ССЫЛКА НА ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ, ПРЕДСТАВЛЕННЫЙ В ЭЛЕКТРОННОМ ВИДЕ
[0001] Официальная копия списка последовательностей предоставляется в электронном виде через EFS-Web в виде списка последовательностей в формате ASCII файлом с именем «SEQ LISTING 18054WO», созданным 27 марта 2018 г. и имеющим размер 175,5 килобайт, и подается одновременно с описанием. Список последовательностей, содержащийся в этом документе в формате ASCII, является частью описания и полностью включен в настоящее описание посредством ссылки.
ОБЛАСТЬ ТЕХНИКИ
[0002] Настоящее изобретение относится к способам защиты плодов свиней от инфицирования вирусом репродуктивного и респираторного синдрома свиней (PRRSV).
УРОВЕНЬ ТЕХНИКИ
[0003] Репродуктивно-респираторный синдром свиней (PRRS) - это наиболее экономически важное заболевание свиней в Северной Америке, Европе и Азии, которое ежегодно обходится североамериканским производителям примерно в 600 миллионов долларов (Holtkamp et al., 2013). Синдромы клинических заболеваний, вызванные инфицированием вирусом репродуктивного и респираторного синдрома свиней (PRRSV), впервые были зарегистрированы в США в 1987 году (Keffaber, 1989), а затем в Европе в 1990 году (Wensvoort et al., 1991). Инфекция PRRSV приводит к респираторным заболеваниям, включая кашель и лихорадку, репродуктивную недостаточность на поздних сроках беременности и снижение показателей роста. Вирус также участвует во множестве взаимодействий синдрома полимикробного заболевания, сохраняя при этом субклиническую инфекцию на протяжении всей жизни (Rowland et al., 2012). Потери являются результатом респираторных заболеваний у молодых свиней, плохой продуктивности роста, репродуктивной недостаточности и внутриутробного развития инфекции (Keffaber, 1989).
[0004] Репродуктивная форма заболевания составляет около 45% потерь в результате абортов, мертвых плодов и респираторных заболеваний у новорожденных. В самой тяжелой форме репродуктивный PRRS может привести к 90% смертности плодов/новорожденных, а также к увеличению смертности свиноматок. Репродуктивная форма PRRS возникает после заражения беременных свиней или свиноматок примерно через 90 дней из 114-дневного периода беременности (Christianson et al., 1993; Rowland, 2010). После начальной фазы репликации в материнских макрофагах вирус проникает через плаценту и начинает продуктивно инфицировать плод. Первоначально вирус поражает лишь небольшое количество плодов, после чего происходит горизонтальная передача вируса от плода к плоду (Wiltdnson et al., 2016). Точный механизм того, как вирус проникает через плаценту, остается неизвестным, но может быть подобным инфицированию макрофага, выступающего в роли «троянского коня» - механизм, который ранее был описан для вируса, повышающего уровень лактатдегидрогеназы (LDV) (Cafruny, 1996). В отличие от альвеолярных макрофагов у взрослых животных, основным местом репликации PRRSV у плода является вилочковая железа (Rowland, 2003). Поскольку плод свиньи становится иммунокомпетентным примерно на 70-й день беременности, инфицирование PRRSV происходит в иммунном окружении плода, содержащем функциональные В- и Т-клетки (Rowland, 2003; Rowland, 2010).
[0005] Свиньи, которые выживают при внутриутробной инфекции, становятся постоянными источниками вируса на последующих этапах производства, что приводит к появлению эндемически инфицированных стад (Rowland, et al., 2003). Самая тяжелая форма репродуктивного заболевания связана с группой высоковирулентных изолятов, называемых атипичными PRRSV (Halbur et al., 1997; Mengeling et al., 1998). Интересно, что многие из атипичных изолятов вируса PRRSV были выращены на фермах, вакцинированных против вируса PRRS (Key et al., 2001). В 2006 году в Китае появился атипичный вирус, получивший название высокопатогенного вируса PRRSV (HP-PRRSV), который продолжает уничтожать популяции свиней в этой стране (Tian et al., 2007). Поскольку в стандартном коммерческом племенном хозяйстве содержится около 5000 свиноматок, вспышка репродуктивного PRRS с высокой смертностью может иметь разрушительные последствия. Для обеспечения устойчивости производства свинины и продовольственной безопасности приоритетными остаются решения по контролю репродуктивного PRRS. Вакцины не смогли контролировать болезнь, в основном из-за генетического разнообразия структурных белков вируса (Shi et al., 2010). На практике интенсивные меры биобезопасности являются единственным средством защиты репродуктивного стада.
[0006] Вирус репродуктивного и респираторного синдрома свиней (PRRSV) принадлежит к семейству Arterividae вместе с вирусом, повышающим уровень лактатдегидрогеназы мышей, вирусом геморрагической лихорадки обезьян и вирусом артериита лошадей. Структурно артеривирусы напоминают тогавирусы, но похожи на коронавирусы, реплицируются через вложенный 3'-ко-концевой набор субгеномных мРНК, которые имеют общий лидер и поли-А-хвост.Артеривирусы обладают важными свойствами, связанными с вирусным патогенезом, включая тропизм к макрофагам и способность вызывать тяжелые заболевания и персистентную инфекцию (Plagemann, 1996). Молекулярные сравнения между североамериканскими и европейскими вирусами относят все изоляты вируса PRRSV к одному из двух генотипов, соответственно. Тип 2 или Тип 1. Несмотря на то, что два генотипа обладают лишь примерно 70% идентичностью на уровне нуклеотидов (Nelsen et al., 1999), оба имеют тропизм к CD163-положительным клеткам, вызывают долгосрочные инфекции и вызывают сходные клинические признаки.
[0007] CD163 представляет собой мембранный белок 130 кДа типа 1, состоящий из девяти доменов богатых цистеином рецепторов фагоцитов (SRCR), и двух спейсерных доменов, а также трансмембранного домена и короткого цито плазматического хвоста (Fabriek et al., 2005). CD163 свиньи содержит 17 экзонов, которые кодируют сигнальную последовательность пептида, за которой следуют девять доменов SRCR, два линкерных домена (также называемые доменами пролин-серин-треонин (PST), расположенные после SRCR 6 и SRCR 9) и цитоплазматический домен, за которым следует короткий цитоплазматический хвост.Поверхностная экспрессия CD163 ограничена клетками моноцитарно-макрофагального происхождения. Помимо функционирования в качестве вирусного рецептора, CD163 выполняет несколько важных функций, связанных с поддержанием нормального гомеостаза. Например, после инфекции или повреждения тканей CD163 действует как молекула-утилизатор, удаляя комплексы гаптоглобин-гемоглобина из крови (Kristiansen et al., 2001). Образующиеся в результате продукты распада гема регулируют связанный воспалительный ответ (Fabriek et al., 2005). Утилизация HbHp является основной функцией CD163 и находится в SRCR 3 (Madsen et al., 2004). Метаболиты, высвобождаемые макрофагами после разложения HbHp, включают билирубин, СО и свободное железо. Одна из важных функций CD163 - предотвращение окислительной токсичности, вызванной свободным гемоглобином (Kristiansen et al., 2001; Scares et al., 2009).
[0008] Другие важные функции С163 включают адгезию эритробластов (SRCR2), функцию рецептора TWEAK (слабый индуктор апоптоза, подобным фактору некроза опухоли) (SRCR 1-4 и 6-9), бактериального рецептора (SRCR5) и рецептора вируса африканской свиньи (Sanchez-Torres et al. 2003). CD163 также может играть роль иммуномодулятора (обсуждается в Van Gorp et al. 2010).
[0009] CD163 как рецептор PRRSV впервые описали Calvert et. al. (2007). Трансфекция непермиссивных клеточных линий кДНК CD163 различных видов, в том числе обезьян, человека, собак и мышей, может сделать клетки чувствительными к инфекции PRRSV (Calvert et al., 2007). Помимо CD163, второй рецепторный белок, CD169 (также известный как сиалоадгезин или SIGLEC1), был идентифицирован как первичный рецептор PRRSV, участвующий в формировании начального взаимодействия с СР5-матриксным (М) гетеродимером, основным белком на поверхности вириона (Delputte et al., 2002). В этой модели последующее взаимодействие между CD163 и гетеротримером GP2, 3, 4 в эндосомном компартменте опосредует освобождение от оболочки и высвобождение вирусного генома в цитоплазму (Van Breedam et al., 2010, Allende et al., 1999). Предыдущая модель, описывающая инфекцию PRRSV альвеолярных макрофагов идентифицировала SIGLEC1 (CD169) в качестве первичного рецептора вируса на поверхности макрофагов; однако, предыдущая работа, использующая свиней SIGLEC1-/-, показала отсутствие различий в репликации вируса по сравнению со свиньями дикого типа (Prather et al., 2013). Эти результаты подтверждают предыдущие исследования in vitro, демонстрирующие, что устойчивые к вирусу PRRSV клеточные линии, лишенные поверхностных CD169 и CD163, поддерживают репликацию вируса после трансфекции плазмидой CD163 (Welch et al., 2010).
[0010] Многие характеристики как патогенеза вируса PRRSV (особенно на молекулярном уровне), так и эпизоотологии плохо изучены, что затрудняет его контроль. В настоящее время производители часто вакцинируют свиней против вируса PRRSV модифицированными живыми аттенуированными штаммами или убитыми вирусными вакцинами, однако существующие вакцины часто не обеспечивают удовлетворительной защиты. Это происходит как из-за вариабельности штаммов, так и из-за недостаточной стимуляции иммунной системы. Помимо опасений по поводу эффективности имеющихся вакцин против вируса PRRSV, существуют убедительные доказательства того, что модифицированная живая вакцина, используемая в настоящее время, может сохраняться в отдельных свиньях и стадах свиней и накапливать мутации (Mengeling et al. 1999), как это было продемонстрировано с вирулентными полевыми изолятами после экспериментального заражения свиней (Rowland et al., 1999). Кроме того, было показано, что вирус вакцины выделяется в сперме вакцинированных хряков (Christopher-Hennings et al., 1997). В качестве альтернативы вакцинации некоторые эксперты рекомендуют в племенных стадах стратегию «испытания и удаления» (Dee et al., 1998). Успешное использование этой стратегии зависит от удаления всех свиней, которые остро или персистентно инфицированы вирусом PRRSV, с последующим строгим контролем для предотвращения повторного введения вируса. Сложность и большая часть затрат, связанных с этой стратегией, заключается в том, что мало известно о патогенезе персистентной инфекции PRRSV, и, следовательно, нет надежных способов для выявления персистентно инфицированных свиней.
[0011] Как можно видеть, в данной области техники существует потребность в разработке стратегий для индукции устойчивости к вирусу PRRSV у животных. Также существует особая потребность в методах защиты плода от инфекции PRRSV во внутриутробном периоде и для предотвращения передачи PRRSV от матери к плоду.
РАСКРЫТИЕ ИЗОБРЕТЕНИЯ
[0012] Предложен способ защиты плода свиньи от заражения вирусом репродуктивного и респираторного синдрома свиней (PRRSV). Способ включает разведение самки свиньи с самцом свиньей. Самка свиньи содержит модифицированные хромосомные последовательности в обоих аллелях своего гена CD163, причем модифицированные хромосомные последовательности снижают чувствительность самки свиньи к инфицированию PRRSV по сравнению с чувствительностью к инфекции PRRSV самки свиньи, которая не содержит никаких модифицированных хромосомных последовательностей в аллелях своего гена CD163. Самец свиньи содержит по меньшей мере один аллель CD163 дикого типа.
[0013] Другие объекты и особенности будут частично очевидны, а частично указаны ниже.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
[0014] Фиг. 1. Для модификации CD163 используются целенаправленные векторы и CRISPR. На панели А показаны экзоны 7, 8 и 9 дикого типа гена CD163, который подвергался модификации с использованием CRISPR. На панели В показан целенаправленный вектор, разработанный для замены экзона 7 свиньи (домен SRCR5 свиньи CD163) ДНК, кодирующей человеческий SRCR8 CD163L. Этот целенаправленный вектор использовали при трансфекциях с отбором по чувствительности к препарату G418. Праймеры для ПЦР для анализа длинных фрагментов на правом плече и на левом плече обозначены стрелками для 1230, 3752, 8791, 7765 и 7775. На панели С изображен целенаправленный вектор, идентичный тому, который показан на панели В, но из которого была удалена кассета Neo. Этот целенаправленный вектор был использован для нацеливания на CD163 в клетках, которые уже были устойчивы к неомицину. Праймеры, используемые в анализах малых делеций, показаны стрелками и помечены как GCD163F и GCD163R. Панель D подчеркивает экзоны, на которые нацелены CRISPR. Расположение CRISPR 10, 131, 256 и 282 показано направленными вниз стрелками на экзоне 7. Числа CRISPR представляют количество пар оснований из соединения интрон-экзон интрона 6 и экзона 7.
[0015] Фиг. 2. Для модификации CD1D использовали целенаправленный вектор и CRISPR. На панели А изображены экзоны 3, 4, 5, 6 и 7 гена CD1D дикого типа, который подвергся модификации с помощью CRISPR. Панель В демонстрирует целенаправленный вектор, предназначенный для замены экзона 3 селективным маркером Neo. Этот целенаправленный вектор использовался в сочетании с CRISPR для модификации CD1D. Праймеры для ПЦР для анализа длинных фрагментов на левом и правом плечах для 3991, 4363, 7373 и 12806 обозначены стрелками. На панели С показаны экзоны; на которые нацелены CRISPR. Расположение CRISPR 4800, 5350, 5620 и 5626 показано направленными вниз стрелками на экзоне 3. Праймеры, используемые в анализах малых делеций, показаны стрелками и помечены как GCD1DF и GCD1DR.
[0016] Фиг. 3. Создание свиней с нокаутом по CD163 и CD1D с помощью CRISPR/Cas9 и SCNT. А) Направленная делеция CD163 в соматических клетках после трансфекции CRISPR/Cas9 и донорной ДНК. Генотип дикого типа дает полосу в 6545 парах оснований (п.н.). Дорожки 16 представляют шесть различных колоний от одной трансфекции CRISPR 10 с Cas9 и донорной ДНК, содержащей Neo. Дорожки 1, 4 и 5 показывают крупную гомозиготную делецию 1500-2000 п. н. Дорожка 2 представляет меньшую гомозиготную делецию. Дорожки 3 и 6 представляют либо аллель дикого типа и мелкую делецию, либо двуаллельную модификацию обоих аллелей. Точные модификации каждой колонии определялись только путем секвенирования колоний, используемых для переноса ядра соматической клетки (somatic cell nuclear transfer SCNT). Слабая полоса дикого типа на некоторых дорожках может свидетельствовать о перекрестном загрязнении фибробластов плода из соседней колонии дикого типа. NTC - безматричный контроль. Б) Направленная делеция CD1D в соматических клетках после трансфекции CRISPR/Cas9 и донорной ДНК. Генотип дикого типа дает полосу 8729 п. н. Дорожки 14 представляют собой колонии с делецией CD1D размером 500-2000 п. н. Дорожка 4, по-видимому, является колонией дикого типа. NTC = безматричный контроль. В) Изображение свиньи с нокаутом CD163, полученной путем SCNT в ходе исследования. Этот поросенок-самец содержит гомозиготную делецию CD163 размером 1506 п.н. Г) Изображение свиней CD1D, полученное в ходе исследования. Эти поросята содержат делецию CD1D размером 1653 п.н. Д) Генотип двух пометов SCNT, содержащих делецию CD163 размером 1506 п. н. Дорожки 1-3 (помет 63) и дорожки 1-4 (помет 64) представляют генотип каждого поросенка из каждого помета. Sow (свиноматка) указывает самку - реципиента эмбрионов SCNT, а дикий тип представляет собой контроль дикого типа. NTC - безматричный контроль. Е) Генотип двух пометов SCNT, содержащих делецию CD1D размером 1653 п. н. Дорожки 1-7 (помет 158) и дорожки 1-4 (помет 159) представляют собой генотип каждого поросенка.
[0017] Фиг. 4. Влияние системы CRISPR/Cas9 на эмбрионы свиньи. А) Частота образования бластоцист после введения различных концентраций системы CRISPR/Cas9 в зиготы. Токсичность системы CRISPR/Cas9 была самой низкой при 10 нг/мкл. Б) Система CRISPR/Cas9 может успешно разрушать экспрессию eGFP в бластоцистах при введении в зиготы. Оригинальное увеличение Х4. В) Типы мутаций eGFV, генерируемые с помощью системы CRISPR/Cas9: генотип дикого типа (SEQ ID NO:16), №1 (SEQ ID NO:17), №2 (SEQ ID NO: 18) и №3 (SEQ ID NO: 19).
[0018] Фиг. 5. Влияние системы CRISPR/Cas9 при направленном воздействии на CD163 в эмбрионах свиней. А) Примеры мутаций, сгенерированных в CD163 системой CRISPR/Cas9: генотип дикого типа (SEQ ID NO:20), №1-1 (SEQ ID NO:21), №1-4 (SEQ ID NO:22) и №2-2 (SEQ ID NO:23). Все эмбрионы, исследованные с помощью секвенирования ДНК, продемонстрировали мутации в CD163 (18/18). CRISPR 131 выделен жирным шрифтом. В) Считывание секвенирования гомозиготной делеции, вызванной системой CRISPR/Cas9. На изображении представлены №1-4 панели А с делецией 2 п. н. CD163.
[0019] Фиг. 6. Воздействие системы CRISPR/Cas9 при введении с двумя типами CRISPR. A) ПЦР-амплификация CD163 в бластоцистах, которым вводили CRISPR/Cas9 в качестве зигот.Дорожки 1, 3, 6 и 12 показывают направленную делецию между двумя разными CRISPR. Б) ПЦР-амплификация CD/D в бластоцистах, которым вводили CRISPR/Cas9 в качестве зигот.По данным гель-электрофореза CD/D имеет более низкую частоту делеции по сравнению с CD163 (3/23); дорожки 1, 8 и 15 показывают очевидные делеции в CD/D. B) Система CRISPR/Cas9 успешно оказала воздействие на два гена, когда система была предоставлена с двумя CRISPR, нацеленными на CD163 и eGFP. Показаны модификации CD163 и eGFP: CD163 дикого типа (SEQ ID NO:24), CD163 №1 (SEQ ID NO:25), CD163 №2 (SEQ ID NO:26), CD163 №3 (SEQ ID NO:27), eGFP дикого типа (SEQ ID NO:28), eGFP №1-1 (SEQ ID NO:29), eGFP №1-2 (SEQ ID NO: 30), eGFP №2 (SEQ ID NO:31), и eGFP №3 (SEQ ID NO:32).
[0020] Фиг. 7. Свиньи с нокаутом CD163, полученные с помощью системы CRISPR/Cas9, введенной в зиготы. А) ПЦР-амплификация CD163 от свиней с нокаутом; явный признак делеции обнаружен в пометах 67-2 и 67-4. Б) Изображение свиней с нокаутом CD163 с суррогатной матерью. Все животные здоровы и не имеют никаких признаков отклонений от нормы. В) Генотип свиней с нокаутом CD163. Последовательность дикого типа показана как SEQ ID NO: 33. Два животных (из пометов 67-1 (SEQ ID NO: 34) и 67-3 (SEQ ID NO: 37)) несут гомозиготную делецию или вставку в CD163. Два других животных (из пометов 67-2 и 67-4) несут двуаллельную модификацию CD163: №67-2 Al (SEQ ID NO:35), №67-2 A2 (SEQ ID NO:36), №67-4 Al (SEQ ID NO:38) и №67-4 a2 (SEQ ID NO:39). Делеция была вызвана введением двух разных CRISPR с системой Cas9. Ни у одного животного, получавшего инъекцию CD163 в зиготу, не было обнаружено мозаичного генотипа.
[0021] Фиг. 8. Свиньи с нокаутом CD1D, полученные с помощью системы CRISPR/Cas9, введенной в зиготы. А) ПЦР-амплификация CD1D от свиней с нокаутом; 166-1 показывает мозаичный генотип CD1D. 166-2, 166-3 и 166-4 не показывают изменения размера ампликона, однако секвенирование ампликона выявило модификации. Дикий тип FF -эмбриональные фибробласты дикого типа. Б) ПЦР-амплификация анализа с длинными фрагментами показала четкую делецию одного аллеля у поросят 166-1 и 166-2. В) Изображение свиней с нокаутом CD1D с суррогатной матерью. D) Данные последовательностей свиней с нокаутом CD1D; дикого типа (SEQ ID NO:40), №166-1.1 (SEQ ID NO: 41), №166-1.2 (SEQ ID NO:42), №166-2 (SEQ ID NO:43), №166-3.1 (SEQ ID NO:44), №166-3.2 (SEQ ID NO:45) и №166-4 (SEQ ID NO:46). Стартовый кодон atg в экзоне 3 выделен жирным шрифтом, а также строчными буквами.
[0022] Фиг. 9. Клинические признаки острой инфекции вирусом PRRSV. Результаты ежедневной оценки наличия респираторных признаков и лихорадки для CD163+/+(n=6) HCD163-/-(n=3).
[0023] Фиг. 10. Гистопатология легких при острой инфекции PRRSV. Репрезентативные микрофотографии тканей, окрашенных гематоксилином-эозином, от свиней дикого типа и свиней-нокаутов. На левой панели показан отек и инфильтрация мононукле арных клеток. Правая панель от свиньи с нокаутом показывает легочную структуру нормального легкого.
[0024] Фиг. 11. Виремия при разных генотипах. Обратите внимание, что данные поросят CD163-/- расположены вдоль оси X.
[0025] Фиг. 12. Продукция антител у свиней с нулевым аллелем, аллелем дикого типа и неохарактеризованным аллелем.
[0026] Фиг. 13. Экспрессия CD163 на клеточной поверхности у отдельных свиней. Линии справа на панелях А неохарактеризованных аллелей, панелях В неохарактеризованных аллелей и панелях CD163+/+, представляют антитело CD163, в то время как линии с левой стороны этих панелей, являются контрольными элементами без антител (фон). Обратите внимание, что у животных CD163-/- окрашивание CD163 перекрывается с фоновым контролем, и что окрашивание CD163 в неохарактеризованных аллелях находится примерно в промежутке между уровнем дикого типа и фоном (также обратите внимание, что это логарифмическая шкала, поэтому менее ~ 10%).
[0027] Фиг. 14. Уровень CD169 на альвеолярных макрофагах от трех репрезентативных свиней и контроля без антител (анти-CD169, меченный FITC).
[0028] Фиг. 15. Виремия при разных генотипах. Обратите внимание, что данные для поросят с аминокислотой Д43 лежат вдоль оси X.
[0029] Фиг. 16. Геномная последовательность экзонов CD163 дикого типа 7-10, используемая в качестве контрольной последовательности (SEQ ID NO: 47). Последовательность включает 3000 п. н. выше экзона 7 до последнего основания экзона 10. Подчеркнутые области показывают расположение, соответственно, экзонов 7, 8, 9 и 10.
[0030] Фиг. 17. Схема модификаций CD163, иллюстрирующая несколько хромосомных модификаций CD163, предсказанный белковый продукт для каждой модификации и относительную экспрессию макрофагов для каждой модификации, измеренную по уровню поверхностного CD163 на альвеолярных макрофагах свиньи (АМС) Черные области указывают на интроны, а белые области - на экзоны. Заштрихованная область указывает на миметик экзона 11 hCD163L1, гомолог экзона 7 свиньи. Серая область указывает на синтезированный интрон с конструкцией PGK Neo.
[0031] Фиг. 18. Схема последовательности белка и гена CD163 свиньи. А) Домены SRCR (овалы) и PST (квадраты) белка CD163 вместе с соответствующими экзонами генов. В) Сравнение CD163 SRCR 5 свиньи (SEQ ID NO: 120) с гомологом человека CD163L1 SRCR 8 (SEQ ID NO: 121).
[0032] Фиг. 19. Типичные результаты поверхностной экспрессии CD163 и CD169 на АМС от свиней дикого типа и свиней с модификацией CD163. Панели А Е показывают результаты для модификаций CD163, как показано на Фиг. 17. Объединенные данные для d7(1467) и d7(1280) показаны на панели D.
[0033] Фиг. 20. Уровни гаптоглобина в сыворотке крови свиней дикого типа и свиней, модифицированных CD163.
[0034] Фиг. 21. Относительная пермиссивность АМС дикого типа и HL11m к инфицированию изолятами PRRSV 2 типа.
[0035] Фиг. 22. Заражение свиней, модифицированных CD 163, изолятами PRRSV 1 и 2 типа.
[0036] Фиг. 23. Вирусная нагрузка для свиней дикого типа и модифицированных CD 163, инфицированных вирусами 2 типа.
[0037] Фиг. 24. Исходы для плода после инфицирования матери PRRSV. Цифры слева обозначают каждую свиноматку («№свиноматки»; см. Таблицу 16 ниже). Под каждым номером свиноматки в скобках указан результат ПЦР для PRRS в сыворотке, измеренный как log10 числа матриц на реакцию. «N» означает отсутствие нуклеиновой кислоты PRRSV (Ct>39). Плоды идентифицируются по количеству и относительному положению в каждом роге матки. Звездочки обозначают образцы ПЦР плода, полученные из брюшной жидкости. Число под каждым плодом представляет собой результат ПЦР PRRS в сыворотке плода (logio числа матриц на реакцию). Число в каждом кружке указывает на наличие анатомической патологии: 1) нормальный плод; 2) маленький плод; 3) изменения плаценты, такие как отслоение плаценты и/или некроз; 4) плод, окрашенный меконием; 5) плод мертвый и некротический. Строчные буквы определяют генотип отдельных плодов (см. Таблицу 16). Ключ: а, А/А; b, С/А; с, В/А; d, Е/А; е, В/С; f, B/D; g, D/C; h, D/D; i, E/C; j, E/D; HO - не определено, так как плод был некротическим; но генотип не был определен.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[0038] Настоящее изобретение направлено на способы защиты плодов свиней от заражения вирусом репродуктивного и респираторного синдрома свиней (PRRSV). Свиньи, имеющие инактивирующие мутации в обоих аллелях гена CD 163, устойчивы к инфицированию PRRSV. В настоящее время неожиданно было обнаружено, что CD163-положительные плоды (например, плоды, которые имеют один или два аллеля CD 163 дикого типа) могут быть защищены от инфекции PRRSV в утробе матери, пока мать обладает инактивирующими мутациями в обоих аллелях своего гена CD163. Так, например, самки, имеющие инактивирующие мутации в обоих аллелях гена CD163, могут быть скрещены с самцами, имеющими два аллеля CD163 дикого типа, и полученные гетерозиготные плоды будут защищены от инфекции PRRSV.
Определения
[0039] При представлении элементов настоящего изобретения или его предпочтительных вариантов воплощения, обозначения в единственном числе и «указанный» предназначены для обозначения того, что существует один или более элементов.
[0040] Термин «и/или» означает любой из элементов, любую комбинацию элементов или все элементы, с которыми этот термин связан.
[0041] Используемый здесь термин «разведение» относится к объединению мужских и женских гамет, так что происходит оплодотворение. Такое соединение может быть осуществлено путем спаривания (совокупления) или искусственными способами in vitro или in vivo. Такие искусственные способы включают, без ограничений, искусственное оплодотворение, искусственное оплодотворение с помощью хирургического вмешательства, оплодотворение in vitro, внутрицитоплазматическую инъекцию сперматозоидов, препарирование вителлинового слоя, культивирование оплодотворенных ооцитов in vitro, перенос яичников и разделение яичников. Используемый здесь термин «разведение» также включает перенос оплодотворенного ооцита в репродуктивный тракт самки.
[0042] Термины «содержащий», «включающий» и «имеющий» предназначены для включения и означают, что могут быть дополнительные элементы, отличные от перечисленных.
[0043] Термин «CRISPR» означает «кластеризированные регулярные промежуточные короткие палиндромные повторы». Системы CRISPR включают системы CRISPR типа I, типа II и типа III.
[0044] Термин «Cas» относится к «белку, связанному с CRISPR». Белки Cas включают, без ограничения, белки-члены семейства Cas9, белки-члены семейства Cas6 (например, Csy4 и Cas6) и белки-члены семейства Cas5.
[0045] Термин «Cas9» обычно может относиться к полипептиду, содержащему по меньшей мере около 5%, 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90% или 100% идентичности последовательности и/или сходства последовательности с полипептидом Cas9 дикого типа (например, Cas9 из S. pyogenes). Иллюстративные последовательности Cas9 представлены SEQ ID NN 1-256 и 795-1346 в Публикации патента США №2016/0046963. SEQ ID NN 1-256 и 795-1346 Публикации патента США №2016/0046963 здесь включены посредством ссылки. «Cas9» может относиться к полипептиду с не более чем примерно 5%, 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90% или 100% идентичностью последовательности и/или сходством последовательности с полипептидом Cas9 дикого типа (например, из S. pyogenes). «Cas9» может относиться к дикому типу или модифицированной форме белка Cas9, который может включать замену аминокислоты, такую как деления, вставка, замена, вариант, мутация, слияние, химера или любую их комбинацию.
[0046] Термин «Cas5» может обычно относиться к полипептиду, содержащему по меньшей мере около 5%, 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90% или 100% идентичности последовательности и/или сходства последовательности с иллюстративным полипептидом Cas5 дикого типа (например, Cas5 из D. vulgaris). Иллюстративные последовательности Cas5 представлены на Фигуре 42 Публикации патента США №2016/0046963. Фиг. 42 Публикации патента США №2016/0046963 включена сюда посредством ссылки. «Cas5» обычно может относиться к полипептиду с максимум примерно 5%, 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90% или 100% идентичности последовательности и/или сходства последовательности с полипептидом Cas5 дикого типа (например, Cas5 из D. vulgaris). «Cas5» может относиться к дикому типу или модифицированной форме белка Cas5, который может включать замену аминокислоты, такую как делеция, вставка, замена, вариант, мутация, слияние, химера или любую их комбинацию.
[0047] Термин «Cas6» может в целом относиться к полипептиду, содержащему по меньшей мере около 5%, 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90% или 100% идентичности последовательности и/или сходства последовательности с иллюстративным полипептидом Cas6 дикого типа (например, Cas6 из Т. thermophilus). Иллюстративные последовательности Cas6 представлены на Фигуре 41 в Публикации патента США №2016/0046963. Фигура 41 публикации патента США №2016/0046963 включена сюда посредством ссылки. «Cas6» в целом может относиться к полипептиду с максимум примерно 5%, 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90% или 100% идентичности последовательности и/или сходства последовательности с полипептидом Cas6 дикого типа (например, из Т. thermophilus). «Cas6» может относиться к дикому типу или модифицированной форме белка Cas6, который может включать замену аминокислоты, такую как делеция, вставка, замена, вариант, мутация, слияние, химера или любую их комбинацию.
[0048] Термины «CRISPR/Cas9» или «система CRISPR/Cas9» относятся к программируемой нуклеазной системе для генной инженерии, которая включает белок Cas9 или его производное, а также одну или более некодирующих РНК, которые могут обеспечивать функцию CRISPR-PHK (crRNA), и транс-активирующую РНК (tracrRNA) для Cas9. CrRNA и tracrRNA можно использовать по отдельности или их можно комбинировать для получения «гидовой РНК» (гРНК). CrRNA или гРНК обеспечивают последовательность, комплементарную геномной мишени.
[0049] Ссылки в данном документе на делецию в нуклеотидной последовательности от нуклеотида x до нуклеотида y означают, что все нуклеотиды в диапазоне были удалены, включая х и у. Так, например, фраза «деления 11 пар оснований от нуклеотида 3137 до нуклеотида 3147 по сравнению с SEQ ID NO: 47» означает, что каждый из нуклеотидов с 3317 по 3147 был удален, включая нуклеотиды 3317 и 3147.
[0050] «Устойчивость животного к заболеванию» - это характеристика животного, при которой животное избегает симптомов заболевания, которые являются результатом взаимодействий между животными и патогенами, таких как взаимодействия между свиньей и PRRSV. То есть предотвращается возможность патогенов вызывать болезни животных и связанные с ними симптомы заболевания или, альтернативно, снижение частоты и/или тяжести клинических признаков или уменьшение клинических симптомов. Специалист в данной области техники поймет, что раскрытые здесь способы могут использоваться с другими композициями и способами, доступными в данной области техники, для защиты животных от атаки патогена.
[0051] Используемые здесь термины «редактирование гена», «редактируемый ген», «генетически отредактированный/модифицированный» и «эффекторы редактирования гена» относятся к использованию хоминг-технологии с природными или искусственно созданными нуклеазами, также именуемыми как «молекулярные ножницы», «хоминг-эндонуклеазы» или «нацеленные эндонуклеазы». Нуклеазы создают специфические двухцепочечные хромосомные разрывы (double-stranded chromosomal breaks - DSB) в желаемых местах генома, которые в некоторых случаях используют эндогенные механизмы клетки для восстановления индуцированного разрыва естественными процессами гомологичной рекомбинации (ГР) и/или негомологичного соединения концов (nonhomologous end-joining - NHEJ). К эффекторам редактирования генов относятся нуклеазы цинковых пальцев (ZFN), эффекторные нуклеазы, подобные активаторам транскрипции (TALEN), системы с кластерными регулярными короткими палиндромными повторами (CRISPR) (например, система CRISPR/Cas9) и мегануклеазы (например, модифицированные мегануклеазы как хоминг-энд о нуклеазы). Термины также включают использование трансгенных процедур и способов, в том числе, например, когда изменение представляет собой делецию или относительно небольшую вставку (обычно менее 20 нуклеотидов) и/или не вводит ДНК от чужеродных видов. Термин также включает животных-потомков, таких как животные, созданные половым скрещиванием или бесполым размножением от животного с отредактированным исходным геном.
[0052] Термины «геномная инженерия», «генная инженерия», «генно-инженерный», «генетически измененный», «генетическое изменение», «модификация генома», «модификация генома» и «генно-модифицированный» могут относиться к изменению генома путем делеции, вставки, мутации или замены определенных последовательностей нуклеиновых кислот. Изменение может быть специфичным для гена или места. Генная инженерия может использовать нуклеазы для разрезания нуклеиновой кислоты, тем самым создавая сайт для изменения. Также рассматривается разработка негеномной нуклеиновой кислоты. Белок, содержащий домен нуклеазы, может связывать и расщеплять нуклеиновую кислоту-мишень, образуя комплекс с нуклеиновой кислотой, нацеленной на нуклеиновую кислоту. В одном примере расщепление может привести к двухцепочечным разрывам в нуклеиновой кислоте-мишени. Нуклеиновая кислота может быть восстановлена, например, с помощью механизма эндогенного негомологичного соединения концов (NHEJ). В следующем примере может быть вставлен фрагмент нуклеиновой кислоты. Модификации нуклеиновых кислот, нацеленных на нуклеиновые кислоты, и сайт-ориентированных полипептидов могут вводить новые функции, которые будут использоваться для геномной инженерии.
[0053] Используемые здесь термины « хоминг-технология ДНК», «хоминг-технология » и «хоминг-эндонуклеаза» включают любые механизмы, которые позволяют нацеливать указанную молекулу на указанную последовательность ДНК, включая белки цинкового пальца (ZF), эффекторы, подобные активаторам транскрипции (TALE) мегануклеазы и системы CRISPR (например, система CRISPR/Cas9).
[0054] Термины «повышенная устойчивость» и «пониженная чувствительность» в данном документе означают, без ограничения, статистически значимое снижение частоты и/или тяжести клинических признаков или клинических симптомов, которые связаны с инфекцией патогеном. Например, «повышенная устойчивость» или «пониженная чувствительность» могут относиться к статистически значимому снижению частоты и/или тяжести клинических признаков или клинических симптомов, которые связаны с инфекцией PRRSV у животного, содержащего модифицированную хромосомную последовательность в белке гена CD163 по сравнению с контрольным животным, имеющим немодифицированную хромосомную последовательность. Термин «статистически значимое уменьшение клинических симптомов» означает, без ограничения, частоту встречаемости по меньшей мере одного клинического симптома в группе модифицированных субъектов, которая после заражения инфекционным агентом по меньшей мере на 10%, предпочтительно по меньшей мере на 20%, более предпочтительно по меньшей мере на 30%, даже более предпочтительно по меньшей мере на 50% и даже более предпочтительно по меньшей мере на 70% ниже, чем в немодифицированной контрольной группе.
[0055] «Нокаут» означает нарушение структуры или регуляторного механизма гена. Нокауты могут быть сгенерированы посредством гомологичной рекомбинации направленных векторов, замещающих векторов или векторов "ударил-убежал" или случайной вставки вектора-ловушки гена, приводящей к полной, частичной или условной потере функции гена.
[0056] Используемый здесь термин «мутация» включает изменения в нуклеотидной последовательности полинуклеотида, например, в гене или кодирующей последовательности ДНК (CDS), по сравнению с последовательностью дикого типа. Термин включает, без ограничения, замены, вставки, сдвиги рамки считывания, делеции, инверсии, транслокации, дупликации, мутации донорных сайтов сплайсинга, точечные мутации и т.п.
[0057] Здесь «уменьшение частоты и/или тяжести клинических признаков» или «уменьшение клинических симптомов» означает, без ограничения, уменьшение количества инфицированных субъектов в группе, уменьшение или исключение количества субъектов с проявлением клинических признаков инфекции или уменьшение тяжести любых клинических признаков, которые присутствуют у одного или более субъектов, по сравнению с инфекцией дикого типа. Например, эти термины охватывают любые клинические признаки инфекции, патологии легких, виремии, продукции антител, снижение нагрузки патогенов, выделение патогенов, снижение передачи патогенов или уменьшение любых клинических признаков, симптоматических для PRRSV. Предпочтительно, эти клинические признаки уменьшаются у одного или более животных по настоящему изобретению по меньшей мере на 10% по сравнению с инфицированными субъектами, не имеющими модификации в гене CD163. Более предпочтительно, клинические признаки уменьшаются у субъектов изобретения по меньшей мере на 20%, предпочтительно по меньшей мере на 30%, более предпочтительно по меньшей мере на 40% и даже более предпочтительно по меньшей мере на 50%.
[0058] «ДНК-связывающий домен TALE» или «TALE» представляет собой полипептид, содержащий один или более повторяющихся доменов/единиц TALE. Домены повторов участвуют в связывании TALE с его родственной последовательностью ДНК-мишени. Одна «повторяющаяся единица» (также называемая «повторением») обычно имеет длину 33-35 аминокислот и демонстрирует по меньшей мере некоторую гомологию последовательности с другими повторяющимися последовательностями TALE в пределах встречающегося в природе белка TALE. Связывающие домены «цинковый палец» и TALE могут быть «сконструированы» для связывания с заранее определенной нуклеотидной последовательностью, например, путем конструирования (изменения одной или более аминокислот) области распознающей спирали встречающихся в природе белков «цинковый палец» или TALE. Следовательно, сконструированные ДНК-связывающие белки (цинковые пальцы или TALE) - это белки, не встречающиеся в природе. Неограничивающими примерами способов конструирования ДНК-связывающих белков являются дизайн и отбор. Сконструированный ДНК-связывающий белок - это белок, не встречающийся в природе, конструкция/состав которого основывается, главным образом, на рациональных критериях. Рациональные критерии для проектирования включают применение правил замены и компьютеризированных алгоритмов для обработки информации в базе данных, в которой хранится информация о существующих проектах ZFP и/или TALE, а также данные о связывании. См., например. Патенты США №№6140081; 6453242 и 6534261; см. также WO 98/53058; WO 98/53059; WO 98/53060; WO 02/016536 и WO 03/016496 и Публикацию США№20110301073.
[0059] «ДНК-связывающий белок цинковые пальцы» (или связывающий домен) представляет собой белок или домен в более крупном белке, который связывает ДНК специфичным для последовательности образом через один или более цинковых пальцев, которые являются участками последовательности аминокислот в связывающем домене, структура которого стабилизируется за счет координации иона цинка. Термин белок цинковые пальцы, связывающий ДНК, часто сокращенно обозначают как «белок цинковые пальцы» или ZFP.
[0060] «Выбранный» белок цинковые пальцы или TALE представляет собой белок, не встречающийся в природе, производство которого происходит, в основном, в результате эмпирического процесса, такого как фаговый дисплей, ловушка взаимодействия или гибридная селекция. См. например. Патент США №5789538; Патент США №5925523; Патент США №6007988; Патент США №6013453; Патент США №6200759; WO 95/19431; WO 96/06166; WO 98/53057; WO 98/54311; WO 00/27878; WO 01/60970; WO 01/88197; WO 02/099084 и Публикацию США №20110301073.
[0061] Различные другие термины определяются ниже.
Способы защиты плодов свиней от инфекции PRRSV
[0062] Предложен способ защиты плода свиньи от заражения вирусом репродуктивного и респираторного синдрома свиней (PRRSV). Способ включает разведение свиньи-самки с свиньей-самцом. Самка свиньи содержит модифицированные хромосомные последовательности в обоих аллелях своего гена CD163, причем модифицированные хромосомные последовательности снижают чувствительность самки свиньи к инфицированию PRRSV по сравнению с чувствительностью к инфекции PRRSV самки свиньи, которая не содержит никаких модифицированных хромосомных последовательностей в аллелях гена CD163. Самец свиньи содержит по меньшей мере один аллель CD163 дикого типа.
[0063] В способах, описанных в данном документе, модифицированные хромосомные последовательности могут быть последовательностями, которые изменены таким образом, что функция белка CD163, связанная с инфекцией PRRSV, нарушается, снижается или устраняется. Таким образом, свиньи-самки, используемые в описанных здесь способах, могут называться «нокаутированными» животными.
[0064] Самец свиньи может содержать два аллеля CD163 дикого типа.
[0065] Термин «аллель CD163 дикого типа», используемый здесь, означает, что последовательность аллеля CD163 представляет собой последовательность, обнаруженную в природе, или что последовательность аллеля CD163 содержит одну или более мутаций (например, вставки, делеции или замены), которые существенно не ухудшают активность CD163. Таким образом, аллели CD163 дикого типа могут содержать полиморфизмы и/или мутации при условии, что эти полиморфизмы или мутации существенно не снижают активность CD163.
[0066] Используя описанные в данном документе способы, плод будет защищен от вирусов PRRSV как типа 1, так и типа 2, включая различные изоляты PRRSV типа 1 и типа 2.
[0067] Таким образом, в способах, описанных в данном документе, модифицированные хромосомные последовательности могут снизить чувствительность самок свиньи к вирусу PRRSV типа 1, PRRSV типа 2 или к вирусам PRRSV типа 1 и типа 2.
[0068] Модифицированные хромосомные последовательности могут снизить чувствительность самок свиней к изоляту PRRSV, выбранному из группы, состоящей из NVSL 97-7895, KS06-72109, Р129, VR2332, С090, AZ25, MLV-ResPRRS, KS62-06274, KS483 (SD23983), С084, SD13-15, Lelystad, 03-1059, 03-1060, SD01-08, 4353PZ и их комбинаций.
[0069] Самка свиньи может включать генетически измененную самку свиньи.
[0070] Генетически модифицированная самка свиньи может быть животным, модифицированным с использованием хоминг-эндонуклеазы. Хоминг-эндонуклеаза может быть природной эндонуклеазой, но предпочтительно представляет собой рационально сконструированную, неприродную хоминг-эндонуклеазу, которая имеет последовательность распознавания ДНК, разработанную таким образом, чтобы эндонуклеаза была нацелена на хромосомную последовательность в гене, кодирующем белок CD163.
[0071] Таким образом, хоминг-эндонуклеаза может быть сконструированной хоминг-эндонуклеазой. Хоминг-эндонуклеаза может включать, например, систему кластерных коротких палиндромных повторов с регулярными интервалами (CRISPR), эффекторную нуклеазу, подобную активатору транскрипции (TALEN), нуклеазу цинкового пальца (ZFN), слитый белок рекомбиназы, мегануклеазу или комбинацию любого из них.
[0072] Хоминг-нуклеаза предпочтительно включает систему CRISPR. Примеры систем CRISPR, которые можно использовать для создания самок свиней для использования в описанных здесь способах, включают, без ограничений, CRISPR/Cas9, CRISPR/Cas5 и CRISPR/Cas6. Использование систем CRISPR для создания генетически модифицированных животных обсуждается ниже.
[0073] В любом из способов, описанных в данном документе, самка свиньи может содержать одинаковую модифицированную хромосомную последовательность в обоих аллелях гена CD163.
[0074] Альтернативно самка свиньи может содержать первую модифицированную хромосомную последовательность в первом аллеле гена CD163 и вторую модифицированную хромосомную последовательность во втором аллеле гена CD163, причем первая и вторая модифицированные хромосомные последовательности отличаются друг от друга.
[0075] В любом из описанных здесь способов каждый аллель гена CD163 самки свиньи может содержать вставку, делецию или их комбинацию.
[0076] Например по меньшей мере один аллель гена CD163 самки свиньи может содержать делецию.
[0077] По меньшей мере один аллель гена CD163 может содержать делецию в рамке считывания.
[0078] По меньшей мере один аллель гена CD163 самки свиньи может содержать вставку.
[0079] В способах, описанных в настоящем документе, модифицированные хромосомные последовательности предпочтительно вызывают снижение продукции или активности белка CD163 по сравнению с продукцией или активностью белка CD163 у самок свиней, у которых отсутствуют модифицированные хромосомные последовательности.
[0080] Предпочтительно, модифицированные хромосомные последовательности приводят к практически отсутствию функционального белка CD163 у самки свиньи. Под «практически без функционального белка CD163» подразумевается, что уровень белка CD163 в животном, потомстве или клетке не определяется или, если его можно обнаружить, он находится на уровне по меньшей мере примерно на 90% ниже, предпочтительно по меньшей мере примерно на 95% ниже, более предпочтительно по меньшей мере примерно на 98%, ниже и даже более предпочтительно по меньшей мере примерно на 99% ниже, чем уровень, наблюдаемый у животного, потомства или клетки, которые не содержат модифицированные хромосомные последовательности.
[0081] Например, в любом из описанных здесь способов самка свиньи не продуцирует белок CD163.
[0082] В любом из описанных здесь способов каждый аллель гена CD163 самки свиньи может включать модификацию экзона 7, модификацию экзона 8, модификацию интрона, примыкающего к экзону 7 или экзону 8, или комбинацию любого из них.
[0083] Например, один или оба аллеля гена CD163 самки свиньи могут включать модификацию экзона 7 гена CD163.
[0084] Модификация в экзоне 7 может включать делецию.
[0085] В любом из описанных здесь способов, где аллель гена CD163 самки свиньи содержит делецию, делеция может включать делецию в рамке считывания.
[0086] Модификация в экзоне 7 может включать вставку.
[0087] В любом из описанных здесь способов модифицированные хромосомные последовательности в одном или обоих аллелях гена CD163 самки свиньи могут привести к неправильному кодированию.
[0088] Если модифицированная хромосомная последовательность приводит к неправильному кодированию аллеля гена CD163, неправильное кодирование может привести к преждевременному стоп-кодону ниже неправильного кодирования в аллеле гена CD163.
[0089] В любом из способов, описанных в данном документе по меньшей мере один из аллелей гена CD163 у самки свиньи содержит модификацию, выбранную из группы, состоящей из: делеции 11 пар оснований от нуклеотида 3137 до нуклеотида 3147 по сравнению с референсной последовательностью SEQ ID NO: 47; вставки 2 пар оснований между нуклеотидами 3149 и 3150 по сравнению с референсной последовательностью SEQ ID NO: 47, с делецией 377 пар нуклеотидов от нуклеотида 2573 до нуклеотида 2949 по сравнению с референсной последовательностью SEQ ID NO: 47 на том же аллеле; делеции 124 пар оснований от нуклеотида 3024 до нуклеотида 3147 по сравнению с референсной последовательностью SEQ ID NO: 47; делеции 123 пар оснований от нуклеотида 3024 до нуклеотида 3146 по сравнению с референсной последовательностью SEQ ID NO: 47; вставки из 1 пары нуклеотидов между нуклеотидами 3147 и 3148 по сравнению с референсной последовательностью SEQ ID NO: 47; делеции 130 пар оснований от нуклеотида 3030 до нуклеотида 3159 по сравнению с референсной последовательностью SEQ ID NO: 47; делеции 132 пар оснований от нуклеотида 3030 до нуклеотида 3161 по сравнению с референсной последовательностью SEQ ID NO: 47; делеции 1506 пар оснований от нуклеотида 1525 до нуклеотида 3030 по сравнению с референсной последовательностью SEQ ID NO: 47; вставки 7 пар оснований между нуклеотидом 3148 и нуклеотидом 3149 по сравнению с референсной последовательностью SEQ ID NO: 47; делеции 1280 пар оснований от нуклеотида 2818 до нуклеотида 4097 по сравнению с референсной последовательностью SEQ ID NO: 47; делеции 1373 пар оснований от нуклеотида 2724 до нуклеотида 4096 по сравнению с референсной последовательностью SEQ ID NO: 47; делеции 1467 пар оснований от нуклеотида 2431 до нуклеотида 3897 по сравнению с референсной последовательностью SEQ ID NO: 47; делеции 1930 пар оснований от нуклеотида 488 до нуклеотида 2,417 по сравнению с референсной последовательностью SEQ ID NO: 47, где удаленная последовательность заменена вставкой из 12 пар оснований, начинающейся с нуклеотида 488, и где имеется дополнительная делеция 129 пар оснований в экзоне 7 от нуклеотида 3044 до нуклеотида 3172 по сравнению с референсной последовательностью SEQ ID NO: 47; делеции 28 пар оснований от нуклеотида 3145 до нуклеотида 3172 по сравнению с референсной последовательностью SEQ ID NO: 47; делеции 1387 пар оснований от нуклеотида 3145 до нуклеотида 4531 по сравнению с референсной последовательностью SEQ ID NO: 47; делеции 1382 пар оснований от нуклеотида 3113 до нуклеотида 4494 по сравнению с референсной последовательностью SEQ ID NO: 47, где удаленная последовательность заменена вставкой из 11 пар оснований, начинающейся с нуклеотида 3113; делеции 1720 пар оснований от нуклеотида 2440 до нуклеотида 4160 по сравнению с референсной последовательностью SEQ ID NO: 47; делеции 452 пар оснований от нуклеотида 3015 до нуклеотида 3466 по сравнению с референсной последовательностью SEQ ID NO: 47; и комбинаций любого из них.
[0090] По меньшей мере один из аллелей гена CD163 у самки свиньи может содержать делецию 11 пар оснований от нуклеотида 3137 до нуклеотида 3147 по сравнению с референсной последовательностью SEQ ID NO: 47.
[0091] По меньшей мере один из аллелей гена CD163 у самки свиньи может содержать вставку 2 пар оснований между нуклеотидами 3149 и 3150 по сравнению с референсной последовательностью SEQ ID NO: 47, с делецией 377 пар оснований от нуклеотида 2573 до нуклеотида 2949 по сравнению с референсной последовательностью SEQ ID NO: 47 на том же аллеле.
[0092] Если модификация включает вставку 2 пар оснований между нуклеотидами 3149 и 3150 по сравнению с референсной последовательностью SEQ ID NO: 47, с делецией 377 пар оснований от нуклеотида 2573 до нуклеотида 2949 по сравнению с референсной последовательностью SEQ ID NO: 47 на том же аллеле, и вставка из 2 пар оснований может содержать динуклеотид AG.
[0093] По меньшей мере один из аллелей гена CD163 у самки свиньи может содержать делецию 124 пар оснований от нуклеотида 3024 до нуклеотида 3147 по сравнению с референсной последовательностью SEQ ID NO: 47.
[0094] По меньшей мере один из аллелей гена CD163 у самки свиньи может содержать делецию 123 пар оснований от нуклеотида 3024 до нуклеотида 3146 по сравнению с референсной последовательностью SEQ ID NO: 47.
[0095] По меньшей мере один из аллелей гена CD163 у самки свиньи может содержать вставку 1 пары оснований между нуклеотидами 3147 и 3148 по сравнению с референсной последовательностью SEQ ID NO: 47.
[0096] Если модификация включает вставку 1 пары оснований между нуклеотидами 3147 и 3148 по сравнению с референсной последовательностью SEQ ID NO: 47, вставка 1 пары оснований может содержать один остаток аденина.
[0097] По меньшей мере один из аллелей гена CD163 у самки свиньи может содержать делецию 130 пар оснований от нуклеотида 3030 до нуклеотида 3159 по сравнению с референсной последовательностью SEQ ID NO: 47.
[0098] По меньшей мере один из аллелей гена CD163 у самки свиньи может содержать делецию 132 пар оснований от нуклеотида 3030 до нуклеотида 3161 по сравнению с референсной последовательностью SEQ ID NO: 47.
[0099] По меньшей мере один из аллелей гена CD163 у самки свиньи может содержать делецию 1506 пар оснований от нуклеотида 1525 до нуклеотида 3030 по сравнению с референсной последовательностью SEQ ID NO: 47.
[00100] По меньшей мере один из аллелей гена CD163 у самки свиньи может содержать вставку из 7 пар оснований между нуклеотидом 3148 и нуклеотидом 3149 по сравнению с референсной последовательностью SEQ ID NO: 47.
[00101] Если модификация включает вставку из 7 пар оснований между нуклеотидом 3148 и нуклеотидом 3149 по сравнению с референсной последовательностью SEQ ID NO: 47, вставка из 7 пар оснований может содержать последовательность ТАСТАСТ (SEQ ID NO:115).
[00102] По меньшей мере один из аллелей гена CD163 у самки свиньи может содержать делецию 1280 пар оснований от нуклеотида 2818 до нуклеотида 4097 по сравнению с референсной последовательностью SEQ ID NO: 47.
[00103] По меньшей мере один из аллелей гена CD163 у самки свиньи может содержать делецию 1373 пар оснований от нуклеотида 2724 до нуклеотида 4096 по сравнению с референсной последовательностью SEQ ID NO: 47.
[00104] По меньшей мере один из аллелей гена CD163 у самки свиньи может содержать делецию 1467 пар оснований от нуклеотида 2431 до нуклеотида 3897 по сравнению с референсной последовательностью SEQ ID NO: 47.
[00105] По меньшей мере один из аллелей гена CD163 у самки свиньи может содержать делецию 1930 пар оснований от нуклеотида 488 до нуклеотида 2417 по сравнению с референсной последовательностью SEQ ID NO: 47, где удаленная последовательность заменена вставкой из 12 пар оснований; начинающейся с нуклеотида 488, и при этом имеется дополнительная делеция 129 пар оснований в экзоне 7 от нуклеотида 3044 до нуклеотида 3172 по сравнению с референсной последовательностью SEQ ID NO: 47.
[00106] Если модификация включает делецию 1930 пар оснований от нуклеотида 488 до нуклеотида 2417 по сравнению с референсной последовательностью SEQ ID NO: 47, где удаленная последовательность заменена вставкой из 12 пар оснований, начинающейся с нуклеотида 488, и при этом имеется дополнительная делеция 129 пар оснований в экзоне 7 от нуклеотида 3044 до нуклеотида 3172 по сравнению с референсной последовательностью SEQ ID NO: 47, вставка из 12 пар оснований может содержать последовательность TGTGGAGAATTC (SEQ ID NO: 116).
[00107] По меньшей мере один из аллелей гена CD163 у самки свиньи может содержать делецию 28 пар оснований от нуклеотида 3145 до нуклеотида 3172 по сравнению с референсной последовательностью SEQ ID NO: 47.
[00108] По меньшей мере один из аллелей гена CD163 у самки свиньи может содержать делецию 1387 пар оснований от нуклеотида 3145 до нуклеотида 4531 по сравнению с референсной последовательностью SEQ ID NO: 47.
[00109] По меньшей мере один из аллелей гена CD163 у самки свиньи может содержать делецию 1382 пар оснований от нуклеотида 3113 до нуклеотида 4494 по сравнению с референсной последовательностью SEQ ID NO: 47, где удаленная последовательность заменена вставкой из 11 пар оснований, начинающейся с нуклеотида 3113.
[00110] Если модификация включает делецию 1382 пар оснований от нуклеотида 3113 до нуклеотида 4494 по сравнению с референсной последовательностью SEQ ID NO: 47, где удаленная последовательность заменена вставкой из 11 пар оснований, начинающейся с нуклеотида 3113, вставка из 11 пар оснований может содержать последовательность AGCCAGCGTGC (SEQ ID NO: 117).
[00111] По меньшей мере один из аллелей гена CD163 у самки свиньи может содержать делецию 1720 пар оснований от нуклеотида 2440 до нуклеотида 4160 по сравнению с референсной последовательностью SEQ ID NO: 47.
[00112] По меньшей мере один из аллелей гена CD163 у самки свиньи может содержать делецию 452 пар оснований от нуклеотида 3015 до нуклеотида 3466 по сравнению с референсной последовательностью SEQ ID NO: 47.
[00113] Например по меньшей мере один из аллелей гена CD163 у самки свиньи содержит модификацию, выбранную из группы, состоящей из вставки из 7 пар оснований между нуклеотидом 3148 и нуклеотидом 3149 по сравнению с референсной последовательностью SEQ ID NO: 47; вставки 2 пар оснований между нуклеотидами 3149 и 3150 по сравнению с референсной последовательностью SEQ ID NO: 47, с делецией 377 пар оснований от нуклеотида 2573 до нуклеотида 2949 по сравнению с референсной последовательностью SEQ ID NO: 47 на том же аллеле; делеции 11 пар оснований от нуклеотида 3137 до нуклеотида 3147 по сравнению с референсной последовательностью SEQ ID NO: 47; делеции 1382 пар оснований от нуклеотида 3113 до нуклеотида 4494 по сравнению с референсной последовательностью SEQ ID NO: 47, где удаленная последовательность заменена вставкой из 11 пар оснований, начинающейся с нуклеотида 3113; и комбинаций любого из них.
[00114] Ген CD163 у самки свиньи может содержать любую комбинацию модифицированных хромосомных последовательностей, описанных в данном документе.
[00115] Например, самка свиньи может содержать вставку 7 пар оснований между нуклеотидом 3148 и нуклеотидом 3149 по сравнению с референсной последовательностью SEQ ID NO: 47 в одном аллеле гена CD163; и вставку 2 пар оснований между нуклеотидами 3149 и 3150 по сравнению с референсной последовательностью SEQ ID NO: 47, с делецией 377 пар оснований от нуклеотида 2573 до нуклеотида 2949 по сравнению с референсной последовательностью SEQ ID NO: 47, в другом аллеле гена CD163.
[00116] Самка свиньи может содержать делецию 1382 пар оснований от нуклеотида 3113 до нуклеотида 4494 по сравнению с референсной последовательностью SEQ ID NO: 47, где удаленная последовательность заменена вставкой из 11 пар оснований, начинающейся с нуклеотида 3113, в одном аллеле гена CD163; и вставки из 2 пар оснований между нуклеотидами 3149 и 3150 по сравнению с референсной последовательностью SEQ ID NO: 47, с делецией 377 пар оснований от нуклеотида 2573 до нуклеотида 2949 по сравнению с референсной последовательностью SEQ ID NO: 47, в другом аллеле гена CD163.
[00117] Самка свиньи может содержать вставку из 7 пар оснований между нуклеотидом 3148 и нуклеотидом 3149 по сравнению с референсной последовательностью SEQ ID NO: 47 в одном аллеле гена CD163; и делецию 11 пар оснований от нуклеотида 3137 до нуклеотида 3147 по сравнению с референсной последовательностью SEQ ID NO: 47 в другом аллеле гена CD163.
[00118] Самка свиньи может содержать делецию 1382 пар оснований от нуклеотида 3113 до нуклеотида 4494 по сравнению с референсной последовательностью SEQ ID NO: 47, где удаленная последовательность заменена вставкой из 11 пар оснований, начинающейся с нуклеотида 3113, в одном аллеле гена CD163; и делецию 11 пар оснований от нуклеотида 3137 до нуклеотида 3147 по сравнению с референсной последовательностью SEQ ID NO: 47 в другом аллеле гена CD163.
[00119] В любом из способов, описанных в данном документе, аллели гена CD163 самки свиньи могут содержать хромосомную последовательность, имеющую по меньшей мере 80% идентичности последовательности с SEQ ID NO: 47 в областях указанной хромосомной последовательности за пределами вставки или делеции.
[00120] Аллели гена CD163 самки свиньи могут содержать хромосомную последовательность, имеющую по меньшей мере 85% идентичности последовательности с SEQ ID NO: 47 в областях указанной хромосомной последовательности вне вставки или делеции.
[00121] Аллели гена CD163 самки свиньи могут содержать хромосомную последовательность, имеющую по меньшей мере 90% идентичности последовательности с SEQ ID NO: 47 в областях указанной хромосомной последовательности вне вставки или делеции.
[00122] Аллели гена CD163 самки свиньи могут содержать хромосомную последовательность, имеющую по меньшей мере 95% идентичности последовательности с SEQ ID NO: 47 в областях указанной хромосомной последовательности вне вставки или делеции.
[00123] Аллели гена CD163 самки свиньи могут содержать хромосомную последовательность, имеющую по меньшей мере 98% идентичности последовательности с SEQ ID NO: 47 в областях указанной хромосомной последовательности вне вставки или делеции.
[00124] Аллели гена CD163 самки свиньи могут содержать хромосомную последовательность, имеющую по меньшей мере 99% идентичности последовательности с SEQ ID NO: 47 в областях указанной хромосомной последовательности вне вставки или делеции.
[00125] Аллели гена CD163 самки свиньи могут содержать хромосомную последовательность, имеющую по меньшей мере 99,9% идентичности последовательности с SEQ ID NO: 47 в областях указанной хромосомной последовательности вне вставки или делеции.
[00126] Аллели гена CD163 самки свиньи могут содержать хромосомную последовательность, имеющую 100% идентичности последовательности с SEQ ID NO: 47 в областях указанной хромосомной последовательности вне вставки или делеции.
[00127] В любом из описанных здесь способов самка свиньи может содержать хромосомную последовательность, включающую SEQ ID NO: 98, 99, 100, 101, 102, 103, 104, 105, 106, 107, 108, 109, 110, 111, 112, 113, 114 или 119 в одном или в обоих аллелях гена CD163.
[00128] Например, самка свиньи может содержать хромосомную последовательность, включающую SEQ ID NO: 99, 102, 103 или 113 в одном или в обоих аллелях гена CD163.
[00129] Аллели гена CD163 у самки свиньи могут содержать любою комбинацию хромосомных последовательностей, включающую SEQ ID NO: 98, 99, 100, 101, 102, 103, 104, 105, 106, 107, 108, 109, 110, 111, 112, 113, 114 или 119. Таким образом, самка свиньи может содержать хромосомную последовательность, включающую любую из SEQ ID NO: 98, 99, 100, 101, 102, 103, 104, 105, 106, 107, 108, 109, 110, 111, 112, 113, 114 или 119 водном аллеле гена CD163 и хромосомную последовательность, включающую любую из SEQ ID NO: 98, 99, 100, 101, 102, 103, 104, 105, 106, 107, 108, 109, 110, 111, 112, 113, 114 или 119 в другом аллеле гена CD163.
[00130] Например, самка свиньи может содержать хромосомную последовательность, включающую SEQ ID NO: 99 в одном аллеле гена CD163, и хромосомную последовательность, включающую SEQ ID NO: 103 в другом аллеле гена CD163.
[00131] Самка свиньи может содержать хромосомную последовательность, включающую SEQ ID NO: 113 в одном аллеле гена CD163, и хромосомную последовательность, включающую SEQ ID NO: 99 в другом аллеле гена CD163.
[00132] Самка свиньи может содержать хромосомную последовательность, включающую SEQ ID NO: 99 в одном аллеле гена CD163, и хромосомную последовательность, включающую SEQ ID NO: 102 в другом аллеле гена CD163.
[00133] Самка свиньи может содержать хромосомную последовательность, включающую SEQ ID NO: 113 в одном аллеле гена CD163, и хромосомную последовательность, включающую SEQ ID NO: 102 в другом аллеле гена CD163.
[00134] В любом из описанных здесь способов разведение будет производить один или более плодов, которые содержат модифицированную хромосомную последовательность в единственном аллеле гена CD163. Поскольку свинья-самка, используемая в способах, описанных в настоящем документе, содержит модифицированные хромосомные последовательности в обоих аллелях ее гена CD163, скрещивание свиньи-самки с самцом свиньи, содержащим по меньшей мере один аллель CD163 дикого типа, даст плоды, унаследовавшие аллель CD163, содержащий модифицированную хромосомную последовательность от самки свиньи и аллель CD163 дикого типа от самца свиньи. Таким образом, в результате разведения будут получены плоды, гетерозиготные по модифицированной хромосомной последовательности CD163. Если самец содержит два аллеля CD163 дикого типа, все плоды, полученные в результате разведения, будут гетерозиготными по модифицированной хромосомной последовательности CD163.
[00135] Плоды, полученные в результате разведения, будут иметь пониженную чувствительность к инфекции PRRSV в утробе матери по сравнению с зародышами в утробе матери свиньи дикого типа.
[00136] В любом из описанных здесь способов разведение может включать спаривание (совокупление) самки свиньи с самцом свиньи.
[00137] В любом из описанных здесь способов разведение может включать искусственное оплодотворение самки животного спермой, полученной от самца животного.
[00138] В любом из описанных здесь способов разведение может включать перенос оплодотворенного ооцита в репродуктивный тракт самки свиньи.
[00139] В способах, в которых разведение включает перенос оплодотворенного ооцита в репродуктивный тракт самки свиньи, оплодотворенный ооцит может быть получен путем оплодотворения ооцита in vitro спермой, полученной от самца свиньи.
[00140] Оплодотворение in vitro может включать внутрицитоплазматическую инъекцию ооцита со спермой, полученной от самца свиньи.
[00141] Когда разведение включает перенос оплодотворенного ооцита в репродуктивный тракт самки свиньи, ооцит может быть ооцитом, полученным от самки свиньи, так что ооцит содержит модифицированные хромосомные последовательности в обоих аллелях гена CD163. В качестве альтернативы ооцит может быть ооцитом, полученным от другой самки свиньи, содержащей модифицированные хромосомные последовательности в обоих аллелях своего гена CD163, при этом модифицированные хромосомные последовательности снижают чувствительность животного к инфекции PRRSV по сравнению с чувствительностью к инфекции PRRSV самки свиньи, которая не содержит модифицированных хромосомных последовательностей в аллелях своего гена CD163. Таким образом, например, можно использовать любой ооцит, имеющий модифицированные хромосомные последовательности в обоих аллелях ее гена CD163 (например, нокауты обоих аллелей гена С163).
[00142] Однако, когда разведение включает перенос оплодотворенного ооцита в репродуктивный тракт самки свиньи, ооцит не обязательно должен содержать какие-либо модифицированные хромосомные последовательности в аллелях гена CD163. Можно использовать ооцит, содержащий два аллеля CD163 дикого типа. Ооцит, содержащий два аллеля CD163 дикого типа, может быть оплодотворен спермой, полученной от самца свиньи, где сперма содержит два аллеля CD163 дикого типа, для создания оплодотворенного ооцита, содержащего два аллеля CD163 дикого типа. Если такой оплодотворенный ооцит (содержащий два аллеля CD163 дикого типа) переносится в репродуктивный тракт самки свиньи, содержащей модифицированные хромосомные последовательности обоих аллелей своего гена CD163, полученный в результате плод будет защищен от инфекции PRRSV.
[00143] В качестве альтернативы, когда разведение включает перенос оплодотворенного ооцита в репродуктивный тракт самки свиньи, оплодотворенный ооцит может содержать модифицированную хромосомную последовательность в единственном аллеле ее гена CD163. Такие оплодотворенные ооциты также будут производить плоды, которые защищены от инфекции PRRSV, после переноса оплодотворенного ооцита в репродуктивный тракт самки свиньи, содержащей модифицированные хромосомные последовательности в обоих аллелях своего гена CD163.
[00144] Таким образом, когда разведение включает перенос оплодотворенного ооцита в репродуктивный тракт самки свиньи, оплодотворенный ооцит может содержать модифицированные хромосомные последовательности в обоих аллелях ее гена CD163 (например, нокауты обоих аллелей гена CD163), может содержать модифицированную хромосомную последовательность только в одном аллеле ее гена CD163 или может содержать только аллели CD163 дикого типа.
Аффинные метки
[00145] «Аффинная метка» может быть либо пептидной аффинной меткой, либо аффинной меткой нуклеиновой кислоты. Термин «аффинная метка» обычно относится к последовательности белка или нуклеиновой кислоты, которая может быть связана с молекулой (например, связана с небольшой молекулой, белком или связана ковалентной связью). Аффинная метка может быть ненативной последовательностью. Пептидная аффинная метка может включать пептид. Пептидная аффинная метка может быть частью расщепленной системы (например, два неактивных пептидных фрагмента могут объединяться вместе в транс-форму для образования активной аффинной метки). Аффинная метка нуклеиновой кислоты может содержать нуклеиновую кислоту. Аффинная метка нуклеиновой кислоты может представлять собой последовательность, которая может избирательно связываться с известной последовательностью нуклеиновой кислоты (например, посредством гибридизации). Аффинная метка нуклеиновой кислоты может представлять собой последовательность, которая может избирательно связываться с белком. Аффинная метка может быть слита с нативным белком. Аффинная метка может быть слита с нуклеотидной последовательностью.
[00146] Иногда одна, две или несколько аффинных меток могут быть слиты с нативным белком или нуклеотидной последовательностью. Аффинную метку можно ввести в нуклеиновую кислоту, нацеленную на нуклеиновую кислоту, с использованием способов транскрипции in vitro или in vivo. Аффинные метки нуклеиновой кислоты могут включать, например, химическую метку, последовательность связывания РНК-связывающего белка, последовательность связывания ДНК-связывающего белка, последовательность, гибридизуемую с полинуклеотидом с аффинной меткой, синтетический аптамер РНК или синтетический аптамер ДНК. Примеры химических аффинных меток нуклеиновых кислот могут включать, без ограничения, рибонуклеотрифосфаты, содержащие биотин, флуоресцентные красители и дигоксегинин. Примеры белок-связывающих аффинных меток нуклеиновых кислот могут включать, без ограничения, связывающую последовательность MS2, связывающую последовательность U1A, последовательности белка, связывающего «петлю-на-стебле», последовательность boxB, последовательность eIF4A или любую последовательность, распознаваемую РНК-связывающим белком. Примеры олигонуклеотидов с аффинной меткой нуклеиновых кислот могут включать, без ограничения, биотинилированные олигонуклеотиды, 2,4-динитрофенил олигонуклеотиды, олигонуклеотиды флуоресцеина и олигонуклеотиды, конъюгированные с первичным амином.
[00147] Аффинная метка нуклеиновой кислоты может представлять собой РНК-аптамер. Аптамеры могут включать аптамеры, которые связываются с теофиллином, стрептавидином, декстраном В512, аденозином, гуанозином, гуанином/ксантином, 7-метил-GTP, аминокислотными аптамерами, такими как аптамеры, которые связываются с аргинином, цитруллином, валином, триптофаном, цианокобаламином, N-метилмезопорфирином IX, флавином, НАД, и аптамеры антибиотиков, такие как аптамеры, которые связываются с тобрамицином, неомицином, ливидомицином, канамицином, стрептомицином, виомицином и хлорамфениколом.
[00148] Аффинная метка нуклеиновой кислоты может содержать последовательность РНК, которая может быть связана с сайт-направленным полипептидом. Сайт-направленный полипептид может быть условно ферментативно неактивным. Последовательность РНК может содержать последовательность, которая может быть связана с членом систем CRISPR типа I, типа II и/или типа III. Последовательность РНК может быть связана с белком - членом семейства RAMP. Последовательность РНК может быть связана с белком - членом семейства Cas9, белком - членом семейства Cas6 (например, Csy4, Cas6). Последовательность РНК может быть связана с белком - членом семейства Cas5 (например, Cas5). Например, Csy4 может связываться с высокой аффинностью с определенной последовательностью шпильки РНК (Кд ~ 50 пМ) и может расщеплять РНК на участке 3' шпильки.
[00149] Аффинная метка нуклеиновой кислоты может содержать последовательность ДНК, которая может быть связана с сайт-направленным полипептидом. Сайт-направленный полипептид может быть условно ферментативно неактивным. Последовательность ДНК может включать последовательность, которая может быть связана с членом систем CRISPR типа I, типа II и/или типа III. Последовательность ДНК может быть связана с белком аргонавта. Последовательность ДНК может быть связана с белком, содержащим домен цинкового пальца, домен TALE или любой другой ДНК-связывающий домен.
[00150] Аффинная метка нуклеиновой кислоты может содержать последовательность рибозима. Подходящие рибозимы могут включать пептидилтрансферазу 23 SrRNA, РНКазу Р, интроны группы I, интроны группы II, разветвленный рибозим GIR1, Leadzyme, рибозимы шпильки, рибозимы в форме головки молотка, рибозимы HDV, рибозимы СРЕВ3, рибозимы VS, рибозим glmS, рибозим СоТС и синтетические рибозимы.
[00151] Пептидные аффинные метки могут содержать метки, которые можно использовать для отслеживания или очистки (например, флуоресцентный белок, такой как зеленый флуоресцентный белок (GFP), YFP, RFP, CFP, mCherry, tdTomato; метка His, (например, метка 6XHis); метка гемагглютинина (НА); метка FLAG; метка Мус; метка GST; метка МВР; метка белка, связывающего хитин; метка кальмодулина; метка V5; метка, связывающая стрептавидин; и т.п.).
[00152] Как нуклеиновые кислоты, так и пептидные аффинные метки могут содержать метки с небольшими молекулами, такие как биотин или дигитоксин, и метки флуоресцентных меток, такие как, например, флуоресцеин, родамин, флуоресцентные красители Alexa, краситель Cyanine3, краситель Cyanine5.
[00153] Аффинные метки нуклеиновой кислоты могут быть расположены на 5'-конце нуклеиновой кислоты (например, нуклеиновой кислоты, нацеленной на нуклеиновую кислоту). Аффинные метки нуклеиновой кислоты могут быть расположены на 3'-конце нуклеиновой кислоты. Аффинные метки нуклеиновой кислоты могут быть расположены на 5'- и на 3'-конце нуклеиновой кислоты. Аффинные метки нуклеиновой кислоты могут быть расположены внутри нуклеиновой кислоты. Пептидные аффинные метки могут быть расположены на N-конце полипептидной последовательности. Пептидные аффинные метки могут быть расположены на С-конце полипептидной последовательности. Пептидные аффинные метки могут быть расположены на N-конце и С-конце полипептидной последовательности. Множество аффинных меток могут быть слиты с нуклеиновой кислотой и/или полипептидной последовательностью.
Агенты захвата
[00154] Используемый здесь термин «агент захвата» может обычно относиться к агенту, который может очищать полипептид и/или нуклеиновую кислоту. Агент захвата может быть биологически активной молекулой или материалом (например, любым биологическим веществом, встречающимся в природе или синтетическим, и включает, без ограничения, клетки, вирусы, субклеточные частицы, белки, в частности, антитела, иммуноглобулины, антигены, липопротеины, гликопротеины, пептиды, полипептиды, белковые комплексы, комплексы (стрепт)авидин-биотин, лиганды, рецепторы или небольшие молекулы, аптамеры, нуклеиновые кислоты, ДНК, РНК, пептидные нуклеиновые кислоты, олигосахариды, полисахариды, липополисахариды, клеточные метаболиты, гаптены, фармакологически активные вещества, алкалоиды, стероиды, витамины, аминокислоты и сахара). В некоторых вариантах воплощения агент захвата может содержать аффинную метку. В некоторых вариантах воплощения агент захвата может предпочтительно связываться с целевым полипептидом или представляющей интерес нуклеиновой кислотой. Агенты захвата могут свободно плавать в смеси. Агенты захвата могут быть связаны с частицей (например, шариком, микросферой, наночастицей). Агенты захвата могут быть связаны с твердой или полужидкой поверхностью. В некоторых случаях агенты захвата необратимо связаны с мишенью. В других случаях агенты захвата обратимо связаны с мишенью (например, если мишень может быть элюирована, или с помощью химического вещества, такого как имидазол).
ДНК-связывающие полипептиды
[00155] Сайт-специфическая интеграция может быть достигнута с использованием факторов, которые способны распознавать и связываться с конкретными нуклеотидными последовательностями, например, в геноме организма-хозяина. Например, многие белки содержат полипептидные домены, которые способны распознавать ДНК и связываться с ней сайт-специфическим образом. Последовательность ДНК, которая распознается ДНК-связывающим полипептидом, может называться «целевой» последовательностью. Полипептидные домены, которые способны распознавать ДНК и связываться с ней сайт-специфическим образом, обычно правильно складываются и функционируют независимо, связывая ДНК сайт-специфическим образом, даже когда экспрессируются в полипептиде, отличном от белка, из которого был первоначально выделен домен. Подобным образом, последовательности-мишени для распознавания и связывания ДНК-связывающими полипептидами обычно могут распознаваться и связываться такими полипептидами, даже если они присутствуют в больших структурах ДНК (например, хромосоме), особенно когда сайт, где расположена последовательность-мишень, находится в месте, о котором известно, что оно является доступным для растворимых клеточных белков (например, в гене).
[00156] Хотя ДНК-связывающие полипептиды, идентифицированные из белков, которые существуют в природе, обычно связываются с конкретной нуклеотидной последовательностью или мотивом (например, с консенсусной распознающей последовательностью), существуют и известны в данной области техники способы модификации многих таких ДНК-связывающих полипептидов с целью узнавания другой нуклеотидной последовательности или мотива. ДНК-связывающие полипептиды включают, например, и без ограничения: ДНК-связывающие домены цинкового пальца; лейциновые «застежки-молнии»; ДНК-связывающие домены UFA; GAL4; TAL; LexA; репрессоры Тет; LacI; и рецепторы стероидных гормонов.
[00157] Например, ДНК-связывающий полипептид может быть цинковым пальцем. Индивидуальные мотивы цинковых пальцев могут быть сконструированы для нацеливания и специфического связывания с любым из большого диапазона участков ДНК. Канонические Cys2His2 (а также неканонические Cys3His) полипептиды цинковых пальцев связывают ДНК, вставляя а-спираль в большую бороздку двойной спирали ДНК-мишени. Распознавание ДНК цинковым пальцем модульное; каждый палец контактирует в первую очередь с тремя последовательными парами оснований в мишени, и несколько ключевых остатков в полипептиде опосредуют распознавание. Путем включения нескольких ДНК-связывающих доменов цинкового пальца в хоминг-эндонуклеазу можно дополнительно повысить ДНК-связывающую специфичность хоминг-эндонуклеазы (и, следовательно, специфичность любых регулирующих эффектов гена, связанных с этим, также может быть увеличена). См., например, Umov et al. (2005) Nature 435:646-51. Таким образом, один или более ДНК-связывающих полипептидов цинковых пальцев можно сконструировать и использовать таким образом, чтобы хоминг-эндонуклеаза, введенная в клетку-хозяин, взаимодействовала с последовательностью ДНК, которая в геноме клетки-хозяина является уникальной.
[00158] Предпочтительно, белок цинкового пальца не является встречающимся в природе, поскольку он сконструирован таким образом, чтобы связываться с избранным сайтом-мишенью. См., например, Beerli et al. (2002) Nature Biotechnol. 20:135-141; Pabo et al. (2001) Ann. Rev. Biochem. 70:313-340; Isalan et al. (2001) Nature Biotechnol. 19:656-660; Segal et al. (2001) Curr. Opin. Biotechnol. 12:632-637; Choo et al. (2000) Curr. Opin. Struct. Biol. 10:411-416; Патенты США №№6453242; 6534261; 6599692; 6503717; 6689558; 7030215; 6794136; 7067317; 7262054; 7070934; 7361635; 7253273; и Публикации патентов США №№2005/0064474; 2007/0218528; 2005/0267061.
[00159] Сконструированный связывающий домен цинкового пальца может иметь новую специфичность связывания по сравнению с природным белком цинкового пальца. К инженерным способам относятся, помимо прочего, рациональный дизайн и различные типы отбора. Рациональный дизайн включает, например, использование баз данных, содержащих триплетные (или квадруплетные) нуклеотидные последовательности и отдельные аминокислотные последовательности цинковых пальцев, в которых каждая триплетная или квадруплетная нуклеотидная последовательность связана с одной или более аминокислотных последовательностей цинковых пальцев, которые связывают конкретную триплетную или квадруплетную последовательность. См., например, Патенты США №№6453242 и 6534261.
[00160] Типичные способы отбора, включая фаговый дисплей и двухгибридные системы, раскрыты в Патентах США №№5789538; 5925523; 6007988; 6013453; 6410248; 6140466; 6200759; и 6242568; а также WO 98/37186; WO 98/53057; WO 00/27878; WO 01/88197 и GB 2338237. Кроме того, повышение специфичности связывания для связывающих доменов цинкового пальца описано, например, в WO 02/077227.
[00161] Кроме того, как описано в этих и других ссылках, домены цинковых пальцев и/или белки цинковых пальцев с множеством пальцев могут быть связаны вместе с использованием любых подходящих линкерных последовательностей, включая, например, линкеры длиной 5 или более аминокислот. В отношении типичных линкерных последовательностей длиной 6 или более аминокислот см., также Патенты США №№6479626; 6903185; и 7153949. Описанные здесь белки могут включать любую комбинацию подходящих линкеров между отдельными цинковыми пальцами белка.
[00162] Выбор сайтов-мишеней: ZFP и способы дизайна и конструирования слитых белков (и полинуклеотидов, кодирующих их) известны специалистам в данной области техники и подробно описаны в Патентах США №№61400815; 789538; 6453242; 6534261; 5925523; 6007988; 6013453; 6200759; WO 95/19431; WO 96/06166; WO 98/53057; WO 98/54311; WO 00/27878; WO 01/60970; WO 01/88197; WO 02/099084; WO 98/53058; WO 98/53059; WO 98/53060; WO 02/016536 и WO 03/016496.
[00163] Кроме того, как раскрыто в этих и других ссылках, домены «цинковые пальцы» и/или белки с «цинковыми пальцами» с множеством пальцев могут быть связаны вместе с использованием любых подходящих линкерных последовательностей, включая, например, линкеры из 5 или более аминокислот в длину. См. также Патенты США №№6479626; 6903185; и 7153949 для типичных линкерных последовательностей длиной 6 или более аминокислот. Описанные здесь белки могут включать любую комбинацию подходящих линкеров между отдельными цинковыми пальцами белка.
[00164] Если самка свиньи для использования в способах, описанных в настоящем документе, была генетически модифицирована с использованием нуклеазы цинкового пальца, самка животного может быть создана с использованием процесса, включающего введение в эмбрион или клетку по меньшей мере одной молекулы РНК, кодирующей целевую нуклеазу цинкового пальца и, необязательно по меньшей мере один вспомогательный полинуклеотид. Способ дополнительно включает инкубацию эмбриона или клетки для обеспечения экспрессии нуклеазы цинкового пальца, при этом двухцепочечный разрыв, введенный в целевую хромосомную последовательность нуклеазой цинкового пальца, восстанавливается с помощью подверженного ошибкам процесса репарации ДНК с негомологичным соединением концов или процесса репарации ДНК, направленного на гомологию. Способ редактирования хромосомных последовательностей, кодирующих белок, связанный с развитием зародышевой линии, с использованием целевой технологии нуклеазы цинковых пальцев является быстрым, точным и высокоэффективным.
[00165] В качестве альтернативы ДНК-связывающий полипептид представляет собой ДНК-связывающий домен из GAL4. GAL4 - это модульный трансактиватор в Saccharomyces cerevisiae, но он также действует как трансактиватор во многих других организмах. См., например, Sadowski et al. (1988) Nature 335:563-4. В этой регуляторной системе экспрессия генов, кодирующих ферменты метаболического пути галактозы у S. cerevisiae, строго регулируется доступным источником углерода. Johnston (1987) Microbiol. Rev. 51:458-76. Транскрипционный контроль этих метаболических ферментов опосредуется взаимодействием между положительным регуляторным белком, GAL4 и симметричной последовательностью ДНК 17 п. н., с которой GAL4 специфически связывается (восходящая последовательность активации - upstream activation sequence (UAS)).
[00166] Нативный GAL4 состоит из 881 аминокислотного остатка с молекулярной массой 99 кДа. GAL4 включает функционально автономные домены, совокупная активность которых определяет активность GAL4 in vivo. Ma and Ptashne (1987) Cell 48:847-53); Brent and Ptashne (1985) Cell 43(3 Pt 2):729-36. 65 N-концевых аминокислот GAL4 составляют ДНК-связывающий домен GAL4. Keegan et al. (1986) Science 231:699-704; Johnston (1987) Nature 328:353-5. Специфичное для последовательности связывание требует присутствия двухвалентного катиона, координированного шестью остатками Cys, присутствующими в ДНК-связывающем домене. Координированный катион-содержащий домен взаимодействует с консервативным триплетом CCG на каждом конце UAS из 17 п. н. и распознает его посредством прямых контактов с большой бороздкой спирали ДНК. Marmorstein et al. (1992) Nature 356:408-14. ДНК-связывающая функция белков позиционирует С-концевые активирующие транскрипцию домены вблизи промотора, так что активирующие домены могут направлять транскрипцию.
[00167] Дополнительные ДНК-связывающие полипептиды, которые можно использовать, включают, например, но без ограничения, связывающую последовательность из гена, индуцируемого AVRBS3; консенсусную связывающую последовательность из гена, индуцируемого AVRBS3, или сконструированную из нее синтетическую связывающую последовательность (например, ДНК-связывающий домен UPA); TAL; LexA (см., например, Brent & Ptashne (1985), выше); LacR (см., например, Labow et al. (1990) Mol. Cell. Biol. 10:3343-56; Bairn et al. (1991) Proc. Natl. Acad. Sci. USA 88(12):5072-6); рецептор стероидных гормонов (Elliston et al. (1990) J. Biol. Chern. 265:11517-121); penpeccop Tet (Патент США №6271341) и мутантный репрессор Tet, который связывается с последовательностью оператора tet в присутствии, но не в отсутствие тетрациклина (Tc); ДНК-связывающий домен NF-kappaB; и компоненты регуляторной системы, описанные в Wang et al. (1994) Proc. Natl. Acad. Sci. USA 91(17):8180-4, которая использует слияние GAL4, рецептора гормона и VP16.
[00168] ДНК-связывающий домен одной или более нуклеаз, используемых в способах и композициях, описанных в данном документе, может включать природный или сконструированный (не встречающийся в природе) эффекторный ДНК-связывающий домен TAL. См., например. Публикация патента США №2011/0301073.
[00169] В качестве альтернативы нуклеаза может включать систему CRISPR. Например, нуклеаза может содержать систему CRISPR/Cas.
[00170] Система (связанная с CRISPR) развивалась у бактерий и археев как адаптивная иммунная система для защиты от вирусной атаки. При воздействии вируса короткие сегменты вирусной ДНК интегрируются в локус CRISPR. РНК транскрибируется из части локуса CRISPR, которая включает вирусную последовательность. Эта РНК, которая содержит последовательность, комплементарную вирусному геному, опосредует нацеливание белка Cas (например, белка Cas9) на последовательность в вирусном геноме. Белок Cas расщепляет и тем самым отключает вирусную мишень. Недавно система CRISPR/Cas была адаптирована для редактирования генома в эукариотических клетках. Введение сайт-специфических двухцепочечных разрывов (site-specific double strand breaks - DSB) позволяет изменять последовательность-мишень с помощью одного из двух эндогенных механизмов репарации ДНК - либо негомологичного концевого соединения (non-homologous end-joining - NHEJ), либо гомологически-направленной репарации (homology-directed repair - HDR). Система CRISPR/Cas также использовалась для регуляции генов, включая репрессию и активацию транскрипции без изменения целевой последовательности. Нацеленная регуляция генов на основе системы CRISPR/Cas может, например, использовать ферментативно неактивный Cas9 (также известный как каталитически мертвый Cas9).
[00171] Системы CRISPR/Cas включают локус CRISPR (сгруппированные с регулярными интервалами короткие палиндромные повторы), который кодирует компоненты РНК системы, и локус Cas (связанный с CRISPR), который кодирует белки. (Jansen et al., 2002. Mol. Microbiol. 43: 1565-1575; Makarova et al., 2002. Nucleic Acids Res. 30: 482-496; Makarova et al., 2006. Biol. Direct 1: 7; Haft et al., 2005. PLoS Comput. Biol. 1: e60). Локусы CRISPR в микробных хозяевах содержат комбинацию генов Cas, а также некодирующие элементы РНК, способные программировать специфичность CRISPR-опосредованного расщепления нуклеиновой кислоты.
[00172] CRISPR типа II является одной из наиболее хорошо изученных систем и выполняет в природе целевой двухцепочечный разрыв ДНК в четыре последовательных этапа. На первом этапе две некодирующие РНК, матрица пре-crRNA и tracrRNA, транскрибируются из локуса CRISPR. На втором этапе tracrRNA гибридизуется с повторяющимися областями пре-crRNA и опосредует процессинг пре-crRNA в зрелые crRNA, содержащие индивидуальные спейсерные последовательности. На третьем этапе зрелый комплекс crRNA: tracrRNA направляет Cas9 к целевой ДНК посредством спаривания оснований Уотсона-Крика между спейсером на crRNA и протоспейсером на целевой ДНК рядом с соседним мотивом протоспейсера (АМС), что является дополнительным требованием для распознавания цели. Наконец, Cas9 опосредует расщепление целевой ДНК с образованием двухцепочечного разрыва внутри протоспейсера.
[00173] Для использования системы CRISPR/Cas для создания целевых вставок и делеций две некодирующие РНК (crRNA и TracrRNA) можно заменить одной РНК, называемой гидовой РНК (гРНК). Активность системы CRISPR/Cas состоит из трех этапов: (i) вставка экзогенных последовательностей ДНК для предотвращения будущих атак в матрицу CRISPR в процессе, называемом «адаптацией», (ii) экспрессия соответствующих белков, а также экспрессия и обработка матрицы с последующей (iii) РНК-опосредованной интерференцией с чужеродной нуклеиновой кислотой. В бактериальной клетке несколько белков Cas участвуют в естественной функции системы CRISPR/Cas и выполняют такие функции, как вставка чужеродной ДНК и т.д.
[00174] Белок Cas может быть «функциональным производным» встречающегося в природе белка Cas. «Функциональное производное» полипептида с нативной последовательностью представляет собой соединение, обладающее качественным биологическим свойством, общим с полипептидом с нативной последовательностью. «Функциональные производные» включают, без ограничения, фрагменты нативной последовательности и производные полипептида с нативной последовательностью и его фрагментов при условии, что они обладают биологической активностью, общей с соответствующим полипептидом с нативной последовательностью. Рассматриваемая здесь биологическая активность представляет собой способность функционального производного гидролизировать субстрат ДНК на фрагменты. Термин «производное» охватывает как варианты аминокислотной последовательности полипептида, так и их ковалентные модификации и слияния. Подходящие производные полипептида Cas или его фрагмента включают, без ограничения, мутанты, слияния, ковалентные модификации белка Cas или его фрагмента. Белок Cas, который включает белок Cas или его фрагмент, а также производные белка Cas или его фрагмента, может быть получен из клетки или синтезирован химически или с помощью комбинации этих двух процедур. Клетка может быть клеткой, которая естественным образом продуцирует белок Cas, или клеткой, которая естественным образом продуцирует белок Cas и генетически сконструирована для производства эндогенного белка Cas с более высоким уровнем экспрессии или для производства белка Cas из экзогенно введенной нуклеиновой кислоты, которая кодирует Cas, являющийся таким же или отличным от эндогенного Cas. В некоторых случаях клетка естественным образом не вырабатывает белок Cas и генетически сконструирована для производства белка Cas.
[00175] Если самка свиньи для использования в способах, описанных в настоящем документе, была генетически модифицирована с использованием системы CRISPR, система CRISPR/Cas9 может быть использована для создания самки свиньи. Чтобы использовать Cas9 для редактирования геномных последовательностей, белок можно доставить прямо в клетку. В качестве альтернативы, мРНК, кодирующая Cas9, может быть доставлена в клетку, или ген, который обеспечивает экспрессию мРНК, кодирующей Cas9, может быть доставлен в клетку. Помимо этого, либо специфичная для мишени crRNA, либо tracrRNA могут быть доставлены непосредственно в клетку, или специфичная(ые) для мишени гРНК может(ут) быть доставлена(ы) в клетку (эти РНК в качестве альтернативы могут продуцироваться геном, сконструированным для экспрессии этих РНК). Выбор сайтов-мишеней и конструирование crRNA/гРНК хорошо известен в данной области техники. Обсуждение конструкции и клонирования гРНК можно найти на http://www.genome-engineering.org/crispr/wp-content/uploads/2014/05/CRISPR-Reagent-Description-Rev20140509.pdf.
[00176] ДНК-связывающий полипептид может специфически распознавать и связываться с целевой нуклеотидной последовательностью, содержащейся в геномной нуклеиновой кислоте организма-хозяина. В некоторых примерах в геноме хозяина можно найти любое количество дискретных примеров целевой нуклеотидной последовательности. Нуклеотидная последовательность-мишень может быть редкой в геноме организма (например, в геноме могут существовать менее примерно 10, примерно 9, примерно 8, примерно 7, примерно 6, примерно 5, примерно 4, примерно 3, примерно 2 или примерно 1 копии(ий) целевой последовательности). Например, целевая нуклеотидная последовательность может быть расположена в уникальном сайте в геноме организма. Нуклеотидные последовательности-мишени, например и без ограничения, могут быть случайным образом распределены по геному относительно друг друга; расположены в разных группах сцепления в геноме; находиться в той же группе связей; расположены на разных хромосомах; расположены на одной хромосоме; расположены в геноме в сайтах, которые экспрессируются в аналогичных условиях в организме (например, под контролем тех же или по существу функционально идентичных регуляторных факторов); и расположены близко друг к другу в геноме (например, последовательности-мишени могут содержаться в нуклеиновых кислотах, интегрированных как конкатемеры в геномных локусах).
Хоминг-эндонуклеазы
[00177] ДНК-связывающий полипептид, который специфически распознает и связывается с целевой нуклеотидной последовательностью, может содержаться в химерном полипептиде с тем, чтобы обеспечивать специфическое связывание с целевой последовательностью химерного полипептида. В примерах такой химерный полипептид может содержать, например, но без ограничения, полипептиды нуклеазы, рекомбиназы и/или лигазы согласно вышеописанным полипептидам. Химерные полипептиды, содержащие ДНК-связывающий полипептид и полипептид нуклеазы, рекомбиназы и/или лигазы, могут также включать другие функциональные полипептидные мотивы и/или домены, такие как, например, и без ограничения: спейсерная последовательность, расположенная между функциональными полипептидами в химерном белке; лидерный пептид; пептид, который нацеливает гибридный белок на органеллу (например, ядро); полипептиды, которые расщепляются клеточным ферментом; пептидные метки (например, Мус, His и т.д.); и другие аминокислотные последовательности, которые не влияют на функцию химерного полипептида.
[00178] Функциональные полипептиды (например, ДНК-связывающие полипептиды и полипептиды нуклеазы) в химерном полипептиде могут быть функционально связаны. Функциональные полипептиды химерного полипептида могут быть функционально связаны посредством их экспрессии из одного полинуклеотида, кодирующего по меньшей мере функциональные полипептиды, лигированные друг с другом в рамке считывания, с тем, чтобы создать химерный ген, кодирующий химерный белок. Альтернативно, функциональные полипептиды химерного полипептида могут быть функционально связаны другими способами, такими как поперечная сшивка независимых экспрессируемых полипептидов.
[00179] ДНК-связывающий полипептид или гидовая РНК, которая специфически распознает и связывается с целевой нуклеотидной последовательностью, может содержаться в составе природного изолированного белка (или его мутанта), где природный изолированный белок или его мутант также содержит полипептид нуклеазы (и может также содержать полипептид рекомбиназы и/или лигазы). Примеры таких изолированных белков включают TALEN, рекомбиназы (например, рекомбиназы Cre, Hin, Tre и FLP), CRISPR/Cas9, управляемый РНК, и мегануклеазы.
[00180] В контексте настоящего описания термин «хоминг-эндонуклеаза» относится к природным или созданным изолированным белкам и их мутантам, которые содержат ДНК-связывающий полипептид или гидовую РНК и полипептид нуклеазы, а также к химерным полипептидам, содержащим ДНК-связывающий полипептид или гидовую РНК и нуклеазу. Может использоваться любая хоминг-эндонуклеаза, содержащая ДНК-связывающий полипептид или гидовую РНК, которая специфически распознает и связывается с целевой нуклеотидной последовательностью, содержащейся в локусе CD 163 (например, либо потому; что целевая последовательность содержится в нативной последовательности в локусе, или потому, что целевая последовательность была введена в локус, например, путем рекомбинации).
[00181] Некоторые примеры подходящих химерных полипептидов включают, без ограничения, комбинации следующих полипептидов: ДНК-связывающие полипептиды «цинковые пальцы»; полипептид нуклеазы FokI; домены TALE; лейциновые «застежки-молнии»; ДНК-связывающие мотивы фактора транскрипции; и домены узнавания и/или расщепления ДНК, выделенные, например, но без ограничения, из TALEN, рекомбиназы (например, рекомбиназы Cre, Hin, RecA, Tre и FLP), управляемого РНК CRISPR/Cas9, мегануклеазы; и другие полипептиды, известные специалистам в данной области техники. Конкретные примеры включают химерный белок, содержащий сайт-специфический ДНК-связывающий полипептид и полипептид нуклеазы. Химерные полипептиды могут быть сконструированы способами, известными специалистам в данной области техники, для изменения последовательности распознавания ДНК-связывающего полипептида, содержащегося в химерном полипептиде, таким образом, чтобы нацелить химерный полипептид на конкретную представляющую интерес нуклеотидную последовательность.
[00182] Химерный полипептид может содержать ДНК-связывающий домен (например, цинковый палец, TAL-эффекторный домен и т.д.) и домен нуклеазы (расщепления). Домен расщепления может быть гетерологичным ДНК-связывающему домену, например ДНК-связывающему домену цинкового пальца и домену расщепления из нуклеазы, или ДНК-связывающему домену TALEN и домену расщепления, или ДНК-связывающему домену мегануклеазы и домену расщепления из другой нуклеазы. Гетерологичные домены расщепления можно получить из любой эндонуклеазы или экзонуклеазы. Примеры эндонуклеаз, из которых может быть получен домен расщепления, включают, без ограничения, эндонуклеазы рестрикции и хоминг-эндонуклеазы. См., например, 2002-2003 Catalogue, New England Biolabs, Beverly, Mass.; and Belfort et al. (1997) Nucleic Acids Res. 25:3379-3388. Известны дополнительные ферменты, расщепляющие ДНК (например, нуклеаза 51; нуклеаза маша; панкреатическая ДНКаза I; микрококковая нуклеаза; дрожжевая эндонуклеаза НО; см. также Linn et al. (eds.) Nucleases, Cold Spring Harbor Laboratory Press, 1993). В качестве источника доменов расщепления и полудоменов расщепления можно использовать один или более из этих ферментов (или их функциональных фрагментов).
[00183] Аналогичным образом, полудомен расщепления может быть получен, как указано выше, из любой нуклеазы или ее части, которая требует димеризации для активности расщепления. В целом, если слитые белки содержат полудомены расщепления, для расщепления требуются два слитых белка. В качестве альтернативы можно использовать один белок, содержащий два полудомена расщепления. Два полудомена расщепления могут происходить из одной и той же эндонуклеазы (или ее функциональных фрагментов), или каждый полудомен расщепления может происходить из другой эндонуклеазы (или ее функциональных фрагментов). Кроме того, сайты-мишени для двух слитых белков расположены предпочтительно по отношению друг к другу, так что связывание двух слитых белков с их соответствующими сайтами-мишенями помещает полудомены расщепления в пространственную ориентацию друг к другу, что позволяет полудоменам расщепления образовывать функциональный домен расщепления, например, путем димеризации. Таким образом, ближние края сайтов-мишеней могут быть разделены 5-8 нуклеотидами или 15-18 нуклеотидами. Однако между двумя сайтами-мишенями может находиться любое целое количество нуклеотидов или пар нуклеотидов (например, от 2 до 50 пар нуклеотидов или более). Обычно сайт расщепления находится между целевыми сайтами.
[00184] Эндонуклеазы рестрикции (рестрикционные ферменты) присутствуют у многих видов и способны к последовательному связыванию с ДНК (в сайте узнавания) и расщеплению ДНК в сайте связывания или рядом с ним, например, так что один или более экзогенных последовательностей (доноры/трансгены) интегрируются в сайты связывания (мишени) или рядом с ними. Некоторые рестрикционные ферменты (например, тип IIS) расщепляют ДНК в сайтах, удаленных от сайта узнавания, и имеют отдельные домены связывания и расщепления. Например, фермент типа US Fok I катализирует двухцепочечное расщепление ДНК на расстоянии 9 нуклеотидов от своего сайта узнавания на одной цепи и 13 нуклеотидов от своего сайта распознавания на другой цепи. См., например, Патенты США №№5356802; 5436150 и 5487994; а также Li et al. (1992) Proc. Natl. Acad. Sci. USA 89:4275-4279; Li et al. (1993) Proc. Natl. Acad. Sci. USA 90:2764-2768; Kirn et al. (1994a) Proc. Natl. Acad. Sci. USA 91:883-887; Kirn et al. (1994b) J. Biol. Chem. 269:31,978-31,982. Таким образом, слитые белки могут включать домен расщепления (или полудомен расщепления) по меньшей мере из одного рестрикционного фермента типа IIS и одного или более связывающих доменов с цинковыми пальцами, которые могут быть или не быть сконструированными.
[00185] Примером рестрикционного фермента типа IIS, домен расщепления которого отделяется от связывающего домена, является Fok I. Этот конкретный фермент активен в виде димера. Bitinaite et al. (1998) Proc. Natl. Acad. Sci. USA 95: 10570-10575. Соответственно, для целей настоящего изобретения часть фермента Fok I, используемая в описанных слитых белках, считается полудоменом расщепления. Таким образом; для целенаправленного двухцепочечного расщепления и/или целевой замены клеточных последовательностей с использованием слияния цинковый палец-Fok I, для восстановления каталитически активного домена расщепления могут быть использованы два слитых белка, каждый из которых содержит полудомен расщепления FokI. В качестве альтернативы также можно использовать одну молекулу полипептида, содержащую ДНК-связывающий домен и два полудомена расщепления FokI.
[00186] Домен расщепления или полудомен расщепления может представлять собой любую часть белка, которая сохраняет активность расщепления или которая сохраняет способность к мультимеризации (например, димеризации) с образованием функционального домена расщепления.
[00187] Типичные рестрикционные ферменты типа IIS описаны в Публикации патента США №2007/0134796. Дополнительные ферменты рестрикции также содержат разделяемые домены связывания и расщепления, и они рассматриваются в настоящем описании. См., например, Roberts et al. (2003) Nucleic Acids Res. 31:418-420.
[00188] Домен расщепления может содержать один или более сконструированных полудоменов расщепления (также называемых мутантами домена димеризации), которые минимизируют или предотвращают гомодимеризацию, как описано, например, в Публикациях патентов США №№2005/0064474; 2006/0188987 и 2008/0131962.
[00189] В качестве альтернативы, нуклеазы могут быть собраны in vivo в целевом сайте нуклеиновой кислоты с использованием так называемой технологии «расщепленного фермента» (см., например, Публикацию патента США№20090068164). Компоненты таких расщепляющихся ферментов могут быть экспрессированы либо в отдельных экспрессионных конструкциях, либо могут быть связаны в одной открытой рамке считывания, где отдельные компоненты разделены, например, с помощью самоотщепляющегося пептида 2А или последовательности IRES. Компонентами могут быть отдельные связывающие домены с цинковыми пальцами или домены связывающего домена мегануклеазы и нуклеиновой кислоты.
Нуклеазы «цинковые пальцы»
[00190] Химерный полипептид может содержать специально разработанную нуклеазу цинкового пальца (ZFN), которая может быть сконструирована для доставки целевого сайт-специфичного разрыва двухцепочечной ДНК, в который может быть интегрирована экзогенная нуклеиновая кислота или донорная ДНК (см. Публикацию патента США 2010/0257638). ZFN представляют собой химерные полипептиды, содержащие неспецифический домен отщепления от эндонуклеазы рестрикции (например, FokI) и полипептид ДНК-связывающего домена цинкового пальца. См., например, Huang et al. (1996) J. Protein Chem. 15:481-9; Kirn et al. (1997a) Proc. Natl. Acad. Sci. USA 94:3616-20; Kirn et al. (1996) Proc. Natl. Acad. Sci. USA 93:1156-60; Kirn et al. (1994) Proc Natl. Acad. Sci. USA 91:883-7; Kirn et al. (1997b) Proc. Natl. Acad. Sci. USA 94:12875-9; Kirn et al. (1997 с) Gene 203:43-9; Kirn et al. (1998) Biol. Chem. 379:489-95; Nahon and Raveh (1998) Nucleic Acids Res. 26:1233-9; Smith et al. (1999) Nucleic Acids Res. 27:674-81. ZFN могут содержать неканонические ДНК-связывающие домены с цинковыми пальцами (см. Публикацию патента США 2008/0182332). Эндонуклеаза рестрикции Fokl должна димеризоваться через нуклеазный домен, чтобы расщепить ДНК и ввести двухцепочечный разрыв. Следовательно, ZFN, содержащие домен нуклеазы из такой эндонуклеазы, также требуют димеризации домена нуклеазы для расщепления целевой ДНК. Mani et al. (2005) Biochem. Biophys. Res. Commun. 334:1191-7; Smith et al. (2000) Nucleic Acids Res. 28:3361-9. Димеризации ZFN могут способствовать два соседних, противоположно ориентированных ДНК-связывающих сайта. Там же.
[00191] Способ сайт-специфической интеграции экзогенной нуклеиновой кислоты по меньшей мере в один локус CD163 хозяина может включать введение в клетку хозяина ZFN, где ZFN распознает и связывается с целевой нуклеотидной последовательностью, где целевая нуклеотидная последовательность содержится в по меньшей мере одном локусе CD163 хозяина. В некоторых примерах целевая нуклеотидная последовательность не содержится в геноме хозяина в любом другом положении, кроме по меньшей мере одного локуса CD163. Например, ДНК-связывающий полипептид ZFN может быть сконструирован для распознавания и связывания с целевой нуклеотидной последовательностью, идентифицированной по меньшей мере в одном локусе CD163 (например, путем секвенирования локуса CD163). Способ сайт-специфической интеграции экзогенной нуклеиновой кислоты по меньшей мере в один функциональный локус CD163 хозяина, который включает введение в клетку хозяина ZFN, может также включать введение в клетку экзогенной нуклеиновой кислоты, при этом рекомбинации экзогенной нуклеиновой кислоты в нуклеиновую кислоту хозяина, содержащую по меньшей мере один локус CD163, способствует сайт-специфическое распознавание и связывание ZFN с последовательностью-мишенью (и последующее расщепление нуклеиновой кислоты, включающей локус CD163).
Необязательные экзогенные нуклеиновые кислоты для интеграции в локус CD163
[00192] Экзогенные нуклеиновые кислоты для интеграции в локус CD163 включают: экзогенную нуклеиновую кислоту для сайт-специфической интеграции по меньшей мере в одном локусе CD163, например, без ограничения, ORF; нуклеиновую кислоту, содержащую нуклеотидную последовательность, кодирующую хоминг-эндонуклеазу; и вектор, содержащий по меньшей мере один из любого или обоих из вышеизложенного. Таким образом, конкретные нуклеиновые кислоты включают нуклеотидные последовательности, кодирующие полипептид, структурные нуклеотидные последовательности и/или сайты узнавания и связывания ДНК-связывающего полипептида.
Необязательные экзогенные молекулы нуклеиновой кислоты для сайт-специфической интеграции
[00193] Как отмечено выше, вставка экзогенной последовательности (также называемая «донорной последовательностью», «донором» или «трансгеном») предусмотрена, например, для экспрессии полипептида, коррекции мутантного гена или для повышенной экспрессии гена дикого типа. Совершенно очевидно, что донорная последовательность обычно не идентична геномной последовательности, в которую она помещена. Донорная последовательность может содержать негомологичную последовательность, фланкированную двумя областями гомологии, чтобы обеспечить эффективную гомологически направленную репарацию (HDR) в представляющем интерес месте. Кроме того, донорные последовательности могут включать векторную молекулу, содержащую последовательности, которые не гомологичны представляющей интерес области в клеточном хроматине. Молекула-донор может содержать несколько прерывистых областей гомологии с клеточным хроматином. Например, для направленной вставки последовательностей, обычно не присутствующих в представляющей интерес области, указанные последовательности могут присутствовать в молекуле донорной нуклеиновой кислоты и фланкированы областями гомологии с последовательностью в представляющей интерес области.
[00194] Донорный полинуклеотид может быть ДНК или РНК, одноцепочечным или двухцепочечным и может быть введен в клетку в линейной или кольцевой форме. См., например, Публикации патентов США №№2010/0047805, 2011/0281361, 2011/0207221 и 2013/0326645. При введении в линейной форме концы донорной последовательности можно защитить (например, от экзонуклеолитической деградации) способами, известными специалистам в данной области техники. Например, один или более дидезоксинуклеотидных остатков добавляются к 3'-концу линейной молекулы и/или самокомплементарные олигонуклеотиды лигируются с одним или обоими концами. См., например, Chang et al. (1987) Proc. Natl. Acad. Sci. USA 84:4959-4963; Nehls et al. (1996) Science 272:886-889. Дополнительные способы защиты экзогенных полинуклеотидов от деградации включают, помимо прочего, добавление концевой(ых) аминогрупп(аминогрупп) и использование модифицированных межнуклеотидных связей, таких как, например, фосфоротиоаты, фосфорамидаты и остатки O-метилибозы или дезоксирибозы.
[00195] Полинуклеотид можно вводить в клетку как часть векторной молекулы, имеющей дополнительные последовательности, такие как, например, точки начала репликации, промоторы и гены, кодирующие устойчивость к антибиотикам. Более того, донорные полинуклеотиды могут быть введены в виде голой нуклеиновой кислоты, в виде комплекса нуклеиновой кислоты с агентом, таким как липосома или полоксамер, или могут быть доставлены вирусами (например, аденовирусом, AAV, вирусом герпеса, ретровирусом, лентивирусом и лентивирусом, дефектным по интегразе (IDLV)).
[00196] Донор обычно интегрирован, так что его экспрессия управляется эндогенным промотором в сайте интеграции, а именно промотором, который управляет экспрессией эндогенного гена, в который интегрирован донор (например, CD163). Однако будет очевидно, что донор может включать промотор и/или энхансер, например конститутивный промотор или индуцибельный или тканеспецифический промотор.
[00197] Кроме того, хотя и не требуются для экспрессии, экзогенные последовательности могут также включать транскрипционные или трансляционные регуляторные последовательности, например, промоторы, энхансеры, инсуляторы, внутренние сайты входа в рибосомы, последовательности, кодирующие пептиды 2А и/или сигналы полиаденилирования.
[00198] Экзогенные нуклеиновые кислоты, которые могут быть интегрированы сайт-специфическим образом по меньшей мере в один локус CD163, чтобы модифицировать локус CD163, включают, например, но без ограничения, нуклеиновые кислоты, содержащие нуклеотидную последовательность, кодирующую представляющий интерес полипептид; нуклеиновые кислоты, содержащие агрономический ген; нуклеиновые кислоты, содержащие нуклеотидную последовательность, кодирующую молекулу РНКи; или нуклеиновые кислоты, разрушающие ген CD163.
[00199] Экзогенная нуклеиновая кислота может быть интегрирована в локус CD163 с тем, чтобы модифицировать локус CD163, где нуклеиновая кислота содержит нуклеотидную последовательность, кодирующую представляющий интерес полипептид, так что нуклеотидная последовательность экспрессируется в хозяине из локуса CD163. В некоторых примерах представляющий интерес полипептид (например, чужеродный белок) экспрессируется из нуклеотидной последовательности, кодирующей представляющий интерес полипептид в коммерческих количествах. В таких примерах представляющий интерес полипептид может быть экстрагирован из клетки-хозяина, ткани или биомассы.
Молекулы нуклеиновых кислот, содержащие нуклеотидную последовательность, кодирующую хоминг-эндонуклеазу
[00200] Нуклеотидная последовательность, кодирующая хоминг-эндонуклеазу, может быть сконструирована путем манипулирования (например, лигирования) нативных нуклеотидных последовательностей, кодирующих полипептиды, содержащиеся в хоминг-эндонуклеазе. Например, нуклеотидная последовательность гена, кодирующего белок, содержащий ДНК-связывающий полипептид, может быть исследована для идентификации нуклеотидной последовательности гена, который соответствует ДНК-связывающему полипептиду, и эта нуклеотидная последовательность может быть использована в качестве элемента нуклеотидной последовательности, кодирующей хоминг-эндонуклеазу, содержащую ДНК-связывающий полипептид. В качестве альтернативы, аминокислотная последовательность хоминг-эндонуклеазы может быть использована для вывода нуклеотидной последовательности, кодирующей хоминг-эндонуклеазу, например, в соответствии с вырожденностью генетического кода.
[00201] В примерных молекулах нуклеиновых кислот, содержащих нуклеотидную последовательность, кодирующую хоминг-эндонуклеазу, последний кодон первой полинуклеотидной последовательности, кодирующей полипептид нуклеазы, и первый кодон второй полинуклеотидной последовательности, кодирующей ДНК-связывающий полипептид, могут быть разделены любым количеством триплетов нуклеотидов, например, без кодирования интрона или «СТОП». Подобным образом, последний кодон нуклеотидной последовательности, кодирующей первую полинуклеотидную последовательность, кодирующую ДНК-связывающий полипептид, и первый кодон второй полинуклеотидной последовательности, кодирующей полипептид нуклеазы, могут быть разделены любым количеством нуклеотидных триплетов. Последний кодон (т.е. самый 3'-концевой в последовательности нуклеиновой кислоты) первой полинуклеотидной последовательности; кодирующей полипептид нуклеазы, и вторая полинуклеотидная последовательность, кодирующая ДНК-связывающий полипептид, могут быть слиты в фазовом регистре с первым кодоном дополнительной полинуклеотидной кодирующей последовательности, непосредственно примыкающей к ней, или отделенной от нее не более чем короткой пептидной последовательностью, такой как последовательность, кодируемая синтетическим нуклеотидным линкером (например, нуклеотидным линкером, который мог быть использован для достижения слияния). Примеры таких дополнительных полинуклеотидных последовательностей включают, например, но без ограничения, метки, нацеливающие пептиды и сайты ферментативного расщепления. Аналогичным образом, первый кодон из самого 5'-конца (в последовательности нуклеиновой кислоты) первой и второй полинуклеотидных последовательностей может быть слит в фазовом регистре с последним кодоном дополнительной полинуклеотидной кодирующей последовательности, непосредственно прилегающей к нему, или отделен от него не более чем короткой пептидной последовательностью.
[00202] Последовательность, разделяющая полинуклеотидные последовательности, кодирующие функциональные полипептиды в хоминг-эндонуклеазе (например, ДНК-связывающий полипептид и полипептид нуклеазы), может, например, состоять из любой последовательности, так что кодируемая аминокислотная последовательность вряд ли будет существенно изменять трансляцию целевой эндонуклеазы. Из-за автономной природы известных полипептидов нуклеаз и известных ДНК-связывающих полипептидов, промежуточные последовательности не будут влиять на соответствующие функции этих структур.
Другие способы нокаута
[00203] Для инактивации генов могут быть использованы различные другие способы, известные в данной области техники, с тем, чтобы получить животных с нокаутом и/или для введения конструкций нуклеиновых кислот животным для получения животных-основателей и для создания линий животных, в которых конструкция с нокаутом или нуклеиновой кислотой интегрирована в геном. Такие способы включают, без ограничения, микроинъекции в пронуклеус (Патент США №4873191), опосредованный ретровирусом перенос генов в зародышевые линии (Van der Putten et al. (1985) Proc. Natl. Acad. Sci. USA 82, 6148-1652), нацеливание гена в эмбриональные стволовые клетки (Thompson et al. (1989) Cell 56, 313-321), электропорацию эмбрионов (Lo (1983) Mol. Cell. Biol. 3, 1803-1814), опосредованную спермой передачу гена (Lavitrano et al. (2002) Proc. Natl. Acad. Sci. USA 99, 14230-14235; Lavitrano et al. (2006) Reprod. Fert. Develop.18, 19-23) и трансформацию in vitro соматических клеток, таких как кумулюсные клетки или клетки молочной железы, или взрослые, фетальные или эмбриональные стволовые клетки, с последующей трансплантацией ядра (Wilmut et al. (1997) Nature 385, 810-813; и Wakayama et al. (1998) Nature 394, 369-374). Особенно полезными способами являются пронуклеарная микроинъекция, перенос генов, опосредованный спермой, и перенос ядра соматических клеток. Генно-модифицированное животное - это животное, все клетки которого имеют модификацию, включая клетки зародышевой линии. Когда используются способы, позволяющие получить животное, которое в своей модификации является мозаичным, животные можно подвергнуть инбридингу, и отобрать потомство, которое является генно-модифицированным. Клонирование, например, может быть использовано для создания мозаичного животного, если его клетки модифицированы в состоянии бластоцисты, или геномная модификация может иметь место при модификации отдельной клетки. Животные, модифицированные таким образом, чтобы они не достигли половой зрелости, могут быть гомозиготными или гетерозиготными по модификации, в зависимости от используемого конкретного подхода. Если конкретный ген инактивирован нокаут-модификацией, обычно требуется гомозиготность. Если конкретный ген инактивирован РНК-интерференцией или доминантно-негативной стратегией, то часто бывает достаточно гетерозиготности.
[00204] Обычно при микроинъекции эмбриона/зиготы конструкцию нуклеиновой кислоты или мРНК вводят в оплодотворенную яйцеклетку; одна или две оплодотворенные яйцеклетки используются в качестве ядерной структуры, содержащей генетический материал из головки сперматозоида и яйцеклетки, и видны внутри протоплазмы. Оплодотворенные яйцеклетки в пронуклеарной стадии могут быть получены in vitro или in vivo (т.е. хирургическим путем извлечены из яйцепровода животных-доноров). Яйцеклетки, оплодотворенные in vitro, могут быть получены следующим образом. Например, яичники свиней могут быть собраны на бойне и поддерживаться при температуре 22-28°С во время транспортировки. Яичники можно промыть и изолировать для аспирации фолликулов, а фолликулы размером 4-8 мм можно аспирировать под вакуумом в конические центрифужные пробирки объемом 50 мл с использованием игл 18 калибра. Фолликулярная жидкость и аспирированные ооциты можно промыть через предварительные фильтры с коммерческим TL-HEPES (Minitube, Verona, Wis.). Могут быть отобраны ооциты, окруженные компактной кумулюсной массой, и помещены в ТСМ-199 OOCYTE MATURATION MEDIUM (Minitube, Verona, Wis.) с добавлением 0,1 мг/мл цистеина, 10 нг/мл эпидермального фактора роста, 10% фолликулярной жидкости свиней, 50 мкМ 2-меркаптоэтанола, 0,5 мг/мл цФМФ, по 10 МЕ/мл гонадотропина сыворотки крови беременных кобыл (pregnant mare serum gonadotropin- PMSG) и хорионического гонадотропина человека (ХГЧ) на приблизительно 22 часа в увлажненном воздухе при 38,7 °С и 5% CO2. Впоследствии ооциты можно перенести в свежую среду созревания ТСМ-199, которая не будет содержать цАМФ, PMSG или ХГЧ, и инкубировать в течение дополнительных 22 часов. Созревшие ооциты могут быть отделены от кумулюсных клеток путем встряхивания в 0,1% гиалуронидазе в течение 1 минуты.
[00205] Для свиней зрелые ооциты можно оплодотворять в 500 мкл микропробирках системы Minitube PORCPRO IVF MEDIUM SYSTEM (Minitube, Verona, Wis.) в 5-луночных планшетах для оплодотворения Minitube. При подготовке к экстракорпоральному оплодотворению (ЭКО) свежесобранную или замороженную сперму хряка можно промыть и ресуспендировать в среде PORCPRO IVF до 400000 сперматозоидов. Концентрацию спермы можно проанализировать с помощью компьютерного анализа спермы (SPERMVISION, Minitube, Verona, Wis.). Окончательное оплодотворение in vitro может быть выполнено в объеме 10 мкл при конечной концентрации приблизительно 40 подвижных сперматозоидов/ооцит, в зависимости от хряка. Все оплодотворяющиеся ооциты можно инкубировать при 38,7°С в атмосфере с 5,0% CO2 в течение шести часов. Через шесть часов после осеменения предполагаемые зиготы можно дважды промыть в NCSU-23 и перенести в 0,5 мл той же среды. Эта система может регулярно производить 20-30% бластоцист у большинства хряков с 10-30% степенью полиспермического осеменения.
[00206] Линеаризованные конструкции нуклеиновых кислот или мРНК можно вводить в один из пронуклеусов или в цитоплазму. Затем инъецированные яйцеклетки могут быть перенесены в самку-реципиента (например, в яйцеводы самки-реципиента) и можно дать им развиться в самке-реципиенте для получения трансгенных животных или животных, подвергнутых редактированию генов. В частности, оплодотворенные in vitro эмбрионы можно центрифугировать при 15000 х g в течение 5 минут для осаждения липидов, что позволяет визуализировать пронуклеус. Эмбрионы можно вводить с помощью инжектора EppendorfFEMTOJET и культивировать до образования бластоцисты. Можно регистрировать скорость дробления эмбриона, образование и качество бластоцисты.
[00207] Эмбрионы могут быть перенесены хирургическим путем в матки асинхронных реципиентов. Как правило, 100-200 (например, 150-200) эмбрионов могут быть помещены в соединение ампула-перешеек яйцевода с использованием 5,5-дюймового катетера ТОМСАТ®. После операции можно провести ультразвуковое исследование беременности в режиме реального времени.
[00208] При переносе ядра соматической клетки трансгенная клетка или клетка с отредактированным геномом, такая как эмбриональный бластомер, фибробласт плода, фибробласт уха взрослого человека или гранулезная клетка, которая включает конструкцию нуклеиновой кислоты, описанную выше, может быть введена в энуклеированный ооцит для образования комбинированной клетки. Ооциты могут быть энуклеированы путем частичного рассечения зоны около полярного тельца с последующим выдавливанием цитоплазмы в области рассечения. Обычно для инъекции трансгенной клетки или клетки с отредактированным геномом в энуклеированный ооцит, остановленный в мейозе 2, используется инъекционная пипетка с острым скошенным кончиком. В некоторых условных обозначениях ооциты, задержанные в мейозе-2, называются яйцеклетками. После получения эмбриона свиньи или крупного рогатого скота (например, путем слияния и активации ооцита) эмбрион переносится в яйцеводы самки-реципиента примерно через 20-24 часа после активации. См., например, Cibelli et al. (1998) Science 280, 1256-1258 и Патенты США №№6548741, 7547816, 7989657 или 6211429. У свиней самок-реципиентов можно проверить на беременность примерно через 20-21 день после переноса эмбрионов.
[00209] Для создания животных, гомозиготных по инактивированному гену, можно использовать стандартные способы разведения от исходных гетерозиготных животных-основателей. Однако гомозиготность может не требоваться. Описанные здесь свиньи с генетической модификацией могут быть скрещены с другими представляющими интерес свиньями.
[00210] После создания животных с отредактированными генами можно оценить инактивацию эндогенной нуклеиновой кислоты с использованием стандартных методик. Первоначальный скрининг может быть проведен с помощью саузерн-блоттинга для того, чтобы определить, произошла ли инактивация. Для описания саузерн-блоттинга см. разделы 9.37-9.52 в Sambrook et al., 1989, Molecular Cloning, A Laboratory Manual, second edition. Cold Spring Harbor Press, Plainview; N.Y. В начальном скрининге также могут использоваться методики полимеразной цепной реакции (ПЦР). ПЦР относится к процедуре или методике, в которой нуклеиновые кислоты-мишени амплифицируются. В целом, информация о последовательности от концов представляющей интерес области или за ее пределами используется для конструирования олигонуклеотидных праймеров, которые идентичны или сходны по последовательности с противоположными цепями матрицы, подлежащей амплификации. ПЦР можно использовать для амплификации конкретных последовательностей ДНК, а также РНК, включая последовательности общей геномной ДНК или общей клеточной РНК. Праймеры обычно имеют длину от 14 до 40 нуклеотидов, но могут иметь длину от 10 нуклеотидов до сотен нуклеотидов. ПЦР описывается, например, в примере праймера ПЦР: A Laboratory Manual, ed. Dieffenbach and Dveksler, Cold Spring Harbor Laboratory Press, 1995. Нуклеиновые кислоты также могут быть амплифицированы с помощью лигазной цепной реакции, амплификации смещения цепи, самоподдерживающейся репликации последовательности или амплификации на основе последовательности нуклеиновой кислоты. См., например, Lewis (1992) Genetic Engineering News 12,1; Guatelli et al. (1990) Proc. Natl. Acad. Sci. USA 87:1874; и Weiss (1991) Science 254:1292. На стадии бластоцисты эмбрионы могут быть индивидуально обработаны для анализа с помощью ПЦР, гибридизации по Саузерну и ПЦР с использованием сплинкеров (см., например, Dupuy et al. Proc Natl Acad Sci USA (2002) 99:4495).
Интерферирующие РНК
[00211] Известно множество систем интерферирующих РНК (РНКи). Двухцепочечная РНК (дцРНК) индуцирует последовательную деградацию транскриптов гомологичных генов. РНК-индуцированный комплекс сайленсинга (RISC) метаболизирует дцРНК до малых 21-23-нуклеотидных малых интерферирующих РНК (миРНК). RISC содержит двухцепочечную РНКазу (дцРНКазу, например. Dicer) и оцРНКазу (например, Argonaut 2 или Ago2). RISC использует антисмысловую цепь в качестве ориентира для поиска расщепляемой мишени. Известны как миРНК, так и микроРНК (микРНК). Способ инактивации гена у генетически модифицированного животного включает индукцию РНК-интерференции против целевого гена и/или нуклеиновой кислоты, так что экспрессия целевого гена и/или нуклеиновой кислоты снижается.
[00212] Например, экзогенная последовательность нуклеиновой кислоты может индуцировать РНК-интерференцию в отношении нуклеиновой кислоты, кодирующей полипептид. Например, для снижения экспрессии этой ДНК может использоваться двухцепочечная малая интерферирующая РНК (миРНК) или коротко шпилечная РНК (кшРНК), гомологичная целевой ДНК. Конструкции для миРНК могут быть получены, как описано, например, в Fire et al. (1998) Nature 391:806; Romano and Masino (1992) Mol. Microbiol. 6:3343; Cogoni et al. (1996) EMBO J. 15:3153; Cogoni and Masino (1999) Nature 399:166; Misquitta and Paterson (1999) Proc. Natl. Acad. Sci. USA 96:1451; и Kennerdell and Carthew (1998) Cell 95:1017. Конструкции для кшРНК могут быть получены, как описано Mchityre and Fanning (2006) BMC Biotechnology 6:1. В целом, кшРНК транскрибируются как одноцепочечные молекулы РНК, содержащие комплементарные участки, которые могут отжигаться и образовывать короткие шпильки.
[00213] Вероятность обнаружения единичной индивидуальной функциональной кшРНК или миРНК, направленной на конкретный ген, высока. Например, предсказуемость конкретной последовательности миРНК составляет около 50%, но может быть сделан ряд интерферирующих РНК с хорошей уверенностью в том, что по меньшей мере одна из них будет эффективной.
[00214] Можно использовать клетки in vitro, клетки in vivo или генетически модифицированное животное, такое как сельскохозяйственное, которое экспрессирует РНКи, направленную против гена, кодирующего CD 163. РНКи может быть, например, выбрана из группы, состоящей из миРНК, кшРНК, дцРНК, RISC и микРНК.
Индуцибельные системы
[00215] Для инактивации гена CD163 может использоваться индуцибельная система. Известны различные индуцибельные системы, позволяющие контролировать инактивацию гена в пространстве и во времени. Было доказано, что некоторые из них функционируют in vivo у свиней.
[00216] Примером индуцибельной системы является система промотора тетрациклин (tet)-OH, которую можно использовать для регулирования транскрипции нуклеиновой кислоты. В этой системе мутированный репрессор Tet (TetR) сливается с доменом активации трансактиваторного белка VP 16 вируса простого герпеса для создания контролируемого тетрациклином активатора транскрипции (tTA), который регулируется tet или доксициклином (dox). В отсутствие антибиотика транскрипция минимальна, в то время как в присутствии tet или dox транскрипция индуцируется. Альтернативные индуцируемые системы включают системы экдизона или рапамицина. Экдизон - это гормон линьки насекомых, производство которого контролируется гетеродимером рецептора экдизона и продуктом гена ультраспиракл (USP). Экспрессия индуцируется обработкой экдизоном или аналогом экдизона, таким как муристерон А. Агент, который вводят животному для запуска индуцибельной системы, называют индукционным агентом.
[00217] Система, индуцируемая тетрациклином, и система рекомбиназы Cre/loxP (конститутивная или индуцибельная) относятся к числу наиболее часто используемых индуцибельных систем. Система, индуцируемая тетрациклином, включает трансактиватор, контролируемый тетрациклином (tTA)/обратный tTA (rtTA). Способ использования этих систем in vivo включает создание двух линий генетически модифицированных животных. Одна линия животных экспрессирует активатор (рекомбиназу tTA, rtTA или Cre) под контролем выбранного промотора. Другая линия животных экспрессирует акцептор, в котором экспрессия представляющего интерес гена (или гена, который должен быть изменен) находится под контролем целевой последовательности для трансактиваторов tTA/rtTA (или фланкируется последовательностями loxP). Спаривание двух животных обеспечивает контроль экспрессии генов.
[00218] Тетрациклин-зависимые регуляторные системы (tet-системы) основаны на двух компонентах, а именно, трансактиваторе, контролируемом тетрациклином (tTA или rtTA), и tTA/rtTA-зависимом промоторе, который контролирует экспрессию нижестоящей кДНК тетрациклин-зависимым образом. В отсутствие тетрациклина или его производных (таких как доксициклин) tTA связывается с последовательностями tetO, разрешая транскрипционную активацию tTA-зависимого промотора. Однако в присутствии доксициклина tTA не может взаимодействовать со своей мишенью и транскрипция не происходит. Система tet, в которой используется tTA, называется tet-OFF, потому что тетрациклин или доксициклин допускают подавление транскрипции. Введение тетрациклина или его производных позволяет контролировать экспрессию трансгена in vivo во времени. rtTA представляет собой вариант tTA, который не функционирует в отсутствие доксициклина, но требует присутствия лиганда для трансактивации. Эта система tet поэтому называется tet-ON. Системы tet были использованы in vivo для индуцируемой экспрессии нескольких трансгенов, кодирующих, например, репортерные гены, онкогены или белки, участвующие в сигнальном каскаде.
[00219] Система Cre/lox использует рекомбиназу Cre, которая катализирует сайт-специфическую рекомбинацию путем кроссовера между двумя удаленными последовательностями распознавания Cre, то есть сайтами loxP. Последовательность ДНК, введенная между двумя последовательностями loxP (называемая ДНК, фланкированная loxP), вырезается с помощью Cre-опосредованной рекомбинации. Контроль экспрессии Cre у трансгенного животного и/или животного с отредактированным геном с использованием либо пространственного контроля (с тканевым или клеточно-специфическим промотором), либо временного контроля (с индуцибельной системой) приводит к контролю вырезания ДНК между двумя сайтами loxP. Одно применение предназначено для условной инактивации гена (условного нокаута). Другой подход заключается в сверхэкспрессии белка, когда фланкированный loxP стоп-кодон вставляют между промоторной последовательностью и представляющей интерес ДНК. Генетически модифицированные животные не экспрессируют трансген до тех пор, пока не будет экспрессирован Cre, что приведет к удалению фланкированного loxP стоп-кодона. Эта система была применена для тканеспецифического онкогенеза и контролируемой экспрессии антигенных рецепторов в В-лимфоцитах. Также были разработаны индуцируемые рекомбиназы Cre. Индуцибельная рекомбиназа Cre активируется только при введении экзогенного лиганда. Индуцибельные рекомбиназы Cre представляют собой слитые белки, содержащие исходную рекомбиназу Cre и специфический лиганд-связывающий домен. Функциональная активность рекомбиназы Cre зависит от внешнего лиганда, который способен связываться с этим специфическим доменом в слитом белке.
[00220] Можно использовать клетки in vitro, клетки in vivo или генетически модифицированное животное, такое как сельскохозяйственное животное, которое содержит ген CD163 под контролем индуцибельной системы. Хромосомная модификация животного может быть геномной или мозаичной. Индуцируемая система может быть, например, выбрана из группы, состоящей из Tet-On, Tet-Off, Cre-lox и Hif1 alpha.
Векторы и нуклеиновые кислоты
[00221] В клетки могут быть введены различные нуклеиновые кислоты для целей нокаута, для инактивации гена, для получения экспрессии гена или для других целей. Используемый здесь термин «нуклеиновая кислота» включает ДНК, РНК и аналоги нуклеиновых кислот, а также нуклеиновые кислоты, которые являются двухцепочечными или одноцепочечными (то есть смысловой или антисмысловой одноцепочечной). Аналоги нуклеиновой кислоты могут быть модифицированы по основному фрагменту, сахарному фрагменту или фосфатному остову для улучшения, например, стабильности, гибридизации или растворимости нуклеиновой кислоты. Модификации основного фрагмента включают дезоксиуридин для дезокситимидина и 5-метил-2'-дезоксицитидин и 5-бром-2'-доксицитидин для дезоксицитидина. Модификации сахарного фрагмента включают модификацию 2'-гидроксила сахара рибозы с образованием 2'-O-метил или 2'-O-аллильных сахаров. Дезоксирибозно-фосфатный остов может быть модифицирован для получения морфолинуклеиновых кислот, в которых каждый основной фрагмент связан с шестичленным морфолино-кольцом или пептидными нуклеиновыми кислотами, в которых дезоксифосфатный остов заменен псевдопептидным остовом, а четыре основания сохранены. См. Summerton and Weller (1997) Antisense Nucleic Acid Drug Dev. 7(3): 187; и Hyrup et al. (1996) Bioorgan. Med. Chem. 4:5. Кроме того, дезоксифосфатный остов может быть заменен, например, фосфоротиоатным или фосфородитиоатным остовом, фосфорамидитом или алкилфосфотриэфирным остовом.
[00222] Целевая последовательность нуклеиновой кислоты-мишени может быть функционально связана с регуляторной областью, такой как промотор. Регуляторные области могут быть регуляторными областями свиней или могут происходить от других видов. В контексте настоящего описания термин «функционально связанный» относится к расположению регуляторной области относительно последовательности нуклеиновой кислоты таким образом, чтобы разрешить или облегчить транскрипцию нуклеиновой кислоты-мишени.
[00223] Промотор любого типа может быть функционально связан с последовательностью нуклеиновой кислоты-мишени. Примеры промоторов включают, без ограничения, тканеспецифические промоторы, конститутивные промоторы, индуцибельные промоторы и промоторы, реагирующие или не реагирующие на конкретный стимул. Подходящие тканеспецифические промоторы могут приводить к преимущественной экспрессии транскрипта нуклеиновой кислоты в бета-клетках и включают, например, промотор человеческого инсулина. Другие тканеспецифические промоторы могут приводить к преимущественной экспрессии, например, в гепатоцитах или ткани сердца и могут включать, соответственно, промоторы тяжелой цепи альбумина или альфа-миозина. Может быть использован промотор, который способствует экспрессии молекулы нуклеиновой кислоты без значительной тканевой или временной специфичности (т.е. конститутивный промотор). Например, можно использовать промотор бета-актина, такой как промотор гена бета-актина курицы, промотор убиквитина, промотор miniCAGs, промотор глицеральдегид-3-фосфатдегидрогеназы (GAPDH) или промотор 3-фосфоглицераткиназы (PGK), а также вирусные промоторы, такие как промотор тимидинкиназы вируса простого герпеса (HSV-TK), промотор SV40 или промотор цитомегаловируса (CMV). Например, в качестве промотора можно использовать гибрид промотора гена бета-актина курицы и энхансера CMV. См., например, Xu et al. (2001) Hum. Gene Ther. 12:563; и Kiwaki et al. (1996) Hum. Gene Ther. 7:821.
[00224] Дополнительные регуляторные области, которые могут быть полезны в конструкциях нуклеиновых кислот, включают, без ограничения, последовательности полиаденилирования, последовательности контроля трансляции (например, внутренний сегмент входа в рибосомы, IRES), энхансеры, индуцибельные элементы или интроны. Такие регуляторные области могут не быть необходимыми, хотя они могут увеличивать экспрессию, влияя на транскрипцию, стабильность мРНК, эффективность трансляции и т.п.Такие регуляторные области могут быть включены в конструкцию нуклеиновой кислоты, если желательно получить оптимальную экспрессию нуклеиновых кислот в клетке(ах). Однако иногда можно получить достаточную экспрессию без таких дополнительных элементов.
[00225] Может использоваться конструкция нуклеиновой кислоты, которая кодирует сигнальные пептиды или селектируемые маркеры. Сигнальные пептиды можно использовать таким образом, чтобы кодируемый полипептид был направлен в конкретное место клетки (например, на поверхность клетки). Неограничивающие примеры селектируемых маркеров включают пуромицин, ганцикловир, аденозиндезаминазу (ADA), аминогликозид фосфотрансферазу (neo, G418, АРН), дигидрофолатредуктазу (DHFR), гигромицин-В-фосфотрансферазу, тимидин-киназу (ТК) и ксантин-гуанин фосфорибозилтрансферазу (XGPRT). Такие маркеры полезны для отбора стабильных трансформантов в культуре. Другие селектируемые маркеры включают флуоресцентные полипептиды, такие как зеленый флуоресцентный белок или желтый флуоресцентный белок.
[00226] Последовательность, кодирующая селектируемый маркер, может быть фланкирована последовательностями распознавания для рекомбиназы, такой как, например, Cre или Flp.Например, селектируемый маркер может быть фланкирован сайтами узнавания loxP (сайты узнавания длиной 34 п. н., распознаваемые рекомбиназой Cre) или сайтами узнавания FRT, так что селектируемый маркер может быть вырезан из конструкции. См. в Orban, et al., Proc. Natl. Acad. Sci. (1992) 89:6861 обзор технологии Cre/lox и Brand and Dymecki, Dev. Cell (2004) 6:7. Транспозон, содержащий Cre- или Flp-активируемый трансген, прерванный геном селектируемого маркера, также может быть использован для получения животных с условной экспрессией трансгена. Например, промотор, управляющий экспрессией маркера/трансгена, может быть либо убиквитарным, либо тканеспецифичным, что приведет к убиквитарной или тканеспецифичной экспрессии маркера у животных F0 (например, свиней). Тканеспецифичная активация трансгена может быть достигнута, например, путем скрещивания свиньи, которая убиквитарно экспрессирует прерванный маркером трансген, со свиньей, экспрессирующей Cre или Flp тканеспецифическим образом, или путем скрещивания свиньи, которая экспрессирует прерванный маркером трансген тканеспецифическим образом со свиньей, которая убиквитарно экспрессирует рекомбиназу Cre или Flp. Контролируемая экспрессия трансгена или контролируемое удаление маркера разрешает экспрессию трансгена.
[00227] Экзогенная нуклеиновая кислота может кодировать полипептид. Последовательность нуклеиновой кислоты, кодирующая полипептид, может включать последовательность метки, которая кодирует «метку», предназначенную для облегчения последующего манипулирования кодируемым полипептидом (например, для облегчения локализации или обнаружения). Последовательности метки могут быть вставлены в последовательность нуклеиновой кислоты, кодирующую полипептид, так, чтобы кодируемая метка располагалась либо на карбоксильном, либо на амино-конце полипептида. Неограничивающие примеры закодированных тегов включают глутатион S-трансферазу (GST) и метку FLAG™ (Kodak, New Haven, Conn.).
[00228] Конструкции нуклеиновых кислот могут быть метилированы с использованием метилазы SssI CpG (New England Biolabs, Ipswich, Mass.). В общем, конструкция нуклеиновой кислоты может быть инкубирована с S-аденозилметионином и SssI CpG-метилазой в буфере при 37°С. Гиперметилирование можно подтвердить, путем инкубирования конструкции с одной единицей эндонуклеазы HinP1I в течение 1 часа при 37°С и анализа путем электрофореза в агарозном геле.
[00229] Конструкции нуклеиновых кислот могут быть с использованием различных методик введены в клетки эмбриона, плода или взрослого животного любого типа, включая, например, половые клетки, такие как ооцит или яйцеклетка, клетка-предшественница, взрослая или эмбриональная стволовая клетка, первичная половая клетка, почечная клетка, такая как клетка РК-15, островковая клетка, бета-клетка, клетка печени или фибробласт, такой как дермальный фибробласт.Неограничивающие примеры способов включают использование систем транспозонов, рекомбинантных вирусов, которые могут инфицировать клетки, или липосом, или других невирусных способов, таких как электропорация, микроинъекция или осаждение фосфатом кальция, которые способны доставлять нуклеиновые кислоты в клетки.
[00230] В системах транспозонов транскрипционная единица конструкции нуклеиновой кислоты, т.е. регуляторная область, функционально связанная с экзогенной последовательностью нуклеиновой кислоты, фланкирована инвертированным повтором транспозона. Несколько систем транспозонов, в том числе, например, Sleeping Beauty (см. Патент США №6613752 и Публикацию патента США №2005/0003542); Frog Prince (Miskey et al. (2003) Nucleic Acids Res. 31:6873); Tol2 (Kawakami (2007) Genome Biology 8(Suppl.l):S7; Minos (Pavlopoulos et al. (2007) Genome Biology 8(Suppl.l):S2); Hsmarl (Miskey et al. (2007)) Mol Cell Biol. 27:4589); и Passport были разработаны для введения нуклеиновых кислот в клетки, включая клетки мышей, человека и свиней. Особенно полезен транспозон Sleeping Beauty. Транспозаза может быть доставлена в виде белка, кодируемого той же конструкцией нуклеиновой кислоты, что и экзогенная нуклеиновая кислота, может быть введена в отдельную конструкцию нуклеиновой кислоты или предоставлена в виде мРНК (например, транскрибируемой и кэпированной in vitro мРНК).
[00231] Также могут быть включены в конструкцию нуклеиновой кислоты инсуляторные элементы для поддержания экспрессии экзогенной нуклеиновой кислоты и ингибирования нежелательной транскрипции генов-хозяев. См., например, Публикацию патента США №2004/0203158. Обычно инсуляторный элемент фланкирует каждую сторону транскрипционной единицы и является внутренним по отношению к инвертированному повтору транспозона. Неограничивающие примеры инсуляторных элементов включают инсуляторы типа области прикрепления матрицы (MAR) и инсуляторные элементы пограничного типа. См., например, Патенты США №№6395549, 5731178, 6100448 и 5610053, и Публикацию патента США №2004/0203158.
[00232] Нуклеиновые кислоты могут быть включены в векторы. Вектор - это широкий термин, который включает любой конкретный сегмент ДНК, который предназначен для перехода от носителя к целевой ДНК. Вектор может называться вектором экспрессии или векторной системой, которая представляет собой набор компонентов, необходимых для встраивания ДНК в геном или другую целевую последовательность ДНК, такую как эписома, плазмида или даже сегмент ДНК вируса/фага. Векторные системы, такие как вирусные векторы (например, ретровирусы, аденоассоциированный вирус и интегрирующие фаговые вирусы) и невирусные векторы (например, транспозоны), используемые для доставки генов животным, имеют два основных компонента: 1) вектор, состоящий из ДНК (или РНК, которая обратно транскрибируется в кДНК) и 2) транспозазу, рекомбиназу или другой фермент интегразы, который распознает как вектор, так и последовательность ДНК-мишени и вставляет вектор в последовательность ДНК-мишени. Векторы чаще всего содержат одну или более кассет экспрессии, которые содержат одну или более последовательностей контроля экспрессии, где последовательность контроля экспрессии представляет собой последовательность ДНК, которая контролирует и регулирует транскрипцию и/или трансляцию другой последовательности, соответственно, ДНК или мРНК.
[00233] Известно много различных типов векторов. Например, известны плазмиды и вирусные векторы, например, ретровирусные векторы. Плазмиды экспрессии млекопитающих обычно имеют точку начала репликации, подходящий промотор и необязательный энхансер, необходимые сайты связывания рибосом, сайт полиаденилирования, донорные и акцепторные сайты сплайсинга, последовательности терминации транскрипции и 5'-фланкирующие нетранскрибируемые последовательности. Примеры векторов включают: плазмиды (которые также могут быть носителями другого типа вектора), аденовирус, аденоассоциированный вирус (AAV), лентивирус (например, модифицированный ВИЧ-1, SIV или FIV), ретровирус (например, ASV, ALV или MoMLV) и транспозоны (например. Sleeping Beauty, Р-элементы, Tol-2, Frog Prince, piggyBac).
[00234] В данном контексте термин нуклеиновая кислота относится как к РНК, так и к ДНК, включая, например, кДНК, геномную ДНК, синтетическую (например, химически синтезированную) ДНК, а также встречающиеся в природе и химически модифицированные нуклеиновые кислоты, например, синтетические основания или альтернативные остовы. Молекула нуклеиновой кислоты может быть двухцепочечной или одноцепочечной (то есть смысловой или антисмысловой одноцепочечной).
[00235] После подробного описания изобретения станет очевидным, что возможны модификации и изменения без выхода за пределы объема изобретения, определенного в прилагаемой формуле изобретения.
ПРИМЕРЫ
[00236] Для дополнительной иллюстрации настоящего изобретения представлены следующие неограничивающие примеры.
Пример 1. Использование системы CRISPR/Cas9 для получения генно-инженерных свиней из ооцитов и эмбрионов, полученных in vitro
[00237] Недавние сообщения, описывающие хоминг-эндонуклеазы, такие как нуклеазы «цинковые пальцы» (ZFN), эффекторные нуклеазы, подобные активатору транскрипции (TALEN), и компоненты системы кластеризированных регулярных промежуточных коротких палиндромных повторов (СРКРК)/связанных с CRISPR (Cas9) предполагают, что генная инженерия (ГЭ) у свиней теперь может быть более эффективной. Направленные хоминг-эндонуклеазы могут вызывать двухцепочечные разрывы (DSB) в определенных местах генома и вызывать либо случайные мутации из-за негомологичного соединения концов (NHEJ), либо, если имеется донорная ДНК, стимуляцию гомологичной рекомбинации (ГР). Целенаправленная модификация генома с помощью ГР может быть достигнута с помощью хоминг-эндонуклеаз, если донорная ДНК предоставляется вместе с целенаправленной нуклеазой. После внесения специфических модификаций в соматические клетки эти клетки использовали для получения ГИ свиней для различных целей с помощью SCNT. Таким образом, хоминг-эндонуклеазы являются полезным инструментом для создания ГИ свиней. Среди различных хоминг-эндонуклеаз предоставляется эффективным подходом система CRISPR/Cas9, адаптированная из прокариот, где она используется в качестве защитного механизма. В природе системе Cas9 требуются три компонента: РНК (~20 оснований), которая содержит область, комплементарную целевой последовательности (цис-репрессированная РНК [crRNA]), РНК, которая содержит область, комплементарную crRNA (трансактивирующая crRNA [tracrRNA]), и Cas9, ферментативный белковый компонент в этом комплексе. Для выполнения функций спаренной по основанию crRNA и tracrRNA может быть сконструирована единая гидовая РНК (гРНК). Комплекс гРНК/белок может сканировать геном и катализировать DSB в областях, которые комплементарны crRNA/гРНК. В отличие от других разработанных нуклеаз, для создания реагентов, необходимых для направленного воздействия на представляющий интерес ген, необходимо разработать только короткий олигомер, тогда как для сборки ZFN и TALEN требуется серия этапов клонирования.
[00238] В отличие от существующих стандартных способов разрушения генов, использование сконструированных нуклеаз дает возможность использовать зиготы в качестве исходного материала для ГИ. Стандартные способы разрушения генов у сельскохозяйственных животных включают ГР в культивируемых клетках и последующую реконструкцию эмбрионов путем переноса ядра соматических клеток (SCNT). Поскольку клонированные животные, полученные с помощью SCNT, иногда проявляют признаки дефектов развития, для исследований обычно используется потомство основателей SCNT/ГИ с тем, чтобы избежать смешения аномалий SCNT и фенотипа, которые могут возникнуть, если в экспериментах используются животные-основатели. Учитывая более длительный период вынашивания у свиней и более высокую стоимость содержания свиней по сравнению с грызунами, сокращение потребности в разведении дает преимущества по времени и стоимости. Недавний отчет продемонстрировал, что прямая инъекция ZFN и TALEN в зиготы свиней может нарушить эндогенный ген и произвести к появлению поросят с желаемыми мутациями. Однако только около 10% поросят продемонстрировали двуаллельную модификацию целевого гена, а у некоторых были представлены мозаичные генотипы. Недавняя статья продемонстрировала, что система CRISPR/Cas9 может вызывать мутации в развивающихся эмбрионах и производить ГИ-свиней с большей эффективностью, чем ZFN или TALEN. Однако ГИ-свиньи, полученные из системы CRISPR/Cas9, также обладали мозаичными генотипами. Кроме того, все вышеупомянутые исследования для экспериментов использовали зиготы, полученные in vivo, которые требуют интенсивных трудозатрат и большого количества свиноматок для получения достаточного количества зигот.
[00239] В настоящем примере описывается эффективный подход к использованию системы CRISPR/Cas9 для создания ГИ-свиней посредством инъекции зигот, полученных in vitro, и модификации соматических клеток с последующей SCNT. Мишенью были два эндогенных гена (CD163 и CD1D) и один трансген (eGFP), и для инъекций SCNT или РНК использовались только, соответственно, ооциты или зиготы, полученные in vitro. CD163, по-видимому, необходим для продуктивного инфицирования вирусом репродуктивного и респираторного синдрома свиней, вирусом, который, как известно, наносит значительный экономический ущерб свиноводству. CD1D считается неклассическим белком главного комплекса гистосовместимости и участвует в презентации липидных антигенов инвариантным естественным Т-клеткам-киллерам. В качестве моделей для сельского хозяйства и биомедицины были созданы свиньи с дефицитом этих генов. Трансген eGFP был использован в качестве мишени для предварительных экспериментов по проверке концепции и оптимизации способов.
МАТЕРИАЛЫ И СПОСОБЫ
[00240] Химические препараты и реагенты. Если не указано иное, все химические вещества, использованные в этом исследовании, были приобретены у Sigma.
Дизайн гРНК для получения специфических CRISPR
[00241] Были сконструированы гидовые РНК для областей в экзоне 7 CD163, которые были уникальными для CD163 дикого типа и не присутствовали в направленном векторе для осуществления обмена доменами (описанном ниже), так что CRISPR будет приводить к DSB в векторе CD163 дикого типа, но не в области направленного вектора для осуществления обмена доменами. Имелось только четыре места, в которых направленный вектор мог вводить однонуклеотидный полиморфизм (SNP), который изменял бы соседний мотив протоспейсера (Spy) S. pyogenes (AMC). Были выбраны все четыре цели, включая:
(SEQ ID NO:1) GGAAACCCAGGCTGGTTGGAgGG (CRISPR 10),
(SEQ ID NO:2) GGAACTACAGTGCGGCACTGtGG (CRISPR 131),
(SEQ ID NO:3) CAGTAGCACCCCGCCCTGACgGG (CRISPR 256) и
(SEQ ID NO:4) TGTAGCCACAGCAGGGACGTcGG (CRISPR 282).
AMC в каждой гРНК можно идентифицировать по жирному шрифту.
[00242] Для мутаций CD1D поиск мишеней CRISPR был произвольно ограничен кодирующей цепью в пределах первых 1000 пар оснований первичного транскрипта. Однако RepeatMasker [26] (библиотека повторов «Pig») идентифицировала повторяющийся элемент, начинающийся с основания 943 первичного транскрипта. Затем поиск мишеней CRISPR был ограничен первыми 942 п.н. первичного транскрипта. Далее поиск был ограничен первыми 873 п. н. первичного транскрипта, поскольку последний Spy AMC расположен в основании 873. Была выбрана первая мишень (CRISPR 4800), поскольку она перекрывалась со стартовым кодоном, расположенным в основании 42 в первичном транскрипте (CCAGCCTCGCCCAGCGACATgGG (SEQ ID NO: 5)). Были выбраны две дополнительные мишени (CRISPR 5620 и 5626), поскольку в произвольно выбранной области они были наиболее удаленными от первого выбора (CTTTCATTTATCTGAACTCAgGG (SEQ ID NO: 6)) и TTATCTGAACTCAGGGTCCCcGG (SEQ ID NO: 7)). Эти мишени перекрываются. По отношению к стартовому кодону наиболее проксимальные Spy AMC были расположены в простой последовательности, содержащей в значительной степени гомополимерную последовательность, как было определено при визуальной оценке. Четвертая мишень (CRISPR 5350) была выбрана потому, что, по сравнению с выбранной первой мишенью, это была самая проксимальная мишень, которая не содержала обширных гомополимерных областей (CAGCTGCAGCATATATTTAAgGG (SEQ ID NO: 8)). Специфичность сконструированных crRNA была подтверждена поиском схожих последовательностей свиней в GenBank. Олигонуклеотиды (Таблица 1) были отожжены и клонированы в вектор р330Х, содержащий две кассеты экспрессии, человеческий кодон-оптимизированный Cas9 S.pyogenes (hSpy), и химерную гидовую РНК. Р330Х был расщеплен при помощи ith BbsI (New England Biolabs) с последующим лабораторным протоколом Zhang (http://www.addgene.org/crispr/zhang/).
[00243] Для направленного воздействия на eGFP были сконструированы две специфические гРНК, нацеленные на кодирующую последовательность eGFP, в первых 60 п.н. стартового кодона eGFP. Обе гРНК, как eGFP1, так и eGFP2 были на антисмысловой цепи, и eGFP1 был непосредственно направлен на стартовый кодон. Последовательность гРНК eGFP1 была следующей: CTCCTCGCCCTTGCTCACCAtGG (SEQ ID NO: 9), и последовательность гРНК eGFP2 была следующей: GACCAGGATGGGCACCACCCcGG (SEQ ID NO: 10).
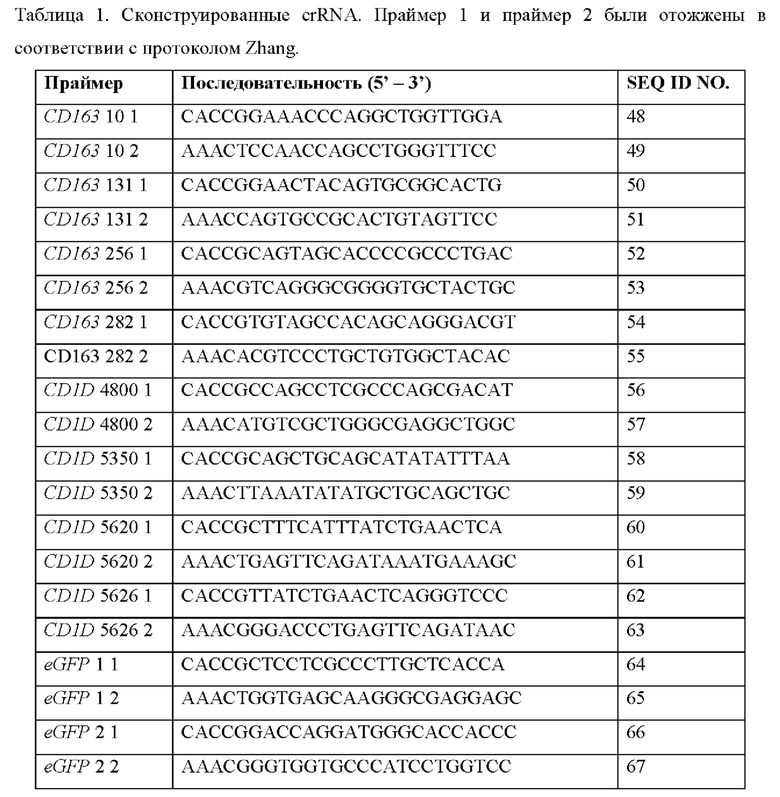
Синтез донорной ДНК для генов CD163 и CD1D
[00244] Оба GD163 и CD1D свиньи были амплифицированы с помощью ПЦР из ДНК, выделенной из фибробластов плода, которые будут использоваться для последующих трансфекций с тем, чтобы гарантировать изогенное соответствие между направленным вектором и трансфицированной клеточной линией. Вкратце, для амплификации фрагмента CD163 из 9538 п.н. использовали метку LA (LA tag) (Clontech), прямой праймер СТСТСССТСАСТСТААССТАСТТ (SEQ ID NO: 11) и обратный праймер TATTTCTCTCACATGGCCAGTC (SEQ ID NO: 12). Фрагмент был подтвержденной последовательностью ДНК и он был использован для построения направленного вектора с заменой домена (Фиг. 1). Этот вектор включал 33 точечные мутации в экзоне 7, так что он кодировал ту же аминокислотную последовательность, что и CD163L человека из экзона 11. Замещенный экзон составлял 315 п. н. Кроме того, последующий интрон был заменен модифицированным интроном миостатина В, который содержал ген селектируемого маркера, который можно было удалить с помощью Cre-рекомбиназы (Cre), и который ранее продемонстрировал нормальный сплайсинг при сохранении сайта loxP (Wells, неопубликованные результаты). Длинное плечо конструкции составляло 3469 п. н. и включало экзон DS с заменой домена. Короткое плечо составляло 1578 п. н. и включало экзоны 7 и 8 (Фиг. 1, панель В). Эта плазмида была использована для попытки заменить кодирующую область экзона 7 в первых экспериментах по трансфекции и она позволила выбрать нацеленные события с помощью селектируемого маркера (G418). Если бы произошло нацеливание, маркер мог быть удален Cre-рекомбиназой. Затем был модифицирован вектор, направленный на DS CD 163 для использования с клеточными линиями, которые уже содержали ген SIGLEC1, разрушенный с помощью Neo, который не мог быть удален при помощи Cre. В этом направленном векторе удаляли кассету Neo, loxP и интрон миостатина В, и в длинном и коротком плече дикого типа оставался только экзон DS (Фиг. 1, панель С).
[00245] Геномную последовательность CD1D свиньи амплифицировали с помощью LA taq с использованием прямого праймера СТСТСССТСАСТСТААССТАСТТ (SEQ ID NO: 13) и обратного праймера GACTGGCCATGTGAGAGAAATA (SEQ ID NO: 14), что в результате дало фрагмент 8729 п. н. ДНК фрагмента секвенировали и использовали для построения направленного вектора, показанного на Фиг. 2. Кассета Neo находится под контролем промотора фосфоглицеринкиназы (PGK) и фланкирована последовательностями loxP, которые были введены для отбора. Длинное плечо конструкции составило 4832 п. н., а короткое плечо составило 3563 п. н. и включало экзоны 6 и 7. Если происходит успешная ГР, экзоны 3,4 и 5 должны были быть удалены и заменены кассетой Neo. Если репарация NHEJ произошла неправильно, экзон 3 будет нарушен.
Получение эмбриональных фибробластов
[00246] Ткань плода свиньи получали на 35 день беременности для создания клеточных линий. Две линии клеток фибробластов плода дикого типа мужского и женского пола были получены от гибрида местной свиньи и свиньи крупной белой породы. В этих исследованиях также использовались фибробласты плода мужского и женского пола, которые ранее были модифицированы для содержания кассеты Neo (SIGLEC1-/- genetics). Эмбриональные фибробласты собирали, как описано, с небольшими изменениями; измельченную ткань каждого плода расщепляли в 20 мл среды для расщепления (среда Игла, модифицированная Дульбекко [DMEM], содержащая L-глутамин и 1 г/л D-глюкозы [Cellgro] с добавлением 200 единиц/мл коллагеназы и 25 единиц Кунитца/мл ДНК-зы I) в течение 5 часов при 38,5°С. После расщепления клетки эмбриональных фибробластов промывали и культивировали с DMEM, 15% эмбриональной бычьей сывороткой (FBS) и 40 мкг/мл гентамицина. После культивирования в течение ночи клетки трипсинизировали и замораживали аликвотами при -80°С в FBS с 10% диметилсульфоксидом и хранили в жидком азоте.
Трансфекция и генотипирование клеток
[00247] Условия трансфекции были в основном такими, как сообщалось ранее. Донорную ДНК всегда использовали в постоянном количестве 1 мкг с различными количествами плазмиды CRISPR/Cas9 (перечислены ниже). Донорную ДНК перед трансфекцией линеаризовали с помощью MLUI (CD163) (NEB) или AFLII (CD1D) (NEB). Пол созданных клеточных линий определяли с помощью ПЦР перед трансфекцией, как описано ранее. Трансфицировали как мужские, так и женские клеточные линии, и данные модификации генома анализировали вместе между трансфекциями. Клеточные линии эмбриональных фибробластов с аналогичным числом пассажей (2-4) культивировали в течение 2 дней и выращивали до 75 85% конфлюэнтности в среде DMEM, содержащей L-глутамин и 1 г/л D-глюкозы (Cellgro) с добавлением 15% FBS, 2,5 нг/мл основного фактора роста фибробластов и 10 мг/мл гентамицина. Клетки фибробластов промывали фосфатно-солевым буфером (PBS) (Life Technologies) и обрабатывали трипсином. Как только клетки отделялись, клетки промывали средой для электр операции (75% солей для клеток [120 мМ KCl, 0,15 мМ CaCl2, 10 мМ K2HPO4, рН 7,6, 5 мМ MgCl2]) и 25% Opti-MEM (Life Technologies). Концентрацию клеток определяли количественно с помощью гемоцитометра. Клетки осаждали при 600 х g в течение 5 минут и ресуспендировали при концентрации 1×106 в среде для электропорации. При каждой электропорации использовалось 200 мкл клеток в кюветах с зазором 2 мм с тремя (1 мс) прямоугольными импульсами, подаваемыми через ВТХ ЕСМ 2001 при 250 В. После электропорации клетки ресуспендировали в DMEM, описанной выше. Для отбора через 24 часа после трансфекции добавляли 600 мкг/мл G418 (Life Technologies), и среду меняли на 7-й день. Колонии собирали на 14-й день после трансфекции. Эмбриональные фибробласты высевали из расчета 10 000 клеток/планшет, если использовался отбор по G418, и из расчета 50 клеток/планшет, если не использовался отбор по G418. Колонии эмбриональных фибробластов собирали с использованием 10-миллиметровых автоклавированных цилиндров для клонирования с герметизированием вокруг каждой колонии с помощью автоклавированной вакуумной смазки. Колонии промывали PBS и собирали с помощью трипсина; затем ресуспендировали в культуральной среде DMEM. Часть (1/3) ресуспендированной колонии переносили в 96-луночный планшет для ПЦР, а оставшиеся (2/3) клетки культивировали в лунках 24-луночного планшета. Осадки клеток ресуспендировали в 6 мкл буфера для лизиса (40 мМ Tris, pH 8,9, 0,9% Triton X-100, 0,4 мг/мл протеиназы K [NEB]), инкубировали при 65°С в течение 30 минут для лизиса клеток с последующим выдерживанием при 85°С в течение 10 минут для инактивации протеиназы K.
ПЦР-скрипит на предмет DS, крупных и мелких делеций
[00248] Выявление репарации, направленной на гомологичную рекомбинацию (ГР). Для идентификации мутаций в CD163 или CD1D применяли ПЦР длинных фрагментов. Для идентификации событий ГР использовались три различных ПЦР-анализа: ПЦР-амплификация областей от последовательностей CD163 или CD1D в донорной ДНК до эндогенных последовательностей CD163 или CD1D с правой или левой стороны, и ПЦР длинных фрагментов, которые амплифицировали большие области CD163 или CD1D, содержащие сконструированные донорные ДНК. Увеличение размера продукта ПЦР на 1,8 т.п.н. (CD1D) или 3,5 т.п.н. (CD163), возникающее в результате добавления экзогенных последовательностей Neo, считалось доказательством направляемой ГР репарации генов. Все условия ПЦР включали начальную денатурацию при 95°С в течение 2 минут, затем 33 цикла по 30 секунд при 94°С, 30 секунд при 50°С и 7-10 минут при 68°С. LA taq использовался для всех анализов в соответствии с рекомендациями производителей. Праймеры представлены в Таблице 2.

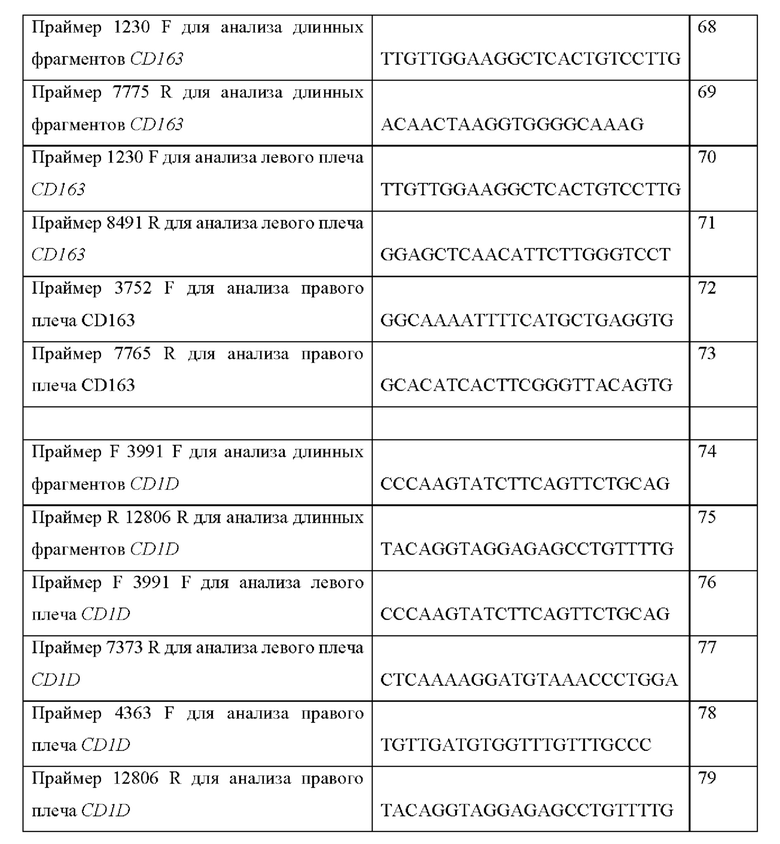
[00249] Анализ мелких делеций (NHEJ). Мелкие делеции определяли с помощью ПЦР-амплификации CD163 или CD1D, фланкирующих предполагаемый участок разреза, введенный системой CRISPR/Cas9. Размер ампликонов составлял 435 п. н. и 1244 п. н. для, соответственно, CD163 и CD1D. Лизаты эмбрионов и фибробластов плода амплифицировали с помощью ПЦР с LA taq. Условиями ПЦР анализов были начальная денатурация при 95°С в течение 2 минут с последующими 33 циклами по 30 секунд при 94°С, 30 секунд при 56°С и 1 минутой при 72°С. Для генотипирования трансфицированных клеток вставки и делеции (INDEL) были идентифицированы путем разделения ампликонов ПЦР с помощью электрофореза в агарозном геле. Для генотипирования эмбрионов полученные продукты ПЦР затем подвергали секвенированию ДНК для выявления мелких делеций с применением прямых праймеров, использованных в ПЦР. Информация о праймерах представлена в Таблице 3.

Перенос ядра соматической клетки (SCNT)
[00250] Для получения эмбрионов путем SCNT использовали либо ооциты свиноматки (ART, Inc.), либо ооциты свиного происхождения с местной бойни. Ооциты, полученные от свиноматок, отправляли на ночь в среду для созревания (ТСМ-199 с 2,9 мМ Hepes, 5 мкг/мл инсулина, 10 нг/мл эпидермального фактора роста [EGF], 0,5 мкг/мл фолликулостимулирующего гормона свиньи [p-FSH], 0,91 мМ пирувата, 0,5 мМ цистеина, 10% свиной фолликулярной жидкости и 25 нг/мл гентамицина) и через 24 часа переносили в свежую среду. После 40-2 часов созревания клетки кумулюса удаляли из ооцитов путем встряхивания в присутствии 0,1% гиалуронидазы. Ооциты, полученные из молодых свиней, созревали, как описано ниже для экстракорпорального оплодотворения (ЭКО). Во время манипуляции ооциты помещали в среду для манипуляции (ТСМ-199 [Life Technologies] с 0,6 мМ NaHCO3, 2,9 мМ Hepes, 30 мМ NaCl, 10 нг/мл гентамицина и 3 мг/мл БСА с осмолярностью 305 мОсм) с добавлением 7,0 мкг/мл цитохалазина В. Полярное тельце вместе с частью прилегающей цитоплазмы, предположительно содержащей пластинку метафазы II, удаляли, и донорскую клетку помещали в перивителлиновое пространство с помощью тонкого стеклянного капилляра. Затем реконструированные эмбрионы были слиты в среде для слияния (0,3 М маннита, 0,1 мМ CaCl2, 0,1 мМ MgCl2 и 0,5 мМ Hepes) с двумя импульсами постоянного тока (интервал 1 секунда) при 1,2 кВ/см в течение 30 секунд с использованием прибора ВТХ Electro Cell Manipulator (Harvard Apparatus). После слияния слитые эмбрионы полностью активировали в темноте при помощи 200 мкМ тимеросала в течение 10 минут и 8 мМ дитиотреитола в течение 30 минут.Затем эмбрионы инкубировали в модифицированной зиготной среде свиней PZM3-MU1 с 0,5 мкМ Scriptaid (S7817; Sigma-Aldrich), ингибитором гистоновой деацетилазы, в течение 14-16 часов, как описано ранее.
Экстракорпоралъное оплодотворение (ЭКО)
[00251] Для ЭКО яичники препубертатных свиней получали с бойни (Farmland Foods Inc.). Незрелые ооциты аспирировали из фолликулов среднего размера (36 мм) с помощью иглы для подкожных инъекций №18, прикрепленной к шприцу на 10 мл. Затем для созревания отбирали ооциты с равномерно темной цитоплазмой и неповрежденными окружающими кумулюсными клетками. Около 50 комплексов кумулюсных ооцитов помещали в лунку, содержащую 500 мкл среды созревания, ТСМ-199 (Invitrogen) с 3,05 мМ глюкозы, 0,91 мМ пирувата натрия, 0,57 мМ цистеина, 10 нг/мл EGF, 0,5 мкг/мл лютеинизирующего гормона (LH), 0,5 мкг/мл ФСГ, 10 нг/мл гентамицина (АРР Pharm) и 0,1% поливиниловый спирт, на 42-44 часа при 38,5°С, 5% CO2 и при увлажненном воздухе. В конце созревания окружающие кумулюсные клетки удаляли из ооцитов путем встряхивания в течение 3 минут в присутствии 0,1% гиалуронидазы. Затем созревшие in vitro ооциты помещали в капли по 50 мкл среды для ЭКО (модифицированная трис-буферная среда, содержащая 113,1 мМ NaCl, 3 мМ KCl, 7,5 мМ CaCl2, 11 мМ глюкозы, 20 мМ Трис, 2 мМ кофеина, 5 мМ пирувата натрия и 2 мг/мл бычьего сывороточного альбумина (БСА)) в группах по 25-30 ооцитов. Один замороженный осадок спермы объемом 100 мкл размораживали в 3 мл Dulbecco PBS с добавлением 0,1% БСА. Замороженную сперму дикого типа либо свежую сперму eGFP промывали центрифугированием в 60% перколле в течение 20 минут при 650 3 g и в модифицированной среде с трис-буфером в течение 10 минут. В некоторых случаях свежесобранная сперма, гетерозиготная по ранее описанному трансгену eGFP, промывалась трижды в PBS. Затем осадок спермы ресуспендировали в среде для ЭКО до 0,5×106 клеток/мл. В капельки с ооцитами вводили пятьдесят микролитров суспензии спермы. Гаметы совместно инкубировали на воздухе в течение 5 часов при 38,5°С в атмосфере с 5% CO2. После оплодотворения эмбрионы инкубировали на воздухе в PZM3-MU1 при 38,5°С и 5% CO2.
Перенос эмбрионов
[00252] Эмбрионы, созданные для получения генетически модифицированных свиней CD163 или CD ID, переносили суррогатным матерям либо на 1-й день (SCNT), либо на 6-й день (инъецированная зигота) после первой активной течки. Для переноса на 6 день зиготы культивировали в течение пяти дополнительных дней в PZM3-MU1 в присутствии 10 нг/мл ps48 (Stemgent, Inc.). Эмбрионы переносили хирургическим путем в ампульно-перешеечное соединение яйцевода суррогатной матери.
Синтез РНК. in vitro для системы CRISPR/Cas9
[00253] Матричную ДНК для транскрипции in vitro амплифицировали с помощью ПЦР (Таблица 4). Матрицей для ПЦР служила плазмида CRISPR/Cas9, используемая для экспериментов по трансфекции клеток. С целью экспрессии Cas9 в зиготах для получения мРНК Cas9 использовали набор mMESSAGE mMACHINE Ultra Kit (Ambion). Затем к мРНК Cas9 добавляли сигнал поли-А с использованием набора Poly (A) tailing kit (Ambion). Гидовые РНК CRISPR были произведены с помощью MEGAshortscript (Ambion). Качество синтезированных РНК визуализировали на 1,5% агарозном геле, а затем разбавляли до конечной концентрации 10 нг/мкл (как гРНК, так и Cas9) и распределяли на аликвоты по 3 мкл.

Микроинъекции сконструированной системы CRISPR/Cas9 в зиготы
[00254] Информационную РНК, кодирующуя Cas9 и гРНК, вводили в цитоплазму оплодотворенных ооцитов через 14 часов после оплодотворения (предполагаемые зиготы) с использованием микроинжектора FemtoJet (Eppendorf). Микроинъекцию проводили в среде для манипуляции на столике с подогревом инвертированного микроскопа Nikon (Nikon Corporation; Tokyo, Japan). Затем инъецированные зиготы переносили в PZM3-MU1 с 10 нг/мл ps48 до дальнейшего использования.
Статистический анализ
[00255] Количество колоний с модифицированным геномом было классифицировано как 1, а колонии без модификации генома были классифицированы как 0. Различия определяли с использованием PROC GLM (SAS) с Р-значением 0,05, которое рассматривается как значимое. Средние значения рассчитывались как средние значения наименьших квадратов. Данные представлены в виде числовых средних значений ± SEM (стандартная ошибка среднего).
РЕЗУЛЬТАТЫ
Нокаут CD163 и CD1D в соматических клетках, обусловленный CRISPR/Cas9
[00256] Была протестирована эффективность четырех различных плазмид CRISPR (гиды 10, 131, 256 и 282), нацеленных на CD163, при количестве 2 мкг/мкл донорной ДНК (Таблица 5). CRISPR 282 привел к значительно большему среднему образованию колоний, чем обработка CRISPR 10 и 256 (Р<0,05). Из анализа ПЦР длинных фрагментов, описанного выше, вместо DS были обнаружены крупные делеции в диапазоне от 503 пар оснований до 1506 пар оснований, образованные путем ГР, как и предполагалось изначально (Фиг. 3, панель А). Это было неожиданным, поскольку предыдущие отчеты с другими системами редактирования ДНК продемонстрировали гораздо меньшие делеции 6 333 п. н. с использованием ZFN у свиней. CRISPR 10 и смесь всех четырех CRISPR привели к большему количеству колоний с модифицированным геномом, чем CRISPR 256 и 282 (таблица 5, Р<0,002). Трансфекция при помощи CRISPR 10 и плазмиды, содержащей Neo, но не имеющей гомологии с CD163, не привела к образованию колоний с крупной делецией. Интересно, что одна моноаллельнаяделеция также была обнаружена тогда, когда донорнаяДНК была введена без какого-либо CRISPR. Этот анализ, вероятно, представляет собой заниженную оценку частоты мутаций, поскольку при секвенировании не проводился скрининг на любые потенциальные мелкие делении, которые нельзя обнаружить на агарозном геле в трансфицированных соматических клетках.
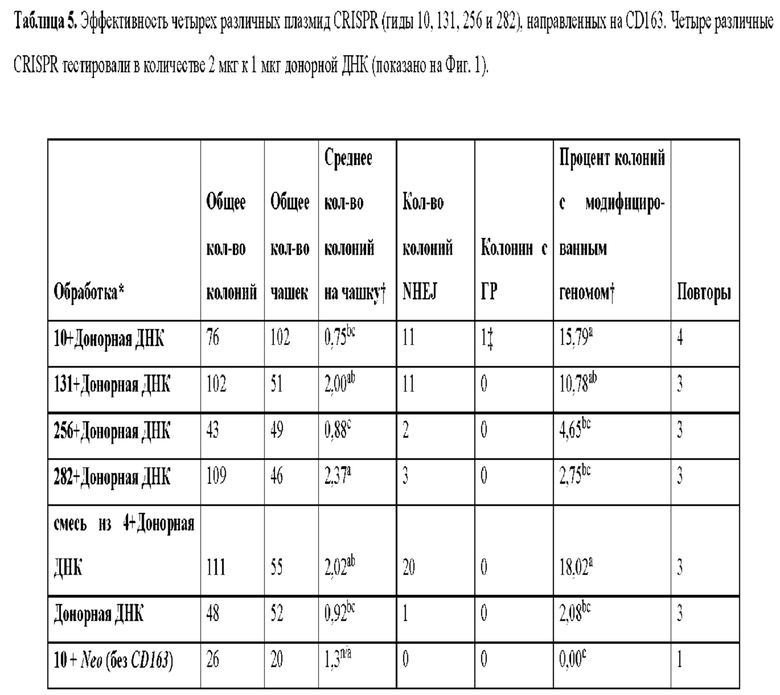
* Смесь из 4 + Донорная ДНК представляет собой смешивание равных количеств по 0,5 мкг каждого CRISPR с 1 мкг донорной ДНК. Обработка донорной ДНК служила в качестве контроля без CRISPR, и обработка 10 + Neo иллюстрирует то, что крупные делении, наблюдаемые при обработке CRISPR, присутствовали только тогда, когда также имелся CD163 донорной ДНК.
 ANOVA был проведен для сравнения среднего количества колоний на чашку для оценки токсичности CRISPR и процента колоний с модифицированным геномом. Р-значения составили, соответственно, 0,025 и 0,0002. н/п = для этой обработки не было повторов, поэтому статистический анализ не проводился.
ANOVA был проведен для сравнения среднего количества колоний на чашку для оценки токсичности CRISPR и процента колоний с модифицированным геномом. Р-значения составили, соответственно, 0,025 и 0,0002. н/п = для этой обработки не было повторов, поэтому статистический анализ не проводился.
 Единственная колония с ГР представляет собой событие частичной ГР.
Единственная колония с ГР представляет собой событие частичной ГР.
a-c Буквы в верхнем индексе указывают на значительную разницу между обработками как для среднего числа колоний на чашку, так и для процента колоний с модифицированным геномом (Р<0,05)
[00257] Первоначальной целью было получение события, направленного на обмен доменами (domain swap - DS) путем ГР для CD163, однако CRISPR не увеличивали эффективность направленного воздействия на CD163. Следует отметить, что различные комбинации этого направленного вектора использовались для модификации CD163 традиционными трансфекциями с помощью ГР, и после скрининга 3399 колоний они привели к 0 целевым событиям (Whitworth and Prather, неопубликованные результаты). Были получены две свиньи с полным DS в результате ГР, который содержал все 33 мутации, которые пытались ввести путем трансфекции с помощью CRISPR 10 и DS-направленного вектора в качестве донорной ДНК.
[00258] Затем была протестирована эффективность CRISPR/Cas9-индуцированных мутаций без селекции препаратами; линия клеток эмбриональных фибробластов, использованная в этом исследовании, уже имела интеграцию устойчивой кассеты Neo и нокаут SIGLEC1. Также проверялось, будет ли соотношение CRISPR/Cas9 и донорной ДНК увеличивать модификацию генома или вызывать токсический эффект при высокой концентрации. Для этого тестирования был избран CRISPR 131, поскольку в предыдущем эксперименте он привел к общему количеству колоний и увеличению процента колоний, обладающих измененным геномом. Увеличение количества ДНК CRISPR 131 с3:1 до 20:1 не оказало значительного влияния на выживаемость фибробластов плода. Процент колоний с геномом, модифицированным NHEJ, существенно не отличался при различных концентрациях CRISPR, однако при соотношении 10:1 наблюдалось наибольшее количество NHEJ (Таблица 6, Р=0,33). Даже при самом высоком соотношении ДНК CRISPR к донорной ДНК (20:1) не отмечалось ГР.
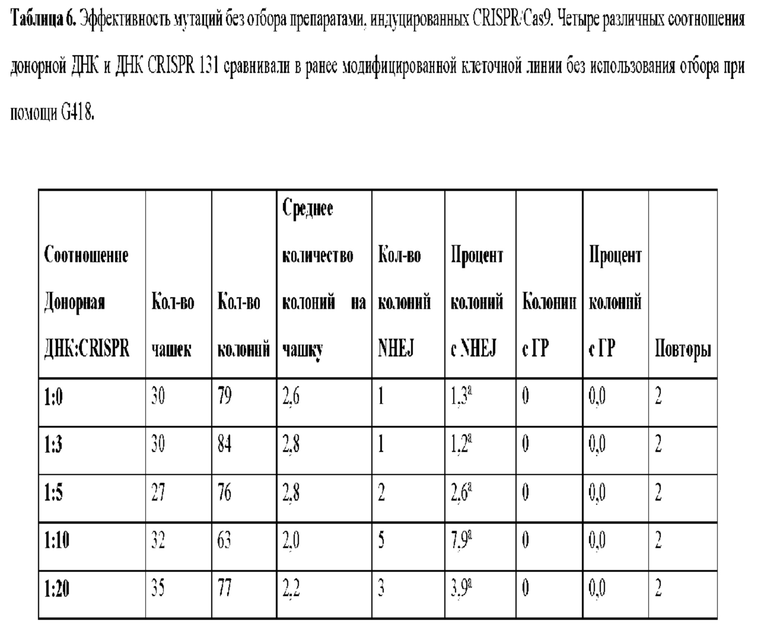
а Значимое различие между обработками для процента колоний с NHEJ-репарацией (Р>0,05).
b Не наблюдалось значимого различия в количестве колоний с модифицированным геномом с повышением концентрации RISPR (P>0,33).
[00259] На основании этого опыта была предпринята попытка целенаправленного разрушения CD1D в соматических клетках. Были разработаны и протестированы как на мужских, так и на женских клетках четыре различных CRISPR. Модификации CD1D можно было обнаружить из трех примененных CRISPR, но использование CRISPR 5350 не привело к модификации CD1D с делецией, достаточно большой для обнаружения с помощью электрофореза в агарозном геле (Таблица 7). Интересно, что в результате ГР не было получено никаких генетических изменений, несмотря на то, что была предоставлена донорная ДНК. Однако наблюдались крупные делеции, подобные тем, что выявлялись в экспериментах с нокаутом CD163 (Фиг. 3, панель В). Если CRISPR/Cas9 не использовался с донорной ДНК, не обнаруживалось никакой целевой модификации CD1D с крупной делецией. Модификация CD1D в результате направленного воздействия под управлением CRISPR/Cas9 составила 4/121 и 3/28 в, соответственно, мужских и женских колониях клеток. В данные по трансфекции были включены только INDEL, обнаруживаемые при электрофорезе в агарозном геле.
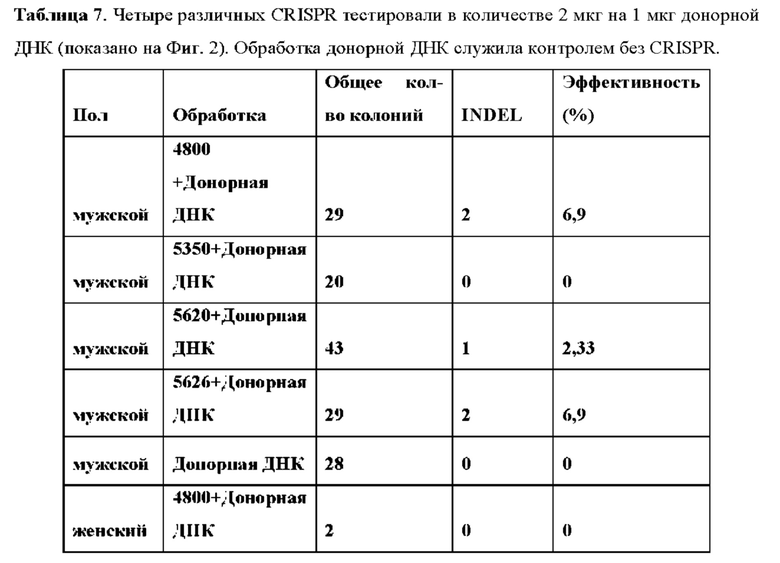
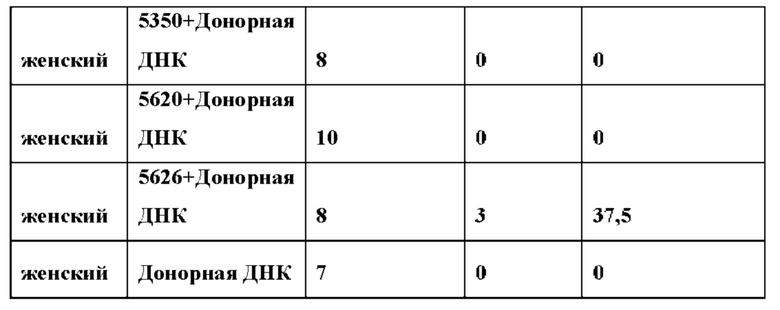
Получение свиней CD163 и CD1D путем SCNT с использованием генетически отредактированных клеток
[00260] Клетки с модификацией CD163 или CD1D использовали для SCNT для получения свиней с нокаутом по CD163 и CD1D (Фиг. 3). Было выполнено семь переносов эмбрионов (CD163, Таблица 8), шесть переносов эмбрионов (CD163-no Neo) и пять переносов эмбрионов (CD1D) в молодые свиньи-реципиентки с эмбрионами SCNT из фибробластов мужского и женского пола, трансфицированных системами CRISPR/Cas9. Шесть (CD163), две (CD163-no Neo) и четыре (CD1D) (Таблица 9) молодых свиней-реципиенток выносили и родили в срок, в результате чего процент наступления беременности составил, соответственно, 85,7%, 33,3% и 80%. Из реципиенток CD163 пять родили здоровых поросят путем кесарева сечения. Одна (О044) опоросилась естественным путем. Размер помета варьировал от одного до восьми. Четыре свиньи были усыплены из-за того, что после рождения они перестали развиваться. Один поросенок был умерщвлен из-за тяжелой волчьей пасти. Все оставшиеся поросята выглядели здоровыми (Фиг. 3, панель С). Два помета поросят-самцов, полученные из фибробластов плода, трансфицированных CRISPR 10 и донорной ДНК, описанные на Фиг. 3, панель В имели делецию 30 п. н. в экзоне 7, соседнем с CRISPR 10, и дополнительную делецию 1476 п. н. предыдущего интрона, таким образом произошло удаление соединения нитрона 6/экзона 7 CD163 (Фиг. 3, панель Е). Генотипы и предсказанные трансляции приведены в Таблице 10. Один поросенок-самец и один помет самок (4 поросенка) были получены в результате трансфекции CD163-no Neo клеток, ранее модифицированных SIGLEC1. Все пятеро поросят были дважды нокаутированы по SIGLEC1 и CD163. Поросенок-самец имел двуаллельную модификацию CD163 с делецией 28 п. н. в экзоне 7 на одном аллеле и делецией 1387 п. н. на другом аллеле, которая включала частичную делецию экзона 7 и полную делецию экзона 8 и последующего интрона, таким образом, происходило удаление соединений экзонов интрона. Поросята-самки имели двуаллельную мутацию CD163, включая делецию 1382 п. н. со вставкой 11 п. н. на одном аллеле и делецию 1720 п. н. CD163 на другом аллеле. Резюме модификаций CD 163 и предсказанных трансляций можно найти в Таблице 10. Резюме модификаций CD1D и предсказанных трансляций с помощью модификации CRISPR можно найти в Таблице 11. Вкратце, родился один помет самок и два помета самцов, в результате чего было получено 13 поросят. Один поросенок умер сразу после рождения. Двенадцать из 13 поросят имели либо двуаллельную, либо гомозиготную делецию CD1D (Фиг. 3, панель F). Один поросенок был дикого типа.
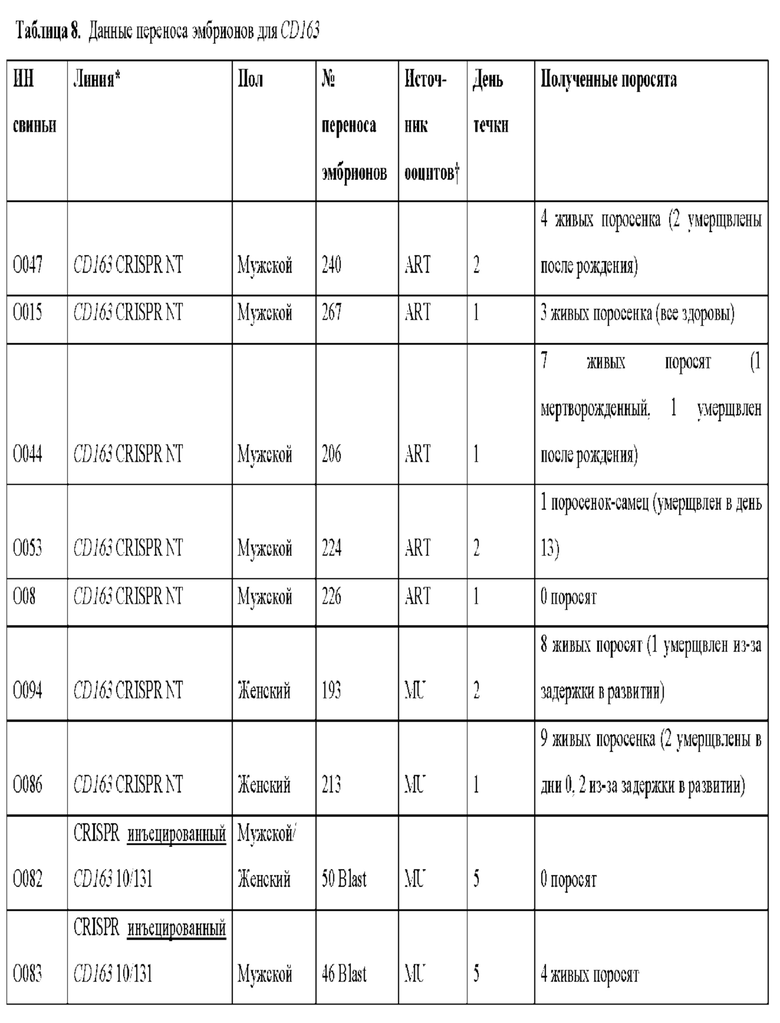
*Линия CD163 CRISPR NT представляет собой эмбрионы, созданные путем переноса ядра (NT) с линией эмбриональных фибробластов, модифицированных путем трансфекции. Эмбрионы с введенными CRISPR представляли собой ЭКО эмбрионы, инъецированные на стадии клеток 1 гидовой РНК CD163 с РНК CAS9. Эмбриональная линия CD163 CRISPR NT-no Neo представляет собой эмбрионы, созданные путем ядерного переноса с предварительно модифицированными эмбриональными фибробластами, которые уже были линией, резистентной kAb, модифицированной путем трансфекции без использования селективного маркера.
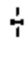 MU относится к ооцитам молодых свиней, которые были аспирированы и созревали в Университете Миссури, как описано в разделе ЭКО Материалов и способов. ART относится к ооцитам свиней, которые были приобретены и созревали, как описано в разделе SCNT Материалов и способов.
MU относится к ооцитам молодых свиней, которые были аспирированы и созревали в Университете Миссури, как описано в разделе ЭКО Материалов и способов. ART относится к ооцитам свиней, которые были приобретены и созревали, как описано в разделе SCNT Материалов и способов.
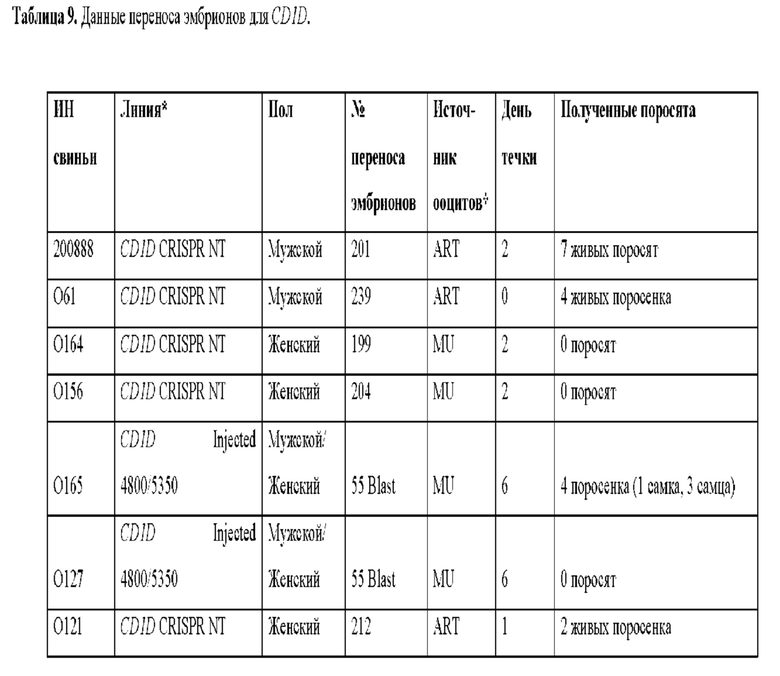
* Линия CD1D CRISPR NT представляет собой эмбрионы, созданные путем переноса ядра (NT) с линией эмбриональных фибробластов, модифицированных путем трансфекции. Эмбрионы с введенными CRISPR представляли собой ЭКО эмбрионы, инъецированные на стадии клеток 1 гидовой РНК CD1D с РНК CAS9.
 MU относится к ооцитам молодых свиней, которые были аспирированы и созревали в Университете Миссури, как описано в разделе ЭКО Материалов и способов. ART относится к ооцитам свиней, которые были приобретены и созревали, как описано в разделе SCNT Материалов и способов.
MU относится к ооцитам молодых свиней, которые были аспирированы и созревали в Университете Миссури, как описано в разделе ЭКО Материалов и способов. ART относится к ооцитам свиней, которые были приобретены и созревали, как описано в разделе SCNT Материалов и способов.
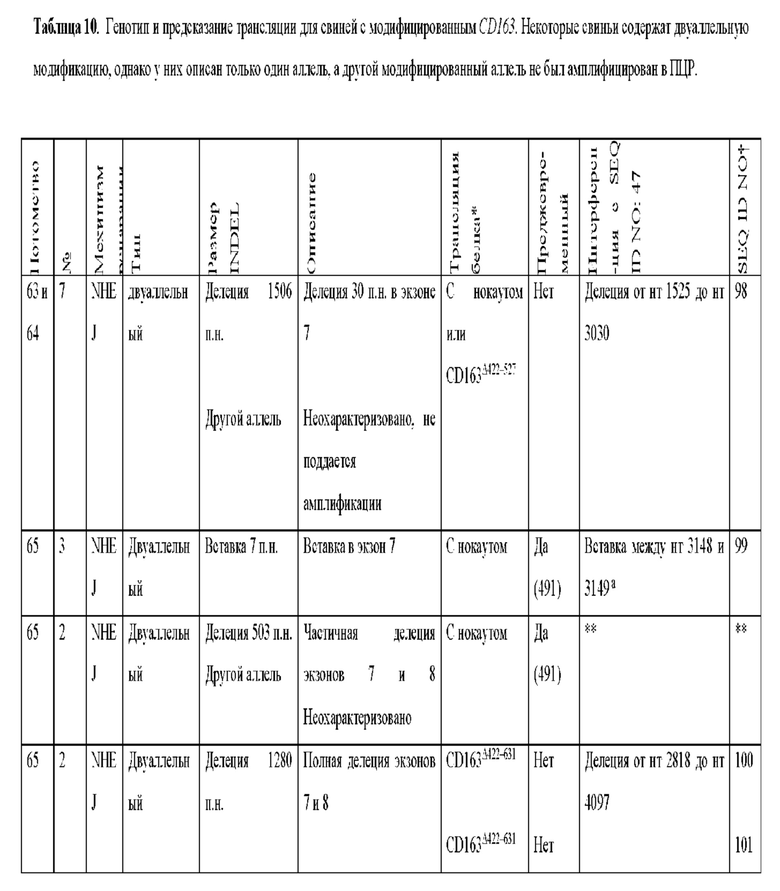
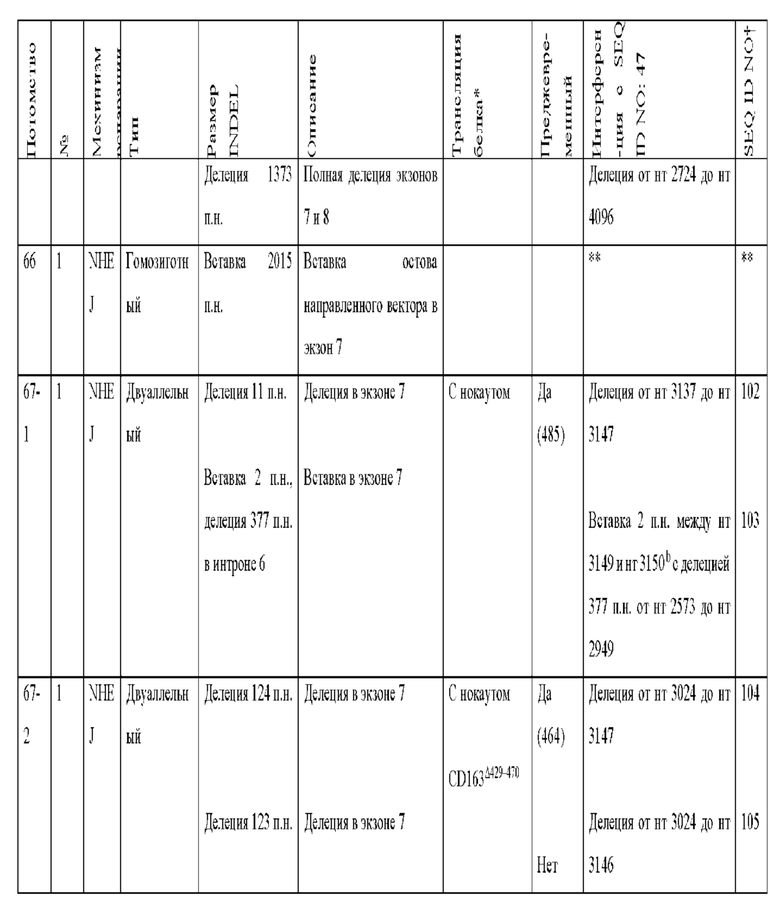
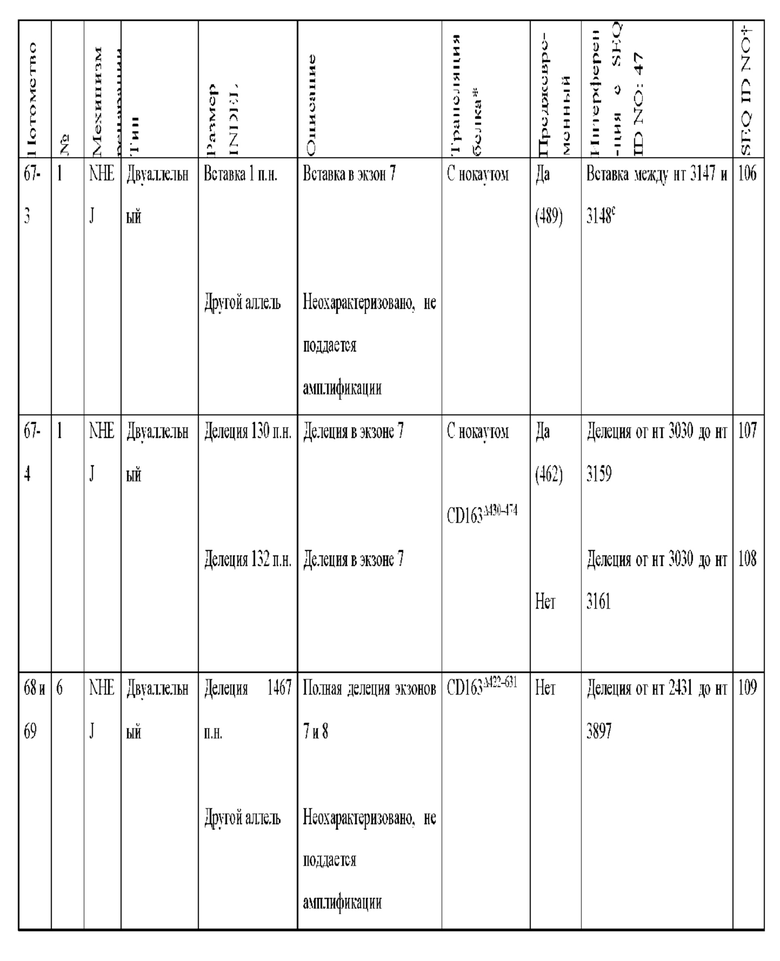
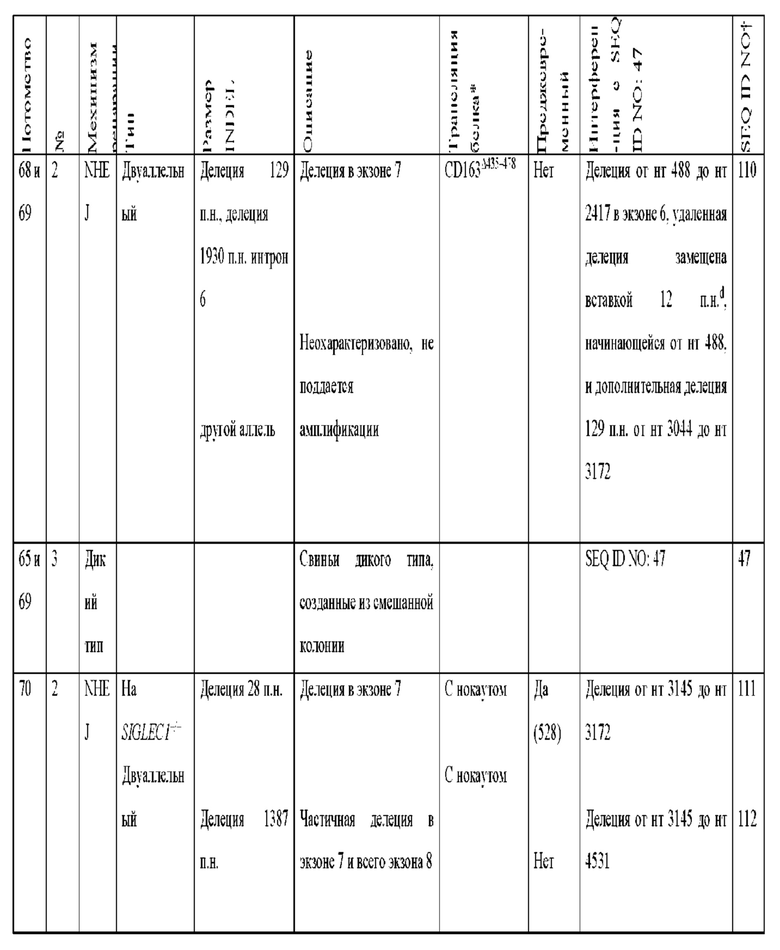
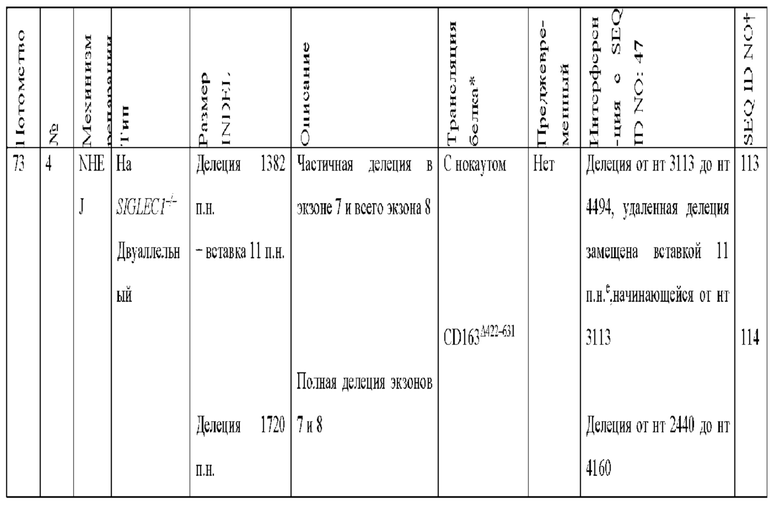
*KO, нокаут
**Не включено, поскольку свиньи были умерщвлены.
 SEQ ID NOs. в этой колонке относится к SEQ ID NOs. для последовательностей, демонстрирующих INDEL по сравнению с SEQ ID NO:47.
SEQ ID NOs. в этой колонке относится к SEQ ID NOs. для последовательностей, демонстрирующих INDEL по сравнению с SEQ ID NO:47.
а Введенной последовательностью была ТАСТАСТ (SEQ ID NO:115)
b Введенной последовательностью была AG.
c Введенной последовательностью был единичный остаток аденина (А),
d Введенной последовательностью была TGTGGAGAATTC (SEQ ID NO:116).
е Введенной последовательностью была AGCCAGCGTGC (SEQ ID NO:117).
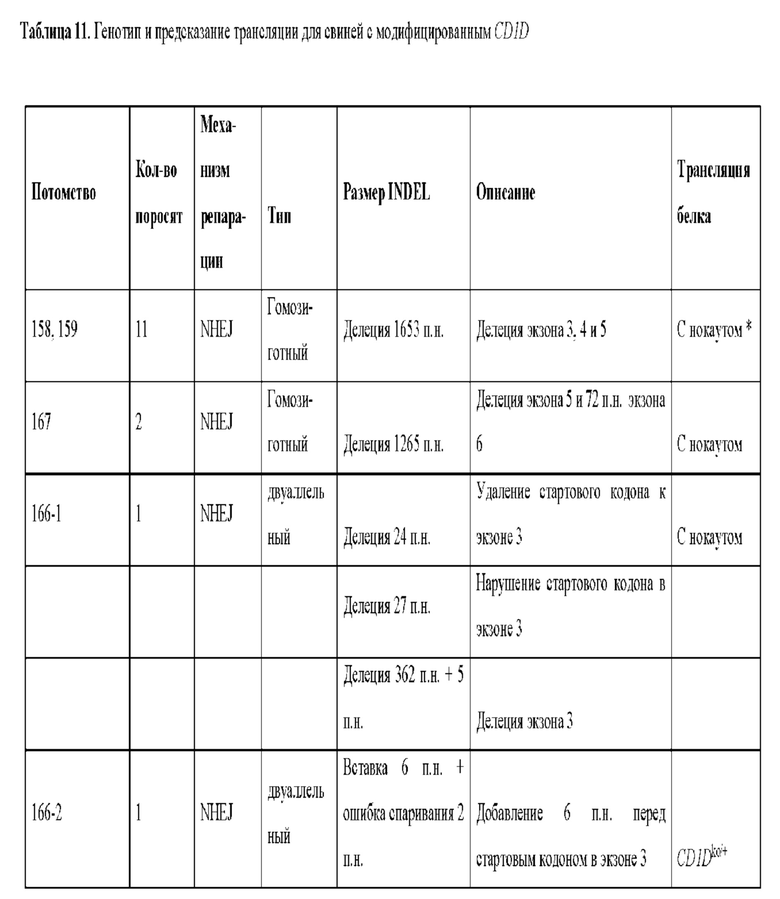
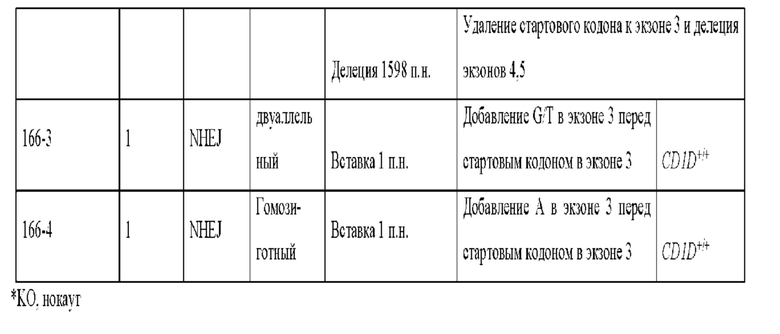 Эффективность системы CRISPR/Cas9 в зиготах свиньи
Эффективность системы CRISPR/Cas9 в зиготах свиньи
[00261] Этот подход, основанный на целенаправленном разрушении CD163 и CD1D в соматических клетках с использованием системы CRISPR/Cas9, был применен к эмбриогенезу свиней. Сначала была протестирована эффективность системы CRISPR/ Cas9 в развивающихся эмбрионах. Система CRISPR/Cas9, направленная на eGFP, была введена в зиготы, оплодотворенные спермой хряка, гетерозиготного по трансгену eGFP. После инъекции наблюдали за последующими эмбрионами, экспрессирующими eGFP. Были протестированы различные концентрации системы CRISPR/Cas9 и исследована цитотоксичность доставленной системы CRISPR/Cas9 (Фиг. 4, панель А); развитие эмбриона после инъекции CRISPR/Cas9 было снижено по сравнению с контролем. Однако все исследованные концентрации CRISPR/Cas9 были эффективны для получения модификации eGFP, поскольку в группе, получавшей инъекцию CRISPR/Cas9, не было обнаружено эмбрионов с экспрессией eGFP (Фиг. 4, панель В); 67,7% контрольных эмбрионов без инъекции окрашивались в зеленый цвет, свидетельствующий об экспрессии eGFP. Когда были генотипированы отдельные бластоцисты, можно было идентифицировать небольшие мутации рядом с сайтами связывания CRISPR (Фиг. 4, панель С). На основе токсичности и эффективности для следующих экспериментов использовали по 10 нг/мкл гРНК и мРНК Cas9.
[00262] Когда в предполагаемые зиготы были введены компоненты CRISPR/Cas9, предназначенные для направленного воздействия на CD163, наблюдалось целевое редактирование генов в последующих бластоцистах. Когда отдельные бластоцисты были генотипированы в отношении мутации CD163, специфические мутации были обнаружены у всех эмбрионов (эффективность редактирования генов 100%). Что еще более важно, в то время, как эмбрионы могли быть обнаружены с гомозиготными или двуаллельными модификациями (соответственно, 8/18 и 3/18) (Фиг. 5), были обнаружены также мозаичные (моноаллельные модификации) генотипы (4/18 эмбрионов). Некоторым эмбрионам из пула (8/10) вводили 2 нг/мкл Cas9 и 10 нг/мкл CRISPR, и различия в эффективности мутагенеза не было обнаружено. Затем, основываясь на результатах in vitro, были введены два CRISPR, представляющие разные гРНК с целью нарушения CD163 или CD1D во время эмбриогенеза с тем, чтобы вызвать специфическую делецию генов-мишеней. В результате отмечена возможность успешно вызывать запланированную делецию CD163 и CD1D путем введения двух гидов. Спланированная делеция определяется как делеция, которая удаляет геномную последовательность между двумя введенными гидами. Среди эмбрионов, получивших два CRISPR, нацеленных на CD165, у всех, кроме одного эмбриона, произошла целевая модификация CD163. Кроме того, было обнаружено, что 5/13 эмбрионов имели спроектированную делецию в CD163 (Фиг. 6, панель А), а 10/13 эмбрионов, по-видимому, имели модификацию CD163 либо гомозиготным, либо двуаллельным образом. Нацеливание на CD1D с помощью двух CRISPR также было эффективным, поскольку все эмбрионы (23/23) продемонстрировали модификацию CD1D. Однако, спроектированная делеция CD1D могла быть обнаружена только у двух эмбрионов (2/23) (Фиг. 6, панель В). Также были обнаружены пять из двадцати трех эмбрионов, обладающих мозаичными генотипами, но остальные эмбрионы имели либо гомозиготную, либо двуаллельную модификацию CD1D. Наконец, проверяли, может ли система CRISPR Cas9 направленно воздействовать на несколько генов в одном и том же эмбрионе. Для этой цели направленное воздействие как на CD163, так и на eGFP проводили в зиготах, оплодотворенных гетерозиготной спермой eGFP. Когда бластоцисты из инъецированных эмбрионов были генотипированы по CD163 и eGFP, было обнаружено, что CD163 и eGFP во время эмбриогенеза были успешно поражены. Результаты секвенирования продемонстрировали, что несколько генов могут быть поражены путем введения нескольких CRISPR с Cas9 (Фиг. 6, панель С).
Получение мутантов по CD163 и CD1D из зигот, инъецированных CRISPR/Cas9
[00263] Основываясь на успехе предыдущего исследования in vitro, было получено несколько зигот, инъецированных CRISPR/Cas9, и 46-55 бластоцист были перенесены реципиенту (поскольку было показано, что это количество эффективно для получения свиней из полученных in vitro эмбрионов). Было выполнено четыре переноса эмбрионов, по два для CD163 и CD1D, и была получена беременность для каждой модификации. Были получены четыре здоровых поросенка, несущие модификации CD163 (Таблица 8). Все поросята в помете 67 от свиноматки-реципиента ID О083 продемонстрировали либо гомозиготную, либо двуаллельную модификацию CD163 (Фиг. 7). У двух поросят была обнаружена спроектированная делеция CD163 при помощи двух доставленных CRISPR. Все поросята были здоровы. В отношении CD1D - одна беременность также дала четырех поросят (помет 166 от свиноматки-реципиента с идентификационным номером 0165): одна самка и три самца (Таблица 9). Один поросенок (166-1) действительно нес мозаичную мутацию CD1D, включая делецию 362 п. н., которая полностью удалила экзон 3, содержащий стартовый кодон (Фиг. 8). Один поросенок содержал вставку 6 п.н. с ошибкой спаривания 2 п.н. на одном аллеле, с большой делецией на другом аллеле. Два остальных поросенка имели двуаллельную вставку одной п.н. Для CD163 мозаичных мутаций обнаружено не было.
ОБСУЖДЕНИЕ
[00264] Повышение эффективности получения свиней с модифицированным геномом может иметь большое значение за счет увеличения количества свиней с отредактированным геномом для сельского хозяйства и биомедицины. Описанные выше данные показывают, что с помощью системы CRISPR/Cas9 с высокой эффективностью можно получать свиней с отредактированным геномом свиней, несущих специфические мутации. Система CRISPR/Cas9 успешно применялась для редактирования генов как в соматических клетках, так и в доимплантационных эмбрионах.
[00265] Когда систему CRISPR/Cas9 вводили в соматические клетки, она успешно индуцировала целенаправленное разрушение целевых генов с помощью NHEJ, но не увеличивала способность к направленному воздействию с помощью редактирования генов. Эффективность таргетинга отдельных CRISPR/Cas9 в соматических клетках варьировалась, что указывает на то, что на эффективность таргетинга может влиять дизайн гида. В частности, при введении CRISPR 5350 и Cas9 в соматические клетки целевую модификацию CD1D получить не удалось. Это свидетельствует о том, что было бы полезно разработать несколько гРНК и проверить их эффективность перед получением свиней. Причина отсутствия направленной на генное редактирование репарации в присутствии донорной ДНК до сих пор неясна. После скрининга 886 колоний (как CD163, так и CD1D), трансфицированных CRISPR и донорной ДНК, только одна колония имела доказательства частичного события редактирования генов. Результаты продемонстрировали, что система CRISPR/Cas9 работала с введенной донорной ДНК, вызывая неожиданные крупные делеции на целевых генах, но не увеличивала эффективность редактирования генов для этих двух конкретных направленных векторов. Однако конкретный механизм того, почему наблюдались крупные делеции, неизвестен. Предыдущие отчеты нашей группы предполагали, что донорная ДНК может эффективно использоваться с ZFN для индукции репарации, направленной на редактирование генов. Аналогичным образом, повышение эффективности нацеливания наблюдалось при использовании донорной ДНК с системой CRISPR/Cas9, но полной репарации, направленной на редактирование генов, отмечено не было. В предыдущем исследовании с использованием ZFN было отмечено, что целенаправленная модификация может происходить посредством комбинации редактирования генов и NHEJ, исходя из того, что была обнаружена частичная рекомбинация введенной донорной ДНК после DSB, индуцированных с помощью ZFN. Одно из объяснений может заключаться в том, что пути редактирования генов и NHEJ не являются независимыми, но могут действовать вместе с тем, чтобы завершить процесс репарации после DSB, индуцированных хоминг-эндонуклеазами. Более высокие концентрации CRISPR могут повысить эффективность направленного воздействия на соматические клетки, хотя в этих результатах экспериментов статистического различия обнаружено не было. Это может указывать на то, что CRISPR является ограничивающим фактором в системе CRISPR/Cas9, но для этого предположения необходима дальнейшая проверка. Клетки-мишени были успешно использованы для производства свиней с отредактированными генами путем SCNT, что указывает на то, что применение CRISPR/Cas9 не влияет на способность клеток к клонированию. Несколько поросят были умерщвлены из-за проблем со здоровьем; однако это не является редкостью у поросят, полученных путем SCNT.
[00266] Когда систему CRISPR/Cas9 вводили в развивающиеся эмбрионы с помощью инъекции зиготы, почти 100% эмбрионов и свиней содержали INDEL в целевом гене, тем самым демонстрируя, что это технология весьма эффективна во время эмбриогенеза. Эффективность, наблюдаемая во время этого исследования, превосходит частоты, описанные в других исследованиях с использованием хоминг-эндонуклеаз во время эмбриогенеза. Уменьшение количества эмбрионов, достигающих стадии бластоцисты, свидетельствует о том, что концентрация CRISPR/Cas9, введенная в этом исследовании, может быть токсичной для эмбрионов. Дальнейшая оптимизация системы доставки может повысить выживаемость эмбрионов и, таким образом, повысить общую эффективность процесса. Наблюдаемый здесь уровень мутагенеза, составляющий почти 100%, отличался от предыдущего сообщения об опосредованном CRISPR/Cas9 нокауте у свиней; однако различие в эффективности между исследованиями может быть связано с выбранной комбинацией гида и мишени. В настоящем исследовании более низкие концентрации CRISPR/Cas9 (по 10 нг/мкл каждая) были эффективны для генерации мутаций в развивающихся эмбрионах и продуцировании свиней с отредактированными генами. Концентрация в зиготах свиней была ниже по сравнению с той, о которой сообщалось ранее (125 нг/мкл Cas9 и 12,5 нг/мкл CRISPR). Более низкая концентрация компонентов CRISPR/Cas9 может быть полезной для развивающихся эмбрионов, поскольку введение избыточных количеств нуклеиновой кислоты в развивающиеся эмбрионы может быть токсичным. В анализах in vitro наблюдались некоторые мозаичные генотипы у эмбрионов, инъецированных CRISPR/Cas9; однако только один поросенок, полученный с помощью этого подхода, имел мозаичный генотип. Потенциально инъекция компонентов CRISPR/Cas9 может быть более эффективной, чем введение других хоминг-эндонуклеаз, поскольку мозаичный генотип считался основным препятствием для использования системы CRISPR/Cas9 в зиготах. Еще одно преимущество использования системы CRISPR/Cas9, продемонстрированное настоящими результатами, заключается в том, что не было утрачено ни одной свиньи с нокаутом по CD163, выращенной из зигот, полученных в результате ЭКО и которые были инъецированы системой CRISPR/Cas9, тогда как несколько поросят, полученных в результате SCNT, были умерщвлены через несколько дней. Это свидетельствует о том, что технология может не только обойти необходимость SCNT в создании нокаутных свиней, но также может решить общие проблемы со здоровьем, связанные с SCNT. Теперь, когда инъекция мРНК CRISPR/Cas9 в зиготы оптимизирована, будущие эксперименты будут также включать совместную инъекцию донорной ДНК.
[00267] Настоящее исследование демонстрирует, что введение двух CRISPR с Cas9 в зиготы может вызывать хромосомные делеции в развивающихся эмбрионах и давать свиней с заданной делецией, то есть специфической делецией между двумя гидами CRISPR. Эта спроектированная делеция может быть полезной, потому что можно указать размер делеции, а не полагаться на случайные события, вызванные NHEJ. В частности, в случае, если имеется вставка/делеция нуклеотидов в количестве, кратном трем, вызванная хоминг-эндонуклеазой, мутация скорее может привести к гипоморфной мутации, потому что сдвига рамки не произойдет. Однако, вводя два CRISPR, можно вызвать более крупные делеции, которые будут иметь более высокий шанс образования нефункционального белка. Интересно, что CDID CRISPR были сконструированы в большей части генома, чем CD 163; расстояние между CD163 CRISPR 10 и 131 составляло 124 п. н., тогда как расстояние между CRISPR 4800 и 5350 для CD1D составляло 550 п. н. Как показано в исследовании, большее расстояние между CRISPR не очень эффективно для создания делеции. Однако, поскольку настоящее исследование включало только ограниченное количество наблюдений и не было необходимости исследовать эффективность отдельных CRISPR, которые здесь не рассматриваются, необходимы дальнейшие исследования для того, чтобы проверить взаимосвязь между расстоянием между CRISPR и вероятностью возникновения предполагаемых делеции.
[00268] Система CRISPR/Cas9 также была эффективна в нацеливании на два гена одновременно в одном и том же эмбрионе, при этом единственным дополнительным этапом было введение одного дополнительного CRISPR с crRNA. Это иллюстрирует легкость разрушения множественных генов по сравнению с другими хоминг-эндонуклеазами. Эти результаты предполагают, что данную технологию можно использовать для направленного воздействия на кластеры генов или семейства генов, которые могут иметь компенсаторный эффект, что затрудняет определение роли отдельных генов в случае, если не нарушены все гены. Результаты демонстрируют, что технология CRISPR/Cas9 может быть применена для создания свиней с отредактированным геномом за счет повышения эффективности направленного воздействия генов в соматических клетках и путем прямой инъекции в зиготу.
Пример 2: Повышенная резистентность к PRRSV у свиней, имеющих модифицированную хромосомную последовательность в гене, кодирующем белок CD163
[00269] За последнюю четверть века вирус репродуктивного и респираторного синдрома свиней (PRRSV) разорил свиноводство. Предположения о способе проникновения вируса включают как SIGLEC1, так и CD163. Хотя нокаут SIGLEC1 не повлиял на реакцию на вирусную инфекцию, в настоящем примере показано, что животные с нулевым CD163 не проявляют клинических признаков инфекции, патологии легких, виремии или продукции антител, которые являются признаками инфекции PRRSV. Был подтвержден не только посредник PRRSV на входе; но если бы животным, созданным таким же путем, разрешили стать пищевым ресурсом, тогда можно было бы описать стратегию предотвращения значительных экономических потерь и страданий животных.
МАТЕРИАЛЫ И СПОСОБЫ
Генотипирование
[00270] Генотипирование было основано как на секвенировании ДНК, так и на секвенировании мРНК. В генотипе самца-производителя была делеция 11 п. н. в одном аллеле, которая при трансляции предусматривала 45 аминокислот в домене 5, что приводило к преждевременному стоп-кодону у аминокислоты 64. В другом аллеле было добавление 2 п.н. в экзоне 7 и делеция 377 п.н. в интроне перед экзоном 7, что при трансляции предусматривало первые 49 аминокислот домена 5, приводящие к преждевременной остановке кода на аминокислоте 85. У одной свиноматки было добавлено 7 п.н. в одном аллеле, который при трансляции предусматривал первые 48 аминокислот домена 5, что привело к преждевременному стоп-кодону на аминокислоте 70. Другой аллель не был охарактеризован (А), поскольку не было полосы от экзона 7, полученной как с помощью ПЦР, так и с помощью ПЦР длинных фрагментов в 6,3 т.п. Остальные 3 свиноматки были клонами и имели делецию 129 п.н. в экзоне 7, что, как предусматривается, приводит к делеции 43 аминокислот из домена 5. Другой аллель не был охарактеризован (В).
Выращивание PRRSV в культуре и получение вирусного инокулюма для заражения свиней описано в соответствии с заявкой 973, утвержденной IBC
[00271] Типовой штамм PRRSV, изолят NVSL 97-7895 (GenBank №AF325691 2001-02-11), был выращен, как описано в утвержденном протоколе 973 IBC. Этот лабораторный изолят использовался в экспериментальных исследованиях около 20 лет (Ladinig и др., 2015). Второй изолят, KS06-72109, был использован для 2-го испытания, как описано ранее (Prather et al., 2013).
Заражение свиней PRRSV
[00272] Для заражения свиней использовали стандартизованный протокол инфицирования PRRSV. Трехнедельных поросят инокулировали приблизительно 104 TCID50 вируса PRRS, который вводили внутримышечным (IM) и интраназальным (IN) путями. За свиньями ежедневно наблюдали, и тех, у кого проявлялись симптомы болезни, лечили в соответствии с рекомендациями ветеринаров CMG. Свиней, находящихся в тяжелом состоянии и имеющих риск заражения, гуманно умерщвляли и собирали их образцы. Персонал и ветеринары не имели информации о генетическом статусе свиней с тем, чтобы исключить предвзятость при оценке или лечении. PRRSV присутствует в жидкостях организма во время инфекции; поэтому образцы крови собирали и хранили при -80°С до проведения измерений для определения количества или степени виремии у каждой свиньи. В конце эксперимента свиней взвешивали и гуманно усыпляли, ткани собирали и фиксировали в 10% забуференном формалине, заливали в парафин и обрабатывали для проведения гистопатологических исследований сертифицированным патологом.
Оценка фенотипа зараженных свиней
[00273] Фенотип свиней оценивали слепым способом ежедневно следующим образом: Каково пищевое поведение у свиньи? Оценка поведения: 0: наилучшее, 1: под вопросом, 2: слегка подавленное, 3: подавленное, 4: умирающее животное. Какое состояние тела у свиньи? Оценка состояния тела: 1: истощение, 2: худощавое, 3: идеальное, 4: жирное, 5: избыточное/ожирение. Какая ректальная температура у свиньи? Нормальная температура тела: 101,6 103,6°F (считается, что лихорадка это выше >104°F). Есть ли хромота (степень)? Какая конечность? Обследуйте конечности на предмет отека суставов и повреждений копыт (проверьте низ и стороны копыта). Оценка хромоты: 1: нет хромоты, 2: слегка неравномерно при ходьбе, кажется жестким в некоторых суставах, но нет хромоты, 3: легкая хромота, небольшая хромота при ходьбе, 4: умеренная хромота, явная хромота, включая хромоту от прикосновения к пальцам ног, 5: сильная хромота, конечности не опираются на вес, нуждаются в поощрении, чтобы стоять/ ходить. Есть ли затрудненное дыхание (степень)? Есть ли дыхание открытым ртом? Есть ли выделения из носа (выделения цвета, количество выделений: легкие/средние/тяжелые)? Вы замечали, что животное кашляет? Есть выделения из глаз? Респираторная оценка: 0: норма, 1: легкая одышка и/или тахипноэ при стрессе (при прикосновении), 2: легкая одышка и/или тахипноэ в покое, 3: умеренная одышка и/или тахипноэ при стрессе (при прикосновении), 4: умеренная одышка и/или тахипноэ в покое, 5: сильная одышка и/или тахипноэ при стрессе (при манипуляциях), 6: сильная одышка и/или тахипноэ в покое. Есть ли признаки диареи (степень тяжести) или рвоты? Есть ли кровь или слизь? Оценка диареи: 0: кал не отмечен, 1: стул нормальный, 2: стул мягкий, но образованный (консистенция мягкого йогурта, образует коровий пирог), 3: жидкая диарея коричневого/желто-коричневого цвета с мелкими частицами фекалий, 4: жидкий понос коричневый/желто-коричневый цвет без твердых частиц фекалий, 5: жидкая диарея, похожая на воду.
[00274] Эта бальная система была разработана Dr. Megan Niederwerder в KSU и базируется на следующих публикациях (Halbur et al., 1995; Merck; Miao et al., 2009; Patience and Thacker. 1989; Winckler and Willen. 2001). Баллы и температуры были проанализированы с использованием ANOVA с разделением на основе генотипов в качестве обработок.
Измерение виремии PRRSV
[00275] Виремию определяли при помощи двух подходов. Титрование вируса выполняли путем добавления серийных разведений сыворотки 1:10 к конфлюэнтным клеткам MARC-145 в 96-луночном планшете. Сыворотку разводили в минимальной эссенциальной среде Игла с добавлением 8% эмбриональной бычьей сыворотки, пенициллина, стрептомицина и амфотерицина В, как описано ранее (Prather et al., 2013). Клетки исследовали через 4 дня инкубации на наличие цитопатического эффекта с помощью микроскопа. Как конечную точку титрования оценивали наибольшее разведение, показывающее цитопатический эффект. Общую РНК выделяли из сыворотки с использованием набора для выделения вирусной РНК Life Technologies MagMAX-96 для измерения вирусной нуклеиновой кислоты. Полимеразную цепную реакцию с обратной транскрипцией проводили с использованием набора EZ-PRRSV МРХ 4.0 от Tetracore на системе ПЦР в реальном времени CFX-96 (Bio-Rad) в соответствии с инструкциями производителя. Каждая реакция (25 мкл) содержала РНК из 5,8 мкл сыворотки. Стандартная кривая была построена путем подготовки серийных разведений контрольной РНК, поставляемой в наборе (Tetracore). Результаты представляются в виде количества матриц на ПЦР.
Окрашивание клеток АМС на SIGLEC1 и CD163
[00276] Альвеолярные макрофаги свиньи (АМС) собирали путем иссечения легких и заполнения их ~ 100 мл холодного фосфатно-солевого буфера. После извлечения промывные клетки с фосфатным буфером осаждали и ресуспендировали в 5 мл холодного физиологического раствора с фосфатным буфером и хранили на льду. Приблизительно 107 АМС инкубировали в 5 мл различных антител (против CD169 свиньи (клон 3В11/11; AbD Serotec); против CD163 свиньи (клон 2А10/11; AbD Serotec)), разбавленных в забуференном фосфатом физиологическом растворе с 5% эмбриональной бычьей сывороткой и 0,1% азидом натрия, в течение 30 минут на льду. Клетки отмывали и ресуспендировали в разведении 1/100 в флуоресцеин изотиоцианате (FITC), конъюгированным с антимышиным IgG (life Technologies), разведенном в буфере для окрашивания, и инкубировали в течение 30 минут на льду. По меньшей мере было проанализировано 104 клеток путем использования проточного цитофотометра FACSCalibur и программного обеспечения Cell Quest (Becton Dickinson).
Измерение PRRSV-специфического Ig
[00277] Для измерения PRRSV-специфического Ig рекомбинантный PRRSV-белок N был экспрессирован в бактериях (Trible et al., 2012) и конъюгирован с магнитными шариками Luminex с использованием набора (Luminex Corporation). Гранулы, связанные с N-белком, разбавляли в фосфатно-солевом буфере, содержащем 10% козьей сыворотки, до 2500 гранул/50 мкл и помещали в лунки 96-луночного круглодонного полистирольного планшета. Сыворотку разводили 1:400 в фосфатно-солевом буфере, содержащем 10% козьей сыворотки, и 50 мкл добавляли в лунки в двух повторах и инкубировали в течение 30 минут при осторожном встряхивании при комнатной температуре. Затем планшет промывали (3 раза) забуференным фосфатом физиологическим раствором, содержащим 10% козьей сыворотки и 50 мкл меченного конъюгированного SP-биотином, аффинно очищенного козьего вторичного антитела против свиней (IgG, Jackson ImmunoResearch) или меченного биотином, аффинно очищенного анти-свиного IgM козы (KPL), разведенного до 2 мкг/мл, в забуференном фосфатом физиологическом растворе, содержащем 10% козьей сыворотки. После 30 минут инкубации планшеты промывали (3 раза), а затем добавляли 50 мкл фикоэритрина, конъюгированного со стрептавидином (2 мкг/мл (Moss, Inc.) в фосфатно-солевом буфере, содержащем 10% козьей сыворотки. Планшеты промывали через 30 минут и микросферы ресуспендировали в 100 мкл забуференного фосфатом физиологического раствора, содержащего 10% козью сыворотку, и анализировали с использованием MAGPIX и программного обеспечения Luminex xPONENT 4.2. Результаты представлены в виде средней интенсивности флуоресценции (mean fluorescence intensity MFI).
РЕЗУЛЬТАТЫ
[00278] Мутации в CD163 были созданы с использованием технологии CRISPR/Cas9, как описано выше в Примере 1. Несколько животных-основателей были получены путем инъекции зиготы и переноса ядра соматической клетки. Некоторые из этих основателей были повязаны, при этом создав потомство для изучения. Единственный самец-основатель был повязан с самками с двумя генотипами. Самец-основатель (67-1) обладал делецией 11 п.н. в экзоне 7 на одном аллеле и добавлением 2 п.н. в экзоне 7 (и делецией 377 п. н. в предыдущем интроне) другого аллеля и был определен как нулевое животное (CD163-/-). Одна самка-основательница (65-1) имела добавление 7 п.н. в экзоне 7 в одном аллеле и неохарактеризованный соответствующий аллель, и, таким образом, для нокаута была предопределена гетерозиготность. (CD163-/?). Второй генотип самки-основателя (3 животных, которые были клонами) содержал пока неохарактеризованный аллель и аллель с делецией 129 п. н. в экзоне 7. Предполагается, что эта делеция приведет к делеции 43 аминокислот в домене 5. В результате спаривания между ними все поросята унаследовали нулевой аллель от хряка и либо делецию 43 аминокислот, либо один из неохарактеризованных аллелей от свиноматок. В дополнение к поросятам дикого типа, которые служили положительным контролем для вирусного заражения, это дало 4 дополнительных генотипа (Таблица 12).
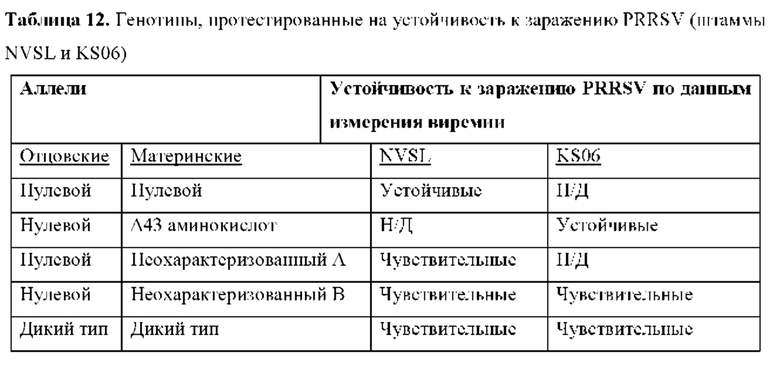
[00279] При отъеме модифицированные поросята и поросята дикого типа соответствующего возраста были доставлены в Университет штата Канзас для заражения PRRSV. Заражение PRRSV проводили, как описано ранее (Prather et al., 2013). Поросята трехнедельного возраста были помещены в помещение для заражения и содержались как одна группа. Все эксперименты были инициированы после одобрения институциональных комитетов по использованию животных и биобезопасности. После акклиматизации свиней заражали изолятом PRRSV NVSL 97-7895 (Ladinig et al., 2015), размноженным на клетках MARC-145 (Kim et al., 1993). Свиней заражали приблизительно 105 TCID50 вируса. Половина инокулюма была введена внутримышечно, а оставшаяся часть - интраназально. Всех инфицированных свиней содержали как одну группу, что позволяло непрерывно подвергаться воздействию вируса от инфицированных собратьев по загону. Образцы крови собирали в различные дни до 35 дней после заражения и в конце в день 35. Свиней вскрывали, ткани фиксировали в 10% забуференном формалине, заливали парафином и обрабатывали для гистопатологии. Связанные с PRRSV клинические признаки, зарегистрированные во время инфекции, включали респираторный дистресс, отсутствие аппетита, летаргию и лихорадку. Результаты по клиническим признакам за период исследования представлены на Фиг. 9. Как и ожидалось, у свиней дикого типа (CD163+/+) были обнаружены ранние признаки инфекции PRRSV, которые достигли пика между 5 и 14 днями и сохранялись в группе в течение оставшейся части исследования. Процент свиней с лихорадкой достиг пика примерно на 10 день. Напротив, у нулевых (CD163 -/-) поросят не было клинических признаков в течение всего периода исследования. Респираторные признаки во время острой инфекции PRRSV отражаются в значительных гистопатологических изменениях в легких (Таблица 9). Инфекция свиней дикого типа продемонстрировала гистопатологию, соответствующую PRRS, включая интерстициальный отек с инфильтрацией мононуклеарными клетками (Фиг. 10). В отличие от этого, не было доказательств легочных изменений у нулевых (CD163-/-) свиней. Размер выборки для различных генотипов невелик; тем не менее, средние оценки были 3,85 (n=7) для дикого типа, 1,75 (n=4) для неохарактеризованного А, 1,33 (n=3) для неохарактеризованного В и 0 (n=3) и для нулевого (CD 163-/-) генотипа.

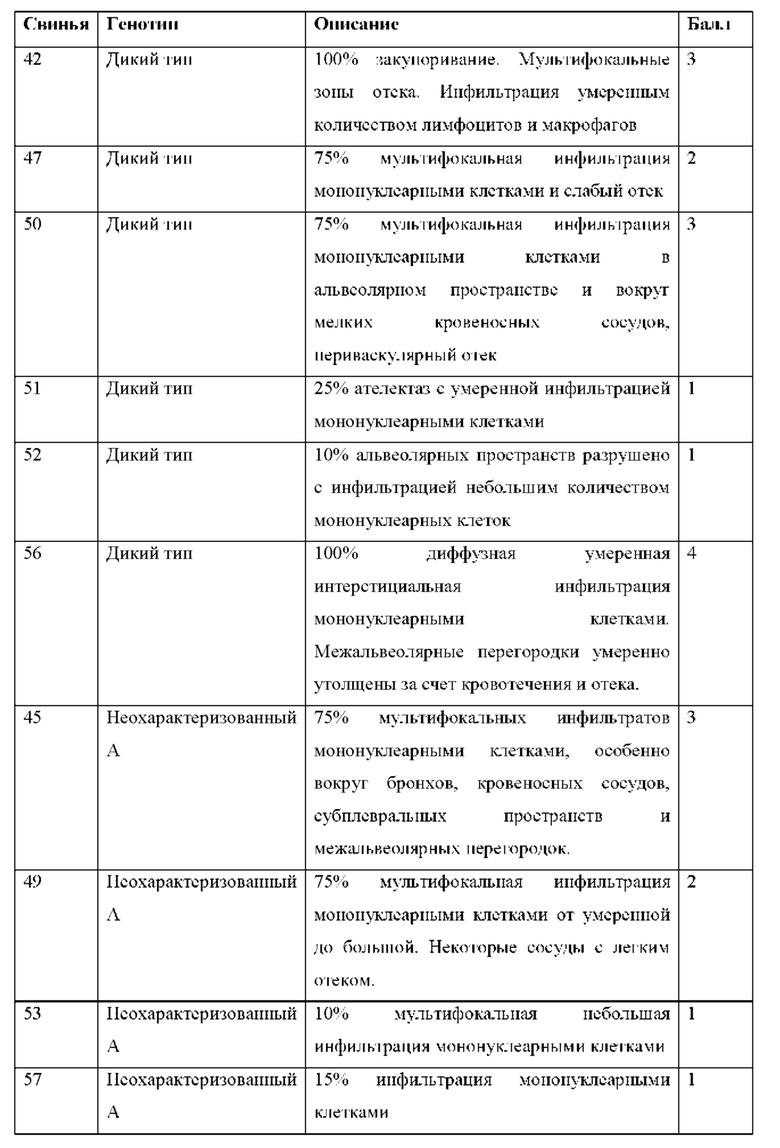
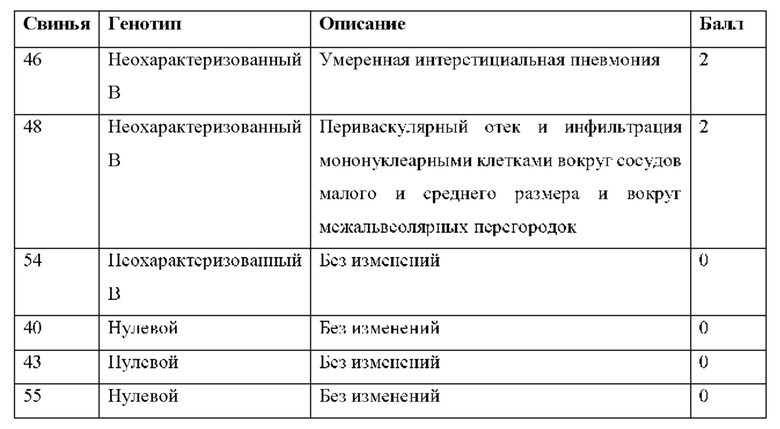
[00280] Пиковые клинические признаки коррелировали с уровнями PRRSV в крови. Измерение вирусной нуклеиновой кислоты проводили путем выделения общей РНК из сыворотки с последующей амплификацией РНК PRRSV с использованием коммерческого ПЦР-теста PRRSV в реальном времени с обратной транскриптазой (Tetracore, Rockville, MD). Стандартная кривая была построена путем подготовки серийных разведений контрольной РНК PRRSV, поставляемой в комплекте для ПЦР в реальном времени, и результаты были стандартизированы в виде количества матриц на 50 мкл реакции ПЦР. Изолят PRRSV соответствовал течению виремии PRRSV у свиней дикого типа CD163+/+(Фиг. 11). Виремия была выражена на четвертый день, достигла пика на 11 день и снизилась до конца исследования. Напротив, вирусная РНК не была обнаружена у свиней CD163-/- в любой момент времени в течение периода исследования. В соответствии с виремией продукция антител свиньями с нулевым и неохарактеризованным аллелем выявлялась к 14 дню и возрастала к 28 дню. У нулевых животных продукции антител отмечено не было (Фиг. 12). Вместе эти данные показывают, что свиньи дикого типа поддерживают репликацию PRRSV с появлением клинических признаков, согласующихся с PRRSV. Напротив, у нокаутных свиней не было ни виремии, ни клинических признаков, хотя свиньи были инокулированы и постоянно контактировали с инфицированными собратьями по загону.
[00281] В конце исследования альвеолярные макрофаги свиньи выделяли с помощью лаважа легких и окрашивали для выявления поверхностной экспрессии SIGLEC1 (CD169, клон 3В11/11) и CD163 (клон 2А10/11), как описано ранее (Prather et al., 2013). Относительно высокий уровень экспрессии CD163 был обнаружен у животных дикого типа CD 163+/+(Фиг. 13). В отличие от этого, свиньи CD163-/- продемонстрировали только фоновые уровни окрашивания анти-CD163, что подтверждает фенотип нокаута. Уровни экспрессии другого маркера макрофагов CD169 были сходными как у свиней дикого типа, так и у свиней с нокаутом (Фиг. 14). Другие поверхностные маркеры макрофагов, включая МНС II и CD 172, были одинаковыми для обоих генотипов (данные не показаны).
[00282] Хотя размер выборки был небольшим, свиньи дикого типа имели тенденцию набирать меньший вес в ходе эксперимента (средний дневной привес 0,81 кг±0,33, n=7) по сравнению со свиньями трех других генотипов (неохарактеризованный А 1,32 кг±0,17, n=4; неохарактеризованный В 1,20 кг±0,16, n=3; нулевой 1,21 кг±0,16, n=3).
[00283] Во втором исследовании 6 свиней дикого типа, 6 свиней с Δ43 аминокислот и 6 свиней с неохарактеризованным аллелем (В) заражали, как описано выше, за исключением того, что для инокуляции поросят использовали KS06-72109. Подобно данным с NVSL, у поросят дикого типа и у неохарактеризованных поросят В развивалась виремия. Однако у свиней с Δ43 аминокислот KS06 не вызывал виремию (Фиг. 15; Таблица 7).
РАССУЖДЕНИЯ И ВЫВОД
[00284] Наиболее клинически значимым заболеванием в свиноводстве является PRRS. Несмотря на то, что программы вакцинации были успешными для предотвращения или ослабления большинства патогенов свиней, PRRCV оказался более сложной задачей. Здесь CD163 идентифицирован как медиатор входа для этого штамма вируса. Хряк-основатель был создан путем инъекции CRISPR/Cas9 в зиготы (Whitworth et al., 2014), и, вследствие этого, трансген отсутствует. Кроме того, один из аллелей свиноматки (также созданный с помощью CRISPR/Cas9) не содержит трансгена. Таким образом, поросенок №40 несет добавление 7 п. н. в одном аллеле и делецию 11 п. н. в другом аллеле, но не имеет трансгена. Эти аллели CD163 устойчивости к вирусу представляют собой незначительные редактирования генома, учитывая тот факт, что геном свиньи насчитывает около 2,8 миллиарда пар оснований (Groenen et al., 2012). Если бы животные, созданные аналогичным образом, были введены в качестве пищевого ресурса, значительные экономические потери можно было бы предотвратить.
Пример 3: Повышенная устойчивость к репродуктивным вирусам свиней генотипа 1 и вирусам PRRS у свиней с заменой домена 5 CD163 SRCR на домен 8 SRCR - человеческий CD163-подобный гомолог
[00285] CD163 считается основным рецептором вируса репродуктивного и респираторного синдрома свиней (PRRSV). В этом исследовании свиней генетически модифицировали (GE) с тем, чтобы они обладали одним из следующих генотипов: полный нокаут (КО) CD163, делеции в домене 5 CD163 богатого цистеином рецептора фагоцитов (SRCR), или замена (обмен доменами) домена SRCR 5 синтезированным экзоном, кодирующим гомолог человеческого CD163-подобного (hCD163Ll) домена SRCR 8. Иммунофенотипирование альвеолярных макрофагов свиней (АМС) показало, что свиньи с нокаутом или делециями домена 5 SRCR не экспрессировали CD163, а АМС не поддерживали инфекцию PRRSV. АМС от свиней, которые обладали гомологом домена hCD163Ll 8, экспрессировали CD163 и поддерживали репликацию вирусов генотипа типа 2, но не типа 1. Заражение свиней с модифицированными CD 163 типичными вирусами типа 1 и типа 2 дало аналогичные результаты. Несмотря на то, что вирусы типа 1 и типа 2 считаются генетически и фенотипически подобными на нескольких уровнях, включая потребность в CD163 в качестве рецептора, результаты демонстрируют явное различие между генотипами PRRSV в распознавании молекулы CD 163.
МАТЕРИАЛЫ И СПОСОБЫ
Геномные модификации гена свиньи CD163
[00286] Эксперименты с участием животных и вирусов проводились в соответствии с Руководством Федерации обществ зоотехников по уходу и использованию сельскохозяйственных животных в исследованиях и обучении, Законом Министерства сельского хозяйства США о защите животных и Положениями о защите животных, и они были одобрены Институциональными комитетами по уходу и использованию животных и Институциональными комитетами по биобезопасности Университета штата Канзас и Университета Миссури. Мутации в CD163, использованные в этом исследовании, были созданы с применением технологии CRISPR/Cas9, как описано выше в предыдущих примерах. Мутации показаны схематически на Фиг. 17. Схема геномной области, показанная на Фиг. 17, охватывает последовательность от интрона 6 до интрона 8 гена CD163 свиньи. Интроны и экзоны схематически изображены на Фиг. 17 не в масштабе. Прогнозируемый белковый продукт показан справа от каждой геномной структуры. Относительная экспрессия макрофагов, измеренная по уровню поверхностного CD163 на АМС, показана в крайнем правом углу Фиг. 17. Черные области обозначают интроны; белые области обозначают экзоны; заштрихованная область указывает миметик экзона 11 hCD163Ll, гомолог экзона 7 свиньи; а серая область указывает синтезированный интрон с конструкцией PGK Neo, как показано на Фиг. 17.
[00287] Конструкция гена CD163 KO-d7(11), показанная на Фиг. 17, имеет делецию 11 пар оснований в экзоне 7 от нуклеотида 3137 до нуклеотида 3147. Конструкция гена CD163 KO-i7(2) имеет вставку из 2 пар оснований в экзоне 7 между нуклеотидами 3149 и 3150, а также делецию 377 пар оснований в интроне перед экзоном 7, от нуклеотида 2573 до нуклеотида 2949. Предполагается, что эти изменения вызовут мутации сдвига рамки считывания и преждевременные стоп-кодоны, что приведет только к частичной трансляции SRCR 5 и фенотипа KO. Три другие мутации вызывают делеции в экзоне 7. Первая, d7(129), имеет делецию 129 пар оснований в экзоне 7 от нуклеотида 3044 до нуклеотида 3172. Конструкция d7(129) также имеет делецию от нуклеотида 488 до нуклеотида 2,417 в экзоне 6, при этом удаленная последовательность заменена вставкой 12 п. н. Две другие делеционные конструкции, d7(1467) и d7(1280), имеют полные делеции экзонов 7 и 8, как показано на Фиг. 17. d7(1467) имеет делецию 1467 пар оснований от нуклеотида 2431 до нуклеотида 3897, a d7(1280) имеет делецию 1280 пар оснований от нуклеотида 2818 до нуклеотида 4097. Для этих делеционных конструкций остальные экзоны CD 163 остались интактными.
[00288] Последняя конструкция, показанная на Фиг. 17, HL11m, была получена с использованием события направленного воздействия, которое удалило экзон 7 и заменило его синтезированным экзоном, кодирующим гомолог SRCR 8 человеческого CD163-подобного белка 1 (домен 8 hCD163Ll кодируется экзоном 11 hCD163L1). Последовательность пептида SRCR 8 была создана путем изменения 33 нуклеотидов в последовательности экзона 7 свиньи. Кассета неомицина была включена в синтезированный экзон для обеспечения возможности скрининга модификации. SEQ ID NO: 118 представляет собой нуклеотидную последовательность для конструкции HL11m в области, соответствующей той же области в референсной последовательности SEQ ID NO: 47.
[00289] Схема свиного белка CD 163 и его гена представлена на Фиг. 18. Домены белка SCRC CD163 (овалы) и PST (квадраты) вместе с соответствующими экзонами гена показаны на панели А Фиг. 18. Сравнение пептидных последовательностей свиного CD163 SRCR 5 (SEQ ID NO: 120) и гомолога человеческого CD163 SRCR 8 (SEQ ID NO: 121) показано на панели В Фиг. 18. Фигура основана на номерах доступа GenBank AJ311716 (CD163 свиньи) и GQ397482 (hCD163-Ll).
Вирусы
[00290] Панель вирусов, использованных в этом примере, приведена в Таблице 14.
Изоляты размножали и титровали на клетках MARC-145 (Kim et al., 1993). Для титрования каждый вирус серийно разводили 1:10 в MEM с добавлением 7% FBS, Pen-Strep (соответственно, 80 единиц/мл и 80 мкг/мл), 3 мкг мл FUNGIZONE (амфотерицин В) и 25 мМ HEPES. Разбавленные образцы добавляли в четырех повторах к конфлюэнтным клеткам MARC-145 в 96-луночном планшете до конечного объема 200 мкл на лунку и инкубировали в течение четырех дней при 37°С с 5% СО2. Конечная точка титрования была определена как последняя лунка с цитопатическим эффектом (ЦПЭ). Инфекционная доза для культуры ткани в 50% (ТСГО50/МЛ) была рассчитана с использованием описанного ранее способа (Reed and Muench 1938).
**НД, Нет данных.
Инфицирование альвеолярных макрофагов
[00291] Приготовление и инфицирование макрофагов выполняли, как описано ранее (Gaudreault, et al., 2009 и Patton, et al., 2008). У умерщвленных свиней удаляли легкие и промывали, выливая в трахею 100 мл холодного фосфатно-солевого буфера (PBS). Трахеи зажимали, а легкие осторожно массировали. Альвеолярное содержимое выливали в 50 мл центрифужные пробирки и хранили на льду. Альвеолярные макрофаги свиней (АМС) осаждали центрифугированием при 1200×g в течение 10 минут при 4°С. Осадки ресуспендировали и один раз промывали холодным стерильным PBS. Осадки клеток ресуспендировали в замораживающей среде, содержащей 45% RPMI 1640, 45% эмбриональной телячьей сыворотки (FBS) и 10% диметилсульфоксида (ДМСО), и хранили в жидком азоте до использования. Замороженные клетки размораживали на льду, подсчитывали и доводили до 5×105 клеток/мл в среде (RPMI 1640 с добавлением 10% FBS, PenStrep и FUNGIZONE; RPMI-FBS). В 96-луночные планшеты добавляли примерно 103 АМС на лунку и инкубировали в течение ночи при 37°С в 5% СО2. Клетки осторожно промывали для удаления неприкрепившихся клеток. В лунки в трех повторах добавляли серийные 1:10 разведения вируса. После инкубации в течение ночи клетки промывали PBS и фиксировали в течение 10 минут 80% ацетоном. После высушивания лунки окрашивали моноклональными антителами SDOW-17, специфичными для N-белка PRRSV (Rural Technologies Inc.), разведенными 1: 1000 в PBS с 1% рыбьим желатином (PBS-FG; Sigma Aldrich). После 30-минутной инкубации при 37°С клетки промывали PBS и окрашивали меченным ALEXAFLUOR 488 антимышиным IgG (Thermofisher Scientific), разведенным 1: 200 в PBS-FG. Планшеты инкубировали в течение 30 минут в темноте при 37°С, промывали PBS и просматривали под флуоресцентным микроскопом. Инфекционная доза в культуре ткани 50% (TCID50)/мл была рассчитана в соответствии с описанным ранее способом (Reed and Muench 1938).
Измерение поверхностной экспрессии CD 169 и CD 163 на АМС
[00292] Окрашивание на поверхностную экспрессию CD 169 и CD 163 проводили, как описано ранее (Prather et al., 2013). Приблизительно 1×106 АМС помещали в пробирки для проточной цитометрии из полистирола (FACS) размером 12 мм × 75 мм и инкубировали в течение 15 минут при комнатной температуре в 1 мл PBS с 10% нормальной мышиной сывороткой для блокирования Fc рецепторов. Клетки осаждали центрифугированием и ресуспендировали в 5 мкл FITC-конъюгированных мышиных mAb против свиного CD169 (клон 3В11/11; AbD Serotec) и 5 мкл РЕ-конъюгированных мышиных mAb против свиного CD163 (клон: 2А10/11, AbD Serotec). После 30 минут инкубации клетки дважды промывали PBS, содержащим 1% бычий сывороточный альбумин (BSA Fraction V; Hyclone), и сразу же анализировали на проточном цитометре BD LSR Fortessa (BD Biosciences) с программным обеспечением FCS Express 5 (De Novo Software). Для каждого образца было проанализировано не менее 10000 клеток.
Измерение виремии PRRS
[00293] РНК выделяли из 50 мкл сыворотки с использованием набора для выделения вирусов MagMAX 96 Ambion (Applied Biosystems) в соответствии с инструкциями производителя. РНК PRRSV определяли количественно с использованием реагентов специфичных для мишени Target-Specific Reagents (Tetracore), для ОТ-ПЦР в реальном времени EZ-PRRSV МРХ 4.0 в соответствии с инструкциями производителя. Каждый планшет содержал стандарты количественного определения Tetracore и контрольные наборы, предназначенные для использования с реагентами ОТ-ПЦР. ПЦР проводили на системе обнаружения ПЦР в реальном времени CFX96 Touch (Bio-Rad) в 96-луночном формате с использованием рекомендуемых параметров цикла. Результаты ПЦР были представлены как logio числа копий РНК PRRSV на 50 мкл реакционного объема, что приблизительно соответствует числу копий на мл сыворотки. Площадь под кривой (AUC) для виремии с течением времени рассчитывалась с помощью GraphPad Prism версия 6.00 для Windows.
Измерение антител к PRRSV
[00294] Иммунофлуоресцентный анализ с использованием микросфер (FMIA) для обнаружения антител против белка нуклеокапсида (N) вируса PRRSV выполняли, как описано ранее (Stephenson et al., 2015). Рекомбинантный белок N PRRSV связывали с гранулами карбоксилированных полистирольных микросфер Luminex MAGPLEX в соответствии с инструкциями производителя. Для FMIA примерно 2500 покрытых антигеном гранул, суспендированных в 50 мкл PBS с 10% козьей сыворотки (PBS-GS), помещали в каждую лунку 96-луночного полистирольного круглодонного планшета. Сыворотки разбавляли 1:400 в PBS-GS и добавляли по 50 мкл в каждую лунку. Планшет заворачивали в фольгу и инкубировали 30 минут при комнатной температуре при осторожном встряхивании. Планшет помещали на магнит, и шарики трижды промывали 190 мкл PBS-GS. Для обнаружения IgG 50 мкл конъюгированных с биотином-SP аффинно очищенных вторичных антител козы против свиньи (IgG, Jackson ImmunoResearch) разводили до 2 мкг/мл в PBS-GS и добавляли по 100 мкл в каждую лунку. Планшет инкубировали при комнатной температуре в течение 30 минут и трижды промывали с последующим добавлением 50 мкл фикоэритрина, конъюгированного со стрептавидином (2 мкг/мл в PBS-GS; SAPE). Через 30 минут микросферы промывали, ресуспендировали в 100 мкл PBS-GS и анализировали с использованием прибора MAGPIX (LUMINEX) и программного обеспечения LUMINEX xPONENT 4.2. Рассчитывали среднюю интенсивность флуоресценции (MFI) минимум для 100 микросфер.
Измерение гаптоглобина (HP)
[00295] Количество Hp в сыворотке измеряли с использованием набора Hp ELISA для свиньи (Genway Biotech Inc.) и выполняли этапы в соответствии с инструкциями производителя. Образцы сыворотки разбавляли 1:10000 в 1-кратном растворе разбавителя и пипетировали в двух повторах в 96-луночный планшет для ELISA, предварительно покрытый антителами против свиного Hp, инкубировали при комнатной температуре в течение 15 минут, затем промывали три раза. В каждую лунку добавляли конъюгат анти-Нр-пероксидаза хрена (HRP) и инкубировали в темноте при комнатной температуре в течение 15 минут. Планшет промывали и в каждую лунку добавляли 100 мкл раствора хромогена-субстрата. После инкубации в темноте в течение 10 минут в каждую лунку добавляли 100 мкл стоп-раствора. Планшет считывали при 450 нм на считывающем устройстве для микропланшетов на основе фильтра Fluostar Omega (BMG Labtech).
РЕЗУЛЬТАТЫ
Фенотипические свойства АМС от CD163-модифицированных свиней
[00296] Для охвата мононуклеарной субпопуляции клеток в материале лаважа легких использовали свойство клеток к прямому и боковому рассеянию. Типичные результаты окрашивания CD169 и CD163 для различных хромосомных модификаций, показанных на Фиг. 17, представлены на Фиг. 19. В репрезентативном примере, представленном на панели А Фиг. 19, более 91% АМС от свиней дикого типа были положительными как по CD169, так и по CD163. Результаты для 12 свиней дикого типа, использованных в этом исследовании, показали в среднем 85+/- 8% дважды положительных клеток. Как показано на панели В Фиг. 19, АМС от свиней CD163 с нокаутом не продемонстрировали признаки CD163, но сохранили нормальные поверхностные уровни CD169. Хотя было предсказано, что полипептиды CD163, полученные от генотипов с делециями (17(1467) и (17(1280), должны продуцировать модифицированные полипептиды CD163, прикрепленные к поверхности АМС, результаты иммуноокрашивания не показали поверхностной экспрессии CD163 (см. Фиг. 19, панель D). Поскольку MAb 2А10 распознает эпитоп, расположенный в первых трех доменах SRCR, отсутствие выявления не стало результатом делеции иммунореактивного эпитопа. Генотип d7(129), как предполагается, обладает делецией 43 аминокислоты в SRCR 5 (см. Фиг. 17). В примере, представленном на панели С Фиг. 19, только 2,4% клеток попали в дважды положительный квадрант. Анализ АМС от девяти свиней (17(129), использованных в этом исследовании, показал процент двойных положительных клеток в диапазоне от 0% до 3,6% (среднее значение = 0,9%). Поверхностная экспрессия CD 169 оставалась аналогичной АМС дикого типа. Для целей данного исследования все свиньи, обладающие генотипами с нокаутом, d7(1467), d7(1280) и d7(129), были отнесены к категории обладающих фенотипом с нулевым CD 163.
[00297] Модификация CD163, содержащая пептидную последовательность HL11m домена 8 hCD163Ll, продемонстрировала двойную экспрессию CD163+и CD169+на АМС (панель Е Фиг. 19). Однако у всех свиней HL11m, проанализированных в этом исследовании, поверхностная экспрессия CD163 была заметно снижена по сравнению с АМС дикого типа. Уровни CD163 попадали в континуум экспрессии, начиная от не обнаруживаемого CD163 до свиней, обладающих умеренными уровнями CD163. В примере, показанном на панели Е Фиг. 19, примерно 60% клеток находились в дважды положительном квадранте, в то время как 40% клеток окрашивались только на CD169. Анализ АМС от 24 свиней HL11m показал, что 38+/- 12% клеток АМС были положительными только на CD169, а 54+/- 14% были дважды положительными (CD169+CD163+).
Уровни циркулирующего гаптоглобина у свиней дикого типа и CD 163-модифщированных свиней
[00298] В качестве поглощающей молекулы CD163 отвечает за удаление комплексов HbHp из крови (Fabriek, et al., 2005; Kristiansen et al., 2001; и Madsen et al., 2004). Уровень Hp в сыворотке обеспечивает удобный способ определения общих функциональных свойств макрофагов, экспрессирующих CD163. Уровни Hp в сыворотке от свиней дикого типа, HLllm и нулевых свиней без CD 163 измеряли в возрасте трех-четырех недель, непосредственно перед инфицированием вирусом PRRSV. Результаты, представленные на Фиг. 20, показали, что в сыворотке от свиней дикого типа было наименьшее количество Hb (среднее значение А450=23+/- 0,18, n=10). Среднее значение и стандартное отклонение для каждой группы составили для дикого типа - 0,23+/- 0,18, n=10; для HL11m - 1,63+/-0,8, n=11; и для нулевой группы - 2,06+/- 0,57, n=9. Нулевая группа состояла из генотипов, которые не экспрессировали CD163 (свиньи с нулевым фенотипом CD 163). Измерения Hp проводили на одном планшете для ELISA. Группы с одинаковой буквой существенно не различались (р>0,05, однофакторный дисперсионный анализ Краскела-Уоллиса с последующим тестом Даннета). Среднее значение А450 для свиней дикого типа значительно отличалось от значения для свиней с нулевым уровнем HL11m и CD163 (р<0,05). Хотя среднее значение А450 было ниже для группы HLllm по сравнению с группой без CD163 (А450=1,6+/- 0,8 против 2,1+/- 0,6), разница не была статистически значимой. Поскольку взаимодействие между HbHp и CD163 происходит через SRCR 3 (Madsen et al., 2004), повышенное количество циркулирующего Hp у свиней HLllm по сравнению со свиньями дикого типа, вероятно, не было следствием снижения сродства CD 163 к Hb/Hp, а было результатом снижения количества макрофагов CD163+ наряду со сниженной экспрессией CD 163 на оставшихся макрофагах (см. панель Е Фиг. 19).
Инфицирование АМС вирусами типа 1 и типа 2
[00299] Пермиссивность CD165-модифицированных свиней к PRRSV первоначально оценивали путем инфицирования клеток АМС in vitro с помощью панели из шести изолятов PRRSV типа 1 и девяти изолятов PRRSV типа 2 (список вирусов см. в Таблице 14). Вирусы в панели представляют разные генотипы, а также различия в нуклеотидных и пептидных последовательностях, патогенезе и годе выделения. Данные, представленные в Таблице 15, показывают результаты экспериментов с использованием АМС от трех свиней для каждой группы генотипа CD163. Перечисленные вирусы соответствуют изолятам PRRSV, приведенным в Таблице 14. Результаты представлены как среднее+/- стандартное отклонение процента инфицированных АМС. СD163-нулевые АМС были получены от свиней, экспрессирующих аллель d7(129) (см. Фигуры 17 и 19 для, соответственно, генных конструкций CD163 и экспрессии CD163 на АМС).

[00300] Как и ожидалось, АМС дикого типа инфицировались всеми вирусами. Напротив, свиньи с CD163-нулевым фенотипом были отрицательными в отношении инфицирования всеми вирусами. Заметное различие наблюдалась в ответах АМС от свиней HL11m. Ни один из вирусов типа 1 не смог инфицировать АМС HL11m; тогда как все вирусы в панели типа 2 инфицировали АМС HL11m, хотя и в гораздо более низком проценте по сравнению с АМС дикого типа.
[00301] Пермиссивность также оценивали путем сравнения конечных точек титрования вирусов между АМС дикого типа и HL11m для одних и тех же вирусов типа 2. Результаты показаны для двух свиней дикого типа и двух свиней HL11m (Фиг. 21). Значения log10 TCID50 были рассчитаны на основании заражения культур макрофагов тем же образцом вируса. Результаты заражения представляют двух разных свиней каждого генотипа. Вирусы, использованные для заражения, перечислены в Таблице 14. Значения log10 TCID50 для АМС от свиней HL11m были на 1-3 log ниже по сравнению с АМС дикого типа, инфицированными тем же вирусом. Единственным исключением было заражение вакцинным штаммом модифицированного живого вируса. В совокупности, результаты показывают, что АМС от свиней HL11m обладают пониженной чувствительностью или пермиссивностью к заражению вирусами 2 типа.
Инфицирование CD 163-модифицированных свиней вирусами Типа 1 и Типа 2
[00302] Свиньи дикого типа (кружки), HL11m (квадраты) и CD163-нулевые (треугольники) были инфицированы репрезентативными вирусами типа 1 (SD13-15) (Фиг. 22, панель А, левый график) и типа 2 (NVSL 97- 7895) (Фиг. 22, панель А, правый график). Свиньи с нулевым фенотипом произошли от аллелей с нокаутом и d(1567) (см. Фиг. 17). Свиней трех генотипов, инокулированных одним и тем же вирусом, смешивали в одном загоне, что позволяло непрерывно подвергать CD163-модифицированных свиней вирусам, выделенным из загонов с животными дикого типа. Количество свиней, инфицированных репрезентативным вирусом типа 1, составляло: дикий тип (n=4), HL11m (n=5) и нулевые (n=3); и вирусом типа 2: дикий тип (n=4), HL11m (n=4) и нулевые (n=3). Как показано на Фиг. 22, свиньи с нулевым CD163, инфицированные вирусом 1 или 2 типа, были отрицательными по виремии во все моменты времени и не имели сероконверсии. Как и ожидалось, свиньи дикого типа были продуктивно инфицированы, имея средний уровень виремии, приближающийся к 106 матрицам на 50 мкл реакции ПЦР через 7 дней после заражения обоими вирусами. К 14 дню все свиньи дикого типа имели сероконверсию (см. Фиг. 22, панель В). В соответствии с результатами инфицирования АМС (Таблица 15), пять свиней HL11m, инфицированных вирусом Типа 1, не продемонстрировали признаков виремии или антител к PRRSV. Все свиньи HL11m, инфицированные изолятом Типа 2, NVSL, поддерживали инфекцию и имели сероконверсию (Фиг. 22, панель В). Наличие пониженной пермиссивности свиней HL11m неясно. Средняя виремия для трех из четырех свиней HL11m была аналогична свиньям дикого типа. Однако у одной свиньи HL11m, №101 (светлые квадраты на Фиг. 22, панель А правый график), виремия была значительно снижена по сравнению с другими свиньями в группе генотипа HL11m. Объяснение снижения уровня виремии свиньи №101 на 3-4- log не было ясным, но предполагало, что некоторые свиньи HL11m могут быть менее чувствительны к PRRSV, наблюдение, подтверждающее результаты инфекции АМС in vitro (Таблица 15). Поскольку все свиньи были заражены одинаковым количеством вируса и оставались смешанными со свиньями дикого типа, более низкая виремия у свиньи №101 не была результатом получения меньшего количества вируса или меньшего воздействия вируса. Проточная цитометрия макрофагов показала, что экспрессия CD163 у свиньи №101 была сопоставима с экспрессией других свиней HL11 т (данные не представлены). Не наблюдалось различий в последовательности миметической последовательности экзона 11.
[00303] Дополнительные исследования с инфицированием вирусами были проведены с использованием двух вирусов, NVSL 97-7895 и KS06-72109. Результаты показаны на Фиг. 23. За свиньями наблюдали в течение 35 дней после заражения, и данные представляли как площадь под кривой (AUC) для измерений виремии, проведенных через 3, 7, 11, 14, 21, 28 и 35 дней после заражения. Как показано на Фиг. 23, для NVSL среднее значение AUC для семи свиней дикого типа, инфицированных NVSL, составило 168+/- 8 против 165+/- 15 для семи свиней HL11m. Для KS06 средние значения AUC для шести свиней дикого типа и шести свиней HL11m составили, соответственно, 156+/- 9 и 163+/- 13. Для обоих вирусов не было статистически значимой разницы между свиньями дикого типа и HL11m (р>0,05). При общем рассмотрении результаты показали, что свиньи HL11m не могли поддерживать инфекцию вирусом PRRSV 1 Типа, но сохранили способность заражения вирусами Типа 2. Несмотря на то, что наблюдалось снижение пермиссивности для PRRSV в АМС от свиней HL11m, инфицированных in vitro изолятами Типа 2, это различие не отражалось на свиньях. В отношении результатов, показанных на Фиг. 23: здесь вирусную нагрузку определяли путем расчета площади под кривой (AUC) для каждой свиньи в течение 35-дневного периода инфицирования. Расчет AUC проводили с использованием log10 ПЦР-виремий, полученных в дни 0, 4, 7, 10, 14, 21, 28 и 35 после заражения. Горизонтальные линии показывают среднее и стандартное отклонение. Ключевые слова: дикий тип = свиньи дикого типа, HL11 = свиньи генотипа HL11m; Нулевой = CD163-нулевой генотип.
ОБСУЖДЕНИЕ
[00304] CD163 представляет собой поверхностный белок макрофагов, важный для удаления избыточного гемоглобина из крови и модуляции воспаления в ответ на повреждение ткани. Он также действует как вирусный рецептор. CD163 участвует как в про-, так и в противовоспалительном ответе (Van Gorp et al., 2010). CD163-положительные макрофаги классифицируются в альтернативно активируемой группе макрофагов М2, которые обычно описываются как высоко фагоцитирующие и противовоспалительные. Макрофаги М2 участвуют в очистке и восстановлении после механического повреждения ткани или инфекции (Stein et al., 1992). Для противовоспалительных свойств экспрессия CD163 активируется противовоспалительными белками, такими как IL-10 (Sulahian, et al., 2002). Во время воспаления CD163 уменьшает воспаление за счет уменьшения окислительного процесса путем удаления циркулирующего гема из крови. Продукты распада гема, такие как биливердин, билирубин и окись углерода, являются мощными противовоспалительными молекулами (Soares and Bach, 2009 и Jeney et al., 2002). Что касается провоспалительного действия, сшивание CD163 на поверхности макрофагов антителом к CD163 или бактериями приводит к локализованному высвобождению провоспалительных цитокинов, включая IL-6, GM-CSF, TNFα и IL-1β (Van den Heuvel et al., 1999 и Fabriek et al., 2009).
[00305] GE свиньи, у которых отсутствует CD163, не могут поддерживать репликацию изолята PRRSV Типа 2 (Whitworth et al., 2016). В этом исследовании изучение инфицирования in vitro демонстрируют устойчивость макрофагов с нулевым фенотипом CD163 к обширной панели изолятов PRRSV Типа 1 и Типа 2, что дополнительно увеличивает устойчивость и потенциально включает все изоляты PRRSV (Таблица 15). Устойчивость макрофагов CD163-нулевого фенотипа к вирусам Типа 1 и Типа 2 была подтверждена in vivo (Фиг. 22 и Фиг. 23). На основании этих результатов можно исключить вклад других рецепторов PRRSV, ранее описанных в литературе (Zhang and Yoo, 2015). Например, Shanmukhappa et al. (2007) показали, что непермиссивные клетки BHK, трансфицированные плазмидой CD151, приобрели способность поддерживать репликацию PRRSV, а инкубация с поликлональным анти-CD151 антителом, как было показано, значительно снижает инфицирование клеток MARC-145. Кроме того, ранее было показано, что линия клеток обезьян, SJPL, первоначально разработанная для использования в размножении вирусов свиного гриппа, поддерживает репликацию PRRSV (Provost, et al., 2012). Важные свойства клеточной линии SJPL включали присутствие CD151 и отсутствие сиалоадгезина и CD163. Взятые вместе, эти данные предоставили убедительные доказательства того, что одного CD151 достаточно для поддержки репликации PRRSV. Результаты этого исследования, показывающие отсутствие инфекции PRRSV у макрофагов и свиней, обладающих нулевым фенотипом CD163, показывают, что CD151 как альтернативный рецептор для PRRSV не является биологически значимым.
[00306] Вирусные белки GP2a и GP4, которые образуют часть комплекса гетеротримеров GP2a, GP3, GP4 на поверхности PRRSV, могут быть осаждены совместно с CD 163 в анализах методом афинной адсорбции из клеток, трансфицированных плазмидами GP2 и GP4 (Das, et al., 2009). Предположительно, GP2 и GP4 образуют взаимодействие с одним или более доменов CD 163 SRCR. Анализы на инфекционность in vitro, включающие остов кДНК CD163 свиньи, содержащий обмен доменами между SRCR 5 свиньи и гомологом из hCD163-Ll SRCR 8, дополнительно локализовали область в SRCR 5, используемую вирусами Типа 1 (Van Gorp, et al., 2010). Интересно предположить, что стабильное взаимодействие между GP2/GP4 и CD163 происходит через SRCR 5. Дополнительные вирусные гликопротеины, такие как GP3 и GP5, могут дополнительно стабилизировать комплекс вирус-рецептор или могут действовать как молекулы корецептора. Потребность в SRCR 5 изучалась в этом исследовании путем заражения макрофагов и свиней, обладающих аллелем HL11m, которые воссоздали обмен доменами CD163L1 SRCR 8 путем замены 33 п.н. в экзоне 7 свиньи. Аллель HL11m также включал кассету неомицина для отбора клеток, положительных в отношении генетического изменения (Фиг. 17). Свиньи HL11m экспрессировали CD163 на АМС, хотя и в меньшей степени по сравнению с АМС дикого типа (Фиг. 19, сравните панели А и Е). Снижение экспрессии, вероятно, было связано с наличием кассеты неомицина, которая располагалась между миметиком экзона 11 и следующим интроном. Свиньи HL11m не были пермиссивны в отношении заражения вирусом Типа 1, что подтверждает важность SRCR 5. Однако макрофаги HL11m и свиньи HL11m действительно поддерживали инфицирование вирусами Типа 2. Основываясь на результатах титрования вируса и процента инфицирования АМС от свиней HL11m, было показано общее снижение проницаемости для вируса по сравнению с макрофагами дикого типа (Таблица 15 и Фиг. 17). Снижение пермиссивности может быть связано со снижением уровней CD163 на макрофагах HL11m в сочетании со сниженным сродством вируса к модифицированному белку CD 163. Высказывая предположение, что вирусы Типа 2 нуждаются в SRCR 5, и что L1 SRCR 8 может функционировать как подходящий заменитель, более низкое сродство можно объяснить различием пептидных последовательностей между SRCR 8 человека и SRCR 5 свиньи (см. Фиг. 18, панель В). Однако сниженная пермиссивность АМС не транслировалась на свиней. Средняя виремия для свиней HL11 т существенно не отличалась по сравнению со свиньями дикого типа (Фиг. 23). Помимо АМС, в виремию вносит вклад инфицирование PRRSV внутрисосудистых, септальных макрофагов и макрофагов из лимфоузлов (Lawson et al., 1997 и Morgan et al., 2014). Потенциальный вклад этих и других популяций CD163-положительных клеток в поддержание общей вирусной нагрузки у свиней HL11m заслуживает дальнейшего изучения.
[00307] Несмотря на то, что плазмиды CD163 с делециями доменов SRCR, стабильно экспрессируются в клетках НЕК (Van Gorp et al., 2010), делеция экзонов 7 и 8 в (17(1467) и d7(1280) привела к отсутствию обнаруживаемой поверхностной экспрессии CD 163 (Фиг. 19, панель D). Поскольку mAB 2А10, используемое для проточной цитометрии, распознает три N-концевых домена SRCR (Van Gorp et al., 2010) и, возможно, 7-й и 8-й домены (Sanchez, et al., 1999), отсутствие обнаружения не было связано с удалением эпитопа 2А10 в мутированных белках. В то время как на АМС от некоторых свиней d7(129) можно было обнаружить небольшое количество экспрессии CD163 (см. Фиг. 19, панель С), количество экспрессированного белка было недостаточным для поддержки инфекции PRRSV в АМС или у свиней. Отсутствие экспрессии CD163 в мутантах с делециями экзона 7 и 8 до конца не изучено, но, вероятно, оно является результатом деградации мРНК и/или белка.
[00308] В 2003 году CD163 был идентифицирован как рецептор вируса африканской чумы свиней (ASFV; Sanchez-Torres et al., 2003). Этот вывод был основан на наблюдении, что инфицированные макрофаги обладают зрелым CD163-позитивным фенотипом, а антитела к CD163, такие как 2А10, блокируют инфицирование макрофагов вирусом АЧС in vitro. Остается определить, устойчивы ли свиньи с нулевым CD163 к инфекции вирусом АЧС.
[00309] Модели клеточных культур, включающие модификации рецептора PRRSV, дали ценную информацию о механизмах проникновения, репликации и патогенеза PRRSV. Одним из уникальных аспектов этого исследования было проведение параллельного эксперимента in vivo на свиньях с модифицированными рецепторами. Это исследование имеет важное значение для возможности разработки профилактических методов лечения одного из самых серьезных заболеваний, с которыми когда-либо сталкивалась мировая свиноводческая отрасль.
Пример 4: Нокаут материнского CD163 защищает плоды от инфицирования вирусом репродуктивного и респираторного синдрома свиней (PRRSV).
[00310] Примеры с 1 по 3 выше демонстрируют, что свиньи, имеющие полный нокаут (KO) гена CD163, лишены экспрессии CD163 на макрофагах и не способны поддерживать инфекцию PRRSV (см. также Whitworth et al., 2016; Wells et al., 2017). Поскольку экспрессия CD163 является доминантным признаком и наследуется классическим способом по Менделю, потомство, обладающее нормальной экспрессией и функцией CD163, может быть получено путем скрещивания самки свиньи с нокаутом CD163-/- с самцом CD163+/+ дикого типа. Для этого исследования молодые свиньи CD163 с нокаутом были скрещены с хряками дикого типа с получением гетерозиготных плодов CD163+/- с тем, чтобы определить, будет ли присутствие генотипа CD163 с нокаутом у матери достаточным для защиты плодов после заражения матери PRRSV. В этом исследовании у свиноматок, обладающих полным нокаутом рецептора CD163, CD163-позитивные плоды, полученные между 109 днями беременности или 20 днями после заражения матери, были полностью защищены от PRRSV. Результаты демонстрируют практические средства устранения репродуктивного заболевания, связанного с PRRSV, которое является основным источником экономических трудностей для сельского хозяйства.
Материалы и способы
[00311] Редактирование гена CD163. Способы CRISPR/Cas9, используемые для создания всех аллелей с нокаутом, подробно описаны в Примерах 1-3 выше. В экспериментах, описанных в этом примере, использовали животных дикого типа и животных с нокаутом или гетерозиготных животных, полученных, как описано в Примерах 1-3 и имеющих аллели, описанные в Таблице 16. Каждый из этих аллелей описан в Примерах 1-3 и в публикации РСТ №WO 2017/023570, которая полностью включена в настоящий документ посредством ссылки. Специфические изменения для аллелей В, D и Е также описаны в Whitworth et al., 2014, а специфические изменения в аллеле С (вставка 2 п.н.) описаны в Whitworth et al., 2016. Все аллели, описанные в Таблице 16, были идентифицированы на основе секвенирования ДНК. Нокаутный генотип подтверждали отсутствием экспрессии CD163, которую измеряли окрашиванием альвеолярных макрофагов моноклональными антителами против CD163, 2А10, как описано выше в Примере 2.
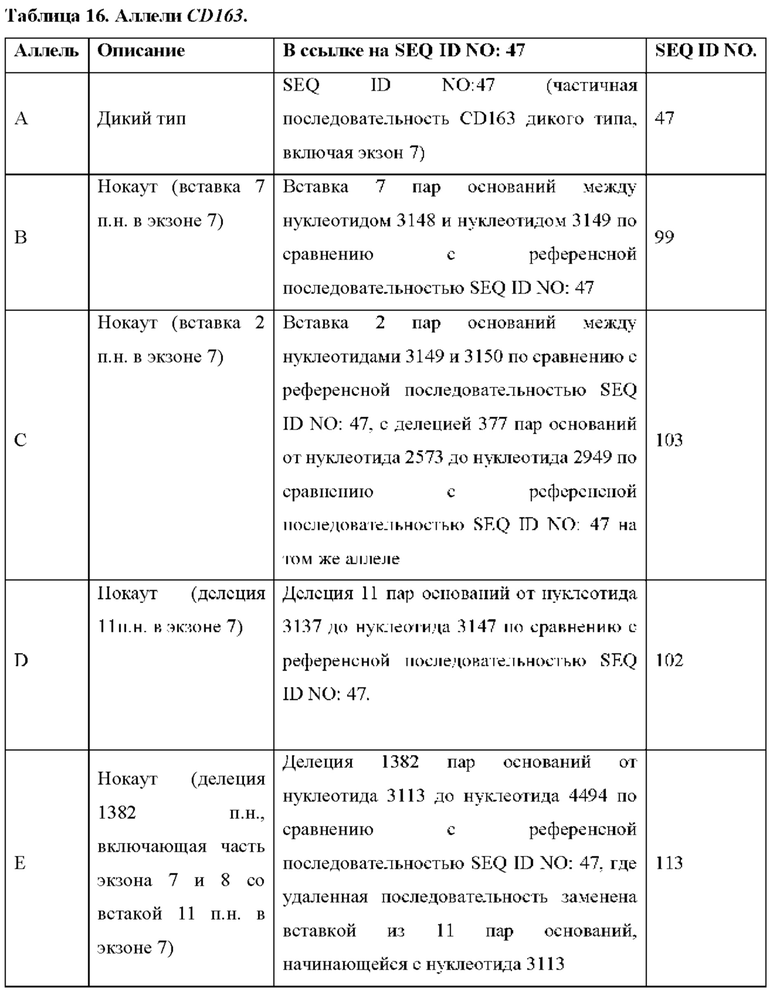
[00312] Инфекция PRRSV. Штамм PRRSV, использованный в этом исследовании, NVSL 97-7895 (NVSL), представляет собой лабораторный штамм, выделенный в 1997 году из стада в Юго-восточной Айове, США, которое переживало бурю абортов, вызванных PRRSV (Halbur et al., 1997). Вирус, поддерживаемый как изолят с низким числом пассажей, размножали и титровали на клетках MARC-145. На 89-91 день беременности молодых свиней инокулировали 105 TCID50 вируса, разведенного в 5 мл культуральной среды. Половину посевного материала вводили внутримышечно, а оставшуюся часть вводили интраназально. Все молодые свиньи содержались в среде, которая позволяла непрерывно подвергаться воздействию вируса, выделяемого инфицированными собратьями по загону. Образцы крови отбирали у молодых свиней до заражения, через семь дней после инокуляции (дпи) и во время эвтаназии. Нуклеиновую кислоту PRRSV измеряли путем выделения общей РНК из сыворотки с последующей ПЦР PRRSV в реальном времени с обратной транскриптазой (Tetracore, Rockville, MD). Стандартная кривая была построена с использованием стандартов количественного определения, поставляемых в комплекте ОТ-ПЦР. Результаты представлены в виде log10 матриц на 25 мкл реакции, что приблизительно соответствует количеству матриц вирусной РНК на мл крови.
РЕЗУЛЬТАТЫ
[00313] Подробное описание нокаутных аллелей, использованных в этом исследовании, показано в Таблице 16 выше. Каждый нокаутный аллель обладал мутацией в экзоне 7, которая, как было предсказано, приводила к сдвигу рамки считывания кодона с последующим преждевременным стоп-кодоном в мРНК. Спаривания между родителями дикого типа и CD163 с нокаутом суммированы в Таблице 17. Первой группой из трех свиноматок, которые служили в качестве положительного инфекционного контроля, были самки CD163+/+, несущие CD163+/+ плоды (группа++/++). Вторую группу (- -/+-) составляли CD163-/- свиноматки, несущие CD163+/- плоды. В этой группе CD 163-/- свиноматки были неспособны поддерживать репликацию PRRS, в то время как CD163+/- плоды сохраняют чувствительность к PRRS-инфекции. И, наконец, третью группу (- -/- -) составили CD163-/- свиноматки, несущие CD 163"" плоды. В последней группе и свиноматки, и плоды должны были быть устойчивыми к инфекции.

[00314] Клинические признаки у инфицированных свиноматок дикого типа включали летаргию и временное отсутствие аппетита. Свиноматки с нокаутом не имели клинических признаков. В течение периода исследования у одной свиноматки дикого типа, №139, беременность прервалась на 106 день (15 дпи). Нуклеиновая кислота PRRSV, измеренная на 7 день после инокуляции, показала уровень виремии для Свиноматки №139, равный 5,5 log10 матриц на реакцию, что свидетельствует о наличии продуктивной инфекции PRRSV. Между 15 и 20 днями после инокуляции все оставшиеся свиноматки были подвергнуты эвтаназии и были немедленно удалены рога матки. Начиная с кончика каждого рога, плоды и плаценты удаляли и оценивали на предмет наличия анатомической патологии. У каждого плода был взят образец крови. Если кровь не могла быть получена, образец жидкости был взят из брюшной полости. Количество плодов, извлеченных от каждой свиноматки, указано в Таблице 17. Для CD163 дикого типа (группа++/++) (включая абортировавшую свиноматку) количество плодов составляло 16, 14 и 12 (среднее значение=14,0). У свиноматок CD163 с нокаутом, несущих CD163+/- плоды (группа - -/+-), было получено 14, 17 и 11 плодов (среднее значение = 13,6). Для самок CD163 с нокаутом, несущих плоды CD163 с нокаутом (группа - -/- -), количество плодов составляло было 7 и 9. Результаты для виремии плода и макропатологии представлены на Фиг. 24 и в Таблице 18.
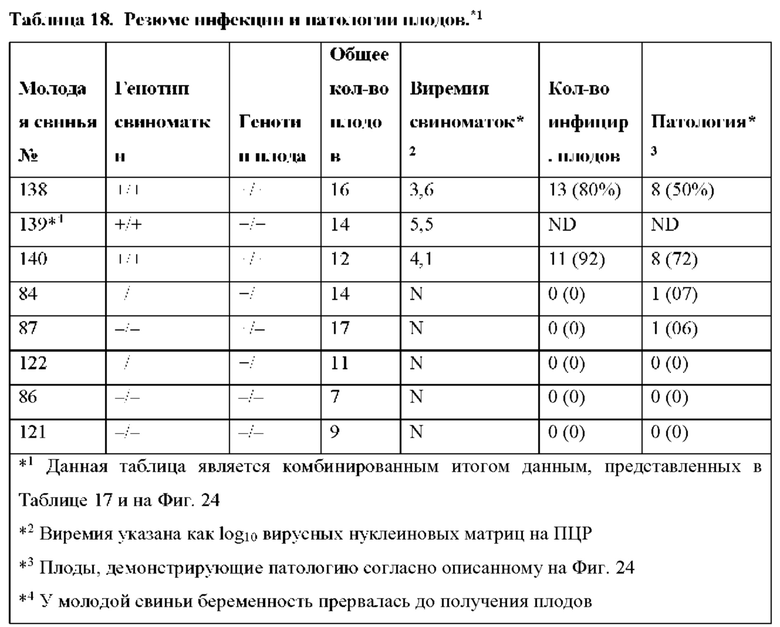
[00315] Фиг. 24 изображает каждый исход для плода после инфицирования матери PRRSV. Цифры слева идентифицируют каждую свиноматку. Под каждой свиноматкой в скобках указан результат ПЦР PRRS в сыворотке, измеренный как log10 матриц на реакцию. «N» означает негативный результат по нуклеиновой кислоте PRRSV (Ct>39). Плоды идентифицируются по количеству и относительному положению в каждом роге матки. Звездочки обозначают образцы ПЦР плода, полученные из абдоминальной жидкости. Число под каждым плодом представляет собой результат ПЦР PRRS в сыворотке плода (log10 матриц на реакцию). Цифра в каждом кружке указывает на наличие анатомической патологии: 1) нормальный плод; 2) маленький плод; 3) изменения плаценты, такие как отслоение плаценты и/или некроз; 4) плод, окрашенный меконием; 5) плод мертвый и некротический. Строчные буквы обозначают генотип отдельных плодов (см. Таблицу 17). Обозначения: а, А/А; b, С/А; с, В/А; d, Е/А; е, В/С; f, B/D; g, D/C; h, D/D; i, E/C; j, E/D; ND, не определено, поскольку плод некротизирован; nd, генотип не был определен.
[00316] На анатомическом уровне у 50% и 72% плодов, полученных от двух свиноматок CD163 дикого типа (++/++), №138 и №140, была обнаружена некоторая степень патологии, включая более мелкие плоды по сравнению с нормой (11% всех плодов), плоды с отслоившейся или некротической плацентой (14%), окрашенные меконием (7%), а также мертвые и некротические плоды (25%). Наблюдения патологии типичны для репродуктивного PRRS. Те же пометы показали высокий уровень инфицирования PRRSV, при этом 92% плодов дали положительный результат на наличие нуклеиновой кислоты PRRSV. Результаты ПЦР для плодов от свиноматок дикого типа иллюстрируют два важных свойства инфицирования плода PRRSV. Во-первых, между плодами отмечалось большое различие в концентрации вируса, обнаруживаемого в сыворотке крови, в результате заражения плодов в разное время. Во-вторых, не всегда уровень виремии коррелировал с патологией. Например, плод №5 от свиноматки №138 обладал высоким уровнем виремии (7,3 log10 матриц на реакцию), но, тем не менее, плод не был поражен. Причина несоответствия между виремией и патологией неясна. Одна из возможностей заключается в том, что патология плода является результатом повреждения тканей со стороны матери и не связана с уровнем виремии плода. В полевых условиях эти нормальные, но инфицированные новорожденные поросята, могут действовать как «суперраспространители», что способствует быстрому распространению PRRSV по производственной системе. Для группы - -/+- (свиноматки №84, 87 и 122) все плоды выглядели нормальными, за незначительным исключением двух плодов, которые были меньше других однопометников. Размер меньше нормального, вероятно, является следствием скученности в роге матки, что уменьшает площадь поверхности плаценты, тем самым ограничивая рост развивающегося плода. Все матери и плоды в группе - -/+- были отрицательными на присутствие нуклеиновой кислоты PRRSV. В последней группе - -/- -не было видимой патологии, и все свиноматки (№86 и 121) и плоды были отрицательными по нуклеиновой кислоте PRRSV.
[00317] Результаты этого исследования ясно демонстрируют, что отсутствие CD163 у свиноматки достаточно для защиты плода, чувствительного к PRRSV. Хотя CD163-позитивное потомство, полученное от свиноматок CD163 с нокаутом, чувствительно к вирусу сразу после рождения, защита от PRRSV in utero обеспечивает средства для устранения основного источника экономических потерь и страданий животных.
[00318] Примеры, раскрытые в данном документе, приведены в качестве иллюстрации и не предназначены для ограничения объема изобретения.
[00319] Принимая во внимание вышеизложенное, будет видно, что достигнуты несколько целей изобретения и достигнуты другие полезные результаты.
[00320] Поскольку в вышеуказанные продукты и способы могут быть внесены различные изменения без отклонения от объема изобретения, предполагается, что все материалы, содержащиеся в приведенном выше описании и показанные на прилагаемом(ых) чертеже(ах), должны интерпретироваться как иллюстративные и не в ограничительном смысле.
ССЫЛКИ
Allende R, et al., 1999. North American and European porcine reproductive and respiratory syndrome viruses differ in non-structural protein coding regions. J. Gen. Virol. 80: 307 315. Bauer BK, et al., Arginine supplementation in vitro increases porcine embryo development and affects mRN A transcript expression. Reprod Fertil Dev 2011; 23:107.
Beaton BP, et al., Compound transgenics: recombinase- mediated gene stacking. In: Pinkert CA (ed.). Transgenic Animal Technology: A Laboratory Handbook, 3rd ed. Waltham, MA: Elsevier; 2014:565-578.
Benfield DA, et al., 1992. Characterization of swine infertility and respiratory syndrome (SIRS) virus (Isolate ATCC VR-2332). J Vet Diag Invest 4:127-133.
Boddicker, et al., 2014. Genome-wide association and genomic prediction for host response to porcine reproductive and respiratory syndrome virus infection. Genetics, selection, evolution: GSE46, 18.
Borg NA, et al., CD1d-1ipid- antigen recognition by the semi-invariant NKT T-cell receptor. Nature 2007; 448:44^19.
Brinster RL, et al., Factors affecting the efficiency of introducing foreign DNA into mice by microinjecting eggs. Proc Natl Acad Sci U S A 1985; 82:4438-1442.
Cafruny, WA, et al, Trojan Horse macrophages: studies with the murine lactate dehydrogenase-elevating virus and implications for sexually transmitted virus infection. J. Gen. Virol. 77(12): 3005-3012(1996).
Calvert JG, et al., 2007. CD163 expression confers susceptibility to porcine reproductive and respiratory syndrome viruses. J Virol. 81:7371-7379.
Carter DB, et al., Phenotyping of transgenic cloned piglets. Cloning Stem Cells 2002; 4:131-145. Cong L, et al., Multiplex genome engineering using CRISPR/Cas systems. Science 2013; 339:819 823.
Christianson, W. T. et al. Experimental reproduction of swine infertility and respiratory syndrome in pregnant sows. American Journal of Veterinary Research 53, 485-488 (1993). Christopher-Hennings, et al., Effects of a modified-live virus vaccine against porcine reproductive and respiratory syndrome in boars, American J. Veterinary Research 58(1), 40 45 (1997). Dai Y, et al., Targeted disruption of the alphal,3-galactosyltransferase gene in cloned pigs. Nat Biotechnol 2002; 20:251-255.
Das PB, et al., 2009. The minor envelope glycoproteins GP2a and GP4 of porcine reproductive and respiratory syndrome virus interact with the receptor CD 163. J Virol. 84:1731-40. Dee et al., Elimination of porcine reproductive and respiratory syndrome virus using a test and removal process, The Veterinary Record 143, 474-476 (1998).
Delputte PL, et al., 2002. Involvement of the matrix protein in attachment of porcine reproductive and respiratory syndrome virus to a heparin-like receptor on porcine alveolar macrophages. J Virol. 76:4312-4320.
Etzerodt, A., et al., 2013. Plasma clearance of hemoglobin and haptoglobin in mice and effect of CD163 gene targeting disruption. Antioxidants & redox signaling 18, 2254-2263. Etzerodt, A., et al., 2013. CD163 and inflammation: biological, diagnostic, and therapeutic aspects. Antioxidants & redox signaling 18, 2352-2363.
Fabriek BO, et al., 2005. The macrophage scavenger receptor CD163. Immunobiology. 210:153-160.
Fabriek BO, et al., 2009. The macrophage scavenger receptor CD163 functions as an innate immune sensor for bacteria. Blood. 113:887-892.
Gaj T, et al., ZFN, TALEN, and CRISPR/Cas- based methods for genome engineering. Trends Biotechnol 2013; 31: 397^105.
Gaudreault N, et al., 2009. Factors affecting the permissiveness of porcine alveolar macrophages for porcine reproductive and respiratory syndrome virus. Archiv Virol. 154:133-136. Graversen, J.H., et al., 2012. Targeting the hemoglobin scavenger receptor CD163 in macrophages highly increases the anti-inflammatory potency of dexamethasone. Molecular therapy: the journal of the American Society of Gene Therapy 20, 1550-1558.
Groenen, M.A., et al., 2012. Analyses of pig genomes provide insight into porcine demography and evolution. Nature 491, 393-398.
Hai T, et al., One-step generation of knockout pigs by zygote injection of CRISPR/Cas system. Cell Res 2014; 24: 372-375.
Halbur, P.G., et al., 1995. Comparison of the pathogenicity of two US porcine reproductive and respiratory syndrome virus isolates with that of the Lelystad virus. Veterinary pathology 32, 648-660.
Halbur, P. & Bush, E. B. Update on abortion storms and sow mortality. Journal of Swine Health and Production 5, 73 (1997).
Hammer RE, et al., Production of transgenic rabbits, sheep and pigs by microinjection. Nature 1985;315:680-683.
Hauschild J, et al., Efficient generation of a biallelic knockout in pigs using zinc-finger nucleases. Proc Natl Acad Sci USA 2011; 108:12013-12017.
Holfkamp, D.J., et al., 2013. Assessment of the economic impact of porcine reproductive and respiratory syndrome virus on United States pork producers. Journal of Swine Health and Production 21, 72-84.
Hwang WY, et al., Efficient genome editing in zebrafish using a CRISPR-Cas system. Nat Biotechnol 2013; 31:227-229.
Jeney V, et al., 2002. Pro-oxidant and cytotoxic effects of circulating heme. Blood. 100:879-887. Keffaber, K.K., 1989. Reproductive failure of unknown etiology. American Association of Swine Practitioners Newsletter 1, 1-10.
Key, K. F. et al. Genetic variation and phylogenetic analyses of the ORF5 gene of acute porcine reproductive and respiratory syndrome virus isolates. Veterinary Microbiology 83, 249-263 (2001).
Kim, H.S., et al.,1993. Enhanced replication of porcine reproductive and respiratory syndrome (PRRS) virus in a homogeneous subpopulation of MA-104 cell line. Arch Virol 133, 477-483. Kristiansen M, et al., 2001. Identification of the haemoglobin scavenger receptor. Nature. 409:198-201.
Kwon DN, et al., Production of biallelic CMP-Neu5Ac hydroxylase knock-out pigs. Sci Rep 2013; 3:1981.
Ladinig, A., et al., 2015. Pathogenicity of three type 2 porcine reproductive and respiratory syndrome virus strains in experimentally inoculated pregnant gilts. Virus Res 203, 24-35. Lai L, et al., Production of alpha-1,3-galactosyltransferase knockout pigs by nuclear transfer cloning. Science 2002; 295:1089-1092.
Lai L, et al., Creating genetically modified pigs by using nuclear transfer. Reprod Biol Endocrinol 2003; 1:82.
Lai L, et al., Production of cloned pigs by using somatic cells as donors. Cloning Stem Cells 2003; 5:233-241.
Lawson SR, et al.,1997. Porcine reproductive and respiratory syndrome virus infection of gnotobiotic pigs: sites of virus replication and co-localization with MAC-387 staining at 21 days post-infection. Virus Res. 51:105-113.
Lee K, et al., Engraftment of human iPS cells and allogeneic porcine cells into pigs with inactivated RAG2 and accompanying severe combined immunodeficiency. Proc Natl Acad Sci USA 2014; 111:7260-7265.
Lee K, et al., Piglets produced from cloned blastocysts cultured in vitro with GM-CSF. Mol Reprod Dev2013; 80: 145-154.
Li D, et al., Heritable gene targeting in the mouse and rat using a CRISPR-Cas system. Nat Biotechnol 2013; 31:681 683.
Lillico SG, et al., Live pigs produced from genome edited zygotes. Sci Rep 2013; 3:2847. Machaty Z, et al., Complete activation of porcine oocytes induced by the sulfhydryl reagent, thimerosal. Biol Reprod 1997; 57:1123 1127.
Madsen M, et al., 2004. Molecular characterization of the haptoglobin-hemoglobin receptor CD 163. Ligand binding properties of the scavenger receptor cysteine-rich domain region. J Biol Chem. 279:51561-51567.
Mengeling, W. L. et al. Clinical consequences of exposing pregnant gilts to strains of porcine reproductive and respiratory syndrome (PRRS) virus isolated from field cases of "atypical" PRRS. American Journal of Veterinary Research. 59, 1540-1544(1998).
Merck, The Merck Veterinary Manual.
http://www.merckmanuals.com/vet/appendixes/reference guides/normal rectal temperature ran ges.html.
Miao, Y.L., et al., 2009. Centrosome abnormalities during porcine oocyte aging. Environmental and molecular mutagenesis 50, 666-671.
Morgan SB, et al., 2014. Pathology and Virus Distribution in the Lung and Lymphoid Tissues of Pigs Experimentally Inoculated with Three Distinct Type 1 PRRS Virus Isolates of Varying Pathogenicity. Transbound Emerg Dis. Nov 10. pp 1-11.
Nelsen CJ, et al., 1999. Porcine reproductive and respiratory syndrome virus comparison: divergent evolution on two continents. J. Virol. 73, 270-280.
Neumann EJ, et al., Assessment of the economic impact of porcine reproductive and respiratory syndrome on swine production in the United States. J Am Vet Med Assoc 2005; 227:385-392. Niu Y, et al., Generation of gene-modified Cynomolgus monkey via Cas9/RNA-mediated gene targeting in one-cell embryos. Cell 2014; 156:836-843.
Patience, J.F., et al.,1989. Swine Nutrition Guide, in: Center, P.S. (Ed.), University of Saskatchewan, Saskatoon, CA, pp.149-171.
Patton JP, et al., 2008. Modulation of CD 163 receptor expression and replication of porcine reproductive and respiratory syndrome virus in porcine macrophages. Virus Res. 140: 161-171. Plagemann, P. G. W. Lactate dehydrogenase-elevating virus and related viruses. In Fields Virology, 3rd Ed, Fields, B. ed. Lippincott- Raven, Philadelphia, pp 1105-1120 (1996). Prather RS, et al., An intact sialoadhesin (Sn/SIGLEC1/CD169) is not required for attachment/ internalization of the porcine reproductive and respiratory syndrome virus. J Virol 2013; 87:9538 9546.
Prather, R.S., et al., 2013. An intact sialoadhesin (Sn/SIGLEC1/CD169) is not required for attachment/internalization of the porcine reproductive and respiratory syndrome virus. J Virol 87, 9538-9546.
Provost С, et al., 2012. Identification of a new cell line permissive to porcine reproductive and respiratory syndrome virus infection and replication which is phenotypically distinct from M ARC-145 cell line. Virol J. 9:267.
Reed JL, et al., 1938. A simple method of estimating fifty percent endpoints. The American Journal of Hygiene 27:493-197.
Ross JW, et al., Optimization of square-wave electroporation for transfection of porcine fetal fibroblasts. Transgenic Res 2010; 19:611-620.
Rowland, R.R.R., et al., The evolution of porcine reproductive and respiratory syndrome virus: quasispecies and emergence of a virus subpopulation during infection of pigs with VR-2332, Virology 259, 262-266 (1999).
Rowland, R. R.R. et al. Lymphotropism of porcine reproductive and respiratory syndrome virus replication during persistent infection of pigs originally exposed to virus in utero. Veterinary Microbiology 96, 219-235 (2003).
Rowland, R. R.R. The interaction between PRRSV and the late gestation pig fetus. Virus Research 154, 114-122 (2010).
Rowland, R.R., et al., 2012. Control of porcine reproductive and respiratory syndrome (PRRS) through genetic improvements in disease resistance and tolerance. Frontiers in genetics 3, 260. Sanchez C, et al., 1999. The porcine 2A10 antigen is homologous to human CD163 and related to macrophage differentiation. Journal of Immunology 162:5230-5237.
Sanchez-Torres C, et al., 2003. Expression of porcine CD163 on monocytes/macrophages correlates with permissiveness to African swine fever infection. Arch Virol. 148:2307-2323. Schaer, C. A., et al., 2006a. Constitutive endocytosis of CD163 mediates hemoglobin-heme uptake and determines the noninflammatory and protective transcriptional response of macrophages to hemoglobin. Circulation research 99, 943-950.
Schaer, D.J., et al., 2006b. CD163 is the macrophage scavenger receptor for native and chemically modified hemoglobins in the absence of haptoglobin. Blood 107, 373-380.
Schaer, D.J., et al., 2006c. Hemophagocytic macrophages constitute a major compartment of heme oxygenase expression in sepsis. European journal of haematology 77, 432-436.
Shanmukhappa, K, et al., 2007. Role of CD151, A tetraspanin, in porcine reproductive and respiratory syndrome virus infection. Virol J. 4:62.
Shimozawa N, et al., Abnormalities in cloned mice are not transmitted to the progeny. Genesis 2002; 34:203-207.
Shi, M. et al. Phylogeny-based evolutionary, demographical, and geographical dissection of North American type 2 porcine reproductive and respiratory syndrome viruses. Journal of Virology 84, 8700-8711 (2010).
Smit, AFA, et al., RepeatMasker Open-3.0. 1996-2010. Current Version: open-4.0.5 (RMLib: 20140131 and Dfam: 1.2). http:// www.repeatmasker.org>. CD163: accessed July 25, 2014; CDID: accessed August 27, 2013.
Soares MP, et al., 2009. Heme oxygenase-1: from biology to therapeutic potential. Trends Mol Med. 15:50-58.
Stein M, et al.,1992. Interleukin 4 potently enhances murine macrophage mannose receptor activity: a marker of alternative immunologic macrophage activation. J Exp Med. 176:287-292. Stephenson R, et al., 2015. Multiplex serology for common viral infections in feral pigs in Hawaii between 2007 and 2010. Accepted for publication. J Wildlife Dis 51:239-2343. Sulahian TH1, et al., 2002. 2002. Human monocytes express CD163, which is upregulated by IL-10 and identical to p155. Cytokine. 2000 12:1312-1321.
Terns MP, et al., CRISPR-based adaptive immune systems. Curr Opin Microbiol 2011; 14:321 327.
Tian, K. et al. Emergence of fatal PRRSV variants: unparalleled outbreaks of atypical PRRS in
China and molecular dissection of the unique hallmark. PLoS One. 2, e526 (2007).
Trible, B.R., et al., 2012. Recognition of the different structural forms of the capsid protein determines the outcome following infection with porcine circovirus type 2. J Virol 86, 13508-13514.
U.S. Department of Agriculture and U.S. Department of Health and Human Services. Dietary Guidelines for Americans, 2010. 7th Edition,
Van Breedam W, et al., Porcine reproductive and respiratory syndrome virus entry into the porcine macrophage. J Gen Virol 2010; 91:1659 1667.
Van Breedam, W., et al., 2010. Porcine reproductive and respiratory syndrome virus entry into the porcine macrophage. J Gen Virol 91, 1659-1667.
Van den Heuvel MM, et al., 1999. Regulation of CD 163 on human macrophages: cross-linking of CD163 induces signaling and activation. J Leukoc Biol. 66:858-866.
van den Hoff MJ, et al., Electroporation in 'intracellular' buffer increases cell survival. Nucleic Acids Res 1992; 20:2902.
Van Gorp, H., et al., 2010. Scavenger receptor CD163, a Jack-of-all-trades and potential target for cell-directed therapy. Mol Immunol 47, 1650-1660.
Van Gorp H, et al., 2010. Identification of the CD 163 protein domains involved in infection of the porcine reproductive and respiratory syndrome virus. J Virol. 84:3101-3105.
Walters EM, et al., Advancing swine models for human health and diseases. Mo Med 2013; 110:212-215.
Wang H, et al., One-step generation of mice carrying mutations in multiple genes by CRISPR/Cas-mediated genome engineering. Cell 2013; 153: 910-918.
Welch SK, et al., 2010. A brief review of CD163 and its role in PRRSV infection. Virus Res. 154:98-103.
Wells, K.D., et al., 2014. Use of the CRISPR/Cas9 system to produce genetically engineered pigs from in vitro-derived oocytes and embryos. Biol Reprod 91, 78.
Wells, K.D. et al. Substitution of porcine CD163 SRCR domain 5 with a CD163-like homolog confers resistance of pigs to genotype1 but not genotype 2 porcine reproductive and respiratory syndrome (PRRS) viruses. Journal of Virology In press (2017).
Wensvoort, G., et al., 1991. Mystery swine disease in The Netherlands: the isolation of Lelystad virus. Veterinary Quarterly 13, 121-130.
Whitworth KM, et al., 2016. CD163 facilitates both entry and replication of porcine reproductive and respiratory syndrome virus. Nature Biotech. 34:20-22.
Whitworth KM, et al., Method of oocyte activation affects cloning efficiency in pigs. Mol Reprod Dev 2009;76:490-500.
Whitworth et al. Use of the CRISPR/Cas9 system to produce genetically engineered pigs from in vitro-derived oocytes and embryos, Biology of Reproduction 91(3): 1 13 (2014). Whitworth KM, et al., Activation method does not alter abnormal placental gene expression and development in cloned pigs. Mol Reprod Dev 2010; 77:1016-1030.
Whitworth KM, et al., Scriptaid corrects gene expression of a few aberrantly reprogrammed transcripts in nuclear transfer pig blastocyst stage embryos. Cell Reprogram 2011; 13:191-204. Whitworth, K.M., et al., 2014. Use ofthe CRISPR/Cas9 system to produce genetically engineered pigs from in vitro-derived oocytes and embryos. Biol Reprod 91, 78.
Whyte JJ, et al., Genetic modifications of pigs for medicine and agriculture. Mol Reprod Dev 2011;78:879-891.
Whyte JJ, et al., Gene targeting with zinc finger nucleases to produce cloned eGFP knockout pigs. Mol Reprod Dev 2011; 78:2.
Wiedenheft B, et al., RNA-guided genetic silencing systems in bacteria and archaea. Nature 2012; 482:331-338.
Wilkinson et al. Genome-wide analysis of the transcriptional response to porcine reproductive and respiratory syndrome virus infection at the maternal/fetal interface and in the fetus. BMC Genomics 17, 383 (2016).
Whitworth, K.M. et al. Gene-edited pigs are protected from porcine reproductive and respiratory syndrome virus. Nature Biotechnology 34, 20-22 (2016).
Winckler, С, et al., 2001. The reliability and repeatability of a lameness scoring system for use as an indicator of welfare in dairy cattle. Acta Agricultura Scandinavica Section A. Animal Science, 103-107.
Yang D, et al., Generation of PPARgamma mono-allelic knockout pigs via zinc-finger nucleases and nuclear transfer cloning. Cell Res 2011; 21:979-982.
Yoshioka K, et al., Birth of piglets derived from porcine zygotes cultured in a chemically defined medium. Biol Reprod 2002; 66:112-119.
Zhang Q, et al., 2015. PRRS virus receptors and their role for pathogenesis. Vet Microbiol. 177:229-241.
Zhao J, et al., Histone deacetylase inhibitors improve in vitro and in vivo developmental competence of somatic cell nuclear transfer porcine embryos. Cell Reprogram 2010; 12:75 83. Zhao J, et al., Significant improvement in cloning efficiency of an inbred miniature pig by histone deacetylase inhibitor treatment after somatic cell nuclear transfer. Biol Reprod 2009; 81:525 530.

--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Curators of the University of Missouri
Prather, Randall
Wells, Kevin
Whitworth, Kristin
<120> СПОСОБЫ ЗАЩИТЫ ПЛОДОВ СВИНЕЙ ОТ ИНФИЦИРОВАНИЯ ВИРУСОМ РЕПРОДУКТИВНОГО
И РЕСПИРАТОРНОГО СИНДРОМА СВИНЕЙ (PRRSV)
<130> UMO 18054.WO
<160> 121
<170> Патентна версия 3.5
<210> 1
<211> 23
<212> ДНК
<213> Sus scrofa
<400> 1
ggaaacccag gctggttgga ggg 23
<210> 2
<211> 23
<212> ДНК
<213> Sus scrofa
<400> 2
ggaactacag tgcggcactg tgg 23
<210> 3
<211> 23
<212> ДНК
<213> Sus scrofa
<400> 3
cagtagcacc ccgccctgac ggg 23
<210> 4
<211> 23
<212> ДНК
<213> Sus scrofa
<400> 4
tgtagccaca gcagggacgt cgg 23
<210> 5
<211> 23
<212> ДНК
<213> Sus scrofa
<400> 5
ccagcctcgc ccagcgacat ggg 23
<210> 6
<211> 23
<212> ДНК
<213> Sus scrofa
<400> 6
ctttcattta tctgaactca ggg 23
<210> 7
<211> 23
<212> ДНК
<213> Sus scrofa
<400> 7
ttatctgaac tcagggtccc cgg 23
<210> 8
<211> 23
<212> ДНК
<213> Sus scrofa
<400> 8
cagctgcagc atatatttaa ggg 23
<210> 9
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 9
ctcctcgccc ttgctcacca tgg 23
<210> 10
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 10
gaccaggatg ggcaccaccc cgg 23
<210> 11
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 11
ctctccctca ctctaaccta ctt 23
<210> 12
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 12
tatttctctc acatggccag tc 22
<210> 13
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 13
ctctccctca ctctaaccta ctt 23
<210> 14
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 14
gactggccat gtgagagaaa ta 22
<210> 15
<211> 27767
<212> ДНК
<213> Sus scrofa
<400> 15
atacaagtgc cttttacaga caatctgcac aagttatttg ttagacatat ttgattatag 60
aattaatatt aaaaggggtt ataacaatca agcattgata atttaattat gtttgcctat 120
tttactttag ttttttgaca taactgtgta actattgcga tttttttatt cctaatgtaa 180
ttagttcaaa acaaagtgca gaaatttaaa atattcaatt caacaacagt atataagtca 240
atattccccc cttaaatttt tacaaatctt tagggagtgt ttctcaattt ctcaatttct 300
ttggttgttt catgtcccat atggaagaaa acatgggtgt gaaagggaag cttactcttt 360
tgattacttc ccttttctgg ttgactccac ctccattatg aagcctttct gtatttttgt 420
ggaagtgaaa tgatttttag aattcttagt ggttctcttc ttcaggagaa catttctagg 480
taataataca agaagattta aatggcataa aaccttggaa tggacaaact cagaatggtg 540
ctacatgaaa actctggatc tgcaggtaaa atcttctcat ttattctata tttacctttt 600
aaatagagtg tagcaatatt ccgacagtca atcaatctga tttaatagtg attggcatct 660
ggagaagaag taacagggaa aaaggcaata agctttataa ggggaacttt tatcttccat 720
agactcaaaa ttgaagacgt gactagaaga ttgctagatt tggcatcagt tttgtaaaat 780
tgctgaggtg aaattaagta agggatgaaa attaactaaa ttgtgttgag tatgaaacta 840
gtagttgtta gaaaagatag aacatgaagg aatgaatatt gattgaaagt tgatgaccta 900
gaggacattt agactaacac ctctgagtgt caaagtctaa tttatgattt acatcgatgc 960
gttaaactca tttaacattc ttactttttt cccctcaagc atttaagctg aagtataaca 1020
tttcacatga aagcctggat tataaatgca cagttcagtg acctatctca gaggagtgac 1080
tgccatagca ttttttttgt ctttttgcct tcagagccac agcaacgcgg gatccgaagc 1140
cgcgtctgcg acccacacca cagctcacgg caatgccgga tctttaaccc actgagcgag 1200
gccggggatc gaacccgcag tctcatggtt cctagtagga ttcgttaacc actgcgccac 1260
gacgggaact cctaccatag catttttact tttaagttac tgttggttta gagtaagaag 1320
gagaaatgag agtgatggag cgtttgctat atttggagac aaggtcctat attggaggtt 1380
ctcaaatata aattttgtcg ctttttcctc caatgtattg ttcaactact atttagcagg 1440
ccactgtgcc aggtactggt gaaactggtg aacatgatag atgtaattca ttccctcatg 1500
gaactttcca tctaacaatg tggatcaggt aggcttggag atgagaatgc cagtggttga 1560
ctatgactct gtggctgaag ggagagctac tcacttcgta gtttcatcaa tgtctttttg 1620
gttttccagg ttttaagccc tgctcttgca attcttttcc cttctccaac tttcttctaa 1680
tttctcaccc ctaggatgcc tataaacatg agtattttca aagctacttc actgaggtta 1740
tatgatcctg gtgtgaattt ttcctgcctg acttgccatt tagaaggaag tgtttcctgg 1800
aatttccatt gtggcttggt ggttaaagac cctgcattgt ctctgtgagg atgtgggttc 1860
aatctctggc ctcattcagt gagtgggtta aggatctggt gtcgctgcaa gctgtggcta 1920
agatcccaca ttgccatgtc tgtggtgtag actggcacct ggagctctga tttgaccaca 1980
atcttaggaa cttcagatgt ggccataaaa aggaaaaaaa agttaggaag ggttttctgt 2040
cttgtttgga ccttcgttaa tctcaaacct ttggaaccat ctctcctcca aaacctcctt 2100
tgggtaagac tgtatgtttg ccctctctct tcttttcgca gactttagaa gatgttctgc 2160
ccatttaagt tccttcactt tggctgtagt cgctgttctc agtgcctgct tggtcactag 2220
ttctcttggt gagtactttg acaaatttac ttgtaaccga gcccaactgt gacaagaaac 2280
actgaaaagc aaataattgc tcctgaagtc tagatagcat ctaaaaacat gcttcatggt 2340
ttcaaggatc atatattgaa accccaggga tcctctagag tcgacctgca gcatgcaggg 2400
gggggggggg gggggggggg gggggggggg gggggggggg gggggggggg gggggggggg 2460
gggggggggg gggggggggg gggggggggg gggggggggg gtgcataagg aaagactatc 2520
tcaacgtctt attcctcagc ttacattaga tttgaaactc tagtcaccta aaatgcaaat 2580
ctcatttact taccatcaga gatattaatg acctatagaa ttcagcataa ataaagtttc 2640
atgtatggat attagcttat ggttctagtc actgctaatt gaaacctgtg atattgctgt 2700
ttgttttgac tcctatgaaa taacattctc ccattgtacc atggatgggt ccagaaacat 2760
ttctcaaatc ctggcttgaa aaaataaata agtaatctaa agaataataa ttctctactt 2820
gctctttgaa tcttgaccaa ttgctgcatt tacctattgt tacaggagga aaagacaagg 2880
agctgaggct aacgggtggt gaaaacaagt gctctggaag agtggaggtg aaagtgcagg 2940
aggagtgggg aactgtgtgt aataatggct gggacatgga tgtggtctct gttgtttgta 3000
ggcagctggg atgtccaact gctatcaaag ccactggatg ggctaatttt agtgcaggtt 3060
ctggacgcat ttggatggat catgtttctt gtcgagggaa tgagtcagct ctctgggact 3120
gcaaacatga tggatgggga aagcataact gtactcacca acaggatgct ggagtaacct 3180
gctcaggtaa gacatacaca aataagtcaa gcctatacat gaaatgcttt gtgggaaaaa 3240
atgtatagat gagttaaaaa caaaaaggaa ccagttttct ataagtcatc tagtccatgt 3300
ataaaattac ccaatccatt actaaaagac cacttctggt attttacaca tgacaaagcc 3360
catattaaaa aaaaaaaatt cagaagagat tctgaatgct ataataaatg agcaagtgac 3420
tagcttcaat tttatattag gtcattctac cttctacttc tacatgaaaa tatcataatg 3480
tctaagttaa ttccttgtcc cctttcccaa taaagcactg ctttcatgca ctggcctatg 3540
aatcatgaac tttttgccct ttaactgatg atcaacttac caaatcaaga aataaatatt 3600
cttagcactg atcctttttt gttgttgttg gaggaagaat gttttgcaaa gtagaattgc 3660
ttttttctgt ttaacagtgc tattcatttc atttacatgg tcgttttaat ttataaaaca 3720
tttcataagt ttcacctcat atgcccttac aataactcag gaagttatat gttagacctt 3780
tctgctgaca aatcccagag tcatgtttct gacccagttc agattccttg gcttcccatt 3840
tctctttgct catgtcattg acctttatgc agccctctta cctcccacct ttctattaca 3900
gaccatctcc tccataggac tggtgttaga aagtactaat ctctacccag gcattgtggt 3960
gcaatgtggg cagcacaggc tggtatctag aaaaatgctg aagtgaattc cagctcagct 4020
gctcgttaat actatcgttt taagtaagct gttcaatcct ttgaaattca ctttctgagc 4080
actcagtgat ataataaatg tagagctact ggtacactgt ctggtatgta ataggtgtta 4140
ccaattaacc ttagtttcct catgggtcac tggttctcat tacctagaca actcatttct 4200
ctttcttcct ctttctcttt ctccattctc ctcctccttc ttcctcttct tcttgtctgt 4260
tattgttata tcattttgct gagaaagtta agaaataaca actctaacct ctacatcgac 4320
cacctagagc aaagttaaaa ataataataa accttgccag actcttacta taattgttgc 4380
tgtctataga gttgactgtt taagttaaga catcagtata tatttttaat ttttgtgttt 4440
tttttttcat acttttacat gaggatcctt tatataagga tgagttaaac aaacttgatt 4500
tttgaagttt atacccctga ggctcaactg cataataata gaaagggatc catagcctct 4560
caaggactta actagtttca tgagttttca gaatctgaat ttctgagatt ctccacccca 4620
attaaagctc aagcctcaga acatatatcc ttctcttggt aaattctatt cttatcacat 4680
gcgtaataat aaaaaagaga gatgttggag acagattttt ttcctcacat tctgtctcta 4740
ctgttttcta ggtgtttgat tctgtgttat ttaacctcag tttgcttatc tgtgaagtag 4800
ggattatggt aataacatat aatgctttat gttgtaaaga ctaaagaaga tagcatatgt 4860
aacacatttg gaacagggaa tgcatatttt gattgtgagc tcttattatt attaccattc 4920
agccctaata aaaatcttgg taagtggaag gctttggatt tcagaacttt taaaatctaa 4980
ttactttttc aaaaaagaac ttcttagggt tttttttttt taaccacaaa gtgtttctat 5040
tttttaggtg tcccaaaatt tcgttccaaa tatctttttc tcagatattt tagtcctcat 5100
agaacaccta gggatagtgg atagagaaaa ttttctttat taaaaagctg ttctttgcta 5160
aaaattgtag caggtacttt tgggaggggg gaaaactttg attcagaaac tgctaagaca 5220
tggagtgttt tgactaattt ttcctcaatt tttaatgttt tttataccat agggtacttt 5280
tgcaaactat tatgcatact tatatatttt tacttttttc ctgtctttta acttccaaat 5340
tcaacttcag acaattattc atgcactaaa ctgtttgtag taagaaagat taaaattaaa 5400
aaattaacca ttcaacaaat gactggtttg ccatttttac tactttgttg tatgaacaat 5460
ttttttttct acaaatgaat actttgagtc tgatttatcc attcctacat aaaagttttt 5520
actatatctt agtattggaa ggaaacaaaa caaaacacaa tgtaaatttt aatctataaa 5580
ttttgggggg gtaaatatac atagatgaaa gtcttaacca ttaattagag tcaaaagatt 5640
aaaattctcc aatatgtgaa cttaggctgc atccaaaatg aagcatcatt tttaaggaca 5700
gcatcaaaag tgaccagagg aattttactt tctttctttt tttttttttt gaattttagt 5760
ttctaaactc acttctgaat aaatacaact tctaaattct cgtcttttct ctactctaga 5820
tggatctgat ttagagatga ggctggtgaa tggaggaaac cggtgcttag gaagaataga 5880
agtcaaattt caaggaacgt ggggaacagt gtgtgatgat cacttcaaca taaatcatgc 5940
ttctgtggtt tgtaaacaac ttgaatgtgg aagtgctgtc agtttctctg gttcagctaa 6000
ttttggagaa ggttctggac caatctggtt tgatgatctt gtatgcaatg gaaatgagtc 6060
agctctctgg aactgcaaac atgaaggatg gggaaagcac aattgcgatc atgctgagga 6120
tgctggagtg atttgcttaa gtaaggactg acctgggttt gttctgttct ccatgagagg 6180
gcaaaaaaag gggagtaaaa gtcttaaaag ctcaaactgt taaaaacata atgatgattg 6240
cttcttttat catcttatta ttatctaatt tcaggtcgaa attctagtac ctgtgcagtt 6300
ttttacctta actgaaatta agataaatag gatagggagg aaggatgagc agtgacattt 6360
aggtccaagt catgaggtta gaaggaaatg ttcagagaat agcccattcc ctcagccctc 6420
aaagaaagaa agaaagaaaa agaaaaaaaa aaagaaagct taactagaaa attttgttct 6480
ctggatgttt tagaggcaaa ccatcccttt atcattccta cctacaaagc cttctcttaa 6540
tcacattacc caccctttcc tactatagtc aggggggggg gggggggggg gggggggggg 6600
gggggggggg gggggggggg gggggggggg gggggggggg gggggggggg gggggggggg 6660
gggggggggg gtgaaaaaag aaccaaacaa tttcaacaaa aaaccaaaca attccaacaa 6720
aattggtcca ataagcaaac ctctagataa atttcagtgc cctggatgtt ttgttaggaa 6780
ctcttcctac aatgcgtgct ttccattctg aaaagtccta tctacttgcc tgatccactt 6840
ctccttccat cctaaacgat tttcagtggt agtatattac tgttgtctct gtctctactt 6900
atatatcttc cccttttcac tcactcctct caggtacagc tcttcagttt gcccttattc 6960
ttgtttcctt gtcaatgact tgttttgtgt ccctcttaca gatggagcag acctgaaact 7020
gagagtggta gatggagtca ctgaatgttc aggaagattg gaagtgaaat tccaaggaga 7080
atggggaaca atctgtgatg atggctggga tagtgatgat gccgctgtgg catgtaagca 7140
actgggaggt ccaactgctg tcactgccat tgtcgagtta acgccagtga gggactggac 7200
acattggctc acacacatac agccatgaca cgatctgctc tatggtccga tgattaaagg 7260
gggggggggg gggggggggg gggggggggg gggggggggg gggggggggg gggggggggg 7320
gggggggggg gggggggggg gggggggggg ggggggggag aagagctggt ggacatttct 7380
ggaaaggaac caaaacccgg aagggccttg ttcttcagga tttgggatgg attggggagg 7440
gagaaaattg tttctaatat ttcttggtgg gaattctttt acagttgtga caaatctttc 7500
acatattctt catttgagta gtttggaggg ttgtctgact gttttctata ataaatgtcc 7560
caagtgctat gaggtaccac atttcaaatt ctaattctac ctgaagctcc aaaaagacaa 7620
aatgttatag gtcttttctt tatatctaat ttgcttatgg tttttagcca ttgacaattt 7680
ttttttctta actcttgaaa ctataaccct atttctaacc aaattcatgt tctatactgg 7740
ctcttcaaaa acccaggaga tgggaaagcc agaatctcca gtgtttcagc ttctgggaag 7800
gagcaagttt ttaaaaatac cctctgggag ctaaattcca catgtatcta tggcctaagt 7860
gtatgtttat tttgcagatg gatcagatct ggaactgaga cttaaaggtg gaggcagcca 7920
ctgtgctggg acagtggagg tggaaattca gaaactggta ggaaaagtgt gtgatagaag 7980
ctggggactg aaagaagctg atgtggtttg caggcagctg ggatgtggat ctgcactcaa 8040
aacatcatat caagtttatt ccaaaaccaa ggcaacaaac acatggctgt ttgtaagcag 8100
ctgtaatgga aatgaaactt ctctttggga ctgcaagaat tggcagtggg gtggacttag 8160
ttgtgatcac tatgacgaag ccaaaattac ctgctcaggt aagaatttca atcaatgtgt 8220
taggaaattg cattctactt tcttttacat gtagctgtcc agttttccca gcaccacttg 8280
ttgaagagac tgtcttttct tcatcatata gtcctacatc ctttgtcata aattaattga 8340
ccataggtgt gtgggtttat atctgggctc tctattctgt tcctttgatc tatatgtctg 8400
tttttatgcc agcaccatgc tgttttgatt actatagctt tgtagtatca tctgaagtca 8460
ggaaacatga ttcctccagc tttgttcttc tttctcaaga ttgttttgtc tattcagagt 8520
ttatgttccc atgcagattt aatttttaaa tttatttaat ttttattttt tatttttaat 8580
ttaaattaat ttaaattttt tatttcccaa cgtacagcca agggggccag ggtaaccttt 8640
acatgtatac attaaaaatt tcaggttttt cccccaccca tttctttctg ttggcaagta 8700
aatttttgaa caaagtttcc caatgctttt taaggggaat tcccttgggg gggggggggg 8760
gggggggggg gggggggggg gggggggggg gggggggggg gggggggggg gggggggggg 8820
gggggggggg gggggggggg gggggggggg gggggggggg agacgaaatt gactatattt 8880
tctttgttgg gaatctttta cagttgtgac aaatctttca catattcttc atttgagtag 8940
tttggagggt tgtctgactg ttttctataa taaatgtccc aagtgctatg aggtaccaca 9000
tttcaaattc taattctacc tgaagctcca aaaagacaaa atgttatagg tcttttcttt 9060
atatctaatt tgcttatggt ttttagccat tgacaatttt tttttcttaa ctcttgaaac 9120
tataacccta tttctaacca aattcatgtt ctatactggc tcttcaaaaa cccaggagat 9180
gggaaagcca gaatctccag tgtttcagct tctgggaagg agcaagtttt taaaaatacc 9240
ctctgggagc taaattccac atgtatctat ggcctaagtg tatgtttatt ttgcagatgg 9300
atcagatctg gaactgagac ttaaaggtgg aggcagccac tgtgctggga cagtggaggt 9360
ggaaattcag aaactggtag gaaaagtgtg tgatagaagc tggggactga aagaagctga 9420
tgtggtttgc aggcagctgg gatgtggatc tgcactcaaa acatcatatc aagtttattc 9480
caaaaccaag gcaacaaaca catggctgtt tgtaagcagc tgtaatggaa atgaaacttc 9540
tctttgggac tgcaagaatt ggcagtgggg tggacttagt tgtgatcact atgacgaaac 9600
caaaattacc tgctcaggta agaatttcaa tcaatgtgtt aggaaaattg cattctactt 9660
tcttttacat gtagctgtcc agttttccca gcaccacttg ttgaaaaaac tgtctttttc 9720
ttcatcatat agtcctacat cccttggcca taaattaatt gaccataagg ggtgtgggtt 9780
taatatccgg ggctcctcaa ttcgggtccc ttggatccta aaagccggtt ttataacccg 9840
acacatggcc tgtttttgac taaataaaac ctttggaaaa caatcccgaa ggtcgaggaa 9900
catggaatcc ccccaacaaa ggaccttctt tccccaaaaa tgcggctcag ccaactcaaa 9960
aagattttat gaatcacaaa ccgcacatta tcttcctaaa attactattc ctatgtttta 10020
atttgcaaag tcattccgat atagttggcg cagagtaact catttagata tccaccccac 10080
cagttcctca ctcaagtaag gggggggggg gggggggggg gggggggggg gggggggggg 10140
gggggggggg gggggggggg gggggggggg gggggggggg gggggggggg gggggggggc 10200
ccccatgtga gattttgtgt gtcctttaag agtggagtct ctatttccca ctgctctctg 10260
gttctcccca aagtaagccc tgctggcttt caaaacttct gggagcttgc cttcttggta 10320
taggactcct gggctaggga gtctaatgtt tggcttagac cccttactgc ttgggaagaa 10380
tctctgcaac tgtaatgaat tatcttccta tttgtgggtt gctgaggata tggtcttaac 10440
tgttctgtgt tctacccctc ctatccatct tgttgtggtt ccttctttat atctttagtt 10500
gtagaaaagt ttttcttatc aacagttgct ctgtaaattg taacttgggt gtacacctag 10560
taggaggtga gctcagggtc ttcctactct gccatcttgg ccatgtcctc taaacatttt 10620
ggtgtatttc actgcaacct ttttaaaaat ctcaaaagtg agctgtgatt ggctagtctt 10680
gtggataatc tctagcattt gatgctaatc atatttatac aaatactttg ttgaaaagtg 10740
atgccttttt aactattatt aaaaaacgta ttgacataac tattgctatt atactgaaaa 10800
gaaagacctt agagaaaata gcataagagc aaaaccatta aacatggaga catctagtca 10860
tagggtggaa attttatgtg gtccatatcc cctaaccagt ggctttacac caggcacatc 10920
ctaactaaga tctgctccca agtgtcttcc ctgatgcttt aaattgtgtt acatggaaac 10980
tatcctttga tgaagaaatg caacctttta aaatacaaca ttgaaacttt tgtgctttaa 11040
ttttgctttt caacattttt tctttttaaa agaagaaatt tatttgtttt tttaaatttt 11100
aatggccacg gcatatggaa gttctcaggc cagggataga attcaagcca caggtgcgac 11160
ccatgccaca actgctgcaa caccagatcc tttaacccac tgcaccaggc cagggattga 11220
agccttgcct tactgacaat ctgagccact tcagtcagat aaagaaattt cttcattaag 11280
cagagtattc acatggttta aacttcaaaa tattaaagtg taaactcttt ccccaccact 11340
gtccccagct caccaactct acttaccaca gacaactgat gtggttaggg tatttaaata 11400
gtaaatccaa gaaaatataa acaaatccgt atatataggt ttcaccccat tttattatcc 11460
taatgttgca tatcatataa actatactgt cccttgggta ttcacttagt aaaatatttt 11520
gatcataatt tcctatcagt atttaaagag ctttctgaaa ttatttctgt ataacatttc 11580
ttttctcatc ggtagggggg gggggggggg gggggggggg gggggggggg gggggggggg 11640
gggggggggg gggggggggg gggggggggg gggggggggg gggggggggg ggggaatggg 11700
aagaaaaaac caccatggtt aatttttttt atccctctac acccgggaaa attacccttg 11760
gggccacact tttctataga aaggggatta tttaaaaggg tctgaaaaag aatttttttt 11820
tcgaaagggg aaatatttgg cctaacttag tcacataagc catgttctct ggcaagttag 11880
gtaacataca tttttgtcat tgggggcaac aaaaacaatt ttccttttgg accttttggg 11940
actccgcatt ggttagggaa ggggaagtat attggaattc ggaaaattcc ttccaaatta 12000
aaaaaggttt gttattttca tattaaccta tttcatatta attagcatga attccagcgc 12060
cattaaaagg gaaaacacct ggagtggtaa gaaaaaagtt tttttttctc tttttttttt 12120
ttttttttta atggccacat ctgtggcatg tgaagttccc aggctagggg tcgaatagga 12180
gctacagctg ccagcttgca ccacagccac aacaatgcca gagccaagcc tcatctgcga 12240
cctataccac aactcatggc aatgctggtt ccttaacccc ctgagtgagg cctggggtca 12300
aacccacatc ctcatggata ctaaccggct ttgttaccgc tgagccatga gggaaactcc 12360
ctttttctca ttgaaaataa gtcaaataga taagcagctt aaggctgttt gggtgattct 12420
gtggtccagt aattatcaaa tcctactgga caagaataga gaatgtgcaa atgagggaac 12480
gtgttggtga gatcaggctc tgcccactga gctatcctct gtcatgggcc ctgtgctgtt 12540
ctcagagctg tacttcctag ggcattgttc tcatttcaat tctgagttca gtgtggagag 12600
tatacgtgtg tgggggctgc acgcttttca caacccactt tctgctgata ctgatttagg 12660
gatccttgga ttgctttaca gttgagtcat cattaactag tgtcacttgc cttcaaagtc 12720
agcaaaataa ttgtctccaa actagtaggc ttctagtgta tttgctttaa tccaatgcca 12780
tgtgaaagta acatggtcaa agaataagtt atataccttg acctaccctg tgaccaggct 12840
cttcctctta atttattgac cactgcctta aggtcatttg aaaccatggg tttgggagga 12900
aggcaaggcc taaatcccgt ctttgttgga aggctcactg tccttgtctt tagagcatca 12960
ttttttttta aactggggta cagtttattt acagtgttgt gtcaatttct gctgtacagc 13020
acagtgaccc agtcatacac atacatacat tctttttctc atactatctt caattttatt 13080
ttctgctaag tctgccattt tatcatcacc tcagtttgaa ggacaggata tttagagttt 13140
gttttttttt tccccccaat cctgcaattt ctaaattata agactctcaa ttagccgtat 13200
ataacagctg caggcacagg atgtctccct cacaaaattg gtatttttcc ttccatttct 13260
tcttgcagtt tggctatttc ttgtctgagt tcatctctct ttttaagtgt taaaaagggc 13320
aaggaggatt catgctatgt caacattatg attttttctt ttctatactt gataagagta 13380
tacttttccc aaatgtcatc caacttttca gcatcagttt ggacatggtt ttcttttcaa 13440
ggtggtattt ctctaatgtc acttgaataa caagactcgt tagttctcca ggctacaata 13500
tcctagtctg agtatattct gcatgttaat tctattcagc cacatccata atttaggttt 13560
tattcctgga acacctcact tttttttttt tttttggtct ttttatagcc ataaccatgg 13620
catatggagg ttcccaggct aggggtctaa tctgagcttt agccactggc ccatgccaca 13680
gccacagcca tgccacatct gagccacatc tgtgaccttt tccacagctc acagaaacac 13740
cagatcccta acccactgag tgaggccagg ggtcaaacct gtaacccttc catggttcct 13800
agtcagattc gttcctctgt accacgatgg gaattcctaa tacctcactt atgataacac 13860
attctgaatt atttaggatt ctattatact gcatgtaata gaaatcccaa atagcaaaat 13920
ttgcaactta aggcaggttc ctgtctttac aaaatcatgt tttcctttgc tatatgtgca 13980
ctttgctttc ctctgtgaat tccctttttt gttatatttc tatagctttt ggaaacactt 14040
ttacttattt gggggggcct agatttttaa ccctctcctt gtttttctag aaatagagtt 14100
tataatttta tttcttcatt tacttgatac tttcaagaga ttcccaggaa aaaaattatg 14160
gaaatactgt ctctgtgcct gccaagttca aactaagaat tgtataatct gttttaattc 14220
ttaagcattt atagatgaca aggctttgtg tctgataggg gccagcgaac tcagtaaaga 14280
gggaagatga gaaagataat ggcaagaatt tatccctgaa gtgtagtttt gacaaaccag 14340
tcacaaagag gtctaagaaa ttttggtcac aaagttgttt tgaatcccag gcattttatt 14400
tgcaatgatt gcatatgttc tggaaaggac atctgaacct aagaaatagt tcatttgcat 14460
tgtgttatat tttactaagg tctgagaaat aatcttgaga tgagaatgaa ctctacttct 14520
tcagagtctg gaaggaataa attatgaaaa tgtattaatg cttctttaaa ccatattgta 14580
tatttatcta ttactaaaca aaaagaagta gctctattta tttatttatt tatttattta 14640
tttatgtctt ttgtctcttt agggccacac ctgtggcata tggaggttcc caggctagag 14700
gtccaattgg agatgtagca gccagcctat gccagagcca ccgcaacacg ggatctgagc 14760
cacgtctgtg acttacacca cagctcacag caacgcctga tcctcaaccc actgagcgag 14820
gccagggatc gaacccatgt cctcatggat gctagttggg ttcgttaact gctgagccat 14880
gatgggaact ccaaattaat tatttcttat atttgttctt catatattca tttctataga 14940
aagaaataaa tacagattca gttaatgatg gcaggtaaaa gcttaactta ttaatcaaag 15000
gagttaatcc aggcacaaaa attcaattca tggctctctg ttaaaattta ggtataggtt 15060
tagcaggaag aaaaggttag tagatgcaga ctattacatt tagaatggat ggacaatgaa 15120
gtcctactat acagcacagg gaactatatc caatctcttg ggatagaata tgatggaaga 15180
caaaatcaga acaagagagt atatatatat gtgtgtgtgt gtgtgtgtgt gtgtgtgtgt 15240
gtgtgtgtgt gtgtgactgg gtcaccctgc ggcacagcag aaattggcag aacattgtaa 15300
atcaactata ctttaatagg aaaaatactt ttaagggcta aatttccaat attctaacca 15360
tgtacacaga gtaaatgtca taaggatgcc agtctgtgta gagattgatg tgttactagc 15420
agattcatga aataaaggct gaggatgtag tccccaagtc acttctgagt ggaagaattt 15480
ctcctttgtc ctggactcaa atattttagg ataaaggaaa aaagaagata tttatagaag 15540
ggacttgttt tcaagtactt gacaaaattt caccattaaa gagaaatttg tgggagttcc 15600
catcgtggct cagtggaaac aaatccaact aggaaccatg aggttgtggg tttgatccct 15660
ggcctcactc agtgggttaa ggatccggtg ttgccgtgag ctgtggtgta ggttgcagac 15720
acggttctga tcctgcgttg ctgtggctgt ggctgtggtg taggccagca gcaaacagct 15780
ctgattagac ccctagcctg gaaacctcca tatgccacag gtgcagccct aaaaagacaa 15840
aaaaagagaa aagacaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaag 15900
aacccccaga ggtatttatt tgtttttgcc ttttttcact gactgttctt tgtttgtttg 15960
tttgagactg atctagaaga ctagagatta caagaaatat ggatttggct cactctaaga 16020
aactgctttc attccaaggt ttgggtctat ccaaaagtgg aatagaatca tatgaatact 16080
agtttatgag tatttagtga gaggaatttc aagctcaaat aatgattcag caagattaaa 16140
ttaaggaggg aattttcctt gtggctgagt gggttaagga cccaatgttg tctctgtgag 16200
gatgtaggtt ccatcctggg ctttgctcat taggttaagg atctggcatt gctgcagctc 16260
agacccagtg ctgccctggt tgtggcttag gccaaagctg cagctccaat tcaatctctg 16320
gcctgggaac ctccatgtgc tacaaggtgc ggccttaaaa ggaaaaaaaa aaaattaaat 16380
caaggactca agagtctttc attatttgtg ttgtggaagc tatatttgtt ttaaagtctt 16440
agttgtgttt agaaagcaag atgttcttca actcaaattt gggagggaac ttgtttcata 16500
catttttaat ggataagtgg caaaattttc atgctgaggt gatctatagt gttgtaatgc 16560
agaatatagt cagatcttga acattttagg aagttggtga gggccaattg tgtatctgtg 16620
ccatgctgat aagaatgtca agggatcaca agaattcgtg ttatttgaca gcagtcatct 16680
ttaaaaggca tttgagaaag tccaatttca aatgcatttc ctttctttaa aagataaatt 16740
gaagaaaata agtctttatt tcccaagtaa attgaattgc ctctcagtct gttaaaagaa 16800
actcttacct tgatgattgc gctcttaacc tggcaaagat tgtctttaaa atctgagctc 16860
catgtcttct gctttatttc tggtgtgcct ttgactccag attacagtaa atggaggact 16920
gagtataggg ctaaaaagta gagagaatgg atgcatatta tctgtggtct ccaatgtgat 16980
gaatgaagta ggcaaatact caaaggaaag agaaagcatg ctccaagaat tatgggttcc 17040
agaaggcaaa gtcccagaat tgtctccagg gaaggacagg gaggtctaga atcggctaag 17100
cccactgtag gcagaaaaac caagaggcat gaatggcttc cctttctcac ttttcactct 17160
ctggcttact cctatcatga aggaaaatat tggaatcata ttctccctca ccgaaatgct 17220
atttttcagc ccacaggaaa cccaggctgg ttggagggga cattccctgc tctggtcgtg 17280
ttgaagtaca acatggagac acgtggggca ccgtctgtga ttctgacttc tctctggagg 17340
cggccagcgt gctgtgcagg gaactacagt gcggcactgt ggtttccctc ctggggggag 17400
ctcactttgg agaaggaagt ggacagatct gggctgaaga attccagtgt gaggggcacg 17460
agtcccacct ttcactctgc ccagtagcac cccgccctga cgggacatgt agccacagca 17520
gggacgtcgg cgtagtctgc tcaagtgaga cccagggaat gtgttcactt tgttcccatg 17580
ccatgaagag ggtagggtta ggtagtcaca gacatctttt taaagccctg tctccttcca 17640
ggatacacac aaatccgctt ggtgaatggc aagaccccat gtgaaggaag agtggagctc 17700
aacattcttg ggtcctgggg gtccctctgc aactctcact gggacatgga agatgcccat 17760
gttttatgcc agcagcttaa atgtggagtt gccctttcta tcccgggagg agcacctttt 17820
gggaaaggaa gtgagcaggt ctggaggcac atgtttcact gcactgggac tgagaagcac 17880
atgggagatt gttccgtcac tgctctgggc gcatcactct gttcttcagg gcaagtggcc 17940
tctgtaatct gctcaggtaa gagaataagg gcagccagtg atgagccact catgacggtg 18000
ccttaagagt gggtgtacct aggagttccc attgtggctc agtggtaaca aactcgactg 18060
gtatccatga gggtatgggt ttgatccctg gccttgctca atgggttaag gatccagcat 18120
tgctgtgagc tgtggtatag gttgcagact ctgctcaggt cccatgttgc tgtgattgtg 18180
gtgtaggctg actgctgcag cttcaatttg acccctagcc cgggaatttc cataggccac 18240
acgtgcagca ctaaggaagg aaaaaaagaa aaaaaaaaaa aaagagtggg tgtgcctata 18300
gtgaagaaca gatgtaaaag ggaagtgaaa gggattcccc cattctgagg gattgtgaga 18360
agtgtgccag aatattaact tcatttgact tgttacaggg aaagtaaact tgactttcac 18420
ggacctccta gttacctggt gcttactata tgtcttctca gagtacctga ttcattccca 18480
gcctggttga cccatccccc tatctctatg gctatgttta tccagagcac atctatctaa 18540
cactccagct gatcttcctg acacagctgt ggcaaccctg gatcctttaa ccaactgtgc 18600
caggctggag atcaaaccta agcctctgca gcaacccaag ctgctgcagt cagattttta 18660
accccctgtg ccactgtggg tatctccgat attttgtatc ttctgtgact gagtggtttg 18720
ctgtttgcag ggaaccagag tcagacacta tccccgtgca attcatcatc ctcggaccca 18780
tcaagctcta ttatttcaga agaaaatggt gttgcctgca taggtgagaa tcagtgacca 18840
acctatgaaa atgatctcaa tcctctgaaa tgcattttat tcatgtttta tttcctcttt 18900
gcagggagtg gtcaacttcg cctggtcgat ggaggtggtc gttgtgctgg gagagtagag 18960
gtctatcatg agggctcctg gggcaccatc tgtgatgaca gctgggacct gaatgatgcc 19020
catgtggtgt gcaaacagct gagctgtgga tgggccatta atgccactgg ttctgctcat 19080
tttggggaag gaacagggcc catttggctg gatgagataa actgtaatgg aaaagaatct 19140
catatttggc aatgccactc acatggttgg gggcggcaca attgcaggca taaggaggat 19200
gcaggagtca tctgctcggg taagttctgc acatcacttc gggttacagt gatttaagaa 19260
acaactaagg tggggcaaag ggtagtgagg catatccatc agagcaaatt ccttgaaata 19320
cggactcaga gggaaccatt gtgagattga ggttcccaga ggtgtggatt taatgaatta 19380
gtgttacctc atgtacaagg tagtatacta ccagaaagat aaaaattcag aagcgagttt 19440
gcagcaaaac tcatagggag aacttctttt ataaataata tgaagctgga tatttagtgc 19500
accacctgat gaccacttta ttaataaata aagagttcct gttgtggcgc agcggaaatg 19560
aatccgacaa ataatcatga gtttgcgggt ttgatccctg acctcgctca gtgggttggg 19620
gatctggtgt tgccatgagc tgtggtgtag gtcgcagatg ctgcttggat cctgctttgc 19680
tgtggctgtg gtataggctt gtggctacag ctccgatttg accgctagcc tgggaacctc 19740
catatgctgc gggggtggcc ctcaaaagaa aaataaataa ataagtaaat aaataagtag 19800
tttaaaaagg acaagaagaa atatatttgg tgttatattc tacagagaca aagataatca 19860
ccatgcccga ttgatttttc aaggcatata aatgagacgt catgggagca aaaatggtca 19920
taatacaatg cccttgtttt gtgtacatgg taagatttta gaaagcattg tgaggtaaaa 19980
aagtgtactc agttataata tattggggaa aacagtacta tgagaagtaa aaaaatctac 20040
atgccggaag ttattttttt aatgtctctt ttagagtcgc acatgcggca tatggaggtt 20100
cccaggctag gggtcgaatc agagctatag ccactggctt atggcacagc cacaacaaca 20160
ctagatctga gccacatcag cgacctatac tatagctcat ggcaatgcca gatccttaac 20220
ctactgagcc aagccatggg tcaaatccag gtcctcacgg atcctaggca aattcatttc 20280
tgctgagcca cgaagggaac tcctcagaag tgattttgat gttactttct tttcatgaca 20340
aatctggtaa agtacataca catagaaact gaagtgtcag aaagggaaat atttcatttt 20400
aaggtaatgt atacaaaaca gtggttttac catctgagta tctcgctaaa ttttaactat 20460
caaggacaat tgccaaaaaa aaaaaaaaaa gagagagaga gagaacagaa tagggttatg 20520
aagctaaaat cacagggtta tgaagctaaa atcacagtaa tttagggaga aaaaaatcca 20580
aagcatgtaa ttgataaaag gttctgagcc tttgtttgag atttagaatt caacttagaa 20640
ataccggtgg tattttaaag cagtccataa gtataaaatc caaggctaaa aaaccagaag 20700
gtatttgtag aacaaatata ttttaataag ctctaccaag tcatccagaa gctattaaag 20760
aattactggt cactgacata gtgtacctgt tttcaaggcc attcttacat cagaataaag 20820
ggagagcacc ctctgaatct tcagaaaaga tgtgaaagtg ctaattctct atttcatccc 20880
agagttcatg tctctgagac tgatcagtga aaacagcaga gagacctgtg cagggcgcct 20940
ggaagttttt tacaacggag cttggggcag cgttggcaag aatagcatgt ctccagccac 21000
agtgggggtg gtatgcaggc agctaggctg tgcagacaga ggggacatca gccctgcatc 21060
ttcagacaag acagtgtcca ggcacatgtg ggtggacaat gttcagtgtc ctaaaggacc 21120
tgacacacta tggcagtgcc catcatctcc atggaagaag agactggcca gcccctcaga 21180
ggagacatgg atcacatgtg ccagtgagta tccattcttt agcgccactg ttatcttctg 21240
atctacctaa gcagaagttt tataatctgt agttaatccc tattctacct ggatgatggg 21300
attcattctg tttaatttgg tgtgcaggta ttcagcatca gtgatcattt tcccaaagac 21360
catcatgctc tgatggtctt ctcaaaagtt ctaatcagtt gcttcctccg tgaacagttg 21420
aggagcagag aatatgtaat tcagaatttg actattgaat catcccattt ttctttcaca 21480
tagtcttttg ttgcactgaa tataaggaga gaagcagtca gaaagatcaa tcctgaatta 21540
tttctccatt ctacatctgt tttaaatttc aaaaaaaaaa attgttatag gtgatttaca 21600
atgtctgtca atttctgctc tacagcaaag tgacccagtt atttacatat acattctgtt 21660
tctcatattt ttaaaccagg agatttctat ctgcctggcg gtttgaggga atttaacatt 21720
atgcatttat gttaacttta ttcacctgat gttttctaag tcatactgag attcttatgc 21780
ccaggatgga atacacctgg tttgctggaa agacatgtgc tttcataaag acgaattttg 21840
gaaaaaatat aaaatttaaa aggcccatta aataagcaaa gttttaagag atttcaaaaa 21900
aaatttcatc tctctctttt cctctttgac ctcttgggca cgttcatctt ctcaaatatg 21960
atcttggtgt ttctgacttt tcagacaaaa taagacttca agaaggaaac actaattgtt 22020
ctggacgtgt ggagatctgg tacggaggtt cctggggcac tgtgtgtgac gactcctggg 22080
accttgaaga tgctcaggtg gtgtgccgac agctgggctg tggctcagct ttggaggcag 22140
gaaaagaggc cgcatttggc caggggactg ggcccatatg gctcaatgaa gtgaagtgca 22200
aggggaatga aacctccttg tgggattgtc ctgccagatc ctggggccac agtgactgtg 22260
gacacaagga ggatgctgct gtgacgtgtt caggtgaggg cagagagtct ggattgagct 22320
tggaagctct ggcagcaaag agagggtggg cggtgacctg cattgggtaa agattggaag 22380
gtccagccta aggatctggt ggtgggggga gacatgatgt ttcagtctga agaatgatga 22440
aaacctgtgt ggttacgcat gggccttcgc cgaggaaagg gacataacta ccatgtatcc 22500
tcctgcagag ggaggaagaa ctaggggatt ctagttttgt gtgggaagga gcagtttact 22560
tggctcagga ggcactaaag gctcagatag gaaacagaga tctgttccat tcttactccc 22620
agaactgatt ctcttctctt ttctcctaca gaaattgcaa agagccgaga atccctacat 22680
gccacaggta tatcaaaaag tttaagaaca tgggacccat tgtctgcatt ttgtggaatc 22740
cctcttatta agacattctg ggtcagaagt tctgaggatt tgacatttac ttcagctatc 22800
tgttatctta cccaagagag ggatggtaac taggaaccca ggtcttttag ctaagacatt 22860
atcacctctt gtgatgttta cttgttctca ggtcgctcat cttttgttgc acttgcaatc 22920
tttggggtca ttctgttggc ctgtctcatc gcattcctca tttggactca gaagcgaaga 22980
cagaggcagc ggctctcagg tctgaacaaa attacggtct ctctaatgtt tctatgggag 23040
aagaagcctc tctggataat aaaacaaaaa aattacattc aagtatcagt tggccagaaa 23100
gagggaacct agaagaggtt taagcagttt ctccgaaaca gggaacaaga attcagagaa 23160
gaaaaggcac attggctgta ctgatgatac ctgcactcgc tatgtatgtt taatggggga 23220
cagtagagaa ttgatagttt agaaggagta tgcttatatg gttctggatg aatcctgtat 23280
ccccccaaac atttattttc tcttactata tacttattac taatttaact cttctgtcaa 23340
gccatgtgct aggttctgaa gatggttcag acttggataa ccaagtgctt ttgttttcat 23400
ggaatttcca gtttagtgga agagataaat atgtaaacaa ataaatgggg gggggggggg 23460
gggggggggg gggggggggg gggggggggg gggggggggg gggggggggg gggggggggg 23520
gggggggggg gggggggggg gggggggggg gggggggggg gggggggggg gggggggggg 23580
gggggggggg gggggggggg gggggggggg gggggggggg gggggggggg gggggggggg 23640
gggggggggg gggggggggg gggggggggg gggggggggg gggggggggg gggggggggg 23700
gggggggggg gggggggggg gggggggggg gggggggggg gggggggggg gggggggggg 23760
gggggggggg gggggggggg gggggggggg gggggggggg gggggggggg gggggggggg 23820
gggggggggg gggggggggg gggggggggg gggggggggg gggggggggg gggggggggg 23880
gggggggggg gggggggggg gggggggggg gggggggggg gggggggggg gggggggggg 23940
gggggggggg gggggggggg gggggggggg gggggggggg gggggggggg gggggggggg 24000
gggggggggg gggggggggg gggggggggg gggggggggg ggggggggtt ggcgggcccc 24060
cctcgaggtc gacggtatcg ataagcttga tatcgaattc gtgagccaga ggacgagact 24120
agagatggat gatgactacg ttatgcttgc actgctgggg aaaagcacac atagggaggg 24180
aacgttttat tatgacccag tccctaacct atgacctctg ttatcagttt tctcaggagg 24240
agagaattct gtccatcaaa ttcaataccg ggagatgaat tcttgcctga aagcagatga 24300
aacggatatg ctaaatccct caggtccgtg ggttctttga ggggctgtag ccctggggtt 24360
cagatcagca gctgcagttg aggttgaggc atgctacttt gcatagcagt agaaagaaat 24420
ctcaactgta ataggaagct tgggatgcat atgaggaaga aaggcaagaa tgaactacaa 24480
attattctta gggaagataa aaattgcagt catggggaga cctctggctg agagggccgt 24540
gattatttct gacagaggga ttatggagta gaatatgatg gcttggacct tttttcacta 24600
aaacaagtca gtcttctcaa aggtagttta gcttttcata tatctttcac agtttcttcc 24660
attcccattt cctgccattt tcctttctct aacttttatt tattatattt tttcctaaaa 24720
gtttaaattt tctatatctt tatcccttca gaagccatcc ctagtcacag gactagtctc 24780
atttcccatt atgtaatgct tctttctctg tctgttgact tctatttaga accagtgcac 24840
taaatctgcc tttaggaaca tacctctgct aggttgcaag aaatatccca ttccccactc 24900
actctgtgaa gactcaatgc ttctcaatat tccttacctc ctgagaggga cttgcctcac 24960
ttctttaatc caagggactc gatttttgcc aaaactaagt caggaaaacc tacataagac 25020
ataggaaaga cttgctgtgc ttcttaaacc ccactgtttg ttttcctaat tgtgaacagt 25080
atttttaaag ttaacaagag agcttctaag gcacttgagg ggagatctga tttatttccc 25140
agtaattatt ttcttccttt cagaaaattc cactgaataa gatggtttta acggatgtgg 25200
gactaatttt tgtgtctaaa tctcttccta tttctggatg aaaaaaagga gaccactctg 25260
aagtacaatg aaaaggaaaa tgggaattat aacctggtga ggtgagtagg aagaatttat 25320
tcatcattgc tgaaaacagg tacattcctt ttgaaagttg agaactcctc tggtattaga 25380
aaaaaaaaaa gaacgtatat acacatatat ttccatgtct atgtttatgt ttgtaaatcc 25440
atattcagaa tatgcaacaa ctttttataa ctatgacttc agtccatctt ttagttacat 25500
atatattcta aacaacaact attgctaaga gaagctgggt aagtaaatgt gaataaatct 25560
tctaaagata ttacaggaag ttcctgctgc ggctcagtgg gttaaagact tgatgtcttt 25620
gtgaagatga gggctcgagc cctggcctca ctcagtgagt taaggatcta gcattgctgt 25680
aagctgcagc gtaggttgca gatagggctc agatccagtg ttgctgtggc tgtggcctca 25740
gttgcagctc tgattcaacc cttaggcgag gaacttccat atgcagcaaa tgtggccatt 25800
aaaaaaaaaa aaaacattat aggagtcatt tcataaaaga gataagacgt ttctatagtt 25860
atatagtgca tactctggta aagatagtat aggatactat aggaatatag aaagcttgcc 25920
tatgaaaatt tgggaagatt gtggaaaaga catctcaaaa tatggcatag aaaagaatca 25980
tatctttgag gaacagtaag tttttcattc aaaaccgtgt attgaacata cttgtggtga 26040
caagtggtgt cctgagtact aaaaattcag tgataaaaga tgctcttgac aaagacatgg 26100
ctgttgaata gaaggtctca ctgtcaatgt gtgggaatta tggacagcct atgtggacac 26160
agggaataga tgagactcta ggctggaagg ctgcattgag cccaataatg aatggtcctg 26220
tctgatatat ttcatgctca tattttattt tagggactat tggggaggtg gtgggttttg 26280
gaagattaag ctgaggcaag acacaatcag attgcctttt ataatttact ttcaggagga 26340
aagtctaact aaaaaaagaa ttcgatatca agcttatcga taccgtcgac ctcgaggagg 26400
cccgcctgcc cttttggggg gggggggggg gggggggggg gggggggggg gggggggggg 26460
gggggggggg gggggggggg gggggggggg gggggggggg gggggggggg gggggcagca 26520
ccaattttat tattggcggg aataaagaga aaaatgtaat ttcaaagatt gctgttggaa 26580
atgaggggtg tggtagcttt tggagaaagc attctggaga cttctattaa tttttttttt 26640
ttaagtgctt caaagatcct ttgatccaac aattctactc ctaaaaattt cttccataca 26700
gataaagcca tttgtctgta tataacaaat agaagagaat tcctttttgc agccttgtta 26760
gtagtgcccc caaactggaa acaaagtgaa tatcagtcag tggggtagcg gctggaaaaa 26820
ttttagtgca cccaaccaac aaagaaaaac catgcacaaa aattcaataa atatcatctc 26880
acttttgtgt tcatgttatt gaatataatt aaacataatg tttacatcta taaaattatc 26940
atatgtatac atgtaaagaa acattaaaac atttttaaca gactgtaaac ttgaggactg 27000
tgaatgactt ttgattgata atctcaaaca tatggatact attctgatgt aataaataat 27060
gattaaattt tttccctaaa gagtaatcac tactgaatcg ttgcctcaga atcatatgga 27120
ggtgctttta aaaaaggcat ttctgcactg ttgttctctg gaatagaagt aattcttatg 27180
tacactgaag tttgaaaatc attgcattta agtgttctgt tcaggaaagt agtgtgcttt 27240
ttaatatttg tgagtgaatg agtaacacaa tacattatat cacattttaa tgtaattcta 27300
cacatgtgca tatgaagaga aaagtaacat ttttttctat ttatgtcttt agttcagcct 27360
ttaagatacc ttgatgaaga cctggactat tgaatgagca agaatctgcc tcttacactg 27420
aagattacaa tacagtcctc tgtctcctgg tattccaaag actgctgttg aatttctaaa 27480
aaatagattg gtgaatgtga ctactcaaag ttgtatgtaa gactttcaag ggcattaaat 27540
aaaaaagaat attgctgatt cttgttcttg attttctgaa tttctgaatc tcttattggg 27600
cttctaattt aaaaaaaaat atctgggcgc ccgcagatat cgaactcttg ggcagtgtga 27660
ccaaacgaag acatatccaa tcaagcatgc aaatggacca gcccactgta ctagcacgct 27720
gtggcagcca atctgaccga gaaagcagac aaccgcaggg agcaacg 27767
<210> 16
<211> 55
<212> ДНК
<213> Sus scrofa
<400> 16
ggtcgccacc atggtgagca agggcgagga gctgttcacc ggggtggtgc ccatc 55
<210> 17
<211> 54
<212> ДНК
<213> Sus scrofa
<400> 17
ggtcgccacc atggccatga gcaagggcga ggagctgttc accggggtgg tgcc 54
<210> 18
<211> 48
<212> ДНК
<213> Sus scrofa
<400> 18
ggtcgccacc atggtgagca agggcgagga gctgttcacc ggggtggt 48
<210> 19
<211> 55
<212> ДНК
<213> Sus scrofa
<400> 19
ggtcgccacc atggttgagc aagggcgagg agctgttcac cggggtggtg cccat 55
<210> 20
<211> 43
<212> ДНК
<213> Sus scrofa
<400> 20
tgcagggaac tacagtgcgg cactgtggtt tccctcctgg ggg 43
<210> 21
<211> 60
<212> ДНК
<213> Sus scrofa
<400> 21
tgcagggaac tacagtgcgg cactgtaaac cactactact gtggtttccc tcctgggggg 60
<210> 22
<211> 41
<212> ДНК
<213> Sus scrofa
<400> 22
tgcagggaac tacagtgcgg ctgtggtttc cctcctgggg g 41
<210> 23
<211> 46
<212> ДНК
<213> Sus scrofa
<400> 23
tgcagggaac tacagtgcgg aactactgtg gtttccctcc tggggg 46
<210> 24
<211> 31
<212> ДНК
<213> Sus scrofa
<400> 24
gaaacccagg ctggttggag gggacattcc c 31
<210> 25
<211> 24
<212> ДНК
<213> Sus scrofa
<400> 25
gaaacccagg ctggggacat tccc 24
<210> 26
<211> 13
<212> ДНК
<213> Sus scrofa
<400> 26
aggggacatt ccc 13
<210> 27
<211> 13
<212> ДНК
<213> Sus scrofa
<400> 27
gaaacccatt ccc 13
<210> 28
<211> 31
<212> ДНК
<213> Sus scrofa
<400> 28
ggtcgccacc atggtgagca agggcgagga g 31
<210> 29
<211> 32
<212> ДНК
<213> Sus scrofa
<400> 29
ggtcgccacc atggctgagc aagggcgagg ag 32
<210> 30
<211> 29
<212> ДНК
<213> Sus scrofa
<400> 30
ggtcgccacc atggtgagag ggcgaggag 29
<210> 31
<211> 32
<212> ДНК
<213> Sus scrofa
<400> 31
ggtcgccacc atggttgagc aagggcgagg ag 32
<210> 32
<211> 48
<212> ДНК
<213> Sus scrofa
<400> 32
ggtcgccacc atggtgagca agggcgagga gaacccaggc tggttgga 48
<210> 33
<211> 49
<212> ДНК
<213> Sus scrofa
<400> 33
tgctgtgcag ggaactacag tgcggcactg tggtttccct cctgggggg 49
<210> 34
<211> 38
<212> ДНК
<213> Sus scrofa
<400> 34
tgctgtgcag ggaactctgt ggtttccctc ctgggggg 38
<210> 35
<211> 22
<212> ДНК
<213> Sus scrofa
<400> 35
ctgtggtttc cctcctgggg gg 22
<210> 36
<211> 23
<212> ДНК
<213> Sus scrofa
<400> 36
actgtggttt ccctcctggg ggg 23
<210> 37
<211> 50
<212> ДНК
<213> Sus scrofa
<400> 37
tgctgtgcag ggaactacag tgcggcaact gtggtttccc tcctgggggg 50
<210> 38
<211> 10
<212> ДНК
<213> Sus scrofa
<400> 38
tcctgggggg 10
<210> 39
<211> 8
<212> ДНК
<213> Sus scrofa
<400> 39
ctgggggg 8
<210> 40
<211> 52
<212> ДНК
<213> Sus scrofa
<400> 40
agagagcaga gccagcgact cgcccagcga catggggtac ctgccgtttg tg 52
<210> 41
<211> 33
<212> ДНК
<213> Sus scrofa
<400> 41
agagagcaga gccagcgact cgcccagcga gat 33
<210> 42
<211> 30
<212> ДНК
<213> Sus scrofa
<400> 42
agagagcaga gccagcgact cgcccagcga 30
<210> 43
<211> 50
<212> ДНК
<213> Sus scrofa
<400> 43
agagccagcc tcgcccagca ggggtaccat ggggtacctg ccgtttgtgt 50
<210> 44
<211> 53
<212> ДНК
<213> Sus scrofa
<400> 44
agagagcaga gccagcgact cgcccagcga gcagtgggta cctgccgttt gtg 53
<210> 45
<211> 53
<212> ДНК
<213> Sus scrofa
<400> 45
agagagcaga gccagcgact cgcccagcga tcagtgggta cctgccgttt gtg 53
<210> 46
<211> 53
<212> ДНК
<213> Sus scrofa
<400> 46
agagagcaga gccagcgact cgcccagcga acatggggta cctgccgttt gtg 53
<210> 47
<211> 4990
<212> ДНК
<213> Sus scrofa
<400> 47
tatagatgac aaggctttgt gtctgatagg ggccagcgaa ctcagtaaag agggaagatg 60
agaaagataa tggcaagaat ttatccctga agtgtagttt tgacaaacca gtcacaaaga 120
ggtctaagaa attttggtca caaagttgtt ttgaatccca ggcattttat ttgcaatgat 180
tgcatatgtt ctggaaagga catctgaacc taagaaatag ttcatttgca ttgtgttata 240
ttttactaag gtctgagaaa taatcttgag atgagaatga actctacttc ttcagagtct 300
ggaaggaata aattatgaaa atgtattaat gcttctttaa accatattgt atatttatct 360
attactaaac aaaaagaagt agctctattt atttatttat ttatttattt atttatgtct 420
tttgtctctt tagggccaca cctgtggcat atggaggttc ccaggctaga ggtccaattg 480
gagatgtagc agccagccta tgccagagcc accgcaacac gggatctgag ccacgtctgt 540
gacttacacc acagctcaca gcaacgcctg atcctcaacc cactgagcga ggccagggat 600
cgaacccatg tcctcatgga tgctagttgg gttcgttaac tgctgagcca tgatgggaac 660
tccaaattaa ttatttctta tatttgttct tcatatattc atttctatag aaagaaataa 720
atacagattc agttaatgat ggcaggtaaa agcttaactt attaatcaaa ggagttaatc 780
caggcacaaa aattcaattc atggctctct gttaaaattt aggtataggt ttagcaggaa 840
gaaaaggtta gtagatgcag actattacat ttagaatgga tggacaatga agtcctacta 900
tacagcacag ggaactatat ccaatctctt gggatagaat atgatggaag acaaaatcag 960
aacaagagag tatatatata tgtgtgtgtg tgtgtgtgtg tgtgtgtgtg tgtgtgtgtg 1020
tgtgtgactg ggtcaccctg cggcacagca gaaattggca gaacattgta aatcaactat 1080
actttaatag gaaaaatact tttaagggct aaatttccaa tattctaacc atgtacacag 1140
agtaaatgtc ataaggatgc cagtctgtgt agagattgat gtgttactag cagattcatg 1200
aaataaaggc tgaggatgta gtccccaagt cacttctgag tggaagaatt tctcctttgt 1260
cctggactca aatattttag gataaaggaa aaaagaagat atttatagaa gggacttgtt 1320
ttcaagtact tgacaaaatt tcaccattaa agagaaattt gtgggagttc ccatcgtggc 1380
tcagtggaaa caaatccaac taggaaccat gaggttgtgg gtttgatccc tggcctcact 1440
cagtgggtta aggatccggt gttgccgtga gctgtggtgt aggttgcaga cacggttctg 1500
atcctgcgtt gctgtggctg tggctgtggt gtaggccagc agcaaacagc tctgattaga 1560
cccctagcct ggaaacctcc atatgccaca ggtgcagccc taaaaagaca aaaaaagaga 1620
aaagacaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa gaacccccag 1680
aggtatttat ttgtttttgc cttttttcac tgactgttct ttgtttgttt gtttgagact 1740
gatctagaag actagagatt acaagaaata tggatttggc tcactctaag aaactgcttt 1800
cattccaagg tttgggtcta tccaaaagtg gaatagaatc atatgaatac tagtttatga 1860
gtatttagtg agaggaattt caagctcaaa taatgattca gcaagattaa attaaggagg 1920
gaattttcct tgtggctgag tgggttaagg acccaatgtt gtctctgtga ggatgtaggt 1980
tccatcctgg gctttgctca ttaggttaag gatctggcat tgctgcagct cagacccagt 2040
gctgccctgg ttgtggctta ggccaaagct gcagctccaa ttcaatctct ggcctgggaa 2100
cctccatgtg ctacaaggtg cggccttaaa aggaaaaaaa aaaaattaaa tcaaggactc 2160
aagagtcttt cattatttgt gttgtggaag ctatatttgt tttaaagtct tagttgtgtt 2220
tagaaagcaa gatgttcttc aactcaaatt tgggagggaa cttgtttcat acatttttaa 2280
tggataagtg gcaaaatttt catgctgagg tgatctatag tgttgtaatg cagaatatag 2340
tcagatcttg aacattttag gaagttggtg agggccaatt gtgtatctgt gccatgctga 2400
taagaatgtc aagggatcac aagaattcgt gttatttgac agcagtcatc tttaaaaggc 2460
atttgagaaa gtccaatttc aaatgcattt cctttcttta aaagataaat tgaagaaaat 2520
aagtctttat ttcccaagta aattgaattg cctctcagtc tgttaaaaga aactcttacc 2580
ttgatgattg cgctcttaac ctggcaaaga ttgtctttaa aatctgagct ccatgtcttc 2640
tgctttattt ctggtgtgcc tttgactcca gattacagta aatggaggac tgagtatagg 2700
gctaaaaagt agagagaatg gatgcatatt atctgtggtc tccaatgtga tgaatgaagt 2760
aggcaaatac tcaaaggaaa gagaaagcat gctccaagaa ttatgggttc cagaaggcaa 2820
agtcccagaa ttgtctccag ggaaggacag ggaggtctag aatcggctaa gcccactgta 2880
ggcagaaaaa ccaagaggca tgaatggctt ccctttctca cttttcactc tctggcttac 2940
tcctatcatg aaggaaaata ttggaatcat attctccctc accgaaatgc tatttttcag 3000
cccacaggaa acccaggctg gttggagggg acattccctg ctctggtcgt gttgaagtac 3060
aacatggaga cacgtggggc accgtctgtg attctgactt ctctctggag gcggccagcg 3120
tgctgtgcag ggaactacag tgcggcactg tggtttccct cctgggggga gctcactttg 3180
gagaaggaag tggacagatc tgggctgaag aattccagtg tgaggggcac gagtcccacc 3240
tttcactctg cccagtagca ccccgccctg acgggacatg tagccacagc agggacgtcg 3300
gcgtagtctg ctcaagtgag acccagggaa tgtgttcact ttgttcccat gccatgaaga 3360
gggtagggtt aggtagtcac agacatcttt ttaaagccct gtctccttcc aggatacaca 3420
caaatccgct tggtgaatgg caagacccca tgtgaaggaa gagtggagct caacattctt 3480
gggtcctggg ggtccctctg caactctcac tgggacatgg aagatgccca tgttttatgc 3540
cagcagctta aatgtggagt tgccctttct atcccgggag gagcaccttt tgggaaagga 3600
agtgagcagg tctggaggca catgtttcac tgcactggga ctgagaagca catgggagat 3660
tgttccgtca ctgctctggg cgcatcactc tgttcttcag ggcaagtggc ctctgtaatc 3720
tgctcaggta agagaataag ggcagccagt gatgagccac tcatgacggt gccttaagag 3780
tgggtgtacc taggagttcc cattgtggct cagtggtaac aaactcgact ggtatccatg 3840
agggtatggg tttgatccct ggccttgctc aatgggttaa ggatccagca ttgctgtgag 3900
ctgtggtata ggttgcagac tctgctcagg tcccatgttg ctgtgattgt ggtgtaggct 3960
gactgctgca gcttcaattt gacccctagc ccgggaattt ccataggcca cacgtgcagc 4020
actaaggaag gaaaaaaaga aaaaaaaaaa aaaagagtgg gtgtgcctat agtgaagaac 4080
agatgtaaaa gggaagtgaa agggattccc ccattctgag ggattgtgag aagtgtgcca 4140
gaatattaac ttcatttgac ttgttacagg gaaagtaaac ttgactttca cggacctcct 4200
agttacctgg tgcttactat atgtcttctc agagtacctg attcattccc agcctggttg 4260
acccatcccc ctatctctat ggctatgttt atccagagca catctatcta acactccagc 4320
tgatcttcct gacacagctg tggcaaccct ggatccttta accaactgtg ccaggctgga 4380
gatcaaacct aagcctctgc agcaacccaa gctgctgcag tcagattttt aaccccctgt 4440
gccactgtgg gtatctccga tattttgtat cttctgtgac tgagtggttt gctgtttgca 4500
gggaaccaga gtcagacact atccccgtgc aattcatcat cctcggaccc atcaagctct 4560
attatttcag aagaaaatgg tgttgcctgc ataggtgaga atcagtgacc aacctatgaa 4620
aatgatctca atcctctgaa atgcatttta ttcatgtttt atttcctctt tgcagggagt 4680
ggtcaacttc gcctggtcga tggaggtggt cgttgtgctg ggagagtaga ggtctatcat 4740
gagggctcct ggggcaccat ctgtgatgac agctgggacc tgaatgatgc ccatgtggtg 4800
tgcaaacagc tgagctgtgg atgggccatt aatgccactg gttctgctca ttttggggaa 4860
ggaacagggc ccatttggct ggatgagata aactgtaatg gaaaagaatc tcatatttgg 4920
caatgccact cacatggttg ggggcggcac aattgcaggc ataaggagga tgcaggagtc 4980
atctgctcgg 4990
<210> 48
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 48
caccggaaac ccaggctggt tgga 24
<210> 49
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 49
aaactccaac cagcctgggt ttcc 24
<210> 50
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 50
caccggaact acagtgcggc actg 24
<210> 51
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 51
aaaccagtgc cgcactgtag ttcc 24
<210> 52
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 52
caccgcagta gcaccccgcc ctgac 25
<210> 53
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 53
aaacgtcagg gcggggtgct actgc 25
<210> 54
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 54
caccgtgtag ccacagcagg gacgt 25
<210> 55
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 55
aaacacgtcc ctgctgtggc tacac 25
<210> 56
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 56
caccgccagc ctcgcccagc gacat 25
<210> 57
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 57
aaacatgtcg ctgggcgagg ctggc 25
<210> 58
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 58
caccgcagct gcagcatata tttaa 25
<210> 59
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 59
aaacttaaat atatgctgca gctgc 25
<210> 60
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 60
caccgctttc atttatctga actca 25
<210> 61
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 61
aaactgagtt cagataaatg aaagc 25
<210> 62
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 62
caccgttatc tgaactcagg gtccc 25
<210> 63
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 63
aaacgggacc ctgagttcag ataac 25
<210> 64
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 64
caccgctcct cgcccttgct cacca 25
<210> 65
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 65
aaactggtga gcaagggcga ggagc 25
<210> 66
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 66
caccggacca ggatgggcac caccc 25
<210> 67
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 67
aaacgggtgg tgcccatcct ggtcc 25
<210> 68
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 68
ttgttggaag gctcactgtc cttg 24
<210> 69
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 69
acaactaagg tggggcaaag 20
<210> 70
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 70
ttgttggaag gctcactgtc cttg 24
<210> 71
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 71
ggagctcaac attcttgggt cct 23
<210> 72
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 72
ggcaaaattt tcatgctgag gtg 23
<210> 73
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 73
gcacatcact tcgggttaca gtg 23
<210> 74
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 74
cccaagtatc ttcagttctg cag 23
<210> 75
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 75
tacaggtagg agagcctgtt ttg 23
<210> 76
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 76
cccaagtatc ttcagttctg cag 23
<210> 77
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 77
ctcaaaagga tgtaaaccct gga 23
<210> 78
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 78
tgttgatgtg gtttgtttgc cc 22
<210> 79
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 79
tacaggtagg agagcctgtt ttg 23
<210> 80
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 80
ggaggtctag aatcggctaa gcc 23
<210> 81
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 81
ggctacatgt cccgtcaggg 20
<210> 82
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 82
gcaggccact aggcagatga a 21
<210> 83
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 83
gagctgacac ccaagaagtt cct 23
<210> 84
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 84
ggctctagag cctctgctaa cc 22
<210> 85
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 85
ggacttgaag aagtcgtgct gc 22
<210> 86
<211> 44
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 86
taatacgact cactataggg agaatggact ataaggacca cgac 44
<210> 87
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 87
gcgagctcta ggaattctta c 21
<210> 88
<211> 40
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 88
ttaatacgac tcactatagg ctcctcgccc ttgctcacca 40
<210> 89
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 89
aaaagcaccg actcggtgcc 20
<210> 90
<211> 38
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 90
ttaatacgac tcactatagg aaacccaggc tggttgga 38
<210> 91
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 91
aaaagcaccg actcggtgcc 20
<210> 92
<211> 38
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 92
ttaatacgac tcactatagg aactacagtg cggcactg 38
<210> 93
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 93
aaaagcaccg actcggtgcc 20
<210> 94
<211> 40
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 94
ttaatacgac tcactatagg ccagcctcgc ccagcgacat 40
<210> 95
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 95
aaaagcaccg actcggtgcc 20
<210> 96
<211> 40
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 96
ttaatacgac tcactatagg cagctgcagc atatatttaa 40
<210> 97
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 97
aaaagcaccg actcggtgcc 20
<210> 98
<211> 3484
<212> ДНК
<213> Sus scrofa
<400> 98
tatagatgac aaggctttgt gtctgatagg ggccagcgaa ctcagtaaag agggaagatg 60
agaaagataa tggcaagaat ttatccctga agtgtagttt tgacaaacca gtcacaaaga 120
ggtctaagaa attttggtca caaagttgtt ttgaatccca ggcattttat ttgcaatgat 180
tgcatatgtt ctggaaagga catctgaacc taagaaatag ttcatttgca ttgtgttata 240
ttttactaag gtctgagaaa taatcttgag atgagaatga actctacttc ttcagagtct 300
ggaaggaata aattatgaaa atgtattaat gcttctttaa accatattgt atatttatct 360
attactaaac aaaaagaagt agctctattt atttatttat ttatttattt atttatgtct 420
tttgtctctt tagggccaca cctgtggcat atggaggttc ccaggctaga ggtccaattg 480
gagatgtagc agccagccta tgccagagcc accgcaacac gggatctgag ccacgtctgt 540
gacttacacc acagctcaca gcaacgcctg atcctcaacc cactgagcga ggccagggat 600
cgaacccatg tcctcatgga tgctagttgg gttcgttaac tgctgagcca tgatgggaac 660
tccaaattaa ttatttctta tatttgttct tcatatattc atttctatag aaagaaataa 720
atacagattc agttaatgat ggcaggtaaa agcttaactt attaatcaaa ggagttaatc 780
caggcacaaa aattcaattc atggctctct gttaaaattt aggtataggt ttagcaggaa 840
gaaaaggtta gtagatgcag actattacat ttagaatgga tggacaatga agtcctacta 900
tacagcacag ggaactatat ccaatctctt gggatagaat atgatggaag acaaaatcag 960
aacaagagag tatatatata tgtgtgtgtg tgtgtgtgtg tgtgtgtgtg tgtgtgtgtg 1020
tgtgtgactg ggtcaccctg cggcacagca gaaattggca gaacattgta aatcaactat 1080
actttaatag gaaaaatact tttaagggct aaatttccaa tattctaacc atgtacacag 1140
agtaaatgtc ataaggatgc cagtctgtgt agagattgat gtgttactag cagattcatg 1200
aaataaaggc tgaggatgta gtccccaagt cacttctgag tggaagaatt tctcctttgt 1260
cctggactca aatattttag gataaaggaa aaaagaagat atttatagaa gggacttgtt 1320
ttcaagtact tgacaaaatt tcaccattaa agagaaattt gtgggagttc ccatcgtggc 1380
tcagtggaaa caaatccaac taggaaccat gaggttgtgg gtttgatccc tggcctcact 1440
cagtgggtta aggatccggt gttgccgtga gctgtggtgt aggttgcaga cacggttctg 1500
atcctgcgtt gctgtggctg tggcacattc cctgctctgg tcgtgttgaa gtacaacatg 1560
gagacacgtg gggcaccgtc tgtgattctg acttctctct ggaggcggcc agcgtgctgt 1620
gcagggaact acagtgcggc actgtggttt ccctcctggg gggagctcac tttggagaag 1680
gaagtggaca gatctgggct gaagaattcc agtgtgaggg gcacgagtcc cacctttcac 1740
tctgcccagt agcaccccgc cctgacggga catgtagcca cagcagggac gtcggcgtag 1800
tctgctcaag tgagacccag ggaatgtgtt cactttgttc ccatgccatg aagagggtag 1860
ggttaggtag tcacagacat ctttttaaag ccctgtctcc ttccaggata cacacaaatc 1920
cgcttggtga atggcaagac cccatgtgaa ggaagagtgg agctcaacat tcttgggtcc 1980
tgggggtccc tctgcaactc tcactgggac atggaagatg cccatgtttt atgccagcag 2040
cttaaatgtg gagttgccct ttctatcccg ggaggagcac cttttgggaa aggaagtgag 2100
caggtctgga ggcacatgtt tcactgcact gggactgaga agcacatggg agattgttcc 2160
gtcactgctc tgggcgcatc actctgttct tcagggcaag tggcctctgt aatctgctca 2220
ggtaagagaa taagggcagc cagtgatgag ccactcatga cggtgcctta agagtgggtg 2280
tacctaggag ttcccattgt ggctcagtgg taacaaactc gactggtatc catgagggta 2340
tgggtttgat ccctggcctt gctcaatggg ttaaggatcc agcattgctg tgagctgtgg 2400
tataggttgc agactctgct caggtcccat gttgctgtga ttgtggtgta ggctgactgc 2460
tgcagcttca atttgacccc tagcccggga atttccatag gccacacgtg cagcactaag 2520
gaaggaaaaa aagaaaaaaa aaaaaaaaga gtgggtgtgc ctatagtgaa gaacagatgt 2580
aaaagggaag tgaaagggat tcccccattc tgagggattg tgagaagtgt gccagaatat 2640
taacttcatt tgacttgtta cagggaaagt aaacttgact ttcacggacc tcctagttac 2700
ctggtgctta ctatatgtct tctcagagta cctgattcat tcccagcctg gttgacccat 2760
ccccctatct ctatggctat gtttatccag agcacatcta tctaacactc cagctgatct 2820
tcctgacaca gctgtggcaa ccctggatcc tttaaccaac tgtgccaggc tggagatcaa 2880
acctaagcct ctgcagcaac ccaagctgct gcagtcagat ttttaacccc ctgtgccact 2940
gtgggtatct ccgatatttt gtatcttctg tgactgagtg gtttgctgtt tgcagggaac 3000
cagagtcaga cactatcccc gtgcaattca tcatcctcgg acccatcaag ctctattatt 3060
tcagaagaaa atggtgttgc ctgcataggt gagaatcagt gaccaaccta tgaaaatgat 3120
ctcaatcctc tgaaatgcat tttattcatg ttttatttcc tctttgcagg gagtggtcaa 3180
cttcgcctgg tcgatggagg tggtcgttgt gctgggagag tagaggtcta tcatgagggc 3240
tcctggggca ccatctgtga tgacagctgg gacctgaatg atgcccatgt ggtgtgcaaa 3300
cagctgagct gtggatgggc cattaatgcc actggttctg ctcattttgg ggaaggaaca 3360
gggcccattt ggctggatga gataaactgt aatggaaaag aatctcatat ttggcaatgc 3420
cactcacatg gttgggggcg gcacaattgc aggcataagg aggatgcagg agtcatctgc 3480
tcgg 3484
<210> 99
<211> 4997
<212> ДНК
<213> Sus scrofa
<400> 99
tatagatgac aaggctttgt gtctgatagg ggccagcgaa ctcagtaaag agggaagatg 60
agaaagataa tggcaagaat ttatccctga agtgtagttt tgacaaacca gtcacaaaga 120
ggtctaagaa attttggtca caaagttgtt ttgaatccca ggcattttat ttgcaatgat 180
tgcatatgtt ctggaaagga catctgaacc taagaaatag ttcatttgca ttgtgttata 240
ttttactaag gtctgagaaa taatcttgag atgagaatga actctacttc ttcagagtct 300
ggaaggaata aattatgaaa atgtattaat gcttctttaa accatattgt atatttatct 360
attactaaac aaaaagaagt agctctattt atttatttat ttatttattt atttatgtct 420
tttgtctctt tagggccaca cctgtggcat atggaggttc ccaggctaga ggtccaattg 480
gagatgtagc agccagccta tgccagagcc accgcaacac gggatctgag ccacgtctgt 540
gacttacacc acagctcaca gcaacgcctg atcctcaacc cactgagcga ggccagggat 600
cgaacccatg tcctcatgga tgctagttgg gttcgttaac tgctgagcca tgatgggaac 660
tccaaattaa ttatttctta tatttgttct tcatatattc atttctatag aaagaaataa 720
atacagattc agttaatgat ggcaggtaaa agcttaactt attaatcaaa ggagttaatc 780
caggcacaaa aattcaattc atggctctct gttaaaattt aggtataggt ttagcaggaa 840
gaaaaggtta gtagatgcag actattacat ttagaatgga tggacaatga agtcctacta 900
tacagcacag ggaactatat ccaatctctt gggatagaat atgatggaag acaaaatcag 960
aacaagagag tatatatata tgtgtgtgtg tgtgtgtgtg tgtgtgtgtg tgtgtgtgtg 1020
tgtgtgactg ggtcaccctg cggcacagca gaaattggca gaacattgta aatcaactat 1080
actttaatag gaaaaatact tttaagggct aaatttccaa tattctaacc atgtacacag 1140
agtaaatgtc ataaggatgc cagtctgtgt agagattgat gtgttactag cagattcatg 1200
aaataaaggc tgaggatgta gtccccaagt cacttctgag tggaagaatt tctcctttgt 1260
cctggactca aatattttag gataaaggaa aaaagaagat atttatagaa gggacttgtt 1320
ttcaagtact tgacaaaatt tcaccattaa agagaaattt gtgggagttc ccatcgtggc 1380
tcagtggaaa caaatccaac taggaaccat gaggttgtgg gtttgatccc tggcctcact 1440
cagtgggtta aggatccggt gttgccgtga gctgtggtgt aggttgcaga cacggttctg 1500
atcctgcgtt gctgtggctg tggctgtggt gtaggccagc agcaaacagc tctgattaga 1560
cccctagcct ggaaacctcc atatgccaca ggtgcagccc taaaaagaca aaaaaagaga 1620
aaagacaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa gaacccccag 1680
aggtatttat ttgtttttgc cttttttcac tgactgttct ttgtttgttt gtttgagact 1740
gatctagaag actagagatt acaagaaata tggatttggc tcactctaag aaactgcttt 1800
cattccaagg tttgggtcta tccaaaagtg gaatagaatc atatgaatac tagtttatga 1860
gtatttagtg agaggaattt caagctcaaa taatgattca gcaagattaa attaaggagg 1920
gaattttcct tgtggctgag tgggttaagg acccaatgtt gtctctgtga ggatgtaggt 1980
tccatcctgg gctttgctca ttaggttaag gatctggcat tgctgcagct cagacccagt 2040
gctgccctgg ttgtggctta ggccaaagct gcagctccaa ttcaatctct ggcctgggaa 2100
cctccatgtg ctacaaggtg cggccttaaa aggaaaaaaa aaaaattaaa tcaaggactc 2160
aagagtcttt cattatttgt gttgtggaag ctatatttgt tttaaagtct tagttgtgtt 2220
tagaaagcaa gatgttcttc aactcaaatt tgggagggaa cttgtttcat acatttttaa 2280
tggataagtg gcaaaatttt catgctgagg tgatctatag tgttgtaatg cagaatatag 2340
tcagatcttg aacattttag gaagttggtg agggccaatt gtgtatctgt gccatgctga 2400
taagaatgtc aagggatcac aagaattcgt gttatttgac agcagtcatc tttaaaaggc 2460
atttgagaaa gtccaatttc aaatgcattt cctttcttta aaagataaat tgaagaaaat 2520
aagtctttat ttcccaagta aattgaattg cctctcagtc tgttaaaaga aactcttacc 2580
ttgatgattg cgctcttaac ctggcaaaga ttgtctttaa aatctgagct ccatgtcttc 2640
tgctttattt ctggtgtgcc tttgactcca gattacagta aatggaggac tgagtatagg 2700
gctaaaaagt agagagaatg gatgcatatt atctgtggtc tccaatgtga tgaatgaagt 2760
aggcaaatac tcaaaggaaa gagaaagcat gctccaagaa ttatgggttc cagaaggcaa 2820
agtcccagaa ttgtctccag ggaaggacag ggaggtctag aatcggctaa gcccactgta 2880
ggcagaaaaa ccaagaggca tgaatggctt ccctttctca cttttcactc tctggcttac 2940
tcctatcatg aaggaaaata ttggaatcat attctccctc accgaaatgc tatttttcag 3000
cccacaggaa acccaggctg gttggagggg acattccctg ctctggtcgt gttgaagtac 3060
aacatggaga cacgtggggc accgtctgtg attctgactt ctctctggag gcggccagcg 3120
tgctgtgcag ggaactacag tgcggctact actactgtgg tttccctcct ggggggagct 3180
cactttggag aaggaagtgg acagatctgg gctgaagaat tccagtgtga ggggcacgag 3240
tcccaccttt cactctgccc agtagcaccc cgccctgacg ggacatgtag ccacagcagg 3300
gacgtcggcg tagtctgctc aagtgagacc cagggaatgt gttcactttg ttcccatgcc 3360
atgaagaggg tagggttagg tagtcacaga catcttttta aagccctgtc tccttccagg 3420
atacacacaa atccgcttgg tgaatggcaa gaccccatgt gaaggaagag tggagctcaa 3480
cattcttggg tcctgggggt ccctctgcaa ctctcactgg gacatggaag atgcccatgt 3540
tttatgccag cagcttaaat gtggagttgc cctttctatc ccgggaggag caccttttgg 3600
gaaaggaagt gagcaggtct ggaggcacat gtttcactgc actgggactg agaagcacat 3660
gggagattgt tccgtcactg ctctgggcgc atcactctgt tcttcagggc aagtggcctc 3720
tgtaatctgc tcaggtaaga gaataagggc agccagtgat gagccactca tgacggtgcc 3780
ttaagagtgg gtgtacctag gagttcccat tgtggctcag tggtaacaaa ctcgactggt 3840
atccatgagg gtatgggttt gatccctggc cttgctcaat gggttaagga tccagcattg 3900
ctgtgagctg tggtataggt tgcagactct gctcaggtcc catgttgctg tgattgtggt 3960
gtaggctgac tgctgcagct tcaatttgac ccctagcccg ggaatttcca taggccacac 4020
gtgcagcact aaggaaggaa aaaaagaaaa aaaaaaaaaa agagtgggtg tgcctatagt 4080
gaagaacaga tgtaaaaggg aagtgaaagg gattccccca ttctgaggga ttgtgagaag 4140
tgtgccagaa tattaacttc atttgacttg ttacagggaa agtaaacttg actttcacgg 4200
acctcctagt tacctggtgc ttactatatg tcttctcaga gtacctgatt cattcccagc 4260
ctggttgacc catcccccta tctctatggc tatgtttatc cagagcacat ctatctaaca 4320
ctccagctga tcttcctgac acagctgtgg caaccctgga tcctttaacc aactgtgcca 4380
ggctggagat caaacctaag cctctgcagc aacccaagct gctgcagtca gatttttaac 4440
cccctgtgcc actgtgggta tctccgatat tttgtatctt ctgtgactga gtggtttgct 4500
gtttgcaggg aaccagagtc agacactatc cccgtgcaat tcatcatcct cggacccatc 4560
aagctctatt atttcagaag aaaatggtgt tgcctgcata ggtgagaatc agtgaccaac 4620
ctatgaaaat gatctcaatc ctctgaaatg cattttattc atgttttatt tcctctttgc 4680
agggagtggt caacttcgcc tggtcgatgg aggtggtcgt tgtgctggga gagtagaggt 4740
ctatcatgag ggctcctggg gcaccatctg tgatgacagc tgggacctga atgatgccca 4800
tgtggtgtgc aaacagctga gctgtggatg ggccattaat gccactggtt ctgctcattt 4860
tggggaagga acagggccca tttggctgga tgagataaac tgtaatggaa aagaatctca 4920
tatttggcaa tgccactcac atggttgggg gcggcacaat tgcaggcata aggaggatgc 4980
aggagtcatc tgctcgg 4997
<210> 100
<211> 3710
<212> ДНК
<213> Sus scrofa
<400> 100
tatagatgac aaggctttgt gtctgatagg ggccagcgaa ctcagtaaag agggaagatg 60
agaaagataa tggcaagaat ttatccctga agtgtagttt tgacaaacca gtcacaaaga 120
ggtctaagaa attttggtca caaagttgtt ttgaatccca ggcattttat ttgcaatgat 180
tgcatatgtt ctggaaagga catctgaacc taagaaatag ttcatttgca ttgtgttata 240
ttttactaag gtctgagaaa taatcttgag atgagaatga actctacttc ttcagagtct 300
ggaaggaata aattatgaaa atgtattaat gcttctttaa accatattgt atatttatct 360
attactaaac aaaaagaagt agctctattt atttatttat ttatttattt atttatgtct 420
tttgtctctt tagggccaca cctgtggcat atggaggttc ccaggctaga ggtccaattg 480
gagatgtagc agccagccta tgccagagcc accgcaacac gggatctgag ccacgtctgt 540
gacttacacc acagctcaca gcaacgcctg atcctcaacc cactgagcga ggccagggat 600
cgaacccatg tcctcatgga tgctagttgg gttcgttaac tgctgagcca tgatgggaac 660
tccaaattaa ttatttctta tatttgttct tcatatattc atttctatag aaagaaataa 720
atacagattc agttaatgat ggcaggtaaa agcttaactt attaatcaaa ggagttaatc 780
caggcacaaa aattcaattc atggctctct gttaaaattt aggtataggt ttagcaggaa 840
gaaaaggtta gtagatgcag actattacat ttagaatgga tggacaatga agtcctacta 900
tacagcacag ggaactatat ccaatctctt gggatagaat atgatggaag acaaaatcag 960
aacaagagag tatatatata tgtgtgtgtg tgtgtgtgtg tgtgtgtgtg tgtgtgtgtg 1020
tgtgtgactg ggtcaccctg cggcacagca gaaattggca gaacattgta aatcaactat 1080
actttaatag gaaaaatact tttaagggct aaatttccaa tattctaacc atgtacacag 1140
agtaaatgtc ataaggatgc cagtctgtgt agagattgat gtgttactag cagattcatg 1200
aaataaaggc tgaggatgta gtccccaagt cacttctgag tggaagaatt tctcctttgt 1260
cctggactca aatattttag gataaaggaa aaaagaagat atttatagaa gggacttgtt 1320
ttcaagtact tgacaaaatt tcaccattaa agagaaattt gtgggagttc ccatcgtggc 1380
tcagtggaaa caaatccaac taggaaccat gaggttgtgg gtttgatccc tggcctcact 1440
cagtgggtta aggatccggt gttgccgtga gctgtggtgt aggttgcaga cacggttctg 1500
atcctgcgtt gctgtggctg tggctgtggt gtaggccagc agcaaacagc tctgattaga 1560
cccctagcct ggaaacctcc atatgccaca ggtgcagccc taaaaagaca aaaaaagaga 1620
aaagacaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa gaacccccag 1680
aggtatttat ttgtttttgc cttttttcac tgactgttct ttgtttgttt gtttgagact 1740
gatctagaag actagagatt acaagaaata tggatttggc tcactctaag aaactgcttt 1800
cattccaagg tttgggtcta tccaaaagtg gaatagaatc atatgaatac tagtttatga 1860
gtatttagtg agaggaattt caagctcaaa taatgattca gcaagattaa attaaggagg 1920
gaattttcct tgtggctgag tgggttaagg acccaatgtt gtctctgtga ggatgtaggt 1980
tccatcctgg gctttgctca ttaggttaag gatctggcat tgctgcagct cagacccagt 2040
gctgccctgg ttgtggctta ggccaaagct gcagctccaa ttcaatctct ggcctgggaa 2100
cctccatgtg ctacaaggtg cggccttaaa aggaaaaaaa aaaaattaaa tcaaggactc 2160
aagagtcttt cattatttgt gttgtggaag ctatatttgt tttaaagtct tagttgtgtt 2220
tagaaagcaa gatgttcttc aactcaaatt tgggagggaa cttgtttcat acatttttaa 2280
tggataagtg gcaaaatttt catgctgagg tgatctatag tgttgtaatg cagaatatag 2340
tcagatcttg aacattttag gaagttggtg agggccaatt gtgtatctgt gccatgctga 2400
taagaatgtc aagggatcac aagaattcgt gttatttgac agcagtcatc tttaaaaggc 2460
atttgagaaa gtccaatttc aaatgcattt cctttcttta aaagataaat tgaagaaaat 2520
aagtctttat ttcccaagta aattgaattg cctctcagtc tgttaaaaga aactcttacc 2580
ttgatgattg cgctcttaac ctggcaaaga ttgtctttaa aatctgagct ccatgtcttc 2640
tgctttattt ctggtgtgcc tttgactcca gattacagta aatggaggac tgagtatagg 2700
gctaaaaagt agagagaatg gatgcatatt atctgtggtc tccaatgtga tgaatgaagt 2760
aggcaaatac tcaaaggaaa gagaaagcat gctccaagaa ttatgggttc cagaagggaa 2820
agggattccc ccattctgag ggattgtgag aagtgtgcca gaatattaac ttcatttgac 2880
ttgttacagg gaaagtaaac ttgactttca cggacctcct agttacctgg tgcttactat 2940
atgtcttctc agagtacctg attcattccc agcctggttg acccatcccc ctatctctat 3000
ggctatgttt atccagagca catctatcta acactccagc tgatcttcct gacacagctg 3060
tggcaaccct ggatccttta accaactgtg ccaggctgga gatcaaacct aagcctctgc 3120
agcaacccaa gctgctgcag tcagattttt aaccccctgt gccactgtgg gtatctccga 3180
tattttgtat cttctgtgac tgagtggttt gctgtttgca gggaaccaga gtcagacact 3240
atccccgtgc aattcatcat cctcggaccc atcaagctct attatttcag aagaaaatgg 3300
tgttgcctgc ataggtgaga atcagtgacc aacctatgaa aatgatctca atcctctgaa 3360
atgcatttta ttcatgtttt atttcctctt tgcagggagt ggtcaacttc gcctggtcga 3420
tggaggtggt cgttgtgctg ggagagtaga ggtctatcat gagggctcct ggggcaccat 3480
ctgtgatgac agctgggacc tgaatgatgc ccatgtggtg tgcaaacagc tgagctgtgg 3540
atgggccatt aatgccactg gttctgctca ttttggggaa ggaacagggc ccatttggct 3600
ggatgagata aactgtaatg gaaaagaatc tcatatttgg caatgccact cacatggttg 3660
ggggcggcac aattgcaggc ataaggagga tgcaggagtc atctgctcgg 3710
<210> 101
<211> 3617
<212> ДНК
<213> Sus scrofa
<400> 101
tatagatgac aaggctttgt gtctgatagg ggccagcgaa ctcagtaaag agggaagatg 60
agaaagataa tggcaagaat ttatccctga agtgtagttt tgacaaacca gtcacaaaga 120
ggtctaagaa attttggtca caaagttgtt ttgaatccca ggcattttat ttgcaatgat 180
tgcatatgtt ctggaaagga catctgaacc taagaaatag ttcatttgca ttgtgttata 240
ttttactaag gtctgagaaa taatcttgag atgagaatga actctacttc ttcagagtct 300
ggaaggaata aattatgaaa atgtattaat gcttctttaa accatattgt atatttatct 360
attactaaac aaaaagaagt agctctattt atttatttat ttatttattt atttatgtct 420
tttgtctctt tagggccaca cctgtggcat atggaggttc ccaggctaga ggtccaattg 480
gagatgtagc agccagccta tgccagagcc accgcaacac gggatctgag ccacgtctgt 540
gacttacacc acagctcaca gcaacgcctg atcctcaacc cactgagcga ggccagggat 600
cgaacccatg tcctcatgga tgctagttgg gttcgttaac tgctgagcca tgatgggaac 660
tccaaattaa ttatttctta tatttgttct tcatatattc atttctatag aaagaaataa 720
atacagattc agttaatgat ggcaggtaaa agcttaactt attaatcaaa ggagttaatc 780
caggcacaaa aattcaattc atggctctct gttaaaattt aggtataggt ttagcaggaa 840
gaaaaggtta gtagatgcag actattacat ttagaatgga tggacaatga agtcctacta 900
tacagcacag ggaactatat ccaatctctt gggatagaat atgatggaag acaaaatcag 960
aacaagagag tatatatata tgtgtgtgtg tgtgtgtgtg tgtgtgtgtg tgtgtgtgtg 1020
tgtgtgactg ggtcaccctg cggcacagca gaaattggca gaacattgta aatcaactat 1080
actttaatag gaaaaatact tttaagggct aaatttccaa tattctaacc atgtacacag 1140
agtaaatgtc ataaggatgc cagtctgtgt agagattgat gtgttactag cagattcatg 1200
aaataaaggc tgaggatgta gtccccaagt cacttctgag tggaagaatt tctcctttgt 1260
cctggactca aatattttag gataaaggaa aaaagaagat atttatagaa gggacttgtt 1320
ttcaagtact tgacaaaatt tcaccattaa agagaaattt gtgggagttc ccatcgtggc 1380
tcagtggaaa caaatccaac taggaaccat gaggttgtgg gtttgatccc tggcctcact 1440
cagtgggtta aggatccggt gttgccgtga gctgtggtgt aggttgcaga cacggttctg 1500
atcctgcgtt gctgtggctg tggctgtggt gtaggccagc agcaaacagc tctgattaga 1560
cccctagcct ggaaacctcc atatgccaca ggtgcagccc taaaaagaca aaaaaagaga 1620
aaagacaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa gaacccccag 1680
aggtatttat ttgtttttgc cttttttcac tgactgttct ttgtttgttt gtttgagact 1740
gatctagaag actagagatt acaagaaata tggatttggc tcactctaag aaactgcttt 1800
cattccaagg tttgggtcta tccaaaagtg gaatagaatc atatgaatac tagtttatga 1860
gtatttagtg agaggaattt caagctcaaa taatgattca gcaagattaa attaaggagg 1920
gaattttcct tgtggctgag tgggttaagg acccaatgtt gtctctgtga ggatgtaggt 1980
tccatcctgg gctttgctca ttaggttaag gatctggcat tgctgcagct cagacccagt 2040
gctgccctgg ttgtggctta ggccaaagct gcagctccaa ttcaatctct ggcctgggaa 2100
cctccatgtg ctacaaggtg cggccttaaa aggaaaaaaa aaaaattaaa tcaaggactc 2160
aagagtcttt cattatttgt gttgtggaag ctatatttgt tttaaagtct tagttgtgtt 2220
tagaaagcaa gatgttcttc aactcaaatt tgggagggaa cttgtttcat acatttttaa 2280
tggataagtg gcaaaatttt catgctgagg tgatctatag tgttgtaatg cagaatatag 2340
tcagatcttg aacattttag gaagttggtg agggccaatt gtgtatctgt gccatgctga 2400
taagaatgtc aagggatcac aagaattcgt gttatttgac agcagtcatc tttaaaaggc 2460
atttgagaaa gtccaatttc aaatgcattt cctttcttta aaagataaat tgaagaaaat 2520
aagtctttat ttcccaagta aattgaattg cctctcagtc tgttaaaaga aactcttacc 2580
ttgatgattg cgctcttaac ctggcaaaga ttgtctttaa aatctgagct ccatgtcttc 2640
tgctttattt ctggtgtgcc tttgactcca gattacagta aatggaggac tgagtatagg 2700
gctaaaaagt agagagaatg gattgaaagg gattccccca ttctgaggga ttgtgagaag 2760
tgtgccagaa tattaacttc atttgacttg ttacagggaa agtaaacttg actttcacgg 2820
acctcctagt tacctggtgc ttactatatg tcttctcaga gtacctgatt cattcccagc 2880
ctggttgacc catcccccta tctctatggc tatgtttatc cagagcacat ctatctaaca 2940
ctccagctga tcttcctgac acagctgtgg caaccctgga tcctttaacc aactgtgcca 3000
ggctggagat caaacctaag cctctgcagc aacccaagct gctgcagtca gatttttaac 3060
cccctgtgcc actgtgggta tctccgatat tttgtatctt ctgtgactga gtggtttgct 3120
gtttgcaggg aaccagagtc agacactatc cccgtgcaat tcatcatcct cggacccatc 3180
aagctctatt atttcagaag aaaatggtgt tgcctgcata ggtgagaatc agtgaccaac 3240
ctatgaaaat gatctcaatc ctctgaaatg cattttattc atgttttatt tcctctttgc 3300
agggagtggt caacttcgcc tggtcgatgg aggtggtcgt tgtgctggga gagtagaggt 3360
ctatcatgag ggctcctggg gcaccatctg tgatgacagc tgggacctga atgatgccca 3420
tgtggtgtgc aaacagctga gctgtggatg ggccattaat gccactggtt ctgctcattt 3480
tggggaagga acagggccca tttggctgga tgagataaac tgtaatggaa aagaatctca 3540
tatttggcaa tgccactcac atggttgggg gcggcacaat tgcaggcata aggaggatgc 3600
aggagtcatc tgctcgg 3617
<210> 102
<211> 4979
<212> ДНК
<213> Sus scrofa
<400> 102
tatagatgac aaggctttgt gtctgatagg ggccagcgaa ctcagtaaag agggaagatg 60
agaaagataa tggcaagaat ttatccctga agtgtagttt tgacaaacca gtcacaaaga 120
ggtctaagaa attttggtca caaagttgtt ttgaatccca ggcattttat ttgcaatgat 180
tgcatatgtt ctggaaagga catctgaacc taagaaatag ttcatttgca ttgtgttata 240
ttttactaag gtctgagaaa taatcttgag atgagaatga actctacttc ttcagagtct 300
ggaaggaata aattatgaaa atgtattaat gcttctttaa accatattgt atatttatct 360
attactaaac aaaaagaagt agctctattt atttatttat ttatttattt atttatgtct 420
tttgtctctt tagggccaca cctgtggcat atggaggttc ccaggctaga ggtccaattg 480
gagatgtagc agccagccta tgccagagcc accgcaacac gggatctgag ccacgtctgt 540
gacttacacc acagctcaca gcaacgcctg atcctcaacc cactgagcga ggccagggat 600
cgaacccatg tcctcatgga tgctagttgg gttcgttaac tgctgagcca tgatgggaac 660
tccaaattaa ttatttctta tatttgttct tcatatattc atttctatag aaagaaataa 720
atacagattc agttaatgat ggcaggtaaa agcttaactt attaatcaaa ggagttaatc 780
caggcacaaa aattcaattc atggctctct gttaaaattt aggtataggt ttagcaggaa 840
gaaaaggtta gtagatgcag actattacat ttagaatgga tggacaatga agtcctacta 900
tacagcacag ggaactatat ccaatctctt gggatagaat atgatggaag acaaaatcag 960
aacaagagag tatatatata tgtgtgtgtg tgtgtgtgtg tgtgtgtgtg tgtgtgtgtg 1020
tgtgtgactg ggtcaccctg cggcacagca gaaattggca gaacattgta aatcaactat 1080
actttaatag gaaaaatact tttaagggct aaatttccaa tattctaacc atgtacacag 1140
agtaaatgtc ataaggatgc cagtctgtgt agagattgat gtgttactag cagattcatg 1200
aaataaaggc tgaggatgta gtccccaagt cacttctgag tggaagaatt tctcctttgt 1260
cctggactca aatattttag gataaaggaa aaaagaagat atttatagaa gggacttgtt 1320
ttcaagtact tgacaaaatt tcaccattaa agagaaattt gtgggagttc ccatcgtggc 1380
tcagtggaaa caaatccaac taggaaccat gaggttgtgg gtttgatccc tggcctcact 1440
cagtgggtta aggatccggt gttgccgtga gctgtggtgt aggttgcaga cacggttctg 1500
atcctgcgtt gctgtggctg tggctgtggt gtaggccagc agcaaacagc tctgattaga 1560
cccctagcct ggaaacctcc atatgccaca ggtgcagccc taaaaagaca aaaaaagaga 1620
aaagacaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa gaacccccag 1680
aggtatttat ttgtttttgc cttttttcac tgactgttct ttgtttgttt gtttgagact 1740
gatctagaag actagagatt acaagaaata tggatttggc tcactctaag aaactgcttt 1800
cattccaagg tttgggtcta tccaaaagtg gaatagaatc atatgaatac tagtttatga 1860
gtatttagtg agaggaattt caagctcaaa taatgattca gcaagattaa attaaggagg 1920
gaattttcct tgtggctgag tgggttaagg acccaatgtt gtctctgtga ggatgtaggt 1980
tccatcctgg gctttgctca ttaggttaag gatctggcat tgctgcagct cagacccagt 2040
gctgccctgg ttgtggctta ggccaaagct gcagctccaa ttcaatctct ggcctgggaa 2100
cctccatgtg ctacaaggtg cggccttaaa aggaaaaaaa aaaaattaaa tcaaggactc 2160
aagagtcttt cattatttgt gttgtggaag ctatatttgt tttaaagtct tagttgtgtt 2220
tagaaagcaa gatgttcttc aactcaaatt tgggagggaa cttgtttcat acatttttaa 2280
tggataagtg gcaaaatttt catgctgagg tgatctatag tgttgtaatg cagaatatag 2340
tcagatcttg aacattttag gaagttggtg agggccaatt gtgtatctgt gccatgctga 2400
taagaatgtc aagggatcac aagaattcgt gttatttgac agcagtcatc tttaaaaggc 2460
atttgagaaa gtccaatttc aaatgcattt cctttcttta aaagataaat tgaagaaaat 2520
aagtctttat ttcccaagta aattgaattg cctctcagtc tgttaaaaga aactcttacc 2580
ttgatgattg cgctcttaac ctggcaaaga ttgtctttaa aatctgagct ccatgtcttc 2640
tgctttattt ctggtgtgcc tttgactcca gattacagta aatggaggac tgagtatagg 2700
gctaaaaagt agagagaatg gatgcatatt atctgtggtc tccaatgtga tgaatgaagt 2760
aggcaaatac tcaaaggaaa gagaaagcat gctccaagaa ttatgggttc cagaaggcaa 2820
agtcccagaa ttgtctccag ggaaggacag ggaggtctag aatcggctaa gcccactgta 2880
ggcagaaaaa ccaagaggca tgaatggctt ccctttctca cttttcactc tctggcttac 2940
tcctatcatg aaggaaaata ttggaatcat attctccctc accgaaatgc tatttttcag 3000
cccacaggaa acccaggctg gttggagggg acattccctg ctctggtcgt gttgaagtac 3060
aacatggaga cacgtggggc accgtctgtg attctgactt ctctctggag gcggccagcg 3120
tgctgtgcag ggaactctgt ggtttccctc ctggggggag ctcactttgg agaaggaagt 3180
ggacagatct gggctgaaga attccagtgt gaggggcacg agtcccacct ttcactctgc 3240
ccagtagcac cccgccctga cgggacatgt agccacagca gggacgtcgg cgtagtctgc 3300
tcaagtgaga cccagggaat gtgttcactt tgttcccatg ccatgaagag ggtagggtta 3360
ggtagtcaca gacatctttt taaagccctg tctccttcca ggatacacac aaatccgctt 3420
ggtgaatggc aagaccccat gtgaaggaag agtggagctc aacattcttg ggtcctgggg 3480
gtccctctgc aactctcact gggacatgga agatgcccat gttttatgcc agcagcttaa 3540
atgtggagtt gccctttcta tcccgggagg agcacctttt gggaaaggaa gtgagcaggt 3600
ctggaggcac atgtttcact gcactgggac tgagaagcac atgggagatt gttccgtcac 3660
tgctctgggc gcatcactct gttcttcagg gcaagtggcc tctgtaatct gctcaggtaa 3720
gagaataagg gcagccagtg atgagccact catgacggtg ccttaagagt gggtgtacct 3780
aggagttccc attgtggctc agtggtaaca aactcgactg gtatccatga gggtatgggt 3840
ttgatccctg gccttgctca atgggttaag gatccagcat tgctgtgagc tgtggtatag 3900
gttgcagact ctgctcaggt cccatgttgc tgtgattgtg gtgtaggctg actgctgcag 3960
cttcaatttg acccctagcc cgggaatttc cataggccac acgtgcagca ctaaggaagg 4020
aaaaaaagaa aaaaaaaaaa aaagagtggg tgtgcctata gtgaagaaca gatgtaaaag 4080
ggaagtgaaa gggattcccc cattctgagg gattgtgaga agtgtgccag aatattaact 4140
tcatttgact tgttacaggg aaagtaaact tgactttcac ggacctccta gttacctggt 4200
gcttactata tgtcttctca gagtacctga ttcattccca gcctggttga cccatccccc 4260
tatctctatg gctatgttta tccagagcac atctatctaa cactccagct gatcttcctg 4320
acacagctgt ggcaaccctg gatcctttaa ccaactgtgc caggctggag atcaaaccta 4380
agcctctgca gcaacccaag ctgctgcagt cagattttta accccctgtg ccactgtggg 4440
tatctccgat attttgtatc ttctgtgact gagtggtttg ctgtttgcag ggaaccagag 4500
tcagacacta tccccgtgca attcatcatc ctcggaccca tcaagctcta ttatttcaga 4560
agaaaatggt gttgcctgca taggtgagaa tcagtgacca acctatgaaa atgatctcaa 4620
tcctctgaaa tgcattttat tcatgtttta tttcctcttt gcagggagtg gtcaacttcg 4680
cctggtcgat ggaggtggtc gttgtgctgg gagagtagag gtctatcatg agggctcctg 4740
gggcaccatc tgtgatgaca gctgggacct gaatgatgcc catgtggtgt gcaaacagct 4800
gagctgtgga tgggccatta atgccactgg ttctgctcat tttggggaag gaacagggcc 4860
catttggctg gatgagataa actgtaatgg aaaagaatct catatttggc aatgccactc 4920
acatggttgg gggcggcaca attgcaggca taaggaggat gcaggagtca tctgctcgg 4979
<210> 103
<211> 4615
<212> ДНК
<213> Sus scrofa
<400> 103
tatagatgac aaggctttgt gtctgatagg ggccagcgaa ctcagtaaag agggaagatg 60
agaaagataa tggcaagaat ttatccctga agtgtagttt tgacaaacca gtcacaaaga 120
ggtctaagaa attttggtca caaagttgtt ttgaatccca ggcattttat ttgcaatgat 180
tgcatatgtt ctggaaagga catctgaacc taagaaatag ttcatttgca ttgtgttata 240
ttttactaag gtctgagaaa taatcttgag atgagaatga actctacttc ttcagagtct 300
ggaaggaata aattatgaaa atgtattaat gcttctttaa accatattgt atatttatct 360
attactaaac aaaaagaagt agctctattt atttatttat ttatttattt atttatgtct 420
tttgtctctt tagggccaca cctgtggcat atggaggttc ccaggctaga ggtccaattg 480
gagatgtagc agccagccta tgccagagcc accgcaacac gggatctgag ccacgtctgt 540
gacttacacc acagctcaca gcaacgcctg atcctcaacc cactgagcga ggccagggat 600
cgaacccatg tcctcatgga tgctagttgg gttcgttaac tgctgagcca tgatgggaac 660
tccaaattaa ttatttctta tatttgttct tcatatattc atttctatag aaagaaataa 720
atacagattc agttaatgat ggcaggtaaa agcttaactt attaatcaaa ggagttaatc 780
caggcacaaa aattcaattc atggctctct gttaaaattt aggtataggt ttagcaggaa 840
gaaaaggtta gtagatgcag actattacat ttagaatgga tggacaatga agtcctacta 900
tacagcacag ggaactatat ccaatctctt gggatagaat atgatggaag acaaaatcag 960
aacaagagag tatatatata tgtgtgtgtg tgtgtgtgtg tgtgtgtgtg tgtgtgtgtg 1020
tgtgtgactg ggtcaccctg cggcacagca gaaattggca gaacattgta aatcaactat 1080
actttaatag gaaaaatact tttaagggct aaatttccaa tattctaacc atgtacacag 1140
agtaaatgtc ataaggatgc cagtctgtgt agagattgat gtgttactag cagattcatg 1200
aaataaaggc tgaggatgta gtccccaagt cacttctgag tggaagaatt tctcctttgt 1260
cctggactca aatattttag gataaaggaa aaaagaagat atttatagaa gggacttgtt 1320
ttcaagtact tgacaaaatt tcaccattaa agagaaattt gtgggagttc ccatcgtggc 1380
tcagtggaaa caaatccaac taggaaccat gaggttgtgg gtttgatccc tggcctcact 1440
cagtgggtta aggatccggt gttgccgtga gctgtggtgt aggttgcaga cacggttctg 1500
atcctgcgtt gctgtggctg tggctgtggt gtaggccagc agcaaacagc tctgattaga 1560
cccctagcct ggaaacctcc atatgccaca ggtgcagccc taaaaagaca aaaaaagaga 1620
aaagacaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa gaacccccag 1680
aggtatttat ttgtttttgc cttttttcac tgactgttct ttgtttgttt gtttgagact 1740
gatctagaag actagagatt acaagaaata tggatttggc tcactctaag aaactgcttt 1800
cattccaagg tttgggtcta tccaaaagtg gaatagaatc atatgaatac tagtttatga 1860
gtatttagtg agaggaattt caagctcaaa taatgattca gcaagattaa attaaggagg 1920
gaattttcct tgtggctgag tgggttaagg acccaatgtt gtctctgtga ggatgtaggt 1980
tccatcctgg gctttgctca ttaggttaag gatctggcat tgctgcagct cagacccagt 2040
gctgccctgg ttgtggctta ggccaaagct gcagctccaa ttcaatctct ggcctgggaa 2100
cctccatgtg ctacaaggtg cggccttaaa aggaaaaaaa aaaaattaaa tcaaggactc 2160
aagagtcttt cattatttgt gttgtggaag ctatatttgt tttaaagtct tagttgtgtt 2220
tagaaagcaa gatgttcttc aactcaaatt tgggagggaa cttgtttcat acatttttaa 2280
tggataagtg gcaaaatttt catgctgagg tgatctatag tgttgtaatg cagaatatag 2340
tcagatcttg aacattttag gaagttggtg agggccaatt gtgtatctgt gccatgctga 2400
taagaatgtc aagggatcac aagaattcgt gttatttgac agcagtcatc tttaaaaggc 2460
atttgagaaa gtccaatttc aaatgcattt cctttcttta aaagataaat tgaagaaaat 2520
aagtctttat ttcccaagta aattgaattg cctctcagtc tgttaaaaga aagaaggaaa 2580
atattggaat catattctcc ctcaccgaaa tgctattttt cagcccacag gaaacccagg 2640
ctggttggag gggacattcc ctgctctggt cgtgttgaag tacaacatgg agacacgtgg 2700
ggcaccgtct gtgattctga cttctctctg gaggcggcca gcgtgctgtg cagggaacta 2760
cagtgcggca cagtgtggtt tccctcctgg ggggagctca ctttggagaa ggaagtggac 2820
agatctgggc tgaagaattc cagtgtgagg ggcacgagtc ccacctttca ctctgcccag 2880
tagcaccccg ccctgacggg acatgtagcc acagcaggga cgtcggcgta gtctgctcaa 2940
gtgagaccca gggaatgtgt tcactttgtt cccatgccat gaagagggta gggttaggta 3000
gtcacagaca tctttttaaa gccctgtctc cttccaggat acacacaaat ccgcttggtg 3060
aatggcaaga ccccatgtga aggaagagtg gagctcaaca ttcttgggtc ctgggggtcc 3120
ctctgcaact ctcactggga catggaagat gcccatgttt tatgccagca gcttaaatgt 3180
ggagttgccc tttctatccc gggaggagca ccttttggga aaggaagtga gcaggtctgg 3240
aggcacatgt ttcactgcac tgggactgag aagcacatgg gagattgttc cgtcactgct 3300
ctgggcgcat cactctgttc ttcagggcaa gtggcctctg taatctgctc aggtaagaga 3360
ataagggcag ccagtgatga gccactcatg acggtgcctt aagagtgggt gtacctagga 3420
gttcccattg tggctcagtg gtaacaaact cgactggtat ccatgagggt atgggtttga 3480
tccctggcct tgctcaatgg gttaaggatc cagcattgct gtgagctgtg gtataggttg 3540
cagactctgc tcaggtccca tgttgctgtg attgtggtgt aggctgactg ctgcagcttc 3600
aatttgaccc ctagcccggg aatttccata ggccacacgt gcagcactaa ggaaggaaaa 3660
aaagaaaaaa aaaaaaaaag agtgggtgtg cctatagtga agaacagatg taaaagggaa 3720
gtgaaaggga ttcccccatt ctgagggatt gtgagaagtg tgccagaata ttaacttcat 3780
ttgacttgtt acagggaaag taaacttgac tttcacggac ctcctagtta cctggtgctt 3840
actatatgtc ttctcagagt acctgattca ttcccagcct ggttgaccca tccccctatc 3900
tctatggcta tgtttatcca gagcacatct atctaacact ccagctgatc ttcctgacac 3960
agctgtggca accctggatc ctttaaccaa ctgtgccagg ctggagatca aacctaagcc 4020
tctgcagcaa cccaagctgc tgcagtcaga tttttaaccc cctgtgccac tgtgggtatc 4080
tccgatattt tgtatcttct gtgactgagt ggtttgctgt ttgcagggaa ccagagtcag 4140
acactatccc cgtgcaattc atcatcctcg gacccatcaa gctctattat ttcagaagaa 4200
aatggtgttg cctgcatagg tgagaatcag tgaccaacct atgaaaatga tctcaatcct 4260
ctgaaatgca ttttattcat gttttatttc ctctttgcag ggagtggtca acttcgcctg 4320
gtcgatggag gtggtcgttg tgctgggaga gtagaggtct atcatgaggg ctcctggggc 4380
accatctgtg atgacagctg ggacctgaat gatgcccatg tggtgtgcaa acagctgagc 4440
tgtggatggg ccattaatgc cactggttct gctcattttg gggaaggaac agggcccatt 4500
tggctggatg agataaactg taatggaaaa gaatctcata tttggcaatg ccactcacat 4560
ggttgggggc ggcacaattg caggcataag gaggatgcag gagtcatctg ctcgg 4615
<210> 104
<211> 4866
<212> ДНК
<213> Sus scrofa
<400> 104
tatagatgac aaggctttgt gtctgatagg ggccagcgaa ctcagtaaag agggaagatg 60
agaaagataa tggcaagaat ttatccctga agtgtagttt tgacaaacca gtcacaaaga 120
ggtctaagaa attttggtca caaagttgtt ttgaatccca ggcattttat ttgcaatgat 180
tgcatatgtt ctggaaagga catctgaacc taagaaatag ttcatttgca ttgtgttata 240
ttttactaag gtctgagaaa taatcttgag atgagaatga actctacttc ttcagagtct 300
ggaaggaata aattatgaaa atgtattaat gcttctttaa accatattgt atatttatct 360
attactaaac aaaaagaagt agctctattt atttatttat ttatttattt atttatgtct 420
tttgtctctt tagggccaca cctgtggcat atggaggttc ccaggctaga ggtccaattg 480
gagatgtagc agccagccta tgccagagcc accgcaacac gggatctgag ccacgtctgt 540
gacttacacc acagctcaca gcaacgcctg atcctcaacc cactgagcga ggccagggat 600
cgaacccatg tcctcatgga tgctagttgg gttcgttaac tgctgagcca tgatgggaac 660
tccaaattaa ttatttctta tatttgttct tcatatattc atttctatag aaagaaataa 720
atacagattc agttaatgat ggcaggtaaa agcttaactt attaatcaaa ggagttaatc 780
caggcacaaa aattcaattc atggctctct gttaaaattt aggtataggt ttagcaggaa 840
gaaaaggtta gtagatgcag actattacat ttagaatgga tggacaatga agtcctacta 900
tacagcacag ggaactatat ccaatctctt gggatagaat atgatggaag acaaaatcag 960
aacaagagag tatatatata tgtgtgtgtg tgtgtgtgtg tgtgtgtgtg tgtgtgtgtg 1020
tgtgtgactg ggtcaccctg cggcacagca gaaattggca gaacattgta aatcaactat 1080
actttaatag gaaaaatact tttaagggct aaatttccaa tattctaacc atgtacacag 1140
agtaaatgtc ataaggatgc cagtctgtgt agagattgat gtgttactag cagattcatg 1200
aaataaaggc tgaggatgta gtccccaagt cacttctgag tggaagaatt tctcctttgt 1260
cctggactca aatattttag gataaaggaa aaaagaagat atttatagaa gggacttgtt 1320
ttcaagtact tgacaaaatt tcaccattaa agagaaattt gtgggagttc ccatcgtggc 1380
tcagtggaaa caaatccaac taggaaccat gaggttgtgg gtttgatccc tggcctcact 1440
cagtgggtta aggatccggt gttgccgtga gctgtggtgt aggttgcaga cacggttctg 1500
atcctgcgtt gctgtggctg tggctgtggt gtaggccagc agcaaacagc tctgattaga 1560
cccctagcct ggaaacctcc atatgccaca ggtgcagccc taaaaagaca aaaaaagaga 1620
aaagacaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa gaacccccag 1680
aggtatttat ttgtttttgc cttttttcac tgactgttct ttgtttgttt gtttgagact 1740
gatctagaag actagagatt acaagaaata tggatttggc tcactctaag aaactgcttt 1800
cattccaagg tttgggtcta tccaaaagtg gaatagaatc atatgaatac tagtttatga 1860
gtatttagtg agaggaattt caagctcaaa taatgattca gcaagattaa attaaggagg 1920
gaattttcct tgtggctgag tgggttaagg acccaatgtt gtctctgtga ggatgtaggt 1980
tccatcctgg gctttgctca ttaggttaag gatctggcat tgctgcagct cagacccagt 2040
gctgccctgg ttgtggctta ggccaaagct gcagctccaa ttcaatctct ggcctgggaa 2100
cctccatgtg ctacaaggtg cggccttaaa aggaaaaaaa aaaaattaaa tcaaggactc 2160
aagagtcttt cattatttgt gttgtggaag ctatatttgt tttaaagtct tagttgtgtt 2220
tagaaagcaa gatgttcttc aactcaaatt tgggagggaa cttgtttcat acatttttaa 2280
tggataagtg gcaaaatttt catgctgagg tgatctatag tgttgtaatg cagaatatag 2340
tcagatcttg aacattttag gaagttggtg agggccaatt gtgtatctgt gccatgctga 2400
taagaatgtc aagggatcac aagaattcgt gttatttgac agcagtcatc tttaaaaggc 2460
atttgagaaa gtccaatttc aaatgcattt cctttcttta aaagataaat tgaagaaaat 2520
aagtctttat ttcccaagta aattgaattg cctctcagtc tgttaaaaga aactcttacc 2580
ttgatgattg cgctcttaac ctggcaaaga ttgtctttaa aatctgagct ccatgtcttc 2640
tgctttattt ctggtgtgcc tttgactcca gattacagta aatggaggac tgagtatagg 2700
gctaaaaagt agagagaatg gatgcatatt atctgtggtc tccaatgtga tgaatgaagt 2760
aggcaaatac tcaaaggaaa gagaaagcat gctccaagaa ttatgggttc cagaaggcaa 2820
agtcccagaa ttgtctccag ggaaggacag ggaggtctag aatcggctaa gcccactgta 2880
ggcagaaaaa ccaagaggca tgaatggctt ccctttctca cttttcactc tctggcttac 2940
tcctatcatg aaggaaaata ttggaatcat attctccctc accgaaatgc tatttttcag 3000
cccacaggaa acccaggctg gttctgtggt ttccctcctg gggggagctc actttggaga 3060
aggaagtgga cagatctggg ctgaagaatt ccagtgtgag gggcacgagt cccacctttc 3120
actctgccca gtagcacccc gccctgacgg gacatgtagc cacagcaggg acgtcggcgt 3180
agtctgctca agtgagaccc agggaatgtg ttcactttgt tcccatgcca tgaagagggt 3240
agggttaggt agtcacagac atctttttaa agccctgtct ccttccagga tacacacaaa 3300
tccgcttggt gaatggcaag accccatgtg aaggaagagt ggagctcaac attcttgggt 3360
cctgggggtc cctctgcaac tctcactggg acatggaaga tgcccatgtt ttatgccagc 3420
agcttaaatg tggagttgcc ctttctatcc cgggaggagc accttttggg aaaggaagtg 3480
agcaggtctg gaggcacatg tttcactgca ctgggactga gaagcacatg ggagattgtt 3540
ccgtcactgc tctgggcgca tcactctgtt cttcagggca agtggcctct gtaatctgct 3600
caggtaagag aataagggca gccagtgatg agccactcat gacggtgcct taagagtggg 3660
tgtacctagg agttcccatt gtggctcagt ggtaacaaac tcgactggta tccatgaggg 3720
tatgggtttg atccctggcc ttgctcaatg ggttaaggat ccagcattgc tgtgagctgt 3780
ggtataggtt gcagactctg ctcaggtccc atgttgctgt gattgtggtg taggctgact 3840
gctgcagctt caatttgacc cctagcccgg gaatttccat aggccacacg tgcagcacta 3900
aggaaggaaa aaaagaaaaa aaaaaaaaaa gagtgggtgt gcctatagtg aagaacagat 3960
gtaaaaggga agtgaaaggg attcccccat tctgagggat tgtgagaagt gtgccagaat 4020
attaacttca tttgacttgt tacagggaaa gtaaacttga ctttcacgga cctcctagtt 4080
acctggtgct tactatatgt cttctcagag tacctgattc attcccagcc tggttgaccc 4140
atccccctat ctctatggct atgtttatcc agagcacatc tatctaacac tccagctgat 4200
cttcctgaca cagctgtggc aaccctggat cctttaacca actgtgccag gctggagatc 4260
aaacctaagc ctctgcagca acccaagctg ctgcagtcag atttttaacc ccctgtgcca 4320
ctgtgggtat ctccgatatt ttgtatcttc tgtgactgag tggtttgctg tttgcaggga 4380
accagagtca gacactatcc ccgtgcaatt catcatcctc ggacccatca agctctatta 4440
tttcagaaga aaatggtgtt gcctgcatag gtgagaatca gtgaccaacc tatgaaaatg 4500
atctcaatcc tctgaaatgc attttattca tgttttattt cctctttgca gggagtggtc 4560
aacttcgcct ggtcgatgga ggtggtcgtt gtgctgggag agtagaggtc tatcatgagg 4620
gctcctgggg caccatctgt gatgacagct gggacctgaa tgatgcccat gtggtgtgca 4680
aacagctgag ctgtggatgg gccattaatg ccactggttc tgctcatttt ggggaaggaa 4740
cagggcccat ttggctggat gagataaact gtaatggaaa agaatctcat atttggcaat 4800
gccactcaca tggttggggg cggcacaatt gcaggcataa ggaggatgca ggagtcatct 4860
gctcgg 4866
<210> 105
<211> 4867
<212> ДНК
<213> Sus scrofa
<400> 105
tatagatgac aaggctttgt gtctgatagg ggccagcgaa ctcagtaaag agggaagatg 60
agaaagataa tggcaagaat ttatccctga agtgtagttt tgacaaacca gtcacaaaga 120
ggtctaagaa attttggtca caaagttgtt ttgaatccca ggcattttat ttgcaatgat 180
tgcatatgtt ctggaaagga catctgaacc taagaaatag ttcatttgca ttgtgttata 240
ttttactaag gtctgagaaa taatcttgag atgagaatga actctacttc ttcagagtct 300
ggaaggaata aattatgaaa atgtattaat gcttctttaa accatattgt atatttatct 360
attactaaac aaaaagaagt agctctattt atttatttat ttatttattt atttatgtct 420
tttgtctctt tagggccaca cctgtggcat atggaggttc ccaggctaga ggtccaattg 480
gagatgtagc agccagccta tgccagagcc accgcaacac gggatctgag ccacgtctgt 540
gacttacacc acagctcaca gcaacgcctg atcctcaacc cactgagcga ggccagggat 600
cgaacccatg tcctcatgga tgctagttgg gttcgttaac tgctgagcca tgatgggaac 660
tccaaattaa ttatttctta tatttgttct tcatatattc atttctatag aaagaaataa 720
atacagattc agttaatgat ggcaggtaaa agcttaactt attaatcaaa ggagttaatc 780
caggcacaaa aattcaattc atggctctct gttaaaattt aggtataggt ttagcaggaa 840
gaaaaggtta gtagatgcag actattacat ttagaatgga tggacaatga agtcctacta 900
tacagcacag ggaactatat ccaatctctt gggatagaat atgatggaag acaaaatcag 960
aacaagagag tatatatata tgtgtgtgtg tgtgtgtgtg tgtgtgtgtg tgtgtgtgtg 1020
tgtgtgactg ggtcaccctg cggcacagca gaaattggca gaacattgta aatcaactat 1080
actttaatag gaaaaatact tttaagggct aaatttccaa tattctaacc atgtacacag 1140
agtaaatgtc ataaggatgc cagtctgtgt agagattgat gtgttactag cagattcatg 1200
aaataaaggc tgaggatgta gtccccaagt cacttctgag tggaagaatt tctcctttgt 1260
cctggactca aatattttag gataaaggaa aaaagaagat atttatagaa gggacttgtt 1320
ttcaagtact tgacaaaatt tcaccattaa agagaaattt gtgggagttc ccatcgtggc 1380
tcagtggaaa caaatccaac taggaaccat gaggttgtgg gtttgatccc tggcctcact 1440
cagtgggtta aggatccggt gttgccgtga gctgtggtgt aggttgcaga cacggttctg 1500
atcctgcgtt gctgtggctg tggctgtggt gtaggccagc agcaaacagc tctgattaga 1560
cccctagcct ggaaacctcc atatgccaca ggtgcagccc taaaaagaca aaaaaagaga 1620
aaagacaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa gaacccccag 1680
aggtatttat ttgtttttgc cttttttcac tgactgttct ttgtttgttt gtttgagact 1740
gatctagaag actagagatt acaagaaata tggatttggc tcactctaag aaactgcttt 1800
cattccaagg tttgggtcta tccaaaagtg gaatagaatc atatgaatac tagtttatga 1860
gtatttagtg agaggaattt caagctcaaa taatgattca gcaagattaa attaaggagg 1920
gaattttcct tgtggctgag tgggttaagg acccaatgtt gtctctgtga ggatgtaggt 1980
tccatcctgg gctttgctca ttaggttaag gatctggcat tgctgcagct cagacccagt 2040
gctgccctgg ttgtggctta ggccaaagct gcagctccaa ttcaatctct ggcctgggaa 2100
cctccatgtg ctacaaggtg cggccttaaa aggaaaaaaa aaaaattaaa tcaaggactc 2160
aagagtcttt cattatttgt gttgtggaag ctatatttgt tttaaagtct tagttgtgtt 2220
tagaaagcaa gatgttcttc aactcaaatt tgggagggaa cttgtttcat acatttttaa 2280
tggataagtg gcaaaatttt catgctgagg tgatctatag tgttgtaatg cagaatatag 2340
tcagatcttg aacattttag gaagttggtg agggccaatt gtgtatctgt gccatgctga 2400
taagaatgtc aagggatcac aagaattcgt gttatttgac agcagtcatc tttaaaaggc 2460
atttgagaaa gtccaatttc aaatgcattt cctttcttta aaagataaat tgaagaaaat 2520
aagtctttat ttcccaagta aattgaattg cctctcagtc tgttaaaaga aactcttacc 2580
ttgatgattg cgctcttaac ctggcaaaga ttgtctttaa aatctgagct ccatgtcttc 2640
tgctttattt ctggtgtgcc tttgactcca gattacagta aatggaggac tgagtatagg 2700
gctaaaaagt agagagaatg gatgcatatt atctgtggtc tccaatgtga tgaatgaagt 2760
aggcaaatac tcaaaggaaa gagaaagcat gctccaagaa ttatgggttc cagaaggcaa 2820
agtcccagaa ttgtctccag ggaaggacag ggaggtctag aatcggctaa gcccactgta 2880
ggcagaaaaa ccaagaggca tgaatggctt ccctttctca cttttcactc tctggcttac 2940
tcctatcatg aaggaaaata ttggaatcat attctccctc accgaaatgc tatttttcag 3000
cccacaggaa acccaggctg gttactgtgg tttccctcct ggggggagct cactttggag 3060
aaggaagtgg acagatctgg gctgaagaat tccagtgtga ggggcacgag tcccaccttt 3120
cactctgccc agtagcaccc cgccctgacg ggacatgtag ccacagcagg gacgtcggcg 3180
tagtctgctc aagtgagacc cagggaatgt gttcactttg ttcccatgcc atgaagaggg 3240
tagggttagg tagtcacaga catcttttta aagccctgtc tccttccagg atacacacaa 3300
atccgcttgg tgaatggcaa gaccccatgt gaaggaagag tggagctcaa cattcttggg 3360
tcctgggggt ccctctgcaa ctctcactgg gacatggaag atgcccatgt tttatgccag 3420
cagcttaaat gtggagttgc cctttctatc ccgggaggag caccttttgg gaaaggaagt 3480
gagcaggtct ggaggcacat gtttcactgc actgggactg agaagcacat gggagattgt 3540
tccgtcactg ctctgggcgc atcactctgt tcttcagggc aagtggcctc tgtaatctgc 3600
tcaggtaaga gaataagggc agccagtgat gagccactca tgacggtgcc ttaagagtgg 3660
gtgtacctag gagttcccat tgtggctcag tggtaacaaa ctcgactggt atccatgagg 3720
gtatgggttt gatccctggc cttgctcaat gggttaagga tccagcattg ctgtgagctg 3780
tggtataggt tgcagactct gctcaggtcc catgttgctg tgattgtggt gtaggctgac 3840
tgctgcagct tcaatttgac ccctagcccg ggaatttcca taggccacac gtgcagcact 3900
aaggaaggaa aaaaagaaaa aaaaaaaaaa agagtgggtg tgcctatagt gaagaacaga 3960
tgtaaaaggg aagtgaaagg gattccccca ttctgaggga ttgtgagaag tgtgccagaa 4020
tattaacttc atttgacttg ttacagggaa agtaaacttg actttcacgg acctcctagt 4080
tacctggtgc ttactatatg tcttctcaga gtacctgatt cattcccagc ctggttgacc 4140
catcccccta tctctatggc tatgtttatc cagagcacat ctatctaaca ctccagctga 4200
tcttcctgac acagctgtgg caaccctgga tcctttaacc aactgtgcca ggctggagat 4260
caaacctaag cctctgcagc aacccaagct gctgcagtca gatttttaac cccctgtgcc 4320
actgtgggta tctccgatat tttgtatctt ctgtgactga gtggtttgct gtttgcaggg 4380
aaccagagtc agacactatc cccgtgcaat tcatcatcct cggacccatc aagctctatt 4440
atttcagaag aaaatggtgt tgcctgcata ggtgagaatc agtgaccaac ctatgaaaat 4500
gatctcaatc ctctgaaatg cattttattc atgttttatt tcctctttgc agggagtggt 4560
caacttcgcc tggtcgatgg aggtggtcgt tgtgctggga gagtagaggt ctatcatgag 4620
ggctcctggg gcaccatctg tgatgacagc tgggacctga atgatgccca tgtggtgtgc 4680
aaacagctga gctgtggatg ggccattaat gccactggtt ctgctcattt tggggaagga 4740
acagggccca tttggctgga tgagataaac tgtaatggaa aagaatctca tatttggcaa 4800
tgccactcac atggttgggg gcggcacaat tgcaggcata aggaggatgc aggagtcatc 4860
tgctcgg 4867
<210> 106
<211> 4991
<212> ДНК
<213> Sus scrofa
<400> 106
tatagatgac aaggctttgt gtctgatagg ggccagcgaa ctcagtaaag agggaagatg 60
agaaagataa tggcaagaat ttatccctga agtgtagttt tgacaaacca gtcacaaaga 120
ggtctaagaa attttggtca caaagttgtt ttgaatccca ggcattttat ttgcaatgat 180
tgcatatgtt ctggaaagga catctgaacc taagaaatag ttcatttgca ttgtgttata 240
ttttactaag gtctgagaaa taatcttgag atgagaatga actctacttc ttcagagtct 300
ggaaggaata aattatgaaa atgtattaat gcttctttaa accatattgt atatttatct 360
attactaaac aaaaagaagt agctctattt atttatttat ttatttattt atttatgtct 420
tttgtctctt tagggccaca cctgtggcat atggaggttc ccaggctaga ggtccaattg 480
gagatgtagc agccagccta tgccagagcc accgcaacac gggatctgag ccacgtctgt 540
gacttacacc acagctcaca gcaacgcctg atcctcaacc cactgagcga ggccagggat 600
cgaacccatg tcctcatgga tgctagttgg gttcgttaac tgctgagcca tgatgggaac 660
tccaaattaa ttatttctta tatttgttct tcatatattc atttctatag aaagaaataa 720
atacagattc agttaatgat ggcaggtaaa agcttaactt attaatcaaa ggagttaatc 780
caggcacaaa aattcaattc atggctctct gttaaaattt aggtataggt ttagcaggaa 840
gaaaaggtta gtagatgcag actattacat ttagaatgga tggacaatga agtcctacta 900
tacagcacag ggaactatat ccaatctctt gggatagaat atgatggaag acaaaatcag 960
aacaagagag tatatatata tgtgtgtgtg tgtgtgtgtg tgtgtgtgtg tgtgtgtgtg 1020
tgtgtgactg ggtcaccctg cggcacagca gaaattggca gaacattgta aatcaactat 1080
actttaatag gaaaaatact tttaagggct aaatttccaa tattctaacc atgtacacag 1140
agtaaatgtc ataaggatgc cagtctgtgt agagattgat gtgttactag cagattcatg 1200
aaataaaggc tgaggatgta gtccccaagt cacttctgag tggaagaatt tctcctttgt 1260
cctggactca aatattttag gataaaggaa aaaagaagat atttatagaa gggacttgtt 1320
ttcaagtact tgacaaaatt tcaccattaa agagaaattt gtgggagttc ccatcgtggc 1380
tcagtggaaa caaatccaac taggaaccat gaggttgtgg gtttgatccc tggcctcact 1440
cagtgggtta aggatccggt gttgccgtga gctgtggtgt aggttgcaga cacggttctg 1500
atcctgcgtt gctgtggctg tggctgtggt gtaggccagc agcaaacagc tctgattaga 1560
cccctagcct ggaaacctcc atatgccaca ggtgcagccc taaaaagaca aaaaaagaga 1620
aaagacaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa gaacccccag 1680
aggtatttat ttgtttttgc cttttttcac tgactgttct ttgtttgttt gtttgagact 1740
gatctagaag actagagatt acaagaaata tggatttggc tcactctaag aaactgcttt 1800
cattccaagg tttgggtcta tccaaaagtg gaatagaatc atatgaatac tagtttatga 1860
gtatttagtg agaggaattt caagctcaaa taatgattca gcaagattaa attaaggagg 1920
gaattttcct tgtggctgag tgggttaagg acccaatgtt gtctctgtga ggatgtaggt 1980
tccatcctgg gctttgctca ttaggttaag gatctggcat tgctgcagct cagacccagt 2040
gctgccctgg ttgtggctta ggccaaagct gcagctccaa ttcaatctct ggcctgggaa 2100
cctccatgtg ctacaaggtg cggccttaaa aggaaaaaaa aaaaattaaa tcaaggactc 2160
aagagtcttt cattatttgt gttgtggaag ctatatttgt tttaaagtct tagttgtgtt 2220
tagaaagcaa gatgttcttc aactcaaatt tgggagggaa cttgtttcat acatttttaa 2280
tggataagtg gcaaaatttt catgctgagg tgatctatag tgttgtaatg cagaatatag 2340
tcagatcttg aacattttag gaagttggtg agggccaatt gtgtatctgt gccatgctga 2400
taagaatgtc aagggatcac aagaattcgt gttatttgac agcagtcatc tttaaaaggc 2460
atttgagaaa gtccaatttc aaatgcattt cctttcttta aaagataaat tgaagaaaat 2520
aagtctttat ttcccaagta aattgaattg cctctcagtc tgttaaaaga aactcttacc 2580
ttgatgattg cgctcttaac ctggcaaaga ttgtctttaa aatctgagct ccatgtcttc 2640
tgctttattt ctggtgtgcc tttgactcca gattacagta aatggaggac tgagtatagg 2700
gctaaaaagt agagagaatg gatgcatatt atctgtggtc tccaatgtga tgaatgaagt 2760
aggcaaatac tcaaaggaaa gagaaagcat gctccaagaa ttatgggttc cagaaggcaa 2820
agtcccagaa ttgtctccag ggaaggacag ggaggtctag aatcggctaa gcccactgta 2880
ggcagaaaaa ccaagaggca tgaatggctt ccctttctca cttttcactc tctggcttac 2940
tcctatcatg aaggaaaata ttggaatcat attctccctc accgaaatgc tatttttcag 3000
cccacaggaa acccaggctg gttggagggg acattccctg ctctggtcgt gttgaagtac 3060
aacatggaga cacgtggggc accgtctgtg attctgactt ctctctggag gcggccagcg 3120
tgctgtgcag ggaactacag tgcggcaact gtggtttccc tcctgggggg agctcacttt 3180
ggagaaggaa gtggacagat ctgggctgaa gaattccagt gtgaggggca cgagtcccac 3240
ctttcactct gcccagtagc accccgccct gacgggacat gtagccacag cagggacgtc 3300
ggcgtagtct gctcaagtga gacccaggga atgtgttcac tttgttccca tgccatgaag 3360
agggtagggt taggtagtca cagacatctt tttaaagccc tgtctccttc caggatacac 3420
acaaatccgc ttggtgaatg gcaagacccc atgtgaagga agagtggagc tcaacattct 3480
tgggtcctgg gggtccctct gcaactctca ctgggacatg gaagatgccc atgttttatg 3540
ccagcagctt aaatgtggag ttgccctttc tatcccggga ggagcacctt ttgggaaagg 3600
aagtgagcag gtctggaggc acatgtttca ctgcactggg actgagaagc acatgggaga 3660
ttgttccgtc actgctctgg gcgcatcact ctgttcttca gggcaagtgg cctctgtaat 3720
ctgctcaggt aagagaataa gggcagccag tgatgagcca ctcatgacgg tgccttaaga 3780
gtgggtgtac ctaggagttc ccattgtggc tcagtggtaa caaactcgac tggtatccat 3840
gagggtatgg gtttgatccc tggccttgct caatgggtta aggatccagc attgctgtga 3900
gctgtggtat aggttgcaga ctctgctcag gtcccatgtt gctgtgattg tggtgtaggc 3960
tgactgctgc agcttcaatt tgacccctag cccgggaatt tccataggcc acacgtgcag 4020
cactaaggaa ggaaaaaaag aaaaaaaaaa aaaaagagtg ggtgtgccta tagtgaagaa 4080
cagatgtaaa agggaagtga aagggattcc cccattctga gggattgtga gaagtgtgcc 4140
agaatattaa cttcatttga cttgttacag ggaaagtaaa cttgactttc acggacctcc 4200
tagttacctg gtgcttacta tatgtcttct cagagtacct gattcattcc cagcctggtt 4260
gacccatccc cctatctcta tggctatgtt tatccagagc acatctatct aacactccag 4320
ctgatcttcc tgacacagct gtggcaaccc tggatccttt aaccaactgt gccaggctgg 4380
agatcaaacc taagcctctg cagcaaccca agctgctgca gtcagatttt taaccccctg 4440
tgccactgtg ggtatctccg atattttgta tcttctgtga ctgagtggtt tgctgtttgc 4500
agggaaccag agtcagacac tatccccgtg caattcatca tcctcggacc catcaagctc 4560
tattatttca gaagaaaatg gtgttgcctg cataggtgag aatcagtgac caacctatga 4620
aaatgatctc aatcctctga aatgcatttt attcatgttt tatttcctct ttgcagggag 4680
tggtcaactt cgcctggtcg atggaggtgg tcgttgtgct gggagagtag aggtctatca 4740
tgagggctcc tggggcacca tctgtgatga cagctgggac ctgaatgatg cccatgtggt 4800
gtgcaaacag ctgagctgtg gatgggccat taatgccact ggttctgctc attttgggga 4860
aggaacaggg cccatttggc tggatgagat aaactgtaat ggaaaagaat ctcatatttg 4920
gcaatgccac tcacatggtt gggggcggca caattgcagg cataaggagg atgcaggagt 4980
catctgctcg g 4991
<210> 107
<211> 4860
<212> ДНК
<213> Sus scrofa
<400> 107
tatagatgac aaggctttgt gtctgatagg ggccagcgaa ctcagtaaag agggaagatg 60
agaaagataa tggcaagaat ttatccctga agtgtagttt tgacaaacca gtcacaaaga 120
ggtctaagaa attttggtca caaagttgtt ttgaatccca ggcattttat ttgcaatgat 180
tgcatatgtt ctggaaagga catctgaacc taagaaatag ttcatttgca ttgtgttata 240
ttttactaag gtctgagaaa taatcttgag atgagaatga actctacttc ttcagagtct 300
ggaaggaata aattatgaaa atgtattaat gcttctttaa accatattgt atatttatct 360
attactaaac aaaaagaagt agctctattt atttatttat ttatttattt atttatgtct 420
tttgtctctt tagggccaca cctgtggcat atggaggttc ccaggctaga ggtccaattg 480
gagatgtagc agccagccta tgccagagcc accgcaacac gggatctgag ccacgtctgt 540
gacttacacc acagctcaca gcaacgcctg atcctcaacc cactgagcga ggccagggat 600
cgaacccatg tcctcatgga tgctagttgg gttcgttaac tgctgagcca tgatgggaac 660
tccaaattaa ttatttctta tatttgttct tcatatattc atttctatag aaagaaataa 720
atacagattc agttaatgat ggcaggtaaa agcttaactt attaatcaaa ggagttaatc 780
caggcacaaa aattcaattc atggctctct gttaaaattt aggtataggt ttagcaggaa 840
gaaaaggtta gtagatgcag actattacat ttagaatgga tggacaatga agtcctacta 900
tacagcacag ggaactatat ccaatctctt gggatagaat atgatggaag acaaaatcag 960
aacaagagag tatatatata tgtgtgtgtg tgtgtgtgtg tgtgtgtgtg tgtgtgtgtg 1020
tgtgtgactg ggtcaccctg cggcacagca gaaattggca gaacattgta aatcaactat 1080
actttaatag gaaaaatact tttaagggct aaatttccaa tattctaacc atgtacacag 1140
agtaaatgtc ataaggatgc cagtctgtgt agagattgat gtgttactag cagattcatg 1200
aaataaaggc tgaggatgta gtccccaagt cacttctgag tggaagaatt tctcctttgt 1260
cctggactca aatattttag gataaaggaa aaaagaagat atttatagaa gggacttgtt 1320
ttcaagtact tgacaaaatt tcaccattaa agagaaattt gtgggagttc ccatcgtggc 1380
tcagtggaaa caaatccaac taggaaccat gaggttgtgg gtttgatccc tggcctcact 1440
cagtgggtta aggatccggt gttgccgtga gctgtggtgt aggttgcaga cacggttctg 1500
atcctgcgtt gctgtggctg tggctgtggt gtaggccagc agcaaacagc tctgattaga 1560
cccctagcct ggaaacctcc atatgccaca ggtgcagccc taaaaagaca aaaaaagaga 1620
aaagacaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa gaacccccag 1680
aggtatttat ttgtttttgc cttttttcac tgactgttct ttgtttgttt gtttgagact 1740
gatctagaag actagagatt acaagaaata tggatttggc tcactctaag aaactgcttt 1800
cattccaagg tttgggtcta tccaaaagtg gaatagaatc atatgaatac tagtttatga 1860
gtatttagtg agaggaattt caagctcaaa taatgattca gcaagattaa attaaggagg 1920
gaattttcct tgtggctgag tgggttaagg acccaatgtt gtctctgtga ggatgtaggt 1980
tccatcctgg gctttgctca ttaggttaag gatctggcat tgctgcagct cagacccagt 2040
gctgccctgg ttgtggctta ggccaaagct gcagctccaa ttcaatctct ggcctgggaa 2100
cctccatgtg ctacaaggtg cggccttaaa aggaaaaaaa aaaaattaaa tcaaggactc 2160
aagagtcttt cattatttgt gttgtggaag ctatatttgt tttaaagtct tagttgtgtt 2220
tagaaagcaa gatgttcttc aactcaaatt tgggagggaa cttgtttcat acatttttaa 2280
tggataagtg gcaaaatttt catgctgagg tgatctatag tgttgtaatg cagaatatag 2340
tcagatcttg aacattttag gaagttggtg agggccaatt gtgtatctgt gccatgctga 2400
taagaatgtc aagggatcac aagaattcgt gttatttgac agcagtcatc tttaaaaggc 2460
atttgagaaa gtccaatttc aaatgcattt cctttcttta aaagataaat tgaagaaaat 2520
aagtctttat ttcccaagta aattgaattg cctctcagtc tgttaaaaga aactcttacc 2580
ttgatgattg cgctcttaac ctggcaaaga ttgtctttaa aatctgagct ccatgtcttc 2640
tgctttattt ctggtgtgcc tttgactcca gattacagta aatggaggac tgagtatagg 2700
gctaaaaagt agagagaatg gatgcatatt atctgtggtc tccaatgtga tgaatgaagt 2760
aggcaaatac tcaaaggaaa gagaaagcat gctccaagaa ttatgggttc cagaaggcaa 2820
agtcccagaa ttgtctccag ggaaggacag ggaggtctag aatcggctaa gcccactgta 2880
ggcagaaaaa ccaagaggca tgaatggctt ccctttctca cttttcactc tctggcttac 2940
tcctatcatg aaggaaaata ttggaatcat attctccctc accgaaatgc tatttttcag 3000
cccacaggaa acccaggctg gttggagggt cctgggggga gctcactttg gagaaggaag 3060
tggacagatc tgggctgaag aattccagtg tgaggggcac gagtcccacc tttcactctg 3120
cccagtagca ccccgccctg acgggacatg tagccacagc agggacgtcg gcgtagtctg 3180
ctcaagtgag acccagggaa tgtgttcact ttgttcccat gccatgaaga gggtagggtt 3240
aggtagtcac agacatcttt ttaaagccct gtctccttcc aggatacaca caaatccgct 3300
tggtgaatgg caagacccca tgtgaaggaa gagtggagct caacattctt gggtcctggg 3360
ggtccctctg caactctcac tgggacatgg aagatgccca tgttttatgc cagcagctta 3420
aatgtggagt tgccctttct atcccgggag gagcaccttt tgggaaagga agtgagcagg 3480
tctggaggca catgtttcac tgcactggga ctgagaagca catgggagat tgttccgtca 3540
ctgctctggg cgcatcactc tgttcttcag ggcaagtggc ctctgtaatc tgctcaggta 3600
agagaataag ggcagccagt gatgagccac tcatgacggt gccttaagag tgggtgtacc 3660
taggagttcc cattgtggct cagtggtaac aaactcgact ggtatccatg agggtatggg 3720
tttgatccct ggccttgctc aatgggttaa ggatccagca ttgctgtgag ctgtggtata 3780
ggttgcagac tctgctcagg tcccatgttg ctgtgattgt ggtgtaggct gactgctgca 3840
gcttcaattt gacccctagc ccgggaattt ccataggcca cacgtgcagc actaaggaag 3900
gaaaaaaaga aaaaaaaaaa aaaagagtgg gtgtgcctat agtgaagaac agatgtaaaa 3960
gggaagtgaa agggattccc ccattctgag ggattgtgag aagtgtgcca gaatattaac 4020
ttcatttgac ttgttacagg gaaagtaaac ttgactttca cggacctcct agttacctgg 4080
tgcttactat atgtcttctc agagtacctg attcattccc agcctggttg acccatcccc 4140
ctatctctat ggctatgttt atccagagca catctatcta acactccagc tgatcttcct 4200
gacacagctg tggcaaccct ggatccttta accaactgtg ccaggctgga gatcaaacct 4260
aagcctctgc agcaacccaa gctgctgcag tcagattttt aaccccctgt gccactgtgg 4320
gtatctccga tattttgtat cttctgtgac tgagtggttt gctgtttgca gggaaccaga 4380
gtcagacact atccccgtgc aattcatcat cctcggaccc atcaagctct attatttcag 4440
aagaaaatgg tgttgcctgc ataggtgaga atcagtgacc aacctatgaa aatgatctca 4500
atcctctgaa atgcatttta ttcatgtttt atttcctctt tgcagggagt ggtcaacttc 4560
gcctggtcga tggaggtggt cgttgtgctg ggagagtaga ggtctatcat gagggctcct 4620
ggggcaccat ctgtgatgac agctgggacc tgaatgatgc ccatgtggtg tgcaaacagc 4680
tgagctgtgg atgggccatt aatgccactg gttctgctca ttttggggaa ggaacagggc 4740
ccatttggct ggatgagata aactgtaatg gaaaagaatc tcatatttgg caatgccact 4800
cacatggttg ggggcggcac aattgcaggc ataaggagga tgcaggagtc atctgctcgg 4860
<210> 108
<211> 4858
<212> ДНК
<213> Sus scrofa
<400> 108
tatagatgac aaggctttgt gtctgatagg ggccagcgaa ctcagtaaag agggaagatg 60
agaaagataa tggcaagaat ttatccctga agtgtagttt tgacaaacca gtcacaaaga 120
ggtctaagaa attttggtca caaagttgtt ttgaatccca ggcattttat ttgcaatgat 180
tgcatatgtt ctggaaagga catctgaacc taagaaatag ttcatttgca ttgtgttata 240
ttttactaag gtctgagaaa taatcttgag atgagaatga actctacttc ttcagagtct 300
ggaaggaata aattatgaaa atgtattaat gcttctttaa accatattgt atatttatct 360
attactaaac aaaaagaagt agctctattt atttatttat ttatttattt atttatgtct 420
tttgtctctt tagggccaca cctgtggcat atggaggttc ccaggctaga ggtccaattg 480
gagatgtagc agccagccta tgccagagcc accgcaacac gggatctgag ccacgtctgt 540
gacttacacc acagctcaca gcaacgcctg atcctcaacc cactgagcga ggccagggat 600
cgaacccatg tcctcatgga tgctagttgg gttcgttaac tgctgagcca tgatgggaac 660
tccaaattaa ttatttctta tatttgttct tcatatattc atttctatag aaagaaataa 720
atacagattc agttaatgat ggcaggtaaa agcttaactt attaatcaaa ggagttaatc 780
caggcacaaa aattcaattc atggctctct gttaaaattt aggtataggt ttagcaggaa 840
gaaaaggtta gtagatgcag actattacat ttagaatgga tggacaatga agtcctacta 900
tacagcacag ggaactatat ccaatctctt gggatagaat atgatggaag acaaaatcag 960
aacaagagag tatatatata tgtgtgtgtg tgtgtgtgtg tgtgtgtgtg tgtgtgtgtg 1020
tgtgtgactg ggtcaccctg cggcacagca gaaattggca gaacattgta aatcaactat 1080
actttaatag gaaaaatact tttaagggct aaatttccaa tattctaacc atgtacacag 1140
agtaaatgtc ataaggatgc cagtctgtgt agagattgat gtgttactag cagattcatg 1200
aaataaaggc tgaggatgta gtccccaagt cacttctgag tggaagaatt tctcctttgt 1260
cctggactca aatattttag gataaaggaa aaaagaagat atttatagaa gggacttgtt 1320
ttcaagtact tgacaaaatt tcaccattaa agagaaattt gtgggagttc ccatcgtggc 1380
tcagtggaaa caaatccaac taggaaccat gaggttgtgg gtttgatccc tggcctcact 1440
cagtgggtta aggatccggt gttgccgtga gctgtggtgt aggttgcaga cacggttctg 1500
atcctgcgtt gctgtggctg tggctgtggt gtaggccagc agcaaacagc tctgattaga 1560
cccctagcct ggaaacctcc atatgccaca ggtgcagccc taaaaagaca aaaaaagaga 1620
aaagacaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa gaacccccag 1680
aggtatttat ttgtttttgc cttttttcac tgactgttct ttgtttgttt gtttgagact 1740
gatctagaag actagagatt acaagaaata tggatttggc tcactctaag aaactgcttt 1800
cattccaagg tttgggtcta tccaaaagtg gaatagaatc atatgaatac tagtttatga 1860
gtatttagtg agaggaattt caagctcaaa taatgattca gcaagattaa attaaggagg 1920
gaattttcct tgtggctgag tgggttaagg acccaatgtt gtctctgtga ggatgtaggt 1980
tccatcctgg gctttgctca ttaggttaag gatctggcat tgctgcagct cagacccagt 2040
gctgccctgg ttgtggctta ggccaaagct gcagctccaa ttcaatctct ggcctgggaa 2100
cctccatgtg ctacaaggtg cggccttaaa aggaaaaaaa aaaaattaaa tcaaggactc 2160
aagagtcttt cattatttgt gttgtggaag ctatatttgt tttaaagtct tagttgtgtt 2220
tagaaagcaa gatgttcttc aactcaaatt tgggagggaa cttgtttcat acatttttaa 2280
tggataagtg gcaaaatttt catgctgagg tgatctatag tgttgtaatg cagaatatag 2340
tcagatcttg aacattttag gaagttggtg agggccaatt gtgtatctgt gccatgctga 2400
taagaatgtc aagggatcac aagaattcgt gttatttgac agcagtcatc tttaaaaggc 2460
atttgagaaa gtccaatttc aaatgcattt cctttcttta aaagataaat tgaagaaaat 2520
aagtctttat ttcccaagta aattgaattg cctctcagtc tgttaaaaga aactcttacc 2580
ttgatgattg cgctcttaac ctggcaaaga ttgtctttaa aatctgagct ccatgtcttc 2640
tgctttattt ctggtgtgcc tttgactcca gattacagta aatggaggac tgagtatagg 2700
gctaaaaagt agagagaatg gatgcatatt atctgtggtc tccaatgtga tgaatgaagt 2760
aggcaaatac tcaaaggaaa gagaaagcat gctccaagaa ttatgggttc cagaaggcaa 2820
agtcccagaa ttgtctccag ggaaggacag ggaggtctag aatcggctaa gcccactgta 2880
ggcagaaaaa ccaagaggca tgaatggctt ccctttctca cttttcactc tctggcttac 2940
tcctatcatg aaggaaaata ttggaatcat attctccctc accgaaatgc tatttttcag 3000
cccacaggaa acccaggctg gttggagggc tggggggagc tcactttgga gaaggaagtg 3060
gacagatctg ggctgaagaa ttccagtgtg aggggcacga gtcccacctt tcactctgcc 3120
cagtagcacc ccgccctgac gggacatgta gccacagcag ggacgtcggc gtagtctgct 3180
caagtgagac ccagggaatg tgttcacttt gttcccatgc catgaagagg gtagggttag 3240
gtagtcacag acatcttttt aaagccctgt ctccttccag gatacacaca aatccgcttg 3300
gtgaatggca agaccccatg tgaaggaaga gtggagctca acattcttgg gtcctggggg 3360
tccctctgca actctcactg ggacatggaa gatgcccatg ttttatgcca gcagcttaaa 3420
tgtggagttg ccctttctat cccgggagga gcaccttttg ggaaaggaag tgagcaggtc 3480
tggaggcaca tgtttcactg cactgggact gagaagcaca tgggagattg ttccgtcact 3540
gctctgggcg catcactctg ttcttcaggg caagtggcct ctgtaatctg ctcaggtaag 3600
agaataaggg cagccagtga tgagccactc atgacggtgc cttaagagtg ggtgtaccta 3660
ggagttccca ttgtggctca gtggtaacaa actcgactgg tatccatgag ggtatgggtt 3720
tgatccctgg ccttgctcaa tgggttaagg atccagcatt gctgtgagct gtggtatagg 3780
ttgcagactc tgctcaggtc ccatgttgct gtgattgtgg tgtaggctga ctgctgcagc 3840
ttcaatttga cccctagccc gggaatttcc ataggccaca cgtgcagcac taaggaagga 3900
aaaaaagaaa aaaaaaaaaa aagagtgggt gtgcctatag tgaagaacag atgtaaaagg 3960
gaagtgaaag ggattccccc attctgaggg attgtgagaa gtgtgccaga atattaactt 4020
catttgactt gttacaggga aagtaaactt gactttcacg gacctcctag ttacctggtg 4080
cttactatat gtcttctcag agtacctgat tcattcccag cctggttgac ccatccccct 4140
atctctatgg ctatgtttat ccagagcaca tctatctaac actccagctg atcttcctga 4200
cacagctgtg gcaaccctgg atcctttaac caactgtgcc aggctggaga tcaaacctaa 4260
gcctctgcag caacccaagc tgctgcagtc agatttttaa ccccctgtgc cactgtgggt 4320
atctccgata ttttgtatct tctgtgactg agtggtttgc tgtttgcagg gaaccagagt 4380
cagacactat ccccgtgcaa ttcatcatcc tcggacccat caagctctat tatttcagaa 4440
gaaaatggtg ttgcctgcat aggtgagaat cagtgaccaa cctatgaaaa tgatctcaat 4500
cctctgaaat gcattttatt catgttttat ttcctctttg cagggagtgg tcaacttcgc 4560
ctggtcgatg gaggtggtcg ttgtgctggg agagtagagg tctatcatga gggctcctgg 4620
ggcaccatct gtgatgacag ctgggacctg aatgatgccc atgtggtgtg caaacagctg 4680
agctgtggat gggccattaa tgccactggt tctgctcatt ttggggaagg aacagggccc 4740
atttggctgg atgagataaa ctgtaatgga aaagaatctc atatttggca atgccactca 4800
catggttggg ggcggcacaa ttgcaggcat aaggaggatg caggagtcat ctgctcgg 4858
<210> 109
<211> 3523
<212> ДНК
<213> Sus scrofa
<400> 109
tatagatgac aaggctttgt gtctgatagg ggccagcgaa ctcagtaaag agggaagatg 60
agaaagataa tggcaagaat ttatccctga agtgtagttt tgacaaacca gtcacaaaga 120
ggtctaagaa attttggtca caaagttgtt ttgaatccca ggcattttat ttgcaatgat 180
tgcatatgtt ctggaaagga catctgaacc taagaaatag ttcatttgca ttgtgttata 240
ttttactaag gtctgagaaa taatcttgag atgagaatga actctacttc ttcagagtct 300
ggaaggaata aattatgaaa atgtattaat gcttctttaa accatattgt atatttatct 360
attactaaac aaaaagaagt agctctattt atttatttat ttatttattt atttatgtct 420
tttgtctctt tagggccaca cctgtggcat atggaggttc ccaggctaga ggtccaattg 480
gagatgtagc agccagccta tgccagagcc accgcaacac gggatctgag ccacgtctgt 540
gacttacacc acagctcaca gcaacgcctg atcctcaacc cactgagcga ggccagggat 600
cgaacccatg tcctcatgga tgctagttgg gttcgttaac tgctgagcca tgatgggaac 660
tccaaattaa ttatttctta tatttgttct tcatatattc atttctatag aaagaaataa 720
atacagattc agttaatgat ggcaggtaaa agcttaactt attaatcaaa ggagttaatc 780
caggcacaaa aattcaattc atggctctct gttaaaattt aggtataggt ttagcaggaa 840
gaaaaggtta gtagatgcag actattacat ttagaatgga tggacaatga agtcctacta 900
tacagcacag ggaactatat ccaatctctt gggatagaat atgatggaag acaaaatcag 960
aacaagagag tatatatata tgtgtgtgtg tgtgtgtgtg tgtgtgtgtg tgtgtgtgtg 1020
tgtgtgactg ggtcaccctg cggcacagca gaaattggca gaacattgta aatcaactat 1080
actttaatag gaaaaatact tttaagggct aaatttccaa tattctaacc atgtacacag 1140
agtaaatgtc ataaggatgc cagtctgtgt agagattgat gtgttactag cagattcatg 1200
aaataaaggc tgaggatgta gtccccaagt cacttctgag tggaagaatt tctcctttgt 1260
cctggactca aatattttag gataaaggaa aaaagaagat atttatagaa gggacttgtt 1320
ttcaagtact tgacaaaatt tcaccattaa agagaaattt gtgggagttc ccatcgtggc 1380
tcagtggaaa caaatccaac taggaaccat gaggttgtgg gtttgatccc tggcctcact 1440
cagtgggtta aggatccggt gttgccgtga gctgtggtgt aggttgcaga cacggttctg 1500
atcctgcgtt gctgtggctg tggctgtggt gtaggccagc agcaaacagc tctgattaga 1560
cccctagcct ggaaacctcc atatgccaca ggtgcagccc taaaaagaca aaaaaagaga 1620
aaagacaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa gaacccccag 1680
aggtatttat ttgtttttgc cttttttcac tgactgttct ttgtttgttt gtttgagact 1740
gatctagaag actagagatt acaagaaata tggatttggc tcactctaag aaactgcttt 1800
cattccaagg tttgggtcta tccaaaagtg gaatagaatc atatgaatac tagtttatga 1860
gtatttagtg agaggaattt caagctcaaa taatgattca gcaagattaa attaaggagg 1920
gaattttcct tgtggctgag tgggttaagg acccaatgtt gtctctgtga ggatgtaggt 1980
tccatcctgg gctttgctca ttaggttaag gatctggcat tgctgcagct cagacccagt 2040
gctgccctgg ttgtggctta ggccaaagct gcagctccaa ttcaatctct ggcctgggaa 2100
cctccatgtg ctacaaggtg cggccttaaa aggaaaaaaa aaaaattaaa tcaaggactc 2160
aagagtcttt cattatttgt gttgtggaag ctatatttgt tttaaagtct tagttgtgtt 2220
tagaaagcaa gatgttcttc aactcaaatt tgggagggaa cttgtttcat acatttttaa 2280
tggataagtg gcaaaatttt catgctgagg tgatctatag tgttgtaatg cagaatatag 2340
tcagatcttg aacattttag gaagttggtg agggccaatt gtgtatctgt gccatgctga 2400
taagaatgtc aagggatcac aagaattcgt gagctgtggt ataggttgca gactctgctc 2460
aggtcccatg ttgctgtgat tgtggtgtag gctgactgct gcagcttcaa tttgacccct 2520
agcccgggaa tttccatagg ccacacgtgc agcactaagg aaggaaaaaa agaaaaaaaa 2580
aaaaaaagag tgggtgtgcc tatagtgaag aacagatgta aaagggaagt gaaagggatt 2640
cccccattct gagggattgt gagaagtgtg ccagaatatt aacttcattt gacttgttac 2700
agggaaagta aacttgactt tcacggacct cctagttacc tggtgcttac tatatgtctt 2760
ctcagagtac ctgattcatt cccagcctgg ttgacccatc cccctatctc tatggctatg 2820
tttatccaga gcacatctat ctaacactcc agctgatctt cctgacacag ctgtggcaac 2880
cctggatcct ttaaccaact gtgccaggct ggagatcaaa cctaagcctc tgcagcaacc 2940
caagctgctg cagtcagatt tttaaccccc tgtgccactg tgggtatctc cgatattttg 3000
tatcttctgt gactgagtgg tttgctgttt gcagggaacc agagtcagac actatccccg 3060
tgcaattcat catcctcgga cccatcaagc tctattattt cagaagaaaa tggtgttgcc 3120
tgcataggtg agaatcagtg accaacctat gaaaatgatc tcaatcctct gaaatgcatt 3180
ttattcatgt tttatttcct ctttgcaggg agtggtcaac ttcgcctggt cgatggaggt 3240
ggtcgttgtg ctgggagagt agaggtctat catgagggct cctggggcac catctgtgat 3300
gacagctggg acctgaatga tgcccatgtg gtgtgcaaac agctgagctg tggatgggcc 3360
attaatgcca ctggttctgc tcattttggg gaaggaacag ggcccatttg gctggatgag 3420
ataaactgta atggaaaaga atctcatatt tggcaatgcc actcacatgg ttgggggcgg 3480
cacaattgca ggcataagga ggatgcagga gtcatctgct cgg 3523
<210> 110
<211> 3603
<212> ДНК
<213> Sus scrofa
<400> 110
tatagatgac aaggctttgt gtctgatagg ggccagcgaa ctcagtaaag agggaagatg 60
agaaagataa tggcaagaat ttatccctga agtgtagttt tgacaaacca gtcacaaaga 120
ggtctaagaa attttggtca caaagttgtt ttgaatccca ggcattttat ttgcaatgat 180
tgcatatgtt ctggaaagga catctgaacc taagaaatag ttcatttgca ttgtgttata 240
ttttactaag gtctgagaaa taatcttgag atgagaatga actctacttc ttcagagtct 300
ggaaggaata aattatgaaa atgtattaat gcttctttaa accatattgt atatttatct 360
attactaaac aaaaagaagt agctctattt atttatttat ttatttattt atttatgtct 420
tttgtctctt tagggccaca cctgtggcat atggaggttc ccaggctaga ggtccaattg 480
gagatgttgt ggagaattcc acaagaattc gtgttatttg acagcagtca tctttaaaag 540
gcatttgaga aagtccaatt tcaaatgcat ttcctttctt taaaagataa attgaagaaa 600
ataagtcttt atttcccaag taaattgaat tgcctctcag tctgttaaaa gaaactctta 660
tatagatgac aaggctttgt gtctgatagg ggccagcgaa ctcagtaaag agggaagatg 720
agaaagataa tggcaagaat ttatccctga agtgtagttt tgacaaacca gtcacaaaga 780
ggtctaagaa attttggtca caaagttgtt ttgaatccca ggcattttat ttgcaatgat 840
tgcatatgtt ctggaaagga catctgaacc taagaaatag ttcatttgca ttgtgttata 900
ttttactaag gtctgagaaa taatcttgag atgagaatga actctacttc ttcagagtct 960
ggaaggaata aattatgaaa atgtattaat gcttctttaa accatattgt atatttatct 1020
attactaaac aaaaagaagt agctctattt atttatttat ttatttattt atttatgtct 1080
tttgtctctt tagggccaca cctgtggcat atggaggttc ccaggctaga ggtccaattg 1140
gagatgttgt ggagaattcc acaagaattc gtgttatttg acagcagtca tctttaaaag 1200
gcatttgaga aagtccaatt tcaaatgcat ttcctttctt taaaagataa attgaagaaa 1260
ataagtcttt atttcccaag taaattgaat tgcctctcag tctgttaaaa gaaactctta 1320
ccttgatgat tgcgctctta acctggcaaa gattgtcttt aaaatctgag ctccatgtct 1380
tctgctttat ttctggtgtg cctttgactc cagattacag taaatggagg actgagtata 1440
gggctaaaaa gtagagagaa tggatgcata ttatctgtgg tctccaatgt gatgaatgaa 1500
gtaggcaaat actcaaagga aagagaaagc atgctccaag aattatgggt tccagaaggc 1560
aaagtcccag aattgtctcc agggaaggac agggaggtct agaatcggct aagcccactg 1620
taggcagaaa aaccaagagg catgaatggc ttccctttct cacttttcac tctctggctt 1680
actcctatca tgaaggaaaa tattggaatc atattctccc tcaccgaaat gctatttttc 1740
agcccacagg aaacccaggc tggttggagg ggacattccc tgctctcact ttggagaagg 1800
aagtggacag atctgggctg aagaattcca gtgtgagggg cacgagtccc acctttcact 1860
ctgcccagta gcaccccgcc ctgacgggac atgtagccac agcagggacg tcggcgtagt 1920
ctgctcaagt gagacccagg gaatgtgttc actttgttcc catgccatga agagggtagg 1980
gttaggtagt cacagacatc tttttaaagc cctgtctcct tccaggatac acacaaatcc 2040
gcttggtgaa tggcaagacc ccatgtgaag gaagagtgga gctcaacatt cttgggtcct 2100
gggggtccct ctgcaactct cactgggaca tggaagatgc ccatgtttta tgccagcagc 2160
ttaaatgtgg agttgccctt tctatcccgg gaggagcacc ttttgggaaa ggaagtgagc 2220
aggtctggag gcacatgttt cactgcactg ggactgagaa gcacatggga gattgttccg 2280
tcactgctct gggcgcatca ctctgttctt cagggcaagt ggcctctgta atctgctcag 2340
gtaagagaat aagggcagcc agtgatgagc cactcatgac ggtgccttaa gagtgggtgt 2400
acctaggagt tcccattgtg gctcagtggt aacaaactcg actggtatcc atgagggtat 2460
gggtttgatc cctggccttg ctcaatgggt taaggatcca gcattgctgt gagctgtggt 2520
ataggttgca gactctgctc aggtcccatg ttgctgtgat tgtggtgtag gctgactgct 2580
gcagcttcaa tttgacccct agcccgggaa tttccatagg ccacacgtgc agcactaagg 2640
aaggaaaaaa agaaaaaaaa aaaaaaagag tgggtgtgcc tatagtgaag aacagatgta 2700
aaagggaagt gaaagggatt cccccattct gagggattgt gagaagtgtg ccagaatatt 2760
aacttcattt gacttgttac agggaaagta aacttgactt tcacggacct cctagttacc 2820
tggtgcttac tatatgtctt ctcagagtac ctgattcatt cccagcctgg ttgacccatc 2880
cccctatctc tatggctatg tttatccaga gcacatctat ctaacactcc agctgatctt 2940
cctgacacag ctgtggcaac cctggatcct ttaaccaact gtgccaggct ggagatcaaa 3000
cctaagcctc tgcagcaacc caagctgctg cagtcagatt tttaaccccc tgtgccactg 3060
tgggtatctc cgatattttg tatcttctgt gactgagtgg tttgctgttt gcagggaacc 3120
agagtcagac actatccccg tgcaattcat catcctcgga cccatcaagc tctattattt 3180
cagaagaaaa tggtgttgcc tgcataggtg agaatcagtg accaacctat gaaaatgatc 3240
tcaatcctct gaaatgcatt ttattcatgt tttatttcct ctttgcaggg agtggtcaac 3300
ttcgcctggt cgatggaggt ggtcgttgtg ctgggagagt agaggtctat catgagggct 3360
cctggggcac catctgtgat gacagctggg acctgaatga tgcccatgtg gtgtgcaaac 3420
agctgagctg tggatgggcc attaatgcca ctggttctgc tcattttggg gaaggaacag 3480
ggcccatttg gctggatgag ataaactgta atggaaaaga atctcatatt tggcaatgcc 3540
actcacatgg ttgggggcgg cacaattgca ggcataagga ggatgcagga gtcatctgct 3600
cgg 3603
<210> 111
<211> 4962
<212> ДНК
<213> Sus scrofa
<400> 111
tatagatgac aaggctttgt gtctgatagg ggccagcgaa ctcagtaaag agggaagatg 60
agaaagataa tggcaagaat ttatccctga agtgtagttt tgacaaacca gtcacaaaga 120
ggtctaagaa attttggtca caaagttgtt ttgaatccca ggcattttat ttgcaatgat 180
tgcatatgtt ctggaaagga catctgaacc taagaaatag ttcatttgca ttgtgttata 240
ttttactaag gtctgagaaa taatcttgag atgagaatga actctacttc ttcagagtct 300
ggaaggaata aattatgaaa atgtattaat gcttctttaa accatattgt atatttatct 360
attactaaac aaaaagaagt agctctattt atttatttat ttatttattt atttatgtct 420
tttgtctctt tagggccaca cctgtggcat atggaggttc ccaggctaga ggtccaattg 480
gagatgtagc agccagccta tgccagagcc accgcaacac gggatctgag ccacgtctgt 540
gacttacacc acagctcaca gcaacgcctg atcctcaacc cactgagcga ggccagggat 600
cgaacccatg tcctcatgga tgctagttgg gttcgttaac tgctgagcca tgatgggaac 660
tccaaattaa ttatttctta tatttgttct tcatatattc atttctatag aaagaaataa 720
atacagattc agttaatgat ggcaggtaaa agcttaactt attaatcaaa ggagttaatc 780
caggcacaaa aattcaattc atggctctct gttaaaattt aggtataggt ttagcaggaa 840
gaaaaggtta gtagatgcag actattacat ttagaatgga tggacaatga agtcctacta 900
tacagcacag ggaactatat ccaatctctt gggatagaat atgatggaag acaaaatcag 960
aacaagagag tatatatata tgtgtgtgtg tgtgtgtgtg tgtgtgtgtg tgtgtgtgtg 1020
tgtgtgactg ggtcaccctg cggcacagca gaaattggca gaacattgta aatcaactat 1080
actttaatag gaaaaatact tttaagggct aaatttccaa tattctaacc atgtacacag 1140
agtaaatgtc ataaggatgc cagtctgtgt agagattgat gtgttactag cagattcatg 1200
aaataaaggc tgaggatgta gtccccaagt cacttctgag tggaagaatt tctcctttgt 1260
cctggactca aatattttag gataaaggaa aaaagaagat atttatagaa gggacttgtt 1320
ttcaagtact tgacaaaatt tcaccattaa agagaaattt gtgggagttc ccatcgtggc 1380
tcagtggaaa caaatccaac taggaaccat gaggttgtgg gtttgatccc tggcctcact 1440
cagtgggtta aggatccggt gttgccgtga gctgtggtgt aggttgcaga cacggttctg 1500
atcctgcgtt gctgtggctg tggctgtggt gtaggccagc agcaaacagc tctgattaga 1560
cccctagcct ggaaacctcc atatgccaca ggtgcagccc taaaaagaca aaaaaagaga 1620
aaagacaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa gaacccccag 1680
aggtatttat ttgtttttgc cttttttcac tgactgttct ttgtttgttt gtttgagact 1740
gatctagaag actagagatt acaagaaata tggatttggc tcactctaag aaactgcttt 1800
cattccaagg tttgggtcta tccaaaagtg gaatagaatc atatgaatac tagtttatga 1860
gtatttagtg agaggaattt caagctcaaa taatgattca gcaagattaa attaaggagg 1920
gaattttcct tgtggctgag tgggttaagg acccaatgtt gtctctgtga ggatgtaggt 1980
tccatcctgg gctttgctca ttaggttaag gatctggcat tgctgcagct cagacccagt 2040
gctgccctgg ttgtggctta ggccaaagct gcagctccaa ttcaatctct ggcctgggaa 2100
cctccatgtg ctacaaggtg cggccttaaa aggaaaaaaa aaaaattaaa tcaaggactc 2160
aagagtcttt cattatttgt gttgtggaag ctatatttgt tttaaagtct tagttgtgtt 2220
tagaaagcaa gatgttcttc aactcaaatt tgggagggaa cttgtttcat acatttttaa 2280
tggataagtg gcaaaatttt catgctgagg tgatctatag tgttgtaatg cagaatatag 2340
tcagatcttg aacattttag gaagttggtg agggccaatt gtgtatctgt gccatgctga 2400
taagaatgtc aagggatcac aagaattcgt gttatttgac agcagtcatc tttaaaaggc 2460
atttgagaaa gtccaatttc aaatgcattt cctttcttta aaagataaat tgaagaaaat 2520
aagtctttat ttcccaagta aattgaattg cctctcagtc tgttaaaaga aactcttacc 2580
ttgatgattg cgctcttaac ctggcaaaga ttgtctttaa aatctgagct ccatgtcttc 2640
tgctttattt ctggtgtgcc tttgactcca gattacagta aatggaggac tgagtatagg 2700
gctaaaaagt agagagaatg gatgcatatt atctgtggtc tccaatgtga tgaatgaagt 2760
aggcaaatac tcaaaggaaa gagaaagcat gctccaagaa ttatgggttc cagaaggcaa 2820
agtcccagaa ttgtctccag ggaaggacag ggaggtctag aatcggctaa gcccactgta 2880
ggcagaaaaa ccaagaggca tgaatggctt ccctttctca cttttcactc tctggcttac 2940
tcctatcatg aaggaaaata ttggaatcat attctccctc accgaaatgc tatttttcag 3000
cccacaggaa acccaggctg gttggagggg acattccctg ctctggtcgt gttgaagtac 3060
aacatggaga cacgtggggc accgtctgtg attctgactt ctctctggag gcggccagcg 3120
tgctgtgcag ggaactacag tgcgtcactt tggagaagga agtggacaga tctgggctga 3180
agaattccag tgtgaggggc acgagtccca cctttcactc tgcccagtag caccccgccc 3240
tgacgggaca tgtagccaca gcagggacgt cggcgtagtc tgctcaagtg agacccaggg 3300
aatgtgttca ctttgttccc atgccatgaa gagggtaggg ttaggtagtc acagacatct 3360
ttttaaagcc ctgtctcctt ccaggataca cacaaatccg cttggtgaat ggcaagaccc 3420
catgtgaagg aagagtggag ctcaacattc ttgggtcctg ggggtccctc tgcaactctc 3480
actgggacat ggaagatgcc catgttttat gccagcagct taaatgtgga gttgcccttt 3540
ctatcccggg aggagcacct tttgggaaag gaagtgagca ggtctggagg cacatgtttc 3600
actgcactgg gactgagaag cacatgggag attgttccgt cactgctctg ggcgcatcac 3660
tctgttcttc agggcaagtg gcctctgtaa tctgctcagg taagagaata agggcagcca 3720
gtgatgagcc actcatgacg gtgccttaag agtgggtgta cctaggagtt cccattgtgg 3780
ctcagtggta acaaactcga ctggtatcca tgagggtatg ggtttgatcc ctggccttgc 3840
tcaatgggtt aaggatccag cattgctgtg agctgtggta taggttgcag actctgctca 3900
ggtcccatgt tgctgtgatt gtggtgtagg ctgactgctg cagcttcaat ttgaccccta 3960
gcccgggaat ttccataggc cacacgtgca gcactaagga aggaaaaaaa gaaaaaaaaa 4020
aaaaaagagt gggtgtgcct atagtgaaga acagatgtaa aagggaagtg aaagggattc 4080
ccccattctg agggattgtg agaagtgtgc cagaatatta acttcatttg acttgttaca 4140
gggaaagtaa acttgacttt cacggacctc ctagttacct ggtgcttact atatgtcttc 4200
tcagagtacc tgattcattc ccagcctggt tgacccatcc ccctatctct atggctatgt 4260
ttatccagag cacatctatc taacactcca gctgatcttc ctgacacagc tgtggcaacc 4320
ctggatcctt taaccaactg tgccaggctg gagatcaaac ctaagcctct gcagcaaccc 4380
aagctgctgc agtcagattt ttaaccccct gtgccactgt gggtatctcc gatattttgt 4440
atcttctgtg actgagtggt ttgctgtttg cagggaacca gagtcagaca ctatccccgt 4500
gcaattcatc atcctcggac ccatcaagct ctattatttc agaagaaaat ggtgttgcct 4560
gcataggtga gaatcagtga ccaacctatg aaaatgatct caatcctctg aaatgcattt 4620
tattcatgtt ttatttcctc tttgcaggga gtggtcaact tcgcctggtc gatggaggtg 4680
gtcgttgtgc tgggagagta gaggtctatc atgagggctc ctggggcacc atctgtgatg 4740
acagctggga cctgaatgat gcccatgtgg tgtgcaaaca gctgagctgt ggatgggcca 4800
ttaatgccac tggttctgct cattttgggg aaggaacagg gcccatttgg ctggatgaga 4860
taaactgtaa tggaaaagaa tctcatattt ggcaatgcca ctcacatggt tgggggcggc 4920
acaattgcag gcataaggag gatgcaggag tcatctgctc gg 4962
<210> 112
<211> 3603
<212> ДНК
<213> Sus scrofa
<400> 112
tatagatgac aaggctttgt gtctgatagg ggccagcgaa ctcagtaaag agggaagatg 60
agaaagataa tggcaagaat ttatccctga agtgtagttt tgacaaacca gtcacaaaga 120
ggtctaagaa attttggtca caaagttgtt ttgaatccca ggcattttat ttgcaatgat 180
tgcatatgtt ctggaaagga catctgaacc taagaaatag ttcatttgca ttgtgttata 240
ttttactaag gtctgagaaa taatcttgag atgagaatga actctacttc ttcagagtct 300
ggaaggaata aattatgaaa atgtattaat gcttctttaa accatattgt atatttatct 360
attactaaac aaaaagaagt agctctattt atttatttat ttatttattt atttatgtct 420
tttgtctctt tagggccaca cctgtggcat atggaggttc ccaggctaga ggtccaattg 480
gagatgtagc agccagccta tgccagagcc accgcaacac gggatctgag ccacgtctgt 540
gacttacacc acagctcaca gcaacgcctg atcctcaacc cactgagcga ggccagggat 600
cgaacccatg tcctcatgga tgctagttgg gttcgttaac tgctgagcca tgatgggaac 660
tccaaattaa ttatttctta tatttgttct tcatatattc atttctatag aaagaaataa 720
atacagattc agttaatgat ggcaggtaaa agcttaactt attaatcaaa ggagttaatc 780
caggcacaaa aattcaattc atggctctct gttaaaattt aggtataggt ttagcaggaa 840
gaaaaggtta gtagatgcag actattacat ttagaatgga tggacaatga agtcctacta 900
tacagcacag ggaactatat ccaatctctt gggatagaat atgatggaag acaaaatcag 960
aacaagagag tatatatata tgtgtgtgtg tgtgtgtgtg tgtgtgtgtg tgtgtgtgtg 1020
tgtgtgactg ggtcaccctg cggcacagca gaaattggca gaacattgta aatcaactat 1080
actttaatag gaaaaatact tttaagggct aaatttccaa tattctaacc atgtacacag 1140
agtaaatgtc ataaggatgc cagtctgtgt agagattgat gtgttactag cagattcatg 1200
aaataaaggc tgaggatgta gtccccaagt cacttctgag tggaagaatt tctcctttgt 1260
cctggactca aatattttag gataaaggaa aaaagaagat atttatagaa gggacttgtt 1320
ttcaagtact tgacaaaatt tcaccattaa agagaaattt gtgggagttc ccatcgtggc 1380
tcagtggaaa caaatccaac taggaaccat gaggttgtgg gtttgatccc tggcctcact 1440
cagtgggtta aggatccggt gttgccgtga gctgtggtgt aggttgcaga cacggttctg 1500
atcctgcgtt gctgtggctg tggctgtggt gtaggccagc agcaaacagc tctgattaga 1560
cccctagcct ggaaacctcc atatgccaca ggtgcagccc taaaaagaca aaaaaagaga 1620
aaagacaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa gaacccccag 1680
aggtatttat ttgtttttgc cttttttcac tgactgttct ttgtttgttt gtttgagact 1740
gatctagaag actagagatt acaagaaata tggatttggc tcactctaag aaactgcttt 1800
cattccaagg tttgggtcta tccaaaagtg gaatagaatc atatgaatac tagtttatga 1860
gtatttagtg agaggaattt caagctcaaa taatgattca gcaagattaa attaaggagg 1920
gaattttcct tgtggctgag tgggttaagg acccaatgtt gtctctgtga ggatgtaggt 1980
tccatcctgg gctttgctca ttaggttaag gatctggcat tgctgcagct cagacccagt 2040
gctgccctgg ttgtggctta ggccaaagct gcagctccaa ttcaatctct ggcctgggaa 2100
cctccatgtg ctacaaggtg cggccttaaa aggaaaaaaa aaaaattaaa tcaaggactc 2160
aagagtcttt cattatttgt gttgtggaag ctatatttgt tttaaagtct tagttgtgtt 2220
tagaaagcaa gatgttcttc aactcaaatt tgggagggaa cttgtttcat acatttttaa 2280
tggataagtg gcaaaatttt catgctgagg tgatctatag tgttgtaatg cagaatatag 2340
tcagatcttg aacattttag gaagttggtg agggccaatt gtgtatctgt gccatgctga 2400
taagaatgtc aagggatcac aagaattcgt gttatttgac agcagtcatc tttaaaaggc 2460
atttgagaaa gtccaatttc aaatgcattt cctttcttta aaagataaat tgaagaaaat 2520
aagtctttat ttcccaagta aattgaattg cctctcagtc tgttaaaaga aactcttacc 2580
ttgatgattg cgctcttaac ctggcaaaga ttgtctttaa aatctgagct ccatgtcttc 2640
tgctttattt ctggtgtgcc tttgactcca gattacagta aatggaggac tgagtatagg 2700
gctaaaaagt agagagaatg gatgcatatt atctgtggtc tccaatgtga tgaatgaagt 2760
aggcaaatac tcaaaggaaa gagaaagcat gctccaagaa ttatgggttc cagaaggcaa 2820
agtcccagaa ttgtctccag ggaaggacag ggaggtctag aatcggctaa gcccactgta 2880
ggcagaaaaa ccaagaggca tgaatggctt ccctttctca cttttcactc tctggcttac 2940
tcctatcatg aaggaaaata ttggaatcat attctccctc accgaaatgc tatttttcag 3000
cccacaggaa acccaggctg gttggagggg acattccctg ctctggtcgt gttgaagtac 3060
aacatggaga cacgtggggc accgtctgtg attctgactt ctctctggag gcggccagcg 3120
tgctgtgcag ggaactacag tgcgattcat catcctcgga cccatcaagc tctattattt 3180
cagaagaaaa tggtgttgcc tgcataggtg agaatcagtg accaacctat gaaaatgatc 3240
tcaatcctct gaaatgcatt ttattcatgt tttatttcct ctttgcaggg agtggtcaac 3300
ttcgcctggt cgatggaggt ggtcgttgtg ctgggagagt agaggtctat catgagggct 3360
cctggggcac catctgtgat gacagctggg acctgaatga tgcccatgtg gtgtgcaaac 3420
agctgagctg tggatgggcc attaatgcca ctggttctgc tcattttggg gaaggaacag 3480
ggcccatttg gctggatgag ataaactgta atggaaaaga atctcatatt tggcaatgcc 3540
actcacatgg ttgggggcgg cacaattgca ggcataagga ggatgcagga gtcatctgct 3600
cgg 3603
<210> 113
<211> 3619
<212> ДНК
<213> Sus scrofa
<400> 113
tatagatgac aaggctttgt gtctgatagg ggccagcgaa ctcagtaaag agggaagatg 60
agaaagataa tggcaagaat ttatccctga agtgtagttt tgacaaacca gtcacaaaga 120
ggtctaagaa attttggtca caaagttgtt ttgaatccca ggcattttat ttgcaatgat 180
tgcatatgtt ctggaaagga catctgaacc taagaaatag ttcatttgca ttgtgttata 240
ttttactaag gtctgagaaa taatcttgag atgagaatga actctacttc ttcagagtct 300
ggaaggaata aattatgaaa atgtattaat gcttctttaa accatattgt atatttatct 360
attactaaac aaaaagaagt agctctattt atttatttat ttatttattt atttatgtct 420
tttgtctctt tagggccaca cctgtggcat atggaggttc ccaggctaga ggtccaattg 480
gagatgtagc agccagccta tgccagagcc accgcaacac gggatctgag ccacgtctgt 540
gacttacacc acagctcaca gcaacgcctg atcctcaacc cactgagcga ggccagggat 600
cgaacccatg tcctcatgga tgctagttgg gttcgttaac tgctgagcca tgatgggaac 660
tccaaattaa ttatttctta tatttgttct tcatatattc atttctatag aaagaaataa 720
atacagattc agttaatgat ggcaggtaaa agcttaactt attaatcaaa ggagttaatc 780
caggcacaaa aattcaattc atggctctct gttaaaattt aggtataggt ttagcaggaa 840
gaaaaggtta gtagatgcag actattacat ttagaatgga tggacaatga agtcctacta 900
tacagcacag ggaactatat ccaatctctt gggatagaat atgatggaag acaaaatcag 960
aacaagagag tatatatata tgtgtgtgtg tgtgtgtgtg tgtgtgtgtg tgtgtgtgtg 1020
tgtgtgactg ggtcaccctg cggcacagca gaaattggca gaacattgta aatcaactat 1080
actttaatag gaaaaatact tttaagggct aaatttccaa tattctaacc atgtacacag 1140
agtaaatgtc ataaggatgc cagtctgtgt agagattgat gtgttactag cagattcatg 1200
aaataaaggc tgaggatgta gtccccaagt cacttctgag tggaagaatt tctcctttgt 1260
cctggactca aatattttag gataaaggaa aaaagaagat atttatagaa gggacttgtt 1320
ttcaagtact tgacaaaatt tcaccattaa agagaaattt gtgggagttc ccatcgtggc 1380
tcagtggaaa caaatccaac taggaaccat gaggttgtgg gtttgatccc tggcctcact 1440
cagtgggtta aggatccggt gttgccgtga gctgtggtgt aggttgcaga cacggttctg 1500
atcctgcgtt gctgtggctg tggctgtggt gtaggccagc agcaaacagc tctgattaga 1560
cccctagcct ggaaacctcc atatgccaca ggtgcagccc taaaaagaca aaaaaagaga 1620
aaagacaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa gaacccccag 1680
aggtatttat ttgtttttgc cttttttcac tgactgttct ttgtttgttt gtttgagact 1740
gatctagaag actagagatt acaagaaata tggatttggc tcactctaag aaactgcttt 1800
cattccaagg tttgggtcta tccaaaagtg gaatagaatc atatgaatac tagtttatga 1860
gtatttagtg agaggaattt caagctcaaa taatgattca gcaagattaa attaaggagg 1920
gaattttcct tgtggctgag tgggttaagg acccaatgtt gtctctgtga ggatgtaggt 1980
tccatcctgg gctttgctca ttaggttaag gatctggcat tgctgcagct cagacccagt 2040
gctgccctgg ttgtggctta ggccaaagct gcagctccaa ttcaatctct ggcctgggaa 2100
cctccatgtg ctacaaggtg cggccttaaa aggaaaaaaa aaaaattaaa tcaaggactc 2160
aagagtcttt cattatttgt gttgtggaag ctatatttgt tttaaagtct tagttgtgtt 2220
tagaaagcaa gatgttcttc aactcaaatt tgggagggaa cttgtttcat acatttttaa 2280
tggataagtg gcaaaatttt catgctgagg tgatctatag tgttgtaatg cagaatatag 2340
tcagatcttg aacattttag gaagttggtg agggccaatt gtgtatctgt gccatgctga 2400
taagaatgtc aagggatcac aagaattcgt gttatttgac agcagtcatc tttaaaaggc 2460
atttgagaaa gtccaatttc aaatgcattt cctttcttta aaagataaat tgaagaaaat 2520
aagtctttat ttcccaagta aattgaattg cctctcagtc tgttaaaaga aactcttacc 2580
ttgatgattg cgctcttaac ctggcaaaga ttgtctttaa aatctgagct ccatgtcttc 2640
tgctttattt ctggtgtgcc tttgactcca gattacagta aatggaggac tgagtatagg 2700
gctaaaaagt agagagaatg gatgcatatt atctgtggtc tccaatgtga tgaatgaagt 2760
aggcaaatac tcaaaggaaa gagaaagcat gctccaagaa ttatgggttc cagaaggcaa 2820
agtcccagaa ttgtctccag ggaaggacag ggaggtctag aatcggctaa gcccactgta 2880
ggcagaaaaa ccaagaggca tgaatggctt ccctttctca cttttcactc tctggcttac 2940
tcctatcatg aaggaaaata ttggaatcat attctccctc accgaaatgc tatttttcag 3000
cccacaggaa acccaggctg gttggagggg acattccctg ctctggtcgt gttgaagtac 3060
aacatggaga cacgtggggc accgtctgtg attctgactt ctctctggag gcagccagcg 3120
tgctttgcag ggaaccagag tcagacacta tccccgtgca attcatcatc ctcggaccca 3180
tcaagctcta ttatttcaga agaaaatggt gttgcctgca taggtgagaa tcagtgacca 3240
acctatgaaa atgatctcaa tcctctgaaa tgcattttat tcatgtttta tttcctcttt 3300
gcagggagtg gtcaacttcg cctggtcgat ggaggtggtc gttgtgctgg gagagtagag 3360
gtctatcatg agggctcctg gggcaccatc tgtgatgaca gctgggacct gaatgatgcc 3420
catgtggtgt gcaaacagct gagctgtgga tgggccatta atgccactgg ttctgctcat 3480
tttggggaag gaacagggcc catttggctg gatgagataa actgtaatgg aaaagaatct 3540
catatttggc aatgccactc acatggttgg gggcggcaca attgcaggca taaggaggat 3600
gcaggagtca tctgctcgg 3619
<210> 114
<211> 3270
<212> ДНК
<213> Sus scrofa
<400> 114
tatagatgac aaggctttgt gtctgatagg ggccagcgaa ctcagtaaag agggaagatg 60
agaaagataa tggcaagaat ttatccctga agtgtagttt tgacaaacca gtcacaaaga 120
ggtctaagaa attttggtca caaagttgtt ttgaatccca ggcattttat ttgcaatgat 180
tgcatatgtt ctggaaagga catctgaacc taagaaatag ttcatttgca ttgtgttata 240
ttttactaag gtctgagaaa taatcttgag atgagaatga actctacttc ttcagagtct 300
ggaaggaata aattatgaaa atgtattaat gcttctttaa accatattgt atatttatct 360
attactaaac aaaaagaagt agctctattt atttatttat ttatttattt atttatgtct 420
tttgtctctt tagggccaca cctgtggcat atggaggttc ccaggctaga ggtccaattg 480
gagatgtagc agccagccta tgccagagcc accgcaacac gggatctgag ccacgtctgt 540
gacttacacc acagctcaca gcaacgcctg atcctcaacc cactgagcga ggccagggat 600
cgaacccatg tcctcatgga tgctagttgg gttcgttaac tgctgagcca tgatgggaac 660
tccaaattaa ttatttctta tatttgttct tcatatattc atttctatag aaagaaataa 720
atacagattc agttaatgat ggcaggtaaa agcttaactt attaatcaaa ggagttaatc 780
caggcacaaa aattcaattc atggctctct gttaaaattt aggtataggt ttagcaggaa 840
gaaaaggtta gtagatgcag actattacat ttagaatgga tggacaatga agtcctacta 900
tacagcacag ggaactatat ccaatctctt gggatagaat atgatggaag acaaaatcag 960
aacaagagag tatatatata tgtgtgtgtg tgtgtgtgtg tgtgtgtgtg tgtgtgtgtg 1020
tgtgtgactg ggtcaccctg cggcacagca gaaattggca gaacattgta aatcaactat 1080
actttaatag gaaaaatact tttaagggct aaatttccaa tattctaacc atgtacacag 1140
agtaaatgtc ataaggatgc cagtctgtgt agagattgat gtgttactag cagattcatg 1200
aaataaaggc tgaggatgta gtccccaagt cacttctgag tggaagaatt tctcctttgt 1260
cctggactca aatattttag gataaaggaa aaaagaagat atttatagaa gggacttgtt 1320
ttcaagtact tgacaaaatt tcaccattaa agagaaattt gtgggagttc ccatcgtggc 1380
tcagtggaaa caaatccaac taggaaccat gaggttgtgg gtttgatccc tggcctcact 1440
cagtgggtta aggatccggt gttgccgtga gctgtggtgt aggttgcaga cacggttctg 1500
atcctgcgtt gctgtggctg tggctgtggt gtaggccagc agcaaacagc tctgattaga 1560
cccctagcct ggaaacctcc atatgccaca ggtgcagccc taaaaagaca aaaaaagaga 1620
aaagacaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa gaacccccag 1680
aggtatttat ttgtttttgc cttttttcac tgactgttct ttgtttgttt gtttgagact 1740
gatctagaag actagagatt acaagaaata tggatttggc tcactctaag aaactgcttt 1800
cattccaagg tttgggtcta tccaaaagtg gaatagaatc atatgaatac tagtttatga 1860
gtatttagtg agaggaattt caagctcaaa taatgattca gcaagattaa attaaggagg 1920
gaattttcct tgtggctgag tgggttaagg acccaatgtt gtctctgtga ggatgtaggt 1980
tccatcctgg gctttgctca ttaggttaag gatctggcat tgctgcagct cagacccagt 2040
gctgccctgg ttgtggctta ggccaaagct gcagctccaa ttcaatctct ggcctgggaa 2100
cctccatgtg ctacaaggtg cggccttaaa aggaaaaaaa aaaaattaaa tcaaggactc 2160
aagagtcttt cattatttgt gttgtggaag ctatatttgt tttaaagtct tagttgtgtt 2220
tagaaagcaa gatgttcttc aactcaaatt tgggagggaa cttgtttcat acatttttaa 2280
tggataagtg gcaaaatttt catgctgagg tgatctatag tgttgtaatg cagaatatag 2340
tcagatcttg aacattttag gaagttggtg agggccaatt gtgtatctgt gccatgctga 2400
taagaatgtc aagggatcac aagaattcgt gttatttgac ttgttacagg gaaagtaaac 2460
ttgactttca cggacctcct agttacctgg tgcttactat atgtcttctc agagtacctg 2520
attcattccc agcctggttg acccatcccc ctatctctat ggctatgttt atccagagca 2580
catctatcta acactccagc tgatcttcct gacacagctg tggcaaccct ggatccttta 2640
accaactgtg ccaggctgga gatcaaacct aagcctctgc agcaacccaa gctgctgcag 2700
tcagattttt aaccccctgt gccactgtgg gtatctccga tattttgtat cttctgtgac 2760
tgagtggttt gctgtttgca gggaaccaga gtcagacact atccccgtgc aattcatcat 2820
cctcggaccc atcaagctct attatttcag aagaaaatgg tgttgcctgc ataggtgaga 2880
atcagtgacc aacctatgaa aatgatctca atcctctgaa atgcatttta ttcatgtttt 2940
atttcctctt tgcagggagt ggtcaacttc gcctggtcga tggaggtggt cgttgtgctg 3000
ggagagtaga ggtctatcat gagggctcct ggggcaccat ctgtgatgac agctgggacc 3060
tgaatgatgc ccatgtggtg tgcaaacagc tgagctgtgg atgggccatt aatgccactg 3120
gttctgctca ttttggggaa ggaacagggc ccatttggct ggatgagata aactgtaatg 3180
gaaaagaatc tcatatttgg caatgccact cacatggttg ggggcggcac aattgcaggc 3240
ataaggagga tgcaggagtc atctgctcgg 3270
<210> 115
<211> 7
<212> ДНК
<213> Sus scrofa
<400> 115
tactact 7
<210> 116
<211> 12
<212> ДНК
<213> Sus scrofa
<400> 116
tgtggagaat tc 12
<210> 117
<211> 11
<212> ДНК
<213> Sus scrofa
<400> 117
agccagcgtg c 11
<210> 118
<211> 8532
<212> ДНК
<213> Sus scrofa
<400> 118
tatagatgac aaggctttgt gtctgatagg ggccagcgaa ctcagtaaag agggaagatg 60
agaaagataa tggcaagaat ttatccctga agtgtagttt tgacaaacca gtcacaaaga 120
ggtctaagaa attttggtca caaagttgtt ttgaatccca ggcattttat ttgcaatgat 180
tgcatatgtt ctggaaagga catctgaacc taagaaatag ttcatttgca ttgtgttata 240
ttttactaag gtctgagaaa taatcttgag atgagaatga actctacttc ttcagagtct 300
ggaaggaata aattatgaaa atgtattaat gcttctttaa accatattgt atatttatct 360
attactaaac aaaaagaagt agctctattt atttatttat ttatttattt atttatgtct 420
tttgtctctt tagggccaca cctgtggcat atggaggttc ccaggctaga ggtccaattg 480
gagatgtagc agccagccta tgccagagcc accgcaacac gggatctgag ccacgtctgt 540
gacttacacc acagctcaca gcaacgcctg atcctcaacc cactgagcga ggccagggat 600
cgaacccatg tcctcatgga tgctagttgg gttcgttaac tgctgagcca tgatgggaac 660
tccaaattaa ttatttctta tatttgttct tcatatattc atttctatag aaagaaataa 720
atacagattc agttaatgat ggcaggtaaa agcttaactt attaatcaaa ggagttaatc 780
caggcacaaa aattcaattc atggctctct gttaaaattt aggtataggt ttagcaggaa 840
gaaaaggtta gtagatgcag actattacat ttagaatgga tggacaatga agtcctacta 900
tacagcacag ggaactatat ccaatctctt gggatagaat atgatggaag acaaaatcag 960
aacaagagag tatatatata tgtgtgtgtg tgtgtgtgtg tgtgtgtgtg tgtgtgtgtg 1020
tgtgtgactg ggtcaccctg cggcacagca gaaattggca gaacattgta aatcaactat 1080
actttaatag gaaaaatact tttaagggct aaatttccaa tattctaacc atgtacacag 1140
agtaaatgtc ataaggatgc cagtctgtgt agagattgat gtgttactag cagattcatg 1200
aaataaaggc tgaggatgta gtccccaagt cacttctgag tggaagaatt tctcctttgt 1260
cctggactca aatattttag gataaaggaa aaaagaagat atttatagaa gggacttgtt 1320
ttcaagtact tgacaaaatt tcaccattaa agagaaattt gtgggagttc ccatcgtggc 1380
tcagtggaaa caaatccaac taggaaccat gaggttgtgg gtttgatccc tggcctcact 1440
cagtgggtta aggatccggt gttgccgtga gctgtggtgt aggttgcaga cacggttctg 1500
atcctgcgtt gctgtggctg tggctgtggt gtaggccagc agcaaacagc tctgattaga 1560
cccctagcct ggaaacctcc atatgccaca ggtgcagccc taaaaagaca aaaaaagaga 1620
aaagacaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa gaacccccag 1680
aggtatttat ttgtttttgc cttttttcac tgactgttct ttgtttgttt gtttgagact 1740
gatctagaag actagagatt acaagaaata tggatttggc tcactctaag aaactgcttt 1800
cattccaagg tttgggtcta tccaaaagtg gaatagaatc atatgaatac tagtttatga 1860
gtatttagtg agaggaattt caagctcaaa taatgattca gcaagattaa attaaggagg 1920
gaattttcct tgtggctgag tgggttaagg acccaatgtt gtctctgtga ggatgtaggt 1980
tccatcctgg gctttgctca ttaggttaag gatctggcat tgctgcagct cagacccagt 2040
gctgccctgg ttgtggctta ggccaaagct gcagctccaa ttcaatctct ggcctgggaa 2100
cctccatgtg ctacaaggtg cggccttaaa aggaaaaaaa aaaaattaaa tcaaggactc 2160
aagagtcttt cattatttgt gttgtggaag ctatatttgt tttaaagtct tagttgtgtt 2220
tagaaagcaa gatgttcttc aactcaaatt tgggagggaa cttgtttcat acatttttaa 2280
tggataagtg gcaaaatttt catgctgagg tgatctatag tgttgtaatg cagaatatag 2340
tcagatcttg aacattttag gaagttggtg agggccaatt gtgtatctgt gccatgctga 2400
taagaatgtc aagggatcac aagaattcgt gttatttgac agcagtcatc tttaaaaggc 2460
atttgagaaa gtccaatttc aaatgcattt cctttcttta aaagataaat tgaagaaaat 2520
aagtctttat ttcccaagta aattgaattg cctctcagtc tgttaaaaga aactcttacc 2580
ttgatgattg cgctcttaac ctggcaaaga ttgtctttaa aatctgagct ccatgtcttc 2640
tgctttattt ctggtgtgcc tttgactcca gattacagta aatggaggac tgagtatagg 2700
gctaaaaagt agagagaatg gatgcatatt atctgtggtc tccaatgtga tgaatgaagt 2760
aggcaaatac tcaaaggaaa gagaaagcat gctccaagaa ttatgggttc cagaaggcaa 2820
agtcccagaa ttgtctccag ggaaggacag ggaggtctag aatcggctaa gcccactgta 2880
ggcagaaaaa ccaagaggca tgaatggctt ccctttctca cttttcactc tctggcttac 2940
tcctatcatg aaggaaaata ttggaatcat attctccctc accgaaatgc tatttttcag 3000
cccacaggaa acccaggctg gttggagctg acatgccctg ctctggtcgt gttgaagtaa 3060
aacatgcaga cacgtggggc tccgtctgtg attctgactt ctctctgcat gcggccaacg 3120
tgctgtgcag ggaactaaat tgcggcgatg cgatttccct ctcggtggga gatcactttg 3180
gaaaaggaaa tggactgacc tgggctgaaa aattccagtg tgaggggagc gagacccacc 3240
ttgcactctg cccaatagta caacaccctg aagacacatg tatccacagc agggaagtcg 3300
gcgtagtctg ctcaagtaag agtttactga aaataacact cttaaaatct tgttatgttt 3360
ttattcataa tgtgaatgag tagtagtgga aaataactac cagtttccta agcttataac 3420
ttcgtatagc atacattata cgaagttata agcctgcagg aattctaccg ggtaggggag 3480
gcgcttttcc caaggcagtc tggagcatgc gctttagcag ccccgctggg cacttggcgc 3540
tacacaagtg gcctctggcc tcgcacacat tccacatcca ccggtaggcg ccaaccggct 3600
ccgttctttg gtggcccctt cgcgccacct tctactcctc ccctagtcag gaagttcccc 3660
cccgccccgc agctcgcgtc gtgcaggacg tgacaaatgg aagtagcacg tctcactagt 3720
ctcgtgcaga tggacagcac cgctgagcaa tggaagcggg taggcctttg gggcagcggc 3780
caatagcagc tttgctcctt cgctttctgg gctcagaggc tgggaagggg tgggtccggg 3840
ggcgggctca ggggcgggct caggggcggg gcgggcgccc gaaggtcctc cggaggcccg 3900
gcattctgca cgcttcaaaa gcgcacgtct gccgcgctgt tctcctcttc ctcatctccg 3960
ggcctttcga cctgcagcca ccatggccat gattgaacag gatggcctgc atgcaggttc 4020
tccagctgcc tgggtggaga gactgtttgg ctatgactgg gcacagcaga ccattggttg 4080
ctctgatgca gcagtgttca gactgtcagc ccagggcagg ccagtcctgt ttgtgaagac 4140
agacctcagt ggggctctca atgagctgca ggatgaggct gccagactct cctggctggc 4200
aacaactggg gtcccctgtg cagctgtcct ggatgtggtc acagaagctg gaagggactg 4260
gctcctgctg ggtgaggtgc ctgggcagga cctcctgtcc tctcacctgg ctccagctga 4320
gaaagtgtca atcatggctg atgccatgag aagactccac accctggacc cagccacctg 4380
cccctttgac caccaggcca agcacaggat tgagagggcc agaaccagga tggaggctgg 4440
cctggtggac caggatgacc tggatgaaga acaccagggc ctggcccctg ctgaactgtt 4500
tgccaggctc aaggcatcca tgccagatgg tgaggacctg gtggtgactc atggggatgc 4560
ctgcctgccc aacatcatgg tggaaaatgg aaggttctct ggcttcattg actgtggcag 4620
gctgggagtg gctgacaggt accaggacat tgccctggca accagggaca ttgcagaaga 4680
gctgggggga gaatgggcag acaggttcct ggtgctctat ggcattgcag cccctgactc 4740
ccagagaatt gccttctaca gactgctgga tgagttcttc taaatcgatt ataatcagcc 4800
ataccacatt tgtagaggtt ttacttgctt taaaaaacct cccacacctc cccctgaacc 4860
tgaaacataa aatgaatgca atgttgttgt taaacttgtt tattgcagct tataatggtt 4920
acaaataaag caatagcatc acaaatttca caaataaagc atttttttca ctgcattcta 4980
gttgtggttt gtccaaactc atcaatgtgg aaaataacta ccagtttcct aagcttataa 5040
cttcgtatag catacattat acgaagttat aagctagaca aaagtatctt accccaatgg 5100
tagccctgta cccaataaaa gtaggtgttc agtttcatat cctatgaaat accctcttga 5160
tacttttact ttgcatgagg atttagaaga aaaaagtttt actataatcc ttaacttagg 5220
aaattctttt gaattggaaa tgaaacacaa attgcttttc attgatatgc catatgatta 5280
tatgaataaa acatgaaatc ttcatattgg attctagtat atacccaagt aaatattttt 5340
tccctagaag agtgccaagt gtgttaaaac cttttggttt aataaagcag aaaaaaataa 5400
actctaaaaa tcataattaa aaatgaaatg cttttattta tagcaattaa ctacaacatg 5460
tttagactta catactatta aatataatat atttaagatc ccctcatgat aaatatgttc 5520
attattttgt aggctgttga tgcactaata tgtatgtaga ttactttgtg aattgccctt 5580
aataaaattt aaaactttag gctagtaaac ctgtaacact caacttagtt ctgaactatc 5640
tcactattct tttgcaagaa tttacttagg taatgccaac taatttattc caaggccaaa 5700
aagatgacaa tgtcttatat attataaaaa ctaataaaaa ccattttaaa acctagtata 5760
aatttaaagg tacttgctct tctggttcat ctcttctttt gtttacttct gctttcaaaa 5820
acttatttat tgtgaccata ttctttactt ccatttattg ttataattta taagatacta 5880
tacttgcaag caataaatgt tatcttttta gcttttaaat ggtctcattt gaaaagaata 5940
tataattagt aagtcatagc tactttaaat aaaaacttat tctttaagag attaaacact 6000
tctccaagtg atctgttttt ctttaattaa aacgttatta actcccaaaa tgatgttatt 6060
gtttttttat aatcttaaat accaataatt accaggtcta ttttgatttt gatacaggat 6120
aaaaactact attaattact taagaatgtg ttctttttta tatgtaccat tttcatgatc 6180
aaagttggtg atatgactga ggttttgatt attattaaac agatagttaa tatgatatat 6240
tcctcatttt tccaaatgaa aggaaaaatg tcttatatgg aggaaaagat tggggcaggg 6300
ggattagtaa attattactt aaatatctga ataggaggat ttttcaatga aaggataaag 6360
gaagaatgat tgtatcatct gaatctttcc ctccctttcc tggagtttgt cctttcaacc 6420
cagtatacct accactccct tcatcaccta ctttcccatt acagtcccta tgtgttgggt 6480
ggtaactatt ttgttttggt gttaatatcc aagtttccct taataacacc tagtgaatgg 6540
aggaaggatg agcataccta cccatcagac atatttagcc accatattta atcaacaagc 6600
atgaagaaag gaagctagcc tctccccttc ctttcctcct gcctctctct ctcttctctg 6660
tcctcgctcc ctttcttccc atcaatattt tcagagcacc tcttatgcgc caggcattgg 6720
gatactcaaa ctggaggaaa caagaaaaaa aaaaaaaaaa ggcgaagacc tcagggaaat 6780
ttatattgct gctatatttt tttgagccta gtgtaaatta aaattcctta atgctgtgcc 6840
ttttaaaaac acaaataagc aaaatagttt atttcttcaa cagttaaatc cttagggtag 6900
gaaagtgatt caggatctat tgctactatt aactcttctt tcattttcac acaggataca 6960
cacaaatccg cttggtgaat ggcaagaccc catgtgaagg aagagtggag ctcaacattc 7020
ttgggtcctg ggggtccctc tgcaactctc actgggacat ggaagatgcc catgttttat 7080
gccagcagct taaatgtgga gttgcccttt ctatcccggg aggagcacct tttgggaaag 7140
gaagtgagca ggtctggagg cacatgtttc actgcactgg gactgagaag cacatgggag 7200
attgttccgt cactgctctg ggcgcatcac tctgttcttc agggcaagtg gcctctgtaa 7260
tctgctcagg taagagaata agggcagcca gtgatgagcc actcatgacg gtgccttaag 7320
agtgggtgta cctaggagtt cccattgtgg ctcagtggta acaaactcga ctggtatcca 7380
tgagggtatg ggtttgatcc ctggccttgc tcaatgggtt aaggatccag cattgctgtg 7440
agctgtggta taggttgcag actctgctca ggtcccatgt tgctgtgatt gtggtgtagg 7500
ctgactgctg cagcttcaat ttgaccccta gcccgggaat ttccataggc cacacgtgca 7560
gcactaagga aggaaaaaaa gaaaaaaaaa aaaaaagagt gggtgtgcct atagtgaaga 7620
acagatgtaa aagggaagtg aaagggattc ccccattctg agggattgtg agaagtgtgc 7680
cagaatatta acttcatttg acttgttaca gggaaagtaa acttgacttt cacggacctc 7740
ctagttacct ggtgcttact atatgtcttc tcagagtacc tgattcattc ccagcctggt 7800
tgacccatcc ccctatctct atggctatgt ttatccagag cacatctatc taacactcca 7860
gctgatcttc ctgacacagc tgtggcaacc ctggatcctt taaccaactg tgccaggctg 7920
gagatcaaac ctaagcctct gcagcaaccc aagctgctgc agtcagattt ttaaccccct 7980
gtgccactgt gggtatctcc gatattttgt atcttctgtg actgagtggt ttgctgtttg 8040
cagggaacca gagtcagaca ctatccccgt gcaattcatc atcctcggac ccatcaagct 8100
ctattatttc agaagaaaat ggtgttgcct gcataggtga gaatcagtga ccaacctatg 8160
aaaatgatct caatcctctg aaatgcattt tattcatgtt ttatttcctc tttgcaggga 8220
gtggtcaact tcgcctggtc gatggaggtg gtcgttgtgc tgggagagta gaggtctatc 8280
atgagggctc ctggggcacc atctgtgatg acagctggga cctgaatgat gcccatgtgg 8340
tgtgcaaaca gctgagctgt ggatgggcca ttaatgccac tggttctgct cattttgggg 8400
aaggaacagg gcccatttgg ctggatgaga taaactgtaa tggaaaagaa tctcatattt 8460
ggcaatgcca ctcacatggt tgggggcggc acaattgcag gcataaggag gatgcaggag 8520
tcatctgctc gg 8532
<210> 119
<211> 4538
<212> ДНК
<213> Sus scrofa
<400> 119
tatagatgac aaggctttgt gtctgatagg ggccagcgaa ctcagtaaag agggaagatg 60
agaaagataa tggcaagaat ttatccctga agtgtagttt tgacaaacca gtcacaaaga 120
ggtctaagaa attttggtca caaagttgtt ttgaatccca ggcattttat ttgcaatgat 180
tgcatatgtt ctggaaagga catctgaacc taagaaatag ttcatttgca ttgtgttata 240
ttttactaag gtctgagaaa taatcttgag atgagaatga actctacttc ttcagagtct 300
ggaaggaata aattatgaaa atgtattaat gcttctttaa accatattgt atatttatct 360
attactaaac aaaaagaagt agctctattt atttatttat ttatttattt atttatgtct 420
tttgtctctt tagggccaca cctgtggcat atggaggttc ccaggctaga ggtccaattg 480
gagatgtagc agccagccta tgccagagcc accgcaacac gggatctgag ccacgtctgt 540
gacttacacc acagctcaca gcaacgcctg atcctcaacc cactgagcga ggccagggat 600
cgaacccatg tcctcatgga tgctagttgg gttcgttaac tgctgagcca tgatgggaac 660
tccaaattaa ttatttctta tatttgttct tcatatattc atttctatag aaagaaataa 720
atacagattc agttaatgat ggcaggtaaa agcttaactt attaatcaaa ggagttaatc 780
caggcacaaa aattcaattc atggctctct gttaaaattt aggtataggt ttagcaggaa 840
gaaaaggtta gtagatgcag actattacat ttagaatgga tggacaatga agtcctacta 900
tacagcacag ggaactatat ccaatctctt gggatagaat atgatggaag acaaaatcag 960
aacaagagag tatatatata tgtgtgtgtg tgtgtgtgtg tgtgtgtgtg tgtgtgtgtg 1020
tgtgtgactg ggtcaccctg cggcacagca gaaattggca gaacattgta aatcaactat 1080
actttaatag gaaaaatact tttaagggct aaatttccaa tattctaacc atgtacacag 1140
agtaaatgtc ataaggatgc cagtctgtgt agagattgat gtgttactag cagattcatg 1200
aaataaaggc tgaggatgta gtccccaagt cacttctgag tggaagaatt tctcctttgt 1260
cctggactca aatattttag gataaaggaa aaaagaagat atttatagaa gggacttgtt 1320
ttcaagtact tgacaaaatt tcaccattaa agagaaattt gtgggagttc ccatcgtggc 1380
tcagtggaaa caaatccaac taggaaccat gaggttgtgg gtttgatccc tggcctcact 1440
cagtgggtta aggatccggt gttgccgtga gctgtggtgt aggttgcaga cacggttctg 1500
atcctgcgtt gctgtggctg tggctgtggt gtaggccagc agcaaacagc tctgattaga 1560
cccctagcct ggaaacctcc atatgccaca ggtgcagccc taaaaagaca aaaaaagaga 1620
aaagacaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa gaacccccag 1680
aggtatttat ttgtttttgc cttttttcac tgactgttct ttgtttgttt gtttgagact 1740
gatctagaag actagagatt acaagaaata tggatttggc tcactctaag aaactgcttt 1800
cattccaagg tttgggtcta tccaaaagtg gaatagaatc atatgaatac tagtttatga 1860
gtatttagtg agaggaattt caagctcaaa taatgattca gcaagattaa attaaggagg 1920
gaattttcct tgtggctgag tgggttaagg acccaatgtt gtctctgtga ggatgtaggt 1980
tccatcctgg gctttgctca ttaggttaag gatctggcat tgctgcagct cagacccagt 2040
gctgccctgg ttgtggctta ggccaaagct gcagctccaa ttcaatctct ggcctgggaa 2100
cctccatgtg ctacaaggtg cggccttaaa aggaaaaaaa aaaaattaaa tcaaggactc 2160
aagagtcttt cattatttgt gttgtggaag ctatatttgt tttaaagtct tagttgtgtt 2220
tagaaagcaa gatgttcttc aactcaaatt tgggagggaa cttgtttcat acatttttaa 2280
tggataagtg gcaaaatttt catgctgagg tgatctatag tgttgtaatg cagaatatag 2340
tcagatcttg aacattttag gaagttggtg agggccaatt gtgtatctgt gccatgctga 2400
taagaatgtc aagggatcac aagaattcgt gttatttgac agcagtcatc tttaaaaggc 2460
atttgagaaa gtccaatttc aaatgcattt cctttcttta aaagataaat tgaagaaaat 2520
aagtctttat ttcccaagta aattgaattg cctctcagtc tgttaaaaga aactcttacc 2580
ttgatgattg cgctcttaac ctggcaaaga ttgtctttaa aatctgagct ccatgtcttc 2640
tgctttattt ctggtgtgcc tttgactcca gattacagta aatggaggac tgagtatagg 2700
gctaaaaagt agagagaatg gatgcatatt atctgtggtc tccaatgtga tgaatgaagt 2760
aggcaaatac tcaaaggaaa gagaaagcat gctccaagaa ttatgggttc cagaaggcaa 2820
agtcccagaa ttgtctccag ggaaggacag ggaggtctag aatcggctaa gcccactgta 2880
ggcagaaaaa ccaagaggca tgaatggctt ccctttctca cttttcactc tctggcttac 2940
tcctatcatg aaggaaaata ttggaatcat attctccctc accgaaatgc tatttttcag 3000
cccacaggaa acccagctca acattcttgg gtcctggggg tccctctgca actctcactg 3060
ggacatggaa gatgcccatg ttttatgcca gcagcttaaa tgtggagttg ccctttctat 3120
cccgggagga gcaccttttg ggaaaggaag tgagcaggtc tggaggcaca tgtttcactg 3180
cactgggact gagaagcaca tgggagattg ttccgtcact gctctgggcg catcactctg 3240
ttcttcaggg caagtggcct ctgtaatctg ctcaggtaag agaataaggg cagccagtga 3300
tgagccactc atgacggtgc cttaagagtg ggtgtaccta ggagttccca ttgtggctca 3360
gtggtaacaa actcgactgg tatccatgag ggtatgggtt tgatccctgg ccttgctcaa 3420
tgggttaagg atccagcatt gctgtgagct gtggtatagg ttgcagactc tgctcaggtc 3480
ccatgttgct gtgattgtgg tgtaggctga ctgctgcagc ttcaatttga cccctagccc 3540
gggaatttcc ataggccaca cgtgcagcac taaggaagga aaaaaagaaa aaaaaaaaaa 3600
aagagtgggt gtgcctatag tgaagaacag atgtaaaagg gaagtgaaag ggattccccc 3660
attctgaggg attgtgagaa gtgtgccaga atattaactt catttgactt gttacaggga 3720
aagtaaactt gactttcacg gacctcctag ttacctggtg cttactatat gtcttctcag 3780
agtacctgat tcattcccag cctggttgac ccatccccct atctctatgg ctatgtttat 3840
ccagagcaca tctatctaac actccagctg atcttcctga cacagctgtg gcaaccctgg 3900
atcctttaac caactgtgcc aggctggaga tcaaacctaa gcctctgcag caacccaagc 3960
tgctgcagtc agatttttaa ccccctgtgc cactgtgggt atctccgata ttttgtatct 4020
tctgtgactg agtggtttgc tgtttgcagg gaaccagagt cagacactat ccccgtgcaa 4080
ttcatcatcc tcggacccat caagctctat tatttcagaa gaaaatggtg ttgcctgcat 4140
aggtgagaat cagtgaccaa cctatgaaaa tgatctcaat cctctgaaat gcattttatt 4200
catgttttat ttcctctttg cagggagtgg tcaacttcgc ctggtcgatg gaggtggtcg 4260
ttgtgctggg agagtagagg tctatcatga gggctcctgg ggcaccatct gtgatgacag 4320
ctgggacctg aatgatgccc atgtggtgtg caaacagctg agctgtggat gggccattaa 4380
tgccactggt tctgctcatt ttggggaagg aacagggccc atttggctgg atgagataaa 4440
ctgtaatgga aaagaatctc atatttggca atgccactca catggttggg ggcggcacaa 4500
ttgcaggcat aaggaggatg caggagtcat ctgctcgg 4538
<210> 120
<211> 101
<212> Белок
<213> Sus scrofa
<400> 120
Pro Arg Leu Val Gly Gly Asp Ile Pro Cys Ser Gly Arg Val Glu Val
1 5 10 15
Gln His Gly Asp Thr Trp Gly Thr Val Cys Asp Ser Asp Phe Ser Leu
20 25 30
Glu Ala Ala Ser Val Leu Cys Arg Glu Leu Gln Cys Gly Thr Val Val
35 40 45
Ser Leu Leu Gly Gly Ala His Phe Gly Glu Gly Ser Gly Gln Ile Trp
50 55 60
Ala Glu Glu Phe Gln Cys Glu Gly His Glu Ser His Leu Ser Leu Cys
65 70 75 80
Pro Val Ala Pro Arg Pro Asp Gly Thr Cys Ser His Ser Arg Asp Val
85 90 95
Gly Val Val Cys Ser
100
<210> 121
<211> 100
<212> Белок
<213> Homo sapiens
<400> 121
Leu Arg Leu Val Asp Gly Asp Ser Arg Cys Ala Gly Arg Val Glu Ile
1 5 10 15
Tyr His Asp Gly Phe Trp Gly Thr Ile Cys Asp Asp Gly Trp Asp Leu
20 25 30
Ser Asp Ala His Val Val Cys Gln Lys Leu Gly Cys Gly Val Ala Phe
35 40 45
Asn Ala Thr Val Ser Ala His Phe Gly Glu Gly Ser Gly Pro Ile Trp
50 55 60
Leu Asp Asp Leu Asn Cys Thr Gly Met Glu Ser His Leu Trp Gln Cys
65 70 75 80
Pro Ser Arg Gly Trp Gly Gln His Asp Cys Arg His Lys Glu Asp Ala
85 90 95
Gly Val Ile Cys
100
<---
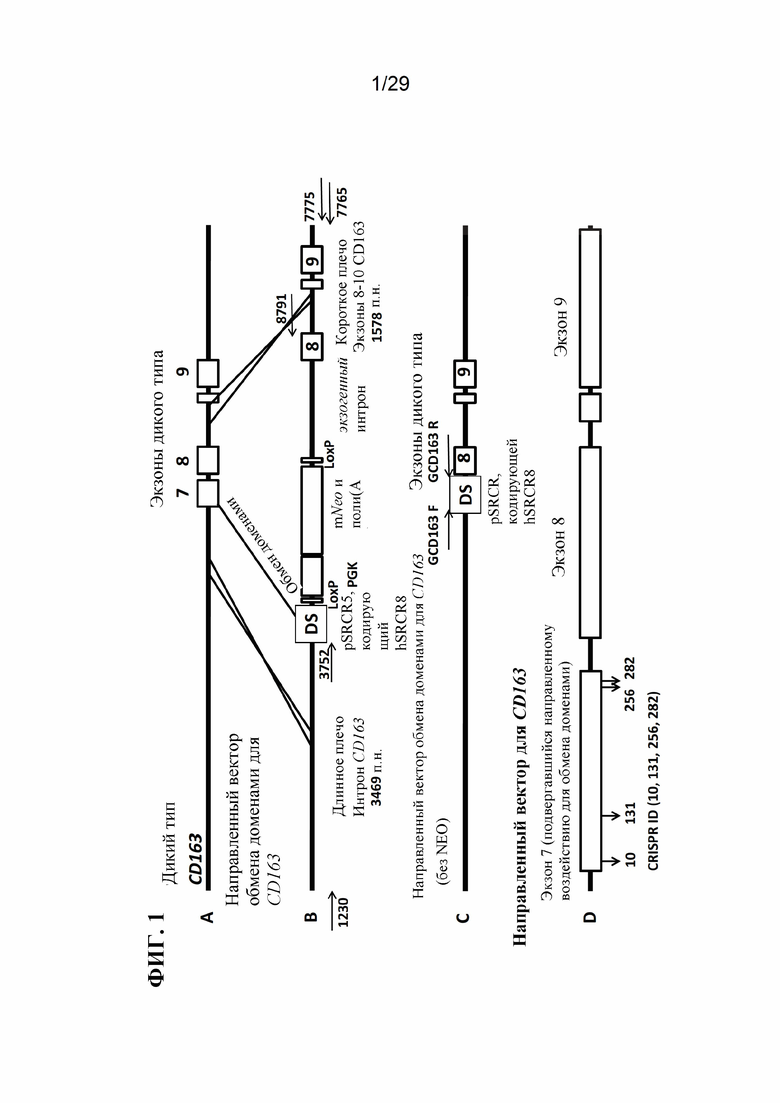
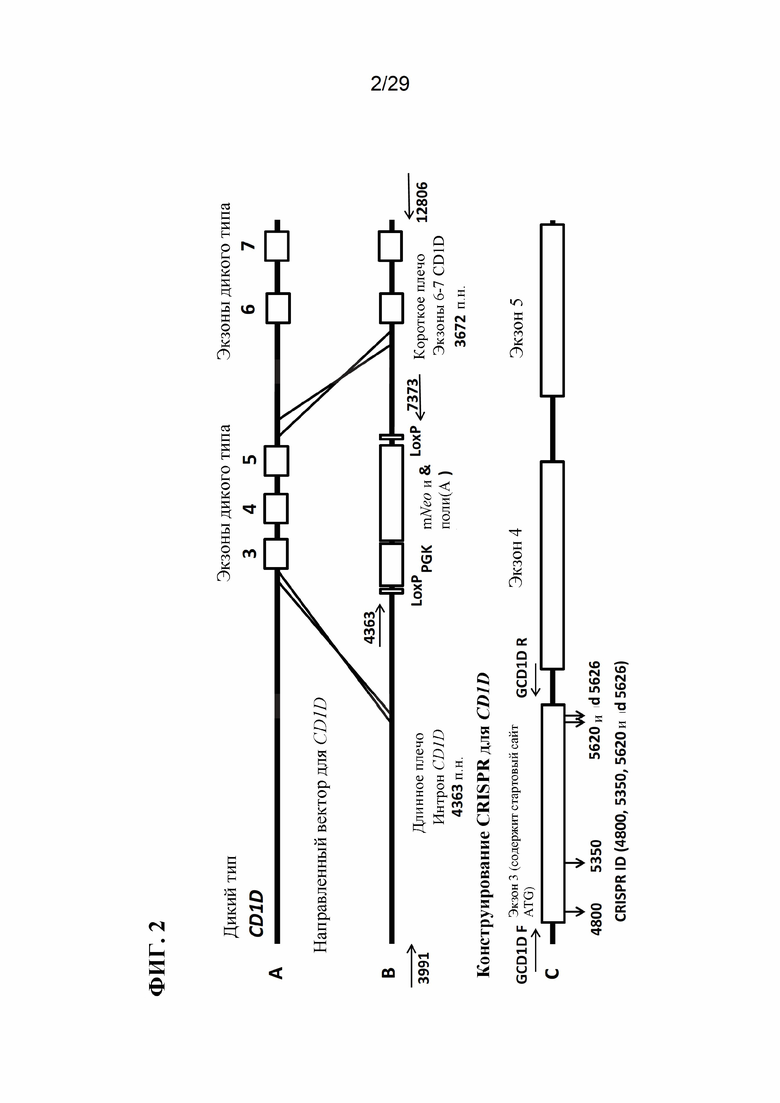

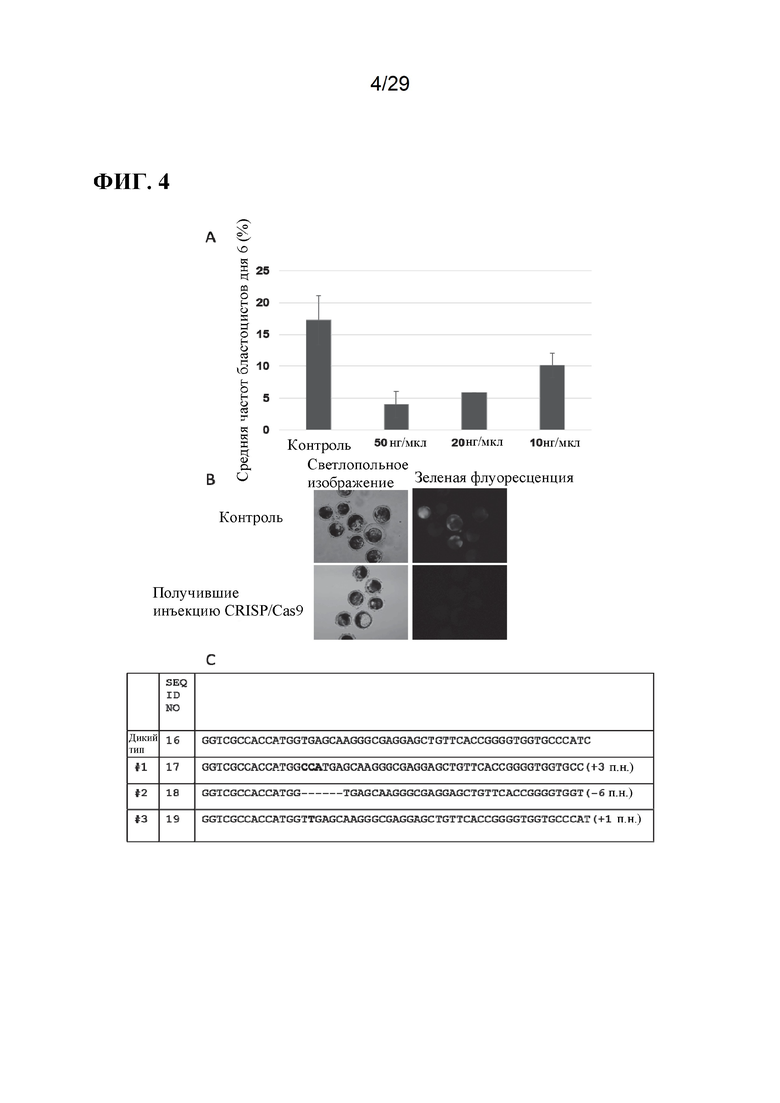
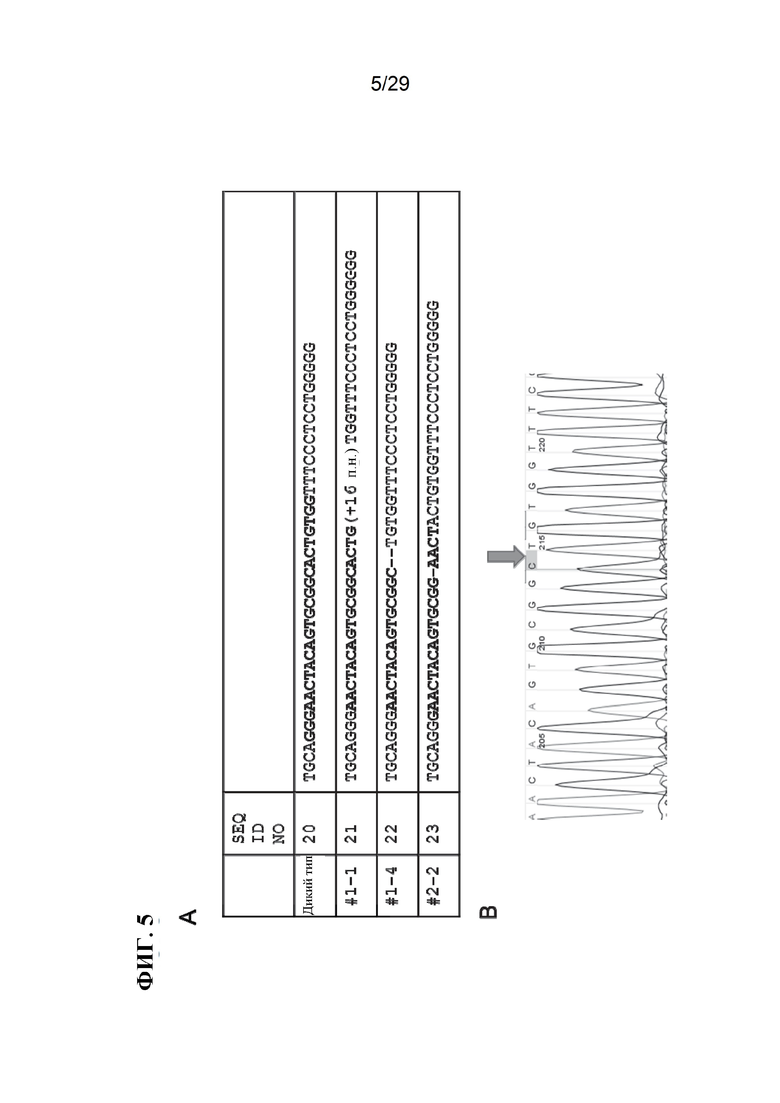
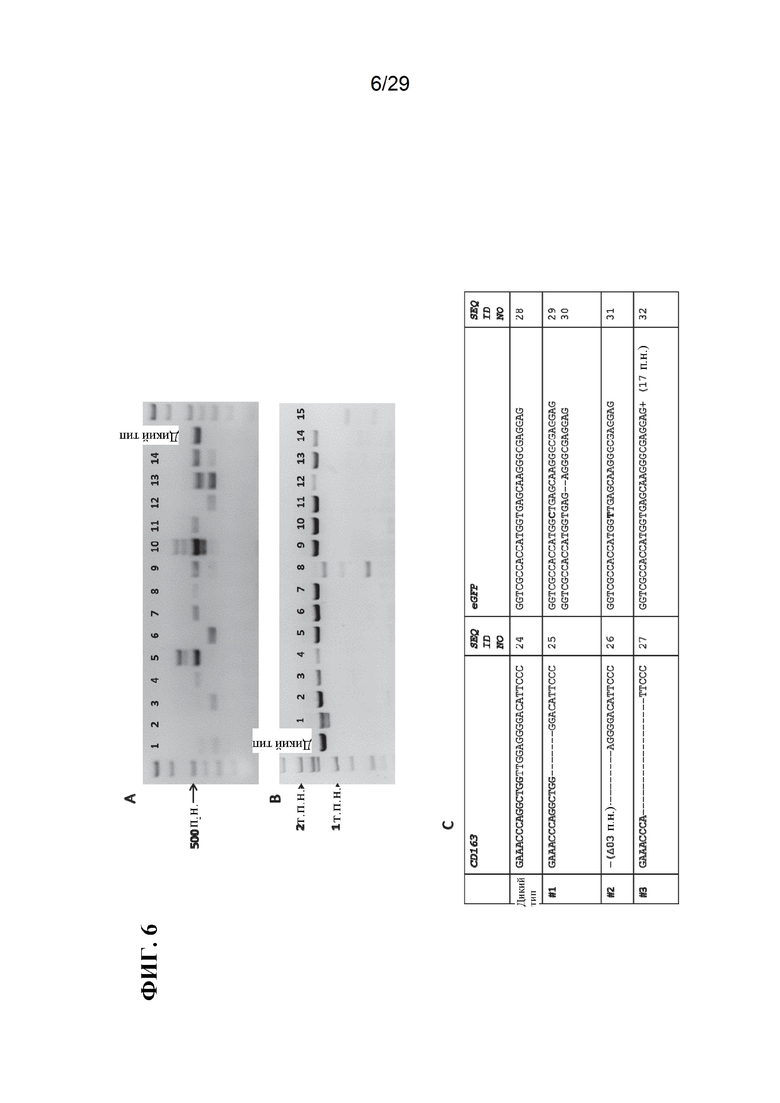


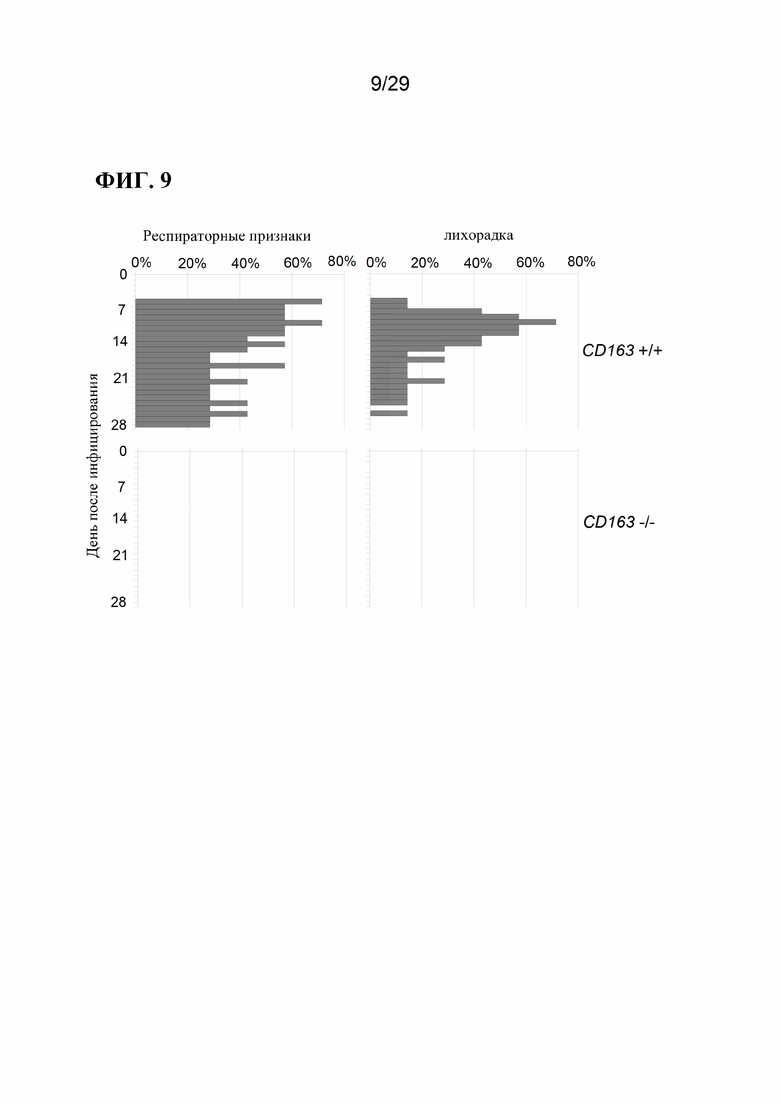

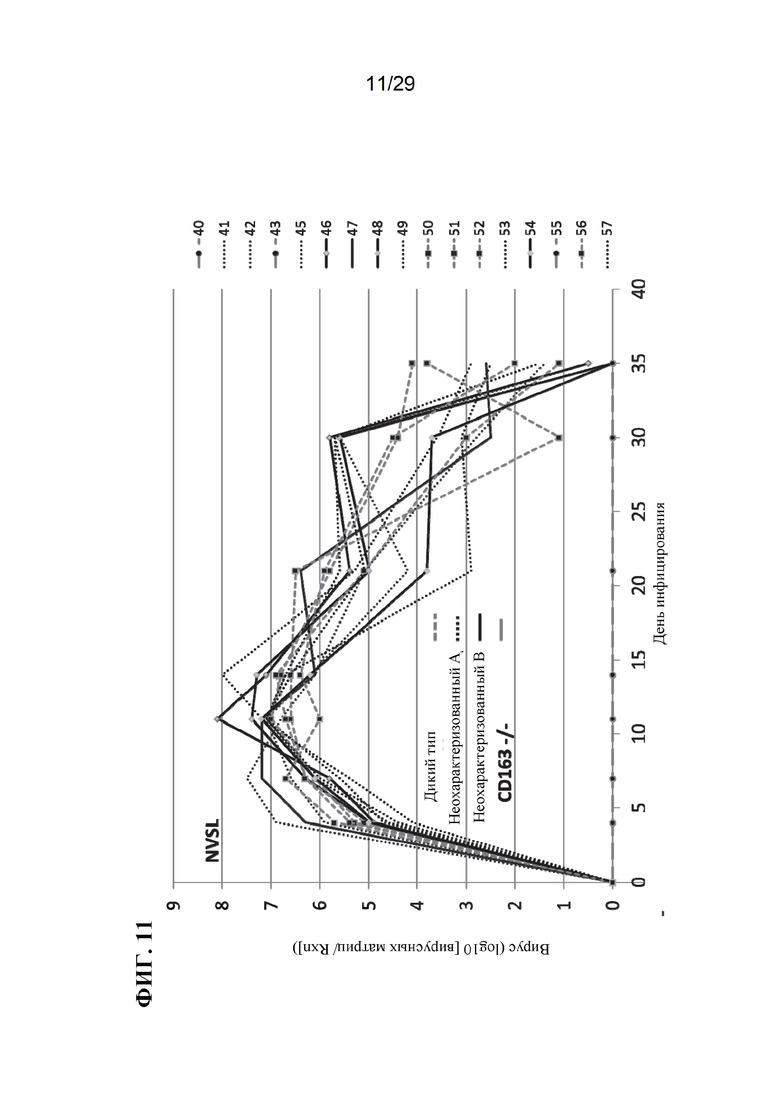
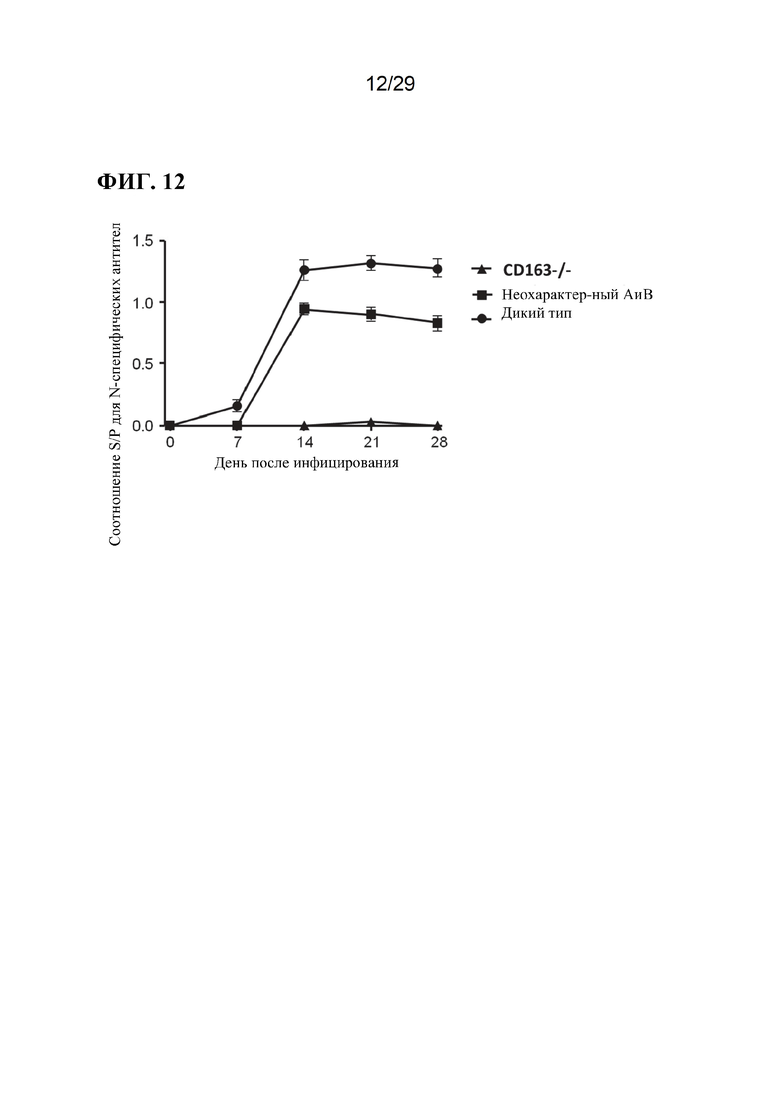
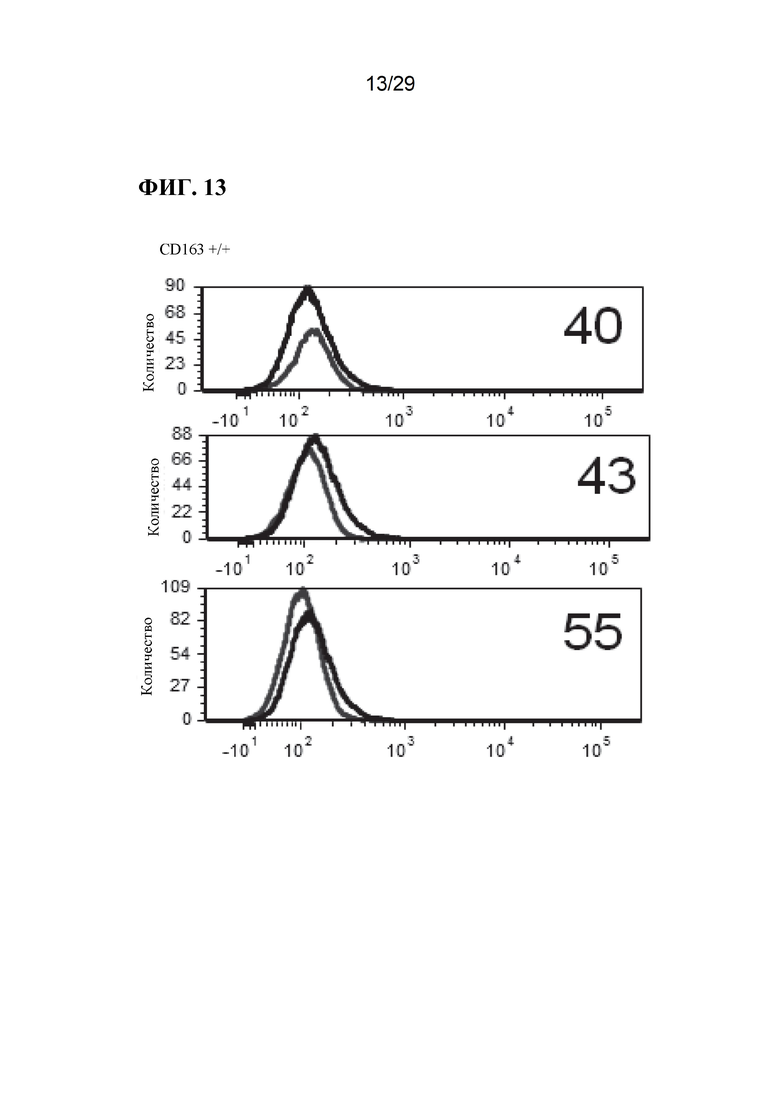
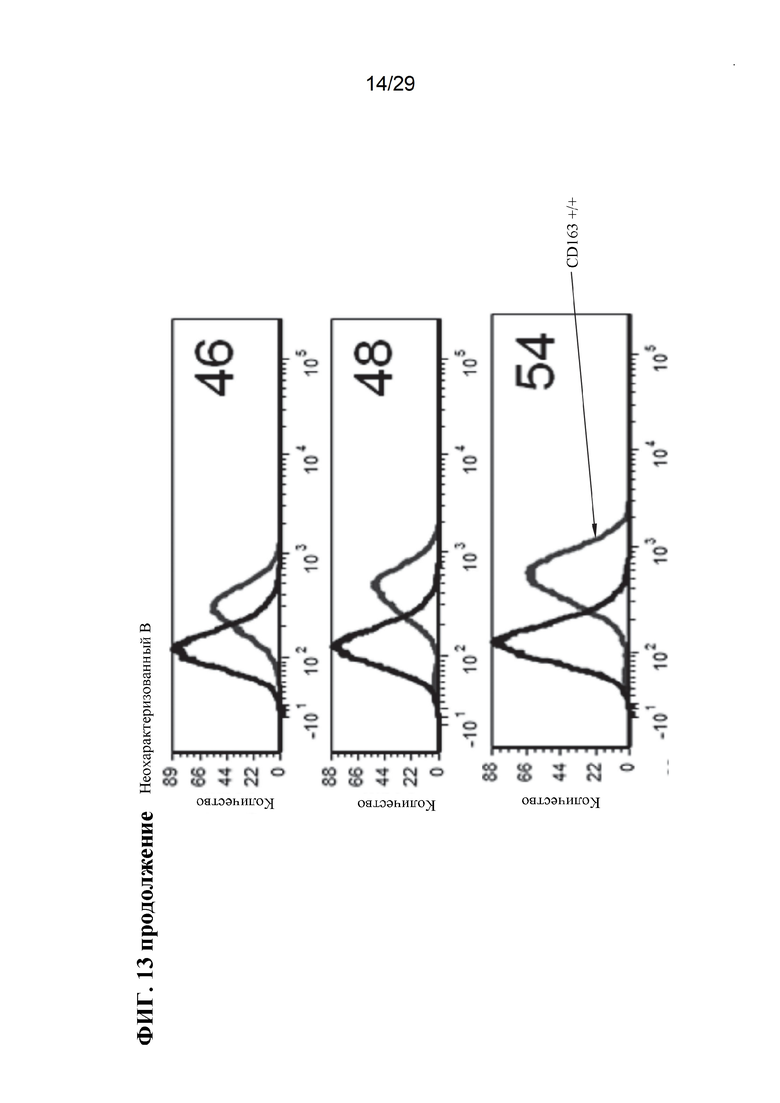
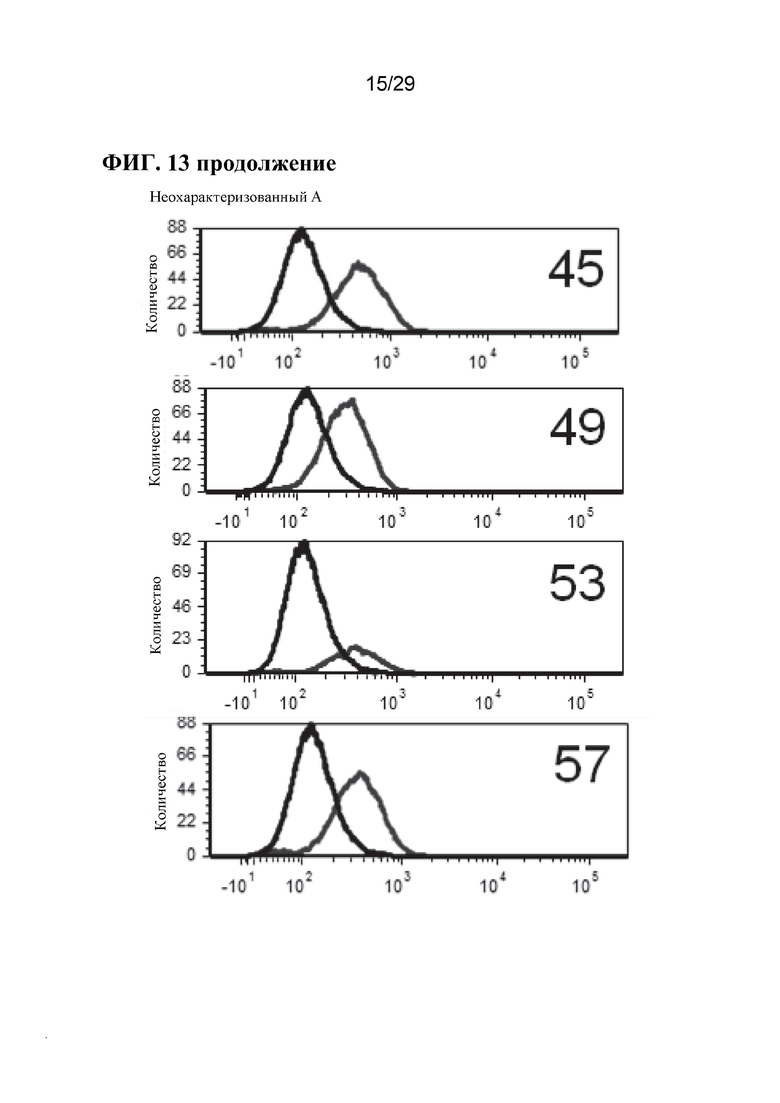
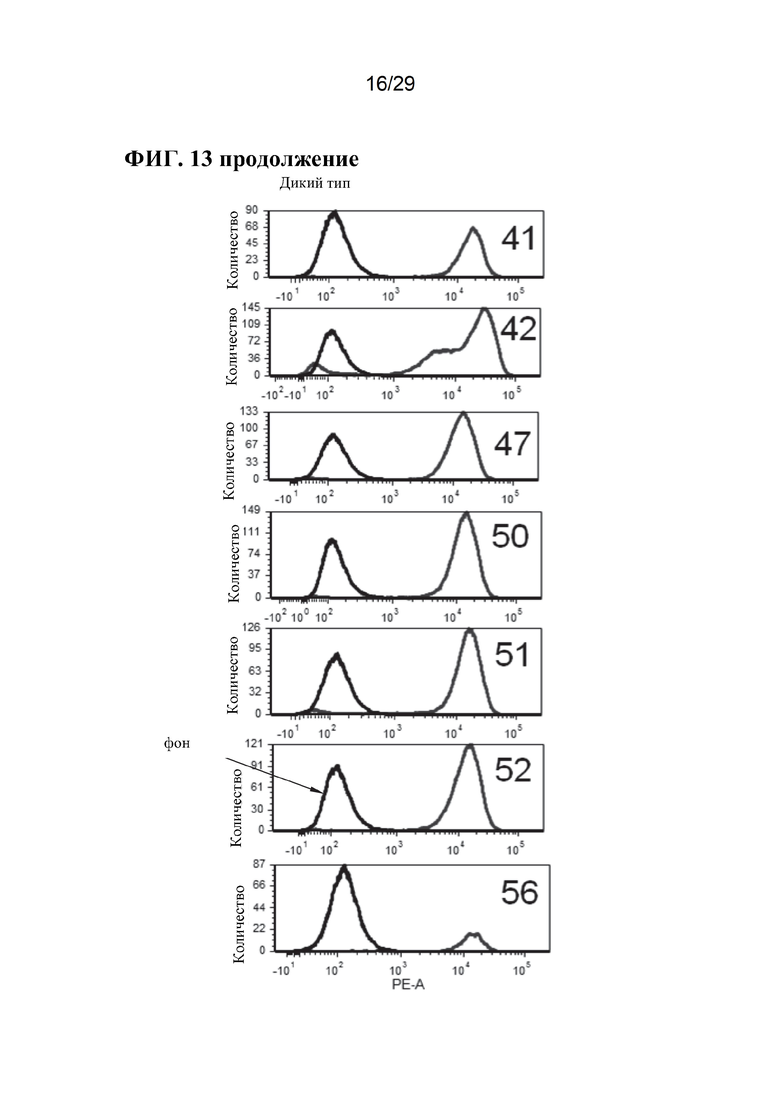
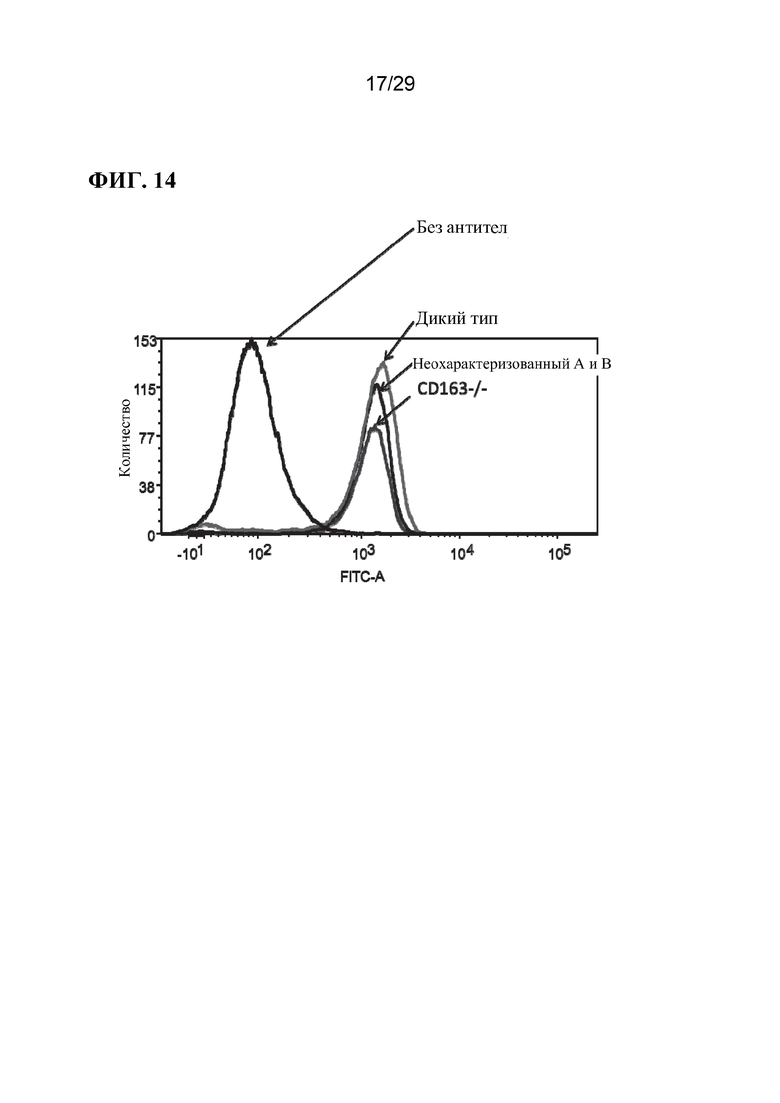
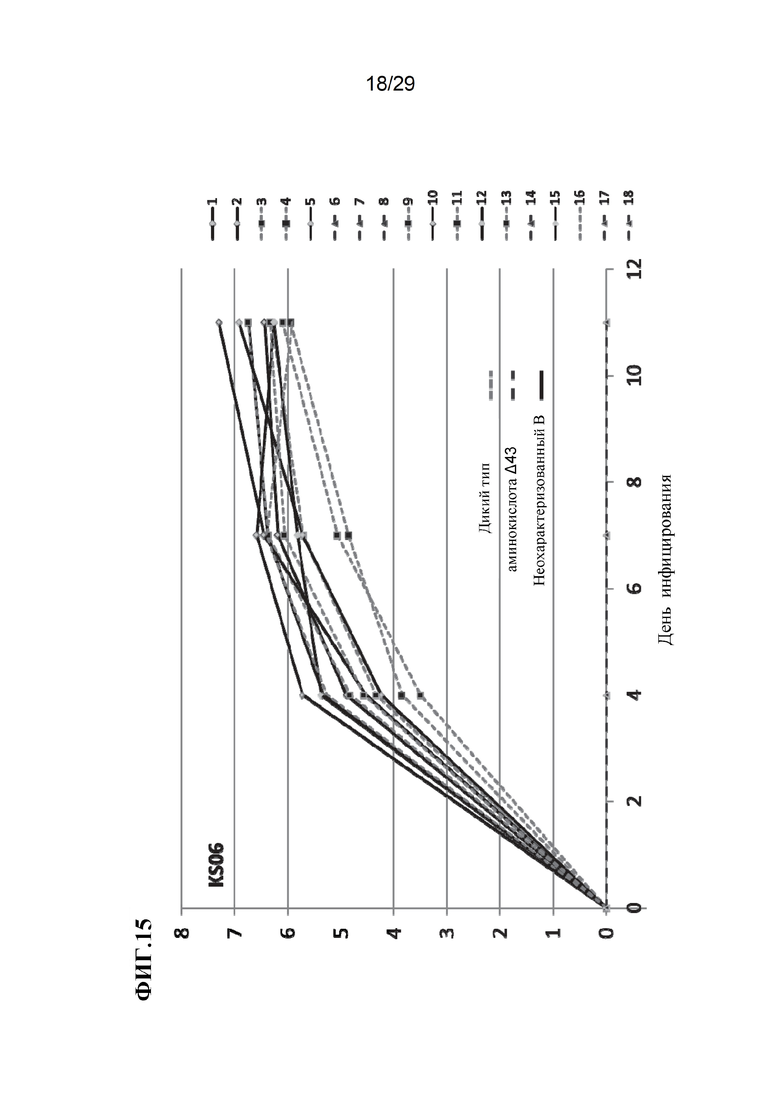


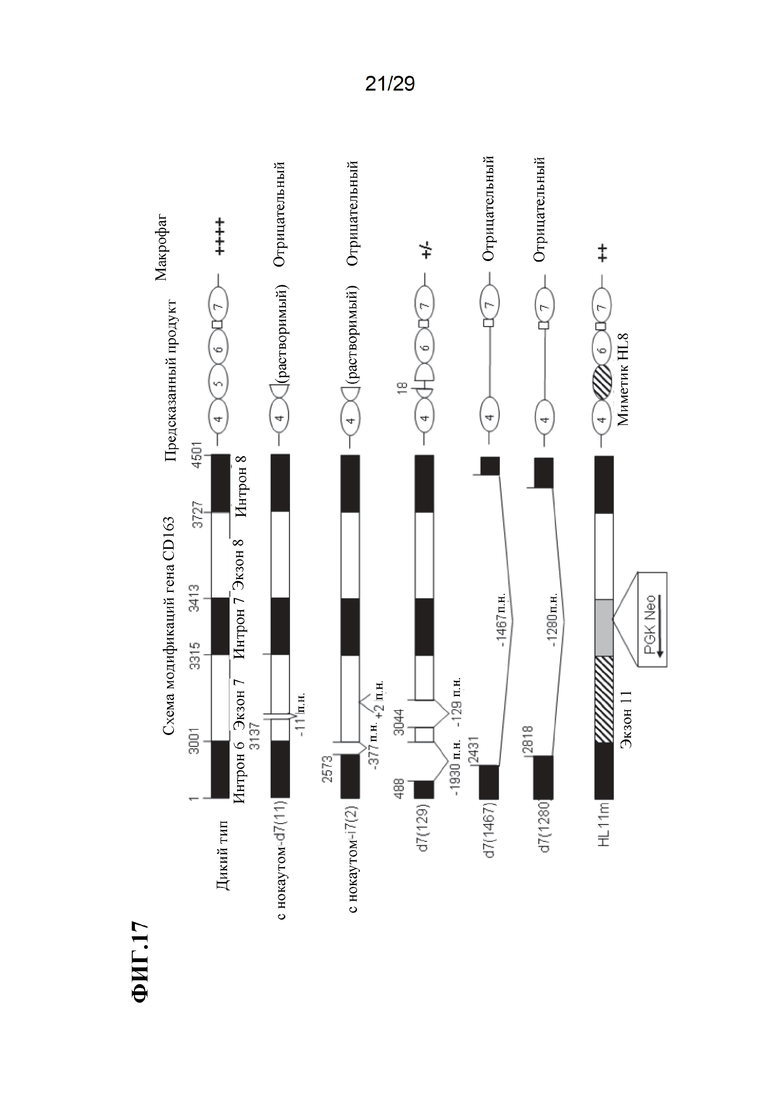
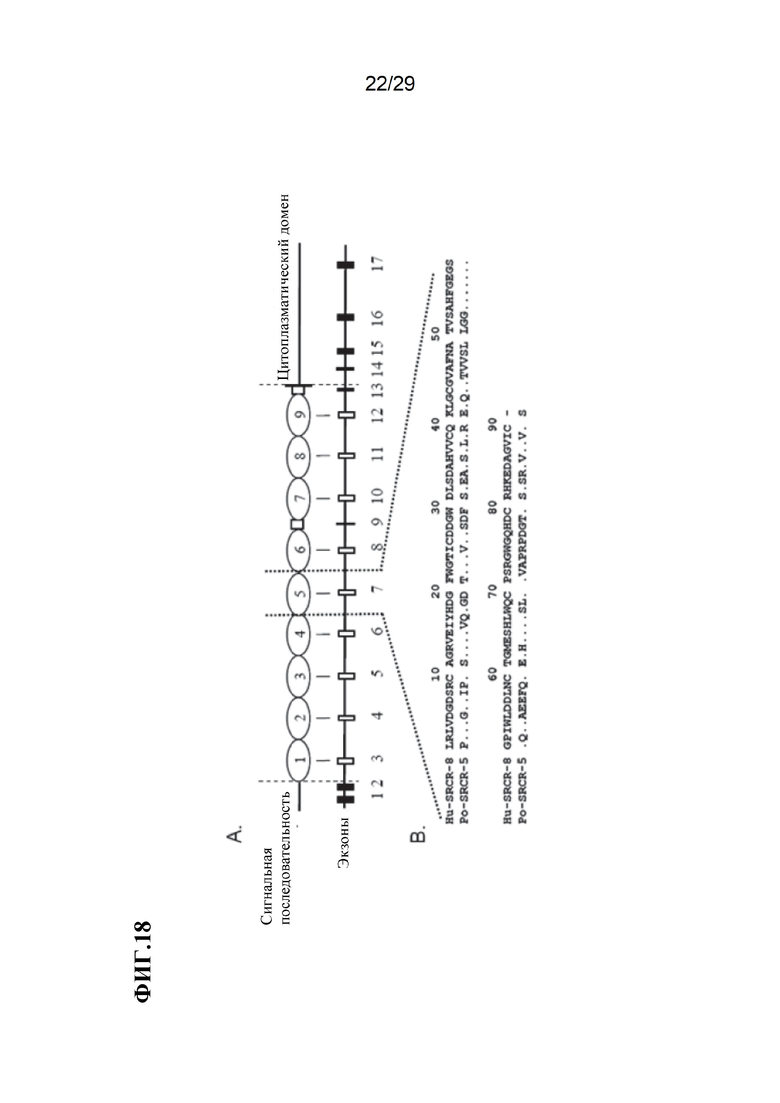
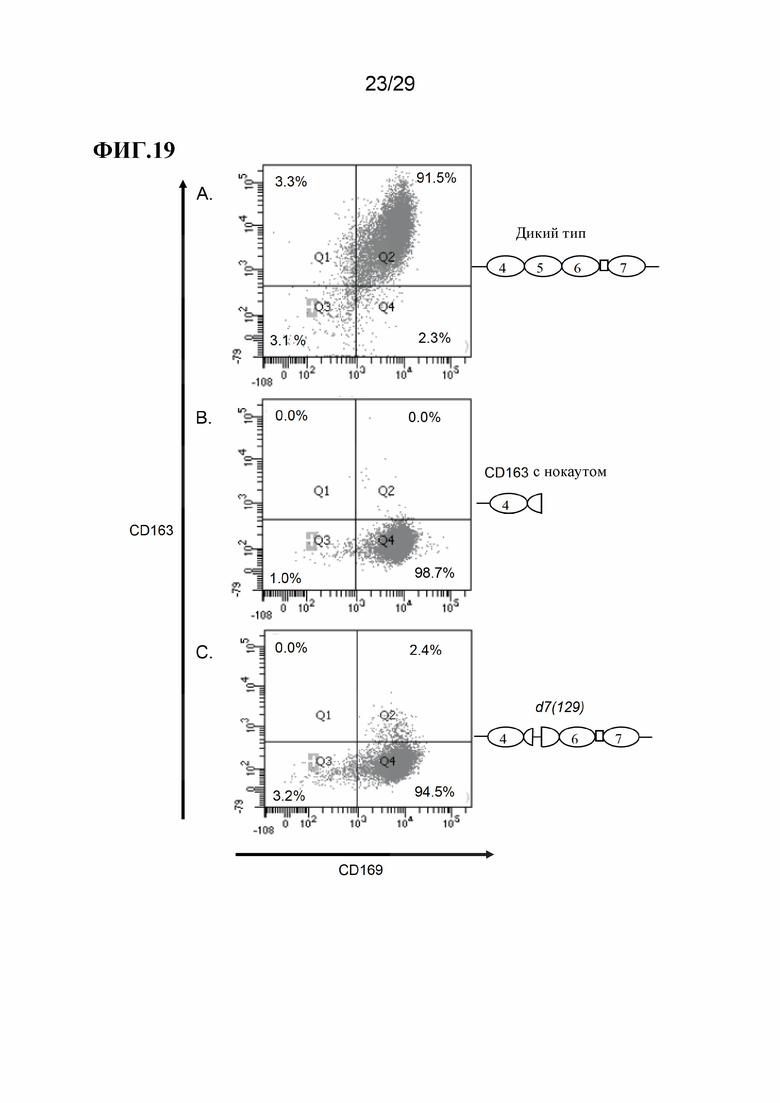
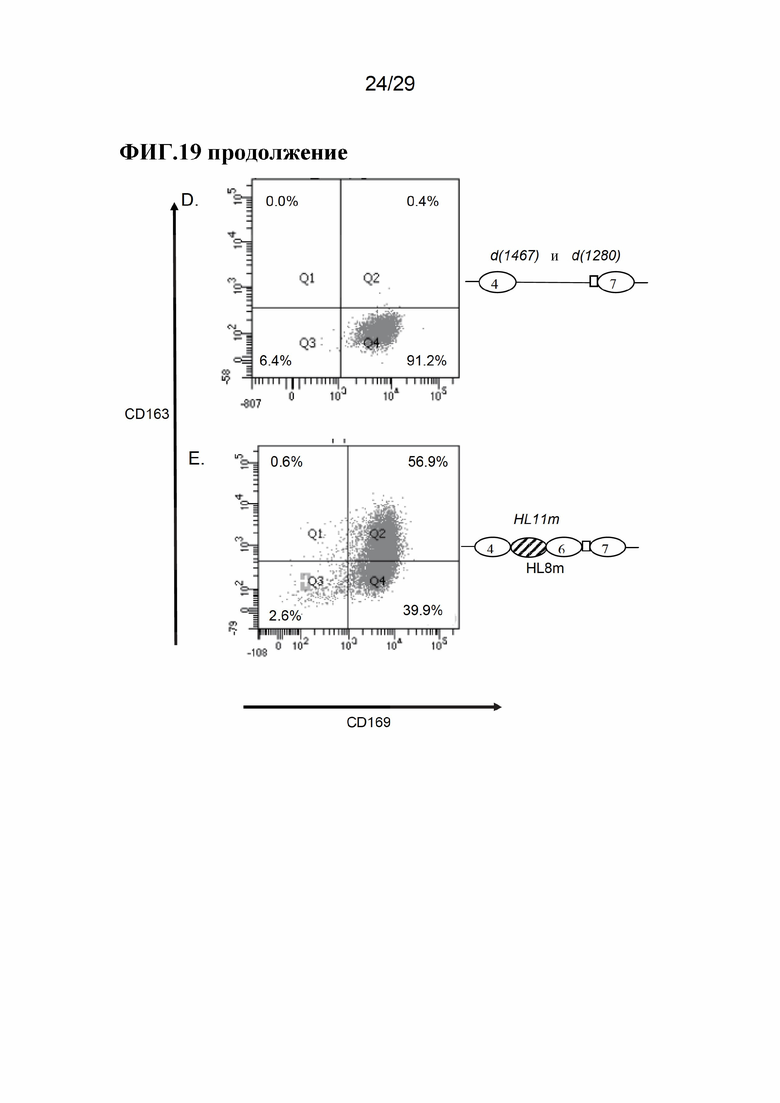
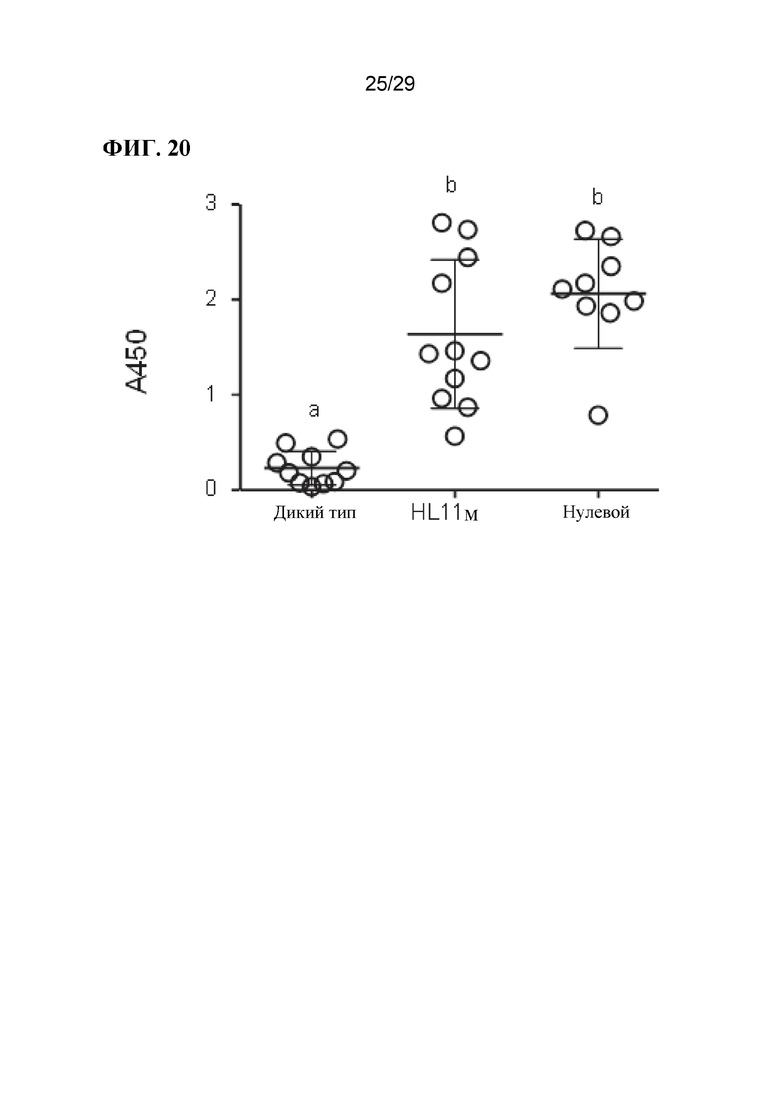
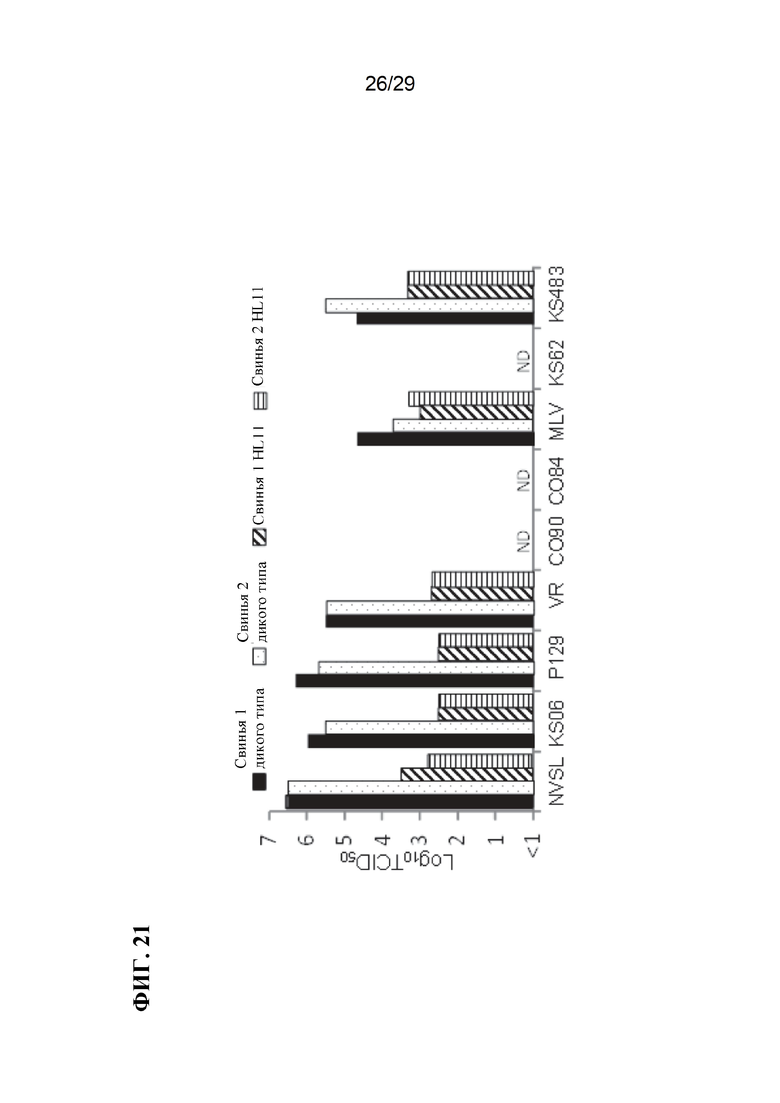
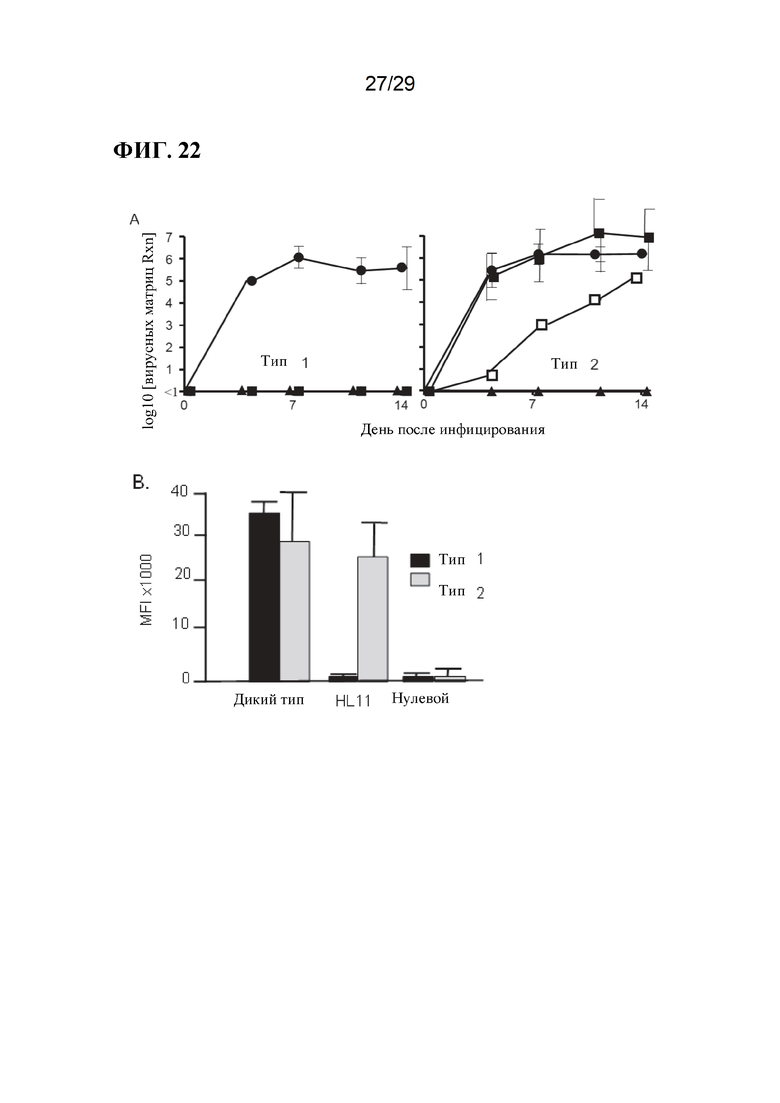
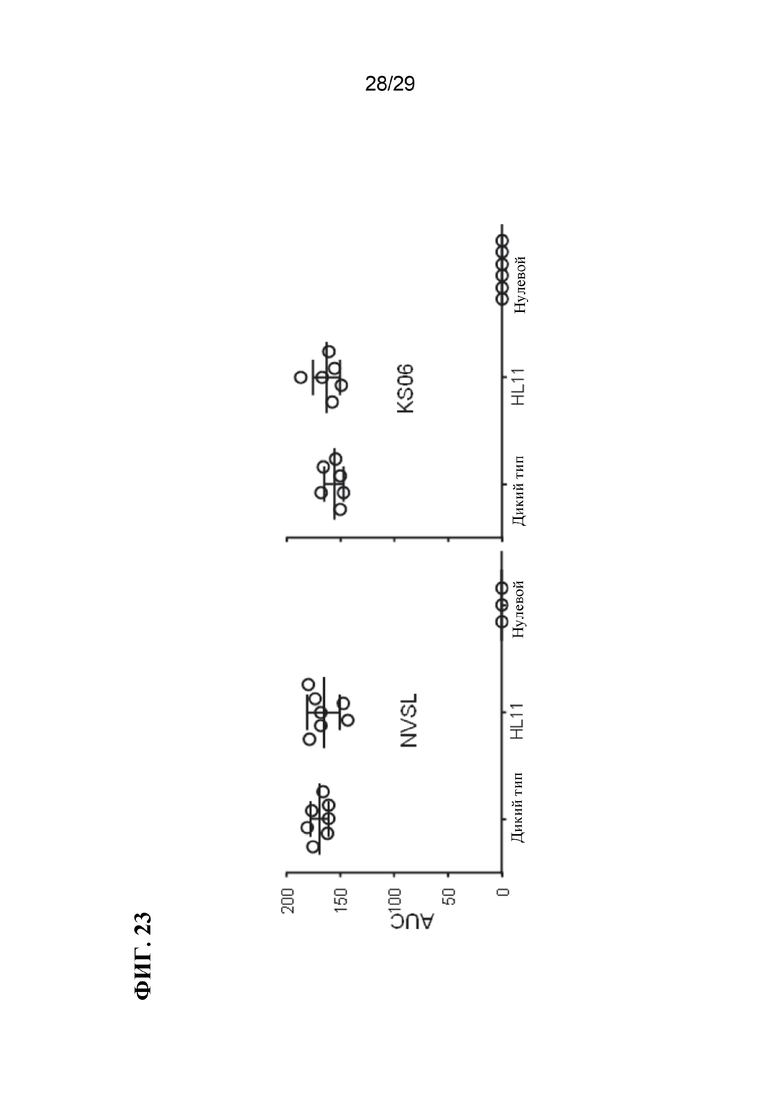
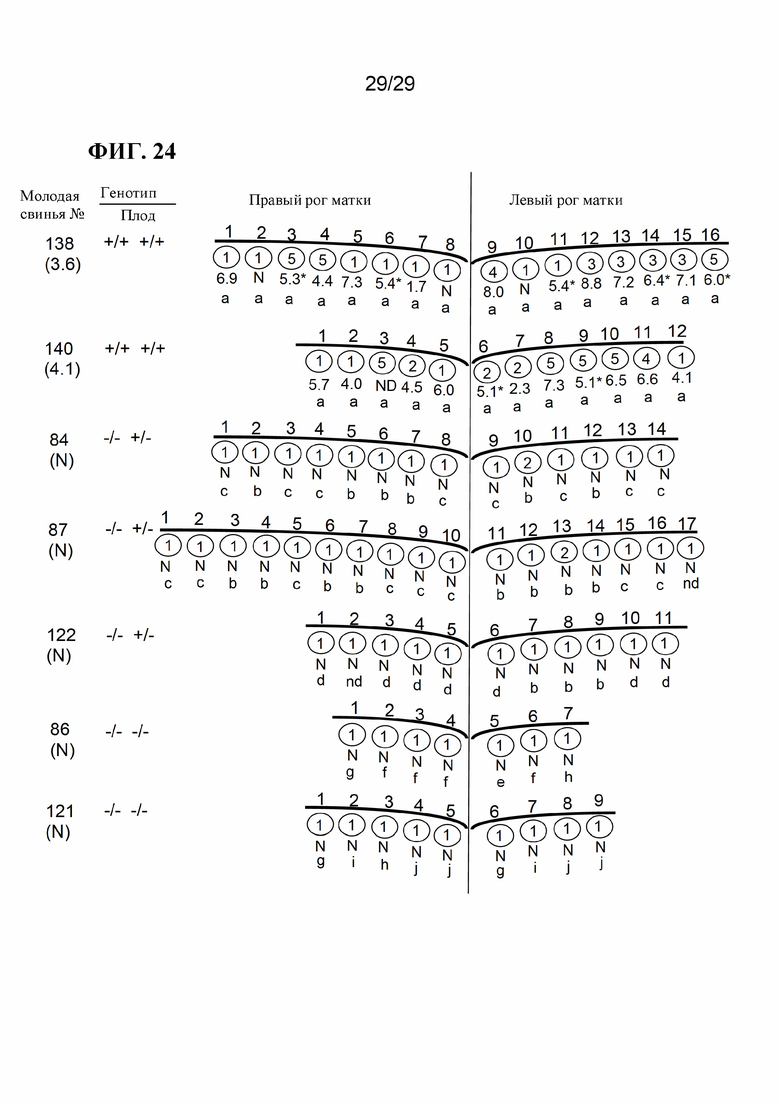
Изобретение относится к области биотехнологии, в частности к способу защиты плода свиньи от заражения вирусом репродуктивного и респираторного синдрома свиней (PRRSV), включающему разведение самки свиньи с самцом свиньи. При этом самка свиньи содержит модифицированные хромосомные последовательности в обоих аллелях гена CD163, причем модифицированная хромосомная последовательность представляет собой вставку в гене, кодирующем белок CD163, делецию в гене, кодирующем белок CD163, или их комбинацию. Изобретение эффективно для снижения чувствительности самки свиньи к заражению PRRSV. 30 з.п. ф-лы, 24 ил., 18 табл., 4 пр.
1. Способ защиты плода свиньи от заражения вирусом репродуктивного и респираторного синдрома свиней (PRRSV), включающий разведение самки свиньи с самцом свиньи, при этом:
самка свиньи содержит модифицированные хромосомные последовательности в обоих аллелях гена CD163, причем модифицированная хромосомная последовательность представляет собой вставку в гене, кодирующем белок CD163, делецию в гене, кодирующем белок CD163, или их комбинацию,
причем вставка выбрана из вставки из 7 пар оснований между нуклеотидом 3148 и нуклеотидом 3149 по сравнению с референсной последовательностью SEQ ID NO: 47, и вставки из 2 пар оснований между нуклеотидами 3149 и 3150 по сравнению с референсной последовательностью SEQ ID NO: 47; и вставки 1 пары оснований между нуклеотидами 3147 и 3148 по сравнению с референсной последовательностью SEQ ID NO: 47, и
причем делеция выбрана из делеции 11 пар оснований от нуклеотида 3137 до нуклеотида 3147 по сравнению с референсной последовательностью SEQ ID NO: 47, делеции 1280 пар оснований от нуклеотида 2818 до нуклеотида 4097 по сравнению с референсной последовательностью SEQ ID NO: 47, делеции 1930 пар оснований от нуклеотида 488 до нуклеотида 2417 по сравнению с референсной последовательностью SEQ ID NO: 47, которая заменена вставкой из 12 пар оснований, начинающейся с нуклеотида 488, делеции 129 пар оснований в экзоне 7 от нуклеотида 3044 до нуклеотида 3172 по сравнению с референсной последовательностью SEQ ID NO: 47, делеции 1467 пар оснований от нуклеотида 2431 до нуклеотида 3897 по сравнению с референсной последовательностью SEQ ID NO: 47, делеции в рамке считывания в экзоне 7 гена, кодирующего белок CD163, делеции экзонов 7 и 8 гена, кодирующего белок CD163, делеции в экзоне 7 гена, кодирующего белок CD163, что приводит к введению стоп-кодона в экзон 7, делеции 123 пар оснований от нуклеотида 3024 до нуклеотида 3146 по сравнению с референсной последовательностью SEQ ID NO: 47, делеции 1373 пар оснований от нуклеотида 2724 до нуклеотида 4096 по сравнению с референсной последовательностью SEQ ID NO: 47, делеции 1720 пар оснований от нуклеотида 2440 до нуклеотида 4160 по сравнению с референсной последовательностью SEQ ID NO: 47, делеции 124 пар оснований от нуклеотида 3024 до нуклеотида 3147 по сравнению с референсной последовательностью SEQ ID NO: 47, делеции 130 пар оснований от нуклеотида 3030 до нуклеотида 3159 по сравнению с референсной последовательностью SEQ ID NO: 47, делеции 132 пар оснований от нуклеотида 3030 до нуклеотида 3161 по сравнению с референсной последовательностью SEQ ID NO: 47, делеции 28 пар оснований от нуклеотида 3145 до нуклеотида 3172 по сравнению с референсной последовательностью SEQ ID NO: 47,
причем модифицированные хромосомные последовательности снижают чувствительность самки свиньи к заражению PRRSV по сравнению с чувствительностью к заражению PRRSV самки свиньи, которая не содержит любые модифицированные хромосомные последовательности в аллелях гена CD163; и
самец свиньи содержит по меньшей мере один аллель CD163 дикого типа.
2. Способ по п. 1, отличающийся тем, что самец свиньи содержит два аллеля CD163 дикого типа.
3. Способ по п. 1 или 2, отличающийся тем, что модифицированные хромосомные последовательности снижают чувствительность плода свиньи к вирусу PRRSV Типа 1, PRRSV Типа 2 или к обоим вирусам PRRSV Типа 1 и Типа 2.
4. Способ по п. 3, отличающийся тем, что модифицированные хромосомные последовательности снижают чувствительность самки свиньи к изоляту PRRSV, выбранному из группы, состоящей из NVSL 97-7895, KS06-72109, P129, VR2332, CO90, AZ25, MLV-ResPRRS, KS62-06274, KS483 (SD23983), CO84, SD13-15, Lelystad, 03-1059, 03-1060, SD01-08, 4353PZ и их комбинаций.
5. Способ по любому из пп. 1–4, отличающийся тем, что самка свиньи включает генетически модифицированную самку свиньи.
6. Способ по п. 5, отличающийся тем, что самка свиньи генетически модифицирована с применением хоминг-эндонуклеазы.
7. Способ по п. 6, отличающийся тем, что хоминг-эндонуклеаза включает сконструированную хоминг-эндонуклеазу.
8. Способ по п. 6 или 7, отличающийся тем, что хоминг-эндонуклеаза включает систему кластерных коротких палиндромных повторов с регулярными интервалами (CRISPR), эффекторную нуклеазу, подобную активатору транскрипции (TALEN), нуклеазу цинковых пальцев (ZFN), слитый белок рекомбиназы, мегануклеазу или комбинацию любых из них.
9. Способ по любому из пп. 1–8, отличающийся тем, что самка свиньи генетически модифицирована с применением системы CRISPR.
10. Способ по любому из пп. 1–9, отличающийся тем, что самка свиньи содержит первую модифицированную хромосомную последовательность в первом аллеле гена CD163 и вторую модифицированную хромосомную последовательность во втором аллеле гена CD163, причем первая и вторая модифицированные хромосомные последовательности отличаются друг от друга.
11. Способ по любому из пп. 1–10, отличающийся тем, что модифицированные хромосомные последовательности вызывают снижение продукции или активности белка CD163 по сравнению с продукцией или активностью белка CD163 у самки свиньи, у которой отсутствуют модифицированные хромосомные последовательности.
12. Способ по любому из пп. 1–11, отличающийся тем, что модифицированные хромосомные последовательности приводят к практически отсутствию продуцирования функционального белка CD163 у самки свиньи.
13. Способ по любому из пп. 1–12, отличающийся тем, что самка свиньи не продуцирует белок CD163.
14. Способ по любому из пп. 1–13, отличающийся тем, что модифицированные хромосомные последовательности в одном или обоих аллелях гена CD163 самки свиньи приводят к неправильному кодированию.
15. Способ по п. 14, отличающийся тем, что неправильное кодирование приводит к преждевременному стоп-кодону ниже неправильного кодирования в аллеле гена CD163.
16. Способ по любому из пп. 1-15, отличающийся тем, что:
модификация включает вставку из 2 пар оснований между нуклеотидами 3149 и 3150 по сравнению с референсной последовательностью SEQ ID NO: 47, с делецией 377 пар оснований от нуклеотида 2573 до нуклеотида 2949 по сравнению с референсной последовательностью SEQ ID NO: 47 на том же аллеле, и вставка из 2 пар оснований включает динуклеотид AG;
модификация включает вставку из 1 пары оснований между нуклеотидами 3147 и 3148 по сравнению с референсной последовательностью SEQ ID NO: 47, и вставка из 1 пары оснований содержит единичный остаток аденина;
модификация включает вставку из 7 пар оснований между нуклеотидом 3148 и нуклеотидом 3149 по сравнению с референсной последовательностью SEQ ID NO: 47, и вставка из 7 пар оснований содержит последовательность TACTACT (SEQ ID NO: 115);
модификация включает делецию 1930 пар оснований от нуклеотида 488 до нуклеотида 2417 по сравнению с референсной последовательностью SEQ ID NO: 47, где удаленная последовательность заменена вставкой из 12 пар оснований, начинающейся с нуклеотида 488, и при этом имеется дополнительная делеция 129 пар оснований в экзоне 7 от нуклеотида 3044 до нуклеотида 3172 по сравнению с референсной последовательностью SEQ ID NO: 47, и где вставка из 12 пар оснований содержит последовательность TGTGGAGAATTC (SEQ ID NO: 116); или
модификация включает делецию 1382 пар оснований от нуклеотида 3113 до нуклеотида 4494 по сравнению с референсной последовательностью SEQ ID NO: 47, где удаленная последовательность заменена вставкой из 11 пар оснований, начинающейся с нуклеотида 3113, и вставка из 11 пар оснований включает последовательность AGCCAGCGTGC (SEQ ID NO: 117).
17. Способ по п. 16, отличающийся тем, что по меньшей мере один из аллелей гена CD163 самки свиньи содержит модификацию, выбранную из группы, состоящей из:
вставки из 7 пар оснований между нуклеотидом 3148 и нуклеотидом 3149 по сравнению с референсной последовательностью SEQ ID NO: 47;
вставки из 2 пар оснований между нуклеотидами 3149 и 3150 по сравнению с референсной последовательностью SEQ ID NO: 47, с делецией 377 пар оснований от нуклеотида 2573 до нуклеотида 2949 по сравнению с референсной последовательностью SEQ ID NO: 47 на том же аллеле;
делеции 11 пар оснований от нуклеотида 3137 до нуклеотида 3147 по сравнению с референсной последовательностью SEQ ID NO: 47;
делеции 1382 пар оснований от нуклеотида 3113 до нуклеотида 4494 по сравнению с референсной последовательностью SEQ ID NO: 47, где удаленная последовательность заменена вставкой из 11 пар оснований, начинающейся с нуклеотида 3113;
и комбинаций любых из них.
18. Способ по п. 17, отличающийся тем, что самка свиньи содержит:
(а) вставку из 7 пар оснований между нуклеотидом 3148 и нуклеотидом 3149 по сравнению с референсной последовательностью SEQ ID NO: 47 в одном аллеле гена CD163; и
вставку из 2 пар оснований между нуклеотидами 3149 и 3150 по сравнению с референсной последовательностью SEQ ID NO: 47, с делецией 377 пар оснований от нуклеотида 2573 до нуклеотида 2949 по сравнению с референсной последовательностью SEQ ID NO: 47, в другом аллеле гена CD163;
(б) делецию 1382 пар оснований от нуклеотида 3113 до нуклеотида 4494 по сравнению с референсной последовательностью SEQ ID NO: 47, где удаленная последовательность заменена вставкой из 11 пар оснований, начинающейся с нуклеотида 3113, в одном аллеле гена CD163; и
вставку из 2 пар оснований между нуклеотидами 3149 и 3150 по сравнению с референсной последовательностью SEQ ID NO: 47, с делецией 377 пар оснований от нуклеотида 2573 до нуклеотида 2949 по сравнению с референсной последовательностью SEQ ID NO: 47, в другом аллеле гена CD163;
(в) вставку из 7 пар оснований между нуклеотидом 3148 и нуклеотидом 3149 по сравнению с референсной последовательностью SEQ ID NO: 47 в одном аллеле гена CD163; и
делецию 11 пар оснований от нуклеотида 3137 до нуклеотида 3147 по сравнению с референсной последовательностью SEQ ID NO: 47 в другом аллеле гена CD163; или
(г) делецию 1382 пар оснований от нуклеотида 3113 до нуклеотида 4494 по сравнению с референсной последовательностью SEQ ID NO: 47, где удаленная последовательность заменена вставкой из 11 пар оснований, начинающейся с нуклеотида 3113, в одном аллеле гена CD163; и
делецию 11 пар оснований от нуклеотида 3137 до нуклеотида 3147 по сравнению с референсной последовательностью SEQ ID NO: 47 в другом аллеле гена CD163.
19. Способ по п. 18, отличающийся тем, что самка свиньи содержит:
(а) вставку из 7 пар оснований между нуклеотидом 3148 и нуклеотидом 3149 по сравнению с референсной последовательностью SEQ ID NO: 47 в одном аллеле гена CD163; и
вставку из 2 пар оснований между нуклеотидами 3149 и 3150 по сравнению с референсной последовательностью SEQ ID NO: 47, с делецией 377 пар оснований от нуклеотида 2573 до нуклеотида 2949 по сравнению с референсной последовательностью SEQ ID NO: 47, в другом аллеле гена CD163; или
(б) делецию 1382 пар оснований от нуклеотида 3113 до нуклеотида 4494 по сравнению с референсной последовательностью SEQ ID NO: 47, где удаленная последовательность заменена вставкой из 11 пар оснований, начинающейся с нуклеотида 3113, в одном аллеле гена CD163; и
вставку из 2 пар оснований между нуклеотидами 3149 и 3150 по сравнению с референсной последовательностью SEQ ID NO: 47, с делецией 377 пар оснований от нуклеотида 2573 до нуклеотида 2949 по сравнению с референсной последовательностью SEQ ID NO: 47, в другом аллеле гена CD163.
20. Способ по любому из пп.1-19, отличающийся тем, что аллели гена CD163 самки свиньи содержат хромосомную последовательность, имеющую по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98%, по меньшей мере 99%, по меньшей мере 99,9% или 100% идентичность последовательности SEQ ID NO: 47 в областях указанной хромосомной последовательности вне вставки или делеции.
21. Способ по любому из пп. 1–20, отличающийся тем, что самка свиньи содержит хромосомную последовательность, включающую SEQ ID NO: 98, 99, 100, 101, 102, 103, 104, 105, 106, 107, 108, 109, 110, 111, 112, 113, 114 или 119 в одном или в обоих аллелях гена CD163.
22. Способ по п. 21, отличающийся тем, что самка свиньи содержит хромосомную последовательность, включающую SEQ ID NO: 99, 102, 103 или 113 в одном или в обоих аллелях гена CD163.
23. Способ по п. 22, отличающийся тем, что самка свиньи содержит:
(а) хромосомную последовательность, включающую SEQ ID NO: 99 в одном аллеле гена CD163, и хромосомную последовательность, включающую SEQ ID NO: 103 в другом аллеле гена CD163;
(б) хромосомную последовательность, включающую SEQ ID NO: 113 в одном аллеле гена CD163, и хромосомную последовательность, включающую SEQ ID NO: 99 в другом аллеле гена CD163;
(в) хромосомную последовательность, включающую SEQ ID NO: 99 в одном аллеле гена CD163, и хромосомную последовательность, включающую SEQ ID NO: 102 в другом аллеле гена CD163; или
(г) хромосомную последовательность, включающую SEQ ID NO: 113 в одном аллеле гена CD163, и хромосомную последовательность, включающую SEQ ID NO: 102 в другом аллеле гена CD163.
24. Способ по п. 23, отличающийся тем, что самка свиньи содержит:
(a) хромосомную последовательность, включающую SEQ ID NO: 99 в одном аллеле гена CD163, и хромосомную последовательность, включающую SEQ ID NO: 103 в другом аллеле гена CD163; или
(б) хромосомную последовательность, включающую SEQ ID NO: 113 в одном аллеле гена CD163, и хромосомную последовательность, включающую SEQ ID NO: 99 в другом аллеле гена CD163.
25. Способ по любому из пп. 1–24, отличающийся тем, что в результате разведения получают один или более плодов, которые содержат модифицированную хромосомную последовательность в единичном аллеле гена CD163.
26. Способ по п. 25, отличающийся тем, что плоды имеют пониженную чувствительность к заражению PRRSV в утробе матери по сравнению с плодами в утробе матери - самки свиньи дикого типа.
27. Способ по любому из пп. 1–26, отличающийся тем, что разведение включает скрещивание самки свиньи с самцом свиньи.
28. Способ по любому из пп. 1–26, отличающийся тем, что разведение включает искусственное оплодотворение самки спермой, полученной от самца.
29. Способ по любому из пп. 1–26, отличающийся тем, что разведение включает перенос оплодотворенного ооцита в репродуктивный тракт самки свиньи.
30. Способ по п. 29, отличающийся тем, что оплодотворенный ооцит был получен путем оплодотворения ооцита in vitro спермой, полученной от самца свиньи.
31. Способ по п. 30, отличающийся тем, что оплодотворение in vitro включает внутрицитоплазматическую инъекцию ооцита со спермой, полученной от самца свиньи.
| US2018027783 A1, 01.02.2018 | |||
| WO2017023570 A1, 09.02.2017 | |||
| YANG H | |||
| et al., CD163 knockout pigs are fully resistant to highly pathogenic porcine reproductive and respiratory syndrome virus, Antiviral Research, 2018, doi: 10.1016/j.antiviral.2018.01.004 | |||
| JAY G | |||
| CALVERT et al., CD163 Expression Confers Susceptibility to Porcine Reproductive and |

