Изобретение относится к области биотехнологии. Может быть использовано при бактериологических исследованиях по идентификации бактерий комплекса Acinetobacter calcoaceticus - Acinetobacter baumannii (комплекс ACB), а также в производстве питательных сред для этих исследований.
Известен способ фенотипической идентификации видов комплекса АСВ, в том числе A.seifertii (Nemec A., Krizova L., Maixnerova M., Sedo О., Brisse S., Higgins P.G. Acinetobacter seifertii sp.nov., a member of the Acinetobacter calcoaceticus - Acinetobacter baumannii complex, isolated from human clinical specimens. Int. J.Sys. Evol. Microbiol. 2015, vol. 65, no.3, pp. 934-942. https://doi.org/10.1099/ijs.0.000043). Чистые культуры бактерий комплекса АСВ изучали тестами на рост при 30°С, 37°С, 41°С, 44°С в сердечно-мозговом бульоне (Оксоид). Окисление и кислотообразование на глюкозе исследовали на среде Хью-Лейфсона. Цитохромоксидазу выявляли 1% раствором тетраметилпарафенилендиамина. Гемолиз изучали на кровяном агаре с 5% крови овец и человека. Тесты утилизации выполняли в жидкой минеральной среде с солями аммония и добавлением 0,1% (масс/об) источника углерода при 30°С. Использовали 33 источника углерода: ацетат, L-аргинин, L-аспартат, 4-амино-бутират, бета-аланин, L-арабиноза, транс-аконитат, азелат, бензоат, 2,3-бутандиол, L-глутамат, глюконат, D-глюкозу, гистамин, глутарат, гентизат, L-гистидин, 4-гидроксибензоат, L- лейцин, левулинат, D-малат, малонат, L-орнитин, путресцин, D-рибозу, L-тартрат, трикарбаллилат, тригонеллин. фенилацетат, L-фенилаланин, цитрат, цитраконат, этанол. Тесты учитывали через 6 суток. Фенотипический анализ также проводили с использованием системы API 20 NE (bio Merieux) в соответствии с инструкцией производителя. Фенотипические характеристики A.seifertii в целом соответствовали свойствам других представителей комплекса АСВ и дополнительно продемонстрировали известные трудности в фенотипической дифференциации между видами комплекса АСВ. Несмотря на отсутствие однозначных диагностических признаков между видами можно отметить некоторые свойства или их комбинации, которые были типичными для A.seifertii, но редко встречаются у других видов. Большинство штаммов A.seifertii росли при 41°С, но не росли при 44°С; ни один из штаммов A.seifertii не рос на L-арабинозе, тогда как все штаммы вида A. nosocomialis и большинство штаммов A.baumannii (84%), A.pittii (85%), A.calcoaceticus (27%) утилизировали этот субстрат. Таким образом, точная фенотипическая идентификация бактерий A.seifertii традиционными метаболичесими и физиологическими тестами остается ненадежной.
Известен также способ идентификации видов бактерий, в том числе A.seifertii, с использованием фенотипического метода матрично-активиро-ванной лазерной десорбции/ионизации с время пролетной масс-спектрометрией((MALDI-TOF/MS). Объектом исследования является чистая культура микроорганизмов (часть колонии бактерий), которую исследуют согласно инструкции производителя, например, масс-спектрометром Microflex с базой данных MALDI Byotyper (Bruker Daltonics Inc., Германия).Уровни идентификации бактерий трактуются по критериям, указанным в инструкции: score 2300-3000 высокая вероятность идентификации вида; 2000-2299 надежная идентификация рода, вероятная идентификация вида; 1700-1999 вероятная идентификация рода; 0000-1699 ненадежная идентификация. При наличии в базе данных представителей A.baumannii, A.pittii, A.nosocomialis, A.calcoaceticus обеспечивается быстрая и точная идентификация этих видов.В связи с открытием двух новых видов A.seifertii, A.dijkshoorniae, включенных в группу Acinetobacter baumannii (Ab), было проведено исследование этих бактерий для расчета средних спектров видов (MSP) и создания моделей распознавания образов. Видоспецифические пики были идентифицированы для штаммов всех видов и MSP, полученная из трех штаммов A.seifertii, двух штаммов A.dijkshoorniae, и двух штаммов A.nosocomialis были добавлены в базу данных таксономии Braker, что позволило успешно идентифицировать все изоляты с использованием спектров любого бактериального экстракта или колонии. Своевременное уточнение и пополнение базы данных для метода обеспечивает точную идентификацию бактерий (Mari-Almirall М., Cosgaya С, Higgins P.G., Van Assche A., Telli М., Huys G., Lievens В., Seifert M., Dijkshoorn L., Roca I., Vila J. MALDI-TOF/MS identification of species from the Acinerobacter baumannii (Ab) group revisited: inclusion of the novel A.seifertii and A.dijkshooriae species. Clin. Microbiol. Infect. 2017, Mar; vol. 23, no. 3, pp. 210.e1-210.e9. https://doi:10.1016/j.cmi.2016.11.020).
Прототипом заявляемого способа нами избран способ фенотипической идентификации видов комплекса АСВ, в том числе A.seifertii по статье (Nemec A., Krizova L., Maixnerova М., Sedo О., Brisse S., Higgins P.G. Acinetobacter seifertii sp.nov., a member of the Acinetobacter calcoaceticus - Acinetobacter baumannii complex, isolated from human clinical specimens. Int. J.Sys. Evol. Microbiol. 2015, vol. 65, no. 3, pp. 934-942. https://doi.org/10.1099/ijs.0.000043), так как он содержит полную характеристику фенотипических признаков A.seifertii и других видов комплекса АСВ, а также наиболее близок заявляемому способу по совокупности существенных признаков.
Недостатком прототипного способа является недостаточная точность фенотипической идентификации бактерий вида A.seifertii.
Недостатком аналога, основанного на применении фенотипического метода MALDI-TOF масс-спектрометрии, является необходимость сложного прибора и недостаточная точность, в некоторых случаях, идентификации A.seifertii при несвоевременном обновлении базы данных.
Целью изобретения является повышение точности исследования по фенотипической идентификации бактерий вида A.seifertii.
В соответствии с изобретением решение указанной технической задачи достигается тем, что после предварительного отбора материала, подлежащего исследованию, - чистой культуры грамотрицательных коккобактерий, растущих при 30°С, 37°С, 41°С, аэробов, неподвижных, оксидазоотрицательных. каталазоположительных, не ферментирующих, но окисляющих D-глюкозу с образованием кислоты, не имеющих нитратредуктазы; утилизирующих в качестве единственного источника углерода ацетат натрия (контроль отсутствия ауксотрофности), этанол, трикарбаллиловую кислоту; не утилизирующих D-глюкозу; суспендирукт исследуемую суточную агаровую культуру бактерий по половине петли диаметром 2 мм в лунке планшета в 0,1 мл 0,85% раствора хлорида натрия; засевают по одной петле суспензии бактерий штрихом на поверхность сектора плотной питательной среды, содержащей источник углерода D-ксилозу (CAS 58-86-6), NH4Cl, NH4NO3, Na2SO4, K2HPO4< KH2PO4< MgSO4; бромтимоловый синий 1,6% водный раствор, уплотнитель агар бактериологический, дистиллированную воду при следующем содержании ингредиентов: D-ксилоза (CAS 58-86-6), 1,0-3,0 г; NH4Cl 5,0 г; NH4NO3 1,0 г; Na2SO4 2,0 г; K2HPO4 3,0 г; KH2PO4 1,0 г; MgSO4 0,1 г; бромтимоловый синий (1,6%) водный раствор) 4 мл: агар бактериологический 14,0-16,0 г; вода дистиллированная 1 л, рН 7,2±0,2; стерилизация среды при 112°С в течение 20 мин, а также засевают одну петлю суспензии бактерий на поверхность питательной среды такого же состава, но без источника углерода D-ксилозы (контроль основы среды);инкубируют посевы при 30°С в аэробных условиях в течение 24-48 ч; определяют принадлежность исследуемых бактерий к виду A.seifertii по отсутствию их роста на питательной среде с D-ксилозой и питательной среде без источника углерода D-ксилозы при наличии роста на питательной среде такого же состава с ацетатом натрия в ходе предварительного отбора культур. Примечание. Бактериологический контроль качества питательной среды заявляемого способа проводят при изготовлении среды: контрольные штаммы - клинический штамм A.baumannii (положительный контроль), клинический штамм A.seifertii (отрицательный контроль); суспендируют суточные агаровые культуры контрольных штаммов по половине петли диаметром 2 мм в лунках планшета в 0,1 мл 0,85% растворе хлорида натрия и засевают каждый штамм по одной петле суспензии бактерий штрихом на поверхность питательной среды с D-ксилозой, такой же среды без D-ксилозы (контроль основы среды),такой же среды с ацетатом натрия вместо D-ксилозы (контроль отсутствия ауксотрофности); инкубируют посевы при 30°С в аэробных условиях 24 ч, затем учитывают результат - питательная среда пригодна к использованию при наличии на среде с D-ксилозой по штриху посева роста газона бактерий A.baumannii и отсутствия роста A.seifertii; при отсутствии на питательной среде без D-ксилозы роста бактерий A.baumannii и A.seifertii; при наличии на питательной среде с ацетатом натрия роста бактерий A.baumannii и A.seifertii.
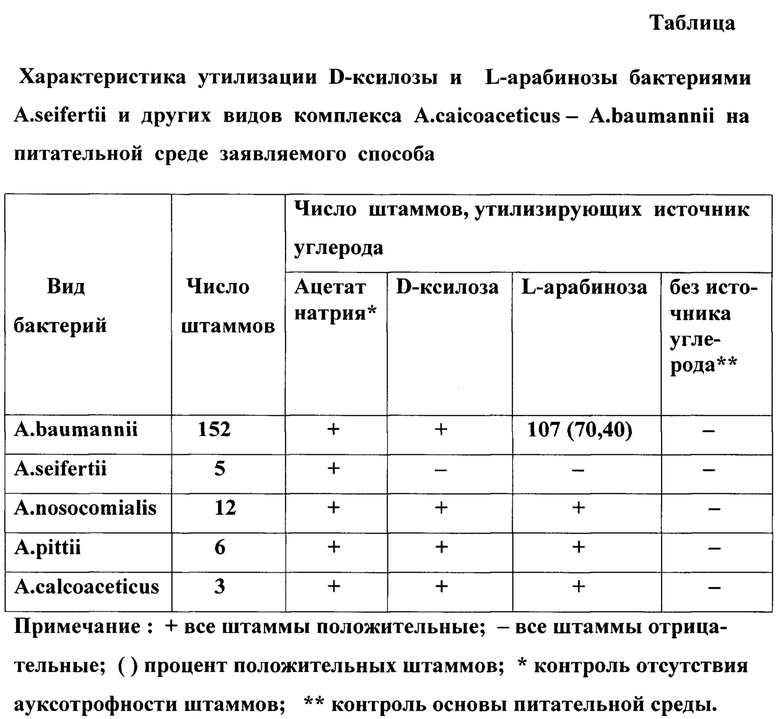
Первый отличительный существенный признак способа - использование в питательной среде заявляемого способа D-ксилозы (CAS 58-86-6) в качестве единственного источника углерода. Этот признак не применялся в прототипе и аналогах. Более того, тест на утилизацию D-ксилозы не использовался при фенотипической характеристике всех видов ацинетобактеров комплекса A.calcoaceticus- A.baumannii. Его пригодность для идентификации A.seifertii не очевидна и требовала сравнительного изучения всех видов ацинетобактеров комплекса АСВ. Нами были изучены 5 штаммов A.seifertii, из них 3 клинических изолята, выделенных в Военно-медицинской академии в 2019, 2023, 2024 годах, и 2 штамма из коллекции культур Санкт-Петербургского института имени Пастера. Также были изучены клинические изоляты: 152 штамма A.baumannii, в том числе 37 штаммов A.baumannii bv. tryptophandestruens, 12 штаммов A.nosocomialis, 6 штаммов A.pittii и 3 штамма A.calcoaceticus, выделенных из реки Невы. Вид всех штаммов был идентифицирован методом MALDI-TOF масс-спектрометрии при их выделении. Сведения о штаммах A.dijkshoorniae были получены по описанию этого вида (https://doi.org/10.1099/ijsem.0.001318). Было установлено, что все штаммы A.seifertii однозначно не утилизировали D-ксилозу в качестве единственного источника углерода, а все штаммы прочих изученных видов ацинетобактеров группы АСВ однозначно утилизировали D-ксилозу (табл.). Учитывая,что в прототипе среди тестов утилизации 33 субстратов лучший результат по дифференциации бактерий A.seifertii показал тест утилизации L-арабинозы, нами было проведено сравнительное исследование тестов утилизации L-арабинозы и D-ксилозы по методике заявляемого спосо6а. Было установлено, что утилизация L-арабинозы однозначно отрицательная у всех штаммов A.seifertii и однозначно положительная у всех штаммов видов A.pittii, A,nosocomialis, A calcoaceticus, но часто отрицательная у штаммов вида A.baumannii. Из 152 штаммов A.baumannii только 107(70,40%) утилизировали L-арабинозу (табл.). Не утилизировали L-арабинозу 45 штаммов A.baumannii, которые включали все штаммы биовара A.baumannii bv. tryptophandestruens (37 штаммов). Высокая частота этого биовара у бактерий вида A.baumannii не позволяет использовать тест утилизации L арабинозы для надежной идентификации A.seifertii. Отличительный существенный признак непосредственно определяет решение поставленной технической задачи повышения точности исследования по фенотипической идентификации бактерий вида A.seifertii, так как только тест утилизации D-ксилозы позволяет точно отличить все штаммы A.seifertii от всех штаммов прочих изученных видов ацинетобактеров комплекса АСВ, что ранее было невозможно фенотипическими тестами без применения приборов метода MALDI-TOF MS. Механизм достижения этого технического результата не известен из уровня техники и впервые представлен только в материалах нашей заявки.
Второй отличительный существенный признак способа - использование в питательной среде заявляемого способа уплотнителя агар бактериологический. Плотная питательная среда создает оптимальные аэробные условия для аэробных ацинетобактеров, позволяет точно визуально определять наличие или отсутствие роста бактерий по штриху посева, сокращает время исследования. Агар не утилизируется большинством бактерий, поэтому его используют в составе синтетических питательных сред, например, цитратный агар Симмонса, агар Козера.
Существенным признаком способа является предварительный отбор материала, подлежащего исследованию, - чистой культуры грамотрицательных бактерий, растущих при 30°С, 37°С, 41°С, аэробов, неподвижных, оксидазоотрицательных, каталазоположительных; не ферментирующих, но окисляющих D-глюкозу с образованием кислоты; не имеющих нитратредуктазы; утилизирующих в качестве единственного источника углерода ацетат натрия (контроль отсутствия ауксотрофности), этанол, трикарбаллиловую кислоту; не утилизирующих D-глюкозу, что устанавливает принадлежность изучаемых культур к ацинетобактерам комплекса АСВ и исключает от дальнейшего исследования ауксотрофные штаммы бактерий.
Существенным признаком также является использование суточных агаровых культур бактерий, их суспендирование в 0,1 мл 0,85% раствора хлорида натрия, посев одной петли суспензии бактерий штрихом па поверхность питательной среды с единственным источником углерода, что позволяет исключить занос на среду посторонних органических субстратов.
Существенным признаком является состав питательной среды заявляемого способа: комплекс солей (г/л) NH4Cl 5,0; NH4NO3 1,0; Na2SO4 2,0; K2HPO4 3,0; KH2PO4 1,0; MgSO4 0,1, который определяет синтетический состав среды, содержит минеральные источники азота, но не содержит источников углерода, что направляет метаболизм бактерий на утилизацию изучаемого органического субстрата как единственного источника углерода; обеспечивает оптимальный рН среды 7,2±0,2; бромтимоловый синий (1,6% водный раствор) позволяет визуально контролировать рН сред. Существенные признаки также оптимальная температура культивирования 30°С, аэробные условия, продолжительность инкубации посевов 24-48 ч.
Существенный признак - посев одной петли суспензии исследуемых бактерий штрихом на поверхность питательной среды без источника углерода (контроль основы среды), что позволяет более точно выявлять отсутствие роста бактерий на питательной среде с источником углерода (D-ксилоза) путем сравнительной визуальной оценки штрихов посевов после их инкубации.
Примеры, подтверждающие возможность осуществления способа
Пример 1. Исследуемый материал - чистая культура бактерий штамма №844, выделенная из мокроты больного, у которой предварительно выявлена совокупность признаков: грамотрицательные коккобактерии, растут при 30°С, 37°С, 41°С, аэробы, неподвижные, оксидазоотрицательные, каталазоположительные; не ферментируют, но окисляют D-глюкозу с образованием кислоты; не имеют нитратредуктазы; утилизируют в качестве единственного источника углерода ацетат натрия (контроль отсутствия ауксотрофности), этанол, трикарбаллиловую кислоту; не утилизируют D-глюкозу. Суспендируют исследуемую суточную агаровую культуру по половине петли диаметром 2 мм в лунке планшета с 0,1 мл 0,85% раствора хлорида натрия, засевают по одной петле суспензии бактерий штрихом на сектор питательной среды заявляемого способа, содержащей D-ксилозу 1,0 г/л и агар бактериологический 14,0 г/л и остальные ингредиенты по Примечанию к примеру 1, а также на сектор такой же среды без D-ксилозы (контроль основы среды); инкубируют посевы при 30°С в аэробных условиях в течение 24 ч, после чего учитывают результат .Исследуемый штамм №844 не растет на питательных средах с D-ксилозой и без D-ксилозы при наличии их роста на питательной среде с ацетатом натрия при предварительном отборе культур, что указывает на отсутствие у них ауксотрофности и принадлежность к виду A.seifertii по признаку отсутствия утилизации D-ксилозы. Контрольная видовая идентификация штамма №844 методом MALDI_TOF масс-спектрометрии с использованием микробиологического анализатора Вас-toSCREEN (НПФ «Литех», Россия) на базе масс-спектрометра Laser LT2 Plus (Scientific Analysis Instruments, Великобритания) подтвердила принадлежность его к виду A.seifertii.
Примечание к примеру 1. Методика приготовления и контроля питательной среды заявляемого способа. В 1 л дистиллированной воды вносят, г/л: D-ксилоза (CAS 58-86-6)? 1,0; NH4Cl 5,0; NH4NO3 1,0; Na2SO4 2,0; K2HPO4 3,0; KH2PO4 1,0; MgSO4 0,1; бромтимоловый синий 1,6% водный раствор 4 мл; агар бактериологический 14,0; растворяют все ингредиенты при нагревании, устанавливают рН 7,2±0,2, стерилизуют при 112°С в течение 20 мин, разливают в чашки Петри. Питательная среда прозрачная, зеленого цвета, пригодна к использованию в течение 30 суток при хранении от 4°С до 8°С. Бактериологический контроль питательной среды проводят при ее изготовлении используя контрольные клинические штаммы бактерий A.baumannii (положительный контроль) и A.seifertii (отрицательный контроль). Суточные агаровые культуры контрольных штаммов суспендируют по половине петли диаметом 2 мм в 0,1 мл 0,85% растворе хлорида натрия в лунке планшета, засевают по петле суспензии бактерий штрихом на сектор питательной среды, инкубируют посевы в аэробных условиях при 30°С в течение 24-48 ч, затем учитывают результат: питательная среда пригодна к использованию при наличии роста по штриху посева газона бактерий A.baumannii и отсутствии роста по штриху посева бактерий A.seifertii. Другие варианты питательной среды заявляемого способа - среда без D-ксилозы (для контроля основы среды) и среда с ацетатом натрия (для контроля отсутствия ауксотрофности) изготавливаются таким же образом. Их бактериологический контроль проводят теми же контрольными штаммами по такой же методике. Питательная среда для контроля основы среды пригодна к использованию, если на ней отсутствует рост бактерий A.baumannii и A.seifertii; питательная среда для контроля отсутствия ауксотрофности бактерий пригодна к использованию, если на ней растут бактерии A.baumannii и A.seifertii.
Пример 2. Исследуемый материал - чистая культура штамма №8957, выделенная из мазка с зева больного, у которой предварительно выявлена совокупность признаков: грамотрицательные коккобактерии, растущие при 30°С, 37°С, 41°С, аэробы, неподвижные, оксидазоотрицательные, каталазоположительные; не ферментируют, но окисляют D-глюкозу с образованием кислоты; не имеют нитратредуктазы; утилизируют в качестве единственного источника углерода ацетат натрия (контроль отсутствия ауксотрофности), этанол, трикарбаллиловую кислоту; не утилизируют D-глюкозу. Суспендируют исследуемую суточную агаровую культуру по половине петли диаметром 2 мм в лунке планшета в 0,1 мл 0,855 раствора хлорида натрия, засевают одну петлю суспензии бактерий штрихом на сектор среды заявляемого способа, содержащей D-ксилозу 3,0 г/л и агар бактериологический 16,0 г/л и остальные ингредиенты среды по Примечанию к примеру 1, а также на сектор такой же среды без D-ксилозы (контроль основы среды); инкубируют посевы при 30°С в аэробных условиях в течение 48 ч. затем учитывают результат. Бактерии штамма №8957 не растут на поверхности питательных сред с D-ксилозой и без D-ксилозы при наличии их роста на питательной среде с ацетатом натрия, что указывает на отсутствие у них ауксотрофности и приннадлежность их к виду A.seifertii по признаку отсутствия утилизации D-ксилозы. Контрольная видовая идентификация штамма №8957 методом MALDI-TOF масс-спектрометрии с использованием микробиологического анализатора BactoSCREEN (НПФ «Литех», Россия) на базе масс-спектрометра Laser LT2 Plus (Scientific Analysis Instruments, Великобритания) подтвердила принадлежность его к виду A.seifertii.
Пример 3. Исследуемый материал - чистая культура штамма A.baumannii №3459, выделенного в 2019 году из клинического материала и идентифицированная при выделении методом MALDI-TOF масс-спектрометрии с применением необновленной базы данных MALDI Biotyp-ег масс-спектрометром Microflex (Bruker Daltonix, Германия). Этот штамм был единственным среди штаммов A.baumannuu по необычному сочетанию наличия быстрой уреазы и отсутствия утилизации L-арабинозы. Суточную агаровую культуру штамма суспендируют по половине петли диаметром 2 мм в лунке планшета в 0,1 мл 0,85% раствора хлорида натрия, засевают по одной петле суспензии бактерий штрихом на сектор питательной среды заявляемого способа, содержащей D-ксилозу 2,0 г/л и агар бактериологический 15 г/л и остальные ингредиенты по Примечанию к примеру 1, а также на сектор такой же среды без D-ксилозы (контроль основы среды) и сектор такой же среды с ацетатом натрия вместо D-ксилозы (контроль отсутствия ауксотрофности бактерий), инкубируют посевы при 30°С в аэробных условиях в течение 48 ч затем учитывают результат. Исследуемый штамм A.baumannii №3459 не растет на питательной среде с D-ксилозой и питательной среде без D-ксилозы при наличии его роста на питательной среде с ацетатом натрия, что указывает на отсутствие ауксотрофности и принадлежность штамма №3459 к виду A.seifertii по признаку отсутствия утилизации D-ксилозы. Контрольная видовая идентификация штамма №3459 методом MALDI-TOF масс-спектрометрии с использованием микробиологического анализатора BactoSCREEN (НПФ «Литех», Россия) на базе масс-спектрометра Laser LT2 Plus (Scientific Analysis Instruments, Великобритания) подтвердила принадлежность его к виду A.seifertii.
Таким образом, при всех предельных концентрациях основных ингредиентов питательной среды заявляемого способа получены точные результаты идентификации бактерий A.seifertii, что свидетельствуют о достижении заявленным способом поставленной технической задачи повышения точности исследования по фенотипической идентификации бактерий вида Acinetobacter seifertii.
Изобретение относится к биотехнологии. Предложен способ фенотипической идентификации бактерий Acinetobacter seifertii, предусматривающий предварительный отбор культуры бактерий с совокупностью признаков комплекса A.calcoaceticus-A.baumannii и отсутствием ауксотрофности; суспендирование агаровой культуры в 0,85% растворе хлорида натрия; посев суспензии штрихом на питательную среду, содержащую единственный источник углерода D-ксилозу, NH4Cl, NH4NO3, Na2SO4, K2HPO4, KH2PO4, MgSO4, бромтимоловый синий 1,6% раствор, агар бактериологический и дистиллированную воду в заданных соотношениях, а также посев суспензии бактерий на такую же среду без D-ксилозы. Проводят аэробную инкубацию при 30°С в течение 24-48 ч с последующей идентификацией A.seifertii по отсутствию роста на питательной среде с D-ксилозой и среде без D-ксилозы, но наличию роста бактерий на среде с ацетатом натрия при предварительном отборе культуры. Изобретение позволяет повысить точность фенотипической идентификации бактерий Acinetobacter seifertii. 1 табл., 3 пр.
Способ фенотипической идентификации бактерий Acinetobacter seifertii, включающий предварительный отбор материала, подлежащего исследованию, - чистой культуры грамотрицательных коккобактерий, растущих при 30°С, 37°С, 41°С, аэробов, неподвижных, оксидазоотрицательных, каталазоположительных; не ферментирующих, но окисляющих D-глюкозу с образованием кислоты; не имеющих нитратредуктазы; утилизирующих в качестве единственного источника углерода ацетат натрия -контроль отсутствия ауксотрофности, этанол, трикарбаллиловую кислоту; не утилизирующих D-глюкозу; суспендирование суточной агаровой культуры бактерий по половине петли диаметром 2 мм в лунке планшета в 0,1 мл 0,85% раствора хлорида натрия; посев одной петли суспензии бактерий штрихом на поверхность сектора плотной питательной среды, содержащей источник углерода, NH4Cl, NH4NO3, Na2SO4, K2HPO4, KH2PO4, MgSO4; бромтимоловый синий 1,6% водный раствор, уплотнитель среды, дистиллированную воду; а также посев одной петли суспензии бактерий штрихом на поверхность питательной среды этого же состава без источника углерода -контроль основы среды, инкубирование посевов при 30°С в аэробных условиях в течение 24-48 ч; определение принадлежности бактерий к виду A.seifertii по отсутствию их роста на поверхности питательной среды с источником углерода и без источника углерода, но наличию роста на питательной среде с ацетатом натрия при предварительном отборе культур, при этом питательная среда содержит в качестве единственного источника углерода D-ксилозу CAS 58-86-6, а также уплотнитель агар бактериологический при следующем содержании ингредиентов:
при этом питательную среду стерилизуют при 112° в течение 20 мин; рН 7,2±0,2.
| NEMEC ALEXANDR et al | |||
| Acinetobacter seifertii sp | |||
| nov., a member of the Acinetobacter calcoaceticus - Acinetobacter baumannii complex isolated from human clinical specimens; International journal of systematic and evolutionary microbiology, 2015, N 65, p | |||
| Накаливаемый катод для вакуумных приборов | 1924 |
|
SU934A1 |
| СИВОЛОДСКИЙ Е.П., ЗУЕВА Е.П | |||
| Таксономическое и прикладное значение профилей | |||

