[1] Данная заявка испрашивает приоритет по заявке на патент Китая № 2019109413286, поданной 30 сентября 2019 г., содержание которой включено в данный документ во всей своей полноте.
Область техники
[2] Настоящее изобретение относится к области биофармацевтических препаратов, в частности к антителу, нацеливающемуся на CD3, биспецифическому антителу и их применению.
Уровень техники
[3] Т-лимфоциты представляют собой важный класс клеток, участвующих в адаптивном иммунном ответе, и при этом Т-клетки распознают антигены посредством Т-клеточного рецептора (TCR). TCR не распознает поверхностные эпитопы антигена непосредственно, однако специфически распознает молекулярные комплексы антиген-пептид-MHC (pMHC), представленные на поверхности антигенпрезентирующих клеток (APC) или клеток-мишеней. Специфичность Т-клеточного ответа опосредована распознаванием pMHC молекулярными комплексами TCR и CD3. TCR представляет собой гетеродимер, состоящий из двух различных трансмембранных полипептидных цепей, представленных четырьмя пептидными цепями, включающими α, β, γ и δ; причем в соответствии с различными сочетаниями пептидных цепей TCR подразделяют на TCRαβ и TCRγδ. CD3 содержит различные трансмембранные полипептидные цепи, т.е. γ, δ, ε и ζ, которые взаимодействуют с образованием гомодимеров или гетеродимеров в составе комплексов TCR-CD3. Например, комплексы TCR-CD3 предусматривают димер TCRαβ, димер CD3γε, димер CD3δε и димер CD3ζζ. Поскольку цитоплазматическая область пептидной цепи TCR очень короткая, то обычно предполагается, что сигнал активации, генерируемый при распознавании TCR антигена, передается в Т-клетку пептидной цепью CD3.
[4] Ввиду важной роли CD3 в инициировании иммунного ответа средства, обеспечивающие передачу сигналов, нацеливающиеся на сигнальный путь TCR-CD3, в частности моноклональные антитела, нацеливающиеся на CD3, считаются эффективными средствами, которые могут модулировать иммунный процесс и применяться для лечения воспалительных или аутоиммунных заболеваний. Фактически, антитело к CD3 ортоклон OKT3 являлось первым одобренным терапевтическим антителом. OKT3 было впервые одобрена FDA США в 1985 году для лечения острого отторжения после трансплантации органов. Хотя иммуносупрессивная способность, возникающая в результате повторного введения OKT3, обеспечивала эффективное лечение отторжения после трансплантации почки, его применение ограничивалось синдромом первичной токсической реакции на дозу; представляющим собой синдром, который, как полагают, связан с опосредованной OKT3 активацией Т-клеток и высвобождением цитокинов. Впоследствии, в 2010 году, OKT3 было отозвано с рынка ввиду индуцирования серьезного цитокинового шторма и проблем с иммуногенностью, ассоциированных с мышиными антителами, помимо других факторов.
[5] Другая проблема, связанная с антителами к CD3, заключается в том, что многие антитела к CD3 оказались видоспецифическими, например, OKT3 реагирует с CD3 шимпанзе, но не с CD3 других приматов, как например гомологи CD3 макаков или мышиные гомологи CD3. Видовая специфичность моноклональных антител к CD3 является серьезным препятствием для их разработки в качестве лекарственных средств на основе антител для лечения заболеваний у человека. Любое новое лекарственное средство-кандидат должно пройти тщательную доклиническую проверку, прежде чем его можно будет применять в клинических испытаниях с участием людей. Цель доклинического тестирования заключается в том, чтобы подтвердить, что лекарственное средство-кандидат обладает требуемой активностью и, что наиболее важно, что лекарственное средство-кандидат является безопасным. Доклинические тестирование в отношении безопасности включают введение лекарственного средства-кандидата представителям представляющих интерес видов, предпочтительно приматам, отличным от человека. Однако высшие приматы, особенно шимпанзе, считаются видами, находящимися по угрозой исчезновения, и использование таких животных для тестирования безопасности лекарственных средств строго ограничено. Видами, описанными в уровне техники, подходящими для тестирования в отношении оценки безопасности, могут являться макаки, в частности яванские макаки. Однако трудно предоставить достоверные данные по доклинической оценке безопасности антител к CD3, у которых отсутствует специфичная для видов приматов перекрестная реактивность. Среди известных антител, которые связываются с CD3 человека, SP34 является одним из очень немногих, которые могут связываться с несколькими CD3 приматов (например, CD3 человека и яванского макака) (см. Salmeron, A. et al, J Immunol 147 (1991) 3047-3052; Conrad M.L., et. al, Cytometry A 71 (2007) 925-933).
[6] Хотя моноклональные антитела к CD3 прошли клиническую проверку в отношении своей эффективности для определенных групп заболеваний, тем не менее, в последние годы антитела к CD3 все чаще используются в разработке лекарственных средств на основе биспецифических антител. В настоящее время во всем мире на долю антител, представляющих собой биспецифический рекрутер T-клеток на основе CD3 (BsTCE), приходится более чем половина программ, предусматривающих использование биспецифических антител на клинической или доклинической стадиях. Биспецифические антитела к CD3 BsTCE, с одной стороны, демонстрируют столь же высокую эффективность, как и средства терапии на основе CAR-T-клеток, а с другой стороны, их можно получать и коммерциализировать, также как и традиционные моноклональные антитела. Среди биспецифических антител, одобренных в настоящее время для продажи во всем мире, самыми первыми были одобрены катумаксомаб (одобренный Европейским агентством по лекарственным средствам (EMA) в 2009 г. и отозванный из США в 2013 г.) и блинатумомаб (одобренный Управлением по контролю качества пищевых продуктов и лекарственных средств (FDA) в 2014 г.), оба из которых являются BsTCE. Антитело к CD3 является важным компонентом в конструировании BsTCE. Биспецифическое антитело BsTCE может связываться с двумя мишенями одновременно: один конец распознает опухолеассоциированный антиген (ТАА) на поверхности опухолевых клеток, тогда как другой конец связывается с молекулой CD3 на Т-клетках. В присутствии опухолевых клеток связывание биспецифического антитела BsTCE с поверхностью опухолевых клеток может обеспечивать рекрутинг и активацию Т-клеток, находящихся вблизи опухолевых клеток, что в свою очередь обеспечивает уничтожение опухолевых клеток. При проектировании и конструировании различных структур биспецифических антител BsTCE выбор и оптимизация антител к CD3 имеют первостепенную важность. Во-первых, крайне важна видоспецифичность моноклональных антител к CD3, особенно применительно к перекрестной реакции у нечеловекообразных обезьян. Во-вторых, также крайне важна аффинность антитела к CD3 по отношению к комплексу CD3; при этом антитело к CD3 с высокой аффинностью может характеризоваться удерживанием антитела в селезенке и других участках, что затрудняет его контактное взаимодействие с опухолью; и высокая аффинность может также обуславливать чрезмерное стимулирование Т-клеток, приводя в результате к высокому уровню высвобождения цитокинов. В третьих, валентные связи, участвующие в связывании антител к CD3, также играют важную роль, при этом ранее было обнаружено, что многовалентные формы биспецифических антител к CD3 могут вызывать побочные эффекты путем активации Т-клеток без связывания опухолеассоциированных антигенов, и, таким образом, подавляющее большинство исследуемых биспецифических антител к CD3 находится в форме моновалентных антител к CD3.
[7] В дополнение к антителам к CD3 крайне важной является также схема структуры биспецифических антител BsTCE. Существуют различные структуры биспецифических антител BsTCE, которые можно разделить на две основные категории: IgG-подобные структуры, содержащие Fc, и структуры на основе фрагментов антител без Fc. Например, блинатумомаб представляет собой структуру с одной полипептидной цепью, состоящую из двух последовательно соединенных одноцепочечных фрагментов вариабельной области антитела (scFv), однако эта структура имеет короткий период полужизни, требует непрерывной внутривенной перфузии и очень неудобна в применении. Таким образом, Fc-содержащие структуры используются во многих биспецифических антителах BsTCE для улучшения молекулярной стабильности и фармакокинетических свойств. Однако, поскольку для CD3-связывающего домена в BsTCE обычно требуется моновалентная форма, то Fc-содержащие структуры часто бывают асимметричными. В случае этих асимметричных структур, содержащих Fc, имеется множество необходимых к преодолению технических трудностей, таких как проблема гомодимеризации тяжелых цепей в асимметричной структуре, проблема несоответствия легких цепей, молекулярное перекрестное связывание, вызванное рецептором Fcγ, и функциональные эффекты, такие как ADCC или CDC и т.д. Для конструирования биспецифических антител BsTCE из антител IgG к TAA и антител IgG к CD3 можно выбирать различные асимметричные структуры [фигура 16 (A)], при этом одной из широко используемых структур является IgG-подобная структура, которая сохраняет два независимых Fab-домена, которая при этом содержит четыре различающиеся полипептидные цепи [две различающиеся тяжелые цепи и две различающиеся легкие цепи, структура показана на фигуре 16 (B)], с молекулярной массой, примерно соответствующей молекулярной массе обычного моноклонального антитела; при этом данная структура может приводить к образованию побочных продуктов, содержащих несколько комбинаций, обусловленных наличием множества различающихся полипептидных цепей, что создает большую трудность для очистки при экспрессии и осуществления способа получения антитела. Если Fab антитела к CD3 подвергнут модифицированию с получением структуры scFv, то «четырехцепочечная» структура может быть превращена в «трехцепочечную» структуру [показана на фигуре 16 (C)], что дополнительно снижает число комбинаций побочных продуктов, а следовательно сложность его получения. Для того, чтобы сконструировать биспецифическое антитело BsTCE, авторы настоящего изобретения предприняли попытку преобразования мышиного IgG-антитела SP34 в scFv, однако независимо от того, какой был выбран способ расположения (VH/VL) или в какой степени была изменена длина связывающего пептида, стабильный scFv получить не удавалось; таким образом, в данной области техники имеется острая потребность в стабильном моноклональном антителе к CD3, особенно в стабильной структуре scFv, полученной на его основе.
[8] Таким образом, в данной области техники существует острая потребность в антителе к CD3, которое способно связываться с CD3 приматов, имеет подходящую способность связывания CD3 и имеет стабильную одноцепочечную структуру scFv.
Содержание настоящего изобретения
[9] Техническая проблема, которая должна быть решена в данной области, состоит в том, чтобы преодолеть недостаток, связанный с отсутствием низкоантигенных, эффективных и безопасных антител к CD3 и биспецифических антител с асимметричными структурами; при этом настоящее изобретение предусматривает антитело, нацеливающееся на CD3, биспецифическое антитело и их применение.
[10] Для решения вышеупомянутой технической проблемы техническим решением, обеспечиваемым первым аспектом настоящего изобретения, является антитело, нацеливающееся на CD3, содержащее вариабельную область легкой цепи (VL) и вариабельную область тяжелой цепи (VH); где аминокислотная последовательность VL представлена под SEQ ID NO: 56 или ее мутантным вариантом, VH представлена мутантным вариантом аминокислотной последовательности, представленной под SEQ ID NO: 42, предусматривающей одну или более мутаций в положениях 30, 73, 76, 78, 93 и 94 (в соответствии со схемой нумерации по Chothia). Мутация может вызывать добавление, делецию или замену одного или более аминокислотных остатков в исходной аминокислотной последовательности. Антитело, нацеливающееся на CD3, по настоящему изобретению изменяет способность связывания с Т-клетками и снижает уровень высвобождения цитокинов, и вследствие этого ожидается снижение токсичности, ассоциированной с синдромом высвобождения цитокинов.
[11] В предпочтительном примере VH характеризуется наличием мутаций в положениях, выбранных из следующих групп:
[12] (a) положение 30;
[13] (b) положения 30, 73 и 76;
[14] (c) положения 30, 93 и 94;
[15] (d) положения 30, 73 и 93;
[16] (e) положения 30 и 93;
[17] (f) положения 30, 76 и 78;
[18] (g) положения 73, 76, 93 и 94;
[19] (h) положения 76, 78 и 93;
[20] (i) положения 30, 73, 76, 93 и 94;
[21] (j) положения 30, 76, 78 и 93.
[22] В предпочтительном примере VH характеризуется наличием мутаций, выбранных из следующих групп:
[23] (a) N30S;
[24] (b) N30S, D73N и S76N;
[25] (c) N30S, V93A и R94K;
[26] (d) N30S, D73N и V93A;
[27] (e) N30S и V93T;
[28] (f) N30S, S76N и L78A;
[29] (g) D73N, S76N, V93A и R94K;
[30] (h) S76N, L78A и V93T;
[31] (i) N30S, D73N, S76N, V93A и R94K;
[32] (j) N30S, S76N, L78A и V93T.
[33] При условии, что VH антитела характеризуется наличием определенных выше мутаций, антитело по настоящему изобретению является дополнительно мутированным по аминокислотной последовательности VL, представленной под SEQ ID NO: 56, или по аминокислотной последовательности VH, представленной под SEQ ID NO: 42, и полученная в результате аминокислотная последовательность характеризуется 80%, 85%, 90%, 95%, 98%, 99% или большей идентичностью с исходной аминокислотной последовательностью, и аминокислотные последовательности, которые поддерживают или улучшают функцию антитела, также входят в объем правовой охраны настоящего изобретения.
[34] В предпочтительном примере аминокислотная последовательность VH представлена под любым из SEQ ID NO: 43-55, и/или аминокислотная последовательность VL представлена под любым из SEQ ID NO: 57-60.
[35] В предпочтительном примере
[36] аминокислотная последовательность VH представлена под SEQ ID NO: 44, и аминокислотная последовательность VL представлена под SEQ ID NO: 58; или
[37] аминокислотная последовательность VH представлена под SEQ ID NO: 51, и аминокислотная последовательность VL представлена под SEQ ID NO: 58; или
[38] аминокислотная последовательность VH представлена под SEQ ID NO: 44, и аминокислотная последовательность VL представлена под SEQ ID NO: 60; или
[39] аминокислотная последовательность VH представлена под SEQ ID NO: 51, и аминокислотная последовательность VL представлена под SEQ ID NO: 60; или
[40] аминокислотная последовательность VH представлена под SEQ ID NO: 45, и аминокислотная последовательность VL представлена под SEQ ID NO: 58; или
[41] аминокислотная последовательность VH представлена под SEQ ID NO: 52, и аминокислотная последовательность VL представлена под SEQ ID NO: 58; или
[42] аминокислотная последовательность VH представлена под SEQ ID NO: 43, и аминокислотная последовательность VL представлена под SEQ ID NO: 58; или
[43] аминокислотная последовательность VH представлена под SEQ ID NO: 43, и аминокислотная последовательность VL представлена под SEQ ID NO: 60; или
[44] аминокислотная последовательность VH представлена под SEQ ID NO: 50, и аминокислотная последовательность VL представлена под SEQ ID NO: 58; или
[45] аминокислотная последовательность VH представлена под SEQ ID NO: 47, и аминокислотная последовательность VL представлена под SEQ ID NO: 58; или
[46] аминокислотная последовательность VH представлена под SEQ ID NO: 48, и аминокислотная последовательность VL представлена под SEQ ID NO: 58; или
[47] аминокислотная последовательность VH представлена под SEQ ID NO: 49, и аминокислотная последовательность VL представлена под SEQ ID NO: 58; или
[48] аминокислотная последовательность VH представлена под SEQ ID NO: 53, и аминокислотная последовательность VL представлена под SEQ ID NO: 58; или
[49] аминокислотная последовательность VH представлена под SEQ ID NO: 54, и аминокислотная последовательность VL представлена под SEQ ID NO: 58; или
[50] аминокислотная последовательность VH представлена под SEQ ID NO: 43, и аминокислотная последовательность VL представлена под SEQ ID NO: 57; или
[51] аминокислотная последовательность VH представлена под SEQ ID NO: 44, и аминокислотная последовательность VL представлена под SEQ ID NO: 57; или
[52] аминокислотная последовательность VH представлена под SEQ ID NO: 43, и аминокислотная последовательность VL представлена под SEQ ID NO: 59; или
[53] аминокислотная последовательность VH представлена под SEQ ID NO: 44, и аминокислотная последовательность VL представлена под SEQ ID NO: 59; или
[54] аминокислотная последовательность VH представлена под SEQ ID NO: 51, и аминокислотная последовательность VL представлена под SEQ ID NO: 57; или
[55] аминокислотная последовательность VH представлена под SEQ ID NO: 55, и аминокислотная последовательность VL представлена под SEQ ID NO: 58; или
[56] аминокислотная последовательность VH представлена под SEQ ID NO: 46, и аминокислотная последовательность VL представлена под SEQ ID NO: 58.
[57] В предпочтительном примере антитело предусматривает одноцепочечный вариабельный фрагмент антитела (scFv), предусматривающий VL-линкер-VH или VH-линкер-VL; предпочтительно линкер (т.е. линкерный пептид) представляет собой (GGGGS)n [сокращенно (G4S)n] или его вариант, где n представляет собой отличное от нуля натуральное число, предпочтительно от 1 до 20, более предпочтительно аминокислотная последовательность линкера представлена под SEQ ID NO: 65, SEQ ID NO: 66 или SEQ ID NO: 67; более предпочтительно аминокислотная последовательность scFv представлена под SEQ ID NO: 73, SEQ ID NO: 74, SEQ ID NO: 75, SEQ ID NO: 78, SEQ ID NO: 79 или SEQ ID NO: 80; еще более предпочтительно антитело дополнительно содержит кристаллизуемый фрагмент (Fc), при этом Fc связан с scFv посредством шарнира.
[58] В предпочтительном примере антитело дополнительно содержит константную область, предпочтительно человеческую константную область; предпочтительно человеческая константная область предусматривает константную область человеческой легкой цепи и константную область человеческой тяжелой цепи, и при этом константная область человеческой легкой цепи предпочтительно представляет собой константную область человеческой легкой цепи κ, приведенную под SEQ ID NO: 61, или константную область человеческой легкой цепи λ, приведенную под SEQ ID NO: 62; более предпочтительно константная область человеческой тяжелой цепи представлена изотипами hIgG1, hIgG2, hIgG3, hIgG4 или их вариантом, предпочтительно константной областью тяжелой цепи, приведенной под SEQ ID NO: 63 или SEQ ID NO: 64.
[59] Для решения вышеупомянутой технической проблемы техническим решением, предусмотренным вторым аспектом настоящего изобретения, является биспецифическое антитело. Биспецифическое антитело по настоящему изобретению имеет трехцепочечную структуру, что может уменьшать число комбинаций побочных продуктов, а следовательно сложность его получения; однако невозможно разработать биспецифическое антитело посредством незначительной модификации антитела согласно существующей технологии. Как описано в уровне техники, чтобы сконструировать биспецифическое антитело BsTCE авторы настоящего изобретения предприняли попытку преобразования мышиного IgG-антитела SP34 в scFv, однако независимо от того, какой был выбран способ расположения (VH/VL) или в какой степени была изменена длина линкерного пептида, стабильный scFv получить не удавалось. После разработки нескольких схем мутаций и осуществления процедур проверки авторы настоящего изобретения обнаружили, что только некоторые из этих мутаций могут обеспечивать для scFv сохранение стабильной структуры. Биспецифическое антитело по настоящему изобретению содержит первую функциональную область белка и вторую функциональную область белка, где первая функциональная область белка предусматривает антитело, нацеливающееся на CD3, согласно первому аспекту настоящего изобретения; при этом предпочтительно биспецифическое антитело содержит следующие три цепи: (1) VL1-линкер-VH1-шарнир-CH2-CH3 («выступ») или VH1-линкер-VL1-шарнир-CH2-CH3 («выступ»), относящиеся к первой функциональной области белка, (2) VH2-CH1-шарнир-CH2-CH3 («впадина»), относящуюся ко второй функциональной области белка, и (3) VL2-CL, относящуюся ко второй функциональной области белка; при этом вторая функциональная область белка представляет собой антитело, не нацеливающееся на CD3, предпочтительно антитело, нацеливающееся на B7H4, или антитело, нацеливающееся на ROR1, и линкер предпочтительно представляет собой (G4S)n, где n представляет собой отличное от нуля натуральное число, предпочтительно от 1 до 20, и более предпочтительно аминокислотная последовательность линкера представлена под SEQ ID NO: 65, SEQ ID NO: 66 или SEQ ID NO: 67; более предпочтительно биспецифическое антитело содержит VL1-линкер-VH1-шарнир-CH2-CH3 («выступ»), приведенную под SEQ ID NO: 88, VH2-CH1-шарнир-CH2-CH3 («впадина»), приведенную под SEQ ID NO: 86, и VL2-CL, приведенную под SEQ ID NO: 83, или VL1-линкер-VH1-шарнир-CH2-CH3 («выступ»), приведенную под SEQ ID NO: 88, VH2-CH1-шарнир-CH2-CH3 («впадина»), приведенную под SEQ ID NO: 87, и VL2-CL, приведенную под SEQ ID NO: 85. Биспецифическое антитело по настоящему изобретению обеспечивает преодоление недостатка, связанного с нестабильностью плеча одноцепочечного антитела, нацеливающегося на CD3, которое при этом является стабильным и обладает способностью связываться с Т-клетками. Получение биспецифического антитела, содержащего только три цепи, характеризуется легкостью осуществления, и при этом сложность его получения является сниженной.
[60] Для решения вышеупомянутой технической проблемы техническим решением, предусмотренным третьим аспектом настоящего изобретения, является выделенная нуклеиновая кислота, кодирующая антитело, нацеливающееся на CD3, согласно первому аспекту настоящего изобретения или биспецифическое антитело согласно второму аспекту настоящего изобретения.
[61] Для решения вышеупомянутой технической проблемы техническим решением, предусмотренным четвертым аспектом настоящего изобретения, является вектор экспрессии, содержащий выделенную нуклеиновую кислоту согласно третьему аспекту настоящего изобретения; предпочтительно вектор экспрессии выбран из ретровирусного вектора, лентивирусного вектора, аденовирусного вектора и вектора на основе аденоассоциированного вируса.
[62] Для решения вышеупомянутой технической проблемы техническим решением, предусмотренным пятым аспектом настоящего изобретения, является генетически модифицированная клетка, трансфицированная вектором экспрессии согласно четвертому аспекту настоящего изобретения; предпочтительно генетически модифицированная клетка представляет собой эукариотическую клетку.
[63] Для решения вышеупомянутой технической проблемы техническим решением, предусмотренным шестым аспектом настоящего изобретения, является фармацевтическая композиция, содержащая антитело, нацеливающееся на CD3, согласно первому аспекту настоящего изобретения, биспецифическое антитело согласно второму аспекту настоящего изобретения, генетически модифицированную клетку согласно пятому аспекту настоящего изобретения и фармацевтически приемлемый носитель; предпочтительно фармацевтическая композиция дополнительно содержит антитело, направленное против иммунной контрольной точки.
[64] Для решения вышеупомянутой технической проблемы техническим решением, предусмотренным седьмым аспектом настоящего изобретения, является применение антитела, нацеливающегося на CD3, согласно первому аспекту настоящего изобретения, биспецифического антитела согласно второму аспекту настоящего изобретения, выделенной нуклеиновой кислоты согласно третьему аспекту настоящего изобретения, вектора экспрессии согласно четвертому аспекту настоящего изобретения, генетически модифицированной клетки согласно пятому аспекту настоящего изобретения или фармацевтической композиции согласно шестому аспекту настоящего изобретения в изготовлении лекарственного препарата для лечения опухоли.
[65] Для решения вышеупомянутой технической проблемы техническим решением, предусмотренным восьмым аспектом настоящего изобретения, является комбинация наборов, предусматривающая набор A и набор B; при этом набор A содержит антитело, нацеливающееся на CD3, согласно первому аспекту настоящего изобретения, биспецифическое антитело согласно второму аспекту настоящего изобретения, генетически модифицированную клетку согласно пятому аспекту настоящего изобретения или фармацевтическую композицию согласно шестому аспекту настоящего изобретения; набор B содержит другие антитела, биспецифические антитела, генетически модифицированные клетки или фармацевтические композиции, другие антитела, биспецифические антитела, генетически модифицированные клетки или фармацевтические композиции, нацеливающиеся на CD3, B7H4, ROR1 или другую мишень. Набор A и набор B можно использовать в любом порядке, при этом набор A можно использовать перед набором B, или набор B можно использовать перед набором A. Лекарственное средство в наборе A присутствует в инъекционной форме, такой как инъекция, и лекарственное средство в наборе B присутствует в инъекционной форме, такой как инъекция, или в форме для проглатывания, такой как таблетка или пилюля.
[66] Для лечения соответствующей опухоли пациенту можно вводить антитело, нацеливающееся на CD3, согласно первому аспекту настоящего изобретения, биспецифическое антитело согласно второму аспекту настоящего изобретения, генетически модифицированную клетку согласно пятому аспекту настоящего изобретения, фармацевтическую композицию согласно шестому аспекту настоящего изобретения или комбинацию наборов согласно восьмому аспекту настоящего изобретения.
[67] Основываясь на общеизвестных знаниях в данной области техники упомянутые выше предпочтительные условия можно произвольно объединять с получением предпочтительных примеров настоящего изобретения.
[68] Все реагенты и исходные материалы, используемые в настоящем изобретении, являются коммерчески доступными.
[69] Положительные и прогрессивные эффекты, обеспечиваемые настоящим изобретением, являются следующими:
[70] 1. Моноклональное антитело по настоящему изобретению изменяет способность связывания с Т-клетками и снижает уровень высвобождения цитокинов, и вследствие этого ожидается снижение токсичности, ассоциированной с синдромом высвобождения цитокинов.
[71] 2. Биспецифическое антитело, полученное на его основе, обеспечивает преодоление недостатка, связанного с нестабильностью плеча одноцепочечного антитела, нацеливающегося на CD3, которое при этом является стабильным и обладает способностью связываться с Т-клетками.
[72] 3. Получение биспецифического антитела, содержащего только три цепи, характеризуется легкостью осуществления, и при этом сложность его получения является сниженной.
Краткое описание графических материалов
[73] На фиг. 1 показаны результаты HPLC-SEC одноцепочечного антитела к CD3 после одностадийной очистки: (A) PR000275, (B) PR000276, (C) PR000307 и (D) PR000308.
[74] На фиг. 2 показано выравнивание последовательностей гуманизированных мутантных вариантов VH SP34.
[75] На фиг. 3 показано выравнивание последовательностей гуманизированных мутантных вариантов VL SP34.
[76] На фиг. 4 показаны различия в значимых участках различных мутантных последовательностей VH/VL, где на (A) показана мутантная последовательность VH, и на (B) показана мутантная последовательность VL.
[77] На фиг. 5 показаны (A) результаты SDS-PAGE и (B) результаты HPLC-SEC одноцепочечного антитела к CD3 PR000510 после одностадийной очистки.
[78] На фиг. 6 показана способность связывания антитела к CD3 PR000260 с (A) рекомбинантными клетками CHOK1, сверхэкспрессирующими CD3 человека, и (B) рекомбинантными клетками CHOK1, сверхэкспрессирующими CD3 яванского макака.
[79] На фиг. 7 показана способность связывания антитела к CD3 с Т-клетками человека, в том числе кривая связывания и относительная интенсивность MFI (интенсивность флуоресценции MFI для антитела, связывающегося с Т-клетками человека при определенных концентрациях, и относительный показатель по сравнению с исходным антителом PR000260 (SP34)) или максимальная MFI, где (A) PR000511, PR000512, PR000513, PR000514 и PR000260 связываются с Т-клетками человека, (B) PR001848, PR001849 и PR000260 связываются с Т-клетками человека, (C) PR002467, PR002468, PR002469, PR002470, PR002471, PR002472, PR001848 и PR000260 связываются с Т-клетками человека, (D) PR001848, PR002742, PR002743 и PR000260 связываются с Т-клетками человека, (E) PR002833, PR002834, PR002835, PR002836, PR002837, PR002742, PR001848, PR002469 и PR000260 связываются с Т-клетками человека, (F) PR003886, PR001848 и PR002742 связываются с Т-клетками человека, (G) PR001848, PR002469 и PR004616 связываются с Т-клетками человека.
[80] На фиг.8 показана способность связывания одноцепочечного антитела к CD3 с Т-клетками человека, в том числе кривая связывания и относительная интенсивность MFI (интенсивность флуоресценции MFI для антитела, связывающегося с Т-клетками человека при определенной концентрации, и относительный показатель по сравнению с исходным антителом PR000260 (SP34)), где (A) PR000510, PR000624, PR000627 и PR000260 связываются с Т-клетками человека, (B) PR001850 и PR000260 связываются с Т-клетками человека.
[81] На фиг. 9 показана способность связывания антитела к CD3 с Т-клетками яванского макака.
[82] На фиг. 10 показана способность антитела к CD3 активировать Т-клетки человека для обеспечения продуцирования цитокина IFN-γ, где (A) PR000511, PR000512, PR000513, PR000514 и PR000260 активируют Т-клетки, (B) PR001848, PR001849 и PR000260 активируют Т-клетки, (C) PR002468, PR002469, PR002471 и PR001848 активируют Т-клетки, (D) PR002742, PR001848 и PR000260 активируют Т-клетки, (E) PR002833, PR002834, PR002835, PR002836, PR002837 и PR000260 активируют Т-клетки, (F) PR003886, PR001848 и PR002742 активируют Т-клетки, (G) PR001848, PR002469 и PR004616 активируют Т-клетки.
[83] На фиг. 11 показана способность одноцепочечного антитела к CD3 (PR000510, PR000623, PR000624, PR000627 и PR000260) активировать Т-клетки человека для обеспечения продуцирования цитокина IFN-γ.
[84] На фиг. 12 показаны результаты SDS-PAGE образцов биспецифического антитела (A) PR002883 и (B) PR002885, полученных путем одностадийной очистки.
[85] На фиг. 13 показана способность связывания моноклонального антитела и биспецифического антитела с (A) клетками SK-BR-3 и (B) Т-клетками человека.
[86] На фиг. 14 показана способность к уничтожению клеток-мишеней, опосредованная биспецифическим антителом PR002883 in vitro; где на (A) показано уничтожение клеток SK-BR-3, а на (B) показаны уровни высвобождения IFN-γ.
[87] На фиг. 15 показана способность связывания моноклонального антитела и биспецифического антитела с (A) клетками Panc-1 и (B) Т-клетками человека.
[88] На фиг. 16 показана структура моноклонального антитела или биспецифического антитела; (A) IgG-структура, (B) асимметричная «четырехцепочечная» структура, (C) асимметричная «трехцепочечная» структура, предусматривающие одноцепочечное антитело.
Подробное описание предпочтительного варианта осуществления
[89] Настоящее изобретение дополнительно проиллюстрировано ниже с помощью примеров, однако при этом настоящее изобретение не ограничивается объемом описанных примеров. Экспериментальные способы, для которых в нижеследующих примерах не указаны конкретные условия, были выбраны в соответствии с обычными способами и условиями или в соответствии с техническими требованиями.
[90] В данной заявке термин «антитело» обычно относится к белку, содержащему фрагмент, который связывается с антигеном и необязательно обеспечивает возможность фрагменту, который связывается с антигеном, придавать фрагменту, являющемуся каркасом или остовом, конформацию, которая способствует связыванию антитела с антигеном. Как правило, антитело может содержать вариабельную область легкой цепи антитела (VL), вариабельную область тяжелой цепи антитела (VH) или и то и другое. VH- и VL-области можно дополнительно разделить на гипервариабельные области, называемые областями, определяющими комплементарность (CDR), которые рассредоточены в пределах более консервативных областей, называемых каркасными областями (FR). Каждая из VH и VL может состоять из трех областей CDR и четырех областей FR, которые могут располагаться в следующем порядке от амино-конца к карбокси-концу: FR1, CDR1, FR2, CDR2, FR3, CDR3 и FR4. Вариабельные области тяжелой и легкой цепей содержат связывающие домены, взаимодействующие с антигеном. Примеры антител включают без ограничения антитело, антигенсвязывающий фрагмент (Fab, Fab', F(ab)2, Fv-фрагмент, F(ab')2, scFv, ди-scFv и/или dAb), иммуноконъюгат, мультиспецифическое антитело (например, биспецифическое антитело), фрагмент антитела, производное антитела, аналог антитела или слитый белок и т. п., при условии, что они проявляют требуемую антигенсвязывающую активность.
[91] В настоящей заявке термин «вариабельный» обычно относится к тому факту, что определенные части последовательности вариабельного домена антитела существенно различаются, что обуславливает связывание различных специфических антител с их конкретным антигеном и специфичность по отношению к нему. Однако вариабельность не распределена равномерно по всей вариабельной области антитела. Она сконцентрирована в трех сегментах вариабельных областей легкой и тяжелой цепей, известных как область, определяющая комплементарность (CDR), или область высокой вариабельности (HVR). Часть вариабельного домена, являющаяся консервативной в более высокой степени, известна как каркас (FR). Каждый из доменов с вариабельной структурой из природных тяжелой и легкой цепей содержит четыре области FR, большинство из которых принимают β-складчатую конформацию и соединены тремя CDR, которые образуют петлевую связь, а в некоторых случаях образуют часть β-складчатой структуры. CDR в каждой цепи находятся в непосредственной близости друг от друга, являясь разделенными областью FR, и образуют антигенсвязывающий участок антитела вместе с CDR из другой цепи, при этом константные области не участвуют непосредственно в связывании антитела с антигеном, однако проявляют различные эффекторные функции, такие как участие в антителозависимой цитотоксичности, опосредуемой антителом. В настоящее время в уровне техники CDR антитела можно определять посредством различных способов, таких как схема определения согласно Kabat на основе вариабельности последовательностей (см. Kabat et al., Sequences of proteins of immunological interest, Fifth Edition, National Institutes of Health, Bethesda, Maryland (1991)) и схема определения согласно Chothia на основе расположения структурных петлевых областей (см. Al-Lazikani et al., JMol Biol 273:927-48, 1997). В настоящей заявке комбинированная схема определения, предусматривающая определение согласно Kabat и определение согласно Chothia, также используется для идентификации аминокислотных остатков в последовательностях вариабельных доменов и последовательностях полноразмерных антител (таблица 1).
Таблица 1. Способы определения CDR антитела, предусматриваемые в настоящей заявке (см. http://bioinf.org.uk/abs/)
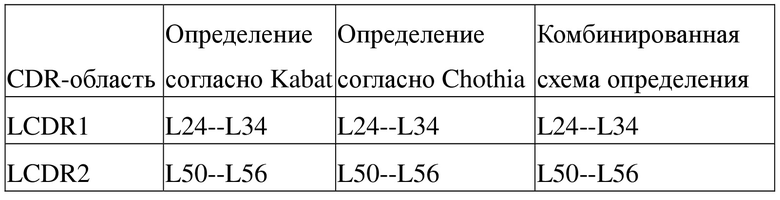
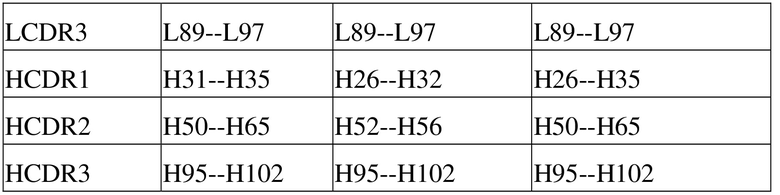
[92] При этом Laa-Lbb может относиться к аминокислотной последовательности от положения аа (схема нумерации согласно Chothia) до положения bb (схема нумерации согласно Chothia), начиная с N-конца легкой цепи антитела; Haa-Hbb может относиться к аминокислотной последовательности от положения аа (схема кодирования согласно Chothia) до положения bb (схема нумерации согласно Chothia), начиная с N-конца тяжелой цепи антитела. Например, L24-L34 может относиться к аминокислотной последовательности от положения 24 до положения 34, начиная с N-конца легкой цепи антитела в соответствии со схемой нумерации согласно Chothia; H26-H32 может относиться к аминокислотной последовательности от положения 26 до положения 32, начиная с N-конца тяжелой цепи антитела в соответствии со схемой нумерации согласно Chothia.
[93] Эффекторные функции, опосредованные доменом Fc антитела, такие как ADCC и CDC, также являются крайне важными биологическими функциями, причем различные изоформы IgG характеризуются различными функциями применительно к ADCC или CDC, например, IgG1 и IgG3 характеризуются сильными эффектами ADCC и CDC, в то время как IgG2 и IgG4 характеризуются относительно слабыми эффектами. Кроме того, первоначальная эффекторная функция Fc также может быть подвергнута модулированию с помощью аминокислотных мутаций или модификаций для изменения способности связывания Fc с рецептором Fc. Например, двойная мутация «LALA» (L234A/L235A) в IgG1 значительно снижает аффинность к FcγRIIIA (CD16A) и, таким образом, снижает эффект ADCC. Кроме того, мутация P329G значительно снижает связывание IgG1 нескольких рецепторов Fcγ (см. Schlothauer T, Herter S, Koller CF, et al. Protein Eng Des Sel. 2016 Oct; 29(10): 457-466). В настоящей заявке, чтобы уменьшить связывание антител к CD3 с рецептором Fcγ, в Fc этих антител к CD3 вводили двойную мутацию «LALA» (L234A/L235A) или тройную мутацию «LALAPG» (L234A/L235A/P329G).
Пример 1. Получение и анализ рекомбинантного антитела с определением его характеристик
1.1. Получение рекомбинантного антитела класса IgG
[94] После получения последовательностей вариабельных доменов легкой и тяжелой цепей, кодирующих молекулу антитела, молекулу рекомбинантного антитела можно получить путем экспрессии последовательностей вариабельных доменов легкой и тяжелой цепей, слитых с соответствующими последовательностями константных доменов легкой и тяжелой цепей человеческого антитела, с применением традиционных методик рекомбинантной ДНК. В данном примере последовательность вариабельного домена тяжелой цепи антитела (VH) генетически синтезировали и встраивали в плазмидный вектор для экспрессии в клетках млекопитающего, кодирующий последовательность константного домена тяжелой цепи человеческого антитела изотипа IgG1, с получением кодируемой полноразмерной тяжелой цепи антитела изотипа IgG1, и при этом двойную мутацию «LALA» (L234A/L234A) (SEQ ID NO: 63) или тройную мутацию «LALAPG» (L234A/L235A/P329G) (SEQ ID NO: 64) вводили в константную область тяжелой цепи IgG1 для уменьшения уровня связывания антитела с рецептором Fcγ. Последовательность вариабельного домена легкой цепи антитела (VL) генетически синтезировали и встраивали в плазмидный вектор для экспрессии в клетках млекопитающего, кодирующий последовательность константного домена легкой цепи κ человеческого антитела (SEQ ID NO: 61), с получением кодируемой полноразмерной легкой цепи κ антитела; в качестве альтернативы VL генетически синтезировали и встраивали в плазмидный вектор для экспрессии в клетках млекопитающего, кодирующий последовательность константного домена легкой цепи λ человеческого антитела (SEQ ID NO: 62), с получением полноразмерной легкой цепи λ, получая кодируемое антитело.
[95] Посредством котрансфекции клетки-хозяина млекопитающего (например, клетки эмбриональной почки человека HEK293) плазмидами, кодирующими тяжелую цепь антитела, и плазмидами, кодирующими легкую цепь антитела, можно получать очищенное рекомбинантное антитело в виде комплекса правильно спаренных легких и тяжелых цепей с применением обычных методик экспрессии и очистки рекомбинантных белков. В частности, клетки HEK293 размножали в среде для экспрессии FreeStyle™ F17 Expression Medium (Thermo, № A1383504). Перед осуществлением транзиентной трансфекции концентрацию клеток доводили до 6-8×105 клеток/мл и их инкубировали в шейкере при 37°C, 8% CO2, в течение 24 часов при концентрации клеток 1,2×106 клеток/мл. Получали 30 мл культивируемых клеток. Плазмиду, кодирующую тяжелую цепь, и плазмиду, кодирующую легкую цепь, смешивали в соотношении 2:3, в общей сложности 30 мкг плазмид растворяли в 1,5 мл среды с уменьшенным содержанием сыворотки крови Opti-MEM (Thermo, № 31985088) и фильтровали через мембрану с порами 0,22 мкм. Затем 1,5 мл Opti-MEM растворяли в 120 мкл PEI (Polysciences, № 23966-2) в концентрации 1 мг/мл и выдерживали в течение 5 минут. К плазмиде медленно добавляли PEI, после чего осуществляли инкубирование в течение 10 минут при комнатной температуре. Смешанный раствор плазмиды и PEI медленно добавляли по каплям в колбу для культивирования, при этом колбу для культивирования подвергали встряхиванию. Трансфицированные клетки инкубировали при 37°C, 8% CO2, в шейкере в течение 5 дней. Жизнеспособность клеток определяли через 5 дней. Затем культуры собирали центрифугированием при 3300 g в течение 10 минут для обеспечения сбора надосадочной жидкости. Примеси в надосадочной жидкости удаляли центрифугированием на высокой скорости. Колонку для гравитационного элюирования (Bio-Rad, № 7311550), содержащую MabSelect™ (GE Healthcare Life Science, № 71-5020-91 AE), уравновешивали с помощью PBS (pH 7,4) и промывали количеством PBS, равным 2-5-кратному объему колонки. В колонку загружали образец надосадочной жидкости и ее промывали количеством PBS, равным 5-10-кратному объему колонки. Затем целевой белок элюировали с помощью 0,1 М глицина при рН 3,5, затем доводили до нейтрального рН с помощью трис-HCl при рН 8,0, окончательно концентрировали с использованием пробирки для ультрафильтрации (Millipore, № UFC901024), а затем осуществляли замену на буфер PBS и получали очищенный раствор рекомбинантных антител. Наконец, измеряли концентрацию с помощью NanoDrop (Thermo Scientific™ NanoDrop™ One), дозировали и сохраняли раствор очищенных рекомбинантных антител для использования в качестве резервного.
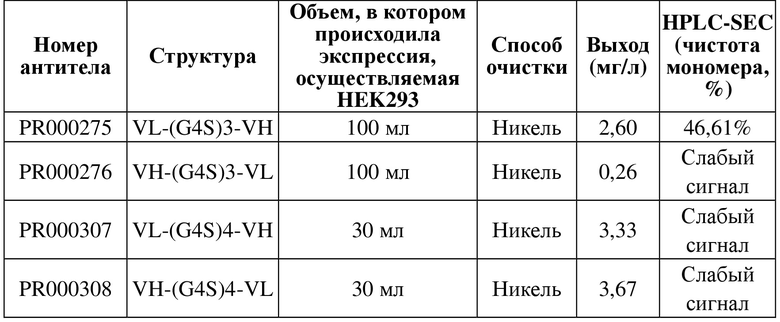
1.2. Получение моновалентного рекомбинантного антитела scFv-his
[96] Последовательности VH и VL антитела связывали гибким пептидом (линкером) с получением одной полипептидной цепи, кодирующей как VH, так и VL, т.е. фрагмента, представленного вариабельной областью одноцепочечного антитела (scFv). Если выбран линкерный пептид подходящей длины, такой как (G4S)3 (SEQ ID NO: 65) или (G4S)4 (SEQ ID NO: 66), то VH и VL могут надлежащим образом подвергаться укладке и сборке в функциональные антитела. Различные структуры scFv (VH-линкер-VL или VL-линкер-VH) можно конструировать в зависимости от различных расположений VH и VL и различий в линкерном пептиде. Одиночный scFv содержит антигенсвязывающую область, состоящую из пары VH и VL, которая обычно связывает только одну молекулу антигена и поэтому называется моновалентной связывающей молекулой.
[97] Для облегчения очистки в данном примере С-конец scFv сливали с His-меткой, состоящей из 6-гистидина. Плазмиду, кодирующую scFv и His-метку, генетически синтезировали и встраивали в плазмидный вектор для экспрессии в клетках млекопитающих с получением плазмиды, кодирующей scFv-his, которую трансфицировали в клетку-хозяина млекопитающего (например, клетку эмбриональной почки человека HEK293), а затем рекомбинантный белок можно подвергать очистке с применением обычных методик экспрессии и очистки рекомбинантных белков. В частности, клетки HEK293 размножали в среде для экспрессии FreeStyle™ F17 Expression Medium (Thermo, № A1383504). Перед осуществлением транзиентной трансфекции концентрацию клеток доводили до 6-8×105 клеток/мл и их инкубировали в шейкере при 37°C, 8% CO2, в течение 24 часов при концентрации клеток 1,2×106 клеток/мл. Получали 30 мл культивируемых клеток. Растворяли 30 мкг плазмиды в 1,5 мл среды с уменьшенным содержанием сыворотки крови Opti-MEM (Thermo, № 31985088) и фильтровали через мембрану с порами 0,22 мкм. Затем 1,5 мл Opti-MEM растворяли в 120 мкл PEI (Polysciences, № 23966-2) в концентрации 1 мг/мл и выдерживали в течение 5 минут. К плазмиде медленно добавляли PEI, после чего осуществляли инкубирование в течение 10 минут при комнатной температуре. Смешанный раствор плазмиды и PEI медленно добавляли по каплям в колбу для культивирования, при этом колбу для культивирования подвергали встряхиванию. Трансфицированные клетки инкубировали при 37°C, 8% CO2, в шейкере в течение 5 дней. Жизнеспособность клеток определяли через 5 дней. Затем культуры собирали центрифугированием при 3300 g в течение 10 минут для обеспечения сбора надосадочной жидкости. Примеси в надосадочной жидкости удаляли центрифугированием на высокой скорости. Колонку для гравитационного элюирования (Bio-Rad, № 7311550), содержащую сефарозный гель Ni Sepharose excel (GE Healthcare Life Science, № 17-3712-01), Ni Sepharose excel (GE Healthcare Life Science, № 17-3712-01), уравновешивали с помощью PBS (pH 7,4) и промывали количеством PBS, равным 2-5-кратному объему колонки. В колонку загружали образец надосадочной жидкости и ее промывали количеством PBS, равным 5-10-кратному объему колонки. Сначала элюировали неспецифически адсорбированные гетеропротеины с помощью буфера А (содержащего 20 мМ имидазола, 150 мМ фосфата, рН 8,0), а затем целевой белок с помощью буфера В (содержащего 500 мМ имидазола, 150 мМ фосфата, рН 8,0), наконец, использовали пробирки для ультрафильтрации (Millipore, № UFC901024) для концентрирования и замены раствора на буфер PBS и для получения раствора очищенных рекомбинантных антител. Наконец, измеряли концентрацию с помощью NanoDrop (Thermo Scientific™ NanoDrop™ One), дозировали и сохраняли раствор очищенных рекомбинантных антител для использования в качестве резервного.
1.3. Получение двухвалентного рекомбинантного антитела scFv-Fc
[98] В данном примере рекомбинантную молекулу scFv-Fc конструировали посредством слияния последовательности Fc константной области IgG1 человека (Glu216-Lys447, содержащей шарнирную область, домен СН2 и домен СН3) с scFv на его С-конце, и при этом посредством гомодимеризации Fc образовывалась двухвалентная молекула димера scFv-Fc, которая способна была связывать две молекулы антигена одновременно. При этом также вводили двойную мутацию «LALA» (L234A/L235A) или тройную мутацию «LALAPG» (L234A/L235A/P329G) в Fc для уменьшения связывания антитела с рецептором Fcγ. Полипептидную последовательность, кодирующую scFv-Fc, генетически синтезировали и встраивали в плазмидный вектор для экспрессии в клетках млекопитающих с получением плазмиды, кодирующей scFv-Fc, которую затем трансфицировали в клетки-хозяева млекопитающего (например, клетку эмбриональной почки человека HEK293), а затем применяли способ экспрессии и очистки белка, описанный в примере 1.1, с получением очищенного рекомбинантного белка.
1.4. Анализ чистоты белка с использованием HPLC-SEC
[99] Чистоту и агрегатную форму образцов белка анализировали с помощью эксклюзионной хроматографии (SEC) с разделением молекул по размеру. Аналитическую колонку TSKgel G3000SWxl (Tosoh Bioscience, № 08541, 5 мкм, 7,8 мм×30 см) соединяли с жидкостным хроматографом высокого давления (HPLC) (Agilent Technologies, Agilent 1260 Infinity II) и уравновешивали буфером PBS при комнатной температуре в течение по меньшей мере 1 часа. В систему вводили соответствующее количество образца белка (по меньшей мере 10 мкг), профильтрованного через мембрану с порами 0,22 мкм, и задавали следующую программу HPLC: колонку загружали образцом с буфером PBS при скорости потока 1,0 мл/мин в течение максимум 20 минут. В ходе HPLC генерировались аналитические отчеты HPLC, в которых при их генерации сообщалось время удерживания компонентов с различным размером молекул в образце.
Пример 2. Рекомбинантная экспрессия мышиного-человеческого химерного антитела к CD3, представляющего собой антитело SP34.
[100] SP34 представляет собой антитело к CD3e человека мышиного происхождения, которое связывает ряд различных CD3 приматов и обладает функцией активирования Т-клеток. Последовательности вариабельных областей VH и VL SP34 раскрыты в WO 2016071004A1. В настоящей заявке аминокислотная последовательность VH SP34 представлена под SEQ ID NO: 42, и ее соответствующий мышиный ген V зародышевого типа представляет собой IGHV10-1; аминокислотная последовательность VL SP34 представлена под SEQ ID NO: 56, и ее соответствующий мышиный ген V зародышевого типа представляет собой IGLV1. В данном примере последовательность VH SP34 подвергали слиянию с последовательностью константного домена тяжелой цепи человеческого антитела изотипа IgG1 (SEQ ID NO: 63), предусматривающей двойную мутацию «LALA» (L234A/L235A), с получением полноразмерной тяжелой цепи мышиного-человеческого химерного антитела SP34 изотипа IgG1; аминокислотную последовательность VL SP34 сливали с последовательностью константного домена легкой цепи λ человеческого антитела (SEQ ID NO: 62) с получением полноразмерной легкой цепи λ мышиного-человеческого химерного антитела SP34.
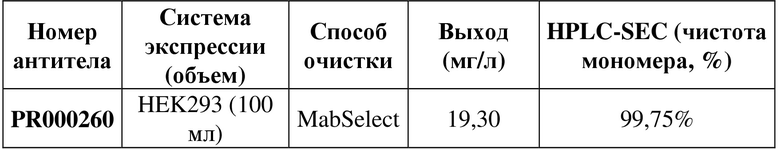
[101] Мышиное-человеческое химерное рекомбинантное антитело SP34 PR000260 получали в соответствии со способом из примера 1.1. В нижеследующей таблице 2 показаны данные по рекомбинантной экспрессии PR000260.
Таблица 2. Экспрессия и очистка рекомбинантного антитела PR000260
Пример 3. Преобразование антитела к CD3, представляющего собой мышиное антитело SP34, в рекомбинантное антитело scFv
[102] Последовательность VH (SEQ ID NO: 42) и последовательность VL (SEQ ID NO: 56) SP34 связывались гибким пептидом (линкером) с получением одной полипептидной цепи, кодирующей как VH, так и VL, т.е. фрагмента, представленного вариабельной областью одноцепочечного антитела (scFv). В зависимости от различных расположений VH и VL и различных длин линкерных пептидов (SEQ ID NO: 65, SEQ ID NO: 66) обеспечивалась возможность конструирования различных структур scFv, и при этом His-метку, состоящую из 6-гистидина, сливали с С-концом scFv для обеспечения очистки. Для конструирования scFv согласно настоящей заявке также мог использоваться линкерный пептид, приведенный под SEQ ID NO: 67.
[103] В данном примере четыре молекулы рекомбинантного антитела scFv (PR000275, PR000276, PR000307, PR000308) получали в соответствии со способом из примера 1.2. Номера последовательностей этих четырех молекул рекомбинантных антител scFv перечислены в таблице 3 ниже; в нижеследующей таблице 4 показаны данные по экспрессии этих четырех рекомбинантных молекул; и результаты HPLC-SEC для этих четырех молекул после одностадийной очистки показаны на фигуре 1, где на (A) показан результат для PR000275, на (B) показан результат для PR000276, на (C) показан результат для PR000307 и на (D) показан результат для PR000308. Можно увидеть, что при использовании последовательностей VH и VL SP34 для конструирования scFv, независимо от того, какая использовалась модель выравнивания (VH/VL) или в какой степени была изменена длина линкерного пептида, стабильный scFv получить не удавалось.
Таблица 3. Структура и номер последовательности для молекул четырех рекомбинантных антител
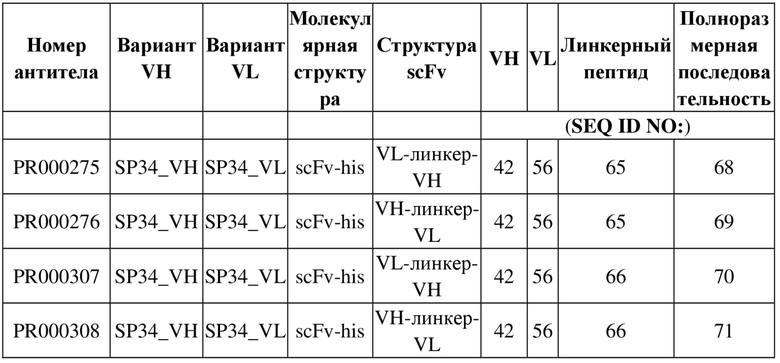
Таблица 4. Экспрессия и очистка молекул рекомбинантных антител scFv
Пример 4. Оптимизация последовательности SP34
4.1. Гуманизация последовательностей вариабельной области и мутация каркасной области
[104] В данном примере для гуманизации последовательности применяли способ «трансплантации CDR», т.е. трансплантации CDR VH мышиного антитела в каркасную область VH человеческого антитела и трансплантация CDR VL мышиного антитела в каркасную область VL человеческого антитела. Последовательность каркасной области VH или VL человеческого антитела можно получать из последовательностей человеческих генов зародышевого типа или последовательностей антител, которые были перегруппированы с помощью V(D)J, или консенсусных последовательностей конкретного семейства генов VH или VL человеческого антитела. В данном примере последовательности каркасной области, обеспечиваемые последовательностями человеческих генов зародышевого типа, использовались в качестве гуманизированных матричных последовательностей, т.е. фрагмент человеческого гена V зародышевого типа обеспечивал последовательности каркасных областей FR1, FR2 и FR3, а фрагмент человеческого гена J зародышевого типа обеспечивал последовательность каркасной области FR4. Наконец, последовательности гуманизированной вариабельной области (VH или VL) конструировали в виде (человеческая)FR1-(мышиная)CDR1-(человеческая)FR2-(мышиная)CDR2-(человеческая)FR3-(мышиная)CDR3-(человеческая)FR4.
[105] В данном примере последовательность фрагмента человеческого гена V зародышевого типа IGHV3-73*01 или фрагмента человеческого гена V зародышевого типа IGHV3-23*01 использовали в качестве гуманизированной матрицы в комбинации с последовательностью фрагмента человеческого гена J зародышевого типа IGHJ1*01 с получением последовательности каркасной области. Аминокислотные мутации вводили в один или более сайтов в положение 30, положение 73, положение 76, положение 78, положение 93 или положение 94 (в соответствии со схемой нумерации согласно Chothia) с получением нескольких различных мутантных последовательностей VH.
[106] В данном примере последовательность фрагмента человеческого гена V зародышевого типа IGLV7-46*02 в комбинации с последовательностью фрагмента человеческого гена J зародышевого типа IGLJ2*01 или последовательностью фрагмента человеческого гена V зародышевого типа IGKV1-39*01 в комбинации с последовательностью фрагмента человеческого гена J зародышевого типа IGKJ4*01 использовали в качестве гуманизированной матрицы с получением последовательности каркасной области. Аминокислотные мутации вводили в нулевое или более высокое количество сайтов в положение 2, положение 36, положение 46, положение 49, положение 66, положение 69, положение 71 или положение 87 (в соответствии со схемой нумерации согласно Chothia) с получением нескольких различных мутантных последовательностей VL.
[107] В нижеследующей таблице 5 перечислены номера последовательностей вариабельной области антитела, ее оптимизированных мутантных последовательностей (FV) и последовательностей областей CDR и FR, определенных согласно Chothia.
Таблица 5. Вариабельная область антитела SP34 и ее оптимизированные мутантные последовательности (FV), а также перечень последовательностей областей CDR и FR, определенных согласно Chothia
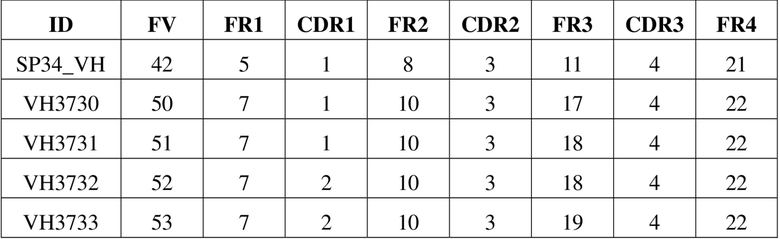
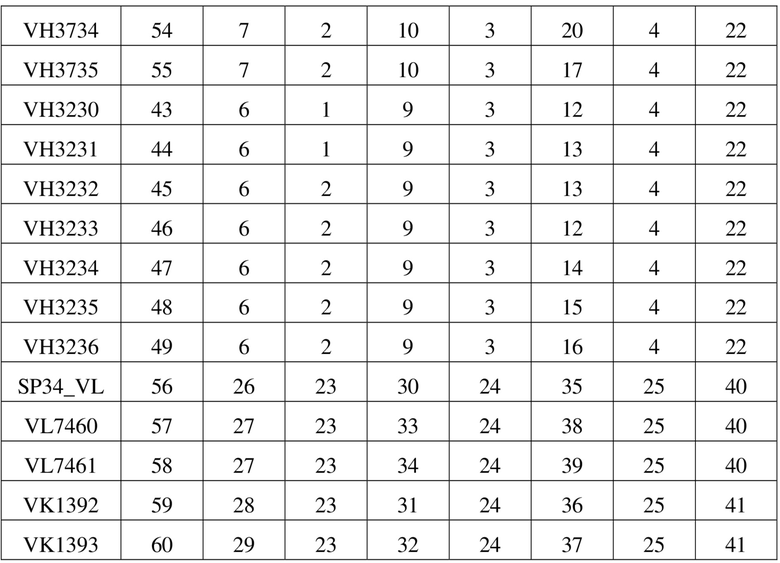
[108] На фигуре 2 представлен список сравниваемых мутантных последовательностей VH. На фигуре 3 представлен список сравниваемых мутантных последовательностей VL. Различия в последовательностях мутантных вариантов VH и мутантных вариантов VL в значимых сайтах перечислены в (A) и (B) на фигуре 4 соответственно. Как видно на фигурах 2-4, мутации антитела по настоящему изобретению в VH имели место в одном или более сайтов, выбранных из положения 30, положения 73, положения 76, положения 78, положения 93 или положения 94 аминокислотной последовательности, приведенной под SEQ ID NO: 42. Мутации в VL происходили в положении 2, положении 36, положении 46, положении 49, положении 66, положении 69, положении 71 и/или положении 87 последовательности, приведенной под SEQ ID NO: 56. Более подробную информацию о мутациях можно найти в таблице 5 для характеристик последовательностей VH3730, VH3731, VH3732, VH3733, VH3734, VH3735, VH3230, VH3231, VH3232, VH3233, VH3234, VH3235, VH3236, VL7460, VL7461, VK1392 и VK1393.
4.2. Молекулы рекомбинантных антител, содержащие мутантные варианты с оптимизированными последовательностями
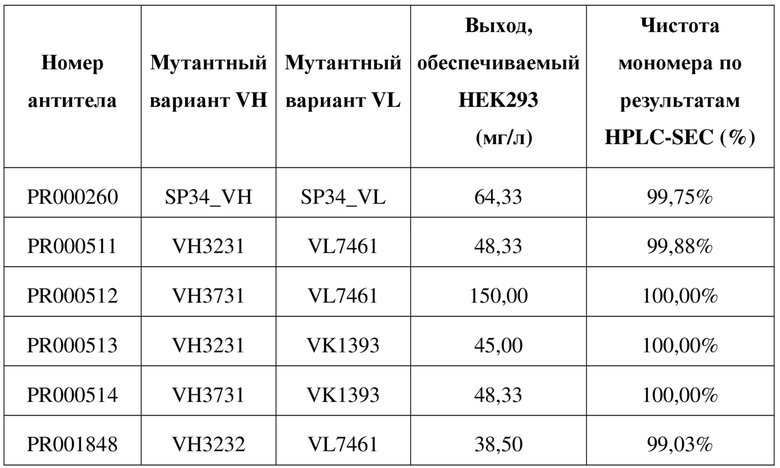
[109] Последовательности мутантных вариантов VH и мутантных вариантов VL, полученные в примере 4.1, спаривали и объединяли, а рекомбинантное антитело IgG конструировали в соответствии со способом, описанным в примере 1.1, где двойную мутацию «LALA» или тройную мутацию «LALAPG» вводили в константную область тяжелой цепи IgG1 для снижения эффекторной функции Fc. В таблице 6 перечислены последовательности молекул рекомбинантных антител, которые были оптимизированы в отношении последовательности. В таблице 7 приведены данные в отношении экспрессии рекомбинантных антител. За исключением трех молекул IgG, сконструированных с помощью мутантного варианта VH, представляющего собой VH3230, которые характеризовались очень низкими значениями выхода при экспрессии, все другие молекулы IgG характеризовались приемлемыми значениями выхода при экспрессии.
Таблица 6. Список последовательностей химерного антитела SP34 или антитела с оптимизированной последовательностью
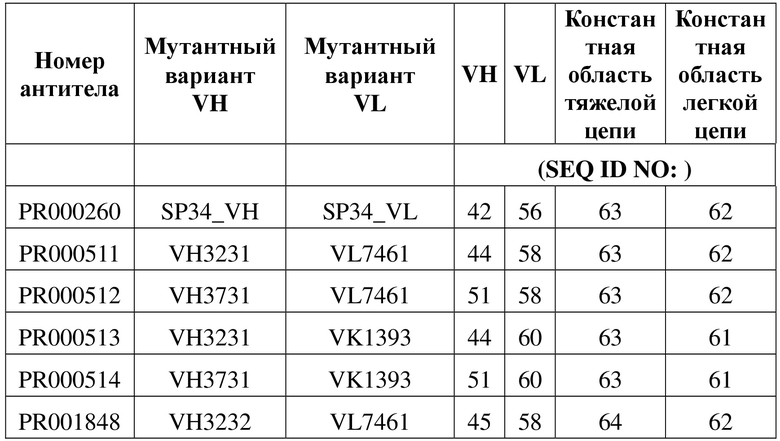
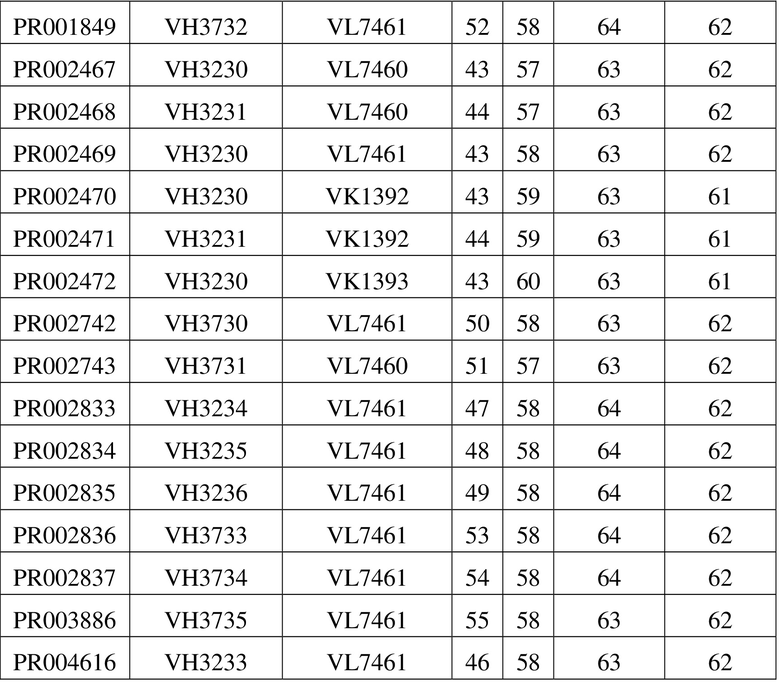
Таблица 7. Выход при экспрессии и чистота рекомбинантного антитела
4.3. Рекомбинантные молекулы scFv, содержащие мутантные варианты с оптимизированными последовательностями
[110] Последовательности мутантных вариантов VH и мутантных вариантов VL, полученные в примере 4.1, спаривали и объединяли, и множество молекул рекомбинантного двухвалентного антитела scFv получали в соответствии со способом, описанным в примере 1.3. В следующих таблицах 8, 9 соответственно перечислена информация относительно последовательностей и экспрессии белка scFv. Как видно из таблицы 9, PR000510 и PR000627 представляют собой молекулы, характеризующиеся особенно улучшенными показателями экспрессии и стабильности. На фигуре 5 показаны результаты (A) SDS-PAGE и (B) HPLC-SEC PR000510, на которых видно, что они характеризуются хорошей мономерную чистоту без наличия явных агрегатов.
Таблица 8. Информация относительно структуры и последовательности молекул scFv, сконструированных на основе мутантных вариантов с оптимизированными последовательностями
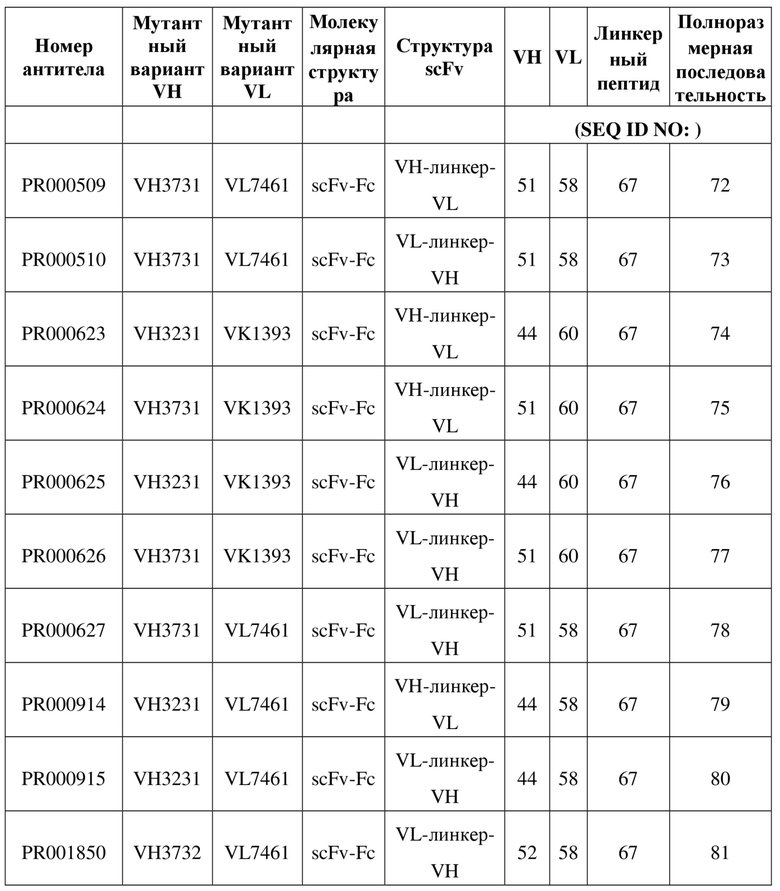
Таблица 9. Данные относительно экспрессии антитела scFv после оптимизации последовательности
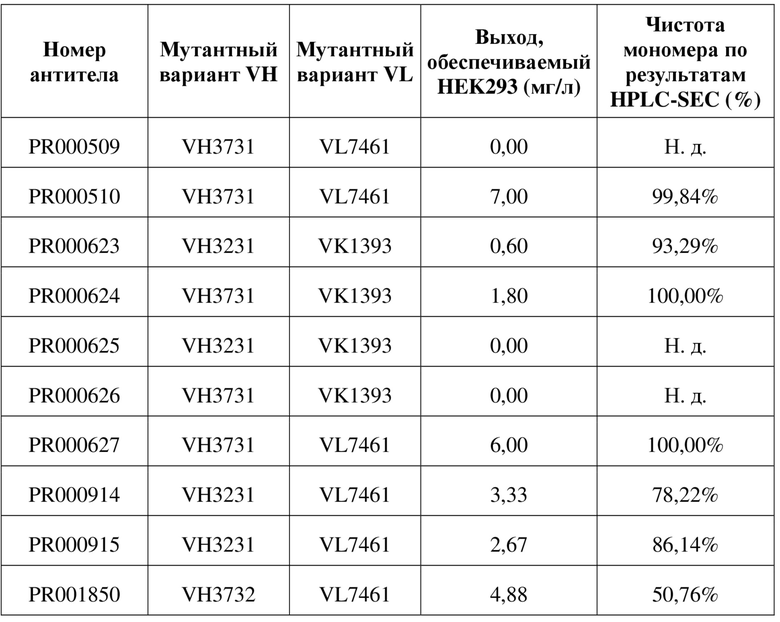
Пример 5. Использование FACS для определения связывающей способности антитела к CD3 и клеток, экспрессирующих CD3
[111] Способ проточного анализа FACS применяли для анализа связывания антитела к CD3 с CD3-экспрессирующими клетками, при этом CD3-экспрессирующие клетки могли представлять собой: клетки CHOK1, сверхэкспрессирующие CD3 человека, или клетки HEK293 (плазмидами, кодирующими ORF цепей γ, δ, ε и ζ CD3 человеческого происхождения, и плазмидами, кодирующими ORF цепей α и β TCR человека, котрансфицировали клетки-хозяева CHOK1 (ATCC, CCL-61) или HEK293 (ATCC, CRL-1573) для конструирования стабильных клеточных линий, экспрессирующих структуру, представленную комплексом TCR/CD3 человека); клетки CHOK1 или HEK293, сверхэкспрессирующие CD3 яванского макака; Т-клетки человека всех типов (выделенные с помощью набора для выделения Т-клеток человека всех типов (Miltenyi, № 130-096-535) из РВМС); Т-клетки яванского макака всех типов. В частности, собранные клетки дважды промывали с помощью PBS, содержащего 2% FBS (буфер FACS), и ресуспендировали в буфере для FACS, распределяли по 96-луночным планшетам по 1×105 клеток на лунку, центрифугировали при 500 g в течение 5 минут. Надосадочную жидкость удаляли и добавляли 100 мкл предварительно подвергнутого градиентному разбавлению антитела к CD3, затем инкубировали в течение 1 часа при комнатной температуре и дважды промывали буфером для FACS. Клетки ресуспендировали в буфере для FACS, в котором разбавляли вторичное антитело Alexa Fluor 488 AffiniPure, представляющее собой антитело козы к IgG человека, являющееся специфичным по отношению к фрагменту Fcγ (Jackson ImmunoResearch, № 109-545-098), затем инкубировали при комнатной температуре в темноте в течение 30 минут. Клетки дважды промывали буфером для FACS и ресуспендировали в 200 мкл буфера для FACS. Значение сигнала флуоресцентной люминесценции считывали посредством проточной цитометрии (BD FACS CANTOII или ACEA NovoCyte) и полученные данные обрабатывали и анализировали с помощью программного обеспечения FlowJo версии 10 (FlowJo, LLC). Программное обеспечение GraphPad Prism 8 использовали для обработки данных и графического анализа, и параметры, такие как кривые связывания и значения EC50, могли быть получены с помощью четырехпараметрической нелинейной аппроксимации.
[112] На фигуре 6 показана способность связывания антитела к CD3, полученного в примере 2, с рекомбинантными клетками CHOK1, сверхэкспрессирующими CD3 человека (фигура 6 (A)), и рекомбинантными клетками CHOK1, сверхэкспрессирующими CD3 яванского макака (фигура 6 (B)). Результаты указали на то, что химерное антитело SP34 PR000260 обладало высокой способностью связывания как с CD3 человека, так и с CD3 яванского макака.
[113] На (A) - (G) фигуры 7 показана способность связывания антител к CD3, полученных в примере 4.2 (в том числе PR000260 и его мутантных вариантов), с Т-клетками человека всех типов соответственно. Интенсивность флуоресценции MFI для антитела к CD3, связывающегося с Т-клетками человека всех типов, и относительный показатель по отношению к исходному антителу PR000260 рассчитывали при концентрации антитела, составляющей 7,4 или 10 мкг/мл. В частности, после оптимизации последовательности антитела SP34 изотипа IgG PR000512, PR000513, PR001849 и PR002837 характеризовались способностью связывания, сравнимой с PR000260 (т.е. химерным антителом SP34); в то время как PR000514 характеризовалось несколько более высокой способностью связывания, чем PR000260; PR000511, PR001848, PR002469, PR002472, PR002742, PR002833, PR002834, PR002835, PR002836, PR003886 и PR004616 характеризовались более низкой способностью связывания с Т-клетками; PR002467, PR002468, PR002470, PR002471 и PR002743, с другой стороны, почти не связывали Т-клетки (или сигнал не мог быть выявлен при текущей концентрации антител). Приведенные выше результаты указывают на то, что в рамках настоящего изобретения посредством оптимизации последовательности антитела к CD3 было осуществлено получение нескольких новых антител, которые обладают различающимися способностями связываться с Т-клетками человека и могут использоваться в различных сценариях применения.
[114] На (A) и (B) фигуры 8 показана связывающая способность одноцепочечного антитела к CD3, представленного в формате scFv-Fc, полученного в примере 4.3, с Т-клетками человека всех типов. Интенсивность флуоресценции MFI для антитела к CD3, связывающегося с Т-клетками человека всех типов, и относительный показатель по отношению к исходному антителу PR000260 рассчитывали при концентрации антитела, составляющей 7,4 или 10 мкг/мл. В частности, после оптимизации гуманизации представленного в формате scFv антитела SP34 PR000624 имело сравнимую или немного более высокую способность связывания при сравнении с PR000260; PR000510 и PR000627 имели сравнимую или немного меньшую способность связывания при сравнении с PR000260; и PR001850 имело значительно более низкую способность связывания с Т-клетками при сравнении с PR000260. Приведенные выше результаты указывают на то, что в рамках настоящего изобретения также получали несколько стабильных одноцепочечных антител в форме scFv посредством оптимизации последовательности антитела к CD3, которые способны связываться с Т-клетками человека и подходят для применения в таких сценариях, как конструирование биспецифических антител.
[115] На фигуре 9 показана способность некоторых антител к CD3, полученных в примере 4.2, связываться с Т-клетками яванского макака всех типов. Можно видеть, что различные молекулы обладали различной способностью связывания с Т-клетками яванского макака всех типов и положительно коррелировали с их способностью связывания с Т-клетками человека всех типов; т.е. молекулы, которые характеризовались сильным связыванием с Т-клетками человека всех типов, также характеризовались сильным связыванием с Т-клетками яванского макака всех типов, и наоборот.
Пример 6. Определение активации, индуцированной антителом к CD3 на Т-клетках человека
[116] Градиентными разбавлениями антител к CD3 (например, 50, 10, 5, 1, 0,5, 0,05 мкг/мл) покрывали 96-луночные планшеты для культивирования клеток, при этом предусматривали лунки в трех повторностях для каждой концентрации и по 50 мкл раствора на лунку, инкубирование осуществляли в течение ночи при 4°C. Плотность клеток РВМС человека (MiaoTong Biology) или Т-клеток человека всех типов (выделенных с помощью набора для выделения Т-клеток человека всех типов (Miltenyi, № 130-096-535) из РВМС) доводили до 7,5×105/мл и добавляли человеческое антитело к CD28 в концентрации 1 мкг/мл, после чего в планшет для культивирования клеток добавляли по 200 мкл на лунку полученной в результате смеси и осуществляли инкубирование в инкубаторе с CO2. После 72 часов инкубации отбирали надосадочную жидкость и определяли содержание в ней IFN-γ с использованием набора для ELISA IFN-γ (Thermo, № 88-7316-77). Для обработки данных и графического анализа использовали программное обеспечение GraphPad Prism 8.
[117] На (A) - (G) фигуры 10 показана способность каждого антитела к CD3 (в том числе химерного антитела SP34), полученного в примере 4.2, активировать Т-клетки человека соответственно. В случае если концентрация антител составляла 1 мкг/мл, PR000511, PR000512, PR000513 и PR000514 обеспечивали продуцирование значительно более низких уровней IFN-γ после активации Т-клеток, чем PR000260; в случае если концентрация антитела составляла 10 мкг/мл, PR000512, PR000513 и PR000514 активировали клетки с обеспечением несколько более низких уровней IFN-γ, чем PR000260 (фигура 10 (A)). В случае если концентрация антитела составляла 0,5 мкг/мл и 5 мкг/мл, уровень IFN-γ, обеспечиваемый при активации клеток с помощью PR001848, был значительно ниже, чем уровень для PR000260 (фигура 10 (B)). Кроме того, также выявили эффект активации Т-клеток антителами PR002468, PR002469, PR002471, PR002742, PR002833, PR002834, PR002835, PR002836, PR002837, PR001848, PR003886 и PR004616 ((C) - (G) на фигуре 10) в концентрациях 0,5 мкг/мл, 5 мкг/мл и 50 мкг/мл, при этом результаты показали, что уровень IFN-γ, продуцируемого Т-клетками, активированными этими антителами, был намного ниже, чем уровни, обеспечиваемые при активации с помощью PR000260, при этом не выявляли высвобождения IFN-γ Т-клетками, активированными PR002468 и PR002471, и выявляли только слабый уровень IFN-γ при использовании 50 мкг/мл PR002469 и PR002835; уровень IFN-γ, продуцируемого Т-клетками, активированными PR002742 и PR003886, был сопоставимым и немного более низким, чем в случае PR001848; уровень IFN-γ, продуцируемого Т-клетками, активированными PR002469 и PR004616, был сопоставимым и значительно более низким, чем в случае PR001848. Приведенные выше результаты указывают на то, что в рамках настоящего изобретения посредством оптимизации последовательности антитела к CD3 было осуществлено получение нескольких новых антител, которые обладают различающимися способностями активации в Т-клетках человека, могут обеспечивать контроль высвобождения различных уровней цитокинов и могут использоваться в различных сценариях применения.
[118] На фигуре 11 показана способность антитела к CD3, представленного в формате scFv-Fc, полученного в примере 4.3, активировать Т-клетки человека. PR000510, PR000623, PR000624 и PR000627 при концентрациях 1 мкг/мл и 10 мкг/мл продемонстрировали более низкие уровни IFN-γ, чем PR000260, однако выше, чем антитело изотипического контроля, что указывало на то, что эти четыре молекулы ограничивали высвобождение цитокинов, регулируя уровни активации Т-клеток. Приведенные выше результаты указывают на то, что в рамках настоящего изобретения также получали несколько стабильных одноцепочечных антител в форме scFv посредством оптимизации последовательности антитела к CD3, которые характеризуются более слабой активацией Т-клеток человека, демонстрируя более низкие уровни высвобождения цитокинов, и подходят для применения в таких сценариях, как конструирование биспецифических антител.
Пример 7. Биспецифическое антитело, нацеливающееся на B7H4, содержащее антитело к CD3, представленное в формате scFv
[119] B7H4, член семейства трансмембранных белков B7, характеризуется высоким уровнем экспрессии в различных солидных опухолевых тканях, как например при раке молочной железы, яичника и эндометрия, в то время как он не характеризуется экспрессией или характеризуется крайне слабой экспрессией в нормальных тканях, что делает B7H4 очень специфичным опухоль-ассоциированным антигеном-мишенью. Конструировали молекулу биспецифического антитела, нацеливающегося как на B7H4, так и на CD3, которая способна избирательно активировать Т-клетки вблизи опухолевых клеток посредством нацеливания на B7H4 и связывания с ним на поверхности опухолевых клеток, обеспечивая тем самым специфическое уничтожение опухолевых клеток.
7.1. Получение антитела к B7H4
[120] Последовательность вариабельной области антитела к B7H4 может быть получена из WO 2016040724, а рекомбинантное антитело PR000014 изотипа IgG, связывающее мишень B7H4, конструировали в соответствии со способами из примера 1.1. В нижеследующей таблице 10 приведена информация относительно последовательности антитела к B7H4 PR000014.
Таблица 10. Информация относительно последовательности легкой и тяжелой цепей антитела к B7H4 PR000014

7.2. Получение биспецифического антитела, нацеливающегося на B7H4, содержащего антитело к CD3, представленное в формате scFv
[121] Использовали последовательность антитела к B7H4 PR000014, полученного в примере 7.1, и последовательность одноцепочечного антитела к CD3 PR000627, полученного в примере 4.3, для конструирования молекулы биспецифического антитела PR002883, нацеливающейся на B7H4 × CD3, которая содержала три полипептидные цепи: тяжелую цепь, содержащую scFv одноцепочечного антитела к CD3 (SEQ ID NO: 88), тяжелую цепь, содержащую VH антитела к B7H4 (SEQ ID NO: 86), и легкую цепь, содержащую VL антитела к B7H4 (SEQ ID NO: 83). Структура показана на фигуре 16 (C). Поскольку молекула имела особую асимметричную структуру, то в константные области двух тяжелых цепей вводили различные аминокислотные мутации, чтобы уменьшить образование гомологичных димеров тяжелых цепей. В то же время вводили тройную мутацию «LALAPG» (L234A/L235A/P329G) в константную область тяжелой цепи для предупреждения перекрестного связывания и снижения эффекторной функции, обусловленной связыванием рецептора Fcγ.
[122] Рекомбинантный белок биспецифического антитела PR002883 получали с применением способа, описанного в примере 1.1, в комбинации с плазмидами в некотором соотношении (например, 1:1:1 или других соотношениях) и последующей одностадийной аффинной очисткой. Последовательность биспецифического антитела PR002883 представлена в таблице 11; экспрессия биспецифического антитела представлена в таблице 12.
Таблица 11. Цепи биспецифического антитела и соответствующая информация относительно последовательности

Таблица 12. Экспрессия биспецифического антитела

[123] На фигуре 12 (А) показаны результаты для биспецифического антитела PR002883 после одностадийной очистки посредством анализа SDS-PAGE. Он показал, что его основными побочными продуктами являются неполностью собранные молекулы с небольшим количеством высокополимерных компонентов, количество которых можно уменьшить за счет оптимизации стадии очистки или оптимизации показателя уровня трансфекции плазмидами.
7.3. Связывание с опухолевыми клетками, экспрессирующими B7H4
[124] В данном примере исследовали способность биспецифического антитела связывать опухолевые клетки SK-BR-3 (ATCC, HTB-30), экспрессирующие B7H4 человека. В частности, собирали суспензию клеток SK-BR-3, доводили плотность клеток до 1×106/мл и инокулировали их по 100 мкл/лунка в 96-луночный планшет с V-образным дном (Corning, № 3894); затем добавляли подлежащее тестированию антитело в 2-кратной концентрации от конечной концентрации, полученной с помощью 3-кратного градиентного разбавления, в объеме 100 мкл/лунка. Клетки инкубировали при 4°С в темноте в течение 2 часов. После этого клетки дважды промывали с помощью 100 мкл/лунка предварительно охлажденного PBS, центрифугировали при 500 g, 4°C, в течение 5 минут. Надосадочную жидкость удаляли. Затем добавляли 100 мкл/лунка вторичного антитела Alexa Fluor 488 AffiniPure, представляющего собой антитело козы к IgG человека, являющегося специфичным по отношению к фрагменту Fcγ (Jackson ImmunoResearch, № 109-545-098), и клетки инкубировали в условиях отсутствия света при 4°C в темноте в течение 1 часа. Затем клетки дважды промывали с помощью 100 мкл/лунка предварительно охлажденного PBS, центрифугировали при 500 g в течение 5 минут. Надосадочную жидкость удаляли. Наконец, клетки ресуспендировали с помощью 200 мкл/лунка предварительно охлажденного PBS. Значение сигнала флуоресцентной люминесценции считывали посредством проточной цитометрии (BD FACS CANTOII или ACEA NovoCyte) и полученные данные обрабатывали и анализировали с помощью программного обеспечения FlowJo версии 10 (FlowJo, LLC). Программное обеспечение GraphPad Prism 8 использовали для обработки данных и графического анализа, и параметры, такие как кривые связывания и значения EC50, могли быть получены с помощью четырехпараметрической нелинейной аппроксимации.
[125] На фигуре 13 (А) показана способность связывания моноклонального антитела, полученного в примере 7.1, и биспецифического антитела, полученного в примере 7.2, с клеткой SK-BR-3. Можно видеть, что биспецифическое антитело PR002883 имело сравнимую или даже лучшую способность связывания по сравнению с моноклональным антителом PR000014.
7.4. Связывание Т-клеток человека
[126] Способность биспецифического антитела PR002883 связываться с Т-клетками человека всех типов определяли посредством способа, описанного в примере 5. Как показано на фигуре 13 (В), PR002883 способно связываться с Т-клетками человека всех типов.
7.5. Уничтожение in vitro клеток линии SK-BR-3, характеризующихся высоким уровнем экспрессии B7H4, посредством биспецифического антитела и высвобождение цитокинов
[127] Для исследования способности уничтожать in vitro клетки-мишени, опосредованной биспецифическим антителом B7H4 × CD3, в качестве эффекторных клеток использовали РВМС человека, а клетки линии SK-BR-3, характеризующиеся высоким уровнем экспрессии B7H4 (ATCC, HTB-30), использовали в качестве клеток-мишеней для анализов уничтожения in vitro и выявления высвобождения цитокинов. В частности, в каждую лунку E-планшета (ACEA Biosciences Inc., № 05232368001) добавляли по 50 мкл среды RPMI1640/10% FBS, после чего осуществляли уравновешивание в инкубаторе, содержащем 5% CO2, при 37°C в течение 30 минут, а затем Е-планшет помещали в прибор xCELLigence RTCA (ACEA Biosciences) для тестирования в отношении нормальности. Плотность SK-BR-3 доводили до 0,4×106 клеток/мл с помощью среды RPMI1640/10% FBS, затем инокулировали в E-планшет по 50 мкл клеток/лунка, а затем E-планшет помещали на xCELLigence RTCA на ночь для выявления клеточного индекса. Плотность РВМС доводили до 4×106 клеток/мл с помощью среды RPMI1640/10% FBS и инокулировали в E-планшет по 50 мкл клеток/лунка, затем подлежащее тестированию антитело, представленное в 4-кратной концентрации относительно конечной концентрации, полученной посредством 5-кратного градиентного разбавления, добавляли при 50 мкл/лунка, при этом самая высокая конечная концентрация антитела составляла 0,2 нМ, и при этом имелось 7 концентраций для каждого антитела, конечное соотношение эффекторных клеток и клеток-мишеней составляло 10: 1, причем предусматривали две повторности. В то же время в планшет вносили холостой контроль: SKBR3 + РВМС + среда RPMI1640/10% FBS; E-планшет инкубировали при 37°C в инкубаторе, содержащем 5% CO2, в течение 24 часов. После инкубации E-планшет помещали на прибор xCELLigence RTCA для выявления клеточного индекса.
[128] Эффект специфического уничтожения, оказываемый антителом, рассчитывали с применением определенного в ходе анализа клеточного индекса в следующей формуле:
% уничтожения клеток = (1 - тестовый образец/холостой контроль) * 100%.
[129] Надосадочную жидкость клеточной культуры собирали для выявления высвобождения цитокина IFN-γ. См. инструкции по эксплуатации набора IFN-γ (набор с планшетами без покрытия, предназначенный для ELISA IFN-гамма человека IFN gamma Human Uncoated ELISA Kit, Thermo, № 88-7316-77) для выявления с помощью ELISA.
[130] Как показано на (A) и (B) фигуры 14, биспецифическое антитело PR002883 могло активировать Т-клетки с высвобождением цитокинов, таких как IFN-γ, и эффективно уничтожать опухолевые клетки SK-BR-3. Почти 100% опухолевых клеток подвергались уничтожению, когда концентрация биспецифического антитела составляла 0,01 мкг/мл (фигура 14 (А)).
Пример 8. Биспецифическое антитело, нацеливающееся на ROR1, содержащее антитело к CD3, представленное в формате scFv
[131] ROR1 представлял собой неактивный трансмембранный белок тирозинпротеинкиназы, который сверхэкспрессируется во многих опухолях, но фактически не экспрессируется в нормальных тканях. ROR1 способствует пролиферации и миграции клеток при хроническом лимфоцитарном лейкозе посредством передачи сигнала по пути передачи сигналов Wnt после взаимодействия с Wnt5a, выступая в качестве рецептора, и способствует эпителиально-мезенхимальной трансформации (EMT) в солидных опухолях. Опухолеспецифическая экспрессия ROR1 делает его подходящим опухолеассоциированным антигеном-мишенью для разработки терапевтических средств. Конструирование молекулы биспецифического антитела, нацеливающегося как на ROR1, так и на CD3, может обеспечивать избирательную активацию Т-клеток вблизи опухолевых клеток посредством нацеливания на ROR1 на поверхности опухолевых клеток и связывания с ним, обеспечивая тем самым специфическое уничтожение опухолевых клеток.
8.1. Получение антитела к ROR1
[132] Последовательность вариабельной области антитела к ROR1 могла быть получена из WO 2016094873, а конструкция рекомбинантного антитела PR000374 класса IgG, нацеливающегося на ROR1, - в соответствии со способами из примера 1.1. В нижеследующей таблице 13 приведена информация относительно последовательности антитела к ROR1, представляющего собой PR000374.
Таблица 13. Перечень последовательностей антитела к ROR1, представляющего собой PR000374

8.2. Получение биспецифического антитела, нацеливающегося на ROR1, содержащего антитело к CD3, представленное в формате scFv
[133] Конструирование молекулы биспецифического антитела PR002885, нацеливающегося на ROR1 × CD3, осуществляли с использованием последовательности антитела к ROR1, представляющего собой PR000374, полученного в примере 8.1, и последовательности одноцепочечного антитела к CD3, представляющего собой PR000627, полученного в примере 4.3, которое содержало три полипептидные цепи: тяжелую цепь, содержащую scFv одноцепочечного антитела к CD3 (SEQ ID NO 88), тяжелую цепь, содержащую VH антитела к ROR1 (SEQ ID NO: 87), и легкую цепь, содержащую VL антитела к ROR1 (SEQ ID NO: 85). Структура показана на фигуре 16 (C). Поскольку молекула имела особую асимметричную структуру, то в константные области двух тяжелых цепей вводили различные аминокислотные мутации, чтобы уменьшить образование гомологичных димеров тяжелых цепей. В то же время вводили тройную мутацию «LALAPG» (L234A/L235A/P329G) в константную область тяжелой цепи для предупреждения перекрестного связывания и снижения эффекторной функции, обусловленной связыванием рецептора Fcγ.
[134] Рекомбинантный белок биспецифического антитела PR002885 получали с применением способа, описанного в примере 1.1, в комбинации с плазмидами в некотором соотношении (например, 1:1:1 или других соотношениях) и последующей одностадийной аффинной очисткой. Последовательность биспецифического антитела PR002885 представлена в таблице 14; экспрессия биспецифического антитела представлена в таблице 15.
Таблица 14. Цепи биспецифического антитела и номер соответствующей последовательности
Таблица 15. Экспрессия биспецифического антитела
[135] На фигуре 12 (B) показаны результаты для биспецифического антитела PR002885 после одностадийной очистки посредством анализа SDS-PAGE. Он показал, что его основными побочными продуктами являются неполностью собранные молекулы с небольшим количеством высокополимерных компонентов, количество которых можно уменьшить за счет оптимизации стадии очистки или оптимизации показателя уровня трансфекции плазмидами.
8.3. Связывание с опухолевыми клетками, экспрессирующими ROR1
[136] В данном примере исследовали способность биспецифического антитела связывать опухолевые клетки Panc-1 (ATCC, CRL-1469), экспрессирующие ROR1 человека. В частности, собирали суспензию клеток Panc-1, доводили плотность клеток до 1×106/мл и инокулировали их по 100 мкл/лунка в 96-луночный планшет с V-образным дном (Corning, № 3894); затем добавляли подлежащее тестированию антитело в 2-кратной концентрации от конечной концентрации, полученной с помощью 3-кратного градиентного разбавления, в объеме 100 мкл/лунка. Клетки инкубировали при 4°С в темноте в течение 2 часов. После этого клетки дважды промывали с помощью 100 мкл/лунка предварительно охлажденного PBS, центрифугировали при 500 g, 4°C в течение 5 минут. Надосадочную жидкость удаляли. Затем добавляли 100 мкл/лунка флуоресцентного вторичного антитела Alexa Fluor 488 AffiniPure, представляющего собой антитело козы к IgG человека, являющегося специфичным по отношению к фрагменту Fcγ (Jackson ImmunoResearch, № 109-545-098), и клетки инкубировали в условиях отсутствия света при 4°C в темноте в течение 1 часа. Затем клетки дважды промывали с помощью 100 мкл/лунка предварительно охлажденного PBS, центрифугировали при 500 g в течение 5 минут. Надосадочную жидкость удаляли. Наконец, клетки ресуспендировали с помощью 200 мкл/лунка предварительно охлажденного PBS. Значение сигнала флуоресцентной люминесценции считывали посредством проточной цитометрии (BD FACS CANTOII или ACEA NovoCyte) и полученные данные обрабатывали и анализировали с помощью программного обеспечения FlowJo версии 10 (FlowJo, LLC). Программное обеспечение GraphPad Prism 8 использовали для обработки данных и графического анализа, и параметры, такие как кривые связывания и значения EC50, могли быть получены с помощью четырехпараметрической нелинейной аппроксимации.
[137] На фигуре 15 (А) показана способность связывания моноклонального антитела, полученного в примере 8.1, и биспецифического антитела, полученного в примере 8.2, с клеткой Panc-1. Можно видеть, что как биспецифическое антитело PR002885, так и моноклональное антитело PR000374 могут связывать Panc-1.
8.4. Связывание Т-клеток человека
[138] Способность биспецифического антитела PR002885 связываться с Т-клетками человека всех типов определяли посредством способа, описанного в примере 5. Как показано на фигуре 15 (В), PR002885 способно связываться с Т-клетками человека всех типов.
| название | год | авторы | номер документа |
|---|---|---|---|
| БИСПЕЦИФИЧЕСКОЕ АНТИТЕЛО | 2019 |
|
RU2796019C2 |
| PSMA×CD3 БИСПЕЦИФИЧЕСКОЕ ОДНОЦЕПОЧЕЧНОЕ АНТИТЕЛО С МЕЖВИДОВОЙ СПЕЦИФИЧНОСТЬЮ | 2009 |
|
RU2559531C2 |
| Терапевтические антитела и их применения | 2016 |
|
RU2722381C2 |
| АНТИ-СD3-АНТИТЕЛО И МОЛЕКУЛА, СОДЕРЖАЩАЯ ЭТО АНТИТЕЛО | 2017 |
|
RU2790326C2 |
| МУЛЬТИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА И СПОСОБЫ ИХ ПОЛУЧЕНИЯ И ПРИМЕНЕНИЯ | 2018 |
|
RU2811477C2 |
| АНТИТЕЛА, СВЯЗЫВАЮЩИЕ CD3 | 2017 |
|
RU2807216C2 |
| Биспецифическое антитело и его применение | 2019 |
|
RU2785292C2 |
| БИСПЕЦИФИЧЕСКОЕ ОДНОЦЕПОЧЕЧНОЕ АНТИТЕЛО К PSMAxCD3 С МЕЖВИДОВОЙ СПЕЦИФИЧНОСТЬЮ | 2011 |
|
RU2617942C2 |
| PSCAXCD3, CD19XCD3, C-METXCD3, ЭНДОСИАЛИНXCD3, EPCAMXCD3, IGF-1RXCD3 ИЛИ FAP-АЛЬФАXCD3 БИСПЕЦИФИЧЕСКОЕ ОДНОЦЕПОЧЕЧНОЕ АНТИТЕЛО С МЕЖВИДОВОЙ СПЕЦИФИЧНОСТЬЮ | 2009 |
|
RU2547600C2 |
| АНТИТЕЛА, СВЯЗЫВАЮЩИЕ CD3 | 2017 |
|
RU2790288C2 |

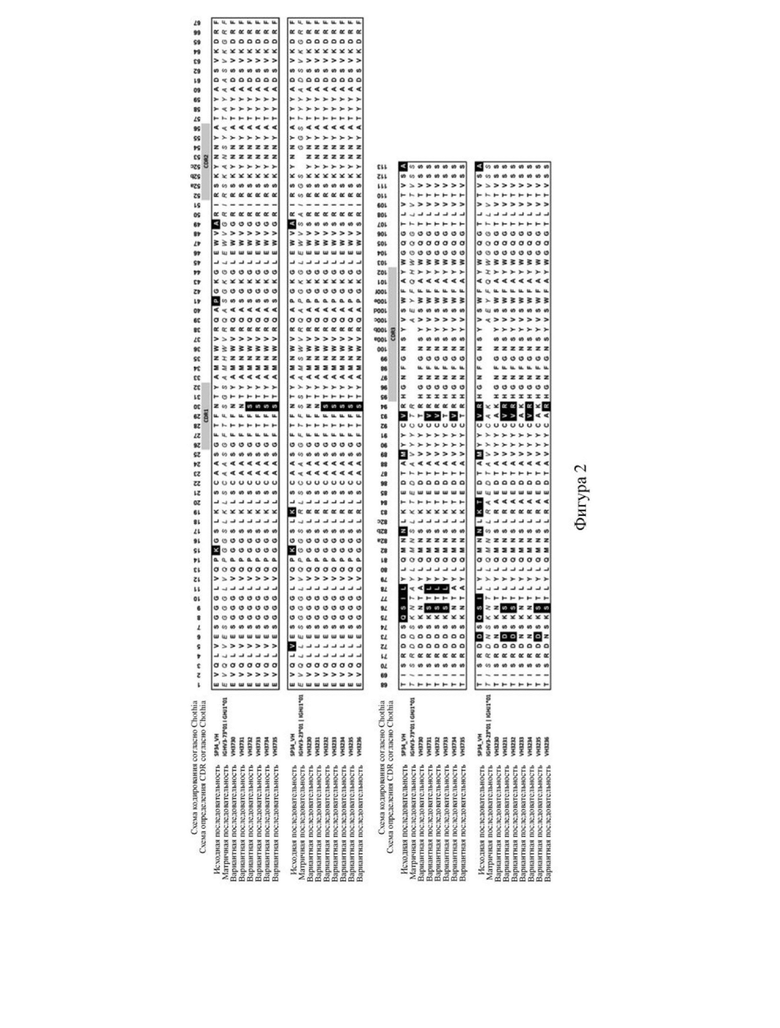
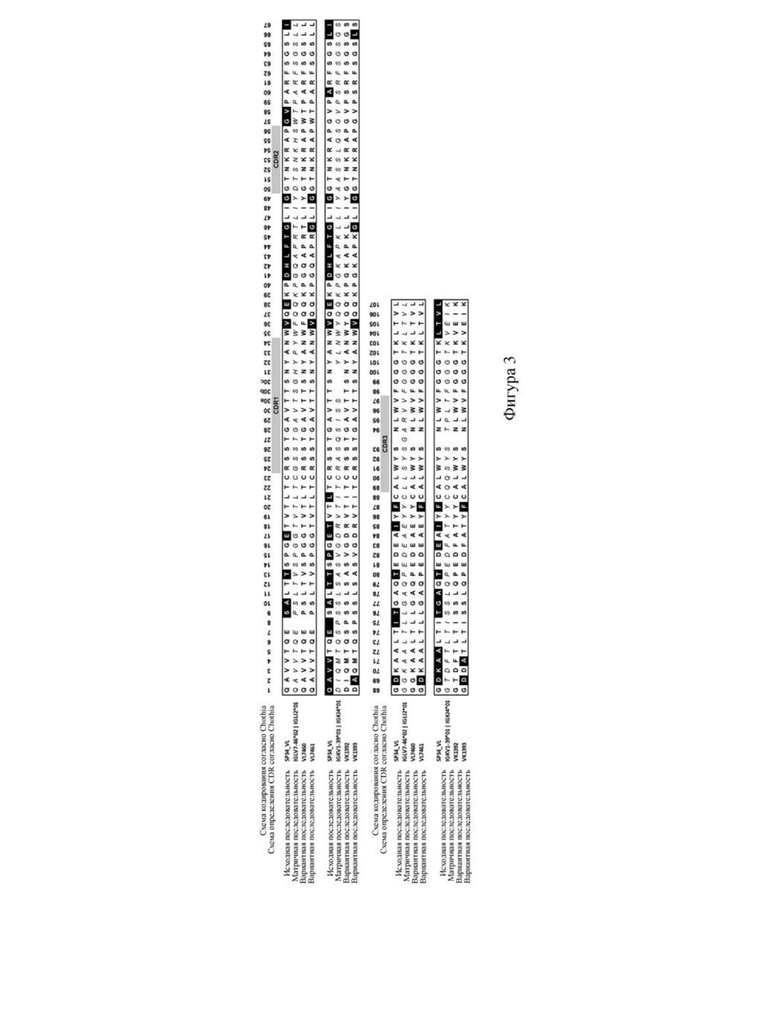
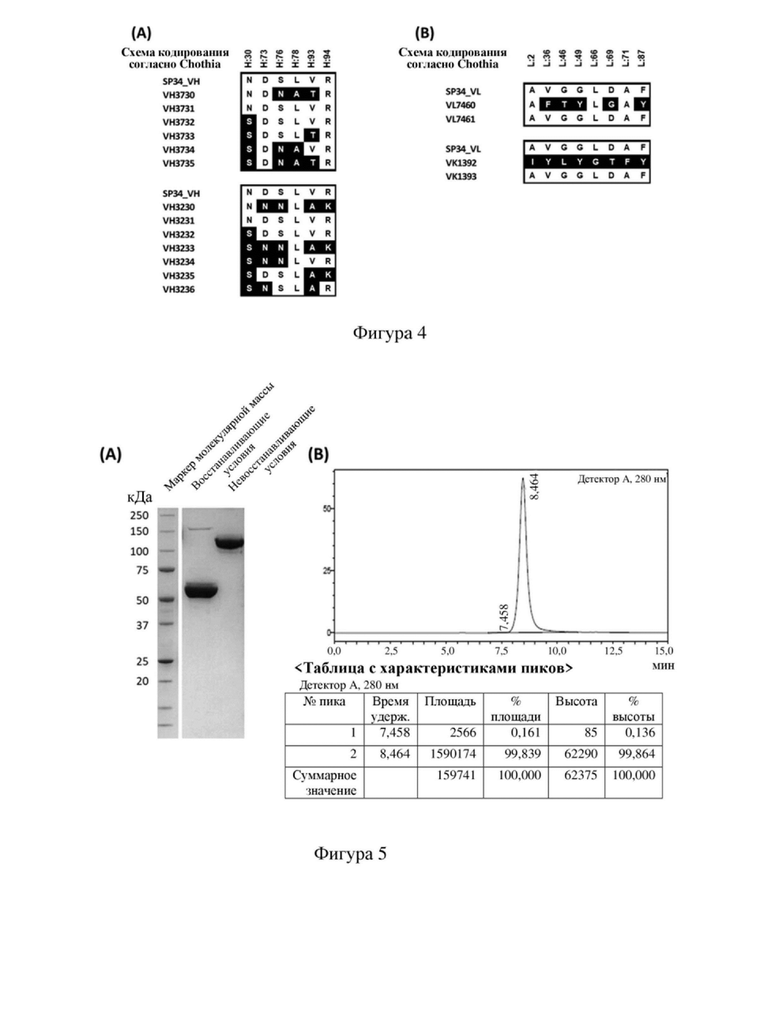
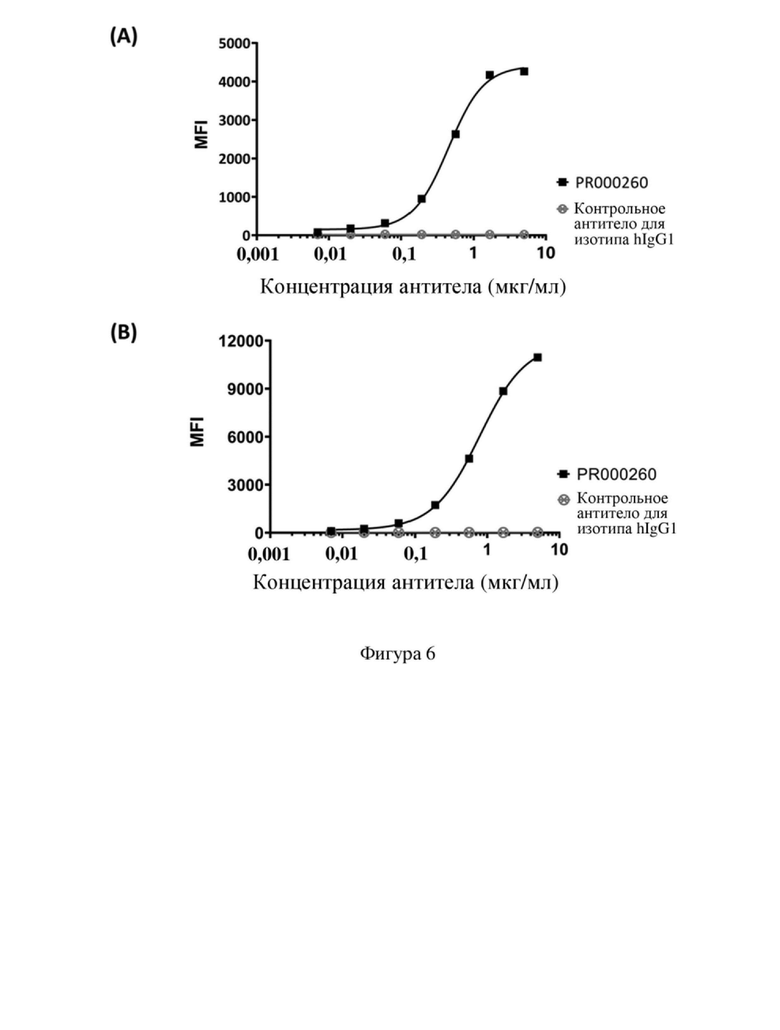
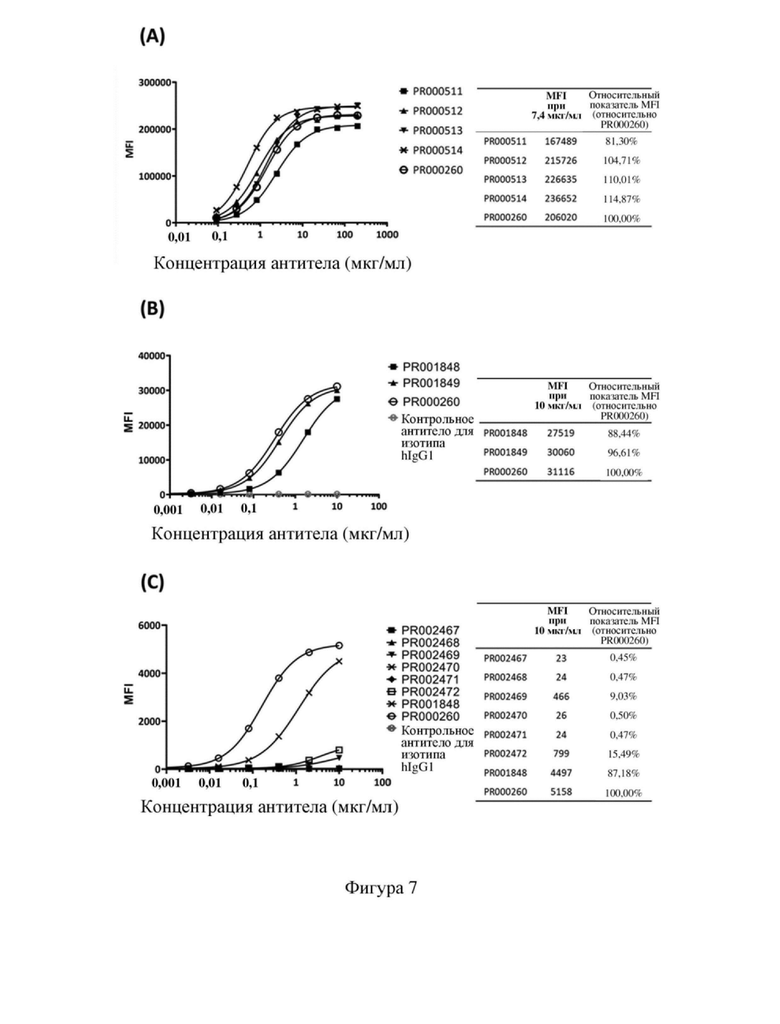
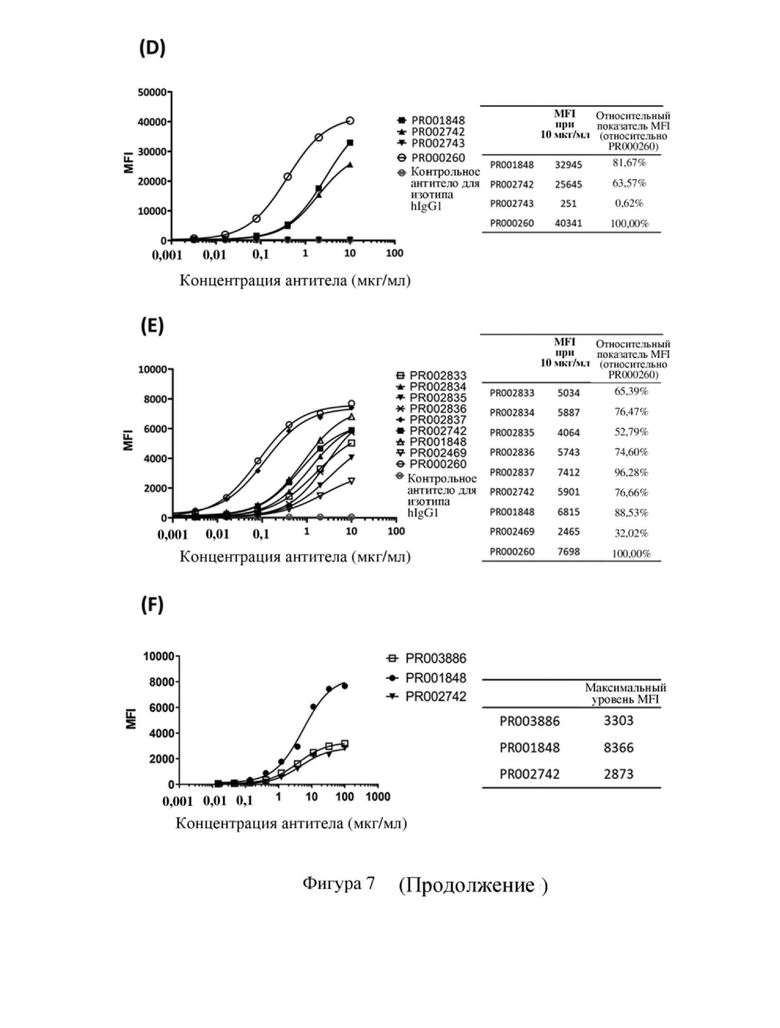
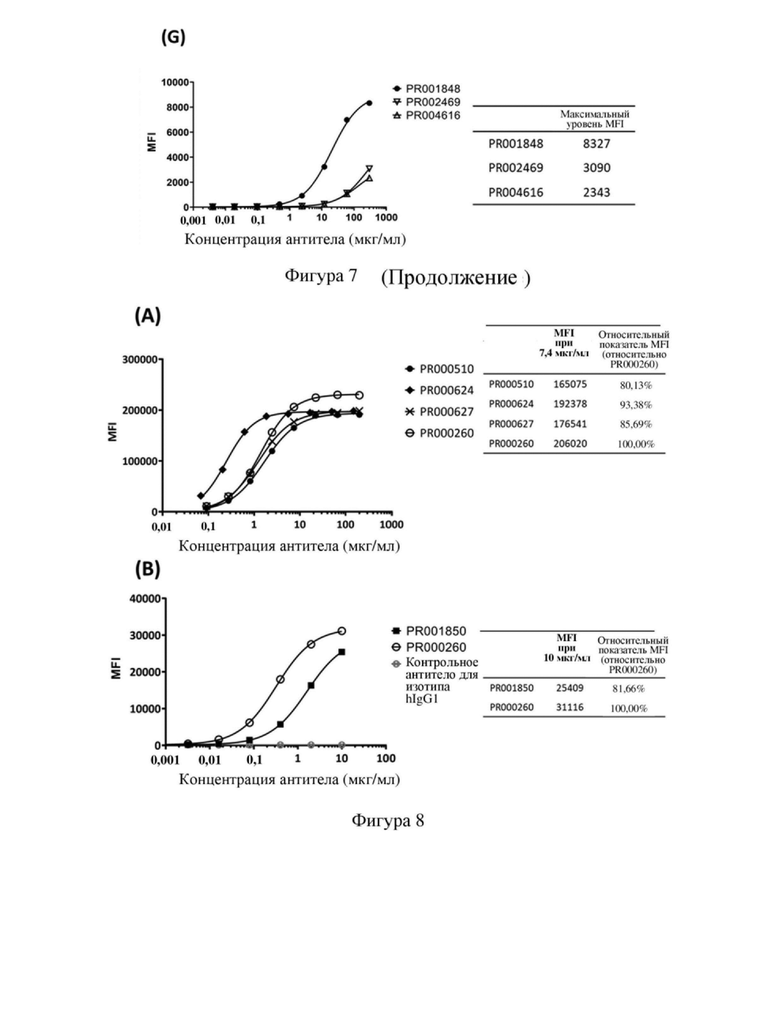
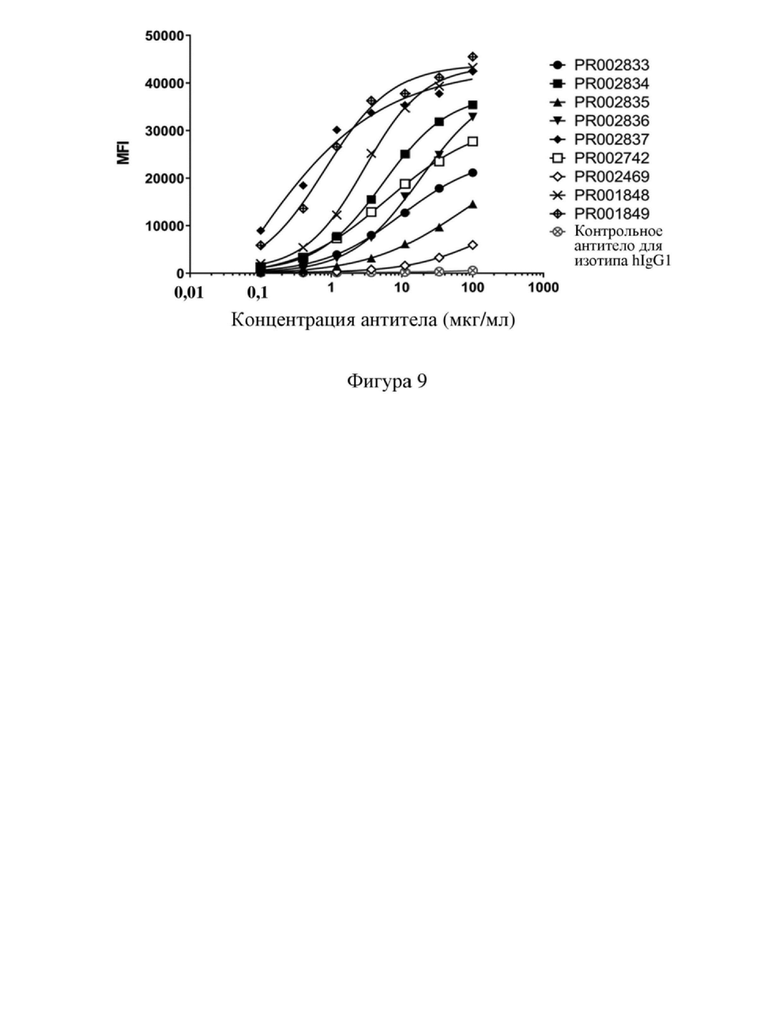
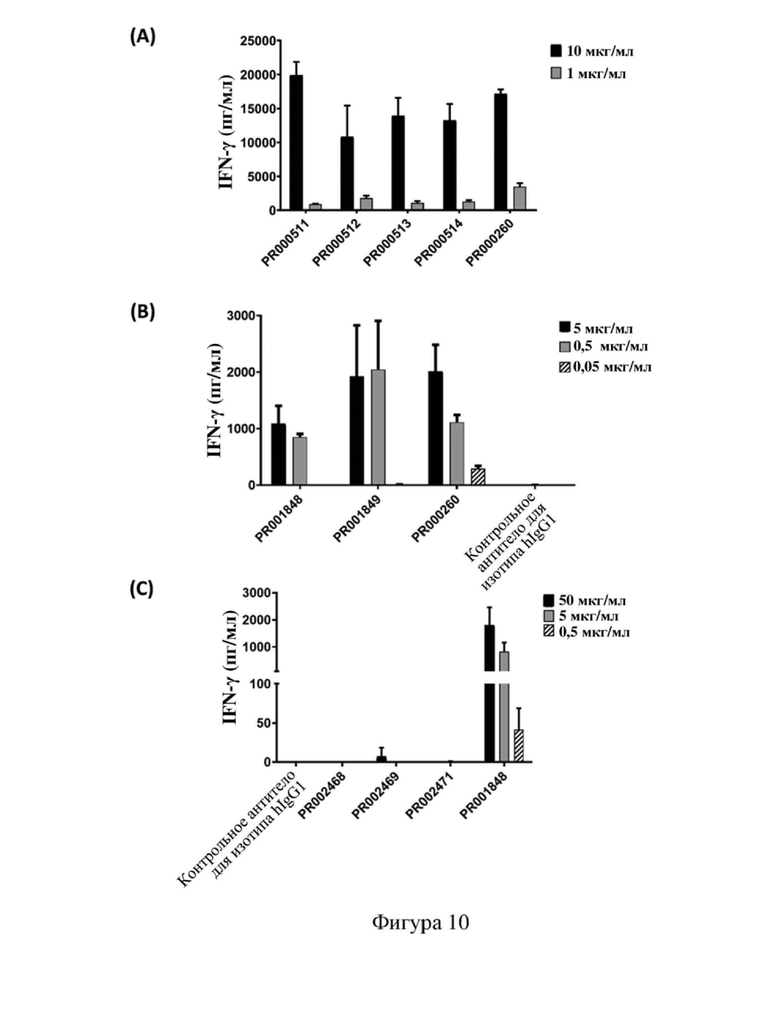
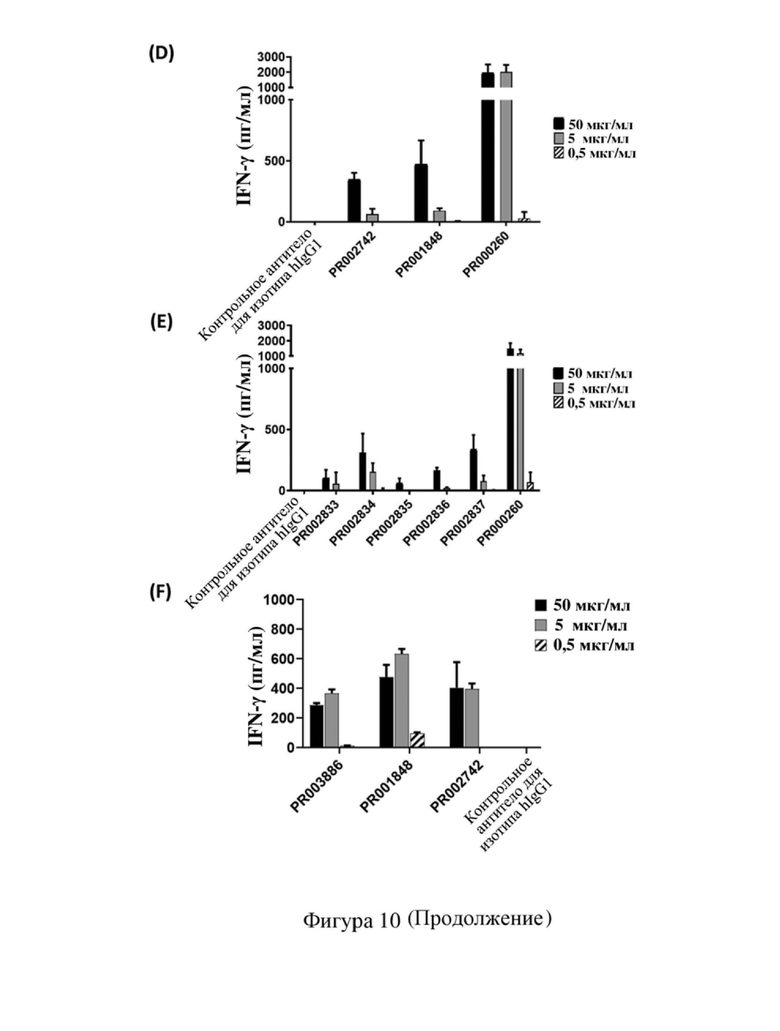
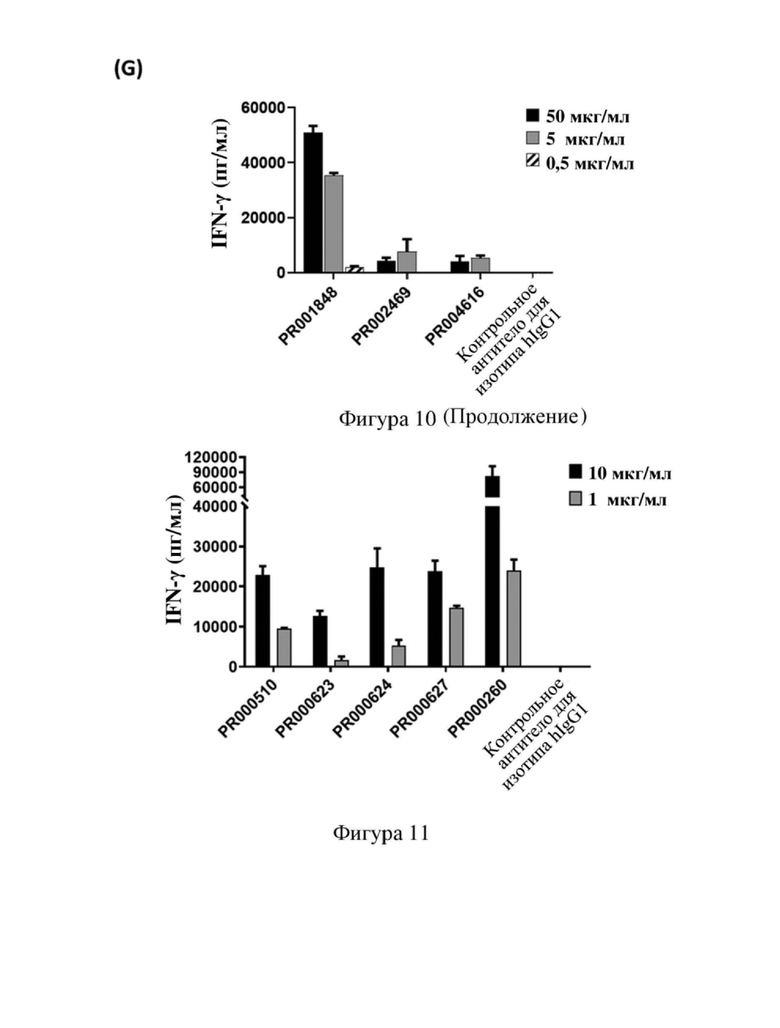
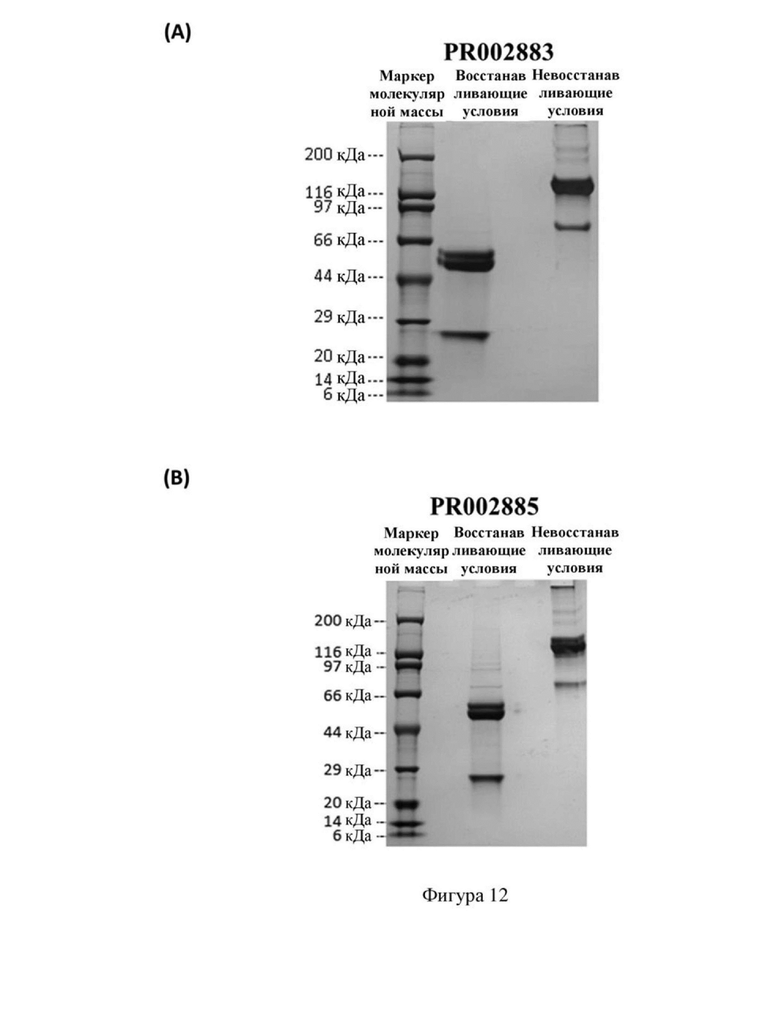
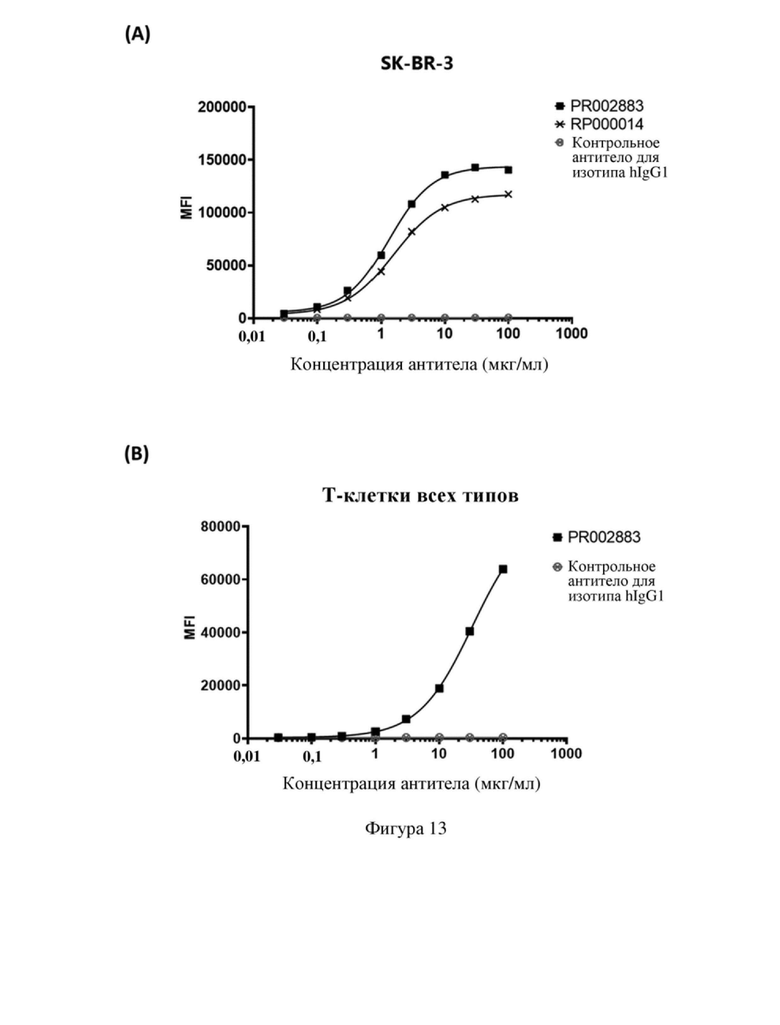
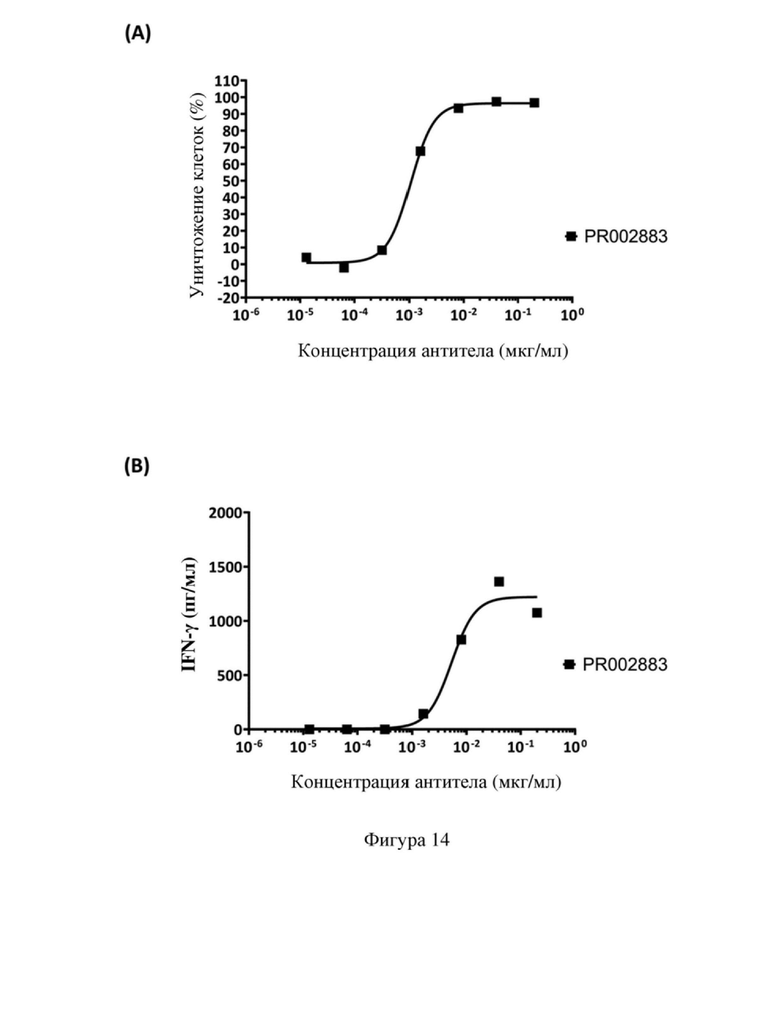
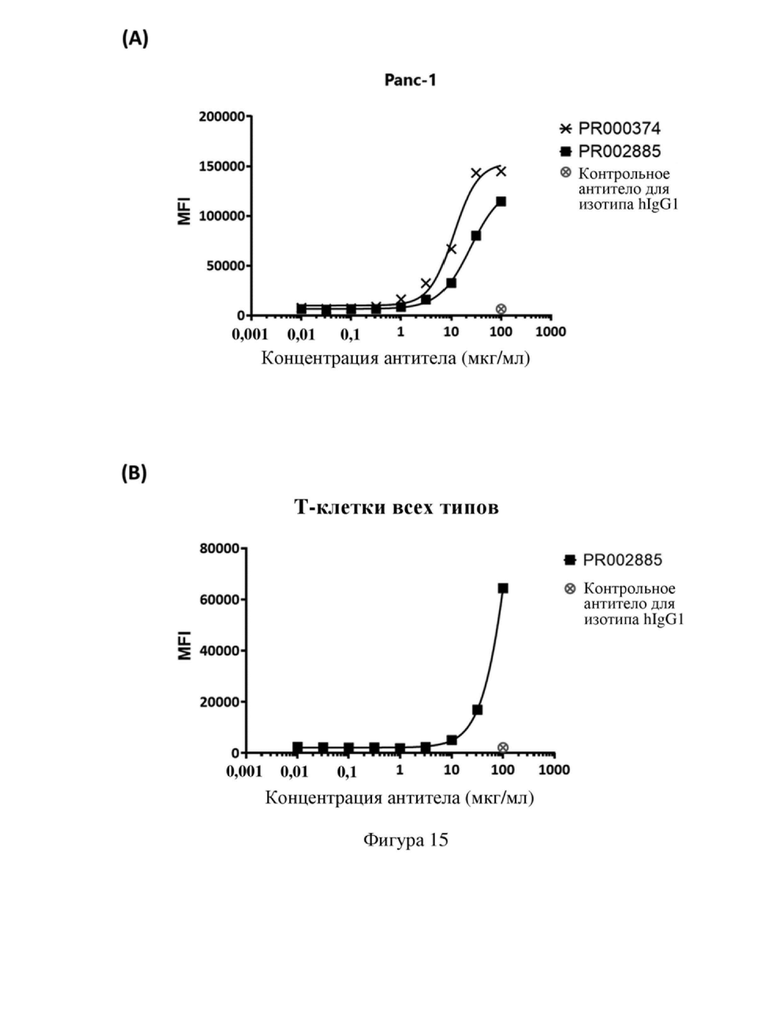
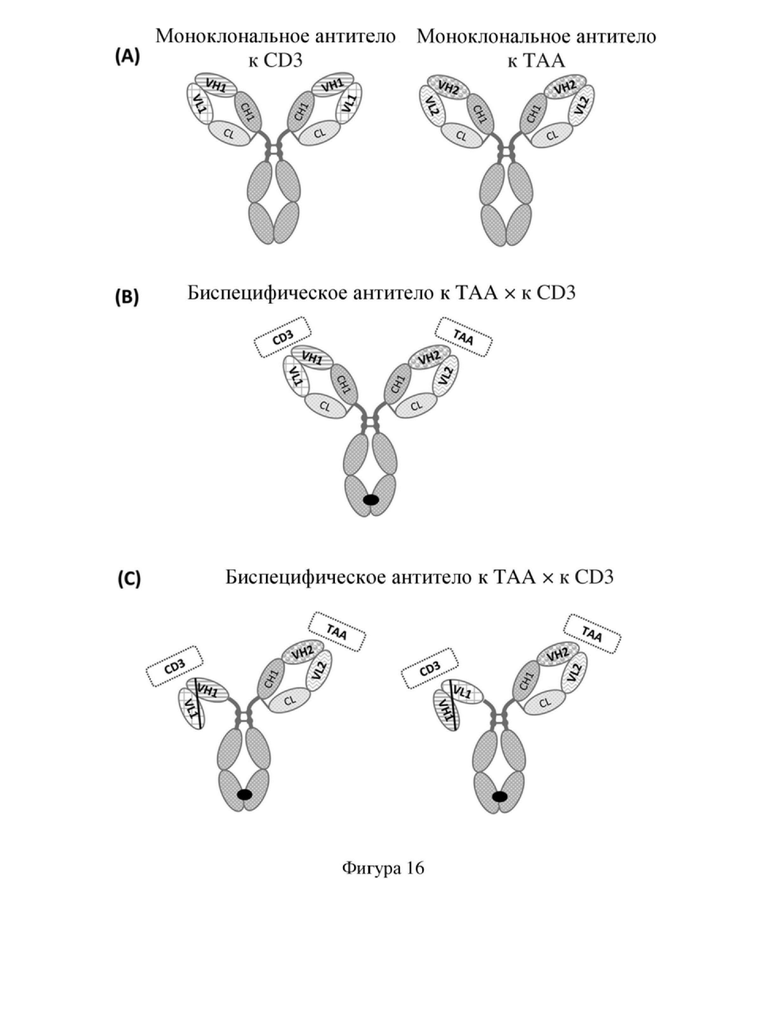
Группа изобретений относится к области биофармацевтических препаратов, в частности к антителу, нацеливающемуся на CD3, биспецифическому антителу, их получению и применению. Антитело, нацеливающееся на CD3, содержит аминокислотную последовательность VH, представленную под любым из SEQ ID NO: 43-55, и аминокислотную последовательность VL, представленную под любым из SEQ ID NO: 57-60. Биспецифическое антитело содержит первую функциональную область белка, предусматривающую антитело, нацеливающееся на CD3, и вторую функциональную область белка. Также предложены выделенная нуклеиновая кислота, вектор экспрессии, генетически модифицированная клетка для получения антитела, фармацевтическая композиция и их применение в изготовлении лекарственного препарата для лечения опухоли с CD3 в качестве мишени. Антитело способно связываться с CD3 приматов, обеспечивает снижение токсичности, вызванной синдромом высвобождения цитокинов, а полученное на его основе биспецифическое антитело является стабильным и обладает способностью связываться с Т-клетками. 8 н. и 16 з.п. ф-лы, 16 ил., 15 табл., 8 пр.
1. Антитело, нацеливающееся на CD3, содержащее вариабельную область легкой цепи (VL) и вариабельную область тяжелой цепи (VH); где
аминокислотная последовательность VH представлена под SEQ ID NO: 44, и аминокислотная последовательность VL представлена под SEQ ID NO: 58; или
аминокислотная последовательность VH представлена под SEQ ID NO: 51, и аминокислотная последовательность VL представлена под SEQ ID NO: 58; или
аминокислотная последовательность VH представлена под SEQ ID NO: 44, и аминокислотная последовательность VL представлена под SEQ ID NO: 60; или
аминокислотная последовательность VH представлена под SEQ ID NO: 51, и аминокислотная последовательность VL представлена под SEQ ID NO: 60; или
аминокислотная последовательность VH представлена под SEQ ID NO: 45, и аминокислотная последовательность VL представлена под SEQ ID NO: 58; или
аминокислотная последовательность VH представлена под SEQ ID NO: 52, и аминокислотная последовательность VL представлена под SEQ ID NO: 58; или
аминокислотная последовательность VH представлена под SEQ ID NO: 43, и аминокислотная последовательность VL представлена под SEQ ID NO: 58; или
аминокислотная последовательность VH представлена под SEQ ID NO: 43, и аминокислотная последовательность VL представлена под SEQ ID NO: 60; или
аминокислотная последовательность VH представлена под SEQ ID NO: 50, и аминокислотная последовательность VL представлена под SEQ ID NO: 58; или
аминокислотная последовательность VH представлена под SEQ ID NO: 47, и аминокислотная последовательность VL представлена под SEQ ID NO: 58; или
аминокислотная последовательность VH представлена под SEQ ID NO: 48, и аминокислотная последовательность VL представлена под SEQ ID NO: 58; или
аминокислотная последовательность VH представлена под SEQ ID NO: 49, и аминокислотная последовательность VL представлена под SEQ ID NO: 58; или
аминокислотная последовательность VH представлена под SEQ ID NO: 53, и аминокислотная последовательность VL представлена под SEQ ID NO: 58; или
аминокислотная последовательность VH представлена под SEQ ID NO: 54, и аминокислотная последовательность VL представлена под SEQ ID NO: 58; или
аминокислотная последовательность VH представлена под SEQ ID NO: 44, и аминокислотная последовательность VL представлена под SEQ ID NO: 57; или
аминокислотная последовательность VH представлена под SEQ ID NO: 44, и аминокислотная последовательность VL представлена под SEQ ID NO: 59; или
аминокислотная последовательность VH представлена под SEQ ID NO: 51, и аминокислотная последовательность VL представлена под SEQ ID NO: 57; или
аминокислотная последовательность VH представлена под SEQ ID NO: 55, и аминокислотная последовательность VL представлена под SEQ ID NO: 58; или
аминокислотная последовательность VH представлена под SEQ ID NO: 46, и аминокислотная последовательность VL представлена под SEQ ID NO: 58.
2. Антитело по п. 1, где антитело предусматривает одноцепочечное антитело (scFv), предусматривающее VL-линкер-VH или VH-линкер-VL.
3. Антитело по п. 2, где линкер представляет собой (G4S)n или его вариант, где n представляет собой отличное от нуля натуральное число.
4. Антитело по п. 3, где аминокислотная последовательность линкера представлена под SEQ ID NO: 65, SEQ ID NO: 66 или SEQ ID NO: 67.
5. Антитело по п. 4, где аминокислотная последовательность scFv представлена под SEQ ID NO: 73, SEQ ID NO: 74, SEQ ID NO: 75, SEQ ID NO: 78, SEQ ID NO: 79 или SEQ ID NO: 80.
6. Антитело по п. 5, где антитело дополнительно содержит Fc, при этом Fc связан с scFv посредством шарнира.
7. Антитело по п. 1, где антитело дополнительно содержит константную область.
8. Антитело по п. 7, где константная область представляет собой человеческую константную область, при этом человеческая константная область предусматривает константную область человеческой легкой цепи и константную область человеческой тяжелой цепи.
9. Антитело по п. 8, где константная область человеческой легкой цепи представляет собой константную область человеческой легкой цепи κ, приведенную под SEQ ID NO: 61, или константную область человеческой легкой цепи λ, приведенную под SEQ ID NO: 62.
10. Антитело по п. 9, где константная область человеческой тяжелой цепи представлена изотипами hIgG1, hIgG2, hIgG3, hIgG4 или их вариантом.
11. Антитело по п. 10, где константная область человеческой тяжелой цепи представляет собой константную область тяжелой цепи, приведенной под SEQ ID NO: 63 или SEQ ID NO: 64.
12. Биспецифическое антитело для лечения опухоли с CD3 в качестве мишени, содержащее первую функциональную область белка и вторую функциональную область белка, где первая функциональная область белка предусматривает антитело, нацеливающееся на CD3, по любому из пп. 1-11.
13. Биспецифическое антитело по п. 12, где биспецифическое антитело содержит следующие три цепи: (1) VL1-линкер-VH1-шарнир-CH2-CH3 («выступ») или VH1-линкер-VL1-шарнир-CH2-CH3 («выступ»), относящиеся к первой функциональной области белка, (2) VH2-CH1-шарнир-CH2-CH3 («впадина»), относящуюся ко второй функциональной области белка, и (3) VL2-CL, относящуюся ко второй функциональной области белка; при этом вторая функциональная область белка представляет собой антитело, не нацеливающееся на CD3.
14. Биспецифическое антитело по п. 13, где вторая функциональная область белка представляет собой антитело, нацеливающееся на B7H4, или антитело, нацеливающееся на ROR1.
15. Биспецифическое антитело по п. 13 или 14, где линкер представляет собой (G4S)n, где n представляет собой отличное от нуля натуральное число.
16. Биспецифическое антитело по п. 15, где аминокислотная последовательность линкера представлена под SEQ ID NO: 65, SEQ ID NO: 66 или SEQ ID NO: 67.
17. Биспецифическое антитело по любому из пп. 12-16, где биспецифическое антитело содержит VL1-линкер-VH1-шарнир-CH2-CH3 («выступ»), приведенную под SEQ ID NO: 88, VH2-CH1-шарнир-CH2-CH3 («впадина»), приведенную под SEQ ID NO: 86, и VL2-CL, приведенную под SEQ ID NO: 83, или VL1-линкер-VH1-шарнир-CH2-CH3 («выступ»), приведенную под SEQ ID NO: 88, VH2-CH1-шарнир-CH2-CH3 («впадина»), приведенную под SEQ ID NO: 87, и VL2-CL, приведенную под SEQ ID NO: 85.
18. Выделенная нуклеиновая кислота для получения антитела, кодирующая антитело, нацеливающееся на CD3, по любому из пп. 1-11.
19. Выделенная нуклеиновая кислота для получения антитела, кодирующая биспецифическое антитело по любому из пп. 12-17.
20. Вектор экспрессии для получения антитела, содержащий выделенную нуклеиновую кислоту по п. 18 или 19.
21. Генетически модифицированная клетка для получения антитела, трансфицированная вектором экспрессии по п. 20.
22. Фармацевтическая композиция для лечения опухоли с CD3 в качестве мишени, содержащая антитело, нацеливающееся на CD3, по любому из пп. 1-11, биспецифическое антитело по любому из пп. 12-17, генетически модифицированную клетку по п. 21 и фармацевтически приемлемый носитель.
23. Фармацевтическая композиция по п. 22, где фармацевтическая композиция дополнительно содержит антитело, направленное против иммунной контрольной точки.
24. Применение антитела, нацеливающегося на CD3, по любому из пп. 1-11, биспецифического антитела по любому из пп. 12-17, выделенной нуклеиновой кислоты по п. 18 или 19, вектора экспрессии по п. 20, генетически модифицированной клетки по п. 21 или фармацевтической композиции по п. 22 или 23 в изготовлении лекарственного препарата для лечения опухоли с CD3 в качестве мишени.
| WO 2016071355 A1, 12.05.2016 | |||
| WO 2017136659 A2, 10.08.2017 | |||
| WO 2019075405 A1, 18.04.2019 | |||
| WO 2015001085 A1, 08.01.2015 | |||
| EP 3492591 A1, 05.06.2019 | |||
| WO 2019008379 A1, 10.01.2019 | |||
| CD19XCD3-СПЕЦИФИЧЕСКИЕ ПОЛИПЕПТИДЫ И ИХ ПРИМЕНЕНИЕ | 1999 |
|
RU2228202C2 |
| RU 2016132863 A, 20.02.2018. | |||

