Область техники, к которой относится изобретение
Настоящее изобретение относится к области опухолевой терапии и молекулярной иммунологии и касается антител против PDL-1, их фармацевтических композиций и их применения. В частности, настоящее изобретение касается моноклонального антитела против PDL-1.
УРОВЕНЬ ТЕХНИКИ
Сигнальный путь PD-1/PDL-1 играет важную роль в регуляции иммунотолерантности, при микробных инфекциях и иммунном уклонении опухолей. PD-1 (белок-1 запрограммированной смерти клеток) в основном экспрессируется на T-клетках и других иммунных клетках, а его лиганд PDL-1 сильно экспрессирован во многих типах опухолей человека. Присутствие белка PDL-1 было продемонстрировано иммуногистохимическим методом при раке молочной железы человека, раке легких, раке желудка, колоректальном раке, раке пищевода, раке яичников, раке шейки матки, почечноклеточной карциноме, раке мочевого пузыря, раке поджелудочной железы, глиоме и меланоме. Более того, уровень экспрессии PDL-1 тесно связан с клиническим лечением и прогнозом пациентов.
Блокирование сигнального пути PD-1/PDL-1 может активировать заторможенные T-клетки и индуцировать активированные T-клетки на борьбу с раковыми клетками. Блокирование передачи сигналов PD-1/PDL-1 может усилить пролиферацию специфичных к опухолевым антигенам T-клеток, которые играют роль в уничтожении опухолевых клеток, а затем ингибируют рост локальных опухолей (Julie R et al., 2012, N Engl J Med. 366: 2455-2465). Моноклональные антитела к PDL-1 могут усиливать секрецию IFN-γ инфильтрирующими опухоль T-клетками CD8+, указывая на то, что блокировка сигнального пути PD-1/PDL-1 играет роль в иммунном ответе опухолевых клеток с тем, чтобы вызвать иммунный ответ (Blank C et al., 2006, Int. J. Cancer, 119: 317-327).
Кроме того, PDL-1 также может связываться с B7-1 in vivo. Исследования показали, что комплекс PDL-1/B7-1 является отрицательным сигналом для активации T-клеток, а само взаимодействие может привести к снижению поверхностных маркеров активации T-клеток и ингибировать пролиферацию T-клеток.
IL-2 (интерлейкин-2) является своеобразным лимфокином, который секретируется Th-клетками и обладает широким спектром иммунной активности: 1) он стимулирует пролиферацию и дифференцировку T-клеток; 2) стимулирует образование цитотоксических T-лимфоцитов; 3) стимулирует пролиферацию и дифференцировку NK-клеток и повышает активность NK-клеток; 4) стимулирует образование активируемых лимфокином клеток-киллеров (клеток LAK), которые являются разновидностью уничтожающих опухоли иммунных клеток, трансформирующихся из лимфоцитов при стимуляции IL-2 в течение 3-6 дней in vitro. IFN-γ (гамма-интерферон) вырабатывается T-клетками и может ингибировать пролиферацию опухолевых клеток, усиливать презентацию антигена под действием MHC, стимулировать экспрессию фактора некроза опухолей и предотвращать ангиогенез опухолей. Недавние исследования показали, что IFN-γ может подавлять способность опухолевых клеток уклоняться от атак иммунной системы путем регулирования экспрессии Fas/FasL у опухолевых клеток и повышения чувствительности опухолевых клеток к опосредованному Fas апоптозу, что ведет к торможению клеток злокачественных опухолей.
Сейчас вообще считается, что антитела, нацеленные на путь PDL-1, приведут к прорыву в лечении различных опухолей, включая немелкоклеточный рак легких, почечноклеточную карциному, рак яичников, меланому (Homet M.B., Parisi G., et al. Anti-PD1 Therapy in Melanoma. Semin Oncol. 2015 Jun;42(3):466-473), лейкемию и анемию (Held SA, Heine A, et al. Advances in immunotherapy of chronic myeloid leukemia CML. Curr Cancer Drug Targets 2013 Sep;13(7):768-74).
В настоящее время по-прежнему необходимо разрабатывать новые антитела против PDL-1 с лучшим сродством связывания и большей эффективностью блокирования (PDL-1 с PD-1) для активации T-лимфоцитов.
CУЩНОСТЬ ИЗОБРЕТЕНИЯ
В результате глубоких исследований и творческой работы авторы изобретения использовали рекомбинантный PDL-1, экспрессируемый клетками млекопитающих, в качестве антигена для иммунизации мышей, и собирали клетки селезенки у мышей и сливали с клетками миеломы, получая гибридомы. При скрининге большого числа гибридом был получен следующий штамм клеток гибридомы: LT005, который был депонирован в Китайском центре коллекций типовых культур (CCTCC) 4 августа 2015 г. с номером доступа CCTCC No. C2015133.
Авторы изобретения неожиданно обнаружили, что штамм LT005 клеток гибридомы секретирует моноклональное антитело (названное 5C10), которое может специфически связываться с PDL-1 и эффективно блокировать связывание PDL-1 с PD-1. Кроме того, авторы изобретения также открыли два других моноклональных антитела, названных 5F10 и 9F6, которые блокируют связывание PDL-1 с PD-1.
Кроме того, авторы изобретения творчески получили гуманизированные антитела против PDL-1, названные 5C10H1L1, 5C10H1L2, 5C10H2L1 и 5C10H2L2, соответственно.
Кроме того, авторы изобретения также творчески мутировали константную область 5C10H2L2 и получили антитело 5C10H2L2-IgG1mt, у которого была эффективно снижена ADCC (опосредованная антителами клеточная цитотоксичность) и/или CDC (зависимая от комплемента цитотоксичность).
Кроме того, авторы изобретения также обнаружили, что антитела по изобретению, особенно 5C10, 5C10H1L1, 5C10H1L2, 5C10H2L1, 5C10H2L2, 5F10, 9F6 и 5C10H2L2-IgG1mt, могут эффективно связывать и активировать T-клетки человека, индуцируя секрецию IFN-γ и IL-2, что указывает на возможность профилактики и лечения рака легких, меланомы, почечных опухолей, рака яичников, лейкемии и анемии.
Таким образом, было совершено настоящее изобретение.
В одном аспекте изобретение касается моноклонального антитела или его антиген-связывающего фрагмента, причем:
данное моноклональное антитело содержит вариабельную область тяжелой цепи, включающую участки CDR, приведенные в SEQ ID NO: 15-17, и/или содержит вариабельную область легкой цепи, включающую участки CDR, приведенные в SEQ ID NO: 18-20;
или
данное моноклональное антитело содержит вариабельную область тяжелой цепи, включающую участки CDR, приведенные в SEQ ID NO: 29-31, и/или содержит вариабельную область легкой цепи, включающую участки CDR, приведенные в SEQ ID NO: 32-34;
или
данное моноклональное антитело содержит вариабельную область тяжелой цепи, включающую участки CDR, приведенные в SEQ ID NO: 35-37, и/или содержит вариабельную область легкой цепи, включающую участки CDR, приведенные в SEQ ID NO: 38-40.
Антитела 5C10, 5C10H1L1, 5C10H1L2, 5C10H2L1 или 5C10H2L2 имеют одни и те же следующие аминокислотные последовательности участков CDR:
HCDR1: GFSLSNYD (SEQ ID NO: 15)
HCDR2: IWTGGAT (SEQ ID NO: 16)
HCDR3: VRDSNYRYDEPFTY (SEQ ID NO: 17)
LCDR1: QSIGTN (SEQ ID NO: 18)
LCDR2: YAS (SEQ ID NO: 19)
LCDR3: QQSNSWPYT (SEQ ID NO: 20).
Антитело 5F10 имеет следующие аминокислотные последовательности участков CDR:
HCDR1: GFDIKDTY (SEQ ID NO: 29)
HCDR2: IDPADGNT (SEQ ID NO: 30)
HCDR3: ARGLGAWFAS (SEQ ID NO: 31)
LCDR1: QDITNS (SEQ ID NO: 32)
LCDR2: YTS (SEQ ID NO: 33)
LCDR3: QQGHTLPPT (SEQ ID NO: 34).
Антитело 9F6 имеет следующие аминокислотные последовательности участков CDR:
HCDR1: GFNIKDTY (SEQ ID NO: 35)
HCDR2: IDPANGNT (SEQ ID NO: 36)
HCDR3: SRGPPGGIGEYIYAMDY (SEQ ID NO: 37)
LCDR1: SSVSSSY (SEQ ID NO: 38)
LCDR2: STS (SEQ ID NO: 39)
LCDR3: HQYHRSPPT (SEQ ID NO: 40).
Вышеуказанные CDRs могут быть получены с помощью технических подходов, известных специалистам в данной области. Например, путем анализа аминокислотной последовательности вариабельной области тяжелой цепи или легкой цепи по системе IMGT в базе данных VBASE2.
В некоторых воплощениях представлено такое моноклональное антитело или его антигенсвязывающий фрагмент, у которого:
вариабельная область тяжелой цепи имеет аминокислотную последовательность, выбранную из SEQ ID NO: 2, SEQ ID NO: 6 и SEQ ID NO: 10, и/или вариабельная область легкой цепи имеет аминокислотную последовательность, выбранную из SEQ ID NO: 4, SEQ ID NO: 8 и SEQ ID NO: 12;
или
вариабельная область тяжелой цепи имеет аминокислотную последовательность по SEQ ID NO: 21 и/или вариабельная область легкой цепи имеет аминокислотную последовательность по SEQ ID NO: 23;
или
вариабельная область тяжелой цепи имеет аминокислотную последовательность по SEQ ID NO: 25 и/или вариабельная область легкой цепи имеет аминокислотную последовательность по SEQ ID NO: 27.
В некоторых воплощениях моноклональное антитело выбирают из числа следующих (1)–(7):
(1) аминокислотная последовательность вариабельной области тяжелой цепи приведена в SEQ ID NO: 2, а аминокислотная последовательность вариабельной области легкой цепи приведена в SEQ ID NO: 4 (5C10);
(2) аминокислотная последовательность вариабельной области тяжелой цепи приведена в SEQ ID NO: 6, а аминокислотная последовательность вариабельной области легкой цепи приведена в SEQ ID NO: 8 (5C10H1L1);
(3) аминокислотная последовательность вариабельной области тяжелой цепи приведена в SEQ ID NO: 10, а аминокислотная последовательность вариабельной области легкой цепи приведена в SEQ ID NO: 12 (5C10H2L2 или 5C10H2L2-IgG1mt);
(4) аминокислотная последовательность вариабельной области тяжелой цепи приведена в SEQ ID NO: 6, а аминокислотная последовательность вариабельной области легкой цепи приведена в SEQ ID NO: 12 (5C10H1L2);
(5) аминокислотная последовательность вариабельной области тяжелой цепи приведена в SEQ ID NO: 10, а аминокислотная последовательность вариабельной области легкой цепи приведена в SEQ ID NO: 8 (5C10H2L1);
(6) аминокислотная последовательность вариабельной области тяжелой цепи приведена в SEQ ID NO: 21, а аминокислотная последовательность вариабельной области легкой цепи приведена в SEQ ID NO: 23 (5F10);
(7) аминокислотная последовательность вариабельной области тяжелой цепи приведена в SEQ ID NO: 25, а аминокислотная последовательность вариабельной области легкой цепи приведена в SEQ ID NO: 27 (9F6).
В некоторых воплощениях моноклональное антитело или его антиген-связывающий фрагмент выбирают из Fab, Fab′, F(ab′)2, Fd, Fv, dAb, фрагментов с определяющими комплементарность участками, одноцепочечных антител (напр., scFv), гуманизированных антител, химерных антител или диател.
В некоторых воплощениях моноклональное антитело или его антиген-связывающий фрагмент связывается с PDL-1 со значением EC50 менее 100 нМ, к примеру, менее 10 нМ, 1 нМ, 0,9 нМ, 0,8 нМ, 0,7 нМ, 0,6 нМ, 0,5 нМ, 0,4 нМ, 0,3 нМ, 0,2 нМ, 0,1 нМ или меньше, причем предпочтительно EC50 определяется непрямым методом ELISA.
В некоторых воплощениях представлено моноклональное антитело или его антигенсвязывающий фрагмент, причем моноклональное антитело включает не содержащую CDR область, при этом не содержащая CDR область происходит из другого вида, чем мышь, к примеру, из антител человека.
Предпочтительно константная область моноклонального антитела выбирается из константных областей IgG1, IgG2, IgG3 или IgG4 человека.
Предпочтительно константная область моноклонального антитела представлена мутантной константной областью IgG1 человека; более предпочтительно мутантная константная область IgG1 человека содержит 1, 2 или 3 мутации в положении 234, 235 и 237 по системе нумерации EU, причем мутации выбраны из: L234A, L235A и G237A.
В некоторых воплощениях представлено моноклональное антитело или его антигенсвязывающий фрагмент, причем моноклональное антитело вырабатывается клетками гибридомы штамма LT005, а клетки гибридомы штамма LT005 депонированы в Китайском центре коллекций типовых культур (CCTCC) под номером доступа CCTCC No. C2015133.
Другой аспект настоящего изобретения касается выделенной молекулы нуклеиновой кислоты A, которая включает последовательность нуклеотидов, кодирующую вариабельную область тяжелой цепи антитела, причем:
данное антитело содержит вариабельную область тяжелой цепи, включающую участки CDR, приведенные в SEQ ID NOs: 15-17;
предпочтительно тяжелая цепь данного антитела имеет аминокислотную последовательность по SEQ ID NO: 2, SEQ ID NO: 6 или SEQ ID NO: 10;
более предпочтительно данная молекула нуклеиновой кислоты имеет нуклеотидную последовательность по SEQ ID NO: 1, SEQ ID NO: 5 или SEQ ID NO: 9.
В другом воплощении изобретения данное антитело содержит вариабельную область тяжелой цепи, включающую участки CDR, приведенные в SEQ ID NOs: 29-31,
предпочтительно тяжелая цепь данного антитела имеет аминокислотную последовательность по SEQ ID NO: 21,
более предпочтительно данная молекула нуклеиновой кислоты имеет нуклеотидную последовательность по SEQ ID NO: 22.
В другом воплощении изобретения данное антитело содержит вариабельную область тяжелой цепи, включающую участки CDR, приведенные в SEQ ID NOs: 35-37,
предпочтительно тяжелая цепь данного антитела имеет аминокислотную последовательность по SEQ ID NO: 25,
более предпочтительно данная молекула нуклеиновой кислоты имеет нуклеотидную последовательность по SEQ ID NO: 26.
Следующий аспект настоящего изобретения касается выделенной молекулы нуклеиновой кислоты B, которая включает последовательность нуклеотидов, кодирующую вариабельную область легкой цепи антитела, причем:
данное антитело содержит вариабельную область легкой цепи, включающую участки CDR, приведенные в SEQ ID NOs: 18-20;
предпочтительно легкая цепь данного антитела имеет аминокислотную последовательность по SEQ ID NO: 4, SEQ ID NO: 8 или SEQ ID NO: 12;
более предпочтительно данная молекула нуклеиновой кислоты имеет нуклеотидную последовательность по SEQ ID NO: 3, SEQ ID NO: 7 или SEQ ID NO: 11.
В другом воплощении изобретения данное антитело содержит вариабельную область легкой цепи, включающую участки CDR, приведенные в SEQ ID NOs: 32-34,
предпочтительно легкая цепь данного антитела имеет аминокислотную последовательность по SEQ ID NO: 23,
более предпочтительно данная молекула нуклеиновой кислоты имеет нуклеотидную последовательность по SEQ ID NO: 24.
В другом воплощении изобретения данное антитело содержит вариабельную область легкой цепи, включающую участки CDR, приведенные в SEQ ID NOs: 38-40,
предпочтительно легкая цепь данного антитела имеет аминокислотную последовательность по SEQ ID NO: 27,
более предпочтительно данная молекула нуклеиновой кислоты имеет нуклеотидную последовательность по SEQ ID NO: 28.
Следующий аспект настоящего изобретения касается выделенной молекулы нуклеиновой кислоты C, которая включает предыдущую молекулу нуклеиновой кислоты A и молекулу нуклеиновой кислоты B; необязательно молекула нуклеиновой кислоты C также включает последовательность линкера для соединения молекулы нуклеиновой кислоты A и молекулы нуклеиновой кислоты B.
Следующий аспект настоящего изобретения касается вектора, который содержит выделенную молекулу нуклеиновой кислоты A, выделенную молекулу нуклеиновой кислоты B или выделенную молекулу нуклеиновой кислоты C.
Следующий аспект настоящего изобретения касается клетки хозяина, которая содержит выделенную молекулу нуклеиновой кислоты A, выделенную молекулу нуклеиновой кислоты B или выделенную молекулу нуклеиновой кислоты C либо вектор.
Что касается термина “выделенная молекула нуклеиновой кислоты A”, “выделенная молекула нуклеиновой кислоты B” или “выделенная молекула нуклеиновой кислоты C”, то буквы A, B или C использовались только для большей ясности или для различения, а сама буква не имеет особого значения.
Следующий аспект настоящего изобретения касается способа получения моноклонального антитела или его антигенсвязывающего фрагмента, описанных выше, который включает следующие стадии: культивирование клеток хозяина по изобретению в подходящих условиях и извлечение моноклонального антитела или его антиген-связывающего фрагмента из клеточных культур.
Следующий аспект настоящего изобретения касается клеток гибридомы штамма LT005, который депонирован в Китайском центре коллекций типовых культур (CCTCC) под номером доступа CCTCC No. C2015133.
Другой аспект настоящего изобретения касается моноклонального антитела или его антигенсвязывающего фрагмента, который способен конкурентно связываться с антигенным эпитопом антитела или фрагментом, секретируемым клетками гибридомы штамма LT005. Предпочтительно антитело или его антигенсвязывающий фрагмент обладает какой-либо из следующих активностей:
препарата, блокирующего связывание PDL-1 с PD-1 или B7-1,
препарата, регулирующего (напр., уменьшающего) активность PDL-1 или уровень PDL-1,
препарата, устраняющего иммуносупрессию организма под действием PD-1 или PDL-1, или
препарата, усиливающего экспрессию IFN-γ и/или IL-2 в T-лимфоцитах.
Следующий аспект настоящего изобретения касается конъюгата, включающего моноклональное антитело или его антиген-связывающий фрагмент и соединительную часть, причем моноклональное антитело представляет собой любое из моноклональных антител или их антигенсвязывающих фрагментов, описанных в изобретении, а соединительная часть представляет собой детектируемую метку, предпочтительно она представляет собой радиоактивный изотоп, флуоресцентное вещество, люминесцентное вещество, окрашенное вещество или фермент.
Следующий аспект настоящего изобретения касается набора, содержащего моноклональное антитело или его антигенсвязывающий фрагмент или конъюгат, описанные выше.
Предпочтительно набор дополнительно содержит вторичное антитело, которое специфически распознает моноклональное антитело или его антигенсвязывающий фрагмент; необязательно вторичное антитело помечено детектируемой меткой типа радиоактивного изотопа, флуоресцентного вещества, люминесцентного вещества, окрашенного вещества или фермента.
Следующий аспект настоящего изобретения касается применения данного моноклонального антитела или его антигенсвязывающего фрагмента либо конъюгата при изготовлении набора, причем данный набор применяется для выявления наличия или уровня PDL-1 в образце.
Следующий аспект настоящего изобретения касается фармацевтической композиции, содержащей моноклональное антитело или его антигенсвязывающий фрагмент либо конъюгат настоящего изобретения и необязательно также фармацевтически приемлемый носитель и/или наполнитель.
Следующий аспект настоящего изобретения касается применения моноклонального антитела или его антигенсвязывающего фрагмента или конъюгата настоящего изобретения при изготовлении лекарственного средства для профилактики и/или лечения и/или вспомогательного лечения и/или диагностики опухолей или анемии; предпочтительно такие опухоли выбирают из рака молочной железы, рака легких типа немелкоклеточного рака легких, рака печени, рака желудка, колоректального рака типа рака толстой кишки или рака прямой кишки, рака пищевода, рака яичников, рака шейки матки, рака почек, рака простаты, рака мочевого пузыря, рака поджелудочной железы, глиомы, меланомы и лейкемии.
Следующий аспект настоящего изобретения касается применения моноклонального антитела или его антигенсвязывающего фрагмента или конъюгата настоящего изобретения при изготовлении:
препарата для блокировки связывания PDL-1 с PD-1 или с B7-1,
препарата для регулирования (напр., уменьшения) активности PDL-1 или уровня PDL-1,
препарата для устранения иммуносупрессии под действием PD-1 или PDL-1, или
препарата для усиления экспрессии IFN-γ и/или IL-2 в T-лимфоцитах.
Следующий аспект настоящего изобретения касается способа in vivo или in vitro, который включает стадию введения в клетки эффективного количества моноклонального антитела или его антигенсвязывающего фрагмента или конъюгата по изобретению, причем данный способ является:
способом блокировки связывания PDL-1 с PD-1 или с B7-1,
способом регулирования (напр., уменьшения) активности PDL-1 или уровня PDL-1,
способом устранения иммуносупрессии под действием PD-1 или PDL-1, или
способом усиления экспрессии IFN-γ и/или IL-2 в T-лимфоцитах.
В одном воплощении настоящего изобретения данный способ не предназначается для терапевтического применения.
Следующий аспект настоящего изобретения касается способа лечения и/или профилактики опухолей или анемии, включающего стадию введения субъекту эффективного количества моноклонального антитела или его антигенсвязывающего фрагмента либо конъюгата по изобретению; предпочтительно опухоли выбирают из рака молочной железы, рака легких типа немелкоклеточного рака легких, рака печени, рака желудка, колоректального рака типа рака толстой кишки или рака прямой кишки, рака пищевода, рака яичников, рака шейки матки, рака почек, рака простаты, рака мочевого пузыря, рака поджелудочной железы, глиомы, меланомы и лейкемии.
РАСКРЫТИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
В настоящем изобретении, если не указано иначе, научные и технические термины, используемые в этом изобретении, должны иметь значения, которые обычно понимаются специалистами в данной области. Кроме того, лабораторные процедуры, используемые в данном изобретении, связанные с культивированием клеток, молекулярной генетикой, химией нуклеиновых кислот и иммунологией, представляет собой общие процедуры, которые используются в соответствующих областях. В то же время, чтобы лучше понять изобретение, ниже приводятся определения и интерпретации соответствующих терминов.
В настоящем изобретении, при упоминании аминокислотной последовательности белка PDL-1 (лиганда-1 запрограммированной смерти, NCBI GenBank ID: NP_054862,1), включая полноразмерный белок PDL-1 или внеклеточный домен PDL-1 (PDL-1ECD) или фрагменты, содержащие PDL-1ECD; это также охватывает слитые белки PDL-1ECD, к примеру, фрагменты, слитые с Fc IgG мыши или человека (mFc или hFc). Кроме того, как это понятно специалистам в данной области, белок PDL-1 также включает такие белки, в которые естественным образом или искусственно введены мутации аминокислотной последовательности (включая, без ограничения, замены, делеции и/или добавления), не влияющие на биологические функции. Таким образом, в настоящем изобретении термин “белок PDL-1” должен охватывать все такие последовательности, включая приведенный выше список последовательностей, а также его естественные или искусственные варианты. Кроме того, при упоминании фрагмента последовательности белка PDL-1 это означает не только вышеуказанные фрагменты последовательности, но также и соответствующие фрагменты последовательностей естественных или искусственных вариантов.
Термин “EC50” в настоящем изобретении служит для обозначения концентрации для достижения 50% от максимального эффекта, то есть концентрации, которая может вызвать 50% от максимального эффекта.
Термин “антитело” в настоящем изобретении служит для обозначения молекулы иммуноглобулина, которая обычно состоит из двух пар полипептидных цепей (каждая пара содержит “легкую” (L) цепь и “тяжелую” (H) цепь). Легкие цепи антител можно классифицировать как κ- и λ-цепи. Тяжелые цепи можно классифицировать как: μ, δ, γ, α или ε, а соответствующие антитела определяются как IgM, IgD, IgG, IgA и IgE, соответственно. У легких и тяжелых цепей вариабельная и константная область соединяются участком “J” примерно в 12 или больше аминокислот, а тяжелая цепь также содержит участки “D” примерно из 3 или больше аминокислот. Каждая тяжелая цепь состоит из вариабельной области тяжелой цепи (VH) и константной области тяжелой цепи (CH). Константная область тяжелой цепи состоит из 3 доменов (CH1, CH2 и CH3). Каждая легкая цепь состоит из вариабельной области легкой цепи (VL) и константной области легкой цепи (CL). Константная область легкой цепи состоит из одного домена (CL). Константная область антитела может опосредовать связывание иммуноглобулинов с тканями или факторами хозяина, включая различные клетки иммунной системы (напр., эффекторные клетки) и первый компонент (C1q) классической системы комплемента. Области VH и VL можно еще подразделить на сильно изменчивые участки (которые называют участками, определяющими комплементарность, т.е. CDR) и консервативные участки, называемые каркасными (FR), которые разбросаны между участками CDR. Каждый VH и VL состоит из 3 CDR и 4 FR, расположенных в следующем порядке: FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4, с N-конца до C-конца. Вариабельные области (VH и VL) тяжелой цепи и легкой цепи образуют антигенсвязывающий сайт. Отнесение аминокислот к участкам или доменам соответствует определению в Kabat sequences of proteins of immunological interest (National Institutes of Health, Bethesda, Md. (1987 и 1991)) или Chothia & Lesk (J. Mol. Biol., 196: 901-917 (1987); Chothia et al., Nature 342: 878-883 (1989)). Термин “антитело” не ограничивается каким-либо конкретным методом получения антител. К примеру, он включает, в частности, рекомбинантные антитела, моноклональные антитела и поликлональные антитела. Антитела могут относиться к различным изотипам или субизотипам, напр., антитела типа IgG (напр., подтипа IgG1, IgG2, IgG3 или IgG4), IgA1, IgA2, IgD, IgE или IgM.
Термин “антиген-связывающий фрагмент” антитела служит для обозначения полипептида, содержащего фрагмент полноразмерного антитела, который сохраняет способность к специфическому связыванию с тем же антигеном, что и полноразмерное антитело, и/или конкурировать с полноразмерным антителом за специфическое связывание антигена, а также известен как “антиген-связывающая часть”. Поскольку он часто встречается в тексте Fundamental Immunology, Ch. 7 (Paul W., ed., second edition, Raven Press, N.Y. (1989)), то он включен в настоящее изобретение путем ссылки на все случаи. Антиген-связывающие фрагменты можно получить по технологии рекомбинантной ДНК или путем ферментативного или химического расщепления интактных антител. В некоторых случаях антигенсвязывающие фрагменты включают фрагменты Fab, Fab′, F(ab′)2, Fd, Fv, dAb и определяющих комплементарность участков (CDR), фрагменты одноцепочечных антител (напр., scFv), химерных антител, диател и тому подобные полипептиды, содержащие по меньшей мере такую часть полипептида, которая достаточна для придания антителу способности к специфическому связыванию антигена.
В настоящем изобретении термин “Fd-фрагмент” означает фрагмент антитела, состоящий из доменов VH и CH1; термин “Fv-фрагмент” означает фрагмент антитела, состоящий из одноцепочечных доменов VL и VH антитела; термин “dAb-фрагмент” означает фрагмент антитела, состоящий из домена VH (Ward et al., Nature 341: 544-546 (1989)); термин “Fab-фрагмент” означает фрагмент антитела, состоящий из доменов VL, VH, CL и CH1; термин “F(ab′)2-фрагмент” означает фрагмент антитела, содержащий два Fab-фрагмента, соединенные мостиками из дисульфидных связей в шарнирной области.
В некоторых случаях антигенсвязывающий фрагмент антитела представляет собой одноцепочечное антитело (напр., scFv), у которого домены VL и VH образуют моновалентные молекулы при помощи линкера с образованием одной полипептидной цепи (напр. см. Bird et al., Science 242: 423-426 (1988); Huston et al., Proc. Natl. Acad. Sci. USA, 85: 5879-5883 (1988)). Такие молекулы scFv могут иметь общую структуру типа NH2-VL-линкер-VH-COOH или NH2-VH-линкер-VL-COOH. Соответствующая новейшая технология линкера состоит из повторяющейся аминокислотной последовательности GGGGS или ее вариантов. Например, (GGGGS)4, но можно использовать и его варианты (Holliger et al. (1993), Proc. Natl. Acad. Sci. USA, 90: 6444-6448). Другие линкеры, которые можно использовать в данном изобретении, описаны в Alfthan et al. (1995), Protein Eng. 8: 725-731; Choi et al. (2001) Eur. J. Immunol. 31: 94-106; Hu et al. (1996), Cancer Res. 56: 3055-3061; Kipriyanov et al. (1999), J. Mol. Biol. 293: 41-56; и Roovers et al. (2001), Cancer Immunol.
В некоторых случаях антигенсвязывающий фрагмент антитела представляет собой диатело, то есть бивалентное антитело, у которого VH и VL экспрессируются в одной полипептидной цепи, однако, используется очень короткий линкер для предотвращения спаривания двух доменов из одной и той же цепи, при этом домен вынужден спариваться с комплементарным доменом из другой цепи, и образуются два сайта связывания антигена (напр., см. Holliger P. et al., Proc. Natl. Acad. Sci. USA, 90: 6444 -6448 (1993); и Poljak RJ et al., Structure 2: 1121-1123 (1994)).
В других случаях антигенсвязывающий фрагмент антитела представляет собой “биспецифичное антитело”, что определяется как первое антитело (фрагмент) и второе антитело (фрагмент) или миметик антитела, связанные соединительным плечом, причем способы связывания включают, без ограничения, химические реакции, слияние генов и ферментативные реакции. Антигенсвязывающие фрагменты антител могут представлять собой “мультиспецифичные антитела”, как-то триспецифичные антитела и тетраспецифичные антитела, причем первые могут специфически связываться с тремя различными антигенами, а последние могут специфически связываться с 4 различными антигенами. Например, искусственные белки анкириновых повторов (DARPin), соединенные или связанные с антителами типа IgG, scFv с Fc-фрагментами антител, как в CN 104341529 A; финомер против-IL-17a с антителом против IL-6R, как в WO 2015/141862 A1.
В других случаях антигенсвязывающий фрагмент антитела представляет собой “биспецифичный конъюгат антитела”, что определяется как первое антитело (фрагмент) и второй фрагмент с биологической функцией (не антитела и не его миметика), связанные соединительным плечом, причем методы связывания включают, без ограничения, химические реакции, слияние генов и ферментативные реакции, а вторые фрагменты с биологической функцией включают пептиды со связывающей активностью, белки, полиэтиленгликоль (ПЭГ), радионуклиды, нуклеиновые кислоты, низкомолекулярные токсины, рецепторы или лиганды и т.д., при этом конъюгат сохраняет активности каждого фрагмента, приобретая тем самым двойные функции/биспецифичность.
Антигенсвязывающие фрагменты (напр., описанные выше фрагменты антител) можно получить из данного антитела (напр., 5C10, 5C10H1L1, 5C10H1L2, 5C10H2L1 и 5C10H2L2 в настоящем изобретении) стандартными методами, известными специалистам в данной области (к примеру, методами рекомбинантной ДНК либо ферментативного или химического расщепления), причем к антигенсвязывающим фрагментам применимы те же методы специфического скрининга, что и к интактным антителам.
В настоящем изобретении, если специально не указано, термин “антитело” охватывает не только интактные антитела, но и антигенсвязывающие фрагменты антител.
Термин “mAb” и “моноклональное антитело” служит для обозначения фрагментов антител или молекул антител из очень гомологичной группы, то есть группы идентичных молекул антител, если не произойдет естественная мутация. Моноклональные антитела очень специфичны к единственному эпитопу на антигене. Поликлональные антитела отличаются от моноклональных тем, что поликлональные антитела обычно содержат по меньшей мере 2 или несколько различных антител, которые распознают разные эпитопы на одном и том же антигене. Моноклональные антитела обычно получают по гибридомной технологии, о которой впервые сообщали Kohler et al. (Nature, 256: 495, 1975), но могут быть получены и по технологии рекомбинантной ДНК (см. также USP 4,816,567).
Термин “химерное антитело” служит для обозначения антител, у которых часть легкой цепи и/или тяжелой цепи происходит из одного антитела (которое может происходить из определенного вида или относится к определенному классу или подклассу антител), а другая часть легкой цепи и/или тяжелой цепи происходит из другого антитела (которое может происходить из одного и того же или другого вида или относится к одному или нескольким классам или подклассам антител). Тем не менее, они по-прежнему сохраняют активность связывания с антигеном мишени (USP 4,816,567 на Cabilly et al.; Morrison et al., Proc. Natl. Acad. Sci. USA, 81: 6851, 6855 (1984)).
Термин “гуманизированное антитело” служит для обозначения иммуноглобулина человека (акцепторное антитело), у которого все или часть участков CDR заменены на участки CDR из антитела не человека (донорного антитела), причем донорным антителом может быть антитело не человека (напр., мыши, крысы или кролика) с ожидаемой специфичностью, сродством или реакционной способностью. Кроме того, аминокислотные остатки каркасной области (FR) акцепторного антитела могут быть заменены на аминокислоты из антител не человека или аминокислоты из других антител для дальнейшего улучшения характеристик антитела. Более подробную информацию о гуманизированных антителах см., к примеру, в Jones et al., Nature, 321:522-525 (1986); Reichmann et al., Nature, 332:323-329 (1988); Presta, Curr. Op. Struct. Biol., 2:593-596 (1992); и Clark, Immunol. Today 21: 397-402 (2000).
Методы гуманизации основываются на сочетании одного или нескольких общепринятых методов гуманизации. Например, применяются методы, описанные ниже.
Гуманизация может осуществляться путем пересадки CDR. В этом методе сначала определяют участки CDR у антитела мыши, а затем пересаживают 6 участков CDR из тяжелой цепи и легкой цепи мыши на человеческую матрицу с высокой гомологией к области FR мыши. Человеческая матрица может быть выбрана из исходной гаметной последовательности (зародышевой линии) типа гаметной последовательности, полученной из базы данных IMGT, или же из последовательностей зрелых антител типа последовательности антитела, полученной из базы данных GenBank. В антитело с привитыми CDR можно вводить обратные мутации. Некоторые аминокислоты матрицы человека можно подвергнуть обратной мутации на аминокислоты матрицы мыши с тем, чтобы улучшить сродство антитела.
Гуманизация также может осуществляться путем пересадки SDR. В этом методе сначала нужно определить участки SDR у антитела мыши. Участки SDR можно определить такими методами, как сканирование по аланину. Затем участки SDR мыши пересаживают на человеческую матрицу с высокой гомологией к матрице мыши. Человеческая матрица может быть выбрана из исходной гаметной последовательности (зародышевой линии) типа гаметной последовательности, полученной из базы данных IMGT, или же из последовательностей зрелых антител типа последовательности антитела, полученной из базы данных GenBank. В антитело с привитыми SDR можно вводить обратные мутации. Некоторые аминокислоты матрицы человека можно подвергнуть обратной мутации на аминокислоты матрицы мыши с тем, чтобы улучшить сродство антитела. Tamura M., D.E. Milenic, M. Iwahashi, E. Padlan, J. Schlom & S.V. Kashmiri. Structural correlates of an anti-carcinoma antibody: identification of specificity determining residues (SDRs) and development of a minimally immunogenic antibody variant by retention of SDRs only. J. Immunol., 164, 1432-41 (2000).
Гуманизация также может осуществляться путем переделки поверхности. В этом методе создается модель антитела мыши путем компьютерного моделирования гомологии или анализа кристаллической структуры белка. В соответствии с моделью определяются аминокислоты на поверхности антитела, и эти аминокислоты подвергают мутации на соответствующие аминокислоты антитела человека. Можно выбрать те аминокислоты антител человека, которые встречаются с высокой частотой в одном и том же месте. Padlan E.A. A possible procedure for reducing the immunogenicity of antibody variable domains while preserving their ligand-binding properties. Mol. Immunol., 28, 489-98 (1991).
Гуманизация также может осуществляться путем гипергуманизации. В этом методе сначала определяют участки CDR у антитела мыши, а затем в качестве матрицы выбирают последовательность человека с высокой гомологией к этим 6 участкам CDR, на которые пересаживают 6 участков CDR мыши. Человеческая матрица может быть выбрана из исходной гаметной последовательности (зародышевой линии) типа гаметной последовательности, полученной из базы данных IMGT, или же из последовательностей зрелых антител типа последовательности антитела, полученной из базы данных GenBank. В антитело с привитыми CDR можно вводить обратные мутации. Некоторые аминокислоты матрицы человека можно подвергнуть обратной мутации на аминокислоты матрицы мыши с тем, чтобы улучшить сродство антитела. Tan P., D.A. Mitchell, T.N. Buss, M.A. Holmes, C. Anasetti & J. Foote. “Superhumanized” antibodies: reduction of immunogenic potential by complementarity determining region grafting with human germline sequences: application to an anti-CD28. J. Immunol., 169, 1119-25 (2002).
Термин “выделенный” или “выделение” относится к получению чего-либо искусственным путем в естественном состоянии. Если какие-то вещества или ингредиенты в природе являются “выделенными”, то либо изменилось естественное окружение, в котором находились данные вещества или ингредиенты, либо они были выделены в естественном окружении, или же имели место оба случая. Например, у живых животных естественные полинуклеотиды или полипептиды не являются выделенными, а процесс, при котором достигается высокая чистота тех же самых полинуклеотидов или полипептидов, выделенных в этом естественном состоянии, называется выделением. Термин “выделенный” или “выделение” не исключает присутствия искусственных или синтетических веществ и не исключает наличия других примесей, не влияющих на активность.
Термин “вектор” служит для обозначения носителя из нуклеиновой кислоты, в который может быть вставлен полинуклеотид. Экспрессирующий вектор представляет собой такой вектор, который может экспрессировать белок, кодируемый вставленным полинуклеотидом. Вектор может быть введен в клетки хозяина путем трансформации, трансдукции или трансфекции для экспрессии переносимых генетических элементов в клетках хозяина. Векторы хорошо известны специалистам в данной области, включая, без ограничения: плазмиды; фагемиды; космиды; искусственные хромосомы типа искусственных хромосом дрожжей (YAC), бактериальные искусственные хромосомы (BAC) или искусственные хромосомы из P1 (PAC); фаги типа фага λ или фага M13; и вирусы животных. Вирусы животных, которые можно использовать в качестве векторов, включают, без ограничения, ретровирусы (в том числе лентивирусы), аденовирусы, аденоассоциированные вирусы, герпесвирусы (типа вируса простого герпеса), поксвирусы, бакуловирусы, папилломавирусы, паповавирусы (типа SV40). Вектор может содержать различные контролирующие экспрессию элементы, включая, без ограничения, последовательности промоторов, последовательности инициации транскрипции, последовательности энхансеров, селекционные элементы и гены-репортеры. Кроме того, вектор также может содержать начало репликации.
Термин “клетка хозяина” служит для обозначения клеток, которые могут использоваться для введения векторов, включая, без ограничения, прокариотические клетки типа Escherichia coli или Bacillus subtilis, клетки грибов типа дрожжевых клеток или Aspergillus, клетки насекомых типа клеток S2 или Sf9 дрозофилы или клетки животных, как-то фибробласты, клетки CHO, клетки COS, клетки NS0, клетки HeLa, клетки BHK, клетки HEK 293, или клетки человека.
Термин “специфическое связывание” служит для обозначения реакции неслучайного связывания между двумя молекулами типа реакции между антителом и его антигеном-мишенью. В некоторых воплощениях специфическое связывание антитела с антигеном (или антитела со специфичностью к антигену) означает то, что антитело связывается с антигеном со сродством связывания (KD) менее 10−5 М, к примеру, менее 10−6 M, 10−7 M, 10−8 M, 10−9 M, 10−10 М или даже меньше.
Термин “KD” служит для обозначения равновесной константы диссоциации специфического взаимодействия антитело-антиген, которая применяется для описания сродства связывания между антителом и антигеном. Чем меньше равновесная константа диссоциации, тем прочнее связывание антиген-антитело и тем выше сродство между антителом и антигеном. Обычно антитело (к примеру, моноклональное антитело 5C10, 5C10H1L1, 5C10H1L2, 5C10H2L1 или 5C10H2L2 настоящего изобретения) связывается с антигеном (напр., с белком PDL-1) со значением KD менее 10−5 M, например, менее 10−6 M, 10−7 M, 10−8 M, 10−9 M, 10−10 M или даже меньше при измерении на детекторе молекулярных взаимодействий Fortebio.
Термины “моноклональное антитело” и “mAb” имеют одинаковые значения и применяются взаимозаменяемо; термины “поликлональное антитело” и “pAb” имеют одинаковые значения и применяются взаимозаменяемо; термины “полипептид” и “белок” имеют одинаковые значения и применяются взаимозаменяемо. Кроме того, в настоящем изобретении аминокислоты обозначаются однобуквенными или трехбуквенными сокращениями, принятыми в этой области. Например, аланин может обозначаться как A или Ala.
Термины “гибридома” и “линия клеток гибридомы” применяются взаимозаменяемо, а при упоминании терминов “гибридома” и “линия клеток гибридомы” они также включают субклоны и клетки потомства гибридомы. Например, при упоминании клеток гибридомы штамма LT005 это также относится к субклонам и потомству клеток гибридомы штамма LT005.
Термин “фармацевтически приемлемый носитель и/или наполнитель” служит для обозначения таких носителей и/или эксципиентов, которые совместимы с реципиентами и активными ингредиентами по фармакологии и/или физиологии, что хорошо известно в данной области техники (напр., см. Remington’s Pharmaceutical Sciences. Edited by Gennaro AR, 19th ed. Pennsylvania: Mack Publishing Company, 1995), включая, без ограничения: модуляторы рН, поверхностно-активные вещества, адъюванты, усилители ионной силы. Например, рН-модуляторы включают, без ограничения, физраствор на фосфатном буфере; поверхностно-активные вещества включают, без ограничения, катионные, анионные или неионные поверхностно-активные вещества типа Tween-80; усилители ионной силы включают, без ограничения, хлорид натрия.
Термин “адъювант” служит для обозначения неспецифических иммуностимулирующих средств, которые усиливают иммунный ответ на антиген либо изменяют тип иммунного ответа при введении в организм до или вместе с антигеном. Существует много различных адъювантов, включая, без ограничения, алюминиевые адъюванты (напр., гидроксид алюминия), адъювант Фрейнда (напр., полный адъювант Фрейнда и неполный адъювант Фрейнда), Corynebacterium parvum, липополисахарид, цитокины и т.д. В экспериментах на животных чаще всего применяется адъювант Фрейнда. В клинических испытаниях чаще применяется адъювант из гидроксида алюминия.
Термин “эффективная доза” служит для обозначения количества, достаточного для получения требуемого эффекта или по крайней мере части требуемого эффекта. Например, эффективная доза для профилактики таких заболеваний, как опухоли, означает такое количество, которое достаточно для предотвращения, ингибирования или задержки возникновения заболевания типа опухоли; эффективная доза для лечения заболевания означает такое количество, которое достаточно для излечения или по крайней мере частичной защиты пациента от заболевания и его осложнений. Определение такой эффективной дозы входит в сферу компетенции специалистов данной области техники. Например, эффективная доза для лечения заболевания будет зависеть от тяжести заболевания, общего состояния иммунной системы пациента, общего состояния пациента, как-то возраста, веса и пола, принимаемых лекарств, а также от другого лечения, применяемого в то же самое время.
Преимущества настоящего изобретения
Моноклональное антитело 5C10 в настоящем способно специфически связываться с PDL-1, очень эффективно блокируя взаимодействие PDL-1 с PD-1, эффективно устраняя иммунносупрессию иммунной системы под действием PDL-1 и активируя T-лимфоциты.
КРАТКОЕ ОПИСАНИЕ ФИГУР
Фиг. 1. Анализ слитого белка PDL-1ECD-mFc методом SDS-PAGE. Образцы и их количество при нанесении, слева направо: маркер (10 мкл); образец, нанесенный на хроматографическую колонку (10 мкл); проходная фракция (10 мкл); элюирование (10 мкл).
Фиг. 2. Анализ слитого белка PD-1-hFc методом SDS-PAGE. Образцы и их количество при нанесении, слева направо: образец, нанесенный на хроматографическую колонку (10 мкл); маркер (10 мкл).
Фиг. 3. Анализ слитого белка B7-1-hFc методом SDS-PAGE. Образцы и их количество при нанесении, слева направо: образец, нанесенный на хроматографическую колонку (10 мкл); маркеры (10 мкл).
Фиг. 4. Анализ антитела 5C10 методом SDS-PAGE. Образцы и их количество при нанесении, слева направо: маркеры (10 мкл); восстановленный образец белка (1 мкг); проходная фракция из хроматографической колонки; невосстановленный образец белка (1 мкг).
Фиг. 5. Анализ антитела 5C10H1L1 (гуманизированное 5C10) методом SDS-PAGE. Образцы и их количество при нанесении, слева направо: маркеры (10 мкл); элюирование из хроматографической колонки (10 мкл); проходная фракция (10 мкл); образец, нанесенный на хроматографическую колонку (10 мкл).
Фиг. 6. Анализ антитела 5C10H1L2 (гуманизированное 5C10) методом SDS-PAGE. Образцы и их количество при нанесении, слева направо: маркеры (10 мкл); элюирование из хроматографической колонки (10 мкл); проходная фракция (10 мкл); образец, нанесенный на хроматографическую колонку (10 мкл).
Фиг. 7. Анализ антитела 5C10H2L1 (гуманизированное 5C10) методом SDS-PAGE. Образцы и их количество при нанесении, слева направо: маркеры (10 мкл); элюирование из хроматографической колонки (10 мкл); проходная фракция (10 мкл); образец, нанесенный на хроматографическую колонку (10 мкл).
Фиг. 8. Анализ антитела 5C10H2L2 (гуманизированное 5C10) методом SDS-PAGE. Образцы и их количество при нанесении, слева направо: маркеры (10 мкл); элюирование из хроматографической колонки (10 мкл); проходная фракция (10 мкл); образец, нанесенный на хроматографическую колонку (10 мкл).
Фиг. 9. Кинетические параметры связывания антитела 5C10H2L2 с PDL-1.
Фиг. 10. Кинетические параметры связывания антитела HpLp с PDL-1.
Фиг. 11. Кинетические параметры связывания антитела PCAB с PDL-1.
Фиг. 12. Блокировка взаимодействия PDL-1 с PD-1 антителом 5C10, 5C10H2L2 или HpLp (Fortebio).
Фиг. 13. Связывание 5C10H1L1 с положительными по PDL-1 клетками 293T.
Фиг. 14. Связывание 5C10H1L2 с положительными по PDL-1 клетками 293T.
Фиг. 15. Связывание 5C10H2L1 с положительными по PDL-1 клетками 293T.
Фиг. 16. Связывание 5C10H2L2 с положительными по PDL-1 клетками 293T.
Фиг. 17. Связывание HpLp с положительными по PDL-1 клетками 293T.
Фиг. 18. Связывание PCAB с положительными по PDL-1 клетками 293T.
Фиг. 19. Связывание 5C10H1L1, 5C10H1L2, 5C10H2L2 или 5C10H2L1 с рекомбинантным белком PDL-1 человека по данным непрямого метода ELISA.
Фиг. 20. Связывание 5C10H2L2 или HpLp с рекомбинантным белком PDL-1 обезьяны по данным непрямого метода ELISA.
Фиг. 21. Связывание 5C10H2L2 с рекомбинантным белком PDL-1 человека, PDL-2 человека или PDL-1 мыши по данным ELISA.
Фиг. 22. 5C10H1L1, 5C10H1L2, 5C10H2L2 или 5C10H2L1 конкурируют с PD-1 за связывание с PDL-1 (конкурентный метод ELISA).
Фиг. 23. 5C10H1L2 конкурирует с B7-1 за связывание с PDL-1 (конкурентный метод ELISA).
Фиг. 24. 5C10H1L2 повышает секрецию IFN-γ, блокируя взаимодействие PDL-1 с PD-1.
Фиг. 25. 5C10H1L2 повышает секрецию IL-2, блокируя взаимодействие PDL-1 с PD-1.
Фиг. 26A. Сродство и кинетические константы связывания 5C10H2L2-IgG1mt с FcγRIIIa по данным Biacore.
Фиг. 26B. Сродство и кинетические константы связывания Tecentriq® с FcγRIIIa по данным Biacore.
Фиг. 27A. Сродство и кинетические константы связывания 5C10H2L2-IgG1mt с C1q по данным Biacore.
Фиг. 27B. Сродство и кинетические константы связывания Tecentriq® с C1q по данным Biacore.
Фиг. 28. Лечебное действие 5C10H2L2-IgG1mt на клетки немелкоклеточного рака легких.
Описание хранения биологического материала
Клетки гибридомы LT005 были депонированы в Китайском центре коллекций типовых культур (CCTCC) при Уханьском университете (почтовый индекс: 430072) 4 августа 2015 г. с номером доступа CCTCC No. C2015133.
Примеры
Нижеследующие примеры приводятся с тем, чтобы обеспечить специалистам в данной области полное раскрытие и описание способов получения и применения способов и композиций по изобретению и не должны ограничивать объем того, что авторы изобретения считают своим изобретением. Методы или условия, не указанные в примерах, могут выполняться в соответствии с литературой или спецификациями на препараты. Реактивы или инструменты, изготовители которых не указаны, являются стандартными препаратами, которые можно приобрести на рынке.
Мыши BALB/c, используемые в настоящем изобретении, приобретали в Медицинском центре лабораторных животных Guangdong. T-клетки, используемые в настоящем изобретении, получали от Akeso Biopharma Co., Ltd.
Препаративный пример 1. Рекомбинантный белок PDL-1ECD-mFc
1. Синтез гена PDL-1ECD-mFc
Химерный ген конструировали так, чтобы он включал внеклеточный домен PDL-1 (лиганда-1 белка-1 запрограммированной смерти клеток, NCBI GenBank ID:NP_054862.1), именуемый PDL-1ECD, и Fc-фрагмент IgG мыши (mFc). Для улучшения эффективности экспрессии целевого гена в клетках 293F последовательность нуклеотидов была оптимизирована по кодонам и синтезирована на GenScript Biotech Co., Ltd. В научной литературе обозначения PDL-1 и PD-L1 могут применяться взаимозаменяемым образом, а в настоящем изобретении принято единое обозначение PDL-1.
2. Получение плазмиды pUC57simple-PDL-1ECD-mFc
Слитый ген PDL-1ECD-mFc, синтезированный на GenScript Biotech Co., Ltd., клонировали в экспрессирующий вектор pUC57simple (полученный от GenScript Biotech Co., Ltd.), получая плазмиду pUC57simple-PDL-1ECD-mFc.
3. Конструирование рекомбинантной плазмиды pcDNA3.1-PDL-1ECD-mFc
Плазмиду pUC57simple-PDL-1-ECD-mFc подвергали расщеплению ферментами (XbaI и BamHI), а затем проводили электрофорез. Выделенный фрагмент гена PDL-1ECD-mFc клонировали в экспрессирующий вектор pcDNA3.1 (приобретенный у Invitrogen) посредством лигирования, получая плазмиду pcDNA3.1-PDL-1ECD-mFc. Затем продукты лигирования трансформировали в клетки E. coli штамма DH5a (приобретенные у Tiangen Biotech Co. Ltd.) в соответствии с инструкциями. Положительные на pcDNA3.1-PDL-1ECD-mFc колонии получали путем скрининга, а затем амплифицировали стандартным методом. Рекомбинантную плазмиду экстрагировали с помощью набора DP103-03 (Tiangen Biotech (Beijing) Co., Ltd.) в соответствии с инструкциями к набору.
Рекомбинантную плазмиду pcDNA3.1-PDL-1-ECD-mFc трансфецировали в клетки 293F (Invitrogen) с помощью набора для трансфекции липофектамином (Invitrogen).
Трансфецированные клетки 293F культивировали в течение 7 дней. Затем супернатанты культур, содержащие рекомбинантный белок, подвергали очистке при помощи высокоскоростного центрифугирования, вакуумной фильтрации через микропористую мембрану и колонки HiTrap-protein A-HP, получая слитый белок PDL-1ECD-mFc, который подвергали анализу методом электрофореза SDS-PAGE с добавлением рабочего буфера для электрофореза в восстановительных условиях. Как видно из фиг. 1, молекулярный вес целевого белка составляет около 53 кД.
Препаративный пример 2. Получение рекомбинантного белка PD-1-hFc
1. Синтез гена PD-1-hFc
Химерный ген конструировали так, чтобы он включал внеклеточный домен PD-1 (белка-1 запрограммированной смерти клеток, NCBI GenBank ID: NP_005009.2), именуемый PD-1ECD, и Fc-фрагмент IgG человека (hFc). Для улучшения эффективности экспрессии целевого гена в клетках 293F последовательность нуклеотидов была оптимизирована по кодонам и синтезирована на GenScript Biotech Co., Ltd.
2. Получение плазмиды pUC57simple-PD-1ECD-TEV-hFc
Ген PD-1ECD-TEV-hFc клонировали в экспрессирующий вектор pUC57simple (полученный от GenScript Biotech Co., Ltd.), получая плазмиду pUC57simple-PD-1ECD-TEV-hFc.
3. Конструирование плазмиды pcDNA3.1-PD-1ECD-TEV-hFc
Плазмиду pUC57simple-PD-1ECD-TEV-hFc подвергали расщеплению ферментами (XbaI и BamHI). Очищенный фрагмент гена PD-1ECD-TEV-hFc клонировали в экспрессирующий вектор pcDNA3.1 (приобретенный у Invitrogen), получая pcDNA3.1-PD-1ECD-TEV-hFc, которую трансформировали в клетки E. coli штамма DH5a (приобретенные у Tiangen). Получали положительные колонии путем скрининга, которые затем размножали по стандартной методике культивирования E. coli DH5a и экстрагировали рекомбинантную плазмиду по инструкции к набору DP103-03 (Tiangen Biotech (Beijing) Co., Ltd.).
Рекомбинантную плазмиду pcDNA3.1-PD-1ECD-TEV-hFc трансфецировали в клетки 293F (Invitrogen) по инструкции к набору для трансфекции липофектамином (Invitrogen).
Клетки 293F трансфецировали с помощью pcDNA3.1-PD-1ECD-TEV-hFc и культивировали в течение 7 дней. Затем супернатанты культуры, содержащие рекомбинантный белок, очищали при помощи высокоскоростного центрифугирования, вакуумной фильтрации через микропористую мембрану и колонки Mabselect SuRe, получая слитый белок PD-1ECD-TEV-hFc, который подвергали анализу методом электрофореза SDS-PAGE с добавлением рабочего буфера для электрофореза в восстановительных условиях (фиг. 2).
Препаративный пример 3. Получение рекомбинантного белка B7-1-hFc
1. Синтез гена B7-1-hFc
Химерный ген конструировали так, чтобы он включал внеклеточный домен B7-1 (кластера дифференцировки 80 (также CD80 и B7-1), NCBI GenBank ID: NP_005182.1), именуемый B7-1ECD, и Fc-фрагмент IgG человека (hFc). Для улучшения эффективности экспрессии целевого гена в клетках 293F последовательность нуклеотидов была оптимизирована по кодонам и синтезирована на GenScript Biotech Co., Ltd.
2. Получение плазмиды pUC57simple-B7-1ECD-hFc
Ген B7-1ECD-hFc клонировали в экспрессирующий вектор pUC57simple (полученный от GenScript Biotech Co., Ltd.), получая плазмиду pUC57simple-B7-1ECD-hFc.
3. Конструирование плазмиды pcDNA3.1-PD-1ECD-TEV-hFc
Плазмиду pUC57simple-B7-1ECD-hFc подвергали расщеплению ферментами (XbaI и BamHI). Очищенный фрагмент гена B7-1ECD-hFc клонировали в экспрессирующий вектор pcDNA3.1 (приобретенный у Invitrogen) посредством лигирования, получая плазмиду pcDNA3.1-B7-1ECD-hFc. Затем продукты лигирования трансформировали в клетки E. coli штамма DH5a (приобретенные у Tiangen). Проводили скрининг колоний и получали положительные по pcDNA3.1-B7-1ECD-hFc клоны, которые затем размножали по стандартной методике культивирования E. coli DH5a и экстрагировали рекомбинантную плазмиду по инструкции к набору DP103-03 (Tiangen Biotech (Beijing) Co., Ltd.).
Рекомбинантную плазмиду pcDNA3.1-B7-1ECD-hFc трансфецировали в клетки 293F (Invitrogen) по инструкции к набору для трансфекции липофектамином (Invitrogen).
Клетки 293F трансфецировали с помощью плазмиды pcDNA3.1-B7-1ECD-hFc и культивировали в течение 7 дней. Затем супернатанты культуры, содержащие рекомбинантный белок, очищали при помощи высокоскоростного центрифугирования, вакуумной фильтрации через микропористую мембрану и колонки Mabselect SuRe, получая слитый белок B7-1ECD-hFc, который подвергали анализу методом электрофореза SDS-PAGE с добавлением рабочего буфера для электрофореза в восстановительных условиях (фиг. 3).
Пример 1. Получение клеток гибридомы штамма LT005 и моноклональных антител 5C10, 5F10 и 9F6
Рекомбинантный белок PDL-1ECD-mFc, экспрессированный в клетках млекопитающих, использовали в качестве иммуногена для иммунизации мышей. У иммунизированных мышей извлекали спленоциты и сливали с клетками миеломы, получая клетки гибридомы. После скрининга большого количества образцов получали клетки гибридомы штамма LT005, вырабатывающие моноклональное антитело 5C10, которое может специфически связываться с PDL-1. Также в настоящем изобретении были получены два других моноклональных антитела 5F10 и 9F6.
1. Получение клеток гибридомы
Рекомбинантный слитый белок PDL-1ECD-mFc, полученный в Препаративном примере 1, использовали в качестве иммуногена для иммунизации мышей BALB/c (Медицинский центр лабораторных животных Guangdong). У иммунизированных мышей извлекали спленоциты и сливали их с клетками миеломы мыши, получая клетки гибридомы в соответствии с общими методами (напр., Stewart S.J., “Monoclonal Antibody Production”, in Basic Methods in Antibody Production and Characterization, Eds. G.C. Howard and D.R. Bethell, Boca Raton: CRC Press, 2000).
Для получения клеток гибридомы, вырабатывающих антитела, способные специфически связываться с PDL-1-ECD-mFc, проводили анализ непрямым методом ELISA, используя в качестве покрывающего антигена PDCD-1-ECD-mFc. После этого клетки гибридомы подвергали конкурентному методу ELISA и отбирали клетки, секретирующие моноклональные антитела, конкурирующие с PD-1 (PD-1-hFc, полученным в Препаративном примере 2) при связывании с PDL-1. А затем получали стабильные клетки гибридомы штамма LT005, вырабатывающие моноклональное антитело 5C10, методом серийных разведений.
Клетки гибридомы штамма LT005 депонировали в Китайском центре коллекций типовых культур (CCTCC) при Уханьском университете (почтовый индекс: 430072) 4 августа 2015 г. с номером доступа CCTCC No. C2015133.
Таким же образом получали еще два штамма клеток гибридомы, вырабатывающих антитела мыши (именуемые 5F10 и 9F6, соответственно).
2. Получение моноклональных антител 5C10, 5F10 и 9F6
Клетки гибридомы штамма PDL-1-5C10 культивировали в среде, содержащей 10% (низкое содержание IgG) фетальной бычьей сыворотки (FBS) в течение 7 дней, а затем собирали супернатанты клеточной культуры и очищали, получая антитело 5C10.
Таким же образом получали антитела 5F10, 9F6 в соответствии с описанным выше способом.
3. Анализ антитела 5C10 методом SDS-PAGE
Образцы очищенного белка вносили в восстановительный рабочий буфер и в невосстановительный рабочий буфер, соответственно. Вместе с проходной фракцией после очистки все образцы кипятили и наносили на гель SDS-PAGE для анализа. Результаты показали, что молекулярная масса восстановленного белка составляет примерно 50 кДа и 25 кДа, а невосстановленного белка – примерно 150 кД (фиг. 4)
4. Определение сродства, конкурентного сродства и клеточного сродства у мышиных антител 5C10, 5F10 и 9F6
Сродство антител определяли методом ELISA и конкурентным методом ELISA по методикам, описанным в Примерах 9 и 13, соответственно, а сродство к клеткам определяли методом FACS по методике, описанной в Примере 8.
Результаты представлены в табл. 1.
Таблица 1. Сродство, конкурентное сродство и клеточное сродство у мышиных
антител 5C10, 5F10 и 9F6
Результаты показали, что три мышиных антитела не уступают эталонному антителу PCAB (полученному в Примере 5) в отношении сродства и конкурентного сродства. Антитело 5C10 проявляло наилучшие показатели по клеточному сродству и положительному ответу. Клеточное сродство у 5F10 было лучше, чем у PCAB, а содержание положительных у 9F6 было лучше, чем у PCAB.
Пример 2. Получение последовательностей тяжелых цепей и легких цепей моноклональных антител 5C10, 5F10 и 9F6
Из полученных в Примере 1 клеток гибридомы штамма LT005 экстрагировали общую мРНК с помощью набора для выделения РНК (Tiangen, DP430) в соответствии с инструкцией производителя.
Синтезировали кДНК с помощью набора для синтеза первой нити кДНК TransScript SuperMix фирмы TransGen Biotech в соответствии с инструкцией производителя и амплифицировали методом ПЦР. На продуктах ПЦР проводили TA-клонирование в соответствии с инструкциями из набора для клонирования pEASY-T1 (TransGen, CT101). Продукты TA-клонирования подвергали секвенированию и получили следующие результаты.
Нуклеотидная последовательность, кодирующая VH антитела 5C10 (360 п.о.):
CAGGTGCAACTGAAGGAGTCAGGACCTGGCCTGGTGGCGCCCTCACAGAACCTGTCCATTACCTGCACTGTCTCTGGGTTCTCATTAAGCAACTATGATATAAGCTGGATTCGCCAGCCACCAGGAAAGGGTCTGGAGTGGCTCGGAGTAATATGGACTGGTGGAGCCACAAATTATAATTCAGCTTTCATGTCCAGACTGAGCATCAGTAGGGACAACTCCAAGAGCCAAGTTTTCTTAAAAATGAACAGTCTGCAAACTGATGACACAGCCATATATTACTGTGTGAGAGATTCGAACTATAGGTACGACGAGCCGTTTACTTACTGGGGCCAAGGGACTCTGGTCACTGTCTCTGCA (SEQ ID NO: 1).
Аминокислотная последовательность VH антитела 5C10 (120 а.к.):
QVQLKESGPGLVAPSQNLSITCTVSGFSLSNYDISWIRQPPGKGLEWLGVIWTGGATNYNSAFMSRLSISRDNSKSQVFLKMNSLQTDDTAIYYCVRDSNYRYDEPFTYWGQGTLVTVSA (SEQ ID NO: 2).
Нуклеотидная последовательность, кодирующая VL антитела 5C10 (318 п.о.):
GACATCTTGCTGACTCAGTCTCCAGCCATCCTGTCTGTGAGTCCAGGAGAAAGAGTCAGTCTCTCCTGCAGGGCCAGTCAGAGCATTGGCACAAACATACACTGGTTTCAGCAAAGAACAAATGGTTCTCCAAGGCTTCTCATAAAGTATGCTTCTGAGTCTATCTCTGGGATCCCTTCCAGGTTTAGTGGCAGTGGATCAGGGACAGATTTTACTCTTAGCATCAACAGTGTGGAGTCTGAAGATATTGCAGATTACTACTGTCAACAAAGTAATAGCTGGCCGTACACGTTCGGAGGGGGGACCAAGCTGGAAATA (SEQ ID NO: 3).
Аминокислотная последовательность VL антитела 5C10 (106 а.к.):
DILLTQSPAILSVSPGERVSLSCRASQSIGTNIHWFQQRTNGSPRLLIKYASESISGIPSRFSGSGSGTDFTLSINSVESEDIADYYCQQSNSWPYTFGGGTKLEI (SEQ ID NO: 4).
Таким же образом были получены последовательности легких и тяжелых цепей моноклональных антител 9F6 и 5F10.
Аминокислотная последовательность VH антитела 5F10 (117 а.к.):
EVQLQQSGAELVKPGASVKLSCTASGFDIKDTYIHWVKQRPEQGLEWIGRIDPADGNTRYDPKFQDKTTITTDTSSNTAHLQLSSLTSEDTAVYYCARGLGAWFASWGQGTLVTVSA (SEQ ID NO: 21).
Нуклеотидная последовательность, кодирующая VH антитела 5F10 (351 п.о.):
GAGGTTCAGCTGCAGCAGTCTGGGGCAGAGCTTGTGAAGCCAGGGGCCTCAGTCAAGTTGTCCTGCACAGCTTCTGGCTTCGACATTAAAGACACCTATATCCACTGGGTGAAGCAGAGGCCTGAACAGGGCCTGGAGTGGATTGGAAGGATTGATCCTGCGGACGGTAATACTAGGTATGACCCGAAGTTCCAGGACAAGACCACTATAACAACCGACACATCCTCCAACACAGCCCACCTGCAGCTCAGCAGCCTGACATCTGAGGACACTGCCGTCTATTACTGTGCTAGAGGCCTCGGAGCTTGGTTTGCTTCCTGGGGCCAAGGGACTCTGGTCACTGTCTCTGCA (SEQ ID NO: 22).
Аминокислотная последовательность VL антитела 5F10 (106 а.к.):
DIQMTQTTSSLSASLGDRVTISCRASQDITNSLNWYQQKPDGTVKLLIHYTSRLHSGVPSRFSGSGSGTDYSLTISNLEQEDIATYFCQQGHTLPPTFGGGTKLEI (SEQ ID NO: 23).
Нуклеотидная последовательность, кодирующая VL антитела 5F10 (318 п.о.):
GATATCCAGATGACACAGACTACATCCTCCCTGTCTGCCTCTCTGGGAGACAGAGTCACCATCAGTTGCAGGGCAAGTCAGGACATTACCAATTCCTTAAACTGGTATCAGCAGAAACCAGATGGAACTGTTAAACTCCTGATCCACTACACATCAAGATTACACTCAGGAGTCCCATCAAGGTTCAGTGGCAGTGGGTCTGGAACAGATTATTCTCTCACCATTAGCAACCTGGAGCAAGAAGATATTGCCACTTACTTTTGCCAACAGGGTCATACGCTTCCTCCGACGTTCGGTGGAGGCACCAAGCTGGAAATC (SEQ ID NO: 24).
Аминокислотная последовательность VH антитела 9F6 (124 а.к.):
EVQLQQSGAELVKPGASVKLSCTASGFNIKDTYMYWVKQRPEQGLEWIGRIDPANGNTKYDPKFQGKATITADTSANTAYLQLSSLTSEDTAVYYCSRGPPGGIGEYIYAMDYWGQGTSVTVSS (SEQ ID NO: 25).
Нуклеотидная последовательность, кодирующая VH антитела 9F6 (372 п.о.):
GAGGTTCAGCTGCAGCAGTCTGGGGCAGAGCTTGTGAAGCCAGGGGCCTCAGTCAAGTTGTCCTGCACAGCTTCTGGCTTCAACATTAAAGACACCTATATGTACTGGGTGAAGCAGAGGCCTGAACAGGGCCTGGAGTGGATTGGAAGGATTGATCCTGCGAATGGTAATACTAAATATGACCCGAAGTTCCAGGGCAAGGCCACTATAACAGCAGACACATCCGCCAACACAGCCTACCTGCAGCTCAGCAGCCTGACATCTGAGGACACTGCCGTCTATTACTGTTCTAGAGGCCCTCCAGGAGGTATCGGCGAGTATATCTATGCTATGGACTACTGGGGTCAAGGAACCTCAGTCACCGTCTCCTCA (SEQ ID NO: 26).
Аминокислотная последовательность VL антитела 9F6 (107 а.к.):
QIVLTQSPAIMSASLGERVTMTCTASSSVSSSYLHWYQQKPGSSPKLWIYSTSNLASGVPARFSGSGSGTSYSLTISSMEAEDAATYYCHQYHRSPPTFGGGTKLEI (SEQ ID NO: 27).
Нуклеотидная последовательность, кодирующая VL антитела 9F6 (321 п.о.):
CAAATTGTTCTCACCCAGTCTCCAGCAATCATGTCTGCATCTCTAGGGGAACGGGTCACCATGACCTGCACTGCCAGCTCAAGTGTAAGTTCCAGTTACTTGCACTGGTACCAGCAGAAGCCAGGATCCTCCCCCAAACTCTGGATTTATAGCACATCCAACCTGGCTTCTGGAGTCCCAGCTCGCTTCAGTGGCAGTGGGTCTGGGACCTCTTACTCTCTCACAATCAGCAGCATGGAGGCTGAAGATGCTGCCACTTATTACTGCCACCAGTATCATCGTTCCCCACCCACGTTCGGTGGAGGCACCAAGCTGGAAATC (SEQ ID NO: 28).
Пример 3. Разработка последовательностей легких и тяжелых цепей гуманизированных антител 5C10H1L1, 5C10H1L2, 5C10H2L1 и 5C10H2L2
Для конструирования мутаций использовали трехмерную кристаллическую структуру белка PDL-1 (PDB Code 3BIK, Lin DY et al., PNAS USA 105(8): 3011-6 (2008)) и структуру 5C10, полученную при компьютерном моделировании на основе последовательности в Примере 2, а затем составляли мутантные последовательности вариабельной области антител 5C10H1L1, 5C10H1L2, 5C10H2L1, 5C10H2L2 (константная область тяжелой цепи: C-область тяжелой цепи IgG1, номер доступа: P01857; константная область легкой цепи: C-область κ-цепи Ig, номер доступа: P01834). Последовательности вариабельной области представлены ниже.
1. Последовательности легкой и тяжелой цепи гуманизированного моноклонального антитела 5C10H1L1
Нуклеотидная последовательность, кодирующая VH антитела 5C10H1L1 (360 п.о.):
CAGGTCCAGCTGCAGGAGTCAGGCCCCGGCCTGGTGAAGCCCAGTGAGAACCTGTCAATCACCTGCACAGTCTCTGGCTTCTCACTGAGCAATTACGACATCAGTTGGATTCGACAGCCCCCTGGAAAGGGCCTGGAATGGCTGGGCGTGATCTGGACAGGCGGGGCAACTAACTATAATCCAGCCTTTAAAAGCCGGCTGACCATTTCCAGAGACAACTCCAAGTCTCAGGTGTCTCTGAAAATGAGCTCCCTGCAGGCCGCTGATACCGCTGTGTACTATTGTGTCAGGGACAGCAATTACCGCTATGATGAGCCCTTCACATACTGGGGGCAGGGAACTCTGGTGACCGTCTCTAGT (SEQ ID NO: 5).
Аминокислотная последовательность VH антитела 5C10H1L1 (120 а.к.):
QVQLQESGPGLVKPSENLSITCTVSGFSLSNYDISWIRQPPGKGLEWLGVIWTGGATNYNPAFKSRLTISRDNSKSQVSLKMSSLQAADTAVYYCVRDSNYRYDEPFTYWGQGTLVTVSS (SEQ ID NO: 6).
Нуклеотидная последовательность, кодирующая VL антитела 5C10H1L1 (321 п.о.):
GAAATCGTGCTGACACAGAGCCCTGACACACTGAGCGTGACTCCCAAGGAGAAAGTCACCCTGACATGCCGGGCATCACAGAGCATCGGAACAAACATTCACTGGTTCCAGCAGAGACCAGGCCAGAGCCCCAAGCTGCTGATCAAATACGCCTCCGAATCTATCAGTGGCATTCCTTCCCGATTCTCAGGCAGCGGGTCCGGAACCGACTTTACTCTGACCATTAACTCTGTGGAGGCTGAAGATGCCGCTACATACTATTGCCAGCAGTCTAATAGTTGGCCTTATACCTTCGGCCAGGGGACAAAGCTGGAGATCAAA (SEQ ID NO: 7).
Аминокислотная последовательность VL антитела 5C10H1L1 (107 а.к.):
EIVLTQSPDTLSVTPKEKVTLTCRASQSIGTNIHWFQQRPGQSPKLLIKYASESISGIPSRFSGSGSGTDFTLTINSVEAEDAATYYCQQSNSWPYTFGQGTKLEIK (SEQ ID NO: 8).
2. Последовательности легкой и тяжелой цепи гуманизированного моноклонального антитела 5C10H2L2
Нуклеотидная последовательность, кодирующая VH антитела 5C10H2L2 (360 п.о.):
CAGGTCCAGCTGCAGGAGTCCGGCCCCGGCCTGGTGAAGCCCTCCGAGACACTGTCTATCACCTGCACAGTCAGCGGCTTCTCACTGAGCAACTACGACATCTCCTGGATTCGACAGCCCCCTGGAAAGGGCCTGGAATGGCTGGGCGTGATCTGGACAGGCGGGGCAACTAACTATAATCCAGCCCTGAAATCTCGGCTGACTATTAGTAGAGACAACTCAAAGAATCAGGTGTCCCTGAAAATGAGCTCCGTCACCGCCGCTGATACAGCTGTGTACTATTGTGTCAGGGACAGCAATTACCGCTATGATGAGCCCTTTACCTACTGGGGGCAGGGAACTCTGGTGACCGTCTCTAGT (SEQ ID NO: 9).
Аминокислотная последовательность VH антитела 5C10H2L2 (120 а.к.):
QVQLQESGPGLVKPSETLSITCTVSGFSLSNYDISWIRQPPGKGLEWLGVIWTGGATNYNPALKSRLTISRDNSKNQVSLKMSSVTAADTAVYYCVRDSNYRYDEPFTYWGQGTLVTVSS (SEQ ID NO: 10).
Нуклеотидная последовательность, кодирующая VL антитела 5C10H2L2 (321 п.о.):
GAAATCGTGCTGACACAGTCTCCTGATACCCTGAGCGTGACTCCCAAGGAGAAAGTCACCCTGACATGCAGGGCATCACAGAGCATCGGAACAAACATTCACTGGTTCCAGCAGAAGCCAGGCCAGAGCCCCAAGCTGCTGATCAAATACGCCTCCGAATCTATTAGTGGAGTGCCTTCCCGCTTCTCAGGCAGCGGGTCCGGAACCGACTTTACTCTGACCATCAACTCTGTGGAGGCTGAAGATGCCGCTACATACTATTGCCAGCAGTCTAATAGTTGGCCTTATACCTTCGGCCAGGGGACAAAGCTGGAGATCAAA (SEQ ID NO: 11).
Аминокислотная последовательность VL антитела 5C10H2L2 (107 а.к.):
EIVLTQSPDTLSVTPKEKVTLTCRASQSIGTNIHWFQQKPGQSPKLLIKYASESISGVPSRFSGSGSGTDFTLTINSVEAEDAATYYCQQSNSWPYTFGQGTKLEIK (SEQ ID NO: 12).
3. Последовательности легкой и тяжелой цепи гуманизированного моноклонального антитела 5C10H1L2
Нуклеотидная последовательность, кодирующая VH антитела 5C10H1L2:
SEQ ID NO: 5.
Аминокислотная последовательность VH антитела 5C10H1L2: SEQ ID NO: 6.
Нуклеотидная последовательность, кодирующая VL антитела 5C10H1L2:
SEQ ID NO: 11.
Аминокислотная последовательность VL антитела 5C10H1L2: SEQ ID NO: 12.
4. Последовательности легкой и тяжелой цепи гуманизированного моноклонального антитела 5C10H2L1
Нуклеотидная последовательность, кодирующая VH антитела 5C10H2L1:
SEQ ID NO: 9.
Аминокислотная последовательность VH антитела 5C10H2L1: SEQ ID NO: 10.
Нуклеотидная последовательность, кодирующая VL антитела 5C10H2L1:
SEQ ID NO: 7.
Аминокислотная последовательность VL антитела 5C10H2L1: SEQ ID NO: 8.
Пример 4. Получение гуманизированных из 5C10 антител 5C10H1L1, 5C10H1L2, 5C10H2L1, 5C10H2L2 и их анализ методом SDS-PAGE
Клонировали кДНК тяжелой цепи (VH: SEQ ID NO: 5 или SEQ ID NO: 9; CH: hIgG1) и легкой цепи (последовательность VL: SEQ ID NO: 7 или SEQ ID NO: 11; CL: каппа-цепь человека) для 5C10H1L1, 5C10H1L2, 5C10H2L1 и 5C10H2L2 в вектор pUC57simple (GenScript Biotech Co., Ltd), получая плазмиды pUC57simple-5C10H1, pUC57simple5C10L1, pUC57simple-5C10H2 и pUC57simple-5C10L2.
Далее эти последовательности клонировали в вектор pcDNA3.1 по методике, описанной в Препаративном примере 1. Экстрагировали рекомбинантные плазмиды и совместно трансфецировали в клетки 293F. После культивирования в течение 7 дней супернатанты культур подвергали очистке при помощи высокоскоростного центрифугирования, вакуумной фильтрации через микропористую мембрану и колонки HiTrap-protein A-HP.
Образцы очищенного белка вносили с восстановительный и невосстановительный рабочий буфер по отдельности, все образцы кипятили и наносили на гель SDS-PAGE для анализа. Результаты представлены на фиг. 5, фиг. 6, фиг. 7 и фиг. 8, причем восстановленный образец содержит две полосы на геле, соответствующие полосам в 50 кД и 25 кД по белковых маркерам, соответственно, а невосстановленный белок содержит одну полосу в 150 кДа.
Пример 5. Анализ кинетических параметров связывания у гуманизированного антитела 5C10H2L2
Кинетические параметры связывания гуманизированного антитела 5C10H2L2 с антигеном PDL-1 (NCBI GenBank ID: NP_054862.1, нуклеотидная последовательность: SEQ ID NO: 13, аминокислотная последовательность: SEQ ID NO: 14) определяли методом Fortebio.
1. Приготовление образцов
(1) Белок PDL-1-mFc получали по методике, описанной в Препаративном примере 1, а затем белок PDL-1-mFc расщепляли протеазой TEV и очищали методом колоночной хроматографии, получая антиген PDL-1.
Последовательность ДНК PDL-1 (870 п.о.):
ATGAGGATTTTCGCCGTCTTTATCTTTATGACCTACTGGCATCTGCTGAACGCTTTTACTGTGACCGTCCCCAAGGATCTGTATGTGGTGGAGTACGGAAGCAACATGACTATCGAGTGCAAGTTCCCCGTGGAAAAACAGCTGGACCTGGCCGCTCTGATTGTCTATTGGGAGATGGAAGATAAGAATATCATTCAGTTTGTGCACGGCGAGGAAGACCTGAAAGTCCAGCATAGCTCCTACAGGCAGCGCGCCCGACTGCTGAAGGATCAGCTGTCCCTGGGGAACGCAGCCCTGCAGATCACCGACGTGAAACTGCAGGATGCTGGAGTCTACAGGTGCATGATCTCTTACGGCGGGGCTGATTATAAGCGCATTACAGTGAAAGTCAATGCACCTTATAACAAGATCAATCAGAGAATTCTGGTGGTCGACCCAGTGACCAGTGAGCACGAACTGACATGTCAGGCTGAGGGCTACCCCAAGGCAGAAGTGATCTGGACCTCTAGTGATCATCAGGTCCTGTCAGGGAAAACCACAACTACCAACAGCAAGCGAGAGGAAAAACTGTTCAATGTGACATCCACTCTGAGGATCAACACAACTACCAATGAGATTTTCTATTGCACTTTTCGGAGACTGGACCCTGAGGAAAACCACACCGCAGAGCTGGTCATCCCAGAACTGCCACTGGCACACCCACCTAATGAGCGAACACACCTGGTCATCCTGGGAGCCATTCTGCTGTGCCTGGGCGTCGCTCTGACTTTCATTTTTCGGCTGAGAAAGGGGCGGATGATGGACGTGAAAAAGTGTGGCATTCAGGATACTAACTCAAAAAAGCAGTCCGATACCCATCTGGAAGAAACC (SEQ ID NO: 13).
Аминокислотная последовательность PDL-1 (290 а.к.):
MRIFAVFIFMTYWHLLNAFTVTVPKDLYVVEYGSNMTIECKFPVEKQLDLAALIVYWEMEDKNIIQFVHGEEDLKVQHSSYRQRARLLKDQLSLGNAALQITDVKLQDAGVYRCMISYGGADYKRITVKVNAPYNKINQRILVVDPVTSEHELTCQAEGYPKAEVIWTSSDHQVLSGKTTTTNSKREEKLFNVTSTLRINTTTNEIFYCTFRRLDPEENHTAELVIPELPLAHPPNERTHLVILGAILLCLGVALTFIFRLRKGRMMDVKKCGIQDTNSKKQSDTHLEET
(SEQ ID NO: 14).
(2) Получение антител положительного контроля HpLp и PCAB
В настоящем изобретении в качестве положительного контроля выбрано HpLp или PCAB, причем HpLp имеется в продаже как Atezolizumab (торговая марка Tecentriq®), а PCAB – антитело к PDL-1 в клинических испытаниях.
Atezolizumab (торговая марка Tecentriq®) приобретали у Roche. Способ получения HpLp (также известно как KF025HpLp) приведен в US 2010/0203056 A1 (напр., в примере 10), причем последовательность VH антитела HpLp описана в SEQ ID NO: 20, а последовательность VL описана в SEQ ID NO: 21.
Способ получения PCAB приведен в US 7,943,743 B2 (напр., в примере 1), причем последовательность VH антитела описана в SEQ ID NO: 1, а последовательность VL описана в SEQ ID NO: 11.
2. Методы
Для определения сродства 5C10H2L2, HpLp и PCAB к антигену PDL-1 на поверхности сенсора AR2G фиксировали антиген PDL-1 (5 мкг/мл) путем связывания по аминогруппам с последующей блокировкой этаноламином. После уравновешивания в ФСБТ проводили связывание между антигеном, захваченным на биосенсоре, и антителом, причем делали серийные разведения антител при 3-кратном разведении (исходная концентрация: 200 нМ) в ФСБТ (10 мМ). Сродство связывания 5C10H2L2, HpLp and PCAB с антигеном PDL-1 анализировали с помощью Fortebio Data Analysis 7.0.
3. Результаты
Сродство и кинетические константы связывания 5C10H2L2, HpLp и PCAB с PDL-1 представлены в табл. 2 и на фиг. 9-11.
Таблица 2. Сродство и кинетические константы связывания 5C10H2L2,
HpLp и PCAB с PDL-1
KD: константа диссоциации; kon: константа скорости ассоциации; koff: константа скорости диссоциации; KD = koff/kon.
Результаты показали, что все три антитела связываются с антигеном с очень высоким сродством. При этом сродство связывания с антигеном у 5C10H2L2 и HpLp было выше, чем у PCAB.
Пример 6. Блокирование взаимодействия PDL-1 с PD-1 антителами 5C10, 5C10H2L2 и HpLp (Fortebio)
Для выявления способности 5C10, 5C10H2L2 и HpLp блокировать взаимодействие PDL-1 с PD-1 на поверхности сенсора AR2G фиксировали антиген PDL-1 (5 мкг/мл) путем связывания по аминогруппам, а затем блокировали этаноламином. После уравновешивания в ФСБТ проводили связывание между антигеном, захваченным на биосенсоре, и антителом, причем делали серийные разведения антител при 3-кратном разведении (исходная концентрация: 33,33 нМ) в ФСБТ (10 мМ). Затем кончики биосенсора погружали в раствор, содержащий белок PD-1 (10 мкг/мл), на 420 сек.
Как видно из фиг. 12, каждое антитело было способно эффективно ингибировать связывание PDL-1 человека с PD-1 зависимым от дозы образом, а интенсивность флуоресценции при каждой дозе и значения EC50 представлены в табл. 3.
Таблица 3. Блокировка взаимодействия PDL-1 с PD-1 антителами
5C10, 5C10H2L2 и HpLp
Результаты показали, что все три антитела способны эффективно ингибировать связывание PDL-1 человека с PD-1 зависимым от дозы образом.
Пример 7. Блокирование взаимодействия PDL-1 с PD-1 антителами 5C10H2L2 и HpLp
Сравнивали способность 5C10H2L2 блокировать взаимодействие PD1/PDL-1 с HpLp методом HTRF с помощью набора для анализа связывания PD1/PDL-1 (CISBIO, 63ADK000CPLPEH). Готовили серийные разведения антител 5C10H2L2 и HpLp при 3-кратном разведении (исходная концентрация: 100 мкг/мл, 10 градиентов) в буфере для разведения. В растворы вносили 2 мкл образца, 4 мкл PDL-1-EuK и 4 мкл Tag-PD1 с последующим кратковременным центрифугированием и инкубацией (20 мин при комнатной температуре). Затем в растворы добавляли 10 мкл антитела Tag-XL665 с последующим кратковременным центрифугированием и инкубацией (2 часа при комнатной температуре). Наконец, считывали показания на приборе PHERA Star Fs (BMG) и проводили анализ данных с помощью Graph Prism.
Результаты показали, что HpLp и 5C10H2L2 обладают близкой способностью к блокированию взаимодействия PD1/PDL-1 (67,29 нг/мл и 68,97 нг/мл, соответственно). Оба антитела могут эффективно ингибировать связывание PDL-1 человека с PD-1.
Пример 8. Связывание гуманизированных антител против PDL-1 с клетками, экспрессирующими PDL-1, при определении методом FACS
Сначала конструировали клетки 293Т, экспрессирующие PDL-1, а затем эти клетки метили гуманизированными антителами 5C10H1L1, 5C10H1L2, 5C10H2L1, 5C10H2L2 и антителами положительного контроля (HpLp и PCAB). Специфическое связывание антител 5C10H1L1, 5C10H1L2, 5C10H2L1, 5C10H2L2 и антител положительного контроля (HpLp и PCAB) с антигеном анализировали методом FACS.
1. Конструирование клеток 293Т, экспрессирующих PDL-1
Содержащий PDL-1 вектор pLenti6.3-PDL-1 (приобретенный у фирмы Invitrogen) трансфецировали в клетки 293T в соответствии с руководством к набору для трансфекции липофектамином (фирмы Invitrogen). После скрининга получали клетки, стабильно экспрессирующие PDL-1.
2. Мечение антителами и анализ методом FACS
Собирали клетки 293T после стандартной обработки трипсином, а количество клеток на 1 пробирку для сбора составляло 2×105. Готовили разведения каждого из антител в PBS (1% BSA) в концентрации 50 нМ, 20 нМ, 10 нМ, 3 нМ, 1 нМ, 0,1 нМ, 0,01 нМ и 0 нМ, соответственно. Затем разведения антител инкубировали с экспрессирующими PDL-1 клетками 293T на льду в течение 2 часов, после чего отмывали 3 раза с помощью PBS. Разводили FITC-антитело козы против IgG человека (1:100) в PBS и вносили в каждую пробирку по 100 мкл, а затем инкубировали 1 час на льду. После 3-кратной отмывки в PBS клетки ресуспендировали в 300 мкл PBS и детектировали сигналы флуоресценции через настроенные на FITC каналы проточного цитометра.
3. Результаты
Связывание антител 5C10H1L1, 5C10H1L2, 5C10H2L1, 5C10H2L2 и антител для положительного контроля (HpLp и PCAB) с экспрессирующими PDL-1 клетками 293T представлено на фиг. 13-18.
Результаты показали, что все антитела способны эффективно связываться с PDL-1 на поверхности клеток 293T зависимым от дозы образом. После количественного анализа флуоресценции у связавшихся антител строили кривые связывания по стандартной модели и рассчитывали эффективность связывания в виде EC50 для каждого антитела, как показано в табл. 4.
Таблица 4. Анализ интенсивности флуоресценции при связывании антител
5C10H1L1, 5C10H1L2, 5C10H2L2, 5C10H2L1, HpLp, PCAB с антигеном PDL-1 на поверхности клеток 293T методом FACS
Результаты показали, что все антитела могут эффективно связываться с целевым белком (PDL-1) на поверхности клеток 293T зависимым от дозы образом.
Пример 9. Определение сродства связывания гуманизированных антител против PDL-1 с PDL-1 непрямым методом ELISA
Для оценки сродства связывания 5C10H1L1, 5C10H1L2, 5C10H2L2, 5C10H2L1 и антител положительного контроля (HpLp и PCAB) с PDL-1 человека проводили анализ непрямым твердофазным иммуноферментным методом (ELISA). Планшет для ELISA покрывали PDL-1 человека и инкубировали при 4°C в течение ночи, а затем блокировали 1% BSA при 37°C в течение 2 ч. Затем в каждую лунку добавляли антитела и инкубировали при 37°C в течение 30 мин. Добавляли вторичное антитело – конъюгированное с HRP козье антитело против IgG (H+L) человека (Jackson, 109-035-088). Добавляли субстрат TMB (Neogen, 308177) для хромогенной реакции и инкубировали 5 минут. Считывали поглощение при 450 нм.
Результаты представлены на фиг. 19. Как видно из неё, 5C10H1L1, 5C10H1L2, 5C10H2L2, 5C10H2L1, HpLp и PCAB могут эффективно связываться с PDL-1 человека зависимым от дозы образом. В табл. 5 приведена интенсивность флуоресценции при каждой дозе и расчетная эффективность связывания, представленная значением EC50 после построения кривой.
Таблица 5. Связывание антител 5C10H1L1, 5C10H1L2, 5C10H2L2, 5C10H2L1,
HpLp и PCAB с PDL-1 человека (непрямой метод ELISA)
Результаты показали, что антитела по изобретению могут эффективно связываться с PDL-1 человека зависимым от дозы образом.
Пример 10. Сродство связывания антитела 5C10H2L2 с PDL-1 обезьяны при определении непрямым методом ELISA
Учитывая, что эксперименты по фармакокинетике и токсикологии проводятся на экспериментальных животных, целью этого эксперимента является определение того, может ли антитело 5C10H2L2 связываться с PDL-1 обезьяны; если антитело 5C10H2L2 может связываться с PDL-1 обезьяны, то обезьян можно использовать для экспериментов по фармакокинетике и токсикологии.
Связывание 5C10H2L2 и положительного контроля – HpLp с PDL-1 обезьяны измеряли непрямым методом ELISA. Планшет для ELISA покрывали PDL-1 обезьяны и инкубировали при 4°C в течение ночи, а затем блокировали 1% BSA в PBS при 37°C в течение 2 ч. Затем в каждую лунку добавляли антитела и инкубировали при 37°C в течение 30 мин. Добавляли вторичное антитело – конъюгированное с HRP козье антитело против IgG (H+L) человека (Jackson, 109-035-088). Добавляли субстрат TMB (Neogen, 308177) для хромогенной реакции и инкубировали 5 минут. Считывали поглощение при 450 нм.
Результаты по активности связывания 5C10H2L2 и HpLp с PDL-1 обезьяны представлены на фиг. 20. Как видно из фиг. 20, 205C10H2L2 и HpLp эффективно связываются с PDL-1 обезьяны зависимым от дозы образом. В табл. 6 приведена интенсивность флуоресценции при каждой дозе и расчетная эффективность связывания, представленная значением EC50 после построения кривой.
Таблица 6. Связывание антител 5C10H2L2 и HpLp с PDL-1 обезьяны
(непрямой метод ELISA)
Результаты показали, что оба антитела – 5C10H2L2 и HpLp могут эффективно связываться с PDL-1 обезьяны зависимым от дозы образом, причем связывающая способность выше у 5C10H2L2, чем у HpLp.
Пример 11. Связывание 5C10H2L2 с PDL-1 человека, PD-L2 человека и PDL-1 мыши при определении непрямым методом ELISA
Связывание 5C10H2L2 с PDL-1 человека, PD-L2 человека (Sino Biological Inc., кат. № 10292-H08H) и PDL-1 мыши (Sino Biological Inc., кат. № 50010-M08H) анализировали непрямым методом ELISA. На планшет для ELISA по отдельности вносили PDL-1 человека, PD-L2 человека или PDL-1 мыши при 0,5 мкг/мл по 100 мкл на лунку и инкубировали при 4°C в течение ночи. Делали серийные разведения антител, начиная с 1 мкг/мл (3-кратное разведение, 11 градиентов). Лунки блокировали 1% BSA при 37°C в течение 2 часов. Добавляли вторичное антитело – конъюгированное с HRP козье антитело против IgG (H+L) человека (Jackson, 109-035-088) в разведении 1:20000. Добавляли субстрат TMB (Neogen, 308177) для хромогенной реакции и инкубировали 5 минут. Считывали поглощение при 450 нм.
Результаты по связыванию 5C10H2L2 с PDL-1 человека, PD-L2 человека и PDL-1 мыши представлены на фиг. 21. Как видно из фиг. 21, 5C10H2L2 может эффективно связываться с PDL-1 человека зависимым от дозы образом. По интенсивности флуоресценции при каждой дозе рассчитывали эффективность связывания после построения кривой, получая значение EC50 = 9,16 нг/мл, в то же время оно не связывается с PD-L2 человека и PDL-1 мыши.
В заключение, 5C10H2L2 специфически связывается с PDL-1 человека, тогда как атезолизумаб может связываться с PDL-1 мыши (Tecentriq® Pharmacology Review, FDA, номер заявки 761034Orig1s000). Эти данные показывают, что антитело 5C10H2L2 обладает отличной специфичностью.
Пример 12. Определение активности конкурентного связывания гуманизированных антител к PDL-1 с PD-1 конкурентным методом ELISA
Способность 5C10H1L1, 5C10H1L2, 5C10H2L2, 5C10H2L1 и положительных контролей (HpLp и PCAB) конкурировать с PD-1 за связывание с PDL-1 человека оценивали конкурентным методом ELISA. Планшет для ELISA покрывали рецептором и инкубировали при 4°C в течение ночи. Блокировали лунки 1% BSA при 37°C в течение 2 ч. После этого смешивали антитело и PDL-1-mFc и инкубировали при комнатной температуре в течение 15 мин, а затем вносили в каждую лунку и инкубировали при 37°C в течение 30 мин. Добавляли вторичное антитело – конъюгированное с HRP козье антитело против IgG (H+L) мыши (Jackson, 109-035-062). Добавляли субстрат TMB (Neogen, 308177) для хромогенной реакции и инкубировали 5 минут. Считывали поглощение при 450 нм.
Результаты по активности связывания этих антител с PDL-1 представлены на фиг. 22. Видно, что 5C10H1L1, 5C10H1L2, 5C10H2L2, 5C10H2L1, HpLp и PCAB могут конкурировать с PD-1 за связывание с PDL-1 зависимым от дозы образом. В табл. 7 приведена интенсивность флуоресценции при каждой дозе и расчетная эффективность связывания, представленная значением EC50 после построения кривой.
Таблица 7. Конкурентное связывание антител 5C10H1L1, 5C10H1L2, 5C10H2L2, 5C10H2L1, HpLp и PCAB с PDL-1 (конкурентный метод ELISA)
Результаты показали, что все исследованные антитела могут конкурентно и эффективно связываться с антигеном PDL-1 зависимым от дозы образом.
Пример 13. Определение активности конкурентного связывания антител к PDL-1 5C10H2L2 и B7-1 конкурентным методом ELISA
Способность 5C10H2L2 и антител положительного контроля (HpLp и PCAB) конкурировать за связывание PDL-1 с B7-1 (B7-1-hFc, получено в Препаративном примере 3) определяли конкурентным методом ELISA. Планшет для ELISA покрывали PDL-1 и инкубировали при 4°C в течение ночи. Блокировали лунки 1% BSA при 37°C в течение 2 ч. После этого смешивали антитело и PDL-1-mFc и инкубировали при комнатной температуре в течение 15 мин, а затем вносили в каждую лунку и инкубировали при 37°C в течение 30 мин. Добавляли вторичное антитело – конъюгированное с HRP козье антитело против IgG (H+L) мыши (Jackson, 109-035-062). Добавляли субстрат TMB (Neogen, 308177) для хромогенной реакции и инкубировали 5 минут. Считывали поглощение при 450 нм.
Результаты по конкуренции 5C10H2L2 с B7-1 за связывание PDL-1 представлены на фиг. 23, которые показали, что 5C10H2L2, HpLp и PCAB могут эффективно конкурировать с B7-1 за связывание с PDL-1. В табл. 8 приведена интенсивность флуоресценции при каждой дозе и расчетная эффективность связывания, представленная значением EC50 после построения кривой.
Таблица 8. Результаты по конкуренции антител 5C5C10H2L2, HpLp, PCAB и B7-1
за связывание с PDL-1 конкурентным методом ELISA
* Неполная конкуренция. Следует отметить, что окно данных по HpLp в эксперименте по конкурентному связыванию было меньше, у двух других антител. Как видно из табл. 7, снижение OD не проявляется при возрастании концентрации HpLp, начиная с разведения 1:27 (как-то 1:9, 1:3, 3 мкг/мл).
Результаты показывают, что все исследованные антитела могут конкурировать с B7-1 за связывание PDL-1, причем активность конкурентного связывания у 5C10H2L2 сильнее, чем у PCAB, а значение EC50 у 5C10H2L2 почти наполовину меньше, чем у PCAB. Активность конкурентного связывания у HpLp повышается незначительно при повышении концентрации.
Пример 14. Анализ клеточной биологической активности 5C10H2L2 и антител положительного контроля (HpLp и PCAB)
Для исследования влияния моноклонального антитела 5C10H2L2 и положительного контроля (HpLp и PCAB) на секрецию IL-2 и IFN-γ в мононуклеарных клетках периферической крови (МКПК) использовали Ficoll-Paque Plus (GE Healthcare, серия No. 171440-02) для выделения МКПК. Для индукции к МКПК добавляли IL-4 (Peprotech K2513, 1000 ед./мл) и GM-CSF (Peprotech H1513, 1000 ед./мл) на 6 дней. После этого для индукции к МКПК и клеткам еще добавляли TNF-α (Peprotech G1513, 200 ед./мл) на 3 дня, получая клетки DC.
Выделяли T-клетки из МКПК. Смешивали клетки DC с T-клетками в соотношении 1:10 и культивировали их. После 5-6-дневной инкубации при различных концентрациях антитела 5C10H2L2 (hIgG в качестве контроля) проводили ELISA для оценки секреции IFN-γ (набор фирмы Dakewe Biotech Inc.) и IL-2 (набор фирмы Dakewe Biotech Inc.).
Результаты по секреции IFN-γ и IL-2 из смеси DC и T-клеток представлены на фиг. 24 и фиг. 25, соответственно. Результаты показывают, что 5C10H2L2, HpLp и PCAB могут эффективно индуцировать секрецию IFN-γ и IL-2 зависимым от дозы образом.
Пример 15. Разработка и получение моноклонального антитела 5C10H2L2-IgG1mt с модифицированной константной областью IgG1
В настоящем изобретении константная область тяжелой цепи представляет собой C-область тяжелой цепи IgG1, номер доступа: P01857; константная область легкой цепи представляет собой C-область κ-цепи Ig, номер доступа: P01834. Аминокислоты в положениях 234, 235 и 237 по системе нумерации EU подвергали мутации следующим образом: L234A, L235A и G237A. Мутантное антитело было названо 5C10H2L2-IgG1mt, а его получали по методике из Примера 4.
Пример 16. Динамическое сродство 5C10H2L2-IgG1mt к FcγRIIIa и C1q при определении методом ForteBio
1. Сродство и кинетику связывания 5C10H2L2-IgG1mt и Tecentriq® с FcγRIIIa характеризовали методом ForteBio (фирмы Pall, кат. № Octet, Qke) следующим образом.
Очищенный FcγRIIIa-биотин сажали на покрытый стрептавидином SA-чип путем связывания биотина со стрептавидином по стандартной методике с помощью наборов фирмы ForteBio при фиксированных условиях (1 мкг/мл FcγRIIIa-Biotin, 300 сек). Проводили связывание чипа с антителом при концентрации 4000 нМ в течение 120 сек с последующей инкубацией в ФСБТ (pH 7,4) в течение 180 сек для диссоциации. Кривые связывания и диссоциации анализировали с помощью программного обеспечения Octet.
Результаты, представленные на фиг. 26А и 26В, показали, что 5C10H2L2-IgG1mt и Tecentriq® не связываются с FcγRIIIa, свидетельствуя, что оба они не обладают активностью ADCC.
2. Сродство и кинетику связывания 5C10H2L2-IgG1mt и Tecentriq® с C1q (фирмы Fitzgerald, кат. № 32R-AC049) характеризовали методом ForteBio следующим образом.
Очищенное антитело сажали на покрытый стрептавидином SA-чип путем связы-вания биотина со стрептавидином по стандартной методике с помощью наборов фирмы ForteBio при фиксированных условиях (20 мкг/мл антитела-биотина, 300 сек). Затем проводили связывание чипа с C1q при концентрации 200 нМ (2-кратное разведение) в течение 120 сек с последующей инкубацией в ФСБТ (pH 7,4) в течение 180 сек для диссоциации. Кривые связывания и диссоциации анализировали с помощью программного обеспечения Octet. Для того, чтобы свести к минимуму влияние сродства на оценку константы связывания, учитывали только сегмент данных, соответствующий началу фаз связывания и диссоциации. Значения KD, kon и koff представлены в табл. 9, а кривые приведены на фиг. 27A и 27B.
Таблица 9. Динамическое сродство 5C10H2L2-IgG1mt и Tecentriq® к C1q
Результаты показывают, что 5C10H2L2-IgG1mt обладает меньшим динамическим сродством к C1q, чем Tecentriq®.
Пример 17. Активность CDC у 5C10H2L2-IgG1mt
Положительные по PDL-1 опухолевые клетки HCC1954 (приобретали у ATCC, кат. № CRL-2338) культивировали в соответствующей среде (RPMI1640 + 10% бычьей сыворотки). Готовили серийные разведения 5C10H2L2-IgG1mt в среде (RPMI1640 + 10% сыворотки человека), начиная со 10000 мкг/мл (5-кратные разведение, 10 градиентов). Вышеприведенные опухолевые клетки обрабатывали трипсином, собирали клетки в несколько пробирок и ресуспендировали в соответствующей среде (RPMI1640 + 10% сыворотки человека), а затем вносили в 96-луночный планшет (10000 клеток на лунку) с различными разведениями антител для инкубации на 5 часов. После этого в каждую лунку добавляли 20 мкл реагента CCK8 (фирмы Dongren Chemical Technology Co., Ltd., кат. № CK04, серия: JJ744) для инкубации на 3 часа. Считывали поглощение при 450 нм на считывающем устройстве (Molecular Devices, Model: SpectraMax M2). Активность дегидрогеназы в митохондриях отражает цитотоксичность антитела для клеток HCC1954.
Результаты показывают, что 5C10H2L2-IgG1mt не вызывает эффекта CDC на клетках HCC1954.
Пример 18. Ингибирование развития опухолей in vivo при раке толстой кишки
1. Образцы
5C10H2L2-IgG1mt, Tecentriq® и IgG человека получали из Sichuan Kelun Pharmaceutical Research Institute Co., Ltd. Tecentriq® приобретали у Roche, а IgG человека приобретали у Chengdu Rongsheng Pharmaceutical Co., Ltd.
Приготовление: три образца разбавляли физраствором, содержащим 0,1% BSA, до требуемых концентраций.
Клетки и животные
Клетки MC-38/H-11 происходят из клеток MC-38 рака толстой кишки мыши (фирмы Cobioer, кат. № CB606025), у которых эндогенный PDL-1 мыши был “вырублен” при помощи CRISPR/Cas9, а в клетки трансфецирован PDL-1 человека. Таким образом, клетки MC-38/H-11 будут экспрессировать только белок PDL-1 человека.
Мышей C57BL/6, 7-8-недельных, самок, приобретали у Shanghai Slac Laboratory Animal Co., Ltd.
2. Процедура
Каждой мыши вводили подкожно 1×105 клеток MC-38/H-11 и случайным образом разбивали на группы для внутрибрюшинного введения (IP) образцов через день (Q2D), начиная со второго дня от инокуляции опухолей (D0). Вводили следующие дозы: IgG человека (15 мг/кг), 5C10H2L2-IgG1mt (1,5, 5, 15 мг/кг) и Tecentriq® (15 мг/кг). В каждой группе было по 10 мышей, а вводимый объем составлял 0,1 мл/10 г массы тела.
3. Экспериментальные показатели
Влияние препаратов на рост опухолей отмечали по T/C% или по ингибированию роста опухолей (TGI) (%).
Диаметр опухолей измеряли два раза в неделю с помощью штангенциркуля. Объем опухолей (V) рассчитывали как:
V = 1/2 × a × b2, где a и b означают длину и ширину, соответственно.
T/C% = T/C × 100, где C означает объем опухолей или вес опухолей в контрольной группе, а T означает объем опухолей или вес опухолей в опытной группе.
Ингибирование роста опухолей (TGI) (%) = (C − T)/C × 100, где C означает объем опухолей или вес опухолей в контрольной группе, а T означает объем опухолей или вес опухолей в опытной группе.
4. Результаты
Результаты представлены ниже в табл. 10.
Таблица 10. Действие 5C10H2L2-IgG1mt (1,5 мг/кг, 5 мг/кг и 15мг/кг) и Tecentriq® на подкожные ксенотрансплантаты MC-38/H-11 рака толстой кишки
(1,5 мг/кг)
(5 мг/кг)
(15 мг/кг)
Примечание. Мышей разбивали на группы случайным образом, а первое введение было в D0. D27 означает 27-й день после первого введения.
Степень TGI при 1,5, 5 и 15 мг/кг 5C10H2L2-IgG1mt у подкожных ксенотрансплантатов МС-38/Н-11 составляла 63,9%, 75,8% и 68,6%, соответственно, исходя из среднего объема опухолей. Учитывая, что индивидуальные вариации размера опухолей в каждой группе были очень большими, было целесообразно использовать медианный объем опухолей для расчета ингибирования роста опухолей. При этом условии степень TGI составляла 100%, 100% и 100%. Контрольный препарат Tecentriq® (15 мг/кг) давал TGI 93,8% (при расчетах по медианному объему опухолей). Степень TGI у МС-38/Н-11 при 5C10H2L2-IgG1mt (1,5, 5, 15 мг/кг) составляла 100%, 100% и 100% при расчетах по медианному весу опухолей. Tecentriq® (15 мг/кг) давал TGI 93,7%. Значения TGI при расчетах по медианному объему опухолей и медианному весу опухолей были очень близкими, что свидетельствует о надежности вычислений по медианному объему опухолей. 5C10H2L2-IgG1mt (1,5, 5, 15 мг/кг) не только ингибировал рост опухолей, но также ингибировал онкогенез. Под конец эксперимента (D27) встречаемость опухолей при 1,5, 5, 15 мг/кг 5C10H2L2-IgG1mt составляла 40, 40 и 40%, соответственно. В группе Tecentriq® встречаемость составляла 50%. Все препараты хорошо переносились несущими опухоли мышами, не отмечалось значительных потерь веса или других симптомов. 5C10H2L2-IgG1mt (1,5, 5, 15 мг/кг) по сравнению с Tecentriq® оказывал более сильное противоопухолевое действие на модели подкожной трансплантации клеток MC-38/H-11 рака толстой кишки.
Пример 19. Ингибирование развития опухолей in vivo при раке легких
Постановка модели на животных. Мышам NOG подкожно инокулировали клетки HCC827 немелкоклеточного рака легких (приобретали у ATCC, кат. № CRL-2868). Когда объем опухолей достигал 100 мм3, мышам сразу вводили внутривенно активированные РВМСs человека, имитируя иммунную систему человека перед введением.
Схема дозирования. 10 мг/кг, внутривенная инъекция, один раз в два дня, всего 4 раза. Объем опухолей измеряли два раза в неделю после введения. Мышей разбивали на три группы: контроль (IgG), 5C10H2L2-IgG1mt и Tecentriq®, по 6 мышей в каждой группе.
Кривая роста опухолей представлена на фиг. 28.
Результаты показали, что объем опухолей в группе 5C10H2L2-IgG1mt был явно меньше, чем в группе Tecentriq® и в контрольной группе IgG, начиная с 4-го дня. Рост опухолей в группе 5C10H2L2-IgG1mt почти полностью подавлялся. Напротив, опухоли в группе Tecentriq® и в контрольной группе IgG продолжали расти. Результаты показали, что антитело по настоящему изобретению проявляет более сильный противоопухолевый эффект in vivo, чем Tecentriq®.
Конкретные воплощения настоящего изобретения были описаны подробно, что должно быть понятно специалистам в данной области. Согласно всем деталям, которые были раскрыты, в них могут вноситься различные модификации и замены, которые, тем не менее, должны находиться в пределах охраны настоящего изобретения. Полный объем изобретения изложен в прилагаемой формуле изобретения и любых её эквивалентах.











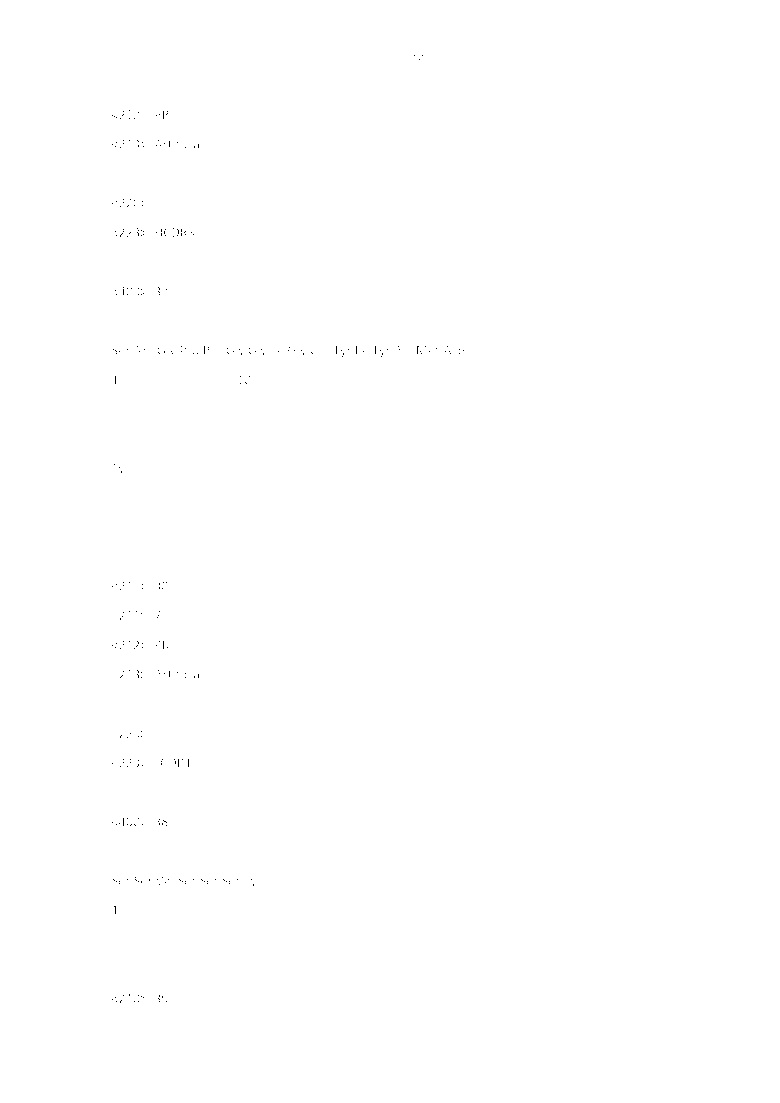

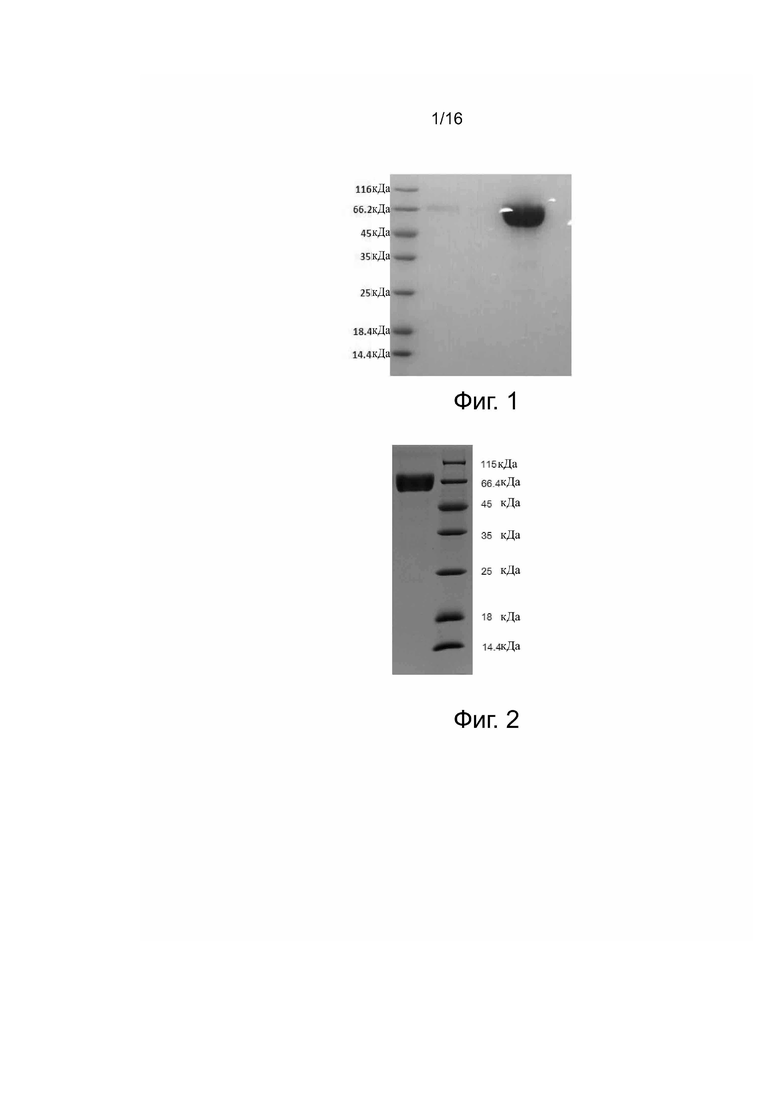
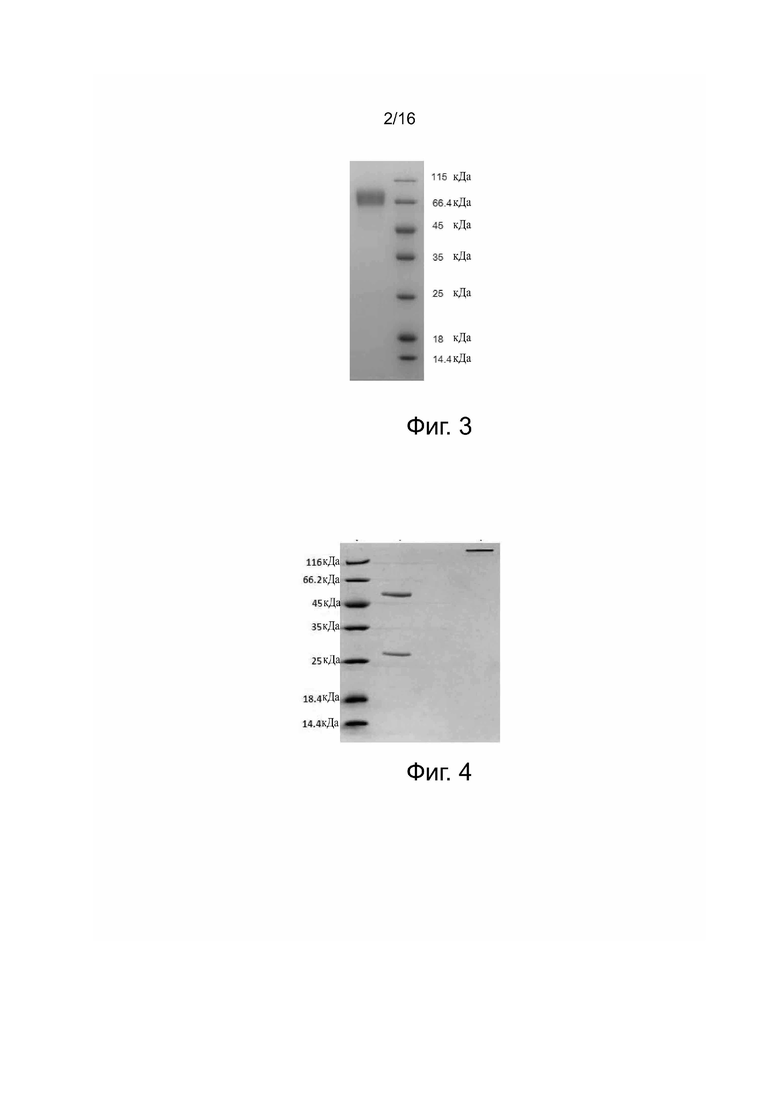

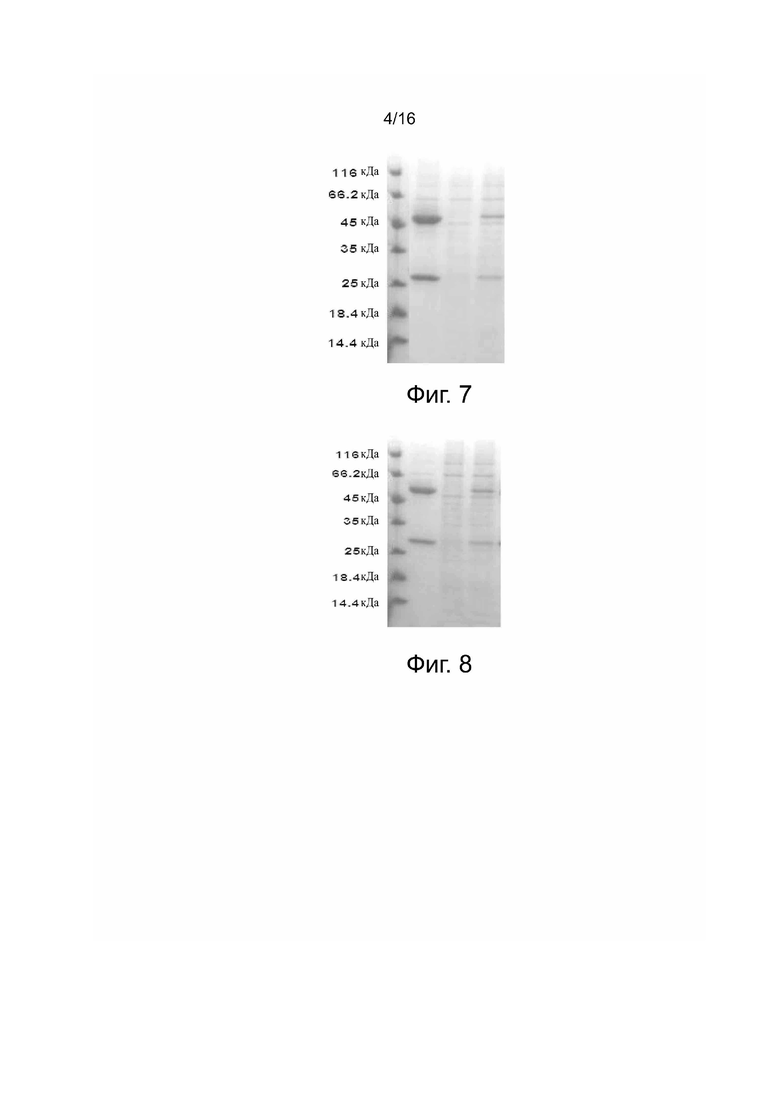
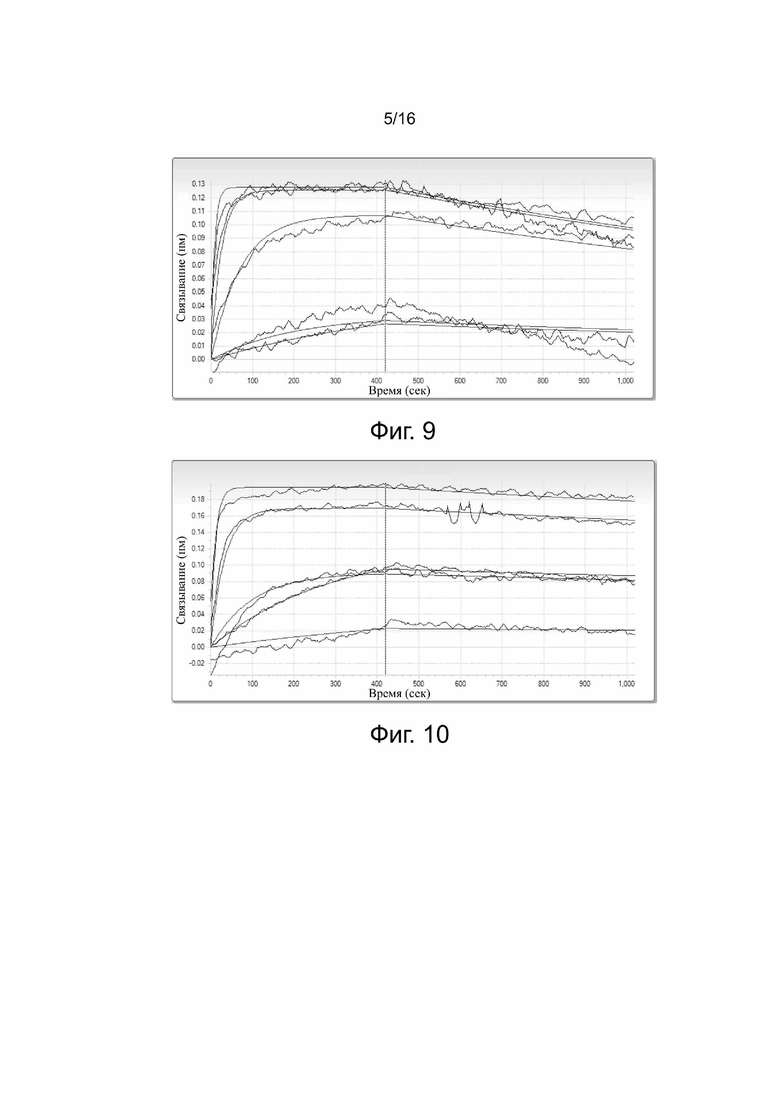
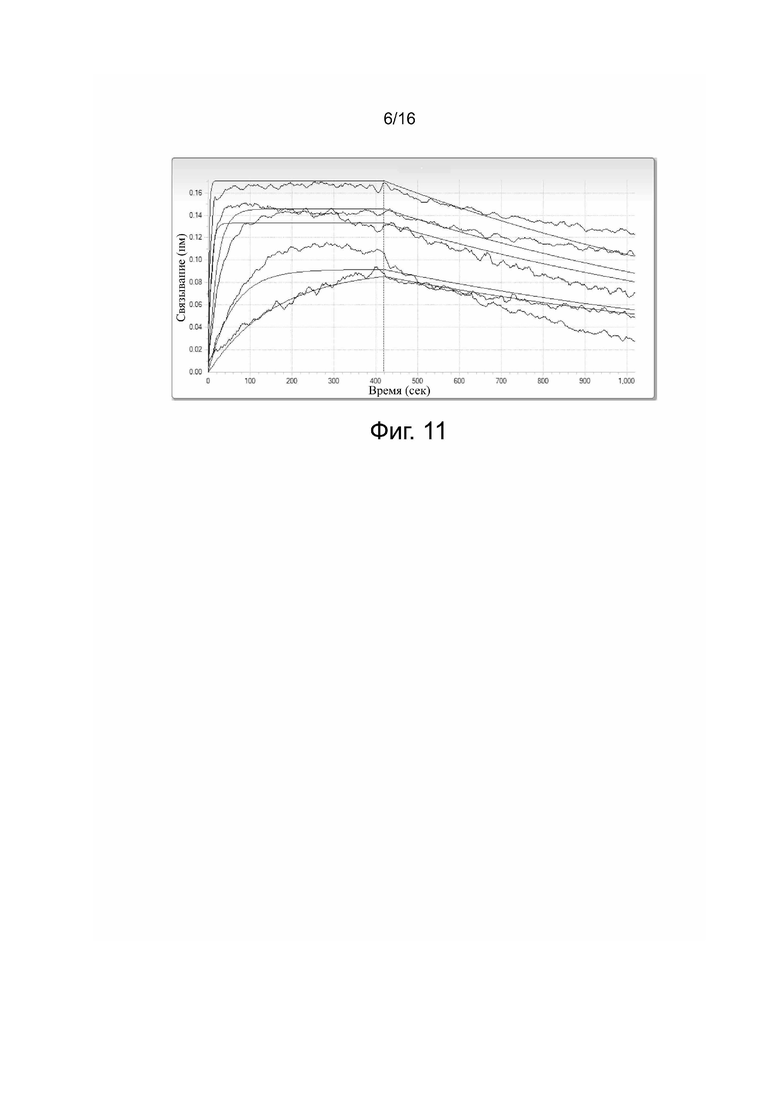

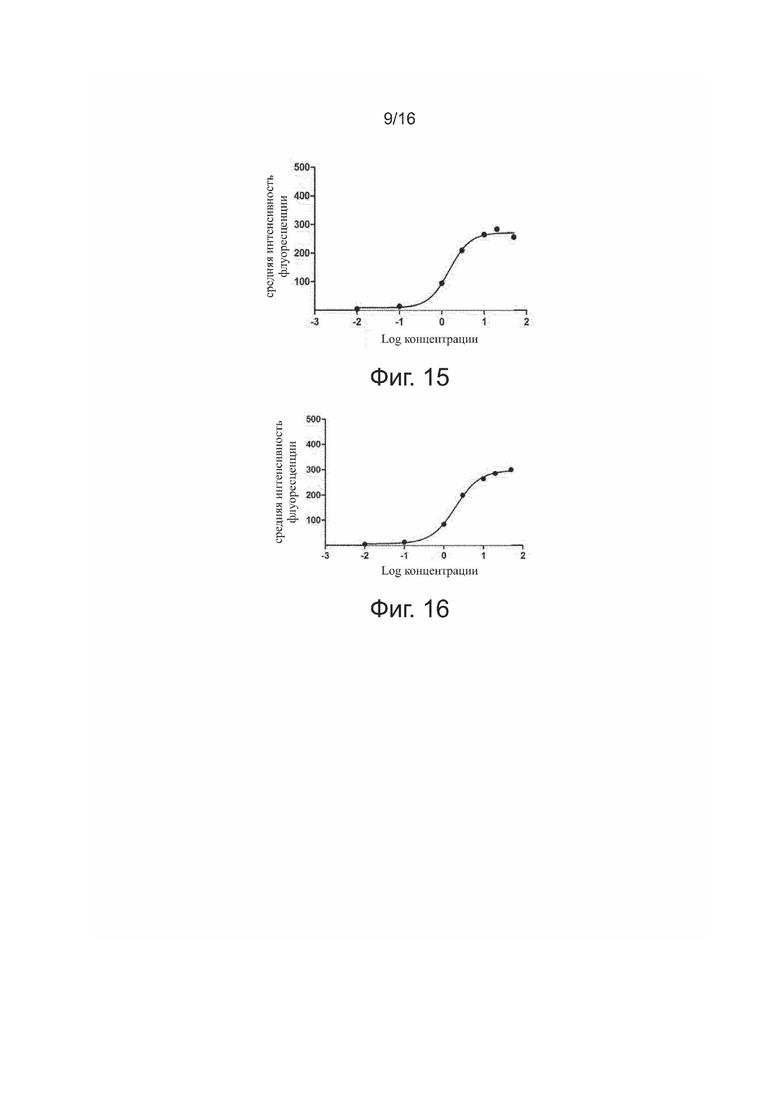
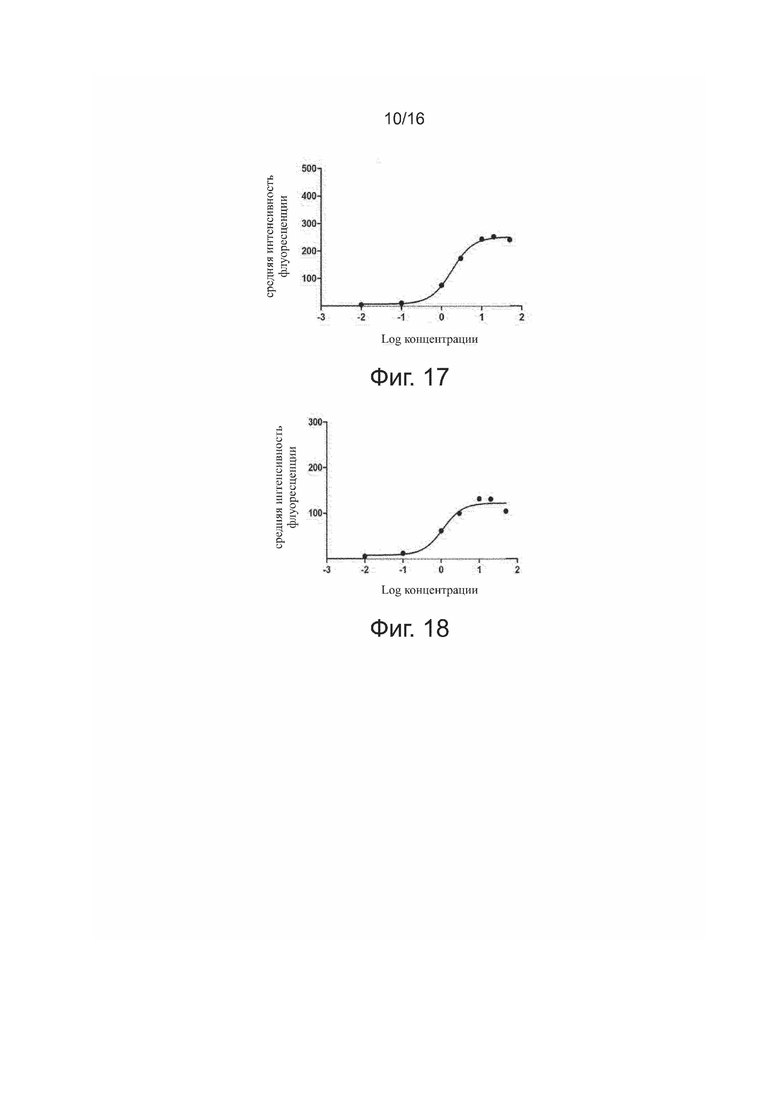
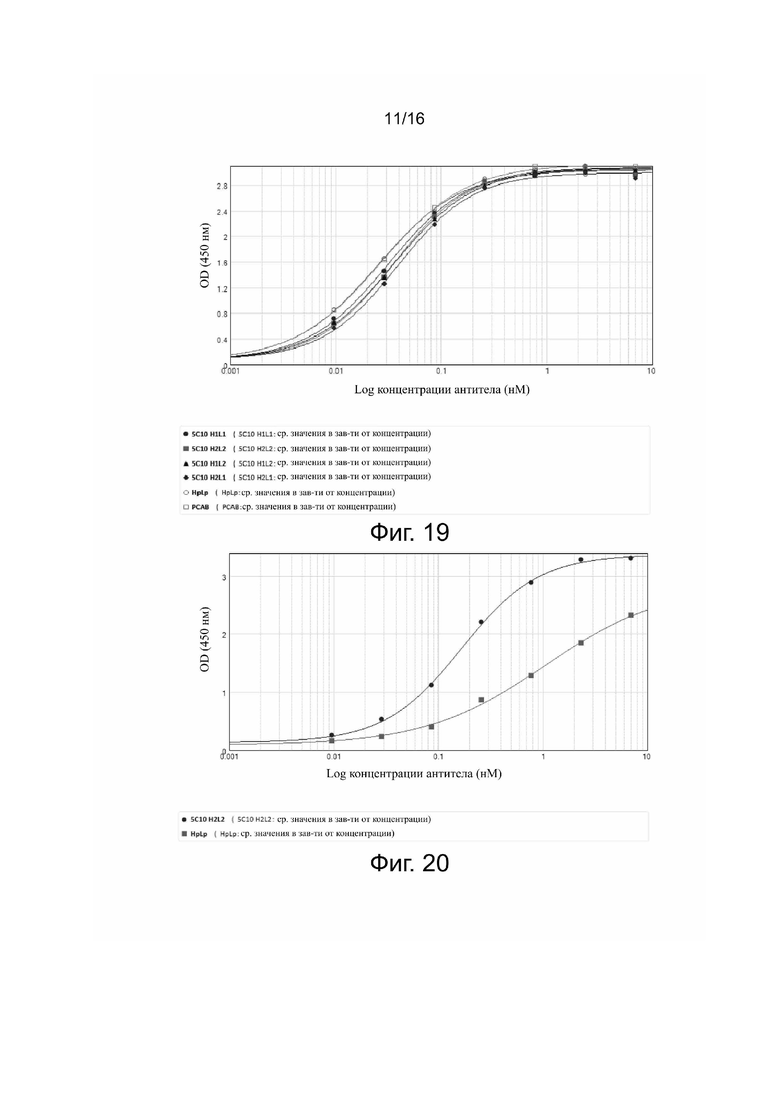
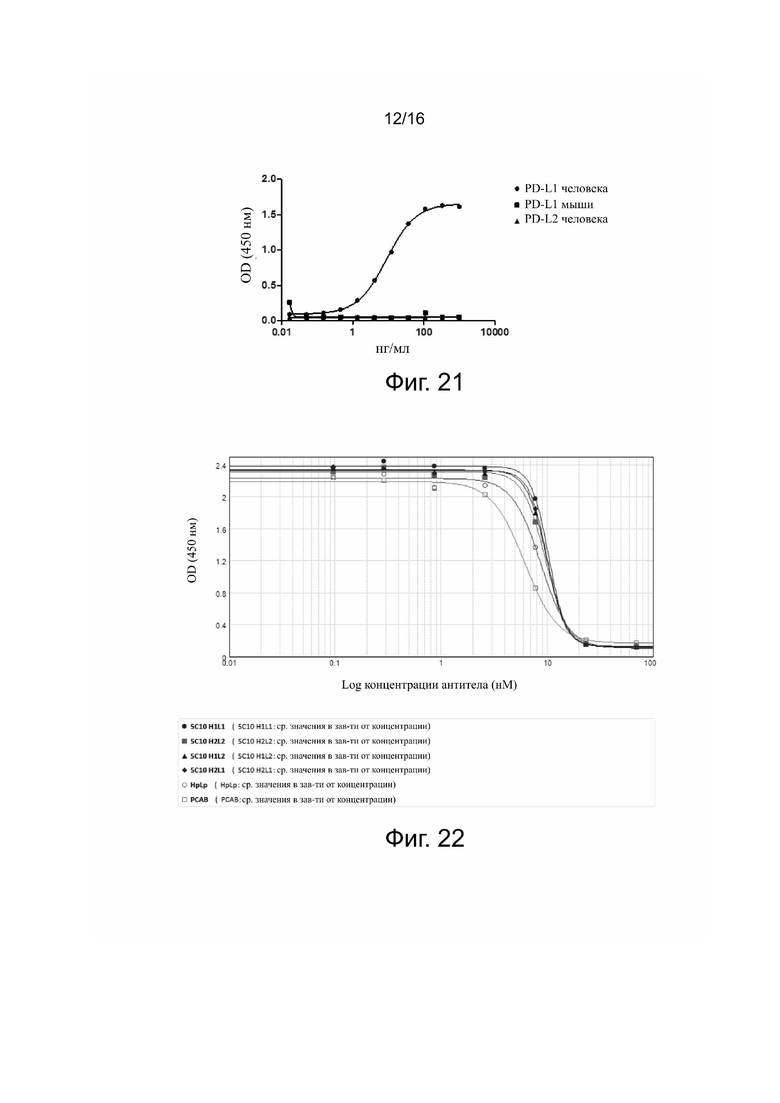
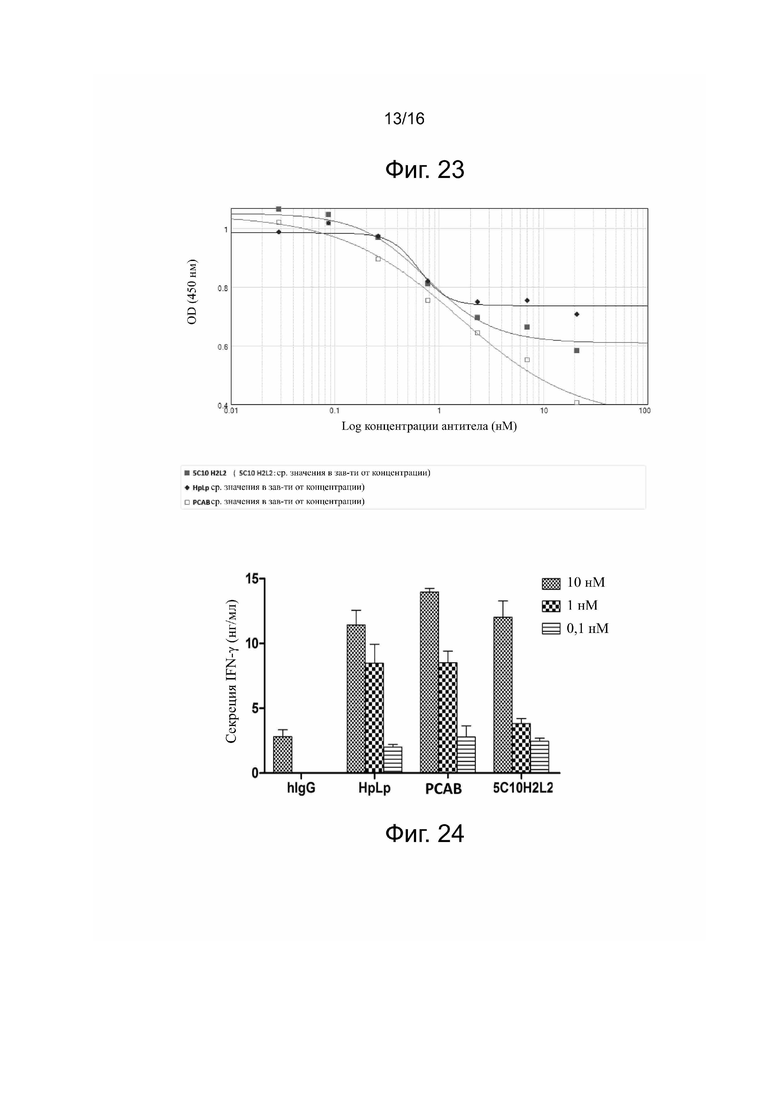
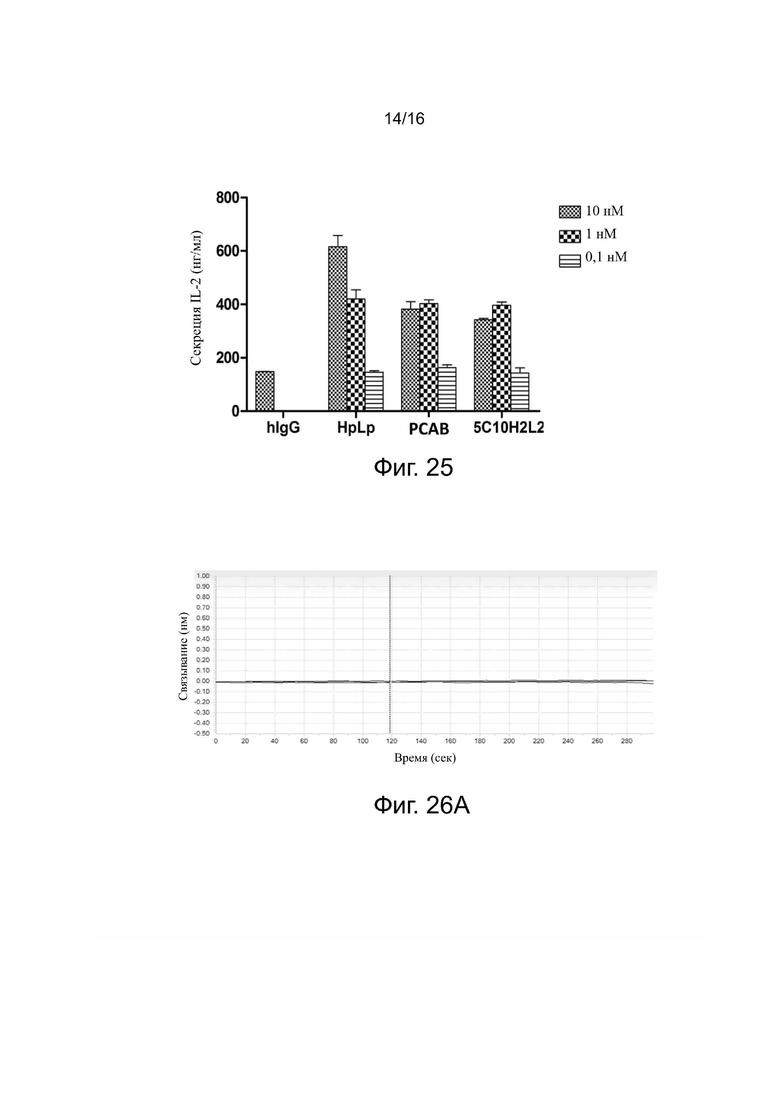
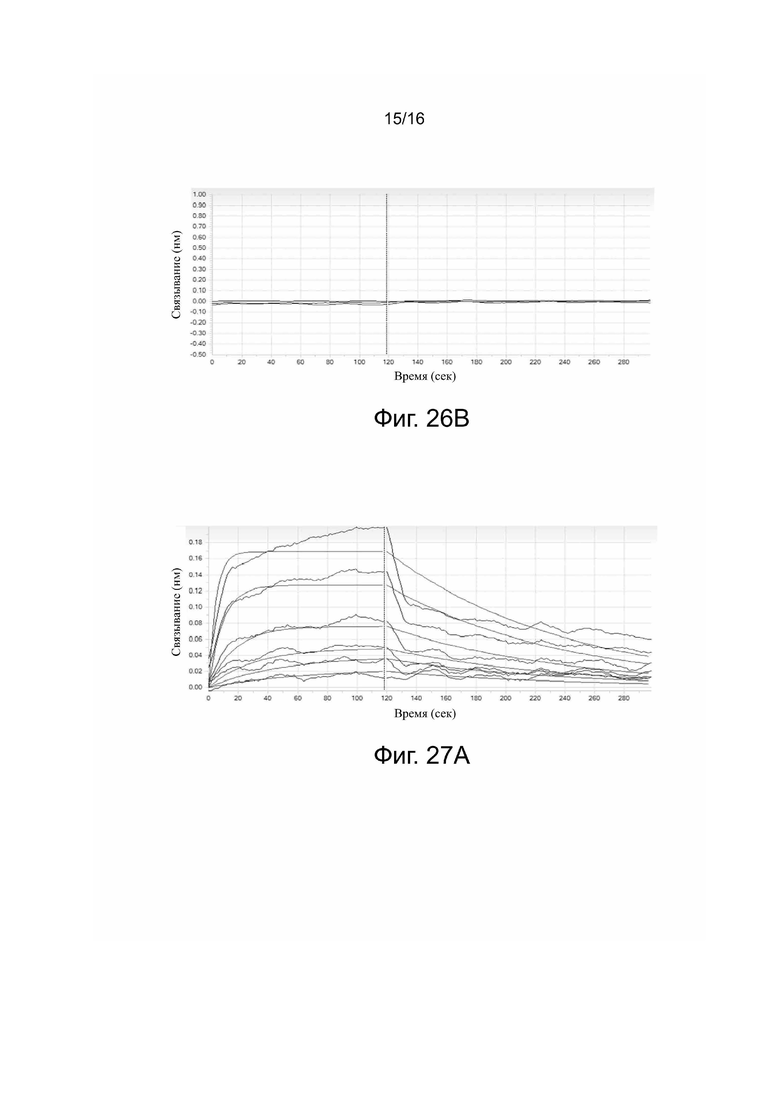
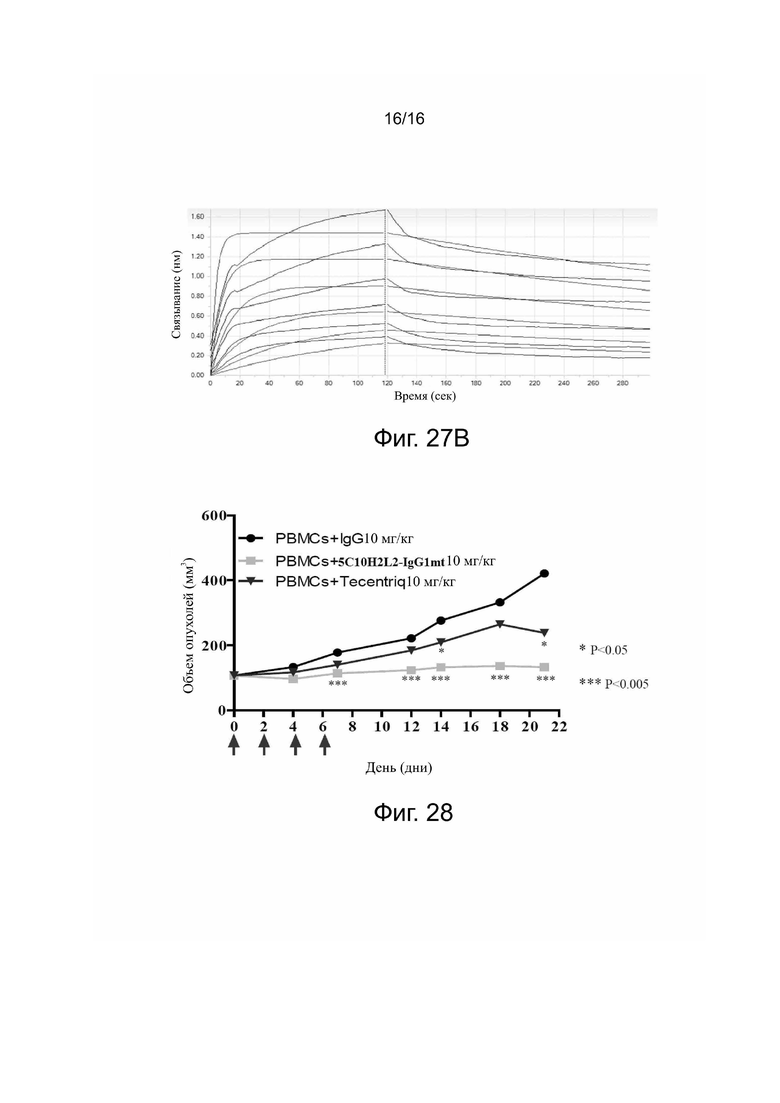
Настоящее изобретение относится к области иммунологии. Предложены анти-PDL-1 моноклональные антитела или их антигенсвязывающие фрагменты. Также рассмотрена молекула нуклеиновой кислоты, экспрессирующий вектор, клетка-хозяин, способ получения антитела и штамм клеток гибридомы. Кроме того, предложены конъюгат, бифункциональный конъюгат и мультиспецифичное антитело для связывания PDL-1, набор для обнаружения наличия или уровня PDL-1 и фармацевтическая композиция для лечения и/или профилактики. Также описаны применения анти-PDL-1 антитела, способ блокировки связывания и способ лечения и/или профилактики. Данное изобретение может найти дальнейшее применение в терапии опухолей или анемии. 18 н. и 39 з.п. ф-лы, 30 ил., 10 табл., 19 пр.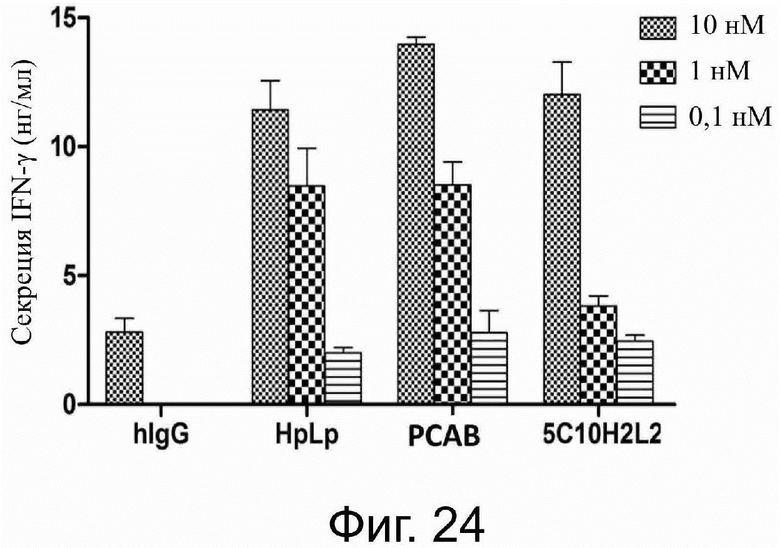
1. Анти-PDL-1 моноклональное антитело или его антигенсвязывающий фрагмент, причем:
данное анти-PDL-1 моноклональное антитело имеет вариабельную область тяжелой цепи, включающую HCDR1, HCDR2 и HCDR3, как представлено в SEQ ID NO: 2, SEQ ID NO: 6 или SEQ ID NO: 10, и имеет вариабельную область легкой цепи, содержащую LCDR1, LCDR2 и LCDR3, как представлено в SEQ ID NO: 4, SEQ ID NO: 8 или SEQ ID NO: 12;
или
данное анти-PDL-1 моноклональное антитело имеет вариабельную область тяжелой цепи, содержащую HCDR1, HCDR2 и HCDR3, как представлено в SEQ ID NO: 21, и имеет вариабельную область легкой цепи, содержащую LCDR1, LCDR2 и LCDR3, как представлено в SEQ ID NO: 23;
или
данное анти-PDL-1 моноклональное антитело имеет вариабельную область тяжелой цепи, содержащую HCDR1, HCDR2 и HCDR3, как представлено в SEQ ID NO: 25, и имеет вариабельную область легкой цепи, содержащую LCDR1, LCDR2 и LCDR3, как представлено в SEQ ID NO: 27.
2. Анти-PDL-1 моноклональное антитело или его антигенсвязывающий фрагмент, причем:
данное анти-PDL-1 моноклональное антитело имеет вариабельную область тяжелой цепи, включающую HCDR1, как представлено в SEQ ID NO: 15, HCDR2, как представлено в SEQ ID NO:16, и HCDR3, как представлено в SEQ ID NO:17, и содержит вариабельную область легкой цепи, включающую LCDR1, как представлено в SEQ ID NO: 18, LCDR2, как представлено в SEQ ID NO:19, и LCDR3, как представлено в SEQ ID NO:20;
или
данное анти-PDL-1 моноклональное антитело имеет вариабельную область тяжелой цепи, включающую HCDR1, как представлено в SEQ ID NO: 29, HCDR2, как представлено в SEQ ID NO: 30, и HCDR3, как представлено в SEQ ID NO:31, и содержит вариабельную область легкой цепи, включающую LCDR1, как представлено в SEQ ID NO: 32, LCDR2, как представлено в SEQ ID NO: 33, и LCDR3, как представлено в SEQ ID NO: 34;
или
данное анти-PDL-1 моноклональное антитело имеет вариабельную область тяжелой цепи, включающую HCDR1, как представлено в SEQ ID NO: 35, HCDR2, как представлено в SEQ ID NO: 36, и HCDR3, как представлено в SEQ ID NO: 37, и имеет вариабельную область легкой цепи, включающую LCDR1, как представлено в SEQ ID NO: 38, LCDR2, как представлено в SEQ ID NO: 39, и LCDR3, как представлено в SEQ ID NO: 40,
где HCDR1, HCDR2, HCDR3, LCDR1, LCDR2 и LCDR3 получены с помощью определения по IMGT в базе данных VBASE2.
3. Анти-PDL-1 моноклональное антитело или его антигенсвязывающий фрагмент по п. 2, при этом:
вариабельная область тяжелой цепи имеет аминокислотную последовательность, выбранную из SEQ ID NO: 2, SEQ ID NO: 6 и SEQ ID NO: 10, и вариабельная область легкой цепи имеет аминокислотную последовательность, выбранную из SEQ ID NO: 4, SEQ ID NO: 8 и SEQ ID NO: 12;
или
вариабельная область тяжелой цепи имеет аминокислотную последовательность по SEQ ID NO: 21 и вариабельная область легкой цепи имеет аминокислотную последовательность по SEQ ID NO: 23;
или
вариабельная область тяжелой цепи имеет аминокислотную последовательность по SEQ ID NO: 25 и вариабельная область легкой цепи имеет аминокислотную последовательность по SEQ ID NO: 27.
4. Анти-PDL-1 моноклональное антитело или его антигенсвязывающий фрагмент по п. 3, где
вариабельная область тяжелой цепи имеет аминокислотную последовательность SEQ ID NO: 2, а вариабельная область легкой цепи имеет аминокислотную последовательность SEQ ID NO: 4;
вариабельная область тяжелой цепи имеет аминокислотную последовательность SEQ ID NO: 6, а вариабельная область легкой цепи имеет аминокислотную последовательность SEQ ID NO: 8;
вариабельная область тяжелой цепи имеет аминокислотную последовательность SEQ ID NO: 6, а вариабельная область легкой цепи имеет аминокислотную последовательность SEQ ID NO: 12;
вариабельная область тяжелой цепи имеет аминокислотную последовательность SEQ ID NO: 10, а вариабельная область легкой цепи имеет аминокислотную последовательность SEQ ID NO: 8; или же
вариабельная область тяжелой цепи имеет аминокислотную последовательность SEQ ID NO: 10, а вариабельная область легкой цепи имеет аминокислотную последовательность SEQ ID NO: 12.
5. Анти-PDL-1 моноклональное антитело или его антигенсвязывающий фрагмент по п. 2 или 3, при этом антитело или его антигенсвязывающий фрагмент выбраны из Fab, Fab′, F(ab′)2, Fd, Fv, одноцепочечных антител, гуманизированных антител, химерных антител или диател.
6. Анти-PDL-1 моноклональное антитело или его антигенсвязывающий фрагмент по п.5, где одноцепочечное антитело представляет собой scFv.
7. Анти-PDL-1 моноклональное антитело или его антигенсвязывающий фрагмент по п. 2 или 3, при этом антитело или его антигенсвязывающий фрагмент связывается с PDL-1 со значением EC50 менее 100 нМ.
8. Анти-PDL-1 моноклональное антитело или его антигенсвязывающий фрагмент по п. 2 или 3, при этом анти-PDL-1 моноклональное антитело или его антигенсвязывающий фрагмент связывается с PDL-1 со значением EC50 менее 10 нМ, 1 нМ, 0,9 нМ, 0,8 нМ, 0,7 нМ, 0,6 нМ, 0,5 нМ, 0,4 нМ, 0,3 нМ, 0,2 нМ, 0,1 нМ или меньше.
9. Анти-PDL-1 моноклональное антитело или его антигенсвязывающий фрагмент по п. 7 или 8, причем EC50 определяется непрямым методом ELISA.
10. Анти-PDL-1 моноклональное антитело или его антигенсвязывающий фрагмент по п. 2 или 3, при этом анти-PDL-1 моноклональное антитело включает не содержащую CDR область, причем не содержащая CDR область происходит из другого вида, чем мышь.
11. Анти-PDL-1 моноклональное антитело или его антигенсвязывающий фрагмент по п. 2 или 3, при этом анти-PDL-1 моноклональное антитело включает не содержащую CDR область, причем не содержащая CDR область происходит из антитела человека.
12. Анти-PDL-1 моноклональное антитело или его антигенсвязывающий фрагмент по п. 2 или 3, при этом константная область моноклонального антитела выбрана из константных областей IgG1, IgG2, IgG3 или IgG4 человека.
13. Анти-PDL-1 моноклональное антитело или его антигенсвязывающий фрагмент по п. 2 или 3, при этом константная область моноклонального антитела представлена мутантной константной областью IgG1 человека.
14. Анти-PDL-1 моноклональное антитело или его антигенсвязывающий фрагмент по п. 13, при этом мутантная константная область IgG1 человека содержит мутацию N297A в константной области тяжелой цепи по системе нумерации EU.
15. Анти-PDL-1 моноклональное антитело или его антигенсвязывающий фрагмент по п. 13 или 14, при этом мутантная константная область тяжелой цепи IgG1 человека содержит по меньшей мере одну мутацию в положении 234, 235 или 237 по системе нумерации EU и после мутации снижается динамическое сродство антитела или его антигенсвязывающего фрагмента к FcγRIIIa и/или C1q.
16. Анти-PDL-1 моноклональное антитело или его антигенсвязывающий фрагмент по п. 13 или 14, при этом мутантная константная область IgG1 человека содержит 1, 2 или 3 мутации в положении 234, 235 и 237 по системе нумерации EU, причем мутации выбраны из L234A, L235A и G237A.
17. Анти-PDL-1 моноклональное антитело или его антигенсвязывающий фрагмент по любому из пп. 13-16, при этом мутация снижает активность ADCC и/или активность CDC антитела или его антигенсвязывающего фрагмента.
18. Анти-PDL-1 моноклональное антитело или его антигенсвязывающий фрагмент по п. 2 или 3, при этом анти-PDL-1 моноклональное антитело вырабатывается клетками гибридомы штамма LT005, а клетки гибридомы штамма LT005 депонированы в Китайском центре коллекций типовых культур (CCTCC) под номером доступа CCTCC No. C2015133.
19. Выделенная молекула нуклеиновой кислоты, кодирующая моноклональное антитело против PDL-1 по любому из пп. 2-17, которая включает молекулу нуклеиновой кислоты, кодирующую вариабельную область тяжелой цепи моноклонального антитела, и молекулу нуклеиновой кислоты, кодирующую вариабельную область легкой цепи моноклонального антитела, причем:
указанная вариабельная область тяжелой цепи указанного моноклонального антитела содержит HCDR1, как представлено в SEQ ID NO: 15, HCDR2, как представлено в SEQ ID NO: 16, и HCDR3, как представлено в SEQ ID NO: 17; и указанная вариабельная область легкой цепи указанного моноклонального антитела содержит LCDR1, как представлено в SEQ ID NO: 18, LCDR2, как представлено в SEQ ID NO: 19, и LCDR3, как представлено в SEQ ID NO: 20;
или
указанная вариабельная область тяжелой цепи указанного моноклонального антитела содержит HCDR1, как представлено в SEQ ID NO: 29, HCDR2, как представлено в SEQ ID NO: 30, и HCDR3, как представлено в SEQ ID NO: 31; и указанная вариабельная область легкой цепи указанного моноклонального антитела содержит LCDR1, как представлено в SEQ ID NO: 32, LCDR2, как представлено в SEQ ID NO: 33, и LCDR3, как представлено в SEQ ID NO: 34;
или
указанная вариабельная область тяжелой цепи указанного моноклонального антитела содержит HCDR1, как представлено в SEQ ID NO: 35, HCDR2, как представлено в SEQ ID NO: 36, и HCDR3, как представлено в SEQ ID NO: 37; и указанная вариабельная область легкой цепи указанного моноклонального антитела содержит LCDR1, как представлено в SEQ ID NO: 38, LCDR2, как представлено в SEQ ID NO: 39, и LCDR3, как представлено в SEQ ID NO: 40,
где HCDR1, HCDR2, HCDR3, LCDR1, LCDR2 и LCDR3 получены с помощью определения по IMGT в базе данных VBASE2.
20. Выделенная молекула нуклеиновой кислоты по п. 19, в которой:
вариабельная область тяжелой цепи указанного моноклонального антитела имеет аминокислотную последовательность SEQ ID NO: 2, SEQ ID NO: 6 или SEQ ID NO: 10; и вариабельная область легкой цепи указанного моноклонального антитела имеет аминокислотную последовательность SEQ ID NO: 4, SEQ ID NO: 8 или SEQ ID NO: 12;
или
вариабельная область тяжелой цепи указанного моноклонального антитела имеет аминокислотную последовательность SEQ ID NO: 21, а вариабельная область легкой цепи указанного моноклонального антитела имеет аминокислотную последовательность SEQ ID NO: 23;
или
вариабельная область тяжелой цепи указанного моноклонального антитела имеет аминокислотную последовательность SEQ ID NO: 25, а вариабельная область легкой цепи указанного моноклонального антитела имеет аминокислотную последовательность SEQ ID NO: 27.
21. Выделенная молекула нуклеиновой кислоты по п. 19, где:
молекула нуклеиновой кислоты, кодирующая вариабельную область тяжелой цепи моноклонального антитела, имеет нуклеотидную последовательность SEQ ID NO: 1, SEQ ID NO: 5 или SEQ ID NO: 9; и молекула нуклеиновой кислоты, кодирующая вариабельную область легкой цепи указанного моноклонального антитела, имеет нуклеотидную последовательность SEQ ID NO: 3, SEQ ID NO: 7 или SEQ ID NO: 11;
или же
молекула нуклеиновой кислоты, кодирующая вариабельную область тяжелой цепи моноклонального антитела, имеет нуклеотидную последовательность SEQ ID NO: 22; и молекула нуклеиновой кислоты, кодирующая вариабельную область легкой цепи указанного моноклонального антитела, имеет нуклеотидную последовательность SEQ ID NO: 24;
или же
молекула нуклеиновой кислоты, кодирующая вариабельную область тяжелой цепи моноклонального антитела, имеет нуклеотидную последовательность SEQ ID NO: 26; и молекула нуклеиновой кислоты, кодирующая вариабельную область легкой цепи указанного моноклонального антитела, имеет нуклеотидную последовательность SEQ ID NO: 28.
22. Выделенная молекула нуклеиновой кислоты по любому из пп. 19-21, которая дополнительно содержит линкер для соединения молекулы нуклеиновой кислоты, кодирующей вариабельную область тяжелой цепи моноклонального антитела, и молекулы нуклеиновой кислоты, кодирующей вариабельную область легкой цепи моноклонального антитела.
23. Экспрессирующий вектор, который включает выделенную молекулу нуклеиновой кислоты по любому из пп. 19-22.
24. Клетка-хозяин для получения анти-PDL-1 моноклонального антитела по любому из пп.2-17, которая включает выделенную молекулу нуклеиновой кислоты по любому из пп. 19-22 или экспрессирующий вектор по п. 23.
25. Способ получения анти-PDL-1 моноклонального антитела или его антигенсвязывающего фрагмента по любому из пп. 2-17, который включает следующие стадии: культивирование клетки-хозяина по п. 24 в подходящих условиях и извлечение анти-PDL-1 моноклонального антитела или его антигенсвязывающего фрагмента из культуры клетки-хозяина.
26. Штамм клеток гибридомы LT005 для получения анти-PDL-1 моноклонального антитела по п.18, которые были депонированы в Китайском центре коллекций типовых культур (CCTCC) под номером доступа CCTCC No. C2015133.
27. Конъюгат для связывания PDL-1, который включает анти-PDL-1 моноклональное антитело или его антигенсвязывающий фрагмент и соединительную часть, причем указанное анти-PDL-1 моноклональное антитело представляет собой анти-PDL-1 моноклональное антитело или его антигенсвязывающий фрагмент по любому из пп. 2-17, а соединительная часть представляет собой детектируемую метку.
28. Конъюгат по п. 27, в котором соединительная часть представляет собой радиоактивный изотоп, флуоресцентное вещество, люминесцентное вещество, окрашенное вещество или фермент.
29. Бифункциональный конъюгат антитела для связывания PDL-1, который включает анти-PDL-1 моноклональное антитело или его антигенсвязывающий фрагмент и конъюгированную часть, причем указанное антитело представляет собой анти-PDL-1 моноклональное антитело или его антигенсвязывающий фрагмент по любому из пп. 2-17, а конъюгированная часть представляет собой второй биологически функциональный фрагмент.
30. Бифункциональный конъюгат антитела по п. 29, в котором второй биологически функциональный фрагмент обладает связывающей активностью и представляет собой белок, полиэтиленгликоль (ПЭГ), нуклид, нуклеиновую кислоту, низкомолекулярный токсин, рецептор или лиганд.
31. Мультиспецифичное антитело для связывания PDL-1, которое получено при конъюгировании первого антитела или его фрагмента с другим антителом или его фрагментом или с миметиком антитела, причем каждое антитело или его фрагмент либо миметик антитела сохраняет исходную специфичность связывания, а первое антитело или его фрагмент представляет собой анти-PDL-1 моноклональное антитело или его антигенсвязывающий фрагмент по любому из пп. 2-17.
32. Мультиспецифичное антитело по п. 31, в котором мультиспецифичное антитело представляет собой биспецифичное антитело, или триспецифичное антитело, или тетраспецифичное антитело.
33. Набор для обнаружения наличия или уровня PDL-1 в образце, который включает анти-PDL-1 моноклональное антитело или его антигенсвязывающий фрагмент по любому из пп. 2-17, или же включает конъюгат по п. 27 или 28, или же включает конъюгат бифункционального антитела по п. 29 или 30, или же включает мультиспецифичное антитело по п. 31 или 32, где набор дополнительно включает вторичное антитело, которое специфически распознает анти-PDL-1 моноклональное антитело или его антигенсвязывающий фрагмент.
34. Набор по п. 33, где вторичное антитело помечено детектируемой меткой.
35. Набор по п. 34, где детектируемая метка представляет собой радиоактивный изотоп, флуоресцентное вещество, люминесцентное вещество, окрашенное вещество или фермент.
36. Применение анти-PDL-1 моноклонального антитела или его антигенсвязывающего фрагмента по любому из пп. 2-17, либо конъюгата по п. 27 или 28, либо конъюгата бифункционального антитела по п. 29 или 30, либо мультиспецифичного антитела по п. 31 или 32 при изготовлении набора, причем набор применяется для выявления наличия или уровня PDL-1 в образце.
37. Фармацевтическая композиция для лечения и/или профилактики опухоли или анемии, сопровождаемых иммуносупрессией под действием сигнального пути PD-1/PDL-1, которая включает анти-PDL-1 моноклональное антитело или его антигенсвязывающий фрагмент по любому из пп. 2-17, либо конъюгат по п. 27 или 28, либо конъюгат бифункционального антитела по п. 29 или 30, либо мультиспецифичное антитело по п. 31 или 32, и где фармацевтическая композиция также включает фармацевтически приемлемый носитель и/или наполнитель.
38. Применение анти-PDL-1 моноклонального антитела или его антигенсвязывающего фрагмента по любому из пп. 2-17, либо конъюгата по п. 27 или 28, либо конъюгата бифункционального антитела по п. 29 или 30, либо мультиспецифичного антитела по п. 31 или 32 при изготовлении лекарственного средства для профилактики, и/или лечения, и/или вспомогательного лечения опухоли или анемии, сопровождаемых иммуносупрессией под действием сигнального пути PD-1/PDL-1.
39. Применение по п. 38, в котором такая опухоль выбрана из рака молочной железы, рака легких, рака печени, рака желудка, колоректального рака, рака пищевода, рака яичников, рака шейки матки, рака почек, рака простаты, рака мочевого пузыря, рака поджелудочной железы, глиомы, меланомы и лейкемии.
40. Применение по п. 39, в котором указанный рак легкого представляет собой немелкоклеточный рак легкого.
41. Применение по п. 39, где указанным колоректальным раком является рак толстой кишки или рак прямой кишки.
42. Применение моноклонального антитела против PDL-1 или его антигенсвязывающего фрагмента по любому из пп. 2-17, или конъюгата по п. 27 или 28, или конъюгата бифункционального антитела по п. 29 или 30, или мультиспецифичного антитела по п. 31 или 32 при изготовлении лекарственного средства для диагностики опухоли или анемии, сопровождаемых иммуносупрессией под действием сигнального пути PD-1/PDL-1.
43. Применение по п. 42, в котором указанная опухоль выбрана из рака молочной железы, рака легкого, рака печени, рака желудка, колоректального рака, рака пищевода, рака яичника, рака шейки матки, рака почки, рака простаты, рака мочевого пузыря, рака поджелудочной железы, глиомы, меланомы и лейкемии.
44. Применение по п. 43, где указанный рак легкого представляет собой немелкоклеточный рак легкого.
45. Применение по п. 43, где указанным колоректальным раком является рак толстой кишки или рак прямой кишки.
46. Применение анти-PDL-1 моноклонального антитела или его антигенсвязывающего фрагмента по любому из пп. 2-17, либо конъюгата по п. 27 или 28, либо конъюгата бифункционального антитела по п. 29 или 30, либо мультиспецифичного антитела по п. 31 или 32 при изготовлении препарата для блокировки связывания PDL-1 с PD-1 или с B7-1.
47. Применение по п. 46, где указанный препарат предназначен для уменьшения активности PDL-1 или уровня PDL-1.
48. Применение по п. 46, где указанный препарат предназначен для устранения иммуносупрессии под действием PD-1 или PDL-1.
49. Применение по п. 46, где указанный препарат предназначен для усиления экспрессии IFN-γ и/или IL-2 в T-лимфоцитах.
50. Способ in vivo или in vitro, который включает стадию введения в клетки эффективного количества анти-PDL-1 моноклонального антитела или его антигенсвязывающего фрагмента по любому из пп. 2-17, либо конъюгата по п. 27 или 28, либо конъюгата бифункционального антитела по п. 29 или 30, либо мультиспецифичного антитела по п. 31 или 32, и представляет собой способ блокировки связывания PDL-1 с PD-1 или с B7-1.
51. Способ по п. 50, где указанный способ предназначен для уменьшения активности PDL-1 или уровня PDL-1.
52. Способ по п. 50, где указанный способ предназначен для устранения иммуносупрессии под действием PD-1 или PDL-1.
53. Способ по п. 50, где указанный способ предназначен для усиления экспрессии IFN-γ и/или IL-2 в T-лимфоцитах.
54. Способ лечения и/или профилактики опухолей или анемии, сопровождаемых иммуносупрессией под действием сигнального пути PD-1/PDL-1, включающий стадию введения субъекту эффективного количества анти-PDL-1 моноклонального антитела или его антигенсвязывающего фрагмента по любому из пп. 2-17, либо конъюгата по п. 27 или 28, либо конъюгата бифункционального антитела по п. 29 или 30, либо мультиспецифичного антитела по п. 31 или 32.
55. Способ по п. 54, в котором такая опухоль выбрана из рака молочной железы, рака легких, рака печени, рака желудка, рака кишечника, рака пищевода, рака яичников, рака шейки матки, рака почек, рака простаты, рака мочевого пузыря, рака поджелудочной железы, глиомы, меланомы и лейкемии.
56. Способ по п. 55, в котором указанный рак легкого представляет собой немелкоклеточный рак легкого.
57. Способ по п. 55, в котором указанный колоректальный рак представляет собой рак толстой кишки или рак прямой кишки.
| Устройство для закрепления лыж на раме мотоциклов и велосипедов взамен переднего колеса | 1924 |
|
SU2015A1 |
| Станок для изготовления деревянных ниточных катушек из цилиндрических, снабженных осевым отверстием, заготовок | 1923 |
|
SU2008A1 |
| CA 2954868 A1, 14.01.2016 | |||
| Резиновая заготовка для подошв | 1929 |
|
SU19344A1 |
| Способ приготовления лака | 1924 |
|
SU2011A1 |

