Настоящее изобретение относится к растениям розы, таким как срезочные розы, садовые розы, горшечные розы и подвои роз, содержащим по меньшей мере два гена, обеспечивающих устойчивость к болезнетворному организму, вызывающему мучнистую росу. Конкретно, настоящее изобретение относится к растениям розы, устойчивым к мучнистой росе, вызываемой болезнетворным организмом Podosphaera pannosa, также известным как Sphaerotheca pannosa var. rosae. Настоящее изобретение также относится к методам селекции растений роз по настоящему изобретению, устойчивых к мучнистой росе.
Мучнистая роса (МР) является основной болезнью листьев у срезочной, горшечной и садовой розы и вызывается облигатным биотрофным аскомицетом Podosphaera pannosa, также известным как Sphaerotheca pannosa. Симптомы заражения мучнистой росой включают замедленный рост растений и деформацию листьев из-за образования белых мучнистых конидий, которые появляются на поверхностях растения, таких как листья, цветы, стебли и бутоны. Не существует официальных показателей потерь при производстве розы в результате мучнистой росы, но борьба с мучнистой росой на трех основных тепличных культурах: огурцах, томатах и розах - в Канаде обходится в C$ 6000/га.
С болезнью мучнистой росой можно бороться с помощью химических препаратов, но применение фунгицидов является дорогостоящим, трудоемким и вредным для окружающей среды. Правительствами во всем мире устанавливаются все более жесткие правила по применению химических препаратов в садоводстве, что в комбинации с вышеуказанными факторами означает, что создание разновидностей розы, устойчивой к мучнистой росе, является основным фактором для управления и избегания финансовых потерь из-за массового появления мучнистой росы.
Хотя Podosphaera pannosa способен заражать широкий ряд реципиентов, включая Prunus cerasus и Prunus avium, взаимодействие между Podosphaera pannosa и розой характеризуется высокой степенью специфичности. Например, некоторые патотипы гриба описаны полвека назад, и более поздний анализ для проверки совместимости между 8 монокондиальными изолятами и 18 генотипами реципиентов показал высокий уровень специфичности и видовое разнообразие.
Срезочные розы восприимчивы к мучнистой росе (МР). Для того, чтобы вывести срезочные розы, устойчивые к МР, необходимо идентифицировать гены устойчивости. Некоторые публикации показывают данные для генетической основы устойчивости к мучнистой росе в роду Rosa, причем устойчивость распространяется от количественной до качественной устойчивости. Однако коммерчески важные срезочные розы (Rosa hybrida), как правило, тетраплоидные, и локусы количественных признаков (ЛКП (QTL)) для устойчивости к мучнистой росе показаны главным образом в других видах или разновидностях, чем Rosa hybrida.
Например, с использованием диплоидной гибридной BC1 популяции Rosa multiflora (n=117), созданной скрещиванием устойчивой линии (88/124-46) с восприимчивой линией (82/78-1) и возвратным скрещиванием гибрида F1 95/13-90 с 82/78-1, обнаружено, что ген Rpp1 является главным действующим геном в группе сцепления 3, придающей устойчивость к МР до 10 дней после инокуляции с действием по доминантному моногенному типу. С использованием другой диплоидной гибридной популяции R. multiflora F1 (n=270), созданной скрещиванием устойчивой линии (95/13-39) с восприимчивой линией (Sp3 или 82/78-1) из той же программы селекции перекрестным опылением, которая предполагает интрогрессию генов из тетраплоидных садовых роз в Rosa multiflora, картированы ЛКП устойчивости в шести различных средах. В целом обнаружено 28 различных ЛКП в группах сцепления 1, 2, 3, 4, 6 и 7, причем сильный кластер ЛКП образуется в группах сцепления 3 и 4, что в совокупности является показателем полигенного механизма устойчивости. Существенная пропорция наблюдаемой фенотипической вариансы в отношении устойчивости (31%) унаследована от восприимчивого родителя, и, что интересно, вышеуказанное действие гена Rpp1 в этой популяции не видно.
При диплоидном скрещивании (n=90) между розой сорта «Yesterday» и Rosa wichurana 9 ЛКП для устойчивости до 10 дней после инокуляции против 2 рас обнаружены в группах сцепления 2, 3, 5 и 6, причем каждый объясняет от 15% до 74% фенотипической вариансы. Из 9 ЛКП только один обнаружен с использованием обеих рас. Хотя большинство аллелей устойчивости происходит из Rosa wichurana, оба родителя вносят вклад в изменение устойчивости.
Единственный пример обеспечивающих устойчивость ЛКП в тетраплоидной срезочной розе - стебли популяции К5 популяции F1, которая создана скрещиванием двух культурных сортов P540 и P867, которые оба являются только частично устойчивыми. Использовали два различных изолята моноспорового гриба, и устойчивость оценивали с использованием трех различных оценок болезни: оценку болезни через 11 дней после инокуляции (dpi), латентный период и скорость развития симптомов. Для каждой из трех оценок болезни показано от 16 до 28 маркеров, перекрывающих все 7 групп сцепления, ассоциированных с мучнистой росой, хотя строгое статистическое обоснование не дается. Хотя ряд маркеров ассоциируется с различными оценками болезни, ни один из маркеров не обнаружен для обоих изолятов. Для каждой из шести комбинаций изолята и оценки болезни затем выполняли несколько регрессий с использованием 4 или 5 из наиболее значимых маркеров. Фенотипическая варианса, объясняемая несколькими регрессиями, колеблется от 10,4 до 22,3%, показывая, что устойчивость в этом гибриде является количественной и регулируется несколькими генами с только минимальным действием каждого.
Таким образом, хотя опубликовано несколько исследований ЛКП для устойчивости розы к мучнистой росе, общей темой является то, что имеется мало последовательности и совпадения в опубликованных результатах. Например, устойчивость колеблется от моногенной до количественной. Кроме того, ЛКП не участвуют в различных исследованиях, что предполагает, что может иметь место гетерогенность в генетических механизмах, лежащих в основе узнавания белков гриба и, таким образом, устойчивости к мучнистой росе. Частично такая гетерогенность может являться результатом разнообразия генов среди и даже внутри изолятов, которое в комбинации с гипотетической моделью взаимодействия ген-для-гена объясняет, что локации и размеры действия ЛКП могут зависеть от используемого изолята.
С другой стороны, отсутствие подтверждения ставит под сомнение надежность сделанных выводов из-за нередко малого размера образца или малого покрытия маркера или их комбинации. Диплоидную популяцию F1 Rosa roxburghii (сорт Guinong №. 6 × сорт Guinong № 5) оценивали при естественных нагрузках болезни, 4 аналога гена устойчивости (RGA), которые не могут быть приписаны к группе сцепления, ассоциируются с CRPM1 - главным R локусом, который не приписан к LG, что объясняет 72% фенотипического изменения при устойчивости к мучнистой росе.
Устойчивость на основе локуса устойчивости к мучнистой росе (MLO) сначала идентифицировали у ячменя (Hordeum vulgare) как рецессивный аллель, обеспечивающий надежную устойчивость против всех изолятов милдью (сем. Blumeria graminis, sp. horde). Устойчивость на основе MLO не является уникальной для ячменя, так как с тех пор ортологи MLO открыты во многих растениях других родов, таких как Pisum, Arabidopsis и Solanum, где показано, что утрата мутаций в функциональных генах MLO ведет к широкому спектру устойчивости к мучнистой росе. Семейство белков MLO состоит из 7 кладов, и при всей распространенности в наземных растениях, функция большинства генов MLO неизвестна, и все гены MLO, которые, как показано, ассоциируются с устойчивостью к мучнистой росе, обнаружены в кладе V. У яблони (Malus domestica) - члена семейства Rosaceae, из 21 гена MLO только три гомолога MLO, из которых два принадлежат к кладу V, активируются после заражения мучнистой росой. Это предполагает, что обнаружение гомологов MLO само по себе не является достаточным для идентификации гена, вовлеченного в устойчивость к мучнистой росе.
Однако, несмотря на надежность, устойчивость на основе MLO часто имеет разрушительное плейотропное действие, такое как некротическая пятнистость листьев и пониженная энергия, создавая таким образом внушительные препятствия при планировании стратегии селекции. У розы (диплоидные гибриды Rosa multiflora и тетраплоидный Rosa hybrida, сорт Pariser Charme) четыре гена MLO, созданные RhMLO1 до 4, идентифицированы на основе гомологии их последовательностей с хорошо охарактеризованными последовательностями, полученными у ячменя и Arabidopsis. Каждый ген, размещенный между двумя и шестью аллелями, состоит из 15 экзонов, и полная длина кодирующей последовательности составляла во всех случаях приблизительно 1700 п.о.. Гены MLO распределены по нескольким группам сцепления, причем rhMLO3 и rhMLO4 группируются вместе в LG1 (между 40 и 45 cM), rhMLO2 обнаружен в LG3 (ca. в 35 cM), и rhMLO1 обнаружен в LG5 около 60 cM.
На сегодня известно, что нет потери функциональных мутантов, но показано, что все четыре гомолога MLO принадлежат кладу V - единственному кладу, для которого известно, что гены MLO могут играть роль в устойчивости к PM. Только один из четырех генов MLO, обнаруженных в розе, функционально связан с устойчивостью к мучнистой росе, так как трансформация Rosa multiflora "Baiyu" антисмысловым rhMLO1 дала растения с повышенными (но не абсолютными) уровнями устойчивости к мучнистой росе (до 15 дней после инокуляции) по сравнению с нетрансгенными контрольными растениями.
Учитывая вышеизложенное, в технике существует потребность в дополнительном предоставлении генов устойчивости к мучнистой росе.
Целью настоящего изобретения, среди других целей, является удовлетворение указанной выше потребности в технике.
Согласно настоящему изобретению, указанная выше цель, среди других целей, удовлетворяется предоставлением растений розы путем, изложенным в прилагаемой формуле изобретения.
Конкретно, указанная выше цель, среди других целей, удовлетворяется путем предоставления растений розы, устойчивых к мучнистой росе и включающих в своем ядерном геноме по меньшей мере одну нуклеотидную последовательность, представленную SEQ ID No. 1, и по меньшей мере одну нуклеотидную последовательность, представленную SEQ ID No. 2, причем совместное присутствие SEQ ID No. 1 и SEQ ID No. 2 в указанном ядерном геноме обеспечивает устойчивость к мучнистой росе.
Авторы настоящего изобретения неожиданно обнаружили, что комбинация SEQ ID No. 1 и SEQ ID No. 2 обеспечивает прочную устойчивость к мучнистой росе. Согласно изобретению, существенно, что объединяются оба гена устойчивости к мучнистой росе, поскольку SEQ ID No. 1, в отсутствие SEQ ID No. 2, предоставляет неподдающуюся детекции устойчивость розы к мучнистой росе, в то время как SEQ ID No. 2 в отсутствие SEQ ID No. 1 обеспечивает слабую устойчивость к мучнистой росе. Авторы настоящего изобретения также неожиданно обнаружили, что гены устойчивости по настоящему изобретению являются доминантными генами, т.е. присутствие по одной копии обоих генов является достаточным для обеспечения устойчивости к мучнистой росе.
Согласно предпочтительному воплощению, настоящее изобретение относится к устойчивости к мучнистой росе против болезнетворного для растений организма аскомицета Podosphaera pannosa, также известного как Sphaerotheca pannosa var. rosae. Для розы болезнетворный для растений организм аскомицет Podosphaera pannosa является основным болезнетворным организмом, вызывающим мучнистую росу.
Согласно другому предпочтительному воплощению, растениями розы по настоящему изобретению являются растения Rosa hybrida, и ядерный геном по настоящему изобретению является геномом тетраплоида. В частности, в геномах полиплоидов, таких как геномы диплоидов, триплоидов, тетраплоидов, гексаплоидов или октаплоидов, наличие доминантных генов устойчивости к мучнистой росе обеспечивает основное преимущество избегания множества промежуточных фенотипов мучнистой росы, зависящего от числа копий присутствующих генов устойчивости. Геномы полиплоидов по настоящему изобретению, такие как геномы триплоидов, тетраплоидов, гексаплоидов или октаплоидов, можно соответственно получить прямо или косвенно через удваивание генома, включающего SEQ ID No. 1 и 2. Например, тетраплоид, гексаплоид и октаплоид можно легко получить из генома диплоида, включающего SEQ ID No. 1 и 2, и геном гексаплоида также можно получить путем удваивания генома триплоидного растения.
Согласно еще одному предпочтительному воплощению, настоящее изобретение относится к устойчивым к мучнистой росе растениям розы, включающим в своем ядерном геноме по меньшей мере одну, предпочтительно две, предпочтительнее три, даже предпочтительнее четыре нуклеотидные последовательности, представленные SEQ ID No. 1, например, 2, 3, 4, 5 или 6 в случае генома гексаплоида или, например, 2, 3, 4, 5, 6, 7 или 8 в случае генома октаплоида, и/или по меньшей мере одну, предпочтительно две, предпочтительнее три, даже предпочтительнее четыре нуклеотидные последовательности, представленные SEQ ID No. 2, например, 2, 3, 4, 5 или 6 в случае генома гексаплоида или, например, 2, 3, 4, 5, 6, 7 или 8 в случае генома октаплоида.
Согласно еще одному предпочтительному воплощению, настоящее изобретение относится к устойчивым к мучнистой росе растениям розы, включающим в своем ядерном геноме по меньшей мере одну, предпочтительно три, предпочтительнее четыре нуклеотидные последовательности, например, 2, 3, 4, 5 или 6 в случае генома гексаплоида или, например, 2, 3, 4, 5, 6, 7 или 8 в случае генома октаплоида, представленные SEQ ID No. 1, или по меньшей мере одну, предпочтительно две, предпочтительнее три и даже предпочтительнее четыре нуклеотидные последовательности, представленные SEQ ID No. 2, например, 2, 3, 4, 5 или 6 в случае генома гексаплоида или, например, 2, 3, 4, 5, 6, 7 или 8 в случае генома октаплоида.
Согласно еще одному предпочтительному воплощению, настоящее изобретение относится к устойчивым к мучнистой росе растениям розы, включающим в своем ядерном геноме по меньшей мере одну, предпочтительно две, предпочтительнее три и даже предпочтительнее четыре нуклеотидные последовательности, представленные SEQ ID No. 1, например, 2, 3, 4, 5 или 6 в случае генома гексаплоида или, например, 2, 3, 4, 5, 6, 7 или 8 в случае генома октаплоида, и по меньшей мере одну, предпочтительно две, предпочтительнее три и даже предпочтительнее четыре нуклеотидные последовательности, представленные SEQ ID No. 2, например, 2, 3, 4, 5 или 6 в случае генома гексаплоида или, например, 2, 3, 4, 5, 6, 7 или 8 в случае генома октаплоида.
Согласно особенно предпочтительному воплощению, устойчивые к мучнистой росе растения розы по настоящему изобретению выбирают из группы, включающей срезочную розу, горшечную розу, подвой розы и садовую розу, предпочтительно срезочную розу.
Согласно еще одному особенно предпочтительному воплощению, устойчивые к мучнистой росе растения розы по настоящему изобретению обнаруживают доминантный фенотип.
С учетом благоприятных свойств устойчивости к мучнистой росе, обеспеченной синергетическим эпистатическим действием между двумя доминантными генами, настоящее изобретение также относится к способам селекции устойчивого к мучнистой росе растения розы, указанного выше, включающему стадии:
а) выделения ДНК ядерного генома из растения розы;
b) установления присутствия SEQ ID No. 1 и SEQ ID No. 2 в выделенной ДНК ядерного генома;
с) установления фенотипа указанного растения розы в отношении мучнистой росы, причем присутствие SEQ ID No. 1 и SEQ ID No. 2 указывает на устойчивый к мучнистой росе фенотип.
Настоящее изобретение далее будет описываться подробнее в следующем примере. В примере делается ссылка на фигуры, описанные далее.
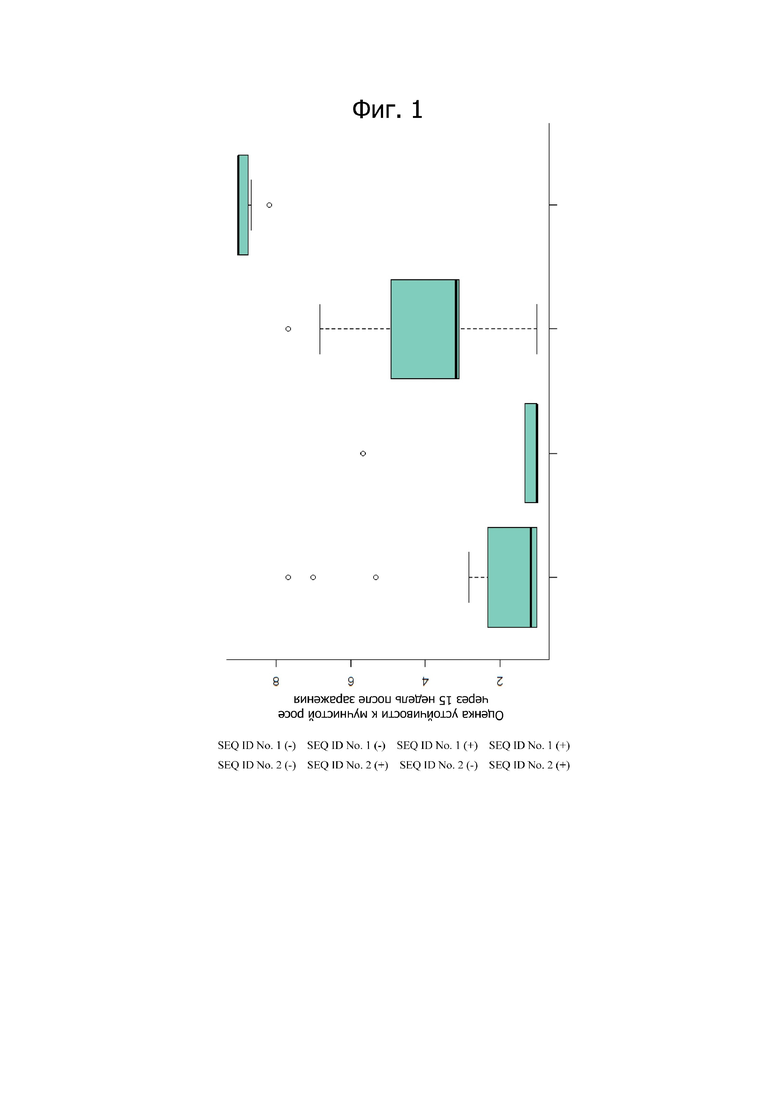
Фигура 1 представляет прямоугольную диаграмму, показывающую действие аллелей устойчивости SEQ ID No. 1 и SEQ ID No. 2 по отдельности и в тандеме. Присутствие аллеля устойчивости показано знаком «+» и нулевого аллеля устойчивости показано знаком «-». Растения с обоими аллелями устойчивости являются высоко устойчивыми.
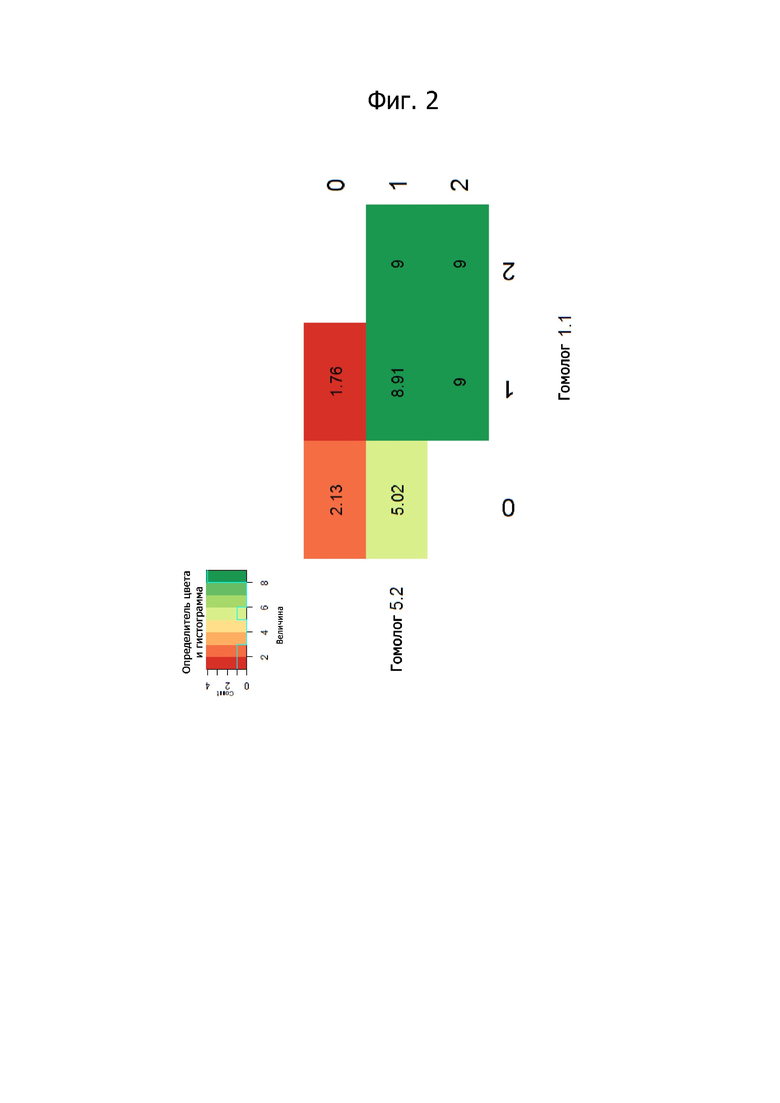
Фигура 2 представляет число копий SEQ ID No. 1 и SEQ ID No. 2, требуемое для обеспечения устойчивости к мучнистой росе по настоящему изобретению у растений розы.
Пример
Введение
Авторы изобретения в данном случае исследовали число, масштаб действия и генетические позиции ЛКТ, лежащие в основе устойчивости к РМ у популяции тетраплоидной розы (Rosa hybrida). Авторы представляют полученные данные, которые показывают отчетливое основное действие ЛКТ в группе сцепления 1 и группе сцепления 5, поясняющие 20% и 90% фенотипической вариансы в устойчивости к РМ, соответственно. Авторы также показывают, что действие ЛКТ видно только тогда, когда аллели устойчивости присутствуют в обоих ЛКТ, так как растения, имеющие аллели устойчивости в обоих ЛКТ, являются высоко устойчивыми до 15 недель после инокуляции, в то время как растения только с одним или нулевым аллелем устойчивости в указанной временной рамке показывают симптомы РМ.
Способ
Создают популяцию тетраплоидного гибрида F1 Rosa hybrida ручным опылением тетраплоидной срезочной розы RS-1183 ("Avalanche", далее называемой P1) пыльцой тетраплоидной садовой розы. Один из полученных отпрысков F1 самоопыляют для создания популяции F2. Родителей растений отпрыска 235 F1 и 42 F2 скринируют на устойчивость против PM (Podosphaera pannosa). Изолят первоначально выделяют из зараженных роз в садовой теплице, и получают инокулят по предварительному анализу на PM. Инокуляцию выполняют по схеме блоков, и шесть черенков каждого сорта рандомизируют в 6 блоков. Биоанализ осуществляют в условиях длинного дня при температуре, установленной на 20°C и 23°C для дня и ночи, соответственно. Относительная влажность чередуется от 60% в течение дня до 85% ночью. Для каждого растения популяции F1 уровни заражения оценивают через 1, 3, 6, 9, 12 и 15 недель после заражения, для каждого растения популяции F2 уровни заражения оценивают через 6 и 12 недель после заражения, и уровни заражения растений обеих популяций оценивают по шкале от 1 до 9, где 1 представляет наиболее чувствительные особи, и 9 представляет полностью устойчивые особи.
Все растения генотипируют с использованием матрицы WagRhSNP Axiom SNP. Этот чип содержит 68893 SNP, которые таргетируются двумя зондами с каждого направления. Контроль качества выполняют с использованием R пакета FitPoly, и 67779 маркеров сохраняются для 51685 SNP. После удаления SNP с более 5% потерянных данных остается 42143 маркера. В целом успешно генотипированы 232 особи F1, из которых 3 удалены, так как они были генетическими выбросами, и одна из-за потерянных фенотипических данных. Дополнительный контроль качества выполняют путем контролирования воспроизводимости генотипов родителей, неожиданной сегрегации, генотипических выбросов, асимметричных маркеров и нулевых аллелей, а также различий между растениями.
Полученную ранее генетическую карту (с использованием популяции К5) используют для картирования этих коррелированных SNP для группы сцепления (LG) и генетической позиции. Все ассоциированные SNP сегрегированы, следуя программе, где устойчивый родитель является симплексом, и восприимчивый родитель является нуллиплексом. Для хромосом, где обнаружен ЛКП, карты сцепления сконструированы в JoinMap с использованием маркеров, которые являлись симплексом в P2 и нуллиплексом в P1, и анализы ЛКП выполняют в MapQTL.
Для каждого участка генома, который значительно ассоциирован с устойчивостью к РМ, создают праймеры KASP, таргетирующие наиболее значительно ассоциированные SNP, а также один SNP с любой стороны. Праймеры KASP создают с использованием фланкирующих последовательностей зондов, таргетирующих ассоциированные SNP на матрице WagRhSNP Axiom SNP.
Родителей и в целом 48 произвольно выбранных растений F1 генотипируют со всеми SNP с использованием анализов KASP. Генотипы оценивают как число аллелей устойчивости, имеющихся у особи. Так как устойчивый родитель имеет одну копию аллеля устойчивости в каждом ассоциированном SNP, а восприимчивый родитель ноль, доза генотипа в F1 ограничивается до 0 (нуллиплекс для аллеля устойчивости) и 1 (симплекс для аллеля устойчивости).
Результаты
В целом 267 маркеров имеют корреляцию с устойчивостью к РМ >0,35. Все маркеры с высокой корреляцией найдены в группе сцепления 1 и 5 на генетической карте, полученной с использованием популяции К5. Для маркеров, которые включены в генетическую карту по описанию, а также карту, полученную с использованием популяции К5, порядок консервативный, подтверждающий, что конструкция карты сцепления для этих двух групп сцепления является успешной.
Через три недели после инокуляции SNP M23333_428 у гомолога 5.2 объясняет до 90% фенотипической вариансы (LOD = 114,2). Второй SNP G54183_559 обнаружен через 15 недель после инокуляции у гомолога LG1 1.1 (LOD =23,1) при 60,5 cM, что объясняет 20,3% фенотипической вариансы. Одновременный анализ ЛКП с использованием модели множественных ЛКП показывает, что ЛКП в гомологах 5.2 и 1.1 необходимы для абсолютной устойчивости через 15 недель. В целом с использованием анализов KASP генотипированы 86 растений с данными устойчивости к PM, из которых 48 являются потомством F1, 29 являются растениями F2 (чистое потомство одного самоопыленного растения F1), и 4 являются P1 и P2 (включая дубликаты обоих).
Авторы сначала анализировали ассоциацию между генотипами SNP и устойчивостью к PM в популяции F1. Данные генотипирования изменялись от 87% (для G8670_490) до 100%. При рассмотрении ассоциации между генотипами по KASP и устойчивостью к PM отчетливым показателем устойчивости к РМ через 15 недель после инокуляции являлось присутствие аллелей устойчивости в большинстве прочно ассоциированных SNP в обоих гомологах. Все растения с такой комбинацией генотипа показывают оценку PM больше 8 (высокая устойчивость, фиг. 1), в то время, как растения с генотипами устойчивости в одном локусе или при отсутствии таковых никогда не были высокоустойчивыми, а преимущественно высоковосприимчивыми (фиг. 2). Анализ с использованием ANOVA показывает, что синергетический эпистатический эффект является весьма значимым (таблица 1). Также геномный анализ растений розы показывает, что SEQ ID No 1 и 2 непосредственно связаны с генами устойчивости, что лежит в основе устойчивости по настоящему изобретению.
Таблица 1. Параметрические оценки влияния SEQ ID No. 1: TTTGTTCATTATAAACTCATTCCTCGCTTCCTCAACCTTCTCTGAAACGACC) и SEQ ID No. 2: GGCTTTTCGCCCTGCGTCTTGCTCTCCAAAAACTCACTACTAATTTGTCA на устойчивость к мучнистой росе (15 недель после инокуляции) из линейной модели. Положительные параметрические оценки показывают, что устойчивые генотипы более устойчивы, чем восприимчивые генотипы. Положительный член, характеризующий взаимодействие, показывает синергетическое эпистатическое действие: эффект присутствия устойчивого генотипа в одном гомологе более сильный, если устойчивый генотип также присутствует в другом гомологе.
После демонстрации того, что для придания устойчивости необходимо присутствие генов устойчивости в обоих локусах, авторы изобретения затем проверили, является ли тип действия в каждом локусе полностью доминантным, иными словами, нет ли различия в устойчивости к РМ между растениями, имеющими один аллель устойчивости в каждом локусе, и растениями, которые имеют несколько аллелей устойчивости в каждом локусе. Для этого объединили данные для популяции F2 с данными для родителей и популяции F1. Популяции F2 являются самоопыляемыми популяциями, полученными путем самоопыления растения F1 с одним аллелем устойчивости в каждом локусе, причем таким образом предполагая полисомное наследование, авторы ожидали растения с 0, 1 и 2 копиями в каждом локусе в полученной совокупности данных. Устойчивость к РМ в популяции F2 оценивали только до 12 недель после инокуляции. Устойчивость к РМ через 12 недель после инокуляции сильно коррелирует с устойчивостью к РМ через 15 недель после инокуляции (r=0,98), что означает, что ограничение анализов 12 неделями после инокуляции не оказывает существенного влияния на выводы авторов изобретения.
И действительно, ясно показано, что одного аллеля устойчивости в каждом локусе достаточно для придания абсолютной устойчивости через 12 недель после инокуляции, и присутствие нескольких генов устойчивости на локус не придает существенной дополнительной устойчивости (фиг. 2), что является четким доказательством, что аллели устойчивости являются доминантными над аллелями восприимчивости.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> ДЮММЕН ГРУП Б.В.
<120> РОЗА, УСТОЙЧИВАЯ К МУЧНИСТОЙ РОСЕ
<130> 4/2XH16/22
<150> NL2019209
<151> 2017-07-10
<160> 2
<170> BiSSAP 1.3.6
<210> 1
<211> 52
<212> DNA
<213> Rosa
<400> 1
tttgttcatt ataaactcat tcctcgcttc ctcaaccttc tctgaaacga cc 52
<210> 2
<211> 50
<212> DNA
<213> Rosa
<400> 2
ggcttttcgc cctgcgtctt gctctccaaa aactcactac taatttgtca 50
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| РАСТЕНИЯ ОГУРЦА, УСТОЙЧИВЫЕ К ЗАБОЛЕВАНИЯМ | 2006 |
|
RU2418405C2 |
| НОВЫЕ ГЕНЫ УСТОЙЧИВОСТИ К ИНФЕКЦИЯМ AGROBACTERIUM TUMEFACIENS У РОЗЫ | 2019 |
|
RU2824669C1 |
| УСТОЙЧИВАЯ К ЛОЖНОЙ МУЧНИСТОЙ РОСЕ КАПУСТА И СПОСОБ ЕЕ СЕЛЕКЦИИ | 2018 |
|
RU2805675C2 |
| РАСТЕНИЯ ТОМАТА С УЛУЧШЕННОЙ УСТОЙЧИВОСТЬЮ К БОЛЕЗНЯМ | 2015 |
|
RU2721952C2 |
| РАСТЕНИЯ САЛАТ-ЛАТУК, ИМЕЮЩИЕ РЕЗИСТЕНТНОСТЬ ПРОТИВ NASONOVIA RIBISNIGRI БИОТИПА 1 | 2015 |
|
RU2731639C2 |
| МОЛЕКУЛЯРНЫЕ МАРКЕРЫ, АССОЦИИРОВАННЫЕ С УСТОЙЧИВОСТЬЮ ПОДСОЛНЕЧНИКА К OROBANCHE | 2018 |
|
RU2776361C2 |
| QTL УРОЖАЯ В РАСТЕНИЯХ ОГУРЦА | 2015 |
|
RU2723586C2 |
| МАРКЕРЫ, СЦЕПЛЕННЫЕ С УСТОЙЧИВОСТЬЮ РАСТЕНИЙ СОИ К SCN | 2011 |
|
RU2620973C2 |
| ГЕН-ВОССТАНОВИТЕЛЬ RF4 ДЛЯ ЦИТОПЛАЗМАТИЧЕСКОЙ МУЖСКОЙ СТЕРИЛЬНОСТИ (CMS) C-ТИПА КУКУРУЗЫ, МОЛЕКУЛЯРНЫЕ МАРКЕРЫ И ИХ ПРИМЕНЕНИЕ | 2011 |
|
RU2603005C2 |
| QTL УРОЖАЯ В РАСТЕНИЯХ ОГУРЦА | 2015 |
|
RU2723581C2 |
Изобретение относится к области биохимии, в частности к устойчивому к мучнистой росе трансгенному растению розы. Также раскрыт способ идентификации устойчивого к мучнистой росе растения розы. Изобретение позволяет эффективно бороться с мучнистой росой. 2 н. и 7 з.п. ф-лы, 2 ил., 1 табл., 1 пр.
1. Устойчивое к мучнистой росе трансгенное растение розы, включающее в своем ядерном геноме одну нуклеотидную последовательность, представленную SEQ ID No. 1, и одну нуклеотидную последовательность, представленную SEQ ID No. 2, причем совместное присутствие SEQ ID No. 1 и SEQ ID No. 2 в указанном ядерном геноме обеспечивает устойчивость к мучнистой росе.
2. Устойчивое к мучнистой росе трансгенное растение розы по п. 1, причем указанная устойчивость к мучнистой росе является устойчивостью против болезнетворного для растений организма аскомицета Podosphaera pannosa.
3. Устойчивое к мучнистой росе трансгенное растение розы по п. 1 или 2, причем указанное растение розы представляет собой Rosa hybrida, и указанный ядерный геном является геномом тетраплоида, геномом гексаплоида, геномом октаплоида или геномом диплоида.
4. Устойчивое к мучнистой росе трансгенное растение розы по любому из пп. 1-3, включающее в своем ядерном геноме две нуклеотидные последовательности, представленные SEQ ID No. 1, и/или две нуклеотидные последовательности, представленные SEQ ID No. 2.
5. Устойчивое к мучнистой росе трансгенное растение розы по п. 3 или 4, включающее в своем ядерном геноме три нуклеотидные последовательности, представленные SEQ ID No. 1, и/или три нуклеотидные последовательности, представленные SEQ ID No. 2.
6. Устойчивое к мучнистой росе трансгенное растение розы по любому из пп. 3-5, включающее в своем ядерном геноме четыре нуклеотидные последовательности, представленные SEQ ID No. 1, и/или четыре нуклеотидные последовательности, представленные SEQ ID No. 2.
7. Устойчивое к мучнистой росе трансгенное растение розы по любому из пп. 1-6, причем указанное растение розы выбирают из группы, включающей срезочную розу, горшечную розу, садовую розу и подвой розы.
8. Устойчивое к мучнистой росе трансгенное растение розы по любому из пп. 1-7, причем указанная устойчивость к мучнистой росе является доминантной устойчивостью.
9. Способ идентификации устойчивого к мучнистой росе растения розы по любому из пп. 1-8, включающий стадии:
а) выделения ДНК ядерного генома из растения розы;
b) установления присутствия SEQ ID No. 1 и SEQ ID No. 2 в выделенной ДНК ядерного генома;
с) установления фенотипа указанного растения розы в отношении мучнистой росы, причем присутствие SEQ ID No. 1 и SEQ ID No. 2 указывает на устойчивый к мучнистой росе фенотип.
| RU 2015112578 A, 27.10.2016 | |||
| МОЛЕКУЛЯРНЫЙ МАРКЕР FR_ER1 И ЕГО ИСПОЛЬЗОВАНИЕ ДЛЯ СЕЛЕКЦИИ НОВЫХ СОРТОВ ГОРОХА, УСТОЙЧИВЫХ К МУЧНИСТОЙ РОСЕ | 2013 |
|
RU2593691C2 |
| СПОСОБ СОЗДАНИЯ ЛИНИЙ ЯРОВОЙ МЯГКОЙ ПШЕНИЦЫ С УДЛИНЕННЫМ СРОКОМ КОЛОШЕНИЯ И С КОМПЛЕКСНОЙ УСТОЙЧИВОСТЬЮ К ГРИБНЫМ БОЛЕЗНЯМ | 2013 |
|
RU2535985C1 |
| RU 2012147570 A, 10.09.2014. | |||

