Сведения о родственных заявках
Настоящая заявка испрашивает приоритет по предварительной заявке на патент США № 61/844,168, поданной 9 июля 2013 г. и включённой в данный документ в виде ссылки во всей своей полноте для всех целей.
Декларация государственного интереса
Данное изобретение было создано при правительственной поддержке по грантам DE-FG02-02ER63445 от Министерства энергетики, NSF-SynBERC от Национального научного фонда и SA5283-11210 от Национального научного фонда. Правительство имеет определённые права на данное изобретение.
Предпосылки создания изобретения
Бактериальные и архейные CRISPR/Cas-системы используют короткие направляющие РНК (гидРНК) в комплексе с белками Cas для направленной деградации комплементарных последовательностей, содержащихся во вторгшейся в клетку чужеродной нуклеиновой кислоте. См. Deltcheva, E. et al. CRISPR RNA maturation by trans-encoded small RNA and host factor RNase III [Созревание CRISPR РНК, обеспечиваемое транс-кодированной малой РНК и фактором хозяина РНКазой III]. Nature 471, 602-607 (2011); Gasiunas, G, Barrangou, R., Horvath, P. & Siksnys, V. Cas9-crRNA ribonucleoprotein complex mediates specific DNA cleavage for adaptive immunity in bacteria [Рибонуклеопротеиновый комплекс Cas9-crРНК опосредует специфическое расщепление ДНК для адаптивного иммунитета у бактерий]. Proceedings of the National Academy of Sciences of the United States of America 109, E2579-2586 (2012); Jinek, M. et al. A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity [Программируемая двойная-РНК-направляемая эндонуклеаза ДНК в адаптивном иммунитете бактерий]. Science 337, 816-821 (2012); Sapranauskas, R. et al. The Streptococcus thermophilus CRISPR/Cas system provides immunity in Escherichia coli [CRISPR/Cas-система бактерий Streptococcus thermophilus обеспечивает иммунитет у Escherichia coli]. Nucleic acids research 39, 9275-9282 (2011); и Bhaya, D., Davison, M. & Barrangou, R. CRISPR-Cas systems in bacteria and archaea: versatile small RNAs for adaptive defense and regulation [CRISPR/Cas-системы бактерий и архей: универсальные малые РНК для адаптивной защиты и регуляции]. Annual review of genetics 45, 273-297 (2011). Недавно проведенное воссоздание CRISPR-системы II типа бактерий S. pyogenes in vitro показало, что crРНК (CRISPR РНК), слитой со стандартной транс-кодированной tracrRNA (транс-активирующей CRISPR РНК), достаточно для того, чтобы направить белок Cas9 к месту последовательность-специфического расщепления ДНК-мишени, соответствующих crРНК. Экспрессирование гидРНК, гомологичной сайту-мишени, приводит к привлечению Cas9 и деградации ДНК-мишени. См. H. Deveau et al., Phage response to CRISPR-encoded resistance in Streptococcus thermophilus [Ответ фага на CRISPR-кодируемую устойчивость у Streptococcus thermophilus]. Journal of Bacteriology 190, 1390 (Feb, 2008).
Краткое описание сущности изобретения
Аспекты настоящего изобретения направлены на мультиплексную (множественную) модификацию ДНК в клетке, где используется одна или несколько направляющих РНК (рибонуклеиновых кислот) для того, чтобы направить экспрессируемый клеткой фермент, обладающий нуклеазной активностью, например, ДНК-связывающий белок, обладающий нуклеазной активностью, к месту целевой локализации на ДНК (дезоксирибонуклеиновой кислоте), где фермент разрезает ДНК и экзогенная донорная нуклеиновая кислота вставляется в ДНК, например, гомологичной рекомбинацией. Аспекты настоящего изобретения включают проведение нескольких циклов или повторение этапов модификации ДНК в клетке для создания клетки, имеющей множественные модификации ДНК. Модификации могут включать инсерцию экзогенных донорных нуклеиновых кислот.
Множественные инсерции экзогенных нуклеиновых кислот могут осуществляться в одну стадию, путем введения в клетку, которая экспрессирует фермент, нуклеиновых кислот, кодирующих множество РНК, и множество экзогенных донорных нуклеиновых кислот, например, котрансформацией, при этом РНК экспрессируются, и при этом каждая РНК из указанного множества направляет фермент к конкретному участку ДНК, фермент разрезает ДНК и одна из множества экзогенных нуклеиновых кислот вставляется в ДНК на место разреза. Согласно этому аспекту, множество изменений или модификаций ДНК в клетке создаются за один цикл.
Множественные инсерции экзогенных нуклеиновых кислот могут осуществляться в клетке путем проведения нескольких повторных стадий или циклов введения в клетку, которая экспрессирует фермент, одной или нескольких нуклеиновых кислот, кодирующих одну или несколько РНК либо множество РНК, и одну или несколько экзогенных нуклеиновых кислот либо множество экзогенных нуклеиновых кислот, при этом РНК экспрессируется и направляет фермент к конкретному участку ДНК, фермент разрезает ДНК и экзогенная нуклеиновая кислота вставляется в ДНК на место разреза, что приводит к получению клетки, имеющей множественные изменения или инсерции экзогенной ДНК в ее ДНК. Согласно одному аспекту, клетка, экспрессирующая фермент, может представлять собой клетку, которая экспрессирует фермент естественным путём, или клетку, которая была генетически изменена таким образом, чтобы она могла экспрессировать фермент, например, введением в клетку нуклеиновой кислоты, кодирующей фермент, которая может экспрессироваться клеткой. Таким образом, аспекты настоящего изобретения включают цикл стадий введения РНК в клетку, которая экспрессирует фермент; введение экзогенной донорной нуклеиновой кислоты в клетку, экспрессирующую РНК; образование колокализуемого комплекса из РНК, фермента и ДНК; ферментативное расщепление ДНК ферментом и инсерцию донорной нуклеиновой кислоты в ДНК. Повторение цикла или повторение вышеуказанных стадий приводит к мультиплексной генетической модификации клетки во множестве локусах, т.е. клетки, имеющей множественные генетические модификации.
Согласно некоторым аспектам изобретения, предлагается способ увеличения частоты гомологичной рекомбинации за счёт циклически повторяющейся методики, описанной выше. В одном варианте осуществления способа направляемое разрезание геномной ДНК с помощью Cas9 стимулирует экзогенную ДНК за счёт резкого увеличения частоты гомологичной рекомбинации. Согласно одному из дополнительных аспектов, экзогенная донорная нуклеиновая кислота включает гомологичные последовательности или “плечи”, фланкирующие участок разреза. Согласно одному из дополнительных аспектов, экзогенная донорная нуклеиновая кислота включает последовательность, удаляющую разрезаемую последовательность. Согласно одному из дополнительных аспектов, экзогенная донорная нуклеиновая кислота включает гомологичные последовательности или “плечи”, фланкирующие участок разреза, и последовательность для удаления сайта разреза. Таким образом, Cas9 может использоваться в качестве негативной селекции против клеток, которые не содержат экзогенную донорную ДНК. Соответственно предлагается метод отрицательной селекции для идентификации клеток, имеющих высокую частоту рекомбинации.
Согласно некоторым аспектам, ДНК-связывающие белки или ферменты в контексте настоящего изобретения включают белок, который образует комплекс с гидРНК, направляющей комплекс к последовательности двухцепочечной ДНК, при этом комплекс связывается с последовательностью ДНК. Согласно одному аспекту, фермент может представлять собой РНК-направляемый ДНК-связывающий белок, такой как РНК-направляемый ДНК-связывающий белок системы CRISPR II типа, который связывается с ДНК и направляется РНК. Согласно одному аспекту, РНК-направляемым ДНК-связывающим белком является белок Cas9.
Этот аспект настоящего изобретения может рассматриваться как колокализация РНК и ДНК-связывающего белка на или с двухцепочечной ДНК. Таким образом, комплекс ДНК-связывающий белок/гидРНК может использоваться для разрезания множества участков двухцепочечной ДНК с целью создания клетки с множественными генетическими модификациями, такими как множественные вставки экзогенной донорной ДНК.
Согласно некоторым аспектам, предлагается способ внесения множества изменений в ДНК-мишень в клетке, экспрессирующей фермент, который образует колокализуемый комплекс с РНК, комплементарной ДНК-мишени, и который расщепляет ДНК-мишень сайт-специфическим образом, включающий (a) введение в клетку первой чужеродной нуклеиновой кислоты, кодирующей одну или несколько РНК, комплементарных ДНК-мишени, которая(ые) направляет(ют) фермент к ДНК-мишени, где одна или несколько РНК и фермент являются членами колокализуемого комплекса для ДНК-мишени; введение в клетку второй чужеродной нуклеиновой кислоты, кодирующей одну или несколько последовательностей донорной нуклеиновой кислоты, в котором одна или несколько РНК и одна или несколько последовательностей донорной нуклеиновой кислоты экспрессируются, где одна или несколько РНК и одна или несколько последовательностей донорной нуклеиновой кислоты колокализуются на ДНК-мишени, фермент расщепляет ДНК-мишень и донорная нуклеиновая кислота вставляется в ДНК-мишень с получением изменённой ДНК в клетке; и повторение стадии (a) множество раз для внесения множественных изменений в ДНК в клетке.
Согласно одному аспекту, клетка является эукариотической клеткой. Согласно одному аспекту, клетка является дрожжевой клеткой, растительной клеткой или животной клеткой. Согласно одному аспекту, клетка является клеткой млекопитающего.
Согласно одному аспекту, РНК содержит примерно от 10 до 500 нуклеотидов. Согласно одному аспекту, РНК содержит примерно от 20 до 100 нуклеотидов.
Согласно одному аспекту, одна или несколько РНК является направляющей РНК (гидРНК). Согласно одному аспекту, одна или несколько РНК представляет собой слияние tracrРНК-crРНК.
Согласно одному аспекту, ДНК является геномной ДНК, митохондриальной ДНК, вирусной ДНК или экзогенной ДНК.
Дополнительные отличительные признаки и преимущества некоторых вариантов осуществления настоящего изобретения будут более очевидны из нижеследующего описания вариантов и фигур и формулы изобретения.
Краткое описание фигур
Вышеизложенные и другие отличительные признаки и преимущества настоящих вариантов осуществления будут более понятны из нижеследующего подробного описания иллюстративных вариантов осуществления вкупе с прилагаемыми чертежами, на которых:
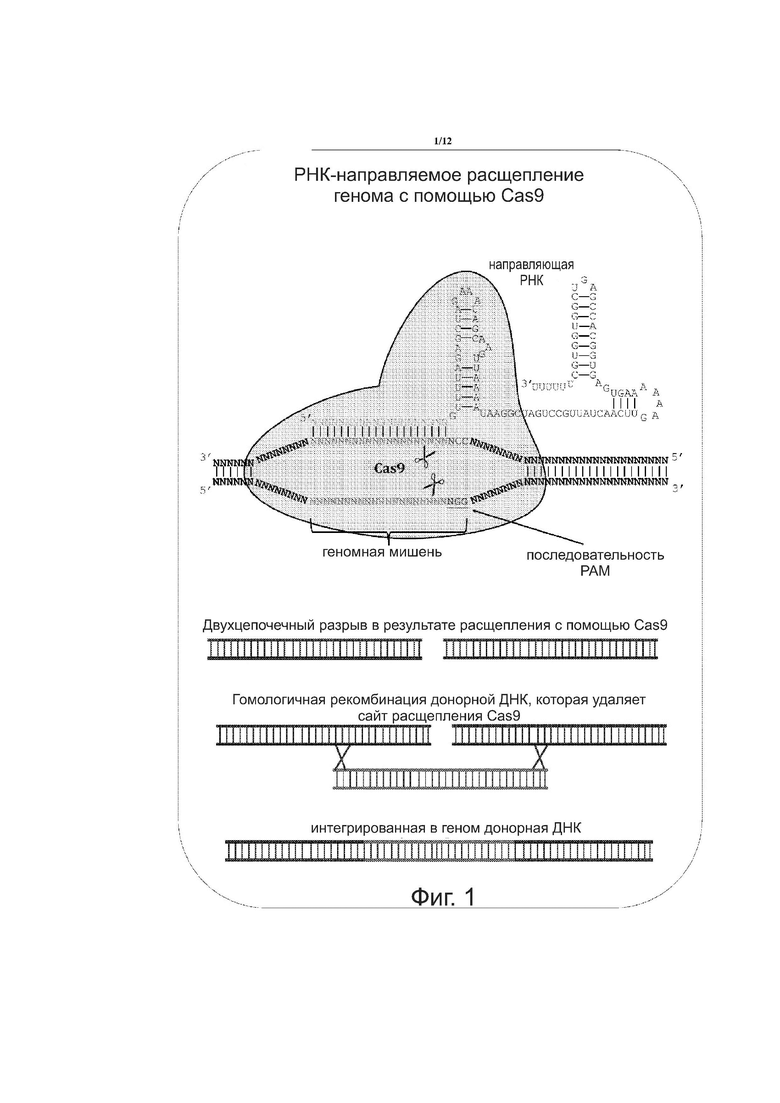
на фиг. 1 показана схема РНК-направляемого расщепления генома с помощью Cas9;
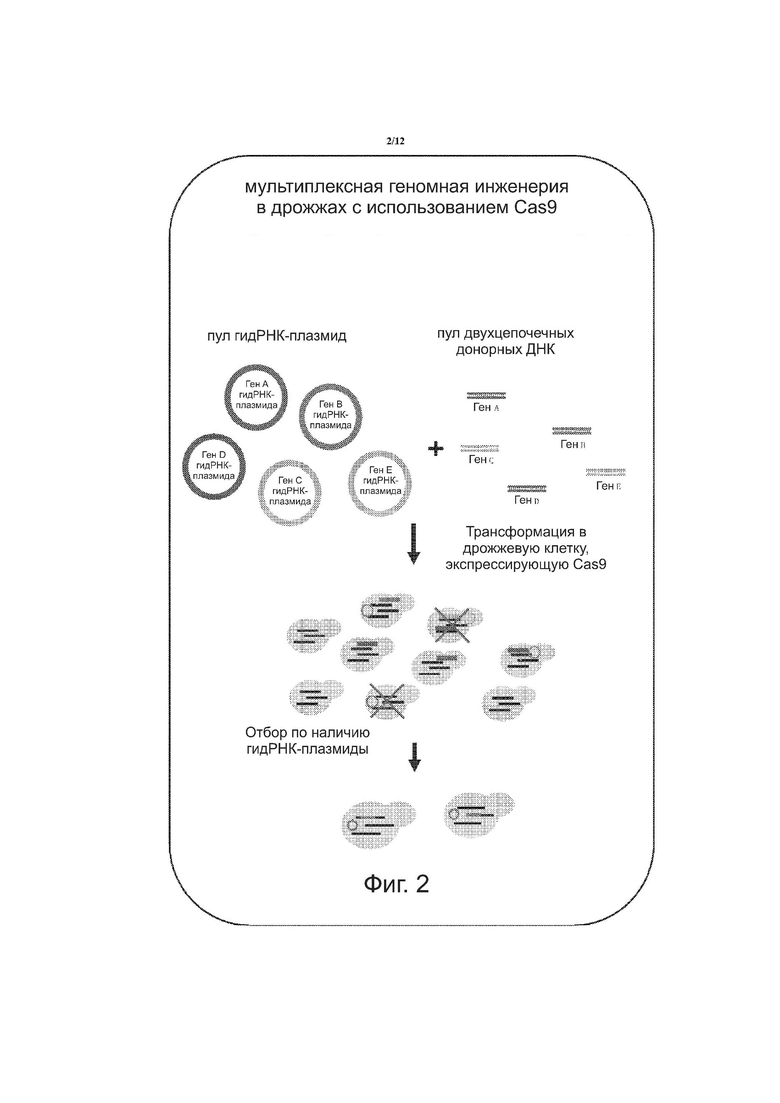
на фиг. 2 схематически показана мультиплексная геномная инженерия в дрожжах с использованием Cas9;
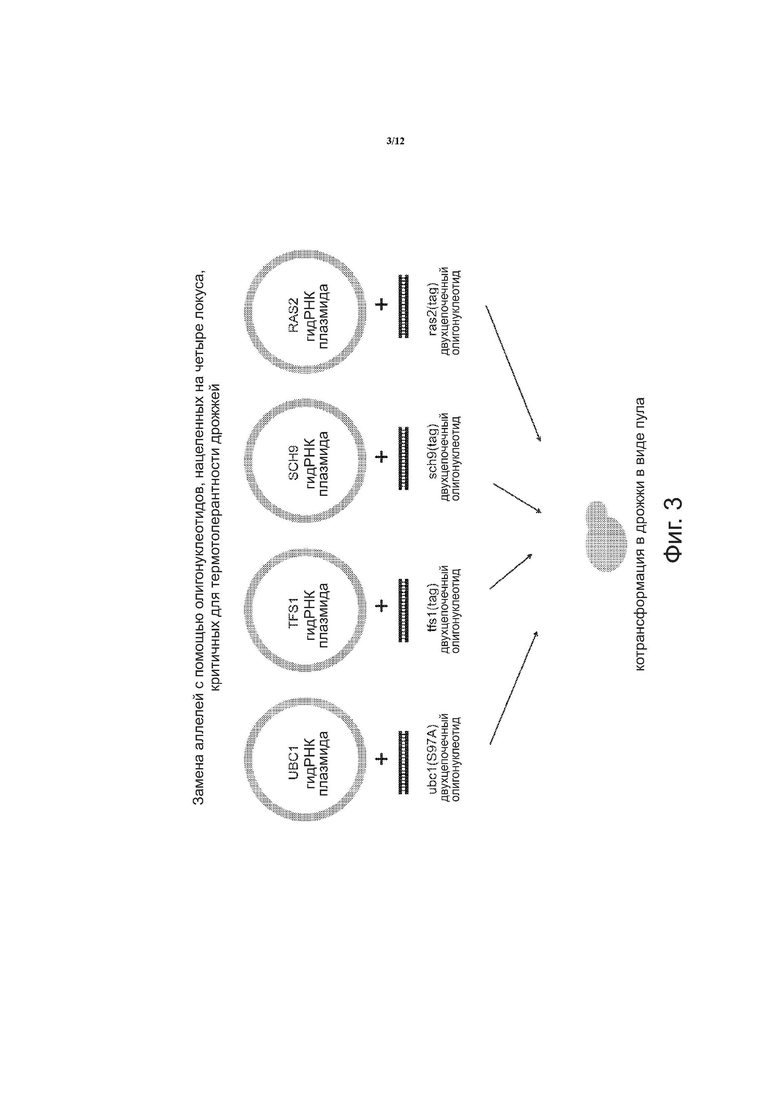
на фиг. 3 схематически показана замена аллелей с использованием олигонуклеотидов, нацеленных на четыре локуса, критических для термотолерантности дрожжей;
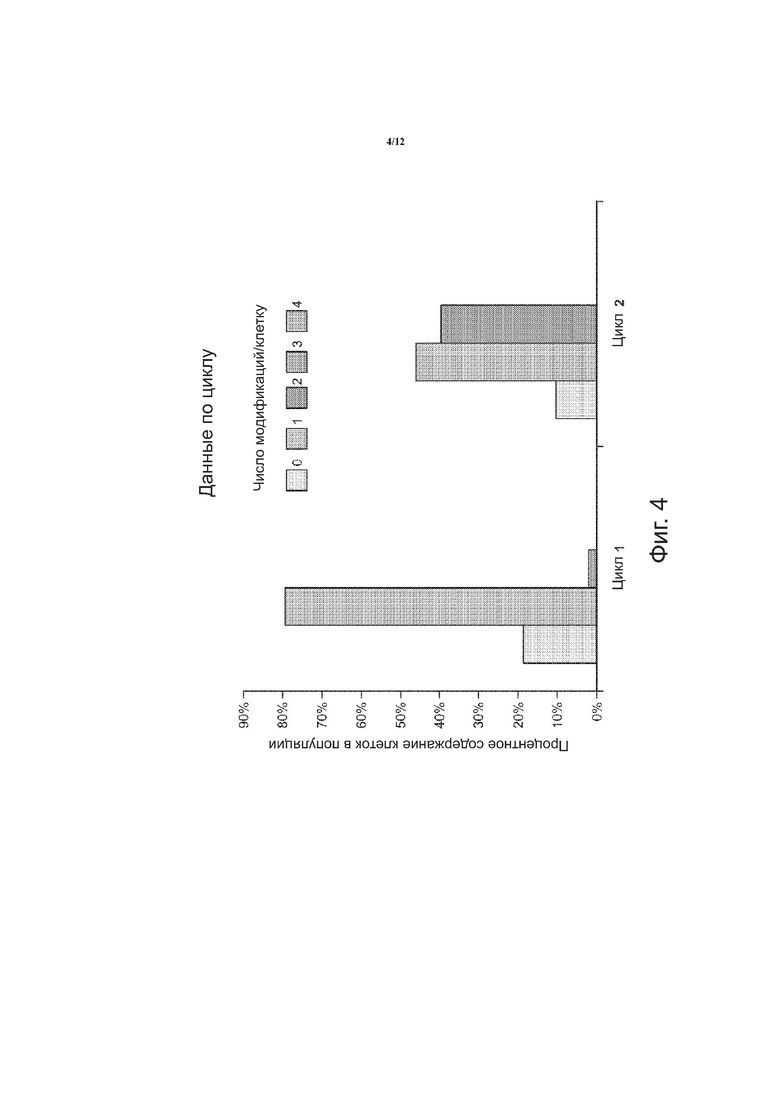
на фиг. 4 приводится диаграмма, показывающая число модификаций на клетку после одного цикла и после двух циклов;
на фиг. 5А приводится таблица штаммов, имеющих мутации. Фиг. 5B показывает термотолерантность при тепловом шоке у различных штаммов;
на фиг. 6A приводятся графические данные по частоте трансформации. На фиг. 6В приводятся графические данные по частоте отдельных рекомбинаций. На фиг. 6C приводятся графические данные по частоте совместных рекомбинаций в локусах can1 и KanMX;
на фиг. 7 приводятся графические данные по мультиплексному линейному введению кассеты в два локуса;
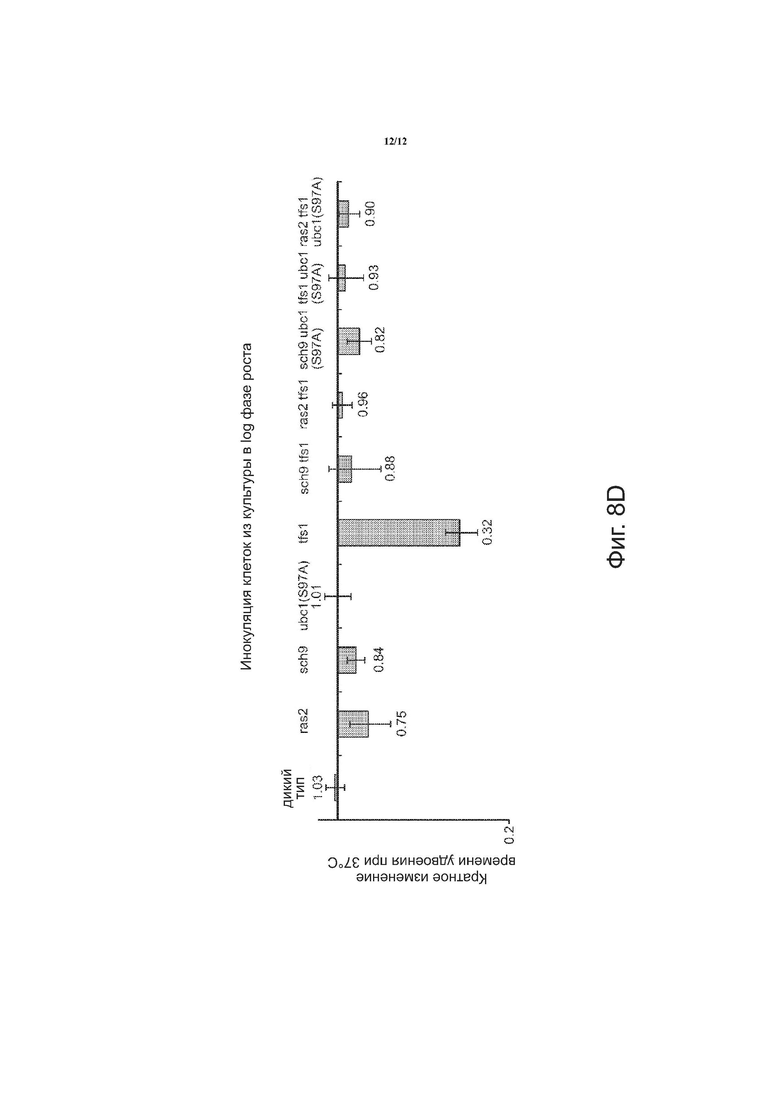
на фиг. 8A приводятся графические данные по кратному изменению времени удвоения при 30°С. На фиг. 8B приводятся графические данные по кратному изменению времени удвоения при 37°С. На фиг. 8C приводятся графические данные по кратному изменению времени удвоения при 42°С для клеток, инокулированных из культуры в поздней стационарной фазе роста. На фиг. 8D приводятся графические данные по кратному изменению времени удвоения при 42°C для клеток, инокулированных из культуры в поздней log фазе роста.
Подробное описание
Варианты осуществления настоящего изобретения основываются на многократном применении экзогенной ДНК, ферментов нуклеаз, таких как ДНК-связывающие белки, и направляющих РНК (гидРНК), которые колокализуются на ДНК и расщепляют или разрезают ДНК с инсерцией экзогенной ДНК, например, гомологичной рекомбинацией. Такие ДНК-связывающие белки хорошо известны специалистам в данной области техники как связывающиеся с ДНК для различных целей. Такие ДНК-связывающие белки могут быть ДНК-связывающими белками, встречающимися в природе. ДНК-связывающие белки, входящие в объём настоящего изобретения, включают те из них, которые могут направляться РНК, называемой в описании гидРНК. Согласно этому аспекту, гидРНК и РНК-направляемый ДНК-связывающий белок образуют колокализационный комплекс на ДНК. Эти ДНК-связывающие белки, обладающие нуклеазной активностью, известны специалистам в данной области техники и включают встречающиеся в природе ДНК-связывающие белки, обладающие нуклеазной активностью, такие как белки Cas9, присутствующие, например, в системах CRISPR II типа. Эти белки Cas9 и белки систем CRISPR II типа подробно описаны в литературе. См. Makarova et al., Nature Reviews, Microbiology, Vol. 9, June 2011, pp. 467-477, включая всю дополнительную информацию, которая в полном объёме включена в виде ссылок в данный документ.
Функция типичных ДНК-связывающих белков, обладающих нуклеазной активностью, состоит во внесении одноцепочечного надреза (ника) или разрезании двухцепочечной ДНК. Указанная нуклеазная активность проявляется у ДНК-связывающего белка, имеющего одну или несколько полипептидных последовательностей, проявляющих нуклеазную активность. Типичные ДНК-связывающие белки могут иметь два отдельных нуклеазных домена, при этом каждый из доменов ответственен за разрезание или надрезание конкретной цепи двухцепочечной ДНК. Типичные полипептидные последовательности, обладающие нуклеазной активностью, известные специалистам в данной области техники, включают McrA-HNH-связанный нуклеазный домен и RuvC-подобный нуклеазный домен. Таким образом, типичные ДНК-связывающие белки – это белки, которые от природы содержат один или более из McrA-HNH-связанного нуклеазного домена и RuvC-подобного нуклеазного домена.
Примером ДНК-связывающего белка является РНК-направляемый ДНК-связывающий белок системы CRISPR II типа. Примером ДНК-связывающего белка является белок Cas9.
В S. pyogenes Cas9 создает двухцепочечный разрыв с тупыми концами на 3по слева (выше) от прилежащего к протоспейсеру мотива (PAM) в результате процесса, опосредованного двумя каталитическими доменами в белке: доменом HNH, который расщепляет комплементарную нить ДНК, и RuvC-подобным доменом, который расщепляет некомплементарную нить. См. Jinke et al., Science 337, 816-821 (2012); включён в полном объёме в настоящий документ в виде ссылки. Известно, что белки Cas9 присутствуют во многих CRISPR-системах II типа, включая нижеследующие как идентифицированные в дополнительной информации к Makarova et al., Nature Reviews, Microbiology, Vol. 9, June 2011, pp. 467-477: Methanococcus maripaludis C7; Corynebacterium diphtheriae; Corynebacterium efficiens YS-314; Corynebacterium glutamicum ATCC 13032 Kitasato; Corynebacterium glutamicum ATCC 13032 Bielefeld; Corynebacterium glutamicum R; Corynebacterium kroppenstedtii DSM 44385; Mycobacterium abscessus ATCC 19977; Nocardia farcinica IFM10152; Rhodococcus erythropolis PR4; Rhodococcus jostii RHAl; Rhodococcus opacus B4 uid36573; Acidothermus cellulolyticus 11B; Arthrobacter chlorophenolicus A6; Kribbella flavida DSM 17836 uid43465; Thermomonospora curvata DSM 43183; Bifidobacterium dentium Bdl; Bifidobacterium longum DJO10A; Slackia heliotrinireducens DSM 20476; Persephonella marina EX HI; Bacteroides fragilis NCTC 9434; Capnocytophaga ochracea DSM 7271; Flavobacterium psychrophilum JIP02 86; Akkermansia muciniphila ATCC BAA 835; Roseiflexus castenholzii DSM 13941; Roseiflexus RSI; Synechocystis PCC6803; Elusimicrobium minutum Peil91; некультивируемые бактерии Termite group 1, филотип Rs D17; Fibrobacter succinogenes S85; Bacillus cereus ATCC 10987; Listeria innocua; Lactobacillus casei; Lactobacillus rhamnosus GG; Lactobacillus salivarius UCC118; Streptococcus agalactiae A909; Streptococcus agalactiae NEM316; Streptococcus agalactiae 2603; Streptococcus dysgalactiae equisimilis GGS 124; Streptococcus equizooepidemicus MGCS10565; Streptococcus gallolyticus UCN34 uid46061; Streptococcus gordonii Challis ssp. CHI; Streptococcus mutans NN2025 uid46353; Streptococcus mutans; Streptococcus pyogenes Ml GAS; Streptococcus pyogenes MGAS5005; Streptococcus pyogenes MGAS2096; Streptococcus pyogenes MGAS9429; Streptococcus pyogenes MGAS 10270; Streptococcus pyogenes MGAS6180; Streptococcus pyogenes MGAS315; Streptococcus pyogenes SSI-1; Streptococcus pyogenes MGAS 10750; Streptococcus pyogenes NZ131; Streptococcus thermophilus CNRZ1066; Streptococcus thermophilus LMD-9; Streptococcus thermophilus LMG 18311; Clostridium botulinum A3 Loch Maree; Clostridium botulinum B Eklund 17B; Clostridium botulinum Ba4 657; Clostridium botulinum F Langeland; Clostridium cellulolyticum H10; Finegoldia magna ATCC 29328; Eubacterium rectale ATCC 33656; Mycoplasma gallisepticum; Mycoplasma mobile 163K; Mycoplasma penetrans; Mycoplasma synoviae 53; Streptobacillus moniliformis DSM 12112; Bradyrhizobium BTAil; Nitrobacter hamburgensis X14; Rhodopseudomonas palustris BisB 18; Rhodopseudomonas palustris BisB5; Parvibaculum lavamentivorans DS-1; Dinoroseobacter shibae DFL 12; Gluconacetobacter diazotrophicus Pal 5 FAPERJ; Gluconacetobacter diazotrophicus Pal 5 JGI; Azospirillum B510 uid46085; Rhodospirillum rubrum ATCC 11170; Diaphorobacter TPSY uid29975; Verminephrobacter eiseniae EF01-2; Neisseria meningitides 053442; Neisseria meningitides alphal4; Neisseria meningitides Z2491; Desulfovibrio salexigens DSM 2638; Campylobacter jejuni doylei 269 97; Campylobacter jejuni 81116; Campylobacter jejuni; Campylobacter lari RM2100; Helicobacter hepaticus; Wolinella succinogenes; Tolumonas auensis DSM 9187; Pseudoalteromonas atlantica T6c; Shewanella pealeana ATCC 700345; Legionella pneumophila Paris; Actinobacillus succinogenes 130Z; Pasteurella multocida; Francisella tularensis novicida U112; Francisella tularensis holarctica; Francisella tularensis FSC 198; Francisella tularensis tularensis; Francisella tularensis WY96-3418; и Treponema denticola ATCC 35405. Таким образом, аспекты настоящего изобретения направлены на белок Cas9, присутствующий в CRISPR-системе II типа.
В литературе белок Cas9 может упоминаться специалистами в данной области техники как Csnl. Последовательность белка Cas9 в S. pyogenes, которая является предметом описанных в настоящем документе экспериментов, приводится ниже. См. Deltcheva et al., Nature 471, 602-607 (2011); включён в полном объёме в настоящий документ в виде ссылки.

Согласно одному аспекту, РНК-направляемый ДНК-связывающий белок включает гомологи и ортологи Cas9, которые сохраняют способность белка к связыванию с ДНК, направляются РНК и разрезают ДНК. Согласно одному аспекту, белок Cas9 содержит приведенную выше последовательность встречающегося в природе Cas9 из S. pyogenes, или белковую последовательность, имеющую по меньшей мере 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95%, 98% или 99% гомологии с ней, которая является ДНК-связывающим белком, например, РНК-направляемым ДНК-связывающим белком.
Согласно одному аспекту, предлагается сконструированная система Cas9-гидРНК, которая обеспечивает при необходимости сайт-специфическое РНК-направляемое разрезание генома, и модификацию генома инсерцией экзогенных донорных нуклеиновых кислот. Направляющие РНК (гидРНК) комплементарны сайтам-мишеням или локусам-мишеням на ДНК. Направляющие РНК могут быть химерами crРНК-tracrРНК. Cas9 связывается на или вблизи геномной ДНК-мишени. Один или более направляющих РНК связываются на или вблизи геномной ДНК-мишени. Cas9 разрезает геномную ДНК-мишень, и на место разреза в ДНК вставляется экзогенная донорная ДНК.
Таким образом, способы направлены на использование направляющей РНК (гидРНК) с белком Cas9 и экзогенной донорной нуклеиновой кислотой для множественных инсерций экзогенных донорных нуклеиновых кислот в ДНК в клетке, экспрессирующей Cas9, путём цикличного введения нуклеиновой кислоты, кодирующей РНК, и экзогенной донорной нуклеиновой кислоты, экспрессии РНК; колокализации РНК, Cas9 и ДНК таким образом, чтобы инициировать разрезание ДНК; и инсерцию экзогенной донорной нуклеиновой кислоты. Стадии способа могут осуществляться в циклическом режиме с проведением требуемого количества циклов для достижения желательного числа модификаций ДНК. Соответственно способы настоящего изобретения направлены на редактирование генов-мишеней с использованием описанных в настоящем документе белков Cas9 и гидРНК для осуществления мультиплексной генетической и эпигенетической инженерии клеток.
Прочие аспекты настоящего изобретения направлены на использование ДНК-связывающих белков или систем в целом для мультиплексной вставки экзогенных донорных нуклеиновых кислот в ДНК, например, в геномную ДНК клетки, такой как человеческая клетка. Специалист в данной области техники сможет легко идентифицировать типичные ДНК-связывающие системы на основе настоящего изобретения.
Клетки согласно настоящему изобретению включают любую клетку, в которую могут вводиться и экспрессироваться, как описано выше, чужеродные нуклеиновые кислоты. Само собой разумеется, что описанные здесь основные концепции настоящего изобретения не ограничиваются видом клетки. Клетки по настоящему изобретению включают эукариотические клетки, прокариотические клетки, клетки животных, клетки растений, клетки грибов, клетки архей, клетки эубактерий и т.п. Клетки включают эукариотические клетки, такие как клетки дрожжей, клетки растений и клетки животных. Специфические клетки включают клетки млекопитающего, например, клетки человека. Помимо этого, клетки включают любые клетки, в которых целесообразно или желательно модифицировать ДНК.
Нуклеиновые кислоты-мишени включают любую нуклеинокислотную последовательность, которые может быть полезно надрезать или разрезать используя колокализационный комплекс, как описано выше. Нуклеиновые кислоты-мишени включают гены. Для целей настоящего изобретения ДНК, например, двухцепочечная ДНК может включать -нуклеиновую кислоту-мишень, а колокализационный комплекс может связываться с ней или же колокализоваться на ней иным образом в области нуклеиновой кислоты-мишени или либо вблизи нее, причём таким образом, чтобы колокализационный комплекс мог оказывать желательное воздействие на нуклеиновую кислоту-мишень. Такие нуклеиновые кислоты-мишени могут включать эндогенные (или встречающиеся в природе) нуклеиновые кислоты и экзогенные (или чужеродные) нуклеиновые кислоты. Специалист на основе настоящего изобретения сможет легко идентифицировать или создать направляющие РНК (гидРНК) и белки Cas9, которые колокализуются на ДНК, включающей нуклеиновую кислоту-мишень. Специалист сможет также идентифицировать регулирующие транскрипцию белки или домены, которые подобным образом колокализуются на ДНК, включающей нуклеиновую кислоту-мишень. ДНК включает геномную ДНК, митохондриальную ДНК, вирусную ДНК или экзогенную ДНК. Согласно одному аспекту, материалы и методы, применяемые при осуществлении настоящего изобретения на практике, включают те из них, какие описаны в Di Carlo, et al., Nucleic Acids Research, 2013, vol. 41, No. 7, p. 4336-4343; включён в настоящий документ в полном объёме в виде ссылки для всех целей, включая типичные штаммы и питательные среды, плазмидную конструкцию, трансформацию плазмид, электропорацию транзиентной гидРНК-кассеты и донорных нуклеиновых кислот, трансформацию гидРНК-плазмиды донорной ДНК в Cas9-экспрессирующих клетках, индукцию галактозой Cas9, идентификацию CRISPR-Cas мишеней в геноме дрожжей и др. Дополнительные ссылки, включающие информацию, материалы и методы, полезные для специалиста при реализации изобретения, см. в Mali, P, Yang, L., Esvelt, K.M., Aach, J., Guell, M., DiCarlo, J.E., Norville, J.E. и Church, G.M. (2013) RNA-Guided human genome engineering via Cas9 [РНК-направляемая инженерия генома человека с использованием Cas9]. Science, 10.1126fscience.l232033; Storici, F., Durham, C.L., Gordenin, D.A. и Resnick, M.A. (2003) Chromosomal site-specific double-strand breaks are efficiently targeted for repair by oligonucleotides in yeast [Хромосомные сайт-специфические двухцепочечные разрывы могут эффективно служить мишенью для репарации олигонуклеотидами в дрожжах]. PNAS, 100, 14994-14999; Jinek, M., Chylinski, K., Fonfara, I., Hauer, M., Doudna, J.A. и Charpentier, E. (2012) A programmable dual-RNA-Guided DNA endonuclease in adaptive bacterial immunity [Программируемая двойная-РНК-направляемая эндонуклеаза ДНК в адаптивном иммунитете бактерий]. Science, 337, 816-821, каждый из который включён в полном объёме в настоящую заявку в виде ссылки для всех целей.
Чужеродные нуклеиновые кислоты (т.е. те, которые не являются частью природного нуклеинокислотного состава клетки) могут вводиться в клетку любым пригодным для такого введения методом, известным специалистам в данной области техники. Такие методы включают трансфекцию, трансдукцию, вирусную трансдукцию, микроинъекцию, липофекцию, нуклеофекцию, бомбардировку наночастицами, трансформацию, конъюгацию и т.п. Специалист в данной области техники легко поймёт и адаптирует при необходимости такие методы, используя доступные литературные источники.
Нижеследующие примеры приводятся в качестве репрезентативных для настоящего изобретения. Эти примеры не имеют целью ограничить объём настоящего изобретения, поэтому приведенные и другие эквивалентные варианты осуществления изобретения будут очевидны из описания настоящего изобретения, фигур и приложенной формулы изобретения.
Пример I
Общий способ мультиплексированного редактирования генов в дрожжах с использованием CRISPR-Cas9
Cas9 из иммунной системы CRISPR бактерий Streptococcus pyogenes используется для стимуляции гомологичной рекомбинации и отбора против клеток, которые не рекомбинируют трансформированную ДНК в Saccharomyces cerevisiae. Общий способ РНК-направляемого расщепления ДНК с использованием Cas9 представлен на фиг.1. Между Cas9, гидРНКом и ДНК-мишенью образуется колокализационный комплекс. Двухцепочечный разрыв в ДНК-мишень вносит Cas9. Затем донорная ДНК вставляется в ДНК путём гомологичной рекомбинации. Донорная ДНК содержит фланкирующие последовательности с обеих сторон от места разреза и последовательность, которая удаляет сайт расщепления ферментом Cas9. Результатом этого является интеграция (включение) донорной ДНК в ДНК, которая может быть геномной ДНК.
Предлагается следующий общий способ высокочастотной рекомбинации донорной ДНК с помощью мультиплексированной инженерии ДНК в дрожжах с использованием Cas9 (см. фиг. 2). Клетки, не имеющие от природы РНК-направляемой эндонуклеазы Cas9, можно трансформировать ДНК таким образом, чтобы клетка могла экспрессировать РНК-направляемую эндонуклеазу Cas9. Выращиваются клетки, которые экспрессируют РНК-направляемую эндонуклеазу Cas9. Конструируется плазмида, включающая одну или несколько нуклеиновых кислот, кодирующих одну или несколько гидРНК, и селективный маркёр, известный специалистам в данной области техники, для введения в клетку и экспрессии одной или нескольких гидРНК. На фиг. 2 показан пул плазмид (каждая с нуклеиновой кислотой, кодирующей гидРНК), используемых для различных генов, вставляемых в геномную ДНК клетки, т.е. для гена A, гена B, гена C, гена D и гена E. Создаётся также пул донорных ДНК, включающих двухцепочечные донорные ДНК для гена A, гена B, гена C, гена D и гена E.
Клетки промываются и кондиционируются с ацетатом лития. Клетки могут также промываться и смешиваться с пулом экзогенных донорных нуклеиновых кислот, таких как двухцепочечные олигонуклеотиды, например, ДНК-кассета, и плазмидами, включающими нуклеиновые кислоты, кодирующие гидРНК. Как показано на фиг. 2, клетки трансформируются экзогенными донорными нуклеиновыми кислотами и плазмидами с использованием PEG 3350 и ацетата лития.
Как показано на фиг. 2, клетки отбираются по наличию одной или более гидРНК с помощью селективного маркёра. Отобранные клетки экспрессируют одну или несколько гидРНК. Между гидРНК, РНК-направляемой эндонуклеазой Cas9 и ДНК в клетке образуются один или несколько колокализационных комплексов. Эндонуклеаза разрезает ДНК и донорная нуклеиновая кислота вставляется в клетку путём рекомбинации, например, гомологичной рекомбинации. Затем клетки избавляют от плазмиды и впоследствии необязательно подвергаются одному или дополнительным циклам вышеописанных стадий. Может проводиться множество циклов. Клетка, подвергнутая множеству циклов, проявляет высокую частоту рекомбинации. Альтернативно, клетки деселектируются для сохранения плазмиды или же помещаются в питательные среды с последующим отбором против клеток с плазмидой. Затем процесс повторяется, начиная со стадии выращивания клеток. Таким образом, способы включают цикл стадий с клетками, уже модифицированными на предыдущем цикле, или отбор клеток из предыдущего цикла, которые не были модифицированы, и последующий цикл стадий с немодифицированными клетками для осуществления модификации ДНК, как описано в настоящем документе.
Пример II
Подробный протокол циклов
Клетки выращиваются (ауксотрофы по урацилу, с постоянной экспрессией Cas9) до оптической плотности от 0,8 до 1,0 в 5 мл дрожжевых сред SC или SC + FOA (фтороротовая кислота) (100 мкг/мл). Клетки осаждаются при 2250g в течение 3 минут и промываются один раз 10 мл воды; затем клетки высушиваются на солнце и ресуспендируются в 1 мл 100 мМ раствора ацетата лития. Осажденные клетки ресуспендируются в 500 мкл 100 мМ раствора ацетата лития. Создаётся трансформационная смесь путём добавления в следующем порядке (на 50 мкл клеток): смеси ДНК, включающей 1 нмоль пула двухцепочечных олигонуклеотидов и по 5 мкг каждой из гидРНК (вектор p426 с урациловым маркёром) с последующим добавлением воды до 70 мкл для достижения требуемого конечного объёма; 240 мкл 50%-го раствора PEG 3350 и 36 мкл 1 M раствора ацетата лития. Смесь инкубируется при 30°C в течение 30 минут. Затем смесь перемешивается на вортексе, и клетки подвергаются тепловому шоку путём инкубирования смеси при 42°C в течение 20 минут. После этого клетки осаждаются, супернатант отбрасывается. Клетки инокулируют в 5 мл среды SC-урацил с последующим отбором клеток, содержащих гидРНК-плазмиду, по генам урацила,. Клеткам дают возможность восстановиться в течение 2 суток. Спустя двое суток проводят инокуляцию 100 мкл клеточной культуры в 5 мл свежеприготовленной среды SC и оставляют для роста на 12 часов с последующей деселекцией для сохранения плазмиды. Затем проводят инокуляцию 100 мкл клеточных культур в 5 мл питательных сред SC и SC + FOA (100мкг/мл) для отбора против клеток с плазмидой. Этим завершается один цикл процесса. Процесс повторяется для любого числа желательных циклов. Общий процесс может включать 1 цикл, 2 цикла, 3 цикла, 4 цикла, 5 циклов, 6 циклов, 7 циклов, 8 циклов, 9 циклов, 10 циклов, 15 циклов, 20 циклов, 25 циклов и т.д.
Пример III
Термотолерантность к тепловому шоку при отборе мутантов
С помощью способов, описанных в настоящем документе, была показана термотолерантность к тепловому шоку в отборе мутантов. Гены, которые, как известно, увеличивают термотолерантность в дрожжах при их нокауте или точечной мутации, служили мишенью для описанной выше системы направляющая РНК-Cas9. Для мутации были выбраны четыре гена: UBC1, SCH9, TFS1 и RAS2. SCH9 – это протеинкиназа, которая регулирует гены осмостресса, стресса, вызываемого нарушением питания, и стресса, вызываемого изменением внешней среды. TFS1 ингибирует карбоксипептидазы Y и Ira2p, подавляет GAP-активность Ras и реагирует на репликативный стресс ДНК. RAS2 является GTP-связывающим белком, который регулирует азотное голодание и вовлекается в проводящие пути стресс-ответа. Для каждого из SCH9, TFS 1 и RAS2 была создана донорная ДНК, которая представляет собой аллель, содержащий мутацию серин → аланин в кодирующей области. UBC1-E2 – это убиквитин-конъюгирующий фермент. Была создана донорная ДНК, включающая точечную мутацию, которая удаляет сайт фосфорилирования, что приводит к возникновению термотолерантности.
С помощью способов, описанных в настоящем документе, на гены нацелили направляющую РНК (гидРНК), сконструированную таким образом, чтобы она могла направить Cas9- расщепление в локусах генов, вместе с двухцепочечным олигонуклеотидом для внесения изменений. Как показано на фиг. 3, замена аллелей была достигнута с помощью олигонуклеотидов, нацеленных на четыре локуса, ответственных за термотолерантность в дрожжах. Согласно схеме, были созданы четыре плазмиды, каждая из которых содержала нуклеиновую кислоту, кодирующую направляющую РНК, для одного из генов: UBC1 gРНК плазмида, TFS1 gРНК плазмида, SCH9 gРНК плазмида и RAS2 gРНК плазмида. Каждая плазмида имела соответствующий двухцепочечный донорный олигонуклеотид: двухцепочечный олигонуклеотид ubcl (S97A), двухцепочечный олигонуклеотид tfsl (tag), двухцепочечный олигонуклеотид sch9 (tag) и двухцепочечный олигонуклеотид ras (tag). Плазмиды и соответствующие двухцепочечные донорные олигонуклеотиды котрансформировались в дрожжах в виде пула. Были проведены два цикла; и число модификаций на клетку как функция процентного содержания клеток в клеточной популяции показано на фиг. 4. Значительное число клеток после цикла 2 включало одну или две модификации. Удалось выделить один тройной мутант (данные не показаны).
На фиг. 5A приведена таблица штаммов, полученных описанными в настоящем документе способами, в том числе штаммов, трансформированных одним донорным олигонуклеотидом, штаммов, трансформированных двумя донорными олигонуклеотидами, и штамма, трансформированного тремя донорными олигонуклеотидами. На фиг. 5B показано влияние инкубации при 42°C в течение трёх часов по сравнению с вариантом без инкубации и незначительное сокращение числа клеток дикого типа. На фиг. 5B также показано влияние инкубации при 55°C в течение двух часов по сравнению с вариантом без инкубации. Мутантами, наиболее толерантными к тепловому шоку при 55°C, оказались sch9, sch9 tfsl и tfsl ubcl(s97a).
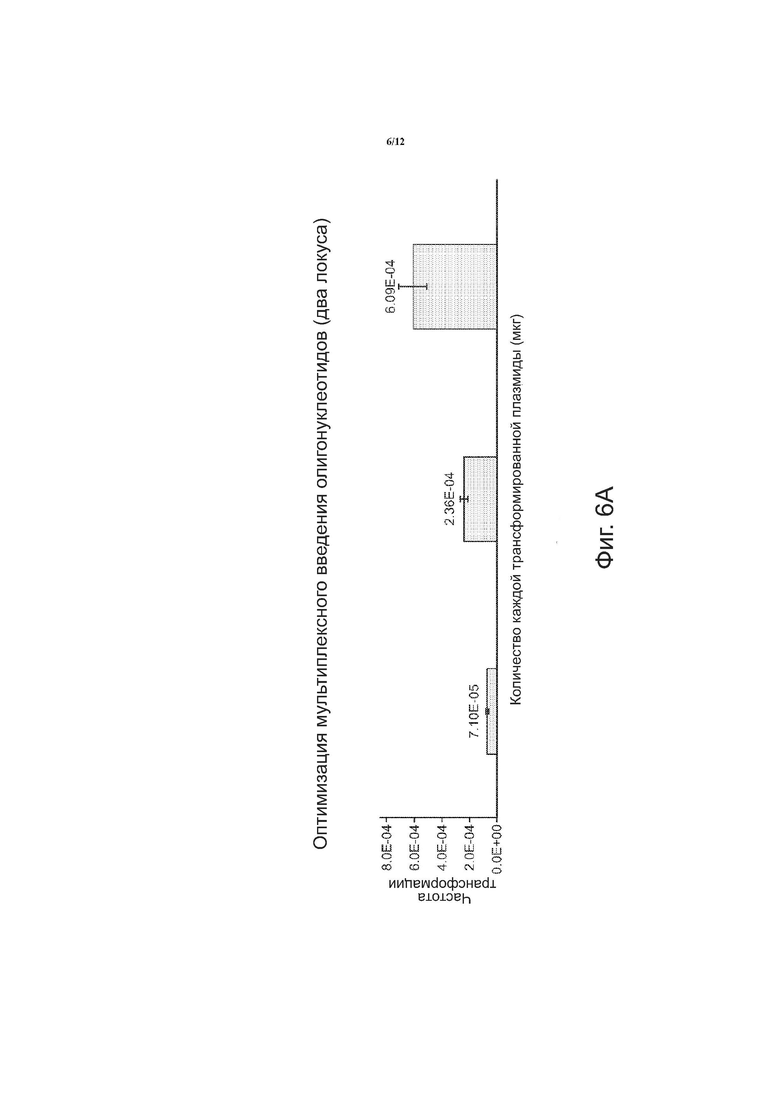
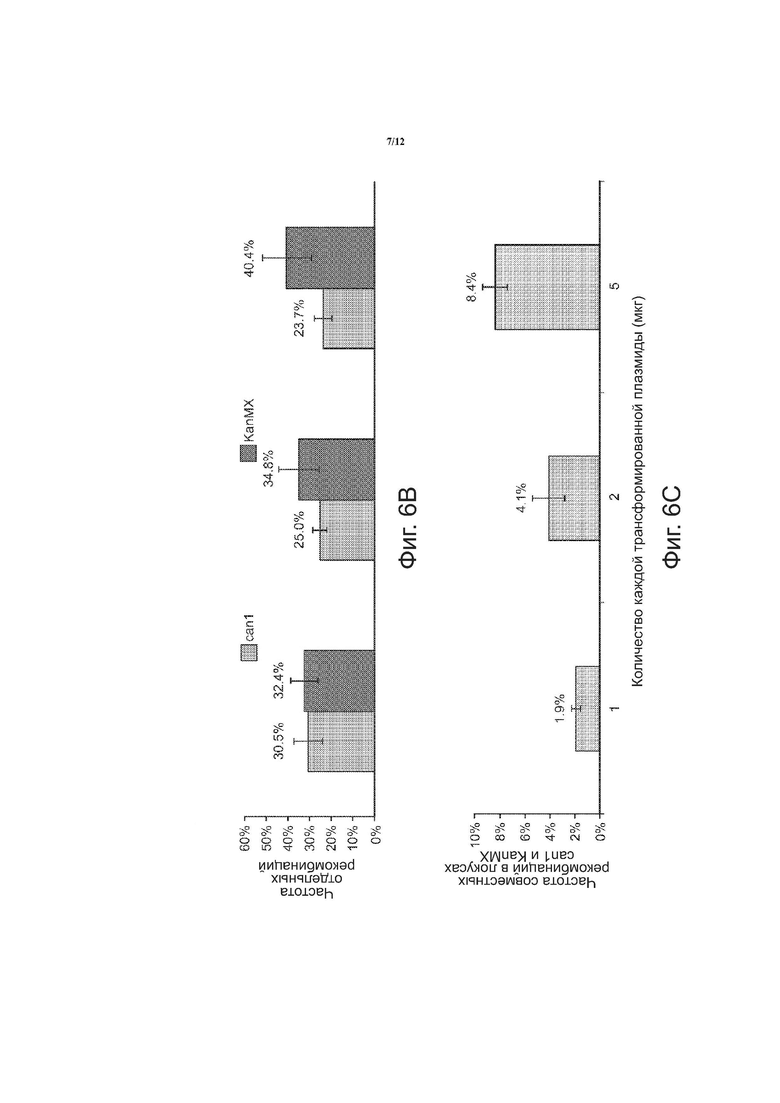
На фиг. 6 в целом приведена графическая информация по оптимизации мультиплексного введения олигонуклеотидов в два локуса. На фиг. 6A показана зависимость частоты трансформации от количества каждой трансформированной плазмиды (мкг). На фиг. 6B показана зависимость частоты отдельных рекомбинаций от количества каждой трансформированной плазмиды (мкг). На фиг. 6C показана зависимость частоты совместной рекомбинации в локусах canl и KanMX от количества каждой трансформированной плазмиды (мкг).
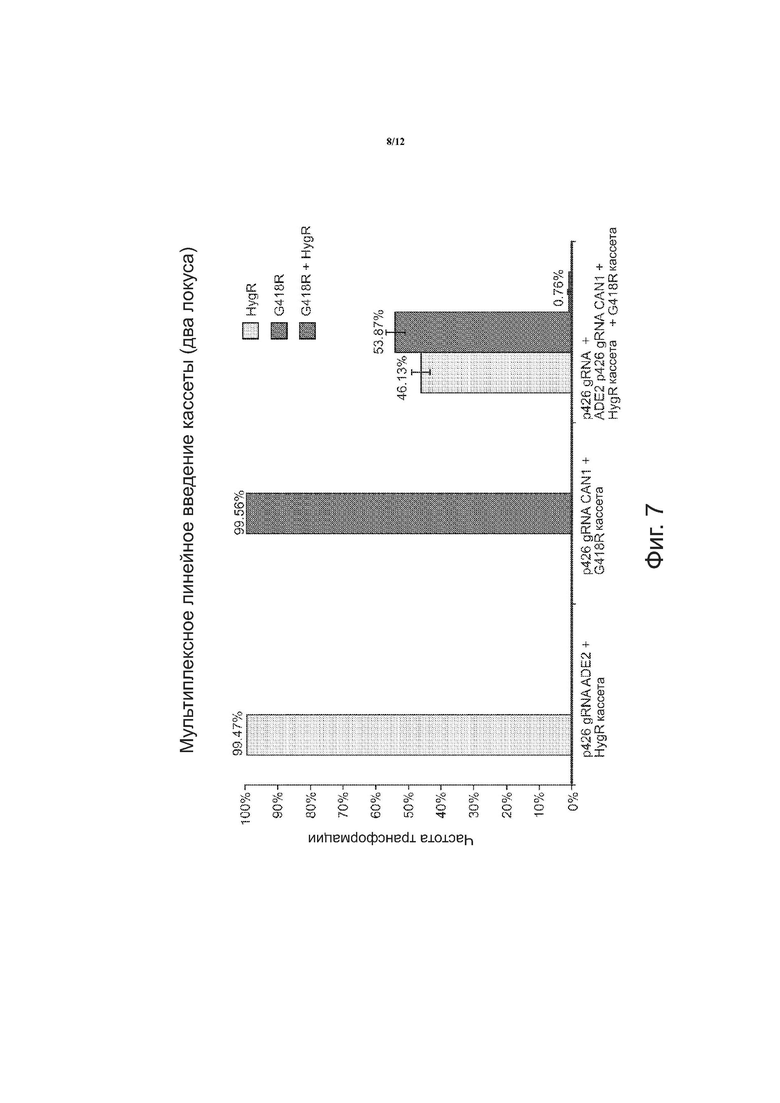
На фиг. 7 в целом приведена графическая информация по мультиплексному линейному введению кассеты в два локуса. Первый слева большой столбец показывает частоту трансформации для p426 gРНК ADE2 + HygR кассеты; следующий столбец – частоту трансформации для p426 gРНК CAN1 + G418R кассеты; следующие три столбца – частоту трансформации для p426 gРНК + ADE2 p426 gРНК CAN1 + HygR кассеты + G418R кассета.
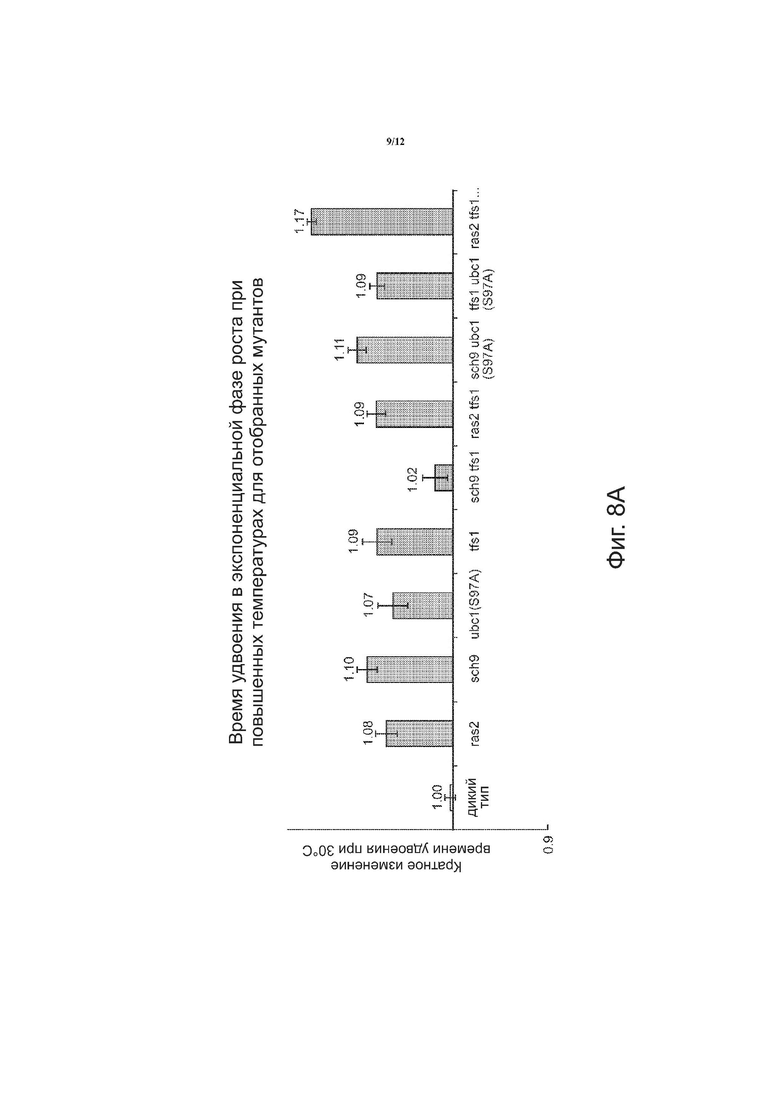
На фиг. 8 в целом приведены результаты анализа скорости роста, показывающие время удвоения в экспоненциальной фазе роста при повышенных температурах для отбора мутантов. На фиг. 8A приведена диаграмма кратного изменения времени удвоения при 30°C для клеток дикого типа и идентифицированных мутантов. На фиг. 8B приведена диаграмма кратного изменения времени удвоения при 37°C для клеток дикого типа и идентифицированных мутантов. На фиг. 8C приведена диаграмма кратного изменения времени удвоения при 42°C для дикого типа и идентифицированных мутантов при инокуляции клеток из культуры в поздней стационарной фазе роста. На фиг. 8D приведена диаграмма кратного изменения времени удвоения при 42°C для клеток дикого типа и идентифицированных мутантов при инокуляции клеток из культуры в log фазе роста. Графические данные показывают меньшее время удвоения при 37°C для sch9 tfsl и tfsl ubcl(S97A). Графические данные показывают меньшее время удвоения при 42°C для ras2 tfsl, sch9 ubcl(S97A), tfsl ubcl(S97A) и ras2 tfsl ubcl(S97A).
| название | год | авторы | номер документа |
|---|---|---|---|
| МУЛЬТИПЛЕКСНАЯ ГЕНОМНАЯ ИНЖЕНЕРИЯ, НАПРАВЛЯЕМАЯ РНК | 2014 |
|
RU2706562C2 |
| НАПРАВЛЯЕМАЯ РНК РЕГУЛЯЦИЯ ТРАНСКРИПЦИИ | 2014 |
|
RU2690935C2 |
| ГЕНОМНАЯ ИНЖЕНЕРИЯ | 2014 |
|
RU2688462C2 |
| НАПРАВЛЯЕМАЯ РНК РЕГУЛЯЦИЯ ТРАНСКРИПЦИИ | 2014 |
|
RU2756865C2 |
| ОРТОГОНАЛЬНЫЕ БЕЛКИ Cas9 ДЛЯ РНК-НАПРАВЛЯЕМОЙ РЕГУЛЯЦИИ И РЕДАКТИРОВАНИЯ ГЕНОВ | 2014 |
|
RU2748433C2 |
| ОРТОГОНАЛЬНЫЕ БЕЛКИ CAS9 ДЛЯ РНК-НАПРАВЛЯЕМОЙ РЕГУЛЯЦИИ И РЕДАКТИРОВАНИЯ ГЕНОВ | 2014 |
|
RU2704981C2 |
| ОРТОГОНАЛЬНЫЕ БЕЛКИ CAS9 ДЛЯ РНК-НАПРАВЛЯЕМОЙ РЕГУЛЯЦИИ И РЕДАКТИРОВАНИЯ ГЕНОВ | 2021 |
|
RU2771583C1 |
| ГЕНОМНАЯ ИНЖЕНЕРИЯ | 2021 |
|
RU2812848C2 |
| РНК-НАПРАВЛЯЕМАЯ ИНЖЕНЕРИЯ ГЕНОМА ЧЕЛОВЕКА | 2013 |
|
RU2699523C2 |
| ГЕНОМНАЯ ИНЖЕНЕРИЯ | 2014 |
|
RU2764757C2 |



Изобретение относится к области биотехнологии, в частности к эукариотической клетке, постоянно экспрессирующей направляемый РНК ДНК-связывающий белок системы CRISPR типа II, который образует комплекс колокализации с направляющей РНК, комплементарной ДНК-мишени, и которая расщепляет ДНК-мишень сайт-специфическим образом. Указанная клетка включает множество направляющих РНК, комплементарных различным сайтам ДНК, где направляющая РНК представляет собой слияние tracrРНК-crРНК, и множество последовательностей экзогенных донорных нуклеиновых кислот. Изобретение эффективно для введения множества модификаций в целевую ДНК. 10 з.п. ф-лы, 8 ил., 3 пр.
1. Эукариотическая клетка для мультиплексной геномной инженерии, постоянно экспрессирующая направляемый РНК ДНК-связывающий белок системы CRISPR типа II, который образует комплекс колокализации с направляющей РНК, комплементарной ДНК-мишени, и которая расщепляет ДНК-мишень сайт-специфическим образом, включающая
(а) множество направляющих РНК, комплементарных различным сайтам ДНК, где направляющая РНК представляет собой слияние tracrРНК-crРНК, и
(б) множество последовательностей экзогенных донорных нуклеиновых кислот.
2. Эукариотическая клетка по п.1, отличающаяся тем, что содержит несколько вставок экзогенных донорных нуклеиновых кислот в указанную ДНК.
3. Эукариотическая клетка по п.1, в которой фермент представляет собой Cas9.
4. Эукариотическая клетка по п.1, отличающаяся тем, что представляет собой дрожжевую клетку, растительную клетку или клетку животного.
5. Эукариотическая клетка по п.1, в которой направляющая РНК содержит от около 10 до около 500 нуклеотидов.
6. Эукариотическая клетка по п.1, в которой направляющая РНК содержит от около 20 до около 100 нуклеотидов.
7. Эукариотическая клетка по п.1, в которой указанная ДНК представляет собой геномную ДНК, митохондриальную ДНК, вирусную ДНК или экзогенную ДНК.
8. Эукариотическая клетка по п.1, в которой направляющая РНК присутствует в плазмиде.
9. Эукариотическая клетка по п.1, в которой направляющая РНК и последовательность экзогенной нуклеиновой кислоты присутствуют в плазмиде.
10. Эукариотическая клетка по п.1, в которой последовательность экзогенной нуклеиновой кислоты включает гомологические последовательности или плечо, фланкирующее участок разреза.
11. Эукариотическая клетка по п.1, в которой последовательность экзогенной нуклеиновой кислоты включает последовательность для удаления сайта разреза.
| JIANG, WENYAN, et al | |||
| RNA-guided editing of bacterial genomes using CRISPR-Cas systems, Nature biotechnology, 2013 (published online 29 January 2013), 31(3): 233-239 | |||
| DICARLO J.E | |||
| et al | |||
| Genome engineering in Saccharomyces cerevisiae using CRISPR-Cas systems, Nucleic Acids Res., 2013 (Published online 4 March 2013), 41(7): 4336-4343 | |||
| MALI P | |||
| et |

