Область техники
Изобретение относится к области биотехнологии и касается способа получения целевых биологически активных рекомбинантных белков и полипептидов (протеинов, белков-мишеней), производимых с использованием микробиологического синтеза.
Уровень техники
Традиционные способы микробиологического синтеза биологически активных рекомбинантных белков и полипептидов (протеинов, белков-мишеней) предусматривают создание и использование генетических конструкций, штаммов и методов их культивирования, а также, как правило, включают сложные комбинации хроматографических методов очистки целевых продуктов. Разработка и применение этих способов обычно требует значительных экономических и временных затрат.В этой связи большую актуальность приобретают универсальные способы, позволяющие ускорить разработку технологий получения белков-мишеней и снизить себестоимость конечных продуктов [обзоры Yang X., Pistolozzi М, Lin Zh. New trends in aggregating tags for therapeutic protein purification. Biotechnology Letters, 2018, 40, 745-753; Gomari M.M., Saraygord-Afshari N., Farsimadan M., Rostami N., Aghamiri S., Farajollahi M.M. Opportunities and challenges of the tag-assisted protein purification techniques: Applications in the pharmaceutical industry. Biotechnol Adv, 2020, 45, 107653].
Их разработка требует решения, по крайней мере, двух взаимосвязанных технологических задач: получения целевых белков-мишеней в биологически активном состоянии и их хотя бы первичного отделения/очистки от основной массы примесных клеточных белков. При этом успешное решение первой задачи, например, путем биосинтеза белков-мишеней с использованием развитой технологии растворимых гибридных белков, включающих универсальные аффинные метки, как правило, приводит к необходимости применения дорогостоящих сорбентов на стадии аффинной очистки. С другой стороны, удешевление очистки путем применения центрифугирования или фильтрации целевых белков, получаемых в нерастворимом виде в тельцах включения (ТВ) в клетках E.coli, сопровождается необходимостью разрабатывать трудоемкие и не до конца эффективные методы восстановления биологически активного состояния (корректной конформации) белков-мишеней с применением процедур денатурации/ренатурации.
В этой связи актуальность приобретают компромиссные способы получения биологически активных белков-мишеней, не требующие применения процедур денатурации/ренатурации и при этом позволяющие для очистки/отделения целевых продуктов, по крайней мере, первичной, использовать недорогие методы центрифугирования или фильтрования.
Из предшествующего уровня техники известна группа способов получения белков-мишеней с помощью растворимых SUMO-фьюжнов, включающих последовательности белков SUMO и пептидных тэгов, опосредующих аффинную очистку [Butt T.R., Edavettal S.C., Hall J.P., Mattern M.R. SUMO fusion technology for difficult-to-express proteins. Protein Expr Purif, 2005, 43(1), 1 -9; Marblestone J.G., Edavettal S.C., Lim Y., Lim P., Zuo X., Butt T.R. Comparison of SUMO fusion technology with traditional gene fusion systems: Enhanced expression and solubility with SUMO. Protein Set, 2006, 15(1), 182-189]. Они также обеспечивают получение белков-мишеней в биологически активном состоянии без использования ренатурации и позволяют упростить их очистку за счет использования универсальных аффинных сорбентов. Для их реализации наиболее часто используют белок SUMO дрожжей Saccharomyces cerevisiae [Johnson et al., 1997; Muller et al., 1998], кодируемый геном SMT3 и состоящий из 98 аминокислотных остатков [MossessovaE., Lima CD. Ulp1-SUMO crystal structure and genetic analysis reveal conserved interactions and a regulatory element essential for cell growth in yeast. Mol Cell, 2000, 5(5), 865-876]. Использование SUMO является ключевым элементом и залогом высокой результативности указанной группы способов. Присутствие SUMO обеспечивает высокие показатели растворимости и процессинга фьюжнов, а также стимулирует корректную свертку белков-мишеней [Malakhov М.Р., Mattern M.R., Malakhova О.А., Drinker M., Weeks S.D., Butt T.R. SUMO fusions and SUMO-specific protease for efficient expression and purification of proteins. J Struct Funct Genomics, 2004, 5(1-2), 75-86; Butt T.R., Edavettal S.C., Hall J.P., Mattern M.R. SUMO fusion technology for difficult-to-express proteins. Protein Expr Purif, 2005, 43(1), 1-9; Marblestone J.G., Edavettal S.C., Lim Y., Lim P., Zuo X., Butt T.R. Comparison of SUMO fusion technology with traditional gene fusion systems: Enhanced expression and solubility with SUMO. Protein Set, 2006, 15(1), 182-189].
При реализации указанной группы способов захват и первичные стадии очистки белков-мишеней (из клеточных лизатов) проводят без разрушения SUMO-фьюжнов. Высвобождение белков-мишеней из состава SUMO-фьюжнов осуществляют на завершающих стадиях очистки с использованием ферментативного гидролиза (процессинга), осуществляемого под действием производных SUMO-специфичной протеиназы Ulp1, например, ULP275 [RU 2451076] или других вариантов [Li S.-J., Hochstrasser М. The Ulp1 SUMO isopeptidase: distinct domains required for viability, nuclear envelope localization, and substrate specificity. J Cell Biol, 2003, 160, 1069-1081; Butt T.R., Edavettal S.C., Hall J.P., Mattern M.R. SUMO fusion technology for difficult-to-express proteins. Protein Expr Purif, 2005, 43(1), 1-9].
Возможность использовать производные протеиназы Ulp1 относят к преимуществам указанной группы способов, поскольку:
1) действие протеиназ, направленное на пептидную или изопептидную связь, соединяющую консервативный С-концевой дипептид Гли-Гли SUMO и следующий с ним аминокислотный остаток белка-мишени, обладает исключительной высокой селективностью и практически не зависит от природы этого аминокислотного остатка (исключая пролин), который, в том числе, может быть представлен целым структурным элементом, включающим дисульфидную связь [RU 2441072];
2) протеиназы обладают чрезвычайно высокой активностью и обеспечивают эффективный гидролиз SUMO-фьюжнов в соотношении фермента к субстрату 1: 5000 и более [Malakhov М.Р., Mattern M.R., Malakhova О.A., Drinker М., Weeks S.D., Butt T.R. SUMO fusions and SUMO-specific protease for efficient expression and purification of proteins. J Struct Fund Genomics, 2004, 5(1-2), 75-86; Butt T.R., Edavettal S.C., Hall J.P., Mattern M.R. SUMO fusion technology for difficult-to-express proteins. Protein Expr Purif, 2005, 43(1), 1-9; Marblestone J.G., Edavettal S.C., Lim Y, Lim P., Zuo X., Butt T.R. Comparison of SUMO fusion technology with traditional gene fusion systems: Enhanced expression and solubility with SUMO. Protein Sci., 2006, 15(1), 182-189];
3) протеиназы обладают чрезвычайно высокой специфичностью и способны безошибочно распознавать SUMO в составе любых SUMO-фьюжнов, поскольку взаимодействуют не с отдельным сайтом, а с трехмерной структурой SUMO [Wilkinson K.D. Regulation of ubiquitin-dependent processes by deubiquitinating enzymes. FASEB J, 1997, 11, 1245-1256; Chung C.H., Baek S.H. Deubiquitinating enzymes: their diversity and emerging roles. Biochem Biophys Res Commun, 1999,266, 633-640; Mossessova E., Lima CD. Ulp1-SUMO crystal structure and genetic analysis reveal conserved interactions and a regulatory element essential for cell growth in yeast. Mol Cell, 2000, 5(5), 865-876; Butt T.R., Edavettal S.C., Hall J.P., Mattern M.R. SUMO fusion technology for difficult-to-express proteins. Protein Expr Purif, 2005, 43(1), 1-9];
4) протеиназы обладают низкой чувствительности к условиям ферментативной реакции [Malakhov M.R, Mattern M.R., Malakhova О.А., Drinker M., Weeks S.D., Butt T.R. SUMO fusions and SUMO-specific protease for efficient expression and purification of proteins. J Struct Funct Genomics, 2004, 5(1-2), 75-86; Butt T.R., Edavettal S.C., Hall J.P., Mattern M.R. SUMO fusion technology for difficult-to-express proteins. Protein Expr Purif, 2005, 43(1), 1-9].
В совокупности уникальные физико-химические характеристики SUMO и уникальные ферментативные характеристики производных протеиназы Ulp1 обеспечивают их эффективное применение для решения широкого круга задач: (1) увеличения экспрессии белков-мишеней в клетках эукариот и прокариот; (2) увеличения растворимости и стимулирования корректной свертки белков-мишеней в составе SUMO-фьюжнов в ходе их биосинтеза; (3) упрощения очистки белков-мишеней; (4) получения белков-мишеней с заданными N-концевыми последовательностями [Malakhov M.R, Mattern M.R., Malakhova О.А., Drinker M., Weeks S.D., Butt T.R. SUMO fusions and SUMO-specific protease for efficient expression and purification of proteins. J Struct Funct Genomics, 2004, 5(1-2), 75-86; Marblestone J.G., Edavettal S.C., Lim Y., Lim P., Zuo X., Butt T.R. Comparison of SUMO fusion technology with traditional gene fusion systems: Enhanced expression and solubility with SUMO. Protein Sci, 2006, 15(1), 182-189].
В то же время уникальные свойства и высокая эффективность их применения для организации биосинтеза и процессинга растворимых SUMO-содержащих фьюжнов, ни в коей мере не являются обоснованием возможности и эффективности их применения в отношение фьюжнов, синтезируемых в форме нерастворимых ТВ. Сведений о процессинге SUMO-фьюжнов в составе нерастворимых ТВ в источниках научно-технической информации не обнаружено.
Наиболее близким к заявляемому изобретению является способ биосинтеза биологически активных белков-мишеней в составе нерастворимых интеин-содержащих гибридных белков (фьюжнов), содержащих самоассоциирующие тэги (CAT). В процессе биосинтеза такие САТ-содержащие фьюжны самоассоциируют с образованием нерастворимых агрегатов, называемых активными тельцами включения (АТВ) [US 9200306]. От обычных телец включения (ТВ) АТВ отличает присутствие в их составе белков-мишеней в корректно свернутом биологически активном состоянии. Высвобождение белков-мишеней из состава АТВ осуществляется в процессе очистки в результате автокаталитического интеин-зависимого гидролиза (процессинга) фьюжнов [обзор Lin Zh., Zhao Q., Xing L., Zhou В., Wang X. Aggregating tags for column-free protein purification. Biotechnol. J., 2015, 10, 1877-1886]. Данный способ эффективно используют для получения рекомбинантного глюкагоноподобного пептида и других белков [US 9200306; Xing L., Xu W., Zhou В., Chen Y., Lin Z. Facile expression and purification of the antimicrobial peptide histatin 1 with a cleavable self-aggregating tag (cSAT) in Escherichia coli. Protein Expr. Purif., 2013, 88, 248-253; Zhao Q., Xu W, Xing L., Lin Z. Recombinant production of medium-to large-sized peptides in Escherichia coli using a cleavable self-aggregating tag. Microb Cell Fact, 2016, 15, 136; Zhao Q., Zhou В., Gao X., Xing L., Wang X., Lin Z. A cleavable self-assembling tag strategy for preparing proteins and peptides with an authentic N-terminus. Biotechnol J, 2017, 12:1600656; Lin Z., Ndengue P.P.A., Jing Y, Zhao L., Yang X. Facile expression and purification of active human growth hormone in E.coli by a cleavable self-aggregating tag scheme. Protein Expr Purif, 2021, 188, 105974].
Ключевым технологическим преимуществом указанного способа является обегченное разделение целевых белков и примесных клеточных белков с использованием центрифугирования (осаждения) или фильтрации, снижающее трудозатраты и себестоимость производимых протеинов. Реализация способа включает три этапа. На первом из них осаждение/фильтрация используется для отделения нерастворимых АТВ от растворимых клеточных белков. На втором этапе осуществляют автокаталитический интеин-зависимый процессинг фьюжнов и высвобождение белков-мишеней из состава нерастворимых АТВ. На третьем этапе осаждение/фильтрация используют для отделения растворимых белков-мишеней, высвобожденных из состава АТВ, от остатков нерастворимых АТВ. Результирующие препараты белков-мишеней характеризуются высокой степенью первичной очистки [US 9200306; Xing L., Xu W., Zhou В., Chen Y., Lin Z. Facile expression and purification of the antimicrobial peptide histatin 1 with a cleavable self-aggregating tag (cSAT) in Escherichia coli. Protein Expr. Purif., 2013, 88, 248-253; Zhao Q., Xu W., Xing L., Lin Z. Recombinant production of medium-to large-sized peptides in Escherichia coli using a cleavable self-aggregating tag. Microb Cell Fact, 2016, 15, 136; Zhao Q., Zhou В., Gao X., Xing L., Wang X., Lin Z. A cleavable self-assembling tag strategy for preparing proteins and peptides with an authentic N-terminus. Biotechnol J, 2017, 12:1600656; Lin Z., Ndengue P.P.A., Jing Y, Zhao L., Yang X. Facile expression and purification of active human growth hormone in E.coli by a cleavable self-aggregating tag scheme. Protein Expr Purif, 2021, 188, 105974], при необходимости их дальнейшая очистка может проводиться с использованием стандартных методов, известных из уровня техники.
Эффективность указанного способа снижают потери, возникающие вследствие недостаточно эффективной свертки отдельных компонентов фьюжна, неполного и/или преждевременного высвобождения белка-мишени из состава фьюжна [Zhao Q., Zhou В., Gao X., Xing L., Wang X., Lin Z. A cleavable self-assembling tag strategy for preparing proteins and peptides with an authentic N-terminus. Biotechnol J, 2017, 12:1600656], наличия структурных особенностей фьюжна, снижающих автокаталитическую активность интеинов [Mathys S., Evans Т.С., Chute I.C., Wu H., Chong S., Benner J., Liu X.Q., Xu M.Q. Characterization of a self-splicing mini-intein and its conversion into autocatalytic N- and C-terminal cleavage elements: facile production of protein building blocks for protein ligation. Gene, 1999, 231(1-2), 1-13], деградации белка-мишени в ходе продолжительного процессинга. К числу ограничений способа может относиться также предпочтительная локализация белка-мишени на N-конце синтезируемого фьюжна, поскольку именно такая конфигурация фьюжна используется в большинстве работ. Известна лишь одна работа, в которой синтезируемый фьюжн содержал белок-мишень в С-концевом положении [Zhao Q., Zhou В., Gao X., Xing L., Wang X., Lin Z. A cleavable self-assembling tag strategy for preparing proteins and peptides with an authentic N-terminus. Biotechnol J, 2017, 12:1600656].
Технической проблемой, на решение которой направлено заявляемое изобретение, является расширение арсенала средств для получения биологически активных рекомбинантных белков-мишеней.
Раскрытие сущности изобретения
Техническим результатом заявляемого изобретения является получение биологически активных рекомбинантных белков-мишеней, производимых с использованием микробиологического синтеза без применения процедур денатурации/ренатурации.
Для достижения технического результата предложен способ получения биологически активных рекомбинантных протеинов, содержащих до 4 дисульфидных связей, заключающийся в разработке генетической конструкции, кодирующей предшественник целевого белка-мишени, содержащий в своем составе расположенные в порядке перечисления от N-конца к С-концу последовательность самоассоциирующего пептида L6KD, заключающего следующие друг за другом 6 остатков лейцина, последовательность белка SUMO - продукта гена Smt3 дрожжей Saccharomyces cerevisiae и последовательность целевого белка-мишени, первый N-концевой остаток, следующий за последовательностью белка SUMO, отличен от пролина, и использования созданной генетической конструкции для получения целевого белка-мишени путем трансформации этой конструкцией подходящего реципиентного штамма, культивирования трансформированного штамма в условиях, обеспечивающих экспрессию генетической конструкции и микробиологический синтез предшественника целевого белка-мишени в форме нерастворимых телец включения, с последующим выделением синтезированных телец включения из клеток штамма-продуцента, высвобождением из их состава растворимого целевого белка-мишени с помощью подходящей производной SUMO-специфичной протеиназы, отделением остатков нерастворимых телец включения с помощью центрифугирования и дальнейшей очисткой высвобожденного белка-мишени с использованием методов молекулярной биологии, известных из уровня техники, без применения ренатурации.
Для осуществления биосинтеза SUMO-фьюжна, заключающего последовательность целевого белка-мишени разрабатывают генетическую конструкцию, кодирующую целевой SUMO-фьюжн, и получают рекомбинантный штамм-продуцент, в клетках которого осуществляют экспрессию генетической конструкции. Предпочтительно для биосинтеза SUMO-фьюжна создают штаммы-продуценты на основе клеток бактерий или дрожжей.
Присутствие в составе SUMO-фьюжна последовательности белка SUMO стимулирует синтез целевого белка-мишени в корректной свернутом биологически активном состоянии, не требующем ренатурации. В этой связи, синтезированные ТВ относятся к категории АТВ.
- синтезированные АТВ извлекают из клеток штамма-продуцента, для чего, применяя методы, известные из уровня техники, вскрывают клетки штамма-продуцента и с помощью центрифугирования или фильтрации отделяют нерастворимые АТВ от растворимых клеточных белков, после чего суспендируют АТВ в подходящем буфере и получают суспензию нерастворимых АТВ для дальнейшей работы;
- суспензию АТВ обрабатывают подходящей SUMO-специфичной протеиназой, обеспечивающей гидролиз пептидной связи, соединяющей последовательность белка SUMO с последовательностью целевого белка-мишени в составе SUMO-фьюжна, и высвобождение целевого белка-мишени в растворимую фракцию;
- высвобожденный из состава АТВ растворимый целевой белок-мишень отделяют от остатков нерастворимых АТВ с использованием центрифугирования или фильтрации и получают препарат целевого биологически активного белка-мишени. При этом ни на одном из этапов осуществления заявляемого способа не применяют процедуру ренатурации целевого белка-мишени или его предшественника.
При этом АТВ, формируемые в клетках штамма-продуцента в результате самоассоциации SUMO-фьюжнов, содержащих в своем составе последовательность белка SUMO в сочетании самоассоциирующим пептидом, обеспечивают доступ SUMO-специфичной протеиназы к сайтам протеолиза и, как результат, высокую эффективность процессинга, высвобождение и последующее отделение целевого белка-мишени в форме растворимого белка от остатков нерастворимых АТВ. В результате ферментативного процессинга суспензии нерастворимых АТВ получают препарат растворимого целевого белка-мишени, который обладает биологической активностью и, как правило, имеет уровень чистоты, превышающий 85%. При необходимости доочистка полученного препарата может быть проведена с использованием методов, известных из уровня техники, но без применения ренатурации.
Таким образом, заявляемый способ обеспечивает получение и полностью контролируемое высвобождение биологически активного белка-мишени, как в способе-прототипе на основе использования SUMO. В то же время благодаря использованию самоассоциирующего пептида согласно заявляемому способу первичную очистку целевых SUMO-фьюжнов осуществляют в облеченном режиме с помощью центрифугирования или фильтрации, как по способу-прототипу на основе фьюжнов с интеином.
Было обнаружено, что фьюжны белка SUMO, известного своей высокой растворимостью [Malakhov M.R, Mattern M.R., Malakhova O.A., Drinker M., Weeks S.D., Butt T.R. SUMO fusions and SUMO-specific protease for efficient expression and purification of proteins. J Struct Funct Genomics, 2004, 5(1-2), 75-86; Marblestone J.G., Edavettal S.C., Lim Y, Lim R, Zuo X., Butt T.R. Comparison of SUMO fusion technology with traditional gene fusion systems: Enhanced expression and solubility with SUMO. Protein ScL, 2006, 15(1), 182-189], содержащие на N-конце самоассоциирующий пептид L6KD, способны эффективно синтезироваться в форме нерастворимых ТВ.
Было обнаружено, что в составе целевых SUMO-фьюжнов, синтезируемых в форме нерастворимых ТВ, белок SUMO стимулирует корректную свертку белков-мишеней и тем самым обеспечивает их биосинтез в биологически активном состоянии, что позволяет называть синтезируемые ТВ активными тельцами включения (АТВ).
Было обнаружено, что каталитически-активные производные SUMO-специфичной протеиназы Ulp1 дрожжей S. cerevisiae способны осуществлять специфический ферментативный гидролиз (процессинг) SUMO-фьюжнов, синтезированных в форме нерастворимых АТВ, и высвобождать из их состава растворимые белки-мишени, пригодные для дальнейшей очистки с использованием методов, известных из уровня техники, без применения ренатурации.
Характерной особенностью заявляемого способа является С-концевое расположение белков-мишеней в составе целевых SUMO-фьюжнов, синтезируемых в форме нерастворимых АТВ.
Краткое описание чертежей
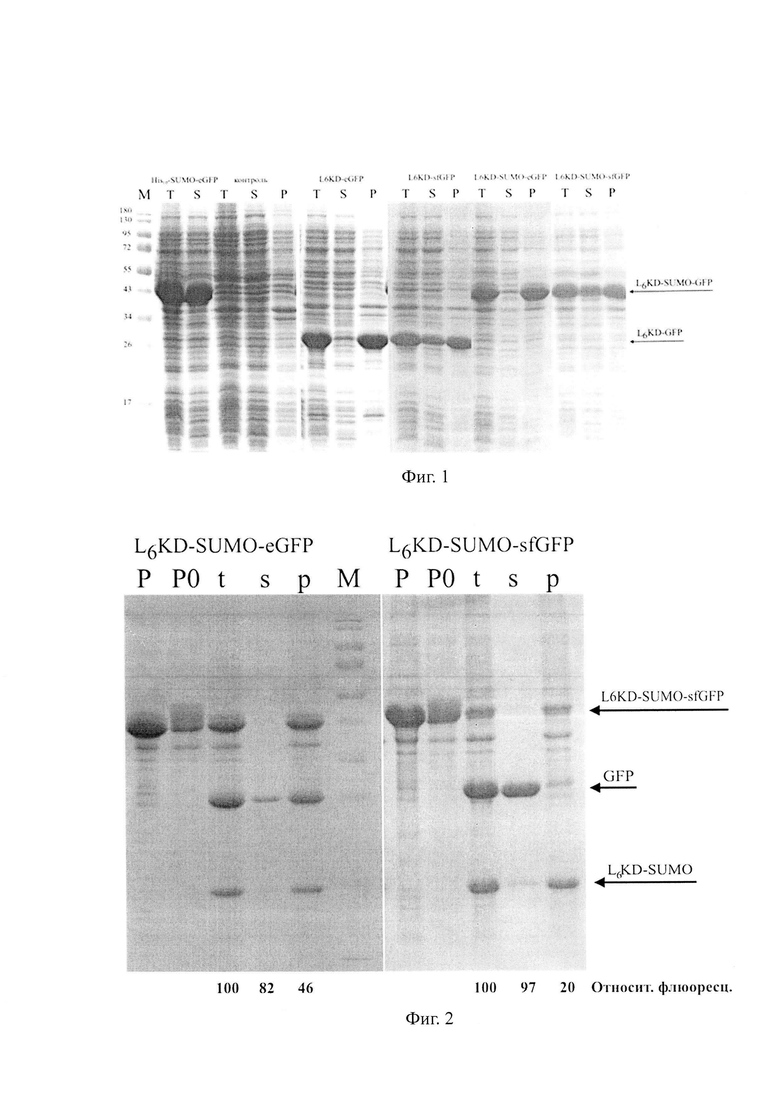
На фиг. 1 показан анализ растворимости целевых SUMO-фьюжнов, содержащих флюоресцентные белки-мишени eGFP и sfGFP, синтезируемых в клетках E.coli. Состав целевых фьюжнов указан над дорожками. На дорожки в пропорциональном количестве наносили образцы фракций суммарных (Т), растворимых (S) и нерастворимых (Р) клеточных белков. Положение полос, соответствующих различным целевым фьюжнам, показано стрелками. М-маркеры молекулярного веса М (кДа).
На фиг. 2 показан анализ процессинга нерастворимых целевых фьюжнов, содержащих флюоресцентные белки-мишени eGFP и sfGFP. Состав целевых фьюжнов указан над дорожками. На дорожки в пропорциональном количестве наносили: исходный образец нерастворимых клеточных белков (Р), контрольный образец нерастворимых клеточных белков, не обрабатывавшийся протеиназой Ulp275 (РО), а также образцы суммарных (t), растворимых (s) и нерастворимых (р) белков фракции нерастворимых клеточных белков, обработанных протеиназой Ulp275. Образцы были обработаны SUMO-протеиназой Ulp275 в пропорции 1:250 к количеству целевого фьюжна в образце в течение 22 часов при температуре 30°С. Стрелками показано положение полос, соответствующих указанным белкам. М - маркеры молекулярного веса белков. Приведены данные относительной флюоресценции соответствующих образцов флюоресцентных белков-мишеней eGFP и sfGFP. За 100% принята флюоресценция образцов t.
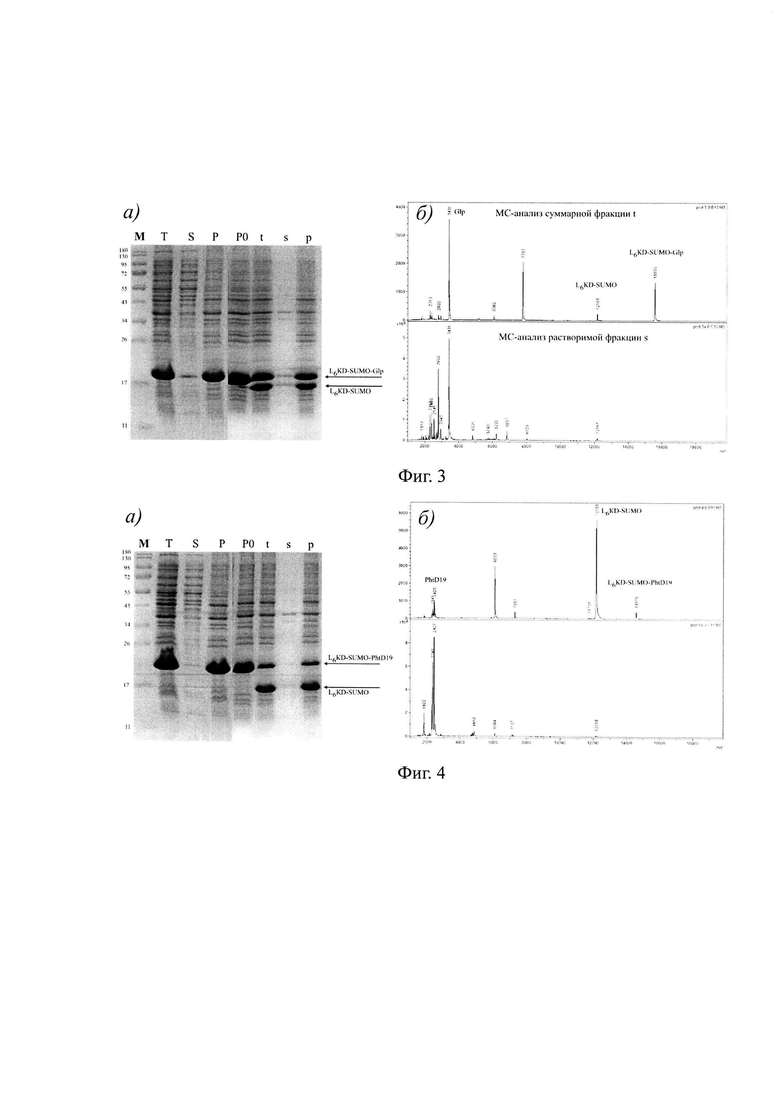
На фиг. 3 показан: а) анализ биосинтеза, растворимости и процессинга целевого фьюжна L6KD-SUMO-Glp.На дорожки в пропорциональном количестве наносили препараты фракций суммарного (Т), растворимого (S) и нерастворимого (Р) клеточных белков, контрольный образец нерастворимых клеточных белков, не подвергавшийся обработке протеиназой Ulp275 (РО), а также образцы суммарных (t), растворимых (s) и нерастворимых (р) белков фракции нерастворимых клеточных белков, обработанных протеиназой Ulp275. Стрелками показано положение полос, соответствующих исходному фьюжну LeKD-SUMO-Glp и его процессированному фрагменту L6KD-SUMO. М - маркеры молекулярного веса белков, б) масс-спектрометрический анализ образцов суммарных (t, сверху) и растворимых (s, снизу) белков фракции нерастворимых клеточных белков, обработанных протеиназой Ulp275. Показано положение пиков, соответствующих исходному целевому фьюжну LeKD-SUMO-Glp и продуктам его процессинга, белку L6KD-SUMO и целевому белку-мишени Glp.
На фиг. 4 показан: а) анализ биосинтеза, растворимости и процессинга целевого фьюжна L6KD-SUMO-PhtD19. На дорожки в пропорциональном количестве наносили препараты фракций суммарного (Т), растворимого (S) и нерастворимого (Р) клеточных белков, контрольный образец нерастворимых клеточных белков, не подвергавшийся обработке протеиназой U1p275 (РО), а также образцы суммарных (t), растворимых (s) и нерастворимых (р) белков фракции нерастворимых клеточных белков, обработанных протеиназой Ulp275. Стрелками показано положение полос, соответствующих исходному фьюжну L6KD-SUMO-PhtD19 и его процессированному фрагменту L6KD-SUMO. М - маркеры молекулярного веса белков, б) масс-спектрометрический анализ образцов суммарных (t, сверху) и растворимых (s, снизу) белков фракции нерастворимых клеточных белков, обработанных протеиназой U1p275. Показано положение пиков, соответствующих исходному целевому фьюжну L6KD-SUMO-PhtD19 и продуктам его процессинга, белку L6KD-SUMO и целевому белку-мишени PhtD19.
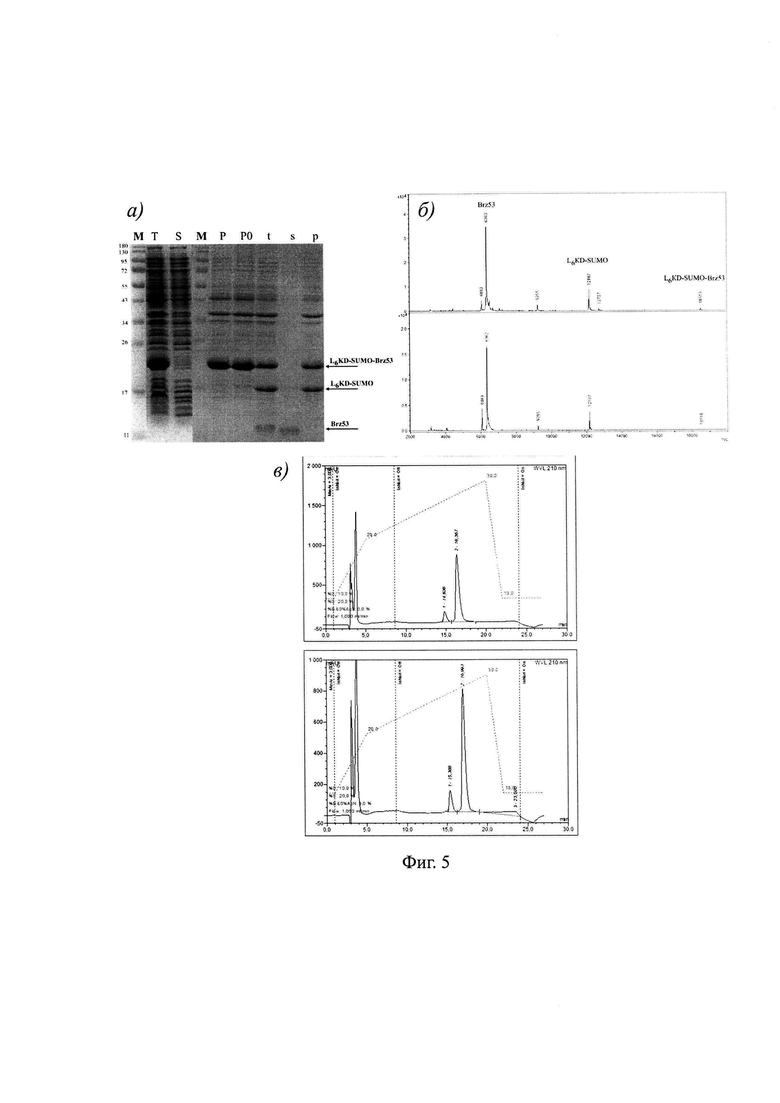
На фиг. 5 показан: а) анализ биосинтеза, растворимости и процессинга целевого фьюжна L6KD-SUMO-Brz53. На дорожки в пропорциональном количестве наносили препараты фракций суммарного (Т), растворимого (S) и нерастворимого (Р) клеточных белков, контрольный образец нерастворимых клеточных белков, не подвергавшийся обработке протеиназой Ulp275 (РО), а также образцы суммарных (t), растворимых (s) и нерастворимых (р) белков фракции нерастворимых клеточных белков, обработанных протеиназой Ulp275. Стрелками показано положение полос, соответствующих исходному фьюжну L6KD-SUMO-Brz53, его процессированному фрагменту L6KD-SUMO и зрелому целевому белку-мишени - браззеину Brz53. М - маркеры молекулярного веса белков, б) масс-спектрометрический анализ образцов суммарных (t, сверху) и растворимых (s, снизу) белков фракции нерастворимых клеточных белков, обработанных протеиназой Ulp275. Показано положение пиков, соответствующих исходному целевому фьюжну L6KD-SUMO-Brz53 и продуктам его процессинга, белку LeKD-SUMO и целевому белку-мишени Brz53; в) ВЭЖХ анализ образца целевого белка-мишени Brz53, полученного с использованием заявляемого способа (сверху), и контрольного образца браззеина Brz53, секретированного в клетках дрожжей (снизу).
Осуществление изобретения
1) С использованием методов генетической инженерии получают генетическую конструкцию, кодирующую рекомбинантную молекулу целевого SUMO-фьюжна, который включает три принципиальные части:
N-концевую часть, заключающую последовательность самоассоциирующего пептида L6KD, отвечающего за биосинтез SUMO-фьюжна в форме нерастворимых АТВ;
- центральную часть, заключающую последовательность белка SUMO дрожжей S. cerevisiae, стимулирующего корректную свертку целевого белка-мишени и его высвобождение из состава нерастворимых АТВ под действием SUMO-специфичной протеиназы;
- С-концевую часть, заключающую последовательность целевого белка-мишени.
2) Конструируют штамм-продуцент, обеспечивающий биосинтез SUMO-фьюжна, заключающего последовательность целевого белка-мишени. С этой целью в клетки подходящего реципиентного штамма, например -реципиентного штамма Е. coli BL21(DE3), путем трансформации вводят созданную генетическую конструкцию, кодирующую целевой SUMO-фьюжн, заключающий последовательность белка-мишени.
3) Разрабатывают и осуществляют культивирование штамма-продуцента, обеспечивающее эффективный биосинтез и накопление целевого SUMO-фьюжна в форме нерастворимых АТВ. Для этого посевной материал клеток штамма-продуцента культивируют при подходящей температуре в диапазоне от 18°С до 40°С, например, при 37°С в питательной среде, включающей источники углерода, азота и минеральные соли, обычно используемые для выращивания клеток E.coli [Маниатис Т., Фрич Э., Сэмбрук Дж. Методы генетической инженерии. Молекулярное клонирование. М.: Мир, 1984. - 479 с.]. Культивирование осуществляют с использованием шейкеров или в биореакторах (ферментерах). Индукцию экспрессии генетической конструкции, обеспечивающей биосинтез целевого SUMO-фьюжна, осуществляют путем внесения индуктора в среду культивирования. В качестве индуктора используют изопропил-P-D-1-тиогалактопиранозид (ИПТГ) в концентрации от 0,1 до 2 мМ или лактозу в концентрации от 0,5% до 3%. Индуктор вносят на стадии роста культуры, предшествующей стационарной фазе роста. Индукцию осуществляют на протяжении не менее 1 ч. В процессе индукции клетки штамма-продуцента синтезируют и накапливают целевой SUMO-фьюжн в форме нерастворимых АТВ в количестве до 40% от суммарного белка клеток. По окончании культивирования, например, с использованием центрифугирования производят отделение биомассы клеток штамма-продуцента, содержащих целевой SUMO-фьюжн, от культуральной среды.
4) Полученную биомассу клеток штамма-продуцента подвергают дезинтеграции. Для дезинтеграции используют подходящее ультразвуковое оборудование или Френч-пресс, после чего, применяя центрифугирование или фильтрацию, из полученного клеточного лизата отделяют фракцию нерастворимых клеточных белков, среди которых с помощью электрофореза обнаруживают целевой SUMO-фьюжн в форме нерастворимых АТВ в количестве до 85%) от суммы белков выделенной фракции.
5) Выделенные АТВ суспендируют в буфере подходящего состава, например, фосфатно-солевом или трис-буфере, после чего полученную суспензию инкубируют в течение от 0,01 часа до 48 часов при температуре от 4°С до 37°С в присутствие каталитически-активного фрагмента SUMO-специфичной протеиназы Ulp1 дрожжей Saccharomyces cerevisiae, например, Ulp275 [RU 2451076], применяемого в пропорции от 1:10000 до 1:10 (по весу) относительно количества целевого SUMO-фьюжна. В процессе инкубации осуществляют ферментативный гидролиз от 10% до 100% SUMO-фьюжна, присутствующего в образце, и высвобождают из его состава растворимый целевой белок-мишень. При необходимости, используя специфические методы, известные из уровня техники, оценивают активность процессированного белка-мишени.
6) По окончании ферментативного гидролиза SUMO-фьюжна из состава целевого препарата с эффективностью до 90% удаляют остатки нерастворимых АТВ. С этой целью смесь растворимого белка-мишени и остатков нерастворимых АТВ вновь подвергают центрифугированию или фильтрации. В результате получают препарат растворимого целевого белка-мишени с чистотой свыше 90%.
7) При необходимости с использованием методов, известных из уровня техники, производят доочистку полученного растворимого белка-мишени без применения ренатурации и получают конечный препарат, содержащий целевой биологически активный рекомбинантный белок-мишень (протеин).
Пример 1. Конструирование плазмиды pET28S-His10-eGFP для биосинтеза фьюжна His10-eGFP
С целью получения вспомогательной плазмиды проводят ПЦР-амплификацию гена eGFP. Матрицей для амплификации служит плазмида pEGFP (Clontech). Для амплификации используют праймеры: 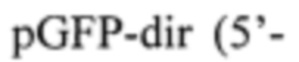

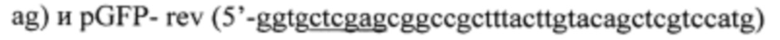
В составе праймеров подчеркнуты последовательности сайтов узнавания рестриктаз NcoI, BglII и BamIII (pGFP-dir), а также XhoI (pGFP-rev).
Амплифицированный фрагмент ДНК очищают из геля, используя с этой целью набор Qiagen (Qiagen, cat.№28706) и после открывания концов с использованием рестриктаз NcoI и XhoI клонируют в лабораторном векторе pET28S, расщепленном по таким же сайтам. Вектор pET28S, являющийся производным стандартного вектора pET28b+(Novagen), содержит мутацию, вместо уникального сайта BglII вектор содержит последовательность сайта Sail.
В результате клонирования получают плазмиду pET28S-His10-eGFP. В составе этой плазмиды клонированный фрагмент ДНК заключает структурный ген eGFP прецизионно слитый с 5'-концевой последовательностью ДНК, кодирующей 10 остатков гистидина, находящейся в окружении сайтов узнавания рестриктаз BglII и BamIII (см. структуру праймера pGFP-dir). Плазмиду pET28S-His10-eGFP используют для биосинтеза и анализа экспрессии фьюжна Hism-eGFP, а также для клонирования других фрагментов ДНК.
Пример 2. Клонирование гена SMT3, кодирующего белок SUMO дрожжей Saccharomyces cerevisiae
Ген SMT3 амплифицируют в ходе ПЦР с использованием в качестве матрицы хромосомной ДНК штамма S.cerevisiae Y618 [Kartasheva et al., 1996, Yeast 12: 1297-1300]. Амплификацию проводят в две стадии. Сначала амплифицируют два перекрывающихся фрагмента ДНК, для чего используют следующие пары праймеров:
Фрагмент 1 размером 129 п.о.:
N450 (5'-atatccatggaaaagagatctgactcagaagtcaatcaagaa)
N454 (5'-cttgaagaaaatctctgaa)
Фрагмент 2 размером 180 п.о.:
N453 (5'-ttcagagattttcttcaag)
N452 (5'-atatcaattggatccaccaatctgttctctgtga).
В составе праймеров подчеркнуты последовательности сайтов узнавания рестриктаз NcoI и BglII (N450), а также BamIII (N452).
Амплифицированные фрагменты ДНК элюируют из агарозного геля как в примере 1, и их смесь используют для ПЦР-лигирования. Д ля этого проводят ПЦР-амплификацию на смеси фрагментов 1 и 2 в качестве матрицы. Праймерами для амплификации служат N450 и N468. Полученный в результате ПЦР фрагмент ДНК размером 310 п. о. элюируют из агарозного геля, обрабатывают рестриктазами NcoI и BamIII и клонируют в плазмиде pET28S-His10-eGFP (Пример 1), расщепленной по таким же сайтам. В результате клонирования получают плазмиду pET28S-SUMO-eGFP, в составе которой нуклеотидную последовательность клонированного гена подтверждают секвенированием.
Полученную плазмиду pET28S-SUMO-eGFP используют для конструирования других плазмид в качестве источника BglII/XhoI фрагмента ДНК, кодирующего слитый белок SUMO-eGFP.
Пример 3. Конструирование плазмиды pET28S-HIS10-SUMO-EGFP ДЛЯ БИОСИНТЕЗА И АНАЛИЗА СВОЙСТВ ФЬЮЖНА HIS10-SUMO-EGFP
Плазмиду pET28S-His10-SUMO-eGFP конструируют путем замещения уникального BamIII/XhoI фрагмента ДНК плазмиды pET28S-His10-eGFP, кодирующего eGFP, на уникальный BglII/XhoI фрагмент ДНК плазмиды pET28S-SUMO-eGFP, кодирующий фьюжн SUMO-eGFP.
Полученную плазмиду pET28S-His10-SUMO-eGFP используют для биосинтеза и анализа свойств контрольного фьюжна His10-SUMO-eGFP, включающего последовательность белка SUMO, но не включающего самоассоциирующий пептид LeKD.
Пример 4. Конструирование плазмиды pET28-L6KD-eGFP для биосинтеза и анализа свойств АТВ, содержащих фьюжн L6KD-eGFP
Для получения фрагмента ДНК, кодирующего слитый белок L6KD-eGFP, используют ПЦР. Матрицей для амплификации служит плазмида pET28S-His10-eGFP. Амплификацию проводят в две стадии. Сначала получают базовый фрагмент ДНК, для амплификации которого используют праймеры:
N1472(5'-tataccatggtgctgttactgctgttgaaggatggatccatggtgagcaagggc)
N619(5'-ggtggcagcagccaactcagctt)
Затем, используя базовый фрагмент в качестве матрицы, получают целевой фрагмент ДНК, который амплифицируют с помощью праймеров N1473 (5'-tataccatgggtttgctgttactgctgttgaa) и N619.
В составе праймеров подчеркнуты последовательности сайтов узнавания рестриктаз NcoI (N1472 и N1473) и BamIII (N1472).
Амплифицированный фрагмент ДНК элюируют из агарозного геля, обрабатывают рестриктазами NcoI и XhoI и клонируют в плазмиде рЕТ28b+(Novagen), расщепленной по таким же сайтам. В результате клонирования получают плазмиду pET28-L6KD-eGFP, в составе которой нуклеотидную последовательность клонированного гена подтверждают секвенированием.
Полученную плазмиду pET28-L6KD-eGFP используют для биосинтеза и анализа свойств нерастворимых АТВ, содержащих фьюжн L6KD-eGFP, не включающий последовательность белка SUMO.
Пример 5. Конструирование плазмиды pET28-L6KD-SUMO-eGFP для биосинтеза и анализа свойств АТВ, содержащих фьюжн L6KD-SUMO-eGFP
Плазмиду pET28-L6KD-SUMO-eGFP конструируют путем замещения уникального ВаmIII/XhoI фрагмента ДНК плазмиды pET28-L6KD-eGFP, кодирующего белок-мишень eGFP, на уникальный BgHI/XhoI фрагмент ДНК плазмиды pET28S-SUMO-eGFP, кодирующий слитый белок SUMO-eGFP.
Сконструированную плазмиду pET28-L6KD-SUMO-eGFP используют для биосинтеза и анализа свойств нерастворимых АТВ, содержащих фьюжн L6KD-SUMO-eGFP, в составе которого последовательность белка-мишени eGFP слита с последовательностью белка L6KD-SUMO.
Пример 6. Конструирование плазмиды pET28-L6KD-sfGFP для биосинтеза и анализа свойств АТВ, содержащих фьюжн L6KD-sfGFP
Известен аналог белка eGFP - белок sfGFP - «superfolder GFP», обладающий собственной улучшенной эффективностью свертки [Aronson D.E., Costantini L.M., Snapp E.L. Superfolder GFP is fluorescent in oxidizing environments when targeted via the Sec translocon. Traffic, 2011, 12(5), 543-8].
Фрагмент ДНК, кодирующий белок sfGFP, получают с помощью ПЦР. Матрицей для ПЦР служит плазмида pAAV-CAG-NES-GAF-CaMP2-sfGFP [Subach О.М. and Subach F.V. GAF-CaMP3-sfGFP, an enhanced version of the near-infrared genetically encoded positive phytochrome-based calcium indicator for the visualization of neuronal activity. Int J Mol Scl, 2020, 21(18), 6883], для ПЦР используют праймеры pGFP-dir и pGFP- rev (Пример 1). В результате амплификации получают целевой фрагмент ДНК, заключающий ген sfGFP, который элюируют из агарозного геля, обрабатывают рестриктазами ВаmIII. и XhoI и используют для замещения исходного ВаmIII/XhoI фрагмента ДНК в плазмиде pET28-L6KD-eGFP (Пример 4). В результате замещения получают плазмиду pET28-L6KD-sfGFP, в составе которой нуклеотидную последовательность клонированного гена подтверждают секвенированием.
Полученную плазмиду pET28-L6KD-sfGFP используют для биосинтеза и анализа свойств нерастворимых АТВ, содержащих фьюжн L6KD-sfGFP, включающий последовательность эффективно сворачиваемого варианта белка sfGFP и не включающий последовательность белка SUMO.
Пример 7. Конструирование плазмиды pET28-L6KD-SUMO-sfGFP для биосинтеза и анализа свойств АТВ, содержащих фьюжн L6KD-SUMO-sfGFP
Конструирование проводят, как описано в примере 6 за исключением того, что для замещения исходного фрагмента ДНК используют плазмиду pET28-L6KD-SUMO-eGFP (Пример 5). В результате замещения получают плазмиду pET28-L6KD-SUMO-sfGFP, которую используют для биосинтеза и анализа свойств нерастворимых АТВ, содержащих фьюжн L6KD-SUMO-sfGFP, в составе которого последовательность эффективно сворачиваемого варианта белка-мишени sfGFP слита с последовательностью белка L6KD-SUMO.
Пример 8. Конструирование плазмиды pET28-L6KD-SUMO-Glp для биосинтеза и анализа свойств АТВ, содержащих фьюжн L6KD-SUMO-Glp
Подготавливают вспомогательную плазмиду pET28-Glp, содержащую фрагмент ДНК, кодирующий модифицированный глюкагоно-подобный пептид 1 человека Glp. Плазмиду pET28-Glp получают из плазмиды рЕТ28-Glp20 [RU 2642260] путем замещения уникального NcoI/BamIII фрагмента ДНК, содержащего ген Glp20, на фрагмент «В», имеющий липкие концы NcoI и ВаmIII, кодирующий модифицированный глюкагоно-подобный пептид 1 человека [RU 2642260]. В полученной плазмиде pET28-Glp последовательность ДНК, кодирующая ген Glp, содержит уникальный сайт узнавания рестриктазы KpnI и заканчивается терминирующим кодоном, расположенным сразу после сайта ВаmIII.
Фрагмент ДНК, кодирующий фьюжн L6KD-SUMO, получают с помощью ПЦР, матрицей для ПЦР служит плазмида pET28-L6KD-SUMO-eGFP. Амплификацию проводят с использованием праймеров N685 (5'-taatacgactcactataggg) и N1790 (5'-taaaggtaccttcaccatgtccaccaatctgttctctgt). В составе праймера N1790 подчеркнута последовательность сайта узнавания рестриктазы KpnI.
Амплифицированный фрагмент ДНК обрабатывают рестриктазами NcoI и KpnI и полученный NcoIlKpnI фрагмент ДНК используют для замещения NcoIlKpnI фрагмента ДНК в плазмиде pET28-Glp.В результате получают плазмиду pET28-L6KD-SUMO-Glp, которую используют для биосинтеза и анализа свойств нерастворимых АТВ, содержащих фьюжн L6KD-SUMO-Glp, в составе которого последовательность белка-мишени Glp слита с последовательностью белка LeKD-SUMO.
Пример 9. Конструирование плазмиды pET28-L6KD-SUMO-PhtD19 для биосинтеза и анализа свойств АТВ, содержащих Фьюжн L6KD-SUMO-PhtD19
Известен пептид PhtD_19, называемый здесь пептидом PhtD19, являющийся фрагментом pneumococcal histidine triad protein D fragment (a.a. 200-219) [Papastamatiou T, Routsias JG, Koutsoni O, Dotsika E, Tsakris A, Spoulou V. Evaluation of protective efficacy of selected immunodominant B-cell epitopes within virulent surface proteins of Streptococcus pneumoniae. Infect. Immun., 2018, 86(3), е00673-17]. Пептид является иммунодоминантным B-клеточным эпитопом белка, расположенного на поверхности клеток Streptococcus pneumoniae, и представляет интерес в качестве кандидатного компонента соответствующей вакцины.
Фрагмент ДНК, кодирующий пептид PhtD19, получают с помощью ПЦР, который проводят, используя смесь праймеров N-PhtD19 (5'-ggcgtagaggatcgagatct), N1762 (5'-ggtaatgatcaccatgcggcacaatatatgcatcgccagatctcgatcctctacgcc) и N1763 (5'-tatactcgagttacagttcgttcttcggaatgtaatggtaatgatcaccatgcggc). Амплифицированный фрагмент ДНК обрабатывают рестриктазами BglUII и XhoI и полученный BglII/XhoI фрагмент ДНК используют для замещения ВаmIII/XhoI фрагмента ДНК в плазмиде pET28-L6KD-SUMO-eGFP. В результате получают плазмиду pET28-L6KD-SUMO-PhtD19, которую используют для биосинтеза и анализа свойств нерастворимых АТВ, содержащих фьюжн L6KD-SUMO-PhtD19, в составе которого последовательность белка-мишени PhtD19 слита с последовательностью белка L6KD-SUMO.
Пример 10. Конструирование плазмиды pET28-L6KD-SUMO-Brz53 для биосинтеза и анализа свойств АТВ, содержащих фьюжн L6KD-SUMO-Brz53
Известен белок браззеин, обладающий сладким вкусом и относящийся к семейству дефензин-подобных белков (Defensin-like protein) Pentadiplandra brazzeana (GeneBank P56552). Одна из зрелых более сладких форм белка, называемая здесь Brz53, имеет размер 53 аминокислотных остатка и содержит 4 дисульфидные связи [Hellekant G., Danilova V. Brazzein a small, sweet protein: discovery and physiological overview. Chem Senses, 2005, 30 Suppl 1, i88-9].
Структура праймеров, используемых для конструирования фрагмента ДНК, кодирующего белок Brz53, приведена ниже.


Фрагмент ДНК Brzz, кодирующий белок Brz53, получают с использованием ПЦР-амплификации, проводимой на смеси олигонуклеотидов b3 и и4 с использованием праймеров N1840 и b2. Структура получаемого синтетического гена и кодируемого ею белка приведены ниже:

Параллельно с использованием праймеров N450 (Пример 2) и N1841 (5'-tccaccaatctgttctctgtg) амплифицируют фрагмент ДНК SUMO-F, кодирующий белок SUMO. Матрицей для амплификации служит ДНК плазмиды pET28S-SUMO-eGFP.
На завершающем этапе смесь амплифицированных фрагментов ДНК Brzz и SUMO-F используют для ПЦР-лигирования. С этой целью проводят ПЦР-амплификацию на смеси фрагментов ДНК и праймеров N450 и b2. Результирующий ПЦР фрагмент ДНК размером очищают из агарозного геля, обрабатывают рестриктазами ЕсоШ и XhoI и клонируют в плазмиде рЕТ28-L6KD)-SUMO-eGFP (Пример 5), расщепленной по таким же сайтам. В результате клонирования получают плазмиду pET28-L6KD-SUMO-Brz53, в составе которой нуклеотидную последовательность клонированного гена подтверждают секвенированием.
Плазмиду pET28-L6KD-SUMO-Brz53 используют для биосинтеза и анализа свойств нерастворимых АТВ, содержащих фьюжн L6KD-SUMO-Brz53, в составе которого последовательность белка-мишени Brz53 слита с последовательностью белка L6KD-SUMO.
Пример 11. Получение контрольного образца браззеина Brz53, секретированного в клетках дрожжей Pichia pastoris
Для получения контрольного образца браззеина используют синтетический ген браззеина Brz53 и конструируют штамм-продуцент дрожжей Pichia pastoris, аналогично тому, как описано в работе [Poirier N., Roudnitzky N., Brockhoff A., Belloir C., Maison M., Thomas-Danguin Т., Meyerhof W., Briand L. Efficient production and characterization of the sweet-tasting brazzein secreted by the yeast Pichia pastoris. J Agric Food Chem., 2012, 60(39), 9807-9814]. Для конструирования штамма-продуцента применяют ранее разработанный молекулярно-генетический инструментарий [RU 2522479]. Очистку контрольного образца браззеина Brz53 проводят, как описано в работе [Poirier N., Roudnitzky N., Brockhoff A., Belloir C., Maison M., Thomas-Danguin Т., Meyerhof W., Briand L. Efficient production and characterization of the sweet-tasting brazzein secreted by the yeast Pichia pastoris. J Agric Food Chem., 2012, 60(39), 9807-9814].
Полученный контрольный образец браззеина используют в качестве образца сравнения для выполнения ВЭЖХ анализа браззеина Brz53, произведенного с использованием заявляемого способа.
Пример 12. Получение штамма-продуцента целевого фьюжна (варианты)
Штамм - продуцент целевого фьюжна (варианты) получают путем трансформации реципиентного штамма E.coli BL21(DE3) (Novagen) - ВКПМ В-10189. Трансформацию осуществляют путем введения в клетки реципиентного штамма плазмиды для биосинтеза целевого фьюжна (примеры 3-10) с применением реактива СаСl2 [Маниатис Т., Фрич Э., Сэмбрук Дж. Методы генетической инженерии. Молекулярное клонирование. М.: Мир, 1984. - 479 с.]. Колонии трансформированного штамма отбирают на селективной среде, содержащей антибиотик канамицин. В результате трансформации получают трансформированный штамм - продуцент, который содержит плазмиду для биосинтеза целевого фьюжна и в ответ на внесение в среду культивирования индуктора ИПТГ или лактозы осуществляет биосинтез целевого фьюжна.
Пример 13. Биосинтез белка-мишени в составе целевого фьюжна (варианты)
Биосинтез целевого фьюжна осуществляют следующим образом. Сначала получают посевную культуру штамма-продуцента. Для этого одну колонию штамма-продуцента засевают в пробирку, содержащую 3 мл среды YTS следующего состава (мас. %): дрожжевой экстракт - 1, триптон бакто - 2, NaCl - 1, канамицин - 0,003, вода - остальное. Посевную культуру штамма-продуцента выращивают на ротационном шейкере со скоростью вращения 250 об/мин в течение 4 часов при температуре 37°С.
На втором этапе посевную культуру переносят в среду для индукции. С этой целью 0,5 мл посевной культуры переносят в колбу, содержащую 50 мл среды TRB следующего состава (мас. %): дрожжевой экстракт - 2,4; триптон бакто - 1,2; одномолярный фосфатный буфер (рН7) - 10; одномолярный раствор сульфата магния - 0,2; лактоза - 0,5; глицерин - 0,5; канамицин - 0,009; вода - остальное. Культивирование в среде для индукции продолжают в течение 20 часов на ротационном шейкере со скоростью вращения 250 об/мин при оптимальной температуре в тех же условиях.
Для биосинтеза целевых фьюжнов с использованием плазмид, сконструированных как описано в примерах 3-9, оптимальной температурой является 30°С.
Для биосинтеза целевого фьюжна с использованием плазмиды, сконструированной как описано в примере 10, оптимальной температурой является 22°С.
По окончании культивирования измеряют оптическую плотность клеточной культуры при длине волны 600 нм, используют подходящий спектрофотометр. Затем в отдельную пробирку отбирают объем клеточной культуры (предварительно хорошо перемешанной на вортексе), соответствующий 20 о.е. Клетки осаждают центрифугированием в течение 2 минут со скоростью 13200 об/мин, надосадочную жидкость тщательно удаляют с использованием водоструйного насоса. Полученный клеточный осадок промывают. С этой целью осадок суспендируют в 1 мл буфера PBS и вновь центрифугируют в том же режиме, надосадочную жидкость удаляют.
В результате получают образец клеток штамма-продуцента, который используют для анализа результатов биосинтеза целевого фьюжна.
Пример 14. Методы анализа белка-мишени в составе целевого фьюжна (варианты)
Дезинтеграция клеток штамма-продуцента
Для проведения анализа целевого фьюжна проводят дезинтеграцию клеток штамма-продуцента. С этой целью образец клеток штамма-продуцента ресуспендируют в 1 мл подходящего буфера, например, буфера PBS или буфера 50 мМ Tris-HCl, рН 7.0. Суспензию клеток хорошо перемешивают на вортексе, охлаждают на льду подвергают дезинтеграции, например, с использованием дезинтегратора ультразвукового лабораторного Bioblock Scientific Vibra cell 75043, в режиме: 5 секунд пульс, 10 секунд пауза, 40 секунд (то есть 9 микроциклов). Отбирают 100 мкл пробы Т суммарного лизата.
Остальной объем лизата центрифугируют 10 минут при 13200 об/мин при температуре 4°С. Отбирают 100 мкл пробы S осветленного лизата, после чего надосадочную жидкость тщательно удаляют с использованием водоструйного насоса. Фракцию осадка ресуспендируют в 0,9 мл PBS, перемешивая на вортексе до гомогенного состояния и получают пробу Р нерастворимых клеточных белков.
Определение флюоресценции
Образцы фракций клеточного лизата штамма-продуцента, синтезировавшего флюоресцентный белок-мишень (eGFP или sfGFP) в составе целевого фьюжна, используют для измерения флюоресценции.
Показатель флюоресценции отражает количество корректно свернутого биологически активного белка eGFP в образце.
В лунки плашки для измерения флюоресценции вносят по 160 мкл буфера PBS. Затем в эти лунки вносят отобранные пробы Т, S и Р по 40 мкл в лунку. Конечный объем жидкости во всех лунках составляет 200 мкл. Образцы в лунках тщательно перемешивают с использованием горизонтального шейкера. Измерения проводят с использованием подходящего оборудования, например, с помощью микропланшетного анализатора Turner Biosystems Modulus II с использованием фильтра с длинами волн: ех 525 нм, em 580-640 нм. Полученные значения флюоресценции делят на 4 для расчета удельной величины, соответствующей 1 о.е. клеток.
Полученные значения флюоресценции образцов подвергают математической и статистической обработке и результирующие значения заносят в итоговую таблицу.
Оценка уровня биосинтеза и растворимых свойств целевого фьюжна
Образцы белков отобранных проб Т, S и Р подвергают анализу с использованием гель-электрофорореза в 15% полиакриламидном геле в денатурирующих восстанавливающих условиях с окрашиванием Кумасси G-250 по стандартной процедуре.
Обнаружив на геле на дорожке, соответствующей образцу Т, S или Р, полосу целевого фьюжна, используют экспертную оценку и сопоставляют ее интенсивность с интенсивностью остальных белковых полос на этой же дорожке. В результате получают оценочный показатель относительного содержания целевого фьюжна в соответствующей фракции клеточного лизата. Оценочный показатель образца Т характеризует уровень биосинтеза целевого фьюжна в клетках штамма-продуцента.
Сопоставляя оценочные показатели интенсивности полос целевого фьюжна на дорожках, соответствующих образцам Т, S и Р, делают заключение о растворимых свойствах целевого фьюжна и о его биосинтезе в форме нерастворимых телец включения в клетках штамма-продуцента.
Процессинг нерастворимых АТВ с помощью SUMO-специфичной протеазы Анализируют процессинг белков, являющихся составной частью пробы Р.
100 мкл пробы Р используют для измерения концентрации белка. С этой целью образец пробы Р центрифугируют 10 минут при 13200 об/мин при температуре 4°С, надосадочную жидкость удаляют водоструйным насосом, осадок растворяют в 100 мкл 8 М мочевины и используют для измерения концентрации белка по методу Брэдфорда [Bradford М.М. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 1976, 72, 248-254].
Оставшиеся 800 мкл суспензии нерастворимого белка пробы Р подвергают осаждению с помощью центрифугирования в прежнем режиме, полученный осадок суспендируют в 800 мкл буфера 10 мМ Tris-HCl, рН 8.0 и разделяют на аликвоты по 200 мкл в каждой.
В соответствии с результатами определения концентрации белка по Брэдфорду в полученные аликвоты объемом 200 мкл вносят препарат очищенного фермента SUMO-протеиназы Ulp275 [RU 2451076] в соотношении 1/100 относительно количества целевого фьюжна в каждой аликвоте. Разницу в объемах полученных реакционных смесей выравнивают с помощью буфера. Реакцию проводят при 30°С в течении 4 часов.
По завершении реакции содержимое каждой реакционной смеси перемешивают и отбирают пробу "t" суммарного белка для анализа с использованием гель-электрофореза. Оставшуюся часть образцов подвергают центрифугированию в течение 10 минут при 13200 об/мин при 4°С. После центрифугирования отбирают образец "s" надосадочной жидкости, содержащей фракцию растворимых белков реакционной смеси. Остальную надосадочную жидкость тщательно удаляют с помощью водоструйного насоса. Полученный осадок суспендируют в 200 мкл исходного буфера и отбирают образец "р" нерастворимых белков.
Анализ отобранных образцов t, s и р проводят с помощью гель-электрофореза.
В результате анализа оценивают эффективность процессинга целевого фьюжна под действием SUMO-протеиназы Ulp275 и растворимые свойства белка-мишени, высвобождаемого из состава целевого фьюжна под действием протеиназы. В случае, показатель относительного содержания белка-мишени в образце s превосходит аналогичный показатель образца t, считают, что белок-мишень обладает растворимыми свойствами.
Растворимость коротких белков-мишеней, полученных в результате процессинга АТВ с помощью SUMO-специфичной протеиназы и не разрешаемых с помощью гель-электрофореза, оценивают с использованием масс-спектрометрического анализа продуктов процессинга. Считают, что белок-мишень обладает растворимыми свойствами, если по данным масс-спектрометрии показатель относительного содержания белка-мишени в образце s превосходит аналогичный показатель образца t.
Масс-спектрометрический анализ продуктов процессинга
Массспектрометрический анализ проводят на MALDI-времяпролетном масс-спектрометре UltrafleXtreme ("Bruker Daltonics", Германия), оснащенном УФ-лазером (Nd). Масс-спектры получают в линейной моде режима положительных ионов с использованием рефлектрона. Точность измеренных средних масс составляет 3 Да. Для определения расчетных значений масс белков используют программный пакет Vector NTI ("Thermo Fisher Scientific", США).
Сравнительный ВЭЖХ-анализ браззеина Brz53
Сравнительный анализ браззеина проводят с помощью офВЭЖХ, например, с использованием высокоэффективного жидкостного хроматографа UltiMate 3000 HPLC (Thermo Fisher Scientific, США) и колонки YMC-Pack ODS-A (С18), 5 мкм, 300А, 4,6 × 250 мм (YMC CO., LTD, Япония). Разделение компонентов осуществляют путем градиентного элюирования, в качестве подвижной фазы используют: буфер А - 0,2%-ная трифторуксусная кислота (Sigma Aldrich, США), буфер Б - 80% ацетонитрил (J.T. Baker, Нидерланды) в буфере А. Скорость потока - 1,0 мл/мин. Температура колонки - 35°С. Детектирование проводят при длине волны 210 нм. Объем вводимой пробы - 20 мкл.
Пример 15. Применение заявляемого способа для получения белков-мишеней eGFP и sfGFP
Целевые белки-мишени eGFP и sfGFP различаются между собой эффективностью сворачивания, белок eGFP обладает низкой, a sfGFP («superfolder GFP») - намного более высокой эффективностью корректного сворачивания [Aronson D.E., Costantini L.M., Snapp E.L. Superfolder GFP is fluorescent in oxidizing environments when targeted via the Sec translocon. Traffic, 2011, 12(5), 543-8].
Как показывают результаты, синтез белков-мишеней eGFP и sfGFP в составе целевых фьюжнов L6KD-eGFP, L6KE, -SUMO-eGFP, L6KD-sfGFP и L6KD-SUMO-sfGFP, независимо от присутствия SUMO в их составе, осуществляется преимущественно во фракции (Р) нерастворимых клеточных белков и характеризуется высоким уровнем синтеза, их доля во фракции суммарных (Т) клеточных белков превышает 20% (фиг.1). В этих же условиях контрольный фьюжн His10-SUMO-eGFP, не содержащий L6KD), накапливается преимущественно во фракции растворимых (S) клеточных белков (фиг. 1).
Анализ флюоресцирующей активности (Табл. 1) свидетельствует о том, что предшественники целевых белков-мишеней eGFP и sfGFP, синтезируемые в составе нерастворимых ТВ, обнаруживаемых во фракции Р клеточных лизатов, обладают флюоресцентной (биологической) активностью, то есть -формируют АТВ.
При этом включение SUMO в состав предшественника eGFP, обладающего низкой эффективностью сворачивания, увеличивает синтез активного eGFP в клетках с 4% до 20% относительно контрольного уровня, также с 2% до 96% возрастает доля активных eGFP в составе нерастворимых АТВ. Включение SUMO в состав предшественника sfGFP, обладающего высокой эффективностью сворачивания, не влияет на показатели уровня синтеза активного sfGFP в клетках и доли активного sfGFP в составе нерастворимых АТВ (табл.1).
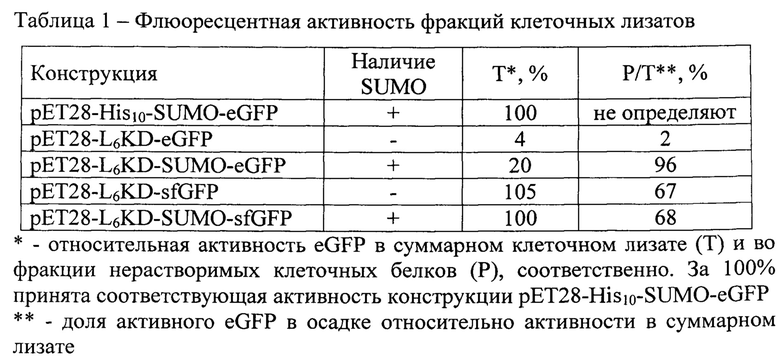
Таким образом, представленные результаты доказывают, что:
1) применение заявляемого способа обеспечивает синтез предшественников целевых биологически активных белков-мишеней eGFP и sfGFP в форме нерастворимых АТВ;
2) включение белка SUMO в состав предшественников целевых белков eGFP и sfGFP оказывает положительное (eGFP) или нейтральное (sfGFP) влияние на уровень синтеза и способность предшественника активного белка-мишени формировать нерастворимые АТВ.
Результаты, представленные на Фиг. 2 свидетельствуют о том, что действие SUMO-специфичной протеиназы Ulp275 на АТВ, содержащие целевые фьюжны L6KD-SUMO-eGFP и L6KD-SUMO-sfGFP, обеспечивает эффективное высвобождение зрелых биологически активных белков-мишеней eGFP и sfGFP в растворимую фракцию. При этом центрифугирование процессированных образцов обеспечивает очистку целевых растворимых белков-мишеней eGFP и sfGFP от остатков нерастворимых АТВ (Фиг. 2).
В совокупности представленные результаты доказывают высокую эффективность получения биологически активных белков-мишеней eGFP и sfGFP с использованием заявляемого способа.
Пример 16. Применение заявляемого способа для получения белков-мишеней Brz53. Glp и PhtD19
Целевые белки-мишени Brz53, Glp и PhtD19 отличаются друг от друга и от белков-мишеней eGFP и sfGFP биологической активностью, третичной структурой, молекулярной массой, эффективностью сворачивания, наличием дисульфидных связей и др.
Результаты применения заявляемого способа для получения белков-мишеней Glp, PhtD19 и Brz53 в составе целевых фьюжнов LeKD-SUMO-Glp, L6KD-SUMO-PhtD19 и L6KD-SUMO-Brz53 представлены на Фиг. 3а, 4а и 5а, соответственно. Представленные результаты показывают, что каждый из целевых фьюжнов синтезируется в клетках штамма-продуцента в составе ТВ во фракции нерастворимых клеточных белков (Фиг. 3а, 4а и 5а). Выделение из клеток и последующая обработка нерастворимых ТВ с помощью SUMO-специфичной протеиназы Ulp275 обеспечивает высвобождение целевых белков Glp, PhtD19 и Brz53 в растворимую фракцию и осаждение нерастворимых остатков исходных ТВ с использованием центрифугирования (Фиг. 3а, 4а и 5а).
Биологическую активность белков-мишеней Glp, PhtD19 и Brz53, высвобождаемых из состава ТВ, непосредственное измерение которой затруднено, доказывают путем определения ключевых показателей, характерных для биологически активных форм этих белков. Таким показателем является молекулярный вес указанных белков-мишеней.
Результаты масс-спектрометрического анализа молекулярного веса белков, получаемых путем процессинга АТВ с использованием протеиназы Ulp275, приведены на Фиг. 3б, 4б и 5б и отражены в таблице 2. Как хорошо видно, измеренные показатели целевых белков-мишеней Glp, PhtD19 и Brz53 совпадают с расчетными величинами. Таким образом, заявляемый способ обеспечивает получение белков-мишеней Glp и PhtD19, обладающих целевыми характеристиками. С другой стороны, соответствие измеренного и расчетного показателей молекулярного веса доказывает наличие в структуре белка Brz53 4 дисульфидных связей. Анализ с использованием обращенно-фазовой ВЭЖХ (Фиг. 5в) также подтверждает идентичность белка-мишени Brz53, полученного с использованием заявляемого способа, и контрольного образца биологически активного браззеина Brz53, содержащего в своем составе 4 дисульфидные связи, полученного путем секреции в дрожжах.
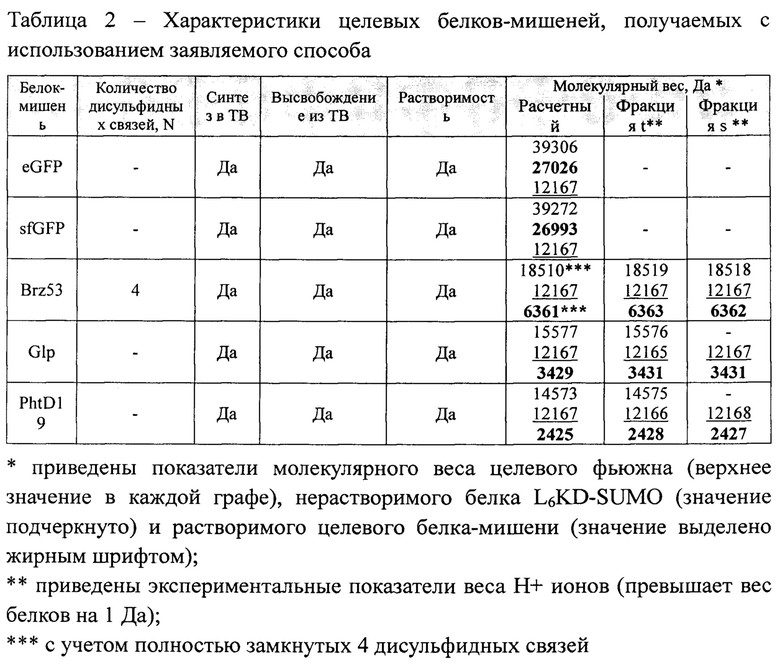
Таким образом, результаты анализов доказывают, что заявляемый способ обеспечивает высокую эффективность получения целевых белков-мишеней Glp, PhtD19 и Brz53, обладающих характеристиками биологически активных форм этих белков. При этом, как и ранее в отношение белков-мишеней eGFP и sfGFP, для получения целевых белков-мишеней Glp, PhtD19 и Brz53 процедура денатурации/ренатурации не применялась.
Таким образом, совокупность представленных данных доказывает, что заявляемый способ обеспечивает высокую эффективность получения целевых белков-мишеней, имеющих отличия в биологической активности, третичной структуре, молекулярной массе, эффективности сворачивания, наличии дисульфидных связей и др. При этом получаемые с использованием заявляемого способа целевые белки-мишени демонстрируют биологическую активность непосредственно или обладают физико-химические характеристиками, свойственными их биологически активным формам.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ ПРОТЕИНАЗЫ Ulp275 | 2011 |
|
RU2451076C1 |
| ГИБРИДНЫЙ БЕЛОК (ВАРИАНТЫ), ШТАММ БАКТЕРИЙ ESCHERICHIA COLI - ПРОДУЦЕНТ ГИБРИДНОГО БЕЛКА (ВАРИАНТЫ) И СПОСОБ ПОЛУЧЕНИЯ БЕЗМЕТИОНИНОВОГО ИНТЕРФЕРОНА АЛЬФА-2 ЧЕЛОВЕКА | 2011 |
|
RU2453604C1 |
| ГИБРИДНЫЙ БЕЛОК, ШТАММ БАКТЕРИЙ ESCHERICHIA COLI - ПРОДУЦЕНТ ГИБРИДНОГО БЕЛКА И СПОСОБ ПОЛУЧЕНИЯ БЕЗМЕТИОНИНОВОГО ИНТЕРФЕРОНА АЛЬФА-2b ЧЕЛОВЕКА ИЗ ЭТОГО ГИБРИДНОГО БЕЛКА | 2010 |
|
RU2441072C1 |
| Протеиновая платформа для получения антиген-презентирующих наночастиц | 2023 |
|
RU2814986C1 |
| Генетическая конструкция, кодирующая предшественник белка YB-1 человека, штамм Escherichia coli - продуцент предшественника белка YB-1 человека, способ микробиологического синтеза этого предшественника | 2019 |
|
RU2728237C1 |
| ГИБРИДНЫЙ БЕЛОК НА ОСНОВЕ L-АСПАРАГИНАЗЫ Wolinella succinogenes, ШТАММ Escherichia coli - ПРОДУЦЕНТ ГИБРИДНОГО БЕЛКА (ВАРИАНТЫ) И СПОСОБ ПОЛУЧЕНИЯ ГИБРИДНОГО БЕЛКА, ОБЛАДАЮЩЕГО ПРОТИВООПУХОЛЕВОЙ АКТИВНОСТЬЮ | 2014 |
|
RU2562166C1 |
| Температурочувствительный мутантный интеин для нерастворимой экспрессии предшественника целевого белка | 2015 |
|
RU2619217C1 |
| Полипептид для понижения уровня сахара в крови на основе глюкагоноподобного пептида-1 человека, рекомбинантный штамм-продуцент E. coli и способ получения этого полипептида | 2016 |
|
RU2642260C1 |
| СИСТЕМА ЭКСПРЕССИИ И СПОСОБ ПОЛУЧЕНИЯ НЕМОДИФИЦИРОВАННЫХ РЕКОМБИНАНТНЫХ БЕЛКОВ В Escherichia coli С ЕЁ ИСПОЛЬЗОВАНИЕМ | 2015 |
|
RU2604796C1 |
| СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО БЕЛКА ПАУТИНЫ, СЛИТЫЙ БЕЛОК, РЕКОМБИНАНТНАЯ ДНК, ВЕКТОР ЭКСПРЕССИИ, КЛЕТКА-ХОЗЯИН И ШТАММЫ-ПРОДУЦЕНТЫ | 2010 |
|
RU2451023C1 |
Способ получения биологически активных рекомбинантных протеинов, содержащих до 4 дисульфидных связей. Способ предусматривает разработку генетической конструкции, кодирующей предшественник целевого белка-мишени, содержащий в своем составе расположенные в порядке перечисления от N-конца к С-концу последовательность самоассоциирующего пептида L6KD, включающего следующие друг за другом 6 остатков лейцина, последовательность белка SUMO - продукта гена Smt3 дрожжей Saccharomyces cerevisiae и последовательность целевого белка-мишени, первый N-концевой остаток которого, следующий за последовательностью белка SUMO, отличен от пролина. Созданную генетическую конструкцию используют для получения целевого белка-мишени путем трансформации этой конструкцией подходящего реципиентного штамма, культивирования трансформированного штамма в условиях, обеспечивающих экспрессию генетической конструкции и микробиологический синтез предшественника целевого белка-мишени в форме нерастворимых телец включения, с последующим выделением синтезированных телец включения из клеток штамма-продуцента, высвобождением из их состава растворимого целевого белка-мишени с помощью подходящей производной SUMO-специфичной протеиназы, отделением остатков нерастворимых телец включения с помощью центрифугирования и дальнейшей очисткой высвобожденного белка-мишени с использованием методов молекулярной биологии, без применения ренатурации. Изобретение обеспечивает эффективное получение биологически активных рекомбинантных белков-мишеней, производимых с использованием микробиологического синтеза без применения процедур денатурации/ренатурации. 5 ил., 2 табл., 16 пр.
Способ получения биологически активных рекомбинантных протеинов, содержащих до 4 дисульфидных связей, заключающийся в разработке генетической конструкции, кодирующей предшественник целевого белка-мишени, содержащий в своем составе расположенные в порядке перечисления от N-конца к С-концу последовательность самоассоциирующего пептида L6KD, заключающего следующие друг за другом 6 остатков лейцина, последовательность белка SUMO - продукта гена Smt3 дрожжей Saccharomyces cerevisiae и последовательность целевого белка-мишени, первый N-концевой остаток, следующий за последовательностью белка SUMO, отличен от пролина, и использования созданной генетической конструкции для получения целевого белка-мишени путем трансформации этой конструкцией подходящего реципиентного штамма, культивирования трансформированного штамма в условиях, обеспечивающих экспрессию генетической конструкции и микробиологический синтез предшественника целевого белка-мишени в форме нерастворимых телец включения, с последующим выделением синтезированных телец включения из клеток штамма-продуцента, высвобождением из их состава растворимого целевого белка-мишени с помощью подходящей производной SUMO-специфичной протеиназы, отделением остатков нерастворимых телец включения с помощью центрифугирования и дальнейшей очисткой высвобожденного белка-мишени с использованием методов молекулярной биологии, известных из уровня техники, без применения ренатурации.
| US 9200306 B2, 01.12.2015 | |||
| СПОСОБ ПОЛУЧЕНИЯ ПРОТЕИНАЗЫ Ulp275 | 2011 |
|
RU2451076C1 |
| ГИБРИДНЫЙ БЕЛОК, ШТАММ БАКТЕРИЙ ESCHERICHIA COLI - ПРОДУЦЕНТ ГИБРИДНОГО БЕЛКА И СПОСОБ ПОЛУЧЕНИЯ БЕЗМЕТИОНИНОВОГО ИНТЕРФЕРОНА АЛЬФА-2b ЧЕЛОВЕКА ИЗ ЭТОГО ГИБРИДНОГО БЕЛКА | 2010 |
|
RU2441072C1 |
| YANG X | |||
| ET AL | |||
| Cleavable Self-Aggregating Tags (cSAT) for Therapeutic Peptide Expression and Purification | |||
| In: Garcia Fruitоs, E., Aris Giralt, A | |||
| (eds) Insoluble Proteins | |||
| Methods in Molecular Biology, 2022, vol 2406 | |||
| Humana, New York, NY | |||
| PP | |||
| Способ получения продукта конденсации бетанафтола с формальдегидом | 1923 |
|
SU131A1 |

