ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
По настоящей заявке испрашивается приоритет предварительной заявки США № 62/482368, поданной 6 апреля 2017 года под названием «МОЛЕКУЛЯРНЫЕ МАРКЕРЫ, АССОЦИИРОВАННЫЕ С УСТОЙЧИВОСТЬЮ ПОДСОЛНЕЧНИКА К OROBANCHE», содержание которой в полном объеме включено в качестве ссылки.
ССЫЛКА НА СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ, ПОДАННЫХ В ЭЛЕКТРОННОМ ВИДЕ
Официальная копия списка последовательностей подана в электронном виде посредством EFS-Web в виде списка последовательностей в формате ASCII с файлом, который называется «74816-US-PSP-20170406_ST25», создан 08/13/2018, имеет размер 7,41 килобайт, и был подан одновременно с описанием. Список последовательностей, который содержится в этом документе в формате ASCII, является частью описания и включен в настоящий документ в качестве ссылки в полном объеме.
ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к способам, полезным при отборе растений подсолнечника по повышенной устойчивости к Orobanche.
ПРЕДШЕСТВУЮЩИЙ ИЗОБРЕТЕНИЮ УРОВЕНЬ ТЕХНИКИ
Orobanche (или заразиха) представляет собой род из более чем 200 видов паразитических травянистых растений в семействе Orobanchaceae, в основном, родом из умеренного Северного полушария. Orobanche cumana (заразиха подсолнечная) является облигатным нефотосинтезирующим корневым паразитическим растением на растении подсолнечника (Helianthus annuus L.). Заразиха является маленьким растением, от 10 до 60 см в высоту в зависимости от вида. Лучше всего она распознается по стеблям от желтого до соломенного цветов, полностью лишенным хлорофилла, с белыми, желтыми или голубоватто-желтыми цветами, похожими на львиный зев. Цветочные побеги чешуйчатые, с плотным концевым шипом, несущим от десяти до двадцати цветов у большинства видов, хотя у O. uniflora они одиночные. Листья представляют собой просто треугольные чешуйки. Семена мелкие, коричневато-коричневые и чернеют с возрастом. Эти растения в основном цветут с конца зимы до поздней весны. Когда они не цветут, ни одна часть растений не видна над поверхностью почвы.
Поскольку у них нет хлорофилла, они полностью зависят от других растений относительно питательных веществ. Семена заразихи остаются в состоянии покоя в почве, часто в течение многих лет, пока их не стимулируют к прорастанию определенные соединения, которые производятся корнями живого растения. Рассада заразихи выбрасывает корнеобразный росток, который прикрепляется к корням ближайших хозяев. После прикрепления к хозяину, заразиха лишает хозяина воды и питательных веществ.
Каково ее экономическое влияние? Этот паразитический сорняк является серьезной угрозой в Европе, особенно в странах вокруг Черного моря и в Испании (Molinero-Ruiz et al, 2013). В России воздействие О. cumana связано с выращиванием H. annuus (подсолнечника) и задокументировано еще в первые годы двадцатого века. В последнее время проблема О. cumana на подсолнечнике возросла в Турции, Болгарии, Испании, Греции, Румынии, Венгрии, Израиле и Сербии (Parker and Riches, 1993; ter Borg, 1994; Garcia-Torres et al., 1995). Во всех этих странах серьезные потери происходят на циклической основе, поскольку О. cumana развила новую вирулентность, преодолевая любое сопротивление, которое растениеводы-селекционеры сумели внедрить в культуру. Зараженные растения имеют меньший диаметр головки, и можно ожидать, что восприимчивые сорта потеряют по меньшей мере до 50% своей урожайности, и были зарегистрированы потери в 100%. Уровни из 4, 6, 8 и 25 O. cumana на растение-хозяина могут привести к потерям 20, 52, 82 и 90% соответственно (Shalom et al., 1988). Шиндрова и соавт. (1998) отметили, что пораженные подсолнухи были короче, с меньшим диаметром головки и меньшей урожайностью на головку.
Генетическая устойчивость к O. cumana является, как правило, качественной или вертикальной, и, таким образом, популяции O. cumana обычно делятся на расы (Vranceanu et al, 1980). Эти расы периодически преодолевают все доступные источники устойчивости. До настоящего времени были описаны восемь рас, A-H, и очень часто сообщают о расах F, G и H (Kaya, 2014). В качестве возможных причин быстрого появления новых рас O. cumana были названы генетическое разнообразие, мутации и селекция в определенных совокупностях генов и поток генов между популяциями дикой и сорной O. cumana (Pineda-Martos et al 2013). Несмотря на то, что использование гербицидов, таких как имидазолинон, имеет некоторую степень эффективности, а растворы для управления урожаем, в том числе соляризация почвы, могут помочь при заражениях, селекция по генетической устойчивости остается наиболее эффективным способом борьбы с заражением Orobanche.
Настоящее изобретение дополнительно относится к способам борьбы с сорняками или нежелательной растительностью в непосредственной близости с растением подсолнечника по изобретению. Один способ включает нанесение эффективного количества AHAS-ингибирующего гербицида, в частности, гербицида имидазолинона или сульфонилкарбамида, на сорняки и растение подсолнечника. Другой способ включает контакт семян подсолнечника по настоящему изобретению до посева и/или после предварительного проращивания с эффективным количеством AHAS-ингибирующего гербицида, в частности гербицида имидазолинона или сульфонилкарбамида. Настоящее изобретение дополнительно относится к семенам подсолнечника по настоящему изобретению, обработанным эффективным количеством AHAS-ингибирующего гербицида. Растения и семена подсолнечника для применения в этих способах содержат в своих геномах первый аллель AHASL1 и второй аллель AHASL1. Первый аллель AHASL1 кодирует белок подсолнечника AHASL1, содержащий аминокислотную замену A122T. Второй аллель AHASL1 кодирует белок подсолнечника AHASL1, содержащий аминокислотную замену A205V или аминокислотную замену P197L.
Настоящее изобретение дополнительно относится к способам борьбы с сорняками-паразитами Orobanche cumana и Orobanche cernua, также известными как заразиха, на зараженных растениях подсолнечника. Способ включает нанесение эффективного количества гербицида имидазолинон на сорняки и на устойчивое к гербициду растение подсолнечника по настоящему изобретению, в частности, растение подсолнечника, содержащее два аллеля A122T или растение подсолнечника, содержащее один аллель A122T AHASL1 и один аллель A205V AHASL1.
Селекция для придания подсолнечнику устойчивости к Orobanche является основным и наиболее эффективным способом для борьбы с потерей урожая, связанной с заражением Orobanche. Разработка молекулярных генетических маркеров облегчило картирование и отбор агрономически важных признаков у подсолнечников, и были идентифицированы локусы количественных признаков (QTL) для устойчивости к Orobanche. QTL, придающие устойчивость к Orobanche, ранее были идентифицированы на LG4, 6, 9, 10, 11, 13, 15, 16 и 17 (Louarn et al, 2016). Однако, из-за высокой скорости, с которой Orobanche преодолевает устойчивость, крайне необходимы дополнительные источники устойчивости.
Интрогрессия QTL посредством использования молекулярных маркеров, связанных с устойчивостью к Orobanche, увеличит скорость и точность перемещения устойчивости к Orobanche в элитные гибриды подсолнечника, таким образом, улучшая уровень устойчивости в субтропической зародышевой плазме. Включение устойчивости к Orobanche в элитную зародышевую плазму подсолнечника может предотвратить распространение вирусного заболевания капусты в неэндемичные регионы.
Несмотря на то, что информация о QTL для устойчивости к Orobanche доступна в данной области, немногие родословные могут быть классифицированы как высокоустойчивые, и имеется мало доказательств какой-либо сильной устойчивости к устойчивости к Orobanche у коммерчески доступных гибридов. Существует необходимость в коммерчески приемлемых гибридах, устойчивых к Orobanche, и в способе для разработки и отслеживания устойчивых инбридов и гибридов подсолнечника при помощи скрещивания с использованием маркера.
В настоящем документе предлагается способ картирования QTL устойчивости к Orobanche в популяции DH с использованием подхода двухродительского картирования QTL. Настоящее изобретение позволяет проводить отбор потомства, которое содержит геномную основу агрономически желаемого родителя и геномный признак устойчивости к Orobanche устойчивого родителя-донора. Настоящее изобретение также позволяет отслеживать QTL устойчивости к Orobanche для того чтобы интрогрессировать признак устойчивости к Orobanche в новые растения путем обычного скрещивания.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Одним из вариантов осуществления настоящего изобретения является способ отбора растения с повышенной устойчивостью к Orobanche. Способ включает этапы: a) детекции по меньшей мере одной маркерной нуклеиновой кислоты; и, b) отбора растения, содержащего маркерную нуклеиновую кислоту, и, таким образом, отбора растения с повышенной устойчивостью к Orobanche. Растение предпочтительно представляет собой растение подсолнечника.
В вариантах осуществления изобретения, маркерная нуклеиновая кислота выбрана из группы, состоящей из DHAI000240, DHAG000732, DHAI007796 и DHAI007334. В дополнительных вариантах осуществления изобретения выбрана по меньшей мере одна маркерная нуклеиновая кислота, и предпочтительно, выбраны по меньшей мере две маркерные нуклеиновые кислоты.
В другом варианте осуществления изобретения предлагается способ отбора растения подсолнечника с повышенной устойчивостью к Orobanche, способ, включающий: a) детекцию по меньшей мере одной маркерной нуклеиновой кислоты, где по меньшей мере одна маркерная нуклеиновая кислота выбрана из группы, состоящей из DHAI000240, DHAG000732, DHAI007796 и DHAI007334; и, b) отбор растения, содержащего по меньшей мере одну маркерную нуклеиновую кислоту, и, таким образом, отбор растения подсолнечника с повышенной устойчивостью к Orobanche. Растения подсолнечника, полученные при помощи способов, описываемых в настоящем документе, также включены в настоящее изобретение.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ И СПИСКОВ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
Изобретение можно понять более полно из последующего подробного описания и прилагаемых чертежей и списка последовательностей, которые являются частью заявки. Список последовательностей содержит однобуквенный код для символов нуклеотидной последовательности и трехбуквенные коды для аминокислот, как определено в соответствии со стандартами IUPAC-IUBMB, описанными в Nucleic Acids Research 13:3021-3030 (1985) и в Biochemical Journal 219 (No. 2): 345-373 (1984), которые включены в настоящий документ в качестве ссылки в полном объеме. Символы и формат, использованные для данных с нуклеотидными и аминокислотными последовательностями, удовлетворяют правилам, изложенным в 37 C.F.R. §1822.
На фигуре 1 представлен QTL, устойчивый к Orobanche, OrDeb-2, картированный в группе сцепления 4.
На фигуре 2 представлен пиковый LOD-балл и процент изменчивости, объясненной за счет QTL в группе сцепления 4.
SEQ ID NO:1 представляет собой последовательность, из которой был разработан анализ DHAI000240 KASP™.
SEQ ID NO:2 представляет собой праймер для аллеля 1 для анализа DHAI000240 KASP™.
SEQ ID NO:3 представляет собой праймер для аллеля 2 для анализа DHAI000240 KASP™.
SEQ ID NO:4 представляет собой общий обратный праймер для анализа DHAI000240 KASP™.
SEQ ID NO:5 представляет собой последовательность, из которой был разработан анализ DHAI009612 KASP™.
SEQ ID NO:6 представляет собой праймер для аллеля 1 для анализа DHAI009612 KASP™.
SEQ ID NO:7 представляет собой праймер для аллеля 2 для анализа DHAI009612 KASP™.
SEQ ID NO:8 представляет собой общий обратный праймер для анализа DHAI009612 KASP™.
SEQ ID NO:9 представляет собой последовательность, из которой был разработан анализ DHAI000732 KASP™.
SEQ ID NO:10 представляет собой праймер для аллеля 1 для анализа DHAG000732 KASP™.
SEQ ID NO:11 представляет собой праймер для аллеля 2 для анализа DHAG000732 KASP™.
SEQ ID NO:12 представляет собой общий обратный праймер для анализа DHAG000732 KASP™.
SEQ ID NO:13 представляет собой последовательность, из которой был разработан анализ DHAI009613 KASP™.
SEQ ID NO:14 представляет собой праймер для аллеля 1 для анализа DHAI009613 KASP™.
SEQ ID NO:15 представляет собой праймер для аллеля 2 для анализа DHAI009613 KASP™.
SEQ ID NO:16 представляет собой общий обратный праймер для анализа DHAI009613 KASP™.
SEQ ID NO:17 представляет собой последовательность, из которой был разработан анализ DHAI007334 KASP™.
SEQ ID NO:18 представляет собой праймер для аллеля 1 для анализа DHAI007334 KASP™.
SEQ ID NO:19 представляет собой праймер для аллеля 2 для анализа DHAI007334 KASP™.
SEQ ID NO:20 представляет собой общий обратный праймер для анализа DHAI007334 KASP™.
SEQ ID NO:21 представляет собой последовательность, из которой был разработан анализ DHAI000243 KASP™.
SEQ ID NO:22 представляет собой праймер для аллеля 1 для анализа DHAI000243 KASP™.
SEQ ID NO:23 представляет собой праймер для аллеля 2 для анализа DHAI000243 KASP™.
SEQ ID NO:24 представляет собой общий обратный праймер для анализа DHAI000243 KASP™.
SEQ ID NO:25 представляет собой последовательность, из которой был разработан анализ DHAI009614 KASP™.
SEQ ID NO:26 представляет собой праймер для аллеля 1 для анализа DHAI009614 KASP™.
SEQ ID NO:27 представляет собой праймер для аллеля 2 для анализа DHAI009614 KASP™.
SEQ ID NO:28 представляет собой общий обратный праймер для анализа DHAI009614 KASP™.
SEQ ID NO:29 представляет собой последовательность, из которой был разработан анализ DHAI007796 KASP™.
SEQ ID NO:30 представляет собой праймер для аллеля 1 для анализа DHAI007796 KASP™.
SEQ ID NO:31 представляет собой праймер для аллеля 2 для анализа DHAI007796 KASP™.
SEQ ID NO:32 представляет собой общий обратный праймер для анализа DHAI007796 KASP™.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к способам выявления и отбора растений подсолнечника с повышенной устойчивостью к Orobanche. Последующие определения предоставлены с целью понимания изобретения.
Термин «аллель» относится к одной из двух или более различных нуклеотидных последовательностей, которые встречаются в специфическом локусе.
«Ампликон» представляет собой амплифицированную нуклеиновую кислоту, например, нуклеиновую кислоту, которую получают путем амплификации матричной нуклеиновой кислоты любым доступным способом амплификации (например, ПЦР, ЛЦР, транскрипция или т.п.).
Термин «амплификация» в контексте амплификации нуклеиновой кислоты представляет собой любой процесс, в котором производят дополнительные копии выбранной нуклеиновой кислоты для ее транскрибированной формы. Типичные способы амплификации включают различные способы репликации на основе полимеразы, в том числе полимеразную цепную реакцию (ПЦР), способы, опосредованные лигазой, такие как лигазная цепная реакция (ЛЦР) и способы амплификации на основе РНК-полимеразы (например, путем транскрипции).
Термин «сборка» применяют к бактериальным искусственным клонам (BAC) и их склонностям собираться вместе и формировать непрерывные фрагменты ДНК. «Сборки» BAC в контиг основаны на выравнивании последовательностей, если BAC секвенирован, или происходят путем выравнивания фингерпринта BAC с фингерпринтами других BAC.
Аллель «ассоциирован с» признаком, когда он сцеплен с ним, и когда присутствие аллеля является указанием на то, что желаемый признак или форма признака будут встречаться у растения, содержащего аллель.
«Бактериальная искусственная хромосома (BAC)» представляет собой клонирующий вектор, который получен из природного F-фактора Escherichia coli. BAC может принимать большие вставки из последовательности ДНК. У подсолнечника, ряд BAC, или бактериальных искусственных хромосом, каждая содержащая большую вставку геномной ДНК подсолнечника, были собраны в контиги (перекрывающиеся смежные генетические фрагменты, или «смежные ДНК»).
«Обратное скрещивание» относится к способу, при котором гибридное потомство повторно скрещивается с одним из родителей. В схеме «обратного скрещивания», «донорный» родитель относится к родительскому растению с желаемым геном или локусом, который предполагается интрогрессировать. «Реципиентный» родитель (использован один раз или более одного раза) или «рекуррентный» родитель (использован два раза или более двух раз) относится к родительскому растению, в которое предполагается интрогрессировать ген или локус. Например, см. Ragot, M. et al. (1995) Marker-assisted backcrossing: a practical example, in Techniques et Utilisations des Marqueurs Moleculaires Les Colloques, Vol. 72, pp. 45-56, и Openshaw et al., (1994) Marker-assisted Selection in Backcross Breeding, Analysis of Molecular marker Data, pp. 41-43. Исходное скрещивание приводит к поколению F1: термин "BC1" тогда относится ко второму использованию рекуррентного родителя, "BC2" относится к третьему использованию рекуррентного родителя, и так далее.
Сантиморганида («cM») представляет собой единицу измерения частоты рекомбинации. Одна cM равна вероятности 1%, что маркер в одном генетическом локусе будет отделен от маркера во втором генетическом локусе из-за кроссинговера в одном поколении.
«Хромосомный интервал» обозначает непрерывную линейную последовательность геномной ДНК, которая у растений расположена на одной хромосоме. Генетические элементы или гены, расположенные в одном хромосомном интервале, физически связаны. Размер хромосомного интервала конкретно не ограничен. В некоторых аспектах, генетические элементы, расположенные внутри одного хромосомного интервала, генетически связаны, как правило, с расстоянием генетической рекомбинации, например, менее чем или равным 20 cM, или альтернативно, менее чем или равным 10 cM. То есть, два генетических элемента внутри одного хромосомного интервала подвергаются рекомбинации с частотой менее чем или равной 20% или 10%, соответственно.
Термин «хромосомный интервал» обозначает все и любые интервалы, определяемые любыми маркерами, перечисленными в настоящем изобретении. Предлагается хромосомный интервал, который коррелирует с устойчивостью к Orobanche. Этот интервал, расположенный на LG4, включает и фланкирован DHAI0000240 и DHAI007796. Существует по меньшей мере три подинтервала в хромосомном интервале DHAI0000240 и DHAI007796: DHAI0000240 и DHAG000732, DHA000732 и DHAI007334, и DHAI007334 и DHAI007796.
Термин «комплементарный» относится к нуклеотидной последовательности, которая комплементарна указанной нуклеотидной последовательности, т.е., последовательности связываются по правилам спаривания оснований.
Термин "смежные ДНК" относится к перекрывающимся смежным генетическим фрагментам.
Термин «скрещенный» или «скрещиваться» означает слияние гамет посредством опыления для получения потомства (например, клеток, семян или растений). Термин относится как к половому скрещиванию (опыление одного растения другим), так и к самооплодотворению (самоопыление, например, когда пыльца и яйцеклетка от одного растения). Термин «скрещивание» относится к акту слияния гамет посредством опыления для получения потомства.
Термин «элитная линия» относится к любой линии, которая получена в результате скрещивания и отбора для превосходных агрономических характеристик. Элитное растение представляет собой любое растение из элитной линии.
«Благоприятный аллель» представляет собой аллель в определенном локусе, который придает желаемый фенотип или вносит вклад в желаемый фенотип, например, преобладающую повышенную устойчивость к Orobanche, или альтернативно, представляет собой аллель, который позволяет идентифицировать растения с пониженной устойчивостью к Orobanche, которые можно удалить из программы скрещивания или высадки селекции («обратный отбор»). Благоприятным аллелем маркера является маркерный аллель, который сегрегирует с благоприятным фенотипом, или альтернативно, сегрегирует с неблагоприятным фенотипом растения, таким образом, обеспечивая преимущество идентификации растений.
«Фрагмент» предназначен для обозначения части нуклеотидной последовательности. Фрагменты можно использовать в качестве зондов для гибридизации или ПЦР-праймеров при использовании способов, описываемых в настоящем документе.
«Генетическая карта» представляет собой описание генетических взаимосвязей между локусами на одной или нескольких хромосомах (или хромосомы) данного вида, как правило, изображенное в форме диаграммы или таблицы. Для каждой генетической карты, расстояния между локусами измеряют посредством частот рекомбинации между ними, а рекомбинации между локусами можно выявлять с использованием ряда молекулярных генетических маркеров (также называемых молекулярные маркеры). Генетическая карта является продуктом картируемой популяции, типов используемых маркеров, и полиморфного потенциала каждого маркера между различными популяциями. Порядок и генетическое расстояние между локусами могут различаться на разных генетических картах. Однако, информацию, такую как положение и порядок маркеров, можно коррелировать между картами, определяя физическое расположение маркеров на одной из групп сцепления подсолнечника, в референсном геноме сборки Ha412HO bronze, который общедоступен в интернет. Специалист в данной области может использовать общедоступный геномный браузер для того, чтобы определить физическое расположение маркеров на хромосоме.
Термин «генетический маркер» относится к любому типу маркера на основе нуклеиновой кислоты, включая в качестве неограничивающих примеров, полиморфизм длин рестрикционных фрагментов (ПДРФ) (Botstein et al, 1998), простую повторяющуюся последовательность (SSR) (Jacob et al., 1991), случайно амплифицируемую полиморфную ДНК (RAPD) (Welsh et al., 1990), рестрикционный полиморфизм амплифицированных последовательностей (CAPS) (Rafalski and Tingey, 1993, Trends in Genetics 9:275-280), полиморфизм длин амплифицированных фрагментов (AFLP) (Vos et al, 1995, Nucleic Asids Res. 23:4407-4414), однонуклеотидный полиморфизм (SNP) (Brookes, 1999, Gene 234:177-186), амплифицированную область, охарактеризованную нуклеотидной последовательностью (SCAR) (Pecan and Michelmore, 1993, Theor. Appl. Genet, 85:985-993), маркирующий последовательность участок (STS) (Onozaki et al. 2004, Euphytica 138:255-262), одноцепочечный конформационный полиморфизм (SSCP) (Orita et al., 1989, Proc Natl Aced Sci USA 86:2766-2770), повтор между простыми последовательностями (ISR) (Blair et al. 1999, Theor. Appl. Genet. 98:780-792), полиморфизм амплифицированных последовательностей между ретротранспозонами (IRAP), амплифицированный полиморфизм ретротранспозон-микросателлит (REMAP) (Kalendar et al., 1999, Theor. Appl. Genet 98:704-711), продукт расщепления РНК (такой как метка Lynx) и т.п.
«Частота генетической рекомбинации» представляет собой частоту событий кросинговера (рекомбинация) между двумя генетическими локусами. Частоту рекомбинации можно наблюдать по сегрегации маркеров и/или признаков после мейоза.
«Геном» относится к общей ДНК или полному набору генов, расположенном на хромосоме или наборе хромосом.
Термин «генотип» представляет собой генетическую конституцию индивидуума (или группы индивидуумов) по одному или нескольким генетическим локусам, в отличие от наблюдаемого признака (фенотипа). Генотип определяется аллелем/аллелями одного или нескольких известных локусов, которые индивидуум унаследовал от его родителей. Термин «генотип» можно использовать для ссылки на генетическую конституцию индивидуума по одному локусу, по нескольким локусам, или, в более общем смысле, термин «генотип» можно использовать для ссылки на генетическую характеристику индивидуума по всем генам в его геноме.
«Зародышевая плазма» относится к генетическому материалу индивидуума или от индивидуума (например, растения), группы индивидуумов (например, линии, сорта или семейства растений), или клона, полученного из линии, сорта, вида или культуры растений. Зародышевая плазма может быть частью организма или клетки или может быть отделена от организма или клетки. В основном, зародышевая плазма предоставляет генетический материал со специфической молекулярной характеристикой, которая обеспечивает физическую основу для некоторых или всех наследственных качеств организма или клеточной культуры. Как применяют в настоящем документе, зародышевая плазма включает клетки, семена или ткани, из которых можно выращивать новые растения, или части растений, такие как листья, побеги, пыльцу, или клетки, которые можно культивировать в целое растение.
Референсный геном подсолнечника «сборка Ha415HO bronze» является общедоступным на «Sunflower Genome Database». «Sunflower Genome Database» также предлагает ряд других геномных ресурсов, генетические и физические карты с высокой плотностью, а также данные о последовательностях и транскриптоме для различных анализов диких и культивированных генотипов.
«Гаплотип» представляет собой генотип индивидуума по множеству генетических локусов, т.е. комбинацию аллелей. Как правило, генетические локусы, описываемые гаплотипом, связаны физически и генетически, т.е., находятся на одном и том же сегменте хромосомы. Термин «гаплотип» может относиться к полиморфизмам последовательности в конкретном локусе, таком как локус одного маркера, или полиморфизмам последовательности в нескольких локусах по хромосомному сегменту указанного генома. Последний вариант также может быть обозначен как «маркерные гаплотипы» или «маркерные аллели», и последние из них можно также обозначать как «гаплотипы длинных фрагментов».
«Наследуемость (h2)» признака в популяции представляет собой долю наблюдаемых различий по признаку между индивидуумами внутри популяции, при этом различия вызваны генетическими различиями. Величина h2 для QTL представляет собой процент изменчивости, который объясняется генетикой, а не окружающей средой.
«Гетерозисная группа» включает набор генотипов, которые хорошо проявляются при скрещивании с генотипами из другой гетерозисной группы (Hallauer at al. (1998) Corn breeding, p. 463-564. In G. F. Sprague and J. W. Dudley (ed) Corn and sunflower improvement). Инбредные линии делятся на гетерозисные группы, и дополнительно подразделяются на семейства внутри гетерозисных групп на основании нескольких критериев, таких как поколение, ассоциации на основе молекулярных маркеров и проявление в комбинациях гибридов (Smith at al. (1990) Theor. Appl. Gen. 80:833-840).
Термин «гетерозиготный» означает генетическое состояние, где различные аллели располагаются в соответствующем локусе на гомологичных хромосомах.
Термин «гомозиготный» означает генетическое состояние, где идентичные аллели располагаются в соответствующем локусе на гомологичных хромосомах.
«Гибридизация» или «гибридизация нуклеиновых кислот» относится к спариванию коплементарных цепей РНК и ДНК, а также к спариванию комплементарных одиночных цепей ДНК.
Термин «гибридизироваться» означает образование пар оснований между комплементарными областями цепей нуклеиновой кислоты.
Термин «вставка/делеция» относится к вставке или делеции, где одна линия может быть обозначена как имеющая вставку относительно второй линии, или вторая линия может быть обозначена как имеющая делецию относительно первой линии.
Термин «интрогрессия» или «интрогрессирование» относится к переносу желаемого аллеля генетического локуса из одного генетического окружения в другое. Например, интрогрессия желаемого аллеля конкретного локуса может быть передана по меньшей мере одному потомку посредством полового скрещивания между двумя родителями одного и того же вида, где по меньшей мере один из родителей имеет желаемый аллель в геноме. Альтернативно, например, перенос аллеля может происходить путем рекомбинации между двумя донорскими геномами, например, в слитом протопласте, где по меньшей мере один из донорских протопластов имеет желаемый аллель в геноме. Желаемый аллель может быть, например, выбранным аллелем маркера, QTL, трансгеном или т.п. В любом случае, с потомством, содержащим желаемый аллель, можно повторно проводить обратное скрещивание с линией с желаемым генетическим окружением и отбирать по желаемому аллелю, чтобы получить аллель, закрепленный в выбранном генетическом окружении. Например, локус группы сцепления 4, описываемый в настоящем документе, можно интрогрессировать в рекуррентного родителя, который восприимчив к устойчивости к Orobanche. Линия рекурентного родителя с интрогрессированным геном или локусом тогда имеет повышенную устойчивость к Orobanche.
Как применяют в настоящем документе, термин «сцепление» применяют для описания степени, с которой один маркерный локус ассоциирован с другим маркерным локусом или каким-то другим локусом (например, локусом устойчивости с Orobanche). Связь сцепления между молекулярным маркером и фенотипом дается как «вероятность» или «скорректированная вероятность». Сцепление можно выражать в виде желаемого предела или диапазона. Например, в некоторых вариантах осуществления любой маркер сцеплен (генетически и физически) с любым другим маркером, когда маркеры разделены менее чем 50, 40, 30, 25, 20, или 15 единицами картирования на cM). В некоторых аспектах, выгодно определить заключенный в скобки диапазон сцепления, например, между 10 и 20 cM, между 10 и 30 cM, или между 10 и 40 cM. Чем ближе маркер связан со вторым локусом, тем лучшим индикатором для второго локуса становится этот маркер. Таким образом, «близко сцепленные локусы», такие как маркерный локус и второй локус демонстрируют частоту рекомбинации между локусами 10% или меньше, предпочтительно приблизительно 9% или меньше, still более предпочтительно приблизительно 8% или меньше, еще более предпочтительно приблизительно 7% или меньше, еще более предпочтительно приблизительно 6% или меньше, еще более предпочтительно приблизительно 5% или меньше, еще более предпочтительно приблизительно 4% или меньше, еще более предпочтительно приблизительно 3% или меньше, и еще более предпочтительно приблизительно 2% или меньше. В высокопредпочтительных вариантах осуществления, релевантные локусы демонстрируют частоту рекомбинации приблизительно 1% или меньше, например, приблизительно 0,75% или меньше, более предпочтительно приблизительно 0,5% или меньше, или еще более предпочтительно приблизительно 0,25% или меньше. Два локуса, которые расположены на одной хромосоме, и на таком расстоянии, что рекомбинация между двумя локусами происходит с частотой менее чем 10% (например, приблизительно 9%, 8%, 7%, 6%, 5%, 4%, 3%, 2%, 1%, 0,75%, 0,5%, 0,25%, или меньше) также являются «проксимальными» друг для друга. Поскольку одна cM представляет собой расстояние между двумя маркерами, которое показывает 1% частоты рекомбинации, любой маркер является близко сцепленным (генетически и физически) с любым другим маркером, который находится в непосредственной близости, например, на расстоянии 10 cM или менее. Два близко сцепленных маркера на одной хромосоме могут быть расположены друг от друга в 9, 8, 7, 6, 5, 4, 3, 2, 1, 0,75, 0,5 или 0,25 cM или меньше.
Термин «неравновесие по сцеплению» относится к неслучайной сегрегации генетических локусов или признаков для обоих локусов. В любом случае неравновесное сцепление подразумевает, что соответствующие локусы находятся в достаточной физической близости по длине хромосомы, так что они сегрегируются вместе с большей, чем случайная (т. е. неслучайной) частотой (в случае совместного сегрегирования признаков, локусы, лежащие в основе этих признаков, находятся в достаточной близости друг к другу). Маркеры, которые показывают неравновесие по сцеплению, считаются сцепленными. Сцепленные локусы совместно сегрегируют более чем в 50% случаев, например, приблизительно от 51% до приблизительно 100% случаев. Другими словами, два маркера, которые совместно сегрегируют, имеют частоту рекомбинации менее 50% (и, по определению, разделены менее чем 50 сМ на одной и той же хромосоме). Как применяют в настоящем документе, сцепление может быть между двумя маркерами, или альтернативно между маркером и фенотипом. Маркерный локус может быть «ассоциирован с» (сцеплен с) признаком, например, повышенной устойчивостью к Orobanche. Степень сцепления молекулярного маркера с фенотипическим признаком измеряют, например, как статистическую вероятность совместной сегрегации этого молекулярного маркера с фенотипом.
Неравновесие по сцеплению наиболее часто оценивают при помощи параметра r2, который рассчитывают с использованием формулы, описанной Hill, W. G. и Robertson, A, Theor Appl. Genet 38:226-231 (1988). Когда r2=1, между двумя маркерными локусами существует полное LD, что означает, что маркеры не разделяются рекомбинацией и имеют одинаковую частоту аллелей. Значений r2 выше 1/3 указывают на LD достаточно сильное, чтобы быть полезным для картирования (Ardlie at al., Nature Reviews Genetics 3:299-309 (2002)). Таким образом, аллели находятся в неравновесии по сцеплению, когда значения r2 между попарно сравниваемыми маркерными локусами больше или равны 0,33, 0,4, 0,5, 0,6, 0,7, 0,8, 0,9 или 1,0.
Как применяют в настоящем документе, «неравновесие по сцеплению» описывает ситуацию, в которой два маркера сегрегируют независимо, т.е., распределяются среди потомства случайным образом. Маркеры, которые показывают неравновесие по сцеплению, считаются несцепленными (лежат они или нет на одной хромосоме).
«Величину логарифма соотношения шансов (LOD)» или «LOD-балл» (Risch, Science 255:803-804 (1992)) применяют для интервального картирования для описания степени сцепления между двумя маркерными локусами. LOD-балл, равный трем, между двумя маркерами указывает на то, что вероятность присутствия сцепления в 1000 раз выше, чем его отсутствие, в то время как LOD-балл, равный двум, указывает на то, что вероятность сцепления в 100 раз выше, чем его отсутствие. LOD-балл, больше или равный двум, можно использовать для выявления сцепления.
«Локус» представляет собой положение на хромосоме, где расположен ген или маркер.
Термин «растение подсолнечника» включает в себя целое растение подсолнечника, растительные клетки подсолнечника, протопласт растения подсолнечника, растительную клетку подсолнечника или тканевые культуры подсолнечника, из которых можно регенерировать растения подсолнечника, растительные клетки подсолнечника, которые интактны в растениях подсолнечника или частях растения подсолнечника, такие как семена подсолнечника, головки подсолнечника, цветы подсолнечника, семядоли подсолнечника, листья подсолнечника, стебли подсолнуха, почки подсолнечника, корни подсолнуха, верхушки корней подсолнечника и т.п.
«Orobanche (Orobanche spp)» представляют собой виды паразитических сорняков, которые приводят к разрушительным потерям урожая у производителей подсолнечника.
«Маркер» представляет собой нуклеотидную последовательность или кодированный ею продукт (например, белок), используемый в качестве ориентира. Для того чтобы маркеры были полезны при обнаружении рекомбинаций, они должны выявлять различия или полиморфизмы в популяции, подвергаемой мониторингу. Для молекулярных маркеров это означает различия на уровне ДНК из-за различий в полинуклеотидной последовательности (например, SSR, RFLP, AFLP, SNP). Геномная изменчивость может иметь любое происхождение, например, вставки, делации, дупликации, повторяющиеся элементы, точечные мутации, рекомбинационные события или наличие и последовательность мобильных генетических элементов. Молекулярные маркеры могут быть получены из геномных или экспрессированных нуклеиновых кислот (например, EST) и могут также относиться к нуклеиновым кислотам, используемым в качестве зондов или пар праймеров, способных амплифицировать последовательности фрагментов с помощью способов на основе ПЦР. Большое количество молекулярных маркеров подсолнечника известно в данной области и опубликовано или доступно из различных источников.
Маркеры, соответствующие генетическим полиморфизмам между представителями популяции, могут быть обнаружены способами, хорошо известными в данной области. Эти способы включают, например, секвенирование ДНК, способы специфической амплификации последовательности на основе ПЦР, детекцию RFLP, детекцию изозимовых маркеров, детекцию полинуклеотидных полиморфизмов с помощью аллель-специфической гибридизации (ASH), детекцию амплифицированных вариабельных последовательностей генома растения, детекцию само-поддерживаемой репликации последовательности, детекцию SSR, детекцию SNP или детекцию AFLP. Хорошо известны также способы обнаружения маркеров экспрессируемых последовательностей (EST) и маркеров SSR, полученных на основе EST и RAPD.
«Маркерный аллель», альтернативно «аллель маркерного локуса», может относиться к одной из множества полиморфных нуклеотидных последовательностей, обнаруженных в маркерном локусе в популяции, которая полиморфна по маркерному локусу.
«Селекция при помощи маркера» (MAS) представляет собой способ, при котором фенотипы выбирают на основании маркерных генотипов.
«Обратная селекция при помощи маркера» представляет собой способ, при котором используют маркерные генотипы для выявления растений, которые не должны быть отобраны, что позволяет удалить их из программы скрещивания или высаживания.
«Маркерный локус» представляет собой специфическое местоположение на хромосоме в геноме вида, где можно найти специфический маркер. Маркерный локус можно использовать для отслеживания присутствия второго маркерного локуса, например, сцепленного локуса, который кодирует фенотипический признак или вносит вклад в экспрессию фенотипического признака. Например, маркерный локус можно использовать для мониторинга сегрегации аллелей в локусе, таком как a QTL или одиночный ген, который генетически или физически сцеплен с маркерным локусом.
«Маркерный зонд» представляет собой последовательность или молекулу нуклеиновой кислоты, которую можно использовать для выявления присутствия маркерного локуса путем гибридизации нуклеиновых кислот, например, зонд из нуклеиновой кислоты, который комплементарен последовательности маркерного локуса. Для гибридизации нуклеиновых кислот использовать маркерные зонды, содержащие 30 или более смежных нуклеотидов маркерного локуса («всю или часть» последовательности маркерного локуса). Альтернативно, в некоторых аспектах, маркерный зонд относится к зонду любого типа, который способен различать конкретный аллель (т.е. генотип), присутствующий в маркерном локусе.
Термин "молекулярный маркер" можно использовать для обозначения генетического маркера, как определено выше, или кодируемого им продукта (например, белка), используемого в качестве ориентира при идентификации сцепленного локуса. Маркер может быть получен из геномных нуклеотидных последовательностей или из экспрессированных нуклеотидных последовательностей (например, из сплайсированной РНК, кДНК и т.д.) или из кодированного полипептида. Термин также относится к последовательностям нуклеиновых кислот, которые комплементарны маркерным последовательностям или фланкируют маркерные последовательности, таким как нуклеиновые кислоты, используемые в качестве зондов или пар праймеров, способных амплифицировать маркерную последовательность. «Зонд с молекулярным маркером» представляет собой последовательность или молекулу нуклеиновой кислоты, которую можно использовать для определения присутствия маркерного локуса, например, зонд из нуклеиновой кислоты, который комплементарен последовательности маркерного локуса. Альтернативно, в некоторых аспектах маркерный зонд относится к зонду любого типа, который способен различать конкретный аллель (т.е. генотип), присутствующий в маркерном локусе. Нуклеиновые кислоты являются «комплементарными», когда они специфически гибридизуются в растворе, например, согласно правилам спаривания оснований по Уотсону-Крику. Некоторые из описанных в данном документе маркеров также обозначены как гибридизационные маркеры, когда они расположены в области вставки/делеции, такой как неколлинеарная область, описываемая в данном документе. Это потому, что область вставки представляет собой, по определению, полиморфизм, видимый через растение без вставки. Таким образом, маркер должен только указывать, присутствует ли область вставки/делеции или отсутствует. Для выявления таких гибридизационных маркеров можно использовать любую подходящую технологию детекции, например, в примерах, предоставленных в настоящем документе, применяют технологию SNP.
«Нуклеотидная последовательность», «полинуклеотид», «последовательность нуклеиновой кислоты», и «фрагмент нуклеиновой кислоты» используются взаимозаменяемо и относятся к полимеру РНК или ДНК, который является одно- или двухцепочечным, необязательно содержащим синтетические, неприродные или измененные нуклеотидные основания. «Нуклеотид» представляет собой мономерную единицу, из которой конструируют полимеры ДНК или РНК, и состоит из пуринового или пиримидинового основания, пентозы и группы фосфорной кислоты. Нуклеотиды (как правило, находящиеся в их 5'-монофосфатной форме) обозначают при помощи однобуквенного обозначения следующим образом: «A» для аденилата или дезоксиаденилата (для РНК или ДНК, соответственно), «C» для цитидилата или дезоксицитидилата, «G» для гуанилата или дезоксигуанилата, «U» для уридилата, «T» для дезокситимидилата, «R» для пуринов (A или G), «Y» for пиримидины (C или T), «K» для G или T, «H» для A или C или T, «I» для инозина, и «N» для любого нуклеотида.
Термины «фенотип», или «фенотипический признак», или «признак» относятся к одному или нескольким признакам организма. Фенотип можно наблюдать невооруженным глазом, или любыми другими способами оценки, известными в данной области, например, микроскопией, биохимическим анализом или электромеханическим анализом. В некоторых случаях, фенотип напрямую контролируется одним геном или генетическим локусом, т.е., «признак, контролируемый одним геном». В остальных случаях, фенотип представляет собой результат нескольких генов.
«Физическая карта» генома представляет собой карту, показывающую линейный порядок опознаваемых ориентиров (включая гены, маркеры и т.д.) на ДНК хромосомы. Однако, в отличие от генетических карт, расстояние между ориентирами является абсолютным (например, измерено в парах оснований или выделенных и перекрывающихся смежных генетических фрагментах) и не основано на генетической рекомбинации.
«Растение» может быть целым растением, любой его частью или клеткой или тканевой культурой, полученной из растения. Таким образом, термин «растение» может относиться к любому из: целое растение, компоненты или органы растения (например, листья, стебли, корни и т.д.), ткани растения, семена, растительные клетки и/или их потомство. Растительная клетка представляет собой клетку растения, взятую из растения или полученную путем культивирования из клетки, взятой из растения.
«Полиморфизм» представляет собой изменение в ДНК, которое встречается слишком часто, чтобы быть результатом просто новой мутации. Полиморфизм должен иметь частоту в популяции по меньшей мере 1%. Полиморфизм может быть однонуклеотидным полиморфизмом, или SNP, или полиморфизмом типа вставка/делеция, также называемым в настоящем документе как «indel».
«Значение вероятности» или «p-значение» представляет собой статистическую вероятность того, что конкретная комбинация фенотипа и наличия или отсутствия определенного маркерного аллеля является случайной. Таким образом, чем ниже показатель вероятности, тем выше вероятность того, что фенотип и конкретный маркер будут совместно сегрегировать. В некоторых аспектах показатель вероятности считается «значимым» или «незначимым». В некоторых вариантах осуществления значение вероятности 0,05 (р=0,05 или 5% вероятности) случайной сортировки считают значимым показателем совместной сегрегации. Однако приемлемой вероятностью может быть любая вероятность менее 50% (р=0,5). Например, значимая вероятность может быть менее чем 0,25, менее чем 0,20, менее чем 0,15, менее чем 0,1, менее чем 0,05, менее чем 0,01 или менее чем 0,001.
Термин «потомство» относится к поколению, полученному после скрещивания.
«Растение-потомок» получают при скрещивании двух растений.
«Референсная последовательность» представляет собой определенную последовательность, которую используют в качестве основы для сравнения последовательностей. Референсную последовательность получают путем генотипирования ряда линий по локусу, выравнивания нуклеотидных последовательностей в программе для выравнивания последовательностей (например, Sequencher), а затем получения консенсусной последовательности выравнивания.
«Однонуклеотидный полиморфизм (SNP)» представляет собой аллельное однонуклеотидное изменение - A, T, C или G - в последовательности ДНК, представляющей один локус по меньшей мере от двух индивидуумов одного вида. Например, два секвенированных фрагмента ДНК, представляющие один и от же локус по меньшей мере от двух индивидуумов одного вида, AAGCCTA и AAGCTTA, содержат различие по одному нуклеотиду.
Фраза «при жестких условиях» относится к условиям, при которых зонд или полинуклеотид будет гибридизироваться с конкретной последовательностью нуклеиновой кислоты, как правило, в сложной смеси нуклеиновых кислот, но по существу не с другими последовательностями. Жесткие условия зависимы от последовательности и будут различными при различных обстоятельствах.
Выравнивания последовательностей и расчет процента идентичности можно определять при помощи ряда способов сравнения, разработанных для детекции гомологичных последовательностей, включая в качестве неограничивающих примеров, программу MEGALIGN® из биоинформатической компьютерной оболочки LASERGENE® (DNASTAR® Inc., Madison, Wis.). Если не указано иное, множественное сравнение последовательностей, представленных в настоящем документе проводили при помощи способа выравнивания Clustal V (Higgins и Sharp, CABIOS. 5:151-153 (1989)) с параметрами по умолчанию (GAP PENALTY=10, GAP LENGTH PENALTY=10), Параметры по умолчанию для попарного выравнивания и расчета процента идентичности белковых последовательностей при помощи способа Clustal V представляют собой KTUPLE=1, GAP PENALTY=3, WINDOW=5 и DIAGONALS SAVED=5. Для нуклеиновых кислот эти параметры представляют собой KTUPLE=2, GAP PENALTY=5, WINDOW=4 и DIAGONALS SAVED=4. После выравнивания последовательностей при помощи программы Clustal V, возможно получить значения «процента идентичности» и «расхождения», рассматривая таблицу «расстояния между последовательностями» в этой же программе; если не указано иное, проценты идентичности и расхождения, предоставленные и заявленные в настоящем документе, рассчитывали таким образом.
Перед подробным описанием настоящего изобретения следует понимать, что это изобретение не ограничено конкретными вариантами осуществления. Также следует понимать, что терминология, используемая в данном документе, предназначена для описания конкретных вариантов осуществления и не предназначена для ограничения. Как применяют в настоящем документе и в прилагаемой формуле изобретения, термины в единственном числе и формах единственного числа включают ссылки на множественное число, если содержание явно не указывает на иное. Таким образом, например, ссылка на «растение» также включает в себя множество растений. В зависимости от контекста использование термина «растение» может также включать генетически сходное или идентичное потомство этого растения. Использование термина «нуклеиновая кислота» необязательно включает в себя множество копий этой молекулы нуклеиновой кислоты.
Генетическое картирование
В течение уже достаточно долгого времени известно, что определенные генетические локусы, коррелирующие с конкретными фенотипами, могут быть картированы в геноме организма. Растениевод-селекционер может эффективно использовать молекулярные маркеры для идентификации желаемых индивидуумов, выявляя маркерные аллели, которые показывают статистически значимую вероятность совместной сегрегации с желаемым фенотипом, что проявляется как неравновесие по сцеплению. Идентифицируя молекулярный маркер или кластеры молекулярных маркеров, которые совместно сегрегируют с интересующим признаком, селекционер может быстро отобрать желаемый фенотип, выбирая подходящий аллель молекулярного маркера (способ, называемый селекция при помощи маркера, или MAS).
Ряд способов хорошо известен в данной области для обнаружения молекулярных маркеров или кластеров молекулярных маркеров, которые сегрегируют совместно с интересующим признаком. Основная идея, лежащая в основе этих способов, заключается в обнаружении маркеров, для которых альтернативные генотипы (или аллели) имеют значительно отличающиеся средние фенотипы. Таким образом, сравниваются маркерные локусы по величине различий между альтернативными генотипами (или аллелями) или по уровню значимости этих различий. Предполагается, что гены признаков расположены ближе всего к маркеру/маркерам, которые имеют наибольшее ассоциированное генотипическое различие.
Два таких способа, используемых для обнаружения интересующих локусов признаков, представляют собой: 1) анализ ассоциаций на основе популяции и 2) классический анализ сцепления. В популяционном анализе ассоциаций, линии получают из ранее существовавших групп популяций с несколькими основателями, например, элитные селекционные линии. Популяционный анализ ассоциаций основывается на распаде неравновесия по сцеплению (LD) и идее, что в неструктурированной популяции только корреляции между генами, контролирующими интересующий признак, и маркерами, тесно связанными с этими генами, останутся после стольких поколений случайного спаривания. В действительности, большинство ранее существовавших групп популяций имеют подструктуру популяции. Таким образом, использование подхода структурированной ассоциации помогает контролировать структуру популяции путем распределения индивидуумов по популяциям, используя данные, полученные от маркеров, случайно распределенных по геному, тем самым сводя к минимуму неравновесие из-за структуры популяции в отдельных популяциях (также называемых субпопуляциями). Фенотипические значения сравнивают с генотипами (аллелями) у каждого маркерного локуса для каждой линии в субпопуляции. Значительная ассоциация маркер-признак указывает на дозу близости между маркерным локусом и одним или несколькими генетическими локусами, которые участвуют в проявлении этого признака.
Те же самые принципы лежат в основе классического анализа сцепления; однако, LD получают путем создания популяции из небольшого числа основателей. Основатели выбраны, чтобы максимизировать уровень полиморфизма в пределах построенной популяции, а полиморфные участки оценивают по их уровню ко-сегрегации с данным фенотипом. Был использован ряд статистических способов для выявления значимых ассоциаций маркер-признак. Одним из таких способов является подход интервального картирования (Lander and Botstein, Genetics 121: 185-199 (1989), в котором каждую из многих позиций на генетической карте (скажем, с интервалами в 1 cM) тестируют на вероятность того, что ген, который контролирует интересующий признак, находится в этой позиции. Данные генотипов/фенотипов применяют для расчета LOD-балла (логарифмического отношения правдоподобия) для каждого тестового положения. Когда показатель LOD превышает пороговое значение, это является значимым доказательством для местоположения гена, который контролирует интересующий признак, в этой позиции на генетической карте (который попадет между двумя определенными маркерными локусами).
Маркеры, ассоциированные с устойчивостью к Orobanche
В настоящем документе идентифицированы маркеры, ассоциированные с повышенной устойчивостью к Orobanche. Способы включают выявление по меньшей мере одного маркерного аллеля, ассоциированного с улучшенной устойчивостью, в зародышевой плазме растения подсолнечника. Маркерный локус можно выбирать из любых маркерных локусов, приведенных в таблице 1, включая DHAI000240, DHAG000732, DHAI007796 и DHAI007334 и любых других маркеров, сцепленных с этими маркерами.
Генетические элементы или гены, расположенные на непрерывном линейном участке геномной ДНК на одной хромосоме, физически сцеплены. DHAI000240, DHAG000732, DHAI007796 и DHAI007334, тесно связанные с устойчивостью к Orobanche, определяют QTL устойчивости к Orobanche. Любой полинуклеотид, который собирается в непрерывную ДНК между и включая SEQ ID NO:1 (референсная последовательность для DHAI0000240), или нуклеотидная последовательность, которая на 95% идентична SEQ ID NO:1 на основании способа выравнивания Clustal V, и SEQ ID NO:29 (референсная последовательность для DHAI007796), или нуклеотидная последовательность, которая на 95% идентична SEQ ID NO:29 на основании способа выравнивания Clustal V, может содержать маркерные локусы, которые ассоциированы с устойчивостью к Orobanche.
Генетические элементы или гены, расположенные на непрерывном линейном участке геномной ДНК на нескольких группах сцепления, физически сцеплены для субинтервала DHAI0000240 и DHAG000732. DHAI0000240 и DHAG000732, тесно связанные с устойчивостью к Orobanche, определяют QTL устойчивости к Orobanche. Любой полинуклеотид, который собирается в непрерывную ДНК между и включая SEQ ID NO:1 (референсная последовательность для DHAI0000240), или нуклеотидная последовательность, которая на 95% идентична SEQ ID NO:1 на основании способа выравнивания Clustal V, и SEQ ID NO:9 (референсная последовательность для DHAG000732), или нуклеотидная последовательность, которая на 95% идентична SEQ ID NO:9 на основании способа выравнивания Clustal V, может содержать маркерные локусы, которые ассоциированы с устойчивостью к Orobanche.
Генетические элементы или гены, расположенные на непрерывном линейном участке геномной ДНК на одной хромосоме, физически сцеплены для субинтервала DHAG000732 и DHAI007334. DHAG000732 и DHAI007334, тесно связанные с устойчивостью к Orobanche, определяют QTL устойчивости к Orobanche. Любой полинуклеотид, который собирается в непрерывную ДНК между и включая SEQ ID NO:9 (референсная последовательность для DHAG000732), или нуклеотидная последовательность, которая на 95% идентична SEQ ID NO:9 на основании способа выравнивания Clustal V, и SEQ ID NO:17 (референсная последовательность для DHAI007334), или нуклеотидная последовательность, которая на 95% идентична SEQ ID NO:17 на основании способа выравнивания Clustal V, может содержать маркерные локусы, которые ассоциированы с устойчивостью к Orobanche.
Генетические элементы или гены, расположенные на непрерывном линейном участке геномной ДНК на одной хромосоме, физически сцеплены для субинтервала DHAI007334 и DHAI007796. DHAI007334 и DHAI007796, тесно связанные с устойчивостью к Orobanche, определяют QTL устойчивости к Orobanche. Любой полинуклеотид, который собирается в непрерывную ДНК между и включая SEQ ID NO:17 (референсная последовательность для DHAI007334), или нуклеотидная последовательность, которая на 95% идентична SEQ ID NO:17 на основании способа выравнивания Clustal V, и SEQ ID NO:29 (референсная последовательность для DHAI007796), или нуклеотидная последовательность, которая на 95% идентична SEQ ID NO:29 на основании способа выравнивания Clustal V, может содержать маркерные локусы, которые ассоциированы с устойчивостью к Orobanche.
Распространенной мерой сцепления является частота, с которой признаки ко-сегрегируют. Ее можно выражать в процентах от ко-сегрегации (частота рекомбинации) или в санти-Морганидах (сМ). cM представляет собой единицу измерения частоты генетической рекомбинации. Одна cM равна 1% вероятности того, что признак в одном генетическом локусе будет отделен от признака в другом локусе из-за кроссинговера в одном поколении (что означает, что признаки сегрегируют вместе в 99% случаев). Поскольку хромосомное расстояние приблизительно пропорционально частоте событий кроссинговера между признаками, существует приблизительное физическое расстояние, которое коррелирует с рекомбинационной частотой.
Маркерные локусы сами по себе являются признаками и их можно оценивать в соответствии со стандартным анализом сцепления, отслеживая маркерные локусы во время сегрегации. Таким образом, одна cM равна 1% вероятности того, что маркерный локус будет отделен от другого локуса из-за кросинговера в одном поколении.
Можно использовать другие маркеры, сцепленные с маркерами, перечисленными в таблице 1, для предсказания устойчивости к Orobanche у растения подсолнечника. Такие маркеры включают любые маркеры в пределах 50 cM от DHAI000240, DHAG000732, DHAI007796 и DHAI007334, маркеров, ассоциированных с устойчивостью к Orobanche. Чем ближе маркер расположен к гену, контролирующему интересующий признак, тем более эффективным и полезным будет этот маркер в качестве индикатора желаемого признака. Близко сцепленные локусы демонстрируют частоту кросинговера между локусами приблизительно 10% или меньше, предпочтительно приблизительно 9% или меньше, еще более предпочтительно приблизительно 8% или меньше, еще более предпочтительно приблизительно 7% или меньше, еще более предпочтительно приблизительно 6% или меньше, еще более предпочтительно приблизительно 5% или меньше, еще более предпочтительно приблизительно 4% или меньше, еще более предпочтительно приблизительно 3% или меньше, и еще более предпочтительно приблизительно 2% или меньше. В высоко предпочтительных вариантах осуществления, релевантные локусы (например, маркерный локус и локус-мишень) демонстрируют частоту рекомбинации приблизительно 1% или меньше, например, приблизительно 0,75% или меньше, более предпочтительно приблизительно 0,5% или меньше, или еще более предпочтительно приблизительно 0,25% или меньше. Таким образом, локусы расположены приблизительно на 10 cM, 9 cM, 8 cM, 7 cM, 6 cM, 5 cM, 4 cM, 3 cM, 2 cM, 1 cM, 0,75 cM, 0,5 cM или 0,25 cM или меньше друг от друга. Другими словами, два локуса, которые расположены на одной хромосоме и на таком расстоянии, что рекомбинация между двумя локусами и происходит с частотой менее чем 10% (например, приблизительно 9%, 8% 7%, 6%, 5%, 4%, 3%, 2% 1%, 0,75%, 0,5%, 0,25% или меньше) «проксимальны» друг к другу.
Хотя конкретные маркерные аллели могут демонстрировать совместную сегрегацию с повышенной устойчивостью к Orobanche, важно отметить, что маркерный локус не обязательно ответственен за проявление фенотипа устойчивости к Orobanche. Например, не является обязательным, чтобы маркерная полинуклеотидная последовательность была частью гена, который придает повышенную устойчивость к Orobanche (например, была частью открытой рамки считывания гена). Ассоциация между конкретным маркерным аллелем и фенотипом повышенной устойчивости к Orobanche обусловлена исходной фазой «сопряженного» сцепления между маркерным аллелем и аллелем в предковой линии подсолнечника, из которой произошел аллель. В конце концов, при повторной рекомбинации, события кросинговера между маркерным и генетическим локусом могут изменить эту ориентацию. По этой причине благоприятный маркерный аллель может меняться в зависимости от фазы сцепления, которая существует в «устойчивом» родителе, используемом для создания сегрегационных популяций. Это не меняет того факта, что маркер можно использовать для мониторинга сегрегации фенотипа. Изменяется только то, какой маркерный аллель считается эффективным в данной сегрегирующей популяции.
Термин «хромосомный интервал» обозначает любые и все интервалы, определенные любым из набора маркеров, изложенных в настоящем изобретении. Предлагается хромосомный интервал, который коррелирует с устойчивостью к Orobanche. Этот интервал, расположенный в группе сцепления 4, включает и фланкирован DHAI0000240 и DHAI007796. Существуют по меньшей мере три подинтервала этого хромосомного интервала DHAI0000240 и DHAI007796: DHAI0000240 и DHAG000732, DHA000732 и DHAI007334, и DHAI007334 и DHAI007796.
Для выявления хромосомных интервалов доступен ряд способов, хорошо известных в данной области. Границы таких хромосомных интервалов очерчены так, чтобы охватить маркеры, которые будут сцеплены с геном, контролирующим интересующий признак. Другими словами, хромосомный интервал нарисован так, что любой маркер, который находится в пределах этого интервала (включая конечные маркеры, которые определяют границы интервала), можно использовать в качестве маркера для устойчивости к Orobanche. Описанный выше интервал охватывает кластер маркеров, которые сегрегируют совместно с устойчивостью к Orobanche. Кластеризация маркеров происходит в относительно небольших доменах на хромосомах, что указывает на присутствие гена, контролирующего интересующий признак в этих хромосомных регионах.
Интервал был очерчен, чтобы охватить маркеры, которые сегрегируют совместно с устойчивостью к Orobanche. Интервал охватывает маркеры, которые картируются в пределах интервала, а также маркеры, которые определяют конечные точки. Например, DHAI0000240 и DHAI007796, разделенные 3906661 п.н. на основании последовательности LG4 (из сборки Ha412HO bronze, находящейся в общедоступной базе данных генома подсолнечника), определяют хромосомный интервал, включающий кластер маркеров, которые сегрегируют совместно с устойчивостью к Orobanche. Второй пример включает подинтервал DHAI0000240 и DHAG000732, разделенный 1522089 п.н. на основании последовательности LG4, который определяет хромосомный интервал, охватывающий кластер маркеров, которые сегрегируют совместно с устойчивостью к Orobanche. Третий пример включает подинтервал DHAG000732 и DHAI007334, разделенный 1564876 п.н. на основании последовательности LG4, который определяет хромосомный интервал, охватывающий кластер маркеров, которые сегрегируют совместно с устойчивостью к Orobanche. Четвертый пример включает в себя подинтервал DHAI007334 и DHAI007796, разделенные 819532 п.н. на основании последовательности LG4, который определяет хромосомный интервал, охватывающий кластер маркеров, которые сегрегируют совместно с устойчивостью к Orobanche. Интервал, описываемый концевыми маркерами, которые определяют конечные точки интервала, будет включать в себя концевые маркеры и любые маркеры, локализующиеся в пределах этого хромосомного домена, независимо от того, известны ли эти маркеры в настоящее время или неизвестны.
Хромосомные интервалы могут определяться по маркерам, которые сцеплены (показывают неравновесие по сцеплению) с представляющим интерес маркером, и являются обычной мерой неравновесия по сцеплению (LD) в отношении исследований ассоциации. Если значение r2 для LD между любым маркерным локусом LG4, лежащим в интервале DHAI0000240 и DHAI007796, или одном из трех подинтервалов DHAI0000240 и DHAI007796: DHAI0000240 и DHAG000732, DHA000732 и DHAI007334, и DHAI0073396 и/или любом другом подинтервале DHAI0000240 и DHAI007796, и идентифицированным маркером в пределах этого интервала, у которого аллель связан с повышенной устойчивостью к Orobanche, больше чем 1/3 (Ardlie et al. Nature Reviews Genetics 3: 299-309 (2002)), локусы сцеплены.
Маркер по изобретению также может быть комбинацией аллелей в маркерных локусах, иначе известной как гаплотип. Специалисты в данной области ожидают, что могут быть дополнительные полиморфные сайты в маркерных локусах на хромосоме и около 2 маркеров, идентифицированных в указанном документе, где один или несколько полиморфных сайтов находятся в неравновесном сцеплении (LD) с аллелем, связанным с повышенной устойчивостью к Orobanche. Говорят, что два конкретных аллеля в разных полиморфных сайтах находятся в LD, если присутствие аллеля в одном из сайтов имеет тенденцию предсказывать присутствие аллеля в другом сайте на той же хромосоме (Stevens, Mol. Diag. 4: 309-17 (1999)).
Селекция при помощи маркера
Молекулярные маркеры можно использовать в ряде способов для селекции растений (например, см. Staub et al. (1996) Hortscience 729-741; Tanksley (1983) Plant Molecular Biology Reporter 1: 3-8). Одной из основных областей, представляющих интерес, является повышение эффективности обратного скрещивания и интрогрессирования гены с помощью селекции при помощи маркера (MAS). Молекулярный маркер, который демонстрирует связь с локусом, влияющим на желаемый фенотипический признак, предоставляет полезный инструмент для селекции признака в популяции растений. Это особенно верно в отношении признаков, которые трудно фенотипировать из-за их зависимости от условий окружающей среды. В эту категорию входят признаки, связанные с устойчивостью к биотическим и абиотическим стрессам. Эта категория также включает признаки, которые очень дороги для фенотипа из-за трудоемкой искусственной инокуляции или поддержания управляемых стрессовых условий среды. Другая категория признаков включает те, которые ассоциированы с разрушением растения как такового. Разрушительное фенотипирование было узким местом для реализации MAS для качественных признаков семян. Поскольку анализы ДНК-маркеров не зависят от окружающей среды, являются надежными, удобными, менее трудоемкими, менее дорогостоящими и занимают меньше физического пространства, чем фенотипирование в полевых условиях, можно анализировать гораздо большие популяции, увеличивая шансы найти рекомбинант с целевым сегментом из линии донора, перенесенный в линию реципиента. Чем ближе сцепление, тем полезнее маркер, так как рекомбинация с меньшей вероятностью происходит между маркером и геном, вызывающим признак, что может привести к ложно-положительным результатам. Наличие фланкирующих маркеров уменьшает вероятность того, что ложноположительная селекция, поскольку потребуется двойная рекомбинация. Идеальная ситуация - иметь маркер в самом гене, чтобы между маркером и геном не происходила рекомбинация. Такой маркер называется «идеальный маркер».
Когда ген интрогрессируют путем MAS, вводится не только ген, но и фланкирующие области (Gepts. (2002). Crop Sci; 42: 1780-1790). Это обозначают как «перемещение сцепления». В тех случаях, когда донорское растение не является родственным для растения-реципиента, эти фланкирующие области несут дополнительные гены, которые могут кодировать агрономически нежелательные признаки. Это «перемещение сцепления» может также привести к снижению урожайности или другим отрицательным агрономическим характеристикам даже после нескольких циклов обратного скрещивания в элитную линию подсолнечника. Это также иногда обозначают как «перемещение урожайности». Размер фланкирующей области может быть уменьшен путем дополнительного обратного скрещивания, хотя это не всегда успешно, так как у заводчиков нет контроля над размером региона или контрольными точками рекомбинации (Young et al. (1998) Genetics 120: 579-585). В классическом скрещивании, как правило, является случайностью, что выбраны рекомбинации, которые способствуют уменьшению размера донорского сегмента (Tanksley et al. (1989). Biotechnology 7: 257-264).
Даже после 20 обратных скрещиваний в обратных скрещиваниях этого типа можно ожидать, что найдется значительная часть донорской хромосомы, еще сцепленная с выбранным геном. С помощью маркеров, однако, можно выбрать тех редких индивидуумов, которые подверглись рекомбинации рядом с интересующим геном. У 150 растений с обратным скрещиванием существует 95% вероятность того, что по меньшей мере одно растение перенесет кроссинговер в пределах 1 сМ от гена, на основании расстояния на генетической карте одного мейоза. Маркеры позволят провести однозначную идентификацию таких индивидуумов. При одном дополнительном обратном скрещивании из 300 растений вероятность 95% кросинговера в пределах расстояния в 1 сМ на генетической карте одного мейоза с другой стороны гена будет равна 95%, что приведет к образованию сегмента вокруг целевого гена менее 2 сМ на основании расстояния на генетической карте одного мейоза. Это может быть выполнено в двух поколениях с маркерами, в то время как это потребовало бы в среднем 100 поколений без маркеров (См. Tanksley et al., выше). Когда известно точное местоположение гена, фланкирующие маркеры, окружающие ген, могут быть использованы для отбора рекомбинаций в популяциях разных размеров. Например, в меньших по размеру популяциях можно ожидать рекомбинации дальше от гена, поэтому для обнаружения рекомбинации потребуются более дистальные фланкирующие маркеры.
Ключевыми компонентами для реализации MAS являются: (i) определение популяции, в рамках которой будет определена ассоциация маркер-признак, которая может быть сегрегирующей популяцией или случайной или структурированной популяцией; (ii) мониторинг сегрегации или ассоциации полиморфных маркеров относительно признака и определение сцепления или ассоциации с использованием статистических способов; (iii) определение набора желательных маркеров на основе результатов статистического анализа, и (iv) использование и/или экстраполяция этой информации к текущему набору селекционной зародышевой плазмы, чтобы сделать возможными селекционные решения на основе маркеров. Маркеры, описанные в этом изобретении, а также другие типы маркеров, такие как SSR и FLP, можно использовать в протоколах селекции при помощи маркеров.
SSR можно определить как относительно короткие серии тандемных повторов ДНК длиной 6 п.н. или меньше (Tautz (1989) Nucleic Acid Research 17: 6463-6471; Wang et al. (1994) Theoretical and Applied Genetics, 88: 1-6). Полиморфизмы возникают из-за различий в количестве повторяющихся звеньев, вероятно, вызванных проскальзыванием во время репликации ДНК (Levinson and Gutman (1987) Mol Biol Evol 4: 203-221). Изменение длины повторов может быть обнаружено путем конструирования ПЦР-праймеров для консервативной неповторяющейся фланкирующей области (Weber и май (1989) Am J Hum Genet. 44: 388-396), SSR очень подходят для картирования и MAS, поскольку они мультиаллельные, кодоминантные, воспроизводимые и поддаются высокопроизводительной автоматизации (Rafalski et al. (1996). Generating and using DNA markers in plants. In Non-mammalian genomic analysis: a practical guide. Academic Press, стр. 75-135).
Можно получать различные виды SSR-маркеров, а профили SSR из устойчивых линий можно получать путем электрофореза продуктов амплификации в геле. Оценка маркерного генотипа основана на размере амплифицированного фрагмента.
Также могут быть созданы различные типы FLP-маркеров. Чаще всего амплификационные праймеры применяются для полиморфизмов длин фрагментов. Такие маркерные FLP во многом похожи на маркерные SSR, за исключением того, что амплифицируемая праймерами область не является, как правило, областью с высоким содержанием повторов. Кроме того, амплифицированная область, или ампликон, будет иметь достаточную вариабельность среди зародышевой плазмы, часто из-за вставок или делеций, так что фрагменты, генерируемые амплификационными праймерами, могут различаться среди полиморфных индивидуумов.
SNP-маркеры выявляют нуклеотидные замены одной пары оснований. Из всех типов молекулярных маркеров SNPs являются наиболее распространенными, поэтому обладают потенциалом для обеспечения самого высокого разрешения генетической карты (Bhattramakki et al. 2002 Plant Molecular Biology 48: 539-547). SNP можно анализировать с еще более высокой пропускной способностью, чем SSR, так называемым «ультра-высокопроизводительным» способом, поскольку они не требуют большого количества ДНК, и анализ автоматизации может быть простым. SNP также обещают быть относительно недорогими системами. Эти три фактора вместе делают SNP очень привлекательными для применения в МАС. Для генотипирования SNP доступно несколько способов, включая гибридизацию, удлинение праймера, расщепление олигонуклеотида, расщепление нуклеазой, минисеквенирование и кодируемые сферы. Такие способы были рассмотрены в: Gut (2001) Hum Mutat 17 pp, 475-492: Shi (2001) Clin Chem 47, pp. 164-172; Kwok (2000) Pharmacogenomics 1, pp. 95-100: Bhattramakki and Rafalski (2001) Discovery and application of single nucleotide polymorphism markers in plants. В кн .: R, J Henry, Ed, Plant Genotyping: The DNA Fingerprinting of Plants, CABI Publishing, VVallingford. Широкий спектр коммерчески доступных технологий использует эти и другие способы получения информации о SNP, включая Masscode™. (Qiagen), Invader® (Third Wave Technologies), SnapShot® (Applied Biosystems), Taqman® (Applied Biosystems) и Beadarrays™ (Illumina).
Ряд SNP вместе в пределах последовательности, или вдоль сцепленны последовательностей можно использовать для описания гаплотипа для любого конкретного генотипа (Ching et al. (2002), BMC Genet. 3:19 pp. Gupta et al. 2001, Rafalski (2002b), Plant Science 162: 329-333). Гаплотипы могут быть более информативными, чем отдельные SNP и могут быть более описательными для любого конкретного генотипа. Например, одиночный SNP может представлять собой аллель T для конкретной линии или сорта с повышенной устойчивостью к Orobanche, но аллель T может также встречаться в селекционной популяции подсолнечника, используемой в качестве рекуррентных родителей. В этом случае гаплотип, например, комбинация аллелей в сцепленных SNP-маркерах, может быть более информативным. Как только уникальный гаплотип был присвоен хромосомной области донора, этот гаплотип можно использовать в этой популяции или любой ее подгруппе, чтобы определить, есть ли у индивида определенный ген. См., например, WO 2003054229. Использование автоматизированных высокопроизводительных платформ для детекции маркеров, известных специалистам в данной области, делает этот процесс высокопроизводительным и эффективным.
Последовательности, указанные в таблице 1, могут быть легко использованы для получения дополнительных полиморфных SNP (и других маркеров) в пределах интервала QTL, указанного в этом описании. Маркеры в пределах описанной области карты могут быть гибридизованы с BAC или другими геномными библиотеками, или электронно выровнены с геномными последовательностями, чтобы найти новые последовательности в том же приблизительном местоположении, что и описанные маркеры.
В дополнение к SSR, FLP и SNP, как указано выше, также широко используются другие типы молекулярных маркеров, в том числе в качестве неограничивающих примеров, маркеры, полученные из EST, RAPD и другие маркеры на основе нуклеиновой кислоты.
Профили изозимов и связанные морфологические характеристики могут, в некоторых случаях, также косвенно использоваться в качестве маркеров. Несмотря на то, что они напрямую не определяют различия ДНК, на них часто влияют специфические генетические различия. Однако маркерн, которые обнаруживают вариации ДНК, гораздо более многочисленны и полиморфны, чем изозим или морфологические маркеры (Tanksley (1983) Plant Molecular Biology Reporter 1: 3-8).
Выравнивание последовательностей или контигов также можно использовать для поиска последовательности вверх или вниз по хромосоме от конкретных маркеров, перечисленных в документе. Эти новые последовательности, близкие к маркерам, описанным в документе, затем используют для обнаружения и разработки функционально эквивалентных маркеров. Например, различные физические и/или генетические карты выравнивают, чтобы локализовать эквивалентные маркеры, не описанные в этом изобретении, но находящиеся в аналогичных областях. Эти карты могут быть в пределах видов подсолнечника или даже среди других видов, чьи геномы имеют некоторый уровень коллинеарности на макро- и микроуровне с подсолнухом, такие как рис и сорго.
В основном, MAS использует полиморфные маркеры, которые были идентифицированы как имеющие значительную вероятность совместной сегрегации с устойчивостью к Orobanche. Предполагается, что такие маркеры картируются рядом с локусами количественных признаков (QTL), дают растению свой устойчивый к Orobanche фенотип, и считаются индикаторами, или маркерами, для желаемого признака. Маркеры проверяет растение подсолнечника на наличие желаемого аллеля, и те, которые содержат желаемый генотип в одном или нескольких локусах, как ожидается, передадут желаемый генотип вместе с желаемым фенотипом своему потомству. Способы идентификации растений подсолнечника, которые имеют повышенную устойчивость к Orobanche, путем идентификации растений, имеющих указанный аллель в любом из маркерных локусов, описываемых в документе, включая DHAI000240, DHAG000732, DHAI007796 и DHAI007334, представлены в настоящем документе.
Интервал, представленный в этом документе, находит применение в MAS для селекции растений, которые демонстрируют повышенную устойчивость к Orobanche. Любой маркер, который картируется в интервале LG4, определенном и включающем DHAI000240 и DHAI007796, можно использовать для этой цели. Кроме того, гаплотипы, включающие аллели в одном или нескольких маркерных локусах в пределах интервалов LG4, определяемых и включающих в себя DHAI000240 и DHAG000732, DHAG000732 и DHAI007334, или DHAI007334 и DHAI007796, можно использовать для введения повышенной устойчивости к Orobanche в линии или сорта подсолнечника. Любой аллель или гаплотип, который находится в неравновесном сцеплении с аллелем, связанным с повышенной устойчивостью к Orobanche, можно использовать в MAS для селекции растений с повышенной устойчивостью к Orobanche.
ПРИМЕРЫ
Следующие примеры предлагаются для иллюстрации, но не для ограничения прилагаемой формулы изобретения. Следует понимать, что примеры и варианты осуществления, описываемые в настоящем документе, предназначены только для иллюстративных целей и что специалисты в данной области знают различные реагенты или параметры, которые могут быть изменены, без выхода за пределы сущности или объема прилагаемой формулы изобретения.
Пример 1: Картируемая популяция и фенотипические данные
Картируемая популяция F2 для определения устойчивости к Orobanche получали от скрещивания восприимчивой запатентованной B-линии DAS с устойчивой линией Deb-2, полученной от Consejo Superior de Investigaciones Científicas (CSIC). Deb-2 переносит ген (-ы) устойчивости к расе G Orobanche от Helianthus debilis. Картируемая популяция состояла из 289 F2 индивидуумов в дополнение к родителям.
Популяцию проверяли на устойчивость к Orobanche к расе G. Семена подсолнечника проращивали в горшках в отсутствие Orobanche. Через два дня после прорастания проросшие семена были высажены в почву, зараженную Orobanche расы G. Полив растений был остановлен после 38-го дня и начался процесс оценки. На 45-й день почву и растения удаляли из горшка, и оценивали растения на основе появления растений Orobanche в категории «устойчивые (без появления растений Orobanche)» или «восприимчивые (1 или более растений Orobanche). Оценивали пять растений для каждого F2.
Пример 2: Генотипирование популяции F2 и картирование QTL
Сто восемь индивидуумов из популяции F2 генотипировали с патентованным DAS чипом с 9000 SNP подсолнечника (Illumina, San Diego, CA). Получали карту геномного сцепления, состоящую из 807 SNP-маркеров в 20 группах сцепления. Основной QTL устойчивости, OrDeb-2, был картирован в группе сцепления 4 генома подсолнечника (LG 4; Фиг. 1). QTL объяснил 64,4% от общего фенотипического отклонения в популяции картирования F2 и имел показатель LOD 40,4 (фиг. 2). Пик QTL пришелся на интервал в 3906780 п.н. между SNP-маркерами DHAI000240 и DHAI007796.
JoinMap 4.0 (Van Ooijen, JW et al, 2006) использовали для создания генетической карты сцепления. Для JoinMap 4.0 требуется только один входной файл от популяции, обозначенный как файл локусов генотипов. В файле локусов генотипов популяции DH, восприимчивые родительские аллели были названы «A», устойчивые родительские аллели были названы «B». Недостающие данные были представлены тире (-) в файле локусов генотипов. MapQTL 6.0 (Van Ooijen, JW et al, 2008) использовали для сопоставления QTL. MapQTL 6.0 требуется три входных файла, включая файл локусов генотипов, файл карты и файл количественных данных. Файл локусов генотипов (файл loc) содержит коды генотипов для всех локусов сегрегирующей популяции, как описано выше. Файл карты содержит данные от JoinMap и содержит предполагаемые положения на карте для всех локусов. Файл количественных данных (файл qua) содержал оценки Orobanche для каждой линии картируемой популяции. Использовали способ интервального картирования. Когда показатель LOD превышал порог значимости (1000 результатов теста перестановки) в группе сцепления, выявляли сегрегационный QTL; положение с наибольшим LOD в группе сцепления было расчетным положением QTL на карте.
Маркеры, идентифицированные в интервале QTL с использованием MapQTL 6.0, сравнивали в программе BLAST с общедоступным эталонным геномом подсолнечника, сборкой Ha412HO bronze, для определения физического местоположения маркеров. Физическое местонахождение маркеров описано в таблице 1.
Пример 3: Дизайн анализа конкурентной аллель-специфической ПЦР (KASPar™)
Анализы KASPar™ были разработаны с помощью менеджера документооборота Kraken™ (LGC Genomics, Beverly, MA) с использованием последовательностей, сконструированных для анализа, которые представляли собой геномные последовательности, фланкирующие SNP. Маркеры в интервале QTL, связанные с ними последовательности и аллели устойчивости описаны в таблице 1.
Генотипирование SNP проводили с использованием анализов KASP™ для SNP. ДНК, объемом 2 мкл (1,5-2,5 нг/мкл), разливали в 384-луночные ПЦР-планшеты. Планшеты центрифугировали в течение 1 минуты при 3000 об./мин. и сушили в течение 2 часов при 65°C. Реакционную смесь KASP™ получали с использованием 1× мастер-микса KASP™ (LGC Genomics, Beverly, MA), как описано производителем, и добавляли 4,0 мкл реакционной смеси добавлены в ПЦР-планшеты. Планшеты были запечатаны лазером оптически прозрачной постоянной пленкой и центрифугированы в течение 5 минут при 3000 об./мин. Термоциклирование проводили с использованием программы для ступенчатой ПЦР в Hydrocycler™ (LGC Genomics) со следующими условиями: 94°С в течение 15 минут, затем 10 циклов при 94°С в течение 20 секунд и ступенчатый спуск до 65-57°С в течение 60 секунд (падение 0,8°С за цикл). За этим последовали 29 циклов при 94°C в течение 20 секунд и отжиг при 57°C в течение 60 секунд. После ПЦР планшеты центрифугировали при 2500 об./мин. в течение 1 минуты, и полученные продукты анализировали с использованием микроспектрофотометра PHERAStar для чтения планшетов (BMG Labtech, Cary, NC), используя химию красителей FAM и HEX, чтобы различать генотипы. Затем данные оценивали с использованием приложения KlusterCaller в системе управления лабораторной информацией Kraken (LGC Genomics).
После завершения KASP™ ПЦР и считывания флуоресценции, необработанные данные интенсивности флуоресценции непосредственно из спектрофотометра для чтения планшетов анализировали в Kraken Workflow Manager™ (LGC Genomics). Получали график с ОФУ (единица относительной флуоресценции) FAM в качестве оси x и VIC или HEX в качестве оси y. Определения зиготности были сделаны на основе разделения кластера в режиме кластера.
Таблица 1: Анализ последовательностей KASP™ по SNP-маркерам, расположенным в интервале QTL LG 4 для OrDeb-2. Физическое положение маркеров основано на сборке Ha412HO bronze.
(SEQ ID NO)
(SEQ ID NO)
(SEQ ID NO)
Пример 4: Валидация маркеров
Проводили скрининг маркеров, ассоциированных с QTL Orobanche на LG 4, против второй популяции с источником устойчивости Deb-2, скрещенной с восприимчивой элитной линией. Исследовали фенотип родителей и 180 потомков, как описано ранее, а затем генотипировали по DHAI000240, DHAG000732, DHAI007796 и DHAI007334. 128 потомков с фенотипами устойчивости также имели устойчивые или гетерозиготные генотипы. 52 потомка с восприимчивыми фенотипами также имели генотипы восприимчивости (Таблица 2).
Таблица 2: Результаты валидации маркеров на второй популяции с устойчивостью, связанной с Deb-2. Представлено число побегов Orobanche и распознавание фенотипа и генотипа, где S=восприимчивый, R=устойчивый, A=восприимчивый аллель, B=устойчивый аллель, и H=гетерозиготный.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Dow AgroSciences, LLC
Gao, Wenxiang
Ripley, Van
Channabasavaradhya, Chandra
Meyer, David
Velasco, Leonardo
Benson, Robert
Perez Vich, Begona
Erickson, Angela
Fernandez Martinez, Jose Maria
Ren, Ruihua
Avery, Milan
<120> МОЛЕКУЛЯРНЫЕ МАРКЕРЫ, АССОЦИИРОВАННЫЕ С УСТОЙЧИВОСТЬЮ
ПОДСОЛНЕЧНИКА К OROBANCHE
<130> 74816-US-PSP
<160> 32
<170> PatentIn version 3.5
<210> 1
<211> 120
<212> ДНК
<213> Helianthus annuus
<220>
<221> прочий признак
<222> (58)..(58)
<223> n является a, t, c, or g
<400> 1
agtgattgtg ggagcatggt taggtgccgc tagatcgttg gacaaacagt tcaccgcnct 60
raggcaagag gaggagctcg agaaggagat ggaaagagcc gcggttaaaa taccgattgt 120
<210> 2
<211> 41
<212> ДНК
<213> Искусственная последовательность
<220>
<223> DHAI000240_A1
<400> 2
gaaggtgacc aagttcatgc tcgagctcct cctcttgcct t 41
<210> 3
<211> 41
<212> ДНК
<213> Искусственная последовательность
<220>
<223> DHAI000240_A2
<400> 3
gaaggtcgga gtcaacggat tcgagctcct cctcttgcct c 41
<210> 4
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> DHAI000240_C
<400> 4
ccgctagatc gttggacaaa cagtt 25
<210> 5
<211> 120
<212> ДНК
<213> Helianthus annuus
<220>
<221> прочий признак
<222> (65)..(65)
<223> n является a, t, c or g
<400> 5
ttctaaaatc cacgtaaaag tacagtattt cgatgttaca caagatcgta actgggaccg 60
ygganttaaa agtgctaaat atcccggagt tccgtacacg ttctttggtc agagaaacgg 120
<210> 6
<211> 44
<212> ДНК
<213> Искусственная последовательность
<220>
<223> DHAI009612_A1
<400> 6
gaaggtgacc aagttcatgc tcacaagatc gtaactggga ccgt 44
<210> 7
<211> 43
<212> ДНК
<213> Искусственная последовательность
<220>
<223> DHAI009612_A2
<400> 7
gaaggtcgga gtcaacggat tacaagatcg taactgggac cgc 43
<210> 8
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> DHAI009612_C
<400> 8
cggaactccg ggatatttag cactt 25
<210> 9
<211> 99
<212> ДНК
<213> Helianthus annuus
<220>
<221> прочий признак
<222> (97)..(97)
<223> n является a, t, c or g
<400> 9
gagaaacgga tgtcgggttt cgttatatca agatgcgcac gtccccgatg gtttcattcc 60
yaaaataccc cttgctaacg gggaatttta cgagccnca 99
<210> 10
<211> 45
<212> ДНК
<213> Искусственная последовательность
<220>
<223> DHAG000732_A1
<400> 10
gaaggtgacc aagttcatgc ttccccgtta gcaaggggta tttta 45
<210> 11
<211> 44
<212> ДНК
<213> Искусственная последовательность
<220>
<223> DHAG000732_A2
<400> 11
gaaggtcgga gtcaacggat tccccgttag caaggggtat tttg 44
<210> 12
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> DHAG000732_C
<400> 12
cgcacgtccc cgatggtttc at 22
<210> 13
<211> 119
<212> ДНК
<213> Helianthus annuus
<220>
<221> прочий признак
<222> (99)..(99)
<223> n является a, t, c or g
<400> 13
gatmccccgt ttcattcgct tttccgaacg cttgataccg cccaccatga cgatttccay 60
cagccgaact acaccggcgc ttccataacc aaaggcggnc cccgcgaacc gtggcacga 119
<210> 14
<211> 41
<212> ДНК
<213> Искусственная последовательность
<220>
<223> DHAI009613_A1
<400> 14
gaaggtgacc aagttcatgc tcgccggtgt agttcggctg a 41
<210> 15
<211> 40
<212> ДНК
<213> Искусственная последовательность
<220>
<223> DHAI009613_A2
<400> 15
gaaggtcgga gtcaacggat tgccggtgta gttcggctgg 40
<210> 16
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> DHAI009613_C
<400> 16
ttgataccgc ccaccatgac gattt 25
<210> 17
<211> 251
<212> ДНК
<213> Helianthus annuus
<400> 17
ttctgtcgga gttacctggg taattgttcg tttttgtatt cgattgttct accaaaaaga 60
aacatcatca tcatcggcat ccatagaacg tgatgtgttt aataggtata atgtggcraa 120
actctaacag ttgattcgtc caggttcgtt cgtggatgct tacatctctt tagattcgta 180
tgacgatcaa caaaacgcca ggattcgatt agtagctaag aagctgattg ttgattctaa 240
ctgaagggtt t 251
<210> 18
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> DHAI007334_A1
<400> 18
gaaggtgacc aagttcatgc tgtgatgtgt ttaataggta taatgtggca 50
<210> 19
<211> 48
<212> ДНК
<213> Искусственная последовательность
<220>
<223> DHAI007334_A2
<400> 19
gaaggtcgga gtcaacggat tgatgtgttt aataggtata atgtggcg 48
<210> 20
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> DHAI007334_C
<400> 20
aacctggacg aatcaactgt tagagttt 28
<210> 21
<211> 86
<212> ДНК
<213> Helianthus annuus
<400> 21
tcaaaggatg tctttgacca gaaagcyagt aataagtact ggattgatgt ccagttgcgt 60
tggggtgatt atgattctca tgatat 86
<210> 22
<211> 47
<212> ДНК
<213> Искусственная последовательность
<220>
<223> DHAI000243_A1
<400> 22
gaaggtgacc aagttcatgc tcaaaggatg tctttgacca gaaagct 47
<210> 23
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> DHAI000243_A2
<400> 23
gaaggtcgga gtcaacggat taaaggatgt ctttgaccag aaagcc 46
<210> 24
<211> 29
<212> ДНК
<213> Искусственная последовательность
<220>
<223> DHAI000243_C
<400> 24
gcaactggac atcaatccag tacttatta 29
<210> 25
<211> 120
<212> ДНК
<213> Helianthus annuus
<220>
<221> прочий признак
<222> (94)..(95)
<223> n является a, t, c or g
<400> 25
gggtcaaaag cgtcttggtc aacttgccaa gtggaaaaca gcagaagaag tggctgctct 60
kgttcgttcc ttacccgttg aagaacaacc caanncaaat cattgtgaca cgtaaaggaa 120
<210> 26
<211> 44
<212> ДНК
<213> Искусственная последовательность
<220>
<223> DHAI009614_A1
<400> 26
gaaggtgacc aagttcatgc tcagcagaag aagtggctgc tctt 44
<210> 27
<211> 43
<212> ДНК
<213> Искусственная последовательность
<220>
<223> DHAI009614_A2
<400> 27
gaaggtcgga gtcaacggat tagcagaaga agtggctgct ctg 43
<210> 28
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> DHAI009614_C
<400> 28
gggttgttct tcaacgggta aggaa 25
<210> 29
<211> 203
<212> ДНК
<213> Helianthus annuus
<400> 29
ctgctgttgc ctcccctgct actagtttct tgctgatcag gttcagtatg tgatgtatgt 60
caatctttta ttatgytgct gttgatctta taatatgatc tgtttggtat gtgtgttgat 120
tatgcgtata tagtcaaatt ttaacatggc ccgtttcgac ccgtacctat tattattatc 180
aaaagttcaa aacccatttt gac 203
<210> 30
<211> 52
<212> ДНК
<213> Искусственная последовательность
<220>
<223> DHAI007796_A1
<400> 30
gaaggtgacc aagttcatgc tcaaacagat catattataa gatcaacagc aa 52
<210> 31
<211> 51
<212> ДНК
<213> Искусственная последовательность
<220>
<223> DHAI007796_A2
<400> 31
gaaggtcgga gtcaacggat taaacagatc atattataag atcaacagca g 51
<210> 32
<211> 29
<212> ДНК
<213> Искусственная последовательность
<220>
<223> DHAI007796_C
<400> 32
gatcaggttc agtatgtgat gtatgtcaa 29
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| МОЛЕКУЛЯРНЫЕ МАРКЕРЫ НИЗКОГО СОДЕРЖАНИЯ ПАЛЬМИТИНОВОЙ КИСЛОТЫ В ПОДСОЛНЕЧНИКЕ (HELIANTHUS ANNUS) И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2013 |
|
RU2670517C2 |
| SNP-МАРКЕРЫ И ОТБОР В ОТНОШЕНИИ НИЗКОГО СОДЕРЖАНИЯ КЛЕТЧАТКИ В ПРЕДСТАВИТЕЛЯХ РОДА BRASSICA | 2019 |
|
RU2834673C2 |
| ГЕНЕТИЧЕСКИЕ ЛОКУСЫ, СВЯЗАННЫЕ С УСТОЙЧИВОСТЬЮ К ПЫЛЬНОЙ ГОЛОВНЕ У МАИСА | 2009 |
|
RU2562864C2 |
| ГЕНЕТИЧЕСКИЙ ЛОКУС, АССОЦИИРОВАННЫЙ С ГНИЛЬЮ КОРНЯ И СТЕБЛЯ, ОБУСЛОВЛЕННОЙ PHYTOPHTHORA, У СОИ | 2016 |
|
RU2748688C2 |
| РАСТЕНИЯ ОГУРЦА, УСТОЙЧИВЫЕ К ЗАБОЛЕВАНИЯМ | 2006 |
|
RU2418405C2 |
| РОЗА, УСТОЙЧИВАЯ К МУЧНИСТОЙ РОСЕ | 2018 |
|
RU2769284C2 |
| МОЛЕКУЛЯРНЫЕ МАРКЕРЫ ГЕНА RLM4 РЕЗИСТЕНТНОСТИ К ЧЕРНОЙ НОЖКЕ BRASSICA NAPUS И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2014 |
|
RU2718584C2 |
| РАСТЕНИЕ, УСТОЙЧИВОЕ К НАСЕКОМЫМ | 2008 |
|
RU2522477C2 |
| МОЛЕКУЛЯРНЫЕ МАРКЕРЫ ГЕНА RLM2 РЕЗИСТЕНТНОСТИ К ЧЕРНОЙ НОЖКЕ BRASSICA NAPUS И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2014 |
|
RU2717017C2 |
| РАСТЕНИЯ КУКУРУЗЫ, ОТЛИЧАЮЩИЕСЯ ЛОКУСАМ КОЛИЧЕСТВЕННЫХ ПРИЗНАКОВ QTL | 2008 |
|
RU2560599C2 |
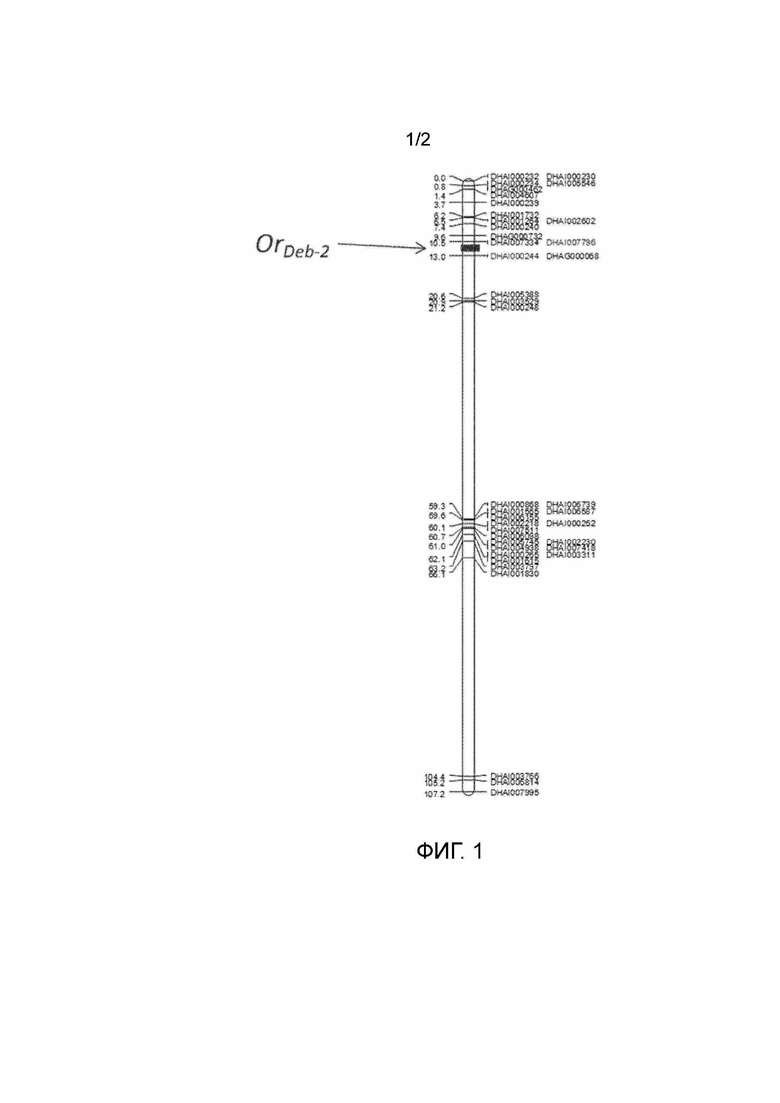
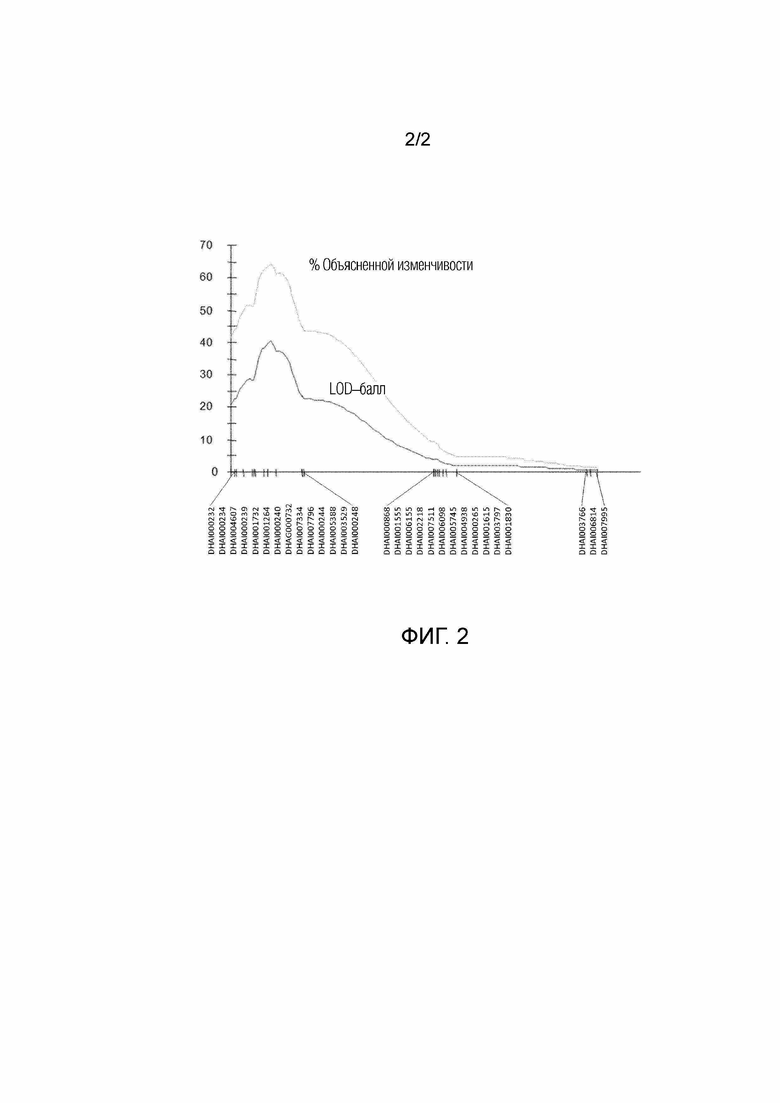
Изобретение относится к области биохимии, в частности к способу отбора растения подсолнечника, которое демонстрирует повышенную устойчивость к Orobanche. Изобретение позволяет отбирать растения подсолнечника с повышенной устойчивостью к Orobanche. 8 з.п. ф-лы, 2 ил., 2 табл., 4 пр.
1. Способ отбора растения подсолнечника, которое демонстрирует повышенную устойчивость к Orobanche, включающий стадии: a) выявления хромосомного интервала, содержащего один или более маркеров однонуклеотидного полиморфизма (SNP), представленных в SEQ ID NO: 1, SEQ ID NO: 9, SEQ ID NO: 17 или SEQ ID NO: 29, связанных с устойчивостью к Orobanche; и b) отбора растения, содержащего маркер SNP, и, таким образом, отбора растения, которое демонстрирует повышенную устойчивость к Orobanche.
2. Способ по п. 1, в котором хромосомный интервал, содержащий один или более маркеров SNP, представленных в SEQ ID NO: 1, SEQ ID NO: 9, SEQ ID NO: 17 или SEQ ID NO: 29, выявляется с помощью секвенирования ДНК, амплификации последовательности на основе ПЦР, аллель-специфической гибридизации или множества однонуклеотидных полиморфизмов (SNP).
3. Способ по п. 1, в котором выбраны по меньшей мере две маркера SNP.
4. Способ по п. 1, в котором выбраны по меньшей мере три маркера SNP.
5. Способ по п. 1, в котором выбраны по меньшей мере четыре маркера SNP.
6. Способ по п. 1, отличающийся тем, что один или более маркеров SNP выявляют с помощью множества SNP.
7. Способ по п. 1, отличающийся тем, что один или более маркеров SNP выявляют с помощью секвенирования ДНК.
8. Способ по п. 1, отличающийся тем, что один или более маркеров SNP выявляют с помощью конкурентной аллель-специфичной ПЦР.
9. Способ по п. 8, в котором одну или несколько маркерных нуклеиновых кислот определяют с использованием аллель-специфических праймеров SEQ ID NO: 2 или SEQ ID NO: 3 для SNP с SEQ ID NO: 1; SEQ ID NO: 10 или SEQ ID NO: 11 для SNP с SEQ ID NO: 9; SEQ ID NO: 18 или SEQ ID NO: 19 для SNP с SEQ ID NO: 17; или SEQ ID NO: 30 или SEQ ID NO: 31 для SNP с SEQ ID NO: 29.
| Приспособление для очистки, охлаждения, смешивания, поглощения или т.п. воздуха, газов и паров жидкостями | 1926 |
|
SU23910A1 |
| ГЕНЕТИЧЕСКИЕ МАРКЕРЫ, АССОЦИИРОВАННЫЕ С ЗАСУХОУСТОЙЧИВОСТЬЮ КУКУРУЗЫ | 2010 |
|
RU2575605C2 |
| RU 2015121354 A, 27.12.2016. | |||

