Изобретение относится к области биотехнологии, а именно к генно-инженерным препаратам медицинского назначения и технологии их получения, точнее к технологии получения безметионинового интерферона альфа-2b.
Интерфероны альфа-2 (IFN-α2) человека относятся к классу цитокинов, обладающих противовирусной активностью, и являются представителями семейства альфа-интерферонов, секретируемых практически всеми типами вирус-инфицированных клеток человека [Pfeffer et al., 1998, Cancer Research 58:2489-2499].
Рекомбинантные IFN-α2 применяют для терапии вирусных и опухолевых заболеваний [Samuel, 2001, Clinical Microbiology Reviews, 14:778-809; Schadendorf et al., 2009, Annals of Oncology, 20 (Supplement 6): vi41-vi50; Pfeffer et al., 1998, Cancer Research 58:2489-2499], в том числе для лечения твердых опухолей, таких как рак мочевого пузыря, рак почки, ВИЧ-индуцированная саркома Капоши и др. [Torti, 1988, J. Clin. Oncol., 6:476-483; Vugrin et al., 1985, Cancer Treat. Rep., 69:817-820; Rios et al., 1985, J. Clin. Oncol., 3:506-512], а также для лечения хронических форм гепатитов В и С и иных вирусных заболеваний [Clark V., Nelson D.R., 2009, Clin Liver Dis, 13:351-363].
N-концевым аминокислотным остатком природных IFN-α2 является цистеин, связанный дисульфидной связью с 98 цистеином белка [Bodo G., Fogy I., 1985, The Interferon System, pp. 23-27; Wetzel et al., 1981, J. Interfer. Res., 1:381-90]. В то же время у некоторой части молекул рекомбинантных IFN-α2, выделяемых из клеток Escherichia coli (Е. coli), на N-конце остается дополнительный остаток метионина (формилметионина), что делает рекомбинантный IFN-α2 модифицированным белком.
Избыточный N-концевой остаток метионина или формилметионина может влиять на стабильность и иммуногенность белков [Ben-Bassat A., Bauer K., 1987, Nature 326, 315], а также в ряде случаев приводит к потере их биологической активности [Rabbani et al., 1988, J Biol Chem, 263:1307-1313]. В связи с тем, что с 2005 г Европейская Фармакопея ограничила производство и использование на территории европейских стран препаратов, содержащих модифицированный IFN-α2 [Eur. Pharm., 5th ed.: 2004], проблема удаления N-концевого метионина из состава IFN-α2 бактериального происхождения приобрела еще большее значение.
В клетках Е. coli, остающихся одними из основных продуцентов рекомбинантных белков, удаление (процессинг) N-концевого метионина детерминируется радиусом бокового радикала, следующего за метионином аминокислотного остатка синтезируемого белка [Hirel et al., 1989, Proc Nati Acad Sci USA, 86(21):8247-51; Dalboge et al., FEBS Lett. 1990; 266(1-2):1-3]. Однако такой процессинг часто оказывается неполным в случае гиперэкспрессии белка [Yasueda et al., 1991, Appl Microbiol Biotechnol, 36:211-215; Hwang et al., 1999, Biochem J, 338:335-342].
В качестве ферментативных способов удаления N-концевого метионина из состава рекомбинантных белков используют:
- удаление метионина с использованием метионинаминопептидазы [Shapiro et al., 1988, Anal. Biochem, 175:450-461; Solbiati et al., 1999, J Mol Biol, 290:607-14; Liao et al., 2004, Protein Science, 13:1802-1810];
- способы, основанные на биосинтезе удлиненного белка, между N-концевым метионином и зрелой частью которого введена короткая последовательность сайта узнавания какой-либо протеиназы. Последующая протеиназная обработка удлиненного белка приводит к удалению введенной последовательности вместе с N-концевым метионином [US 5665566; Belagaje et al., 1997, Protein Sci, 6:1953-1962; Hosfield Т., Lu Q., 1999, Anal Biochem, 269:10-6; Jenny R.J. et al., 2003, Protein Express Purif, 31:1-11].
К интерферону альфа-2 человека относят два аллельных варианта интерферона: интерферон альфа-2а и интерферон альфа-2b, отличающиеся в кодирующей области одиночной нуклеотидной заменой в позиции 137 (2а:А, 2b:G) [Kaluz et al., 1993 Acta Virol. 37:97-100; Kaluz et al., 1994, Acta Virol. 38:101-4; Lee et al., 1995, J Interferon Cytokine Res.15:341-9], которая приводит к аминокислотной замене (2a:Lys, 2b:Arg).
В настоящее время известен способ получения безметионинового интерферона альфа-2b человека (RU 2432401, 2011), включающий в себя микробиологический синтез белка-предшественника путем культивирования бактерий Escherichia coli, содержащих плазмиды с геном интерферона альфа-2b человека и нуклеотидной последовательностью, кодирующей сайт протеолиза энтеропептидазой Asp-Asp-Asp-Asp-Lys, расположенный перед нуклеотидной последовательностью, кодирующей зрелую часть интерферона альфа-2b человека, с последующим выделением телец включения синтезированного предшественника, частичную ренатурацию выделенного предшественника в присутствии препятствующего замыканию дисульфидных связей дитиоэритритола, гидролиз предшественника ферментом энтеропептидазой с образованием безметионинового интерферона альфа-2b человека, а после завершения реакции гидролиза предшественника полную ренатурацию интерферона альфа-2b человека в присутствии пары соединений цистин и цистеин и очистку полученного безметионинового интерферона альфа-2b человека методом хроматографии на КМ-сефарозе.
Так же известен способ [KR 100327971, 2000-11-15] получения безметионинового интерферона альфа-2b человека из синтезированного в клетках штамма Е. coli КССМ 10053 белка-предшественника, являющегося метионинсодержащим интерфероном альфа-2b человека, с помощью его обработки ферментом метионинаминопептидазой.
Основным недостатком способа является необходимость проведения длительной (12-20 часов) ферментативной обработки белка-предшественника. Дополнительные трудности может вызывать последующий перевод безметионинового белка в биологически активное состояние в условиях, необходимых для формирования дисульфидных связей [Morehead et al., 1984, Biochemistry, 23:2500-7], а также процесс отделения целевого безметионинового интерферона от метионинсодержащего предшественника вследствие незначительности различий в их физико-химических свойствах.
Известен способ получения безметионинового интерферона альфа-2 человека [RU 2453604, 2012], включающий микробиологический синтез и выделение белка-предшественника безметионинового интерферона альфа-2 человека с последующим ферментативным выщеплением из его состава безметионинового интерферона, причем в качестве продуцента белка-предшественника используют бактериальный штамм Escherichia coli ВКПМ В-10414 - продуцент гибридного белка предшественник безметионинового интерферона альфа-2, а для ферментативного выщепления безметионинового интерферона используют протеиназу ULP275.
Недостатком данной технологии способа является низкая активность полученного интерферона альфа-2b (2,0×108 МЕ/мг).
Известен штамм Е. coli BDEES4 и рекомбинантная плазмида pES4-4, содержащая промотор PT7, усилитель трансляции гена 10 фага Т7, последовательность искусственного гена рекомбинантного интерферона альфа 2b, последовательность гена β-лактамазы, детерминирующего устойчивость к антибиотику ампициллин, и терминатор транскрипции фага Т7 [RU 2258081, 2005]. Уровень экспрессии интерферона составляет 20-30% от всех клеточных белков.
Основным недостатком технологии является использование в плазмиде селективного гена β-лактамазы, детерминирующего устойчивость штамма к антибиотику ампициллин. При культивировании штамма β-лактомаза секретируется из бактериальной клетки и разрушает антибиотик. В результате чего в процессе ферментации происходит постоянное неконтролируемое снижение селективного давления для плазмидосодержащих клеток штамма, накопление безплазмидных клеток и, как следствие, снижение выхода интерферона. Кроме того, штамм-продуцент, содержащий плазмиду pES4-4, синтезирует интерферон альфа-2b с дополнительной аминокислотой метионин на N-конце молекулы.
Наиболее близким по технической сущности к предлагаемому изобретению является группа изобретений, в состав которой входят рекомбинантная плазмида pFM-IFN-17, обеспечивающая экспрессию интерферона альфа-2b человека, рекомбинантная плазмида pFM-AP, обеспечивающая экспрессию фермента метионинаминопептидазы Е. coli, биплазмидный штамм Escherichia coli FM-IFN-AP (pFM-IFN-17, pFM-AP) - продуцент безметионинового рекомбинантного интерферона альфа-2b человека [RU 2610173, 2017]. Способ получения интерферона включает в себя получение телец включения, их растворение смесью гуанидин гидрохлорида и дитиотреитола в трис-буферном растворе, инкубирование 2 час при комнатной температуре, отделение осадка центрифугированием и фильтрованием. Полученный раствор интерферона ренатурируют, фильтруют и подвергают диафильтрации. Затем добавляют концентрированный раствор ацетата натрия, и инкубируют 10-12 часов при 40°С. Раствор интерферона фильтруют через мембранный фильтр и очищают последовательно гидрофобной хроматографией на катионообменной смоле CM Sepharose FF, гидрофобной смоле Phenyl Sepharose HP (GE Healthcare), а затем с помощью обратно-фазной хроматографии на сорбенте С18. Далее белок очищают на катионообменном сорбенте MacroPrep High и подвергают колоночной гель-фильтрации на смоле Sephacryl S200 или Superdex 75 (GE Healthcare).
Недостатком данного изобретения является сложная многостадийная технология получения конечного продукта, в результате чего выход целевого продукта с 200 г биомассы, полученной при культивировании штамма, составляет 1-1,5 г субстанции.
Задачей, решаемой авторами являлась разработка более простой технологии получения рекомбинантного безметионинового интерферона альфа-2b, позволяющей получить целевой продукт с более высоким выходом.
Для решения поставленной задачи была создана новая последовательности ДНК, кодирующая метионинаминопептидазу Escherichia coli, на основе которой получены плазмидный экспрессионный вектор pRh15A и штамм бактерий Escherichia coli BL21 Star (DE3) [pAYC-ET-(hIFN-α2b)-lacI, pRh15A], Использование этого штамма позволило осуществлять более простую технологию выделения и обеспечить повышенный выход интерферона.
Технический результат достигался созданием группы изобретений, включающей в себя:
- последовательность ДНК (Seq ID No 1), кодирующая метионинаминопептидазу Escherichia coli;
- плазмидный экспрессионный вектор pRh15A;
- штамм бактерий Escherichia coli BL21 Star (DE3) [pAYC-ET-(hIFN-α2b)-lacI, pRh15A];
- способ получения интерферона альфа-2b человека, включающий в себя культивирование штамма Escherichia coli BL21 Star (DE3) [pAYC-ET-(hIFN-α2b)-lacI], отделение интерферонсодержащей биомассы, с последующим выделением интерферона путем последовательной хроматографической очистки.
Оптимальным режимом очистки при этом является последовательная четырехстадийная хроматографическая очистка на Фенил-сефарозе, Солозе КГ 20/30, Q-сефарозе и SP-целлюлозе.
Оптимальными условиями очистки являлись:
- на стадии очистки на Фенил-сефарозе - рН 5,2, использование в качестве растворителя 0,1 М трис-ацетатного буферного раствора, содержавшего 0,26 М натрия хлорида, 1 mM этилен-диаминотетра-уксусной кислоты (ЭДТА) и 0,1 масс. % метионина;
- на стадии очистки на Солозе КГ 20/30 - рН 5,2, колонка, содержащая сорбент, уравновешивается 0,25 М трис-ацетатным буферным раствором, содержавшим 1 mM ЭДТА и 0,1 мас. % метионина, отмывка уравновешивающим буферным раствором, элюцию проводят 0,05 М трисовым буферным раствором с 1 mM ЭДТА и 0,1 масс. % метионина;
- на стадии очистки на Q-сефарозе - рН 6,5; колонка, содержащая сорбент, уравновешивается 0,25 М трис-ацетатным буферным раствором, содержавшим, 1 mM ЭДТА и 0,1 масс. % метионина; отмывка уравновешивающим буферного раствора, а элюцию проводят 0,25 М трис-ацетатным буферным раствором с 1 mM ЭДТА и 0,1 масс. % метионина при рН 5,9;
- на стадии очистки на Sp-целлюлозе: рН 5,2, колонка, содержащая сорбент уравновешивается 0,25 М трис-ацетатным буферным раствором, содержащим 1 mM ЭДТА и 0,1 масс. % метионина; отмывается использованием 0,05 М натрий- ацетатного буферного раствора, а элюцию проводят 1000 мл 0,05 М натрий- ацетатным буферным раствором с добавлением 0,15 М натрия.
Все стадии хроматографии оптимально проводить в присутствии 2% метионина, что гарантирует исключение вероятности образования окисленной формы интерферона альфа-2b.
Сущность изобретения иллюстрируется следующими последовательностями и рисунками.
Фиг. 1. Нуклеотидная последовательность SEQ ID No1 синтетической ДНК, продуцирующей метионинаминопептидазу.
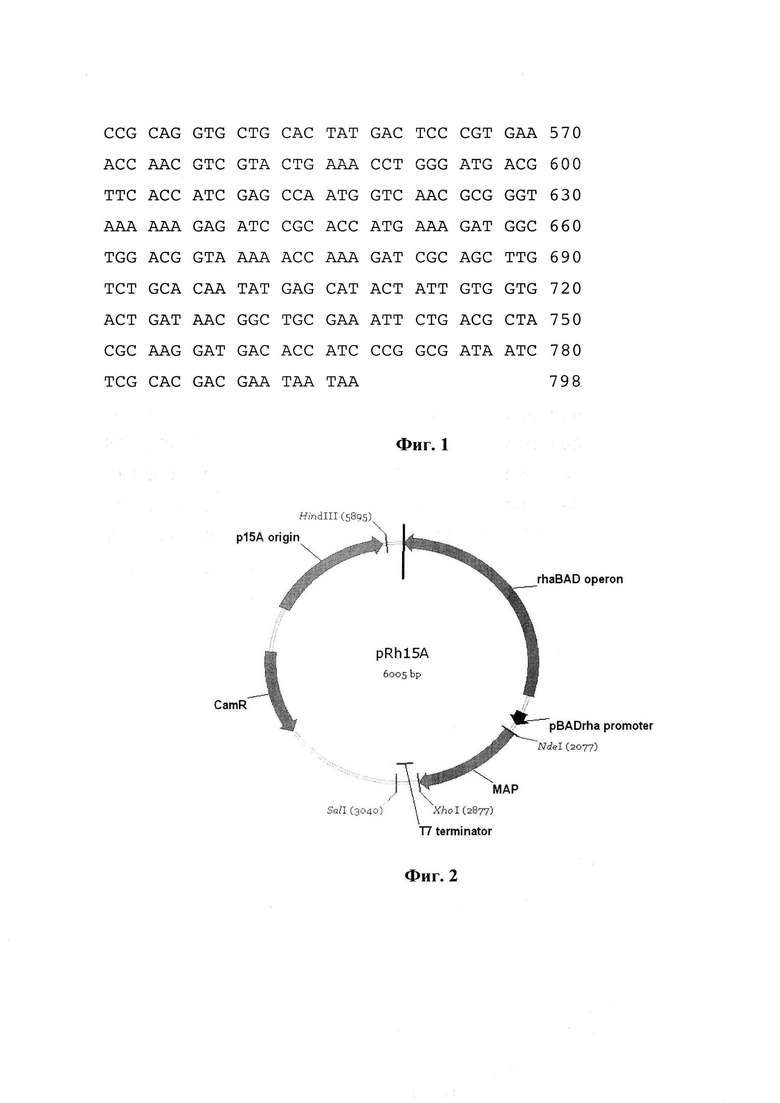
Фиг. 2. Схема плазмидного вектора pRh15A, несущего последовательность гена метионинаминопептидазы, где
MAP - ген метионинаминопептидазы Е. coli.
pBAD promoter - прокариотический промотор, служащий сигналом инициации транскрипции генов MAP
Т7 terminator - терминаторы транскрипции генов MAP
rhaBAD operon - регуляторная последовательность rhaBAD оперона, необходимая для регуляции экспрессии генов, находящихся под контролем промотора pBADrha.
р15А origin - сайт начала репликации плазмид
CamR - последовательность гена хлорамфеникол ацетилтрансферазы, обеспечивающей устойчивость бактерий к хлорамфениколу
SalI, HindIII, XhoI, NdeI - рестрикционные сайты, необходимые для объединения отдельных фрагментов в векторе
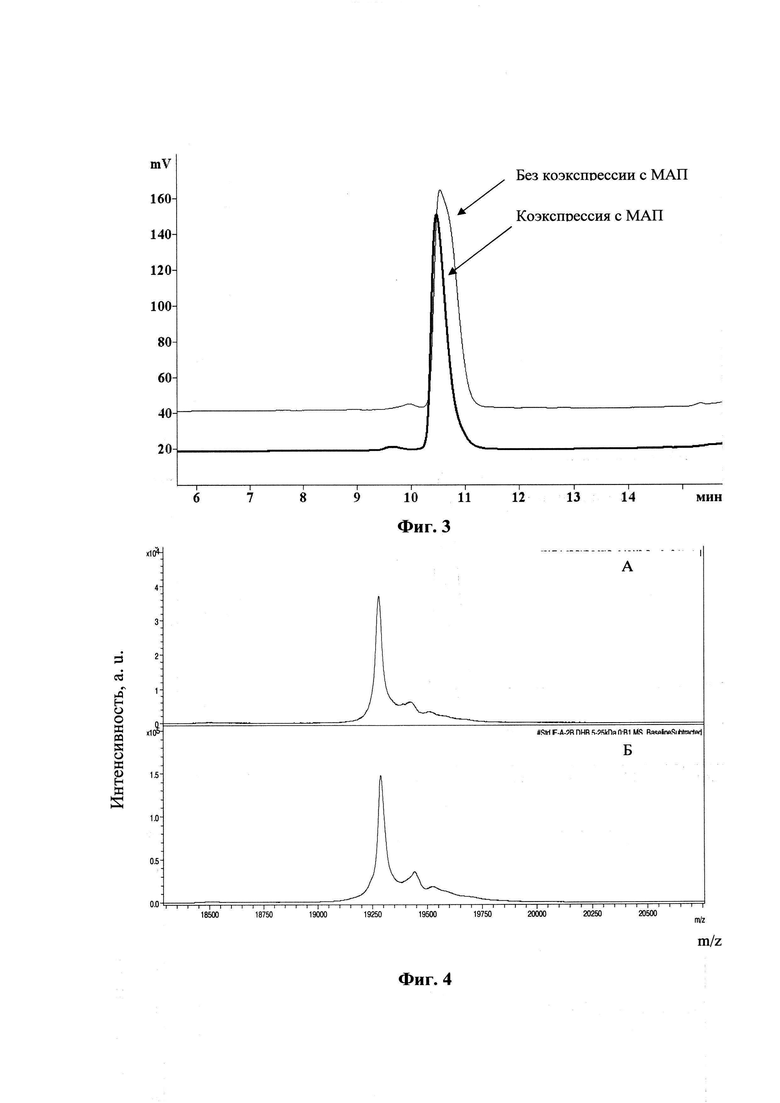
Фиг. 3. Наложение хроматограмм ОФ-ВЭЖХ разделения образцов интерферона альфа-2b человека, полученных при коэкспресии с МАП и без коэкспресии с МАП.
Фиг. 4. Спектрограмма полученного безметионинового интерферона альфа- 2b (А) и стандарта, рекомендованного европейской фармакопеей (Б)
Последовательность ДНК, кодирующая метионинаминопептидазу Е. coli, была амплифицирована с использованием в качестве матрицы ДНК из культуры бактерий штамма Escherichia coli BL21Star[DE3]. По концам к гену были добавлены олигонуклеотиды, формирующие сайты рестрикции NdeI и XhoI. (Полученная последовательность ДНК, кодирующая метионинаминопептидазу Escherichia coli с добавленными олигонуклеотидами, формирующими сайты рестрикции NdeI и XhoI приведена на SEQ ID No 1 -фиг1.
Ген был обработан вышеуказанными рестриктазами и клонирован в подготовленный аналогичным образом вектор pRhD(-), полученный из вектора pET22b(+) (Novagen, USA) с помощью следующих праймеров:
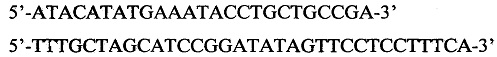
Для получения экспрессионного вектора pRhD(-)/MAP использовали плазмиду pRhD(-).
Последовательность гена метионинаминопептидазы была амплифицирована с геномной ДНК клеток Е. coli штамма BL21(DE3) с помощью следующих праймеров:

Амплифицированный фрагмент ДНК размером 800 п. н., концы которого были открыты с помощью рестриктаз NdeI и XhoI, клонировали в вектор pRhD(-), линеаризованный с помощью этих же рестриктаз.
В результате клонирования была получена плазмида pRhD(-)/MAP. Корректность сборки была доказана секвенированием.
В полученной плазмиде сайт репликации pBR322 и ген беталактамазы были заменены на сайт репликации р15А и ген устойчивости к хлорамфениколу из плазмиды pACYC184 (NEB). Данная модификация необходима для эффективной коэкспрессии двух белков с различных плазмид внутри одной клетки. Полученный плазмидный вектор получил название pRhD(-)/MAP.
Фрагмент последовательности ДНК, содержащий ген метионинаминопептидазы и регулирующие его экспрессию последовательности, был амплифицирован с плазмиды pRhD(-)/MAP с помощью следующих праймеров:
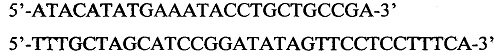
Амплифицированный фрагмент ДНК размером 3146 п. н., концы которого были открыты с помощью рестриктаз HindIII и SalI, был клонирован в вектор pACYC184, линеаризованный с помощью этих же рестриктаз. В результате клонирования был получен вектор pRh15A, содержащий ген метионинаминопептидазы под контролем регулируемого рамнозного промотора, сайт репликации р15А и ген устойчивости к хлорамфениколу для проведения селекции. Корректность сборки плазмиды была доказана секвенированием. Полученный экспрессионный плазмидный вектор получил название pRh15A.
Схема экспрессионного плазмидного вектора pRh15A приведена на фиг. 2.
Полученный штамм бактерий Escherichia coli BL21Star[DE3](pAYC-ET-(hIFN-α2b)-lacI, pRh15A) - продуцент безметионинового интерферона альфа-2b человека, был получен трансформацией бактериальной клетки двумя экспрессионными плазмидами - экспрессионной плазмидой, содержащей последовательность ДНК, кодирующей ген интерферона альфа-2b человека и экспрессионной плазмидой, содержащей последовательность ДНК, кодирующую метионинаминопептидазу Е. coli.
Полученный штамм Escherichia coli BL21Star[DE3](pAYC-ET-(hIFN-α2b)-lacI, pRh15A) характеризуется следующими культурально-морфологическими и физико-биохимическими свойствами:
Культурально-морфологические особенности штамма. Грам-отрицательные прямые палочки, размером 1,1-1,5×2,0-3,0 мкм, одиночные, спор и капсул не образуют. Каталазоположительные. Оксидазоотрицательные. Факультативные анаэробы. Клетки хорошо растут на простых питательных средах, содержащих и не содержащих стрептомицин и хлорамфеникол, например, на среде LB. На агаризованной среде - колонии гладкие, круглые, слабо выпуклые, с ровным краем. В жидких средах образуют равномерную светорассеивающую суспензию, при хранении без перемешивания оседают на дно. Клетки растут в интервале температур от 8°С до 43°С, интервал для культивирования - 28-38°С, оптимум роста при 37°С. Интервал рН 5-8. Катализируют D-глюкозу и некоторые другие углеводы с образованием кислоты и газа, не сбраживают галактозу. Реакция Фогес-Проскауэра отрицательная, не образуют H2S, гидролизуют мочевину.
Характеристики полезного вещества, синтезируемого штаммами. Рекомбинантный белок - интерферон альфа-2b человека с отщепленным N-концевым остатком метионина (безметиониновый интерферон альфа-2b человека).Активность штамма. Продуктивность штаммов - выход рекомбинантного интерферона альфа-2b человека составляет не менее 10% интерферона альфа-2b от суммамарных клеточных белков (по данным компьютерного денситометрирования сканированного геля) при культивировании в условиях индукции.
Условия хранения штамма:
- лиофильно высушенным в среде, содержащей 17-18% сахарозы, 3,5% желатина, 19,6% обезжиренного молока (до 2-х лет),
- замороженным при -70°С (до 12 месяцев) в 15%-ном растворе глицерина (или 7% растворе диметилсульфоксида) в питательной среде LB (Бакто-триптон - 10 г/л, дрожжевой экстракт - 5 г/л, хлористый натрий - 10 г/л, рН 6.8),
- криоконсервированным при -196°С в жидком азоте (бессрочно): суспензию клеток в 15%-ном растворе глицерина с высокой плотностью (109 - 1010 клеток в 1 мл) разливают в криопробирки и помещают в холодильник с температурой -70°С на 2 часа, а затем переносят в жидкий азот, где и хранят при -196°С. Оттаивание замороженных клеток должно быть как можно более быстрым: криопробирки погружают на 2 мин в водяную баню с температурой 35-45°С. Клетки из криопробирок высеивают на богатые питательные среды, например 2xLB.
Способ получения безметионинового интерферона альфа-2b из биомассы штамма-продуцента включает в себя получение биомассы (лизирование клеток, отмывку телец включения (ТВ), денатурацию ТВ, ренатурацию, осаждение сульфатом аммония, рН-фракционирование) и хроматографическую очистку.
Лизирование проводится с использованием буферного раствора. Для полноты лизиса суспензия лизата подвергалась двукратному замораживанию.
После размораживания для расщепления нуклеиновых кислот к лизату добавлялся раствор магния хлорида и ДНК-аза. Лизат центрифугировали, после чего проводили отмывку телец включения последовательной обработкой следующими растворами: раствором 4 М мочевины раствором 40% изопропилового спирта и водой.
Затем проводили процесс первичной денатурации в буферном растворе 8М гуанидингидрохлорида, содержащим 0,05-0.15 масс. % метионина и вторичной денатурации дитиотреитолом. Процесс ренатурации осуществлялся аргинином, а высаливание интерферона альфа-2b сульфатом аммония. Полученный продукт подвергали рН-фракционирование уксусной кислотой до значения рН 5,2.
Хроматографическая очистка проводится последовательно на фенил-сефарозе, Солозе КГ 20/30, Q-сефарозе и Sp-целлюлозе.
Оптимальными условиями очистки являлись:
- на стадии очистки на фенил-сефарозе - 700 мл сорбента, рН 5,2 с использованием в качестве растворителя 3000 мл 0,1 М трис-ацетатного буферного раствора, содержавшего 0,26 М натрия хлорида, 1 mM этилен-диаминотетрауксусной кислоты (ЭДТА) и 0,1 масс. % метионина;
- на стадии очистки на солозе КГ 20/30 - колонка, содержащая 500 мл сорбента, уравновешивается 2500 мл 0,25 М трис-ацетатным буферным раствором, содержавшим, 1 mM ЭДТА и 0,1 мас. % метионина, рН5,2. Отмывается с использованием 1500 мл уравновешивающего буферного раствора, а элюцию проводят 2000 мл 0,05 М трис-буферным раствором с 1 mM ЭДТА и 0,1 масс. % метионина.
- на Стадии очистки на Q-сефарозе - колонка, содержащая 1000 мл сорбента, уравновешивается 8000 мл 0,25 М трис-ацетатным буферным раствором, содержавшим, 1 mM ЭДТА и 0,1 мас. % метионина, рН6,5. Отмывается с использованием 1500 мл уравновешивающего буферного раствора, а элюцию проводят 5000 мл 0,25 М трис-ацетатным буферным раствором с 1 mM ЭДТА и 0,1 мас. % метионина, рН 5,9.
- на стадии очистки на Sp-целлюлозе: колонка, содержащая 100 мл сорбента уравновешивается 500 мл 0,25 М трис-ацетатным буферным раствором, содержащим 1 mM ЭДТА и 0,1 мас. % метионина; отмывается использованием 500 мл 0,05 М натрий-ацетатного буферного раствора, рН 5,2 а элюцию проводят 1000 мл 0,05 М натрий- ацетатным буферным раствором с добавлением 0,15 М натрия хлорида при рН 5,2.
После всех стадий очистки получаемый интерферон альфа-2b имеет чистоту >95% по ОФ-ВЭЖХ.
Сущность и преимущества заявляемой группы изобретений иллюстрируются следующими примерами.
Пример 1. Конструирование плазмиды pRhD(-)
С целью создания плазмиды pRhD(-) была амплифицирована последовательность полилинкера на матрице плазмиды pET22b(+) (Novagen, USA) с помощью следующих праймеров:
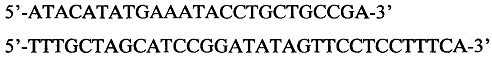
Допустимо использование праймеров в интервале концентраций от 5 до 25 мкМ, однако лучший результат достигается при использовании праймеров в концентрации 10 мкМ. Амплифицированный фрагмент полилинкера размером 305 п. н., концы которого были открыты с помощью рестриктаз NdeI и NheI, был клонирован в плазмиду pET302/NT-His (Thermo Fisher Scientific, USA), линеаризованную с помощью этих же рестриктаз, с помощью лигазы фага Т4. Количество добавляемых рестриктаз может варьироваться от 0.1 до 1 ед., тем не менее, оптимальное количество - 0.25 ед. каждой рестриктазы на 1 реакцию. Количество добавляемой лигазы может варьироваться от 0.5 до 5 ед., тем не менее, оптимальное количество - 1 ед. на реакцию. В результате был получен вектор рЕТ322.
Затем на матрице генома Е. coli штамма DH10B (Invitrogen, USA) был амплифицирован фрагмент (продукт 2030 п. н.), содержащий регуляторную часть rhaBAD-оперона. Для этого использовали следующую пару праймеров:
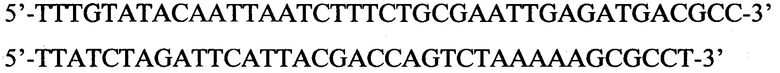
Допустимо использование праймеров в интервале концентраций от 5 до 25 мкМ, однако лучший результат достигается при использовании праймеров в концентрации 10 мкМ. Амплифицированный фрагмент размером 2030 п. н., концы которого были открыты с помощью рестриктаз AccI и XbaI, клонировали в плазмиду рЕТ322, линеаризованную с помощью этих же рестриктаз, с помощью лигазы фага Т4. Количество добавляемых рестриктаз может варьироваться от 0.1 до 1 ед., тем не менее, оптимальное количество - 0.25 ед. каждой рестриктазы на 1 реакцию. Количество добавляемой лигазы может варьироваться от 0.5 до 5 ед., тем не менее, оптимальное количество - 1 ед. на реакцию. В результате был получен вектор pRhD(-). Корректность сборки была доказана секвенированием.
Полученная плазмида pRhD(-) содержала в своем составе ген бета-лактамазы, обеспечивающий устойчивость к антибиотикам бета-лактамной группы, сайт репликации pBR322, а также регулирующие элементы и промотер rhaBAD-оперона, что позволяет индуцировать и регулировать экспрессию целевого белка посредством концентрации рамнозы в среде.
Конструирование экспрессионного вектора pRhD(-)/MAP
Для получения экспрессионных векторов использовали плазмиду pRhD(-).
На первом этапе последовательность гена метионинаминопептидазы была амплифицирована с геномной ДНК клеток Е. coli штамма BL21(DE3) с помощью следующих праймеров:

Допустимо использование праймеров в интервале концентраций от 5 до 25 мкМ, однако лучший результат достигается при использовании праймеров в концентрации 10 мкМ. Амплифицированный фрагмент ДНК размером 800 п. н., концы которого были открыты с помощью рестриктаз NdeI и XhoI, клонировали в вектор pRhD(-), линеаризованный с помощью этих же рестриктаз, с помощью лигазы фага Т4. Количество добавляемых рестриктаз может варьироваться от 0.1 до 1 ед., тем не менее, оптимальное количество - 0.25 ед. каждой рестриктазы на 1 реакцию. Количество добавляемой лигазы может варьироваться от 0.5 до 5 ед., тем не менее, оптимальное количество - 1 ед. на реакцию.
В результате клонирования была получена плазмида pRhD(-)/MAP. Корректность сборки плазмиды была доказана секвенированием.
На втором этапе в полученной плазмиде сайт репликаций pBR322 и ген бета-лактамазы были заменены на сайт репликации р15А и ген устойчивости к хлорамфениколу из плазмиды pACYC184 (NEB). Данная модификация была осуществлена для эффективной коэкспрессии двух белков с различных плазмид внутри одной клетки.
Конструирование экспрессионного вектора pRh15A
Фрагмент последовательности ДНК, содержащий ген метионинаминопептидазы и регулирующие его экспрессию последовательности, был амплифй-цирован с плазмиды pRhD(-)/MAP с помощью следующих праймеров:
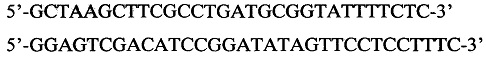
Допустимо использование праймеров в интервале концентраций от 5 до 25 мкМ, однако лучший результат достигается при использовании праймеров в концентрации 10 мкМ. Амплифицированный фрагмент ДНК размером 3146 п. н., концы которого были открыты с помощью рестриктаз HindIII и SalI, был клонирован в вектор pACYC184, линеаризованный с помощью этих же рестриктаз, с помощью лигазы фага Т4. Количество добавляемых рестриктаз может варьироваться от 0.1 до 1 ед., тем не менее, оптимальное количество - 0.25 ед. каждой рестриктазы на 1 реакцию. Количество добавляемой лигазы может варьироваться от 0.5 до 5 ед., тем не менее, оптимальное количество - 1 ед. на реакцию.
В результате клонирования был получен вектор pRh15A, содержащий ген метионинаминопептидазы под контролем регулируемого рамнозного промотора, сайт репликации р15А и ген устойчивости к хлорамфениколу для проведения селекции. Корректность сборки плазмиды была доказана секвенированием.
Пример 2. Получение штамма культивируемых клеток - продуцента безметионинового интерферона альфа-2b человека
Для экспрессии гена интерферона альфа-2b использовали созданный в соответствий с примером 1 плазмидный экспрессионный вектор pRh15A. Электрокомпетентные клетки Е. coli BL21 [DE3] трансформировали смесью плазмид pRhl5A и pAYC-ET-(hIFN-α2b)-lacI в эквимолярном соотношении. Трансформацию проводили с помощью аппарата Bio-Rad MicroPulser (Bio-Rad, США) согласно инструкции производителя. Положительные трансформанты отбирали на агаризованной среде, содержащей смесь двух антибиотиков (стрептомицин в конечной концентрации от 50 до 125 мкг/мл, предпочтительно 100 мкг/мл; и хлорамфеникол в конечной концентрации от 20 до 40 мкг/мл, предпочтительно 25 мкг/мл), что гарантировало содержание обеих плазмид в отобранных колониях. Наличие в трансформантах обеих плазмид также подтверждали путем ПЦР с использованием двух пар праймеров, специфических для гена интерферона альфа-2b и гена МАП, соответственно. Полученный штамм получил название Escherichia coli BL21 Star (DE3) [pAYC-ET-(hIFN-α2b)-lacI, pRh15A].
Пример 3. Анализ чистоты и гомогенности полученного интерферона альфа-2b
Для определения количества белковых примесей в полученном образце интерферона альфа-2b пробы разделяли методом ВЭЖХ в обратной фазе на колонке Phenomenex Jupiter С18. 10 мкг (±2 мкг) ИЛ-36РА в 20 мкл (±4 мкл) раствора наносили на колонку в 50 мМ (±5 мМ) фосфатном буфере, рН 6,0 (допустимый интервал рН 5,8-6,2). Элюцию проводили градиентом ацетонитрила с 30% до 70% в 0,1% ТФУ, рН 2,5 (±0,2), за 20 минут (±5 мин) при скорости потока 1,5 мл/мин и температуре +35°С (±2°С).
Детектирование проводили на УФ-детекторе при длине волны 220 нм. Типовая хроматограмма ОФ-ВЭЖХ разделения интерферона альфа-2b по этой методике представлена на Фиг. 3.
Для определения примеси формы интерферона альфа-2b, содержащей неотщепленный N-концевой остаток метионина, проводили ОФ-ВЭЖХ по методу, описанному выше. При этом элюцию проводили градиентом ацетонитрила с 39,6% до 40,8%, в остальном условия были такими же. Типовая хроматограмма ОФ-ВЭЖХ разделения интерферона альфа-2b по этой методике представлена на Фиг. 4
После всех стадий очистки по результатам анализов ВЭЖХ получаемый интерферон альфа-2b имеет чистоту >95%.
Для доказательства полноты отщепления N-концевого остатка метионина, проводили MALDI масс-спектрометрию в с помощью масс-спектрометра с источником ионизации ultrafleXtreme (Bruker, Германия). При этом ионизацию образцов (стандарта, рекомендованного европейской фармакопеей и полученного безметионинового интерферона альфа-2b) проводили с матрицы DHB. Спектрограммы стандарта, рекомендованного европейской фармакопеей (Б) и полученного безметионинового интерферона альфа-2b (А), представлены на Фиг. 5.
По результатам анализов ВЭЖХ и MALDI масс-спектрометрии получаемый безметиониновый интерферон альфа-2b имеет чистоту >95% и является аналогом стандартного образца безметионинового интерферона альфа-2b. При этом N-концевой остаток метионина отщепляется in vivo и целевой белок не требует дальнейшей обработки ферментами, что приводит к сокращению технологии и экономических затрат.
Пример 4. Способ получения безметилированного (Met-) интерферона альфа-2b человека из биомассы штамма Escherichia coli BL21 Star (DE3) [pAYC-ET-(hIFN-α2b)-lacI, pRh15A].
Культивирование штамма проводили в ферментере Bioflo с рабочим объемом 10 л. Состав среды: кислотный гидролизат казеина - 15 г/л; дрожжевой экстракт - 7,5 г/л, хлорид натрия 5 г/л, стрептомицин - 50 мг/л, хлорамфеникол - 30 мг/л.
Начальные параметры культивирования: рН - 7,0; температура - 37оС; насыщение кислородом - 100%. Далее, по мере роста культуры, уровень насыщения среды кислородом поддерживался автоматически на 40% от полного насыщения при данной температуре. рН среды до конца культивирования поддерживался на уровне 6,9 с помощью 10% раствора ортофосфорной кислоты. Начальная оптическая плотность культуры 0,3-0,5ед. при длине волны 540 нм. Через 4-4.5 часа культивирования, по достижении оптической плотности культуры 6-8 ед. при длине волны 540 нм и исчерпании глюкозы была введена рамноза (индуктор МАП) до конечной концентрации 0,02%. Через 0,5 часа был введен индуктор интерферона альфа-2b IPTG до конечной концентрации 0,5 мМ. Продолжительность индукции 4 часа. Конечная оптическая плотность составляла 20-22 ед. при длине волны 540 нм. Количество биомассы с влажностью 85% после сепарации при 10000 об\мин составляло 250 грамм.
Переработку биомассы через тельца включения проводили по стандартной схеме (US Patent 5789551, RU 2165455, RU 2242516, RU 2610173 A. Beldarrain, Y. Cruz, O. Cruz, M. Navarro, M.Gil., "Purification and conformational properties of human interferon alfa-2b produced in E.coli"., Biotechnol. Appl. Biochem., 2001., 33., 173-182).
Хроматографическую очистку проводили последовательно на фенил-сефарозе, Солозе КГ 20/30, Q- сефарозе и Sp-целлюлозе. Все стадии хроматографии проводили в присутствии 2% метионина, что гарантировало исключение вероятности образования окисленной формы интерферона альфа-2b
Условиями очистки являлись:
- на стадии очистки на фенил-сефарозе - 700 мл сорбента, рН 5,2 с использованием в качестве растворителя 3000 мл 0,1 М трис-ацетатного буферного раствора, содержавшего 0,26 М натрия хлорида, 1 mM этилен-диаминотетрауксусной кислоты (ЭДТА) и 0,1 масс. % метионина;
- на стадии очистки на Солозе КГ 20/30 - колонка, содержащая 500 мл сорбента, уравновешивается 2500 мл 0,25 М трис-ацетатным буферным раствором, содержавшим, 1 mM ЭДТА и 0,1 мас. % метионина, рН5,2. Отмывается с использованием 1500 мл уравновешивающего буферного раствора, а элюцию проводят 2000 мл 0,05 М трисовым буферным раствором с 1 mM ЭДТА и 0,1 масс. % метионина;
- на стадии очистки на Q-сефарозе - колонка, содержащая 1000 мл сорбента, уравновешивается 8000 мл 0,25 М трис-ацетатным буферным раствором, содержавшим, 1 mM ЭДТА и 0,1 мас. % метионина, рН 6,5. Отмывка проводилась с использованием 1500 мл уравновешивающего буферного раствора, а элюция - 5000 мл 0,25 М трис-ацетатным буферным раствором с 1 mM ЭДТА и 0,1 масс. % метионина, рН 5,9;
- на стадии очистки на Sp-целлюлозе: колонка, содержащая 100 мл сорбента уравновешивается 500 мл 0,25 М трис-ацетатным буферным раствором, содержащим 1 mM ЭДТА и 0,1 масс. % метионина; отмывается использованием 500 мл 0,05 М натрий-ацетатного буферного раствора, рН 5,2 а элюцию проводят 1000 мл 0,05 М натрий- ацетатным буферным раствором с добавлением 0,15 М натрия хлорида при рН 5,2.
После всех стадий очистки количество получаемого безметионинового интерферона альфа-2b составило 2,1 г с чистотой 95,8% по ОФ-ВЭЖХ.
В результате реализации заявляемой группы изобретений был получен интерферон альфа-2b с чистотой более 95% по ОФ-ВЭЖХ и выходом на 30-50%, превышающим выход при реализации ближайшего аналога.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ БЕЗМЕТИОНИНОВОГО ИНТЕРФЕРОНА-АЛЬФА2b ЧЕЛОВЕКА | 2009 |
|
RU2432401C2 |
| ГИБРИДНЫЙ БЕЛОК, ШТАММ БАКТЕРИЙ ESCHERICHIA COLI - ПРОДУЦЕНТ ГИБРИДНОГО БЕЛКА И СПОСОБ ПОЛУЧЕНИЯ БЕЗМЕТИОНИНОВОГО ИНТЕРФЕРОНА АЛЬФА-2b ЧЕЛОВЕКА ИЗ ЭТОГО ГИБРИДНОГО БЕЛКА | 2010 |
|
RU2441072C1 |
| ГИБРИДНЫЙ БЕЛОК (ВАРИАНТЫ), ШТАММ БАКТЕРИЙ ESCHERICHIA COLI - ПРОДУЦЕНТ ГИБРИДНОГО БЕЛКА (ВАРИАНТЫ) И СПОСОБ ПОЛУЧЕНИЯ БЕЗМЕТИОНИНОВОГО ИНТЕРФЕРОНА АЛЬФА-2 ЧЕЛОВЕКА | 2011 |
|
RU2453604C1 |
| СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО ИНТЕРФЕРОНА АЛЬФА-2b И ИНТЕРФЕРОНСОДЕРЖАЩИЙ ПРЕПАРАТ (ВАРИАНТЫ) | 2005 |
|
RU2294372C1 |
| СПОСОБ ПОЛУЧЕНИЯ БЕЗМЕТИОНИНОВОГО РЕЦЕПТОРНОГО АНТАГОНИСТА ИНТЕРЛЕЙКИНА-36, ШТАММ БАКТЕРИЙ ESCHERICHIA COLI BL21[DE3] - ПРОДУЦЕНТ БЕЗМЕТИОНИНОВОГО РЕЦЕПТОРНОГО АНТАГОНИСТА ИНТЕРЛЕЙКИНА-36 (ВАРИАНТЫ), СПОСОБ ИХ ПОЛУЧЕНИЯ, ПЛАЗМИДНЫЕ ВЕКТОРЫ pBAD15A и pET15A И ПОСЛЕДОВАТЕЛЬНОСТЬ ДНК ДЛЯ ИХ ПОЛУЧЕНИЯ | 2017 |
|
RU2672816C9 |
| Штамм бактерий Escherichia Coli - продуцент рекомбинантного белка IL-29 | 2020 |
|
RU2760585C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pSS5, КОДИРУЮЩАЯ СИНТЕЗ РЕКОМБИНАНТНОГО ЧЕЛОВЕЧЕСКОГО АЛЬФА-2b ИНТЕРФЕРОНА, ШТАММ ESCHERICHIA COLI SS5 - ПРОДУЦЕНТ РЕКОМБИНАНТНОГО ЧЕЛОВЕЧЕСКОГО АЛЬФА-2b ИНТЕРФЕРОНА И СПОСОБ ПОЛУЧЕНИЯ ИНТЕРФЕРОНА АЛЬФА-2b | 1999 |
|
RU2165455C1 |
| РАСТВОР ДЛЯ ИНЪЕКЦИЙ, РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pSX50, КОДИРУЮЩАЯ СИНТЕЗ РЕКОМБИНАНТНОГО ЧЕЛОВЕЧЕСКОГО АЛЬФА-2b ИНТЕРФЕРОНА, ШТАММ Escherichia coli SX50 - ПРОМЫШЛЕННЫЙ ШТАММ-ПРОДУЦЕНТ РЕКОМБИНАНТНОГО ЧЕЛОВЕЧЕСКОГО АЛЬФА-2b ИНТЕРФЕРОНА И СПОСОБ ПРОМЫШЛЕННОГО ПОЛУЧЕНИЯ ИНТЕРФЕРОНА АЛЬФА-2b | 2006 |
|
RU2319502C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pIF TREN, КОДИРУЮЩАЯ ПОЛИПЕПТИД ИНТЕРФЕРОНА АЛЬФА-2b ЧЕЛОВЕКА, И ШТАММ БАКТЕРИЙ Escherichia coli-ПРОДУЦЕНТ ПОЛИПЕПТИДА ИНТЕРФЕРОНА АЛЬФА-2b ЧЕЛОВЕКА | 2006 |
|
RU2326168C1 |
| Рекомбинантная плазмида pFM-IFN-17, обеспечивающая экспрессию интерферона альфа-2b человека, рекомбинантная плазмида pFM-АР, обеспечивающая экспрессию фермента метионинаминопептидазы E. coli, биплазмидный штамм Escherichia coli FM-IFN-АР (pFM-IFN-17, pFM-АР) - продуцент (Met-) рекомбинантного интерферона альфа-2b человека | 2016 |
|
RU2610173C1 |

Группа изобретений относится к области биотехнологии. Предложены плазмидный экспрессионный вектор pRh15A для получения безметионинового интерферона альфа-2b человека, штамм - продуцент безметионинового интерферона альфа-2b человека и способ получения безметионинового интерферона альфа-2b человека. Плазмидный экспрессионный вектор pRh15A содержит ген метионинаминопептидазы Escherichia coli, рамнозный промотор (pBADrha), rhaBAD оперон, терминатор транскрипции генов фага Т7, ориджин репликации плазмид р15А, ген хлорамфеникол ацетилтрансферазы (CamR) и рестрикционные сайты, необходимые для объединения отдельных фрагментов в векторе. Предложенный штамм представляет собой штамм Escherichia coli BL21Star[DE3], содержащий плазмидный экспрессионный вектор pAYC-ET-(hIFN-a2b)-lacI и вектор pRh15A. Способ получения безметионинового интерферона альфа-2b человека, включающий в себя культивирование указанного штамма, отделение интерферонсодержащей биомассы с последующей хроматографической очисткой целевого продукта. Группа изобретений обеспечивает получение полностью безметионинового интерферона альфа-2b с высоким выходом. 3 н.п. ф-лы, 4 ил., 4 пр.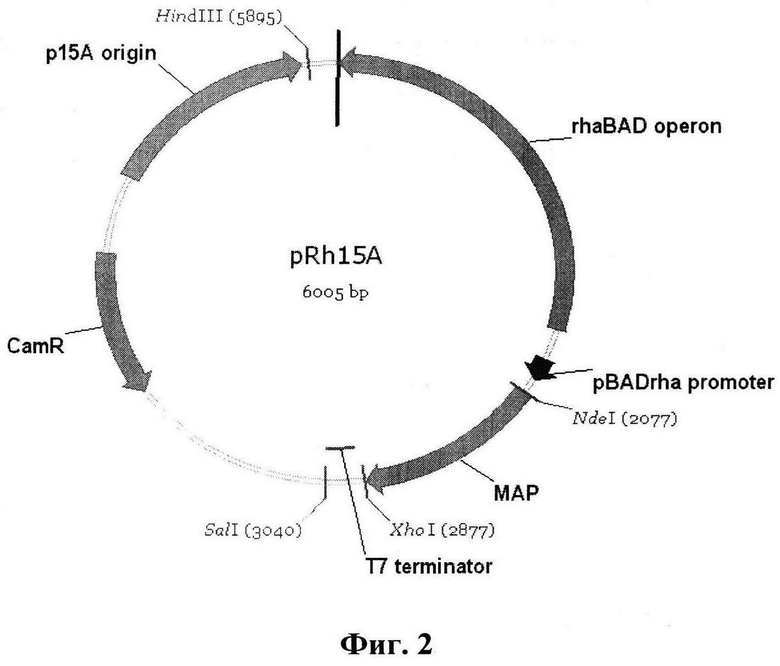
1. Плазмидный экспрессионный вектор pRh15A для получения безметионинового интерферона альфа-2b человека, содержащий:
- ген метионинаминопептидазы Escherichia coli,
- рамнозный промотор (pBADrha),
- rhaBAD оперон,
- терминатор транскрипции генов фага Т7,
- ориджин репликации плазмид р15А,
- ген хлорамфеникол ацетилтрансферазы (CamR) и
- рестрикционные сайты, необходимые для объединения отдельных фрагментов в векторе, карта которого приведена на Фиг. 2.
2. Штамм - продуцент безметионинового интерферона альфа-2b человека, представляющий собой штамм Escherichia coli BL21Star[DE3], содержащий плазмидный экспрессионный вектор pAYC-ET-(hIFN-a2b)-lacI и вектор по п.1.
3. Способ получения безметионинового интерферона альфа-2b человека, включающий в себя культивирование штамма Escherichia coli - продуцента безметионинового интерферона альфа-2b человека, отделение интерферонсодержащей биомассы с последующей хроматографической очисткой целевого продукта, отличающийся тем, что культивируют штамм по п.2, а хроматографическую очистку проводят в несколько этапов:
- на фенил-сефарозе при рН 5,2 с использованием в качестве растворителя 0,1 М трис-ацетатного буферного раствора, содержащего 0,26 NaCl, 1 мМ этилен-диаминотетрауксусной кислоты (ЭДТА) и 0,1% метионина,
- затем хроматографию проводят последовательно на сорбентах Солоза КГ 20/30, Q-сефароза и SP-целлюлоза, при этом интерферон альфа-2b человека сорбируется в 0,25 М трис-ацетатном буферном растворе, содержащем 1 мМ ЭДТА и 0,1% метионина,
- на Солозе КГ 20/30 при рН 5,2, элюцию осуществляют 0,05 М трис-буферным раствором с 1 мМ ЭДТА и 0,1% метионина,
- на Q-сефарозе при рН 6,5, а элюцию проводят 0,25 М трис-ацетатным буферным раствором, содержащим 1 мМ ЭДТА и 0,1% метионина при рН 5,9,
- на SP-целлюлозе при рН 5,2, отмывку проводят 0,05 М натрий-ацетатным буферным раствором, а элюцию проводят 0,05 М натрий-ацетатным буферным раствором с добавлением 0,15 М натрия хлорида.
| Рекомбинантная плазмида pFM-IFN-17, обеспечивающая экспрессию интерферона альфа-2b человека, рекомбинантная плазмида pFM-АР, обеспечивающая экспрессию фермента метионинаминопептидазы E. coli, биплазмидный штамм Escherichia coli FM-IFN-АР (pFM-IFN-17, pFM-АР) - продуцент (Met-) рекомбинантного интерферона альфа-2b человека | 2016 |
|
RU2610173C1 |
| CN 102367441 A, 07.03.2012 | |||
| СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО ИНТЕРФЕРОНА АЛЬФА-2b И ИНТЕРФЕРОНСОДЕРЖАЩИЙ ПРЕПАРАТ (ВАРИАНТЫ) | 2005 |
|
RU2294372C1 |
| ШТАММ Escherichia coli BL21 (DE3)[pAYC-ET-(hIFN-α2b)-IacI]-ПРОДУЦЕНТ РЕКОМБИНАНТНОГО ЧЕЛОВЕЧЕСКОГО АЛЬФА-2b ИНТЕРФЕРОНА И СПОСОБ ЕГО КУЛЬТИВИРОВАНИЯ | 2005 |
|
RU2303063C2 |
| Вертикальный гидравлический пресс | 1980 |
|
SU946981A1 |

