Изобретение относится к области медицинской биотехнологии, а именно к пептидам, обладающим антибактериальной активностью, и может быть использовано в клинической медицине и ветеринарии.
В последние десятилетия во всем мире наблюдается быстрое распространение штаммов патогенных бактерий, обладающих множественной лекарственной устойчивостью (МЛУ). [Antimicrobial Resistance Collaborators. Global burden of bacterial antimicrobial resistance in 2019: a systematic analysis. Lancet. 2022. Vol. 399. P. 629-655. doi: 10.1016/SO140-6736(21)02724-0.].
Мутации, обуславливающие лекарственную устойчивость, включают в себя модификацию мишени антибиотика, уменьшение проникновения антибиотика в клетку, активацию механизмов удаления антибиотика из клетки, изменения метаболизма бактерий, ферментативное разрушение антибиотика. Например, у грамотрицательных бактерий основным механизмом резистентности к β-лактамным антибиотикам является их разрушение β-лактамазами, в то время, как у грамположительных бактерий - модификация пенициллин-связывающих белков.
Гены, определяющие резистентность, легко передаются между клетками бактерий с помощью мобильных генетических элементов (плазмид, транспозонов, умеренных бактериофагов) и в результате накопления нескольких таких генов в одной бактерии возникают штаммы патогенов, резистентные одновременно к нескольким антибиотикам, называемые штаммами с множественной, лекарственной устойчивостью (МЛУ) [Munita J. М., Arias С.A. Mechanisms of Antibiotic Resistance // Microbiol. Spectr. 2016. Vol. 4. P. 10.1128/microbiolspec.VMBF-0016-2015. doi: 10.1128/microbiolspec.VMBF-0016-2015.].
В частности, известны МЛУ-штаммы Klebsiella pneumonia, Acinetobacter baumanii и др., устойчивые к цефалоспрорину, карбапенемам, полимиксинам [Tang М., Kong X., Нао J., et al. Epidemiological Characteristics and Formation Mechanisms of Multidrug-Resistant Hypervirulent Klebsiella pneumonia // Front Microbiol. 2020. Vol. 11. P. 581543. doi: 10.3389/fmicb.2020.581543.; Ibrahim S., Al-Saryi N., Al-Kadmy I.M.S., et al. Multidrug-resistant Acinetobacter baumannii as an emerging concern in hospitals // Mol. Biol. Rep.2021.Vol. 48. P. 6987-6998. doi: 10.1007/sl 1033-021-06690-6.; Matson H.H., Jones B.M., Wagner J.L., et al. Growing resistance in Stenotrophomonas maltophilia? //Am. J. Health Syst. Pharm. 2019. Vol. 76. P. 2004-2005. doi: 10.1093/ajhp/zxz247.]. Борьба с МЛУ-патогенами является серьезной проблемой для клинической медицины.
Для подавления роста патогенных бактерий представляется перспективным использование антимикробных пептидов [Pini A., Falciani С, Mantengoli Е., et al. A novel tetrabranched antimicrobial peptide that neutralizes bacterial lipopolysaccharide and prevents septic shock in vivo // FASEB J. 2010. Vol. 24. P. 1015-1022. doi: 10.1096/fj09-145474.].
Например, известен искусственный (не встречающийся в природе) пептид RFRRFRLPFPvRPPIRIH-NH2, подавляющий рост ряда патогенных бактерий, в том числе МЛУ-штаммов Acinetobacter baumannii и Klebsiella pneumoniae, при этом минимальная ингибирующая рост данных штаммов концентрация пептида (МИК50%) составила 8 мкМ [RU 2771493, дата регистрации 05.05.2022].
Целью изобретения является расширение спектра антимикробных пептидов, подавляющих рост патогенных бактерий.
Для достижения указанной цели разработаны новые антимикробные пептиды мицецилин и рицецидин по настоящему изобретению. Данные пептиды, в частности, подавляют рост патогенных микроорганизмов, в том числе МЛУ-штаммов Escherichia coli, Klebsiella pneumoniae, Acinetobacter baumanii, Pseudomonas aeruginosa, Staphylococcus aureus.
Известно, что у кольчатых червей, аннелид, защита от патогенов обусловлена исключительно функционированием системы врожденного иммунитета. В частности, у данных организмов открыто несколько высокоактивных антимикробных пептидов, которые первоначально синтезируются в виде более крупных про-белков. Данные пептиды располагаются на С-концах про-белков, вслед за глобулярным доменом BRICHOS, и между зрелым антимикробным пептидом и BRICHOS-доменом располагается линкер с сайтом протеолиза фуриновой протеиназы.
Первыми открытыми BRICHOS антимикробными пептидами аннелид являются ареницины 1-3, изолированные из Arenicola marina [https://doi.Org/10.1016/j.febslet.2004.10.012] [ПАТЕНТ WO 2007/023163 А1].
Позднее были идентифицированы еще несколько BRICHOS антимикробных пептидов аннелид, в частности пептиды алвинеллацин из Alvinella pompejana [https://doi.org/10.1371/journal.pone.0095737] [ПАТЕНТ ЕР2516453 В1], два пептида никомицин 1 и 2 из Nicomache minor [https://doi.org/10.3390/md16110401] [ПАТЕНТ RU 2721273] и пептид капителлацин из Capitella teleta [https://doi.org/10.3390/md18120620]. Практически все известные антимикробные пептиды из аннелид с BRICHOS про-белком имеют структуру Р-шпильки, стабилизированную дисульфидными связями.
Для идентификации новых высокоактивных антимикробных пептидов со структурой Р-шпильки был проведен биоинформатический поиск кодирующих белки последовательностей, содержащих одиночный домен BRICHOS с С-концевым пептидом. Для поиска использована аминокислотная последовательность BRICHOS-домена прекурсора ареницина 1 (UniprotID: Q5SC60, аминокислотные остатки 26-181).
Поиск гомологов среди эукариотических белков выполнен при помощи BLAST с использованием матрицы BLOSUM-62 и выравниванием без пробелов. Из найденных гомологов выделены те, в которых на С-конце домена BRICHOS присутствует дополнительный участок длиной не более 50 аминокислотных остатков.
Проведенный нами анализ выявил несколько кодирующих последовательностей в геномах амфибий Microcaecilia unicolor и Rhinatrema bivittatum, наиболее интересными из которых являются последовательность гена LOCI 15457007 из генома Microcaecilia unicolor базы данных NCBI (сборка генома NCBI RefSeq assembly: GCF_901765095.1) и последовательность гена LOCI 15075475 из генома Rhinatrema bivittatum базы данных NCBI (сборка генома NCBI RefSeq assembly: GCF_901001135.1). Предположительные сайты фуриновой протеиназы, состоящие из двух остатков аргинина (RR), были обнаружены в линкерах кодируемых указанными генами белков (белковые последовательности ХР 030042296 и ХР_029431717), расположенных между BPJCHOS-доменами и С-концевыми короткими амфифильным цистеин-богатыми пептидами. С-концевые фрагменты, следующие за предположительными сайтами протеолиза, обладают высоким содержанием гидрофобных аминокислот и аргинина, а также симметричным расположением цистеинов, что позволило идентифицировать их как катионные антимикробные пептиды.
Уникальность идентифицированных пептидов обусловлена несколькими факторами.
Во-первых, они являются первыми антимикробными пептидами с BRICHOS-доменом за пределами типа кольчатых червей и, соответственно, первыми антимикробными пептидами данного класса у позвоночных. Во-вторых, про-белки идентифицированных пептидов обладают уникальным строением, которое сочетает наличие гомолога белка легочного сурфактанта SP-C на N-конце и антимикробного пептида на С-конце.
Таким образом, идентифицированные белки в своем строении обладают сходством как с BRICHOS антимикробными пептидами аннелид, так и с легочным сурфактантом млекопитающих. В-третьих, паттерн расположения остатков цистеина и формируемых ими дисульфидных связей в идентифицированных, а именно СХ5СХ4СХ5С, где С обозначает остатки цистеина, а X обозначает любые другие аминокислотные остатки, отличается от всех известных антимикробных пептидов со структурой (3-шпильки.
Для идентифицированных антимикробных пептидов было проведено моделирование с помощью алгоритма AlphaFold v.2, которое предсказало пространственную структуру антимикробных пептидов в виде антипараллельной бета-шпильки с правой закрученностью и близким расположением четырех цистеинов, что, в свою очередь, предполагает стабилизацию структуры данных пептидов двумя внутримолекулярными S-S связями.
Идентифицированные антимикробные пептиды были названы мицецилином и рицецилином, для пептида из Microcaecilia unicolor и Rhinatrema bivittatum, соответственно. Аминокислотные последовательности мицецилина и рицецилина представлены на фиг. 1. Цистеины, которые могут образовывать дисульфидные связи, отмечены желтым цветом. Сайт протеолиза отмечен пунктирной линией.
Изобретение поясняется следующими примерами:
Пример 1. Синтез новых антимикробных пептидов
Пептиды мицецилин (МС) и рицецилин (RC) были синтезированы посредством твердофазного синтеза из Fmoc-аминокислот с помощью автоматического синтезатора пептидов Symphony X (Protein Technologies, США). После завершения синтеза и снятия со смолы пептиды были очищены методом обращенно-фазовой жидкостной хроматографии на хроматографе NGC Bio-Rad system (Hercules, USA), колонка Symmetry С18, 19 x 300 mm, 5 μm (Waters, Germany).
Чистота пептидов, по данным аналитической хроматографии, выполненной на колонке Luna (С-18, 250x4,6 мм, 5 мкм (Phenimenex, США), составляла не менее 95%. Пептиды были рефолдированы в окисляющих условиях для формирования дисульфидных связей. Молекулярные массы были подтверждены данными масс-спектрометрического анализа MALDI TOF (Matrix-assisted laser desorption/ionization Time-of Fligft) на приборе Ultraflex (Bruker, США).
Пример 2. Исследование антимикробных свойств пептидов
Для исследования использовались изолированные из клинического материала штаммы бактерий Acinetobacter baumannii - 2 штамма, Escherichia coli, Klebsiella pneumoniae, Pseudomonas aeruginosa и Staphylococcus aureus - no 1 штамму, а также один лабораторный штамм Escherichia coli ML 35p, устойчивой только к ампициллину. Устойчивость выделенных штаммов бактерий к антибиотикам определяли диск-диффузионным методом (таблица 1).
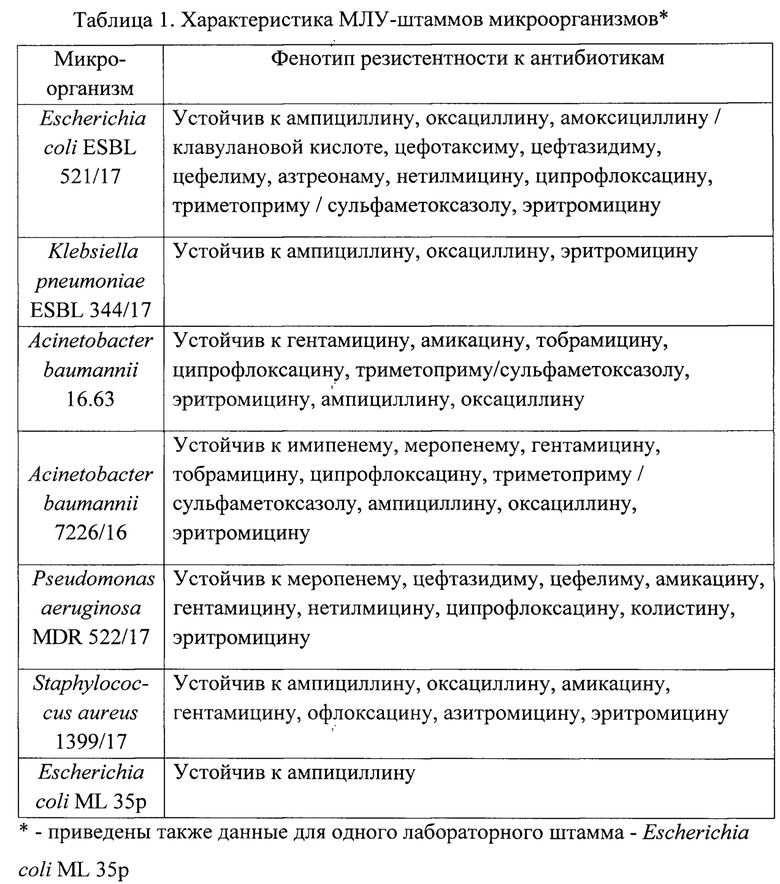
Антимикробную активность пептидов определяли методом серийных разведений в жидкой питательной среде, содержащей микроорганизмы, по стандартной методике, оптимизированной ранее для пептидных препаратов [Tossi A., Scocchi М., Zanetti М., et al. An approach combining cDNA amplification and chemical synthesis for the identification of novel, cathelicidin-derived, antimicrobial peptides // Antibacterial Peptide Protocols. Methods In Molecular Biology, vol. 78 / W. Shafer (Ed.). Totowa, NJ: Humana Press Inc., 1997. P. 133-151. doi: 10.1385/0-89603-408-9:133], рассчитывая величины их минимальных ингибирующих концентраций (МИК), как наименьших концентраций, в присутствии которых визуально не наблюдался рост микроорганизмов.
Микроорганизмы культивировали в 2,1% бульоне Мюллера-Хинтона М391 (HiMedia, Индия). Серийные разведения веществ в 10 мМ натрий-фосфатном буфере рН 7,4, содержащем 0,1% БСА, производили в стерильных 60-луночных микрокамерах, ячейки которых предварительно инкубировали в течение часа с 0,1% раствором БСА для предотвращения неспецифического связывания пептидов со стенками лунок, после чего раствор БСА из лунок удаляли.
В экспериментальные и контрольные (содержащие буфер без антимикробных веществ) лунки вносили суспензию бактерий в логарифмической фазе роста в 2,1% бульоне Мюллера-Хинтона и равный объем раствора пептида. Конечное содержание бактерий составило 5×105 КОЕ/мл [Wiegand I., Hilpert К., Hancock R.E.W. Agar and broth dilution methods to determine the minimal inhibitory concentration (MIC) of antimicrobial substances // Nat. Protocols. 2008. Vol.3. P. 163-175. doi: 10.1038/nprot.2007.521.]. Использовали по три параллели контрольных и опытных проб. Итоговое значение МИК рассчитывали как медиану на основании 3-5 независимых экспериментов.
Результаты проведенных экспериментов представлены в таблице 2. В данной работе в качестве параметра, характеризующего минимальную ингибирующую концентрацию (МИК), использовали также среднее геометрическое минимальных ингибирующих рост микроорганизмов концентраций (GMHK). Результаты представлены в таблице 2.
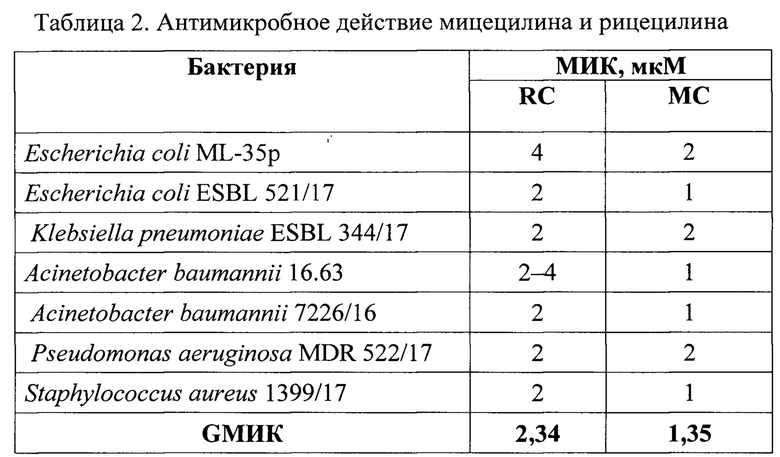
Таким образом, мицецилин и рицецилин подавляют рост МЛУ-штаммов бактерий в концентрациях от 1 до 4 мкМ.
Пример 3. Исследование гемолитической и цитотоксической активности мицецилина и рицецилина
3.1. Определение гемолитической активности
Проводили двукратные серийные разведения исследуемых антимикробных пептидов и добавляли по 3 мкл раствора каждого пептида в забуференном физиологическом растворе (ЗФР) к 27 мкл подготовленной суспензии эритроцитов, выделенных из донорской крови человека стандартным способом. После инкубации в течение 30 мин при +37°С с периодическим перемешиванием реакцию останавливали, внося в каждую микропробирку по 90 мкл охлажденного ЗФР. Пробы центрифугировали при 8000 g в течение 4 минут, после чего по 100 мкл надосадочной жидкости переносили в плоскодонный 96-луночный планшет для последующей регистрации оптической плотности (OD). В качестве отрицательного контроля на спонтанный гемолиз, в пробы вместо раствора пептида вносили аналогичный объем стерильного ЗФР. Положительным контролем, соответствующим полному лизису эритроцитов, служили пробы, в которых вместо пептида к суспензии эритроцитов добавляли 10% водный раствор детергента Triton Х-100. Все пробы, контрольные и опытные, готовили в трех повторностях. Поглощение проб определяли при длине волны 540 нм, близкой к одному из максимумов поглощения гемоглобина; с использованием планшетного спектрофотометра SpectraMax 250 (Molecular Devices, США). Процент гемолиза в экспериментальных пробах рассчитывали относительно положительного (100% гемолиз) и отрицательного (0% гемолиз) контролей по формуле:
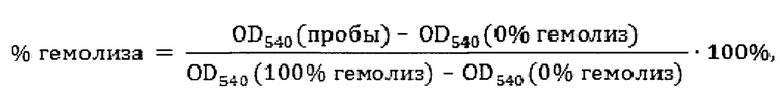
где OD540 пробы, 0% гемолиза и 100% гемолиза - величины оптической плотности содержащей пептид пробы, а также отрицательного и положительного контролей соответственно, измеренные при длине волны 540 нм.
Эксперимент повторяли трижды, результаты представляли, как «среднее±среднеквадратичное отклонение». Построение графиков зависимости интенсивности гемолиза от концентрации осуществляли с использованием программы SigmaPlot 12.0 (Systat Software Inc., США).
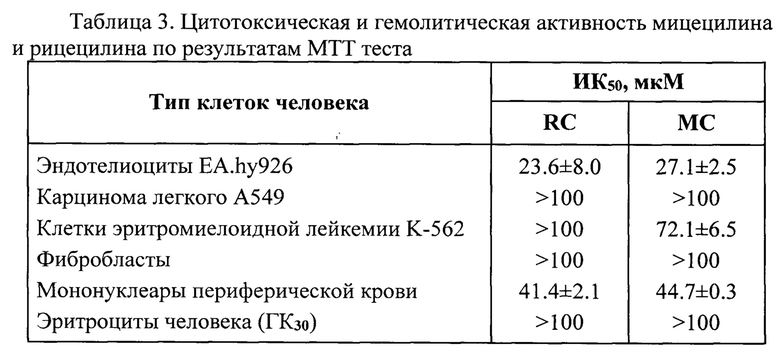
Гемолиз эритроцитов в присутствии 20-100 мкМ мицецилина и рицецилина не отличался от спонтанного гемолиза в отсутствие пептидов. Между тем, в присутствии 100 мкМ известного пептида PG-1 в данных условиях происходил лизис не менее 85% эритроцитов. Таким образом, гемолитические концентрации данных пептидов превышают 100 мкМ. Гемолитические концентрации пептидов ГК30, при которых происходит разрушение 30% эритроцитов, также превышают 100 мкМ (таблица 3).
3.2. Определение цитотоксической активности
Исследовали цитотоксическую активность пептидов по отношению к следующим клеткам человека: мононуклеарным клеткам периферической крови здоровых доноров, культивируемым фибробластам человека, клеткам эритромиелоидной лейкемии человека К562, клеткам карциномы легкого человека А549, клеткам эндотелиальной линии ЕА.Ну926 путем постановки МТТ-теста, позволяющего определять суммарную активность дыхательных ферментов митохондрий по восстановлению неокрашенных форм МТТ (3-4,5-диметилтиазол-2-ил-2,5-дифенилтетразола бромида) до голубого кристаллического формазана [Mosmann, Т. Rapid Colorimetric Assay for Cellular Growth and Survival: Application to Proliferation and Cytotoxicity Assays // J. Immunol. Methods. 1983. Vol. 65. P. 55-63.].
К 90 мкл клеточной суспензии в бессывороточной питательной среде, помещенной в лунки 96-луночных планшетов, добавляли 10 мкл раствора пептида различной концентрации (серийные двукратные разведения) в трех параллелях. В контрольные лунки добавляли по 10 мкл среды. Планшеты далее помещали в СО2 инкубатор на 20 часов. За четыре часа до окончания срока инкубации в лунки планшетов добавляли по 10 мкл раствора МТТ (5 мг/мл в ЗФР). По окончании инкубации в лунки вносили по 100 мкл изопропанола с 0,04 М НС1, перемешивали и проводили измерение оптической плотности раствора в лунках планшета при длине волны 540 нм (вычитая величину оптической плотности при 690 нм, как фоновую) на спектрофотометре SpectraMax250 (США). Для расчета медианной (полумаксимальной) ингибирующей концентрации пептидов (ИК50) использовали программу Sigma Plot 12. Результаты представлены в табл. 3.
Пример 4. Подсчет индексов селективности мицецилина и рицецилина
С целью объективной оценки антимикробной, гемолитической и цитотоксической активности пептидов были подсчитаны индексы специфичности. Индекс ИСг/б представляет собой отношение ГК30 (эффективной гемолитической концентрации, вызывающую гемолиз 30% эритроцитов) к GMИK
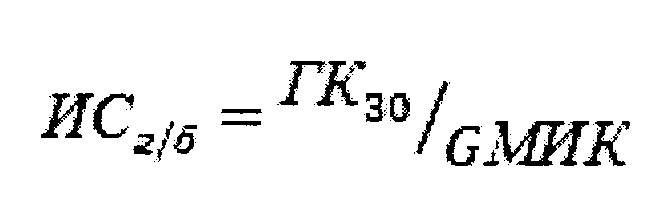
Индекс ИСэу/б является отношением среднего геометрического полумаксимальных эффективных цитотоксических концентраций (ИК50), вызывающих гибель нормальных эукариотических клеток (мононуклеаров периферической крови и фибробластов кожи человека), к GMИK

Результаты представлены в таблице 4.
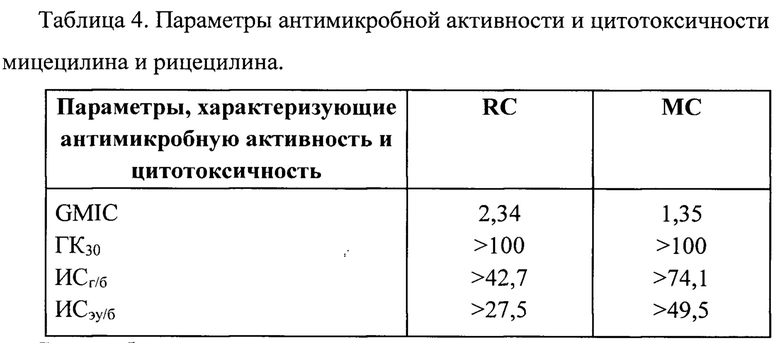
Таким образом, рицецилин и мицецилин имеют высокую антимикробную активность и низкую токсичность для клеток человека, что свидетельствует о перспективности их использования для лечения инфекций, в том числе инфекций, вызванных бактериями с множественной лекарственной устойчивостью.
--->
<?xml version="1.0" encoding="UTF-8"?>
<!DOCTYPE ST26SequenceListing PUBLIC "-//WIPO//DTD Sequence Listing
1.3//EN" "ST26SequenceListing_V1_3.dtd">
<ST26SequenceListing originalFreeTextLanguageCode="ru"
dtdVersion="V1_3" fileName="Ricecilin and Micecilin.xml"
softwareName="WIPO Sequence" softwareVersion="2.3.0"
productionDate="2024-03-20">
<ApplicationIdentification>
<IPOfficeCode>RU</IPOfficeCode>
<ApplicationNumberText>2023132474</ApplicationNumberText>
<FilingDate>2023-12-04</FilingDate>
</ApplicationIdentification>
<ApplicantFileReference>624</ApplicantFileReference>
<EarliestPriorityApplicationIdentification>
<IPOfficeCode>RU</IPOfficeCode>
<ApplicationNumberText>2023132474</ApplicationNumberText>
<FilingDate>2023-12-04</FilingDate>
</EarliestPriorityApplicationIdentification>
<ApplicantName languageCode="ru">Федеральное государственное
бюджетное научное учреждение "Институт экспериментальной
медицины"</ApplicantName>
<ApplicantNameLatin>Federal State Budgetary Scientific Institution
"Institute of Experimental Medicine"</ApplicantNameLatin>
<InventorName languageCode="ru">Шамова Ольга Валерьевна
</InventorName>
<InventorNameLatin>Shamova Olga</InventorNameLatin>
<InventionTitle languageCode="ru">Синтетические аналоги природных
пептидов, обладающих антибактериальной активностью</InventionTitle>
<SequenceTotalQuantity>2</SequenceTotalQuantity>
<SequenceData sequenceIDNumber="1">
<INSDSeq>
<INSDSeq_length>20</INSDSeq_length>
<INSDSeq_moltype>AA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..20</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>protein</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q2">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>ACLRIRVCNRYYCYVFLRCF</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="2">
<INSDSeq>
<INSDSeq_length>20</INSDSeq_length>
<INSDSeq_moltype>AA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..20</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>protein</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q4">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>FCIRVRVCNHYFCFYALRCF</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
</ST26SequenceListing>
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| Бета-шпилечный пептид, обладающий антимикробной активностью в отношении бактерий с множественной лекарственной устойчивостью | 2023 |
|
RU2812977C1 |
| Пептид, обладающий антибактериальной активностью в отношении микроорганизмов с множественной лекарственной устойчивостью | 2021 |
|
RU2771493C1 |
| Пептид, проявляющий антибактериальные и противоопухолевые свойства | 2019 |
|
RU2702661C1 |
| СПОСОБ ПОЛУЧЕНИЯ АНТИМИКРОБНЫХ ПЕПТИДОВ С ПОНИЖЕННОЙ ГЕМОЛИТИЧЕСКОЙ АКТИВНОСТЬЮ | 2006 |
|
RU2316336C1 |
| ПЕПТИДЫ ЛАТАРЦИНЫ, ПРОЯВЛЯЮЩИЕ АНТИМИКРОБНУЮ АКТИВНОСТЬ | 2006 |
|
RU2319745C1 |
| БЕТА-ШПИЛЕЧНЫЙ ПОЛИПЕПТИД, ОБЛАДАЮЩИЙ АНТИМИКРОБНОЙ АКТИВНОСТЬЮ | 2015 |
|
RU2624020C2 |
| ПЕПТИДЫ ЛАТАРЦИНЫ, ПРОЯВЛЯЮЩИЕ АНТИМИКРОБНУЮ АКТИВНОСТЬ | 2005 |
|
RU2302467C1 |
| ПЕПТИДЫ, ПРОЯВЛЯЮЩИЕ АНТИМИКРОБНУЮ АКТИВНОСТЬ | 2005 |
|
RU2302425C2 |
| ПЕПТИДЫ ЛАТАРЦИНЫ, ПРОЯВЛЯЮЩИЕ АНТИМИКРОБНУЮ АКТИВНОСТЬ | 2006 |
|
RU2306148C1 |
| ПЕПТИДЫ ЛАТАРЦИНЫ, ПРОЯВЛЯЮЩИЕ АНТИМИКРОБНУЮ АКТИВНОСТЬ | 2005 |
|
RU2302466C1 |

Изобретение относится к области биотехнологии, конкретно к пептидам, обладающим антибактериальной активностью, и может быть использовано в медицине в антибактериальной терапии. Предложены синтетические аналоги природных пептидов – мицецилин и рицецилин, которые разработаны на основе идентифицированных антимикробных пептидов из Microcaecilia unicolor и из Rhinatrema bivittatum соответственно. Изобретение обеспечивает получение пептидов, которые подавляют рост штаммов бактерий с множественной лекарственной устойчивостью. 2 н.п. ф-лы, 1 ил., 4 табл., 4 пр.
1. Синтетический аналог природного пептида рицецилин формулы ACLRIRVCNRYYCYVFLRCF, обладающий антибактериальной активностью и характеризующийся аминокислотной последовательностью SEQ ID No: 1.
2. Синтетический аналог природного пептида мицецилин формулы FCIRVRVCNHYFCFYALRCF, обладающий антибактериальной активностью и характеризующийся аминокислотной последовательностью SEQ ID No: 2.
| ШАМОВА О.В., Пептидомика - медицине: ожидания и реальность, Актовая речь, 15.12.2022, Санкт-Петербург, 29с., найдено в интернет [31/07/2024] по адресу: https://iemspb.ru/wp-content/uploads/2022/12/%D0%90%D0%BA%D1%82%D0%BE%D0%B2%D0%B0%D1%8F-%D1%80%D0%B5%D1%87%D1%8C-%D0%A8%D0%B0%D0%BC%D0%BE%D0%B2%D0%B0-%D0%9E.%D0%92.pdf; | |||
| БД "NCBI Reference |

