ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
[001] Настоящая заявка испрашивает приоритет согласно § 119(3) раздела 35 Свода законов США по предварительным заявкам на патент США №№62/685203 (поданной 14 июня 2018 года), 62/702206 (поданной 23 июля 2018 года) и 62/812580 (поданной 1 марта 2019 года), каждая из которых включена в данный документ посредством ссылки во всей своей полноте.
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
[002] Официальная копия перечня последовательностей подана в электроном виде через EFS-Web в виде перечня последовательностей в формате ASCII под названием "10347_ST25.txt", который был создан 13 июня 2019 года, имеет размер приблизительно 50 килобайт и подан одновременно с описанием. Перечень последовательностей, содержащийся в данном документе в формате ASCII, является частью описания и включен в данный документ посредством ссылки во всей своей полноте.
УРОВЕНЬ ТЕХНИКИ
[003] Продукты на основе моноклональных антител революционизировали биофармацевтическую индустрию и достигли значительного прогресса в лечении нескольких заболеваний. Во многих из таких продуктов на основе моноклональных антител использовались природные свойства молекул антител (т.е. традиционные генные сегменты иммуноглобулинов) и, в некоторых случаях, были включены другие свойства, такие как мечение (например, пегилирование, мечение радиоактивным изотопом) или конъюгация с другими лекарственными средствами. При современном темпе одобрения примерно 70 продуктов на основе моноклональных антител ожидаются на рынке к 2020 году. Несмотря на такой прогресс и знания, полученные при применении моноклональных антител для терапевтического применения, остаются заболевания, связанные с мишенями, представляющими сложность для связывания и/или доступа моноклональных антител, что подчеркивает необходимость в различных подходах для разработки эффективных способов лечения.
КРАТКОЕ ОПИСАНИЕ
[004] Настоящее изобретение охватывает признание того, что желательным является получение с помощью генной инженерии животных, отличных от человека (например, грызуна (например, крысы, например, мыши), для создания дополнительных in vivo систем для идентификации и разработки новых терапевтических средств на основе антител и, в некоторых вариантах осуществления, средств, представляющих собой антитела (например, моноклональные антитела и/или их фрагменты), которые можно применять для лечения различных заболеваний. Более того, настоящее изобретение также охватывает признание того, что желательным является получение с помощью генной инженерии животных, отличных от человека, имеющих сконструированный определяющий разнообразие кластер (DH) тяжелой цепи (или сконструированную DH-область) в пределах вариабельной области тяжелой цепи иммуноглобулина (например, гетерологичной вариабельной области тяжелой цепи иммуноглобулина, например, вариабельной области тяжелой цепи человеческого иммуноглобулина), содержащей один или более DH-сегментов, сконструированных таким образом, чтобы они функционально связывались с последовательностью сигнала рекомбинации и позволяли перегруппировку DH-DH, которая может приводить к повышению экспрессии антител, содержащих определяющую комплементарность область три (CDR3), которая характеризуется большей длиной аминокислотной последовательности по сравнению с CDR3 дикого типа (или эталоном), и которая благодаря разнообразию в некоторых вариантах осуществления направляет связывание с конкретными антигенами. В некоторых вариантах осуществления животные, отличные от человека, описанные в данном документе, обеспечивают in vivo системы для разработки антител и/или терапевтических средств на основе антител для введения людям.
[005] В данном документе описаны нуклеотидные молекулы, содержащие по меньшей мере один определяющий разнообразие генный сегмент тяжелой цепи иммуноглобулина (DH), функционально связанный с 23-мерной последовательностью сигнала рекомбинации (RSS), где необязательно генный сегмент DH может быть локализован в пределах сконструированной DH-области, например, вариабельной области тяжелой цепи иммуноглобулина (например, вариабельной области тяжелой цепи человеческого или гуманизированного иммуноглобулина). Таким образом, в некоторых вариантах осуществления нуклеотидная молекула, описанная в данном документе, содержит сконструированную определяющую разнообразие область (DH) тяжелой цепи иммуноглобулина, которая содержит по меньшей мере один генный сегмент DH, функционально связанный с 23-мером (RSS). В некоторых вариантах осуществления сконструированная DH-область содержит (i) по меньшей мере один генный сегмент DH, функционально связанный с 23-мерной RSS, и (ii) неперегруппированный генный сегмент DH, фланкированный с одной стороны 12-мерной RSS и с другой стороны другой 12-мерной RSS, где (i) по меньшей мере один генный сегмент DH, функционально связанный с 23-мерной RSS, и (ii) генный сегмент DH зародышевой линии, фланкированный с одной стороны 12-мерной RSS и с другой стороны другой 12-мерной RSS, функционально связаны таким образом, что (i) и (ii) могут соединиться при событии рекомбинации DH-DH в соответствии с правилом 12/23. Также предусмотрены нацеливающиеся векторы, животные, отличные от человека (например, грызуны (например, крысы или мыши)) и клетки животного, отличного от человека (например, клетки грызуна (например, клетки крысы или клетки мыши), содержащие нуклеотидную молекулу, описанную в данном документе, способы применения нуклеотидной молекулы, описанной в данном документе, и т.д.
[006] В некоторых вариантах осуществления генный сегмент DH, функционально связанный с 23-мерной RSS, содержит человеческий генный сегмент DH, функционально связанный с 23-мерной RSS. В некоторых вариантах осуществления человеческий генный сегмент DH содержит по меньшей мере 19 нуклеотидов и/или кодирует два цистеина. В некоторых вариантах осуществления человеческий генный сегмент DH содержит по меньшей мере 20 нуклеотидов и/или кодирует два цистеина. В некоторых вариантах осуществления человеческий генный сегмент DH содержит по меньшей мере 23 нуклеотида и/или кодирует два цистеина. В некоторых вариантах осуществления человеческий генный сегмент DH содержит по меньшей мере 28 нуклеотидов и/или кодирует два цистеина. В некоторых вариантах осуществления человеческий генный сегмент DH содержит по меньшей мере 31 нуклеотид и/или кодирует два цистеина. В некоторых вариантах осуществления человеческий генный сегмент DH содержит по меньшей мере 37 нуклеотидов и/или кодирует два цистеина. В некоторых вариантах осуществления человеческий генный сегмент DH, функционально связанный с 23-мерной RSS, содержит человеческий генный сегмент DH2, функционально связанный с 23-мерной RSS. В некоторых вариантах осуществления генный сегмент DH2 содержит по меньшей мере 30 нуклеотидов и/или кодирует два цистеина. В некоторых вариантах осуществления человеческий генный сегмент DH, функционально связанный с 23-мерной RSS, содержит генный сегмент DH, выбранный из группы, состоящей из человеческого генного сегмента DH3-3, генного сегмента DH3-9, генного сегмента DH3-10, генного сегмента DH3-16, генного сегмента DH3-22, человеческого генного сегмента DH2-2, человеческого генного сегмента DH2-8 или человеческого генного сегмента DH2-15. В некоторых вариантах осуществления человеческий генный сегмент DH, функционально связанный с 23-мерной RSS, содержит человеческий генный сегмент DH, выбранный из группы, состоящей из человеческого генного сегмента DH3-3, человеческого генного сегмента DH2-2, человеческого генного сегмента DH2-8 и человеческого генного сегмента DH2-15. В некоторых вариантах осуществления человеческий генный сегмент DH, функционально связанный с 23-мерной RSS, содержит человеческий генный сегмент DH, выбранный из группы, состоящей из человеческого генного сегмента DH3-3, человеческого генного сегмента DH2-2, человеческого генного сегмента DH2-8 и человеческого генного сегмента DH2-15. В некоторых вариантах осуществления человеческий генный сегмент DH содержит человеческий генный сегмент DH3-3. В некоторых вариантах осуществления человеческий генный сегмент DH содержит человеческий генный сегмент DH2-2. В некоторых вариантах осуществления человеческий генный сегмент DH содержит человеческий генный сегмент DH2-8. В некоторых вариантах осуществления человеческий генный сегмент DH содержит человеческий генный сегмент DH2-15.
[007] В некоторых вариантах осуществления генный сегмент DH, функционально связанный с 23-мерной RSS, содержит: (а) человеческий генный сегмент DH3-3, функционально связанный с 23-мерной RSS, где необязательно 23-мерная RSS является смежной с 5'-концом человеческого генного сегмента DH3-3, (b) человеческий генный сегмент DH2-2, функционально связанный с 23-мерной RSS, где необязательно 23-мерная RSS является смежной с 3'-концом человеческого генного сегмента DH2-2, (с) человеческий генный сегмент DH2-8, функционально связанный с 23-мерной RSS, где необязательно 23-мерная RSS является смежной с 3'-концом человеческого генного сегмента DH2-8, (d) человеческий генный сегмент 2-15, функционально связанный с 23-мерной RSS, где необязательно 23-мерная RSS является смежной с 3'-концом человеческого генного сегмента DH2-15 или (е) любую комбинацию из (a)-(d).
[008] В некоторых вариантах осуществления нуклеотидная молекула, описанная в данном документе (например, генный сегмент DH, функционально связанный с 23-мерной RSS, сконструированная DH-область, вариабельная область тяжелой цепи иммуноглобулина и т.д.), содержит нуклеотидную последовательность, содержащую последовательность, представленную под SEQ ID NO: 52.
[009] В некоторых вариантах осуществления нуклеотидная молекула, описанная в данном документе (например, генный сегмент DH, функционально связанный с 23-мерной RSS, сконструированная DH-область, вариабельная область тяжелой цепи иммуноглобулина и т.д.), содержит нуклеотидную последовательность, содержащую последовательность, представленную под SEQ ID NO:61.
[0010] В некоторых вариантах осуществления нуклеотидная молекула, описанная в данном документе (например, генный сегмент DH, функционально связанный с 23-мерной RSS, сконструированная DH-область, вариабельная область тяжелой цепи иммуноглобулина и т.д.), содержит нуклеотидную последовательность, содержащую последовательность, представленную под SEQ ID NO:70.
[0011] В некоторых вариантах осуществления нуклеотидная молекула, описанная в данном документе (например, генный сегмент DH, функционально связанный с 23-мерной RSS, сконструированная DH-область, вариабельная область тяжелой цепи иммуноглобулина и т.д.), содержит нуклеотидную последовательность, содержащую последовательность, представленную под SEQ ID NO:71.
[0012] В некоторых вариантах осуществления нуклеотидная молекула, описанная в данном документе (например, генный сегмент DH, функционально связанный с 23-мерной RSS, сконструированная DH-область, вариабельная область тяжелой цепи иммуноглобулина и т.д.), содержит нуклеотидную последовательность, содержащую последовательность, представленную под SEQ ID NO:72.
[0013] В некоторых вариантах осуществления генный сегмент DH, функционально связанный с 23-мерной RSS, содержит от 5'-конца к 3'-концу 23-мерную RSS и генный сегмент DH, например, содержит от 5'-конца к 3'-концу 23-мерную RSS, (человеческий) генный сегмент DH и 12-мерную RSS, например, 23-мерная RSS является смежной с 5'-концом генного сегмента DH, например, генный сегмент DH функционально связан с 23-мерной RSS 5'-конца. В некоторых вариантах осуществления генный сегмент DH, функционально связанный с 23-мерной RSS, содержит человеческий генный сегмент DH3-3, функционально связанный с 23-мерной RSS 5'-конца, например, нуклеотидная молекула содержит от 5'-конца к 3'-концу 23-мерную RSS, человеческий генный сегмент DH3-3 и 12-мерную RSS.
[0014] Сконструированные DH-области, содержащие по меньшей мере один генный сегмент DH с 23-мерной RSS 5'-конца, могут дополнительно содержать неперегруппированный генный сегмент DH, фланкированный с одной стороны 12-мерной RSS и с другой стороны другой 12-мерной RSS (например, неперегруппированный генный сегмент DH, который фланкирован с одной стороны 12-мерной RSS и с другой стороны другой 12-мерной RSS, содержит генный сегмент DH зародышевой линии, например, генный сегмент DH в его конфигурации зародышевой линии и т.д.), где неперегруппированный генный сегмент DH располагается выше и функционально связан с по меньшей мере одним генным сегментом DH, функционально связанным с 23-мером 5'-конца. В некоторых вариантах осуществления неперегруппированный генный сегмент DH, фланкированный с одной стороны 12-мерной RSS и с другой стороны другой 12-мерной RSS, содержит неперегруппированный человеческий генный сегмент DH, фланкированный с одной стороны 12-мерной RSS и с другой стороны другой 12-мерной RSS.
[0015] В некоторых вариантах осуществления генный сегмент DH, функционально связанный с 23-мерной RSS, содержит от 5'-конца к 3'-концу генный сегмент DH и 23-мерную RSS, например, содержит от 5'-конца к 3'-концу 12-мерную RSS, (человеческий) генный сегмент DH и 23-мерную RSS, например, 23-мерная RSS является смежной с 3'-концом генного сегмента DH, например, генный сегмент DH функционально связан с 23-мером 3'-конца и т.д. В некоторых вариантах осуществления генный сегмент DH, функционально связанный с 23-мерной RSS 3'-конца, содержит человеческий генный сегмент DH2, функционально связанный с 23-мерной RSS 3'-конца, где генный сегмент DH2 выбран из группы, состоящей из человеческого генного сегмента DH2-2, человеческого генного сегмента DH2-8 и человеческого генного сегмента DH2-15. В некоторых вариантах осуществления генный сегмент DH, функционально связанный с 23-мерной RSS, содержит от 3'-конца к 3'-концу: человеческий генный сегмент DH2-2, функционально связанный с 23-мерной RSS 3'-конца, человеческий генный сегмент DH2-8, функционально связанный с 23-мерной RSS 3'-конца, и человеческий генный сегмент DH2-15, функционально связанный с 23-мерной RSS 3'-конца. В некоторых вариантах осуществления генный сегмент DH, функционально связанный с 23-мерной RSS 3'-конца, содержит от 5' к 3': первую смежную нуклеотидную последовательность, содержащую 12-мерную RSS, человеческий генный сегмент DH2-2 и 23-мерную RSS, вторую смежную нуклеотидную последовательность, содержащую 12-мерную RSS, человеческий генный сегмент DH2-8 и 23-мерную RSS, и третью смежную нуклеотидную последовательность, содержащую 12-мерную RSS, человеческий генный сегмент DH2-15 и 23-мерную RSS.
[0016] Сконструированные DH-области, содержащие по меньшей мере один генный сегмент DH, функционально связанный с 23-мерной RSS 3'-конца, могут дополнительно содержать неперегруппированный генный сегмент DH, который фланкирован с одной стороны 12-мерной RSS и с другой стороны другой 12-мерной RSS (например, неперегруппированный генный сегмент DH, фланкированный с одной стороны 12-мерной RSS и с другой стороны другой 12-мерной RSS, может содержать генный сегмент DH зародышевой линии, например, генный сегмент DH в его конфигурации зародышевой линии и т.д.), где неперегруппированный генный сегмент DH, фланкированный с одной стороны 12-мерной RSS и с другой стороны другой 12-мерной RSS, расположен ниже и функционально связан с по меньшей мере одним генным сегментом DH, функционально связанным с 23-мерной RSS 3'-конца. В некоторых вариантах осуществления неперегруппированный генный сегмент DH, фланкированный с одной стороны 12-мерной RSS и с другой стороны другой 12-мерной RSS, содержит неперегруппированный человеческий генный сегмент DH, фланкированный с одной стороны 12-мерной RSS и с другой стороны другой 12-мерной RSS.
[0017] В некоторых вариантах осуществления сконструированная DH-область, описанная в данном документе, содержит (i) один или более неперегруппированных человеческих генных сегментов DH, где каждый из одного или более человеческих генных сегментов DH фланкирован на 5'- и 3'-концах 12-мерной RSS, и (ii) по меньшей мере один генный сегмент DH, функционально связанный с 23-мерной RSS, содержит человеческий генный сегмент DH, например, человеческий генный сегмент DH3-3, функционально связанный на своем 5'-конце с 23-мерной RSS. В некоторых вариантах осуществления сконструированная DH-область, описанная в данном документе, содержит от 5'-конца к 3'-концу (i) по меньшей мере один генный сегмент DH, функционально связанный с 23-мерной RSS, например, по меньшей мере один человеческий генный сегмент DH2, функционально связанный на своем 3'-конце с 23-мерной RSS, где необязательно по меньшей мере один человеческий генный сегмент DH2, функционально связанный на своем 3'-конце с 23-мерной RSS, содержит человеческий генный сегмент DH2-2, функционально связанный на своем 3'-конце с 23-мерной RSS, человеческий генный сегмент DH2-8, функционально связанный на своем 3'-конце с 23-мерной RSS, человеческий генный сегмент DH2-15, функционально связанный на своем 3'-конце с 23-мерной RSS, или любую их комбинацию, и (ii) один или множество человеческих генных сегментов DH, где каждый из указанных одного или множества человеческих генных сегментов DH фланкирован на своих 5'- и 3'-концах 12-мерной RSS.
[0018] В некоторых вариантах осуществления сконструированная DH-область, описанная в данном документе, содержит только человеческие генные сегменты DH.
[0019] В некоторых вариантах осуществления нуклеотидная молекула, описанная в данном документе (например, сконструированные DH-области, вариабельные области тяжелой цепи иммуноглобулина и т.д.), содержит генный сегмент DH, функционально связанный с 23-мерной RSS, и неперегруппированный генный сегмент DH, фланкированный с одной стороны 12-мерной RSS и с другой стороны другой 12-мерной RSS, где генный сегмент DH, функционально связанный с 23-мерной RSS, и неперегруппированный генный сегмент DH, фланкированный с одной стороны 12-мерной RSS и с другой стороны другой 12-мерной RSS, не подвергались рекомбинации с (i) другим генным сегментом DH, (ii) генным сегментом VH, (iii) генным сегментом JH или (iv) любой их комбинацией. В некоторых вариантах осуществления нуклеотидные молекулы, описанные в данном документе, включают молекулы, содержащие сконструированные DH-области, содержащие один или более генных сегментов DH, подвергшихся рекомбинации с (i) другим генным сегментом DH, (ii) генным сегментом VH, (iii) генным сегментом JH или любой их комбинацией, например, нуклеотидные молекулы, содержащие перегруппированную VDJ- или VDDJ-кодирующую последовательность, которая кодирует вариабельную область тяжелой цепи иммуноглобулина.
[0020] Соответственно, в некоторых вариантах осуществления нуклеотидная молекула, описанная в данном документе, содержащая сконструированную DH-область, описанную в данном документе, дополнительно содержит в функциональной связи: (а) по меньшей мере один неперегруппированный вариабельный (VH) генный сегмент тяжелой цепи иммуноглобулина (например, неперегруппированный человеческий генный сегмент VH6-1), который располагается выше и функционально связан со сконструированной DH-областью, (b) по меньшей мере один неперегруппированный соединяющий (JH) генный сегмент тяжелой цепи иммуноглобулина (например, неперегруппированный человеческий JH6), который располагается ниже и функционально связан со сконструированной DH-областью, или комбинацию (а) и (b).
[0021] В некоторых вариантах осуществления по меньшей мере один неперегруппированный генный сегмент VH содержит полный репертуар функциональных неперегруппированных человеческих генных сегментов VH, охватывающих и содержащих неперегруппированные человеческие генные сегменты VH3-74 и VH1-6. В некоторых вариантах осуществления по меньшей мере один неперегруппированный генный сегмент VH содержит полный репертуар функциональных неперегруппированных человеческих генных сегментов VH, охватывающих и содержащих неперегруппированные человеческие генные сегменты VH3-74 и VH1-6 в конфигурации зародышевой линии. В некоторых вариантах осуществления по меньшей мере один неперегруппированный генный сегмент JH содержит неперегруппированный человеческий генный сегмент JH4, неперегруппированный человеческий генный сегмент JH5 и неперегруппированный человеческий генный сегмент JH6. В некоторых вариантах осуществления по меньшей мере один неперегруппированный генный сегмент JH содержит неперегруппированный человеческий генный сегмент JH1, неперегруппированный человеческий генный сегмент JH2, неперегруппированный человеческий генный сегмент JH3, неперегруппированный человеческий генный сегмент JH4, неперегруппированный человеческий генный сегмент JH5 и неперегруппированный человеческий генный сегмент JH6. В некоторых вариантах осуществления по меньшей мере один неперегруппированный генный сегмент JH содержит неперегруппированный человеческий генный сегмент JH1, неперегруппированный человеческий генный сегмент JH2, неперегруппированный человеческий генный сегмент JH3, неперегруппированный человеческий генный сегмент JH4, неперегруппированный человеческий генный сегмент JH5 и неперегруппированный человеческий генный сегмент JH6 в конфигурации зародышевой линии.
[0022] В некоторых вариантах осуществления нуклеотидная молекула, описанная в данном документе, содержит вариабельную (VH) область тяжелой цепи иммуноглобулина, содержащую сконструированную DH-область, описанную в данном документе, например, содержит в функциональной связи от 5'-конца к 3'-концу:
(a) по меньшей мере один неперегруппированный вариабельный (VH) генный сегмент тяжелой цепи иммуноглобулина,
(b) сконструированную DH-область, содержащую по меньшей мере один генный сегмент DH, функционально связанный с 23-мерной RSS, и
(c) по меньшей мере один неперегруппированный соединяющий (JH) генный сегмент тяжелой цепи иммуноглобулина.
[0023] В некоторых вариантах осуществления
(а) по меньшей мере один неперегруппированный генный сегмент VH содержит (i) неперегруппированный человеческий генный сегмент VH6-1,
(ii) неперегруппированный человеческий генный сегмент VH2- 1 и неперегруппированный человеческий генный сегмент VH6-1 и/или
(iii) все функциональные неперегруппированные человеческие генные сегменты VH, охватывающие и содержащие человеческие генные сегменты от неперегруппированного VH3-74 до неперегруппированного VH6-1, например, все функциональные неперегруппированные человеческие генные сегменты VH в конфигурации зародышевой линии, где необязательно ген Adam6 грызуна заменяет псевдоген между неперегруппированными человеческими генными сегментами VH2- 1 и VH6-1;
(b) сконструированная DH-область содержит от 5'-конца к 3'-концу:
(i) один или более, например, множество неперегруппированных человеческих генных сегментов DH, где каждый из множества неперегруппированных человеческих генных сегментов DH фланкирован на его 5'- и 3'-концах 12-мерной RSS, и человеческий генный сегмент DH, функционально связанный на своем 3'-конце с 23-мерной RSS, где необязательно множество неперегруппированных человеческих генных сегментов DH содержит неперегруппированные человеческие генные сегменты DH, охватывающие и содержащие неперегруппированный человеческий генный сегмент DH1-1 и неперегруппированный человеческий генный сегмент DH1-26 в конфигурации зародышевой линии и/или где человеческий генный сегмент, функционально связанный на своем 5'-конце с 23-мерной RSS, представляет собой человеческий генный сегмент DH3-3, например, где сконструированный DH содержит полный репертуар неперегруппированных человеческих генных сегментов DH в конфигурации зародышевой линии, за исключением неперегруппированного человеческого генного сегмента DH7-27, который заменен на генный сегмент DH3-3, функционально связанный с 23-мерной RSS 5'-конца;
(ii) по меньшей мере один человеческий генный сегмент DH, функционально связанный на своем 3'-конце с 23-мерной RSS, и один или более, например, множество человеческих генных сегментов DH, где каждый из одного или множества человеческих генных сегментов DH фланкирован на своих 5'- и 3'-концах 12-мерной RSS, где необязательно по меньшей мере один человеческий генный сегмент DH, функционально связанный на своем 3'-конце с 23-мерной RSS, содержит генный сегмент DH2-2, функционально связанный на своем 3'-конце с 23-мерной RSS, человеческий генный сегмент DH2-8, функционально связанный на своем 3'-конце с 23-мерной RSS, человеческий генный сегмент DH2-15, функционально связанный на своем 3'-конце с 23-мерной RSS, и/или где один или множество человеческих генных сегментов DH содержат генные сегменты DH, охватывающие и содержащие неперегруппированный человеческий генный сегмент DH1-1 и неперегруппированный человеческий генный сегмент DH7-27, где необязательно сконструированная DH содержит полный репертуар неперегруппированных человеческих генных сегментов DH в конфигурации зародышевой линии, за исключением того, что неперегруппированные человеческие генные сегменты DH2-2, DH2-8 и DH2-15 соответственно заменены на генный сегмент DH2-2, функционально связанный на своем 3'-конце с 23-мерной RSS, человеческий генный сегмент DH2-8, функционально связанный на своем 3'-конце с 23-мерной RSS, и человеческий генный сегмент DH2-15, функционально связанный на своем 3'-конце с 23-мерной RSS;
(iii) или комбинацию (b)(i) и (b)(ii); и
(с) по меньшей мере один неперегруппированный генный сегмент JH содержит
(i) неперегруппированный человеческий генный сегмент JH6,
(ii) неперегруппированный человеческий генный сегмент JH4, неперегруппированный человеческий генный сегмент JH5 и неперегруппированный человеческий генный сегмент JH6 и/или
(iii) полный репертуар неперегруппированных человеческих генных сегментов JH, например, неперегруппированный человеческий генный сегмент JHI, неперегруппированный человеческий генный сегмент JH2, неперегруппированный человеческий генный сегмент JH3, неперегруппированный человеческий генный сегмент JH4, неперегруппированный человеческий генный сегмент JH5 и неперегруппированный человеческий генный сегмент JH6, где необязательно неперегруппированные человеческие генные сегменты JHI, JH2, JH3, JH4, JH5, и JH6 находятся в конфигурации зародышевой линии. В некоторых вариантах осуществления VH-область иммуноглобулина (содержащая по меньшей мере один функциональный генный сегмент VH, сконструированную DH-область и по меньшей мере один функциональный генный сегмент JH) представляет собой VH-область человеческого иммуноглобулина, например, каждый генный сегмент (например, каждый генный сегмент VH, DH и JH в нем), включая генный сегмент DH, функционально связанный с 23-мерной RSS, представляет собой человеческий (VH, DH или JH) генный сегмент.
[0024] В некоторых вариантах осуществления нуклеотидная молекула, содержащая VH-область иммуноглобулина, описанную в данном документе, содержит в функциональной связи от 5'-конца к 3'-концу
(a) по меньшей мере неперегруппированный человеческий генный сегмент VH6-1, например, все или часть всех функциональных неперегруппированных человеческих генных сегментов VH, охватывающих и содержащих человеческие генные сегменты от неперегруппированного VH3-74 до неперегруппированного VH6-1, например, полный репертуар функциональных неперегруппированных человеческих генных сегментов VH, необязательно содержащих ген Adam6 грызуна между двумя неперегруппированными человеческими генными сегментами VH, например, (например, где ген Adam6 грызуна расположен между человеческим генным сегментом VH1-2 и человеческим генным сегментом VH6-1)
(b) человеческую сконструированную DH-область, содержащую от 5'-конца к 3'-концу неперегруппированные человеческие генные сегменты DH, охватывающие и содержащие неперегруппированный человеческий генный сегмент DH1-1 и неперегруппированный человеческий генный сегмент DH1-26 в конфигурации зародышевой линии и неперегруппированный человеческий генный сегмент DH3-3, функционально связанный с 23-мерной RSS 5'-конца, например, полный репертуар неперегруппированных человеческих генных сегментов DH в конфигурации зародышевой линии, за исключением неперегруппированного человеческого генного сегмента DH7-27, который заменен на неперегруппированный человеческий генный сегмент DH3-3, функционально связанный с 23-мерной RSS в области 5'-конца, и
(c) по меньшей мере неперегруппированный человеческий генный сегмент JH6.
[0025] В некоторых вариантах осуществления нуклеотидная молекула, содержащая VH-область иммуноглобулина, описанную в данном документе, содержит в функциональной связи от 5'-конца к 3'-концу:
(a) по меньшей мере неперегруппированный человеческий генный сегмент VH6-1, например, все или часть всех функциональных неперегруппированных человеческих генных сегментов VH, охватывающих и содержащих человеческие генные сегменты от неперегруппированного VH3-74 до неперегруппированного VH6-1, например, полный репертуар функциональных неперегруппированных человеческих генных сегментов VH, необязательно содержащих ген Adam6 грызуна между двумя неперегруппированными человеческими генными сегментами VH, например, (например, где ген Adam6 грызуна расположен между человеческим генным сегментом VH1-2 и человеческим генным сегментом VH6-1)
(b) человеческую сконструированную DH-область, содержащую от 5'-конца к 3'-концу неперегруппированные человеческие генные сегменты DH, охватывающие и содержащие неперегруппированный человеческий генный сегмент DH1-1 и неперегруппированный человеческий генный сегмент DH1-26 в конфигурации зародышевой линии и неперегруппированный человеческий генный сегмент DH3-3, функционально связанный с 23-мерной RSS 5'-конца, например, полный репертуар неперегруппированных человеческих генных сегментов DH в конфигурации зародышевой линии, за исключением неперегруппированного человеческого генного сегмента DH7-27, который заменен на неперегруппированный человеческий генный сегмент DH3-3, функционально связанный с 23-мерной RSS в области 5'-конца, и
(c) неперегруппированный человеческий генный сегмент JH4, неперегруппированный человеческий генный сегмент JH5 и неперегруппированный человеческий генный сегмент JH6.
[0026] В некоторых вариантах осуществления нуклеотидная молекула, содержащая VH-область иммуноглобулина, описанную в данном документе, содержит в функциональной связи от 5'-конца к 3'-концу
(а) по меньшей мере неперегруппированный человеческий генный сегмент VH6-1, например, все или часть всех функциональных неперегруппированных человеческих генных сегментов VH, охватывающих и содержащих человеческие генные сегменты от неперегруппированного VH3-74 до неперегруппированного VH6-1, например, полный репертуар функциональных неперегруппированных человеческих генных сегментов VH, необязательно содержащих ген Adam6 грызуна между двумя неперегруппированными человеческими генными сегментами VH, например, (например, где ген Adam6 грызуна расположен между человеческим генным сегментом VH1-2 и человеческим генным сегментом VH6-1)
(b) человеческую сконструированную DH-область, содержащую от 5'-конца к 3'-концу неперегруппированный человеческий генный сегмент DH1-1, генный сегмент DH2-2, функционально связанный на своем 3'-конце с 23-мерной RSS, человеческий генный сегмент DH2-8, функционально связанный на своем 3'-конце с 23-мерной RSS, человеческий генный сегмент DH2-15, функционально связанный на своем 3'-конце с 23-мерной RSS, и неперегруппированные человеческие генные сегменты DH, охватывающие и содержащие неперегруппированный человеческий генный сегмент DH3-16 и неперегруппированный человеческий генный сегмент DH7-27, где необязательно сконструированная DH-область содержит полный репертуар неперегруппированных человеческих генных сегментов DH в конфигурации зародышевой линии, за исключением того, что неперегруппированные человеческие генные сегменты DH2-2, DH2-8 и DH2-15 соответственно заменены на генный сегмент DH2-2, функционально связанный на своем 3'-конце с 23-мерной RSS, человеческий генный сегмент DH2-8, функционально связанный на своем 3'-конце с 23-мерной RSS и человеческий генный сегмент DH2-15, функционально связанный на своем 3'-конце с 23-мерной RSS, и
(c) полный репертуар неперегруппированных человеческих генных сегментов JH, например, неперегруппированный человеческий генный сегмент JHI, неперегруппированный человеческий генный сегмент JH2, неперегруппированный человеческий генный сегмент JH3, неперегруппированный человеческий генный сегмент JH4, неперегруппированный человеческий генный сегмент JH5 и неперегруппированный человеческий генный сегмент JH6, где необязательно неперегруппированные человеческие генные сегменты JHI, JH2, JH3, JH4, JH5 и JH6 находятся в конфигурации зародышевой линии.
[0027] В некоторых вариантах осуществления нуклеотидная молекула, описанная в данном документе, содержит от 5'-конца к 3'-концу (а) вариабельную (VH) область тяжелой цепи (человеческого) иммуноглобулина, содержащую по меньшей мере один (человеческий) генный сегмент VH, сконструированную (человеческую) dh-область, описанную в данном документе, и по меньшей мере один (человеческий) генный сегмент JH, функционально связанный с (b) константной областью (CH) тяжелой цепи иммуноглобулина или ее частью, где необязательно CH представляет собой CH грызуна, которая содержит область интронного энхансера грызуна, ген IgM грызуна, ген IgD грызуна, ген IgG грызуна, ген IgA грызуна, ген IgE грызуна или любую их комбинацию. В некоторых вариантах осуществления нуклеотидная молекула, описанная в данном документе, содержит от 5'-конца к 3'-концу (а) VH-область тяжелой цепи человеческого иммуноглобулина, содержащую по меньшей мере один человеческий генный сегмент VH, человеческую сконструированную DH-область, описанную в данном документе, и по меньшей мере один человеческий генный сегмент JH, функционально связанный с (b) CH-областью грызуна, содержащей по меньшей мере область интронного энхансера грызуна и необязательно ген IgM грызуна. В некоторых вариантах осуществления нуклеотидная молекула, описанная в данном документе, содержит от 5'-конца к 3'-концу (а) человеческую VH-область, содержащую по меньшей мере один человеческий генный сегмент VH, человеческую сконструированную DH-область, описанную в данном документе, и по меньшей мере один человеческий генный сегмент JH, функционально связанный с (b) CH-областью грызуна, содержащей по меньшей мере область интронного энхансера грызуна и ген IgM грызуна. В некоторых вариантах осуществления нуклеотидная молекула, описанная в данном документе, содержит от 5'-конца к 3'-концу (а) человеческую VH-область, содержащую по меньшей мере один человеческий генный сегмент VH, человеческую сконструированную DH-область, описанную в данном документе, и по меньшей мере один человеческий генный сегмент JH, функционально связанный с (b) эндогенной CH-областью грызуна, например, в эндогенном локусе тяжелой цепи иммуноглобулина грызуна. В некоторых вариантах осуществления грызун может представлять собой крысу. В некоторых вариантах осуществления грызун может представлять собой мышь.
[0028] В некоторых вариантах осуществления нуклеотидная молекула, описанная в данном документе, дополнительно содержит ген Adam6 грызуна. В некоторых вариантах осуществления ген Adam6 грызуна расположен между человеческими генными сегментами VH2-1 и VH6-1, например, заменяет человеческий ген Adam6, расположенный между человеческими генными сегментами VH2-1 и VH6-1 в конфигурации зародышевой линии. В некоторых вариантах осуществления грызун может представлять собой крысу. В некоторых вариантах осуществления грызун может представлять собой мышь.
[0029] В некоторых вариантах осуществления нуклеотидная молекула, описанная в данном документе, содержит одну или более кассет для отбора по устойчивости к лекарственному средству, например, ген устойчивости к лекарственному средству, фланкированный одним или более сайтами сайт-специфической рекомбинации, например, ген устойчивости к лекарственному средству неомицину, фланкированный сайтом распознавания сайт-специфический рекомбинации loxP, где по меньшей мере одна из одной или более кассет устойчивости к лекарственному средству необязательно располагается непосредственно выше по меньшей мере одного генного сегмента DH, функционально связанного с 23-мерной RSS. В некоторых вариантах осуществления нуклеотидная молекула содержит последовательность, проиллюстрированную на фигуре 2.
[0030] В данном документе также описаны нацеливающиеся векторы, например, для модифицирования генома (например, генома зародышевой линии) животного, отличного от человека (например, грызуна, такого как крыса или мышь) для того, чтобы он содержал сконструированную DH-область, описанную в данном документе. Как правило, нацеливающийся вектор, описанный в данном документе, содержит любую из нуклеотидных молекул, описанных в данном документе, и необязательно содержит 5'- и 3'-плечи гомологии для гомологичной рекомбинации в пределах вариабельной области тяжелой цепи иммуноглобулина, где необязательно вариабельная область тяжелой цепи иммуноглобулина представляет собой вариабельную область тяжелой цепи человеческого или гуманизированного иммуноглобулина. В некоторых вариантах осуществления нацеливающийся вектор, описанный в данном документе, содержит 5'-плечо гомологии, содержащее неперегруппированный человеческий генный сегмент, например, генный сегмент VH6-1 и/или 3'-плечо гомологии, содержащее CH-область грызуна (например, мыши) или ее часть, например, CH-область интронного энхансера грызуна (мыши) и/или ген IgM грызуна (мыши).
[0031] В некоторых вариантах осуществления нацеливающийся вектор содержит нуклеотидную молекулу, описанную в данном документе, и 5'- и 3'-плечи гомологии, расположенные в конфигурации для создания условий для гомологичной рекомбинации с последовательностью тяжелой цепи иммуноглобулина, при этом последовательность тяжелой цепи иммуноглобулина необязательно может быть расположена в эндогенном локусе тяжелой цепи иммуноглобулина грызуна и/или содержит вариабельную область тяжелой цепи человеческого или гуманизированного иммуноглобулина. В некоторых вариантах осуществления нацеливающийся вектор, описанный в данном документе, содержащий 5'-плечо гомологии, который содержит неперегруппированный человеческий генный сегмент (например, генный сегмент VH6-1), человеческую сконструированную DH-область, по меньшей мере один неперегруппированный человеческий генный сегмент JH и 3'-плечо гомологии, содержащее CH-область интронного энхансера грызуна (мыши), и/или грызун (мышь) может быть применим для конструирования DH-области у грызуна, содержащей гуманизированный локус тяжелой цепи иммуноглобулина, например, мыши, содержащий замену вариабельных последовательностей мышиного иммуноглобулина на вариабельные последовательности человеческого иммуноглобулина, например, мыши линии VELOCIMMUNE®, которые могут необязательно содержать функциональный ген ADAM6, который восстанавливает их фертильность, модифицированную последовательность эндогенного гена константной области тяжелой цепи, содержащую интактный эндогенный ген IgM и другой эндогенный модифицированный ген (например, IgG) константной области для получения обратных химерных антител, отличных от IgM, в которых отсутствует функциональный СН1-домен, последовательность, кодирующую обратную химерную гуманизированную общую легкую цепь, последовательность, кодирующую обратную химерную гуманизированную легкую каппа-цепь, последовательность, кодирующую обратную химерную гуманизированную легкую лямбда-цепь, последовательность, кодирующую гибридную каппа/лямбда или легкую каппа/лямбда-цепь, неперегруппированные человеческие генные сегменты тяжелой цепи зародышевой линии и/или (не)перегруппированные генные сегменты легкой цепи зародышевой линии, модифицированные с помощью кодона гистидина для экспрессии вариабельных доменов, которые содержат аминокислоты, представляющие собой гистидин, и могут демонстрировать чувствительное к рН связывание антигена, и/или терминальную дезоксинуклеотидилтрансферазу (TdT) для повышения разнообразия антигенных рецепторов. См., например, патенты США №№9035128; 9066502; 9163092; 9150662; 9334333; 9850462;9844212;9029628;9006511;9394373;9206261; 9206262; 9206263; 9226484; 9399683;9540452;9012717;9796788,8697940;8754287; 9334334; 9801362; 9332742; 9969814; публикации заявок на патент США 2011/0195454, 2012/0021409, 2012/0192300, 2013/0185821, 2013/0302836, 2013/0045492 и 2018/0125043; РСТ-публикации №№ WO 2017/210586 и WO 2019/113065, каждый из которых включен в данный документ посредством ссылки во всей своей полноте.
[0032] Таким образом, в данном документе описаны способы конструирования dh-области для рекомбинации DH-DH, например, у грызуна. В некоторых вариантах осуществления способ включает модифицирование DH-области, содержащей один или множество неперегруппированных генных сегментов DH, таким образом, чтобы она содержала по меньшей мере один генный сегмент DH, функционально связанный с 23-мерной RSS. В некоторых вариантах осуществления способ модифицирования вариабельной области тяжелой цепи иммуноглобулина для конструирования рекомбинации DH-DH включает получение вариабельной области тяжелой цепи иммуноглобулина, содержащей DH-область, содержащую один или более неперегруппированных генных сегментов DH, где каждый из неперегруппированных генных сегментов DH фланкирован с одной стороны 12-мерной RSS и с другой стороны другой 12-мерной RSS, и дальнейшее модифицирование DH-области таким образом, чтобы она дополнительно содержала по меньшей мере один генный сегмент DH, функционально связанный с 23-мерной RSS. В некоторых вариантах осуществления DH-область представляет собой человеческую DH-область, содержащую один или множество неперегруппированных человеческих генных сегментов DH, например, где множество неперегруппированных человеческих генных сегментов DH необязательно содержит все функциональные неперегруппированные человеческие генные сегменты DH, охватывающие и содержащие генные сегменты DH1-1 и DH7-27, например, в конфигурации зародышевой линии. В некоторых вариантах осуществления модифицирование включает замену одного или более из одного или множества неперегруппированных генных сегментов DH, фланкированных с одной стороны 12-мерной RSS и с другой стороны другой 12-мерной RSS, например, функционального неперегруппированного человеческого генного сегмента DH, на по меньшей мере один генный сегмент DH, функционально связанный с 23-мерной RSS. В некоторых вариантах осуществления ближайший к 3'-концу неперегруппированный генный сегмент DH Эн-области, фланкированный с одной стороны 12-мерной RSS и с другой стороны другой 12-мерной RSS, заменен на генный сегмент DH, функционально связанный с 23-мерной RSS, где генный сегмент DH, функционально связанный с 23-мерной RSS, содержит от 5'-конца к 3'-концу 23-мерную RSS, генный сегмент DH и 12-мерную RSS. В некоторых вариантах осуществления неперегруппированный генный сегмент DH, фланкированный с одной стороны 12-мерной RSS и с другой стороны другой 12-мерной RSS, заменен на соответствующий генный сегмент DH, сконструированный таким образом, чтобы он был функционально связан с 23-мерной RSS. В некоторых вариантах осуществления, где DH-область содержит неперегруппированный человеческий генный сегмент DH7-27 (например, генный сегмент DH7-27 зародышевой линии), неперегруппированный человеческий генный сегмент DH7-27 заменен на человеческий генный сегмент DH (например, на неперегруппированный человеческий генный сегмент DH3-3), функционально связанный с 5'-концом 23-мерной RSS. В некоторых вариантах осуществления, где DH-область содержит неперегруппированный генный сегмент DH2-2, неперегруппированный генный сегмент DH2-8 и/или неперегруппированный генный сегмент DH2-15 (например, где сконструированная DH содержит полный репертуар неперегруппированных человеческих генных сегментов DH в конфигурации зародышевой линии), неперегруппированный генный сегмент DH2-2, неперегруппированный генный сегмент DH2-8 и/или неперегруппированный генный сегмент DH2-15 соответственно заменены на генный сегмент DH2-2, функционально связанный на своем 3'-конце с 23-мерной RSS, человеческий генный сегмент DH2-8, функционально связанный на своем 3'-конце с 23-мерной RSS и/или человеческий генный сегмент DH2-15, функционально связанный на своем 3'-конце с 23-мерной RSS. В некоторых вариантах осуществления модифицирование включает замену одной из двух 12-мерных RSS, фланкирующих неперегруппированный генный сегмент DH (например, человеческий генный сегмент DH зародышевой линии), на 23-мерную RSS. В некоторых вариантах осуществления вариабельная область тяжелой цепи иммуноглобулина, подлежащая модификации, содержит, в дополнение к DH-области, JH-область (необязательно содержащую полный репертуар человеческих генных сегментов JH зародышевой линии, содержащий человеческий генный сегмент JH1 зародышевой линии, человеческий генный сегмент JH2 зародышевой линии, человеческий генный сегмент JH3 зародышевой линии, человеческий генный сегмент JH4 зародышевой линии, человеческий генный сегмент JH5 зародышевой линии и человеческий генный сегмент JH6 зародышевой линии, где необязательно человеческие генные сегменты JH1, JH2, JH3, JH4, JH5 и JH6 зародышевой линии находятся в конфигурации зародышевой линии), функционально связанную с DH-областью, и замена неперегруппированного генного сегмента DH, фланкированного с одной стороны 12-мерной RSS и с другой стороны другой 12-мерной RSS, включает делецию по меньшей мере одного неперегруппированного генного сегмента JH, содержащегося в JH-области, например, приводит к делеции неперегруппированных человеческих генных сегментов JH1, JH2, JH3, JH4 и/или JH5, смежных с DH-областью. В некоторых вариантах осуществления делеция по меньшей мере одного генного сегмента JH зародышевой линии, содержащегося в JH-области, включает делецию неперегруппированных человеческих генных сегментов JH1, JH2 и JH3 и необязательно дополнительную делецию генных сегментов JH4 и JH5. В некоторых вариантах осуществления замена одного или более из всех функциональных генных сегментов DH, например, замена генного сегмента DH7-27 на генный сегмент DH, функционально связанный с 23-мерной RSS 5'-конца, например, на генный сегмент DH3-3, функционально связанный с 23-мерной RSS 5'-конца, приводит к делеции неперегруппированных человеческих генных сегментов JH1, JH2 и JH3, смежных с DH-областью. В некоторых вариантах осуществления замена одного или более из всех функциональных генных сегментов DH, например, замена генного сегмента DH7-27 на генный сегмент DH, функционально связанный с 23-мерной RSS 5'-конца, например, на генный сегмент DH3-3, функционально связанный с 23-мерной RSS 5'-конца, приводит к делеции неперегруппированных человеческих генных сегментов JH1, JH2, JH3, JH4 и JH5, смежных с DH-областью. В некоторых вариантах осуществления DH-область модифицирована таким образом, что она содержит по меньшей мере один генный сегмент DH, функционально связанный с 23-мерной RSS, где по меньшей мере один генный сегмент DH, функционально связанный с 23-мерной RSS, содержит (а) человеческий генный сегмент DH3-3, функционально связанный с 23-мерной RSS, где необязательно 23-мерная RSS является смежной с 5'-концом генного сегмента DH3-3, (b) человеческий генный сегмент DH2-2, функционально связанный с 23-мерной RSS, где необязательно 23-мерная RSS является смежной с З'-концом генного сегмента DH2-2, (с) человеческий генный сегмент DH2-8, функционально связанный с 23-мерной RSS, где необязательно 23-мерная RSS является смежной с З'-концом генного сегмента DH2-8, (d) человеческий генный сегмент DH2-15, функционально связанный с 23-мерной RSS, где необязательно 23-мерная RSS является смежной с З'-концом генного сегмента DH2-15, или (е) любую комбинацию (a)-(d).
[0033] Такие способы могут привести к получению вариабельной области тяжелой цепи иммуноглобулина, содержащей сконструированную DH-область, например, нуклеотидную молекулу, содержащую вариабельную область тяжелой цепи (человеческого) иммуноглобулина, описанную в данном документе, где (не)перегруппированная область тяжелой цепи (человеческого) иммуноглобулина может быть необязательно связана с константной областью тяжелой цепи иммуноглобулина, отличного от человеческого, или ее частью, например, константной областью тяжелой цепи иммуноглобулина грызуна, содержащей по меньшей мере CH-область интронного энхансера грызуна и/или ген IgM грызуна, например, необязательно в эндогенном локусе тяжелой цепи иммуноглобулина, отличного от человеческого. В некоторых вариантах осуществления после рекомбинации такой локус тяжелой цепи иммуноглобулина, например, эндогенный локус тяжелой цепи иммуноглобулина, содержит кодирующую последовательность перегруппированной вариабельной области тяжелой цепи иммуноглобулина, которая кодирует вариабельный домен тяжелой цепи иммуноглобулина, например, вариабельный домен тяжелой цепи иммуноглобулина, содержащий определяющую комплементарность область 3 (CDR3) длиной более 20 аминокислот, где длина может представлять собой результат рекомбинации VH(DHA-DHB)JH, VHDHJH6 или VH(DHA-DHB)JH6. Соответственно, в данном документе предусмотрены грызуны, клетки грызунов, локусы и/или нуклеотидные молекулы, содержащие перегруппированную последовательность VH(DHA-DHB)JH, VH(DH)JH6 или VH(DHA-DHB)JH6 тяжелой цепи иммуноглобулина, кодирующую вариабельный домен тяжелой цепи иммуноглобулина, содержащий CDR3, где длина CDR3 составляет по меньшей мере 20 аминокислот.
[0034] Также в данном документе описаны животные, отличные от человека, например, грызуны, содержащие сконструированную DH-область по настоящему изобретению, например, нуклеиновые кислоты, нацеливающиеся векторы и/или локусы тяжелых цепей иммуноглобулинов, описанные в данном документе, например, в своем геноме, например, в геноме зародышевой линии. Также в данном документе описаны геномы таких животных, отличных от человека, например, геномы грызунов. В некоторых вариантах осуществления в данном документе описан грызун, геном зародышевой линии которого содержит, или геном зародышевой линии грызуна, содержащий вариабельную область тяжелой цепи иммуноглобулина, описанную в данном документе, где вариабельная область тяжелой цепи иммуноглобулина содержит: (i) по меньшей мере один неперегруппированный вариабельный (VH) генный сегмент тяжелой цепи, (ii) сконструированную определяющую разнообразие область (DH) вариабельной области тяжелой цепи, где сконструированная DH-область содержит один или более неперегруппированных генных сегментов DH, каждый из которых фланкирован с одной стороны 12-мерной RSS и с другой стороны другой 12-мерной RSS, и один или более генных сегментов DH, каждый из которых функционально связан с 23-мерной последовательностью сигнала рекомбинации (RSS), и (iii) по меньшей мере один неперегруппированный соединяющий (JH) генный сегмент тяжелой цепи, где (i)-(iii) находятся в функциональной связи таким образом, что после рекомбинации вариабельная область тяжелой цепи иммуноглобулина содержит перегруппированную последовательность вариабельной области тяжелой цепи, кодирующую вариабельный домен тяжелой цепи иммуноглобулина, где необязательно перегруппированная последовательность вариабельной области тяжелой цепи образуется после события рекомбинации VH(DH-DH)JH, где необязательно по меньшей мере один из одного или более генных сегментов DH, каждый из которых функционально связан с а 23-мерной последовательностью сигнала рекомбинации (RSS), соединяется с одним из одного или более неперегруппированных генных сегментов DH, каждый из которых фланкирован с одной стороны 12-мерной RSS и с другой стороны другой 12-мерной RSS, во время события рекомбинации VH(DH-DH). В некоторых вариантах осуществления один или более сегментов DH, функционально связанных с 23-мерной RSS, содержат генный сегмент DH, функционально связанный с 23-мерной RSS 3'-конца. В некоторых вариантах осуществления один или более сегментов DH, функционально связанных с 23-мерной RSS, содержат генный сегмент DH, функционально связанный с 23-мерной RSS 5'-конца. В некоторых вариантах осуществления вариабельная область тяжелой цепи иммуноглобулина представляет собой вариабельную область тяжелой цепи человеческого иммуноглобулина, содержащую только человеческие генные сегменты VH, DH и JH. В некоторых вариантах осуществления вариабельная область тяжелой цепи человеческого иммуноглобулина функционально связана с константной областью тяжелой цепи иммуноглобулина. В некоторых вариантах осуществления константная область тяжелой цепи иммуноглобулина представляет собой эндогенную константную область тяжелой цепи иммуноглобулина животного, отличного от человека, или генома животного, отличного от человека, например, расположена в эндогенном локусе тяжелой цепи иммуноглобулина. В некоторых вариантах осуществления вариабельная область тяжелой цепи иммуноглобулина содержит человеческие генные сегменты VH, охватывающие от VH3-74 до VH6-1 в конфигурации зародышевой линии. В некоторых вариантах осуществления вариабельная область тяжелой цепи иммуноглобулина содержит человеческий генный сегмент JH6. В некоторых вариантах осуществления вариабельная область тяжелой цепи иммуноглобулина содержит одну или более нуклеотидных молекул, кодирующих один или более полипептидов Adam6 грызуна (например, ген Adam6 грызуна), которые необязательно могут быть вставлены между двумя человеческими генными сегментами VH (например, могут быть вставлены между человеческим генным сегментом VH1-2 и человеческим генным сегментом VH6-1) и/или вставлены вместо человеческого псевдогена Adam6. В некоторых вариантах осуществления животное, отличное от человека, представляет собой грызуна. В некоторых вариантах осуществления геном грызуна является гетерозиготным по сконструированной DH-области. В некоторых вариантах осуществления геном грызуна является гомозиготным по сконструированной DH-области. В некоторых вариантах осуществления грызун может представлять собой крысу. В некоторых вариантах осуществления грызун может представлять собой мышь. В некоторых вариантах осуществления грызун, геном грызуна или клетка грызуна представляют собой крысу, мышь, геном крысы, геном мыши, клетку крысы или клетку мыши, например, эмбриональную стволовую клетку грызуна (крысы или мыши).
[0035] В некоторых вариантах осуществления грызун, геном грызуна или клетка грызуна, описанные в данном документе, дополнительно содержат перегруппированную VDJ тяжелой цепи и/или кодирующие последовательности VH(DHA-DHB)JH, кодирующие вариабельные домены тяжелой цепи иммуноглобулина. В некоторых вариантах осуществления в данном документе описано животное, отличное от человека, например, грызун, например, крыса или мышь, которое содержит (1) в своем геноме зародышевой линии, например, в зародышевой клетке, локус тяжелой цепи иммуноглобулина, содержащий сконструированную DH-область, содержащую генный сегмент DH, функционально связанный с 23-мерной RSS, и (2) в своем соматическом геноме, например, в В-клетке, перегруппированную кодирующую последовательность VH(DHA-DHB)JH вариабельной области тяжелой цепи, где первый или второй генный сегмент DH (DHA или DHB соответственно) получен из генного сегмента DH, функционально связанного с 23-мерной RSS, или его части (например, содержит последовательность, идентичную по меньшей мере части генного сегмента DH, функционально связанного с 23-мерной RSS, его варианту, подвергнутому соматической гипермутации, и/или его вырожденному варианту). В некоторых вариантах осуществления, где В-клетка представляет собой наивную В-клетку, и/или перегруппированная последовательность, кодирующая вариабельную область тяжелой цепи, функционально связана с последовательностью константной области IgM, по меньшей мере один из генных сегментов DHA и DHB содержит по меньшей мере 9 последовательных нуклеотидов, которые выравниваются с нуклеотидной молекулой, кодируемой генным сегментом DH, функционально связанным с 23-мерной RSS, и каждый из генных сегментов DHA и DHB содержит по меньшей мере 5 последовательных нуклеотидов, которые выравниваются с нуклеотидной молекулой генного сегмента DH зародышевой линии. В некоторых вариантах осуществления, где В клетка представляет собой плазматическую В-клетку или В-клетку памяти, и/или перегруппированная последовательность, кодирующая вариабельную область тяжелой цепи, подвергнута соматической гипермутации и/или функционально связана с последовательностью константной области иммуноглобулина, отличного от IgM (например, IgG, IgA, IgE и т.д.), каждый из DHA и DHB соответственно демонстрирует 40% идентичность с первым и вторым генными сегментами DH зародышевой линии, с максимально 1 нуклеотидом, подвергнутым мутации. В некоторых вариантах осуществления по меньшей мере 95% от всех перегруппированных последовательностей VDJ и/или VH(DHA-DHB)JH, кодирующих тяжелую цепь у грызуна, имеют длину CDR3, составляющую по меньшей мере 10 аминокислот, где необязательно по меньшей мере 70% от всех перегруппированных последовательностей VDJ и/или VH(DHA-DHB)JH, кодирующих тяжелую цепь у грызуна, имеют длину CDR3, составляющую по меньшей мере 11 аминокислот, необязательно по меньшей мере 15% от всех перегруппированных последовательностей VDJ и/или VH(DHA-DHB)JH, кодирующих тяжелую цепь у грызуна, имеют длину CDR3, составляющую по меньшей мере 14 аминокислот. В некоторых вариантах осуществления совокупность кодирующих последовательностей VDJ и/или VH(DHA-DHB)JH у грызуна имеет длину CDR3, составляющую по меньшей мере 15 аминокислот, необязательно по меньшей мере 16 аминокислот, необязательно по меньшей мере 17 аминокислот и необязательно по меньшей мере 18 аминокислот. В некоторых вариантах осуществления в данном документе описан грызун или клетка грызуна, которая экспрессирует, или нуклеиновая кислота или локус иммуноглобулина, которые содержат перегруппированную кодирующую последовательность VH(DHA-DHB)JH тяжелой цепи, где второй генный сегмент DH зародышевой линии представляет собой DH3-3, где необязательно перегруппированная кодирующая последовательность VH(DHA-DHB)JH тяжелой цепи кодирует CDR3, имеющий длину, составляющую более 20 аминокислот. В некоторых вариантах осуществления в данном документе описан грызун или клетка грызуна, которая экспрессирует, или нуклеиновая кислота или локус иммуноглобулина, которые содержат перегруппированную кодирующую последовательность VH(DHA-DHB)JH тяжелой цепи, где первый генный сегмент DH зародышевой линии представляет собой DH2-2, DH2-8 или DH2-15, где необязательно перегруппированная кодирующая последовательность VH(DHA-DHB)JH тяжелой цепи кодирует CDR3, имеющий длину, составляющую более 20 аминокислот. В некоторых вариантах осуществления в данном документе описан грызун или клетка грызуна, которая экспрессирует перегруппированную кодирующую последовательность VH(DHA-DHB)JH6 тяжелой цепи, кодирующую CDR3, имеющий длину, составляющую более 20 аминокислот.
[0036] В некоторых вариантах осуществления предусмотрены геном грызуна, нуклеиновая кислота или локус иммуноглобулина, содержащие перегруппированную кодирующую последовательность VH(DHA-DHB)JH тяжелой цепи человеческого иммуноглобулина, функционально связанную с последовательностью константной области тяжелой цепи иммуноглобулина грызуна. В некоторых вариантах осуществления генный сегмент DHB получен из человеческого генного сегмента DH3-3 зародышевой линии. В некоторых вариантах осуществления генный сегмент DHA получен из человеческого генного сегмента DH2-2, DH2-8 или DH2-15 зародышевой линии. В некоторых вариантах осуществления перегруппированная тяжелая цепь определена как кодирующая последовательность VH(DHA-DHB)JH, с учетом того, что каждый из DHA и DHB соответственно демонстрирует 40% идентичность с первым и вторым генными сегментами DH зародышевой линии, с максимально 1 нуклеотидом, подвергнутым мутации. В некоторых вариантах осуществления перегруппированная кодирующая последовательность VH(DHA-DHB)JH тяжелой цепи кодирует CDR3, имеющий длину, составляющую более 20 аминокислот. В некоторых вариантах осуществления генный сегмент JH получен из человеческого генного сегмента JH6 зародышевой линии. В некоторых вариантах осуществления предусмотрен грызун или клетка грызуна, содержащая кодирующую последовательность VH(DHA-DHB)JH. В некоторых вариантах осуществления грызун представляет собой крысу или мышь, или клетка грызуна представляет собой клетку крысы или клетку мыши. В некоторых вариантах осуществления клетка грызуна представляет собой В-клетку грызуна. В некоторых вариантах осуществления предусмотрена гибридома, содержащая В-клетку грызуна, экспрессирующую перегруппированную кодирующую последовательность VH(DHA-DHB)JH тяжелой цепи, слитую с клеткой миеломы. В некоторых вариантах осуществления последовательность VH(DHA-DHB)JH подтверждена как результат события рекомбинации DH-DH, когда каждая из последовательностей, определенных как DHA и DHB, демонстрирует 40% идентичность с первым и вторым генными сегментами DH зародышевой линии соответственно, с максимально 1 нуклеотидом, подвергнутым мутации.
[0037] В некоторых вариантах осуществления предусмотрено животное, отличное от человека, или клетка, отличная от человеческой, геном которых содержит вариабельную область тяжелой цепи иммуноглобулина, которая содержит сконструированную DH-область, где сконструированная DH-область содержит по меньшей мере один сегмент DH, который функционально связан с первой и второй последовательностями сигнала рекомбинации (RSS). В некоторых вариантах осуществления первая RSS представляет собой 23-мерную RSS, и вторая RSS представляет собой 12-мерную RSS. В некоторых вариантах осуществления первая RSS представляет собой 12-мерную RSS, и вторая RSS представляет собой 23-мерную RSS.
[0038] В некоторых вариантах осуществления предусмотрено животное, отличное от человека, геном которого содержит вариабельную область тяжелой цепи иммуноглобулина, которая содержит сконструированную DH-область, где сконструированная DH-область содержит один или более сегментов DH, каждый из которых функционально связан с 23-мерной последовательностью сигнала рекомбинации (RSS).
[0039] В некоторых вариантах осуществления предусмотрена клетка, отличная от человеческой, или ткань, геном которых содержит вариабельную область тяжелой цепи иммуноглобулина, которая содержит сконструированную DH-область, где сконструированная DH-область содержит один или более сегментов DH, каждый из которых функционально связан с 23-мерной последовательностью сигнала рекомбинации (RSS). В некоторых вариантах осуществления клетка происходит из лимфоидной или миелоидной линии дифференцировки. В некоторых вариантах осуществления клетка представляет собой лимфоцит. В некоторых вариантах осуществления клетка выбрана из В-клетки, дендритной клетки, макрофага, моноцита и Т-клетки. В некоторых вариантах осуществления ткань выбрана из жировой ткани, ткани мочевого пузыря, головного мозга, молочной железы, костного мозга, глаза, сердца, кишечника, почки, печени, легкого, лимфатического узла, мышцы, поджелудочной железы, плазмы крови, сыворотки крови, кожи, селезенки, желудка, тимуса, яичка, яичника или любой их комбинации.
[0040] В некоторых вариантах осуществления предусмотрена иммортализованная клетка, полученная из клетки, отличной от человеческой, описанной в данном документе, например, клетка гибридомы, полученная посредством слияния В-клетки, выделенной из животного, отличного от человека, как описано в данном документе, с клеткой миеломы.
[0041] В некоторых вариантах осуществления клетка, отличная от человеческой, представляет собой эмбриональную стволовую (ES) клетку, отличную от человеческой. В некоторых вариантах осуществления эмбриональная стволовая клетка, отличная от человеческой, представляет собой эмбриональную стволовую клетку грызуна. В некоторых вариантах осуществления эмбриональная стволовая клетка грызуна представляет собой эмбриональную стволовую клетку мыши и получена из линии 129, линии C57BL или их смеси. В некоторых вариантах осуществления эмбриональная стволовая клетка грызуна представляет собой эмбриональную стволовую клетку мыши и представляет собой смесь линий 129 и C57BL.
[0042] В некоторых вариантах осуществления предусмотрено применение эмбриональной стволовой клетки, отличной от человеческой, описанной в данном документе, для получения животного, отличного от человека. В некоторых вариантах осуществления эмбриональная стволовая клетка, отличная от человеческой, представляет собой эмбриональную стволовую клетку мыши и ее применяют для получения мыши, содержащей вариабельную область тяжелой цепи иммуноглобулина, которая включает сконструированную DH-область, описанную в данном документе. В некоторых вариантах осуществления эмбриональная стволовая клетка, отличная от человеческой, представляет собой эмбриональную стволовую клетку крысы и ее применяют для получения крысы, содержащей вариабельную область тяжелой цепи иммуноглобулина, которая включает сконструированную DH-область, описанную в данном документе. В некоторых вариантах осуществления неограничивающий иллюстративный способ получения крысы, содержащей вариабельную область тяжелой цепи иммуноглобулина, которая включает сконструированную DH-область, может включать способы, раскрытые в US 20140309487, включенном в данный документ посредством ссылки во всей своей полноте.
[0043] В некоторых вариантах осуществления предусмотрен эмбрион, отличный от человеческого, содержащий описанную в данном документе эмбриональную стволовую клетку, отличную от человеческой, созданный из нее, полученный из нее или сгенерированный из нее. В некоторых вариантах осуществления эмбрион, отличный от человеческого, представляет собой эмбрион грызуна; в некоторых вариантах осуществления - эмбрион мыши; в некоторых вариантах осуществления - эмбрион крысы.
[0044] В некоторых вариантах осуществления предусмотрено применение эмбриона, отличного от человеческого, описанного в данном документе, для получения животного, отличного от человека. В некоторых вариантах осуществления эмбрион, отличный от человеческого, представляет собой эмбрион мыши, и его применяют для получения мыши, содержащей вариабельную область тяжелой цепи иммуноглобулина, которая включает сконструированную DH-область, описанную в данном документе. В некоторых вариантах осуществления эмбрион, отличный от человеческого, представляет собой эмбрион крысы, и его применяют для получения крысы, содержащей вариабельную область тяжелой цепи иммуноглобулина, которая включает сконструированную DH-область, описанную в данном документе.
[0045] В данном документе предусмотрены способы получения грызуна, геном которого содержит сконструированную DH-область, способ, включающий (а) модифицирование генома эмбриональной стволовой клетки грызуна таким образом, чтобы он содержал фрагмент ДНК, содержащий один или более сегментов DH, каждый из которых функционально связан с 23-мерной RSS, например, где фрагмент ДНК содержит нуклеотидную молекулу, нацеливающийся вектор и/или сконструированную DH-область, описанную в данном документе, и (b) получение грызуна с применением модифицированной эмбриональной стволовой клетки грызуна, полученной из (а). В некоторых вариантах осуществления способы включают модифицирование неперегруппированной DH-области вариабельной области тяжелой цепи иммуноглобулина таким образом, чтобы она содержала по меньшей мере один сегмент DH, функционально связанный с 23-мерной RSS, где неперегруппированная DH-область дополнительно содержит один или более неперегруппированных генных сегментов DH, каждый из которых фланкирован с одной стороны 12-мерной RSS и с другой стороны другой 12-мерной RSS, с получением таким образом указанного грызуна. В некоторых вариантах осуществления модифицирование включает замену одного или более неперегруппированных человеческих генных сегментов DH и необязательно замену или делецию одного или более генных сегментов JH, при этом по меньшей мере один сегмент DH функционально связан с 23-мерной RSS. В некоторых вариантах осуществления вариабельная область тяжелой цепи иммуноглобулина представляет собой вариабельную область тяжелой цепи человеческого иммуноглобулина, например, где вариабельная область тяжелой цепи человеческого иммуноглобулина содержит неперегруппированный генный кластер VH тяжелой цепи человеческого иммуноглобулина, содержащий по меньшей мере один генный сегмент VH, неперегруппированную DH-область тяжелой цепи человеческого иммуноглобулина, содержащую один или более неперегруппированных человеческих генных сегментов DH, и неперегруппированный генный кластер JH тяжелой цепи человеческого иммуноглобулина, содержащий по меньшей мере один неперегруппированный человеческий генный сегмент JH, где неперегруппированная DH-область тяжелой цепи человеческого иммуноглобулина модифицирована таким образом, что она содержит по меньшей мере один сегмент DH, функционально связанный с 23-мерной RSS. В некоторых вариантах осуществления стадия модифицирования приводит к получению вариабельной области тяжелой цепи иммуноглобулина, содержащей (i) полный репертуар функциональных человеческих генных сегментов VH, например, всех функциональных генных сегментов VH, охватывающих и содержащих сегменты от VH3-74 до VH6-1, (ii) полный репертуар неперегруппированных человеческих генных сегментов DH, за исключением DH7-27, замененного на по меньшей мере один генный сегмент DH, функционально связанный с 23-мерной RSS, и (iii) по меньшей мере неперегруппированный человеческий генный сегмент JH6 и необязательно по меньшей мере неперегруппированный генный сегмент JH4, неперегруппированный генный сегмент JH5 и неперегруппированный генный сегмент JH6. В некоторых вариантах осуществления по меньшей мере один генный сегмент DH, функционально связанный с 23-мерной RSS, содержит генный сегмент DH3-3, функционально связанный с 23-мерной RSS 5'-конца. В некоторых вариантах осуществления стадия модифицирования приводит к получению вариабельной области тяжелой цепи иммуноглобулина, содержащей (i) полный репертуар функциональных человеческих генных сегментов VH, например, всех функциональных генных сегментов VH, охватывающих и содержащих сегменты с VH3-74 по VH6-1, (ii) полный репертуар неперегруппированных человеческих генных сегментов DH, за исключением неперегруппированных человеческих генных сегментов DH2-2, DH2-8 и DH2-15, соответственно замененных на генный сегмент DH2-2, функционально связанный на своем 3'-конце с 23-мерной RSS, человеческий генный сегмент DH2-8, функционально связанный на своем 3'-конце с 23-мерной RSS, и человеческий генный сегмент DH2-15, функционально связанный на своем 3'-конце с 23-мерной RSS, и (iii) полный репертуар неперегруппированных человеческих генных сегментов JH, например, неперегруппированный человеческий генный сегмент JH1, неперегруппированный человеческий генный сегмент JH2, неперегруппированный человеческий генный сегмент JH3, неперегруппированный человеческий генный сегмент JH4, неперегруппированный человеческий генный сегмент JH5 и неперегруппированный человеческий генный сегмент JH6. В некоторых вариантах осуществления вариабельная область тяжелой цепи иммуноглобулина (а) дополнительно содержит один или более генов Adam6 грызуна, где необязательно один или более генов Adam6 грызуна расположены между двумя неперегруппированными генными сегментами VH, например, между неперегруппированным человеческим генным сегментом VH1-2 и неперегруппированным человеческим генным сегментом VH6-1, и/или (b) функционально связана с константной областью тяжелой цепи иммуноглобулина, где необязательно константная область тяжелой цепи иммуноглобулина представляет собой эндогенную константную область тяжелой цепи иммуноглобулина грызуна, например, эндогенную константную область тяжелой цепи иммуноглобулина грызуна в эндогенном локусе тяжелой цепи иммуноглобулина. В некоторых вариантах осуществления грызун представляет собой крысу или мышь.
[0046] В некоторых вариантах осуществления предусмотрен набор, содержащий животное, отличное от человека, описанное в данном документе, клетку или ткань, отличную от человеческой, описанную в данном документе, иммортализованную клетку, описанную в данном документе, эмбриональную стволовую клетку, отличную от человеческой, описанную в данном документе, или эмбрион, отличный от человеческого, описанный в данном документе.
[0047] В некоторых вариантах осуществления предусмотрен набор, описанный в данном документе, для применения для изготовления и/или разработки лекарственного средства (например, антитела или его антигенсвязывающего фрагмента) для терапии или диагностики. В некоторых вариантах осуществления предусмотрен набор, описанный в данном документе, для применения для изготовления и/или разработки лекарственного средства (например, антитела или его антиген связывающего фрагмента) для лечения, предупреждения или снижения интенсивности проявления заболевания, нарушения или состояния.
[0048] В некоторых вариантах осуществления предусмотрены трансген, конструкция нуклеиновой кислоты, ДНК-конструкция или нацеливающийся вектор, описанные в данном документе. В некоторых вариантах осуществления трансген, конструкция нуклеиновой кислоты, ДНК-конструкция или нацеливающийся вектор содержат сконструированную DH-область, описанную в данном документе. В некоторых вариантах осуществления трансген, конструкция нуклеиновой кислоты, ДНК-конструкция или нацеливающийся вектор содержат фрагмент ДНК, который содержит один или более сегментов DH, функционально связанных с 23-мерной RSS. В некоторых вариантах осуществления трансген, конструкция нуклеиновой кислоты, ДНК-конструкция или нацеливающийся вектор дополнительно содержат один или более селективных маркеров. В некоторых вариантах осуществления трансген, конструкция нуклеиновой кислоты, ДНК-конструкция или нацеливающийся вектор дополнительно содержат один или более сайтов сайт-специфической рекомбинации (например, loxP, Frt или их комбинации). В некоторых вариантах осуществления трансген, конструкция нуклеиновой кислоты, ДНК-конструкция или нацеливающийся вектор изображены на фигуре 2.
[0049] В некоторых вариантах осуществления предусмотрено применение трансгена, конструкции нуклеиновой кислоты, ДНК-конструкции или нацеливающегося вектора, описанных в данном документе, для получения животного, отличного от человека, клетки, отличной от человеческой, эмбриональной стволовой клетки, отличной от человеческой, и/или эмбриона, отличного от человеческого.
[0050] В некоторых вариантах осуществления каждый из одного или более сегментов DH функционально связан с 23-мерной RSS 3'-конца. В некоторых вариантах осуществления каждый из одного или более сегментов DH функционально связан с 23-мерной RSS 5'-конца.
[0051] В некоторых вариантах осуществления сконструированная DH-область содержит один сегмент DH, функционально связанный с 23-мерной RSS 5'-конца. В некоторых вариантах осуществления сконструированная DH-область содержит один сегмент DH, функционально связанный с RSS 3'-конца. В некоторых вариантах осуществления из одного сегмента DH, функционально связанного с 23-мерной RSS 5'-конца, один сегмент DH представляет собой синтетический сегмент DH; в некоторых вариантах осуществления синтетический человеческий сегмент DH; в некоторых вариантах осуществления синтетический человеческий сегмент DH, характеризующийся последовательностью, которая идентична или практически идентична человеческому сегменту DH3-3. В некоторых вариантах осуществления из одного сегмента DH, функционально связанного с 23-мерной RSS 3'-конца, один сегмент DH представляет собой синтетический сегмент DH; в некоторых вариантах осуществления синтетический человеческий сегмент DH; в некоторых вариантах осуществления синтетический человеческий сегмент DH, характеризующийся последовательностью, которая идентична или практически идентична человеческому сегменту DH3-3.
[0052] В некоторых вариантах осуществления сконструированная DH-область содержит три сегмента DH, каждый из которых функционально связан с 23-мерной RSS 5'-конца. В некоторых вариантах осуществления из трех сегментов DH, каждый из которых функционально связан с 23-мерной RSS 5'-конца, три сегмента DH представляют собой синтетические сегменты DH. В некоторых вариантах осуществления из трех сегментов DH, каждый из которых функционально связан с 23-мерной RSS 5'-конца, три сегмента DH представляют собой человеческие сегменты семейства DH2. В некоторых вариантах осуществления из трех сегментов DH, каждый из которых функционально связан с 23-мерной RSS 5'-конца, три сегмента DH выбраны из человеческого DH2-2, человеческого DH2-8, человеческого DH2-15, человеческого DH2-21 и их комбинаций. В некоторых вариантах осуществления из трех сегментов DH, каждый из которых функционально связан с 23-мерной RSS 3'-конца, три сегмента DH представляют собой человеческий DH2-2, человеческий DH2-8 и человеческий DH2-15.
[0053] В некоторых вариантах осуществления сконструированная DH-область содержит три сегмента DH, каждый из которых функционально связан с 23-мерной RSS 3'-конца. В некоторых вариантах осуществления из трех сегментов DH, каждый из которых функционально связан с 23-мерной RSS 3'-конца, три сегмента DH представляют собой синтетические сегменты DH. В некоторых вариантах осуществления из трех сегментов DH, каждый из которых функционально связан с 23-мерной RSS 3'-конца, три сегмента DH представляют собой человеческие сегменты семейства DH2. В некоторых вариантах осуществления из трех сегментов DH, каждый из которых функционально связан с 23-мерной RSS 3'-конца, три сегмента DH выбраны из человеческого DH2-2, человеческого DH2-8, человеческого DH2-15, человеческого DH2-21 и их комбинаций. В некоторых вариантах осуществления из трех сегментов DH, каждый из которых функционально связан с 23-мерной RSS 3'-конца, три сегмента DH представляют собой человеческий DH2-2, человеческий DH2-8 и человеческий DH2-15.
[0054] В некоторых вариантах осуществления сконструированная DH-область, описанная в данном документе, содержит множество человеческих сегментов DH, где по меньшей мере один из множества человеческих генных сегментов DH функционально связан с RSS 5'-конца или 3'-конца; в некоторых вариантах осуществления с 23-мерной RSS 5'-конца. В некоторых вариантах осуществления сконструированная DH-область, описанная в данном документе, содержит множество человеческих сегментов DH, где по меньшей мере три из множества человеческих генных сегментов DH функционально связаны с RSS 5'-конца или 3'-конца; в некоторых вариантах осуществления с 23-мерной RSS 3'-конца.
[0055] В некоторых вариантах осуществления в геноме предусмотренного животного, отличного от человека, клетки, отличной от человеческой, или ткани, отличной от человеческой, отсутствует один или более сегментов DH дикого типа. В некоторых вариантах осуществления в геноме предусмотренного животного, отличного от человека, клетки, отличной от человеческой, или ткани, отличной от человеческой, отсутствуют все или практически все сегменты DH дикого типа. В некоторых вариантах осуществления геном предусмотренного животного, отличного от человека, клетки, отличной от человеческой, или ткани, отличной от человеческой, содержит только человеческие сегменты DH.
[0056] В некоторых вариантах осуществления вариабельная область тяжелой цепи иммуноглобулина представляет собой вариабельную область тяжелой цепи человеческого иммуноглобулина. В некоторых вариантах осуществления вариабельная область тяжелой цепи человеческого иммуноглобулина функционально связана с константной областью тяжелой цепи иммуноглобулина. В некоторых вариантах осуществления константная область тяжелой цепи иммуноглобулина представляет собой эндогенную (например, отличную от человеческой) константную область тяжелой цепи иммуноглобулина.
[0057] В некоторых вариантах осуществления вариабельная область тяжелой цепи человеческого иммуноглобулина содержит человеческие генные сегменты VH от VH3-74 до VH6-1. В некоторых вариантах осуществления вариабельная область тяжелой цепи человеческого иммуноглобулина содержит по меньшей мере человеческий генный сегмент JH JH6. В некоторых вариантах осуществления вариабельная область тяжелой цепи человеческого иммуноглобулина содержит по меньшей мере человеческие генные сегменты JH JH4, JH5 и JH6. В некоторых вариантах осуществления вариабельная область тяжелой цепи человеческого иммуноглобулина содержит человеческие генные сегменты JH JH1, JH2, JH3, JH4, JH5 и JH6.
[0058] В некоторых вариантах осуществления в геноме животного, отличного от человека, клетки, отличной от человеческой, или ткани, отличной от человеческой, отсутствует эндогенный ген Adam6. В некоторых вариантах осуществления геном животного, отличного от человека, клетки, отличной от человеческой, или ткани, отличной от человеческой, дополнительно содержит вставку одной или более нуклеотидных последовательностей, кодирующих один или более полипептидов Adam6 грызуна; в некоторых вариантах осуществления одна или более нуклеотидных последовательностей вставлены между первым и вторым человеческими генными сегментами VH; в некоторых вариантах осуществления одна или более нуклеотидных последовательностей вставлены вместо человеческого псевдогена Adam6; в некоторых вариантах осуществления одна или более нуклеотидных последовательностей вставлены между человеческим генным сегментом VH и человеческим генным сегментом DH. В некоторых вариантах осуществления первый человеческий генный сегмент VH представляет собой человеческий VH1-2, и второй человеческий генный сегмент VH представляет собой человеческий VH6- 1.
[0059] В некоторых вариантах осуществления предусмотренное животное, отличное от человека, клетка, отличная от человеческой, или ткань, отличная от человеческой, являются гомозиготными, гетерозиготными или гемизиготными по сконструированной DH-области, описанной в данном документе. В некоторых вариантах осуществления предусмотренное животное, отличное от человека, клетка, отличная от человеческой, или ткань, отличная от человеческой, являются трансгенными по сконструированной DH-области, описанной в данном документе.
[0060] В некоторых вариантах осуществления предусмотрен способ получения животного, отличного от человека, геном которого содержит вариабельную область тяжелой цепи иммуноглобулина, которая содержит сконструированную DH-область, при этом способ включает (а) осуществление вставки фрагмента ДНК в эмбриональную стволовую клетку, отличную от человеческой, где указанный фрагмент ДНК содержит один или более сегментов DH, каждый из которых функционально связан с 23-мерной RSS; (b) получение эмбриональной стволовой клетки, отличной от человеческой, образованной на стадии (а); и (с) создание животного, отличного от человека, с применением эмбриональной стволовой клетки, отличной от человеческой, из стадии (b).
[0061] В некоторых вариантах осуществления фрагмент ДНК содержит один или более сегментов DH, каждый из которых функционально связан с 23-мерной RSS 3'-конца. В некоторых вариантах осуществления фрагмент ДНК содержит один сегмент DH, функционально связанный с 23-мерной RSS 3'-конца. В некоторых вариантах осуществления фрагмент ДНК содержит один синтетический человеческий сегмент DH, функционально связанный с 23-мерной RSS 3'-конца. В некоторых вариантах осуществления фрагмент ДНК содержит один синтетический человеческий сегмент DH3-3, функционально связанный с 23-мерной RSS 3'-конца. В некоторых вариантах осуществления фрагмент ДНК содержит три сегмента DH, каждый из которых функционально связан с 23-мерной RSS 3'-конца. В некоторых вариантах осуществления фрагмент ДНК содержит три человеческих сегмента DH, каждый из которых функционально связан с 23-мерной RSS 3'-конца. В некоторых вариантах осуществления фрагмент ДНК содержит три человеческих сегмента DH, каждый из которых функционально связан с 23-мерной RSS 3'-конца, где человеческие сегменты DH представляют собой человеческий DH2-2, человеческий DH2-8 и человеческий DH2-15.
[0062] В некоторых вариантах осуществления фрагмент ДНК содержит один или более сегментов DH, каждый из которых функционально связан с 23-мерной RSS 5'-конца. В некоторых вариантах осуществления фрагмент ДНК содержит один сегмент DH, функционально связанный с 23-мерной RSS 5'-конца. В некоторых вариантах осуществления фрагмент ДНК содержит один синтетический человеческий сегмент DH, функционально связанный с 23-мером 5'-конца. В некоторых вариантах осуществления фрагмент ДНК содержит один синтетический человеческий сегмент DH3-3, функционально связанный с 23-мерной RSS 5'-конца. В некоторых вариантах осуществления фрагмент ДНК содержит один синтетический человеческий сегмент DH3-3, функционально связанный с 23-мерной RSS 5'-конца, где синтетический человеческий сегмент DH3-3 расположен в человеческой DH-области на месте человеческого сегмента DH7-27.
[0063] В некоторых вариантах осуществления фрагмент ДНК содержит один или более селективных маркеров. В некоторых вариантах осуществления фрагмент ДНК содержит один или более сайтов сайт-специфической рекомбинации.
[0064] В некоторых вариантах осуществления предусмотрен способ получения животного, отличного от человека, геном которого содержит вариабельную область тяжелой цепи иммуноглобулина, которая содержит сконструированную DH-область, при этом способ включает стадию модифицирования генома животного, отличного от человека, или клетки животного, отличного от человека, таким образом, чтобы они содержали вариабельную область тяжелой цепи иммуноглобулина, которая содержит сконструированную DH-область, где сконструированная DH-область содержит один или более сегментов DH, каждый из которых функционально связан с 23-мерной RSS, с получением таким образом указанного животного, отличного от человека.
[0065] В некоторых вариантах осуществления получения животного, отличного от человека, геном животного, отличного от человека, или клетки животного, отличного от человека, модифицирован таким образом, что он содержит один или более сегментов DH, каждый из которых функционально связан 23-мерной RSS 5'-конца. В некоторых вариантах осуществления получения животного, отличного от человека, геном животного, отличного от человека, или клетки животного, отличного от человека, модифицирован таким образом, что он содержит один или более сегментов DH, каждый из которых функционально связан 23-мерной RSS 3'-конца.
[0066] В некоторых вариантах осуществления предусмотрен способ получения антитела с помощью животного, отличного от человека. В некоторых вариантах осуществления способ получения антитела или получения нуклеиновой кислоты, кодирующей его, включает иммунизацию животного, отличного от человека (например, грызуна (например, крысы или мыши), с помощью антигена, где грызун содержит геном зародышевой линии, содержащий сконструированную DH-область, которая содержит один или более сегментов DH, каждый из которых функционально связан с 23-мерной RSS, и обеспечение у грызуна индукции иммунного ответа на антиген, включая образование антитела, которое связывает антиген, или нуклеиновой кислоты, кодирующей его. В некоторых вариантах осуществления способ дополнительно включает извлечение антитела или нуклеиновой кислоты, кодирующей его, из грызуна или клетки грызуна, например, из В-клетки или гибридомы. В некоторых вариантах осуществления каждый из одного или более сегментов DH функционально связан с 23-мерной RSS 5'-конца. В некоторых вариантах осуществления каждый из одного или более сегментов DH функционально связан с 23-мерной RSS 3'-конца.
[0067] В некоторых вариантах осуществления способ включает стадии (а) иммунизации животного, отличного от человека, с помощью антигена, где указанное животное, отличное от человека, содержит геном, содержащий вариабельную область тяжелой цепи иммуноглобулина, которая содержит сконструированную DH-область, где сконструированная DH-область содержит один или более сегментов DH, каждый из которых функционально связан с 23-мерной RSS; (b) содержания грызуна в условиях, достаточных для того, чтобы у животного, отличного от человека, индуцировался иммунный ответ на антиген; и (с) извлечения антитела, которое связывает антиген, из животного, отличного от человека, или клетки животного, отличного от человека. В некоторых вариантах осуществления клетка животного, отличного от человека, представляет собой В-клетку. В некоторых вариантах осуществления клетка животного, отличного от человека, представляет собой гибридому.
[0068] В некоторых вариантах осуществления предусмотрено животное, отличное от человека, геном которого содержит вариабельную область тяжелой цепи человеческого иммуноглобулина, которая содержит один или более человеческих генных сегментов VH, сконструированную DH-область, которая содержит по меньшей мере один человеческий сегмент DH, функционально связанный с 23-мерной RSS, и по меньшей мере один человеческий генный сегмент JH, где вариабельная область тяжелой цепи человеческого иммуноглобулина функционально связана с одним или более эндогенными генами константной области иммуноглобулина таким образом, что грызун характеризуется тем, что когда он иммунизирован с помощью антигена, он образует антитела, содержащие вариабельные домены человеческой тяжелой цепи, которые содержат области CDR3, полученные при рекомбинации человеческих DH-DH и/или усиленной рекомбинации с генным сегментом JH6, и где антитела демонстрируют специфическое связывание с антигеном. В некоторых вариантах осуществления по меньшей мере один человеческий сегмент DH, функционально связанный с 23-мерной RSS, расположен на месте человеческого сегмента DH7-27. В некоторых вариантах осуществления вариабельная область тяжелой цепи человеческого иммуноглобулина содержит менее чем все шесть человеческих генных сегментов JH. В некоторых вариантах осуществления вариабельная область тяжелой цепи человеческого иммуноглобулина содержит только один из шести человеческих генных сегментов JH. В некоторых вариантах осуществления в вариабельной области тяжелой цепи человеческого иммуноглобулина содержится человеческий генный сегмент JH6 и отсутствует функциональный генный сегмент JH1, отсутствует функциональный генный сегмент JH2, отсутствует функциональный генный сегмент JH3, отсутствует функциональный генный сегмент JH4 и отсутствует функциональный генный сегмент JH5. В некоторых вариантах осуществления вариабельная область тяжелой цепи человеческого иммуноглобулина содержит только три из шести человеческих генных сегментов JH. В некоторых вариантах осуществления в вариабельной области тяжелой цепи человеческого иммуноглобулина содержится только человеческий генный сегмент JH4, человеческий генный сегмент JH5 и человеческий генный сегмент JH6, и отсутствует функциональный генный сегмент JH1, отсутствует функциональный генный сегмент JH2 и отсутствует функциональный генный сегмент JH3. В некоторых вариантах осуществления, где животное, отличное от человека, геном которого содержит вариабельную область тяжелой цепи человеческого иммуноглобулина, содержащую менее всех шести человеческих генных сегментов JH, например, вариабельную область тяжелой цепи человеческого иммуноглобулина, которая содержит только один из шести человеческих генных сегментов JH (например, вариабельную область тяжелой цепи человеческого иммуноглобулина, в которой содержится человеческий генный сегмент JH6 и отсутствует функциональный генный сегмент JHI, отсутствует функциональный генный сегмент JH2, отсутствует функциональный генный сегмент JH3, отсутствует функциональный генный сегмент JH4 и отсутствует функциональный генный сегмент JH5), проявляет усиленную рекомбинацию с генным сегментом JH6 по сравнению с контрольным животным, отличным от человека, геном которого содержит вариабельную область тяжелой цепи человеческого иммуноглобулина, содержащую все шесть человеческих генных сегментов JH, например, животное, отличное от человека, содержит более высокий процент перегруппированных последовательностей тяжелой цепи иммуноглобулина, содержащих последовательность генного сегмента JH6 или ее часть, и/или кодирующих CDR3, имеющую длину, составляющую по меньшей мере 20 аминокислот, чем контрольное животное, отличное от человека. В некоторых вариантах осуществления, где животное, отличное от человека, геном которого содержит вариабельную область тяжелой цепи человеческого иммуноглобулина, содержащую менее всех шести человеческих генных сегментов JH, например, вариабельную область тяжелой цепи человеческого иммуноглобулина, которая содержит только три из шести человеческих генных сегментов JH (например, вариабельную область тяжелой цепи человеческого иммуноглобулина, в которой содержится человеческий генный сегмент JH4, человеческий генный сегмент JH5 и человеческий генный сегмент JH6, и отсутствует функциональный генный сегмент JHI, отсутствует функциональный генный сегмент JH2 и отсутствует функциональный генный сегмент JH3), проявляет усиленную рекомбинацию с генным сегментом JH6 по сравнению с контрольным животным, отличным от человека, геном которого содержит вариабельную область тяжелой цепи человеческого иммуноглобулина, содержащую все шесть человеческих генных сегментов JH, например, животное, отличное от человека, содержит более высокий процент перегруппированных последовательностей тяжелой цепи иммуноглобулина, содержащих последовательность генного сегмента JH6 или ее часть, и/или кодирующих CDR3, длина которой составляет по меньшей мере 20 аминокислот, по сравнению с контрольным животным, отличным от человека.
[0069] В некоторых вариантах осуществления предусмотрено животное, отличное от человека, геном которого содержит вариабельную область тяжелой цепи человеческого иммуноглобулина, которая содержит один или более человеческих генных сегментов VH, сконструированную DH-область, которая содержит по меньшей мере один генный сегмент DH, фланкированный на своем 3'-конце 23-мерной RSS, и по меньшей мере два человеческих генных сегмента JH, где вариабельная область тяжелой цепи человеческого иммуноглобулина функционально связана с одним или более эндогенными генами константной области иммуноглобулина таким образом, что грызун характеризуется тем, что когда он иммунизирован с помощью антигена, он образует антитела, содержащие вариабельные домены человеческой тяжелой цепи, которые содержат области CDR3, полученные при рекомбинации человеческих DH-DH, и где антитела демонстрируют специфическое связывание с антигеном. В некоторых вариантах осуществления сконструированная DH-область содержит по меньшей мере три человеческих сегмента DH, каждый из которых фланкирован на своем 3'-конце 23-мерной RSS. В некоторых вариантах осуществления сконструированная DH-область содержит три человеческих сегмента DH, каждый из которых фланкирован на своем 3'-конце 23-мерной RSS, где указанные три человеческих сегмента DH представляют собой человеческий DH2-2, человеческий DH2-8 и человеческий DH2-15. В некоторых вариантах осуществления вариабельная область тяжелой цепи человеческого иммуноглобулина содержит человеческие генные сегменты VH от VH3-74 до VH6-1.
[0070] В некоторых вариантах осуществления антиген является патогеном, например, патогеном бактериальной, грибковой или вирусной природы. В некоторых вариантах осуществления иммунизация животного, отличного от человека, описанная в данном документе, включает инфицирование животного, отличного от человека, с помощью патогена, например, патогена бактериальной, грибковой или вирусной природы. В некоторых вариантах осуществления иммунизация животного, отличного от человека, описанная в данном документе, включает введение животному, отличному от человека, геномного или белкового материала, выделенного из патогена, например, патогена бактериальной, грибковой или вирусной природы. В некоторых вариантах осуществления антиген представляет собой рецептор (например, рецептор комплемента, рецептор хемокина и т.д.) или его часть. В некоторых вариантах осуществления антиген представляет собой нуклеиновую кислоту, кодирующую рецептор, или ее часть. В некоторых вариантах осуществления антиген представляет собой клетку, экспрессирующую рецептор, или ее часть. В некоторых вариантах осуществления антиген представляет собой ионный канал или его часть. В некоторых вариантах осуществления антиген представляет собой нуклеиновую кислоту, кодирующую ионный канал, или ее часть. В некоторых вариантах осуществления антиген представляет собой клетку, экспрессирующую ионный канал, или ее часть.
[0071] В некоторых вариантах осуществления предусмотренные животное, отличное от человека, клетка, отличная от человеческой, или ткань, отличная от человеческой, содержат геном, который дополнительно содержит вставку одного или более человеческих генных сегментов VL и одного или более человеческих генных сегментов JL в эндогенном локусе легкой цепи. В некоторых вариантах осуществления человеческие сегменты VL и JL представляют собой генные сегменты Vκ и Jκ и вставлены в эндогенный κ локус легкой цепи. В некоторых вариантах осуществления человеческие генные сегменты Vκ и Jκ функционально связаны с геном Сκ грызуна (например, геном Сκ мыши или крысы). В некоторых вариантах осуществления человеческие сегменты VL и JL представляют собой генные сегменты Vλ и Jλ и вставлены в эндогенный λ локус легкой цепи. В некоторых вариантах осуществления человеческие генные сегменты Vλ и Jλ функционально связаны с геном Сλ грызуна (например, с геном Сλ мыши или крысы).
[0072] В некоторых вариантах осуществления предусмотрено применение животного, отличного от человека, клетки, отличной от человеческой, или ткани, отличной от человеческой, описанных в данном документе, для изготовления и/или разработки лекарственного средства или вакцины для применения в медицине, например, для применения в качестве лекарственного препарата. В некоторых вариантах осуществления предусмотрено применение животного, отличного от человека, клетки, отличной от человеческой, или ткани, отличной от человеческой, описанных в данном документе, для изготовления и/или разработки антитела для введения людям. В некоторых вариантах осуществления предусмотрено применение животного, отличного от человека, клетки, отличной от человеческой, или ткани, отличной от человеческой, описанных в данном документе, для изготовления лекарственного препарата для лечения, предупреждения или снижения интенсивности проявления заболевания, нарушения или состояния.
[0073] В некоторых вариантах осуществления предусмотрено животное, отличное от человека, клетка, отличная от человеческой, или ткань, отличная от человеческой, описанные в данном документе, для применения для изготовления и/или разработки лекарственного средства для терапии или диагностики. В некоторых вариантах осуществления предусмотрено животное, отличное от человека, клетка, отличная от человеческой, или ткань, отличная от человеческой, описанные в данном документе, для применения для изготовления лекарственного препарата для лечения, предупреждения или снижения интенсивности проявления заболевания, нарушения или состояния.
[0074] В некоторых вариантах осуществления животное, отличное от человека, предусмотренное в данном документе, является грызуном; в некоторых вариантах осуществления - мышью; в некоторых вариантах осуществления - крысой. Во многих вариантах осуществления клетка животного, отличного от человека, предусмотренная в данном документе, представляет собой клетку грызуна; в некоторых вариантах осуществления клетку мыши; в некоторых вариантах осуществления клетку крысы. Во многих вариантах осуществления ткань животного, отличного от человека, предусмотренная в данном документе, представляет собой ткань грызуна; в некоторых вариантах осуществления - ткань мыши; в некоторых вариантах осуществления - ткань крысы.
[0075] В некоторых вариантах осуществления 23-мерная RSS содержит нуклеотидную последовательность, содержащую последовательность, представленную под SEQ ID NO: 151.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
[0076] Графические материалы, включенные в данный документ, которые состоят из последующих фигур, приведены только в иллюстративных целях, а не для ограничения.
[0077] На фигуре 1 без соблюдения масштаба показана общая иллюстрация вариантов осуществления настоящего изобретения, демонстрирующая порядок сборки генных сегментов при событии рекомбинации DJ в случае генных сегментов вариабельной области тяжелой цепи иммуноглобулина с немодифицированной DH-областью (верхняя панель) и сконструированной dh-областью (нижняя панель). 12-мерные последовательности сигнала рекомбинации (RSS) изображены в виде незакрашенных треугольников. 23-мерные RSS изображены в виде треугольников с вертикальной штриховкой. Иллюстративные немодифицированные генные сегменты VH (незакрашенные прямоугольники) и генные сегменты DH (закрашенные прямоугольники) соответственно изображены в виде незакрашенных и закрашенных прямоугольников и обозначены с использованием общей номенклатуры с использованием букв алфавита. Генные сегменты D (например, DH3-3), функционально связанные с 23-мерной RSS, и немодифицированные генные сегменты JH соответственно изображены в виде прямоугольников с горизонтальной штриховкой и незакрашенных прямоугольников и обозначены с применением их надлежащей номенклатуры. Также показан промотор μ0 (μ 0 pro). Не показана рекомбинация генного сегмента VH с рекомбинированным генным сегментом DJ.
[0078] На фигуре 2 без соблюдения масштаба показаны иллюстрации иллюстративных вариантов осуществления нацеливающихся векторов в соответствии с примером 1. Пунктирными линиями обозначены генные сегменты DH, которые включены в нацеливающийся вектор, но подробно не изображены.
[0079] На фигуре 3 без соблюдения масштаба показана иллюстрация неограничивающего иллюстративного варианта осуществления вставки нацеливающегося вектора 23:DH3-3:12/JH6 в локус вариабельной области тяжелой цепи гуманизированного иммуноглобулина в геноме ES-клеток мыши посредством электропорации (ЕР). Пунктирными линиями обозначены генные сегменты VH или генные сегменты DH, которые включены в локус вариабельной области тяжелой цепи, но подробно не изображены.
[0080] На фигуре 4 без соблюдения масштаба показана иллюстрация неограничивающего иллюстративного варианта осуществления Cre-опосредованной делеции кассет для отбора в локусах тяжелой цепи гуманизированного иммуноглобулина после электропорации и интеграции нацеливающегося вектора 23:DH3-3:12/JH6, как описано в примерах 1 и 2. Пунктирными линиями обозначены генные сегменты VH или генные сегменты DH, которые включены в локус вариабельной области тяжелой цепи, но подробно не изображены.
[0081] На фигуре 5 без соблюдения масштаба показана иллюстрация неограничивающего иллюстративного варианта осуществления вставки нацеливающегося вектора 23:DH3-3:12/JH4-6 в локус вариабельной области тяжелой цепи гуманизированного иммуноглобулина в геноме ES-клеток мыши посредством электропорации (ЕР). Пунктирными линиями обозначены генные сегменты VH или генные сегменты DH, которые включены в локус вариабельной области тяжелой цепи, но подробно не изображены.
[0082] На фигуре 6 без соблюдения масштаба показана иллюстрация неограничивающего иллюстративного варианта осуществления Cre-опосредованной делеции кассет для отбора в локусах тяжелой цепи гуманизированного иммуноглобулина после электропорации и интеграции нацеливающегося вектора 23:DH3-3:12/JH4-6, как описано в примерах 1 и 2. Пунктирными линиями обозначены генные сегменты VH или генные сегменты DH, которые включены в локус вариабельной области тяжелой цепи, но подробно не изображены.
[0083] На фигуре 7 без соблюдения масштаба показана иллюстрация неограничивающего иллюстративного варианта осуществления вставки нацеливающегося вектора 12:DH2-2:23|12:DH2-8:23|12:DH2-15:23/JH1-6 в локус вариабельной области тяжелой цепи гуманизированного иммуноглобулина в геноме ES-клеток мыши посредством электропорации (ЕР). Пунктирными линиями обозначены генные сегменты VH или генные сегменты DH, которые включены в локус вариабельной области тяжелой цепи, но подробно не изображены. Псевдогены изображены в виде закрашенных треугольников.
[0084] На фигуре 8 без соблюдения масштаба показана иллюстрация иллюстративного неограничивающего варианта осуществления Cre-опосредованной делеции кассет для отбора в локусах тяжелой цепи гуманизированного иммуноглобулина после электропорации и интеграции нацеливающегося вектора 12:DH2-2:23|12:DH2-8:23|12:DH2-15:23, как описано в примерах 1 и 2. Пунктирными линиями обозначены генные сегменты VH или генные сегменты DH, которые включены в локус вариабельной области тяжелой цепи, но подробно не изображены.
[0085] На фигуре 9А показаны результаты применительно к варианту осуществления настоящего изобретения, демонстрирующие доли в процентах (оси у) всех функциональных считываний для иммуноглобулинов (Ig), полученных в результате события рекомбинации DH-DH (нижняя панель), содержащих CDR3 с определенной аминокислотной длиной (оси х), выделенных из животных, модифицированных с помощью нацеливающегося вектора 12:DH2-2:23|12:DH2-8:23|12:DH2-15:23. "S1," "S2" и "S3" использованы для обозначения каждой отдельной экспериментальной мыши.
[0086] На фигуре 9В показаны результаты применительно к варианту осуществления настоящего изобретения, демонстрирующие доли в процентах (оси у) всех считываний Ig, полученных в результате события рекомбинации DH-DH (нижняя панель), содержащих CDR3 с определенной аминокислотной длиной (оси х), выделенных из животных, модифицированных с помощью нацеливающегося вектора 12:DH2-2:23|12:DH2-8:23|12:DH2-15:23. "S1," "S2" и "S3" использованы для обозначения каждой отдельной экспериментальной мыши.
[0087] На фигуре 10А показаны результаты применительно к варианту осуществления настоящего изобретения, демонстрирующие доли в процентах (оси у) всех функциональных считываний для иммуноглобулинов (Ig), полученных в результате события рекомбинации DH-DH, содержащих CDR3, содержащую определенное количество остатков цистеина (оси х), выделенных из животных, модифицированных с помощью нацеливающегося вектора 12:DH2-2:23|12:DH2-8:23|12:DH2-15:23. "S1," "S2" и "S3" использованы для обозначения каждой отдельной экспериментальной мыши.
[0088] На фигуре 10В показаны результаты применительно к варианту осуществления настоящего изобретения, демонстрирующие доли в процентах (оси у) всех считываний для иммуноглобулинов (Ig), полученных в результате события рекомбинации DH-DH, содержащих CDR3, содержащую определенное количество остатков цистеина (оси х), выделенных из животных, модифицированных с помощью нацеливающегося вектора 12:DH2-2:23|12:DH2-8:23|12:DH2-15:23. "S1," "S2" и "S3" использованы для обозначения каждой отдельной экспериментальной мыши.
[0089] На фигуре 11 показаны результаты применительно к варианту осуществления настоящего изобретения, демонстрирующие титры иллюстративных антител (ось у) у отдельных мышей с локусами к вариабельных областей тяжелой и легкой цепей гуманизированного иммуноглобулина (VI; см., например, патенты США №№8697940 и 8642835; каждый из которых включен в данный документ посредством ссылки во всей своей полноте) и мышей, модифицированных с помощью нацеливающегося вектора 23:DH3-3: 12/JH6, описанного в данном документе (V(DD)J), обе когорты которых были иммунизированы с применением ДНК-иммуногена, кодирующего рецептор, сопряженный с G-белком (GPCR). Титры антител определяли с помощью анализа связывания клеток линии MSD с применением 293 клеток, сконструированных для экспрессии GPCR (GPCR; ось х), и с применением клеток, которые не экспрессируют GPCR (контроль; ось х).
[0090] На фигуре 12А показаны результаты применительно к варианту осуществления настоящего изобретения, демонстрирующие доли в процентах (%; оси у) перегруппированных последовательностей VHDHJH гена тяжелой цепи иммуноглобулина и последовательностей гена, предположительно являющихся результатом перегруппировки VHDHA-DHBJH в соответствии со строгим критерием, представленным в таблице 7, выделенных из костного мозга (ВМ) или селезенки мышей, модифицированных с помощью нацеливающегося вектора 23:DH3-3: 12/JH6, и кодирующих CDR3 (HCDR3) тяжелой цепи, имеющую аминокислотную (АА) длину, составляющую от 5 до 30 аминокислот (оси х). Количества клеток ВМ и селезенки не нормализованы относительно друг друга. n=1.
[0091] На фигуре 12В показаны результаты применительно к варианту осуществления настоящего изобретения, демонстрирующие увеличенную диаграмму, показанную на панели на фигуре 12А. Количества клеток ВМ и селезенки не нормализованы относительно друг друга. n=1.
[0092] На фигуре 12С показаны результаты применительно к варианту осуществления настоящего изобретения, демонстрирующие доли в процентах для считываний с CDR3 длиной более 21 аминокислотного остатка как в костном мозге, так и в селезенке мышей, модифицированных с помощью нацеливающегося вектора 23:DH3-3:12/JH6. Количества клеток ВМ и селезенки не нормализованы относительно друг друга. n=1.
[0093] На фигуре 13 показаны результаты применительно к варианту осуществления настоящего изобретения, демонстрирующие доли в процентах (%; оси у) предположительно перегруппированных последовательностей VHDHA-DHBJH гена тяжелой цепи иммуноглобулина (в соответствии со строгим критерием, представленным в таблице 7), выделенных из костного мозга (ВМ) или селезенки мышей, модифицированных с помощью нацеливающегося вектора 23:DH3-3:12/JH6, и кодирующих CDR3 (HCDR3) тяжелой цепи с аминокислотной (АА) длиной, длина каждой из которых превышала 20 аминокислот (оси х). Количества клеток ВМ и селезенки не нормализованы относительно друг друга. n=1.
ОПРЕДЕЛЕНИЯ
[0094] Объем настоящего изобретения определяется формулой изобретения, прилагаемой к настоящему документу, и не ограничивается конкретными вариантами осуществления, описанными в данном документе; специалисты в данной области техники в ходе ознакомления с настоящим изобретением будут осведомлены о различных модификациях, которые могут быть эквивалентны таким описанным вариантам осуществления или иным образом находящимся в пределах объема формулы изобретения. В целом терминология соответствует общепринятому значению в данной области техники, если явно не указано иное. Точные определения определенных терминов приведены в данном документе ниже; значения этих и других терминов в конкретных случаях в рамках этого описания будут понятны специалистам в данной области техники из контекста. Дополнительные определения для следующих терминов и других терминов изложены по всему описанию. Ссылки, приведенные в данном описании, или соответствующие их части включены в данный документ посредством ссылки во всей своей полноте.
[0095] Порядковые термины, такие как «первый», «второй», «третий» и т.д., используемые в формуле изобретения для модификации элемента формулы изобретения, сами по себе не означают какой-либо приоритет, предшествование или порядок расположения одного элемента формулы изобретения перед другим или временной порядок, в котором выполняются действия способа, но используются исключительно в качестве отметок для проведения различий между одним элементом формулы изобретения, имеющим определенное название, и другим элементом, имеющим такое же название (за исключением используемого порядкового термина), для проведения различий между элементами формулы изобретения.
[0096] Форму единственного числа в описании и формуле изобретения, если четко не указано противоположное, следует понимать как охватывающие определяемые объекты в форме множественного числа. Признаки, раскрытые в пунктах формулы изобретения или частях описания, в которых содержится «или» между одним или более членами группы, считаются охваченными, если один, более одного или все члены группы присутствуют в указанном продукте или способе, используются в них или иным образом имеют отношение к ним, если не указано противоположное или иное не очевидно из контекста. Настоящее изобретение охватывает варианты осуществления, в которых только один член группы присутствует в указанном продукте или способе, используется в них или иным образом имеет отношение к ним. Настоящее изобретение также охватывает варианты осуществления, в которых более одного или все члены группы присутствуют в указанном продукте или способе, используются в них или иным образом имеют отношение к ним. Кроме того, следует понимать, что настоящее изобретение охватывает все варианты, комбинации и перестановки, в которых одно или более ограничений, элементов, условий, описательных терминов и т.д. из одного или более перечисленных пунктов формулы изобретения входят в другой пункт формулы изобретения, зависимый от того же основного пункта формулы изобретения (или, в зависимости от того, что применимо, любого другого пункта формулы изобретения), если не указано иное или если специалисту в данной области не будет очевидным возникновение противоречия или несоответствия. В тех случаях, когда элементы представлены в виде перечней (например, в виде группы Маркуша или в аналогичном формате), следует понимать, что также раскрыта каждая подгруппа элементов, и любой(-ые) элемент(-ы) можно исключить из группы. Следует понимать, что, как правило, в тех случаях, когда настоящее изобретение или аспекты настоящего изобретения рассматриваются как включающие конкретные элементы, признаки и т.д., варианты осуществления настоящего изобретения или аспекты настоящего изобретения состоят из таких элементов, признаков и т.д. или состоят главным образом из них. С целью упрощения такие варианты осуществления не были в каждом случае специально прямо изложены в данном документе. Также следует понимать, что любой вариант осуществления или аспект настоящего изобретения может быть явным образом исключен из формулы изобретения независимо от упоминания конкретного исключения в описании.
[0097] Используемые в данной заявке термины «приблизительно» и «примерно» применяются в качестве эквивалентов. Предполагается, что все числа, используемые в настоящей заявке, вместе с термином «приблизительно/примерно» или без него охватывают любые нормальные отклонения, например, +/- 5%, понятные специалисту в соответствующей области техники.
[0098] «Введение» относится к введению композиции субъекту или в живую систему (например, в клетку, орган, ткань, организм или их соответствующий компонент или набор компонентов). Специалисты в данной области техники поймут, что путь введения может варьироваться, например, в зависимости от субъекта или системы, которым будет вводиться композиция, природы композиции, цели введения и т.д.
[0099] Например, в некоторых вариантах осуществления введение субъекту-животному (например, человеку или грызуну) может быть бронхиальным (в том числе посредством бронхиальной инсталляции), трансбуккальным, энтеральным, внутрикожным, внутриартериальным, интрадермальным, внутрижелудочным, интрамедуллярным, внутримышечным, интраназальным, внутрибрюшинным, интратекальным, внутривенным, интравентрикулярным, чресслизистым, назальным, пероральным, ректальным, подкожным, подъязычным, местным, трахеальным (в том числе посредством интратрахеальной инстилляции), трансдермальным, вагинальным и/или витреальным. В некоторых вариантах осуществления введение может подразумевать прерывистое введение доз. В некоторых вариантах осуществления введение может предусматривать непрерывное введение доз (например, перфузия) в течение по меньшей мере выбранного периода времени. В некоторых вариантах осуществления антитело, полученное с помощью животного, отличного от человека, раскрытого в данном документе, может быть введено субъекту (например, субъекту человеку или грызуну). В некоторых вариантах осуществления фармацевтическая композиция содержит антитело, полученное с помощью животного, отличного от человека, раскрытого в данном документе. В некоторых вариантах осуществления фармацевтическая композиция может содержать буфер, разбавитель, вспомогательное вещество или любую их комбинацию. В некоторых вариантах осуществления фармацевтическая композиция, содержащая антитело, полученное с помощью животного, отличного от человека, раскрытого в данном документе, может быть включена в контейнер для хранения или введения, например, флакон, шприц (например, шприц для внутривенной инфузии) или пакет (например, пакет для внутривенной инфузии).
[00100] Термин «биологически активный»: обозначает характеристику любого средства, обладающего активностью в биологической системе in vitro или in vivo (например, в организме). Например, средство, которое в случае присутствия в организме оказывает биологическое воздействие в пределах этого организма, считается биологически активным.
[00101] В конкретных вариантах осуществления, в которых белок или полипептид являются биологически активными, часть этого белка или полипептида, обладающая по меньшей мере одним общим с данным белком или полипептидом видом биологической активности, обычно называется «биологически активной» частью.
[00102] Термин «сравнимый» относится к двум или более средствам, объектам, ситуациям, наборам условий и т.д., которые могут не быть идентичны друг другу, но которые являются в достаточной степени похожими для обеспечения возможности их сравнения, чтобы на основании наблюдаемых различий или сходств можно было сделать вполне обоснованные заключения. Специалисту в данной области техники будет понятно из контекста, какая степень идентичности необходима в любых заданных обстоятельствах для двух или более таких средств, объектов, ситуаций, наборов условий и т.д., чтобы считать их сравнимыми.
[00103] Термин «консервативный» относится к консервативной аминокислотной замене, т.е. к замене аминокислотного остатка на другой аминокислотный остаток, содержащий R-группу боковой цепи с аналогичными химическими свойствами (например, зарядом или гидрофобностью). Как правило, консервативная аминокислотная замена практически не будет изменять представляющие интерес функциональные свойства белка, например способность рецептора связываться с лигандом. Примеры групп аминокислот, которые имеют боковые цепи с аналогичными химическими свойствами, включают: алифатические боковые цепи, такие как у глицина, аланина, валина, лейцина и изолейцина; алифатические боковые цепи с гидроксильными группами, такие как у серина и треонина; амид-содержащие боковые цепи, такие как у аспарагина и глутамина; ароматические боковые цепи, такие как у фенилаланина, тирозина и триптофана; основные боковые цепи, такие как у лизина, аргинина и гистидина; кислотные боковые цепи, такие как у аспарагиновой кислоты и глутаминовой кислоты; и серосодержащие боковые цепи, такие как у цистеина и метионина. Группы консервативных аминокислотных замен включают, например, валин/лейцин/изолейцин, фенилаланин/тирозин, лизин/аргинин, аланин/валин, глутамат/аспартат и аспарагин/глутамин.
[00104] B некоторых вариантах осуществления консервативная аминокислотная замена может представлять собой замену любого нативного остатка в белке аланином, используемую, например, в аланин-сканирующем мутагенезе. В некоторых вариантах осуществления проводят консервативную замену, которая имеет положительное значение в матрице логарифмической функции правдоподобия РАМ250, раскрытой в Gonnet, G.Н. et al., 1992, Science 256:1443-1445, включенной в данный документ посредством ссылки во всей своей полноте. В некоторых вариантах осуществления замена представляет собой умеренно консервативную замену, где замена имеет неотрицательное значение в матрице логарифмической функции правдоподобия РАМ250.
[00105] Выражение «контроль»: относится к понятному в данной области значению слова «контроль», подразумевающему стандарт, с которым сравнивают результаты. Как правило, контроли применяют для повышения целостности в экспериментах путем разделения переменных для того, чтобы сделать вывод об этих переменных. В некоторых вариантах осуществления контроль представляет собой реакцию или анализ, проводимые одновременно с тестовыми реакцией или анализом для обеспечения объекта сравнения. «Контроль» может относиться к «контрольному животному». «Контрольное животное» может иметь модификацию, описанную в данном документе, модификацию, отличную от описанной в данном документе, или не иметь модификации (т.е. обозначать животное дикого типа). В одном эксперименте используют «тест» (т.е. тестируемую переменную). Во втором эксперименте, «контрольном», тестируемую переменную не используют. Контроль может представлять собой положительный контроль или отрицательный контроль.
[00106] B некоторых вариантах осуществления контроль представляет собой исторический контроль (т.е. тест или анализ, проведенный ранее, или количество или результат, известные заранее). В некоторых вариантах осуществления контроль представляет собой или включает напечатанную или иным образом сохраненную запись.
[00107] «Разрушение» обозначает результат события гомологичной рекомбинации с молекулой ДНК (например, с эндогенной гомологичной последовательностью, такой как ген или локус гена).
[00108] B некоторых вариантах осуществления разрушение может приводить к вставке, делеции, замене, замещению, миссенс-мутации или сдвигу рамки считывания в последовательности(ях) ДНК или любой их комбинации или может быть представлено такими. Вставки могут включать вставку целых генов, фрагментов генов, например, экзонов, которые по происхождению могут быть отличны от эндогенной последовательности (например, являются гетерологичной последовательностью), или кодирующих последовательностей, полученных или выделенных из конкретного представляющего интерес гена. В некоторых вариантах осуществления разрушение может приводить к повышению экспрессии и/или активности гена или продукта гена (например, белка, кодируемого геном). В некоторых вариантах осуществления разрушение может приводить к снижению уровня экспрессии и/или активности гена или продукта гена. В некоторых вариантах осуществления разрушение может приводить к изменению последовательности гена или кодируемого продукта гена (например, кодируемого белка). В некоторых вариантах осуществления разрушение может приводить к изменению последовательности хромосомы или положения хромосомы в геноме. В некоторых вариантах осуществления разрушение может приводить к усечению или фрагментации гена или кодируемого продукта гена (например, кодируемого белка). В некоторых вариантах осуществления разрушение может приводить к удлинению гена или кодируемого продукта гена. В некоторых таких вариантах осуществления за счет разрушения может обеспечиваться сборка слитого белка. В некоторых вариантах осуществления разрушение может влиять на уровень экспрессии, но не на активность гена или продукта гена. В некоторых вариантах осуществления разрушение может влиять на активность, но не на уровень экспрессии гена или продукта гена. В некоторых вариантах осуществления разрушение может не оказывать значительного воздействия на уровень экспрессии гена или продукта гена. В некоторых вариантах осуществления разрушение может не оказывать значительного воздействия на активность гена или продукта гена. В некоторых вариантах осуществления разрушение может не оказывать значительного воздействия ни на уровень экспрессии, ни на активность гена или продукта гена. В некоторых вариантах осуществления значительное воздействие может быть измерено с помощью, например, но без ограничения, t-критерия Стьюдента.
[00109] «Эндогенный локус» или «эндогенный ген» относится к генетическому локусу, находящемуся в исходном или эталонном организме (или клетке) до введения изменения, разрушения, делеции, вставки, модификации, замещения или замены, описанных в данном документе.
[00110] B некоторых вариантах осуществления эндогенный локус содержит последовательность, частично или полностью, встречающуюся в природе. В некоторых вариантах осуществления эндогенный локус представляет собой локус дикого типа. В некоторых вариантах осуществления эталонный организм представляет собой организм дикого типа. В некоторых вариантах осуществления эталонный организм представляет собой сконструированный организм. В некоторых вариантах осуществления эталонный организм представляет собой организм, разводимый в лаборатории (или дикого типа, или сконструированный).
[00111] «Эндогенный промотор» относится к промотору, который в естественных условиях, например, в организме дикого типа, ассоциирован с эндогенным геном или генетическим локусом.
[00112] «Сконструированный» обозначает, в целом, аспект проведения манипуляций человеком. Как понятно специалистам в данной области техники и что является для них общепринятой практикой, потомство сконструированного полинуклеотида или клетки, как правило, по-прежнему называют «сконструированным», даже если фактическую манипуляцию проводили на предшествующем объекте. Кроме того, как будет понятно специалистам в данной области техники, доступно множество методик, посредством которых можно осуществлять «конструирование», описанное в данном документе.
[00113] B некоторых вариантах осуществления полинуклеотид может считаться «сконструированным» в том случае, если две или более последовательности, которые не соединены друг с другом в этом порядке в природе, подвергаются манипуляции человеком для непосредственного соединения друг с другом в сконструированном полинуклеотиде. В некоторых вариантах осуществления сконструированный с помощью генной инженерии полинуклеотид может содержать регуляторную последовательность, которая обнаруживается в природе в функциональной связи с первой кодирующей последовательностью, но не в функциональной связи со второй кодирующей последовательностью, соединенную человеком таким образом, чтобы она была функционально связанной со второй кодирующей последовательностью. В вариантах осуществления в настоящем документе, описывающих сконструированный с помощью генной инженерии генный сегмент DH, генный сегмент DH подвергают манипуляции человеком таким образом, чтобы он был функционально связан (например, был фланкирован по меньшей мере с одной стороны, был смежным, граничил, непосредственно прилегал) с 23-мерной RSS. В некоторых вариантах осуществления генный сегмент DH, сконструированный таким образом, что он функционально связан с 23-мерной RSS, получают из другого генного сегмента DH, например, генный сегмент DH, функционально связанный с 23-мерной RSS, содержит нуклеотидную последовательность, идентичную нуклеотидной последовательности другого генного сегмента DH, с отличиями, обусловленными лишь вырожденностью генетического кода и/или заменой 12-мерной RSS на 23-мерную RSS. Другой генный сегмент DH и сконструированный генный сегмент DH, полученный из него (например, генный сегмент DH, функционально связанный с 23-мерной RSS), могут считаться соответствующими генными сегментами. Например человеческий генный сегмент DH3-3, функционально связанный с 23-мерной RSS, может считаться соответствующим генному сегменту DH3-3, фланкированному с одной стороны 12-мерной RSS и с другой стороны другой 12-мерной RSS, где генный сегмент DH3-3, функционально связанный с 23-мерной RSS, генный сегмент DH3-3, фланкированный с одной стороны 12-мерной RSS и с другой стороны другой 12-мерной RSS, имеют идентичную нуклеотидную последовательность, за исключением отличий, обусловленных вырожденностью генетического кода и/или заменой одной из двух 12-мерных RSS на 23-мерную RSS. В качестве альтернативы или дополнительно, в некоторых вариантах осуществления первая и вторая последовательности нуклеиновых кислот, каждая из которых кодирует полипептидные элементы или домены, которые в природе не связаны друг с другом, могут быть связаны друг с другом в единый сконструированный полинуклеотид. Аналогично, в некоторых вариантах осуществления клетка или организм могут считаться «сконструированными» в том случае, если они были подвергнуты манипуляции таким образом, чтобы их генетическая информация была изменена (например, был введен новый генетический материал, который ранее отсутствовал, или ранее присутствовавший генетический материал был изменен или удален).
[00114] B некоторых вариантах осуществления «конструирование» может включать отбор или разработку (например, последовательностей нуклеиновых кислот, полипептидных последовательностей, клеток, тканей и/или организмов) путем использования компьютерных систем, запрограммированных для проведения анализа или сравнения, или в иных случаях для обеспечения анализа, рекомендации в отношении и/или отбора, последовательностей, изменений и т.д. В качестве альтернативы или дополнительно в некоторых вариантах осуществления «конструирование» может включать применение методологий химического синтеза in vitro и/или методик рекомбинантных нуклеиновых кислот, таких как, например, амплификация нуклеиновой кислоты (например, посредством полимеразной цепной реакции), гибридизация, мутация, трансформация, трансфекция и т.д. и/или любой из множества методологий контролируемого размножения. Как будет понятно специалистам в данной области, множество таких общепринятых методик (например, для рекомбинантной ДНК, синтеза олигонуклеотидов и культивирования и трансформации тканей (например, электропорация, липофекция и т.д.)) хорошо известны из уровня техники и описаны в различных общих и более специфических литературных источниках, которые цитируются и/или обсуждаются во всем настоящем описании. См., например, Sambrook et al., Molecular Cloning: A Laboratory Manual (2d ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, Нью-Йорк, 1989); включен в данный документ посредством ссылки во всей своей полноте).
[00115] «Ген» относится к последовательности ДНК в хромосоме, которая кодирует продукт (например, РНК-продукт и/или полипептидный продукт). С целью внесения ясности, термин «ген», как правило, относится к части нуклеиновой кислоты, которая кодирует полипептид; при этом данный термин может необязательно охватывать регуляторные последовательности, что будет понятно из контекста специалистам в данной области техники. Это определение не подразумевает исключение применения термина «ген» к не кодирующим белок единицам экспрессии, а скорее поясняет то, что в большинстве случаев термин, используемый в данном документе, относится к нуклеиновой кислоте, кодирующей полипептид.
[00116] B некоторых вариантах осуществления ген содержит кодирующую последовательность (т.е. последовательность, которая кодирует конкретный продукт). В некоторых вариантах осуществления ген содержит некодирующую последовательность. В некоторых вариантах осуществления ген может содержать как кодирующую (например, экзонную), так и некодирующую (например, интронную) последовательность. В некоторых вариантах осуществления ген может содержать одну или более регуляторных последовательностей (например, промоторы, энхансеры и т.д.) и/или интронных последовательностей, которые, например, могут контролировать или влиять на один или более аспектов экспрессии гена (например, специфическую для типа клеток экспрессию, индуцируемую экспрессию и т.д.).
[00117] Выражение «гетерологичный» относится к средству или объекту из другого источника. Например, при использовании в отношении полипептида, гена или продукта гена, представленного в конкретной клетке или организме, термин поясняет, что релевантный полипептид или его фрагмент, ген или его фрагмент, или продукт гена или его фрагмент (1) был сконструирован человеком; (2) был введен в клетку или организм (или в их предшественника) человеком (например, посредством генной инженерии) и/или (3) в естественных условиях не вырабатывается соответствующей клеткой или организмом (например, соответствующим типом клетки или типом организма) или отсутствует в них. Другой пример включает полипептид или его фрагмент, ген или его фрагмент, или продукт гена или его фрагмент, которые обычно присутствуют в конкретной нативной клетке или организме, но были модифицированы, например, за счет мутации или помещения под контроль не связанных в естественных условиях и, в некоторых вариантах осуществления, не являющихся эндогенными регуляторных элементов (например, промотора).
[00118] «Клетка-хозяин» относится к клетке, в которую были введены гетерологичные (например, экзогенные) нуклеиновая кислота или белок. При ознакомлении с данным изобретением специалистам в данной области техники будет понятно, что такие термины обозначают не только конкретную рассматриваемую клетку, но также используются для обозначения потомства такой клетки. Поскольку в последующих поколениях вследствие влияния мутаций или окружающей среды могут происходить определенные модификации, такое потомство в действительности может не быть идентичным родительской клетке, но по-прежнему включается в объем термина «клетка-хозяин».
[00119] B некоторых вариантах осуществления клетка-хозяин представляет собой или включает прокариотическую или эукариотическую клетку. В вариантах осуществления клетка-хозяин представляет собой или включает клетку млекопитающего. Как правило, клетка-хозяин представляет собой любую клетку, подходящую для приема и/или продуцирования гетерологичных нуклеиновой кислоты или белка, независимо от того, к какому царству живой природы относится клетка. Иллюстративные клетки предусматривают клетки прокариот и эукариот (одноклеточных или многоклеточных), бактериальные клетки (например, штаммы Escherichia coli, Bacillus spp., Streptomyces spp. и т.д.), клетки микобактерий, клетки грибов, клетки дрожжей (например, Saccharomyces cerevisiae, Schizosaccharomyces pombe, Pichiapastoris, Pichia methanolica, и т.д.), клетки растений, клетки насекомых (например, SF-9, SF-21, клетки насекомых, зараженные бакуловирусами, Trichoplusiani, и т.д.), клетки животных, отличных от человека, клетки человека или продукты слияния клеток, такие как, например, гибридомы или квадромы.
[00120] B некоторых вариантах осуществления клетка представляет собой клетку человека, обезьяны, человекообразной обезьяны, хомяка, крысы или мыши. В некоторых вариантах осуществления клетка является эукариотической и выбрана из следующих клеток: СНО (например, СНО K1, DXB-11 СНО, Veggie-CHO), COS (например, COS-7), клетки сетчатки, Vero, CV1, клетки почки (например, HEK293, 293 EBNA, MSR293, MDCK, HaK, BHK), HeLa, HepG2, WI38, MRC 5, Colo205, НВ 8065, HL-60 (например, BHK21), Jurkat, Daudi, А431 (эпидермальные), CV-1, U937, 3Т3, L-клетки, клетки С127, SP2/0, NS-0, ММТ 060562, клетки Сертоли, клетки BRL 3А, клетки НТ1080, клетки миеломы, опухолевые клетки и линии клеток, полученные из вышеупомянутых клеток. В некоторых вариантах осуществления клетка содержит один или более вирусных генов, например, клетка сетчатки, которая экспрессирует вирусный ген (например, клетка PER.C6®). В некоторых вариантах осуществления клетка-хозяин представляет собой или предусматривает выделенную клетку. В некоторых вариантах осуществления клетка-хозяин представляет собой часть ткани. В некоторых вариантах осуществления клетка-хозяин представляет собой часть организма.
[00121] «Гуманизированный» относится к молекуле (например, нуклеиновой кислоте, белку и т.д.), которая не являлась человеческой по происхождению и в которой часть была заменена на соответствующую часть соответствующей человеческой молекулы, за счет чего модифицированная (например, гуманизированная) молекула сохраняет свою биологическую функцию и/или сохраняет структуру, которая осуществляет сохраненную биологическую функцию. В отличие от этого, термин «человеческий» и т.п. охватывает молекулы, характеризующиеся исключительно человеческим происхождением, например человеческие нуклеотиды или белок, содержащие исключительно человеческие нуклеотидные и аминокислотные последовательности соответственно. Термин «человеческий (гуманизированный)» используют для отображения того, что человеческая (гуманизированная) молекула может быть (а) человеческой молекулой или (b) гуманизированной молекулой.
[00122] «Идентичность» применительно к сравнению последовательностей относится к идентичности, определенной с помощью ряда различных алгоритмов, известных из уровня техники, которые можно применять для измерения идентичности нуклеотидных и/или аминокислотных последовательностей.
[00123] B некоторых вариантах осуществления значения идентичности, описываемые в данном документе, определяют с применением выравнивания ClustalW версии 1.83 (медленного), в котором используются штраф за открытие гэпа 10,0, штраф за продление гэпа 0,1, а также с применением матрицы сходства Gonnet (MACVECTOR™ 10.0.2, Mac Vector Inc., 2008).
[00124] «In vitro» обозначает события, которые происходят в искусственной окружающей среде, например, в пробирке или реакционном сосуде, в культуре клеток и т.д., а не в пределах многоклеточного организма.
[00125] «In vivo» означает события, которые происходят в пределах многоклеточного организма, такого как человек и/или животное, отличное от человека. В контексте клеточных систем данный термин может использоваться в отношении событий, которые происходят в пределах живой клетки (по сравнению, например, с in vitra-системами).
[00126] «Выделенный» относится к веществу и/или объекту, которые были (1) отделены по меньшей мере от некоторых компонентов, с которыми они были связаны при первоначальном получении (либо в природе, либо в экспериментальных условиях), и/или (2) сконструированы, продуцированы, получены и/или изготовлены в результате действий человека. Выделенные вещества и/или объекты могут быть отделены от приблизительно 10 или более других компонентов, с которыми они были изначально связаны. В некоторых вариантах осуществления выделенные средства характеризуются чистотой, составляющей по меньшей мере приблизительно 80% или больше. Вещество является «чистым», если оно практически не содержит других компонентов. В некоторых вариантах осуществления, что будет понятно специалистам в данной области, вещество может по-прежнему считаться «выделенным» или даже «чистым» после объединения с определенными другими компонентами, такими как, например, один или более носителей или наполнителей (например, с буфером, растворителем, водой и т.д.); в таких вариантах осуществления процент выделения или чистоты вещества рассчитывают без учета таких носителей или наполнителей.
[00127] B качестве лишь одного примера, в некоторых вариантах осуществления биологический полимер, такой как полипептид или полинуклеотид, встречающийся в природе, считается «выделенным», если: (а) он ввиду своего происхождения или источника получения не связан с некоторыми или всеми компонентами, сопровождающими его в его нативном состоянии в природе; (b) он практически не содержит других полипептидов или нуклеиновых кислот того же вида из вида, который вырабатывает их в природе; или (с) он экспрессируется или иным образом связан с компонентами в клетке или другой системе экспрессии, не принадлежащей к виду, который вырабатывает его в природе. Таким образом, например, в некоторых вариантах осуществления полипептид, синтезируемый химическим путем или синтезируемый в клеточной системе, отличной от той, которая продуцирует его в природе, считается «выделенным» полипептидом. В качестве альтернативы или дополнительно, в некоторых вариантах осуществления полипептид, который был подвергнут очистке с помощью одной или более методик, может считаться «выделенным» полипептидом в той степени, в которой он был отделен от других компонентов: а) с которыми он связан в природе и/или b) с которыми он был связан при первоначальном получении.
[00128] «Животное, отличное от человека», относится к любому позвоночному живому существу, которое не является человеком.
[00129] B некоторых вариантах осуществления животное, отличное от человека, является круглоротым, костной рыбой, хрящевой рыбой (например, акула или скат), земноводным, пресмыкающимся, млекопитающим и птицей. В некоторых вариантах осуществления млекопитающее, отличное от человека, представляет собой примата, козу, овцу, свинью, собаку, корову или грызуна. В некоторых вариантах осуществления животное, отличное от человека, представляет собой грызуна, такого как крыса или мышь.
[00130] Термин «нуклеиновая кислота» в его наиболее широком смысле относится к любому соединению и/или веществу, которое включено или может быть включено в состав олигонуклеотидной цепи и как правило является взаимозаменяемым по отношению к молекуле нуклеиновой кислоты, последовательности нуклеиновой кислоты, нуклеотиду, нуклеотидной молекуле, где указанные термины также являются взаимозаменяемыми по отношению друг к другу.
[00131] B некоторых вариантах осуществления «нуклеиновая кислота» представляет собой соединение и/или вещество, которое включено или может быть включено в состав олигонуклеотидной цепи посредством фосфодиэфирной связи. Как будет очевидно из контекста, в некоторых вариантах осуществления термин «нуклеиновая кислота» относится к отдельным остаткам нуклеиновой кислоты (например, нуклеотидам и/или нуклеозидам); в некоторых вариантах осуществления термин «нуклеиновая кислота» относится к олигонуклеотидной цепи, содержащей отдельные остатки нуклеиновой кислоты. В некоторых вариантах осуществления «нуклеиновая кислота» представляет собой или содержит РНК; в некоторых вариантах осуществления «нуклеиновая кислота» представляет собой или содержит ДНК. В некоторых вариантах осуществления «нуклеиновая кислота» представляет собой, содержит или состоит из одного или более остатков природной нуклеиновой кислоты. В некоторых вариантах осуществления «нуклеиновая кислота» представляет собой, содержит или состоит из одного или более аналогов нуклеиновой кислоты. В некоторых вариантах осуществления аналог нуклеиновой кислоты отличается от «нуклеиновой кислоты» тем, что в нем не используется фосфодиэфирный остов. Например, в некоторых вариантах осуществления «нуклеиновая кислота» представляет собой, содержит или состоит из одной или более «пептидных нуклеиновых кислот», которые известны из уровня техники и имеют в остове пептидные связи вместо фосфодиэфирных связей и считаются находящимися в пределах объема настоящего изобретения. В качестве альтернативы или дополнительно в некоторых вариантах осуществления «нуклеиновая кислота» содержит одну или более фосфоротиоатных и/или 5'-N-фосфорамидитных связей вместо фосфодиэфирных связей. В некоторых вариантах осуществления «нуклеиновая кислота» представляет собой, содержит или состоит из одного или более природных нуклеозидов (например, аденозина, тимидина, гуанозина, цитидина, уридина, дезоксиаденозина, дезокситимидина, дезоксигуанозина и дезоксицитидина). В некоторых вариантах осуществления «нуклеиновая кислота» представляет собой, содержит или состоит из одного или более аналогов нуклеозидов (например, 2-аминоаденозина, 2-тиотимидина, инозина, пирролопиримидина, 3-метиладенозина, 5-метилцитидина, С5-пропинилцитидина, С5-пропинилуридина, 2-аминоаденозина, С5-бромуридина, С5-фторуридина, С5-йодуридина, С5-пропинилуридина, С5-пропинилцитидина, С5-метилцитидина, 2-аминоаденозина, 7-деазааденозина, 7-деазагуанозина, 8-оксоаденозина, 8-оксогуанозина, O(6)-метилгуанина, 2-тиоцитидина, метилированных оснований, интеркалированных оснований и их комбинаций). В некоторых вариантах осуществления «нуклеиновая кислота» содержит один или более модифицированных сахарных остатков (например, 2'-фтор-рибозу, рибозу, 2'-дезоксирибозу, арабинозу и гексозу) по сравнению с таковыми в природных нуклеиновых кислотах. В некоторых вариантах осуществления «нуклеиновая кислота» имеет нуклеотидную последовательность, которая кодирует функциональный продукт гена, такой как РНК или белок. В некоторых вариантах осуществления «нуклеиновая кислота» имеет нуклеотидную последовательность, которая кодирует фрагмент полипептида (например, пептид). В некоторых вариантах осуществления «нуклеиновая кислота» содержит один или более интронов. В некоторых вариантах осуществления «нуклеиновая кислота» содержит один или более экзонов. В некоторых вариантах осуществления «нуклеиновая кислота» содержит одну или более кодирующих последовательностей. В некоторых вариантах осуществления «нуклеиновую кислоту» получают посредством одного или более из выделения из природного источника, ферментативного синтеза путем полимеризации на основе комплементарной матрицы (in vivo или in vitro), воспроизведения в рекомбинантной клетке или системе и химического синтеза. В некоторых вариантах осуществления «нуклеиновая кислота» имеет длину, составляющую по меньшей мере 3 или более остатков. В некоторых вариантах осуществления «нуклеиновая кислота» является однонитевой; в некоторых вариантах осуществления «нуклеиновая кислота» является двухнитевой. В некоторых вариантах осуществления «нуклеиновая кислота» имеет нуклеотидную последовательность, содержащую по меньшей мере один элемент, который кодирует полипептид или его фрагмент, или комплементарна последовательности, которая кодирует полипептид или его фрагмент. В некоторых вариантах осуществления «нуклеиновая кислота» обладает ферментативной активностью.
[00132] «Функционально связанный» относится к контактному положению, при котором описанные компоненты находятся во взаимосвязи, позволяющей им функционировать надлежащим образом.
[00133] B некоторых вариантах осуществления функционально связанные нуклеотидные последовательности являются смежными друг с другом, например, последовательность нуклеиновой кислоты, содержащая генный сегмент иммуноглобулина, функционально связанный с RSS, содержит нуклеотидную последовательность генного сегмента иммуноглобулина, смежную с нуклеотидной последовательностью RSS, например, генный сегмент иммуноглобулина фланкирован по меньшей мере с одной стороны (например, примыкает) нуклеотидной последовательностью RSS непрерывным образом, чтобы генный сегмент иммуноглобулина непосредственно прилегал к нуклеотидной последовательности RSS.
[00134] B других вариантах осуществления функциональная связь не требует смежного положения. Например, неперегруппированные генные сегменты вариабельной области «функционально связанные» друг с другом, способны к перегруппировке с образованием перегруппированного гена вариабельной области, где указанные неперегруппированные генные сегменты вариабельной области могут необязательно быть смежными друг с другом. Неперегруппированные генные сегменты вариабельной области, функционально связанные друг с другом и со смежным геном константной области, способны к перегруппировке с образованием перегруппированного гена вариабельной области, который экспрессируется в сочетании с геном константной области в виде полипептидной цепи антиген связывающего белка. Контролирующая последовательность, «функционально связанная» с кодирующей последовательностью, лигирована таким образом, что экспрессия кодирующей последовательности достигается в условиях, подходящих для функционирования контролирующих последовательностей. «Функционально связанные» последовательности включают как последовательности, контролирующие экспрессию, смежные с геном, представляющим интерес, так и последовательности, контролирующие экспрессию, действующие по транс-типу или на некотором расстоянии, осуществляя контроль над геном, представляющим интерес.
[00135] Термин «последовательность, контролирующая экспрессию», относится к полинуклеотидным последовательностям, которые являются необходимыми для осуществления воздействия на экспрессию и процессинг кодирующих последовательностей, с которыми они лигированы. «Последовательности, контролирующие экспрессию», включают соответствующие последовательности для инициации и терминации транскрипции, промоторные и энхансерные последовательности; сигналы для эффективного процессинга РНК, такие как сигналы сплайсинга и полиаденилирования; последовательности, стабилизирующие цитоплазматическую мРНК; последовательности, повышающие эффективность трансляции (т.е. консенсусную последовательность Козак); последовательности, повышающие стабильность белка; и при необходимости последовательности, усиливающие секрецию белка. Природа таких контролирующих последовательностей различается в зависимости от организма-хозяина. Например, у прокариот такие контролирующие последовательности обычно предусматривают промотор, участок связывания рибосом и последовательность для терминации транскрипции, тогда как у эукариот обычно такие контролирующие последовательности предусматривают промоторы и последовательность для терминации транскрипции. Термин «контролирующие последовательности» подразумевает включение компонентов, наличие которых является существенным для экспрессии и процессинга, и также может включать дополнительные компоненты, наличие которых является преимущественным, например, лидерные последовательности и последовательности-партнеры по слиянию.
[00136] Как правило, каждый неперегруппированный генный сегмент V, генный сегмент D или генный сегмент J иммуноглобулина функционально связан (например, ассоциирован, фланкирован с одной или с двух сторон, является смежным и т.д.) с последовательностью сигнала рекомбинации (RSS), которая может представлять собой 12-мерную RSS или 23-мерную RSS. Любой генный сегмент, фланкированный с каждой стороны RSS (включая генный сегмент DH, функционально связанный с 23-мерной RSS), не подвергался рекомбинации, и, таким образом, может считаться "неперегруппированным" генным сегментом. В некоторых вариантах осуществления в данном документе неперегруппированный генный сегмент может содержать генный сегмент в своей конфигурации зародышевой линии (например, дикого типа), например, как генный сегмент VH зародышевой линии, так и генный сегмент JH зародышевой линии фланкированы с обеих сторон 23-мерной RSS. Напротив, генный сегмент DH зародышевой линии, например, неперегруппированный генный сегмент DH, фланкирован с каждой стороны 12-мерной RSS. Генный сегмент DH, функционально связанный с 23-мерной RSS, например, сконструированный генный сегмент DH, также не подвергался рекомбинации и, таким образом, содержит (i) 23-мерную RSS и (ii) 12-мерную RSS. Сконструированный генный сегмент DH (например, генный сегмент DH, функционально связанный с 23-мерной RSS), способен к перегруппировке с другим генным сегментом DH, функционально связанным с 12-мерной RSS, в соответствии с правилом рекомбинации 12/23.
[00137] Термин «физиологические условия» имеет понятное в данной области техники значение, обозначающее условия, при которых клетки или организмы живут и/или размножаются. В некоторых вариантах осуществления данный термин относится к условиям внешней или внутренней окружающей среды, которые могут возникать в природе относительно организма или системы клеток. В некоторых вариантах осуществления физиологические условия представляют собой условия, присутствующие в пределах организма человека или животного, отличного от человека, в частности, те условия, которые присутствуют в месте и/или в пределах области хирургического вмешательства. Физиологические условия, как правило, включают, например, температурный диапазон, составляющий 20-40°С, атмосферное давление, составляющее 1, показатель рН, составляющий 6-8, концентрацию глюкозы, составляющую 1-20 мМ, концентрацию кислорода при уровнях, равным атмосферным, и силу тяжести, которая наблюдается на земном шаре. В некоторых вариантах осуществления условия в лаборатории регулируют и/или поддерживают при физиологических условиях. В некоторых вариантах осуществления физиологические условия наблюдаются в организме (например, в животном, отличном от человека).
[00138] Выражение «полипептид» относится к любой полимерной цепи из аминокислот.
[00139] B некоторых вариантах осуществления полипептид имеет аминокислотную последовательность, встречающуюся в природе. В некоторых вариантах осуществления полипептид имеет аминокислотную последовательность, не встречающуюся в природе. В некоторых вариантах осуществления полипептид имеет аминокислотную последовательность, которая содержит части, встречающиеся в природе отдельно друг от друга (т.е. от двух или более различных организмов, например части, принадлежащие организму человека и организму, отличному от человеческого). В некоторых вариантах осуществления полипептид имеет аминокислотную последовательность, которая является сконструированной в том смысле, что она разработана и/или получена в результате действий человека. В некоторых вариантах осуществления полипептид может содержать или состоять из нескольких фрагментов, каждый из которых встречается в том же исходном полипептиде в другом пространственном порядке относительно друг друга, чем порядок, встречающийся в представляющем интерес полипептиде (например, фрагменты, которые напрямую связаны в исходном полипептиде, могут быть пространственно разделены в представляющем интерес полипептиде или наоборот, и/или фрагменты могут присутствовать в представляющем интерес полипептиде в другом порядке, чем порядок в исходном), таким образом, что представляющий интерес полипептид является производным своего исходного полипептида.
[00140] Термин «рекомбинантный» относится к полипептидам, которые разработаны, сконструированы, получены, экспрессированы, созданы или выделены с помощью рекомбинантных способов, таким как полипептиды, экспрессированные с использованием рекомбинантного вектора экспрессии, введенного путем трансфекции в клетку-хозяина, полипептиды, выделенные из комбинаторной библиотеки рекомбинантных человеческих полипептидов (Hoogenboom Н.R., 1997 TIB Tech. 15:62-70; Hoogenboom Н., and Chames P., 2000, Immunology Today 21: 371-378; Azzazy H., and Highsmith W.E., 2002, Clin. Biochem. 35: 425-445; Gavilondo J.V., and Larrick J.W., 2002, BioTechniques 29: 128-145), антитела, выделенные из животного (например, мыши), трансгенного по генам человеческих иммуноглобулинов (см., например, Taylor, L.D., et al., 1992, Nucl. Acids Res. 20: 6287-6295; Little M. et al., 2000, Immunology Today 21: 364-370; Kellermann S.A. and Green L.L., 2002, Current Opinion in Biotechnology 13: 593-597; Murphy, A.J., et al., 2014, Proc. Natl. Acad. Sci. U. S. A. 111 (14): 5153-5158, каждый из которых включен в данный документ посредством ссылки во всей своей полноте), или полипептиды, которые получены, экспрессированы, созданы или выделены любыми другими способами, предусматривающими сплайсинг выбранных элементов последовательности друг с другом.
[00141] B некоторых вариантах осуществления один или более из таких выбранных элементов последовательностей обнаруживаются в природе. В некоторых вариантах осуществления один или более из таких выбранных элементов последовательности разработаны in silico. В некоторых вариантах осуществления один или более из таких выбранных элементов последовательностей получают в результате мутагенеза (например, in vivo или in vitro) элемента известной последовательности, например, из природного или синтетического источника. Например, в некоторых вариантах осуществления рекомбинантный полипептид содержит последовательности, встречающиеся в геноме (или полипептиде) представляющего интерес организма-источника (например, человека, мыши и т.д.). В некоторых вариантах осуществления рекомбинантный полипептид содержит последовательности, которые встречаются в природе отдельно друг от друга (т.е. из двух или более разных организмов, например, часть, принадлежащая человеку, и часть, отличная от человеческой) в двух разных организмах (например, человека и организме, отличном от человеческого). В некоторых вариантах осуществления рекомбинантный полипептид имеет аминокислотную последовательность, полученную в результате мутагенеза (например, in vitro или in vivo, например, в животном, отличном от человека) таким образом, что аминокислотные последовательности рекомбинантных полипептидов представляют собой последовательности, которые, несмотря на то, что они происходят из полипептидных последовательностей и являются родственными им, в естественных условиях могут не существовать in vivo в пределах генома животного, отличного от человека.
[00142] «Эталон» относится к стандартному или контрольному средству, животному, группе, индивидууму, популяции, образцу, последовательности или значению, с которым сравнивают представляющие интерес средство, животное, группу, индивидуума, популяцию, образец, последовательность или значение. «Эталон» может относиться к «эталонному животному». «Эталонное животное» может иметь модификацию, описанную в данном документе, модификацию, отличную от описанной в данном документе, или не иметь модификации (т.е. представлять собой животное дикого типа). Как будет понятно специалистам в данной области техники, эталонное средство, животное, когорту, индивидуума, популяцию, образец, последовательность или значение, как правило, определяют или характеризуют в условиях, сравнимых с условиями, используемыми для определения или получения характеристик средства, животного (например, млекопитающего), когорты, индивидуума, популяции, образца, последовательности или значения, представляющих интерес.
[00143] B некоторых вариантах осуществления эталонное средство, животное, группу, индивидуума, популяцию, образец, последовательность или значение тестируют и/или определяют практически одновременно с тестированием или определением представляющего интерес средства, животного, группы, индивидуума, популяции, образца, последовательности или значения. В некоторых вариантах осуществления эталонное средство, животное, группа, индивидуум, популяция, образец, последовательность или значение представляют собой исторический эталон, необязательно воплощенный в материальной форме. В некоторых вариантах осуществления эталон может относиться к контролю.
[00144] Выражение «практически» относится к качественному состоянию проявления полной или практически полной меры или степени выраженности представляющих интерес характеристики или свойства. Специалисту в области биологии будет понятно, что биологические и химические явления редко, если это вообще происходит, продолжаются до окончания и/или протекают до завершения или достигают или избегают абсолютного результата. Таким образом, термин «практически» используется в данном документе для охвата потенциального отсутствия завершенности, присущей многим биологическим и химическим явлениям.
[00145] Выражение «практически гомологичный» относится к сравнению аминокислотных последовательностей или последовательностей нуклеиновых кислот. Как будет понятно специалистам в данной области техники, две последовательности обычно считаются «практически гомологичными», если они содержат гомологичные остатки в соответствующих положениях. Гомологичные остатки могут являться идентичными остатками. В качестве альтернативы, гомологичные остатки могут представлять собой неидентичные остатки с соответственно сходными структурными и/или функциональными характеристиками. Например, как хорошо известно специалистам в данной области техники, определенные аминокислоты, как правило, классифицируют как «гидрофобные» или «гидрофильные» аминокислоты и/или как имеющие «полярные» или «неполярные» боковые цепи. Замена одной аминокислотой на другую аминокислоту того же типа часто может считаться «гомологичной» заменой. Типичное распределение аминокислот по категориям приведено ниже:



[00146] Как хорошо известно из уровня техники, аминокислотные последовательности или последовательности нуклеиновых кислот можно сравнивать с помощью любого из множества алгоритмов, в том числе таковых, доступных в коммерческих компьютерных программах, таких как BLASTN для нуклеотидных последовательностей и BLASTP, BLAST с гэпами и PSI-BLAST для аминокислотных последовательностей. Такие иллюстративные программы описаны в Altschul, S.F. et al., 1990, J. Mol. Biol., 215 (3): 403-410; Altschul, S.F. et al., 1996, Methods in Enzymol. 266: 460-80; Altschul, S.F. et al., 1997, Nucleic Acids Res., 25: 3389-402; Baxevanis, A.D., and B.F.F. Ouellette (eds.) Bioinformatics: A Practical Guide to the Analysis of Genes and Proteins, Wiley, 1998; and Misener et al. (eds.) Bioinformatics Methods and Protocols (Methods in Molecular Biology, Vol. 132), Humana Press, 1998. В дополнение к идентификации гомологичных последовательностей, программы, упомянутые выше, как правило, дают представление о степени гомологии.
[00147] B некоторых вариантах осуществления две последовательности считаются практически гомологичными, если по меньшей мере 95% или более их соответствующих остатков являются гомологичными на протяжении рассматриваемого фрагмента, состоящего из остатков. В некоторых вариантах осуществления рассматриваемый фрагмент представляет собой полную последовательность. В некоторых вариантах осуществления рассматриваемый фрагмент содержит по меньшей мере 9 или более остатков. В некоторых вариантах осуществления рассматриваемый фрагмент содержит смежные остатки вдоль полной последовательности. В некоторых вариантах осуществления рассматриваемый фрагмент содержит расположенные с промежутками остатки вдоль полной последовательности, например расположенные с промежутками остатки, сводимые вместе за счет складчатой конформации полипептида или его части. В некоторых вариантах осуществления рассматриваемый фрагмент содержит по меньшей мере 10 или больше остатков.
[00148] Выражение «практически идентичный» относится к сравнению аминокислотных последовательностей или последовательностей нуклеиновых кислот. Как будет понятно рядовым специалистам в данной области техники, две последовательности обычно считаются «практически идентичными», если они содержат идентичные остатки в соответствующих положениях. Как хорошо известно из уровня техники, аминокислотные последовательности или последовательности нуклеиновых кислот можно сравнивать с помощью любого из множества алгоритмов, в том числе таковых, доступных в коммерческих компьютерных программах, таких как BLASTN для нуклеотидных последовательностей и BLASTP, BLAST с гэпами и PSI-BLAST для аминокислотных последовательностей. Такие иллюстративные программы описаны в Altschul, S. F. et al., 1990, J. Mol. Biol., 215(3): 403-410; Altschul, S. F. et al., 1996, Methods in Enzymol. 266:460-80; Altschul, S. F. et al., 1997, Nucleic Acids Res., 25:3389-3402; Baxevanis, A.D., and B. F. F. Ouellette (eds.) Bioinformatics: A Practical Guide to the Analysis of Genes and Proteins, Wiley, 1998; and Misener et al. (eds.) Bioinformatics Methods and Protocols (Methods in Molecular Biology, Vol.132), Humana Press, 1998. В дополнение к идентификации идентичных последовательностей, программы, упомянутые выше, как правило, дают представление о степени идентичности.
[00149] B некоторых вариантах осуществления две последовательности считаются практически идентичными, если по меньшей мере 95% или более их соответствующих остатков являются идентичными на протяжении рассматриваемого фрагмента, состоящего из остатков. В некоторых вариантах осуществления рассматриваемый фрагмент представляет собой полную последовательность. В некоторых вариантах осуществления рассматриваемый фрагмент содержит по меньшей мере 10 или больше остатков.
[00150] «Нацеливающийся вектор» или «нацеливающаяся конструкция» относится к полинуклеотидной молекуле, содержащей нацеливающийся участок. Нацеливающийся участок содержит последовательность, которая идентична или практически идентична последовательности в целевой клетке, ткани или животном, и обеспечивает интеграцию нацеливающейся конструкции в положение в пределах генома клетки, ткани или животного посредством гомологичной рекомбинации. Также предусмотрены нацеливающиеся участки, которые нацеливаются с помощью сайтов распознавания сайт-специфической рекомбиназы (например, сайтов loxP или Frt).
[00151] B некоторых вариантах осуществления нацеливающаяся конструкция, описанная в данном документе, дополнительно содержит последовательность нуклеиновой кислоты или ген, представляющие особенный интерес, селектируемый маркер, контрольные и/или регуляторные последовательности, а также другие последовательности нуклеиновой кислоты, обеспечивающие возможность рекомбинации, опосредованной добавлением экзогенных белков, которые способствуют или содействуют рекомбинации с участием таких последовательностей. В некоторых вариантах осуществления нацеливающаяся конструкция, описанная в данном документе, дополнительно содержит представляющий интерес полный или неполный ген, где представляющий интерес ген является гетерологичным геном, который кодирует полный или неполный полипептид, имеющий функцию, сходную с таковой у белка, кодируемого эндогенной последовательностью. В некоторых вариантах осуществления нацеливающаяся конструкция, описанная в данном документе, дополнительно содержит представляющий интерес гуманизированный полный или неполный ген, где представляющий интерес гуманизированный ген кодирует полный или неполный полипептид, имеющий функцию, сходную с таковой у полипептида, кодируемого эндогенной последовательностью. В некоторых вариантах осуществления нацеливающаяся конструкция (или нацеливающийся вектор) могут содержать последовательность нуклеиновой кислоты, в отношении которой проведена манипуляция человеком. Например, в некоторых вариантах осуществления нацеливающаяся конструкция (или нацеливающийся вектор) могут быть сконструированы таким образом, что они содержат сконструированный или рекомбинантный полинуклеотид, который содержит две или более последовательности, которые не связаны друг с другом в таком порядке в природе, пока не подвергаются манипуляции человеком для связывания напрямую друг с другом в сконструированном или рекомбинантном полинуклеотиде.
[00152] «Трансген» или «трансгенная конструкция» относится к последовательности нуклеиновой кислоты (кодирующей, например, представляющий интерес полный или неполный полипептид), которую человек ввел в клетку с помощью таких способов, которые описаны в, для трансгенного животного или клетки, в которые он введен. Трансген может данном документе. Трансген может быть частично или полностью гетерологичным, т.е. чужеродным содержать одну или более последовательностей регуляции транскрипции и любую другую нуклеиновую кислоту, такую как интроны или промоторы, которые могут быть необходимы для экспрессии выбранной последовательности нуклеиновой кислоты. Трансген может содержать один или более селектируемых маркеров, которые обеспечивают возможность для последовательного отбора потомства (например, клеток), которое получило трансген.
[00153] Термины «трансгенное животное», «трансгенное животное, отличное от человека» или Tg+ используются взаимозаменяемо в данном документе и обозначают любое не встречающееся в природе животное, отличное от человека, у которого одна или более клеток животного, отличного от человека, содержат гетерологичную нуклеиновую кислоту и/или ген, кодирующие представляющий интерес полный или неполный полипептид.
[00154] B некоторых вариантах осуществления гетерологичную последовательность нуклеиновой кислоты и/или ген вводят в клетку напрямую или опосредованно, путем введения в клетку-предшественника, путем преднамеренной генетической манипуляции, например с помощью микроинъекции или путем инфицирования рекомбинантным вирусом. Термин «генетическая манипуляция» не включает классические методики скрещивания, а скорее относится к введению молекулы(молекул) рекомбинантной ДНК. Данная молекула может быть интегрирована в хромосому или может представлять собой внехромосомно реплицирующуюся ДНК. Термин «Tg+» включает животных, которые являются гетерозиготными или гомозиготными по гетерологичной нуклеиновой кислоте и/или гену, и/или животных, которые имеют одну или множество копий гетерологичной нуклеиновой кислоты и/или гена.
[00155] «Вектор» относится к молекуле нуклеиновой кислоты, способной транспортировать другую нуклеиновую кислоту, с которой она связана.
[00156] B некоторых вариантах осуществления векторы способны к внехромосомной репликации и/или экспрессии нуклеиновых кислот, с которыми они соединены, в клетке-хозяине, такой как эукариотическая и/или прокариотическая клетка. Векторы, способные управлять экспрессией функционально связанных генов, называются в данном документе «векторами экспрессии».
[00157] Термин «дикий тип» включает его понятное в данной области техники значение, которое обозначает объект, обладающий структурой и/или активностью, встречающимися природе в «нормальном» (в противоположность мутантному, болезненному, измененному и т.д.) состоянии или окружении. Специалистам в данной области техники будет понятно, что гены и полипептиды дикого типа часто существуют в нескольких различных формах (например, аллелях).
[00158] Другие признаки, цели и преимущества настоящего изобретения очевидны из нижеследующего подробного описания некоторых вариантов осуществления. Следует, однако, понимать, что подробное описание, хоть и указывает на некоторые варианты осуществления настоящего изобретения, приведено только в целях иллюстрации, а не ограничения. Различные изменения и модификации в пределах объема настоящего изобретения станут очевидны специалистам в данной области техники из подробного описания.
ПОДРОБНОЕ ОПИСАНИЕ ОПРЕДЕЛЕННЫХ ВАРИАНТОВ ОСУЩЕСТВЛЕНИЯ
[00159] B настоящем изобретении предусмотрены, среди прочих объектов, трансгенные или полученные с помощью генетической инженерии животные, отличные от человека, содержащие гетерологичный генетический материал, кодирующий одну или более частей (функциональных фрагментов, связывающих частей и т.д.) человеческих иммуноглобулинов, где указанный гетерологичный генетический материал встроен в локус вариабельной области тяжелой цепи иммуноглобулина таким образом, что гетерологичный генетический материал функционально связан с генами константных областей (CH) тяжелой цепи. Предполагается, что такие животные, отличные от человека, проявляют способность вырабатывать антитела, кодируемые перегруппированными последовательностями V(DD)J. Также предполагается, что у таких животных, отличных от человека, имеется популяция антител, характеризующихся вариабельными участками тяжелой цепи с повышенным разнообразием CDR3 по сравнению с популяцией антител с разнообразием CDR3 вариабельной области тяжелой цепи иммуноглобулина, полученной за счет локуса вариабельной области тяжелой цепи иммуноглобулина дикого типа (или локуса вариабельной области тяжелой цепи иммуноглобулина, который встречается в природе). Таким образом, предусмотренные животные, отличные от человека, особенно применимы для разработки терапевтических средств на основе антител, которые связывают определенные антигены, в частности, антигены, ассоциированные со сниженной и/или слабой иммуногенностью, или антигены, которые характеризуются одним или более эпитопами, которые являются труднодоступными для связывания традиционными (или дикого типа) антителами. В частности, настоящее изобретение охватывает введение 23-мерной последовательности сигнала рекомбинации, примыкающей (например, на 5'-конце или 3'-конце) к одному или более сегментам DH DH-о6ласти в вариабельной области тяжелой цепи иммуноглобулина, что приводит к возможности DH-DH-перегруппировки сегментов в ходе рекомбинации VDJ и экспрессии антител, содержащих вариабельные области тяжелой цепи и, в частности, области CDR3, которые могут характеризоваться большей аминокислотной длиной по сравнению с антителами, полученными при рекомбинации VDJ с участием отдельных сегментов DH. Такие трансгенные животные, отличные от человека, обеспечивают in vivo систему для идентификации и разработки антител и/или терапевтических средств на основе антител, связывающих мишени заболеваний, которая превосходит нацеливающие способности общепринятых технологий поиска новых лекарственных средств. Кроме того, такие трансгенные животные, отличные от человека, обеспечивают применимую систему на основе модели на животном для разработки антител и/или терапевтических средств на основе антител, фокусирующихся или сконструированных для работы с нарушениями взаимодействий белок-белок, которые являются ключевыми при различных заболеваниях и/или патологических состояниях, поражающих человека.
[00160] Как показано на верхней панели на фигуре 1, рекомбинация между генными сегментами иммуноглобулина следует правилу, часто называемому правилом 12/23, согласно которому генные сегменты, фланкированные последовательностями сигнала рекомбинации (RSS), соединяются в ходе упорядоченного процесса. Каждая RSS состоит из консервативного блока из семи нуклеотидов (гептамера; 5'-CACAGTG-3'; SEQ ID NO: 144), который является смежным с кодирующей последовательностью (например, с сегментом VH, DH или JH), после которого расположена неконсервативная область, известная как спейсер, длина которой составляет 12 п.о. или 23 п.о., и второй консервативный блок, состоящий из девяти нуклеотидов (нонамер; 5'-АСАААААСС-3'; SEQ ID NO: 145). Последовательность спейсера может варьироваться, но его консервативная длина соответствует одному или двум оборотам двойной спирали ДНК. Таким образом, последовательности гептамера и нонамера располагаются на одной стороне спирали ДНК, где они могут быть связаны комплексом белков, катализирующим рекомбинацию. RSS со спейсером длиной 23 п.о. представляет собой 23-мерную RSS, и RSS со спейсером длиной 12 п.о. представляет собой 12-мерную RSS. Согласно правилу рекомбинации 12/23 обычно рекомбинация возможна между 12-мерной RSS и 23-мерной RSS и невозможна между 23-мерной RSS и другой 23-мерной RSS или между 12-мерной RSS и другой 12-мерной RSS, например, невозможна прямая рекомбинация VH зародышевой линии с JH зародышевой линии (т.е. соединение 23-мера с 23-мером) или рекомбинация DH зародышевой линии с DH зародышевой линии (т.е. соединение 12-мера с 12-мером), хотя сообщалось об исключениях.
[00161] B некоторых вариантах осуществления животные, отличные от человека, описанные в данном документе, содержат вариабельную область тяжелой цепи иммуноглобулина, содержащую сконструированный кластер разнообразия (т.е. сконструированную DH-область), характеризующуюся наличием одного или более сегментов DH, каждый из которых функционально связан с 23-мерной RSS и соответственно способен к рекомбинации DH-DH. В некоторых вариантах осуществления антитела, содержащие CDR3, полученные в результате такой рекомбинации, могут характеризоваться как обладающие повышенным разнообразием, что является результатом способности молекулы с большей аминокислотной длиной к прямому связыванию с определенными антигенами (например, вирусами, мембранными каналами и т.д.). В некоторых вариантах осуществления животные, отличные от человека, описанные в данном документе, содержат вариабельный (VH) и соединяющий (JH) человеческие генные сегменты тяжелой цепи, функционально связанные со сконструированной с помощью генной инженерии DH-областью таким образом, что между указанными VH, JH и более чем одним DH-сегментом происходит рекомбинация VDJ с последующим получением вариабельной области тяжелой цепи, которая связывает антиген, представляющий интерес. В некоторых вариантах осуществления сконструированная DH-область, описанная в данном документе, содержит один или более (например, 1, 2, 3, 4, 5, 10 или более) человеческих сегментов DH, сконструированных для обеспечения возможности (или способствования) рекомбинации DH-DH с более высокой частотой по сравнению с эталонным локусом вариабельной области тяжелой цепи иммуноглобулина. В некоторых вариантах осуществления животные, отличные от человека, описанные в данном документе, содержат несколько генных сегментов VH и JH, функционально связанных с одним или более сегментами DH, каждый из которых функционально связан с 23-мерной последовательностью сигнала рекомбинации (RSS) в вариабельной области тяжелой цепи иммуноглобулина в геноме животного, отличного от человека. Во многих вариантах осуществления сегменты VH и JH представляют собой человеческие генные сегменты VH и JH.
[00162] B некоторых вариантах осуществления животные, отличные от человека, описанные в данном документе, дополнительно содержат локус легкой цепи человеческого или гуманизированного иммуноглобулина (например, κ и/или λ), и таким образом животные, отличные от человека, продуцируют антитела, содержащие человеческие вариабельные участки (т.е. тяжелой и легкой цепи) и константные участки, отличные от человеческих. В некоторых вариантах осуществления указанный локус легкой цепи человеческого или гуманизированного иммуноглобулина содержит генные сегменты VL и JL, функционально связанные с константной областью легкой цепи грызуна (например, Сκ или Сλ грызуна). В некоторых вариантах осуществления животные, отличные от человека, описанные в данном документе, дополнительно содержат локус легкой цепи иммуноглобулина, как описано в патентах США №№9796788; 9969814; публикациях заявок на патент США №№2011/0195454 А1, 2012/0021409 А1, 2012/0192300 А1, 2013/0045492 А1, 2013/0185821 А1, 2013/0302836 А1, 2018/0125043; публикациях международных заявок на патент №№ WO 2011/097603, WO 2012/148873, WO 2013/134263, WO 2013/184761, WO 2014/160179, WO 2014/160202 и WO2019/113065, каждая из которых включена в данный документ посредством ссылки во всей своей полноте).
[00163] Различные аспекты настоящего изобретения подробно описаны в следующих разделах. Использование разделов не подразумевает ограничения вариантами осуществления, описанными в данном документе. Каждый раздел может быть применен к любому аспекту или варианту осуществления, описанным в данном документе. В настоящей заявке использование «или» означает «и/или», если не указано иное.
Рекомбинация VDJ
[00164] Гены, отвечающие за синтез иммуноглобулинов, присутствуют во всех клетках животного и сгруппированы в генные сегменты, последовательно расположенные на хромосоме. Конфигурация наследуемых человеческих генных сегментов, например, конфигурация зародышевой линии человеческих генных сегментов, например, порядок расположения человеческих генных сегментов в геноме зародышевой линии (например, в геноме, передаваемом следующему поколению) человека, может быть найдена в Lefranc, М.-Р., Exp.Clin. Immunogenet, 18, 100-116 (2001), включенном в данный документ посредством ссылки во всей своей полноте, в котором также показаны функциональные генные сегменты и псевдогены, находящиеся в пределах локуса тяжелой цепи человеческого иммуноглобулина в конфигурации зародышевой линии. Серия событий рекомбинации при участии нескольких генетических компонентов служит для сборки иммуноглобулинов из упорядоченной группы генных сегментов (например, V, D и J). Известно, что данная сборка генных сегментов является неточной и, таким образом, разнообразие иммуноглобулинов достигается как вследствие комбинации разных генных сегментов, так и образования уникальных соединений за счет неточного соединения. Дополнительная вариативность создается за счет процесса, известного как соматическая гипермутация, при котором последовательность вариабельного участка иммуноглобулинов изменяется с повышением аффинности и специфичности в отношении антигена. Молекула иммуноглобулина представляет собой полипептид Y-образной формы, состоящий из двух идентичных легких и тяжелых цепей, каждая из которых имеет два структурных компонента: один вариабельный домен и один константный домен. Именно вариабельные домены тяжелой и легкой цепей образованы за счет сборки генных сегментов, тогда как константные домены сливаются с вариабельными доменами в процессе сплайсинга РНК. Несмотря на то, что механизм сборки (или связывания) генных сегментов в тяжелой и легкой цепях является схожим, в случае легких цепей требуется только одно событие соединения (т.е. V с J), тогда как в случае тяжелых цепей требуется два события соединения (т.е. D с J и V с DJ).
[00165] Сборка генных сегментов вариабельных областей для тяжелой и легкой цепей (называемая соответственно рекомбинацией VDJ и рекомбинацией VJ) направляется консервативными последовательностями некодирующей ДНК, фланкирующими каждый генный сегмент, которые называются последовательностями сигнала рекомбинации (RSS), что обеспечивает перегруппировки ДНК в точных местоположениях относительно кодирующих последовательностей V, D и J (см., например, Ramsden, D.A. et al., 1994, Nuc. Acids Res. 22(10): 1785-96). Иллюстративное схематическое изображение последовательностей, участвующих в рекомбинации VDJ генных сегментов тяжелой цепи, понятное специалисту в данной области техники, представлено на фигуре 1. Каждая RSS состоит из консервативного блока, состоящего из семи нуклеотидов (гептамера), который является смежным с кодирующей последовательностью (например, сегментом V), после которой расположен спейсер (состоящий либо из 12 п.о., либо из 23 п.о.), и второго консервативного блока, состоящего из девяти нуклеотидов (нонамера). Несмотря на то, что значительная дивергенция последовательностей в пределах спейсера длиной 12 п.о. или 23 п.о. у разных индивидуумов допустима, длина данных последовательностей, как правило, не варьируется. Рекомбинация между генными сегментами иммуноглобулинов следует правилу, часто называемому правилом 12/23, согласно которому генные сегменты, фланкированные RSS со спейсером длиной 12 п.о. (или 12-мером), как правило, соединяются с генным сегментом, фланкированным спейсером длиной 23 п.о. (или 23-мером; см., например, Hiom, K. and М. Gellert, 1998, Mol. Cell. 1(7): 1011-9, включенный в данный документ посредством ссылки во всей своей полноте). Сообщалось, что последовательность RSS оказывает влияние на эффективность и/или частоту рекомбинации с конкретным генным сегментом (см., например, Ramsden, D.A and G.E. Wu, 1991, Proc. Natl. Acad. Sci. U.S.A. 88:10721-5; Boubnov, N.V. et al., 1995, Nuc. Acids Res. 23:1060-7; Ezekiel, U.R. et al., 1995, Immunity 2:381-9; Sadofsky, M. et al., 1995, Genes Dev. 9:2193-9; Cuomo, C.A. et al., 1996, Mol. Cell Biol. 16:5683-90; Ramsden, D.A. et al., 1996, EMBO J 15:3197-3206, каждый из которых включен в данный документ посредством ссылки во всей своей полноте). Действительно, во многих сообщениях отмечается чрезвычайно смещенное и вариабельное использование генных сегментов, в частности, сегментов DH, у разных индивидуумов. Если не указано иное или если специалисту в данной области не будет очевидным возникновение противоречия или несоответствия, предполагается, что неперегруппированный генный сегмент, если нет указания на содержание одной RSS, содержит две RSS, с которыми данный генный сегмент связан в естественных условиях, например, фланкирован ими, функционально связан с ними и т.д. В некоторых вариантах осуществления в данном документе неперегруппированный генный сегмент может содержать генный сегмент в конфигурации зародышевой линии (например, дикого типа), например, как генный сегмент VH зародышевой линии, так и генный сегмент JH зародышевой линии фланкированы с обеих сторон 23-мерной RSS. Напротив, генный сегмент DH зародышевой линии, например, неперегруппированный генный сегмент DH, фланкирован с каждой стороны 12-мерной RSS.
[00166] Таким образом, неперегруппированный генный сегмент может также относиться к генному сегменту в конфигурации зародышевой линии, включая любую RSS, ассоциированную с такой конфигурацией зародышевой линии. Более того, несколько генных сегментов в своей конфигурации зародышевой линии, как правило, относятся не только к каждому отдельному генному сегменту, находящемуся в своей конфигурации зародышевой линии (например, неперегруппированному), но также к порядку и/или расположению функциональных генных сегментов. См., например, Lefranc, М.-Р., Exp.Clin. Immunogenet, 18, 100-116 (2001); включенный в данный документ посредством ссылки во всей своей полноте, в качестве источника информации о конфигурации зародышевой линии человеческих генных сегментов V, D и J.
[00167] Сборка генных сегментов с образованием вариабельных областей тяжелой и легкой цепей приводит к образованию антигенсвязывающих областей (или сайтов) иммуноглобулинов. Такие антигенсвязывающие области характеризуются, помимо всего прочего, присутствием гипервариабельных областей, которые обычно обозначаются как области, определяющие комплементарность (CDR). В тяжелой и легкой цепях находится по три CDR (т.е. в общей сложности шесть CDR), причем и CDR1, и CDR2 полностью кодируются генным сегментом V. Однако CDR3 кодируется последовательностью, образующейся в результате соединения сегментов V и J в случае легких цепей и сегментов V, D и J в случае тяжелых цепей. Таким образом, дополнительный генный сегмент, используемый в ходе рекомбинации для получения кодирующей последовательности вариабельной области тяжелой цепи, существенно повышает разнообразие антигенсвязывающих сайтов тяжелых цепей. Таким образом, у предусмотренных животных, отличных от человека, содержащих сконструированную DH-область, описанную в данном документе, генерируется разнообразие CDR3, характеризующихся рекомбинацией DH-DH и большей аминокислотной длиной по сравнению с областями CDR3 вариабельной области тяжелой цепи, полученными в ходе традиционной рекомбинации VDJ.
[00168] Без ограничения какой-либо теорией есть основания полагать, что достижение дополнительного разнообразия в репертуаре CDR3 тяжелой цепи возможно посредством увеличения количества соединений, образующих последовательность гена перегруппированной вариабельной области тяжелой цепи, и/или увеличения длины области CDR3. Один механизм увеличения количества соединений и/или увеличения длины области CDR3 может состоять в соединении первого сегмента DH (который может иметь общее обозначение DH"A") с другим сегментом DH (который может иметь общее обозначение DH"B") при рекомбинации DH-DH до последовательных событий рекомбинации DH-JH и V-DHJH, приводящих к образованию генной последовательности VH(DHA-DHB)JH. Долгое время считалось, что в естественных условиях события рекомбинации DH-DH были невозможны согласно правилу 12/23 (Alt, F.W. et al., 1984, EMBO J. 3(6): 1209-19; включен в данный документ посредством ссылки во всей своей полноте). Однако, оказывается, что события рекомбинации DH-DH происходят с очень низкой частотой у людей в обход правила 12/23, (в 1 из 800 или ~0,125% наивных В-клеток; см., например, Ollier, P.J. et al. 1985, EMBO J. 4(13B):3681-88; Milner, E.C.B. et al. 1986, Immunol. Today 7:36-40; Liu, Z. et al., 1987, Nucleic Acids Res. 15(11):4688; Liu, Z. et al., 1987, Nucleic Acids Res. 15(15):6296; Meek, K.D. et al., 1989, J. Exp.Med. 169(2):519-33; Meek, K.D. et al., 1989, J. Exp.Med. 170:39-57; Baskin, B. et al. 1998, Clin. Exp.Immunol. 112:44-7; Briney, B.S. et al., 2012 Immunol. 137: 56-64; каждый из которых включен в данный документ посредством ссылки во всей своей полноте) и считаются основным механизмом образования необычно длинных CDR3, наблюдающихся в некоторых тяжелых цепях (Janeway's Immunobiology. 9th Edition. Kenneth Murphy, Casey Weaver. Chapter 5, 2017; включен посредством ссылки во всей своей полноте). Однако, некоторые потенциальные терапевтические мишени (например без ограничения вирусы, рецепторы клеточной поверхности, трансмембранные белки типа IV, такие как GPCR, ионные каналы) имеют замаскированные или глубоко расположенные эпитопы, которые являются недоступными для нормальных антител, но могу быть распознаны антителами с длинными последовательностями HCDR3. Например, антитела с очень длинными HCDR3 часто обнаруживаются у пациентов с хроническими вирусными инфекциями и в некоторых случаях характеризуются нейтрализующей активностью широкого спектра (например нейтрализующие антитела широкого спектра к HIV-1 или вирусу гриппа). Для отбора антител, способных достигать этих скрытых эпитопов, было бы полезно повысить частоту образования тяжелых цепей с очень длинными HCDR3. Без ограничения какой-либо конкретной теорией, одним способом для достижения этого является увеличение частоты перегруппировок VH(DHA-DHB)JH тяжелой цепи посредством конструирования сегментов DH таким образом, чтобы они содержали 23-мерную RSS, и/или увеличение частоты рекомбинации генного сегмента DH с более длинным генным сегментом JH6. Предполагается, что у мышей соединение DH-JH происходит в виде двух упорядоченных стадий: (1) первичной перегруппировки проксимального сегмента DQ52 (обозначаемого DH7-27 у людей) с одним из ближайших сегментов JH (JH1 или JH2). Сильный промотор μ0, расположенный выше DQ52, затем обеспечивает возможность оставшимся сегментам JH (JH3 и JH4) стать более доступными для генов, активизирующих рекомбинацию (RAG); (2) вторичной перегруппировки дистального сегмента DH с одним из оставшихся сегментов JH (JH3 или JH4). См. фигуру 1, верхняя панель. Это согласуется с более частым использованием нижележащих сегментов JH, наблюдающимся как у мышей (JH3-JH4), так и у людей (JH4-JH6) (Nitschke et al. 2001 J. Immunol. 166:2540-52, включен в данный документ посредством ссылки во всей своей полноте). Без ограничения какой-либо конкретной теорией, предполагается, что замена DH7-27 и JH1-JH3 (или JH1-JH5) на синтетический ген DH (например, синтетический сегмент DH3-3), содержащий 23-мерную RSS 5'-конца и 12-мерную RSS 3'-конца, приведет к высокой частоте рекомбинации VH(DH-DH)JH и может происходить в виде механизма, состоящего из трех стадий: (1) перегруппировки 23-DH-12 с JH4, JH5 или JH6 с получением 23(DH)JH, (2) перегруппировки дистальных DH (DH1-1 - DH1-26) с 23(DH)JH с получением 12(DH-DH)JH (перегруппировка VH с 23(DH)JH была бы невозможна согласно правилу 12/23), (3) перегруппировки VH с 12(DH-DH)JH с получением кодирующей последовательности VDDJ, кодирующей вариабельный домен тяжелой цепи иммуноглобулина. См., например, фигуру 1, нижняя панель. Также предполагается, что замена генных сегментов DH2-2, DH2-8 и DH2-15, которые являются длинными генными сегментами DH (содержащих более 31 нуклеотидов, кодирующих два цистеина, которые способны к образованию дисульфидной связи, предположительно стабилизирующей области HCDR3 [см., например, Wang et al. 2013 Cell 153:1379-93, включен в данный документ посредством ссылки во всей своей полноте], соответственно на те генные сегменты, функционально связанные с 12-мерной RSS 5'-конца и с 23-мерной RSS 3'-конца, также приведет к высокой частоте рекомбинации VH(DH-DH)JH, которая может происходить по механизму, состоящему из трех стадий: (1) перегруппировки 12:DH-12 с JH4, JH5 или JH6 с получением 12(DH)JH, (2) перегруппировки дистальных 12:DH2-2:23, 12:DH2-8:23 или 12:DH2-15-23 с 12(DH)JH с получением 12(DH-DH)JH и (3) перегруппировки VH с 12(DH-DH)JH с получением кодирующей последовательности VDDJ, кодирующей вариабельный домен тяжелой цепи иммуноглобулина. Как описано в данном документе, сконструированную DH-область конструировали с помощью размещения 23-мерного спейсера во фланкирующее положение на 5'- или 3'-конце, смежное с одним или более сегментами DH, тем самым создавая возможность для рекомбинации DH с DH в локусе вариабельной области тяжелой цепи гуманизированного иммуноглобулина.
[00169] B некоторых вариантах осуществления животные, отличные от человека, описанные в данном документе, содержат локус вариабельной области тяжелой цепи иммуноглобулина, который демонстрирует рекомбинацию VDJ, которая происходит без соблюдения правила 12/23 по сравнению с эталонным животным, отличным от человека. В некоторых вариантах осуществления описанные в данном документе животные, отличные от человека, содержат одну или более RSS, смежных или фланкирующих один или более сегментов DH, являющихся модифицированными по сравнению с сегментами DH дикого типа. В некоторых вариантах осуществления каждый из одного или более генных сегментов DH вариабельной области тяжелой цепи иммуноглобулина животного, отличного от человека, описанного в данном документе, функционально связан с 23-мерной RSS 5'-конца или 3'-конца таким образом, что частота рекомбинации DH-DH повышается у животного, отличного от человека, по сравнению с эталонным животным, отличным от человека. В некоторых вариантах осуществления эффективность и/или частоту рекомбинации можно определить по частоте использования генных сегментов в популяции последовательностей антител (например, от индивидуума или группы индивидуумов; см., например, Arnaout, R. et al., 2011, PLoS One 6(8):e22365; Glanville, J. et al., 2011, Proc. Natl. Acad. Sci. U.S.A. 108(50):20066-71; каждый из которых включен в данный документ посредством ссылки во всей своей полноте). Таким образом, животные, отличные от человека, описанные в данном документе, могут в некоторых вариантах осуществления содержать один или более сегментов DH, каждый из которых функционально связан или фланкирован 23-мерной RSS таким образом, что рекомбинация DH-DH происходит с повышенной частотой по сравнению с рекомбинацией DH-DH у эталонного животного, отличного от человека. В некоторых вариантах осуществления грызун, содержащий сконструированную DH-область, описанную в данном документе, демонстрирует по меньшей мере 3-кратное повышение частоты рекомбинации DH-DH по сравнению с рекомбинацией DH-DH у эталонного животного, отличного от человека. В некоторых вариантах осуществления грызун, содержащий сконструированную DH-область, описанную в данном документе, демонстрирует по меньшей мере 4-кратное повышение частоты рекомбинации DH-DH по сравнению с рекомбинацией DH-DH у эталонного животного, отличного от человека. В некоторых вариантах осуществления грызун, содержащий сконструированную DH-область, описанную в данном документе, демонстрирует по меньшей мере 5-кратное повышение частоты рекомбинации DH-DH по сравнению с рекомбинацией DH-DH у эталонного животного, отличного от человека. В некоторых вариантах осуществления грызун, содержащий сконструированную DH-область, описанную в данном документе, демонстрирует по меньшей мере 10-кратное повышение частоты рекомбинации DH-DH по сравнению с рекомбинацией DH-DH у эталонного животного, отличного от человека. В некоторых вариантах осуществления грызун, содержащий сконструированную DH-область, описанную в данном документе, демонстрирует по меньшей мере 20-кратное повышение частоты рекомбинации DH-DH по сравнению с рекомбинацией DH-DH у эталонного животного, отличного от человека. В некоторых вариантах осуществления грызун, содержащий сконструированную DH-область, описанную в данном документе, демонстрирует по меньшей мере 30-кратное повышение частоты рекомбинации DH-DH по сравнению с рекомбинацией DH-DH у эталонного животного, отличного от человека. В некоторых вариантах осуществления грызун, содержащий сконструированную DH-область, описанную в данном документе, демонстрирует по меньшей мере 40-кратное повышение частоты рекомбинации DH-DH по сравнению с рекомбинацией DH-DH у эталонного животного, отличного от человека. В некоторых вариантах осуществления грызун, содержащий сконструированную DH-область, описанную в данном документе, демонстрирует по меньшей мере 50-кратное повышение частоты рекомбинации DH-DH по сравнению с рекомбинацией DH-DH у эталонного животного, отличного от человека.
Предусмотренные in vivo системы
[00170] Настоящее изобретение основано на признании того, что определенные антигены ассоциированы с низкой и/или слабой иммуногенностью и, следовательно, являются неудовлетворительными мишенями для терапевтических средств на основе антител. Действительно, многие мишени заболеваний (например, вирусы, белки каналов) характеризуются как с трудом поддающиеся лечению или не поддающиеся медикаментозной терапии. Таким образом, настоящее изобретение основано на создании in vivo системы для разработки антител и терапевтических средств на основе антител, которые устраняют недостатки, связанные с общепринятыми технологиями и/или подходами к поиску новых лекарственных средств. В некоторых вариантах осуществления в настоящем изобретении предусмотрена in vivo система, характеризующаяся наличием локусов иммуноглобулина, в частности, локусов вариабельной области тяжелой цепи гуманизированного иммуноглобулина, которые содержат сконструированную DH-область, в которой одна или более RSS DH изменены, модифицированы или сконструированы из формата 12-мер-DH-12-мер в формат 12-мер-DH-23-мер или 23-мер-DH-12-мер, тем самым создавая возможность для рекомбинации DH-DH с повышенной частотой по сравнению с рекомбинацией DH-DH, наблюдаемой в эталонной in vivo системе. Настоящее изобретение конкретно демонстрирует получение с помощью генной инженерии трансгенного грызуна, геном которого содержит вариабельную область тяжелой цепи иммуноглобулина, которая содержит сконструированную DH-область, где указанная сконструированная DH-область содержит один или более сегментов DH, каждый из которых функционально связан с 23-мерной RSS, расположенной относительно каждого из одного или более сегментов DH таким образом, чтобы создавать условия для повышения частоты рекомбинации DH-DH. Способы, описанные в данном документе, могут быть адаптированы для получения любого количества сегментов DH (например, традиционных или синтетических), каждый из которых функционально связан с 23-мерной RSS 5'- или 3'-конца для создания сконструированной DH-области, описанной в данном документе. После интеграции в вариабельную область тяжелой цепи иммуноглобулина (т.е. помещения в функциональную связь с генными сегментами VH и JH и/или одной или более константными областями) сконструированная DH-область создает условия для рекомбинации генных сегментов VH и JH с более чем одним сегментом DH, например, для получения антител, характеризующихся тяжелыми цепями с дополнительным разнообразием (т.е. длинными CDR3, например, где по меньшей мере 95% последовательностей CDR3 тяжелой цепи имеют длину, составляющую по меньшей мере 14 аминокислотных остатков) для прямого связывания с определенными антигенами. В некоторых вариантах осуществления такие вариабельные области тяжелой цепи обладают способностью достигать труднодоступные эпитопы вирусов, белков каналов, GPCR и т.д.
[00171] Без ограничения какой-либо конкретной теорией, авторами отмечается, что данные, представленные в данном документе, демонстрируют, что в некоторых вариантах осуществления грызуны, геном которых содержит вариабельный локус тяжелой цепи иммуноглобулина, содержащий сконструированную с помощью генной инженерии DH-область, характеризующуюся вставкой сегмента DH, функционально связанного с 23-мерной RSS 5'-конца, эффективно образуют антитела, получаемые при рекомбинации V(DD)J. Авторами также отмечается, что данные, представленные в данном документе, демонстрируют, что в некоторых вариантах осуществления грызуны, геном которых содержит вариабельный локус тяжелой цепи иммуноглобулина, содержащий сконструированную с помощью генной инженерии DH-область, характеризующуюся вставкой трех сегментов DH, каждый из которых функционально связан с 23-мерной RSS 3'-конца, эффективно образуют антитела, получаемые при рекомбинации V(DD)J. Также в данном документе отмечается, что делеция некоторых или всех пяти генных сегментов JH, расположенных выше генного сегмента JH6, приводит к предпочтительной рекомбинации с участием генного сегмента JH6. Примечательно, что генный сегмент JH6 содержит 63 нуклеотида, например, на 10 нуклеотидов больше, чем другие генные сегменты JH1, JH2, JH3, JH4 и JH5, которые соответственно содержат 52, 53, 50, 40 и 51 нуклеотид. Таким образом, настоящее изобретение по меньшей мере в некоторых вариантах осуществления охватывает разработку in vivo системы для получения антител и/или терапевтических средств на основе антител для мишеней заболеваний, которые с трудом поддаются лечению, например, посредством получения грызунов, которые образуют перегруппированные последовательности генов вариабельной области тяжелой цепи иммуноглобулина, например, VHDHJH и VH(DHA-DHB)JH, например, VH(DHA-DHB)JH6, последовательности, кодирующие вариабельные домены тяжелой цепи с CDR3, длина которых составляет по меньшей мере 20 аминокислот, например, с CDR3, длина которых составляет 20-30 аминокислот. В некоторых вариантах осуществления по меньшей мере 8-10% перегруппированных генных последовательностей вариабельной области тяжелой цепи иммуноглобулина, например, генные последовательности VHDHJH и VH(DHA-DHB)JH животного, отличного от человека, описанного в данном документе, кодируют области CDR3, длина которых составляет по меньшей мере 21 аминокислоту.
[00172] B некоторых вариантах осуществления в данном документе описывается животное, отличное от человека, например, грызун, например, крыса или мышь, которое содержит (1) в своем геноме зародышевой линии, например, в зародышевой клетке, локус тяжелой цепи иммуноглобулина, содержащий сконструированную с помощью генной инженерии DH-область, содержащую генный сегмент DH, функционально связанный с 23-мерной RSS, и (2) в своем соматическом геноме, например, в В-клетке, перегруппированную кодирующую последовательность тяжелой цепи VH(DHA-DHB)JH, где первый или второй генный сегмент DH (т.е. DHA или DHB из кодирующей последовательности VH(DHA-DHB)JH соответственно) содержит генный сегмент DH, функционально связанный с 23-мерной RSS, или ее частью, например, где первый или второй генный сегмент DH содержит по меньшей мере 9 последовательных нуклеотидов, совпадающих при выравнивании с генным сегментом DH, функционально связанным с 23-мерной RSS, и где и первый и второй генные сегменты DH содержат по меньшей мере 5 последовательных нуклеотидов, совпадающих при выравнивании с соответствующим генным сегментом DH зародышевой линии, независимо от какого-либо перекрывания.
[00173] B некоторых вариантах осуществления предусмотренные животные, отличные от человека, содержат локус тяжелой цепи иммуноглобулина, характеризующийся наличием нескольких человеческих генных сегментов VH, DH и JH, сгруппированных в конфигурации зародышевой линии и функционально связанных с генами константной области тяжелой цепи иммуноглобулина, энхансерами и регуляторными областями, отличными от человеческих. В некоторых вариантах осуществления предусмотренные животные, отличные от человека, содержат один или более человеческих генных сегментов VH, один или более человеческих генных сегментов DH и один или более человеческих генных сегментов JH, функционально связанных с константной областью тяжелой цепи иммуноглобулина, отличного от человеческого.
[00174] B некоторых вариантах осуществления предусмотренные животные, отличные от человека, содержат по меньшей мере человеческие генные сегменты VH VH3-74, VH3-73, VH3-72, VH2-70, VH1-69, VH3-66, VH3-64, VH4-61, VH4-59, VH1-58, VH3-53, VH5-51, VH3-49, VH3-48, VH1-46, VH1-45, VH3-43, VH4-39, VH4-34, VH3-33, VH4-31, VH3-30, VH4-28, VH2-26, VH1-24, VH3-23, VH3-21, VH3-20, VH1-18, VH3-15, VH3-13, VH3-11, VH3-9, VH1-8, VH3-7, VH2-5, VH7-4-1, VH4-4, VH1-3, VH1-2 и VH6-1.
[00175] B некоторых вариантах осуществления предусмотренные животные, отличные от человека, содержат человеческие генные сегменты DH DH1-1, Dh2-2, DH3-3, DH4-4, DH5-5, DH6-6, DH1-7, DH2-8, DH3-9, DH3-10, DH5-12, DH6-13, DH2-15, DH3-16, DH4-17, DH5-18, DH6-19, DH1-20, DH2-21, DH3-22, DH6-25 и DH1-26. В некоторых вариантах осуществления предусмотренные животные, отличные от человека, дополнительно содержат DH1-14, DH4-11, DH4-23, DH5-24 или их комбинацию. В некоторых вариантах осуществления, предусмотренные животные, отличные от человека, дополнительно содержат человеческий DH7-27.
[00176] В некоторых вариантах осуществления в данном документе описано животное, отличное от человека, например, грызун, например, крыса или мышь, которое содержит (1) в своем геноме зародышевой линии, например, в зародышевой клетке, локус тяжелой цепи иммуноглобулина, содержащий сконструированную DH-область, содержащую генный сегмент DH, функционально связанный с 23-мерной RSS, и (2) в своем соматическом геноме, например, в В-клетке, перегруппированную кодирующую последовательность тяжелой цепи VH(DHa-DHb)JH, где первый или второй генный сегмент DH (т.е. DHA или DHB из кодирующей последовательности VH(DHa-DHb)JH соответственно) содержит генный сегмент DH, функционально связанный с 23-мерной RSS или ее частью, например, где первый или второй генный сегмент DH содержит по меньшей мере 9 последовательных нуклеотидов, совпадающих при выравнивании с генным сегментом DH, функционально связанным с 23-мерной RSS, и где и первый и второй генные сегменты DH содержат по меньшей мере 5 последовательных нуклеотидов, совпадающих при выравнивании с соответствующим генным сегментом DH зародышевой линии, независимо от какого-либо перекрывания. В некоторых вариантах осуществления предусмотренные животные, отличные от человека, содержат человеческие генные сегменты DH, функционально связанные с 23-мерной RSS. В некоторых вариантах осуществления человеческий генный сегмент DH, функционально связанный с 23-мерной RSS, содержит два кодона цистеина. В некоторых вариантах осуществления человеческий генный сегмент DH, функционально связанный с 23-мерной RSS, содержит по меньшей мере 37 нуклеотидов. В некоторых вариантах осуществления генный сегмент DH человека, функционально связанный с 23-мерной RSS, содержит по меньшей мере 19 нуклеотидов. В некоторых вариантах осуществления генный сегмент DH человека, функционально связанный с 23-мерной RSS, содержит по меньшей мере 20 нуклеотидов. В некоторых вариантах осуществления генный сегмент DH человека, функционально связанный с 23-мерной RSS, содержит по меньшей мере 23 нуклеотида. В некоторых вариантах осуществления генный сегмент DH человека, функционально связанный с 23-мерной RSS, содержит по меньшей мере 28 нуклеотидов. В некоторых вариантах осуществления генный сегмент DH человека, функционально связанный с 23-мерной RSS, содержит по меньшей мере 31 нуклеотид. В некоторых вариантах осуществления генный сегмент DH человека, функционально связанный с 23-мерной RSS, содержит по меньшей мере 37 нуклеотидов.
[00177] В некоторых вариантах осуществления человеческий генный сегмент DH, функционально связанный с 23-мерной RSS, содержит человеческий генный сегмент DH1. В некоторых вариантах осуществления человеческий генный сегмент DH, функционально связанный с 23-мерной RSS, содержит человеческий генный сегмент DH2. В некоторых вариантах осуществления человеческий генный сегмент DH, функционально связанный с 23-мерной RSS, содержит человеческий генный сегмент DH3. В некоторых вариантах осуществления человеческий генный сегмент DH, функционально связанный с 23-мерной RSS, содержит человеческий генный сегмент DH4. В некоторых вариантах осуществления человеческий генный сегмент DH, функционально связанный с 23-мерной RSS, содержит человеческий генный сегмент DH5. В некоторых вариантах осуществления человеческий генный сегмент DH, функционально связанный с 23-мерной RSS, содержит человеческий генный сегмент DH6. В некоторых вариантах осуществления генный сегмент DH человека, функционально связанный с 23-мерной RSS, содержит генный сегмент DH7 человека.
[00178] В некоторых вариантах осуществления генный сегмент DH человека, функционально связанный с 23-мерной RSS, содержит генный сегмент DH1-1 человека. В некоторых вариантах осуществления генный сегмент DH человека, функционально связанный с 23-мерной RSS, содержит генный сегмент DH2-2 человека. В некоторых вариантах осуществления человеческий генный сегмент DH, функционально связанный с 23-мерной RSS, содержит человеческий генный сегмент DH3-3. В некоторых вариантах осуществления человеческий генный сегмент DH, функционально связанный с 23-мерной RSS, содержит человеческий генный сегмент DH4-4. В некоторых вариантах осуществления человеческий генный сегмент DH, функционально связанный с 23-мерной RSS, содержит человеческий генный сегмент DH5-5. В некоторых вариантах осуществления человеческий генный сегмент DH, функционально связанный с 23-мерной RSS, содержит человеческий генный сегмент DH6-6. В некоторых вариантах осуществления человеческий генный сегмент DH, функционально связанный с 23-мерной RSS, содержит человеческий генный сегмент DH1-7. В некоторых вариантах осуществления человеческий генный сегмент DH, функционально связанный с 23-мерной RSS; содержит человеческий генный сегмент DH2-8. В некоторых вариантах осуществления человеческий генный сегмент DH, функционально связанный с 23-мерной RSS, содержит человеческий генный сегмент DH3-9. В некоторых вариантах осуществления человеческий генный сегмент DH, функционально связанный с 23-мерной RSS, содержит человеческий генный сегмент DH3-10. В некоторых вариантах осуществления человеческий генный сегмент DH, функционально связанный с 23-мерной RSS, содержит человеческий генный сегмент DH5-12. В некоторых вариантах осуществления человеческий генный сегмент DH, функционально связанный с 23-мерной RSS, содержит человеческий генный сегмент DH6-13. В некоторых вариантах осуществления человеческий генный сегмент DH, функционально связанный с 23-мерной RSS, содержит человеческий генный сегмент DH2-15. В некоторых вариантах осуществления человеческий генный сегмент DH, функционально связанный с 23-мерной RSS, содержит человеческий генный сегмент DH3-16. В некоторых вариантах осуществления человеческий генный сегмент DH, функционально связанный с 23-мерной RSS, содержит человеческий генный сегмент DH4-17. В некоторых вариантах осуществления человеческий генный сегмент DH, функционально связанный с 23-мерной RSS, содержит человеческий генный сегмент DH5-18. В некоторых вариантах осуществления человеческий генный сегмент DH, функционально связанный с 23-мерной RSS, содержит человеческий генный сегмент DH6-19. В некоторых вариантах осуществления человеческий генный сегмент DH, функционально связанный с 23-мерной RSS, содержит человеческий генный сегмент DH1-20. В некоторых вариантах осуществления человеческий генный сегмент DH, функционально связанный с 23-мерной RSS, содержит человеческий генный сегмент DH2-21. В некоторых вариантах осуществления человеческий генный сегмент DH, функционально связанный с 23-мерной RSS, содержит человеческий генный сегмент DH3-22. В некоторых вариантах осуществления человеческий генный сегмент DH, функционально связанный с 23-мерной RSS, содержит человеческий генный сегмент DH6-25. В некоторых вариантах осуществления человеческий генный сегмент DH, функционально связанный с 23-мерной RSS, содержит человеческий генный сегмент DH1-26. В некоторых вариантах осуществления человеческий генный сегмент DH, функционально связанный с 23-мерной RSS, содержит человеческий генный сегмент DH1-14. В некоторых вариантах осуществления человеческий генный сегмент DH, функционально связанный с 23-мерной RSS; содержит человеческий генный сегмент DH4-11. В некоторых вариантах осуществления человеческий генный сегмент DH, функционально связанный с 23-мерной RSS, содержит человеческий генный сегмент DH4-23. В некоторых вариантах осуществления человеческий генный сегмент DH, функционально связанный с 23-мерной RSS, содержит человеческий генный сегмент DH5-24. В некоторых вариантах осуществления человеческий генный сегмент DH, функционально связанный с 23-мерной RSS, содержит человеческий генный сегмент DH7-27.
[00179] В некоторых вариантах осуществления предусмотренные животные, отличные от человека, содержат полный или практически полный репертуар человеческих генных сегментов DH, как правило сгруппированных в порядке, наблюдаемом в неперегруппированном вариабельном локусе человеческого генома, где один из человеческих генных сегментов DH полного или практически полного репертуара заменен на генный сегмент DH, сконструированный таким образом, что он функционально связан с 23-мерной RSS. В некоторых вариантах осуществления один из человеческих генных сегментов DH полного или практически полного репертуара заменен на соответствующий генный сегмент DH, сконструированный таким образом, что он функционально связан с 23-мерной RSS, например, генный сегмент DH2-2 дикого типа заменен на генный сегмент DH2-2, сконструированный таким образом, что он функционально связан с 23-мерной RSS. В некоторых вариантах осуществления один из человеческих генных сегментов DH полного или практически полного репертуара заменен на другой генный сегмент DH, сконструированный таким образом, что он функционально связан с 23-мерной RSS, например, генный сегмент DH 7-27 дикого типа заменен на генный сегмент DH3-3, сконструированный таким образом, что он функционально связан с 23-мерной RSS. В некоторых вариантах осуществления генный сегмент DH1-1 полного или практически полного репертуара человеческих генных сегментов DH, как правило сгруппированных в порядке, наблюдаемом в неперегруппированном вариабельном локусе человеческого генома, заменен на соответствующий или другой генный сегмент DH, сконструированный таким образом, что он функционально связан с 23-мерной RSS, например, с 23-мерной RSS 3'-конца. В некоторых вариантах осуществления генный(генные) сегмент(сегменты) DH1-1, DH2-2, DH2-8, DH2-15, DH2-21 или любая их комбинация полного или практически полного репертуара человеческих генных сегментов DH, как правило сгруппированных в порядке, наблюдаемом в неперегруппированном вариабельном локусе человеческого генома, заменен(заменены) на соответствующий или другой генный сегмент DH, сконструированный таким образом, что он функционально связан с 23-мерной RSS, например, с 23-мерной RSS 3'-конца. В некоторых вариантах осуществления каждый из генных сегментов DH2-2, DH2-8 и DH2-15 полного или почти полного репертуара человеческих генных сегментов DH, как правило сгруппированных в порядке, наблюдаемом в неперегруппированном вариабельном локусе человеческого генома, заменен на соответствующий или другой генный сегмент DH, сконструированный таким образом, что он функционально связан с 23-мерной RSS, например, с 23-мерной RSS 3'-конца.
[00180] В некоторых вариантах осуществления предусмотренные животные, отличные от человека, содержат по меньшей мере человеческий генный сегмент JH JH6. В некоторых вариантах осуществления предусмотренные животные, отличные от человека, содержат по меньшей мере человеческие генные сегменты JH JH4, JH5 и JH6. В некоторых вариантах осуществления предусмотренные животные, отличные от человека, содержат по меньшей мере человеческие генные сегменты JH JH1, JH2, JH3, JH4, JH5 и JH6.
[00181] В некоторых вариантах осуществления константная область тяжелой цепи иммуноглобулина, отличного от человеческого, содержит один или более генов константной области тяжелой цепи иммуноглобулина, отличного от человеческого, такого как, например, иммуноглобулин М (IgM), иммуноглобулин D (IgD), иммуноглобулин G (IgG), иммуноглобулин Е (IgE) и иммуноглобулин A (IgA). В некоторых вариантах осуществления константная область тяжелой цепи иммуноглобулина, отличного от человеческого, содержит гены константной области IgM грызуна, IgD грызуна, IgG3 грызуна, IgG1 грызуна, IgG2b грызуна, IgG2a грызуна, IgE грызуна и IgA грызуна. В некоторых вариантах осуществления указанные человеческие генные сегменты VH, DH и JH функционально связаны с одним или более энхансеров тяжелой цепи иммуноглобулина, отличного от человеческого (т.е., энхансерными последовательностями или энхансерными областями). В некоторых вариантах осуществления указанные человеческие генные сегменты VH, DH и JH функционально связаны с одной или более регуляторными областями тяжелой цепи иммуноглобулина, отличного от человеческого (или регуляторными последовательностями). В некоторых вариантах осуществления указанные человеческие генные сегменты VH, DH и JH функционально связаны с одним или более энхансеров (или энхансерными последовательностями) тяжелой цепи иммуноглобулина, отличного от человеческого, и одной или более регуляторными областями (или регуляторными последовательностями) тяжелой цепи иммуноглобулина, отличного от человеческого.
[00182] В некоторых вариантах осуществления предусмотренные животные, отличные от человека, не содержат (или у них отсутствует) эндогенный ген Adam6. В некоторых вариантах осуществления предусмотренные животные, отличные от человека, не содержат или у них отсутствует эндогенный ген Adam6 (или последовательность, кодирующая Adam6) в том же положении в геноме зародышевой линии, в каком он находится в геноме зародышевой линии у животного, отличного от человека, дикого типа того же вида. В некоторых вариантах осуществления предусмотренные животные, отличные от человека, не содержат или у них отсутствует псевдоген Adam6 человека. В некоторых вариантах осуществления предусмотренные животные, отличные от человека, содержат вставку по меньшей мере одной нуклеотидной последовательности, которая кодирует один или более (например, грызуна) полипептидов Adam6, отличных от человеческих. Указанная вставка может находиться вне сконструированного локуса тяжелой цепи иммуноглобулина, описанного в данном документе (например, выше генного сегмента VH, расположенного наиболее близко к 5'-концу), в пределах сконструированного локуса тяжелой цепи иммуноглобулина или где-либо еще в геноме зародышевой линии животного, отличного от человека (например, произвольным образом введенная последовательность, кодирующая Adam6, отличный от человеческого), клетки или ткани, отличных от человеческих. В некоторых вариантах осуществления предусмотренные животные, отличные от человека, не содержат или у них отсутствует функциональный эндогенный псевдоген Adam6.
[00183] В некоторых вариантах осуществления предусмотренное животное, отличное от человека, клетка, отличная от человеческой, или ткань, отличная от человеческой, описанные в данном документе, не экспрессируют на выявляемом уровне полную или неполную эндогенную VH-область, отличную от человеческой, в молекуле антитела. В некоторых вариантах осуществления предусмотренное животное, отличное от человека, клетка, отличная от человеческой, или ткань, отличная от человеческой, описанные в данном документе, не содержат (или характеризуются отсутствием или делецией) одной или более нуклеотидных последовательностей, которые кодируют полную или неполную эндогенную VH-область, отличную от человеческой (например, VH, DH и/или JH) в молекуле антитела. В некоторых вариантах осуществления предусмотренное животное, отличное от человека, клетка, отличная от человеческой, или ткань, отличная от человеческой, описанные в данном документе, обладают геномом зародышевой линии, который содержит делецию эндогенных генных сегментов VH, DH и JH, отличных от человеческих, полностью или частично. В некоторых вариантах осуществления предусмотренное животное, отличное от человека, является фертильным.
[00184] В некоторых вариантах осуществления предусмотренные животные, отличные от человека, дополнительно содержат сконструированный локус легкой цепи иммуноглобулина κ, характеризующийся наличием нескольких человеческих генных сегментов Vκ и Jκ, сгруппированных в конфигурации зародышевой линии и вставленных выше, а также функционально связанных с геном Сκ, отличным от человеческого. В некоторых вариантах осуществления сконструированный локус легкой цепи иммуноглобулина κ содержит по меньшей мере человеческие генные сегменты Vκ, которые находятся в проксимальном вариабельном кластере (или проксимальном плече, или проксимальной дупликации) локуса легкой цепи человеческого иммуноглобулина κ. В некоторых вариантах осуществления сконструированный локус легкой цепи иммуноглобулина κ содержит по меньшей мере человеческие генные сегменты Vκ Vκ2-40, Vκ1-39, Vκ1-33, Vκ2-30, Vκ2-8, Vκ1-27, Vκ2-24, Vκ6-21, Vκ3-20, Vκ1-17, Vκ1-16, Vκ3-15, Vκ1-12, Vκ3-11, Vκ1-9, Vκ1-8, Vκ1-6, Vκ1-5, Vκ5-2 и Vκ4-1. В некоторых вариантах осуществления сконструированный локус легкой цепи иммуноглобулина κ содержит по меньшей мере 1, 2, 3, 4, 5, 10 или 15 человеческих генных сегментов Vκ, выбранных из Vκ2-40, Vκ1-39, Vκ1-33, Vκ2-30, Vκ2-8, Vκ1-27, Vκ2-24, Vκ6-21, Vκ3-20, Vκ1-17, Vκ1-16, Vκ3-15, Vκ1-12, Vκ3-11, Vκ1-9, Vκ1-8, Vκ1-6, Vκ1-5, Vκ5-2 и Vκ4-1. В некоторых вариантах осуществления сконструированный локус легкой цепи иммуноглобулина κ содержит человеческие генные сегменты Jκ Jκ1, Jκ2, Jκ3, Jκ4 и Jκ5. В некоторых вариантах осуществления сконструированный локус легкой цепи иммуноглобулина κ содержит по меньшей мере 1, 2, 3 или 4 человеческих генных сегмента Jκ, выбранных из Jκ1, Jκ2, Jκ3, Jκ4 и Jκ5.
[00185] Bo многих вариантах осуществления указанные человеческие генные сегменты Vκ и Jκ функционально связаны с одним или более энхансерами (т.е. энхансерными последовательностями или энхансерными областями) легкой цепи иммуноглобулина κ, отличного от человеческого. В некоторых вариантах осуществления указанные человеческие генные сегменты Vκ и Jκ функционально связаны с интронной энхансерной областью легкой цепи мышиного Igκ (Igκ Ei или Eiκ). Во многих вариантах осуществления указанные человеческие генные сегменты Vκ и Jκ функционально связаны с одной или более регуляторными областями (или регуляторными последовательностями) легкой цепи иммуноглобулина κ, отличного от человеческого. В некоторых вариантах осуществления указанные человеческие генные сегменты Vκ и Jκ функционально связаны с энхансерной областью 3'-конца легкой цепи мышиного Igκ (Igκ 3'Е или 3'Еκ). В некоторых вариантах осуществления указанные человеческие генные сегменты vκ и Jκ функционально связаны с мышиным Eiκ и функционально связаны с мышиным 3'Еκ. В некоторых вариантах осуществления указанные человеческие генные сегменты Vκ и Jκ функционально связаны с одним или более энхансерами (или энхансерными последовательностями, или энхансерными областями) легкой цепи иммуноглобулина κ, отличного от человеческого, и одной или более регуляторными областями (или регуляторными последовательностями) легкой цепи иммуноглобулина κ, отличного от человеческого. В некоторых вариантах осуществления сконструированный локус легкой цепи иммуноглобулина κ содержит такие же энхансерные области (или энхансерные последовательности) легкой цепи иммуноглобулина κ, отличного от человеческого, какие встречаются в локусе легкой цепи иммуноглобулина κ дикого типа. В некоторых вариантах осуществления сконструированный локус легкой цепи иммуноглобулина κ содержит такие же энхансерные области (или энхансерные последовательности) легкой цепи Igκ, отличного от человеческого, какие встречаются в локусе легкой цепи иммуноглобулина κ дикого типа другого вида (например, другого вида грызунов).
[00186] В некоторых вариантах осуществления сконструированный локус легкой цепи иммуноглобулина κ, описанный в данном документе, не содержит (т.е. характеризуется отсутствием) человеческого гена VpreB (или последовательности, кодирующей ген VpreB).
[00187] В некоторых вариантах осуществления ген Сκ, отличный от человеческого, сконструированного с помощью генной инженерии локуса легкой цепи иммуноглобулина κ включает ген Сκ грызуна, такой как, например, ген Сκ мыши или ген Сκ крысы. В некоторых вариантах осуществления ген Сκ, отличный от человеческого, сконструированного с помощью генной инженерии локуса легкой цепи иммуноглобулина κ представляет собой или содержит ген Сκ мыши из генетического фона, который включает линию 129, линию BALB/c, линию C57BL/6, помесную линию 129xC57BL/6 или их комбинации.
[00188] В некоторых вариантах осуществления предусмотренное животное, отличное от человека, клетка, отличная от человеческой, или ткань, отличная от человеческой, описанные в данном документе, не экспрессируют на выявляемом уровне полную или неполную эндогенную Vκ-область, отличную от человеческой, в молекуле антитела. В некоторых вариантах осуществления предусмотренное животное, отличное от человека, клетка, отличная от человеческой, или ткань, отличная от человеческой, описанные в данном документе, не содержат (или характеризуются отсутствием или делецией) одной или более нуклеотидных последовательностей, которые кодируют полную или неполную эндогенную Vκ-область, отличную от человеческой, в молекуле антитела. В некоторых вариантах осуществления предусмотренное животное, отличное от человека, клетка, отличная от человеческой, или ткань, отличная от человеческой, описанные в данном документе, обладают геномом зародышевой линии, который содержит делецию эндогенных генных сегментов Vκ и Jκ, отличных от человеческих, полностью или частично.
[00189] В некоторых вариантах осуществления предусмотренные животные, отличные от человека, дополнительно содержат локус легкой цепи иммуноглобулина дикого типа или инактивированный (например, с помощью нацеливания на ген) локус легкой цепи иммуноглобулина λ.
[00190] В некоторых вариантах осуществления предусмотренное животное, отличное от человека, клетка, отличная от человеческой, или ткань, отличная от человеческой, описанные в данном документе, не экспрессируют на выявляемом уровне полную или неполную эндогенную Vλ-область, отличную от человеческой, в молекуле антитела. В некоторых вариантах осуществления предусмотренное животное, отличное от человека, клетка, отличная от человеческой, или ткань, отличная от человеческой, описанные в данном документе, не содержат (или характеризуются отсутствием или делецией) одной или более нуклеотидных последовательностей, которые кодируют полную или неполную эндогенную Vλ-область, отличную от человеческой, в молекуле антитела. В некоторых вариантах осуществления предусмотренное животное, отличное от человека, клетка, отличная от человеческой, или ткань, отличная от человеческой, описанные в данном документе, обладают геномом зародышевой линии, который содержит делецию эндогенных генных сегментов Vλ и Jλ, отличных от человеческих, полностью или частично. В некоторых вариантах осуществления предусмотренное животное, отличное от человека, клетка, отличная от человеческой, или ткань, отличная от человеческой, описанные в данном документе, обладают геномом зародышевой линии, который содержит делецию эндогенных генных сегментов Vλ, Jλ и Cλ, отличных от человеческих, полностью или частично.
[00191] Руководство для получения нацеливающих векторов, клеток, отличных от человеческих и животных, отличных от человека, несущих такие сконструированные с помощью генной инженерии локусы иммуноглобулина, можно найти в патентах США №№8642835, 8697940, 9006511, 9012717, 9029628, 9035128, 9066502, 9150662 и 9163092, которые включены в данный документ посредством ссылки во всей их полноте. Специалисты в данной области знают ряд технологий, известных из уровня техники, для осуществления такого конструирования с помощью генной инженерии и/или манипуляции в отношении геномов, отличных от человеческих (например, млекопитающего), или для получения, обеспечения или производства иным образом таких последовательностей, предназначенных для введения в геном зародышевой линии животных, отличных от человека.
ДНК-конструкции
[00192] Как правило, молекулу полинуклеотида, содержащую генные сегменты иммуноглобулина, описанные в данном документе, в частности, один или более генных сегментов DH, каждый из которых функционально связан в области 5'- или 3'-конца с 23-мерной RSS, встраивают в вектор, предпочтительно ДНК-вектор, чтобы реплицировать молекулу полинуклеотида в подходящей клетке-хозяине.
[00193] Благодаря их размеру, сегменты DH можно клонировать напрямую из геномных источников, доступных от коммерческих поставщиков, или конструировать in silico на основании опубликованных последовательностей, доступных в GenBank. В качестве альтернативы последовательности иммуноглобулина могут быть получены из библиотек бактериальных искусственных хромосом (ВАС). Библиотеки ВАС содержат вставки со средним размером 100-150 т.н., и они способны нести вставки размером до 300 т.н. (Shizuya, et al., 1992, Proc. Natl. Acad. Sci., USA 89:8794-8797; Swiatek, et al., 1993, Genes and Development 7:2071-2084; Kim, et al., 1996, Genomics 34 213-218; включенные в данный документ посредством ссылки во всей своей полноте). Например, геномные библиотеки ВАС для человека и мыши были сконструированы и являются коммерчески доступными (например, Invitrogen, Карлсбад, Калифорния). Геномные библиотеки ВАС также могут служить источником последовательностей иммуноглобулина, а также областей транскрипционного контроля.
[00194] В качестве альтернативы последовательности иммуноглобулина можно выделять, клонировать и/или переносить из искусственных хромосом дрожжей (YAC). Полный локус иммуноглобулина или его часть можно клонировать и помещать в одну или более YAC. Если используют несколько YAC, и при этом они содержат перекрывающиеся гомологичные области, их можно рекомбинировать внутри штаммов дрожжей-хозяев с получением единой конструкции, представляющей полный локус. Плечи YAC можно дополнительно модифицировать с помощью кассет для отбора для млекопитающих путем их усовершенствования таким образом, чтобы содействовать введению конструкций в эмбриональные стволовые клетки или эмбрионы с помощью способов, известных из уровня техники и/или описанных в данном документе.
[00195] ДНК-конструкции можно получать с помощью способов, известных из уровня техники. Например, ДНК-конструкцию можно получить в виде части большей плазмиды. Такое получение позволяет эффективно клонировать и отбирать правильные конструкции, как это известно из уровня техники. Фрагменты ДНК, содержащие один или более DH-сегментов, каждый из которых функционально связан в области 5'- или 3'-конца с 23-мерной RSS, как описано в данном документе, могут быть расположены между подходящими сайтами рестрикции в плазмиде таким образом, чтобы их можно было легко выделить из остальных последовательностей плазмиды для включения в состав генома требуемого животного.
[00196] В некоторых вариантах осуществления способы, применяемые для получения плазмид и трансформации организмов-хозяев, известны из уровня техники. Другие подходящие системы экспрессии, подходящие как для прокариотических, так и для эукариотических клеток, а также общие процедуры рекомбинации см. в Molecular Cloning: A Laboratory Manual, 2nd Ed., ed. by Sambrook, J. et al., Cold Spring Harbor Laboratory Press: 1989; включенный в данный документ посредством ссылки во всей своей полноте.
Получение животных, отличных от человека
[00197] Предусмотрены животные, отличные от человека, которые экспрессируют антитела, обладающие вариативностью CDR3 тяжелой цепи, характеризующейся большей аминокислотной длиной, являющейся результатом интеграции 23-мерных RSS, смежных с одним или более DH-сегментами, что обеспечивает возможность для DH-DH-рекомбинации в пределах вариабельной области тяжелой цепи иммуноглобулина в геноме животного, отличного от человека. Подходящие примеры, описанные в данном документе, включают грызунов, в частности, мышей. Один или более DH-сегментов во многих вариантах осуществления содержат гетерологичные DH-сегменты (например, DH-сегменты человека). Также предусмотрены животные, отличные от человека, эмбрионы, отличные от человеческих, клетки, отличные от человеческих и нацеливающие конструкции, отличные от человеческих, для получения животных, отличных от человека, эмбрионов, отличных от человеческих, и клеток, отличных от человеческих, содержащих указанные DH-сегменты, способные к DH-DH-рекомбинации с повышенной частотой по сравнению с животными дикого типа или эталонными животными, отличными от человека.
[00198] В некоторых вариантах осуществления один или более DH-сегментов модифицированы таким образом, чтобы быть смежными (или функционально связанными) в области 5'- или 3'-конца с 23-мерной RSS в пределах кластера вариативности (т.е. DH-области) вариабельной области тяжелой цепи иммуноглобулина в геноме животного, отличного от человека. В некоторых вариантах осуществления DH-область (или ее часть) вариабельной области тяжелой цепи иммуноглобулина не подвергают делеции (т.е. она является интактной). В некоторых вариантах осуществления DH-область (или ее часть) вариабельной области тяжелой цепи иммуноглобулина изменяют, разрушают, подвергают делеции, конструируют с помощью генной инженерии или заменяют одним или более DH-сегментами, каждый из которых функционально связан в области 5'- или 3'-конца с 23-мерной RSS. В некоторых вариантах осуществления всю или практически всю DH-область заменяют одним или более синтетическими DH-сегментами, каждый из которых функционально связан в области 5'- или 3'-конца с 23-мерной RSS; в некоторых вариантах осуществления один или более стандартных генных сегментов DH в DH-области вариабельной области тяжелой цепи иммуноглобулина не подвергают делеции или замене. В некоторых вариантах осуществления DH-область содержит как стандартные сегменты DH (т.е. один или более сегментов DH, каждый из которых функционально связан с 12-мерной RSS 5'-конца и с 12-мерной RSS 3'-конца), так и сконструированные генные сегменты DH (т.е. один или более сегментов DH, каждый из которых функционально связан с 23-мерной RSS 5'- или 3'-конца. В некоторых вариантах осуществления DH-область, описанная в данном документе, представляет собой синтетическую DH-область. В некоторых вариантах осуществления DH-область представляет собой DH-область человека. В некоторых вариантах осуществления DH-область представляет собой DH-область мыши. В некоторых вариантах осуществления сконструированную с помощью генной инженерии DH-область (или ее часть), описанную в данном документе, встраивают в вариабельную область тяжелой цепи иммуноглобулина таким образом, чтобы указанная сконструированная с помощью генной инженерии DH-область (или ее часть) была функционально связана с одним или более генными сегментами VH и/или одним или более генный сегментами JH. В некоторых вариантах осуществления указанную сконструированную с помощью генной инженерии DH-область встраивают в одну из двух копий вариабельной области тяжелой цепи иммуноглобулина, в результате получая животное, отличное от человека, которое является гетерозиготным по отношению к указанной сконструированной с помощью генной инженерии DH-области. В некоторых вариантах осуществления предусмотрено животное, отличное от человека, которое является гомозиготным по сконструированной с помощью генной инженерии DH-области. В некоторых вариантах осуществления предусмотрено животное, отличное от человека, которое является гетерозиготным по сконструированной с помощью генной инженерии DH-области.
[00199] В некоторых вариантах осуществления животное, отличное от человека, описанное в данном документе, содержит интегрированную произвольным образом вариабельную область тяжелой цепи иммуноглобулина человека, которая содержит DH-область, которая содержит один или более DH-сегментов, каждый из которых функционально связан в области 5'- или 3'-конца с 23-мерной RSS в пределах его генома. Таким образом, такие животные, отличные от человека, могут быть описаны как имеющие трансген тяжелой цепи иммуноглобулина человека, содержащий сконструированную с помощью генной инженерии DH-область. Сконструированная с помощью генной инженерии DH-бласть может быть выявлена с помощью разнообразных способов, включающих, например, ПЦР, вестерн-блоттинг, Саузерн-блоттинг, определение полиморфизма длины рестрикционных фрагментов (RFLP) или анализа приобретения аллеля (GOA) или потери аллеля (LOA). В некоторых таких вариантах осуществления животное, отличное от человека, описанное в данном документе, является гетерозиготным по отношению к сконструированной с помощью генной инженерии DH-области, описанной в данном документе. В некоторых таких вариантах осуществления животное, отличное от человека, описанное в данном документе, является гомозиготным по отношению к сконструированной с помощью генной инженерии DH-область, описанной в данном документе. В некоторых таких вариантах осуществления животное, отличное от человека, описанное в данном документе, является гемизиготным по отношению к сконструированной с помощью генной инженерии DH-области, описанной в данном документе. В некоторых таких вариантах осуществления животное, отличное от человека, описанное в данном документе, содержит одну или более копий сконструированной с помощью генной инженерии DH-области, описанной в данном документе.
[00200] В некоторых вариантах осуществления сконструированная с помощью генной инженерии DH-область животного, отличного от человека, описанного в данном документе, содержит по меньшей мере один DH-сегмент, который ассоциирован (или функционально связан) в области 5'-конца с 23-мерной RSS. В некоторых вариантах осуществления сконструированная с помощью генной инженерии DH-область животного, отличного от человека, описанного в данном документе, содержит по меньшей мере один DH-сегмент, который ассоциирован (или функционально связан) в области 3'-конца с 23-мерной RSS. В некоторых вариантах осуществления сконструированная с помощью генной инженерии DH-область животного, отличного от человека, описанного в данном документе, содержит по меньшей мере DH3-3-сегмент человека, который ассоциирован (или функционально связан) в области 5'-конца с 23-мерной RSS. В некоторых вариантах осуществления сконструированная DH-область животного, отличного от человека, описанного в данном документе, содержит по меньшей мере человеческий сегмент DH2, который ассоциирован (или функционально связан) с 23-мерной RSS 3' конца, где человеческий сегмент DH2 выбран из группы, состоящей из человеческого DH2-2, человеческого DH2-8, человеческого DH2-15 и человеческого DH2-21.
[00201] В некоторых вариантах осуществления сконструированная DH-область животного, отличного от человека, описанного в данном документе, содержит более одного сегмента DH, каждый из которых ассоциирован (или функционально связан) с 23-мерной RSS 5'-конца. В некоторых вариантах осуществления сконструированная с помощью генной инженерии DH-область животного, отличного от человека, описанного в данном документе, содержит более одного DH-сегмента, каждый из которых ассоциирован (или функционально связан) на своем 3'-конце с 23-мерной RSS. В некоторых вариантах осуществления сконструированная DH-область животного, отличного от человека, описанного в данном документе, содержит человеческие сегменты DH2-2, DH2-8 и DH2-15, каждый из которых ассоциирован (или функционально связан) с 23-мерной RSS 3'-конца.
[00202] Предусмотрены композиции и способы получения животных, отличных от человека, геном которых содержит вариабельную область тяжелой цепи иммуноглобулина, которая содержит сконструированную с помощью генной инженерии DH-область, где сконструированная с помощью генной инженерии DH-область содержит один или более DH-сегментов, каждый из которых ассоциирован (или функционально связан) в области 3'- или 5'-конца с 23-мерной RSS, включая композиции и способы получения животных, отличных от человека, которые экспрессируют антитела, содержащие вариабельную область тяжелой цепи, которая содержит CDR3-область, имеющую аминокислотную последовательность, кодирующуюся более чем одним DH-сегментом локуса тяжелой цепи иммуноглобулина, который содержит генные сегменты VH и JH человека, функционально связанные с одним или более генами константной области тяжелой цепи, отличными от человеческих. В некоторых вариантах осуществления также предусмотрены композиции и способы получения животных, отличных от человека, которые экспрессируют такие антитела под контролем эндогенного(-ых) энхансер(-ов) и/или эндогенной(-ых) регуляторной(-ых) последовательность(-ей). В некоторых вариантах осуществления также предусмотрены композиции и способы для получения животных, отличных от человека, которые экспрессируют такие антитела под контролем гетерологичного(-ых) энхансера(ов) и/или гетерологичной(-ых) регуляторной(-ых) последовательности(ей). Способы включают вставку одного или более DH-сегментов и других последовательностей, которые обеспечивают возможность DH-DH-рекомбинации с повышенной частотой по сравнению с DH-сегментами дикого типа, в геном животного, отличного от человека таким образом, чтобы происходила экспрессия антитела, которое содержит тяжелые цепи иммуноглобулина, полученного в результате рекомбинации V(DD)J.
[00203] В некоторых вариантах осуществления способы включают вставку ДНК, которая содержит генный сегмент DH с 23-мерной RSS 5'-конца и 12-мерной RSS 3'-конца, функционально связанный с одним или более генными сегментами JH. Как описано в данном документе, генный сегмент DH располагается ниже μ0-промоторной последовательности. В некоторых вариантах осуществления способы включают вставку DH-сегмента с 23-мерной RSS на своем 5'-конце и 12-мерной RSS на своем 3'-конце в положение относительно μ0-промоторной последовательности таким образом, чтобы указанный генный сегмент JH являлся доступным для генов RAG (например, RAG-1 и/или RAG-2) во время рекомбинации. Генетический материал, который содержит DH-сегмент и фланкирующие RSS, описанные выше, может быть вставлен в геном животного, отличного от человека, за счет чего создается животное, отличное от человека, содержащее сконструированную DH-область, которая содержит указанный DH-сегмент и необходимые RSS для обеспечения условий для рекомбинации со смежным генным сегментом DH и генными сегментами VH и JH.
[00204] В некоторых вариантах осуществления способы включают вставку ДНК, которая содержит три генных сегмента DH, каждый из которых связан с 12-мерной RSS на своем 5'-конце и 23-мерной RSS на своем 3'-конце, функционально связанными с шестью генными сегментами JH. Как описано в данном документе, генные сегменты DH располагаются среди нескольких генных сегментов DH, каждый из которых связан с 12-мерной RSS на своем 5'- и 3'-конце. В некоторых вариантах осуществления способы включают вставку генных сегментов DH2-2, DH2-8 и DH2-15, каждый из которых связан с 12-мерной RSS на своем 5'-конце и 23-мерной RSS на своем 3'-конце в кластер разнообразия, содержащий несколько других генных сегментов DH, каждый из которых связан со стандартной RSS или RSS дикого типа. Генетический материал, который содержит генные сегменты DH и фланкирующие RSS, описанные выше, может быть вставлен в геном животного, отличного от человека, за счет чего создается животное, отличное от человека, содержащее сконструированную DH-область, которая содержит указанные DH-сегменты и необходимые RSS для обеспечения условий для рекомбинации со смежными генными сегментами DH и генными сегментами VH и JH.
[00205] При необходимости последовательности, соответствующие (или кодирующие) DH-сегментам могут быть модифицированы таким образом, чтобы включать кодоны, оптимизированные для экспрессия в животном, отличном от человека (например, см. патенты США №№5670356 и 5874304; каждый из которых включен в данный документ посредством ссылки во всей своей полноте). Кодон-оптимизированные последовательности представляют собой синтетические последовательности, и они предпочтительно кодируют идентичный полипептид (или биологически активный фрагмент полноразмерного полипептида, обладающий практически такой же активностью, что и полноразмерный полипептид), кодируемый исходным полинуклеотидом, не являющимся кодон-оптимизированным. В некоторых вариантах осуществления последовательности соответствующие (или кодирующие) DH-сегментам могут предусматривать измененную последовательность, чтобы оптимизировать частоту использования кодонов для конкретного типа клетки (например, клетки грызуна). Например, кодоны последовательностей, соответствующих DH-сегментам, которые должны быть встроены в геном животного, отличного от человека (например, грызуна), могут быть оптимизированы для экспрессии в клетке животного, отличного от человека. Такая последовательность может быть описана как кодон-оптимизированная последовательность.
[002 06] Вставка DH-сегментов, функционально связанных с 23-мерной RSS на своем 5'- или 3'-конце, в DH-область таким образом, чтобы указанные DH-сегменты были функционально связаны с генными сегментами VH и JH (например, несколькими из генных сегментов VH и JH), требует относительно минимальной модификации генома и приводит к экспрессии антител, содержащих тяжелые цепи, характеризующиеся наличием CDR3 с большими значениями аминокислотной длины.
[00207] Способы получения трансгенных животных, отличных от человека, в том числе с помощью нокаутов и нокинов, хорошо известны из уровня техники (см., например, Gene Targeting: A Practical Approach, Joyner, ed., Oxford University Press, Inc. (2000); включенный в данный документ посредством ссылки во всей своей полноте). Например, получение трансгенных грызунов может необязательно предусматривать разрушение генетических локусов одного или более эндогенных генов грызуна (или генных сегментов) и введение одного или более DH-сегментов, каждый из которых функционально связан с 23-мерной RSS, в геном грызуна, в некоторых вариантах осуществления, в то же место, что и эндогенный ген (или генные сегменты) грызуна. В некоторых вариантах осуществления один или более DH-сегментов, каждый из которых функционально связан с 23-мерной RSS, вводят в DH-область вставленного произвольным образом локуса тяжелой цепи иммуноглобулина в геноме грызуна. В некоторых вариантах осуществления один или более DH-сегментов, каждый из которых функционально связан с 23-мерной RSS, вводят в DH-область эндогенного локуса тяжелой цепи иммуноглобулина в геноме грызуна; в некоторых вариантах осуществления эндогенный локус тяжелой цепи иммуноглобулина изменен, модифицирован или сконструирован таким образом, чтобы содержать генные сегменты человека (например, V и/или J), функционально связанные с одним или более генами константной области (например, человека или мышиные).
[00208] Схематическое изображение (без соблюдения масштаба) иллюстративных нацеливающих векторов для конструирования сконструированной DH-области и интеграции в эмбриональные стволовые (ES) клетки грызуна для получения грызуна, геном которого содержит вариабельную область тяжелой цепи иммуноглобулина, которая включает сконструированный кластер разнообразия (т.е. DH-область), где указанный кластер разнообразия содержит один или более DH-сегментов, каждый из которых функционально связан с 23-мерной RSS на своем 5'-или 3'-конце, представлено на фигуре 2. Иллюстративные стратегии и способы встраивания таких векторов в вариабельные области тяжелой цепи иммуноглобулина в геноме ES-клеток грызуна представлены на фигурах 3-8. На каждой из фигур 2-8 в соответствующих местах указаны сайты распознавания рестрикционного фермента NotI, указаны названия и примерное расположение (короткая пунктирная линия) различных наборов праймеров/зондов (см. таблицу 4) для различных изображенных (без соблюдения масштаба) аллелей и, если не указано иное, незакрашенные символы и полосы обозначают последовательность человека, в то время как закрашенные символы и темные полосы обозначают последовательность мыши. Для каждой из фигур применяются следующие сокращения: spec: ген устойчивости к спектиномицину; neo: ген устойчивости к неомицину; hyg: ген устойчивости к гигромицину; lp: сайт распознавания сайт-специфической рекомбинации loxP; Ei: мышиный интронный энхансер тяжелой цепи; IgM: мышиный ген константной области иммуноглобулина М; L: последовательность сайта loxP; Frt: целевая последовательность для распознавания флиппазой; μ0 pro: последовательность промотора μ0.
[00209] Как показано на фигуре 2, фрагменты ДНК, содержащие несколько DH-сегментов, содержащих один DH-сегмент, функционально связанный с 23-мерной RSS на своем 5'-конце (фигура 2, верхняя и средняя часть), или содержащих три DH-сегмента, каждый из которых функционально связан с 23-мерной RSS на своем 3'-конце (фигура 2, нижняя часть), получены с применением технологии VELOCIGENE® (см., например, патент США №6586251 и Valenzuela et al., 2003, Nature Biotech. 21(6): 652-659; включенные в данный документ посредством ссылки во всей своей полноте) и методик молекулярной биологии, известных из уровня техники. На фигуре 2, если не указано иное, незакрашенные символы и полосы обозначают последовательность человека, в то время как закрашенные символы и темные полосы обозначают последовательность мыши. Изображено относительное расположение, без соблюдения масштаба, генных сегментов VH6-1, DH2-2, DH2-8, DH2-15, DH3-3 и JH1, JH2, JH3, JH4, JH5 и JH6 человека при их наличии, и любой генный сегмент без названия представляет собой генный сегмент DH. Как правило DH область, обозначенная диагональными линиями, содержит полный репертуар неперегруппированных генных сегментов DH человека (см., например, www.imgt.org/IMGTrepertoire/index.php?section=LocusGeries&repertoire=locus&species=human&group=IGH; включенный в данный документ посредством ссылки во всей своей полноте) с учетом следующих исключений: в составе двух верхних нацеливающих векторов отсутствует последний ген DH7-27, который заменен последовательностью, содержащей генный сегмент DH3-3, фланкированный 23-мерной RSS на своем 5'-конце и 12-мерной RSS на своем 3'-конце (изображенный в виде незакрашенной стрелки); в составе нижнего нацеливающего вектора отсутствует неперегруппированный генный сегмент DH2-2 человека, неперегруппированный генный сегмент DH2-8 человека и генные сегменты DH2-15, которые соответственно заменены генным сегментом DH2-2, фланкированным 12-мерной RSS на своем 5'-конце и 23-мерной RSS на своем 3'-конце, генным сегментом DH2-8, фланкированным 12-мерной RSS на своем 5'-конце и на своем 3'-конце 23-мерной RSS, и генным сегментом DH2-15, фланкированным 12-мерной RSS на своем 5'-конце и 23-мерной RSS на своем 3'-конце (где каждый сконструированный генный сегмент DH2-2, DH2-8 и DH2-15 изображен в виде незакрашенной стрелки). В виде незакрашенных вертикальных треугольников изображены места расположения уникальных искусственных 40-меров, гомологичных праймерам и зондам последовательности, помещенным в нацеливающий вектор 12:DH2-2:23|12:DH2-8:23|12:DH2-15:23 для обеспечения гарантии правильной модификации нацеливающего вектора/ES-клетки. Последовательности уникальных искусственных 40-меров, обозначенные «1», «2», «10», «16», «8» и «18», представлены под SEQ ID NO:73, SEQ ID NO:74, SEQ ID NO:75, SEQ ID NO:76, SEQ ID NO:77 и SEQ ID NO:78 соответственно.
[00210] Фрагменты ДНК собирают с применением плеч гомологии для точной прицельной вставки в локус вариабельной области тяжелой цепи гуманизированного иммуноглобулина (фигуры 3, 5, 7). Кассета для отбора (например, по устойчивости к неомицину), фланкированная сайтами распознавания сайт-специфической рекомбинации (например, loxP), вставлена в нацеливающие векторы для обеспечения проверки надлежащей вставки в клоны ES-клеток и может быть удалена с помощью временной экспрессии рекомбиназы (например, Cre) в положительных клонах ES-клеток (фигуры 4, 6, 8). Фрагменты ДНК содержат последовательности, необходимые для надлежащей сборки (т.е. рекомбинации), транскрипции и экспрессии вариабельных областей тяжелой цепи после внедрения в локус тяжелой цепи иммуноглобулина. Нацеливающие векторы конструируют таким образом, чтобы сконструированная DH-область была фланкирована на своем 5'-конце геномной ДНК VH человека и на своем 3'-конце геномной ДНК JH человека и геномной ДНК константной области тяжелой цепи (например, интронного энхансера и гена константной области IgM), отличной от человеческой (например, грызуна). Окончательные нацеливающие векторы для встраивания в геном клетки, отличной от человеческой (например, эмбриональной стволовой клетки грызуна), содержат геномную ДНК VH (например, содержат один или более генных сегментов VH), сконструированную DH-область, геномную ДНК JH на своем 3'-конце и геномную ДНК константной области тяжелой цепи, отличную от человеческой (например, грызуна), все из которых функционально связаны для обеспечения условий для рекомбинации между генными сегментами VH, сконструированной DH-областью и генным сегментом JH после интеграции в геном животного, отличного от человека. После сборки нацеливающие векторы линеаризируют и вводят с помощью электропорации в ES-клетки грызуна.
[00211] Нацеливающие векторы вводят в эмбриональные стволовые клетки грызуна (например, мыши) таким образом, чтобы последовательность, содержащаяся в нацеливающем векторе (т.е. сконструированная DH-область) приводила к способности клетки, отличной от человеческой, или животного, отличного от человека (например, мышь), экспрессировать антитела, которые содержат CDR3, имеющие аминокислоты, кодируемые более чем одним DH-сегментом.
[00212] Как описано в данном документе, получают трансгенных грызунов, у которых сконструированная DH-область была введена в локус тяжелой цепи иммуноглобулина генома грызуна (например, локус тяжелой цепи иммуноглобулина, сконструированный таким образом, чтобы содержать генные сегменты вариабельной области человека, который может представлять собой эндогенный локус тяжелой цепи иммуноглобулина сконструированный указанным образом).
[00213] Локусы иммуноглобулина, содержащие генные сегменты вариабельной области человека известны из уровня техники и могут быть найдены, например, в патентах США №№5633425; 5770429; 5814318; 6075181; 6114598; 6150584; 6998514; 7795494; 7910798; 8232449; 8502018; 8697940; 8703485; 8754287;8791323; 8809051; 8907157; 9035128; 9145588; 9206263; 9447177; 9551124; 9580491 и 9475559, каждый из которых включен в данный документ посредством ссылки во всей своей полноте, а также в публикациях заявок на патент США №№20100146647, 20110195454, 20130167256, 20130219535, 20130326647, 20130096287 и 2015/0113668, каждая из которых включена в данный документ посредством ссылки во всей своей полноте, и в публикациях согласно РСТ №№WO2007117410, WO2008151081, WO2009157771, WO2010039900, WO2011004192, WO2011123708 и WO2014093908, каждая из которых включена в данный документ посредством ссылки во всей своей полноте.
[00214] B некоторых вариантах осуществления животные, отличные от человека, раскрытые в данном документе, содержат экзогенные трансгены полностью человеческого иммуноглобулина, содержащие сконструированную DH-область, которые способны перестраиваться в предшественниках В-клеток у мышей (Alt et al., 1985, Immunoglobulin genes in transgenic mice, Trends Genet 1:231-236; включенная в данный документ посредством ссылки во всей своей полноте). В таких вариантах осуществления трансгены полностью человеческого иммуноглобулина, содержащие сконструированную DH-область, могут быть вставлены (произвольным образом) и эндогенные гены иммуноглобулина также могут быть нокаутированы (Green et al., 1994, Antigen-specific human monoclonal antibodies from mice engineered with human Ig heavy and light chain YACs, Nat Genet 7:13-21; Lonberg et al., 1994, Antigen-specific human antibodies from mice comprising four distinct genetic modifications, Nature 368:856-859; Jakobovits et al., 2007, From XenoMouse technology to panitumumab, the first fully human antibody product from transgenic mice, Nat Biotechnol 25:1134-1143; где каждая из публикаций включена посредством ссылки во всей своей полноте), например, при этом эндогенные локусы тяжелой цепи и легкой κ-цепи иммуноглобулина инактивируют, например путем направленной делеции небольших, но важных частей каждого эндогенного локуса, с последующим введением человеческих локусов гена иммуноглобулина в виде произвольно встроенных больших трансгенов или минихромосом (Tomizuka et al., 2000, Double trans-chromosomic mice: maintenance of two individual human chromosome fragments containing Ig heavy and kappa loci and expression of fully human antibodies, PNAS USA 97:722-727; включенная посредством ссылки во всей своей полноте).
[00215] B некоторых вариантах осуществления локусы тяжелой и легкой цепей человеческого или гуманизированного иммуноглобулина, содержащие сконструированную DH-область, находятся в эндогенных локусах тяжелой и легкой цепей иммуноглобулина соответственно. Ранее был описан способ осуществления масштабной генетической замены in situ для локусов генов вариабельных областей иммуноглобулина зародышевой линии мыши локусами генов вариабельных областей иммуноглобулина зародышевой линии человека с сохранением способности мышей давать потомство. См., например, патенты США №№6596541 и 8697940, каждый из которых включен в данный документ посредством ссылки во всей своей полноте. В частности, описана точная замена шести мегабаз локусов генов вариабельных областей как мышиной тяжелой цепи, так и мышиной легкой κ-цепи иммуноглобулина их человеческими аналогами, при этом константные области мыши оставались интактными. В результате получали мышей, у которых имелась точная замена всего репертуара вариабельных областей иммуноглобулина зародышевой линии эквивалентными последовательностями вариабельных областей иммуноглобулина зародышевой линии человека при сохранении константных областей мыши. Вариабельные области человека связаны с константными областями мыши с образованием химерных человеческих-мышиных локусов иммуноглобулина, которые перегруппировываются и экспрессируются на физиологически приемлемых уровнях. Экспрессируемые антитела представляют собой «обратные химеры», т.е. они содержат человеческие последовательности вариабельной области и мышиные последовательности константной области. Этих мышей, имеющих гуманизированные вариабельные области иммуноглобулина, у которых экспрессируются антитела, имеющие человеческие или гуманизированные вариабельные области и мышиные константные области, называют мышами VELOCIMMUNE®.
[00216] Гуманизированные мыши VELOCIMMUNE® демонстрируют наличие полностью функциональной гуморальной иммунной системы, которая практически неотличима от таковой у мышей дикого типа. Они демонстрируют нормальные клеточные популяции на всех стадиях развития В-клеток. У них продемонстрирована нормальная морфология лимфоидных органов. Последовательности антител мышей VELOCIMMUNE® демонстрируют нормальную перегруппировку V(D)J и нормальные значения частоты соматических гипермутаций. Популяции антител у этих мышей отображают распределения изотипов, которые возникают в результате нормального переключения класса (например, нормального цис-переключения изотипа). Иммунизация мышей VELOCIMMUNE® приводит к устойчивым гуморальным иммунным ответам, при которым вырабатывается большой, разнообразный репертуар антител, имеющих человеческие вариабельные домены иммуноглобулина, подходящие для применения в качестве терапевтических кандидатов. Эта платформа обеспечивает богатый источник человеческих последовательностей вариабельной области иммуноглобулина с естественным образом созревшей аффинностью для получения фармацевтически приемлемых антител и других антиген связывающих белков. Также было показано, что замена даже одного эндогенного генного сегмента VH на генный сегмент VH человека может приводить к иммунному ответу, предусматривающему гуманизированный вариабельный домен иммуноглобулина. См., например, Tien et al. (2016) Cell 166:1471-84; включенную в данный документ посредством ссылки во всей своей полноте. Получение мышей VELOCIMMUNE® обеспечивает точная замена мышиных вариабельных последовательностей иммуноглобулина человеческими вариабельными последовательностями иммуноглобулина, таким образом, чтобы человеческие вариабельные последовательности иммуноглобулина являлись функционально связанными с эндогенной(-ыми) отличной(-ыми) от человеческой(-их) последовательностью(-ями) гена константной области обратным химерным способом.
[00217] Мыши, модифицированные обратным химерным способом, включают мышей, модифицированных таким образом, чтобы они содержали в эндогенном локусе иммуноглобулина человеческую(гуманизированную) вариабельную область (например, содержащую (D), J и один или более генных сегментов V человека), функционально связанную с эндогенной константной областью, например
(а) в эндогенном локусе тяжелой цепи:
(i) неперегруппированную человеческую (гуманизированную) вариабельную область тяжелой цепи иммуноглобулина в функциональной связи с эндогенной константной областью тяжелой цепи, при этом неперегруппированная человеческая (гуманизированная) вариабельная область тяжелой цепи иммуноглобулина содержит несколько неперегруппированных человеческих генных сегментов VH вариабельной области тяжелой цепи (например, все функциональные неперегруппированные человеческие генные сегменты VH), один или более неперегруппированных генных сегментов DH тяжелой цепи иммуноглобулина и один или более неперегруппированных генных сегментов JH тяжелой цепи иммуноглобулина,
при этом необязательно один или более неперегруппированных генных сегментов DH тяжелой цепи иммуноглобулина и один или более неперегруппированных генных сегментов JH тяжелой цепи иммуноглобулина представляют собой один или более неперегруппированных человеческих генных сегментов DH тяжелой цепи иммуноглобулина (например, все функциональные человеческие генные сегменты DH) и/или один или более неперегруппированных человеческих генных сегментов JH тяжелой цепи иммуноглобулина (например, все функциональные человеческие генные сегменты JH);
(ii) рестриктированную неперегруппированную человеческую (гуманизированную) вариабельную область тяжелой цепи в функциональной связи с эндогенной константной областью тяжелой цепи, при этом рестриктированная неперегруппированная человеческая (гуманизированная) вариабельная область тяжелой цепи состоит по сути из одного неперегруппированного человеческого генного сегмента VH вариабельной области тяжелой цепи, функционально связанного с одним или несколькими неперегруппированными генными сегментами DH тяжелой цепи иммуноглобулина и одним или несколькими неперегруппированными генными сегментами JH тяжелой цепи иммуноглобулина, при этом необязательно один или более неперегруппированных генных сегментов DH тяжелой цепи иммуноглобулина и один или более неперегруппированных генных сегментов JH тяжелой цепи иммуноглобулина представляют собой один или более неперегруппированных человеческих генных сегментов DH тяжелой цепи иммуноглобулина и/или один или более неперегруппированных человеческих генных сегментов JH тяжелой цепи иммуноглобулина соответственно;
(iii) модифицированную гистидином неперегруппированную человеческую (гуманизированную) вариабельную область тяжелой цепи в функциональной связи с эндогенной константной областью тяжелой цепи, при этом модифицированная гистидином неперегруппированная человеческая (гуманизированная) вариабельная область тяжелой цепи содержит неперегруппированную последовательность гена вариабельной области тяжелой цепи иммуноглобулина, содержащую в последовательности, кодирующей определяющую комплементарность область 3 (CDR3), замену по меньшей мере одного негистидинового кодона гистидиновым кодоном или вставку по меньшей мере одного гистидинового кодона; или
(iv) последовательность, кодирующую иммуноглобулин, состоящий только из тяжелых цепей, содержащую неперегруппированную человеческую (гуманизированную) вариабельную область тяжелой цепи в функциональной связи с эндогенной константной областью тяжелой цепи, при этом эндогенная константная область тяжелой цепи содержит (1) интактный эндогенной ген IgM, который кодирует изотип IgM, который соединяется с легкой цепью, и (2) отличный от IgM ген, например ген IgG, не содержащий последовательность, которая кодирует функциональный домен СН1, при этом отличный от IgM ген кодирует отличный от IgM изотип, не содержащий домен СН1, способный ковалентно связываться с константным доменом легкой цепи; и/или
(b) в эндогенном локусе легкой цепи:
(i) неперегруппированную человеческую (гуманизированную) вариабельную область легкой цепи иммуноглобулина в функциональной связи с эндогенной константной областью легкой цепи, при этом неперегруппированная человеческая (гуманизированная) вариабельная область легкой цепи иммуноглобулина содержит несколько неперегруппированных человеческих генных сегментов VL вариабельной области легкой цепи (например, все функциональные человеческие неперегруппированные генные сегменты VL человека) и один или более неперегруппированных генных сегментов JL легкой цепи иммуноглобулина,
при этом необязательно один или более неперегруппированных генных сегментов JL легкой цепи иммуноглобулина представляют собой один или более неперегруппированных человеческих генных сегментов JL легкой цепи иммуноглобулина (например, все функциональные человеческие генные сегменты JHL),
при этом необязательно эндогенный локус легкой цепи иммуноглобулина представляет собой эндогенный локус легкой каппа-цепи (κ) иммуноглобулина, неперегруппированная человеческая (гуманизированная) вариабельная область легкой цепи иммуноглобулина содержит человеческие вариабельные κ (Vκ) и соединяющие κ (Jκ) генные сегменты, и при этом эндогенная константная область легкой цепи представляет собой эндогенную последовательность константной области κ-цепи, и/или при этом эндогенный локус легкой цепи иммуноглобулина представляет собой эндогенную легкую лямбда-цепь (λ) иммуноглобулина, неперегруппированная человеческая (гуманизированная) вариабельная область легкой цепи иммуноглобулина содержит человеческие вариабельные генные сегменты λ (Vλ) и соединяющие генные сегменты λ (Jλ) и эндогенная константная область легкой цепи представляет собой эндогенную последовательность константной области λ-цепи, при этом необязательно эндогенный локус легкой λ-цепи иммуноглобулина содержит (а) один или более человеческих генных сегментов Vλ, (b) один или более человеческих генных сегментов Jλ и (с) один или более человеческих генных сегментов Cλ, при этом (а) и (b) функционально связаны с (с) и константным генным сегментом легкой цепи иммуноглобулина грызуна (Cλ), и при этом эндогенный локус легкой λ-цепи иммуноглобулина дополнительно содержит один или более энхансеров легкой λ-цепи иммуноглобулина грызуна (Eλ), и один или несколько энхансеров легкой λ-цепи (Eλ) иммуноглобулина человека, с необязательным содержанием трех человеческих Eλ;
(ii) последовательность, кодирующую общую легкую цепь, содержащую перегруппированную человеческую (гуманизированную) последовательность вариабельной области легкой цепи в функциональной связи с эндогенной константной областью легкой цепи, при этом перегруппированная человеческая (гуманизированная) последовательность вариабельной области легкой цепи содержит человеческий генный сегмент VL, перегруппированный с генным сегментом JL легкой цепи иммуноглобулина;
(iii) рестриктированную неперегруппированную человеческую (гуманизированную) вариабельную область легкой цепи в функциональной связи с эндогенной константной областью легкой цепи, при этом рестриктированная неперегруппированная человеческая (гуманизированная) вариабельная область легкой цепи содержит не более двух неперегруппированных человеческих вариабельных (VL) генных сегментов легкой цепи иммуноглобулина, функционально связанных с одним или несколькими неперегруппированными человеческими соединяющими генными сегментами легкой цепи иммуноглобулина (JL);
(iv) модифицированную гистидином неперегруппированную человеческую (гуманизированную) вариабельную область легкой цепи в функциональной связи с эндогенной константной областью легкой цепи, при этом модифицированная гистидином неперегруппированная человеческая (гуманизированная) вариабельная область легкой цепи содержит неперегруппированную человеческую (гуманизированную) последовательность гена вариабельной области легкой цепи иммуноглобулина, содержащую в последовательности, кодирующей определяющую комплементарность область 3 (CDR3), замену по меньшей мере одного негистидинового кодона гистидиновым кодоном или вставку по меньшей мере одного гистидинового кодона; или
(v) модифицированную гистидином перегруппированную человеческую (гуманизированную) вариабельную область легкой цепи в функциональной связи с эндогенной константной областью легкой цепи, при этом модифицированная гистидином перегруппированная человеческая (гуманизированная) вариабельная область легкой цепи содержит перегруппированную человеческую (гуманизированную) последовательность гена вариабельной области легкой цепи иммуноглобулина, содержащую в последовательности, кодирующей определяющую комплементарность область 3 (CDR3), замену по меньшей мере одного негистидинового кодона гистидиновым кодоном или вставку по меньшей мере одного гистидинового кодона,
при этом необязательно мышь дополнительно содержит
(i) локус человеческой (гуманизированной) тяжелой цепи иммуноглобулина, содержащий функциональный ген ADAM6, за счет чего мышь проявляет фертильность дикого типа животного, отличного от человека; и/или
(ii) экзогенный ген терминальной дезоксинуклеотидилтрансферазы (TdT) для обеспечения повышенной вариативности антигенного рецептора, необязательно такой, что по меньшей мере 10% перегруппированных генов вариабельной области содержат добавления, не соответствующие матрице, где указанные мыши были описаны ранее. См., например, патенты США №№8697940; 8754287; 9204624; 9334334; 9801362; 9332742 и 9516868; публикации заявок на патент США №№20110195454, 20120021409, 20120192300, 20130045492; 20150289489; 20180125043; 20180244804; публикации согласно РСТ №№WO2019/113065, WO2017210586 и WO2011163314; Lee et al. (2014) Nature Biotechnology 32:356, каждый из которых включен в данный документ посредством ссылки во всей своей полноте.
[00218] B некоторых вариантах осуществления настоящее изобретение включает генетически модифицированное животное, отличное от человека, чей геном, например геном зародышевой линии, содержит:
эндогенный локус иммуноглобулина, содержащий вариабельную область тяжелой цепи иммуноглобулина, содержащую генный сегмент VH человека, сконструированную DH-область человека по настоящему изобретению и генный сегмент JH человека, где указанная вариабельная область тяжелой цепи иммуноглобулина функционально связана с константной областью, и/или
эндогенный локус цепи, содержащий вариабельную область легкой цепи иммуноглобулина, содержащую генный сегмент VL человека и генные сегменты JL человека, где указанная вариабельная область легкой цепи иммуноглобулина функционально связана с константной областью.
[00219]B некоторых вариантах осуществления животное, отличное от человека, например грызун, например крыса или мышь, содержит в своем геноме замену одного или нескольких эндогенных сегментов VH, DH и JH в эндогенном локусе тяжелой цепи иммуноглобулина одним или более сегментами VH, DH и JH человека, где указанные один или более сегментов VH, DH и JH человека содержат генный сегмент DH человека, функционально связанный с 23-мерной RSS, и функционально связаны с эндогенным геном тяжелой цепи иммуноглобулина; и необязательно неперегруппированный или перегруппированный человеческий сегмент VL и JL, функционально связанный с геном константной области легкой цепи (CL) иммуноглобулина, отличного от человеческого, например грызуна, например мыши или крысы, или человека, например, в эндогенном локусе легкой цепи, отличной от человеческой.
[00220] B определенных вариантах осуществления генетически модифицированные животные, отличные от человека содержат в своем геноме, например, например, в геноме зародышевой линии, локус (экзогенный или эндогенный) иммуноглобулина, содержащий вариабельную область иммуноглобулина, содержащую один или более неперегруппированных генных сегментов вариабельной области иммуноглобулина человека, содержащих сконструированную DH-область и константную область иммуноглобулина, содержащую ген константной области иммуноглобулина, и в котором один или более неперегруппированных генных сегментов вариабельной области иммуноглобулина человека функционально связаны с геном константной области иммуноглобулина.
[00221] Как правило, генетически модифицированный локус иммуноглобулина содержит вариабельную область иммуноглобулина (содержащую генные сегменты вариабельной области иммуноглобулина), функционально связанную с константной областью иммуноглобулина. В некоторых вариантах осуществления генетически модифицированный локус иммуноглобулина содержит один или более неперегруппированных генных сегментов вариабельной области тяжелой цепи иммуноглобулина человека, содержащих сконструированную DH-область, функционально связанных с геном тяжелой цепи константной области. В некоторых вариантах осуществления генетически модифицированный локус иммуноглобулина содержит неперегруппированные генные сегменты вариабельной κ-области иммуноглобулина человека, функционально связанные с геном константной области κ-цепи. В некоторых вариантах осуществления генетически модифицированный локус иммуноглобулина содержит неперегруппированные генные сегменты вариабельной области λ иммуноглобулина человека, функционально связанные с геном константной области κ-цепи. В некоторых вариантах осуществления генетически модифицированный локус иммуноглобулина содержит неперегруппированные генные сегменты вариабельной области λ иммуноглобулина человека, функционально связанные с геном константной области λ-цепи.
[00222] B определенных вариантах осуществления животное, отличное от человека, содержит в эндогенном локусе тяжелой цепи неперегруппированную человеческую (гуманизированную) вариабельную область тяжелой цепи иммуноглобулина, содержащую сконструированную DH-область, в функциональной связи с эндогенной константной областью тяжелой цепи, при этом вариабельная область иммуноглобулина содержит один или более неперегруппированных генных сегментов вариабельной области тяжелой цепи Ig человека. В некоторых вариантах осуществления один или более неперегруппированных генных сегментов вариабельной области Ig человека содержат по меньшей мере один вариабельный сегмент тяжелой цепи (VH) иммуноглобулина человека, один или более вариативных сегментов тяжелой цепи иммуноглобулина (DH) (например, один или более неперегруппированных сегментов тяжелой цепи иммуноглобулина человека DH, функционально связанных с 23-мерной RSS), и один или более соединяющих сегментов тяжелой цепи иммуноглобулина (JH) (необязательно один или более неперегруппированных сегментов JH человека). В некоторых вариантах осуществления неперегруппированные генные сегменты вариабельной области Ig человека содержат несколько неперегруппированных сегментов VH человека, один или более неперегруппированных сегментов DH (человека) (например, один или более неперегруппированных сегментов DH (человека), функционально связанных с 23-мерной RSS), и один или более неперегруппированных сегментов JH (человека). В некоторых вариантах осуществления неперегруппированные генные сегменты вариабельной области Ig человека содержат по меньшей мере 3 генных сегмента VH, по меньшей мере 18 генных сегментов VH, по меньшей мере 20 генных сегментов VH, по меньшей мере 30 генных сегментов VH, по меньшей мере 40 генных сегментов VH, по меньшей мере 50 генных сегментов VH, по меньшей мере 60 генных сегментов VH, по меньшей мере 70 генных сегментов VH или по меньшей мере 80 генных сегментов VH. В некоторых вариантах осуществления неперегруппированные генные сегменты Ig человека содержат все из функциональных генных сегментов DH человека, где по меньшей мере один из функциональных генных сегментов DH человека модифицируют таким образом, чтобы он был функционально связан с 23-мерной RSS. В некоторых вариантах осуществления неперегруппированные генные сегменты Ig человека содержат все из функциональных генных сегментов JH человека. Иллюстративные вариабельные области, содержащие генные сегменты тяжелой цепи Ig, представлены, например, в Macdonald et al, Proc. Natl. Acad. Sci. USA 111:5147-52 и в дополнительной информации, которые включены в данный документ посредством ссылки во всей своей полноте.
[00223] B некоторых вариантах осуществления животные, отличные от человека, предусмотренные в данном документе, содержат в эндогенном локусе тяжелой цепи рестриктированную неперегруппированную человеческую (гуманизированную) вариабельную область тяжелой цепи в функциональной связи с эндогенной константной областью тяжелой цепи, содержащей по меньшей мере ген IgM, отличный от человеческого, при этом рестриктированная неперегруппированная человеческая (гуманизированная) вариабельная область тяжелой цепи характеризуется наличием единственного генного сегмента VH человека, нескольких генных сегментов DH (например, генных сегментов DH человека, содержащих один или более неперегруппированный сегментов DH (человека), функционально связанных с 23-мерной RSS), и несколько генных сегментов JH (например геннных сегментов JH человека), при этом рестриктированный локус тяжелой цепи иммуноглобулина способен перегруппировываться и образовывать несколько отдельных перегруппировок, при этом каждая перегруппировка получена из единственного генного сегмента VH человека, одного из сегментов DH и одного из сегментов JH, и при этом каждая перегруппировка кодирует отличающийся вариабельный домен тяжелой цепи (например, как описано в публикации заявки на патент США №20130096287, которая включена в данный документ посредством ссылки во всей своей полноте). В некоторых вариантах осуществления единственный генный сегмент VH человека представляет собой VH1-2 или VH1-69.
[00224] B определенных вариантах осуществления животное, отличное от человека, содержит в эндогенном локусе легкой цепи неперегруппированную человеческую (гуманизированную) вариабельную область легкой цепи иммуноглобулина в функциональной связи с эндогенной константной областью легкой цепи. В некоторых вариантах осуществления неперегруппированная человеческая (гуманизированная) вариабельная область легкой цепи иммуноглобулина содержит неперегруппированные генные сегменты вариабельной κ-области Ig человека. В некоторых вариантах осуществления неперегруппированная (гуманизированная) вариабельная область иммуноглобулина человека содержит несколько неперегруппированных сегментов Vκ человека и один или более неперегруппированных сегментов JK человека. В некоторых вариантах осуществления неперегруппированные генные сегменты вариабельной области иммуноглобулина человека включают все из сегментов Jκ человека. В некоторых вариантах осуществления генные сегменты вариабельной области иммуноглобулина содержат четыре функциональных сегмента Vκ все из сегментов Jκ человека. В некоторых вариантах осуществления генные сегменты вариабельной области иммуноглобулина содержат 16 функциональных сегментов Vκ и все сегменты Jκ человека (например, все функциональные сегменты Vκ и сегменты Jκ человека). В некоторых вариантах осуществления неперегруппированные генные сегменты вариабельной области иммуноглобулина человека включают все из сегментов Vκ человека и все сегменты Jκ человека. Иллюстративные вариабельные области, содержащие генные сегменты κ Ig, представлены, например, в Macdonald et al., Proc. Natl. Acad. Sci. USA 111:5147-52 и в дополнительной информации, которые включены в данный документ посредством ссылки во всей своей полноте.
[00225] B некоторых вариантах осуществления рестриктированная неперегруппированная человеческая (гуманизированная) вариабельная область легкой цепи в функциональной связи с эндогенной константной областью легкой цепи характеризуется тем, что неперегруппированная человеческая (гуманизированная) вариабельная область легкой цепи содержит не более двух генных сегментов VL человека и несколько генных сегментов JL (например, двойную легкую цепь мыши или DLC, как описано в патенте США №9796788, который включен в данный документ посредством ссылки во всей своей полноте). В некоторых вариантах осуществления генные сегменты VL представляют собой генные сегменты Vκ. В некоторых вариантах осуществления генные сегменты VL представляют собой генные сегменты Vλ. В некоторых вариантах осуществления генные сегменты Vκ представляют собой генные сегменты IGKV3-20 и IGKV1-39. В некоторых вариантах осуществления животное, отличное от человека, содержит ровно два неперегруппированных генных сегмента Vκ человека и пять неперегруппированных генных сегмента Jκ человека, функционально связанных с мышиной константной областью легкой цепи, в эндогенных локусах легкой κ-цепи мыши, при этом необязательно ровно два неперегруппированных генных сегмента Vκ человека представляют собой генный сегмент Vκ1-39 человека и генный сегмент Vκ3-20 человека, при этом пять неперегруппированных генных сегмента Jκ человека представляют собой генный сегмент Jκ1 человека, генный сегмент Jκ2 человека, генный сегмент Jκ3 человека, генный сегмент Jκ4 человека и генный сегмент Jκ5 человека, при этом неперегруппированные генные сегменты легкой каппа-цепи человека способны перегруппировываться и кодировать вариабельные домены антитела человека, и, кроме того, при этом необязательно животное, отличное от человека, не содержит эндогенный генный сегмент Vκ, который способен перегруппировываться с образованием вариабельной области легкой цепи иммуноглобулина.
[00226] В определенных вариантах осуществления неперегруппированная человеческая (гуманизированная) вариабельная область легкой цепи иммуноглобулина в функциональной связи с эндогенной константной областью легкой цепи содержит неперегруппированные генные сегменты вариабельной области Igλ человека. В некоторых вариантах осуществления неперегруппированные генные сегменты вариабельной области иммуноглобулина человека включают несколько сегментов Vλ человека и один или более сегментов Jλ человека. В некотором варианте осуществления неперегруппированные генные сегменты вариабельной области иммуноглобулина человека включают один или более сегментов Vλ человека, один или более сегментов Jλ человека и одну или более последовательностей константной области Сλ человека. В некоторых вариантах осуществления неперегруппированные генные сегменты вариабельной области человека содержат все из сегментов Vλ человека. В некоторых вариантах осуществления неперегруппированные генные сегменты вариабельной области человека содержат все из сегментов Jλ человека. Иллюстративные вариабельные области, содержащие генные сегменты к Ig, представлены, например, в патентах США №№9035128 и 6998514, каждый из которых включен в данный документ посредством ссылки во всей своей полноте. В некоторых вариантах осуществления неперегруппированная человеческая (гуманизированная) вариабельная область легкой цепи иммуноглобулина в функциональной связи с эндогенной константной областью легкой цепи содержит (а) один или более генных сегментов Vλ человека, (b) один или более генных сегментов Jλ человека и (с) один или более генных сегментов Сλ человека, при этом (а) и (b) функционально связаны с (с) и эндогенным (например, грызуна) генным сегментом Сλ, и при этом эндогенный локус легкой λ-цепи иммуноглобулина дополнительно содержит один или более энхансеров легкой λ-цепи иммуноглобулина (Eλ) грызуна и один или более энхансеров легкой λ-цепи иммуноглобулина человека (Еλ), с необязательным содержанием трех Еλ человека.
[00227] B определенных вариантах осуществления неперегруппированная человеческая (гуманизированная) вариабельная область легкой цепи иммуноглобулина в функциональной связи с эндогенной константной областью легкой цепи содержит неперегруппированные генные сегменты вариабельной области Igλ человека, функционально связанные с эндогенным (например, грызуна, например крысы или мыши) геном Сκ, так что у животного, отличного от человека, экспрессируется легкая цепь иммуноглобулина, которая содержит человеческую последовательность вариабельного домена κ, полученную из генных сегментов Vλ и Jλ, слитых с эндогенным константным κ-доменом, см., например, патент США №9226484, включенный в данный документ посредством ссылки во всей своей полноте.
[00228] B некоторых вариантах осуществления вариабельная область иммуноглобулина, содержащая неперегруппированные генные сегменты вариабельной области иммуноглобулина человека, также содержит межгенные последовательности вариабельной области иммуноглобулина человека. В некоторых вариантах осуществления вариабельная область иммуноглобулина содержит межгенные последовательности вариабельной области Ig, отличного от человеческого (например, грызуна, крысы, мыши). В некоторых вариантах осуществления межгенная последовательность имеет эндогенное видовое происхождение.
[00229] B некоторых вариантах осуществления вариабельная область иммуноглобулина представляет собой перегруппированную вариабельную область легкой цепи (универсальную вариабельную область легкой цепи). В некоторых вариантах осуществления перегруппированный ген вариабельной области легкой цепи Ig представляет собой перегруппированный ген вариабельной области легкой цепи Ig человека. Иллюстративные перегруппированные вариабельные области легкой цепи Ig представлены, например, в патентах США №№9969814; 10130181 и 10143186, и публикациях заявок на патент США №№20120021409, 20120192300, 20130045492, 20130185821, 20130302836 и 20150313193, каждая из которых включена в данный документ посредством ссылки во всей своей полноте. В некоторых вариантах осуществления для получения биспецифических антител применяют организм, отличный от человеческого (организм с «универсальной легкой цепью»), содержащий универсальную вариабельную область легкой цепи. В некоторых вариантах осуществления последовательность, кодирующая общую легкую цепь, содержит одну перегруппированную человеческую последовательность Vκ/Jκ легкой цепи иммуноглобулина, функционально связанную с эндогенной константной областью легкой цепи, при этом одна перегруппированная человеческая последовательность Vκ/Jκ легкой цепи иммуноглобулина представляет собой либо (i) последовательность Vκ1-39/Jκ5 человека, содержащую генный сегмент Vκ1-39 человека, слитый с генным сегментом Jκ5 человека, либо (ii) последовательность Vκ3-20/Jκ1 человека, содержащую генный сегмент Vκ3-20 человека, слитый с генным сегментом Jκ1 человека.
[00230] B некоторых вариантах осуществления вариабельная область иммуноглобулина представляет собой вариабельную область легкой цепи и/или тяжелой цепи иммуноглобулина, которая содержит вставки и/или замены гистидиновых кодонов, предназначенных для придания рН-зависимых свойств связывания антителам, вырабатываемым в таком организме, отличном от человеческого. В некоторых из таких вариантов осуществления гистидиновые кодоны вставляют и/или вставляют посредством замещения в последовательности нуклеиновой кислоты, кодирующие CDR3. Различные такие локусы легкой и/или тяжелой цепи иммуноглобулина представлены в патентах США №№9301510; 9334334 и 9801362 и в публикации заявки на патент США №20140013456, каждый из которых включен в данный документ посредством ссылки во всей своей полноте. В некоторых вариантах осуществления модифицированная гистидином перегруппированная человеческая (гуманизированная) вариабельная область легкой цепи в функциональной связи с эндогенной константной областью легкой цепи содержит одну перегруппированную человеческую последовательность гена вариабельной области легкой цепи иммуноглобулина, содержащую последовательности сегментов Vκ и Jκ человека, при этом необязательно последовательность сегмента Vκ получена из генного сегмента Vκ1-39 или Vκ3-20 человека, и при этом одна перегруппированная человеческая последовательность гена вариабельной области легкой цепи иммуноглобулина содержит замену по меньшей мере одного негистидинового кодона последовательности сегмента Vκ гистидиновым кодоном, который экспрессируется в положении, выбранном из группы, состоящей из 105, 106, 107, 108, 109, 111 и их комбинации (согласно нумерации IMGT). В некоторых вариантах осуществления модифицированная гистидином неперегруппированная человеческая (гуманизированная) вариабельная область тяжелой цепи в функциональной связи с эндогенной константной областью тяжелой цепи содержит неперегруппированную человеческую (гуманизированную) последовательность гена вариабельной области тяжелой цепи иммуноглобулина, содержащую в последовательности, кодирующей определяющую комплементарность область 3 (CDR3) (например, в (человеческом) генном сегменте DH, модифицированном таким образом, чтобы быть функционально связанным с 23-мерной RSS), замену по меньшей мере одного негистидинового код она гистидиновым кодоном или вставку по меньшей мере одного гистидинового кодона. В некоторых вариантах осуществления неперегруппированная человеческая (гуманизированная) последовательность гена вариабельной области тяжелой цепи иммуноглобулина содержит следующие генные сегменты: неперегруппированный генный сегмент VH человека, неперегруппированный генный сегмент DH человека или синтетический генный сегмент DH, содержащий генный сегмент DH, функционально связанный с 23-мерной RSS, и неперегруппированный генный сегмент JH человека, где необязательно неперегруппированный генный сегмент DH человека, или синтетический генный сегмент DH, или генный сегмент DH, функционально связанный с 23-мерной RSS, содержит замену по меньшей мере одного негистидинового кодона гистидиновым кодоном или вставку по меньшей мере одного гистидинового кодона. В некоторых вариантах осуществления модифицированная гистидином неперегруппированная человеческая (гуманизированная) вариабельная область легкой цепи в функциональной связи с эндогенной константной областью тяжелой цепи содержит неперегруппированный генный сегмент VL и неперегруппированный генный сегмент JL. В некоторых вариантах осуществления модифицированная гистидином неперегруппированная человеческая (гуманизированная) вариабельная область легкой цепи содержит не более двух неперегруппированных генных сегментов VL человека (например, не более двух генных сегментов Vκ) и один или более неперегруппированных генных сегментов JL человека (например, Jκ), при этом каждый из не более двух генных сегментов VL человека содержит в последовательности, кодирующей CDR3, замену по меньшей мере одного негистидинового кодона гистидиновым кодоном или вставку по меньшей мере одного гистидинового кодона. В некоторых вариантах осуществления не более двух неперегруппированных генных сегментов Vκ человека представляют собой генные сегменты Vκ1-39 и Vκ3-20 человека, при этом каждый содержит одну или несколько замен негистидинового кодона гистидиновым кодоном, и при этом генные сегменты Vκ и Jκ человека способны перегруппировываться, и генные сегменты Vκ и Jκ человека кодируют человеческий вариабельный домен легкой цепи, содержащий один или более остатков гистидина в положении, выбранном из группы, состоящей из 105, 106, 107, 108, 109, 111 (согласно нумерации IGMT) и их комбинации, при этом один или более остатков гистидина получают в результате одной или более замен.
[00231] B некоторых вариантах осуществления константная область иммуноглобулина предусматривает ген константной области тяжелой цепи. В некоторых вариантах осуществления ген константной области тяжелой цепи представляет собой ген константной области тяжелой цепи человека. В некоторых вариантах осуществления ген константной области тяжелой цепи имеет эндогенное видовое происхождение. В некоторых вариантах осуществления ген константной области тяжелой цепи представляет собой ген константной области мыши или ген константной области крысы. В некоторых вариантах осуществления ген константной области представляет собой смесь человеческой последовательности и последовательности, отличной от человеческой. Например, в некоторых вариантах осуществления ген константной области кодирует область СН1 человека и область СН2 и/или СН3, отличную от человеческой (например, эндогенного видового происхождения, мыши, крысы). В некоторых вариантах осуществления ген константной области тяжелой цепи представляет собой ген константной области Cμ, Сδ, Сγ (Cγ1, Сγ2, Сγ3, Сγ4), Сα или Сε. В некоторых вариантах осуществления ген константной области представляет собой эндогенный ген константной области. В некоторых вариантах осуществления ген константной области кодирует мутантную область СН1 таким образом, что у животного, отличного от человека, экспрессируются антитела, состоящие только из тяжелых цепей (см., например, патент США №8754287, публикацию заявки на патент США №2015/0289489, каждый из которых включен в данный документ посредством ссылки во всей своей полноте). В некоторых вариантах осуществления, например, где цель заключается в выработке тяжелых цепей для получения биспецифических антител (например, в организмах с универсальной или двойной легкой цепью), Fc-домены тяжелых цепей содержат модификации для облегчения образования гетеродимера тяжелых цепей и/или для подавления образования гомодимера тяжелых цепей. Такие модификации представлены, например, в патентах США №№5731168; 5807706; 5821333; 7642228 и 8679785 и в публикации заявки на патент США №2013/0195849, каждый из которых включен в данный документ посредством ссылки во всей своей полноте.
[00232] B некоторых вариантах осуществления константная область иммуноглобулина предусматривает ген константной области легкой цепи. В некоторых вариантах осуществления ген константной области легкой цепи представляет собой ген константной области κ. В некоторых вариантах осуществления ген константной области легкой цепи представляет собой ген константной области λ. В некоторых вариантах осуществления ген константной области легкой цепи имеет эндогенное видовое происхождение. В некоторых вариантах осуществления ген константной области легкой цепи представляет собой ген константной области мыши или ген константной области крысы. В некоторых вариантах осуществления ген константной области легкой цепи представляет собой смесь человеческой последовательности и последовательности, отличной от человеческой.
[00233] B некоторых вариантах осуществления вариабельная область иммуноглобулина предусматривает генные сегменты вариабельной области человека и ген константной области иммуноглобулина, с которым функционально связаны генные сегменты вариабельной области, расположенные в эндогенном локусе иммуноглобулина. В некоторых вариантах осуществления эндогенный локус иммуноглобулина представляет собой эндогенный локус тяжелой цепи. В некоторых вариантах осуществления эндогенный локус иммуноглобулина представляет собой эндогенный локус κ. В некоторых вариантах осуществления эндогенный локус иммуноглобулина представляет собой эндогенный локус λ. В некоторых вариантах осуществления ген константной области, с которым функционально связаны генные сегменты вариабельной области человека, представляет собой эндогенный ген константной области.
[00234] B некоторых вариантах осуществления один или более эндогенных локусов иммуноглобулина или часть одного или более эндогенных локусов (например, вариабельная область и/или константная область) в геноме животного, отличного от человека, предусмотренного в данном документе, являются инактивированными. Эндогенные локусы гена вариабельной области иммуноглобулина и их части могут быть инактивированы с применением любого способа, известного из уровня техники, в том числе без ограничения делеции локуса или его части из генома организма, замены локуса или его части отличающейся последовательностью нуклеиновой кислоты, инверсии части локуса и/или перемещения части локуса в другое положение в геноме организма, отличного от человеческого. В некоторых вариантах осуществления инактивация локуса является только частичной инактивацией. В некоторых вариантах осуществления локус вариабельной области инактивирован, но константная область остается функциональной (например, поскольку она функционально связана с генными сегментами вариабельной области, отличными от эндогенных).
[00235] B некоторых вариантах осуществления генетически модифицированное животное, отличное от человека, содержит инактивированный эндогенный локус тяжелой цепи иммуноглобулина. В некоторых вариантах осуществления эндогенный локус тяжелой цепи иммуноглобулина или его часть инактивированы с помощью делеции, замены, перемещения и/или инверсии по меньшей мере части эндогенной вариабельной области эндогенного локуса тяжелой цепи. В некоторых вариантах осуществления по меньшей мере часть вариабельной области эндогенного локуса тяжелой цепи, которая подвергается делеции, замене, перемещению и/или инверсии, содержит сегменты J вариабельной области. В некоторых вариантах осуществления эндогенный локус тяжелой цепи иммуноглобулина или его часть инактивированы с помощью делеции, замены, перемещения и/или инверсии по меньшей мере части эндогенной константной области эндогенного локуса тяжелой цепи. В некоторых вариантах осуществления по меньшей мере часть константной области эндогенного локуса тяжелой цепи, которая подвергается делеции, замене, перемещению и/или инверсии, содержит ген Oμ эндогенной константной области.
[00236] B некоторых вариантах осуществления генетически модифицированное животное, отличное от человека, содержит инактивированный эндогенный локус κ-цепи иммуноглобулина. В некоторых вариантах осуществления эндогенный локус κ-цепи иммуноглобулина или его часть инактивированы с помощью делеции, замены, перемещения и/или инверсии по меньшей мере части эндогенной вариабельной области эндогенного локуса κ-цепи. В некоторых вариантах осуществления по меньшей мере часть вариабельной области эндогенного локуса κ-цепи, которая подвергается делеции, замене, перемещению и/или инверсии, содержит сегменты J вариабельной области. В некоторых вариантах осуществления эндогенный локус κ-цепи иммуноглобулина или его часть инактивированы с помощью делеции, замены, перемещения и/или инверсии по меньшей мере части эндогенной константной области эндогенного локуса κ-цепи. В некоторых вариантах осуществления по меньшей мере часть константной области эндогенного локуса κ-цепи, которая подвергается делеции, замене, перемещению и/или инверсии, содержит ген Сκ эндогенной константной области.
[00237] B некоторых вариантах осуществления генетически модифицированное животное, отличное от человека, содержит инактивированный эндогенный локус λ-цепи иммуноглобулина. В некоторых вариантах осуществления эндогенный локус λ-цепи иммуноглобулина или его часть инактивированы с помощью делеции, замены, перемещения и/или инверсии по меньшей мере части эндогенной вариабельной области эндогенного локуса λ-цепи. В некоторых вариантах осуществления по меньшей мере часть по меньшей мере одного генного кластера V-J-C в эндогенном локусе λ-цепи подвергается делеции, замене, перемещению и/или инверсии. В некоторых вариантах осуществления эндогенный локус λ-цепи иммуноглобулина или его часть инактивированы с помощью делеции, замены, перемещения и/или инверсии по меньшей мере части эндогенной константной области эндогенного локуса λ-цепи. В некоторых вариантах осуществления по меньшей мере часть константной области эндогенного локуса λ-цепи, которая подвергается делеции, замене, перемещению и/или инверсии, содержит ген С эндогенной константной области.
[00238] B различных вариантах осуществления модификации локуса иммуноглобулина не влияют на фертильность животного, отличного от человека. В некоторых вариантах осуществления локус тяжелой цепи содержит функциональный, например, эндогенный ген ADAM6a, ген ADAM6b или оба, и генетическая модификация не влияет на экспрессию и/или функцию эндогенного гена ADAM6a, гена ADAM6b или обоих. В некоторых вариантах осуществления геном генетически модифицированного животного, отличного от человека, дополнительно содержит эктопически расположенный функциональный, например, эндогенный ген ADAM6a, ген ADAM6b или оба. Иллюстративные животные, отличные от человека, экспрессирующие экзогенный ADAM6a и/или ADAM6b, описаны в патентах США №№8642835 и 8697940, каждый из которых включен в данный документ посредством ссылки во всей своей полноте.
[00239] B некоторых вариантах осуществления генетически модифицированное животное, отличное от человека, дополнительно содержит и экспрессирует экзогенную терминальную дезоксинуклеотидилтрансферазу (TdT) для обеспечения повышенной вариативности антигенного рецептора. Иллюстративные животные, отличные от человека, экспрессирующие экзогенный TdT, описаны в публикации согласно РСТ WO 2017210586, которая включена в данный документ посредством ссылки во всей своей полноте.
[00240] B некоторых вариантах осуществления геном предусмотренного животного, отличного от человека, дополнительно содержит один или более генов тяжелой и/или легкой цепей иммуноглобулина человека (см., например, патент США №8502018; патент США №8642835; патент США №8697940; патент США №8791323 и публикации заявок на патент США №2013/0096287 А1 и 2018/0125043 А1 и публикацию согласно РСТ № WO 2019/113065, каждая из которых включена в данный документ посредством ссылки во всей своей полноте). В качестве альтернативы, сконструированную DH-область можно вводить в эмбриональную стволовую клетку другой модифицированной линии, такой как, например, линия VELOCIMMUNE® (см., например, патент США №8502018 или патент США №8642835; включенные в данный документ посредством ссылки во всей своей полноте). В некоторых вариантах осуществления животных, отличных от человека, описанных в данном документе, можно получить путем введения целенаправленно воздействующего вектора, описанного в данном документе, в клетку из модифицированной линии. В качестве лишь одного примера, нацеливающий вектор, описанный в данном документе, может быть введен в животное, отличное от человека, описанное в патентах США №№8642835 и 8697940; включенных в данный документ посредством ссылки во всей своей полноте, где указанное животное, отличное от человека, экспрессирует антитела, которые содержат полностью человеческие вариабельные области и константные области мыши. В некоторых вариантах осуществления животных, отличных от человека, описанных в данном документе, получают таким образом, чтобы они дополнительно содержали гены иммуноглобулина человека (гены вариабельной и/или константной области). В некоторых вариантах осуществления животные, отличные от человека, описанные в данном документе, содержат сконструированную DH-область, описанную в данном документе, и генетический материал от гетерологичного вида (например, человека), где генетический материал кодирует, полностью или частично, один или более вариабельных участков тяжелой и/или легкой цепей человека.
[00241] Животных, отличных от человека, описанных в данном документе, можно получить, как описано выше, или с использованием способов, известных из уровня техники, таким образом, чтобы они содержали дополнительные человеческие или гуманизированные гены, зачастую в зависимости от предполагаемого применения животного, отличного от человека. Генетический материал таких дополнительных человеческих или гуманизированных генов можно вводить посредством дальнейшего изменения генома клеток (например, эмбриональных стволовых клеток), имеющих генетические модификации, описанные выше, или при необходимости с помощью методик скрещивания, известных из уровня техники, с другими генетически модифицированными линиями, которые описаны.
[00242] Например, как описано в данном документе, животные, отличные от человека, содержащие сконструированную DH-область, могут дополнительно содержать (например, посредством применения стратегий кроссбридинга или множественного генного нацеливания) одну или более модификаций, описанных в публикациях заявок на патент США №№2011-0195454 А1, 2012-0021409 А1, 2012-0192300 А1, 2013-0045492 А1, 2013-0185821 А1, 2013-0198880 А1, 2013-0302836 А1, 2015-0059009 А1; публикациях международных заявок на патент №№ WO 2011/097603, WO 2012/148873, WO 2013/134263, WO 2013/184761, WO 2014/160179, WO 2014/160202; все из которых включены в данный документ посредством ссылки во всей своей полноте.
[00243] Трансгенное животное-родоначальник, отличное от человека, может быть идентифицировано на основании наличия сконструированной DH-области в его геноме и/или экспрессии антител, которые содержат CDR3-области, содержащие аминокислоты, полученные в результате DH-DH-рекомбинации в тканях или клетках животного, отличного от человека. Трансгенное животное-родоначальник, отличное от человека, затем может применяться для разведения дополнительных животных, отличных от человека, несущих сконструированную DH-область, таким образом получая серию животных, отличных от человека, при этом каждое несет одну или более копий сконструированной DH-области. Более того, трансгенных животных, отличных от человека, несущих сконструированную DH-область, при необходимости, можно дополнительно скрещивать с другими трансгенными животными, отличными от человека, несущими другие трансгены (например, гены иммуноглобулина человека), которые описаны.
[00244] Можно также получать трансгенных животных, отличных от человека, содержащих выбранные системы, которые позволяют регулировать экспрессию трансгена или управлять ею. Иллюстративные системы содержат систему рекомбиназы Cre/loxP из бактериофага Р1 (см., например, Lakso, M. et al., 1992, Proc. Natl. Acad. Sci. USA 89:6232-6236, включенный посредством ссылки во всей своей полноте) и систему рекомбиназы FLP/Frt из S. cerevisiae (O'Gorman, S. et al, 1991, Science 251:1351-1355; каждый из которых включен посредством ссылки во всей своей полноте). Таких животных можно получать путем конструирования «двойных» трансгенных животных, например, путем спаривания двух трансгенных животных, одно из которых содержит трансген, содержащий выбранную модификацию (например, сконструированную DH-область), а другое содержит трансген, кодирующий рекомбиназу (например, рекомбиназу Cre).
[00245] Хотя в данном документе подробно обсуждаются варианты осуществления, в которых сконструированная dh-область используется в мыши (т.е. мышь со сконструированной DH-областью, функционально связанной с генными сегментами VH и JH человека, все из которых функционально связаны с одним или более генами константной области тяжелой цепи мыши), также предусмотрены другие животные, отличные от человека, которые содержат сконструированную DH-область. В некоторых вариантах осуществления, такие животные, отличные от человека, содержат сконструированную DH-область, функционально связанную с эндогенными генными сегментами VH и JH. В некоторых вариантах осуществления, такие животные, отличные от человека, содержат сконструированную DH-область, функционально связанную с гуманизированными генными сегментами VH и JH. Такие животные, отличные от человека, подразумевают любых животных, которые могут быть генетически модифицированы таким образом, чтобы экспрессировать антитела, содержащие CDR3, которые содержат аминокислоты, полученные в результате DH-DH-рекомбинации с повышенной частотой по сравнению с локусом тяжелой цепи иммуноглобулина дикого типа, как раскрыто в данном документе, включающих, например, млекопитающих, например мышь, крысу, кролика, свинью, животных подсемейства бычьих (например, корову, быка, буйвола), оленя, овцу, козу, курицу, кошку, собаку, хорька, примата (например, игрунку, макаку-резуса) и т.д. Например, для тех животных, отличных от человека, для которых подходящие генетически модифицируемые ES-клетки являются труднодоступными, используют другие способы для получения животного, отличного от человека, содержащего генетическую модификацию. Такие способы предусматривают, например, модификацию генома клеток, отличных от клеток ES (например, фибробласта или индуцированной плюрипотентной клетки), и использование переноса ядер соматических клеток (SCNT) для переноса генетически модифицированного генома в подходящую клетку, например, в энуклеированный ооцит, и вынашивание модифицированной клетки (например, модифицированного ооцита) животным, отличным от человека, в подходящих условиях с образованием эмбриона.
[00246] Способы модификации генома животного, отличного от человека (например, свиньи, коровы, грызуна, курицы и т.д.), включают, например, использование нуклеазы с «цинковыми пальцами» (ZFN), эффекторной нуклеазы, подобной активатору транскрипции (TALEN), или Cas-белка (т.е. системы CRISPR/Cas) для модификации генома таким образом, чтобы он содержал сконструированную DH-область, описанную в данном документе. Руководство по способам модификации генома зародышевой линии животного, отличного от человека, можно найти, например, в публикациях заявок на патент США №№2015-0376628 А1, US 2016-0145646 А1 и US 2016-0177339 А1; включенных в данный документ посредством ссылки во всей своей полноте.
[00247] B некоторых вариантах осуществления животное, отличное от человека, описанное в данном документе, представляет собой млекопитающее. В некоторых вариантах осуществления животное, отличное от человека, описанное в данном документе, представляет собой мелкое млекопитающее, например млекопитающее надсемейства Dipodoidea или Muroidea. В некоторых вариантах осуществления генетически модифицированное животное, описанное в данном документе, представляет собой грызуна. В некоторых вариантах осуществления грызун, описанный в данном документе, выбран из мыши, крысы и хомяка. В некоторых вариантах осуществления грызун, описанный в данном документе, выбран из животных надсемейства Muroidea. В некоторых вариантах осуществления генетически модифицированное животное, описанное в данном документе, относится к семейству, выбранному из Calomyscidae (например, мышевидные хомячки), Cricetidae (например, хомяки, крысы и мыши Нового света, полевки), Muridae (настоящие мыши и крысы, карликовые песчанки, иглистые мыши, косматые хомяки), Nesomyidae (лазающие мыши, скальные мыши, белохвостые крысы, малагасийские крысы и мыши), Platacanthomyidae (например, колючие сони) и Spalacidae (например, слепыши, бамбуковые крысы и цокоры). В некоторых вариантах осуществления генетически модифицированный грызун, описанный в данном документе, выбран из настоящей мыши или крысы (семейство Muridae), песчанки, иглистой мыши и косматого хомяка. В некоторых вариантах осуществления генетически модифицированная мышь, описанная в данном документе, является представителем семейства Muridae. В некотором варианте осуществления животное, отличное от человека, описанное в данном документе, представляет собой грызуна. В некоторых вариантах осуществления грызун, описанный в данном документе, выбран из мыши и крысы. В некоторых вариантах осуществления животное, отличное от человека, описанное в данном документе, представляет собой мышь.
[00248] B некоторых вариантах осуществления животное, отличное от человека, описанное в данном документе, представляет собой грызуна, который представляет собой мышь линии C57BL, выбранной из C57BL/A, C57BL/An, C57BL/GrFa, C57BL/KaLwN, C57BL/6, C57BL/6J, C57BL/6ByJ, C57BL/6NJ, C57BL/10, C57BL/10ScSn, C57BL/10Cr и C57BL/Ola. В некоторых вариантах осуществления мышь по настоящему изобретению представляет собой мышь из линии 129, выбранной из группы, состоящей из линий, которые представляют собой 129Р1, 129Р2, 129Р3, 129X1, 129S1 (например, 129S1/SV, 129S1/SvIm), 129S2, 129S4, 129S5, 129S9/SvEvH, 129/SvJae, 129S6 (129/SvEvTac), 129S7, 129S8, 129T1, 129T2 (см., например, Festing et al., 1999, Mammalian Genome 10:836; Auerbach, W. et al., 2000, Biotechniques 29(5): 1024-1028, 1030, 1032; включенный в данный документ посредством ссылки во всей своей полноте). В некоторых вариантах осуществления описанная в данном документе генетически модифицированная мышь представляет собой помесь вышеупомянутой линии 129 и вышеупомянутой линии C57BL/6. В некоторых вариантах осуществления мышь, описанная в данном документе, представляет собой помесь вышеупомянутых линий 129 или помесь вышеупомянутых линий BL/6. В некоторых вариантах осуществления линия 129 в описанной в данном документе помеси представляет собой линию 129S6 (129/SvEvTac). В некоторых вариантах осуществления мышь, описанная в данном документе, принадлежит к линии BALB, например, к линии BALB/c. В некоторых вариантах осуществления мышь, описанная в данном документе, представляет собой помесь линии BALB и другой вышеупомянутой линии.
[00249] B некоторых вариантах осуществления животное, отличное от человека, описанное в данном документе, представляет собой крысу. В некоторых вариантах осуществления крыса, описанная в данном документе, выбрана из крысы линии Wistar, линии LEA, линии Sprague Dawley, линии Fischer, F344, F6 и Dark Agouti. В некоторых вариантах осуществления линия крыс, описанная в данном документе, представляет собой помесь двух или более линий, выбранных из группы, состоящей из Wistar, LEA, Sprague Dawley, Fischer, F344, F6 и Dark Agouti.
Способы
[00250] Было разработано несколько in vitro и in vivo методик для получения терапевтических средств на основе антител. В частности, характерной особенностью in vivo методик является получение трансгенных животных (т.е. грызунов), содержащих гены иммуноглобулина человека, случайным образом встроенные в геном животного (например, см. патент США №5569825, включенный в данный документ посредством ссылки во всей своей полноте) или точно помещенные в эндогенный локус иммуноглобулина в функциональной связи с эндогенными константными областями иммуноглобулина животного (например, см. патенты США №№8502018; 8642835; 8697940 и 8791323, каждый из которых включен в данный документ посредством ссылки во всей своей полноте). Оба подхода были эффективны в получении перспективных кандидатов терапевтических антител для применения у человека. Кроме того, оба подхода имеют преимущество по сравнению с in vitro подходами в том, что кандидаты антител выбирают из репертуаров антител, полученных in vivo, что включает отбор по признаку аффинности и специфичности к антигену в пределах внутренней среды иммунной системы хозяина. При таком способе антитела связываются с естественным образом презентируемым антигеном (в пределах релевантных биологических эпитопов и поверхностей), а не с искусственными средами или с использованием in silico прогнозирований, которые могут сопутствовать in vitro методикам. Несмотря на то, что с помощью in vivo технологий получают репертуары функциональных антител, создание антител к сложным (например, к вирусам, полипептидам каналов) или цитоплазматическим антигенам остается затруднительным. Кроме того, получение антител к полипептидам, которые характеризуются высокой степенью идентичности последовательности у разных видов (например, человека и мыши), остается проблемой вследствие иммунной толерантности.
[00251] Таким образом, настоящее изобретение, среди прочего, основывается на признании того, что конструирование in vivo системы, характеризующейся производством антител, имеющих дополнительное разнообраие CDR, в частности CDR3, полученных в результате нестандартной перегруппировки генного сегмента (т.е. DH-DH-рекомбинации), может осуществляться с применением одного или более DH-сегментов, каждый из которых функционально связан с 23-мерной RSS на своем 5'-или 3'-конце. При наличии 23-мерной RSS на 5- или 3'-конце достигается повышение частоты рекомбинации между сегментами DH по сравнению с локусом тяжелой цепи иммуноглобулина, в котором отсутствуют такие сконструированные генные сегменты DH. Такое дополнительное разнообразие может управлять связыванием с конкретными антигенами (например, вирусами, полипептидами каналов). После рекомбинации с участием генного сегмента VH по меньшей мере двух генных сегментов DH и генного сегмента JH формируется последовательность, кодирующая вариабельную области тяжелой цепи, которая содержит CDR3-область, содержащую аминокислотную последовательность, полученную в результате DH-DH-рекомбинации. Кроме того, данная CDR3, полученная в результате DH-DH-рекомбинации, содержит дополнительное разнообразие, связанное с увеличенной аминокислотной длиной. Кроме того, такая CDRS-область обладает способностью к прямому связыванию с определенным антигеном (или эпитопом), в других случаях являющихся недоступными для связывания антителом, полученным в результате стандартной VDJ-рекомбинации.
[00252] Предусмотренные животные, отличные от человека, могут быть использованы для получения человеческого антитела, при этом человеческое антитело содержит вариабельные домены, полученные из одной или более последовательностей нуклеиновой кислоты вариабельного участка, кодируемых генетическим материалом из клетки животного, отличного от человека, описываемого в данном документе. Например, предусмотренное животное, отличное от человека, иммунизируют с применением антигена, представляющего интерес (например, полипептида вируса или канала, полного или его части) при условиях и в течение времени, достаточных для того, чтобы у животного, отличного от человека, выработался иммунный ответ на указанный антиген, представляющий интерес. Антитела выделяют из животного, отличного от человека (или из одной или более клеток, например, из одной или более В-клеток), и характеризуют посредством проведения различных анализов, с помощью которых измеряют, например, аффинность, специфичность, проводят картирование эпитопов, определяют способность к блокированию взаимодействия лиганд-рецептор, подавлению активации рецепторов и т.д. В некоторых вариантах осуществления антитела, вырабатываемые предусмотренными животными, отличными от человека, содержат один или более человеческих вариабельных доменов, которые получают из одной или более человеческих нуклеотидных последовательностей вариабельной области, выделенных из животного, отличного от человека. В некоторых вариантах осуществления у предусмотренных животных, отличных от человека, может быть индуцирована выработка антител к лекарственным средствам (например, антиидиотипического антитела).
[00253] Животные, отличные от человека, описанные в данном документе, представляют улучшенную in vivo систему и источник биологических материалов (например, клеток) для получения человеческих антител, применимых для различных анализов. В некоторых вариантах осуществления предусмотренных животных, отличных от человека, применяют для разработки терапевтических средств, которые нацеливаются на один или более вирусов и/или модулируют активность вируса и/или модулируют взаимодействие вируса с другими партнерами по связыванию (например, поверхностным клеточным рецептором). В некоторых вариантах осуществления предусмотренных животных, отличных от человека, применяют для разработки терапевтических средств, которые нацеливаются на один или более белков каналов и/или модулируют активность белков каналов и/или модулируют взаимодействие белков каналов с другими партнерами по связыванию. В некоторых вариантах осуществления предусмотренных животных, отличных от человека, применяют для идентификации, скрининга и/или разработки кандидатных терапевтических средств (например, антител, siRNA и т.д.), которые связывают один или более полипептидов вирусов, каналов или сопряженного с G-белком рецептора (GPCR). В некоторых вариантах осуществления предусмотренных животных, отличных от человека, применяют для скрининга и разработки кандидатных терапевтических средств (например, антител, siRNA и т.д.), которые блокируют активность одного или более вирусных полипептидов, одного или более полипептидов каналов человека или одного или более полипептидов GPCR человека. В некоторых вариантах осуществления предусмотренных животных, отличных от человека, применяют для определения профиля связывания антагонистов и/или агонистов одного или более полипептидов GPCR человека или одного или более полипептидов каналов человека. В некоторых вариантах осуществления предусмотренных животных, отличных от человека, применяют для определения эпитопа или эпитопов для одного или более кандидатных терапевтических антител, которые связывают один или более полипептидов GPCR человека или которые связывают один или более полипептидов каналов человека.
[00254] B некоторых вариантах осуществления предусмотренных животных, отличных от человека, применяют для определения фармакокинетических профилей антител. В некоторых вариантах осуществления, каждое из одного или более предусмотренных животных, отличных от человека, и одного или более контрольных или эталонных животных, отличных от человека, подвергают воздействию одного или более кандидатных терапевтических средств на основе антител в различных дозах (например, 0,1 мг/кг, 0,2 мг/кг, 0,3 мг/кг, 0,4 мг/кг, 0,5 мг/кг, 1 мг/кг, 2 мг/кг, 3 мг/кг, 4 мг/кг, 5 мг/кг, 7,5 мг/кг, 10 мг/кг, 15 мг/кг, 20 мг/кг, 25 мг/кг, 30 мг/кг, 40 мг/кг или 50 мг/кг или более). Кандидатные терапевтические антитела могут быть введены дозами посредством любого желаемого пути введения, в том числе парентерального и отличных от парентерального путей введения. Парентеральные пути включают, например, внутривенный, внутриартериальный, интрапортальный, внутримышечный, подкожный, внутрибрюшинный, интраспинальный, интратекальный, интрацеребровентрикулярный, внутричерепной, интраплевральный или другие пути инъекции. Пути, отличные от парентеральных, включают, например, пероральный, интраназальный, трансдермальный, внутрилегочный, ректальный, трансбуккальный, вагинальный, внутриглазной. Введение также можно осуществлять путем непрерывной инфузии, местного применения, замедленного высвобождения из имплантатов (гелей, мембран и т.п.) и/или внутривенной инъекции, например, с применением пакета для жидкостей для внутривенной инфузии. У животных, отличных от человека (гуманизированных и контрольных), производят забор крови в различные моменты времени (например, 0 ч., 6 ч., 1 день, 2 дня, 3 дня, 4 дня, 5 дней, 6 дней, 7 дней, 8 дней, 9 дней, 10 дней, 11 дней или вплоть до 30 или более дней). Могут проводиться различные анализы для определения фармакокинетических профилей вводимых кандидатных терапевтических антител с использованием образцов, полученных от животных, отличных от человека, описанных в данном документе, в том числе без ограничения анализы уровня общего IgG, гуморального ответа на терапевтические средства, агглютинации и т.д.
[00255] B некоторых вариантах осуществления предусмотренных животных, отличных от человека, применяют для измерения терапевтического эффекта блокирования или модулирования активности целевого антигена и действия на экспрессию гена в результате клеточных изменений или плотности целевого антигена на клеточной поверхности (в случае с поверхностным клеточным рецептором) клеток животных, отличных от человека, описанных в данном документе. В некоторых вариантах осуществления предусмотренное животное, отличное от человека, или клетки, выделенные из него, подвергают воздействию кандидатного терапевтического средства, которое связывает антиген, представляющий интерес, и по истечении последовательного периода времени анализируют в отношении эффектов, производимых на целевые антиген-зависимые процессы (или взаимодействия), например, взаимодействия лиганд-рецептор или передачу сигнала с помощью антигена.
[00256] B некоторых вариантах осуществления предусмотренных животных, отличных от человека, применяют для измерения терапевтического эффекта блокирования или модуляции активности канала (или передачи сигнала с помощью канала, или взаимодействий, опосредованных каналом, или потенциалов действия канала) и эффекта в отношении генной экспрессии в результате клеточных изменений или плотности каналов в клетках животных, отличных от человека, описываемых в данном документе. В некоторых вариантах осуществления предусмотренное животное, отличное от человека, или клетки, выделенные из него, подвергают воздействию кандидатного терапевтического средства, которое связывает полипептид канала человека (или его часть) и по истечении последовательного периода времени анализируют в отношении эффектов, производимых на канал-зависимые процессы (или взаимодействия), например, взаимодействия лиганд-рецептор или потенциалы действия канала.
[00257] B некоторых вариантах осуществления предусмотренные животные, отличные от человека, экспрессируют антитела, таким образом, можно создавать клетки, линии клеток и культуры клеток, служащие в качестве источника антител для применения в анализах, основанных на связывании, и функциональных анализах, например, для анализа связывания или функции антагониста или агониста, в частности, где антагонист или агонист является специфичным к последовательности полипептида человека или эпитопу, или в качестве альтернативы специфичным к последовательности полипептида человека или эпитопу, который принимает участие во взаимодействии (связывании) «лиганд-рецептор». В некоторых вариантах осуществления эпитопы, связываемые кандидатными терапевтическими антителами или siRNA, могут определяться с применением клеток, выделенных из предусмотренных животных, отличных от человека.
[00258] B некоторых вариантах осуществления клетки из предусмотренных животных, отличных от человека, могут быть выделены и использованы для определенных целей или могут поддерживаться в культуре в течение многих поколений. В некоторых вариантах осуществления клетки от предусмотренного животного, отличного от человека, подвергают иммортализации (например, посредством использования вируса) и поддерживают в культуре в течение неопределенного периода времени (например, в последовательных культурах).
[00259] B некоторых вариантах осуществления животные, отличные от человека, описанные в данном документе, обеспечивают in vivo систему для получения вариантов антител, которые связывают целевой антиген человека. Такие варианты включают антитела, обладающие требуемой функциональностью, специфичностью, низкой перекрестной реактивностью с общим эпитопом, который встречается у двух или более целевых антигенов человека. В некоторых вариантах осуществления предусмотренных животных, отличных от человека, применяют для создания панелей антител для получения серии вариантов антител, которые подвергают скринингу в отношении необходимой или улучшенной функциональности.
[00260] B некоторых вариантах осуществления животные, отличные от человека, описанные в данном документе, обеспечивают in vivo систему для получения библиотек антител. Такие библиотеки обеспечивают источник последовательностей вариабельной области тяжелой и легкой цепей, которые могут быть привиты на разные Fc-участки, исходя из необходимой эффекторной функции, и/или использоваться в качестве источника для созревания аффинности у последовательности вариабельной области, с использованием методик, известных из уровня техники (например, сайт-направленный мутагенез, ПЦР с внесением ошибок и т.д.).
[00261] B некоторых вариантах осуществления животные, отличные от человека, описанные в данном документе, обеспечивают in vivo систему для анализа и тестирования лекарственного средства или вакцины. В некоторых вариантах осуществления кандидатное лекарственное средство или вакцину можно вводить одному или более предусмотренным животным, отличным от человека, с последующим наблюдением за животными, отличными от человека, для определения одного или более иммунных ответов на лекарственное средство или вакцину, профиля безопасности лекарственного средства или вакцины или эффекта в отношении заболевания или состояния и/или одного или более симптомов заболевания или состояния. Иллюстративные способы, используемые для определения профиля безопасности, включают измерение токсичности, оптимальной концентрации дозы, ответа с участием антитела (т.е. антитела к лекарственному средству), эффективности лекарственного средства или вакцины и возможных факторов риска. Такие лекарственные средства или вакцины можно улучшать и/или разрабатывать с применением таких животных, отличных от человека.
[00262] Эффективность вакцины можно определять с помощью ряда способов. Вкратце, животных, отличных от человека, описанных в данном документе, вакцинируют с помощью способов, известных из уровня техники, и затем заражают вакциной, или вакцину вводят уже инфицированному животному, отличному от человека. Ответ животного(-ых), отличного(-ых) от человека, на вакцину можно измерять путем мониторинга животного(-ых), отличного(-ых) от человека (или выделенных из него(них) клеток), и/или проведения одного или более анализов в их отношении для определения эффективности вакцины. Ответ животного(-ых), отличного(-ых) от человека, на вакцину затем сравнивают с ответом контрольных животных, используя один или более показателей, известных из уровня техники и/или описанных в данном документе.
[00263] Эффективность вакцины можно дополнительно определять с помощью анализов, основанных на нейтрализации вируса. Вкратце, животных, отличных от человека, описанных в данном документе, иммунизируют и производят забор сыворотки крови в разные дни после иммунизации. Серийные разведения сыворотки крови предварительно инкубируют в присутствии вируса и в ходе этого периода времени антитела в сыворотке крови, специфичные к вирусу, связываются с ним. Смесь вирус/сыворотка крови затем добавляют к пермиссивным клеткам для определения инфицирующей способности с помощью анализа бляшкообразования или анализа микронейтрализации. Если антитела в сыворотке крови нейтрализуют вирус, то наблюдается меньшее количество бляшек или меньшее количество относительных люциферазных единиц по сравнению с контрольной группой.
[00264] Животные, отличные от человека, описанные в данном документе, обеспечивают улучшенную in vivo систему для разработки и определения характеристик терапевтических средств на основе антител для применения в лечении рака и/или воспалительных заболеваний. Воспаление в течение длительного времени связывали с раком (см., например, обзор в Grivennikov, S.I. et al., 2010, Cell 140:883-99; Rakoff-Nahoum, S., 2006, Yale J. Biol. Med. 79:123-30, каждый из которых включен в данный документ посредством ссылки во всей своей полноте). Действительно, окружение развивающейся опухоли в некоторой степени характеризуется инфильтрацией различными медиаторами воспаления. Также хроническое воспаление может приводить к более высокой вероятности развития рака. Таким образом, в некоторых вариантах осуществления описанное в данном документе животное, отличное от человека, обеспечивает in vivo систему для разработки и/или идентификации противоопухолевых и/или противовоспалительных терапевтических средств. В некоторых вариантах осуществления предусмотренным животным, отличным от человека, или контрольным животным, отличным от человека (например, имеющим генетическую модификацию, отличную от описанной в данном документе, или не имеющим генетической модификации, т.е. дикого типа), может быть имплантирована опухоль (или опухолевые клетки) с последующим введением одного или более кандидатных терапевтических средств. В некоторых вариантах осуществления кандидатные терапевтические средства могут содержать полиспецифическое антитело (например, биспецифическое антитело) или смесь антител. В некоторых вариантах осуществления кандидатные терапевтические средства предусматривают комбинированную терапию, такую как, например, введение доз двух или более моноспецифических антител последовательно или одновременно. Перед введением одного или более кандидатных терапевтических средств может быть предоставлено достаточное время для развития опухоли в одном или более местоположениях в пределах организма животного, отличного от человека. Пролиферацию, рост, выживаемость и т.д. опухолевых клеток можно измерять как перед введением, так и после введения кандидатного терапевтического средства (средств). При необходимости у животного, отличного от человека, также можно измерять цитотоксичность кандидатных терапевтических средств.
Наборы
[00265] Настоящее изобретение дополнительно предусматривает пакет или набор, содержащие один или более контейнеров, заполненных по меньшей мере одним животным, отличным от человека, клеткой, отличной от человеческой, фрагментом ДНК и/или вектором для целенаправленного воздействия, описанными в данном документе. Наборы можно использовать в любом применимом способе (например, в способе исследования). Необязательно связанным с таким(-ими) контейнером(-ами) может быть уведомление в форме, предписанной правительственным органом, регулирующим производство, применение или продажу фармацевтических препаратов или биологических продуктов, при этом данное уведомление отражает (а) одобрение органом, регулирующим производство, применение или продажу, для введения человеку, (b) указания по применению и/или договор, который регулирует передачу материалов и/или биологических продуктов (например, животного, отличного от человека, или клеток, отличных от человеческих, описанных в данном документе) между двумя или более организациями, или как первое, так и второе.
[00266] Другие признаки настоящего изобретения станут очевидными из следующего описания иллюстративных вариантов осуществления, которые приведены в иллюстративных целях и не подразумеваются как ограничивающие его.
ПРИМЕРЫ
[00267] Следующие примеры приведены для того, чтобы обеспечить специалистов в данной области описанием того, как создавать и применять способы и композиции по настоящему изобретению, и не предназначены для ограничения объема того, что авторы настоящего изобретения рассматривают в качестве своего изобретения. Если не указано иное, температура указана в градусах Цельсия, а давление является атмосферным или близким к нему.
Пример 1. Разработка и конструирование нацеливающего вектора
[00268] Данный пример иллюстрирует конструирование нацеливающих векторов для вставки в геном животного, отличного от человека, такого как грызун (например, мышь). В частности, в способах, описанных в данном примере, продемонстрировано получение нацеливающих векторов для вставки в геном эмбриональных стволовых (ES) клеток грызуна (например, мыши) для получения грызуна, геном которого содержит вариабельную область тяжелой цепи иммуноглобулина, которая включает сконструированную область разнообразия (DH) тяжелой цепи, где указанная сконструированная DH-область содержит один или более DH-сегментов, каждый из которых функционально связан с 23-мерной RSS на своем 5'- или 3'-конце. В данном примере описаны три нацеливающих вектора: первый нацеливающий вектор, содержащий сконструированную DH-область, которая содержит расположенный проксимально (или с 3'-конца) DH-сегмент, функционально связанный с 23-мерной RSS на своем 5'-конце и функционально связанный с одним JH-сегментом человека, второй нацеливающий вектор, содержащий сконструированную DH-область, которая содержит расположенный проксимально (или с 3'-конца) DH-сегмент, функционально связанный с 23-мерной RSS на своем 5'-конце и функционально связанный с тремя JH-сегментами человека, и третий нацеливающий вектор, содержащий сконструированную DH-область, которая содержит три DH-сегмента, каждый из которых связан с 23-мерной RSS на своем 3'-конце и функционально связан с шестью JH-сегментами человека (фигура 2). Каждый из этих нацеливающих векторов получали и по отдельности вставляли в гуманизированную вариабельную область тяжелой цепи иммуноглобулина (фигуры 3, 5, 7). Как описано ниже, сконструированную DH-область располагали в функциональной связи с вариабельным сегментом тяжелой цепи (VH) и соединяющим сегментом тяжелой цепи (JH) таким образом, чтобы после VDJ-рекомбинации экспрессировались антитела, содержащие CDR3, полученные в результате DH-DH-рекомбинации.
[00269] Нацеливающие векторы, содержащие один или более DH-сегментов человека, каждый из которых связан с 23-мерной RSS на своем 5'- или 3'-конце, для вставки в вариабельную область тяжелой цепи иммуноглобулина создавали с использованием технологии VELOCIGENE® (см., например, патент США №6586251 и Valenzuela et al., 2003, Nature Biotech. 21(6):652-659; включенные в данный документ посредством ссылки во всей своей полноте) и методик молекулярной биологии, известных из уровня техники. Способы, описанные в данном примере, могут при необходимости применяться для использования любого DH-сегмента, набора DH-сегментов или комбинации DH-сегментов.
А. Нацеливающий вектор 23:DH3-3:12/JH6 (фигура 2)
[00270] Вкратце, первый нацеливающий вектор конструировали с применением геномного фрагмента ДНК, содержащего несколько DH-сегментов человека, и синтетического человеческого DH3-3-сегмента, расположенного на месте DH7-27 человека. Донора для модификации in vitro Cas9/GA получали посредством синтеза de novo (Blue Heron Bio) и он содержал от 5'-конца к 3'-концу: (а) плечо гомологии длиной 100 п. о. начинающееся с 350 п. о. выше 12-мерной RSS на 5'-конце DH7-27, (b) сайты AgeI и XhoI для вставки кассеты устойчивости к неомицину, (с) область длиной 250 п.о. выше 12-мерной RSS на 5'-конце DH7-27, (d) синтетический человеческий DH3-3-сегмент, сконструированный с 23-мерной RSS на своем 5'-конце (от JH4) и 12-мерной RSS на своем 3'-конце (от DH3-3), и (3) блок гомологии длиной 100 п. о., начинающийся с 3 п. о. ниже JH5. Кассету loxp-UbC-Em7-Neo-loxp, например, ген устойчивости к неомицину, фланкированный сайтами распознавания сайт-специфической рекомбинации lοxΡ, располагали на ~250 п. о. выше синтетического DH-сегмента и в нисходящем направлении посредством лигирования в сайты AgeI и XhoI для обеспечения условий для отбора в Е. coli и ES-клетках мыши. Данного донора 23:DH3-3: 12/JH6 (SEQ ID NO: 61), содержащего кассету устойчивости к неомицину, применяли для модификации ВАС с помощью in vitro Cas9/GA с применением двух комплексов Cas9:gRNA. Перед модификацией ВАС была идентична по последовательности химерному локусу IgH мыши VELOCIMMUNE® (см., например, фигуры 3 и 4, демонстрирующие примерный неограничивающий эндогенный локус тяжелой цепи иммуноглобулина мыши VELOCIMMUNE®, являющейся гетерозиготной по аллелю 6394, содержащему гуманизированную вариабельную область, в составе которой отсутствовали генные сегменты JH и которая функционально связана с эндогенной последовательностью константной области тяжелой цепи иммуноглобулина мыши, и аллелю 1460, содержащему гуманизированную вариабельную область, функционально связанную с эндогенной последовательностью константной области тяжелой цепи иммуноглобулина мыши). В частности, ВАС была идентична аллелю 1460, начиная от наиболее проксимально расположенного гена VH человека (VH6-1), и содержала все 27 генов DH человека, все 6 генов JH человека, интронный энхансер (Εμ) IgH мыши, область переключения (8μ) IgM мыши и первые 4 экзона гена IgM мыши. ВАС также содержала кассету устойчивости к спектиномицину (spec) и ~29 т. н. межгенной последовательности человека выше гена VH6-1. Соединение «человек-мышь» составляло 222 п. о. от 3'-конца гена JH6 человека и 490 п. о. от 5'-конца энхансера Εμ мыши. Вставка донора 23:D3-3: 12/JH6 в ВАС путем GA привела к получению окончательного нацеливающего вектора 23:D3-3: 12/JH6, содержащего замену генного сегмента DH7-7 сконструированным сегментом 23:D3-3:12, и делеции генных сегментов JH1, JH2, JH3, JH4 и JH5 (фигура 2). В таблице 1 представлена последовательность gRNA, праймеров и зондов, применявшихся для определения точности конструирования нацеливающего вектора 23:DH3-3: 12/JH6.


[00271] Нацеливающий вектор 23:D3-3:12/JH6 линеаризировали с NotI и вводили с помощью электропорации в эмбриональные стволовые клетки мыши, обладающие геномом, гетерозиготным по гуманизированной вариабельной области тяжелой цепи иммуноглобулина (т.е. первому аллелю тяжелой цепи (1460het), содержащему несколько VH, DH и JH-сегментов человека, функционально связанных с константной областью тяжелой цепи иммуноглобулина грызуна, содержащей энхансеры и регуляторные области тяжелой цепи грызуна, и содержащих вставленную нуклеотидную последовательность, кодирующую один или более мышиных генов Adam6 (например, патенты США №№8642835 и 8697940, каждый из которых включен в данный документ посредством ссылки во всей своей полноте); и второму аллелю тяжелой цепи (6394het), содержащему несколько VH и DH-сегментов человека, делецию JH-области и константную область тяжелой цепи иммуноглобулина грызуна, содержащую энхансеры и регуляторные области тяжелой цепи грызуна, и содержащему вставленную нуклеотидную последовательность, кодирующую один или более мышиных генов Adam6 (например, патенты США №№8642835 и 8697940, каждый из которых включен в данный документ посредством ссылки во всей своей полноте)) и гомозиготным (НО) по гуманизированному эндогенному κ-локусу, содержащему полный репертуар генных сегментов Vκ и Jκ легкой цепи иммуноглобулина человека, функционально связанных с эндогенной Сκ-областью тяжелой цепи иммуноглобулина мыши (1293) (см. фигуру 3). Аллель 6799 получали после электропорации и надлежащей гомологичной рекомбинации аллеля 1460 с нацеливающим вектором 23:D3-3: 12/JH6, содержащим кассету устойчивости к неомицину. (Фигура 3). Такие сконструированные ES-клетки мыши, гетерозиготные по аллелям 6394 и 679, применяли для обеспечения эффективного скрининга положительных ES-клонов (см. ниже) и последовательного cre-опосредованного удаления кассет устойчивости к лекарственному средству, при этом после указанной делеции аллели 6394 и 6799 соответственно назывались 6643 и 6800 (фигура 4).
В. Нацеливающий вектор 23:DH3-3:12/JH4-6 (фигура 2)
[00272] Сходным образом конструировали другой нацеливающий вектор (23:DH3-3: 12/JH4-6) с применением такого же геномного фрагмента ДНК, содержащего несколько DH-сегментов человека и синтетического человеческого DH3-3-сегмента, расположенного на месте DH7-27 человека. Однако этот второй нацеливающий вектор включал три из шести JH-сегментов человека (т.е. JH4, JH5, JH6). Для создания окончательного нацеливающего вектора 23:DH3-3: 12/JH4-6, применяли донора для модификации ВАС, идентичной по последовательности химерному локусу IgH мыши VELOCIMMUNE® (см., например, фигуры 3 и 4, демонстрирующие примерный неограничивающий эндогенный локус тяжелой цепи иммуноглобулина мыши VELOCIMMUNE®, являющейся гетерозиготной по аллелю 6394, содержащему гуманизированную вариабельную область, в составе которой отсутствуют генные сегменты JH и которая функционально связана с эндогенной последовательностью константной области тяжелой цепи иммуноглобулина мыши, и аллелю 1460, содержащему гуманизированную вариабельную область, функционально связанную с эндогенной последовательностью константной области тяжелой цепи иммуноглобулина мыши). Донор содержал от 5'-конца к 3'-концу: (а) блок гомологии длиной 100 п. о., начинающийся с 350 п. о. выше 12-мерной RSS на 5'-конце D7-27, (b) сайты AgeI и XhoI для вставки кассеты устойчивости к неомицину, (с) область длиной 250 п. о. выше 12-мерной RSS на 5'-конце DH7-27, (d) синтетический человеческий DH3-3-сегмент, фланкированный на своем 5'-конце 23-мерной RSS (от JH4) и на своем 3'-конце 12-мерной RSS (от DH3-3), и (е) блок гомологии длиной 100 п. о., начинающийся с 3 п. о. ниже JH3. Кассету loxp-UbC-Em7-Neo-loxp, например, ген устойчивости к неомицину, фланкированный сайтами распознавания сайт-специфической рекомбинации loxΡ располагали на ~250 п. о. выше синтетического DH-сегмента посредством лигирования в сайты AgeI и XhoI для обеспечения условий для отбора в Е. coli и ES-клетках мыши. Данного донора 23:D3-3: 12/JH4-6, содержащего нуклеотидную последовательность, представленную под SEQ ID NO: 52, включающего кассету устойчивости к неомицину, применяли для модификации ВАС, идентичной по последовательности аллелю 1460 (фигуры 3 и 4). В частности, ВАС была идентична аллелю 1460, начиная от наиболее проксимально расположенного гена VH человека (VH6-1), и содержала все 27 генов DH человека, все 6 генов JH человека, интронный энхансер (Εμ) IgH мыши, область переключения (8μ) IgM мыши и первые 4 экзона гена IgM мыши. ВАС также содержала кассету устойчивости к спектиномицину (spec) и -29 т.н. межгенной последовательности человека выше гена VH6-1. Соединение «человек-мышь» составляло 222 п. о. от 3'-конца гена JH6 человека и 490 п. о. от 5'-конца энхансера Εμ мыши. Вставка донора 23:D3-3: 12/JH4-6 в ВАС с помощью in vitro Cas9/GA с применением двух комплексов Cas9:gRNA привела к получению окончательного нацеливающего вектора 23:D3-3: 12/JH4-6 (фигура 2), содержащего замену генного сегмента DH7-7 сконструированным сегментом 23:D3-3:12, и делеции генных сегментов JH1, JH2, и JH3. В таблице 2 представлена последовательность gRNA, праймеров и зондов, применявшихся для определения точности конструирования нацеливающего вектора 23:DH3-3:12/JH6.


[00273] Нацеливающий вектор 23:D3-3: 12/JH4-6 также линеаризировали с NotI и вводили с помощью электропорации в эмбриональные стволовые клетки мыши, обладающие геномом, гетерозиготным по гуманизированной вариабельной области тяжелой цепи иммуноглобулина (т.е. первому аллелю тяжелой цепи (1460het), содержащему несколько VH, DH, и JH-сегментов человека, функционально связанных с константной областью тяжелой цепи иммуноглобулина грызуна, содержащей энхансеры и регуляторные области тяжелой цепи грызуна, и содержащих вставленную нуклеотидную последовательность, кодирующую один или более мышиных генов Adam6 [например, патенты США №№8642835 и 8697940, каждый из которых включен в данный документ посредством ссылки во всей своей полноте] ; и второму аллелю тяжелой цепи (6394het), содержащему несколько сегментов VH и DH человека, делецию JH-области и константную область тяжелой цепи иммуноглобулина грызуна, содержащую энхансеры и регуляторные области тяжелой цепи грызуна, и содержащему вставленную нуклеотидную последовательность, кодирующую один или более мышиных генов Adam6 [например, патенты США №№8642835 и 8697940, каждый из которых включен в данный документ посредством ссылки во всей своей полноте] ) и гомозиготным (НО) по гуманизированному эндогенному к-локусу, содержащему полный репертуар генных сегментов Vκ и Jκ легкой цепи иммуноглобулина человека, функционально связанных с эндогенной Ск-областью тяжелой цепи иммуноглобулина мыши (1293) (см. фигуру 5). Аллель 6797 получали после электропорации и надлежащей гомологичной рекомбинации аллеля 1460 с нацеливающим вектором 23:D3-3: 12/JH4-6, содержащим кассету устойчивости к неомицину. (Фигура 5). Такие сконструированные ES-клетки мыши, гетерозиготные по аллелям 6394 и 6797, применяли для обеспечения эффективного скрининга положительных ES-клонов (см. ниже) и последовательного сге-опосредованного удаления кассет устойчивости к лекарственному средству, при этом после указанной делеции аллели 6394 и 6797 соответственно назывались 6643 и 6798 (фигура 6).
С. Нацеливающий вектор 12:DH2-2:23|12:DH2-8:23|12:DH2-15:23 /JH1-6 (фигура 2)
[00274] Вкратце, третий нацеливающий вектор конструировали с применением фрагмента ДНК, содержащего несколько DH-сегментов человека, которые содержали три генных сегмента семейства DH2 (DH2-2, DH2-8 и DH2-15), при этом каждый имел 12-мерную RSS на своем 5'-конце и 23-мерную RSS на своем 3'-конце. Прежде всего, ВАС была идентичной по последовательности химерному локусу IgH мыши VELOCIMMUNE® (см., например, фигуры 3 и 4, демонстрирующие примерный неограничивающий эндогенный локус тяжелой цепи иммуноглобулина мыши VELOCIMMUNE®, являющейся гетерозиготной по аллелю 6394, содержащему гуманизированную вариабельную область, в составе которой отсутствуют генные сегменты JH и которая функционально связана с эндогенной последовательностью константной области тяжелой цепи иммуноглобулина мыши, и аллелю 1460, содержащему гуманизированную вариабельную область, функционально связанную с эндогенной последовательностью константной области тяжелой цепи иммуноглобулина мыши). В честности, ВАС, которая была идентична аллелю 1460, начиная от наиболее проксимально расположенного гена VH человека (VH6-1), и содержала все 27 генов DH человека, все 6 генов JH человека, интронный энхансер (Εμ) IgH мыши, область переключения (8μ) IgM мыши и первые 4 экзона гена IgM мыши, модифицировали с применением бактериальной гомологичной рекомбинации (BHR) с целью осуществления вставки кассеты loxp-UbC-Em7-Neo-loxp, например, гена устойчивости к неомицину, фланкированного сайтами распознавания сайт-специфической рекомбинации loxΡ, на 512 п. о. выше первого DH-сегмента человека (DH1-1) для обеспечения условий для отбора в Е. coli и ES-клетках мыши. Эту полученную ВАС последовательно модифицировали с использованием трех доноров посредством in vitro Cas9/GA.
1. Первый донор для модификации DH2-2 содержал от 5'-конца к 3'-концу: блок гомологии длиной 50 п. о., начинавшийся с 1419 п. о. выше 12-мерной RSS на 5'-конце DH2-2, сайт множественного клонирования (MreI-NsiI-EcoRI-Kpni-MreI), модифицированный ген D2-2 с 12-мерной RSS на своем 3'-конце, замененной оптимизированной 23-мерной RSS на своем 3'-конце, полученной из VH1-69 человека (CACAGTGTGA АААСССАСАТ CCTGAGAGTG АСАСАААСС; Т->А, G->C; SEQ ID NO: 151), и блок гомологии длиной 50 п. о., заканчивающийся на 863 п. о. ниже 23-мерной RSS на 3'-конце D2-2. Нуклеотидная последовательность этого первого донора для модификации DH2-2 представлена под SEQ ID NO: 70. Учитывая, что DH-область состояла из четырех прямых повторов длиной -10 т.н., проектирование уникальных праймеров и зондов для скрининга было крайне трудным. Для решения данной задачи две уникальных последовательности длиной 40 п. о. вставляли следующим образом: одну на 74 п. о. ниже блока гомологии на 5'-конце и одну на 40 п. о. выше блока гомологии на 3'-конце. Такие уникальные 40-меры применяли в качестве сайтов связывания для праймеров для проведения ПЦР/секвенирования и зондов Taqman, они были изображены в виде незакрашенных вертикальных треугольников «1» и «2» на фигуре 2 и содержали нуклеотидную последовательность, представленную под SEQ ID NO: 73 и SEQ ID NO: 74 соответственно.
2. Второй донор для модификации DH2-8 содержал от 5'-конца к 3'-концу: блок гомологии длиной 50 п. о., начинавшийся с 552 п. о. выше 12-мерной RSS на 5'-конце DH2-8, сайт множественного клонирования (MreI-NsiI-EcoRI-KpnI-MreI), модифицированный ген D2-8 с 12-мерной RSS на своем 3'-конце, замененной оптимизированной 23-мерной RSS на своем 3'-конце, полученной из VH1-69 человека (CACAGTGTGA АААСССАСАТ CCTGAGAGTG АСАСАААСС; Т->А, G->C SEQ ID NO: 151), и блок гомологии длиной 50 п. о., заканчивающийся на 867 п. о. ниже 23-мерной RSS на 3'-конце D2-8. Нуклеотидная последовательность этого второго донора для модификации DH2-8 представлена под SEQ ID NO: 71. Учитывая, что DH-область состояла из четырех прямых повторов длиной ~10 т. н., проектирование уникальных праймеров и зондов для скрининга было крайне трудным. Для решения данной задачи две уникальных последовательности длиной 40 п. о. вставляли следующим образом: одну на 74 п. о. ниже блока гомологии на 5'-конце и одну на 40 п. о. выше блока гомологии на 3'-конце. Такие уникальные 40-меры применяли в качестве сайтов связывания для праймеров для проведения ПЦР/секвенирования и зондов Taqman, они были изображены в виде незакрашенных вертикальных треугольников «10» и «16» на фигуре 2 и содержали нуклеотидную последовательность, представленную под SEQ ID NO: 75 и SEQ ID NO: 76 соответственно.
3. Третий донор для модификации DH2-15 содержал от 5'-конца к 3'-концу: блок гомологии длиной 50 п. о., начинавшийся с 391 п. о. выше 12-мерной RSS на 5'-конце DH2-15, сайт множественного клонирования (MreI-NsiI-EcoRI-KpnI-MreI), модифицированный ген D2-15 с 12-мерной RSS на своем 3'-конце, замененной оптимизированной 23-мерной RSS на своем 3'-конце, полученной из VHl-69 человека (CACAGTGTGA АААСССАСАТ CCTGAGAGTG АСАСАААСС; Т->А, G->C SEQ ID NO: 151), и блок гомологии длиной 50 п. о., заканчивающийся на 867 п. о. ниже 23-мерной RSS на 3'-конце D2-15. Нуклеотидная последовательность этого третьего донора для модификации DH2-2 представлена под SEQ ID NO: 72. Учитывая, что DH-область состояла из четырех прямых повторов длиной -10 т.н., проектирование уникальных праймеров и зондов для скрининга было крайне трудным. Для решения данной задачи две уникальных последовательности длиной 40 п. о. вставляли следующим образом: одну на 74 п. о. ниже блока гомологии на 5'-конце и одну на 40 п. о. выше блока гомологии на 3'-конце. Такие уникальные 40-меры применяли в качестве сайтов связывания для праймеров для проведения ПЦР/секвенирования и зондов Taqman, они были изображены в виде незакрашенных вертикальных треугольников «8» и «18» на фигуре 2 и содержали нуклеотидную последовательность, представленную под SEQ ID NO:77 и SEQ ID NO: 78 соответственно.
После модификации in vitro Cas9/GA с применением трех доноров, окончательный нацеливающий вектор содержал от 5'-конца к 3'-концу: плечо гомологии на своем 5'-конце длиной 49 т.н., содержавшее ДНК вариабельной области человека, кассету устойчивости к неомицину, сконструированную DH-область человека, включавшую три DH-сегмента человека, каждый из которых был фланкирован 12-мерной RSS на своем 5'-конце и 23-мерной RSS на своем 3'-конце, шесть JH-сегментов человека и плечо гомологии на своем 3'-конце длиной 24 т.н., содержавшее интронный энхансер (Ei) тяжелой цепи мыши и ген константной области IgM мыши. (Фигура 2). В таблице 3 представлена последовательность gRNA, праймеров и зондов, применявшихся для определения точности конструирования нацеливающего вектора 12:DH2-2:23|12:DH2-8:23|12:DH2-15:23 /JH1-6.



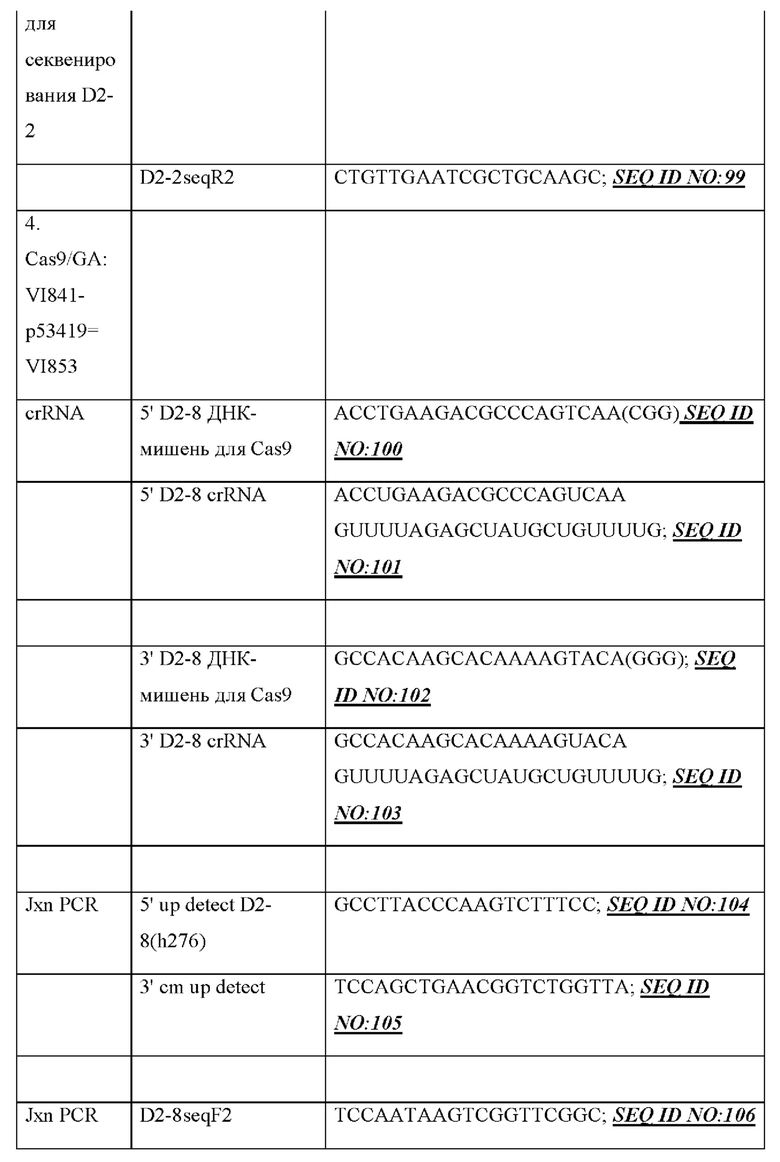

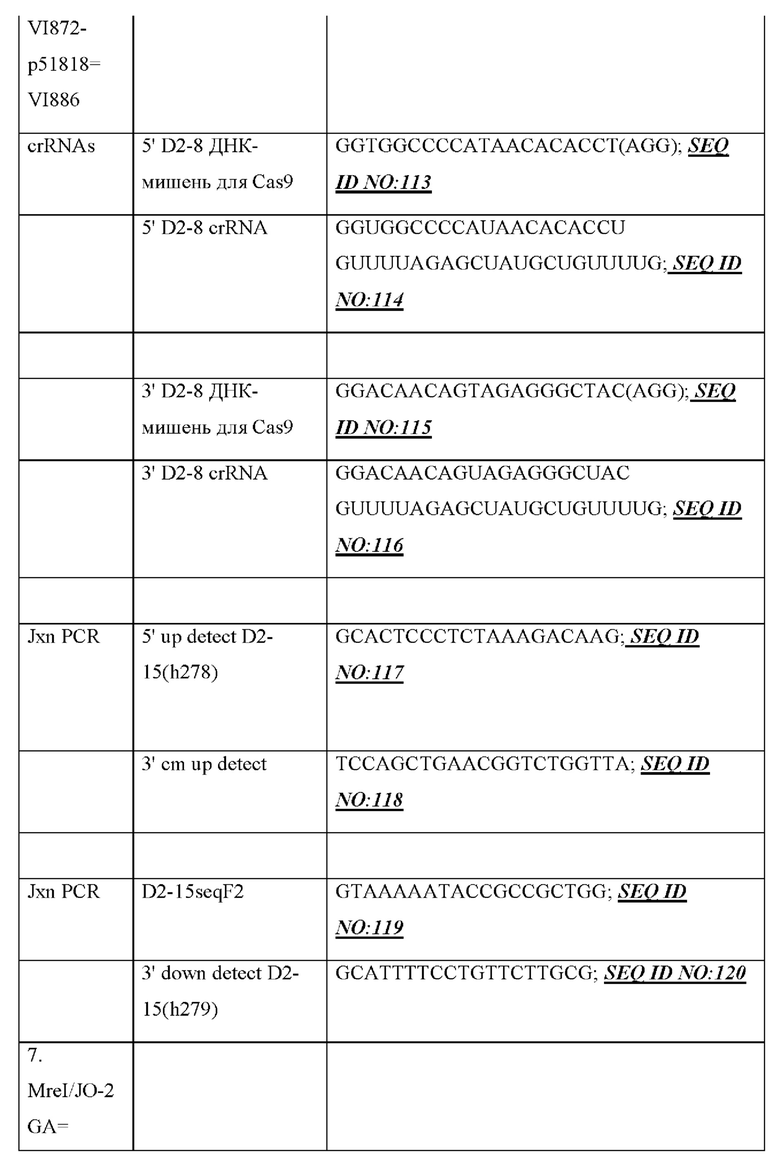
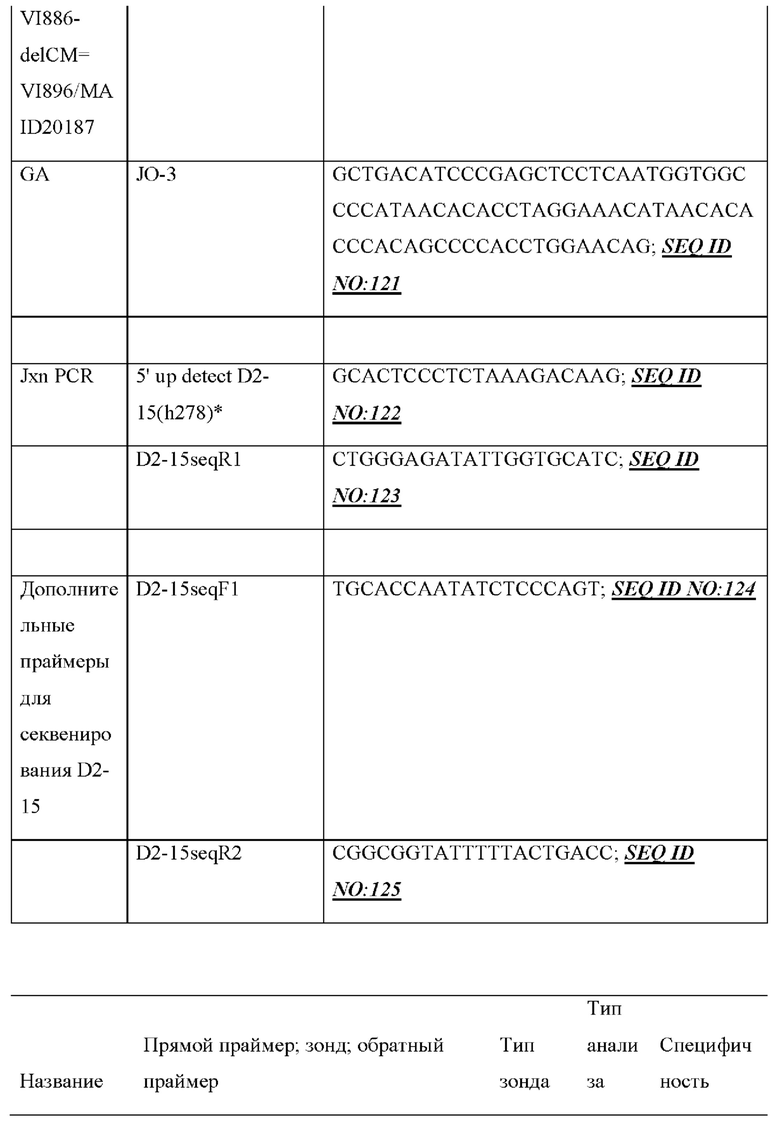

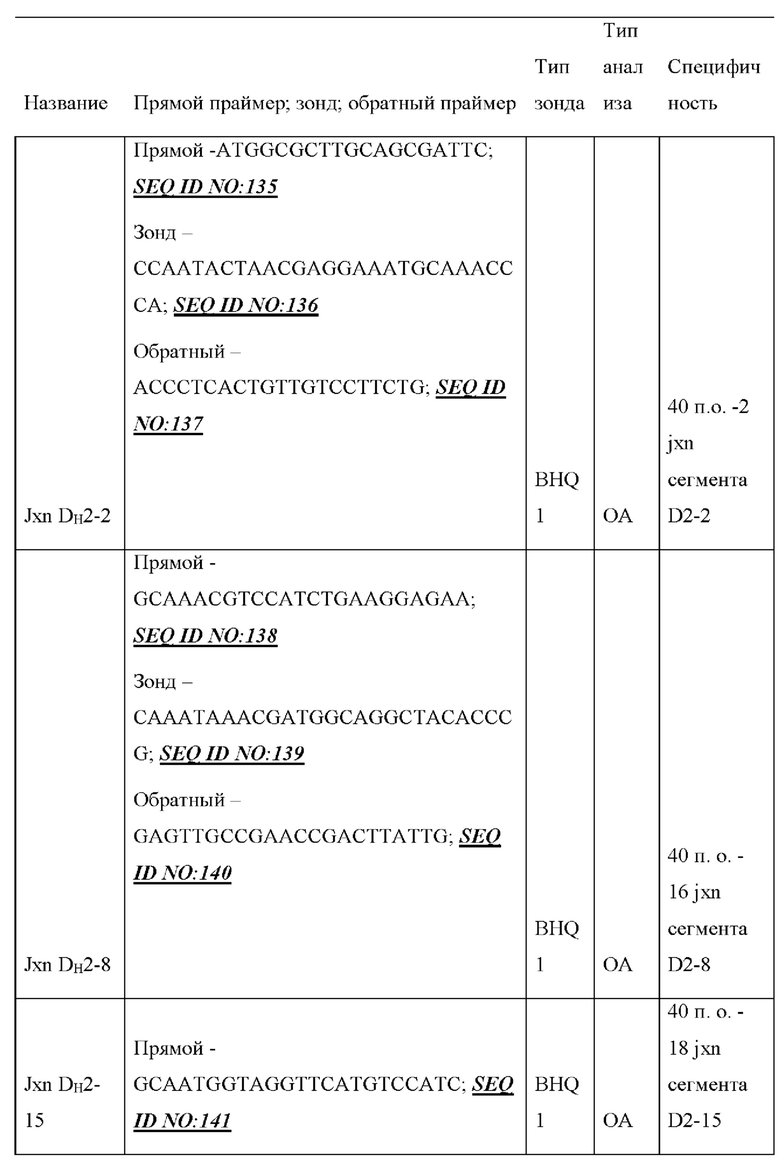

[00275] Нацеливающий вектор 12:DH2-2:23:|12:DH2-8:23|12:DH2-15:23×3/JH1-6 линеаризировали с NotI и вводили с помощью электропорации в эмбриональные стволовые клетки мыши, обладающие геномом, гомозиготным по гуманизированной вариабельной области тяжелой цепи иммуноглобулина, которая содержала полный репертуар функциональных генных сегментов VH человека, делецию DH-области за исключением DH7-27 и полный репертуар функциональных генных сегментов JH человека (6011), и который был гомозиготным (НО) по гуманизированному эндогенному κ-локусу, содержащему полный репертуар генных сегментов Vκ и Jκ легкой цепи иммуноглобулина человека, функционально связанных с эндогенной Сκ-областью тяжелой цепи иммуноглобулина мыши (1293) (см. фигуру 7). Аллель 20187 получали после электропорации и надлежащей гомологичной рекомбинации аллеля 6011 с нацеливающим вектором 12:DH2-2:23|12:DH2-8:23|12:DH2-15:23/JH1-6, содержащим кассету устойчивости к неомицину. (Фигура 7). Такие сконструированные ES-клетки мыши применяли для обеспечения эффективного скрининга положительных ES-клонов (см. ниже) и последовательного cre-опосредованного удаления кассеты устойчивости к неомицину, при этом после указанной делеции аллель 20187 соответственно называли 20188 (фигура 8).
Пример 2. Скрининг ES-клеток
[00276] Данный пример демонстрирует получение животных, отличных от человека (например, грызунов), геном которых содержит вариабельную область тяжелой цепи иммуноглобулина, содержащую сконструированную область разнообразия тяжелой цепи (DH), где сконструированная DH-область содержит один или более DH-сегментов, каждый из которых функционально связан с 23-мерной RSS.
[00277] Правильность сборки нацеливающих векторов, описанных в примере 1, и целенаправленной вставки фрагментов ДНК в кластер разнообразия гуманизированного локуса тяжелой цепи иммуноглобулина, содержащегося в ДНК-клонах ВАС, подтверждали с помощью секвенирования и проведения полимеразной цепной реакции на всем протяжении конструирования каждого нацеливающего вектора. Подвергнутую нацеливанию ДНК ВАС, подтвержденную полимеразной цепной реакцией, затем вводили в эмбриональные стволовые (ES) клетки мыши посредством электропорации с последующим культивированием в среде для отбора. Геном ES-клеток, применявшийся для электропорации каждого нацеливающего вектора, изображен на фигурах 3, 5 и 7 соответственно. Колонии, устойчивые к лекарственному средству, отбирали через 10 дней после электропорации и подвергали скринингу с использованием TAQMAN™ и кариотипированию в отношении точности нацеливания, как было описано ранее (Valenzuela et al., выше; Frendewey, D. et al., 2010, Methods Enzymol. 476:295-307, которые включены в данный документ посредством ссылки во всей своей полноте), с применением наборов праймер/зонд, с помощью которых определяли надлежащую интеграцию сконструированных генных сегментов DH (таблица 4; F: прямой праймер, Р: зонд, R: обратный праймер).
[00278] Применяли способ VELOCIMOUSE (DeChiara, Т.М. et al., 2010, Methods Enzymol. 476:285-294; DeChiara, T.M., 2009, Methods Mol. Biol. 530:311-324; Poueymirou et al., 2007, Nat. Biotechnol. 25:91-99, которые включены в данный документ посредством ссылки во всей своей полноте), при котором подвергнутые нацеливанию ES-клетки вводили в эмбрионы мышей Swiss Webster на стадии 8 бластомеров, не прошедших компактизацию, для получения здоровых мышей поколения F0, происходящих полностью из ES-клеток, гетерозиготных по сконструированной DH-области и в которых экспрессируются антитела, содержащие вариабельные области тяжелой цепи, которые содержат области CDR3, полученные в результате DH-DH-рекомбинации. Гетерозиготного самца поколения F0 скрещивали с самками C57B16/NTac для получения гетерозигот F1, которых скрещивали между собой с получением гомозигот поколения F2 и мышей дикого типа.
[00279] Кассету для отбора по устойчивости к лекарственному средству необязательно можно удалить с помощью последовательного добавления рекомбиназы (например, путем обработки с помощью Cre) или путем скрещивания с линией мышей Cre-deleter (см., например, патентную публикацию международной заявки №WO 2009/114400, включенную в данный документ посредством ссылки во всей своей полноте), чтобы удалить любую выбранную кассету, фланкированную lox, которая была введена с помощью нацеливающей конструкции, которую не удаляют, например, на стадии ES-клеток или в эмбрионе. У мышей необязательно сохраняли кассету для отбора. Кассеты для отбора, встроенные с помощью генной инженерии в нацеливающие векторы, описанные в данном документе, удаляли в положительных ES-клетках с помощью временной экспрессии рекомбиназы Cre (см. фигуры 4, 6 и 8 соответственно).




Пример 3. Определение характеристик мышей, модифицированных с применением нацеливающих векторов 23:DH3-3:12/JH6, 23:DH3-3:12/JH4-6 или 12:DH2-2:23|12:DH2-8:23|12:DH2-15:23/JH1-6
[00280] Иммунофенотипирование
[00281] Иммунофенотипический анализ животных, модифицированных с применением нацеливающих векторов 23:DH3-3:12/JH6 или 23:DH3-3:12/JH4-6, проводили с использованием мышей, гетерозиготных по соответствующим модифицированным аллелям 6800 или 6795 (см. фигуры 4 и 6). Иммунофенотипический анализ животных, модифицированных с применением нацеливающего вектора 12:DH2-2:23|12:DH2-8:2-23|12:DH2-15:23/JH1-6, проводили с использованием мышей, которых скрещивали с получением гомозиготности по модифицированному аллелю 20188. Развитие В-клеток анализировали у таких животных с помощью сортировки флуоресцентно-активированных клеток (FACS). Вкратце, забор селезенки и костей нижних конечностей (бедренной кости и большеберцовой кости) производили от:
мышей VELOCIMMUNE® (n=3, 26% C57BL/6 23% 129S6/SvEvTac 51% Balb/cAnNTac; см., например, патенты США №№8502018 и 8642835),
мышей 6643het/6800het//1293ho (25% C57BL/6NTac 25% 129S6/SvEvTac 50% Balb/cAnNTac, модифицированных с применением нацеливающего вектора 23:DH3-3:12/JH6; см. фигуру 4; n=3;
мышей 6643het/6798het//1293ho (25% C57BL/6NTac 25% 129S6/SvEvTac 50% Balb/cAnNTac, модифицированных с применением нацеливающего вектора 23:DH3-3:12/JH4-6; см. фигуру 6; n=3) или
мышей 20188ho/1293ho (25% C57BL/6NTac 25% 129S6/SvFvTac 50% Balb/cAnNTac, модифицированных с применением нацеливающего вектора 12:DH2-2:23|12:DH2-8:23|12:DH2.1523/JH1-6; n=3).
Костный мозг собирали из бедренных костей с помощью центрифугирования. Эритроциты из препаратов селезенки и костного мозга лизировали с помощью буфера для лизиса ACK (Gibco) с последующей промывкой с помощью 1xPBS с 2% FBS. Выделенные клетки (1×106) инкубировали с выбранными смесями антител в течение 30 мин. при +4°С: окрашивание 1: антитело крысы к CD43 мыши, конъюгированное с FITC (Biolegend 121206, клон 1 В11), антитело крысы к с-kit мыши, конъюгированное с РЕ (Biolegend 105808, клон 2 В8), антитело крысы к IgM мыши, конъюгированное с РеСу (eBiosciences 25-5790-82, клон П/41), антитело крысы KlgD мыши, конъюгированное с PerCP-Cy5.5 (Biolegend 405710, клон 11-26 с. 2а), антитело крысы к CD3 мыши, конъюгированное с РВ (Biolegend 100214, клон 17-А2), антитело крысы к В220 мыши, конъюгированное с АРС (eBiosciences 17-0452-82, клон RA3-6B2), и антитело крысы к CD19 мыши, конъюгированное с АРС-Н7 (BD 560143 клон 1D3); окрашивание 2: антитело крысы к легкой каппа-цепи мыши, конъюгированное с FITC (BD 550003, клон 187.1), антитело крысы к легкой лямбда-цепи мыши, конъюгированное с РЕ (Biolegend 407308, клон RML-42), антитело крысы к IgM мыши, конъюгированное с РеСу (eBiosciences 25-5790-82, клон II/41), антитело крысы к IgD мыши, конъюгированное с PerCP-Cy5.5 (Biolegend 405710, клон 11-26 с. 2а), антитело крысы к CD3 мыши, конъюгированное с РВ (Biolegend 100214, клон 17-А2), антитело крысы к В220 мыши, конъюгированное с АРС (eBiosciences 17-0452-82, клон RA3-6B2), и антитело крысы к CD19 мыши, конъюгированное с АРС-Н7 (BD 560143 клон 1D3). После окрашивания клетки промывали и затем фиксировали в 2% формальдегиде. Получение данных осуществляли на проточном цитометре BD LSRFORTESSA™ (BD Biosciences) и анализировали с помощью программного обеспечения FLOWJO™ (BD Biosciences).
[00282] У мышей, гетерозиготных по аллелям 6800 или 6798, общее число В-клеток снизилось как в селезенке, так и в костном мозге и наблюдалось повышение числа про-В-клеток в костном мозге по сравнению с контрольными мышами VELOCIMMUNE® (данные не представлены). В целом, В-клетки характеризовались одинаковым использованием κ и λ (данные не представлены). У мышей, гомозиготных по аллелю 20188, наблюдался немного более высокий уровень λ+В-клеток в селезенке (данные не представлены). Ни одно из отличий, наблюдаемых в популяциях В-клеток у мышей, экспрессирующих аллели 6800, 6798 или 20188, по сравнению с контрольными мышами VELOLCIMMUNE® не считали влияющим на развитие В-клеток этих животных (данные не представлены).
[00283] Частота DH-DH-перегруппировок и определение их характеристик
[00284] Перегруппировки генов у неиммунизированных наивных мышей, модифицированных с применением нацеливающего вектора 23:DH3-3:12/JH6, нацеливающего вектора 23:DH3-3:12/JH4-6 или нацеливающего вектора 12:DH2-2:23|12:DH2-8:23 12:DH2-15:23 анализировали с помощью секвенирования репертуара IgM. Библиотеку репертуара IgM В-клеток селезенки получали с помощью набора для амплификации cDNA SMARTer™ RACE (быстрая амплификация концов cDNA) (Clontech) с применением праймеров, специфичных по отношению к константной области IgM мыши (таблица 5, с помощью «nnnnnn» обозначены последовательности с шагом 6 п. о. для обеспечения мультиплексирования образцов для секвенирования). Полученную библиотеку репертуара затем секвенировали при помощи секвенатора MiSeq (Illumina).
[00285] Последовательности со спаренными концами Illumina MiSeq (2×300 циклов) объединяли и фильтровали на основании качества и полного совпадения с праймером константной области тяжелой цепи IgM. Перегруппированные последовательности тяжелой цепи выравнивали с эталонными базами данных для V, D и J (IMGT) зародышевой линии. Последующим анализам подвергали только продуктивные перегруппировки, называемые последовательностями без стоп-кодонов, в пределах открытой рамки считывания и VDJ-соединения внутри рамки считывания.

[00286] Как ожидалось, у неиммунизированных животных, модифицированных с применением нацеливающего вектора 23:DH3-3:12/JH6 или нацеливающего вектора 23:DH3-3:12/JH4-6, не определялась экспрессия DH7-27 и использование JH соответственно ограничивалось JH6 или JH4, JH5 и JH6 (данные не представлены). У неиммунизированных животных, модифицированных с применением нацеливающего вектора 12:DH2-2:23|12:DH2-8:23|12:DH2-15:23, определялась рекомбинация с участием всех генных сегментов JH1-6, однако рекомбинация с участием JH4 определялась с более высокой частотой (приблизительно на 50%) как у модифицированных, так и у контрольных животных (данные не представлены). У всех модифицированных животных использовались различные генные сегменты VH (например, без ограничения VH4-39, VH3-23 и т.д.) и различные генные сегменты DH (например, без ограничения DH1-7, DH3-10 и DH5-12) рекомбинировались со сконструированными с помощью генной инженерии генными сегментпми DH. Интересно, что использование генных сегментов DH2-2, DH2-8 и DH2-15 было более высоким у животных, модифицированных с применением нацеливающего вектора 12:DH2-2:23|12:DH2-8:23|12:DH2-15:23 (2,41%; 6,34%; 6,91%; n=3) по сравнению с контрольными животными (2,57%, 3,44%; 3,58%; n=3). (Таблица 6).
[00287]Функциональные последовательности, содержащие как минимум два последовательных DH-сегмента, определявшиеся в пределах области соединения, затем подтверждали как полученные в результате события рекомбинации DH-DH с применением следующих критериев. Для последовательностей, выделенных от мышей, модифицированных с применением нацеливающего вектора 23:DH3-3: 12/JH6 или нацеливающего вектора 23:DH3-3: 12/JH4-6, событие рекомбинации DH-DH считали подтвержденным, когда (1) определенный как «успешный» или DH-сегмент на 3'-конце при выравнивании совпадал минимум по 9 последовательным парам оснований с последовательностью зародышевой линии генного сегмента DH IGHD3-3, и (2) оба определенных лидерных на 5'-конце и успешных на 3'-конце генных сегмента DH совпадали при выравнивании минимум по 5 последовательным парам оснований с генным сегментом DH зародышевой линии без учета какого-либо перекрытия. Для последовательностей, выделенных от мышей, модифицированных с применением нацеливающего вектора 12:DH2-2:23|12:DH2-8:23|12:DH2-15:23, событие рекомбинации DH-DH считали подтвержденным, когда (1) определенный как «лидерный» или DH-сегмент на 5'-конце при выравнивании совпадал минимум по 9 последовательным парам оснований с последовательностью зародышевой линии генного сегмента DH IGHD2-2, генного сегмента DH IGHD2-8 или генного сегмента H IGHD2-15, и (2) оба определенных лидерных на 5'-конце и успешных на 3'-конце генных сегмента DH совпадали при выравнивании минимум по 5 последовательным нуклеотидам с генным сегментом DH зародышевой линии без учета какого-либо перекрытия.
[00288] B таблице 6 представлено число функциональных считываний для Ig в виде абсолютной величины и доли в процентах от всех считываний, доли в процентах от функциональных считываний, полученных для генного сегмента DH2-2, генного сегмента DH2-8 или генного сегмента DH2-15, и доли в процентах от всех считываний, полученных для генного сегмента DH2-2, генного сегмента DH2-8 или генного сегмента DH2-15, удовлетворяющих критериям для подтверждения в качестве результата события рекомбинации DH-DH.

*Слияние DH-DH для каждого из этих контрольных животных подтверждали с помощью тех же критериев, какие были указаны для животных, модифицированных с применением нацеливающего вектора 12:DH2-2:23|12:DH2-8:23|12:DH2-15:23.
[00289] Как показано в таблице 6, у всех неиммунизированных модифицированных животных демонстрировалось повышение числа событий рекомбинации DH-DH по сравнению с контрольными животными. У животных, модифицированных с применением сконструированных генных сегментов 2-2, 2-8 и 2-15, например, гомозиготных по аллелю 20188, на события рекомбинации DH-DH приходилось от 1,69% до 4,16% всех перегруппировок, содержащих один из трех сконструированных с помощью генной инженерии генных сегментов DH2-2, 2-8, 2-15. В отличие от этого, у контрольных животных на события рекомбинации DH-DH приходилось менее 0,2% всех перегруппировок с участием генного сегмента 2-2, 2-8 или 2-15. В отдельном эксперименте частота подтвержденных событий рекомбинации DH-DH с участием сконструированного генного сегмента DH3-3 у мышей, гетерозиготных по аллелю 6800 (фигура 4) определялась в диапазоне от 0,6 до 1,2% всех функциональных считываний, полученных для генного сегмента DH3-3.
[00290]У всех модифицированных животных перегруппированные VH(DHA-DHB)JH перегруппировки кодировали CDR3 с большей длиной (например, без ограничения CDR3 с длиной, составляющей приблизительно 21 аминокислоту) и содержащие больше цистеиновых остатков (например, фигуры 9 и 10).
[00291] Таким образом, мыши, модифицированные с применением сконструированного с помощью генной инженерии генного сегмента DH, демонстрировали относительно нормальный фенотип в отношении В-клеток и повышение частоты событий рекомбинации DH-DH за счет использования сконструированного с помощью генной инженерии генного сегмента DH. Более того, рекомбинированные последовательности DH-DH кодировали CDR3 с большей длиной и более высокой частотой внедрения цистеина.
Пример 4. Получение антител
[00292] B данном примере продемонстрировано получение антител у грызуна, геном которого содержит вариабельную область тяжелой цепи иммуноглобулина, которая включает сконструированную с помощью генной инженерии DH-область, описанную в данном документе. При необходимости, для иммунизации грызунов, содержащих сконструированную DH-область, описанную в данном документе, можно применять способы, описанные в данном примере, или способы иммунизации, хорошо известные из уровня техники, с использованием полипептидов или их фрагментов (например, пептиды, полученные из требуемого эпитопа) или комбинации полипептидов или их фрагментов.
[00293] Вкратце, когортам мышей, содержащих сконструированную DH-область, описываемую в данном документе, вводили представляющий интерес антиген с использованием способов иммунизации, известных из уровня техники. Мониторинг гуморального иммунного ответа (т.е. определение титра в сыворотке) осуществляли с помощью иммунологического анализа ELISA и/или полученных клеток почредством MSD. При достижении требуемого иммунного ответа собирали спленоциты (и/или другую лимфатическую ткань) и подвергали их слиянию с клетками миеломы мыши для сохранения их жизнеспособности и образования иммортализированных линий клеток гибридомы. Проводили скрининг (например, с помощью анализа ELISA или MSD) и отбор линий клеток гибридомы для идентификации линий клеток гибридомы, которые продуцировали антиген-специфические антитела. При необходимости, гибридомы можно дополнительно характеризовать в отношении относительной аффинности связывания и изотипу. С использованием данной методики получали несколько антиген-специфических химерных антител (т.е. антител, имеющих вариабельные домены человека и константные домены грызуна).
[00294] ДНК, кодирующую вариабельные участки тяжелой цепи и легких цепей, можно выделять и связывать с требуемыми изотипами (константными областями) тяжелой цепи и легкой цепи для получения полностью человеческих антител. Такое антитело может продуцироваться в клетке, такой как клетка СНО. Полностью человеческие антитела затем характеризуют в отношении относительной аффинности связывания и/или активности нейтрализации представляющего интерес антигена.
[00295] ДНК, кодирующую антиген-специфические химерные антитела или вариабельные домены легкой и тяжелой цепей, можно выделять напрямую из антиген-специфических лимфоцитов. Изначально выделяли химерные антитела с высокой аффинностью, имеющие вариабельную область человека и константную область грызуна, и характеризовали и подвергали отбору в отношении требуемых характеристик, включающих без ограничения аффинность, селективность, эпитоп и т.д. Константные области грызуна (например, без ограничения крысы, мыши и т.д.) заменяли на требуемые константные области человека для получения полностью человеческих антител. Не смотря на то, что отобранная константная область может быть различной в зависимости от конкретного применения, характеристики, касающиеся высокой аффинности связывания антигена или специфичности к мишени, присущи вариабельной области. Антиген-специфические антитела также выделяли напрямую из антиген-положительных В-клеток (из иммунизированных мышей) без слияния с клетками миеломы, как описано, например, в патенте США №7582298, включенном в данный документ посредством ссылки в полном объеме. С помощью данного способа получали несколько полностью человеческих антиген-специфических антител (т.е. антител, обладающих человеческими вариабельными доменами и человеческими константными доменами).
[00296] B одном эксперименте контрольных мышей (например, мышей VELOCIMMUNE® с гуманизированными локусами вариабельной области тяжелой и к легкой цепи иммуноглобулина (n=4-6; см., например, патенты США №№8697940 и 8642835; каждый из которых включен в данный документ посредством ссылки во всей своей полноте)) и тестируемых мышей (например, мышей, модифицированных таким образом, чтобы они содержали сконструированные области 23:DH3-3:12/JH4-6 или 23:DH3-3: 12/JH6, описанные в данном документе (n=4-6)) иммунизировали с помощью разных антигенов и/или нуклеиновых кислот, их кодирующих (рецептора сопряженного с G-белком (GPCR), ионного канала, рецептора хемокина или патогена).
[00297] Мыши могли вырабатывать иммунный ответ к каждому из тестируемых антигенов, при этом выраженность ответа варьировала в диапазоне от низких титров до высоких специфичных титров (данные не предоставлены). Мыши со сконструированной областью 23:DH3-3:12/JH6, описанной в данном документе, демонстрировали титры к клеточной линии экспрессирующей GPCR, сравнимые с контрольными мышами с гуманизированными локусами вариабельной области тяжелой и к легкой цепи иммуноглобулина (фигура 11; закрашенными кругами обозначена клеточная линия, полученная с помощью генной инженерии, которая экспрессировала GPCR; незакрашенными кругами обозначены контрольные клетки, которые не экспрессировали GPCR).
[00298] Более того, 100% из 23 тестируемых антител, специфичных к рецептору, сопряженному с G-белком (GPCR), которые вырабатывались у мышей, содержавших сконструированную с помощью генной инженерии DH-область, имели CDR3 тяжелой цепи длиной 10 или более аминокислот; 20% имели CDR3 длиной 14 аминокислот и 1 антитело имело CDR3 длиной 18 аминокислот (данные не представлены). В отличие от этого, только приблизительно 60% из 8 тестируемых антител, специфичных к GPCR, которые вырабатывались у мышей с гуманизированными локусами вариабельной области тяжелой и к легкой цепи иммуноглобулина, но без сконструированной DH-области, имели CDR3 длиной 10 аминокислот или более, ни одно из них не имело CDR3 длиной 14 или более (данные не представлены).
[00299] Несколько противовирусных антител выделяли от мыши со сконструированной областью 23:DH3-3: 12/JH4-6, где по меньшей мере одно демонстрировало нейтрализующую активность выше по сравнению с молекулой сравнения (данные не предоставлены).
[00300] Антитела от мыши со сконструированной 23:DH3-3: 12/JH6 и иммунизированной при помощи ионного канала, содержали популяцию антител, имеющую HCDR3 длиной более 21 аминокислоты, полученных в результате как перегруппировки VHDHJH, так и перегруппировки VHDHA-DHBJH (как определялось согласно жестким критериям, описанным для данного примера), содержащих последовательности, кодирующие HCDR3 с большей длиной, большее количество которых отмечалось в костном мозге по сравнению с селезенкой (фигура 12). Анализ длин CDR3, кодировавшихся последовательностями, которые определяли как полученные в результате перегруппировки VHDHA-DHBJH в соответствии с жесткими критериями по данному примеру (согласно жестким критериям по данному примеру каждый определяющийся генный сегмент DH должен демонстрировать как минимум 40% идентичность по отношению к генному сегменту D зародышевой линии, генный сегмент DH на 3'-конце должен быть получен из генного сегмента DH3-3, и в каждом генном сегменте DH должна быть максимум 1 нуклеотидная мутация) представлен на фигуре 13. В таблице 7 ниже представлено сравнение показателей частоты рекомбинации VHDHA-DHBJH в костном мозге или селезенке с применением жестких критериев, описанных в данном примере, или нежестких критериев по данному примеру (где согласно нежестким критериям по данному примеру определяемый генный сегмент DH должен демонстрировать идентичность по как минимум 5 нуклеотидам по отношению к генному сегменту D зародышевой линии, генный сегмент DH на 3'-конце должен быть получен из генного сегмента DH3-3).
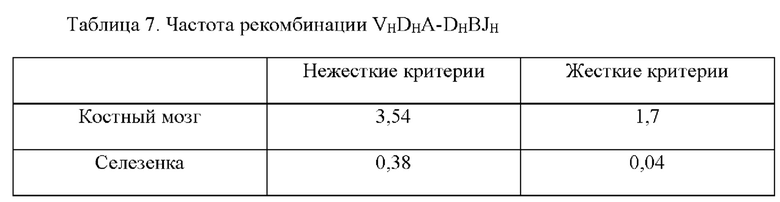
ЭКВИВАЛЕНТЫ
[00301] Рассмотрев описанные таким образом некоторые аспекты по меньшей мере одного варианта осуществления настоящего изобретения, специалистам в данной области будет понятно, что различные изменения, модификации и улучшения будут очевидны специалистам в данной области. Подразумевается, что такие изменения, модификации и улучшения являются частью настоящего изобретения, и подразумевается, что они находятся в пределах сути и объема настоящего изобретения. Соответственно, вышеприведенные описание и графические материалы представлены исключительно в качестве примера, и настоящее изобретение подробно описано в нижеследующей формуле изобретения.
[00302] Специалистам в данной области будут понятны типичные среднеквадратические отклонения или ошибки, присущие значениям, получаемым в анализах или других процессах, описанных в данном документе. Публикации, веб-сайты и другие справочные материалы, упоминаемые в данном документе для описания предпосылок настоящего изобретения и для предоставления дополнительных подробностей, относящихся к его практическому осуществлению, включены в данный документ посредством ссылки во всей своей полноте.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Regeneron Pharmaceuticals, Inc.
MURPHY, Andrew J.
MACDONALD, Lynn
GUO, Chunguang
MCWHIRTER, John
VORONINA, Vera
<120> ЖИВОТНЫЕ, ОТЛИЧНЫЕ ОТ ЧЕЛОВЕКА, СПОСОБНЫЕ К
DH-DH-ПЕРЕГРУППИРОВКЕ, И ПУТИ ИХ ПРИМЕНЕНИЯ
<130> 009108.347/10347
<150> 62685203
<151> 14.06.2018
<150> 62702206
<151> 23.07.2018
<150> 62812580
<151> 01.03.2019
<160> 151
<170> Версия PatentIn 3.5
<210> 1
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> hIgHJ2
<400> 1
gaggctgtgc tactggtact tc
22
<210> 2
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> hIgHJ2
<400> 2
ccgtggcacc ctggtcactg t
21
<210> 3
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> hIgHJ2
<400> 3
tggtgcctgg acagagaag
19
<210> 4
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VI741-42h1
<400> 4
tttttgtgca ccccttaatg g
21
<210> 5
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VI741-42h1
<400> 5
atgtggtatt acgatttttg ga
22
<210> 6
<211> 27
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VI741-42h1
<400> 6
ctctgtgaca ctgtgggtat aataacc
27
<210> 7
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VI742h2
<400> 7
tgtcacagag tccatcaaaa acct
24
<210> 8
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VI742h2
<400> 8
caccaccccc tctc
14
<210> 9
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VI742h2
<400> 9
cctgagaccc tggcaagct
19
<210> 10
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Neo
<400> 10
ggtggagagg ctattcggc
19
<210> 11
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Neo
<400> 11
tgggcacaac agacaatcgg ctg
23
<210> 12
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Neo
<400> 12
gaacacggcg gcatcag
17
<210> 13
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Jxn DH2-2
<400> 13
atggcgcttg cagcgattc
19
<210> 14
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Jxn DH2-2
<400> 14
ccaatactaa cgaggaaatg caaaccca
28
<210> 15
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Jxn DH2-2
<400> 15
accctcactg ttgtccttct g
21
<210> 16
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Jxn DH2-8
<400> 16
gcaaacgtcc atctgaagga gaa
23
<210> 17
<211> 27
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Jxn DH2-8
<400> 17
caaataaacg atggcaggct acacccg
27
<210> 18
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Jxn DH2-8
<400> 18
gagttgccga accgacttat tg
22
<210> 19
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Jxn DH2-15
<400> 19
gcaatggtag gttcatgtcc atc
23
<210> 20
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Jxn DH2-15
<400> 20
accgccgctg gtactccaat
20
<210> 21
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Jxn DH2-15
<400> 21
ccctggctgt ctgggtttg
19
<210> 22
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> hIgHJ6
<400> 22
cagcagaggg ttccatgaga a
21
<210> 23
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> hIgHJ6
<400> 23
caggacaggg ccacggacag tc
22
<210> 24
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> hIgHJ6
<400> 24
gccacccaga gaccttctgt
20
<210> 25
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> hIgH31
<400> 25
atcacactca tcccatcccc
20
<210> 26
<211> 29
<212> ДНК
<213> Искусственная последовательность
<220>
<223> hIgH31
<400> 26
cccttcccta agtaccacag agtgggctc
29
<210> 27
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> hIgH31
<400> 27
cacagggaag caggaactgc
20
<210> 28
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> mIgHp2
<400> 28
gccatgcaag gccaagc
17
<210> 29
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> mIgHp2
<400> 29
ccaggaaaat gctgccagag cctg
24
<210> 30
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> mIgHp2
<400> 30
agttcttgag ccttagggtg ctag
24
<210> 31
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> mADAM6-1
<400> 31
ctgagcatgg tgtcccatct
20
<210> 32
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> mADAM6-1
<400> 32
tgcagataag tgtcatctgg gcaa
24
<210> 33
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> mADAM6-1
<400> 33
ccagagtact gtagtggctc taag
24
<210> 34
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> hIgk5
<400> 34
ccccgtcctc ctcctttttc
20
<210> 35
<211> 32
<212> ДНК
<213> Искусственная последовательность
<220>
<223> hIgk5
<400> 35
tcatgtccat taacccattt accttttgcc ca
32
<210> 36
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> hIgk5
<400> 36
tgcaagtgct gccagcaag
19
<210> 37
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> hIgK21
<400> 37
catttggcta catatcaaag ccg
23
<210> 38
<211> 27
<212> ДНК
<213> Искусственная последовательность
<220>
<223> hIgK21
<400> 38
cctgagccag ggaacagccc actgata
27
<210> 39
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> hIgK21
<400> 39
acatggctga ggcagacacc
20
<210> 40
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> mIgKd2
<400> 40
gcaaacaaaa accactggcc
20
<210> 41
<211> 37
<212> ДНК
<213> Искусственная последовательность
<220>
<223> mIgKd2
<400> 41
ctgttcctct aaaactggac tccacagtaa atggaaa
37
<210> 42
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> mIgKd2
<400> 42
ggccacattc catgggttc
19
<210> 43
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> hyg
<400> 43
tgcggccgat cttagcc
17
<210> 44
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> hyg
<400> 44
acgagcgggt tcggcccatt c
21
<210> 45
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> hyg
<400> 45
ttgaccgatt ccttgcgg
18
<210> 46
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> hIgHD-J.1
<400> 46
tggcagctgt tcaaccatgt
20
<210> 47
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> hIgHD-J.1
<400> 47
atccactgtc ccagacagca cc
22
<210> 48
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> hIgHD-J.1
<400> 48
ggcggtttgt ggagtttcct
20
<210> 49
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> hIgHD-J.2
<400> 49
tgttgcctcc ctgcttctag
20
<210> 50
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> hIgHD-J.2
<400> 50
cccagaccct cccttgttcc tga
23
<210> 51
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> hIgHD-J.2
<400> 51
ggcccacgca ttgttcag
18
<210> 52
<211> 3263
<212> ДНК
<213> Искусственная последовательность
<220>
<223> >p46458 (донор 23-D3-3-12_delJ1-3]
<400> 52
tacgtaccct ggccaccgcc acccgggcct ctctgctgct gggaaaacaa gtcagaaagc
60
aagtggatga gaggtggcgt gacagaccca gcttcagatc tgctctaccg gtataacttc
120
gtataatgta tgctatacga agttatatgc atggcctccg cgccgggttt tggcgcctcc
180
cgcgggcgcc cccctcctca cggcgagcgc tgccacgtca gacgaagggc gcagcgagcg
240
tcctgatcct tccgcccgga cgctcaggac agcggcccgc tgctcataag actcggcctt
300
agaaccccag tatcagcaga aggacatttt aggacgggac ttgggtgact ctagggcact
360
ggttttcttt ccagagagcg gaacaggcga ggaaaagtag tcccttctcg gcgattctgc
420
ggagggatct ccgtggggcg gtgaacgccg atgattatat aaggacgcgc cgggtgtggc
480
acagctagtt ccgtcgcagc cgggatttgg gtcgcggttc ttgtttgtgg atcgctgtga
540
tcgtcacttg gtgagtagcg ggctgctggg ctggccgggg ctttcgtggc cgccgggccg
600
ctcggtggga cggaagcgtg tggagagacc gccaagggct gtagtctggg tccgcgagca
660
aggttgccct gaactggggg ttggggggag cgcagcaaaa tggcggctgt tcccgagtct
720
tgaatggaag acgcttgtga ggcgggctgt gaggtcgttg aaacaaggtg gggggcatgg
780
tgggcggcaa gaacccaagg tcttgaggcc ttcgctaatg cgggaaagct cttattcggg
840
tgagatgggc tggggcacca tctggggacc ctgacgtgaa gtttgtcact gactggagaa
900
ctcggtttgt cgtctgttgc gggggcggca gttatggcgg tgccgttggg cagtgcaccc
960
gtacctttgg gagcgcgcgc cctcgtcgtg tcgtgacgtc acccgttctg ttggcttata
1020
atgcagggtg gggccacctg ccggtaggtg tgcggtaggc ttttctccgt cgcaggacgc
1080
agggttcggg cctagggtag gctctcctga atcgacaggc gccggacctc tggtgagggg
1140
agggataagt gaggcgtcag tttctttggt cggttttatg tacctatctt cttaagtagc
1200
tgaagctccg gttttgaact atgcgctcgg ggttggcgag tgtgttttgt gaagtttttt
1260
aggcaccttt tgaaatgtaa tcatttgggt caatatgtaa ttttcagtgt tagactagta
1320
aattgtccgc taaattctgg ccgtttttgg cttttttgtt agacgtcgag ctagcctaga
1380
ttgggaaccc gggtctctcg aattgttgac aattaatcat cggcatagta tatcggcata
1440
gtataatacg acaaggtgag gaactaaacc atccaccatg attgaacaag atggattgca
1500
cgcaggttct ccggccgctt gggtggagag gctattcggc tatgactggg cacaacagac
1560
aatcggctgc tctgatgccg ccgtgttccg gctgtcagcg caggggcgcc cggttctttt
1620
tgtcaagacc gacctgtccg gtgccctgaa tgaactgcag gacgaggcag cgcggctatc
1680
gtggctggcc acgacgggcg ttccttgcgc agctgtgctc gacgttgtca ctgaagcggg
1740
aagggactgg ctgctattgg gcgaagtgcc ggggcaggat ctcctgtcat ctcaccttgc
1800
tcctgccgag aaagtatcca tcatggctga tgcaatgcgg cggctgcata cgcttgatcc
1860
ggctacctgc ccattcgacc accaagcgaa acatcgcatc gagcgagcac gtactcggat
1920
ggaagccggt cttgtcgatc aggatgatct ggacgaagag catcaggggc tcgcgccagc
1980
cgaactgttc gccaggctca aggcgcgcat gcccgacggc gaggatctcg tcgtgaccca
2040
tggcgatgcc tgcttgccga atatcatggt ggaaaatggc cgcttttctg gattcatcga
2100
ctgtggccgg ctgggtgtgg cggaccgcta tcaggacata gcgttggcta cccgtgatat
2160
tgctgaagag cttggcggcg aatgggctga ccgcttcctc gtgctttacg gtatcgccgc
2220
tcccgattcg cagcgcatcg ccttctatcg ccttcttgac gagttcttct gaagatccgc
2280
tgtaagtctg cagaaattga tgatctatta aacaataaag atgtccacta aaatggaagt
2340
ttttcctgtc atactttgtt aagaagggtg agaacagagt acctacattt tgaatggaag
2400
gattggagct acgggggtgg gggtggggtg ggattagata aatgcctgct ctttactgaa
2460
ggctctttac tattgcttta tgataatgtt tcatagttgg atatcataat ttaaacaagc
2520
aaaaccaaat taagggccag ctcattcctc ccactcatga tctatagatc tatagatctc
2580
tcgtgggatc attgtttttc tcttgattcc cactttgtgg ttctaagtac tgtggtttcc
2640
aaatgtgtca gtttcatagc ctgaagaacg agatcagcag cctctgttcc acatacactt
2700
cattctcagt attgttttgc caagttctaa ttccatcaga cctcgacctg cagccggcgc
2760
gccataactt cgtataatgt atgctatacg aagttatgtc gacctcgaga atttacaaaa
2820
gaaaaggaaa aacacacttg gcagccttca gcactctaat gattcttaac agcagcaaat
2880
tattggcaca agactccaga gtgactggca gggttgaggg ctggggtctc ccacgtgttt
2940
tggggctaac agcggaaggg agagcactgg caaaggtgct gggggcccct ggacccgacc
3000
cgccctggag accgcagcca catcagcccc cagccccaca ggccccctac cagccgcagg
3060
gtttttgtgc accccttaat ggggcctccc acaatgtggt attacgattt ttggagtggt
3120
tattataccc acagtgtcac agagtccatc aaaaaccaag atggctttcc ttctgcctcc
3180
tttctctggg cccagcgtcc tctgtcctgg agctgggaga taatgtccgg gggctccttg
3240
gtctgcgctg ggccatgtac gta
3263
<210> 53
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Сайт распознавания ДНК
<400> 53
tcagaaagca agtggatgag agg
23
<210> 54
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 5' D7.27 gRNA
<400> 54
ucagaaagca aguggaugag guuuuagagc uaugcuguuu ug
42
<210> 55
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Сайт распознавания ДНК
<400> 55
gctccaggac agaggacgct ggg
23
<210> 56
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 3' JH1-3 del gRNA
<400> 56
gcuccaggac agaggacgcu guuuuagagc uaugcuguuu ug
42
<210> 57
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 5' up detect D7-27
<400> 57
gtgaacaggt ggaaccaac
19
<210> 58
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 3' ub pro-200
<400> 58
ccagtgccct agagtcaccc a
21
<210> 59
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 5' neo detect
<400> 59
ctcccactca tgatctatag a
21
<210> 60
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 3' down detect JH1-3 del
<400> 60
gtcccagttc ccaaagaaag
20
<210> 61
<211> 3263
<212> ДНК
<213> Искусственная последовательность
<220>
<223> >p46459 (донор 23-D3-3-12_delJ1-5)
<400> 61
tacgtaccct ggccaccgcc acccgggcct ctctgctgct gggaaaacaa gtcagaaagc
60
aagtggatga gaggtggcgt gacagaccca gcttcagatc tgctctaccg gtataacttc
120
gtataatgta tgctatacga agttatatgc atggcctccg cgccgggttt tggcgcctcc
180
cgcgggcgcc cccctcctca cggcgagcgc tgccacgtca gacgaagggc gcagcgagcg
240
tcctgatcct tccgcccgga cgctcaggac agcggcccgc tgctcataag actcggcctt
300
agaaccccag tatcagcaga aggacatttt aggacgggac ttgggtgact ctagggcact
360
ggttttcttt ccagagagcg gaacaggcga ggaaaagtag tcccttctcg gcgattctgc
420
ggagggatct ccgtggggcg gtgaacgccg atgattatat aaggacgcgc cgggtgtggc
480
acagctagtt ccgtcgcagc cgggatttgg gtcgcggttc ttgtttgtgg atcgctgtga
540
tcgtcacttg gtgagtagcg ggctgctggg ctggccgggg ctttcgtggc cgccgggccg
600
ctcggtggga cggaagcgtg tggagagacc gccaagggct gtagtctggg tccgcgagca
660
aggttgccct gaactggggg ttggggggag cgcagcaaaa tggcggctgt tcccgagtct
720
tgaatggaag acgcttgtga ggcgggctgt gaggtcgttg aaacaaggtg gggggcatgg
780
tgggcggcaa gaacccaagg tcttgaggcc ttcgctaatg cgggaaagct cttattcggg
840
tgagatgggc tggggcacca tctggggacc ctgacgtgaa gtttgtcact gactggagaa
900
ctcggtttgt cgtctgttgc gggggcggca gttatggcgg tgccgttggg cagtgcaccc
960
gtacctttgg gagcgcgcgc cctcgtcgtg tcgtgacgtc acccgttctg ttggcttata
1020
atgcagggtg gggccacctg ccggtaggtg tgcggtaggc ttttctccgt cgcaggacgc
1080
agggttcggg cctagggtag gctctcctga atcgacaggc gccggacctc tggtgagggg
1140
agggataagt gaggcgtcag tttctttggt cggttttatg tacctatctt cttaagtagc
1200
tgaagctccg gttttgaact atgcgctcgg ggttggcgag tgtgttttgt gaagtttttt
1260
aggcaccttt tgaaatgtaa tcatttgggt caatatgtaa ttttcagtgt tagactagta
1320
aattgtccgc taaattctgg ccgtttttgg cttttttgtt agacgtcgag ctagcctaga
1380
ttgggaaccc gggtctctcg aattgttgac aattaatcat cggcatagta tatcggcata
1440
gtataatacg acaaggtgag gaactaaacc atccaccatg attgaacaag atggattgca
1500
cgcaggttct ccggccgctt gggtggagag gctattcggc tatgactggg cacaacagac
1560
aatcggctgc tctgatgccg ccgtgttccg gctgtcagcg caggggcgcc cggttctttt
1620
tgtcaagacc gacctgtccg gtgccctgaa tgaactgcag gacgaggcag cgcggctatc
1680
gtggctggcc acgacgggcg ttccttgcgc agctgtgctc gacgttgtca ctgaagcggg
1740
aagggactgg ctgctattgg gcgaagtgcc ggggcaggat ctcctgtcat ctcaccttgc
1800
tcctgccgag aaagtatcca tcatggctga tgcaatgcgg cggctgcata cgcttgatcc
1860
ggctacctgc ccattcgacc accaagcgaa acatcgcatc gagcgagcac gtactcggat
1920
ggaagccggt cttgtcgatc aggatgatct ggacgaagag catcaggggc tcgcgccagc
1980
cgaactgttc gccaggctca aggcgcgcat gcccgacggc gaggatctcg tcgtgaccca
2040
tggcgatgcc tgcttgccga atatcatggt ggaaaatggc cgcttttctg gattcatcga
2100
ctgtggccgg ctgggtgtgg cggaccgcta tcaggacata gcgttggcta cccgtgatat
2160
tgctgaagag cttggcggcg aatgggctga ccgcttcctc gtgctttacg gtatcgccgc
2220
tcccgattcg cagcgcatcg ccttctatcg ccttcttgac gagttcttct gaagatccgc
2280
tgtaagtctg cagaaattga tgatctatta aacaataaag atgtccacta aaatggaagt
2340
ttttcctgtc atactttgtt aagaagggtg agaacagagt acctacattt tgaatggaag
2400
gattggagct acgggggtgg gggtggggtg ggattagata aatgcctgct ctttactgaa
2460
ggctctttac tattgcttta tgataatgtt tcatagttgg atatcataat ttaaacaagc
2520
aaaaccaaat taagggccag ctcattcctc ccactcatga tctatagatc tatagatctc
2580
tcgtgggatc attgtttttc tcttgattcc cactttgtgg ttctaagtac tgtggtttcc
2640
aaatgtgtca gtttcatagc ctgaagaacg agatcagcag cctctgttcc acatacactt
2700
cattctcagt attgttttgc caagttctaa ttccatcaga cctcgacctg cagccggcgc
2760
gccataactt cgtataatgt atgctatacg aagttatgtc gacctcgaga atttacaaaa
2820
gaaaaggaaa aacacacttg gcagccttca gcactctaat gattcttaac agcagcaaat
2880
tattggcaca agactccaga gtgactggca gggttgaggg ctggggtctc ccacgtgttt
2940
tggggctaac agcggaaggg agagcactgg caaaggtgct gggggcccct ggacccgacc
3000
cgccctggag accgcagcca catcagcccc cagccccaca ggccccctac cagccgcagg
3060
gtttttgtgc accccttaat ggggcctccc acaatgtggt attacgattt ttggagtggt
3120
tattataccc acagtgtcac agagtccatc aaaaacctcc tcaccacccc ctctctgagt
3180
ccacttaggg agactcagct tgccagggtc tcagggtcag agtcttggag gcattttgga
3240
ggtcaggaaa gaaagcctac gta
3263
<210> 62
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Сайт распознавания ДНК
<400> 62
tcagaaagca agtggatgag agg
23
<210> 63
<211> 42
<212> DNA
<213> Искусственная последовательность
<220>
<223> 5' D7.27 crRNA
<400> 63
ucagaaagca aguggaugag guuuuagagc uaugcuguuu ug
42
<210> 64
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Сайт распознавания ДНК
<400> 64
ttagggagac tcagcttgcc agg
23
<210> 65
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 3' JH1-5 del crRNA
<400> 65
uuagggagac ucagcuugcc guuuuagagc uaugcuguuu ug
42
<210> 66
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 5' up detect D7-27
<400> 66
gtgaacaggt ggaaccaac
19
<210> 67
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 3' ub pro-200
<400> 67
ccagtgccct agagtcaccc a
21
<210> 68
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 5' neo detect
<400> 68
ctcccactca tgatctatag a
21
<210> 69
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 3' down detect JH1-5 del
<400> 69
acgcaatcat cacgacagc
19
<210> 70
<211> 3345
<212> ДНК
<213> Искусственная последовательность
<220>
<223> >p51816 (донор 12-D2-2-23)
<400> 70
actagtagaa gataagctcc agacccccaa gactgggacc tgccttcctg ccaccgcgcc
60
ggcgatgcat ccgggtaggg gaggcgcttt tcccaaggca gtctggagtt gacaattaat
120
catcggcata gtatatcggc atagtataat acgacaaggt gaggaactaa accatggaga
180
aaaaaatcac tggatatacc accgttgata tatcccaatg gcatcgtaaa gaacattttg
240
aggcatttca gtcagttgct caatgtacct ataaccagac cgttcagctg gatattacgg
300
cctttttaaa gaccgtaaag aaaaataagc acaagtttta tccggccttt attcacattc
360
ttgcccgcct gatgaatgct catccggaat tccgtatggc aatgaaagac ggtgagctgg
420
tgatatggga tagtgttcac ccttgttaca ccgttttcca tgagcaaact gaaacgtttt
480
catcgctctg gagtgaatac cacgacgatt tccggcagtt tctacacata tattcgcaag
540
atgtggcgtg ttacggtgaa aacctggcct atttccctaa agggtttatt gagaatatgt
600
ttttcgtctc agccaatccc tgggtgagtt tcaccagttt tgatttaaac gtggccaata
660
tggacaactt cttcgccccc gttttcacca tgggcaaata ttatacgcaa ggcgacaagg
720
tgctgatgcc gctggcgatt caggttcatc atgccgtttg tgatggcttc catgtcggca
780
gaatgcttaa tgaattacaa cagtactgcg atgagtggca gggcggggcg taaggatcct
840
gccatttcat tacctctttc tccgcacccg acatagataa agcttatcga taccgtcgac
900
ctcgaggggg ggcccggtac ccgccggcgc ttgtagctcc agacctccgt gcctcccccg
960
accacttact gttagatagg cattgatcta ggcacggggc gcgatgttga cacgggccag
1020
ggagctgttc cacaaagatc aaccccaaac cgggaccgcc tggcactcgc gccgctgcca
1080
cttccctctc catttgttcc cagcacctct gtgctccctc cctcctccct ccttcagggg
1140
aacagcctgt gcagcccctc cctgcacccc acaccctggg gaggcccaac cctgcctcca
1200
gccctttctc ccccgctgct cttcctgccc atccagacaa ccctggggtc ccatccctgc
1260
agcctacacc ctggtctcca cccagacccc tgtctctccc tccagacacc cctcccaggc
1320
caaccctgca cacgcaggcc ctcccctttt ctgctcccag agcctcaccc tctaccctct
1380
gtgcctaccc cctgcctcct cctgcccaca actcgagctc ttcctctcct ggggcccctg
1440
agccatggca ctgaccgtgc actcccaccc ccacactgcc catgccctca ccttcctcct
1500
ggacactctg accccgctcc cctcttggac ccagccctgg tatttccagg acaaaggctc
1560
acccaagtct tccccatgca ggcccttgcc ctcactgccc ggttacacgg cagcctcctg
1620
tgcacagaag cagggagctc agcccttcca caggcagaag gcactgaaag aaatcggcct
1680
ccagcaccct gatgcacgtc cgcctgtgtc tctcactgcc cgcacctgca gggaggctcg
1740
gcactccctg taaagacgag ggatccaggc agcaacatca tgggagaatg cagggctccc
1800
agacagccca gccctctcgc aggcctctcc tgggaagaga cctgcagcca ccactgaaca
1860
gccacggagc ccgctggata gtaactgagt cagtgaccga cctggagggc aggggagcag
1920
tgaaccggag cccagaccat agggacagag accagccgct gacatcccga gcccctcact
1980
ggcggcccca gaacaccgcg tggaaacaga acagacccac attcccacct ggaaaagggc
2040
agacactgct gagcccccag caccagccct gagaaacacc aggcaacggc atcagagggg
2100
gctcctgaga aagaaaggag gggaggtctc cttcaccagc aagtacttcc cttgaccaaa
2160
aacagggtcc acgcaactcc cccaggacaa aggaggagcc ccctgtacag cactgggctc
2220
agagtcctct cccacacacc ctgagtttca gacaaaaacc ccctggaaat catagtatca
2280
gcaggagaac tagccagaga cagcaagagg ggactcagtg actcccgcgg ggacaggagg
2340
attttgtggg ggctcgtgtc actgtgagga tattgtagta gtaccagctg ctatacccac
2400
agtgtgaaaa cccacatcct gagagtgaca caaacccctg ctgtaaacgc ttccacttct
2460
ggagctgagg ggctgggggg agcgtctggg aagtagggcc taggggtggc catcaatgcc
2520
caaaacgcac cagactcccc cccagacatc accccactgg ccagtgagca gagtaaacag
2580
aaaatgagaa gcagctggga agcttgcaca ggccccaagg aaagagcttt ggcgggtgtg
2640
caagagggga tgcgggcaga gcctgagcag ggccttttgc tgtttctgct ttcctgtgca
2700
gatagttcca taaactggtg ttcaagatcg atggctggga gtgagcccag gaggacagtg
2760
tgggaagggc acagggaagg agaagcagcc gctatcctac actgtcatct ttcaagagtt
2820
tgccctgtgc ccacaatgct gcatcatggg atgcttaaca gctgatgtag acacagctaa
2880
agagagaatc agtgaaatgg atttgcagca cagatctgaa taaattctcc agaatgtgga
2940
gccacacaga agcaagcaca aggaaagtgc ctgatgcaag ggcaaagtac agtgtgtacc
3000
ttcaggctgg gcacagacac tctgaaaagc cttggcagga actccctgca acaaagcaga
3060
gccctgcagg caatgccagc tccagagccc tccctgagag cctcatgggc aaagatgtgc
3120
acaacaggtg tttctcatag ccccaaactg agaatgaagc aaacagccat ctgaaggaaa
3180
acaggcaaat aaacgatggc aggttcatgg cgcttgcagc gattcaacag ctaaccaata
3240
ctaacgagga aatgcaaacc cagacagcca gaaggacaac agtgagggtt acaggtgact
3300
ctgtggttga gttcatgaca atgctgagta attggagtaa ctagt
3345
<210> 71
<211> 2484
<212> ДНК
<213> Искусственная последовательность
<220>
<223> >p53419 (донор 12-D2-8-23)
<400> 71
actagtccca gtcctctcac aggcctctcc tgggaagaga cctgaagacg cccagtcgcc
60
ggcgatgcat ccgggtaggg gaggcgcttt tcccaaggca gtctggagtt gacaattaat
120
catcggcata gtatatcggc atagtataat acgacaaggt gaggaactaa accatggaga
180
aaaaaatcac tggatatacc accgttgata tatcccaatg gcatcgtaaa gaacattttg
240
aggcatttca gtcagttgct caatgtacct ataaccagac cgttcagctg gatattacgg
300
cctttttaaa gaccgtaaag aaaaataagc acaagtttta tccggccttt attcacattc
360
ttgcccgcct gatgaatgct catccggaat tccgtatggc aatgaaagac ggtgagctgg
420
tgatatggga tagtgttcac ccttgttaca ccgttttcca tgagcaaact gaaacgtttt
480
catcgctctg gagtgaatac cacgacgatt tccggcagtt tctacacata tattcgcaag
540
atgtggcgtg ttacggtgaa aacctggcct atttccctaa agggtttatt gagaatatgt
600
ttttcgtctc agccaatccc tgggtgagtt tcaccagttt tgatttaaac gtggccaata
660
tggacaactt cttcgccccc gttttcacca tgggcaaata ttatacgcaa ggcgacaagg
720
tgctgatgcc gctggcgatt caggttcatc atgccgtttg tgatggcttc catgtcggca
780
gaatgcttaa tgaattacaa cagtactgcg atgagtggca gggcggggcg taaggatcct
840
gccatttcat tacctctttc tccgcacccg acatagataa agcttatcga taccgtcgac
900
ctcgaggggg ggcccggtac ccgccggcgc aacggagtct aacaccaaac ctccctggag
960
gccgatgggc gagcaaaatc cgacacgtaa ctacaagtag ggcctgatct agtaacggag
1020
tcattgccag acctggaggc aggggagcag tgagcccgag cccacaccat agggccagag
1080
ggcacagcaa ctgacatccc aagccactca ctggtggtcc cacaacaccc catggaaaga
1140
ggacagaccc acagtcccac ctggaccagg gcagagactg ctgagaccca gcaccagaac
1200
caaccaagaa acaccaggca acagcatcag agggggctct ggcagaacag aggaggggag
1260
gtctccttca ccagcaggcg cttcccttga ccgaagacag gatccatgca actcccccag
1320
gacaaaggag gagccccttg ttcagcactg ggctcagagt cctctccaag acacccagag
1380
tttcagacaa aaaccccctg gaatgcacag tctcagcagg agagccagcc agagccagca
1440
agatggggct cagtgacacc cgcagggaca ggaggatttt gtgggggctc gtgtcactgt
1500
gaggatattg tactaatggt gtatgctata cccacagtgt gaaaacccac atcctgagag
1560
tgacacaaac ccctactgca aacgcattcc acttctgggg ctgaggggct gggggagcgt
1620
ctgggaaata gggctcaggg gtgtccatca atgcccaaaa cgcaccagac tcccctccat
1680
acatcacacc caccagccag cgagcagagt aaacagaaaa tgagaagcaa gctggggaag
1740
cttgcacagg ccccaaggaa agagctttgg cgggtgtgta agaggggatg cgggcagagc
1800
ctgagcaggg ccttttgctg tttctgcttt cctgtgcaga gagttccata aactggtgtt
1860
cgagatcaat ggctgggagt gagcccagga ggacagcgtg ggaagagcac agggaaggag
1920
gagcagccgc tatcctacac tgtcatcttt caaaagtttc ccttgtgccc acattgctgc
1980
atcatgggat gcttaagagc tgatgtagac acagctaaag agagaatcag tgagatggat
2040
ttgcagcaca gatctgaata aattctccag aatgtggagc agcacagaag caagcacaca
2100
gaaagtgcct gatgcaagga caaagttcag tgggcacctt caggcattgc tgctgggcac
2160
agacattctg aaaagccctg gcaggaactc cctgtgacaa agcagaaccc tcaggcaatg
2220
ccagccccag agccctccct gagagcctca tgggcaaaga tgtgcacaac aggtgtttct
2280
catagcccca aactgagagc aaagcaaacg tccatctgaa ggagaacagg caaataaacg
2340
atggcaggct acacccgtgc atccaataag tcggttcggc aactcaactt catgaaatgc
2400
aaacccagac agccacaagc acaaaagtac agggttataa gcgactctgg ttgagttcat
2460
gacaatgctg agtaattgac tagt
2484
<210> 72
<211> 2321
<212> ДНК
<213> Искусственная последовательность
<220>
<223> >p51818 (донор 12-D2-15-23)
<400> 72
actagtgaga ccagccgctg acatcccgag ctcctcaatg gtggccccat aacacacgcc
60
ggcgatgcat ccgggtaggg gaggcgcttt tcccaaggca gtctggagtt gacaattaat
120
catcggcata gtatatcggc atagtataat acgacaaggt gaggaactaa accatggaga
180
aaaaaatcac tggatatacc accgttgata tatcccaatg gcatcgtaaa gaacattttg
240
aggcatttca gtcagttgct caatgtacct ataaccagac cgttcagctg gatattacgg
300
cctttttaaa gaccgtaaag aaaaataagc acaagtttta tccggccttt attcacattc
360
ttgcccgcct gatgaatgct catccggaat tccgtatggc aatgaaagac ggtgagctgg
420
tgatatggga tagtgttcac ccttgttaca ccgttttcca tgagcaaact gaaacgtttt
480
catcgctctg gagtgaatac cacgacgatt tccggcagtt tctacacata tattcgcaag
540
atgtggcgtg ttacggtgaa aacctggcct atttccctaa agggtttatt gagaatatgt
600
ttttcgtctc agccaatccc tgggtgagtt tcaccagttt tgatttaaac gtggccaata
660
tggacaactt cttcgccccc gttttcacca tgggcaaata ttatacgcaa ggcgacaagg
720
tgctgatgcc gctggcgatt caggttcatc atgccgtttg tgatggcttc catgtcggca
780
gaatgcttaa tgaattacaa cagtactgcg atgagtggca gggcggggcg taaggatcct
840
gccatttcat tacctctttc tccgcacccg acatagataa agcttatcga taccgtcgac
900
ctcgaggggg ggcccggtac ccgccggcgc ctaggaaaca taacacaccc acagccccac
960
ctggaacaga tagatgcacc aatatctccc agtaagctca agctcgcggg gcagagactg
1020
ctgagccccc agcaccagcc ccaagaaaca ccaggcaaca gtatcagagg gggctcccga
1080
gaaagagagg aggggagatc tccttcacca tcaaatgctt cccttgacca aaaacagggt
1140
ccacgcaact cccccaggac aaaggaggag ccccctatac agcactgggc tcagagtcct
1200
ctctgagaca ccctgagttt cagacaacaa cccgctggaa tgcacagtct cagcaggaga
1260
acagaccaaa gccagcaaaa gggacctcgg tgacaccagt agggacagga ggattttgtg
1320
ggggctcgtg tcactgtgag gatattgtag tggtggtagc tgctactccc acagtgtgaa
1380
aacccacatc ctgagagtga cacaaacccc tactgcaaac acccccactc ctggggctga
1440
ggggctaggg gagcgtctgg gaagtagggt ccaggggtgt ctatcaatgt ccaaaatgca
1500
ccagactccc cgccaaacac caccccacca gccagcgagc agggtaaaca gaaaatgaga
1560
ggctctggga agcttgcaca ggccccaagg aaagagcttt ggcgggtgtg caagagggga
1620
tgcaggcaga gcctgagcag ggccttttgc tgtttctgct ttcctgtgca gagagttcca
1680
taaactggtg ttcaagatca gtggctggga atgagcccag gagggcagtc tgtgggaaga
1740
gcacagggaa ggaggagcag ccgctatcct acactgtcat ctttcaaaag tttgccttgt
1800
gaccacacta ttgcatcatg ggatgcttaa gagctgatgt agacacagct aaagagagaa
1860
tcagtgagat gaatttgcag catagatctg aataaactct ccagaatgtg gagcagtaca
1920
gaagcaaaca cacagaaagt gcctgatgca aggacaaagt tcagtgggca ccttcaggca
1980
ttgctgctgg gcacagacac tctgaaaagc cctggcagga tctccctgcg acaaagcaga
2040
accctcaggc aatgccagcc ccagagccct ccctgagagc gtcatgggga aagatgtgca
2100
gaacagctga ttatcataga ctcaaactga gaacagagca aacgtccatc tgaagaacag
2160
tcaaataagc aatggtaggt tcatgtccat ctcatcggtc agtaaaaata ccgccgctgg
2220
tactccaatg caaacccaga cagccagggg acaacagtag agggctacag gcggctttgc
2280
ggttgagttc atgacaatgc tgagtaattg gagtaactag t
2321
<210> 73
<211> 40
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 1 P51816(D2-2)
<400> 73
tgttagatag gcattgatct aggcacgggg cgcgatgttg
40
<210> 74
<211> 40
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 2 P51816(D2-2)
<400> 74
gcgcttgcag cgattcaaca gctaaccaat actaacgagg
40
<210> 75
<211> 40
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 10 P53419(D2-8)
<400> 75
cgagcaaaat ccgacacgta actacaagta gggcctgatc
40
<210> 76
<211> 40
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 16 P53419(D2-8)
<400> 76
ctacacccgt gcatccaata agtcggttcg gcaactcaac
40
<210> 77
<211> 40
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 8 P51818(D2-15)
<400> 77
atagatgcac caatatctcc cagtaagctc aagctcgcgg
40
<210> 78
<211> 40
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 18 P51818(D2-15)
<400> 78
tccatctcat cggtcagtaa aaataccgcc gctggtactc
40
<210> 79
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 5' hIgHD HU2(h268)
<400> 79
gcagtaaccc tcaggaagca
20
<210> 80
<211> 31
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 3' hIgHD HU2 AgeI(h269)
<400> 80
cttccaccgg tacagccaca ccaaggtcat c
31
<210> 81
<211> 31
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 5' hIgHD HD2 XhoI(h271)
<400> 81
cttccctcga ggaacactgt cagctcccac a
31
<210> 82
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 3' hIgHD HD2(h272)
<400> 82
agatcctcca tgcgtgctg
19
<210> 83
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 5' hIgHD HU2 detect(h270)
<400> 83
tctcctctcg tcgcctctac
20
<210> 84
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 3' ub pro-200
<400> 84
ccagtgccct agagtcaccc a
21
<210> 85
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 5' neo detect
<400> 85
ctcccactca tgatctatag a
21
<210> 86
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 3' hIgHD HD2 detect(h273)
<400> 86
gggtgcttgg gtcctgttag
20
<210> 87
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 5' D2-2 Cas9 ДНК-мишень
<400> 87
aggtctggag ctacaagcgg tgg
23
<210> 88
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 5' D2-2 gRNA
<400> 88
aggucuggag cuacaagcgg guuuuagagc uaugcuguuu ug
42
<210> 89
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 3' D2-2 ДНК-мишень для Cas9
<400> 89
aggacaacag tgagggttac agg
23
<210> 90
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 3' D2-2 gRNA
<400> 90
aggacaacag ugaggguuac guuuuagagc uaugcuguuu ug
42
<210> 91
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 5' up detect D2-2(h274)
<400> 91
gtcaaaggtg gaggcagtg
19
<210> 92
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 3' cm up detect
<400> 92
tccagctgaa cggtctggtt a
21
<210> 93
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> D2-2seqF2
<400> 93
agcgattcaa cagctaacc
19
<210> 94
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 3' down detect D2-2(h275)
<400> 94
ccagtagaag taacgaccac
20
<210> 95
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> JO-1
<400> 95
ctccagaccc ccaagactgg gacctgcctt cctgccaccg cttgtagctc cagacctccg
60
tgcctccccc gaccacttac
80
<210> 96
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 5' up detect D2-2(h274)
<400> 96
gtcaaaggtg gaggcagtg
19
<210> 97
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> D2-2seqR1
<400> 97
cccgtgccta gatcaatgc
19
<210> 98
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> D2-2seqF1
<400> 98
aggcattgat ctaggcacg
19
<210> 99
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> D2-2seqR2
<400> 99
ctgttgaatc gctgcaagc
19
<210> 100
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 5' D2-8 ДНК-мишень для Cas9
<400> 100
acctgaagac gcccagtcaa cgg
23
<210> 101
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 5' D2-8 crRNA
<400> 101
accugaagac gcccagucaa guuuuagagc uaugcuguuu ug
42
<210> 102
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 3' D2-8 ДНК-мишень для Cas9
<400> 102
gccacaagca caaaagtaca ggg
23
<210> 103
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 3' D2-8 crRNA
<400> 103
gccacaagca caaaaguaca guuuuagagc uaugcuguuu ug
42
<210> 104
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 5' up detect D2-8(h276)
<400> 104
gccttaccca agtctttcc
19
<210> 105
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 3' cm up detect
<400> 105
tccagctgaa cggtctggtt a
21
<210> 106
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> D2-8seqF2
<400> 106
tccaataagt cggttcggc
19
<210> 107
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 3' down detect D2-8(h277)
<400> 107
gcagaagtaa ccaccactg
19
<210> 108
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> JO-2
<400> 108
tcacaggcct ctcctgggaa gagacctgaa gacgcccagt caacggagtc taacaccaaa
60
cctccctgga ggccgatggg
80
<210> 109
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 5' up detect D2-8(h276)
<400> 109
gccttaccca agtctttcc
19
<210> 110
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> D2-8seqR1
<400> 110
gccctacttg tagttacgtg
20
<210> 111
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> D2-8seqF1
<400> 111
gcaaaatccg acacgtaac
19
<210> 112
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> D2-8seqR2
<400> 112
cgaaccgact tattggatgc
20
<210> 113
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 5' D2-8 ДНК-мишень для Cas9
<400> 113
ggtggcccca taacacacct agg
23
<210> 114
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 5' D2-8 crRNA
<400> 114
gguggcccca uaacacaccu guuuuagagc uaugcuguuu ug
42
<210> 115
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 3' D2-8 ДНК-мишень для Cas9
<400> 115
ggacaacagt agagggctac agg
23
<210> 116
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 3' D2-8 crRNA
<400> 116
ggacaacagu agagggcuac guuuuagagc uaugcuguuu ug
42
<210> 117
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 5' up detect D2-15(h278)
<400> 117
gcactccctc taaagacaag
20
<210> 118
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 3' cm up detect
<400> 118
tccagctgaa cggtctggtt a
21
<210> 119
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> D2-15seqF2
<400> 119
gtaaaaatac cgccgctgg
19
<210> 120
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 3' down detect D2-15(h279)
<400> 120
gcattttcct gttcttgcg
19
<210> 121
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> JO-3
<400> 121
gctgacatcc cgagctcctc aatggtggcc ccataacaca cctaggaaac ataacacacc
60
cacagcccca cctggaacag
80
<210> 122
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 5' up detect D2-15(h278)*
<400> 122
gcactccctc taaagacaag
20
<210> 123
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> D2-15seqR1
<400> 123
ctgggagata ttggtgcatc
20
<210> 124
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> D2-15seqF1
<400> 124
tgcaccaata tctcccagt
19
<210> 125
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> D2-15seqR2
<400> 125
cggcggtatt tttactgacc
20
<210> 126
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 23D3.312-1
<400> 126
tttttgtgca ccccttaatg g
21
<210> 127
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 23D3.312-1
<400> 127
atgtggtatt acgatttttg ga
22
<210> 128
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 23D3.312-1
<400> 128
tctgtgacac tgtgggtata ataacc
26
<210> 129
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 23D3.312-2
<400> 129
cccacagtgt cacagagtcc at
22
<210> 130
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 23D3.312-2
<400> 130
caagatggct ttccttctgc ctcc
24
<210> 131
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 23D3.312-2
<400> 131
tcccagctcc aggacagag
19
<210> 132
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 23D3.312-3
<400> 132
tgtcacagag tccatcaaaa acct
24
<210> 133
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 23D3.312-3
<400> 133
caccaccccc tctc
14
<210> 134
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 23D3.312-3
<400> 134
cctgagaccc tggcaagct
19
<210> 135
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Jxn DH2-2
<400> 135
atggcgcttg cagcgattc
19
<210> 136
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Jxn DH2-2
<400> 136
ccaatactaa cgaggaaatg caaaccca
28
<210> 137
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Jxn DH2-2
<400> 137
accctcactg ttgtccttct g
21
<210> 138
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Jxn DH2-8
<400> 138
gcaaacgtcc atctgaagga gaa
23
<210> 139
<211> 27
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Jxn DH2-8
<400> 139
caaataaacg atggcaggct acacccg
27
<210> 140
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Jxn DH2-8
<400> 140
gagttgccga accgacttat tg
22
<210> 141
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Jxn DH2-15
<400> 141
gcaatggtag gttcatgtcc atc
23
<210> 142
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Jxn DH2-15
<400> 142
accgccgctg gtactccaat
20
<210> 143
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Jxn DH2-15
<400> 143
ccctggctgt ctgggtttg
19
<210> 144
<211> 7
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Искусственный
<400> 144
cacagtg
7
<210> 145
<211> 9
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Искусственный
<400> 145
acaaaaacc
9
<210> 146
<211> 29
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотид для смены матрицы
<400> 146
agcagtggta tcaacgcaga gtacatggg
29
<210> 147
<211> 54
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймеры к константной области IgM для 1-го цикла ПЦР
<400> 147
acactctttc cctacacgac gctcttccga tctgggaaga catttgggaa ggac
54
<210> 148
<211> 57
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймеры к PE2-PIIA для 1-го цикла ПЦР
<400> 148
gtgactggag ttcagacgtg tgctcttccg atctaagcag tggtatcaac gcagagt
57
<210> 149
<211> 68
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймеры для 2-го цикла ПЦР
<220>
<221> другой_признак
<222> (30)..(35)
<223> n представляет собой a, c, g или t
<400> 149
aatgatacgg cgaccaccga gatctacacn nnnnnacact ctttccctac acgacgctct
60
tccgatct
68
<210> 150
<211> 64
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймеры для 2-го цикла ПЦР
<220>
<221> другой_признак
<222> (25)..(30)
<223> n представляет собой a, c, g или t
<400> 150
caagcagaag acggcatacg agatnnnnnn gtgactggag ttcagacgtg tgctcttccg
60
atct
64
<210> 151
<211> 39
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 23-мерная RSS из VH1-69
<400> 151
cacagtgtga aaacccacat cctgagagtg acacaaacc
39
<---


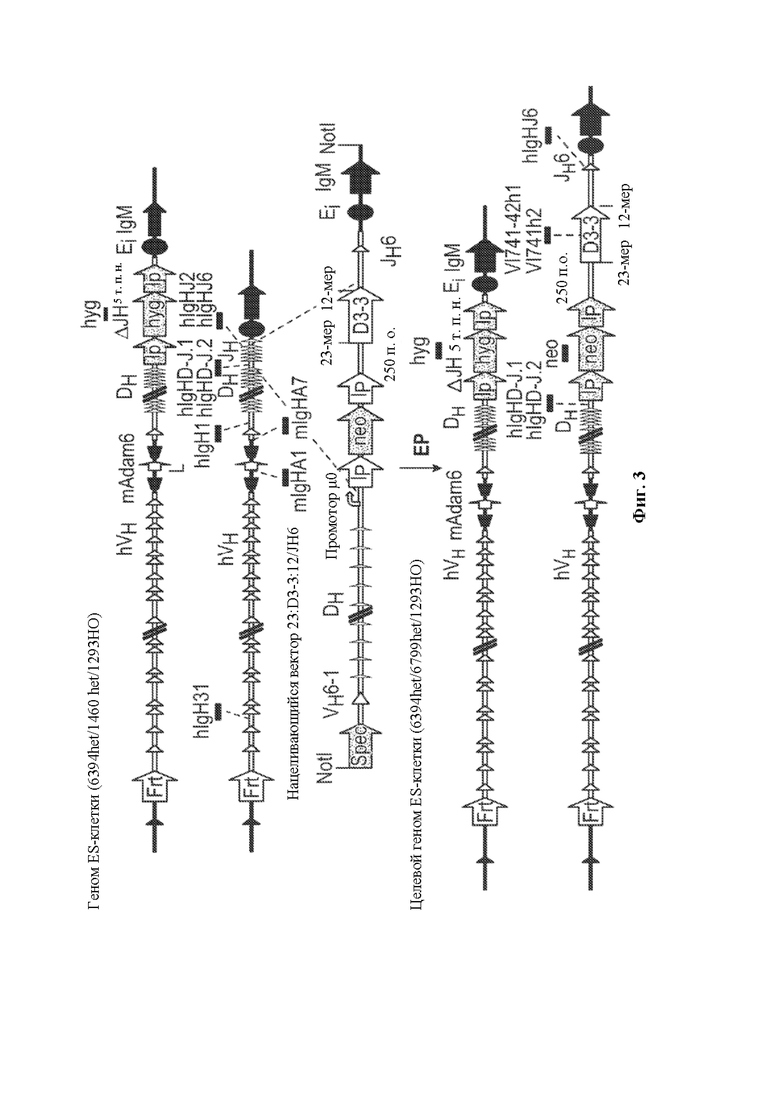
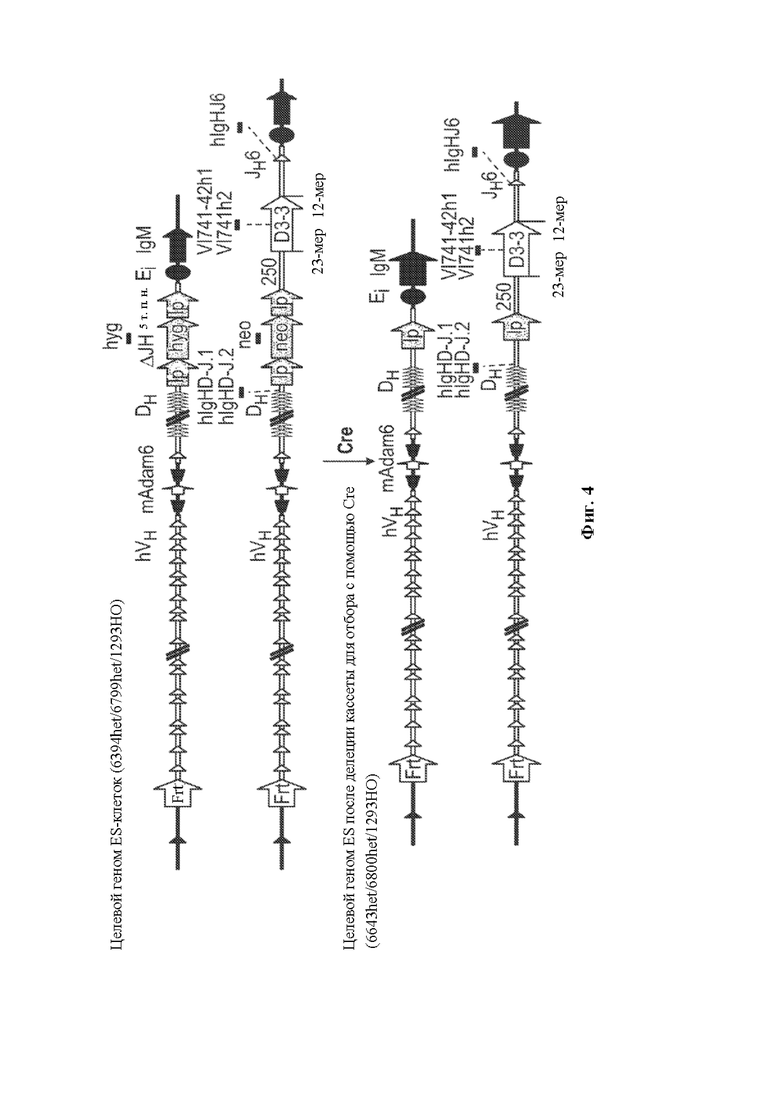
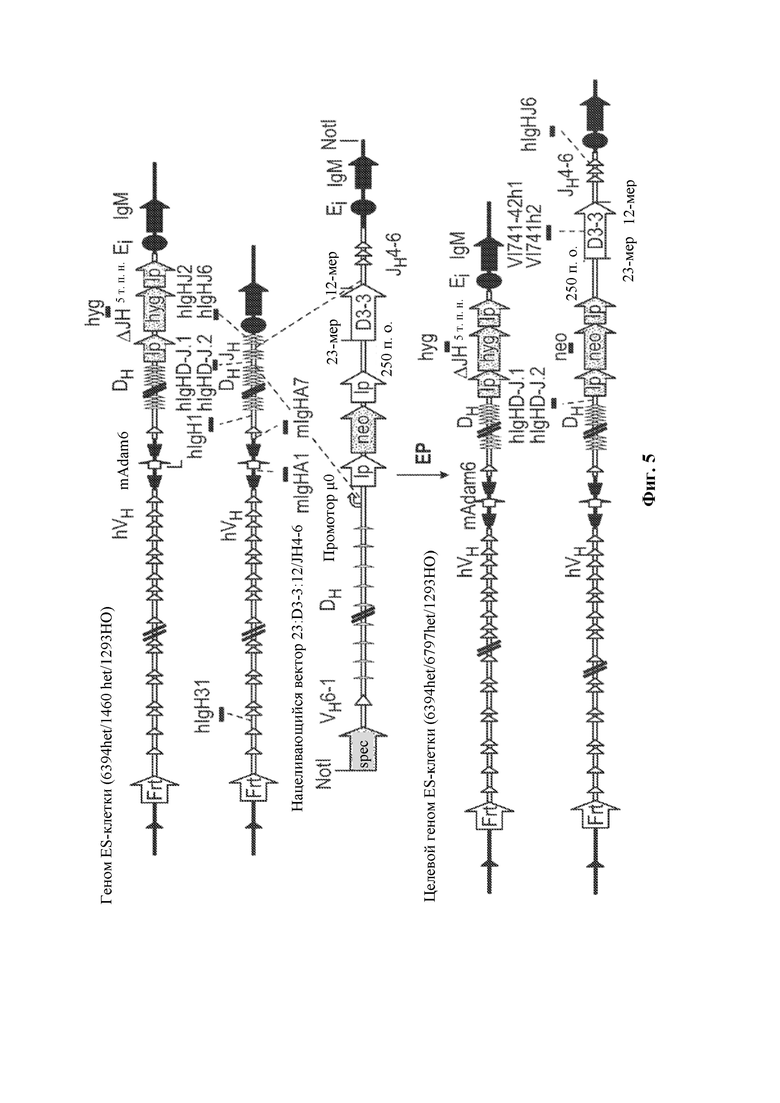


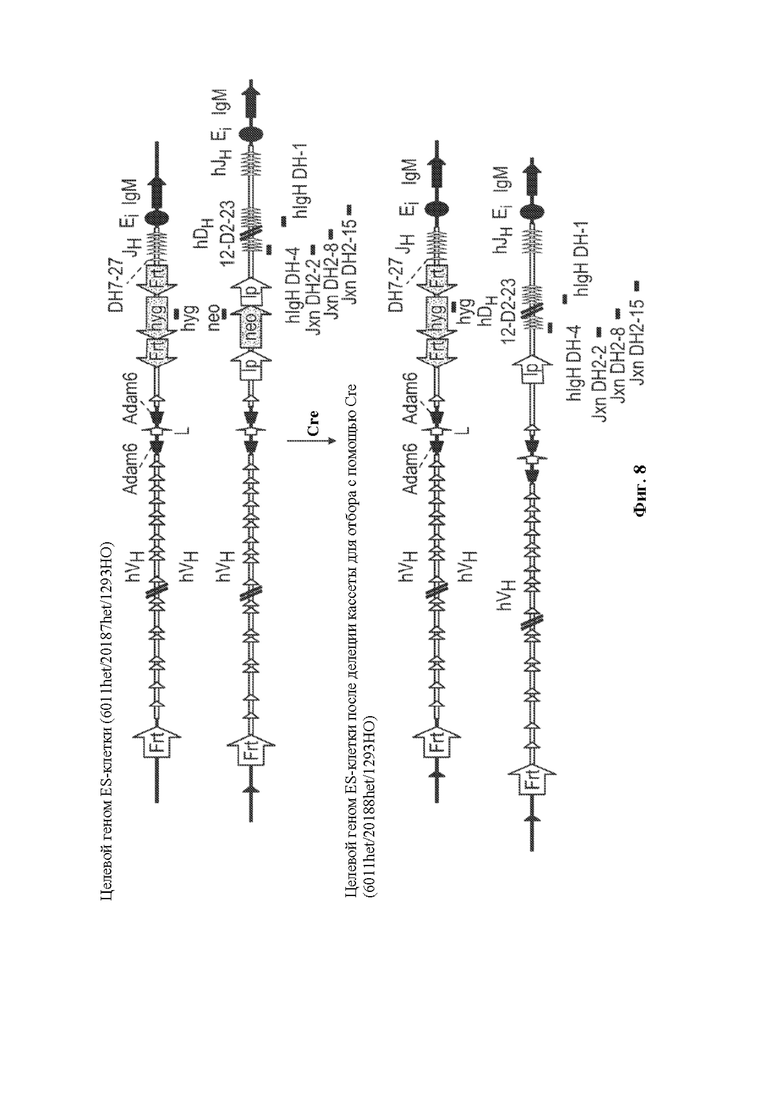

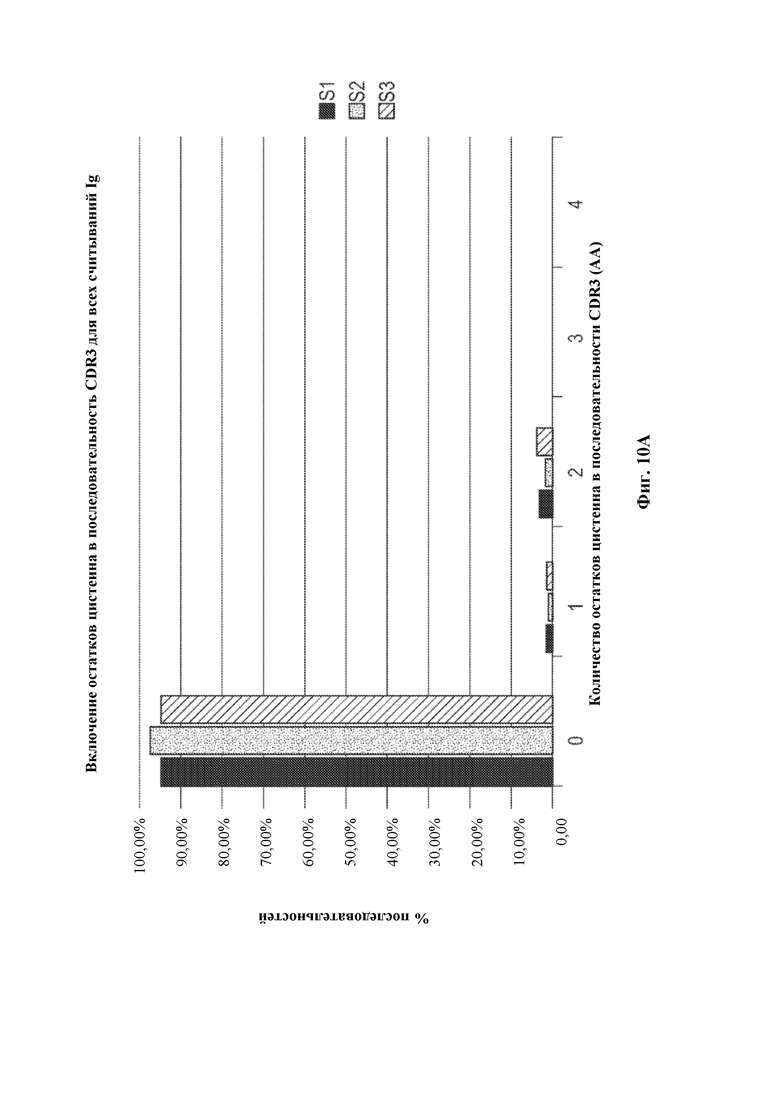
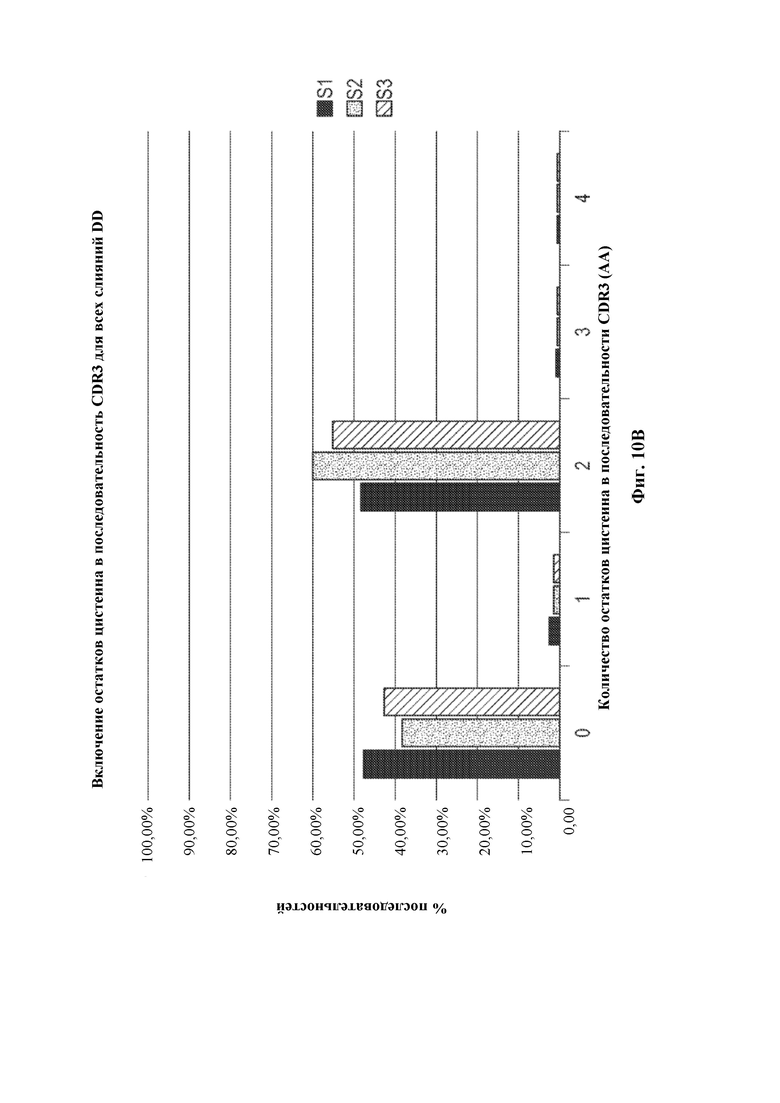
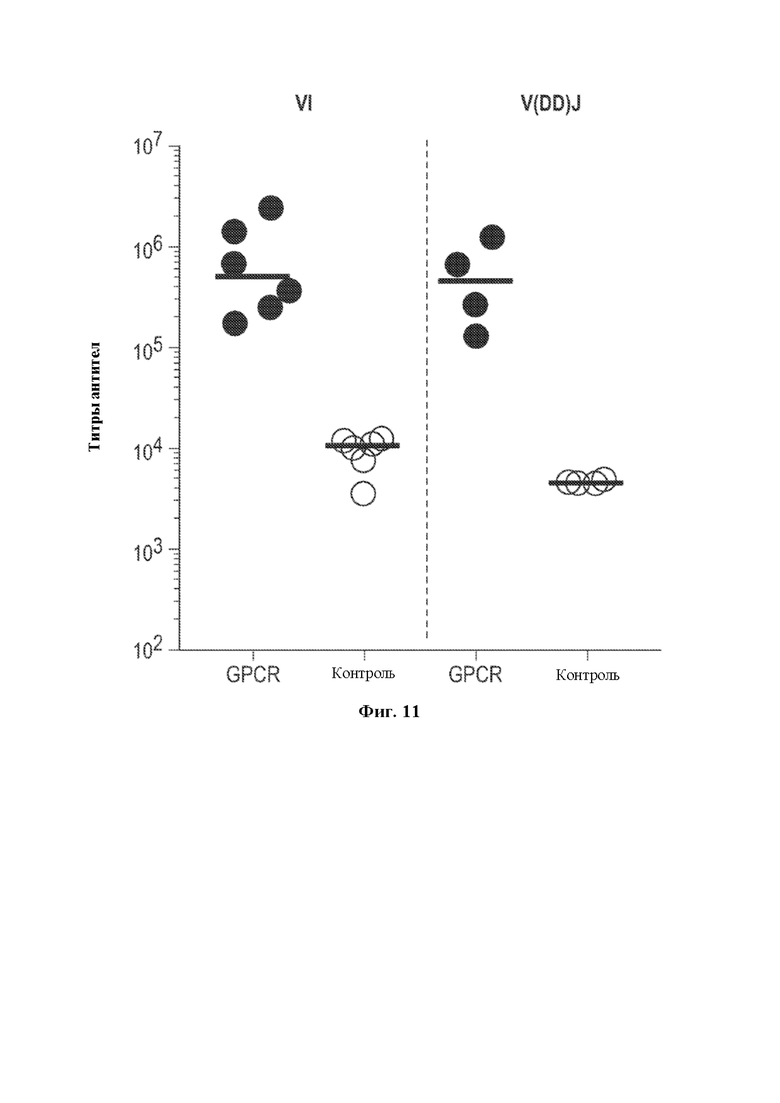


Изобретение относится к области биотехнологии. Описана группа изобретений, включающая нуклеотидную молекулу для конструирования рекомбинации DH-DH, нацеливающий вектор, способ модифицирования вариабельной области тяжелой цепи иммуноглобулина для конструирования рекомбинации DH-DH у животного, нуклеиновую кислоту тяжелой цепи иммуноглобулина для конструирования рекомбинации DH-DH, нуклеиновую кислоту грызуна для конструирования рекомбинации DH-DH, клетку грызуна для конструирования рекомбинации DH-DH, грызуна для конструирования рекомбинации DH-DH, эмбрион грызуна для конструирования рекомбинации DH-DH, способ получения грызуна, геном которого содержит сконструированную DH-область (варианты), способ получения антитела и способ получения нуклеиновой кислоты, кодирующей антитело. Изобретение расширяет арсенал средств для конструирования рекомбинации DH-DH. 12 н. и 66 з.п. ф-лы, 17 ил., 7 табл., 4 пр.
1. Нуклеотидная молекула, содержащая сконструированную определяющую вариабельность область тяжелой цепи (DH) иммуноглобулина, для конструирования рекомбинации DH-DH, где сконструированная DH-область содержит:
(i) сконструированный генный сегмент DH, содержащий генный сегмент DH, непосредственно прилегающий к 23-мерной последовательности сигнала рекомбинации (RSS), и
(ii) неперегруппированный генный сегмент DH, фланкированный со своего 5’-конца первой 12-мерной RSS и со своего 3’-конца второй 12-мерной RSS, и
где (i) сконструированный генный сегмент DH и (ii) неперегруппированный генный сегмент DH функционально связаны.
2. Нуклеотидная молекула по п. 1, где нуклеотидная молекула дополнительно содержит:
(a) по меньшей мере один неперегруппированный вариабельный (VH) генный сегмент тяжелой цепи иммуноглобулина, который находится выше и функционально связан со сконструированной DH-областью,
(b) по меньшей мере один неперегруппированный соединяющий (JH) генный сегмент тяжелой цепи иммуноглобулина, который находится ниже и функционально связан со сконструированной DH-областью, или
(c) комбинацию (a) и (b).
3. Нуклеотидная молекула по п. 2, где
(a) по меньшей мере один неперегруппированный генный сегмент VH содержит неперегруппированный человеческий генный сегмент VH6-1,
(b) сконструированная DH-область содержит только человеческий генный сегмент DH,
(c) по меньшей мере один неперегруппированный генный сегмент JH содержит неперегруппированный человеческий генный сегмент JH6 или
(d) любую комбинацию (a), (b) и (c).
4. Нуклеотидная молекула по п. 2 или 3, где по меньшей мере один неперегруппированный генный сегмент JH содержит неперегруппированный человеческий генный сегмент JH4, неперегруппированный человеческий генный сегмент JH5 и неперегруппированный человеческий генный сегмент JH6.
5. Нуклеотидная молекула по любому из пп. 2-4, где по меньшей мере один неперегруппированный генный сегмент JH содержит неперегруппированный человеческий генный сегмент JH1, неперегруппированный человеческий генный сегмент JH2, неперегруппированный человеческий генный сегмент JH3, неперегруппированный человеческий генный сегмент JH4, неперегруппированный человеческий генный сегмент JH5 и неперегруппированный человеческий генный сегмент JH6.
6. Нуклеотидная молекула по любому из пп. 2-5, где по меньшей мере один неперегруппированный генный сегмент VH содержит полный репертуар функциональных неперегруппированных человеческих генных сегментов VH.
7. Нуклеотидная молекула по любому из пп. 1-6, где нуклеотидная молекула дополнительно содержит один или более функциональных генов Adam6 грызуна.
8. Нуклеотидная молекула по любому из пп. 1-7, где сконструированный генный сегмент DH содержит 23-мерную RSS, непосредственно прилегающую к 5’-концу генного сегмента DH.
9. Нуклеотидная молекула по любому из пп. 1-7, где сконструированный генный сегмент DH содержит 23-мерную RSS, непосредственно прилегающую к 3’-концу генного сегмента DH.
10. Нуклеотидная молекула по любому из пп. 1-9, где сконструированный генный сегмент DH содержит:
(a) человеческий генный сегмент DH3-3, непосредственно прилегающий к 23-мерной RSS;
(b) человеческий генный сегмент DH2-2, непосредственно прилегающий к 23-мерной RSS;
(c) человеческий генный сегмент DH2-8, непосредственно прилегающий к 23-мерной RSS;
(d) человеческий генный сегмент DH2-15, непосредственно прилегающий к 23-мерной RSS; или
(e) любую комбинацию (a)-(d).
11. Нуклеотидная молекула по любому из пп. 1-10, содержащая нуклеотидную последовательность, представленную под SEQ ID NO: 52.
12. Нуклеотидная молекула по любому из пп. 1-11, содержащая нуклеотидную последовательность, представленную под SEQ ID NO: 61.
13. Нуклеотидная молекула по любому из пп. 1-12, содержащая нуклеотидную последовательность, представленную под SEQ ID NO: 70.
14. Нуклеотидная молекула по любому из пп. 1-13, содержащая нуклеотидную последовательность, представленную под SEQ ID NO: 71.
15. Нуклеотидная молекула по любому из пп. 1-14, содержащая нуклеотидную последовательность, представленную под SEQ ID NO: 72.
16. Нуклеотидная молекула по любому из пп. 1-5, содержащая в функциональной связи от 5’-конца к 3’-концу:
(a) по меньшей мере один неперегруппированный человеческий генный сегмент VH;
(b) сконструированную DH-область, где сконструированный генный сегмент DH содержит человеческий генный сегмент DH3-3, непосредственно прилегающий к 23-мерной RSS, где 23-мерная RSS непосредственно прилегает к 5’-концу человеческого генного сегмента DH3-3; и
(c) по меньшей мере один неперегруппированный человеческий генный сегмент JH.
17. Нуклеотидная молекула по любому из пп. 1-15, содержащая в функциональной связи от 5’-конца к 3’-концу:
(a) по меньшей мере один неперегруппированный человеческий генный сегмент VH;
(b) сконструированную DH-область, где
(i) сконструированный генный сегмент DH содержит человеческий генный сегмент DH2, непосредственно прилегающий к 23-мерной RSS, где 23-мерная RSS непосредственно прилегает к 3’-концу человеческого генного сегмента DH2;
(ii) сконструированный генный сегмент DH содержит человеческий генный сегмент DH2-2, непосредственно прилегающий к 23-мерной RSS;
(iii) сконструированный генный сегмент DH содержит человеческий генный сегмент DH2-8, непосредственно прилегающий к 23-мерной RSS;
(iv) сконструированный генный сегмент DH содержит человеческий генный сегмент DH2-15, непосредственно прилегающий к 23-мерной RSS; или
(v) любая их комбинация, и
(c) по меньшей мере один неперегруппированный человеческий генный сегмент JH.
18. Нуклеотидная молекула по п. 16 или 17, где по меньшей мере один неперегруппированный человеческий генный сегмент VH содержит от 5’-конца к 3’-концу неперегруппированный человеческий генный сегмент VH1-2 и неперегруппированный человеческий генный сегмент VH6-1, где нуклеотид дополнительно содержит один или более функциональных генов Adam6 грызуна между неперегруппированным человеческим генным сегментом VH1-2 и неперегруппированным человеческим генным сегментом VH6-1 и где неперегруппированный человеческий генный сегмент VH1-2, один или более функциональных генов Adam6 грызуна и неперегруппированный человеческий генный сегмент VH6-1 являются смежными.
19. Нуклеотидная молекула по любому из пп. 16-18, где по меньшей мере один неперегруппированный человеческий генный сегмент VH содержит полный репертуар функциональных неперегруппированных человеческих генных сегментов VH.
20. Нуклеотидная молекула по любому из пп. 16-19, где по меньшей мере один неперегруппированный человеческий генный сегмент JH содержит неперегруппированный человеческий генный сегмент JH6.
21. Нуклеотидная молекула по п. 20, где по меньшей мере один неперегруппированный человеческий генный сегмент JH содержит неперегруппированный человеческий генный сегмент JH4, неперегруппированный человеческий генный сегмент JH5 и неперегруппированный человеческий генный сегмент JH6.
22. Нуклеотидная молекула по п. 21, где по меньшей мере один неперегруппированный человеческий генный сегмент JH содержит неперегруппированный человеческий генный сегмент JH1, неперегруппированный человеческий генный сегмент JH2, неперегруппированный человеческий генный сегмент JH3, неперегруппированный человеческий генный сегмент JH4, неперегруппированный человеческий генный сегмент JH5 и неперегруппированный человеческий генный сегмент JH6.
23. Нуклеотидная молекула по любому из пп. 1-22, которая дополнительно содержит на своем 3’-конце константную (CH) область тяжелой цепи иммуноглобулина или ее часть.
24. Нуклеотидная молекула по п. 23, где CH-область или ее часть представляет собой CH-область грызуна или ее часть.
25. Нуклеотидная молекула по п. 24, где при этом CH-область грызуна или ее часть содержит область интронного энхансера грызуна и ген IgM грызуна.
26. Нуклеотидная молекула по любому из пп. 1-25, дополнительно содержащая кассету для отбора по устойчивости к лекарственному средству, которая расположена выше сконструированного генного сегмента DH.
27. Нуклеотидная молекула по любому из пп. 1-25, где (i) сконструированный генный сегмент DH и (ii) неперегруппированный генный сегмент DH могут соединиться при событии рекомбинации DH-DH в соответствии с правилом 12/23.
28. Нацеливающийся вектор, содержащий нуклеотидную молекулу по любому из пп. 1-26 и 5’- и 3’-плечи гомологии, расположенные в конфигурации для создания условий для гомологичной рекомбинации с последовательностью тяжелой цепи иммуноглобулина в эндогенном локусе тяжелой цепи иммуноглобулина грызуна.
29. Способ модифицирования вариабельной области тяжелой цепи иммуноглобулина для конструирования рекомбинации DH-DH у животного, отличного от человека, при этом способ включает:
(а) получение последовательности вариабельной цепи тяжелой цепи иммуноглобулина, содержащей DH-область, содержащую один или более неперегруппированных генных сегментов DH, где каждый неперегруппированный генный сегмент DH фланкирован на своем 5’-конце первой 12-мерной последовательностью сигнала рекомбинации (RSS) и на своем 3’-конце второй 12-мерной RSS, и
(b) конструирование DH-области таким образом, чтобы она дополнительно содержала по меньшей мере один сконструированный генный сегмент DH, где сконструированный генный сегмент DH содержит генный сегмент DH, непосредственно прилегающий к 23-мерной RSS,
где в полученной сконструированной DH-области сконструированный генный сегмент DH и по меньшей мере один из одного или более неперегруппированных генных сегментов DH функционально связаны и могут соединиться при событии рекомбинации DH-DH в соответствии с правилом 12/23.
30. Способ по п. 29, где один или более неперегруппированных генных сегментов DH содержат по меньшей мере один неперегруппированный человеческий генный сегмент DH.
31. Способ по п. 30, где по меньшей мере один из неперегруппированных человеческих генных сегментов DH содержит все функциональные человеческие генные сегменты DH.
32. Способ по любому из пп. 29-31, где DH-область, содержащая один или более неперегруппированных генных сегментов DH, содержит два или более неперегруппированных генных сегмента DH, каждый из которых фланкирован на своем 5’-конце первой 12-мерной RSS и на своем 3’-конце второй 12-мерной RSS, а конструирование включает:
(a) замену по меньшей мере одного из двух или более неперегруппированных генных сегментов DH на сконструированный генный сегмент DH,
(b) замену первой 12-мерной RSS или второй 12-мерной RSS по меньшей мере одним из двух или более неперегруппированных генных сегментов DH на 23-мерную RSS или
(c) комбинацию (a) и (b).
33. Способ по п. 32, где последовательность вариабельной цепи тяжелой цепи иммуноглобулина дополнительно содержит JH-область, содержащую по меньшей мере один неперегруппированный генный сегмент JH,
где JH-область функционально связана с DH-областью и
где замена по меньшей мере одного из двух или более неперегруппированных генных сегментов DH на сконструированный генный сегмент DH включает делецию по меньшей мере одного неперегруппированного генного сегмента JH.
34. Способ по п. 33, где JH-область содержит полный репертуар человеческих генных сегментов JH зародышевой линии, содержащий неперегруппированный человеческий генный сегмент JH1, неперегруппированный человеческий генный сегмент JH2, неперегруппированный человеческий генный сегмент JH3, неперегруппированный человеческий генный сегмент JH4, неперегруппированный человеческий генный сегмент JH5 и неперегруппированный человеческий генный сегмент JH6, и
где делеция по меньшей мере одного генного сегмента JH зародышевой линии включает делецию
(i) неперегруппированного человеческого генного сегмента JH1, неперегруппированного человеческого генного сегмента JH2 и неперегруппированного человеческого генного сегмента JH3 или
(ii) неперегруппированного человеческого генного сегмента JH1, неперегруппированного человеческого генного сегмента JH2, неперегруппированного человеческого генного сегмента JH3, неперегруппированного человеческого генного сегмента JH4 и неперегруппированного человеческого генного сегмента JH5.
35. Способ по любому из пп. 29-34, где сконструированный генный сегмент DH содержит:
(a) человеческий генный сегмент DH3-3, непосредственно прилегающий к 23-мерной RSS,
(b) человеческий генный сегмент DH2-2, непосредственно прилегающий к 23-мерной RSS,
(c) человеческий генный сегмент DH2-8, непосредственно прилегающий к 23-мерной RSS,
(d) человеческий генный сегмент DH2-15, непосредственно прилегающий к 23-мерной RSS, или
(e) любую комбинацию (a)-(d).
36. Способ по любому из пп. 32-35, где конструирование включает замену ближайшего к 3’-концу неперегруппированного генного сегмента DH DH-области на сконструированный генный сегмент DH.
37. Способ по любому из пп. 32-36, где конструирование включает замену по меньшей мере одного из одного или более неперегруппированных генных сегментов DH, который соответствует сконструированному генному сегменту DH на сконструированный генный сегмент DH.
38. Нуклеиновая кислота тяжелой цепи иммуноглобулина для конструирования рекомбинации DH-DH, содержащая нуклеотидную молекулу по любому из пп. 1-27 или нацеливающийся вектор по п. 28.
39. Нуклеиновая кислота грызуна для конструирования рекомбинации DH-DH, содержащая нуклеотидную молекулу по любому из пп. 1-27, нацеливающийся вектор по п. 28 или нуклеиновую кислоту тяжелой цепи иммуноглобулина по п. 38.
40. Нуклеиновая кислота грызуна по п. 39, где нуклеиновая кислота грызуна содержит вариабельную область тяжелой цепи иммуноглобулина, которая содержит:
(i) неперегруппированные генные сегменты VH,
(ii) сконструированную DH-область и
(iii) неперегруппированные генные сегменты JH,
где (i)-(iii) находятся в функциональной связи таким образом, чтобы после рекомбинации вариабельная область тяжелой цепи иммуноглобулина содержала перегруппированную последовательность вариабельной области тяжелой цепи, кодирующую вариабельный домен тяжелой цепи иммуноглобулина.
41. Нуклеиновая кислота грызуна по п. 40, где сконструированный генный сегмент DH содержит генный сегмент DH, непосредственно прилегающий к 23-мерной RSS 3’-конца.
42. Нуклеиновая кислота грызуна по п. 40 или 41, где сконструированный генный сегмент DH содержит генный сегмент DH, непосредственно прилегающий к 23-мерной RSS 5’-конца.
43. Нуклеиновая кислота грызуна по любому из пп. 41, 42, где вариабельная область тяжелой цепи иммуноглобулина представляет собой вариабельную область тяжелой цепи человеческого иммуноглобулина, содержащую только человеческие генные сегменты VH, человеческие генные сегменты DH и человеческие генные сегменты JH.
44. Нуклеиновая кислота грызуна по п. 43, где вариабельная область тяжелой цепи человеческого иммуноглобулина функционально связана с CH-областью.
45. Нуклеиновая кислота грызуна по п. 44, где CH-область представляет собой эндогенную CH-область.
46. Нуклеиновая кислота грызуна по любому из пп. 40-45, где неперегруппированные генные сегменты VH содержат полный репертуар функциональных неперегруппированных человеческих генных сегментов VH.
47. Нуклеиновая кислота грызуна по любому из пп. 40-46, где неперегруппированные генные сегменты JH содержат человеческий генный сегмент JH6.
48. Нуклеиновая кислота грызуна по любому из пп. 40-47, где в нуклеиновой кислоте грызуна отсутствует эндогенный ген Adam6.
49. Нуклеиновая кислота грызуна по любому из пп. 40-47, дополнительно содержащая один или более генов Adam6 грызуна.
50. Нуклеиновая кислота грызуна по п. 49, где один или более генов Adam6 грызуна вставлены между двумя человеческими генными сегментами VH.
51. Нуклеиновая кислота грызуна по п. 50, где один или более генов Adam6 грызуна вставлены между неперегруппированным человеческим генным сегментом VH1-2 и неперегруппированным человеческим генным сегментом VH6-1.
52. Нуклеиновая кислота грызуна по п. 49, где нуклеиновая кислота не содержит человеческий псевдоген Adam6.
53. Нуклеиновая кислота грызуна по п. 49, где один или более генов Adam6 грызуна вставлены между неперегруппированным человеческим генным сегментом VH и неперегруппированным человеческим генным сегментом DH.
54. Нуклеиновая кислота грызуна по любому из пп. 39-53, дополнительно содержащая ген терминальной дезоксинуклеотидилтрансферазы.
55. Нуклеиновая кислота грызуна по любому из пп. 39-54, характеризующаяся тем, что она является гомозиготной по сконструированной DH-области.
56. Нуклеиновая кислота грызуна по любому из пп. 39-54, характеризующаяся тем, что она является гетерозиготной по сконструированной DH-области.
57. Нуклеиновая кислота грызуна по любому из пп. 39-56, где нуклеиновая кислота грызуна представляет собой нуклеиновую кислоту крысы или нуклеиновую кислоту мыши.
58. Клетка грызуна для конструирования рекомбинации DH-DH, содержащая нуклеиновую кислоту грызуна по любому из пп. 39-57.
59. Клетка грызуна по п. 58, где клетка грызуна представляет собой клетку крысы или клетку мыши.
60. Клетка грызуна по п. 58 или 59, где клетка грызуна представляет собой эмбриональную стволовую клетку грызуна.
61. Грызун для конструирования рекомбинации DH-DH, полученный из эмбриональной стволовой клетки грызуна по п. 60, где грызун содержит нуклеиновую кислоту грызуна в геноме своей зародышевой линии.
62. Грызун по п. 61, дополнительно содержащий соматический геном, который содержит перегруппированную кодирующую последовательность VH(DHA-DHB)JH тяжелой цепи,
где перегруппированная кодирующая последовательность VH(DHA-DHB)JH тяжелой цепи функционально связана с последовательностью гена константной области IgM и/или где DHA или DHB получены из сконструированного генного сегмента DH или его части и по меньшей мере 9 пар оснований из DHA или DHB являются идентичными по меньшей мере 9 парам оснований из сконструированного генного сегмента DH.
63. Грызун по п. 61 или 62, где:
(i) по меньшей мере 95% от всех перегруппированных кодирующих последовательностей тяжелой цепи VDJ у грызуна имеют длину CDR3, составляющую по меньшей мере 10 аминокислот;
(ii) по меньшей мере 70% от всех перегруппированных кодирующих последовательностей тяжелой цепи VDJ у грызуна имеют длину CDR3, составляющую по меньшей мере 11 аминокислот;
(iii) по меньшей мере 15% от всех перегруппированных кодирующих последовательностей тяжелой цепи VDJ у грызуна имеют длину CDR3, составляющую по меньшей мере 14 аминокислот или
(iv) любая комбинация из (i), (ii) и (iii).
64. Эмбрион грызуна для конструирования рекомбинации DH-DH, полученный из эмбриональной стволовой клетки грызуна по п. 60, где эмбрион грызуна содержит нуклеиновую кислоту грызуна в своем геноме.
65. Способ получения грызуна, геном которого содержит сконструированную DH-область, при этом способ включает получение грызуна из эмбриональной стволовой клетки по п. 60.
66. Способ получения грызуна, геном которого содержит сконструированную DH-область, при этом способ включает
(a) модифицирование генома эмбриональной стволовой клетки грызуна таким образом, чтобы он содержал фрагмент ДНК, содержащий один или более сконструированных генных сегментов DH, каждый из которых содержит генный сегмент DH, непосредственно прилегающий к 23-мерной RSS, где фрагмент ДНК содержит нуклеотидную молекулу по любому из пп. 1-27, нацеливающийся вектор по п. 28 или нуклеиновую кислоту тяжелой цепи иммуноглобулина по п. 38, и
(b) получение грызуна с применением модифицированной эмбриональной стволовой клетки грызуна из стадии (a).
67. Способ по п. 65 или 66, где грызун представляет собой крысу или мышь.
68. Способ получения антитела, включающий
иммунизацию грызуна с помощью антигена, где указанный грызун содержит в геноме своей зародышевой линии нуклеиновую кислоту грызуна в соответствии с любым из пп. 39-57, и
обеспечение индукции у грызуна иммунного ответа на антиген, включая образование антитела, которое связывает антиген.
69. Способ по п. 68, дополнительно включающий извлечение антитела из грызуна или клетки грызуна.
70. Способ по п. 68, где клетка грызуна представляет собой В-клетку или гибридому.
71. Способ по любому из пп. 68-70, где один или более сконструированных генных сегментов DH, каждый из которых непосредственно прилегает к 23-мерной RSS, содержит генный сегмент DH, функционально связанный с 23-мерной RSS 5’-конца.
72. Способ по любому из пп. 68-70, где один или более сконструированных генных сегментов DH, каждый из которых непосредственно прилегает к 23-мерной RSS, содержит генный сегмент DH, функционально связанный с 23-мерной RSS 3’-конца.
73. Способ по любому из пп. 67-72, где грызун представляет собой крысу или мышь.
74. Способ получения нуклеиновой кислоты, кодирующей антитело, включающий:
иммунизацию грызуна антигеном, при этом грызун содержит в геноме своей зародышевой линии нуклеиновую кислоту грызуна по любому из пп. 39-57, и
обеспечение индукции у грызуна иммунного ответа на антиген, включая образование нуклеиновой кислоты, кодирующей антитело, которое связывает антиген.
75. Способ по п. 74, дополнительно включающий извлечение нуклеиновой кислоты, кодирующей антитело, из грызуна или клетки грызуна.
76. Способ по п. 75, где клетка грызуна представляет собой В-клетку или гибридому.
77. Способ по любому из пп. 74-76, где один или более сконструированных генных сегментов DH, каждый из которых непосредственно прилегает к 23-мерной RSS, содержит генный сегмент DH, функционально связанный с 23-мерной RSS 5’-конца.
78. Способ по любому из пп. 74-76, где один или более сконструированных генных сегментов DH, каждый из которых непосредственно прилегает к 23-мерной RSS, содержит генный сегмент DH, функционально связанный с 23-мерной RSS 3’-конца.
| TUAILLON N | |||
| et al., VHD rearrangements in human immunoglobulin heavy chain minilocus transgenic mice, European Journal of Immunology, 2000, V | |||
| Способ обработки медных солей нафтеновых кислот | 1923 |
|
SU30A1 |
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| JUNG D | |||
| et al., Mechanism and control of V (D) J recombination at the immunoglobulin heavy chain locus, Annu | |||
| Rev | |||
| Immunol, 2006, V | |||
| Пишущая машина для тюркско-арабского шрифта | 1922 |
|
SU24A1 |
| ЛАДА А | |||
| Г | |||
| и др., ВОПЛОЩЕНИЕ | |||
