ГОСУДАРСТВЕННАЯ ПОДДЕРЖКА
[0001] Данное изобретение было выполнено при поддержке Правительства согласно договору номер (USDA/ARS) 58-1940-8-868, представленной Департаментом Сельского Хозяйства. Правительство обладает определенными правами на изобретение.
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
[0002] В данной заявке содержится Перечень Последовательностей, озаглавленный «UMO 11053. WO SEQ_ST25», созданный 15 мая 2012, который включен в данный документ во всей полноте посредством ссылки.
ОБЛАСТЬ ТЕХНИКИ
[0003] Настоящее изобретение, в общем плане, относится к генетически модифицированной свинье, у которой по меньшей мере один аллель гена SIGLEC1 был инактивирован и/или был инактивирован по меньшей мере один аллель гена CD163. Также предложены способы получения таких трансгенных свиней.
УРОВЕНЬ ТЕХНИКИ
[0004] Репродуктивно-Респираторный Синдром Свиней (РРСС) является одной из наиболее экономически значимых болезней свиней. Впервые это заболевание было выявлено в США в 1987 (Keffaber 1989), а в Европе - в 1990 (Wensvoort et al. 1991). Молекулярный анализ прототипных вирусов РРСС (ВРРСС) VR-2332 и Lelystad (американский и европейский изоляты, соответственно) позволил предположить, что на двух континентах почти одновременно появились два дивергентных штамма, вероятно, по причине похожих изменений в работе со свиньями (Murtaugh et al. 1995; Nelsen et al. 1999). После возникновения этот вирус распространился во всем мире и ВРРСС Европейского генотипа был выявлен в американских стадах свиней (Ropp et al. 2004). РРСС характеризуется тяжелой и иногда смертельной дыхательной недостаточностью и репродуктивной недостаточностью, а также предрасполагает инфицированных свиней к бактериальным патогенам, а также иным вирусным патогенам (Benfield et al. 1992), и является ключевым элементом экономически значимого Комплекса Респираторных Заболеваний Свиней (КРЗС). Наиболее постоянными патологическими повреждениями, вызываемыми ВРРСС при острой инфекции, являются интерстициальная пневмония и легкий лимфоцитарный энцефалит (Plagemann 1996). После острой фазы ВРРСС-инфекции, обычно характеризующейся виремией и клиническим заболеванием, множество свиней полностью выздоравливают, хотя и являются носителями вируса с низкой нагрузкой в течение длительного периода времени. Эти свиньи-носители постоянно инфицированы ВРРСС и периодически или постоянно выделяют вирус, поэтому могут инфицировать интактных свиней при прямом или косвенном контакте. Хроническая инфекция ВРРСС была хорошо задокументирована в экспериментальных условиях (Albina et al. 1994; Allende et al. 2000; Benfield et al. 1998; Christopherhennings et al. 1995; Sur et al. 1996; Yoon et al. 1993). В частности, инфекционный вирус обнаруживался до 157 суток после инфекции (Wills et al. 1997). Основными клетками-мишенями и при острой, и при хронической инфекции являются макрофаги и моноциты (Molitor et al. 1997), хотя было установлено, что также поражаются пневмоциты и стволовые клетки эпителия яичек (Sur et al. 1996; Suretal. 1997).
[0005] Этиологическим агентом РРСС является оболочечный вирус, содержащий РНК положительной полярности, принадлежащий к семейству Arteriviridae отряда Nidovirales. Другие члены семейства Arteriviridae включают вирус подъема уровня лактатдегидрогеназы (LDV) мышей, вирус артериита лошадей (EAV) и вирус геморрагической лихорадки обезьян (SHFV). Анализ данных о геномной последовательности выявил наличие большого разнообразия между штаммами ВРРСС, а также существование высококонсервативных доменов (Andreyev et al. 1997; Meng 2000; Meng et al. 1995). Организация генома ВРРСС аналогична с другими Артеривирусами, геномная РНК функционирует как информационная РНК для репликазных белков ORF1a (Plagemann 1996). ORF1a и 1b включают приблизительно 80% вирусного генома и кодируют РНК-зависимую РНК-полимеразу, а также полипротеины, которые подвергаются процессингу в неструктурные белки (Snijder, Meulenberg 1998). С помощью вируса Lelystad, было выявлено, что ORF 2-7 кодируют вирусные структурные белки. Белок, закодированный ORF 5 (GP5) и М (Van Breedam et al. 2010b) может играть роль в индуцировании апоптоза, вызываемого ВРРСС (Suarez et al. 1996; Sur et al. 1997) и, как предполагается, является белком прикрепления в близкородственном Вирусе подъема уровня лактатдегиддрогеназы. Малые оболочечные гликопротеины GP2a и GP4 ВРРСС взаимодействуют с CD163 (Das et al. 2010). Имеются данные, исходя из которых можно предположить, что связывание с SIGLEC1 (иммуноглобулиноподобный лектин 1, связывающий сиаловую кислоту) необходимо для проникновения в клетки и, фактически, для вирусной инфекции необходимо двойное связывание и с SIGLEC1, и с CD163 (Van Gorp et al. 2008).
[0006] Многие характеристики патогенеза (особенно на молекулярном уровне) и эпизоотиологии ВРРСС плохо установлены, поэтому попытки его контроля затруднительны. Чтобы лучше его изучить, были разработаны инфекционные клоны ВРРСС (Nielsen et al. 2003). В настоящее время производители часто вакцинируют свиней против ВРРСС вакцинами с модифицированными живыми ослабленными штаммами или с убитыми вирусами. Однако доступные в настоящее время вакцины часто не обеспечивают удовлетворительной защиты, зачастую по причине вариабельности вируса и недостаточной стимуляции иммунной системы. Защитный иммунный ответ возможен, поскольку было показано, что предшествующее воздействие может обеспечить полную защиту при нагрузке свиней гомологичным штаммом ВРРСС (Lager et al. 1999). Однако при нагрузке гетерологическими штаммами защитный иммунитет не был ни разу достоверно продемонстрирован. Помимо неудовлетворительной эффективности имеющихся вакцин против ВРРСС, имеются убедительные свидетельства в пользу того, что используемая модифицированная живая вакцина может персистировать в отдельных стадах свиней и накапливать мутации (Mengeling et al. 1999), как это было показано с вирулентными полевыми изолятами после экспериментального инфицирования свиней (Rowland et al. 1999). Помимо этого, было показано, что вирус из вакцины выделяется со спермой вакцинированных хряков (Christopherhermings et al. 1997). Вместо вакцинации, некоторые авторы выступают в пользу стратегии «проверки и удаления» в племенных стадах (Dee, Molitor 1998). Успешность применения этой стратегии зависит от удаления всех свиней и с острой, и с хронической инфекцией ВРРСС с последующим строгим контролем для профилактики повторного внедрения вируса. Сложность и большая часть издержек, связанных с этой стратегией, заключаются в том, что о патогенезе хронической ВРРСС-инфекции мало известно и поэтому нет надежных способов для идентификации хронически инфицированных свиней.
[0007] Предполагаемый клеточный рецептор ВРРСС, который был идентифицирован при помощи моноклональных антител, очищен и секвенирован (Vanderheijden et al. 2003; Wissink et al. 2003), получил наименование SIGLEC1. Эта молекула обладает сходством с сиалоадгезинами и, как было показано, опосредует проникновение ВРРСС в невосприимчивые клетки, однако рекомбинантная клеточная линия, экспрессирующая этот рецептор, не смогла поддержать продуктивную репликацию ВРРСС (Vanderheijden et al. 2003). Существенно, что молекулы сиаловой кислоты, присутствующие на поверхности ВРРСС, как было показано, необходимы для инфицирования альвеолярных макрофагов. Следом за связыванием вируса с этим рецептором, происходит проникновение ВРРСС по механизму опосредованного рецептором эндоцитоза (Nauwynck et al. 1999). Ключевая роль сиалоадгезина при проникновении ВРРСС была установлена в экспериментах, в которых был продемонстрирован вирусный захват клетками РК15 (клеточная линия, не позволяющая репликацию ВРРСС), трансфицированными свиным сиалоадгезином, но не нетрансфицированными контрольными клетками РК15 (Vanderheijden et al. 2003). Дальнейшие исследования этой группой показали, что взаимодействия между сиаловой кислотой на поверхности вириона ВРРСС и молекулой сиалоадгезина были необходимы для инфицирования ВРРСС альвеолярных макрофагов (Delputte, Nauwynck 2004). Обратная стратегия, а именно, удаление сиаловой кислоты с поверхности ВРРСС или предварительное инкубирование со специфическими для сиаловой кислоты лектинами также привело к блокированию инфекции (Delputte et al. 2004; Delputte, Nauwynck 2004; Van Breedam et al. 2010b). Независимо от специфических исследований проникновения ВРРСС, использовался сайт-направленный мутагенез, чтобы идентифицировать шесть ключевых аминокислотных остатков, необходимых для связывания сиаловой кислоты с мышиным сиалоадгезином (Vinson et al. 1996), который на 69% идентичен свиному сиалоадгезину. Заслуживает внимания то, что шесть аминокислот, идентифицированных в мышином сиалоадгезине, сохранены и в свиной молекуле.
[0008] SIGLEC1 является трансмембранным рецептором семейства иммуноглобулиноподобных лектинов, связывающихся с сиаловой кислотой. Вначале он был описан как рецептор мышиных макрофагов, связывающий овечьи эритроциты (Crocker and Gordon 1986). Он экспрессируется макрофагами в гемопоэтической и лимфоидной тканях. SIGLEC-гены состоят из N-терминального домена из V-набора, содержащего сайт связывания сиаловой кислоты, последующего переменного количества доменов из С2-набора, трансмембранного домена и цитоплазматического хвоста. В отличие от других SIGLEC, SIGLEC1 не имеет тирозинового мотива в цитоплазматическом хвосте (Oetke et al. 2006). Сайт связывания сиаловой кислоты был картирован в N-терминальном иммуноглобулиноподобном домене V-набора (Nath et al. 1995). Остаток R116, судя по всему, является одной из аминокислот, необходимых для связывания с сиаловой кислотой (Crocker et al. 1999; Delputte et al. 2007). Таким образом, интактный N-терминальный домен необходим и достаточен для связывания ВРРСС с SIGLEC1 (Van Breedam et al. 2010a). Сообщалось о SIGLEC 1-нокаутных мышах, которые были жизнеспособны, фертильны и без аномалий развития (Oetke et al. 2006). Однако у этих мышей наблюдались небольшие изменения популяций В- и Т-клеток и понижение уровня иммуноглобулина М.
[0009] Процесс инфекции ВРРСС начинается с первичного связывания с гепарансульфатом на поверхности альвеолярного макрофага. Затем происходит прочное связывание с сиалоадгезином (SIGLEC1, также обозначается CD169 или SN). Затем вирус интернализуется по клатрин-опосредованному эндоцитозу. Затем другая молекула - CD163, способствует обнажению вируса в эндосоме (Van Breedam et al. 2010a). Геном вируса высвобождается и происходит инфицирование клетки.
[0010] В CD163 имеется 17 экзонов, белок состоит из внеклеточной области с 9 цистеин-богатыми (SRCR) доменами фагоцитарных рецепторов, трансмембранного сегмента и короткого цитоплазматического хвоста. При различном сплайсинге одного гена получается несколько разных вариантов (Ritter et al. 1999а; Ritter et al. 1999b). В значительной степени это варьирование обусловлено длиной цитоплазматического хвоста.
[0011] CD163 выполняет ряд важных функций, включая действие в качестве фагоцитарного рецептора к гаптоглобину-гемоглобину. Элиминация свободного гемоглобина из крови является важной функцией CD163, поскольку группа гема может быть очень токсичной (Kristiansen et al. 2001). CD163 имеет цитоплазматический хвост, облегчающий эндоцитоз. Мутация этого хвоста приводит к снижению захвата комплекса гептоглобин-гемоглобин (Nielsen et al. 2006). Другие функции С163 включают адгезию эритробластов (SRCR2), работу в качестве рецептора TWEAK (SRCR 1-4 и 6-9), бактериального рецептора (SRCR5), рецептора вируса африканских свиней (Sanchez-Torres et al. 2003), а также возможную роль в качестве иммуномодулятора (обсуждалось в (Van Gorp et al. 2010a)).
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
[0012] В одном аспекте, настоящее изобретение представляет собой генетически модифицированную свинью, у которой по меньшей мере инактивирован один аллель гена SIGLEC1 и/или инактивирован по меньшей мере один аллель гена CD163, причем инактивация аллеля CD163 приводит к белку CD163, неспособному связывать и/или обнажать вирус репродуктивно-респираторного синдрома свиней (ВРРСС).
[0013] Другой аспект настоящего изобретения представлен генетически модифицированной свиньей, у которой инактивирован по меньшей мере один аллель гена SIGLEC1, полученного по способу, включающему энуклеацию ооцита свиньи; слияние ооцита с фибробластной клеткой донорской свиньи, содержащей в геноме фибробластной клетки по меньшей мере, один инактивированный аллель гена SIGLEC1; и активацию ооцита для получения эмбриона.
[0014] Настоящее изобретение также относится к генетически модифицированной свинье, у которой инактивирован по меньшей мере один аллель гена CD163, причем инактивация аллеля CD163 приводит к белку CD163, неспособному к связыванию и/или обнажению вируса репродуктивно-респираторного синдрома свиней (ВРРСС), полученного по способу, включающему энуклеацию ооцита свиньи; слияние ооцита с фибробластной клеткой донорской свиньи, содержащей в геноме фибробластной клетки по меньшей мере, один инактивированный аллель гена CD163; и активацию ооцита для получения эмбриона.
[0015] В другом аспекте, настоящее изобретение является генетически модифицированной свиньей, у которой инактивированы оба аллеля гена SIGLEC1, полученного по способу, включающему спаривание самца генетически модифицированной свиньи, у которого инактивирован по меньшей мере один аллель гена SIGLEC1, с самкой генетически модифицированной свиньи, у которой инактивирован по меньшей мере один аллель гена SIGLEC1, с получением F1-потомства, и скрининг F1-потомства для выявления генетически модифицированных свиней, у которых инактивированы оба аллеля гена SIGLEC1.
[0016] Другой аспект настоящего изобретения заключается в получении генетически модифицированной свиньи, у которой инактивированы оба аллеля гена CD163, полученного по способу, включающему спаривание самца генетически модифицированной свиньи, у которого инактивирован по меньшей мере, один аллель гена CD163, с самкой генетически модифицированной свиньи, у которой инактивирован по меньшей мере один аллель гена CD163, с получением F1-потомства, и скрининг F1-потомства для выявления генетически модифицированных свиней, у которых инактивированы оба аллеля гена CD163.
[0017] Настоящее изобретение также относится к генетически модифицированной свинье, полученной по любому из трех способов, у которой инактивированы оба аллеля гена S1GLEC1 и оба аллеля гена CD163, причем инактивация аллеля CD163 приводит к белку CD163, неспособному связывать и/или обнажать вирус репродуктивно-респираторного синдрома свиней (ВРРСС). Первый такой способ включает спаривание генетически модифицированной свиньи, у которой инактивирован по меньшей мере один аллель гена SIGLEC1, с генетически модифицированной свиньей, у которой инактивирован по меньшей мере один аллель гена CD163, с получением F1-потомства, и скрининг F1-потомства для выявления генетически модифицированных свиней, у которых инактивирован по меньшей мере один аллель гена SIGLEC1, и инактивирован по меньшей мере один аллель гена CD163. Этот способ дополнительно включает спаривание друг с другом генетически модифицированных свиней, у которых инактивирован по меньшей мере один аллель гена S1GLEC1, и инактивирован по меньшей мере один аллель гена CD163, с получением Р2-потомства, и скрининг Р2-потомства для выявления генетически модифицированных свиней, у которых инактивированы оба аллеля гена SIGLEC1 и оба аллеля гена CD163.
[0018] Второй такой способ включает спаривание генетически модифицированной свиньи, у которой инактивированы оба аллеля гена S1GLEC1, с генетически модифицированной свиньей, у которой инактивированы оба аллеля гена CD163, с получением F1-потомства, спаривание F1-потомства, чтобы получить Р2-потомство и скрининг Р2-потомства для выявления генетически модифицированных свиней, у которых оба аллеля гена SIGLEC1 и оба аллеля гена CD163 инактивированы.
[0019] Третий такой способ включает спаривание генетически модифицированной свиньи, у которой инактивированы по меньшей мере один аллель гена SIGLEC1 и по меньшей мере один аллель гена CD163, с другой генетически модифицированной свиньей, у которой инактивированы по меньшей мере один аллель гена SIGLEC1 и по меньшей мере один аллель гена CD163, с получением F1-потомства, и скрининг F1-потомства для выявления генетически модифицированных свиней, у которых оба аллеля гена SIGLEC1 и оба аллеля гена CD163 инактивированы.
[0020] Настоящее изобретение также относится к потомству любых описанных выше генетически модифицированных свиней, у которых: (1) по меньшей мере, один аллель гена SIGLEC1 был инактивирован; (2) по меньшей мере, один аллель гена CD163 был инактивирован; (3) по меньшей мере, один аллель гена SIGLEC1 и один аллель гена CD163 были инактивированы; или (4) оба аллеля гена SIGLEC1 и оба аллеля гена CD163 были инактивированы. В таком потомстве, у которого один или оба аллеля гена CD163 были инактивированы, инактивация приводит к белку CD163, неспособному связывать и/или обнажать вирус репродуктивно-респираторного синдрома свиней (ВРРСС).
[0021] Настоящее изобретение также направлено на способ получения генетически модифицированной свиньи, у которой по меньшей мере один аллель гена SIGLEC1 был инактивирован. Способ включает энуклеацию ооцита свиньи; слияние ооцита с фибробластной клеткой донорской свиньи, содержащей в геноме фибробластной клетки по меньшей мере один инактивированный аллель гена SIGLEC1; и активацию ооцита для получения эмбриона.
[0022] В еще одном аспекте, настоящее изобретение является способом получения генетически модифицированной свиньи, у которой инактивирован по меньшей мере один аллель гена CD163, причем инактивация аллеля CD163 приводит к белку CD163, неспособному связывать и/или обнажать вирус репродуктивно-респираторного синдрома свиней (ВРРСС). Этот способ включает энуклеацию ооцита свиньи; слияние ооцита с фибробластной клеткой донорской свиньи, содержащей в геноме фибробластной клетки по меньшей мере один инактивированный аллель гена CD 163; и активацию ооцита для получения эмбриона.
[0023] Настоящее изобретение также направлено на способ получения генетически модифицированной свиньи, у которой по меньшей мере оба аллеля гена SIGLEC1 были инактивированы. Способ включает спаривание самки генетически модифицированной свиньи, у которой инактивирован по меньшей мере один аллель гена SIGLEC1, с самцом генетически модифицированной свиньи, у которого инактивирован по меньшей мере один аллель гена SIGLEC1, с получением F1-потомства, и скрининг F1-потомства для выявления генетически модифицированных свиней, у которых инактивированы оба аллеля гена SIGLEC1.
[0024] Настоящее изобретение также направлено на способ получения генетически модифицированной свиньи, у которой инактивированы оба аллеля гена CD163, причем инактивация аллеля CD163 приводит к белку CD163, неспособному к связыванию и/или обнажению ВРРСС. Этот способ включает спаривание самки генетически модифицированной свиньи, у которой инактивирован по меньшей мере один аллель гена CD163, с самцом генетически модифицированной свиньи, у которого инактивирован по меньшей мере один аллель гена CD 163, с получением F1-потомства, и скрининг F1-потомства для выявления генетически модифицированных свиней, у которых инактивированы оба аллеля гена CD163.
[0025] В еще одном аспекте, настоящее изобретение является способом получения генетически модифицированной свиньи, у которой инактивированы оба аллеля гена SIGLEC1 и оба аллеля гена CD163, причем инактивация аллелей CD163 приводит к белку CD163, неспособному связывать и/или обнажать вирус репродуктивно-респираторного синдрома свиней (ВРРСС). Способ включает спаривание генетически модифицированной свиньи, у которой инактивирован по меньшей мере один аллель гена S1GLEC1, с генетически модифицированной свиньей, у которой инактивирован по меньшей мере один аллель гена CD163, с получением F1-потомства, и скрининг F1-потомства для выявления генетически модифицированных свиней, у которых инактивирован по меньшей мере один аллель гена S1GLEC1, и инактивирован по меньшей мере один аллель гена CD163. Способ дополнительно включает спаривание друг с другом генетически модифицированных свиней, у которых инактивирован по меньшей мере один аллель гена S1GLEC1, и инактивирован по меньшей мере один аллель гена CD163, с получением Р2-потомства, и скрининг Р2-потомства для выявления генетически модифицированных свиней, у которых инактивированы оба аллеля гена S1GLEC1 и оба аллеля гена CD163.
[0026] Настоящее изобретение также относится к другому способу получения генетически модифицированной свиньи, у которой инактивированы оба аллеля гена SIGLEC1 и оба аллеля гена CD163, причем инактивация гена CD163 приводит к белку CD163, неспособному связывать и/или обнажать вирус репродуктивно-респираторного синдрома свиней (ВРРСС). Этот способ включает спаривание генетически модифицированной свиньи, у которой инактивированы оба аллеля гена SIGLEC1, с генетически модифицированной свиньей, у которой инактивированы оба аллеля гена CD163, с получением F1-потомства, спаривание F1-потомства, чтобы получить Р2-потомство и скрининг F2-потомства для выявления генетически модифицированных свиней, у которых оба аллеля гена SIGLEC1 и оба аллеля гена СD163 инактивированы.
[0027] Настоящее изобретение также направлено на еще один способ получения генетически модифицированной свиньи, у которой инактивированы оба аллеля гена SIGLEC 1 и оба аллеля гена CD163, причем инактивация гена CD163 приводит к белку CD163, неспособному связывать и/или обнажать вирус репродуктивно-респираторного синдрома свиней (ВРРСС). Способ включает спаривание генетически модифицированной свиньи, у которой инактивированы по меньшей мере один аллель гена SIGLEC1 и по меньшей мере один аллель гена CD163, с другой генетически модифицированной свиньей, у которой инактивированы по меньшей мере один аллель гена SIGLEC1 и по меньшей мере один аллель гена CD163, с получением F1-потомства, и скрининг F1-потомства для выявления генетически модифицированных свиней, у которых оба аллеля гена SIGLEC1 и оба аллеля гена CD163 инактивированы.
[0028] В других аспектах, настоящее изобретение относится к потомству генетически модифицированной свиньи, полученному по любому из способов выше, у которого инактивированы один или оба аллеля гена SIGLEC1 и/или инактивированы один или оба аллеля гена CD163, причем инактивация аллелей CD 163приводит к белку CD163, неспособному связывать и/или обнажать вирус репродуктивно-респираторного синдрома свиней (ВРРСС).
[0029] Другие цели и характеристики будут частично понятны и частично отмечены ниже.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
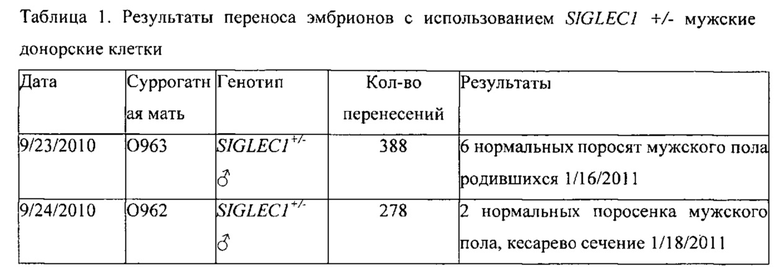
[0030] На Фигуре 1 показано строение гена сиалоадгезина и схема нацеленного вектора. Фигуры 1А и 1В являются схематическими диаграммами, показывающими, что гены человеческого (Фиг. 1А) и мышиного (Фиг. 1В) сиалоадгезина состоят из 21 экзона и простираются приблизительно на 20 кб. На Фигуре 1С представлен мутационный анализ экзона 2 (ДНК-последовательность экзона 2 показана на Фигуре 1С под SEQ ID NO: 7, а аминокислотная последовательность, кодируемая экзоном 2, показана на Фигуре 1С под SEQ ID NO: 8). Мутационный анализ выявил 6 аминокислот, придающих связывание сиалоадгезина с лигандом (выделены обведенным/жирным текстом). На Фигуре 1D представлена схема нацеленного вектора, используемого для замены части экзона 1 и экзонов 2 и 3 SIGLEC1 на стоп-кодоны. В вектор также включена неомициновая кассета селекции (neo), управляемая промотором ФГК.
[0031] На Фигуре 2 показаны схема нацеленного вектора, строение гена сиалоадгезина и строение измененного гена сиалоадгезина.
[0032] На Фигуре 3 представлена фотография геля, показывающая ПЦР-скрининг для идентификации гена сиалоадгезина.
[0033] На Фигуре 4 представлена фотография геля, показывающая ПЦР-скрининг для идентификации равных количеств аллелей сиалоадгезина дикого типа и целевого типа.
[0034] На Фигуре 5 представлена структурная организация CD163 дикого типа (слева), содержащего 9 внеклеточных SRCR-доменов, 2 богатых пролином, серином и треонином (PST) домена, трансмембранную область и внутриклеточный цитоплазматический хвост. Генетически модифицированный CD163 представлен справа. Структурная организация домена остается такой же, за исключением того, что SRCR-домен 5 был заменен на SRCR-домен 8 из лиганда CD163 (CD163L).
[0035] На Фигуре 6 представлен вектор нацеливания CD163, в котором плечи вектора являются участками ДНК с последовательностью, идентичной природному CD163 или дикого типа, что позволяет отжиг вектора с CD163, уже присутствующим в клетках. Модифицированная ДНК, которая находится между двумя плечами CD163 затем может быть внедрена в клетки ДНК посредством гомологической рекомбинации.
ОПРЕДЕЛЕНИЯ
[0036] «Нокаутная свинья» представляет собой генетически модифицированную свинью, у которой было изменено функционирование одного или обоих аллелей гена, к примеру, путем частичной или полной делеции гена. Если один аллель гена нокаутирован, то свинья гетерозиготна по нокаутированному гену; если нокаутированы оба аллеля, то свинья гомозиготна по нокаутированному гену.
[0037] Термин «донорская клетка» обозначает клетку, от которой происходит ядро или материал хроматина, используемые для ядерной передачи. Как описано в других местах данного документа, ядерная передача может включать перенос ядра или хроматина, выделенных из донорской клетки, или перенос всей донорской клетки, включая ядро или материал хроматина.
[0038] Термин «генная модификация» обозначает одно или более изменений генной последовательности (включая кодирующие последовательности и некодирующие последовательности, такие как последовательности интронов, промоторов и 5' и 3'-нетранслируемые последовательности), которые изменяют экспрессию или активность гена. Такие модификации включают, к примеру, инсерции (например, гетерологических последовательностей, таких как селектируемые маркеры и/или сигналы останова), делеции, мутации сдвига рамки считывания, бессмысленные мутации, миссенс-мутации, точечные мутации или их комбинации.
[0039] Термин «реципиентная клетка» обозначает клетку, в которую вводится донорская клетка, ядро донорской клетки или хроматин донорской клетки. Реципиентные клетки перед ядерной передачей энуклеируются подходящим способом. Примеры реципиентных клеток включают ооциты, зиготы и клетки двухклеточных эмбрионов.
[0040] «Малые интерферирующие РНК» (миРНК) обозначают двухнитевые молекулы РНК, обладающие способностью специфически влиять на экспрессию белка. миРНК обычно имеют от приблизительно 10 до приблизительно 30 нуклеотидов в длину. Длина молекулы миРНК основывается на длине антисмысловой цепи молекулы миРНК.
ОПИСАНИЕ ПРЕДПОЧТИТЕЛЬНЫХ ВОПЛОЩЕНИЙ
[0041] Настоящее изобретение направлено на генетически модифицированных свиней, резистентных к инфекции вирусом репродуктивно-респираторного синдрома свиней (ВРРСС). Инфекционная способность ВРРСС зависит от трех специфических медиаторов проникновения: (1) первоначальное связывание с гепарансульфатом, (2) связывание/интернализация сиалоадгезином (SIGLEC1), и (3) интернализация/обнажение вируса, оказываемые CD163. Таким образом, предотвращение взаимодействия между ВРРСС и SIGLEC1 и/или ВРРСС и CD163 приводит к невозможности ВРРСС инфицировать хозяина. В связи с этим, настоящее изобретение направлено на генетически модифицированную свинью, у которой по меньшей мере инактивирован один аллель гена SIGLEC1 и/или у которой инактивирован по меньшей мере один аллель гена CD163, причем инактивация аллеля CD163 приводит к белку CD163, неспособному связывать и/или обнажать вирус репродуктивно-респираторного синдрома свиней (ВРРСС). Эти свиньи также могут называться нокаутными по SIGLEC 1 и/или CD163 свиньями.
[0042] Изобретение включает свиней, у которых только один аллель целевого гена (SIGLEC 1 и/или CD163) был инактивирован, а другой остался неизменным. Эти животные, называемые в данном документе «гетерозиготными» или «гемизиготными» животными могут использоваться в подходах к разведению для получения гомозиготных мутантов. Также в изобретение включены гомозиготные мутантные свиньи, у которых инактивированы оба аллеля целевого гена по одному или разным способам. Соответственно, настоящее изобретение включает генетически модифицированных свиней, у которых: (1) один аллель гена SIGLEC1 был инактивирован; (2) один аллель гена CD163 был инактивирован; (3) оба аллеля гена SIGLEC1 были инактивированы; (4) оба аллеля гена CD163 были инактивированы; (5) оба аллеля гена SIGLEC1 и один аллель гена CD163 были инактивированы; (6) один аллель гена SIGLEC 1 и оба аллеля гена CD163 были инактивированы; (7) один аллель гена SIGLEC1 и один аллель гена CD163 были инактивированы; или (8) оба аллеля гена SIGLEC1 и оба аллеля гена CD163 были инактивированы. В каждом из этих случаев и в общем контексте настоящей заявки, инактивация аллеля(ей) CD163 приводит к белку CD163, неспособному связывать и/или обнажать вирус репродуктивно-респираторного синдрома свиней (ВРРСС).
[0043] Нацеливание на ген с целью получения животных по изобретению может привести к инактивации гена посредством разрушения, удаления, изменения или перемещения последовательностей целевого гена. Способы инактивации гена хорошо известны в данной области техники. К примеру, целевой ген может быть инактивирован внедрением гетерологической последовательности (такой как селектируемый маркер и/или стоп-кодон) в целевой ген, делецией части гена или всего гена, изменением гена (например, мутацией рамки считывания, бессмысленной мутацией, миссенс-мутацией, точечной мутацией, заменой части гена или всего гена на другую нуклеиновокислотную последовательность) или комбинацией любого из вышеперечисленного.
[0044] Внедренные последовательности могут заменить ранее существующие последовательности в гене или могут быть добавлены к таким последовательностям, в зависимости от схемы целевой конструкции. Схема целевых конструкций может быть изменена, в зависимости от того, необходимо ли полностью нокаутировать функцию гена или поддерживать сниженный уровень функционирования. В случае SIGLEC1, желателен полный нокаут функции. В качестве примера и, не ограничиваясь описанным, ген SIGLEC1 может быть нокаутирован делецией части экзона 1 и всех экзонов 2 и 3, к примеру, заменой части экзона 1 и всех экзонов 2 и 3 на неомицин-селектируемую кассету. В некоторых воплощениях модифицирование может также включать добавление LoxP-сайтов с любой стороны последовательности мутируемого гена SIGLEC1. В некоторых случаях нацеленная конструкция может содержать оба loxP-сайта, фланкирующие внедряемую последовательность и CRE-рекомбиназу. В других случаях может использоваться двухвекторная система, в которой нацеленная конструкция включает loxP-сайты, фланкирующие внедряемую последовательность, а второй вектор включает трансген, кодирующий CRE-рекомбиназу. Трансген для CRE-рекомбиназы может располагаться под влияние тканеспецифического промотора, так чтобы его экспрессия отображала ген SIGLEC1, который нефункционален в конкретных клеточных линиях. Аналогичным образом, антибиотик-селектируемая кассета может быть фланкирована loxP-сайтами так, чтобы она могла быть удалена на более позднем этапе при помощи Cre-рекомбиназы.
[0045] В случае CD163, желательно инактивировать только его функцию ВРРСС-связывания и/или его обнажения, оставляя другие функции CD163 минимально затронутыми или незатронутыми. Не ограничиваясь какой-либо конкретной теорией, считается, что полностью нокаутные по CD163 особи могут быть нежизнеспособными или серьезно скомпрометированными по причине роли, которую играет CD163 в связывании и интернализации комплексов гемоглобин-гаптоглобин. Соответственно, CD163 может быть деактивирован разрушением пятого N-терминального фагоцитарного богатого цистеином (SRCR) домена CD163, для которого было показано, что он играет роль в ВРРСС-инфекции (Van Gorp et al., 2010), оставляя остальные домены незатронутыми. SRCR домен 5 может быть генетически модифицирован, например, внедрением точечных мутаций, которые изменяют структуру этого домена или обменом этого домена на другой. К примеру, SRCR домен 5 может быть замещен на SRCR домен 8 из CD163 лиганда (CD163L), поскольку этот «обмен» домена, как было показано, снижает относительную инфекционность ВРРСС до 0% в культивированных клетках (Van Gorp et al 2010).
[0046] По другим подходам, кодирующие последовательности целевого гена не изменяются или изменяются минимально, а взамен нацеливание производится на последовательности, влияющие на экспрессию целевого гена, такие как промоторные последовательности. В любом случае, инсерция селектируемого маркера часто желательна для облегчения идентификации клеток, в которых произошло нацеливание. При необходимости, такие маркеры или иные внедренные последовательности могут быть позднее удалены, например, при помощи Cre-Lox или аналогичных систем.
[0047] При модифицировании целевого гена используются нуклеиновокислотные конструкции с участками гомологии с целевым геном (например, SIGLEC1 или CD163) или с фланкирующими областями целевого гена, так чтобы интегрирование конструкций в геном изменяло экспрессию гена, изменением последовательности гена и/или изменением уровней экспрессии гена. Таким образом, для изменения гена нацеленная конструкция обычно разрабатывается так, чтобы она включала три основные области: (i) первую область, которая гомологична целевому локусу, на который производится нацеливание (например, ген SIGLEC1 или CD163, либо их фланкирующая последовательность), (ii) вторую область, представляющую собой гетерологическую полиуклеотидную последовательность (например, кодирующую селектируемый маркер, такой как белок резистентности к антибиотику), которая специфически замещает часть локуса нацеливания или внедрена в локус нацеливания и (iii) третью область, которая, подобно первой области, гомологична локусу нацеливания, но обычно не смежна с первой областью конструкции. Гомологическая рекомбинация между нацеленной конструкцией и локусом нацеливания дикого типа приводит к делеции любых локусных последовательностей между двумя областями гомологии, отраженными в нацеленном векторе и замещению этой последовательности на гетерологическую последовательность (например, гетерологической последовательности, кодирующей селектируемый маркер) или внедрению ее внутрь той последовательности. Типичные конструкция и вектор для выполнения такого нацеленного модифицирования описаны в Примере 1; тем не менее, известны другие векторы, которые могут применяться по таким подходам, и которые с легкостью могут быть адаптированы для использования по изобретению.
[0048] Чтобы облегчить гомологическую рекомбинацию, первая и третья области нацеленных векторов (см. выше) включают последовательности, которые проявляют значительную идентичность последовательности с генами, на которые они нацелены (или фланкирующими областями). К примеру, первая и третья области нацеленных векторов могут иметь последовательности, которые по меньшей мере приблизительно на 80% по меньшей мере приблизительно на 90% по меньшей мере приблизительно на 95% по меньшей мере приблизительно на 98% по меньшей мере приблизительно на 99% или приблизительно на 100% идентичны целевым генам или фланкирующим областям. Идентичность последовательности обычно измеряют при помощи BLAST® (Средство поиска основного локального выравнивания) или BLAST®2 с параметрами по умолчанию, указанными там (см. Altschul et al., J. Mol. Biol. 215:403-410, 1990; Tatiana et al., FEMS Microbiol. Lett. 174:247-250, 1999). Таким образом, последовательности, которые по меньшей мере приблизительно на 80% по меньшей мере приблизительно на 90% по меньшей мере приблизительно на 95% по меньшей мере приблизительно на 98%) по меньшей мере приблизительно на 99% или даже приблизительно на 100%) идентичны последовательностью локусам целевых генов могут использоваться по изобретению для облегчения гомологической рекомбинации.
[0049] Общий размер двух гомологических областей (т.е., первой и третьей областей, описанных выше) может составлять, к примеру, от приблизительно 2 килобаз (кб) до приблизительно 25 кб (например, от приблизительно 4 кб до приблизительно 20 кб, от приблизительно 5 кб до приблизительно 15 кб, или от приблизительно 6 кб до приблизительно 10 кб). Размер области, заменяющей часть целевого локуса или внедряемой в целевой локус (вторую область, описанную выше), может составлять, к примеру, от приблизительно 0,5 килобаз (кб) до приблизительно 5 кб (например, от приблизительно 1 кб до приблизительно 4 кб или от приблизительно 3 кб до приблизительно 4 кб).
[0050] Могут применяться различные способы доставки нацеленной конструкции. Для доставки нацеленной конструкции могут использоваться способы клеточной трансфекции, включая кальций-фосфатную, липофекцию, электропорацию и инъекцию ядра. Если ген является транскрипционно активным в используемом типе клетки, то может применяться стратегия беспромоторного селектируемого маркера, так, чтобы устойчивость к антибиотику формировалась только у клеток, претерпевших событие рекомбинации в транскрибируемой единице. Альтернативно, если ген транскрипционно неактивен в используемом типе клетки, то могут применяться промоторы, которые индуцируемы, тканеспецифичны или содержат изоляторы, такие как области присоединения матрицы (MAR).
[0051] Альтернативно, чтобы «заглушить» транскрипцию SIGLEC1, может применяться технология миРНК. Антисмысловая технология хорошо известна в данной области техники. Короче говоря, применяется нуклеотидная последовательность, содержащая от приблизительное до приблизительно 29 нуклеотидов, которая является комплементарной по отношению к смысловой последовательности иРНК SIGLEC1. Степень комплементарности обычно находится в пределах от приблизительно 70% до приблизительно 100%. Предпочтительно, комплементарность больше, чем приблизительно 80%, более предпочтительно, больше, чем приблизительно 90%, и еще более предпочтительно, больше, чем приблизительно 95%. Области иРНК SIGLEC1, которые пригодны для нацеливания миРНК, легко могут быть определены путем сравнения эффективности некоторых антисмысловых последовательностей, выполненных таким образом, чтобы быть комплементарными по отношению к различным областям иРНК SIGLEC1 для предотвращения продукции белка SIGLEC1. Такие эксперименты могут быть легко выполнены без необоснованного экспериментирования по любой из техник, известных в данной области техники.
[0052] Векторы, используемые для экспрессии миРНК, хорошо известны в данной области техники. Вектор может быть круговой или линейной ДНК любой длины, либо встраивающейся в геном хозяина, либо поддерживающей себя в эписомальной форме. В общем, кассеты экспрессии миРНК могут быть лигированы в вектор ДНК-трансфекции, такой как плазмида или лентивирусный, аденовирусный, альфавирусный, ретровирусный или иной вирусный вектор. Типичные вирусные векторные системы для млекопитающих включают аденовирусные векторы; векторы адено-ассоциированного вируса типа 1 («AAV-1») или адено-ассоциированного вируса типа 2 («AAV-2»); векторы гепатита дельта; живые, ослабленные дельта-вирусы; герпесвирусные векторы; альфавирусные векторы; или ретровирусные векторы (включая лентивирусные векторы).
[0053] Трансформация клеток млекопитающих может выполняться по стандартным методикам, известным в данной области техники. Может использоваться любая из хорошо известных процедур по внедрению чужеродных нуклеотидных последовательностей в клетки-хозяева, если они успешно встраивают в клетку-хозяина по меньшей мере миРНК-конструкцию. Эти процедуры включают использование вирусной трансдукции (к примеру, путем использования любого из вирусных векторов, указанных в предыдущем параграфе, например, ретровирусной инфекцией, необязательно, в присутствии полибрена для усиления эффективности инфекции), кальций-фосфатной трансфекции, электропорации, биолистической системы доставки частиц (т.е., генных пушек), липосом, микроинъекции и любого из других известных способов введения клонированной геномной ДНК, кДНК, синтетической ДНК или иного чужеродного генетического материала в клетку-хозяина. По настоящему изобретению, миРНК против SIGLEC1 вводится в донорскую фибробластную клетку свиньи, которую затем используют для получения трансгенно модифицированной свиньи по настоящему изобретению.
[0054] Трансгенные животные по изобретению могут быть получены с использованием следующей общей процедуры переноса ядра соматической клетки. Вкратце, геном соматической клетки свиньи (например, фетальный фибробласт) генетически модифицируют генным нацеливанием, как описано выше, чтобы получить донорскую клетку. Затем ядро такой генетически модифицированной донорской клетки (или донорскую клетка полностью, включая ядро) переносят в реципиентную клетку, к примеру, в энуклеированный ооцит. Донорская клетка может быть слита с энуклеированным ооцитом или донорское ядро донорской клетки само по себе может быть инъецировано в реципиентную клетку или инъецировано в перивителлиновое пространство рядом с мембраной ооцита.
[0055] Таким образом, после получения соматических клеток, в которых было произведено нацеливание на целевой ген (на один или оба аллеля, как описано выше), может быть выполнен ядерный перенос. Необязательно, генетически модифицированные донорские клетки могут быть криоконсервированы до ядерного переноса. Пригодные к использованию реципиентные клетки включают ооциты, оплодотворенные зиготы или клетки двуклеточных эмбрионов, все из которых могут быть или могут не быть энуклеированы.
[0056] Реципиентные ооциты могут быть получены по способам, известным в данной области техники или же могут быть получены из коммерческих источников (например, BoMed Inc., Мэдисон, Висконсин). Ооцит может быть получен от «подсвинка», т.е. от нерожавшей самки свиньи, либо от «свиноматки», т.е. от рожавшей самки свиньи.
[0057] Соответственно, генетически модифицированная свинья, у которой инактивирован по меньшей мере один аллель гена SIGLEC1, может быть получена посредством энуклеации ооцита свиньи; слияния ооцита с фибробластной клеткой донорской свиньи, содержащей в геноме фибробластной клетки по меньшей мере один инактивированный аллель гена SIGLEC1; и активации ооцита для получения эмбриона. Аналогичным образом, генетически модифицированная свинья, у которой инактивирован по меньшей мере один аллель гена CD163, причем инактивация аллеля CD163 приводит к белку CD163, неспособному к связыванию и/или обнажению вируса репродуктивно-респираторного синдрома свиней (ВРРСС), может быть получена посредством энуклеации ооцита свиньи; слияния ооцита с фибробластной клеткой донорской свиньи, содержащей в геноме фибробластной клетки по меньшей мере один инактивированный аллель гена CD163; и активации ооцита для получения эмбриона. Оба способа могут дополнительно включать этап переноса эмбриона в репродуктивный тракт суррогатной свиньи, отличающейся тем, что у суррогатной свиньи начался эструс, но не завершилась овуляция; и отличающейся тем, что беременность и роды в срок дают генетически модифицированную свинью, геном которой содержит по меньшей мере один инактивированный аллель SIGLEC1 и/или CD163. Настоящее изобретение также относится к потомству такой генетически модифицированной свиньи, у которого по меньшей мере инактивирован один аллель гена SIGLEC1 и/или у которого инактивирован по меньшей мере один аллель гена CD163, причем инактивация аллеля CD163 приводит к белку CD163, неспособному связывать и/или обнажать вирус репродуктивно-респираторного синдрома свиней (ВРРСС).
[0058] Способы энуклеации ооцитов свиньи известны в данной области техники, и энуклеации можно добиться по любому из стандартных способов. К примеру, энуклеации ооцита можно добиться при помощи микропипетки в микроманипуляционной среде.
[0059] Введение окруженного мембраной ядра от донорской клетки свиньи в энуклеированный реципиентный ооцит с образованием ооцита, содержащего донорское ядро, может выполняться путем слияния мембраны окруженного мембраной ядра из донорской клетки млекопитающего с мембраной энуклеированного реципиентного ооцита млекопитающего с образованием ооцита, содержащего ядро донорской клетки млекопитающего. Альтернативно, такое введение может выполняться путем микроинъекции окруженного мембраной ядра из донорской клетки млекопитающего в энуклеированный реципиентный ооцит млекопитающего с образованием ооцита, содержащего ядро донорской клетки млекопитающего. К примеру, можно вводить донорскую клетку (или ядро) в пространство под блестящей оболочкой или в перивителлиновое пространство энуклеированного реципиентного ооцита и впоследствии выполнять слияние мембраны с получением ооцита, содержащего в цитоплазме донорское ядро. В способах, представленных в данном документе, можно использовать все варианты введения донорского ядерного материала в энуклеированный реципиентный ооцит млекопитающего, которые известны обычному специалисту в данной области техники.
[0060] К примеру, этап слияния может выполняться в среде для слияния. Альтернативно, для облегчения слияния могут применяться инактивированный вирус или фузогенный агент, такой как полиэтиленгликоль (ПЭГ). См., например, Graham (1969) Wistar Inst. Symp. Monogr. 9:19-33 и Mc Grath et al. (1983) Science 220:1300-1302 об использовании вирусов; Fisher et al. (1981) Tech. Cell. Physiol. 1:1-36 о химически индуцированном слиянии клеток; и Berg (1982) Bioelectrochem. Bioenerg. 9:223-228, а также Robl et al. (1987) J. Anim. Sci. 64:642-647 об электрическом индуцировании слияния клеток.
[0061] В US 6211429 В1, содержание которого включено в данный документ посредством ссылки, описаны способы развития активированных ооцитов in vitro и in vivo. Термин «активированный» или «активация» обозначает способность неоплодотворенного ооцита развиваться до по меньшей мере пронуклеарного этапа или до последующих этапов, после обработки ооцит-модифицирующим агентом и восстанавливающим агентом. Говоря в общем, пронуклеарный этап достигается приблизительно через три-семь часов после такой обработки. Термин «ооцит-модифицирующий агент» обозначает агент, который может реагировать с субстратом на ооците или в нем, к примеру, с тиоловой (-SH) группой, которая может быть белковой тиоловой группой; эффект от этой реакции при последующей обработке ооцита восстанавливающим агентом по способам, представленным в US 6211429 В1, приводит к активации ооцитов млекопитающих.
[0062] При совместном использовании -SH или ооцит-модифицирующего агента, такого как тимеросал и -SH восстанавливающего агента, такого дитиотреитол, возможна полная активация ооцитов млекопитающих. Сочетание короткой обработки тимеросалом с только одним кальциевым переходом до обработки восстанавливающим агентом, таким как ДТТ, особенно эффективно для достижения активации. Тимеросал запускает серию Са2+ выбросов в ооцитах млекопитающих, которая с последующим инкубированием с восстанавливающим агентом, таким как ДТТ, может стимулировать образование пронуклеуса. Так, после вымывания тимеросала, добавляют ДТТ, чтобы обратить действие тимеросала с последующим вымыванием, чтобы позволить эмбриону продолжить развитие. Комбинированная обработка тимеросал/ДТТ также индуцирует экзоцитоз кортикальной гранулы, последующее уплотнение блестящей оболочки и развитие активированных ооцитов до этапа бластоцисты.
[0063] Помимо тимеросала, могут применяться другие ооцит-модифицирующие агенты, такие как трет-бутилгидропероксид; тиомочевина; фенилэфрин; N-алкилмалеимиды, такие как N-этилмалеимид; окисленный глутатион; альфа-галокислоты, такие как иодацетат, хлорацетат и бромацетат; иодацетамид; п-меркурибензоат; п-хлормеркурибензоат;
5,5'-дитиобис(2-нитробензойная кислота) (DTNB);
(2-триметиламмоний)этилметантиосульфонат (MTSET); и (2-сульфонатоэтил)метантиосульфонат (MTSES). Пригодные восстанавливающие агенты, такие как восстановители тиоловой (-SH) группы, помимо ДТТ, включают, но, не ограничиваясь перечисленным, дитиоэритритол (ДТЭ); бета-меркаптоэтанол; цистеин; восстановленный глутатитон; восстановленную тиомочевину; тиогликолят; и аскорбиновую кислоту.
[0064] Период времени, в течение которого ооциты контактируют с ооцит-модифицирующим агентом является периодом, эффективным для активации ооцитов при последующей обработке восстанавливающим агентом. Такой период времени может быть в пределах от приблизительно 5 минут до приблизительно 20 минут, предпочтительно от приблизительно 5 минут до приблизительно 15 минут, или, более предпочтительно, от приблизительно 5 минут до приблизительно 12 минут. Период времени, в течение которого ооциты контактируют с восстанавливающим агентом должен быть достаточно долгим, чтобы привести к активации ооцитов при предшествующей обработке ооцит-модифицирующим агентом. Такой период времени может быть в пределах от приблизительно 5 минут до приблизительно 1 часа, предпочтительно от приблизительно 10 минут до приблизительно 45 минут, более предпочтительно, от приблизительно 20 минут до приблизительно 40 минут, еще более предпочтительно, приблизительно 30 минут.
[0065] Контактирование энуклеированного ооцита с восстанавливающим агентом после контактирования с ооцит-модифицирующим агентом может происходить почти сразу же после или в течение периода времени в пределах от приблизительно 5 секунд до приблизительно 5 минут после воздействия ооцит-модифицирующего агента на ооцит. Ооцит, обработанный ооцит-модифицирующим агентом, может переноситься в среду, содержащую восстанавливающий агент, в отсутствие промежуточного промывочного этапа. Альтернативно, ооцит, обработанный ооцит-модифицирующим агентом, может промываться контрольной или содержащей восстанавливающий агент средой для почти полного удаления ооцит-модифицирующего агента перед культивированием ооцита в среде, содержащей восстанавливающий агент. В качестве другой альтернативы, восстанавливающий агент может добавляться непосредственно к ооциту, в то время как последний все еще находится в среде, содержащей ооцит-модифицирующий агент.
[0066] Альтернативно, активация ооцита также может достигаться при ряде химических обработок, не включающих кальций, например, ингибированием протеинкиназы (Mayes et al. (1995) Biol. Reprod. 53:270-275) или ингибированием синтеза белка (Nussbaum et al. (1995) Mol. Reprod. Dev. 41:70-75).
[0067] После активации ооцит обычно недолго культивируют in vitro. Затем полученный эмбрион переносят в суррогатную самку, и развитие эмбриона продолжается суррогатно. К примеру, эмбрионы могут культивироваться в течение приблизительно недели, а затем трансфицироваться хирургически или нехирургически в репродуктивный тракт суррогата. Эмбрионы могут трансфицироваться в фаллопиеву трубу через яичниковые бахромки суррогата. Альтернативно, эмбрионы могут трансфицироваться в фаллопиеву трубу при помощи катетера, проникающего через стенку фаллопиевой трубы. Другой способ переноса эмбрионов включает культивирование их до этапа бластоцисты с последующим введением в репродуктивный тракт суррогатной свиньи. Эти способы хорошо известны в данной области техники и могут с легкостью применяться для получения генетически модифицированных свиней по настоящему изобретению.
[0068] Дополнительные способы для получения генетически модифицированных свиней и других крупных животных известны в данной области техники и могут использоваться по настоящему изобретению (см., к примеру, US 2005/0120400 A1; Патент США Nо. 5995577; WO 95/16670; WO 96/07732; WO 97/00669; WO 97 00668; WO 2005/104835; Lai et al., Reproductive Biology and Endocrinology 1:82, 2003; Hao et al., Transgenic Res. 15:739-750, 2006; Li et al., Biology of Reproduction 75:226-230, 2006; Lai et al., Nature Biotechnology 24(4):435-436, 2006; Lai et al., Methods in Molecular Biology 254(2): 149-163, 2004; Lai et al., Cloning and Stem Cells 5(4):233-241, 2003; Park et al., Animal Biotechnology 12(2):173-181, 2001; Lai et al., Science 295:1089-1092, 2002; Park et al., Biology of Reproduction 65:1681-1685,2001; содержимое которых включено в данный документ посредством ссылки).
[0069] Другие способы, пригодные для получения генетически модифицированных свиней включают инъекцию или трансдукцию нуклеаз (нуклеазы с цинковыми пальцами или Tal-нуклеазы, которые были бы нацелены на интересующий ген) в соматические клетки, с последующим ядерным переносом соматической клетки и переносом эмбриона в суррогат. Помимо этого также может выполняться сперма-опосредованное или опосредованное внутрицитоплазматической инъекцией спермия (ICSI) генетическое модифицирование. Вкратце, при ICSI-опосредованном модифицировании, нацеленную конструкцию смешивают со спермием и инъецируют оба компонента в ооцит. При сперма-опосредованном модифицировании конструкцию смешивают со спермой и для того, чтобы суррогатная мать забеременела, применяют оплодотворение in vitro (IVF) или осеменение. Опытный специалист может с легкостью использовать эти способы для получения нокаутных свиней по настоящему изобретению. Хотя они и не развиваются полностью в свинью, эмбриональные стволовые клетки или индуцированные плюрипотентные клетки, по технологии, разработанной для мышей, могут быть генетически модифицированы и использоваться либо в качестве донорских клеток для переноса ядра соматической клетки, либо для получения химерных животных.
[0070] Как указывалось выше, настоящее изобретение также направлено на генетически модифицированную свинью, у которой (1) инактивированы оба аллеля гена SIGLEC1, (2) инактивированы оба аллеля гена CD163, причем инактивация аллелей CD163 приводит к белку CD163, неспособному связывать и/или обнажать вирус репродуктивно-респираторного синдрома свиней (ВРРСС), или (3) инактивированы оба аллеля гена SIGLEC1 и оба аллеля гена CD163. Генетически модифицированные свиньи, которые гомозиготны по инактивации гена, могут быть получены путем спаривания гетерозиготных по инактивации гена свиней и скринингом потомства для идентификации животных, гомозиготных по инактивации гена(ов). Настоящее изобретение также направлено на потомство такой генетически модифицированной свиньи, у которой инактивированы один или оба аллеля гена SIGLEC1 и/или инактивированы один или оба аллеля гена CD163, причем инактивация аллелей CD163 приводит к белку CD163, неспособному связывать и/или обнажать вирус репродуктивно-респираторного синдрома свиней (ВРРСС).
[0071] Соответственно, настоящее изобретение направлено на способ получения генетически модифицированной свиньи, у которой инактивированы оба аллеля гена SIGLEC1, посредством спаривания самки генетически модифицированной свиньи, у которой инактивирован по меньшей мере один аллель гена SIGLEC1, с самцом генетически модифицированной свиньи, у которого инактивирован по меньшей мере один аллель SIGLEC1, с получением F1-потомства, и скринингом F1-потомства для выявления генетически модифицированных свиней, у которых инактивированы оба аллеля гена SIGLEC1. Аналогично, настоящее изобретение направлено на способ получения генетически модифицированной свиньи, у которой инактивированы оба аллеля гена CD163, посредством спаривания самки генетически модифицированной свиньи, у которой инактивирован по меньшей мере один аллель гена CD163, с самцом генетически модифицированной свиньи, у которого инактивирован по меньшей мере один аллель CD163, с получением F1-потомства, и скринингом F1-потомства для выявления генетически модифицированных свиней, у которых инактивированы оба аллеля гена CD163.
[0072] По настоящему изобретению также предлагаются способы получения генетически модифицированной свиньи, у которой инактивированы оба аллеля гена SIGLEC1 и оба аллеля гена CD163. Один такой способ включает спаривание генетически модифицированной свиньи, у которой инактивирован по меньшей мере один аллель гена SIGLEC1, с генетически модифицированной свиньей, у которой инактивирован по меньшей мере один аллель гена CD163 с получением F1-потомства; скрининг F1-потомства для идентификации генетически модифицированной свиньи, у которой инактивирован по меньшей мере один аллель гена SIGLEC1 иинактивирован по меньшей мере один аллель гена CD163; спаривание друг с другом генетически модифицированных свиней, у которых инактивирован по меньшей мере один аллель гена SIGLEC1, и инактивирован по меньшей мере один аллель гена CD163, с получением Р2-потомства, и скрининг Р2-потомства для выявления генетически модифицированных свиней, у которых инактивированы оба аллеля гена SIGLEC1 и оба аллеля гена CD163.
[0073] Способы получения генетически модифицированной свиньи, у которой по меньшей мере один аллель гена SIGLEC1 был инактивирован и/или по меньшей мере один аллель гена CD163 был инактивирован, представлены в разделах выше. Скрининг потомства может выполняться так, как принято стандартно в данной области техники, например, использованием ПЦР или Саузерн-блоттингом.
[0074] Другой способ получения генетически модифицированной свиньи, у которой инактивированы оба аллеля гена SIGLEC1, и инактивированы оба аллеля гена CD163, включает спаривание генетически модифицированной свиньи, гомозиготной по инактивации SIGLECI, с генетически модифицированной свиньи, гомозиготной по инактивации CD163, с получением F1-потомства, спаривание F1-потомства, чтобы получить Р2-потомство и скрининг Р2-потомства для выявления животных, гомозиготных и по инактивации SIGLEC1, и инактивации CD163.
[0075] Еще один способ получения генетически модифицированной свиньи, у которой инактивированы оба аллеля гена SIGLEC1 и оба аллеля гена CD163, включает спаривание генетически модифицированной свиньи, у которой инактивированы по меньшей мере один аллель гена SIGLEC1 и по меньшей мере один аллель гена CD163, с другой генетически модифицированной свиньей, у которой инактивированы по меньшей мере один аллель гена SIGLEC1 и по меньшей мере один аллель гена CD163, с получением F1-потомства, и скрининг F1-потомства для выявления генетически модифицированных свиней, у которых оба аллеля гена SIGLEC1 и оба аллеля гена CD165 инактивированы.
[0076] Настоящее изобретение также относится к потомству генетически модифицированной свиньи, полученному по любому из способов выше, у которого инактивированы один или оба аллеля гена SIGLEC1 и/или инактивированы один или оба аллеля гена CD163, причем инактивация гена CD163 приводит к белку CD163, неспособному связывать и/или обнажать вирус репродуктивно-респираторного синдрома свиней (ВРРСС).
[0077] Помимо того, что гомозиготных мутантных животных можно получать по подходам разведения, в которых используются гетерозиготные животные, их также можно получать по подходу, в котором клетка (например, фетальный фибробласт) с мутацией в одном аллеле, такая как клетка, выделенная от животного, полученного по способам, описанным выше, подвергается генному нацеливанию посредством гомологической рекомбинации, чтобы достичь модифицирования оставшегося аллеля. Затем полученная донорская клетка может использоваться в качестве источника модифицированного ядра для ядерного переноса в реципиентную клетку, такую как энуклеированный ооцит, приводя к образованию гомозиготного мутантного эмбриона, который при имплантации в суррогатную самку, развивается в гомозиготное мутантное животное. Генетически модифицированная свинья, у которой оба аллеля гена(ов) SIGLEC1 и/или СD163 были инактивированы, также может быть получена посредством инъекции или трансфекции цинк-пальцевых нуклеаз или Tal-нуклеаз (которые могут нацеливаться на оба аллеля гена одновременно) в соматических клетках, с последующим переносом ядра соматической клетки (ПЯСК) и переносом эмбриона суррогатной матери для получения такой свиньи. Настоящее изобретение также относится к потомству такой генетически модифицированной свиньи, у которого инактивированы один или оба аллеля гена SIGLEC1 и/или инактивированы один или оба аллеля гена CD163, причем инактивация гена CD163 приводит к белку CD163, неспособному связывать и/или обнажать вирус репродуктивно-респираторного синдрома свиней (ВРРСС).
[0078] После детального описания изобретения будет очевидно, что возможны его модификации и варианты без отступания от охвата изобретения, определенного в прилагаемой формуле изобретения.
ПРИМЕРЫ
[0100] Следующие неограничивающие примеры представлены для дальнейшей демонстрации настоящего изобретения.
ПРИМЕР 1
[0101] Модифицирование гена SIGLEC1
[0102] В подходе, используемом для усечения гена сиалоадгезина, применялась гомологическая рекомбинация для удалеения экзонов, кодирующих белок и введения преждевременных остановов в оставшейся кодирующей последовательности гена сиалоадгезина. Ген сиалоадгезина свиньи (SIGLEC1, референсная последовательность NCBI NM_214346) кодирует 210 кДа белок с транскрипта иРНК в 5193 баз (Vanderheijden et al. 2003). Последовательность генома свиньи в районе гена сиалоадгезина (Номер доступа в Genbank CU 467609) использовалась для получения олигонуклеотидов для усиления фрагментов генома при помощи высокоточной ПЦР [AccuTaq (Invitrogen)] для получения нацеленной конструкции. Один фрагмент («верхнее плечо») включал первый кодирующий экзон и 3304 пар оснований вверх от начала трансляции. Второй («нижнее плечо») фрагмент имел длину 4753 пар оснований и включал большинство интронов вниз после третьего кодирующего экзона, вплоть до 6-го интрона (включая 4-й, 5-й и 6-й кодирующие экзоны). Исходя из сравнения с геномными последовательностями мышиного и человеческого сиалоадгезина, прогнозировалось, что ген сиалоадгезина свиньи состоит из 21 экзона (Фиг. 1А, 1В). Экзон 2 консервативен для свиньи, мыши и человека. Аминокислотное выравнивание экзона 2 выявило, что шесть аминокислот мышиного сиалоадгезина, для которых известно, что они ассоциированы с активностью в связывании сиаловой кислоты, сохранены в свинном сиалоадгезине (Фиг. 1С). Стратегия изначального нацеливания фокусировалась на создании изменений в гене сиалоадгезина так, чтобы из мутантного гена не ожидалось получения функционального белка. Другие стратегии инактивации могут включать нацеленную модификацию выбранных остатков в экзоне 2 гена сиалоадгезина или изменение доменов иммуноглобулина так, чтобы избежать связывания вируса РРСС (возможно, изменением их порядка или замещением их на похожие домены от других видов). Дополнительные модификации могут включать фланкирование неомициновой кассеты или одного иммуноглобулин-подобных доменов сайтами loxP чтобы при желании позволить индуцируемое или тканеспецифическое извлечение.
[0103] При нарушении текущего гена, часть экзона 1 и экзоны 2 и 3 полностью заменялись неомиционовой кассетой селекции с использованием вектора заместительного типа (Фиг. 1D) (Mansour et al. 1988). В плазмидной конструкции промотор фосфоглицеролкиназы (ФГК) использовался для управления экспрессией неомициновой кассеты, чтобы позволить положительную селекцию трансфицированных колоний.
[0104] Подготовка Донорской Клетки
[0105] Первичная клеточная линия мужских фетальных фибробластов 35 суток беременности была выделена от приобретенных крупных белых свиней (Ландрас). Клетки были культивированы и выращены в течение 48 часов до 80%-й конфлюэтности в среде Игла модификации Дульбекко (DMEM), содержащей 5 мМ глутамина, натрия бикарбонат (3,7 г/л), пенициллин-стрептомицин и 1 г/л d-глюкозы с добавкой 15% фетальной телячьей сыворотки Hyclone, 10 мкг/мл гентамицина и 2,5 нг/мл главного фактора роста фибробластов (Sigma). За четыре часа до трансфекции среду удаляли и заменяли на свежую. Фибробластные клетки были промыты 10 мл фосфатного солевого буферного раствора (DPBS; Invitrogen) и помещены в колбу вместимостью 75 см с 1 мл 0,05% трипсина-ЭДТА (Invitrogen). Клетки были ресуспендированы в DMEM и собраны путем центрифугирования при 600 xg в течение 10 минут. Клетки были промыты Opti-MEM (Invitrogen) и вновь центрифугированы при 600 xg в течение 10 минут. Для ресуспендирования осадка использовались цитосоли (75% цитосолей [120 мМ KCl, 0,15 мМ CaCl2, 10 мМ K2НРO4; рН 7,6, 5 мМ MgCl2]) и 25% Opti-Mem (van den Hoff et al. 1992). Клетки были подсчитаны в гемоцитометре и их концентрация была доведена до 1×106 клеток на мл. Электропорацию клеток выполняли с 4 мкг однонитевой нацеленной ДНК (получали тепловой денатурацией) в 200 мкл трансфекционной среды, состоящей из 1x106 клеток/мл. Клетки были подвергнуты элетропорации в электрической ячеке-манипуляторе ВТХ ЕСМ 2001 с тремя импульсами 250 В длительностью по 1 мс. Электропорированные клетки были разбавлены DMEM/FBS/ФРФ при 10000 клеток на 13 см чашку. Электропорированные клетки культивировались в течение ночи без селективного давления. На следующий день среду заменяли питательной средой, содержащей G418 (0,6 мг/мл). Спустя 10 дней селекции, были выделены G418-резистентные колонии, которые были перенесены в 24-луночные планшеты для экспансии. После выращивания в 24-луночных планшетах, клетки были разделены (~половину использовали для выделения геномной ДНК) в 6-луночные планшеты. Для определения успешности нацеливания на ген сиалоадгезина использовали ПЦР. В реакции использовались олигонуклеотиды, которые отжигались в фосфоглюкокиназной (ФГК) кассете (включающей Neo), спаренной с теми, которые отжигались с геномной ДНК в сиалоадгезиновом локусе, расположенном за пределами целевых плеч (см. Фиг. 2). Таким образом, было выявлено успешное нацеливание на оба «плеча». Был идентифицирован нацеленный клон фибробластов (4-18) и некоторые из клеток культуры использовались для ядерного переноса (см. ниже), а остальные были заморожены для последующего применения. Для дополнительного подтверждения успешности нацеливания, выполнялась проверка гомологической рекомбинации способом Саузерн-блоттинга.
[0106] Сбор ооцитов и созревание In Vitro(WM)
[0107] Ооциты свиней были приобретены у ART Inc (Мэдисон, Висконсин) и доведены до зрелости по инструкциям поставщика. Спустя 42-44 ч созревания in vitro, ооциты были отделены от кумулюсных клеток путем осторожного перемешивания в 0,5 мг/мл гиалуронидазе. После удаления кумулюсных клеток, выбирались ооциты с хорошей морфологией и видимым полярным тельцем (метафаза II), которые содержались в микроманипуляционной среде при 38,5°C до ядерного переноса.
[0108] Перенос ядра соматической клетки. Слияние/Активация Комплексных Ооцитов после Ядерного Переноcа, и Развитие в Культуре In Vitro
[0109] Под микроскопом бескумулюсный ооцит захватывали в удерживающую микропипетку в каплях микроманипуляционной среды с добавкой 7,5 мкг/мл цитохалазина В и покрытых минеральным маслом. Блестящую оболочку протыкали тонкой стеклянной инъекционной микропипеткой рядом с первым полярным тельцем, и первое полярное тельце с расположенной рядом цитоплазмой, содержащей хромосомы метафазы II, втягивали в пипетку, пипетку вытаскивали и содержимое выливали. Выбирали одну округлую и яркую донорскую клетку с гладкой поверхностью, которую переносили в перивителлиновое пространство рядом с мембраной ооцита (Lai et al. 2006; Lai et al. 2002).
[0110] Комплекс ядерного переноса (ооцит + фибробласт) сливали в среде для слияния с низким содержанием кальция (0,3 М маннитола, 0,1 мМ СаС12⋅2Н2O,0,1 мМ MgCl2⋅6H2O и 0,5 мМ HEPES). Затем слитые ооциты подвергали активации путем обработки 200 мкМ тимеросала в течение 10 минут в темноте, с последующим ополаскиванием и обработкой 8 мМ дитиотреитола (ДТТ) в течение 30 минут; затем ооциты вновь ополаскивались для удаления ДТТ (Machaty and Prather 2001; Machaty et al. 1997). После сливания/активации ооциты были промыты трижды средой для свиных зигот 3 (PZM3) с добавкой 4 мг/мл БСА (Im et al. 2004), и культивировались при 38,5°C в увлажненной атмосфере с 5% О2, 90% N2 и 5% СО2 в течение 30 минут. Успешно слитые комплексы культивировались 15-21 часов до хирургического переноса эмбриона в суррогат.
[0111] Подготовка Суррогата. Перенос Эмбриона. Диагностика Беременности и Роды
[0112] Суррогатные подсвинки были синхронизированы путем введения 18-20 мг REGU-MATE (0,22% раствор альтреногеста) (Intervet, Миллсборо, Делавер), который подмешивали в корм в течение 14 дней по схеме, в зависимости от этапа эстрального цикла. После последнего введения REGU-MATE (105 часов), внутримышечно вводили 1000 единиц чХГ, чтобы вызвать эструс. В суррогатный пул также включали других суррогатов, которые естественным образом подходили по циклу к соответствующей дате. Использовались суррогаты, у которых отмечался день начала (день 0) эструса или первый день после него (Lai et al. 2002). Суррогаты подготавливались асептически и производился каудально-вентральный разрез для обнажения репродуктивного тракта. Эмбрионы помещались в одну фаллопиеву трубу через бахромки яичника. Суррогатные матери проверялись на беременность путем ультразвукового исследования приблизительно на 30 сутки, а затем еще раз спустя неделю беременности. Свиньи обычно рожают на 114 сутки беременности.
[0113] После трансфекции и скрининга фибробластных клеток, были отобраны кандидатные донорские клетки, которые использовались для переноса ядра соматической клетки (ПЯСК). Шестьсот шестьдесят шесть ПЯСК-эмбрионов были перенесены двум суррогатным матерям. Одна принесла 6 нормальных поросят мужского пола на 115 сутки беременности, у другой суррогатной матери выполнили кесарево сечение на 117 сутки беременности, получив два нормальных поросенка мужского пола, как показано в Таблице 1.
[0114] На фигуре 2 показано строение гена сиалоадгезина (SIGLEC1), нацеленного вектора и ожидаемый после рекомбинации генотип. На верхней вкладке Фигуры 2 показана нацеленная конструкция, применявшаяся для гомологической рекомбинации. Как описано выше, «верхний» фрагмент ДНК, использовавшийся для создания нацеленной конструкции, содержал ~3,5 кб вверх от экзона 1 и включал часть экзона 1 (после стартового кодона). «Нижний» фрагмент ДНК начинался в пределах интрона 3 и включал экзоны 4, 5, 6 и часть экзона 7. Большая часть экзона 1 и экзоны 2 и 3 полностью были замещены на неомициновую (neo) кассету; тимидинкиназная (ТК) кассета была введена сразу же после нижнего плеча при необходимости для использования в качестве маркера отрицательной селекции. На нижней вкладке Фигуры 2 показан мутантный ген сиалоадгезина после гомологической рекомбинации. Стрелками показаны олигонуклеотиды сайтов связывания, использованные для ПЦР-скрининга нацеленных клеточных линий и клонированных свиней. Нацеливание верхнего и нижнего плеча выполнялось путем олигонуклеотидного отжига, как показано стрелками, обозначающими праймеры «верхний, нацеленный на сиало C» и «обратный ФГК поли A» (SEQ ID NO: 1 и 3), «прямой промотор ФГК» и «7Rw1» (SEQ ID NO: 4 и 6), соответственно. Олигонуклеотидные последовательности перечислены в Таблице 2 ниже. При дополнительной проверке ПЦР использовались олигонуклеотиды, которые фланкировали район усечения/кассету Neo (праймеры «Экзон 1 конец ck» и «Интрон 3 ck обратный» (SEQ ID NO: 2 и 5, соответственно)). Продукта поврежденного аллеля и аллеля дикого типа отличались по длине приблизительно на 500 пар оснований.
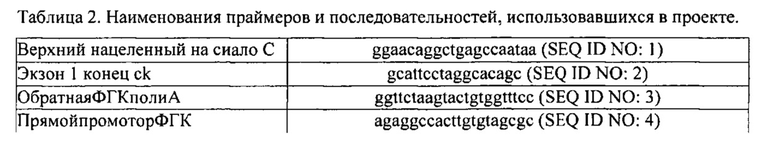

Праймерам из Таблицы 2 были присвоены идентификационные номера последовательностей исходя из положения стрелок в нижней части Фигуры 2 слева направо. Таким образом, самая левая стрелка в нижней части Фигуры 2 указывает положение праймера«верхний, нацеленный на сиало C» (SEQ ID NO: 1), следующая стрелка справа указывает положение праймера «Экзон 1 конец ck» (SEQ ID NO: 2), следующая стрелка справа от той указывает положение праймера «обратная ФГК поли A» (SEQ ID NO: 3), и так далее.
[0115] Скрининг на предмет инактивации SIGLEC1
[0116] Геномная ДНК была выделена от поросят и использовалась для подтверждения событий нацеливания. Успешность нацеливания на ген SIGLEC1 подтверждалась при помощи ПЦР с олигонуклеотидами, которые отжигались с кассетой Neo, спаренными с нуклеотидами, которые отжигались с геномной ДНК SIGLEC1, находившейся за областью, содержавшейся в нацеленной конструкции. На верхней вкладке Фигуры 3, нацеливание «верхнего» плеча изучалось при помощи олигонуклеотидов «обратная ФГК поли A» и «верхний нацеленный на сиало C» (SEQ ID NOs. 3 и 1, соответственно; показано на Фигуре 2). Был получен продукт ожидаемого размера (~4500 пар оснований). На нижней вкладке показано, что успешность нацеливания «нижнего» плеча определяли при помощи нуклеотидов «прямой промотор ФГК» и «7Rw1» (SEQ ID NOs: 4 и 6, соответственно; показано на Фигуре 2). Был получен продукт ожидаемого размера (~5000 пар оснований). Контрольное «нижнее плечо плазмиды» представляло собой частичную конструкцию, включающую кассету Neo с фрагментом гена сиалоадгезина, содержащим большую часть интрона 3 и большую часть экзона 7. Олигонуклеотид 7Rw1 был способен к отжигу с последовательностью экзона 7, присутствующей в плазмиде, и вместе с олигонуклеотидом прямого промотора ФГК, он был способен образовывать продукт, идентичный тому, который бы получался при успешном нацеливании. На обеих вкладках показаны реакции ПЦР нацеливания, выполненные на геномной ДНК, экстрагированной из восьми клонов поросят, полученных от линии нацеленных фетальных фибробластов 4-18.
[0117] Детектирование аллелей сиалоадгезина дикого и нацеленного типа выполнялось с использованием ПЦР с олигонуклеотидами, которые отжигались с ДНК, фланкирующей нацеленную область гена сиалоадгезина. Использовались олигонуклеотиды «Экзон 1 конец ck» и «Интрон 3 ck обратный» (SEQ ID NO: 2 и 5, соответственно; показано на Фигуре 2). Полученные продукты имели ~2400 пар оснований для аллеля дикого типа и ~2900 пар оснований для нацеленного аллеля. На Фигуре 4, вкладке слева (Фиг. 4А) показаны тестовые реакции, выполненные с геномной ДНК дикого типа, нацеленной плазмидой, использовавшейся для трансфекций, геномной ДНК (4-18) из успешно нацеленного клона фибробластов (обращают на себя внимание две полосы) и геномная ДНК (4-3) из ненацеленного клона фибробластов. На вкладке справа (Фиг. 4В) показана реакция, выполненная на геномной ДНК, экстрагированной из восьми клонов поросят, полученных от линии нацеленных фетальных фибробластов 4-18. В каждой полосе отмечалось два продукта ПЦР с ожидаемыми размерами, в то время как ДНК дикого типа и шаблонные нацеленные плазмиды давали только по одной линии. Таким образом, все поросята, полученные посредством ПЯСК, были гетерозиготны по предполагаемой мутации, а именно SIGLEC +/-.
[0118] Получение Гомозиготных Животных
[0119] Самцы генетически модифицированных свиней, идентифицированные как гетерозиготные по инактивации SIGLEC1, использовались как родительские особи (поколение F0), и спаривались с самками дикого типа для получения самцов и самок с одним инактивированным аллелем SIGLEC1 (F1). Самцы F1 затем спаривались с самками F1 для получения Р2-потомства с двумя инактивированными аллелями SIGLEC1. Такие животные могут быть идентифицированы после рождения посредством ПЦР, описанной выше и направленной на SIGLEC1 или, иначе, Саузерн-блоттингом.
ПРИМЕР 2
[0120] Получение Конструкции, Нацеленной на CD163
[0121] Как было установлено, делеция цитоплазматического домена CD163 приводит к неспособности ВРРСС к инфицированию; аналогично действует делеция или модификация SRCR домена 5. Поскольку некоторые из SRCR доменов CD163 обладают важными функциями для выживания животного, например, для удаления гемоглобина, модифицирование гена при условии сохранения других функций является хорошей стратегией для получения свиней, резистентных к ВРРСС. Предшествующие исследования также показали, что замена домена SRCR5 на домен CD163L 8 также блокирует инфекционную способность (Van Gorp et al. 2010b). Таким образом, нацеленная конструкция может быть сконструирована так, как показано на Фигуре 6, с заменой SRCR5 в CD163 на SRCR-домен CD163L.
[0122] После изготовления нацеленной конструкции, генетически модифицированная свинья, гетерозиготная по инактивации CD163, причем инактивация аллеля CD163 приводит к белку CD163, неспособному связывать и/или обнажать вирус репродуктивно-респираторного синдрома свиней (ВРРСС), может быть получена по способам, которые в общих чертах аналогичны описанным в Примере 1. Вначале нацеленный вектор вводят в донорскую клетку, в которой он может рекомбинировать с эндогенным геном CD163. Затем отбираются донорские клетки с этой специфической модификацией, которые используются для переноса ядра соматической клетки с получением генетически модифицированного эмбриона. Затем эмбрион переносят суррогатной матери для вынашивания. После идентификации у трансгенной свиньи инактивированного аллеля CD163 посредством ПЦР или Саузерн-блоттинга, животных доращивают до половозрелости и используют для естественного размножения с передачей гена потомству.
[0123] ЛИТЕРАТУРА
[0124] Albina Е, Madec F, Cariolet R, Torrison J. 1994. Immune Response and Persistence of the Porcine Reproductive and Respiratory Syndrome Virus in Infected Pigs and Farm Units. Veterinary Record 134(22):567-573.
[0125] Allende R, Laegreid WW, Kutish GF, Galeota JA, Wills RW, Osorio FA. 2000. Porcine reproductive and respiratory syndrome virus: Description of persistence in individual pigs upon experimental infection. Journal of Virology 74(22): 10834-10837.
[0126] Andreyev VG, Wesley RD, Mengeling WL, Vorwald AC, Lager KM. 1997. Genetic Variation and Phylogenetic Relationships of 22 Porcine Reproductive and Respiratory Syndrome Virus (Prrsv) Field Strains Based on Sequence Analysis of Open Reading Frame 5. Archives of Virology 142(5):993-1001.
[0127] Benfield DA, Nelson E, Collins JE, Harris L, Goyal SM, Robison D, Christianson WT, Morrison RB, Gorcyca D, Chladek D. 1992. Characterization of swine infertility and respiratory syndrome (SIRS) virus (isolate ATCC VR-2332). Journal of Veterinary Diagnostic Investigation 4:127-133.
[0128] Benfield DA, Nelson J, Rossow KD, Rowland RR, Lawson SR, Steffen M, Collins M. 1998. Pathogenesis and persistence of PRRS. Proceedings, Allen D. Leman Swine Conference: 169-171.
[0129] Christopherhennings J, Nelson EA, Hines RJ, Nelson JK, Swenson SL, Zimmerman JJ, Chase CCL, Yaeger MJ, Benfield DA. 1995. Persistence of Porcine Reproductive and Respiratory Syndrome Virus in Serum and Semen of Adult Boars. Journal of Veterinary Diagnostic Investigation 7(4):456-464.
[0130] Christopherhennings J, Nelson EA, Nelson JK, Benfield DA. 1997. Effects of a Modified-Live Virus Vaccine against Porcine Reproductive and Respiratory Syndrome in Boars. American Journal of Veterinary Research 58(1):40-45.
[0131] Crocker PR, Gordon S. 1986. Properties and distribution of a lectin-like hemagglutinin differentially expressed by murine stromal tissue macrophages. J Exp Med 164(6):1862-1875.
[0132] Crocker PR, Vinson M, Kelm S, Drickamer K. 1999. Molecular analysis of sialoside binding to sialoadhesin by NMR and site-directed mutagenesis. Biochemical Journal 341 (Part 2):355-361.
[0133] Das PB, Dinh PX, Ansari IH, de Lima M, Osorio FA, Pattnaik AK. 2010. The minor envelope glycoproteins GP2a and GP4 of porcine reproductive and respiratory syndrome virus interact with the receptor CD163. J Virol 84(4):1731-1740.
[0134] Dee SA, Molitor TW. 1998. Elimination of Porcine Reproductive and Respiratory Syndrome Virus Using a Test and Removal Process. Veterinary Record 143(17):474-476.
[0135] Delputte PL, Meerts P, Costers S, Nauwynck HJ. 2004. Effect of virus-specific antibodies on attachment, internalization and infection of porcine reproductive and respiratory syndrome virus in primary macrophages. Vet Immunol Immunopathol 102(3): 179-188.
[0136] Delputte PL, Nauwynck HJ. 2004. Porcine arterivirus infection of alveolar macrophages is mediated by sialic acid on the virus. Journal of Virology 78(15):8094-8101.
[0137] Delputte PL, Van Breedam W, Delrue I, Oetke C, Crocker PR, Nauwynck HJ. 2007. Porcine arterivirus attachment to the macrophage-specific receptor sialoadhesin is dependent on the sialic acid-binding activity of the N-terminal immunoglobulin domain of sialoadhesin. J Virol 81(17):9546-9550.
[0138] Im G-S, Lai L, Liu Z, Hao Y, Prather RS. 2004. In vitro development of preimplantation porcine nuclear transfer embryos cultured in different media and gas atmospheres.
Theriogenology 61:1125-1135.
[0139] Keffaber KK. 1989. Reproductive failure of unknown etiology. American Association of Swine Practitioners Newsletter 1:1-9.
[0140] Kristiansen M, Graversen JH, Jacobsen C, Sonne O, Hoffman HJ, Law SK, Moestrup SK. 2001. Identification of the haemoglobin scavenger receptor. Nature 409(6817):198-201.
[0141] Lager KM, Mengeling WL, Brockmeier SL. 1999. Evaluation of protective immunity in gilts inoculated with the NADC-8 isolate of porcine reproductive and respiratory syndrome virus (PRRSV) and challenge-exposed with an antigenically distinct PRRSV isolate. American Journal of Veterinary Research 60(8):1022-1027.
[0142] Lai L, Kang JX, Li R, Wang J, Witt W, Yong HY, Hao Y, Wax D, Murphy CN, Rieke A, Samuel M, Linville ML, Korte SW, Evans R, Starzl ТЕ, Prather RS, Dai Y. 2006. Generation of cloned transgenic pigs rich in omega-3 fatty acids. Nature Biotechnology 24:435-437.
[0143] Lai LX, Kolber-Simonds D, Park KW, Cheong HT, Greenstein JL, Im GS, Samuel M, Bonk A, Rieke A, Day BN, Murphy CN, Carter DB, Hawley RJ, Prather RS. 2002. Production of alpha-1,3-galactosyltransferase knockout pigs by nuclear transfer cloning. Science 295(5557):1089-1092.
[0144] Machaty Z, Prather RS. 2001. Complete activation of mammalian oocytes. US Patent Issued April 17: #6,211,429.
[0145] Machaty Z, Wang WH, Day BN, Prather RS. 1997. Complete Activation of Porcine Oocytes Induced by the Sulfhydryl Reagent, Thimerosal. Biology of Reproduction 57(5):1123-1127.
[0146] Mansour SL, Thomas ICR, Capecchi MR. 1988. Disruption of the proto-oncogene int-2 in mouse embryo-derived stem cells: a general strategy for targeting mutations to non- selectable genes. Nature 336:348-352.
[0147] Meng XJ. 2000. Heterogeneity of porcine reproductive and respiratory syndrome virus: implications for current vaccine efficacy and future vaccine development [Review], Veterinary Microbiology 74(4):309-329.
[0148] Meng XJ, Paul PS, Halbur PG, Morozov I. 1995. Sequence Comparison of Open Reading Frames 2 to 5 of Low and High Virulence United States Isolates of Porcine Reproductive and Respiratory Syndrome Virus. Journal of General Virology 76(Part 12):3181-3188.
[0149] Mengeling WL, Vorwald AC, Lager KM, Clouser DF, Wesley RD. 1999. Identification and clinical assessment of suspected vaccine-related field strains of porcine reproductive and respiratory syndrome virus. American Journal of Veterinary Research 60(3):334-340.
[0150] Molitor TW, Bautista EM, Choi CS. 1997. Immunity to Prrsv - Double-Edged Sword. Veterinary Microbiology 55(l-4):265-276.
[0151] Murtaugh MP, Elam MR, Kakach LT. 1995. Comparison of the Structural Protein Coding Sequences of the Vr-2332 and Lelystad Virus Strains of the Prrs Virus. Archives of Virology 140(8):1451-1460.
[0152] Nath D, van der Merwe PA, Kelm S, Bradfield P, Crocker PR. 1995. The amino-terminal immunoglobulin-like domain of sialoadhesin contains the sialic acid binding site. Comparison with CD22. J Biol Chem 270(44):26184-26191.
[0153] Nauwynck HJ, Duan X, Favoreel HW, Van Oostveldt P, Pensaert MB. 1999. Entry of porcine reproductive and respiratory syndrome virus into porcine alveolar macrophages via receptor-mediated endocytosis. Journal of General Virology 80(Part 2):297-305.
[0154] Nelsen CJ, Murtaugh MP, Faaberg KS. 1999. Porcine reproductive and respiratory syndrome virus comparison: Divergent evolution on two continents. Journal of Virology 73(1):270-280.
[0155] Nielsen HS, Liu G, Nielsen J, Oleksiewicz MB, Botner A, Storgaard T, Faaberg KS. 2003. Generation of an infectious clone of VR-2332, a highly virulent North American type isolate of porcine reproductive and respiratory syndrome virus. Journal of Virology 77(6):3702- 3711.
[0156] Nielsen MJ, Madsen M, Moller HJ, Moestrup SK. 2006. The macrophage scavenger receptor CD163: endocytic properties of cytoplasmic tail variants. J Leukoc Biol 79(4):837-845.
[0157] Oetke C, Vinson MC, Jones C, Crocker PR. 2006. Sialoadhesin-deficient mice exhibit subtle changes in B- and T-cell populations and reduced immunoglobulin M levels. Mol Cell Biol 26(4):1549-1557.
[0158] Plagemann PGW. 1996. Lactate dehydrogenase-elevating virus and related viruses. In Fields Virology, 3rd Ed, Fields, В et al., ed: 1105-1120.
[0159] Ritter M, Buechler C, Langmann T, Orso E, Klucken J, Schmitz G. 1999a. The scavenger receptor CD163: regulation, promoter structure and genomic organization. Pathobiology 67(5-6):257-261.
[0160] Ritter M, Buechler C, Langmann T, Schmitz G. 1999b. Genomic organization and chromosomal localization of the human CD163 (M1 30) gene: a member of the scavenger receptor cysteine-rich superfamily. Biochem Biophys Res Commun 260(2):466-474.
[0161] Ropp SL, Wees СЕМ, Fang Y, Nelson EA, Rossow KD, Bien M, Arndt B, Preszler S, Steen P, Christopher-Hennings J, Collins JE, Benfield DA, Faaberg KS. 2004. Characterization of emerging European-like porcine reproductive and respiratory syndrome virus isolates in the United States. Journal of Virology 78(7):3684-3703.
[0162] Rowland RR, Steffen M, Ackerman T, Benfield DA. 1999. The evolution of porcine reproductive and respiratory syndrome virus: quasispecies and emergence of a virus subpopulation during infection of pigs with VR-2332. Virology 259:262-266.
[0163] Sanchez-Torres C, Gomez-Puertas P, Gomez-del-Moral M, Alonso F, Escribano JM, Ezquerra A, Dominguez J. 2003. Expression of porcine CD163 on monocytes/macrophages correlates with permissiveness to African swine fever infection. Arch Virol 148(12):2307-2323.
[0164] Snijder EJ, Meulenberg JJM. 1998. THE MOLECULAR BIOLOGY OF ARTERIVIRUSES [Review], Journal of General Virology 79(Part 5):961-979.
[0165] Suarez P, Diazguerra M, Prieto C, Esteban M, Castro JM, Nieto A, Ortin J. 1996. Open Reading Frame 5 of Porcine Reproductive and Respiratory Syndrome Virus as a Cause of Virus-Induced Apoptosis. Journal of Virology 70(5):2876-2882.
[0166] Sur JH, Cooper VL, Galeota JA, Hesse RA, Doster AR, Osorio FA. 1996. In Vivo Detection of Porcine Reproductive and Respiratory Syndrome Virus Rna by in Situ Hybridization at Different Times Postinfection. Journal of Clinical Microbiology 34(9):2280-2286.
[0167] Sur JH, Doster AR, Christian JS, Galeota JA, Wills RW, Zimmerman JJ, Osorio FA. 1997. Porcine Reproductive and Respiratory Syndrome Virus Replicates in Testicular Germ Cells, Alters Spermatogenesis, and Induces Germ Cell Death by Apoptosis. Journal of Virology 71(12):9170-9179.
[0168] Van Breedam W, Delputte PL, Van Gorp H, Misinzo G, Vanderheijden N, Duan X, Nauwynck HJ. 2010a. Porcine reproductive and respiratory syndrome virus entry into the porcine macrophage. J Gen Virol 91(Pt 7):1659-1667.
[0169] Van Breedam W, Van Gorp H, Zhang JQ, Crocker PR, Delputte PL, Nauwynck HJ. 2010b. The M/GP(5) glycoprotein complex of porcine reproductive and respiratory syndrome virus binds the sialoadhesin receptor in a sialic acid-dependent manner. PLoS Pathog 6(1):e1000730.
[0170] van den Hoff MJ, Moorman AF, Lamers WH. 1992. Electroporation in 'intracellular' buffer increases cell survival. Nucleic Acids Res 20(11):2902.
[0171] Van Gorp H, Delputte PL, Nauwynck HJ. 2010a. Scavenger receptor CD163, a Jack-of-all-trades and potential target for cell-directed therapy. Mol Immunol 47(7-8):1650-1660.
[0172] Van Gorp H, Van Breedam W, Delputte PL, Nauwynck HJ. 2008. Sialoadhesin and CD163 join forces during entry of the porcine reproductive and respiratory syndrome virus. J Gen Virol 89(Pt 12):2943-2953.
[0173] Van Gorp H, Van Breedam W, Van Doorsselaere J, Delputte PL, Nauwynck HJ. 2010b. Identification of the CD163 protein domains involved in infection of the porcine reproductive and respiratory syndrome virus. J Virol 84(6):3101-3105.
[0174] Vanderheijden N, Delputte PL, Favoreel HW, Vandekerckhove J, Van Damme J, van Woensel PA, Nauwynck HJ. 2003. Involvement of sialoadhesin in entry of porcine reproductive and respiratory syndrome virus into porcine alveolar macrophages. Journal of Virology 77(15):8207-8215.
[0175] Vinson M, Vandermerwe PA, Kelm S, May A, Jones EY, Crocker PR. 1996. Characterization of the Sialic Acid-Binding Site in Sialoadhesin by Site-Directed Mutagenesis. Journal of Biological Chemistry 271(16):9267-9272.
[0176] Welch SKW, Calvert JG. 2010. A brief review of CD163 and its role in PRRSV infection. Virus Research 154:98-103.
[0177] Wensvoort G, Terpstra C, Pol JMA, ter Laak EA, Bloemrad M, de Kluyer EP, Kragten C, van Buiten L, den Besten A, Wagenaar F, Broekhuijsen JM, Moonen PLJM, Zetstra T, de Boer EA, Tibben HJ, de Jong MF, van't Veld P, Groenland GJR, van Gennep JA, Voets MTH, Verheijden JHM, Braamskamp J. 1991. Mystery swine disease in The Netherlands: the isolation of Lelystad virus. Veterinary Quarterly 13:121-130.
[0178] Wills RW, Zimmerman JJ, Yoon KJ, McGinley MJ, Hill HT, Piatt KB, Christopherhennings J, Nelson EA. 1997. Porcine Reproductive and Respiratory Syndrome Virus - a Persistent Infection. Veterinary Microbiology 55(1-4):231-240.
[0179] Wissink EHJ, van Wijk HAR, Pol JMA, Godeke GJ, van Rijn PA, Rottier PJM, Meulenberg JJM. 2003. Identification of porcine alveolar macrophage glycoproteins involved in infection of porcine respiratory and reproductive syndrome virus. Archives of Virology 148(1):177-187.
[0180] Yoon IL, Joo HS, Christianson WE, Morrison RB, Dial GD. 1993. Persistent and contact infection in nursery pigs experimentally infected with PRRS virus. Swine Health and Production 1:5-8.
[0181] При введении элементов настоящего изобретения или его предпочтительного(ых) воплощения(й), существительные в единственном числе и слово «упомянутый» обозначают, что подразумеваются один или больше элементов. Термины «содержащий», «включающий» и «имеющий» имеют включающий характер и подразумевается, что могут быть дополнительные элементы, отличные от перечисленных элементов.
[0182] В свете сказанного выше, будет понятно, что достигнут ряд целей изобретения и иные заслуживающие внимания результаты.
[0183] Поскольку в указанные выше продукты и способы могут быть внесены различные изменения без отступления от охвата изобретения, подразумевается, что все сведения в описании выше и в сопроводительных чертежах должны интерпретироваться как демонстративные, а не ограничительные.
Соответствие Исправленных Пунктов Пунктам в РСТ-заявке




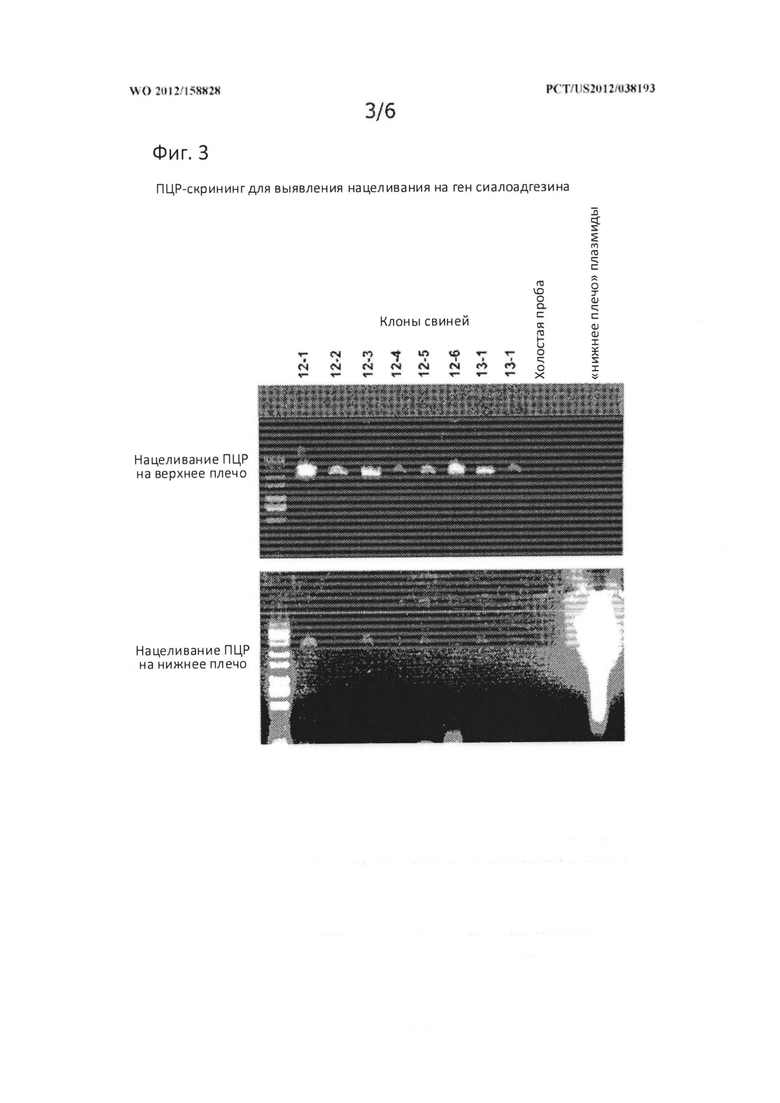
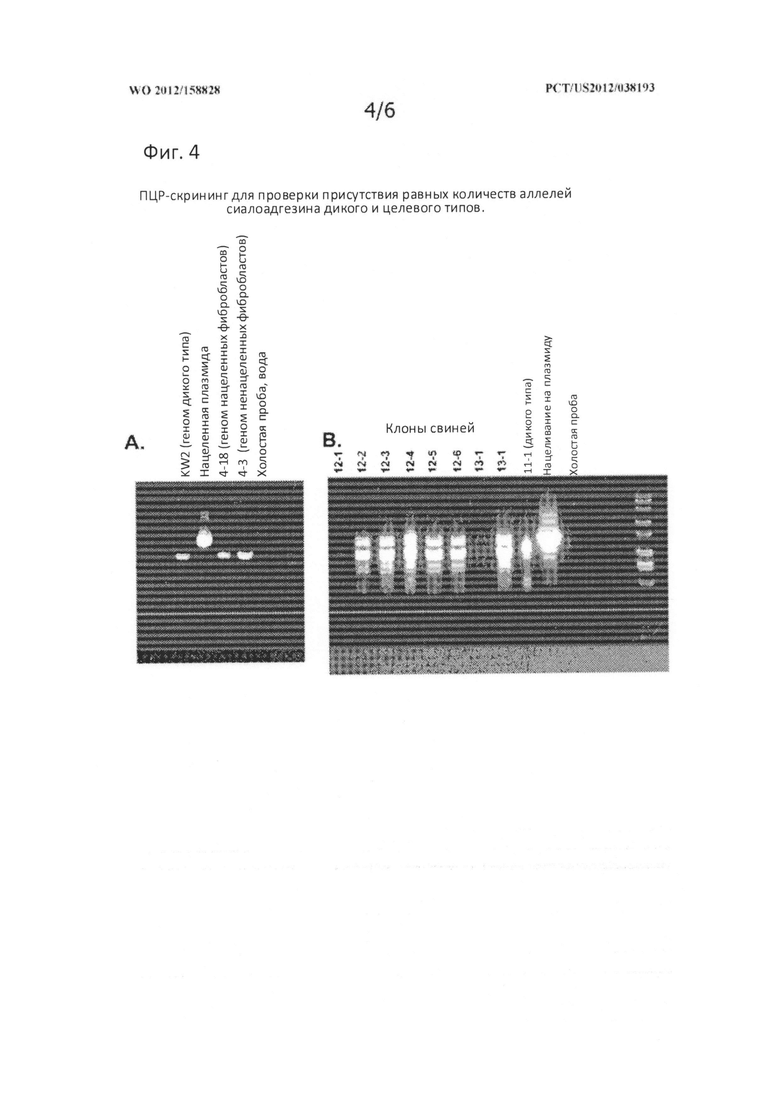
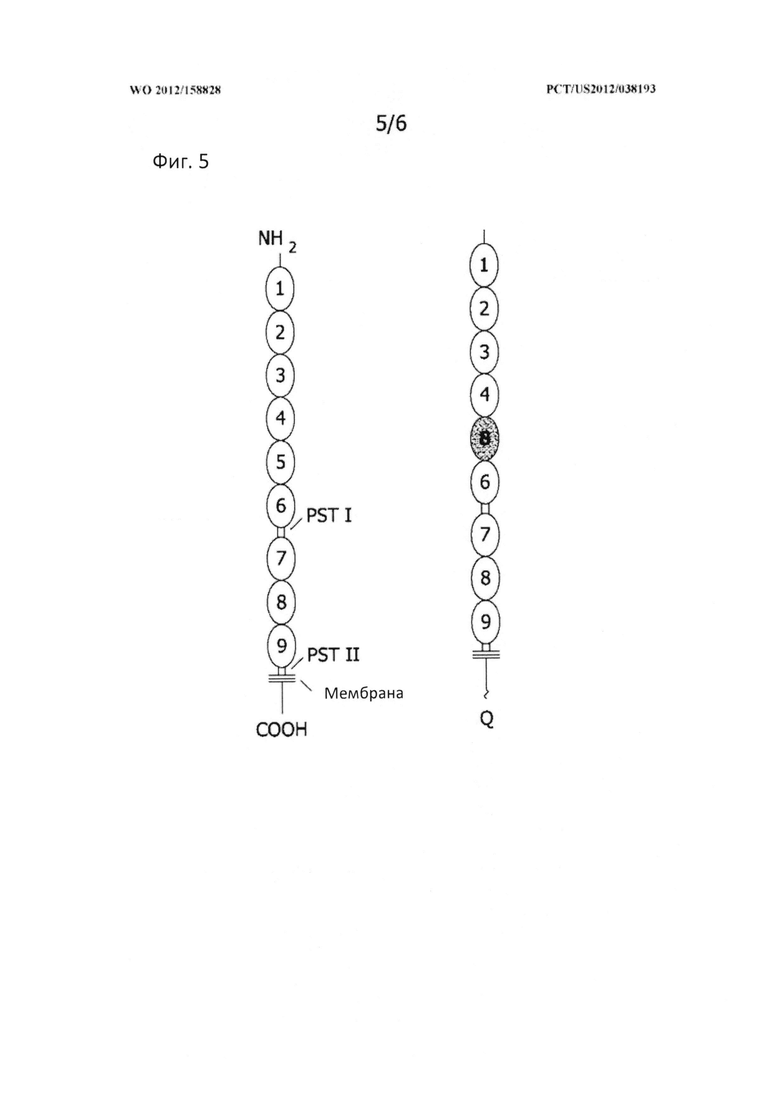
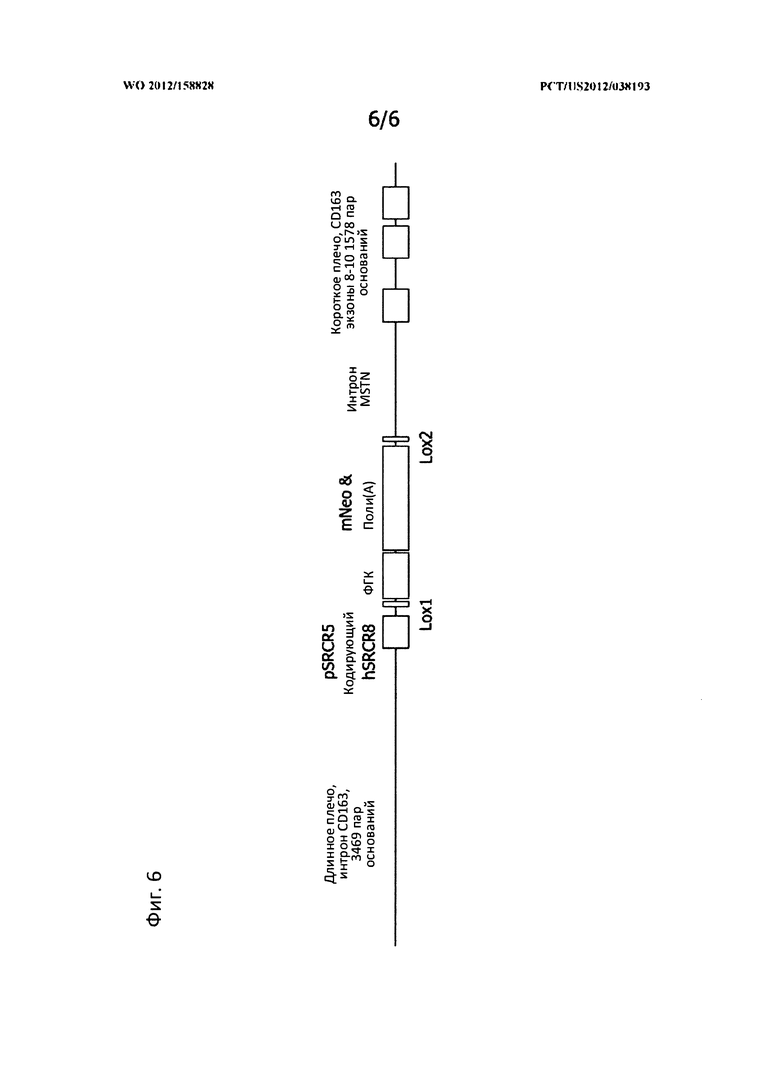
Изобретение относится к генетической инженерии, в частности к генетически модифицированной свинье, которая является резистентной к инфекции вирусом репродуктивно-респираторного синдрома свиней (ВРРСС). Указанная генетически модифицированная свинья содержит инактивирующую мутацию в обоих аллелях гена CD163. Такая инактивирующая мутация приводит к получению белка CD163, который не может связывать ВРРСС, то есть мутация изменяет активность CD163. Указанная генетически модифицированная свинья может дополнительно содержать инактивирующую мутацию в обоих аллелях гена SIGLEC1. Также предложены способы получения таких генетически модифицированных свиней. Настоящее изобретение позволяет получать генетически модифицированных свиней, резистентных к ВРРСС. 5 н. и 19 з.п. ф-лы, 6 ил., 2 пр., 2 табл.
1. Генетически модифицированная свинья, содержащая инактивирующую мутацию в обоих аллелях гена CD163, где указанная свинья является резистентной к инфекции вирусом репродуктивно-респираторного синдрома свиней (ВРРСС).
2. Генетически модифицированная свинья по п. 1, где указанная инактивирующая мутация в аллеле гена CD163 приводит к получению белка CD163, который не может связывать и/или обнажать ВРРСС.
3. Генетически модифицированная свинья по п. 1, где указанные мутации включают инсерции, частичные делеции, полные делеции, мутации со сдвигом рамки считывания, бессмысленные мутации, миссенс-мутации, точечные мутации или их комбинацию.
4. Генетически модифицированная свинья по п. 1, где указанные мутации включают частичные делеции, полные делеции или их комбинацию.
5. Генетически модифицированная свинья по п. 1, где указанная мутация изменяет экспрессию или активность CD163.
6. Генетически модифицированная свинья по п. 1 или 2, где аллель CD163 был инактивирован путем замены экзона 7 на богатый цистеином домен 8 фагоцитарного рецептора (SRCR) из CD163L.
7. Генетически модифицированная свинья по любому из пп. 1-5, где указанная свинья дополнительно содержит инактивирующую мутацию, по меньшей мере, в одном аллеле гена SlGLECl.
8. Генетически модифицированная свинья по любому из пп. 1-5, где указанная свинья дополнительно содержит инактивирующую мутацию в обоих аллелях гена SIGLEC1.
9. Способ получения генетически модифицированной свиньи, содержащей инактивирующие мутации в обоих аллелях гена CD163, где указанная свинья является резистентной к инфекции ВРРСС, где способ включает:
энуклеацию ооцита свиньи;
слияние ооцита с фибробластной клеткой донорской свиньи, где геном фибробластной клетки содержит по меньшей мере, один инактивированный аллель CD163;
активацию ооцита для получения эмбриона;
перенос эмбриона в репродуктивный тракт суррогатной свиньи, у которой начался эструс, но не завершилась овуляция; и при этом беременность и роды в срок дают генетически модифицированную свинью, геном которой содержит, по меньшей мере, один инактивированный аллель CD163;
спаривание самки генетически модифицированной свиньи, геном которой содержит, по меньшей мере, один инактивированный аллель CD163, с самцом генетически модифицированной свиньи, геном которого содержит, по меньшей мере, один инактивированный аллель CD163, для получения F1-потомства;
скрининг F1-потомства для идентификации генетически модифицированной свиньи, у которой оба аллеля гена CD163 были инактивированы.
10. Способ по п. 9, где указанные мутации включают инсерции, частичные делеции, полные делеции, мутации со сдвигом рамки считывания, бессмысленные мутации, миссенс-мутации, точечные мутации или их комбинацию.
11. Способ по п. 9, где указанные мутации включают частичные делеции, полные делеции или их комбинацию.
12. Способ по п. 9, где указанная мутация изменяет экспрессию или активность CD163.
13. Способ по п. 9, где указанные аллели были инактивированы посредством использования рекомбинационной системы Cre-lox.
14. Способ по п. 9, где аллель CD163 инактивирован путем замены экзона 7 на богатый цистеином домен 8 фагоцитарного рецептора (SRCR) из CD163L.
15. Способ по любому из пп. 9-14, где энуклеацию выполняют в микроманипуляционной среде при помощи микропипетки.
16. Способ по любому из пп. 9-14, где слияние выполняют в среде для слияния.
17. Способ по любому из пп. 9-14, где активация ооцита включает инкубирование ооцита в присутствии тимеросала.
18. Способ по любому из пп. 9-14, где перенос включает перенос эмбриона в фаллопиеву трубу через яичниковые бахромки суррогатной матери.
19. Способ получения генетически модифицированной свиньи, которая является резистентной к инфекции ВРРСС, у которой оба аллеля гена SIGLEC1 и оба аллеля гена CD163 инактивированы, где способ включает:
спаривание самки генетически модифицированной свиньи, содержащей инактивирующую мутацию, по меньшей мере, в одном аллеле гена SIGLEC1, с самцом генетически модифицированной свиньи, содержащим инактивирующую мутацию, по меньшей мере, в одном аллеле гена CD163, или спаривание самца генетически модифицированной свиньи, содержащего инактивирующую мутацию, по меньшей мере, в одном аллеле гена SIGLEC1, с самкой генетически модифицированной свиньи, содержащей инактивирующую мутацию, по меньшей мере, в одном аллеле гена CD163, для получения F1-потомства;
скрининг F1-потомства для идентификации генетически модифицированной свиньи, у которой, по меньшей мере, один аллель гена SIGLEC1 инактивирован и, по меньшей мере, один аллель гена CD163 инактивирован;
спаривание генетически модифицированных свиней, у которых, по меньшей мере, один аллель гена SIGLEC1 инактивирован и, по меньшей мере, один аллель гена CD163 инактивирован, друг с другом для получения F2-потомства; и
скрининг F2-потомства для идентификации генетически модифицированной свиньи, у которой оба аллеля гена SIGLEC1 и оба аллеля гена CD163 инактивированы.
20. Способ получения генетически модифицированной свиньи, которая является резистентной к инфекции ВРРСС, у которой оба аллеля гена SIGLEC1 и оба аллеля гена CD163 инактивированы, где способ включает:
спаривание самки генетически модифицированной свиньи, содержащей инактивирующие мутации в обоих аллелях гена SIGLEC1, с самцом генетически модифицированной свиньи, содержащим инактивирующие мутации в обоих аллелях гена CD 163, или спаривание самца генетически модифицированной свиньи, содержащего инактивирующие мутации в обоих аллелях гена SIGLEC1, с самкой генетически модифицированной свиньи, содержащей инактивирующие мутации в обоих аллелях гена
CD163, для получения F1-потомства;
спаривание F1-потомства для получения F2-потомства; и
скрининг F2-потомства для идентификации генетически модифицированной свиньи, у которой оба аллеля гена SIGLEC1 и оба аллеля гена CD163 инактивированы.
21. Способ получения генетически модифицированной свиньи, которая является резистентной к инфекции ВРРСС, у которой оба аллеля гена SIGLEC1 и оба аллеля гена CD163 инактивированы, где способ включает:
спаривание генетически модифицированной свиньи, у которой инактивированы, по меньшей мере, один аллель гена SIGLEC1 и, по меньшей мере, один аллель гена CD163, с другой генетически модифицированной свиньей, у которой инактивированы, по меньшей мере, один аллель гена SIGLEC1 и, по меньшей мере, один аллель гена CD163, с получением F1-потомства; и
скрининг F1-потомства для идентификации генетически модифицированной свиньи, у которой оба аллеля гена SIGLEC1 и оба аллеля гена CD163 инактивированы.
22. Способ по любому из пп. 19-21, отличающийся тем, что при скрининге используются полимеразная цепная реакция (ПЦР) или Саузерн-блот.
23. Способ по любому из пп. 9-14 и 19-21, где указанная свинья является резистентной к инфекции вирусом репродуктивно-респираторного синдрома свиней (ВРРСС).
24. Способ по любому из пп. 9-14 и 12-21, где указанная инактивирующая мутация в аллеле гена CD163 приводит к получению белка CD163, который не может связывать и/или обнажать вирус репродуктивно-респираторного синдрома свиней (ВРРСС).
| US 2011016546 А1, 20.01.2011 | |||
| US 2011016543 А1, 20.01.2011 | |||
| VANDERHEIJDEN N | |||
| et al., Involvement of sialoadhesin in entry of porcine reproductive and respiratory syndrome virus into porcine alveolar macrophages, J.Virol., 2003, Aug, 77(15), pp.8207-8215 | |||
| VAN GORP H | |||
| et al., Sialoadhesin and CD163 join forces during entry of the porcine reproductive and respiratory syndrome virus, J | |||
| Gen | |||
| Virol., 2008, 89, pp.2943-2953 | |||
| VAN GORP H | |||
| et al., Identification of the CD163 protein domains involved in infection of the porcine reproductive and respiratory syndrome virus, J | |||
| Virol., 2010, 84(6), pp.3101-3105 | |||
| РЫБЧИН В.Н., Основы генетической инженерии, Санкт-Петербург, Издательство СПбГТУ, 2002, с.411-418. |

