48
Leu His Thr Leu Leu Asp Leu Gln Val Phe Gly Asn
1 5 10
<210> 49
<211> 27
<212> ДНК
<213> Homo sapiens
<400> 49
gaggttagcc agcatctaga ctatgcc 27
<210> 50
<211> 9
<212> БЕЛОК
<213> Homo sapiens
<400> 50
Glu Val Ser Gln His Leu Asp Tyr Ala
1 5
<210> 51
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированная RSS
<220>
<221> иной_признак
<222> (10)..(21)
<223> n представляет собой a, c, g или t
<400> 51
acaaaaaccn nnnnnnnnnn ncacagtg 28
<210> 52
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированная RSS
<220>
<221> иной_признак
<222> (8)..(19)
<223> n представляет собой a, c, g или t
<400> 52
cacagtgnnn nnnnnnnnna caaaaacc 28
<210> 53
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированная RSS
<220>
<221> иной_признак
<222> (10)..(21)
<223> n представляет собой a, c, g или t
<400> 53
ggwtwytgwn nnnnnnnnnn ncactgtg 28
<210> 54
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированная RSS
<220>
<221> иной_признак
<222> (8)..(19)
<223> n представляет собой a, c, g или t
<400> 54
cacagtgnnn nnnnnnnnnb cmrmaacy 28
<210> 55
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированная RSS
<220>
<221> иной_признак
<222> (10)..(21)
<223> n представляет собой a, c, g или t
<400> 55
gatttttgtn nnnnnnnnnn ntactgtg 28
<210> 56
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированная RSS
<220>
<221> иной_признак
<222> (8)..(19)
<223> n представляет собой a, c, g или t
<400> 56
cacagtgnnn nnnnnnnnna caaaaacc 28
<210> 57
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированная RSS
<220>
<221> иной_признак
<222> (10)..(21)
<223> n представляет собой a, c, g или t
<400> 57
ggattttgtn nnnnnnnnnn ncactgtg 28
<210> 58
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированная RSS
<220>
<221> иной_признак
<222> (8)..(19)
<223> n представляет собой a, c, g или t
<400> 58
cacagtgnnn nnnnnnnnnt caaaaacc 28
<210> 59
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированная RSS
<400> 59
ggattttgta cagccccgag tcactgtg 28
<210> 60
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированная RSS
<400> 60
cacagtgaga aaaactgtgt caaaaacc 28
<210> 61
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированная RSS
<400> 61
ggattttgta cagccccgag tcactgtg 28
<210> 62
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированная RSS
<400> 62
cacagtgaga aaagcttcgt caaaaacc 28
<210> 63
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированная RSS
<400> 63
ggattttgta cagccccgag tcactgtg 28
<210> 64
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированная RSS
<400> 64
cacagtgaga atagctacgt caaaaacc 28
<210> 65
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированная RSS
<400> 65
ggattttgta cagccccgag tcactgtg 28
<210> 66
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированная RSS
<400> 66
cacagtgaga aaaactgtgt caaaaacc 28
<210> 67
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированная RSS
<400> 67
ggattttgta cagccccgag tcactgtg 28
<210> 68
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированная RSS
<400> 68
cacagtgaga aaagctatgt caaaaacc 28
<210> 69
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированная RSS
<400> 69
ggattttgtg ggggctcgtg tcactgtg 28
<210> 70
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированная RSS
<400> 70
cacagtgaca cagccccatt caaaaacc 28
<210> 71
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированная RSS
<400> 71
ggattttgtg ggggctcgtg tcactgtg 28
<210> 72
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированная RSS
<400> 72
cacagtgaca cagccccatt caaaaacc 28
<210> 73
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированная RSS
<400> 73
ggattttgtg ggggctcgtg tcactgtg 28
<210> 74
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированная RSS
<400> 74
cacagtgaca cagacccatt caaaaacc 28
<210> 75
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированная RSS
<400> 75
ggattttgtg ggggttcgtg tcactgtg 28
<210> 76
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированная RSS
<400> 76
cacagtgaca caaccccatt caaaaacc 28
<210> 77
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированная RSS
<400> 77
ggattttgtt gaggtctgtg tcactgtg 28
<210> 78
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированная RSS
<400> 78
cacagtgtca cagagtccat caaaaacc 28
<210> 79
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированная RSS
<400> 79
ggattttgtt gaggtctgtg tcactgtg 28
<210> 80
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированная RSS
<400> 80
cacagtgtca cagagtccat caaaaacc 28
<210> 81
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированная RSS
<400> 81
ggattttgtt gaggtctgtg tcactgtg 28
<210> 82
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированная RSS
<400> 82
cacagtgtca cagagtccat caaaaacc 28
<210> 83
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированная RSS
<400> 83
ggattttgtt gaggtctgtg tcactgtg 28
<210> 84
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированная RSS
<400> 84
cacagtgtca cacggtccat caaaaacc 28
<210> 85
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированная RSS
<400> 85
ggattttgtt gaggtctgtg tcactgtg 28
<210> 86
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированная RSS
<400> 86
cacagtgtca cagagtccat caaaaacc 28
<210> 87
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированная RSS
<400> 87
ggattttgtg aagggtcctc ccactgtg 28
<210> 88
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированная RSS
<400> 88
cacagtgatg aacccagcat caaaaacc 28
<210> 89
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированная RSS
<400> 89
ggattttgtg aagggccctc ccactgtg 28
<210> 90
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированная RSS
<400> 90
cacagtgatg aacccagtgt caaaaacc 28
<210> 91
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированная RSS
<400> 91
ggattttgtg aaggcccctc ccactgtg 28
<210> 92
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированная RSS
<400> 92
cacagtgatg aaactagcat caaaaacc 28
<210> 93
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированная RSS
<400> 93
ggattttgtg aagggccctc ccactgtg 28
<210> 94
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированная RSS
<400> 94
cacagtgatg aaaccagcat caaaaacc 28
<210> 95
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированная RSS
<400> 95
ggattttgtc agggggtgtc acactgtg 28
<210> 96
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированная RSS
<400> 96
cacagtggtg ctgcccatat caaaaacc 28
<210> 97
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированная RSS
<400> 97
ggattttgtc aggcgatgtc acactgtg 28
<210> 98
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированная RSS
<400> 98
cacagtggtg ccgcccatat caaaaacc 28
<210> 99
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированная RSS
<400> 99
ggattttgtc agggggtgtc acactgtg 28
<210> 100
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированная RSS
<400> 100
cacagtggtg ctgcccatat caaaaacc 28
<210> 101
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированная RSS
<400> 101
ggattttgtc agggggtgcc acactgtg 28
<210> 102
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированная RSS
<400> 102
cacagtggtg ccgcccatat caaaaacc 28
<210> 103
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированная RSS
<400> 103
ggattttgta ggtgtctgtg tcactgtg 28
<210> 104
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированная RSS
<400> 104
cacagtgaca ctcgccaggt caaaaacc 28
<210> 105
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированная RSS
<400> 105
ggattttgta ggtgtctgtg tcactgtg 28
<210> 106
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированная RSS
<400> 106
cacagtgaca ctcacccagt caaaaacc 28
<210> 107
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированная RSS
<400> 107
ggattttgta gctgtctgta tcactgtg 28
<210> 108
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированная RSS
<400> 108
cacagtgaca ctcgccaggt caaaaacc 28
<210> 109
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированная RSS
<400> 109
ggtttctgaa gctgtctgtg tcacagtc 28
<210> 110
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированная RSS
<400> 110
cacaatgaca ctgggcagga cagaaacc 28
<210> 111
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированная RSS
<400> 111
gggtttggct gagctgagaa ccactgtg 28
<210> 112
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированная RSS
<400> 112
cacagtgatt ggcagctcta caaaaacc 28
<210> 113
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер и/или зонд
<400> 113
cacaggaaac agctatgacc 20
<210> 114
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер и/или зонд
<400> 114
ccagtgccct agagtcaccc a 21
<210> 115
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер и/или зонд
<400> 115
ctcccactca tgatctatag a 21
<210> 116
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер и/или зонд
<400> 116
ctggggctcg ctttagttg 19
<210> 117
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер и/или зонд
<400> 117
tgatagccgt tgtattcagc 20
<210> 118
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер и/или зонд
<400> 118
ccagtgccct agagtcaccc a 21
<210> 119
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер и/или зонд
<400> 119
gccatttctg tctgcattcg 20
<210> 120
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер и/или зонд
<400> 120
ggttcatcat gccgtttgtg a 21
<210> 121
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер и/или зонд
<400> 121
tgggctcgta gtttgacgtg 20
<210> 122
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер и/или зонд
<400> 122
ttacccacac ttcacgcacg 20
<210> 123
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер и/или зонд
<400> 123
ttaaacgacg cctcgaatg 19
<210> 124
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер и/или зонд
<400> 124
gcaaccattc gttgtagtag 20
<210> 125
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер и/или зонд
<400> 125
ctaacgcagt catgtaatgc 20
<210> 126
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер и/или зонд
<400> 126
gactgtcacc cagcattac 19
<210> 127
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер и/или зонд
<400> 127
cgtcgcctct acgggaaatc 20
<210> 128
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер и/или зонд
<400> 128
ccagtgccct agagtcaccc a 21
<210> 129
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер и/или зонд
<400> 129
gccatttctg tctgcattcg 20
<210> 130
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер и/или зонд
<400> 130
aaacaccacg taggatttac gc 22
<210> 131
<211> 10213
<212> ДНК
<213> Искусственная последовательность
<220>
<223> D6-DH1166
<400> 131
tacgtagccg tttcgatcct cccgaattga ctagtgggta ggcctggcgg ccgctgccat 60
ttcattacct ctttctccgc acccgacata gataccggtg gattcgaatt ctccccgttg 120
aagctgacct gcccagaggg gcctgggccc accccacaca ccggggcgga atgtgtacag 180
gccccggtct ctgtgggtgt tccgctaact ggggctccca gtgctcaccc cacaactaaa 240
gcgagcccca gcctccagag cccccgaagg agatgccgcc cacaagccca gcccccatcc 300
aggaggcccc agagctcagg gcgccggggc ggattttgta cagccccgag tcactgtgcg 360
gaaggatgct gtggttagct ttggcaaagt tttcctgcca ccacagtgag aaaaactgtg 420
tcaaaaaccg tctcctggcc cctgctggag gccgcgccag agaggggagc agccgccccg 480
aacctaggtc ctgctcagct cacacgaccc ccagcaccca gagcacaacg gagtccccat 540
tgaatggtga ggacggggac cagggctcca gggggtcatg gaaggggctg gaccccatcc 600
tactgctatg gtcccagtgc tcctggccag aactgaccct accaccgaca agagtccctc 660
agggaaacgg gggtcactgg cacctcccag catcaacccc aggcagcaca ggcataaacc 720
ccacatccag agccgactcc aggagcagag acaccccagt accctggggg acaccgaccc 780
tgatgactcc ccactggaat ccaccccaga gtccaccagg accaaagacc ccgcccctgt 840
ctctgtccct cactcaggac ctgctgcggg gcgggccatg agaccagact cgggcttagg 900
gaacaccact gtggccccaa cctcgaccag gccacaggcc cttccttcct gccctgcggc 960
agcacagact ttggggtctg tgcagagagg aatcacagag gccccaggct gaggtggtgg 1020
gggtggaaga cccccaggag gtggcccact tcccttcctc ccagctggaa cccaccatga 1080
ccttcttaag ataggggtgt catccgaggc aggtcctcca tggagctccc ttcaggctcc 1140
tccccggtcc tcactaggcc tcagtcccgg ctgcgggaat gcagccacca caggcacacc 1200
aggcagccca gacccagcca gcctgcagtg cccaagccca cattctggag cagagcaggc 1260
tgtgtctggg agagtctggg ctccccaccg cccccccgca caccccaccc acccctgtcc 1320
aggccctatg caggagggtc agagcccccc atggggtatg gacttagggt ctcactcacg 1380
tggctcccct cctgggtgaa ggggtctcat gcccagatcc ccacagcaga gctggtcaaa 1440
ggtggaggca gtggccccag ggccaccctg acctggaccc tcaggctcct ctagccctgg 1500
ctgccctgct gtccctggga ggcctggact ccaccagacc acaggtccag ggcaccgccc 1560
ataggtgctg cccacactca gttcacagga agaagataag ctccagaccc ccaagactgg 1620
gacctgcctt cctgccaccg cttgtagctc cagacctccg tgcctccccc gaccacttac 1680
acacgggcca gggagctgtt ccacaaagat caaccccaaa ccgggaccgc ctggcactcg 1740
ggccgctgcc acttccctct ccatttgttc ccagcacctc tgtgctccct ccctcctccc 1800
tccttcaggg gaacagcctg tgcagcccct ccctgcaccc cacaccctgg ggaggcccaa 1860
ccctgcctcc agccctttct cccccgctgc tcttcctgcc catccagaca accctggggt 1920
cccatccctg cagcctacac cctggtctcc acccagaccc ctgtctctcc ctccagacac 1980
ccctcccagg ccaaccctgc acatgcaggc cctccccttt tctgctgcca gagcctcagt 2040
ttctaccctc tgtgcctacc ccctgcctcc tcctgcccac aactcgagct cttcctctcc 2100
tggggcccct gagccatggc actgaccgtg cactcccacc cccacactgc ccatgccctc 2160
accttcctcc tggacactct gaccccgctc ccctcttgga cccagccctg gtatttccag 2220
gacaaaggct cacccaagtc ttccccatgc aggcccttgc cctcactgcc cggttacacg 2280
gcagcctcct gtgcacagaa gcagggagct cagcccttcc acaggcagaa ggcactgaaa 2340
gaaatcggcc tccagcaccc tgatgcacgt ccgcctgtgt ctctcactgc ccgcacctgc 2400
agggaggctc ggcactccct gtaaagacga gggatccagg cagcaacatc atgggagaat 2460
gcagggctcc cagacagccc agccctctcg caggcctctc ctgggaagag acctgcagcc 2520
accactgaac agccacggag cccgctggat agtaactgag tcagtgaccg acctggaggg 2580
caggggagca gtgaaccgga gcccagacca tagggacaga gaccagccgc tgacatcccg 2640
agcccctcac tggcggcccc agaacaccgc gtggaaacag aacagaccca cattcccacc 2700
tggaacaggg cagacactgc tgagccccca gcaccagccc tgagaaacac caggcaacgg 2760
catcagaggg ggctcctgag aaagaaagga ggggaggtct ccttcaccag caagtacttc 2820
ccttgaccaa aaacagggtc cacgcaactc ccccaggaca aaggaggagc cccctgtaca 2880
gcactgggct cagagtcctc tcccacacac cctgagtttc agacaaaaac cccctggaaa 2940
tcatagtatc agcaggagaa ctagccagag acagcaagag gggactcagt gactcccgcg 3000
gggacaggag gattttgtgg gggctcgtgt cactgtgctg cataccctgc tggacctgca 3060
agtattcggc aactgtgagg ttagccagca tctagactat gcccacagtg acacagcccc 3120
attcaaaaac ccctgctgta aacgcttcca cttctggagc tgaggggctg gggggagcgt 3180
ctgggaagta gggcctaggg gtggccatca atgcccaaaa cgcaccagac tcccccccag 3240
acatcacccc actggccagt gagcagagta aacagaaaat gagaagcagc tgggaagctt 3300
gcacaggccc caaggaaaga gctttggcgg gtgtgcaaga ggggatgcgg gcagagcctg 3360
agcagggcct tttgctgttt ctgctttcct gtgcagatag ttccataaac tggtgttcaa 3420
gatcgatggc tgggagtgag cccaggagga cagtgtggga agggcacagg gaaggagaag 3480
cagccgctat cctacactgt catctttcaa gagtttgccc tgtgcccaca atgctgcatc 3540
atgggatgct taacagctga tgtagacaca gctaaagaga gaatcagtga aatggatttg 3600
cagcacagat ctgaataaat tctccagaat gtggagccac acagaagcaa gcacaaggaa 3660
agtgcctgat gcaagggcaa agtacagtgt gtaccttcag gctgggcaca gacactctga 3720
aaagccttgg caggaactcc ctgcaacaaa gcagagccct gcaggcaatg ccagctccag 3780
agccctccct gagagcctca tgggcaaaga tgtgcacaac aggtgtttct catagcccca 3840
aactgagaat gaagcaaaca gccatctgaa ggaaaacagg caaataaacg atggcaggtt 3900
catgaaatgc aaacccagac agccagaagg acaacagtga gggttacagg tgactctgtg 3960
gttgagttca tgacaatgct gagtaattgg agtaacaaag gaaagtccaa aaaatacttt 4020
caatgtgatt tcttctaaat aaaatttaca gccggcaaaa tgaactatct tcttaaggga 4080
taaactttcc actaggaaaa ctataaggaa aatcaagaaa aggatgatca cataaacaca 4140
gtggtcgtta cttctactgg ggaaggaaga gggtatgaac tgagacacac agggttggca 4200
agtctcctaa caagaacaga acaaatacat tacagtacct tgaaaacagc agttaaaatt 4260
ctaaattgca agaagaggaa aatgcacaca gctgtgttta gaaaattctc agtccagcac 4320
tgttcataat agcaaagaca ttaacccagg ttggataaat aaacgatgac acaggcaatt 4380
gcacaatgat acagacatac attcagtata tgagacattg atgatgtatc cccaaagaaa 4440
tgactttaaa gagaaaaggc ctgatatgtg gtggcactca cctccctggg catccccgga 4500
caggctgcag gcacactgtg tggcagggca ggctggtacc tgctggcagc tcctggggcc 4560
tgatgtggag caggcacaga gccgtatccc cccgaggaca tataccccca aggacggcac 4620
agttggtaca ttccggagac aagcaactca gccacactcc caggccagag cccgagaggg 4680
acgcccatgc acagggaggc agagcccagc tcctccacag ccagcagcac ccgtgcaggg 4740
gccgccatct ggcaggcaca gagcatgggc tgggaggagg ggcagggaca ccaggcaggg 4800
ttggcaccaa ctgaaaatta cagaagtctc atacatctac ctcagccttg cctgacctgg 4860
gcctcacctg acctggacct cacctggcct ggacctcacc tggcctagac ctcacctctg 4920
ggcttcacct gagctcggcc tcacctgact tggaccttgc ctgtcctgag ctcacatgat 4980
ctgggcctca cctgacctgg gtttcacctg acctgggctt cacctgacct gggcctcatc 5040
tgacctgggc ctcactggcc tggacctcac ctggcctggg cttcacctgg cctcaggcct 5100
catctgcacc tgctccaggt cttgctggaa cctcagtagc actgaggctg caggggctca 5160
tccagggttg cagaatgact ctagaacctc ccacatctca gctttctggg tggaggcacc 5220
tggtggccca gggaatataa aaagcctgaa tgatgcctgc gtgatttggg ggcaatttat 5280
aaacccaaaa ggacatggcc atgcagcggg tagggacaat acagacagat atcagcctga 5340
aatggagcct cagggcacag gtgggcacgg acactgtcca cctaagccag gggcagaccc 5400
gagtgtcccc gcagtagacc tgagagcgct gggcccacag cctcccctcg gtgccctgct 5460
acctcctcag gtcagccctg gacatcccgg gtttccccag gcctggcggt aggattttgt 5520
tgaggtctgt gtcactgtgc atggcagcta ctgccagccc gcagccactg gctactgagg 5580
atgccgattc tgagaatagc agcttctact actatgacta cctggatgaa gtagctttca 5640
tgctctgccg gaaggatgct gtggttagct ttggcaaagt tttcctgcca ccacagtgtc 5700
acagagtcca tcaaaaaccc atccctggga accttctgcc acagccctcc ctgtggggca 5760
ccgccgcgtg ccatgttagg attttgactg aggacacagc accatgggta tggtggctac 5820
cgcagcagtg cagcccgtga cccaaacaca cagggcagca ggcacaacag acaagcccac 5880
aagtgaccac cctgagctcc tgcctgccag ccctggagac catgaaacag atggccagga 5940
ttatcccata ggtcagccag acctcagtcc aacaggtctg catcgctgct gccctccaat 6000
accagtccgg atggggacag ggctggccca cattaccatt tgctgccatc cggccaacag 6060
tcccagaagc ccctccctca aggctgggcc acatgtgtgg accctgagag ccccccatgt 6120
ctgagtaggg gcaccaggaa ggtggggctg gccctgtgca ctgtccctgc ccctgtggtc 6180
cctggcctgc ctggccctga cacctgggcc tctcctgggt catttccaag acagaagaca 6240
ttcccaggac agctggagct gggagtccat catcctgcct ggccgtcctg agtcctgcgc 6300
ctttccaaac ctcacccggg aagccaacag aggaatcacc tcccacaggc agagacaaag 6360
accttccaga aatctctgtc tctctcccca gtgggcaccc tcttccaggg cagtcctcag 6420
tgatatcaca gtgggaaccc acatctggat cgggactgcc cccagaacac aagatggccc 6480
acagggacag ccccacagcc cagcccttcc cagaccccta aaaggcgtcc caccccctgc 6540
atctgcccca gggctcaaac tccaggagga ctgactcctg cacaccctcc tgccagacat 6600
cacctcagcc cctcctggaa gggacaggag cgcgcaaggg tgagtcagac cctcctgccc 6660
tcgatggcag gcggagaaga ttcagaaagg tctgagatcc ccaggacgca gcaccactgt 6720
caatgggggc cccagacgcc tggaccaggg cctgcgtggg aaaggcctct gggcacactc 6780
agggggattt tgtgaagggt cctcccactg tgcatggcag ctactgccag cccgcagcca 6840
ctggctactg aggatgccga ttctgagaat agcagcttct actactatga ctacctggat 6900
gaagtagctt tcatgctctg ccggaaggat gctgtggtta gctttggcaa agttttcctg 6960
ccactgcata ccctgctgga cctgcaagta ttcggcaact gtgaggttag ccagcatcta 7020
gactatgccc acagtgatga acccagcatc aaaaaccgac cggactccca aggtttatgc 7080
acacttctcc gctcagagct ctccaggatc agaagagccg ggcccaaggg tttctgccca 7140
gaccctcggc ctctagggac atcttggcca tgacagccca tgggctggtg ccccacacat 7200
cgtctgcctt caaacaaggg cttcagaggg ctctgaggtg acctcactga tgaccacagg 7260
tgccctggcc ccttccccac cagctgcacc agaccccgtc atgacagatg ccccgattcc 7320
aacagccaat tcctggggcc aggaatcgct gtagacacca gcctccttcc aacacctcct 7380
gccaattgcc tggattccca tcccggttgg aatcaagagg acagcatccc ccaggctccc 7440
aacaggcagg actcccacac cctcctctga gaggccgctg tgttccgtag ggccaggctg 7500
cagacagtcc ccctcacctg ccactagaca aatgcctgct gtagatgtcc ccacctggaa 7560
aataccactc atggagcccc cagccccagg tacagctgta gagagagtct ctgaggcccc 7620
taagaagtag ccatgcccag ttctgccggg accctcggcc aggctgacag gagtggacgc 7680
tggagctggg cccatactgg gccacatagg agctcaccag tgagggcagg agagcacatg 7740
ccggggagca cccagcctcc tgctgaccag aggcccgtcc cagagcccag gaggctgcag 7800
aggcctctcc agggggacac tgtgcatgtc tggtccctga gcagcccccc acgtccccag 7860
tcctgggggc ccctggcaca gctgtctgga ccctctctat tccctgggaa gctcctcctg 7920
acagccccgc ctccagttcc aggtgtggat tttgtcaggg ggtgtcacac tgtgcagctt 7980
cttgtgcaag cacagtggtg ctgcccatat caaaaaccag gccaagtaga caggcccctg 8040
ctgtgcagcc ccaggcctcc agctcacctg cttctcctgg ggctctcaag gctgctgttt 8100
tctgcactct cccctctgtg gggagggttc cctcagtggg agatctgttc tcaacatccc 8160
acggcctcat tcctgcaagg aaggccaatg gatgggcaac ctcacatgcc gcggctaaga 8220
tagggtgggc agcctggcgg ggacaggaca tcctgctggg gtatctgtca ctgtgcctag 8280
tggggcactg gctcccaaac aacgcagtcc ttgccaaaat ccccacggcc tcccccgcta 8340
ggggctggcc tgatctcctg cagtcctagg aggctgctga cctccagaat ggctccgtcc 8400
ccagttccag ggcgagagca gatcccaggc cggctgcaga ctgggaggcc accccctcct 8460
tcccagggtt cactgcaggt gaccagggca ggaaatggcc tgaacacagg gataaccggg 8520
ccatccccca acagagtcca ccccctcctg ctctgtaccc cgcacccccc aggccagccc 8580
atgacatccg acaaccccac accagagtca ctgcccggtg ctgccctagg gaggacccct 8640
cagcccccac cctgtctaga ggactgggga ggacaggaca cgccctctcc ttatggttcc 8700
cccacctggc tctggctggg acccttgggg tgtggacaga aaggacgctt gcctgattgg 8760
cccccaggag cccagaactt ctctccaggg accccagccc gagcaccccc ttacccagga 8820
cccagccctg cccctcctcc cctctgctct cctctcatca ccccatggga atccagaatc 8880
cccaggaagc catcaggaag ggctgaggga ggaagtgggg ccactgcacc accaggcagg 8940
aggctctgtc tttgtgaacc cagggaggtg ccagcctcct agagggtatg gtccaccctg 9000
cctatggctc ccacagtggc aggctgcagg gaaggaccag ggacggtgtg ggggagggct 9060
cagggccccg cgggtgctcc atcttggatg agcctatctc tctcacccac ggactcgccc 9120
acctcctctt caccctggcc acacgtcgtc cacaccatcc taagtcccac ctacaccaga 9180
gccggcacag ccagtgcaga cagaggctgg ggtgcagggg ggccgactgg gcagcttcgg 9240
ggagggagga atggaggaag gggagttcag tgaagaggcc cccctcccct gggtccagga 9300
tcctcctctg ggacccccgg atcccatccc ctccaggctc tgggaggaga agcaggatgg 9360
gagaatctgt gcgggaccct ctcacagtgg aatacctcca cagcggctca ggccagatac 9420
aaaagcccct cagtgagccc tccactgcag tgctgggcct gggggcagcc gctcccacac 9480
aggatgaacc cagcaccccg aggatgtcct gccaggggga gctcagagcc atgaaggagc 9540
aggatatggg acccccgata caggcacaga cctcagctcc attcaggact gccacgtcct 9600
gccctgggag gaaccccttt ctctagtccc tgcaggccag gaggcagctg actcctgact 9660
tggacgccta ttccagacac cagacagagg ggcaggcccc ccagaaccag ggatgaggac 9720
gccccgtcaa ggccagaaaa gaccaagttg cgctgagccc agcaagggaa ggtccccaaa 9780
caaaccagga ggattttgta ggtgtctgtg tcactgtgca aacccatgaa aaccccaagg 9840
gagtttggaa ctgccatgcc gatttcggcg ggcatggcac catttggaag ctcttcctcc 9900
ggttccagca gaacctgcta ccacagtgac actcgccagg tcaaaaaccc catcccaagt 9960
cagcggaatg cagagagagc agggaggaca tgtttaggat ctgaggccgc acctgacacc 10020
caggccagca gacgtctcct gtccacggca ccctgccatg tcctgcattt ctggaagaac 10080
aagggcaggc tgaagggggt ccaggaccag gagatgggtc cgctctaccc agagaaggag 10140
ccaggcagga cacaagcccc cacgcgtggg ctcgtagttt gacgtgcgtg aagtgtgggt 10200
aagaaagtac gta 10213
<210> 132
<211> 9828
<212> ДНК
<213> Искусственная последовательность
<220>
<223> D6-D17613
<400> 132
gcggccgctg ccatttcatt acctctttct ccgcacccga catagattac gtaacgcgtg 60
ggctcgtagt ttgacgtgcg tgaagtgtgg gtaagaaagt ccccattgag gctgacctgc 120
ccagagggtc ctgggcccac ccaacacacc ggggcggaat gtgtgcaggc ctcggtctct 180
gtgggtgttc cgctagctgg ggctcacagt gctcacccca cacctaaaac gagccacagc 240
ctccggagcc cctgaaggag accccgccca caagcccagc ccccacccag gaggccccag 300
agcacagggc gccccgtcgg attttgtaca gccccgagtc actgtgcagc ttcttgcaca 360
gtgagaaaag cttcgtcaaa aaccgtctcc tggccacagt cggaggcccc gccagagagg 420
ggagcagcca ccccaaaccc atgttctgcc ggctcccatg accccgtgca cctggagccc 480
cacggtgtcc ccactggatg ggaggacaag ggccgggggc tccggcgggt cggggcaggg 540
gcttgatggc ttccttctgc cgtggcccca ttgcccctgg ctggagttga cccttctgac 600
aagtgtcctc agagagtcag ggatcagtgg cacctcccaa catcaacccc acgcagccca 660
ggcacaaacc ccacatccag ggccaactcc aggaacagag acaccccaat accctggggg 720
accccgaccc tgatgactcc cgtcccatct ctgtccctca cttggggcct gctgcggggc 780
gagcacttgg gagcaaactc aggcttaggg gacaccactg tgggcctgac ctcgagcagg 840
ccacagaccc ttccctcctg ccctggtgca gcacagactt tggggtctgg gcagggagga 900
acttctggca ggtcaccaag cacagagccc ccaggctgag gtggccccag ggggaacccc 960
agcaggtggc ccactaccct tcctcccagc tggaccccat gtcttcccca agataggggt 1020
gccatccaag gcaggtcctc catggagccc ccttcaggct cctctccaga ccccactggg 1080
cctcagtccc cactctagga atgcagccac cacgggcaca ccaggcagcc caggcccagc 1140
caccctgcag tgcccaagcc cacaccctgg aggagagcag ggtgcgtctg ggaggggctg 1200
ggctccccac ccccaccccc acctgcacac cccacccacc cttgcccggg ccccctgcag 1260
gagggtcaga gcccccatgg gatatggact tagggtctca ctcacgcacc tcccctcctg 1320
ggagaagggg tctcatgccc agatcccccc agcagcgctg gtcacaggta gaggcagtgg 1380
ccccagggcc accctgacct ggcccctcag gctcctctag ccctggctgc cctgctgtcc 1440
ctgggaggcc tgggctccac cagaccacag gtctagggca ccgcccacac tggggccgcc 1500
cacacacagc tcacaggaag aagataagct ccagaccccc aggcccggga cctgccttgc 1560
tgctacgact tcctgcccca gacctcgttg ccctcccccg tccacttaca cacaggccag 1620
gaagctgttc ccacacagac caaccccaga cggggaccac ctggcactca ggtcactgcc 1680
atttccttct ccattcactt ccaatgcctc tgtgcttcct ccctcctcct tccttcgggg 1740
gagcaccctg tgcagctcct ccctgcagtc cacaccctgg ggagacccga ccctgcagcc 1800
cacaccctgg ggagacctga ccctcctcca gccctttctc ccccgctgct cttgccaccc 1860
accaagacag ccctggggtc ctgtccctac agcccccacc cagttctcta cctagacccg 1920
tcttcctccc tctaaacacc tctcccaggc caaccctaca cctgcaggcc ctcccctcca 1980
ctgccaaaga ccctcagttt ctcctgcctg tgcccacccc cgtgctcctc ctgcccacag 2040
ctcgagctct tcctctccta gggcccctga gggatggcat tgaccgtgcc ctcgcaccca 2100
cacactgccc atgccctcac attcctcctg gccactccag ccccactccc ctctcaggcc 2160
tggctctggt atttctggga caaagcctta cccaagtctt tcccatgcag gcctgggccc 2220
ttaccctcac tgcccggtta cagggcagcc tcctgtgcac agaagcaggg agctcagccc 2280
ttccacaggc agaaggcact gaaagaaatc ggcctccagc gccttgacac acgtctgcct 2340
gtgtctctca ctgcccgcac ctgcagggag gctcggcact ccctctaaag acgagggatc 2400
caggcagcag catcacagga gaatgcaggg ctaccagaca tcccagtcct ctcacaggcc 2460
tctcctggga agagacctga agacgcccag tcaacggagt ctaacaccaa acctccctgg 2520
aggccgatgg gtagtaacgg agtcattgcc agacctggag gcaggggagc agtgagcccg 2580
agcccacacc atagggccag aggacagcca ctgacatccc aagccactca ctggtggtcc 2640
cacaacaccc catggaaaga ggacagaccc acagtcccac ctggaccagg gcagagactg 2700
ctgagaccca gcaccagaac caaccaagaa acaccaggca acagcatcag agggggctct 2760
ggcagaacag aggaggggag gtctccttca ccagcaggcg cttcccttga ccgaagacag 2820
gatccatgca actcccccag gacaaaggag gagccccttg ttcagcactg ggctcagagt 2880
cctctccaag acacccagag tttcagacaa aaaccccctg gaatgcacag tctcagcagg 2940
agagccagcc agagccagca agatggggct cagtgacacc cgcagggaca ggaggatttt 3000
gtgggggctc gtgtcactgt gctgcatacc ctgctggacc tgcaagtatt cggcaacagt 3060
gaggttagcc agcatctaga ctatgcccac agtgacacag ccccattcaa aaacccctac 3120
tgcaaacgca ttccacttct ggggctgagg ggctggggga gcgtctggga aatagggctc 3180
aggggtgtcc atcaatgccc aaaacgcacc agactcccct ccatacatca cacccaccag 3240
ccagcgagca gagtaaacag aaaatgagaa gcaagctggg gaagcttgca caggccccaa 3300
ggaaagagct ttggcgggtg tgtaagaggg gatgcgggca gagcctgagc agggcctttt 3360
gctgtttctg ctttcctgtg cagagagttc cataaactgg tgttcgagat caatggctgg 3420
gagtgagccc aggaggacag cgtgggaaga gcacagggaa ggaggagcag ccgctatcct 3480
acactgtcat ctttcgaaag tttgccttgt gcccacactg ctgcatcatg ggatgcttaa 3540
cagctgatgt agacacagct aaagagagaa tcagtgagat ggatttgcag cacagatctg 3600
aataaattct ccagaatgtg gagcagcaca gaagcaagca cacagaaagt gcctgatgca 3660
aggacaaagt tcagtgggca ccttcaggca ttgctgctgg gcacagacac tctgaaaagc 3720
cctggcagga actccctgtg acaaagcaga accctcaggc aatgccagcc ccagagccct 3780
ccctgagagc ctcatgggca aagatgtgca caacaggtgt ttctcatagc cccaaactga 3840
gagcaaagca aacgtccatc tgaaggagaa caggcaaata aacgatggca ggttcatgaa 3900
atgcaaaccc agacagccac aagcacaaaa gtacagggtt ataagcgact ctggttgagt 3960
tcatgacaat gctgagtaat tggagtaaca aagtaaactc caaaaaatac tttcaatgtg 4020
atttcttcta aataaaattt acaccctgca aaatgaactg tcttcttaag ggatacattt 4080
cccagttaga aaaccataaa gaaaaccaag aaaaggatga tcacataaac acagtggtgg 4140
ttacttctgc tggggaagga agagggtatg aactgagata cacagggtgg gcaagtctcc 4200
taacaagaac agaacgaata cattacagta ccttgaaaac agcagttaaa cttctaaatt 4260
gcaagaagag gaaaatgcac acagttgtgt ttagaaaatt ctcagtccag cactgttcat 4320
aatagcaaag acattaaccc aggtcggata aataagcgat gacacaggca attgcacaat 4380
gatacagaca tatatttagt atatgagaca tcgatgatgt atccccaaat aaacgacttt 4440
aaagagataa agggctgatg tgtggtggca ttcacctccc tgggatcccc ggacaggttg 4500
caggctcact gtgcagcagg gcaggcgggt acctgctggc agttcctggg gcctgatgtg 4560
gagcaagcgc agggccatat atcccggagg acggcacagt cagtgaattc cagagagaag 4620
caactcagcc acactcccca ggcagagccc gagagggacg cccacgcaca gggaggcaga 4680
gcccagcacc tccgcagcca gcaccacctg cgcacgggcc accaccttgc aggcacagag 4740
tgggtgctga gaggaggggc agggacacca ggcagggtga gcacccagag aaaactgcag 4800
acgcctcaca catccacctc agcctcccct gacctggacc tcactggcct gggcctcact 4860
taacctgggc ttcacctgac cttggcctca cctgacttgg acctcgcctg tcccaagctt 4920
tacctgacct gggcctcaac tcacctgaac gtctcctgac ctgggtttaa cctgtcctgg 4980
aactcacctg gccttggctt cccctgacct ggacctcatc tggcctgggc ttcacctggc 5040
ctgggcctca cctgacctgg acctcatctg gcctggacct cacctggcct ggacttcacc 5100
tggcctgggc ttcacctgac ctggacctca cctggcctcg ggcctcacct gcacctgctc 5160
caggtcttgc tggagcctga gtagcactga gggtgcagaa gctcatccag ggttggggaa 5220
tgactctaga agtctcccac atctgacctt tctgggtgga ggcagctggt ggccctggga 5280
atataaaaat ctccagaatg atgactctgt gatttgtggg caacttatga acccgaaagg 5340
acatggccat ggggtgggta gggacatagg gacagatgcc agcctgaggt ggagcctcag 5400
gacacaggtg ggcacggaca ctatccacat aagcgaggga tagacccgag tgtccccaca 5460
gcagacctga gagcgctggg cccacagcct cccctcagag ccctgctgcc tcctccggtc 5520
agccctggac atcccaggtt tccccaggcc tggcggtagg attttgttga ggtctgtgtc 5580
actgtgcatg gcagctactg ccagcccgca gccactggct actgaggatg ccgattctga 5640
gaatagcagc ttctactact atgactacct ggatgaagta gctttcatgc tccacagtgt 5700
cacagagtcc atcaaaaacc catgcctgga agcttcccgc cacagccctc cccatggggc 5760
cctgctgcct cctcaggtca gccccggaca tcccgggttt ccccaggctg ggcggtagga 5820
ttttgttgag gtctgtgtca ctgtgccaaa cccatgaaaa ccccaaggga gtttggaaca 5880
gccatgccga tttcggcggg catggcacca tttggaagct cttcctccgg ttccagcaga 5940
acctgctacc cacagtgtca cagagtccat caaaaaccca tccctgggag cctcccgcca 6000
cagccctccc tgcaggggac cggtacgtgc catgttagga ttttgatcga ggagacagca 6060
ccatgggtat ggtggctacc acagcagtgc agcctgtgac ccaaacccgc agggcagcag 6120
gcacgatgga caggcccgtg actgaccacg ctgggctcca gcctgccagc cctggagatc 6180
atgaaacaga tggccaaggt caccctacag gtcatccaga tctggctccg aggggtctgc 6240
atcgctgctg ccctcccaac gccagtccaa atgggacagg gacggcctca cagcaccatc 6300
tgctgccatc aggccagcga tcccagaagc ccctccctca aggctgggca catgtgtgga 6360
cactgagagc cctcatatct gagtaggggc accaggaggg aggggctggc cctgtgcact 6420
gtccctgccc ctgtggtccc tggcctgcct ggccctgaca cctgagcctc tcctgggtca 6480
tttccaagac agaagacatt cctggggaca gccggagctg ggcgtcgctc atcctgcccg 6540
gccgtcctga gtcctgctca tttccagacc tcaccgggga agccaacaga ggactcgcct 6600
cccacattca gagacaaaga accttccaga aatccctgcc tctctcccca gtggacaccc 6660
tcttccagga cagtcctcag tggcatcaca gcggcctgag atccccagga cgcagcaccg 6720
ctgtcaatag gggccccaaa tgcctggacc agggcctgcg tgggaaaggc ctctggccac 6780
actcggggat tttgtgaagg gccctcccac tgtgccagct tcttgagcca tgccgatttc 6840
ggcgggcatg gcaccatttg gaagctcttc ctccggttcc agcagaacct gctaccacag 6900
tgatgaaccc agtgtcaaaa accggctgga aacccagggg ctgtgtgcac gcctcagctt 6960
ggagctctcc aggagcacaa gagccgggcc caaggatttg tgcccagacc ctcagcctct 7020
agggacacct gggtcatctc agcctgggct ggtgccctgc acaccatctt cctccaaata 7080
ggggcttcag agggctctga ggtgacctca ctcatgacca caggtgacct ggcccttccc 7140
tgccagctat accagaccct gtcttgacag atgccccgat tccaacagcc aattcctggg 7200
accctgaata gctgtagaca ccagcctcat tccagtacct cctgccaatt gcctggattc 7260
ccatcctggc tggaatcaag aaggcagcat ccgccaggct cccaacaggc aggactcccg 7320
cacaccctcc tctgagaggc cgctgtgttc cgcagggcca ggccctggac agttcccctc 7380
acctgccact agagaaacac ctgccattgt cgtccccacc tggaaaagac cactcgtgga 7440
gcccccagcc ccaggtacag ctgtagagac agtcctcgag gcccctaaga aggagccatg 7500
cccagttctg ccgggaccct cggccaggcc gacaggagtg gacgctggag ctgggcccac 7560
actgggccac ataggagctc accagtgagg gcaggagagc acatgccggg gagcacccag 7620
cctcctgctg accagaggcc cgtcccagag cccaggaggc tgcagaggcc tctccaggga 7680
gacactgtgc atgtctggta cctaagcagc cccccacgtc cccagtcctg ggggcccctg 7740
gctcagctgt ctgggccctc cctgctccct gggaagctcc tcctgacagc cccgcctcca 7800
gttccaggtg tggattttgt caggcgatgt cacactgtgc agcttcttga gcaagcacag 7860
tggtgccgcc catatcaaaa accaggccaa gtagacaggc ccctgctgcg cagccccagg 7920
catccacttc acctgcttct cctggggctc tcaaggctgc tgtctgtcct ctggccctct 7980
gtggggaggg ttccctcagt gggaggtctg tgctccaggg cagggatgat tgagatagaa 8040
atcaaaggct ggcagggaaa ggcagcttcc cgccctgaga ggtgcaggca gcaccacgga 8100
gccacggagt cacagagcca cggagccccc attgtgggca tttgagagtg ctgtgccccc 8160
ggcaggccca gccctgatgg ggaagcctgt cccatcccac agcccgggtc ccacgggcag 8220
cgggcacaga agctgccagg ttgtcctcta tgatcctcat ccctccagca gcatcccctc 8280
cacagtgggg aaactgaggc ttggagcacc acccggcccc ctggaaatga ggctgtgagc 8340
ccagacagtg ggcccagagc actgtgagta ccccggcagt acctggctgc agggatcagc 8400
cagagatgcc aaaccctgag tgaccagcct acaggaggat ccggccccac ccaggccact 8460
cgattaatgc tcaaccccct gccctggaga cctcttccag taccaccagc agctcagctt 8520
ctcagggcct catccctgca aggaaggtca agggctgggc ctgccagaaa cacagcaccc 8580
tccctagccc tggctaagac agggtgggca gacggctgtg gacgggacat attgctgggg 8640
catttctcac tgtcacttct gggtggtagc tctgacaaaa acgcagaccc tgccaaaatc 8700
cccactgcct cccgctaggg gctggcctgg aatcctgctg tcctaggagg ctgctgacct 8760
ccaggatggc tccgtcccca gttccagggc gagagcagat cccaggcagg ctgtaggctg 8820
ggaggccacc cctgcccttg ccggggttga atgcaggtgc ccaaggcagg aaatggcatg 8880
agcacaggga tgaccgggac atgccccacc agagtgcgcc ccttcctgct ctgcaccctg 8940
caccccccag gccagcccac gacgtccaac aactgggcct gggtggcagc cccacccaga 9000
caggacagac ccagcaccct gaggaggtcc tgccaggggg agctaagagc catgaaggag 9060
caagatatgg ggcccccgat acaggcacag atgtcagctc catccaggac cacccagccc 9120
acaccctgag aggaacgtct gtctccagcc tctgcaggtc gggaggcagc tgacccctga 9180
cttggacccc tattccagac accagacaga ggcgcaggcc ccccagaacc agggttgagg 9240
gacgccccgt caaagccaga caaaaccaag gggtgttgag cccagcaagg gaaggccccc 9300
aaacagacca ggaggatttt gtaggtgtct gtgtcactgt gcatggcagc tactgccagc 9360
ccgcagccac tggctactga ggatgccgat tctgagaata gcagcttcta ctactatgac 9420
tacctggatg aagtagcttt catgctcagc cggaaggatg ctgtggttag ctttggcaaa 9480
gttttcctgc cacccacagt gacactcacc cagtcaaaaa ccccattcca agtcagcgga 9540
agcagagaga gcagggagga cacgtttagg atctgagact gcacctgaca cccaggccag 9600
cagacgtctc ccctccaggg caccccaccc tgtcctgcat ttctgcaaga tcaggggcgg 9660
cctgaggggg ggtctagggt gaggagatgg gtcccctgta caccaaggag gagttaggca 9720
ggtcccgagc actcttaatt aaacgacgcc tcgaatggaa ctactacaac gaatggttgc 9780
tctacgtaat gcattcgcta ccttaggacc gttatagtta ggcgcgcc 9828
<210> 133
<211> 9894
<212> ДНК
<213> Искусственная последовательность
<220>
<223> D6-DH114619
<400> 133
tacgtattaa ttaaacgacg cctcgaatgg aactactaca acgaatggtt gctctcccca 60
ttgaggctga cctgcccaga gagtcctggg cccaccccac acaccggggc ggaatgtgtg 120
caggcctcgg tctctgtggg tgttccgcta gctggggctc acagtgctca ccccacacct 180
aaaatgagcc acagcctccg gagcccccgc aggagacccc gcccacaagc ccagccccca 240
cccaggaggc cccagagctc agggcgcccc gtcggatttt gtacagcccc gagtcactgt 300
gcaaacccat gaaaacccca agggagtttg gaaccacagt gagaatagct acgtcaaaaa 360
ccgtccagtg gccactgccg gaggccccgc cagagagggc agcagccact ctgatcccat 420
gtcctgccgg ctcccatgac ccccagcacg cggagcccca cagtgtcccc actggatggg 480
aggacaagag ctggggattc cggcgggtcg gggcaggggc ttgatcgcat ccttctgccg 540
tggctccagt gcccctggct ggagttgacc cttctgacaa gtgtcctcag agagacaggc 600
atcaccggcg cctcccaaca tcaaccccag gcagcacagg cacaaacccc acatccagag 660
ccaactccag gagcagagac accccaatac cctgggggac cccgaccctg atgacttccc 720
actggaattc gccgtagagt ccaccaggac caaagaccct gcctctgcct ctgtccctca 780
ctcaggacct gctgccgggc gaggccttgg gagcagactt gggcttaggg gacaccagtg 840
tgaccccgac cttgaccagg acgcagacct ttccttcctt tcctggggca gcacagactt 900
tggggtctgg gccaggagga acttctggca ggtcgccaag cacagaggcc acaggctgag 960
gtggccctgg aaagacctcc aggaggtggc cactcccctt cctcccagct ggaccccatg 1020
tcctccccaa gataagggtg ccatccaagg caggtgctcc ttggagcccc attcagactc 1080
ctccctggac cccactgggc ctcagtccca gctctgggga tgaagccacc acaagcacac 1140
caggcagccc aggcccagcc accctgcagt gcccaagcac acactctgga gcagagcagg 1200
gtgcctctgg gaggggctga gctccccacc ccacccccac ctgcacaccc cacccacccc 1260
tgcccagcgg ctctgcagga gggtcagagc cccacatggg gtatggactt agggtctcac 1320
tcacgtggct cccatcatga gtgaaggggc ctcaagccca ggttcccaca gcagcgcctg 1380
tcgcaagtgg aggcagaggc ccgagggcca ccctgacctg gtccctgagg ttcctgcagc 1440
ccaggctgcc ctgctgtccc tgggaggcct gggctccacc agaccacagg tccagggcac 1500
cgggtgcagg agccacccac acacagctca caggaagaag ataagctcca gacccccagg 1560
gccagaacct gccttcctgc tactgcttcc tgccccagac ctgggcgccc tcccccgtcc 1620
acttacacac aggccaggaa gctgttccca cacagaacaa ccccaaacca ggaccgcctg 1680
gcactcaggt ggctgccatt tccttctcca tttgctccca gcgcctctgt cctccctggt 1740
tcctccttcg ggggaacagc ctgtgcagcc agtccctgca gcccacaccc tggggagacc 1800
caaccctgcc tggggccctt ccaaccctgc tgctcttact gcccacccag aaaactctgg 1860
ggtcctgtcc ctgcagtccc taccctggtc tccacccaga cccctgtgta tcactccaga 1920
cacccctccc aggcaaaccc tgcacctgca ggccctgtcc tcttctgtcg ctagagcctc 1980
agtttctccc ccctgtgccc acaccctacc tcctcctgcc cacaactcta actcttcttc 2040
tcctggagcc cctgagccat ggcattgacc ctgccctccc accacccaca gcccatgccc 2100
tcaccttcct cctggccact ccgaccccgc cccctctcag gccaagccct ggtatttcca 2160
ggacaaaggc tcacccaagt ctttcccagg caggcctggg ctcttgccct cacttcccgg 2220
ttacacggga gcctcctgtg cacagaagca gggagctcag cccttccaca ggcagaaggc 2280
actgaaagaa atcggcctcc agcaccttga cacacgtccg cccgtgtctc tcactgcccg 2340
cacctgcagg gaggctccgc actccctcta aagacaaggg atccaggcag cagcatcacg 2400
ggagaatgca gggctcccag acatcccagt cctctcacag gcctctcctg ggaagagacc 2460
tgcagccacc accaaacagc cacagaggct gctggatagt aactgagtca atgaccgacc 2520
tggagggcag gggagcagtg agccggagcc cataccatag ggacagagac cagccgctga 2580
catcccgagc tcctcaatgg tggccccata acacacctag gaaacataac acacccacag 2640
ccccacctgg aacagggcag agactgctga gcccccagca ccagccccaa gaaacaccag 2700
gcaacagtat cagagggggc tcccgagaaa gagaggaggg gagatctcct tcaccatcaa 2760
atgcttccct tgaccaaaaa cagggtccac gcaactcccc caggacaaag gaggagcccc 2820
ctatacagca ctgggctcag agtcctctct gagacaccct gagtttcaga caacaacccg 2880
ctggaatgca cagtctcagc aggagaacag accaaagcca gcaaaaggga cctcggtgac 2940
accagtaggg acaggaggat tttgtggggg ctcgtgtcac tgtgcaaacc catgaaaacc 3000
ccaagggagt ttggaactgc aagcacagtg acacagaccc attcaaaaac ccctactgca 3060
aacacaccca ctcctggggc tgaggggctg ggggagcgtc tgggaagtag ggtccagggg 3120
tgtctatcaa tgtccaaaat gcaccagact ccccgccaaa caccacccca ccagccagcg 3180
agcagggtaa acagaaaatg agaggctctg ggaagcttgc acaggcccca aggaaagagc 3240
tttggcgggt gtgcaagagg ggatgcaggc agagcctgag cagggccttt tgctgtttct 3300
gctttcctgt gcagagagtt ccataaactg gtgttcaaga tcagtggctg ggaatgagcc 3360
caggagggca gtctgtggga agagcacagg gaaggaggag cagccgctat cctacactgt 3420
catctttcaa aagtttgcct tgtgaccaca ctattgcatc atgggatgct taagagctga 3480
tgtagacaca gctaaagaga gaatcagtga gatgaatttg cagcatagat ctgaataaac 3540
tctccagaat gtggagcagt acagaagcaa acacacagaa agtgcctgat gcaaggacaa 3600
agttcagtgg gcaccttcag gcattgctgc tgggcacaga cactctgaaa agccttggca 3660
ggatctccct gcgacaaagc agaaccctca ggcaatgcca gccccagagc cctccctgag 3720
agcgtcatgg ggaaagatgt gcagaacagc tgattatcat agactcaaac tgagaacaga 3780
gcaaacgtcc atctgaagaa cagtcaaata agcaatggta ggttcatgca atgcaaaccc 3840
agacagccag gggacaacag tagagggcta caggcggctt tgcggttgag ttcatgacaa 3900
tgctgagtaa ttggagtaac agaggaaagc ccaaaaaata cttttaatgt gatttcttct 3960
aaataaaatt tacaccaggc aaaatgaact gtcttcttaa gggataaact ttcccctgga 4020
aaaactacaa ggaaaattaa gaaaacgatg atcacataaa cacagttgtg gttacttcta 4080
ctggggaagg aagagggtat gagctgagac acacagagtc ggcaagtctc caagcaagca 4140
cagaacgaat acattacagt accttgaata cagcagttaa acttctaaat cgcaagaaca 4200
ggaaaatgca cacagctgtg tttagaaaat tctcagtcca gcactattca taatagcaaa 4260
gacattaacc caggttggat aaataaatga tgacacaggc aattgcacaa tgatacagac 4320
atacatttag tacatgagac atcgatgatg tatccccaaa gaaatgactt taaagagaaa 4380
aggcctgatg tgtggtggca ctcacctccc tgggatcccc ggacaggttg caggcacact 4440
gtgtggcagg gcaggctggt acatgctggc agctcctggg gcctgatgtg gagcaagcgc 4500
agggctgtat acccccaagg atggcacagt cagtgaattc cagagagaag cagctcagcc 4560
acactgccca ggcagagccc gagagggacg cccacgtaca gggaggcaga gcccagctcc 4620
tccacagcca ccaccacctg tgcacgggcc accaccttgc aggcacagag tgggtgctga 4680
gaggaggggc agggacacca ggcagggtga gcacccagag aaaactgcag aagcctcaca 4740
catccacctc agcctcccct gacctggacc tcacctggtc tggacctcac ctggcctggg 4800
cctcacctga cctggacctc acctggcctg ggcttcacct gacctggacc tcacctggcc 4860
tccggcctca cctgcacctg ctccaggtct tgctggaacc tgagtagcac tgaggctgca 4920
gaagctcatc cagggttggg gaatgactct ggaactctcc cacatctgac ctttctgggt 4980
ggaggcatct ggtggccctg ggaatataaa aagccccaga atggtgcctg cgtgatttgg 5040
gggcaattta tgaacccgaa aggacatggc catggggtgg gtagggacat agggacagat 5100
gccagcctga ggtggagcct caggacacag ttggacgcgg acactatcca cataagcgag 5160
ggacagaccc gagtgttcct gcagtagacc tgagagcgct gggcccacag cctcccctcg 5220
gtgccctgct gcctcctcag gtcagccctg gacatcccgg gtttccccag gccagatggt 5280
aggattttgt tgaggtctgt gtcactgtgc atggcagcta ctgccagccc gcagccactg 5340
gctactgagg atgccgattc tgagaatagc agcttctact actatgacta cctggatgaa 5400
gtagctttca tgctctgcga ggttagccag catctagact atgcccacag tgtcacacgg 5460
tccatcaaaa acccatgcca cagccctccc cgcaggggac cgccgcgtgc catgttacga 5520
ttttgatcga ggacacagcg ccatgggtat ggtggctacc acagcagtgc agcccatgac 5580
ccaaacacac agggcagcag gcacaatgga caggcctgtg agtgaccatg ctgggctcca 5640
gcccgccagc cccggagacc atgaaacaga tggccaaggt caccccacag ttcagccaga 5700
catggctccg tggggtctgc atcgctgctg ccctctaaca ccagcccaga tggggacaag 5760
gccaacccca cattaccatc tcctgctgtc cacccagtgg tcccagaagc ccctccctca 5820
tggctgagcc acatgtgtga accctgagag caccccatgt cagagtaggg gcagcagaag 5880
ggcggggctg gccctgtgca ctgtccctgc acccatggtc cctcgcctgc ctggccctga 5940
cacctgagcc tcttctgagt catttctaag atagaagaca ttcccgggga cagccggagc 6000
tgggcgtcgc tcatcccgcc cggccgtcct gagtcctgct tgtttccaga cctcaccagg 6060
gaagccaaca gaggactcac ctcacacagt cagagacaaa gaaccttcca gaaatccctg 6120
tctcactccc cagtgggcac cttcttccag gacattcctc ggtcgcatca cagcaggcac 6180
ccacatctgg atcaggacgg cccccagaac acaagatggc ccatggggac agccccacaa 6240
cccaggcctt cccagacccc taaaaggcgt cccaccccct gcacctgccc cagggctaaa 6300
aatccaggag gcttgactcc cgcataccct ccagccagac atcacctcag ccccctcctg 6360
gaggggacag gagcccggga gggtgagtca gacccacctg ccctcgatgg caggcgggga 6420
agattcagaa aggcctgaga tccccaggac gcagcaccac tgtcaatggg ggccccagac 6480
gcctggacca gggcctgcgt gggaaaggcc gctgggcaca ctcaggggga ttttgtgaag 6540
gcccctccca ctgtgcagct tcttgtgcaa gcaaacccat gaaaacccca agggagtttg 6600
gaactgccat gccgatttcg gcgggcatgg caccatttgg aagctcttcc tccggttcca 6660
gcagaacctg ctaccacagt gatgaaacta gcatcaaaaa ccggccggac acccagggac 6720
catgcacact tctcagcttg gagctctcca ggaccagaag agtcaggtct gagggtttgt 6780
agccagaccc tcggcctcta gggacaccct ggccatcaca gcggatgggc tggtgcccca 6840
catgccatct gctccaaaca ggggcttcag agggctctga ggtgacttca ctcatgacca 6900
caggtgccct ggccccttcc ccgccagcta caccgaaccc tgtcccaaca gctgccccag 6960
ttccaacagc caattcctgg ggcccagaat tgctgtagac accagcctcg ttccagcacc 7020
tcctgccaat tgcctggatt cacatcctgg ctggaatcaa gagggcagca tccgccaggc 7080
tcccaacagg caggactccc gcacaccctc ctctgagagg ccgctgtgtt ccgcagggcc 7140
aggccctgga cagttcccct cacctgccac tagagaaaca cctgccattg tcgtccccac 7200
ctggaaaaga ccactcgtgg agcccccagc cccaggtaca gctgtagaga gactccccga 7260
gggatctaag aaggagccat gcgcagttct gccgggaccc tcggccaggc cgacaggagt 7320
ggacactgga gctgggccca cactgggcca cataggagct caccagtgag ggcaggagag 7380
cacatgccgg ggagcaccca gcctcctgct gaccagaggc ccgtcccaga gcccaggagg 7440
ctgcagaggc ctctccaggg ggacactgtg catgtctggt ccctgagcag ccccccacgt 7500
ccccagtcct gggggcccct ggcacagctg tctggaccct ccctgttccc tgggaagctc 7560
ctcctgacag ccccgcctcc agttccaggt gtggattttg tcagggggtg tcacactgtg 7620
ctgcataccc tgctggacct gcaagtattc ggcaactgtc ggaaggatgc tgtggttagc 7680
tttggcaaag ttttcctgcc accacagtgg tgctgcccat atcaaaaacc aggccaagta 7740
gacaggcccc tgctgtgcag ccccaggcct ccacttcacc tgcttctcct ggggctctca 7800
aggtcactgt tgtctgtact ctgccctctg tggggagggt tccctcagtg ggaggtctgt 7860
tctcaacatc ccagggcctc atgtctgcac ggaaggccaa tggatgggca acctcacatg 7920
ccgcggctaa gatagggtgg gcagcctggc gggggacagt acatactgct ggggtgtctg 7980
tcactgtgcc tagtggggca ctggctccca aacaacgcag tcctcgccaa aatccccaca 8040
gcctcccctg ctaggggctg gcctgatctc ctgcagtcct aggaggctgc tgacctccag 8100
aatgtctccg tccccagttc cagggcgaga gcagatccca ggccggctgc agactgggag 8160
gccaccccct ccttcccagg gttcactgga ggtgaccaag gtaggaaatg gccttaacac 8220
agggatgact gcgccatccc ccaacagagt cagccccctc ctgctctgta ccccgcaccc 8280
cccaggccag tccacgaaaa ccagggcccc acatcagagt cactgcctgg cccggccctg 8340
gggcggaccc ctcagccccc accctgtcta gaggacttgg ggggacagga cacaggccct 8400
ctccttatgg ttcccccacc tgcctccggc cgggaccctt ggggtgtgga cagaaaggac 8460
acctgcctaa ttggccccca ggaacccaga acttctctcc agggacccca gcccgagcac 8520
ccccttaccc aggacccagc cctgcccctc ctcccctctg ctctcctctc atcaccccat 8580
gggaatccgg tatccccagg aagccatcag gaagggctga aggaggaagc ggggccgtgc 8640
accaccgggc aggaggctcc gtcttcgtga acccagggaa gtgccagcct cctagagggt 8700
atggtccacc ctgcctgggg ctcccaccgt ggcaggctgc ggggaaggac cagggacggt 8760
gtgggggagg gctcagggcc ctgcgggtgc tcctccatct tcggtgagcc tcccccttca 8820
cccaccgtcc cgcccacctc ctctccaccc tggctgcacg tcttccacac catcctgagt 8880
cctacctaca ccagagccag caaagccagt gcagacaaag gctggggtgc aggggggctg 8940
ccagggcagc ttcggggagg gaaggatgga gggaggggag gtcagtgaag aggccccctt 9000
cccctgggtc caggatcctc ctctgggacc cccggatccc atcccctcct ggctctggga 9060
ggagaagcag gatgggagaa tctgtgcggg accctctcac agtggaatat ccccacagcg 9120
gctcaggcca gacccaaaag cccctcagtg agccctccac tgcagtcctg ggcctgggta 9180
gcagcccctc ccacagagga cagacccagc accccgaaga agtcctgcca gggggagctc 9240
agagccatga aagagcagga tatggggtcc ccgatacagg cacagacctc agctccatcc 9300
aggcccaccg ggacccacca tgggaggaac acctgtctcc gggttgtgag gtagctggcc 9360
tctgtctcgg accccactcc agacaccaga cagaggggca ggccccccaa aaccagggtt 9420
gagggatgat ccgtcaaggc agacaagacc aaggggcact gaccccagca agggaaggct 9480
cccaaacaga cgaggaggat tttgtagctg tctgtatcac tgtgcagctt cttgtgccat 9540
gccgatttcg gcgggcatgg caccatttgg aagctcttcc tccggttcca gcagaacctg 9600
ctaccacagt gacactcgcc aggtcaaaaa ccccgtccca agtcagcgga agcagagaga 9660
gcagggagga cacgtttagg atctgaggcc gcacctgaca cccagggcag cagacgtctc 9720
ccctccaggg caccctccac cgtcctgcgt ttcttcaaga ataggggcgg cctgaggggg 9780
tccagggcca ggcgataggt cccctctacc ccaaggagga gccaggcagg acccgagcac 9840
cgatgcatct aacgcagtca tgtaatgctg ggtgacagtc agttcgccta cgta 9894
<210> 134
<211> 11977
<212> ДНК
<213> Искусственная последовательность
<220>
<223> D6-DH120126
<400> 134
tacgtaatgc atctaacgca gtcatgtaat gctgggtgac agtcagttcg cctccccatt 60
gaggctgacc tgcccagacg ggcctgggcc caccccacac accggggcgg aatgtgtgca 120
ggccccagtc tctgtgggtg ttccgctagc tggggccccc agtgctcacc ccacacctaa 180
agcgagcccc agcctccaga gccccctaag cattccccgc ccagcagccc agcccctgcc 240
cccacccagg aggccccaga gctcagggcg cctggtcgga ttttgtacag ccccgagtca 300
ctgtgcatgc cgatttcggc gggcatggca ccatttggaa gctcttcctc cggttccagc 360
agaacctgct accacagtga gaaaaactgt gtcaaaaacc gactcctggc agcagtcgga 420
ggccccgcca gagaggggag cagccggcct gaacccatgt cctgccggtt cccatgaccc 480
ccagcaccca gagccccacg gtgtccccgt tggataatga ggacaagggc tgggggctcc 540
ggtggtttgc ggcagggact tgatcacatc cttctgctgt ggccccattg cctctggctg 600
gagttgaccc ttctgacaag tgtcctcaga aagacaggga tcaccggcac ctcccaatat 660
caaccccagg cagcacagac acaaacccca catccagagc caactccagg agcagagaca 720
ccccaacact ctgggggacc ccaaccgtga taactcccca ctggaatccg ccccagagtc 780
taccaggacc aaaggccctg ccctgtctct gtccctcact cagggcctcc tgcagggcga 840
gcgcttggga gcagactcgg tcttagggga caccactgtg ggccccaact ttgatgaggc 900
cactgaccct tccttccttt cctggggcag cacagacttt ggggtctggg cagggaagaa 960
ctactggctg gtggccaatc acagagcccc caggccgagg tggccccaag aaggccctca 1020
ggaggtggcc actccacttc ctcccagctg gaccccaggt cctccccaag ataggggtgc 1080
catccaaggc aggtcctcca tggagccccc ttcagactcc tcccgggacc ccactggacc 1140
tcagtccctg ctctgggaat gcagccacca caagcacacc aggaagccca ggcccagcca 1200
ccctgcagtg ggcaagccca cactctggag cagagcaggg tgcgtctggg aggggctaac 1260
ctccccaccc cccacccccc atctgcacac agccacctac cactgcccag accctctgca 1320
ggagggccaa gccaccatgg ggtatggact tagggtctca ctcacgtgcc tcccctcctg 1380
ggagaagggg cctcatgccc agatccctgc agcactagac acagctggag gcagtggccc 1440
cagggccacc ctgacctggc atctaaggct gctccagccc agacagcact gccgttcctg 1500
ggaagcctgg gctccaccag accacaggtc cagggcacag cccacaggag ccacccacac 1560
acagctcaca ggaagaagat aagctccaga ccccagggcg ggacctgcct tcctgccacc 1620
acttacacac aggccaggga gctgttccca cacagatcaa ccccaaaccg ggactgcctg 1680
gcactagggt cactgccatt tccctctcca ttccctccca gtgcctctgt gctccctcct 1740
tctggggaac accctgtgca gcccctccct gcagcccaca cgctggggag accccaccct 1800
gcctcgggcc ttttctacct gctgcacttg ccgcccaccc aaacaaccct gggtacgtga 1860
ccctgcagtc ctcaccctga tctgcaacca gacccctgtc cctccctcta aacacccctc 1920
ccaggccaac tctgcacctg caggccctcc gctcttctgc cacaagagcc tcaggttttc 1980
ctacctgtgc ccacccccta acccctcctg cccacaactt gagttcttcc tctcctggag 2040
cccttgagcc atggcactga ccctacactc ccacccacac actgcccatg ccatcacctt 2100
cctcctggac actctgaccc cgctcccctc cctctcagac ccggccctgg tatttccagg 2160
acaaaggctc acccaagtct tccccatgca ggcccttgcc ctcactgcct ggttacacgg 2220
gagcctcctg tgcgcagaag cagggagctc agctcttcca caggcagaag gcactgaaag 2280
aaatcagcct ccagtgcctt gacacacgtc cgcctgtgtc tctcactgcc tgcacctgca 2340
gggaggctcc gcactccctc taaagatgag ggatccaggc agcaacatca cgggagaatg 2400
cagggctccc agacagccca gccctctcgc aggcctctcc tgggaagaga cctgcagcca 2460
ccactgaaca gccacggagg tcgctggata gtaaccgagt cagtgaccga cctggagggc 2520
aggggagcag tgaaccggag cccataccat agggacagag accagccgct aacatcccga 2580
gcccctcact ggcggcccca gaacaccccg tggaaagaga acagacccac agtcccacct 2640
ggaacagggc agacactgct gagcccccag caccagcccc aagaaacact aggcaacagc 2700
atcagagggg gctcctgaga aagagaggag gggaggtctc cttcaccatc aaatgcttcc 2760
cttgaccaaa aacagggtcc acgcaactcc cccaggacaa aggaggagcc ccctgtacag 2820
cactgggctc agagtcctct ctgagacagg ctcagtttca gacaacaacc cgctggaatg 2880
cacagtctca gcaggagagc caggccagag ccagcaagag gagactcggt gacaccagtc 2940
tcctgtaggg acaggaggat tttgtggggg ttcgtgtcac tgtgcaaacc catgaaaacc 3000
ccaagggagt ttggaacagc aagcacagtg acacaacccc attcaaaaac ccctactgca 3060
aacgcaccca ctcctgggac tgaggggctg ggggagcgtc tgggaagtat ggcctagggg 3120
tgtccatcaa tgcccaaaat gcaccagact ctccccaaga catcacccca ccagccagtg 3180
agcagagtaa acagaaaatg agaagcagct gggaagcttg cacaggcccc aaggaaagag 3240
ctttggcagg tgtgcaagag gggatgtggg cagagcctca gcagggcctt ttgctgtttc 3300
tgctttcctg tgcagagagt tccataaact ggtattcaag atcaatggct gggagtgagc 3360
ccaggaggac agtgtgggaa gagcacaggg aaggaggagc agccgctatc ctacactgtc 3420
atcttttgaa agtttgccct gtgcccacaa tgctgcatca tgggatgctt aacagctgat 3480
gtagacacag ctaaagagag aatcagtgaa atggatttgc agcacagatc tgaataaatc 3540
ctccagaatg tggagcagca cagaagcaag cacacagaaa gtgcctgatg ccaaggcaaa 3600
gttcagtggg caccttcagg cattgctgct gggcacagac actctgaaaa gcactggcag 3660
gaactgcctg tgacaaagca gaaccctcag gcaatgccag ccctagagcc cttcctgaga 3720
acctcatggg caaagatgtg cagaacagct gtttgtcata gccccaaact atggggctgg 3780
acaaagcaaa cgtccatctg aaggagaaca gacaaataaa cgatggcagg ttcatgaaat 3840
gcaaactagg acagccagag gacaacagta gagagctaca ggcggctttg cggttgagtt 3900
catgacaatg ctgagtaatt ggagtaacag aggaaagccc aaaaaatact tttaatgtga 3960
tttcttctaa ataaaattta cacccggcaa aatgaactat cttcttaagg gataaacttt 4020
cccctggaaa aactataagg aaaatcaaga aaacgatgat cacataaaca cagtggtggt 4080
tacttctact ggggaaggaa gagggtatga gctgagacac acagagtcgg caagtctcct 4140
aacaagaaca gaacaaatac attacagtac cttgaaaaca gcagttaaac ttctaaatcg 4200
caagaagagg aaaatgcaca cacctgtgtt tagaaaattc tcagtccagc actgttcata 4260
atagcaaaga cattaaccca ggttggataa ataagcgatg acacaggcaa ttgcacaatg 4320
atacagacat acattcagta tatgagacat cgatgatgta tccccaaaga aatgacttta 4380
aagagaaaag gcctgatgtg tggtggcaat cacctccctg ggcatccccg gacaggctgc 4440
aggctcactg tgtggcaggg caggcaggca cctgctggca gctcctgggg cctgatgtgg 4500
agcaggcaca gagctgtata tccccaagga aggtacagtc agtgcattcc agagagaagc 4560
aactcagcca cactccctgg ccagaaccca agatgcacac ccatgcacag ggaggcagag 4620
cccagcacct ccgcagccac caccacctgc gcacgggcca ccaccttgca ggcacagagt 4680
gggtgctgag aggaggggca gggacaccag gcagggtgag cacccagaga aaactgcaga 4740
agcctcacac atccctcacc tggcctgggc ttcacctgac ctggacctca cctggcctcg 4800
ggcctcacct gcacctgctc caggtcttgc tggagcctga gtagcactga ggctgtaggg 4860
actcatccag ggttggggaa tgactctgca actctcccac atctgacctt tctgggtgga 4920
ggcacctggt ggcccaggga atataaaaag ccccagaatg atgcctgtgt gatttggggg 4980
caatttatga acccgaaagg acatggccat ggggtgggta gggacagtag ggacagatgt 5040
cagcctgagg tgaagcctca ggacacaggt gggcatggac agtgtccacc taagcgaggg 5100
acagacccga gtgtccctgc agtagacctg agagcgctgg gcccacagcc tcccctcggg 5160
gccctgctgc ctcctcaggt cagccctgga catcccgggt ttccccaggc ctggcggtag 5220
gattttgttg aggtctgtgt cactgtgcat ggcagctact gccagcccgc agccactggc 5280
tactgaggat gccgattctg agaatagcag cttctactac tatgactacc tggatgaagt 5340
agctttcatg ctcagcgagg ttagccagca tctagactat gcccacagtg tcacagagtc 5400
catcaaaaac ccatgcctgg gagcctccca ccacagccct ccctgcgggg gaccgctgca 5460
tgccgtgtta ggattttgat cgaggacacg gcgccatggg tatggtggct accacagcag 5520
tgcagcccat gacccaaaca cacggggcag cagaaacaat ggacaggccc acaagtgacc 5580
atgatgggct ccagcccacc agccccagag accatgaaac agatggccaa ggtcacccta 5640
caggtcatcc agatctggct ccaaggggtc tgcatcgctg ctgccctccc aacgccaaac 5700
cagatggaga cagggccggc cccatagcac catctgctgc cgtccaccca gcagtcccgg 5760
aagcccctcc ctgaacgctg ggccacgtgt gtgaaccctg cgagcccccc atgtcagagt 5820
aggggcagca ggagggcggg gctggccctg tgcactgtca ctgcccctgt ggtccctggc 5880
ctgcctggcc ctgacacctg agcctctcct gggtcatttc caagacattc ccagggacag 5940
ccggagctgg gagtcgctca tcctgcctgg ctgtcctgag tcctgctcat ttccagacct 6000
caccagggaa gccaacagag gactcacctc acacagtcag agacaacgaa ccttccagaa 6060
atccctgttt ctctccccag tgagagaaac cctcttccag ggtttctctt ctctcccacc 6120
ctcttccagg acagtcctca gcagcatcac agcgggaacg cacatctgga tcaggacggc 6180
ccccagaaca cgcgatggcc catggggaca gcccagccct tcccagaccc ctaaaaggta 6240
tccccacctt gcacctgccc cagggctcaa actccaggag gcctgactcc tgcacaccct 6300
cctgccagat atcacctcag ccccctcctg gaggggacag gagcccggga gggtgagtca 6360
gacccacctg ccctcaatgg caggcgggga agattcagaa aggcctgaga tccccaggac 6420
gcagcaccac tgtcaatggg ggccccagac gcctggacca gggcctgtgt gggaaaggcc 6480
tctggccaca ctcaggggga ttttgtgaag ggccctccca ctgtggaggt tagccagcat 6540
ctagactatg cccacagtga tgaaaccagc atcaaaaacc gaccggactc gcagggttta 6600
tgcacacttc tcggctcgga gctctccagg agcacaagag ccaggcccga gggtttgtgc 6660
ccagaccctc ggcctctagg gacacccggg ccatcttagc cgatgggctg atgccctgca 6720
caccgtgtgc tgccaaacag gggcttcaga gggctctgag gtgacttcac tcatgaccac 6780
aggtgccctg gtcccttcac tgccagctgc accagaccct gttccgagag atgccccagt 6840
tccaaaagcc aattcctggg gccgggaatt actgtagaca ccagcctcat tccagtacct 6900
cctgccaatt gcctggattc ccatcctggc tggaatcaag agggcagcat ccgccaggct 6960
cccaacaggc aggactccca cacaccctcc tctgagaggc cgctgtgttc cgcagggcca 7020
ggccgcagac agttcccctc acctgcccat gtagaaacac ctgccattgt cgtccccacc 7080
tggcaaagac cacttgtgga gcccccagcc ccaggtacag ctgtagagag agtcctcgag 7140
gcccctaaga aggagccatg cccagttctg ccgggaccct cggccaggcc gacaggagtg 7200
gacgctggag ctgggcccac actgggccac ataggagctc accagtgagg gcaggagagc 7260
acatgccggg gagcacccag cctcctgctg accagagacc cgtcccagag cccaggaggc 7320
tgcagaggcc tctccagggg gacacagtgc atgtctggtc cctgagcagc ccccaggctc 7380
tctagcactg ggggcccctg gcacagctgt ctggaccctc cctgttccct gggaagctcc 7440
tcctgacagc cccgcctcca gttccaggtg tggattttgt cagggggtgc cacactgtgc 7500
tgcataccct gctggacctg caagtattcg gcaacagtcg gaaggatgct gtggttagct 7560
ttggcaaagt tttcctgcca ccacagtggt gccgcccata tcaaaaacca ggccaagtag 7620
acagacccct gccacgcagc cccaggcctc cagctcacct gcttctcctg gggctctcaa 7680
ggctgctgtc tgccctctgg ccctctgtgg ggagggttcc ctcagtggga ggtctgtgct 7740
ccagggcagg gatgactgag atagaaatca aaggctggca gggaaaggca gcttcccgcc 7800
ctgagaggtg caggcagcac cacagagcca tggagtcaca gagccacgga gcccccagtg 7860
tgggcgtgtg agggtgctgg gctcccggca ggcccagccc tgatggggaa gcctgccccg 7920
tcccacagcc caggtcccca ggggcagcag gcacagaagc tgccaagctg tgctctacga 7980
tcctcatccc tccagcagca tccactccac agtggggaaa ctgagccttg gagaaccacc 8040
cagccccctg gaaacaaggc ggggagccca gacagtgggc ccagagcact gtgtgtatcc 8100
tggcactagg tgcagggacc acccggagat ccccatcact gagtggccag cctgcagaag 8160
gacccaaccc caaccaggcc gcttgattaa gctccatccc cctgtcctgg gaacctcttc 8220
ccagcgccac caacagctcg gcttcccagg ccctcatccc tccaaggaag gccaaaggct 8280
gggcctgcca ggggcacagt accctccctt gccctggcta agacagggtg ggcagacggc 8340
tgcagatagg acatattgct ggggcatctt gctctgtgac tactgggtac tggctctcaa 8400
cgcagaccct accaaaatcc ccactgcctc ccctgctagg ggctggcctg gtctcctcct 8460
gctgtcctag gaggctgctg acctccagga tggcttctgt ccccagttct agggccagag 8520
cagatcccag gcaggctgta ggctgggagg ccacccctgt ccttgccgag gttcagtgca 8580
ggcacccagg acaggaaatg gcctgaacac agggatgact gtgccatgcc ctacctaagt 8640
ccgccccttt ctactctgca acccccactc cccaggtcag cccatgacga ccaacaaccc 8700
aacaccagag tcactgcctg gccctgccct ggggaggacc cctcagcccc caccctgtct 8760
agaggagttg gggggacagg acacaggctc tctccttatg gttcccccac ctggctcctg 8820
ccgggaccct tggggtgtgg acagaaagga cgcctgccta attggccccc aggaacccag 8880
aacttctctc cagggacccc agcccgagca cccccttacc caggacccag ccctgcccct 8940
cctcccctct gctctcctct catcactcca tgggaatcca gaatccccag gaagccatca 9000
ggaagggctg aaggaggaag cggggccgct gcaccaccgg gcaggaggct ccgtcttcgt 9060
gaacccaggg aagtgccagc ctcctagagg gtatggtcca ccctgcctgg ggctcccacc 9120
gtggcaggct gcggggaagg accagggacg gtgtggggga gggctcaggg ccctgcaggt 9180
gctccatctt ggatgagccc atccctctca cccaccgacc cgcccacctc ctctccaccc 9240
tggccacacg tcgtccacac catcctgagt cccacctaca ccagagccag cagagccagt 9300
gcagacagag gctggggtgc aggggggccg ccagggcagc tttggggagg gaggaatgga 9360
ggaaggggag gtcagtgaag aggcccccct cccctgggtc taggatccac ctttgggacc 9420
cccggatccc atcccctcca ggctctggga ggagaagcag gatgggagat tctgtgcagg 9480
accctctcac agtggaatac ctccacagcg gctcaggcca gatacaaaag cccctcagtg 9540
agccctccac tgcagtgcag ggcctggggg cagcccctcc cacagaggac agacccagca 9600
ccccgaagaa gtcctgccag ggggagctca gagccatgaa ggagcaagat atggggaccc 9660
caatactggc acagacctca gctccatcca ggcccaccag gacccaccat gggtggaaca 9720
cctgtctccg gcccctgctg gctgtgaggc agctggcctc tgtctcggac ccccattcca 9780
gacaccagac agagggacag gccccccaga accagtgttg agggacaccc ctgtccaggg 9840
cagccaagtc caagaggcgc gctgagccca gcaagggaag gcccccaaac aaaccaggag 9900
gtttctgaag ctgtctgtgt cacagtcggg catagccacg gctaccacaa tgacactggg 9960
caggacagaa accccatccc aagtcagccg aaggcagaga gagcaggcag gacacattta 10020
ggatctgagg ccacacctga cactcaagcc aacagatgtc tcccctccag ggcgccctgc 10080
cctgttcagt gttcctgaga aaacaggggc agcctgaggg gatccagggc caggagatgg 10140
gtcccctcta ccccgaggag gagccaggcg ggaatcccag ccccctcccc attgaggcca 10200
tcctgcccag aggggcccgg acccacccca cacacccagg cagaatgtgt gcaggcctca 10260
ggctctgtgg gtgccgctag ctggggctgc cagtcctcac cccacaccta aggtgagcca 10320
cagccgccag agcctccaca ggagacccca cccagcagcc cagcccctac ccaggaggcc 10380
ccagagctca gggcgcctgg gtggattttg tacagccccg agtcactgtg ctgcataccc 10440
tgctggacct gcaagtattc ggcaaccaca gtgagaaaag ctatgtcaaa aaccgtctcc 10500
cggccactgc tggaggccca gccagagaag ggaccagccg cccgaacata cgaccttccc 10560
agacctcatg acccccagca cttggagctc cacagtgtcc ccattggatg gtgaggatgg 10620
gggccggggc catctgcacc tcccaacatc acccccaggc agcacaggca caaaccccaa 10680
atccagagcc gacaccagga acacagacac cccaataccc tgggggaccc tggccctggt 10740
gacttcccac tgggatccac ccccgtgtcc acctggatca aagaccccac cgctgtctct 10800
gtccctcact cagggcctgc tgaggggcgg gtgctttgga gcagactcag gtttaggggc 10860
caccattgtg gggcccaacc tcgaccagga cacagatttt tctttcctgc cctggggcaa 10920
cacagacttt ggggtctgtg cagggaggac cttctggaaa gtcaccaagc acagagccct 10980
gactgaggtg gtctcaggaa gacccccagg agggggcttg tgccccttcc tctcatgtgg 11040
accccatgcc ccccaagata ggggcatcat gcagggcagg tcctccatgc agccaccact 11100
aggcaactcc ctggcgccgg tccccactgc gcctccatcc cggctctggg gatgcagcca 11160
ccatggccac accaggcagc ccgggtccag caaccctgca gtgcccaagc ccttggcagg 11220
attcccagag gctggagccc acccctcctc atccccccac acctgcacac acacacctac 11280
cccctgccca gtccccctcc aggagggttg gagccgccca tagggtgggg gctccaggtc 11340
tcactcactc gcttcccttc ctgggcaaag gagcctcgtg ccccggtccc ccctgacggc 11400
gctgggcaca ggtgtgggta ctgggcccca gggctcctcc agccccagct gccctgctct 11460
ccctgggagg cctgggcacc accagaccac cagtccaggg cacagcccca gggagccgcc 11520
cactgccagc tcacaggaag aagataagct tcagaccctc agggccggga gctgccttcc 11580
tgccacccct tcctgcccca gacctccatg ccctccccca accacttaca cacaagccag 11640
ggagctgttt ccacacagtt caaccccaaa ccaggacggc ctggcactcg ggtcactgcc 11700
atttctgtct gcattcgctc ccagcgcccc tgtgttccct ccctcctccc tccttccttt 11760
cttcctgcat tgggttcatg ccgcagagtg ccaggtgcag gtcagccctg agcttggggt 11820
cacctcctca ctgaaggcag cctcagggtg cccaggggca ggcagggtgg gggtgaggct 11880
tccagctcca accgcttcgc taccttagga ccgttatagt taggcgcgcc gtcgaccaat 11940
tctcatgttt gacagcttat catcgaattt ctacgta 11977
<210> 135
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер и/или зонд
<400> 135
tgcggccgat cttagcc 17
<210> 136
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер и/или зонд
<400> 136
acgagcgggt tcggcccatt c 21
<210> 137
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер и/или зонд
<400> 137
ttgaccgatt ccttgcgg 18
<210> 138
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер и/или зонд
<400> 138
cgggtcactg ccatttctg 19
<210> 139
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер и/или зонд
<400> 139
tctgcattcg ctcccagcgc 20
<210> 140
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер и/или зонд
<400> 140
tctgcggcat gaacccaat 19
<210> 141
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер и/или зонд
<400> 141
tggccagaac tgaccctac 19
<210> 142
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер и/или зонд
<400> 142
accgacaaga gtccctcagg 20
<210> 143
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер и/или зонд
<400> 143
ggagtcggct ctggatgtg 19
<210> 144
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер и/или зонд
<400> 144
ggagccaggc aggacaca 18
<210> 145
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер и/или зонд
<400> 145
tgggctcgta gtttgacgt 19
<210> 146
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер и/или зонд
<400> 146
gggactttct tacccacact tca 23
<210> 147
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер и/или зонд
<400> 147
ggtcccgagc actcttaatt aaac 24
<210> 148
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер и/или зонд
<400> 148
cctcgaatgg aactac 16
<210> 149
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер и/или зонд
<400> 149
gggagagcaa ccattcgttg t 21
<210> 150
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер и/или зонд
<400> 150
ccgagcaccg atgcatcta 19
<210> 151
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер и/или зонд
<400> 151
cgcagtcatg taatgc 16
<210> 152
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер и/или зонд
<400> 152
gggaggcgaa ctgactgtca 20
<210> 153
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер и/или зонд
<400> 153
ggtggagagg ctattcggc 19
<210> 154
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер и/или зонд
<400> 154
tgggcacaac agacaatcgg ctg 23
<210> 155
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер и/или зонд
<400> 155
gaacacggcg gcatcag 17
<210> 156
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер и/или зонд
<400> 156
cagtcccgtt gatccagcc 19
<210> 157
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер и/или зонд
<400> 157
cccatcaggg attttgtatc tctgtggacg 30
<210> 158
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер и/или зонд
<400> 158
ggatatgcag cactgtgcca c 21
<210> 159
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер и/или зонд
<400> 159
tcctccaacg acaggtccc 19
<210> 160
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер и/или зонд
<400> 160
tccctggaac tctgccccga caca 24
<210> 161
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер и/или зонд
<400> 161
gatgaactga cgggcacagg 20
<210> 162
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер и/или зонд
<400> 162
atcacactca tcccatcccc 20
<210> 163
<211> 29
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер и/или зонд
<400> 163
cccttcccta agtaccacag agtgggctc 29
<210> 164
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер и/или зонд
<400> 164
cacagggaag caggaactgc 20
<210> 165
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер и/или зонд
<400> 165
gccatgcaag gccaagc 17
<210> 166
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер и/или зонд
<400> 166
ccaggaaaat gctgccagag cctg 24
<210> 167
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер и/или зонд
<400> 167
agttcttgag ccttagggtg ctag 24
<210> 168
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер и/или зонд
<400> 168
ccccacagca aatcacaacc 20
<210> 169
<211> 27
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер и/или зонд
<400> 169
atgcagttgt cacccttgag gccattc 27
<210> 170
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер и/или зонд
<400> 170
tgtttcccag gcgtcactg 19
<210> 171
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер и/или зонд
<400> 171
ctcagtgatt ctggccctgc 20
<210> 172
<211> 32
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер и/или зонд
<400> 172
tgctccacag ctacaaaccc cttcctataa tg 32
<210> 173
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер и/или зонд
<400> 173
ggatgatggc tcagcacaga g 21
<210> 174
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер и/или зонд
<400> 174
tggtcacctc caggagcctc 20
<210> 175
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер и/или зонд
<400> 175
agtctctgct tcccccttgt ggctatgagc 30
<210> 176
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер и/или зонд
<400> 176
gctgcagggt gtatcaggtg c 21
<210> 177
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер и/или зонд
<400> 177
gtgtcacagt cgggcata 18
<210> 178
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер и/или зонд
<400> 178
ccacggctac cacaatgaca ctgg 24
<210> 179
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер и/или зонд
<400> 179
ccttcggctg acttgggatg 20
<210> 180
<211> 66
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность нуклеиновой кислоты мю-конотоксина или
токсина тарантула ProTxII
<400> 180
gagagaagct gcaatggcag acgcggctgc agcagcagat ggagccgcga tcatagcagg 60
tgctgc 66
<210> 181
<211> 22
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность мю-конотоксина или токсина
тарантула ProTxII
<400> 181
Glu Arg Ser Cys Asn Gly Arg Arg Gly Cys Ser Ser Arg Trp Ser Arg
1 5 10 15
Asp His Ser Arg Cys Cys
20
<210> 182
<211> 31
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность нуклеиновой кислоты мю-кононтоксина или
токсина тарантула ProTxII
<400> 182
aggatattgt agcagcagat ggtgctatac c 31
<210> 183
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность мю-конотоксина или токсина
тарантула ProTxII
<400> 183
Gly Tyr Cys Ser Ser Arg Trp Cys Tyr Thr
1 5 10
<210> 184
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность нуклеиновой кислоты мю-конотоксина
или токсина тарантула ProTxII
<400> 184
gtattacgat tgcaactgca gcagatggcg cgaccatagc aggtgctgct attatacc 58
<210> 185
<211> 19
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность мю-конотоксина или токсина
тарантула ProTxII
<400> 185
Tyr Tyr Asp Cys Asn Cys Ser Arg Trp Arg Asp His Ser Arg Cys Cys
1 5 10 15
Tyr Tyr Thr
<210> 186
<211> 66
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность нуклеиновой кислоты мю-конотоксина
или токсина тарантула ProTxII
<400> 186
gagaggctta gctgtggctt ccctaagagc tgccgcagca ggcaaagcaa gcctcacaga 60
tgctgc 66
<210> 187
<211> 22
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность мю-конотоксина или токсина
тарантула ProTxII
<400> 187
Glu Arg Leu Ser Cys Gly Phe Pro Lys Ser Cys Arg Ser Arg Gln Ser
1 5 10 15
Lys Pro His Arg Cys Cys
20
<210> 188
<211> 90
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность нуклеиновой кислоты мю-конотоксина
или токсина тарантула ProTxII
<400> 188
tacagccaga agtggatgtg gacttgcgat agtgagagga agtgcagtga gggtatggta 60
tgccggctgt ggtgtaagaa gaagctctgg 90
<210> 189
<211> 30
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность мю-конотоксина или токсина
тарантула ProTxII
<400> 189
Tyr Ser Gln Lys Trp Met Trp Thr Cys Asp Ser Glu Arg Lys Cys Ser
1 5 10 15
Glu Gly Met Val Cys Arg Leu Trp Cys Lys Lys Lys Leu Trp
20 25 30
<210> 190
<211> 48
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность нуклеиновой кислоты мю-конотоксина
или токсина тарантула ProTxII
<400> 190
agctgcaact gcagcagcaa atggagccgc gaccatagca ggtgctgc 48
<210> 191
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность мю-конотоксина или токсина
тарантула ProTxII
<400> 191
Ser Cys Asn Cys Ser Ser Lys Trp Ser Arg Asp His Ser Arg Cys Cys
1 5 10 15
<210> 192
<211> 57
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность нуклеиновой кислоты мю-конотоксина
или токсина тарантула ProTxII
<400> 192
gagagatgca atggcagacg cggctgcagc agatggcgcg atcatagcag gtgctgc 57
<210> 193
<211> 19
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность мю-конотоксина или токсина
тарантула ProTxII
<400> 193
Glu Arg Cys Asn Gly Arg Arg Gly Cys Ser Arg Trp Arg Asp His Ser
1 5 10 15
Arg Cys Cys
<210> 194
<211> 43
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность нуклеиновой кислоты мю-конотоксина
или токсина тарантула ProTxII
<400> 194
aggatattgt actaatcgga gcaggcaggg tgtatgctat acc 43
<210> 195
<211> 14
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность мю-конотоксина или токсина
тарантула ProTxII
<400> 195
Gly Tyr Cys Thr Asn Arg Ser Arg Gln Gly Val Cys Tyr Thr
1 5 10
<210> 196
<211> 61
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность нуклеиновой кислоты мю-конотоксина
или токсина тарантула ProTxII
<400> 196
gtattacgat tgcaactgca gcagatgggc tcgcgaccat agcaggtgct gctattataa 60
c 61
<210> 197
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность мю-конотоксина или токсина
тарантула ProTxII
<400> 197
Tyr Tyr Asp Cys Asn Cys Ser Arg Trp Ala Arg Asp His Ser Arg Cys
1 5 10 15
Cys Tyr Tyr Asn
20
<210> 198
<211> 76
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность нуклеиновой кислоты мю-конотоксина
или токсина тарантула ProTxII
<400> 198
gtattactat gagagatgca atggcagacg cggctgcagc agatggcgcg atcatagcag 60
gtgctgctat tataac 76
<210> 199
<211> 25
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность мю-конотоксина или токсина
тарантула ProTxII
<400> 199
Tyr Tyr Tyr Glu Arg Cys Asn Gly Arg Arg Gly Cys Ser Arg Trp Arg
1 5 10 15
Asp His Ser Arg Cys Cys Tyr Tyr Asn
20 25
<210> 200
<211> 57
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность нуклеиновой кислоты мю-конотоксина
или токсина тарантула ProTxII
<400> 200
gagaggcttt gtggcttccc taagagctgc agcaggcaaa agcctcacag atgctgc 57
<210> 201
<211> 19
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность мю-конотоксина или токсина
тарантула ProTxII
<400> 201
Glu Arg Leu Cys Gly Phe Pro Lys Ser Cys Ser Arg Gln Lys Pro His
1 5 10 15
Arg Cys Cys
<210> 202
<211> 90
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность нуклеиновой кислоты мю-конотоксина
или токсина тарантула ProTxII
<400> 202
tactgccaga agtggatgtg gactagcgat agtgagagga agtgctgtga gggtatggta 60
agccggctgt ggtgtaagaa gaagctctgg 90
<210> 203
<211> 30
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность мю-конотоксина или токсина
тарантула ProTxII
<400> 203
Tyr Cys Gln Lys Trp Met Trp Thr Ser Asp Ser Glu Arg Lys Cys Cys
1 5 10 15
Glu Gly Met Val Ser Arg Leu Trp Cys Lys Lys Lys Leu Trp
20 25 30
<210> 204
<211> 38
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность нуклеиновой кислоты мю-конотоксина
или токсина тарантула ProTxII
<400> 204
tgcaactgca gcagatggcg cgaccatagc aggtgctg 38
<210> 205
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность мю-конотоксина или токсина
тарантула ProTxII
<400> 205
Cys Asn Cys Ser Arg Trp Arg Asp His Ser Arg Cys
1 5 10
<210> 206
<211> 66
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность нуклеиновой кислоты мю-конотоксина
или токсина тарантула ProTxII
<400> 206
gagagatgct gcaatggcag acgcggctgc agcagcagat ggtgccgcga tcatagcagg 60
tgctgc 66
<210> 207
<211> 22
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность мю-конотоксина или токсина
тарантула ProTxII
<400> 207
Glu Arg Cys Cys Asn Gly Arg Arg Gly Cys Ser Ser Arg Trp Cys Arg
1 5 10 15
Asp His Ser Arg Cys Cys
20
<210> 208
<211> 43
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность нуклеиновой кислоты мю-конотоксина
или токсина тарантула ProTxII
<400> 208
aggatattgt agtggtagca gcagatgggg tagctgctac tcc 43
<210> 209
<211> 14
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность мю-конотоксина или токсина
тарантула ProTxII
<400> 209
Gly Tyr Cys Ser Gly Ser Ser Arg Trp Gly Ser Cys Tyr Ser
1 5 10
<210> 210
<211> 88
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность нуклеиновой кислоты мю-конотоксина
или токсина тарантула ProTxII
<400> 210
gtattatgat tacgagagag cttgcaatgg cagacgcggc tgcagcagat gggctcgcga 60
tcatagcagg tgctgctatc gttatacc 88
<210> 211
<211> 29
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность мю-конотоксина или токсина
тарантула ProTxII
<400> 211
Tyr Tyr Asp Tyr Glu Arg Ala Cys Asn Gly Arg Arg Gly Cys Ser Arg
1 5 10 15
Trp Ala Arg Asp His Ser Arg Cys Cys Tyr Arg Tyr Thr
20 25
<210> 212
<211> 63
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность нуклеиновой кислоты мю-конотоксина
или токсина тарантула ProTxII
<400> 212
gagaggcttg cttgtggctt ccctaagagc tgcagcaggc aagctaagcc tcacagatgc 60
tgc 63
<210> 213
<211> 21
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность мю-конотоксина или токсина
тарантула ProTxII
<400> 213
Glu Arg Leu Ala Cys Gly Phe Pro Lys Ser Cys Ser Arg Gln Ala Lys
1 5 10 15
Pro His Arg Cys Cys
20
<210> 214
<211> 90
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность нуклеиновой кислоты мю-конотоксина
или токсина тарантула ProTxII
<400> 214
tactgccaga agtggatgtg gacttgcgat agtgagagga agagctgtga gggtatggta 60
tgccggctgt ggagtaagaa gaagctctgg 90
<210> 215
<211> 30
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность мю-конотоксина или токсина
тарантула ProTxII
<400> 215
Tyr Cys Gln Lys Trp Met Trp Thr Cys Asp Ser Glu Arg Lys Ser Cys
1 5 10 15
Glu Gly Met Val Cys Arg Leu Trp Ser Lys Lys Lys Leu Trp
20 25 30
<210> 216
<211> 41
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность нуклеиновой кислоты мю-конотоксина
или токсина тарантула ProTxII
<400> 216
tgcaactgca gcagatgggc tcgcgaccat agcaggtgct g 41
<210> 217
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность мю-конотоксина или токсина
тарантула ProTxII
<400> 217
Cys Asn Cys Ser Arg Trp Ala Arg Asp His Ser Arg Cys
1 5 10
<210> 218
<211> 63
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность нуклеиновой кислоты мю-конотоксина
или токсина тарантула ProTxII
<400> 218
gagagagctt gcaatggcag acgcggctgc agcagatggg ctcgcgatca tagcaggtgc 60
tgc 63
<210> 219
<211> 21
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность мю-конотоксина или токсина
тарантула ProTxII
<400> 219
Glu Arg Ala Cys Asn Gly Arg Arg Gly Cys Ser Arg Trp Ala Arg Asp
1 5 10 15
His Ser Arg Cys Cys
20
<210> 220
<211> 31
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность нуклеиновой кислоты мю-конотоксина
или токсина тарантула ProTxII
<400> 220
agcatattgt cggagcaggc agtgctattc c 31
<210> 221
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность мю-конотоксина или токсина
тарантула ProTxII
<400> 221
Ala Tyr Cys Arg Ser Arg Gln Cys Tyr Ser
1 5 10
<210> 222
<211> 82
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность нуклеиновой кислоты мю-конотоксина
или токсина тарантула ProTxII
<400> 222
gtattactat gagaggcttg cttgtggctt ccctaagagc tgcagcaggc aagctaagcc 60
tcacagatgc tgctattact ac 82
<210> 223
<211> 27
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность мю-конотоксина или токсина
тарантула ProTxII
<400> 223
Tyr Tyr Tyr Glu Arg Leu Ala Cys Gly Phe Pro Lys Ser Cys Ser Arg
1 5 10 15
Gln Ala Lys Pro His Arg Cys Cys Tyr Tyr Tyr
20 25
<210> 224
<211> 66
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность нуклеиновой кислоты мю-конотоксина
или токсина тарантула ProTxII
<400> 224
gagaggcttt gctgtggctt ccctaagagc tgccgcagca ggcaatgcaa gcctcacaga 60
tgctgc 66
<210> 225
<211> 22
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность мю-конотоксина или токсина
тарантула ProTxII
<400> 225
Glu Arg Leu Cys Cys Gly Phe Pro Lys Ser Cys Arg Ser Arg Gln Cys
1 5 10 15
Lys Pro His Arg Cys Cys
20
<210> 226
<211> 90
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность нуклеиновой кислоты мю-конотоксина
или токсина тарантула ProTxII
<400> 226
tactgccaga agtggatgtg gacttgcgat agtgagagga agtgctgtga gggtatggta 60
tgccggctgt ggtgtaagaa gaagctctgg 90
<210> 227
<211> 30
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность мю-конотоксина или токсина
тарантула ProTxII
<400> 227
Tyr Cys Gln Lys Trp Met Trp Thr Cys Asp Ser Glu Arg Lys Cys Cys
1 5 10 15
Glu Gly Met Val Cys Arg Leu Trp Cys Lys Lys Lys Leu Trp
20 25 30
<210> 228
<211> 48
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность нуклеиновой кислоты мю-конотоксина
или токсина тарантула ProTxII
<400> 228
tgctgcaact gcagcagcaa atggtgccgc gaccatagca ggtgctgc 48
<210> 229
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность мю-конотоксина или токсина
тарантула ProTxII
<400> 229
Cys Cys Asn Cys Ser Ser Lys Trp Cys Arg Asp His Ser Arg Cys Cys
1 5 10 15
<210> 230
<211> 77
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность нуклеиновой кислоты мю-конотоксина
или токсина тарантула ProTxII
<400> 230
ggtatagtgg ggagaggctt tgtggcttcc ctaagagctg cagcaggcaa aagcctcaca 60
gatgctgcag ctactac 77
<210> 231
<211> 25
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность мю-конотоксина или токсина
тарантула ProTxII
<400> 231
Tyr Ser Gly Glu Arg Leu Cys Gly Phe Pro Lys Ser Cys Ser Arg Gln
1 5 10 15
Lys Pro His Arg Cys Cys Ser Tyr Tyr
20 25
<210> 232
<211> 9975
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Tx-DH1166
<400> 232
tacgtagccg tttcgatcct cccgaattga ctagtgggta ggcctggcgg ccgctgccat 60
ttcattacct ctttctccgc acccgacata gataccggtg gattcgaatt ctccccgttg 120
aagctgacct gcccagaggg gcctgggccc accccacaca ccggggcgga atgtgtacag 180
gccccggtct ctgtgggtgt tccgctaact ggggctccca gtgctcaccc cacaactaaa 240
gcgagcccca gcctccagag cccccgaagg agatgccgcc cacaagccca gcccccatcc 300
aggaggcccc agagctcagg gcgccggggc ggattttgta cagccccgag tcactgtgga 360
gagaagctgc aatggcagac gcggctgcag cagcagatgg agccgcgatc atagcaggtg 420
ctgccacagt gagaaaaact gtgtcaaaaa ccgtctcctg gcccctgctg gaggccgcgc 480
cagagagggg agcagccgcc ccgaacctag gtcctgctca gctcacacga cccccagcac 540
ccagagcaca acggagtccc cattgaatgg tgaggacggg gaccagggct ccagggggtc 600
atggaagggg ctggacccca tcctactgct atggtcccag tgctcctggc cagaactgac 660
cctaccaccg acaagagtcc ctcagggaaa cgggggtcac tggcacctcc cagcatcaac 720
cccaggcagc acaggcataa accccacatc cagagccgac tccaggagca gagacacccc 780
agtaccctgg gggacaccga ccctgatgac tccccactgg aatccacccc agagtccacc 840
aggaccaaag accccgcccc tgtctctgtc cctcactcag gacctgctgc ggggcgggcc 900
atgagaccag actcgggctt agggaacacc actgtggccc caacctcgac caggccacag 960
gcccttcctt cctgccctgc ggcagcacag actttggggt ctgtgcagag aggaatcaca 1020
gaggccccag gctgaggtgg tgggggtgga agacccccag gaggtggccc acttcccttc 1080
ctcccagctg gaacccacca tgaccttctt aagatagggg tgtcatccga ggcaggtcct 1140
ccatggagct cccttcaggc tcctccccgg tcctcactag gcctcagtcc cggctgcggg 1200
aatgcagcca ccacaggcac accaggcagc ccagacccag ccagcctgca gtgcccaagc 1260
ccacattctg gagcagagca ggctgtgtct gggagagtct gggctcccca ccgccccccc 1320
gcacacccca cccacccctg tccaggccct atgcaggagg gtcagagccc cccatggggt 1380
atggacttag ggtctcactc acgtggctcc cctcctgggt gaaggggtct catgcccaga 1440
tccccacagc agagctggtc aaaggtggag gcagtggccc cagggccacc ctgacctgga 1500
ccctcaggct cctctagccc tggctgccct gctgtccctg ggaggcctgg actccaccag 1560
accacaggtc cagggcaccg cccataggtg ctgcccacac tcagttcaca ggaagaagat 1620
aagctccaga cccccaagac tgggacctgc cttcctgcca ccgcttgtag ctccagacct 1680
ccgtgcctcc cccgaccact tacacacggg ccagggagct gttccacaaa gatcaacccc 1740
aaaccgggac cgcctggcac tcgggccgct gccacttccc tctccatttg ttcccagcac 1800
ctctgtgctc cctccctcct ccctccttca ggggaacagc ctgtgcagcc cctccctgca 1860
ccccacaccc tggggaggcc caaccctgcc tccagccctt tctcccccgc tgctcttcct 1920
gcccatccag acaaccctgg ggtcccatcc ctgcagccta caccctggtc tccacccaga 1980
cccctgtctc tccctccaga cacccctccc aggccaaccc tgcacatgca ggccctcccc 2040
ttttctgctg ccagagcctc agtttctacc ctctgtgcct accccctgcc tcctcctgcc 2100
cacaactcga gctcttcctc tcctggggcc cctgagccat ggcactgacc gtgcactccc 2160
acccccacac tgcccatgcc ctcaccttcc tcctggacac tctgaccccg ctcccctctt 2220
ggacccagcc ctggtatttc caggacaaag gctcacccaa gtcttcccca tgcaggccct 2280
tgccctcact gcccggttac acggcagcct cctgtgcaca gaagcaggga gctcagccct 2340
tccacaggca gaaggcactg aaagaaatcg gcctccagca ccctgatgca cgtccgcctg 2400
tgtctctcac tgcccgcacc tgcagggagg ctcggcactc cctgtaaaga cgagggatcc 2460
aggcagcaac atcatgggag aatgcagggc tcccagacag cccagccctc tcgcaggcct 2520
ctcctgggaa gagacctgca gccaccactg aacagccacg gagcccgctg gatagtaact 2580
gagtcagtga ccgacctgga gggcagggga gcagtgaacc ggagcccaga ccatagggac 2640
agagaccagc cgctgacatc ccgagcccct cactggcggc cccagaacac cgcgtggaaa 2700
cagaacagac ccacattccc acctggaaca gggcagacac tgctgagccc ccagcaccag 2760
ccctgagaaa caccaggcaa cggcatcaga gggggctcct gagaaagaaa ggaggggagg 2820
tctccttcac cagcaagtac ttcccttgac caaaaacagg gtccacgcaa ctcccccagg 2880
acaaaggagg agccccctgt acagcactgg gctcagagtc ctctcccaca caccctgagt 2940
ttcagacaaa aaccccctgg aaatcatagt atcagcagga gaactagcca gagacagcaa 3000
gaggggactc agtgactccc gcggggacag gaggattttg tgggggctcg tgtcactgtg 3060
aggatattgt agcagcagat ggtgctatac ccacagtgac acagccccat tcaaaaaccc 3120
ctgctgtaaa cgcttccact tctggagctg aggggctggg gggagcgtct gggaagtagg 3180
gcctaggggt ggccatcaat gcccaaaacg caccagactc ccccccagac atcaccccac 3240
tggccagtga gcagagtaaa cagaaaatga gaagcagctg ggaagcttgc acaggcccca 3300
aggaaagagc tttggcgggt gtgcaagagg ggatgcgggc agagcctgag cagggccttt 3360
tgctgtttct gctttcctgt gcagatagtt ccataaactg gtgttcaaga tcgatggctg 3420
ggagtgagcc caggaggaca gtgtgggaag ggcacaggga aggagaagca gccgctatcc 3480
tacactgtca tctttcaaga gtttgccctg tgcccacaat gctgcatcat gggatgctta 3540
acagctgatg tagacacagc taaagagaga atcagtgaaa tggatttgca gcacagatct 3600
gaataaattc tccagaatgt ggagccacac agaagcaagc acaaggaaag tgcctgatgc 3660
aagggcaaag tacagtgtgt accttcaggc tgggcacaga cactctgaaa agccttggca 3720
ggaactccct gcaacaaagc agagccctgc aggcaatgcc agctccagag ccctccctga 3780
gagcctcatg ggcaaagatg tgcacaacag gtgtttctca tagccccaaa ctgagaatga 3840
agcaaacagc catctgaagg aaaacaggca aataaacgat ggcaggttca tgaaatgcaa 3900
acccagacag ccagaaggac aacagtgagg gttacaggtg actctgtggt tgagttcatg 3960
acaatgctga gtaattggag taacaaagga aagtccaaaa aatactttca atgtgatttc 4020
ttctaaataa aatttacagc cggcaaaatg aactatcttc ttaagggata aactttccac 4080
taggaaaact ataaggaaaa tcaagaaaag gatgatcaca taaacacagt ggtcgttact 4140
tctactgggg aaggaagagg gtatgaactg agacacacag ggttggcaag tctcctaaca 4200
agaacagaac aaatacatta cagtaccttg aaaacagcag ttaaaattct aaattgcaag 4260
aagaggaaaa tgcacacagc tgtgtttaga aaattctcag tccagcactg ttcataatag 4320
caaagacatt aacccaggtt ggataaataa acgatgacac aggcaattgc acaatgatac 4380
agacatacat tcagtatatg agacattgat gatgtatccc caaagaaatg actttaaaga 4440
gaaaaggcct gatatgtggt ggcactcacc tccctgggca tccccggaca ggctgcaggc 4500
acactgtgtg gcagggcagg ctggtacctg ctggcagctc ctggggcctg atgtggagca 4560
ggcacagagc cgtatccccc cgaggacata tacccccaag gacggcacag ttggtacatt 4620
ccggagacaa gcaactcagc cacactccca ggccagagcc cgagagggac gcccatgcac 4680
agggaggcag agcccagctc ctccacagcc agcagcaccc gtgcaggggc cgccatctgg 4740
caggcacaga gcatgggctg ggaggagggg cagggacacc aggcagggtt ggcaccaact 4800
gaaaattaca gaagtctcat acatctacct cagccttgcc tgacctgggc ctcacctgac 4860
ctggacctca cctggcctgg acctcacctg gcctagacct cacctctggg cttcacctga 4920
gctcggcctc acctgacttg gaccttgcct gtcctgagct cacatgatct gggcctcacc 4980
tgacctgggt ttcacctgac ctgggcttca cctgacctgg gcctcatctg acctgggcct 5040
cactggcctg gacctcacct ggcctgggct tcacctggcc tcaggcctca tctgcacctg 5100
ctccaggtct tgctggaacc tcagtagcac tgaggctgca ggggctcatc cagggttgca 5160
gaatgactct agaacctccc acatctcagc tttctgggtg gaggcacctg gtggcccagg 5220
gaatataaaa agcctgaatg atgcctgcgt gatttggggg caatttataa acccaaaagg 5280
acatggccat gcagcgggta gggacaatac agacagatat cagcctgaaa tggagcctca 5340
gggcacaggt gggcacggac actgtccacc taagccaggg gcagacccga gtgtccccgc 5400
agtagacctg agagcgctgg gcccacagcc tcccctcggt gccctgctac ctcctcaggt 5460
cagccctgga catcccgggt ttccccaggc ctggcggtag gattttgttg aggtctgtgt 5520
cactgtggta ttacgattgc aactgcagca gatggcgcga ccatagcagg tgctgctatt 5580
atacccacag tgtcacagag tccatcaaaa acccatccct gggaaccttc tgccacagcc 5640
ctccctgtgg ggcaccgccg cgtgccatgt taggattttg actgaggaca cagcaccatg 5700
ggtatggtgg ctaccgcagc agtgcagccc gtgacccaaa cacacagggc agcaggcaca 5760
acagacaagc ccacaagtga ccaccctgag ctcctgcctg ccagccctgg agaccatgaa 5820
acagatggcc aggattatcc cataggtcag ccagacctca gtccaacagg tctgcatcgc 5880
tgctgccctc caataccagt ccggatgggg acagggctgg cccacattac catttgctgc 5940
catccggcca acagtcccag aagcccctcc ctcaaggctg ggccacatgt gtggaccctg 6000
agagcccccc atgtctgagt aggggcacca ggaaggtggg gctggccctg tgcactgtcc 6060
ctgcccctgt ggtccctggc ctgcctggcc ctgacacctg ggcctctcct gggtcatttc 6120
caagacagaa gacattccca ggacagctgg agctgggagt ccatcatcct gcctggccgt 6180
cctgagtcct gcgcctttcc aaacctcacc cgggaagcca acagaggaat cacctcccac 6240
aggcagagac aaagaccttc cagaaatctc tgtctctctc cccagtgggc accctcttcc 6300
agggcagtcc tcagtgatat cacagtggga acccacatct ggatcgggac tgcccccaga 6360
acacaagatg gcccacaggg acagccccac agcccagccc ttcccagacc cctaaaaggc 6420
gtcccacccc ctgcatctgc cccagggctc aaactccagg aggactgact cctgcacacc 6480
ctcctgccag acatcacctc agcccctcct ggaagggaca ggagcgcgca agggtgagtc 6540
agaccctcct gccctcgatg gcaggcggag aagattcaga aaggtctgag atccccagga 6600
cgcagcacca ctgtcaatgg gggccccaga cgcctggacc agggcctgcg tgggaaaggc 6660
ctctgggcac actcaggggg attttgtgaa gggtcctccc actgtggaga ggcttagctg 6720
tggcttccct aagagctgcc gcagcaggca aagcaagcct cacagatgct gccacagtga 6780
tgaacccagc atcaaaaacc gaccggactc ccaaggttta tgcacacttc tccgctcaga 6840
gctctccagg atcagaagag ccgggcccaa gggtttctgc ccagaccctc ggcctctagg 6900
gacatcttgg ccatgacagc ccatgggctg gtgccccaca catcgtctgc cttcaaacaa 6960
gggcttcaga gggctctgag gtgacctcac tgatgaccac aggtgccctg gccccttccc 7020
caccagctgc accagacccc gtcatgacag atgccccgat tccaacagcc aattcctggg 7080
gccaggaatc gctgtagaca ccagcctcct tccaacacct cctgccaatt gcctggattc 7140
ccatcccggt tggaatcaag aggacagcat cccccaggct cccaacaggc aggactccca 7200
caccctcctc tgagaggccg ctgtgttccg tagggccagg ctgcagacag tccccctcac 7260
ctgccactag acaaatgcct gctgtagatg tccccacctg gaaaatacca ctcatggagc 7320
ccccagcccc aggtacagct gtagagagag tctctgaggc ccctaagaag tagccatgcc 7380
cagttctgcc gggaccctcg gccaggctga caggagtgga cgctggagct gggcccatac 7440
tgggccacat aggagctcac cagtgagggc aggagagcac atgccgggga gcacccagcc 7500
tcctgctgac cagaggcccg tcccagagcc caggaggctg cagaggcctc tccaggggga 7560
cactgtgcat gtctggtccc tgagcagccc cccacgtccc cagtcctggg ggcccctggc 7620
acagctgtct ggaccctctc tattccctgg gaagctcctc ctgacagccc cgcctccagt 7680
tccaggtgtg gattttgtca gggggtgtca cactgtgtac agccagaagt ggatgtggac 7740
ttgcgatagt gagaggaagt gcagtgaggg tatggtatgc cggctgtggt gtaagaagaa 7800
gctctggcac agtggtgctg cccatatcaa aaaccaggcc aagtagacag gcccctgctg 7860
tgcagcccca ggcctccagc tcacctgctt ctcctggggc tctcaaggct gctgttttct 7920
gcactctccc ctctgtgggg agggttccct cagtgggaga tctgttctca acatcccacg 7980
gcctcattcc tgcaaggaag gccaatggat gggcaacctc acatgccgcg gctaagatag 8040
ggtgggcagc ctggcgggga caggacatcc tgctggggta tctgtcactg tgcctagtgg 8100
ggcactggct cccaaacaac gcagtccttg ccaaaatccc cacggcctcc cccgctaggg 8160
gctggcctga tctcctgcag tcctaggagg ctgctgacct ccagaatggc tccgtcccca 8220
gttccagggc gagagcagat cccaggccgg ctgcagactg ggaggccacc ccctccttcc 8280
cagggttcac tgcaggtgac cagggcagga aatggcctga acacagggat aaccgggcca 8340
tcccccaaca gagtccaccc cctcctgctc tgtaccccgc accccccagg ccagcccatg 8400
acatccgaca accccacacc agagtcactg cccggtgctg ccctagggag gacccctcag 8460
cccccaccct gtctagagga ctggggagga caggacacgc cctctcctta tggttccccc 8520
acctggctct ggctgggacc cttggggtgt ggacagaaag gacgcttgcc tgattggccc 8580
ccaggagccc agaacttctc tccagggacc ccagcccgag caccccctta cccaggaccc 8640
agccctgccc ctcctcccct ctgctctcct ctcatcaccc catgggaatc cagaatcccc 8700
aggaagccat caggaagggc tgagggagga agtggggcca ctgcaccacc aggcaggagg 8760
ctctgtcttt gtgaacccag ggaggtgcca gcctcctaga gggtatggtc caccctgcct 8820
atggctccca cagtggcagg ctgcagggaa ggaccaggga cggtgtgggg gagggctcag 8880
ggccccgcgg gtgctccatc ttggatgagc ctatctctct cacccacgga ctcgcccacc 8940
tcctcttcac cctggccaca cgtcgtccac accatcctaa gtcccaccta caccagagcc 9000
ggcacagcca gtgcagacag aggctggggt gcaggggggc cgactgggca gcttcgggga 9060
gggaggaatg gaggaagggg agttcagtga agaggccccc ctcccctggg tccaggatcc 9120
tcctctggga cccccggatc ccatcccctc caggctctgg gaggagaagc aggatgggag 9180
aatctgtgcg ggaccctctc acagtggaat acctccacag cggctcaggc cagatacaaa 9240
agcccctcag tgagccctcc actgcagtgc tgggcctggg ggcagccgct cccacacagg 9300
atgaacccag caccccgagg atgtcctgcc agggggagct cagagccatg aaggagcagg 9360
atatgggacc cccgatacag gcacagacct cagctccatt caggactgcc acgtcctgcc 9420
ctgggaggaa cccctttctc tagtccctgc aggccaggag gcagctgact cctgacttgg 9480
acgcctattc cagacaccag acagaggggc aggcccccca gaaccaggga tgaggacgcc 9540
ccgtcaaggc cagaaaagac caagttgcgc tgagcccagc aagggaaggt ccccaaacaa 9600
accaggagga ttttgtaggt gtctgtgtca ctgtgagctg caactgcagc agcaaatgga 9660
gccgcgacca tagcaggtgc tgccacagtg acactcgcca ggtcaaaaac cccatcccaa 9720
gtcagcggaa tgcagagaga gcagggagga catgtttagg atctgaggcc gcacctgaca 9780
cccaggccag cagacgtctc ctgtccacgg caccctgcca tgtcctgcat ttctggaaga 9840
acaagggcag gctgaagggg gtccaggacc aggagatggg tccgctctac ccagagaagg 9900
agccaggcag gacacaagcc cccacgcgtg ggctcgtagt ttgacgtgcg tgaagtgtgg 9960
gtaagaaagt acgta 9975
<210> 233
<211> 9714
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TxDH17613
<400> 233
gcggccgctg ccatttcatt acctctttct ccgcacccga catagattac gtaacgcgtg 60
ggctcgtagt ttgacgtgcg tgaagtgtgg gtaagaaagt ccccattgag gctgacctgc 120
ccagagggtc ctgggcccac ccaacacacc ggggcggaat gtgtgcaggc ctcggtctct 180
gtgggtgttc cgctagctgg ggctcacagt gctcacccca cacctaaaac gagccacagc 240
ctccggagcc cctgaaggag accccgccca caagcccagc ccccacccag gaggccccag 300
agcacagggc gccccgtcgg attttgtaca gccccgagtc actgtggaga gatgcaatgg 360
cagacgcggc tgcagcagat ggcgcgatca tagcaggtgc tgccacagtg agaaaagctt 420
cgtcaaaaac cgtctcctgg ccacagtcgg aggccccgcc agagagggga gcagccaccc 480
caaacccatg ttctgccggc tcccatgacc ccgtgcacct ggagccccac ggtgtcccca 540
ctggatggga ggacaagggc cgggggctcc ggcgggtcgg ggcaggggct tgatggcttc 600
cttctgccgt ggccccattg cccctggctg gagttgaccc ttctgacaag tgtcctcaga 660
gagtcaggga tcagtggcac ctcccaacat caaccccacg cagcccaggc acaaacccca 720
catccagggc caactccagg aacagagaca ccccaatacc ctgggggacc ccgaccctga 780
tgactcccgt cccatctctg tccctcactt ggggcctgct gcggggcgag cacttgggag 840
caaactcagg cttaggggac accactgtgg gcctgacctc gagcaggcca cagacccttc 900
cctcctgccc tggtgcagca cagactttgg ggtctgggca gggaggaact tctggcaggt 960
caccaagcac agagccccca ggctgaggtg gccccagggg gaaccccagc aggtggccca 1020
ctacccttcc tcccagctgg accccatgtc ttccccaaga taggggtgcc atccaaggca 1080
ggtcctccat ggagccccct tcaggctcct ctccagaccc cactgggcct cagtccccac 1140
tctaggaatg cagccaccac gggcacacca ggcagcccag gcccagccac cctgcagtgc 1200
ccaagcccac accctggagg agagcagggt gcgtctggga ggggctgggc tccccacccc 1260
cacccccacc tgcacacccc acccaccctt gcccgggccc cctgcaggag ggtcagagcc 1320
cccatgggat atggacttag ggtctcactc acgcacctcc cctcctggga gaaggggtct 1380
catgcccaga tccccccagc agcgctggtc acaggtagag gcagtggccc cagggccacc 1440
ctgacctggc ccctcaggct cctctagccc tggctgccct gctgtccctg ggaggcctgg 1500
gctccaccag accacaggtc tagggcaccg cccacactgg ggccgcccac acacagctca 1560
caggaagaag ataagctcca gacccccagg cccgggacct gccttgctgc tacgacttcc 1620
tgccccagac ctcgttgccc tcccccgtcc acttacacac aggccaggaa gctgttccca 1680
cacagaccaa ccccagacgg ggaccacctg gcactcaggt cactgccatt tccttctcca 1740
ttcacttcca atgcctctgt gcttcctccc tcctccttcc ttcgggggag caccctgtgc 1800
agctcctccc tgcagtccac accctgggga gacccgaccc tgcagcccac accctgggga 1860
gacctgaccc tcctccagcc ctttctcccc cgctgctctt gccacccacc aagacagccc 1920
tggggtcctg tccctacagc ccccacccag ttctctacct agacccgtct tcctccctct 1980
aaacacctct cccaggccaa ccctacacct gcaggccctc ccctccactg ccaaagaccc 2040
tcagtttctc ctgcctgtgc ccacccccgt gctcctcctg cccacagctc gagctcttcc 2100
tctcctaggg cccctgaggg atggcattga ccgtgccctc gcacccacac actgcccatg 2160
ccctcacatt cctcctggcc actccagccc cactcccctc tcaggcctgg ctctggtatt 2220
tctgggacaa agccttaccc aagtctttcc catgcaggcc tgggccctta ccctcactgc 2280
ccggttacag ggcagcctcc tgtgcacaga agcagggagc tcagcccttc cacaggcaga 2340
aggcactgaa agaaatcggc ctccagcgcc ttgacacacg tctgcctgtg tctctcactg 2400
cccgcacctg cagggaggct cggcactccc tctaaagacg agggatccag gcagcagcat 2460
cacaggagaa tgcagggcta ccagacatcc cagtcctctc acaggcctct cctgggaaga 2520
gacctgaaga cgcccagtca acggagtcta acaccaaacc tccctggagg ccgatgggta 2580
gtaacggagt cattgccaga cctggaggca ggggagcagt gagcccgagc ccacaccata 2640
gggccagagg acagccactg acatcccaag ccactcactg gtggtcccac aacaccccat 2700
ggaaagagga cagacccaca gtcccacctg gaccagggca gagactgctg agacccagca 2760
ccagaaccaa ccaagaaaca ccaggcaaca gcatcagagg gggctctggc agaacagagg 2820
aggggaggtc tccttcacca gcaggcgctt cccttgaccg aagacaggat ccatgcaact 2880
cccccaggac aaaggaggag ccccttgttc agcactgggc tcagagtcct ctccaagaca 2940
cccagagttt cagacaaaaa ccccctggaa tgcacagtct cagcaggaga gccagccaga 3000
gccagcaaga tggggctcag tgacacccgc agggacagga ggattttgtg ggggctcgtg 3060
tcactgtgag gatattgtac taatcggagc aggcagggtg tatgctatac ccacagtgac 3120
acagccccat tcaaaaaccc ctactgcaaa cgcattccac ttctggggct gaggggctgg 3180
gggagcgtct gggaaatagg gctcaggggt gtccatcaat gcccaaaacg caccagactc 3240
ccctccatac atcacaccca ccagccagcg agcagagtaa acagaaaatg agaagcaagc 3300
tggggaagct tgcacaggcc ccaaggaaag agctttggcg ggtgtgtaag aggggatgcg 3360
ggcagagcct gagcagggcc ttttgctgtt tctgctttcc tgtgcagaga gttccataaa 3420
ctggtgttcg agatcaatgg ctgggagtga gcccaggagg acagcgtggg aagagcacag 3480
ggaaggagga gcagccgcta tcctacactg tcatctttcg aaagtttgcc ttgtgcccac 3540
actgctgcat catgggatgc ttaacagctg atgtagacac agctaaagag agaatcagtg 3600
agatggattt gcagcacaga tctgaataaa ttctccagaa tgtggagcag cacagaagca 3660
agcacacaga aagtgcctga tgcaaggaca aagttcagtg ggcaccttca ggcattgctg 3720
ctgggcacag acactctgaa aagccctggc aggaactccc tgtgacaaag cagaaccctc 3780
aggcaatgcc agccccagag ccctccctga gagcctcatg ggcaaagatg tgcacaacag 3840
gtgtttctca tagccccaaa ctgagagcaa agcaaacgtc catctgaagg agaacaggca 3900
aataaacgat ggcaggttca tgaaatgcaa acccagacag ccacaagcac aaaagtacag 3960
ggttataagc gactctggtt gagttcatga caatgctgag taattggagt aacaaagtaa 4020
actccaaaaa atactttcaa tgtgatttct tctaaataaa atttacaccc tgcaaaatga 4080
actgtcttct taagggatac atttcccagt tagaaaacca taaagaaaac caagaaaagg 4140
atgatcacat aaacacagtg gtggttactt ctgctgggga aggaagaggg tatgaactga 4200
gatacacagg gtgggcaagt ctcctaacaa gaacagaacg aatacattac agtaccttga 4260
aaacagcagt taaacttcta aattgcaaga agaggaaaat gcacacagtt gtgtttagaa 4320
aattctcagt ccagcactgt tcataatagc aaagacatta acccaggtcg gataaataag 4380
cgatgacaca ggcaattgca caatgataca gacatatatt tagtatatga gacatcgatg 4440
atgtatcccc aaataaacga ctttaaagag ataaagggct gatgtgtggt ggcattcacc 4500
tccctgggat ccccggacag gttgcaggct cactgtgcag cagggcaggc gggtacctgc 4560
tggcagttcc tggggcctga tgtggagcaa gcgcagggcc atatatcccg gaggacggca 4620
cagtcagtga attccagaga gaagcaactc agccacactc cccaggcaga gcccgagagg 4680
gacgcccacg cacagggagg cagagcccag cacctccgca gccagcacca cctgcgcacg 4740
ggccaccacc ttgcaggcac agagtgggtg ctgagaggag gggcagggac accaggcagg 4800
gtgagcaccc agagaaaact gcagacgcct cacacatcca cctcagcctc ccctgacctg 4860
gacctcactg gcctgggcct cacttaacct gggcttcacc tgaccttggc ctcacctgac 4920
ttggacctcg cctgtcccaa gctttacctg acctgggcct caactcacct gaacgtctcc 4980
tgacctgggt ttaacctgtc ctggaactca cctggccttg gcttcccctg acctggacct 5040
catctggcct gggcttcacc tggcctgggc ctcacctgac ctggacctca tctggcctgg 5100
acctcacctg gcctggactt cacctggcct gggcttcacc tgacctggac ctcacctggc 5160
ctcgggcctc acctgcacct gctccaggtc ttgctggagc ctgagtagca ctgagggtgc 5220
agaagctcat ccagggttgg ggaatgactc tagaagtctc ccacatctga cctttctggg 5280
tggaggcagc tggtggccct gggaatataa aaatctccag aatgatgact ctgtgatttg 5340
tgggcaactt atgaacccga aaggacatgg ccatggggtg ggtagggaca tagggacaga 5400
tgccagcctg aggtggagcc tcaggacaca ggtgggcacg gacactatcc acataagcga 5460
gggatagacc cgagtgtccc cacagcagac ctgagagcgc tgggcccaca gcctcccctc 5520
agagccctgc tgcctcctcc ggtcagccct ggacatccca ggtttcccca ggcctggcgg 5580
taggattttg ttgaggtctg tgtcactgtg gtattacgat tgcaactgca gcagatgggc 5640
tcgcgaccat agcaggtgct gctattataa ccacagtgtc acagagtcca tcaaaaaccc 5700
atgcctggaa gcttcccgcc acagccctcc ccatggggcc ctgctgcctc ctcaggtcag 5760
ccccggacat cccgggtttc cccaggctgg gcggtaggat tttgttgagg tctgtgtcac 5820
tgtggtatta ctatgagaga tgcaatggca gacgcggctg cagcagatgg cgcgatcata 5880
gcaggtgctg ctattataac cacagtgtca cagagtccat caaaaaccca tccctgggag 5940
cctcccgcca cagccctccc tgcaggggac cggtacgtgc catgttagga ttttgatcga 6000
ggagacagca ccatgggtat ggtggctacc acagcagtgc agcctgtgac ccaaacccgc 6060
agggcagcag gcacgatgga caggcccgtg actgaccacg ctgggctcca gcctgccagc 6120
cctggagatc atgaaacaga tggccaaggt caccctacag gtcatccaga tctggctccg 6180
aggggtctgc atcgctgctg ccctcccaac gccagtccaa atgggacagg gacggcctca 6240
cagcaccatc tgctgccatc aggccagcga tcccagaagc ccctccctca aggctgggca 6300
catgtgtgga cactgagagc cctcatatct gagtaggggc accaggaggg aggggctggc 6360
cctgtgcact gtccctgccc ctgtggtccc tggcctgcct ggccctgaca cctgagcctc 6420
tcctgggtca tttccaagac agaagacatt cctggggaca gccggagctg ggcgtcgctc 6480
atcctgcccg gccgtcctga gtcctgctca tttccagacc tcaccgggga agccaacaga 6540
ggactcgcct cccacattca gagacaaaga accttccaga aatccctgcc tctctcccca 6600
gtggacaccc tcttccagga cagtcctcag tggcatcaca gcggcctgag atccccagga 6660
cgcagcaccg ctgtcaatag gggccccaaa tgcctggacc agggcctgcg tgggaaaggc 6720
ctctggccac actcggggat tttgtgaagg gccctcccac tgtggagagg ctttgtggct 6780
tccctaagag ctgcagcagg caaaagcctc acagatgctg ccacagtgat gaacccagtg 6840
tcaaaaaccg gctggaaacc caggggctgt gtgcacgcct cagcttggag ctctccagga 6900
gcacaagagc cgggcccaag gatttgtgcc cagaccctca gcctctaggg acacctgggt 6960
catctcagcc tgggctggtg ccctgcacac catcttcctc caaatagggg cttcagaggg 7020
ctctgaggtg acctcactca tgaccacagg tgacctggcc cttccctgcc agctatacca 7080
gaccctgtct tgacagatgc cccgattcca acagccaatt cctgggaccc tgaatagctg 7140
tagacaccag cctcattcca gtacctcctg ccaattgcct ggattcccat cctggctgga 7200
atcaagaagg cagcatccgc caggctccca acaggcagga ctcccgcaca ccctcctctg 7260
agaggccgct gtgttccgca gggccaggcc ctggacagtt cccctcacct gccactagag 7320
aaacacctgc cattgtcgtc cccacctgga aaagaccact cgtggagccc ccagccccag 7380
gtacagctgt agagacagtc ctcgaggccc ctaagaagga gccatgccca gttctgccgg 7440
gaccctcggc caggccgaca ggagtggacg ctggagctgg gcccacactg ggccacatag 7500
gagctcacca gtgagggcag gagagcacat gccggggagc acccagcctc ctgctgacca 7560
gaggcccgtc ccagagccca ggaggctgca gaggcctctc cagggagaca ctgtgcatgt 7620
ctggtaccta agcagccccc cacgtcccca gtcctggggg cccctggctc agctgtctgg 7680
gccctccctg ctccctggga agctcctcct gacagccccg cctccagttc caggtgtgga 7740
ttttgtcagg cgatgtcaca ctgtgtactg ccagaagtgg atgtggacta gcgatagtga 7800
gaggaagtgc tgtgagggta tggtaagccg gctgtggtgt aagaagaagc tctggcacag 7860
tggtgccgcc catatcaaaa accaggccaa gtagacaggc ccctgctgcg cagccccagg 7920
catccacttc acctgcttct cctggggctc tcaaggctgc tgtctgtcct ctggccctct 7980
gtggggaggg ttccctcagt gggaggtctg tgctccaggg cagggatgat tgagatagaa 8040
atcaaaggct ggcagggaaa ggcagcttcc cgccctgaga ggtgcaggca gcaccacgga 8100
gccacggagt cacagagcca cggagccccc attgtgggca tttgagagtg ctgtgccccc 8160
ggcaggccca gccctgatgg ggaagcctgt cccatcccac agcccgggtc ccacgggcag 8220
cgggcacaga agctgccagg ttgtcctcta tgatcctcat ccctccagca gcatcccctc 8280
cacagtgggg aaactgaggc ttggagcacc acccggcccc ctggaaatga ggctgtgagc 8340
ccagacagtg ggcccagagc actgtgagta ccccggcagt acctggctgc agggatcagc 8400
cagagatgcc aaaccctgag tgaccagcct acaggaggat ccggccccac ccaggccact 8460
cgattaatgc tcaaccccct gccctggaga cctcttccag taccaccagc agctcagctt 8520
ctcagggcct catccctgca aggaaggtca agggctgggc ctgccagaaa cacagcaccc 8580
tccctagccc tggctaagac agggtgggca gacggctgtg gacgggacat attgctgggg 8640
catttctcac tgtcacttct gggtggtagc tctgacaaaa acgcagaccc tgccaaaatc 8700
cccactgcct cccgctaggg gctggcctgg aatcctgctg tcctaggagg ctgctgacct 8760
ccaggatggc tccgtcccca gttccagggc gagagcagat cccaggcagg ctgtaggctg 8820
ggaggccacc cctgcccttg ccggggttga atgcaggtgc ccaaggcagg aaatggcatg 8880
agcacaggga tgaccgggac atgccccacc agagtgcgcc ccttcctgct ctgcaccctg 8940
caccccccag gccagcccac gacgtccaac aactgggcct gggtggcagc cccacccaga 9000
caggacagac ccagcaccct gaggaggtcc tgccaggggg agctaagagc catgaaggag 9060
caagatatgg ggcccccgat acaggcacag atgtcagctc catccaggac cacccagccc 9120
acaccctgag aggaacgtct gtctccagcc tctgcaggtc gggaggcagc tgacccctga 9180
cttggacccc tattccagac accagacaga ggcgcaggcc ccccagaacc agggttgagg 9240
gacgccccgt caaagccaga caaaaccaag gggtgttgag cccagcaagg gaaggccccc 9300
aaacagacca ggaggatttt gtaggtgtct gtgtcactgt gtgcaactgc agcagatggc 9360
gcgaccatag caggtgctgc cacagtgaca ctcacccagt caaaaacccc attccaagtc 9420
agcggaagca gagagagcag ggaggacacg tttaggatct gagactgcac ctgacaccca 9480
ggccagcaga cgtctcccct ccagggcacc ccaccctgtc ctgcatttct gcaagatcag 9540
gggcggcctg agggggggtc tagggtgagg agatgggtcc cctgtacacc aaggaggagt 9600
taggcaggtc ccgagcactc ttaattaaac gacgcctcga atggaactac tacaacgaat 9660
ggttgctcta cgtaatgcat tcgctacctt aggaccgtta tagttaggcg cgcc 9714
<210> 234
<211> 9797
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TxDH114619
<400> 234
tacgtattaa ttaaacgacg cctcgaatgg aactactaca acgaatggtt gctctcccca 60
ttgaggctga cctgcccaga gagtcctggg cccaccccac acaccggggc ggaatgtgtg 120
caggcctcgg tctctgtggg tgttccgcta gctggggctc acagtgctca ccccacacct 180
aaaatgagcc acagcctccg gagcccccgc aggagacccc gcccacaagc ccagccccca 240
cccaggaggc cccagagctc agggcgcccc gtcggatttt gtacagcccc gagtcactgt 300
ggagagatgc tgcaatggca gacgcggctg cagcagcaga tggtgccgcg atcatagcag 360
gtgctgccac agtgagaata gctacgtcaa aaaccgtcca gtggccactg ccggaggccc 420
cgccagagag ggcagcagcc actctgatcc catgtcctgc cggctcccat gacccccagc 480
acgcggagcc ccacagtgtc cccactggat gggaggacaa gagctgggga ttccggcggg 540
tcggggcagg ggcttgatcg catccttctg ccgtggctcc agtgcccctg gctggagttg 600
acccttctga caagtgtcct cagagagaca ggcatcaccg gcgcctccca acatcaaccc 660
caggcagcac aggcacaaac cccacatcca gagccaactc caggagcaga gacaccccaa 720
taccctgggg gaccccgacc ctgatgactt cccactggaa ttcgccgtag agtccaccag 780
gaccaaagac cctgcctctg cctctgtccc tcactcagga cctgctgccg ggcgaggcct 840
tgggagcaga cttgggctta ggggacacca gtgtgacccc gaccttgacc aggacgcaga 900
cctttccttc ctttcctggg gcagcacaga ctttggggtc tgggccagga ggaacttctg 960
gcaggtcgcc aagcacagag gccacaggct gaggtggccc tggaaagacc tccaggaggt 1020
ggccactccc cttcctccca gctggacccc atgtcctccc caagataagg gtgccatcca 1080
aggcaggtgc tccttggagc cccattcaga ctcctccctg gaccccactg ggcctcagtc 1140
ccagctctgg ggatgaagcc accacaagca caccaggcag cccaggccca gccaccctgc 1200
agtgcccaag cacacactct ggagcagagc agggtgcctc tgggaggggc tgagctcccc 1260
accccacccc cacctgcaca ccccacccac ccctgcccag cggctctgca ggagggtcag 1320
agccccacat ggggtatgga cttagggtct cactcacgtg gctcccatca tgagtgaagg 1380
ggcctcaagc ccaggttccc acagcagcgc ctgtcgcaag tggaggcaga ggcccgaggg 1440
ccaccctgac ctggtccctg aggttcctgc agcccaggct gccctgctgt ccctgggagg 1500
cctgggctcc accagaccac aggtccaggg caccgggtgc aggagccacc cacacacagc 1560
tcacaggaag aagataagct ccagaccccc agggccagaa cctgccttcc tgctactgct 1620
tcctgcccca gacctgggcg ccctcccccg tccacttaca cacaggccag gaagctgttc 1680
ccacacagaa caaccccaaa ccaggaccgc ctggcactca ggtggctgcc atttccttct 1740
ccatttgctc ccagcgcctc tgtcctccct ggttcctcct tcgggggaac agcctgtgca 1800
gccagtccct gcagcccaca ccctggggag acccaaccct gcctggggcc cttccaaccc 1860
tgctgctctt actgcccacc cagaaaactc tggggtcctg tccctgcagt ccctaccctg 1920
gtctccaccc agacccctgt gtatcactcc agacacccct cccaggcaaa ccctgcacct 1980
gcaggccctg tcctcttctg tcgctagagc ctcagtttct cccccctgtg cccacaccct 2040
acctcctcct gcccacaact ctaactcttc ttctcctgga gcccctgagc catggcattg 2100
accctgccct cccaccaccc acagcccatg ccctcacctt cctcctggcc actccgaccc 2160
cgccccctct caggccaagc cctggtattt ccaggacaaa ggctcaccca agtctttccc 2220
aggcaggcct gggctcttgc cctcacttcc cggttacacg ggagcctcct gtgcacagaa 2280
gcagggagct cagcccttcc acaggcagaa ggcactgaaa gaaatcggcc tccagcacct 2340
tgacacacgt ccgcccgtgt ctctcactgc ccgcacctgc agggaggctc cgcactccct 2400
ctaaagacaa gggatccagg cagcagcatc acgggagaat gcagggctcc cagacatccc 2460
agtcctctca caggcctctc ctgggaagag acctgcagcc accaccaaac agccacagag 2520
gctgctggat agtaactgag tcaatgaccg acctggaggg caggggagca gtgagccgga 2580
gcccatacca tagggacaga gaccagccgc tgacatcccg agctcctcaa tggtggcccc 2640
ataacacacc taggaaacat aacacaccca cagccccacc tggaacaggg cagagactgc 2700
tgagccccca gcaccagccc caagaaacac caggcaacag tatcagaggg ggctcccgag 2760
aaagagagga ggggagatct ccttcaccat caaatgcttc ccttgaccaa aaacagggtc 2820
cacgcaactc ccccaggaca aaggaggagc cccctataca gcactgggct cagagtcctc 2880
tctgagacac cctgagtttc agacaacaac ccgctggaat gcacagtctc agcaggagaa 2940
cagaccaaag ccagcaaaag ggacctcggt gacaccagta gggacaggag gattttgtgg 3000
gggctcgtgt cactgtgagg atattgtagt ggtagcagca gatggggtag ctgctactcc 3060
cacagtgaca cagacccatt caaaaacccc tactgcaaac acacccactc ctggggctga 3120
ggggctgggg gagcgtctgg gaagtagggt ccaggggtgt ctatcaatgt ccaaaatgca 3180
ccagactccc cgccaaacac caccccacca gccagcgagc agggtaaaca gaaaatgaga 3240
ggctctggga agcttgcaca ggccccaagg aaagagcttt ggcgggtgtg caagagggga 3300
tgcaggcaga gcctgagcag ggccttttgc tgtttctgct ttcctgtgca gagagttcca 3360
taaactggtg ttcaagatca gtggctggga atgagcccag gagggcagtc tgtgggaaga 3420
gcacagggaa ggaggagcag ccgctatcct acactgtcat ctttcaaaag tttgccttgt 3480
gaccacacta ttgcatcatg ggatgcttaa gagctgatgt agacacagct aaagagagaa 3540
tcagtgagat gaatttgcag catagatctg aataaactct ccagaatgtg gagcagtaca 3600
gaagcaaaca cacagaaagt gcctgatgca aggacaaagt tcagtgggca ccttcaggca 3660
ttgctgctgg gcacagacac tctgaaaagc cttggcagga tctccctgcg acaaagcaga 3720
accctcaggc aatgccagcc ccagagccct ccctgagagc gtcatgggga aagatgtgca 3780
gaacagctga ttatcataga ctcaaactga gaacagagca aacgtccatc tgaagaacag 3840
tcaaataagc aatggtaggt tcatgcaatg caaacccaga cagccagggg acaacagtag 3900
agggctacag gcggctttgc ggttgagttc atgacaatgc tgagtaattg gagtaacaga 3960
ggaaagccca aaaaatactt ttaatgtgat ttcttctaaa taaaatttac accaggcaaa 4020
atgaactgtc ttcttaaggg ataaactttc ccctggaaaa actacaagga aaattaagaa 4080
aacgatgatc acataaacac agttgtggtt acttctactg gggaaggaag agggtatgag 4140
ctgagacaca cagagtcggc aagtctccaa gcaagcacag aacgaataca ttacagtacc 4200
ttgaatacag cagttaaact tctaaatcgc aagaacagga aaatgcacac agctgtgttt 4260
agaaaattct cagtccagca ctattcataa tagcaaagac attaacccag gttggataaa 4320
taaatgatga cacaggcaat tgcacaatga tacagacata catttagtac atgagacatc 4380
gatgatgtat ccccaaagaa atgactttaa agagaaaagg cctgatgtgt ggtggcactc 4440
acctccctgg gatccccgga caggttgcag gcacactgtg tggcagggca ggctggtaca 4500
tgctggcagc tcctggggcc tgatgtggag caagcgcagg gctgtatacc cccaaggatg 4560
gcacagtcag tgaattccag agagaagcag ctcagccaca ctgcccaggc agagcccgag 4620
agggacgccc acgtacaggg aggcagagcc cagctcctcc acagccacca ccacctgtgc 4680
acgggccacc accttgcagg cacagagtgg gtgctgagag gaggggcagg gacaccaggc 4740
agggtgagca cccagagaaa actgcagaag cctcacacat ccacctcagc ctcccctgac 4800
ctggacctca cctggtctgg acctcacctg gcctgggcct cacctgacct ggacctcacc 4860
tggcctgggc ttcacctgac ctggacctca cctggcctcc ggcctcacct gcacctgctc 4920
caggtcttgc tggaacctga gtagcactga ggctgcagaa gctcatccag ggttggggaa 4980
tgactctgga actctcccac atctgacctt tctgggtgga ggcatctggt ggccctggga 5040
atataaaaag ccccagaatg gtgcctgcgt gatttggggg caatttatga acccgaaagg 5100
acatggccat ggggtgggta gggacatagg gacagatgcc agcctgaggt ggagcctcag 5160
gacacagttg gacgcggaca ctatccacat aagcgaggga cagacccgag tgttcctgca 5220
gtagacctga gagcgctggg cccacagcct cccctcggtg ccctgctgcc tcctcaggtc 5280
agccctggac atcccgggtt tccccaggcc agatggtagg attttgttga ggtctgtgtc 5340
actgtggtat tatgattacg agagagcttg caatggcaga cgcggctgca gcagatgggc 5400
tcgcgatcat agcaggtgct gctatcgtta tacccacagt gtcacacggt ccatcaaaaa 5460
cccatgccac agccctcccc gcaggggacc gccgcgtgcc atgttacgat tttgatcgag 5520
gacacagcgc catgggtatg gtggctacca cagcagtgca gcccatgacc caaacacaca 5580
gggcagcagg cacaatggac aggcctgtga gtgaccatgc tgggctccag cccgccagcc 5640
ccggagacca tgaaacagat ggccaaggtc accccacagt tcagccagac atggctccgt 5700
ggggtctgca tcgctgctgc cctctaacac cagcccagat ggggacaagg ccaaccccac 5760
attaccatct cctgctgtcc acccagtggt cccagaagcc cctccctcat ggctgagcca 5820
catgtgtgaa ccctgagagc accccatgtc agagtagggg cagcagaagg gcggggctgg 5880
ccctgtgcac tgtccctgca cccatggtcc ctcgcctgcc tggccctgac acctgagcct 5940
cttctgagtc atttctaaga tagaagacat tcccggggac agccggagct gggcgtcgct 6000
catcccgccc ggccgtcctg agtcctgctt gtttccagac ctcaccaggg aagccaacag 6060
aggactcacc tcacacagtc agagacaaag aaccttccag aaatccctgt ctcactcccc 6120
agtgggcacc ttcttccagg acattcctcg gtcgcatcac agcaggcacc cacatctgga 6180
tcaggacggc ccccagaaca caagatggcc catggggaca gccccacaac ccaggccttc 6240
ccagacccct aaaaggcgtc ccaccccctg cacctgcccc agggctaaaa atccaggagg 6300
cttgactccc gcataccctc cagccagaca tcacctcagc cccctcctgg aggggacagg 6360
agcccgggag ggtgagtcag acccacctgc cctcgatggc aggcggggaa gattcagaaa 6420
ggcctgagat ccccaggacg cagcaccact gtcaatgggg gccccagacg cctggaccag 6480
ggcctgcgtg ggaaaggccg ctgggcacac tcagggggat tttgtgaagg cccctcccac 6540
tgtggagagg cttgcttgtg gcttccctaa gagctgcagc aggcaagcta agcctcacag 6600
atgctgccac agtgatgaaa ctagcatcaa aaaccggccg gacacccagg gaccatgcac 6660
acttctcagc ttggagctct ccaggaccag aagagtcagg tctgagggtt tgtagccaga 6720
ccctcggcct ctagggacac cctggccatc acagcggatg ggctggtgcc ccacatgcca 6780
tctgctccaa acaggggctt cagagggctc tgaggtgact tcactcatga ccacaggtgc 6840
cctggcccct tccccgccag ctacaccgaa ccctgtccca acagctgccc cagttccaac 6900
agccaattcc tggggcccag aattgctgta gacaccagcc tcgttccagc acctcctgcc 6960
aattgcctgg attcacatcc tggctggaat caagagggca gcatccgcca ggctcccaac 7020
aggcaggact cccgcacacc ctcctctgag aggccgctgt gttccgcagg gccaggccct 7080
ggacagttcc cctcacctgc cactagagaa acacctgcca ttgtcgtccc cacctggaaa 7140
agaccactcg tggagccccc agccccaggt acagctgtag agagactccc cgagggatct 7200
aagaaggagc catgcgcagt tctgccggga ccctcggcca ggccgacagg agtggacact 7260
ggagctgggc ccacactggg ccacatagga gctcaccagt gagggcagga gagcacatgc 7320
cggggagcac ccagcctcct gctgaccaga ggcccgtccc agagcccagg aggctgcaga 7380
ggcctctcca gggggacact gtgcatgtct ggtccctgag cagcccccca cgtccccagt 7440
cctgggggcc cctggcacag ctgtctggac cctccctgtt ccctgggaag ctcctcctga 7500
cagccccgcc tccagttcca ggtgtggatt ttgtcagggg gtgtcacact gtgtactgcc 7560
agaagtggat gtggacttgc gatagtgaga ggaagagctg tgagggtatg gtatgccggc 7620
tgtggagtaa gaagaagctc tggcacagtg gtgctgccca tatcaaaaac caggccaagt 7680
agacaggccc ctgctgtgca gccccaggcc tccacttcac ctgcttctcc tggggctctc 7740
aaggtcactg ttgtctgtac tctgccctct gtggggaggg ttccctcagt gggaggtctg 7800
ttctcaacat cccagggcct catgtctgca cggaaggcca atggatgggc aacctcacat 7860
gccgcggcta agatagggtg ggcagcctgg cgggggacag tacatactgc tggggtgtct 7920
gtcactgtgc ctagtggggc actggctccc aaacaacgca gtcctcgcca aaatccccac 7980
agcctcccct gctaggggct ggcctgatct cctgcagtcc taggaggctg ctgacctcca 8040
gaatgtctcc gtccccagtt ccagggcgag agcagatccc aggccggctg cagactggga 8100
ggccaccccc tccttcccag ggttcactgg aggtgaccaa ggtaggaaat ggccttaaca 8160
cagggatgac tgcgccatcc cccaacagag tcagccccct cctgctctgt accccgcacc 8220
ccccaggcca gtccacgaaa accagggccc cacatcagag tcactgcctg gcccggccct 8280
ggggcggacc cctcagcccc caccctgtct agaggacttg gggggacagg acacaggccc 8340
tctccttatg gttcccccac ctgcctccgg ccgggaccct tggggtgtgg acagaaagga 8400
cacctgccta attggccccc aggaacccag aacttctctc cagggacccc agcccgagca 8460
cccccttacc caggacccag ccctgcccct cctcccctct gctctcctct catcacccca 8520
tgggaatccg gtatccccag gaagccatca ggaagggctg aaggaggaag cggggccgtg 8580
caccaccggg caggaggctc cgtcttcgtg aacccaggga agtgccagcc tcctagaggg 8640
tatggtccac cctgcctggg gctcccaccg tggcaggctg cggggaagga ccagggacgg 8700
tgtgggggag ggctcagggc cctgcgggtg ctcctccatc ttcggtgagc ctcccccttc 8760
acccaccgtc ccgcccacct cctctccacc ctggctgcac gtcttccaca ccatcctgag 8820
tcctacctac accagagcca gcaaagccag tgcagacaaa ggctggggtg caggggggct 8880
gccagggcag cttcggggag ggaaggatgg agggagggga ggtcagtgaa gaggccccct 8940
tcccctgggt ccaggatcct cctctgggac ccccggatcc catcccctcc tggctctggg 9000
aggagaagca ggatgggaga atctgtgcgg gaccctctca cagtggaata tccccacagc 9060
ggctcaggcc agacccaaaa gcccctcagt gagccctcca ctgcagtcct gggcctgggt 9120
agcagcccct cccacagagg acagacccag caccccgaag aagtcctgcc agggggagct 9180
cagagccatg aaagagcagg atatggggtc cccgatacag gcacagacct cagctccatc 9240
caggcccacc gggacccacc atgggaggaa cacctgtctc cgggttgtga ggtagctggc 9300
ctctgtctcg gaccccactc cagacaccag acagaggggc aggcccccca aaaccagggt 9360
tgagggatga tccgtcaagg cagacaagac caaggggcac tgaccccagc aagggaaggc 9420
tcccaaacag acgaggagga ttttgtagct gtctgtatca ctgtgtgcaa ctgcagcaga 9480
tgggctcgcg accatagcag gtgctgccac agtgacactc gccaggtcaa aaaccccgtc 9540
ccaagtcagc ggaagcagag agagcaggga ggacacgttt aggatctgag gccgcacctg 9600
acacccaggg cagcagacgt ctcccctcca gggcaccctc caccgtcctg cgtttcttca 9660
agaatagggg cggcctgagg gggtccaggg ccaggcgata ggtcccctct accccaagga 9720
ggagccaggc aggacccgag caccgatgca tctaacgcag tcatgtaatg ctgggtgaca 9780
gtcagttcgc ctacgta 9797
<210> 235
<211> 12029
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TxDH120126
<400> 235
tacgtaatgc atctaacgca gtcatgtaat gctgggtgac agtcagttcg cctccccatt 60
gaggctgacc tgcccagacg ggcctgggcc caccccacac accggggcgg aatgtgtgca 120
ggccccagtc tctgtgggtg ttccgctagc tggggccccc agtgctcacc ccacacctaa 180
agcgagcccc agcctccaga gccccctaag cattccccgc ccagcagccc agcccctgcc 240
cccacccagg aggccccaga gctcagggcg cctggtcgga ttttgtacag ccccgagtca 300
ctgtggagag agcttgcaat ggcagacgcg gctgcagcag atgggctcgc gatcatagca 360
ggtgctgcca cagtgagaaa aactgtgtca aaaaccgact cctggcagca gtcggaggcc 420
ccgccagaga ggggagcagc cggcctgaac ccatgtcctg ccggttccca tgacccccag 480
cacccagagc cccacggtgt ccccgttgga taatgaggac aagggctggg ggctccggtg 540
gtttgcggca gggacttgat cacatccttc tgctgtggcc ccattgcctc tggctggagt 600
tgacccttct gacaagtgtc ctcagaaaga cagggatcac cggcacctcc caatatcaac 660
cccaggcagc acagacacaa accccacatc cagagccaac tccaggagca gagacacccc 720
aacactctgg gggaccccaa ccgtgataac tccccactgg aatccgcccc agagtctacc 780
aggaccaaag gccctgccct gtctctgtcc ctcactcagg gcctcctgca gggcgagcgc 840
ttgggagcag actcggtctt aggggacacc actgtgggcc ccaactttga tgaggccact 900
gacccttcct tcctttcctg gggcagcaca gactttgggg tctgggcagg gaagaactac 960
tggctggtgg ccaatcacag agcccccagg ccgaggtggc cccaagaagg ccctcaggag 1020
gtggccactc cacttcctcc cagctggacc ccaggtcctc cccaagatag gggtgccatc 1080
caaggcaggt cctccatgga gcccccttca gactcctccc gggaccccac tggacctcag 1140
tccctgctct gggaatgcag ccaccacaag cacaccagga agcccaggcc cagccaccct 1200
gcagtgggca agcccacact ctggagcaga gcagggtgcg tctgggaggg gctaacctcc 1260
ccacccccca ccccccatct gcacacagcc acctaccact gcccagaccc tctgcaggag 1320
ggccaagcca ccatggggta tggacttagg gtctcactca cgtgcctccc ctcctgggag 1380
aaggggcctc atgcccagat ccctgcagca ctagacacag ctggaggcag tggccccagg 1440
gccaccctga cctggcatct aaggctgctc cagcccagac agcactgccg ttcctgggaa 1500
gcctgggctc caccagacca caggtccagg gcacagccca caggagccac ccacacacag 1560
ctcacaggaa gaagataagc tccagacccc agggcgggac ctgccttcct gccaccactt 1620
acacacaggc cagggagctg ttcccacaca gatcaacccc aaaccgggac tgcctggcac 1680
tagggtcact gccatttccc tctccattcc ctcccagtgc ctctgtgctc cctccttctg 1740
gggaacaccc tgtgcagccc ctccctgcag cccacacgct ggggagaccc caccctgcct 1800
cgggcctttt ctacctgctg cacttgccgc ccacccaaac aaccctgggt acgtgaccct 1860
gcagtcctca ccctgatctg caaccagacc cctgtccctc cctctaaaca cccctcccag 1920
gccaactctg cacctgcagg ccctccgctc ttctgccaca agagcctcag gttttcctac 1980
ctgtgcccac cccctaaccc ctcctgccca caacttgagt tcttcctctc ctggagccct 2040
tgagccatgg cactgaccct acactcccac ccacacactg cccatgccat caccttcctc 2100
ctggacactc tgaccccgct cccctccctc tcagacccgg ccctggtatt tccaggacaa 2160
aggctcaccc aagtcttccc catgcaggcc cttgccctca ctgcctggtt acacgggagc 2220
ctcctgtgcg cagaagcagg gagctcagct cttccacagg cagaaggcac tgaaagaaat 2280
cagcctccag tgccttgaca cacgtccgcc tgtgtctctc actgcctgca cctgcaggga 2340
ggctccgcac tccctctaaa gatgagggat ccaggcagca acatcacggg agaatgcagg 2400
gctcccagac agcccagccc tctcgcaggc ctctcctggg aagagacctg cagccaccac 2460
tgaacagcca cggaggtcgc tggatagtaa ccgagtcagt gaccgacctg gagggcaggg 2520
gagcagtgaa ccggagccca taccataggg acagagacca gccgctaaca tcccgagccc 2580
ctcactggcg gccccagaac accccgtgga aagagaacag acccacagtc ccacctggaa 2640
cagggcagac actgctgagc ccccagcacc agccccaaga aacactaggc aacagcatca 2700
gagggggctc ctgagaaaga gaggagggga ggtctccttc accatcaaat gcttcccttg 2760
accaaaaaca gggtccacgc aactccccca ggacaaagga ggagccccct gtacagcact 2820
gggctcagag tcctctctga gacaggctca gtttcagaca acaacccgct ggaatgcaca 2880
gtctcagcag gagagccagg ccagagccag caagaggaga ctcggtgaca ccagtctcct 2940
gtagggacag gaggattttg tgggggttcg tgtcactgtg agcatattgt cggagcaggc 3000
agtgctattc ccacagtgac acaaccccat tcaaaaaccc ctactgcaaa cgcacccact 3060
cctgggactg aggggctggg ggagcgtctg ggaagtatgg cctaggggtg tccatcaatg 3120
cccaaaatgc accagactct ccccaagaca tcaccccacc agccagtgag cagagtaaac 3180
agaaaatgag aagcagctgg gaagcttgca caggccccaa ggaaagagct ttggcaggtg 3240
tgcaagaggg gatgtgggca gagcctcagc agggcctttt gctgtttctg ctttcctgtg 3300
cagagagttc cataaactgg tattcaagat caatggctgg gagtgagccc aggaggacag 3360
tgtgggaaga gcacagggaa ggaggagcag ccgctatcct acactgtcat cttttgaaag 3420
tttgccctgt gcccacaatg ctgcatcatg ggatgcttaa cagctgatgt agacacagct 3480
aaagagagaa tcagtgaaat ggatttgcag cacagatctg aataaatcct ccagaatgtg 3540
gagcagcaca gaagcaagca cacagaaagt gcctgatgcc aaggcaaagt tcagtgggca 3600
ccttcaggca ttgctgctgg gcacagacac tctgaaaagc actggcagga actgcctgtg 3660
acaaagcaga accctcaggc aatgccagcc ctagagccct tcctgagaac ctcatgggca 3720
aagatgtgca gaacagctgt ttgtcatagc cccaaactat ggggctggac aaagcaaacg 3780
tccatctgaa ggagaacaga caaataaacg atggcaggtt catgaaatgc aaactaggac 3840
agccagagga caacagtaga gagctacagg cggctttgcg gttgagttca tgacaatgct 3900
gagtaattgg agtaacagag gaaagcccaa aaaatacttt taatgtgatt tcttctaaat 3960
aaaatttaca cccggcaaaa tgaactatct tcttaaggga taaactttcc cctggaaaaa 4020
ctataaggaa aatcaagaaa acgatgatca cataaacaca gtggtggtta cttctactgg 4080
ggaaggaaga gggtatgagc tgagacacac agagtcggca agtctcctaa caagaacaga 4140
acaaatacat tacagtacct tgaaaacagc agttaaactt ctaaatcgca agaagaggaa 4200
aatgcacaca cctgtgttta gaaaattctc agtccagcac tgttcataat agcaaagaca 4260
ttaacccagg ttggataaat aagcgatgac acaggcaatt gcacaatgat acagacatac 4320
attcagtata tgagacatcg atgatgtatc cccaaagaaa tgactttaaa gagaaaaggc 4380
ctgatgtgtg gtggcaatca cctccctggg catccccgga caggctgcag gctcactgtg 4440
tggcagggca ggcaggcacc tgctggcagc tcctggggcc tgatgtggag caggcacaga 4500
gctgtatatc cccaaggaag gtacagtcag tgcattccag agagaagcaa ctcagccaca 4560
ctccctggcc agaacccaag atgcacaccc atgcacaggg aggcagagcc cagcacctcc 4620
gcagccacca ccacctgcgc acgggccacc accttgcagg cacagagtgg gtgctgagag 4680
gaggggcagg gacaccaggc agggtgagca cccagagaaa actgcagaag cctcacacat 4740
ccctcacctg gcctgggctt cacctgacct ggacctcacc tggcctcggg cctcacctgc 4800
acctgctcca ggtcttgctg gagcctgagt agcactgagg ctgtagggac tcatccaggg 4860
ttggggaatg actctgcaac tctcccacat ctgacctttc tgggtggagg cacctggtgg 4920
cccagggaat ataaaaagcc ccagaatgat gcctgtgtga tttgggggca atttatgaac 4980
ccgaaaggac atggccatgg ggtgggtagg gacagtaggg acagatgtca gcctgaggtg 5040
aagcctcagg acacaggtgg gcatggacag tgtccaccta agcgagggac agacccgagt 5100
gtccctgcag tagacctgag agcgctgggc ccacagcctc ccctcggggc cctgctgcct 5160
cctcaggtca gccctggaca tcccgggttt ccccaggcct ggcggtagga ttttgttgag 5220
gtctgtgtca ctgtggtatt actatgagag gcttgcttgt ggcttcccta agagctgcag 5280
caggcaagct aagcctcaca gatgctgcta ttactaccac agtgtcacag agtccatcaa 5340
aaacccatgc ctgggagcct cccaccacag ccctccctgc gggggaccgc tgcatgccgt 5400
gttaggattt tgatcgagga cacggcgcca tgggtatggt ggctaccaca gcagtgcagc 5460
ccatgaccca aacacacggg gcagcagaaa caatggacag gcccacaagt gaccatgatg 5520
ggctccagcc caccagcccc agagaccatg aaacagatgg ccaaggtcac cctacaggtc 5580
atccagatct ggctccaagg ggtctgcatc gctgctgccc tcccaacgcc aaaccagatg 5640
gagacagggc cggccccata gcaccatctg ctgccgtcca cccagcagtc ccggaagccc 5700
ctccctgaac gctgggccac gtgtgtgaac cctgcgagcc ccccatgtca gagtaggggc 5760
agcaggaggg cggggctggc cctgtgcact gtcactgccc ctgtggtccc tggcctgcct 5820
ggccctgaca cctgagcctc tcctgggtca tttccaagac attcccaggg acagccggag 5880
ctgggagtcg ctcatcctgc ctggctgtcc tgagtcctgc tcatttccag acctcaccag 5940
ggaagccaac agaggactca cctcacacag tcagagacaa cgaaccttcc agaaatccct 6000
gtttctctcc ccagtgagag aaaccctctt ccagggtttc tcttctctcc caccctcttc 6060
caggacagtc ctcagcagca tcacagcggg aacgcacatc tggatcagga cggcccccag 6120
aacacgcgat ggcccatggg gacagcccag cccttcccag acccctaaaa ggtatcccca 6180
ccttgcacct gccccagggc tcaaactcca ggaggcctga ctcctgcaca ccctcctgcc 6240
agatatcacc tcagccccct cctggagggg acaggagccc gggagggtga gtcagaccca 6300
cctgccctca atggcaggcg gggaagattc agaaaggcct gagatcccca ggacgcagca 6360
ccactgtcaa tgggggcccc agacgcctgg accagggcct gtgtgggaaa ggcctctggc 6420
cacactcagg gggattttgt gaagggccct cccactgtgg agaggctttg ctgtggcttc 6480
cctaagagct gccgcagcag gcaatgcaag cctcacagat gctgccacag tgatgaaacc 6540
agcatcaaaa accgaccgga ctcgcagggt ttatgcacac ttctcggctc ggagctctcc 6600
aggagcacaa gagccaggcc cgagggtttg tgcccagacc ctcggcctct agggacaccc 6660
gggccatctt agccgatggg ctgatgccct gcacaccgtg tgctgccaaa caggggcttc 6720
agagggctct gaggtgactt cactcatgac cacaggtgcc ctggtccctt cactgccagc 6780
tgcaccagac cctgttccga gagatgcccc agttccaaaa gccaattcct ggggccggga 6840
attactgtag acaccagcct cattccagta cctcctgcca attgcctgga ttcccatcct 6900
ggctggaatc aagagggcag catccgccag gctcccaaca ggcaggactc ccacacaccc 6960
tcctctgaga ggccgctgtg ttccgcaggg ccaggccgca gacagttccc ctcacctgcc 7020
catgtagaaa cacctgccat tgtcgtcccc acctggcaaa gaccacttgt ggagccccca 7080
gccccaggta cagctgtaga gagagtcctc gaggccccta agaaggagcc atgcccagtt 7140
ctgccgggac cctcggccag gccgacagga gtggacgctg gagctgggcc cacactgggc 7200
cacataggag ctcaccagtg agggcaggag agcacatgcc ggggagcacc cagcctcctg 7260
ctgaccagag acccgtccca gagcccagga ggctgcagag gcctctccag ggggacacag 7320
tgcatgtctg gtccctgagc agcccccagg ctctctagca ctgggggccc ctggcacagc 7380
tgtctggacc ctccctgttc cctgggaagc tcctcctgac agccccgcct ccagttccag 7440
gtgtggattt tgtcaggggg tgccacactg tgtactgcca gaagtggatg tggacttgcg 7500
atagtgagag gaagtgctgt gagggtatgg tatgccggct gtggtgtaag aagaagctct 7560
ggcacagtgg tgccgcccat atcaaaaacc aggccaagta gacagacccc tgccacgcag 7620
ccccaggcct ccagctcacc tgcttctcct ggggctctca aggctgctgt ctgccctctg 7680
gccctctgtg gggagggttc cctcagtggg aggtctgtgc tccagggcag ggatgactga 7740
gatagaaatc aaaggctggc agggaaaggc agcttcccgc cctgagaggt gcaggcagca 7800
ccacagagcc atggagtcac agagccacgg agcccccagt gtgggcgtgt gagggtgctg 7860
ggctcccggc aggcccagcc ctgatgggga agcctgcccc gtcccacagc ccaggtcccc 7920
aggggcagca ggcacagaag ctgccaagct gtgctctacg atcctcatcc ctccagcagc 7980
atccactcca cagtggggaa actgagcctt ggagaaccac ccagccccct ggaaacaagg 8040
cggggagccc agacagtggg cccagagcac tgtgtgtatc ctggcactag gtgcagggac 8100
cacccggaga tccccatcac tgagtggcca gcctgcagaa ggacccaacc ccaaccaggc 8160
cgcttgatta agctccatcc ccctgtcctg ggaacctctt cccagcgcca ccaacagctc 8220
ggcttcccag gccctcatcc ctccaaggaa ggccaaaggc tgggcctgcc aggggcacag 8280
taccctccct tgccctggct aagacagggt gggcagacgg ctgcagatag gacatattgc 8340
tggggcatct tgctctgtga ctactgggta ctggctctca acgcagaccc taccaaaatc 8400
cccactgcct cccctgctag gggctggcct ggtctcctcc tgctgtccta ggaggctgct 8460
gacctccagg atggcttctg tccccagttc tagggccaga gcagatccca ggcaggctgt 8520
aggctgggag gccacccctg tccttgccga ggttcagtgc aggcacccag gacaggaaat 8580
ggcctgaaca cagggatgac tgtgccatgc cctacctaag tccgcccctt tctactctgc 8640
aacccccact ccccaggtca gcccatgacg accaacaacc caacaccaga gtcactgcct 8700
ggccctgccc tggggaggac ccctcagccc ccaccctgtc tagaggagtt ggggggacag 8760
gacacaggct ctctccttat ggttccccca cctggctcct gccgggaccc ttggggtgtg 8820
gacagaaagg acgcctgcct aattggcccc caggaaccca gaacttctct ccagggaccc 8880
cagcccgagc acccccttac ccaggaccca gccctgcccc tcctcccctc tgctctcctc 8940
tcatcactcc atgggaatcc agaatcccca ggaagccatc aggaagggct gaaggaggaa 9000
gcggggccgc tgcaccaccg ggcaggaggc tccgtcttcg tgaacccagg gaagtgccag 9060
cctcctagag ggtatggtcc accctgcctg gggctcccac cgtggcaggc tgcggggaag 9120
gaccagggac ggtgtggggg agggctcagg gccctgcagg tgctccatct tggatgagcc 9180
catccctctc acccaccgac ccgcccacct cctctccacc ctggccacac gtcgtccaca 9240
ccatcctgag tcccacctac accagagcca gcagagccag tgcagacaga ggctggggtg 9300
caggggggcc gccagggcag ctttggggag ggaggaatgg aggaagggga ggtcagtgaa 9360
gaggcccccc tcccctgggt ctaggatcca cctttgggac ccccggatcc catcccctcc 9420
aggctctggg aggagaagca ggatgggaga ttctgtgcag gaccctctca cagtggaata 9480
cctccacagc ggctcaggcc agatacaaaa gcccctcagt gagccctcca ctgcagtgca 9540
gggcctgggg gcagcccctc ccacagagga cagacccagc accccgaaga agtcctgcca 9600
gggggagctc agagccatga aggagcaaga tatggggacc ccaatactgg cacagacctc 9660
agctccatcc aggcccacca ggacccacca tgggtggaac acctgtctcc ggcccctgct 9720
ggctgtgagg cagctggcct ctgtctcgga cccccattcc agacaccaga cagagggaca 9780
ggccccccag aaccagtgtt gagggacacc cctgtccagg gcagccaagt ccaagaggcg 9840
cgctgagccc agcaagggaa ggcccccaaa caaaccagga ggtttctgaa gctgtctgtg 9900
tcacagtctg ctgcaactgc agcagcaaat ggtgccgcga ccatagcagg tgctgccaca 9960
atgacactgg gcaggacaga aaccccatcc caagtcagcc gaaggcagag agagcaggca 10020
ggacacattt aggatctgag gccacacctg acactcaagc caacagatgt ctcccctcca 10080
gggcgccctg ccctgttcag tgttcctgag aaaacagggg cagcctgagg ggatccaggg 10140
ccaggagatg ggtcccctct accccgagga ggagccaggc gggaatccca gccccctccc 10200
cattgaggcc atcctgccca gaggggcccg gacccacccc acacacccag gcagaatgtg 10260
tgcaggcctc aggctctgtg ggtgccgcta gctggggctg ccagtcctca ccccacacct 10320
aaggtgagcc acagccgcca gagcctccac aggagacccc acccagcagc ccagccccta 10380
cccaggaggc cccagagctc agggcgcctg ggtggatttt gtacagcccc gagtcactgt 10440
gggtatagtg gggagaggct ttgtggcttc cctaagagct gcagcaggca aaagcctcac 10500
agatgctgca gctactacca cagtgagaaa agctatgtca aaaaccgtct cccggccact 10560
gctggaggcc cagccagaga agggaccagc cgcccgaaca tacgaccttc ccagacctca 10620
tgacccccag cacttggagc tccacagtgt ccccattgga tggtgaggat gggggccggg 10680
gccatctgca cctcccaaca tcacccccag gcagcacagg cacaaacccc aaatccagag 10740
ccgacaccag gaacacagac accccaatac cctgggggac cctggccctg gtgacttccc 10800
actgggatcc acccccgtgt ccacctggat caaagacccc accgctgtct ctgtccctca 10860
ctcagggcct gctgaggggc gggtgctttg gagcagactc aggtttaggg gccaccattg 10920
tggggcccaa cctcgaccag gacacagatt tttctttcct gccctggggc aacacagact 10980
ttggggtctg tgcagggagg accttctgga aagtcaccaa gcacagagcc ctgactgagg 11040
tggtctcagg aagaccccca ggagggggct tgtgcccctt cctctcatgt ggaccccatg 11100
ccccccaaga taggggcatc atgcagggca ggtcctccat gcagccacca ctaggcaact 11160
ccctggcgcc ggtccccact gcgcctccat cccggctctg gggatgcagc caccatggcc 11220
acaccaggca gcccgggtcc agcaaccctg cagtgcccaa gcccttggca ggattcccag 11280
aggctggagc ccacccctcc tcatcccccc acacctgcac acacacacct accccctgcc 11340
cagtccccct ccaggagggt tggagccgcc catagggtgg gggctccagg tctcactcac 11400
tcgcttccct tcctgggcaa aggagcctcg tgccccggtc ccccctgacg gcgctgggca 11460
caggtgtggg tactgggccc cagggctcct ccagccccag ctgccctgct ctccctggga 11520
ggcctgggca ccaccagacc accagtccag ggcacagccc cagggagccg cccactgcca 11580
gctcacagga agaagataag cttcagaccc tcagggccgg gagctgcctt cctgccaccc 11640
cttcctgccc cagacctcca tgccctcccc caaccactta cacacaagcc agggagctgt 11700
ttccacacag ttcaacccca aaccaggacg gcctggcact cgggtcactg ccatttctgt 11760
ctgcattcgc tcccagcgcc cctgtgttcc ctccctcctc cctccttcct ttcttcctgc 11820
attgggttca tgccgcagag tgccaggtgc aggtcagccc tgagcttggg gtcacctcct 11880
cactgaaggc agcctcaggg tgcccagggg caggcagggt gggggtgagg cttccagctc 11940
caaccgcttc gctaccttag gaccgttata gttaggcgcg ccgtcgacca attctcatgt 12000
ttgacagctt atcatcgaat ttctacgta 12029
<210> 236
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер и/или зонд
<400> 236
tcttatcaga cagggggctc tc 22
<210> 237
<211> 55
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер и/или зонд
<400> 237
gtgactggag ttcagacgtg tgctcttccg atcttcacca tggactgsac ctgga 55
<210> 238
<211> 56
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер и/или зонд
<400> 238
gtgactggag ttcagacgtg tgctcttccg atctccatgg acacactttg ytccac 56
<210> 239
<211> 57
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер и/или зонд
<400> 239
gtgactggag ttcagacgtg tgctcttccg atcttcacca tggagtttgg gctgagc 57
<210> 240
<211> 59
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер и/или зонд
<400> 240
gtgactggag ttcagacgtg tgctcttccg atctagaaca tgaaacayct gtggttctt 59
<210> 241
<211> 54
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер и/или зонд
<400> 241
gtgactggag ttcagacgtg tgctcttccg atctatgggg tcaaccgcca tcct 54
<210> 242
<211> 57
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер и/или зонд
<400> 242
gtgactggag ttcagacgtg tgctcttccg atctacaatg tctgtctcct tcctcat 57
<210> 243
<211> 54
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер и/или зонд
<400> 243
acactctttc cctacacgac gctcttccga tctgggaaga catttgggaa ggac 54
<210> 244
<211> 64
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер и/или зонд
<220>
<221> nnnnnn
<222> (25)..(30)
<223> 6-нуклеотидный индекс
<220>
<221> иной_признак
<222> (25)..(30)
<223> n представляет собой a, c, g или t
<400> 244
caagcagaag acggcatacg agatnnnnnn gtgactggag ttcagacgtg tgctcttccg 60
atct 64
<210> 245
<211> 68
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер и/или зонд
<220>
<221> nnnnnn
<222> (29)..(35)
<223> 6-нуклеотидный индекс
<220>
<221> иной_признак
<222> (30)..(35)
<223> n представляет собой a, c, g или t
<400> 245
aatgatacgg cgaccaccga gatctacacn nnnnnacact ctttccctac acgacgctct 60
tccgatct 68
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ЖИВОТНЫЕ, ОТЛИЧНЫЕ ОТ ЧЕЛОВЕКА, СПОСОБНЫЕ К DH-DH-ПЕРЕГРУППИРОВКЕ СКОНСТРУИРОВАННЫХ ГЕННЫХ СЕГМЕНТОВ, И ПУТИ ИХ ПРИМЕНЕНИЯ | 2019 |
|
RU2801802C2 |
| ЖИВОТНЫЕ, НЕ ЯВЛЯЮЩИЕСЯ ЧЕЛОВЕКОМ, ИМЕЮЩИЕ СКОНСТРУИРОВАННУЮ ЛЕГКУЮ ЦЕПЬ ЛЯМБДА ИММУНОГЛОБУЛИНА, И ИХ ПРИМЕНЕНИЕ | 2018 |
|
RU2778410C2 |
| ОТЛИЧНЫЕ ОТ ЧЕЛОВЕКА ЖИВОТНЫЕ СО СКОНСТРУИРОВАННЫМ ЛОКУСОМ ЛЕГКОЙ ЛЯМБДА-ЦЕПИ ИММУНОГЛОБУЛИНА | 2017 |
|
RU2757665C2 |
| МЫШИ С ГУМАНИЗИРОВАННОЙ УНИВЕРСАЛЬНОЙ ЛЕГКОЙ ЦЕПЬЮ | 2012 |
|
RU2787781C2 |
| МЫШИ С ГУМАНИЗИРОВАННОЙ ЛЕГКОЙ ЦЕПЬЮ | 2012 |
|
RU2761639C2 |
| МЫШЬ С ОБЩЕЙ ЛЕГКОЙ ЦЕПЬЮ | 2011 |
|
RU2724663C2 |
| МЫШИ, У КОТОРЫХ ВЫРАБАТЫВАЮТСЯ СВЯЗЫВАЮЩИЕ БЕЛКИ, СОДЕРЖАЩИЕ V-ДОМЕНЫ | 2011 |
|
RU2750219C2 |
| МЫШИ ADAM6 | 2012 |
|
RU2722373C2 |
| МЫШЬ С ОБЩЕЙ ЛЕГКОЙ ЦЕПЬЮ | 2020 |
|
RU2826121C2 |
| МЫШИ, ЭКСПРЕССИРУЮЩИЕ ЛЕГКУЮ ЦЕПЬ С ВАРИАБЕЛЬНОЙ ОБЛАСТЬЮ λ ЧЕЛОВЕКА И КОНСТАНТНОЙ ОБЛАСТЬЮ МЫШИ | 2020 |
|
RU2833566C2 |


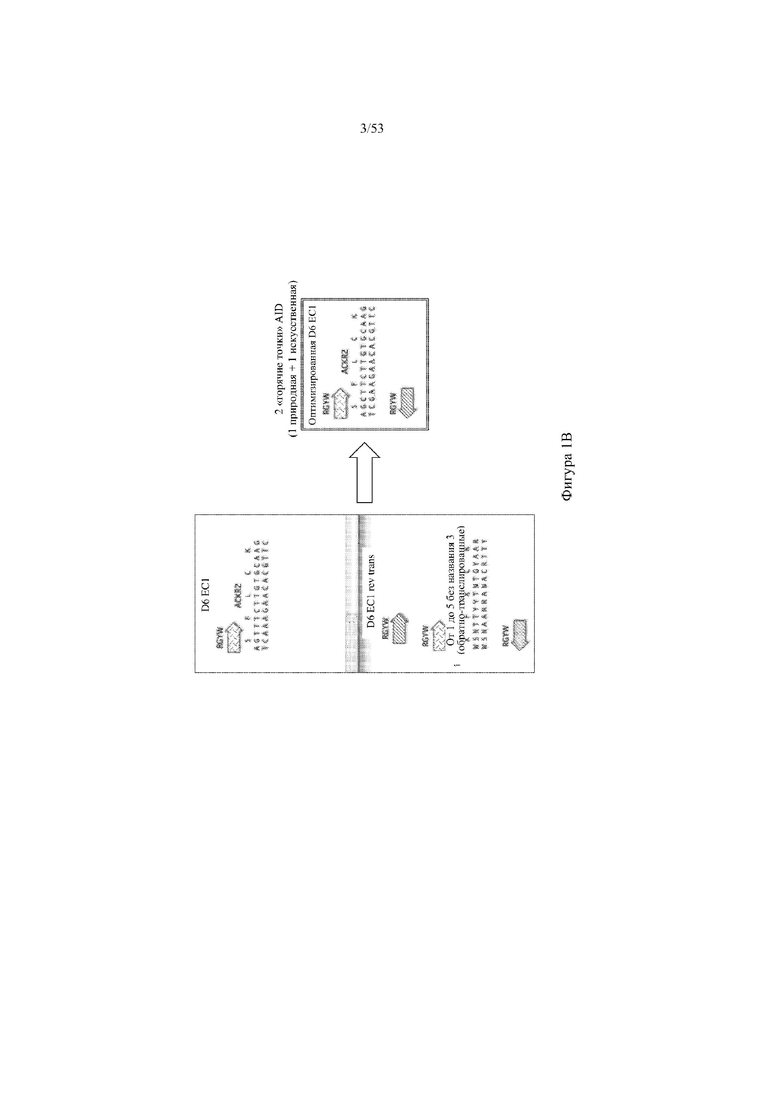
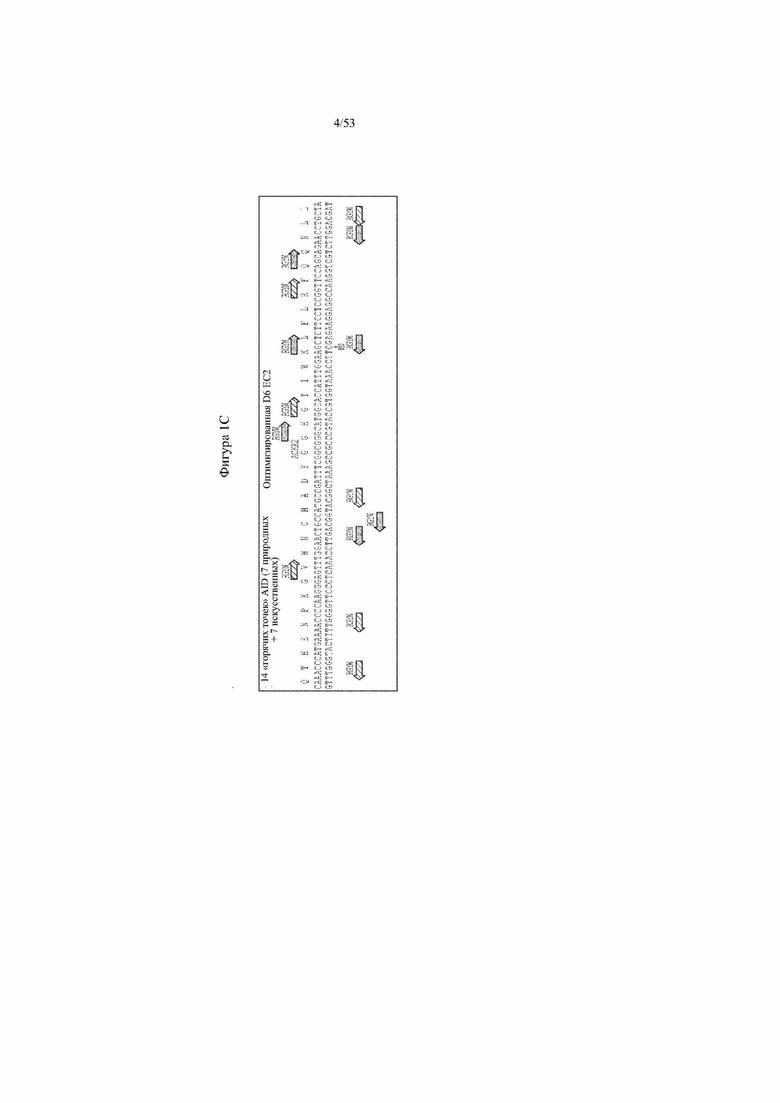

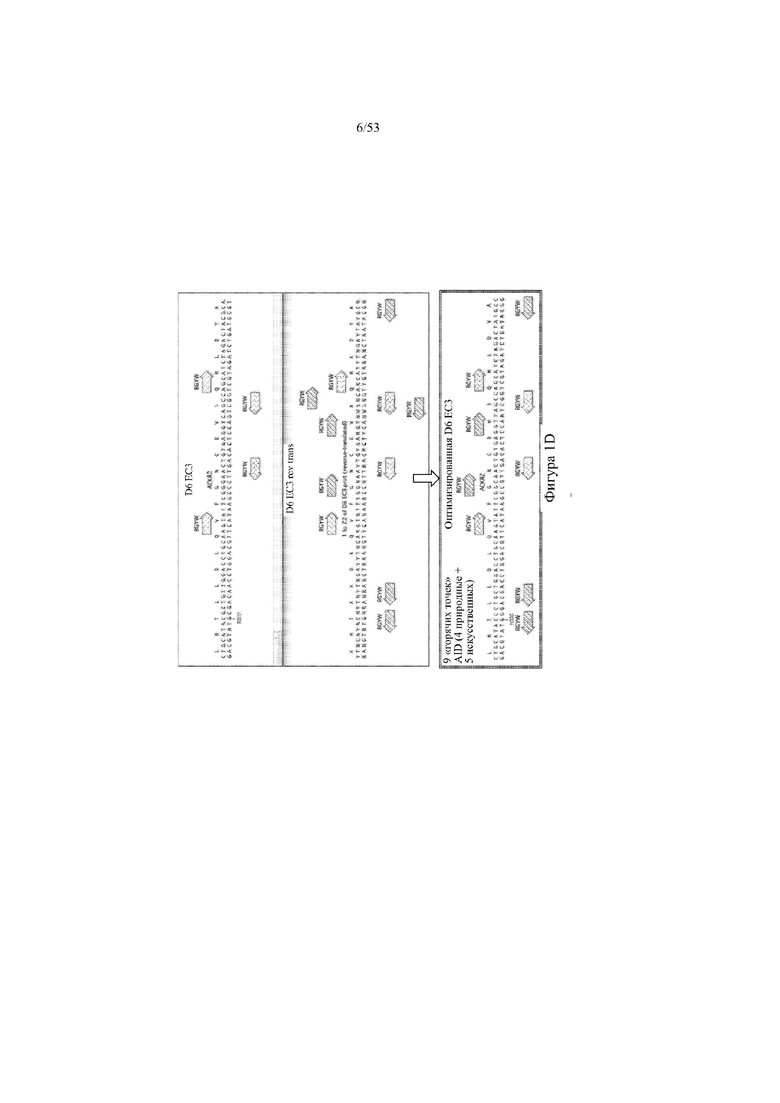
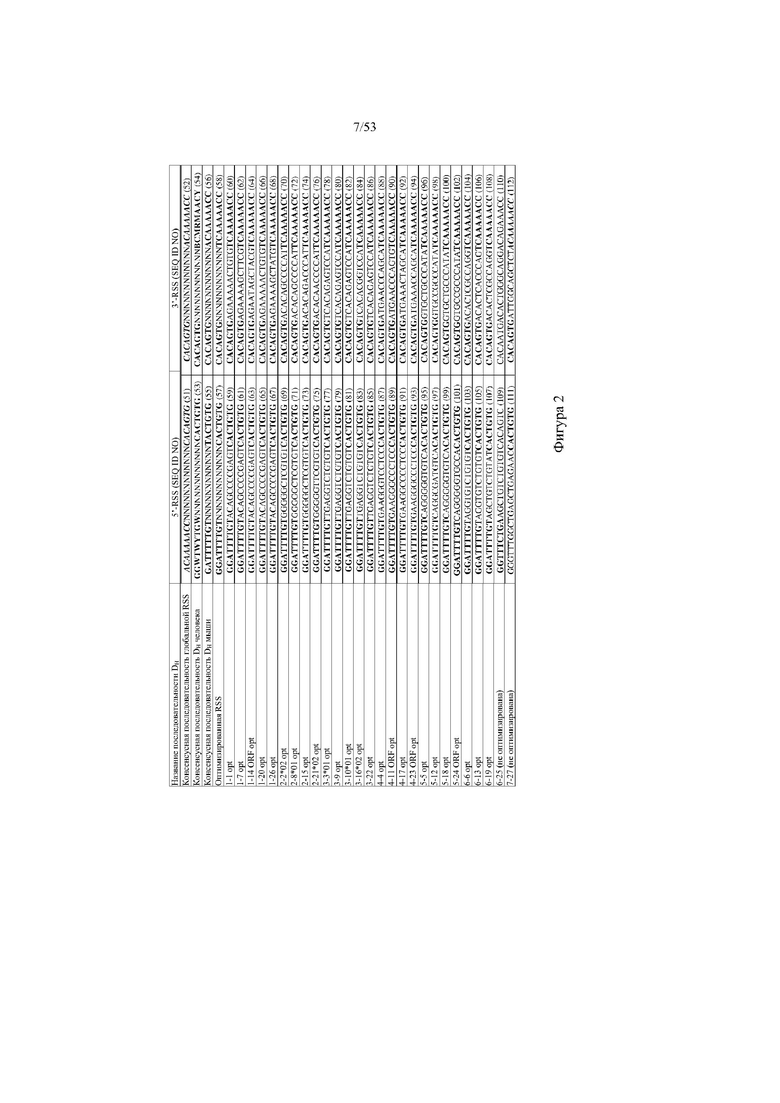
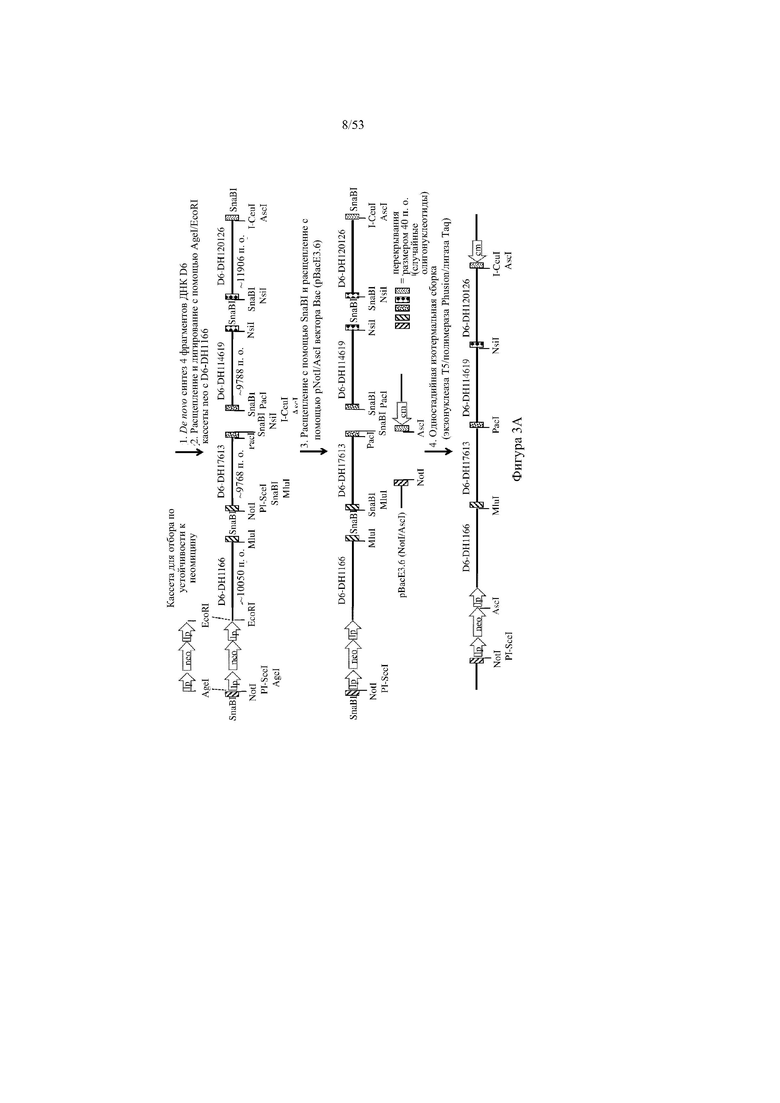
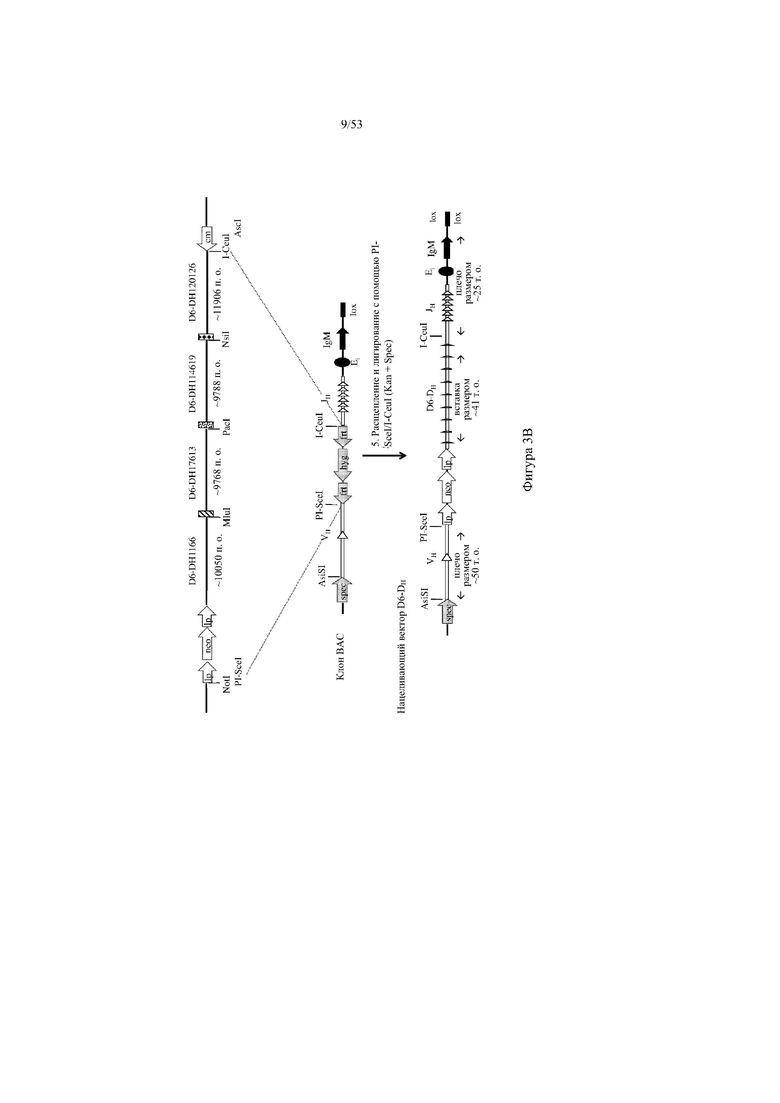

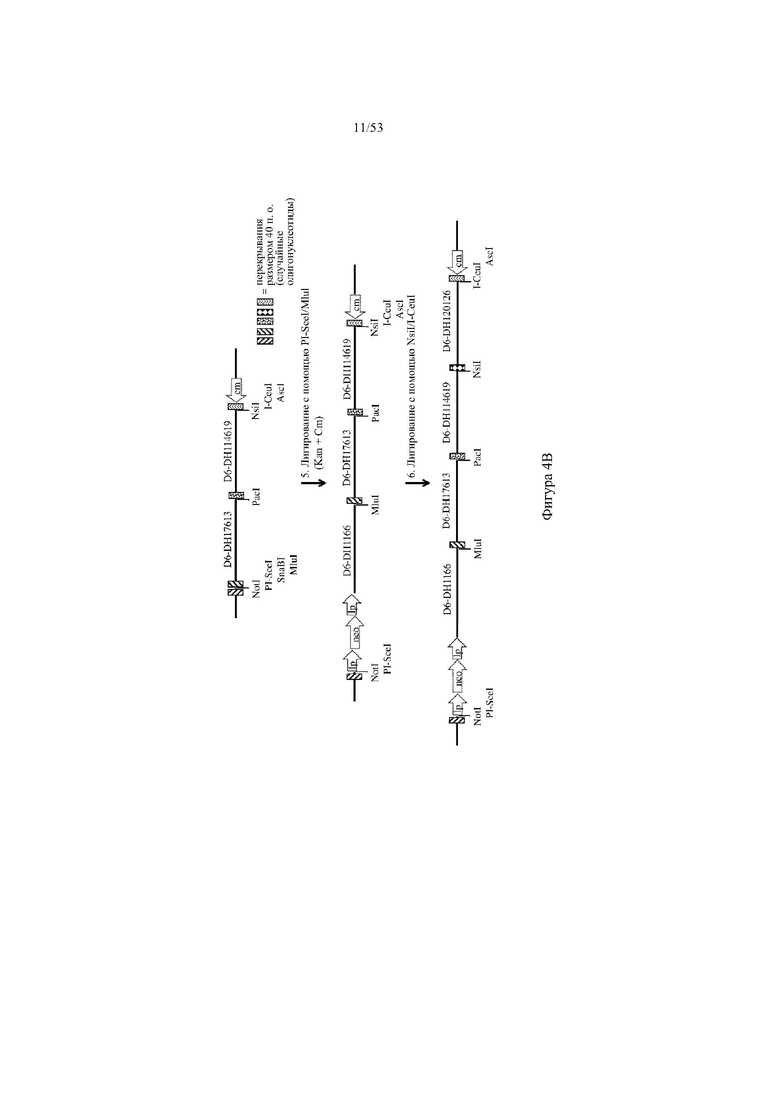

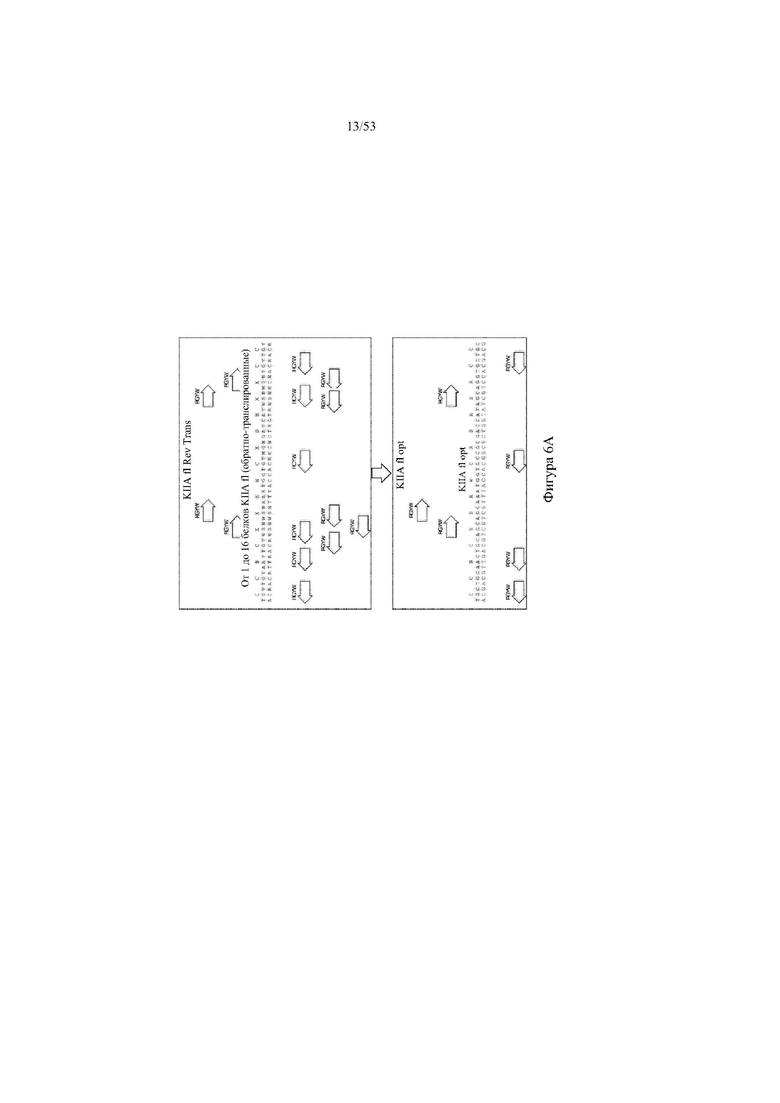
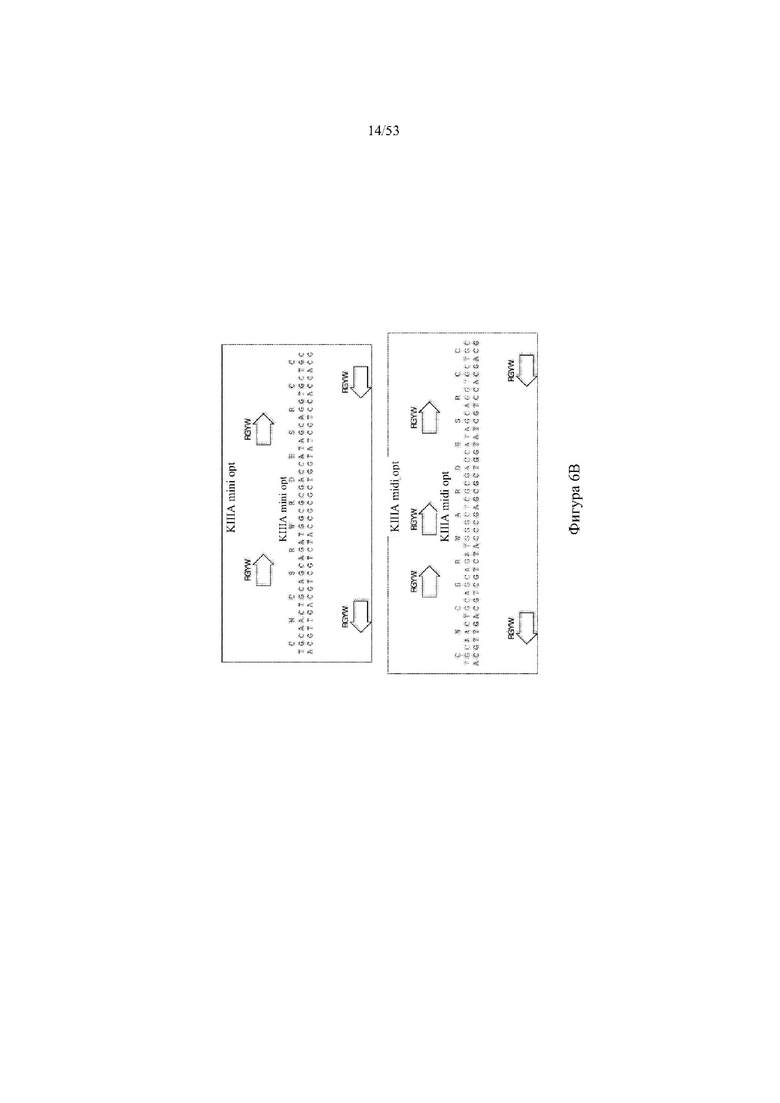
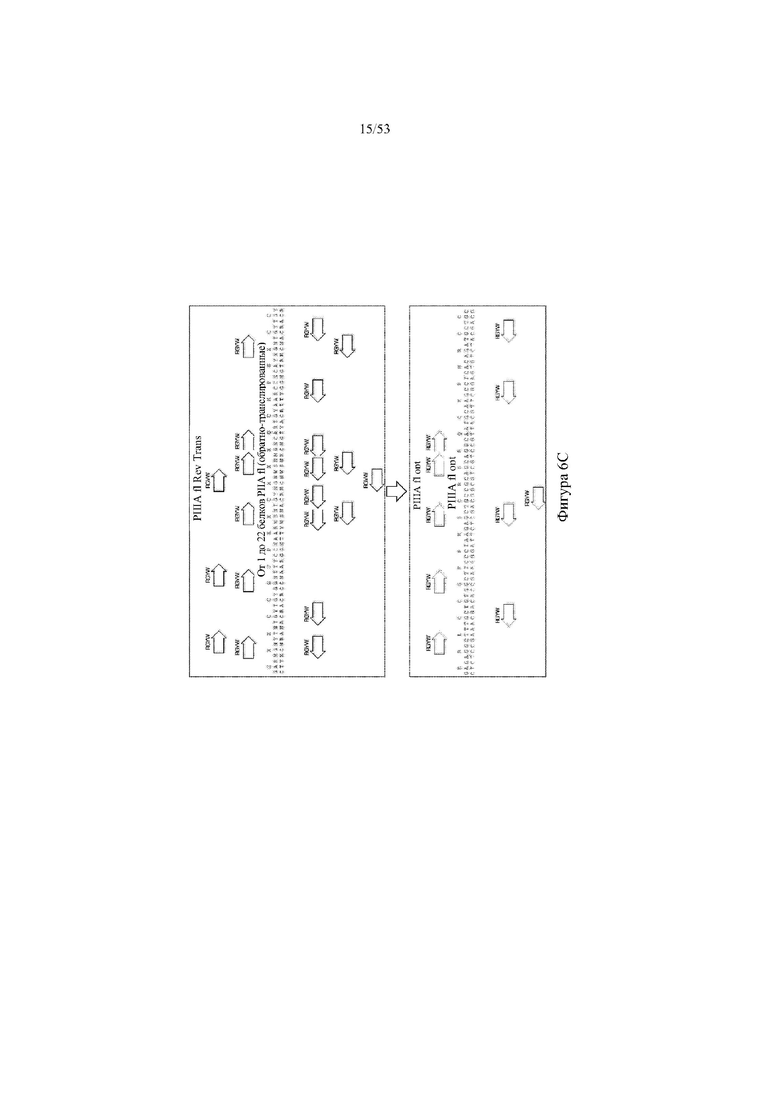
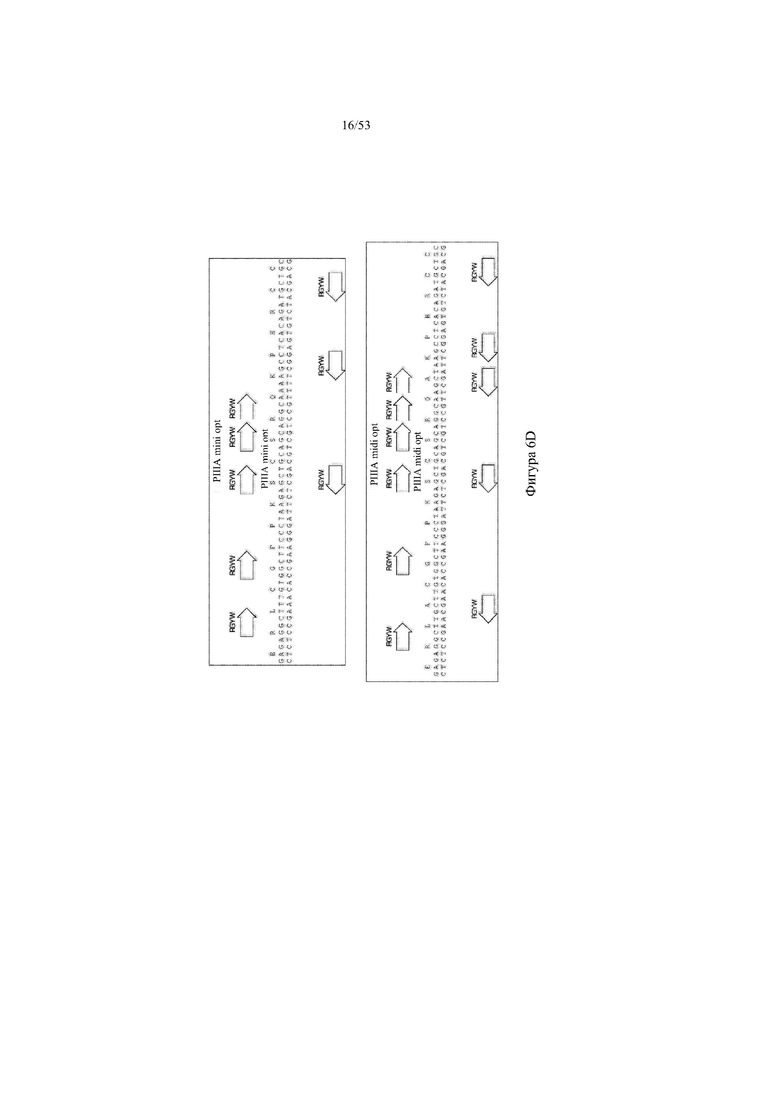
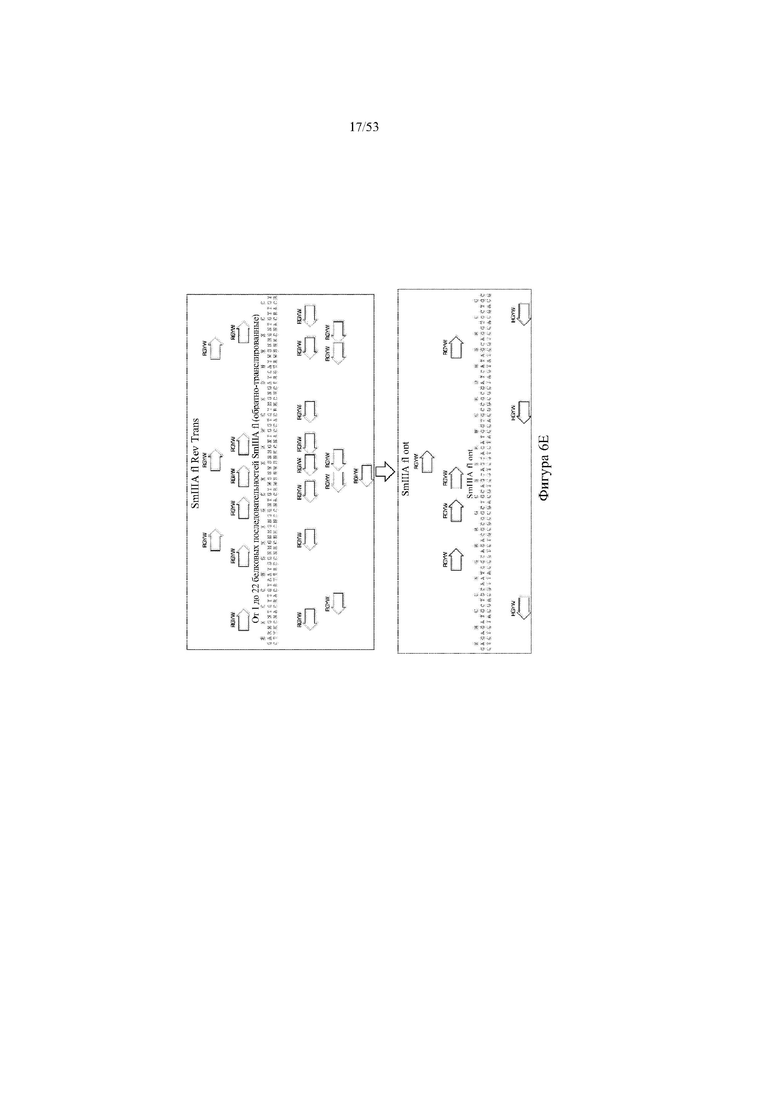
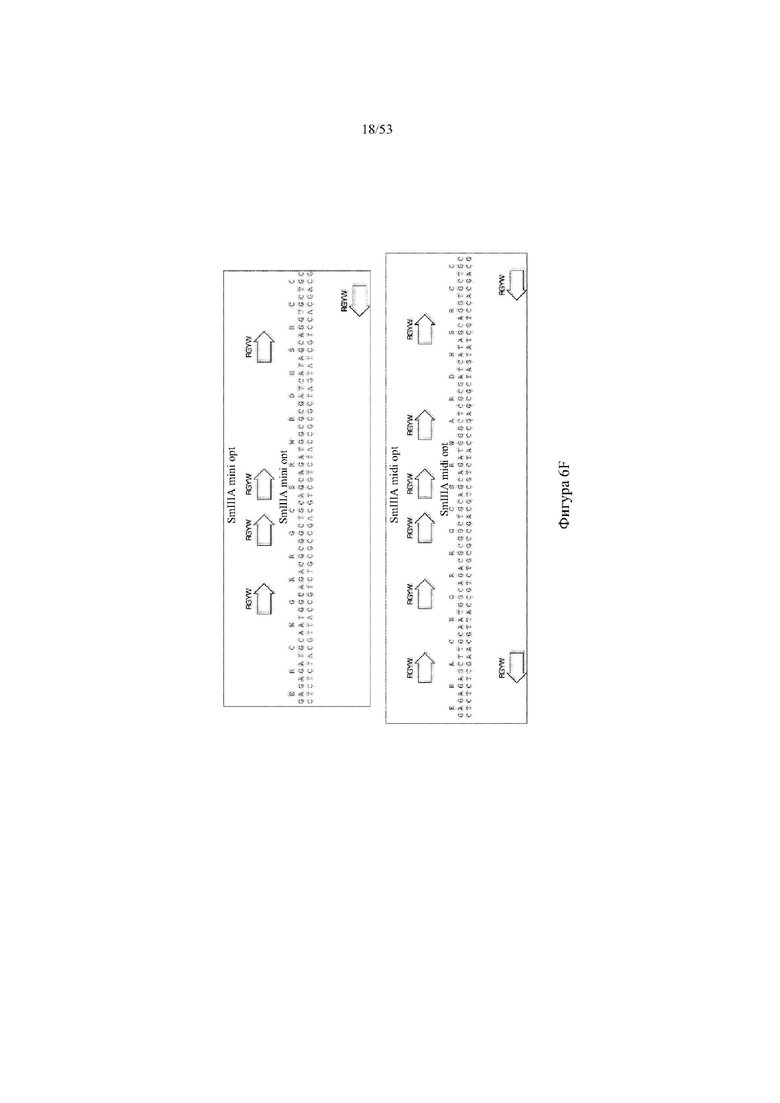
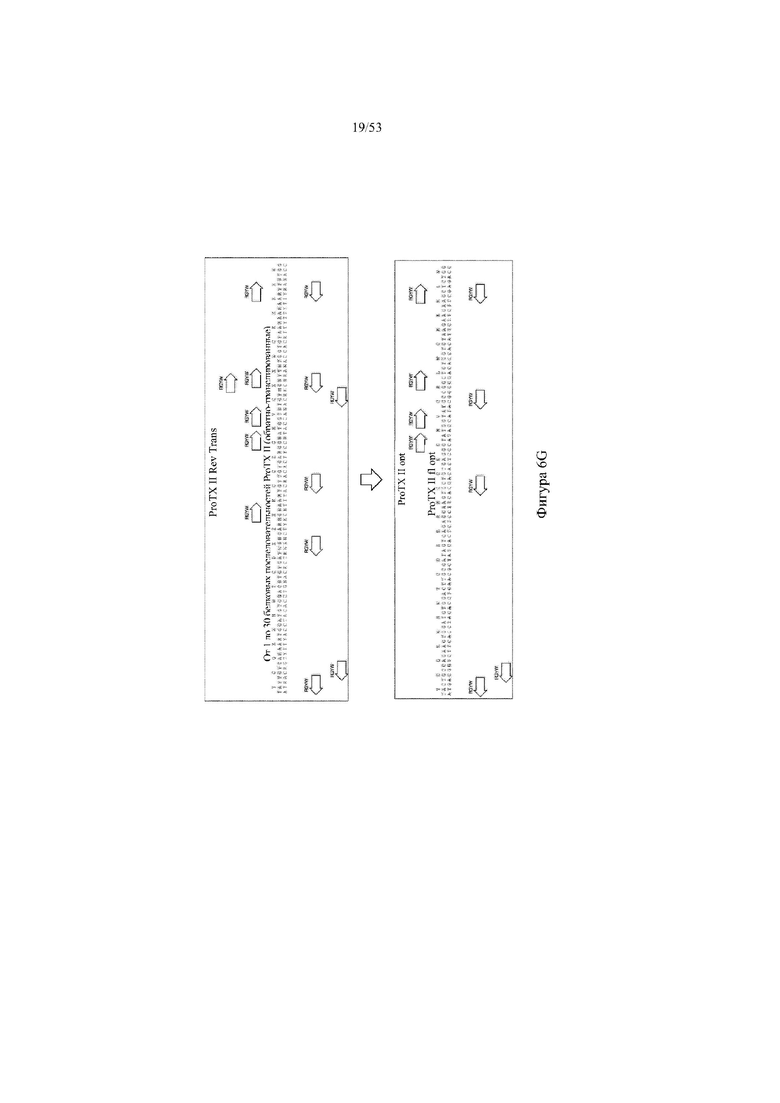
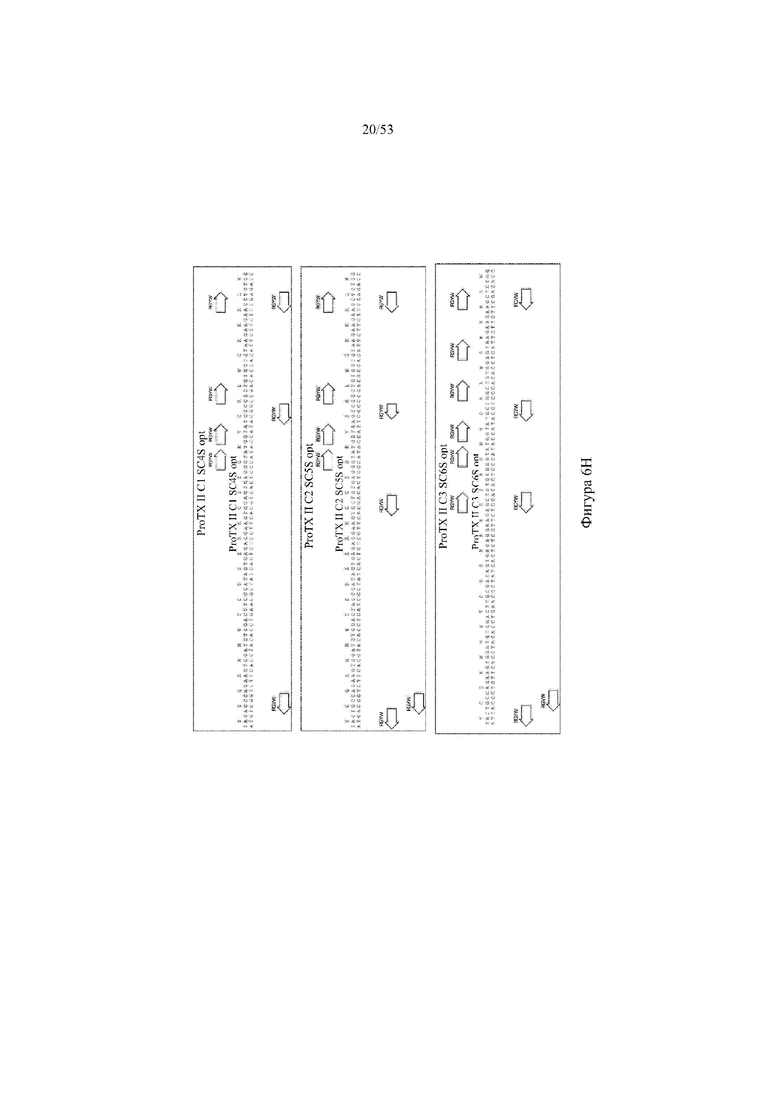
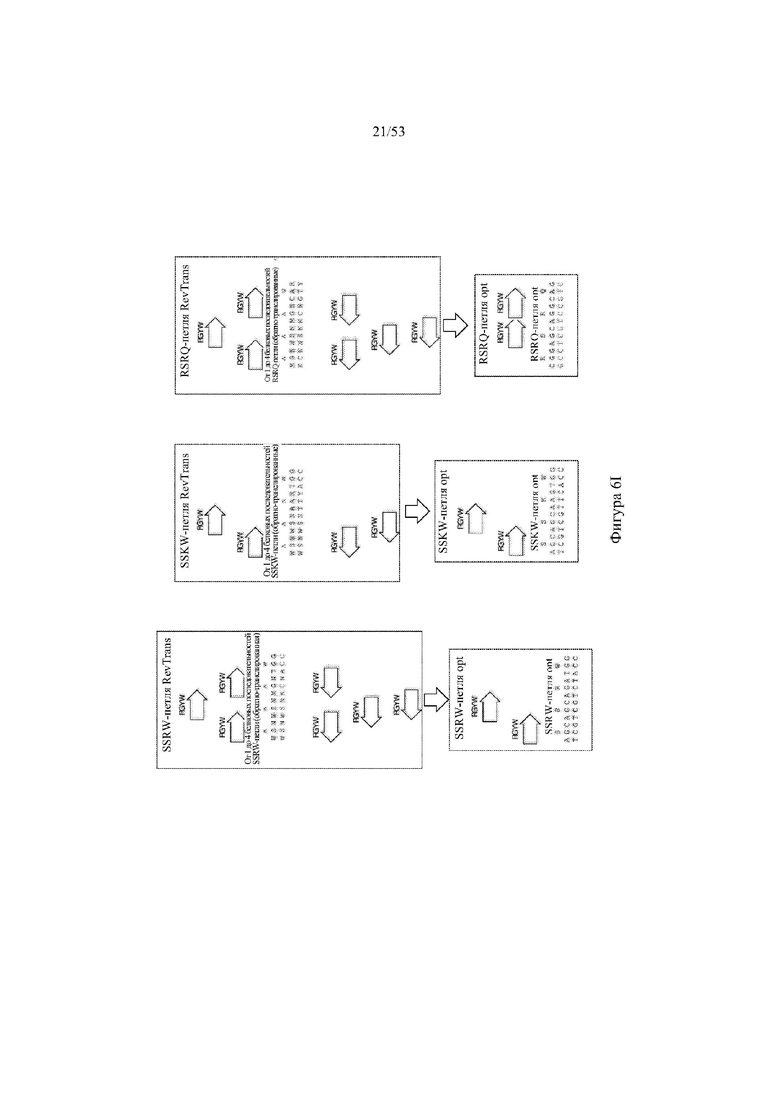
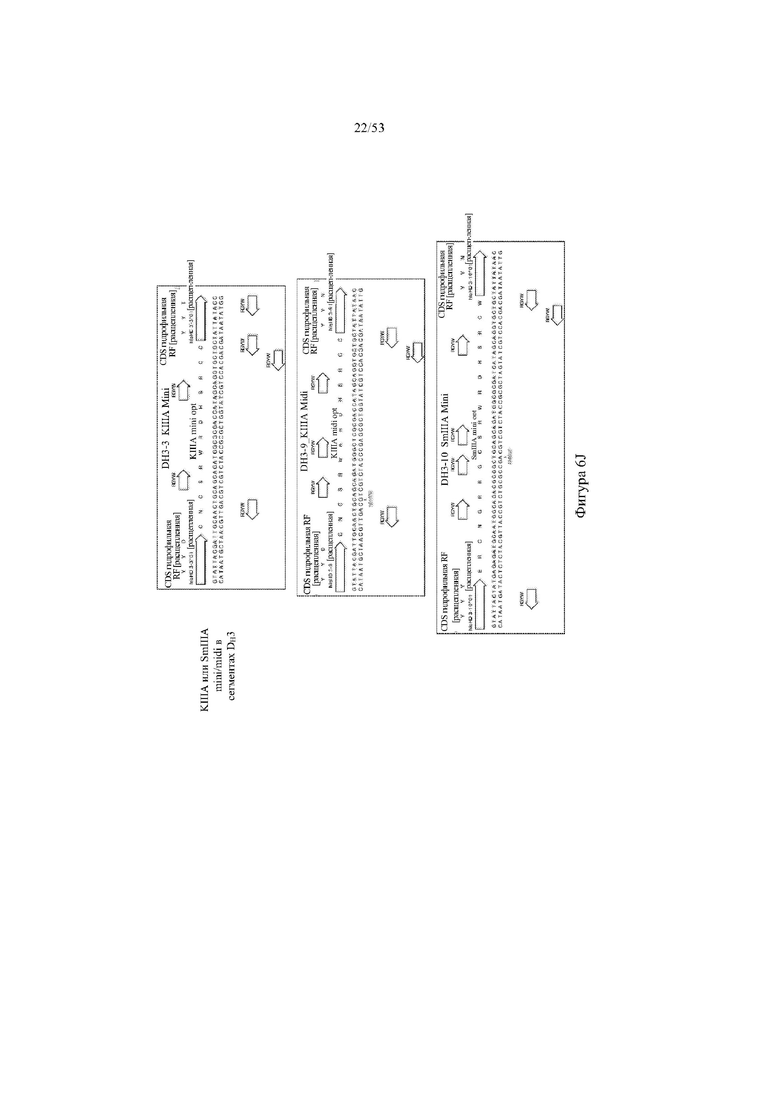
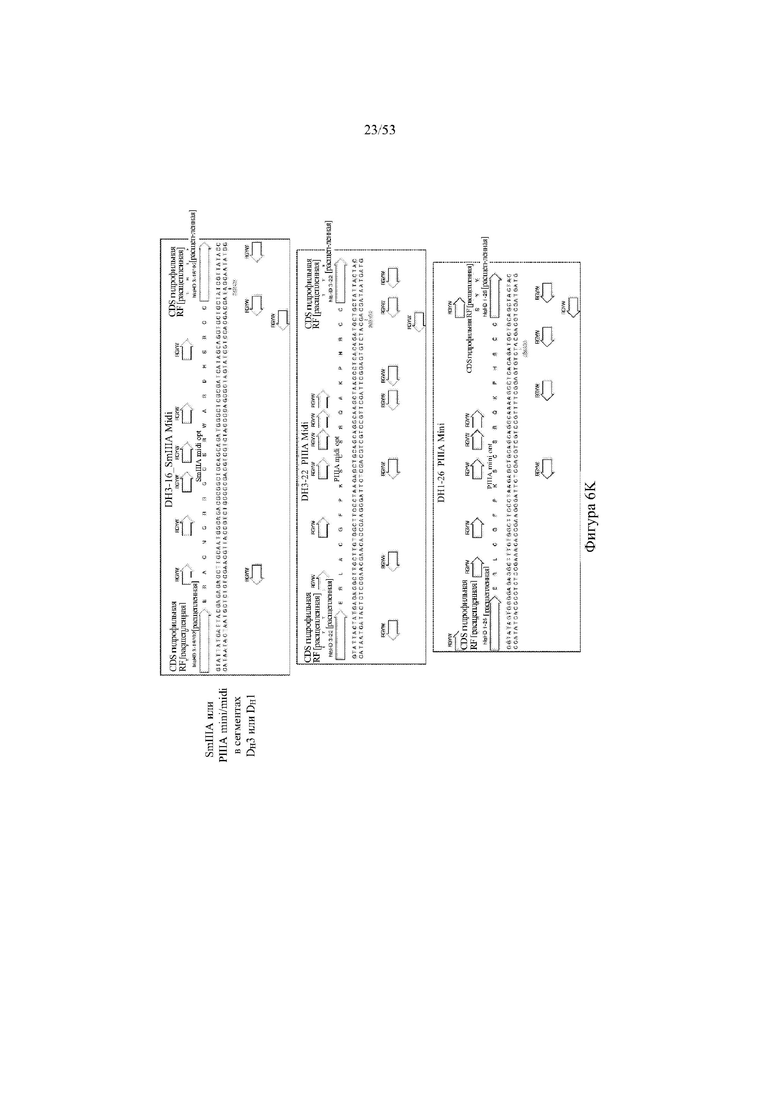
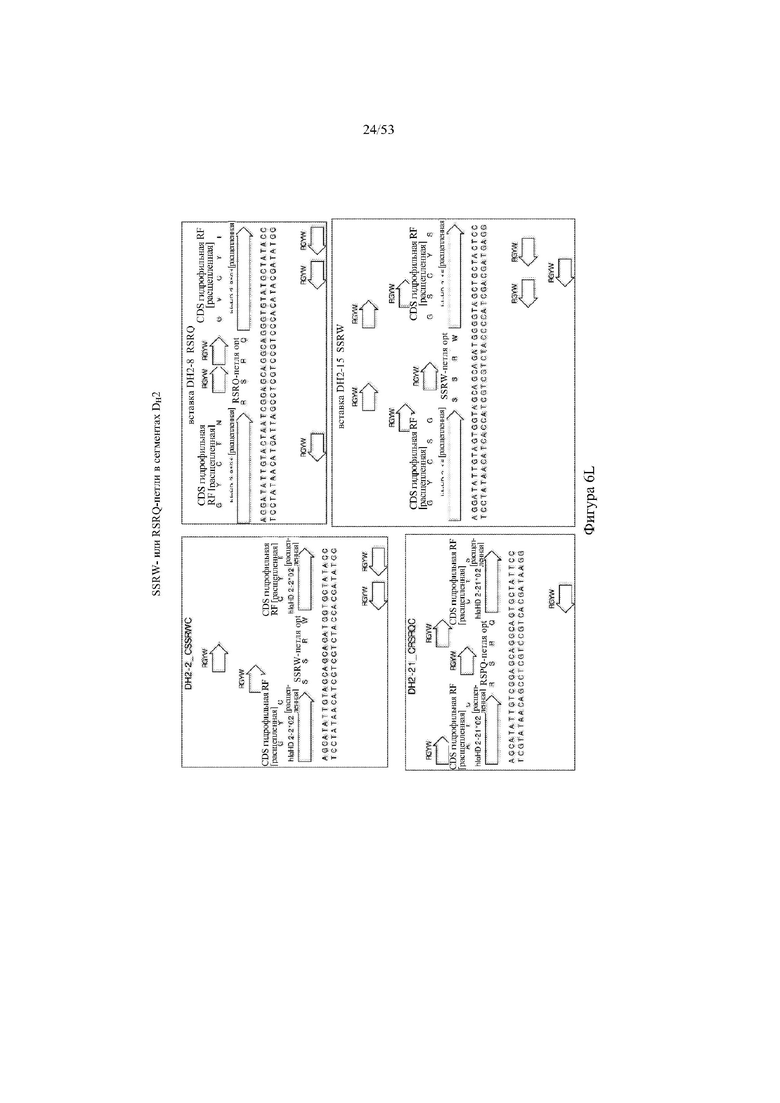
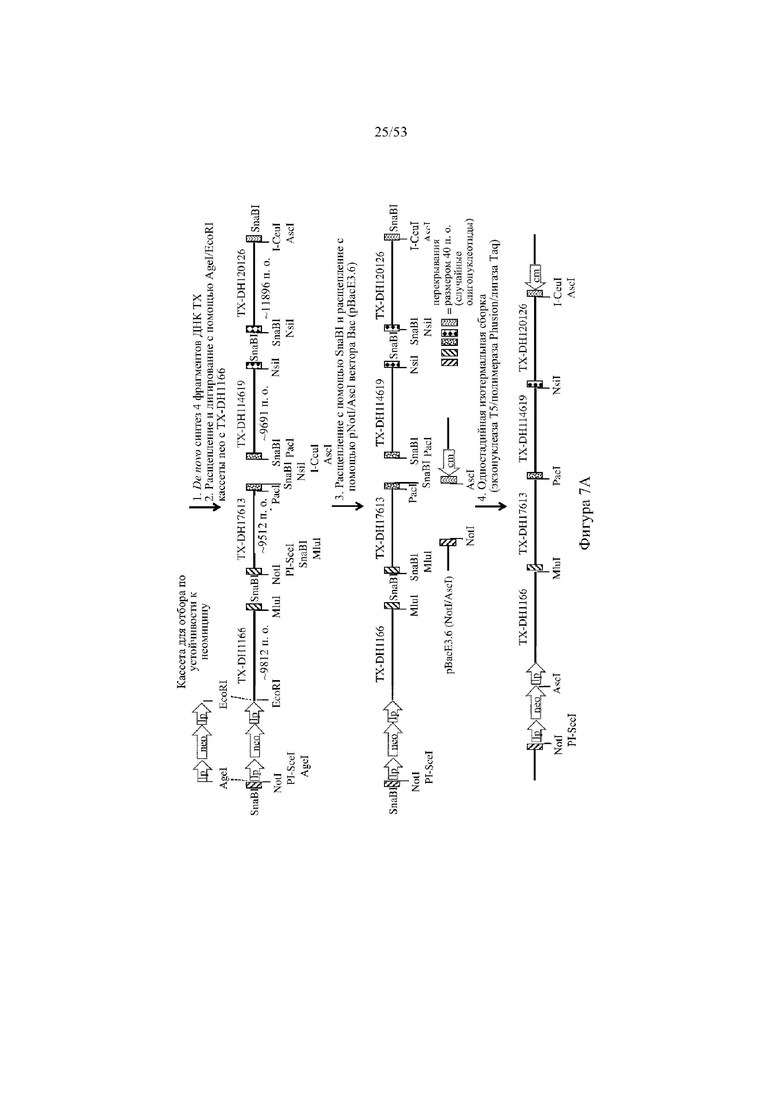
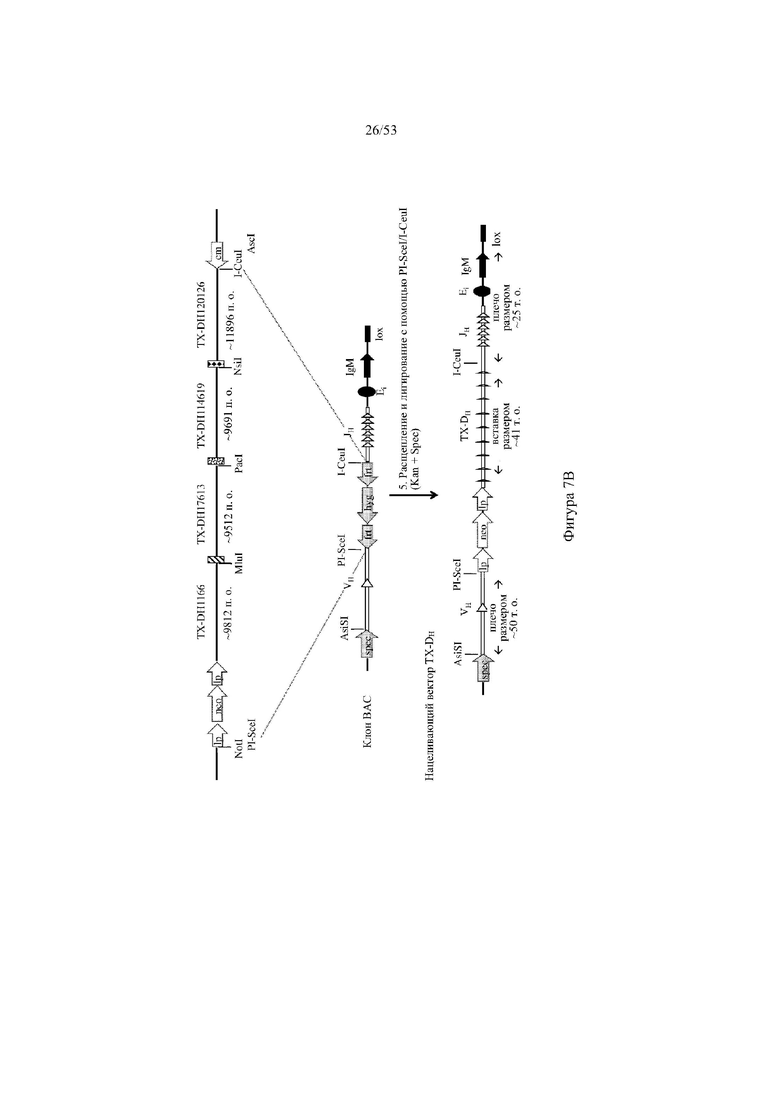

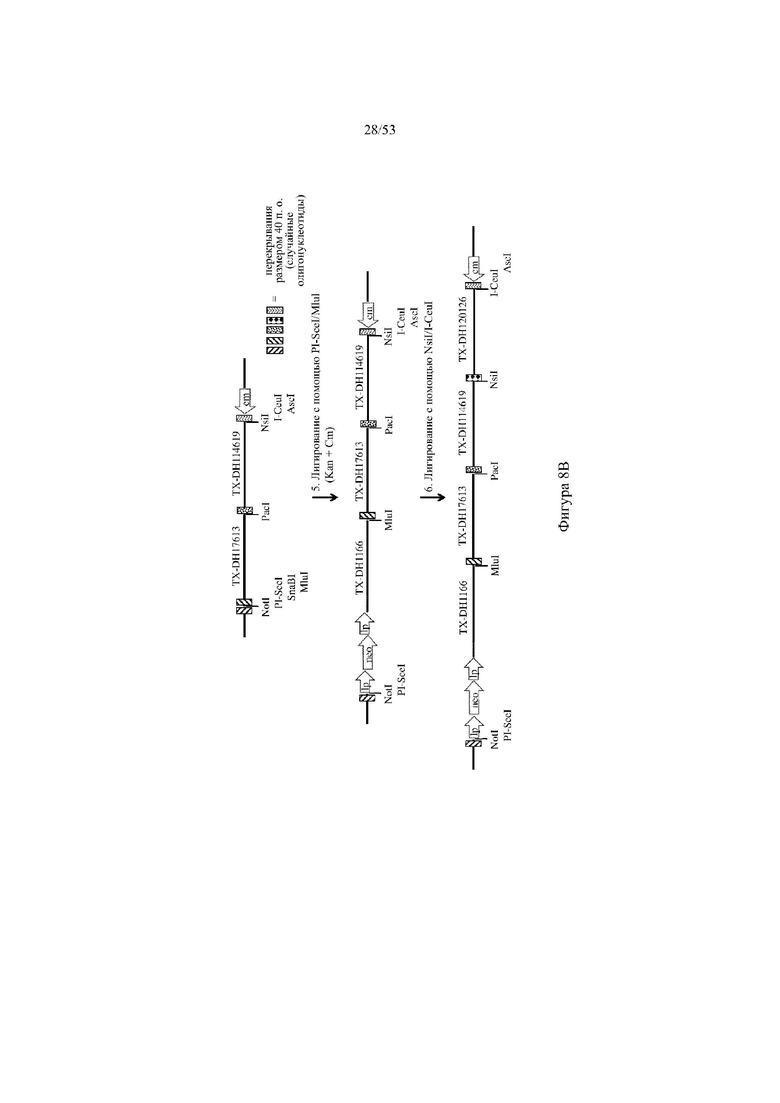
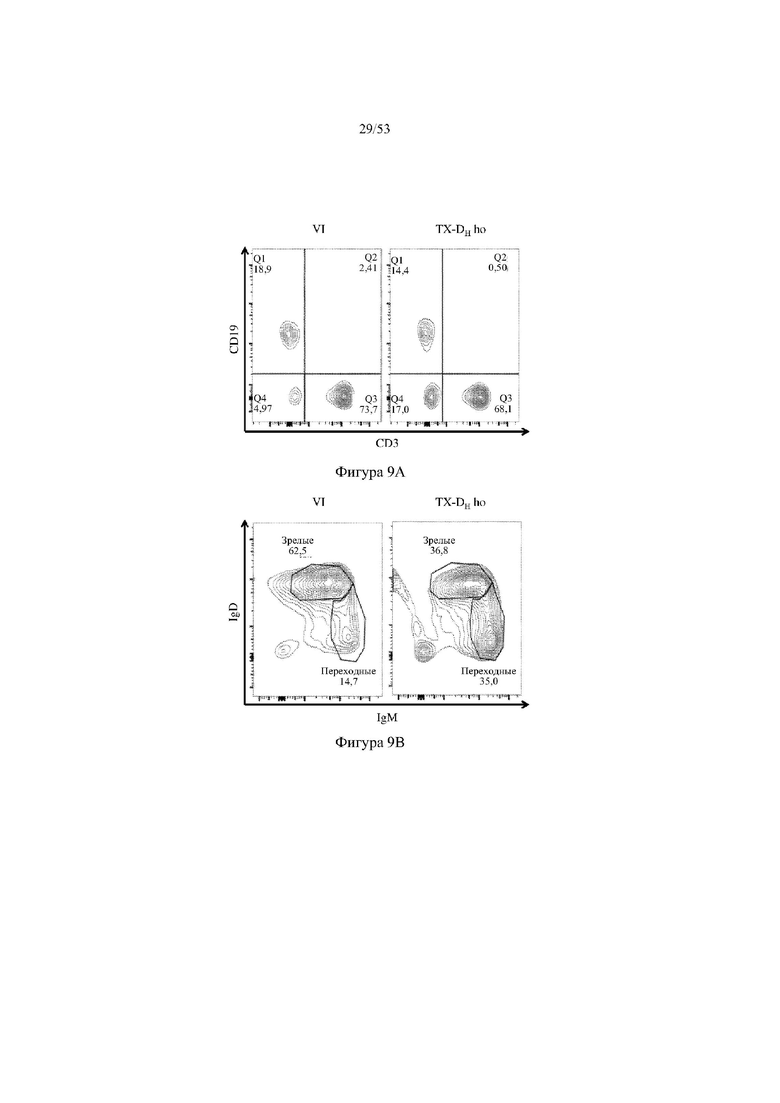
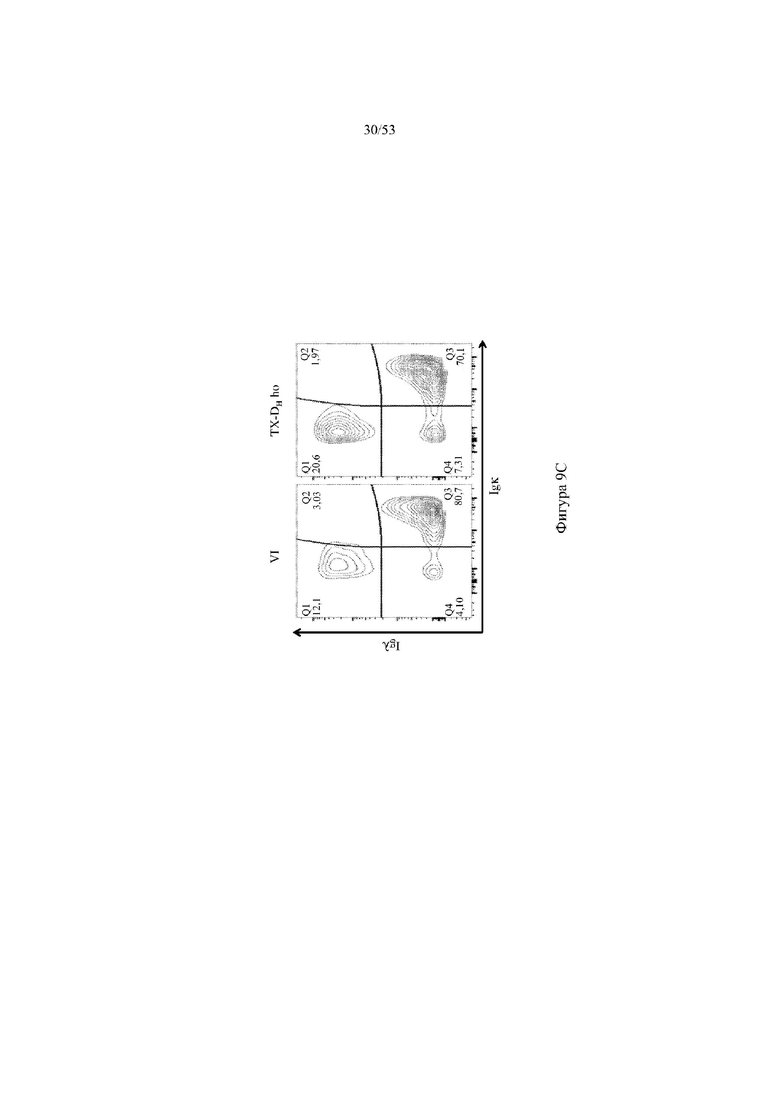


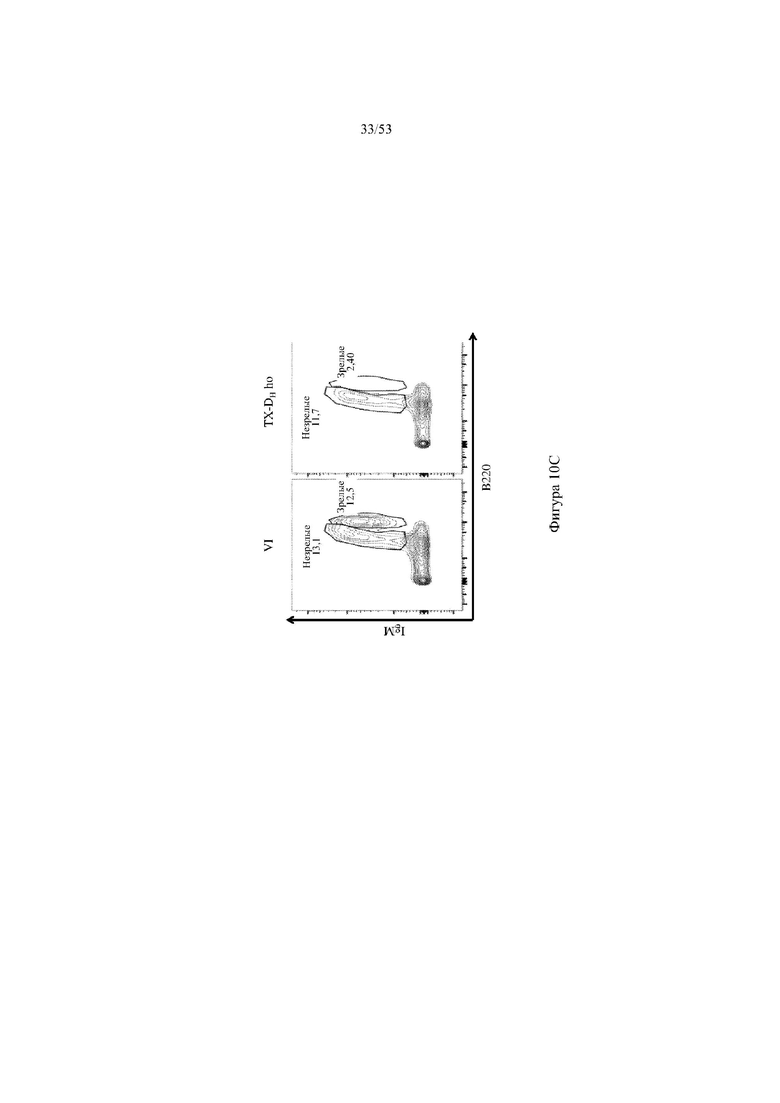
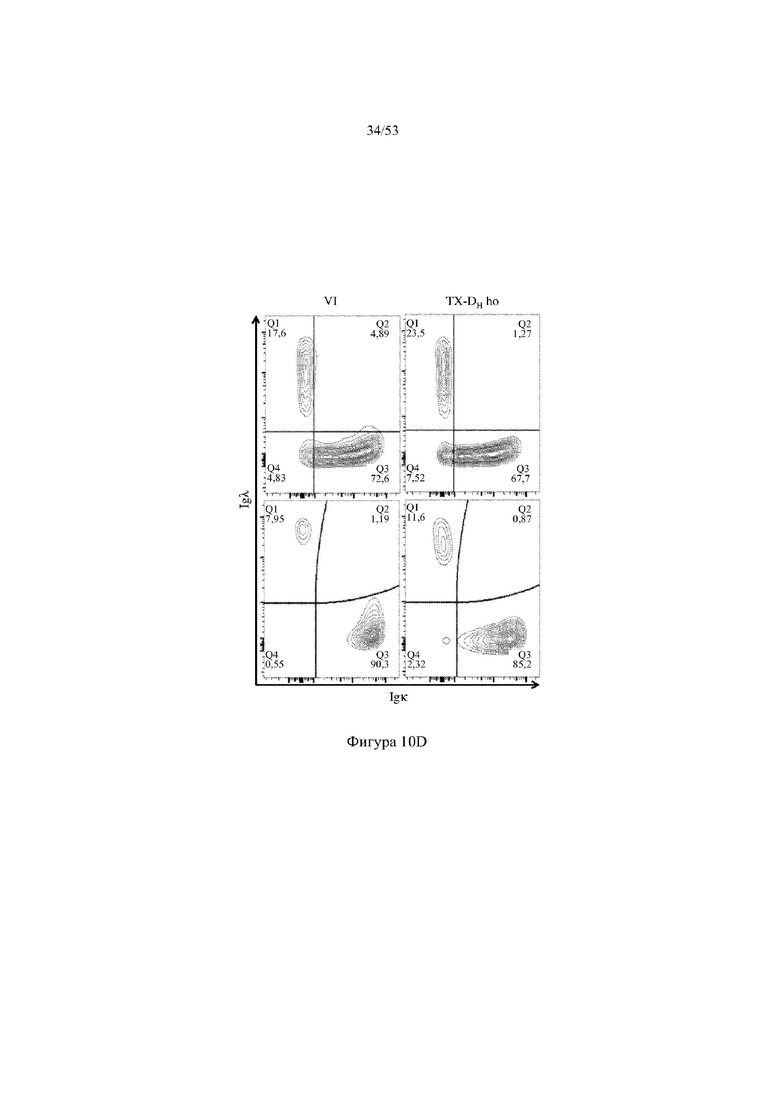
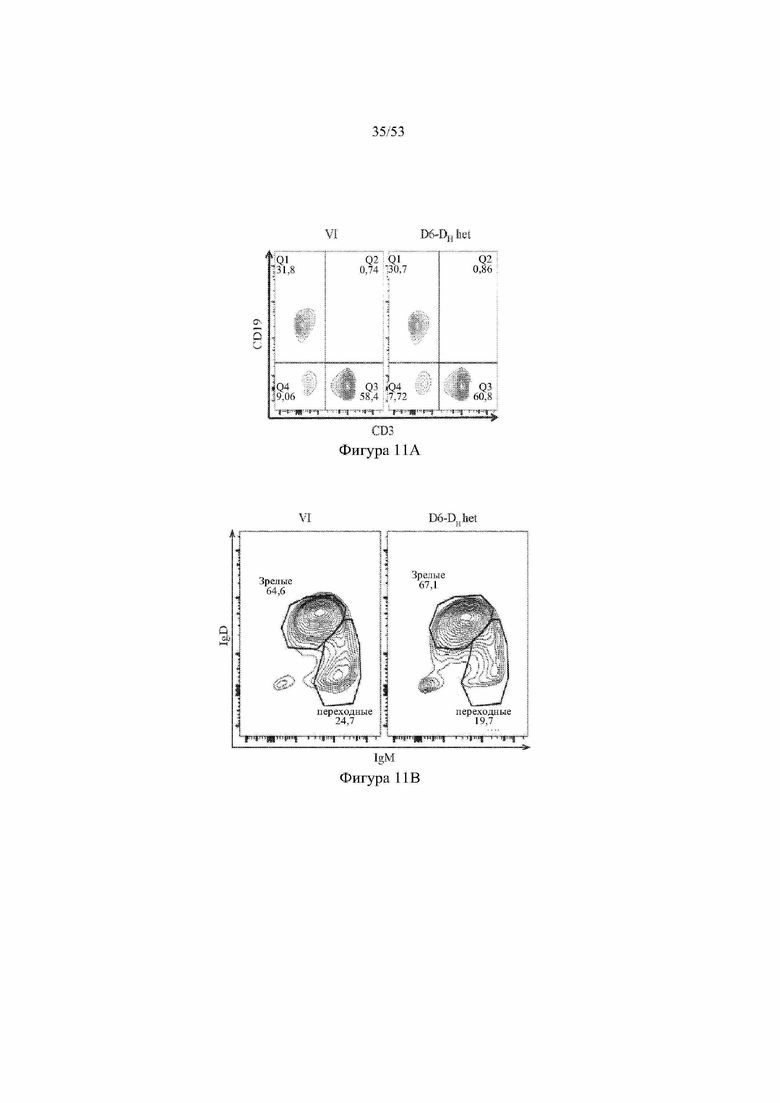
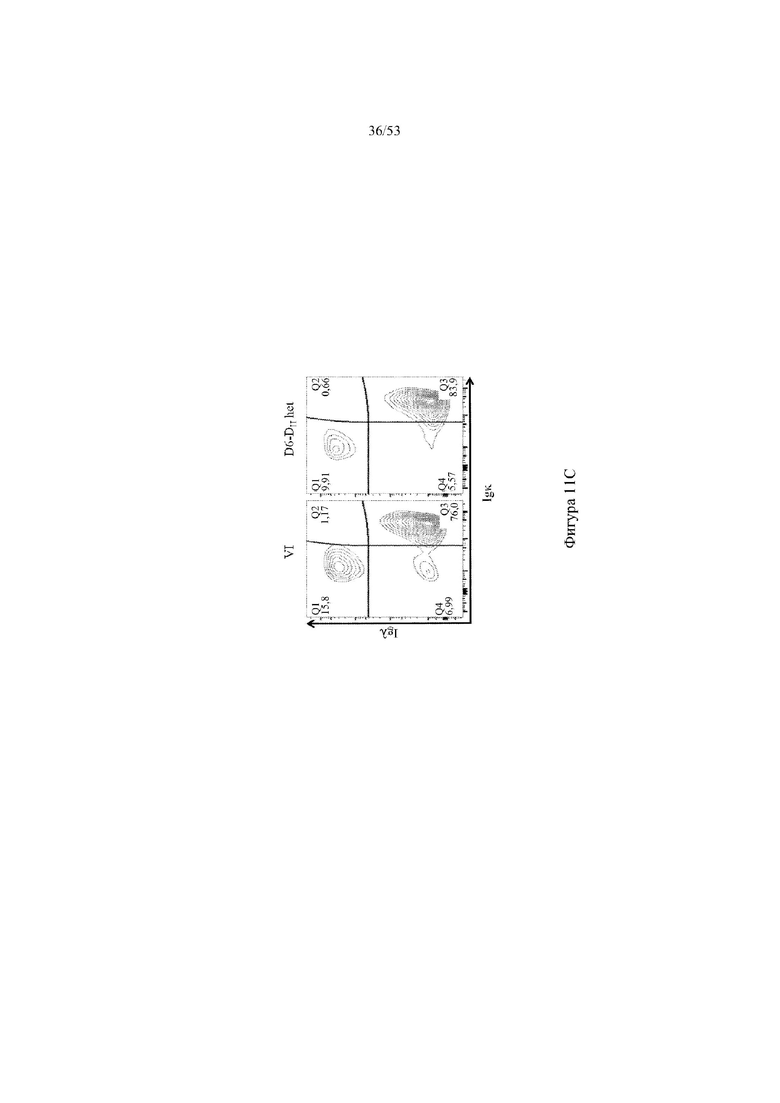
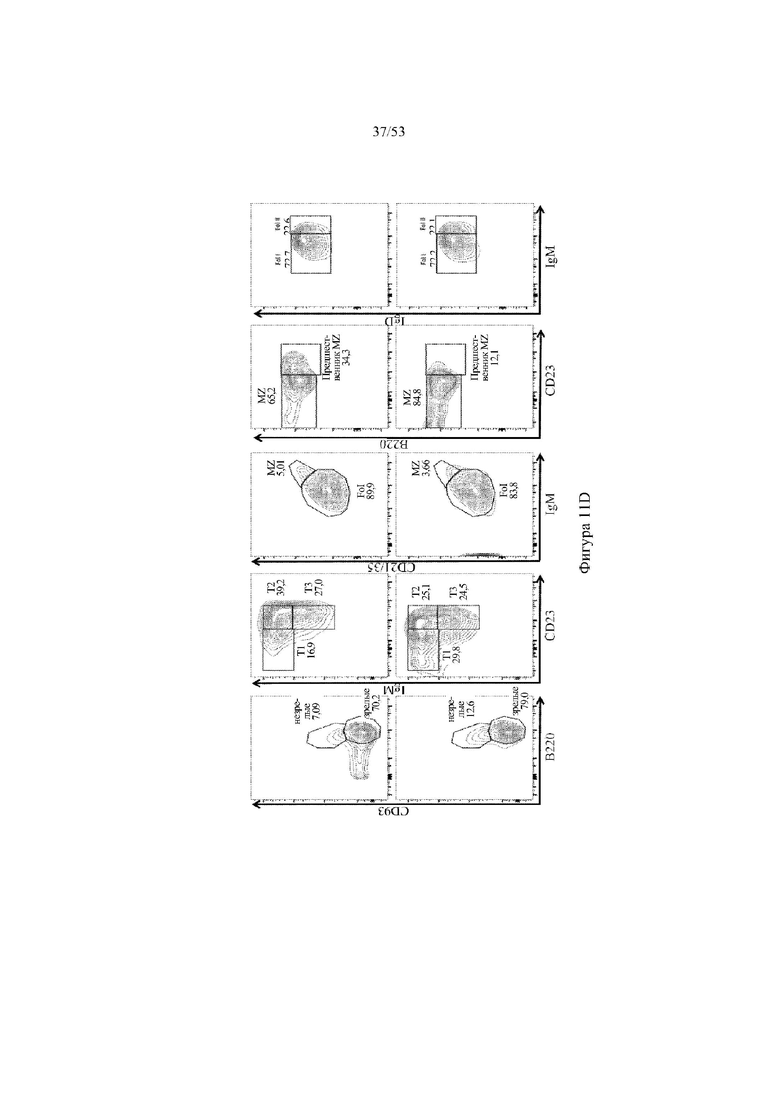
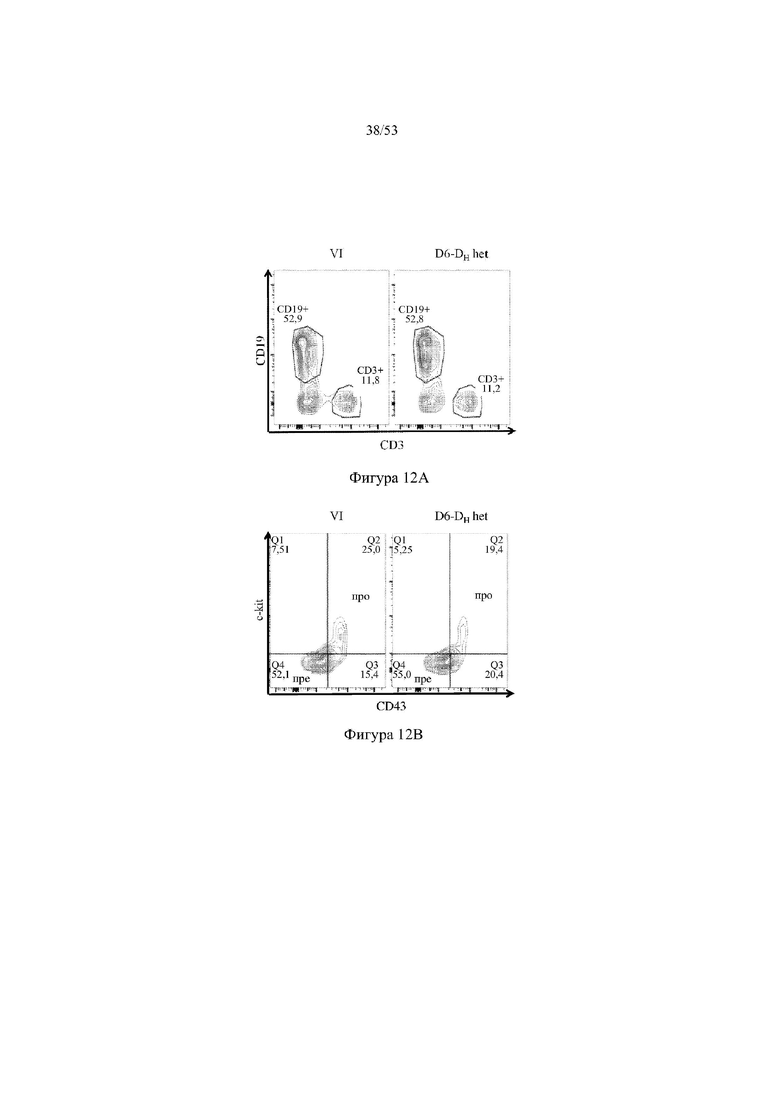

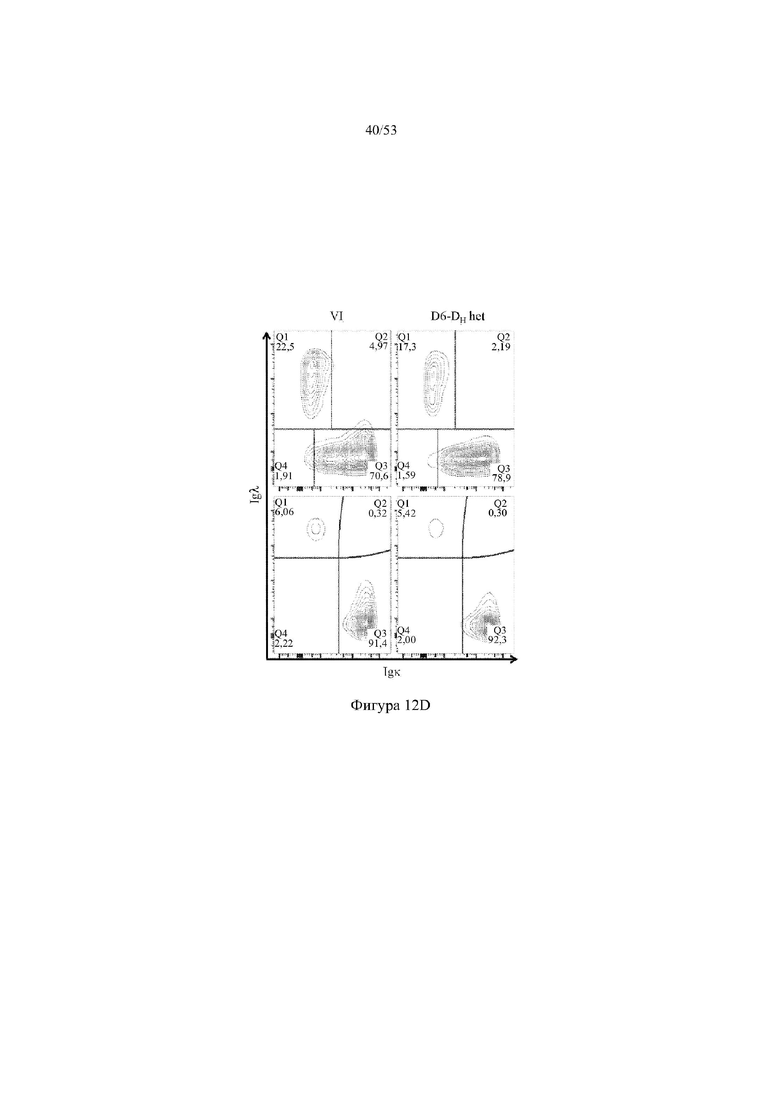
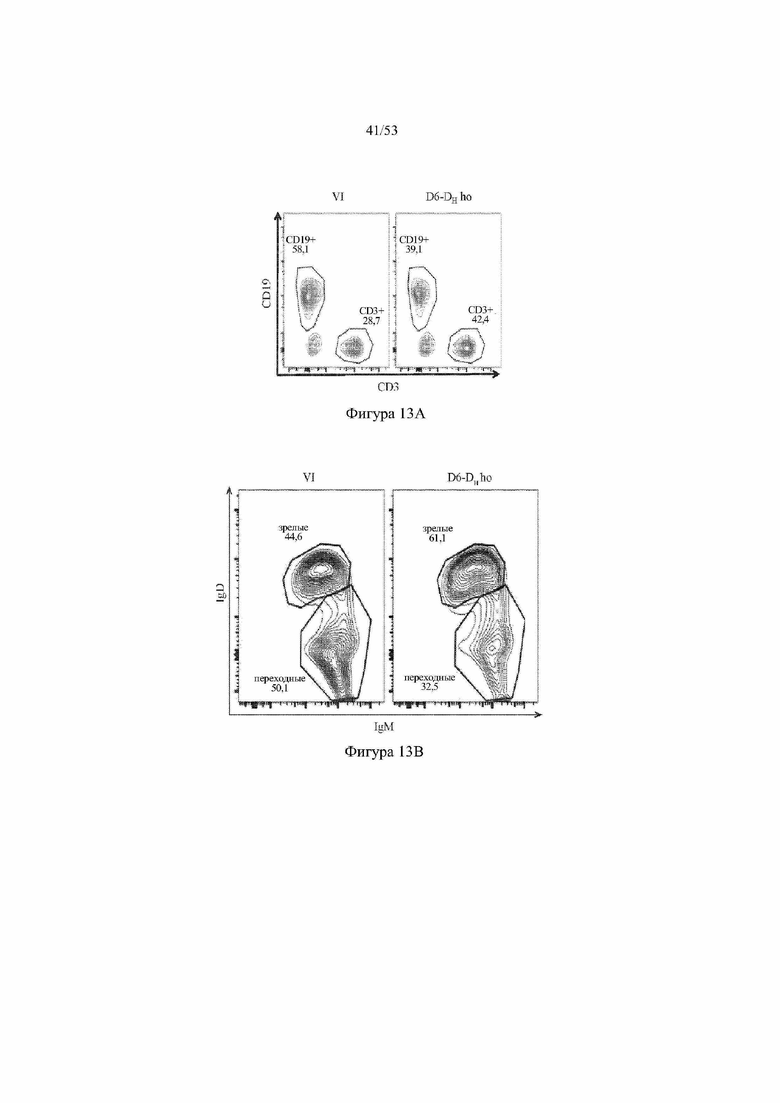
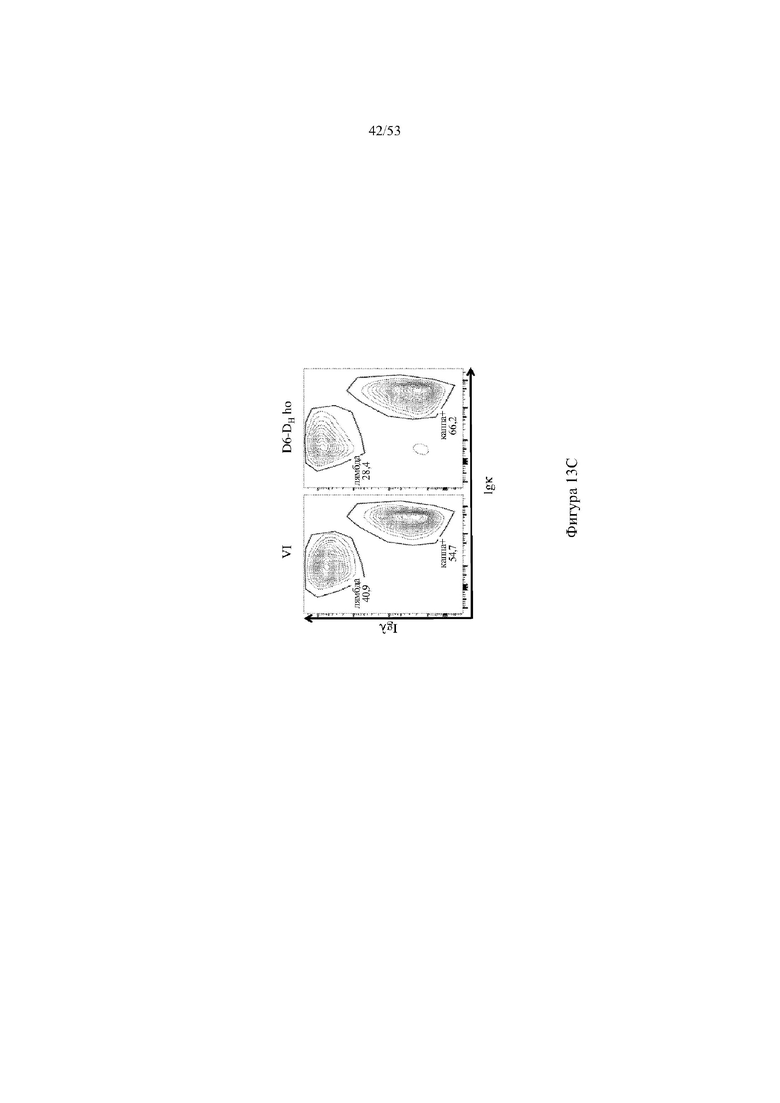

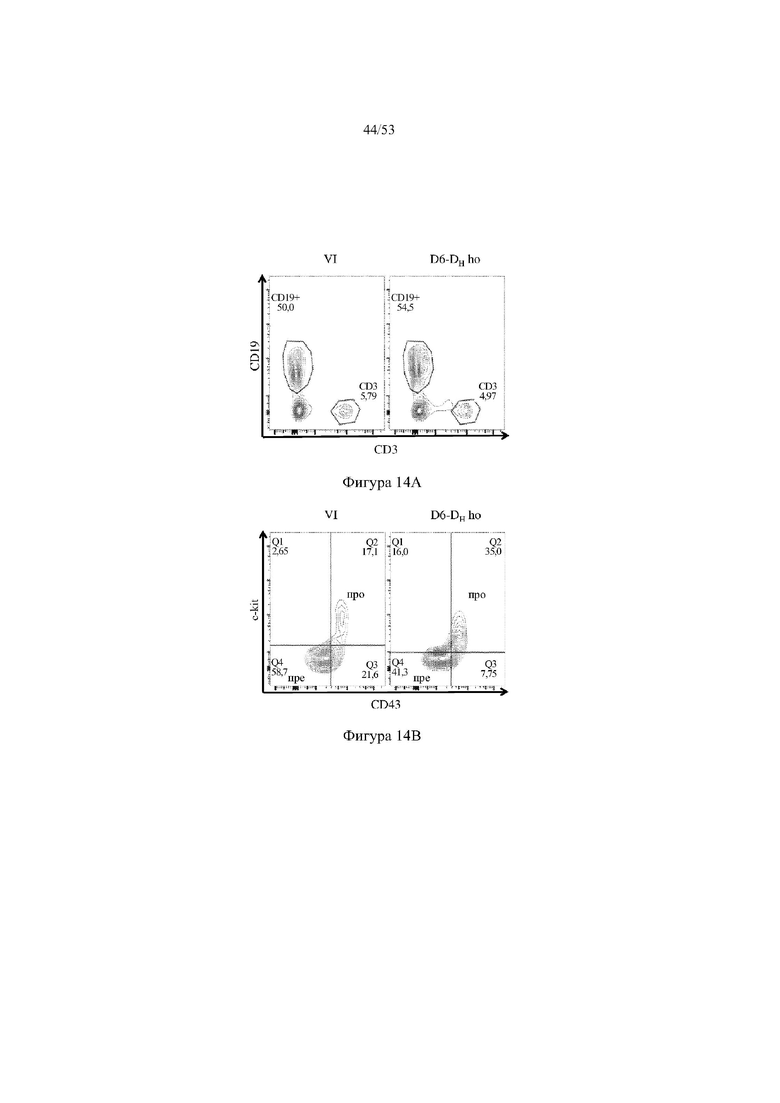

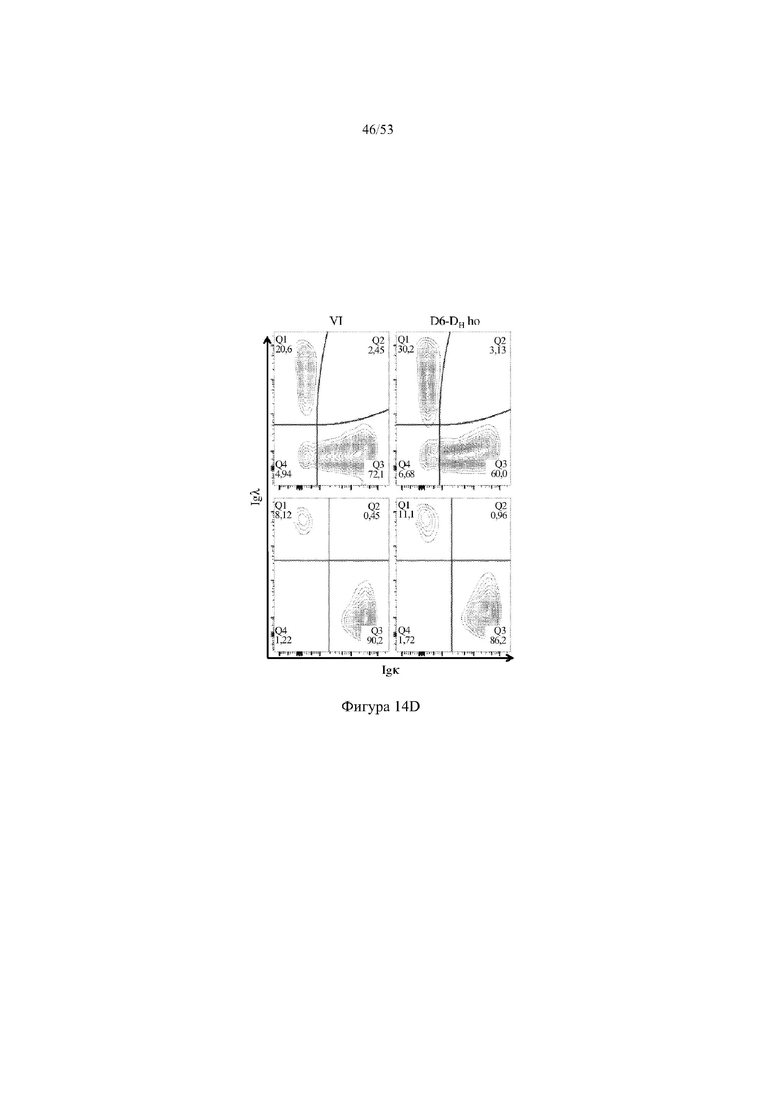


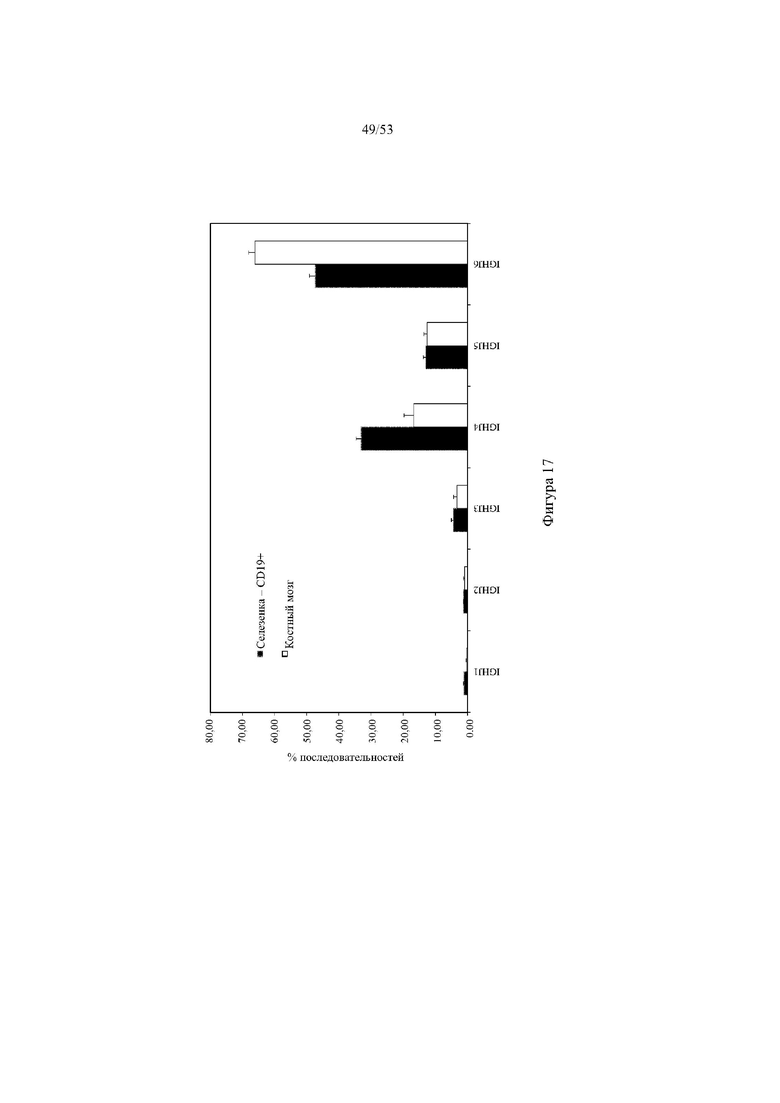
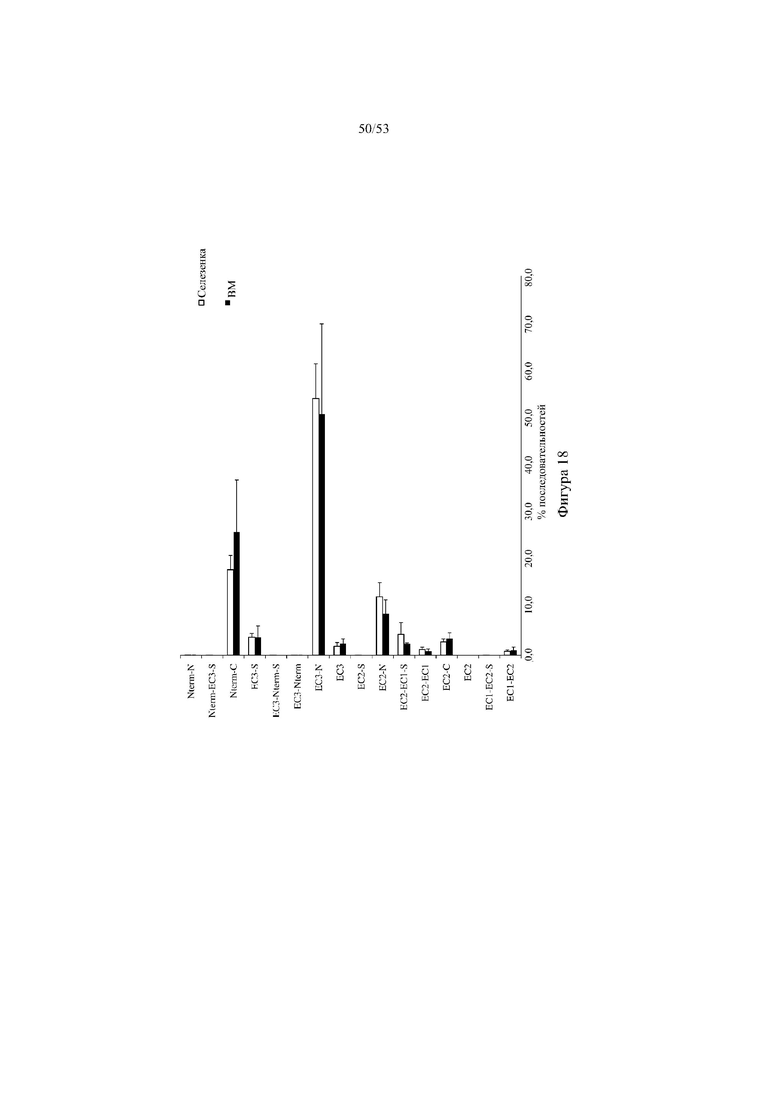
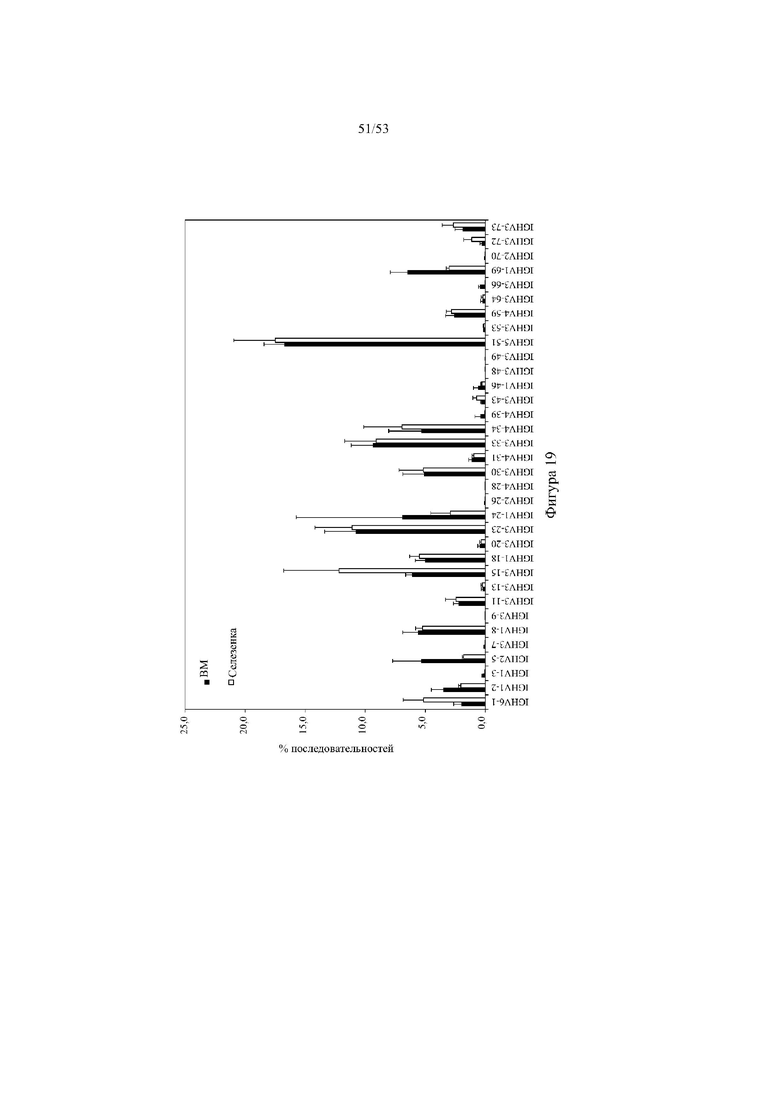

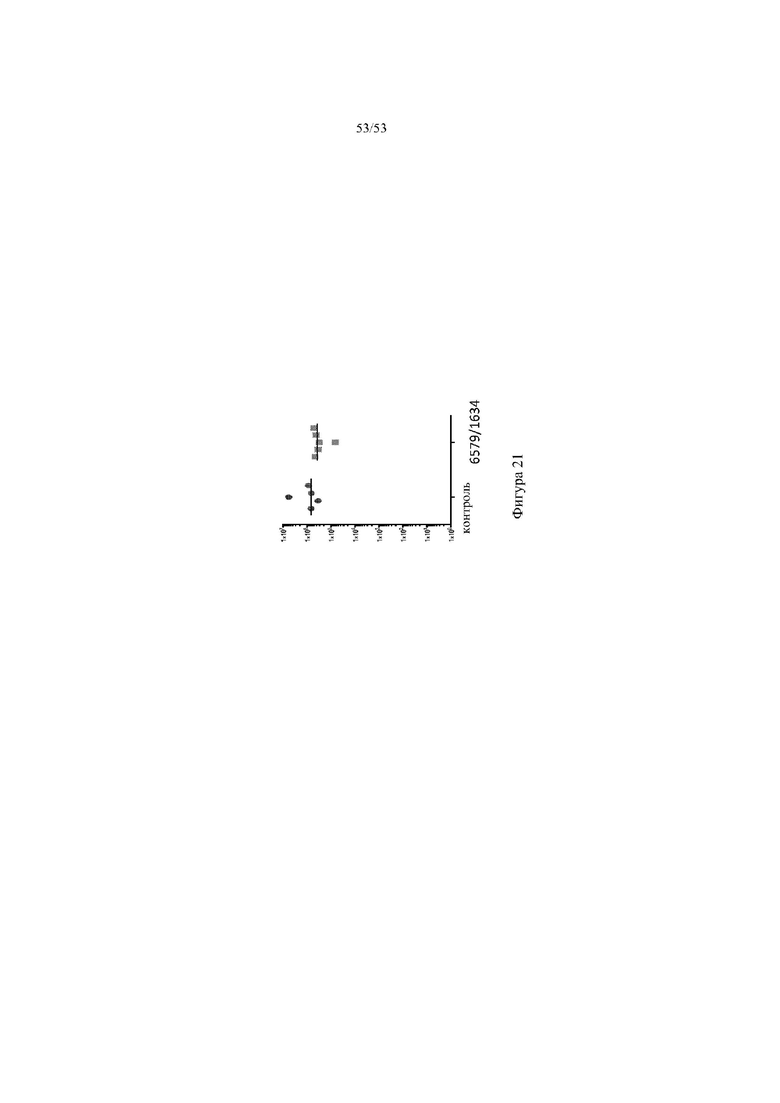
Изобретение относится к области биотехнологии, в частности к крысе или мыши, содержащей зародышевую клетку, которая содержит неперегруппированный вариабельный участок тяжелой цепи иммуноглобулина, функционально связанный с эндогенным константным участком тяжелой цепи иммуноглобулина, и В-клетку, способу ее получения, а также к ее клетке, ткани и эмбриону. Изобретение эффективно для получения антитела. 6 н. и 59 з.п. ф-лы, 21 ил., 12 табл., 8 пр.
1. Крыса или мышь для разработки антител, содержащая
(A) зародышевую клетку, которая содержит в эндогенном локусе тяжелой цепи неперегруппированный вариабельный участок тяжелой цепи иммуноглобулина, функционально связанный с эндогенным константным участком тяжелой цепи иммуноглобулина,
где неперегруппированный вариабельный участок тяжелой цепи иммуноглобулина содержит в функциональной связи:
(i) неперегруппированный генный сегмент VH (hVH) человека,
(ii) сконструированный участок DH, который заменяет один или более эндогенных генных сегментов DH,
где сконструированный участок DH содержит одну или более нуклеотидных последовательностей, каждая из которых
(a) кодирует представляющий интерес полипептид, не являющийся иммуноглобулином, или его часть и
(b) фланкирована первой и второй последовательностью сигнала рекомбинации, и
(iii) неперегруппированный генный сегмент JH (hJH) человека, и
(B) В-клетку,
где неперегруппированный генный сегмент hVH из (i), одна или более нуклеотидных последовательностей из (ii) и неперегруппированный генный сегмент hJH из (iii) рекомбинируют в В-клетке с образованием последовательности перегруппированного вариабельного участка тяжелой цепи hVH/eDH/hJH, функционально связанной с эндогенным константным участком тяжелой цепи иммуноглобулина, и
где в В-клетке экспрессируется перегруппированная последовательность VH/eDH/hJH, функционально связанная с эндогенным константным участком тяжелой цепи иммуноглобулина.
2. Крыса или мышь по п. 1, где представляющий интерес полипептид, не являющийся иммуноглобулином, представляет собой хемокиновый рецептор.
3. Крыса или мышь по п. 2, где хемокиновый рецептор представляет собой атипичный хемокиновый рецептор (ACKR).
4. Крыса или мышь по п. 3, где ACKR представляет собой D6 хемокиновый рецептор-ловушку.
5. Крыса или мышь по п. 4, где сконструированный участок DH включает 5, 10, 15, 20, 25 или больше нуклеотидных последовательностей, каждая из которых кодирует внеклеточную часть D6 хемокинового рецептора-ловушки.
6. Крыса или мышь по п. 5, где сконструированный участок DH включает 25 нуклеотидных последовательностей, каждая из которых кодирует внеклеточную часть D6 хемокинового рецептора-ловушки.
7. Крыса или мышь по п. 5 или 6, где внеклеточная часть D6 хемокинового рецептора-ловушки выбрана из группы, состоящей из N-концевого участка, внеклеточной петли и их комбинаций.
8. Крыса или мышь по п. 6, где каждая из 25 нуклеотидных последовательностей кодирует последовательность, которая по меньшей мере на 95% идентична последовательности, изложенной в SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 5, SEQ ID NO: 7, SEQ ID NO: 9, SEQ ID NO: 11, SEQ ID NO: 13, SEQ ID NO: 15, SEQ ID NO: 17, SEQ ID NO: 19, SEQ ID NO: 21, SEQ ID NO: 23, SEQ ID NO: 25, SEQ ID NO: 27, SEQ ID NO: 29, SEQ ID NO: 31, SEQ ID NO: 33, SEQ ID NO: 35, SEQ ID NO: 37, SEQ ID NO: 39, SEQ ID NO: 41, SEQ ID NO: 43, SEQ ID NO: 45, SEQ ID NO: 47 или SEQ ID NO: 49.
9. Крыса или мышь по п. 6, где каждая из 25 нуклеотидных последовательностей кодирует последовательность, которая практически идентична последовательности, изложенной в SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 5, SEQ ID NO: 7, SEQ ID NO: 9, SEQ ID NO: 11, SEQ ID NO: 13, SEQ ID NO: 15, SEQ ID NO: 17, SEQ ID NO: 19, SEQ ID NO: 21, SEQ ID NO: 23, SEQ ID NO: 25, SEQ ID NO: 27, SEQ ID NO: 29, SEQ ID NO: 31, SEQ ID NO: 33, SEQ ID NO: 35, SEQ ID NO: 37, SEQ ID NO: 39, SEQ ID NO: 41, SEQ ID NO: 43, SEQ ID NO: 45, SEQ ID NO: 47 или SEQ ID NO: 49.
10. Крыса или мышь по п. 6, где каждая из 25 нуклеотидных последовательностей кодирует последовательность, которая идентична последовательности, изложенной в SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 5, SEQ ID NO: 7, SEQ ID NO: 9, SEQ ID NO: 11, SEQ ID NO: 13, SEQ ID NO: 15, SEQ ID NO: 17, SEQ ID NO: 19, SEQ ID NO: 21, SEQ ID NO: 23, SEQ ID NO: 25, SEQ ID NO: 27, SEQ ID NO: 29, SEQ ID NO: 31, SEQ ID NO: 33, SEQ ID NO: 35, SEQ ID NO: 37, SEQ ID NO: 39, SEQ ID NO: 41, SEQ ID NO: 43, SEQ ID NO: 45, SEQ ID NO: 47 или SEQ ID NO: 49.
11. Крыса или мышь по п. 1, где представляющий интерес полипептид, не являющийся иммуноглобулином, представляет собой конотоксин или токсин тарантула.
12. Крыса или мышь по п. 11, где конотоксин выбран из группы, состоящей из α-конотоксина, δ-конотоксина, κ-конотоксина, μ-конотоксина, ω-конотоксина и их комбинаций.
13. Крыса или мышь по п. 12, где конотоксин представляет собой μ-конотоксин.
14. Крыса или мышь по п. 13, где сконструированный участок DH включает 5, 10, 15, 20, 25 или больше нуклеотидных последовательностей, каждая из которых кодирует часть μ-конотоксина, токсина тарантула или их комбинаций.
15. Крыса или мышь по п. 14, где сконструированный участок DH включает 26 нуклеотидных последовательностей, каждая из которых кодирует часть μ-конотоксина и/или токсина тарантула.
16. Крыса или мышь по п. 15, где каждая из 26 нуклеотидных последовательностей кодирует последовательность, которая по меньшей мере на 95% идентична последовательности, изложенной в SEQ ID NO: 180, SEQ ID NO: 182, SEQ ID NO: 184, SEQ ID NO: 186, SEQ ID NO: 188, SEQ ID NO: 190, SEQ ID NO: 192, SEQ ID NO: 194, SEQ ID NO: 196, SEQ ID NO: 198, SEQ ID NO: 200, SEQ ID NO: 202, SEQ ID NO: 204, SEQ ID NO: 206, SEQ ID NO: 208, SEQ ID NO: 210, SEQ ID NO: 212, SEQ ID NO: 214, SEQ ID NO: 216, SEQ ID NO: 218, SEQ ID NO: 220, SEQ ID NO: 222, SEQ ID NO: 224, SEQ ID NO: 226, SEQ ID NO: 228 или SEQ ID NO: 230.
17. Крыса или мышь по п. 15, где каждая из 26 нуклеотидных последовательностей кодирует последовательность, которая практически идентична последовательности, изложенной в SEQ ID NO: 180, SEQ ID NO: 182, SEQ ID NO: 184, SEQ ID NO: 186, SEQ ID NO: 188, SEQ ID NO: 190, SEQ ID NO: 192, SEQ ID NO: 194, SEQ ID NO: 196, SEQ ID NO: 198, SEQ ID NO: 200, SEQ ID NO: 202, SEQ ID NO: 204, SEQ ID NO: 206, SEQ ID NO: 208, SEQ ID NO: 210, SEQ ID NO: 212, SEQ ID NO: 214, SEQ ID NO: 216, SEQ ID NO: 218, SEQ ID NO: 220, SEQ ID NO: 222, SEQ ID NO: 224, SEQ ID NO: 226, SEQ ID NO: 228 или SEQ ID NO: 230.
18. Крыса или мышь по п. 15, где каждая из 26 нуклеотидных последовательностей кодирует последовательность, которая идентична последовательности, изложенной в SEQ ID NO: 180, SEQ ID NO: 182, SEQ ID NO: 184, SEQ ID NO: 186, SEQ ID NO: 188, SEQ ID NO: 190, SEQ ID NO: 192, SEQ ID NO: 194, SEQ ID NO: 196, SEQ ID NO: 198, SEQ ID NO: 200, SEQ ID NO: 202, SEQ ID NO: 204, SEQ ID NO: 206, SEQ ID NO: 208, SEQ ID NO: 210, SEQ ID NO: 212, SEQ ID NO: 214, SEQ ID NO: 216, SEQ ID NO: 218, SEQ ID NO: 220, SEQ ID NO: 222, SEQ ID NO: 224, SEQ ID NO: 226, SEQ ID NO: 228 или SEQ ID NO: 230.
19. Крыса или мышь по любому из предыдущих пунктов, где одна или более нуклеотидных последовательностей содержат последовательность, которая увеличивает частоту возникновения соматической гипермутации в одной или более нуклеотидных последовательностях.
20. Крыса или мышь по любому из предыдущих пунктов, где первая последовательность сигнала рекомбинации содержит последовательность, которая по меньшей мере на 95% идентична первой последовательности сигнала рекомбинации, изложенной в любой из SEQ ID NO: 51-112.
21. Крыса или мышь по любому из предыдущих пунктов, где первая последовательность сигнала рекомбинации содержит последовательность, которая практически идентична первой последовательности сигнала рекомбинации, изложенной в любой из SEQ ID NO: 51-112.
22. Крыса или мышь по любому из предыдущих пунктов, где первая последовательность сигнала рекомбинации содержит последовательность, которая идентична первой последовательности сигнала рекомбинации, изложенной в любой из SEQ ID NO: 51-112.
23. Крыса или мышь по любому из предыдущих пунктов, где вторая последовательность сигнала рекомбинации содержит последовательность, которая по меньшей мере на 95% идентична второй последовательности сигнала рекомбинации, изложенной в любой из SEQ ID NO: 51-112.
24. Крыса или мышь по любому из предыдущих пунктов, где вторая последовательность сигнала рекомбинации содержит последовательность, которая практически идентична второй последовательности сигнала рекомбинации, изложенной в любой из SEQ ID NO: 51-112.
25. Крыса или мышь по любому из предыдущих пунктов, где вторая последовательность сигнала рекомбинации содержит последовательность, которая идентична второй последовательности сигнала рекомбинации, изложенной в любой из SEQ ID NO: 51-112.
26. Крыса или мышь по любому из предыдущих пунктов, где первая и вторая последовательности сигнала рекомбинации выбраны из последовательностей, изложенных в любой из SEQ ID NO: 51-112.
27. Крыса или мышь по любому из предыдущих пунктов, где крыса или мышь сохраняет один или более эндогенных генных сегментов DH дикого типа.
28. Крыса или мышь по любому из предыдущих пунктов, где неперегруппированный генный сегмент VH (hVH) человека содержит множество генных сегментов hVH и/или где неперегруппированный генный сегмент JH (hJH) человека содержит множество генных сегментов hJH.
29. Крыса или мышь по любому из предыдущих пунктов, где:
(i) неперегруппированные генные сегменты VH (hVH) человека заменяют эндогенные генные сегменты VH,
(ii) сконструированный участок DH заменяет эндогенные генные сегменты DH, и
(iii) неперегруппированные генные сегменты JH (hJH) человека заменяют эндогенные генные сегменты JH.
30. Крыса или мышь по любому из предыдущих пунктов, где неперегруппированный вариабельный участок тяжелой цепи иммуноглобулина содержит в функциональной связи:
(i) неперегруппированные генные сегменты VH (hVH) человека, которые заменяют эндогенные генные сегменты VH,
(ii) сконструированный участок DH, который заменяет эндогенные генные сегменты DH, и
(iii) неперегруппированные генные сегменты JH (hJH) человека, которые заменяют все эндогенные генные сегменты JH.
31. Крыса или мышь по любому из предыдущих пунктов, которая представляет собой мышь.
32. Выделенная клетка грызуна для получения грызуна, пригодного для разработки антител, геном которой содержит в эндогенном локусе тяжелой цепи вариабельный участок тяжелой цепи иммуноглобулина, функционально связанный с эндогенным константным участком тяжелой цепи иммуноглобулина,
где неперегруппированный вариабельный участок тяжелой цепи иммуноглобулина содержит:
(i) неперегруппированный генный сегмент VH (hVH) человека,
(ii) сконструированный участок DH, содержащий одну или более нуклеотидных последовательностей, каждая из которых
(a) кодирует представляющий интерес полипептид, не являющийся иммуноглобулином, или его часть и
(b) фланкирована первой и второй последовательностью сигнала рекомбинации, и
(iii) неперегруппированный генный сегмент JH (hJH) человека,
так чтобы неперегруппированный генный сегмент hVH из (i), одна или более нуклеотидных последовательностей из (ii) и неперегруппированный генный сегмент hJH из (iii) рекомбинировали в В-клетке с образованием последовательности перегруппированного вариабельного участка тяжелой цепи hVH/eDH/hJH, функционально связанной с эндогенным константным участком тяжелой цепи иммуноглобулина, и
где клетка грызуна представляет собой клетку крысы или клетку мыши.
33. Выделенная клетка грызуна по п. 32, где клетка представляет собой эмбриональную стволовую клетку или зародышевую клетку.
34. Эмбрион грызуна для получения грызуна, пригодного для разработки антител, содержащий эмбриональные стволовые клетки,
где геном каждой эмбриональной стволовой клетки содержит в эндогенном локусе тяжелой цепи вариабельный участок тяжелой цепи иммуноглобулина, функционально связанный с эндогенным константным участком тяжелой цепи иммуноглобулина,
где неперегруппированный вариабельный участок тяжелой цепи иммуноглобулина содержит:
(i) неперегруппированный генный сегмент VH (hVH) человека,
(ii) сконструированный участок DH, содержащий одну или более нуклеотидных последовательностей, каждая из которых
(a) кодирует представляющий интерес полипептид, не являющийся иммуноглобулином, или его часть и
(b) фланкирована первой и второй последовательностью сигнала рекомбинации, и
(iii) неперегруппированный генный сегмент JH (hJH) человека,
так чтобы неперегруппированный генный сегмент hVH из (i), одна или более нуклеотидных последовательностей из (ii) и неперегруппированный генный сегмент hJH из (iii) рекомбинировали в В-клетке с образованием последовательности перегруппированного вариабельного участка тяжелой цепи hVH/eDH/hJH, функционально связанной с эндогенным константным участком тяжелой цепи иммуноглобулина, и
где эмбрион грызуна представляет собой эмбрион крысы или эмбрион мыши.
35. Выделенная ткань грызуна для разработки антител, содержащая клетку, геном которой содержит в эндогенном локусе тяжелой цепи вариабельный участок тяжелой цепи иммуноглобулина, функционально связанный с эндогенным константным участком тяжелой цепи иммуноглобулина,
где неперегруппированный вариабельный участок тяжелой цепи иммуноглобулина содержит:
(i) неперегруппированный генный сегмент VH (hVH) человека,
(ii) сконструированный участок DH, содержащий одну или более нуклеотидных последовательностей, каждая из которых
(а) кодирует представляющий интерес полипептид, не являющийся иммуноглобулином, или его часть и
(b) фланкирована первой и второй последовательностью сигнала рекомбинации, и
(iii) неперегруппированный генный сегмент JH (hJH) человека,
так чтобы неперегруппированный генный сегмент hVH из (i), одна или более нуклеотидных последовательностей из (ii) и неперегруппированный генный сегмент hJH из (iii) рекомбинировали в В-клетке с образованием последовательности перегруппированного вариабельного участка тяжелой цепи hVH/eDH/hJH, функционально связанной с эндогенным константным участком тяжелой цепи иммуноглобулина, и
где ткань грызуна представляет собой ткань крысы или ткань мыши.
36. Способ получения грызуна, геном которого содержит вариабельный участок тяжелой цепи иммуноглобулина, который включает сконструированный участок DH, при этом способ предусматривает
(a) получение эмбриональной стволовой клетки крысы или мыши, где эмбриональная стволовая клетка крысы или мыши содержит в эндогенном локусе тяжелой цепи иммуноглобулина замену эндогенной последовательности, содержащей эндогенный генный сегмент VH иммуноглобулина, эндогенные генные сегменты DH иммуноглобулина и эндогенный генный сегмент JH иммуноглобулина, на геномную последовательность человека, содержащую неперегруппированный генный сегмент VH человека, неперегруппированные генные сегменты DH иммуноглобулина человека и неперегруппированные генные сегменты JH иммуноглобулина человека,
(b) осуществление замены одного или нескольких неперегруппированных генных сегментов DH иммуноглобулина человека на сконструированный участок DH так, чтобы сконструированный генный сегмент DH был вставлен между неперегруппированным генным сегментом VH человека и неперегруппированными генными сегментами JH человека и в функциональной связи с ними,
где сконструированный участок DH содержит одну или более нуклеотидных последовательностей, каждая из которых
(i) кодирует представляющий интерес полипептид, не являющийся иммуноглобулином, или его часть; и
(ii) фланкирована первой и второй последовательностью сигнала рекомбинации,
так чтобы неперегруппированный генный сегмент VH человека из (i), сконструированный участок DH из (ii) и неперегруппированный генный сегмент JH человека из (iii) были функционально связаны с эндогенным константным участком тяжелой цепи иммуноглобулина и могли рекомбинировать в В-клетке с образованием последовательности перегруппированного вариабельного участка тяжелой цепи hVH/eDH/hJH, которая также функционально связана с эндогенным константным участком тяжелой цепи иммуноглобулина, и
(с) получение крысы или мыши с применением эмбриональной стволовой клетки из (b).
37. Способ по п. 36, где представляющий интерес полипептид, не являющийся иммуноглобулином, представляет собой хемокиновый рецептор.
38. Способ по п. 37, где хемокиновый рецептор представляет собой атипичный хемокиновый рецептор (ACKR).
39. Способ по п. 38, где ACKR представляет собой D6 хемокиновый рецептор-ловушку.
40. Способ по п. 39, где сконструированный участок DH включает 5, 10, 15, 20, 25 или больше нуклеотидных последовательностей, каждая из которых кодирует внеклеточную часть D6 хемокинового рецептора-ловушки.
41. Способ по п. 40, где сконструированный участок DH включает 25 нуклеотидных последовательностей, каждая из которых кодирует внеклеточную часть D6 хемокинового рецептора-ловушки.
42. Способ по п. 40 или 41, где внеклеточная часть D6 хемокинового рецептора-ловушки выбрана из группы, состоящей из N-концевого участка, внеклеточной петли и их комбинаций.
43. Способ по п. 36, где представляющий интерес полипептид, не являющийся иммуноглобулином, представляет собой конотоксин или токсин тарантула.
44. Способ по п. 43, где конотоксин выбран из группы, состоящей из α-конотоксина, δ-конотоксина, κ-конотоксина, μ-конотоксина, ω-конотоксина и их комбинаций.
45. Способ по п. 44, где конотоксин представляет собой μ-конотоксин.
46. Способ по п. 45, где сконструированный участок DH включает 5, 10, 15, 20, 25 или больше нуклеотидных последовательностей, каждая из которых кодирует часть μ-конотоксина, токсина тарантула или их комбинаций.
47. Способ по п. 46, где сконструированный участок DH включает 26 нуклеотидных последовательностей, каждая из которых кодирует часть μ-конотоксина и/или токсина тарантула.
48. Способ по любому из пп. 36-47, где геномная последовательность человека содержит вариабельный участок тяжелой цепи иммуноглобулина человека и где вариабельный участок тяжелой цепи иммуноглобулина человека функционально связан с эндогенным константным участком тяжелой цепи иммуноглобулина.
49. Способ получения антитела от грызуна, при этом способ предусматривает стадии:
(a) иммунизации грызуна, который представляет собой крысу или мышь, антигеном, где грызун содержит зародышевую клетку, которая содержит в эндогенном локусе тяжелой цепи неперегруппированный вариабельный участок тяжелой цепи иммуноглобулина, функционально связанный с эндогенным константным участком тяжелой цепи иммуноглобулина, где вариабельный участок тяжелой цепи иммуноглобулина содержит в функциональной связи
(i) неперегруппированный генный сегмент VH (hVH) человека,
(ii) сконструированный участок DH, где сконструированный участок DH заменяет один или более эндогенных генных сегментов DH и содержит одну или более нуклеотидных последовательностей, каждая из которых
кодирует представляющий интерес полипептид, не являющийся иммуноглобулином, или его часть и
фланкирована первой и второй последовательностью сигнала рекомбинации; и
(iii) неперегруппированный генный сегмент JH (hJH) человека,
(b) содержания грызуна в условиях, подходящих для того, чтобы у грызуна вырабатывался иммунный ответ на антиген; и
(c) выделения из грызуна или клетки грызуна антитела, которое связывает антиген.
50. Способ по п. 49, где клетка грызуна представляет собой В-клетку.
51. Способ по п. 50, дополнительно предусматривающий слияние В-клетки с клеткой миеломы с получением гибридомы.
52. Способ по любому из пп. 49-51, где представляющий интерес полипептид, не являющийся иммуноглобулином, представляет собой хемокиновый рецептор.
53. Способ по п. 52, где хемокиновый рецептор представляет собой атипичный хемокиновый рецептор (ACKR).
54. Способ по п. 53, где ACKR представляет собой D6 хемокиновый рецептор-ловушку.
55. Способ по п. 54, где сконструированный участок DH включает 5, 10, 15, 20, 25 или больше нуклеотидных последовательностей, каждая из которых кодирует внеклеточную часть D6 хемокинового рецептора-ловушки.
56. Способ по п. 55, где сконструированный участок DH включает 25 нуклеотидных последовательностей, каждая из которых кодирует внеклеточную часть D6 хемокинового рецептора-ловушки.
57. Способ по п. 55 или 56, где внеклеточная часть D6 хемокинового рецептора-ловушки выбрана из группы, состоящей из N-концевого участка, внеклеточной петли и их комбинаций.
58. Способ по любому из пп. 49-51, где представляющий интерес полипептид, не являющийся иммуноглобулином, представляет собой конотоксин или токсин тарантула.
59. Способ по п. 58, где конотоксин выбран из группы, состоящей из α-конотоксина, δ-конотоксина, κ-конотоксина, μ-конотоксина, ω-конотоксина и их комбинаций.
60. Способ по п. 59, где конотоксин представляет собой μ-конотоксин.
61. Способ по п. 60, где сконструированный участок DH включает 5, 10, 15, 20, 25 или больше нуклеотидных последовательностей, каждая из которых кодирует часть μ-конотоксина, токсина тарантула или их комбинаций.
62. Способ по п. 61, где сконструированный участок DH включает 26 нуклеотидных последовательностей, каждая из которых кодирует часть μ-конотоксина и/или токсина тарантула.
63. Способ по любому из пп. 49-62, где вариабельный участок тяжелой цепи иммуноглобулина представляет собой вариабельный участок тяжелой цепи иммуноглобулина человека.
64. Способ по любому из пп. 49-63, где вариабельный участок тяжелой цепи иммуноглобулина функционально связан с константным участком тяжелой цепи иммуноглобулина.
65. Способ по п. 64, где константный участок тяжелой цепи иммуноглобулина представляет собой эндогенный константный участок тяжелой цепи иммуноглобулина.
| WO 03084477 A2, 16.10.2003 | |||
| HARRY W SCHROEDER JR et al., Genetic Control of DH Reading Frame and Its Effect on B-cell Development and Antigen-Specifc Antibody Production,Crit Rev Immunol, 2010, Vol.30, N.4, pp.327-344 | |||
| ANDRE M | |||
| VALE et al., The role of evolutionarily conserved germ‐line DH sequence in B‐1 cell development and natural antibody |

