Изобретение относится к биотехнологии, в частности к способу получения пептидов в прокариотах. Оно может быть использовано для получения анальгетического пептида HCRG21 биотехнологическим путем.
HCRG21 - это пептид актинии Heteractis crispa. Ранее было показано, что рекомбинантный аналог HCRG21 обладает пролонгированным анальгетическим действием и может найти применение в качестве активной фармакологической субстанции для изготовления анальгетического лекарственного средства, предназначенного для облегчения острых болевых состояний и боли при хронических патологических процессах, где нарушена нормальная функциональная активность и задействована повышенная активность ионного канала TRPV1. Препарат снимает воспаление, эффективно понижает температуру тела животных и человека [RU 2755206, С1, 14.09.2021]. Для пептида определена основная биологическая мишень - ионный канал TRPV1, который характеризуется своей способностью селективно активироваться капсаицином (жгучий компонент перца), реагировать на изменение значения pH внеклеточной среды, активироваться при превышении критического уровня нагревания (более 43°С) и распознавать множество различных химических молекул. Ингибирование активности TRPV1 в животных тестах приводит к анальгетическому эффекту [Гладких И.Н. и др. Успехи биологической химии, 2021, Т. 61, С. 107-154]. Активность этого пептида как ингибитора ионного канала TRPV1 была определена путем электрофизиологических экспериментов, где было получено значение половинного эффекта ингибирования (IC50) 6,9 мкМ для индуцированного капсаицином тока на ооцитах Xenopus laevis [Monastyrnaya M. et al. Marine Drugs 2016, 14, 229-234].
HCRG21 представляет собой 56-членный полипептид (SEQ ID NO: 1):
RGICSEPKVVGPCTAYFRRFYFDSETGKCTPFIYGGCEGNGNNFETLRACRAICRА
Известен гомологичный анальгетический пептид АРНС3, который можно получить из природного источника [S.A. Kozlov et al. Russian Journal of Bioorganic Chemistry, 2009, vol. 35, No. 6, pp. 711-719], экспрессией в бактериальной системе в составе гибридного белка с тиоредоксином [Yaroslav A. Andreev et al. Marine Drugs. 2013 Dec; 11(12): 5100-5115] или в составе гибридного белка DnaBAPHC3 в виде телец включения, содержащего, наряду с АРНС3, последовательность мини-интеина Ssp DnaB [RU 2619170 C2, 12.05.2017]. Недостатки этих методов заключаются в том, что перенос способов получения одного пептида на другой гомологичный пептид не могут дать гарантировано подобный результат из-за индивидуальных особенностей выделения (из природного сырья) или биосинтеза молекулы внутри клеток продуцентов (биотехнологический синтез).
Другой способ, представленный в патенте RU 2571942 С2, 27.12.2015, позволяет получить рекомбинантный пептидный анальгетик РТ1 с высоким выходом, который обладает модулирующим действием на участвующие в генерации болевого сигнала клеточные рецепторы и может найти применение в медицине и научных исследованиях. Однако этот пептид имеет отличную структуру и биологическую клеточную мишень, а значит, он сам и способ его получения не могут быть использованы для синтеза пептида HCRG21.
Прототип, наиболее близкий к заявляемому для патентования способу получения HCRG21, описан в работе [Monastyrnaya M. et al. Marine Drugs 2016, 14, 229-234], где пептид получали в Escherichia coli (штамм BL21(DE3)) в составе гибридного белка, содержащего целевой пептид HCRG21, фрагмент тиоредоксина и интегрированную аффинную метку - последовательность из шести остатков гистидина.
Недостаток данного метода заключается в необходимости использовать для отделения активного пептида от белка-партнера процедуру гидролиза с применением бромциана, расщепляющего гибридный белок по остатку метионина, дополнительно введенного перед первым N-концевым остатком целевого пептида. Использование в схеме синтеза ядовитых веществ, таких как бромциан, резко снижает круг задач, где можно применять полученную таким способом субстанцию. К тому же выход целевого пептида данным способом производства оказался пониженным - 5,5 мг/л клеточной культуры.
Изобретение решает задачу получения высокопродуктивного бактериального штамма-продуцента, позволяющего получать рекомбинантный анальгетический пептид HCRG21 с высоким выходом и по упрощенной технологии.
Настоящее изобретение относится к гибридному белку SMT3-HCRG21 для получения полипептида HCRG21, имеющему последовательность, состоящую из 176 аминокислотных остатков (SEQ ID NO: 2):
MVKYHHHHHHGSGLVPRGSASMSDSEVNQEAKPEVKPEVKPETHINLKVSDGSSEIFFKIKKTTPLRRLMEAFAKRQGKEMDSLRFLYDGIRIQADQTPEDLDMEDNDIIEAHREQIGGRGICSEPKVVGPCTAYFRRFYFDSETGKCTPFIYGGCEGNGNNFETLRACRAICRA,
и содержащий гексагистидиновую метку (HS), последовательность SMT3, сайт распознавания Ulp1-протеазы и полипептид HCRG21, имеющий аминокислотную последовательность SEQ ID NO: 1.
Настоящее изобретение также относится к рекомбинантной плазмидной ДНК pSMT3_HCRG21 для экспрессии указанного гибридного белка SMT3-HCRG21, имеющей длину 4355 п.о., состоящей из следующих ключевых генетических элементов:
- промотора и терминатора траснкрипции Т7-РНК-полимеразы;
- синтетической нуклеотидной последовательности (SEQ ID NO: 3):
CATATGGTGAAGTATCACCACCATCATCACCATGGCTCAGGCTTAGTTCCTCGTGGCAGTGCCTCCATGAGCGATAGTGAAGTCAACCAAGAAGCGAAACCGGAAGTGAAACCTGAAGTGAAACCGGAGACCCACATCAACCTGAAAGTGAGTGATGGCAGTTCTGAGATCTTCTTCAAGATCAAGAAGACCACTCCGTTACGTCGTCTGATGGAGGCGTTTGCGAAACGCCAAGGCAAAGAAATGGATAGTCTTCGCTTTCTGTATGATGGCATTCGCATTCAAGCTGATCAGACTCCAGAAGATCTGGACATGGAAGACAACGACATCATTGAAGCACATCGTGAACAGATTGGTGGTCGTGGCATCTGCAGCGAACCGAAAGTGGTTGGTCCGTGCACTGCGTACTTTCGTCGCTTCTACTTTGACTCTGAGACTGGCAAGTGCACACCGTTCATCTATGGTGGCTGTGAAGGGAATGGCAACAACTTTGAGACCTTACGTGCGTGTCGTGCGATCTGTCGTGCCTAACTCGAG,
кодирующую гибридный белок согласно настоящему изобретению SMT3-HCRG21, имеющий аминокислотную последовательность SEQ ID NO: 2;
-гена устойчивости к антибиотику канамицину (KanR) для проведения отбора рекомбинантных клеток;
-участка инициации репликации бактериофага f1 (Оri f1).
Настоящее изобретение также относится к штамму Escherichia coli BL21(DE3)/pSMT3_HCRG21, продуцирующему гибридный белок SMT3-HCRG21 (SEQ ID NO: 2), содержащему указанную рекомбинантую плазмиду ДНК pSMT3_HCRG21. Согласно настоящему изобретению штамм Escherichia coli BL21(DE3)/pSMT3_HCRG21 получают путём трансформации клеток штамма Escherichia coli BL 21(DE3) рекомбинантной плазмидой согласно настоящему изобретению.
Предлагаемое изобретение также относится к способу получения полипептида HCRG21, имеющего аминокислотную последовательность SEQ ID NO: 1, включающему стадии:
a) культивирования штамма-продуцента Escherichia coli BL21(DE3)/pSMT3_HCRG21 согласно настоящему изобретению с получением биомассы клеток штамма-продуцента, продуцирующего гибридный белок SMT3-HCRG21 согласно настоящему изобретению;
б) выделения гибридного белка SMT3-HCRG21 согласно настоящему изобретению из биомассы клеток штамма-продуцента, полученных на стадии а);
c) ферментативного расщепления гибридного белка SMT3-HCRG21, полученного на стадии б), с образованием полипептида HCRG21 с аминокислотной последовательностью SEQ ID NO: 1;
д) очистки полипептида HCRG21, полученного на стадии c).
Техническим результатом является образование HCRG21 с последовательностью SEQ ID NO: 1, выход которого в 2 раза превышает выход по способу-прототипу, и достигает 10,2 мг/литр культуры при чистоте препарата 97%, который имеет замкнутые дисульфидные связи по данным масс-спектрометрии и способен ингибировать капсаицин-вызванные токи через ионный канал TRPV1 на модельных клетках. Кроме того, использование в качестве белка партнера фрагмента N-концевой последовательности SMT3, которая распознается и выщепляется специфической протеазой дрожжей Ulp1, позволяет избежать использование в схеме синтеза ядовитого бромциана, что расширяет круг задач, где можно применять полученную таким способом субстанцию.
В составе гибридного белка был использован фрагмент N-концевой последовательности SMT3 для того, чтобы избежать трудностей, связанных с расщеплением рекомбинантного гибридного белка с помощью различных протеаз, таких как TEV, энтерокиназа, фактор X и др., а также с целью удешевления этой стадии. Эта последовательность распознается и выщепляется специфической протеазой дрожжей Ulp1, которая специфична к белкам SMT3 и SUMO-1 [J. Li and M. Hochstrasser, Nature, 1999, 398(6724), 246-251]. Этот подход для синтеза пептидов был уже опробован ранее. Разработчики из НИИ биохимии ФИЦ ФТМ предложили способ получения зрелого аполипопротеина Α-I человека, который является основным белком плазмы крови, участвующим в образовании липопротеинов высокой плотности [RU 2605326 С1, 20.12.2016]. Для получения зрелого белка апоА-I человека был использован рекомбинантный химерный белок SUMO3-apoA-I, включающий с N-конца аминокислотную последовательность убиквитин-подобного белка SUMO3 человека, слитую с С-конца с аминокислотной последовательностью зрелого белка апоА-I человека. В заявке CN110305887A описан слитый белок SUMO-дросомицин и способ его получения. Противогрибковый пептид имеет широкие перспективы применения при приготовлении антибактериальных препаратов, безопасных пищевых продуктов и косметики, а также имеет высокую рыночную стоимость.
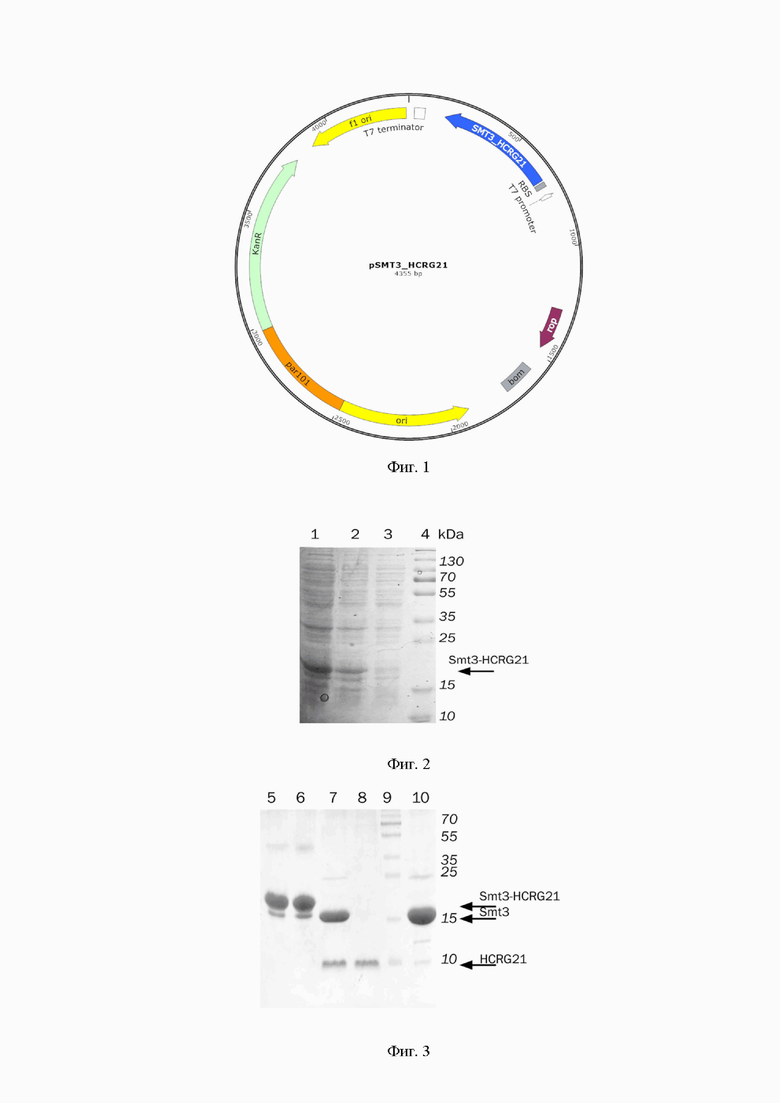
Для заявляемого способа синтезируют ген SMT3-HCRG21 (SEQ ID NO: 2) с учетом оптимизации кодонов для прокариот, далее используют в качестве матрицы плазмиду на основе векторов pET 23a(+) и pET 28a(+), в которую встраивают ген интереса. Полученной экспрессионной плазмидой pSMT3_HCRG21, строение которой приведено на фиг. 1, трансформируют клетки Е.coli BL21 (DE3).
Штамм-продуцент E.coli BL21 (DE3)/pSMT3_HCRG21 выращивают в богатой среде LB, содержащей 50 мг/л канамицина и 0,5-5 г/л глюкозы до достижения максимальной плотности культуры. Индукцию биосинтеза гибридного белка осуществляют лактозой, после чего через час ферментации собирают осадок, отделяют нерастворимый клеточный осадок, ренатурируют гибридный белок и выделяют целевой пептид HCRG21 хроматографически после гидролиза протеазой.
Изобретение иллюстрируют фигуры.
Фиг. 1. Физическая карта плазмиды pSMT3_HCRG21.
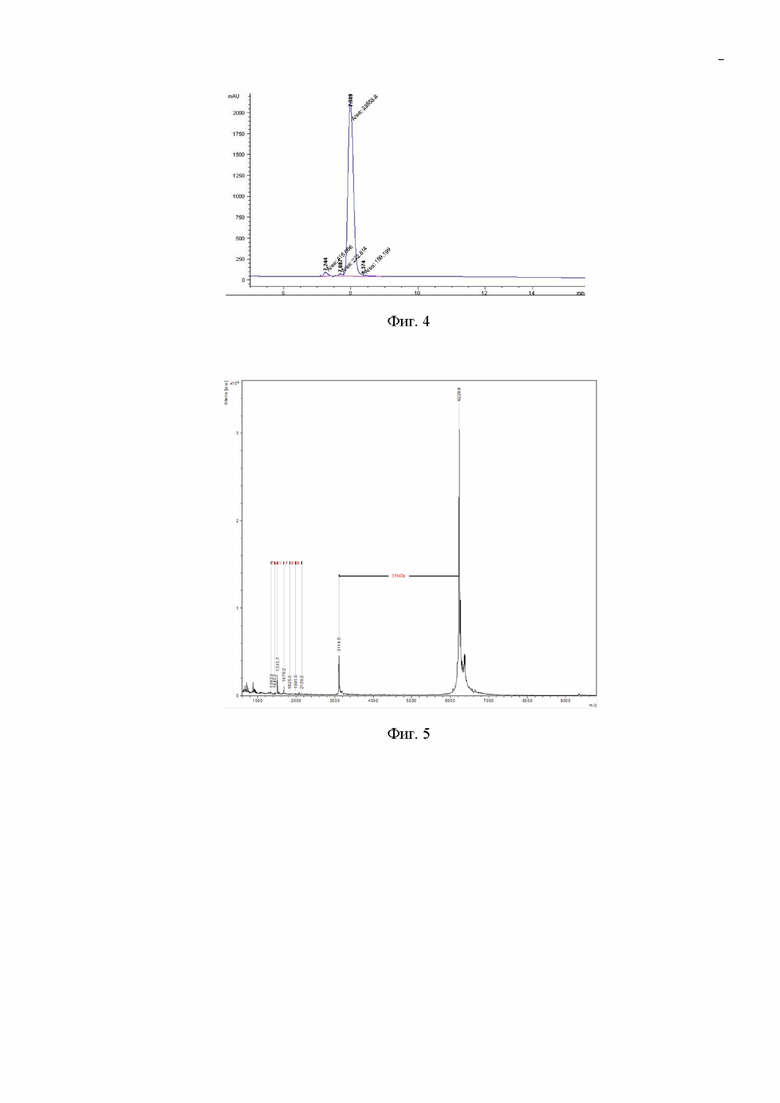
Фиг. 2. Электрофореграмма лизата клеток штамма-продуцента после разрушения биомассы E.coli Bl21(DE3)/pSMT3_HCRG21 в 15% ПААГ. Дорожка 1 - тотальный клеточный лизат; дорожка 2 - клеточный осадок; дорожка 3 - осветленный клеточный лизат; дорожка 4 - маркеры молекулярных масс. Стрелкой указано положение полосы, соответствующее по ожидаемой молекулярной массе гибридному белку SMT3-HCRG21.
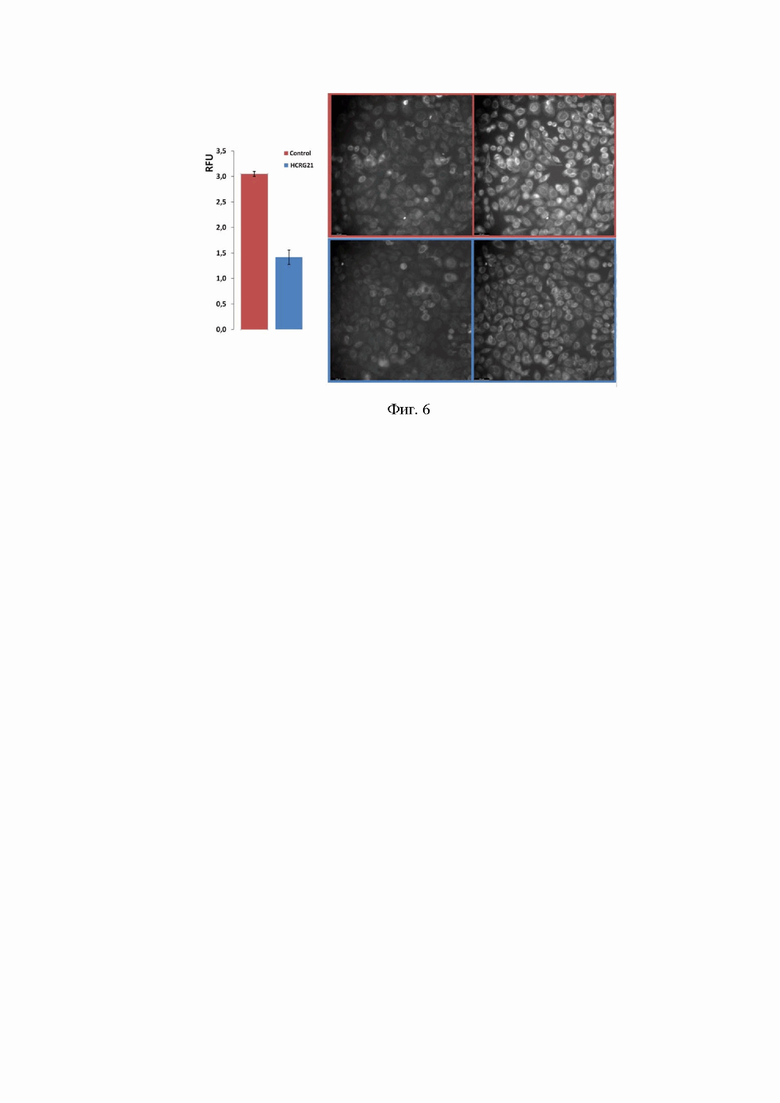
Фиг. 3 Электрофореграмма белков в 12,5% ПААГ. Дорожка 5 - SMT3-HCRG21 до ренатурации, дорожка 6 - SMT3-HCRG21 после ренатурации, дорожка 7 - гидролиз гибридного белка протеазой Ulp-1 (3 часа при комнатной температуре), дорожка 8 - целевой продукт после очистки аффинной хроматографией, дорожка 9 - маркеры молекулярных масс, дорожка 10 - фракция, удерживаемая на второй аффинной хроматографии. Стрелками указаны положения полос, соответствующие по ожидаемой молекулярной массе гибридному белку SMT3-HCRG21, N-концевому фрагменту SMT3 и целевому пептиду HCRG21.
Фиг. 4. Профиль аналитической ВЭЖХ рекомбинантного HCRG21, полученного после очистки методом ОФ-ВЭЖХ. Разделение на колонке VDSpher PUR 100 C18-E (VDS, Германия), 250 x 4,6 мм на скорости 1 мл/мин проводят в градиенте концентрации ацетонитрила от 5 до 90% в 0,1% растворе трифторуксусной кислоты. Расчет площади пиков проводят при длине волны 214 нм.
Фиг. 5. Масс-спектрометрический анализ целевого пептида.
Фиг. 6. Анализ ингибирующей способности целевого пептида методом флуоресцентного имиджинга. Оценивают изменение видимой флуоресценции клеток, экспрессирующих ионный канал TRPV1, после их активации селективным агонистом капсаицином в концентрации 1 мкМ, как разницу между светимостью до добавления агониста (слева) и через 20 секунд после добавления агониста (справа). За 5 минут до начала измерения в клетки добавляют 20 мкл воды (контрольный образец сверху) или 20 мкл раствора HCRG21 до финальной концентрации 5 мкМ (образцы снизу). Расчет относительной флуоресценции по трем независимым лункам проводят по формуле F/F0-1.
Изобретение иллюстрируется примерами.
Пример 1
Получение штамма-продуцента E. coli Bl21(DE3)/pSMT3_HCRG21
Штамм-продуцент Е. coli BL21 (DE3)/pSMT3_HCRG21 получают трансформацией компетентных клеток Е. coli BL21 (DE3) плазмидным вектором pSMT3_HCRG21. К 100 мкл химически компетентных клеток E. coli BL21 (DE3) добавляют 1,5 мкл плазмиды и помещают в лед на 30 минут. Далее клетки помещают в термостат на 45 сек при температуре 42°С. После этого снова помещают клетки в лед на 5 минут. Затем к клеткам добавляют 300 мкл среды SOC и культивируют в течение 1 часа на шейкере (перемешивание 225 об/мин) при 37°С. Содержимое пробирок высеивают на чашки Петри с LB-агаром, содержащим селективный антибиотик канамицин (50 мг/л) и инкубируют в течение 14-16 ч при 37°С. Полученные колонии отбирают для получения маточной культуры и далее нарабатывают культуру в 15 мл пробирках с питательной средой LB, содержащей 50 мг/л канамицина и 0,5% глюкозы (качалка 190 об/мин, температура 37°С, 15-16 часов). Полученные культуры пересевают 2,5%об. в пробирки со свежей питательной средой LB, содержащей 50 мг/л канамицина и 0,5% глюкозы и культивируют до оптической плотности 0,6-0,7 единиц (качалка 190 об/мин, температура 37°С). Далее добавляют глицерин до 15%об и полученный исследовательский банк (музейные культуры) хранят при -70°С.
Пример 2
Получение гибридного белка SMT3-HCRG21
Для приготовления инокулята используют музейную культуру. Инокулируют 0,1% музейной культуры в среде LB, содержащей 50 мг/л канамицина и 5 г/л глюкозы. Через 16 часов инкубации (при 32°С, качалка 180 об/мин) отсутствие посторонней микрофлоры в инокуляте подтверждают микроскопией. Выращивание клеток-продуцентов проводят в 30 литровом ферментере в 20 литрах среды LB при 37°С. Каждый час контролируют культуральную жидкость на значение pH, количество кислорода, оптическую плотность при 600 нм, и концентрацию глюкозы. По достижению OD600 равного 12 понижают расход воздуха c 25 до 15%, вводят в ферментер 0,3 г лактозы и продолжают ферментацию 60 минут при 37°С. Далее собирают влажную биомассу (1 кг) после центрифугирования культуральной среды при 10000 g в течение 20 минут при 10°С.
Полученную биомассу ресуспендируют в 50 мМ Трис-HCl, 20 мМ ЭДТА, pH 8,0 буфере из расчета 10 мл буфера на 1 г биомассы, после чего разрушают на гомогенизаторе. Полученный клеточный лизат центрифугируют при 10000 g в течение 20 минут при 10°С. Полученный клеточный осадок (70 г) повторно ресуспендируют в буфере 50 мМ Трис-HCl, pH 8.0, перемешивают в течение 20 минут и переосаждают при 10000 g в течение 20 минут при 10°С.
Из полученной биомассы, нерастворимого клеточного осадка и осветленного клеточного лизата отбирают пробу в количестве 30 мкл и с красителем бромфеноловым синим нагревают 3 мин при 100°С. Пробы по 4 мкл используют для электрофореза в 15% ДСН-ПААГ. Гель прокрашивают кумасси R-250 по стандартной методике и сканируют с помощью денситометра Shimadzu CS-930 (фиг. 2).
Нерастворимый клеточный осадок промывают 10-20 объемами буфера (50 мМ Трис-HCl, pH 8,0) и растворяют в буфере, содержащем 50 мМ Трис-HCl, pH 8,0 с 8 М мочевиной из расчета 10 мл буфера на 1 г осадка. Экстракцию проводят в течение 30 минут при перемешивании на магнитной мешалке, далее собирают супернатант после центрифугирования при 10000 g в течение 20 минут при 10°С.
Для проведения металл-хелатной хроматографии используют сорбент Ni IMAC SepFast (BioToolomics, England), который предварительно активируют двумя колоночными объемами 0,15 М раствора NiSO4. Рабочую скорость процесса задают 17 мл/мин. После нанесения супернатанта колонку промывают буферным раствором, содержащим 50 мМ Трис-HCl, pH 8,0 и 8 М мочевину, до выхода на плато показателей УФ-поглощения при длине волны 280 нм и проводимости. Элюцию гибридного белка осуществляют линейным градиентом 0-100% буферного раствора, содержащего 50 мМ Трис-HCl, pH 8,0, 8 М мочевину и 500 мМ имидазол, за 120 минут.
Далее проводят очистку SMT3-HCRG21 при помощи анионообменной хроматографии на колонке XK 50 с 200 мл сорбента DEAE Sepharose (GE Healthcare), используя хроматографическую систему Akta Pure. Рабочую скорость процесса задают 17 мл/мин. После нанесения разделяемой фракции колонку промывают буферным раствором, содержащим 50 мМ Трис-HCl, pH 8,0 и 2 М мочевину, до выхода на плато показателей УФ-поглощения при длине волны 280 нм и проводимости. Элюцию осуществляют линейным градиентом 0-100% буферного раствора, содержащего 50 мМ Трис-HCl, pH 8,0, 2 М мочевину и 1 М NaCl, за 80 минут. Детекцию осуществляют по поглощению при длине волны 280 нм. Сбор фракций начинают при достижении показателя УФ-поглощения уровня выше 50 mAU. Фракции анализируют методом белкового электрофореза в ПААГ и те из них, которые содержат более 90% целевого гибридного белка, объединяют для дальнейшей работы.
Пример 3
Получение пептида HCRG21
Очищенный гибридный белок разводят в два раза добавлением равного объёма буферного раствора (50 мМ Трис-HCl, рН 8,5) и оставляют для ренатурации в течение ночи при 4°C и перемешивании (скорость 100 об/мин). После этого к ренатуранту SMT3-HCRG21 добавляют суспензию лиофилизированной протеазы Ulp-1 (1 мг фермента на 100 мг белка) и 1 М раствор дитиотриетола до конечной концентрации 1 мМ. Гидролиз проводят не менее 3 часов при перемешивании при комнатной температуре, после чего реакционную смесь наносят на колонку для проведения металл-хелатной хроматографии.
Для проведения металл-хелатной хроматографии используют сорбент Ni IMAC SepFast (BioToolomics, England), который предварительно активируют двумя колоночными объемами 0,15 М раствора NiSO4. Рабочую скорость процесса задают 17 мл/мин. После нанесения гидролизата, колонку промывают буферным раствором, содержащим 50 мМ Трис-HCl, pH 8,0, и 8 М мочевину, и собирают несвязавшуюся с носителем фракцию до выхода на плато показателей УФ-поглощения при длине волны 280 нм. Элюцию остальных продуктов гидролиза осуществляют линейным градиентом 0-100% буферного раствора, содержащего 50 мМ Трис-HCl, pH 8,0, 8 М мочевину и 500 мМ имидазол, за 20 минут.
На каждой стадии получения отбирают аликвоты растворов в количестве 30 мкл, которые в конце процесса выделения нагревают 3 мин при 100°С с красителем бромфеноловым синим. Пробы по 4 мкл используют для электрофореза в 12,5% ДНС-ПААГ. Гель прокрашивают кумасси R-250 по стандартной методике и сканируют с помощью денситометра Shimadzu CS-930 (фиг. 3).
Для окончательной очистки используют носитель с обращенной фазой ODS-A (YMC, Япония), упакованный в колонку диаметром 1 см, и хроматографическую систему Akta Pure. В качестве мобильной фазы используют раствор А: 0,1% уксусной кислоты, 2% этанола и раствор Б: 0,1% уксусной кислоты, 75% этанола. Колонку уравновешивают раствором А и наносят несвязавшуюся фракцию после второй аффинной хроматографии, после чего колонку промывают раствором А до тех пор, пока УФ-поглощение при длине волны 280 нм и проводимость не достигнут плато. Элюирование проводят в линейном градиенте 0-100% раствора Б в течение 50 минут при скорости потока 2 мл/мин. Собранные фракции анализируют с помощью аналитической ВЭЖХ для определения количественного состава примесей относительно целевого продукта (фиг. 4). Выход целевого продукта составляет 10,2 мг/л клеточной культуры. Фракции с чистотой не менее 97% объединяют и лиофилизируют. Молекулярную массу чистого пептида определяют методом МАЛДИ масс-спектрометрии (фиг. 5).
Пример 4
Тестирование активности HCRG21
Клетки CHO (яичник китайского хомячка), стабильно экспрессирующие ионный канал TRPV1 крысы, высевают в 96-луночный планшет с черными стенками и прозрачным дном и растят при 37°C и 5% CO2 до плотности 40000 клеток на лунку, используя полную среду без антибиотиков. Для индукции экспрессии каналов TRPV1 в лунки вносят тетрациклин до конечной концентрации 1 мкг/мл за 14 часов до начала измерения. Затем клетки загружают флуоресцентным красителем, добавляя по 20 мкл Voltage Sensitive Dye (Molecular Devices, San Jose, USA) (Cat # R8126) на лунку в соответствии с протоколом, разработанным производителем. Через 30 минут инкубации (темнота, 37°C и 5% CO2) в лунки добавляют по 20 мкл испытуемого раствора, для контрольных клеток добавляют 20 мкл воды. Через 5 минут инкубации проводят измерение флуоресценции в видимом поле инвертированного микроскопа Leica DM IL, оснащенного G/R фильтром cube: EX: 225/-450 | DC: 495 595 | EM: 250/-500, производя запись изображения видеокамерой в течение 60 секунд через 40-кратный объектив. Через 5 секунд после начала записи в лунку вносят раствор капсаицина до конечной концентрации 1 мкМ. Относительную флуоресценцию рассчитывают для всего изображения после вычитания фрагмента фона, сравнивая интенсивность сигнала на изображении через 20 секунд после добавления капсаицина (F) с интенсивностью сигнала на изображении до добавления капсаицина (F0) по формуле F/F0-1. Используют три повтора для пептида и три повтора для воды, после чего рассчитывают среднее значение и стандартное отклонение. Определяют, что HCRG21 в концентрации 5 мкМ ингибирует относительную флуоресценцию клеток на 50%, что аналогично ранее опубликованной активности для этого пептида, измеренной для капсаицин активированных токов на ооцитах лягушки.
| название | год | авторы | номер документа |
|---|---|---|---|
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК, КОДИРУЮЩАЯ ГИБРИДНЫЙ БЕЛОК L-HEP, ШТАММ ESCHERICHIA COLI ПРОДУЦЕНТ УКАЗАННОГО БЕЛКА И СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО БЕЛКА | 2019 |
|
RU2716975C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК PTRX-TEVRS-РТН, КОДИРУЮЩАЯ ГИБРИДНЫЙ БЕЛОК, СПОСОБНЫЙ К ПРОТЕОЛИТИЧЕСКОМУ РАСЩЕПЛЕНИЮ С ОБРАЗОВАНИЕМ ФРАГМЕНТА ЭНДОГЕННОГО ЧЕЛОВЕЧЕСКОГО ПАРАТИРЕОИДНОГО ГОРМОНА (1-34), ШТАММ ESCHERICHIA COLI BL21(DE3)/PTRX-TEVRS-РТН - ПРОДУЦЕНТ УКАЗАННОГО БЕЛКА И СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО PTH (1-34) | 2019 |
|
RU2700452C1 |
| СИСТЕМА ЭКСПРЕССИИ И СПОСОБ ПОЛУЧЕНИЯ НЕМОДИФИЦИРОВАННЫХ РЕКОМБИНАНТНЫХ БЕЛКОВ В Escherichia coli С ЕЁ ИСПОЛЬЗОВАНИЕМ | 2015 |
|
RU2604796C1 |
| Способ получения пептида, модулирующего активность пуринергических рецепторов | 2019 |
|
RU2714114C1 |
| Рекомбинантная плазмидная ДНК pER-APHC3, кодирующая гибридный белок, способный к автокаталитическому расщеплению с образованием APHC3, штамм Escherichia coli C3030/pER-APHC3 продуцент указанных белков и способ получения рекомбинантного APCH3 | 2015 |
|
RU2619170C2 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pEst877, ДЕТЕРМИНИРУЮЩАЯ ЭКСПРЕССИЮ ПОЛИПЕПТИДА С АКТИВНОСТЬЮ ЭСТЕРАЗЫ Psychrobacter cryohalolentis К5 НА ПОВЕРХНОСТИ КЛЕТОК ESCHERICHIA COLI, И ШТАММ БАКТЕРИЙ Escherichia coli BL21(DE3)pLysS/pEst877-ПРОДУЦЕНТ ПОЛИПЕПТИДА С АКТИВНОСТЬЮ ЭСТЕРАЗЫ Psychrobacter cryohalolentis К5 НА ПОВЕРХНОСТИ КЛЕТОК | 2013 |
|
RU2526213C1 |
| Рекомбинантный белок, обладающий протективным действием в отношении менингококков (варианты), полинуклеотид, кодирующий рекомбинантный белок, рекомбинантная плазмидная ДНК, содержащая указанный полинуклеотид, клетка-хозяин, содержащая указанную рекомбинантную плазмидную ДНК, способ получения рекомбинантного белка | 2017 |
|
RU2701964C2 |
| ПЛАЗМИДА ДЛЯ ЭКСПРЕССИИ В КЛЕТКАХ БАКТЕРИИ, ПРИНАДЛЕЖАЩЕЙ К РОДУ Escherichia, НЕАКТИВНОГО ПРЕДШЕСТВЕННИКА ДНКазы I ЧЕЛОВЕКА ИЛИ ЕЕ МУТЕИНОВ, БАКТЕРИЯ, ПРИНАДЛЕЖАЩАЯ К РОДУ Escherichia, - ПРОДУЦЕНТ НЕАКТИВНОГО ПРЕДШЕСТВЕННИКА РЕКОМБИНАНТНОЙ ДНКазы I ЧЕЛОВЕКА ИЛИ ЕЕ МУТЕИНА, ПРЕДШЕСТВЕННИК РЕКОМБИНАНТНОЙ ДНКазы I ЧЕЛОВЕКА ИЛИ ЕЕ МУТЕИНА, СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОЙ ДНКазы I ЧЕЛОВЕКА ИЛИ ЕЕ МУТЕИНА, СПОСОБ ПОЛУЧЕНИЯ КОНЪЮГАТОВ ПОЛИЭТИЛЕНГЛИКОЛЯ И РЕКОМБИНАНТНОГО МУТЕИНА ДНКазы I ЧЕЛОВЕКА, ФЕРМЕНТАТИВНО АКТИВНЫЙ КОНЪЮГАТ МУТЕИНА РЕКОМБИНАНТНОЙ ДНКазы I ЧЕЛОВЕКА | 2011 |
|
RU2502803C2 |
| СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО АНАЛЬГЕТИЧЕСКОГО ПЕПТИДА | 2013 |
|
RU2571942C2 |
| Рекомбинантный белок, связывающийся с RBD S-белка SARS-CoV-2 | 2022 |
|
RU2778942C1 |
Изобретение относится к биотехнологии, в частности к получению анальгетических пептидов в прокариотах, и может быть использовано в медицине. Пептид HCRG21 получают путем трансформации компетентных клеток Escherichia coli BL21(DE3) рекомбинантной плазмидной ДНК pSMT3_HCRG21, обеспечивающей синтез гибридного белка SMT3_HCRG21, содержащего целевой пептид, в клетках Escherichia coli. Настоящее изобретение раскрывает штамм Escherichia coli BL21(DE3)/pSMT3_HCRG21, продуцирующий указанный гибридный белок, а также способ получения указанного рекомбинантного пептида HCRG21, предусматривающий культивирование указанного штамма Escherichia coli BL21(DE3)/pSMT3_HCRG21. Технический результат - образование HCRG21, который имеет замкнутые дисульфидные связи по данным масс-спектрометрии и способен ингибировать капсаицин-вызванные токи через ионный канал TRPV1 на модельных клетках. 3 н.п. ф-лы, 6 ил., 4 пр.
1. Рекомбинантная плазмидная ДНК pSMT3_HCRG21, 4355 п. о., для экспрессии гибридного белка SMT3-HCRG21 с аминокислотной последовательностью SEQ ID NO: 2, содержащего пептид HCRG21, имеющий аминокислотную последовательность SEQ ID NO: 1, состоящая из фрагментов ДНК плазмид рЕТ 23а(+) и рЕТ 28а(+), содержащих промотор и терминатор транскрипции Т7-РНК-полимеразы, синтетическую нуклеотидную последовательность SEQ ID NO: 3, кодирующую гибридный белок SMT3-HCRG21, ген устойчивости к антибиотику канамицину для проведения отбора рекомбинантных клеток и участок инициации репликации бактериофага f1.
2. Штамм бактерий Escherichia coli BL21(DE3)/pSMT3_HCRG21 - продуцент анальгетического пептида HCRG21, полученный путем трансформации компетентных клеток Escherichia coli BL21(DE3) рекомбинантной плазмидной ДНК pSMT3_HCRG21 по п. 1.
3. Способ получения рекомбинантного пептида HCRG21, имеющего аминокислотную последовательность SEQ ID NO: 1, включающий трансформацию штамма Escherichia coli BL21(DE3) полученной плазмидной ДHK pSMT3_HCRG21 по п. 1, культивирование полученного штамма-продуцента Escherichia coli BL21(DE3)/pSMT3_HCRG21 по п. 2, выделение гибридного белка из нерастворимого клеточного осадка после разрушения клеток на гомогенизаторе путем экстракции гибридного белка в буферном растворе, содержащем 8 М мочевину, с последующей металл-хелатной и анионообменной хроматографией, дальнейший гидролиз ренатурированного гибридного белка протеазой Ulp-1 и выделение целевого пептида HCRG21 с помощью металл-хелатной и обращеннофазной хроматографии.
| GERON M | |||
| et al | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Разборный с внутренней печью кипятильник | 1922 |
|
SU9A1 |
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| SINTSOVA O | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Разборный с внутренней печью кипятильник | 1922 |
|
SU9A1 |
| Способ восстановления хромовой кислоты, в частности для получения хромовых квасцов | 1921 |
|
SU7A1 |
| СИНЦОВА О | |||
