ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
Данная заявка имеет приоритет по предварительной патентной заявке США № 61/238867, поданной 1-го сентября 2009 г., и по предварительной патентной заявке США № 61/253438, поданной 20-го октября 2009 г., полное описание которых включено в настоящий документ путем ссылки.
ОБЛАСТЬ ТЕХНИКИ
Область данного изобретения в общем относится к способам очистки полипептида, содержащего СН2/СН3-область, включающим связывание полипептида с белком А и элюирование градиентом рН.
ПРЕДПОСЫЛКИ СОЗДАНИЯ ИЗОБРЕТЕНИЯ
Крупномасштабная экономичная очистка белков представляет собой растущую проблему в биотехнологической промышленности. Обычно, белки продуцируют в культурах клеток с использованием либо линий клеток млекопитающих, либо бактериальных клеточных линий, модифицированных методами генной инженерии для продукции целевого белка в результате введения рекомбинатной плазмиды, содержащей ген этого белка. Поскольку используемые клеточные линии представляют собой живые организмы, они должны получать питание из комплексной ростовой среды, содержащей сахара, аминокислоты и ростовые факторы, обычно с добавленными препаратами животной сыворотки. Выделение целевого белка из смеси соединений, используемых для питания клеток, и из побочных продуктов самих клеток до чистоты, достаточной для его использования в качестве терапевтического средства для применения у человека, представляет огромную проблему.
Методики очистки белков из клеточного дебриса исходно зависят от места экспрессии белка. Для некоторых белков можно вызывать их секрецию напрямую из клетки в окружающую ростовую среду; другие синтезируются внутри клетки. В последнем случае, первая стадия способа очистки включает лизис клетки, который можно осуществить рядом способов, включая механическое воздействие, осмотический шок или обработку ферментами. Такое разрушение высвобождает все содержимое клетки в гомогенат и, кроме того, генерирует субклеточные фрагменты, которые сложно удалить вследствие их малого размера. Их обычно удаляют дифференциальным центрифугированием или с помощью фильтрации. Такая же проблема возникает, хотя и в меньшей степени, с секретируемыми напрямую белками вследствие естественной гибели клеток и высвобождения внутриклеточных белков клеток-хозяев в течение процесса продукции белка.
После получения осветленного раствора, содержащего целевой белок, его выделение из других белков, продуцируемых клеткой, обычно пытаются осуществить, используя комбинацию разных хроматографических методик. Для некоторых белков (например, белков для использования в качестве терапевтических средств для применения у людей) обычно используют аффинную хроматографию, в которой задействовано специфичное взаимодействие между очищаемым белком и иммобилизованным захватывающим агентом. Белок А представляет собой адсорбент, пригодный для аффинной хроматографии белков, таких как антитела, которые содержат Fc-область. Белок А представляет собой белок (41 кДа) клеточной стенки Staphylococcus aureas, который связывается с высокой аффинностью (примерно, 10-8 M для IgG человека) с Fc-областью антител. Однако вследствие того, что белки имеют тенденцию к агрегации или некорректному фолдингу, целевой белок (то есть мономер) часто элюируется с таких аффинных колонок вместе с другими примесями, такими как белковые агрегаты, побочные продукты жизнедеятельности самих клеток (то есть, примеси из клеток-хозяев) или загрязнение вирусного фильтра.
Для дополнительного разделения этих примесей и смесей белков на основе их заряда, степени гидрофобности или размера, были разработаны другие методики, такие как ионообменная хроматография, гидрофобная хроматография или эксклюзионная хроматография. Для каждой из этих методик доступны несколько разных хроматографических смол или сорбентов, что позволяет точно подобрать схему очистки для конкретного белка. Основой каждого из этих способов выделения является то, что белки можно заставить либо двигаться с различной скоростью по длинной твердой фазе (например, колонке), достигая физического разделения, которое увеличивается по мере дальнейшего продвижения белков по твердой фазе, либо селективно связываться с разделяющей средой с последующим дифференциальным элюированием разными растворителями. Однако каждый из этих способов требует дополнительных буферов, смол или сорбентов, а также других ресурсов для дополнительной очистки, что, в свою очередь, приводит к более длительному времени обработки и более высокой стоимости. Поэтому, необходимы более эффективные и экономичные способы очистки белковых мономеров.
Способы очистки полипептидов из агрегатов, мультимеров и модифицированных белков с использованием колонки с белком А и элюирования градиентом рН были описаны в патентной заявке США № 12/008,160.
Все публикации, патенты и патентные заявки, цитируемые в настоящем документе, таким образом полностью включены в него путем ссылки для всех целей в такой же степени, как если бы каждая индивидуальная публикация, каждый индивидуальный патент и каждая индивидуальная патентная заявка были бы специально и индивидуально указаны включенными путем ссылки.
КРАТКОЕ ОПИСАНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к способам очистки полипептида, содержащего СН2/СН3-область, в результате связывания полипептида с белком А и элюирования с градиентом рН, начинающимся с низкого рН. Эти способы очистки обеспечивают преимущество достижения лучшего последовательного отделения полипептидов или не-агрегатов от различных примесей, включая примеси из клеток-хозяев, загрязнение вирусного фильтра, вирусные или вирусоподобные частицы, основные варианты полипептида и агрегаты полипептидного мономера, а также обеспечивают более высокую чистоту целевых полипептидных мономеров в очищенной фракции/пуле. Данные способы можно осуществлять с использованием различных хроматографических смол и хроматографических сорбентов с белком А. Эти способы также можно использовать в промышленном или коммерческом процессе, они могут способствовать использованию альтернативных последующих методик очистки помимо колоночной хроматографии.
В одном аспекте изобретение относится к способу очистки полипептида, содержащего СН2/СН3-область, включающему связывание с белком А и элюирование градиентом рН, начинающимся с 5,0 или менее.
В другом аспекте изобретение относится к способу очистки полипептида, содержащего СН2/СН3-область, включающему стадии: (а) связывания полипептида с белком А; и (b) элюирования полипептида градиентом рН, начинающимся с 5,0 или менее, с использованием элюирующего буфера, причем элюирующий буфер включает буфер с высоким рН и буфер с низким рН, а градиент рН образуется в результате регулирования процентного содержания каждого рН-буфера в элюирующем буфере.
В некоторых вариантах осуществления изобретения градиент рН начинается примерно с рН 4,2. В других вариантах осуществления изобретения градиент рН начинается примерно с рН 4,3. В некоторых вариантах осуществления изобретения градиент рН начинается примерно с рН 4,6. В некоторых вариантах осуществления изобретения градиент рН заканчивается на рН 3,0 или более высоком значении. В некоторых вариантах осуществления изобретения градиент рН заканчивается примерно на рН 3,7.
В некоторых вариантах осуществления изобретения буфер с высоким рН имеет значение примерно рН 5,0, причем буфер с низким рН имеет рН примерно 2,7.
В некоторых вариантах осуществления изобретения процентное содержание буфера с низким рН начинается примерно с 35%. В некоторых вариантах осуществления изобретения элюирующий буфер, содержащий примерно 35% буфера с низким рН, включает примерно 16,25 мМ ацетат и примерно 8,25 мМ формиат. В других вариантах осуществления изобретения процентное содержание буфера с низким рН начинается примерно с 25%. В некоторых вариантах осуществления изобретения элюирующий буфер, содержащий примерно 25% буфера с низким рН, включает примерно 18,75 мМ ацетат и примерно 6,25 мМ формиат. В некоторых вариантах осуществления изобретения процентное содержание буфера с низким рН начинается примерно с 40%. В некоторых вариантах осуществления изобретения элюирующий буфер, содержащий примерно 40% буфера с низким рН, включает примерно 15 мМ ацетат и примерно 10 мМ формиат.
В некоторых вариантах осуществления изобретения полипептид наносят с плотностью нагрузки, начинающейся примерно с 14 г/л. В некоторых вариантах осуществления изобретения полипептид наносят с плотностью нагрузки в диапазоне от примерно 14 г/л до примерно 45 г/л.
В некоторых вариантах осуществления изобретения белок А представляет собой смолу с белком А для колоночной хроматографии или хроматографический сорбент с белком А. В некоторых вариантах осуществления хроматографический сорбент с белком А представляет собой мембрану или монолит.
В некоторых вариантах осуществления изобретения белок А представляет собой смолу с белком А для колоночной хроматографии, причем скорость потока для элюирования полипептида варьирует от примерно 5 объемов колонки в час до примерно 25 объемов колонки в час.
В некоторых вариантах осуществления изобретения белок А представляет собой смолу с белком А для колоночной хроматографии, причем очищенная фракция полипептида содержит около или меньше примерно 12 объемов колонки с белком А.
В некоторых вариантах осуществления изобретения примеси из клеток-хозяев отделены от полипептида. В некоторых вариантах осуществления изобретения примеси из клеток-хозяев представляют собой белки клеток яичника китайского хомячка (СНОР).
В некоторых вариантах осуществления изобретения агрегат отделен от полипептида. В других вариантах осуществления изобретения загрязнение вирусного фильтра отделено от полипептида.
В некоторых вариантах осуществления изобретения вирусные частицы или вирусоподобные частицы отделены от полипептида. В некоторых вариантах осуществления изобретения основный вариант полипептида отделен от полипептида.
В некоторых вариантах осуществления изобретения СН2/СН3-область содержит Fc-область иммуноглобулина.
В некоторых вариантах осуществления изобретения полипептид представляет собой антитело. В некоторых вариантах осуществления изобретения антитело является моноклональным антителом, поликлональным антителом, полиспецифичным антителом или фрагментом антитела.
В других вариантах осуществления изобретения полипептид является иммуноадгезином.
В некоторых вариантах осуществления изобретения полипептид имеет чистоту по меньшей мере примерно 98% мономера. В других вариантах осуществления изобретения полипептид имеет чистоту по меньшей мере примерно 99% мономера.
В некоторых вариантах осуществления настоящего изобретения соотношение примесей из клеток-хозяев и очищенного полипептида по меньшей мере примерно на 75% ниже, примерно на 80% ниже, примерно на 85% ниже, примерно на 90% ниже, примерно на 95% ниже, примерно на 96% ниже, примерно на 97% ниже, примерно на 98% ниже или примерно на 99% ниже соотношения в неочищенном полипептиде.
В некоторых вариантах осуществления изобретения соотношение примесей из клеток-хозяев и очищенного полипептида по меньшей мере примерно на 20% ниже соотношения в полипептиде, очищенном способом с использованием ступенчатого элюирования, причем способ с использованием ступенчатого элюирования включает связывание полипептида с белком А и элюирование рН, начиная с рН 3,6 или менее. В некоторых вариантах осуществления изобретения соотношение примесей из клеток-хозяев и очищенного полипептида по меньшей мере примерно на 60% ниже соотношения в полипептиде, очищенном способом с использованием ступенчатого элюирования, причем способ с использованием ступенчатого элюирования включает связывание полипептида с белком А и элюирование рН, начиная с рН 3,6 или менее.
В некоторых вариантах осуществления изобретения очищенный полипептид имеет содержание вирусных или вирусоподобных частиц меньше примерно 15000 частиц/мл. В некоторых вариантах осуществления изобретения очищенный полипептид имеет содержание вирусных или вирусоподобных частиц меньше примерно 12500 частиц/мл, меньше примерно 10000 частиц/мл, меньше примерно 7500 частиц/мл, меньше примерно 5000 частиц/мл, меньше примерно 2500 частиц/мл, меньше примерно 1500 частиц/мл, меньше примерно 1000 частиц/мл, меньше примерно 750 частиц/мл, меньше примерно 500 частиц/мл, меньше примерно 250 частиц/мл, меньше примерно 100 частиц/мл или меньше примерно 50 частиц/мл. В некоторых вариантах осуществления изобретения вирусоподобная частица является ретровирусоподобной частицей.
В некоторых вариантах осуществления изобретения очищенный полипептид имеет степень очистки от вирусных или вирусоподобных частиц по меньшей мере примерно 4 LRV (число порядков снижения количества вируса). В некоторых вариантах осуществления изобретения очищенный полипептид имеет степень очистки от вирусных или вирусоподобных частиц в диапазоне от примерно 4 LRV до примерно 8 LRV. В некоторых вариантах осуществления изобретения очищенный полипептид имеет степень очистки от вирусных или вирусоподобных частиц в диапазоне от примерно 4 LRV до примерно 7 LRV. В некоторых вариантах осуществления изобретения очищенный полипептид имеет степень очистки от вирусных или вирусоподобных частиц примерно 5 LRV, примерно 6 LRV, примерно 7 LRV или примерно 8 LRV. В некоторых вариантах осуществления изобретения вирусоподобная частица является ретровирусоподобной частицей.
В некоторых вариантах осуществления изобретения очищенный полипептид является мономером полипептида.
В некоторых вариантах осуществления изобретения белок А представляет собой модифицированный или немодифицированный белок А-лиганд.
В некоторых вариантах осуществления изобретения очистка является промышленным способом.
В некоторых вариантах осуществления любого из аспектов изобретения способ очистки дополнительно включает при очистке полипептида проведение стадии фильтрации вирусов или стадии ионообменной хроматографии. В некоторых вариантах осуществления изобретения стадию ионообменной хроматографии проводят после стадии очистки.
В некоторых вариантах осуществления любого из аспектов изобретения способ очистки не включает дополнительную стадию удаления агрегатов.
В некоторых вариантах осуществления любого из аспектов изобретения способ очистки не включает дополнительную стадию удаления загрязнения вирусного фильтра.
В некоторых вариантах осуществления любого из аспектов изобретения способ очистки не включает дополнительную стадию удаления основного варианта полипептида. В некоторых вариантах осуществления любого из аспектов изобретения способ очистки не включает дополнительную стадию удаления кислотного варианта полипептида.
В другом аспекте изобретение относится к полипептидному продукту, очищенному описанными в настоящем документе способами.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
На фиг.1 показана хроматограмма ступенчатого градиента рН: по оси х отложен объем в мл от начала элюирования с белка А, а по оси y отложено поглощение (mAU). Также показана отличительная форма кривой УФ-поглощения при 280 нм при элюировании ступенчатым градиентом - большой пик в начале элюирования, затем снижающийся до постоянной высоты и убывающий по мере снижения рН.
На фиг.2 показаны результаты SEC (эксклюзионной хроматографии) фракций анти-VEGF антитела №1. По оси Х отложено время удерживания на SEC-колонке (мин), а на оси Y отложено нормализованное поглощение в УФ (mAU). По мере увеличения номера фракции (то есть, снижения рН по мере прохождения градиентного элюирования) также увеличиваются пики HMWS (высокомолекулярных соединений) и димеров (время удерживания около 12,5 минут и 13,5 минут, соответственно), в то время как пик мономера уменьшается (время удерживания 16 минут). Количественную оценку результатов, полученных из этих кривых, проводили с помощью интегрирования всех пиков (например, HMWS, димера и мономера), сравнивая отдельные относительные площади пиков в виде процентов (например, общую площадь устанавливали как 100%, а профиль интегрирования пиков SEC-кривой (далее профиль SEC-интегрирования) выражали в процентах, как, например, «31% HMWS, 36% димера и 33% мономера»).
На фиг.3 показан график результатов SEC-интегрирования для анти-VEGF антитела №1. На этом графике показано, что для первых девяти элюированных фракций уровень мономера высокий, причем в четырех фракциях он составляет 100%, а уровень димера и HMWS достигает пика позднее в процессе элюирования. Данные результаты демонстрируют, что ступенчатый градиент рН отделяет агрегаты от мономера анти-VEGF антитела №1.
На фиг.4А показан профиль SEC-интегрирования для ряда белковых молекул (анти-CD20 антитела, анти-VEGF антитела №2, анти-MUC16 антитела и анти-CD4 антитела). С помощью ступенчатого градиента рН мономеры были успешно отделены от агрегатов для анти-CD20 антитела, анти-VEGF антитела №2, анти-MUC16 антитела и анти-CD4 антитела.
На фиг.4В показан профиль SEC-интегрирования для негликозилированного анти-Met антитела с одним антиген-связывающим участком, продуцируемого ферментацией бактериальных клеток-хозяев (E. coli). С помощью ступенчатого градиента рН мономеры были успешно отделены от агрегатов для негликозилированного анти-Met антитела с одним антиген-связывающим участком.
На фиг.5 показано параллельное сравнение между стандартным ступенчатым элюированием (контроль; элюирование белков при рН 3,6 или менее без градиента рН) и элюированием ступенчатым градиентом рН анти-CD20 антитела. График элюирования анти-CD20 антитела и СНОР на левой панели показывает уровень СНОР на фракцию в ppm (частей на миллион; показатель, используемый для стандартизации измерения примесей на количество продукта). График элюирования анти-CD20 антитела и агрегатов на правой панели показывает значения интегрирования пиков SEC-кривых на фракцию в течение градиентного элюирования. Вертикальные линии на левой и правой панелях представляют собой символическое объединение содержащихся фракций, в результате которого был бы получен пул элюирования градиентом рН с характеристиками, показанными в таблице внизу слайда.
На фиг.6 показано отделение СНОР от анти-VEGF антитела №1. Уровень СНОР на фракцию выражен в ppm или нг/мл.
На фиг.7 показано отделение СНОР от анти-MUC16 антитела. Уровень СНОР на фракцию выражен в ppm или нг/мл.
Фиг.8 представляет собой наложение хроматограмм на смолах с белком А: MABSELECT™, MABSELECT SURE™, PROSEP® Va, PROSEP® Ultra Plus и POROS® MABCAPTURE™ A.
Фиг.9 представляет собой график Парето, показывающий, что стартовый % В (стартовое значение рН и наклон градиента элюирования) является наиболее важным параметром в определении эффективности отделения агрегатов, за которым следуют плотность нагрузки и время удерживания.
Фиг.10 представляет собой профиль взаимодействия для параметров стартового % В, плотности нагрузки и стартового % В, общей продолжительности элюирования и времени удерживания.
Фиг.11 представляет собой пример элюирования ступенчатым градиентом рН в производственном масштабе.
На фиг.12 показаны результаты исследования по применению ионообменных мембран после элюирования с белка А, в котором использовали два размера катионообменной мембраны и три размера анионообменной мембраны. Мембраны Coin и Nano, обе представляют собой производственный тип в отношении числа слоев мембраны, в то время как мембраны ACRODISC® имеют меньшее число слоев, но являются типичными используемыми лабораторными моделями.
На фиг.13 показан результат сравнения SEC-интегрирования между очисткой на колонке объемом 4,1 л (пилотный производственный масштаб) и колонке 28 мл (лабораторный масштаб).
Фиг.14 представляет собой график падения пропускной способности фильтра VIRESOLVE® Pro для анти-VEGF антитела №1, на котором проведено сравнение элюирования с белка А ступенчатым градиентом рН и стандартного ступенчатого элюирования с белка А в отношении увеличения пропускания большей массы через парвовирусный фильтр VIRESOLVE® Pro. При использовании ступенчатого градиента рН для элюирования с белка наблюдалось 6-кратное увеличение пропускаемой массы.
Фиг.15 представляет собой хроматограмму элюирования полным градиентом рН, показывающую фактические данные, полученные хроматографическим программным обеспечением AKTA UNICORN™ для полного градиента рН при плотности нагрузки 21 г/л. Отсутствие исходного высокого пика при 250 нм в начале градиента указывает на то, что рН в начале элюирования выше необходимого для элюирования продуктов с колонки с белком А.
Фиг.16 представляет собой хроматограмму (полученную с использованием оборудования АКТА) с вариантом градиента рН 5,0-2,7 (0-100%B), указывающую на то, что анти-VEGF антитело №1 элюируется в виде дискретного пика в диапазоне рН 4,6-3,6.
Фиг.17 представляет собой интегрирование пиков, полученных ионообменным анализом вариантов для фракций после элюирования ступенчатым градиентом рН с белка А, которое указывает на отделение основного варианта полипептида в хвостовой части элюирования ступенчатым градиентом.
Фиг.18 представляет собой количество частиц RVLP (ретровирусоподобных частиц) на фракцию, определенное с помощью QPCR-анализа (анализа с помощью количественной полимеразной цепной реакции), выраженное относительно элюирования анти-VEGF антитела №1. Основная масса RVLP элюируется в конце градиента, когда элюируется мало продукта.
Фиг.19 представляет собой LRV (число порядков снижения количества вируса) для каждой фракции при элюировании анти-VEGF антитела №1 с белка А с помощью ступенчатого градиента рН.
На фиг.20 приведены совокупные величины LRV для символических пулов фракций, показывающие, что можно получить более высокое значение LRV в пуле после элюирования с белка А, если исключить более поздние фракции.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к способам очистки полипептида, содержащего СН2/СН3-область, путем связывания полипептида с белком А и элюирования градиентом рН, начинающимся с низкого рН. Авторы изобретения сделали важное открытие, что элюирование полипептидов, содержащих СН2/СН3-область, с белка А градиентом рН при низком рН может обеспечить лучшее последовательное отделение полипептидов от различных примесей, включая примеси из клеток-хозяев, загрязнение вирусного фильтра, вирусные или вирусоподобные частицы, основные варианты полипептида и/или агрегаты полипептида, в очищенной фракции/пуле, а также может обеспечить более высокую чистоту или процентное содержание целевых полипептидных мономеров в очищенной фракции/пуле. Поэтому, изобретение имеет значительные преимущества. Авторы изобретения также открыли, что эти способы можно осуществить, используя различные хроматографические смолы и хроматографические сорбенты с белком А, и что эти способы можно использовать в промышленных или коммерческих процессах, и они могут способствовать применению альтернативных последующих методик очистки помимо колоночной хроматографии (например, мембранных адсорбентов).
Соответственно, один аспект изобретения относится к способу очистки полипептида, содержащего СН2/СН3-область, включающему связывание полипептида с белком А и элюирование градиентом рН, начинающимся с 5,0 или более низкого значения.
В другом аспекте изобретение относится к способу очистки полипептида, содержащего СН2/СН3-область, включающему стадии: (а) связывания полипептида с белком А; и (b) элюирования полипептида градиентом рН, начинающимся с 5,0 или меньшего значения, с использованием элюирующего буфера, причем элюирующий буфер включает буфер с высоким рН и буфер с низким рН, а градиент рН образуется в результате регулирования процентного содержания каждого рН-буфера в элюирующем буфере.
В еще одном аспекте изобретения относится к полипептиду, очищенному способами, описанными в настоящем документе.
В осуществлении настоящего изобретения на практике будут использоваться, если не указано иное, общепринятые методики молекулярной биологии (включая рекомбинатные методики), микробиологии, клеточной биологии, биохимии и иммунологии, которые входят в компетенцию специалиста в данной области. Такие методики полностью описаны в литературе, такой как, Molecular Cloning: A Laboratory Manual, second edition (Sambrook et al., 1989) Cold Spring Harbor Press; Oligonucleotide Synthesis (M.J. Gait, ed., 1984); Methods in Molecular Biology, Humana Press; Cell Biology: A Laboratory Notebook (I.E. Cellis, ed., 1998) Academic Press; Animal Cell Culture (R.I. Freshney, ed., 1987); Introduction to Cell and Tissue Culture (J.P. Mather and P.E. Roberts, 1998) Plenum Press; Cell and Tissue Culture: Laboratory Procedures (A. Doyle, J.B. Griffiths, and D.G. Newell, eds., 1993-1998) J. Wiley and Sons; Methods in Enzymology (Academic Press, Inc.); Handbook of Experimental Immunology (D.M. Weir and C.C. Blackwell, eds.); Gene Transfer Vectors for Mammalian Cells (J.M. Miller and M.P. Calos, eds., 1987); Current Protocols in Molecular Biology (F.M. Ausubel et al., eds., 1987); PCR: The Polymerase Chain Reaction, (Mullis et al., eds., 1994); Current Protocols in Immunology (I.E. Coligan et al., eds., 1991); Short Protocols in Molecular Biology (Wiley and Sons, 1999); Immunobiology (C.A. Janeway and P. Travers, 1997); Antibodies (P. Finch, 1997); Antibodies: a practical approach (D. Catty., ed., IRL Press, 1988-1989); Monoclonal antibodies: a practical approach (P. Shepherd and C. Dean, eds., Oxford University Press, 2000); Using antibodies: a laboratory manual (E. Harlow and D. Lane (Cold Spring Harbor Laboratory Press, 1999); The Antibodies (M. Zanetti and J.D. Capra, eds., Harwood Academic Publishers, 1995).
Определения
Следует понимать, что целевой полипептид в настоящем документе представляет собой полипептид, содержащий СН2/СН3-область, и, поэтому, подходит для очистки с помощью белка А. Термин «СН2/СН3-область» при использовании в настоящем документе относится к таким аминокислотным остаткам в Fc-области молекулы иммуноглобулина, которые взаимодействуют с белком А. В некоторых вариантах осуществления изобретения СН2/СН3-область содержит интактную СН2-область, за которой следует интактная СН3-область, и наиболее предпочтительно содержит Fc-область иммуноглобулина. Примеры белков, содержащих СН2/СН3-область, включают антитела, иммуноадгезины и слитые белки, содержащие целевой белок, слитый или конъюгированный с СН2/СН3-областью.
Термины «полипептид» и «белок» используются в настоящем документе взаимозаменяемо для обозначения полимеров аминокислот любой длины. Полимер может быть линейным или разветвленным, он может содержать модифицированные аминокислоты, и может быть прерван неаминокислотами. Термины также охватывают аминокислотный полимер, модифицированный естественным образом или в результате вмешательства человека; например, модифицированный в результате образования дисульфидных связей, гликозилирования, модификации липидами, ацетилирования, фосфорилирования или любой другой манипуляции или модификации, такой как конъюгация с меткой. Также в определение включены, например, полипептиды, содержащие один или несколько аналогов аминокислот (включая, например, неприродные аминокислоты и т.п.), а также другие модификации, известные в данной области.
Используемый в настоящем документе термин «очищенный полипептид» или «очищенный белок» представляют собой продукт, элюированный после аффинной хроматографии на белке А с использованием способов элюирования рН-градиентом, описанных в настоящем документе. Очищенные полипептиды/белки, предпочтительно содержат мономеры полипептида.
Используемый в настоящем документе термин «неочищенный полипептид», «неочищенный белок» или «белковая нагрузка» представляет собой полипептид или белок в нагружаемом материале или исходном материале до стадии очистки аффинной хроматографией на белке А.
Используемый в настоящем документе термин «примесь» или «примеси» представляет собой материал, отличающийся от целевого полипептидного мономерного продукта. Примеси включают, но не ограничены этим, вариант полипептида (например, кислотный или основный варианты полипептида), фрагмент полипептида, агрегат или производное целевого полипептидного мономера, другой полипептид, липид, нуклеиновую кислоту, эндотоксин, примесь из клеток-хозяев или загрязнение вирусного фильтра.
Используемый в настоящем документе термин «мономер» («мономеры») относится к одной единице полипептида, содержащей СН2/СН3-область. Например, в случае антитела мономер состоит из двух тяжелых цепей и двух легких цепей; в случае антитела с одним антиген-связывающим участком мономер состоит из одной тяжелой цепи и одной легкой цепи.
Используемый в настоящем документе термин «основный вариант полипептида» или «основный вариант» относится к варианту целевого полипептида, который имеет большую основность (например, определенную с помощью катионообменной хроматографии), чем целевой полипептид.
Используемый в настоящем документе термин «кислотный вариант полипептида» или «кислотный вариант» относится к варианту целевого полипептида, который имеет большую кислотность (например, определенную с помощью катионообменной хроматографии), чем целевой полипептид.
Используемый в настоящем документе термин «агрегат» («агрегаты») относится к любым мультимерам полипептида или фрагмента полипептида, содержащим СН2/СН3-область. Например, агрегат может быть димером, тримером, тетрамером или мультимером выше тетрамера, и т.п.
Используемый в настоящем документе термин «примесь из клеток-хозяев» относится к любому белковому загрязнению или побочному продукту, вносимому линией клеток-хозяев, клеточной культуральной жидкостью или клеточной культурой. Примеры включают, но не ограничены этим, белки клеток яичника китайского хомячка (СНОР), белки E. coli, дрожжевые белки, белки клеток COS обезьяны или белки клеток миеломы (например, белки клеток NS0 (клетки плазмацитомы мышей, полученные из мыши линии BALB/c)).
Используемый в настоящем документе термин «загрязнение вирусного фильтра» относится к любой макромолекулярной частице или высокомолекулярным соединениям (HMWS) с гидродинамическим диаметром, аналогичным или выше размера пор парвовирусного фильтра. Загрязнение вирусного фильтра включает, но не ограничено этим, растворимые высокомолекулярные полипептидные агрегаты, и растворимые и/или нерастворимые агрегаты примесей из клеток-хозяев (например, СНОР).
«Клетка-хозяин» включает индивидуальную клетку или клеточную культуру, которая может быть или является реципиентом вектора (вектором) для включения полинуклеотидных вставок для продукции полипептидов. Клетка-хозяин включает потомство одной клетки-хозяина, и потомство может не быть полностью идентично (по морфологии или совпадению геномной ДНК) исходной родительской клетке вследствие природной, случайной или преднамеренной мутации.
«Твердая фаза», используемая в настоящем изобретении, относится к неводному матриксу, к которому может прикрепляться белок А.
«Буфер» представляет собой буферный раствор, который препятствует изменению рН путем действия своих кислотно-основных сопряженных компонентов. Различные буферы, которые можно использовать в зависимости, например, от желаемого рН буфера, описаны в Buffers. A Guide for the Preparation and Use of Buffers in Biological Systems, Gueffroy, D., Ed. Calbiochem Corporation (1975).
«Уравновешивающий буфер» в настоящем документе представляет собой буфер, используемый для подготовки твердой фазы (с иммобилизованным белком А) для нанесения целевого белка.
«Промывочный буфер» в настоящем документе используется для обозначения буфера, который пропускают через твердую фазу (с иммобилизованным белком А) после нанесения и до элюирования целевого белка.
Термин «антитело» используется в максимально широком смысле и определенно охватывает моноклональные антитела (включая полноразмерные моноклональные антитела), поликлональные антитела, полиспецифичные антитела (например, биспецифичные антитела) и фрагменты антител, при условии что они сохраняют СН2/СН3-область или модифицированы, чтобы содержать СН2/СН3-область, описанную в настоящем документе.
«Фрагменты антител» содержат часть полноразмерного антитела, обычно его антиген-связывающую или вариабельную область. Примеры фрагментов антител включают Fab-, Fab'-, F(ab')2- и Fv-фрагменты; молекулы одноцепочечных антител; диантитела; линейные антитела и полиспецифичные антитела, образуемые фрагментами антител. Используемый в настоящем изобретении фрагмент антитела содержит СН2/СН3-область.
Используемый в настоящем документе термин «моноклональное антитело» относится к антителу, полученному из популяции по существу гомогенных антител, то есть индивидуальные антитела, содержащиеся в популяции, являются идентичными за исключением того, что возможные природные мутации могут присутствовать в минорных количествах. Моноклональные антитела являются высоко специфичными, будучи направленными против единственного антигенного сайта. Кроме того, в отличие от стандартных препаратов (поликлональных) антител, которые обычно включают различные антитела, направленные против разных детерминант (эпитопов), каждое моноклональное антитело направлено против единственной детерминанты на антигене. Определитель «моноклональное» указывает на характер антитела, как полученного из по существу гомогенной популяции антител, и его не следует истолковывать так требование получения антитела каким-либо конкретным способом. Например, моноклональные антитела для использования по настоящему изобретению можно изготовить гибридомным способом, впервые описанным Kohleret al, Nature 256:495 (1975), или его можно изготовить методами рекомбинантных ДНК (см., например, патент США № 4816567). «Моноклональные антитела» также можно выделить из фаговых библиотек антител, используя методики, описанные, например, в Clackson et al., Nature 352:624-628 (1991) и Marks et al., J. Mol. Biol. 222:581-597 (1991).
Моноклональные антитела в настоящем документе специально включают «химерные» антитела (иммуноглобулины), в которых часть тяжелой и/или легкой цепи идентична или гомологична соответствующим последовательностям в антителах, полученных из определенных биологических видов или принадлежащих к определенному классу или подклассу антител, в то время как остаток цепи (цепей) идентичен или гомологичен соответствующим последовательностям в антителах, полученных из других биологических видов или принадлежащих другому классу или подклассу антител, а также включают фрагменты таких антител, при условии что они проявляют желаемую биологическую активность (патент США № 4816567 и Morrison et al., Proc. Natl. Acad. Sci. USA 81:6851-6855 (1984)).
Термин «гипервариабельная область» при использовании в настоящем документе относится к аминокислотным остаткам антитела, которые отвечают за связывание антигена. Гипервариабельная область включает аминокислотные остатки из «определяющей комплементарность области» или «CDR» (то есть остатки 24-34 (L1), 50-56 (L2) и 89-97 (L3) в вариабельном домене легкой цепи и остатки 31-35 (H1), 50-65 (H2) и 95-102 (H3) в вариабельном домене тяжелой цепи; Kabat et al., Sequences of Proteins of Immunological Interest, 5 th Ed. Public Health Service, National Institutes of Health, Bethesda, Md. (1991)) и/или остатки из «гипервариабельной петли» (то есть остатки 26-32 (L1), 50-52 (L2) и 91-96 (L3) в вариабельном домене легкой цепи и остатки 26-32 (H1), 53-55 (H2) и 96-101 (H3) в вариабельном домене тяжелой цепи; Chothia and Lesk J. Mol. Biol. 196:901-917 (1987)). «Каркасные» или «FR»-остатки являются аминокислотными остатками вариабельного домена помимо остатков гипервариабельной области, указанных в настоящем документе.
«Гуманизированными» формами антител других биологических видов кроме человека (например, мышиных) являются химерные антитела, которые содержат минимальную последовательность, полученную из иммуноглобулина другого биологического вида кроме человека. Преимущественно гуманизированные антитела являются иммуноглобулинами человека (реципиентным антителом), в которых остатки гипервариабельной области реципиента замещены остатками гипервариабельной области антител других биологических видов кроме человека (донорного антитела), таких как мышь, крыса, кролик или нечеловекообразный примат, имеющими желаемую специфичность, аффинность и связывающую способность. В некоторых случаях, остатки каркасной области (FR) Fv-фрагмента иммуноглобулина человека замещены соответствующими остатками иммуноглобулина другого биологического вида кроме человека. Кроме того, гуманизированные антитела могут содержать остатки, которые не найдены в реципиентном антителе или в донорном антителе. Эти модификации проводят для дополнительного улучшения функционирования антител. В общем, гуманизированное антитело будет содержать по существу все из по меньшей мере одного, и обычно двух, вариабельных доменов, в которых все или по существу все гипервариабельные петли соответствуют петлям иммуноглобулина другого биологического вида кроме человека, и все или по существу все из FR-областей являются FR-областями из последовательности иммуноглобулина человека. Гуманизированное антитело необязательно также будет содержать по меньшей мере часть константной области иммуноглобулина (Fc), обычно, константной области иммуноглобулина человека. Дополнительные подробности см. в Jones et al., Nature 321:522-525 (1986); Riechmann et al., Nature 332:323-329 (1988); и Presta, Curr. Op. Struct. Biol. 2:593-596 (1992).
Используемый в настоящем документе термин «иммуноадгезин» обозначает антителоподобные молекулы, которые объединяют «связывающий домен» гетерологичного «адгезинового» белка (например, рецептора, лиганда или фермента) с эффекторными функциями константного домена иммуноглобулина. Структурно иммуноадгезины включают аминокислотную последовательность адгезина с желаемой специфичностью связывания, которая отличается от сайта узнавания и связывания антигена (антиген-связывающего участка) антитела (то есть является «гетерологичным»), слитую с последовательностью константного домена иммуноглобулина. Предпочтительно, чтобы последовательность константного домена иммуноглобулина в иммуноадгезине была получена из γ1, γ2 или γ4-тяжелых цепей, поскольку иммуноадгезины, содержащие эти области, можно очистить с помощью хроматографии на белке А (Lindmark et al., J. Immunol. Meth. 62: 1-13 (1983)).
Используемый в настоящем документе термин «лиганд-связывающий домен» относится к любому нативному клеточному поверхностному рецептору или его любой области или любому производному, сохраняющим по меньшей мере качественное связывание лиганда соответствующего нативного рецептора. В конкретном варианте осуществления изобретения рецептор относится к клеточному поверхностному полипептиду, имеющему внеклеточный домен, гомологичный члену суперсемейства генов иммуноглобулинов. Другие рецепторы, которые не являются членами суперсемейства генов иммуноглобулинов, но тем не менее ясно предусмотрены в этом определении, являются рецепторами цитокинов, и в частности, рецепторами с тирозинкиназной активностью (тирозинкиназными рецепторами), членами суперсемейств рецепторов гематопоэтина и нейронального ростового фактора и молекулами клеточной адгезии, например, E-, L- и P-селектинами.
Термин «рецептор-связывающий домен» используется для обозначения любого нативного лиганда для рецептора, включая молекулы клеточной адгезии, или любой области или любого производного такого природного лиганда, сохраняющих по меньшей мере качественную способность связывать рецептор соответствующего нативного лиганда. Это определение в том числе определенно включает связывающие последовательности из лигандов вышеуказанных рецепторов.
«Химера антитела и иммуноадгезина» включает молекулу, в которой объединены по меньшей мере один связывающий домен антитела (указанный в настоящем документе) по меньшей мере с одним иммуноадгезином (указанным в настоящей заявке). Примерами химер антител и иммуноадгезина являются химеры CD4-IgG с двойной специфичностью, описанные в Berg et al., PNAS (USA) 88:4723-4727 (1991) и Chamow et al., J. Immunol. 153:4268 (1994).
В настоящем документе, если явно не указано иное, использование единственного числа и т. п. относится к одному или нескольким.
Определение «примерно» относительно значения или параметра в настоящем документе включает (и описывает) варианты осуществления, которые направленны на такое значение или параметр per se. Например, описание, относящееся к «примерно Х» включает описание «Х». Численные диапазоны включают числа, ограничивающие диапазон.
Следует понимать, что если варианты осуществления описаны в настоящем документе выражением «содержащие», прочие аналогичные варианты осуществления, описанные в терминах «состоящие из» и/или «состоящие по существу из», также предусмотрены.
Очистка полипептидов
Способ в настоящем документе включает очистку полипептида, содержащего СН2/СН3-область, от одной или нескольких примесей с помощью аффинной хроматографии на белке А с использованием градиента рН, начинающегося с низкого рН. В одном аспекте полипептид, содержащий СН2/СН3-область, можно очистить способом, включающим связывание полипептида с белком А и элюирование градиентом рН, начинающимся с 5,0 или менее.
В другом аспекте полипептид, содержащий СН2/СН3-область, можно также очистить способом, включающим стадии: (а) связывания полипептида с белком А; и (b) элюирования полипептида градиентом рН, начинающимся с 5,0 или менее, с использованием элюирующего буфера, причем элюирующий буфер включает буфер с высоким рН и буфер с низким рН, а градиент рН образуется в результате регулирования процентного содержания каждого рН-буфера в элюирующем буфере.
Белок А представляет собой модифицированный или немодифицированный белок А-лиганд. Используемый в настоящем изобретении «белок А-лиганд» охватывает природный белок А, белок А, полученный синтетически (например, пептидным синтезом или с помощью рекомбинантных методик), и его варианты, которые сохраняют способность связывать белки, имеющие СН2/СН3-область. Модифицированный белок А-лиганд может быть химически изменен для стабильности в растворах с высоким рН в течение коротких интервалов времени (например, MABSELECT SURE™ (GE Healthcare (Piscataway, NJ)), POROS® MABCAPTURE™ A (Applied Biosystems (Foster City, CA)). Термин «немодифицированный белок А-лиганд», используемый в настоящем документе, охватывает белок А-лиганд, аналогичный белку А, выделенному из природного источника. Немодифицированный белок А-лиганд, например, MABSELECT™, PROSEP™ Va, PROSEP™ Ultra Plus, можно приобрести от GE Healthcare (Piscataway, NJ) или Millipore (Billerica, MA).
Белок А может быть иммобилизован на твердой фазе. Твердой фазой может быть колонка для очистки, дискретная фаза или дискретные частицы, мембрана или фильтр. Примеры материалов для создания твердой фазы включают полисахариды (такие как агароза и целлюлоза) и другие механически стабильные матриксы, такие как кремний (например, стекло с контролируемым размером пор), полистиролдивинилбензол, полиакриламид, керамические частицы и производные любого из вышеуказанного.
Белок А, иммобилизованный на твердой фазе, используют для очистки полипептидов, содержащих СН2/СН3-область. В некоторых вариантах осуществления изобретения твердой фазой является колоночная смола с белком А, включающая смолу на основе стеклянных шариков, смолу на основе кремния или смолу на основе агарозы для иммобилизации белка А. Например, твердой фазой является колонка из стекла с контролируемым размером пор или колонка из кремниевой кислоты. Иногда для предупреждения неспецифического связывания с колонкой колонка покрыта реагентом, таким как глицерин. Колонка PROSEP™ A представляет собой пример колонки из стекла с контролируемым размером пор с белком А, которая покрыта глицерином. В других вариантах осуществления изобретения твердой фазой является сорбент для хроматографии на белке А для иммобилизации белка А. Сорбенты для хроматографии на белке А включают, но не ограничены этим, мембраны (например, мембрану SARTOBIND™ Protein A от компании Sartorius (Goettingen, Germany)) или монолиты (например, монолиты CIM® Protein A HLD от компании BIA Separations (Villach, Austria)).
Твердую фазу для хроматографии на белке А можно уравновесить уравновешивающим буфером, и затем на уравновешенную твердую фазу можно нанести неочищенные полипептиды, содержащие различные примеси (например, собранную клеточную культуральную жидкость). Полипептид можно нанести с буфером для нанесения. Обычно, уравновешивающий буфер для уравновешивания твердой фазы может быть одинаковым с буфером для наноса, но необязательно. По мере прохождения полипептидов через твердую фазу полипептиды и различные примеси адсорбируются на иммобилизованном белке А. Промывающие буферы можно использовать для удаления некоторых примесей, такие как примеси из клеток-хозяев, но не целевых полипептидов.
Уравновешивающий буфер предпочтительно является изотоничным и обычно имеет рН в диапазоне от примерно 6 до примерно 8. Например, уравновешивающий буфер может содержать 25 мМ Tris, 25 мМ NaCl, 5 мМ EDTA и иметь pH 7,1.
Буфер для наноса относится к буферу, который используют для наноса смеси содержащего СН2/СН3-область белка и загрязнений на твердую фазу, на которой иммобилизован белок А. Часто буферы для уравновешивания и наноса являются одинаковыми.
Промывающий буфер может служить для элюирования примесей из клеточной линии или других различных примесей. Проводимость и/или рН промывающего буфера являются такими, что хроматографией на белке А элюируются примеси, а какое-либо значительное количество целевого полипептида не элюируется.
Полипептид, связанный с белком А, можно элюировать градиентом рН, используя один элюирующий буфер или комбинацию элюирующих буферов.
«Элюирующий буфер» используют для элюирования содержащего СН2/СН3-область полипептида с иммобилизованного белка А. Используемый в настоящем изобретении элюирующий буфер содержит буфер с высоким рН и буфер с низким рН, и тем самым образует градиент рН в результате регулирования процентного содержания буфера с высоким рН и буфера с низким рН в элюирующем буфере. В некоторых вариантах осуществления изобретения элюирующий буфер имеет рН в диапазоне от примерно 3 до примерно 5. Значения рН, используемые в настоящем документе, измеряют без присутствия полипептидов. Примеры рН-буферов, которые контролируют рН в этом диапазоне, включают, но не ограничены этим, фосфатный, ацетатный, цитратный буферы, муравьиную кислоту и аммонийный буфер, а также их комбинации. Предпочтительными такими буферами являются ацетатный буфер и муравьиная кислота.
В некоторых вариантах осуществления изобретения градиент рН начинается примерно с 5,0. В некоторых вариантах осуществления изобретения градиент рН начинается со значения ниже 5,0. В некоторых вариантах осуществления изобретения градиент рН начинается со значения в диапазоне от примерно 5,0 до примерно 4,0. В некоторых вариантах осуществления изобретения градиент рН начинается с примерно 4,9, примерно 4,8, примерно 4,7, примерно 4,6, примерно 4,5, примерно 4,4, примерно 4,3, примерно 4,2, примерно 4,1 или примерно 4,0. В некоторых вариантах осуществления изобретения градиент рН начинается с примерно 4,98, примерно 4,96, примерно 4,94, примерно 4,92, примерно 4,90, примерно 4,88, примерно 4,86, примерно 4,84, примерно 4,82, примерно 4,80, примерно 4,78, примерно 4,76, примерно 4,74, примерно 4,72, примерно 4,70, примерно 4,68, примерно 4,66, примерно 4,64, примерно 4,62, примерно 4,60, примерно 4,58, примерно 4,56, примерно 4,54, примерно 4,52, примерно 4,50, примерно 4,48, примерно 4,46, примерно 4,44, примерно 4,42, примерно 4,40, примерно 4,38, примерно 4,36, примерно 4,34, примерно 4,32, примерно 4,30, примерно 4,28, примерно 4,24, примерно 4,22, примерно 4,20, примерно 4,18, примерно 4,16, примерно 4,14, примерно 4,12, примерно 4,10, примерно 4,08, примерно 4,06, примерно 4,04 или примерно 4,02.
В некоторых вариантах осуществления изобретения градиент рН заканчивается примерно на 3,0. В некоторых вариантах осуществления изобретения градиент рН заканчивается на значении выше 3,0. В некоторых вариантах осуществления изобретения градиент рН заканчивается на значении в диапазоне от примерно 3,0 до примерно 4,0. В некоторых вариантах осуществления изобретения градиент рН заканчивается на примерно 3,1, примерно 3,2, примерно 3,3, примерно 3,4, примерно 3,5, примерно 3,6, примерно 3,7, примерно 3,8 или примерно 3,9, В некоторых вариантах осуществления изобретения градиент рН заканчивается на примерно 3,12, примерно 3,14, примерно 3,16, примерно 3,18, примерно 3,20, примерно 3,22, примерно 3,24, примерно 3,26, примерно 3,28, примерно 3,30, примерно 3,32, примерно 3,34, примерно 3,36, примерно 3,38, примерно 3,40, примерно 3,42, примерно 3,44, примерно 3,46, примерно 3,48, примерно 3,50, примерно 3,52, примерно 3,54, примерно 3,56, примерно 3,58, примерно 3,60, примерно 3,61, примерно 3,62, примерно 3,63, примерно 3,64, примерно 3,65, примерно 3,66, примерно 3,67, примерно 3,68, примерно 3,69, примерно 3,70, примерно 3,71, примерно 3,72, примерно 3,73, примерно 3,74, примерно 3,75, примерно 3,76, примерно 3,77, примерно 3,78, примерно 3,79, примерно 3,80, примерно 3,82, примерно 3,84, примерно 3,86, примерно 3,88, примерно 3,9, примерно 3,92, примерно 3,94, примерно 3,96 или примерно 3,98.
В некоторых вариантах осуществления изобретения градиент рН начинается с примерно рН 4,2 и заканчивается на примерно рН 3,7. В некоторых вариантах осуществления изобретения градиент рН начинается с примерно рН 4,24 и заканчивается на примерно рН 3,69. Например, анти-VEGF антитела, анти-CD20 антитела, анти-MUC16 антитела, анти-CD4 антитела и анти-Met антитела с одним антиген-связывающим участком можно очистить с использованием градиента рН, начинающегося с примерно рН 4,24 и заканчивающего на примерно pH 3,69.
В других вариантах осуществления изобретения градиент рН начинается с примерно рН 4,3 и заканчивается на примерно рН 3,7. В некоторых вариантах осуществления изобретения градиент рН (то есть ступенчатый градиент рН) начинается с примерно рН 4,34 и заканчивается на примерно рН 3,69. Например, анти-VEGF антитела, анти-CD20 антитела, анти-MUC16 антитела, анти-CD4 антитела и анти-Met антитела с одним антиген-связывающим участком можно очистить с использованием градиента рН, начинающегося с примерно рН 4,34 и заканчивающего на примерно pH 3,69.
В некоторых вариантах осуществления изобретения градиент рН начинается с примерно рН 4,6 и заканчивается на примерно рН 3,7. В некоторых вариантах осуществления изобретения градиент рН (то есть полный градиент рН) начинается с примерно рН 4,58 и заканчивается на примерно рН 3,69. Например, анти-VEGF антитела, анти-CD20 антитела, анти-MUC16 антитела, анти-CD4 антитела и анти-Met антитела с одним антиген-связывающим участком можно очистить с использованием градиента рН, начинающегося с примерно рН 4,58 и заканчивающего на примерно pH 3,69.
Элюирующий буфер содержит буфер с высоким рН и буфер с низким рН, а градиент рН образуется в результате регулирования процентного содержания каждого рН-буфера в элюирующем буфере. В некоторых вариантах осуществления изобретения буфер с высоким рН имеет рН примерно 5,0, а буфер с низким рН имеет рН примерно 2,7. Например, буфер с высоким рН может представлять собой 25 мМ ацетат и иметь рН 5,0, а буфер с низким рН может представлять собой 25 мМ муравьиную кислоту и иметь рН 2,7.
Регулирование исходного процентного содержания буфера с низким рН может оптимизировать и максимизировать чистоту очищаемого полипептида, а также последующее отделение примесей, включая агрегаты, примеси из клеточной линии, основный вариант полипептида, вирусную частицу, вирусоподобную частицу и загрязнение вирусного фильтра, от мономеров полипептида. Процентное содержание буфера с низким рН в элюирующем буфере может начинаться с примерно 25%, примерно 30%, примерно 35%, примерно 40% или примерно 45%.
В некоторых вариантах осуществления изобретения процентное содержание буфера с низким рН в элюирующем буфере может начинаться примерно с 25%. В некоторых вариантах осуществления изобретения элюирующий буфер, содержащий примерно 25% буфера с низким рН, содержит примерно 19 мМ ацетат, примерно 6 мМ формиат, и имеет буферную проводимость примерно 1140 мкСм/см при pH 4,5-4,6. Например, элюирующий буфер, содержащий примерно 25% буфера с низким рН, содержит 18,75 мМ ацетат, 6,25 мМ формиат и имеет буферную проводимость 1141 мкСм/см при pH 4,58.
В некоторых вариантах осуществления изобретения процентное содержание буфера с низким рН в элюирующем буфере может начинаться примерно с 35%. В некоторых вариантах осуществления изобретения элюирующий буфер, содержащий примерно 35% буфера с низким рН, содержит примерно 16 мМ ацетат, примерно 9 мМ формиат, и имеет буферную проводимость примерно 1040 мкСм/см при pH 4,3-4,4. Например, элюирующий буфер, содержащий примерно 35% буфера с низким рН, содержит 16,25 мМ ацетат, 8,75 мМ формиат, и имеет буферную проводимость 1039 мкСм/см при pH 4,34.
В некоторых вариантах осуществления изобретения процентное содержание буфера с низким рН в элюирующем буфере может начинаться примерно с 40%. В некоторых вариантах осуществления изобретения элюирующий буфер, содержащий примерно 40% буфера с низким рН, содержит примерно 15 мМ ацетат, примерно 10 мМ формиат и имеет буферную проводимость примерно 974 мкСм/см при pH 4,2-4,3. Например, элюирующий буфер, содержащий примерно 40% буфера с низким рН, содержит 15 мМ ацетат, 10 мМ формиат и имеет буферную проводимость 974 мкСм/см при pH 4,24.
В некоторых вариантах осуществления изобретения процентное содержание буфера с низким рН в элюирующем буфере может заканчиваться примерно на 60%. В некоторых вариантах осуществления изобретения элюирующий буфер, содержащий примерно 60% буфера с низким рН, содержит примерно 10 мМ ацетат, примерно 15 мМ формиат и имеет буферную проводимость примерно 763 мкСм/см при pH 3,6-3,7. Например, буфер с низким рН в конце градиента рН может содержать 10 мМ ацетат, 15 мМ формиат и иметь буферную проводимость 763 мкСм/см при pH 3,69.
В некоторых вариантах осуществления изобретения элюирующий буфер имеет буферную проводимость в диапазоне от примерно 1200 мкСм/см до примерно 500 мкСм/см. В некоторых вариантах осуществления изобретения элюирующий буфер имеет буферную проводимость в диапазоне от примерно 1150 мкСм/см до примерно 700 мкСм/см. В некоторых вариантах осуществления изобретения элюирующий буфер имеет буферную проводимость примерно 1145 мкСм/см, примерно 1141 мкСм/см, примерно 1130 мкСм/см, примерно 1120 мкСм/см, примерно 1110 мкСм/см, примерно 1000 мкСм/см, примерно 1039 мкСм/см, примерно 1000 мкСм/см, примерно 974 мкСм/см, примерно 900 мкСм/см, примерно 800 мкСм/см, примерно 763 мкСм/см или примерно 700 мкСм/см.
В некоторых вариантах осуществления изобретения композиция элюирующего буфера представляет собой примерно 9-20 мМ ацетат и 5-15 мМ формиат. В некоторых вариантах осуществления изобретения композиция элюирующего буфера представляет собой примерно 10-19 мМ ацетат и 6-16 мМ формиат.
Регулирование плотности нагрузки полипептида может также оптимизировать и максимизировать чистоту очищаемого полипептида и отделение примесей, включая агрегаты, примеси из клеточной линии, основный вариант полипептида, вирусную частицу, вирусоподобную частицу и загрязнение вирусного фильтра, от мономеров полипептидов.
Термин «плотность нагрузки» или «плотность нанесения» представляет собой количество очищенного полипептида (г) на литр хроматографической смолы или плотность очищенного полипептида на литр объема мембраны/фильтра (л). Плотность нагрузки измеряют в г/л.
В некоторых вариантах полипептид наносят с плотностью нанесения, начинающейся с 14 г/л или примерно с 14 г/л. В некоторых вариантах полипептид наносят с плотностью нанесения в диапазоне от примерно 14 г/л до примерно 5 г/л или 14 г/л до примерно 70 г/л. В некоторых вариантах полипептид наносят с плотностью нанесения примерно 15 г/л, примерно 17 г/л, примерно 19 г/л, примерно 21 г/л, примерно 23 г/л, примерно 25 г/л, примерно 26 г/л, примерно 27 г/л, примерно 28 г/л, примерно 29 г/л, примерно 31 г/л, примерно 33 г/л, примерно 35 г/л, примерно 37 г/л, примерно 39 г/л, примерно 41 г/л, примерно 43 г/л, примерно 45 г/л, примерно 50 г/л, примерно 55 г/л, примерно 60 г/л, примерно 65г/л или примерно 70 г/л.
С помощью регулирования времени удерживания при элюировании полипептида (или скорости элюирующего потока) также можно оптимизировать и максимизировать чистоту полипептида и последующее отделение примесей от мономеров полипептидов. При увеличенной плотности нанесения время удерживания при элюировании полипептида играет гораздо большую роль в способности градиента рН эффективно фракционировать агрегаты. В некоторых вариантах осуществления изобретения полипептид имеет скорость элюирующего потока в диапазоне от примерно 5 объемов колонки в час до примерно 35 объемов колонки в час. В некоторых вариантах осуществления изобретения полипептид элюируют со скоростью потока в диапазоне от примерно 5 объемов колонки в час до примерно 25 объемов колонки в час. В некоторых вариантах осуществления изобретения полипептид элюируют со скоростью потока примерно 5 объемов колонки в час, примерно 7,5 объемов колонки в час, примерно 10 объемов колонки в час, примерно 12,5 объемов колонки в час, примерно 15 объемов колонки в час, примерно 17,5 объемов колонки в час, примерно 20 объемов колонки в час, примерно 22,5 объемов колонки в час, примерно 25 объемов колонки в час, примерно 27,5 объемов колонки в час, примерно 30 объемов колонки в час, примерно 32,5 объемов колонки в час или примерно 35 объемов колонки в час.
Полипептиды, очищенные с использованием способов, описанных в настоящем документе, имеют выход по меньше мере примерно любой из 75% от неочищенного полипептида, 80% от неочищенного полипептида, 85% от неочищенного полипептида, 90% от неочищенного полипептида, 95% от неочищенного полипептида, 96% от неочищенного полипептида, 97% от неочищенного полипептида, 98% от неочищенного полипептида или 99% от неочищенного полипептида.
Выход представляет собой общее количество собранного очищенного полипептида по сравнению с неочищенным полипептидом до аффинной хроматографической очистки на белке А, описанной в настоящем документе, обычно выражаемое в процентах от неочищенного полипептида.
В некоторых вариантах осуществления изобретения соотношение примесей из клеток-хозяев к очищенному полипептиду по меньшей мере примерно на 75% ниже, примерно на 80% ниже, примерно на 85% ниже, примерно на 90% ниже, примерно на 95% ниже, примерно на 96% ниже, примерно на 97% ниже, примерно на 98% ниже или примерно на 99% ниже соотношения в неочищенном полипептиде.
В некоторых вариантах осуществления изобретения соотношение примесей из клеток-хозяев к очищенному полипептиду по меньшей мере примерно на 20% ниже, примерно на 30% ниже, примерно на 40% ниже, примерно на 50% ниже, примерно на 60% ниже или примерно на 70% ниже соотношения в полипептиде, очищенном стадиями с использованием рН, отличающимися от указанных в настоящем изобретении. Например, в общепринятом или типичном ступенчатом способе элюирования с белка А, полипептид очищают путем связывания полипептида с белком А и элюирования полипептида при рН 3,6 или менее без градиента рН. Соответственно, в некоторых вариантах осуществления изобретения соотношение примесей из клеток-хозяев к очищенному полипептиду по меньшей мере примерно на 20% ниже, примерно на 30% ниже, примерно на 40% ниже, примерно на 50% ниже, примерно на 60% ниже или примерно на 70% ниже соотношения в полипептиде, очищенном способом ступенчатого элюирования, причем способ ступенчатого элюирования включает связывание полипептида с белком А и элюирование рН, начиная с 3,6 или менее.
В некоторых вариантах осуществления изобретения соотношение загрязнения вирусного фильтра к очищенному полипептиду по меньшей мере примерно на 75% ниже, меньшей мере примерно на 80% ниже, примерно на 85% ниже, примерно на 90% ниже, примерно на 95% ниже, примерно на 96% ниже, примерно на 97% ниже, примерно на 98% ниже или примерно на 99% ниже соотношения в неочищенном полипептиде.
В некоторых вариантах осуществления изобретения соотношение загрязнения вирусного фильтра к очищенному полипептиду по меньшей мере примерно на 20% ниже, примерно на 30% ниже, примерно на 40% ниже, примерно на 50% ниже, примерно на 60% ниже или примерно на 70% ниже соотношения в полипептиде, очищенном стадиями с использованием рН, отличающимися от указанных в настоящем изобретении. Например, в общепринятом или типичном ступенчатом способе элюирования с белка А, полипептид очищают путем связывания полипептида с белком А и элюирования полипептида при рН 3,6 или менее без градиента рН. Соответственно, в некоторых вариантах осуществления изобретения соотношение загрязнения вирусного фильтра к очищенному полипептиду по меньшей мере примерно на 20% ниже, примерно на 30% ниже, примерно на 40% ниже, примерно на 50% ниже, примерно на 60% ниже или примерно на 70% ниже соотношения в полипептиде, очищенном способом ступенчатого элюирования, причем способ ступенчатого элюирования включает связывание полипептида с белком А и элюирование рН, начиная с 3,6 или меньшего значения.
В некоторых вариантах осуществления изобретения очищенный полипептид имеет количество вирусных частиц или вирусоподобных частиц меньше примерно 15000 частиц/мл. В некоторых вариантах осуществления изобретения очищенный полипептид имеет количество вирусных частиц или вирусоподобных частиц меньше примерно 12500 частиц/мл, меньше примерно 10000 частиц/мл, меньше примерно 7500 частиц/мл, меньше примерно 5000 частиц/мл, меньше примерно 2500 частиц/мл, меньше примерно 1500 частиц/мл, меньше примерно 1000 частиц/мл, меньше примерно 750 частиц/мл, меньше примерно 500 частиц/мл, меньше примерно 250 частиц/мл, меньше примерно 100 частиц/мл или меньше примерно 50 частиц/мл. В некоторых вариантах осуществления изобретения вирусоподобная частица является ретровирусоподобной частицей.
Используемый в настоящем документе термин «вирусная частица» представляет собой вирион, состоящий из ядра с нуклеиновыми кислотами, окруженного защитной белковой оболочкой (капсидом). «Вирусоподобные частицы» представляют собой неинфекционный вирус, который имеет аналогичные морфологические, биохимические или другие свойства. У них поврежден по меньшей мере один из компонентов, необходимых для жизненного цикла вируса. Примером вирусоподобной частицы является ретровирусподобная частица, не способная реплицироваться. Вирусная частица или вирусоподобная частица могут быть эндогенными или экзогенными (привнесенными). Эндогенные вирусная частица или вирусоподобная частица продуцируются линией клеток-хозяев, присутствуют в клетках и клеточной культуральной жидкости и могут рассматриваться как примесь из клеток-хозяев. Экзогенные или привнесенные вирусная или вирусоподобная частицы не получены из линии клеток-хозяев.
В некоторых вариантах осуществления изобретения очищенный полипептид имеет степень очистки от вирусных или вирусоподобных частиц по меньшей мере примерно 4 LRV (число порядков снижения количества вируса). В некоторых вариантах осуществления изобретения очищенный полипептид имеет степень очистки от вирусных или вирусоподобных частиц в диапазоне от примерно 4 LRV до примерно 8 LRV. В некоторых вариантах осуществления изобретения очищенный полипептид имеет степень очистки от вирусных или вирусоподобных частиц в диапазоне от примерно 4 LRV до примерно 7 LRV. В некоторых вариантах осуществления изобретения очищенный полипептид имеет степень очистки от вирусных или вирусоподобных частиц примерно 5 LRV, примерно 6 LRV, примерно 7 LRV или примерно 8 LRV. В некоторых вариантах осуществления изобретения вирусоподобная частица является ретровирусоподобной частицей.
Используемое в настоящем документе LRV представляет собой разницу в числе порядков (общего количества вируса) в неочищенном полипептиде и в очищенном полипептиде.
В некоторых вариантах осуществления изобретения очищенным полипептидом является мономер полипептида.
В некоторых вариантах осуществления изобретения очищенная фракция полипептида содержит примерно или меньше примерно 20 объемов колонки с белком А. В некоторых вариантах осуществления изобретения очищенная фракция полипептида содержит примерно или меньше примерно 15 объемов колонки с белком А. В некоторых вариантах осуществления изобретения очищенная фракция полипептида содержит примерно или меньше примерно 12 объемов колонки с белком А. В некоторых вариантах осуществления изобретения очищенная фракция полипептида содержит примерно или меньше примерно 11, примерно 10, примерно 9, примерно 8, примерно 7, примерно 6, примерно 5,5 или примерно 5,0 объемов колонки с белком А.
В некоторых вариантах осуществления изобретения способы, описанные в настоящем документе, удаляют по меньшей мере две из примесей, описанных в настоящем документе, из целевого полипептидного мономерного продукта. Например, способы удаляют агрегат и примеси из клеток-хозяев, агрегат и загрязнение вирусного фильтра, агрегат и вирусные частицы, агрегат и вирусоподобные частицы, агрегат и основный вариант полипептида, или примесь из линии клеток-хозяев и вирусные частицы и т.п. В некоторых вариантах осуществления изобретения способы, описанные в настоящем документе, удаляют по меньшей мере три из примесей, описанных в настоящем документе, из целевого полипептидного мономерного продукта. Например, способы удаляют агрегат, примесь из клеток-хозяев и загрязнение вирусного фильтра, или агрегат, примесь из клеток-хозяев и вирусные частицы, а также основные варианты полипептида, и т.п. В некоторых вариантах осуществления изобретения способы, описанные в настоящем документе, удаляют по меньшей мере четыре из примесей, описанных в настоящем документе, из целевого полипептидного мономерного продукта. Например, способы удаляют агрегат, примесь из клеток-хозяев, загрязнение вирусного фильтра и вирусные частицы, или агрегат, примесь из клеток-хозяев, загрязнение вирусного фильтра и вирусоподобные частицы. В некоторых вариантах осуществления изобретения способы, описанные в настоящем документе, удаляют по меньшей мере пять из примесей, описанных в настоящем документе, из целевого полипептидного мономерного продукта. Например, способы удаляют агрегат, примесь из клеток-хозяев, загрязнение вирусного фильтра, вирусные частицы и вирусоподобные частицы, и т.п. В некоторых вариантах осуществления изобретения способы, описанные в настоящем документе, удаляют все примеси, описанные в настоящем документе, из целевого полипептидного мономерного продукта.
В некоторых вариантах осуществления изобретения способы, описанные в настоящем документе, не включают дополнительную стадию очистки для удаления агрегата, и очищенные полипептиды имеют чистоту по меньшей мере примерно 98% или примерно 99% мономера. Удаление агрегата, обычно проводимое на отдельной стадии ионообменной хроматографии, не требуется после хроматографии на белке А с использованием градиента рН, описанного в настоящем документе.
В некоторых вариантах осуществления изобретения способы, описанные в настоящем документе, не включают дополнительную стадию очистки для удаления загрязнения вирусного фильтра, и очищенные полипептиды имеют чистоту по меньшей мере примерно 98% или примерно 99% мономера.
В некоторых вариантах осуществления изобретения способы, описанные в настоящем документе, не включают дополнительную стадию очистки для удаления основного или кислотного варианта полипептида.
Полипептид, очищенный с использованием способов, описанных в настоящем документе, можно подвергнуть дополнительным стадиям очистки либо до, либо в течение, либо после стадии хроматографии на белке А. Примеры дополнительных стадий очистки включают, но не ограничены этим, гидроксиаппатитную хроматографию; диализ; аффинную хроматографию с использованием антитела для захвата белка; гидрофобную хроматографию (HIC) (например, фракционирование на HIC); осаждение сульфатом аммония; осаждение полиэтиленгликолем или производным полиэтиленгликоля; анионную и катионную обменную хроматографию; осаждение этанолом; обращенно-фазовую ВЭЖХ; хроматографию на силикагеле; хроматофокусирование; SDS-PAGE; вирусную фильтрацию; гель-фильтрацию и мягкую распределительную хроматографию.
В некоторых вариантах осуществления изобретения полипептиды дополнительно подвергают стадии вирусной фильтрации. Например, парвовирусный фильтр можно использовать на стадии вирусной фильтрации, следующей за стадией хроматографии на белке А с использованием градиента рН, описанной в настоящем документе.
В некоторых вариантах осуществления изобретения полипептиды дополнительно подвергают стадии ионообменной хроматографии. В некоторых вариантах осуществления настоящего изобретения стадия ионообменной хроматографии включает стадию катионообменной хроматографии. В некоторых вариантах осуществления настоящего изобретения стадия ионообменной хроматографии включает стадию анионообменной хроматографии. В некоторых вариантах осуществления настоящего изобретения стадия ионообменной хроматографии включает стадию катионообменной хроматографии и стадию анионообменной хроматографии.
В некоторых вариантах осуществления изобретения стадию ионообменной хроматографии проводят без остановки после стадии хроматографии на белке А, описанной в настоящем документе. Например, после способа хроматографии на белке А, описанного в настоящем документе, можно использовать катионо- и анионобменные хроматографические мембраны вместо стандартных катионо- и/или анионобменных хроматографических колонок для получения очищенных полипептидов с чистотой и выходом сравнимыми с получаемыми способом стандартной хроматографии на белке А без градиента рН с последующими стандартными стадиями катионо- и анионообменной колоночной хроматографии.
В некоторых вариантах осуществления изобретения способы, описанные в настоящем документе, представляют собой промышленные или коммерческие процессы. Используемые в настоящем изобретении промышленные или коммерческие процессы относятся к крупномасштабной очистке белка/полипептида, например, от примерно 1 кг до примерно 25 кг белкового/полипептидного продукта (масштаб ферментера) на процесс очистки.
Полипептиды
Полипептид или белок, очищаемый с использованием способов, описанных в настоящем документе, включает, но не ограничен эти, антитело, иммуноадгезин или полипептид, слитый или конъюгированный с СН2/СН3-областью. Методики получения таких молекул обсуждаются ниже.
Антитела
Антитела в объеме настоящего изобретения включают, но не ограничены этим: анти-CD20 антитела, такие как химерное анти-CD20, «C2B8» в патенте США № 5736137 (РИТУКСАН®); анти-VEGF антитела, включая гуманизированные и/или аффинно-созревшие анти-VEGF антитела, такие как гуманизированное анти-VEGF антитело huA4.6.1 АВАСТИН® (Kim et al., Growth Factors, 7:53-64 (1992), международная публикация № WO 96/30046 и WO 98/45331, опубликованная 15-го октября, 1998) и V3LA; анти-MUC16 антитело; анти-CD4 антитела, такие как антитело cM-7412 (Choy et al. Arthritis Rheum. 39(1):52-56 (1996)) и антитело Ибализумаб (TNX355); анти-MET антитела, такие как анти-С-Met антитело (5D5) с одним антиген-связывающим участком; анти-HER2 антитела Трастузумаб (ГЕРЦЕПТИН®) (Carter et al., Proc. Natl. Acad. Sci. USA, 89:4285-4289 (1992), патент США № 5725856) и гуманизированное 2C4 (WO 01/00245, Adams et al.); химерный или гуманизированный вариант антитела 2Н7 из патента США № 5721108B1, или Тозитумомаб (БЕКСАР®); анти-IL-8 антитела (St John et al., Chest, 103:932 (1993) и международная публикация № WO 95/23865); антитела к антигену стволовых клеток простаты (PSCA) (WO 01/40309); анти-CD40 антитела, включая S2C6 и их гуманизированные варианты (WO 00/75348); анти-CD1 антитела (патент США № 5622700, WO 98/23761, Steppe et al., Transplant Intl. 4:3-7 (1991), и Hourmant et al., Transplantation 58:377-380 (1994)); анти-CD18 (патент США № 5622700, выданный 22-го апреля 1997 г., или WO 97/26912, опубликованная 31-го июля 1997 г.); анти-IgE антитела (включая E25, E26 и E27; патент США № 5714338, выданный 3-го февраля 1998 г., или патент США № 5091313, выданный 25-го февраля 1992 г., WO 93/04173, опубликованная 4-го марта 1993 г., или международная заявка № PCT/US98/13410, поданная 30-го июня 1998 г., патент США № 5714338, Presta et al., J. Immunol. 151:2623-2632 (1993), и международная публикация № WO 95/19181); антитела к Apo-2-рецептору (WO 98/51793, опубликованная 19-го ноября 1998 г.); антитела к TNF-α, включая cA2 (РЕМИКАД®), CDP571 и MAK-195 (см. патент США № 5672347, выданный 30-го сентября 1997 г, Lorenz et al. J. Immunol. 156(4): 1646-1653(1996), и Dhainaut et al. Crit. Care Med. 23(9): 1461-1469 (1995)); антитела к тканевому фактору (TF) (европейский патент № 0420937B1, выданный 9-го ноября, 1994); антитела к интегрину α4β7 человека (WO 98/06248, опубликованная 19-го февраля 1998 г.); антитела к рецептору фактора роста эпидермиса (EGFR) (например, химеризованное или гуманизированное антитело 225, как например, в WO 96/40210, опубликованной 19-го декабря 1996 г.); анти-CD3 антитела, такие как OKT3 (патент США № 4515893, выданный 7-го мая 1985 г.); анти-CD25 или анти-Tac антитела, такие как CHI-621 (СИМУЛЕКТ® и ЗЕНАПАКС® (см. патент США № 5693762, выданный 2-го декабря 1997 г.); анти-CD52 антитела, такие как CAMPATH-1H (Riechmann et al. Nature 332:323-337 (1988)); антитела к Fc-рецептору, такие как антитело М22, направленное против Fсγ RI, как например, в Graziano et al. J. Immunol. 155(10):4996-5002 (1995); антитела к карциноэмбриональному антигену (CEA), такие как hMN-14 (Sharkey et al. Cancer Res. 55(23 Suppl): 5935s-5945s (1995); антитела, направленные против эпителиальных клеток молочной железы, включая huBrE-3, hu-Mc 3 и CHL6 (Ceriani et al. Cancer Res. 55(23): 5852s-5856s (1995); и Richman et al. Cancer Res. 55(23 Supp):5916s-5920s (1995)); антитела, которые связывают клетки карциномы толстой кишки, такие как C242 (Litton et al. Eur J Immunol. 26(1): 1-9 (1996)); ати-CD38 антитела, например, AT 13/5 (Ellis et al. J. Immunol. 155(2):925-937 (1995)); анти-CD33 антитела, такие как Hu M195 (Jurcic et al. Cancer Res 55(23 Suppl):5908s-5910s (1995)) и CMA-676 или CDP771; анти-CD22 антитела, такие как LL2 или Лимфоцид (Juweid et al. Cancer Res 55(23 Suppl):5899s-5907s (1995)); анти-EpCAM антитела, такие как 17-1A (ПАНОРЕКС®); анти-GpIIb/IIIa антитела, такие как абциксимаб или c7E3 Fab (РЕОПРО®); анти-RSV антитела, такие как MEDI-493 (СИНАГИС®); анти-CMV антитела, такие как ПРОТОВИР®; анти-HIV антитела, такие как PRO542; антитела против гепатита, такие как анти-Hep B антитело ОСТАВИР®; анти-CA 125 антитела, такие как Оварекс; антиидиотипическое эпитопу GD3 антитело BEC2; анти-αvβ3 антитела, включая ВИТАКСИН®; анитело к клеткам почечной карциномы человека, такое как ch-G250; ING-1; антитело к антигену 17-1А человека (3622W94); антитело к колоректальной опухоли человека (A33); антитело R24 к меланоме человека, направленное против ганглиозида GD3; антитело к сквамозно-клеточной карциноме человека (SF-25); антитела к лейкоцитарному антигену человека (HLA), такие как Smart ID 10 и анти-HLA DR антитело Онколим (Lym-1).
Помимо антител, специально указанных выше, опытный специалист может создать антитела, направленные против целевого антигена, например, используя методики, описанные ниже.
(i) Выбор и получение антигена
Антитело в настоящем документе направлено против целевого антигена. Предпочтительно, чтобы антиген был биологически важным полипептидом, а введение антитела млекопитающему, страдающему от заболевания или расстройства может приводить к терапевтическому эффекту у этого млекопитающего. Однако также предусмотрены антитела, направленные против неполипептидных антигенов (таких как опухолевые гликолипидные антигены; см. патент США № 5091178). Если антиген является полипептидом, то он может быть трансмембранной молекулой (например, рецептором) или лигандом, таким как ростовой фактор. Примеры антигенов включат белки, описанные в разделе (3) ниже. Примеры молекулярных мишеней антител, охватываемых настоящим изобретением, включают CD-белки, такие как CD3, CD4, CD8, CD19, CD20, CD22 и CD34; членов семейства ErbB-рецепторв, таких как рецепторы EGFR, HER2, HER3 или HER4; молекулы клеточной адгезии, такие как LFA-1, Mac1, p150,95, VLA-4, ICAM-1, VCAM и av/β3-интегрин, включая либо α, либо β его субъединицы (например, анти-CD11a, анти-CD18 или анти-CD11b антитела); ростовые факторы, такие как VEGF; IgE; антигены групп крови; flk2/flt3-рецептор; рецептор ожирения (ОВ); mpl-рецептор; CTLA-4; белок C или любой из других антигенов, указанных в настоящем описании.
Для получения антител в качестве иммуногенов можно использовать растворимые антигены или их фрагменты, необязательно конъюгированные с другими молекулами. В случае трансмембранных молекул, таких как рецепторы, в качестве иммуногена можно использовать их фрагменты (например, внеклеточный домен рецептора). В альтернативном варианте в качестве иммуногена можно использовать клетки, экспрессирующие трансмембранную молекулу. Такие клетки можно получить из природного источника (например, раковых клеточных линий), или они могут представлять собой клетки, трансформированные с помощью рекомбинантных методик для экспрессии трансмембранной молекулы.
Специалистам в данной области будут очевидны другие антигены и их формы, пригодные для получения антител.
(ii) Поликлональные антитела
Поликлональные антитела предпочтительно получать в организме животных в результате многократных подкожных (sc) или внутрибрюшинных (ip) инъекций заданного антигена и адъюванта. Может быть полезно конъюгировать антиген с белком, иммуногенным в иммунизируемых биологических видах, например, с гемоцианином морского блюдечка, сывороточным альбумином, бычьим тиреоглобулином или трипсиновым ингибитором из сои, используя бифункциональный или образующий производные агент, например, малеимидобензоил-сульфосукцинимидный сложный эфир (конъюгация через остатки цистеина), N-гидроксисукцинимид (через остатки лизина), глутаровый альдегид, янтарный ангидрид, SOCl2 или R1N=C=NR, где R и R1 являются различными алкильными группами.
Животных иммунизируют антигеном, иммуногенными конъюгатами или производными, объединяя, например, 100 мкг или 5 мкг белка или конъюгата (для кроликов или мышей, соответственно) с 3 объемами полного адъюванта Фрейнда и вводя раствор с помощью инъекций внутрикожно в несколько участков. Через один месяц животным повторно вводят от 1/5 до 1/10 от исходного количества антигена или конъюгата в полном адъюванте Фрейнда с помощью подкожных инъекций в несколько участков. Через 7-14 дней у животных отбирают кровь, и проводят анализ сыворотки на титр антител. Животным повторно вводят антиген до достижения плато титра. Предпочтительно повторно вводить животным конъюгат одного антигена, но конъюгированный с другим белком и/или посредством другого сшивающего реагента. Конъюгаты также можно получить в рекомбинантной клеточной культуре в виде слитых белков. Также для усиления иммунного ответа соответственно используют агрегирующие агенты, такие как алюмокалиевые квасцы.
(iii) Моноклональные антитела
Моноклональные антитела можно получить с использованием гибридомного способа, впервые описанного Kohler et al., Nature, 256:495 (1975), или можно получить способами рекомбинантных ДНК (патент США № 4816567).
В гибридомном способе мышь или другое подходящее животное-хозяин, такое как хомячок или макака, иммунизируют, как описано выше, для возникновения лимфоцитов, продуцирующих или способных продуцировать антитела, которые будут специфично связывать белок, используемый для иммунизации. В альтернативном варианте, лимфоциты можно иммунизировать in vitro. Затем лимфоциты сливают с миеломными клетками, используя подходящий агент для слияния, такой как полиэтиленгликоль, для образования гибридомной клетки (Goding, Monoclonal Antibodies: Principles and Practice, pp.59-103 (Academic Press, 1986)).
Полученные таким образом клетки гибридомы высевают и растят в подходящей культуральной среде, которая предпочтительно содержит одно или несколько веществ, ингибирующих рост или выживаемость неслитых, родительских миеломных клеток. Например, если у родительских миеломных клеток отсутствует фермент гипоксантин-гуанинфосфорибозилтрансфераза (HGPRT или HPRT), культуральная среда для гибридом обычно будет включать гипоксантин, аминоптерин и тимидин (НАТ-среда), которые препятствуют росту клеток, лишенных HGPRT.
Предпочтительными миеломными клетками являются клетки, которые легко поддаются слиянию, поддерживают на высоком уровне стабильную продукцию антитела отобранными антитело-продуцирующими клетками и чувствительны к среде, такой как НАТ-среда. Из таких клеток предпочтительными миеломными клеточными линиями являются мышиные миеломные линии, такие как полученные из мышиных опухолей MOPC-21 и MPC-11, доступные от Salk Institute Cell Distribution Center, San Diego, Calif. USA, и клетки SP-2 или X63-Ag8-653, доступные из Американской коллекции типовых культур, Rockville, Maryland USA. Для получения моноклональных антител человека также были описаны линии клеток миеломы человека и гетеромиеломы мыши и человека (Kozbor, J. Immunol., 133:3001 (1984); Brodeur et al., Monoclonal Antibody Production Techniques and Applications, pp. 51-63 (Marcel Dekker, Inc., New York, 1987)).
Культуральную среду, в которой выращивают клетки гибридомы, анализируют относительно продукции моноклональных антител, направленных против антигена. Предпочтительно определять специфичность связывания моноклональных антител, продуцируемых клетками гибридомы, с помощью иммунопреципитации или анализом связывания in vitro, таким как радиоиммуноанализ (РИА) или твердофазный иммуноферментный анализ (ELISA).
После идентификации клеток гибридомы, продуцирующих антитела требуемой специфичности, аффинности и/или активности, клоны можно субклонировать с помощью предельного разведения и вырастить стандартными способами (Goding, Monoclonal Antibodies: Principles and Practice, pp.59-103 (Academic Press, 1986)). Подходящие культуральные среды для указанной цели включают, например, среду DMEM или RPMI-1640. Кроме того, клетки гибридомы можно выращивать in vivo в виде асцитных опухолей у животного.
Моноклональные антитела, секретируемые субклонами, соответствующим образом отделяют от культуральной среды, асцитной жидкости или сыворотки обычными способами очистки иммуноглобулинов, такими как, например, хроматография на белке A-сефарозе, на гидроксиапатитах, гель-электрофорез, диализ или аффинная хроматография. Предпочтительно использовать методику аффинной хроматографии на белке А с использованием градиента рН, описанную в настоящем документе.
ДНК, кодирующую моноклональные антитела, без труда выделяют и секвенируют, используя общепринятые методики (например, используя олигонуклеотидные зонды, которые способны специфично связываться с генами, кодирующими тяжелые и легкие цепи моноклональных антител). Клетки гибридомы служат в качестве предпочтительного источника такой ДНК. Для синтеза моноклональных антител в рекомбинантных клетках-хозяевах ДНК после выделения можно поместить в экспрессирующие векторы, которыми затем трансфицируют клетки-хозяева, такие как клетки E. coli, клетки обезьяны (COS), клетки яичника китайского хомячка (CHO) или клетки миеломы, которые в ином случае не продуцируют иммуноглобулин.
ДНК также можно модифицировать, например, замещением кодирующими последовательностями константных доменов тяжелой и легкой цепей человека гомологичных мышиных последовательностей (патент США 4,816,567; Morrison, et al., Proc. Natl. Acad. Sci. USA, 81:6851 (1984)) или ковалентным связыванием с последовательностью, кодирующей иммуноглобулин, всей или части последовательности, кодирующей полипептид неиммуноглобулиновой природы.
Обычно такими полипептидами неиммуноглобулиновой природы заменяют константные домены антитела или ими заменяют вариабельные домены одного антиген-связывающего участка антитела для создания химерного бивалентного антитела, содержащего один антиген-связывающий участок, имеющий специфичность к антигену, и другой антиген-связывающий участок, имеющий специфичность к другому антигену.
Моноклональные антитела можно выделить из фаговых библиотек антител, созданных с использованием методик, описанных в McCafferty et al., Nature, 348: 552-554 (1990). В статьях Clackson et al., Nature, 352: 624-628 (1991) и Marks et al., J. Mol. Biol., 222: 581-597 (1991) описано, соответственно, выделение мышиных антител и антител человека с использованием фаговых библиотек. В последующих публикациях описано получение высокоаффинных (наномолярный диапазон) антител человека с помощью перетасовки цепей (Marks et al., Bio/Technology, 10: 779-783 (1992)), а также с помощью комбинаторной инфекции и рекомбинации in vivo в качестве стратегии для конструирования очень крупных фаговых библиотек (Waterhouse et al., Nuc. Acids. Res., 21: 2265-2266 (1993)). Таким образом, указанные методики являются приемлемой альтернативой традиционным гибридомным методикам получения моноклональных антител.
(iv) Гуманизированные антитела и антитела человека
Гуманизированное антитело имеет один или несколько аминокислотных остатков, введенных в него из источника, который отличен от человека. Указанные аминокислотные остатки из источника, отличного от человека, часто называют «импортируемыми» остатками, которые обычно взяты из «импортируемого» вариабельного домена. Гуманизацию по существу можно выполнить, следуя способу Winter и соавторов (Jones et al., Nature, 321: 522-525 (1986); Riechmann et al., Nature, 332: 323-327 (1988); Verhoeyen et al., Science, 239: 1534-1536 (1988)), замещая CDR или последовательностями CDR грызунов соответствующие последовательности антитела человека. Соответственно такие «гуманизированные» антитела являются химерными антителами (патент США 4816567), в которых существенно меньшая часть, чем интактный вариабельный домен человека, заменена на соответствующую последовательность биологического вида, кроме человека. На практике гуманизированные антитела обычно представляют собой антитела человека, в которых некоторые остатки CDR и возможно некоторые остатки FR замещены остатками из аналогичных участков антител грызунов.
Для снижения иммуногенности очень важен выбор вариабельных доменов человека как легкой, так и тяжелой цепи, используемых для получения гуманизированных антител. По так называемому способу «оптимальной подгонки» проводят скрининг последовательности вариабельного домена антитела грызуна по полной библиотеке известных последовательностей вариабельных доменов человека. Затем в качестве каркаса (FR) человека для гуманизированного антитела принимают последовательность вариабельного домена человека, которая наиболее близка последовательности вариабельного домена грызуна (Sims et al., J. Immunol., 151:2296 (1993)). В другом способе используют определенный каркас, полученный из консенсусной последовательности для всех антител человека конкретной подгруппы легких или тяжелых цепей. Один каркас можно использовать для нескольких разных гуманизированных антител (Carter et al., Proc. Natl. Acad. Sci. USA, 89: 4285 (1992); Presta et al., J. Immunol., 151: 2623 (1993)).
Кроме того, важно, чтобы антитела были гуманизированы с сохранением высокой аффинности к антигену и других полезных биологических свойств. Для достижения указанной цели по предпочтительному способу гуманизированные антитела получают, проводя анализ родительских последовательностей и различных концептуальных гуманизированных продуктов с использованием трехмерных моделей родительских и гуманизированных последовательностей. Трехмерные модели иммуноглобулинов общедоступны и известны специалистам в данной области. Доступны компьютерные программы, которые иллюстрируют и показывают возможные трехмерные конформационные структуры выбранных для исследования иммуноглобулиновых последовательностей. Исследование таких изображений позволяет проанализировать вероятную роль аминокислотных остатков в функционировании выбранной для исследования последовательности иммуноглобулина, т.е. провести анализ остатков, влияющих на способность выбранного для исследования иммуноглобулина связывать свой антиген. Таким образом, можно выбрать и скомбинировать остатки FR из реципиентных и импортируемых последовательностей так, чтобы достичь требуемых характеристик антитела, таких как повышенная аффинность к антигену-мишени (антигенам). Обычно на связывание антигена непосредственно и в наибольшей степени влияют остатки CDR.
В качестве альтернативы, в настоящее время можно получить трансгенных животных (например, мышей), способных после иммунизации продуцировать полный репертуар антител человека в отсутствие продукции эндогенных иммуноглобулинов. Например, было описано, что гомозиготная делеция гена соединительной области тяжелой цепи (JH) антитела у химерных и мутантных по зародышевой линии мышей приводит к полному ингибированию продукции эндогенных антител. Перенос набора генов иммуноглобулинов зародышевой линии человека в таких мутантных по зародышевой линии мышей будет приводить к продукции антител человека при антигенной стимуляции. См., например, Jakobovits et al., Proc. Natl. Acad. Sci. USA, 90: 2551 (1993); Jakobovits et al., Nature, 362: 255-258 (1993); Bruggermann et al., Year in Immuno., 7: 33 (1993); и Duchosal et al. Nature 355: 258 (1992). Антитела человека также можно получить из библиотек фагового дисплея (Hoogenboom et al., J. Mol. Biol., 227: 381 (1991); Marks et al., J. Mol. Biol., 222: 581-597 (1991); Vaughan et al. Nature Biotech 14: 309 (1996)).
(v) Фрагменты антител
Для получения фрагментов антител были разработаны различные методики. Традиционно такие фрагменты получают протеолитическим расщеплением интактных антител (см., например, Morimoto et al., Journal of Biochemical and Biophysical Methods 24: 107-117 (1992) и Brennan et al., Science, 229: 81 (1985)). Однако в настоящее время такие фрагменты могут непосредственно продуцироваться рекомбинантными клетками-хозяевами. Например, фрагменты антител можно выделить из фаговых библиотек антител, обсуждаемых выше. В альтернативном варианте Fab'-SH-фрагменты можно напрямую выделить из клеток E. coli и соединить химическим способом с образованием F(ab')2-фрагментов (Carter et al., Bio/Technology 10:163-167 (1992)). В другом подходе F(ab')2-фрагменты можно выделить напрямую из культуры рекомбинантных клеток-хозяев. Также можно выделить одноцепочечный Fv-фрагмент (scFv). См. WO 93/16185. Другие методики получения фрагментов антител будут известны специалисту в данной области.
(vi) Полиспецифичные антитела
Полиспецифичные антитела специфично связывают по меньшей мере два разных антигена. Несмотря на то, что такие молекулы, как правило, будут связывать только два антигена (т.е. являться биспецифичными антителами, BsAb), антитела с дополнительной специфичностью, такие как триспецифичные антитела, входят в указанное определение при его использовании в настоящем документе.
Способы изготовления биспецифичных антител известны в данной области. Традиционное получение полноразмерных биспецифичных антител основано на совместной экспрессии двух пар иммуноглобулиновых тяжелых и легких цепей, при этом две цепи имеют разные специфичности (Millstein et al., Nature, 305: 537-539 (1983)). Вследствие случайного подбора тяжелых и легких цепей иммуноглобулина такие гибридомы (квадромы) продуцируют возможную смесь из 10 разных молекул антител, из которых только одна имеет правильную биспецифичную структуру. Очистка молекулы с правильной структурой, обычно осуществляемая с использованием аффинной хроматографии, является довольно сложной, а выход продукта низким. Аналогичные методики описаны в WO 93/08829 и в Traunecker et al., EMBO J., 10: 3655-3659 (1991).
Согласно другому подходу, описанному в WO 96/27011, область контакта между парой молекул антител может быть сконструирована таким образом, чтобы максимизировать процент гетеродимеров, выделяемых из рекомбинантной клеточной культуры. Предпочтительная область контакта содержит, по меньшей мере, часть домена CH3-константного домена антитела. В данном способе одну или несколько аминокислот с небольшими боковыми цепями из области контакта первой молекулы антитела заменяют аминокислотами с более крупными боковыми цепями (например, тирозином или триптофаном). В области контакта второй молекулы антитела создают компенсирующие «впадины», имеющие размер, идентичный или сходный с размером большой боковой цепи (цепей), заменяя аминокислоты с большими боковыми цепями аминокислотами с меньшими боковыми цепями (например, на аланином или треонином). Это обеспечивает механизм увеличения выхода гетеродимера по сравнению с другими нежелательными конечными продуктами, такими как гомодимеры.
Биспецифичные антитела включают перекрестно сшитые или «гетероконъюгированные» антитела. Например, одно из антител в гетероконъюгате может быть связано с авидином, а другое с биотином. Такие антитела, например, были предложены для того, чтобы направить клетки иммунной системы к нежелательным клеткам (патент США 4676980), и для лечения ВИЧ-инфекции (WO 91/00360, WO 92/200373 и EP 03089). Гетероконъюгаты антител можно получить, используя любые способы образования перекрестных сшивок. Подходящие сшивающие агенты хорошо известны в данной области и описаны в патенте США 4676980 вместе с рядом методик образования перекрестных сшивок.
Методики создания биспецифичных антител из фрагментов антител также описаны в литературе. Например, биспецифичные антитела можно получить, используя химическую связь. В Brennan et al., Science, 229: 81 (1985) описана процедура, в которой интактные антитела протеолитически расщепляют для получения F(ab')2-фрагментов. Полученные фрагменты восстанавливают в присутствии арсенита натрия, агента, образующего комплексы с дитиольными группами, для того чтобы стабилизировать соседние дитиолы и препятствовать образованию межмолекулярной дисульфидной связи. Затем полученные Fab'-фрагменты превращают в производные тионитробензоата (TNB). Один из Fab'-TNB-производных затем снова превращают в Fab'-тиол восстановлением с использованием меркаптоэтиламина и смешивают с эквимолярным количеством другого Fab'-TNB-производного с образованием биспецифичного антитела. Полученные биспецифичные антитела можно использовать в качестве агентов для избирательной иммобилизации ферментов.
Недавние достижения в данной области обеспечили возможность прямого выделения Fab'-SH-фрагментов из E. coli, которые можно соединить химическим способом с образованием биспецифичных антител. В Shalaby et al., J. Exp. Med., 175: 217-225 (1992) описано получение полностью гуманизированной биспецифичной F(ab')2-молекулы антитела. Каждый Fab'-фрагмент по отдельности секретировали из клеток E. coli и подвергали направленному химическому связыванию in vitro с образованием биспецифичного антитела. Полученное таким образом биспецифичное антитело было способно связываться с клетками, сверхэкспрессирующими ErbB2-рецептор, и здоровыми T-клетками человека, а также запускать литическую активность цитотоксических лимфоцитов человека в отношении опухолей молочной железы человека.
Также описаны различные методики получения и выделения фрагментов биспецифичных антител непосредственно из культуры рекомбинантных клеток. Например, были получены биспецифичные антитела с использованием лейциновых молний. Kostelny et al., J. Immunol., 148(5): 1547-1553 (1992). Пептиды лейциновой молнии из белков Fos и Jun связывали с Fab'-частями двух разных антител с помощью слияния генов. Гомодимеры антител восстанавливали в шарнирной области для образования мономеров, и затем повторно окисляли с образованием гетеродимеров антител. Такой способ также можно использовать для получения гомодимеров антител. Технология «диантител», описанная Hollinger et al., Proc. Natl. Acad. Sci. USA, 90:6444-6448 (1993), предоставляет альтернативный механизм получения фрагментов биспецифичных антител. Фрагменты содержат вариабельный домен тяжелой цепи (VH), связанный с вариабельным доменом легкой цепи (VL) линкером, слишком коротким, чтобы обеспечить возможность образования пар между двумя доменами на одной цепи. Соответственно, VH- и VL-домены одного фрагмента вынуждены образовывать пары с комплементарными VL- и VH-доменами другого фрагмента, образуя при этом два антиген-связывающих участка. Также была опубликована другая стратегия получения фрагментов биспецифичных антител с использованием димеров одноцепочечного Fv-фрагмента (sFv). См. Gruber et al., J. Immunol., 152:5368 (1994). В альтернативном варианте антитела могут представлять собой «линейные антитела», описанные в Zapata et al. Protein Eng. 8(10):1057-1062 (1995). В нескольких словах, данные антитела содержат пару тандемных Fd-сегментов (VH-CH1-VH и VL), которые образуют пару антиген-связывающих областей. Линейные антитела могут быть биспецифичными или моноспецифичными.
Предусмотрены антитела более чем с двумя валентностями. Например, можно получить триспецифичные антитела. Tutt et al. J. Immunol. 147: 60 (1991).
Иммуноадгезины
Наиболее простая и незамысловатая конструкция иммуноадгезина сочетает связывающий домен (связывающие домены) адгезина (например, внеклеточный домен (ECD) рецептора) с шарнирной и Fc областями тяжелой цепи иммуноглобулина. Обычно при получении иммуноадгезинов по настоящему изобретению нуклеиновую кислоту, кодирующую связывающий домен адгезина, сливают на С-конце с нуклеиновой кислотой, кодирующей N-конец последовательности константного домена иммуноглобулина, однако также возможно слияние на N-конце.
Обычно при таком слиянии кодируемый химерный полипептид будет сохранять по меньшей мере функционально активный шарнир, СН2- и CH3-домены константного участка тяжелой цепи иммуноглобулина. Слияние также осуществляют с С-концом Fc-части константного домена или непосредственно на N-конце с СН1 тяжелой цепи или соответствующей областью легкой цепи. Точное место, на котором осуществляют слияние, не является критичным; конкретные сайты хорошо известны, и их можно выбрать для оптимизации биологической активности, секреции или связывающих характеристик иммуноадгезина.
В некоторых вариантах осуществления последовательность адгезина слита с N-концом Fc-домена иммуноглобулина G1 (IgG1). Возможно слияние всей константной области тяжелой цепи с последовательностью адгезина. Однако предпочтительно использовать в слиянии последовательность, которая начинается в шарнирном участке непосредственно выше сайта расщепления папаином, который химически ограничивает Fc-область IgG (т.е. остаток 216, принимая первый остаток константного участка тяжелой цепи как 114), или в аналогичных участках других иммуноглобулинов. В некоторых вариантах осуществления изобретения аминокислотная последовательность адгезина слита с (а) шарнирной областью и/или СН2 и CH3, или (b) CH1, шарниром, СН2 и CH3-доменами тяжелой цепи IgG.
Для создания биспецифичных иммуноадгезинов их собирают в виде мультимеров, и в частности, в виде гетеродимеров или гетеротетрамеров. В общем, эти собранные иммуноглобулины состоят из известных структурных единиц. Основной четырехцепочной структурной единицей является форма, в которой существуют IgG, IgD и IgE. Четырехцепочная единица повторяется в иммуноглобулинах большего молекулярного веса; IgM обычно существует в виде пентамера основных четырехцепочных единиц, соединенных между собой дисульфидными связями. IgA-глобулин и иногда IgG-глобулин также могут находиться в сыворотке в мультимерной форме. В случае мультимера каждая из четырехцепочных единиц может быть одинаковой или разной.
Различные типичные собранные иммуноадгезины в пределах объема изобретения схематически представлены ниже:
(a) ACL-ACL;
(b) ACH-(ACH, ACL-ACH, ACL-VHCH или VLCL-ACH);
(c) ACL-ACH-(ACL-ACH, ACL-VHCH, VLCL-ACH или VLCL-VHCH);
(d) ACL-VHCH-(ACH или ACL-VHCH, или VLCL-ACH);
(e) VLCL-ACH-(ACL-VHCH или VLCL-ACH); и
(f) (A-Y)n-(VLCL-VHCH)2,
в которых, каждый А представляет собой одинаковые или разные аминокислотные последовательности иммуноадгезина;
VL - вариабельный домен легкой цепи иммуноглобулина;
VH - вариабельный домен тяжелой цепи иммуноглобулина:
CL - константный домен легкой цепи иммуноглобулина;
CH - константный домен тяжелой цепи иммуноглобулина;
n - целое число больше 1;
Y - обозначает остаток ковалентно сшивающего агента.
Для краткости в вышеуказанных структурах показаны только ключевые признаки; в них не указаны соединительные (J) или другие домены иммуноглобулинов, а также не показаны дисульфидные связи. Однако когда такие области требуются для связывающей активности, их необходимо создать, чтобы они находились в обычных местах, которые они занимают в молекулах иммуноглобулинов.
В альтернативном варианте последовательности иммуноадгезинов можно встроить между последовательностями тяжелой цепи и легкой цепи иммуноглобулина, получая иммуноглобулин, содержащий химерную тяжелую цепь. В данном варианте осуществления изобретения последовательности иммуноадгезинов слиты с 3'-концом тяжелой цепи иммуноглобулина в каждом плече иммуноглобулина, либо между шарниром и CH2-доменом, либо между CH2 и CH3-доменами. Аналогичные конструкции были опубликованы в Hoogenboom, et al., Mol. Immunol. 28: 1027-1037 (1991).
Хотя присутствие легкой цепи иммуноглобулина не требуется, в иммуноадгезинах по настоящему изобретению легкая цепь иммуноглобулина может присутствовать либо ковалентно связанной с слитым полипептидом адгезина и тяжелой цепью иммуноглобулина, либо непосредственно слитой с адгезином. В первом случае ДНК, кодирующую легкую цепь иммуноглобулина, обычно совместно экспрессирует с ДНК, кодирующей слитый белок адгезина и тяжелой цепи иммуноглобулина. При секреции тяжелая цепь и легкая цепь гибрида будут ковалентно связываться, давая иммуноглобулиноподобную структуру, содержащую связанные дисульфидным мостиком две пары тяжелой цепи и легкой цепи иммуноглобулина. Способы, пригодные для получения таких структур, например, раскрыты в патенте США № 4816567, выданном 28-го марта 1989.
Иммуноадгезины наиболее удобно конструировать путем слияния последовательности кДНК, кодирующей адгезиновую часть с сохранением рамки считывания, с кДНК-последовательностью иммуноглобулина. Однако также можно использовать слияние с геномными фрагментами иммуноглобулинов (см., например, Aruffo et al., Cell 61: 1303-1313 (1990); и Stamenkovic et al., Cell 66: 1133-1144 (1991)). Последний тип слияния требует для экспрессии присутствия регуляторных последовательностей иммуноглобулина. кДНК, кодирующие константные области тяжелой цепи IgG, можно выделить на основе опубликованной последовательности из кДНК-библиотек, полученных из селезенки и лимфоцитов периферической крови, с помощью методик гибридизации или полимеразной цепной реакции (ПЦР). кДНК, кодирующие «адгезиновую» и иммуноглобулиновую части иммуноадгезина, встраивают тандемом в плазмидный вектор, обеспечивающий эффективную экспрессию в выбранных клетках-хозяевах.
Другие полипептиды, содержащие С Н 2/С Н 3-область
Предназначенный для очистки полипептид представляет собой слитый или конъюгированный с СН2/СН3-областью полипептид. Такие слитые полипептиды можно получать для того, чтобы увеличить время полужизни в сыворотке и/или чтобы обеспечить возможность очистки белка с помощью аффинной хроматографии на белке А. Примеры биологически важных белков, которые можно конъюгировать, таким образом включают ренин; гормон роста, включая гормон роста человека и бычий гормон роста; фактор высвобождения гормона роста; паратиреоидный гормон; тироидстимулирующий гормон; липопротеины; альфа-1-антитрипсин; А-цепь инсулина; В-цепь инсулина; проинсулин; фолликулостимулирующий гормон; кальцитонин, лютеинизирующий гормон; глюкагон; факторы свертывания крови, такие как фактор VIIIC, фактор IX, тканевой фактор (TF) и фактор вон Виллебранда; противосвертывающие факторы, такие как белок С; предсердный натрийуретический фактор; легочный сурфактант; активатор плазминогена, такой как урокиназа или из мочи человека, или тканевый активатор плазминогена (t-PA); бомбезин; тромбин; гемопоэтический фактор роста; факторы некроза опухоли альфа и бета; энкефалиназа; RANTES (регулируется при активации и в норме экспрессируется и секретируется Т-клетками); воспалительный белок макрофагов человека (MIP-1-альфа); сывороточный альбумин, такой как сывороточный альбумин человека; ингибирующая субстанция Мюллера; А-цепь релаксина; В-цепь релаксина; прорелаксин; мышиный гонадотропин-ассоциированный пептид; микробный белок, такой как бета-лактамаза; ДНКаза; IgE; цитотоксический Т-лимфоцит-ассоциированный антиген (CTLA), такой как CTLA-4; ингибин; активин; фактор роста эндотелия сосудов (VEGF); рецепторы гормонов или факторов роста; белок А или D; ревматоидные факторы; нейротрофический фактор, такой как костный нейротрофический фактор (BDNF), нейротрофин-3, -4, -5 или -6 (NT-3, NT-4, NT-5 или NT-6) или фактор роста нервов, такой как NGF-β; тромбоцитарный фактор роста (PDGF); фактор роста фибробластов, такой как αFGF и βFGF; эпидермальный фактор роста (EGF); трансформирующий фактор роста (TGF), такой как TGF-альфа и TGF-бета, включая TGF-β1, TGF-β2, TGF-β3, TGF-β4 или TGF-β5; инсулиноподобный фактор роста-I и -II (IGF-I и IGF-II); des(1-3)-IGF-1 (мозговой IGF-I), белки, связывающиеся с инсулиноподобным фактором роста; CD-белки, такие как CD3, CD4, CD8, CD19 и CD20; эритропоэтин; остеоиндуктивные факторы; иммунотоксины; костный морфогенный белок (BMP); интерферон, такой как интерферон α, β и γ; колониестимулирующие факторы (CSF), например, M-CSF, GM-CSF и G-CSF; интерлейкины (IL), например, от IL-1 до IL-10; супероксиддисмутаза; Т-клеточные рецепторы; поверхностные мембранные белки; фактор ускорения распада (CD55); вирусный антиген, такой как, например, часть оболочки ВИЧ; транспортные белки; «домашние» рецепторы; адрессины; регуляторные белки; интегрины, такие как CD11a, CD11b, CD11c, CD18, ICAM, VLA-4 и VCAM; ассоциированный с опухолью антиген, такой как EGFR, HER2-, HER3- или HER4-рецептор, и фрагменты любого из вышеперечисленных полипептидов.
Экспрессия полипептидов
Для очистки с использованием способа, описанного в настоящем документе, полипептид обычно получают, используя рекомбинантные методики. Полипептид также можно получить пептидным синтезом (или другим способом синтеза) или выделить из природного источника.
Для рекомбинантной продукции полипептида кодирующую его нуклеиновую кислоту выделяют и встраивают в реплицирующийся вектор для дальнейшего клонирования (амплификации ДНК) или для экспрессии. Кодирующую полипептид ДНК легко выделить и секвенировать, используя обычные процедуры (например, если полипептид представляет собой антитело, используя олигонуклеотидные пробы, способные специфично связываться с генами, кодирующими тяжелую и легкую цепи антитела). Имеется множество векторов. Компоненты вектора обычно включают без ограничения один или несколько следующих компонентов: сигнальную последовательность, точку инициации репликации, один или несколько маркерных генов, энхансерный элемент, промотор и последовательность терминации транскрипции (например, описанные в патенте США № 5534615, специально включенном в настоящий документ путем ссылки).
Подходящими клетками-хозяевами для клонирования или экспрессии ДНК в векторах по изобретению являются прокариотические клетки, дрожжевые клетки или клетки высших эукариот. Подходящие для указанной цели прокариоты включают эубактерии, такие как грамотрицательные или грамположительные организмы, например, Enterobacteriaceae, такие как Escherichia, например E. coli, Enterobacter, Erwinia, Klebsiella, Proteus, Salmonella, например, Salmonella typhimurium, Serratia, например, Serratia marcescans и Shigella, а также Bacilli, такие как B. subtilis и B. licheniformis (например, B. licheniformis 41P, описанную в DD 266710, опубликованном 12 апреля 1989), Pseudomonas, такие как P. aeruginosa, и Streptomyces. Указанные примеры являются иллюстративными, а не ограничивающими.
Кроме прокариот, подходящими клонирующими или экспрессирующими хозяевами для векторов, кодирующих полипептид, являются эукариотические микроорганизмы, такие как нитчатые грибы или дрожжи. Среди низших эукариотических микроорганизмов-хозяев наиболее широко используются Saccharomyces cerevisiae или обычные пекарские дрожжи. Однако ряд других родов, видов и штаммов являются общедоступными и применимыми в настоящем изобретении, такие как Schizosaccharomyces pombe; хозяева Kluyveromyces, такие как, например, K. lactis, K. fragilis (ATCC 12,424), K. bulgaricus (ATCC 16045), K. wickeramii (ATCC 24178), K. waltii (ATCC 56500), K. drosophilarum (ATCC 36906), K. thermotolerans и K. marxianus; Yarrowia (EP 402226); Pichia pastoris (EP 183070); Candida; Trichoderma reesia (EP 244234); Neurospora crassa; Schwanniomyces, такие как Schwanniomyces occidentalis; и нитчатые грибы, такие как, например, хозяева Neurospora, Penicillium, Tolypocladium и Aspergillus, такие как A. nidulans и A. niger.
Подходящие клетки-хозяева для экспрессии гликозилированных полипептидов получены из многоклеточных организмов. Примерами клеток беспозвоночных являются клетки растений и насекомых. Идентифицированы многочисленные бакуловирусные штаммы и варианты, а также соответствующие пермиссивные клетки-хозяева насекомых, таких как Spodoptera frugiperda (гусеница), Aedes aegypti (комар), Aedes albopictus (комар), Drosophila melanogaster (плодовая мушка) и Bombyx mori. Общедоступно множество вирусных штаммов для трансфекции, например, вариант L-1 NPV Autographa californica и штамм Bm-5 NPV Bombyx mori, и такие вирусы могут быть использованы в качестве вируса по изобретению, в частности, для трансфекции клеток Spodoptera frugiperda. В качестве хозяев также можно использовать культуры растительных клеток хлопчатника, кукурузы, картофеля, сои, петунии, томатов и табака.
Однако наибольший интерес представляли клетки позвоночных, и наращивание клеток позвоночных в культуре (культуре ткани) стало рутинной процедурой. Примерами применимых линий клеток-хозяев млекопитающих являются линия клеток почки обезьяны CV1, трансформированных SV40 (COS-7, ATCC CRL 1651); линия клеток эмбриональной почки человека (клетки 293 или 293, субклонированные для роста в суспензионной культуре, Graham et al., J. Gen Virol. 36: 59 (1977)); клетки почки новорожденного сирийского хомячка (BHK, ATCC CCL 10); клетки яичника китайского хомячка/DHFR (CHO, Urlaub et al., Proc. Natl. Acad. Sci. USA 77:4216 (1980)); клетки Сертоли мышей (TM4, Mather, Biol. Reprod. 23: 243-251 (1980); клетки почки обезьяны (CV1, ATCC CCL 70); клетки почки африканской зеленой обезьяны (VERO-76, ATCC CRL-1587); клетки карциномы шейки матки человека (HELA, ATCC CCL 2); клетки почки собаки (MDCK, ATCC CCL 34); клетки печени серой крысы (BRL 3A, ATCC CRL 1442); клетки легкого человека (W138, ATCC CCL 75); клетки печени человека (Hep G2, HB 8065); клетки опухоли молочной железы мыши (MMT 060562, ATCC CCL51); клетки TRI (Mather et al., Annals N.Y. Acad. Sci. 383: 44-68 (1982)); клетки MRC 5; клетки FS4 и клетки гепатомы человека (Hep G2).
Клетки-хозяева трансформируют вышеописанными экспрессирующими или клонирующими векторами для продукции полипептида и культивируют в обычных питательных средах, соответствующим образом модифицированных для индукции промоторов, отбора трансформантов или амплификации генов, кодирующих целевые последовательности.
Клетки-хозяева, используемые для получения полипептидов, используемых в способах по настоящему изобретению, можно культивировать в различных средах. Доступные в продаже среды, такие как среда Хама F10 (Sigma), минимальная питательная среда ((MEM), Sigma), RPMI-1640 (Sigma), модифицированная по Дульбекко среда Игла (DMEM), Sigma) подходят для культивирования клеток-хозяев. Кроме того, любую из сред, описанных в Ham et al., Meth. Enz. 58:44 (1979), Barnes et al., Anal. Biochem. 102:255 (1980), патентах США №№ 4767704, 4657866, 4927762, 4560655 или 5122469, WO 90/03430, WO 87/00195 или в патенте США № Re. 30985, можно использовать в качестве культуральной среды для клеток-хозяев. Любую из указанных сред можно при необходимости дополнить гормонами и/или другими факторами роста (такими как инсулин, трансферрин или эпидермальный фактор роста), солями (такими как хлорид натрия, кальция, магния и фосфат), буферами (такими как HEPES), нуклеотидами (такими как аденозин и тимидин), антибиотиками (такими как лекарственное соединение ГЕНТАМИЦИН™), микроэлементами (определяемыми как неорганические соединения, обычно присутствующие в конечных концентрациях в микромолярном диапазоне) и глюкозой или эквивалентным источником энергии. Любые другие необходимые добавки также можно включить в соответствующих концентрациях, которые будут известны специалистам в данной области. Условия культивирования, такие как температура, pH и т.п., представляют собой условия, используемые ранее в случае клеток-хозяев, выбранных для экспрессии, и будут известны среднему специалисту в данной области.
При использовании рекомбинантных методик полипептид можно продуцировать внутриклеточно, в периплазматическое пространство, или непосредственно секретировать в среду. Если пептид продуцируется внутриклеточно, на первой стадии удаляют частицы дебриса, либо клетки-хозяева, либо лизированные клетки (например, в результате гомогенизации), например, с помощью центрифугирования или ультрафильтрации. При секреции полипептида в среду супернатанты из таких экспрессионных систем обычно сначала концентрируют, используя доступный в продаже белковый концентрирующий фильтр, например, ультрафильтрующий элемент Amicon или Millipore Pellicon.
Композиции, включающие фармацевтические составы, содержащие полипептиды
Изобретение также включает композиции, такие как фармацевтические составы. Фармацевтический состав, содержащий полипептид, очищенный способами по настоящему изобретению, возможно конъюгированный с гетерологичной молекулой, можно получить смешиванием активного ингредиента, имеющего необходимую степень чистоты, с возможными фармацевтически приемлемыми носителями, вспомогательными веществами или стабилизаторами (Remington's Pharmaceutical Sciences 21st edition (2005)), в виде лиофилизированных составов или водных растворов.
Полипептидный продукт, содержащий СН2/СН3-область, очищенный способами, описанными в настоящем документе, может иметь желаемую степень чистоты по меньшей мере примерно 98% мономера или по меньшей мере примерно 99% мономера.
«Фармацевтически приемлемые» носители, вспомогательные вещества или стабилизаторы являются нетоксичными для реципиента в используемых дозах и концентрациях и включают буферы, как например, фосфат, цитрат и другие органические кислоты; антиоксиданты, включая аскорбиновую кислоту и метионин; консерванты (такие как хлорид октадецилдиметилбензиламмония; хлорид гексаметония; хлорид бензалкония, хлорид бензетония; фенол, бутиловый или бензиловый спирт; алкилпарабены, такие как метил- или пропилпарабен; катехин; резорцин; циклогексанол; 3-пентанол и м-крезол); низкомолекулярный полипептид (примерно меньше 10 остатков); белки, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глутамин, аспарагин, гистидин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включая глюкозу, маннозу или декстрины; хелатирующие агенты, такие как EDTA; сахара, такие как сахароза, маннит, трегалоза или сорбит; солеобразующие противоионы, такие как натрий; комплексы металлов (например, Zn-белковые комплексы); и/или неионогенные поверхностно-активные вещества, такие как TWEEN™, PLURONICS™ или полиэтиленгликоль (ПЭГ).
Состав по изобретению может также содержать более одного действующего соединения, необходимого при конкретном показании, против которого проводится лечение, предпочтительно соединений с дополняющей друг друга активностью, которые не влияют негативно друг на друга. Такие молекулы соответственно присутствуют в комбинации в количестве, эффективном для предполагаемой цели.
Действующие ингредиенты также можно заключить в микрокапсулы, приготовленные, например, методиками коацервации или полимеризации на границе фаз, например, микрокапсулы из гидроксиметилцеллюлозы или желатиновые микрокапсулы и микрокапсулы из полиметилметакрилата, соответственно, в коллоидные системы доставки лекарственных средств (например, липосомы, альбуминовые микросферы, микроэмульсии, наночастицы и нанокапсулы) или в макроэмульсии. Такие методики описаны в Remington's Pharmaceutical Sciences 21st edition (2005).
Составы, предназначенные для введения in vivo, должны быть стерильны. Это легко достигается фильтрацией через стерильные фильтрационные мембраны.
Можно получить препараты с замедленным высвобождением. Подходящие примеры препаратов с замедленным высвобождением включают полупроницаемые матрицы из твердых гидрофобных полимеров, содержащие вариант полипептида, причем такие матрицы имеют определенную форму, например, пленки или микрокапсулы. Примеры матриц для длительного высвобождения включают полиэфиры, гидрогели (например, поли(2-гидроксиэтилметакрилат) или поливиниловый спирт), полилактиды (патент США № 3773919), сополимеры L-глутаминовой кислоты и γ-этил-L-глутамата, недеградируемый этиленвинилацетат, деградируемые сополимеры молочной кислоты и гликолевой кислоты, такие как LUPRON DEPOT™ (инъекционные микросферы, состоящие из сополимера молочной кислоты и гликолевой кислоты и лейпролида ацетата), и поли-D-(-)-3-гидроксимасляную кислоту.
Полипептид, очищенный как описано в настоящем документе, или композицию, содержащую полипептид и фармацевтически приемлемый носитель, затем используют в различных диагностических, терапевтических или других способах применения, известных для таких полипептидов и композиций. Например, полипептид можно использовать для лечения заболевания у млекопитающего путем введения терапевтически эффективного количества полипептида млекопитающему.
Следующие примеры приведены для иллюстрации, а не для ограничения изобретения.
ПРИМЕРЫ
Следует понимать, что примеры и варианты осуществления, описанные в настоящем документе, приведены только в иллюстративных целях, и что различные модификации или изменения в свете приведенного описания будут очевидны для специалистов в данной области и включены в сущность и сферу применения данной заявки.
Пример 1: Хроматография на белке А с элюированием градиентом рН
Проводили очистку шести разных белков, содержащих СН2/СН3-область (анти-VEGF антитела №1, анти-CD20 антитела, анти-VEGF антитела №2, анти-MUC16 антитела, анти-CD4 антитела и анти-Met антитела с одним антиген-связывающим участком), на хроматографической колонке с белком А, используя способ элюирования ступенчатым градиентом рН. Способ кратко описан в таблице 1.
Описание эксперимента
В программе Unicorn был создан файл метода с использованием параметров белковой нагрузки, композиции буферов/рН, перечисленных в таблице 1. Этот файл выполнялся в системе GE Healthcare AKTA (GE Healthcare) Explorer FPLC (жидкостной экспресс-хроматографии белков). FPLC представляет собой настольный инструмент из пластиковых шлангов и насосов, имитирующий процесс промышленной очистки. FPLC создает «градиент рН» в результате смешивания двух буферов в программируемом меняющемся соотношении, используя систему из двух насосов (насоса А и насоса В), в которой поддерживается скорость потока и меняется процент потока от каждого из насосов. В программе Unicorn назначается «% В» относительно другого значения за указанный объем (число объемов колонки).
На стадии уравновешивания колонки, из колонки удаляют раствор для хранения, указанный в таблице 1, и подготавливают ее для нанесения белкового материала. На стадии нанесения белка HCCF (собранную клеточную культуральную жидкость) предварительно фильтруют, используя вакуумный фильтр с размером пор 0,2 микрона, и наносят на колонку с белком А (MABSELECT™, MABSELECT SURE™, POROS® MABCAPTURE™ A, PROSEP® Va или PROSEP® Ultra Plus). Белки наносят с плотностью в диапазоне 14-37 г/л. Большинство экспериментов проводили при нагрузке 21 г/л. На стадии 1-й промывки, использовали промывающий буфер №1 для нанесения на колонку любого белкового материала, оставшегося в шлангах хроматографа AKTA. На стадии 2-й промывки удаляли примеси, такие как CHOP (белки клеток яичника китайского хомячка), с помощью промывающего буфера №2. На стадии 3-й промывки использовали промывающий буфер №3 для удаления промывающего буфера №2 и связанных примесей с колонки для подготовки к стадии элюирования. На стадии элюирования ступенчатым градиентом рН градиент, образуемый точной регуляцией потоков двух рН-буферов с различными значениями рН с помощью двух насосных систем, использовали для постепенного движения от смеси из двух буферов с одним рН до другой смеси, указанной в процентах. Параметр элюирования «35-60% В» соответствует диапазону градиента рН примерно 4,3-3,7. Более конкретно, 35% В соответствует рН элюирующего буфера 4,34, композиции буфера 16,25 мМ ацетата, 8,75 мМ формиата, и проводимости буфера 1039 мкСм/см. 60% В соответствует рН элюирующего буфера 3,69, композиции буфера 10 мМ ацетата, 15 мМ формиата, и проводимости буфера 763 мкСм/см. На данной стадии элюирования в большинстве экспериментов собирали фракции, и анализировали их, используя в качестве метода анализа эксклюзионную ВЭЖХ для определения в ходе элюирования поведения мономера относительно вариантов (HMWS (высокомолекулярных соединений), димера или фрагментов). Для отдельного эксперимента проводили анализ СНОР, и во всех фракциях измеряли концентрацию белка используя УФ-спектрофотометр NanoDrop (Thermo Fischer Scientific, Wilmington, DE). На стадии регенерации использовали регенерирующий буфер для удаления сильно связанных примесей или остатков продукта, для того чтобы минимизировать перенос вещества между экспериментами. На стадии хранения из колонки с белком А удаляли регенерирующий буфер, и хранили колонку в растворе, подобранном для поддержания целостности колонки в нерабочем состоянии.
Длительность стадий и скорость потока измеряли в CV (объемах колонки) и сантиметрах в час, соответственно. Из-за давления скорость потока устанавливали в сантиметрах в час (скорость потока в см/ч делили на высоту слоя колонки для вычисления объемов колонки в час, также стандартной единицы).
Хроматограммы записывали и анализировали с использованием FPLC-системы AKTA (GE Healthcare) и связанного с ней пакета программного обеспечения Unicorn. После проведения очистки на колонке с белком А оценивали и изучали данные УФ-поглощения, рН и проводимости (а также другие измеренные значения или параметры программы/протоколы).
а. Анализ с помощью эксклюзионной хроматографии (SEC)
Анализ с помощью эксклюзионной хроматографии (SEC) проводили на Agilent 1200 series HPLC (Agilent Technologies, USA, кат. № G1329A) и использовали для определения относительного содержания высокомолекулярных соединений (HMWS), димера, мономера и фрагментов в собранных образцах. Использовали колонку TSK G3000SWXL, диаметром 7,8 мм, высотой 300 мм и объемом 14,24 мл (Tosoh Bioscience, Tokyo, Japan, кат. № 08541). Для стандартизации массы, наносимой на аналитическую колонку, каждый образец либо разбавляли приблизительно до концентрации антитела 0,5 г/л, используя HPLC-буфер (фосфат калия/хлорид калия), либо модифицировали введение образцов. Все образцы готовили в стеклянных ампулах Agilent HPLC объемом 1,5 мл. Хроматографию проводили в течение 30 мин при скорости 0,5 мл/мин. Введение образцов регулировали таким образом, чтобы наносить приблизительно 25 нг антитела на образец. Отрицательные контроли, содержащие соответствующие буферы для образцов, анализировали с каждым набором образцов. Кривые УФ-поглощения при 280 нм анализировали либо вручную, используя программное обеспечение ChemStation (Agilent Technologies), либо автоматически, используя программное обеспечение CHROMELEON® (DIONEX, Sunnyvale, CA), для интегрирования отдельных пиков, чтобы получить процентное содержание соединений в образцах. Процентное содержание, полученное в данном анализе, можно умножить на концентрацию (мг/мл) фракции, чтобы получить фактические концентрации или массу для каждого варианта соединения по размеру в образце (например, результат SEC: 4% HMWS, 3% димера, 92% мономера, 1% фрагмента; концентрация образца: 2 г/л; объем образца: 10 мл; концентрация мономера: 1,84 г/л, всего в образце 18,4 мг мономера).
b. Анализ CHOP
Образцы из отдельных экспериментов отбирали в аналитическую группу, для которой проводили стандартный и утвержденный твердофазный иммуноферментный анализ (ELISA) для количественного определения содержания CHOP. Аффинно-очищенные козьи антитела к СНОР иммобилизовали на лунках микротитровочного планшета. Разведения образцов, содержащих СНОР, стандарты и контроли, инкубировали в лунках с последующей инкубацией с козьими анти-СНОР антителами, конъюгированными с пероксидазой хрена. Ферментативную активность пероксидазы детектировали с использованием о-фенилендиаминдигидрохлороида. Количественное определение СНОР проводили, измеряя поглощение при 492 нм в микропланшетном спектрофотометре. Для создания стандартной кривой и автоматического вычисления концентрации СНОР в образце использовали компьютерную программу для аппроксимации кривых. Рабочий диапазон для ELISA обычно составлял от 5 нг/мл до 320 нг/мл. Для сравнения пулов результаты стандартизировали до ppm.
Результаты
Кривая элюирования ступенчатым градиентом показана на фиг.1. Все белки были элюированы до конца снижения рН. Большая часть белков элюировалась с колонки более быстро, но достаточно много белков оставалось на колонке и элюировалось по мере прохождения градиента. С понижением рН возникало разделение между целевым продуктом (диапазон элюирования от примерно рН 4,6 до 3,7 с небольшими вариациями между молекулами) и побочным агрегатом (диапазон элюирования примерно от 3,9 до 3,5).
Для всех шести протестированных белковых молекул (анти-VEGF антитела №1, анти-CD20 антитела, анти-VEGF антитела №2, анти-MUC16 антитела, анти-CD4 антитела и анти-Met антитела с одним антиген-связывающим участком) наблюдалось высокое содержание (~100%) мономера в первых фракциях градиента, а хвостовая часть градиента содержала более высокий уровень агрегированных соединений (>50%). Эти результаты SEC-анализа показаны на фиг.2-4В. Поскольку этот набор тестируемых молекул охватывает большие классы белковых молекул (например, химерные антитела, thioMab (рекомбинантные моноклональные антитела, имеющие точечную мутацию в результате замены одной аминокислоты цистеином), IgG4 и фрагменты антител, продуцируемые E. coli), то данный способ ступенчатого градиента рН предполагает широкое применение для всех полипептидов/белков, содержащих СН2/СН3-область (например, Fc-область).
Пример 2: Отделение СНОР от анти-CD20 антитела, анти-VEGF антитела №1 и анти-MUC16 антитела на хроматографической колонке с белком А с использованием стандартного ступенчатого элюирования и элюирования ступенчатым градиентом рН
При использовании способа элюирования белка ступенчатым градиентом рН, описанного в примере 1, измеряли содержание анти-CD20 антитела на фракцию в мг/мл (по УФ-поглощению при 280 нм на настольном спектрофотометре после хроматографии, которое совпадало с данными УФ-поглощения при 280 нм на AKTA/Unicorn в течение стадии элюирования ступенчатым градиентом, как видно на хроматограмме анти-VEGF антитела №1 на фиг.1А) и содержание СНОР на фракцию в ppm. Как видно из левой панели фиг.5 (элюирование анти-CD20 антитела и СНОР), более поздние фракции при более низких значения рН в ходе элюирования ступенчатым градиентом рН содержали очень мало анти-CD20 антитела по сравнению с количеством СНОР. Данные из контрольного эксперимента, проводимого в то же время, что и элюирование анти-CD20 антитела ступенчатым градиентом рН, показаны в верхней строке таблицы на фиг.5. В контрольном эксперименте использовали такие же условия, как описанные в таблице 1, за исключением того, что не использовали ступенчатый градиент рН на стадии элюирования белка, а белок элюировали при рН 3,6 или менее. В нижней строке показано маленькое, но приемлемое снижение выхода анти-CD20 антитела с использованием ступенчатого градиента рН (следует отметить, что снижение выхода наблюдалось вследствие ожидаемого удаления агрегатов; выход вычисляли, определяя значение титра HCCF, которое включает агрегаты в общее количество продукта, нанесенного на колонку). Примерно меньше 5% уровня агрегатов и меньше примерно половины уровня СНОР наблюдалось по сравнению с контрольным пулом. Результаты указывают на неожиданный эффект увеличения чистоты при использовании элюирования ступенчатым градиентом рН.
Также проводили отделение СНОР от анти-VEGF антитела №1 и анти-MUC16 антитела, используя способ ступенчатого градиента рН, описанный в примере 1. Аналогично ситуации, наблюдаемой на графике содержания СНОР для анти-CD20 антитела, большее количество СНОР элюировалось в конце градиента рН относительно элюирования анти-VEGF антитела, указывая на то, что элюирование ступенчатым градиентом рН отделяет примеси из клеток-хозяев для данной белковой молекулы, также как и для анти-CD20 антитела. См. фиг.6. В случае отделения СНОР от анти-MUC16 антитела, данное антитело имело гораздо более высокое содержание СНОР относительно картины элюирования, наблюдаемой для анти-VEGF антитела №1. См. фиг.7. Соответственно, значительное количество СНОР можно удалить из анти-MUC16 антитела, используя способ ступенчатого градиента рН, описанный выше.
Пример 3: Очистка от вирусных частиц с использованием хроматографии на белке А с элюированием ступенчатым градиентом рН
Оценивали очистку от вирусных частиц анти-VEGF антитела №1 с использованием способа хроматографии на белке А с элюированием ступенчатым градиентом рН, описанным в примере 1.
Хроматография на белке А с элюированием ступенчатым градиентом рН
Все стадии и буферы были такими же, как используемые в примере 1. В фракциях исследовали количество ретровирусоподобных частиц, используя количественную полимеразную цепную реакцию.
а. Количественная полимеразная цепная реакция для определения ретровирусоподобых частиц (RVLP QPCR)
Анализ эндогенных RVLP представляет собой количественный ПЦР в реальном времени. Вирусную РНК выделяли из образцов, используя Qiagen EZ1 (Qiagen, Valencia, CA). Объем образцов составлял 0,4 мл (неразведенную и разведенную 1:10 HCCF, неразведенный пул после элюирования с белка А). Эффективность выделения подтверждали, включая референсный стандартный образец HCCF с известным титром ретровирусных частиц из клеток СНО. Геномную ДНК удаляли расщеплением ДНКазой, обрабатывая выделенный элюат ДНКазой I в концентрации 0,2 ед./мл в течение 30 мин при повышенной температуре. Затем ДНКазу инактивировали при 70°C в течение 15 мин. Отсутствие ретровирусной ДНК подтверждали, анализируя образцы без добавления обратной транскриптазы.
Анализ с помощью ПЦР в реальном времени для количественного определения генома ретровирусов из клеток СНО проводили, как описано в De Wit et al. (Biologicals, 28(3): 137-48 (2000)), но используя новые пробы в расположенной рядом области. Реагенты и методики также были усовершенствованы и улучшены. Последовательности праймеров и проб были разработаны для амплификации фрагмента в высоко консервативной pol-области из генома ретровирусов типа С в клетках СНО. Каждая ретровирусоподобная частица содержит две молекулы геномной РНК. Олигонуклеотидные пробы и праймеры были заказаны в Applied Biosystems (Foster City, CA) и Invitrogen (Carlsbad, CA). Очистку от вирусных частиц выражали как число порядков снижения количества вируса или LRV, которая представляет собой log10 разницы между общим количеством вируса в нанесенном белке (HCCF) и пуле продукта. Общее количество вируса получали из титра вируса (частицы/мл или нейтр.ед./мл) в образцах и объема образца (мл). На фиг.18 показано число эндогенных вирусоподобных частиц для каждой фракции, собранной при элюировании ступенчатым градиентом рН. Некоторое количество вируса элюировалось в начале градиента в большом пике, но большая часть вирусов элюировалась в конце элюирования. Соответственно, отделение данных RVLP от продукта может обеспечить преимущество общей эффективности хроматографии на белке А с элюированием ступенчатым градиентом рН или полным градиентом в отношении очистки от вирусов. На фиг.19 показано LRV для каждой фракции в сравнении с наносом HCCF. График основан на вычислениях с использованием значений с фиг.18. Большое снижение LRV наблюдалось в более поздних элюируемых фракциях, обогащенных агрегатами. LRV было высоким (желаемый эффект) в середине элюирования, во фракциях с высоким содержанием полипептидного мономера.
Пример 4: Очистка от основных вариантов полипептида с использованием хроматографии на белке А с элюированием градиентом рН
Измеряли очистку анти-VEGF антитела №1 от основных вариантов полипептида (или основных вариантов) с использованием способа хроматографии на белке А с элюированием ступенчатым градиентом рН, описанного в примере 1.
Хроматография на белке А с элюированием ступенчатым градиентом рН
Все стадии и буферы были такими же, как используемые в примере 1. Фракции исследовали анализом вариантов с помощью ионообменной хроматографии.
а. Анализ вариантов с помощью ионообменной хроматографии
Аналитический ионообменный хроматографический (IEC) анализ проводили на Agilent 1200 series HPLC (Agilent Technologies, USA, кат. № G1329A) и использовали для определения относительного уровня основного пика к кислотным и основным заряженным вариантам для собранных образцов анти-VEGF антитела №1. Использовали колонку Dionex ProPac WCX-10, 4,6×250 мм (Dionex, кат. № 054993) с градиентом ACES [N-(2-ацетамидо)-2-аминоэтансульфоновой кислоты] и NaCl при повышенной температуре. Подготовка образцов включала смену буфера у образцов на подвижную фазу IEC перед 20-минутным расщеплением карбоксипептидазой (СрВ) при нагревании. В образце на колонку наносили приблизительно 50 мкг анти-VEGF антитела №1. Данные УФ-поглощения при 280 нм получали и интегрировали, используя программное обеспечение ChemStation (Agilent Technologies). Процентное содержание, полученное интегрированием для каждой категории пиков кислотного, основного и главного, анализировали на тенденцию изменения состава заряженных соединений по градиенту. На фиг.17 показан результат интегрирования пиков ионообменного анализа вариантов по 20 фракциям, полученным элюированием ступенчатым градиентом рН с белка А. Процентное содержание основных вариантов, присутствующих во фракциях, значительно повышалось в хвостовой части элюирования ступенчатым градиентом рН с белка А. Данная хвостовая часть элюирования градиентом представляла собой ту же самую часть, в которой наблюдалось повышенное содержание СНОР и агрегатов, описанное в примерах 1 и 2.
Пример 5: Элюирование ступенчатым градиентом рН с использованием различных хроматографических колонок с белком А
Обе смолы, MABSELECT SURE™ и MABSELECT™, тестировали в способе элюирования ступенчатым градиентом рН, описанным выше. Данные две смолы с белком А, несмотря на аналогичное название, имеют присоединенные лиганды с разной аффинностью. MABSELECT™ несет нативный белок А-лиганд, который связывает Fc-части антител. MABSELECT SURE™ несет модифицированный белок А, измененный химически для стабильности в растворах с высоким рН в течение короткого времени. Наложение профилей элюирования АКТА показывает, что кривые элюирования очень похожи для двух смол с белком А. Как видно из профиля SEC-интегрирования для MabSelect на фиг.8, с использованием данной смолы было получено сравнимое отделение агрегатов. Другими смолами, успешно протестированными в отношении отделения агрегатов, были PROSEP® Va, PROSEP® Ultra Plus и POROS® MABCAPTURE™ A. Соответственно, различные аффинные смолы (например, MABSELECT™, MABSELECT SURE™, PROSEP® Va, PROSEP® Ultra Plus и POROS® MABCAPTURE™) можно использовать в способе элюирования ступенчатым градиентом рН для разделения примесей.
Пример 6: Планирование экспериментов (DOE) с использованием различных параметров для способа элюирования градиентом рН
Для анти-VEGF антитела №1 были изучены различные параметры, важные в большинстве хроматографических процессов, с использованием статистически спланированного исследования (35 хроматографических прогонов) в диапазонах, показанных в таблице 2. Параметр «%В в начале элюирования» влияет на стартовый рН стадии элюирования (который играет главную роль в определении формы кривой элюирования). Чем выше начальный %В, тем ниже значение рН при начале элюирования, больше белка элюируется с колонки в первых фракциях, а также тем ниже общий наклон градиента. Параметры изменяли одновременно в дробном факторном эксперименте, разработанном для выявления главных эффектов, а также их взаимодействий. Для всех прогонов в ходе элюирования собирали фракции, и анализировали агрегаты и концентрацию во всех фракциях. Для определения содержания мономеров в символических пулах использовали интерполяционные методы вычисления для пулов, которые будут давать точно 85% выхода (нижний предел целевого выхода стадии), и данные значения использовали для сравнения эффективности отделения мономера от агрегатов для каждого набора параметров прогона.
Диапазоны параметров DOE
Ход эксперимента
Все изменения параметров кроме высоты слоя смолы (который требует набивки различных колонок) проверялись с использование программного обеспечения Unicorn. CV-фракции отбирались на протяжении всех элюирований, анализировались с использованием эксклюзионной ВЭЖХ (описанной выше), и в них измеряли концентрацию белка (УФ-поглощение при 280 нм на УФ-спектрофотометре NanoDrop). Для лучшей стандартизации результатов для сравнения собирали данные из различных наборов фракций и использовали вычисления для интерполяции общего выхода и SEC-профиля пула для каждого прогона.
Использовали программный пакет JMP® (SAS, Cary, NC) для создания плана прогонов дробного факторного исследования параметров. Один из прогонов был выбран из набора в качестве «примера промышленного прогона» благодаря своему высокому выходу, высокому уровню мономера и маленькому размеру пула (фиг.11).
Результаты
График Парето результатов DOE показывает, что «стартовый %В» является наиболее влиятельным параметром, определяющим отделение агрегатов, за которым следует плотность нагрузки (чем меньше, тем лучше) и время удерживания (время удерживания вычисляли делением контролируемых параметров высоты слоя смолы (см/CV) на скорость потока (см/ч), чтобы получить ч/CV). Соответственно, чем ниже стартовый %В, тем выше стартовый рН элюирования и более эффективно отделение агрегатов; чем ниже плотность нагрузки, тем более эффективно отделение агрегатов; и чем больше время удерживания, тем более эффективно отделение агрегатов. См. фиг.9. Кроме того, профили взаимодействия из данного исследования показывают, что существует взаимодействие между параметром плотности нагрузки и стартовым %В, а также существует взаимодействие между временем удерживания и плотностью нагрузки. При увеличенной плотности нагрузки стартовый %В обладает большим эффектом на содержание мономера в символических пулах с 85%-ым выходом, относительно того, если бы прогон проводили с меньшей плотностью нагрузки. Кроме того, при повышенной плотности нагрузки время удерживания играет гораздо большую роль в способности градиента разделять агрегаты более эффективно. При элюировании для получения высокой выработки продукта с использованием способа элюирования ступенчатым градиентом рН необходимо использовать более низкую скорость. См. фиг.10. На фиг.11 показано, что пул из этого прогона имел объем меньше 10 CV (например, 5,4 CV или 6 CV), в то же время внося меньше 1% агрегатов при выходе более 85%.
В дополнение к главным эффектам (график Парето на фиг.9), взаимодействиям (фиг.10) и примеру промышленного прогона (фиг.11) наблюдалось, что общая продолжительность элюирования не влияла на отделение агрегатов, но ее можно менять для снижения размера пула, чтобы получить в результате пул, все еще имеющий высокую чистоту и высокий выход, но в меньшем объеме (что предпочтительно для промышленного масштаба). Также использование ступенчатого градиента рН приводило к меньшим объемам пула с чистотой, практически сравнимой с чистотой, обеспечиваемой полным градиентом (см. пример 8). Также было найдено, что ступенчатый градиент рН устойчив относительно нескольких модификаций способа, включая меньшие изменения в плотности наноса и скорости потока, а также является полностью нечувствительным к высоте слоя смолы.
Пример 7: Очистка белка с использованием хроматографии на ионообменной мембране, следующей за хроматографией на белке А с элюированием ступенчатым градиентом рН
Для того чтобы определить, можно ли исключить последующие стадии колоночной хроматографии из способа очистки или заменить использованием мембран, лабораторный цикл очистки анти-VEGF антитела №1 с помощью хроматографии на белке А с элюированием ступенчатым градиентом рН (пул, содержащий меньше 1% агрегатов, и с высоким выходом) нагружали на следующие далее мембраны. Мембраны MUSTANG® S (Pall corporation) и MUSTANG® Q (Pall corporation) представляют собой катионообменную мембрану и анионообменную мембрану, соответственно. Успех определяли по способности мембран обеспечивать такую же общую чистоту и выход по сравнению с обычным последующим колоночным способом.
Ход эксперимента
Параметры, используемые для определения оптимальных условий нанесения для очистки от СНОР и агрегатов на S- и/или Q-мембранах, были взяты из предшествующих исследований по оптимальным условиям нагрузки для очистки от СНОР на S- и/или Q-мембране с использованием пула после ступенчатого элюирования с белка А в таблицах 3А и 3В. В этих предшествующих исследованиях были показаны перспективные результаты при использовании пула после стандартного ступенчатого элюирования с белка А (контрольная группа, в которой не использовали градиент рН и элюировали белок при рН 3,6 или меньшем значении); однако в этом способе содержание СНОР немного превышало целевой уровень, и этот способ не удалял никаких агрегатов из процесса. В этих предшествующих исследованиях мембраны нагружали в количестве до 5 кг/л мембраны и были найдены оптимальные условия для удаления примесей. Такие же условия использовали для пула после элюирования ступенчатым градиентом рН с белка А для сравнения очистки различных пулов, снятых с белка А, на данных мембранах относительно основных удаляемых примесей, такими как СНОР и агрегаты.
Предшествующие параметры для определения оптимальных условий наноса для очистки от СНОР на S- или Q-мембранах
Предшествующие параметры для определения возможности достижения дополнительной очистки от примесей в результате последовательного использования
S- и Q-мембран
В исходных исследованиях пулы после стандартного ступенчатого элюирования с белка А подводили до определенных значений рН и проводимости и пропускали через лабораторные катионообменную (MUSTANG® S) и анионообменную (MUSTANG Q) мембранные ячейки, соединенные с FPLC-системой очистки AKTA™. Скорость потока поддерживали, и изучали данные по давлению для информации о загрязнении/падении проницаемости (мембраны). Фракции отбирали при различной плотности нагрузки и анализировали содержание СНОР (с помощью ELISA) и концентрацию антител (по УФ-поглощению при 280 нм на УФ-спектрофотометре NanoDrop). Результаты этих анализов сравнивали для различной плотности нагрузки, чтобы найти оптимальные условия нанесения для конечной очистки от СНОР. В последних исследованиях пул после элюирования с белка А ступенчатым градиентом рН подводили до оптимума, найденного в предшествующих исследованиях. Далее сравнивали влияние пула после элюирования ступенчатым градиентом относительно стандартного ступенчатого элюирования с белка А на способность расположенных после мембран удалять СНОР и другие примеси, одновременно сохраняя высокий выход и низкое содержание агрегатов. Для проверки воспроизводимости и масштабируемости результатов также использовали два размера катионообменной мембраны и три размера анионообменной мембраны.
Результаты
При использовании пула после ступенчатого элюирования с белка А в качестве наносимого вещества на последовательность катионообменной и анионообменной мембран наименьший полученный уровень СНОР составлял 15-20 ppm для мембран под нагрузкой 5 кг/л мембраны. Поскольку мембраны не удаляют агрегат, то содержание этих примесей было неприемлемо высоким в конечных пулах. В отличие от этого при наносе пула после элюирования ступенчатым градиентом рН с белка А на такие же мембраны в конечных пулах содержание СНОР составляло 0-15 ppm, а содержание агрегатов было все еще низким, что показывает эффект на весь процесс.
Кроме того, протестированные конечные пулы каждой из трех комбинаций ионообменных мембран имели высокий выход и высокую чистоту. Во всех случаях уровень агрегатов оставался низким при подведении рН и пропускании через мембрану, а содержание СНОР также было ниже относительно наблюдаемого в предшествующих контрольных исследованиях, показывая неожиданный эффект снижения уровня СНОР при использовании для наноса пула с белка А с низким содержанием агрегатов. См. фиг.12. Соответственно, использование ионообменных хроматографических мембран вместо обычных ионообменных хроматографических колонок устраняет необходимость во многих обычных буферах для колоночной хроматографии и другие недостатки, связанные в затратами времени/пространства. Еще более важно то, что поскольку данные заряженные мембраны использовали в режиме перегрузки (то есть вещество пропускают через мембрану при высокой плотности нагрузки без стадий промывки или элюирования, необходимых для получения высоко очищенного пула), мембраны позволяют проводить непрерывную очистку после стадии хроматографии на белке А с элюированием градиентом рН.
Также показано, что профили SEC-интегрирования между пилотным промышленным масштабированием (4,1 л) и лабораторным масштабом (28 мл) являются весьма сходными, что указывает на возможность успешного масштабирования элюирования с белка А ступенчатым градиентом рН, а также указывает на то, что любые результаты маломасштабных экспериментов можно рассматривать как характеристики поведения крупномасштабных экспериментов. См. фиг.13. Эти результаты указывают на неожиданный эффект воспроизводимости и масштабируемости хроматографии на белке А с элюированием ступенчатым градиентом рН.
Пример 8: Эксперименты с парвовирусным фильтром VIRESOLVE® Pro
А. Сравнение падения проницаемости VIRESOLVE® Pro
Сравнивали ступенчатый градиент рН с белка А и стандартное ступенчатое элюирование с белка А (контрольная группа, в которой не использовали градиент рН, и белок элюировали при рН 3,6 или меньшем значении) относительно повышения массы, пропускаемой через парвовирусный фильтр (VIRESOLVE® Pro, Millipore, Inc.). Перед пропусканием через парвовирусный фильтр VIRESOLVE® Pro обычную HCCF очищали как хроматографией с элюированием ступенчатым градиентом рН с белка А, так и хроматографией со стандартным ступенчатым контрольным элюированием с белка А, и пропускали через QSFF (Q Sepharose Fast Flow (анионообменную колонку; GE Healthcare)) в стандартном проточном режиме. Было протестировано несколько условий прогонов через VIRESOLVE® Pro: 1) пул после стандартного ступенчатого элюирования с белка А и прохождения через встроенный стерильный префильтр SHC, 2) пул после стандартного ступенчатого элюирования с белка А и прохождения через катионообменный (СЕХ) мембранный адсорбент в качестве встроенного префильтра, 3) пул после элюирования ступенчатым градиентом рН с белка А и прохождения через SHC-префильтр (незаряженный стерильного уровня фильтр с размером пор 0,2 микрон), и 4) пул после элюирования ступенчатым градиентом рН с белка А и прохождения через СЕХ-мембранный адсорбент в качестве встроенного префильтра. Эксперименты с некоторыми пулами повторяли. VIRESOLVE® Pro использовали в фильтрующем устройстве, в котором используется набор перистальтических насосов, уравновешивающих устройств и датчиков давления для записи данных в электронную таблицу.
Пул после ступенчатого градиента рН и встроенного стерильного фильтра SHC вел себя аналогично пулу после ступенчатого элюирования с катионообменной мембраной, причем оба способа показывали примерно шестикратное увеличение возможной пропускаемой массы через VIRESOLVE® Pro. См. фиг.14. Соответственно, этот результат указывает на неожиданный эффект элюирования с белка А ступенчатым градиентом рН в удалении загрязнений с фильтра VIRESOLVE® Pro без помощи СЕХ-мембранного префильтра. Кроме того, эффективность VIRESOLVE® выигрывала в результате комбинации СЕХ-мембранного адсорбента с пулом после элюирования ступенчатым градиентом рН с белка А, приводя к неожиданному 18-кратному улучшению возможной пропускаемой массы по сравнению с прохождением пула после стандартного ступенчатого элюирования с белка А и SHC через VIRESOLVE® Pro.
В. Анализ СНОР и SEC-анализ после VIRESOLVE® Pro
Образцы отбирали в различные моменты в течение прогона и анализировали на содержание СНОР и с помощью SEC. Последовательность стадий стандартного ступенчатого элюирования с белка А с использованием встроенного SHC-фильтра приводила к тому, что образцы имели достаточно низкий уровень СНОР, но все еще содержали агрегаты. В экспериментах с элюированием с белка А с помощью ступенчатого градиента рН содержание агрегатов оставалось низким на протяжении различных стадий процесса и давало в конечном пуле меньше 1% агрегатов. Этот результат был значительно лучше результата для пула, получаемого после стандартного ступенчатого элюирования. Кроме того, уровень СНОР в пулах, получаемых элюированием ступенчатым градиентом рН в сочетании с SHC-префильтром и фильтром VIRESOLVE® Pro был сравнимым с уровнем СНОР после стандартного ступенчатого элюирования с катионообменной мембраной. Уровни были немного ниже получаемых после стандартной стадии с SHC. См. таблицы 4А и 4В. Эти результаты показывают, что способ с использованием ступенчатого градиента рН не только обеспечивает большую пропускаемую массу по сравнению с пулом после стандартной стадии, но также обеспечивает сравнимую или лучшую чистоту.
Пример 9: Элюирование с белка А полным градиентом рН
Также тестировали элюирование с белка А полным градиентом рН. Все стадии и буферы были такими же, как используемые в способе ступенчатого градиента рН, описанном в примере 1, за исключением того, что градиент 35-60% В, используемый в ступенчатом градиенте рН, был снижен до 25-60% В в начале градиента, что приводило к более высокому рН в начале градиента (начинающемуся примерно с рН 4,6 и заканчивающемуся примерно на 3,7). 25% В соответствует элюирующему буферу с рН 4,58, композиции буфера из 18,75 М ацетата и 6,25 мМ формиата и проводимости буфера 1141 мкСм/см. 60% В соответствует элюирующему буферу с рН 3,69, композиции буфера из 10 М ацетата и 15 мМ формиата и проводимости буфера 763 мкСм/см. См. таблицу 5.
Кривая элюирования полным градиентом показана на фиг.15. Все продукты элюировались до конца снижения рН. Хроматограмма для полного градиента рН была такой же для всех стадий как в случае ступенчатого градиента рН, за исключением того, что элюирование антитела начиналось при более высоком рН. Отсутствие первоначального высокого пика при 280 нм в начале градиента приводило к фракциям с более низкой концентрацией в начале градиента с большим количеством белка, распределенным по остальной стадии элюирования. Это позволяет лучшее разделение веществ, которые элюируются предпочтительно при более высоком рН чем мономер. Таким образом, эта методика может быть одинаково эффективна в очистке от примесей, которые отделяются от целевого продукта при более низких значениях рН (а именно, агрегатов и СНОР, отделяемых с использованием ступенчатого градиента), с единственным недостатком в виде объема конечного пула по сравнению со ступенчатым градиентом рН (более низкие концентрации во фракциях в начала градиента вызывают необходимость объединения большего числа фракций для достижения символического пула с желаемым выходом). Данные SEC-анализа и интегрирования для данного полного градиента также демонстрируют отделение агрегатов от мономера при более низких значениях рН, позволяя предположить, что эффекты и применение ступенчатого градиента рН можно также распространить и на полный градиент.
Пример 10: Образование агрегатов при элюировании с белка А
Для гарантии того, что агрегаты не образуются при низком рН или в хвостовой области элюирования ступенчатым градиентом рН, а также того, что методика элюирования с белка А ступенчатым градиентом рН в действительности отделяет мономер от агрегатов, а не вызывает образование агрегатов, использовали очищенные материалы для того, чтобы показать, что содержание агрегатов наносимого материала не увеличивалось при обработке на белке А с или без компонентов HCCF. См. таблицу 6.
| название | год | авторы | номер документа |
|---|---|---|---|
| ОЧИСТКА АНТИТЕЛ С ПОМОЩЬЮ КАТИОНООБМЕННОЙ ХРОМАТОГРАФИИ | 2008 |
|
RU2498991C2 |
| СПОСОБЫ И КОМПОЗИЦИИ, ВКЛЮЧАЮЩИЕ ОЧИЩЕННЫЕ РЕКОМБИНАНТНЫЕ ПОЛИПЕПТИДЫ | 2014 |
|
RU2671481C2 |
| РАЗДЕЛЕНИЕ БИСПЕЦИФИЧЕСКИХ АНТИТЕЛ И ПОБОЧНЫХ ПРОДУКТОВ ПРОЦЕССА ПОЛУЧЕНИЯ БИСПЕЦИФИЧЕСКИХ АНТИТЕЛ С ПРИМЕНЕНИЕМ ГИДРОКСИАПАТИТНОЙ ХРОМАТОГРАФИИ | 2014 |
|
RU2652911C2 |
| СПОСОБ ВЫДЕЛЕНИЯ ПОВТОРНО СВЕРНУТОГО РЕКОМБИНАНТНОГО БЕЛКА ИЗ КУЛЬТУРЫ ПРОКАРИОТИЧЕСКИХ КЛЕТОК (ВАРИАНТЫ) | 2007 |
|
RU2439076C2 |
| СПОСОБЫ ПОВЫШЕНИЯ ЭФФЕКТИВНОСТИ ЭТАПОВ ОЧИСТКИ БЕЛКА, НАХОДЯЩИХСЯ НИЖЕ ПО ПОТОКУ, С ИСПОЛЬЗОВАНИЕМ МЕМБРАННОЙ ИОНООБМЕННОЙ ХРОМАТОГРАФИИ | 2012 |
|
RU2648999C2 |
| СПОСОБЫ И КОМПОЗИЦИИ, ВКЛЮЧАЮЩИЕ ОЧИЩЕННЫЕ РЕКОМБИНАНТНЫЕ ПОЛИПЕПТИДЫ | 2014 |
|
RU2793783C1 |
| ОСАЖДЕНИЕ И ОЧИСТКА БЕЛКОВ ПОЛИЭЛЕКТРОЛИТАМИ | 2008 |
|
RU2474585C2 |
| СПОСОБ ОЧИСТКИ БЕЛКА СЛИЯНИЯ | 2015 |
|
RU2698654C2 |
| ПОЛУЧЕНИЕ ГЕТЕРОМУЛЬТИМЕРНЫХ БЕЛКОВ | 2011 |
|
RU2624027C2 |
| СПОСОБЫ ОЧИСТКИ ОДНОДОМЕННЫХ АНТИГЕНСВЯЗЫВАЮЩИХ МОЛЕКУЛ | 2009 |
|
RU2553214C2 |

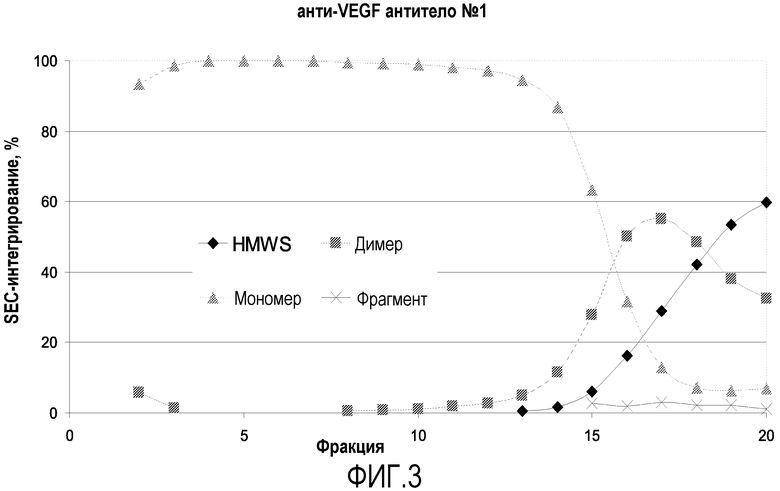
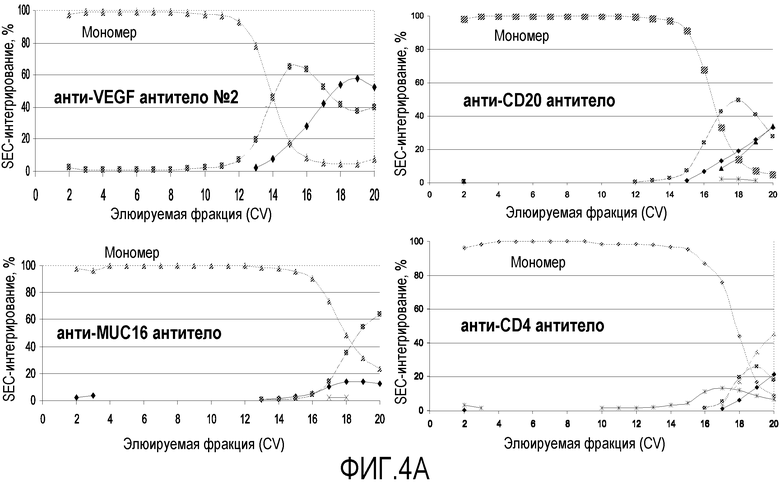
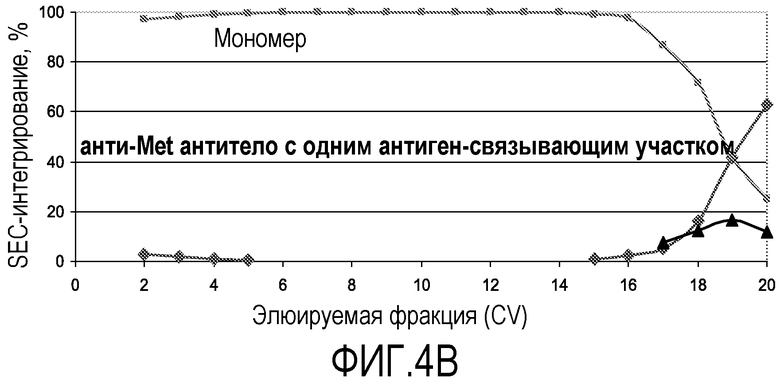
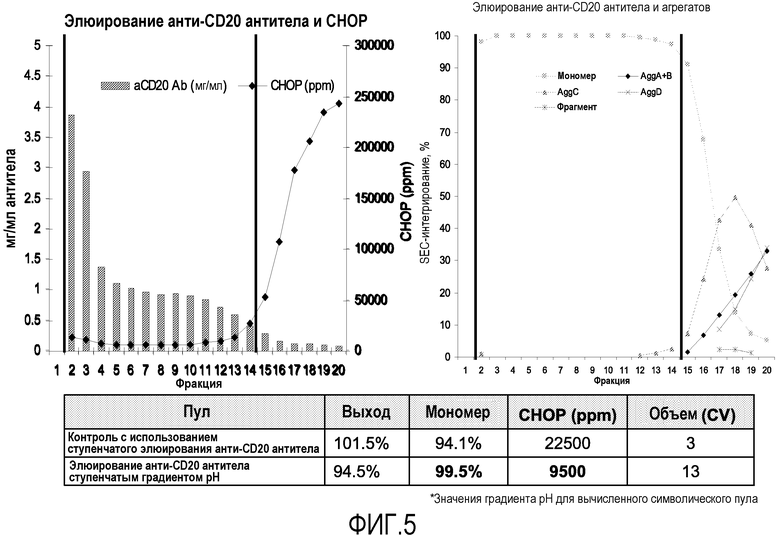
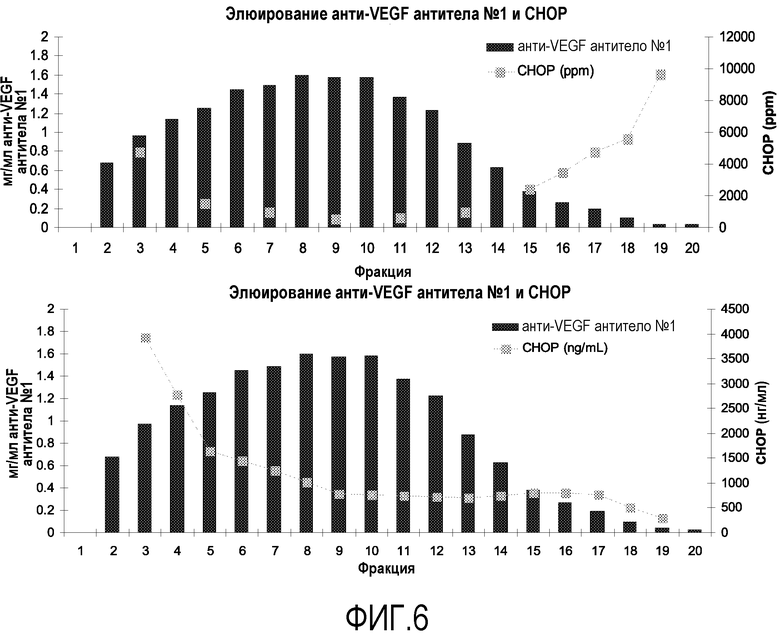
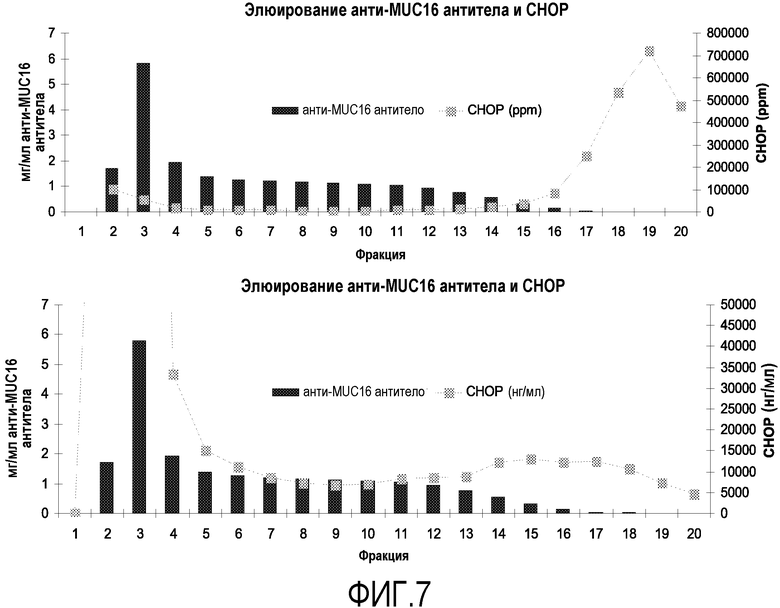
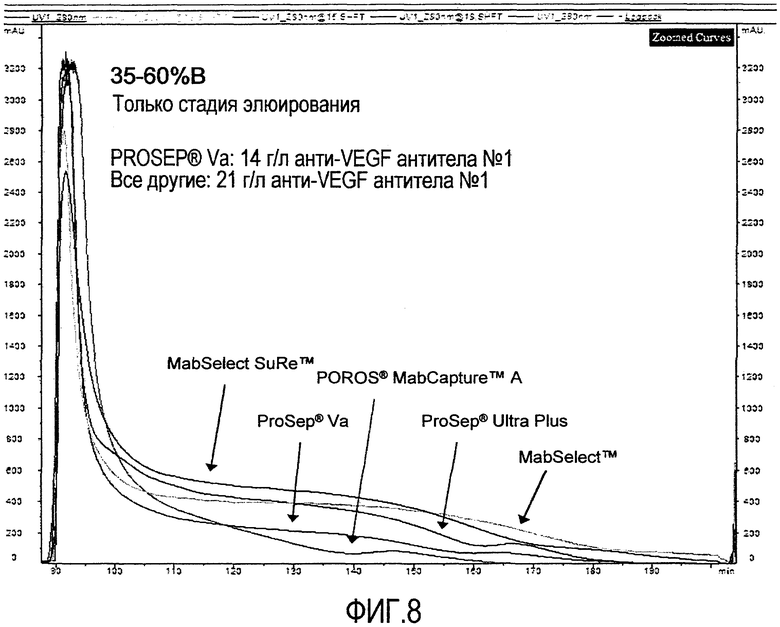
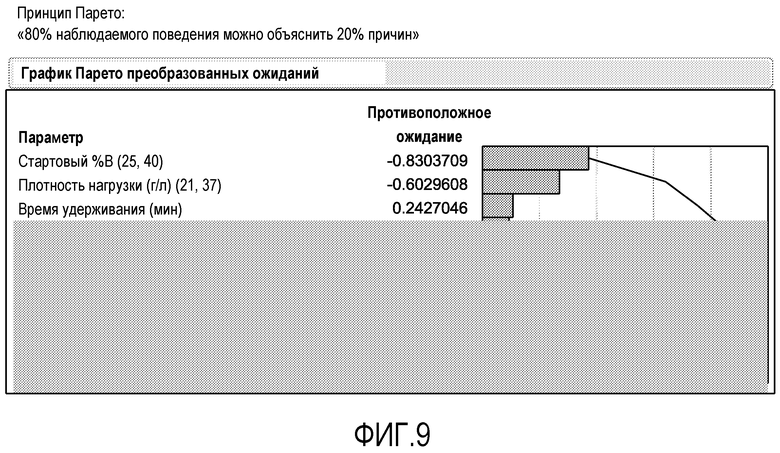
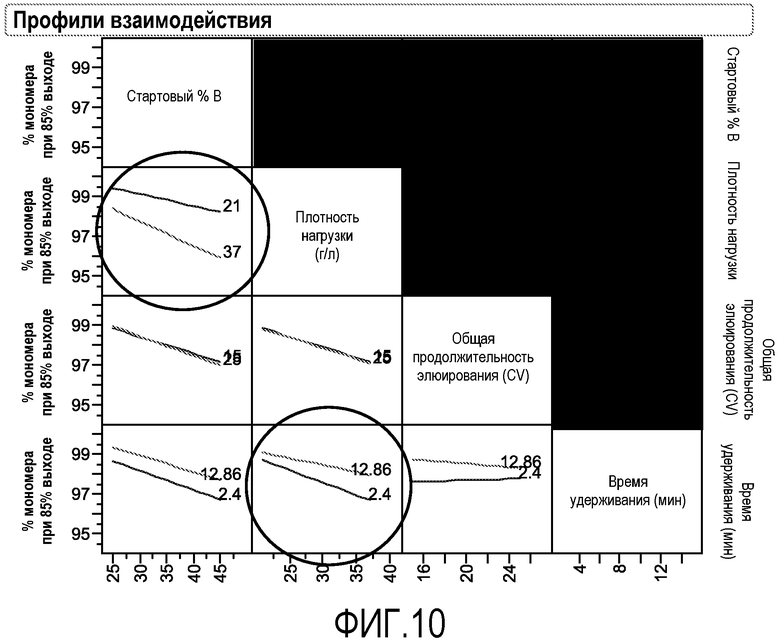
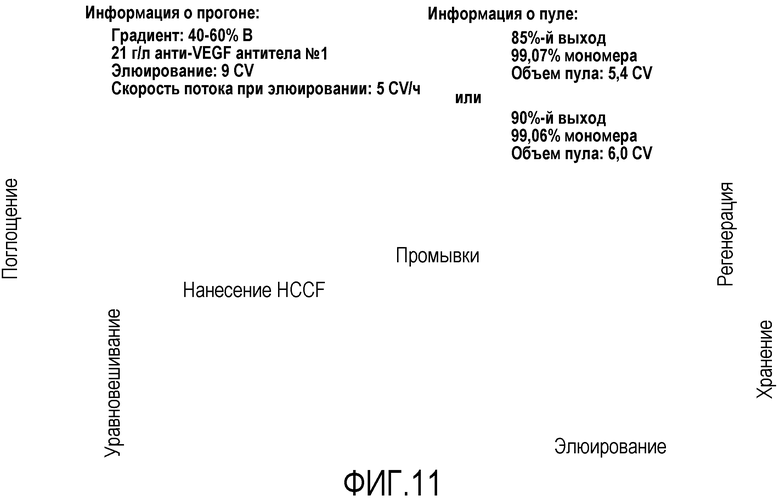
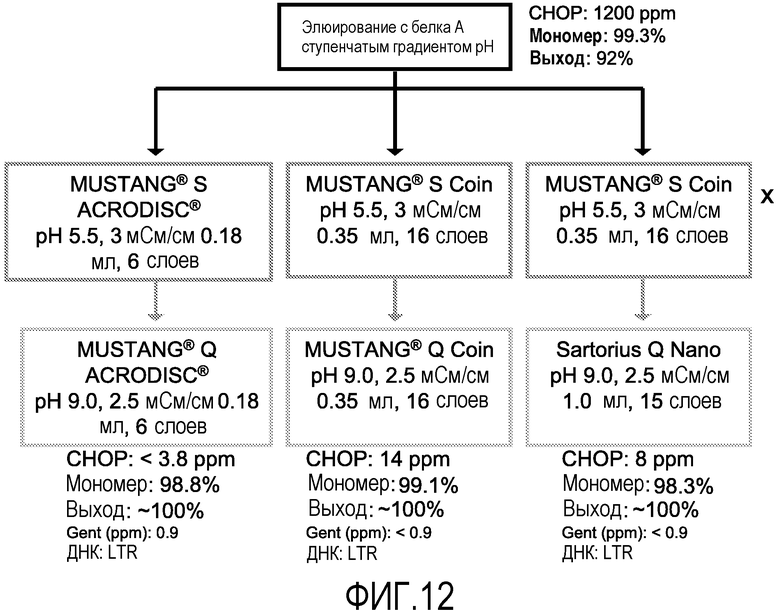
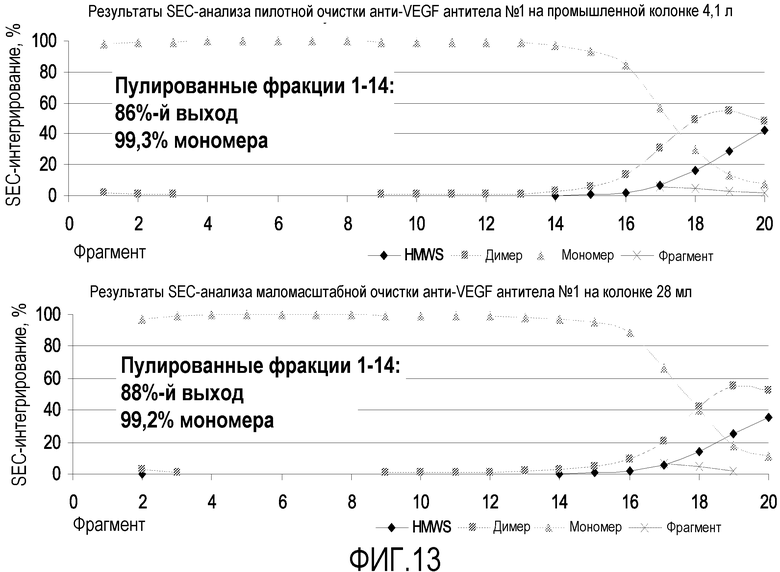
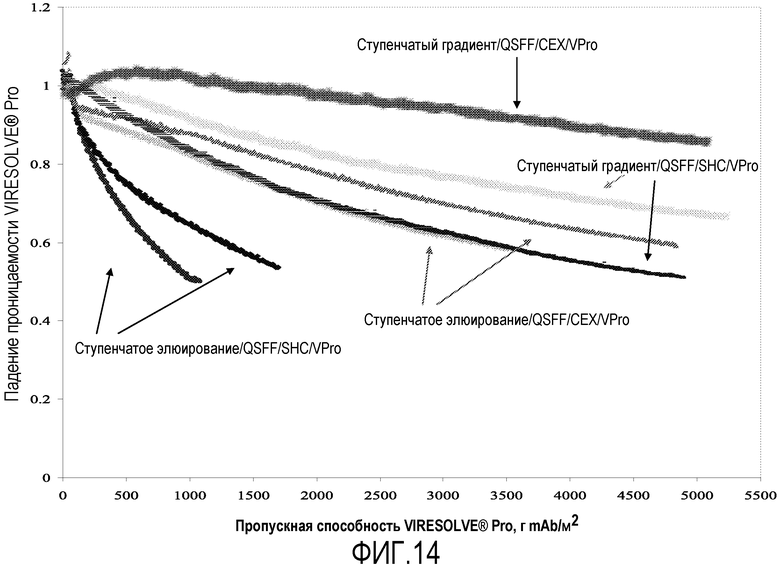

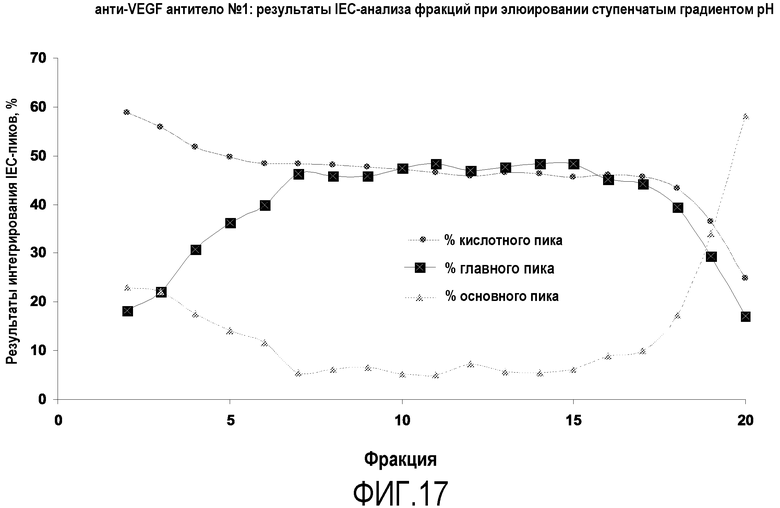
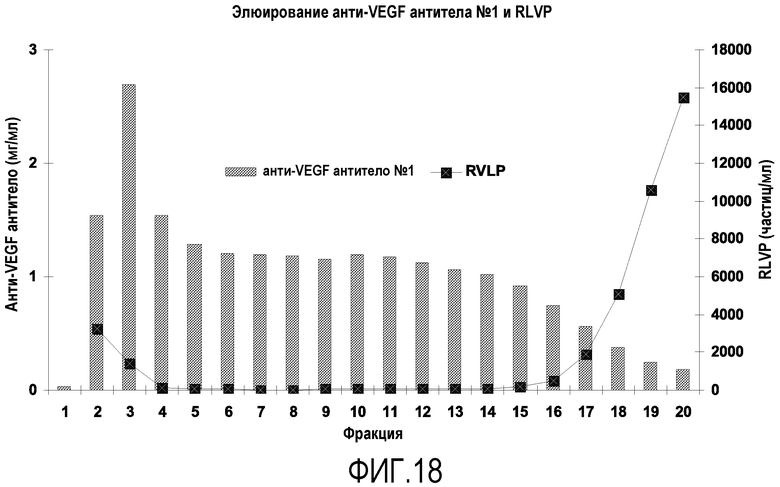
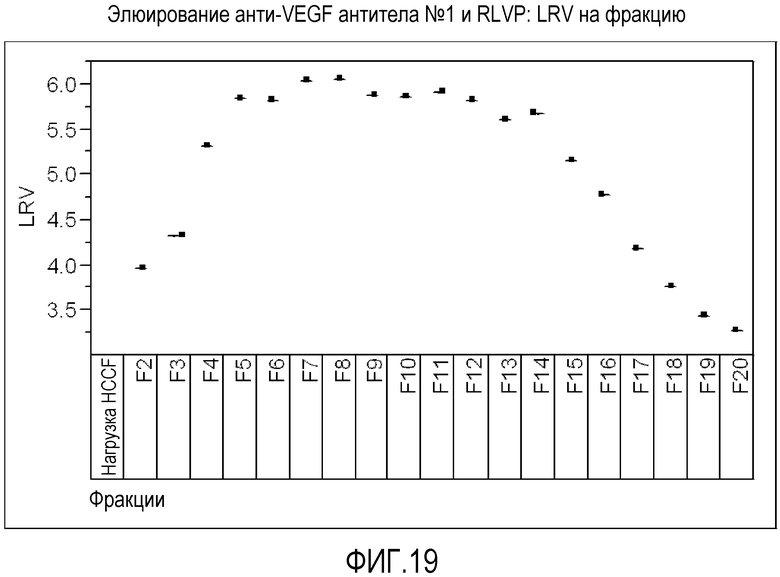
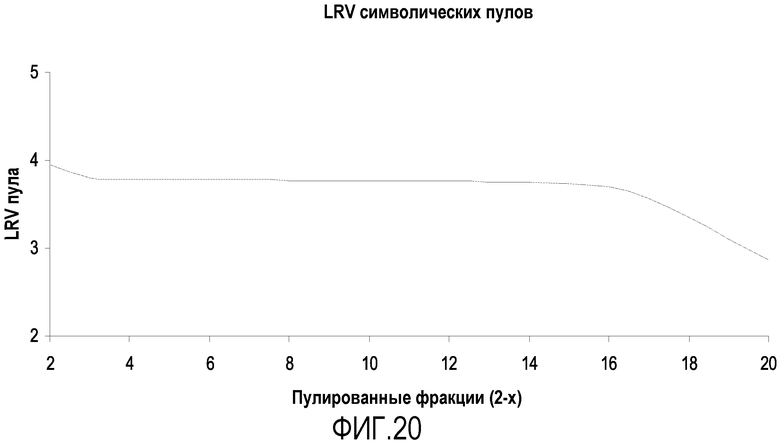
Изобретение относится к биотехнологии, а именно к способу очистки полипептида, содержащего Сн2/Сн3-область. Способ включает связывание полипептида с белком А и элюирование полипептида с градиентом pH, начинающимся с 5,0 или менее, с использованием элюирующего буфера, причем элюирующий буфер включает буфер с высоким pH и буфер с низким pH, а градиент pH образуется в результате регулирования процентного содержания каждого pH-буфера в элюирующем буфере. Буфер с высоким pH имеет pH примерно 5,0. Буфер с низким pH имеет pH примерно 2,7. Изобретение позволяет отделить агрегат, примесь из клеток-хозяев, загрязнение вирусного фильтра, вирусную частицу или вирусоподобную частицу от требуемого полипептида. 20 з.п. ф-лы, 21 ил., 8 табл., 11 пр.
1. Способ очистки полипептида, содержащего Сн2/Сн3-область, включающий стадии:
(a) связывания полипептида с белком А; и
(b) элюирования полипептида с градиентом pH, начинающимся с 5,0 или менее, с использованием элюирующего буфера, причем элюирующий буфер включает буфер с высоким pH и буфер с низким pH, а градиент pH образуется в результате регулирования процентного содержания каждого pH-буфера в элюирующем буфере,
где буфер с высоким pH имеет pH примерно 5,0, и где буфер с низким pH имеет pH примерно 2,7, и где агрегат, примесь из клеток-хозяев, загрязнение вирусного фильтра, вирусную частицу или вирусоподобную частицу отделяют от требуемого полипептида.
2. Способ по п. 1, в котором процентное содержание буфера с низким pH начинается примерно с 35%.
3. Способ по п. 2, в котором элюирующий буфер, содержащий примерно 35% буфера с низким pH, содержит примерно 16,25 мМ ацетат и примерно 8,25 мМ формиат.
4. Способ по п. 1, в котором процентное содержание буфера с низким pH начинается примерно с 25%.
5. Способ по п. 4, в котором элюирующий буфер, содержащий примерно 25% буфера с низким pH, содержит примерно 18,75 мМ ацетат и примерно 6,25 мМ формиат.
6. Способ по п. 1, в котором процентное содержание буфера с низким pH начинается примерно с 40%.
7. Способ по п. 6, в котором элюирующий буфер, содержащий примерно 40% буфера с низким pH, содержит примерно 15 мМ ацетат и примерно 10 мМ формиат.
8. Способ по п. 1, в котором градиент pH начинается примерно с pH 4,6.
9. Способ по любому из пп. 1-8, в котором градиент pH заканчивается на 3,0 или большем значении.
10. Способ по п. 9, в котором градиент pH заканчивается примерно на 3,7.
11. Способ по любому из пп. 1-8 или 10, в котором примесь из клеток-хозяев является белком из клеток яичника китайского хомячка (CHOP).
12. Способ по любому из пп. 1-8, 10, в котором основный вариант полипептида отделяют от полипептида.
13. Способ по любому из пп. 1-8, 10, в котором полипептид представляет собой антитело.
14. Способ по п. 13, в котором антитело является моноклональным антителом, поликлональным антителом, полиспецифичным антителом или фрагментом антитела.
15. Способ по п. 13, где антитело представляет собой анти-VEGF антитело, анти-CD20 антитело, анти-MUC16 антитело, анти-CD4 антитело или анти-МЕТ антитело.
16. Способ по любому из пп. 1-8, 10 или 14, в котором полипептид является иммуноадгезином.
17. Способ по любому из пп. 1-8, 10, или 14, в котором полипептид имеет чистоту по меньшей мере примерно 98% мономера.
18. Способ по п. 1, в котором соотношение примесей из клеток-хозяев и очищенного полипептида по меньшей мере примерно на 20% ниже соотношения в полипептиде, очищенном способом с использованием ступенчатого элюирования, причем способ с использованием ступенчатого элюирования включает связывание полипептида с белком А и элюирование с pH, начиная с 3,6 или менее.
19. Способ по п. 18, в котором соотношение примесей из клеток-хозяев и очищенного полипептида по меньшей мере примерно на 60% ниже соотношения в полипептиде, очищенном способом с использованием ступенчатого элюирования, причем способ с использованием ступенчатого элюирования включает связывание полипептида с белком А и элюирование с pH, начиная с 3,6 или менее.
20. Способ по любому из пп. 18 или 19, в котором очищенный полипептид является мономером полипептида.
21. Способ по любому из пп. 1-8, 10, 14, 18 или 19, в котором очистка является промышленным способом.
| DUHAMEL R.C | |||
| ET AL., pH gradient elution of human IgG1, IgG2 and IgG4 from protein A-Sepharose, JOURNAL OF IMMUNOLOGICAL METHODS, 1979, v | |||
| Способ очистки нефти и нефтяных продуктов и уничтожения их флюоресценции | 1921 |
|
SU31A1 |
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| Способ добывания бензина и иных продуктов из нефти, нефтяных остатков и пр. | 0 |
|
SU211A1 |
| WO2008025747 A1, 06.03.2008 | |||
| WO 2008085988 A1, 17.07.2008 | |||
| US 20080167450 A1, 10.07.2008 | |||
| PAN H | |||
| ET AL., Quantitation of soluble aggregates in recombinant monoclonal antibody cell | |||

