Область изобретения
Данное изобретение относится к композициям для стимуляции иммунного ответа млекопитающих, особенно, Т-клеточного ответа. Изобретение также относится к медицинскому применению таких композиций в иммуноонкологии, включая противоопухолевую терапию, путем промотирования противоопухолевого Т-клеточного ответа у пациента, а также для использования композиций при других заболеваниях и состояниях, когда модулирование баланса между эффекторными Т-клетками и регуляторными Т-клетками в пользу активности эффекторных Т-клеток, например, путем стимуляции эффекторных Т-клеток и/или путем деплетирования регуляторных Т-клеток, создает терапевтически полезный эффект.
Уровень техники
ICOS (индуцибельный Т-клеточный костимулятор) является членом семейства генов CD28, участвующих в регуляции иммунных ответов, в частности, гуморальных иммунных реакций, впервые идентифицированным в 1999 году [1]. Он представляет собой 55 кДа трансмембранный белок, существующий в виде дисульфидно-связанного гомодимера с двумя дифференциально гликозилированными субъединицами. ICOS экспрессируется исключительно на Т-лимфоцитах и присутствует на различных субпопуляциях Т-клеток. Он присутствует на низких уровнях на наивных Т-лимфоцитах, но его экспрессия быстро индуцируется при иммунной активации, усиливаясь в ответ на провоспалительные стимулы, такие как вовлечение TCR и костимуляция с помощью CD28 [2, 3]. ICOS играет определенную роль в поздней фазе активации Т-клеток, формировании Т-клеток памяти и, что важно, в регуляции гуморальных ответов через Т-зависимые В-клеточные ответы [4, 5]. Внутриклеточно ICOS связывает PI3K и активирует киназы фосфатидилинозитол-зависимую киназу 1 (PDK1) и протеинкиназу В (PKB). Активация ICOS предотвращает гибель клеток и усиливает клеточный метаболизм. В отсутствие ICOS (нокаут ICOS) или в присутствии анти-ICOS нейтрализующих антител будет наблюдаться супрессия провоспалительных реакций.
ICOS связывается с лигандом ICOS (ICOSL), экспрессируемым на В-клетках и антигенпрезентирующих клетках (АРС) [6, 7]. В качестве костимулирующей молекулы он участвует в регулировании TCR-опосредованных иммунных ответов и ответов антител на антиген. Экспрессия ICOS на регуляторных Т-клетках может быть важной, поскольку было высказано предположение, что этот тип клеток играет отрицательную роль в иммунологическом надзоре онкологических клеток - имеются свежие данные, подтверждающие это, при раке яичников [8]. Важно отметить, что экспрессия ICOS, как сообщается, является повышенной на внутриопухолевых регуляторных Т-клетках (TRegs) по сравнению с CD4+ и CD8+ эффекторными клетками, присутствующими в микроокружении опухоли. Деплетирование TRegs с использованием антител с Fc-опосредованной клеточной эффекторной функцией продемонстрировало сильную противоопухолевую эффективность в доклинической модели [9]. Все увеличивающийся объем данных указывает на участие ICOS в противоопухолевом эффекте как в животных моделях, так и у пациентов, получающих ингибиторы иммунных контрольных точек. У мышей, дефицитных по ICOS или ICOSL, противоопухолевый эффект анти-CTLA4 терапии снижается [10], тогда как у нормальных мышей лиганд ICOS повышает эффективность анти-CTLA4 лечения при меланоме и раке предстательной железы [11]. Кроме того, ретроспективное исследование пациентов-людей с запущенной меланомой показало повышенные уровни ICOS после лечения ипилимумабом (анти-CTLA4) [12]. Кроме того, экспрессия ICOS повышается у пациентов с раком мочевого пузыря, получавших анти-CTLA4 [13]. Было также отмечено, что у онкологических пациентов, получавших анти-CTLA4 терапию, основная часть опухолеспецифических продуцирующих IFNγ CD4 Т-клеток является ICOS-положительной, а устойчивое повышение ICOS-положительных CD4-Т-клеток коррелирует с выживаемостью [12, 13, 14].
WO 2016/120789 описывает анти-ICOS антитела и предлагает их использование для активации Т-клеток и для лечения рака, инфекционных заболеваний и/или сепсиса. Был получен ряд мышиных анти-ICOS анти-ICOS антител, субпопуляция которых, как сообщается, является агонистами рецептора ICOS человека. Антитело «422.2» было выбрано в качестве оптимизированного прототипа анти-ICOS антитела и было гуманизировано с получением человеческого антитела IgG4PE, обозначенного как «H2L5». Как сообщалось, H2L5 имеет аффинность 1,34 нМ по отношению к человеческому ICOS и 0,95 нМ - к ICOS яванского макака, способно индуцировать продуцирование цитокинов в Т-клетках и увеличивать экспрессию маркеров активации Т-клеток в сочетании со стимуляцией CD3. Однако, сообщалось, что мыши, несущие имплантированные клетки меланомы человека, демонстрируют только минимальные эффекты задержки роста опухоли или увеличения выживаемости при лечении H2L5 hIgG4PE по сравнению с группой, получавшей контрольное лечение. Антитело также не дало значительного дополнительного ингибирования роста опухоли в экспериментах с комбинациями с ипилимумабом (анти-CTLA-4) или пембролизумабом (анти-PD-1) по сравнению с монотерапией ипилимумабом или пембролизумабом. Наконец, у мышей с имплантированными клетками рака толстой кишки (СТ26), низкие дозы мышиного перекрестно-реактивного суррогата H2L5 в сочетании с мышиным суррогатом ипилимумаба или пембролизумаба лишь незначительно улучшали общую выживаемость по сравнению с использованием только анти-CTL4 и анти-PD1 терапии. Аналогичное отсутствие сильного терапевтического эффекта было показано для мышей, несущих имплантированные клетки ЕМТ6.
В WO 2016/154177 описаны дополнительные примеры анти- ICOS анти-ICOS антител. Сообщалось, что эти антитела являются агонистами CD4+ Т-клеток, включая эффекторные CD8+ Т-клетки (TEff), и деплетируют Т-регуляторные клетки (TRegs). Было описано селективное воздействие антител на клетки TEff по сравнению с TReg, причем антитела могут преимущественно деплетировать TRegs, оказывая в то же время минимальное влияние на TEffs, которые экспрессируют более низкий уровень ICOS. Анти-ICOS антитела были предложены для использования при лечении рака, и была описана комбинированная терапия анти-PD-1 или анти-PD-L1 антителами.
Сущность изобретения
Антитело к ICOS, вызывающее повышение активности эффекторных Т-клеток, представляет терапевтический подход в иммуноонкологии и других медицинских контекстах, в которых ответ CD8+ Т-клеток является полезным, включая различные заболевания и состояния, и в схемах вакцинации. При многих заболеваниях и состояниях, включающих иммунный компонент, существует баланс между эффекторными Т-клетками (TEff), которые обеспечивают иммунный ответ CD8+ Т-клеток, и регуляторными Т-клетками (TReg), которые подавляют этот иммунный ответ путем снижения уровня TEffs. Данное изобретение относится к антителам, которые модулируют этот баланс TEff/TReg в пользу активности эффекторных Т-клеток. Антитела, инициирующие деплетирование высокоположительных по ICOS регуляторных Т-клеток, облегчали бы супрессию TEffs и, таким образом, создавали бы суммарный эффект промотирования эффекторного ответа Т-клеток. Дополнительный или комплементарный механизм анти-ICOS антитела заключается в агонистической активности на уровне рецептора ICOS, стимулирующей ответ эффекторных Т-клеток.
Относительная экспрессия ICOS на эффекторных Т-клетках (TEff) по сравнению с регуляторными Т-клетками (TReg) и относительные активности этих клеточных популяций будут влиять на общий эффект анти-ICOS антитела in vivo. Предусматриваемый способ действия сочетает агонизм эффекторных Т-клеток с деплетированием ICOS-положительных регуляторных Т-клеток. Дифференциальные и даже противоположно направленные эффекты воздействия на эти две разные популяции Т-клеток могут быть достигнуты из-за их различных уровней экспрессии ICOS. Двойная модификация вариабельной и постоянной областей, соответственно, анти-ICOS антитела может позволить получить молекулу, которая будет оказывать суммарный положительный эффект на эффекторный Т-клеточный ответ, влияя на соотношение CD8/TReg. Антигенсвязывающий домен агонистического антитела, который активирует рецептор ICOS, может быть объединен с константной областью антитела (Fc), способствующей понижающей регуляции и/или клиренсу высокоэкспрессирующих клеток, с которыми связано антитело. Эффекторную положительную константную область можно использовать для рекрутмента клеточных эффекторных функций против клеток-мишеней (TRegs), например, для стимулирования антителозависимой клеточно-опосредованной цитотоксичности (ADCC) или антителозависимого фагоцитоза клеток (ADCP). Таким образом, антитело может как промотировать активацию эффекторных Т-клеток, так и подавлять иммуносупрессивные Т-регуляторные клетки. Поскольку ICOS экспрессируется на TRegs сильнее, чем на TEffs, может быть достигнут терапевтический баланс, при котором функция Teff промотируется, в то время как TRegs деплетируются, что приводит к итоговому увеличению иммунного ответа Т-клеток (например, противоопухолевому ответу или другому терапевтически полезному Т-клеточному ответу).
Несколько доклинических и клинических исследований показали сильную положительную корреляцию между высоким соотношением эффекторных Т-клеток к клеткам T-reg в микроокружении опухоли (ТМЕ) и общей выживаемостью. Сообщалось, что для пациентов с раком яичников отношение клеток CD8:T-reg является показателем хорошего клинического исхода [15]. Аналогичное наблюдение было сделано у пациентов с метастазирующей меланомой после приема ипилумумаба [16]. В доклинических исследованиях было также показано, что высокое отношение эффекторных клеток:Т-reg в ТМЕ связано с противоопухолевым ответом [43].
Данное изобретение предусматривает антитела, которые связывают ICOS человека. Антитела нацелены на внеклеточный домен ICOS и потому связываются с Т-клетками, экспрессирующими ICOS. Приведены примеры антител, которые были спроектированы так, чтобы они оказывали агонистический эффект на ICOS, тем самым усиливая функцию эффекторных Т-клеток, определяемую по способности увеличивать экспрессию и секрецию IFNγ. Как отмечалось, анти-ICOS антитела также могут быть сконструированы таким образом, чтобы они деплетировали клетки, с которыми связываются, что должно влиять преимущественно на регуляторные Т-клетки, обеспечивающие понижающее регулирование, устраняя супрессорное действие этих клеток на ответ эффекторных Т-клеток и тем самым промотируя, в целом, эффекторный Т-клеточный ответ. Независимо от их механизма действия, эмпирически продемонстрировано, что анти-ICOS антитела в соответствии с данным изобретением действительно стимулируют Т-клеточный ответ и оказывают противоопухолевое действие in vivo, как показано в примерах. За счет выбора пригодных форматов антител, таких как включающие константные области с желательным уровнем эффекторной функции Fc или, при необходимости, отсутствием такой эффекторной функции, анти-ICOS антитела могут быть адаптированы для использования в различных медицинских контекстах, включая лечение заболеваний и состояний, при которых эффекторный Т-клеточный ответ является полезным и/или когда желательна супрессия регуляторных Т-клеток.
Типичные примеры антител включают STIM001, STIM002, STIM002-B, STIM003, STIM004, STIM005, STIM006, STIM007, STIM008 и STIM009, последовательности которых приведены в данном документе.
Антитело в соответствии с данным изобретением может представлять собой антитело, которое конкурирует за связывание с человеческим ICOS с антителом (например, человеческим IgG1 или scFv), содержащим участки, определяющие комплементарность, (CDR) тяжелой и легкой цепей STIM001, STIM002, STIM002-B, STIM003, STIM004, STIM005, STIM006, STIM007, STIM008 или STIM009, необязательно, антителом, содержащим VH- и VL-домены STIM001, STIM002, STIM002-B, STIM003, STIM004, STIM005, STIM006, STIM007, STIM008 или STIM009.
Антитело в соответствии с данным изобретением может содержать один или несколько CDR любого из STIM001, STIM002, STIM002-B, STIM003, STIM004, STIM005, STIM006, STIM007, STIM008 и STIM009 (например, все 6 CDR любого такого антитела, или набор HCDR и/или LCDR) или их варианты, как описано в данном документе.
Антитело может содержать домен VH антитела, содержащий CDRs HCDR1, HCDR2 и HCDR3, и домен VL антитела, содержащий CDRs LCDR1, LCDR2 и LCDR3, причем HCDR3 представляет собой HCDR3 антитела, выбранного из STIM001, STIM002, STIM002-B, STIM003, STIM004, STIM005, STIM006, STIM007, STIM008 и STIM009, или содержит такой HCDR3 с 1, 2, 3, 4 или 5 аминокислотными изменениями. HCDR2 может представлять собой HCDR2 выбранного антитела или может содержать HCDR2 с 1, 2, 3, 4 или 5 аминокислотными изменениями. HCDR1 может представлять собой HCDR1 выбранного антитела или может содержать HCDR1 с 1, 2, 3, 4 или 5 аминокислотными изменениями.
Антитело может содержать домен VL антитела, содержащий CDRs HCDR1, HCDR2 и HCDR3, и домен VL антитела, содержащий CDRs LCDR1, LCDR2 и LCDR3, причем LCDR3 представляет собой LCDR3 антитела, выбранного из STIM001, STIM002, STIM002-B, STIM003, STIM004, STIM005, STIM006, STIM007, STIM008 и STIM009, или содержит LCDR3 с 1, 2, 3, 4 или 5 аминокислотными изменениями. LCDR2 может представлять собой LCDR2 выбранного антитела или может содержать этот LCDR2 с 1, 2, 3, 4 или 5 аминокислотными изменениями. LCDR1 может представлять собой LCDR1 выбранного антитела или может содержать этот LCDR1 с 1, 2, 3, 4 или 5 аминокислотными изменениями.
Антитело может содержать:
домен VH антитела, содержащий участки, определяющие комплементарность, HCDR1, HCDR2 и HCDR3, и
домен VL антитела, содержащий участки, определяющие комплементарность, LCDR1, LCDR2 и LCDR3,
причем участки, определяющие комплементарность, тяжелой цепи принадлежат STIM001, STIM002, STIM002-B, STIM003, STIM004, STIM005, STIM006, STIM007, STIM008 или STIM009, или содержат участки, определяющие комплементарность, тяжелой цепи STIM001, STIM002, STIM002-B, STIM003, STIM004 или STIM005, STIM006, STIM007, STIM008 или STIM009 с 1, 2, 3, 4 или 5 аминокислотными изменениями; и/или
причем участки, определяющие комплементарность, легкой цепи принадлежат STIM001, STIM002, STIM002-B, STIM003, STIM004, STIM005, STIM006, STIM007, STIM008 или STIM009, или содержат участки, определяющие комплементарность, легкой цепи STIM001, STIM002, STIM002-B, STIM003, STIM004, STIM005, STIM006, STIM007, STIM008 или STIM009 с 1, 2, 3, 4 или 5 аминокислотными изменениями.
Антитело может содержать домен VH, содержащий набор гипервариабельных участков тяжелой цепи (HCDR) HCDR1, HCDR2 и HCDR3, причем
HCDR1 представляет собой HCDR1 STIM003,
HCDR2 представляет собой HCDR2 STIM003,
HCDR3 представляет собой HCDR3 STIM003,
или содержащий такой набор HCDR с 1, 2, 3, 4, 5 или 6 аминокислотными изменениями.
Антитело может содержать домен VL, содержащий набор гипервариабельных участков легкой цепи (LCDR) LCDR1, LCDR2 и LCDR3, причем
LCDR1 представляет собой LCDR1 STIM003,
LCDR2 представляет собой LCDR2 STIM003,
LCDR3 представляет собой LCDR3 STIM003,
или содержащий такой набор LCDR с 1, 2, 3 или 4 аминокислотными изменениями. Аминокислотные изменения (например, замены) могут находиться в положении любого остатка CDR. Примеры аминокислотных изменений показаны на Фигуре 35, Фигуре 36 и Фигуре 37, которые показывают выравнивания вариантов последовательностей анти-ICOS антител. Таким образом, аминокислотное изменение в CDR STIM003 может быть заменой остатка, присутствующего в соответствующем положении в антителе CL-74570 или антителе CL-71642, как показано на Фигуре 36.
Примерами аминокислотных изменений в CDR STIM003 являются замены в следующих положениях остатков, определяемых согласно IMGT:
В HCDR1 - замещение в положении 28 IMGT, необязательно, консервативное замещение, например, V28F.
В HCDR2 - замещение в положении 59, 63 и/или 64 IMGT. Необязательно, замещение в положении 59 представляет собой N59I, замещение в положении 63 представляет собой G63D, и/или замещение в положении 64 представляет собой D64N и/или D64S.
В HCDR3 - замещение в положении 106, 108, 109 IMGT и/или 112. Необязательно, замещение в положении 106 представляет собой R106A, замещение в положении 108 представляет собой F108Y, замещение в положении 109 представляет собой Y109F, и/или замещение в положении 112 представляет собой H112N.
В LCDR1, замещение в положении 36, например, R36S.
В LCDR3, замещение в положении 105, 108 и/или 109. Необязательно, замещение в положении 105 представляет собой H105Q, замещение в положении 108 представляет собой D108G, и/или замещение в положении 109 представляет собой M109N или M109S.
Антитела по изобретению могут содержать каркасные области домена VH и/или VL, соответствующие последовательностям генных сегментов зародышевой линии человека. Например, они могут содержать одну или несколько каркасных областей STIM001, STIM002, STIM002-B, STIM003, STIM004, STIM005, STIM006, STIM007, STIM008 или STIM009. Каркасная область или каркасные области могут представлять собой FR1, FR2, FR3 и/или FR4.
Как описано в Примере 12, в Таблице Е12-1 приведены V, D и J генные сегменты человеческой зародышевой линии, которые генерировали VH-домены этих антител посредством рекомбинации, а в Таблице Е12-2 приведены V и J генные сегменты человеческого зародышевой линии, которые генерировали VL домены этих антител посредством рекомбинации. Домены VH и VL антитела по данному изобретению могут быть основаны на таких V(D)J сегментах.
Антитело по изобретению может содержать домен VH антитела, который (i) получен в результате рекомбинации V генного сегмента тяжелой цепи человека, D генного сегмента тяжелой цепи человека и J генного сегмента тяжелой цепи человека, причем
V сегмент представляет собой IGHV1-18 (например, V1-18*01), IGVH3-20 (например, V3-20*d01), IGVH3-11 (например, V3-11*01) или IGVH2-5 (например, V2-5*10);
D генный сегмент представляет собой IGHD6-19 (например, IGHD6-19*01), IGHD3-10 (например, IGHD3-10*01) или IGHD3-9 (например, IGHD3-9*01); и/или
J генный сегмент представляет собой IGHJ6 (например, IGHJ6*02), IGHJ4 (например, IGHJ4*02) или IGHJ3 (например, IGHJ3*02), или (и) содержит каркасные области FR1, FR2, FR3 и FR4, причем
FR1 выравнивается с V генным сегментом зародышевой линии человека IGHV1-18 (например, V1-18*01), IGVH3-20 (например, V3-20*d01), IGVH3-11 (например, V3-11*01) или IGVH2-5 (например, V2-5*10), необязательно, с 1, 2, 3, 4 или 5 аминокислотными изменениями,
FR2 выравнивается с V генным сегментом гена зародышевой линии человека IGHV1-18 (например, V1-18*01), IGVH3-20 (например, V3-20*d01), IGVH3-11 (например, V3-11*01) или IGVH2-5 (например, V2-5*10), необязательно, с 1, 2, 3, 4 или 5 аминокислотными изменениями,
FR3 выравнивается с V генным сегментом зародышевой линии человека IGHV1-18 (например, V1-18*01), IGVH3-20 (например, V3-20*d01), IGVH3-11 (например, V3-11*01) или IGVH2-5 (например, V2-5*10), необязательно, с 1, 2, 3, 4 или 5 аминокислотными изменениями и/или
FR4 выравнивается с J генным сегментом зародышевой линии IGJH6 (например, JH6*02), IGJH4 (например, JH4*02) или IGJH3 (например, JH3*02), необязательно, с 1, 2, 3, 4 или 5 аминокислотными изменениями,
FR1, FR2 и FR3 домена VH обычно совпадают с одним и тем же V генным сегментом зародышевой линии. Так, например, антитело может содержать домен VH, полученный в результате рекомбинации V генного сегмента тяжелой цепи человека IGHV3-20 (например, VH3-20*d01), D генного сегмента тяжелой цепи человека и J генного сегмента тяжелой цепи человека IGJH4 (например, JH4*02). Антитело может содержать каркасные области домена VH FR1, FR2, FR3 и FR4, причем FR1, FR2 и FR3 каждый совпадают с V генным сегментом зародышевой линии IGHV3-20 (например, IGVH3-20*d01) с 1, 2, 3, 4 или 5 аминокислотными изменениями, и FR4, который выравнивается с J генным сегментом зародышевой линии IGHJ4 (например, IGHJ4*02) с 1, 2, 3, 4 или 5 аминокислотными изменениями. Выравнивание может быть точным, но в некоторых случаях один или несколько остатков могут быть мутированы по сравнению с зародышевой линией, поэтому могут присутствовать аминокислотные замещения или, реже, делеции или инсерции.
Антитело по изобретению может содержать домен VL антитела, который (i) получен в результате рекомбинации V генного сегмента легкой цепи человека и J генного сегмента легкой цепи человека, причем
V сегмент представляет собой IGKV2-28 (например, IGKV2-28*01), IGKV3-20 (например, IGKV3-20*01), IGKV1D-39 (например, IGKV1D-39*01) или IGKV3-11 (например, IGKV3-11*01) и/или
J генный сегмент представляет собой IGKJ4 (например, IGKJ4*01), IGKJ2 (например, IGKJ2*04), IGLJ3 (например, IGKJ3*01) или IGKJ1 (например, IGKJ1*01); или (и) содержит каркасные области FR1, FR2, FR3 и FR4, причем
FR1 выравнивается с V генным сегментом зародышевой линии человека IGKV2-28 (например, IGKV2-28*01), IGKV3-20 (например, IGKV3-20*01), IGKV1D-39 (например, IGKV1D-39*01) или IGKV3-11 (например, IGKV3-11*01), необязательно, с 1, 2, 3, 4 или 5 аминокислотными изменениями,
FR2 выравнивается с V генным сегментом зародышевой линии человека IGKV2-28 (например, IGKV2-28*01), IGKV3-20 (например, IGKV3-20*01), IGKV1D-39 (например, IGKV1D-39*01) или IGKV3-11 (например, IGKV3-11*01), необязательно, с 1, 2, 3, 4 или 5 аминокислотными изменениями,
FR3 выравнивается с V генным сегментом зародышевой линии человека IGKV2-28 (например, IGKV2-28*01), IGKV3-20 (например, IGKV3-20*01), IGKV1D-39 (например, IGKV1D-39*01) или IGKV3-11 (например, IGKV3-11*01), необязательно, с 1, 2, 3, 4 или 5 аминокислотными изменениями и/или
FR4 выравнивается с J генным сегментом зародышевой линии человека IGKJ4 (например, IGKJ4*01), IGKJ2 (например, IGKJ2*04), IGKJ3 (например, IGKJ3*01) или IGKJ1 (например, IGKJ1*01), необязательно, с 1, 2, 3, 4 или 5 аминокислотными изменениями.
FR1, FR2 и FR3 домена VL обычно совпадают с одним и тем же V генным сегментом зародышевой линии. Так, например, антитело может содержать домен VL, полученный в результате рекомбинации V генного сегмента легкой цепи человека IGKV3-20 (например, IGKV3-20*01) и J сегмента легкой цепи человека IGKJ3 (например, IGKJ3*01). Антитело может содержать каркасные области домена VL FR1, FR2, FR3 и FR4, причем FR1, FR2 и FR3 каждый совпадают с V генным сегментом зародышевой линии IGKV3-20 (например, IGKV3-20*01) с изменениями до 1, 2, 3, 4 или 5 аминокислот, и FR4, который выравнивается с J генным сегментом зародышевой линии IGKJ3 (например, IGKJ3*01) с изменениями до 1, 2, 3, 4 или 5 аминокислот. Выравнивание может быть точным, но в некоторых случаях один или несколько остатков могут быть мутированы по сравнению с зародышевой линией, поэтому могут присутствовать аминокислотные замещения или, реже, делеции или инсерции.
Антитело в соответствии с изобретением может содержать домен VH антитела, который является доменом VH STIM001, STIM002, STIM002-B, STIM003, STIM004 или STIM005, STIM006, STIM007, STIM008 или STIM009, или который имеет аминокислотную последовательность, по меньшей мере на 90% идентичную последовательности домена VH антитела STIM001, STIM002, STIM002-B, STIM003, STIM004, STIM005, STIM006, STIM007, STIM008 или STIM009. Идентичность аминокислотной последовательности может составлять по меньшей мере 95%.
Антитело может содержать домен VL антитела, который является доменом VL STIM001, STIM002, STIM002-B, STIM003, STIM004 или STIM005, STIM006, STIM007, STIM008 или STIM009, или который имеет аминокислотную последовательность, по меньшей мере, на 90% идентичную последовательности домена VL антитела STIM001, STIM002, STIM002-B, STIM003, STIM004, STIM005, STIM006, STIM007, STIM008 или STIM009. Идентичность аминокислотной последовательности может составлять по меньшей мере 95%.
Домен VH антитела, имеющий HCDR STIM001, STIM002, STIM002-B, STIM003, STIM004, STIM005, STIM006, STIM007, STIM008 или STIM009, или имеющий вариант этих CDR, может быть сопряжен с доменом VL антитела, имеющим LCDRs этого же антитела, или имеющим вариант этих CDR. Аналогично, домен VH любого из STIM001, STIM002, STIM002-B, STIM003, STIM004, STIM005, STIM006, STIM007, STIM008 или STIM009, или вариант этого домена VH может быть сопряжен с доменом VL этого же антитела, или вариантом VL-домена этого же антитела.
Например, антитело может содержать домен VH антитела STIM001 и домен VL STIM001. В другом примере, антитело может содержать домен VH антитела STIM002 и домен VL STIM002. В другом примере антитело может содержать домен VH антитела STIM003 и домен VL STIM003.
Антитела могут включать константные области, необязательно, константные области тяжелой и/или легкой цепи человека. Типичным примером изотипа является IgG, например, IgG1 человека.
Дополнительные аспекты данного изобретения включают молекулы нуклеиновых кислот, кодирующие последовательности антител, описанных в данном документе, клетки-хозяева, содержащие такие нуклеиновые кислоты, и способы получения антител путем культивирования клеток-хозяев и экспрессии и, необязательно, выделения или очистки антител. В результате этого, получают экспрессированное антитело. VH- и VL-домены антител, описанных в данном документе, могут быть получены аналогичным образом и являются аспектами данного изобретения. Также предлагаются фармацевтические композиции, содержащие антитела.
Другие аспекты изобретения относятся к ICOS-нокаутным животным, не являющимся человеком, и их использованию для получения антител к человеческому ICOS. У ICOS-нокаутного животного, ICOS не экспрессируется, например, потому, что кодирующий ICOS ген был инактивирован или удален из генома животного. Такие животные полезны для получения перекрестно-реактивных антител, которые распознают как ICOS человека, так и ICOS видов, не относящихся к человеку. Нормальный процесс иммунной толерантности означает, что лимфоциты, распознающие «собственные» антигены, удаляются или инактивируются для предотвращения аутоиммунных реакций в организме, в то время как отсутствие эндогенного антигена ICOS у не являющегося человеком нокаутного животного означает, что иммунная система животного не должна приобретать толерантность к этому антигену и поэтому может вырабатывать иммунный ответ против ICOS при его инъекции в виде рекомбинантного белка или с использованием клеточных линий или везикул, экспрессирующих ICOS. Иммунный репертуар нокаутного животного должен содержать лимфоциты, способные распознавать белок ICOS, принадлежащий этому виду животных. Подопытное животное, не являющееся человеком (например, мышь), иммунизированное человеческим ICOS, может, таким образом, генерировать антитела, которые связывают как ICOS человека, так и ICOS подопытного животного (например, ICOS мыши).
Это создает как минимум два преимущества. Во-первых, перекрестно-видовое антитело может использоваться для доклинического тестирования на не являющемся человеком подопытном животном до проведения разработок с клиническими испытаниями на людях. Во-вторых, иммунная система нокаутного животного может быть способна распознавать большее количество возможных эпитопов в молекуле ICOS человека по сравнению с распознаваемыми животным, экспрессирующим ICOS, благодаря чему иммунный репертуар нокаутного животного может иметь более высокое функциональное разнообразие антител. Поскольку существует сходство между последовательностями гомологичных молекул ICOS разных видов, иммунная система не являющегося человеком животного обычно может приобретать толерантность к тем областям белка ICOS человека, которые совпадают с ICOS животных, не относящихся к человеку, в то время как у нокаутного животного такая толеризация не происходит.
Возможность использования ICOS-нокаутного животного и его преимущества для получения перекрестно-реактивных антител показаны в примерах. Особенно удивительно, что ICOS-нокаутное животное можно успешно иммунизировать для получения ответа на антитела, поскольку сам ICOS задействован в биологии иммунной системы, такой как образование и поддержание зародышевых центров, и участвует в формировании иммунного ответа посредством его роли на фолликулярных Т-хелперных клетках, которые являются ICOS+ve клетками [37]. С учетом этого, можно предположить, что ICOS-нокаутное животное будет генерировать, в лучшем случае, плохой ответ на антитела. Неожиданно, для ICOS-нокаутных мышей были получены высокие титры, и высокофункциональные антитела были выделены из репертуара антител, в том числе желательные перекрестно-реактивные антитела.
Иллюстративные варианты реализации изобретения изложены в приложенной формуле изобретения.
Краткое описание графических материалов
Некоторые аспекты и варианты реализации изобретения будут далее описаны более подробно со ссылками на прилагаемые чертежи.
Фигура 1: Определение титров сыворотки ICOS-нокаутных (ICOS KO) Kymouse и Kymouse дикого типа против ICOS человека и мыши, экспрессируемых на клетках СНО методом проточной цитометрии. Данные иллюстрируют способность иммуноглобулина в сыворотке (а) ICOS-нокаутных (KO)-мышей (KO) или (b) не-ICOS KO мышей дикого типа (HK или HL), иммунизированных каждая клетками MEF, экспрессирующим и человеческий ICOS, и человеческим белком ICOS, связывать ICOS человека (связывание ICOS человека) или ICOS мыши (связывание ICOS мыши), экспрессируемые на клетках СНО.
Геометрическое среднее является мерой интенсивности флуоресценции связывания иммуноглобулина с клетками, по результатам определения методом проточной цитометрии.
Фигура 2: Анализ нейтрализации лиганда ICOS человека рецептором ICOS человека методом гомогенной флуоресценции с временным разрешением (HTRF). Сравнение профилей нейтрализации анти-ICOS моноклональных антител (mAbs) STIM001-STIM009 в формате человеческого IgG1 с С398.4А и соответствующими изотипическими контролями. Данные представляют четыре эксперимента.
Фигура 3: Анализ нейтрализации методом HTRF мышиного лиганда ICOS мышиным ICOS-рецептором. Сравнение профилей нейтрализации анти-ICOS моноклональных антител (mAbs) STIM001-STIM009 в формате человеческого IgG1 с С398.4А и соответствующими изотипическими контролями. Данные представляют три эксперимента.
Фигура 4: Анализ прямой нейтрализации методом HTRF лиганда ICOS человека рецептором ICOS человека. Сравнение профилей нейтрализации анти-ICOS моноклональных антител (mAbs) STIM001-STIM009 в формате IgG4.PE человека с С398.4А и соответствующими изотипическими контролями. Данные представляют четыре эксперимента.
Фигура 5: Анализ нейтрализации методом HTRF мышиного лиганда ICOS мышиным ICOS-рецептором. Сравнение профилей нейтрализации анти-ICOS моноклональных антител (mAbs) STIM001-STIM009 в формате IgG4.PE человека с С398.4А и соответствующими изотипическими контролями. Данные представляют четыре эксперимента.
Фигура 6а: Исследование концентрационной зависимости опосредованной STIM001 антителозависимой клеточной цитотоксичности (ADCC) на клетках MJ с использованием свежевыделенных NK-клеток в качестве эффекторных клеток. Эффекторные клетки и клетки-мишени (соотношение эффекторы:мишени 5:1) инкубировали вместе с антителом в течение 2 часов. Высвобождение BATDA (бис(ацетоксиметил)-2,2':6',2''-терпиридин-6,6''-дикарбоновой кислоты) лизированными клетками-мишенями измеряли, как описано в инструкции производителя набора. НС - гибридный изотипический контроль.
Фигуры 6b, с, d: Исследование концентрационной зависимости опосредованной STIM001 и STIM003 ADCC на клетках MJ со свежевыделенными NK-клетками в качестве эффекторных клеток. Эффекторные клетки и клетки-мишени (соотношение эффекторы : мишени 5:1) инкубировали вместе с антителом в течение 2 часов. Высвобождение BATDA лизированными клетками-мишенями измеряли, как описано в инструкции производителя набора. НС - гибридный изотипический контроль.
Фигуры 6е, f, g: Исследование концентрационной зависимости опосредованной STIM001 (hIgG1) и STIM003 (hIgG1) ADCC на трансфицированных ICOS клетках CCRF-CEM со свежевыделенными NK-клетками в качестве эффекторных клеток. Эффекторные клетки и клетки-мишени (соотношение эффекторы : мишени 5:1) инкубировали вместе с антителом в течение 4 часов. Высвобождение BATDA лизированными клетками-мишенями измеряли, как описано в инструкции производителя набора. НС - гибридный изотипический контроль.
Фигура 7, Фигура 8, Фигура 9: Анти-ICOS антитело ингибирует рост опухоли СТ26 и улучшает выживаемость при введении в качестве монотерапии или в комбинации с анти-PDL1. STIM001 mIgG2a является более эффективным, чем формат mIgG1. На каждом графике указано число исцеленных животных или животных со стабильным заболеванием.
Фигура 10: 2×2 комбинации СТ26 в in vivo исследованиях эффективности. Каждая группа лечения представлена «паутинной диаграммой» (spider plot), показывающей размеры опухоли отдельных животных (n=10 на группу). В сочетании с анти-PDL1-антителами STIM001 задерживает рост опухоли и улучшает выживаемость получающих лечение животных. Эффективность, наблюдаемая в присутствии STIM001 mIgG2a, выше, чем для STIM001 mIgG1 Наконец, STIM001 mIgG2a в сочетании с анти-PDLI mIgG2a была самой мощной комбинацией, инициирующей противоопухолевый ответ, приводящий к исцелению от болезни 60% животных. Для каждой группы количество животных, вылеченных от их заболевания, указано в верхнем правом углу соответствующих графиков. Введение препаратов осуществлялось в дни 6, 8,10, 13, 15 и 17.
Фигура 11: Графики, показывающие изменение объема опухолей СТ26 со временем у животных, получавших анти-ICOS или анти-PDLI монотерапии или комбинированные терапии. Каждая группа лечения представлена «паутинной диаграммой», показывающей размер опухоли отдельных животных (n=10 на группу). Для каждой группы число животных с размером опухоли менее 100 мм∧3 (стабильное заболевание/исцеление) указано в верхнем правом углу соответствующих графиков. Введение препаратов осуществлялось в дни 6, 8, 10, 13, 15 и 17. Период введения препарата показан заштрихованной областью, (а) Изотипический контроль; (b) Анти-PDL1 mIgG2a AbW; (с) Анти-ICOS STIM003 mIgG1; (d) Анти-ICOS STIM003 mIgG2a; (e) Анти-PDL1 mIgG2a AbW + STIM003 mIgG1; (f) Анти-PDL1 mIgG2a AbW + STIM003 mIgG2a. STIM003 mIgG2 в значительной степени ингибирует рост опухоли СТ26 в сочетании с анти-PDLI (AbW) mIgG2a.
Фигура 12: Тест на активацию MJ-клеток in vitro - иммобилизованные на шариках. Профили стимуляции STIM001, STIM002 и STIM003 анти-ICOS mAbs, иммобилизованных на шариках, по сравнению с анти-ICOS С398.4А и соответствующими изотипическими контролями. Данные представляют среднее для двух экспериментов (n=1 в случае шариков с С398.4А изотипическим контролем).
Фигура 13: Тест на активацию MJ-клеток in vitro - иммобилизованные на планшете. Профили стимуляции STIM001, STIM002, STIM003 и STIM004 анти-ICOS mAbs, иммобилизованных на планшетах, по сравнению с анти-ICOS С398.4А и соответствующими изотипическими контролями. Данные представляют собой среднее для двух экспериментов.
Фигура 14: Анализ связывания STIM001 и STIM003 hIgG1 с активированными Т-клетками методом FACS. (а) показывает типичный эксперимент дозового ответа предварительно меченых антител, связывающихся с активированными Т-клетками, и (b) показывает связывание после дозового ответа «голых» антител с последующим детектированием с помощью вторичного меченого антитела. В таблицах приведены соответствующие значения ЕС50 (М), определенные с помощью программы GraphPad Prism.
Фигура 15: STIM001 и STIM003 показывают изотип-зависимые эффекты на Т-клеточный компартмент в месте опухоли. В общей сложности 1×10Е5 опухолевых клеток СТ-26 имплантировали подкожно самкам мышей Balb/c. На 13-й день и 15-й день после имплантации животным вводили внутрибрюшинно антитела или физиологический раствор (n=10/каждой группе). На 16-й день после имплантации у животных с опухолями извлекают селезенку и опухоли (n=8/каждой группе), диссоциируют и окрашивают для анализа методом FACS. А - процент CD3-клеток, положительных к CD4-клеткам. В -процент CD3-клеток, положительных к клеткам CD8. С - процент CD4-клеток, которые являются Foxp3+ и CD25+. D - процент CD4-клеток в селезенке, положительных к Foxp3+ и CD25+. Е - процент эффекторных CD4-клеток от общего числа CD4-клеток. F -отношение эффекторов CD8 к клеткам T-Reg. G - отношение эффекторов CD4 к клеткам T-Reg. Статистический анализ проводили с использованием программы GraphPad Prism, все группы, получавшие антитело, сравнивали с группой, получавшей физиологический раствор, значения Р указаны в случае их значимости (р<0,05). Значения представляют собой среднее значение + стандартное отклонение (SD) (n=8 мышей/группу). Для F: Значения представляют собой среднее значение + стандартная ошибка среднего (SEM).
Фигура 16: Пример данных исследований концентрационной зависимости эффектов агониста STIM001 (hIgG1) и STIM003 (hIgG1) на выделенных Т-клетках человека, ко-стимулированных с помощью анти-CD3/анти-CD28 магнитных частиц dynabeads в течение 3-х дней в анализе активации Т-клеток 1 (см. Пример 9b). Продуцирование IFN-γ использовалось в качестве показателя агонистического эффекта. STIM001 (hIgG1) и STIM003 (hIgG1) тестировали в иммобилизованных на планшете растворимых или сшитых растворимых (Fc-связанное антитело (Ab)) форматах и сравнивали с гибридным изотипическим контролем (НС hIgG1). Для сравнения иммобилизованный на планшете анализ включал антитело хомячка С398.4А и его изотипический контроль (IgG хомяка). Верхняя панель показывает данные для антител, иммобилизованных на планшете. Нижняя панель показывает данные для антител KgG1 в растворимой и сшитой формах. Левая и правая панели, соответственно, используют Т-клетки от двух независимых доноров-людей.
Фигура 17: Пример набора данных для STIM001 в анализе активации Т-клеток 1 (см. Пример 9). Данные указывают уровни IFN-γ, индуцированные STIM001 (hIgG1), или его гибридным изотипическим контролем (НС IgG1) при одной определенной дозе для Т-клеток от 8 независимых доноров. Иммобилизованное на планшете антитело (Фигура 17а) использовали при концентрации 5 мкг/мл. Растворимые антитела (Фигура 17b) использовали при концентрации 15 мкг/мл. Каждая точка относится к одному донору, обозначенному номером (например, D214). Значимость оценивали с использованием статистического критерия Уилкоксона: *, р<0,05 и **, р<0,01.
Фигура 18: Пример набора данных для STIM003 в анализе активации Т-клеток 1 (см. Пример 9). Данные показывают уровни IFN-γ, индуцированного STIM003 (hIgG1), или его гибридного изотипического контроля (НС hIgG1) в одной заданной дозе для Т-клеток от 8 независимых здоровых доноров-людей. Растворимые антитела (Фиг. 18а) использовали при концентрации 15 мкг/мл. Инактивированное на планшете антитело (Фиг. 18b) использовали при концентрации 5 мкг/мл. Каждая точка относится к одному донору, обозначенному номером (например, D214). Значимость оценивали с использованием статистического критерия Уилкоксона: *, р<0,05 и **, р<0,01.
Фигура 19: Пример данных анализа Т-клеточной активации 2 (см. Пример 9в). Изучение агонистического эффекта STIM001 (hIgG1) и STIM003 (hIgG1) на изолированные Т-клетки человека, стимулированные анти-CD3/анти-CD28 dynabeads в течение 3-х дней, с последующим выдерживанием в среде в течение 3 дней и, наконец, повторной стимуляцией инактивированными на планшете STIM001, STIM003 или С398.4А Ab+/- CD3 Ab. Данные сравнивают уровни IFN-γ (А, В), TNF-α (С, D) и IL-2 (Е, F), индуцированные STIM001, STIM003, с их гибридным контролем IgG1 (А, С, Е), или С398.4А с его контролем IgG хомяка (В, D, F) в одной заданной дозе и в комбинации с CD3 Ab (связывание с TCR). Каждая точка представляет независимого донора, обозначенного номером (например, D190). Статистическую значимость между антителами (Abs) и их изотипическим контролем оценивали с использованием статистического критерия Уилкоксона и указанного значения р. Следует отметить, что концентрация STIM003 несколько отличается от концентрации НС IgG1 (5,4 против 5 мкг/мл).
Фигура 20: График, показывающий процент иммунных клеток (CD8 Т-эффекторные, CD4 Т-эффекторные и CD4/FoxP3 TReg-клетки) в опухолях СТ26 и селезенке опухолевых животных, которые экспрессируют ICOS на своей поверхности. Значения обозначают среднее ± стандартное отклонение (SD) (n=8). Значения Р были рассчитаны с использованием непараметрического критерия Данна для множественных сравнений. NS = незначимые; *** = р<0,001; **** = р<0,0001.
Фигура 21: Относительная экспрессия ICOS на поверхности иммунных клеток - CD8 Т-эффекторных, CD4 Т-эффекторных и CD4/FoxP3 TRegs - по результатам определения средней интенсивности флуоресценции (MFI). Значения обозначают среднее ± стандартное отклонение (SD) (n=8). Значения Р были рассчитаны с использованием непараметрического критерия Данна для множественных сравнений. **** = р<0,0001, ** = р<0,01. Следует обратить внимание на разницу в интенсивности флуоресценции между селезенкой (низкая) и опухолями (высокая).
Фигура 22: Влияние STIM001 и STIM003 на процентное соотношение различных иммунных клеток в микроокружении опухолей СТ26. * = р<0,05.
Фигура 23: Влияние антител STIM001 и STIM003 на процентное содержание регуляторных Т-клеток (CD4+/FoxP3+ клеток) в микроокружении опухолей СТ26. ** = р<0,05, **** = р<0,0001. Значения обозначают среднее ± стандартное отклонение (SD) (n=8). Значения Р были рассчитаны с использованием непараметрического критерия Данна для множественных сравнений.
Фигура 24: STIM001 и STIM003 mIgG2 значительно увеличивают соотношение CD8 эффекторных Т-клеток к TReg и соотношение CD4 эффекторных Т-клеток к TReg в опухолях СТ26. Величину отношения определяли путем деления процента эффекторных клеток в опухоли на процент регуляторных Т-клеток в опухоли.
Фигура 25: Влияние антител на процент иммунных клеток в селезенке животных, несущих опухоль СТ26.
Фигура 26: Влияние антител на процент регуляторных Т-клеток (клеток CD4+/FoxP3+) в селезенке животных, несущих опухоль СТ26.
Фигура 27: (А) Соотношение CD8 эффекторные : Treg и (В) соотношение CD4 : TReg в селезенке животных, несущих опухоль СТ26.
Фигура 28: Поверхностное окрашивание конъюгированных с AF647 STIM001, STIM003 и контроля с гибридом hIgG1 (НС IgG1) на активированной общей популяции Т-клеток яванского макака с Маврикия (Mauritian cynomolgus). Данные анализов с использованием различных доноров-источников Т-клеток показаны в А и В, соответственно. Значения ЕС50 указаны в таблице.
Фигура 29: Кривые Каплана-Мейера для модели СТ26 Balb/C. Затенение показывает период дозирования. LogRank р (значение р для логарифмического рангового критерия) <0,0001.
Фигура 30, Фигура 31, Фигура 32, Фигура 33: Графики, показывающие изменение объемов опухолей А20 со временем у мышей в исследовании, описанном в Примере 20. Каждая группа лечения представлена паутинной диаграммой, показывающей размер опухоли у отдельных животных, n=8 на группу. Для каждой группы в левом нижнем углу графика указано количество животных без признаков опухоли (что указывает на выздоровление). Введение доз проводили в дни 8, 11, 15, 18, 22, 25 и 29 после имплантации опухолевых клеток, а период дозирования обозначен как область, закрашенная в серый цвет. По сравнению с контрольной группой (Фиг. 30) и группой лечения анти-PD-L1 (Фиг. 31), группы лечения STIM001 mIgG2a (Фиг. 32) и STIM003 mIgG2a (Фиг. 33) показали значительное ингибирование роста опухоли А20.
Фигура 34: Данные исследований эффективности СТ26 in vivo, описанных в Примере 11 с, с использованием комбинации антитела анти-PD-L1 mIgG2a с однократной или многократной дозой STIM003 mIgG2a. Каждая группа лечения представлена «паутинной диаграммой», показывающей размеры опухолей отдельных животных (n=8 на группу). Для каждой группы количество животных, излеченных от заболевания, указано в правом нижнем углу соответствующего графика. Дни введения доз каждого антитела обозначены стрелками под соответствующими графиками.
Фигура 35: Аминокислотные последовательности доменов VH (сверху) и VL (снизу) STIM002, показывающие остатки, которые различаются в соответствующих последовательностях STIM001, STIM002B и родственных антител CL-61091, CL-64536, CL-64837, CL-64841 и CL-64912 и/или зародышевой линии человека. Нумерация последовательностей соответствует IMGT.
Фигура 36: Аминокислотные последовательности доменов VH (сверху) и VL (снизу) STIM003, показывающие остатки, которые различаются в соответствующих последовательностях родственных антител CL-71642 и CL-74570 и/или зародышевой линии человека. Нумерация последовательностей соответствует IMGT. Домен VL антитела CL-71642, полученный после секвенирования, показан здесь без N-концевого остатка. Из выравнивания видно, что полная последовательность домена VH будет содержать N-концевую глутаминовую кислоту.
Фигура 37: Аминокислотные последовательности доменов VH (сверху) и VL (снизу) STIM007, показывающие остатки, которые различаются в соответствующих последовательностях STIM008 и/или зародышевой линии человека. Нумерация последовательностей соответствует IMGT.
Фигура 38: Влияние антител STIM003 (анти-ICOS) и AbW (анти-PD-L1) mIgG2a в сингенной модели J558. Каждая группа лечения представлена «паутинной диаграммой», показывающей размеры опухолей отдельных животных (n=10 или n=8 на группу). Монотерапия STIM003 продемонстрировала определенную степень эффективности у 3 из 8 животных, вылеченных от заболевания. Аналогично, анти-PDL1 был эффективным в этой модели у 6 из 8 животных, вылеченных от заболевания к дню 37. В сочетании с анти-PDL1-антителами, STIM003 mIgG2 полностью ингибировал рост опухоли и улучшал выживаемость животных, получающих лечение. Для каждой группы количество животных, излеченных от заболевания, указано в правом нижнем углу соответствующего графика. Дни введения доз обозначены пунктирными линиями (дни 11, 15, 18, 22, 25 и 29).
Фигура 39: Количественная оценка экспрессии ICOS (процент положительных клеток и относительная экспрессия/dMFI (дифференциальная средняя интенсивность флуоресценции)) на разных подтипах OИЛ (опухоль-инфильтрующие лимфоциты) в опухолевой ткани. (А) % подтипов иммунных клеток, положительных к экспрессии ICOS, и (В) dMFI ICOS (относительная экспрессия ICOS на ICOS-положительной клетке) подтипов иммунных клеток животных, обработанных физиологическим раствором или суррогатными антителами анти-PD-L1 или анти-PD-1 Мышам имплантировали 100 мкл 1×106 жизнеспособных клеток/мл в день 0 (n=7 или n=8). Животным вводили внутрибрюшинно (ip) 130 мкг антитела в день 13 и день 15. Образцы тканей выделяли и анализировали на 16-й день. Клетки CD4+/FOXP3+ были включены только для популяции TReg (графики справа) и были исключены из «эффекторных» CD4-клеток (графики слева), которые все являются Foxp3-отрицательными. См. Пример 22.
Фигура 40: Данные исследований эффективности А20 in vivo. Каждая группа лечения представлена «паутинной диаграммой», показывающей размер опухоли отдельных животных (n=10 на группу). Для каждой группы на соответствующем графике указано количество животных, излеченных от их заболевания. Для многократных доз введение проводили в дни 8, 11, 15, 18, 22 и 25, обозначенные пунктирными линиями. Для разовой дозы животные получали инъекцию IP (внутрибрюшинно) только на 8-й день. (А) Физиологический раствор; (В) многократные дозы STIM003 mIgG2a; (С) разовая доза STIM003 mIgG2a. См. Пример 23.
Фигура 41: Кривые Каплана-Мейера для исследований, описанных в Примере 23, с фиксированной дозой 60 мкг STIM003 mIgG2a. SD = разовая доза, день 8. MD = многократные дозы BIW (два раза в неделю) с дня 8.
Фигура 42: Экспрессия ICOS на основных субпопуляциях Т-клеток (T-reg [CD4+/FoxP3+], CD4 Eff [CD4+/FoxP3-] клетки и CD8+) у животных, несущих опухоль СТ26 (n=4 для каждого момента времени), получавших физиологический раствор. Фенотипирование иммунных клеток проводили в дни 1, 2, 3, 4 и 8 после введения и окрашивали для экспрессии ICOS во всех тканях во все моменты времени. A-D показывают процент ICOS-положительных клеток во все моменты времени в четырех разных тканях. Е-Н показывают dMFI ICOS (относительную экспрессию) во все моменты времени во всех четырех разных тканях. См. Пример 24.
Фигура 43: Анализ методом FACS, демонстрирующий деплетирование T-reg в ТМЕ (микроокружение опухоли) в ответ на антитело mIgG2a STIM003. Животным, несущим опухоль СТ-26, вводили однократную дозу (6, 60 или 200 мкг) STIM003 на 12-й день после имплантации опухолевых клеток. Ткани (n=4 для каждого момента времени) собирали для анализа FACS в дни 1, 2, 3, 4 и 8 после введения дозы. Показаны процент клеток Т-reg (CD4+ CD25+ Foxp3+) в опухоли в целом (А) и процент клеток T-reg в крови (В) в разные моменты времени. См. Пример 24.
Фигура 44: Увеличение соотношений CD8 : T-reg и CD4 eff : T-reg в ответ на STIM003 mIgG2a. Опухолевые животные СТ-26 получали разовую дозу (6, 60 или 200 мкг) STIM003 mIgG2a на 12-й день после имплантации опухолевых клеток. Ткани (n=4 для каждого момента времени) собирали для анализа FACS в дни 1, 2, 3, 4 и 8 после введения дозы и рассчитывали отношения Т eff к Т-reg. (А) и (В), отношение CD8:T-reg в опухоли и в крови, (С) и (D) отношение CD4-eff:T-reg в опухоли и в крови. См. Пример 24.
Фигура 45: Лечение STIM003 коррелирует с повышенной дегрануляцией и продуцированием цитокинов Th1 опухоль-инфильтрующими лимфоцитами (ОИЛ). На 8-й день после введения дозы ОИЛ выделяли и проводили анализ методом FACS для определения экспрессии CD107a на CD4 и CD8 Т-клетках (А-В). Параллельно, клетки из диссоциированных опухолей выдерживали в покое в течение 4 часов в присутствии брефельдина-А, клетки окрашивали на маркеры Т-клеток и пермеабилизировали для внутриклеточного окрашивания для детектирования IFN-γ и TNF-α (С-Н). См. Пример 24.
Детальное описание
ICOS (Индуцируемый Т-клеточный ко-стимулятор)
Антитела в соответствии с данным изобретением связывают внеклеточный домен ICOS человека. Таким образом, антитела связывают экспрессирующие ICOS Т-лимфоциты. «ICOS» или «рецептор ICOS», упоминаемые в данном документе, могут быть человеческими ICOS, если из контекста не следует иное. Последовательности ICOS человека, яванского макака и мыши приведены в приложенном перечне последовательностей и доступны в NCBI (Национальный центр биотехнологической информации, США) как человеческий NCBI ID: NP_036224.1, мышиный NCBI ID: NP_059508.2 и яванского макака GenBank ID: ЕНН55098.1.
Перекрестная реактивность
Антитела в соответствии с данным изобретением, предпочтительно, являются перекрестно-реактивными и могут, например, связывать внеклеточный домен ICOS мыши, а также ICOS человека. Антитела могут связывать другие нечеловеческие ICOS, включая ICOS приматов, таких как яванский макак. Анти-ICOS антитело, предназначенное для терапевтического использования у людей, должно связывать ICOS человека, тогда как связывание с ICOS других видов не будет иметь прямой терапевтической значимости в клиническом контексте человека. Тем не менее, приведенные в данном документе данные показывают, что антитела, связывающие ICOS как человека, так и мыши, обладают свойствами, которые делают их особенно пригодными для использования в качестве агонистов и деплетирующих молекул. Это может быть результатом того, что один или несколько конкретных эпитопов являются мишенями перекрестно-реактивных антител. Однако, независимо от основополагающей теории, перекрестно-реактивные антитела имеют высокую ценность и являются превосходными кандидатами в терапевтические молекулы для доклинических и клинических исследований.
Как поясняется в экспериментальных примерах, раскрытые в данном документе антитела STIM были получены с использованием технологии Kymouse™, в которой мыши были подвергнуты генетической модификации с целью обеспечения отсутствия экспрессии ICOS мыши (нокаут ICOS). ICOS-нокаутные трансгенные животные и их использование для получения перекрестно-реактивных антител являются дополнительными аспектами данного изобретения.
Один из способов количественной оценки степени видовой перекрестной реактивности антитела выражается как кратность различия в аффинности к антигену одного вида по сравнению с антигеном другого вида, например, кратность различий в аффинности к человеческому ICOS против ICOS мыши. Аффинность может быть количественно определена как KD, что обозначает равновесную константу диссоциации реакции антитело-антиген, по результатам определения методом SPR, с антителом в формате Fab, как описано в другой части данного документа. Анти-ICOS антитело с видовой перекрестной реактивностью может иметь разницу в аффинности связывания ICOS человека и мыши, составляющую 30-кратную или меньше, 25-кратную или меньше, 20-кратную или меньше, 15-кратную или меньше, 10-кратную или меньше или 5-кратную или меньше. Другими словами, KD связывания внеклеточного домена человеческого ICOS может иметь значение в пределах 30-кратного, 25-кратного, 20-кратного, 15-кратного, 10-кратного или 5-кратного по отношению к KD связывания внеклеточного домена ICOS мыши. Антитела также могут считаться перекрестно-реактивными, если KD для антигена, связывающегося с обоими видами, соответствует пороговому значению, например, если KD связывания человеческого ICOS и KD связывания ICOS мыши оба составляют 10 мМ или меньше, предпочтительно, 5 мМ или меньше, более предпочтительно, 1 мМ или меньше. KD может составлять 10 нМ или меньше, 5 нМ или меньше, 2 нМ или меньше, или 1 нМ или меньше. KD может составлять 0,9 нМ или меньше, 0,8 нМ или меньше, 0,7 нМ или меньше, 0,6 нМ или меньше, 0,5 нМ или меньше, 0,4 нМ или меньше, 0,3 нМ или меньше, 0,2 нМ или меньше, или 0,1 нМ или меньше.
Альтернативной мерой перекрестной реактивности при связывании ICOS человека и ICOS мыши является способность антитела нейтрализовать лиганд ICOS, связывающийся с ICOS-рецептором, например, в анализе методом HTRF (см. Пример 8). Примеры антител с перекрестно-видовой реактивностью представлены в данном документе, включая STIM001, STIM002, STIM002-B, STIM003, STIM005 и STIM006, для каждого из которых была подтверждена способность нейтрализовать связывание человеческого В7-Н2 (лиганда ICOS) с человеческим ICOS и нейтрализовать связывание мышиного В7-Н2 с мышиным ICOS в анализе HTRF. Любое из этих антител или их вариантов может быть выбрано, когда требуется антитело, перекрестно-реактивное к ICOS человека и мыши. Анти-ICOS антитело с перекрестно-видовой реактивностью может иметь значение IC50 ингибирования связывания ICOS человека с рецептором ICOS человека, которое находится в пределах 25-кратного, 20-кратного, 15-кратного, 10-кратного или 5-кратного по отношению к IC50 ингибирования связывания ICOS мыши с мышиным рецептором ICOS, по результатам определения в анализе HTRF. Антитела также могут считаться перекрестно-реактивными, если IC50 ингибирования связывания человеческого ICOS с рецептором ICOS человека и IC50 ингибирования связывания ICOS мыши с рецептором ICOS мыши оба имеют значения, составляющие 1 мМ или меньше, предпочтительно, 0,5 мМ или меньше, например, 30 нМ или меньше, 20 нМ или меньше, 10 нМ или меньше. Значения IC50 могут составлять 5 нМ или меньше, 4 нМ или меньше, 3 нМ или меньше или 2 нМ или меньше. В некоторых случаях IC50 будет составлять по меньшей мере 0,1 нМ, по меньшей мере 0,5 нМ или по меньшей мере 1 нМ.
Специфичность
Антитела в соответствии с данным изобретением, предпочтительно, являются специфическими к ICOS. Это означает, что антитело связывает свой эпитоп на белке-мишени ICOS (ICOS человека и, предпочтительно, ICOS мыши и/или яванского макака, как отмечено выше), но не демонстрирует значительного связывания с молекулами, которые не презентируют такой эпитоп, включая другие молекулы семейства генов CD28. Антитело в соответствии с данным изобретением, предпочтительно, не связывает человеческий CD28. Антитело, предпочтительно, также не связывает CD28 мыши или яванского макака.
CD28 ко-стимулирует ответы Т-клеток при связывании с его лигандами CD80 и CD86 на профессиональных антигенпрезентирующих клетках в контексте распознавания антигена через Т-клеточный рецептор (TCR). Для различных in vivo применений антител, описанных в данном документе, избегание связывания с CD28 считается выгодным. Отсутствие связывание анти-ICOS антитела с CD28 должно позволить CD28 взаимодействовать с его нативными лигандами и генерировать соответствующий костимулирующий сигнал для активации Т-клеток. Кроме того, отсутствие связывания анти-ICOS антитела с CD28 исключает риск суперагонизма. Чрезмерная стимуляция CD28 может индуцировать пролиферацию в покоящихся Т-клетках без нормального требования распознавания родственного антигена через TCR, что потенциально приводит к бесконтрольной (runaway) активации Т-клеток и последующему синдрому высвобождения цитокинов, особенно у людей. Таким образом, нераспознавание CD28 антителами в соответствии с данным изобретением является преимуществом с точки зрения их безопасного клинического применения у людей.
Как обсуждалось в другой части данного документа, данное изобретение распространяется на мультиспецифические антитела (например, биспецифические). Мультиспецифическое (например, биспецифическое) антитело может содержать (i) антигенсвязывающий сайт ICOS антитела и (ii) дополнительный антигенсвязывающий сайт (необязательно, антигенсвязывающий сайт антитела, как описано в данном документе), распознающий другой антиген (например, PD-L1). Специфическое связывание индивидуальных антигенсвязывающих сайтов может быть определено. Таким образом, антитела, которые специфически связывают ICOS, включают антитела, содержащие антигенсвязывающий сайт, который специфически связывает ICOS, причем, необязательно, антигенсвязывающий сайт ICOS содержится в антигенсвязывающей молекуле, которая дополнительно включает один или несколько дополнительных сайтов связывания одного или нескольких других антигенов, например, биспецифическое антитело, которое связывает ICOS и PD-L1.
Аффинность
Аффинность связывания антитела с ICOS может быть определена. Аффинность антитела к его антигену может быть количественно определена в терминах равновесной константы диссоциации KD, отношения Ka/Kd скорости ассоциации или связывания (on-rate) (Ka) и скорости диссоциации или распада (kd) взаимодействия антитело-антиген. Kd, Ka и Kd для связывания антитело-антиген можно измерить с использованием метода поверхностного плазмонного резонанса (SPR).
Антитело в соответствии с данным изобретением может связывать домен ЕС ICOS человека с KD, равной 10 мМ или меньше, предпочтительно, 5 мМ или меньше, более предпочтительно, 1 мМ или меньше. KD может составлять 50 нМ или меньше, 10 нМ или меньше, 5 нМ или меньше, 2 нМ или меньше, или 1 нМ или меньше. KD может составлять 0,9 нМ или меньше, 0,8 нМ или меньше, 0,7 нМ или меньше, 0,6 нМ или меньше, 0,5 нМ или меньше, 0,4 нМ или меньше, 0,3 нМ или меньше, 0,2 нМ или меньше, или 0,1 нМ или меньше. KD может составлять по меньшей мере 0,001 нМ, например, по меньшей мере 0,01 нМ или по меньшей мере 0,1 нМ.
Количественная оценка аффинности может быть выполнена с использованием метода SPR с антителом в формате Fab. Пригодный протокол выглядит следующим образом:
1. Проведение реакции сочетания антитела против IgG человека (или другого антитела с сопоставимой по видам константной областью) с биосенсорным чипом (например, GLM-чипом), такой как реакция сочетания с первичным амином;
2. Экспонирование антитела против IgG человека (или другого подобранного по видам антитела) тестируемым антителом, например, в формате Fab, для захвата тестируемого антитела на чипе;
3. Пропускание тестируемого антигена над поверхностью захвата чипа в диапазоне концентраций, например, при 5000 нМ, 1000 нМ, 200 нМ, 40 нМ, 8 нМ и 2 нМ и при 0 нМ (т.е. только буфер); и
4. Определение аффинности связывания тестируемого антитела с тестируемым антигеном с использованием метода SPR при 25°С. Буфер может иметь рН 7,6, 150 мМ NaCl, 0,05% детергента (например, Р20) и 3 мМ ЭДТА. Буфер может, необязательно, содержать 10 мМ HEPES. HBS-EP может использоваться как подвижный буфер. HBS-EP может быть приобретен у фирмы Teknova Inc (California, номер по каталогу Н8022).
Регенерацию поверхности связывания можно проводить с помощью 10 мМ глицина при рН 1,7. Такая обработка удаляет связанное антитело и позволяет использовать поверхность для другого взаимодействия. Данные связывания могут быть аппроксимированы встроенной моделью 1:1 с использованием стандартных методов, например, с использованием модели, встроенной в программу анализа даанных ProteOn XPR36TM.
Известны различные инструменты SPR, такие как Biacore™, ProteOn XPR36™ (Bio-Rad®) и KinExA® (Sapidyne Instruments, Inc). Рабочие примеры SPR приведены в Примере 7.
Как описано, аффинность может быть определена методом SPR с антителом в формате Fab, с антигеном, связанным с поверхностью чипа, и тестируемым антителом, пропускаемым над чипом в формате Fab в растворе, для определения аффинности взаимодействия мономерного антитела и антигена. Аффинность может быть определена при любом желательном значении рН, например, рН 5,5 или рН 7,6, и любой желательной температуре, например, 25°С или 37°С. Как указано в Примере 7, антитела в соответствии с данным изобретением связывают ICOS человека с кажущейся аффинностью менее 2 нМ, по результатам определения методом SPR с использованием антитела в моновалентном (Fab) формате.
Другие способы измерения связывания антитела с ICOS включают флуоресцентно активируемую сортировку клеток (FACS), например, с использованием клеток (например, клеток СНО) с экзогенной поверхностной экспрессией ICOS или активированных первичных Т-клеток, экспрессирующих эндогенные уровни ICOS. Связывание антитела с ICOS-экспрессирующими клетками, измеренное методом FACS, указывает на то, что антитело способно связывать внеклеточный (ЕС) домен ICOS.
Агонизм рецептора ICOS
Лиганд ICOS (ICOSL, также известный как В7-Н2) представляет собой молекулу, экспрессируемую клеточной поверхностью, которая связывается с рецептором ICOS [17]. Это межклеточное взаимодействие лиганд-рецептор способствует мультимеризации ICOS на поверхности Т-клеток, активируя рецептор и стимулируя дальнейшее прохождение сигнала в Т-клетке. В эффекторных Т-клетках эта активация рецептора стимулирует эффекторный Т-клеточный ответ.
Анти-ICOS антитела могут выступать в качестве агонистов ICOS, имитируя и даже превосходя это стимулирующее действие нативного лиганда ICOS на рецептор. Такой агонизм может быть следствием способности антитела стимулировать мультимеризацию ICOS на Т-клетке. Одним из механизмов этого является то, что антитела образуют межклеточные мостики между ICOS на поверхности Т-клеток и рецепторами на соседней клетке (например, В-клетке, антигенпрезентирующей клетке или другой иммунной клетке), такими как рецепторы Fc. Другой механизм заключается в том, что антитела, имеющие множественные (например, два) антигенсвязывающие сайты (например, две пары доменов VH-VL), соединяют несколько молекул-рецепторов ICOS и, таким образом, способствуют мультимеризации. Может возникать комбинация этих механизмов.
Агонизм можно тестировать в анализах активации Т-клеток in vitro, используя антитело в растворимой форме (например, в формате иммуноглобулина или в другом формате антитела, содержащем два пространственно разделенных антигенсвязывающих сайта, например, две пары VH-VL), включающее или не включающее сшивающий агент, или используя антитело, связанное с твердой поверхностью, для создания иммобилизованной матрицы антигенсвязывающих сайтов. Анализы агонизма могут использовать человеческую ICOS-положительную клеточную линию Т-лимфоцитов, такую как MJ-клетки (АТСС CRL-8294), в качестве Т-клеток-мишеней для активации в таких анализах. Один или несколько показателей активации Т-клеток можно определить для тестового антитела и сравнить с эталонной молекулой или отрицательным контролем, чтобы определить, существует ли статистически значимая (р<0,05) разница в активации Т-клеток, вызываемой тестируемым антителом по сравнению с эталонной молекулой или контролем. Одним из пригодных показателей активации Т-клеток является продуцирование цитокинов, например, IFNγ, TNFα или IL-2. Квалифицированный специалист будет включать, при необходимости, пригодные контроли, стандартизируя условия проведения анализа для тестируемого антитела и контроля. Пригодным отрицательным контролем является антитело в том же формате (например, изотипический контроль), которое не связывает ICOS, например, антитело, специфичное к антигену, не присутствующему в аналитической системе. Существенное различие, наблюдаемое для тестового антитела по сравнению с родственным изотипическим контролем в динамическом диапазоне анализа, указывает на то, что антитело действует как агонист рецептора ICOS в этом анализе.
Агонистическое антитело может быть определено как такое, которое при тестировании в анализе активации Т-клеток:
имеет значительно более низкий ЕС50 для индукции продуцирования IFNγ по сравнению с контрольным антителом;
индуцирует значительно более высокое максимальное продуцирование IFNγ по сравнению с контрольным антителом;
имеет значительно более низкий ЕС50 для индукции продуцирования IFNγ по сравнению с ICOSL-Fc;
индуцирует значительно более высокое максимальное продуцирование IFNγ по сравнению с ICOSL-Fc;
имеет значительно более низкий ЕС50 для индукции продуцирования IFNγ по сравнению с контрольным антителом С398.4А; и/или
индуцирует значительно более высокое максимальное продуцирование IFNγ по сравнению с контрольным антителом С398.4А.
In vitro тесты Т-клеток включают анализ с иммобилизацией на шариках в Примере 13, анализ с иммобилизацией на планшете в Примере 14, и анализ растворимой формы в Примере 15.
Значительно более низкое или значительно более высокое значение может иметь, например, разность до 0,5-кратной, до 0,75-кратной, до 2-кратной, до 3-кратной, до 4-кратной или до 5-кратной по сравнению с эталонным или контрольным значением.
Таким образом, в одном примере антитело в соответствии с данным изобретением имеет значительно более низкий, например, по меньшей мере в 2 раза более низкий ЕС50 для индукции IFNγ в анализе активации клеток MJ с использованием антитела в формате с иммобилизацией на шариках, по сравнению с контролем.
Анализ с иммобилизацией на шариках использует антитело (и, для контрольных или калибровочных экспериментов, контрольное антитело, эталонное антитело или ICOSL-Fc), связанное с поверхностью шариков. Могут использоваться магнитные бусины, различные виды которых являются коммерчески доступными, например, тозил-активированные DYNABEADS М-450 (DYNAL Inc, 5 Delaware Drive, Lake Success, NY 11042 Prod No. 140,03, 140,04). Покрытие на шарики может быть нанесено, как описано в Примере 13 или, обычно, путем растворения материала покрытия в карбонатном буфере (рН 9,6, 0,2 М) или другим способом, известным специалистам в данной области техники. Использование бусинок удобно позволяет определять количество белка, связанного с поверхностью бусин, с хорошей степенью точности. Стандартные методы количественного определения Fc-белка могут быть использованы для количественной оценки белка, связанного на бусинах. Может быть использован любой пригодный метод, со ссылкой на соответствующий стандарт в динамическом диапазоне анализа. Метод DELFIA описан в Примере 13, но могут использоваться ИФА или другие методы.
Агонистическая активность антитела также может быть измерена в первичных Т-лимфоцитах человека ex vivo. Способность антитела индуцировать экспрессию IFNγ в таких Т-клетках указывает на агонизм ICOS. В данном документе описаны два анализа активации Т-клеток с использованием первичных клеток - см. Пример 2, анализ активации Т-клеток 1 и анализ активации Т-клеток 2. Предпочтительно, антитело будет демонстрировать значительную (р<0,05) индукцию IFNγ при 5 мкг/мл, по сравнению с контрольным антителом, в анализе активации Т-клеток 1 и/или анализе активации Т-клеток 2. Как отмечено выше, анти-ICOS антитело может стимулировать активацию Т-клеток в большей степени, чем ICOS-L или С398.4 в таком анализе. Таким образом, антитело может продемонстрировать значительно (р<0,05) большую индукцию IFNγ при 5 мкг/мл по сравнению с контрольным или эталонным антителом в анализе 1 или 2 активации Т-клеток. В качестве альтернативного результата анализа может быть измерена индукция TNFα или IL-2.
Агонизм анти-ICOS антитела может увеличивать его способность изменять баланс между популяциями TReg и TEff клеток in vivo, например, в месте патологии, таком как микроокружение опухоли, в пользу TEff клеток. Способность антитела увеличивать уничтожение опухолевых клеток активированными ICOS-положительными эффекторными Т-клетками может быть определена, как описано в другой части данного документа.
Т-клеточно-зависимое убийство клеток
Эффекторная Т-клеточная функция может быть определена в биологически релевантном контексте с использованием in vitro анализа совместной культивации, в котором опухолевые клетки инкубируют с соответствующими иммунными клетками для инициирования зависимого от иммунных клеток убийства, в котором наблюдается эффект влияния анти-ICOS антитела на убийство опухолевых клеток с помощью TEffs.
Может быть определена способность антитела усиливать уничтожение опухолевых клеток активированными ICOS-положительными эффекторными Т-клетками. Анти-ICOS антитело может стимулировать значительно большее (р<0,05) убийство опухолевых клеток по сравнению с контрольным антителом. Анти-ICOS антитело может стимулировать схожее по величине или большее убийство опухолевых клеток в таком анализе по сравнению с эталонной молекулой, такой как лиганд ICOS или антитело С398.4. Схожая по величине степень убийства опухолевых клеток может быть представлена в виде значения определяемого показателя для тестируемого антитела, менее чем в два раза отличающегося от значения для эталонной молекулы.
Эффективность нейтрализации взаимодействия лиганд-рецептор ICOS
Антитело в соответствии с данным изобретением может представлять собой антитело, которое ингибирует связывание ICOS с его лигандом ICOSL.
Степень, в которой антитело ингибирует связывание рецептора ICOS с его лигандом, называется эффективностью нейтрализации взаимодействия лиганд-рецептор. Эффективность обычно выражается как значение IC50, выраженное в пМ, если не указано иное. В исследованиях связывания лигандов IC50 представляет собой концентрацию, которая снижает связывание рецептора на 50% от максимального уровня специфического связывания. IC50 может быть рассчитана путем построения графика зависимости % специфического связывания рецептора от логарифма концентрации антитела, и использования прикладной программы, такой как Prism (GraphPad), для аппроксимации данных сигмоидальной функцией для определения значений IC50. Эффективность нейтрализации можно определить методом анализа HTRF (гомогенная флуоресценция с временным разрешением). Подробный рабочий пример анализа методом HTRF эффективности нейтрализации взаимодействия лиганд-рецептора приведен в Примере 8.
Значение IC50 может представлять собой среднее для множества измерений. Так, например, значения IC50 могут быть получены по результатам трех повторных экспериментов, что позволяет вычислить среднее значение IC50.
Антитело может иметь значение IC50, равное 1 мМ или меньше, в анализе нейтрализации лиганд-рецептор, например, 0,5 мМ или меньше. IC50 может составлять 30 нМ или меньше, 20 нМ или меньше, 10 нМ или меньше, 5 нМ или меньше, 4 нМ или меньше, 3 нМ или меньше или 2 нМ или меньше. IC50 может составлять по меньшей мере 0,1 нМ, по меньшей мере 0,5 нМ или по меньшей мере 1 нМ.
Антитела
Как описано более подробно в Примерах, мы выделили и охарактеризовали антитела, представляющие особый интерес, обозначенные STIM001, STIM002, STIM002-В, STIM003, STIM004, STIM005, STIM006, STIM007, STIM008 и STIM009. В различных аспектах изобретения, если из контекста не следует иное, антитела могут быть выбраны из любых из этих антител или из подгруппы STIM001, STIM002, STIM003, STIM004 и STIM005. Последовательности каждого из этих антител представлены в прилагаемом перечне последовательностей, причем для каждого антитела приведены следующие последовательности: нуклеотидная последовательность, кодирующая домен VH; аминокислотная последовательность домена VH; аминокислотная последовательность CDR1 VH, аминокислотная последовательность CDR2 VH; аминокислотная последовательность CDR3 VH; нуклеотидная последовательность, кодирующая домен VL; аминокислотная последовательность домена VL; аминокислотная последовательность CDR1 VL; аминокислотная последовательность CDR2 VL; и аминокислотная последовательность CDR3 VL, соответственно. Данное изобретение охватывает анти-ICOS антитела, имеющие последовательности доменов VH и/или VL всех антител, представленные в приложенном перечне последовательностей и/или на чертежах, а также антитела, содержащие HCDR и/или LCDR этих антител, необязательно, имеющие полную аминокислотную последовательность тяжелой цепи и/или легкой цепи.
STIM001 имеет аминокислотную последовательность вариабельной области тяжелой цепи (VH), представленную SEQ ID NO: 366, содержащую аминокислотную последовательность CDRH1, представленную SEQ ID NO: 363, аминокислотную последовательность CDRH2, представленную SEQ ID NO: 364, и аминокислотную последовательность CDRH3, представленную SEQ ID NO: 365. Последовательность нуклеиновой кислоты тяжелой цепи домена VH представлена SEQ ID NO: 367. STIM001 имеет аминокислотную последовательность вариабельной области легкой цепи (VL), представленную SEQ ID NO: 373, содержащую аминокислотную последовательность CDRL1, представленную SEQ ID NO: 370, аминокислотную последовательность CDRL2, представленную SEQ ID NO: 371, и аминокислотную последовательность CDRL3, представленную SEQ ID NO: 372. Последовательность нуклеиновой кислоты легкой цепи домена VL представлена SEQ ID NO: 374. Домен VH может быть объединена с любой из последовательностей константной области тяжелой цепи, описанной в данном документе, например, SEQ ID NO: 193, SEQ ID NO: 195, SEQ ID NO: 197, SEQ ID NO: 199, SEQ ID NO: 201, SEQ ID NO: 203, Seq ID Nos: 205, Seq ID Nos: 340, Seq ID Nos: 524, Seq ID Nos: 526, Seq ID Nos: 528, Seq ID Nos: 530, SEQ ID NO: 532 или SEQ ID NO: 534. Область VL может быть объединена с любой из последовательностей константной области легкой цепи, описанной в данном документе, например, SEQ ID NOs: 207, 209, 211, 213, 215, 217, 219, 221, 223, 225, 227, 229, 231, 233, 235, 237, 536 и 538. Полноразмерная аминокислотная последовательность тяжелой цепи представлена SEQ ID NO: 368 (последовательность нуклеиновой кислоты тяжелой цепи - SEQ ID NO: 369). Полноразмерная аминокислотная последовательность легкой цепи представлена SEQ ID NO: 375 (последовательность нуклеиновой кислоты легкой цепи - SEQ ID NO: 376).
STIM002 имеет аминокислотную последовательность вариабельной области тяжелой цепи (VH), представленную SEQ ID NO: 380, содержащую аминокислотную последовательность CDRH1, представленную SEQ ID NO: 377, аминокислотную последовательность CDRH2, представленную SEQ ID NO: 378, и аминокислотную последовательность CDRH3, представленную SEQ ID NO: 379. Последовательность нуклеиновой кислоты тяжелой цепи домена VH представлена SEQ ID NO: 381. STIM002 имеет аминокислотную последовательность вариабельной области легкой цепи (VL), представленную SEQ ID NO: 387, содержащую аминокислотную последовательность CDRL1, представленную SEQ ID NO: 384, аминокислотную последовательность CDRL2, представленную SEQ ID NO: 385, и аминокислотную последовательность CDRL3, представленную SEQ ID NO: 386. Последовательность нуклеиновой кислоты легкой цепи домена VL представлена SEQ ID NO: 388 или SEQ ID NO: 519. Домен VH может быть объединена с любой из последовательностей константной области тяжелой цепи, описанной в данном документе, например, SEQ ID NO: 193, SEQ ID NO: 195, SEQ ID NO: 197, SEQ ID NO: 199, SEQ ID NO: 201, SEQ ID NO: 203, Seq ID Nos: 205, Seq ID Nos: 340, Seq ID Nos: 524, Seq ID Nos: 526, Seq ID Nos: 528, Seq ID Nos: 530, SEQ ID NO: 532 или SEQ ID NO: 534. Область VL может быть объединена с любой из последовательностей константной области легкой цепи, описанной в данном документе, например, SEQ ID NOs: 207, 209, 211, 213, 215, 217, 219, 221, 223, 225, 227, 229, 231, 233, 235, 237, 536 и 538. Полноразмерная аминокислотная последовательность тяжелой цепи представлена SEQ ID NO: 382 (последовательность нуклеиновой кислоты тяжелой цепи - SEQ ID NO: 383). Полноразмерная аминокислотная последовательность легкой цепи представлена SEQ ID NO: 389 (последовательность нуклеиновой кислоты легкой цепи -SEQ ID NO: 390 или SEQ ID NO: 520).
STIM002-B имеет аминокислотную последовательность вариабельной области тяжелой цепи (VH), представленную SEQ ID NO: 394, содержащую аминокислотную последовательность CDRH1, представленную SEQ ID NO: 391, аминокислотную последовательность CDRH2, представленную SEQ ID NO: 392, и аминокислотную последовательность CDRH3, представленную SEQ ID NO: 393. Последовательность нуклеиновой кислоты тяжелой цепи домена VH представлена SEQ ID NO: 395. STIM002-B имеет аминокислотную последовательность вариабельной области легкой цепи (VL), представленную SEQ ID NO: 401, содержащую аминокислотную последовательность CDRL1, представленную SEQ ID NO: 398, аминокислотную последовательность CDRL2, представленную SEQ ID NO: 399, и аминокислотную последовательность CDRL3, представленную SEQ ID NO: 400. Последовательность нуклеиновой кислоты легкой цепи домена VL представлена SEQ ID NO: 402. Домен VH может быть объединена с любой из последовательностей константной области тяжелой цепи, описанной в данном документе, например, SEQ ID NO: 193, SEQ ID NO: 195, SEQ ID NO: 197, SEQ ID NO: 199, SEQ ID NO: 201, SEQ ID NO: 203, Seq ID Nos: 205, Seq ID Nos: 340, Seq ID Nos: 524, Seq ID Nos: 526, Seq ID Nos: 528, Seq ID Nos: 530, SEQ ID NO: 532 или SEQ ID NO: 534. Область VL может быть объединена с любой из последовательностей константной области легкой цепи, описанной в данном документе, например, SEQ ID NOs: 207, 209, 211, 213, 215, 217, 219, 221, 223, 225, 227, 229, 231, 233, 235, 237, 536 и 538. Полноразмерная аминокислотная последовательность тяжелой цепи представлена SEQ ID NO: 396 (последовательность нуклеиновой кислоты тяжелой цепи - SEQ ID NO: 397). Полноразмерная аминокислотная последовательность легкой цепи представлена SEQ ID NO: 403 (последовательность нуклеиновой кислоты легкой цепи - SEQ ID NO: 404).
STIM003 имеет аминокислотную последовательность вариабельной области тяжелой цепи (VH), представленную SEQ ID NO: 408, содержащую аминокислотную последовательность CDRH1, представленную SEQ ID NO: 405, аминокислотную последовательность CDRH2, представленную SEQ ID NO: 406, и аминокислотную последовательность CDRH3, представленную SEQ ID NO: 407. Последовательность нуклеиновой кислоты тяжелой цепи домена VH представлена SEQ ID NO: 409 или SEQ ID NO: 521. STIM003 имеет аминокислотную последовательность вариабельной области легкой цепи (VL), представленную SEQ ID NO: 415, содержащую аминокислотную последовательность CDRL1, представленную SEQ ID NO: 412, аминокислотную последовательность CDRL2, представленную SEQ ID NO: 413, и аминокислотную последовательность CDRL3, представленную SEQ ID NO: 414. Последовательность нуклеиновой кислоты легкой цепи домена VL представлена SEQ ID NO: 4416. Домен VH может быть объединена с любой из последовательностей константной области тяжелой цепи, описанной в данном документе, например, SEQ ID NO: 193, SEQ ID NO: 195, SEQ ID NO: 197, SEQ ID NO: 199, SEQ ID NO: 201, SEQ ID NO: 203, Seq ID Nos: 205, Seq ID Nos: 340, Seq ID Nos: 524, Seq ID Nos: 526, Seq ID Nos: 528, Seq ID Nos: 530, SEQ ID NO: 532 или SEQ ID NO: 534. Область VL может быть объединена с любой из последовательностей константной области легкой цепи, описанной в данном документе, например, SEQ ID NOs: 207, 209, 211, 213, 215, 217, 219, 221, 223, 225, 227, 229, 231, 233, 235, 237, 536 и 538. Полноразмерная аминокислотная последовательность тяжелой цепи представлена SEQ ID NO: 410 (последовательность нуклеиновой кислоты тяжелой цепи - SEQ ID NO: 411 или SEQ ID NO: 522). Полноразмерная аминокислотная последовательность легкой цепи представлена SEQ ID NO: 417 (последовательность нуклеиновой кислоты легкой цепи -SEQ ID NO: 418).
STIM004 имеет аминокислотную последовательность вариабельной области тяжелой цепи (VH), представленную SEQ ID NO: 422, содержащую аминокислотную последовательность CDRH1, представленную SEQ ID NO: 419, аминокислотную последовательность CDRH2, представленную SEQ ID NO: 420, и аминокислотную последовательность CDRH3, представленную SEQ ID NO: 421. Последовательность нуклеиновой кислоты тяжелой цепи домена VH представлена SEQ ID NO: 423. STIM004 имеет аминокислотную последовательность вариабельной области легкой цепи (VL), представленную SEQ ID NO: 429, содержащую аминокислотную последовательность CDRL1, представленную SEQ ID NO: 426, аминокислотную последовательность CDRL2, представленную SEQ ID NO: 427, и аминокислотную последовательность CDRL3, представленную SEQ ID NO: 428. Последовательность нуклеиновой кислоты легкой цепи домена VL представлена SEQ ID NO: 430 или SEQ ID NO: 431. Домен VH может быть объединена с любой из последовательностей константной области тяжелой цепи, описанной в данном документе, например, SEQ ID NO: 193, SEQ ID NO: 195, SEQ ID NO: 197, SEQ ID NO: 199, SEQ ID NO: 201, SEQ ID NO: 203, Seq ID Nos: 205, Seq ID Nos: 340, Seq ID Nos: 524, Seq ID Nos: 526, Seq ID Nos: 528, Seq ID Nos: 530, SEQ ID NO: 532 или SEQ ID NO: 534. Область VL может быть объединена с любой из последовательностей константной области легкой цепи, описанной в данном документе, например, SEQ ID NOs: 207, 209, 211, 213, 215, 217, 219, 221, 223, 225, 227, 229, 231, 233, 235, 237, 536 и 538. Полноразмерная аминокислотная последовательность тяжелой цепи представлена SEQ ID NO: 424 (последовательность нуклеиновой кислоты тяжелой цепи - SEQ ID NO: 425). Полноразмерная аминокислотная последовательность легкой цепи представлена SEQ ID NO: 432 (последовательность нуклеиновой кислоты легкой цепи -SEQ ID NO: 433 или SEQ ID NO: 434).
STIM005 имеет аминокислотную последовательность вариабельной области тяжелой цепи (VH), представленную SEQ ID NO: 438, содержащую аминокислотную последовательность CDRH1, представленную SEQ ID NO: 435, аминокислотную последовательность CDRH2, представленную SEQ ID NO: 436, и аминокислотную последовательность CDRH3, представленную SEQ ID NO: 437. Последовательность нуклеиновой кислоты тяжелой цепи домена VH представлена SEQ ID NO: 439. STIM005 имеет аминокислотную последовательность вариабельной области легкой цепи (VL), представленную SEQ ID NO: 445, содержащую аминокислотную последовательность CDRL1, представленную SEQ ID NO: 442, аминокислотную последовательность CDRL2, представленную SEQ ID NO: 443, и аминокислотную последовательность CDRL3, представленную SEQ ID NO: 444. Последовательность нуклеиновой кислоты легкой цепи домена VL представлена SEQ ID NO: 446. Домен VH может быть объединена с любой из последовательностей константной области тяжелой цепи, описанной в данном документе, например, SEQ ID NO: 193, SEQ ID NO: 195, SEQ ID NO: 197, SEQ ID NO: 199, SEQ ID NO: 201, SEQ ID NO: 203, Seq ID Nos: 205, Seq ID Nos: 340, Seq ID Nos: 524, Seq ID Nos: 526, Seq ID Nos: 528, Seq ID Nos: 530, SEQ ID NO: 532 или SEQ ID NO: 534. Область VL может быть объединена с любой из последовательностей константной области легкой цепи, описанной в данном документе, например, SEQ ID NOs: 207, 209, 211, 213, 215, 217, 219, 221, 223, 225, 227, 229, 231, 233, 235, 237, 536 и 538. Полноразмерная аминокислотная последовательность тяжелой цепи представлена SEQ ID NO: 440 (последовательность нуклеиновой кислоты тяжелой цепи - SEQ ID NO: 441). Полноразмерная аминокислотная последовательность легкой цепи представлена SEQ ID NO: 447 (последовательность нуклеиновой кислоты легкой цепи - SEQ ID NO: 448).
STIM006 имеет аминокислотную последовательность вариабельной области тяжелой цепи (VH), представленную SEQ ID NO: 452, содержащую аминокислотную последовательность CDRH1, представленную SEQ ID NO: 449, аминокислотную последовательность CDRH2, представленную SEQ ID NO: 450, и аминокислотную последовательность CDRH3, представленную SEQ ID NO: 451. Последовательность нуклеиновой кислоты тяжелой цепи домена VH представлена SEQ ID NO: 453. STIM006 имеет аминокислотную последовательность вариабельной области легкой цепи (VL), представленную SEQ ID NO: 459, содержащую аминокислотную последовательность CDRL1, представленную SEQ ID NO: 456, аминокислотную последовательность CDRL2, представленную SEQ ID NO: 457, и аминокислотную последовательность CDRL3, представленную SEQ ID NO: 458. Последовательность нуклеиновой кислоты легкой цепи домена VL представлена SEQ ID NO: 460. Домен VH может быть объединена с любой из последовательностей константной области тяжелой цепи, описанной в данном документе, например, SEQ ID NO: 193, SEQ ID NO: 195, SEQ ID NO: 197, SEQ ID NO: 199, SEQ ID NO: 201, SEQ ID NO: 203, Seq ID Nos: 205, Seq ID Nos: 340, Seq ID Nos: 524, Seq ID Nos: 526, Seq ID Nos: 528, Seq ID Nos: 530, SEQ ID NO: 532 или SEQ ID NO: 534. Область VL может быть объединена с любой из последовательностей константной области легкой цепи, описанной в данном документе, например, SEQ ID NOs: 207, 209, 211, 213, 215, 217, 219, 221, 223, 225, 227, 229, 231, 233, 235, 237, 536 и 538. Полноразмерная аминокислотная последовательность тяжелой цепи представлена SEQ ID NO: 454 (последовательность нуклеиновой кислоты тяжелой цепи - SEQ ID NO: 455). Полноразмерная аминокислотная последовательность легкой цепи представлена SEQ ID NO: 461 (последовательность нуклеиновой кислоты легкой цепи - SEQ ID NO: 462).
STIM007 имеет аминокислотную последовательность вариабельной области тяжелой цепи (VH), представленную SEQ ID NO: 466, содержащую аминокислотную последовательность CDRH1, представленную SEQ ID NO: 463, аминокислотную последовательность CDRH2, представленную SEQ ID NO: 464, и аминокислотную последовательность CDRH3, представленную SEQ ID NO: 465. Последовательность нуклеиновой кислоты тяжелой цепи домена VH представлена SEQ ID NO: 467. STIM007 имеет аминокислотную последовательность вариабельной области легкой цепи (VL), представленную SEQ ID NO: 473, содержащую аминокислотную последовательность CDRL1, представленную SEQ ID NO: 470, аминокислотную последовательность CDRL2, представленную SEQ ID NO: 471, и аминокислотную последовательность CDRL3, представленную SEQ ID NO: 472. Последовательность нуклеиновой кислоты легкой цепи домена VL представлена SEQ ID NO: 474. Домен VH может быть объединена с любой из последовательностей константной области тяжелой цепи, описанной в данном документе, например, SEQ ID NO: 193, SEQ ID NO: 195, SEQ ID NO: 197, SEQ ID NO: 199, SEQ ID NO: 201, SEQ ID NO: 203, Seq ID Nos: 205, Seq ID Nos: 340, Seq ID Nos: 524, Seq ID Nos: 526, Seq ID Nos: 528, Seq ID Nos: 530, SEQ ID NO: 532 или SEQ ID NO: 534. Область VL может быть объединена с любой из последовательностей константной области легкой цепи, описанной в данном документе, например, SEQ ID NOs: 207, 209, 211, 213, 215, 217, 219, 221, 223, 225, 227, 229, 231, 233, 235, 237, 536 и 538. Полноразмерная аминокислотная последовательность тяжелой цепи представлена SEQ ID NO: 468 (последовательность нуклеиновой кислоты тяжелой цепи - SEQ ID NO: 469). Полноразмерная аминокислотная последовательность легкой цепи представлена SEQ ID NO: 475 (последовательность нуклеиновой кислоты легкой цепи - SEQ ID NO: 476).
STIM008 имеет аминокислотную последовательность вариабельной области тяжелой цепи (VH), представленную SEQ ID NO: 480, содержащую аминокислотную последовательность CDRH1, представленную SEQ ID NO: 477, аминокислотную последовательность CDRH2, представленную SEQ ID NO: 478, и аминокислотную последовательность CDRH3, представленную SEQ ID NO: 479. Последовательность нуклеиновой кислоты тяжелой цепи домена VH представлена SEQ ID NO: 481. STIM008 имеет аминокислотную последовательность вариабельной области легкой цепи (VL), представленную SEQ ID NO: 487, содержащую аминокислотную последовательность CDRL1, представленную SEQ ID NO: 484, аминокислотную последовательность CDRL2, представленную SEQ ID NO: 485, и аминокислотную последовательность CDRL3, представленную SEQ ID NO: 486. Последовательность нуклеиновой кислоты легкой цепи домена VL представлена SEQ ID NO: 488. Домен VH может быть объединена с любой из последовательностей константной области тяжелой цепи, описанной в данном документе, например, SEQ ID NO: 193, SEQ ID NO: 195, SEQ ID NO: 197, SEQ ID NO: 199, SEQ ID NO: 201, SEQ ID NO: 203, Seq ID Nos: 205, Seq ID Nos: 340, Seq ID Nos: 524, Seq ID Nos: 526, Seq ID Nos: 528, Seq ID Nos: 530, SEQ ID NO: 532 или SEQ ID NO: 534. Область VL может быть объединена с любой из последовательностей константной области легкой цепи, описанной в данном документе, например, SEQ ID NOs: 207, 209, 211, 213, 215, 217, 219, 221, 223, 225, 227, 229, 231, 233, 235, 237, 536 и 538. Полноразмерная аминокислотная последовательность тяжелой цепи представлена SEQ ID NO: 482 (последовательность нуклеиновой кислоты тяжелой цепи - SEQ ID NO: 483). Полноразмерная аминокислотная последовательность легкой цепи представлена SEQ ID NO: 489 (последовательность нуклеиновой кислоты легкой цепи - SEQ ID NO: 490).
STIM009 имеет аминокислотную последовательность вариабельной области тяжелой цепи (VH), представленную SEQ ID NO: 494, содержащую аминокислотную последовательность CDRH1, представленную SEQ ID NO: 491, аминокислотную последовательность CDRH2, представленную SEQ ID NO: 492, и аминокислотную последовательность CDRH3, представленную SEQ ID NO: 493. Последовательность нуклеиновой кислоты тяжелой цепи домена VH представлена SEQ ID NO: 495. STIM009 имеет аминокислотную последовательность вариабельной области легкой цепи (VL), представленную SEQ ID NO: 501, содержащую аминокислотную последовательность CDRL1, представленную SEQ ID NO: 498, аминокислотную последовательность CDRL2, представленную SEQ ID NO: 499, и аминокислотную последовательность CDRL3, представленную SEQ ID NO: 500. Последовательность нуклеиновой кислоты легкой цепи домена VL представлена SEQ ID NO: 502. Домен VH может быть объединена с любой из последовательностей константной области тяжелой цепи, описанной в данном документе, например, SEQ ID NO: 193, SEQ ID NO: 195, SEQ ID NO: 197, SEQ ID NO: 199, SEQ ID NO: 201, SEQ ID NO: 203, Seq ID Nos: 205, Seq ID Nos: 340, Seq ID Nos: 524, Seq ID Nos: 526, Seq ID Nos: 528, Seq ID Nos: 530, SEQ ID NO: 532 или SEQ ID NO: 534. Область VL может быть объединена с любой из последовательностей константной области легкой цепи, описанной в данном документе, например, SEQ ID NOs: 207, 209, 211, 213, 215, 217, 219, 221, 223, 225, 227, 229, 231, 233, 235, 237, 536 и 538. Полноразмерная аминокислотная последовательность тяжелой цепи представлена SEQ ID NO: 496 (последовательность нуклеиновой кислоты тяжелой цепи - SEQ ID NO: 497). Полноразмерная аминокислотная последовательность легкой цепи представлена SEQ ID NO: 503 (последовательность нуклеиновой кислоты легкой цепи - SEQ ID NO: 504).
Антитела в соответствии с данным изобретением представляют собой иммуноглобулины или молекулы, содержащие иммуноглобулиновые домены, независимо от того, являются ли они натуральными или частично или полностью синтезированными. Антитела могут быть молекулами IgG, IgM, IgA, IgD или IgE или их антигенспецифическими фрагментами антител (включая, без ограничений, Fab, F(ab')2, Fv, связанные дисульфидными связями Fv, scFv, однодоменное антитело, мультиспецифическое антитело замкнутой конформации, дисульфидсвязанный scfv, диатело (diabody)), независимо от того, получены ли они от любого вида, который естественным образом продуцирует антитело, или созданы по технологии рекомбинантной ДНК; независимо от того, были ли они выделены из сыворотки, В-клеток, гибридом, трансфектом, дрожжей или бактерий. Антитела могут быть гуманизированы с использованием обычных технологий. Термин «антитело» охватывает любой полипептид или белок, содержащий антигенсвязывающий сайт антитела. Антигенсвязывающий сайт (паратоп) является частью антитела, которая связывается с эпитопом антигена-мишени (ICOS) и является комплементарной к нему.
Термин «эпитоп» относится к области антигена, которая связывается антителом. Эпитопы могут быть определены как структурные или функциональные. Функциональные эпитопы обычно являются подмножеством структурных эпитопов и содержат остатки, которые непосредственно влияют на аффинность взаимодействия. Эпитопы также могут быть конформационными, то есть состоящими из нелинейно расположенных аминокислот. В некоторых вариантах реализации, эпитопы могут включать детерминанты, которые являются химически активными поверхностными группировками молекул, таких как аминокислоты, боковые цепи Сахаров, фосфорильные группы или сульфонильные группы, и в некоторых вариантах реализации, могут иметь специфические трехмерные структурные характеристики и/или специфические характеристики заряда.
Антигенсвязывающий сайт представляет собой поли пептид или домен, который содержит один или несколько гипервариабельных участков (CDR) антитела и способен связывать антиген. Например, полипептид содержит CDR3 (например, HCDR3). Например, полипептид содержит CDR 1 и 2 (например, HCDR1 и 2) или CDR 1-3 вариабельного домена антитела (например, HCDRs1-3).
Антигенсвязывающий сайт антитела может быть создан одним или несколькими вариабельными доменами антител. В качестве примера, сайт связывания антитела состоит из одного вариабельного домена, например, вариабельного домена тяжелой цепи (домен VH) или вариабельного домена легкой цепи (домен VL). В другом примере, сайт связывания содержит пару VH/VL или две или более таких пар. Таким образом, антигенсвязывающий сайт антитела может содержать VH и VL.
Антитело может представлять собой цельный иммуноглобулин, включая константные области, или может быть фрагментом антитела. Фрагмент антитела представляет собой часть интактного антитела, например, содержащую антигенсвязывающую и/или вариабельную область интактного антитела. Примеры фрагментов антител включают:
(i) фрагмент Fab, представляющий собой моновалентный фрагмент, состоящий из доменов VL, VH, CL и СН1; (ii) фрагмент F(ab')2, представляющий собой двухвалентный фрагмент, включающий два фрагмента Fab, связанных дисульфидным мостиком в шарнирной области;
(iii) фрагмент Fd, состоящий из доменов VH и СН1;
(iv) фрагмент Fv, состоящий из доменов VL и VH одного плеча антитела,
(v) фрагмент dAb (Ward et al., (1989) Nature 341:544-546, которая в полном объеме включена в данный документ посредством ссылки), состоящий из домена VH или VL; и
(vi) изолированный гипервариабельный участок (CDR), который сохраняет специфическую антигенсвязывающую функциональность.
Другими примерами антител являются Н2-антитела, которые содержат димер тяжелой цепи (5'-\/Н-(необязательный шарнир)-СН2-СН3-3') и лишены легкой цепи.
Обычно используемым фрагментом являются одноцепочечные антитела (например, scFv). Мультиспецифические антитела могут быть образованы из фрагментов антител. Антитело по изобретению, при необходимости, может использовать любой такой формат.
Необязательно, иммуноглобулиновые домены антитела могут быть слиты или конъюгированы с дополнительными полипептидными последовательностями и/или с метками, тэгами, токсинами или другими молекулами. Иммуноглобулиновые домены антитела могут быть слиты или конъюгированы с одной или несколькими различными антигенсвязывающими областями, с образованием молекулы, которая может связывать второй антиген в дополнение к ICOS. Антитело по данному изобретению может быть мультиспецифическим антителом, например, биспецифическим антителом, содержащим (i) антигенсвязывающий сайт антитела к ICOS и (и) дополнительный антигенсвязывающий сайт (необязательно, антигенсвязывающий сайт антитела, как описано в данном документе) который распознает другой антиген (например, PD-L1).
Антитело обычно содержит домен VH и/или VL антитела. Изолированные VH- и VL-домены антител также являются частью изобретения. Вариабельные домены антител представляют собой части легкой и тяжелой цепей антител, которые включают аминокислотные последовательности гипервариабельных участков (CDR, т.е. CDR1, CDR2 и CDR3) и каркасных областей (FR). Таким образом, в каждом домене VH и VL присутствуют CDR и FR. Домен VH содержит набор HCDR, а домен VL содержит набор LCDR. VH относится к вариабельному домену тяжелой цепи. VL относится к вариабельному домену легкой цепи. Каждый VH и VL обычно состоит из трех CDR и четырех FR, расположенных, в направлении от аминоконца к карбоксильному концу, в следующем порядке: FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4. Согласно способам, используемым в данном изобретении, положения аминокислот, отнесенных к CDR и FR, могут быть определены в соответствии с Кабатом (Sequences of Proteins of Immunological Interest (National Institutes of Health, Bethesda, Md., 1987 и 1991)) или в соответствии с номенклатурой IMGT. Антитело может содержать домен VH антитела, содержащий CDR1, CDR2 и CDR3 VH и каркас. Альтернативно или дополнительно, оно может содержать домен VL антитела, содержащий CDR1, CDR2 и CDR3 VL и каркас. Примеры доменов VH и VL и CDR антител в соответствии с данным изобретением приведены в прилагаемом перечне последовательностей, который является частью данного описания. CDR, приведенные в перечне последовательностей, определены в соответствии с системой IMGT [18]. Все последовательности VH и VL, последовательности CDR, наборы CDR и наборы HCDR и наборы LCDR, раскрытые в данном документе, представляют аспекты и варианты реализации изобретения. В используемом в данном документе значении, «набор CDR» содержит CDR1, CDR2 и CDR3. Таким образом, набор HCDR относится к HCDR1, HCDR2 и HCDR3, а набор LCDR относится к LCDR1, LCDR2 и LCDR3. Если не указано иное, «набор CDR» включает HCDR и LCDR.
Антитело по изобретению может содержать один или несколько CDR, как описано в данном документе, например, CDR3 и, необязательно, CDR1 и CDR2, с образованием набора CDR. CDR или набор CDR могут представлять собой CDR или набор CDR любого из STIM001, STIM002, STIM002-B, STIM003, STIM004, STIM005, STIM006, STIM007, STIM008 и STIM009, или могут быть их вариантами, как описано в данном документе.
Изобретение предусматривает антитела, содержащие HCDR1, HCDR2 и/или HCDR3 любого из антител STIM001, STIM002, STIM002-B, STIM003, STIM004, STIM005, STIM006, STIM007, STIM008 и STIM009, и/или LCDR1, LCDR2 и/или LCDR3 любого из этих антител, например, набор CDR. Антитело может содержать набор CDR VH одного из этих антител. Необязательно, оно может также содержать набор CDR VL одного из этих антител, и CDR VL могут принадлежать тому же антителу, что и CDR VH, или другому.
Изобретение также предусматривает домен VH, содержащий раскрытый набор HCDR, и/или домен VL, содержащий раскрытый набор LCDR.
Как правило, домен VH сопряжен с доменом VL для обеспечения антигенсвязывающего сайта антитела, хотя, как описано ниже, для связывания антигена можно использовать только VH- или VL-домен. Домен VH STIM003 может быть сопряжен с доменом VL STIM003, для образования антигенсвязывающего сайта антитела, содержащего как VH, так и VL-домены STIM003. Аналогичные варианты реализации предусмотрены для других доменов VH и VL, раскрытых в данном документе. В других вариантах реализации, VH STIM003 сопряжен с VL-доменом, отличным от VL STIM003. Неоднородность легкой цепи является хорошо известным в данной области фактом. Кроме того, аналогичные варианты реализации предусматриваются изобретением для других доменов VH и VL, раскрытых в данном документе.
Таким образом, VH любого из антител STIM001, STIM002, STIM003, STIM004 и STIM005 может быть соединен с VL любого из антител STIM001, STIM002, STIM003, STIM004 и STIM005. Кроме того, VH любого из антител STIM001, STIM002, STIM002-B, STIM003, STIM004, STIM005, STIM006, STIM007, STIM008 и STIM009 может быть соединен с VL любого из антител STIM001, STIM002, STIM002-B, STIM003, STIM004, STIM005, STIM006, STIM007, STIM008 или STIM009.
Антитело может содержать один или несколько CDR, например, набор CDR, в каркасе антитела. Каркасные области могут представлять собой последовательности генных сегментов зародышевой линии человека. Таким образом, антитело может представлять собой человеческое антитело, имеющее домен VH, содержащий набор HCDR в каркасе зародышевой линии человека. Обычно антитело также имеет домен VL, содержащий набор LCDR, например, в каркасе зародышевой линии человека. «Генный сегмент» антитела, например, VH генный сегмент, D генный сегмент или HJ генный сегмент, относится к олигонуклеотиду, имеющему последовательность нуклеиновой кислоты, из которой получена эта часть антитела, например, VH генный сегмент представляет собой олигонуклеотид, содержащий последовательность нуклеиновой кислоты, которая соответствует домену VH полипептида от FR1 до части CDR3. V, D и J генные сегменты человека рекомбинируют для генерации домена VH, а V и J сегменты человека рекомбинируют для генерации домена VL. Домен или область D относятся к домену или области разнообразия (diversity) цепи антитела. Домен или область J относятся к соединительному домену или области цепи антитела. Соматическая гипермутация может приводить к тому, что домен VH или VL антитела будет имеет каркасные области, которые точно не совпадают или не совмещаются с соответствующими генными сегментами, но выравнивание последовательности может использоваться для идентификации ближайших генных сегментов и, таким образом, определения того, из какой именно комбинации генных сегментов был получен данный конкретный домен VH или VL. При выравнивании последовательностей антител с генными сегментами аминокислотная последовательность антитела может быть выровнена с аминокислотной последовательностью, кодируемой генным сегментом, или нуклеотидная последовательность антитела может быть выровнена непосредственно с нуклеотидной последовательностью генного сегмента.
Выравнивание последовательностей доменов VH и VL антител STIM с родственными антителами и последовательностями зародышевой линии человека показано на Фигуре 35, Фигуре 36 и Фигуре 37.
Антитело по изобретению может представлять собой человеческое антитело или химерное антитело, содержащее вариабельные области человека и нечеловеческие константные области (например, мышиные). Антитело по изобретению, например, имеет вариабельные области человека и, необязательно, также имеет константные области человека.
Таким образом, антитела, необязательно, включают константные области или их части, например, константные области антитела человека или их части. Например, домен VL может быть присоединен своим С-концом к константным доменам легкой цепи каппа или лямбда. Аналогично, домен VH антитела может быть присоединен своим С-концом к полной константной области тяжелой цепи иммуноглобулина или ее части (например, домену СН1 или области Fc), полученной из любого изотипа антитела, например, IgG, IgA, IgE и IgM, и любого из подклассов изотипа, таких как IgG1 или IgG4.
Примеры константных областей тяжелой цепи человека приведены в таблице S1.
Альтернативно, константные области антител по изобретению могут быть нечеловеческими константными областями. Например, когда антитела генерируются у трансгенных животных (примеры которых описаны в другой части данного документа), могут быть получены химерные антитела, содержащие вариабельные области человека и нечеловеческие константные области (животного-хозяина). Некоторые трансгенные животные генерируют полностью человеческие антитела. Другие были сконструированы для получения антител, содержащих химерные тяжелые цепи и полностью человеческие легкие цепи. В тех случаях, когда антитела содержат одну или несколько нечеловеческих константных областей, они могут быть заменены на константные области человека, для получения антител, более пригодных для введения людям в качестве терапевтических композиций, поскольку их иммуногенность в результате этого снижается.
Гидролиз антител ферментом папаином дает два идентичных антигенсвязывающих фрагмента, известных также как фрагменты «Fab», и фрагмент «Fc», не обладающий антигенсвязывающей активности, но способный кристаллизоваться. «Fab» при использовании в данном документе относится к фрагменту антитела, который включает один константный и один вариабельный домен каждой из тяжелой и легкой цепей. Термин «Fc-область» используется в данном документе для определения С-концевой области тяжелой цепи иммуноглобулина, включая Fc-области нативной последовательности и варианты Fc-областей. «Fc-фрагмент» относится к карбоксиконцевым частям обеих Н-цепей, удерживаемых вместе дисульфидными связями. Эффекторные функции антител определяются последовательностями в области Fc-области, которая также распознается Fc-рецепторами (FcR), обнаруженными на определенных типах клеток. Гидролиз антител ферментом пепсином дает фрагмент F(ab')2, в котором два плеча молекулы антитела остаются связанными и содержат два антигенсвязывающих сайта. Фрагмент F(ab')2 обладает способностью сшивать антиген.
«Fv» при использовании в данном документе относится к минимальному фрагменту антитела, которое сохраняет как антигенраспознающие, так и антигенсвязывающие сайты. Эта область состоит из димера вариабельного домена одной тяжелой и одной легкой цепи с сильной нековалентной или ковалентной связью. Именно в этой конфигурации три CDR каждого вариабельного домена взаимодействуют с образованием антигенсвязывающего сайта на поверхности димера VH-VL. В совокупности, шесть CDR обеспечивают антигенсвязывающую специфичность антитела. Однако, даже один вариабельный домен (или половина Fv, содержащего только три CDR, специфичных для антигена) обладает способностью распознавать и связывать антиген, хотя и с более низким сродством, чем полный сайт связывания.
Антитела, раскрытые в данном документе, могут быть модифицированы для увеличения или уменьшения периода полувыведения из сыворотки. В одном варианте реализации, вводят одну или несколько из следующих мутаций: T252L, T254S или T256F, для увеличения биологического периода полувыведения антитела. Биологический период полувыведения также может быть увеличен путем изменения домена CH1 константной области тяжелой цепи или области CL с целью введения эпитопа связывания рецептора реутилизации, взятого из двух петель домена СН2 Fc-области IgG, как описано в патентах США №№5869046 и 6121022, причем раскрытые в этом документе модификации включены в данный документ посредством ссылок. В другом варианте реализации, шарнирную Fc-область антитела или антигенсвязывающего фрагмента по изобретению мутируют для уменьшения биологического периода полувыведения антитела или фрагмента. Одну или несколько аминокислотных мутаций вводят в область интерфейса домена СН2-СН3 фрагмента Fc-шарнира, так чтобы антитело или фрагмент имели нарушенное связывание со стафилококковым (Staphylococcyl) белком A (SpA) по сравнению со связыванием нативного SpA с шарнирным доменом Fc. Специалистам в данной области известны другие способы увеличения периода полувыведения из сыворотки. Так, в одном варианте реализации, антитело или фрагмент пегилируют. В другом варианте реализации, антитело или фрагмент сливают с альбуминсвязывающим доменом, например, альбуминсвязывающее однодоменное антитело (dAb). В другом варианте реализации, антитело или фрагмент ПАСилируют (PASylated) (т.е. проводят сплайсинг (genetic fusion) полипептидных последовательностей, состоящих из PAS (XL-Protein GmbH), которые образует незаряженные структуры неупорядоченных клубков с большим гидродинамическим объемом). В другом варианте реализации, антитело или фрагмент представляют собой XTENylated®/rPEGylated продукты (т.е. проводят сплайсинг пептидной последовательности с неточными повторами (Amunix, Versartis) с терапевтическим пептидом). В другом варианте реализации, антитело или фрагмент представляют собой ELPylated продукты (т.е. проводят сплайсинг с последовательностью с ELP (эластин-подобный полипептид)-повторами (PhaseBio)). Различные такие гибридные продукты, позволяющие увеличить периоды полувыведения, описаны более подробно в Strohl, BioDrugs (2015) 29: 215-239, и эти гибридные продукты, например, приведенные в таблицах 2 и 6, включены в данный документ посредством ссылок.
Антитело может иметь модифицированную константную область, которая повышает стабильность. Так, в одном варианте реализации, константная область тяжелой цепи содержит мутацию Ser228Pro. В другом варианте реализации, раскрытые в данном документе антитела и фрагменты содержат шарнирную область тяжелой цепи, модифицированную с целью изменения числа цистеиновых остатков. Эта модификация может быть использована для облегчения сборки легкой и тяжелой цепей или для повышения или снижения стабильности антитела.
Эффекторные функции Fc - ADCC, ADCP и CDC
Как обсуждалось выше, анти-ICOS антитела могут быть представлены в различных изотипах и с различными константными областями. Примеры последовательностей константной области тяжелой цепи антитела IgG человека показаны в таблице S1. Fc-область антитела в первую очередь определяет его эффекторную функцию в терминах связывания Fc, опосредованной антителами клеточной цитотоксичности (ADCC), комплемент-зависимой цитотоксичности (CDC) и антителозависимого клеточно-опосредованного фагоцитоза (ADCP). Эти «клеточные эффекторные функции», в отличие от эффекторной Т-клеточной функции, включают рекрутирование клеток, несущих рецепторы Fc, к местонахождению клеток-мишеней, приводя к уничтожению связанной с антителом клетки. В дополнение к ADCC и CDC, механизм ADCP [19] представляет собой средство деплетирования связанных с антителом Т-клеток и, таким образом, нацеливания на TRegs с высокой экспрессией ICOS для уничтожения.
Клеточные эффекторные функции ADCC, ADCP и/или CDC также могут проявляться антителами, не имеющими Fc-областей. Антитела могут содержать несколько различных антигенсвязывающих сайтов, один из которых нацелен на ICOS, а другой - на молекулу-мишень, причем взаимодействие с этой молекулой-мишенью индуцирует ADCC, ADCP и/или CDC, например, антитело, содержащее две области scFv, соединенные линкером, причем один scFv может взаимодействовать с клеткой-эффектором.
Антитело в соответствии с данным изобретением может представлять собой антитело, которое проявляет ADCC, ADCP и/или CDC. Альтернативно, у антитела в соответствии с данным изобретением может отсутствовать активность ADCC, ADCP и/или CDC. В любом случае, антитело в соответствии с данным изобретением может содержать или может, необязательно, не содержать Fc-область, которая связывается с одним или несколькими типами рецептора Fc. Использование различных форматов антител и наличие или отсутствие связывания FcR и клеточных эффекторных функций позволяют модифицировать антитело для использования с конкретными терапевтическими целями, как описано в другой части данного документа.
Пригодный формат антител для некоторых терапевтических применений использует константную область человеческого IgG1 дикого типа. Константная область может быть константной областью IgG1 с активированным эффектором, необязательно, обладающей ADCC и/или CDC и/или ADCP активностью. Пригодной последовательностью константной области человеческого IgG1 дикого типа является SEQ ID NO: 340 (IGHG1*01). Дополнительные примеры константных областей IgG1 человека приведены в таблице S1.
Для тестирования терапевтических антител-кандидатов в мышиных моделях заболеваний людей вместо эффектор-положительной константной области человека может быть включена эффектор-положительная константная область мыши, такая как IgG2a мыши (mIgG2a).
Константная область может быть модифицирована для усиления ADCC и/или CDC и/или ADCP.
Величина Fc-опосредованных эффектов может быть усилена путем модификации Fc-домена различными установленными методами. Такие методы повышают аффинность к определенным Fc-рецепторам, тем самым создавая потенциальные разнообразные профили усиления активации. Это может быть достигнуто путем модификации одного или нескольких аминокислотных остатков [20]. Показано, что константные области IgG1 человека, содержащие специфические мутации или измененное гликозилирование в остатке Asn297 (например, N297Q, нумерация по индексу EU), улучшают связывание с Fc-рецепторами. Примеры мутаций включают один или несколько остатков, выбранных из 239, 332 и 330 для константных областей IgG1 человека (или эквивалентных положений в других изотипах IgG). Таким образом, антитело может содержать константную область IgG1 человека, имеющую одну или несколько мутаций, независимо выбранных из N297Q, S239D, I332E и A330L (нумерация по индексу EU). Тройную мутацию (M252Y/S254T/T256E) можно использовать для усиления связывания с FcRn, а другие мутации, влияющие на связывание FcRn, описаны в Таблице 2 [21], причем любая из них может быть использована в данном изобретении.
Повышенная аффинность к Fc-рецепторам может быть также достигнута путем изменения естественного профиля гликозилирования Fc-домена, например, путем генерации вариантов с пониженной степенью фукозилирования или дефукозилированных [22]. Нефукозилированные антитела содержат триманнозильную коровую структуру N-гликанов Fc комплексного типа без остатка фукозы. Эти антитела с модифицированной схемой гликозилирования, не содержащие корового остатка фукозы из N-гликанов Fc, могут проявлять более сильный ADCC, чем фукозилированные эквиваленты, из-за усиления связывающей способности FcγRIIIa. Например, для усиления ADCC, остатки в шарнирной области могут быть изменены для повышения связывания с Fc-гамма RIII [23]. Таким образом, антитело может содержать константную область тяжелой цепи человеческого IgG, которая представляет собой вариант константной области тяжелой цепи человеческого IgG дикого типа, причем вариант константной области тяжелой цепи человеческого IgG связывается с рецепторами Fcγ человека, выбранными из группы, состоящей из FcyRIIB и FcyRIIA, с большей аффинностью, чем при связывании константной области тяжелой цепи человеческого IgG дикого типа с рецепторами Fcγ человека. Антитело может содержать константную область тяжелой цепи IgG человека, которая представляет собой вариант константной области тяжелой цепи человеческого IgG дикого типа, причем вариант константной области тяжелой цепи IgG человека связывается с человеческим FcγRIIB с большей аффинностью, чем при связывании константной области тяжелой цепи человеческого IgG дикого типа с человеческим FcγRIIB. Вариант константной области тяжелой цепи человеческого IgG может быть вариантом IgG1 человека, вариантом IgG2 человека или вариантом константной области тяжелой цепи IgG4 человека. В одном варианте реализации, вариант константной области тяжелой цепи IgG человека содержит одну или несколько аминокислотных мутаций, выбранных из G236D, P238D, S239D, S267E, L328F и L328E (система нумерации по индексу EU). В другом варианте реализации, вариант константной области тяжелой цепи человеческого IgG содержит набор аминокислотных мутаций, выбранных из группы, состоящей из: S267E и L328F; P238D и L328E; P238D и одно или несколько замещений, выбранных из группы, состоящей из E233D, G237D, H268D, P271G и A330R; P238D, E233D, G237D, H268D, P271G и A330R; G236D и S267E; S239D и S267E; V262E, S267E и L328F; и V264E, S267E и L328F (система нумерации по индексу EU). Усиление CDC может быть достигнуто за счет аминокислотных изменений, которые увеличивают аффинность к C1q, первому компоненту классического активационного каскада комплемента [24]. Другим подходом является создание химерного домена Fc, состоящего из сегментов IgG1 человека и IgG3 человека, имеющих более высокую аффинность IgG3 к C1q [25]. Антитела по данному изобретению могут содержать мутированные аминокислоты в остатках 329, 331 и/или 322 для изменения связывания C1q и/или снижения или отмены активности CDC. В другом варианте реализации, антитела или фрагменты антител, раскрытые в данном документе, могут содержать Fc-области с модификациями в остатках 231 и 239, в результате чего аминокислоты заменяют с целью изменения способности антитела фиксировать комплемент. В одном варианте реализации, антитело или фрагмент имеют константную область, содержащую одну или несколько мутаций, выбранных из E345K, E430G, R344D и D356R, в частности, двойную мутацию, содержащую R344D и D356R (система нумерации по индексу EU).
В WO 2008/137915 описаны анти-ICOS антитела с модифицированными Fc-областями, обладающими повышенной эффекторной функцией. Сообщалось, что такие антитела опосредуют усиленную активность ADCC по сравнению с уровнем активности ADCC, опосредованным родительским антителом, содержащим домены VH и VK и Fc-область дикого типа. Антитела в соответствии с данным изобретением могут использовать такие варианты Fc-областей, имеющие эффекторную функцию, как описано в этом документе.
ADCC-активность антитела может быть определена в анализе, как описано в данном документе. ADCC-активность анти-ICOS антитела может быть определена in vitro с использованием ICOS-положительной линии Т-клеток, как описано в Примере 10. ADCC-активность анти-PD-L1 антитела может быть определена in vitro в анализе ADCC с использованием клеток, экспрессирующих PD-L1.
Для некоторых применений (например, в контексте вакцинации) может быть предпочтительным использование антител без эффекторной функции Fc. Могут предусматриваться антитела без константной области или без области Fc - примеры таких форматов антител описаны в другой части данного документа. Альтернативно, антитело может иметь константную область, являющуюся нуль-эффекторной (effector null). Антитело может иметь константную область тяжелой цепи, которая не связывает рецепторы Fcγ, например, константная область может содержать мутацию Leu235Glu (то есть, когда остаток лейцина дикого типа мутируют на остаток глутаминовой кислоты). Другой необязательной мутацией константной области тяжелой цепи является Ser228Pro, которая повышает стабильность. Константной областью тяжелой цепи может быть IgG4, содержащий как мутацию Leu235Glu, так и мутацию Ser228Pro. Эта константная область тяжелой цепи IgG4-PE является нуль-эффекторной.
Альтернативной нуль-эффекторной константной областью человека является инактивированный IgG1. Инактивированная константная область тяжелой цепи IgG1 может содержать аланин в положении 235 и/или 237 (нумерация по индексу EU), например, это может быть последовательность IgG1*01, содержащая мутации L235A и/или G237A («LAGA»).
Вариант константной области тяжелой цепи IgG человека может содержать одну или несколько мутаций аминокислот, которые уменьшают аффинность IgG к человеческому FcγRIIIA, человеческому FcγRIIA или человеческому FcγRI. В одном варианте реализации, FcγRIIB экспрессируется на клетке, выбранной из группы, состоящей из макрофагов, моноцитов, В-клеток, дендритных клеток, эндотелиальных клеток и активированных Т-клеток. В одном варианте реализации, вариант константной области тяжелой цепи человеческого IgG содержит одну или несколько следующих аминокислотных мутаций G236A, S239D, F243L, Т256А, K290A, R292P, S298A, Y300L, V305I, A330L, I332E, Е333А, K334A, А339Т и P396L (система нумерации по индексу EU). В одном варианте реализации, вариант константной области тяжелой цепи человеческого IgG содержит набор аминокислотных мутаций, выбранных из группы, состоящей из: S239D; Т256А; K290A; S298A; I332E; Е333А; K334A; А339Т; S239D и I332E; S239D, A330L и I332E; S298A, Е333А и K334A; G236A, S239D и I332E; и F243L, R292P, Y300L, V305I и P396L (система нумерации по индексу EU). В одном варианте реализации, вариант константной области тяжелой цепи человеческого IgG содержит аминокислотные мутации S239D, A330L или I332E (система нумерации по индексу EU). В одном варианте реализации, вариант константной области тяжелой цепи человеческого IgG содержит аминокислотные мутации S239D и I332E (система нумерации по индексу EU). В одном варианте реализации, вариант константной области тяжелой цепи человеческого IgG представляет собой вариант константной области тяжелой цепи человека IgG1, содержащий аминокислотные мутации S239D и I332E (система нумерации по индексу EU). В одном варианте реализации, антитело или фрагмент содержат афукозилированную Fc-область. В другом варианте реализации, антитело или его фрагмент являются дефукозилированными. В другом варианте реализации, антитело или фрагмент имеют пониженную степень фукозилирования.
Антитело может иметь константную область тяжелой цепи, которая связывает один или несколько типов рецептора Fc, но не индуцирует клеточные эффекторные функции, то есть не опосредует ADCC, CDC или активность ADCP. Такая константная область может быть неспособна связывать специфический Fc-рецептор (рецепторы), ответственный за запуск активности ADCC, CDC или ADCP.
Генерация и модификация антител
Способы идентификации и приготовления антител хорошо известны. Антитела могут быть получены с использованием трансгенных мышей (например, Kymouse™, Velocimouse®, Omnimouse®, Xenomouse®, HuMab Mouse® или MeMo Mouse®), крыс (например, Omnirat®), верблюдовых, акул, кроликов, кур или других животных, не являющихся людьми, иммунизированных ICOS или его фрагментом, или синтетическим пептидом, содержащим представляющий интерес мотив последовательности ICOS, с последующей необязательной гуманизацией константных областей и/или вариабельных областей с получением человеческих или гуманизированных антител. В качестве примера, можно использовать технологии дисплея, такие как дрожжевой, фаговый или рибосомальный дисплей, как будет понятно специалисту. Стандартное созревание аффинности, например, с использованием технологии дисплея, может быть выполнено на следующей стадии после выделения оптимизированного прототипа антитела из трансгенного животного, библиотеки фагового дисплея или другой библиотеки. Типичные примеры пригодных технологий описаны в US 20120093818 (Amgen, Inc), который в полном объеме включен в данное описание посредством ссылки, например, способы, изложенные в параграфах [0309]-[0346].
Иммунизация ICOS-нокаутных не являющихся человеком животных с помощью антигена человеческого ICOS способствует генерации антител, которые распознают как человеческий, так и нечеловеческий ICOS. Как описано в данном документе и проиллюстрировано в примерах, ICOS-нокаутная мышь может быть иммунизирована клетками, экспрессирующими ICOS человека, чтобы стимулировать продуцирование у мыши антител к ICOS человека и мыши, которые могут быть выделены и испытаны на связывание с ICOS человека и ICOS мыши. Таким образом могут быть выбраны перекрестно-реактивные антитела, которые могут быть подвергнуты скринингу на другие желательные свойства, как описано в данном документе. Способы получения антител к антигену (например, антигену человека) путем иммунизации антигеном животных с «нокаутной» экспрессией эндогенного антигена (например, эндогенного мышиного антигена), могут быть осуществлены на животных, способных генерировать антитела, содержащие вариабельные домены человека. Геномы таких животных могут быть модифицированы так, чтобы они содержали локус человеческого или гуманизированного иммуноглобулина, кодирующий генные сегменты вариабельной области человека и, необязательно, эндогенную константную область или константную область человека. Рекомбинация генных сегментов вариабельной области человека генерирует человеческие антитела, которые могут иметь либо нечеловеческую, либо человеческую константную область. Впоследствии нечеловеческие константные области могут быть заменены на константные области человека, причем антитело предназначено для использования in vivo у людей. Такие способы и нокаутные трансгенные животные описаны в WO 2013/061078.
Обычно, Kymouse™, VELOCIMMUNE® или другая мышь или крыса (необязательно, ICOS-нокаутная мышь или крыса, как упомянуто выше), могут быть подвергнуты стимуляции представляющим интерес антигеном, и лимфатические клетки (такие как В-клетки) выделяют у мышей, которые экспрессируют антитела. Лимфатические клетки могут быть слиты с клеточной линией миеломы для получения бессмертных клеточных линий гибридомы, и такие гибридомные клеточные линии подвергают скринингу и селекции для идентификации клеточных линий гибридомы, которые продуцируют антитела, специфичные к представляющему интерес антигену. ДНК, кодирующая вариабельные области тяжелой цепи и легкой цепи, может быть выделена и связана с желательными изотипическими константными областями тяжелой цепи и легкой цепи. Такой белок антитела может продуцироваться в клетке, такой как клетка СНО (яичника китайского хомячка). Альтернативно, ДНК, кодирующая антигенспецифические химерные антитела или вариабельные домены легкой и тяжелой цепей, может быть выделена непосредственно из антиген специфических лимфоцитов.
Вначале, выделяют высокоаффинные химерные антитела, имеющие вариабельную область человека и константную область мыши. Антитела характеризуют и подвергают селекции на желательные характеристики, включая аффинность, селективность, агонизм, зависимое от Т-клеток уничтожение клеток, эффективность нейтрализации, эпитоп и т.д. Константные области мыши необязательно, заменяют на желательную константную область человека для получения полностью человеческого антитела по изобретению, например, дикого типа или модифицированных IgG1 или IgG4 (например, SEQ ID NO: 751, 752, 753 в US 2011/0065902 (который включен в данный документ в полном объеме посредством ссылки). Если выбранная константная область может изменяться в зависимости от конкретного применения, то вариабельная область определяет характеристики высокоаффинного связывания с антигеном и специфичности к мишени.
Таким образом, в другом аспекте, данное изобретение предусматривает трансгенное млекопитающее, отличное от человека, имеющее геном, содержащий человеческий или гуманизированный локус иммуноглобулина, причем млекопитающее не экспрессирует ICOS. Млекопитающее может быть, например, нокаутной мышью или крысой или другими видами лабораторных животных. Трансгенные мыши, такие как Kymouse™, содержат человеческие локусы тяжелых и легких цепей иммуноглобулина, вставленные в соответствующие локусы эндогенных мышиных иммуноглобулинов. Трансгенным млекопитающим в соответствии с данным изобретением может быть такое, которое содержит такие целевые вставки, или оно может содержать локусы тяжелой и легкой цепей иммуноглобулина человека или гены иммуноглобулина, случайным образом вводимые в его геном, которые вставлены в локус, отличный от эндогенного локуса Ig, или находятся на дополнительной хромосоме или хромосомном фрагменте.
Другими аспектами изобретения являются использование таких не являющихся человеком млекопитающих для продуцирования антител к ICOS и способы продуцирования антител или вариабельных доменов тяжелых и/или легких цепей у таких млекопитающих.
Способ продуцирования антитела, которое связывает внеклеточный домен человеческого и нечеловеческого ICOS, может включать создание трансгенного млекопитающего, не являющегося человеком, имеющего геном, содержащий человеческий или гуманизированный локус иммуноглобулина, причем млекопитающее не экспрессирует ICOS, и
(а) иммунизацию млекопитающего антигеном ICOS человека (например, с помощью клеток, экспрессирующих ICOS человека, или очищенным рекомбинантным белком ICOS);
(b) выделение антител, продуцируемых млекопитающим;
(c) тестирование антител на способность связывать человеческий и нечеловеческий ICOS; и
(d) выбор одного или нескольких антител, которые связывают как человеческий, так и нечеловеческий ICOS.
Тестирование на способность связывать ICOS человека и нечеловеческий ICOS, может быть проведено с использованием поверхностного плазмонного резонанса, HTRF, FACS или любого другого способа, описанного в данном документе. Необязательно, определяют аффинности связывания для ICOS человека и мыши. Может быть определена аффинность, или кратность различий в аффинности, для связывания с ICOS человека и ICOS мыши, и таким образом могут быть отобраны антитела, демонстрирующие видовую перекрестную реактивность, (примеры пороговых значений аффинности и кратности различий, которые могут использоваться в качестве критериев отбора, приведены в других частях данного описания). Также, или альтернативно, могут быть определены эффективность нейтрализации активность или кратность различий в эффективности нейтрализации для антитела, предназначенного для ингибирования связывания лиганда ICOS человека и мыши с рецептором ICOS человека и мыши, соответственно, в качестве способа скрининга кросс-реактивных антител, например, в анализе методом HTRF. Также, возможные пороговые значения и кратности различий, которые могут использоваться в качестве критериев при селекции, приведены в других частях данного документа.
Способ может включать тестирование антител на способность связывать нечеловеческие ICOS от животного того же вида, что и иммунизированное млекопитающее, или другого вида. Таким образом, когда трансгенным млекопитающим является мышь (например, Kymouse™), антитела могут быть протестированы на способность связывать ICOS мыши. Если трансгенное млекопитающее является крысой, антитела могут быть испытаны на способность связывать ICOS крысы. Однако, в равной степени полезным может быть определение перекрестной реактивности выделенного антитела к нечеловеческому ICOS, принадлежащему животному другого вида. Таким образом, антитела, образующиеся у коз, могут быть протестированы на связывание с ICOS крысы или мыши. Необязательно, вместо этого или дополнительно, может быть определено связывание с козьим ICOS.
В других вариантах реализации, трансгенное млекопитающее, не являющееся человеком, может быть иммунизировано нечеловеческим ICOS, возможно, ICOS млекопитающего того же вида (например, ICOS-нокатная мышь может быть иммунизирована ICOS мыши) вместо человеческого ICOS. Затем таким же образом определяют аффинность выделенных антител для связывания с ICOS человека и нечеловеческим ICOS, и отбирают антитела, которые связывают как человеческий, так и нечеловеческий ICOS.
Нуклеиновая кислота, кодирующая вариабельный домен тяжелой цепи антитела и/или вариабельный домен легкой цепи отобранного антитела, может быть выделена. Такая нуклеиновая кислота может кодировать всю тяжелую цепь и/или легкую цепь антитела или вариабельный домен (домены) без ассоциированной константной области (областей). Как было отмечено, кодирующие нуклеотидные последовательности могут быть получены непосредственно из клеток мыши, продуцирующих антитела, или В-клетки могут быть иммортализованы или слиты для получения гибридом, экспрессирующих антитело, и кодирования нуклеиновой кислоты, полученной из таких клеток. Необязательно, нуклеиновую кислоту, кодирующую вариабельный домен (домены), затем конъюгируют с нуклеотидной последовательностью, кодирующей константную область тяжелой цепи человека и/или константную область легкой цепи человека, для получения нуклеиновой кислоты, кодирующей тяжелую цепь антитела человека и/или легкую цепь антитела человека, например, кодирующей антитело, содержащее как тяжелую, так и легкую цепь. Как описано в другой части данного документа, эта стадия особенно полезна в тех случаях, когда иммунизированное млекопитающее продуцирует химерные антитела с нечеловеческими константными областями, которые, предпочтительно, заменяют на константные области человека для получения антитела, которое будет менее иммуногенным при введении людям в качестве лекарственного средства. Обеспечение конкретных константных областей изотипа человека также имеет важное значение для определения эффекторной функции антитела, и ряд пригодных константных областей тяжелой цепи описан в данном документе.
Могут быть проведены другие изменения нуклеиновой кислоты, кодирующей вариабельный домен тяжелой и/или легкой цепи антитела, такие как мутация остатков и генерация вариантов, как описано в данном документе.
Выделенная (необязательно, мутантная) нуклеиновая кислота может быть введена в клетки-хозяева, например, клетки СНО, как было описано. Затем клетки-хозяева культивируют в условиях экспрессии антитела или вариабельного домена тяжелой и/или легкой цепи антитела, в любом желательном формате антител. Некоторые возможные форматы антител описаны в данном документе, например, цельные иммуноглобулины, антигенсвязывающие фрагменты и другие конструкты.
Варианты вариабельных доменов аминокислотной последовательности любого из доменов VH и VL или CDR, последовательности которых конкретно раскрыты в данном документе, могут быть использованы в соответствии с данным изобретением, как обсуждалось.
Существует множество причин, по которым может быть желательно создание вариантов, которые включают оптимизацию последовательности антител для крупномасштабного производства, облегчение очистки, повышение стабильности или улучшение пригодности для включения в желательную фармацевтическую композицию. Работы по белковой инженерии могут проводиться на одном или нескольких целевых остатках в последовательности антител, например, с целью замещения какой-либо аминокислоты альтернативной аминокислотой (необязательно, с получением вариантов, содержащих в этом положении все природные аминокислоты, возможно, за исключением Cys и Met), и мониторинг влияния на функцию и экспрессию, чтобы определить наилучшую замену. В некоторых случаях нежелательно замещать остаток на Cys или Met или вводить эти остатки в последовательность, поскольку это может создавать трудности в производстве, например, из-за образования новых внутримолекулярных или межмолекулярных цистеин-цистеиновых связей. При выборе наилучшего прототипа антитела и его оптимизации для производства и клинической разработки, как правило, желательно как можно меньше изменять его антигенсвязывающие свойства или, по крайней мере, сохранять аффинность и эффективность исходной молекулы. Однако, варианты могут также генерироваться для модуляции ключевых характеристик антител, таких как аффинность, перекрестная реактивность или нейтрализующая способность.
Антитело может содержать набор Н и/или L CDR любого из раскрытых антител с одной или несколькими мутациями аминокислот в раскрытом наборе Н и/или L CDR. Мутация может быть аминокислотной заменой, делецией или инсерцией. Так, например, раскрытый набор Н и/или L CDR может иметь одно или несколько аминокислотных замещений. Например, набор Н и/или L CDR может иметь до 12, 11, 10, 9, 8, 7, 6, 5, 4, 3 или 2 мутаций, например, замещений. Например, до 6, 5, 4, 3 или 2 мутаций, например, замещений, может присутствовать в HCDR3, и/или до 6, 5, 4, 3 или 2 мутаций, например, замещений, может присутствовать в LCDR3. Антитело может содержать набор HCDR, LCDR или набор из 6 (Н и L) CDR, указанных для любого антитела STIM в данном документе, или может содержать такой набор CDR с одной или двумя консервативными заменами.
Одна или несколько аминокислотных мутаций могут быть произвольно проведены в каркасных областях домена VH или VL антитела, описанного в данном документе. Например, один или несколько остатков, которые отличаются от соответствующей последовательности сегмента зародышевой линии человека, могут быть возвращены к зародышевой линии. Последовательности генных сегментов зародышевой линии человека, соответствующие доменам VH и VL примеров анти-ICOS антител, указаны в Таблице Е12-1, Таблице Е12-2 и Таблице Е12-3, а выравнивания доменов VH и VL антител с соответствующими последовательностями зародышевой линии показаны на фигурах.
Антитело может содержать домен VH, который имеет по меньшей мере 60, 70, 80, 85, 90, 95, 98 или 99% идентичности аминокислотной последовательности с VH-доменом любого из антител, приведенных в приложенном перечне последовательностей, и/или содержать домен VL, который имеет по меньшей мере 60, 70, 80, 85, 90, 95, 98 или 99% идентичности аминокислотной последовательности с доменом VL любого из этих антител. Алгоритмы, которые могут быть использованы для вычисления % идентичности двух аминокислотных последовательностей, включают, например, BLAST, FASTA или алгоритм Смита-Уотермана (Smith-Waterman), например, с использованием параметров по умолчанию. Конкретные варианты могут включать одно или несколько изменений аминокислотной последовательности (добавление, делеция, замещение и/или инсерция аминокислотного остатка).
Изменения могут быть сделаны в одной или нескольких каркасных областях и/или одном или нескольких CDR. Варианты, необязательно, получают путем мутагенеза CDR. Изменения обычно не приводят к потере функции, поэтому антитело, содержащее измененную таким образом аминокислотную последовательность, может сохранять способность связывать ICOS. Оно может сохранять ту же величину связывающей способности, что и антитело, в котором изменение не проводилось, например, по результатам измерений в анализе, описанном в данном документе. Антитело, содержащее измененную таким образом аминокислотную последовательность, может иметь повышенную способность связывать ICOS.
Изменение может включать замену одного или нескольких аминокислотных остатков неприродной или нестандартной аминокислотой, модификацию одного или нескольких аминокислотных остатков до не встречающейся в природе или нестандартной формы или вставку одной или нескольких неприродных или нестандартных аминокислот в последовательность. Примеры количества и расположения изменений в последовательностях по изобретению описаны в других частях данного документа. Природные аминокислоты включают 20 "стандартных" L-аминокислот, обозначаемых их стандартными однобуквенными кодами G, А, V, L, I, М, Р, F, W, S, Т, N, Q, Y, С, K, R, Н, D, Е. Нестандартные аминокислоты включают любой другой остаток, который может быть включен в полипептидную основную цепь или образуется в результате модификации существующего аминокислотного остатка. Нестандартные аминокислоты могут быть природными или неприродными.
В используемом в данном документе значении, термин «вариант» относится к пептиду или нуклеиновой кислоте, которые отличаются от исходного полипептида или нуклеиновой кислоты делециями, заменами или добавлениями одной или нескольких аминокислот или нуклеиновых кислот, но сохраняют одну или несколько специфических функций или биологических активностей исходной молекулы. Аминокислотные замещения включают изменения, при которых аминокислоту заменяют на другой природный аминокислотный остаток. Такие замещения могут быть классифицированы как «консервативные», и в этом случае аминокислотный остаток, содержащийся в полипептиде, заменяется другой природной аминокислотой аналогичного характера либо по показателю полярности, функциональности или размеру боковой цепи. Такие консервативные замещения хорошо известны в данной области. Замены, охватываемые данным изобретением, также могут быть «неконсервативными», в которых аминокислотный остаток, присутствующий в пептиде, замещен аминокислотой, имеющей отличающиеся свойства, такой как природная аминокислота из другой группы (например, при замене заряженной или гидрофобной аминокислоты аланином) или, альтернативно, в которых природная аминокислота замещена нетрадиционной аминокислотой. В некоторых вариантах аминокислотные замещения являются консервативными. Кроме того, термин вариант, при использовании со ссылкой на полинуклеотид или полипептид, включает ссылки на полинуклеотид или полипептид, которые могут различаться по первичной, вторичной или третичной структуре с эталонным полинуклеотидом или полипептидом, соответственно (например, по сравнению с полинуклеотидом или полипептидом дикого типа).
В некоторых аспектах можно использовать «синтетические варианты», «рекомбинантные варианты» или «химически модифицированные» варианты полинуклеотидов или варианты полипептидов, выделенные или полученные с использованием способов, хорошо известных в данной области техники. «Модифицированные варианты» могут включать консервативные или неконсервативные изменения аминокислот, как описано ниже. Изменения полинуклеотидов могут приводить к аминокислотным заменам, добавлениям, делециям, слияниям и усечениям в полипептиде, кодируемом референсной последовательностью. Некоторые аспекты использования включают варианты вставки, варианты делеции или варианты замещения с заменами аминокислот, включая вставки и замены аминокислот и других молекул), которые обычно не встречаются в пептидной последовательности, являющейся основой варианта, например, без ограничений, инсерцию орнитина, который обычно не встречается в человеческих белках. Термин «консервативная замена» при описании полипептида относится к изменению аминокислотного состава полипептида, которое существенно не влияет на активность полипептида. Например, консервативная замена относится к замещению аминокислотного остатка на другой аминокислотный остаток, обладающий сходными химическими свойствами (например, кислотный, основной, положительно или отрицательно заряженный, полярный или неполярный и т.д.). Консервативные аминокислотные замены включают замещение лейцина на изолейцин или валин, аспартата на глутамат, или треонина на серии. Таблицы консервативных замещений с указанием функционально сходных аминокислот хорошо известны специалистам. Например, каждая из следующих шесть групп включает аминокислоты, являющиеся консервативными заменами друг для друга: 1) аланин (А), серии (S), треонин (Т); 2) аспарагиновая кислота (D), глутаминовая кислота (Е); 3) аспарагин (N), глутамин (Q); 4) аргинин (R), лизин (K); 5) изолейцин (I), лейцин (L), метионин (М), валин (V); и 6) фенилаланин (F), тирозин (Y), триптофан (W). (См. также Creighton, Proteins, W.Н. Freeman and Company (1984), включенна посредством ссылки в полном объеме.) В некоторых вариантах реализации, индивидуальные замены, делеции или добавления, которые изменяют, добавляют или удаляют одну аминокислоту или небольшую часть аминокислот, также могут рассматриваться как «консервативные замещения», если это изменение не снижает активность пептида. Размер вставок или делеций обычно находится в диапазоне от 1 до 5 аминокислот. Выбор консервативных аминокислот может быть осуществлен на основе положения замещаемой аминокислоты в пептиде, например, когда аминокислота расположена на наружной поверхности пептида и подвержена воздействию растворителей, или когда она расположена во внутренней части и не контактирует с растворителями.
Можно выбрать аминокислоту, которая будет заменять имеющуюся аминокислоту на основании положения имеющейся аминокислоты, включая ее контакт с растворителями (т.е., если аминокислота подвергается воздействию растворителей или расположена на внешней поверхности пептида или полипептида, в отличие от локализованных во внутренней части аминокислот, не подверженных воздействию растворителей). Выбор таких консервативных аминокислотных замещений хорошо известен специалистам в данной области, например, как описано в Dordo et al., J. Mol Biol, 1999, 217, 721-739 и Taylor et al., J. Theor. Biol. 119 (1986), 205-218 и S. French and B. Robson, J. Mol. Evol. 19 (1983) 171. Соответственно, можно выбрать консервативные аминокислотные замещения, пригодные для аминокислот, расположенных на внешней поверхности белка или пептида (т.е. аминокислот, контактирующих с растворителем), например, без ограничений, могут быть использованы следующие замещения: замена Y на F, Т на S или K, Р на А, Е на D или Q, N на D или G, R на K, G на N или А, Т на S или K, D на N или Е, I на L или V, F на Y, S на Т или A, R на K, G на N или А, K на R, А на S, K или Р.
В альтернативных вариантах реализации, можно также выбрать консервативные аминокислотные замещения, охватывающие пригодные аминокислоты во внутренней части белка или пептида, например, можно использовать пригодные консервативные замещения аминокислот, находящихся во внутренней части белка или пептида (т.е. аминокислот, не подвергающихся воздействию растворителя), например, без ограничений, можно использовать следующие консервативные замещения: замена Y на F, Т на А или S, I на L или V, W на Y, М на L, N на D, G на А, Т на А или S, D на N, I на L или V, F на Y или L, S на А или Т и А на S, G, Т или V. В некоторых вариантах реализации, термин вариант охватывает также неконсервативные аминокислотные замещения.
Данное изобретение включает способы получения антител, содержащих варианты доменов VH и/или VL, относящиеся к доменам VH и/или VL антитела, представленным в прилагаемом перечне последовательностей. Такие антитела могут быть получены способом, включающим
(i) обеспечение домена VH антитела, представляющего собой вариант аминокислотной последовательности домена VH исходного антитела, путем добавления, делеции, замещения или введения одной или нескольких аминокислот в аминокислотной последовательности домена VH исходного антитела,
причем домен VH исходного антитела является доменом VH любого из антител STIM001, STIM002, STIM002-B, STIM003, STIM004, STIM005, STIM006, STIM007, STIM008 и STIM009, или доменом VH, содержащим участки, определяющие комплементарность, тяжелой цепи любого из этих антител,
(ii) необязательно, объединение полученного таким образом домена VH с доменом VL, для получения комбинации VH/VL, и
(iii) тестирование домена VH или полученной таким образом комбинации доменов VH/VL, для идентификации антитела с одной или несколькими желательными характеристиками.
Желательные характеристики включают связывание с ICOS человека, связывание с ICOS мыши и связывание с другими нечеловеческими ICOS, такими как ICOS яванского макака. Могут быть идентифицированы антитела со сравнимой или более высокой аффинностью к ICOS человека и/или мыши. Другие желательные характеристики включают опосредованное усиление функции эффекторных Т-клеток, путем деплетирования иммуносупрессивных TRegs, или непосредственное, путем активации ICOS-сигнализации на эффекторных Т-клетках. Идентификация антитела с желательной характеристикой может включать идентификацию антитела с описанным в данном документе функциональным признаком, таким как его аффинность, перекрестная реактивность, специфичность, агонизм рецептора ICOS, нейтрализующая активность и/или промотирование зависимого от Т-клеток убийства, любой из которых может быть определен в анализах, как описано в данном документе.
В тех случаях, когда в этот способ включены домены VL, домен VL может быть доменом VL любого из STIM001, STIM002, STIM002-B, STIM003, STIM004, STIM005, STIM006, STIM007, STIM008 или STIM009, или может быть вариантом, полученным путем добавления, делеции, замещения или вставки одной или нескольких аминокислот в аминокислотной последовательности исходного домена VL, причем исходный домен VL является доменом VL любого из STIM001, STIM002, STIM002-B, STIM003, STIM004, STIM005, STIM006, STIM007, STIM008 и STIM009, или доменом VL, содержащим участки, определяющие комплементарность, легкой цепи любого из этих антител.
Способы получения вариантов антител могут, необязательно, включать получение копий антитела или комбинации доменов VH/VL. Способы могут дополнительно включать экспрессию полученного антитела. Можно продуцировать нуклеотидные последовательности, соответствующие желательному VH и/или VL-домену антитела, необязательно, в одном или нескольких векторах экспрессии. Пригодные способы экспрессии, включая рекомбинантную экспрессию в клетках-хозяевах, подробно описаны в данном документе.
Кодирующие нуклеиновые кислоты и способы экспрессии
Может быть получена выделенная нуклеиновая кислота, кодирующая антитела в соответствии с данным изобретением. Нуклеиновая кислота может представлять собой ДНК и/или РНК. Геномная ДНК, кДНК, мРНК или другая РНК синтетического происхождения или любая их комбинация могут кодировать антитело.
Данное изобретение предусматривает конструкты в форме плазмид, векторов, кассет транскрипции или экспрессии, которые содержат по меньшей мере один полинуклеотид, как указано выше. Иллюстративные нуклеотидные последовательности включены в перечень последовательностей. Ссылка на нуклеотидную последовательность, приведенная в данном документе, охватывает молекулу ДНК с указанной последовательностью, и охватывает молекулу РНК с указанной последовательностью, в которой U заменен на Т, если контекст не требует иного.
Данное изобретение также относится к рекомбинантной клетке-хозяину, которая содержит одну или несколько нуклеиновых кислот, кодирующих антитело. Способы получения кодированного антитела могут включать экспрессию из нуклеиновой кислоты, например, путем культивирования рекомбинантных клеток-хозяев, содержащих нуклеиновую кислоту. Таким образом, антитело может быть получено и может быть выделено и/или очищено с использованием любого пригодного метода, а затем использовано при необходимости. Способ получения может включать приготовление композиции продукта, включающей по меньшей мере один дополнительный компонент, такой как фармацевтически приемлемый эксципиент.
Системы клонирования и экспрессии полипептида во множестве различных клеток-хозяев являются общеизвестными. Пригодные клетки-хозяева включают бактерии, клетки млекопитающих, растительные клетки, мицелиальные грибы, дрожжевые и бакуловирусные системы и трансгенные растения и животных.
Экспрессия антител и фрагментов антител в прокариотических клетках хорошо известна в данной области техники. Обычным бактериальным хозяином является Е. coli. Экспрессия в культуре эукариотических клеток также доступна квалифицированным специалистам в данной области как вариант продуцирования. Клеточные линии млекопитающих, доступные специалистам для экспрессии гетерологичного полипептида, включают клетки яичника китайского хомячка (СНО), клетки HeLa, клетки почек детеныша хомяка, клетки меланомы мыши NSO, клетки миеломы крысы YB2/0, клетки почки эмбриона человека, клетки сетчатки эмбриона человека и многие другие.
Векторы могут содержать соответствующие регуляторные последовательности, включая промоторные последовательности, терминаторные последовательности, последовательности полиаденилирования, энхансерные последовательности, маркерные гены и другие последовательности, при необходимости. Нуклеиновая кислота, кодирующая антитело, может быть введена в клетку-хозяина. Нуклеиновую кислоту может быть введена в эукариотические клетки различными способами, включая трансфекцию фосфатом кальция, использование DEAE-декстрана, электропорацию, опосредованную липосомами трансфекцию и трансдукцию с использованием ретровируса или другого вируса, например, коровьей оспы или, для клеток насекомых, бакуловируса. Для введение нуклеиновой кислоты в клетку-хозяина, в частности, эукариотическую клетку, может использоваться вирусная или плазмидная система. Плазмидная система может поддерживаться эписомально или может быть включена в клетку-хозяина или в искусственную хромосому. Включение может осуществляться путем случайной или целенаправленной интеграции одной или нескольких копий в один или несколько локусов. Для бактериальных клеток, пригодные методы включают трансформацию с хлоридом кальция, электропорацию и трансфекцию с использованием бактериофага. Введение может сопровождаться экспрессией нуклеиновой кислоты, например, путем культивирования клеток-хозяев в условиях экспрессии гена, необязательно, с последующим выделением или очисткой антитела.
Нуклеиновая кислота по изобретению может быть интегрирована в геном (например, хромосому) клетки-хозяина. Интеграции может способствовать включение последовательностей, которые промотируют рекомбинацию с геномом, в соответствии со стандартными методами.
Данное изобретение также предусматривает способ, который включает использование нуклеиновой кислоты, описанной в данном документе, в системе экспрессии с целью экспрессии антитела.
Терапевтическое применение
Антитело, описанное в данном документе, может быть использовано в способе лечения организма человека или животного путем терапии. Антитела находят применение при усилении эффекторного Т-клеточного ответа, что создает полезный эффект при ряде заболеваний или состояний, включая лечение различных видов рака или солидных опухолей, и в контексте вакцинации. Усиленный ответ Teff может быть достигнут с использованием антитела, которое модулирует баланс или соотношение между Teffs и Tregs в пользу активности Teff.
Анти-ICOS антитела могут использоваться для деплетирования регуляторных Т-клеток и/или усиления эффекторного ответа Т-клеток у пациента и могут быть введены пациенту для лечения заболевания или состояния, поддающегося лечению, путем деплетирования регуляторных Т-клеток и/или усиления эффекторного ответа Т-клеток.
Антитело по данному изобретению или композиция, содержащая такую молекулу антитела или кодирующую ее нуклеиновую кислоту, могут быть использованы или предоставлены для использования в любом таком способе. Также предусматривается использование антитела, или содержащей его или кодирующую его нуклеиновую кислоту композиции, с целью изготовления лекарственного средства для использования в любом таком способе. Способ типично включает введение антитела или композиции млекопитающему. Пригодные композиции и способы введения описаны в других частях данного документа.
Одним из предусматриваемых терапевтических применений антител является лечение рака. Рак может быть солидной опухолью, например, почечно-клеточным раком (необязательно, карциномой почек, например, светлоклеточным почечно-клеточным раком), раком головы и шеи, меланомой (необязательно, злокачественной меланомой), немелкоклеточным раком легкого (например, аденокарциномой), раком мочевого пузыря, раком яичников, раком шейки матки, раком желудка, раком печени, раком поджелудочной железы, раком молочной железы, герминогенной карциномой яичек, или метастазами твердой опухоли, такой как перечисленные, или он может быть жидкой гематологической опухолью, например, лимфомой (такой как болезнь Ходжкина или неходжкинская лимфома, например, диффузная крупноклеточная В-клеточная лимфома, ДККВЛ) или лейкозом (например, острым миелоидным лейкозом). Анти-ICOS антитело может усилить рассасывание опухоли при меланоме, раке головы и шеи и немелкоклеточном раке легкого и других раках с мутационной нагрузкой от умеренной до высокой [26]. Увеличивая иммунный ответ пациентов на их неопластические поражения, иммунотерапия, использующая анти-ICOS антитело, предлагает перспективу долгосрочного исцеления или долгосрочных ремиссий, потенциально даже в контексте поздней стадии заболевания.
Раки представляют собой неоднородную группу заболеваний, но анти-ICOS антитела предлагают возможность лечения ряда различных видов рака путем использования собственной иммунной системы пациента, которая может убить любую раковую клетку посредством распознавания мутантных или сверхэкспрессируемных эпитопов, которыми раковые клетки отличаются от нормальной ткани. Путем модуляции баланса Teff/Treg, анти-ICOS антитела могут обеспечивать возможность и/или промотировать иммунное распознавание и уничтожение раковых клеток. Хотя анти-ICOS антитела являются, таким образом, полезными терапевтическими средствами для широкого спектра видов рака, существуют определенные категории рака, для которых анти-ICOS терапия является особенно подходящей и/или при которых анти-ICOS терапия может быть эффективной, когда другие терапевтические средства неэффективны.
Одной из таких групп является рак, положительный по экспрессии лиганда ICOS. Раковые клетки могут приобретать способность экспрессировать лиганд ICOS, как было описано для меланомы [27]. Экспрессия лиганда ICOS может обеспечить клеткам избирательное преимущество, поскольку поверхностно-экспрессируемый лиганд связывает ICOS на Tregs, способствуя пролиферации и активации Tregs и, тем самым, подавляя иммунный ответ против рака. Выживания раковых клеток, экспрессирующих лиганд ICOS, может зависеть от такого подавления иммунной системы с помощью Tregs и, таким образом, будут уязвимы для лечения анти-ICOS антителами, нацеленными на Tregs. Это относится также к ракам, развивающимся из клеток, которые естественным образом экспрессируют лиганд ICOS. Продолжающаяся экспрессия лиганда ICOS этими клетками снова обеспечивает преимущество выживания за счет подавления иммунитета. Рак, экспрессирующий лиганд ICOS, может развиваться из антигенпрезентирующих клеток, таких как В-клетки, дендритные клетки и моноциты, и может быть жидкостной гематологической опухолью, такой как указанные в данном документе. Интересно, что эти типы рака, как было показано, также имеют высокую экспрессию ICOS и FOXP3 (данные TCGA (Атлас ракового генома)) - см. Пример 25. Пример 20 данного документа демонстрирует эффективность типичных анти-ICOS антител при лечении опухолей, развивающихся из раковых В-клеток (А20-сингенных клеток), которые экспрессируют лиганд ICOS.
Соответственно, анти-ICOS антитела можно использовать в методах лечения различных видов рака, положительных по экспрессии лиганда ICOS. Кроме того, рак, подлежащий лечению анти-ICOS антителом в соответствии с данным изобретением, может быть положительным по экспрессии ICOS и/или FOXP3 и, необязательно, также экспрессирует лиганд ICOS.
Пациенты могут пройти тестирование, чтобы определить, является ли их рак положительным по экспрессии представляющего интерес белка (например, лиганда ICOS, ICOS и/или FOXP3), например, путем взятия образца для тестирования (например, биопсии опухоли) у пациента и определения экспрессии представляющего интерес белка. Пациентов, рак которых был охарактеризован как положительный по экспрессии одного, двух или всех таких белков, представляющих интерес, отбирают для лечения анти-ICOS антителом. Как было описано в других частях данного документа, анти-ICOS антитело можно использовать в качестве монотерапии или в комбинации с одним или несколькими другими терапевтическими средствами.
Анти-ICOS антитела также дают надежду пациентам, рак которых не поддается лечению антителами или другими лекарственными препаратами, направленными на иммунные контрольные точки, такие как CTLA-4, PD-1, PD-L1, CD137, GITR или CD73. Эти иммунотерапии эффективны против определенных видов рака, но в некоторых случаях рак может не реагировать, или он может перестать реагировать на продолжающееся лечение антителом. Как и антитела к ингибиторам иммунной контрольной точки, анти-ICOS антитела модулируют иммунную систему пациента - тем не менее анти-ICOS антитело может преуспеть там, причем другие такие антитела терпят неудачу. В данном документе показано, что животные, несущие В-клеточные лимфомы А20, можно лечить анти-ICOS антителами для уменьшения роста опухоли, уменьшения размера опухоли и фактического удаления опухоли из организма, в то время как лечение анти-PD-L1 антителом не отличалось от контроля. Сообщалось также, что клеточная линия А20 устойчива к анти-CTLA-4 [28].
Соответственно, анти-ICOS антитела могут быть использованы в способах лечения различных видов рака, которые не поддаются лечению одной или несколькими иммунотерапиями, такими как (любая из или все из) анти-CTLA-4 антитело, анти-PD1 антитело, анти-PD-L1 антитело, анти-CD137 антитело, анти-GITR антитело или анти-CD73 антитело. Рак может быть охарактеризован как невосприимчивый к лечению антителом или другим лекарственным средством, если лечение этим антителом или лекарственным средством существенно не снижает рост рака, например, если опухоль продолжает расти или не уменьшается в размере, или если после периода отклика опухоль повторно инициирует свой рост. Отсутствие ответа на терапевтическое средство может быть определено ex vivo путем тестирования образца (например, образца биопсии опухоли) на уничтожение раковых клеток или ингибирование роста и/или в клинических условиях по результатам наблюдений (например, с использованием технологии визуализации, включая МРТ), показывающим, что пациент, получающий терапию, не реагирует на лечение. Пациентов, у которых рак был охарактеризован как рефрактерный к лечению такой иммунотерапией, отбирают для лечения анти-ICOS антителом.
Кроме того, анти-ICOS антитела могут быть использованы для лечения рака, образующегося из В-клеток, который является устойчивым к лечению анти-CD20 антителом. Анти-ICOS антитела являются способом лечения раковых заболеваний, которые не отвечают или становятся резистентными к терапии анти-CD20-антителами, такими как ритуксимаб. Анти-ICOS антитело можно использовать в качестве терапии второй линии (или дополнительной, или вспомогательной) для таких видов рака. Рак, устойчивый к анти-CD20-антителу, может быть В-клеточным раком, например, В-клеточной лимфомой, такой как диффузная крупноклеточная В-клеточная лимфома. Резистентность рака к анти-CD20 можно определить ex vivo путем тестирования образца (например, образца биопсии опухоли) на уничтожение раковых клеток или ингибирование роста анти-CD20 антителом и/или в клинических условиях на основе наблюдений, показывающих, что пациент, получающий терапию анти-CD20 антителом, не реагирует на лечение. Альтернативно, или дополнительно, рак (например, образец биопсии опухоли) можно тестировать на оценку экспрессии CD20, причем отсутствие или низкий уровень экспрессии CD20 указывают на потерю чувствительности к анти-CD20-антителу.
Таким образом, образцы, полученные от пациентов, могут тестироваться для определения поверхностной экспрессии представляющего интерес белка, например, лиганда ICOS, ICOS, FOXP3 и/или рецептора-мишени, на который нацелен другой терапевтическое средство (например, антирецепторное антитело). Рецептором-мишенью может быть CD20 (на который нацелена терапия анти-CD20 антителом, таким как ритуксимаб), или другой рецептор, такой как PD1, EGFR, HER2 или HER3. Поверхностная экспрессия лиганда ICOS, ICOS, FOXP3 и/или отсутствие или потеря поверхностной экспрессии рецептора-мишени является показателем того, что рак восприимчив к терапии анти-ICOS антителом. Анти-ICOS антитела могут предусматриваться для введения пациенту, чей рак характеризуется поверхностной экспрессией лиганда ICOS, ICOS, FOXP3 и/или отсутствием или потерей поверхностной экспрессии рецептора-мишени, необязательно, в тех случаях, когда пациент ранее получал лечение анти- -CTLA4, анти-PD1, анти-PD-L1 или антителом к рецептору-мишени и не ответил или перестал реагировать на лечение этим антителом, по результатам измерений, указывающим, например, на продолжение или возобновление роста раковых клеток, например, увеличение размер опухоли.
Любой пригодный способ может быть использован для определения того, являются ли раковые клетки положительными в тестах поверхностной экспрессии белка, такого как лиганд ICOS, CD20 или других рецепторов-мишеней, упомянутых в данном документе. Типичным методом является иммуногистохимия, когда образец клеток (например, образец биопсии опухоли) контактируют с антителом к представляющему интерес белку и детектируют связывание антитела с использованием меченого реагента -обычно второго антитела, которое распознает Fc-область первого антитела и несет детектируемую метку, такую как флуоресцентный маркер. Образец может быть объявлен положительным по результатам тестирования, когда по меньшей мере 5% клеток являются мечеными по результатам визуализации путем окрашивания клеток или другим способом детектирования метки. Необязательно, может использоваться более высокий порог отсечения, например, 10% или 25%. Антитело обычно используют в избыточном количестве. Антитела, реагирующие с представляющими интерес молекулами, являются доступными или могут быть получены с помощью простых методов. Для тестирования на лиганд ICOS в настоящее время доступно антитело МАВ1651 фирмы R&D systems в виде мышиного IgG, который распознает лиганд ICOS человека. Для тестирования на экспрессию CD20 можно использовать ритуксимаб. Альтернативным методом является детектирование уровней мРНК лиганда ICOS или рецептора-мишени [27].
Еще одним признаком того, что опухоль будет реагировать на лечение анти-ICOS антителом, является присутствие Tregs в микроокружении опухоли. Активированные Tregs характеризуются высокой поверхностной экспрессией ICOS и Foxp3 (ICOS-high and Foxp3-high surface expression). Присутствие Tregs в опухоли, особенно в повышенных количествах, обеспечивает дополнительное обоснование того, что пациент может быть отобран для лечения анти-ICOS антителом. Tregs могут быть детектированы в образце биопсии опухоли ex vivo, например, методом иммуногистохимии (анализ совместной экспрессии как Foxp3, так и ICOS, с использованием антител к белку-мишени с последующим детектированием меток, как описано выше) или методом диспергирования на отдельные клетки образца для использования в FACS с мечеными антителами к ICOS и Foxp3. Примеры FACS приведены в Примере 17 и Примере 18.
Анти-ICOS антитела могут быть использованы для лечения раковых заболеваний, связанных с инфекционными агентами, таких как вирусно-индуцированные раковые заболевания. К этой категорию относятся плоскоклеточный рак головы и шеи, рак шейки матки, карцинома клеток Меркеля и многие другие. Вирусы, связанные с раком, включают HBV (вирус гепатита В), HCV (вирус гепатита С), HPV (вирус папилломы человека) (рак шейки матки, рак ротоглотки) и EBV (вирус Эпштейна-Барр) (лимфомы Беркитта, рак желудка, лимфома Ходжкина, другие EBV-положительные В-клеточные лимфомы, нозофарингеальная карцинома и посттрансплантационное лимфопролиферативное заболевание). Международное агентство по исследованию рака (International Agency for Research on Cancer) (Монография 100 В) определило следующие основные локализации рака, связанные с инфекционными агентами:
 Желудок/желудочная: Heliobacter pylori
Желудок/желудочная: Heliobacter pylori
Печень: Вирус гепатита В, вирус гепатита С (HCV), Opistorchis viverrini, Clonorchis sinensis
Шейка матки: Вирус папилломы человека (ВПЧ) с ВИЧ или без него
Аногенитальная (пенис, вульва, влагалище, анус): ВПЧ с ВИЧ или без него
Носоглотка: вирус Эпштейна-Барр (EBV)
Ротоглотка: HPV с употреблением табака или алкоголя или без него
Саркома Капоши: Вирус герпеса человека типа 8 с ВИЧ или без него
Неходжкинская лимфома: Н. pylori, EBV с ВИЧ или без него, HCV, лимфотропный вирус Т-клеток человека 1 типа
Лимфома Ходжкина: EBV с ВИЧ или без него
Мочевой пузырь: Schistosoma haematobium.
Антитела в соответствии с данным изобретением могут быть использованы для лечения рака, связанного с или индуцированного любым из этих инфекционных агентов, таких как раковые заболевания, указанные выше.
Стимуляция эффекторного ответа Т-клеток также может способствовать иммунитету против инфекционных заболеваний и/или выздоровлению от инфекционного заболевания у пациента. Таким образом, анти-ICOS антитело можно использовать для лечения инфекционного заболевания путем введения антитела пациенту.
Инфекционные болезни включают те, которые вызваны патогенами, например, бактериальными, грибковыми, вирусными или протозойными патогенами, и лечение может заключаться в том, чтобы стимулировать иммунный ответ у пациента против инфекции возбудителя. Примером бактериального патогена является туберкулез. Примерами вирусных патогенов являются гепатит В и ВИЧ. Примерами протозойных патогенов являются виды Plasmodium, которые вызывают малярию, например, Р. falciparum.
Антитело может быть использовано для лечения инфекций, например, инфицирования любым патогеном, упомянутым в данном документе. Инфекция может быть персистентной или хронической инфекцией. Инфекция может быть локализованной или системной. Длительный контакт между возбудителем и иммунной системой может привести к истощению иммунной системы или развитию толерантности (проявляется, например, за счет увеличения уровней Tregs и нарушения баланса Treg:Teff в пользу Tregs) и/или к иммунному уклонению патогена путем эволюции и модификации выявленных патогенных антигенов. Эти признаки отражают сходные процессы, которые, как считается, происходят при раке. Анти-ICOS антитела представляют терапевтический подход к лечению инфекции патогеном, например, хронической инфекции, посредством модуляции отношения Treg:Teff в пользу Teff и/или других эффектов, описанных в данном документе.
Лечение может быть проведено у пациентов, для которых было диагностировано инфекционное заболевание или инфекция. Альтернативно, лечение может быть превентивным и вводиться пациенту для защиты от заражения болезнью, например, в качестве вакцины, как описано в другой части данного документа.
Также было высказано предположение, что иммунный ответ, особенно IFNγ-зависимый системный иммунный ответ, может быть полезен для лечения болезни Альцгеймера и других патологий ЦНС, объединяющим элементом которых является нейровоспалительный компонент [29]. В WO 2015/136541 предложено лечение болезни Альцгеймера с использованием анти-PD-1 антитела. Анти-ICOS антитела могут использоваться для лечения болезни Альцгеймера или других нейродегенеративных заболеваний, необязательно, в сочетании с одним или несколькими другими иммуномодуляторами (например, антителом к PD-1).
Комбинированная терапия
Лечение иммуномодулирующим антителом, таким как анти-CTLA4, анти-PD1 или анти-PDL1, особенно с эффекторной функцией Fc, может создать среду, в которой дополнительное деплетирование высокоэкспрессирующих иммуносупрессивных клеток ICOS является полезным. Может быть выгодным комбинировать анти-ICOS антитело с таким иммуномодулятором для усиления его терапевтических эффектов.
Пациент, получавший лечение иммуномодулирующим антителом (например, анти-PDL-1, анти-PD-1, анти-CTLA-4), может особенно выиграть от лечения анти-ICOS антителом. Одна из причин этого заключается в том, что иммуномодулирующее антитело может увеличить количество ICOS-положительных Tregs (например, внутриопухолевых Tregs) у пациента. Этот эффект также наблюдается для некоторых других терапевтических средств, таких как рекомбинантный IL-2. Анти-ICOS антитело может уменьшать и/или обращать резкое увеличение или повышение ICOS+ Tregs (например, внутриопухолевых Tregs), возникающие в результате лечения пациента другим терапевтическим агентом. Таким образом, пациент, выбранный для лечения анти-ICOS антителом, возможно, уже получал лечение первым терапевтическим агентом, причем первым терапевтическим агентом является антитело (например, иммуномодулирующее антитело) или другой агент (например, IL-2), которые увеличивают количество ICOS+ Tregs у пациента.
Иммуномодуляторы, с которыми можно комбинировать анти-ICOS антитело, включают антитела к любому из: PDL1 (например, авелумаб), PD-1 (например, пембролизумаб или ниволумаб) или CTLA-4 (например, ипилимумаб или тремелимумаб). Анти-ICOS антитело можно комбинировать с пидилизумабом. В других вариантах реализации, анти-ICOS антитело не вводят в комбинации с анти-CTLA-4 антителом и/или необязательно, вводят в комбинации с терапевтическим антителом, которое не является анти-CTLA-4 антителом.
Например, анти-ICOS антитело может быть использовано в комбинированной терапии анти-PDL1 антителом. Предпочтительно, анти-ICOS антитело является антителом, которое опосредует ADCC, ADCP и/или CDC. Предпочтительно, анти-PDL1 антитело представляет собой антитело, которое опосредует ADCC, ADCP и/или CDC. Примером такой комбинированной терапии является введение анти-ICOS антитела с анти-PDL1 антителом, причем оба антитела имеют эффектор-положительные константные области. Таким образом, анти-ICOS антитело и анти-PDL1 антитело могут оба быть способны опосредовать ADCC, CDC и/или ADCP. Эффекторная функция Fc и выбор константных областей подробно описаны в других частях данного документа, но в качестве одного из примеров, анти-ICOS IgG1 человека может быть скомбинирован с анти-PD-L1 IgG1 человека. Анти-ICOS антитело и/или анти-PD-L1 антитело могут содержать константную область человеческого IgG1 дикого типа. Альтернативно, эффектор-положительная константная область антитела может быть модифицирована с целью повышения эффекторной функции, например, повышения CDC, ADCC и/или ADCP. Примеры константных областей антитела, включая последовательности человеческого IgG1 дикого типа и мутации, изменяющие эффекторную функцию, подробно обсуждаются в других частях данного документа.
Анти-PDL1 -антитела, с которыми может быть скомбинировано анти-ICOS антитело, включают:
Анти-PDL1-антитело, ингибирующее связывание PD-1 с PDL1 и/или ингибирующее PDL1, необязательно, в качестве эффектор-положительного IgG1 человека;
Анти-PD-1 антитело, ингибирующее связывание PD-1 с PDL1 и/или PDL2;
Авелумаб, человеческое IgG1-антитело, которое ингибирует связывание PD-1 с PDL-1. См. WO 2013/079174;
Дурвалумаб (или «MEDI4736»), представляющий собой вариант человеческого IgG1-антитела, имеющий мутации L234A, L235A и 331. См. WO 2011/066389;
Атезолизумаб, вариант человеческого IgG1 антитела, имеющий мутации N297A, D356E и L358M. См. US 2010/0203056;
BMS-936559, человеческое антитело IgG4, содержащее мутацию S228P. См. WO 2007/005874.
В данном документе раскрыты многочисленные дополнительные примеры анти-PDL1 антител, а другие известны в данной области техники. Данные характеризации многих анти-PDL1 антител, упомянутых в данном документе, были опубликованы в US 9567399 и US 9617338, которые включены в данный документ посредством ссылок. Примеры анти-PDL1 антител имеют VH и/или VL-домены, содержащие HCDR и/или LCDR любого из мутантов 1D05, 84G09, мутанта 1D05 НС 1, мутанта 1D05 НС 2, мутанта 1D05 НС 3, мутанта 1D05 НС 4, мутанта 1D05 LC 1, мутанта 1D05 LC 2, мутанта 1D05 LC 3, 411В08, 411С04, 411D07, 385F01, 386Н03, 389А03, 413D08, 413G05, 413F09, 414В06 или 416Е01, как указано в US 9567399 или US 9617338. Антитело может содержать VH- и VL-домен любого из этих антител и может, необязательно, содержать тяжелую и/или легкую цепь, имеющую аминокислотную последовательность тяжелой и/или легкой цепи любого из этих антител. Домены VH и VL этих анти-PDL1 антител дополнительно описаны в других частях данного документа.
Дополнительные примеры анти-PDL1 антител имеют VH и/или VL-домены, содержащие HCDR и/или LCDRs KN-035, СА-170, FAZ-053, М7824, ABBV-368, LY-3300054, GNS-1480, YW243.55.S70, REGN3504, или анти-PD-L1 антитела, раскрытого в любом из WO 2017/034916, WO 2017/020291, WO 2017/020858, WO 2017/020801, WO 2016/111645, WO 2016/197367, WO 2016/061142, WO 2016/149201, WO 2016/000619, WO 2016/160792, WO 2016/022630, WO 2016/007235, WO 2015/179654, WO 2015/173267, WO 2015/181342, WO 2015/109124, WO 2015/112805, WO 2015/061668, WO 2014/159562, WO 2014/165082, WO 2014/100079, WO 2014/055897, WO 2013/181634, WO 2013/173223, WO 2013/079174, WO 2012/145493, WO 2011/066389, WO 2010/077634, WO 2010/036959, WO 2010/089411 и WO 2007/005874. Антитело может содержать VH- и VL-домен любого из этих антител и может, необязательно, содержать тяжелую и/или легкую цепь, имеющую аминокислотную последовательность тяжелой и/или легкой цепи любого из этих антител. Анти-ICOS антитело, которое используется в комбинированной терапии с анти-PD-L1, может быть антителом по данному изобретению, как описано в данном документе. Альтернативно, анти-ICOS антитело может содержать CDR, или домен VH и/или VL, анти-ICOS антитела, раскрытого в любой из следующих публикаций:
WO 2016154177, US 2016304610 - например, любого из антител 7F12, 37А10, 35А9, 36Е10, 16G10, 37A10S713, 37A10S714, 37A10S715, 37A10S716, 37A10S717, 37A10S718, 16G10S71, 16G10S72, 16G10S73, 16G10S83, 35A9S79, 35A9S710 или 35A9S89;
WO 16120789, US 2016215059 - например, антитела, известного как 422.2 и/или H2L5;
WO 14033327, ЕР 2892928, US 2015239978 - например, антитела, известного как 314-8 и/или полученного из гибридомы CNCM I-4180;
WO 12131004, ЕР 2691419, US 9376493, US 20160264666 - например, антитела Icos145-1 и/или антитела, продуцируемого гибридомой CNCM I-4179;
WO 10056804 - например, антитела JMAb 136 или «136»;
WO 9915553, ЕР 1017723 В1, US 7259247, US 7132099, US 7125551, US 7306800, US 7722872, WO 05103086, ЕР 1740617, US 8318905, US 8916155 - например, антитела MIC-944 или 9F3;
WO 983821, US 7932358 B2, US 2002156242, ЕР 0984023, ЕР 1502920, US 7030225, US 7045615, US 7279560, US 7226909, US 7196175, US 7932358, US 8389690, WO 02070010, ЕР 1286668, ЕР 1374901, US 7438905, US 7438905, WO 0187981, ЕР 1158004, US 6803039, US 7166283, US 7988965, WO 0115732, ЕР 1125585, US 7465445, US 7998478 - например, любого JMAb-антитела, например, любого из JMAb-124, JMAb-126, JMAb-127, JMAb-128, JMAb-135, JMAb-136, JMAb-137, JMAb-138, JMAb-139, JMAb-140, JMAb-141, например, JMAb136;
WO 2014/089113 - например, антитела 17G9;
WO 12174338;
US 2016145344;
WO 11020024, EP 2464661, US 2016002336, US 2016024211, US 8840889;
US 8497244.
Анти-ICOS антитело, необязательно, содержит CDRs 37A10S713, как описано в WO 2016154177. Оно может содержать VH- и VL-домены 37A10S713 и может, необязательно, иметь тяжелые и легкие цепи антитела 37A10S713.
Комбинация анти-ICOS антитела с иммуномодулятором может обеспечить повышенный терапевтический эффект по сравнению с монотерапией и может обеспечить достижение терапевтического преимущества при более низкой дозе иммуномодулятора (иммуномодуляторов). Так, например, антитело (например, анти-PD-L1-антитело, необязательно, ипилимумаб), которое используется в комбинации с анти-ICOS антителом, можно вводить с дозировкой 3 мг/кг, а не более обычной дозой 10 мг/кг. Режим введения анти-PD-L1 или другого антитела может включать внутривенное введение в течение 90 минут каждые 3 недели, в общей сложности, 4 дозы.
Анти-ICOS антитело можно использовать для повышения восприимчивости опухоли к лечению анти-PD-L1 антителом, которая может быть определена как уменьшение дозы, при которой анти-PD-L1 антитело оказывает терапевтический эффект. Таким образом, анти-ICOS антитело можно вводить пациенту для снижения дозы анти-PD-L1-антитела, обеспечивающей эффективное лечение рака или опухоли у пациента. Введение анти-ICOS антитела может снизить рекомендуемую или требуемую дозу введения анти-PD-L1-антитела этому пациенту, например, до 75%, 50%, 25%, 20%, 10% или меньше, по сравнению с дозой введения анти-PD-L1 антитела без анти-ICOS. Пациент может получать лечение путем введения анти-ICOS антитела и анти-PD-L1 антитела в комбинированной терапии, как описано в данном документе.
Преимущество сочетания анти-PD-L1 с анти-ICOS может быть распространяться на снижение дозировки каждого агента по сравнению с его использованием в качестве монотерапии. Анти-PD-L1-антитело может быть использовано для уменьшения дозы, при которой анти-ICOS антитело создает терапевтическое преимущество и, таким образом, может быть введено пациенту для снижения дозы анти-ICOS антитела, эффективной для лечения рака или опухоли у пациента. Таким образом, анти-PD-L1 антитело может снижать рекомендуемую или требуемую дозу введения анти-ICOS антитела этому пациенту, например, до 75%, 50%, 25%, 20%, 10% или меньше, по сравнению с дозировкой при введении анти-ICOS антитела без анти-PD-L1. Пациент может получать лечение путем введения анти-ICOS антитела и анти-PD-L1 антитела в комбинированной терапии, как описано в данном документе.
Как описано в Примере 22 данного документа, лечение анти-PD-L1 антителом, особенно, антителом с эффектор-положительным Fc, по-видимому, не увеличивает экспрессию ICOS на клетках Teff. Это выгодно при введении таких антител в сочетании с эффектор-положительными анти-ICOS антителами, когда увеличение экспрессии ICOS на Teffs будет нежелательно увеличивать чувствительность этих клеток к деплетированию анти-ICOS антителом. Таким образом, в сочетании с анти-PD-L1, анти-ICOS терапия может использовать дифференциальную экспрессию ICOS на Teffs по сравнению с Tregs, что позволяет проводить преимущественное деплетирование Tregs с высокими уровнями ICOS. Это, в свою очередь, уменьшает супрессию TEffs и дает суммарный эффект, промотирующий эффекторный ответ Т-клеток у пациента. Эффект нацеливания молекул иммунной контрольной точки на экспрессию ICOS на Т-клетках также изучался ранее - см. Фиг. S6C в работе [30] (дополнительные материалы), причем сообщалось, что лечение анти-CTLA-4 антителом и/или анти-PDL1 антителом увеличивает процент CD4+ Tregs, экспрессирующих ICOS. Эффект терапевтического агента на экспрессию ICOS в Tregs и Teffs может быть фактором при выборе агентов, пригодных для применения в комбинации с анти-ICOS антителами, поскольку эффект анти-ICOS антитела может быть усилен в условиях высокодифференцированной экспрессии ICOS на Tregs и Teffs.
Как описано в данном документе, однократная доза анти-ICOS антитела может быть достаточной для обеспечения терапевтического эффекта, особенно в комбинации с другими терапевтическими средствами, такими как анти-PD-L1 антитело. При опухолевой терапии, обоснованием такого преимущества однократной дозировки может быть то, что анти-ICOS антитело опосредует его действие, по крайней мере частично, путем регулировки или изменения микроокружения опухоли в достаточной степени, чтобы сделать опухоль более чувствительной к иммунной атаке и/или к воздействию других иммуномодуляторов, таких как упомянутые. Регулировка микроокружения опухоли инициируется, например, деплетированием ICOS-положительных опухоль-проникающих T-regs. Так, например, пациент может получить лечение однократной дозой анти-ICOS антитела, за которой следует одна или несколько доз анти-PD-L1 антитела. На протяжении периода лечения, например, шести месяцев или года, анти-ICOS антитело может быть введено в разовой дозе, тогда как другие агенты, например, анти-PD-L1 антитело, необязательно, вводят многократно на протяжении этого периода лечения, причем, предпочтительно, по меньшей мере одну такую дозу вводят после введения анти-ICOS антитела.
Другие примеры комбинированной терапии включают комбинацию анти-ICOS антитела с:
- антагонистом рецептора аденозина А2А («ингибитор A2AR»);
- агонистом CD137 (например, агонистическим антителом);
- антагонистом фермента индоламин-2,3-диоксигеназы, который катализирует разрушение триптофана («ингибитор IDO»). IDO - это иммунная контрольная точка, активируемая в дендритных клетках и макрофагах, участвующая в иммуносупрессии/иммунотолерантности.
Анти-ICOS антитела могут использоваться в комбинированной терапии с IL-2 (например, рекомбинантным IL-2, таким как алдеслейкин). IL-2 можно вводить с высокой дозой (HD). Типичная терапия HD IL-2 включает болюсную инфузию более 500000 МЕ/кг, например, болюсные инфузии 600000 или 720000 МЕ/кг за цикл терапии, причем 10-15 таких болюсных инфузий вводят с интервалом 5-10 часов, например, до 15 болюсных инфузий через каждые 8 часов, с повторением цикла терапии примерно каждые 14-21 дней до 6-8 циклов. Терапия HD IL-2 была успешной при лечении опухолей, особенно меланомы (например, метастазирующей меланомы) и почечно-клеточной карциномы, но ее применение ограничивается высокой токсичностью IL-2, который может вызывать серьезные побочные эффекты.
Было показано, что лечение IL-2 с высокой дозой увеличивает популяцию ICOS-положительных Tregs у больных раком [31]. Сообщалось, что такое увеличение ICOS+ TRegs после первого цикла терапии HD IL-2 коррелировало с худшим клиническим исходом - чем выше число ICOS+ Tregs, тем хуже прогноз. Вариант F42K IL-2 был предложен в качестве альтернативной терапии, чтобы избежать такого нежелательного увеличения ICOS+ Treg-клеток [32]. Однако, другой подход будет заключаться в использовании увеличения ICOS+ Tregs с помощью антитела в соответствии с данным изобретением в качестве терапевтического агента второй линии.
Может быть выгодным комбинировать терапию IL-2 с анти-ICOS антителами, используя способность нацеливания анти-ICOS антител на TRegs с высокой экспрессией ICOS, ингибируя эти клетки и улучшая прогноз для пациентов, получающих терапию IL-2. Совместное введение IL-2 и антитела анти-ICOS может увеличить показатели эффективности ответа, в то же время избегая или уменьшая нежелательные явления в популяции пациентов, получивших лечение. Комбинация может позволить использовать IL-2 в более низкой дозе по сравнению с монотерапией IL-2, снижая риск или уровень побочных эффектов, возникающих в результате терапии IL-2, и в то же время сохраняя или усиливая клиническую пользу (например, снижение роста опухоли, рассасывание твердой опухоли и/или уменьшение метастазирования). Таким образом, добавление анти-ICOS может улучшить лечение пациентов, получающих IL-2, при высокой дозе (HD) или низкой дозе (LD) IL-2.
Соответственно, один из аспектов изобретения предусматривает способ лечения пациента путем введения пациенту анти-ICOS антитела, причем пациент также получает лечение IL-2, например, HD IL-2. Другим аспектом изобретения является анти-ICOS антитело, предназначенное для использования при лечении пациента, причем пациент также получает лечение IL-2, например, HD IL-2. Анти-ICOS антитело можно использовать в качестве терапии второй линии. Таким образом, пациент, возможно, получал лечение IL-2, например, получил по меньшей мере один цикл терапии HD IL-2, и имеет повышенный уровень ICOS+ Treg. Могут проводиться анализы на образцах раковых клеток, например, образцах биопсии опухоли, с использованием иммуногистохимии или FACS, как описано в другой части данного документа, для детектирования клеток, положительных к ICOS, Foxp3, ICOSL и, необязательно, одного или нескольких дополнительных маркеров, представляющих интерес. Способы могут включать определение того, что пациент имеет повышенный уровень ICOS+ Tregs (например, в периферической крови или в биопсии опухоли) после лечения IL-2, причем повышенный уровень указывает на то, что пациенту было бы полезным лечение анти-ICOS антителом. Увеличение Tregs может определяться по отношению к контрольным (не получавшим лечения) особам или к пациенту до проведения терапии IL-2. Такие пациенты с повышенными уровнями Treg представляют собой группу, которой продолжение лечения одним лишь IL-2 может быть бесполезным, но комбинированная терапия анти-ICOS антителом и IL-2 или лечение одним лишь анти-ICOS антителом обеспечивают терапевтический эффект. Таким образом, после подтверждения того, что пациент имеет повышенный уровень ICOS+ Tregs, можно вводить анти-ICOS антитело и/или проводить дополнительную терапию IL-2. Лечение анти-ICOS антителом может обеспечивать селективное нацеливание на, и деплетирование, ICOS+ Tregs по сравнению с другими популяциями Т-клеток у таких пациентов. Это обеспечивает терапевтический эффект, уменьшая иммуносупрессию, опосредуемую этими клетками, и тем самым усиливая активность Teffs против клеток-мишеней, например, опухолевых клеток или инфицированных клеток.
Комбинированную терапию анти-ICOS антителами и IL-2 можно использовать при любом терапевтическом показании, описанном в данном документе и, в частности, для лечения опухоли, например, меланомы, такой как метастазирующая меланома или почечно-клеточная карцинома. Таким образом, в одном примере пациент, получивший лечение анти-ICOS антителом, представляет собой пациента с метастазирующей меланомой, получившего лечение IL-2, например, терапию HD IL-2 или LD IL-2.
В общем, когда анти-ICOS антитело вводят пациенту, получившему лечение первым терапевтическим агентом (например, иммуномодулирующим антителом) или другим агентом (например, IL-2), анти-ICOS антитело можно вводить после минимального периода, равного, например, 24 часам, 48 часам, 72 часам, 1 неделе или 2 неделям после введения первого терапевтического агента. Анти-ICOS антитело может быть введено в течение 2, 3, 4 или 5 недель после введения первого терапевтического агента. Это не исключает дополнительных введений какого-либо из этих агентов в любое время, хотя может быть желательно минимизировать число вводимых доз для облегчения соблюдения режима лечения пациентами и снижения затрат. Предпочтительно, относительное время введения будет выбираться с целью оптимизации их комбинированного эффекта, причем первый терапевтическое средство создает иммунологическую среду (например, повышенные ICOS+ Tregs или высвобождение антигена, как описано ниже), в которой эффект анти-ICOS антитела является особенно полезным. Таким образом, последовательное введение первого терапевтического агента, а затем анти-ICOS антитела, может обеспечить время для действия первого агента, создавая in vivo условия, в которых анти-ICOS антитело сможет проявить свой усиленный эффект. Различные схемы введения, включая одновременное или последовательное комбинированное лечение, описаны в данном документе и могут быть использованы в соответствующих случаях. Когда первым терапевтическим агентом является тот, который увеличивает количество ICOS+ Tregs у пациента, схема лечения пациента может включать определение наличия у пациента увеличенного количества ICOS+ Tregs и затем введение анти-ICOS антитела.
Как было отмечено, использование анти-ICOS антитела комбинированной терапии может обеспечить преимущества снижения эффективной дозы терапевтических средств и/или противодействия неблагоприятным эффектам терапевтических средств, увеличивающих ICOS+ Treg у пациентов. Однако, дополнительные терапевтические преимущества могут быть достигнуты путем выбора первого терапевтического агента, который вызывает высвобождение антигенов клетками-мишенями посредством «гибели иммунологических клеток», и введения первого терапевтического агента в комбинации с анти-ICOS антителом. Как было отмечено, введение анти-ICOS антитела может следовать за введением первого терапевтического агента, причем введение двух агентов разделяется определенным временным окном, как описано выше.
Иммунологическая гибель клеток - признанный способ гибели клеток, отличающийся от апоптоза. Он характеризуется высвобождением АТФ и HMGB1 из клетки и воздействием кальретикулина на плазматическую мембрану [33, 34].
Иммунологическая гибель клеток в ткани-мишени или в клетках-мишенях способствует поглощению клетки антигенпрезентирующей клеткой, что приводит к презентации антигенов клетки-мишени, что, в свою очередь, индуцирует антигенспецифичные клетки Teff. Анти-ICOS антитело может увеличивать величину и/или продолжительность ответа Teff, действуя как агонист ICOS на клетках Teff. Кроме того, если анти-ICOS антитело имеет активированную функцию Fc-эффектора (например, антитело IgG1 человека), то анти-ICOS антитело может вызывать деплетирование антиген-специфических Tregs. Таким образом, благодаря сочетанию любого из или обоих этих эффектов, баланс между клетками Teff и Treg модулируется в пользу усиления активности Teff. Комбинация анти-ICOS антитела с лечением, индуцирующим иммунологическую гибель клеток в ткани-мишени или клеточном типе, например, в опухоли или в раковых клетках, таким образом, стимулирует у пациента иммунный ответ против ткани-мишени или клеток-мишеней, представляя собой форму вакцинации, в которой вакцинный антиген образуется in vivo.
Соответственно, одним из аспектов изобретения является способ лечения рака у пациента путем in vivo вакцинации пациента против его раковых клеток. Другим аспектом изобретения является анти-ICOS антитело для использования в таком способе. Анти-ICOS антитела могут быть использованы в способе, включающем:
лечение пациента терапией, которая вызывает иммунологическую гибель раковых клеток, что приводит к презентации антигена антигенспецифическим эффекторным Т-клеткам и
введение пациенту анти-ICOS антитела, причем анти-ICOS антитело усиливает антигенспецифический эффекторный Т-клеточный ответ против раковых клеток.
Лечение, индуцирующее иммунологическую гибель клеток, включает облучение (например, ионизирующее облучение клеток с использованием ультрафиолетового света спектра С (UVC) или γ-лучей), химиотерапевтические средства (например, оксалиплатин, антрациклины, такие как доксорубицин, идарубицин или митоксантрон, агонисты BK-канала, такие как флоретин или пимаровая кислота, бортезомиб, сердечные гликозиды, циклофосфамид, ингибиторы GADD34/PP1 с митомицином, PDT (фотодинамическая терапия) с гиперицином, полиинозино-полицитидиловую кислоту, 5-фторурацил, гемцитабин, гефитиниб (gefitnib), эрлотиниб или тапсигаргин с цисплатином) и антитела к опухолеассоциированным антигенам. Опухолеассоциированным антигеном может быть любой антиген, который сверхэкспрессируется опухолевыми клетками по сравнению с неопухолевыми клетками этой же ткани, например, HER2, CD20, EGFR. Пригодные антитела включают герцептин (анти-HER2), ритуксимаб (анти-CD20) или цетуксимаб (анти-EGFR).
Таким образом, целесообразно комбинировать анти-ICOS антитело с одним или несколькими такими методами лечения. Необязательно, анти-ICOS антитело вводится пациенту, который уже получил такое лечение. Анти-ICOS антитело можно вводить после периода, например, 24 часа, 48 часов, 72 часа, 1 неделя или 2 недели после проведения лечения, которое индуцирует иммунологическую гибель клеток, например, от 24 до 72 часов после такого лечения. Анти-ICOS антитело можно вводить через 2, 3, 4 или 5 недель после такого лечения. Другие схемы комбинированной терапии описаны в других частях данного документа.
Хотя выше была описана «вакцинация in vivo», можно также проводить обработку опухолевых клеток ex vivo, чтобы индуцировать иммунологическую гибель клеток, после чего клетки могут быть повторно введены пациенту. Вместо того, чтобы вводить агент или проводить лечение, которое индуцирует иммунологическую гибель клеток, непосредственно пациенту, пациенту вводят обработанные опухолевые клетки. Лечение пациента может осуществляться в соответствии со схемами лечения, описанными выше.
Как уже отмечалось, однократная доза анти-ICOS антитела может быть достаточной для обеспечения терапевтического эффекта. Таким образом, в описанных в данном документе способах лечения анти-ICOS антитело, необязательно, вводят в виде разовой дозы. Разовая доза анти-ICOS антитела может деплетировать Tregs у пациента с последующим благоприятным эффектом при таких заболеваниях, как рак. Ранее сообщалось, что транзиентная абляция Tregs оказывает противоопухолевое действие, включая уменьшение прогрессирования опухоли, лечение установленных (established) опухолей и метастазов и продление выживаемости, и может усиливать терапевтический эффект облучения опухолей [35]. Введение разовой дозы анти-ICOS может обеспечить такое деплетирование Treg и может быть использовано для усиления эффектов других терапевтических подходов, используемых в комбинации, таких как лучевая терапия.
Антитела к PD-L1
Антитело к PD-L1 для использования в комбинации с анти-ICOS антителом, будь то в качестве отдельного терапевтического агента или в мультиспецифическом антителе, как описано в данном документе, может содержать антигенсвязывающий сайт любого анти-PD-L1 антитела. В данном документе раскрыты многочисленные примеры анти-PDL1 антител, и другие являются известными в данной области. Данные характеризации многих анти-PDL1 антител, упомянутых в данном документе, были опубликованы в US 9567399 и US 9617338, которые включены в данный документ посредством ссылок.
1D05 имеет аминокислотную последовательность вариабельной области тяжелой цепи (VH), представленную SEQ ID NO: 33, содержащую аминокислотную последовательность CDRH1, представленную SEQ ID NO: 27 (IMGT) или SEQ ID NO: 30 (Кабат), аминокислотную последовательность CDRH2, представленную SEQ ID NO: 28 (IMGT) или SEQ ID NO: 31 (Кабат), и аминокислотную последовательность CDRH3, представленную SEQ ID NO: 29 (IMGT) или SEQ ID NO: 32 (Кабат). Последовательность нуклеиновой кислоты тяжелой цепи домена VH представлена SEQ ID NO: 34. 1D05 имеет аминокислотную последовательность вариабельной области легкой цепи (VL), представленную SEQ ID NO: 43, содержащую аминокислотную последовательность CDRL1, представленную SEQ ID NO: 37 (IMGT) или SEQ ID NO: 40 (Кабат), аминокислотную последовательность CDRL2, представленную SEQ ID NO: 38 (IMGT) или SEQ ID NO: 41 (Кабат), и аминокислотную последовательность CDRL3, представленную SEQ ID NO: 39 (IMGT) или SEQ ID NO: 42 (Кабат). Последовательность нуклеиновой кислоты легкой цепи домена VL представлена SEQ ID NO: 44. Домен VH может быть объединен с любой из последовательностей константной области тяжелой цепи, описанных в данном документе, например, SEQ ID NO: 193, SEQ ID NO: 195, SEQ ID NO: 197, SEQ ID NO: 199, SEQ ID NO: 201, SEQ ID NO: 203, Seq ID Nos: 205, Seq ID Nos: 340, Seq ID Nos: 524, Seq ID Nos: 526, Seq ID Nos: 528, Seq ID Nos: 530, SEQ ID NO: 532 или SEQ ID NO: 534. Область VL может быть объединена с любой из последовательностей константной области легкой цепи, описанной в данном документе, например, SEQ ID NOs: 207, 209, 211, 213, 215, 217, 219, 221, 223, 225, 227, 229, 231, 233, 235, 237, 536 и 538. Полноразмерная аминокислотная последовательность тяжелой цепи представлена SEQ ID NO: 35 (последовательность нуклеиновой кислоты тяжелой цепи - SEQ ID NO: 36). Полноразмерная аминокислотная последовательность легкой цепи представлена SEQ ID NO: 45 (последовательность нуклеиновой кислоты легкой цепи - SEQ ID NO: 46).
84G09 имеет аминокислотную последовательность вариабельной области тяжелой цепи (VH), представленную SEQ ID NO: 13, содержащую аминокислотную последовательность CDRH1, представленную SEQ ID NO: 7 (IMGT) или SEQ ID NO: 10 (Кабат), аминокислотную последовательность CDRH2, представленную SEQ ID NO: 8 (IMGT) или SEQ ID NO: 11 (Кабат), и аминокислотную последовательность CDRH3, представленную SEQ ID NO: 9 (IMGT) или SEQ ID NO: 12 (Кабат). Последовательность нуклеиновой кислоты тяжелой цепи домена VH представлена SEQ ID NO: 14. 84G09 имеет аминокислотную последовательность вариабельной области легкой цепи (VL), представленную SEQ ID NO: 23, содержащую аминокислотную последовательность CDRL1, представленную SEQ ID NO: 17 (IMGT) или SEQ ID NO: 20 (Кабат), аминокислотную последовательность CDRL2, представленную SEQ ID NO: 18 (IMGT) или SEQ ID NO: 21 (Кабат), и аминокислотную последовательность CDRL3, представленную SEQ ID NO: 19 (IMGT) или SEQ ID NO: 22 (Кабат). Последовательность нуклеиновой кислоты легкой цепи домена VL представлена SEQ ID NO: 24. Домен VH может быть объединена с любой из последовательностей константной области тяжелой цепи, описанной в данном документе, например, SEQ ID NO: 193, SEQ ID NO: 195, SEQ ID NO: 197, SEQ ID NO: 199, SEQ ID NO: 201, SEQ ID NO: 203, Seq ID Nos: 205, Seq ID Nos: 340, Seq ID Nos: 524, Seq ID Nos: 526, Seq ID Nos: 528, Seq ID Nos: 530, SEQ ID NO: 532 или SEQ ID NO: 534. Область VL может быть объединена с любой из последовательностей константной области легкой цепи, описанной в данном документе, например, SEQ ID NOs: 207, 209, 211, 213, 215, 217, 219, 221, 223, 225, 227, 229, 231, 233, 235, 237, 536 и 538. Полноразмерная аминокислотная последовательность тяжелой цепи представлена SEQ ID NO: 15 (последовательность нуклеиновой кислоты тяжелой цепи - SEQ ID NO: 16). Полноразмерная аминокислотная последовательность легкой цепи представлена SEQ ID NO: 25 (последовательность нуклеиновой кислоты легкой цепи - SEQ ID NO: 26).
Мутант 1D05 НС 1 имеет аминокислотную последовательность вариабельной области тяжелой цепи (VH), представленную SEQ ID NO: 47, содержащую аминокислотную последовательность CDRH1, представленную SEQ ID NO: 27 (IMGT) или SEQ ID NO: 30 (Кабат), аминокислотную последовательность CDRH2, представленную SEQ ID NO: 28 (IMGT) или SEQ ID NO: 31 (Кабат), и аминокислотную последовательность CDRH3, представленную SEQ ID NO: 29 (IMGT) или SEQ ID NO: 32 (Кабат). Мутант 1 1D05 НС имеет аминокислотную последовательность вариабельной области легкой цепи (VL), представленную SEQ ID NO: 43, содержащую аминокислотную последовательность CDRL1, представленную SEQ ID NO: 37 (IMGT) или SEQ ID NO: 40 (Кабат), аминокислотную последовательность CDRL2, представленную SEQ ID NO: 38 (IMGT) или SEQ ID NO: 41 (Кабат), и аминокислотную последовательность CDRL3, представленную SEQ ID NO: 39 (IMGT) или SEQ ID NO: 42 (Кабат). Последовательность нуклеиновой кислоты легкой цепи домена VL представлена SEQ ID NO: 44. Домен VH может быть объединена с любой из последовательностей константной области тяжелой цепи, описанной в данном документе, например, SEQ ID NO: 193, SEQ ID NO: 195, SEQ ID NO: 197, SEQ ID NO: 199, SEQ ID NO: 201, SEQ ID NO: 203, Seq ID Nos: 205, Seq ID Nos: 340, Seq ID Nos: 524, Seq ID Nos: 526, Seq ID Nos: 528, Seq ID Nos: 530, SEQ ID NO: 532 или SEQ ID NO: 534. Область VL может быть объединена с любой из последовательностей константной области легкой цепи, описанной в данном документе, например, SEQ ID NOs: 207, 209, 211, 213, 215, 217, 219, 221, 223, 225, 227, 229, 231, 233, 235, 237, 536 и 538. Полноразмерная аминокислотная последовательность легкой цепи представлена SEQ ID NO: 45 (последовательность нуклеиновой кислоты легкой цепи - SEQ ID NO: 46).
Мутант 1D05 НС 2 имеет аминокислотную последовательность вариабельной области тяжелой цепи (VH), представленную SEQ ID NO: 48, содержащую аминокислотную последовательность CDRH1, представленную SEQ ID NO: 27 (IMGT) или SEQ ID NO: 30 (Кабат), аминокислотную последовательность CDRH2, представленную SEQ ID NO: 28 (IMGT) или SEQ ID NO: 31 (Кабат), и аминокислотную последовательность CDRH3, представленную SEQ ID NO: 29 (IMGT) или SEQ ID NO: 32 (Кабат). Мутант 2 1D05 НС имеет аминокислотную последовательность вариабельной области легкой цепи (VL), представленную SEQ ID NO: 43, содержащую аминокислотную последовательность CDRL1, представленную SEQ ID NO: 37 (IMGT) или SEQ ID NO: 40 (Кабат), аминокислотную последовательность CDRL2, представленную SEQ ID NO: 38 (IMGT) или SEQ ID NO: 41 (Кабат), и аминокислотную последовательность CDRL3, представленную SEQ ID NO: 39 (IMGT) или SEQ ID NO: 42 (Кабат). Последовательность нуклеиновой кислоты легкой цепи домена VL представлена SEQ ID NO: 44. Домен VH может быть объединена с любой из последовательностей константной области тяжелой цепи, описанной в данном документе, например, SEQ ID NO: 193, SEQ ID NO: 195, SEQ ID NO: 197, SEQ ID NO: 199, SEQ ID NO: 201, SEQ ID NO: 203, Seq ID Nos: 205, Seq ID Nos: 340, Seq ID Nos: 524, Seq ID Nos: 526, Seq ID Nos: 528, Seq ID Nos: 530, SEQ ID NO: 532 или SEQ ID NO: 534. Область VL может быть объединена с любой из последовательностей константной области легкой цепи, описанной в данном документе, например, SEQ ID NOs: 207, 209, 211, 213, 215, 217, 219, 221, 223, 225, 227, 229, 231, 233, 235, 237, 536 и 538. Полноразмерная аминокислотная последовательность легкой цепи представлена SEQ ID NO: 45 (последовательность нуклеиновой кислоты легкой цепи - SEQ ID NO: 46).
Мутант 1D05 НС 3 имеет аминокислотную последовательность вариабельной области тяжелой цепи (VH), представленную SEQ ID NO: 49, содержащую аминокислотную последовательность CDRH1, представленную SEQ ID NO: 27 (IMGT) или SEQ ID NO: 30 (Кабат), аминокислотную последовательность CDRH2, представленную SEQ ID NO: 28 (IMGT) или SEQ ID NO: 31 (Кабат), и аминокислотную последовательность CDRH3, представленную SEQ ID NO: 29 (IMGT) или SEQ ID NO: 32 (Кабат). Мутант 3 1D05 НС имеет аминокислотную последовательность вариабельной области легкой цепи (VL), представленную SEQ ID NO: 43, содержащую аминокислотную последовательность CDRL1, представленную SEQ ID NO: 37 (IMGT) или SEQ ID NO: 40 (Кабат), аминокислотную последовательность CDRL2, представленную SEQ ID NO: 38 (IMGT) или SEQ ID NO: 41 (Кабат), и аминокислотную последовательность CDRL3, представленную SEQ ID NO: 39 (IMGT) или SEQ ID NO: 42 (Кабат). Последовательность нуклеиновой кислоты легкой цепи домена VL представлена SEQ ID NO: 44. Домен VH может быть объединена с любой из последовательностей константной области тяжелой цепи, описанной в данном документе, например, SEQ ID NO: 193, SEQ ID NO: 195, SEQ ID NO: 197, SEQ ID NO: 199, SEQ ID NO: 201, SEQ ID NO: 203, Seq ID Nos: 205, Seq ID Nos: 340, Seq ID Nos: 524, Seq ID Nos: 526, Seq ID Nos: 528, Seq ID Nos: 530, SEQ ID NO: 532 или SEQ ID NO: 534. Область VL может быть объединена с любой из последовательностей константной области легкой цепи, описанной в данном документе, например, SEQ ID NOs: 207, 209, 211, 213, 215, 217, 219, 221, 223, 225, 227, 229, 231, 233, 235, 237, 536 и 538. Полноразмерная аминокислотная последовательность легкой цепи представлена SEQ ID NO: 45 (последовательность нуклеиновой кислоты легкой цепи - SEQ ID NO: 46).
Мутант 1D05 НС 4 имеет аминокислотную последовательность вариабельной области тяжелой цепи (VH), представленную SEQ ID NO: 342, содержащую аминокислотную последовательность CDRH1, представленную SEQ ID NO: 27 (IMGT) или SEQ ID NO: 30 (Кабат), аминокислотную последовательность CDRH2, представленную SEQ ID NO: 28 (IMGT) или SEQ ID NO: 31 (Кабат), и аминокислотную последовательность CDRH3, представленную SEQ ID NO: 29 (IMGT) или SEQ ID NO: 32 (Кабат). Мутант 4 1D05 НС имеет аминокислотную последовательность вариабельной области легкой цепи (VL), представленную SEQ ID NO: 43, содержащую аминокислотную последовательность CDRL1, представленную SEQ ID NO: 37 (IMGT) или SEQ ID NO: 40 (Кабат), аминокислотную последовательность CDRL2, представленную SEQ ID NO: 38 (IMGT) или SEQ ID NO: 41 (Кабат), и аминокислотную последовательность CDRL3, представленную SEQ ID NO: 39 (IMGT) или SEQ ID NO: 42 (Кабат). Последовательность нуклеиновой кислоты легкой цепи домена VL представлена SEQ ID NO: 44. Домен VH может быть объединена с любой из последовательностей константной области тяжелой цепи, описанной в данном документе, например, SEQ ID NO: 193, SEQ ID NO: 195, SEQ ID NO: 197, SEQ ID NO: 199, SEQ ID NO: 201, SEQ ID NO: 203, Seq ID Nos: 205, Seq ID Nos: 340, Seq ID Nos: 524, Seq ID Nos: 526, Seq ID Nos: 528, Seq ID Nos: 530, SEQ ID NO: 532 или SEQ ID NO: 534. Область VL может быть объединена с любой из последовательностей константной области легкой цепи, описанной в данном документе, например, SEQ ID NOs: 207, 209, 211, 213, 215, 217, 219, 221, 223, 225, 227, 229, 231, 233, 235, 237, 536 и 538. Полноразмерная аминокислотная последовательность легкой цепи представлена SEQ ID NO: 45 (последовательность нуклеиновой кислоты легкой цепи - SEQ ID NO: 46).
Мутант 1D05 LC 1 имеет аминокислотную последовательность вариабельной области тяжелой цепи (VH), представленную SEQ ID NO: 33, содержащую аминокислотную последовательность CDRH1, представленную SEQ ID NO: 27 (IMGT) или SEQ ID NO: 30 (Кабат), аминокислотную последовательность CDRH2, представленную SEQ ID NO: 28 (IMGT) или SEQ ID NO: 31 (Кабат), и аминокислотную последовательность CDRH3, представленную SEQ ID NO: 29 (IMGT) или SEQ ID NO: 32 (Кабат). Последовательность нуклеиновой кислоты тяжелой цепи домена VH представлена SEQ ID NO: 34. Мутант 1 1D05 LC имеет аминокислотную последовательность вариабельной области легкой цепи (VL), представленную SEQ ID NO: 50, содержащую аминокислотную последовательность CDRL1, представленную SEQ ID NO: 37 (IMGT) или SEQ ID NO: 40 (Кабат), и аминокислотную последовательность CDRL3, представленную SEQ ID NO: 39 (IMGT) или SEQ ID NO: 42 (Кабат). Последовательность CDRL2 мутанта 1 1D05 LC определяется по системам Кабата или IMGT по последовательности VL, представленной SEQ ID NO: 50. Домен VH может быть объединен с любой из последовательностей константной области тяжелой цепи, описанной в данном документе, например, SEQ ID NO: 193, SEQ ID NO: 195, SEQ ID NO: 197, SEQ ID NO: 199, SEQ ID NO: 201, SEQ ID NO: 203, SEQ ID NO: 205 или SEQ ID NO: 340, Seq ID No: 524, SEQ ID NO: 526, SEQ ID NO: 528, Seq ID No: 530, SEQ ID NO: 532 или SEQ ID NO: 534. Область VL может быть объединена с любой из последовательностей константной области легкой цепи, описанной в данном документе, например, SEQ ID NOs: 207, 209, 211, 213, 215, 217, 219, 221, 223, 225, 227, 229, 231, 233, 235, 237, 536 и 538. Полноразмерная аминокислотная последовательность тяжелой цепи представлена SEQ ID NO: 35 (последовательность нуклеиновой кислоты тяжелой цепи - SEQ ID NO: 36).
Мутант 1D05 LC 2 имеет аминокислотную последовательность вариабельной области тяжелой цепи (VH), представленную SEQ ID NO: 33, содержащую аминокислотную последовательность CDRH1, представленную SEQ ID NO: 27 (IMGT) или SEQ ID NO: 30 (Кабат), аминокислотную последовательность CDRH2, представленную SEQ ID NO: 28 (IMGT) или SEQ ID NO: 31 (Кабат), и аминокислотную последовательность CDRH3, представленную SEQ ID NO: 29 (IMGT) или SEQ ID NO: 32 (Кабат). Последовательность нуклеиновой кислоты тяжелой цепи домена VH представлена SEQ ID NO: 34. Мутант 2 1D05 LC имеет аминокислотную последовательность вариабельной области легкой цепи (VL), представленную SEQ ID NO: 51, содержащую аминокислотную последовательность CDRL1, представленную SEQ ID NO: 37 (IMGT) или SEQ ID NO: 40 (Кабат), аминокислотную последовательность CDRL2, представленную SEQ ID NO: 38 (IMGT) или SEQ ID NO: 41 (Кабат), и аминокислотную последовательность CDRL3, представленную SEQ ID NO: 39 (IMGT) или SEQ ID NO: 42 (Кабат). Домен VH может быть объединена с любой из последовательностей константной области тяжелой цепи, описанной в данном документе, например, SEQ ID NO: 193, SEQ ID NO: 195, SEQ ID NO: 197, SEQ ID NO: 199, SEQ ID NO: 201, SEQ ID NO: 203, Seq ID Nos: 205, Seq ID Nos: 340, Seq ID Nos: 524, Seq ID Nos: 526, Seq ID Nos: 528, Seq ID Nos: 530, SEQ ID NO: 532 или SEQ ID NO: 534. Область VL может быть объединена с любой из последовательностей константной области легкой цепи, описанной в данном документе, например, SEQ ID NOs: 207, 209, 211, 213, 215, 217, 219, 221, 223, 225, 227, 229, 231, 233, 235, 237, 536 и 538. Полноразмерная аминокислотная последовательность тяжелой цепи представлена SEQ ID NO: 35 (последовательность нуклеиновой кислоты тяжелой цепи - SEQ ID NO: 36).
Мутант 1D05 LC 3 имеет аминокислотную последовательность вариабельной области тяжелой цепи (VH), представленную SEQ ID NO: 33, содержащую аминокислотную последовательность CDRH1, представленную SEQ ID NO: 27 (IMGT) или SEQ ID NO: 30 (Кабат), аминокислотную последовательность CDRH2, представленную SEQ ID NO: 28 (IMGT) или SEQ ID NO: 31 (Кабат), и аминокислотную последовательность CDRH3, представленную SEQ ID NO: 29 (IMGT) или SEQ ID NO: 32 (Кабат). Последовательность нуклеиновой кислоты тяжелой цепи домена VH представлена SEQ ID NO: 34. Мутант 3 1D05 LC имеет аминокислотную последовательность вариабельной области легкой цепи (VL), представленную SEQ ID NO: 298, содержащую аминокислотную последовательность CDRL1, представленную SEQ ID NO: 37 (IMGT) или SEQ ID NO: 40 (Кабат), и аминокислотную последовательность CDRL3, представленную SEQ ID NO: 39 (IMGT) или SEQ ID NO: 42 (Кабат). Последовательность CDRL2 мутанта 3 1D05 LC определяется по системам Кабата или IMGT по последовательности VL, представленной SEQ ID NO: 298. Последовательность нуклеиновой кислоты легкой цепи домена VL представлена SEQ ID NO: 44. Домен VH может быть объединен с любой из последовательностей константной области тяжелой цепи, описанной в данном документе, например, SEQ ID NO: 193, SEQ ID NO: 195, SEQ ID NO: 197, SEQ ID NO: 199, SEQ ID NO: 201, SEQ ID NO: 203, SEQ ID NO: 205 или SEQ ID NO: 340, Seq ID No: 524, SEQ ID NO: 526, SEQ ID NO: 528, Seq ID No: 530, SEQ ID NO: 532 или SEQ ID NO: 534. Область VL может быть объединена с любой из последовательностей константной области легкой цепи, описанной в данном документе, например, SEQ ID NOs: 207, 209, 211, 213, 215, 217, 219, 221, 223, 225, 227, 229, 231, 233, 235, 237, 536 и 538. Полноразмерная аминокислотная последовательность тяжелой цепи представлена SEQ ID NO: 35 (последовательность нуклеиновой кислоты тяжелой цепи - SEQ ID NO: 36). Полноразмерная аминокислотная последовательность легкой цепи представлена SEQ ID NO: 45 (последовательность нуклеиновой кислоты легкой цепи - SEQ ID NO: 46).
411В08 имеет аминокислотную последовательность вариабельной области тяжелой цепи (VH), представленную SEQ ID NO: 58, содержащую аминокислотную последовательность CDRH1, представленную SEQ ID NO: 52 (IMGT) или SEQ ID NO: 55 (Кабат), аминокислотную последовательность CDRH2, представленную SEQ ID NO: 53 (IMGT) или SEQ ID NO: 56 (Кабат), и аминокислотную последовательность CDRH3, представленную SEQ ID NO: 54 (IMGT) или SEQ ID NO: 57 (Кабат). Последовательность нуклеиновой кислоты тяжелой цепи домена VH представлена SEQ ID NO: 59. 411В08 имеет аминокислотную последовательность вариабельной области легкой цепи (VL), представленную SEQ ID NO: 68, содержащую аминокислотную последовательность CDRL1, представленную SEQ ID NO: 62 (IMGT) или SEQ ID NO: 65 (Кабат), аминокислотную последовательность CDRL2, представленную SEQ ID NO: 63 (IMGT) или SEQ ID NO: 66 (Кабат), и аминокислотную последовательность CDRL3, представленную SEQ ID NO: 64 (IMGT) или SEQ ID NO: 67 (Кабат). Последовательность нуклеиновой кислоты легкой цепи домена VL представлена SEQ ID NO: 69. Домен VH может быть объединена с любой из последовательностей константной области тяжелой цепи, описанной в данном документе, например, SEQ ID NO: 193, SEQ ID NO: 195, SEQ ID NO: 197, SEQ ID NO: 199, SEQ ID NO: 201, SEQ ID NO: 203, Seq ID Nos: 205, Seq ID Nos: 340, Seq ID Nos: 524, Seq ID Nos: 526, Seq ID Nos: 528, Seq ID Nos: 530, SEQ ID NO: 532 или SEQ ID NO: 534. Область VL может быть объединена с любой из последовательностей константной области легкой цепи, описанной в данном документе, например, SEQ ID NOs: 207, 209, 211, 213, 215, 217, 219, 221, 223, 225, 227, 229, 231, 233, 235, 237, 536 и 538. Полноразмерная аминокислотная последовательность тяжелой цепи представлена SEQ ID NO: 60 (последовательность нуклеиновой кислоты тяжелой цепи - SEQ ID NO: 61). Полноразмерная аминокислотная последовательность легкой цепи представлена SEQ ID NO: 70 (последовательность нуклеиновой кислоты легкой цепи - SEQ ID NO: 71).
411С04 имеет аминокислотную последовательность вариабельной области тяжелой цепи (VH), представленную SEQ ID NO: 78, содержащую аминокислотную последовательность CDRH1, представленную SEQ ID NO: 72 (IMGT) или SEQ ID NO: 75 (Кабат), аминокислотную последовательность CDRH2, представленную SEQ ID NO: 73 (IMGT) или SEQ ID NO: 76 (Кабат), и аминокислотную последовательность CDRH3, представленную SEQ ID NO: 74 (IMGT) или SEQ ID NO: 77 (Кабат). Последовательность нуклеиновой кислоты тяжелой цепи домена VH представлена SEQ ID NO: 79. 411С04 имеет аминокислотную последовательность вариабельной области легкой цепи (VL), представленную SEQ ID NO: 88, содержащую аминокислотную последовательность CDRL1, представленную SEQ ID NO: 82 (IMGT) или SEQ ID NO: 85 (Кабат), аминокислотную последовательность CDRL2, представленную SEQ ID NO: 83 (IMGT) или SEQ ID NO: 86 (Кабат), и аминокислотную последовательность CDRL3, представленную SEQ ID NO: 84 (IMGT) или SEQ ID NO: 87 (Кабат). Последовательность нуклеиновой кислоты легкой цепи домена VL представлена SEQ ID NO: 89. Домен VH может быть объединена с любой из последовательностей константной области тяжелой цепи, описанной в данном документе, например, SEQ ID NO: 193, SEQ ID NO: 195, SEQ ID NO: 197, SEQ ID NO: 199, SEQ ID NO: 201, SEQ ID NO: 203, Seq ID Nos: 205, Seq ID Nos: 340, Seq ID Nos: 524, Seq ID Nos: 526, Seq ID Nos: 528, Seq ID Nos: 530, SEQ ID NO: 532 или SEQ ID NO: 534. Область VL может быть объединена с любой из последовательностей константной области легкой цепи, описанной в данном документе, например, SEQ ID NOs: 207, 209, 211, 213, 215, 217, 219, 221, 223, 225, 227, 229, 231, 233, 235, 237, 536 и 538. Полноразмерная аминокислотная последовательность тяжелой цепи представлена SEQ ID NO: 80 (последовательность нуклеиновой кислоты тяжелой цепи - SEQ ID NO: 81). Полноразмерная аминокислотная последовательность легкой цепи представлена SEQ ID NO: 90 (последовательность нуклеиновой кислоты легкой цепи - SEQ ID NO: 91).
411D07 имеет аминокислотную последовательность вариабельной области тяжелой цепи (VH), представленную SEQ ID NO: 98, содержащую аминокислотную последовательность CDRH1, представленную SEQ ID NO: 92 (IMGT) или SEQ ID NO: 95 (Кабат), аминокислотную последовательность CDRH2, представленную SEQ ID NO: 93 (IMGT) или SEQ ID NO: 96 (Кабат), и аминокислотную последовательность CDRH3, представленную SEQ ID NO: 94 (IMGT) или SEQ ID NO: 97 (Кабат). Последовательность нуклеиновой кислоты тяжелой цепи домена VH представлена SEQ ID NO: 99. 411D07 имеет аминокислотную последовательность вариабельной области легкой цепи (VL), представленную SEQ ID NO: 108, содержащую аминокислотную последовательность CDRL1, представленную SEQ ID NO: 102 (IMGT) или SEQ ID NO: 105 (Кабат), аминокислотную последовательность CDRL2, представленную SEQ ID NO: 103 (IMGT) или SEQ ID NO: 106 (Кабат), и аминокислотную последовательность CDRL3, представленную SEQ ID NO: 104 (IMGT) или SEQ ID NO: 107 (Кабат). Последовательность нуклеиновой кислоты легкой цепи домена VL представлена SEQ ID NO: 109. Домен VH может быть объединена с любой из последовательностей константной области тяжелой цепи, описанной в данном документе, например, SEQ ID NO: 193, SEQ ID NO: 195, SEQ ID NO: 197, SEQ ID NO: 199, SEQ ID NO: 201, SEQ ID NO: 203, Seq ID Nos: 205, Seq ID Nos: 340, Seq ID Nos: 524, Seq ID Nos: 526, Seq ID Nos: 528, Seq ID Nos: 530, SEQ ID NO: 532 или SEQ ID NO: 534. Область VL может быть объединена с любой из последовательностей константной области легкой цепи, описанной в данном документе, например, SEQ ID NOs: 207, 209, 211, 213, 215, 217, 219, 221, 223, 225, 227, 229, 231, 233, 235, 237, 536 и 538. Полноразмерная аминокислотная последовательность тяжелой цепи представлена SEQ ID NO: 100 (последовательность нуклеиновой кислоты тяжелой цепи - SEQ ID NO: 101). Полноразмерная аминокислотная последовательность легкой цепи представлена SEQ ID NO: 110 (последовательность нуклеиновой кислоты легкой цепи - SEQ ID NO: 111).
385F01 имеет аминокислотную последовательность вариабельной области тяжелой цепи (VH), представленную SEQ ID NO: 118, содержащую аминокислотную последовательность CDRH1, представленную SEQ ID NO: 112 (IMGT) или SEQ ID NO: 115 (Кабат), аминокислотную последовательность CDRH2, представленную SEQ ID NO: 113 (IMGT) или SEQ ID NO: 116 (Кабат), и аминокислотную последовательность CDRH3, представленную SEQ ID NO: 114 (IMGT) или SEQ ID NO: 117 (Кабат). Последовательность нуклеиновой кислоты тяжелой цепи домена VH представлена SEQ ID NO: 119. 385F01 имеет аминокислотную последовательность вариабельной области легкой цепи (VL), представленную SEQ ID NO: 128, содержащую аминокислотную последовательность CDRL1, представленную SEQ ID NO: 122 (IMGT) или SEQ ID NO: 125 (Кабат), аминокислотную последовательность CDRL2, представленную SEQ ID NO: 123 (IMGT) или SEQ ID NO: 126 (Кабат), и аминокислотную последовательность CDRL3, представленную SEQ ID NO: 124 (IMGT) или SEQ ID NO: 127 (Кабат). Последовательность нуклеиновой кислоты легкой цепи домена VL представлена SEQ ID NO: 129. Домен VH может быть объединена с любой из последовательностей константной области тяжелой цепи, описанной в данном документе, например, SEQ ID NO: 193, SEQ ID NO: 195, SEQ ID NO: 197, SEQ ID NO: 199, SEQ ID NO: 201, SEQ ID NO: 203, Seq ID Nos: 205, Seq ID Nos: 340, Seq ID Nos: 524, Seq ID Nos: 526, Seq ID Nos: 528, Seq ID Nos: 530, SEQ ID NO: 532 или SEQ ID NO: 534. Область VL может быть объединена с любой из последовательностей константной области легкой цепи, описанной в данном документе, например, SEQ ID NOs: 207, 209, 211, 213, 215, 217, 219, 221, 223, 225, 227, 229, 231, 233, 235, 237, 536 и 538. Полноразмерная аминокислотная последовательность тяжелой цепи представлена SEQ ID NO: 120 (последовательность нуклеиновой кислоты тяжелой цепи - SEQ ID NO: 121). Полноразмерная аминокислотная последовательность легкой цепи представлена SEQ ID NO: 130 (последовательность нуклеиновой кислоты легкой цепи - SEQ ID NO: 131).
386Н03 имеет аминокислотную последовательность вариабельной области тяжелой цепи (VH), представленную SEQ ID NO: 158, содержащую аминокислотную последовательность CDRH1, представленную SEQ ID NO: 152 (IMGT) или SEQ ID NO: 155 (Кабат), аминокислотную последовательность CDRH2, представленную SEQ ID NO: 153 (IMGT) или SEQ ID NO: 156 (Кабат), и аминокислотную последовательность CDRH3, представленную SEQ ID NO: 154 (IMGT) или SEQ ID NO: 157 (Кабат). Последовательность нуклеиновой кислоты тяжелой цепи домена VH представлена SEQ ID NO: 159. 386Н03 имеет аминокислотную последовательность вариабельной области легкой цепи (VL), представленную SEQ ID NO: 168, содержащую аминокислотную последовательность CDRL1, представленную SEQ ID NO: 162 (IMGT) или SEQ ID NO: 165 (Кабат), аминокислотную последовательность CDRL2, представленную SEQ ID NO: 163 (IMGT) или SEQ ID NO: 166 (Кабат), и аминокислотную последовательность CDRL3, представленную SEQ ID NO: 164 (IMGT) или SEQ ID NO: 167 (Кабат). Последовательность нуклеиновой кислоты легкой цепи домена VL представлена SEQ ID NO: 169. Домен VH может быть объединена с любой из последовательностей константной области тяжелой цепи, описанной в данном документе, например, SEQ ID NO: 193, SEQ ID NO: 195, SEQ ID NO: 197, SEQ ID NO: 199, SEQ ID NO: 201, SEQ ID NO: 203, Seq ID Nos: 205, Seq ID Nos: 340, Seq ID Nos: 524, Seq ID Nos: 526, Seq ID Nos: 528, Seq ID Nos: 530, SEQ ID NO: 532 или SEQ ID NO: 534. Область VL может быть объединена с любой из последовательностей константной области легкой цепи, описанной в данном документе, например, SEQ ID NOs: 207, 209, 211, 213, 215, 217, 219, 221, 223, 225, 227, 229, 231, 233, 235, 237, 536 и 538. Полноразмерная аминокислотная последовательность тяжелой цепи представлена SEQ ID NO: 160 (последовательность нуклеиновой кислоты тяжелой цепи - SEQ ID NO: 161). Полноразмерная аминокислотная последовательность легкой цепи представлена SEQ ID NO: 170 (последовательность нуклеиновой кислоты легкой цепи - SEQ ID NO: 171).
389А03 имеет аминокислотную последовательность вариабельной области тяжелой цепи (VH), представленную SEQ ID NO: 178, содержащую аминокислотную последовательность CDRH1, представленную SEQ ID NO: 172 (IMGT) или SEQ ID NO: 175 (Кабат), аминокислотную последовательность CDRH2, представленную SEQ ID NO: 173 (IMGT) или SEQ ID NO: 176 (Кабат), и аминокислотную последовательность CDRH3, представленную SEQ ID NO: 174 (IMGT) или SEQ ID NO: 177 (Кабат). Последовательность нуклеиновой кислоты тяжелой цепи домена VH представлена SEQ ID NO: 179. 389А03 имеет аминокислотную последовательность вариабельной области легкой цепи (VL), представленную SEQ ID NO: 188, содержащую аминокислотную последовательность CDRL1, представленную SEQ ID NO: 182 (IMGT) или SEQ ID NO: 185 (Кабат), аминокислотную последовательность CDRL2, представленную SEQ ID NO: 183 (IMGT) или SEQ ID NO: 186 (Кабат), и аминокислотную последовательность CDRL3, представленную SEQ ID NO: 184 (IMGT) или SEQ ID NO: 187 (Кабат). Последовательность нуклеиновой кислоты легкой цепи домена VL представлена SEQ ID NO: 189. Домен VH может быть объединена с любой из последовательностей константной области тяжелой цепи, описанной в данном документе, например, SEQ ID NO: 193, SEQ ID NO: 195, SEQ ID NO: 197, SEQ ID NO: 199, SEQ ID NO: 201, SEQ ID NO: 203, Seq ID Nos: 205, Seq ID Nos: 340, Seq ID Nos: 524, Seq ID Nos: 526, Seq ID Nos: 528, Seq ID Nos: 530, SEQ ID NO: 532 или SEQ ID NO: 534. Область VL может быть объединена с любой из последовательностей константной области легкой цепи, описанной в данном документе, например, SEQ ID NOs: 207, 209, 211, 213, 215, 217, 219, 221, 223, 225, 227, 229, 231, 233, 235, 237, 536 и 538. Полноразмерная аминокислотная последовательность тяжелой цепи представлена SEQ ID NO: 180 (последовательность нуклеиновой кислоты тяжелой цепи - SEQ ID NO: 181). Полноразмерная аминокислотная последовательность легкой цепи представлена SEQ ID NO: 190 (последовательность нуклеиновой кислоты легкой цепи - SEQ ID NO: 191).
413D08 имеет аминокислотную последовательность вариабельной области тяжелой цепи (VH), представленную SEQ ID NO: 138, содержащую аминокислотную последовательность CDRH1, представленную SEQ ID NO: 132 (IMGT) или SEQ ID NO: 135 (Кабат), аминокислотную последовательность CDRH2, представленную SEQ ID NO: 133 (IMGT) или SEQ ID NO: 136 (Кабат), и аминокислотную последовательность CDRH3, представленную SEQ ID NO: 134 (IMGT) или SEQ ID NO: 137 (Кабат). Последовательность нуклеиновой кислоты тяжелой цепи домена VH представлена SEQ ID NO: 139. 413D08 имеет аминокислотную последовательность вариабельной области легкой цепи (VL), представленную SEQ ID NO: 148, содержащую аминокислотную последовательность CDRL1, представленную SEQ ID NO: 142 (IMGT) или SEQ ID NO: 145 (Кабат), аминокислотную последовательность CDRL2, представленную SEQ ID NO: 143 (IMGT) или SEQ ID NO: 146 (Кабат), и аминокислотную последовательность CDRL3, представленную SEQ ID NO: 144 (IMGT) или SEQ ID NO: 147 (Кабат). Последовательность нуклеиновой кислоты легкой цепи домена VL представлена SEQ ID NO: 149. Домен VH может быть объединена с любой из последовательностей константной области тяжелой цепи, описанной в данном документе, например, SEQ ID NO: 193, SEQ ID NO: 195, SEQ ID NO: 197, SEQ ID NO: 199, SEQ ID NO: 201, SEQ ID NO: 203, Seq ID Nos: 205, Seq ID Nos: 340, Seq ID Nos: 524, Seq ID Nos: 526, Seq ID Nos: 528, Seq ID Nos: 530, SEQ ID NO: 532 или SEQ ID NO: 534. Область VL может быть объединена с любой из последовательностей константной области легкой цепи, описанной в данном документе, например, SEQ ID NOs: 207, 209, 211, 213, 215, 217, 219, 221, 223, 225, 227, 229, 231, 233, 235, 237, 536 и 538. Полноразмерная аминокислотная последовательность тяжелой цепи представлена SEQ ID NO: 140 (последовательность нуклеиновой кислоты тяжелой цепи - SEQ ID NO: 141). Полноразмерная аминокислотная последовательность легкой цепи представлена SEQ ID NO: 150 (последовательность нуклеиновой кислоты легкой цепи - SEQ ID NO: 151).
413G05 имеет аминокислотную последовательность вариабельной области тяжелой цепи (VH), представленную SEQ ID NO: 244, содержащую аминокислотную последовательность CDRH1, представленную SEQ ID NO: 238 (IMGT) или SEQ ID NO: 241 (Кабат), аминокислотную последовательность CDRH2, представленную SEQ ID NO: 239 (IMGT) или SEQ ID NO: 242 (Кабат), и аминокислотную последовательность CDRH3, представленную SEQ ID NO: 240 (IMGT) или SEQ ID NO: 243 (Кабат). Последовательность нуклеиновой кислоты тяжелой цепи домена VH представлена SEQ ID NO: 245. 413G05 имеет аминокислотную последовательность вариабельной области легкой цепи (VL), представленную SEQ ID NO: 254, содержащую аминокислотную последовательность CDRL1, представленную SEQ ID NO: 248 (IMGT) или SEQ ID NO: 251 (Кабат), аминокислотную последовательность CDRL2, представленную SEQ ID NO: 249 (IMGT) или SEQ ID NO: 252 (Кабат), и аминокислотную последовательность CDRL3, представленную SEQ ID NO: 250 (IMGT) или SEQ ID NO: 253 (Кабат). Последовательность нуклеиновой кислоты легкой цепи домена VL представлена SEQ ID NO: 255. Домен VH может быть объединена с любой из последовательностей константной области тяжелой цепи, описанной в данном документе, например, SEQ ID NO: 193, SEQ ID NO: 195, SEQ ID NO: 197, SEQ ID NO: 199, SEQ ID NO: 201, SEQ ID NO: 203, Seq ID Nos: 205, Seq ID Nos: 340, Seq ID Nos: 524, Seq ID Nos: 526, Seq ID Nos: 528, Seq ID Nos: 530, SEQ ID NO: 532 или SEQ ID NO: 534. Область VL может быть объединена с любой из последовательностей константной области легкой цепи, описанной в данном документе, например, SEQ ID NOs: 207, 209, 211, 213, 215, 217, 219, 221, 223, 225, 227, 229, 231, 233, 235, 237, 536 и 538. Полноразмерная аминокислотная последовательность тяжелой цепи представлена SEQ ID NO: 246 (последовательность нуклеиновой кислоты тяжелой цепи - SEQ ID NO: 247). Полноразмерная аминокислотная последовательность легкой цепи представлена SEQ ID NO: 256 (последовательность нуклеиновой кислоты легкой цепи - SEQ ID NO: 257).
413F09 имеет аминокислотную последовательность вариабельной области тяжелой цепи (VH), представленную SEQ ID NO: 264, содержащую аминокислотную последовательность CDRH1, представленную SEQ ID NO: 258 (IMGT) или SEQ ID NO: 261 (Кабат), аминокислотную последовательность CDRH2, представленную SEQ ID NO: 259 (IMGT) или SEQ ID NO: 262 (Кабат), и аминокислотную последовательность CDRH3, представленную SEQ ID NO: 260 (IMGT) или SEQ ID NO: 263 (Кабат). Последовательность нуклеиновой кислоты тяжелой цепи домена VH представлена SEQ ID NO: 265. 413F09 имеет аминокислотную последовательность вариабельной области легкой цепи (VL), представленную SEQ ID NO: 274, содержащую аминокислотную последовательность CDRL1, представленную SEQ ID NO: 268 (IMGT) или SEQ ID NO: 271 (Кабат), аминокислотную последовательность CDRL2, представленную SEQ ID NO: 269 (IMGT) или SEQ ID NO: 272 (Кабат), и аминокислотную последовательность CDRL3, представленную SEQ ID NO: 270 (IMGT) или SEQ ID NO: 273 (Кабат). Последовательность нуклеиновой кислоты легкой цепи домена VL представлена SEQ ID NO: 275. Домен VH может быть объединена с любой из последовательностей константной области тяжелой цепи, описанной в данном документе, например, SEQ ID NO: 193, SEQ ID NO: 195, SEQ ID NO: 197, SEQ ID NO: 199, SEQ ID NO: 201, SEQ ID NO: 203, Seq ID Nos: 205, Seq ID Nos: 340, Seq ID Nos: 524, Seq ID Nos: 526, Seq ID Nos: 528, Seq ID Nos: 530, SEQ ID NO: 532 или SEQ ID NO: 534. Область VL может быть объединена с любой из последовательностей константной области легкой цепи, описанной в данном документе, например, SEQ ID NOs: 207, 209, 211, 213, 215, 217, 219, 221, 223, 225, 227, 229, 231, 233, 235, 237, 536 и 538. Полноразмерная аминокислотная последовательность тяжелой цепи представлена SEQ ID NO: 266 (последовательность нуклеиновой кислоты тяжелой цепи - SEQ ID NO: 267). Полноразмерная аминокислотная последовательность легкой цепи представлена SEQ ID NO: 276 (последовательность нуклеиновой кислоты легкой цепи - SEQ ID NO: 277).
414В06 имеет аминокислотную последовательность вариабельной области тяжелой цепи (VH), представленную SEQ ID NO: 284, содержащую аминокислотную последовательность CDRH1, представленную SEQ ID NO: 278 (IMGT) или SEQ ID NO: 281 (Кабат), аминокислотную последовательность CDRH2, представленную SEQ ID NO: 279 (IMGT) или SEQ ID NO: 282 (Кабат), и аминокислотную последовательность CDRH3, представленную SEQ ID NO: 280 (IMGT) или SEQ ID NO: 283 (Кабат). Последовательность нуклеиновой кислоты тяжелой цепи домена VH представлена SEQ ID NO: 285. 414В06 имеет аминокислотную последовательность вариабельной области легкой цепи (VL), представленную SEQ ID NO: 294, содержащую аминокислотную последовательность CDRL1, представленную SEQ ID NO: 288 (IMGT) или SEQ ID NO: 291 (Кабат), аминокислотную последовательность CDRL2, представленную SEQ ID NO: 289 (IMGT) или SEQ ID NO: 292 (Кабат), и аминокислотную последовательность CDRL3, представленную SEQ ID NO: 290 (IMGT) или SEQ ID NO: 293 (Кабат). Последовательность нуклеиновой кислоты легкой цепи домена VL представлена SEQ ID NO: 295. Домен VH может быть объединена с любой из последовательностей константной области тяжелой цепи, описанной в данном документе, например, SEQ ID NO: 193, SEQ ID NO: 195, SEQ ID NO: 197, SEQ ID NO: 199, SEQ ID NO: 201, SEQ ID NO: 203, Seq ID Nos: 205, Seq ID Nos: 340, Seq ID Nos: 524, Seq ID Nos: 526, Seq ID Nos: 528, Seq ID Nos: 530, SEQ ID NO: 532 или SEQ ID NO: 534. Область VL может быть объединена с любой из последовательностей константной области легкой цепи, описанной в данном документе, например, SEQ ID NOs: 207, 209, 211, 213, 215, 217, 219, 221, 223, 225, 227, 229, 231, 233, 235, 237, 536 и 538. Полноразмерная аминокислотная последовательность тяжелой цепи представлена SEQ ID NO: 286 (последовательность нуклеиновой кислоты тяжелой цепи - SEQ ID NO: 287). Полноразмерная аминокислотная последовательность легкой цепи представлена SEQ ID NO: 296 (последовательность нуклеиновой кислоты легкой цепи - SEQ ID NO: 297).
416Е01 имеет аминокислотную последовательность вариабельной области тяжелой цепи (VH), представленную SEQ ID NO: 349, содержащую аминокислотную последовательность CDRH1, представленную SEQ ID NO: 343 (IMGT) или SEQ ID NO: 346 (Кабат), аминокислотную последовательность CDRH2, представленную SEQ ID NO: 344 (IMGT) или SEQ ID NO: 347 (Кабат), и аминокислотную последовательность CDRH3, представленную SEQ ID NO: 345 (IMGT) или SEQ ID NO: 348 (Кабат). Последовательность нуклеиновой кислоты тяжелой цепи домена VH представлена SEQ ID NO: 350. 416Е01 имеет аминокислотную последовательность вариабельной области легкой цепи (VL), представленную SEQ ID NO: 359, содержащую аминокислотную последовательность CDRL1, представленную SEQ ID NO: 353 (IMGT) или SEQ ID NO: 356 (Кабат), аминокислотную последовательность CDRL2, представленную SEQ ID NO: 354 (IMGT) или SEQ ID NO: 357 (Кабат), и аминокислотную последовательность CDRL3, представленную SEQ ID NO: 355 (IMGT) или SEQ ID NO: 358 (Кабат). Последовательность нуклеиновой кислоты легкой цепи домена VL представлена SEQ ID NO: 360. Домен VH может быть объединена с любой из последовательностей константной области тяжелой цепи, описанной в данном документе, например, SEQ ID NO: 193, SEQ ID NO: 195, SEQ ID NO: 197, SEQ ID NO: 199, SEQ ID NO: 201, SEQ ID NO: 203, Seq ID Nos: 205, Seq ID Nos: 340, Seq ID Nos: 524, Seq ID Nos: 526, Seq ID Nos: 528, Seq ID Nos: 530, SEQ ID NO: 532 или SEQ ID NO: 534. Область VL может быть объединена с любой из последовательностей константной области легкой цепи, описанной в данном документе, например, SEQ ID NOs: 207, 209, 211, 213, 215, 217, 219, 221, 223, 225, 227, 229, 231, 233, 235, 237, 536 и 538. Полноразмерная аминокислотная последовательность тяжелой цепи представлена SEQ ID NO: 351 (последовательность нуклеиновой кислоты тяжелой цепи - SEQ ID NO: 352). Полноразмерная аминокислотная последовательность легкой цепи представлена SEQ ID NO: 361 (последовательность нуклеиновой кислоты легкой цепи - SEQ ID NO: 362).
Конъюгаты антитело-лекарственное средство
Анти-ICOS антитела могут использоваться в качестве носителей цитотоксических агентов для нацеливания на Tregs. Как указано в Примере 18, Tregs, находящиеся в микроокружении опухоли (ТМЕ), сильно экспрессируют ICOS. ICOS  на внутриопухолевых Tregs сильнее, чем на внутриопухолевых Teffs или периферических Tregs. Таким образом, анти-ICOS антитела, меченные токсичным лекарственным средством или пролекарством, могут преимущественно нацеливаться на Tregs в ТМЕ, чтобы доставлять токсичную полезную нагрузку, селективно ингибируя эти клетки. Такое нацеливание цитотоксических агентов обеспечивает дополнительный путь для устранения иммунного супрессивного воздействия Tregs, тем самым изменяя баланс Treg:Teff в пользу активности Teff, и может быть использовано в качестве альтернативы или в сочетании с любым одним или несколькими другими терапевтическими подходами, описанными в данном документе (например, опосредованное Fc-эффекторами ингибирование Tregs, агонизм эффекторных Т-клеток).
на внутриопухолевых Tregs сильнее, чем на внутриопухолевых Teffs или периферических Tregs. Таким образом, анти-ICOS антитела, меченные токсичным лекарственным средством или пролекарством, могут преимущественно нацеливаться на Tregs в ТМЕ, чтобы доставлять токсичную полезную нагрузку, селективно ингибируя эти клетки. Такое нацеливание цитотоксических агентов обеспечивает дополнительный путь для устранения иммунного супрессивного воздействия Tregs, тем самым изменяя баланс Treg:Teff в пользу активности Teff, и может быть использовано в качестве альтернативы или в сочетании с любым одним или несколькими другими терапевтическими подходами, описанными в данном документе (например, опосредованное Fc-эффекторами ингибирование Tregs, агонизм эффекторных Т-клеток).
Соответственно, изобретение предусматривает анти-ICOS антитело, которое конъюгировано с цитотоксическим лекарственным средством или пролекарством. В случае пролекарства, пролекарство способно активироваться в ТМЕ или другом участке-мишени терапевтической активности для создания цитотоксического агента. Активация может быть ответом на инициатор, такой как фотоактивация, например, с использованием инфракрасного света, чтобы активировать конъюгат фотоабсорбирующего вещества [36]. Пространственно-селективная активация пролекарства дополнительно усиливает цитотоксический эффект конъюгата антитело-лекарственное средство, обеспечивая, в сочетании с высокой экспрессией ICOS на внутриопухолевых Tregs, цитотоксический эффект, который является высокоселективным по отношению к этим клеткам.
Для использования в конъюгате антитело-лекарственное средство, цитотоксическое лекарственное средство или пролекарство, предпочтительно, является неиммуногенным и нетоксичным (бездействующим или неактивным) при циркуляции конъюгата антитело-лекарственное средство в крови. Предпочтительно, цитотоксическое лекарственное средство (или пролекарство, в активированном состоянии) является сильнодействующим - например, от двух до четырех молекул лекарственного средства может быть достаточно для уничтожения клетки-мишени. Фотоактивируемым пролекарством является силикафталоцианиновый краситель (IRDye 700 DX), индуцирующий летальное повреждение клеточной мембраны после воздействия инфракрасного света. Цитотоксические лекарственные средства включают антимитотические агенты, такие как монометилауристатин Е и ингибиторы микротрубочек, такие как производные майтанзина, например, мертанзин, DM1, эмтанзин.
Конъюгирование лекарственного средства (или пролекарства) с антителом обычно осуществляется через линкер. Линкер может быть расщепляемым линкером, например, дисульфидной, гидразоновой или пептидной связью. Могут использоваться расщепляемые катепсином линкеры, так чтобы лекарство высвобождалось катепсином в опухолевых клетках. Альтернативно, могут быть использованы нерасщепляемые линкеры, например, тиоэфирная связь. Также могут быть включены дополнительные группы присоединения и/или спейсеры.
Антитело в конъюгате антитело-лекарственное средство может представлять собой фрагмент антитела, такой как Fab'2 или другой антигенсвязывающий фрагмент, как описано в данном документе, так как малый размер таких фрагментов может способствовать их проникновению в участок ткани (например, твердую опухоль).
Анти-ICOS антитело в соответствии с данным изобретением может быть представлено в виде иммуноцитокина. Анти-ICOS антитела также можно вводить вместе с иммуноцитокинами в комбинированной терапии. Ряд примеров антител описан в данном документе для использования в комбинированной терапии с анти-ICOS, и любое из них (например, анти-PD-L1 антитело) может быть представлено в качестве иммуноцитокинов для использования в данном изобретении. Иммуноцитокин содержит молекулу антитела, конъюгированную с цитокином, таким как IL-2. Таким образом, конъюгаты анти-ICOS:IL-2 и конъюгаты анти-PD-L1:IL-2 являются дополнительными аспектами данного изобретения.
Цитокин IL-2 может обладать активностью по отношению к высокоаффинному рецептору (αβγ) IL-2 и/или рецептору IL-2 с промежуточной аффинностью (αβ). IL-2, используемый в иммуноцитокине, может представлять собой IL-2 человека дикого типа или вариант цитокина IL-2, имеющий одну или несколько делеций, замещений или добавок аминокислот, например, IL-2, имеющий делецию от 1 до 10 аминокислот на N-конце. Другие варианты IL-2 включают мутации R38A или R38Q.
Пример анти-PD-L1-иммуноцитокина содержит тяжелую цепь иммуноглобулина и легкую цепь иммуноглобулина, причем тяжелая цепь содержит, в направлении от N- к С-концу:
a) домен VH, содержащий CDRH1, CDRH2 и CDRH3; и
b) константную область тяжелой цепи;
и легкая цепь содержит, в направлении от N- к С-концу:
c) домен VL, содержащий CDRL1, CDRL2 и CDRL3;
d) константную область легкой цепи (CL);
e) необязательно, линкер (L); и
f) цитокин IL-2;
причем домен VH и домен VL состоят из антигенсвязывающего сайта, который специфически связывается с PD-L1 человека; и
причем иммуноцитокин содержит домен VH, который содержит CDRH3, содержащий мотив X1GSGX2YGX3X4FD (SEQ ID NO: 609), причем X1, X2 и X3 независимо представляют собой любую аминокислоту, а Х4 либо присутствует, либо отсутствует, и в случае присутствия может означать любую аминокислоту.
Домены VH и VL могут быть VH- и VL-доменами любого анти- PD-L1 антитела, упомянутого в данном документе, например, доменами VH и VL 1D05.
IL-2 может быть диким типом или вариантом IL-2 человека.
Вакцинация
Анти-ICOS антитела могут быть представлены в вакцинных композициях или могут вводиться совместно с препаратами вакцин. ICOS принимает участие в формировании фолликулярной хелперной клетки и реакции зародышевого центра [37]. Таким образом, антитела-агонисты ICOS обладают потенциальной клинической полезностью в качестве молекулярных адъювантов для повышения эффективности вакцины. Антитела могут использоваться для повышения защитной эффективности многочисленных вакцин, таких как вакцины против гепатита В, малярии, ВИЧ.
В контексте вакцинации, у анти-ICOS антитела, обычно, будет отсутствовать функция Fc-эффектора и, таким образом, оно не опосредует ADCC, CDC или ADCP. Антитело может быть представлено в формате, не имеющем области Fc, или имеющем нуль-эффекторную константную область. Необязательно, анти-ICOS антитело может иметь константную область тяжелой цепи, которая связывает один или несколько типов рецептора Fc, но не индуцирует активность ADCC, CDC или ADCP, или которая проявляет более низкую активность ADCC, CDC и ADCP по сравнению с человеческим IgG1 дикого типа. Такая константная область может быть неспособна связывать, или может связываться с более низкой аффинностью, с конкретным Fc-рецептором (рецепторами), ответственным за запуск активности ADCC, CDC или ADCP. Альтернативно, когда клеточные эффекторные функции являются приемлемыми или желательными в контексте вакцинации, анти-ICOS антитело может содержать константную область тяжелой цепи, положительную по функции Fc-эффектора. Любой из форматов IgG1, IgG4 и IgG4.PE может, например, использоваться для анти-ICOS антител в схемах вакцинации, а другие примеры пригодных изотипов и константных областей антител более подробно изложены в других частях данного документа.
Препараты и введение
Антитела могут быть моноклональными или поликлональными, но предпочтительно представлены в виде моноклональных антител для терапевтического использования. Они могут быть представлены как часть смеси других антител, необязательно, включающей антитела с различной специфичностью связывания.
Антитела в соответствии с изобретением и кодирующая нуклеиновая кислота обычно представлены в изолированной форме. Таким образом, антитела, VH и/или VL-домены и нуклеиновые кислоты могут быть выделены очисткой из их естественной среды или их производственной среды. Выделенные антитела и выделенная нуклеиновая кислота будут не содержать или по существу не содержать материала, с которым они связаны в природных условиях, такого как другие полипептиды или нуклеиновые кислоты, с которыми они встречаются in vivo, или среды, в которой их получают (например, клеточной культуры), когда такое получение осуществляется in vitro по технологии рекомбинантной ДНК. Необязательно, выделенное антитело или нуклеиновая кислота (1) не содержат по меньшей мере некоторых других белков, вместе с которыми оно обычно встречается, (2) по существу не содержат других белков из того же источника, например, того же вида, (3) экспрессируются клеткой другого вида, (4) были отделены от по меньшей мере примерно 50% полинуклеотидов, липидов, углеводов или других материалов, с которыми они связаны в природе, (5) функционально связаны (посредством ковалентного или нековалентного взаимодействия) с полипептидом, с которым они не связаны в природе, или (6) не встречаются в природе.
Антитела или нуклеиновые кислоты могут входить в состав композиций с разбавителями или адъювантами, и при этом фактически оставаться выделенными - например, их можно смешивать с носителями, если они используются для покрытия микротитровального планшета для иммунологических анализов, и могут быть смешаны с фармацевтически приемлемыми носителями или разбавителями при использовании в терапии. Как описано в другой части данного документа, терапевтические препараты могут также включать другие активные ингредиенты. Антитела могут быть гликозилированы либо естественным образом in vivo, либо системами гетерологичных эукариотических клеток, таких как клетки СНО, или они могут быть (например, если они продуцируются экспрессией в прокариотической клетке) негликозилированными. Изобретение охватывает антитела, имеющие модифицированный профиль гликозилирования. В некоторых областях применения, может быть полезна модификация с целью удаления нежелательных сайтов гликозилирования или, например, удаления фрагмента фукозы для повышения функции ADCC [38]. В других областях применения, модификация галактозилирования может быть проведена для модификации CDC.
Как правило, выделенный продукт составляет по меньшей мере примерно 5%, по меньшей мере примерно 10%, по меньшей мере примерно 25%, или по меньшей мере примерно 50% от данного образца. Антитело может по существу не содержать белков или полипептидов или других загрязнителей, присутствующих в его естественной или производственной среде, которые будут препятствовать его терапевтическому, диагностическому, профилактическому, исследовательскому или другому применению.
Антитело может быть идентифицировано, выделено и/или извлечено из компонента его производственной среды (например, естественным образом или рекомбинантными методами). Выделенное антитело может быть свободно от всех других компонентов его производственной среды, например, так, чтобы антитело было очищено до показателей, одобренных FDA, или соответствующих утвержденному стандарту. Загрязняющими компонентами производственной среды, такой как образуемая рекомбинантными трансфицированными клетками, являются материалы, которые обычно препятствуют исследовательскому, диагностическому или терапевтическому применению антител, и могут включать ферменты, гормоны и другие белковые или небелковые растворенные вещества. В некоторых вариантах реализации, антитело будет очищено: (1) до более чем 95% мас. антитела, по результатам определения, например, методом Лоури, а в некоторых вариантах реализации - до более чем 99% мас.; (2) до степени, достаточной для получения по меньшей мере 15 остатков N-концевой или внутренней аминокислотной последовательности с использованием секвенатора с вращающимся стаканом, или (3) до гомогенности, определяемой методом ДСН-ПААГ в невосстановительных или восстановительных условиях с использованием окрашивания кумасси голубым или серебром. Выделенное антитело включает антитело in situ в рекомбинантных клетках, поскольку по меньшей мере один компонент природной среды антитела не будет присутствовать. Однако, обычно выделенное антитело или его кодирующую нуклеиновую кислоту получают с по меньшей мере одной стадией очистки.
Изобретение предусматривает терапевтические композиции, содержащие антитела, раскрытые в данном документе. Также предусматриваются терапевтические композиции, содержащие нуклеиновую кислоту, кодирующую такие антитела. Кодирующие нуклеиновые кислоты более подробно описаны в других частях данного документа и включают ДНК и РНК, например, мРНК. В терапевтических способах, описанных в данном документе, применение нуклеиновой кислоты, кодирующей антитело, и/или клеток, содержащих такую нуклеиновую кислоту, можно использовать в качестве альтернатив (или в дополнение к) композициям, содержащим само антитело. Таким образом, клетки, содержащие нуклеиновую кислоту, кодирующую антитело, причем, необязательно, нуклеиновая кислота стабильно интегрирована в геном, представляют собой медикаменты для терапевтического использования у пациента. Нуклеиновую кислоту, кодирующую анти-ICOS антитело, можно вводить в человеческие В-лимфоциты, необязательно, В-лимфоциты, полученные от предполагаемого пациента и модифицированные ex vivo. Необязательно, используются В-клетки памяти. Введение пациенту клеток, содержащих кодирующую нуклеиновую кислоту, создает резервуар клеток, способных экспрессировать анти-ICOS антитело, которое может обеспечить терапевтическое преимущество в течение более длительного периода времени по сравнению с введением выделенной нуклеиновой кислоты или выделенного антитела.
Композиции могут содержать пригодные носители, эксципиенты и другие агенты, которые включены в композиции для обеспечения улучшенного переноса, доставки, переносимости и т.п. Множество пригодных рецептур можно найти в фармакологическом справочнике, известном всем химикам-фармацевтам: Remington's Pharmaceutical Sciences, Mack Publishing Company, Easton, Pa. Эти рецептуры включают, например, порошки, пасты, мази, желе, воски, масла, липиды, содержащие липиды (катионные или анионные) везикулы (такие как LIPOFECTINT™), конъюгаты ДНК, безводные абсорбционные пасты, эмульсии типа «масло в воде» и «вода в масле», эмульсии карбовакса (полиэтиленгликоли с разной молекулярной массой), полутвердые гели и полутвердые смеси, содержащие карбовакс. См. также Powell et al. "Compendium of excipients for parenteral formulations" PDA (1998) J Pharm Sci Technol 52:238-311. Композиции могут содержать антитело или нуклеиновую кислоту в сочетании с медицинским буфером для инъекций и/или адъювантом.
Композиции антител или кодирующих их нуклеиновых кислот могут быть составлены с учетом желательного способа введения пациенту, например, в жидкости (необязательно, водном растворе) для инъекций. Различные системы доставки известны и могут использоваться для введения фармацевтической композиции по изобретению. Способы введения включают, без ограничений, интрадермальные, внутримышечные, внутрибрюшинные, внутривенные, подкожные, интраназальные, эпидуральные и оральные пути. Составление композиций антител для подкожного введения обычно требует их концентрирование в меньшем объеме по сравнению с внутривенными препаратами. Высокая эффективность антител в соответствии с данным изобретением может позволить использовать их в достаточно низких дозах для того, чтобы сделать подкожную рецептуру практичной, обеспечивая преимущество по сравнению с менее эффективными анти-ICOS антителами.
Композицию можно вводить любым удобным способом, например, путем инфузии или болюсной инъекции, путем абсорбции через эпителиальные или кожно-слизистые оболочки (например, слизистую оболочку полости рта, слизистую оболочку прямой кишки и кишечника и т.д.), и могут вводиться вместе с другими биологически активными агентами. Введение может быть системным или местным.
Фармацевтическая композиция также может быть введена в везикулу, в частности, липосому (см. Langer (1990) Science 249:1527-1533; Treat et al. (1989) в Liposomes in the Therapy of Infectious Disease and Cancer, Lopez Berestein and Fidler (eds.), Liss, New York, pp. 353-365; Lopez-Berestein, ibid., pp. 317-327; для общего ознакомления см. там же).
В определенных ситуациях, фармацевтическая композиция может быть доставлена в системе с контролируемым высвобождением. В одном варианте реализации, может использоваться насос (см. Langer, выше; Sefton (1987) CRC Crit. Ref. Biomed. Eng. 14:201). В другом варианте реализации могут быть использованы полимерные материалы; см. Medical Applications of Controlled Release, Langer and Wise (eds.), CRC Pres., Boca Raton, Fla. (1974). В еще одном варианте реализации, система контролируемого высвобождения может быть размещена вблизи мишени композиции, благодаря чему требуется только часть от системной дозы (см., например, Goodson, в Medical Applications of Controlled Release, supra, vol. 2, pp. 115-138, 1984).
Инъекционные препараты могут включать лекарственные формы для внутривенных, подкожных, внутрикожных и внутримышечных инъекций, капельных инфузий и т.д. Такие препараты для инъекций могут быть приготовлены общеизвестными способами. Например, инъекционные препараты могут быть получены, например, путем растворения, суспендирования или эмульгирования антитела или его соли, описанных выше, в стерильной водной среде или масляной среде, обычно используемой для инъекций. В качестве водной среды для инъекций используются, например, физиологический солевой раствор, изотонический раствор, содержащий глюкозу и другие вспомогательные агенты и т.д., которые могут быть использованы в комбинации с пригодным солюбилизирующим агентом, таким как спирт (например, этанол), многоатомный спирт (например, пропиленгликоль, полиэтиленгликоль), неионогенное поверхностно-активное вещество (например, полисорбат 80, НСО-50 (аддукт полиоксиэтилена (50 моль) и гидрированного касторового масла) и т.д. В качестве масляной среды используют, например, кунжутное масло, соевое масло и т.д., которые могут быть использованы в комбинации с солюбилизирующим агентом, таким как бензилбензоат, бензиловый спирт и т.д. Полученной таким образом инъекцией можно заполнить соответствующую ампулу. Фармацевтическая композиция по данному изобретению может быть доставлена подкожно или внутривенно с использованием стандартной иглы и шприца. Предполагается, что лечение не будет ограничено применением в условиях клиники. Таким образом, будет также полезной подкожная инъекция с использованием безыгольного устройства. В случае подкожной доставки, устройство для доставки типа шприц-ручки может быть легко применено для доставки фармацевтической композиции по данному изобретению. Такое устройство для доставки типа шприц-ручки может предусматривать повторное использование или быть одноразовым. Многоразовое устройство доставки типа шприц-ручки обычно использует сменный картридж, который содержит фармацевтическую композицию. Как только вся фармацевтическая композиция, находящаяся в картридже, будет введена, и картридж опустеет, пустой картридж можно легко выбросить и заменить новым картриджем, содержащим фармацевтическую композицию. Затем устройство для доставки типа шприц-ручки может быть использовано повторно. В одноразовом устройстве для доставки типа шприц-ручки нет сменного картриджа. Вместо этого, одноразовое устройство для доставки типа шприц-ручки заполняют фармацевтической композицией, удерживаемой в резервуаре внутри устройства. Когда вся фармацевтическая композиция из резервуара будет использована, все устройство выбрасывается. Многочисленные повторно используемые устройства для доставки типа шприц-ручки и автоинъектора находят применение для подкожной доставки фармацевтической композиции по данному изобретению. Примеры включают, без ограничений, AUTOPEN™ (Owen Mumford, Inc., Woodstock, UK), шприц-ручку DISETRONIC™ (Disetronic Medical Systems, Burghdorf, Switzerland), шприц-ручку HUMALOG MIX 75/25™, шприц-ручку HUMALOG™, шприц-ручку HUMALIN 70/30™ (Eli Lilly and Co., Indianapolis, Ind.), NOVOPEN™ I, II и III (Novo Nordisk, Copenhagen, Denmark), NOVOPEN JUNIOR™ (Novo Nordisk, Copenhagen, Denmark), шприц-ручку BD™ (Becton Dickinson, Franklin Lakes, NJ), OPTIPENT™, OPTIPEN PRO™, OPTIPEN STARLET™ и OPTICLIKT™ (Sanofi-Aventis, Frankfurt, Germany), в качестве некоторых примеров. Примеры одноразовых устройств для доставки типа шприц-ручки, которые находят применение для подкожной доставки фармацевтической композиции по данному изобретению, включают, без ограничений, шприц-ручку SOLOSTAR™ (Sanofi-Aventis), FLEXPEN™ (Novo Nordisk) и KWIKPEN™ (Eli Lilly).
Преимущественно, фармацевтические композиции для перорального или парентерального применения, описанные выше, готовят в виде лекарственных форм с единичной дозой, соответствующей дозе активных ингредиентов. Такие лекарственные формы с единичной дозой включают, например, таблетки, пилюли, капсулы, инъекции (ампулы), суппозитории и т.д. Количество содержащегося в них вышеуказанного антитела обычно составляет от 5 до 500 мг на лекарственную форму в стандартной дозе; особенно, в форме инъекции, содержание вышеуказанного антитела может составлять от примерно 5 до примерно 100 мг, и от примерно 10 до примерно 250 мг для других лекарственных форм.
Антитело, нуклеиновая кислота или композиция, содержащая их, могут содержаться в медицинском контейнере, таком как флакон, шприц, контейнер для внутривенного введения (IV) или инъекционное устройство. В одном примере, антитело, нуклеиновая кислота или композиция размещаются in vitro и могут находиться в стерильном контейнере. В одном примере, предусматривается набор, содержащий антитело, упаковку и инструкции по применению в терапевтическом методе, как описано в данном документе.
Одним аспектом изобретения является композиция, содержащая антитело или нуклеиновую кислоту по изобретению и один или несколько фармацевтически приемлемых эксципиентов, примеры которых перечислены выше. «Фармацевтически приемлемый» относится к разрешенному или одобренному регулирующим органом федерального правительства США или правительством штата, или включенному в Фармакопею США или другую общепризнанную фармакопею для использования на животных, включая людей. Фармацевтически приемлемый носитель, эксципиент или адъювант можно вводить пациенту вместе с агентом, например, любым антителом или цепью антитела, описанными в данном документе, и не уничтожает его фармакологическую активность, и является нетоксичным при введении в дозах, достаточных для доставки терапевтического количества агента.
В некоторых вариантах реализации, анти-ICOS антитело будет единственным активным ингредиентом в композиции в соответствии с данным изобретением. Таким образом, композиция может состоять из антитела или она может состоять из антитела с одним или несколькими фармацевтически приемлемыми эксципиентами. Однако, композиции в соответствии с данным изобретением, необязательно, включают один или несколько дополнительных активных ингредиентов. Подробное описание агентов, с которыми могут быть объединены анти-ICOS антитела, приведено в других частях данного документа. Необязательно, композиции содержат множество антител (или кодирующих нуклеиновых кислот) в комбинированном препарате, например, одна композиция содержит анти-ICOS антитело и одно или несколько других антител. Другие терапевтические средства, которые могут быть желательными для введения с антителами или нуклеиновыми кислотами в соответствии с данным изобретением, включают анальгетики. Любой такой агент или комбинация агентов могут быть введены в сочетании с, или входить в состав композиций с антителами или нуклеиновыми кислотами в соответствии с данным изобретением, в виде комбинированного или отдельного препарата. Антитело или нуклеиновая кислота в соответствии с данным изобретением могут вводиться отдельно и последовательно или одновременно и, необязательно, в виде комбинированного препарата с другим терапевтическим агентом или агентами, такими как упомянутые.
Анти-ICOS антитела для использования при определенных терапевтических показаниях могут сочетаться с принятым стандартом лечения. Таким образом, для лечения рака, лечение антителом может применяться в схеме лечения, также включающей, например, химиотерапию, хирургию и/или лучевую терапию. Лучевая терапия может проводиться с однократной дозой или дробными дозами, с облучением непосредственно пострадавших тканей либо всего организма.
Несколько композиций можно вводить по отдельности или одновременно. Раздельное введение относится к двум композициям, которые вводят в разное время, например, с промежутком времени между ними по меньшей мере 10, 20, 30 или 10-60 минут друг от друга или 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 12 часов. Можно также вводить композиции с промежутком времени между ними 24 часа или даже больше. Альтернативно, две или более композиций могут вводиться одновременно, например, с промежутком времени между ними менее 10 или меньше 5 минут. Композиции, вводимые одновременно, могут, в некоторых аспектах, вводиться в виде смеси, со схожим механизмом высвобождения во времени для каждого из компонентов или без него, или с разными механизмами.
Антитела и кодирующие их нуклеиновые кислоты могут быть использованы в качестве терапевтических средств. Пациенты в данном документе обычно являются млекопитающими, обычно людьми. Антитело или нуклеиновую кислоту можно вводить млекопитающему, например, любым способом введения, упомянутым в данном документе.
Введение обычно осуществляется в «терапевтически эффективном количестве», которое представляет собой количество, дающее желательный эффект, для которого они вводятся, достаточный для того, чтобы создать полезный для пациента эффект. Точное количество будет зависеть от цели лечения и будет определяться квалифицированным специалистом в данной области техники с использованием известных методов (см., например, Lloyd (1999) The Art, Science and Technology of Pharmaceutical Compounding). Назначение лечения, например, решения о дозировке и т.д., находится в ведении врачей общей практики и других врачей и может зависеть от тяжести симптомов и/или прогрессирования заболевания, лечение которого проводится. Терапевтически эффективное количество или пригодную дозу антитела или нуклеиновой кислоты можно определить, сравнивая ее активность in vitro и активность in vivo на животной модели. Известны способы экстраполяции эффективных доз для мышей и других экспериментальных животных на человека.
Как показали исследования in vivo, описанные в приведенных в данном документе примерах, анти-ICOS антитело может быть эффективным в диапазоне доз. Фармакодинамические исследования описаны в Примере 24.
Анти-ICOS антитела можно вводить в дозах, содержащих количества в одном из следующих диапазонов значений:
от примерно 10 мкг/кг массы тела до примерно 100 мг/кг массы тела,
от примерно 50 мкг/кг массы тела до примерно 5 мг/кг массы тела,
от примерно 100 мкг/кг массы тела до примерно 10 мг/кг массы тела,
от примерно 100 мкг/кг массы тела до примерно 20 мг/кг массы тела,
от примерно 0,5 мг/кг массы тела до примерно 20 мг/кг массы тела, или
около 5 мг/кг массы тела или меньше, например, менее 4, менее 3, менее 2 или менее 1 мг/кг антитела.
Оптимальная терапевтическая доза для человека может составлять от 0,1 до 0,5 мг/кг, например, около 0,1 мг/кг, 0,15 мг/кг, 0,2 мг/кг, 0,25 мг/кг, 0,3 мг/кг, 0,35 мг/кг, 0,4 мг/кг, 0,45 мг/кг или 0,5 мг/кг. При фиксированном дозировании для взрослых, пригодная доза может составлять от 8 до 50 мг или от 8 до 25 мг, например, 15 мг или 20 мг.
В способах лечения, описанных в данном документе, можно вводить одну или несколько доз. В некоторых случаях однократная доза может быть эффективной для достижения долгосрочного полезного эффекта. Таким образом, способ может включать введение одной дозы антитела, его кодирующей нуклеиновой кислоты, или композиции. Альтернативно, можно вводить несколько доз, обычно последовательно и с промежутком в несколько дней, недель или месяцев. Анти-ICOS антитело может вводиться пациенту повторно с интервалами от 4 до 6 недель, например, каждые 4 недели, каждые 5 недель или каждые 6 недель. Необязательно, анти-ICOS антитело можно вводить пациенту один раз в месяц или реже, например, каждые два месяца или каждые три месяца. Соответственно, способ лечения пациента может включать введение пациенту однократной дозы анти-ICOS антитела, без повторного введения в течение по меньшей мере одного месяца, по меньшей мере, двух месяцев, по меньшей мере, трех месяцев и, необязательно, без повторного введения в течение по меньшей мере 12 месяцев.
Как описано в Примере 11с, сопоставимые терапевтические эффекты могут быть получены с использованием однократной или многократных доз анти-ICOS антитела, что может быть результатом того, что однократная доза антитела является эффективной для преобразования микроокружения опухоли. Врачи могут адаптировать схему введения анти-ICOS антитела к заболеванию и пациенту, проходящему терапию, с учетом статуса заболевания и любых других терапевтических средств или терапевтических мер (например, хирургия, лучевая терапия и т.д.), с которыми объединяется анти-ICOS антитело. В некоторых вариантах реализации, эффективную дозу анти-ICOS антитела вводят чаще, чем один раз в месяц, например, раз в три недели, раз в две недели, или раз в неделю. Лечение анти-ICOS антителом может включать несколько доз, вводимых на протяжении по меньшей мере месяца, по меньшей мере шести месяцев, или по меньшей мере одного года.
Используемые здесь термины «лечить», «лечение», «проведение лечения» или «улучшение» относятся к терапевтическому лечению, целью которого является обращение, облегчение, улучшение, подавление, замедление или прекращение прогрессирования или тяжести состояния, связанного с заболеванием или расстройством. Термин «лечение» включает уменьшение или облегчение по меньшей мере одного неблагоприятного эффекта или симптома состояния, заболевания или расстройства. Лечение обычно «эффективно», если снижается один или несколько симптомов или клинических маркеров. Альтернативно, лечение является «эффективным», если прогрессирование заболевания уменьшается или прекращается. Таким образом, «лечение» включает не только улучшение симптомов или маркеров, но также прекращение или, по крайней мере, замедление прогрессирования или ухудшения симптомов по сравнению с тем, что можно было ожидать при отсутствии лечения. Полезные или желательные клинические результаты включают, без ограничений, облегчение одного или нескольких симптомов, уменьшение степени заболевания, стабилизированное (т.е. не ухудшающееся) состояние заболевания, задержку или замедление прогрессирования заболевания, улучшение или ослабление болезненного состояния, ремиссию (частичную или полную) и/или снижение смертности, независимо от того, являются они обнаруживаемыми или не обнаруживаемыми. Термин «лечение» заболевания также включает облегчение от симптомов или побочных эффектов заболевания (включая паллиативное лечение). Эффективное лечение не предусматривает полного исцеления. В некоторых аспектах, способ может также включать исцеление. В контексте изобретения, лечение может быть профилактическим лечением.
Т-клеточная терапия
В WO 2011/097477 описано использование анти-ICOS антител для генерации и пролиферации Т-клеток путем введения в контакт популяции Т-клеток с первым агентом, который обеспечивает первичный сигнал активации (например, анти-CD3-антитело) и вторым агентом, который активирует ICOS (например, анти-ICOS антитело), необязательно, в присутствии агента поляризации Th17, такого как IL-1β, IL-6, нейтрализующего анти-IFNγ и/или анти-IL-4. Анти-ICOS антитела, раскрытые в данном документе, могут быть использованы в таких способах для получения популяций Т-клеток. Могут генерироваться популяции искусственно размноженных в культуре Т-клеток, обладающих терапевтической активностью (например, противоопухолевой активностью). Как описано в WO 2011/097477, такие Т-клетки могут быть использованы в терапевтических целях в методах лечения пациентов с помощью иммунотерапии.
Морфологический анализ анти-ICOS антител в качестве кандидатов терапевтических средств
Было замечено, что при присоединении кандидатов терапевтических анти-ICOS антител к твердой поверхности и введении в контакт с экспрессирующими ICOS Т-клетками, они были способны индуцировать морфологические изменения в клетках. При добавлении ICOS+ Т-клеток в лунки с внутренним покрытием анти-ICOS антителами, наблюдалось изменение начальной округлой формы клеток до формы веретена, с распространением и прилипанием к поверхности, покрытой антителом. Такие морфологические изменение не наблюдались для контрольного антитела. Кроме того, было обнаружено, что эффект является дозозависимым, причем более быстрое и/или более выраженное изменение формы происходит при увеличении концентрации антитела на поверхности. Изменение формы представляет собой суррогатный показатель связывания Т-клеток с ICOS и/или агонизма анти-ICOS антитела. Анализ может быть использован для идентификации антитела, которое способствует мультимеризации ICOS на поверхности Т-клеток. Такие антитела представляют собой терапевтические кандидаты агонистических антител. Удобно, что визуальный показатель, обеспечиваемый этим анализом, является простым методом скрининга антител или клеток, особенно в больших количествах. Анализ может быть автоматизирован для работы в высокопроизводительной системе.
Соответственно, одним аспектом изобретения является анализ для выбора антитела, которое связывает ICOS, необязательно, для выбора агонистического антитела ICOS, причем анализ включает:
обеспечение массива антител, иммобилизованных (присоединенных или прикрепленных) к субстрату в тестовой лунке;
добавление ICOS-экспрессирующих клеток (например, активированных первичных Т-клеток или клеток MJ) в тестовую лунку;
наблюдение морфологии клеток;
детектирование изменения формы клеток в лунке от округлой до сплюснутой к подложке; причем изменение формы указывает, что антитело представляет собой антитело, которое связывает ICOS, необязательно, агонистическое антитело ICOS, и
выбор антитела из тестовой лунки.
Анализ может проводиться с несколькими тестовыми лунками, которые содержат различные антитела для тестирования, необязательно, параллельно, например, в формате 96-луночного планшета. Подложка, предпочтительно, является внутренней поверхностью лунки. Таким образом, обеспечивается двумерная поверхность, на которой может наблюдаться уплощение клеток. Например, дно и/или стенки лунки могут быть покрыты антителом. Связывание антитела с подложкой может осуществляться через константную область антитела.
Может быть включен отрицательный контроль, такой как антитело, достоверно не связывающее ICOS, предпочтительно, антитело, не связывающее антиген на поверхности экспрессирующих ICOS клеток, которые будут использоваться. Анализ может включать количественную оценку степени морфологического изменения и, при тестировании множества антител, выбор антитела, которое вызывает большее морфологическое изменение, чем одно или несколько других тестируемых антител.
Выбор антитела может включать экспрессию нуклеиновой кислоты, кодирующей антитело, присутствующее в тестовой лунке, представляющей интерес, или экспрессию антитела, содержащего CDR или антигенсвязывающий домен этого антитела. Антитело может быть, необязательно, переформатировано, например, для получения антитела, содержащего антигенсвязывающий домен выбранного антитела, например, фрагмента антитела, или антитела, содержащего другую константную область. Выбранное антитело, предпочтительно, имеет константную область IgG1 человека или другую константную область, как описано в данном документе. Дополнительно, может быть составлена композиция выбранного антитела, содержащая один или несколько дополнительных ингредиентов - пригодные фармацевтические композиции описаны в других частях данного документа.
Пункты
Варианты осуществления изобретения изложены в следующих пронумерованных пунктах, которые являются частью описания.
Пункт 1. Выделенное антитело, которое связывает внеклеточный домен ICOS человека и/или мыши, причем антитело содержит домен VH, содержащий аминокислотную последовательность, имеющую по меньшей мере 95% идентичности последовательности с доменом VH STIM003 SEQ ID NO: 408, и домен VL, содержащий аминокислотную последовательность, имеющую по меньшей мере 95% идентичности последовательности с доменом VL STIM003 SEQ ID NO: 415.
Пункт 2. Антитело согласно пункту 1, причем домен VH содержит набор гипервариабельных участков тяжелой цепи (HCDR) HCDR1, HCDR2 и HCDR3, причем
HCDR1 представляет собой HCDR1 STIM003, имеющий аминокислотную последовательность SEQ ID NO: 405,
HCDR2 представляет собой HCDR2 STIM003, имеющий аминокислотную последовательность SEQ ID NO: 406,
HCDR3 представляет собой HCDR3 STIM003, имеющий аминокислотную последовательность SEQ ID NO: 407.
Пункт 3. Антитело согласно пункту 1 или пункту 2, причем домен VL содержит набор гипервариабельных участков легкой цепи (LCDR) LCDR1, LCDR2 и LCDR3, причем
LCDR1 представляет собой STIM003 LCDR1, имеющий аминокислотную последовательность SEQ ID NO: 412,
LCDR2 представляет собой STIM003 LCDR2, имеющий аминокислотную последовательность SEQ ID NO: 413,
LCDR3 представляет собой STIM003 LCDR3, имеющий аминокислотную последовательность SEQ ID NO: 414.
Пункт 4. Антитело согласно пункту 1, причем аминокислотная последовательность домена VH представлена SEQ ID NO: 408 и/или причем аминокислотная последовательность домена VL представлена SEQ ID NO: 415.
Пункт 5. Выделенное антитело, которое связывает внеклеточный домен ICOS человека и/или мыши, содержащее
домен VH антитела, содержащий участки, определяющие комплементарность, (CDR) HCDR1, HCDR2 и HCDR3, и
домен VL антитела, содержащий участки, определяющие комплементарность, LCDR1, LCDR2 и LCDR3, причем
HCDR1 представляет собой HCDR1 STIM001, STIM002, STIM002-B, STIM003, STIM004, STIM005, STIM006, STIM007, STIM008 или STIM009, или содержит такой HCDR1 с 1, 2, 3, 4 или 5 аминокислотными изменениями,
HCDR2 представляет собой HCDR2 STIM001, STIM002, STIM002-B, STIM003, STIM004, STIM005, STIM006, STIM007, STIM008 или STIM009, или содержит такой HCDR2 с 1, 2, 3, 4 или 5 аминокислотными изменениями и/или
HCDR3 представляет собой HCDR3 STIM001, STIM002, STIM002-B, STIM003, STIM004, STIM005, STIM006, STIM007, STIM008 или STIM009, или содержит такой HCDR3 с 1, 2, 3, 4 или 5 аминокислотными изменениями.
Пункт 6. Антитело согласно пункту 5, причем CDR тяжелой цепи антитела принадлежат STIM001, STIM002, STIM002-B, STIM003, STIM004, STIM005, STIM006, STIM007, STIM008 или STIM009, или содержат CDR тяжелой цепи STIM001, STIM002, STIM002-B, STIM003, STIM004, STIM005, STIM006, STIM007, STIM008 или STIM009 с 1, 2, 3, 4 или 5 аминокислотными изменениями.
Пункт 7. Антитело согласно пункту 6, причем домен VH антитела имеет CDR тяжелой цепи STIM003.
Пункт 8. Выделенное антитело, которое связывает внеклеточный домен ICOS человека и/или мыши, содержащее
домен VH антитела, содержащий участки, определяющие комплементарность, HCDR1, HCDR2 и HCDR3, и
домен VL антитела, содержащий участки, определяющие комплементарность, LCDR1, LCDR2 и LCDR3,
причем LCDR1 представляет собой LCDR1 STIM001, STIM002, STIM002-B, STIM003, STIM004 STIM005, STIM006, STIM007, STIM008 или STIM009, или содержит этот LCDR1 с 1, 2, 3, 4 или 5 аминокислотными изменениями,
LCDR2 представляет собой LCDR2 STIM001, STIM002, STIM002-B, STIM003, STIM004, STIM005, STIM006, STIM007, STIM008 или STIM009, или содержит такой LCDR2 с 1, 2, 3, 4 или 5 аминокислотными изменениями и/или
LCDR3 представляет собой LCDR3 STIM001, STIM002, STIM002-B, STIM003, STIM004, STIM005, STIM006, STIM007, STIM008 или STIM009, или содержит такой LCDR3 с 1, 2, 3, 4 или 5 аминокислотными изменениями.
Пункт 9. Антитело согласно любому из пунктов 5-8, причем CDR легкой цепи антитела принадлежат STIM001, STIM002, STIM002-B, STIM003, STIM004, STIM005, STIM006, STIM007, STIM008 или STIM009, или содержат CDR легкой цепи STIM001, STIM002, STIM002-B, STIM003, STIM004, STIM005, STIM006, STIM007, STIM008 или STIM009 с 1, 2, 3, 4 или 5 аминокислотными изменениями.
Пункт 10. Антитело согласно пункту 9, причем домен VL антитела имеет CDR легкой цепи STIM003.
Пункт 11. Антитело согласно любому из пунктов 5-10, содержащее каркасные области домена VH и/или VL генных сегментов зародышевой линии человека.
Пункт 12. Антитело согласно любому из пунктов 5-11, содержащее домен VH, который (i) получен в результате рекомбинации V генного сегмента тяжелой цепи человека, D генного сегмента тяжелой цепи человека и J генного сегмента тяжелой цепи человека, причем
V сегмент представляет собой IGHV1-18 (например, V1-18*01), IGVH3-20 (например, V3-20*d01), IGVH3-11 (например, V3-11*01) или IGVH2-5 (например, V2-5*10);
D генный сегмент представляет собой IGHD6-19 (например, IGHD6-19*01), IGHD3-10 (например, IGHD3-10*01) или IGHD3-9 (например, IGHD3-9*01); и/или
J генный сегмент представляет собой IGHJ6 (например, IGHJ6*02), IGHJ4 (например, IGHJ4*02) или IGHJ3 (например, IGHJ3*02), или
(ii) содержит каркасные области FR1, FR2, FR3 и FR4, причем
FR1 выравнивается с V генным сегментом зародышевой линии человека IGHV1-18 (например, V1-18*01), IGVH3-20 (например, V3-20*d01), IGVH3-11 (например, V3-11*01) или IGVH2-5 (например, V2-5*10), необязательно, с 1, 2, 3, 4 или 5 аминокислотными изменениями,
FR2 выравнивается с V генным сегментом зародышевой линии человека IGHV1-18 (например, V1-18*01), IGVH3-20 (например, V3-20*d01), IGVH3-11 (например, V3-11*01) или IGVH2-5 (например, V2-5*10), необязательно, с 1, 2, 3, 4 или 5 аминокислотными изменениями,
FR3 выравнивается с V генным сегментом зародышевой линии человека IGHV1-18 (например, V1-18*01), IGVH3-20 (например, V3-20*d01), IGVH3-11 (например, V3-11*01) или IGVH2-5 (например, V2-5*10), необязательно, с 1, 2, 3, 4 или 5 аминокислотными изменениями и/или
FR4 выравнивается с J генным сегментом зародышевой линии IGJH6 (например, JH6*02), IGJH4 (например, JH4*02) или IGJH3 (например, JH3*02), необязательно, с 1, 2, 3, 4 или 5 аминокислотными изменениями,
Пункт 13. Антитело согласно любому из пунктов 5-12, содержащее домен VL антитела, который
(i) получен в результате рекомбинации V генного сегмента легкой цепи человека и J генного сегмента легкой цепи человека, причем
V сегмент представляет собой IGKV2-28 (например, IGKV2-28*01), IGKV3-20 (например, IGKV3-20*01), IGKV1D-39 (например, IGKV1D-39*01) или IGKV3-11 (например, IGKV3-11*01) и/или
J генный сегмент представляет собой IGKJ4 (например, IGKJ4*01), IGKJ2 (например, IGKJ2*04), IGLJ3 (например, IGKJ3*01) или IGKJ1 (например, IGKJ1*01); или
(ii) содержит каркасные области FR1, FR2, FR3 и FR4, причем
FR1 выравнивается с V генным сегментом зародышевой линии человека IGKV2-28 (например, IGKV2-28*01), IGKV3-20 (например, IGKV3-20*01), IGKV1D-39 (например, IGKV1D-39*01) или IGKV3-11 (например, IGKV3-11*01), необязательно, с 1, 2, 3, 4 или 5 аминокислотными изменениями,
FR2 выравнивается с V генным сегментом зародышевой линии человека IGKV2-28 (например, IGKV2-28*01), IGKV3-20 (например, IGKV3-20*01), IGKV1D-39 (например, IGKV1D-39*01) или IGKV3-11 (например, IGKV3-11*01), необязательно, с 1, 2, 3, 4 или 5 аминокислотными изменениями,
FR3 выравнивается с V генным сегментом зародышевой линии человека IGKV2-28 (например, IGKV2-28*01), IGKV3-20 (например, IGKV3-20*01), IGKV1D-39 (например, IGKV1D-39*01) или IGKV3-11 (например, IGKV3-11*01), необязательно, с 1, 2, 3, 4 или 5 аминокислотными изменениями и/или
FR4 совпадет с J генным сегментом зародышевой линии человека IGKJ4 (например, IGKJ4*01), IGKJ2 (например, IGKJ2*04), IGKJ3 (например, IGKJ3*01) или IGKJ1 (например, IGKJ1*01), необязательно, с 1, 2, 3, 4 или 5 аминокислотными изменениями.
Пункт 14. Антитело согласно любому из пунктов 5-13, содержащее домен VH антитела, который является доменом VH STIM001, STIM002, STIM002-B, STIM003, STIM004, STIM005, STIM006, STIM007, STIM008 или STIM009, или который имеет аминокислотную последовательность, по меньшей мере на 90% идентичную последовательности домена VH антитела STIM001, STIM002, STIM002-B, STIM003, STIM004, STIM005, STIM006, STIM007, STIM008 или STIM009.
Пункт 15. Антитело согласно любому из пунктов 5-14, содержащее домен VL антитела, который является доменом VL STIM001, STIM002, STIM002-B, STIM003, STIM004, STIM005, STIM006, STIM007, STIM008 или STIM009, или который имеет аминокислотную последовательность, по меньшей мере, на 90% идентичную последовательности домена VL антитела STIM001, STIM002, STIM002-B, STIM003, STIM004, STIM005, STIM006, STIM007, STIM008 или STIM009.
Пункт 16. Антитело согласно пункту 15, содержащее
домен VH антитела, который выбран из домена VH STIM001, STIM002, STIM002-B, STIM003, STIM004, STIM005, STIM006, STIM007, STIM008 или STIM009, или который имеет аминокислотную последовательность, по меньшей мере на 90% идентичную последовательности домена VH антитела STIM001, STIM002, STIM002-B, STIM003, STIM004, STIM005, STIM006, STIM007, STIM008 или STIM009, и
домен VL антитела, который является доменом VL указанного выбранного антитела, или который имеет аминокислотную последовательность, по меньшей мере на 90% идентичную последовательности домена VL указанного выбранного антитела.
Пункт 17. Антитело согласно пункту 16, содержащее домен VH STIM003 и домен VL STIM003.
Пункт 18. Антитело согласно любому из предшествующих пунктов, содержащее константную область антитела.
Пункт 19. Антитело согласно пункту 18, в котором константная область содержит константную область тяжелой и/или легкой цепи человека.
Пункт 20. Антитело согласно пункту 18 или пункту 19, причем константная область является положительной по Fc-эффектору.
Пункт 21. Антитело согласно пункту 20, содержащее Fc-область, которая имеет повышенную функцию ADCC, ADCP и/или CDC по сравнению с нативной Fc-областью человека.
Пункт 22. Антитело согласно любому из пунктов 18-21, причем антитело представляет собой IgG1.
Пункт 23. Антитело согласно пункту 21 или пункту 22, причем антитело является афукозилированным.
Пункт 24. Антитело согласно любому из предшествующих пунктов, которое конъюгировано с цитотоксическим лекарственным средством или пролекарством.
Пункт 25. Антитело согласно любому из предшествующих пунктов, которое является мультиспецифическим антителом.
Пункт 26. Выделенное антитело, которое связывает внеклеточный домен ICOS человека и мыши с аффинностью (KD) менее 50 нМ, по результатам определения методом поверхностного плазменного резонанса.
Пункт 27. Антитело согласно пункту 26, в котором антитело связывает внеклеточный домен ICOS человека и мыши с аффинностью (KD) менее 5 нМ, по результатам определения методом поверхностного плазменного резонанса.
Пункт 28. Антитело согласно пункту 26 или пункту 27, причем KD связывания внеклеточного домена ICOS человека находится в пределах 10-кратного значения KD связывания внеклеточного домена ICOS мыши.
Пункт 29. Композиция, содержащая выделенное антитело согласно любому из предшествующих пунктов и фармацевтически приемлемый эксципиент.
Пункт 30. Композиция, содержащая выделенную нуклеиновую кислоту, кодирующую антитело согласно любому из пунктов 1-28, и фармацевтически приемлемый эксципиент.
Пункт 31. Способ модуляции баланса регуляторных Т-клеток (Tregs) и эффекторных Т-клеток (Teffs) для усиления ответа Teff у пациента, включающий введение пациенту антитела согласно любому из пунктов 1-28 или композиции согласно пункту 29.
Пункт 32. Способ лечения заболевания или состояния, поддающегося терапии путем деплетирования регуляторных Т-клеток (Tregs) и/или усиления ответа эффекторных Т-клеток (Teff) у пациента, включающий введение пациенту антитела согласно любому из пунктов 1-28 или композиции согласно пункту 29.
Пункт 33. Антитело согласно любому из пунктов 1-28 или композиция согласно пункту 29 для использования в методе лечения организма человека путем терапии.
Пункт 34. Антитело или композиция для использования согласно пункту 33, предназначенные для использования в модулировании баланса регуляторных Т-клеток (Tregs) в эффекторных Т-клеток (Teffs) для усиления ответа эффекторных Т-клеток у пациента.
Пункт 35. Антитело или композиция для использования согласно пункту 33, предназначенные для использования при лечении заболевания или состояния, восприимчивого к терапии путем деплетирования регуляторных Т-клеток (Tregs) и/или усиления ответа эффекторных Т-клеток (Teff) у пациента.
Пункт 36. Способ согласно пункту 32 или антитело или композиция для использования согласно пункту 35, причем заболевание представляет собой рак или твердую опухоль.
Пункт 37. Антитело согласно любому из пунктов 1-28 или композиция согласно пункту 29 для использования в способе лечения рака у человека.
Пункт 38. Способ лечения рака у человека, включающий введение пациенту антитела согласно любому из пунктов 1-28 или композиции согласно пункту 29.
Пункт 39. Способ, или антитело, или композиция для использования согласно любому из пунктов 36-38, причем рак представляет собой почечно-клеточный рак, рак головы и шеи, меланому, немелкоклеточный рак легкого или диффузную крупноклеточную В-клеточную лимфому.
Пункт 40. Способ, или антитело, или композиция для использования согласно любому из пунктов 31-39, причем способ включает введение пациенту антитела и другого терапевтического агента и/или проведение лучевой терапии.
Пункт 41. Способ, или антитело, или композиция для использования согласно пункту 40, причем терапевтическое средство представляет собой анти-PD-L1 антитело.
Пункт 42. Способ, или антитело, или композиция для использования согласно пункту 41, причем анти-PD-L1-антитело содержит домен VH, имеющий аминокислотную последовательность SEQ ID NO: 299, и домен VL, имеющий аминокислотную последовательность SEQ ID NO: 300.
Пункт 43. Способ, или антитело, или композиция для использования согласно пункту 41 или пункту 42, причем терапевтическое средство представляет собой иммуноцитокин анти-PD-L1 IL-2.
Пункт 44. Способ, или антитело, или композиция для использования согласно пункту 43, причем анти-PD-L1 антитело представляет собой иммуноцитокин, содержащий дикий тип или вариант IL-2 человека.
Пункт 45. Способ, или антитело, или композиция для использования согласно пункту 44, причем анти-ICOS антитело и анти-PDL1 антитело могут каждое опосредовать ADCC, ADCP и/или CDC.
Пункт 46. Способ, или антитело, или композиция для использования согласно любому из пунктов 41-45, причем анти-ICOS антитело представляет собой антитело IgG1 человека, и анти-PDL1 антитело представляет собой антитело IgG1 человека.
Пункт 47. Способ, или антитело, или композиция для использования согласно пункту 40, причем терапевтическое средство представляет собой анти-PDL1 антитело.
Пункт 48. Способ, или антитело, или композиция для использования согласно пункту 40, причем другим терапевтическим агентом является IL-2.
Пункт 49. Способ, или антитело, или композиция для использования согласно любому из пунктов 40-48, причем способ включает введение анти-ICOS антитела после введения другого терапевтического агента и/или проведения лучевой терапии.
Пункт 50. Способ, или антитело, или композиция для использования согласно любому из пунктов 31-49, причем
анти-ICOS антитело конъюгировано с пролекарством, и причем
способ или применение включают
введение пациенту анти-ICOS антитела и
селективную активацию пролекарства в области ткани-мишени.
Пункт 51. Способ, или антитело, или композиция для использования согласно пункту 50, причем у пациента имеется солидная опухоль, и способ включает селективную активацию пролекарства в опухоли.
Пункт 52. Способ, или антитело, или композиция для использования согласно пункту 50 или пункту 51, включающие селективную активацию пролекарства посредством фотоактивации.
Пункт 53. Комбинация анти-ICOS антитела IgG1 человека и анти-PDL1 антитела IgG1 человека для использования в методе лечения рака у пациента.
Пункт 54. Способ лечения рака у пациента, включающий введение анти-ICOS антитела IgG1 человека и анти-PD-L1 антитела IgG1 человека пациенту.
Пункт 55. Анти-ICOS антитело для использования в способе лечения рака у пациента, причем способ включает введение пациенту анти-ICOS антитела и анти-PD-L1 антитела, причем вводят однократную дозу анти-ICOS-антитела.
Пункт 56. Анти-ICOS антитело для использования согласно пункту 55, причем анти-ICOS антитело представляет собой антитело IgG1 человека, и анти-PD-L1 антитело представляет собой антитело IgG1 человека.
Пункт 57. Комбинация согласно пункту 53, способ согласно пункту 54 или анти-ICOS антитело для использования согласно пункту 55 или пункту 56, причем рак представляет собой почечно-клеточный рак, рак головы и шеи, меланому, немелкоклеточный рак легкого или диффузную крупноклеточную В-клеточную лимфому.
Пункт 58. Способ или антитело, композиция или комбинация для использования согласно любому из пунктов 41-46 или 53-54, причем способ включает введение пациенту анти-ICOS антитела и анти-PD-L1 антитела, причем вводят однократную дозу анти-ICOS антитела.
Пункт 59. Способ или антитело, композиция или комбинация для использования согласно пункту 58, причем способ включает введение однократной дозы антитела анти-ICOS с последующими многократными дозами анти-PD-L1 антитела.
Пункт 60. Способ или антитело, композиция или комбинация для использования согласно любому из пунктов 41-46 или 53-54, причем анти-ICOS антитело и анти-PDL1 антитело представлены в отдельных композициях для введения.
Пункт 61. Способ или антитело, композиция или комбинация для использования согласно любому из пунктов 41-46 или 53-60, причем анти-ICOS антитело и/или анти-PD-L1 антитело содержит константную область IgG1 человека, содержащую аминокислотную последовательность SEQ ID NO: 340.
Пункт 62. Анти-ICOS антитело для использования в способе лечения пациента, включающем введение анти-ICOS антитела пациенту с повышенным уровнем ICOS-положительных регуляторных Т-клеток после лечения другим терапевтическим агентом.
Пункт 63. Способ лечения пациента, включающий введение анти-ICOS антитела пациенту с повышенным уровнем ICOS-положительных регуляторных Т-клеток после лечения другим терапевтическим агентом.
Пункт 64. Анти-ICOS антитело для использования согласно пункту 62 или способ согласно пункту 63, причем способ включает введение пациенту терапевтического агента, определение того, что пациент имеет повышенный уровень ICOS-положительных регуляторных Т-клеток после лечения указанным агентом, и введение анти-ICOS антитела пациенту для снижения уровня регуляторных Т-клеток.
Пункт 65. Анти-ICOS антитело для использования или способ согласно любому из пунктов 62-64, причем терапевтический агент представляет собой IL-2 или иммуномодулирующее антитело (например, анти-PDL-1, анти-PD-1 или анти-CTLA-4).
Пункт 66. Анти-ICOS антитело для использования или способ согласно любому из пунктов 62-65, причем способ включает лечение опухоли, например, меланомы, такой как метастазирующая меланома.
Пункт 67. Анти-ICOS антитело для использования в способе лечения рака у пациента путем in vivo вакцинации пациента против его раковых клеток, причем способ включает
лечение пациента терапией, которая вызывает иммунологическую гибель раковых клеток, что приводит к презентации антигена антигенспецифическим эффекторным Т-клеткам и
введение пациенту анти-ICOS антитела, причем анти-ICOS антитело усиливает антигенспецифический эффекторный Т-клеточный ответ.
Пункт 68. Способ лечения рака у пациента путем in vivo вакцинации пациента против его раковых клеток, причем способ включает
лечение пациента терапией, которая вызывает иммунологическую гибель раковых клеток, что приводит к презентации антигена антигенспецифическим эффекторным Т-клеткам и
введение пациенту анти-ICOS антитела, причем анти-ICOS антитело усиливает антигенспецифический эффекторный Т-клеточный ответ.
Пункт 69. Способ лечения рака у пациента путем in vivo вакцинации пациента против его раковых клеток, причем способ включает введение пациенту анти-ICOS антитела, причем
пациент ранее лечился терапией, которая вызывает иммунологическую гибель раковых клеток, что приводит к презентации антигена антигенспецифическим эффекторным Т-клеткам, и причем
анти-ICOS антитело усиливает антигенспецифический эффекторный Т-клеточный ответ.
Пункт 70. Анти-ICOS антитело для использования или способ согласно любому из пунктов 67-69, причем терапия, которая вызывает иммунологическую гибель клеток, представляет собой облучение раковых клеток, введение химиотерапевтического агента и/или введение антитела, нацеленного на опухолеассоциированный антиген.
Пункт 71. Анти-ICOS антитело для использования или способ согласно пункту 70, причем химиотерапевтический агент представляет собой оксалиплатин.
Пункт 72. Анти-ICOS антитело для использования или способ согласно пункту 70, причем опухолеассоциированный антиген представляет собой HER2 или CD20.
Пункт 73. Анти-ICOS антитело для использования в способе лечения рака у пациента, причем рак является или был охарактеризован как положительный по экспрессии лиганда ICOS и/или FOXP3.
Пункт 74. Способ лечения рака у пациента, причем рак является или был охарактеризован как положительный по экспрессии лиганда ICOS и/или FOXP3, причем способ включает введение пациенту анти-ICOS антитела.
Пункт 75. Анти-ICOS антитело для использования согласно пункту 73 или способ согласно пункту 74, причем способ включает:
тестирование образца от пациента для определения того, экспрессирует ли рак лиганд ICOS и/или FOXP3;
выбор пациента для лечения анти-ICOS антителом; и
введение пациенту анти-ICOS антитела.
Пункт 76. Анти-ICOS антитело для использования согласно пункту 73 или способ согласно пункту 74, причем способ включает введение анти-ICOS антитела пациенту, у которого тестируемый образец показал, что рак является положительным по экспрессии лиганда ICOS и/или FOXP3.
Пункт 77. Анти-ICOS антитело для использования или способ согласно пункту 75 или пункту 76, причем образец представляет собой образец биопсии твердой опухоли.
Пункт 78. Анти-ICOS антитело для использования в способе лечения рака у пациента, причем рак является или был охарактеризован как невосприимчивый к лечению иммуноонкологическим лекарственным средством, например, анти-CTLA-4 антителом, анти-PD1 антителом, анти-PD-L1 антителом, анти-CD137 антителом или анти-GITR антителом.
Пункт 79. Способ лечения рака у пациента, у которого рак является или был охарактеризован как невосприимчивый к лечению иммуноонкологическим лекарственным средством, например, анти-CTLA-4 антителом, анти-PD1 антителом, анти-PD-L1 антителом, анти-CD137 антителом или анти-GITR антителом, причем способ включает введение пациенту анти-ICOS антитела.
Пункт 80. Анти-ICOS антитело для использования согласно пункту 78 или способ согласно пункту 79, причем способ включает:
лечение пациента иммуноонкологическим препаратом;
определение того, что рак не реагирует на препарат;
выбор пациента для лечения анти-ICOS антителом; и
введение пациенту анти-ICOS антитела.
Пункт 81. Анти-ICOS антитело для использования согласно пункту 78 или способ согласно пункту 79, причем способ включает введение анти-ICOS антитела пациенту, у которого рак не отвечает на предшествующее лечение иммуноонкологическим препаратом.
Пункт 82. Анти-ICOS антитело для использования или способ согласно любому из пунктов 73-81, причем рак представляет собой опухоль, образующуюся из клеток, которые приобрели способность экспрессировать лиганд ICOS.
Пункт 83. Анти-ICOS антитело для использования или способ согласно пункту 82, причем рак представляет собой меланому.
Пункт 84. Анти-ICOS антитело для использования или способ согласно любому из пунктов 73-81, причем рак образуется из антигенпрезентирующей клетки, такой как В-лимфоцит (например, В-клеточная лимфома, такая как диффузная крупноклеточная В-клеточная лимфома) или Т-лимфоцит.
Пункт 85. Анти-ICOS антитело для использования или способ согласно любому из пунктов 73-81, причем рак является невосприимчивым к лечению анти-CD20 антителом.
Пункт 86. Анти-ICOS антитело для использования или способ согласно пункту 85, причем рак представляет собой В-клеточную лимфому.
Пункт 87. Анти-ICOS антитело для использования или способ согласно пункту 86, причем анти-CD20 антитело представляет собой ритуксимаб.
Пункт 88. Анти-ICOS антитело для использования или способ согласно любому из пунктов 85-87, причем способ включает лечение пациента анти-CD20 антителом;
определение того, что рак не реагирует на анти-CD20 антитело;
тестирование образца пациента для определения того, экспрессирует ли рак лиганд ICOS;
выбор пациента для лечения анти-ICOS антителом; и
введение пациенту анти-ICOS антитела.
Пункт 89. Анти-ICOS антитело для использования или способ согласно любому из пунктов 85-87, причем способ включает введение анти-ICOS антитела пациенту, у которого рак не отвечает на предшествующее лечение анти-CD20 антителом.
Пункт 90. Анти-ICOS антитело для использования или способ согласно любому из пунктов 67-89, причем рак представляет собой твердую опухоль.
Пункт 91. Анти-ICOS антитело для использования или способ согласно любому из пунктов 67-89, причем рак представляет собой гематологическую жидкую опухоль.
Пункт 92. Анти-ICOS антитело для использования или способ согласно пункту 90 или 91, причем опухоль имеет высокое содержание регуляторных Т-клеток.
Пункт 93. Анти-ICOS антитело для использования или способ согласно любому из пунктов 53-92, причем анти-ICOS антитело является таким, как определено в любом из пунктов 1-28 или представлено в композиции согласно пункту 29.
Пункт 94. Трансгенное млекопитающее, не являющееся человеком, имеющее геном, содержащий человеческий или гуманизированный локус иммуноглобулина, кодирующий генные сегменты вариабельной области человека, причем млекопитающее не экспрессирует ICOS.
Пункт 95. Способ получения антитела, которое связывает внеклеточный домен человеческого и нечеловеческого ICOS, включающий
(a) иммунизацию млекопитающего согласно пункту 94 антигеном ICOS человека;
(b) выделение антител, продуцируемых млекопитающим;
(c) тестирование антител на способность связывать человеческий и нечеловеческий ICOS; и
(d) выбор одного или нескольких антител, которые связывают как человеческий, так и нечеловеческий ICOS.
Пункт 96. Способ согласно пункту 95, включающий иммунизацию млекопитающего клетками, экспрессирующими ICOS человека.
Пункт 97. Способ согласно пункту 95 или пункту 96, включающий
(с) тестирование антител на способность связывать человеческие ICOS и нечеловеческие ICOS с использованием метода поверхностного плазмонного резонанса и определения аффинностей связывания; и
(d) выбор одного или нескольких антител, для которых KD связывания с человеческим ICOS составляет менее 50 нМ, a KD связывания с нечеловеческим ICOS составляет менее 500 нМ.
Пункт 98. Способ согласно пункту 97, включающий
(d) выбор одного или нескольких антител, для которых KD связывания с человеческим ICOS составляет менее 10 нМ, a KD связывания с нечеловеческим ICOS составляет менее 100 нМ.
Пункт 99. Способ согласно любому из пунктов 95-98, включающий
(с) тестирование антител на способность связывать человеческие ICOS и нечеловеческие ICOS с использованием метода поверхностного плазмонного резонанса и определения аффинностей связывания; и
(d) выбор одного или более антител, для которых KD связывания с человеческим ICOS имеет значение в пределах 10-кратного KD связывания с нечеловеческим ICOS.
Пункт 100. Способ согласно пункту 99, включающий
(d) выбор одного или более антител, для которых KD связывания с человеческим ICOS имеет значение в пределах 5-кратного KD связывания с нечеловеческим ICOS.
Пункт 101. Способ согласно любому из пунктов 95-100, включающий тестирование антител на способность связывать нечеловеческие ICOS от того же вида, к которому относится млекопитающие.
Пункт 102. Способ согласно любому из пунктов 95-101, включающий тестирование антител на способность связывать нечеловеческие ICOS от вида, отличного от того, к которому относится млекопитающее.
Пункт 103. Способ согласно любому из пунктов 95-102, в котором млекопитающее представляет собой мышь или крысу.
Пункт 104. Способ согласно любому из пунктов 95-103, в котором нечеловеческий ICOS представляет собой ICOS мыши или ICOS крысы.
Пункт 105. Способ согласно любому из пунктов 95-104, в котором человеческий или гуманизированный локус иммуноглобулина включает генные сегменты вариабельной области человека перед (upstream) эндогенной константной областью.
Пункт 106. Способ согласно пункту 105, включающий
(a) иммунизацию млекопитающего согласно пункту 94 антигеном ICOS человека, причем млекопитающее представляет собой мышь;
(b) выделение антител, продуцируемых мышью;
(c) тестирование антител на способность связывать ICOS человека и ICOS мыши; и
(d) выбор одного или нескольких антител, которые связывают ICOS как человека, так и мыши.
Пункт 107. Способ согласно любому из пунктов 95-106, включающий выделение нуклеиновой кислоты, кодирующей вариабельный домен тяжелой цепи антитела и/или вариабельный домен легкой цепи антитела.
Пункт 108. Способ согласно любому из пунктов 95-107, в котором млекопитающее генерирует антитела посредством рекомбинации генных сегментов вариабельной области человека и эндогенной константной области.
Пункт 109. Способ согласно пункту 107 или пункту 108, включающий конъюгирование нуклеиновой кислоты, кодирующей вариабельный домен тяжелой и/или легкой цепи, с нуклеотидной последовательностью, кодирующей константную область тяжелой цепи человека и/или константную область легкой цепи человека, соответственно.
Пункт 110. Способ согласно любому из пунктов 107-109, включающий введение нуклеиновой кислоты в клетку-хозяина.
Пункт 111. Способ согласно пункту 110, включающий культивирование клетки-хозяина в условиях экспрессии антитела или вариабельного домена тяжелой и/или легкой цепи антитела.
Пункт 112. Антитело или вариабельный домен тяжелой и/или легкой цепи антитела, продуцируемые способом согласно любому из пунктов 95-111.
Пункт 113. Способ выбора антитела, которое связывает ICOS, необязательно, для выбора агонистического антитела ICOS, причем анализ включает:
обеспечение массива антител, иммобилизованных (присоединенных или прикрепленных) к субстрату в тестовой лунке;
добавление ICOS-экспрессирующих клеток (например, активированных первичных Т-клеток или клеток MJ) в тестовую лунку;
наблюдение морфологии клеток;
детектирование изменения формы в лунках от округлой до сплюснутой к подложке внутри лунки; причем изменение формы указывает, что антитело представляет собой антитело, которое связывает ICOS, необязательно, антитело-агонист ICOS;
выбор антитела из тестовой лунки;
экспрессию нуклеиновой кислоты, кодирующей CDR выбранного антитела; и
составление композиции антитела, содержащей один или несколько дополнительных компонентов.
Различные дополнительные аспекты и варианты реализации данного изобретения будут очевидны для специалистов в данной области с учетом описания, приведенного в данном документе. Все документы, упомянутые в этом описании, включая опубликованные американские аналоги любых патентов или патентных заявок, упомянутых в данном документе, включены в данное описание посредством ссылок в полном объеме.
Экспериментальные примеры
В приведенных далее примерах описываются получение, характеризация и эффективность анти-ICOS антител. Антитела генерировали с использованием Kymouse™, трансгенной мышиной платформы, способной генерировать антитела с вариабельными доменами человека. Антитела из Kymouse™ имеют вариабельные домены человека, полученные из V (D) и J сегментов человека, и константных доменов мыши. Эндогенные вариабельные гены мыши были отключены и составляют очень небольшую часть репертуара (менее 0,5% всех вариабельных областей тяжелой цепи имеют мышиное происхождение). Система Kymouse™ описана в Lee et al. 2014 [39], WO 2011/004192, WO 2011/158009 и WO 2013/061098. В этом проекте использовался штамм Kymouse™ НК, в котором локус тяжелой цепи и локус каппа легкой цепи были гуманизированы.
ICOS-нокаутных Kymouse™ иммунизировали либо белком ICOS, либо комбинацией чередующихся бустерных доз белка и клеток, экспрессирующих ICOS человека и мыши.
Были идентифицированы первично отобранные антитела, связывающиеся с человеческим ICOS. Первичными критериями отбора для скрининга были связывание с экспрессируемым клетками ICOS человека (клетки СНО) и связывание с белком ICOS (HTRF). Также оценивали связывание с мышиным белком ICOS и экспрессируемым клетками ICOS мыши (клетки СНО), и принимали во внимание при первичном отборе антител скринингом. Соответствующие этим критериям первично отобранные антитела использовались для вторичного скрининга. Отбор антител во вторичном скрининге подтверждался определением связывания с ICOS человека и мыши, экспрессируемыми на клетках СНО, методом проточной цитометрии.
Из большого количества подвергнутых скринингу антител была идентифицирована небольшая панель, которая связывается с ICOS человека/яванского макака и мыши, по результатам определения методами поверхностного плазмонного резонанса и проточной цитометрии. Эти антитела включали STIM001, STIM002 и его вариант STIM002-B, STIM003, STIM004 и STIM005. Также были отобраны еще четыре антитела STIM006, STIM007, STIM008 и STIM009, имеющие меньшую перекрестную реактивность с ICOS мыши, но демонстрирующие агонизм к рецептору ICOS человека. Данные, представленные в данном документе, указывают на способность анти-ICOS антител выступать в роли агонистов рецептора ICOS в ICOS-положительной CD4+ клеточной линии, а также в анализе на основе первичных Т-клеток, показывают способность убивать клетки в анализе ADCC и способность промотировать противоопухолевый иммунный ответ in vivo.
Пример 1: Создание ICOS-нокаутной мыши
ICOS-нокаутная линия Kymouse™ была создана путем гомологичной рекомбинации в клетках Kymouse™ НК ES. Вкратце, нацеливающий вектор размером 3,5 т.п.н., кодирующий селекцию пуромицином, был введен в ES-клетки. Успешное прицельное введение гена привело к замене небольшого участка (72 п.о.) локуса ICOS мыши на пуромициновую кассету, с разрывом сигнального пептида/стартового кодона гена. Положительные клоны ES были размножены и введены микроинъекциями в мышиные бластоцисты с образованием в результате химер, для получения, в конечном итоге, животных, гомозиготных как по локусам гуманизированных тяжелой цепи и цепи каппа иммуноглобулина, так и по модифицированному нуль-функциональному локусу ICOS.
Пример 2: Получение антигена и клеточной линии
Создание стабильно трансфицированных клеток MEF и CHO-S, экспрессирующих ICOS человека или мыши
Полноразмерные ДНК-последовательности, кодирующие ICOS человека и мыши, были оптимизированы по использованию кодонов для экспрессии у млекопитающих, заказаны в виде синтезированной цепочки ДНК, и клонированы в экспрессионный вектор под контролем промотора CMV с фланкирующими 3'- и 5'-piggyBac-специфическими терминальными повторными последовательностями, способствующими стабильной интеграции в геном клетки (см. [40]). Вектор экспрессии содержал кассету для селекции пуромицином для облегчения создания стабильной клеточной линии. Для создания клеточных линий, экспрессирующих ICOS человека и ICOS мыши, соответственно, человеческую или мышиную экспрессионную плазмиду ICOS ко-трансфицировали с плазмидой, кодирующей транспозазу piggyBac, в клеточную линию эмбриональных фибробластов мыши (MEF) и клетки CHO-S с использованием реагента для трансфекции FreeStyle Max (Invitrogen) в соответствии с инструкциями производителя. Клетки MEF были получены из эмбрионов, полученных от 129S5, скрещенного с самкой мыши C57BL6. Через двадцать четыре часа после трансфекции к среде добавляли пуромицин и выращивали в течение по меньшей мере двух недель для отбора стабильных клеточных линий. Среду клеточной культуры заменяли каждые 3-4 дня. Экспрессию белка ICOS человека или мыши оценивали методом проточной цитометрии с использованием конъюгированных с РЕ антител против человеческого или антимышиного ICOS (eBioscience), соответственно. Полные среды MEF состояли из модифицированной по Дульбекко среды Игла (Gibco), дополненной 10% об./об. фетальной бычьей сыворотки (Gibco). Полная среда CHO-S состояла из среды CD-CHO с добавкой 8 мМ глутамакса (Gibco). Клетки CHO-S представляют собой клеточную линию СНО-3Е7 с включенной системой рТТ5, доступную от Национального научно-исследовательского совета Канады (National Research Council of Canada), но могут быть использованы и другие клеточные линии СНО.
Приготовление клеток MEF для иммунизации мышей
Слой клеточной культуры удаляли и клетки промывали один раз 1х PBS. Клетки обрабатывали в течение 5 минут трипсином для отделения клеток от поверхности тканевой культуры. Клетки собирали и трипсин нейтрализовали добавлением полной среды, содержащей 10% об./об. фетальной бычьей сыворотки (FCS). Клетки затем центрифугировали при 300g в течение 10 минут и промывали 25 мл 1х PBS. Клетки подсчитывали и ресуспендировали при соответствующей концентрации в 1х PBS.
Клонирование и экспрессия рекомбинантных белков
Синтетическую ДНК, кодирующую внеклеточные домены ICOS человека (NCBI ID: NP_036224.1), ICOS мыши (NCBI ID: NP_059508.2) и ICOS яванского макака (GenBank ID: EHH55098.1), клонировали в экспрессионную плазмиду pREP4 (Invitrogen), либо рТТ5 (National Research Council of Canada) с использованием стандартных методов молекулярной биологии. Конструкты также содержали человеческий Fc, мышиный Fc или пептидный мотив FLAG His для облегчения очистки и детектирования. Их добавляли к конструктам ДНК путем амплификации перекрывающихся последовательностей. Все конструкты были секвенированы перед экспрессией для обеспечения правильного состава последовательностей.
Пример 3: Иммунизация
Иммунизировали ICOS-нокаутные HK Kymice™ (см. Пример 1), штамм HK Kymouse™ дикого типа и штамм HL Kymouse™ дикого типа в соответствии со схемами, приведенными в Таблице Е3. Штаммы HK и HL Kymouse™ дикого типа экспрессируют мышиный ICOS дикого типа. В штамме НК локус тяжелой цепи иммуноглобулина и локус легкой цепи каппа гуманизированы, а в штамме HL гуманизированы локус тяжелой цепи иммуноглобулина и локус легкой цепи лямбда.
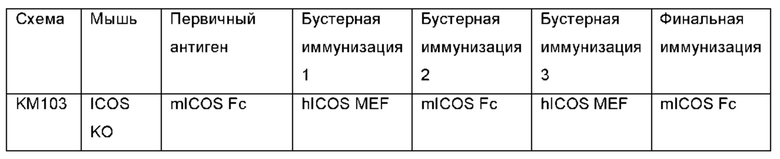
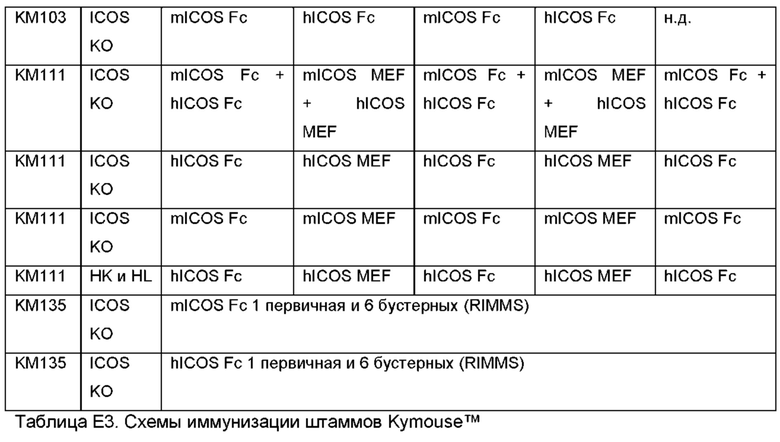
Ключ к таблице:
mICOS Fc = белок ICOS мыши с человеческим Fc
hICOS Fc = белок ICOS человека с человеческим Fc
mICOS MEF = ICOS мыши, экспрессируемый на клетках MEF
hICOS MEF = ICOS человека, экспрессируемый на клетках MEF
mICOS Fc + hICOS Fc = белок ICOS мыши с человеческим Fc + белок ICOS человека с человеческим Fc, вводимые одновременно
mICOS MEF + hICOS MEF=ICOS мыши, экспрессируемый на клетках MEF + ICOS человека, экспрессируемый на клетках MEF, вводимые одновременно
ICOS КО = ICOS-нокаутные HK Kymouse
HK и HL = генотип HK и HL Kymouse дикого типа
RIMMS представляет собой модифицированную процедуру подкожной иммунизации (быстрая иммунизация на нескольких участках); модифицирована в соответствии с Kilpatrick et al. [41]). Схемы иммунизации KM103 и KM111 включали первичную иммунизацию-отдых-бустерную иммунизации при внутрибрюшинном (ip) введении. Система Sigma Adjuvant использовалась для всех иммунизации, а интервалы отдыха обычно составляли от 2 до 3 недель. Финальную иммунизацию проводили внутривенно без адъюванта.
Сыворотки из серийных или терминальных образцов крови анализировали на присутствие специфических антител методом проточной цитометрии, и данные титров использовались (когда это было возможно) для отбора мышей, предназначенных для использования при сортировке В-клеток.
Пример 4: Сравнение титров сыворотки между мышами ICOS КО (нокаутными) и дикого типа
Титры сыворотки иммунизированных ICOS KO и иммунизированных Kymouse дикого типа определяли с использованием проточной цитометрии. У мышей ICOS KO иммунизация антигеном ICOS человека индуцировала иммуноглобулиновый ответ сыворотки со связыванием Ig с ICOS человека и мыши, экспрессируемым на клетках СНО (Фиг. 1а). Напротив, у Kymouse дикого типа (экспрессирующих ICOS мыши) иммунизация тем же самым антигеном ICOS человека продуцировала сыворотки, которые показали заметно сниженное связывание с ICOS мыши по сравнению с связыванием той же самой сыворотки с ICOS человека (Фиг. 1b).
Метод
CHO-S, экспрессирующие ICOS человека или ICOS мыши (см. Пример 2) или нетрансфицированные клетки CHO-S (называемые диким типом (WT)), суспендированные в буфере FACS (PBS+1% мас./об. BSA+0,1% мас./об. азида натрия), распределяли на 96-луночном планшете с V-образными лунками (Greiner) с плотностью 105 клеток на лунку. Титрование сыворотки мыши готовили, разбавляя образцы буфером FACS. Затем добавляли 50 мкл/лунку этого титрования в планшет с клетками. Чтобы определить изменение уровня активности вследствие иммунизации, сыворотку от каждого животного до иммунизации разбавляли до 1/100 в буфере FACS и добавляли 50 мкл/лунку к клеткам. Клетки инкубировали при 4°С в течение 1 часа. Клетки дважды промывали 150 мкл PBS, центрифугируя после каждой стадии промывки и удаляя супернатант аспирацией (центрифугирование при 300×g в течение 3 минут). Для детектирования связывания антител, козий антимышиный IgG АРС (антигенпрезентирующих клеток) (Jackson ImmunoResearch) разбавляли 1/500 в буфере FACS и добавляли к клеткам 50 мкл. В некоторых случаях использовали козий антимышиный IgG AF647 (Jackson ImmunoResearch). Клетки инкубировали 1 час при 4°С в темноте, затем дважды промывали 150 мкл PBS, как описано выше. Для фиксации клеток добавляли 100 мкл 2% об./об. параформальдегида и клетки инкубировали в течение 30 минут при 4°С. Затем клетки осаждали центрифугированием при 300×g и планшеты ресуспендировали в 50 мкл буфера FACS. Интенсивность флуоресцентного сигнала (среднее геометрическое) измеряли методом проточной цитометрии с использованием прибора BD FACS Array.
Пример 5: Сортировка антигенспецифических В-клеток с помощью FACS
В-клетки, экспрессирующие анти-ICOS антитела, были выделены у иммунизированных мышей с использованием способов, по существу, описанных в Примере 1 WO 2015/040401. Вкратце, спленоциты и/или клетки лимфатических узлов, выделенные при проведении схем иммунизации, окрашивали коктейлем антитела, содержащим маркеры для выбора представляющих интерес клеток (CD19), тогда как нежелательные клетки были исключены из конечной сортированной популяции (IgM, IgD, 7AAD). CD19+ В-клетки дополнительно метили флуоресцентно мечеными димерами человеческий ICOS-ECD-Fc и флуоресцентно меченым мышиным ICOS ECD-Fc для детектирования В-клеток, продуцирующих анти-ICOS антитела. Для флуоресцентного мечения ICOS человека и мыши использовали AlexaFluor647 и AlexaFluor488, соответственно - см. Пример 6. Были отобраны клетки, связывающие ICOS человека или ICOS как человека, так и мыши. Эти клетки сортировали по одной методом FACS в буфере для лизиса. Последовательности V-области были выделены с использованием ОТ-ПЦР и двух дополнительных циклов ПЦР, затем были присоединены мостиковыми связями к константной области IgG1 мыши и экспрессированы в клетках HEK293. Супернатанты клеток HEK293 подвергали скринингу на наличие связывания ICOS и функциональных антител. Этот метод далее упоминается как ВСТ.
Пример 6: Скрининг антител из ВСТ
HTRF-скрининг супернатантов ВСТ на связывание с рекомбинантным ICOS-Fc человека и мыши
Супернатанты, собранные из ВСТ в Примере 5, подвергали скринингу на способность секретируемых антител связываться с Fc ICOS человека и Fc ICOS мыши, экспрессируемым в виде рекомбинантных белков. Связывание секретируемых антител с рекомбинантными ICOS человека и мыши идентифицировали методом HTRF® (гомогенная флуоресценция с временным разрешением, Cisbio) с использованием ICOS, меченого FluoProbes®647H (Innova Biosciences) (в данном документе обозначается 647 hICOS или 647 mICOS для ICOS человека и ICOS мыши, меченых FluoProbes®647H, соответственно). 5 мкл супернатанта ВСТ переносили на белый 384-луночный полистирольный планшет с лунками малого объема и несвязывающей поверхностью (Greiner). В каждую лунку добавляли 5 мкл 20 нМ 647 hICOS или 647 mICOS, разбавленных в буфере для анализа HTRF. Для анализа связывания ICOS человека эталонное антитело разбавляли в среде ВСТ (Gibco №А14351-01) до 120 нМ и добавляли 5 мкл в планшет. В лунки отрицательного контроля для анализа связывания ICOS человека (добавляли) 5 мкл мышиного IgG1 (Sigma М9269, в некоторых случаях обозначаемого как СМ7), разбавленного до 120 нМ в среде ВСТ. В случае анализа связывания ICOS мыши эталонное антитело разбавляли в среде ВСТ (Gibco №А14351-01) до 120 нМ и добавляли 5 мкл в планшет. Изотипический контроль IgG2b крысы (R&D Systems) добавляли в лунки отрицательного контроля (R&D Systems) разведенным в среде ВСТ до 120 нМ, и 5 мкл добавляли в планшет. Связывание секретируемых антител к ICOS человека детектировали путем добавления 10 мкл козьего антимышиного IgG (Southern Biotech), непосредственно меченого с помощью криптата европия (Cisbio), разбавленного 1/2000 в буфере для анализа HTRF. В случае анализа связывания ICOS мыши 5 мкл мышиного антитела против IgG2B-UBLB мыши (Southern Biotech) добавляли в лунки положительного и отрицательного контроля и 5 мкл буфера для анализа HTRF добавляли во все остальные лунки планшета. Затем для детектирования связывания добавляли 5 мкл козьего антимышиного IgG (Southern Biotech), непосредственно меченого криптатом европия (Cisbio), разбавленным 1/1000 в буфере для анализа HTRF. Планшет оставляли для инкубации в темноте в течение 2 часов до считывания флуоресценции с временным разрешением при длинах волн излучения 620 нм и 665 нм, 100 вспышек, с использованием планшет-ридера EnVision (Perkin Elmer).
Данные анализировали путем расчета соотношения 665/620 и процента эффекта для каждого образца по уравнению 2 и уравнению 1, соответственно.
Для KM103 и KM11-В1, первичный отбор антител осуществляли на основе эффекта, равного или превышающего 5%, для связывания с ICOS человека и мыши. Для KM135, первичный отбор антител осуществляли на основе эффекта, равного или превышающего 10%, для связывания с ICOS человека и мыши. Для KM111-В2, первичный отбор антител осуществляли на основе эффекта, равного или превышающего 4 процента для связывания с ICOS человека, и эффекта, равного или превышающего 3%, для связывания с ICOS мыши.
Уравнение 1: Расчет процента эффекта при первичном скрининге связывания клеток с помощью Envision и HTRF
Используют величину отношения для лунок (well ratio value) (уравнение 3) или соотношение 665/620 нм (см. уравнение 2) (HTRF)

Неспецифическое связывание = значения для лунок, содержащих изотипический контроль мышиного IgG1
Полное связывание = значения для лунок, содержащих эталонное антитело
Уравнение 2: Расчет отношения 665/620
Отношение 665/620 = (значение 665/620 нм для образца)×10000
Уравнение 3: Расчет соотношения 647/FITC
Данные сначала нормируют по числу клеток путем деления канала mAb (647) на канал FITC (окрашивание клеток) для получения «величины отношения для лунки»:

Скрининг супернатантов ВСТ на связывание с экспрессируемыми клетками ICOS человека и мыши
Супернатанты, собранные из ВСТ в Примере 5, подвергали скринингу на способность секретируемых антител связываться с ICOS человека или мыши, экспрессируемыми на поверхности клеток CHO-S. Для определения связывания человеческого и мышиного ICOS CHO-S, клетки высеивали в 384-луночные обработанные тканевой культурой планшеты с черными стенками и прозрачным дном (Perkin Elmer) при 4×104/лунку в среде F12 (Gibco), дополненной 10% FBS (Gibco), и культивировали в течение ночи. Культуральную среду удаляли из 384-луночных планшетов для анализа. В каждую лунку добавляли по меньшей мере 50 мкл супернатанта ВСТ или 50 мкл эталонного антитела с концентрацией 2 мкг/мл в среде ВСТ, или изотипического контрольного антитела IgG1 (в некоторых случаях обозначается как Cm7, Sigma М9269) при конечной концентрации 2 мкг/мл), разбавленного в среде ВСТ. Планшеты инкубировали в течение 1 часа при 4°С. Супернатант удаляли аспирацией и 50 мкл козьего антимышиного 647 (Jackson immunoresearch) при концентрации 5 мкг/мл с ярко-зеленым (vibrant green) красителем для ДНК (Life Technologies), разведенным 1 к 500 во вторичном буфере для антител (1х PBS+1% BSA+0,1% азида натрия) добавляли для детектирования связывания антитела и визуализации клеток. Планшеты инкубировали в течение 1 ч при 4 градусах. Супернатант удаляли аспирацией, добавляли 25 мкл 4% об./об. параформальдегида и планшеты инкубировали в течение 15 минут при комнатной температуре. Планшеты дважды промывали 100 мкл PBS и затем промывочный буфер полностью удаляли. Интенсивность флуоресценции измеряли с использованием планшет-ридера Envision (Perkin Elmer), измеряющего FITC (возбуждение 494 нм, излучение 520 нм) и alexafluor 647 (возбуждение 650 нм, эмиссия 668 нм). Определяли сигнал анализа, как описано в уравнении 3, и процент эффекта, как в уравнении 1. Полное связывание определяли с использованием эталонного антитела при конечной концентрации анализа 2 мкг/мл. Неспецифическое связывание определяли с использованием изотипического контроля IgG1 мыши (Sigma) при конечной концентрации анализа 2 мкг/мл. Критерии первичного отбора антител были основаны на сигнале анализа и проценте эффекта.
Для KM103, KM111-В1 и KM135, первичный отбор антител осуществлялся на основе эффекта, равного или превышающего 10 процентов. Для KM111-В2, первичный отбор антител осуществлялся на основе эффекта, равного или превышающего 4 процента.
Сводные результаты первичного скрининга
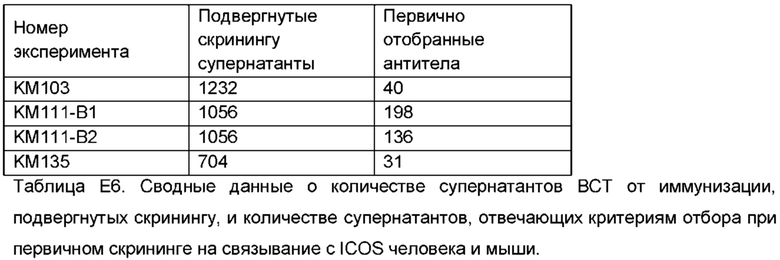
Скрининг методом FACS на связывание с экспрессируемыми клетками ICOS человека и мыши
Супернатанты ВСТ и экспрессированные HEK293 антитела из Примера 5 тестировали на способность связываться с клетками CHO-S, экспрессирующими ICOS человека или мыши.
Клетки CHO-S, экспрессирующие ICOS человека или мыши (см. Пример 2), разводили в буфере FACS (PBS, 1% BSA, 0,1% азида натрия) и распределяли на 96-луночный планшет с V-образными лунками (Greiner) с плотностью 1×105 клеток на лунку. Клетки промывали 150 мкл PBS и центрифугировали при 300 g в течение 3 минут. Для скрининга супернатанта, супернатант отбирали аспирацией и добавляли 150 мкл PBS. Эта стадия промывки повторялась. К промытым клеткам добавляли 30 мкл неразбавленного супернатанта ВСТ или 50 мкл эталонного антитела или контрольного антитела, разбавленного до 5 мкг/мл в среде ВСТ. Клетки инкубировали при 4°С в течение 60 минут. Добавляли 150 мкл буфера FACS и клетки промывали, как описано выше. Для детектирования связывания антител, к клеткам добавляли 50 мкл козьего антимышиного АРС (Jackson ImmunoResearch), разбавленного до 2 мкг/мл в буфере FACS. Клетки инкубировали 4°С в течение 60 минут. Клетки дважды промывали 150 мкл буфера FACS, центрифугировали при 300 g в течение 3 минут после каждой стадии промывки и удаляли супернатант аспирацией. Клетки фиксировали добавлением 25 мкл 4% параформальдегида в течение 20 минут при комнатной температуре. Клетки промывали один раз, как описано выше, и ресуспендировали в буфере FACS для анализа. Интенсивность сигнала АРС (среднее геометрическое) измеряли методом проточной цитометрии с использованием прибора BD FACS Array. Графики данных строили для средних геометрических значений без дальнейших расчетов.
Небольшая подгруппа антител была выбрана как отвечающие более строгим критериям видовой перекрестной реактивности в этом дополнительном скрининге, по сравнению с первичным скринингом. Вкратце:
Из KM103 было отобрано 4 антитела путем определения усредненного значения геометрического среднего для контрольного связывания гибрида с клетками hICOS, mICOS и СНО WT и идентификации антител, имеющих показатели связывания с ICOS мыши и человека, превышающие это значение более чем в 4 раза. Эти 4 антитела были обозначены как STIM001, STIM002-B, STIM007 и STIM009.
Из KM111-В1 было выбрано 4 антитела путем определения усредненного значения геометрического среднего для связывания отрицательного контроля (серый хомячок (Armenian hamster): клон HTK888), с клетками NCOS, mICOS и СНО WT, и идентификации антител, имеющих показатели связывания с ICOS мыши и человека, превышающие это значение более чем в 10 раз.
Из KM111-В2 было выбрано 4 антитела путем определения усредненного значения геометрического среднего для связывания отрицательного контроля (серый хомячок (Armenian hamster): клон HTK888), с клетками NCOS, mICOS и СНО WT, и идентификации антител, имеющих показатели связывания с ICOS мыши и человека, превышающие это значение более чем в 4 раза. Эти 4 антитела включали STIM003, STIM004 и STIM005.
Из KM135 не было выявлено никаких перекрестно-реактивных антител. Из-за технического сбоя метода вторичного скрининга FACS, скрининг также проводили с использованием методов SPR и HTRF, но при этом не было обнаружено антител, соответствующих желательному уровню перекрестной реактивности.
В заключение укажем, что, из различных схем множественной иммунизации, описанных в Примере 3, было подвергнуто скринингу более 4000 супернатантов ВСТ (только для мышей ICOS KO) на связывание с ICOS человека и ICOS мыши, и небольшая панель кандидатов, включающая STIM001, STIM002-B, STIM003, STIM004, STIM005, STIM007 и STIM009, была идентифицирована как имеющие наиболее перспективные характеристики для дальнейшей разработки. Они были использованы для проведения более детальной характеризации.
Отдельно были также выбраны два антитела STIM006 и STIM008, которые не соответствовали критериям видовой перекрестной реактивности, для дополнительной характеризации на основе их способности связывать ICOS человека.
Пример 7: Определение аффинности методом поверхностного плазмонного резонанса (SPR)
Аффинности Fab оптимизированных прототипов антитела ICOS определяли методом SPR с использованием ProteOn XPR3 6 (BioRad). Поверхность захвата антитела против человеческого IgG была создана на чипе биосенсора GLC с использованием реакции сочетания с первичным амином, с иммобилизацией трех антител против человеческого IgG (Jackson Labs 109-005-008,109-006-008 и 309-006-008). ICOS человека (hICOS) и ICOS мыши (mICOS), меченые человеческим Fc, захватывали индивидуально на поверхности антитела против человеческого IgG, и очищенные Fab использовали в качестве аналитов с концентрациями 5000 нМ, 1000 нМ, 200 нМ, 40 нМ и 8 нМ, за исключением STIM003, который использовался с концентрациями 1000 нМ, 200 нМ, 40 нМ, 8 нМ и 2 нМ. Нулевое значение для сенсограмм связывания определяли по двум холостым образцам с инъекцией буфера (т.е. 0 нМ), и данные аппроксимировали с использованием модели 1:1, включенной в программу для анализа данных ProteOn XPR36. Анализ проводили при 25°С и использовали HBS-EP в качестве подвижного буфера.

Кроме того, было проведено сравнение антител: аффинность связывания антигена при разных значениях рН. Как и ранее, димерный белок ICOS человека, представленный в виде внеклеточного домена ICOS, слитого с Fc-областью человека, был захвачен на поверхности захвата против Fc человека, созданной с использованием коктейля 3 антител, иммобилизованных на чипе биосенсора GLC с использованием реакции сочетания первичного амина. SPR-анализ рекомбинантно экспрессированных анти-ICOS Fab проводили в системе ProteOn XPR36 Array (Biorad). Фрагменты Fab использовали в качестве аналита для получения сенсограмм связывания, нулевое значение для которых определяли по двум холостым образцам с впрыскиванием буфера (т.е. 0 нМ). Последующие откалиброванные сенсограммы аппроксимировали моделью 1:1, встроенной в программу анализа данных ProteOn. В таблице Е7-2 представлены аффинность и кинетические данные для антител, все данные определены при 37°С, если не указано иное, с использованием либо HBS-EP при рН 7,4/7,6, либо рН 5,5, как указано. Данные аппроксимировали моделью 1:1. Следует отметить, что данные для STIM002 плохо аппроксимировались моделью 1:1 при рН 7,4 и 5,5 - аффинность этого антитела может быть ниже, чем указано в таблице.
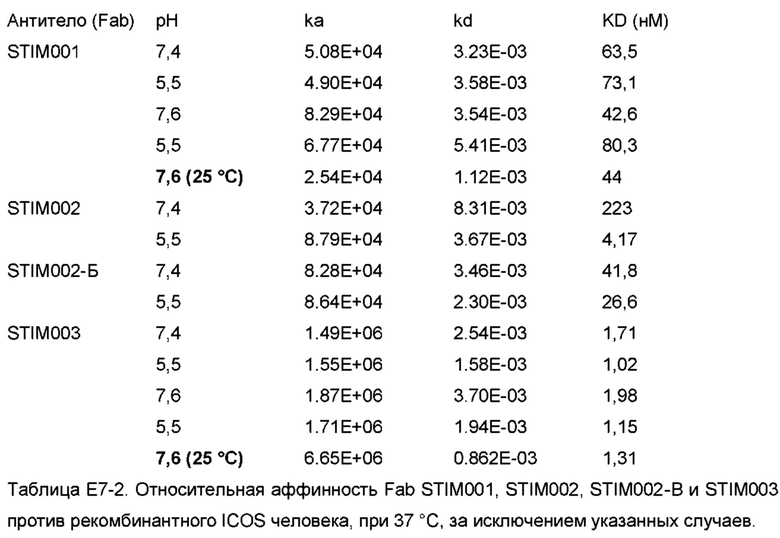
Сравнение данных аффинности при разных значениях рН указывает на то, что антитела сохраняют связывание со своими мишенями в физиологическом диапазоне рН. Микроокружение опухоли может быть относительно кислым по сравнению с кровью, поэтому поддержание аффинности при низком значении рН является потенциальным преимуществом in vivo для улучшения внутриопухолевого деплетирования T-reg.
Пример 8: Нейтрализация связывания лиганда ICOS с рецептором ICOS, по результатам анализа методом HTRF
Отдельные анти-ICOS антитела дополнительно оценивали на их способность нейтрализовать связывание лиганда ICOS (В7-Н2) с ICOS с использованием метода гомогенной флуоресценции с временным разрешением (HTRF). Изотипы моноклональных антител (mAb) IgG1 человека и IgG4.PE человека (нуль-эффекторные) оценивали путем:
- анализа нейтрализации связывания В7-Н2 человека с ICOS человека методом HTRF; и
- анализа нейтрализации связывания В7-Н2 мыши с ICOS мыши методом HTRF.
Для сравнения было включено анти-ICOS антитело С398.4А (в каждом случае - IgG хомяка).
Было обнаружено, что ряд антител обладает высокой нейтрализующей способностью при связывании рецептора-лиганда ICOS человека и/или мыши, и результаты показывают, что некоторые из этих антител проявляют хорошую перекрестную реактивность. Изотип антитела не имел значимого эффекта, различия в результатах между анализами IgG1 и IgG4.PE находились в пределах ошибки эксперимента.
IgG1
В анализах человеческого IgG1, антитело С398.4А дало значение IC50, равное 1,2±0,30 нМ, для нейтрализации лиганда ICOS человека, и IC50, равное 0,14±0,01 нМ, для нейтрализации мышиного лиганда ICOS.
Моноклональные антитела (mAb) IgG1 STIM001, STIM002, STIM003 и STIM005 давали схожие значения IC50 для С398.4А при использовании системы нейтрализации лиганда ICOS человека и также были перекрестно реактивными, нейтрализуя связывание лиганда ICOS мыши с рецептором ICOS мыши.
Два дополнительных перекрестно-реактивных mAb, STIM002-B и STIM004, продемонстрировали более слабую нейтрализацию лиганда ICOS человека и мыши.
STIM006, STIM007, STIM008 и STIM009 показали нейтрализацию человеческого лиганда ICOS, но не продемонстрировали значительной перекрестной реактивности в системе нейтрализации лиганда ICOS мыши. Значения IC50 для нейтрализации мышиного лиганда В7-Н2 не могут быть рассчитаны для этих антител.
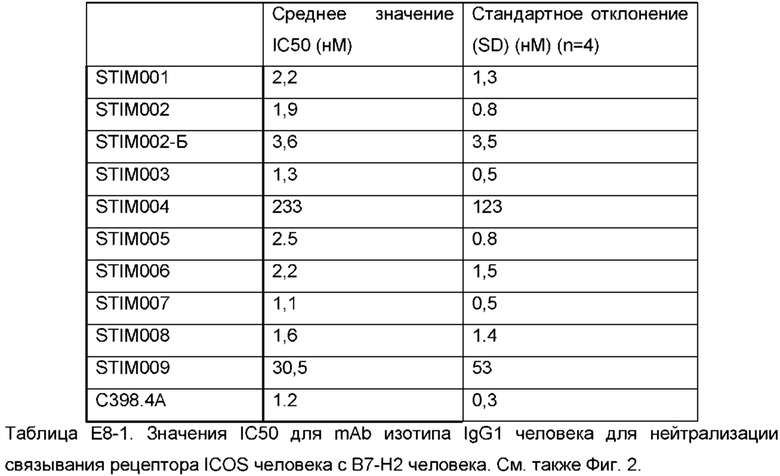
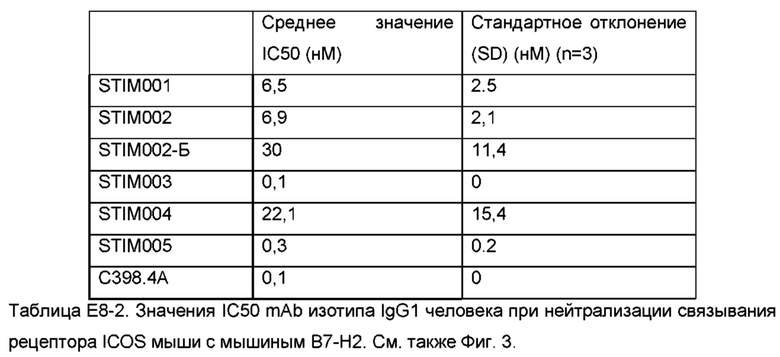
IgG4.PE
Как и ожидалось, mAb IgG4.PE дали результаты, аналогичные полученным для изотипов IgG1.
STIM001, STIM003 и STIM005 показали значения IC50, схожие с С398.4А, при использовании системы нейтрализации лиганда ICOS человека. Эти mAb также были перекрестно-реактивными по нейтрализации лиганда ICOS мыши. STIM002-B и STIM004 дали более слабые значения IC50 для нейтрализации ICOS человека В7-Н2 и мышиным лигандом В7-Н2. STIM007, STIM008 и STIM009 продемонстрировали нейтрализацию связывания лиганда ICOS человека с рецептором ICOS человека, но значения IC50 для нейтрализации мышиного лиганда В7-Н2 не могли быть рассчитаны в этих анализах.
Изотипы IgG4.PE STIM006 и STIM002 не анализировались.
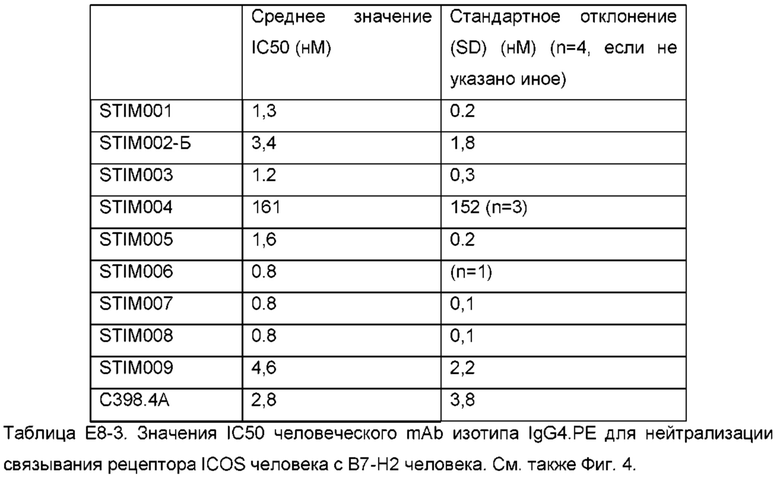

Материалы и методы
Контрольные антитела и изотипический контроль разбавляли в буфере для анализа (0,53 М фторида калия (KF), 0,1% бычьего сывороточного альбумина (BSA) в 1х PBS) от начальной рабочей концентрации, составляющей до 4 мкМ, 1 мкМ, до конечных значений, равных 0,002 нМ, 5,64е-4 нМ, в 11-точечном ряду титрования при разведении 1 к 3. Титрованные антитела добавляли по 5 мкл в 384-луночный (384w) аналитический планшет с белыми стенками (Greiner Bio-One). В лунки положительного контроля добавляли только 5 мкл буфера для анализа.
В нужные лунки добавляли 5 мкл рецептора ICOS (человеческий ICOS-mFc, 20 нМ, конечная концентрация 5 нМ, или мышиный ICOS-mFc 4 нМ, 1 нМ конечная (Chimerigen)). Планшет инкубировали в течение 1 часа при комнатной температуре (RT). После инкубации, 5 мкл лиганда ICOS мыши или человека (В7-Н2, R&D Systems), конъюгированного с Alexa 647 (Innova Bioscience), разбавляли до 32 нМ (8 нМ конечная) для человеческого В7-Н2, или 30 нМ, 7,5 нМ конечная, для мышиного В7-Н2, и добавляли во все лунки аналитического планшета, за исключением лунок отрицательного контроля, в которые вместо этого добавляли 5 мкл буфера для анализа.
Наконец, в каждую лунку добавляли 5 мкл донорного mAb против мышиного IgG (Southern Biotech), меченого криптатом европия (Cis Bio), 40 нМ, 10 нМ конечная концентрация, и анализ оставляли в темноте при комнатной температуре для инкубации еще на 2 часа. После инкубации анализ считывали на планшет-ридере Envision (Perkin Elmer) с использованием стандартного протокола HTRF. Значения каналов 620 нм и 665 нм переносили в Microsoft Excel (Microsoft) и выполняли вычисления % Delta-F и % нейтрализации. Графики кривых титрования и значений IC50 [М] были построены с использованием программы Graphpad (Prism). Значения IC50 вычислялись путем проведения сначала преобразования данных с использованием уравнения X=Log(X). Преобразованные данные затем аппроксимировали с использованием нелинейной регрессии, используя в качестве алгоритма аппроксимации зависимость log(ингибитор) от ответа - переменный наклон (четыре параметра).
Расчет % Delta-F:
Отношение 665/620 нм для преобразования данных измерения соотношения.
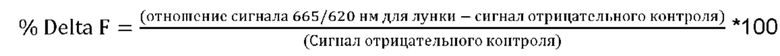
Сигнал отрицательного контроля = среднее значение отношения минимальных сигналов.
% нейтрализации:
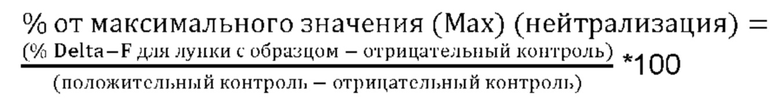
Пример 9а: Активация Т-клеток
Агонистический потенциал воздействия STIM001 и STIM003 на продуцирование цитокинов тестировали в форматах иммобилизации на планшетах и в растворимом состоянии в анализе активации первичных Т-клеток человека, в котором анти-CD3 и анти-CD28 антитела добавляли одновременно к анти-ICOS антителу для индуцирования экспрессии ICOS на эффекторных Т-клетках. Влияние костимуляции ICOS на уровень IFNγ, продуцируемого такими активированными Т-клетками, оценивали методом ИФА через 72 часа после активации.
Материалы и методы
Анализ активации Т-клеток 1:
Выделение мононуклеарных клеток из периферической крови человека (РВМС):
Конусы лейкоцитов собирали у здоровых доноров, и их содержимое разводили до 50 мл фосфатно-солевым буфером (PBS, Gibco) и помещали в верхний слой в 2 центрифужные пробирки на 15 мл Ficoll-Paque (GE Healthcare). РВМС отделяли центрифугированием в градиенте плотности (400 g в течение 40 минут без торможения), переносили в чистую центрифужную пробирку и затем промывали 50 мл PBS, центрифугировали дважды при 300 g в течение 5 мин, и дважды - при 200 g в течение 5 мин. Затем РВМС ресуспендировали в среде R10 (RPMI+10% термоинактивированной фетальной бычьей сыворотки, оба Gibco), и количество клеток и их жизнеспособность оценивали с помощью автоматизированного счетчика клеток EVE™ Automated Cell Counter (NanoEnTek).
Антитела ICOS (Abs): приготовление и разведения:
STIM001 и STIM003 тестировали в трех форматах: иммобилизованными на планшете, растворимыми или растворимыми плюс фрагменты F(ab')2 (109-006-170, Jackson ImmunoResearch), которые сшивают анти-ICOS антитела.
Для формата иммобилизации на планшете: анти-ICOS антитела и их изотипический контроль последовательно разводили 1:3 в PBS для получения конечных концентраций антител в диапазоне от 45 до 0,19 мкг/мл. Покрывали два 96-луночных плоскодонных планшета с высоким связыванием 100 мкл разведенных антител (планшеты для EIA/RIA, Corning) в течение ночи при 4°С. Затем планшет промывали PBS и в каждую лунку добавляли 125 мкл R10.
Для растворимого формата: анти-ICOS антитела и их изотипический контроль серийно разводили 1:3 в среде R10 для получения 2х концентраций маточных растворов антитела в диапазоне значений от 90 до 0,38 мкг/мл. 125 мкл разбавленного антитела пипетировали на два плоскодонных 96-луночных планшета.
Для формата сшитого растворимого антитела: анти-ICOS антитела и их изотипический контроль были смешивали с фрагментами F(ab')2 в отношении 1М к 1М. Смеси антитела/фрагменты F(ab')2 затем последовательно разбавляли 1:3 в среде R10, получая концентрации 2х антитела в диапазоне от 90 до 0,38 мкг/мл для ICOS, и от 60 до 0,24 мкг/мл для фрагментов F(ab')2. 125 мкл разбавленного антитела пипетировали на два плоскодонных 96-луночных планшета.
Выделение Т-клеток, культуры и количественная оценка IFN-γ:
Т-клетки выделяли отрицательным отбором из РВМС с использованием набора для выделения Т-клеток человека EasySep Human Т Cell Isolation Kit (Stemcell Technologies) и ресуспендировали при 2×106/мл в среде R10, дополненной 40 мкл/мл активатора Т-клеток человека Dynabeads Human T-Activator CD3/CD28 (Life Technologies).
125 мкл суспензии Т-клеток добавляли в планшеты, содержащие антитело до конечной концентрации клеток 1×106 клеток/мл и культивировали в течение 72 часов при 37°С и 5% CO2. Затем собирали не содержащие клеток супернатанты и хранили при -20°С до анализа секретированного IFNγ методом ИФА (набор duoset, R&D).
Этот эксперимент повторяли для Т-клеток, выделенных у 6 независимых доноров, и для каждых условий анализа были включены 2 технических реплики.
Анализ активации Т-клеток 2 (анализ STIM-REST-STIM):
Способность агониста STIM001 и STIM003 высвобождать цитокины также тестировали в иммобилизованном на планшете состоянии в анализе Т-клеток человека, причем Т-клетки предварительно стимулировали анти-CD3 и анти-CD28 антителами в течение 3 дней для индуцирования экспрессии ICOS с последующим отдыхом в течение 3 дней, чтобы уменьшить уровни их активации. Экспрессию ICOS подтверждали окрашиванием FACS после стимуляции (день 3) и отдыха (день 6). Эти стимулированные отдохнувшие Т-клетки затем культивировали с STIM001 или STIM003 в присутствии антитела CD3 или без него для оценки потребности в связывании с TCR. Эффекты ко-стимуляции ICOS оценивали через 72 часа по уровням IFNγ, TNFα и IL-2, присутствующих в культуре.
Разбавление и покрытие антителами ICOS:
Анти-человеческий CD3 (клон UCHT1, eBioscience) разбавляли в PBS до 2х концентрации антитела, равной 10 мкг/мл. 50 мкл PBS или 50 мкл разбавленного антитела CD3 пипетировали в 96-луночный плоскодонный планшет с высоким связыванием. STIM001, STIM003 и их изотипический контроль серийно разводили 1:2 в PBS, чтобы получить конечные концентрации 2х антител в диапазоне от 20 до 0,62 мкг/мл. 50 мкл разбавленного анти-ICOS антитела добавляли в лунки, содержащие либо PBS (без связывания с TCR), либо разбавленное антитело CD3 (связывание с TCR). Планшеты покрывали в течение ночи при 4°С.
Выделение Т-клеток, культуры и количественная оценка IFN-γ:
РВМС из конусов лейкоцитов получали, как описано в анализе активации Т-клеток 1. Т-клетки выделяли путем отрицательного отбора из этих РВМС с использованием набора для выделения человеческих Т-клеток EasySep Human Т Cell Isolation Kit (Stemcell Technologies). Т-клетки ресуспендировали при 1×106/мл в среде R10, дополненной 20 мкл/мл активатора человеческих Т-клеток Dynabeads Human T-Activator CD3/CD28 (Life Technologies) и культивировали в течение 3 дней при 37°С и 5% CO2 (стимуляция). На 3-й день из культуры удаляли шарики dynabeads. Т-клетки затем промывали (300 g в течение 5 мин), подсчитывали и ресуспендировали при 1,5×106 мл в среде R10 и культивировали при 37°С и 5% CO2 в течение 3-х дней (фаза отдыха).
На 6-й день стимулированные отдохнувшие Т-клетки промывали (300 g в течение 5 мин), подсчитывали и ресуспендировали при 1×106/мл в среде R10, и 250 мкл суспензии Т-клеток добавляли в планшеты, покрытые антителом ICOS, и культивировали в течение 72 ч при 37°С и 5% CO2. Затем супернатанты, не содержащие клеток, собирали и хранили при -20°С до анализа секретируемых цитокинов на платформе MSD. Этот эксперимент повторяли с Т-клетками, выделенными у 5 независимых доноров, и для каждых условий анализа было включено 3 технических реплики.
Результаты
Как STIM001, так и STIM003 дали положительный результат тестирования индукции экспрессии IFNγ, что демонстрирует агонизм в обоих анализах.
Пример 9b: Данные анализа активации Т-клеток 1
Анализ активации Т-клеток выполняли, как описано в Примере 9а, с использованием Т-клеток, выделенных у 8 независимых доноров, с тестированием каждого из STIM001 и STIM003 в формате человеческом IgG1 Для сравнения было включено анти-ICOS антитело хомячка С398.4А и изотипический контроль антител хомячка. Для каждых условий анализа были включены 2 технические реплики.
Результаты показаны на Фиг. 16, Фиг. 17 и Фиг. 18. Как отмечалось ранее, как STIM001, так и STIM003 дали положительный результат тестирования индуцирования экспрессии IFNγ, что демонстрирует агонистическое действие на первичные Т-клетки человека.
Перекрестно сшитые антитела действовали как агонисты активации Т-клеток, о чем свидетельствует сильное усиление индукции IFNγ в присутствии Fc-связывающих фрагментов F(ab')2 по сравнению с растворимым антителом или с контролем. Экспрессия IFNγ в Т-клетках возрастала с увеличением концентрации сшитых STIM001 или STIM003 (Фиг. 16, нижние панели). Агонизм также наблюдался как для STIM001, так и для STIM003 в формате иммобилизации на планшете и, более слабо, для антитела С398.4А хомячка, о чем свидетельствует увеличение экспрессии IFNγ, наблюдаемое в Т-клетках с ростом концентрации антитела (Фиг. 16, верхние панели).
Величина ответа IFNγ различалась для Т-клеток, полученных от разных доноров, но STIM001 стабильно приводил к увеличению экспрессии IFNγ в Т-клетках по сравнению с экспрессией IFNγ, наблюдаемой с контрольным антителом (НС IgG1). При рассмотрении данных анализов с Т-клетками всех 8 доноров видно, что обработка Т-клеток STIM001 значимо увеличивала экспрессию IFNγ по сравнению с обработкой антителом изотипического контроля, в иммобилизованной на планшете форме, растворимой форме и сшитой форме (Фиг. 17). Таким образом, STIM001 ведет себя как агонист активации Т-клеток во всех трех форматах.
Аналогичные эффекты наблюдались для STIM003 (Фиг. 18). Уровни IFNγ, индуцированные STIM003 hIgG1, сравнивали с уровнями IFNγ, индуцированными его изотипическим контролем (НС IgG1), при определенной дозе антитела в анализе, для 8 независимых доноров. Несмотря на различия между донорами, среднее увеличение уровня IFNγ, индуцированное STIM003, было значимым по сравнению с НС IgG1. Было выдвинуто предположение, что STIM001 и другие антитела STIM, раскрытые в данном документе, обладают потенциалом аналогично промотировать активацию Т-клеток in vivo. Как обсуждалось ранее, агонизм активированных экспрессирующих ICOS Т-клеток может быть опосредуемым связыванием анти-ICOS антитела с, и индуцикцией мультимеризации рецептора ICOS на поверхности Т-клеток. Пример 9с: анализ активации Т-клеток 2
Анализ активации Т-клеток 2 проводили, как описано в Примере 9а.
В отсутствие связывания с TCR (без анти-CD3-антитела) уровни цитокинов, продуцируемых первичными Т-клетками, были низкими, и STIM001 (hIgG1), STIM003 (hIgG1) или антитело С398.4А не индуцировали никакого увеличения даже при самой высокой концентрации, равной 10 мкг/мл. Напротив, когда анти-ICOS антитела были добавлены к Т-клеткам в комбинации с анти-CD3 антителом, STIM001 (hIgG1), STIM003 (hIgG1) и С398.4А все продемонстрировали дозозависимую тенденцию к увеличению экспрессии IFNγ, TNFα и, в меньшей степени, IL-2.
Данные для первичных Т-клеток, обработанных анти-ICOS антителами в условиях связывания с TCR, показаны на Фигуре 19. Хотя заметное увеличение экспрессии цитокинов наблюдалось для каждого из STIM001, STIM003 и С389.4А по сравнению с их соответствующими изотипическими контролями, разница не достигла уровня статистической значимости в этом анализе. Ожидается, что дополнительные реплики анализа с чувствительными первичными Т-клетками от дополнительных доноров позволят получить статистически значимые результаты.
Пример 10а: Анализ ADCC
STIM001 и STIM003, способные убивать (клетки) посредством ADCC, были протестированы в анализе цитотоксичности Delfia BATDA (Perkin Elmer) с использованием первичных NK-клеток человека в качестве эффекторов и клеточной линии MJ с высоким уровнем ICOS (АТСС, CRL-8294) в качестве клеток-мишеней. MJ-клетки представляют собой клетки CD4 Т-лимфоцитов человека, которые экспрессируют высокие уровни белка ICOS.
Этот метод основан на загрузке клеток-мишеней ацетоксиметиловым эфиром лиганда, усиливающего флуоресценцию (BATDA), который быстро проникает через клеточную мембрану. Внутри клетки сложноэфирные связи гидролизуются с образованием гидрофильного лиганда (TDA), который уже не проходит через мембрану. После цитолиза лиганд высвобождается и может детектироваться путем добавления европия, который образует с BATDA высокофлуоресцентный и стабильный хелат (EuTDA). Измеренный сигнал непосредственно коррелирует с количеством лизированных клеток.
Материалы и методы
Мечение клеток-мишеней:
Согласно инструкциям производителя, клетки MJ ресуспендировали при 1×106/мл в среде для анализа (RPMI+10% FBS со сверхнизким содержанием IgG, Gibco) и нагружали 5 мкл/мл реагента Delfia BATDA (Perkin Elmer) в течение 30 минут при 37°С. Затем клетки MJ промывали 3 раза 50 мл PBS (300 g в течение 5 мин) и ресуспендировали при 8×105 мл в среде для анализа, дополненной 2 мМ пробенецида (Life technologies), чтобы уменьшить спонтанное высвобождение BATDA из клеток.
Разбавление антитела ICOS:
STIM001, STIM003 и их изотипический контроль серийно разбавляли 1:4 в среде для анализа + 2 мМ пробенецида для получения 4х конечных концентраций антител в диапазоне до 80 пг/мл.
Выделение и культивация NK-клеток:
РВМС из конусов лейкоцитов получали, как описано в анализе активации Т-клеток 1. NK-клетки выделяли отрицательным отбором от таких РВМС с использованием набора для выделения NK-клеток человека EasySep Human NK Cell Isolation Kit (Stemcell Technologies) и ресуспендировали при 4×106/мл в среде R10+2 мМ пробенецида. Чистота NK-клеток, по результатам проверки путем окрашивания CD3-/CD56+, составляла выше 90%.
В каждую лунку добавляли 50 мкл разбавленного антитела, 50 мкл нагруженных BATDA MJ-клеток, 50 мкл NK-клеток и 50 мкл среды для анализа + 2 мМ пробенецида (конечный объем 200 мкл/лунку), чтобы получить конечную концентрацию антитела в диапазоне с нижним пределом до 20 пг/мл и соотношение эффектор:мишень 5:1. Для определения самопроизвольного и 100%-ного высвобождения BATDA использовали лунки, содержащие только клетки MJ или клетки MJ + буфер для лизиса delfia (Perkin Elmer).
Анализ проводили при 37°С, 5% CO2, в течение 2 ч, после чего переносили 50 мкл супернатанта без клеток в микротитровальные планшеты DELFIA (Perkin Elmer). К супернатантам добавляли 200 мкл раствора европия Delfia Europium (Perkin Elmer) и инкубировали в течение 15 минут при комнатной температуре. Затем флуоресцентный сигнал определяли количественно с помощью многоканального анализатора EnVision Multilabel Reader (PerkinElmer).
Удельное высвобождение, индуцируемое STIM001 и STIM003, рассчитывали в соответствии с приложенными к набору инструкциями. Этот эксперимент повторяли с NK-клетками от независимых доноров и для каждых условий анализа были включены 3 технические реплики.
Результаты
Анти-ICOS антитела STIM001 (hIgG1) и STIM003 (hIgG1) убивают ICOS-положительные MJ-клетки человека в анализе зависимой от первичных NK ADCC (момент времени 2 часа). См. также Фиг. 6а. Субнаномолярные значения ЕС50 были достигнуты в этом анализе для обеих протестированных молекул.

Пример 10b: Анализ ADCC с клетками-мишенями MJ
Эксперимент проводили в соответствии с разделом Материалы и методы Примера 10а. STIM001, STIM003 и изотипическорй контроль серийно разбавляли 1:4 в среде для анализа + 2 мМ пробенецида для получения 4х конечных концентраций антител в диапазоне значений от 40 мкг/мл до 80 пг/мл. В каждую лунку добавляли 50 мкл разбавленного антитела, 50 мкл нагруженных BATDA клеток MJ, 50 мкл NK-клеток и 50 мкл среды для анализа + 2 мМ пробенецида (конечный объем 200 мкл/лунку), чтобы получить конечную концентрацию антитела в диапазоне значений от 10 мкг/мл до 20 пг/мл и соотношение эффектор:мишень 5:1.
Результаты приведены на Фигуре 6 (b-d) и в таблице ниже. STIM001 (hIgG1) и STIM003 (hIgG1) убивали ICOS-положительные клетки MJ человека в анализе ADCC, зависимой от первичных NK, при проведении измерений в момент времени два часа.

Пример 10с: Анализ ADCC с ICOS-трансфицированными клетками-мишенями CCRF-СЕМ
Потенциал STIM001 и STIM003 hIgG1 уничтожения клеток посредством ADCC был дополнительно протестирован в анализе цитотоксичности Delfia BATDA (Perkin Elmer) с использованием первичных NK-клеток человека в качестве эффекторных, и ICOS-трансфицированных клеток CCRF-CEM (АТСС, CRL-119) - в качестве клеток-мишеней, CCRF-CEM представляет собой линию Т-лимфобластов человека, полученную из периферической крови пациента с острым лимфобластным лейкозом. Антитело-опосредованное убийство клеток CCRF-CEM было подтверждено в этом анализе как для STIM001, так и для STIM003.
Материалы и методы
Материалы и методы были такими же, как изложено в Примере 10а, но с использованием в качестве клеток-мишеней клеток CCRF-CEM, полученных от АТСС (АТСС CCL-119), а не MJ-клеток, и с использованием времени инкубации 4 ч.
Клетки CCRF-CEM были трансфицированы ICOS. Синтетическую цепочку ДНК, кодирующую полноразмерный ICOS человека (с сигнальным пептидом, как показано в прилагаемом перечне последовательностей), оптимизированную по использованию кодонов для экспрессии у млекопитающих, клонировали в экспрессионный вектор под контролем промотора CMV и фланкировали специфическими последовательностями концевых повторов 3' и 5' piggyBac, способствующими стабильной интеграции в геном клетки (см. [40]). Вектор экспрессии содержал кассету для селекции пуромицином для облегчения создания стабильной клеточной линии. Экспрессионную плазмиду человеческого ICOS подвергали совместной трансфекции с плазмидой, кодирующей транспозазу piggyBac, в клетки СЕМ CCRF путем электропорации Через 24 часа после трансфекции в среду добавляли пуромицин и выращивали в течение по меньшей мере двух недель для селекции стабильных клеточных линий, при этом среду обменивали каждые 3-4 дня. Экспрессию ICOS человека оценивали методом проточной цитометрии с использованием конъюгированного с РЕ анти-ICOS антитела человека (eBioscience). Полная среда СЕМ состояла из среды Advanced RPMI Medium, содержащей 10% (об./об.) FBS и 2 мМ глутамакса.
STIM001 (hIgG1), STIM003 (hIgG1) и изотипическое контрольное антитело (НС IgG1) серийно разбавляли средой для анализа для получения 4х конечных концентраций антител в диапазоне значений от 20 мкг/мл до 80 пг/мл.
В каждую лунку добавляли 50 мкл разбавленного антитела, 50 мкл нагруженных BATDA ICOS-трансфицированных клеток СЕМ, 50 мкл NK-клеток и 50 мкл среды для анализа (конечный объем 200 мкл/лунку) с получением конечной концентрации антитела в диапазоне от 5 мкг/мл до 20 пг/мл и соотношения эффектор:мишень 5:1.
Результаты
STIM001 (hIgG1) и STIM003 (hIgG1) убивали трансфицированные ICOS клетки CCRF-CEM в анализе ADCC, зависимой от первичных NK, при измерениях в момент времени четыре часа. Результаты приведены на Фигуре 6 (е-g) и в таблице ниже.
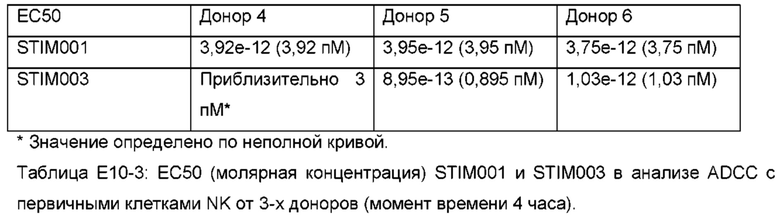
Пример 11а: Сингенная модель СТ26
Улучшенная противоопухолевая эффективность in vivo была продемонстрирована в сингенной модели СТ26 путем комбинирования анти-ICOS (STIM001 mIgG2a, активированный эффектор) с анти-PDLI (10F9G2).
Материалы и методы
Исследования эффективности проводились на мышах Balb/c с использованием модели подкожной карциномы толстой кишки СТ26 (АТСС, CRL-2638). Эта модель слабочувствительна к блокаде PD1/PDL1, и обычно наблюдается только задержка роста опухоли (без стабилизации заболевания или исцеления) в ответ на монотерапии 10F9.G2 (анти-PDLI) и RMT1-14 (анти-PDI). Поэтому эта модель является подходящей для исследований эндогенной резистентности к анти-PDI, анти-PDLI при комбинированной терапии. Все эксперименты in vivo проводились в соответствии с законом Великобритании 1986 г. UK Animal (Scientific Procedures) Act (Акт о животных (научные процедуры)) и Директивой ЕС 86/609 в соответствии с лицензией Министерства внутренних дел Великобритании и были одобрены Наблюдательным советом по защите животных и этическим вопросам Института Бабрахама (Babraham Institute Animal Welfare and Ethical Review Body).
Мыши Balb/c были получены от Charles River UK в возрасте 6-8 недель и с весом более 18 г и размещались в особых условиях без патогенов. В левый бок мышей вводили подкожной инъекцией 1×105 клеток СТ26 (число пересевов менее 20 (Р20)). Если не указано иное, лечение началось на 6-й день после инъекции опухолевых клеток. Клетки СТ26 пассировали in vitro с использованием Accutase (Sigma), дважды промывали в PBS и ресуспендировали в RPMI, дополненной 10% фетальной телячьей сывороткой. Было подтверждено, что жизнеспособность клеток составляла более 90% на момент инъекции опухолевых клеток.
Для исследований in vivo агонист анти-ICOS STIM001 (перекрестно реактивный к белку ICOS мыши) был переформатирован как IgG1 мыши и IgG2a мыши для тестирования в качестве нуль-эффекторной функции и активированной эффекторной функции, соответственно. Анти-PDLI был получен от Biolegend (№ кат.:124325). Гибридные контроли были сгенерированы в Kymab (изотип mIgG2a) или получены из коммерческого источника (изотип HTK888 хомячка, Biolegend (Part No92257, Lot B215504)). Все антитела вводили внутрибрюшинно (IP) в дозе 10 мг/кг (1 мг/мл в 0,9% физиологическом растворе) три раза в неделю с дня 6 (дозирование в течение 2 недель в дни 6-18) в виде монотерапии или путем комбинирования анти-PDLI с анти-ICOS антителами. Вес животных и объем опухоли измеряли 3 раза в неделю начиная с дня инъекции опухолевых клеток. Объем опухоли рассчитывали с использованием модифицированной формулы эллипсоида 1/2(длина × ширина2). Мыши продолжали участвовать в исследованиях до тех пор, пока их опухоли не достигали среднего диаметра 12 мм3 или, в редких случаях, при появлении опухолевых язв (состояние здоровья (welfare)). Эксперимент был остановлен на 50-й день. Статистика выживаемости до конечных точек умерщвления из соображений гуманности (human endpoint) рассчитывалась с использованием метода Каплана-Мейера с помощью программы Prism. Этот подход был использован для определения того, связаны ли конкретные методы лечения с улучшенной выживаемостью.
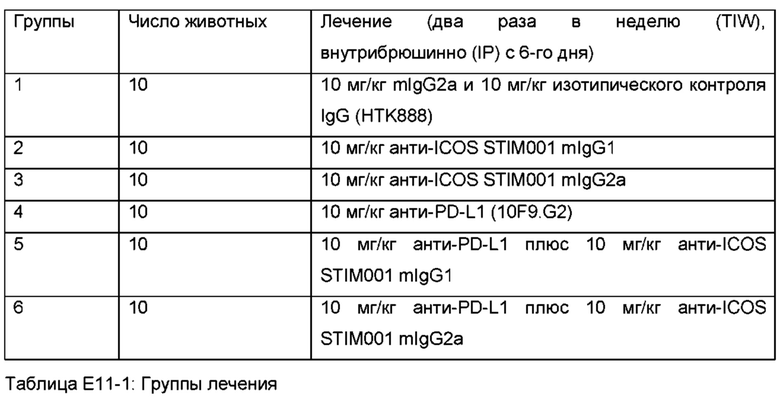
Результаты
Как показано на Фиг. 7, Фиг. 8 и Фиг. 9, агонисты ICOS могут задерживать прогрессирование заболевания и исцелить часть животных от подкожных опухолей СТ-26 либо в виде монотерапии, либо в комбинации с анти-PDLI Монотерапия анти-PDLI индуцировала задержку роста опухоли, но стабилизация заболевания или целебный потенциал не наблюдались. Комбинация была более эффективной при лечении опухолей, чем анти-ICOS монотерапии. Это исследование также показало, что STIM001 в формате IgG2a мыши (активированная эффекторная функция) было более сильнодействующим, чем формат IgG1 мыши (нуль-эффекторный), при инициировании противоопухолевого ответа в этой модели.
Пример 11b: Сильная противоопухолевая эффективность in vivo в сингенной модели СТ26 для комбинации анти-ICOS mIgG2a с анти-PDLI mIgG2a
Были проведены исследования комбинированного применения in vivo STIM001 с мышиным перекрестно-реактивным анти-PDLI антителом человека, обозначенным AbW. Для этой работы in vivo, STIM001 был переформатирован как IgG1 мыши и IgG2a мыши для сравнения его эффективности с низкой эффекторной функцией или в виде молекулы с активированной эффекторной функцией, соответственно. Анти-PDLI AbW было сгенерировано в тех же форматах (IgG1 мыши и IgG2A мыши).
Исследования эффективности проводили на мышах Balb/c с использованием модели подкожной карциномы толстой кишки СТ26 (АТСС, CRL-2638). Мыши Balb/c были получены от Charles River UK в возрасте 6-8 недель и с весом более 18 г и размещались в особых условиях без патогенов. В общей сложности 1×10Е5 клеток СТ26 (число пересевов менее 20 (Р20)) вводили подкожно в правый бок мышей. Если не указано иное, лечение началось на 6-й день после инъекции опухолевых клеток. Клетки СТ26 пассировали in vitro с использованием TrypLE™ Express Enzyme (Thermofisher), дважды промывали в PBS и ресуспендировали в RPMI, дополненной 10% фетальной телячьей сывороткой. Была подтверждена жизнеспособность клеток свыше 90% на момент инъекции опухолевых клеток.
Антитела STIM001 и анти-PDLI вводили одновременно в комбинациях внутрибрюшинно (IP) по 200 мкг каждое (1 мг/мл в 0,9% физиологическом растворе) три раза в неделю с 6-го дня (дозирование в течение 2 недель на протяжении дней 6-17) после имплантации опухолевых клеток, Рост опухоли контролировали и сравнивали с опухолями животных, получавших лечение смесью изотипических контрольных антител (mIgG1 и mIgG2A). Вес животных и объем опухолей измеряли 3 раза в неделю начиная со дня инъекции опухолевых клеток. Объем опухоли рассчитывали с использованием модифицированной формулы эллипсоида 1/2(длина × ширина2). Мыши продолжали участвовать в исследованиях до тех пор, пока их опухоль не достигала среднего диаметра 12 мм3 или, в редких случаях, когда наблюдалось появление опухолевых язв (состояние здоровья). Эксперимент был остановлен на 60-й день. Статистика выживаемости до конечных точек умерщвления из соображений гуманности (human endpoint) рассчитывалась с использованием метода Каплана-Мейера с помощью программы Prism. Этот подход был использован для определения того, связаны ли конкретные методы лечения с улучшенной выживаемостью.

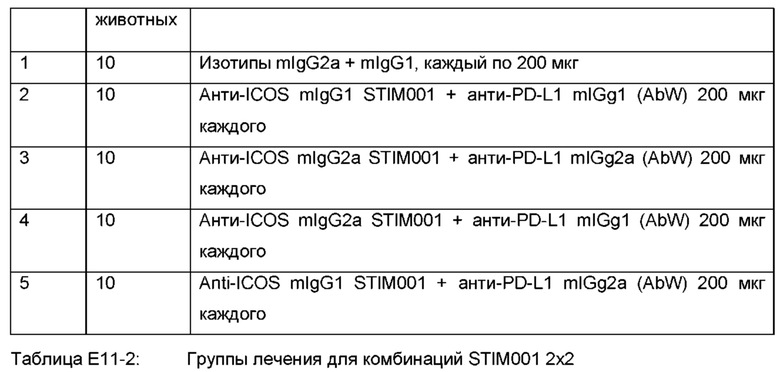
Результаты представлены на Фигуре 10. Все комбинации антител задерживали рост опухоли и увеличивали выживаемость (время достижения конечной точки умерщвления из соображений гуманности (human endpoint)) получавших лечение животных по сравнению с животными, получавшими изотипический контроль. Интересно, что в сочетании с анти-PDL1 (независимо от его формата, mIgG1 или mIgG2a) антитело STIM001 mIgG2a было более эффективным при ингибировании роста опухоли, чем STIM001 в формате mIgG1. Эти данные позволяют предположить, что преимуществом анти-ICOS антитела будет наличие эффекторной функции для максимального увеличения противоопухолевой эффективности. Примечательно, что STIM001 mIgG2a в комбинации с aPD-L1 mIgG2a продемонстрировал самую сильную противоопухолевую эффективность и улучшенную выживаемость (90% животных, продемонстрировавших ответ, и 60% излеченных от болезни на 60-й день).
Аналогично, STIM003 mIgG1 и mIgG2a тестировали как монотерапию или в комбинации с анти-PDL1 (AbW) mIgG2a в тех же моделях опухолей СТ26. STIM003 и анти-PDL1 антитела дозировали животным в виде монотерапии или в комбинации путем внутрибрюшинной инъекции (IP) 200 мкг каждого антитела (1 мг/мл в 0,9% физиологическом растворе) три раза в неделю начиная с 6-го дня (дозирование в течение 2 недель на протяжении дней 6-17) после имплантации опухолевых клеток. В этом эксперименте размеры опухоли контролировали в течение 41 дня. Статистика выживаемости до конечных точек умерщвления из соображений гуманности (human endpoint) рассчитывалась с использованием метода Каплана-Мейера с помощью программы Prism. Этот подход был использован для определения того, связаны ли конкретные методы лечения с улучшенной выживаемостью.


Результаты приведены на Фигуре 11. Монотерапии с использованием aPDL1 (AbW) и STIM003 mIgG2a демонстрировали умеренную противоопухолевую активность (по одному излеченному от болезни животному в каждой группе). Комбинации STIM003 mIgG1 или mIgG2a с aPDL1 (AbW) mIgG2a показали сильную противоопухолевую эффективность. Интересно отметить, что к 41-му дню, в сочетании с aPDL1 mIgG2a, STIM003 mIgG2a был более эффективным при ингибировании роста опухоли, чем STIM003 mIgG1 (60% против 30% животных, излеченных от заболевания, соответственно). Данные также дополнительно подтвердили преимущество эффекторного формата анти-ICOS антител для обеспечения максимальной противоопухолевой эффективности.
В целом, эти данные показывают, что комбинация анти-ICOS антитела STIM001 или STIM003 с анти-PDL1 приводит к наиболее сильному противоопухолевому ответу, когда оба антитела имеют активированную эффекторную функцию. Пригодные соответствующие изотипы антитела человека будут включать человеческий IgG1, необязательно, с дополнительно усиленной эффекторной функцией, например, афукозилированный IgG1.
Кривые Каплана-Мейера для мышей, получавших лечение комбинацией анти-PDL1 mIgG2a и STIM003 mIgG2a и каждым агентом отдельно, показаны на Фигуре 29.
Пример 11 с: Разовая доза антитела STIM003 восстанавливает микроокружение опухоли (ТМЕ) и приводит к сильной противоопухолевой эффективности в сочетании с непрерывным введением анти-PD-L1
В этом исследовании сравнивалось однократное или многократное дозирование STIM003 mIgG2A вместе с многократным дозированием анти-PDL1 антитела (AbW). Данные показывают, что однократная доза анти-ICOS антитела может изменять микроокружение опухоли таким образом, чтобы анти-PD-L1 антитело оказывало больший эффект. Это можно рассматривать как «перезагрузку» микроокружения опухоли (ТМЕ) анти-ICOS антителом.
Как и ранее, эти исследования эффективности проводили на мышах Balb/c с использованием модели подкожной карциномы толстой кишки СТ26 (АТСС, CRL-2638). Мыши Balb/c были получены от Charles River UK в возрасте 6-8 недель и с весом более 18 г и размещались в особых условиях без патогенов. В общей сложности 1×10Е5 клеток СТ26 (число пересевов менее 20 (Р20)) вводили подкожно в правый бок мышей. Если не указано иное, лечение начали на 6-й день после инъекции опухолевых клеток. Клетки СТ26 пассировали in vitro с использованием TrypLE™ Express Enzyme (Thermofisher), дважды промывали в PBS и ресуспендировали в RPMI, дополненной 10% фетальной телячьей сывороткой. Была подтверждена жизнеспособность клеток свыше 90% на момент инъекции опухолевых клеток.
Группы лечения приведены в таблице Е11-4. STIM003 и анти-PDL1 антитела вводили внутрибрюшинно (IP) в дозе 10 мг/кг (1 мг/мл в 0,9% физиологическом растворе). Лечение начали с 6-го дня после имплантации опухолевых клеток. Рост опухоли контролировали и сравнивали с опухолями животных, получавших физиологический раствор. Вес животных и объем опухолей измеряли 3 раза в неделю начиная со дня инъекции опухолевых клеток. Объем опухоли рассчитывали с использованием модифицированной формулы эллипсоида 1/2(длина × ширина2). Мыши продолжали участвовать в исследованиях до тех пор, пока их опухоли не достигали среднего диаметра 12 мм3 или, в редких случаях, при появлении опухолевых язв (состояние здоровья (welfare)). Эксперимент был остановлен на 55-й день.
Данные показаны на Фигуре 34. Параллельное дозирование STIM003 и анти-PDL1 в течение 6 доз начиная с 6-го дня приводило к сильной противоопухолевой эффективности в модели СТ26 у 5 из 8 животных, не имевших опухолей в конце исследований (55-й день). Интересно, что аналогичная противоопухолевая эффективность была достигнута при разовой дозе STIM003 с последующей многократной дозой анти-PDL1 в качестве монотерапии. В комбинации с анти-PD-L1 mIgG2a, наблюдалась аналогичная общая эффективность для однократной дозировки STIM003 (С) и 6 дозами (В). По сравнению с группой, получавшей физиологический раствор (А), в которой только у одного животного наблюдалось спонтанное отторжение опухоли (редкое явление в этой модели), группы, получавшие комбинированные препараты, демонстрировали полное отторжение опухоли у 62,5% животных к концу эксперимента (день 55). Данные свидетельствуют о том, что антитело STIM003 можно использовать для восстановления микроокружения опухоли, и что антитело позволяет резистентным к контрольным точкам опухолям становиться чувствительными к анти-PDL1 Как показано ранее (Пример 11b), линия опухолевых клеток СТ26 не сильно реагирует на анти-PDL1 монотерапию. По-видимому, STIM003 вызывает изменения, которые способствуют противоопухолевой активности анти-PDL1 терапии.
Пример 12: Анализ последовательности антител
Каркасные области антител STIM001, STIM002, STIM002-B, STIM003, STIM004, STIM005, STIM006, STIM007, STIM008 и STIM009 сравнивали с генными сегментами зародышевой линии человека, чтобы идентифицировать ближайшее совпадение. См. Таблицу Е12-1 и Таблицу Е12-2.
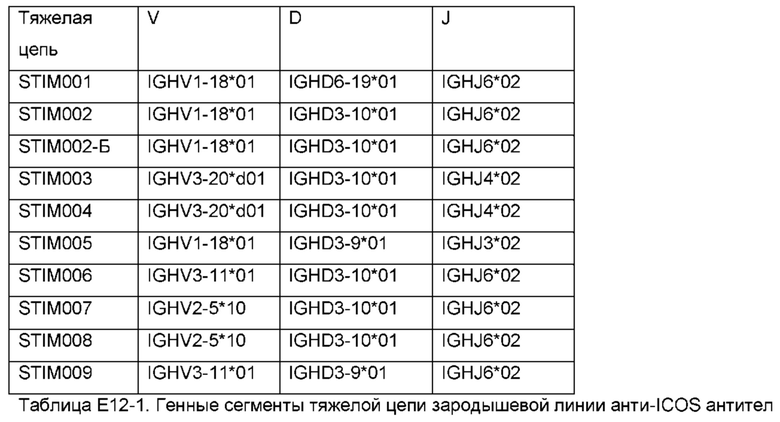
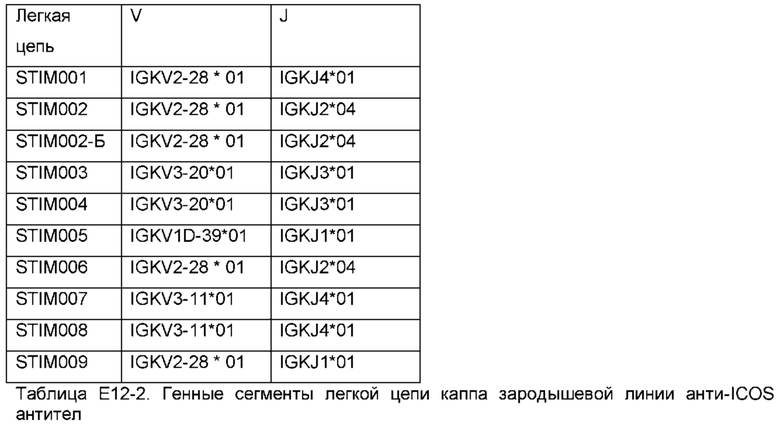
Дополнительные последовательности антител были получены путем секвенирования нового поколения амплификацированной методом ПЦР ДНК антитела из добавочных ICOS-специфических клеток, которые были отсортированы у иммунизированных мышей, как описано в Примере 3. В результате этого был идентифицирован ряд антител, которые можно было сгруппировать в кластеры со STIM001, STIM002 или STIM003 на основе их V и J генных сегментов тяжелой и легкой цепей и длины CDR3. CL-61091 был сгруппирован с STIM001; CL-64536, CL-64837, CL-64841 и CL-64912 были сгруппированы с STIM002; и CL-71642 и CL-74570 были сгруппированы с STIM003. Выравнивания последовательностей доменов VH и VL антитела показаны на Фиг. 35-37.
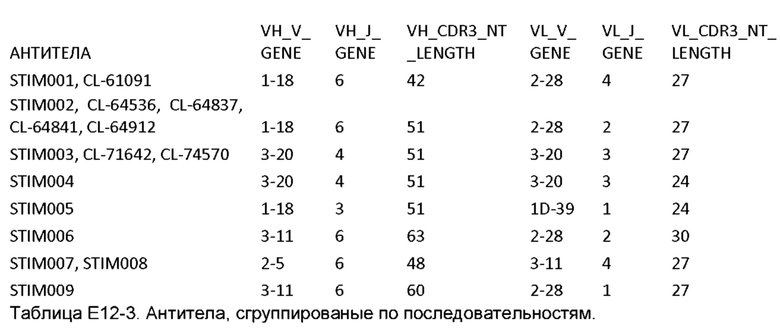
Пример 13: Агонизм ICOS-экспрессирующих клеток MJ с иммобилизованным с шариками антителом
Антитела STIM001, STIM002 и STIM003, анти-ICOS антитело С398.4А и лиганд ICOS (ICOSL-Fc), были все ковалентно связаны с шариками и оценивались по их способности индуцировать экспрессию IFN-γ цитокина клетками MJ, выращенными в культуре. IgG1 человека и изотипические контроли клона С398.4А, иммобилизованные на шариках, анализировали параллельно.
Данные показаны на Фигуре 12 и в Таблице Е13 ниже.
Каждое из анти-ICOS антител продемонстрировало агонизм в этом анализе, стимулируя клетки MJ, по результатам определения с помощью количественного определения IFN-γ, значительно сильнее, чем наблюдалось для их родственных изотипических контролей в динамическом диапазоне анализа.
STIM003 и клон С398.4А давали более низкие значения верхней асимптоты (95% доверительный интервал (CI): от 3,79 до 5,13 и от 3,07 до 4,22, соответственно), но более мощные значения LogEC50 (95% доверительный интервал от -9,40 до -9,11 и от -9,56 до -9,23, соответственно), по сравнению со STIM001 (95% доверительный интервал верхней асимптоты: от 7,21 до 8,88 и 95% доверительный интервал LogEC50: от -8,82 до -8,63) и STIM002 (95% доверительный интервал верхней асимптоты: от 5,38 до 6,95 и 95% доверительный интервал LogEC50 от -9,00 до -8,74). Поскольку для ICOSL-Fc и изотипического контроля клона С398.4А были получены неполные кривые (верхняя часть динамического диапазона анализа), полученные путем аппроксимации значения верхней асимптоты и LogEC50 не считались достоверными. Контроль по гибриду IgG1 человека давал полную кривую, однако площадь под кривой существенно не отличалась от 0, и поэтому он не считался агонистом.

Материалы и методы анализа активации клеток MJ - иммобилизованные на шариках
Связывающие белки для магнитных частиц, представляющие интерес
Анти-ICOS антитела, контрольные антитела и ICOSL-Fc связывали с шариками следующим образом.
Тозил-активированные Dynabeads М-450 (Invitrogen, приблизительно 2×10^8 шариков/образец) инкубировали со 100 мкг каждого образца белка в течение ночи при комнатной температуре при перемешивании. Шарики трижды промывали DPBS (Gibco) и инкубировали с 1 М Трис-HCl, рН 8,0 (UltraPure™, Gibco) в течение 1 часа при комнатной температуре с перемешиванием для блокирования несвязанных реакционноспособных сайтов. Шарики трижды промывали DPBS и, наконец, ресуспендировали в 0,5 мл DPBS/образец.
Затем количество каждого представляющего интерес белка на шариках определяли следующим образом. Черные плоскодонные планшеты с высоким связыванием для ИФА (Greiner) были покрыты антителом против IgG человека (Southern Biotech) или против IgG серого хомячка (Armenian Hamster) (Jackson ImmunoResearch) при 4 мкг/мл в DPBS, 50 мкл/лунку, в течение ночи при 4°С. Затем лунки трижды промывали DPBS + 0,1% Твин (Tween), 200 мкл/лунку, и блокировали 200 мкл/лунку DPBS + 1% BSA в течение 1 часа при комнатной температуре. Лунки снова трижды промывали DPBS + 0,1% Твин. Образцы маточного белка количественно определяли спектрометрически, а шарики подсчитывали на счетчике клеток. Затем инкубировали серийные разбавления образцов белка и шариков на планшетах при 50 мкл/лунку в течение 1 часа при комнатной температуре перед повторным промыванием три раза DPBS + 0,1% Твин. Добавляли 50 мкл/лунку биотинилированного антитела против серого хомячка либо антитела против человеческого IgG-европия в DPBS + 0,1% BSA и инкубировали в течение 1 часа при комнатной температуре. В случае добавления биотинилированного антитела против серого хомячка С398.4А добавляли еще одну стадию инкубации с 50 мкл/лунку стрептавидина-европия (Perkin Elmer), разбавленного 1:500 в буфере для анализа (Perkin Elmer), и инкубировали в течение 1 часа при комнатной температуре. Лунки промывали три раза 200 мкл/лунку TBS + 0,1% Твин перед проявлением анализа путем добавления 50 мкл/лунку усиливающего раствора Delfia (Perkin Elmer), инкубирования в течение 10 минут при комнатной температуре и измерения флуоресценции, испускаемой при 615 нм на многоканальном планшет-ридере Envision Multilabel Plate Reader. Количество белка на шариках определяли экстраполяцией значений сигналов, полученных для известных концентраций несвязанных образцов белка.
Анализ активации клеток MJ in vitro - иммобилизованные на шариках
Клеточную линию MJ [G11] (АТСС CRL-8294) выращивали в IMDM (Gibco или АТСС), дополненной 20%-ной термоинактивированной FBS. Клетки подсчитывали и добавляли 15000 клеток/лунку (50 мкл/лунку) клеточной суспензии в 96-луночные прозрачные плоскодонные полистирольные стерильные микропланшеты, обработанные ТС. Шарики подсчитывали и серийные разбавления 1:2 в диапазоне от 1,5×10^6 шариков/лунку до приблизительно 5860 шариков/лунку (50 мкл/лунку) добавляли к клеткам в двух или трех репликах. Для учета фона, несколько лунок планшета содержали только клетки MJ (100 мкл/лунку). Клетки и гранулы совместно культивировали в планшетах в течение 3 дней при 37°С и 5% CO2, после чего супернатанты собирали центрифугированием и использовали для определения содержания IFN-γ.
Измерение уровней IFN-γ
Содержание IFN-γ в каждой лунке определяли с использованием модификации набора для ИФА Human IFN-gamma DuoSet ELISA (R&D Systems). Черные плоскодонные планшеты с высокими связыванием (Greiner) покрывали захватным антителом (50 мкл/лунку) в течение ночи при 4 мкг/мл в DPBS. Лунки промывали три раза 200 мкл/лунку DPBS + 0,1% Твин. Лунки блокировали 200 мкл/лунку 1% BSA в DPBS (мас./об.), промывали три раза 200 мкл/лунку DPBS + 0,1% Твин, а затем добавляли в каждую лунку 50 мкл/лунку либо стандартных растворов IFN-γ в RPMI или неразведенного клеточного супернатанта и инкубировали в течение 1 часа при комнатной температуре. Лунки промывали три раза 200 мкл/лунку DPBS + 0,1% Твин перед добавлением 50 мкл/лунку биотинилированного антитела обнаружения при 200 нг/мл в DPBS + 0,1% BSA и инкубировали в течение 1 ч при комнатной температуре. Лунки промывали три раза 200 мкл/лунку DPBS + 0,1% Твин перед добавлением 50 мкл/лунку стрептавидина-европия (Perkin Elmer), разбавленного 1:500 в буфере для анализа (Perkin Elmer) и инкубировали в течение 1 ч при RT. Лунки промывали три раза 200 мкл/лунку TBS + 0,1% Твин перед проявлением анализа путем добавления 50 мкл/лунку усиливающего раствора Delfia (Perkin Elmer), и инкубировали в течение 10 мин при комнатной температуре и измеряли флуоресценцию, испускаемую при 615 нм, на многофункциональном планшет-ридере Envision Multilabe Plate Readerl.
Анализ данных
Значения IFN-γ для каждой лунки были интерполированы по калибровочной кривой, с вычетом средних фоновых уровней для лунок, содержащих только клетки. Скорректированные на фон значения затем использовали в программе GraphPad Prism для аппроксимации 4-параметрической логарифмически логистической кривой концентрация-ответ.
Пример 14: Агонизм ICOS-экспрессирующих клеток MJ с иммобилизованным на планшете антителом
Альтернативный анализ агонизма экспрессирующих ICOS Т-клеток использует антитела в иммобилизованном на планшете формате.
Материалы и методы анализа активации клеток MJ - иммобилизованные на планшете
Покрытие антителами: 96-луночные стерильные плоские планшеты с высоким связыванием (Costar) покрывали в течение ночи при 4°С 100 мкл/лунку серийными разведениями 1:2 представляющих интерес белков (анти-ICOS антитела, контрольные антитела и ICOSL-Fc) в DPBS (Gibco) в диапазоне от 10 мкг/мл до 0,02 мкг/мл в двух или трех репликах. Для учета фона, несколько лунок планшета были покрыты только DPBS. Затем планшеты промывали три раза 200 мкл/лунку DPBS перед добавлением клеток.
Стимуляция клеток: клеточную линию MJ [G11] (АТСС CRL-8294) выращивали в IMDM (Gibco или АТСС), дополненной 20%-ной термоинактивированной FBS. Клетки подсчитывали и в планшеты, покрытые белком, добавляли 15000 клеток/лунку (100 мкл/лунку) клеточной суспензии. Клетки культивировали в планшетах в течение 3 дней при 37°С и 5% CO2. Клетки отделяли от среды центрифугированием и супернатанты собирали для определения содержания IFN-γ.
Измерение уровней IFNγ и анализ данных проводили, как описано в Примере 13.
Результаты
Результаты приведены на Фигуре 13 и ниже в Таблице Е14-1. Таким образом, STIM001, STIM002 и STIM003 показали все сильный агонизм, измеряемый по секреции IFN-γ, с близкими значениями LogEC50 (LogEC50 95% Cl: от -7,76 до -7,64, от -7,79 до -7,70 и от -7,82 до -7,73, соответственно) и верхних значений (верхние значения 95% Cl: от 2,06 до 2,54, от 2,44 до 2,93, и от 2,01 до 2,41, соответственно). Клон С398.4А показал схожее значение LogEC50 (LogEC50 95% Cl: от -7.78 до -7.60), но меньшую величину верхнего значения (верхнее значение, 95% Cl: от 1,22 до 1,63), чем у STIM001-STIM003. STIM004 также продемонстрировал агонизм в этом анализе, но был менее активным, достигая умеренной величины верхнего значения (верхнее значение, 95% Cl: от 0,16 до 0,82), при близком значении LogEC50 (LogEC50 95% Cl: от -7,91 до -7,21). STIM001, STIM002 и STIM003 были более сильными агонистами, чем ICOSL-Fc (LogEC50 95% Cl: от -7,85 до -7,31, и верхнее значение, 95% Cl: от 0,87 до 2,45).
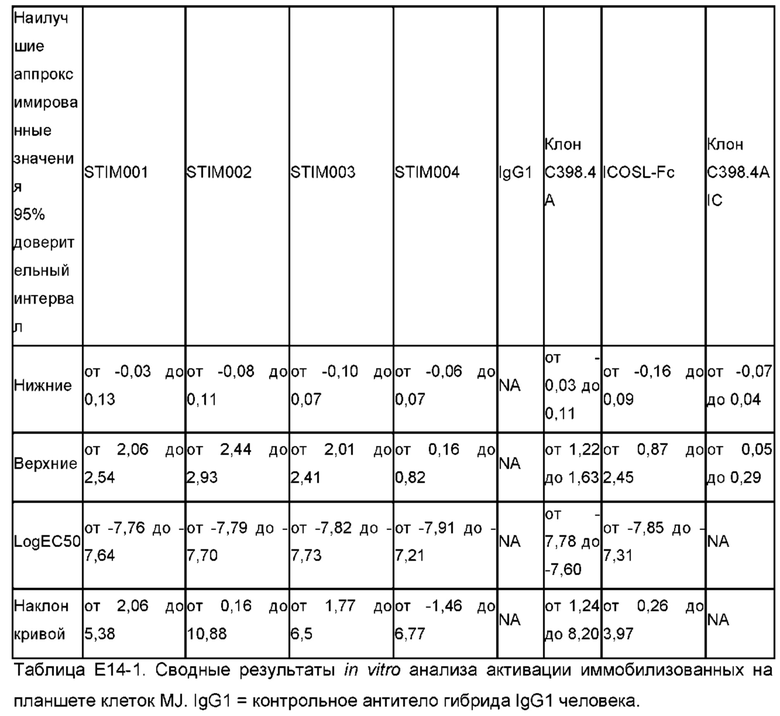
Пример 15: Агонизм ICOS-экспрессирующих клеток MJ к антителу в растворимой форме
В отличие от анализов, описанных в Примере 13 и Примере 14, которые использовали антитело, расположенное на твердой поверхности, этот анализ определяет, обладает ли антитело в растворимой форме агонистическим действием на Т-клетки, экспрессирующие ICOS.
Материалы и методы анализа активации клеток MJ - растворимая форма
Клеточную линию MJ [G11] (АТСС CRL-8294) выращивали в IMDM (Gibco или АТСС), дополненной 20%-ной термоинактивированной FBS. Клетки подсчитывали и добавляли 15000 клеток/лунку (50 мкл/лунку) клеточной суспензии в 96-луночные прозрачные плоскодонные полистирольные стерильные микропланшеты, обработанные ТС. Серийные разведения 1:2 белков, представляющих интерес, в диапазоне от 10 мкг/мл до 0,01953125 мкг/мл, взятых отдельно либо с добавлением сшивающего реагента (AffiniPure F(ab')2 Fragment Goat Anti-Human IgG, Fc Fragment Specific, Jackson ImmunoResearch) были добавлены к клеткам в двух или трех репликах (50 мкл/лунку). Для учета фона, несколько лунок планшета содержали только клетки MJ (100 мкл/лунку). Клетки и гранулы совместно культивировали в планшетах в течение 3 дней при 37°С и 5% CO2, после чего супернатанты собирали центрифугированием и использовали для определения содержания IFN-γ.
Измерение уровней IFNγ и анализ данных проводили, как описано в Примере 13.
Результаты
STIM001 и STIM002 показали значительный агонизм в растворимом состоянии, измеряемый по секреции IFN-γ, по сравнению с контролем гибридом IgG4.PE человека. Сшивание моноклонального антитела через козий фрагмент F(ab')2 Fc против IgG человека увеличивало уровни секретируемого IFN-γ еще сильнее.
Пример 16: Связывание антител с активированными Т-клетками
A. ICOS человека
В этом анализе подтверждается способность анти-ICOS антител распознавать внеклеточный домен ICOS в его нативном контексте на поверхности активированных первичных Т-клеток человека.
Общие Т-клетки (клетки CD3) выделяли и культивировали в течение 3 дней с шариками CD3/CD28 dynabeads (Thermofisher), чтобы индуцировать экспрессию ICOS на их поверхности. Поверхностное окрашивание STIM001, STIM003 и контроля с гибридом hIgG1 (НС IgG1) определяли двумя методами, а именно детектирования после прямого связывания предварительно меченых антител (антител, непосредственно конъюгированных с AF647), или опосредованно с использованием вторичного козьего антитела против Fc человека AF647. Окрашенные клетки анализировали на проточном цитометре Attune, и интенсивность окрашивания была представлена как среднее значение интенсивности флуоресценции (MFI). ЕС50 определяли с использованием программы GraphPad Prism.
Результаты приведены на Фигуре 14. После активации, общие CD3 Т-клетки явным образом окрашивались hIgG1 как STIM001, так и STIM003. Примечательно, что насыщение связывания STIM003 с активированными Т-клетками происходило при более низкой концентрации, чем для STIM001, что указывает на более высокую аффинность STIM003 к ICOS человека. ЕС50 STIM003 была примерно в 100 раз ниже, чем у STIM001 (0,148 нМ против 17 нМ, в непрямом анализе связывания).
В. ICOS приматов, не являющихся человеком
В этом анализе подтверждается способность анти-ICOS антител распознавать внеклеточный домен ICOS в его нативном контексте на поверхности активированных первичных Т-клеток приматов, не являющихся человеком (NHP).
РВМС из цельной крови 2 яванских макаков с о. Маврикия (Wickham Laboratories) выделяли градиентным центрифугированием и культивировали в течение 3 дней с шариками MACSiBeads CD2/CD3/CD28 (Miltenyi) для индуцирования экспрессии ICOS на их поверхности. Поверхностное окрашивание STIM001, STIM003 и контроля с гибридом hIgG1 (НС IgG1) определяли после прямого связывания предварительно меченых антител AF647 (от 80 мкг до 8 пг/мл). Клетки также метили V450-CD3 для оценки окрашивания на субпопуляциях Т-клеток. Окрашенные клетки анализировали на проточном цитометре Attune (Thermofisher), и интенсивность окрашивания была представлена как средняя интенсивность флуоресценции (MFI). ЕС50 определяли с использованием программы GraphPad Prism.
Результаты приведены на Фигуре 28. После активации, Т-клетки явным образом окрашивались hIgG1 как STIM001, так и STIM003. Как было отмечено, при связывании с Т-клетками человека, насыщение связывания STIM003 с активированными Т-клетками NHP происходило при более низкой концентрации, чем для STIM001, что указывает на то, что из этих двух антител ICOS STIM0003 обладает более высокой аффинностью. Значения ЕС50 для связывания с ICOS NHP были аналогичны значениям, полученным для связывания с ICOS человека.

Пример 17: Анализ субпопуляций Т-клеток в опухоль-инфильтрирующих лимфоцитах и периферических Т-клетках
Фармакодинамические исследования показали, что анти-ICOS антитела STIM001 и STIM003 в изотипе mIgG2a существенно деплетируют TRegs, увеличивают процент CD4+ эффекторных клеток и увеличивают соотношение CD4+ эффектор/TReg, а также соотношение CD8+/TReg, в микроокружении опухоли (ТМЕ).
Повышенное отношение CD8+/TReg и увеличенное количество CD4+ эффекторных клеток внутри ТМЕ могут коллективно вносить вклад в клиренс опухоли СТ26, наблюдаемый, когда эти анти-ICOS антитела были введены совместно с анти-PDL1 антителом в исследованиях эффективности STIM001 и STIM003 (Пример 11).
Метод
Исследование фармакодинамики проводили на самках мышей Balb/c, несущих клетки мышиной карциномы толстой кишки СТ-26 (АТСС, CRL-2638). Мышей Balb/c в возрасте 6-8 недель и с весом более 18 г, которых поставляла фирма Charles River UK, размещали в специальных непатогенных условиях. В общей сложности 1×10Е5 опухолевых клеток СТ-26 (число пересевов: Р8) вводили подкожно в правый бок. Все животные, несущие опухоль СТ-26, были разбиты на 6 групп (таблица Е17-1), и индивидуальным мышам вводили дважды (в день 13 и день 15 после имплантации опухолевых клеток) 200 мкг антитела или физиологического раствора. CD3+ Т-клетки животных, несущих опухоль СТ-26, анализировали методом FACS на 16-й день после имплантации опухоли.

Результаты
Животные, получавшие лечение STIM001 и STIM003 в изотипе mIgG2a, показали более низкий процент CD4+ CD3+ CD45+ клеток на участке опухоли по сравнению с группой, получившей физиологический раствор (Фигура 15А), в то время как лечение STIM001 или STIM003 оказывало очень незначительный эффект на процент CD8+ CD3+ CD45+ клеток на участке опухоли (Фиг. 15В). Уменьшение CD4+ Т-клеток можно объяснить сильным снижением процента Т-регуляторных клеток во всех группах, получавших лечение антителами STIM001 и STIM003. Примечательно, что животные, получавшие STIM001 и STIM003 в изотипе mIgG2a, продемонстрировали резкое снижение T-Regs (CD4+ Foxp3+ CD25+ ) в ТМЕ, тогда как STIM001 и STIM003 в изотипе mIgG1 оказывали лишь незначительное влияние на содержание T-Reg в ТМЕ. Кроме того, животные, получавшие STIM001 и STIM003 в изотипе mIgG2a, имели пониженное содержание T-Reg в ТМЕ по сравнению с животными, получавшими коммерческое анти-CTLA-4 антитело (9Н10, Biolegend, № кат. 106208), которое, как известно, деплетирует Т-Reg [42], но этот результат не достигал статистической значимости (Фигура 15С). Эффект STIM001 и STIM003 в изотипах mIgG1 или mIgG2a на компартмент T-Reg был более специфичным по отношению к опухоль-инфильтрующими лимфоцитами (ОИЛ). Деплетирование T-reg не наблюдалось на периферии (как описано выше для анти-CTLA4 [43]) (Фигура 15D). Изменения в содержании T-Reg также приводили к значительному увеличению процентного содержания внутриопухолевых CD4-эффекторных клеток (CD4+ Foxp3- CD25-) (Фигура 15Е), аналогично значительно возрастало соотношение CD4 эффектор/T-Reg и CD8/T-Reg в ТМЕ у животных, получавших STIM001 и STIM003 в mIgG2a (Фигура 15F и Фигура 15G).
Пример 18: Влияние анти-ICOS антитела на уровень экспрессирующих ICOS Т-клеток в опухоли СТ26 и селезенке
Был проведен анализ с целью количественной оценки процента иммунных клеток в опухоли по сравнению с селезенкой, путем анализа общего количества иммунных клеток в тканях опухоли и селезенки после лечения анти-ICOS антителом STIM001 или STIM003. STIM001 и STIM003 mIgG2a вызывали значительное уменьшение Treg в опухоли, но не в селезенке, что указывало на селективный к опухоли эффект. Это деплетирование было селективным для Tregs по сравнению с другими подтипами Т-клеток. Представленные в данном документе результаты помогают понять влияние антител STIM на элементы иммунной системы (immune contexture) и подтверждают, что анти-ICOS антитела с Fc-областями с активированной эффекторной функцией могут сильно деплетировать TRegs.
Материалы и методы
Мышам, несущим опухоли СТ26, вводили две дозы STIM001, STIM003 или анти-CTLA-4 антитела (9Н10). Анти-CTLA4 антитело было включено в качестве положительного контроля деплетирования Treg, поскольку ранее было показано, что анти-CTLA-4 антитело селективно уменьшает Tregs в опухолях [43].
Элементы иммунной системы в опухолях и селезенке получавших лечение животных анализировали методом FACS после дезагрегации ткани.
Характеристики антител для FACS, используемых в этом исследовании, приведены в Таблице Е18. Все антитела для FACS использовались в концентрации, рекомендованной поставщиком. Данные FACS регистрировали с использованием проточного цитометра Attune N×T, и данные были проанализированы с использованием программного обеспечения FlowJo.
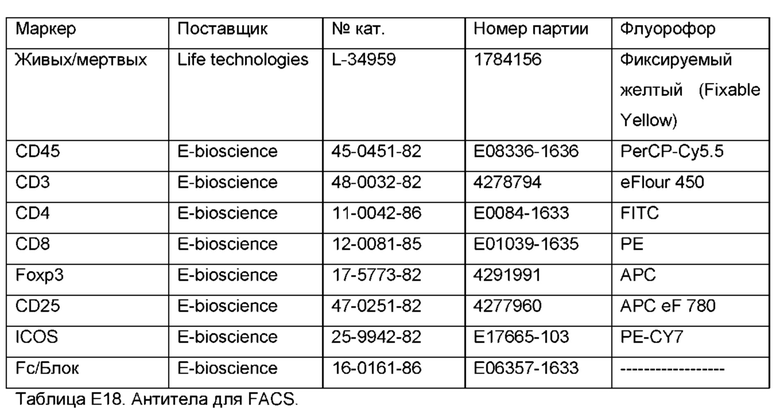
Результаты
Экспрессию ICOS определяли в опухолях СТ26 и в селезенке несущих опухоли животных. Мы наблюдали увеличение содержания инфильтрирующих опухоль иммунных клеток, экспрессирующих белок ICOS (Фиг. 20), что указывает на то, что иммунные клетки в опухолях чаще являются положительными по экспрессии ICOS, чем иммунные клетки на периферии. Почти все (>90%) TRegs в опухоли не получавших лечение животных были положительными по экспрессии ICOS, тогда как CD8+ эффекторные Т-клетки в опухоли не имели таких способностей (ок. 60%). Сравнение субпопуляций Т-клеток (также у не получавших лечения мышей) в опухоли и в селезенке показало, что значительно более высокий (р<0,0001) процент внутриопухолевых Tregs были ICOS-положительными по сравнению с Tregs в селезенке, и значительно более высокий (р<0,001) процент внутриопухолевых клеток CD4+ Teff были ICOS-положительными по сравнению с клетками CD4+ Teff в селезенке.
Также у мышей до лечения уровень экспрессии ICOS на иммунных клетках в микроокружении опухолей СТ26 был значительно выше, чем на иммунных клетках в селезенке (Фиг. 21). Экспрессия ICOS была увеличена на поверхности всех проанализированных субпопуляций иммунных клеток (CD8 Т-эффекторы, CD4 Т-эффекторы и клетки CD4/FoxP3 TReg) в микроокружении опухоли. Следует отметить, что хотя иммунные клетки в опухолях и селезенке экспрессируют ICOS, иммунные клетки в опухоли экспрессируют значительно большее количество ICOS (на что указывают более высокие значения MFI, Фиг. 21), чем клетки в селезенке (на что указывают более низкие значения MFI, Фиг. 21). Важно отметить, что TRegs в опухоли экспрессируют самые высокие уровни ICOS, как сообщалось ранее [11].
Животным, несущим опухоль СТ26, вводили 2 дозы антитела STIM001 или STIM003 и анти-CTLA-4 антитело. Антитела STIM не влияли на общий процент CD45-положительных клеток (маркер иммунных клеток) в опухолях при использовании в формате mIgG1 или mIgG2. Лечение этими антителами также не оказывало существенного влияния на процентное содержание CD8 эффекторных Т-клеток в опухолях СТ26 (Фиг. 22). Лечение STIM001 изотипа mIgG2a приводило к значительному (р<0,05) деплетированию CD4+ эффекторных Т-клеток, но ни один из STIM001 mIgG1, STIM003 mIgG1 и STIM003 mIgG2a не влиял на процентное содержание CD4+ эффекторных Т-клеток.
Лечение анти-CTLA-4 вызывало заметное (хотя и не статистически значимое) увеличение клеток CD45+ и CD8+ эффекторных Т-клеток в ТМЕ, но не влияло на CD4+ эффекторные Т-клетки (Фигура 22).
Антитела STIM существенным образом влияли на регуляторные Т-клетки в опухоли. Как показано на Фигуре 23, STIM001 mIgG2a и STIM003 mIgG2a значительно и селективно деплетировали TRegs (имеющие высокое содержание по экспрессии ICOS) в микроокружении опухоли. Интересно, что анти-CTLA-4 антитело, которое, несмотря на его включение в этот эксперимент в качестве положительного контроля деплетирования TReg, было менее эффективным, чем антитела STIM mIgG2a, по показателям деплетирования TReg.
Это селективное деплетирование TRegs приводило к увеличению отношения CD8 эффекторных Т-клеток к TRegs в опухоли, и к увеличению отношения CD4 эффекторных Т-клеток к TRegs в опухоли, которые оба должны способствовать противоопухолевому иммунному ответу. Данные для соотношений показаны на Фигуре 24.
В отличие от деплетирования внутриопухолевых Tregs под действием STIM001 mIgG2a и STIM003 mIgG2a, для Tregs в селезенке такой эффект не наблюдался (Фиг. 25, Фиг. 26, Фиг. 27). Это указывает на то, что эффекты анти-ICOS антител на деплетирования Tregs не были системными во всех тканях. Такая селективность может быть выгодной для терапевтических анти-ICOS антител при лечении опухолей у пациентов, так как преимущественное деплетирование Tregs в микроокружении опухоли может выборочно снижать супрессию противоопухолевых эффекторных Т-клеток, одновременно минимизируя побочные эффекты в других частях организма. Таким образом, анти-ICOS антитела могут способствовать противоопухолевому ответу в иммунной системе с низким риском нежелательной активации более широкого ответа Т-клеток, который может вызвать препятствующие лечению аутоиммунные нежелательные явления.
Пример 19: Стабильность антитела
Испытывали STIM003 IgG1 человека на стабильность при хранении, замораживании/оттаивании и очистке, и было установлено, что он сохраняет свою стабильность во всех условиях испытаний. % агрегации определяли методом ВЭЖХ.
Не наблюдалось существенного изменения процентного содержания мономера (>99%) после хранения в течение 3 месяцев при 4°С в буфере (10 мМ фосфата натрия, 40 мМ хлорида натрия, рН 7,0).
При испытании на тепловую денатурацию все образцы (n=15) имели одинаковую Tm (без существенной разницы между аликвотами) и сопоставимые кривые тепловой денатурации.
Не наблюдалось значительных изменений Tm (≈70,3°С), процентного содержания мономера или профиля ДСН-ПААГ после 3 циклов замораживания и оттаивания.
Не наблюдалось значительных изменений Tm (≈70,3°С), процентного содержания мономера или профиля ДСН-ПААГ после 7 дней хранения при комнатной температуре.
Выход после очистки белком А составлял 90%.
Пример 20: Монотерапевтическая эффективность анти-ICOS антитела против роста опухоли А20 у мыши
Анти-ICOS антитела STIM001 mIgG2a и STIM003 mIgG2a показали сильную противоопухолевую эффективность при использовании в качестве монотерапий in vivo в сингенной модели мышей А20.
Материалы и методы
Исследование эффективности проводили на мышах BALB/c с использованием модели подкожной саркомы ретикулярных клеток А20 (АТСС, TIB-208). Линия клеток А20 представляет собой линию В-клеточной лимфомы BALB/c, полученную из спонтанного новообразования ретикулярных клеток, обнаруженного у старой мыши BALB/cAnN. Сообщалось, что эта клеточная линия является ICOSL-положительной.
Мыши BALB/c поставлялись фирмой Charles River UK с весом более 18 грамм и размещались в специфических непатогенных условиях. В общей сложности 5×10е5 клеток А20 (число пересевов менее 20 (Р20)) вводили мышам подкожно в правый бок. Клетки А20 пассировали in vitro, дважды промывали в PBS и повторно суспендировали в RPMI с добавкой 10% фетальной телячьей сыворотки. Было подтверждено, что жизнеспособность клеток на момент инъекции опухолевых клеток составляла более 85%. Если не указано иное, введение антител или изотипа начинали с дня 8 после введения опухолевых клеток.
Анти-ICOS антитела STIM001 и STIM003 были получены в формате изотипа IgG2a мыши. Также было приготовлено перекрестно-реактивное анти-PD-L1 антитело мыши (AbW) в том же формате изотипа (IgG2a мыши). Антитела STIM001, STIM003 и анти-PD-L1 вводили внутрибрюшинно (IP), по 200 мкг каждого антитела, два раза в неделю, начиная с 8-го дня (дозирование на протяжении 3 недель между днями 8-29) после имплантации опухолевых клеток. Вес животных и объем опухоли измеряли 3 раза в неделю начиная с дня инъекции опухолевых клеток. Объем опухоли рассчитывали с использованием модифицированной формулы эллипсоида 1/2(длина × ширина2). Мыши продолжали участвовать в исследованиях до тех пор, пока их опухоль не достигла среднего диаметра 12 мм. Эксперимент был остановлен на 43-й день после имплантации опухолевых клеток. Рост опухоли контролировали и сравнивали с опухолями животных, получавших антитело изотипического контроля (mIgG2a). Группы лечения приведены в Таблице Е20 ниже.

Результаты
Монотерапевтическое введение либо STIM001, либо STIM003 (mIgG2a) в модели опухоли А20 привело к полному противоопухолевому ответу (Фиг. 32, Фиг. 33). Все животные, которым вводили STIM001 или STIM003, были излечены от заболевания. Такой результат отличается от полученного в группах изотипического контроля и PD-L1 mIgG2a (Фиг. 30, Фиг. 31). В редких случаях, у некоторых животных наблюдалась регрессия опухолей в группах изотипического контроля (спонтанная регрессия) и анти-PDL-1, но лечение анти-ICOS антителом давало значительно большую эффективность. В конце исследования, 3 из 8 контрольных животных и 2 из 8 животных, получавших анти-PDL-1, не имели опухолей. Однако, все животные, получавшие либо STIM001, либо STIM003, не имели опухолей в конце исследования (8 из 8 мышей в обеих группах), что соответствует 100%-ному исцелению при использовании анти-ICOS антител.
Пример 21: Сильная противоопухолевая эффективность in vivo в сингенной модели миеломы J558 для комбинации анти-ICOS антитела и анти- PD-L1 антитела
Анти-ICOS антитело STIM003 mIgG2a и анти-PD-L1 антитело AbW mIgG2a вводили по отдельности и в комбинации в модели опухоли J558. Это сингенная модель мышиной миеломы. Было обнаружено, что анти-ICOS антитело ингибирует рост опухоли при введении в виде монотерапии или в комбинации с анти-PD-L1.
Материалы и методы
Исследования противоопухолевой эффективности проводили на мышах Balb/c с использованием подкожно клеточной линии плазмацитома : миелома J558 (АТСС, TIB-6). Мыши Balb/c были получены от Charles River UK в возрасте 6-8 недель и с весом более 18 г и размещались в особых условиях без патогенов. В общей сложности 5×106 клеток (число пересевов менее 15 (Р15)) вводили подкожно (в 100 мкл) в правый бок мышей. Если не указано иное, на 11-й день после инъекции опухолевых клеток животные были рандомизированы по размеру опухолей, и было начато лечение. Клетки J558 пассировали in vitro с использованием TrypLE™ Express Enzyme (Thermofisher), дважды промывали в PBS и ресуспендировали в DMEM с добавкой 10% фетальной телячьей сыворотки. Была подтверждена жизнеспособность клеток свыше 90% на момент инъекции опухолевых клеток.
Лечение начинали, когда опухоли достигли среднего объема ок. 140 мм^3. Затем животных распределяли на 4 группы со сходным средним размером опухоли (см. группы дозировки в Таблице Е-21). Оба антитела, которые являются перекрестно-реактивными для мышей, вводили внутрибрюшинно (IP) с 11-го дня (после имплантации опухолевых клеток) два раза в неделю в течение 3 недель (Фиг. 38), за исключением случаев, когда животные требовали исключения из исследований из-за состояния здоровья (welfare) (редко) или размера опухоли. В качестве контроля, группе животных (n=10) вводили в то же самое время физиологический раствор. Для группы комбинированного лечения, оба антитела STIM003 и анти-PDL1 вводили одновременно внутрибрюшинно (IP) в количестве 60 мкг и 200 мкг, соответственно (в 0,9% физиологическом растворе). Рост опухоли контролировали в течение 37 дней и сравнивали с опухолями животных, получавших физиологический раствор. Вес животных и объем опухоли измеряли 3 раза в неделю со дня инъекции опухолевых клеток. Объем опухоли рассчитывали с использованием модифицированной формулы эллипсоида 1/2(длина × ширина2). Мыши продолжали участвовать в исследованиях, пока их опухоль не достигла среднего диаметра 12 мм3 или, в редких случаях, когда наблюдалось появление опухолевых язв (состояние здоровья (welfare)).

Результаты
Сингенные опухоли J558 были очень агрессивными, и все животные в контрольной группе физиологического раствора (n=10) должны были быть исключены из исследований к 21-му дню из-за размера опухоли. Анти-STIM003 mIgG2a и анти-PDL1 mIgG2a продемонстрировали хорошую эффективность в качестве монотерапий в этой модели, с 37,5% и 75% животных, вылеченных от болезни, соответственно. Важно отметить, что сочетание двух антител привело к тому, что у 100% животных опухоли плазмоцитомы были отторгнуты к дню 37. Данные показаны на Фигуре 38.
Пример 22. Введение анти-PD1 увеличивает экспрессию ICOS на ОИЛ значительно сильнее, чем анти-PD-L1 антитело
Фармакодинамическое исследование проводили на животных, имеющих установленные опухоли СТ26, для оценки влияния лечения анти-PD-L1 или анти-PD-1 антителами на экспрессию ICOS на субпопуляциях опухоль-инфильтрующих лимфоцитов (ОИЛ). Сравнивали следующие антитела:
• анти-PD-L1 AbW mIgG1 [ограниченная эффекторная функция]
• анти-PD-L1 AbW mIgG2a [с эффекторной функцией]
• анти-PD-L1 10F9.G2 крысы IgG2b [с эффекторной функцией]
• анти-PD1 антитело RMT1-14 крысы IgG2a [нуль-эффекторное].
Опухоли получивших лечение мышей были выделены, диссоциированы на отдельные клетки и окрашены на CD45, CD3, CD4, CD8, FOXP3 и ICOS.
Материалы и методы
Анти-PD-1 RMP1-14 IgG2a крысы (BioXCell, номер по каталогу: ВЕ0146), анти-PD-L1 10F9.G2 IgG2b крысы (Bio-Legend, номер по каталогу: 124325) и анти-PD-L1 AbW mIgG1 и mIgG2a тестировали в модели опухоли СТ26 путем введения внутрибрюшинно (ip) 130 мкг в дни 13 и 15 после имплантации опухолевых клеток. На 16-й день животных умерщвляли и опухоли мышей собирали для анализа методом FACS. Опухоли гомогенизировали с использованием набора для гомогенизации мышиных опухолей (Miltenyi Biotec) и гомогенизировали. Полученные клеточные суспензии осветляли на 70 мкм фильтрах, собирали осадок клеток и ресуспендировали в буфере FACS при 2 миллионах клеток/лунку в 96-луночном планшете. Клеточные суспензии инкубировали с анти-16/32 моноклональным антителом (eBioscience) и окрашивали антителами для FACS, специфичными к CD3 (17А2), CD45 (30-F11), CD4 (RM4-5), CD8 (53-6,7) и ICOS (7E.17G9), все от eBioscience Ltd. Клетки также окрашивали фиксируемым красителем для определения жизнеспособности LiveDead Yellow (Life technologies). Для внутриклеточного окрашивания Foxp3 образцы фиксировали, пермеабилизировали и окрашивали антителом, специфичным к Foxp3 (eBioscience, FJK-16). Образцы ресуспендировали в PBS и данные регистрировали на проточном цитометре Attune (Invitrogen) и анализировали с использованием прикладной программы FlowJo V10 (Treestar).
Результаты
Лечение анти-PD1 и анти-PD-L1 антителами приводило лишь к незначительному увеличению процента клеток CD8 и Т Regs, экспрессирующих ICOS, в измеренный момент времени. Однако, в ответ на IgG2a анти-PD1 крысы на поверхности клеток ICOS+ve CD8 наблюдалось четкое и значимое (по сравнению с группой, получавшей физиологический раствор) увеличение экспрессии ICOS (повышенная dMFI). Было также отмечено, что экспрессия ICOS повышается на CD4 эффекторных и CD4 Т Reg клетках, хотя этот эффект не достигает уровня статистической значимости. Это анти-PD1 антитело индуцировало заметное увеличение экспрессии ICOS на CD8 эффекторных клетках, которое почти не наблюдалось в случае анти-PD-L1 mIgG2a. Аналогично, при сравнении различных форматов анти-PD-L1 антител, у некоторых получавших лечение животных было обнаружено, что антитело, имеющее самую низкую эффекторную функцию (mIgG1), было ассоциировано с более высокой экспрессией ICOS на эффекторных клетках CD8 и CD4 по сравнению с антителом, имеющем эффекторную функцию (mIgG2a и ratIgG2b), которые редко демонстрировали такой эффект. См. Фиг. 39.
Увеличение экспрессии ICOS на эффекторных CD8/CD4 Т-клетках может создавать эффект повышения чувствительности этих клеток к деплетированию анти-ICOS антителами (например, при лечении мышей с использованием STIM003 mIgG2a). Антитело, демонстрирующее более низкую индукцию ICOS у эффекторных CD8 и CD4 Т-клеток, может быть предпочтительным для использования в комбинации с анти-ICOS антителом. Данные этих исследований показывают, что эффектор-положительное анти-PD-L1 антитело может быть особенно пригодным для комбинации с эффектор-положительным анти-ICOS антителом, что отражает противоопухолевую эффективность, наблюдаемую при комбинировании анти-PDL1 mIgG2a со STIM003 mIgG2a, как описано в других примерах в данном документе.
Пример 23: Сильная противоопухолевая эффективность монотерапии с однократным введением анти-ICOS антитела in vivo в сингенной модели В-лимфомы
Этот эксперимент подтверждает противоопухолевую эффективность STIM003 mIgG2a в качестве монотерапии. Сильная противоопухолевая эффективность была продемонстрирована после кратковременного воздействия STIM003 mIgG2a.
Материалы и методы
Исследования эффективности проводили на мышах BALB/c с использованием модели подкожной саркомы ретикулярных клеток А20 (АТСС №CRL-TIB-208). Мышей BALB/c, поставляемых фирмой Charles River UK в возрасте 6-8 недель и с весом более 18 г, размещали в специальных беспатогенных условиях. В общей сложности 5×10Е5 клеток А20 (число пересевов менее 20 (Р20)) вводили подкожно в правый бок мышей. Лечение начали на 8-й день после инъекции опухолевых клеток, как указано в таблице ниже. Клетки А20 пассировали in vitro с использованием TrypLE™ Express Enzyme (Thermofisher), дважды промывали в PBS и ресуспендировали в RPMI, дополненной 10% фетальной телячьей сывороткой. Было подтверждено, что жизнеспособность клеток на момент инъекции опухолевых клеток составляла более 85%. STIM003 mIgG2a использовали либо в виде разовой дозы (SD) 60 мкг (эквивалент 3 мг/кг для животного весом 20 г), либо в виде многократных доз (MD, два раза в неделю в течение 3 недель) 60 мкг. Противоопухолевую эффективность, наблюдаемую в ответ на две схемы лечения, сравнивали с противоопухолевой эффективностью для животных, получавших физиологический раствор (MD, два раза в неделю в течение 3 недель). Антитела вводили внутрибрюшинно (IP) в виде раствора 1 мг/мл в 0,9% физиологическом растворе. Вес животных и объем опухолей измеряли 3 раза в неделю начиная со дня инъекции опухолевых клеток. Объем опухоли рассчитывали с использованием модифицированной формулы эллипсоида 1/2(длина × ширина2). Мыши продолжали участвовать в исследованиях до тех пор, пока их опухоль не достигала среднего диаметра 12 мм или, реже, до появления опухолевых язв (состояние здоровья).

Результаты
Как многократная, так и разовая доза STIM003 mIgG2a обеспечивала сильную и значительную противоопухолевую эффективность монотерапии, что подтверждается количеством животных без признаков роста опухоли в конечной точке (день 41). Введение разовой дозы (SD) привело к исцелению 7 из 10 животных, тогда как многократная доза вылечила 9 из 10 животных, получивших инъекции В-клеточных лимфобластов А20. Все животные из группы, получавшей физиологический раствор, должны были быть исключены из исследований к 40-му дню из-за размеров опухолей. См. Фиг. 40.
Статистику выживаемости до конечных точек умерщвления из соображений гуманности рассчитывали по кривым Каплана-Мейера (Фиг. 41) с использованием программы GraphPad Prism V7.0. Этот подход был использован для определения того, было ли лечение связано с улучшенной выживаемостью. Значения отношения рисков (Мантеля-Хензеля (Mantel-Haenszel)) и ассоциированные с ними значения Р (логранговые по Мантелю-Коксу (Log-Rank Mantel-Cox)) приведены в таблице ниже.

Пример 24. Временные и дозозависимые эффекты анти-ICOS антитела у животных, несущих опухоль СТ-26
В этом примере представлены результаты фармакодинамических исследований, оценивающих влияние анти-ICOS антитела на иммунные клетки у мышей, несущих опухоли СТ-26. Подтипы Т- и В-клеток из разных тканей анализировали методом FACS после однократной дозы STIM003 mIgG2a.
Методы
Животным, несущим опухоль СТ-26, вводили внутрибрюшинно (i.p.) физиологический раствор или STIM003 в дозе 200 мкг, 60 мкг или 6 мкг на 12-й день после имплантации опухолевых клеток. Опухолевые ткани, кровь, дренирующие опухоль лимфатические узлы (TDLN) и селезенку отбирали в дни 1, 2, 3, 4 и 8 после лечения. Опухоли гомогенизировали для получения одноклеточной суспензии, используя набор для гомогенизации мышиных опухолей (Miltenyi Biotec). Ткань селезенки гомогенизировали с использованием устройства для гомогенизации gentleMACS, эритроциты лизировали с использованием буфера для лизиса эритроцитов (RBC). Дренирующие опухоли лимфоузлы механически измельчали для получения одноклеточных суспензий. Полученные клеточные суспензии осветляли на фильтрах 70 мкм или 40 мкм в зависимости от ткани, затем клетки дважды промывали в полной среде RMPI и, наконец, ресуспендировали в ледяном буфере FACS. Общую кровь собирали в пробирки для плазмы и эритроциты лизировали с использованием буфера для лизиса эритроцитов (RBC), клетки дважды промывали в полной среде RMPI и, наконец, ресуспендировали в ледяном буфере FACS. Одноклеточную суспензию всех тканей распределяли на 96-луночных планшетах с глубокими лунками для анализа методом FACS. Клетки окрашивали красителем для определения жизнеспособности Live Dead Fixable Yellow (Life technologies). Клеточные суспензии инкубировали с анти-CD16/CD32 моноклональным антителом (eBioscience) и окрашивали антителами FACS, специфичным по отношению к CD3 (17А2), CD45 (30-F11), CD4 (RM4-5), CD8 (53-6.7), CD25 (РС61.5), ICOSL (HK5.3), В220 (RA3-6B2), Ki-67 (SoIA15), CD107a (eBio1D4B), IFN-γ (XMG1.2), TNF-α (MP6-XT22), Foxp3 (FJK-16s) и ICOS (7E.17G9), все от фирмы eBioscience Ltd. Для считывания цитокинов методом FACS одноклеточные суспензии из опухолей высевали в 24-луночный планшет на 4 часа в присутствии брефельдина А. Для внутриклеточного окрашивания образцы фиксировали, пермеабилизировали и окрашивали специфическими антителами. Наконец, образцы ресуспендировали в PBS и данные регистрировали с помощью проточного цитометра Attune (Invitrogen) и анализировали с использованием программного обеспечения FlowJo V10 (Treestar).
Результаты и их обсуждение приведены ниже.
Наблюдается высокая экспрессия ICOS на внутриопухолевых T-regs в модели СТ26
Мы продемонстрировали, при сравнении процента опухоль-инфильтрирующих лимфоцитов (ОИЛ), экспрессирующих ICOS, с процентом иммунных клеток в селезенке, крови и дренирующих опухоль лимфоузлах (TDLN), что в микроокружении опухолей СТ-26 большее число иммунных клеток экспрессировало ICOS, чем в других тканях. Что еще более важно, процент ICOS-положительных T-reg клеток во всех тканях и во все моменты времени был выше, чем процент ICOS-положительных CD4 или CD8 эффекторных Т-клеток. Важно отметить, что значения dMFI (относительная экспрессия) для ICOS также имели порядок ранжирования, аналогичный экспрессии внутриопухолевых T-reg, являющихся высокоположительными по экспрессии ICOS по сравнению с другими подтипами ОИЛ. Интересно отметить, что не наблюдалось заметного изменения процентного содержания ICOS+ ОИЛ на протяжении этого эксперимента. Подобные результаты были также получены для селезенки и дренирующих опухоль лимфоузлов (TDLN). С другой стороны, в крови экспрессия ICOS является относительно стабильной на эффекторных Т-клетках, но увеличивается в ходе эксперимента на T-regs. В целом данные показали, что большее число клеток экспрессирует ICOS в микроокружении опухоли, и эти положительные клетки также экспрессируют большее количество молекул ICOS на своей поверхности. Что еще более важно, Т regs в ОИЛ являются очень положительными по ICOS. См. Фиг. 42.
Сильное деплетирование внутриопухолевых T-reg-клеток в ответ на введение STIM003
В ответ на антитело mIgG2a STIM003 наблюдалось сильное и быстрое деплетирование T-reg клеток (CD4+ CD25+ Foxp3) в микроокружении опухоли (ТМЕ). Поскольку T-regs имеют высокую экспрессию ICOS по сравнению с другими субпопуляциями Т-клеток, ожидается, что анти-ICOS антитело с эффекторной функцией будет преимущественным образом деплетировать эти клетки. При более низкой дозе STIM003 (6 мкг, что соответствует 0,3 мг/кг для животного весом 20 г) наблюдалось непрерывное деплетирование T-reg, и к 3-му дню большая часть T-reg в микроокружении опухоли была деплетирована. Интересно, что к 8-му дню клетки T-reg репопулируют микроокружение опухоли, а затем достигают уровня, немного превышающего уровень, наблюдаемый у животных, получающих физиологический раствор. Репопуляция клеток Т-reg при более низких дозах может быть связана с увеличением пролиферирующих CD4 Т-клеток в микроокружении опухоли, о чем свидетельствует наблюдаемое увеличение Ki-67+ CD4 Т-клеток. При дозе более 6 мкг наблюдалось долгосрочное деплетирование клеток T-reg в микроокружении опухоли, на что указывает полное деплетирование Т Reg до последнего момента времени, проанализированного в этих исследованиях (8-й день). В то же время в крови при всех дозах наблюдалось кратковременное деплетирование клеток T-reg. Важно отметить, что к 8-му дню все получавшие лечение животные имели сходный (или более высокий, для дозы 6 мкг) уровень клеток T-reg в крови по сравнению с животными, получавшими физиологический раствор. Данные приведены на Фигуре 43. Примечательно, что, аналогично данным, ранее опубликованным для деплетирующих антител CTLA-4, не наблюдалось существенного изменения процентного содержания клеток T-reg в тканях селезенки или дренирующих опухоль лимфоузлов, что указывает на то, что клетки T-reg могут быть защищены от деплетирования в этих органах.
Таким образом, сильное деплетирование клеток T-reg в микроокружении опухоли было достигнуто в модели СТ-26 при дозе, составляющей всего лишь 6 мкг на животное. Однако, доза 60 мкг приводила к длительному деплетированию до 8 дней после инъекции STIM003 mIgG2a. Этот результат не улучшался при использовании более высокой дозы (200 мкг).
STIM003 mIgG2a увеличивал соотношения CD8:T Reg и CD4:T Reg
Влияние STIM003 на соотношения T-eff:T-reg показано на Фигуре 44.
STIM003 mIgG2a увеличивал отношение CD8:T-reg, а также отношение CD4 eff:T-reg. Хотя все дозы лечения были связаны с увеличением соотношения T-eff к T-reg, промежуточная доза 60 мкг (эквивалент 3 мг/кг для животных весом 20 г) была связана с самым высоким соотношением к 8-му дню после введения препарата.
Интересно отметить, что при дозе 6 мкг соотношения оставались высокими до 4-го дня, но к 8-му дню после лечения они совпадали с показателями для животных, получавших физиологический раствор. Это можно объяснить репопуляцией TRegs, наблюдаемой для этой дозы к 8-му дню после лечения. С другой стороны, при дозе 60 или 200 мкг отношение Teff к T-reg оставалось высоким во все моменты времени. Это объясняется долговременным деплетированием Tregs при этих дозах. Примечательно, что при более высокой дозе (200 мкг), несмотря на долговременное деплетирование Treg, наблюдалось лишь незначительное улучшение отношения к 8-му дню. Это можно объяснить некоторым деплетированием ICOSINT эффекторных клеток при высокой концентрации STIM003.
В целом данные продемонстрировали деплетирование TREG и увеличение соотношения эффекторные клетки : Т reg при всех тестируемых дозах. Однако, оптимальная доза, составляющая 60 мкг (ок. 3 мг/кг) обеспечивала как долгосрочное деплетирование T-reg, так и самые высокие значения соотношения T-eff к T-reg, которые ассоциируются с наиболее благоприятным иммунным контекстом для инициирования противоопухолевого иммунного ответа. Интересно, что аналогичная картина наблюдалась в крови, причем промежуточная доза 60 мкг была связана с самым высоким соотношением T-eff к T-reg. Важно отметить, что в крови улучшение соотношения наблюдалось в более ранний момент времени (между 3-м и 4-м днями).
Активация эффекторных клеток в ответ на STIM003
Поверхностную экспрессию CD107a на опухоль-инфильтрующих Т-эффекторных клетках ранее идентифицировали как надежный маркер для клеток, которые были активированы и проявляют цитотоксическую активность [44]. В данном исследовании этот маркер использовался для подтверждения того, что STIM003, помимо деплетирования Т-regs, может стимулировать цитотоксическую активность эффекторных Т-клеток в микроокружении опухоли. Интересно отметить, что на 8-й день после лечения наблюдалось увеличение поверхностной экспрессии CD107a на компартментах как CD4, так и CD8 Т-клеток, при всех дозах STIM003. Более того, такое повышение экспрессии CD107a на поверхности как CD4, так и CD8 Т-клеток, по-видимому, выходит на плато, когда животным вводят дозу 60 мкг, поскольку при 200 мкг улучшений не наблюдалось.
Чтобы дополнительно продемонстрировать активацию эффекторных клеток в микроокружении опухоли, был проведен анализ высвобождения цитокинов CD4 и CD8 ОИЛ методом FACS. Как и ожидалось, и в соответствии с данными по агонизму in vitro, приведенными в предыдущих Примерах данного документа, STIM003 mIgG2a во всех дозах промотировал продуцирование провоспалительных цитокинов IFN-γ и TNF-α эффекторными CD4 и CD8 Т-клетками. Индукция продуцирования провоспалительных цитокинов оказалась высокой при оптимальной дозе 60 мкг. Действительно, 60 мкг STIM003 значительно увеличивали продуцирование цитокинов CD4 Т-клетками. Аналогичная тенденция наблюдалась для продуцирования провоспалительного цитокина IFN-γ и TNF-α эффекторными CD8 Т-клетками в микроокружении опухоли. Данные приведены на Фигуре 45.
Таким образом, STIM003 во всех дозах приводил к активации Т-клеток в микроокружении опухоли, на что указывает (1) наличие маркера дегрануляции CD107a на их поверхности и (2) продуцирование Th1-цитокинов (IFNγ и TNFα) Т-клетками. Это свидетельствует о том, что STIM003 сильно влияет на иммунный контекст в микроокружении опухоли и играет двойную роль деплетирования клеток Treg и стимулирования активности убийства Т эффекторных клеток.
Определение доз для человека
Основываясь на данных доклинической эффективности, полученных для мышей, можно сделать предварительные прогнозы клинической дозы, пригодной для людей, на основе соответствующей биологической площади поверхности (BSA) [45].
Например, принимая оптимальную дозу анти-ICOS IgG для мышей равной 3 мг/кг (60 мкг), и используя методы работы [45], соответствующая доза для человека составляет 0,25 мг/кг.
По формулам Мостеллера, для индивидуума весом 60 кг и ростом 1,70 м, величина БСА составляет 1,68 м2. Умножение дозы в мг/кг на коэффициент, равный 35,7 (60/1,68) дает фиксированную дозу 15 мг. Для человека весом 80 кг соответствующая фиксированная доза составила бы 20 мг.
Дозы могут быть оптимизированы для терапии человека в клинических испытаниях для определения безопасных и эффективных схем лечения.
Пример 25: Биоинформационный анализ данных для образцов опухолей
Одной из целевых групп различных видов рака в соответствии с данным изобретением являются раковые опухоли, связанные с относительно высоким уровнем ICOS+ иммуносупрессивных Tregs.
Чтобы идентифицировать типы рака, ассоциированные с высоким содержанием Tregs, были получены данные транскриптома из общедоступного массива данных Атлас ракового генома (The Cancer Genome Atlas, TCGA) и проанализированы на уровни экспрессии ICOS и FOXP3. TCGA представляет собой крупномасштабные исследования, в которых каталогизированы геномные и транскриптомные данные, накопленные для различных видов рака, и включает мутации, варианты числа копий, экспрессию гена мРНК и микроРНК (miRNA) и метилирование ДНК, наряду со значительным объемом метаданных образца.
Анализ обогащения набора генов (GSEA) проводили следующим образом. Данные РНК-секвенирования (RNA seq) генной экспрессии, собранные в рамках консорциума TCGA, были загружены из браузера функциональной геномики UCSC Xena Functional Genomics Browser как log2(normalized_count+1). Образцы неопухолевой ткани исключали из набора данных, оставляя данные для 20530 генов из 9732 образцов. Алгоритм из работы [46] и его реализация в [47], позволяющий вычислить оценки обогащения для генов в определенном наборе генов, использовали для транспонирования значений уровня гена в оценку набора генов для каждого образца. Набор генов, представляющий интерес, был определен как содержащий как ICOS, так и FOXP3. Образцы были сгруппированы по первичному заболеванию, а оценки ssGSEA для каждой группы были сопоставлены по 33 группам первичных заболеваний. Было обнаружено, что группами заболеваний, продемонстрировавшими наивысшие медианные показатели, являются лимфоидная неопластическая диффузная крупноклеточная b-клеточная лимфома, тимома, плоскоклеточная карцинома головы и шеи, хотя диффузная крупноклеточная b-клеточная лимфома показала мультимодальное распределение баллов с подмножеством, имеющим высокие показатели, и остальной группой с оценками ниже медианной для группы.
В ранговом порядке от наивысшего до самого низкого значения ssGSEA для экспрессии ICOS и FOXP3, верхними 15 типами рака были:
TGCT (n=156) герминальная опухоль яичников
STAD (n=415) аденокарцинома желудка
CESC (n=305)цервикальная плоскоклеточная карцинома и эндоцервикальная аденокарцинома
LUAD (n=517) аденокарцинома легкого
LAML (n=173) острый миелоидный лейкоз
ESCA (n=185) карцинома пищевода
LUSC (n=502) плоскоклеточная карцинома легкого
LIHC (n=373) гепатоцеллюлярная карцинома печени
Причем n - количество образцов от пациентов с этим типом рака в наборе данных TCGA. Анти-ICOS антитела, раскрытые в данном документе, могут быть использованы для лечения этих и других видов рака.
Раки, которые связаны с относительно высоким уровнем ICOS+ иммуносупрессивных Tregs, и которые дополнительно экспрессируют PD-L1, могут особенно хорошо реагировать на лечение комбинацией анти-ICOS антитела и анти-PD-L1 антитела. Соответствующие схемы лечения и предназначенные для этого антитела были уже подробно описаны в предшествующем описании.
Используя тот же набор данных TCGA, что и раньше, оценки обогащения для ICOS и FOXP3 были скоррелированы с уровнями экспрессии PD-L1 с использованием ранговой корреляции Спирмена, и сгруппированы по признаку первичного заболевания. Были рассчитаны значения Р для каждой группы, а значение p, равное 0,05 (с коррекцией множественного сравнения по Бонферрони), было принято как статистически значимое. Группами заболеваний с наивысшими корреляциями между ICOS/FOXP3 и экспрессией PD-L1 были:
TGCT (n=156) герминогенная опухоль яичников
BLCA (n=407) уротелиальная карцинома мочевого пузыря
CESC (n=305) цервикальная плоскоклеточная карцинома и эндоцервикальная аденокарцинома
STAD (n=415) аденокарцинома желудка
LUAD (n=517) аденокарцинома легкого
Пациенты могут быть отобраны для лечения после анализа, определяющего, что их рак ассоциирован с ICOS+ иммуносупрессивными Tregs и экспрессией PD-L1. Для типов рака, имеющих, как указано выше, высокую степень корреляции, может быть достаточно определения наличия чего-то одного из ICOS+ иммуносупрессивных Tregs и экспрессии PD-L1 (например, выше порогового значения). В этом контексте могут использоваться иммуногистохимические анализы PD-L1.
Пример 26: Оценка дополнительных анти-ICOS антител
Последовательности антител CL-74570 и CL-61091, идентифицированных в Примере 12, были синтезированы и экспрессированы в формате IgG1 в клетках HEK.
Функциональную характеризацию этих антител проводили с использованием анализа методом гомогенной флуоресценции с временным разрешением (HTRF), аналогичного описанному в Примере 6, с модификациями для адаптации анализа к использованию очищенного IgG1, а не супернатанта ВСТ. Вместо супернатанта ВСТ использовали 5 мкл супернатанта, содержащего антитела IgG1 человека, экспрессированные клетками HEK, и общий объем составлял до 20 мкл на лунку, при использовании буфера для HTRF, как было описано ранее. В качестве отрицательного контроля использовали антитело IgG1 человека. Оба антитела проявляли эффект более 5% для связывания с ICOS человека и мыши, при расчете с использованием Уравнения 1, и поэтому подтвердили положительные результаты теста в этом анализе.
Способность этих антител связывать ICOS человека и мыши, экспрессируемый на поверхности клеток CHO-S, была дополнительно подтверждена с использованием анализа Mirrorball. В этом анализе 5 мкл супернатанта, содержащего анти-ICOS Ig-1, переносили во все лунки 384-луночных черных планшетов mirrorball (Corning). Связывание анти-ICOS антител детектировали путем добавления во все лунки 10 мкл козьего анти-человеческого 488 (Jackson Immunoresearch), разбавленного в аналитическом буфере (PBS + 1% BSA + 0,1% азида натрия) в концентрации 0,8 мг/мл.
Для лунок положительного контроля в планшеты добавляли 5 мкл эталонного антитела, разбавленного в среде для анализа до 2,2 мкг/мл. Для лунок отрицательного контроля в планшеты добавляли 5 мкл контрольного гибрида IgG1, разведенного в среде для анализа до 2,2 мкг/мл. 10 мкМ DRAQ5 (Thermoscientific) добавляли к 0,4×106/мл клеткам, ресуспендированным в буфере для анализа, и добавляли 5 мкл во все лунки. Планшеты инкубировали в течение 2 часов при 4 градусах.
Интенсивность флуоресценции измеряли с помощью планшет-ридера Mirrorball (ТТР Labtech), измеряя Alexafluor 488 (возбуждение 493 нм, излучение 519 нм) для популяции 500-700 одиночных клеток. Аналитический сигнал измеряли как медиану средней интенсивности (канал FL2).
Общее связывание определяли с использованием эталонного антитела при концентрации анализа 2,2 мкг/мл. Неспецифическое связывание определяли с использованием контроля с гибридом hIgG1 при концентрации анализа 2,22 мкг/мл. Оба антитела проявляли эффект более 1 процента и поэтому подтвердили положительные результаты теста в этом анализе.

Каждый из CL-74570 и CL-61091 также продемонстрировал связывание с ICOS человека и мыши, экспрессируемым на клетках CHO-S, по результатам определения методом проточной цитометрии. Скрининг методом FACS проводили по способу, аналогичному описанному в Примере 6, с модификациями для адаптации анализа к использованию очищенного IgG1, а не супернатанта ВСТ. Оба антитела продемонстрировали связывание, более чем в 10 раз превышающее среднее значение геометрического среднего (geomean) для связывания отрицательного контроля с клетками hICOS, mICOS и WT СНО.
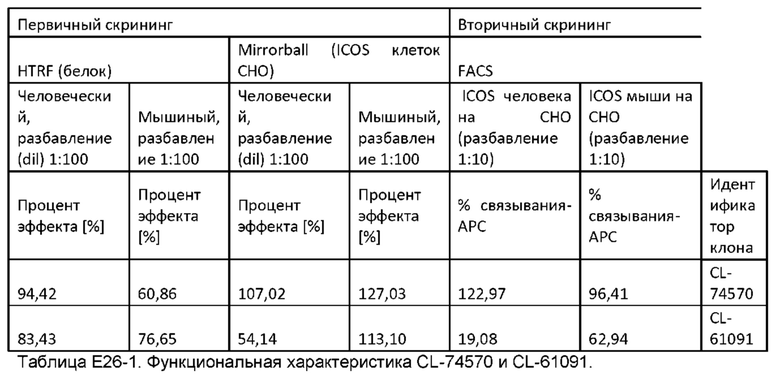
Список литературы
1. Hutloff A, et al. ICOS is an inducible T-cell co-stimulator structurally and functionally related to CD28. Nature. 1999 Jan 21; 397(6716): 263-6.
2. Beier KC, et al. Induction, binding specificity and function of human ICOS. Eur J Immunol. 2000 Dec; 30 (12): 3707-17.
3. Coyle AJ, et al. The CD28-related molecule ICOS is required for effective T cell-dependent immune responses. Immunity. 2000 Jul; 13(1): 95-105.
4. Dong C, et al. ICOS co-stimulatory receptor is essential for T-cell activation and function. Nature. 2001 Jan 4; 409(6816): 97-101.
5. Mak TW, et al.. Costimulation through the inducible costimulator ligand is essential for both T helper and В cell functions in T cell-dependent В cell responses. Nat Immunol. 2003 Aug; 4(8): 765-72.
6. Swallow MM, Wallin JJ, Sha WC. B7h, a novel costimulatory homolog of B7.1 and B7.2, is induced by TNFalpha. Immunity. 1999 Oct; 11(4): 423-32.
7. Wang S, et al. Costimulation of T cells by B7-H2, a B7-like molecule that binds ICOS. Blood. 2000 Oct 15; 96(8):2808-13.
8. Conrad C, Gilliet M. Plasmacytoid dendritic cells and regulatory T cells in the tumor microenvironment: A dangerous liaison. Oncoimmunology. 2013 May 1; 2(5): e2388.
9. Simpson et al., Fc-dependent depletion of tumor-infiltrating regulatory T cells co-defines the efficacy of anti-CTLA-4 therapy against melanoma. J. Exp. Med. 210(9): 1695-1710 2013
10. Fu T, He Q, Sharma P. The ICOS/ICOSL pathway is required for optimal antitumor responses mediated by anti-CTLA-4 therapy. Cancer Res. 2011 Aug 15; 71(16): 5445-54.
11. Fan X, Quezada SA, Sepulveda MA, Sharma P, Allison JP. Engagement of the ICOS pathway markedly enhances efficacy of CTLA-4 blockade in cancer immunotherapy. J Exp Med. 2014 Apr 7; 211(4): 715-25.
12. Carthon, B.C., et al. Preoperative CTLA-4 blockade: Tolerability and immune monitoring in the setting of a presurgical clinical trial. Clin. Cancer Res. 16: 2861-2871.
13. Liakou CI, et al. CTLA-4 blockade increases IFNgamma-producing CD4 + ICOShi cells to shift the ratio of effector to regulatory T cells in cancer patients. Proc Natl Acad Sci USA. 2008 Sep 30; 105(39): 14987-92.
14. Vonderheide, R.H., et al. 2010. Tremelimumab in combination with exemestane in patients with advanced breast cancer and treatment-associated modulation of inducible costimulator expression on patient T cells. Clin. Cancer Res. 16: 3485-3494.
15. Preston CC, et al., The ratios of CD8+ T cells to CD4+ CD25+ FOXP3+ and FOXP3- T cells correlate with poor clinical outcome in human serous ovarian cancer. PLoS One Nov 14; 8(11): e80063.
16. Hodi FS, et al., Immunologic and clinical effects of antibody blockade of cytotoxic T lymphocyte-associated antigen 4 in previously vaccinated cancer patients. PNAS 2008 Feb 26; 105(8): 3005-10
17. Chattopadhyay et al., Structural Basis of Inducible Costimulatory Ligand Function: Determination of the Cell Surface Oligomeric State and Functional Mapping of the Receptor Binding Site of the Protein, J. Immunol. 177 (6): 3920-3929 2006
18. Lefranc MP, IMGT unique numbering for immunoglobulin and T cell receptor variable domains and Ig superfamily V-like domains, Dev Comp Immunol. 27(1): 55-77 2003
19.  et al., "Antibody-Dependent Phagocytosis of Tumor Cells by Macrophages: A Potent Effector Mechanism of Monoclonal Antibody Therapy of Cancer", Cancer Res., 75(23), December 1, 2015
et al., "Antibody-Dependent Phagocytosis of Tumor Cells by Macrophages: A Potent Effector Mechanism of Monoclonal Antibody Therapy of Cancer", Cancer Res., 75(23), December 1, 2015
20. Laza ret al., 2006, Proc. Natl. Acad. Sci. U.S.A., Mar 14; 103(11): 4005-10
21. Dall et al., Immunol 2002; 169: 5171-5180
22. Natsume et al., 2009, Drug Des. Devel. Ther., 3: 7-16, или Zhou Q., Biotechnol. Bioeng., 2008, Feb 15, 99(3): 652-65)
23. Shields et al., 2001, J. Biol. Chem., Mar 2; 276(9): 6591-604)
24. Idusogie et al., J. Immunol., 2001, 166: 2571-2575
25. Natsume et al., 2008, Cancer Res., 68: 3863-3872
26. Alexandrov LB, et al. Signatures of mutational processes in human cancer. Nature. 2013 Aug 22; 500(7463): 415-21
27. Martin-Orozco et al., Melanoma Cells Express ICOS Ligand to Promote the Activation and Expansion of T-Regulatory Cells, Cancer Research 70(23): 9581-9590 2010
28. Houot et al., Therapeutic effect of CD137 immunomodulation in lymphoma and its enhancement by Treg depletion, Blood 114: 3431-3438 2009
29. Baruch K. et al. PD-1 immune checkpoint blockade reduces pathology and improves memory in mouse models of Alzheimer's disease. Nat Med 22(2): 137-137 2016
30. Curran et al., PD01 and CTLA-4 combination blockade expands infiltrating T cells and reduces regulatory T and myeloid cells within B16 melanoma tumours, PNAS 107(9): 4275-4280 2010
31. Sim et al., IL-2 therapy promotes suppressive ICOS+ Treg expansion in melanoma patients, J Clin Invest 2014
32. Sim et al., IL-2 variant circumvents ICOS+ regulatory T cell expansion and promotes NK cell activation, Cancer Immunol Res 2016
33. Kroemer et al. Immunologic Cell Death in Cancer Therapy, Ann Rev Immunol. 31: 51-72 2013
34. Galluzzi, Zitvogel & Kroemer Canc. Imm. Res. 4: 895-902 2016
35. Bos et al., Transient regulatory T cell ablation deters oncogene-driven breast cancer and enhances radiotherapy, J Exp Med 210(11): 2434-2446 2013
36. Sato et al., Spatially selective depletion of tumor-associated regulatory T cells with near-infrared photoimmunotherapy, Science Translational Medicine 8(352) 2016
37. Crotty S. T follicular helper cell differentiation, function, and roles in disease. Immunity. 2014 Oct 16; 41(4): 529-42.
38. Shields et al. (2002) JBC 277: 26733
39. Lee et al, Nature Biotechnology, 32: 6-363, 2014
40. Yusa K, Zhou L, Li MA, Bradley A, Craig NL. A hyperactive piggyBac transposase for mammalian applications, Proc Natl Acad Sci USA. 2011 Jan 25
41. Kilpatrick et al., Rapid development of affinity matured monoclonal antibodies using RIMMS; Hybridoma; 16(4): 381-9 Aug 1997
42. Simpson, T.R. et al., Fc-dependent depletion of tumor-infiltrating regulatory T cells co-defines the efficacy of anti-CTLA-4 therapy against melanoma. The Journal of experimental medicine, 210(9): 1695-710 2013
43. Selby, M.J. et al., Anti-CTLA-4 antibodies of IgG2a isotype enhance antitumor activity through reduction of intratumoral regulatory T cells. Cancer immunology research, 1(1): 32-42 2013.
44. Rubio V., et al. Ex vivo identification, isolation and analysis of tumor-cytolytic T cells. Nat Med. 2003 Nov; 9(11): 1377-82.
45. Nair & Jacob., A simple practice guide for dose conversion between animals and human. J Basic Clin Pharma 2016; 7: 27-31
46. D.A. Barbie, et al., "Systematic RNA interference reveals that oncogenic KRAS-driven cancers require TBK1.," Nature, vol. 462, no. 7269, pp. 108-12, 2009
47. 
 R. Castelo, and J. Guinney, "GSVA: gene set variation analysis for microarray and RNA-Seq data," BMC Bioinformatics, vol. 14, no. 1, p. 7, 2013
R. Castelo, and J. Guinney, "GSVA: gene set variation analysis for microarray and RNA-Seq data," BMC Bioinformatics, vol. 14, no. 1, p. 7, 2013
Последовательности
Антитело STIM001
Нуклеотидная последовательность домена VH: SEQ ID NO: 367

Аминокислотная последовательность домена VH: SEQ ID NO: 366

Аминокислотная последовательность VH CDR1: 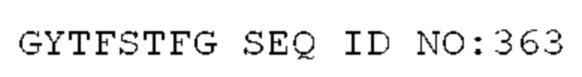
Аминокислотная последовательность VH CDR2: 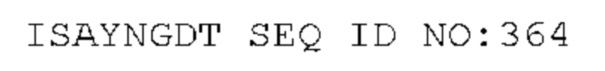
Аминокислотная последовательность VH CDR3: 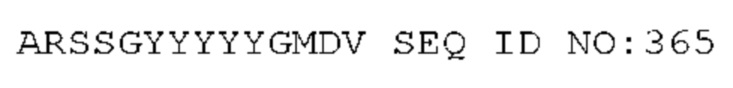
Нуклеотидная последовательность домена VL: SEQ ID NO: 374

Аминокислотная последовательность домена VL: SEQ ID NO: 373
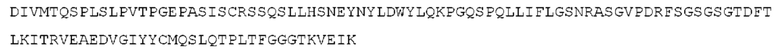
Аминокислотная последовательность VL CDR1: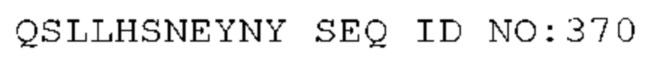
Аминокислотная последовательность VL CDR2: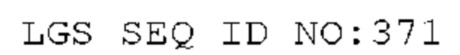
Аминокислотная последовательность VL CDR3: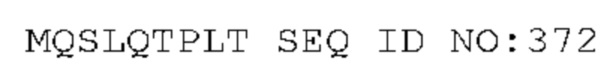
Антитело STIM002
Нуклеотидная последовательность домена VH: SEQ ID NO: 381
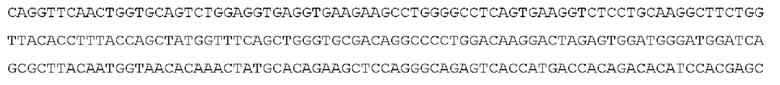
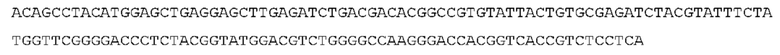
Аминокислотная последовательность домена VH: SEQ ID NO: 380
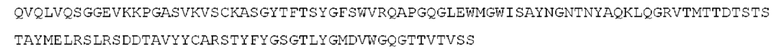
Аминокислотная последовательность VH CDR1: 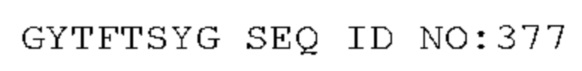
Аминокислотная последовательность VH CDR2: 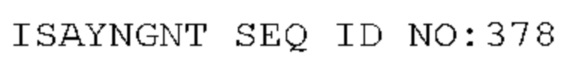
Аминокислотная последовательность VH CDR3: 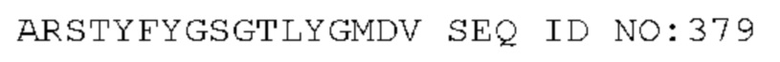
Нуклеотидная последовательность домена VL: 388
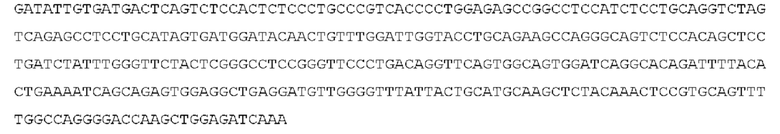
Скорректированная нуклеотидная последовательность домена VL STIM002: SEQ ID NO: 519

Аминокислотная последовательность домена VL: SEQ ID NO: 387
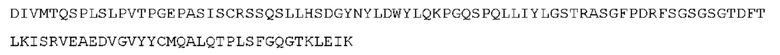
Аминокислотная последовательность VL CDR1: 
Аминокислотная последовательность VL CDR2: 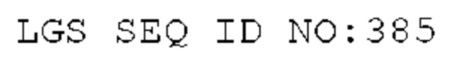
Аминокислотная последовательность VL CDR3: 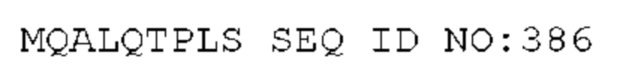
Антитело STIM002-B
Нуклеотидная последовательность домена VH: SEQ ID NO: 395
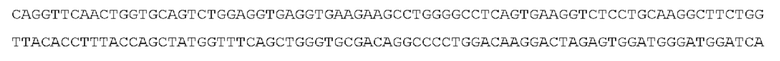
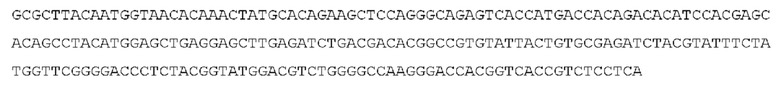
Аминокислотная последовательность домена VH: SEQ ID NO: 394

Аминокислотная последовательность VH CDR1: 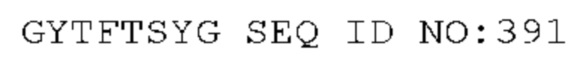
Аминокислотная последовательность VH CDR2: 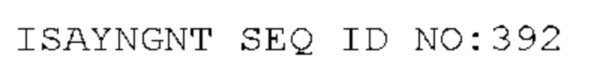
Аминокислотная последовательность VH CDR3: 
Нуклеотидная последовательность домена VL: SEQ ID NO: 402

Аминокислотная последовательность домена VL: SEQ ID NO: 401
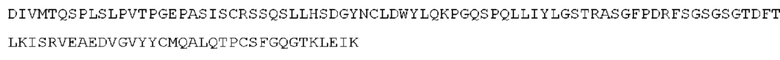
Аминокислотная последовательность VL CDR1: 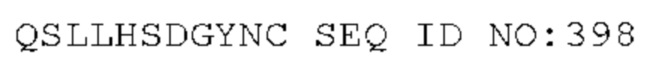
Аминокислотная последовательность VL CDR2: 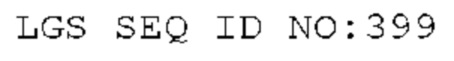
Аминокислотная последовательность VL CDR3: 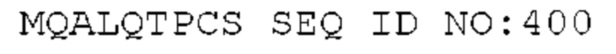
Антитело STIM003
Нуклеотидная последовательность домена VH: SEQ ID NO: 409

Скорректированная нуклеотидная последовательность домена VH STIM003: SEQ ID NO: 521
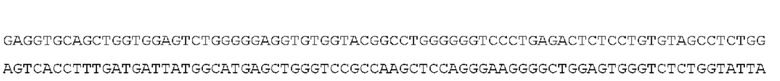
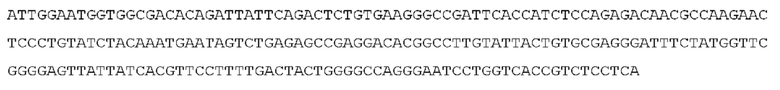
Аминокислотная последовательность домена VH: SEQ ID NO: 408
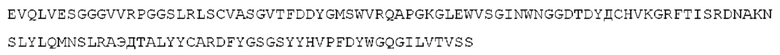
Аминокислотная последовательность VH CDR1: 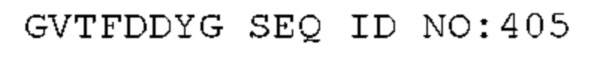
Аминокислотная последовательность VH CDR2: 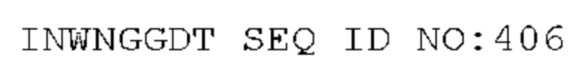
Аминокислотная последовательность VH CDR3: 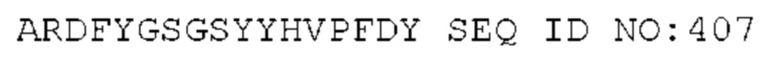
Нуклеотидная последовательность домена VL: SEQ ID NO: 416

Аминокислотная последовательность домена VL: SEQ ID NO: 415

Аминокислотная последовательность VL CDR1: 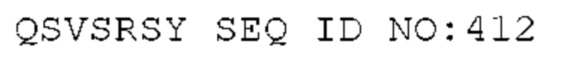
Аминокислотная последовательность VL CDR2: 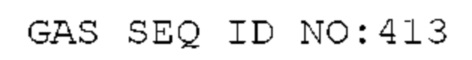
Аминокислотная последовательность VL CDR3: 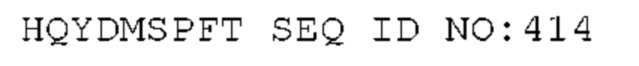
Антитело STIM004
Нуклеотидная последовательность домена VH:

SEQ ID NO: 423
Аминокислотная последовательность домена VH:
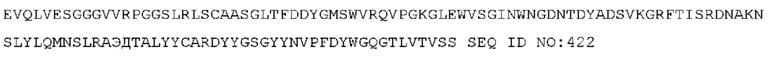
Аминокислотная последовательность VH CDR1: 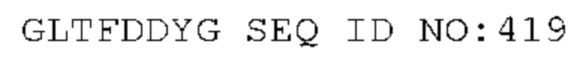
Аминокислотная последовательность VH CDR2: 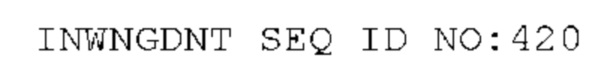
Аминокислотная последовательность VH CDR3: 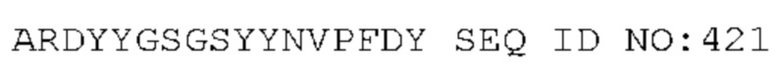
Нуклеотидная последовательность домена VL: SEQ ID NO: 431

Аминокислотная последовательность домена VL, кодируемая вышеуказанной нуклеотидной последовательностью домена VL.
Скорректированная нуклеотидная последовательность домена VL: SEQ ID NO: 430
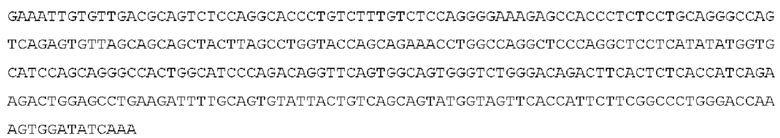
Скорректированная аминокислотная последовательность домена VL: SEQ ID NO: 432
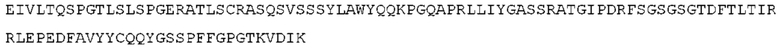
Аминокислотная последовательность VL CDR1: 
Аминокислотная последовательность VL CDR2: 
Аминокислотная последовательность VL CDR3: 
Антитело STIM005
Нуклеотидная последовательность домена VH: SEQ ID NO: 439

Аминокислотная последовательность домена VH: SEQ ID NO: 438
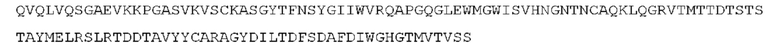
Аминокислотная последовательность VH CDR1: 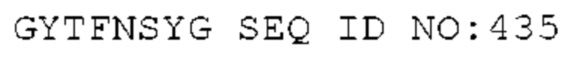
Аминокислотная последовательность VH CDR2: 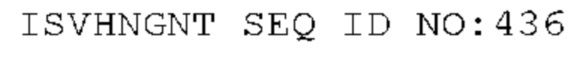
Аминокислотная последовательность VH CDR3: 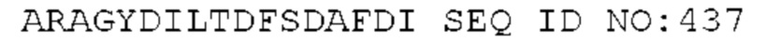
Нуклеотидная последовательность домена VL: SEQ ID NO: 446
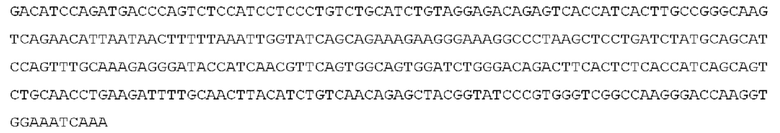
Аминокислотная последовательность домена VL: SEQ ID NO: 445
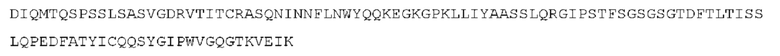
Аминокислотная последовательность VL CDR1: 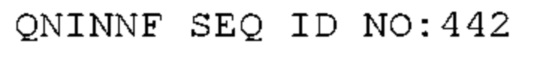
Аминокислотная последовательность VL CDR2: 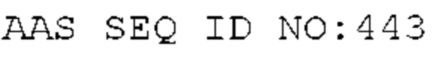
Аминокислотная последовательность VL CDR3: 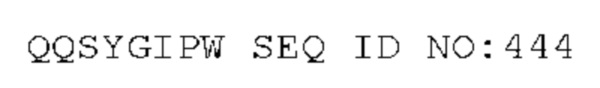
Антитело STIM006
Нуклеотидная последовательность домена VH: SEQ ID NO: 453

Аминокислотная последовательность домена VH: SEQ ID NO: 454

Аминокислотная последовательность VH CDR1: 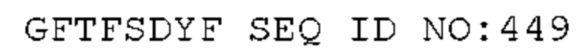
Аминокислотная последовательность VH CDR2: 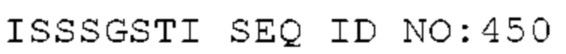
Аминокислотная последовательность VH CDR3: 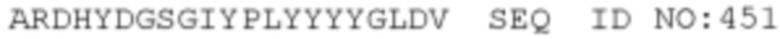
Нуклеотидная последовательность домена VL: SEQ ID NO: 460
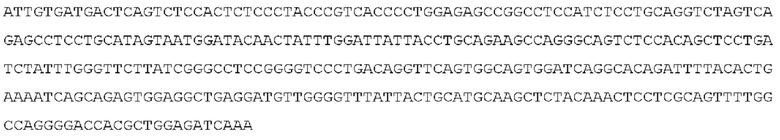
Аминокислотная последовательность домена VL: SEQ ID NO: 459
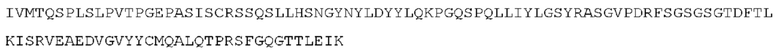
Аминокислотная последовательность VL CDR1: 
Аминокислотная последовательность VL CDR2: 
Аминокислотная последовательность VL CDR3: 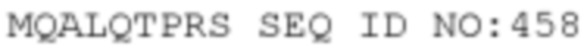
Антитело STIM007
Нуклеотидная последовательность домена VH: SEQ ID NO: 467

Аминокислотная последовательность домена VH: SEQ ID NO: 466
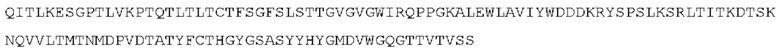
Аминокислотная последовательность VH CDR1: 
Аминокислотная последовательность VH CDR2: 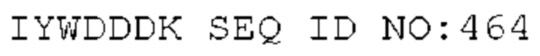
Аминокислотная последовательность VH CDR3: 
Нуклеотидная последовательность домена VL: SEQ ID NO: 474


Аминокислотная последовательность домена VL: SEQ ID NO: 473

Аминокислотная последовательность VL CDR1: 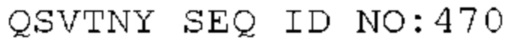
Аминокислотная последовательность VL CDR2: 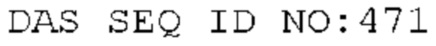
Аминокислотная последовательность VL CDR3: 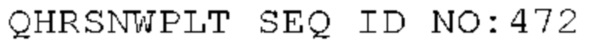
Антитело STIM008
Нуклеотидная последовательность домена VH: SEQ ID NO: 481

Аминокислотная последовательность домена VH: SEQ ID NO: 480
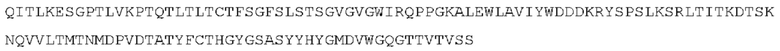
Аминокислотная последовательность VH CDR1: 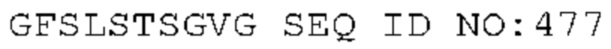
Аминокислотная последовательность VH CDR2: 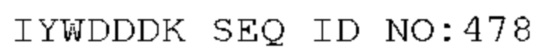
Аминокислотная последовательность VH CDR3: 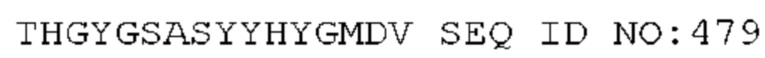
Нуклеотидная последовательность домена VL: SEQ ID NO: 488

Аминокислотная последовательность домена VL: SEQ ID NO: 489
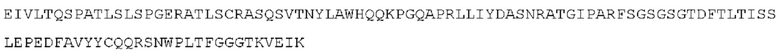
Аминокислотная последовательность VL CDR1: 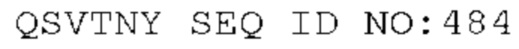
Аминокислотная последовательность VL CDR2: 
Аминокислотная последовательность VL CDR3: 
Антитело STIM009
Нуклеотидная последовательность домена VH: SEQ ID NO: 495

Аминокислотная последовательность домена VH: SEQ ID NO: 494

Аминокислотная последовательность VH CDR1: 
Аминокислотная последовательность VH CDR2: 
Аминокислотная последовательность VH CDR3: 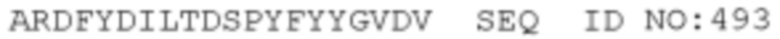
Нуклеотидная последовательность домена VL: SEQ ID NO: 502
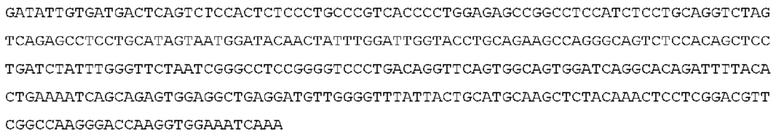
Аминокислотная последовательность домена VL: SEQ ID NO: 501
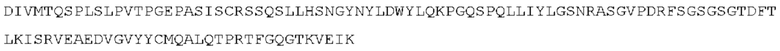
Аминокислотная последовательность VL CDR1: 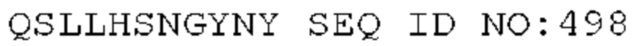
Аминокислотная последовательность VL CDR2: 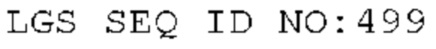
Аминокислотная последовательность VL CDR3: 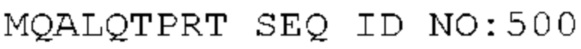
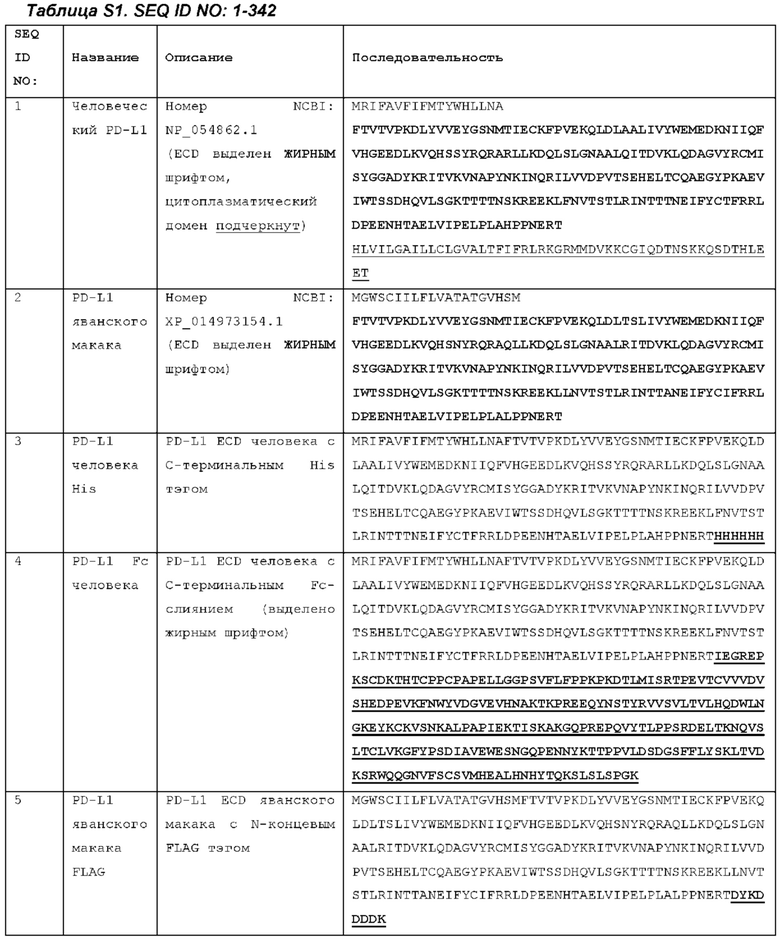

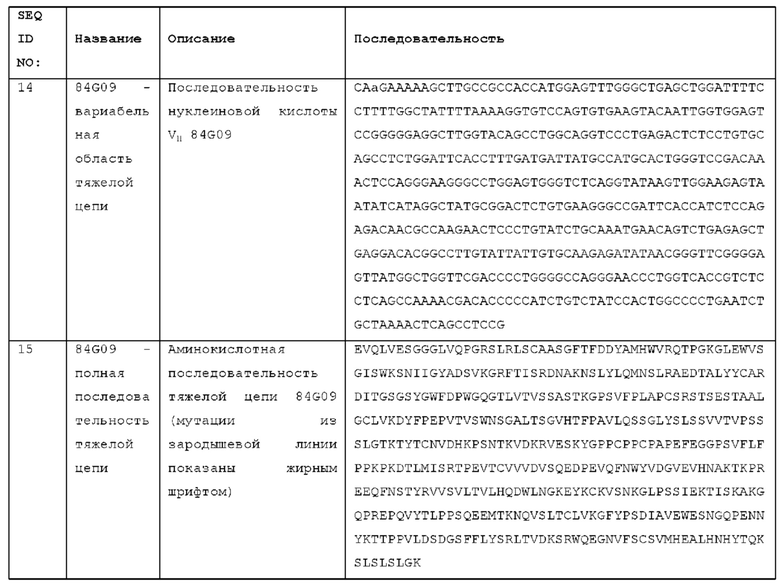
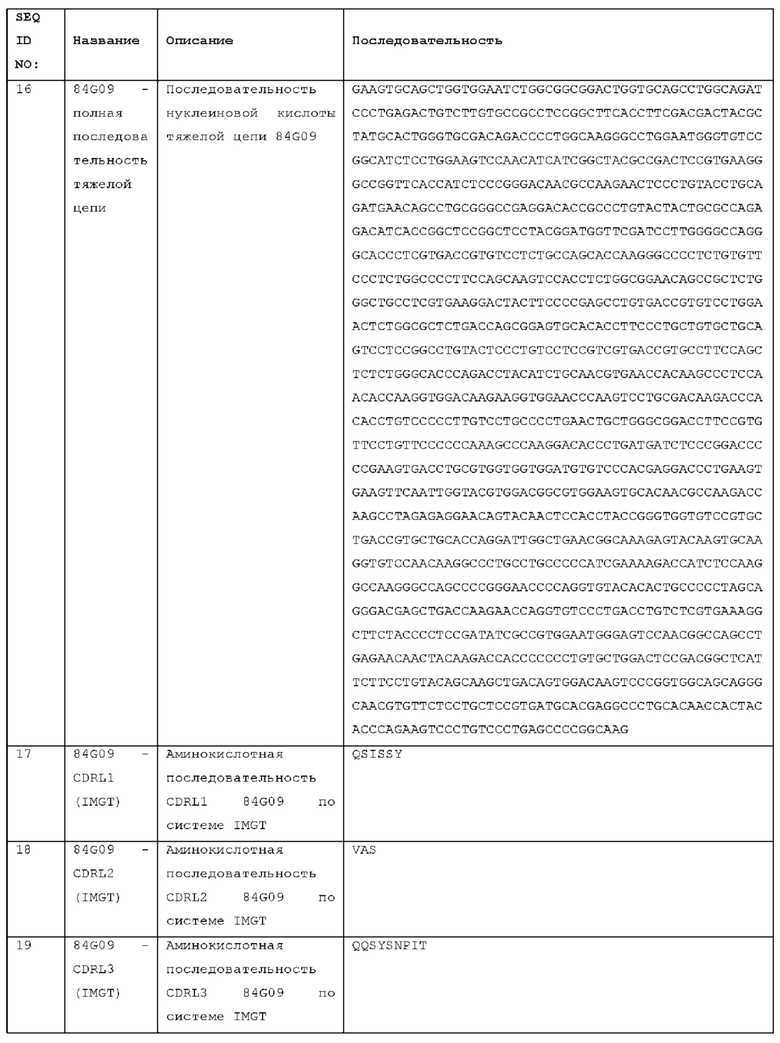
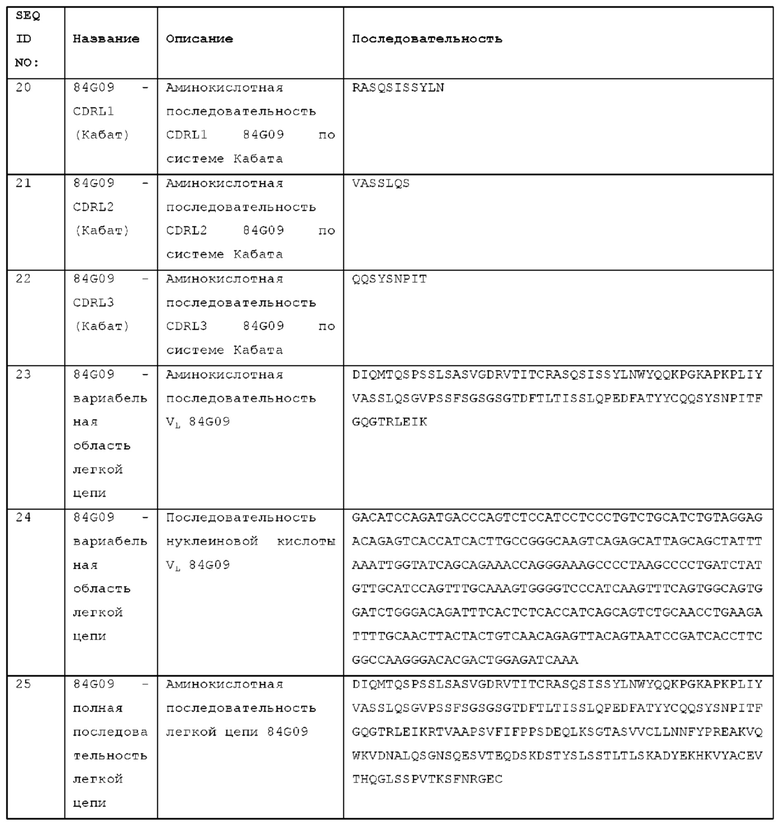
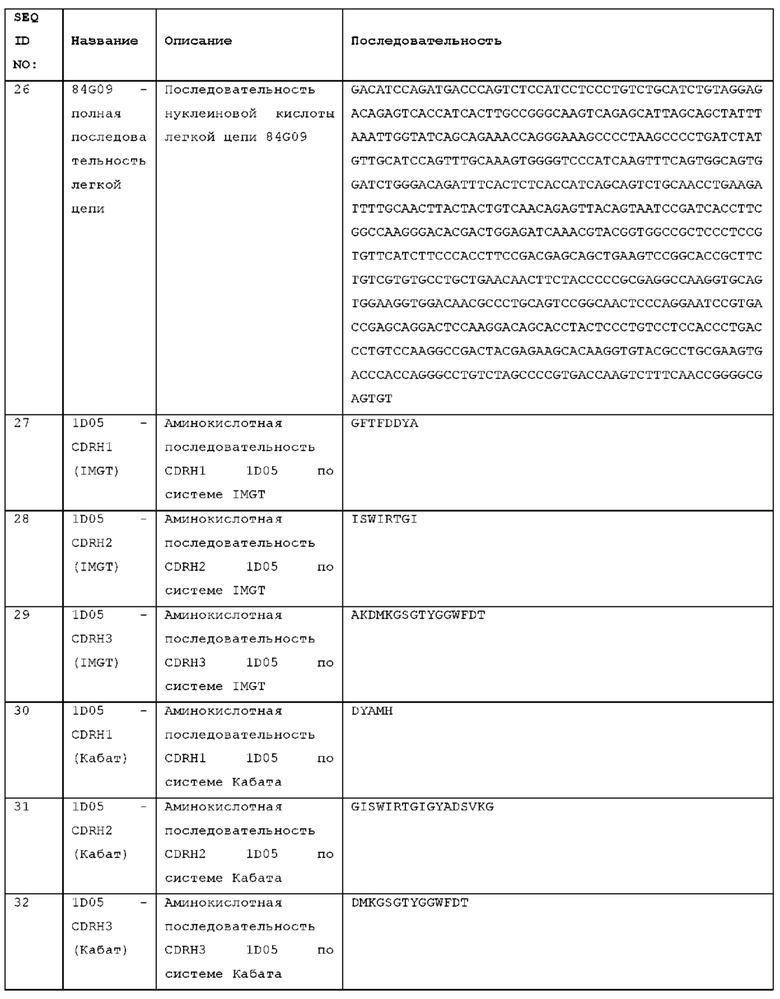
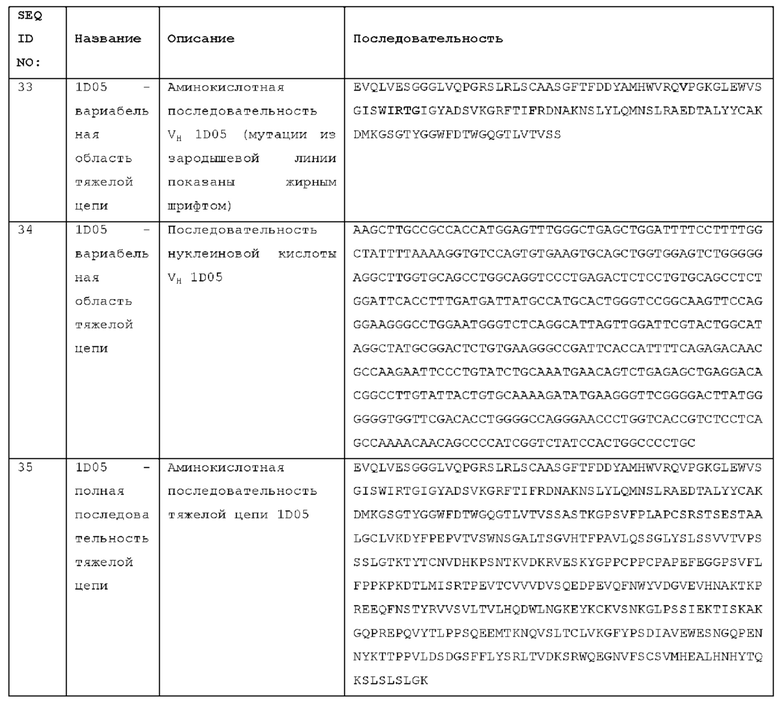
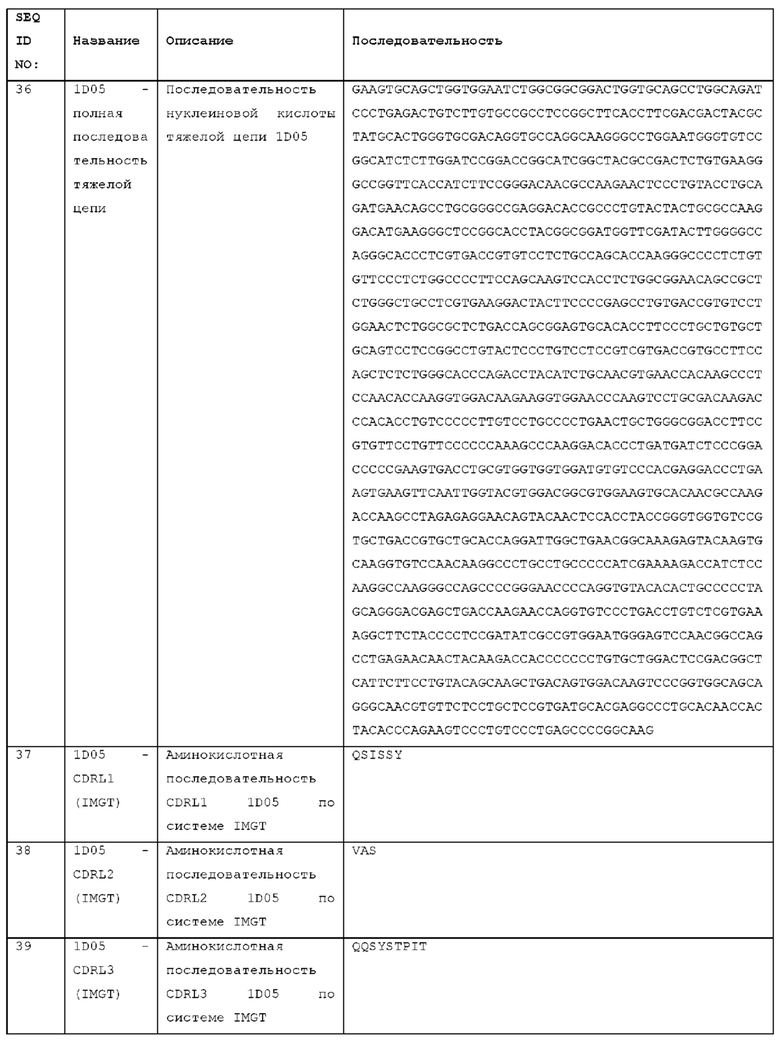
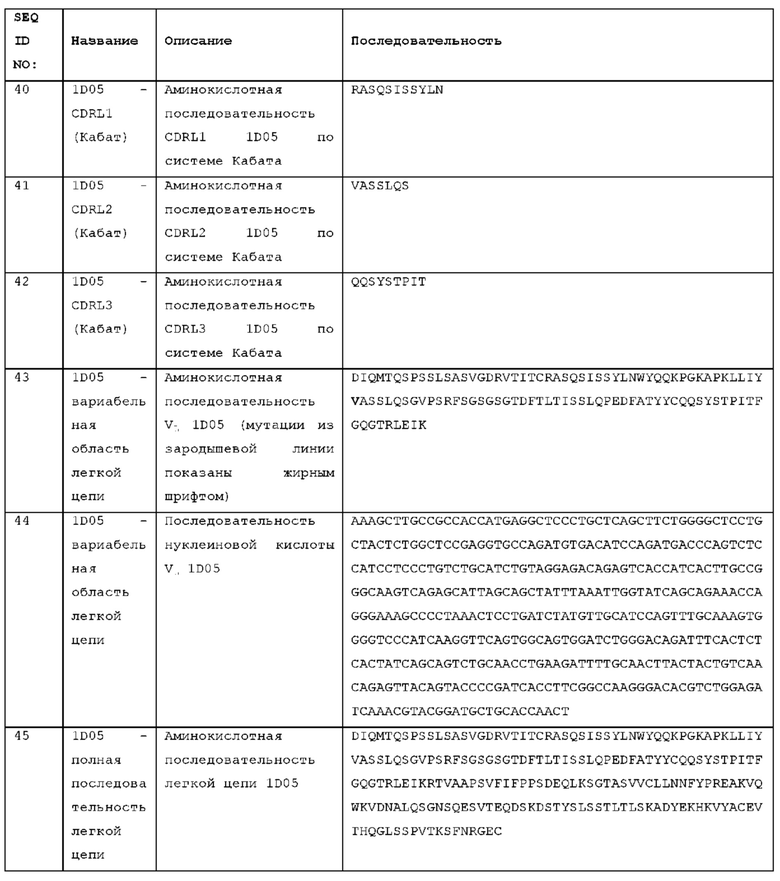
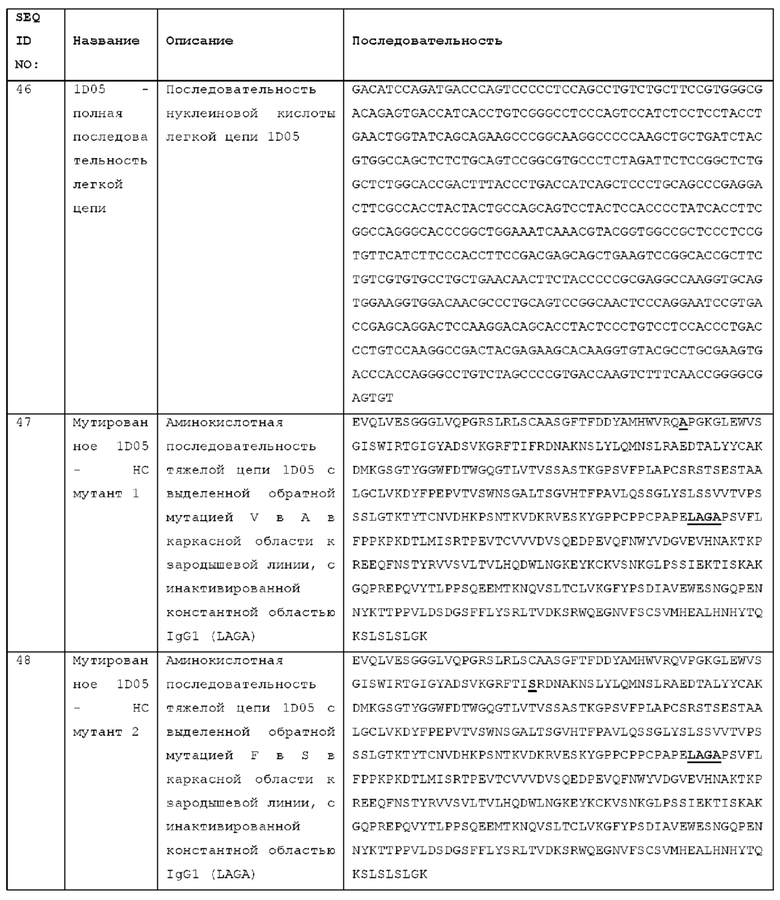
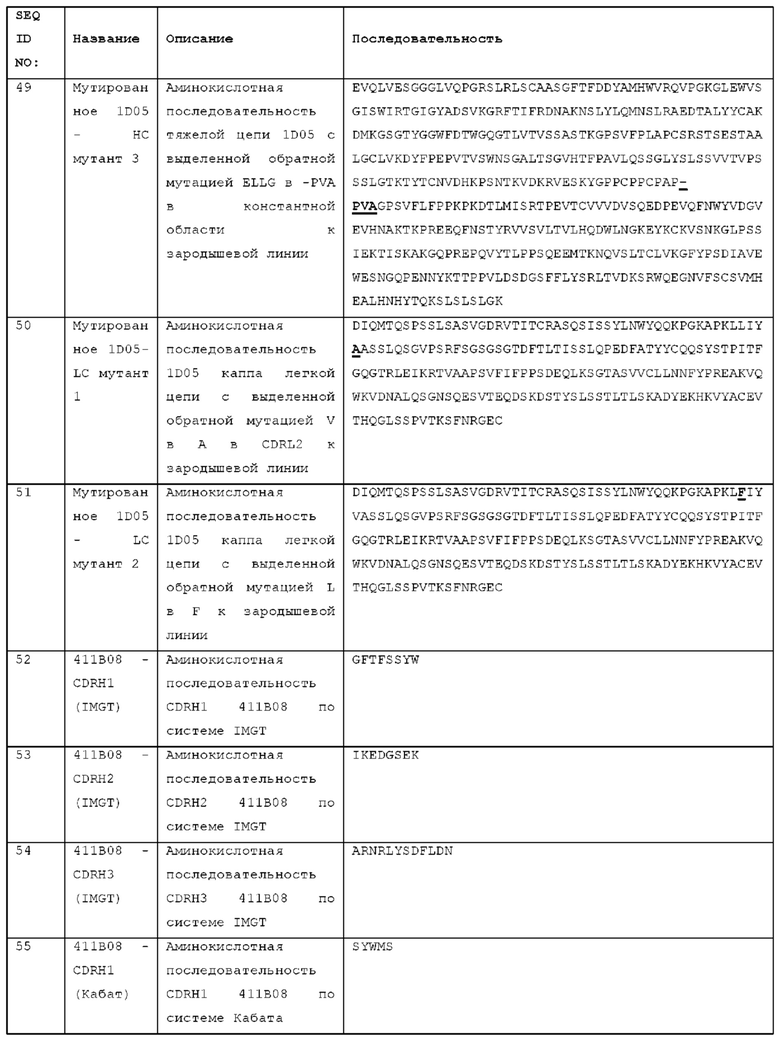
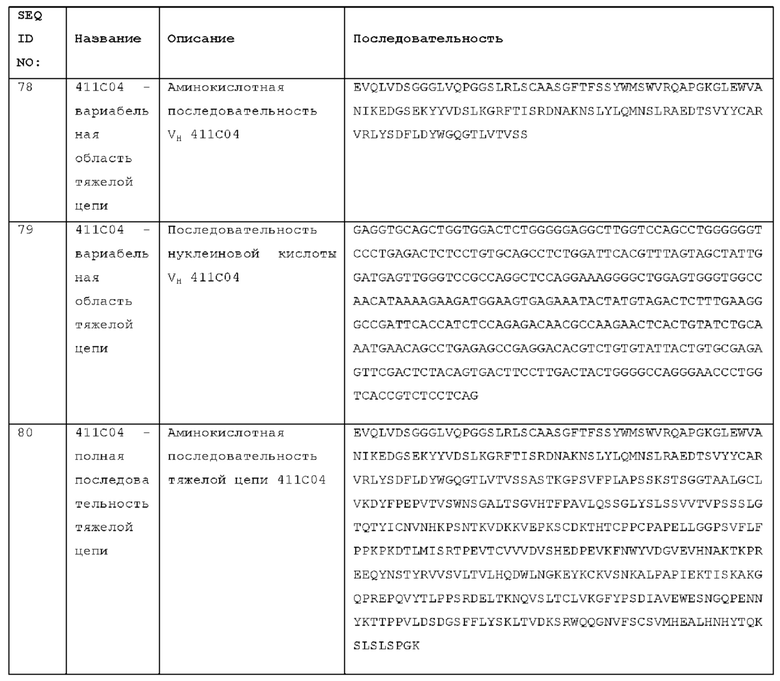
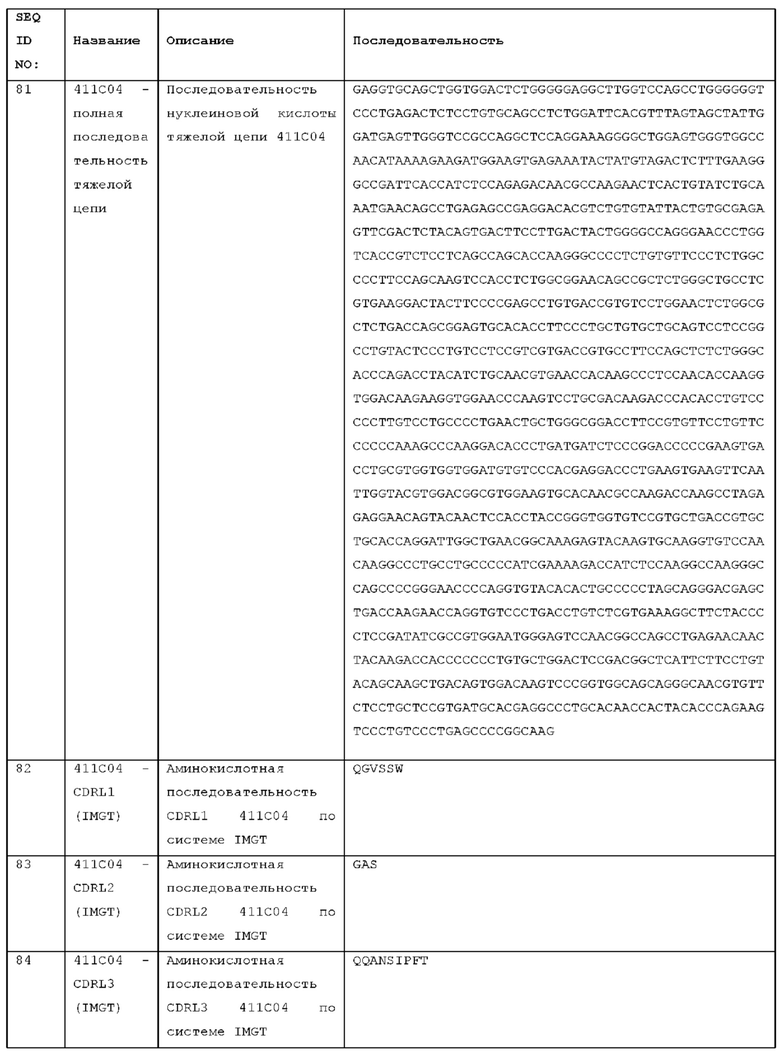
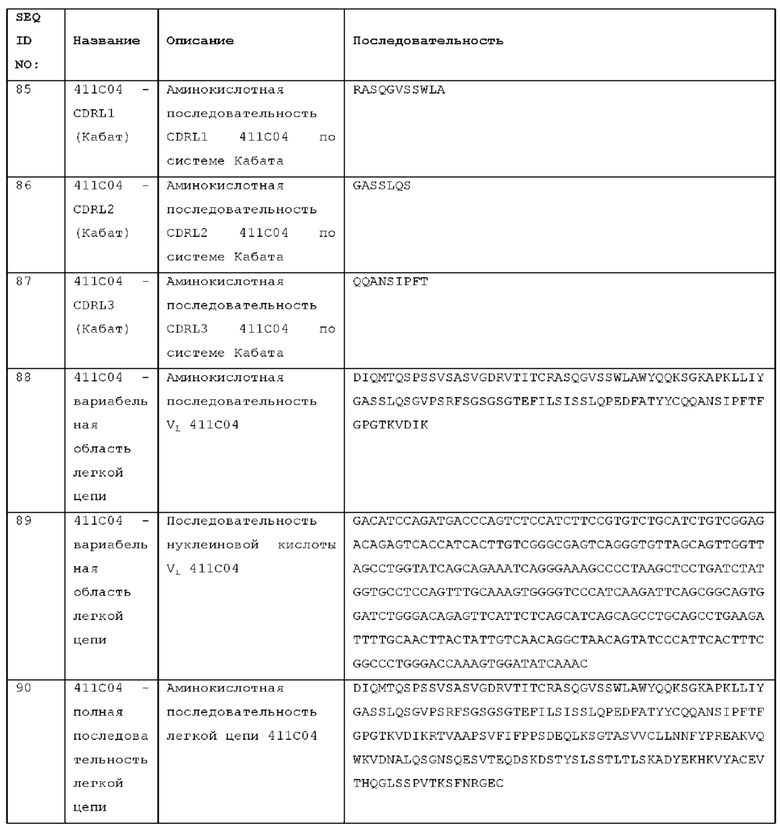
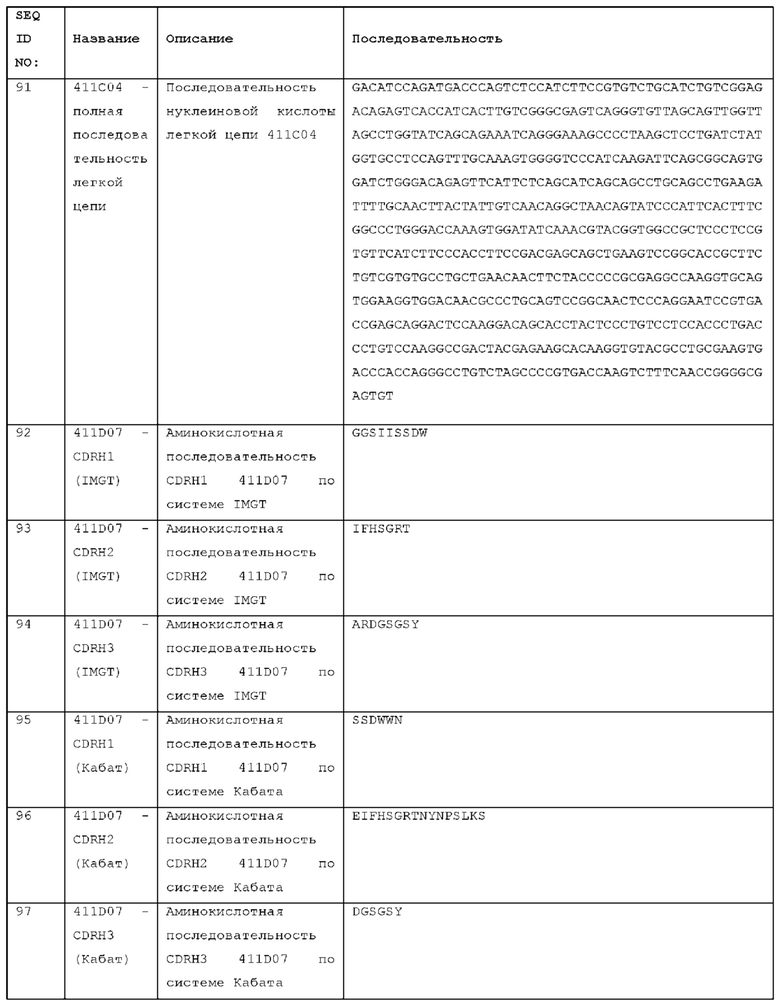



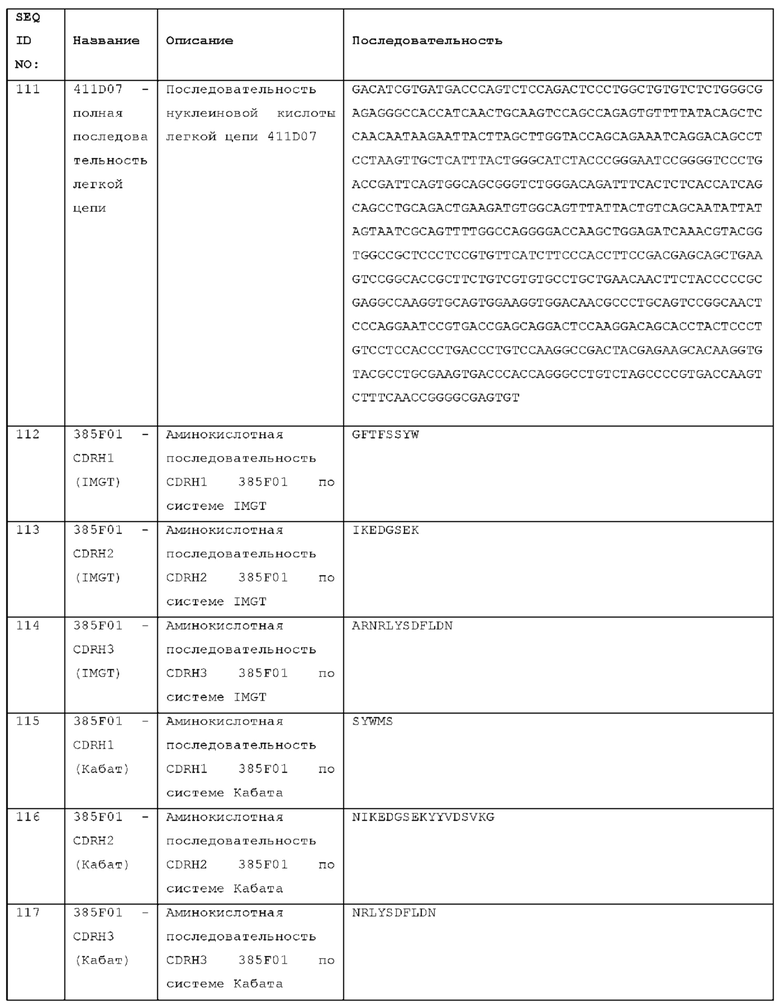
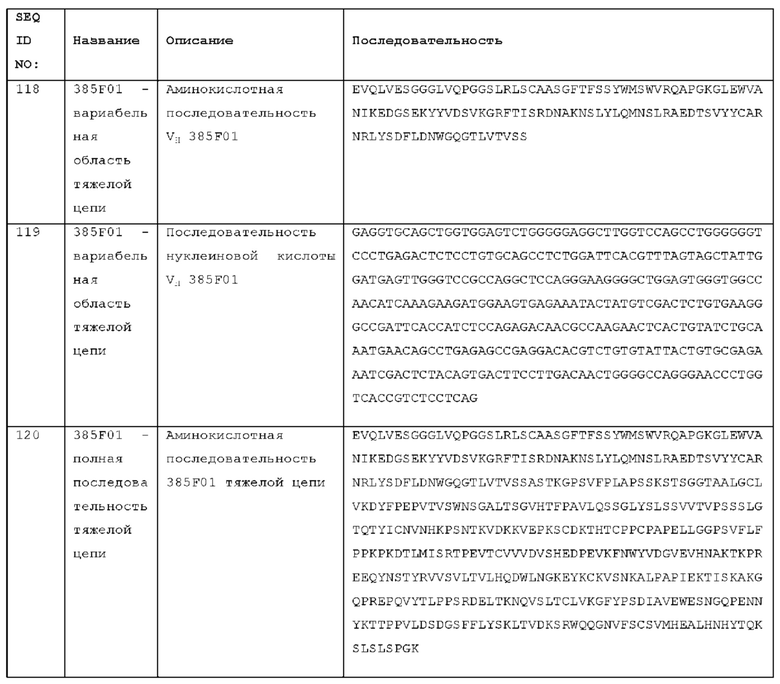
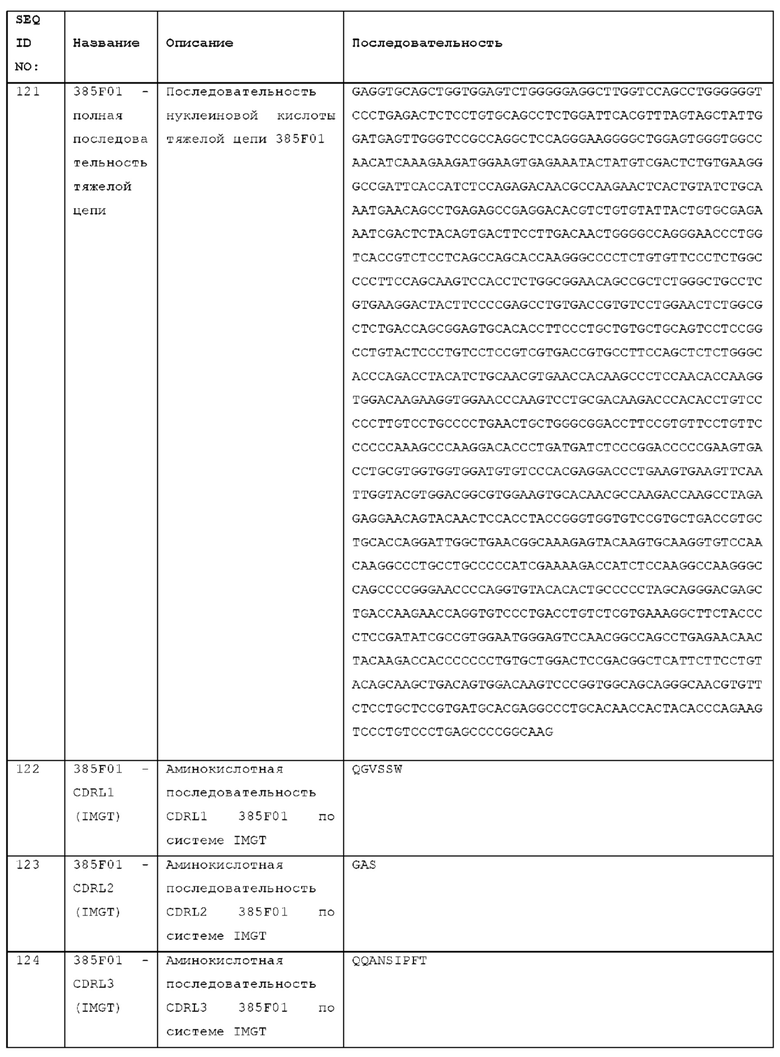
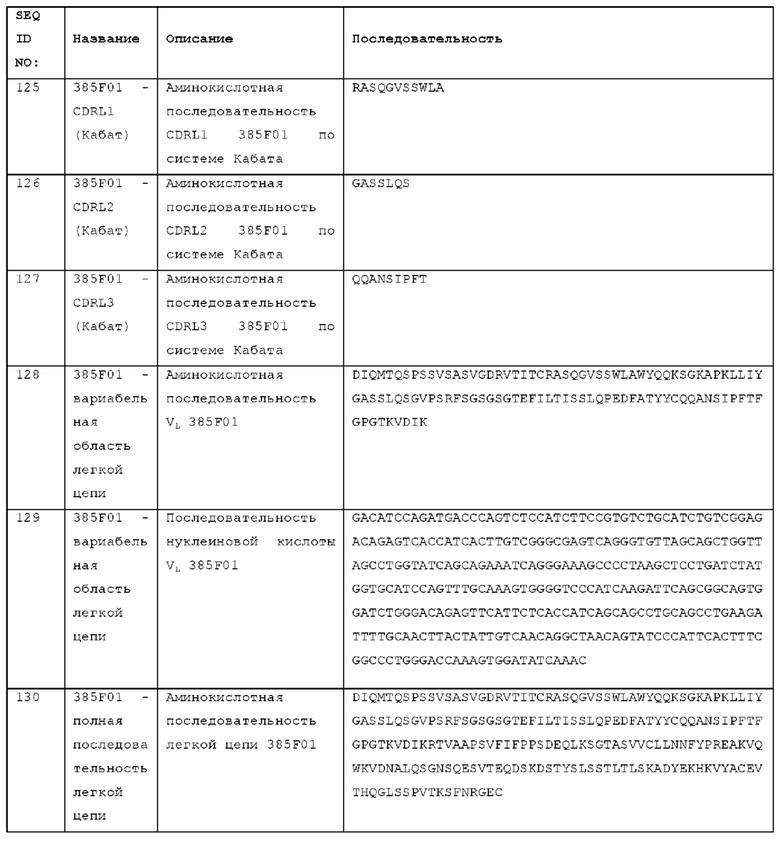
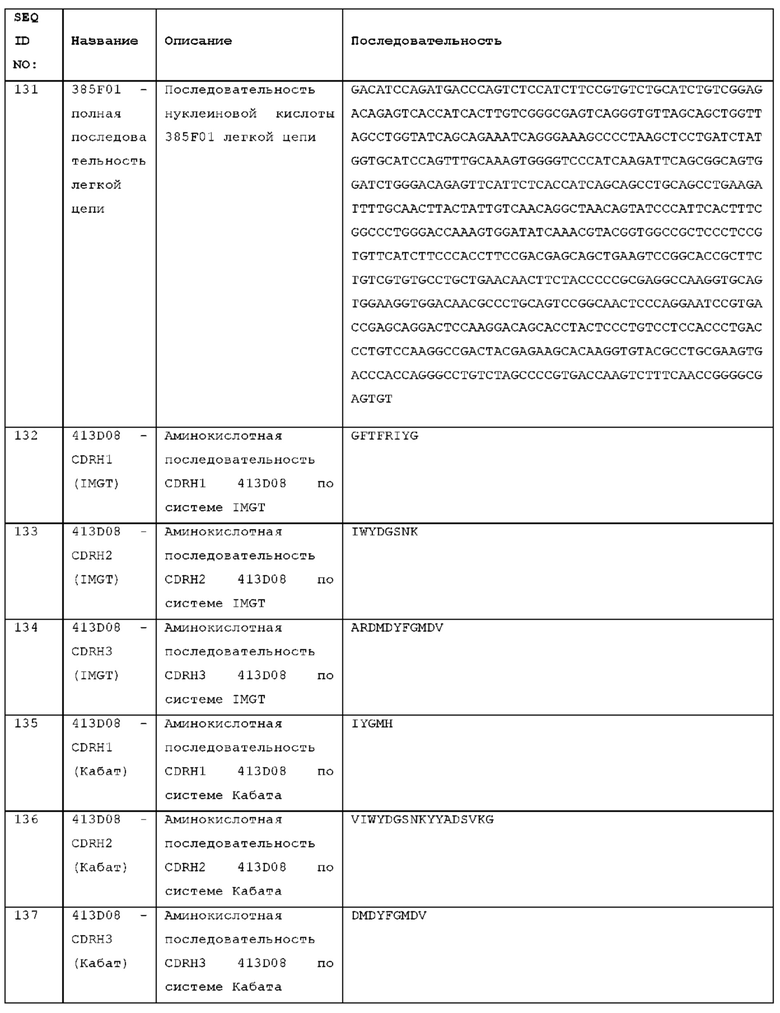
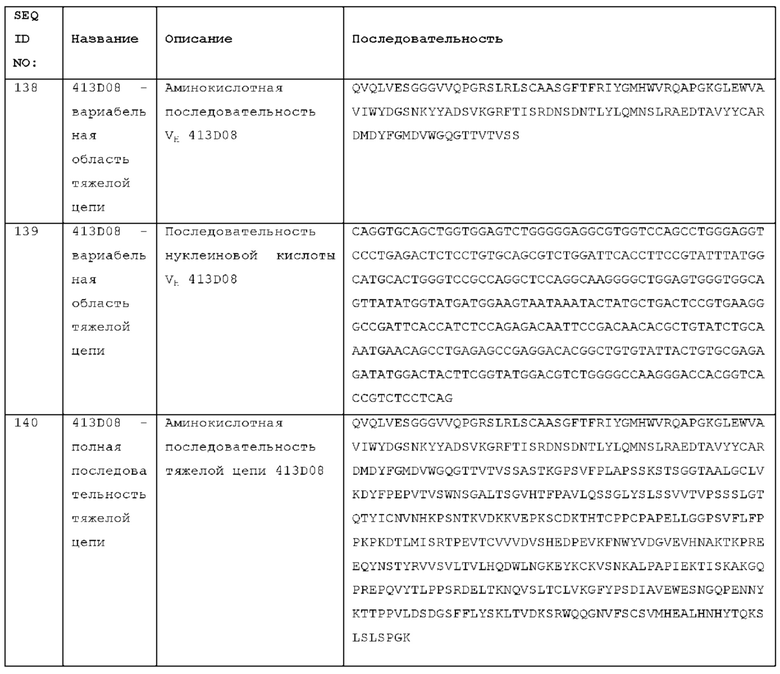
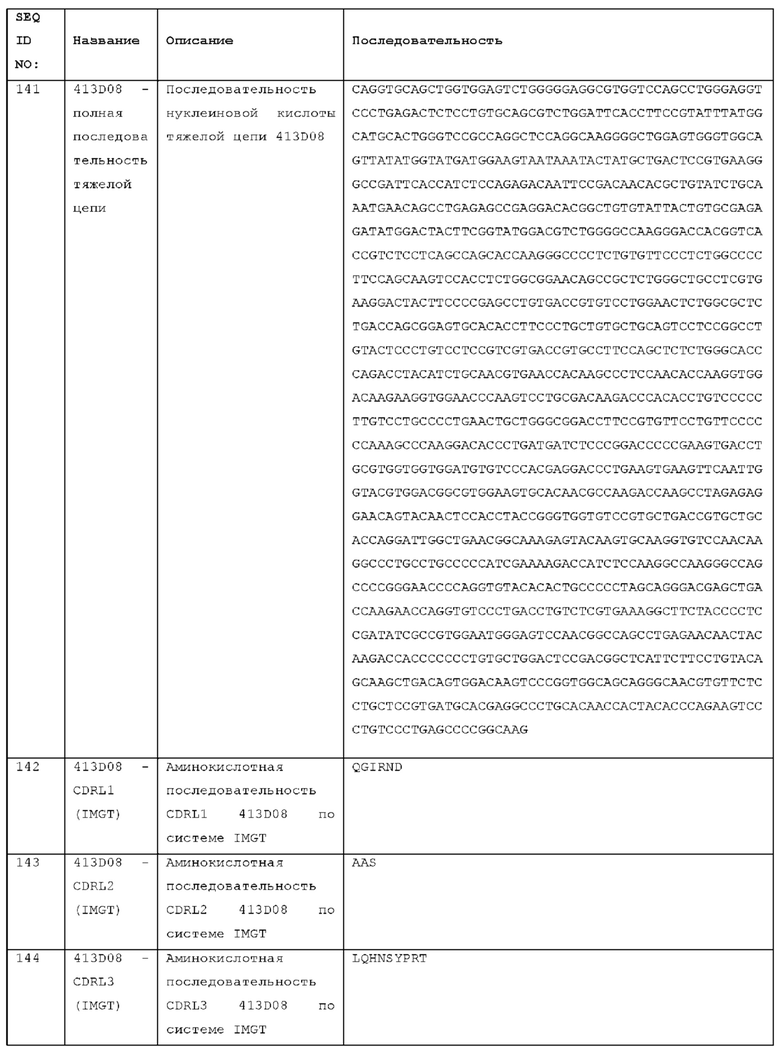
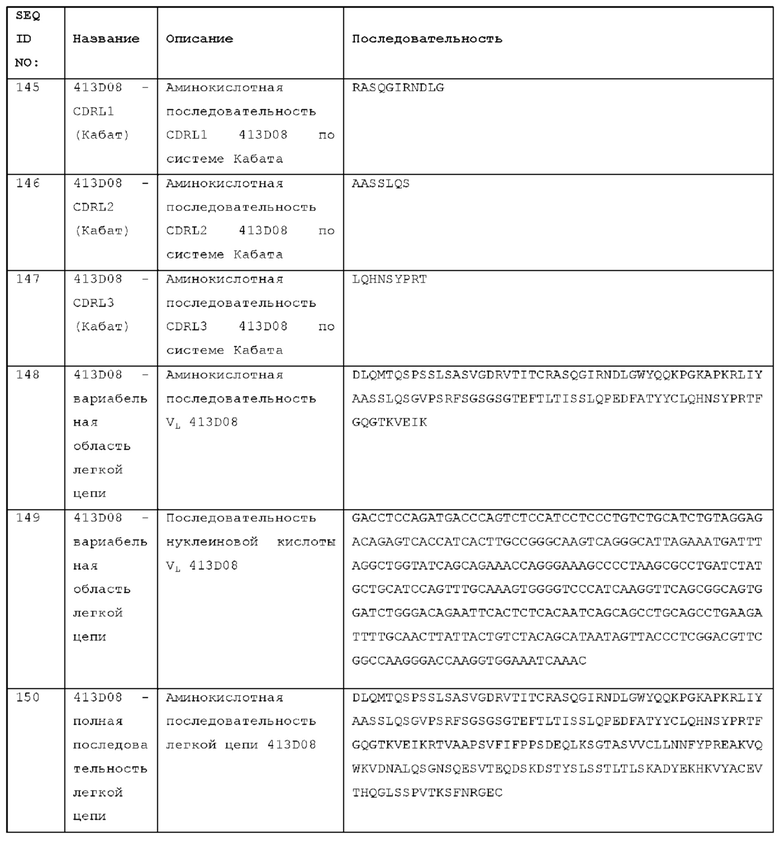

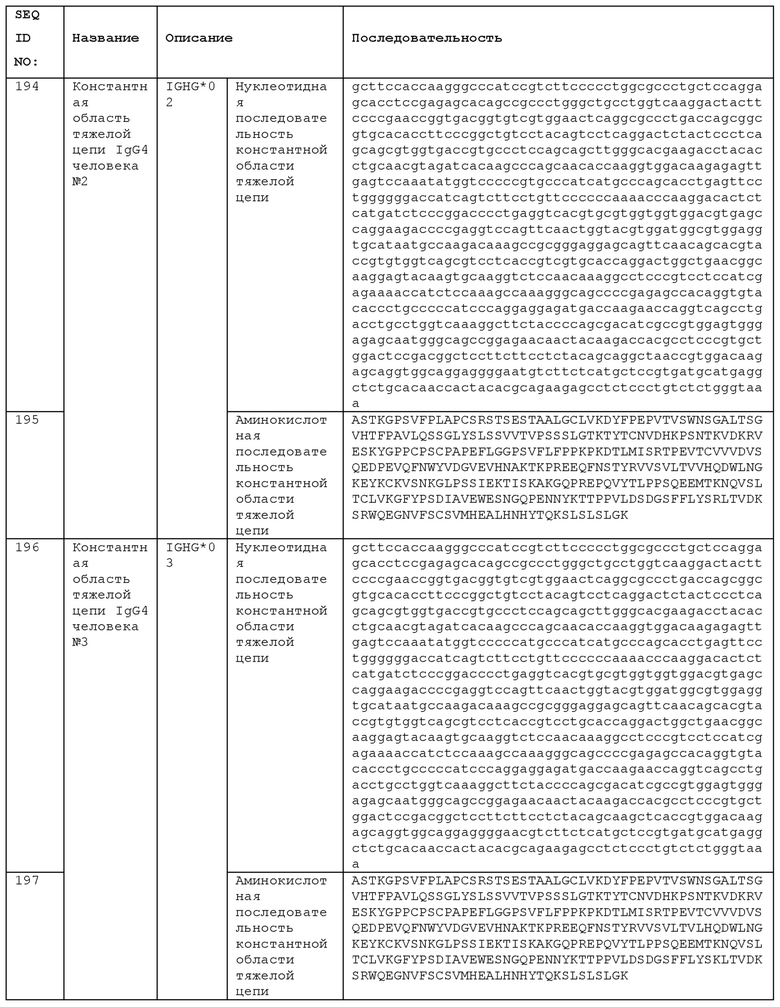
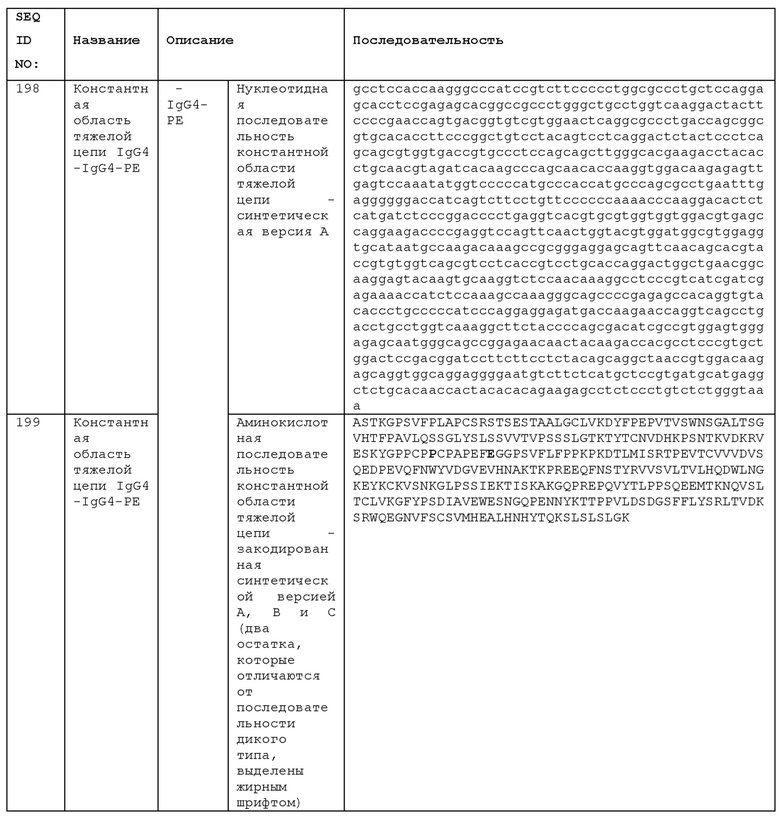
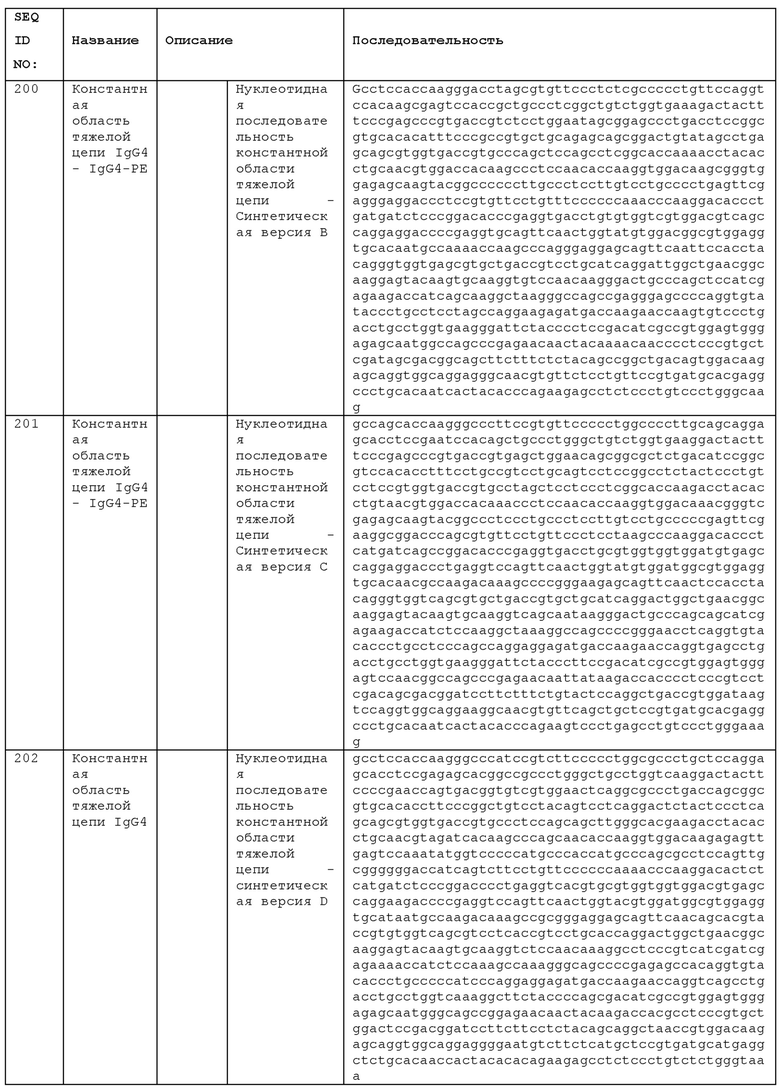
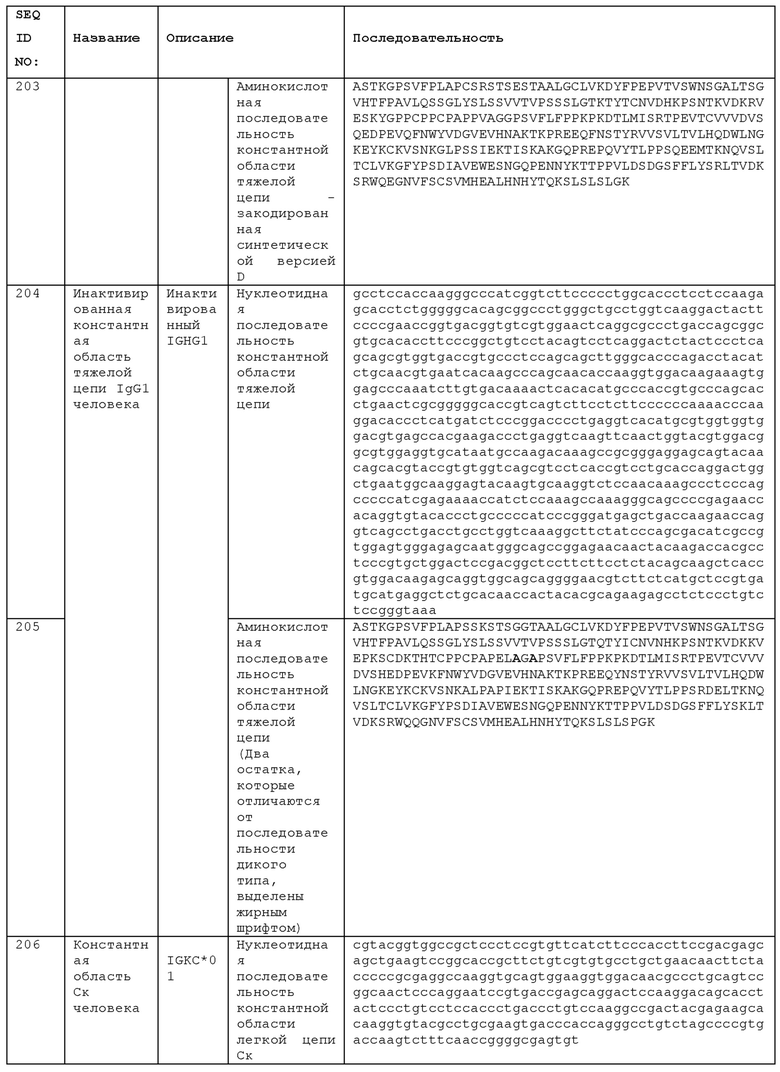
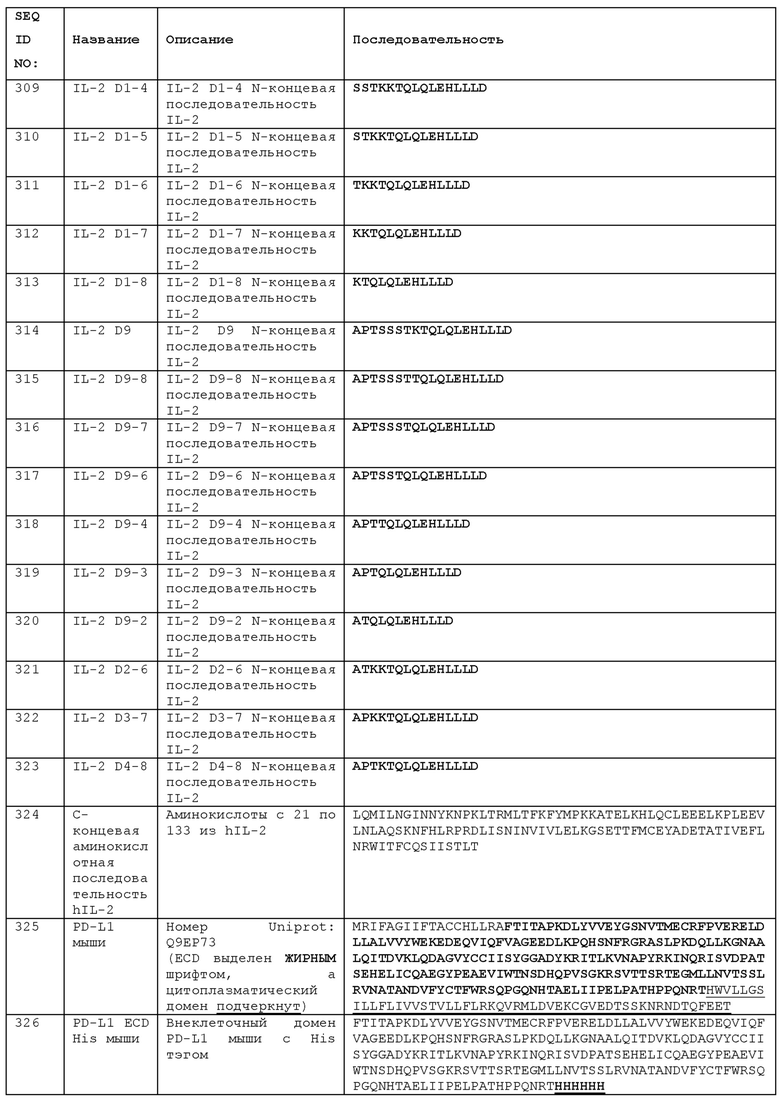
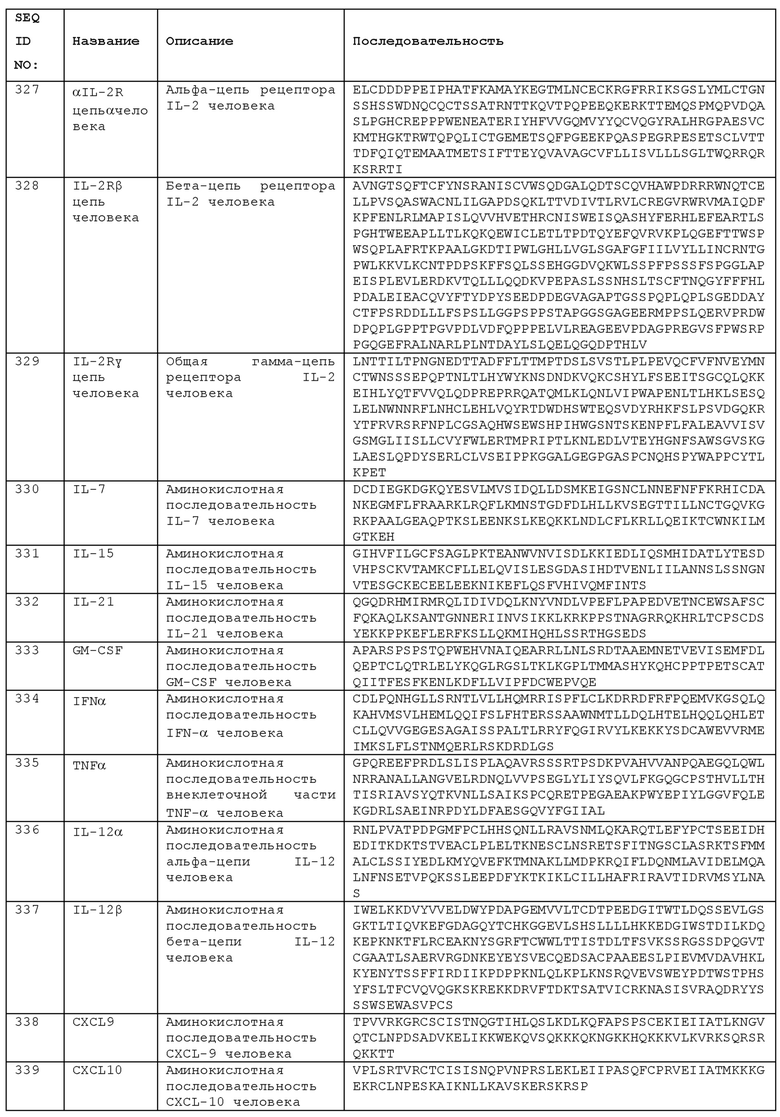
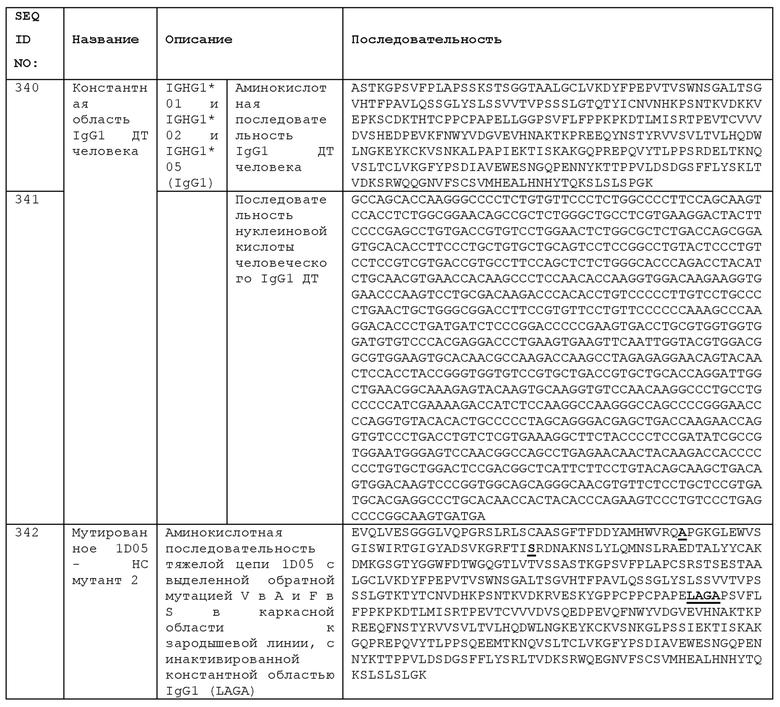

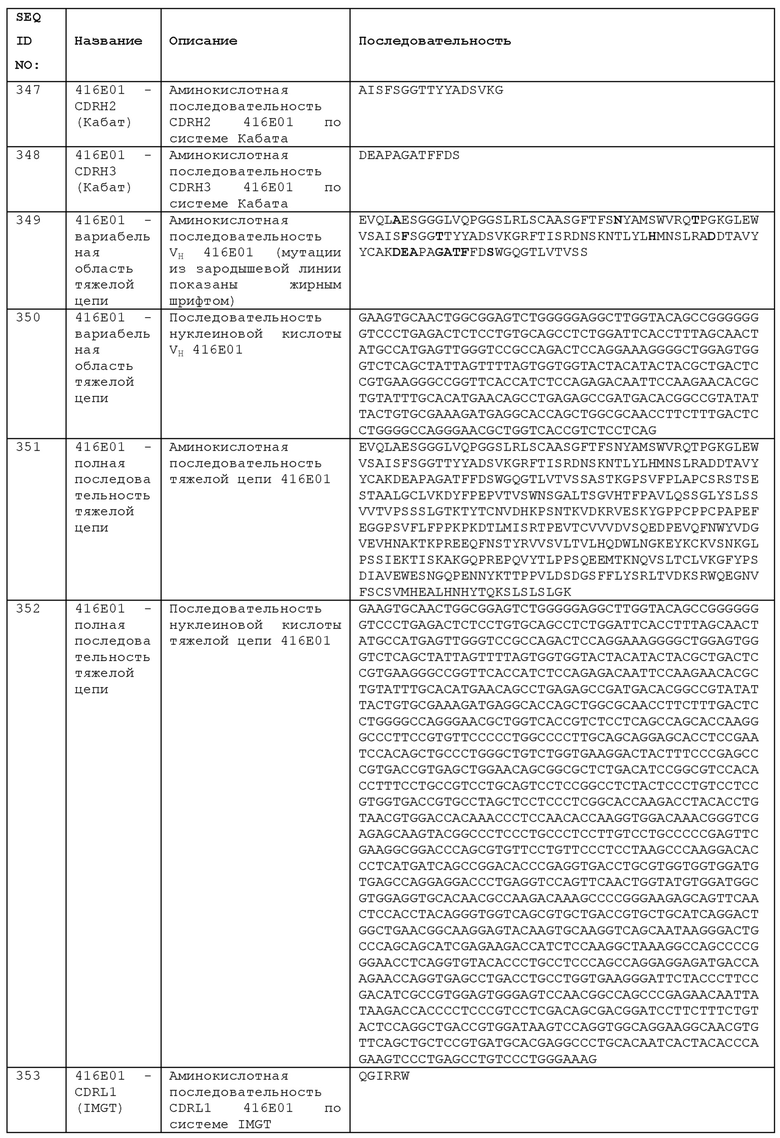
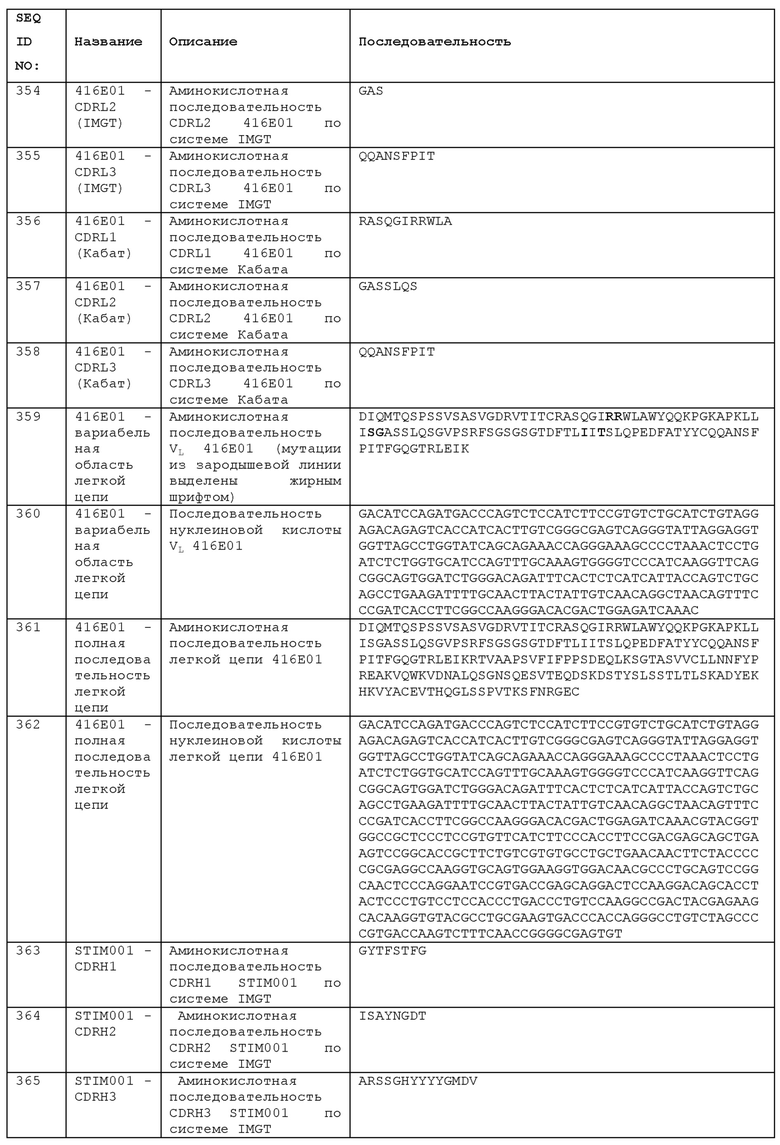

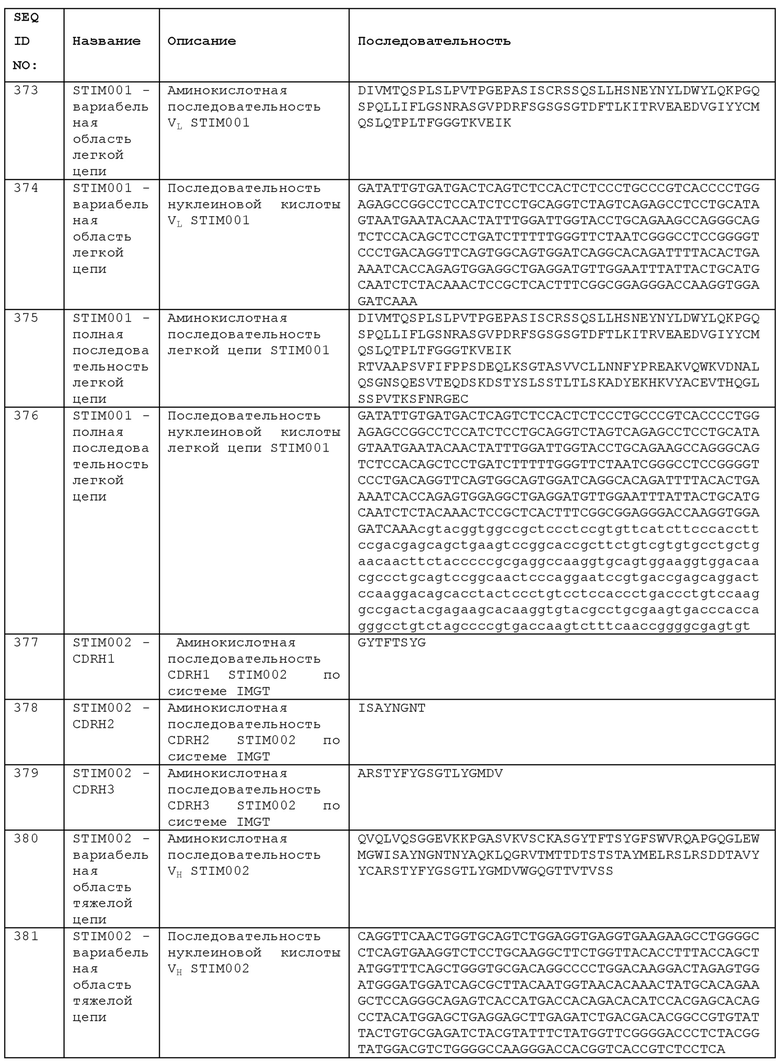

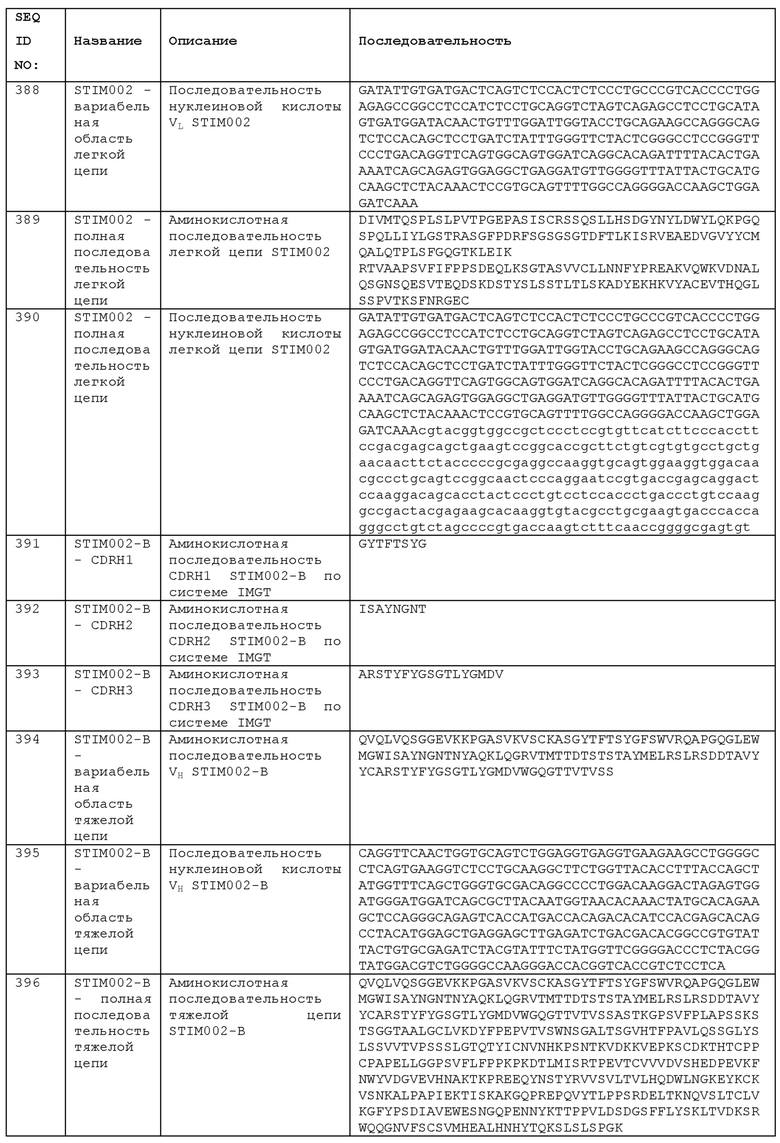
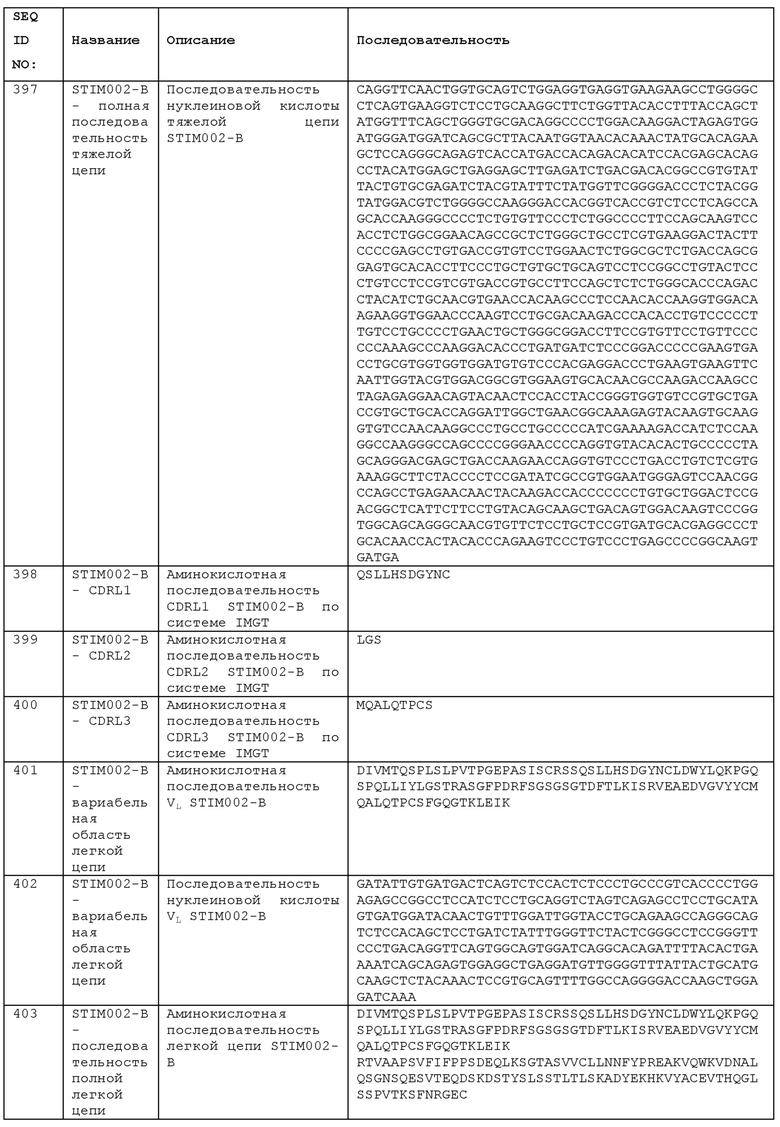
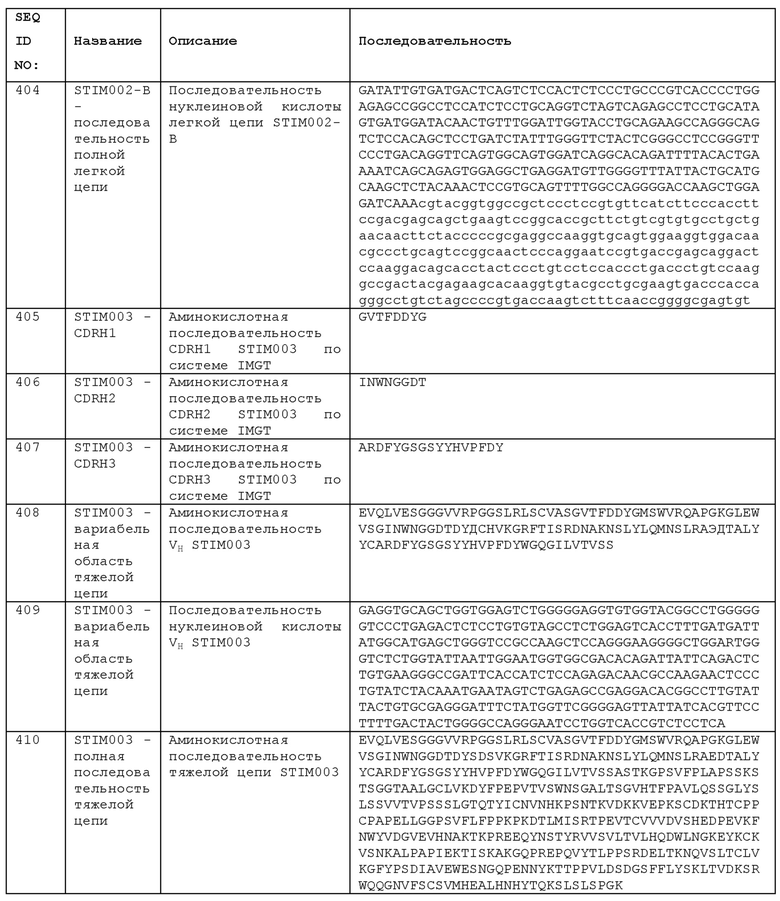
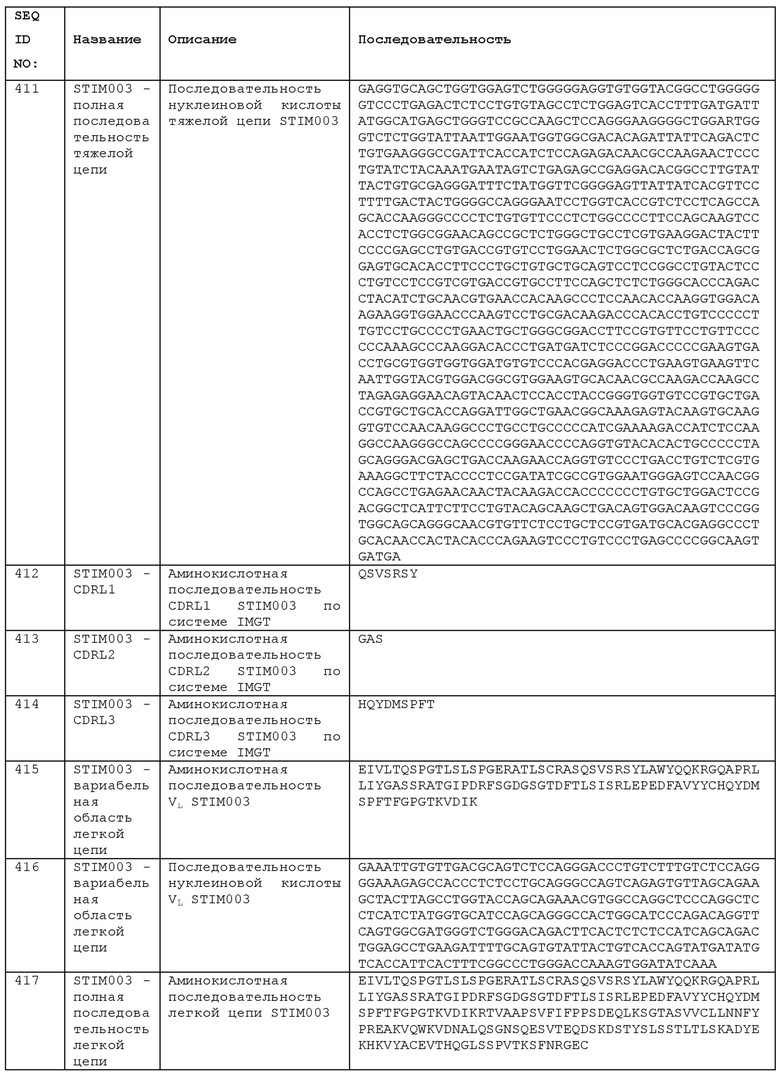
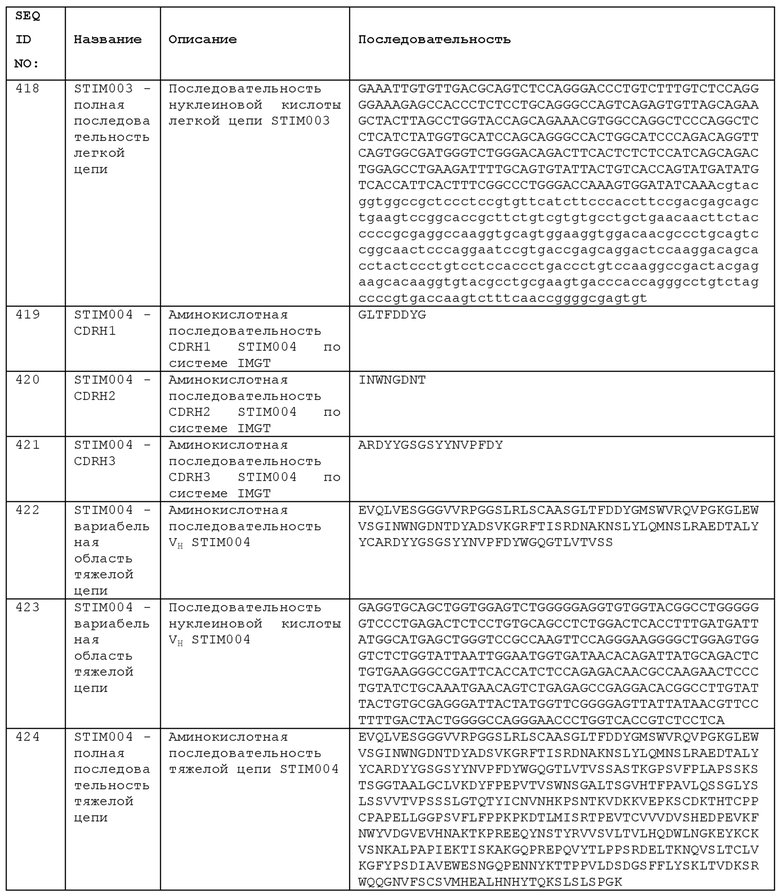
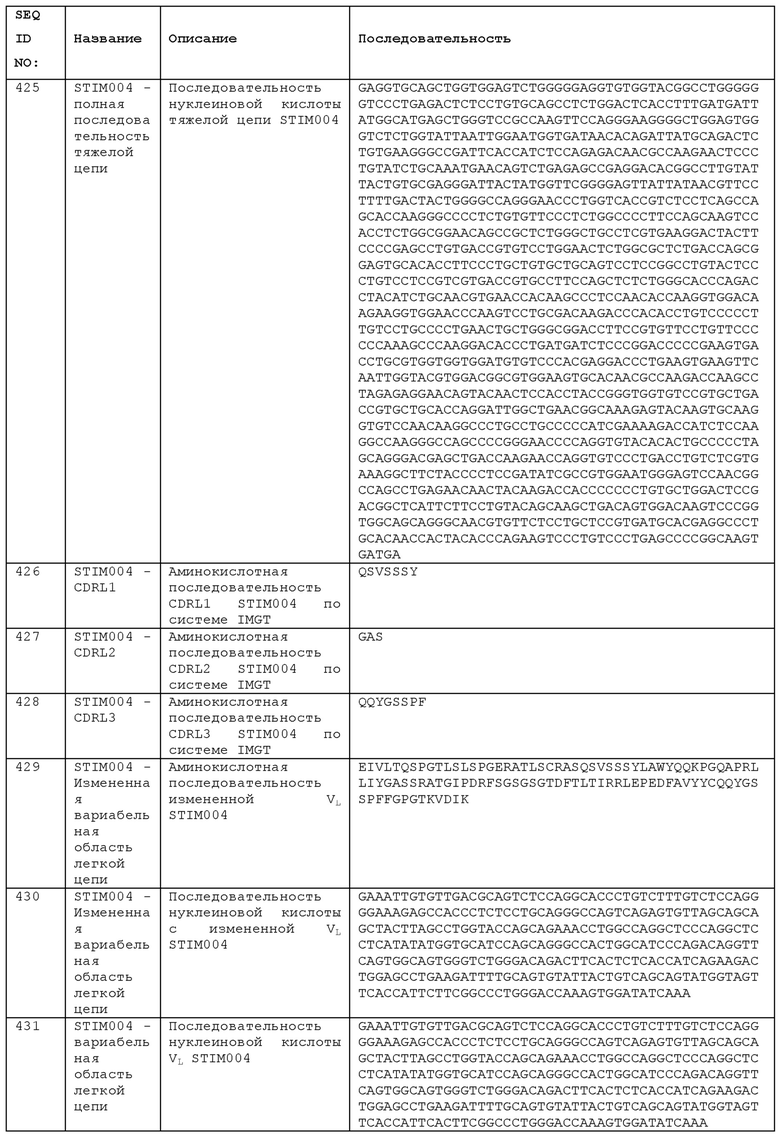

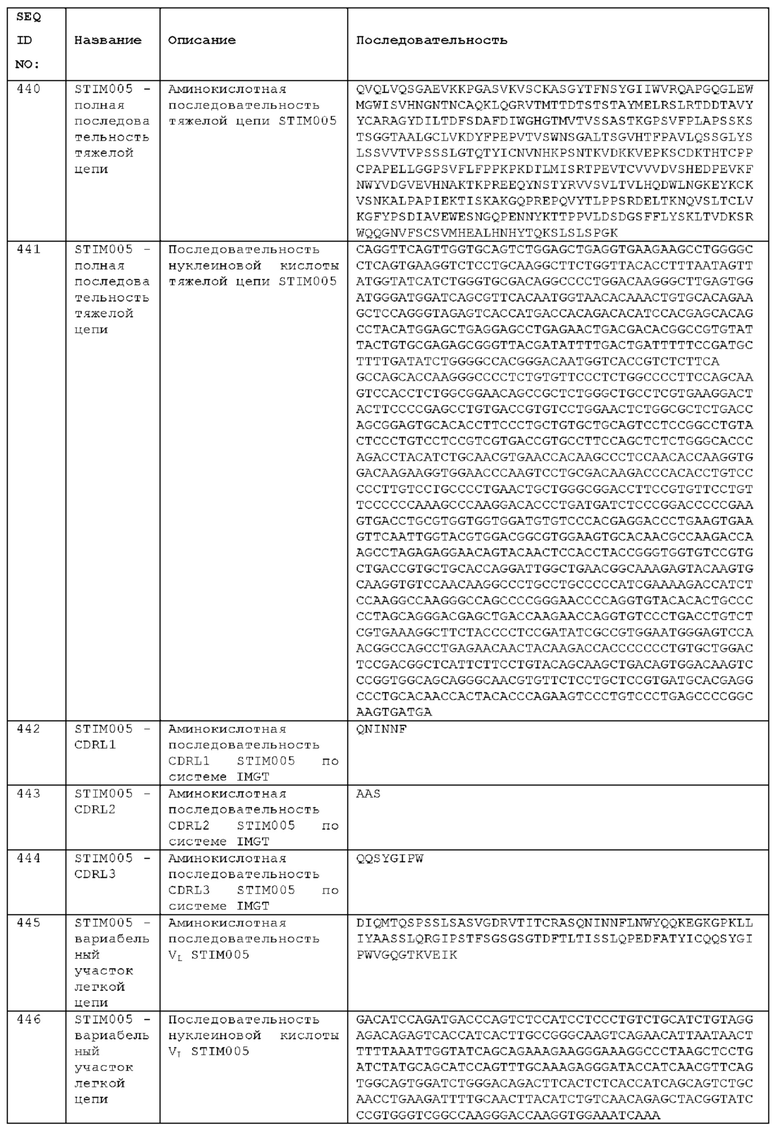
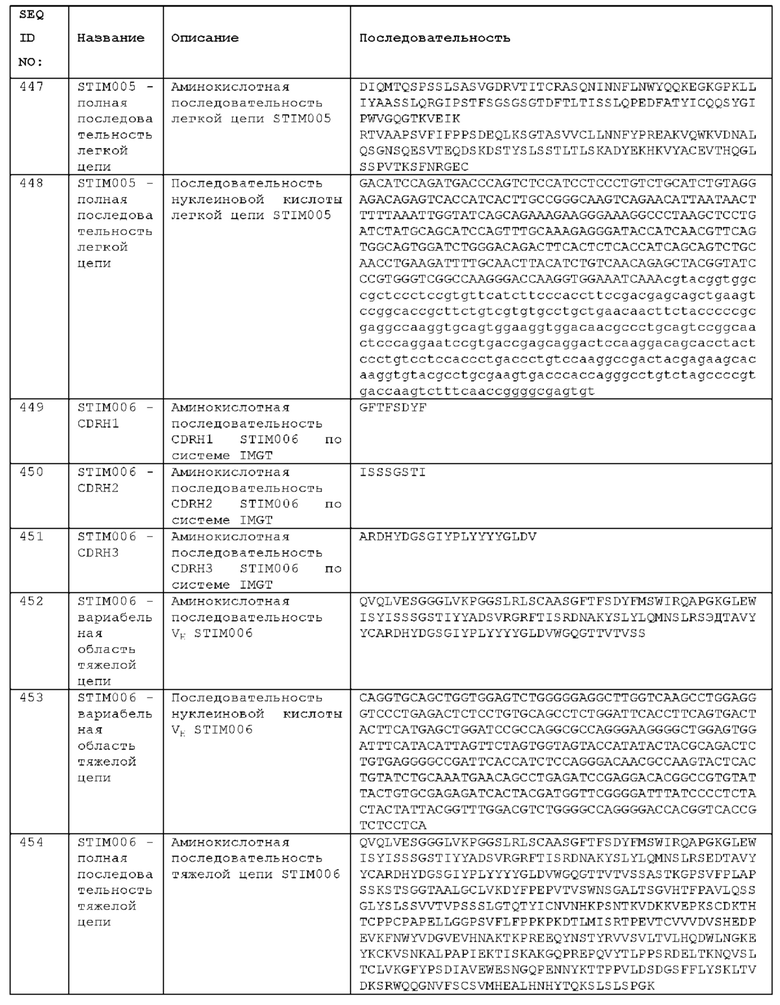
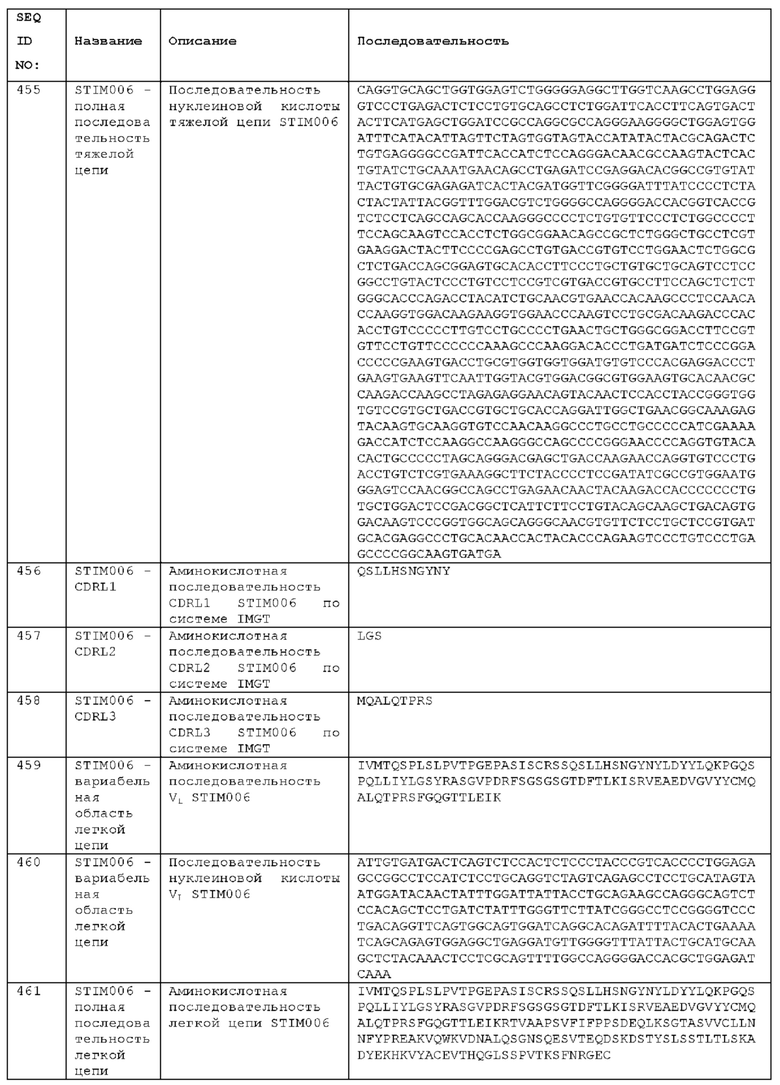
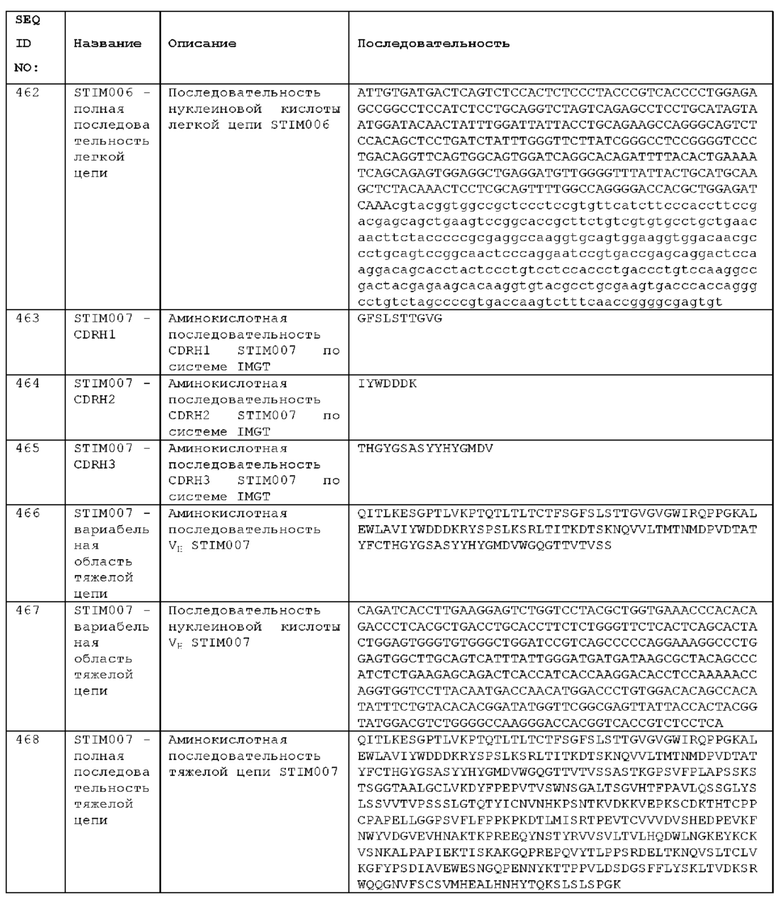
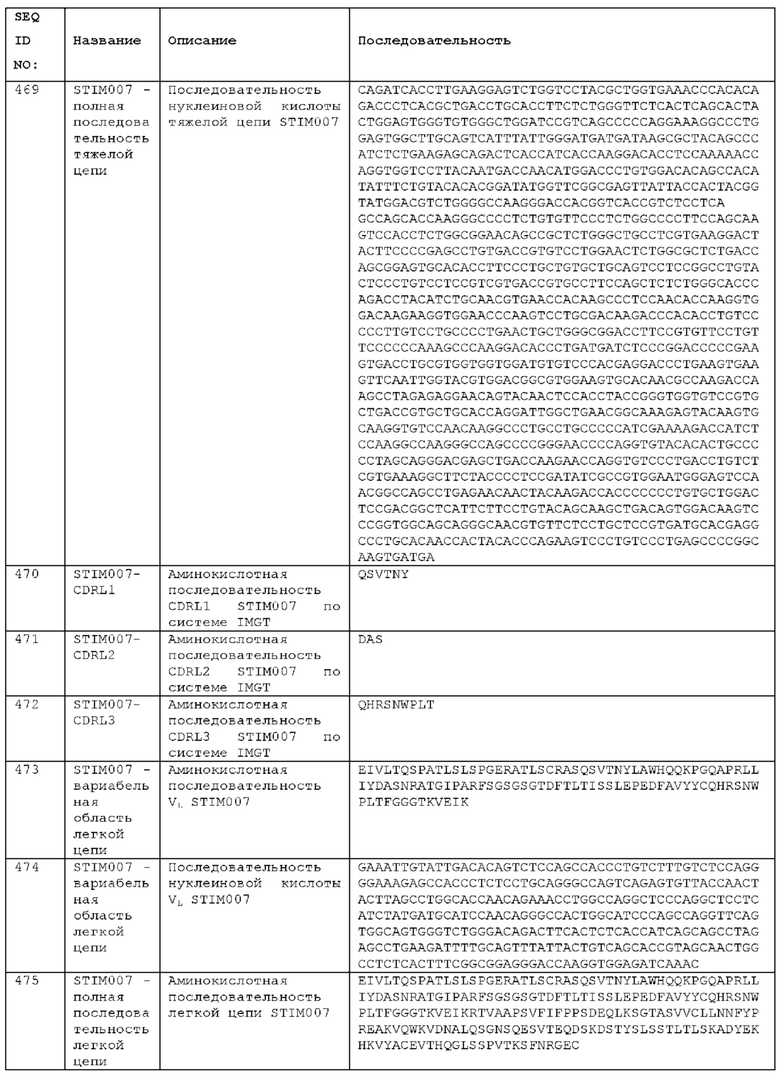
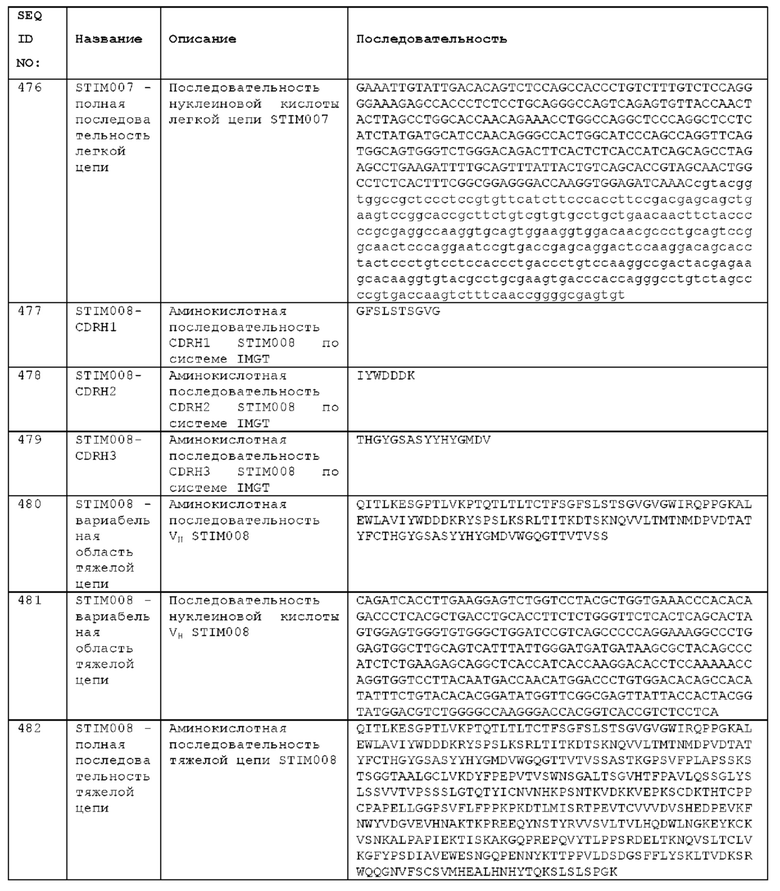

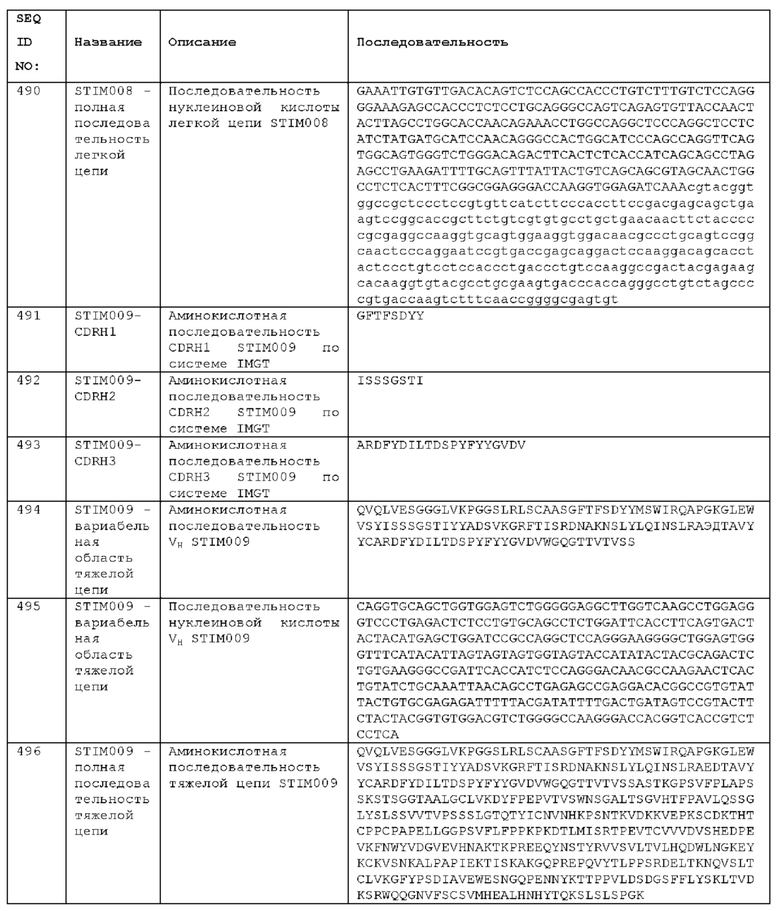
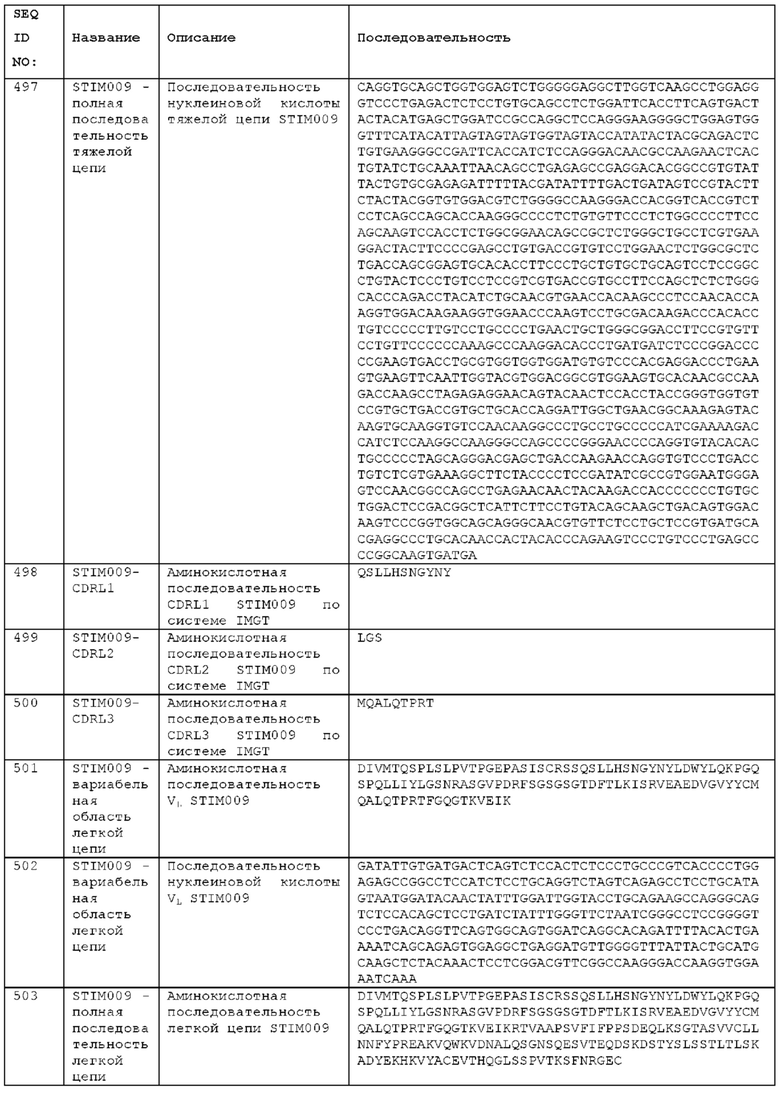
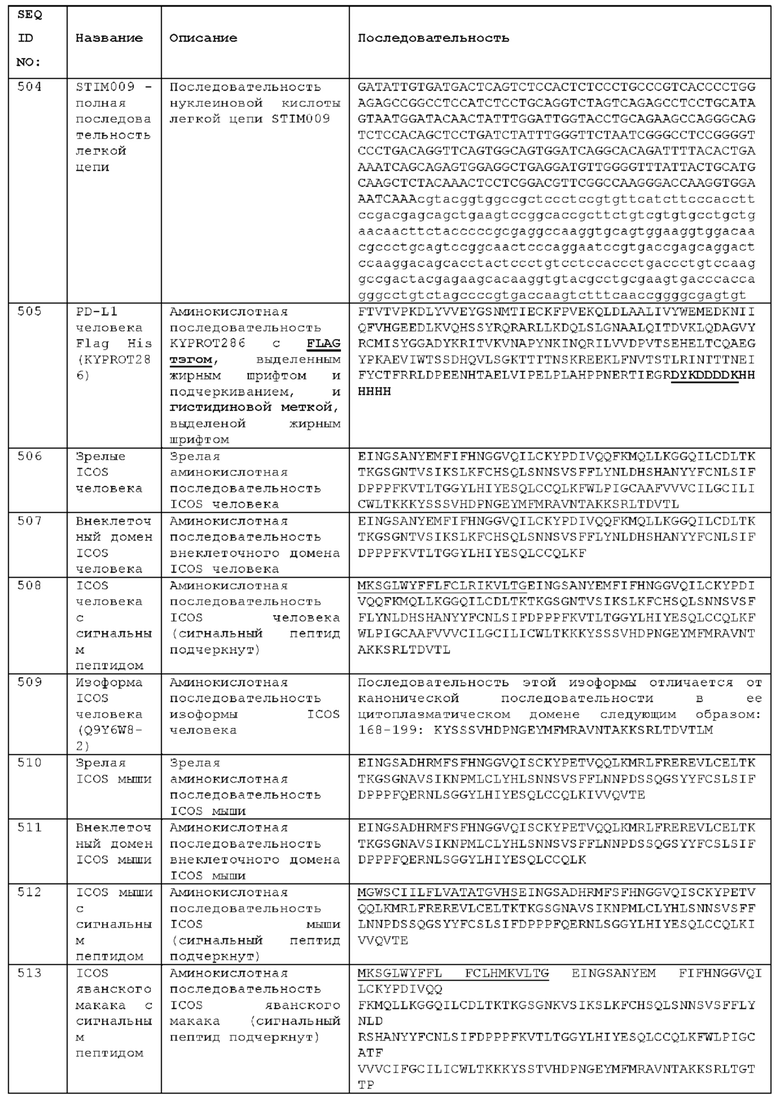
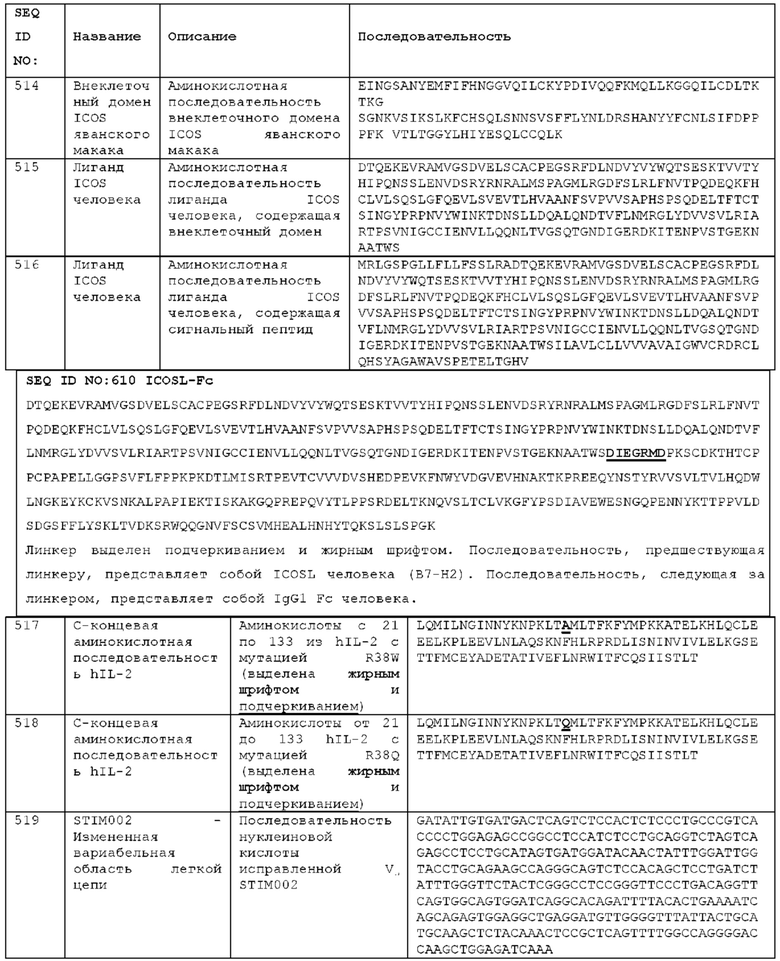
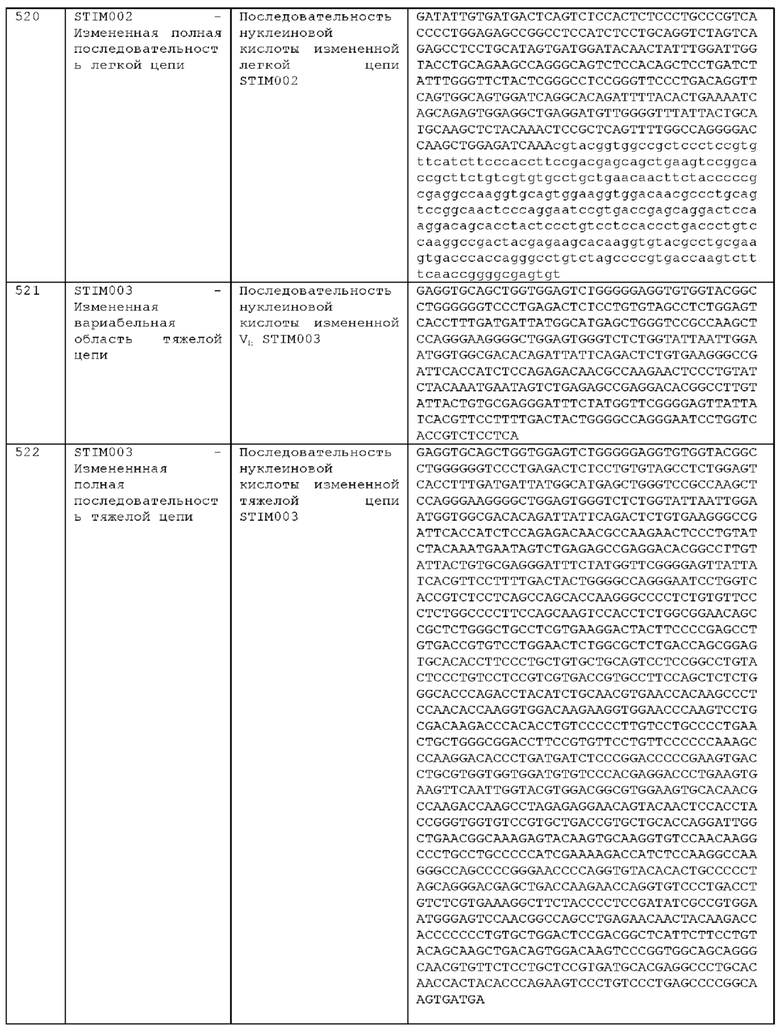

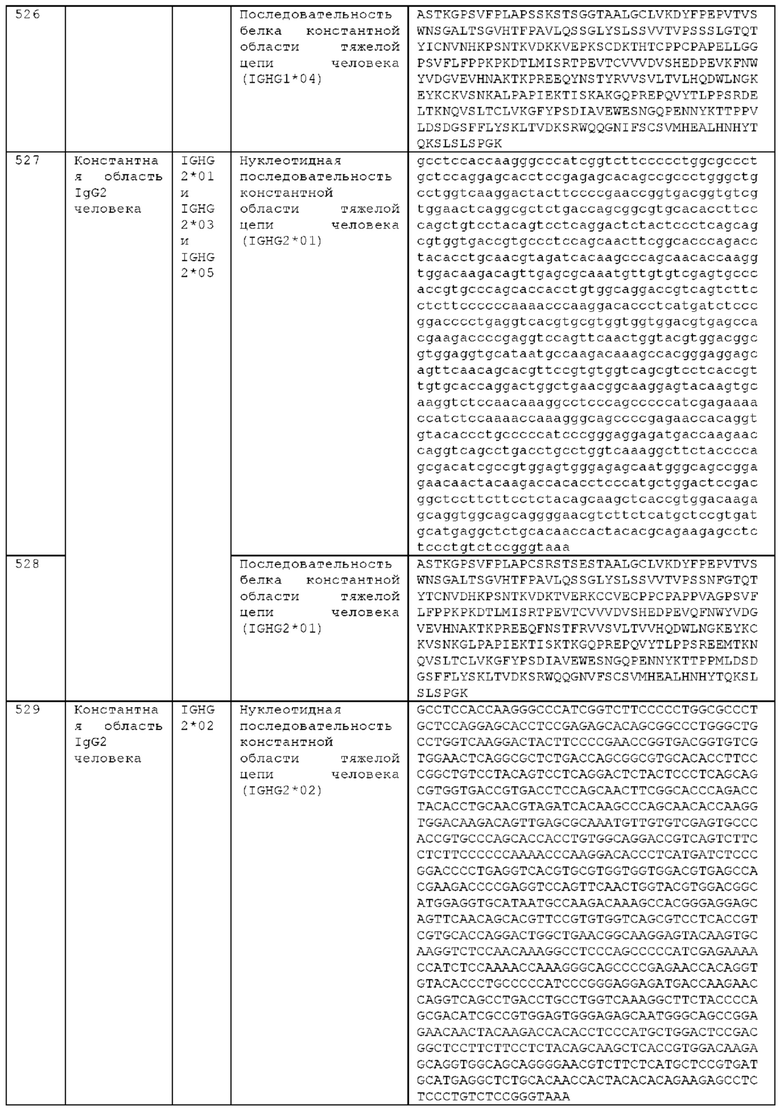
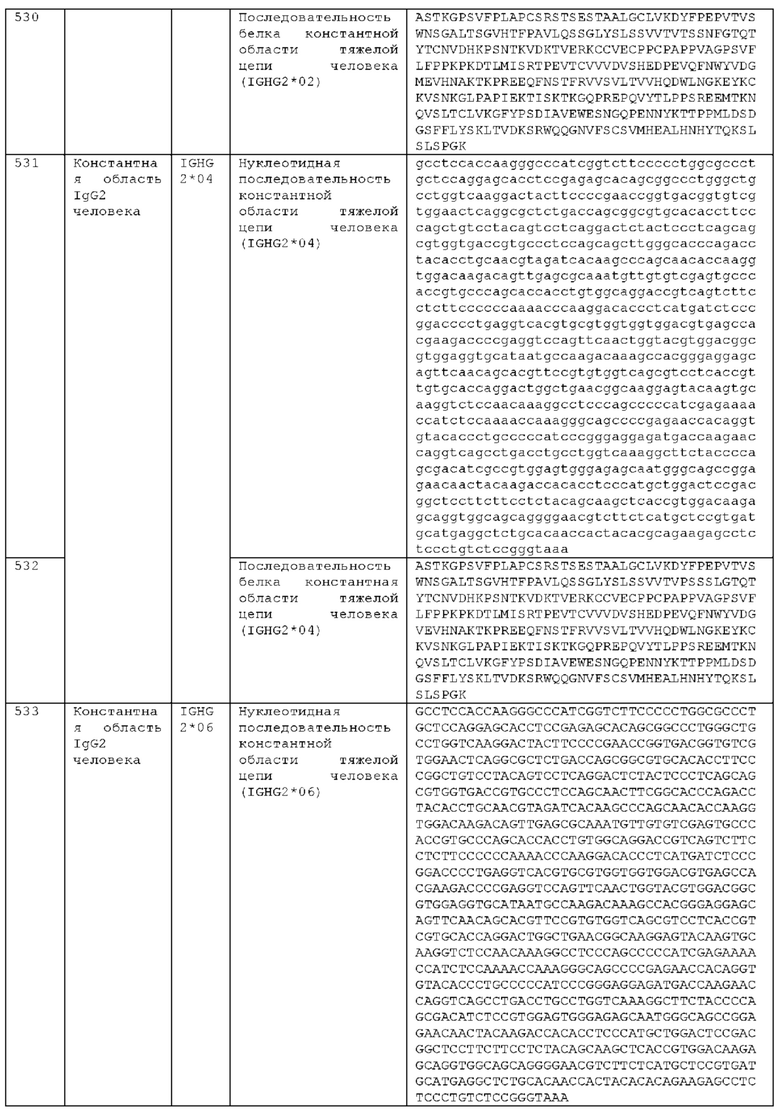

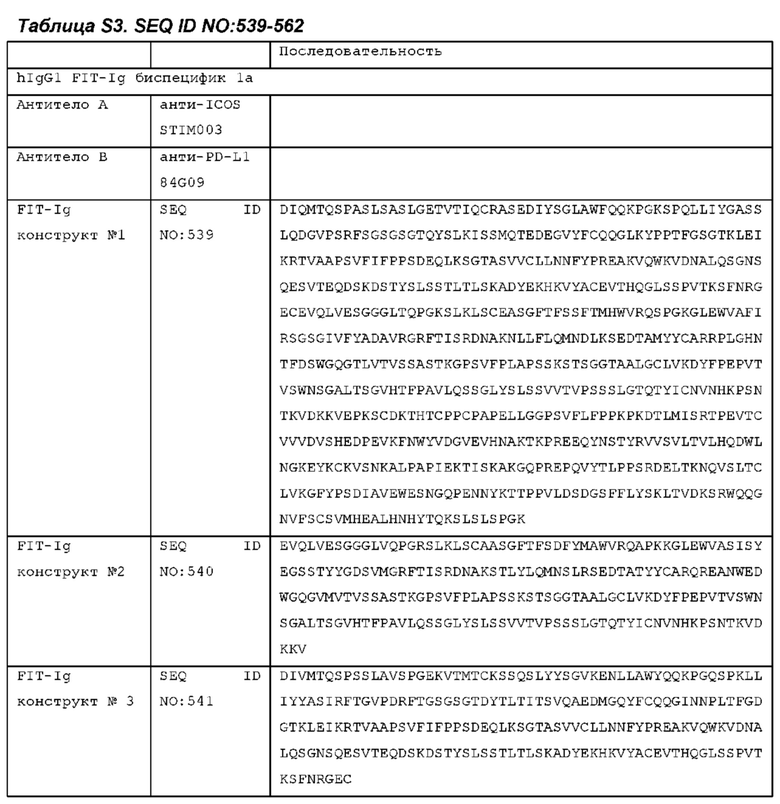
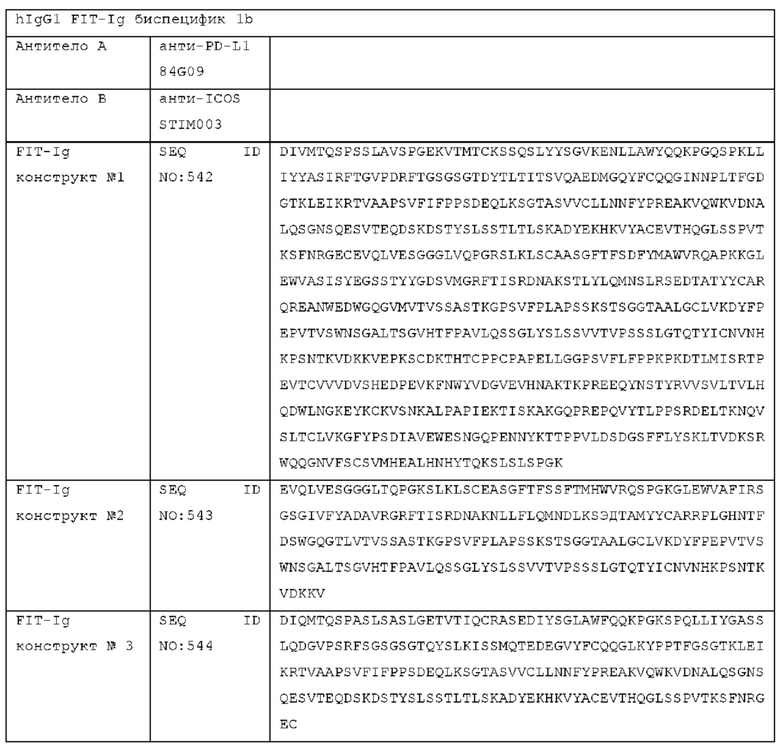
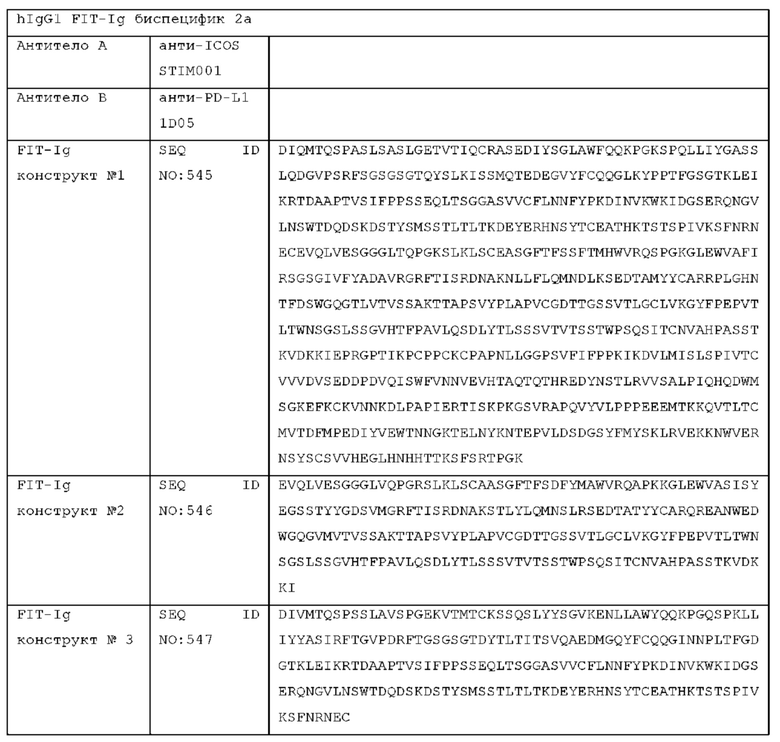

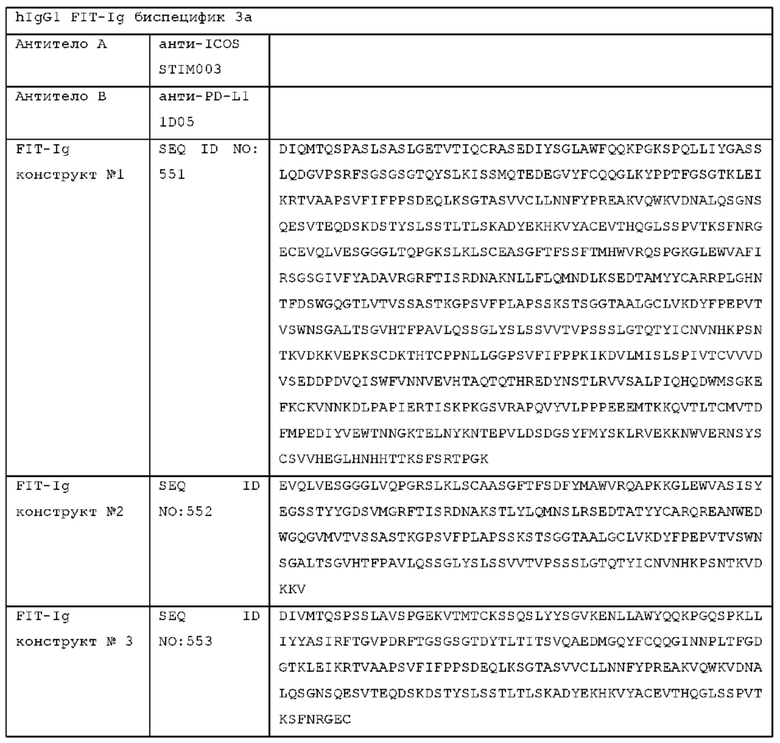
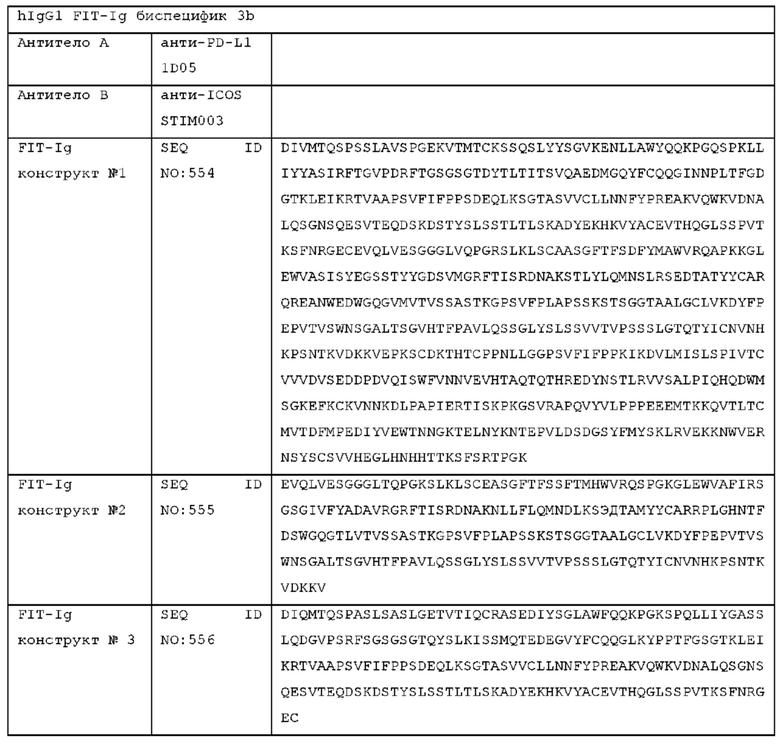
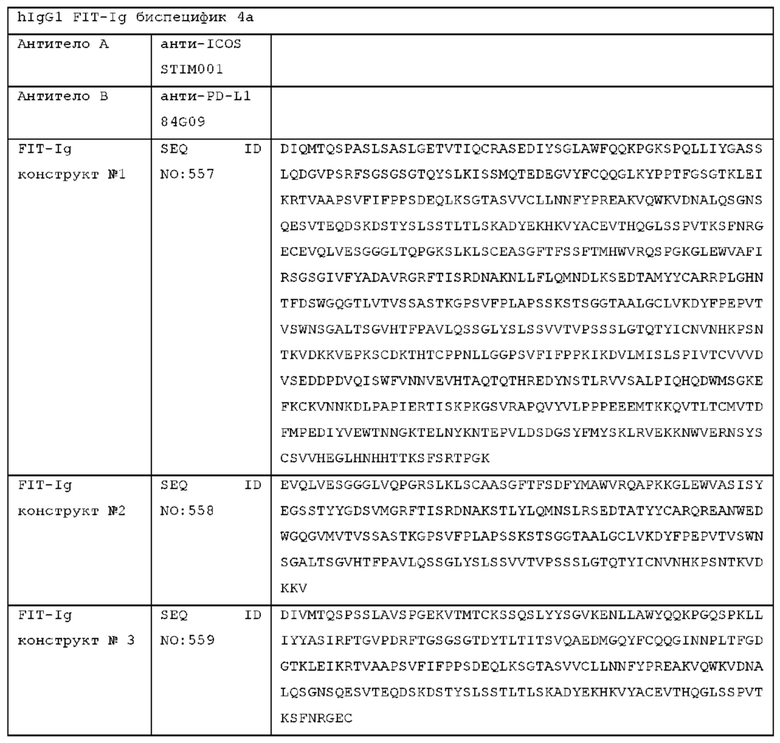
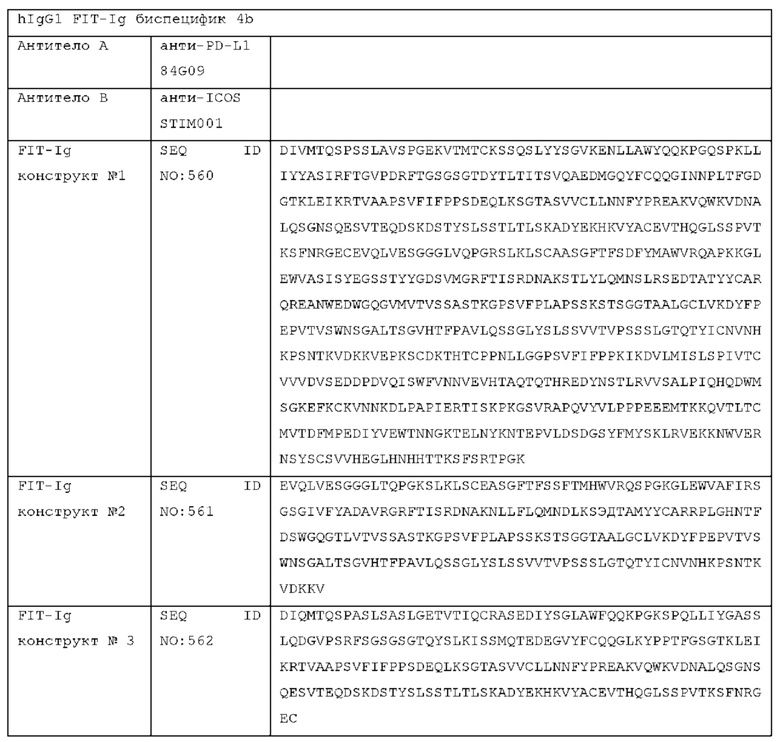
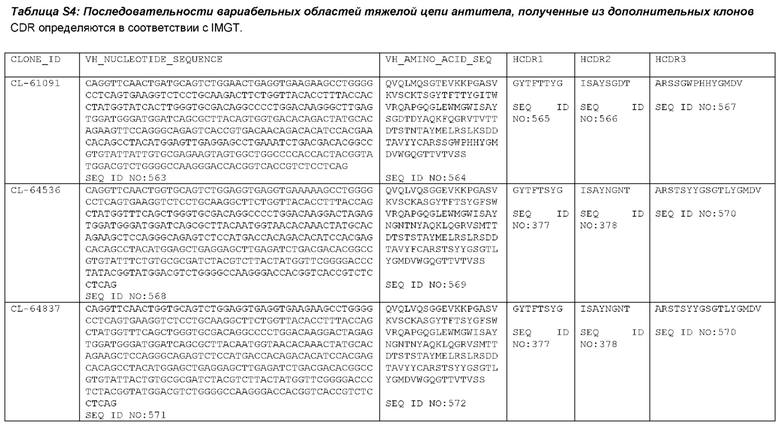
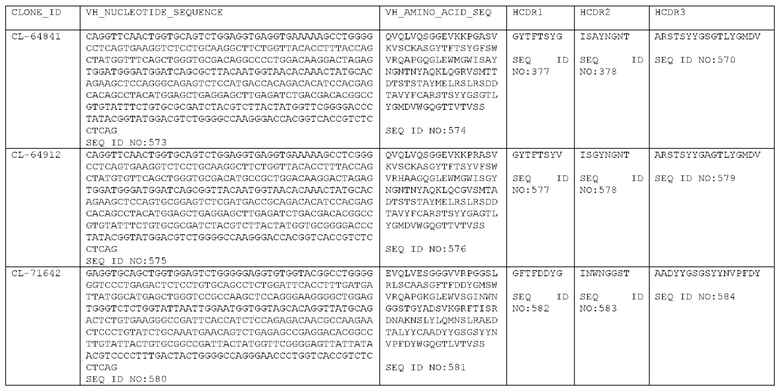


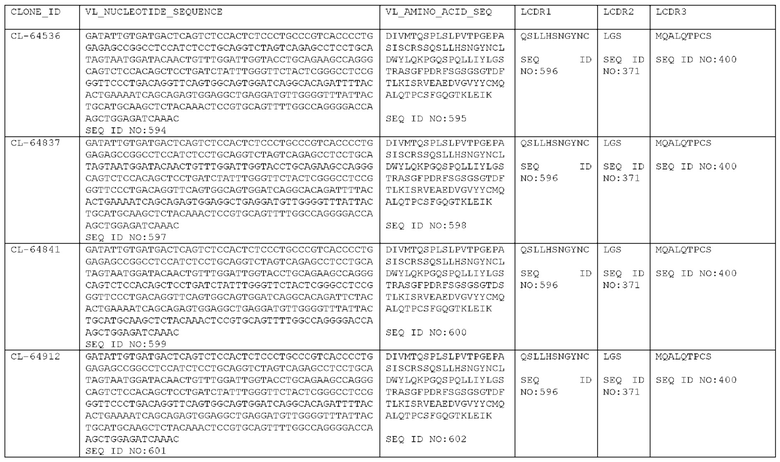

| название | год | авторы | номер документа |
|---|---|---|---|
| PD-L1 специфические антитела | 2017 |
|
RU2756236C2 |
| Анти-PD-L1 и IL-2 цитокины | 2017 |
|
RU2769282C2 |
| АНТИТЕЛА К ICOS | 2016 |
|
RU2742241C2 |
| ГУМАНИЗИРОВАННЫЕ АНТИТЕЛА К OX40 И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2015 |
|
RU2709742C2 |
| АНТИ-TIGIT АНТИГЕНСВЯЗЫВАЮЩИЕ БЕЛКИ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2729379C1 |
| АНТИТЕЛО ПРОТИВ PDL-1, ЕГО ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ И ПРИМЕНЕНИЕ | 2017 |
|
RU2693661C2 |
| АНТИТЕЛА К PD-1 И ИХ ПРИМЕНЕНИЕ | 2017 |
|
RU2761640C2 |
| АНТИТЕЛА ПРОТИВ ОХ40 И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2012 |
|
RU2562874C1 |
| МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА ЧЕЛОВЕКА К БЕЛКУ ПРОГРАММИРУЕМОЙ СМЕРТИ 1 (PD-1) И СПОСОБЫ ЛЕЧЕНИЯ РАКА С ИСПОЛЬЗОВАНИЕМ АНТИ-PD-1-АНТИТЕЛ САМОСТОЯТЕЛЬНО ИЛИ В КОМБИНАЦИИ С ДРУГИМИ ИММУНОТЕРАПЕВТИЧЕСКИМИ СРЕДСТВАМИ | 2013 |
|
RU2599417C2 |
| АНТИТЕЛА ПРОТИВ PD-1 И ИХ ПРИМЕНЕНИЕ | 2018 |
|
RU2788616C2 |
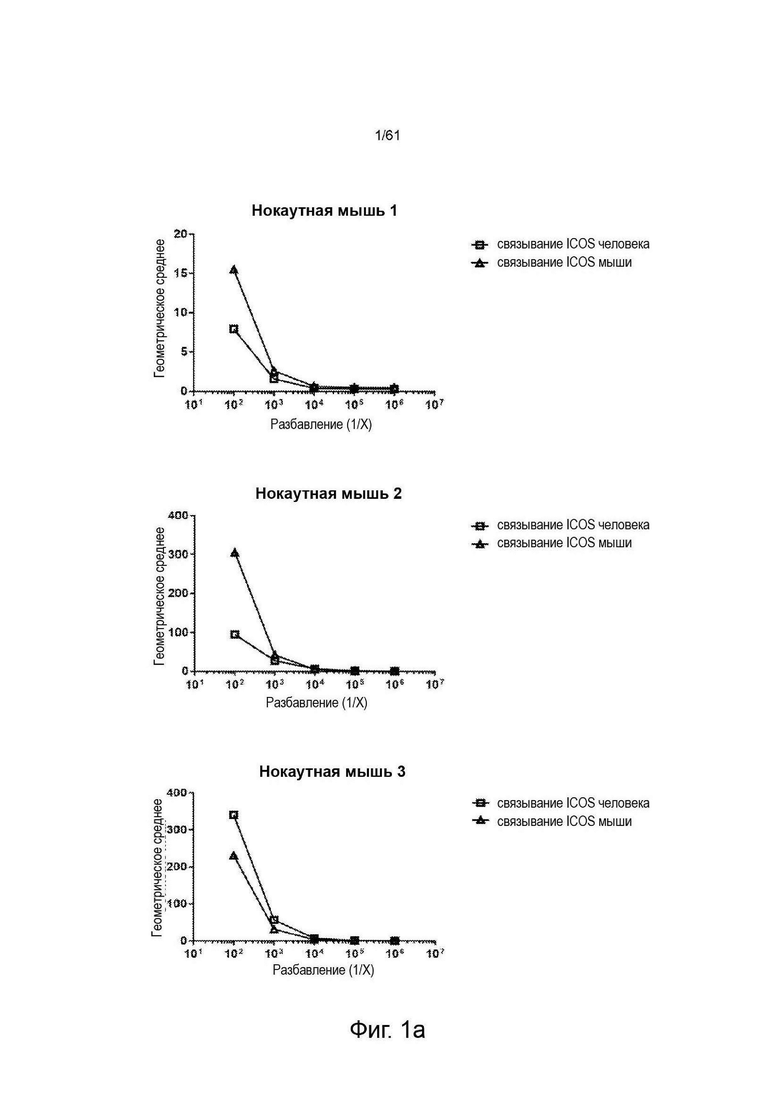
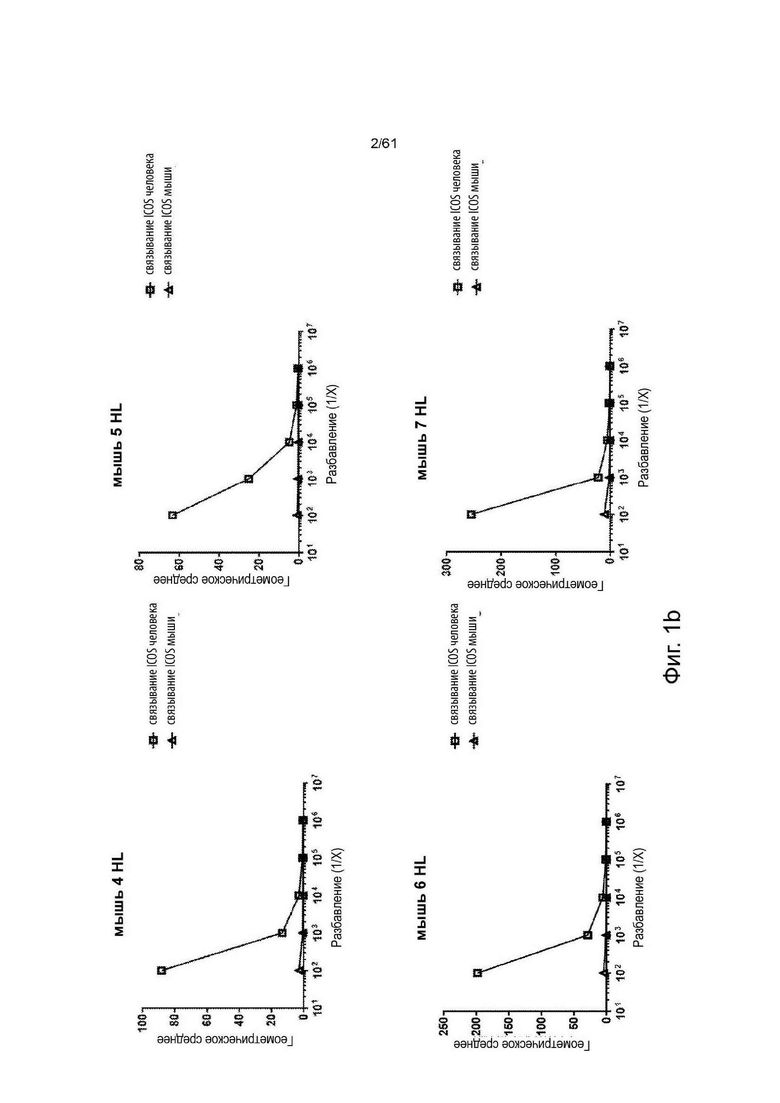
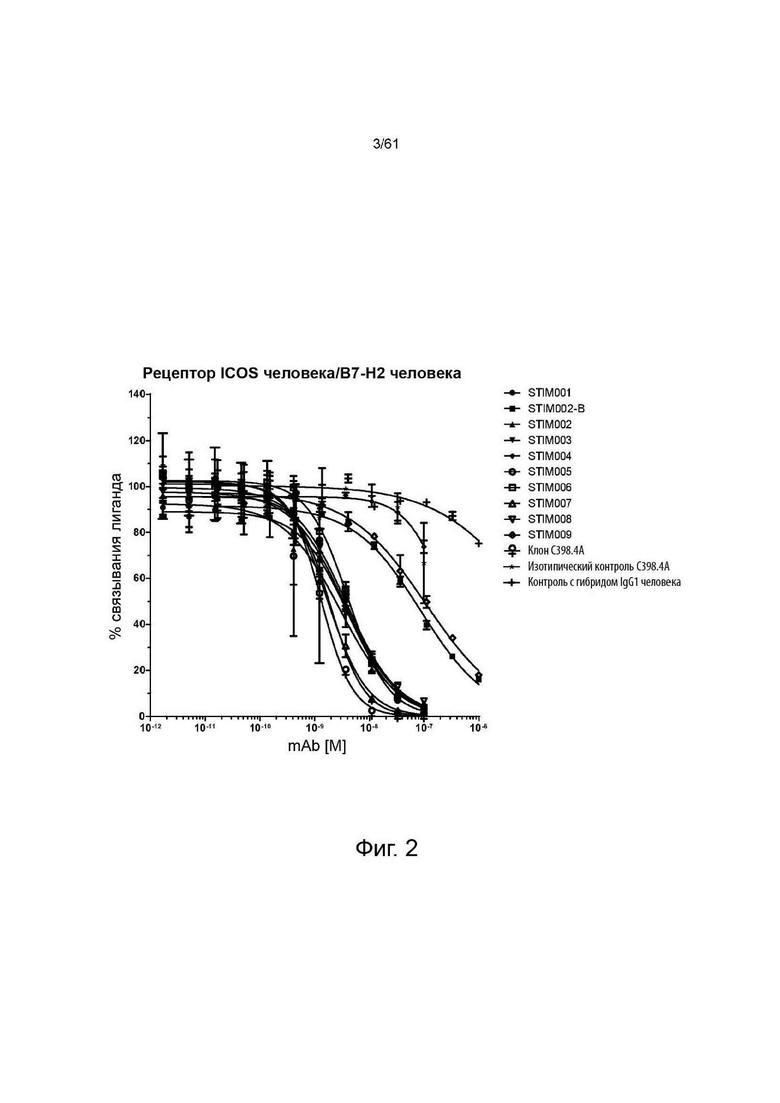
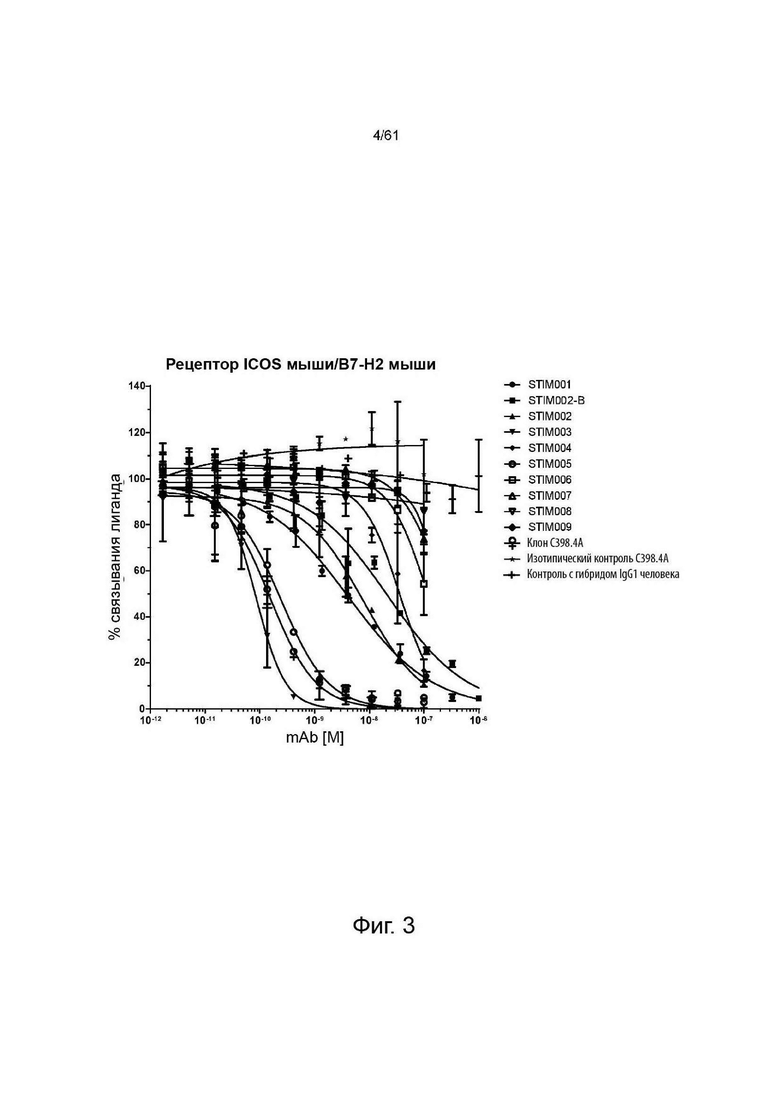

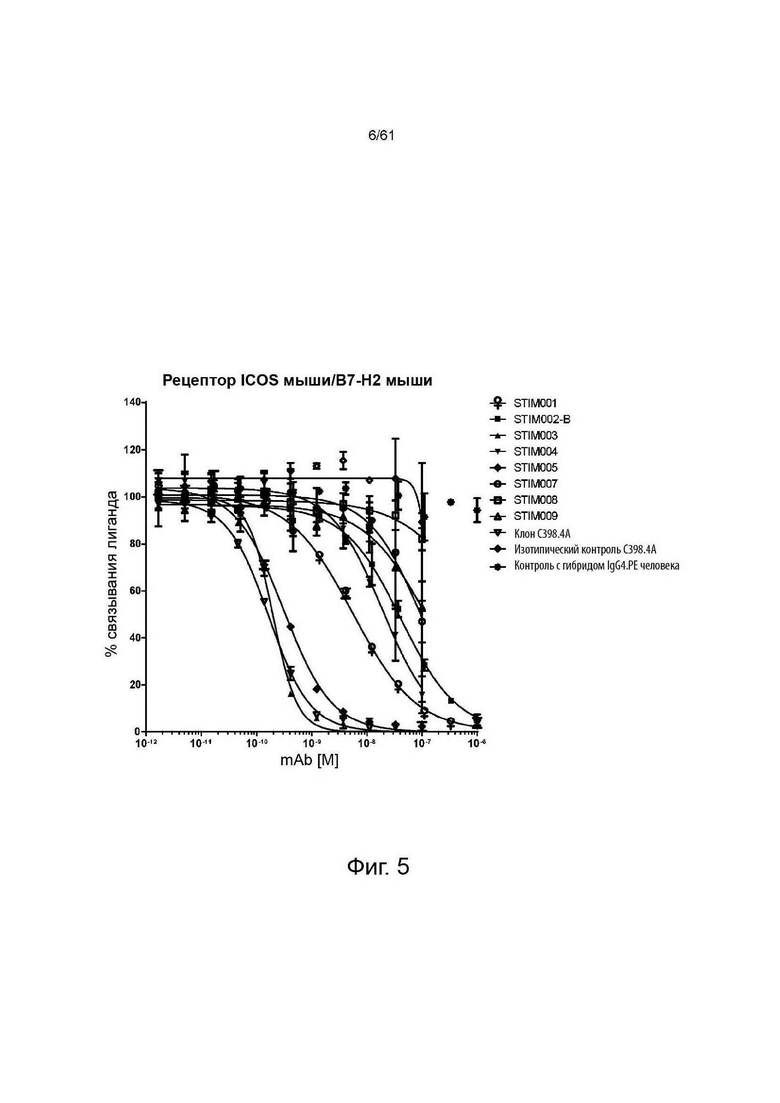
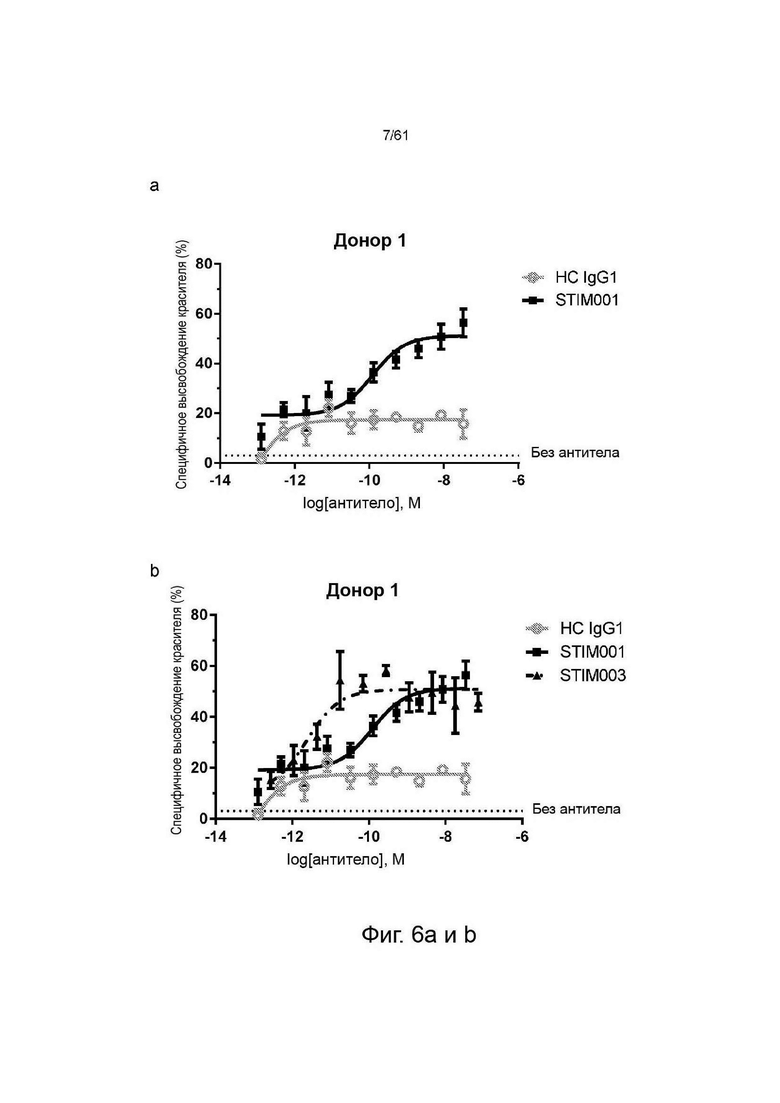
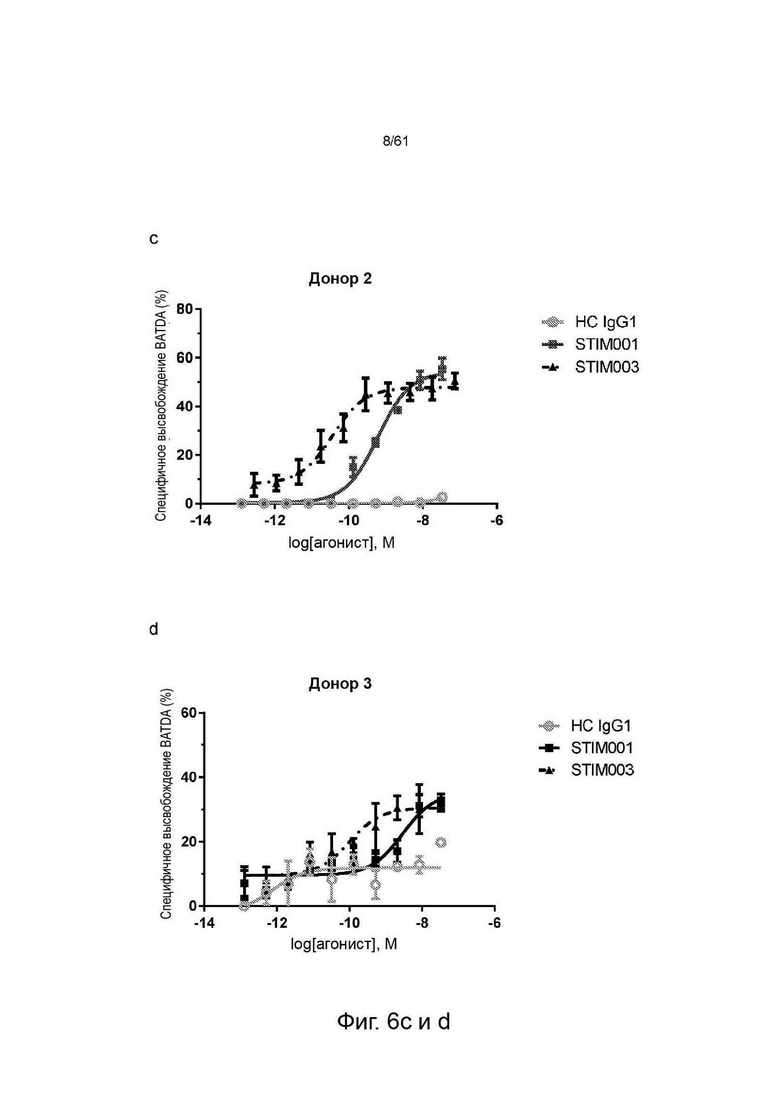
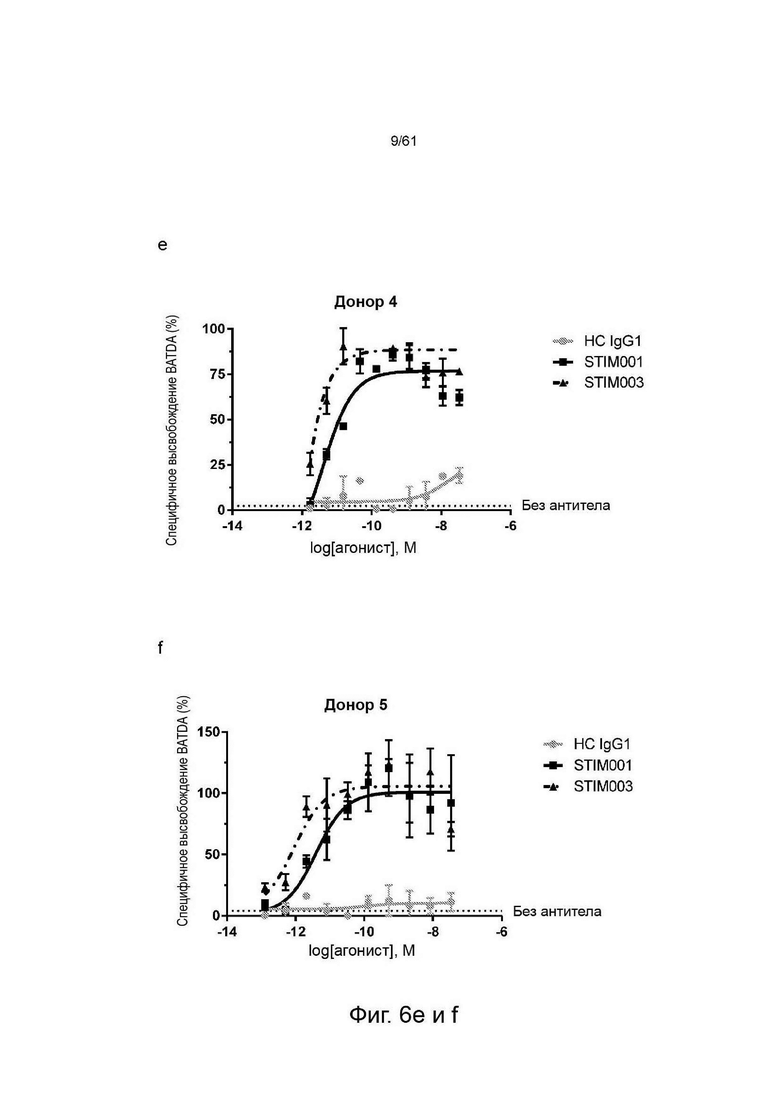
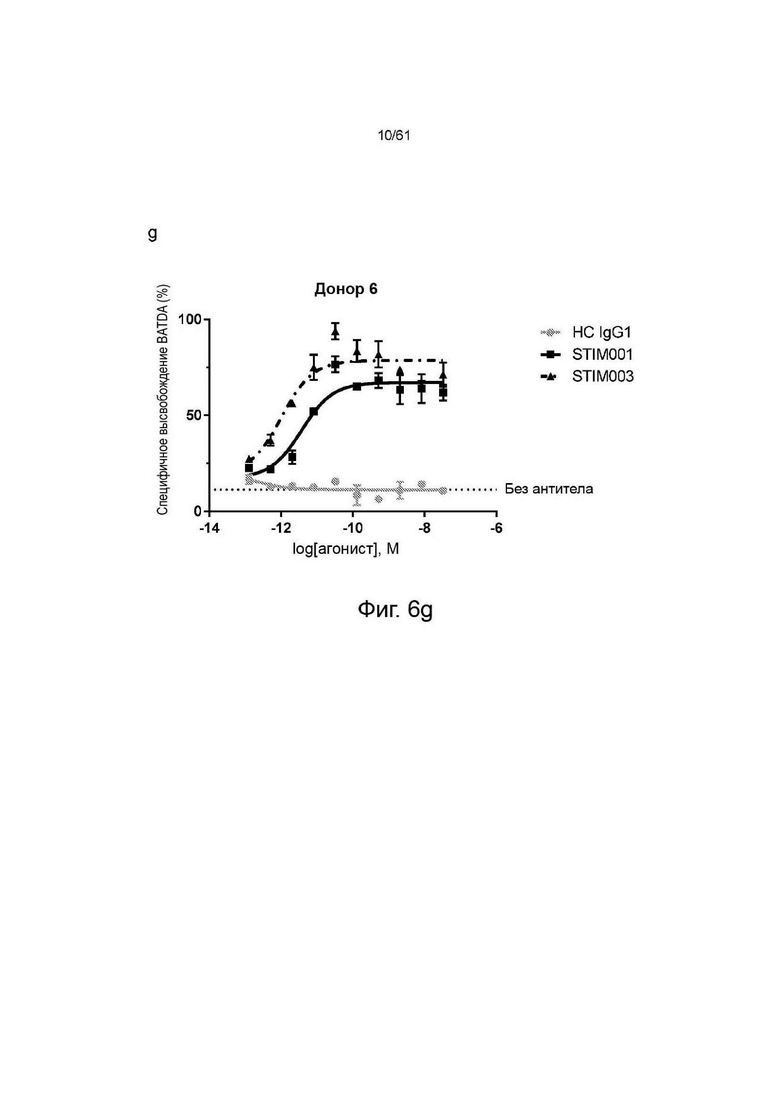
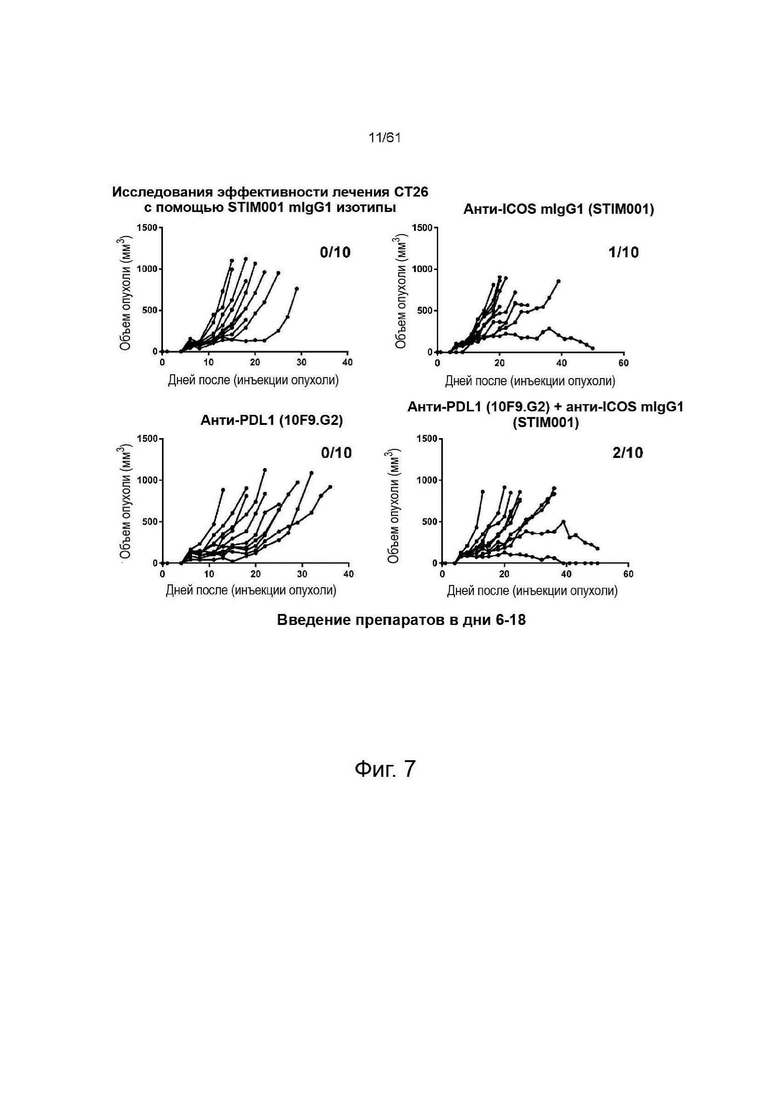
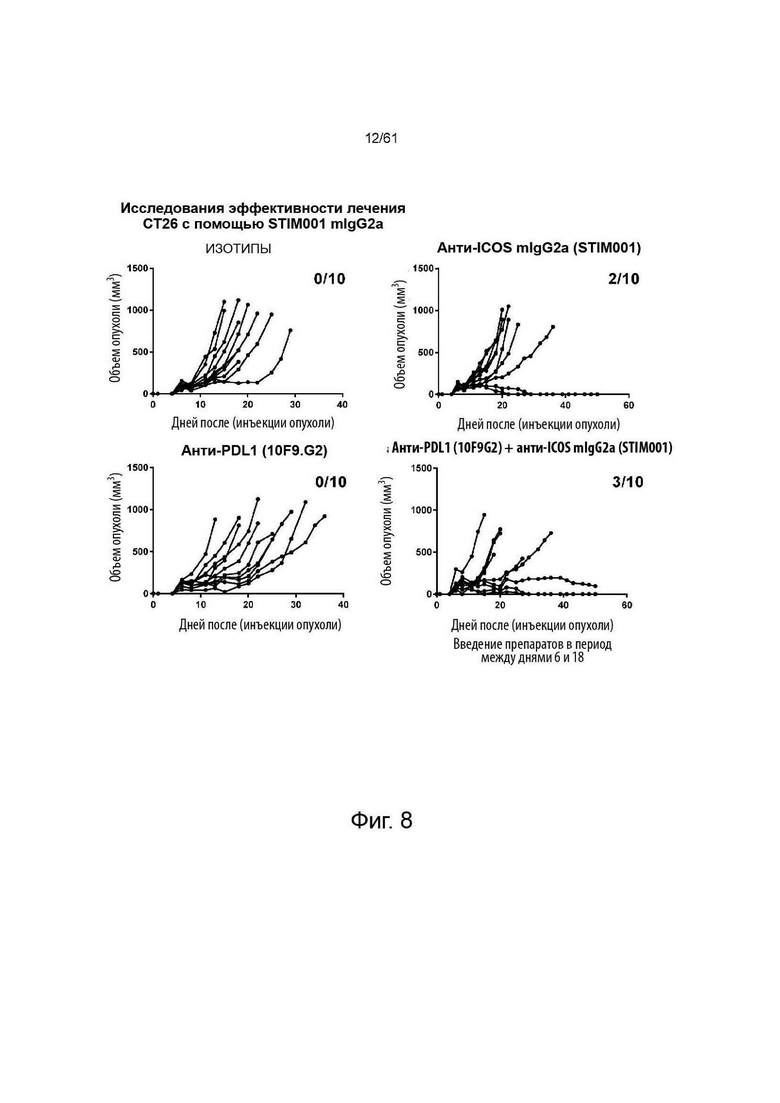

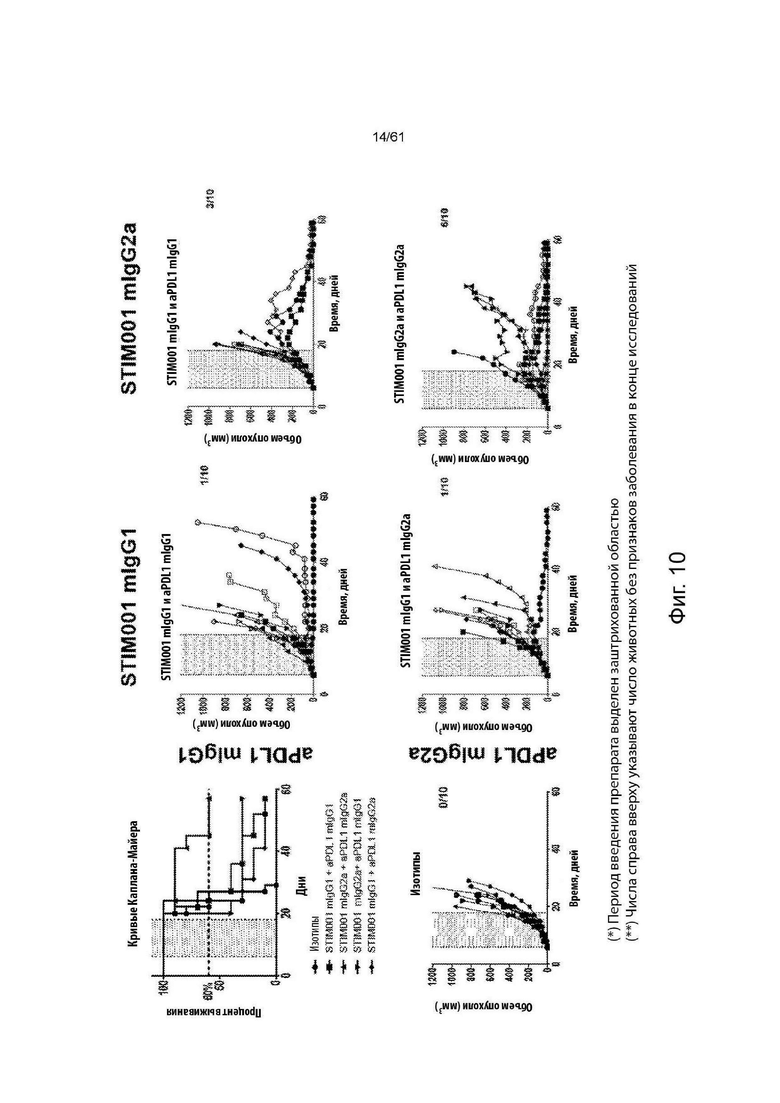
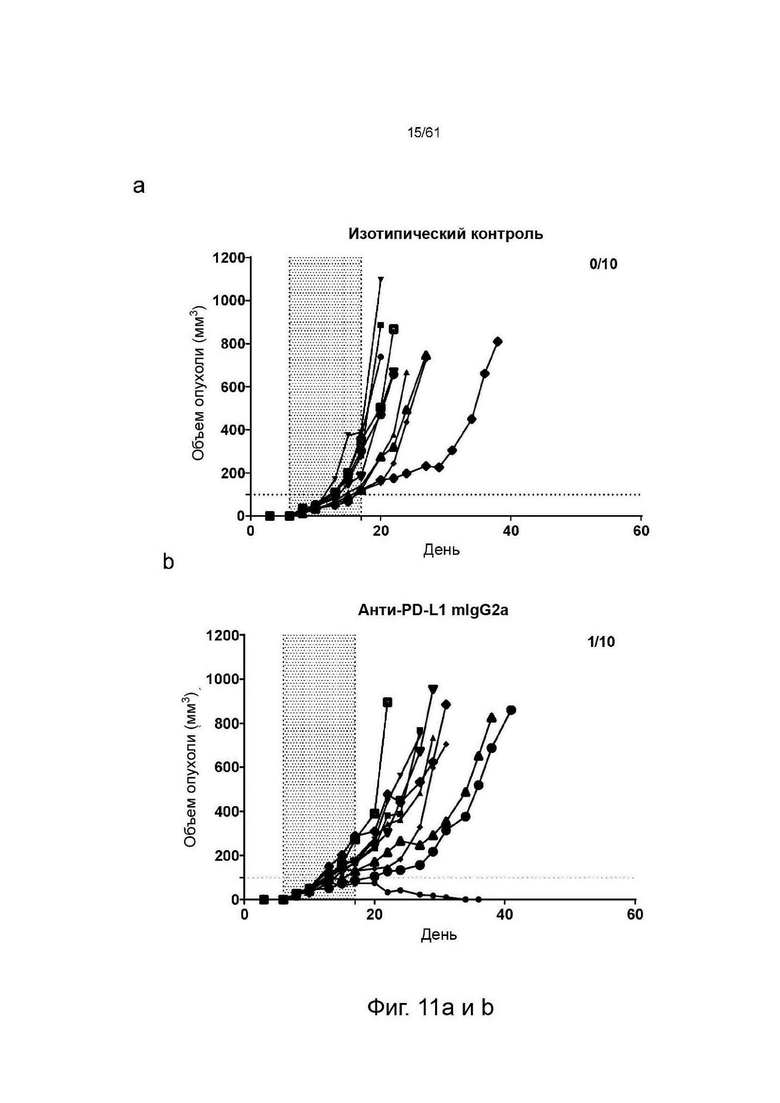
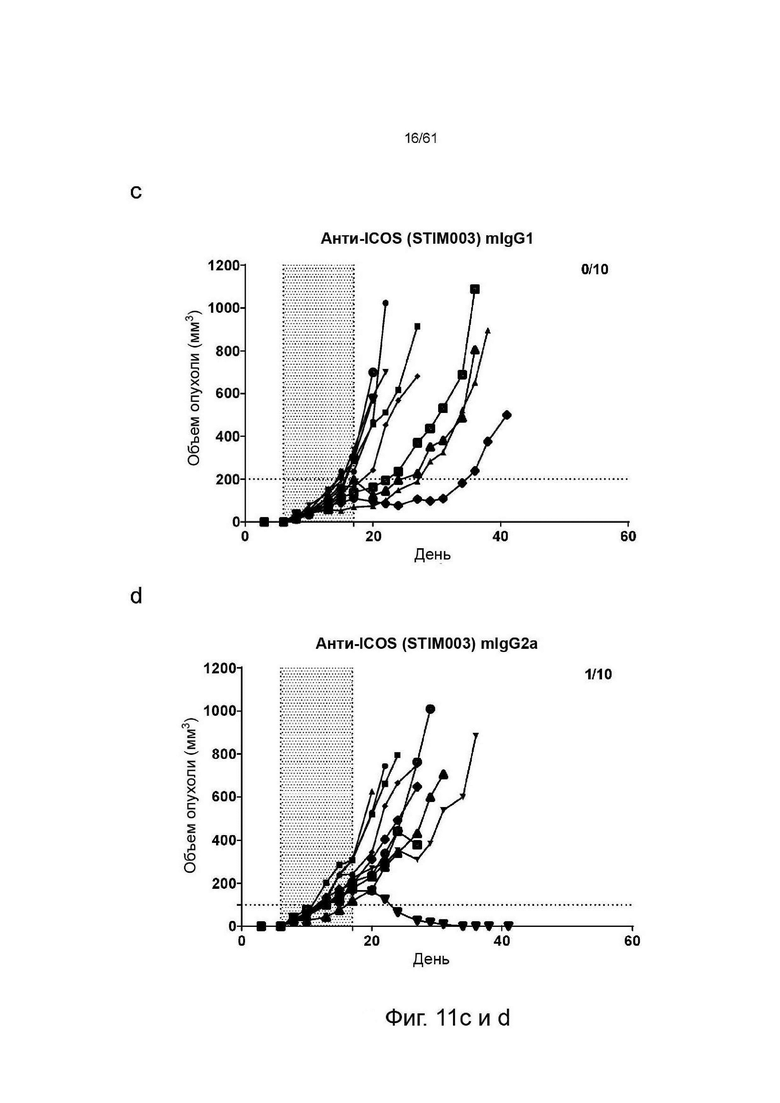
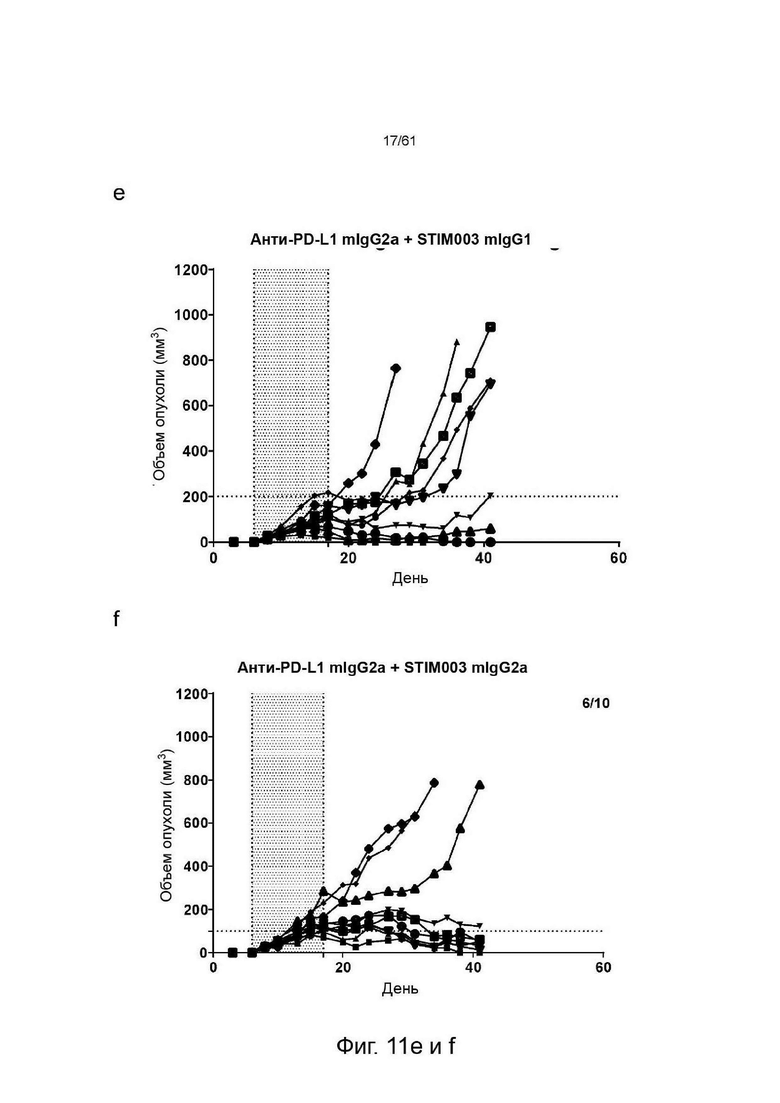
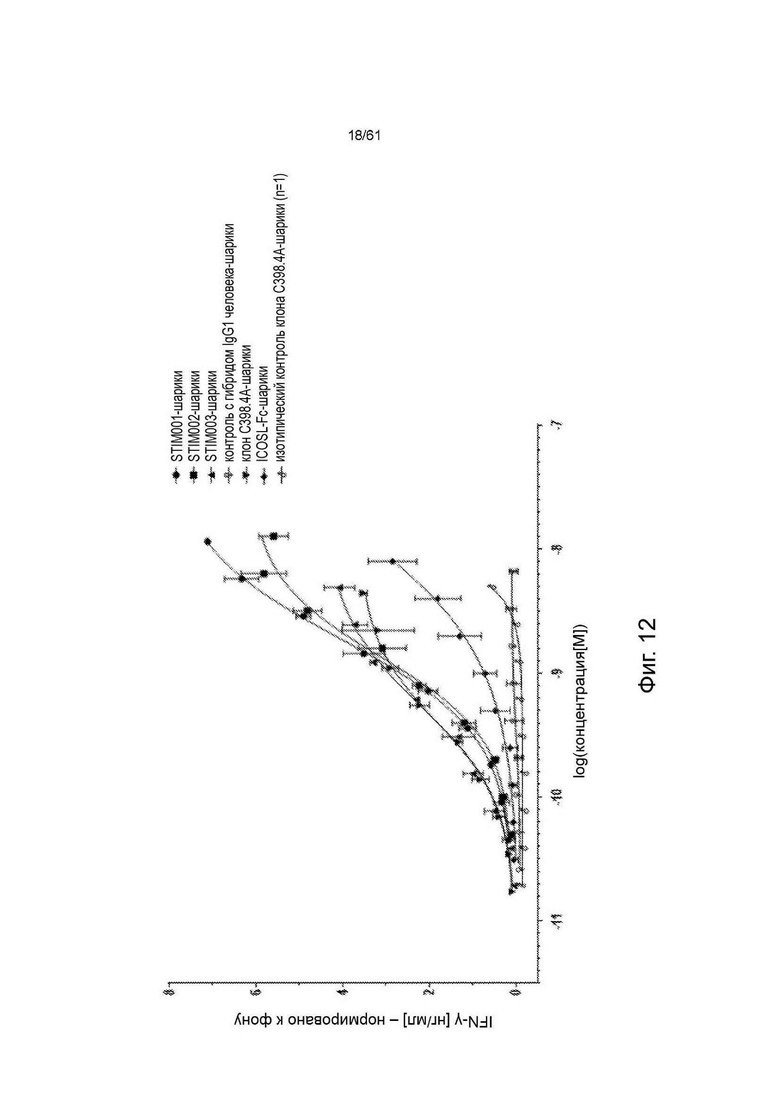
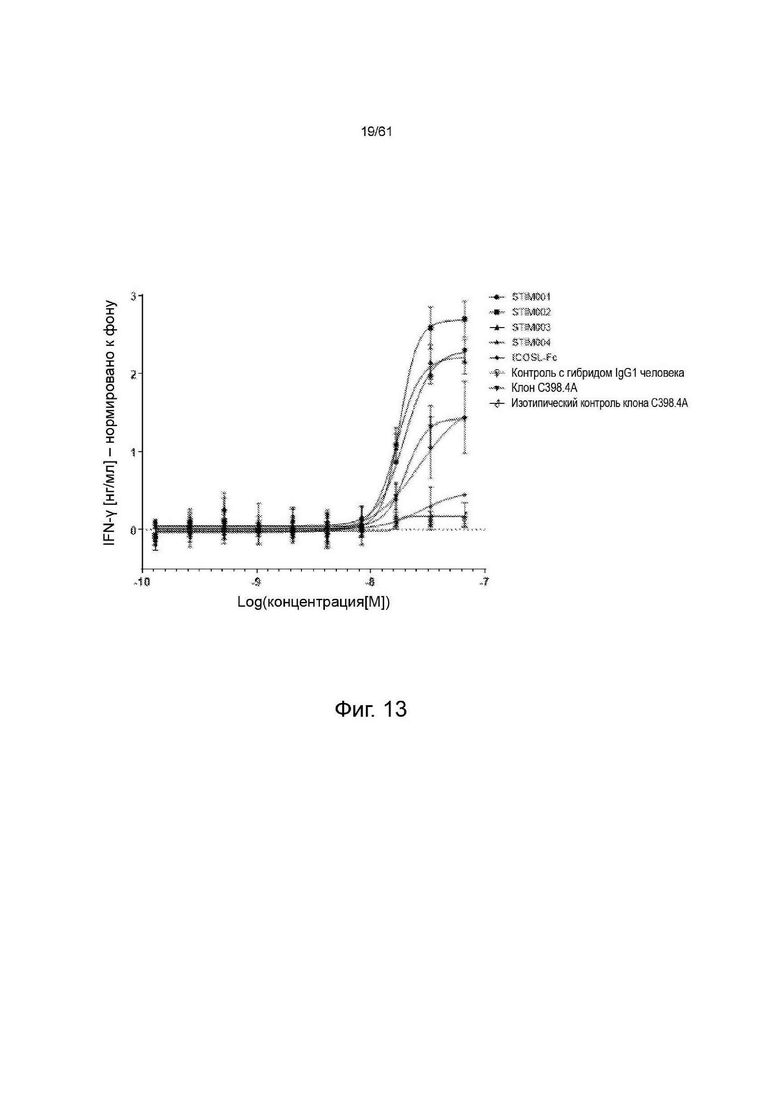
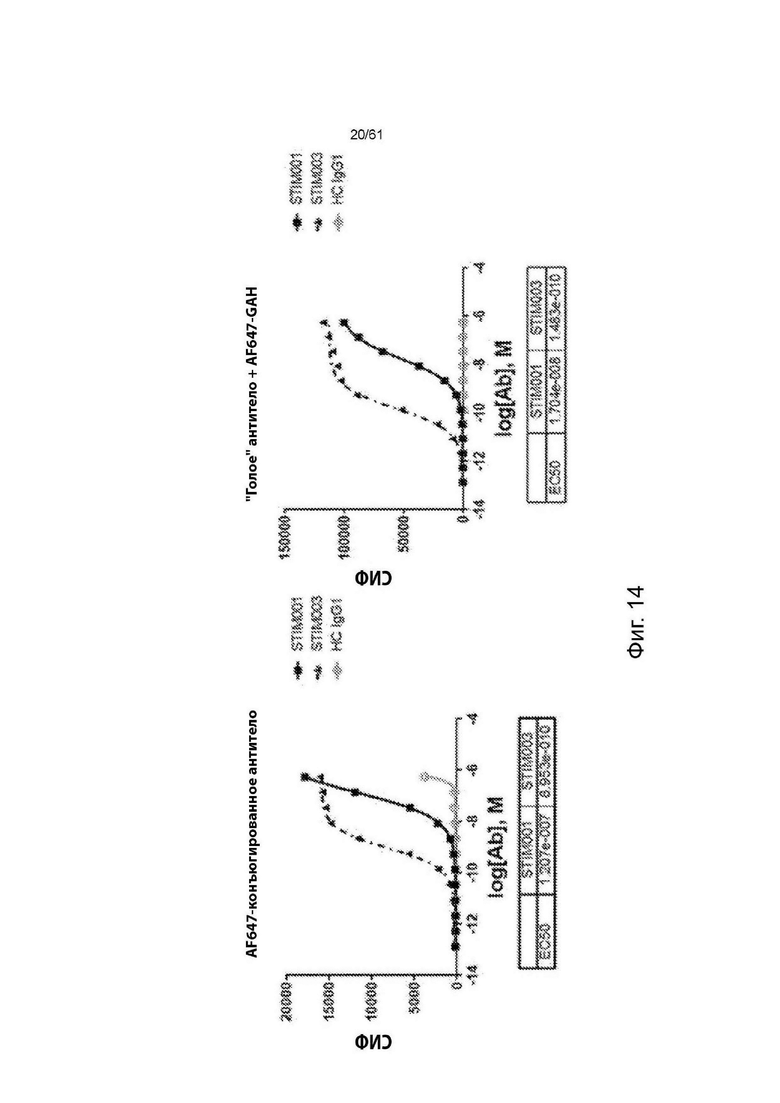
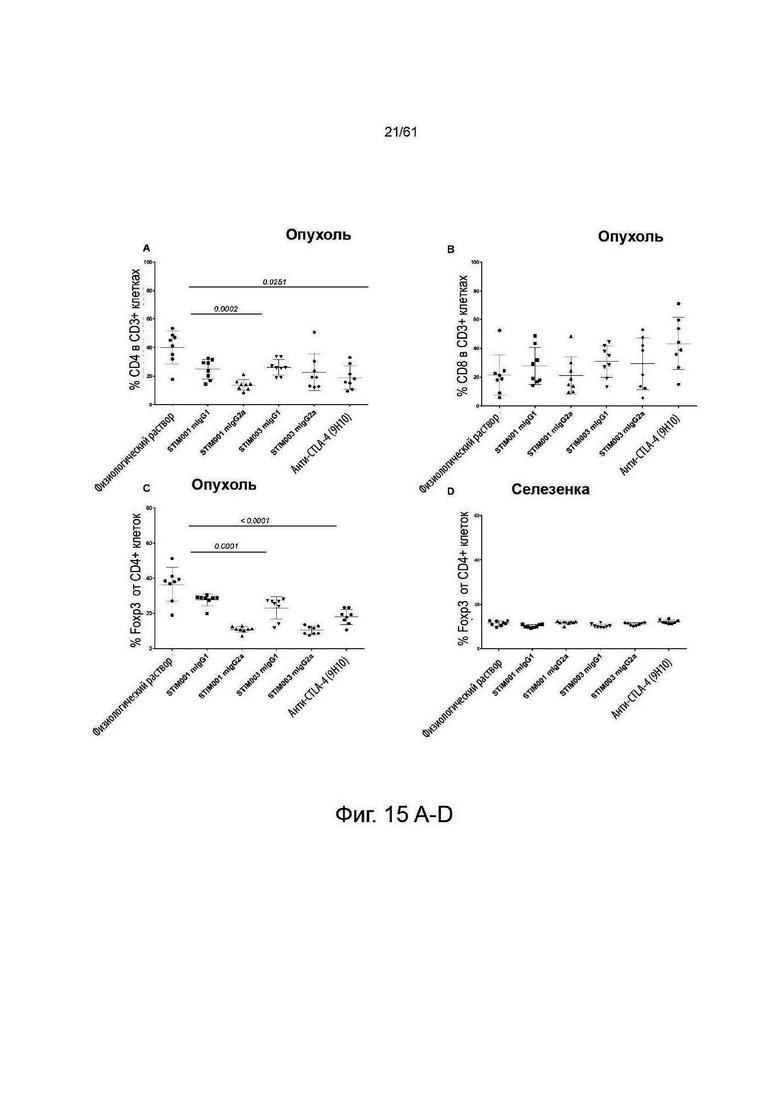
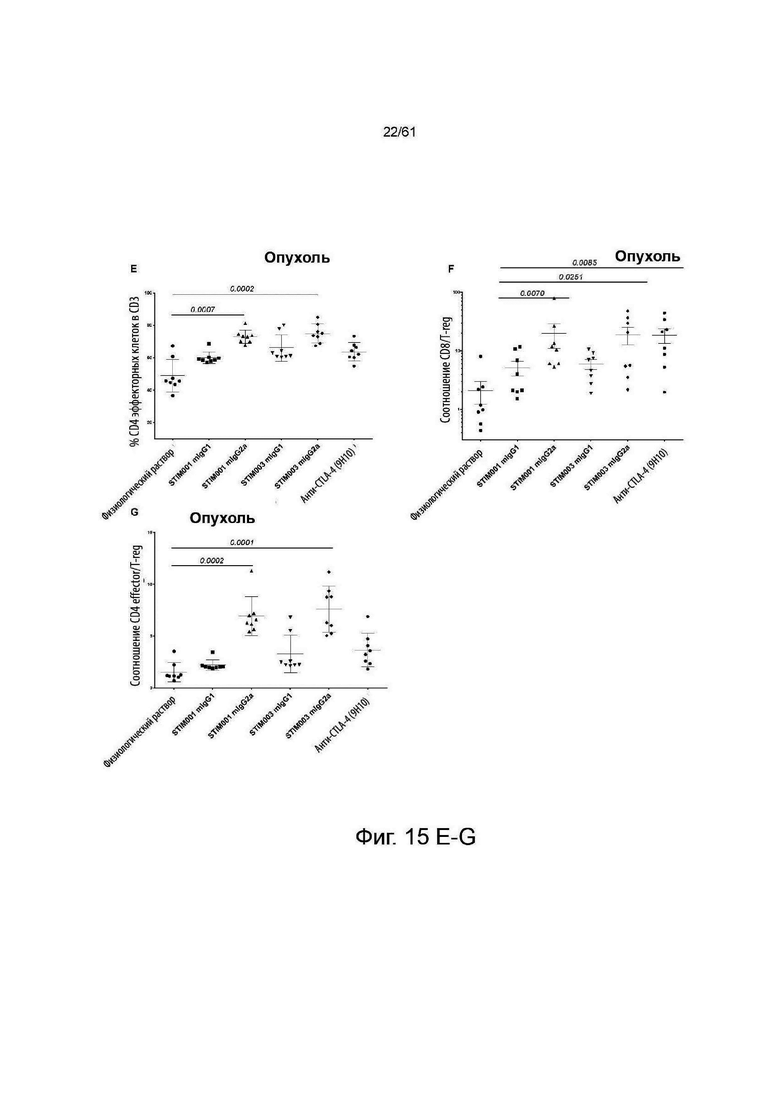
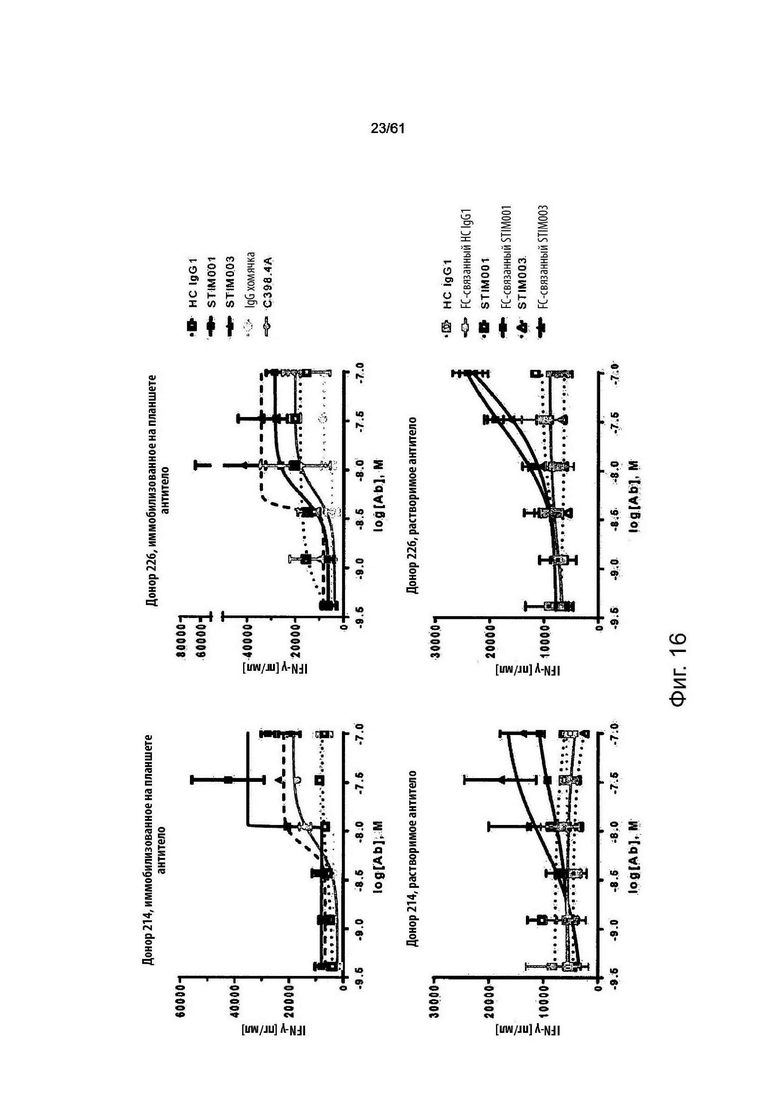
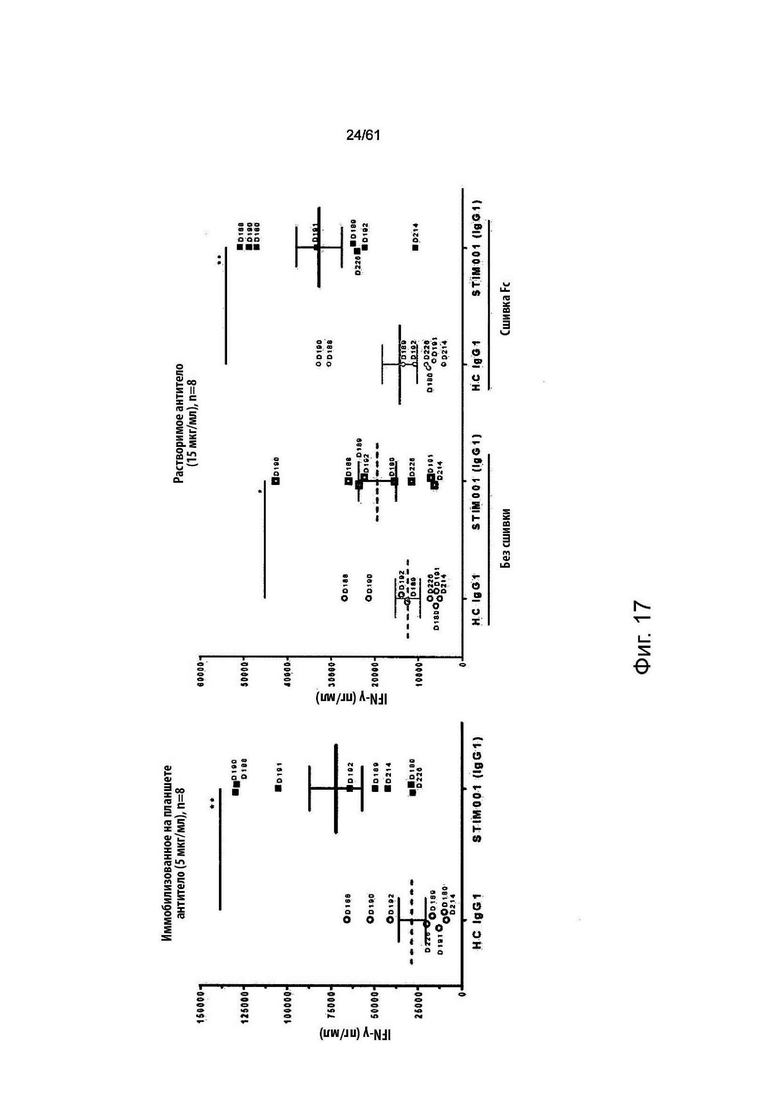

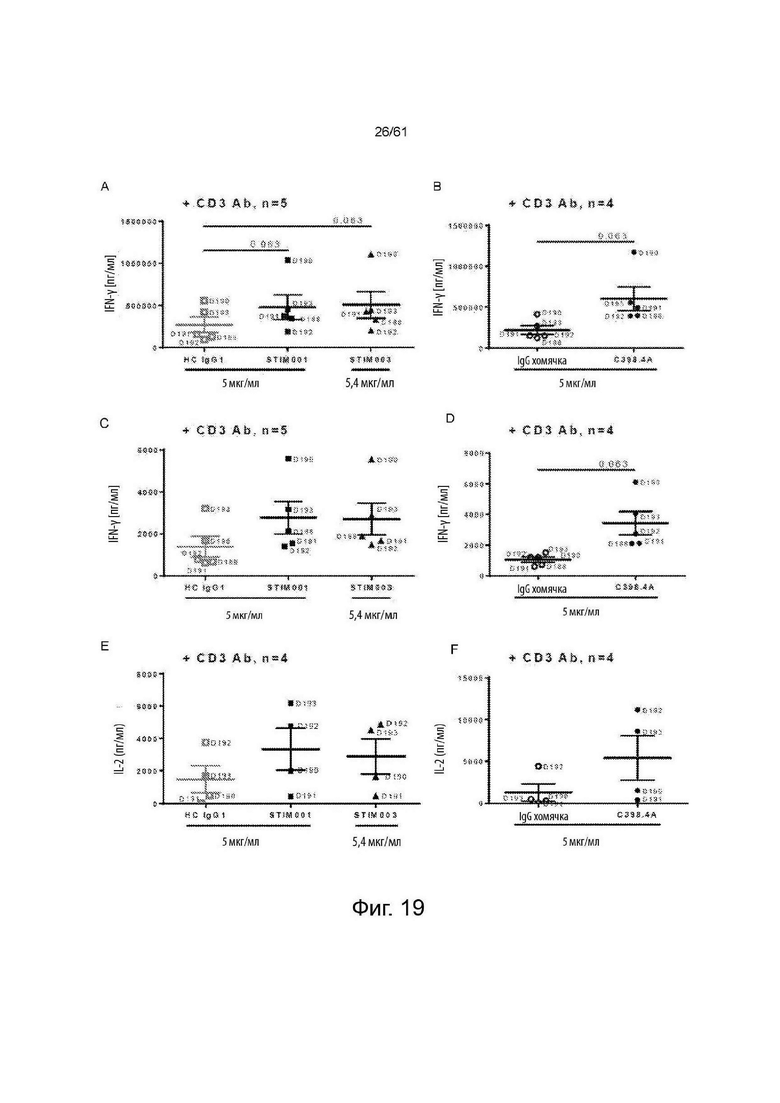
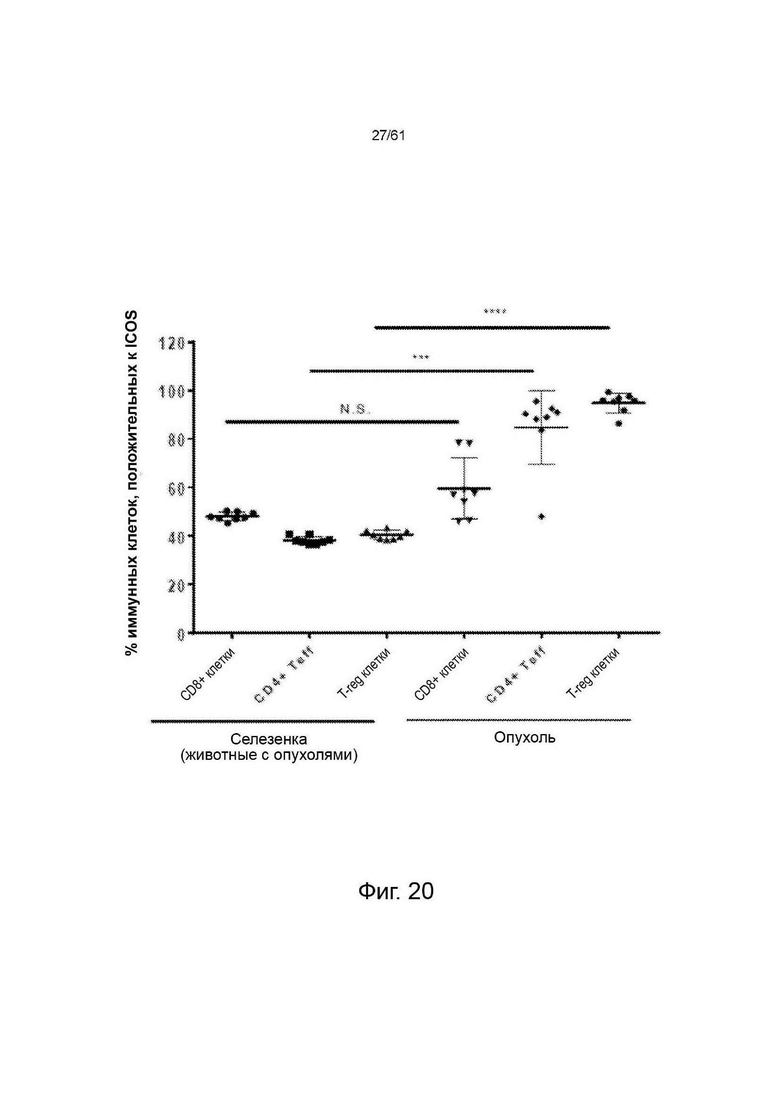
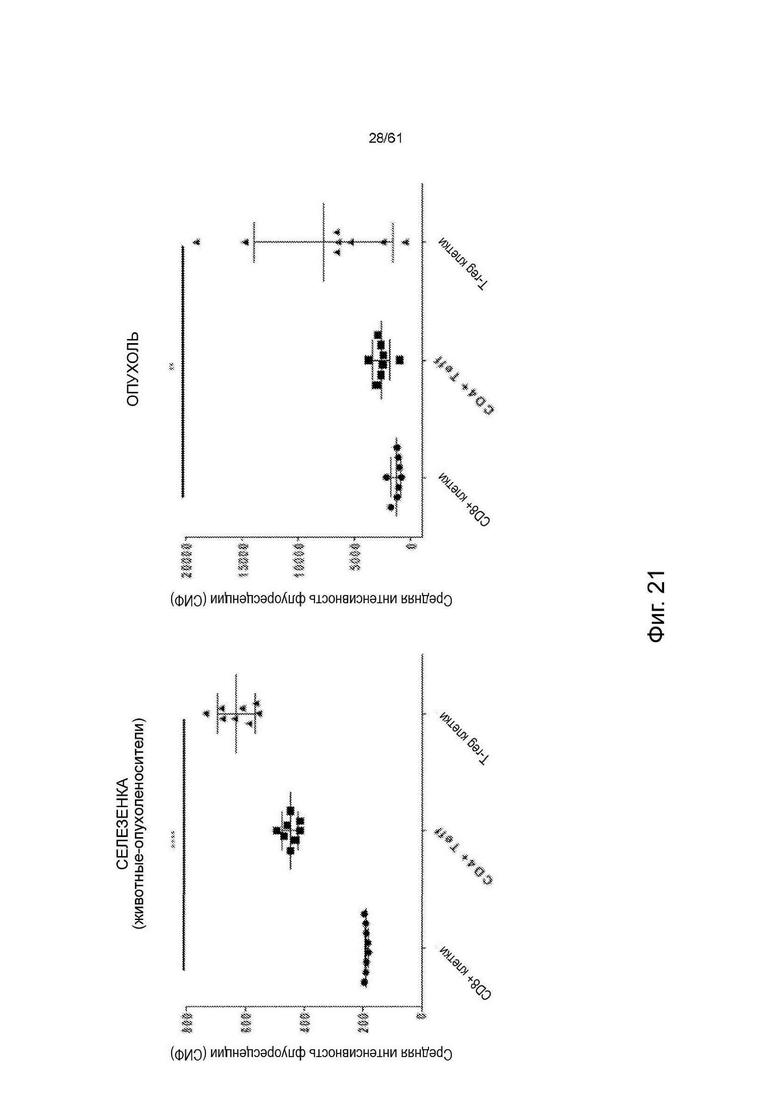

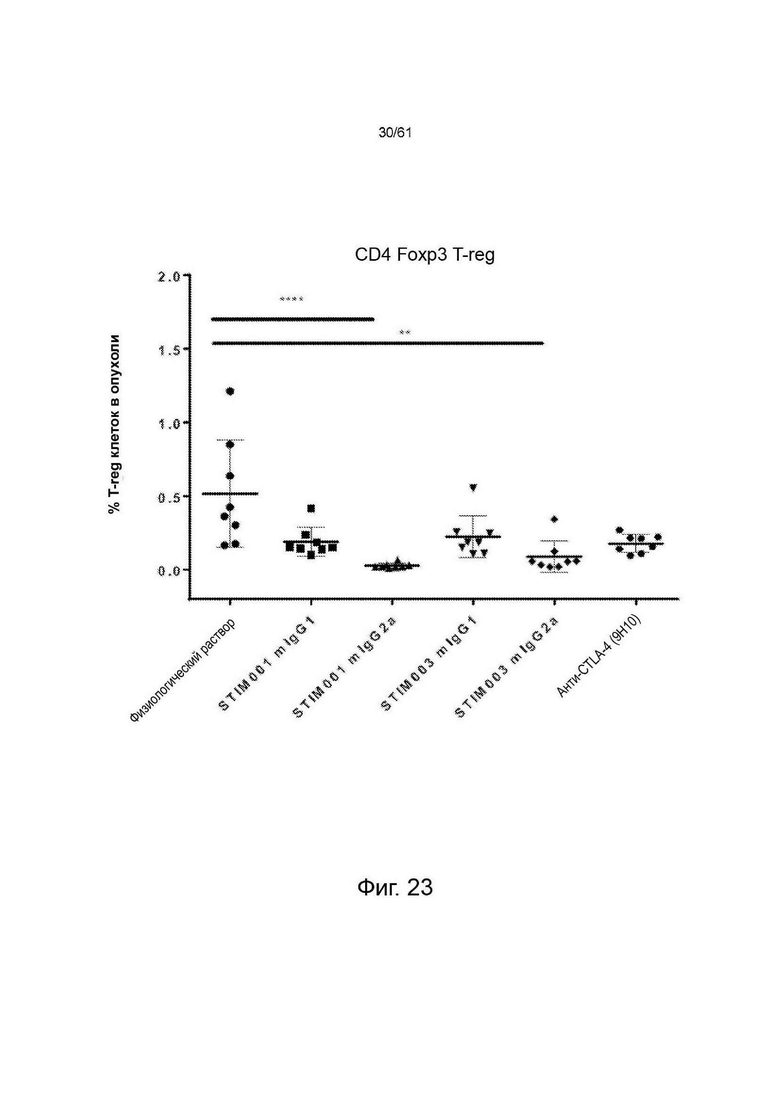
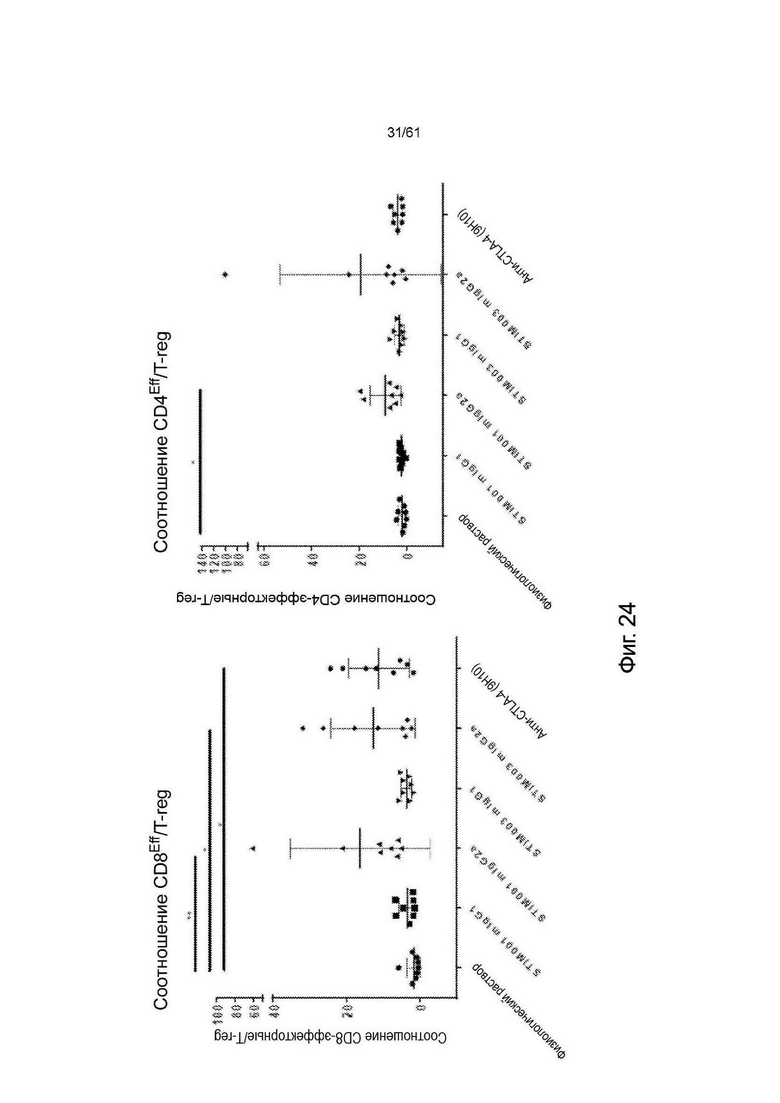

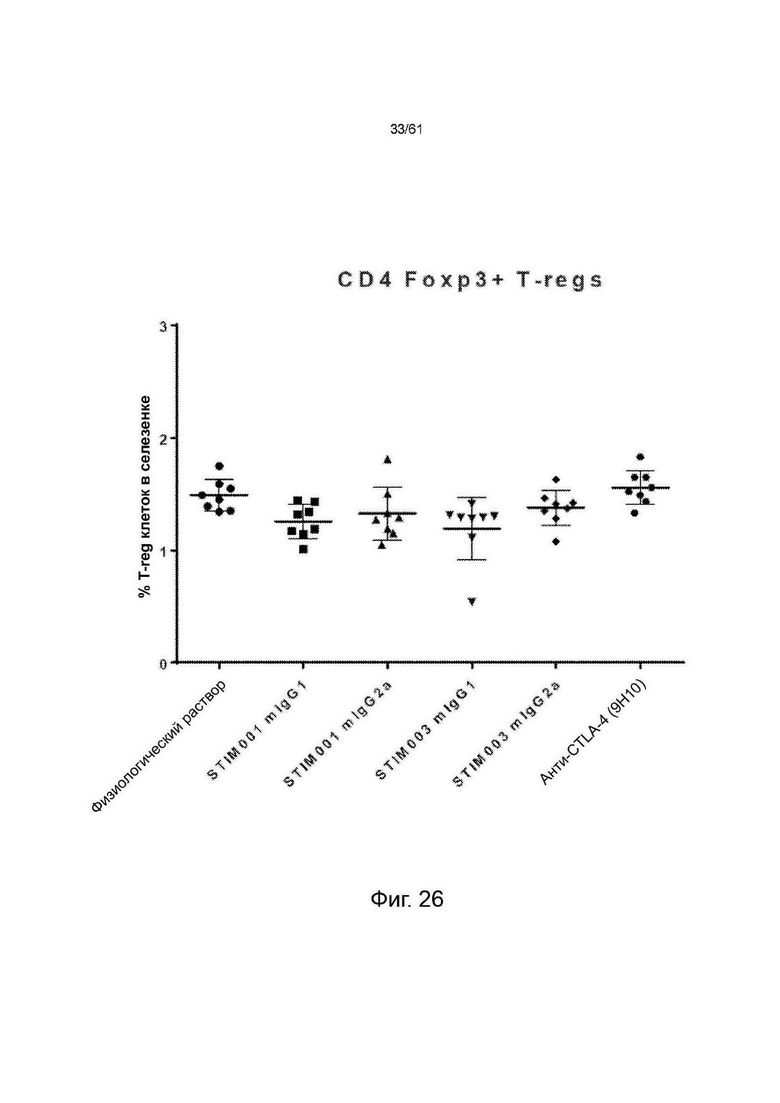
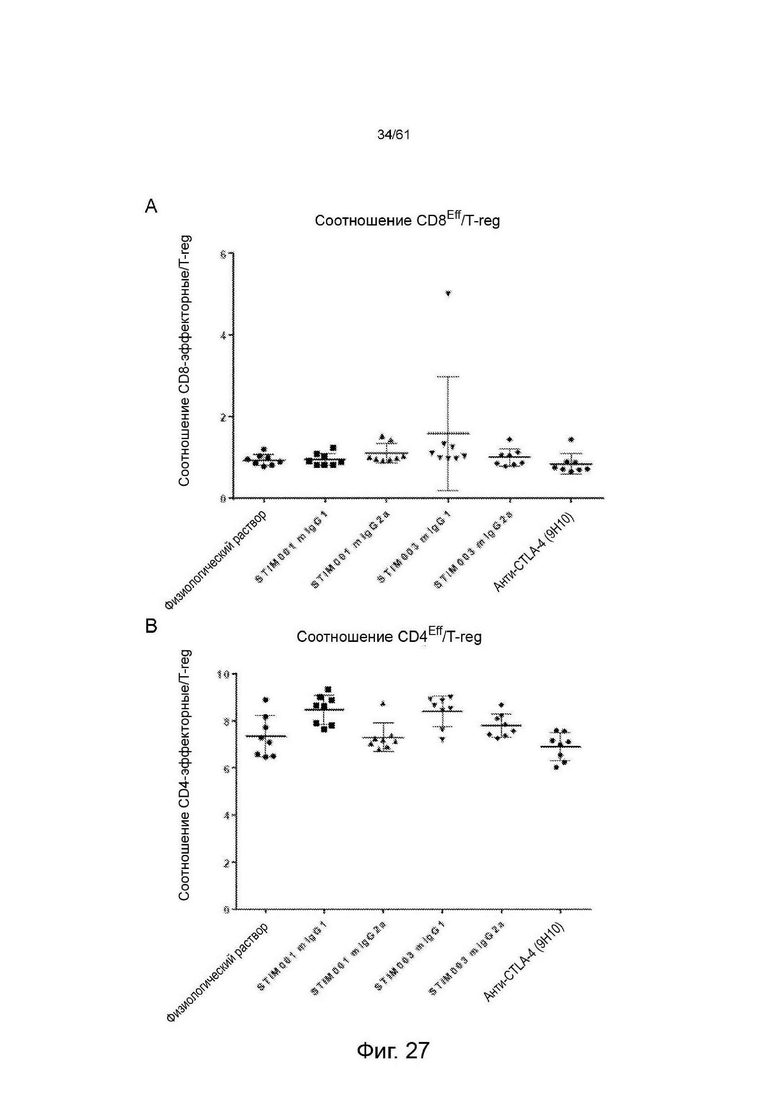
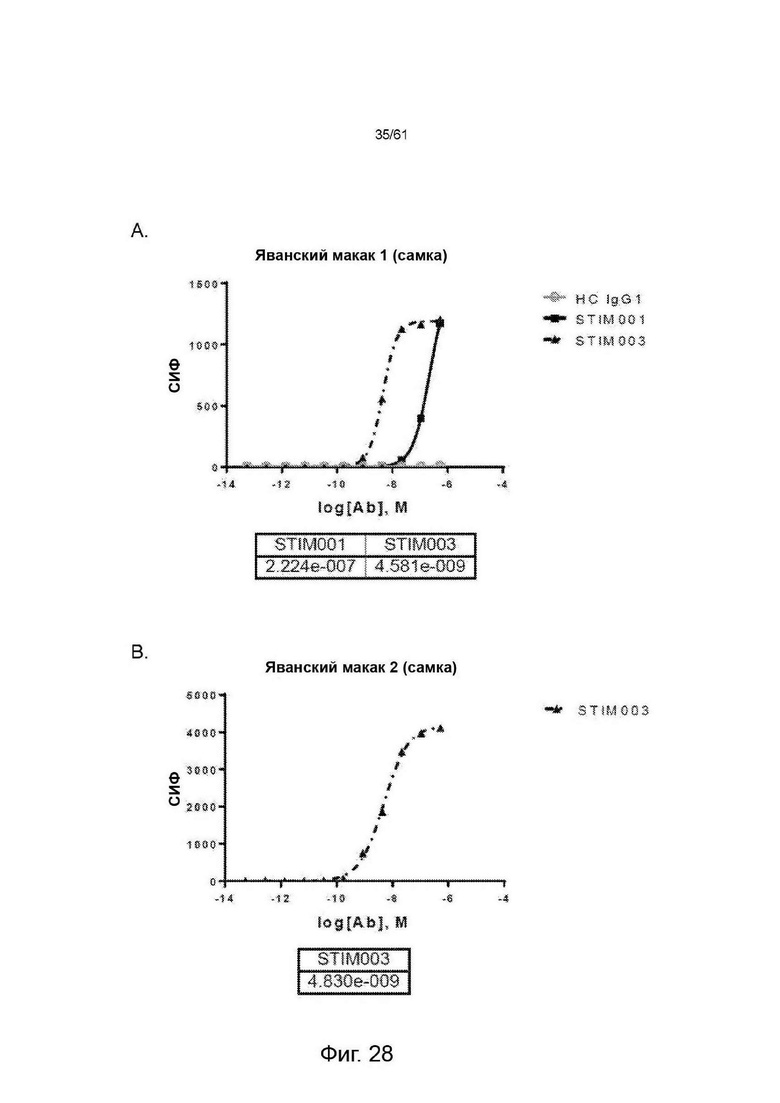
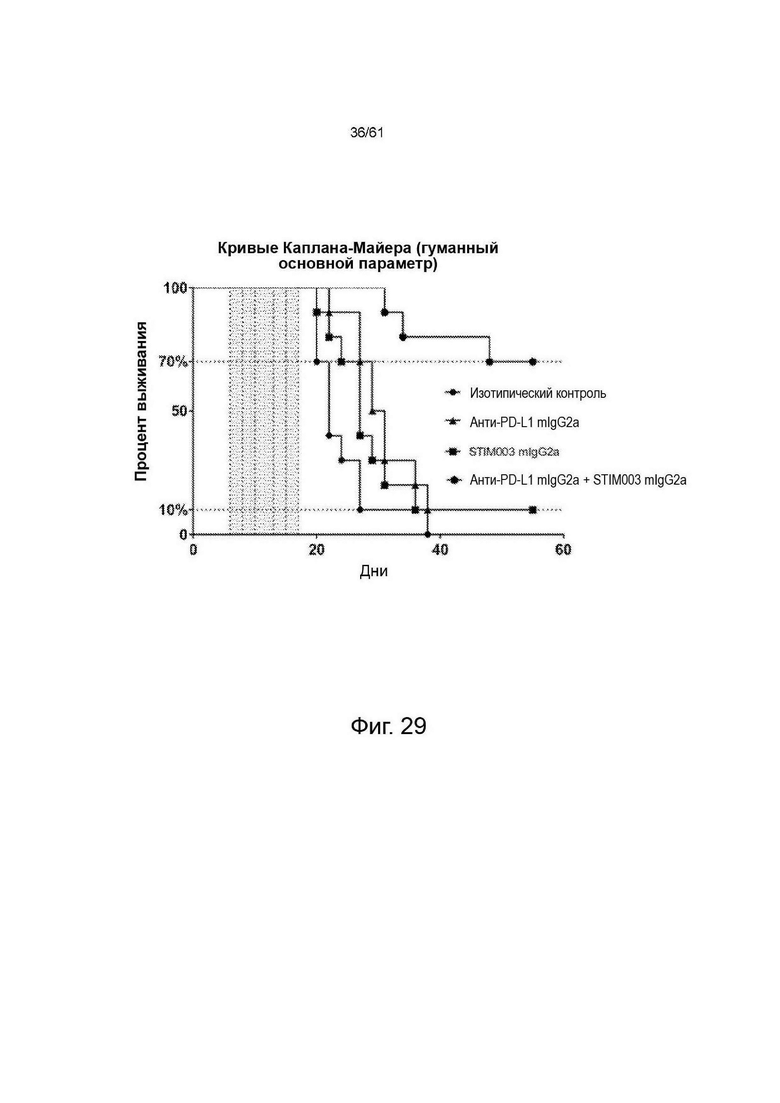
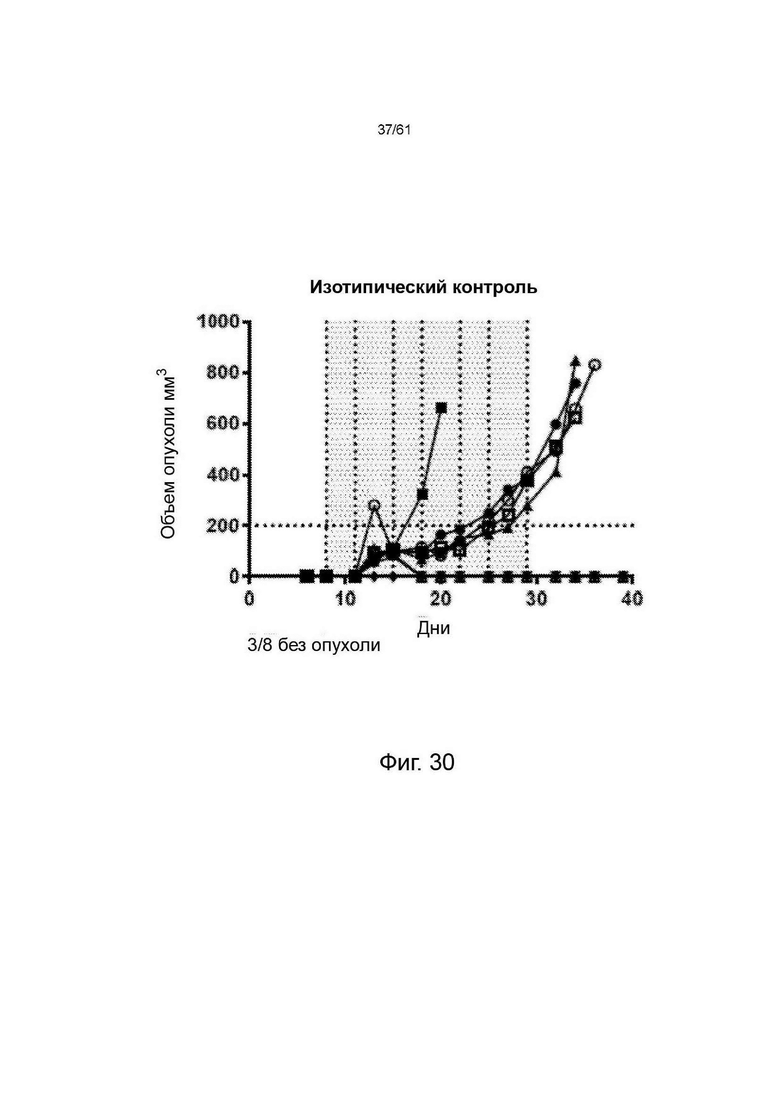

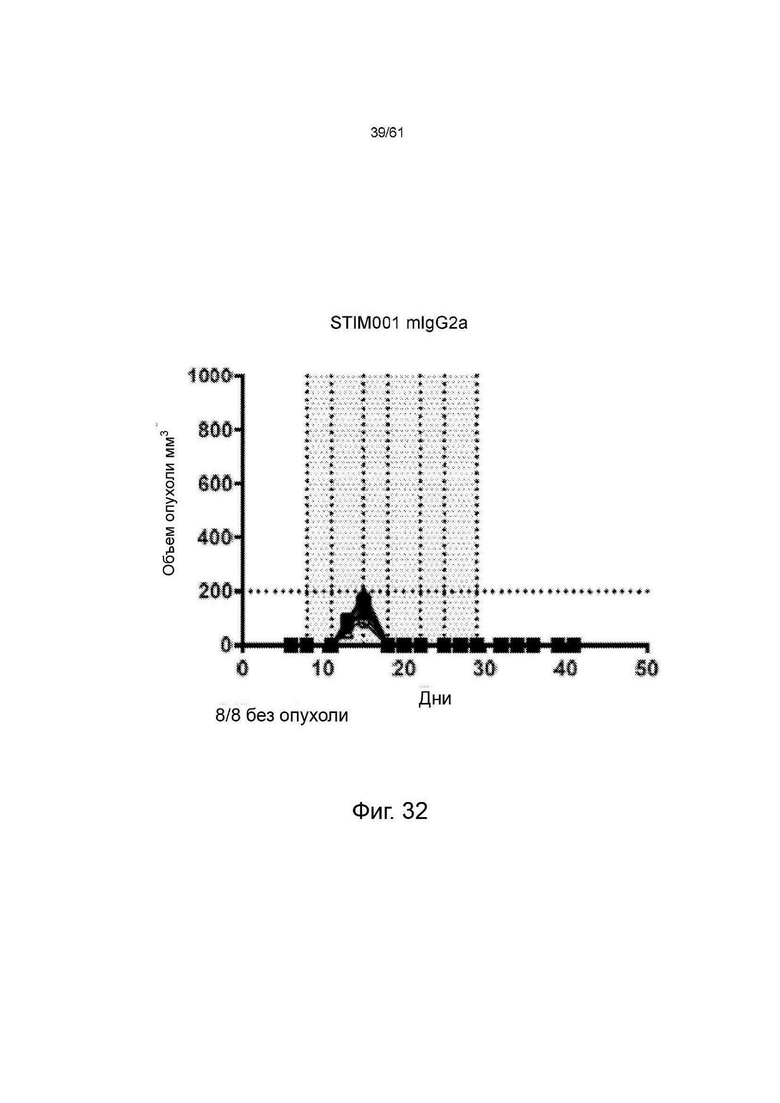

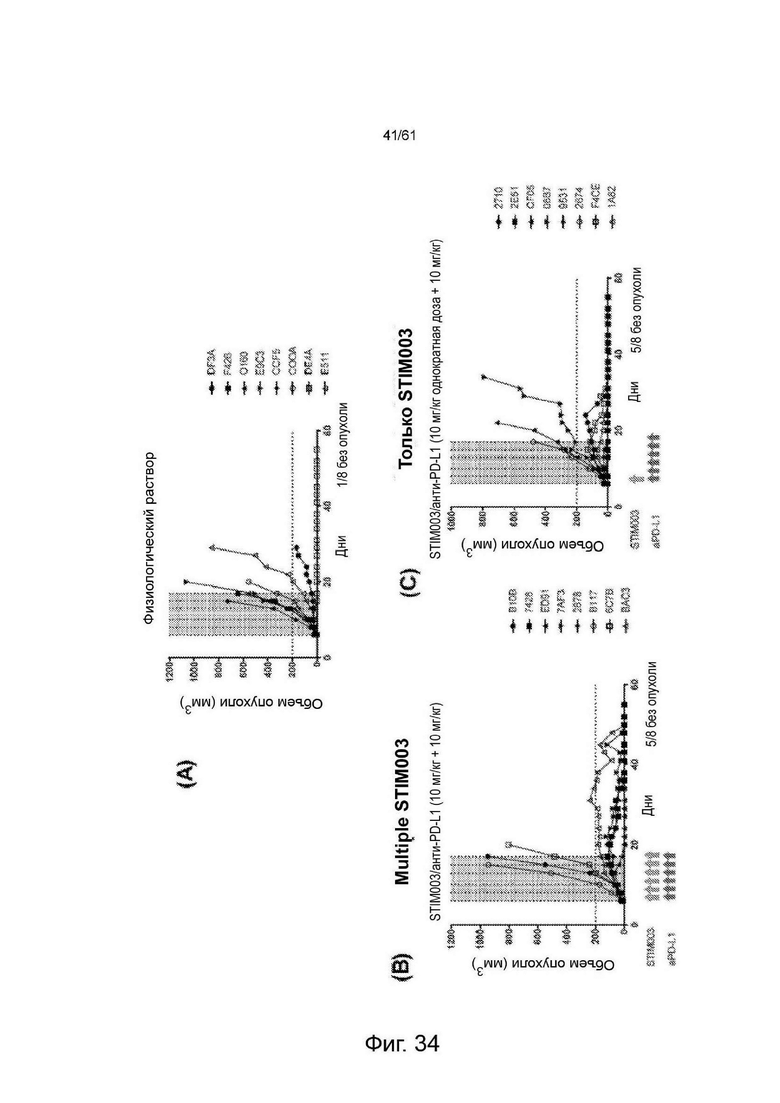
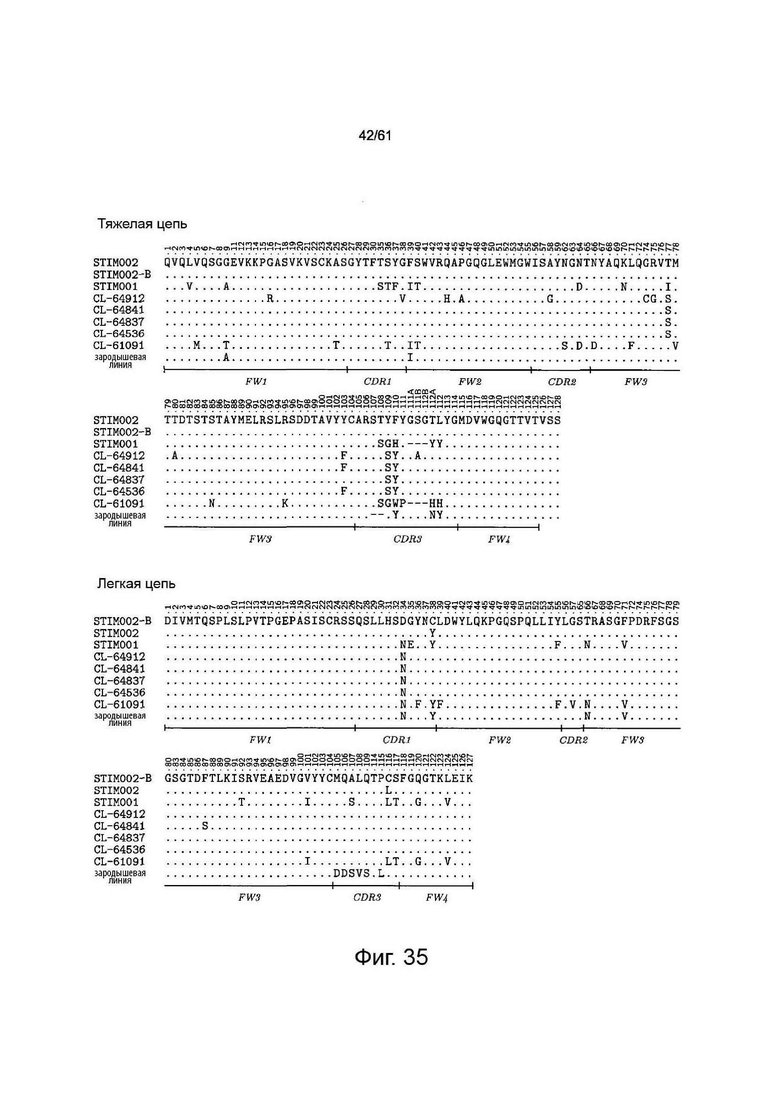




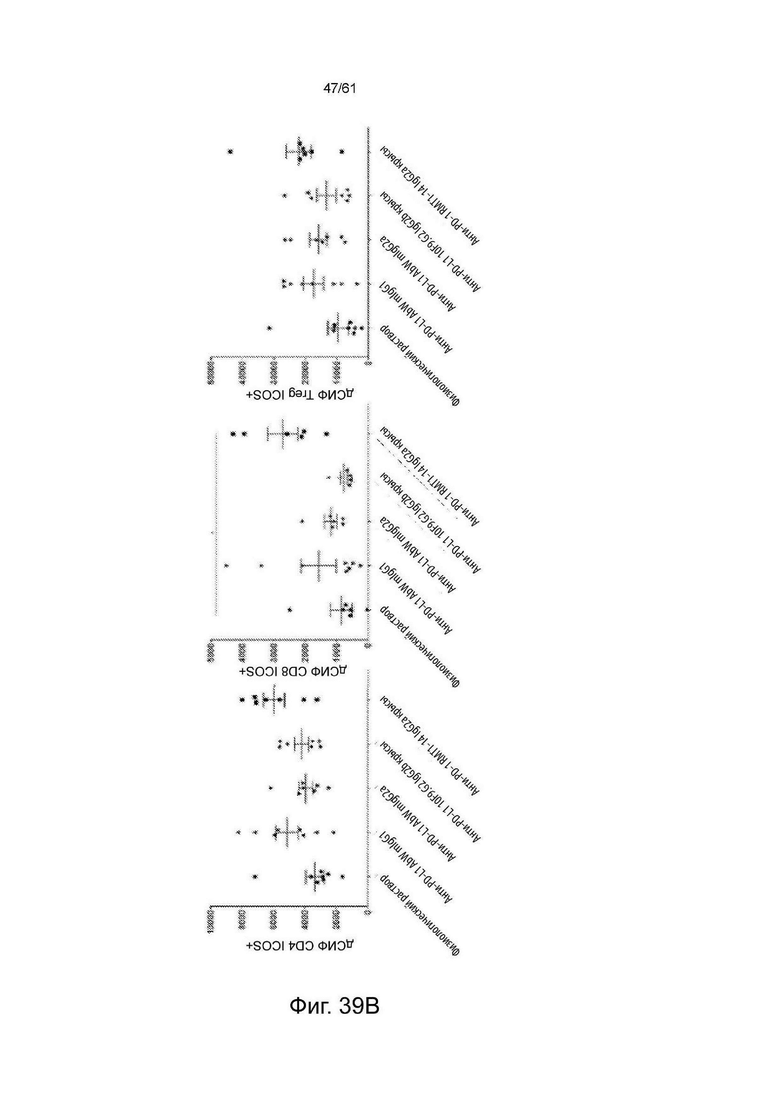

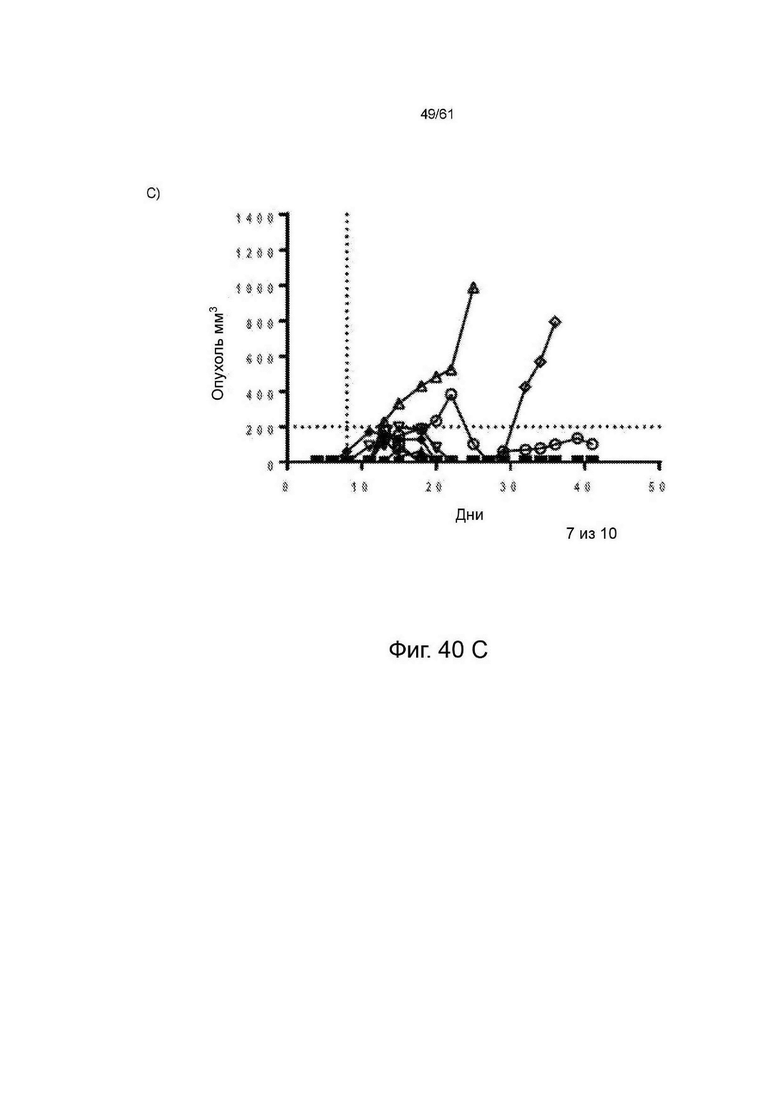

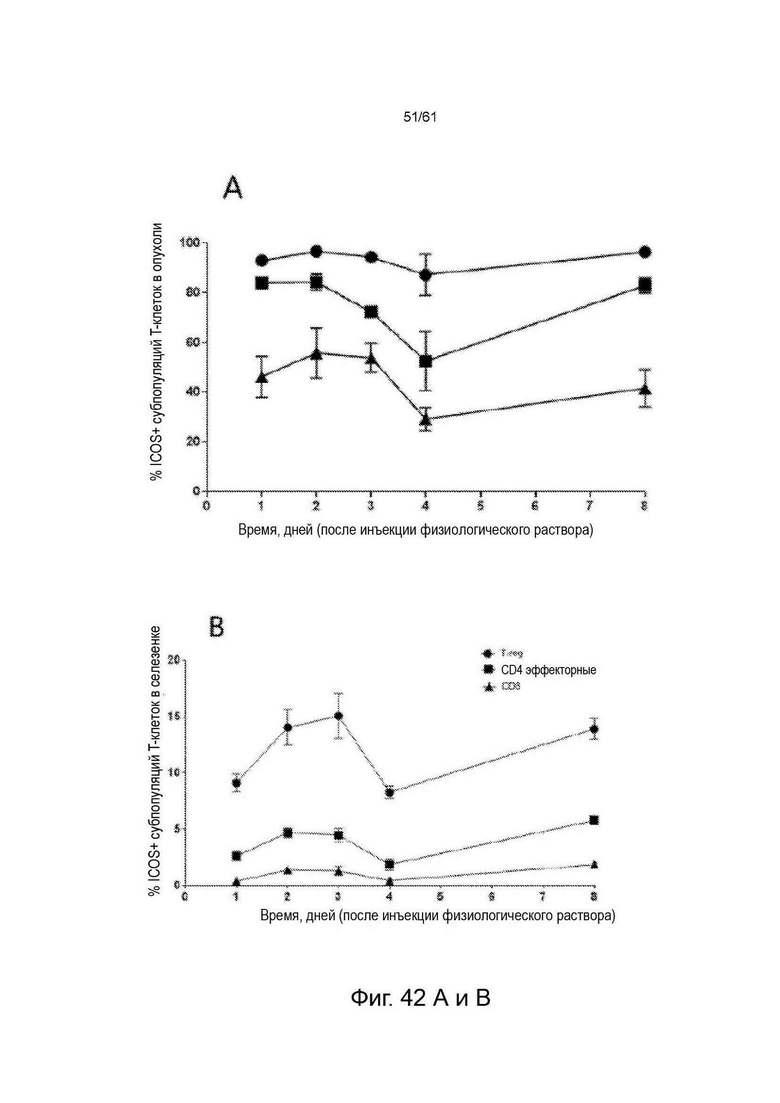

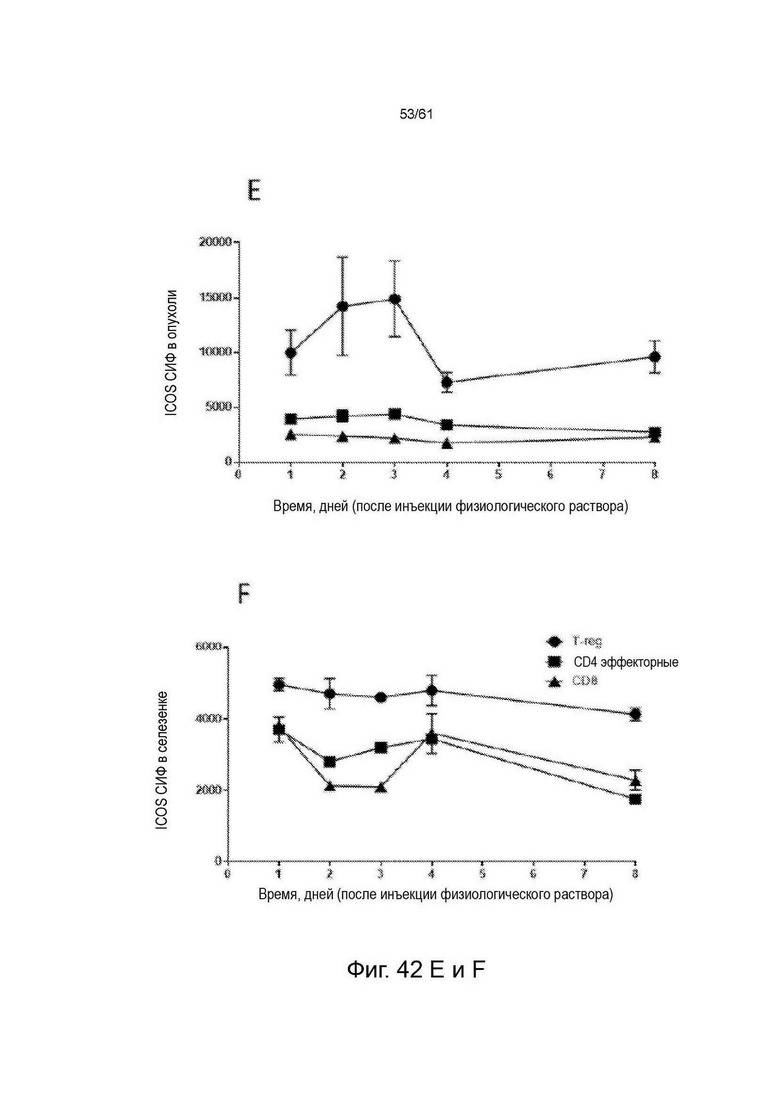
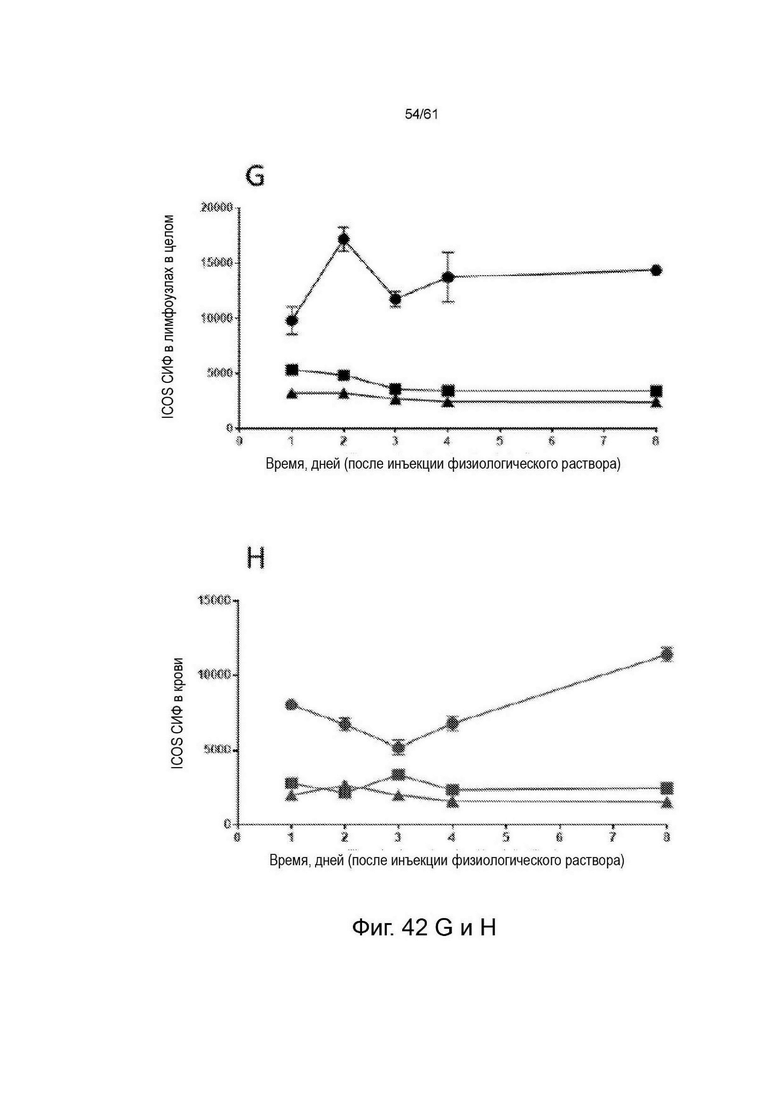

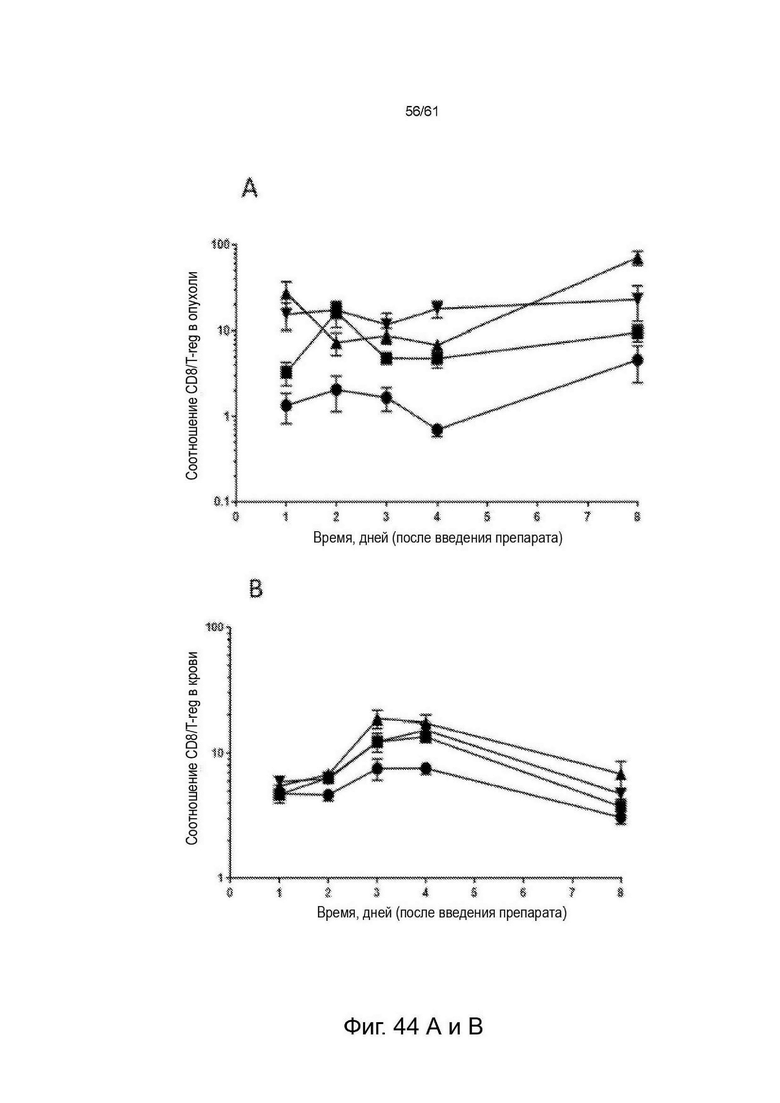
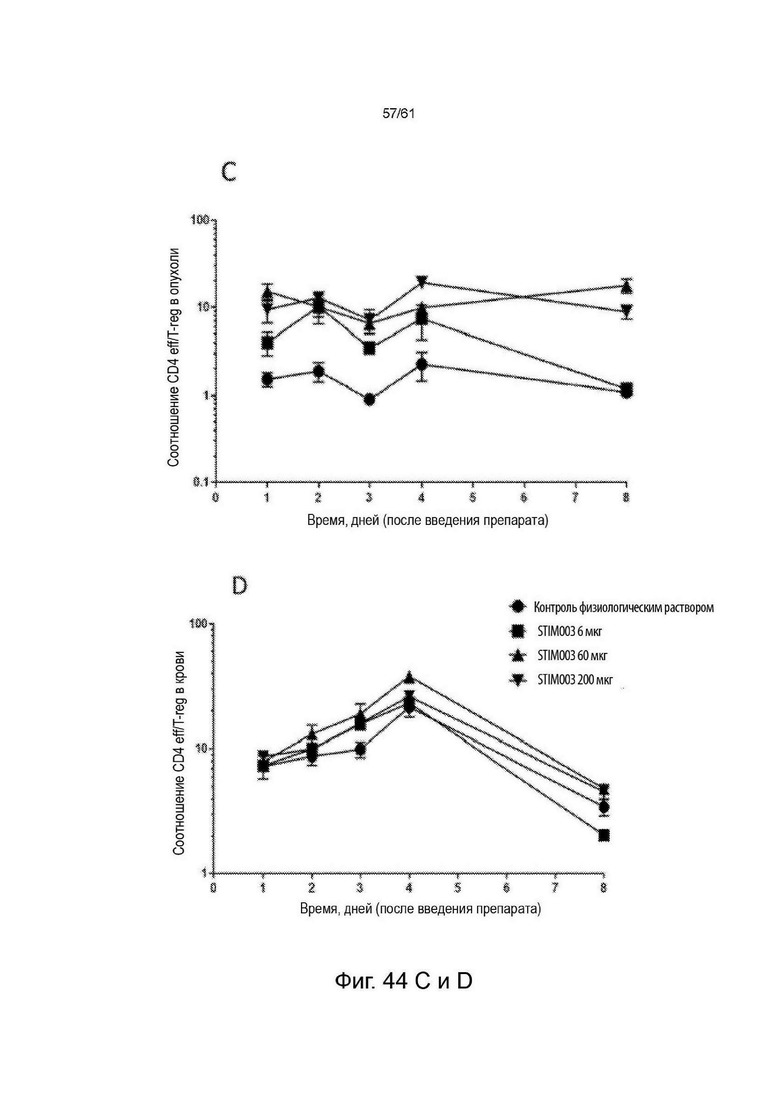
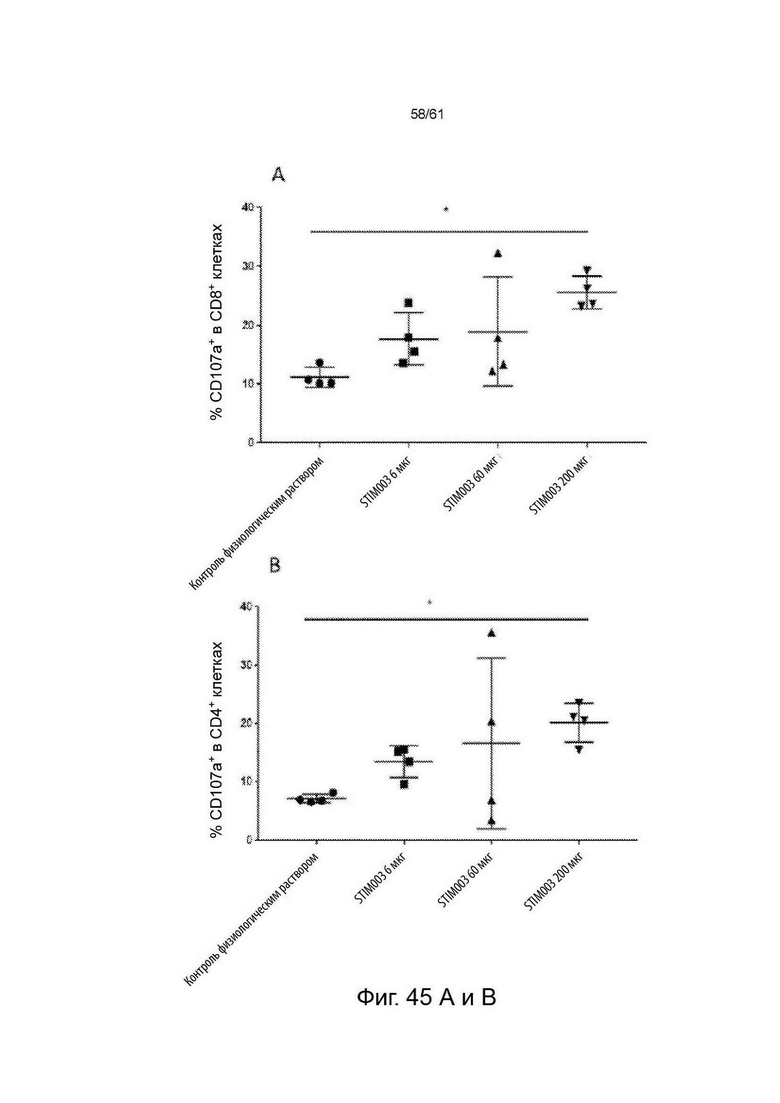
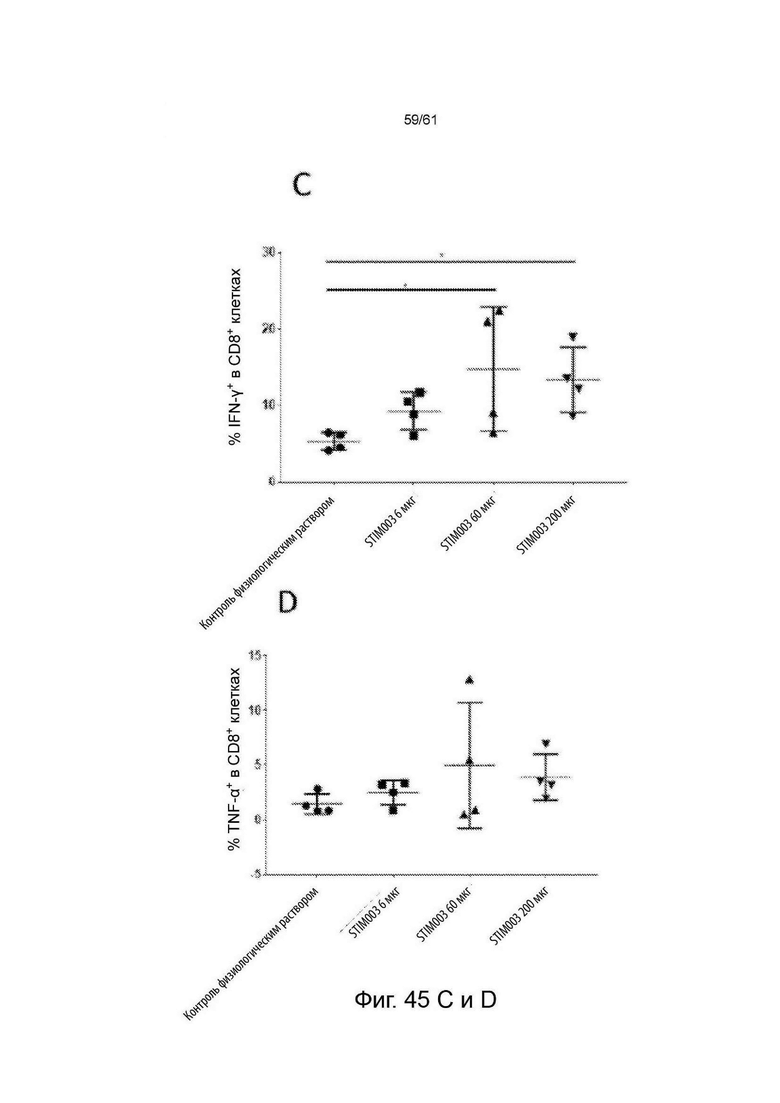
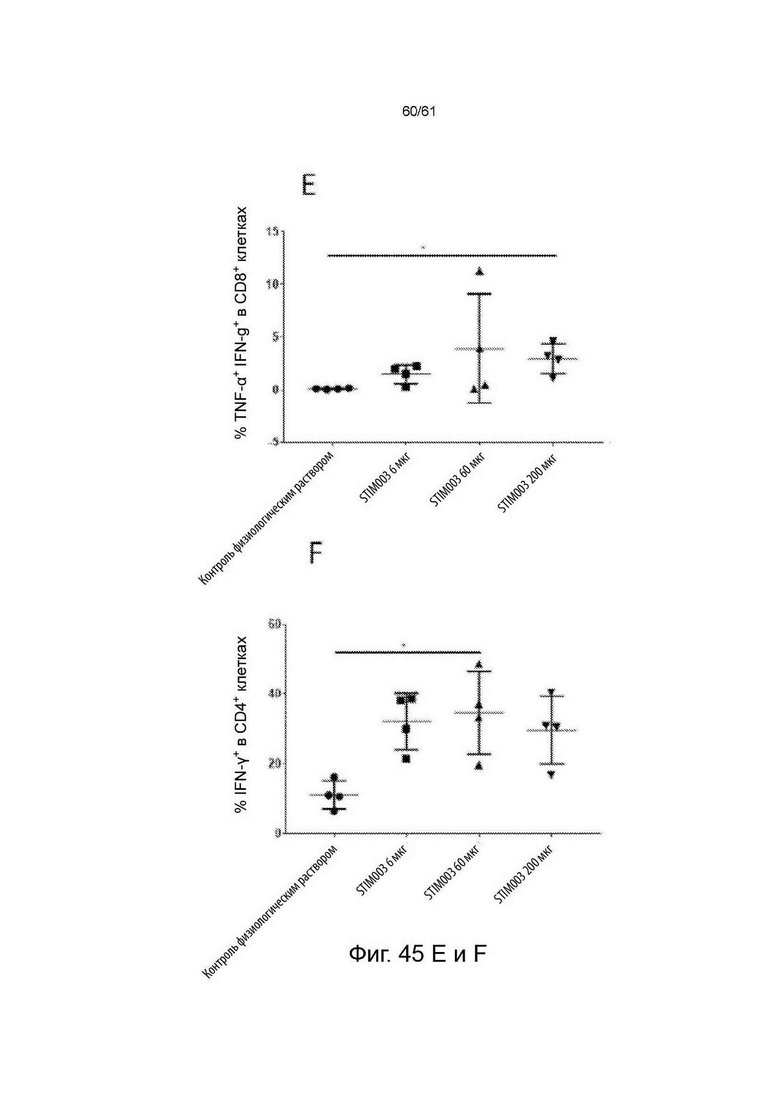
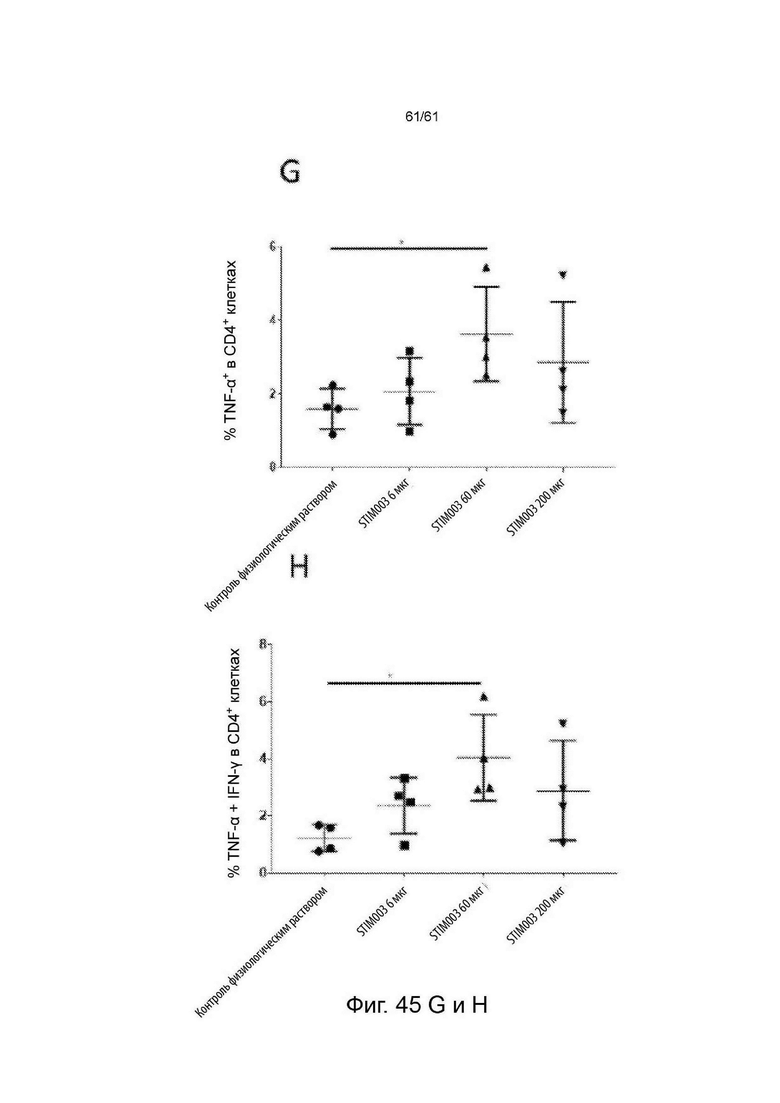
Изобретение относится к области биотехнологии, в частности к выделенному антителу, специфически связывающему внеклеточный домен индуцибельного Т-клеточного костимулятора (ICOS) человека и мыши, а также к содержащей его композиции. Также раскрыта клетка-хозяин in vitro для экспрессии вышеуказанного антитела. Изобретение эффективно для стимуляции Т-клеточного иммуного ответа, а также для лечения рака. 6 н. и 20 з.п. ф-лы, 45 ил., 32 табл., 26 пр.
1. Выделенное антитело, специфически связывающее внеклеточный домен индуцибельного Т-клеточного костимулятора (ICOS) человека и мыши, причем антитело содержит домен VH, содержащий аминокислотную последовательность, имеющую по меньшей мере 95% идентичности последовательности с доменом VH с SEQ ID NO: 408, и домен VL, содержащий аминокислотную последовательность, имеющую по меньшей мере 95% идентичности последовательности с доменом VL с SEQ ID NO: 415.
2. Антитело по п. 1, отличающееся тем, что домен VH содержит набор участков, определяющих комплементарность, тяжелой цепи (HCDR) HCDR1, HCDR2 и HCDR3, причем
HCDR1 представляет собой HCDR1, имеющий аминокислотную последовательность SEQ ID NO: 405,
HCDR2 представляет собой HCDR2, имеющий аминокислотную последовательность SEQ ID NO: 406,
HCDR3 представляет собой HCDR3, имеющий аминокислотную последовательность SEQ ID NO: 407.
3. Антитело по п. 2, отличающееся тем, что домен VL содержит набор участков, определяющих комплементарность, легкой цепи (LCDR) LCDR1, LCDR2 и LCDR3, причем
LCDR1 представляет собой LCDR1, имеющий аминокислотную последовательность SEQ ID NO: 412,
LCDR2 представляет собой LCDR2, имеющий аминокислотную последовательность SEQ ID NO: 413,
LCDR3 представляет собой LCDR3, имеющий аминокислотную последовательность SEQ ID NO: 414.
4. Антитело по п. 3, отличающееся тем, что домен VH содержит набор участков, определяющих комплементарность, тяжелой цепи (HCDR) HCDR1, HCDR2 и HCDR3, причем
HCDR1 представляет собой HCDR1, имеющий аминокислотную последовательность SEQ ID NO: 405,
HCDR2 представляет собой HCDR2, имеющий аминокислотную последовательность SEQ ID NO: 406,
HCDR3 представляет собой HCDR3, имеющий аминокислотную последовательность SEQ ID NO: 407, и где
домен VL содержит набор участков, определяющих комплементарность, легкой цепи (LCDR) LCDR1, LCDR2 и LCDR3, причем
LCDR1 представляет собой LCDR1, имеющий аминокислотную последовательность SEQ ID NO: 412,
LCDR2 представляет собой LCDR2, имеющий аминокислотную последовательность SEQ ID NO: 413,
LCDR3 представляет собой LCDR3, имеющий аминокислотную последовательность SEQ ID NO: 414.
5. Антитело по п. 1, отличающееся тем, что аминокислотная последовательность домена VH представляет собой SEQ ID NO: 408 и аминокислотная последовательность домена VL представляет собой SEQ ID NO: 415.
6. Антитело, специфически связывающее внеклеточный домен индуцибельного Т-клеточного костимулятора (ICOS) человека и мыши, содержащее
домен VH, содержащий набор участков, определяющих комплементарность, тяжелой цепи (HCDR) HCDR1, HCDR2 и HCDR3, причем
HCDR1 представляет собой HCDR1 с SEQ ID NO: 405, необязательно содержащий консервативное замещение в положении 28,
HCDR2 представляет собой HCDR2 с SEQ ID NO: 406, необязательно содержащий замещение в положении 59, положении 63 и/или положении 64, и
HCDR3 представляет собой HCDR3 с SEQ ID NO: 407, необязательно содержащий замещение в положении 108, положении 109 и/или положении 112; и
домен VL, содержащий набор участков, определяющих комплементарность, легкой цепи (LCDR) LCDR1, LCDR2 и LCDR3, причем
LCDR1 представляет собой LCDR1 с SEQ ID NO: 412, необязательно содержащий замещение в положении 36,
LCDR2 представляет собой LCDR2 с SEQ ID NO: 413, и
LCDR3 представляет собой LCDR3 с SEQ ID NO: 414, необязательно содержащий замещение в положении 108 или положении 109.
7. Антитело по п. 6, отличающееся тем, что
замещение в положении 28 HCDR1 представляет собой консервативное замещение, например V28F;
замещение в положении 59 HCDR2 представляет собой N59I;
замещение в положении 63 HCDR2 представляет собой G63D;
замещение в положении 64 HCDR2 представляет собой D64N;
замещение в положении 108 HCDR3 представляет собой F108Y;
замещение в положении 109 HCDR3 представляет собой Y109F; и/или
замещение в положении 112 HCDR3 представляет собой H112N.
8. Антитело по п. 6, отличающееся тем, что
замещение в положении 36 LCDR1 представляет собой R36S;
замещение в положении 108 LCDR3 представляет собой D108G; и/или
замещение в положении 109 LCDR3 представляет собой M109N.
9. Антитело по п. 6, отличающееся тем, что
HCDR1 представляет собой SEQ ID NO: 582,
HCDR2 представляет собой SEQ ID NO: 587,
HCDR3 представляет собой SEQ ID NO: 588,
LCDR1 представляет собой SEQ ID NO: 426,
LCDR2 представляет собой SEQ ID NO: 413, и
LCDR3 представляет собой SEQ ID NO: 608.
10. Антитело по п. 6, отличающееся тем, что аминокислотная последовательность домена VH имеет по меньшей мере 95% идентичности последовательности SEQ ID NO: 586 и аминокислотная последовательность домена VL имеет по меньшей мере 95% идентичности последовательности SEQ ID NO: 607.
11. Антитело по п. 10, отличающееся тем, что аминокислотная последовательность домена VH представляет собой SEQ ID NO: 586 и аминокислотная последовательность домена VL представляет собой SEQ ID NO: 607.
12. Антитело по любому из пп. 1-11, отличающееся тем, что содержит константную область антитела.
13. Антитело по п. 12, отличающееся тем, что константная область содержит константную область тяжелой и/или легкой цепи человека.
14. Антитело по п. 12, отличающееся тем, что константная область является Fc-эффектор-положительной.
15. Антитело по п. 14, отличающееся тем, что антитело представляет собой IgG1.
16. Антитело по п. 1, отличающееся тем, что антитело представляет собой IgG1 человека, содержащее тяжелую цепь SEQ ID NO: 410 и легкую цепь SEQ ID NO: 417.
17. Композиция для стимуляции Т-клеточного иммуного ответа, содержащая эффективное количество выделенного антитела по любому из пп. 1-11 или 16 и фармацевтически приемлемый эксципиент.
18. Композиция для стимуляции Т-клеточного иммуного ответа, содержащая эффективное количество выделенной нуклеиновой кислоты, кодирующей антитело по любому из пп. 1-11 или 16, и фармацевтически приемлемый эксципиент.
19. Клетка-хозяин in vitro для экспрессии антитела, способного к специфическому связыванию внеклеточного домена ICOS, содержащая нуклеиновую кислоту, кодирующую антитело по любому из пп. 1-11 или 16.
20. Способ лечения рака у пациента-человека, включающий введение пациенту антитела по любому из пп. 1-11 или 16.
21. Способ по п. 20, отличающийся тем, что рак представляет собой почечно-клеточный рак, рак головы и шеи, меланому, немелкоклеточный рак легкого, карциному пищевода, рак шейки матки или диффузную крупноклеточную В-клеточную лимфому.
22. Способ по п. 20, отличающийся тем, что способ включает введение пациенту антитела и другого терапевтического агента и/или проведение лучевой терапии.
23. Способ по п. 22, отличающийся тем, что терапевтический агент представляет собой анти-PD-L1 антитело.
24. Способ по п. 23, отличающийся тем, что анти-PD-L1 антитело представляет собой атезолизумаб.
25. Способ по п. 23, отличающийся тем, что анти-ICOS антитело и анти-PDL1 антитело способны каждое опосредовать ADCC, ADCP и/или CDC.
26. Способ по п. 25, отличающийся тем, что анти-ICOS антитело представляет собой антитело IgG1 человека и анти-PDL1 антитело представляет собой антитело IgG1 человека.
| KUTLU ELPEK et al., Abstract A059: Efficacy of anti-ICOS agonist monoclonal antibodies in preclinical tumor models provides a rationale for clinical development as cancer immunotherapeutics, Cancer Immunol Res, 2016;4(1 Suppl):Abstract nr A059 | |||
| CHRISTOPHER HARVEY et al., Efficacy of anti-ICOS agonist monoclonal antibodies in preclinical tumor |

