Область техники
Изобретение относится к области биотехнологии и касается новой платформы для синтеза белков, способных к эффективной самосборке в антиген-презентирующие наночастицы, пригодные для создания вакцинирующих препаратов.
Уровень техники
Вакцинирование остается наиболее экономичным способом предотвращения заболеваний. Пандемия новой коронавирусной инфекции Covid-19 наглядно продемонстрировала потребность в проведении ускоренных разработок и испытания вакцин против новых патогенов. Также стало очевидным, что наличие задела в виде продвинутой платформы способно решительно сократить расходы и продолжительность стадии разработки и снизить риски, связанные с эффективностью создаваемого препапата.
На этом фоне выросла актуальность рекомбинантных платформ, позволяющих разрабатывать препараты, обладающие протективным эффектом, на основе лишь информации об уникальных макромолекулярных паттернах патогена-мишени, но без непосредственного использования природного патогенного материала. Известным примером такой платформы является аденовирусный носитель, применявшийся при создании новой российской вакцины «Спутник V». «Узким местом» подобных платформ является высокая собственная иммуногенность, вызывающая ухудшение протективных свойств вакцинирующих препаратов, разработанных на одной и той же платформе, по мере их применении на одних и тех же пациентах. Причина в том, что уже первое терапевтическое применение вызывает развитие иммунитета, нацеленного на нейтрализацию высокоиммуногенной платформы-носителя, присутствующей во всех изготовленных на ее основе препаратах, пусть даже защищающих от разных патогенов. В этой связи разработчики вакцин во всем мире заинтересованы в недорогих и безопасных биологических платформах, обладающих мощными адъювантными свойствами и при этом низкой, в идеале - нулевой, собственной иммуногенностью. Однако область поиска таких платформ существенно ограничена экономическими соображениями и требованиями безопасности.
На протяжении двух последних десятилетий растет интерес к использованию коротких самоассоциирующих пептидов (САП), называемых тагами агрегации, для получения искусственных биологически активных белковых комплексов и НЧ, способных служить основой для разработки вакцин [Gomes et al, 2017; Negahdaripour et al, 2017; Lei et al, 2019; Tsoras & Champion, 2019; Malonis et al, 2021]. В число таких САП входят и мицеллообразующие пептиды [Trimaille & Verrier, 2015; Hamley, 2021].
В ряде случаев САП, присоединенные к N- или С-концам целевых белков, придают этим белкам свойства самособирающихся фьюжнов (СФ), способных к самоассоциации (самосборке) и формированию НЧ без нарушения корректной свертки и биологической активности. В частности, такими свойствами обладает известный гидрофобный мицеллообразующий САП L6KD, включающий 6 остатков лейцина [Zhou et al, 2012; Zhao et al, 2016; Zhao et al, 2017].
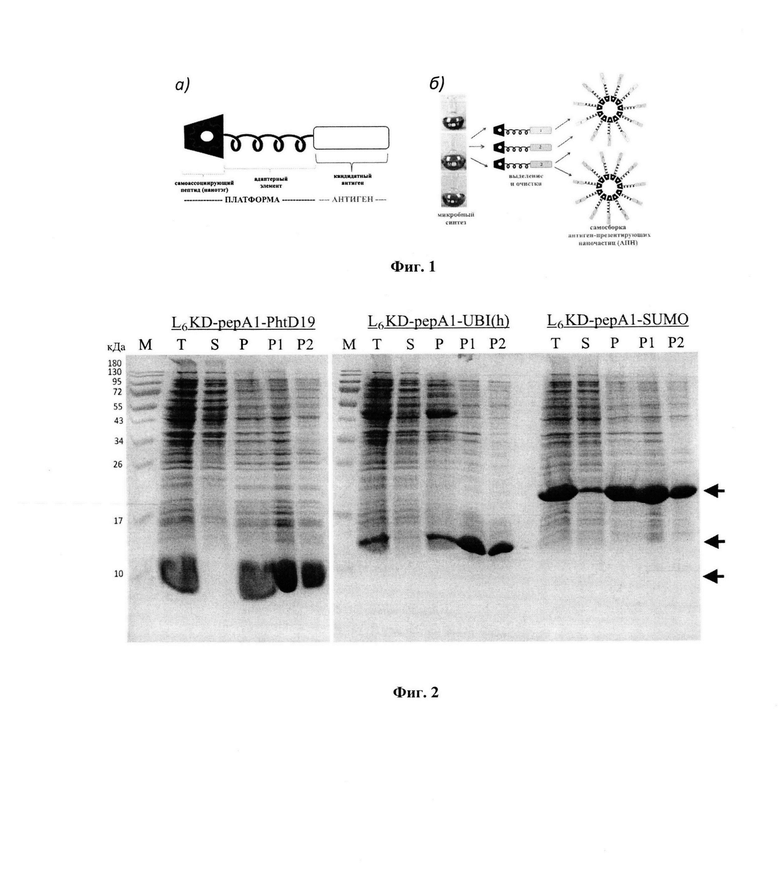
Ближайшим аналогом заявляемой протеиновой платформы является платформа на основе белка SUMO, представляющая собой соединение N-концевого САП L6KD с белком SUMO дрожжей Saccharomyces cerevisiae [Gorbunov et al, 2022]. Платформу-аналог используют для конструирования СФ, содержащих С-концевой вариабельный антиген-мишень. В составе СФ белок SUMO выполняет функцию адаптерного элемента, осуществляющего связь N-концевого САП L6KD С-концевым антигеном-мишенью и одновременно обеспечивающего эффективный биосинтез СФ в тельцах включения в клетках Е. coli. С помощью криоэлектронной микроскопии было показано, что очищенные СФ в процессе ренатурации формируют фибриллярные САПН с упорядоченной структурой, диаметром -17,5 нм и длиной до 500 нм. Показано, что (1) целевой антиген-мишень экспонируется на поверхности САПН (фиг. 1); (2) препараты САПН вызывают сильный антительный ответ у мышей без применения адъювантов, что свидетельствует о наличии у них сильных адъювантных свойств, (3) целевой антиген и платформа обладают близкими показателями иммуногенности [Gorbunov et al, 2022].
От традиционных биосинтетических платформ, например, на основе оболочечных белков различных вирусов, применяемых для получения вирусоподобных частиц наноразмера, платформу-аналог отличает механизм самосборки. В то время как для большинства природных самособирающихся белков формирование ВПЧ/НЧ обеспечивает сложный комплекс многочисленных труднопредсказуемых взаимодействий, ключевым элементом платформы-аналога, отвечающим за формирование САПН, является короткий САП L6KD), процесс самосборки которого относительно прост и подчиняется известным законам мицеллообразования. Это значительно упрощает процесс получения препаратов САПН и расширяет возможности по их применению.
От платформы - ближайшего аналога заявляемую платформу отличает адаптерный элемент, соединяющий N-концевой САП L6KD с С-концевым антигеном-мишенью. В отличие от адаптерного белка SUMO размером 98 аминокислотных остатков адаптерные элементы в составе ПФ имеют размер менее 35 аминокислотных остатков. За счет этого размен пептидной платформы снижен до величины менее 45, а о что выводит ее и абсолютные лидеры в области миниатюризации платформ, используемых для биосинтеза протеинов с целью получения САПН. Вдобавок, как показывают результаты, укороченная пептидная платформа обеспечивает: (а) низкий уровень собственной иммуногенности САПН, не содержащих С-концевой целевой пептидный антиген; (б) мощный антительный ответ на целевой пептидный антиген. В сумме это обеспечивает пептидным САПН улучшенное на порядок соотношение показателей собственной и специфической иммуногенности по сравнению с ближайшим аналогом.
Технической проблемой, на решение которой направлена заявляемая группа изобретений, является расширение арсенала протеиновых платформ, предназначенных для получения самособирающихся антиген-презентирующих наночастиц для разработки на их основе вакцинирующих препаратов.
Раскрытие сущности изобретения
Техническим результатом заявляемого изобретения является создание протеиновой платформы для биосинтеза, самосборки и презентации целевого антигена в составе самособирающихся антиген-презентирующих наночастиц.
Для достижения технического результата предложена протеиновая платформа для получения биосинтетических антиген-презентирующих наночастиц, содержащая в своем составе самоассоциирующий пептид L6KD, соединенный с пептидом pepA1, составной частью которого является последовательность SEQ ID NO: 1 или SEQ ID NO: 2 или SEQ ID NO: 3 или SEQ ID NO: 4.
Заявляемое изобретение раскрывает первичную структуру и неожиданное применение пептида pepA1, составной частью которого является последовательность SEQ ID NO: 1 или SEQ ID NO: 2 или SEQ ID NO: 3 или SEQ ID NO: 4. В частности, раскрывается использование пептида pepA1 в качестве адаптерного элемента в составе новой протеиновой платформы L6KD-pepA1, которое обеспечивает применение этой платформы для конструирования и биосинтеза искусственных антиген-содержащих протеинов в форме телец включения в клетках E.coli, очистки и самосборки этих протеинов в антиген-презентирующие наночастицы (САПН), вызывающие иммунный ответ без использования адъюванта.
Заявляемое изобретение раскрывает также универсальные свойства новой протеиновой платформы, а именно ее способность обеспечивать получение САПН с целевыми антигенами разного размера, имеющими разные аминокислотные последовательности. Универсальные свойства платформы подтверждают, например, с помощью получения препарата САПН с антигенным полипептидом PhtD19 пневмококка размером 19 аминокислотных остатков (а.о.) [Gorbunov et al, 2022], с белком убиквитином человека размером 76 а.о. [GenBank P0CG47] и с белком SUMO (Smt3) дрожжей S. cerevisiae размером 97 а.о [GenBank Q12306]. С помощью методов электронной микроскопии и динамического светорассеяния показано, что, независимо от пазмеоа и свойств целевого антигена, заявляемая платформа обеспечивает его получение в составе САПН.
Из уровня техники известна способность протеиновых наночастиц, независимо от их происхождения, вызывать иммунный ответ, позволяющий использовать их в качестве основы вакцинирующих препаратов [Gomes et al, 2017; Negahdaripour et al, 2017; Lei et al, 2019; Tsoras & Champion, 2019; Malonis et al, 2021]. В связи с тем, что данная способность признается в качестве правила, обобщающего свойства различных протеиновых наночастиц, полученных с использованием разных платформ, заявляемое изобретение подтверждает, сто препараты САПН получаемые на основе новой протеиновой платформы, не являются исключением из данного правила. С этой целью на базе новой протеиновой платформы получают, например, препарат САПН с целевым антигенным полипептидом PhtD19. Анализ биологической активности этого препарата доказывает, что он обладает иммуногенными свойствами и вызывает у животных иммунный ответ против целевого антигена без использования адъюванта, причем показатели этого ответа значительно превосходят показатели, характерные для препарата, получаемого на основе платформы ближайшего аналога. Тем самым, заявляемое изобретение не только подтверждает потенциал, но и раскрывает преимущества использования новой протеиновой платформы для создания вакцинирующих препаратов.
Заявляемое изобретение раскрывает также модификации пептида pepA1, сохраняющие модифицированный пептид пригодным для использования в качестве адаптерного элемента в составе новой протеиновой платформы. В частности, раскрыта информация о допустимых делециях и аминокислотных заменах в С-концевой части пептида pepA1, что позволяет установить минимальные требования к последовательности пептида pepA1, а именно к наличию его в составе последовательности SEQ ID NO: 1 или SEQ ID NO: 2 или SEQ ID NO: 3 или SEQ ID NO: 4.
Тем самым заявляемое изобретение раскрывает новую протеиновую платформу для получения вакцинирующих препаратов.
Краткое описание чертежей
На фиг. 1 представлена принципиальная структура самособирающихся фьюжнов СФ (а) и схема получения моно- и мультивалентных самособирающихся антиген-
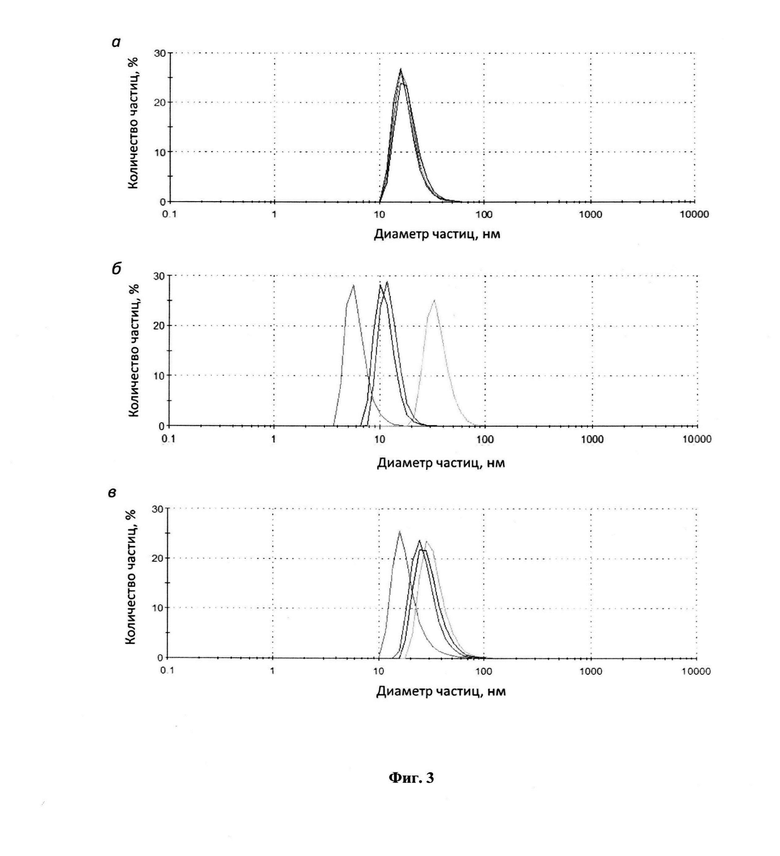
На фиг. 2 представлен сравнительный анализ уровня биосинтеза, растворимости и эффективности ренатурации целевого фьюжна в клетках E.coli. На дорожки нанесены образцы Т, S, Р, Р1 и Р2 клеточных лизатов штаммов-продуцентов E.coli, полученные как описано в примерах 18 и 19. Стрелки в порядке сверху вниз указывают положение целевых фьюжнов L6KD-pepA1-PhtD19, L6KD-pepA1-UBI(h) и L6KD-pepA1-SUMO. М - предокрашенные маркеры молекулярного веса белков (Pierce-26616), молекулярный вес маркеров указан слева.
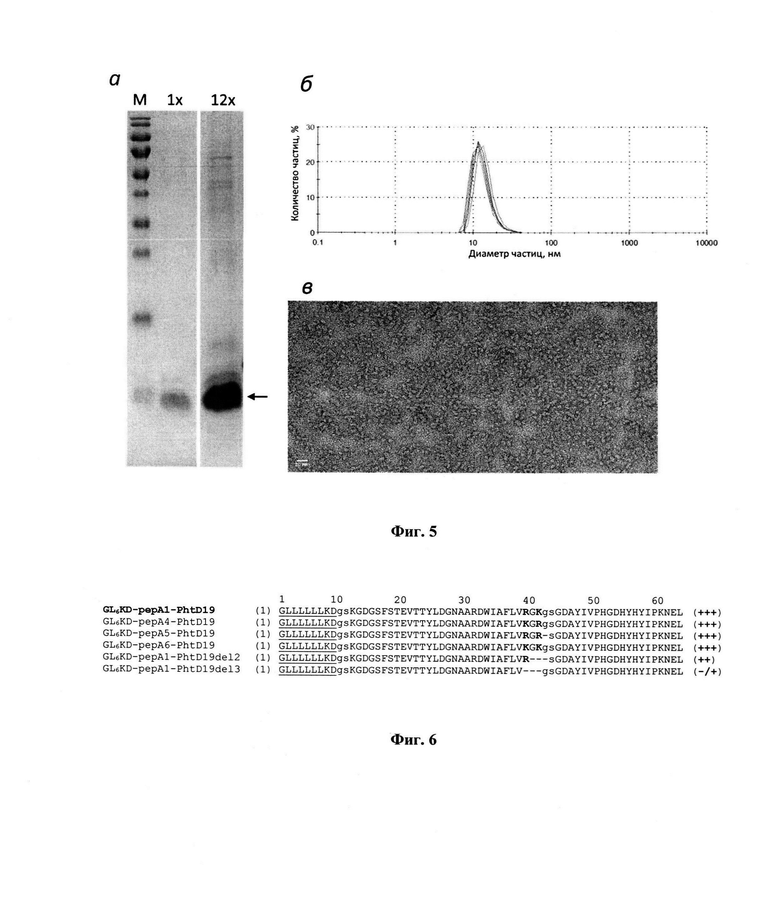
На фиг. 3 показано распределение количества наночастиц от размера в образцах ренатуированных фьюженов а- L6KD-рерА1-PhtD19, 6 - L6KD-pepA1-UBI(h), в - L6KD-pepA1-SUMO. Распределения получены на основе приборной обработки результатов динамического рассеяния света. Каждый образец подвергали 4 независимым сеансам измерения.
На фиг. 4 приведены результаты трансмиссионной электронной микроскопии образцов ренатурированных фьюжнов а - L6KD-рерА1-PhtD19, 6 - L6KD-pepA1-UBI(h), в - L6KD-pepA1-SUMO.
На фиг. 5 показаны результаты анализа физико-химических характеристик высокоочищенного образца САПН, полученного с использованием фьюжна LeKD-pepA1-gs5-PhtD19, сконструированного на базе заявляемой платформы, а - оценка чистоты препарата САПН с помощью гель-электрофореза. На дорожки нанесен препарат САПН в 1-кратном и 12-кратном количестве. Стрелка указывает положение целевого белка. М - предокрашенные маркеры молекулярного веса белков (Pierce-26616), как на фиг. 2; 6 - распределение количества наночастиц от размера. Приведены результаты обработки 5 независимых сеансов динамического рассеяния света; в - трансмиссионная электронная микроскопия образца САПН.
На фиг. 6 приведены первичные последовательности контрольного фьюжна L6KD-рерА1-PhtD19 и его мутантных производных. Серым маркером в N-концевой части фьюжнов выделена последовательность мицеллообразующего пептида L6KD. Серым маркером в С-концевой части фьюжнов выделена последовательность антигенного пептида PhtD19. Заглавными буквами в центральной части фьюжнов показаны последовательности адаптерного пептида pepA1. Аминокислотные замены выделены жирным шрифтом. Строчными буквами выделены линкерные (спейсерные) последовательности. В правой части рисунка в скобках приведены оценочные уровни синтеза соответствующих фьюжнов в клетку E.coli. Условные обозначения + высокий уровень, - низкий уровень.
Осуществление изобретения
Пример 1. Праймеры, используемые для осуществления изобретения

Пример 2. Конструирование вспомогательной плазмиды pET28-L6KD-sfGFP
Для получения целевой плазмиды проводят ПЦР-амплификацию гена sfGFP. В процессе амплификации 5'-конец гена sfGFP удлиняют за счет последовательности, кодирующей самоассоциирующий пептид L6KD. Матрицей для амплификации служит, например, плазмида pAAV-CAG-NES-GAF-CaMP2-sfGFP, любезно предоставленная Ф.В. Субачем (НИЦ «Курчатовский институт»). Для амплификации используют праймеры N1472, N1473 и N1686 (табл. 1).
Амплификацию проводят в две стадии. Сначала ген sfGFP амплифицируют с использованием праймеров N1472 и N1686, а затем полученный фрагмент ДНК реамплифицируют с использованием праймеров N1473 и N1686.
Полученный в результате амплифицированный фрагмент ДНК очищают из геля, используя с этой целью набор Qiagen (Qiagen. cat. №28706) и после открывания концов с использованием рестриктаз Ncol и -А7 го1 клонируют в плазмиде pET28b+(Novagen), расщепленной по этим же сайтам.
В результате клонирования получают плазмиду pET28-L6KD-sfGFP. В составе этой плазмиды клонированный фрагмент ДНК заключает структурный ген sfGFP прецизионно слитый с 5'-концевой последовательностью ДНК, кодирующей самоассоциирующий пептид L6KD, находящейся в окружении сайтов узнавания рестриктаз NcoI и BamHI.
Плазмиду pET28-L6KD-sfGFP используют в качестве вспомогательной для дальнейших этапов конструирования.
Пример 3. Конструктирование вспомогательной плазмиды pET28bi2-L6KD-sfGFP.
Для получения вспомогательной плазмиды pET28bi2-L6KD-sfGFP проводят ПЦР-амплификацию фрагмента ДНК плазмиды pET28b+(Novagen). Для амплификации используют праймеры N619 и N1742 (табл. 1).
Амплифицированный фрагмент ДНК очищают из геля, используя с этой целью набор Qiagen и после открывания концов с использованием рестриктаз BamHI и NcoI клонируют в плазмиде pET28-L6KD-sfGFP (пример 2), расщепленной по уникальным сайтам BglII и NcoI. В результате клонирования получают плазмиду pET28bi2-L6KD-sfGFP, которая в области промотора Т7 вместо сайта BglII содержит уникальный сайт SalI.
Вспомогательную плазмиду pET28bi2-L6KD-sfGFP используют для клонирования других фрагментов ДНК.
Пример 4. Конструирование фрагмента ДНК, кодирующего пептид pepA1
Фрагмент ДНК, кодирующий пептид pepA1 получают с помощью ПНР, которую проводят на смеси трех праймеров N1755, N1756 и N1757 (табл. 1).
Амплифицированный фрагмент ДНК размером 100 пар оснований элюируют из агарозного геля как в примере 2, и после открывания концов с использованием рестриктаз BglII и XhoI клонируют в плазмиде pET28b+(Novagen), расщепленной по таким же сайтам. В результате клонирования получают плазмиду pET28del-pepA1, в составе которой нуклеотидную последовательность клонированного фрагмента ДНК SEQ ID NO: 1, кодирующего пептид pepA1, подтверждают секвенированием.
Плазмиду pET28del-pepA1 используют для получения генетических конструкций, кодирующих самособирающиеся пептидные фьюжны, заключающие в своем составе в качестве адаптерной части пептид pepA1, имеющий последовательность SEQ ID NO: 1.
Пример 5. Клонирование фрагмента ДНК, кодирующего белок SUMO дрожжей S.cerevisiae
Ген SMT3, кодирующий белок SUMO дрожжей S.cerevisiae, амплифицируют в ходе ПЦР с использованием в качестве матрицы хромосомной ДНК штамма S.cerevisiae Y618 [Kartasheva et al., 1996]. Амплификацию проводят в две стадии. Сначала амплифицируют два перекрывающихся фрагмента ДНК. Фрагмент 1 амплифицируют с использованием праймеров N450 и N454 (Табл. 1). Фрагмент 2 амплифицируют с использованием праймеров N453 и N468' (Табл. 1).
Амплифицированные фрагменты ДНК элюируют из агарозного геля как в примере 2, смешивают, и полученную смесь используют для ПЦР-лигирования смешанных фрагментов ДНК. Праймерами для лигирующей ПЦР служат N450 и N468'. Полученный в результате ПЦР-лигирования фрагмент ДНК элюируют из агарозного геля, обрабатывают рестриктазами BglII и XhoI и клонируют в плазмиде pET28b+(Novagen), расщепленной по тем же сайтам. В результате клонирования получают плазмиду pET28del-SUMO, в составе которой нуклеотидную последовательность клонированного фрагмента ДНК подтверждают секвенированием.
Полученная плазмида pET28del-SUMO служит источником уникального BglIIIXhoI фрагмента ДНК, кодирующего белок SUMO дрожжей S. cerevisiae размером 97 а.о.
Пример 6. Конструирование фрагмента ДНК, кодирующего белок убиквитин человека
Согласно данным GenBank аминокислотные последовательности убиквитина человека и убиквитина дрожжей S. cerevisiae отличаются всего тремя заменами. В этой связи с целью получения фрагмента ДНК, кодирующего убиквитин человека, используют ген убиквитина дрожжей, который клонируют, как описано ранее [RU 2489481], в который с помощью ПЦР вносят необходимые замены.
Для внесения замен с помощью ПЦР в качестве матрицы используют ДНК плазмиды pPDX3-GAL1U [RU 2489481]. ПРР проводят в две стадии. Сначала амплифицируют два перекрывающихся фрагмента ДНК. Один фрагмент амплифицируют с использованием праймеров N1747 и N1293 (Табл. 1), а другой - с использованием праймеров N1292 и N1748 (Табл. 1).
Амплифицированные фрагменты ДНК элюируют из агарозного геля как в примере 2, смешивают, и полученную смесь используют для ПЦР-лигирования смешанных фрагментов ДНК. Праймерами для лигирующей ПЦР служат N1747 и N1748. Полученный в результате ПЦР-лигирования фрагмент ДНК элюируют из агарозного геля, обрабатывают рестриктазами BglII и XhoI и клонируют в плазмиде pET28b+(Novagen), расщепленной по тем же сайтам. В результате клонирования получают плазмиду pET28del-UBI(h), в составе которой нуклеотидную последовательность клонированного фрагмента ДНК подтверждают секвенированием.
Полученная плазмида pET28del-UBI(h) служит источником уникального BglII/XhoI фрагмента ДНК, кодирующего белок убиквитин человека размером 76 а.о.
Пример 7. Конструирование фрагмента ДНК, кодирующего пептид PhtD19
Конструирование описано в работе Горбунова с соавт. [Gorbunov et al, 2022]. Для конструирования используют праймеры N1734, N1762 и N1763 (табл. 1). В результате конструирования получают плазмиду pET28del-PhtD19, в составе которой нуклеотидную последовательность клонированного фрагмента ДНК SEQ ID NO: 1, кодирующего пептид PhtD19, подтверждают секвенированием.
Плазмиду pET28del-PhtD19 используют для получения генетических конструкций, кодирующих самособирающиеся пептидные фьюжны, заключающие в своем составе в качестве целевого С-концевого антигена иммуногенный пептид PhtD19, представляющий собой использованный в работе Горбунова с соавт. [Gorbunov et al, 2022] фрагмент (а.о. 200-219) поверхностного белка D гистидиновой триады пневмококка Streptococcus pneumoniae [Papastamatiou et al, 2018].
Пример 8. Конструирование фрагмента ДНК, кодирующего линкерный пептид Glv4Ser(gs5)
Фрагмент ДНК. кодирующий линкерный пептид gs5 получают с помотью ПНР. которую проводят на плазмидной матрице pET28b+с использованием праймеров N1742 и Pr(gs5) (табл. 1).
Амплифицированный фрагмент ДНК гидролизуют по уникальным сайтам NcoIl и XhoI с использованием соответствующих рестриктаз и полученный NcoI/XhoI фрагмент ДНК используют для замещения NcoI/XhoI фрагмента ДНК в плазмиде pET28bi2-GL6KD-sfGFP. В результате клонирования получают плазмиду pET28bi2-gs5, в составе которой нуклеотидную последовательность клонированного фрагмента ДНК, кодирующего пептид gs5, подтверждают секвенированием.
Плазмиду pET28bi2-gs5 используют для получения генетических конструкций кодирующих белковые фьюжны, заключающие в своем составе линкерный пептид gs5.
Пример 9. Конструирование фрагмента ДНК, кодирующего слитый пептид gs5-PhtD19
Плазмиду pET28bi-gs5-PhtD19, содержащую фрагмент ДНК, кодирующий слитый пептид gs5-PhtD19, получают путем замещения уникального MluI/BglII фрагмента ДНК плазмиды pET28del-PhtD19 (пример 7) на MluI/BglII фрагмент ДНК плазмиды pET28bi-gs5 (пример 8).
Плазмиду pET28bi-gs5-PhtD19 используют в качестве вспомогательной для получения последующих конструкций.
Пример 10. Конструирование плазмиды pET28bi-L6KD-рерА1 для синтеза самособирающегося фьюжна L6KD-pepA1
Плазмиду pET28bi2-L6KD-pepA1 конструируют путем замещения уникального MluI/BglII фрагмента ДНК плазмиды pET28del-pepA1 (пример 4) на уникальный MluI/BglII фрагмент ДНК плазмиды pET28bi2-L6KD-sfGFP (пример 3).
Полученная плазмида pET28bi2-L6KD-pepA1 служит источником фрагмента ДНК, кодирующего белок платформы L6KD-рерА1, содержащей N-концевой самоассоциирующий пептид L6KD, слитый с адаптерным пептидом pepA1. Ее используют для конструирования плазмид для синтеза целевых самособирающихся фьюжнов.
Пример 11. Конструирование плазмиды pET28bi2-L6KD-pepA1-gs5-PhtD19 для синтеза самособирающегося фьюжна L6KD-pepA1-gs5-PhtD19
Плазмиду pET28bi2-L6KD-pepA1-gs5-PhtD19 конструируют путем замещения уникального MluI/BglII фрагмента ДНК плазмиды pET28bi-gs5-PhtD19 (пример 9) на уникальный MluI/BglII фрагмент ДНК плазмиды pET28bi2-L6KD-pepA1 (пример 10).
Полученную плазмиду pET28bi2-L6KD-pepA1-gs5-PhtD19 используют для биосинтеза и анализа свойств целевого самособирающегося фьюжна L6KD-pepA1-gs5-PhtD19, заключающего N-концевой самоассоциирующий пептид LeKD, адаптерный пептид pepA1, линкерный пептид gs5 и С-концевой целевой антигенный пептид PhtD19.
Пример 12. Конструирование плазмиды pET28bi2-L6KD-pepA1-PhtD19 для синтеза самособирающегося фьюжна L6KD-рерА1- PhtD19
Плазмиду pET28bi2-L6KD-pepA1-PhtD19 конструируют путем замещения уникального MluI/BglII фрагмента ДНК плазмиды pET28del-PhtD19 (пример 7) на уникальный MluI/BglII фрагмент ДНК плазмиды pET28bi2-L6KD-pepA1 (пример 10).
Полученную плазмиду pET28bi2-L6KD-pepA1-gs5-PhtD19 используют для биосинтеза и анализа свойств целевого самособирающегося фьюжна L6KD-pepA1-PhtD19, заключающего N-концевой самоассоциирующий пептид L6KD, адаптерный пептид pepA1 и С-концевой целевой антигенный пептид PhtD19.
Пептидный фьюжн L6KD-pepA1-PhtD19 отличается от пептидного фьюжна L6KD-pepA1-gs5-PhtD19 отсутствием линкерного элемента gs5.
Пример 13. Конструирование плазмиды pET28bi2-L6KD-pepA1-UBI(h) для биосинтеза самособирающегося фьюжна LeKD-pepA1-UBi(h))
Плазмиду pET28bi2-L6KD-pepA1-UBI(h) конструируют путем замещения уникального MluI/BglII фрагмента ДНК плазмиды pET28del-UBI(h) (пример 6) на уникальный MluI/BglII фрагмент ДНК плазмиды pET28bi-ЬбКХ)-рерА1 (пример 10).
Полученную плазмиду pET28bi2-L6KD-pepA1-UBI(h) используют для биосинтеза и анализа свойств целевого самособирающегося фьюжна L6KD-pepA1-UBI(h), заключающего N-концевой самоассоциирующий пептид L6KD, адаптерный пептид pepA1 и С-концевой целевой антиген - белок убиквитин человека UBI(h).
Пример 14. Конструирование плазмиды pET28bi2-L6KD-pepA1-SUMO для биосинтеза самособирающегося фьюжна LeKD-pepA1-SUMO
Плазмиду pET28bi2-L6KD-pepA1-SUMO конструируют путем замещения уникального MluI/BglII фрагмента ДНК плазмиды pET28del-SUMO (пример 5) на уникальный MluI/BglII фрагмент ДНК плазмиды pET28bi2-L6KD-pepA1 (пример 10).
Полученную плазмиду pET28bi2-L6KD-pepA1-SUMO используют для биосинтеза и анализа свойств целевого самособирающегося фьюжна L6KD-pepA1-SUMO, заключающего N-концевой самоассоциирующий пептид LeKD, адаптерный пептид pepA1 и С-концевой целевой антиген - белок SUMO дрожжей S. cerevisiae.
Пример 15. Конструирование плазмиды pET28bi2-His10-SUMO(gg)-pepA1 для биосинтеза контрольного фьюжна His10-SUMO(gg)-pepA1
Плазмиду pET28bi2-His10-SUMO(gg)-pepA1 конструируют путем замещения уникального MluI/BglII фрагмента ДНК плазмиды pET28del-pepA1 (пример 4) на уникальный MluI/BglII фрагмент ДНК плазмиды pET28s-His10-SUMO(gg)-His6 [Gorbunov et al, 2022].
Полученную плазмиду pET28bi2-His10-SUMO(gg)-pepA1 используют для биосинтеза контрольного фьюжна His10-SUMO(gg)-pepA1, используемого для анализа уровня антительного ответа на пептид pepA1 с помощью ИФА.
Пример 16. Получение штамма E.coli - продуцента целевого самособирающегося фьюжна или контрольного фьюжна
Штамм - пролуцент целевого или контрольного фьюжна получают путем трансформации реципиентного штамма E.coli BL21(DE3) (Novagen) - ВКПМ В-10189. Трансформацию осуществляют путем введения в клетки реципиентного штамма плазмиды для биосинтеза целевого или контрольного фьюжна с применением реактива CaCl2 [Маниатис с соавт., 1984]. Для трансформации используют плазмиды, которые конструируют, как описано в примерах 11-15. Колонии трансформированного штамма отбирают на селективной среде, содержащей антибиотик канамицин. В результате трансформации получают трансформированный штамм, который содержит определенную плазмиду и в ответ на внесение в среду культивирования индуктора ИПТГ или лактозы осуществляет биосинтез целевого или контрольного фьюжна фьюжна.
Пример 17. Биосинтез и очистка контрольного фьюжна His10-SUMO(gg)-pepA1
Биосинтез и очистку образца контрольного фьюжна, содержащего тэг His10, проводят с использованием металл-хелатной хроматографии, как описано [Gorbunov et al, 2022]. Для оиосинтеза используют конструкцию, описанную в примере 15. Продуцент белка получают как описано в примере 16.
Пример 18. Получение целевого самособирающегося фьюжна во фракции нерастворимых клеточных белков
Биосинтез целевого фьюжна осуществляют следующим образом. Сначала получают посевную культуру. Для этого одну колонию штамма-продуцента (пример 17) засевают в пробирку, содержащую 3 мл среды YTS следующего состава (мас. %): дрожжевой экстракт - 1, триптон бакто - 2, NaCl - 1, канамицин - 0,003, вода - остальное. Посевную культуру заявляемого штамма выращивают на ротационном шейкере со скоростью вращения 250 об/мин в течение 4 часов при температуре 37°С.
На втором этапе посевную культуру переносят в среду для индукции. С этой целью 0,5 мл посевной культуры переносят в колбу, содержащую 50 мл среды TRB следующего состава (мас. %): дрожжевой экстракт - 2,4; триптон бакто - 1,2; одномолярный фосфатный буфер (рН7) - 10; одномолярный раствор сульфата магния - 0,2; лактоза - 0,5; глицерин - 0,5; канамицин - 0,009; вода - остальное. Культивирование в среде для индукции продолжают в течение 20 часов на ротационном шейкере со скоростью вращения 250 об/мин при температуре от 18°С до 42°С в тех же условиях.
По окончании культивирования измеряют оптическую плотность клеточной культуры при длине волны 600 нм, используют подходящий спектрофотометр. В микроцентрифужную пробирку объемом 1,5 мл отбирают 20 о.е. клеточной культуры (предварительно хорошо перемешанной на вортексе). Клетки осаждают центрифугированием в течение 2 минут со скоростью 13200 об/мин, надосадочную жидкость тщательно удаляют с использованием водоструйного насоса. Полученный клеточный осадок промывают. С этой целью осадок суспендируют в 1 мл буфера PBS и вновь центрифугируют в том же режиме, надосадочную жидкость удаляют.
На следующем этапе осуществляют дезинтеграцию собранных клеток штамма-продуцента. С этой целью полученный образец клеток штамма-продуцента ресуспендируют в 1 мл подходящего буфера, например, буфера PBS или буфера 50 мМ Tris-HCl, рН 7.0. Суспензию клеток хорошо перемешивают на вортексе, охлаждают на льду и подвергают дезинтеграции, например, с использованием дезинтегратора ультразвукового лабораторного Bioblock Scientific Vibra cell 75043, в режиме: 5 секунд пульс, 10 секунд пауза, 40 секунд (то есть 9 микроциклов). По завершении дезинтеграции отбирают 100 мкл суммарной фракции Т клеточного лизата.
Остальной объем лизата центрифугируют 10 минут при 13200 об/мин при температуре 4°С.Отбирают 100 мкл осветленной надосадочной жидкости, получая образец S фракции растворимых клеточных белков, после чего оставшуюся надосадочную жидкость тщательно удаляют с использованием водоструйного насоса. Оставшийся осадок ресуспендируют в 0,9 мл PBS, перемешивая на вортексе до гомогенного состояния и получают образец Р фракции нерастворимых клеточных белков.
Целевой фьюжн в образце Р нерастворимых клеточных белков используют для ренатурации и последующего анализа, а также для получения высокоочищенных САПН.
Пример 19. Ренатурация целевого фьюжна
Для ренатурации используют образец Р нерастворимых клеточных белков ресуспендированных в 0,9 мл PBS до гомогенного состояния (пример 18). Образец Р содержит смесь всех нерастворимых клеточных белков. 50 мкл образца Р отбирают в чистую пробирку для последующего анализа с помощью гель-электрофореза.
Оставшийся образец Р центрифугируют 10 минут при 13200 об/мин при температуре 4°С, после чего надосадочную жидкость тщательно удаляют с использованием водоструйного насоса. Полученный осадок тщательно растворяют в 85 мкл 8М мочевины. Полученный раствор подвергают центрифугированию в тех же условиях, надосадочную жидкость переносят в чистую микроцентрифужную пробирку и называют образцом Р1.
В соответствие с процедурой получения в 5 мкл образца Р1 содержатся белки, экстрагированных мочевиной из 50 мкл образца Р0 нерастворимых клеточных белков.
Белки образца Р1 подвергают процедуре ренатурации. С этой целью к 80 мкл образца Р1 прибавляют 720 мкл буфера PBS, то есть осуществляют 10-кратное разведение белкового экстракта, снимая денатурирующее действие мочевины и обеспечивая условия, подходящие для ренатурации белков. Разбавленный раствор инкубируют при температуре +4°С в течение 30 минут для завершения процесса ренатурации, после чего вновь центрифугируют его в прежних условиях. Надосадочную жидкость переносят в чистую микроцентрифужную пробирку и называют образцом Р2.
В соответствии с процедурой получения образец Р2 содержит белки, экстрагированных мочевиной из образца Р нерастворимых клеточных белков и успешно ренатурированные.
Пример 20. Получение высокоочищенного препарата САПН
Целевой фьюжн L6KD-pepA1-gs5-PhtD19, синтезированный в клетках штамма-продуцента во фракции Р нерастворимых клеточных белков, как описано в примере 18, подвергают очистке с одновременной ренатурацией и формированием САПН.
С этой целью 3 г биомассы клеток, содержавших целевой фьюжн, суспендируют в 30 мл буфера PBS рН 7,4, содержащего 5 мМ EDTA и 1 мМ PMSF и подвергают дезинтеграции ультразвуком в течение 7 минут при амплитуде 40%, в режиме: pulse on - 5 секунд, pulse off - 12. Тельца включения (ТВ) осаждают центрифугированием в течение 20 минут при 16500 об/мин.
Полученные ТВ дважды промывают 15 мл раствора 1%-го Triton Х-100 в буфере PBS, содержащем 5 мМ EDTA и растворяют в 15 мл 8М мочевины в течение не менее 15 часов, при +4°С. Нерастворившийся клеточный дебрис удаляют центрифугированием при 16500 об/мин в течение 30 минут. В результате получают раствор денатурированного целевого фьюжна.
Ренатурацию проводят путем медленного вливания раствора денатурированного целевого фьюжна к 10-кратному избытку буфера А (20 мМ трис-HCl, 5 мМ EDTA, рН 8,5). По окончании процесса получают ренатурационную смесь. Концентрация целевого фьюжна в ренатурационной смеси составляет около 0,5 мг/мл, концентрация мочевины-0,8 М. Ренатурационную смесь оставляют на 1-3 дня при +4°С без перемешивания для завершения процесса ренатурации фьюжна и одновременного формирования САПН.
Полученный препарат САПН наносят на колонку HiTrapQ FF объемом 5 мл, предварительно уравновешенную буфером А. Элюцию целевого вещества осуществляют с использованием ступенчатого градиента NaCl в буфере А. Целевое вещество элюируют преимущественно при 0,5М NaCl. Полученный препарат обессоливают и концентрируют посредством ультрафильтрации в ячейке Amicon объемом 50 мл, через мембрану с порогом отсечения 100 кДа до концентрации целевого фьюжна в препарате 0,3 мг/мл.
В результате получают препарат целевых САПН. По данным гель-электрофореза чистота целевого фьюжна в препарате САПН превышает 90%.
Пример 21. Методические подходы к анализу свойств целевого фьюжна
Анализ растворимости целевого фьюжна в клетках E.coli
Образцы фракций Т, S и Р клеточного лизата (пример 18) подвергают анализу с использованием гель-электрофорореза в 15% полиакриламидном геле в денатурирующих восстанавливающих условиях с окрашиванием Кумасси G-250 по стандартной процедуре.
Выявив на геле полосу целевого фьюжна и сопоставив ее интенсивность на дорожках, соответствующих образцам Т, S и Р, делают заключение об уровне синтеза целевого фьюжна в клетках E.coli в форме нерастворимого белка (в форме телец включения). Анализ способности целевого фьюжна к ренатурации
Для проведения анализа используют образцы Р, Р1 и Р2, полученные в процессе ренатурации (пример 19). Данные образцы в пропорциональном количестве анализируют с использованием гель-электрофорореза в 15% полиакриламидном геле в денатурирующих восстанавливающих условиях с окрашиванием Кумасси G-250 по стандартной процедуре. Выявив на геле полосу целевого фьюжна и сопоставив ее интенсивность на дорожках, соответствующих образцам Р, Р1 и Р2, делают заключение об эффективности перехода целевого фьюжна из нерастворимого в растворимое состояние в условиях ренатурации, то есть о способности целевого фьюжна к эффективной ренатурации.
Анализ способности целевого фьюжна к формированию САПН в условиях ренатурации
Образец Р2, полученный в результате ренатурации (пример 19), подвергают анализу с использованием методов динамического светорассеяния и трансмиссионной электронной микроскопии с негативным контрастированием, как описано [Gorbunov et al, 2022].
В результате анализа делают заключение о способности целевого фьюжна к самосборке с образованием САПН в условиях ренатурации.
Пример 22. Методические подходы к анализу высокоочищенного препарата САПН Анализ чистоты препарата САПН
Анализ проводят с использованием гель-электрофорореза в 15% полиакриламидном геле в денатурирующих восстанавливающих условиях с окрашиванием Кумасси G-250 по стандартной процедуре. На дорожки наносят неконцентрированный (1х) образец препарата и концентрированный (12х) образец. Выявив на геле полосу целевого фьюжна, сопоставляют ее интенсивность в пробе 1х с интенсивностью примесных полос в пробе 12х и по результатам сопоставления делают заключение о чистоте целевого фьюжна в высокоочищенном образце САПН.
Анализ структурных особенностей высокоочищенных САПН
Структурные свойства высокоочищенных САПН, получаемых как описано в примере 20, оценивают с использованием методов динамического светорассеяния и криогенной или трансмиссионной электронной микроскопии, как описано [Gorbunov et al, 2022]. Полученные результаты анализируют, в том числе путем сопоставления с опубликованными данными, и на основании анализа делают заключение о структурных характеристиках САПН.
Анализ иммунологических свойств высокоочищенных САПН
Иммунологические свойства высокоочищенных САПН определяют, оценивая уровень антительного ответа на препарат САПН в условиях безадъювантной иммунизации животных, например, мышей линии c57b1/6. Иммунизацию и анализ результатов проводят, как описано [Gorbunov et al, 2022]. Для определения показателей иммунного ответа используют иммуно-ферментный анализ (ИФА). В число показателей, определяемых с помощью ИФА входят: количество (доля) серопозитивных животных а также титр антител.
На основание данных ИФА определяют количество (долю) серопозитивных животных, обнаруживших достоверный антительный отклик на введенный препарат, и среднегеометрический показатель титра антител, обнаруженных в крови серопозитивных животных, специфически взаимодействующих с полипептидом платформы pepA1 и с целевым антигеном PhtD19.
Для анализа титра антител, специфически взаимодействующих с полипептидом платформы pepA1, в качестве подложки для ИФА используют препарат белка His10-SUMO(gg)-pepA1, получение которого описано в примере 17.
Для анализа титра антител, специфически взаимодействующих с целевым антигеном PhtD19, в качестве подложки для ИФА используют белок His10-SUMO(gg)-PhtD19 [Gorbunov et al, 2022].
Полученные результаты анализируют, в том числе, путем сопоставления с опубликованными данными [Gorbunov et al, 2022], и на основании анализа делают заключение об иммунологических свойствах иелевых САПН.
Пример 23. Свойства целевых фьюжнов
С целью определения свойств целевых фьюжнов L6KD-pepA1-PhtD19, L6KD-рерА1-UBI(h) и L6KD-pepA1-SUMO, синтезированных в клетках E.coli на базе новой протеиновой платформы проводят сравнительный анализ:
- уровня синтеза и растворимости. Анализ проводят, как описано в примере 21. Результаты анализа показывают (фиг. 2), что целевые фьюжны характеризуются: (1) сравнимо высоким уровнем биосинтеза; (2) преимущественным накоплением во фракции нерастворимых клеточных белков.
- эффективности ренатурации. Анализ проводят как описано в примере 21. Результаты анализа показывают (фиг. 2), что целевые фьюжны: (1) одинаково эффективно экстрагируются мочевиной из фракции нерастворимых клеточных белков; (2) с одинаковой эффективностью переходят из нерастворимого в растворимое состояние в условиях ренатурации, то есть, одинаково эфективно ренатурируют.
- образцов ренатурированных фьюжнов с использованием метода динамического рассеяния света, как описано в примере 21. Результаты анализа показывают (фиг. 3), что: а) независимо от свойств антигена целевые фьюжны ренатурируют с образованием большого количества наночастиц; б) размер получаемых наночастиц зависит от свойств антигена в составе целевых фьюжнов.
- образцов ренатурированных фьюжнов с использованием трансмиссионной электронной микроскопии с негативным контрастированием, как описано в примере 21. Результаты анализа показывают (фиг. 4), что образцы ренатурированных целевых фьюжнов содержат о своем составе значительное количество наночастиц, размер которых зависит от свойств антигена в составе целевого фьюжна. При этом результаты электронной микроскопии хорошо согласуется с результатами анализа с использованием динамического светорассеяния (фиг. 3).
На основание результатов сравнительного анализа заключают, что, независимо от размера и других свойств целевых антигенов, новая протеиновая платформа обеспечивает целевым фьюжнам:
- эффективный синтез в клетках E.coli;
- накопление в клетках E.coli во фракции нерастворимых белков;
- эффективную ренатурацию,
- эффективное формирование САПН в процессе ренатурации.
Учитывая общность свойств целевых фьюжнов и препаратов САПН, получаемых на базе новой протеиновой платформы, для определения иммунологических свойств выбирают произвольный вариант САПН, например, на основе фьюжна L6KD-pepA1-gs5-PhtD19. Высокоочищенный препарат САПН на основе фьюжна L6KD-pepA1-gs5-PhtD19 получают, как описано в примере 20.
Полученный препарат подвергают анализу, результаты которого представлены на фиг. 5. Для проведения анализа используют методы электрофореза в полиакриламидном геле в денатурирующих восстанавливающих условиях (фиг. 5а), динамического рассеяния света (фиг. 56) и трансмиссионной электронной микроскопии с негативным контрастированием (фиг. 5в), как описано в примере 22. В результате анализа определяют, что чистота целевого фьюжна в высокоочищенном препарате САПН превышает 90% (фиг. 5а), при этом наблюдается сходство характеристик высокоочищенного препарата САПН с образцами ренатурированных целевых фьюжнов, полученных на базе заявляемых платформ (сравнить данные фиг. 5б и 5в с данными фиг. 3 и фиг. 4). На основание результатов анализа заключают, что высокоочищенный препарат САПН на основе фьюжна L6KD-pepA1-gs5-PhtD19 обладает свойствами типичными для препаратов САПН, получаемых на базе новой протеиновой платформы.
Для определения иммунологических свойств высокоочищенный препарат САПН используют для иммунизации и получения сывороток крови животных, которые подвергают ИФА, как описано в примере 22.
Для оценки и сравнительного анализа иммунологических свойств препарата САПН на основе фьюжна L6KD-pepA1-gs5-PhtD19 используют иммунологические показатели ближайшего аналога, САПН на основе фьюжна L6KD-SUMO(gg)-PhtD19, определенные в идентичном эксперименте [Gorbunov et al, 2022]. Результаты сравнительного анализа приведены в табл. 2.
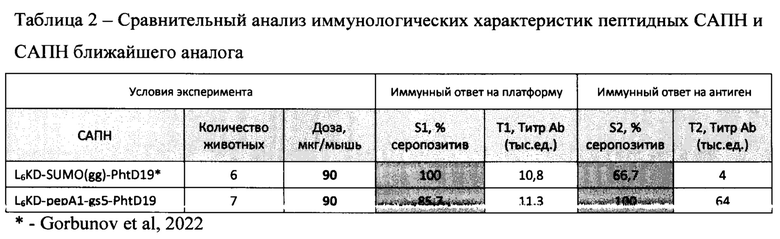
Данные, представленные в табл. 2, показывают, что препарат САПН на платформе ближайшего аналога и на новой протеиновой платформе L6KD)-рерА1 характеризуются сравнимыми показателями иммунного ответа на белки платформы. Доля серопозитивных животных для этих платформ составляет 100% или близко к 100% (85,7%) при титре антител 10,8 и 11,3 тыс.единиц, соответственно. В то же время показатели ответа на целевой антиген у сравниваемых платформ разнятся драматически. Очевидное преимущество демонстрирует новая протеиновая платформа. Так, при использовании новой платформы L6KD-рерА1 доля серопозитивных животных, откликнувшихся на целевой антиген, в 1,5 раза (100% против 66,7%), а титр антител против целевого антигена - в 16 раз (64 тыс.ед против 4 тыс.ед) превышают показатели ближайшего аналога (табл. 2). При этом соотношение величины специфического ответа против целевого антигена к неспецифическому ответу против платформы у заявляемой пептидной платформы примерно в 15 раз превышает показатель ближайшего аналога. Это с очевидностью указывает на то, что относительно ближайшего аналога заявляемая платформа обладает существенно более сильными адъювантными свойствами при существенно более низкой собственной иммуногенности.
Другими словами, сравнительный анализ свидетельствует о существенно большей иммунологической специфичности и эффективности заявляемой пептидной платформы по сравнению с ближайшим аналогом.
Пример 24. Модификации пептида pepA1
Границы и допустимые модификации пептида pepA1, определяющие пригодность модифицированного пептида для использования в качестве основы новой протеиновой платформы, устанавливают с помощью мутантных фьюжнов, содержащих делеции и аминокислотные замены в С-концевой части пептида pepA1. Первичные последовательности мутантных фьюжнов приведены на фиг. 6.
Конструкции для синтеза мутантных фьюжнов получают на основе плазмиды pET28bi2-L6KD-pepA1-PhtD19, используемой для синтеза контрольного самособирающегося фьюжна L6KD-pepA1-PhtD19 (пример 12). Для конструирования делеций и аминокислотных замен в составе фьюжна используют ПЦР.
Получение продуцентов мутантных фьюжнов осуществляют, как описано в примере i6. Синтез, ренатурацию и анализ свойств мутантных фьюжнов проводят, как описано в примерах 18, 19 и 21, соответственно.
В результате анализа свойств мутантных фьюжнов устанавливают, что:
- фьюжны L6KD-pepA1-PhtD19, L6KD-pepA4-PhtD19, UKD-pepA5-PhtD19 и UKD-pepA6-PhtD19, содержащие в С-концевой части пептида pepA1 сочетания остатков аргинина и лизина RGK, как в последовательности SEQ ID NO: 1, или KGR, как в последовательности SEQ ГО NO: 2, или RGR, как в последовательности SEQ ID NO: 3, или KGK, как в последовательности SEQ ID NO:4, соответственно, сохраняют в полном объеме уровень синтеза, растворимые свойства и способность к ренатурации;
- по сравнению с фьюжном L6KD-pepA1-PhtD19 мутантный фьюжн L6KD-рерА1-PhtD19del2, содержащий в С-концевой части пептида pepA1 делецию двух аминокислотных остатков, обладает пониженным уровнем синтеза при сохранении в полном объеме растворимых свойств и способности к ренатурации;
- по сравнению с фьюжном L6KD-pepA1-PhtD19 мутантный фьюжн LeKD-pepA1-PhtD19del3, содержащий в С-концевой части пептида pepA1 делецию трех аминокислотных остатков, практически утрачивает способность к синтезу в клетках E.coli.
На основание полученных данных устанавливают, что в функцию адаптерного элемента в составе новой протеиновой платформы обеспечивает пептид pepA1, составной частью которого является последовательность SEQ ID NO: 1 или SEQ ID NO: 2 или SEQ ID NO: 3 или SEQ ID NO: 4.
Таким образом, примеры, раскрывающие сущность заявляемого изобретения, показывают, что новая протеиновая платформа обеспечивает:
- получение САПН, способных нести целевые антигены (полипептиды и белки) различного размера. При этом платформа детерминирует высокоэффективный синтез целевых фьюжнов в клетках E.coli во фракции нерастворимых белков, а также способность фьюжнов формировать САПН;
- получение значительно более специфичного и сильного иммунного ответа на целевой антиген в составе препарата САПН по сравнению с платформой ближайшего аналога, что свидетельствует о преимуществах использования новой протеиновой платформы для получения вакцинирующих препаратов.
--->
Перечень последовательностей
<110> Федеральное государственное бюджетное учреждение «Национальный
исследовательский центр «Курчатовский институт» (НИЦ «Курчатовский институт»)
<120> Протеиновая платформа для получения антиген-презентирующих наночастиц
<160> 4
<210> 1
<211> 30
<212> PRT
<213> Peptide PepA1
<221> misc_feature
<222> (1)…(30)
<223> Artificial sequence
<400> 1
Lys Gly Asp Gly Ser Phe Ser Thr Glu Val Thr Thr Tyr Leu Asp Gly
1 5 10 15
Asn Ala Ala Arg Asp Trp Ile Ala Phe Leu Val Arg Gly Lys
20 25 30
<210> 2
<211> 30
<212> PRT
<213> Peptide PepA1
<221> misc_feature
<222> (1)…(30)
<223> Artificial sequence
<400> 1
Lys Gly Asp Gly Ser Phe Ser Thr Glu Val Thr Thr Tyr Leu Asp Gly
1 5 10 15
Asn Ala Ala Arg Asp Trp Ile Ala Phe Leu Val Lys Gly Arg
20 25 30
<210> 3
<211> 30
<212> PRT
<213> Peptide PepA1
<221> misc_feature
<222> (1)…(30)
<223> Artificial sequence
<400> 1
Lys Gly Asp Gly Ser Phe Ser Thr Glu Val Thr Thr Tyr Leu Asp Gly
1 5 10 15
Asn Ala Ala Arg Asp Trp Ile Ala Phe Leu Val Arg Gly Arg
20 25 30
<210> 4
<211> 30
<212> PRT
<213> Peptide PepA1
<221> misc_feature
<222> (1)…(30)
<223> Artificial sequence
<400> 1
Lys Gly Asp Gly Ser Phe Ser Thr Glu Val Thr Thr Tyr Leu Asp Gly
1 5 10 15
Asn Ala Ala Arg Asp Trp Ile Ala Phe Leu Val Lys Gly Lys
20 25 30
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ БИОЛОГИЧЕСКИ АКТИВНЫХ РЕКОМБИНАНТНЫХ ПРОТЕИНОВ | 2022 |
|
RU2807615C2 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pET22b(+)/slurp-2, КОДИРУЮЩАЯ БЕЛОК SLURP-2, И ШТАММ БАКТЕРИЙ Escherichia coli BL21(DE3) pET22b(+)/slurp-2- ПРОДУЦЕНТ БЕЛКА SLURP-2 ЧЕЛОВЕКА | 2015 |
|
RU2583307C2 |
| Температурочувствительный мутантный интеин для нерастворимой экспрессии предшественника целевого белка | 2015 |
|
RU2619217C1 |
| Способ получения рекомбинантного экзопротеина А Pseudomonas aeruginosa | 2016 |
|
RU2636346C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pЕТ22b(+)/SLURP-1, КОДИРУЮЩАЯ БЕЛОК SLURP-1, И ШТАММ БАКТЕРИЙ Escherichia coli BL21(DE3)/pET22b(+)/SLURP-1-ПРОДУЦЕНТ БЕЛКА SLURP-1 ЧЕЛОВЕКА | 2010 |
|
RU2453602C2 |
| Способ получения рекомбинантного пептидогликан-ассоциированного липопротеина (PAL) Legionella pneumophila | 2016 |
|
RU2634385C1 |
| СИСТЕМА ЭКСПРЕССИИ И СПОСОБ ПОЛУЧЕНИЯ НЕМОДИФИЦИРОВАННЫХ РЕКОМБИНАНТНЫХ БЕЛКОВ В Escherichia coli С ЕЁ ИСПОЛЬЗОВАНИЕМ | 2015 |
|
RU2604796C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pET22b(+)/вд-ЛИНКС1, КОДИРУЮЩАЯ БЕЛОК СО СВОЙСТВАМИ ЛИНКС1, И ШТАММ БАКТЕРИЙ Escherichia coli BL21(DE3)/pET22b(+)/вд-ЛИНКС1-ПРОДУЦЕНТ БЕЛКА СО СВОЙСТВАМИ ЛИНКС1 ЧЕЛОВЕКА | 2009 |
|
RU2404246C1 |
| ГИБРИДНЫЙ БЕЛОК (ВАРИАНТЫ), ШТАММ БАКТЕРИЙ ESCHERICHIA COLI - ПРОДУЦЕНТ ГИБРИДНОГО БЕЛКА (ВАРИАНТЫ) И СПОСОБ ПОЛУЧЕНИЯ БЕЗМЕТИОНИНОВОГО ИНТЕРФЕРОНА АЛЬФА-2 ЧЕЛОВЕКА | 2011 |
|
RU2453604C1 |
| Рекомбинантная плазмидная ДНК pSMT3_HCRG21, кодирующая гибридный белок SMT3-HCRG21, штамм бактерий Escherichia coli BL21(DE3)/pSMT3_HCRG21 - продуцент анальгетического пептида HCRG21 и способ получения рекомбинантного анальгетического пептида HCRG21 | 2022 |
|
RU2798545C1 |

Изобретение относится к области биотехнологии, а именно к слитому пептиду, содержащему в своем составе расположенные от N-конца к C-концу самоассоциирующий пептид L6KD, пептид pepA1 и антиген. Изобретение эффективно для получения биосинтетических антиген-презентирующих наночастиц. 6 ил., 2 табл., 24 пр.
Слитый пептид для получения биосинтетических антиген-презентирующих наночастиц, содержащий в своем составе расположенные от N-конца к C-концу самоассоциирующий пептид L6KD, пептид pepA1, имеющий последовательность, выбранную из группы SEQ ID NO:1-4, и антиген.
| GORBUNOV A.A | |||
| et al | |||
| Vaccine building kit': Combining peptide bricks to elicit a desired immune response without adding an adjuvant, 2022, Nanomedicine 17(2), DOI:10.2217/nnm-2021-0424 | |||
| MANICA NEGAHDARIPOUR et al | |||
| Harnessing self-assembled peptide nanoparticles in epitope vaccine design, Biotechnol Adv, 2017, 35(5):575-596, doi: |

