Изобретение относится к области фармацевтической биотехнологии, а именно к способам производства терапевтических препаратов, выделения и очистки рекомбинантных белков, в частности к получению высокоочищенного препарата ромиплостим. Изобретение относится к способу получения правильно свернутых белков и к способу хроматографии для разделения правильно свернутых и неправильно свернутых конформаций данного белка. Способ является высокоэффективным для разделения правильно свернутого ромиплостима и неправильно свернутого ромиплостима и контаминирующих примесей с высоким выходом продукта высокой чистоты и с хорошим соотношением правильно свернутого ромиплостима к неправильно свернутому ромиплостиму. Основной сферой применения полученного предлагаемым способом терапевтического рекомбинантного белка ромиплостим является лечение тромбоцитопении у пациентов от одного года и старше, резистентных к другим видам лечения (например, глюкокортикостероидам, иммуноглобулинам) с хронической иммунной (идиопатической) тромбоцитопенической пурпурой (ИТП).
Изобретение решает задачу создания способа получения в промышленных объемах терапевтического препарата ромиплостим, способствует выполнению цели государственной политики РФ по развитию национальной фармацевтической промышленности в сфере обеспечения населения жизненно необходимыми и важнейшими лекарственными препаратами отечественного производства.
ОПИСАНИЕ ЧЕРТЕЖЕЙ
SEQ ID NO 1: Первичная последовательность фрагмента ДНК, заключающая ген белка ромиплостим.
SEQ ID NO 2: Аминокислотная последовательность ТМР пептида (от англ. ТРО Mimetic Peptid, пептид, имитирующий ТРО).
SEQ ID NO 3: Аминокислотная последовательность мономера химерного белка с С-концевой химеризацией плеча тяжелой цепи IgG, составленного двумя пептидными последовательностями из 14 аминокислот (SEQ ID NO: 1) с линкерными вставками из полиглицина, длиною 269 аминокислотных остатков.
Фиг. 1 Механизм действия ромиплостима и элтромбопага.
Активация рецептора тромбопоэтина тромбопоэтином (ТРО) или агонистами рецептора. ТРО или ромиплостим связываются с рецептором тромбопоэтина на поверхности тромбоцитов через домен CRH-2, превращая неактивный рецептор в активный димер, запуская внутриклеточный каскад через фосфорилирование тирозинкиназы JAK2 и последующей активацией транскрипционного фактора STAT, что приводит к пролиферации этих клеток. Элтромбопаг связывается с трансмембранным регионом ТРО рецептора и также запускает внутриклеточный каскад, приводящий к пролиферации тромбоцитов.
Фиг. 2 Распространенность редких болезней из группы «24 нозологий» по нозологиям (80 субъектов Российской Федерации, 93,0% населения страны). Серым цветом выделена распространенность заболеванием идиопатической тромбоцитопенической пурпурой.
Фиг. 3 Структура ромиплостима
Ромиплостим состоит из части IgG1 (Fc домен) с двумя пептидами - миметиками тромбопоэтина (Тромбопоэтин рецептор связывающий домен, 14 а.а.), которые соединены между собой и С-концевой частью Т тяжелой цепи IgG глициновыми линкерами (Полиглицидиновый линкер 5 а.а. и Полиглицидиновый линкер 8 а.а.).
Межцепочечные дисульфидные связи (аминокислотные остатки цистеина в положении С7 и С10) и внутрицепочечные дисульфидные связи СН2 и СН3 (аминокислотные остатки цистеина в положении С42-С102, С148-С206).
Фиг. 4 Строение ромиплостима с димерной последовательностью ТМР миметика. Последовательность ТМР миметика указана с использованием однобуквенного кода.
Фиг. 5 Электрофореграмма образцов отмывных растворов и раствора телец включения. Образцы 1, 2, 3 - отмывные растворы, образец 4 - раствор телец включения.
Фиг. 6 Определение содержания мономерной единицы ромиплостима методом ГФ ВЭЖХ. Кривая 1 - буфер плацебо + дитиотриэтол (ДТТ), 2 - буфер для растворения телец включения, 3 - раствор восстановленного лекарственного препарата (ЛП) Энплейт, 4 - раствор телец включения ромиплостима.
Фиг. 7 Электрофореграмма образцов рефолдируемой смеси и ЛП Энплейт в восстанавливающих и невосстанавливающих условиях. Образец 1 - рефолдируемая смесь в невосстанавливающих условиях, Образец 2 - в восстанавливающих условиях,
Образец 3 - ЛП Энплейт в невосстанавливающих условиях,
Образец 4 - ЛП Энплейт в восстанавливающих условиях.
Фиг. 8 Определение хроматографической чистоты рефолдируемой смеси методом ОФ ВЭЖХ.
Фиг. 9 Электрофореграмма фракций с хроматографий на Q Sepharose FF и MabSelect SuRe в восстанавливающих и невосстанавливающих условиях.
Образец 1 - фракция проскока с MabSelect SuRe; Образец 2 - фракция отмыва с MabSelect SuRe; Образец 3 - элюат с MabSelect SuRe; Образец 4 - элюат с Q Sepharose FF;
Фиг. 10A Профиль хроматографии с MabSelect SuRe.
Пик 1 - отмыв; Пик 2 - элюат 1, серым цветом выделены фракции с содержанием главной формы ромиплостима не менее 60%, формы 1 не более 2%, формы 4 не более 10% по результатам анализа ОФ ВЭЖХ; Пик 3 - элюат2; Пик 4 - регенерация;
Фиг. 10В Определение хроматографической чистоты фракций с хроматографии на MabSelect SuRe методом ОФ ВЭЖХ.
Фиг. 11 Профиль хроматографии с YMC S30-1.
Пик 1 - элюат 1, серым цветом выделены фракции с содержанием главной формы ромиплостима не менее 87%, формы 3 не более 8%, формы 4 не более 4% по результатам анализа ОФ ВЭЖХ; Пик 2 - элюат 2; Пик 3 - регенерация;
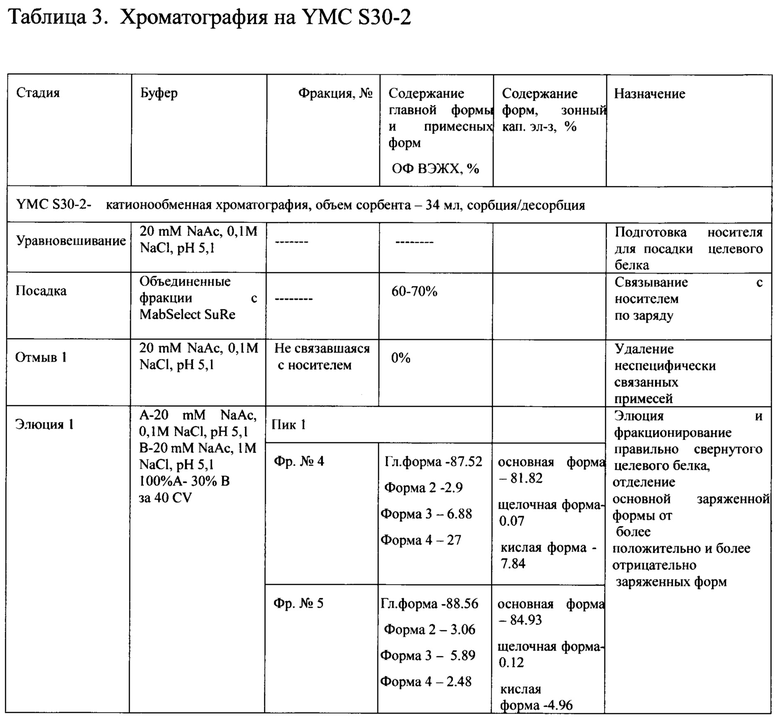
Фиг. 12А Профиль хроматографии с YMC S30-2.
Пик 1 - элюат 1, серым цветом выделены фракции с содержанием главной формы ромиплостима не менее 90% по результатам анализа ОФ ВЭЖХ и не менее 87% по результатам капиллярного зонного электрофореза; Пик 2 - регенерация;
Фиг. 12В Определение хроматографической чистоты фракций с хроматографии на YMC S 30 -2 методом ОФ ВЭЖХ.
Фиг. 12С Определение хроматографической чистоты фракций с хроматографии на YMC S 30 -2 методом капиллярного зонного электрофореза.
Фиг. 13А Определение хроматографической чистоты готовой формы ромиплостима методом ОФ ВЭЖХ.
Фиг. 13В Определение хроматографической чистоты готовой формы ромиплостима методом ГФ ВЭЖХ.
Фиг. 13С Определение хроматографической чистоты готовой формы ромиплостима методом капиллярного зонного электрофореза.
Фиг. 14А Ингибирование связывания тромбопоэтина с ТПО-рецептором различными сериями ГФ ромиплостима.
Фиг. 14В Графики зависимости концентрации фосфорилированной тирозинкиназы JAK2 от логарифма концентрации ромиплостима в разведениях серий ГФ ромиплостима: А-донор №1, В-донор №2, С-донор №3.
Иммунная тромбоцитопения (ИТП) (прежнее название идиопатическая тромбоцитопеническая пурпура) - это заболевание, манифестирующее иммунно-опосредованной тромбоцитопенией. В развитии ИТП ведущую роль играют антитела, направленные против узкого спектра гликопротеинов поверхности тромбоцитов и мегакариоцитов. Чаще всего ИТП встречается у женщин детородного возраста и при беременности (Abir Zainal et al., Immune thrombocytopenic purpura // Journal of community hospital internal medicine perspectives - 2019 - V. 9. - N. 1. - p. 59-61). Однако встречается у лиц обоих полов в любом возрасте. С учетом относительно сниженного уровня тромбопоэтина в крови пациентов с ИТП, а также способности антитромбоцитарных антител вызывать апоптоз мегакариоцитов и нарушать продукцию ими тромбоцитов, было предположено, что активация рецепторов c-Mpl (тромбопоэтиновый рецептор) может приводить к повышению числа тромбоцитов у пациентов с хронической формой ИТП.
За демонстрацией воздействия тромбопоэтина (ТРО) на рост мегакариоцитов и производство тромбоцитов, и подтверждение его роли, как основного фактора роста, регулирующего продукцию тромбоцитов, быстро последовала разработка рекомбинантного человеческого rHuTPO. Гликозилированная полноразмерная молекула с аминокислотной последовательностью, идентичной последовательности нативного ТРО, была получена в культуре клеток млекопитающих. PEG-rHuMGDF (фактор роста и развития мегакариоцитов), пегилированный продукт, содержащий часть нативной последовательности ТРО, был произведен в E.coli и впоследствии модифицирован ковалентным присоединением фрагмента полиэтиленгликоля 20 кД (Archimbaud Е. et al., A randomized, doubleblind, placebo-controlled study with pegylated recombinant human megakaryocyte growth and development factor (PEG-rHuMGDF) as an adjunct to chemotherapy for adults with de novo acute myeloid leukemia // Blood. - 1999. - 94. - p. - 3694-3701). Однако развитие как MGDF, так и rHuTPO было остановлено после того, как было обнаружено, что некоторые пациенты и здоровые добровольцы выработали антитела к PEG-rHuMGDF на повторное введение. Эти антитела перекрестно реагировали с нативным ТРО, что приводило к длительной резистентности к лечению тромбоцитопении (Basser R.L. et al., Development of pancytopenia with neutralizing antibodies to thrombopoietin after multicycle chemotherapy supported by megakaryocyte growth and development factor//Blood. - 2002. - 99. - p. 2599-2602). Исследования продолжались, что привело к разработке следующего поколения средств, стимулирующих тромбопоэз, - пептидов ТРО и не пептидов, миметиков с небольшой структурной гомологией с природным ТРО. Два семейства небольших пептидов, неродственных друг другу по структуре, которые связываются с человеческим рецептором ТРО и конкурируют за связывание с природным ТРО, были идентифицированы из библиотек рекомбинантных пептидов. Затем создавали мутагенизированные библиотеки для дальнейшей оптимизации белков с наилучшей связывающей способностью, что привело к выделению очень активного пептида. Указанный пептид, состоящий из 14 аминокислотных остатков (SEQ ID NO: 2), обозначенный как ТМР (от англ. ТРО Mimetic Peptid, пептид, имитирующий ТРО), не обнаруживал гомологии по аминокислотной последовательности с ТРО (S.Е. Cwirla et al, Peptide agonist of the thrombopoietin receptor as potent as the natural cytokine // Science. - 1997. - 276. - 5319. - p.1696-1699).
Учеными компании Amgen был сконструирован рекомбинантный белок, состоящий из домена Fc-носителя, связанного с несколькими c-Mpl-связывающими доменами, и был назван AMG531 или АМР-2 (Amgen Megakaryopoiesis Protein-2) (David J. Kuter, The biology of thrombopoietin and thrombopoietin receptor agonists // Int J Hematol. - 2013. - 98. - p. 10-23), оригинальное название ромиплостим (Virginia С.Broudy, Nancy L. Lin, AMG531 stimulates megakaryopoiesis in vitro by binding to Mpl // Cytokine - 2004. - 25. - p. 52-60). AMG 531, АМР-2 (ромиплостим) являлся биологическим стимулятором ТРО, разработанным для преодоления проблемы перекрестной реакции антител на ТРО с помощью случайной пептидной последовательности для активации c-Mpl рецептора. (Wojciech Krzyzansk et al, Pharmacokinetic and Pharmacodynamic Modeling of Romiplostim in Animals // Pharm Res. - 2013. - 30. - p. 655-669).
Впоследствии фармакодинамические и фармакокинетические исследования и исследования метаболизма были проведены на мышах, крысах, кроликах, собаках и обезьянах. Ромиплостим хорошо переносился и вызывал дозозависимое повышение уровня тромбоцитов у всех видов животных с широким спектром межвидового различия. (Hartley, С, The novel thrombopoietic agent AMG 531 is effective in pre-clinical models of chemo/radiotherapy induced thrombocytopenia//Proceedings of the American Association for Cancer Research. - 2005. - 46. - (abstract 1233)).
Эффективность ромиплостима была продемонстрирована в клинических исследованиях ромиплостима фазы I - II (Bussel et al, AMG 531, a Thrombopoiesis-Stimulating Protein, for Chronic ITP1 // The New England Journal of Medicine, - 2006. - 355. - р. 1672-1681) и III фазы у пациентов с ИТП (Kuter D.J. et al,. Efficacy of romiplostim in patients with chronic immune thrombocytopenic purpura: a double-blind randomised controlled trial // Lancet. - 2008.-371. - p. 395-403.). В результате, ромиплостим был одобрен для лечения ИТП у взрослых в США и Австралии в 2008 г.
Ромиплостим впервые был лицензирован в ЕС в 2009 г. для использования у пациентов со спленэктомией и без спленэктомии с хронической ИТП, которая не поддается лечению другими методами (например, кортикостероидами, внутривенными иммуноглобулинами, (http://www.ema.europa.eu/docs/en_GB/document_librarv/EPAR_Product_Information/human/000942/WC500039537.pdf). Nplate Summary of Product Characteristics 2018. Accessed May 4, 2018).
В 2014 году показания были расширены, и теперь в них включены пациенты без спленэктомии, которым хирургическое вмешательство противопоказано. Показания были ограничены только взрослыми пациентами во время проведения этого исследования, но в настоящее время ромиплостим одобрен для пациентов старше 1 года. Ромиплостим был впервые рекомендован National Institute for Health and Care Excellence (NICE) в 2011 году. NICE в настоящее время рекомендует ромиплостим в качестве варианта лечения взрослых с хронической ИТП, если их состояние не поддается стандартному активному лечению и спасательной терапии, или они имеют тяжелое заболевание и высокий риск кровотечения, который требует частых курсов спасательной терапии (https://www.nice.org.uk/guidance/ta221/ chapter/1 - Guidance. Romiplostim for the treatment of chronic immune (idiopathic) thrombocytopenic purpura. National Institute for Health and Care Excellence 2014; Technology appraisal guidance (TA221)).
Ромиплостим в настоящее время выпускается компанией Amgen Inc. под торговым наименованием Энплейт® (NPLATE®).
Пациенты с хронической ИТП испытывают снижение качества жизни, связанного со здоровьем (Health-Related Quality of Life (HRQoL), связанное с усталостью, беспокойством по поводу своего внешнего вида из-за синяков, а также из-за нарушения способности выполнять рутинную повседневную деятельность. Действительно, недавно сообщалось, что такие пациенты имеют худшее HRQoL, чем пациенты с гипертонией, артритом или раком (McMillan et al, Self-reported health-related quality of life in adults with chronic immune thrombocytopenic purpur // American Journal of Hematology. - 2008. - 83. - p. 150-154).
Клинические исследования таких стимуляторов тромбопоэза как ромиплостим (Энплэйт®, Амджен) и элтромбопаг (Револейд®, Глаксо Смит Кляйн) показали вероятность достижения ответа у более 80% пациентов при низкой токсичности. Поэтому эти препараты сегодня являются основными в лечении пациентов с тяжелой хронической формой ИТП (Indraraj Umesh Doobaree et al., Primary immune thrombocytopenia (ITP) treated with romiplostim in routine clinical practice: retrospective study from the United Kingdom ITP Registry// Eur J Haematol -2019. - 102. - p.416-423, Drew Provan, Adrian C. Newland, Current Management of Primary Immune Thrombocytopenia//Adv. Ther. - 2015. - 32. - p.875-887). На Фиг. 1. продемонстрирован механизм действия ромиплостима и элтромбопага.
Ромиплостим относится к группе препаратов, предназначенных для патогенетической терапии редких (орфанных) заболеваний. Как правило, это дорогостоящие лекарственные препараты, не имеющие аналоговых замен и являющиеся жизненно необходимыми. Важной особенностью этих лекарственных препаратов является их «таргетность», высокая эффективность (с достижением ответа на терапию, приближающегося к 100%), хороший профиль безопасности и переносимости в большинстве случаев, что немаловажно с учетом длительной пожизненной терапии. Применение именно таких препаратов достоверно и значимо снижает количество необратимых, инвалидизирующих поражений жизненно важных органов и летальность, повышает качество жизни пациентов по сравнению с применяемыми ранее методами наилучшей поддерживающей (симптоматической) терапии, приводит к значительному улучшению медико-социальной реабилитации больных и улучшению показателей долгосрочной выживаемости (вплоть до сравнимой по полу и возрасту со здоровой популяцией).
Для решения задачи расширения доступа к жизненно необходимой терапии больным с редкими заболеваниями авторы изобретения предлагают новый способ получения в промышленных объемах рекомбинантного высокоочищенного препарата ромиплостим. Ромиплостим представляет собой пептидное антитело, состоящее из дисульфидно - связанных константной тяжелой цепи человеского IgG1 и легкой каппа-цепи (фрагмент Fc) с двумя идентичными пептидными последовательностями ковалентно связанных по остатку 228 тяжелой цепи с использованием полиглицина (James В. Bussel et al., AMG 531, a Thrombopoiesis-Stimulating Protein, for Chronic ITP// The new England journal of medicine - 2006. - 355. - p. 1672-81) (Фиг. 3).
Для усиления способности к связыванию была применена стратегия димеризации С-концевых участков по отношению к ТРО миметическому пептиду (ТМР). Было обнаружено, что димер, связанный по С-концевому участку, обладает улучшенной связывающей аффинностью и существенно увеличенной активностью in vitro в исследованиях клеточной пролиферации (William Dower, Stewene Cwirla et al., Peptide Agonists of the Thrombopoietin Receptor,Thrornbopoietin: From Molecule to Medicine // Stem Cells. - 1998. - 16. - p.21-29-). Строение ромиплостима с димерной последовательностью ТМР миметика представлено на Фиг. 4.
Пептиды представляют собой уникальный класс фармацевтических соединений, молекулярно сбалансированных между небольшими молекулами и белками, но все же биохимически и терапевтически отличающихся от обоих. Как собственные сигнальные молекулы для многих физиологических функций, пептиды предоставляют возможность для терапевтического вмешательства, которое тесно имитирует естественные пути. Однако энтузиазм в отношении пептидной терапии сдерживается некоторыми ограничениями нативных пептидов, такими как короткий период полувыведения из плазмы и незначительная пероральная биодоступность. Короткий период полураспада объясняется наличием многочисленных пептидаз и экскреторными механизмами, которые инактивируют пептиды (Jolene L. Lau, Michael K. Dunn, Therapeutic peptides: Historical perspectives, current development trends, and future directions // Bioorganic & Medicinal Chemistry. - 2018. - 26. - p. 2700-2707). Были разработаны различные стратегии расширения стабильности и улучшения фармакокинетических свойств пептидов, одна из этих стратегий состояла в том, чтобы пришить пептиды к области Fc иммуноглобулина G (IgG), тем самым продлевая период полураспада в плазме биологически активных пептидов. Наличие части Fc приводит к увеличению гидродинамического радиуса молекулы выше порога клубочковой фильтрация почками, вследствие увеличения молекулярной массы (около 60 кДа), возможности использования неонатального Fc-рецептора и защите от лизосомальной деградации (Grant Shimamoto, Colin Gegg,Tom Boone and Christophe Queva, Peptibodies A flexible alternative format to antibodies// mAbs. - 2012. - V. 4. - I.5. - p. 586-591).
Fc содержащие полипептиды можно получать с использованием рекомбинантных технологий в клеточных системах дрожжей, бактерий, насекомых, растений и животных (ЕР 2 752 427 A1, Antibodies containing therapeutic peptides).
В настоящее время ромиплостим производится Amgen Inc, Boulder, СО, USA. с помощью технологии рекомбинантной ДНК в Escherichia coli (Е. coli). В коммерческом производственном процессе используются операции ферментации Е. coli и обработки клеток для получения промежуточного продукта (суспензии). Процесс очистки начинается с оттаивания суспензии с последующей солюбилизацией. Ромиплостим очищают с использованием серии стадий хроматографии, концентрирования и диафильтрации для получения очищенного основного вещества (Australian Public Assessment Report (AUSPAR) For Romiplostim//Sample Auspar. - 2008. - p. 1-31).
Существует три основных преимущества производства биологически активных веществ, биосинтез которых осуществляется в E.coli, по сравнению с биосинтезом в клетках млекопитающих или растений: короткое время культивирования, высокий уровень экспрессии, отсутствие посттрансляционных модификаций.
В литературе описаны способы производства E.coli-экспрессируемых пептидных антител, начиная с создания генетической конструкции штамма -продуцента рекомбинантного белка, операций блока ферментации, которые включают в себя культивирование клеток, сбор клеток, дезинтеграцию клеточной биомассы при высоком давлении или ультразвуком с последующим получением и промывкой нерастворимых тел включения, которые содержат денатурированный рекомбинантый белок. Тельца включения образуются в результате неспособности бактериальной клетки-хозяина правильно складывать рекомбинантные белки при высоких уровнях экспрессии, и, как следствие, белки становятся нерастворимыми. Это особенно верно в отношении больших, сложных белков эукариотического происхождения.
Образование неправильно свернутых рекомбинантных белков в некоторой степени ограничивают коммерческую полезность бактериальной ферментации для получения рекомбинантных больших, сложных белков с высокой эффективностью.
С момента появления рекомбинантной экспрессии белков на коммерчески жизнеспособных уровнях в системах экспрессии не млекопитающих, таких как бактерии, были разработаны различные методы для получения правильно свернутых белков из телец включения. Эти методы обычно следуют после процедуры экспрессии белка, который обычно осаждается в тела включения, лизируют клетки, собирают тела включения и затем солюбилизируют их в буфере для солюбилизации, включающем денатурирующий агент или поверхностно-активное вещество и, необязательно, восстановитель, который раскрывает белки и разбирает тела включения на отдельные белковые цепи практически без структуры. Затем белковые цепи разбавляют или промывают буфером для рефолдинга, который поддерживает ренатурацию до биологически активной формы. Когда остатки цистеина присутствуют в первичной аминокислотной последовательности белка, часто необходимо осуществить рефолдинг в среде, которая позволяет правильно образовывать дисульфидные связи (например, используя окислительно-восстановительную пару (Grant Shimamoto, Colin Gegg,Tom Boone and Christophe Queva, Peptibodies A flexible alternative format to antibodies// mAbs. - 2012. - V. 4, - I.5, - p. 586-591, US 6,835,809 B1 Liu et al. Thrombopoietic compounds, WO 00/24770, Liu Chuan -Fa, Thrombopoietic compounds, AU 2004200687 B2, Feige U. et al, Modified peptides as therapeutic agents, Shultz et al. Refolding proteins using a chemically controlled redox state // US 8,952,138 B2). Рефолдированный белок подвергают процессу хроматографической очистки, который оптимизируется для каждого пептидного антитела для обеспечения лучшего выхода продукта при оптимальной очистке от примесей, таких как продукты деградации, мисфолды, аберрантные изомеры, агрегатные, окисленные, деамидированные и другие формы целевого белка, примесей клеточных белков, нуклеиновых кислот, эндотоксинов и липидов. Затем очищенный продукт подвергают концентрированию и переводу в буфер готовой формы диафильтрацией в тангенциальном потоке на мембранах и получают фармацевтическую субстанцию (ФС). Для получения лекарственного препарата ФС лиофилизируют.
Коммерческий производственный процесс получения ромиплостима затрагивается в отчетах (Report on the Deliberation Results. - 2010. - p. - 102, https://www.pmda.go.jp/files/000236304.pdf, Australian Public Assessment Report (AUSPAR) For Romiplostim//Sample Auspar. - 2008. - p. 1-31), но не указываются ни условия процессов ни используемые материалы.
В публикациях компании Amgen: US6,835,809 B1, Liu et al. Trombopoietic compounds, US2006/0234307 A1, Feige et al., Modified peptides as therapeutic agents, EP1144454 B2, Feige et al., Modified peptides as therapeutic agents, US7,169,905 B2, Feige U., Modified peptides as therapeutic agents, US2004/0053845, Feige et al., Modified peptides as therapeutic agents, US2006/0234307, Feige et al., Modified peptides as therapeutic agents, US8044174 B2, Liu et al., Trombopoietic compounds, US9,534,032 B2, Liu et al., Trombopoietic compounds, US2006/0234307 Al, Modified peptides as therapeutic agents, US7,169,905 B2, Feige, Modified peptides as therapeutic agents, описан способ получения вариантов Fc содержащих антител с ТМР пептидами на N или С конце или TMPi-(L)n-TMP2 (Ln - аминокислотные остатки глицина (Gly), где n составляет от 1 до 20, а когда n больше 1, до половины остатков Gly могут быть заменены другой аминокислотой, выбранной из оставшихся 19 природных аминокислот или их стереоизомеров), с использованием стандартных методов для получения генетической конструкции в плазмиде pAMG21 Fc в с экспрессией в Е. coli GM221. Один из вариантов выше указанных соединений, описанный в патенте US6,835,809 B1 Liu et al. Thrombopoietic compounds, и является ромиплостимом. Генетическая конструкция, содержащаяся в плазмиде paMG21 (pAMG21-Fc-TMP-TMP), которая в свою очередь, содержится в штамме-хозяине GM221, зарегистрирована в АТСС под инвентарным номером 98957, с даты депонирования 22 октября 1998 г.
Культуры Е. coli GM221 с pAMG21-Fc-TMP-TMP или Fc-TMP-(L)n-ТМР культивируют в среде LB, содержащей 50 мкг/мл канамицина, инкубируют 3 часа при 37°С перед индукцией. Индукцию генных продуктов Fc-TMP-TMP с LuxPR промотором, осуществляют добавлением синтетического аутоиндуктора N-(3-оксогексаноил)-БЬ гомосеринлактона. Ферментацию проводят в стандартных условиях в масштабе 10 л, в результате чего уровень экспрессии Fc-TMP-TMP был аналогичен уровню, полученному в лабораторном масштабе. Экспрессируемый продукт накапливался в клетках в виде телец включения. Для получения телец включения клеточную биомассу диспергируют в воде в соотношении(1/10) и дезинтегрируют гомогенизацией под высоким давлением (2 прохода при 14000 psi (965,3 бар)). Тельца включения собирают центрифугированием при 4200 rpm в течение 60 минут и солюбилизируют с использованием 6М гуанидинина гидрохлорида в качестве хаотропного агента в восстанавливающих условиях с 8 мМ ДТТ. Ренатурируют белок методом разбавления в 20-25 раз в 50 мМ трис-HCl буферном растворе с 2 М мочевиной, 160 мМ аргинином и 3 mM цистеином при рН 8,5. Смесь перемешивают в течение 16-48 часов при 4°С, а затем концентрируют примерно в 10 раз с помощью ультрафильтрации. Концентрат разбавляют буферным раствором 10 mM Трис с 1,5 М мочевиной, рН 9,0, рН этой смеси доводят до 5,0 с помощью уксусной кислоты. Образовавшийся осадок удаляют центрифугированием. Целевой белок из супернатанта очищают двумя последовательными хроматографиями на SP Sepharose Fast Flow с использованием градиента за 20 колоночных объемов (CV) в натрий -ацетатной буферной системе, рН 5.0.
В патентах US2006/0140934 A1, Gegg et al., Modified Fc molecules, US2007/0269369 A1, Gegg et al, Modified Fc molecules, US7,442,778,Gegg et al, Modified Fc molecules, US7,750,128 B2, Gegg et al., Modified Fc molecules, описан наиболее близкий к заявленному изобретению способ получения пептидных антител, в котором по меньшей мере один биологически активный пептид включен в качестве внутренней последовательности в домен Fc. В другом варианте этого изобретения линейный, свободный пептид, AMP 2 (АМР-2, IEGPTLROWLAARA (SEQ ID NO: 2) был вставлен в домен Fc-петли человеческого IgG1. Конструкция АМР2 Fc-loop обозначена как клон Amgen №6875. Клон Fc-loop AMP 2 (# 6875) был трансформирован в Е. coli обычными методами с экспрессией на высоких уровнях во фракции нерастворимых телец включения. Выделенную фракцию телец включения в соотношении 1/10 солюбилизировали в 6 М гуанидин-HCl, 50 мМ Трис, 8 мМ DTT, рН 9 при комнатной температуре при перемешивании в течение 1 часа. Рефолдинг проводили методом разбавления 1:25 в 2 М мочевине, 50 мМ Трис, 4 мМ цистеине, 1 мМ цистамине, рН 8,5 при 4°С. при перемешивании в течение 48 часов, затем аликвоты оценивали с помощью SDS-PAGE и обращенно-фазовой ВЭЖХ.
Очищали пептидное антитело с помощью 2-х последовательных хроматографий. Первую стадию очистки проводили на колонке с белком А, уравновешенную в буфере 2 М мочевина, 50 мМ Трис с рН 8,5. Фильтрованную реакционную смесь наносили на колонку и затем колонку промывали с 2 объемами уравновешивающего буфера с последующими 2 объемами PBS. Элюировали пептидное антитело 50 мМ натрий-ацетатным буфером (NaOAc), рН 3 и быстро нейтрализовали разбавлением 1: 4 10 мМ NaOAc, 50 мМ NaCl, рН 5.0. Разбавленный элюат снова фильтровали и наносили на катионообменную колонку с SP Sepharose HP (Pharmacia), уравновешенную 10 мМ NaOAc, 50 мМ. NaCl, рН 5. Затем фракции пептидных антител элюировали линейным градиентом 50-500 мМ NaCl. Фракции объединяли и концентрировали примерно до 2 мг/мл.
Для решения задачи получения высокоочищенного и биологически активного препарата ромиплостим в промышленных объемах авторы изобретения предлагают усовершенствованный способ его выделения и очистки. Рекомбинантный белок был получен продукцией в E.coli в виде нерастворимых телец включения. Далее из E.coli выделяли и отмывали тельца включения с последующей их солюбилизацией и рефолдингом белка для получения димерной формы за счет образования 4-х внутрицепочечных и 2-х межцепочечных дисульфидных связей как внутри молекулы каждого мономера, так и между находящимися в растворе разными молекулами мономеров. Далее предлагается проводить хроматографическую очистку белка из рефолдируемой смеси, где первую стадию проводят на сильном анионобменном сорбенте, представляющем собой высокосшитую гранулированную агарозу, в режиме проскока с последующей сорбцией целевого белка на аффинном сорбенте, матрица которого представляет собой агарозу с поперечными сшивками, модифицированную рекомбинантным белком А; с последующими эффективными отмывками и мягкими условиями элюции, и двух катионообменных хроматографий, первая из которых является разделяющей, вторая полирующей и концентрирующей, и последующим переводом в буфер готовой формы гель-фильтрацией с получением готовой формы высокоочищенного препарата ромиплостим, не менее 0,9 г с литра бактериальной культуры.
Основным техническим результатом изобретения является получение в промышленных масштабах высокоочищенного и биологически активного препарата ромиплостим.
Так как продукт синтезируется в бактериальной цитоплазме преимущественно в не растворенном состоянии в виде телец включения при культивировании E.coli ВКПМ В-13251 с уровнем биосинтеза не менее 900 мг/л культуральной жидкости, требуется проведение рефолдинга.
Высокоэффективное извлечение телец включения из клеток достигается в результате процедуры лизиса бактериальных клеток с использованием ультразвукового гомогенизатора или при масштабировании гомогенизатора высокого давления.
В заявленном изобретении продемонстрировано получение фармацевтической субстанции высокоочищенного препарата ромиплостим благодаря разработанной схеме. В результате эффективных промывок получены очищенные от примесей клеточные белки телец включения, после оптимального рефолдинга и последующей хроматографической очистки, которая, в совокупности с применением некоторых стандартных приемов хроматографической очистки белков и введением двух стадий катионообменной хроматографии, первая из которых является разделяющей, а вторая полирующей и концентрирующей, с последующим переводом в буфер готовой формы гель-фильтрацией, является эффективной для разделения правильно свернутого ромиплостима и неправильно свернутого ромиплостима и контаминирующих примесей, таких как продукты деградации, мисфолды, аберрантные изомеры, агрегатные, окисленные, деамидированные и другие формы целевого белка, примесей клеточных белков, нуклеиновых кислот, эндотоксинов и липидов. Результирующий выход продукта способен обеспечивать получение препарата ромиплостима высокой чистоты в отношении правильно свернутого ромиплостима сравнению с неправильно свернутым ромиплостимом и получение готовой формы в количестве не менее 0,9 г с литра бактериальной культуры.
Осуществление изобретения проиллюстрировано на примере получения генно-инженерной конструкции (ГИК) pGNR69-001 и экспрессию целевого белка путем культивирования E.Coli ВКПМ В-13251 в реакторе Biostat Bplus Twin 5L. Процесс культивирования банка клеток ВКПМ В-13251 был ориентирован на получение высокого количества биомассы с целенаправленной локализацией белка ромиплостим в телах включения.
Представленный в изобретении пример очистки белка иллюстрирует лизис клеточной биомассы и получение телец включения, использование отмывок телец включения для удаления контаминирующих примесей клеточных компонентов уже на первой стадии буферными растворами, содержащими 2% Тритон Х-100, 2М мочевину, 2М NaCl и 5 мМ ЭДТА. Отмывки оказались эффективными и удаляли большинство примесных клеточных белков, нуклеиновых кислот, липидов и эндотоксинов. На Фиг. 5 представлена электрофореграмма образцов отмывных растворов и раствор телец включения, полученных с применением отмывных растворов, образцы 1, 2, 3 демонстрируют наличие примесных белков в отмывных растворах и отсутствие целевого белка, образец 4, раствор телец включения, наличие практически свободного от примесей клеточных белков денатурированного мономера химерного белка.
Растворение полученных телец включения может быть осуществлено в соответствующем растворе гуанидина гидрохлорида, обычно на уровне, эквивалентном 7 - 8М гуанидина гидрохлорида, который полностью денатурирует белок. Белок восстанавливали с помощью DTT (дитиотреитол), 5-20 мМ, при рН 8,5, предпочтительнее 10 мМ, и инкубировали при комнатной температуре в течение приблизительно 1,5-2 часов. Представленный в изобретении пример рефолдинга иллюстрирует идентифицированные и оцененные условия рефолдинга, которые оптимизируют выход целевого белка и минимизируют образование аберрантных изомеров, агрегатов и мисфолдов. Прежде всего была подобрана концентрация рефолдируемого белка. Типичные концентрации рефолдинга для сложных молекул, таких как молекулы, содержащие два или более дисульфида, составляют менее 2,0 г/л и более типично 0,001- 0,5 г/л (US20100292447, Pitner WR, et aL. Method and agent for refolding proteins, US7732183, Tomasselli A. et al., Methods for refolding enzymes, Krishnanand Tiwari et al., Refolding of recombinant human granulocyte colony stimulating factor: Effect of cysteine/cystine redox system// Indian Journal of Biochemistry & Biophysics. - 2012. - V. 49, - p. 285-288). Таким образом, рефолдинг больших масс сложного белка, такого как антитело, пептидное антитело или другой Fc-белок в промышленных масштабах представляет значительные ограничения из-за больших объемов, необходимых для рефолдинга белков при этих типичных концентрациях, и является общей проблемой промышленности.
В связи с большими объемами материала и размерами пула, используемыми при работе с производством белка в промышленном масштабе, можно сэкономить значительное время и ресурсы, исключив или упростив один, или несколько этапов процесса, например, увеличив концентрацию белка в рефолдируемой смеси. Хотя рефолдинг белков был ранее продемонстрирован при более высоких концентрациях, рефолдируемые белки были либо значительно меньше по молекулярной массе, либо менее сложными молекулами, содержащими только одну или две дисульфидные связи (см., например, Creighton Т., Kinetic Role of a Meta-stable Native-like Two-disulphide Species in the Folding Transition of Bovine Pancreatic Trypsin Inhibitor // J. Mol.Biol. - 1984. - 179. - p. 497-526, Creighton Т., Renaturation of the Reduced Bovine Pancreatic Trypsin Inhibitor,// J. Mol.Biol. - 1974. - 87. - p. 563-577). Кроме того, в процессах рефолдинга для таких белков использовались детергенты (US 2011/0077384 A1, Yumioka et al., Protein refolding method) или использовалась стратегия ренатурации под высоким давлением (US 7,767,795 В2, Randolph et al., High pressure refolding of protein aggregates and inclusion bodies). Более сложные молекулы, такие как антитела, пептидные антитела и другие крупные белки, часто имеющие более двух дисульфидных связей, от 8 и до 24, в результате описанных выше условий либо не растворяются полностью при использовании детергентов, либо могут образовать гомо- или гетеро-димеры. В патенте US 20100324269 A1, Shultz et al., Refolding proteins using a chemically controlled redox state, - описан способ рефолдинга Fc-содержащих белков молекулярным весом выше 57 к Да и содержащим 8 дисульфидных связей при концентрации 6 г/л, при наличии в буфере для рефолдинга добавок, препятствующих агрегации и способствующих стабильности рефолдируемого белка, таких как 4 М мочевина, 20,9% глицерин, 150 мМ аргинин и в присутствии окислительно/восстановительной пары 2,03 М цистеин/2,75 мМ цистамин. Разбавленную реакционную смесь титровали до щелочного рН (между рН 8 и рН 10) и инкубировали при 5°С в неаэробных условиях 12-72 часа. Описанный процесс продемонстрировал стабильную масштабируемость от 1 L до 2000 L с выходом желаемого продукта приблизительно 27-35% в обоих масштабах.
Изложенный выше способ имеет свои недостатки, высокое содержание мочевины и глицерина мешает дальнейшей хроматографической очистке и предполагает либо разбавление, либо диафильтрацию, а также наличие дополнительного оборудования для создания неаэробных условий, что создает определенные трудности при масштабировании процесса.
В настоящем изобретение предложен масштабируемый способ рефолдинга ромиплостима методом разбавления в 21 раз при концентрации от 0,7 до 1 г/л и установленных значения стабилизирующих концентраций добавок мочевины, аргинина, рН, окислительно/восстановительной пары, температуры и времени рефолдинга. Наилучшие результаты, определенные методом ОФ ВЭЖХ и ГФВЭЖХ, были получены с использованием буфера для рефолдинга, содержащего денатурирующий агент мочевину в концентрации 2,1М, подавитель агрегации аргинин в концентрации 0,168М, окислительно-восстановительные компоненты 3 mM цистин/ 4 mM цистеин в буфере 50 mM Трис-HCl, рН-9.0, при постоянном перемешивании при 4-8°С в аэробных условиях в течение 60-72 часов. Описанный процесс продемонстрировал стабильную масштабируемость от 3 L до 30 L с выходом желаемого продукта приблизительно 40-50% в обоих масштабах.
Описанный технический результат имеет одно из решающих значений для получения качественного целевого белка и способов дальнейшей очистки, так как именно на этой стадии происходит структурная сборка молекулы целевого белка, его димерной формы, за счет образования дисульфидных связей как внутри молекулы каждого мономера так и между находящимися в растворе молекулами мономеров, каждый из которых представляет собой фьюжен аминокислотных последовательностей Fc-части антитела человека изотипа 1 с синтетическим пептидом-миметиком тромбопоэтина на С-конце. Правильно рефолдированный белок должен иметь четыре внутрицепочечные и две межцепочечные дисульфидные связи.
От описанных в отчете по ромиплостиму Kyowa Hakko Kirin Co., Ltd. Australian Public Assessment Report (AUSPAR) for Romiplostim. - 2010. - p.102 и патентах компании Amgen (US 6,835,809 B1, Liu et al. Thrombopoietic compounds, US 6,660,843 В1, Feige et al., Modified peptides as therapeutic agents, ЕР 1 144 454 B2, Feige et al., Modified peptides as therapeutic agents, AU 2004200687, Feige Ulrich Modified peptides as therapeutic agents, US 457169905 B2, Feige Modified peptides as therapeutic agents, US 2004/0053845 Modified peptides as therapeutic agents, US 2006/0234307 Modified peptides as therapeutic agents, US 6,835,809 B1, Liu et al. Thrombopoietic compounds, US 8044174 B2. Liu et al., Thrombopoietic compounds, US 9,534,032 B2, Liu et al. Thrombopoietic compounds, WO 00/24770, Thrombopoietic compounds, US 2006/0234307 A1, Modified peptides as therapeutic agents, 2006. US 7,169,905 B2 Modified peptides as therapeutic agents, 6,117,655 A 9/2000 Capon et al.) кислотной обработки рефолдированного белка с последующими концентрированием и диафильтрацией отказались в виду высоких потерь целевого белка.
Хроматографическая очистка заключалась в проведении анионообменной хроматографии на сильном анионообменном сорбенте в режиме проскока, аффинной хроматографии на сорбенте для моноклональных антител MabSelect SuRe с эффективными отмывками и мягкими условиями элюции, двух катионообменных хроматографий на YMC S30, первая из которых является разделяющей, вторая полирующей и концентрирующей, и последующим переводом в буфер готовой формы гель-фильтрацией на Superdex G25. Конкретные условия проведения каждой стадии хроматографии и используемые марки сорбентов раскрыты в описании, но могут варьировать в известных специалистам пределах. Для первой стадии очистки, исходя из физико-химических свойств целевого белка и по литературным источникам (Fayaz S. et al. Expression, purification and biological activity assessment of romiplostim biosimilar peptibody // DARU J. Pharm. Sci. - 2016. - V. 24. - p. 1-5), разработаны, в частности, условия аффинной хроматографии на MabSelect SuRe, специально созданном аффинном сорбенте на основе агарозной матрицы и лиганда производного белка А, который специфически связывается с Fc-частью антител. Использование такого сорбента позволяет сорбировать на хроматографическую колонку все имеющие сформированную Fc-часть варианты молекул ромиплостима непосредственно из реакционной смеси после рефолдинга. На рынке доступны и другие сорбенты с лигандом протеина А, присоединенным к большому количеству различных матриц, таких как сшитая агароза, пористое стекло с модифицированной поверхностью, полистирол с покрытием, гидрогель, заполненный керамической оболочкой, и другие материалы на основе органических полимеров. Например, сорбент MAbSelect основан на матрице из агарозного геля с высокой степенью поперечных связей, что позволяет выдерживать более высокие скорости потока, сорбент MAbselect Xtra использует тот же химический состав основной цепи, что и MAbSelect, но имеет более широкий размер пор для улучшения массопереноса и, следовательно, способности динамического связывания. Уменьшение площади поверхности за счет более крупных пор компенсируется увеличением плотности лиганда. Смола Protein А на основе стекла с контролируемыми порами от EMD Millipore (ProSep) имеет лучшие характеристики текучести и диффузионные свойства, чем смолы Protein А на основе агарозы. Однако она страдает недостатком уменьшенной площади поверхности для связывания из-за большего диаметра пор. Сорбент MabSelect SuRe состоит из сконструированного белкового лиганда с доменом гомотетрамер-Z, в котором ряд остатков аспарагина был заменен для устранения взаимодействия с вариабельной областью, тем самым уменьшая гетерогенность связывания между антителами и позволяя смоле выдерживать более сильные щелочные условия. Стойкость к щелочам является важным преимуществом, поскольку позволяет многократно использовать 0,1-0,5М NaOH для очистки и дезинфекции. Хотя основные матрицы и плотности лигандов для MabSelect и MabSelect SuRe идентичны, последний обеспечивает большую стабильность в щелочных условиях, используемых в протоколах CIP (протоколы санации и регенерации) (Anurag S. Rathore et al., Re-use of Protein A Resin: Fouling and Economics // BioPharm International. - 2015. - V. 3. - , I. 28. - p.28-33).
Для оптимизации хроматографии на MabSelect SuRe и увеличения выхода целевого белка необходимо было решить несколько проблем: увеличить время жизни дорогостоящего сорбента, полноту посадки и выход белка в элюции, учитывая при этом, что целевой продукт стабилен только в узком диапазоне рН от 4,5 до 5,5.
Стоимость сорбентов с протеином А составляет от 8000 до 15000 долларов США за литр, однако, помимо этого аспекта, еще одной проблемой является загрязнение сорбента из-за неспецифической адсорбции, а так же потеря лиганда в результате протеолиза из-за присутствующих в клеточных лизатах протеаз, вызывающие снижение адсорбционной способности и более короткий срок службы смолы (Ana Mayela Ramos-de-la-Pena et. al, Protein A chromatography: Challenges and progress in the purification of monoclonal antibodies // J. Separation Science. - 2009. - 42. - p. 1816-1827, Anurag S. Rathore et al., Re-use of Protein A Resin: Fouling and Economics // BioPharm International. - 2015. - V. 3. - , I. 28. - p. 28-33). Для решения этой проблемы авторы применили анионообменную хроматографию на Q Sepharose FF в режиме проскока перед аффинной хроматографией на MabSelect SuRe. Были подобраны условия хроматографии, при которых липополисахариды, нуклеиновые кислоты и некоторые клеточные белки сорбировались на Q Sepharose FF, в то время как целевой белок без потерь проходил через колонку и сорбировался на MabSelect SuRe. Для того, чтобы избежать агрегации и обеспечить полноту посадки целевого белка при рН 7,2 и высоком содержании аргинина, который уменьшает специфическое связывание белка с лигандом, раствор рефолдированного белка разводили в 3 раза буфером для уравновешивания для MabSelect SuRe 20 mM натрий фосфат, рН - 7.2, содержащим 1,5М мочевину. Для решения проблем с полнотой элюирования целевого белка и конформационных изменений антител, вызывающих деградацию и агрегацию при низком рН (2,5 - 3,5), необходимым для элюции антител из колонки с белком-А, были разработаны более мягкие условия элюирования. Из литературных источников известно, что аргинин облегчает элюцию антител из колонки с белком-А, не нарушая структуру и поддерживая мономерное состояние (Arakawa Т. et al.6 Elution of antibodies from a Protein-A column by aqueous arginine solutions // Protein Expr. Purif. - 2004. - V. 36. - 2. - P. 244-248). Варьируя рН и концентрацию аргинина в элюирующем буфере, были установлены условия отмывки буферным раствором 30 мМ цитратом натрия, содержащим 0.2М аргинина, рН-4,4 (W1), при которой удалялись родственные примеси, трудно удаляемые при дальнейшей очистке, и условия элюции -линейный градиент за 10 колоночных объемов: от W1 до 100% буфера 30 мМ цитрат натрия, содержащего 0,5 М аргинина, рН-4,0. Элюат собирали по фракциям, в каждую фракцию добавляли 1М Трис непосредственно после получения фракции и доводили рН до 5,0-5,05 раствором 5М NaOH. Отбирали фракции с содержанием основного вещества не менее 60%, и с содержанием примесной формы 1 не более 2%, определяемых методом ОФ ВЭЖХ (Таблица 1, Фиг. 10. А., 10. В.). Применение оптимизированной хроматографии на MabSelect SuRe позволило получить с первой стадии очистки продукт с высоким содержанием структурно правильно фолдированного мономера ромиплостима (более 98,8% по ГФ ВЭЖХ и более 70% по ОФ ВЭЖХ). Для последующих стадий очистки необходимо было подобрать такие сорбенты и условия очистки, чтобы отделить от целевого белка близкородственные мисфолды, аберрантные изомеры, окисленные, деамидированные и другие модифицированные формы. Необходимо было учитывать, что, как было указано выше, белок стабилен только в узком диапазоне рН от 4,5 до 5,5. В качестве наиболее пригодных для этой цели рассматривались катионообменная и гидрофобные хроматографии, и соответственно сорбенты, такие как SP Sepharose FF, Capto S, CM Sepharose FF, Butyl Sepharose, Capto Butyl. Был проведен ряд экспериментов в различных условиях сорбции и элюции. Все эти способы не привели к существенному снижению количества близкородственных примесей в продукте.
Неожиданный прогресс был достигнут при проведении 2-х последовательных катионообменных хроматографий на двух колонках с различным объемом сорбента, представляющего собой гидрофильную полимерную матрицу с размером частиц от 30 мкм до 75 мкм, и размером пор приблизительно 100 нм. В частности, был использован сорбент YMC S30 в условиях сорбции и элюции продолжительным (40-42 колоночных объемов) линейным градиентом до 0,3 - 0,38 М NaCl (Фиг. 11А, Таблицы 2, 3). Первая из двух стадий катионообменной хроматографии является разделяющей, вторая полирующей и концентрирующей.
Для обеих стадий катионобменной хроматографии приемлемым является единый уравновешивающий буфер, рН которого варьирует в диапазоне 4,9-5,2, содержащий 0,1М NaCl. Элюцию на первой катионообменной хроматографии проводят линейным градиентом 0,1М до 0,30-0,38 М NaCl в натрий - ацетатном буфере, имеющим рН в диапазоне 4,9-5,1 за 40 колоночных объемов. На второй катионообменной хроматографии, которая является одновременно полирующей и концентрирующей для следующего процесса - гельфильтрации, целевой белок элюируют линейным градиентом 0,1М до 0,30 -0,3 5М NaCl в натрий -ацетатном буфере, имеющим рН в диапазоне 4,9-5,1 за 41-42 колоночных объема.
Для получения финальной готовой формы ромиплостима были разработаны и изучены два метода: концентрирование и диафильтрация в буфер готовой формы в системе тангенциального потока с использованием мембранной кассеты Pellicon XL Cassete 30 kDa: PES (50 см2) и перевод в буфер готовой формы гель-фильтрационной хроматографией на Superdex G25. Наиболее предпочтительным и удобным для коммерческого производства оказался способ гельфильтрации на колонке с сорбентом Superdex G25, так как при таком способе целевой белок мягко и без потерь переводится в буфер, в котором содержатся все необходимые добавки для поддержания структурной гомогенности, физико-химических свойств и биологической активности получаемой готовой формы. Заключительная стадия гельфильтрационной хроматографии позволяет получить гомогенный препарат в буфере готовой формы.
В результате очистки заявленным способом удалось получить целевой белок с содержанием основного вещества не менее 88%, определяемым методом ОФ ВЭЖХ (Фиг. 13А), с содержанием мономера не менее 98%, определяемым методом ГФ ВЭЖХ (Фиг. 13В) и содержанием основной заряженной формы не менее 70%, определяемой методом капиллярного зонного электрофореза (Фиг. 13С) и обладающий специфической биологической активностью (Фиг. 14А, Фиг. 14В).
Материалы и методы
Синтез белка может проводиться с использованием различных генетических конструкций, авторы для осуществления изобретения создали штамм генно-инженерно-модифицированного микроорганизма E.coli ВКПМ В-13251: реципиентного штамма-хозяина BL21(DE3) плазмидой pGNR-069-001, несущей ген белка Fc-пептидилированного белка ромиплостим. Для трансляции в E.coli оптимизированную последовательность ДНК, кодирующую белок ромиплостим, нуклеотидная последовательность которого приведена в составе SEQ ID NO1, вводили в состав экспрессионного плазмидного вектора, кодирующего мономер химерного белка с С-концевой химеризацией плеча тяжелой цепи IgG, составленного двумя пептидными последовательностями из 14 аминокислот (SEQ ID NO: 2) с линкерными вставками из полиглицина, длиною 269 аминокислотных остатков и молекулярной массой 29,5 кДа (аминокислотная последовательность представлена SEQ ID NO 3, (нуклеотидная последовательность ромиплостима представлена в патенте US 6,835,809 В1, Liu et al., Thrombopoietic compondus)). Последовательность гена ромиплостима состоит из Fc-фрагмента человеческого иммуноглобулина IgG1, в которой каждая одноцепочечная субъединица соединена ковалентной связью на С-конце с дважды повторяющимся синтетическим пептидом, связывающимся с рецептором тромбопоэтина, с линкерными вставками из полиглицина, состоящей из 269 аминокислотных остатков с расчетной молекулярной массой 29,543 кДа (Фиг. 3).
Для экспрессии Ромиплостима получают генно-инженерную конструкцию (ГИК) pGNR69-001. Для получения экспрессионного вектора, была синтезирована оптимизированная для экспрессии в Е. coli. Нуклеотидная последовательность (SEQ ID NO 1), содержащая кодирующую ромиплостим открытую рамку считывания вместе со стартовым кодоном ATG и стоп-сигналом ТАА, фланкированную с 5' и 3' концов сайтами рестрикции NcoI и BamHI соответственно. Синтезированная последовательность была клонирована по вышеуказанным сайтам в вектор, в котором экспрессия белка с клонированной последовательности находится под контролем Т7 промотора. Полученный в результате вектор далее обозначается как pGNR69-001.
Вектором pGNR69-001 был трансформирован штамм E.coli BL21(DE3). Полученный в результате штамм-продуцент BL21(DE3)pGNR69-001 депонирован во Всероссийской коллекции промышленных микроорганизмов (ВКПМ) под инвентарным номером - ВКПМ В-13251.
Для получения целевого белка ромиплостима проводили культивирование E.Coli ВКПМ В-13251 в реакторе Biostat Bplus Tween 5L. Процесс культивирования банка клеток ВКПМ В-13251 был ориентирован на получение высокого количества биомассы с целенаправленной локализацией белка ромиплостима в телах включения и протекал в два этапа. На первом этапе проводили культивирование клеток ВКПМ В-13251 в колбах Эрленмейра в объеме, достаточном для использования на втором этапе производства в качестве посевной культуры. Посевная культура фасовалась в пробирки и замораживалась при -70°С. Одной пробиркой с рабочим банком клеток ВКПМ В-13251 инокулировали 250 мл стерильной среды ЕСМ (Экспериментальная клеточная среда) и растили при 37°С в течение 16 часов при 180 оборотов в минуту.
Второй этап ферментирования включал следующие шаги:
(1) 250 мл стартовой культуры ВКПМ В-13251, выращенная на первом этапе, была загружена в реакторе Biostat Bplus Tween 5L с 3 л среды ЕСМ;
(2) рН в ферментере поддерживали на уровне 7.0 с помощью растворов 12,5% аммиака и 10% ледяной уксусной кислоты;
(3) DO в реакторе установлен на 40% насыщения, который регулировался скоростью оборотов мешалки и изменением процента растворенного кислорода до момента индукции, и затем снижался до 20% в пост-индукционную фазу культивирования;
(4) температура ферментации установлена на 37°С;
(5) добавка питательных растворов (40% дрожжевого экстракта и 10% глицерина) добавлялась экспоненциально после полного потребления глюкозы в среде (обычно спустя 8 часов) из расчета:
V=V0*e0.18t
где V - объем питательного раствора, добавляемый в мл/час;
V0 - 1/100 от стартового объема среды в реакторе в мл;
t - время ферментирования после начала фазы добавления питательных растворов (час).
Экспрессия ромиплостима индуцировалась добавлением ИПТГ до финальной концентрации 1,0 мМ, когда оптическая плотность культуры ВКПМ В-13251 составляла 50-70 оптических единиц. Пост-индукционная фаза ферментирования продолжалась еще 3 часа с добавлением питательных растворов и с почасовым отбором проб для контроля оптической плотности культуры. Конечная оптическая плотность культуры спустя 3 часа после постиндукционной фазы ферментирования составляла 100-150 о.е. Выход биомассы составлял от 80-100 г/л культуры. Белок ромисплостим имеет молекулярную массу размером 29 543 Да, экспрессируется на уровне 20-30% от тотального белка и находится исключительно в форме телец включения. Бактериальную биомассу собирали центрифугированием и хранили при - 70°С.
Замороженную биомассу клеток E.coli суспендировали в буфере для дезинтеграции 20 мМ Tris HCl, 300 мМ NaCl, рН 7,5 с добавлением ингибитора протеаз для ингибирования протеолиза фенил-метил-сульфонил-фторида (PMSF). Для дезинтеграции клеток был использован стандартный метод ультразвуковой дезинтеграции, который при масштабировании был заменен на дезинтеграцию при высоком давлении. Тела включения получали центрифугированием из дезинтегрированной суспензии клеток при 25000xg при 4°С в течение 40 минут. Полученные тела включения последовательно отмывали 50 mM Tris HCl, рН 7,5 буферными растворами, содержащими 2% Triton Х-100, 2 М мочевину, 2 М NaCl и 5 мМ ЭДТА. Отмытые тела включения растворяли в буферном растворе 7М гуанидин гидрохлорид, 50 мМ Tris HCl, 10 мМ DTT, рН 8,2 при комнатной температуре в течение 1,5 -2 часов. Содержание мономерной единицы ромиплостима определяли методом ГФ ВЭЖХ восстановленных тел включения ромиплостима и восстановленного раствора ЛП Энплейт, по которому строилась калибровочная кривая (Фиг. 6). Рефолдинг целевого белка ромиплостим проводили методом разбавления в 21 в буфере 50 мМ Трис-HCl; 2,1 М мочевина; 0,168 М аргинин; 3 мМ цистин; 4 мМ цистеин; рН 9,0, при концентрации белка (от 0,7 до 1 мг/мл) в течение 60-72 часов при 4-8°С при постоянном перемешивании. На Фиг. 7 представлена электрофореграмма образцов рефолдируемой смеси в восстанавливающих и в не восстанавливающих условиях (образец 1, 2) в сравнении с ЛП Энплейт (образец 3,4), препараты демонстрирует практически одинаковые наборы полос на геле, что свидетельствует об отсутствии значимых различий в их организации и указывает о полном прохождении ренатурации. Количественное содержание и качество рефолдированного ромиплостима контролировали методом ГФ ВЭЖХ и ОФ ВЭЖХ (Фиг. 6, Фиг. 8). Выход рефолдированного целевого белка сохранялся при масштабировании процесса с 3 л до 30 л рефолдируемой смеси и составлял от 40% до 50% правильно свернутой димерной формы ромиплостима (Фиг. 8). Дальнейшую очистку ромиплостима производили путем разделения правильно свернутых и неправильно свернутых конформаций данного белка, отделения остатков контаминирующих примесей клеточных белков, липополисахаридов, ДНК из рефолдированного раствора анионообменной хроматографией в режиме проскока на Q Sepharose FF, последующей аффинной хроматографией на MabSelect SuRe в условиях отмывки близкородственных примесей буферным раствором 30 мМ цитратом натрия, содержащим 0.2 М аргинина, рН-4,4 и мягкой элюцией целевого правильно свернутого белка линейным градиентом за 10 колоночных объемов от 100% буфера для отмывки до 100% буфера для элюции 30 мМ цитрата натрия, содержащего 0,5 М аргинина, рН-4,0.
Носители Q Sepharose FF и MabSelect SuRe уравновешиваются одним и тем же 20 мМ натрий фосфатным буфером с 1,5М мочевиной рН-7,2. Элюирование связанных с носителями соединений происходит разными буферами. Элюцию с Q Sepharose FF осуществляли буферным раствором 20 мМ Tris HCl, рН 7,8 с 1М NaCl и наблюдали большой пик оптической плотности в УФ-области, соответствующий в большей степени поглощению ДНК, а также примеси клеточных белков, кроме того на Q Sepharose FF осуществляется отделение бактериальных эндотоксинов. На Фиг. 9 представлена электрофореграмма фракций, полученных с хроматографий на Q Sepharose FF и MabSelect SuRe в восстанавливающих и не восстанавливающих условиях. Образец 4 соответствует образцу элюата с Q Sepharose FF, образец 3 соответствует элюату с сорбента MabSelect SuRe, образец 2 соответствует фракции отмыва, образец 1 - фракции, не связавшейся с носителем. Электрофореграмма наглядно демонстрирует, что целевой белок, не связываясь с сорбентом Q Sepharose FF, полностью задерживается на MabSelect SuRe, на носителе Q Sepharose FF задерживаются только примесные белки, ДНК и липополисахариды. Удаление подобных примесей способствует уменьшению неспецифической сорбции MabSelect SuRe, тем самым усиливая эффективность аффинной хроматографии и увеличивая срок службы дорогостоящего носителя. Элюат с MabSelect SuRe собирали по фракциям, которые анализировались методом ОФ ВЭЖХ.
Анализ полученных фракций с MabSelect SuRe методом ОФ ВЭЖХ указан на Фиг. 10В. На Фиг. 10А представлен профиль хроматографии. Большой пик оптической плотности в УФ-области (пик 1, пик отмыва) наблюдали при рН 4,4 и 0,2 М аргинина, он содержал помимо основного вещества, как менее гидрофобные формы основного вещества, которые хуже удерживались на аффинном сорбенте, чем основное вещество. (Фиг. 10В, формы 1-2), так и более гидрофобные формы (Фиг. 10В, формы 3-4). Так как все эти примеси элюировались с носителя раньше основного вещества, вероятнее всего это неправильно свернутые формы, мономерные единицы, не сформировавшие димер, но также имеющие Fc-домен. При элюировании градиентом рН и аргинина при достижении рН 4,3 и 0,25 М аргинина начинается второй пик, соответствующий основному правильно свернутому веществу (Фиг. 10A., пик 2, серое окрашивание), содержащему правильно сложенный Fc-домен, оставшиеся более сильно связанные с сорбентом формы, имеющие Fc-домен, элюировали при рН 3.2, в присутствии 0,5 М аргинина (Фиг. 10A., пик 3). Более низкий рН, например, рН 3.2, являлся необходимым для элюирования прочно связанных с сорбентом соединений. Тот факт, что эти молекулы связывались с белком А, означает, что они также являлись Fc-слитыми белками. Вследствие того, что эти соединения элюировались при более низком рН по сравнению с правильно свернутым ромиплостимом, они связывались с белком А прочнее. Эти молекулы были более гидрофобными, как показал анализ ОФ ВЭЖХ (Фиг. 10В), и являлись неправильно свернутыми молекулами ромиплостима, скорее всего растворимыми агрегатами, мисфолдами или окисленными формами. Большой пик оптической плотности в УФ-области (Фиг. 10A., пик 4, пик регенерации) наблюдался при регенерации сорбента 0,5 М NaOH. То, что эти соединения настолько прочно связывались с белком А, что удалялись только щелочным раствором довольно высокой молярности, свидетельствует о том, что эти соединения скорее всего нерастворимые агрегаты и нерефолдированный белок. По результатам анализа ОФ ВЭЖХ отбирали фракции с содержанием главной формы не менее 60% и формы 1 не более 2%, формы 4 не более 10% для дальнейшей очистки правильно свернутого целевого белка от оставшихся агрегатов, мисфолдов, окисленных и деамидированных форм.
Сводная таблица по результатам аффинной хроматографии на MabSelect SuRe представлена в таблице 1.
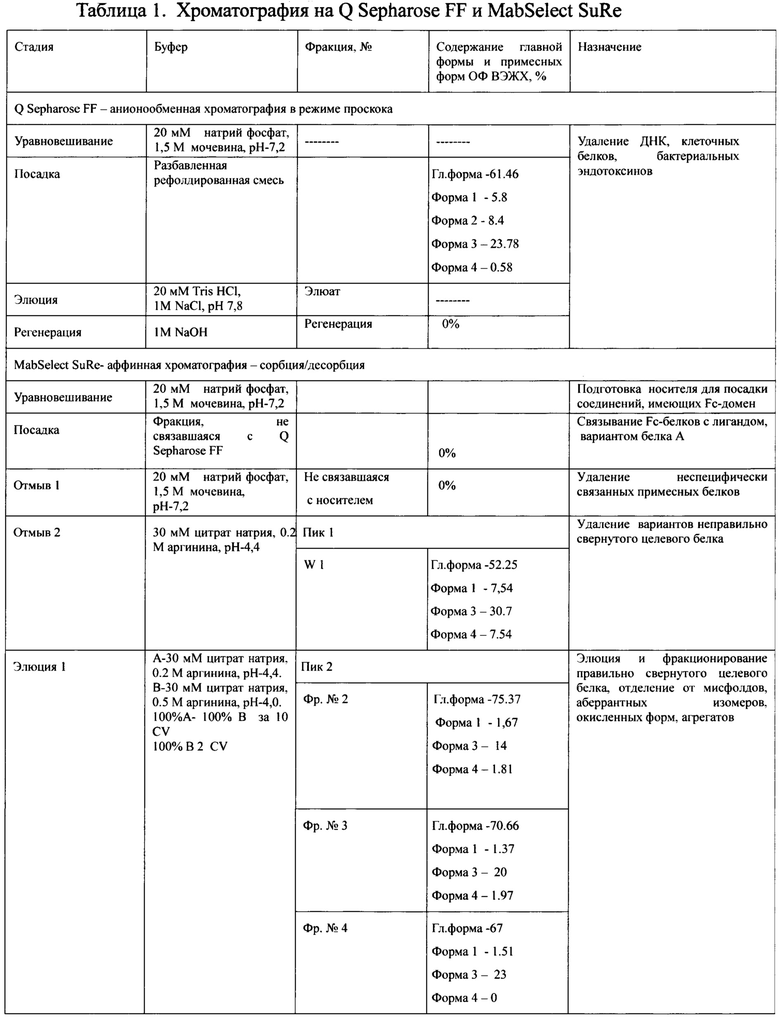
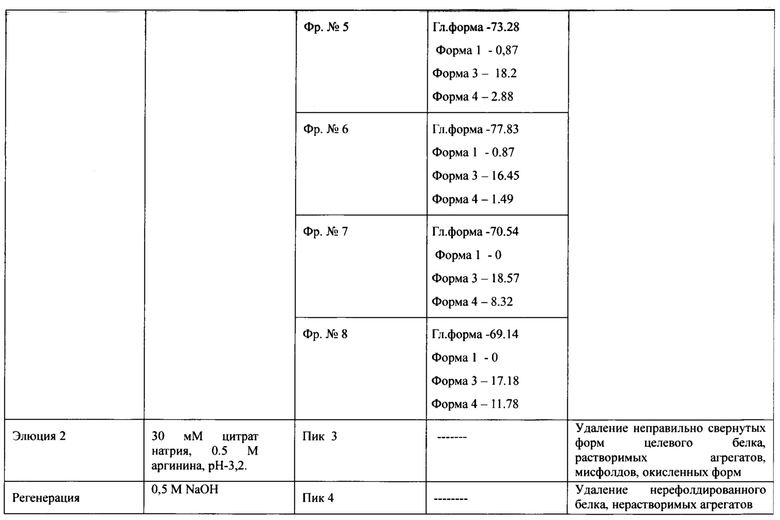
Дальнейшую доочистку производили двумя катионообменными хроматографиями с различным объемом сорбента YMC S30 в условиях сорбции и элюции продолжительным (40 колоночных объемов и более) линейным градиентом NaCl (Фиг. 11А, Фиг. 12А, таблица 2, таблица 3), первая из которых является разделяющей, вторая полирующей и концентрирующей.
Для проведения первой хроматографии на сорбенте YMC S30, фракции, отобранные с MabSelect SuRe, объединяли, разбавляли водой очищенной до электропроводности 9-10 mSm/sm, и наносили на колонку с YMC S30, уравновешенную буфером 20 mM натрий ацетат, 0,1М NaCl, рН 5,1. Элюировали градиентом от буфера 20 mM натрий ацетат, 0,1М NaCl, рН 5,1 до буфера 20 mM натрий ацетат, 0,3 М NaCl, рН 5,1 за 40 колоночных объемов. Элюат собирали по фракциям, которые анализировались методом ОФ ВЭЖХ. На Фиг. 11А представлен профиль хроматографии с YMC S30-1, где можно наблюдать 4 предпика, соответствующих примесным формам и большой пик оптической плотности, соответствующий главной форме целевого белка (пик 1, серое окрашивание) при достижении электропроводности 22-23 mSm/sm. Кроме того, наблюдается высокий узкий пик, соответствующий элюции буфера 20 mM натрий ацетата, 1М NaCl, рН 5,1, в котором элюируются более заряженные формы (пик. 2), и узкий пик регенерации, соответствующий вероятнее всего примесям, имеющим наибольший заряд (пик. 3). Сводная таблица по результатам хроматографии на YMC S30-1 представлена в таблице 2.
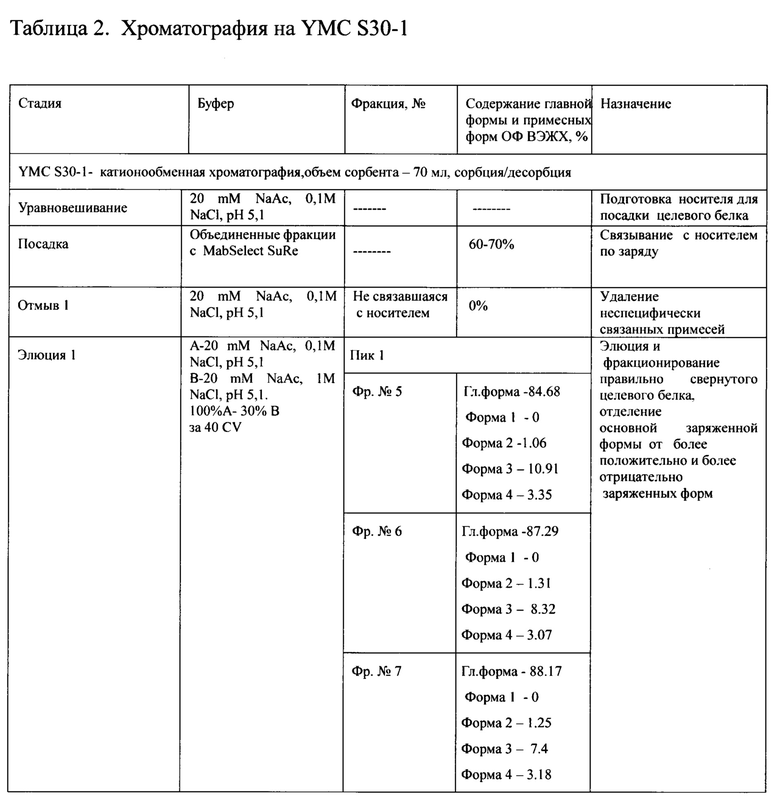
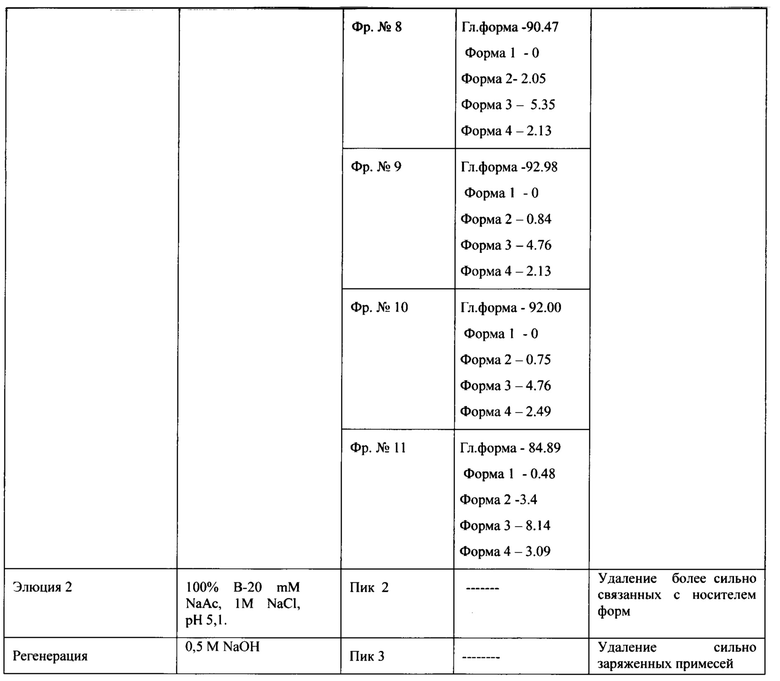
Для дальнейшей очистки целевого белка отбирали фракции по результатам анализа ОФ ВЭЖХ с содержанием главной формы не менее 87% и формы 3 не более 8%, формы 4 не более 4%.
Для проведения второй хроматографии на сорбенте YMC S30, которая является одновременно полирующей и концентрирующей для следующего процесса - гель-фильтрации, фракции отобранные с хроматографии YMC S30-1, объединяли, разбавляли водой очищенной до электропроводности 10 mSm/sm и наносили на колонку с YMC S30, уравновешенную буфером 20 mM натрий ацетат, 0,1М NaCl, рН 5,1. Элюировали градиентом от буфера 20 mM натрий ацетат, 0,1М NaCl, рН 5,1 до буфера 20 mM натрий ацетат, 0,3 М -0,35М NaCl, рН 5,1 за 40-42 колоночных объемов. Элюат собирали по фракциям, которые анализировались методом ОФ ВЭЖХ и методом капиллярного зонного электрофореза на содержание основной изоформы. На Фиг. 12.А. представлен профиль хроматографии, где можно наблюдать один пик оптической плотности, соответствующий главной форме целевого белка при достижении электропроводности 22-23 mSm/sm (пик 1). Пик имеет небольшое растянутое правое плечо, соответствующее примесям кислых форм (Фиг 12.В., Фиг. 12.С, Таблица 3.). Наблюдаются так же и небольшие узкие пики, соответствующие элюции буфера 20 mM натрий ацетат, 1М NaCl, рН 5,1, и регенерации (пики 2, 3).
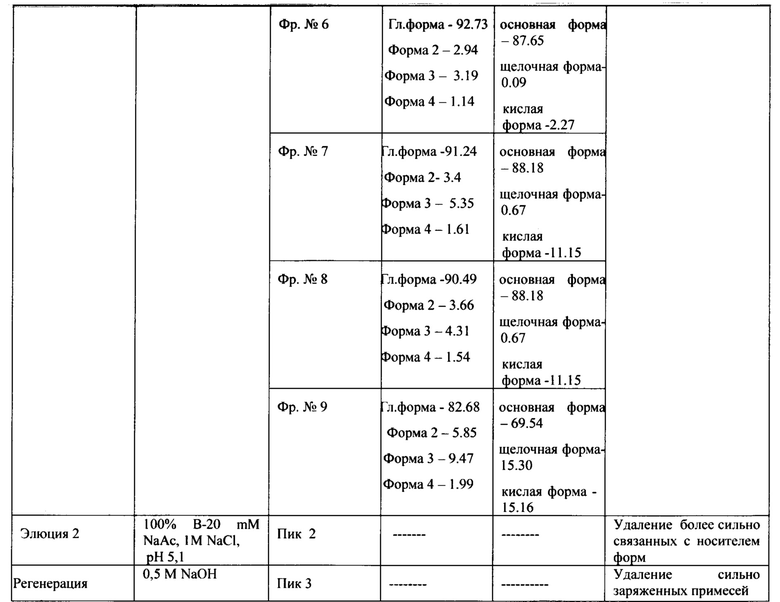
Сводная таблица по результатам хроматографии на YMC S30-2 представлена в таблице 3.
Для перевода целевого белка в буфер готовой формы методом гель-фильтрационной хроматографии на Superdex G25 фракции с хроматографии на YMC S30-2 отбирали по результатам анализа ОФ ВЭЖХ с содержанием главной формы не менее 90%, по результатам капиллярного зонного электрофореза с содержанием основной заряженной формы не менее 87%. Фракции с 6 по 8 объединяли и вносили в колонку с носителем Superdex G25, элюировали буфером готовой формы: 10,0 mM гистидин, 4% маннитол, 2,0% сахароза, 0,0045% полисорбат 20, рН 5.0 при комнатных условиях, результаты хроматографии представлены в таблице 4.

Полученный препарат разбавляли до нужной концентрации, стерильно фильтровали. Выход белка составляет не менее 0,9 г на литр бактериальной культуры.
Измерения параметров различных партий готовой формы целевого белка ромиплостим показали, что в результате очистки заявленным способом получается целевой белок с содержанием основного вещества не менее 88%, определяемым методом ОФ ВЭЖХ (Фиг. 13.А.), с содержанием мономера не менее 98%, определяемым методом ГФ ВЭЖХ (Фиг. 13.В) и содержанием основной заряженной формы не менее 70%, определяемой методом капиллярного зонного электрофореза (Фиг. 13. Специфическая биологическая активность препарата, определяемая методом конкурентного твердофазного иммуноферментного анализа, составила в среднем 95,3±5,3% (результаты представлены на Фиг. 14.А.) и по уровню концентрации высвобожденной из тромбоцитов человека фосфорилированной тирозинкиназы JAK2 составила в среднем 100±6% (результаты представлены на Фиг. 14.В.).
Фармацевтическая субстанция может содержать различные примеси биологической природы связанные с продуцентом: (эндотоксины, остаточная ДНК штамма-продуцента, белки клетки-хозяина). Содержание данных примесей может оказывать влияние на безопасность препарата, поэтому должно быть оценено в очищенном продукте. Содержание бактериальных эндотоксинов, рН и осмоляльность определялись в соответствии с нормами Государственной Фармакопеи РФ (Государственная фармакопея РФ http://femb.ru/femb/pharmacopea.php). Результаты анализа представлены в таблице 5. Количественное определение белков штамма продуцента (E.coli) определяемых методом иммуноферментного анализа в различных партиях готовой формы ромиплостима составило менее 4 нг/мг при норме не более 10 нг/мг белка ("Draft Guidance on Specifications: Test Procedures and Acceptance Criteria for Biotechnological/Biological Products", (FDA, CBER, ICH), 24 July 1998., Jonathan Briggs and Peter R. Panfili, Quantitation of DNA and Protein Impurities in Biopharmaceuticals // Perspective: Analytical Biotechnology, Anal. Chem. - 1991. - 63. - p. 850-859).
Для определения остаточной ДНК использовали метод полимеразной цепной реакции (ГШР) в реальном времени. В его основе лежит амплификации участка ДНК с одновременным измерением возрастания флуоресцентного сигнала. Измеряя количество циклов, необходимых для достижения порогового уровня флуоресценции, можно рассчитать исходное количество ДНК в образце. Для интерпретации результатов измерения используются калибровочные стандарты с известной концентрацией.
При исследовании серий ГФ ромиплостима измеренное значение концентрации ДНК для всех образцов составило величину ниже предела обнаружения (2,1 пг/мл).
Эти значения соответствуют международным рекомендациям, согласно которым содержание остаточной ДНК не должно превышать 10 нг на 1 дозу препарата (WHO. Guidelines on the quality, safety and efficacy of biotherapeutic protein products prepared by recombinant DNA technology. Replacement of annex 3 of WHO technical report series. - 2013. - N.814. http://www.who.int/biologicals/biotherapeutics/rDNA_DB_final_19_Nov_2013.pd f>.
Для лучшего понимания сущности изобретения ниже следуют примеры его осуществления.
Пример 1. Микробиологический синтез белка
Заявляемый способ реализован для штамма-продуцента ВКПМ В-13251 при культивировании в ферментере (Sartorius Biostat В Plus Twin 5L, Германия) в объеме среды 3 л. Весь процесс культивирования ведут при температуре 37°С. Водородный показатель рН среды поддерживают на уровне 7,0-7,1, титрованием 10% раствором уксусной кислоты и 12,5% раствором аммиака. Уровень аэрации 3 л/мин и концентрации растворенного кислорода на уровне 40%, который регулируется каскадом оборотов мешалки.
Выращенный инокулят в объем 250 мл вносят в биореактор со стерильной питательной средой ЕСМ. В качестве стартового субстрата в среду вносят глюкозу в количестве 10 г/л и глицерина 30 г/л.
В период пред-индукционной фазы роста клетки утилизируют глюкозу из среды. Уровень рН падет. Спустя 8-9 часов клетки достигают оптической плотности 50-70 оптических единиц. Экспрессия Ромиплостима индуцируется добавлением ИПТГ до финальной концентрации 1,0 мМ. Далее постиндукционная фаза ферментирования продолжается еще 3 часа с добавлением питательных растворов (40% глицерин, 1% дрожжевого экстракта) и с почасовым отбором проб для контроля оптической плотности культуры. Спустя 3 часа конечная оптическая плотность культуры достигает 100-150 о.е. Процесс ферментирования прекращается. Бактериальную биомассу собирают центрифугированием при 10 000 об/мин в течение 20 мин и помещают на хранение при - 70°С.
Пример 2. Выделение и очистка белка ромиплостим из бактериальной биомассы.
2.1 Разрушение клеток штамма-продуцента, получение телец включения.
Буфер А - 20 мМ Tris HCl, 300 мМ NaCl, 0,001% PMSF рН 7,5;
Осадок биомассы (m=30,0±1,0 г), из холодильника помещают в мерный пластиковый стакан объемом 0,5 л, к осадку клеток доливают 300±0,1 мл свежеприготовленного буферного раствора А. В стакан помещают насадку диспергатора и дисперпергируют в течение 10-15 минут. Стакан с гомогенизированной массой клеток-продуцентов помещают в ледяную баню под зонд ультразвукового дезинтегратора. Обрабатывают ультразвуком на большом щупе в течение 15 минут при амплитуде 25% (цикл: 30 сек - ультразвук, 40 сек - охлаждение). Разливают суспензию в центрифужные стаканы и центрифугируют 25000xg при 4°С в течение 40 мин. После завершения центрифугирования надосадочную жидкость отбрасывают.
2.2 Отмывка телец включения
Буфер В: 50 мМ Tris HCl, 2% Triton Х-100, 5mM EDTA, рН 7,5;
Буфер С: 50 mM Tris HCl, 2М мочевина; 5 mM EDTA, рН 7,5;
Буфер D: 50 mM Tris HCl, 2М NaCl; 5 mM EDTA, рН 7,5;
Буфер Е: 20 mM Tris HCl, 1 mM EDTA, рН 7,5;
Осадок телец включения, помещают в емкость объемом 0,25 л, к осадку доливают 200±0,1 мл буферного раствора В. В стакан помещают насадку диспергатора и диспергируют в течение 3-5 минут. Центрифужный стакан с раствором телец включения центрифугируют при 30 000 х g (14 000 об/мин) при 5°С в течение 35 мин. После завершения центрифугирования надосадочную жидкость отбрасывают.
Выше описанную процедуру последовательно повторяют с буферами C, D, E.
В результате получают не менее - 13,6±2 г отмытых телец включения.
Полученный продукт хранят при температуре минус 80°С.
2.3. Растворение телец включения
Буфер L: 50 mM Tris HCl, 10 mM DTT, 7М гуанидин хлорид, рН 8,2;
Осадок телец включения (m=15,0±0,1 г) из холодильника помещают в мерный пластиковый стакан объемом 0,2 л, к осадку доливают 150±0,1 мл свежеприготовленного раствора L. Диспергируют в течение 5 минут. Стакан помещают на магнитную мешалку и перемешивают при комнатной температуре в течение 90-120 мин. Суспензию центрифугируют 40000 х g при 8°С в течение 120 мин. После завершения центрифугирования осторожно отбирают надосадочную жидкость.
2.4. Рефолдинг.
Буфер R: 2,1 М мочевина; 0,168 М аргинин; 50 mM Трис-HCl; 3 mM цистин; 4 mM цистеин; рН-9.0;
Раствор телец включения объемом 150±10 мл разводят в 20-21 раз охлажденным до 8±1°С буфером R объемом 3000 мл, для чего емкость с буфером R помещают на магнитную мешалку и медленно по каплям при постоянном перемешивании добавляют раствор телец включения, измеряют рН раствора, при отклонении рН от 9,0 корректируют хлористоводородной кислотой или раствором натрия гидроксида. Емкость помещают на мешалку и продолжают процесс фолдирования белка при температуре 4-7±1°С при постоянном перемешивании в течении 70-72 часов.
2.5. Аффинная хроматография на MabSelect SuRe.
Буфер F - 20 мМ натрий фосфат, 1,5 М мочевина, рН-7,2;
Буфер W1 - 30 мМ цитрат натрия, 0.2 М аргинин, рН-4,4.
Буфер G - 30 мМ цитрат натрия, 0,5 М аргинин, рН-4,0;
Буфер Р - 30 мМ цитрат натрия, 0,5 М аргинин, рН-3,2;
Буфера для регенерации колонки:
100 мМ Трис, рН 7,5;
0,5 M NaOH
20% этиловый спирт
Первую стадию очистки целевого белка проводят на последовательно собранных колонках: колонке, заполненной сорбентом Q Sepharose FF объемом 80 мл, и колонке, заполненной сорбентом MabSelect SuRe объемом 60 мл, с использованием хроматографической системы Akta Pure 25 при комнатной температуре.
Колонки промывают 10 колоночными объемами буферного раствора F со скоростью потока 113 см/ч. В емкость, вместимостью 10 л, вносят 6300 мл буфера F и добавляют 3150 мл раствора фолдированного белка, доводят рН раствора до 7,2±0,01 концентрированной фосфорной кислотой и наносят на колонки. После нанесения сорбенты промывают 3 колоночными объемами буферного раствора F, затем колонку с сорбентом Q Sepharose FF отсоединяют и передают на регенерацию, а колонку с сорбентом MabSelect SuRe последовательно промывают 2 колоночными объемами буферного раствора F, 10-12 колоночными объемами буферного раствора W1, при снижении оптической плотности на выходе с колонки до 100 мА для элюции и фракционирования целевого белка и его производных связанных с сорбентом используют линейный градиент от 0% буферного раствора W до 100% буферного раствора G за 10 колоночных объемов. Фракции собирают по 40 мл при повышении оптической плотности раствора, выходящего с колонки до 120 mAU, относительно базовой линии, при снижении оптической плотности ниже 120 mAU сбор фракций прекращают. В каждую фракцию сразу добавляют по 1 мл раствора 1М Трис рН 8,0 и доводят рН до 5,0±0,1 5 М раствором гидроксида натрия. По завершению градиента сорбент промывают 1,8 колоночных объема буферного раствора Р и 1,8 колоночных объема раствора 0,5 М гидроксида натрия и передают на стадию консервации.
Фракции, содержащие целевой белок, с содержанием основного вещества не менее 60% и с содержанием примесного пика №1 не более 2%, определяемых методом ОФ ВЭЖХ, объединяют.
Для консервации колонки с MabSelect SuRe, сорбент последовательно промывают 5 колоночными объемами буферного раствора 01, М Tris HCl, рН 7,5 и 3 колоночными объемами 20% раствора спирта этилового.
Для регенерации колонки с Q Sepharose FF сорбент промывают 3 колоночными объемами раствора 0,5 М гидроксида натрия, выдерживают в этом растворе не менее 60 мин, снова промывают 1,8 колоночными объемами этого же раствора, затем последовательно промывают 5 колоночными объемами буферного раствора 0,1 М Tris HCl, рН 7,5 и 3 колоночными объемами 20% раствора спирта этилового.
2.6. Катионообменная хроматография на YMC S30 - 1
Буфер Н - 20 mM Na ацетат, 0,1 М NaCl, рН 5,0;
Буфер Y - 20 mM Na ацетат, 1,0 М NaCl, рН 5,0;
Вторую стадию очистки целевого белка проводят на колонке, заполненной сорбентом YMC S30 объемом 70 мл, с использованием хроматографической системы Akta Pure 25 при комнатной температуре.
Колонку промывают 3 колоночными объемами буферного раствора Y, затем 10 колоночными объемами буферного раствора Н со скоростью потока 226 см/ч.
Объединенные фракции с хроматографии на MabSelect SuRe разбавляют в 4 раза охлажденной до 4°С водой очищенной, корректируют рН раствора до 5,0±0,01 ледяной уксусной кислотой или 5 М раствором гидроксида натрия, корректируют электропроводность до 8,5±0,5 мСм/см 5М раствором натрия хлорида и наносят на колонку. После нанесения сорбент промывают 5 колоночными объемами буферного раствора Н, для элюции и фракционирования целевого белка и его производных, связанных с сорбентом, используют линейный градиент от 0% буферного раствора Н до 30% буферного раствора Y за 40 колоночных объемов. Фракции собирают при повышении оптической плотности раствора, выходящего с колонки до 136 mAU, относительно базовой линии, при снижении оптической плотности ниже 70 mAU сбор фракций прекращают. Сбор фракций для анализа можно начинать после выхода четырех предпиков. По завершению градиента сорбент промывают 1,8 колоночных объема 100% буферного раствора Y и передают на стадию регенерации.
Фракции, содержащие целевой белок, с содержанием основного вещества не менее 87%, определяемых методом ОФ ВЭЖХ, объединяют.
Для регенерации колонки сорбент последовательно промывают 3 колоночными объемами 0,5 М раствора гидроксида натрия, 5 колоночными объемами буферного раствора 0,1 М Tris HCl, рН 7,5 и 3 колоночными объемами 20% раствора спирта этилового. Хранят колонну с закрытыми входным и выходным отверстиями при температуре 2...8°С.
2.7. Катионообменная хроматография YMC S30 - 2
Третью стадию очистки целевого белка проводят на колонке, заполненной сорбентом YMC S30 объемом 35 мл, с использованием хроматографической системы Akta Pure 25 при комнатной температуре.
Колонку промывают 3 колоночными объемами буферного раствора Y, затем 10 колоночными объемами (для полного уравновешивания) буферного раствора Н со скоростью потока 239 см/ч.
Объединенные фракции с хроматографии на YMC S30 - 1 разбавляют в 2,5 раза охлажденной до 4°С водой очищенной, корректируют рН раствора до 5,0±0,01 ледяной уксусной кислотой или 5 М раствором гидроксида натрия, корректируют электропроводность до 8,5±0,5 мСм/см 5М раствором натрия хлорида, если необходимо, и наносят на колонку. После нанесения сорбент промывают 5 колоночными объемами буферного раствора Н, для элюции и фракционирования целевого белка и его производных, связанных с сорбентом, используют линейный градиент от 0% буферного раствора Н до 30% буферного раствора Y за 40-42 колоночных объема. Фракции собирают при повышении оптической плотности раствора, выходящего с колонки до 60 mAU, относительно базовой линии, при снижении оптической плотности ниже 40 mAU сбор фракций прекращают. По завершению градиента сорбент промывают 1,8 колоночных объема 100% буферного раствора Y и передают на стадию регенерации.
Фракции, содержащие целевой белок с содержанием основного вещества не менее 88%, определяемым методом ОФ ВЭЖХ, с содержанием мономера не менее 98%, определяемым методом ГФ ВЭЖХ и содержанием основной формы не менее 70%, определяемых методом капиллярный зонный электрофорез, объединяют.
Для регенерации колонки сорбент последовательно промывают 3 колоночными объемами 0,5 М раствора гидроксида натрия, 5 колоночными объемами буферного раствора 0,1 М Tris HCl, рН 7,5 и 3 колоночными объемами 20% раствора спирта этилового. Хранят колонну с закрытыми входным и выходным отверстиями при температуре 2…8°С.
2.8 Гель-фильтрационная хроматография на Superdex G 25
Буфер U - 10,0 mM гистидин, 4% маннитол, 2,0% сахароза, 0,0045% полисорбат 20, рН 5.0;
Четвертую стадию очистки целевого белка (замена буфера) проводят на колонке, заполненной сорбентом Superdex G 25 Fine объемом 240 мл, с использованием хроматографической системы Akta Pure 25 при комнатной температуре.
Колонку промывают 2 колоночными объемами буферного раствора U со скоростью потока 20-24 см/ч.
Из объединенной фракции с хроматографии на YMC S30 - 2, отбирают половину раствора и наносят на колонку с сорбентом Superdex G 25 Fine со скоростью потока 24 см/ч, целевой белок выходит при протекании через сорбент 0,3-0,33 колоночного объема буфера U (10±1 мин/ 80±1 мл), целевой белок начинают собирать с ростом оптической плотности выше 65 mAU относительно базовой линии. При снижении оптической плотности ниже 70 mAU сбор белка прекращают. После прохождения через носитель 1,4 колоночного объема буферного раствора U, начинают очистку второй порции. Элюаты объединяют, перемешивают в течение 2-3 минут, измеряют объем.
Для регенерации колонки Superdex G 25 сорбент промывают 3 колоночными объемами 0,5 М гидроксида натрия, 3 колоночными объемами воды очищенной и 3 колоночными объемами 20% раствора спирта этилового.
2.9 Приготовление рабочего раствора готовой формы
Буфер U - 10,0 mM гистидин, 4% маннитол, 2,0% сахароза, 0,0045% полисорбат 20, рН 5.0;
Рассчитывают и добавляют необходимое количество буфера U, чтобы получить концентрацию белка от 0,53 до 0,55 мг/мл. Рабочий раствор белка аккуратно перемешивают и оставляют при температуре 2…8°С на 60 мин. Измеряют объем раствора и содержание белка и добавляют необходимое количество буфера U, чтобы получить концентрацию от 0,53 до 0,55 мг/мл.
2.10 Стерилизующая фильтрация
Стерилизующую фильтрацию рабочего раствора готовой формы проводят в ламинарном потоке воздуха. Для стерилизующей фильтрации используют стерилизующую систему FILTERMAX, 250 ml, PES, с размером пор 0,22 мкм, используя мембранный вакуумный насос.
После фильтрации отбирают пробу для проведения технологического контроля. Полученный раствор является готовой формой ромиплостима.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОЙ ЭНДОНУКЛЕАЗЫ SERRATIA MARCESCENS | 2021 |
|
RU2795623C2 |
| ПРОМЫШЛЕННЫЙ СПОСОБ ПОЛУЧЕНИЯ И ОЧИСТКИ РЕКОМБИНАНТНОГО ГОРМОНА РОСТА ЧЕЛОВЕКА ИЗ ТЕЛЕЦ ВКЛЮЧЕНИЯ | 2011 |
|
RU2473556C1 |
| ПРОМЫШЛЕННЫЙ СПОСОБ ПОЛУЧЕНИЯ И ОЧИСТКИ РЕКОМБИНАНТНОГО ИНТЕРФЕРОНА β-1b ЧЕЛОВЕКА ИЗ ТЕЛЕЦ ВКЛЮЧЕНИЯ | 2011 |
|
RU2473696C1 |
| СПОСОБЫ ОЧИСТКИ БЕЛКОВ, СОДЕРЖАЩИХ ОБЛАСТЬ Fc | 2006 |
|
RU2415865C2 |
| СПОСОБ ОЧИСТКИ РЕКОМБИНАНТНОГО ГРАНУЛОЦИТАРНОГО КОЛОНИЕСТИМУЛИРУЮЩЕГО ФАКТОРА ЧЕЛОВЕКА | 2011 |
|
RU2440417C1 |
| ПЛАЗМИДНЫЙ ВЕКТОР pRh15A ДЛЯ ПОЛУЧЕНИЯ БЕЗМЕТИОНИНОВОГО ИНТЕРФЕРОНА АЛЬФА-2b, ШТАММ БАКТЕРИЙ ESCHERICHIA COLI BL21 DE3 - ПРОДУЦЕНТ БЕЗМЕТИОНИНОВОГО ИНТЕРФЕРОНА АЛЬФА-2b И СПОСОБ ПОЛУЧЕНИЯ БЕЗМЕТИОНИНОВОГО ИНТЕРФЕРОНА АЛЬФА-2b | 2017 |
|
RU2697375C2 |
| Способ получения активного начала вакцины против ротавирусной инфекции белка FliCVP6VP8 | 2017 |
|
RU2649132C1 |
| СПОСОБЫ И КОМПОЗИЦИИ, ВКЛЮЧАЮЩИЕ ОЧИЩЕННЫЕ РЕКОМБИНАНТНЫЕ ПОЛИПЕПТИДЫ | 2014 |
|
RU2671481C2 |
| УЛУЧШЕННАЯ ОЧИСТКА БЕЛКА ПОСРЕДСТВОМ МОДИФИЦИРОВАННОГО ЭЛЮИРОВАНИЯ БЕЛКА А | 2010 |
|
RU2571929C2 |
| СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО ФРАГМЕНТА ПЛАЗМИНОГЕНА ЧЕЛОВЕКА, ОБЛАДАЮЩЕГО АНТИАНГИОГЕННЫМ ДЕЙСТВИЕМ | 2011 |
|
RU2458987C1 |


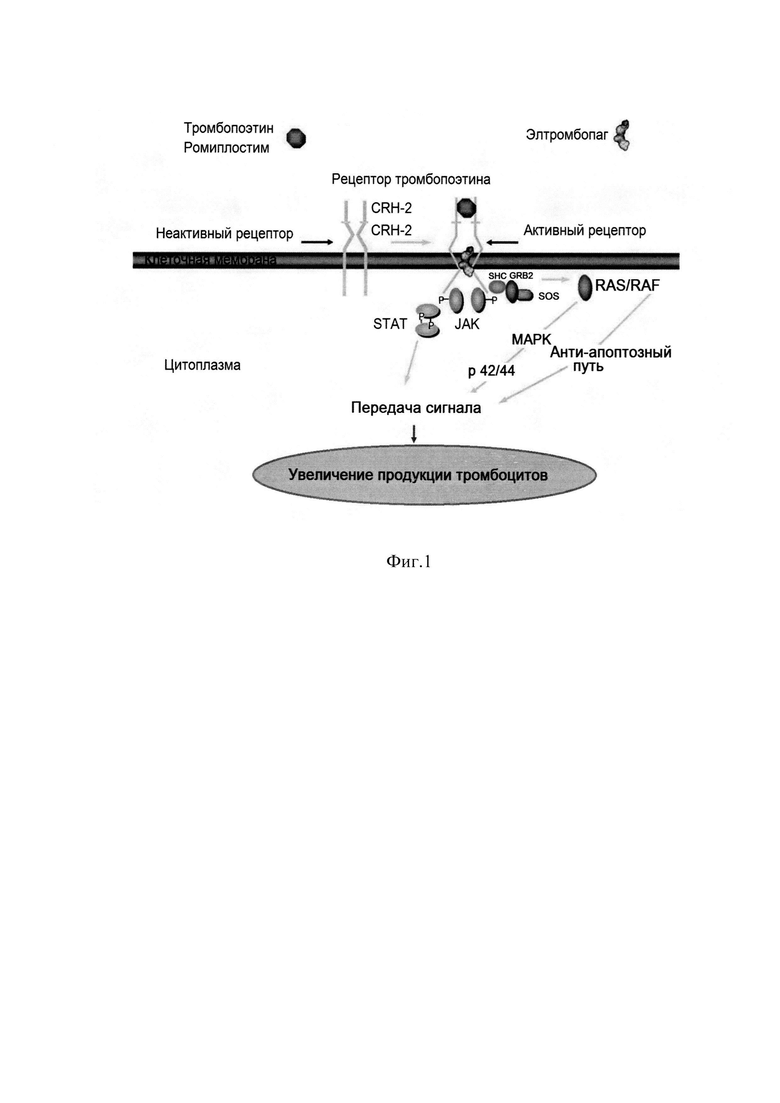
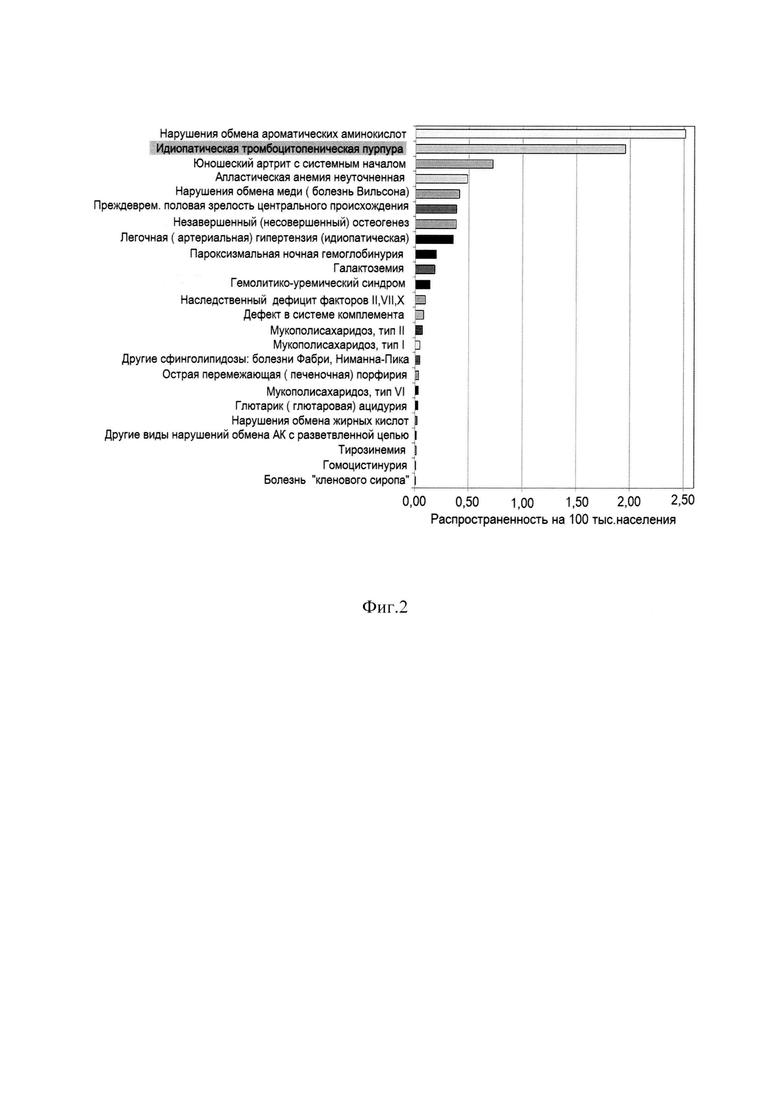
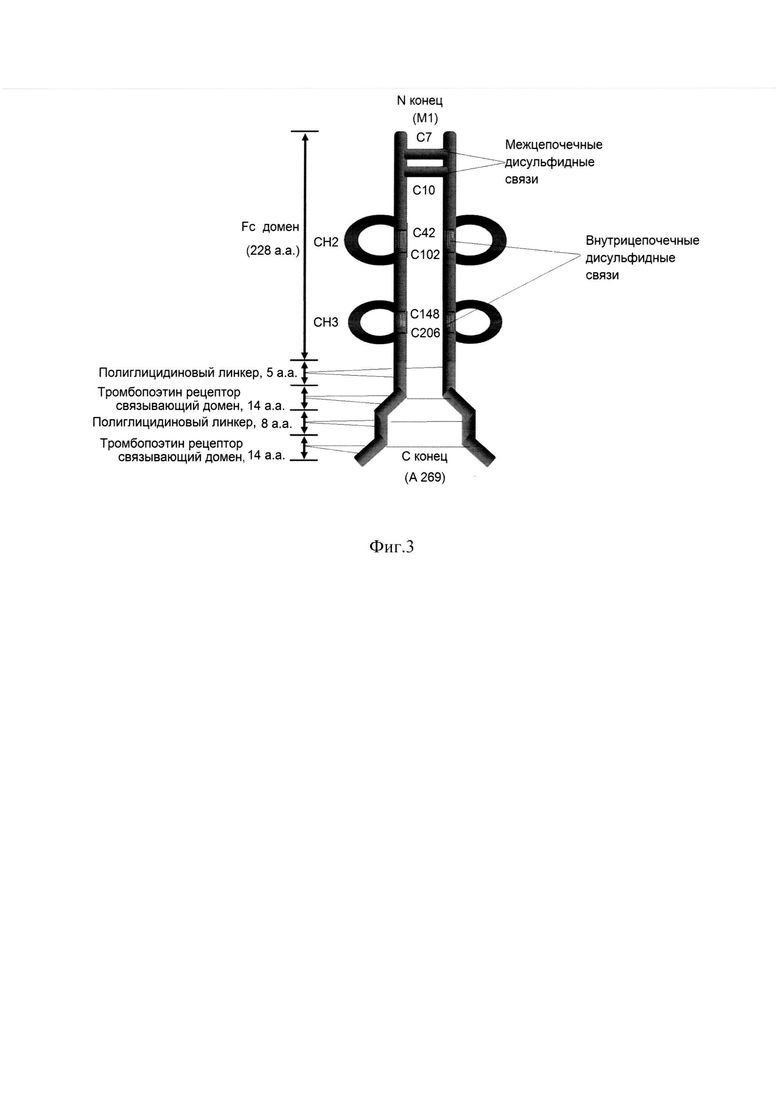

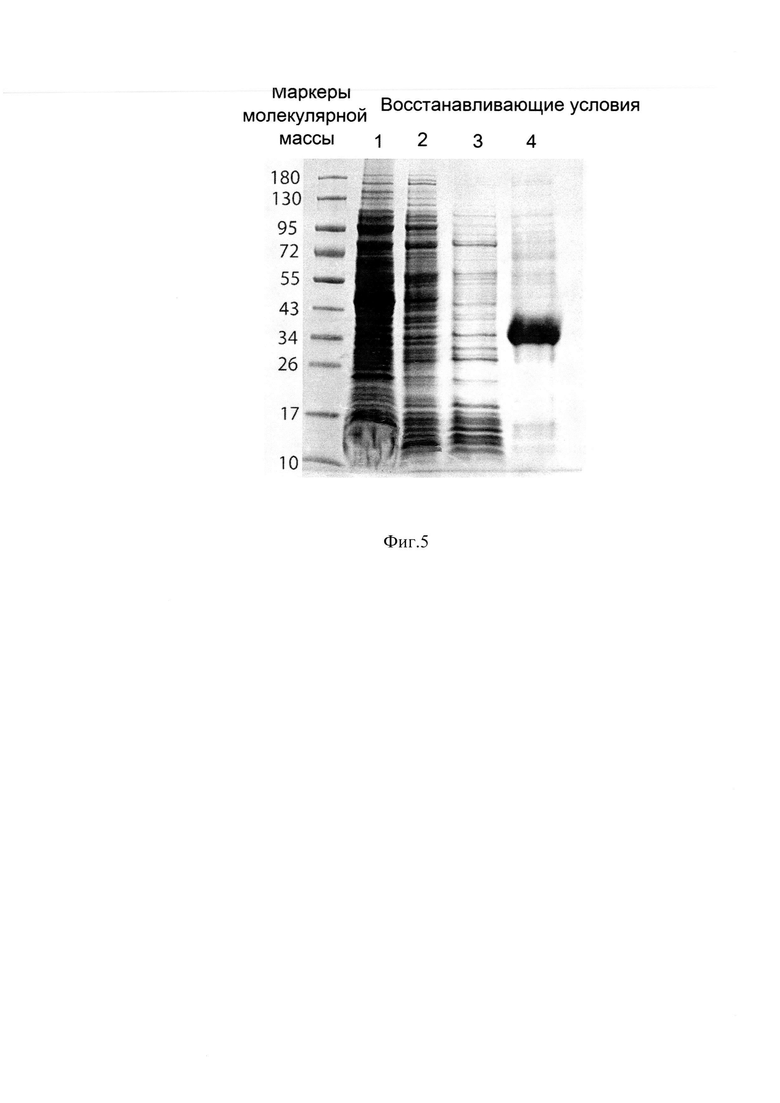
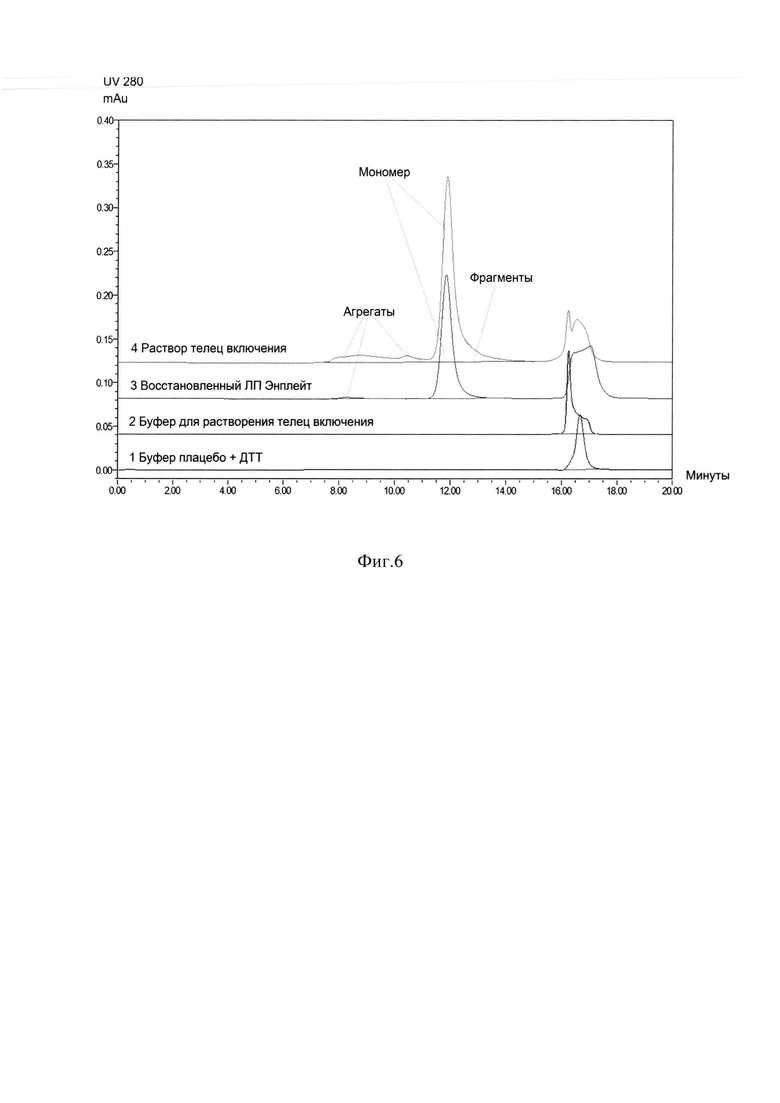
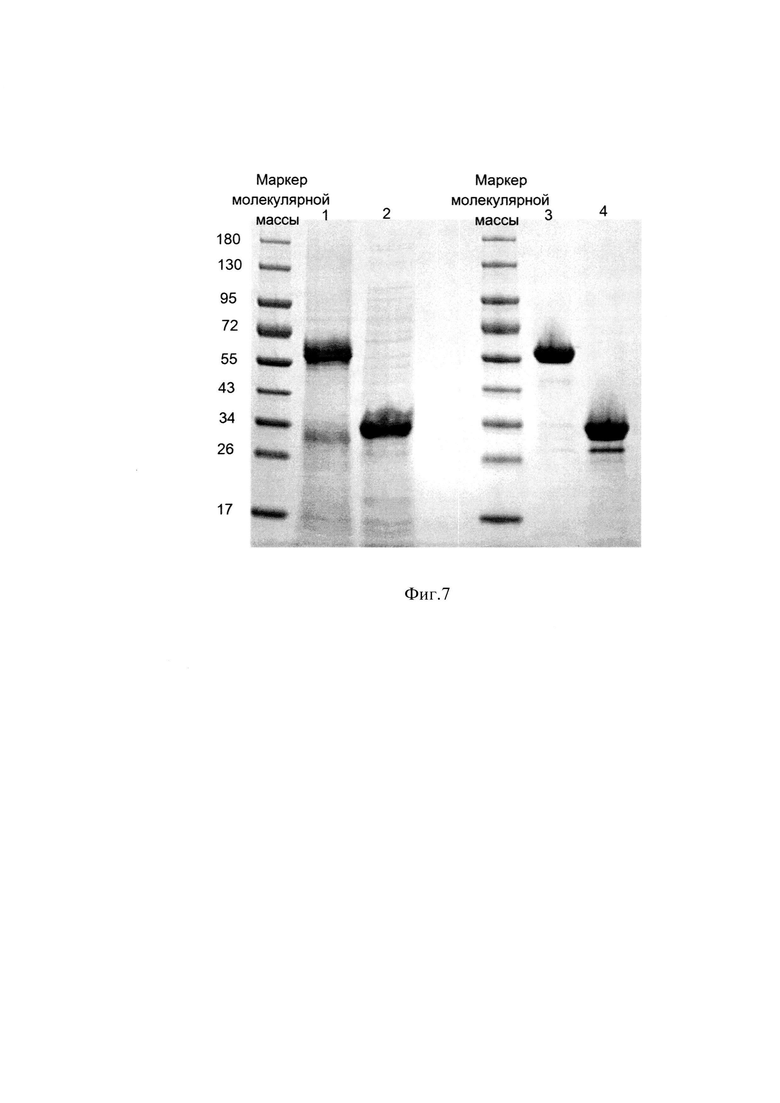
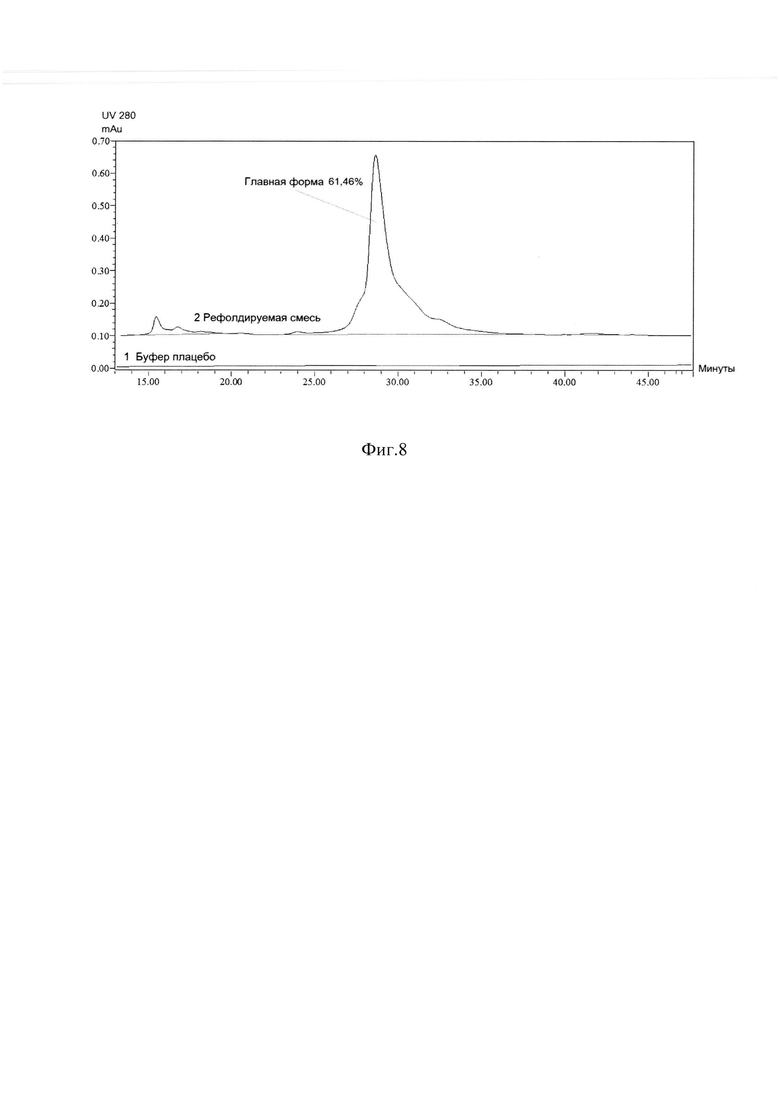
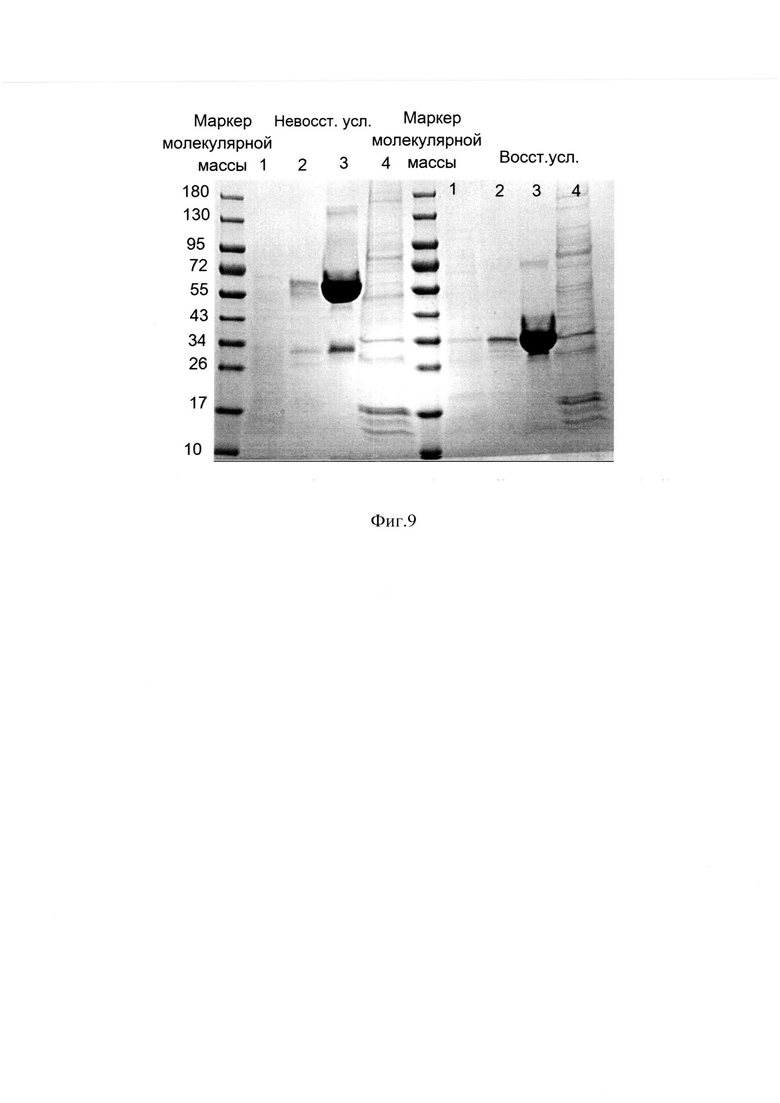
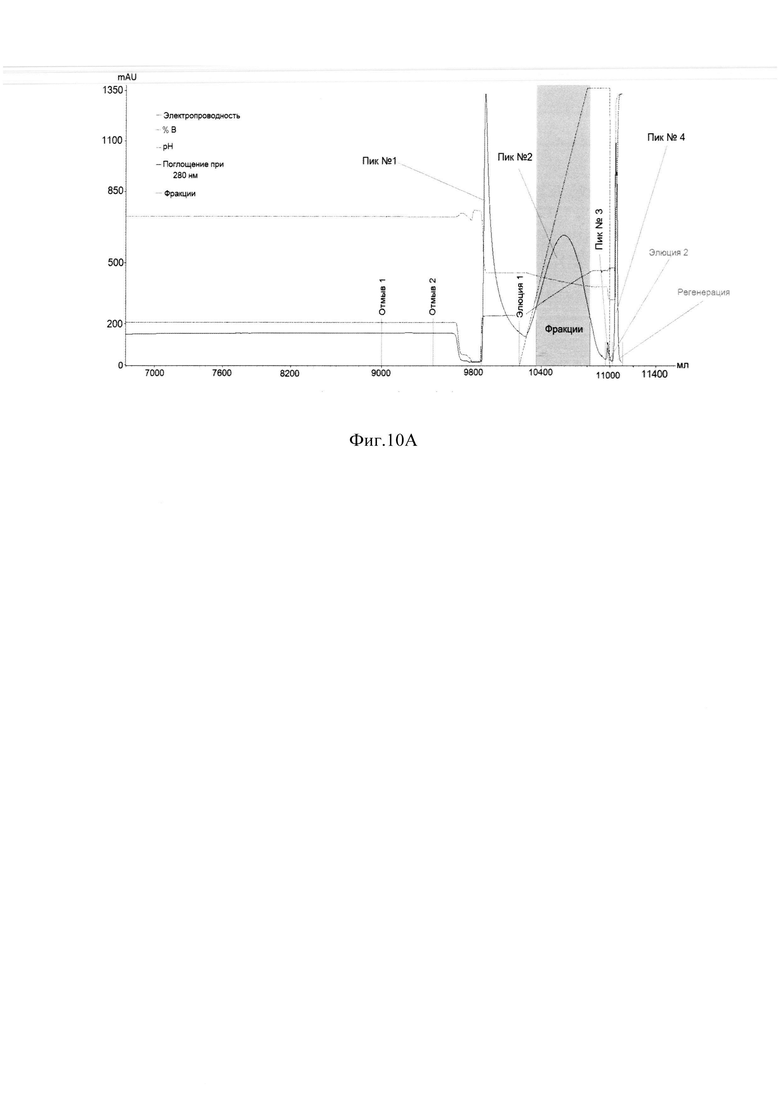
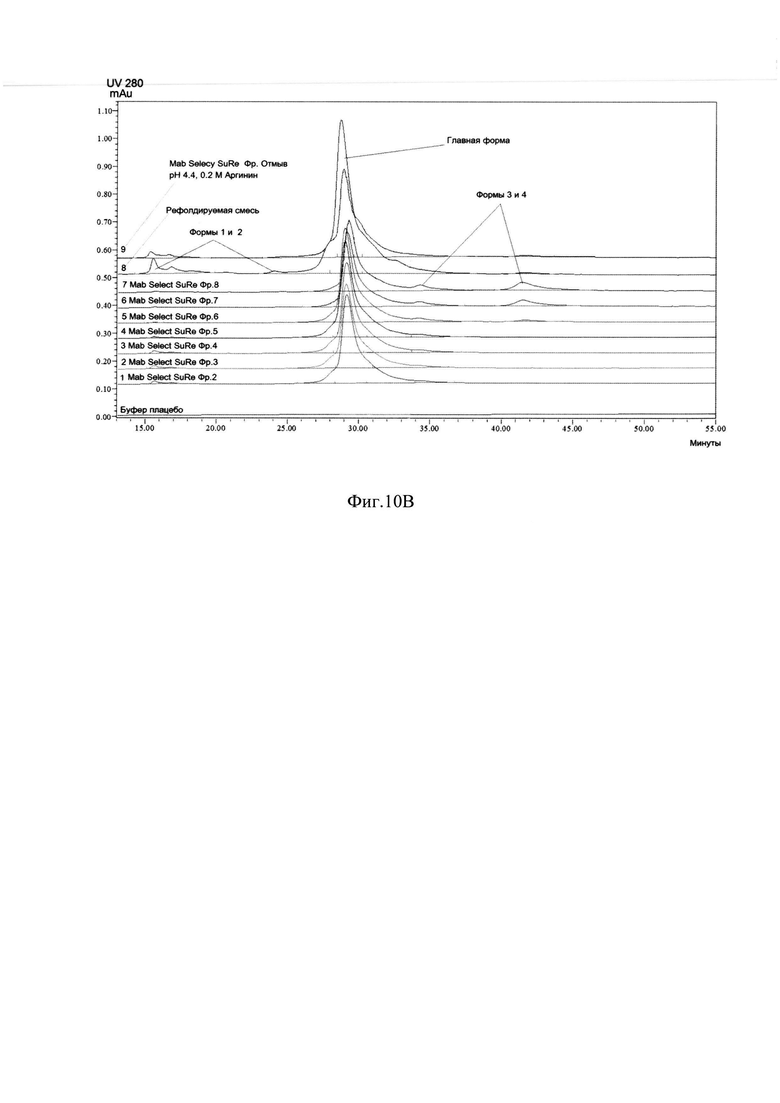
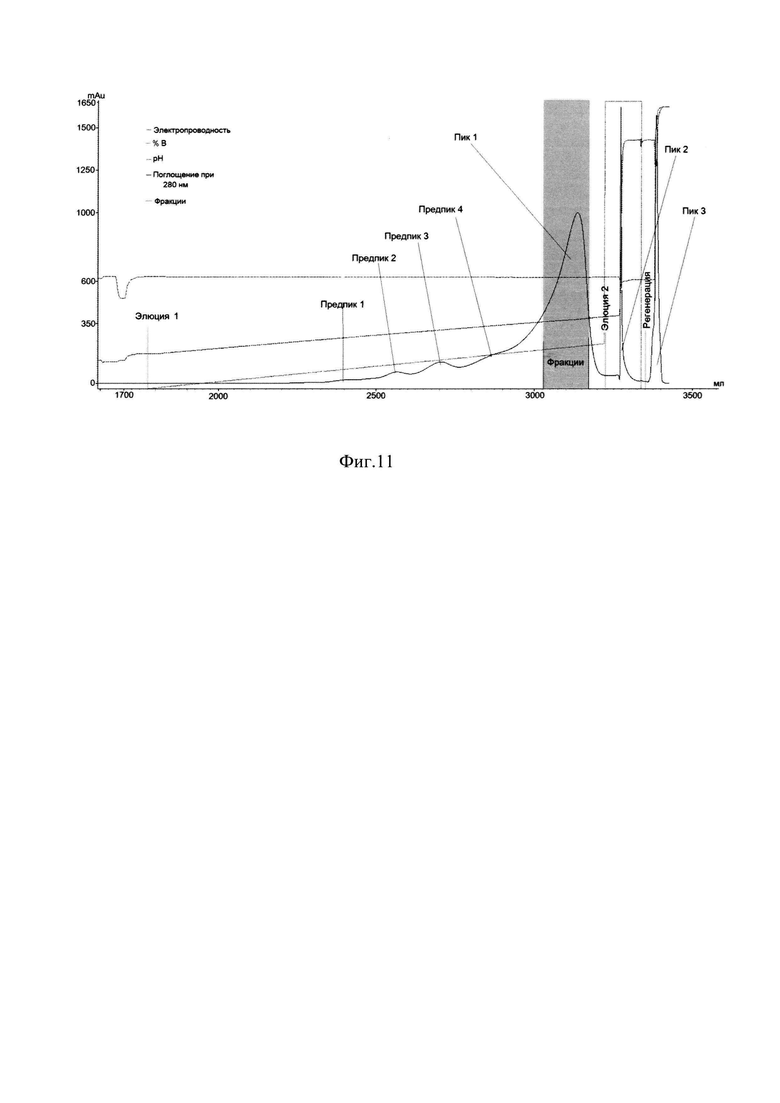
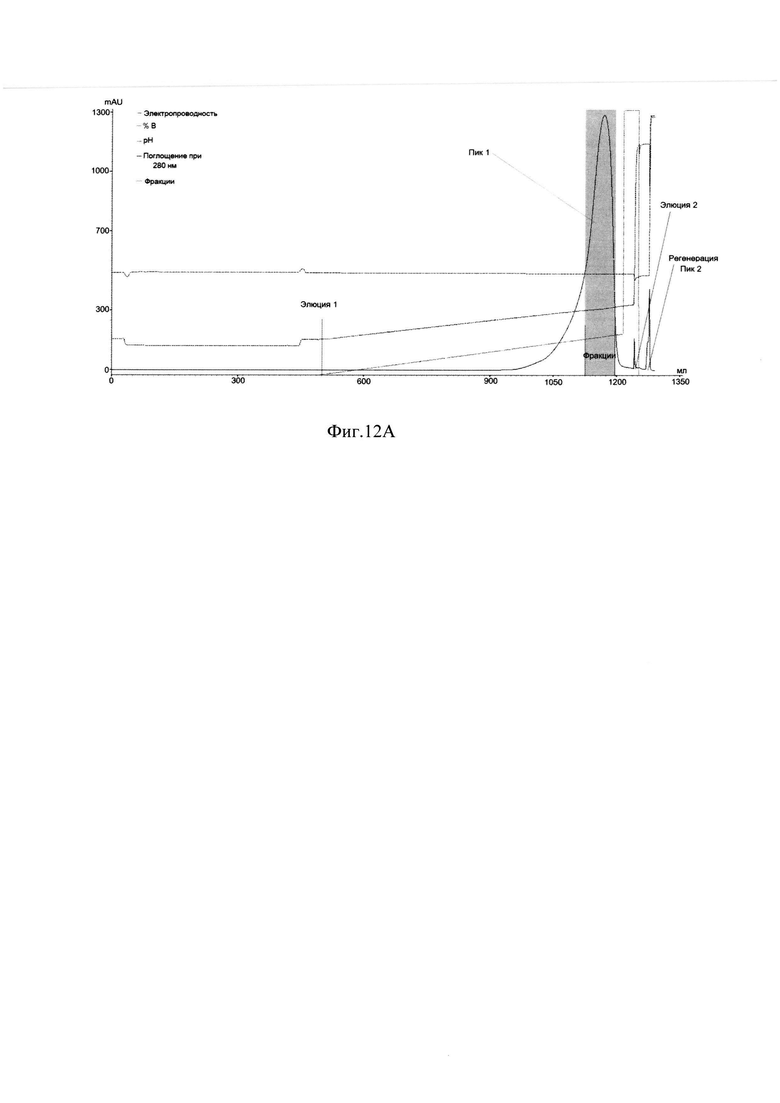
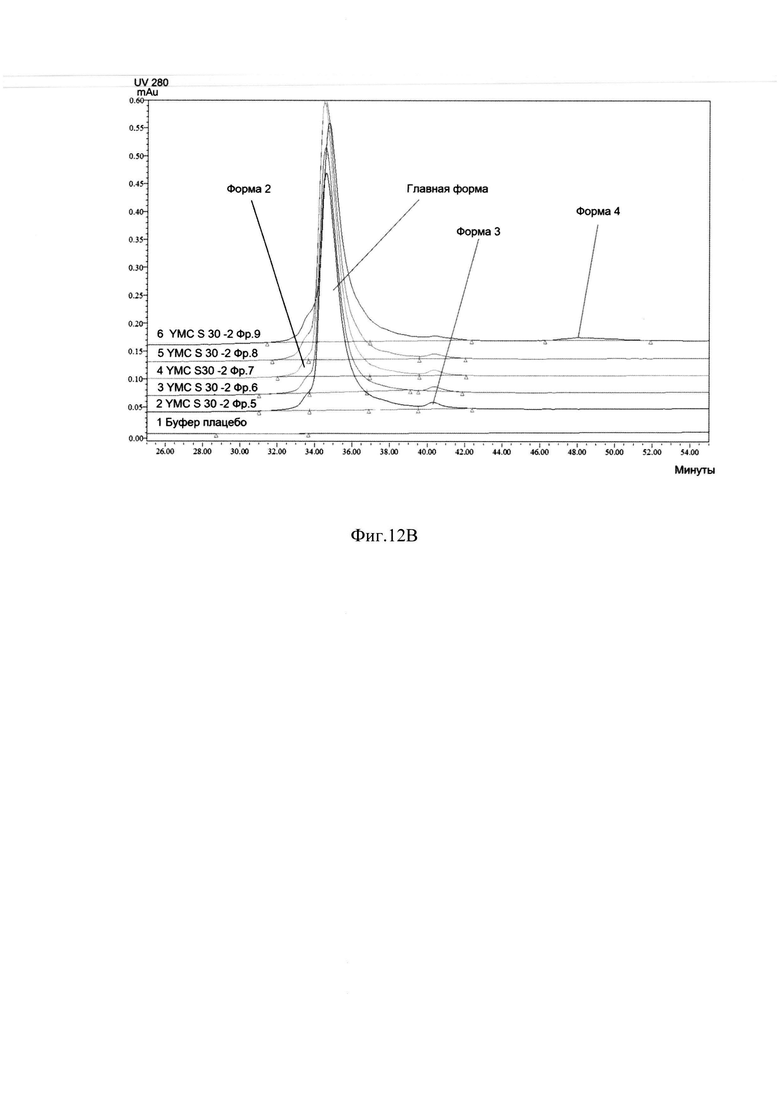
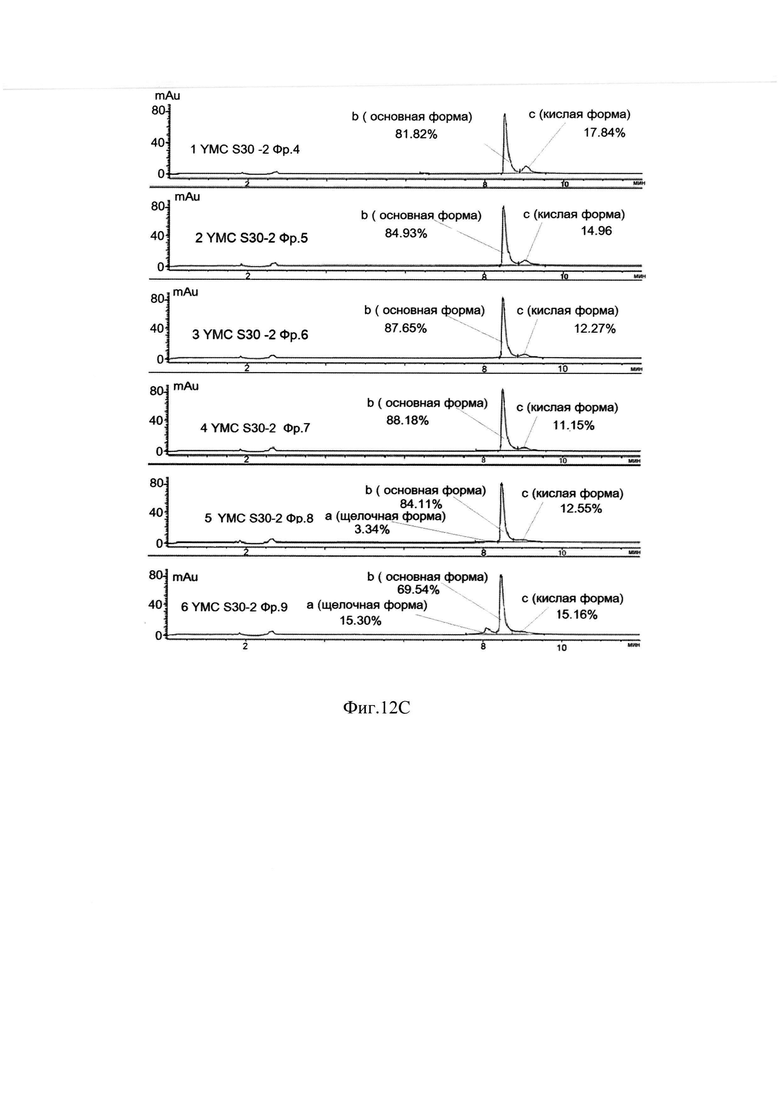
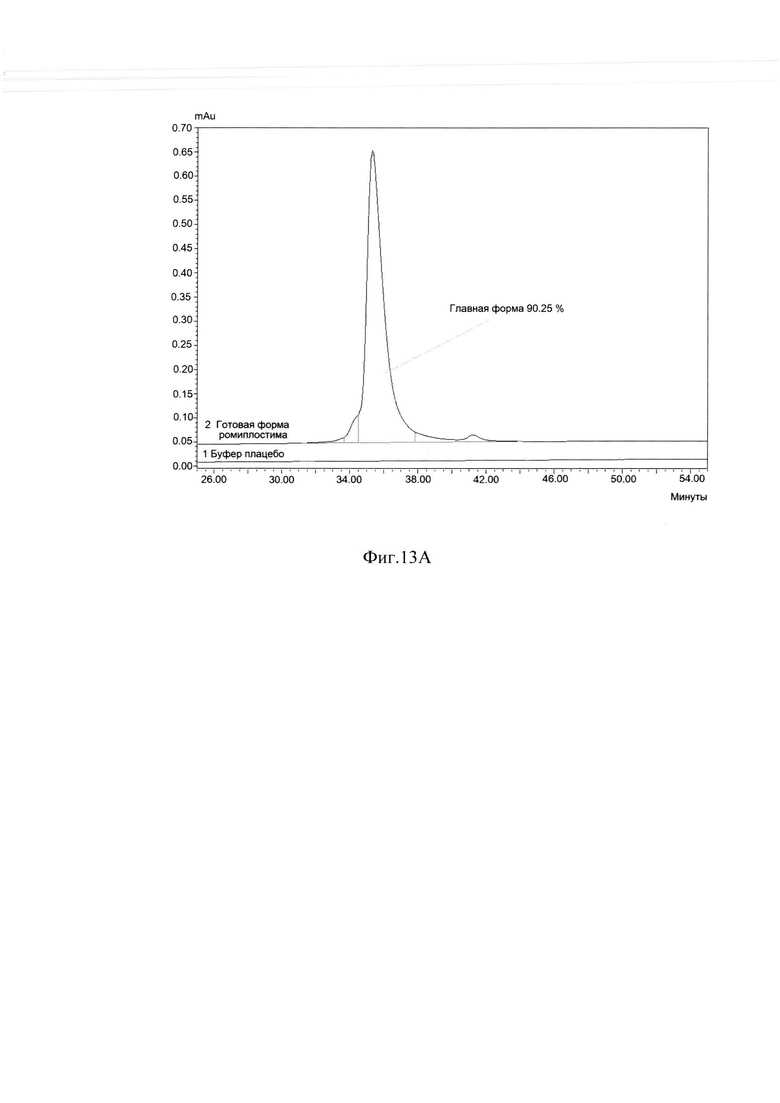
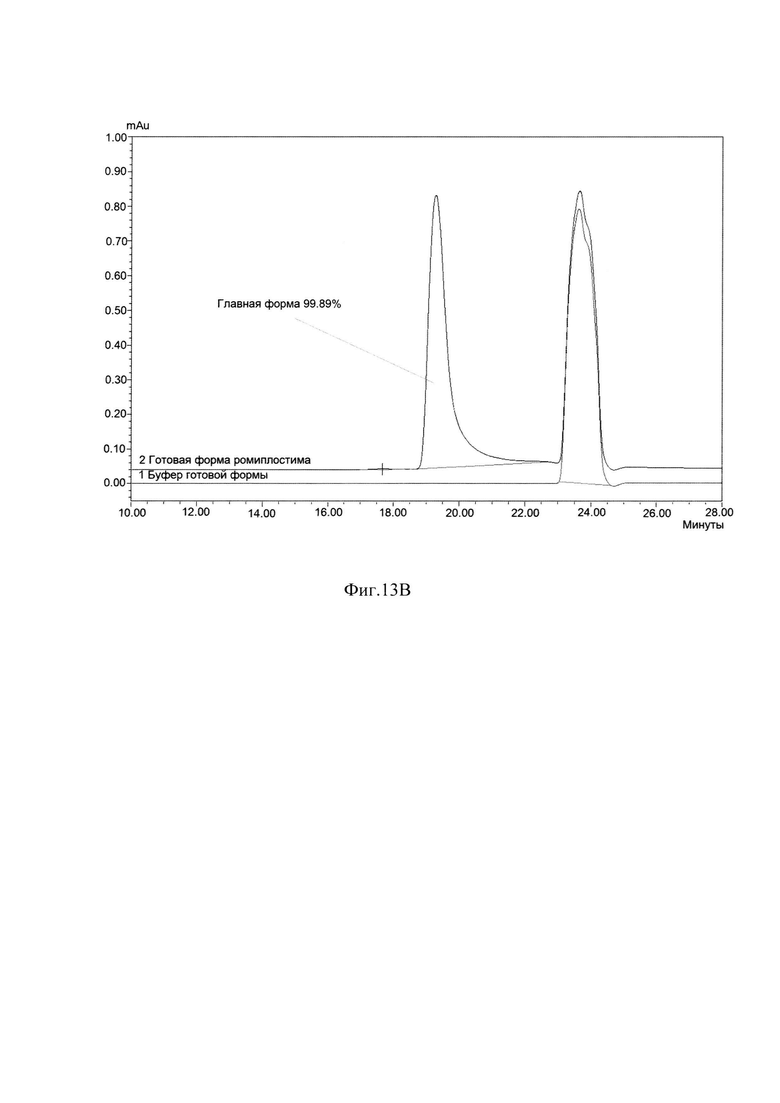
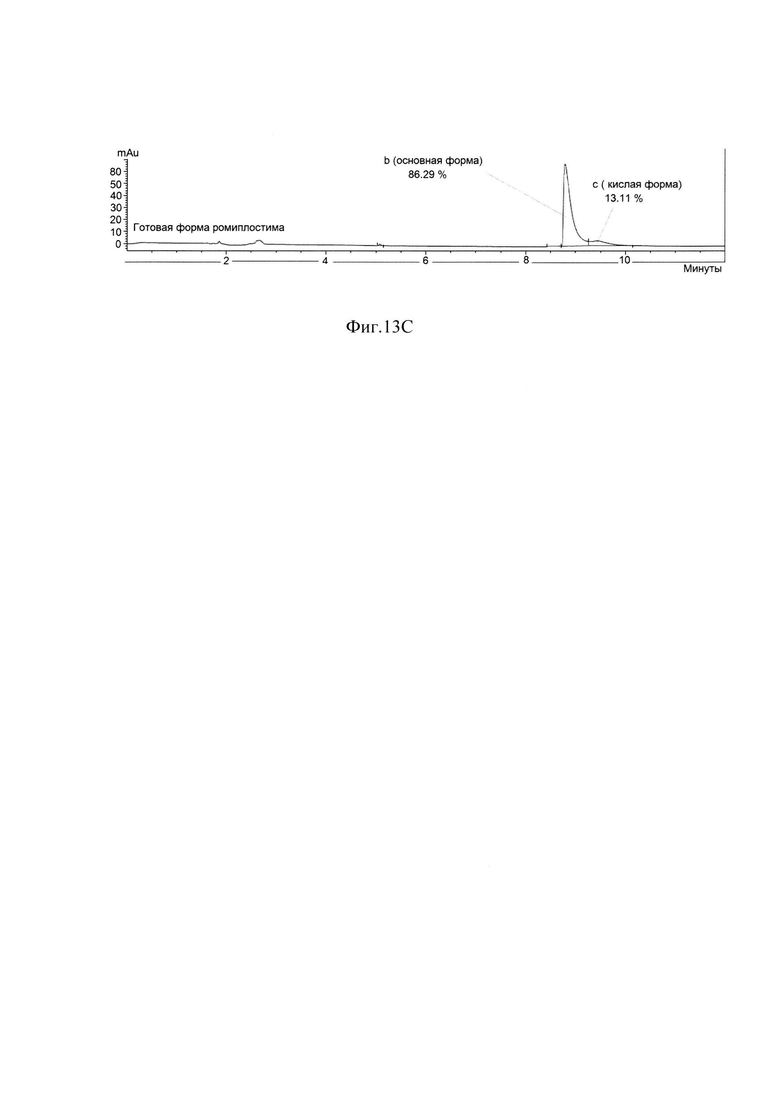
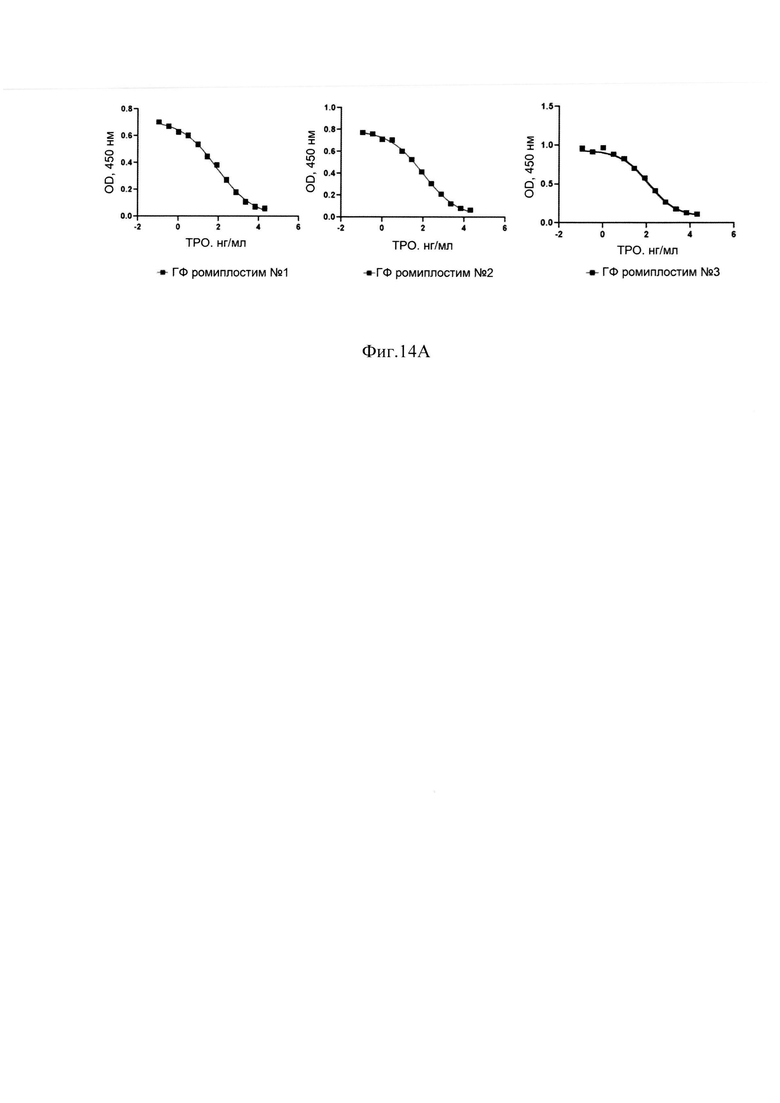
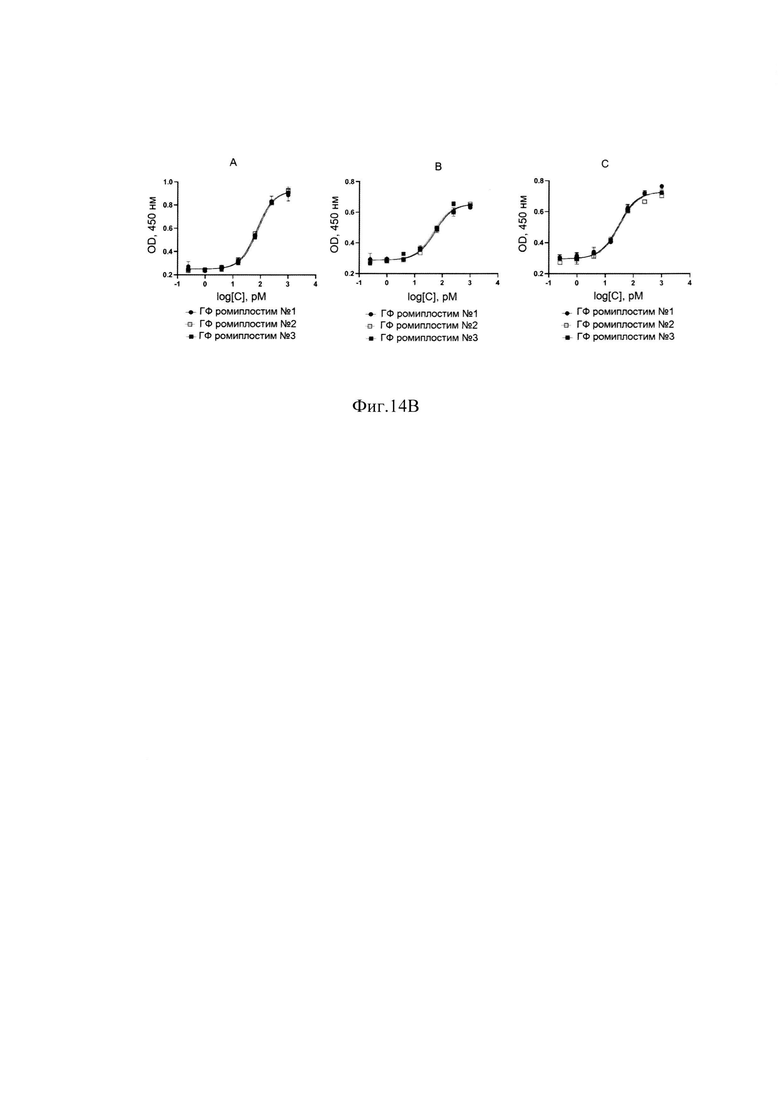
Изобретение относится к области фармацевтической биотехнологии, а именно к способу очистки рекомбинантных белков, а именно к получению высокоочищенного препарата ромиплостим. Изобретение представляет собой способ очистки рекомбинантного ромиплостима, полученного в E. coli, включающий выделение и отмывку телец включения, рефолдинг белка и хроматографическую очистку, при этом основное удаление контаминирующих примесей проводят на стадии выделения телец включения путем их последовательной отмывки, а дополнительное удаление примесей включает проведение 2-х последовательных стадий хроматографии на одном и том же катионообменном сорбенте, где первая стадия является разделяющей, а вторая полирующей и концентрирующей. 6 з.п. ф-лы, 14 ил., 5 табл., 2 пр.
1. Способ очистки рекомбинантного ромиплостима, полученного в E. coli, включающий выделение и отмывку телец включения, рефолдинг белка и хроматографическую очистку, отличающийся тем, что основное удаление контаминирующих примесей клеточных белков, нуклеиновых кислот, липидов и эндотоксинов проводят на стадии выделения телец включения путем их последовательной отмывки, а дополнительное удаление примесей включает проведение 2-х последовательных стадий хроматографии на одном и том же катионообменном сорбенте, где первая стадия является разделяющей, а вторая - полирующей и концентрирующей.
2. Способ по п. 1, где хроматографическую очистку проводят последовательно вначале в режиме проскока на сильном анионообменном сорбенте, с последующей сорбцией целевого белка на аффинном сорбенте, модифицированном рекомбинантным белком А, и далее проводят две стадии хроматографии на катионообменном сорбенте.
3. Способ по п. 2, где катионообменный сорбент представляет собой гидрофильную полимерную матрицу с размером частиц приблизительно от 30 до 75 мкм и размером пор приблизительно 100 нм.
4. Способ по п. 1, где очищенный белок получают переводом в буфер готовой формы гель-фильтрацией.
5. Способ по п. 1, где буфер для отмывки телец включения содержит Triton Х-100, мочевину, NaCl и ЭДТА.
6. Способ по п. 1, где буфер для проведения рефолдинга включает 3 мM цистина и 4 мM цистеина.
7. Способ по п. 1, где рефолдинг проводят при концентрации белка от 0,7 до 1 мг/мл.
| US 20060140934 A1, 24.09.2004 | |||
| ЖИДЕЦКИЙ А.В | |||
| и др | |||
| Кипятильник для воды | 1921 |
|
SU5A1 |
| гос | |||
| ун-та | |||
| Биология, 2017, н.2, стр.58-65 | |||
| EP 1904522 A1, 15.07.2005 | |||
| КИВЕРО А.Д | |||
| и др | |||
| Модификация центрального метаболизма E | |||
| coli для оптимизации процесса биотрансформации | |||
