Область техники
Настоящее изобретение, в общем, относится к терапевтическим микровезикулам¸ полученным из пробиотических бактерий, и их применениям.
Уровень техники
Продовольственная и сельскохозяйственная организация Объединенных Наций определила пробиотики как «живые микроорганизмы, которые при введении в адекватных количествах приносят пользу для здоровья хозяина». Пробиотики влияют на иммунные функции хозяина посредством либо модуляции состава микробиоты, либо метаболической активности, либо даже посредством прямого взаимодействия с компонентами иммунной системы, находящимися в слизистой оболочке пищеварительного тракта. Более 60% иммунных клеток располагаются в слизистой оболочке пищеварительного тракта, и выборка информации о структуре и составе микробиоты преобразуется в местные и системные эффекты через циркулирующие иммунные клетки. После пробиотических взаимодействий иммунные механизмы могут активироваться, что выражается в высвобождении медиаторов иммунных реакций, таких как цитокины, выработке антител и активации лимфоцитов, а также других иммунных клеток. Такие активированные клетки, цитокины и/или соединения, высвобождаемые под действием пробиотиков, будут осуществлять иммуномодулирующие функции в различных участках тела через кровоток. Пробиотики также могут предотвращать или подавлять пролиферацию патогенов и подавлять выработку факторов вирулентности патогенами.
В настоящее время в качестве пробиотиков используется несколько различных бактериальных штаммов, включая бактерии, продуцирующие молочную кислоту, такие как отдельные штаммы Lactobacillus и Bifidobacterium. Эффективность пробиотических бактерий зависит от штамма, и каждый штамм может способствовать улучшению здоровья хозяина посредством различных механизмов.
Существует множество разновидностей пробиотических добавок, но положительное влияние на здоровье варьируется среди бактериальных штаммов, и пока мало что известно о способах контроля и регуляции конкретного пробиотического или биологического эффекта. Каждый бактериальный штамм имеет различные механизмы, с помощью которых опосредуются специфические эффекты для улучшения состояния здоровья и облегчения симптомов, например, желудочно-кишечных расстройств, включая диарею и запор, воспалительное заболевание кишечника (IBD), синдром раздраженного кишечника (IBS) и младенческие колики. Младенческие колики представляют собой состояние, которое может быть крайне стрессовым для семей, где имеет место такая проблема, и значительно ухудшить качество жизни. Кроме того, младенческие колики потенциально могут иметь долгосрочные последствия для младенцев в более позднем возрасте. Было показано, что хорошо изученный пробиотический бактериальный штамм Lactobacillus reuteri DSM 17938 значительно сокращает время плача у младенцев, страдающих коликами. Однако время проявления этого эффекта не является немедленным, и может пройти от одной до трех недель, прежде чем ребенок получит пользу от лечения. Таким образом, существует большая потребность в более быстродействующих вмешательствах, а также в более эффективных, например, для уменьшения периода дискомфорта и плача у детей с коликами.
Сущность изобретения
Основная цель настоящего изобретения заключается в обеспечении терапевтических микровезикул, полученных из пробиотических бактерий.
Данная и другие цели достигаются вариантами осуществления, раскрытыми здесь.
Настоящее изобретение определено в независимых пунктах формулы изобретения. Дополнительные варианты осуществления изобретения определены в зависимых пунктах формулы изобретения.
Аспект вариантов осуществления относится к способу получения терапевтических микровезикул. Способ включает культивирование бактерий пробиотического бактериального штамма в культуральной среде. Пробиотический бактериальный штамм выбран из группы, состоящей из штамма Lactobacillus, штамма Bifidobacterium и их комбинации. Способ также включает подвергание бактерий индуцирующей биотической обработке во время культивирования для индукции продукции терапевтических микровезикул бактериями. Индуцирующая биотическая обработка выбрана из группы, состоящей из сокультивирования бактерий с бактериями другого бактериального штамма, культивирования бактерий в присутствии кондиционированной среды после культивирования бактерий другого бактериального штамма и их комбинации. Другой бактериальный штамм представляет собой штамм Bifidobacterium, и другой бактериальный штамм отличается от пробиотического бактериального штамма.
Другие аспекты вариантов осуществления изобретения относятся к пробиотической композиции, содержащей бактерии пробиотического бактериального штамма и терапевтические микровезикулы, продуцированные пробиотическим бактериальным штаммом или другим пробиотическим бактериальным штаммом, для применения в лечении колик и/или для применения в лечении заболевания, выбранного из группы, состоящей из желудочно-кишечного расстройства или заболевания у младенцев или детей, желудочно-кишечного болевого расстройства, заболевания, связанного с потерей костной массы, заболевания пародонта и их комбинации. Пробиотический бактериальный штамм и другой пробиотический бактериальный штамм выбран из группы, состоящей из штамма Lactobacillus, штамма Bifidobacterium и их комбинации.
Дополнительные аспекты вариантов осуществления относятся к пробиотической композиции для применения в лечении колик и/или для применения в лечении заболевания, выбранного из группы, состоящей из желудочно-кишечного расстройства или заболевания у младенцев или детей, желудочно-кишечного болевого расстройства, заболевания, связанного с потерей костной массы, заболевания пародонта и их комбинации. Пробиотическая композиция включает быстродействующий компонент в виде терапевтических микровезикул, полученных из бактерий пробиотического бактериального штамма. Пробиотический бактериальный штамм выбран из группы, состоящей из штамма Lactobacillus, штамма Bifidobacterium и их комбинации. Пробиотическая композиция также содержит медленнодействующий компонент в виде бактерий пробиотического бактериального штамма или другого пробиотического бактериального штамма. Другой пробиотический бактериальный штамм выбран из группы, состоящей из штамма Lactobacillus, штамма Bifidobacterium и их комбинации. Быстродействующий компонент и медленнодействующий компонент вместе обеспечивают пролонгированный терапевтический эффект при введении субъекту.
Еще одни аспекты вариантов осуществления относятся к терапевтическим микровезикулам, выделенным из бактерий пробиотического бактериального штамма, для применения в лечении колик и/или для применения в лечении заболевания, выбранного из группы, состоящей из желудочно-кишечного расстройства или заболевания у младенцев или детей, желудочно-кишечного болевого расстройства, заболевания, связанного с потерей костной массы, заболевания пародонта и их комбинации. Пробиотический бактериальный штамм выбран из группы, состоящей из штамма Lactobacillus, штамма Bifidobacterium и их комбинации.
Еще один аспект вариантов осуществления изобретения относится к бактериальному штамму, где бактериальный штамм представляет собой Bifidobacterium longum DSM 32947 или Bifidobacterium longum DSM 32948.
Еще один аспект вариантов осуществления изобретения относится к пробиотической композиции, содержащей бактерии штамма Lactobacillus, предпочтительно штамма L. reuteri, и более предпочтительно штамма L. reuteri, выбранного из группы, состоящей из L. reuteri DSM 17938, L. reuteri DSM 32846 и их комбинации. Композиция также содержит бактерии штамма Bifidobacterium longum, выбранного из группы, состоящей из B. longum DSM 32947, B. longum DSM 32948 и их комбинации, или кондиционированную среду после культивирования штамма B. longum.
Терапевтические микровезикулы, продуцированные пробиотическими бактериями, были способны воспроизводить полезные эффекты пробиотических бактерий, как здесь показано. Кроме того, терапевтические микровезикулы фактически были более эффективными, чем пробиотические бактерии, которые их продуцируют, о чем свидетельствует более быстрое начало их специфических полезных эффектов, как показано в примерах.
Краткое описание фигур
Варианты осуществления вместе с их дополнительными целями и преимуществами можно лучше всего понять, обратившись к нижеследующему описанию вместе с прилагаемыми фигурами, где:
На фиг. 1 приведены результаты определения активности 5'-нуклеотидазы в кондиционированной среде после подвергания Lactobacillus reuteri DSM 17938 различным индуцирующим обработкам.
На фиг. 2 показано влияние добавления L. reuteri DSM 17938 (DSM) на фиг. 2A, кондиционированной среды (CM) на фиг. 2B, культуральной среды (бульона) на фиг. 2C, микровезикул, полученных из DSM (микроВ) на фиг. 2D, или кондиционированной среды минус микровезикулы (CM-микроВ) на фиг. 2E, на скорости распространения сократительного комплекса (PCC) на отрезках мышиной тощей кишки in vitro. Верхние панели: гистограммы, показывающие средние значения и стандартные ошибки. Значения P, полученные из анализа парных t-критериев, приведены над горизонтальными полосами. Нижние панели: графики индивидуальных значений разницы (обработка-контрольный раствор Кребса) с 95% доверительными интервалами для каждого соответствующего графика в верхнем ряду. На фиг. 3-7 показаны эти же отношения между верхней и нижней панелями.
На фиг. 3 показано влияние добавления DSM (фиг. 3A), CM (фиг. 3B), бульона (фиг. 3C), микроВ (фиг. 3D) или CM-мкВ (фиг. 3E) на частоту PCC на отрезках мышиной тощей кишки in vitro.
На фиг. 4 показано влияние добавления DSM (фиг. 4A), CM (фиг. 4B), бульона (фиг. 4C), микроВ (фиг. 4D) или CM-мкВ (фиг. 4E) на максимальную амплитуду PCC на отрезках мышиной тощей кишки in vitro.
На фиг. 5 показано влияние добавления DSM (фиг. 5A), CM (фиг. 5B), бульона (фиг. 5C), микроВ (фиг. 5D) или CM-мкВ (фиг. 5E) на скорость PCC на отрезках мышиной ободочной кишки in vitro.
На фиг. 6 показано влияние добавления DSM (фиг. 6A), CM (фиг. 6B), бульона (фиг. 6C), микроВ (фиг. 6D) или CM-микроВ (фиг. 5E) на частоту PCC на отрезках мышиной ободочной кишки in vitro.
На фиг. 7 показано влияние добавления DSM (фиг. 7A), CM (фиг. 7B), бульона (фиг. 7C), микроВ (фиг. 7D) или CM-микроВ (фиг. 7E) на максимальную амплитуду PCC на отрезках мышиной ободочной кишки in vitro.
На фиг. 8 приведены обобщенные результаты, представленные на фиг. 2-7 по скорости, частоте и максимальной амплитуде в тощей кишке и ободочной кишке.
На фиг. 9 показано влияние микровезикул на передачу сигналов TrpV1. На графике показано капсаицин-индуцированная ответная реакция, полученная с использованием микровезикул, выделенных из L. reuteri DSM 17938 (DSM-МВ), и микровезикул, выделенных из бактериального штамма Lactobacillus rhamnosus JB-1 (МВ-JB-1).
На фиг.10 представлен график, показывающий время начала ответной реакции на модели возбуждения брыжеечного нерва с использованием L. reuteri DSM 17938 (DSM) и микровезикул, выделенных из L. reuteri DSM 17938 (МВ).
На фиг.11 показано, что микровезикулы (МВ), полученные из L. reuteri DSM 17938, обладают иммуномодулирующим действием и подавляют ответную секрецию IFN-γ и IL-17A. Оценка иммуномодулирующих эффектов очищенных МВ, полученных из L. reuteri DSM 17938, в культурах PBMC. (Фиг. 11A) PBMC культивировали в течение 48 ч в присутствии L. reuteri (Lr)-МВ в соотношении (МВ:клетки) 500:1, 100:1 и 20:1 с последующим количественным определением уровней секретированного IL-6, IL-10, IL-17A и IFN-γ (n=8). (Фиг.11B) PBMC стимулировали Staphylococcus aureus (Sa)-CFS (2,5%) в присутствии L.r-МВ в соотношении (МВ:клетки) 500:1, 100:1 и 20:1 с последующим количественным определением секретированных уровни IFN-γ IL-17A. Показаны относительные значения, нормализованные к одним S. aureus-CFS (n=8). Прямоугольники включают данные между 25-м и 75-м процентилями с медианными значениями в виде центральной линии и полосами ошибок, показывающими минимальное и максимальное значение. Гистограммы показывают медианное значение с интерквартильным размахом.
На фиг. 12 показано, что микровезикулы, полученные из L. reuteri DSM 17938 и L. reuteri DSM 17938, защищают целостность эпителия от разрушительного действия энтеротоксигенных Escherichia coli (ETEC).
На фиг.13 показана 5'-нуклеотидазная активность в образцах МВ, полученных из L. reuteri DSM 17938 (культивирование L. reuteri DSM 17938 с добавлением 4% супернатанта B. longum ATCC BAA-999 или B. longum DSM 32947 на среде SIM). (DSM 17938+4% DSM 32947 sup. или DSM 17938+4% ATCC BAA-999 sup.) в сравнении с L. reuteri DSM 17938 в SIM (контроль) или L. reuteri DSM 17938 с 4% супернатантом L. paracasei LMG-P-17806 (DSM 17938 на среде SIM или DSM 17938+4% LMG-P-17806 sup.)
На фиг. 14 приведены те же данные, что и на фиг. 13, но которые были нормализованы по отношению к 5'-нуклеотидазной активности и оптической плотности DSM 17938 на среде SIM.
На фиг.15 показана 5'-нуклеотидазная активность в контрольных образцах (DSM 17938 в SIM) по сравнению с образцами, полученными после биотической индуцирующей обработки посредством сокультивирования L. reuteri DSM 17938 с 25% клеток B. longum DSM 32947. Результаты были нормализованы по отношению к 5'-нуклеотидазной активности и оптической плотности DSM 17938 на среде SIM.
На фиг.16 показана 5'-нуклеотидазная активность в образцах МВ после культивирования L. reuteri DSM 32846 с 4% супернатантом B. longum DSM 32947 на среде SIM (DSM 32846+4% DSM 32947 sup.) по сравнению с контрольными L. reuteri DSM 32846 на среде SIM (DSM 32846 на среде SIM). Результаты были нормализованы по отношению к 5'-нуклеотидазной активности и оптической плотности DSM 32846 на среде SIM.
На фиг. 17 показано, что микровезикулы, полученные из L. reuteri DSM 32846, более эффективны в индукции продуцирования IL-6 по сравнению с микровезикулами, выделенными из L. reuteri DSM 17938.
На фиг. 18 и 19 показана индукция секреции IL-6 под действием МВ, выделенных из L. reuteri DSM 32846 и L. reuteri DSM 17938 после культивирования с 4% супернатантом из штаммов B. longum по сравнению с контрольными образцами (DSM 32846 или DSM 17938 соответственно).
На фиг. 20 показана индуцированная продукция IL-6 под действием МВ, полученных из L. reuteri DSM 17938, после сокультивирования с 25% клеток B. longum DSM32947 по сравнению с контрольными образцами (DSM 17938).
На фиг.21 показано, что МВ, полученные из L. reuteri DSM 32846, были частично способны защищать эпителиальный монослой от инфицирования ETEC, вызывающего снижение TEER. На фигуре также показан защитный эффект МВ, полученных из L. reuteri DSM 32846, в отношении повреждения монослоя под действием ETEC в эксперименте с утечкой FITC-декстрана.
На фиг. 22 приведено сравнение между защитным эффектом МВ, полученными из L. reuteri DSM 32846, в эксперименте с утечкой FITC-декстрана и эффектом, полученным с МВ, полученными из L. reuteri DSM 17938. Предварительная обработка монослоев эпителиальных клеток МВ, полученными из L. reuteri DSM 32846, уменьшала утечку FITC-декстрана более эффективно, особенно при более низких концентрациях МВ, по сравнению с МВ, полученными из L. reuteri DSM 17938.
Подробное описание изобретения
Настоящее изобретение, в общем, относится к терапевтическим микровезикулам, полученным из пробиотических бактерий, и их применениям.
Определения
Микровезикулы (МВ, микроВ), также называемые в данной области техники, например, мембранными везикулами, везикулами внешней мембраны, внеклеточными везикулами, представляют собой известную форму коммуникации, которая используется бактериями и эукариотическими клетками. Высвобождение биоактивных МВ с поверхности клетки сохраняется на протяжении всей «жизни» микробов, у бактерий, архей, грибов и паразитов, и продуцирование МВ было показано как in vitro, так и in vivo, что указывает на влияние этих поверхностных органелл на микробную физиологию и патогенез через доставку важных сигнальных молекул, ферментов и токсинов. Бактериальные МВ регулярно продуцируются и выделяются как грамположительными, так и грамотрицательными бактериями, и результаты протеомных исследований показали, что состав таких МВ может отличаться от состава родительских бактерий. МВ могут включать липидные молекулы, молекулы РНК, молекулы ДНК и/или белок. Кроме того, МВ также могут содержать поверхностные компоненты родительских бактерий.
МВ, продуцированные пробиотическими бактериями, как здесь описано, обозначены здесь как терапевтические МВ для указания того, что МВ обладают терапевтическим эффектом. Данный терапевтический эффект МВ может быть таким же или, по меньшей мере, подобен пробиотическому эффекту самих пробиотических бактерий, продуцирующих МВ, при введении субъекту. Следовательно, МВ будут оказывать терапевтический эффект у субъекта, в результате которого можно подавлять, лечить или профилактировать, включая замедление начала развития болезненного состояния, заболевания или расстройства у субъекта, как дополнительно описано в данном документе.
Культуральная среда или питательная среда является стартовой средой, в которой будут культивироваться бактерии.
Кондиционированная среда представляет собой культуральную или питательную среду, в которой культивировались бактерии. Таким образом, такая кондиционированная среда включает любые соединения или агенты, включая МВ, высвобождаемые бактериями в культуральную среду. Бактерии удаляются из культуральной среды и, следовательно, не являются частью кондиционированной среды. Кондиционированная среда может быть получена, например, центрифугированием, седиментацией и/или осаждением культуры бактериальных клеток с получением кондиционированной среды в виде супернатанта.
Клеточная суспензия представляет собой смесь культивированных бактерий и культуральной среды, включая любые соединения или агенты, такие как МВ, высвобождаемые бактериями в культуральную среду, т. е. в кондиционированную среду. Клеточная суспензия представляет собой конечный результат брожения.
Пробиотический бактериальный штамм предпочтительно представляет собой штамм пробиотических бактерий, продуцирующих молочную кислоту, иногда также называемых молочнокислыми бактериями. Бактерии, продуцирующие молочную кислоту, представляют собой группу грамположительных бактерий с низким % содержанием GC в геноме, кислотоустойчивых, обычно не образующих спор, анаэробных, палочковидных или кокковидных бактерий, которые имеют общие метаболические и физиологические характеристики. Данные бактерии продуцируют молочную кислоту в качестве основного конечного продукта метаболизма углеводного брожения. К родам бактерий, продуцирующих молочную кислоту, относятся Lactobacillus, Leuconostoc, Pediococcus, Lactococcus и Streptococcus. Bifidobacterium не относится к традиционным молочнокислым бактериям по причине их генетических отличий, но эти бактерии имеют свойства, которые перекрываются с молочнокислыми бактериями, и у них имеет место метаболизм, в результате которого продуцируется молочная кислота в качестве основного конечного продукта брожения, хотя они продуцируют намного меньше молочной кислоты, чем Lactobacillus. Бифидобактерии являются строго анаэробными, и обычно в большом количестве обнаруживаются в толстом кишечнике.
МВ, продуцированные пробиотическими бактериями, важны, поскольку они представляют собой средства коммуникации между пробиотическими бактериями и окружающими клетками-хозяевами, такими как клетки слизистой оболочки в организме человека, например, клетки слизистой оболочки кишечника в желудочно-кишечной системе, слизистой оболочки ротовой полости или влагалища. Следовательно, МВ могут передавать информацию, такую как пробиотическая информация, от бактериальных клеток хозяину. Следовательно, существует потребность в повышении продукции терапевтических МВ из пробиотических бактерий, которые можно использовать в пробиотических и терапевтических применениях. Экспериментальные данные, представленные здесь, показывают, что МВ, продуцированные пробиотическими бактериями, могут воспроизводить пробиотические или терапевтические эффекты самих пробиотических бактерий, что показано действием выделенных МВ на передачу болевых сигналов и моторику желудочно-кишечного тракта. МВ не только воспроизводили эффекты пробиотических бактерий на передачу болевых сигналов, но и фактически были более эффективными, о чем свидетельствует их способность функционировать быстрее, чем сами пробиотические бактерии, что приводило к более раннему началу наблюдаемого эффекта. Данное открытие было весьма неожиданным. Экспериментальные данные, представленные здесь, также показывают, что терапевтические МВ, выделенные из пробиотического бактериального штамма, обладают иммуностимулирующим действием и способны подавлять специфические цитокины, ассоциированные с аутоиммунными заболеваниями, и что они также защищают целостность эпителиального барьера. Экспериментальные данные, представленные здесь, также показывают, что продукцию МВ можно индуцировать биотической обработкой во время культивирования, например, посредством добавления супернатанта после культивирования другого бактериального штамма к пробиотическому бактериальному штамму или посредством сокультивирования пробиотических бактерий с бактериями другого бактериального штамма.
Таким образом, в настоящем изобретении описаны протоколы, которые можно использовать для получения терапевтических МВ и/ или для повышения присущей им или эндогенной продукции терапевтических МВ.
Аспект вариантов осуществления изобретения включает способ получения терапевтических МВ. Способ включает культивирование бактерий пробиотического бактериального штамма в культуральной среде и подвергание бактерий индуцирующей биотической обработке во время культивирования для индукции продукции терапевтических МВ бактериями. Пробиотический бактериальный штамм выбран из группы, состоящей из штамма Lactobacillus, штамма Bifidobacterium и их комбинации. В данном аспекте индуцирующая биотическая обработка выбрана из группы, состоящей из сокультивирования бактерий пробиотического бактериального штамма с бактериями другого бактериального штамма, культивирования бактерий пробиотического бактериального штамма в присутствии кондиционированной среды после культивирования бактерий другого бактериального штамма и их комбинации. Другой бактериальный штамм представляет собой штамм Bifidobacterium, и другой бактериальный штамм отличается от пробиотического бактериального штамма.
Следовательно, в данном аспекте получение терапевтических МВ включает культивирование бактерий пробиотического бактериального штамма и в то же время стимуляцию бактерий на продуцирование терапевтических МВ посредством подвергания бактерий индуцирующей обработке во время культивирования для индукции продукции терапевтических МВ.
В рамках настоящего изобретения, выражение «индуцировать продукцию терапевтических МВ» включает стимуляцию бактерий на продукцию терапевтических МВ, включая стимулирование, усиление или увеличение продукции терапевтических МВ бактериями. Альтернативно или дополнительно, выражение «индуцировать продукцию терапевтических МВ» включает более эффективное высвобождение терапевтических МВ из бактерий, что приводит к большему количеству высвобождаемых терапевтических МВ бактериями по сравнению с тем, когда бактерии не подвергаются индуцирующей обработке. Альтернативно или дополнительно, выражение «индуцировать продукцию терапевтических МВ» включает продукцию бактериями более сильных или более эффективных терапевтических МВ. В таком случае терапевтические МВ, продуцированные бактериями, подвергнутыми индуцирующей обработке, обладают усиленным терапевтическим эффектом по сравнению с МВ, продуцированными неиндуцированными бактериями, т. е. бактериями, не подвергшимися индуцирующей обработке. Следовательно, индуцирующую обработку согласно вариантам осуществления можно использовать, например, для повышения продукции терапевтических МВ в бактериях пробиотического бактериального штамма, которые уже имеют присущую им или эндогенную продукцию таких терапевтических МВ. В этом случае индуцирующая обработка усиливает эту присущую им или эндогенную продукцию МВ бактериями, давая больше терапевтических МВ при воздействии индуцирующей обработки по сравнению с тем, когда они не подвергаются индуцирующей обработке. Выражение «индуцировать продукцию терапевтических МВ» также включает индуцирование продукции таких терапевтических МВ в бактериях пробиотического бактериального штамма, которые в противном случае не имеют значительной продукции МВ, когда они не подвергаются индуцирующей обработке. Следовательно, выражение «индуцировать продукцию терапевтических МВ» посредством индуцирующей обработки включает в себя как увеличение присущей им или эндогенной продукции терапевтических МВ в бактериях, так и продукцию de novo терапевтических МВ в бактериях.
Индуцирующая обработка, как здесь описано ниже, представляет собой обработку бактерий, которая индуцирует, включая усиление, продукцию бактериями терапевтических МВ. Следовательно, подвергая бактерии, по меньшей мере, одной индуцирующей обработке во время культивирования в соответствии с различными вариантами осуществления, бактерии модифицируются или индуцируются для продукции терапевтических МВ.
Культивирование бактерий может быть выполнено в соответствии с известным протоколом культивирования в подходящем устройстве для культивирования, ферментере или биореакторе, включая, помимо прочего, биореакторы непрерывного действия с механическим перемешиванием, эрлифтные биореакторы, биореакторы с полым волокном и биореакторы с вращающейся системой культивирования клеток (RCCS). Конкретные условия культивирования предпочтительно выбирают с учетом конкретного пробиотического бактериального штамма.
В одном варианте осуществления культуральная среда, содержащая терапевтические МВ и пробиотические бактерии, т. е. клеточная суспензия, подвергается консервации, например, посредством сушки и/или замораживания. Типичные примеры сушки включают распылительную сушку, сублимационную сушку, распылительную сублимационную сушку и вакуумную сушку.
Клеточную суспензию можно необязательно концентрировать до или во время консервации для уменьшения общего объема клеточной суспензии, а также для концентрирования бактериальных клеток, терапевтических МВ и любых других соединений или агентов, присутствующих в ней.
Например, клеточная суспензия может быть сконцентрирована до объема, соответствующего примерно от 5% до 95% от исходного объема, такого как 5, 10, 15, 20, 25, 30, 35, 40, 45, 50, 55, 60, 65, 70, 75, 80, 85, 90 или 95% от исходного объема.
Для концентрирования клеточной суспензии можно использовать различные методы и процессы. Например, клеточная суспензия может быть сконцентрирована удалением воды и, необязательно, других веществ, таких как органические кислоты, сахара и соли, из клеточной суспензии. Можно использовать фильтрующее устройство, которое в основном пропускает воду через фильтрующую мембрану. Данный процесс называется осмосом и может выполняться в различных рабочих режимах, таких как режим обратного осмоса, режим прямого осмоса. В одном варианте осуществления концентрирование также может быть выполнено осаждением образца с использованием химических соединений. Химическое осаждение можно использовать для концентрирования клеточной суспензии с добавлением денатурирующих растворителей или солей. Различные типы хроматографических установок также можно использовать для концентрирования и разделения входных образцов, например, в соответствии с их химическими свойствами и размером. Например, эксклюзионная хроматография функционирует по принципу разделения образцов по размеру, захватывая более мелкие молекулы в маленькие поры и вытесняя более крупные молекулы и частицы. Ионная хроматография помогает разделить молекулы с определенными зарядами. Например, в анионообменной хроматографии может использоваться суммарный отрицательный заряд, находящийся на бактериальных мембранах и МВ, и связывание их с положительно заряженной хроматографической матрицей. Затем их можно элюировать, увеличивая ионную силу окружающей подвижной фазы. Другим метод концентрирования является ультрафильтрация. Ультрафильтрация основана на механическом, а не на химическом взаимодействии. Фильтрующие устройства могут иметь отсечение по молекулярной массе, означая, что все, что превышает определенную молекулярную массу, удерживается на фильтре, в то время как более мелкие молекулы, соли и т. д проходят через фильтр. Обычно используются мембраны с четко определенным размером пор. Данные типы устройств и процессов ультрафильтрации также используются в различных установках, таких как прямая проточная фильтрация (DFF) или тангенциальная проточная фильтрация (TFF). Тангенциальная проточная фильтрация (TFF), также известная как фильтрация поперечным потоком, отличается от других систем фильтрации тем, что жидкость проходит параллельно фильтру, а не проталкивается через мембрану перпендикулярно. Данный метод является предпочтительным за счет его непрерывной фильтрации и воспроизводимых характеристик. Частицы, которые проходят через мембрану, пермеат, отводятся в сторону, а остальная часть, ретентат, возвращается обратно в загрузочный материал.
В одном варианте осуществления бактерии подвергаются индуцирующей обработке во время культивирования для индукции, в том числе, для усиления, продукции терапевтических МВ и высвобождения терапевтических МВ в культуральную среду. Это означает, что терапевтические МВ, продуцированные бактериями пробиотического бактериального штамма после подвергания индуцирующей обработке, высвобождаются из бактерий в культуральную среду. В дополнение или альтернативно, по меньшей мере, некоторые из терапевтических МВ, продуцированные бактериями, могут быть связаны и/или присоединены к клеточной мембране и/или клеточной стенке бактерий.
В одном варианте осуществления способ включает выделение терапевтических МВ из культуральной среды, например, из клеточной суспензии или из кондиционированной среды. В одном варианте осуществления выделение терапевтических МВ включает подвергание культуральной среды, например клеточной суспензии, по меньшей мере, одному центрифугированию при относительной центробежной силе, выбранной в пределах первого диапазона c получением супернатанта, обедненного бактериями, т. е. кондиционированной среды и подвергание кондиционированной среды, по меньшей мере, одному ультрацентрифугированию при относительной центробежной силе, выбранной в пределах второго диапазона, с получением осадка, содержащего МВ. Второй диапазон больше чем первый.
Первое или, по меньшей мере, одно центрифугирование при относительной центробежной силе, выбранной в пределах первого диапазона, выполняется для удаления живых бактерий и крупного дебриса из культуральной среды с образованием тем самым осадка, который отбрасывается, и супернатанта, содержащего терапевтические МВ, обозначенного кондиционированной средой выше. Данная первая стадия процесса выделения может включать в себя одну стадию центрифугирования, но предпочтительно включает, по меньшей мере, две стадии центрифугирования для более эффективного удаления бактерий и крупного дебриса. В случае, по меньшей мере, двух стадий центрифугирования все они могут проводиться при одинаковой относительной центробежной силе. Однако, как правило, более эффективно увеличивать относительную центробежную силу на каждой последующей стадии центрифугирования. Первый диапазон предпочтительно составляет от 100 × g до 50000 × g, например, от 200 × g до 25000 × g, и предпочтительно от 500 × g до 15000 × g. Например, первая стадия центрифугирования может быть при 4000 × g, и вторая стадия центрифугирования - при 10000 × g. В качестве альтернативы можно использовать одну стадию центрифугирования при 600 × g. Супернатант также или альтернативно может быть пропущен через микронный фильтр (от 0,20 мкм до 0,50 мкм, например, 0,45 мкм) для удаления любого дебриса и/или бактерий, оставшихся после центрифугирования.
Кондиционированную среду, содержащую терапевтические МВ, затем подвергают, по меньшей мере, одному ультрацентрифугированию при относительной центробежной силе, выбранной во втором диапазоне, с получением осадка, содержащего МВ. Данная вторая стадия может включать одну или более стадий ультрацентрифугирования. В случае нескольких стадий ультрацентрифугирования все они могут проводиться при одной и той же относительной центробежной силе, или относительная центробежная сила может быть увеличена, как описано выше. Второй диапазон предпочтительно равен или больше 75000 × g, например, равен или выше 85000 × g, предпочтительно равен или выше 100000 × g. Например, можно использовать относительную центробежную силу 118000 × g.
В одном варианте осуществления стадия выделения также включает загрузку кондиционированной среды на градиент сахарозы или сахарозную «подушку» и центрифугирование при относительной центробежной силе, выбранной во втором диапазоне.
В дополнение или альтернативно кондиционированную среду можно профильтровать перед проведением ультрацентрифугирования. В таком случае можно использовать фильтр со средним размером пор, например, от 0,20 мкм до 0,50 мкм.
Выделенные терапевтические МВ можно подвергнуть консервации, например, сушкой, например распылительной сушкой, сублимационной сушкой, распылительной сублимационной сушкой или вакуумной сушкой и/или замораживанием. В одном варианте осуществления изобретения терапевтические МВ являются стабильными после хранения в течение, по меньшей мере, 1 месяца, по меньшей мере, 3 месяцев, по меньшей мере, 5 месяцев или, по меньшей мере, 7 месяцев.
Индуцирующая биотическая обработка выбрана из группы, состоящей из сокультивирования пробиотического бактериального штамма с бактериями другого бактериального штамма, культивирования пробиотического бактериального штамма в присутствии кондиционированной среды после культивирования бактерий другого бактериального штамма и их комбинации. Другой бактериальный штамм предпочтительно представляет собой пробиотический бактериальный штамм. Например, другие бактерии могут относиться к штамму Bifidobacterium, предпочтительно к штамму Bifidobacterium longum, и более предпочтительно к B. longum DSM 32947 и/или DSM 32948 (депонированы BioGaia AB в соответствии с Будапештским договором в Немецкой коллекции микроорганизмов и клеточных культур (Inhoffenstrasse 7B, D-38124 Braunschweig, Германия) 1 ноября 2018 г.). Вышеприведенные примеры штаммов Bifidobacterium особенно пригодны в сочетании с бактериями штамма Lactobacillus, который используется в качестве пробиотического бактериального штамма и, в частности, с бактериями штамма L. reuteri, такими как L. reuteri DSM 17938 (депонированы BioGaia AB в соответствии с Будапештским договором в Немецкой коллекции микроорганизмов и клеточных культур (Inhoffenstrasse 7B, D-38124 Braunschweig, Германия) 30 января 2006 г.) и/или L. reuteri DSM 32846 (депонированы BioGaia AB в соответствии с Будапештским договором в Немецкой коллекции микроорганизмов и клеточных культур (Inhoffenstrasse 7B, D-38124 Braunschweig, Германия) 4 июля 2018 г.).
В конкретном варианте осуществления бактерии пробиотического бактериального штамма можно сокультивировать с бактериями другого бактериального штамма. Альтернативно или дополнительно, кондиционированная среда после культивирования бактерий другого бактериального штамма может быть добавлена к культуральной среде, содержащей бактерии пробиотического бактериального штамма.
В одном варианте осуществления способ также включает подвергание бактерий индуцирующей абиотической обработке во время культивирования для индукции, в том числе, увеличения, продукции терапевтических МВ бактериями. Следовательно, в данном варианте осуществления бактерии подвергаются как индуцирующей биотической обработке, так и индуцирующей абиотической обработке.
Абиотическая обработка относится к обработке «неживыми» химическими и физическими компонентами, которые влияют на живые организмы. Биотическая обработка относится к обработке биотическим материалом, который представляет собой либо живые организмы, либо получен из живых организмов.
В конкретном варианте осуществления абиотическая обработка представляет собой обработку абиотическим стрессором, т.е. абиотическую обработку, которая индуцирует стрессовую реакцию у бактерий пробиотического бактериального штамма при подвергании абиотической обработке во время культивирования. В одном варианте осуществления абиотический стрессор выбран из группы, состоящей из окислительного стресса (обработка кислородом), температурного стресса, рН-стресса, ультрафиолетового (УФ) стресса и их комбинации.
Обработка кислородом означает, что бактерии подвергаются воздействию повышенной концентрации кислорода. В одном варианте осуществления повышенная концентрация кислорода представляет собой нетоксичную концентрацию кислорода. В конкретном варианте осуществления подвергание бактерий кислородной обработке включает подвергание относительные толерантных к кислороду анаэробных бактерий, микроаэрофильных бактерий, аэробных бактерий и/или факультативных анаэробных бактерий воздействию повышенных концентраций кислорода, т. е. повышенных нетоксичных концентраций кислорода, во время культивирования для индукции продукции терапевтических МВ анаэробными бактериями, относительно толерантными к кислороду, микроаэрофильными бактериями, аэробными бактериями и/или факультативными анаэробными бактериями.
В рамках настоящего изобретения, используемая обработка кислородом, не включает добавление каких-либо активных форм кислорода (ROS), таких как пероксиды, включая пероксид водорода, супероксид или гидроксильные радикалы.
Повышенная концентрация кислорода подразумевает концентрацию кислорода в культуральной среде, которая выше, чем (нормальная) концентрация кислорода, которая в противном случае выбрана как оптимальная или, по меньшей мере, подходящая для культивирования бактерий пробиотического бактериального штамма, однако не является токсичной для бактерий. В одном варианте осуществления нетоксичная концентрация кислорода не вызывает значительной гибели бактериальных клеток, означая, что подвергшиеся воздействию бактерии все еще жизнеспособны, по меньшей мере, до 70%, предпочтительно, по меньшей мере, до 75%, более предпочтительно, по меньшей мере, до 80%, например, по меньшей мере до 85% или 90%, или даже выше, чем при воздействии на бактерии нормальной концентрации кислорода. Это увеличение концентрации кислорода может быть достигнуто добавлением (барботированием) кислорода или воздуха к культуральной среде за один или несколько приемов или в течение длительного периода времени. Альтернативно или дополнительно, увеличение концентрации кислорода может быть достигнуто встряхиванием или перемешиванием культуральной среды, содержащей бактерии, включая увеличение количества или уровня встряхивания или перемешивания культуральной среды. Концентрация кислорода, например, нетоксичная концентрация, может варьироваться для разных бактериальных штаммов, но обычно может быть установлена в пределах от 0,1% до 10%. В одном варианте осуществления концентрация кислорода составляет от 0,5% до 2%. В другом варианте концентрация кислорода составляет от 2% до 5%. В еще одном варианте осуществления концентрация кислорода составляет от 5% до 10%.
Конкретный аспект вариантов осуществления изобретения включает способ получения терапевтических МВ. Способ включает культивирование бактерий пробиотического бактериального штамма в культуральной среде и подвергание бактерий окислительной обработке во время культивирования для индукции продуцирования терапевтических МВ бактериями.
Следовательно, в данном аспекте вариантов осуществления продуцирование МВ индуцируется, в том числе усиливается, у бактерий, подверганием их обработке кислородом (окислительным стрессом), но не обязательно в сочетании с подверганием бактерий любой индуцирующей биотической обработке.
В одном варианте осуществления бактерии пробиотического бактериального штамма выбраны из группы, состоящей из анаэробных бактерий с относительной толерантностью к кислороду, микроаэрофильных бактерий, аэробных бактерий и/или факультативных анаэробных бактерий, предпочтительно выбраны из группы, состоящей из аэробных бактерий и факультативных анаэробных бактерий.
В конкретном варианте осуществления бактерии выбраны из группы, состоящей из штамма Lactobacillus, штамма Bifidobacterium и их комбинации. Предпочтительные штаммы Lactobacillus и Bifidobacterium могут быть выбраны среди нижеприведенных иллюстративных примеров предпочтительных бактериальных штаммов.
Температурный стресс может быть вызван повышением температуры культивирования выше нормальной температуры, используемой для культивирования бактерий в биореакторе, т. е. так называемый высокотемпературный стресс. Например, если нормальная температура культивирования составляет 37°C, то температуру можно повысить, по меньшей мере, до 42°C, по меньшей мере, до 43°C или, по меньшей мере, до 44°C, и более предпочтительно, по меньшей мере, до 45°C, например, по меньшей мере, до 46°C, по меньшей мере, до 47°C, по меньшей мере, до 48°C, по меньшей мере, до 49°C или, по меньшей мере, до 50°C. Вместо того, чтобы подвергнуть бактерии высокотемпературному стрессу, бактерии можно подвергнуть низкотемпературному стрессу, т. е. понижением температуры культивирования ниже нормальной температуры культивирования. Например, температура культивирования может быть понижена до 10°C, например, до 8°C или ниже, до 6°C или ниже или до 4°C или ниже.
pH-стресс может быть вызван снижением pH культуральной среды, в которой культивируются бактерии, от нормального или базового pH до кислого или более кислого pH. Альтернативно, бактерии могут быть временно удалены из культуральной среды, а затем подвергнуты pH-стрессу с последующим внесением бактерий, подвергнутых pH-стрессу, в культуральную среду или в свежую культуральную среду. Например, pH можно снизить с нормального диапазона pH от 6,5 до 7 до pH 2 или ниже.
УФ-стресс может быть вызван подверганием бактерий УФ-обработке, например, направлением УФ-света в культуральную среду, содержащую бактерии.
В одном варианте осуществления способ также включает подвергание бактерий воздействию стресс-индуцирующего агента во время культивирования для индукции, в том числе увеличения, продукции терапевтических МВ бактериями. Следовательно, в данном варианте осуществления бактерии подвергаются воздействию как стресс-индуцирующего агента, так и индуцирующей биотической обработке и/или индуцирующей абиотической обработке.
В одном варианте осуществления стресс-индуцирующий агент выбран из группы, состоящей из фруктозы; сахарозы; лизоцима, например, из куриного яйца, также известного как мурамидаза или N-ацетилмурамидгликангидролаза; муцина, например, выделенного из кишечника свиней; β-лактама, например ампициллина, и их комбинации.
В конкретном варианте осуществления стресс-индуцирующий агент представляет собой сахарозу. Сахароза может быть добавлена во время культивирования бактерий для индукции продукции МВ указанными бактериями. Например, сахароза может быть добавлена в культуральную среду для обеспечения концентрации сахарозы в диапазоне от 0,3% до 10% в культуральной среде, такой как 0,3, 0,4, 0,5, 0,6, 0,7, 0,8, 0,9, 1, 1,5, 2, 2,5, 3, 3,5, 4, 4,5, 5, 5,5, 6, 6,5, 7, 7,5, 8, 8,5, 9, 9,5 или 10%.
Вышеописанные примеры индуцирующих обработок можно объединить, например, комбинированием нескольких, т. е., по меньшей мере, двух, абиотических обработок, нескольких биотических обработок, обработок несколькими стресс-индуцирующими агентами, по меньшей мере, одной абиотической обработки и, по меньшей мере, одной биотической обработки, по меньшей мере, одной абиотической обработки и обработки, по меньшей мере, одним стресс-индуцирующим агентом, по меньшей мере, одной биотической обработки и обработки стресс-индуцирующим агентом, или, по меньшей мере, одной абиотической обработки, по меньшей мере одной биотической обработки и обработки, по меньшей мере, одним стресс-индуцирующим агентом.
Продолжительность воздействия индуцирующей обработки может быть выбрана с учетом конкретного типа обработки, конкретного штамма пробиотических бактерий и условий культивирования, таких как тип биореактора. Например, бактерии могут подвергаться абиотической обработке в течение 10 мин, 15 мин, 30 мин, 45 мин, 1 ч, 1,25 ч, 1,5 ч, 1,75 ч, 2 ч, 2,25 ч, 2,5 ч, 2,75 ч, 3 ч, 3,25 ч, 3,5 ч, 3,75 ч, 4 ч, 4,25 ч, 4,5 ч, 4,75 ч, 5 ч или более, в качестве иллюстративных, но не ограничивающих примеров. Также возможны более длительные периоды воздействия абиотического стресса, такие как в течение ночи, 12 ч, 18 ч, 24 ч или даже более. Добавление стресс-индуцирующего агента может включать добавление, по меньшей мере, одного стресс-индуцирующего агента, в культуральную среду один раз или несколько раз, т. е. по меньшей мере, два раза. Добавление бактерий другого бактериального штамма или кондиционированной среды из таких бактерий также можно выполнить один или более раз.
Пробиотический бактериальный штамм предпочтительно представляет собой штамм пробиотических бактерий, продуцирующих молочную кислоту, и, в частности, выбран из Lactobacillus и Bifidobacterium. Lactobacillus включают несколько видов, в том числе, L. acetotolerans, L. acidifarinae, L. acidipiscis, L. acidophilus, L. agilis, L. algidus, L. alimentarius, L. amylolyticus, L. amylophilus, L. amylotrophicus, L. amylovorus, L. animalis, L. antri, L. apodemi, L. aviaries, L. bifermentans, L. brevis, L. buchneri, L. camelliae, L. casei, L. catenaformis, L. ceti, L. coleohominis, L. collinoides, L. composti, L. concavus, L. coryniformis, L. crispatus, L. crustorum, L. curvatus, L. delbrueckii subsp. bulgaricus, L. delbrueckii subsp. delbrueckii, L. delbrueckii subsp. lactis, L. dextrinicus, L. diolivorans, L. equi, L. equigenerosi, L. farraginis, L. farciminis, L. fermentum, L. fornicalis, L. fructivorans, L. frumenti, L. fuchuensis, L. gallinarum, L. gasseri, L. gastricus, L. ghanensis, L. graminis, L. hammesii, L. hamster, L. harbinensis, L. hayakitensis, L. helveticus, L. hilgardii, L. homohiochii, L. iners, L. ingluviei, L. intestinalis, L. jensenii, L. johnsonii, L. kalixensis, L. kefiranofaciens, L. kefiri, L. kimchi, L. kitasatonis, L. kunkeei, L. leichmannii, L. lindneri, L. malefermentans, L. mali, L. manihotivorans, L. mindensis, L. mucosae, L. murinus, L. nagelii, L. namurensis, L. nantensis, L. oligofermentans, L. oris, L. panis, L. pantheris, L. parabrevis, L. parabuchneri, L. paracasei, L. paracollinoides, L. parafarraginis, L. parakefiri, L. paralimentarius, L. paraplantarum, L. pentosus, L. perolens, L. plantarum, L. pontis, L. protectus, L. psittaci, L. rennini, L. reuteri, L. rhamnosus, L. rimae, L. rogosae, L. rossiae, L. ruminis, L. saerimneri, L. sakei, L. salivarius, L. sanfranciscensis, L. satsumensis, L. secaliphilus, L. sharpeae, L. siliginis, L. spicheri, L. suebicus, L. thailandensis, L. ultunensis, L. vaccinostercus, L. vaginalis, L. versmoldensis, L. vini, L. vitulinus, L. zeae и L. zymae. Предпочтительные примеры такого пробиотического бактериального штамма включают Lactobacillus reuteri, Lactobacillus mucosae, Lactobacillus gasseri и Lactobacillus plantarum. В настоящее время предпочтительные примеры такого пробиотического бактериального штамма включают штаммы Lactobacillus reuteri, такие как Lactobacillus reuteri DSM 17938 and Lactobacillus reuteri DSM 32846. Предпочтительными штаммами Bifidobacterium являются B. adolescentis, B. breve, B. longum, B. animalis, B. infantis, B. thermophilum, B. bifidum и B. lactis. Еще одним предпочтительным видом Bifidobacterium является B. longum. В настоящее время предпочтительными штаммами Bifidobacterium являются B. longum DSM 32947 и B. longum DSM 32948.
L. reuteri представляет собой толерантный к кислороду (альтернативно аэротолерантный или относительный толерантный к кислороду) анаэроб, т. е. может генерировать АТФ только брожением.
Дополнительные аспекты вариантов осуществления изобретения относятся к Bifidobacterium longum DSM 32947, Bifidobacterium longum DSM 32948 и композициям, таким как фармацевтические композиции, пищевые композиции, пищевые добавки и пробиотические композиции, содержащие B. longum DSM 32947 и/или B. longum DSM 32948. В конкретном варианте осуществления бактерии бактериального штамма, т. е. B. longum DSM 32947 и/или B. longum DSM 32948, находятся в высушенной или лиофилизированной форме.
Связанные аспекты изобретения включают пробиотические композиции, содержащие бактерии штамма Bifidobacterium longum, выбранные из группы, состоящей из B. longum DSM 32947, B. longum DSM 32948 и их комбинации, и бактерии другого пробиотического бактериального штамма, предпочтительно штамма Lactobacillus, и более предпочтительно штамма L. reuteri и, в частности, штамма L. reuteri, выбранного из группы, состоящей из L. reuteri DSM 17938, L. reuteri DSM 32846 и их комбинации.
Дополнительные аспекты включают пробиотические композиции, содержащие пробиотический бактериальный штамм, предпочтительно штамм Lactobacillus и более предпочтительно штамм L. reuteri, и в частности штамм L. reuteri, выбранный из группы, состоящей из L. reuteri DSM 17938, L. reuteri DSM 32846 и их комбинации, и кондиционированную среду после культивирования штамма Bifidobacterium longum, выбранного из группы, состоящей из Bifidobacterium longum DSM 32947, Bifidobacterium longum DSM 32948 и их комбинации.
Бактерии, содержащиеся в пробиотической композиции согласно вышеизложенному, предпочтительно находятся в пробиотической композиции в высушенном виде, например, лиофилизированные или высушенные замораживанием, высушенные распылительной сушкой или высушенные распылительной лиофильной сушкой или высушенные вакуумной сушкой бактерии.
Бактерии B. longum DSM 32947 или B. longum DSM 32948, находящиеся в вышеописанных пробиотических композициях, могут быть представлены в высушенной форме, например, высушенной замораживанием (лиофилизированной), распылительной сушкой, распылительной лиофильной сушкой или вакуумной сушкой форме. Если композиции также содержат бактерии другого пробиотического бактериального штамма, такого как L. reuteri DSM 17938 и/или L. reuteri DSM 32846, то эти бактерии также могут быть предоставлены в композициях в высушенной форме, такой как высушенная замораживанием (лиофилизированная), высушенная распылительной сушкой, высушенная распылительной лиофильной сушкой или вакуумной сушкой форма.
Бактерии B. longum DSM 32947 и DSM 32948 были модифицированы (адаптированы или эволюционированы) из родительских штаммов посредством многоступенчатого селекционного процесса для улучшения роста и уменьшения проблемы с гетерогенным ростом. Таким образом, B. longum DSM 32947 и DSM 32948 демонстрируют повышенный рост. Эти штаммы не встречаются в природе, поскольку они были вынуждены эволюционировать, т. е. они не являются нативными или встречающимися в природе бактериальными штаммами.
Многоступенчатая селекция Bifidobacterium из клинических образцов включала выделение Bifidobacterium, выделенных из клинических образцов, на чашках с агаром MRS. Для повышения роста и уменьшения проблемы гетерогенного роста, бактерии подвергали следующей процедуре:
1. Посев штрихом на чашки с агаром MRS и после трех суток анаэробного культивирования при 37°C селекцию колонии с хорошим ростом;
2. Инокуляция отобранной колонии в бульон MRS и инкубирование при 37°C в анаэробных условиях;
3. Отбор образца и повторение стадий 1 и 2 до тех пор, пока не будут получены желаемые характеристики; и
4. Суспендирование бактерий в 15% глицерине и хранение при -70°C.
Вышеописанная обработка приводила к повышению роста и снижала проблему с гетерогенной морфологии колоний.
В результате подвергания бактерий, по меньшей мере, одной индуцирующей обработке во время культивирования в соответствии с изобретением, бактерии модифицируются и/или индуцируются для продуцирования, в том числе увеличения, продукции, терапевтических МВ, и они предпочтительно высвобождают данные МВ в культуральную среду. Индуцирующая обработка относится к изменениям в условиях культивирования, которые оказывают сильное влияние на продукцию МВ бактериями по сравнению с бактериями, не подвергавшимися индуцирующей обработке во время культивирования.
Для оценки и/или измерения повышенной продукции МВ или измененной эффективности МВ, которая была индуцирована или вызвана в результате различных индуцирующих обработок, можно применять различные способы. Возможен вариант количественного определения МВ, например, с использованием прибора Nanosight, или с использованием проточной цитометрии, или с использованием флуоресцентного красителя для окрашивания мембраны и тем самым количественного определения количества МВ. Например, флуоресценцию можно измерить (после отмывки), используя планшетный ридер (и сравнивая со стандартной кривой). Более простой способ представляет собой также сравнение размера гранул осадка или осадка после центрифугирования или измерение массы осадка. Осадок большего размера или большей массы означает большее количество МВ. Другими способами оценки эффективности МВ является измерение активности МВ на более сложных моделях in vitro или моделях in vivo, например, описанных в разделе «Примеры».
Важным метаболическим процессом в организме человека является метаболизм пуринов, в котором пурины метаболизируются и расщепляются под действием определенных ферментов. Примером такого фермента является экто-5'-нуклеотидаза (CD73), заякоренная в клеточной мембране 5'-нуклеотидаза, которая считается ключевым ферментом в генерации аденозина. Некоторые пробиотические бактерии содержат ген 5'-нуклеотидазы и продуцируют активный фермент 5'-нуклеотидазу и, следовательно, способны продуцировать аденозин. Активность 5'-нуклеотидазы и, таким образом, продукция аденозина может иметь место внеклеточно, т. е. вне или на поверхности бактерий, и в результате она может, например, находиться в супернатанте или другой внеклеточной жидкости, продуцированной бактериями. Таким образом, активный фермент 5'-нуклеотидаза может находиться на поверхности клетки, например, в форме, заякоренной в клеточной стенке 5'-нуклеотидазы, внеклеточно из бактериальной клетки, например, в супернатанте и/или в виде 5'-нуклеотидазы, ассоциированной с мембраной МВ. Как следствие, в этой группе бактерий образование аденозина и/или активность 5'-нуклеотидазы (EC 3.1.3.5) можно использовать в качестве маркера для определения эффективности продукции МВ.
Другие возможные модели включают, не ограничиваясь этим, модели, которые в настоящее время используются для оценки пробиотического эффекта бактериального штамма. Например, они включает доклинические модели in vitro, на которых можно измерить моторику кишечника или восприятие/передачу болевых сигналов, что демонстрирует, например, типичный дискомфорт, связанный с коликами у младенцев и другими функциональными желудочно-кишечными расстройствами. В настоящем документе используется несколько моделей для оценки потенциального действия на младенческие колики, например, модели, описанные в примерах 1, 3, 4, 9 и 10. Также включены модели клеточной иммуностимуляции, на которых можно оценить уровень выбранных цитокинов. Другой механизм, посредством которого пробиотические бактерии проявляют свое действие, относится к снижению проницаемости слизистой оболочки, т. е. к защите целостности эпителиального барьера. Таким образом, эффективность МВ можно оценить на модели проницаемости эпителия с использованием ETEC (энтеротоксигенной кишечной палочки) in vitro. Также соответствующие модели на животных могут быть использованы для исследования эффектов различных индуцирующих обработок, и также эффекты могут быть дополнительно оценены в клинических испытаниях у людей.
Терапевтические МВ можно, необязательно после консервации, вводить млекопитающему, например, в форме выделенных терапевтических МВ, в виде пробиотической композиции, как здесь дополнительно описано ниже, или в виде обработанной культуральной среды, кондиционированной среды или клеточной суспензии, например в виде высушенной культуральной среды, в том числе высушенной замораживанием (лиофилизированной), высушенной распылительной сушкой, высушенной распылительной лиофильной сушкой или высушенной вакуумной сушкой культуральной среды, кондиционированной среды или клеточной суспензии из бактерий пробиотического бактериального штамма и пробиотического бактериального штамма.
Вариант осуществления изобретения относится к пробиотической композиции, содержащей бактерии пробиотического бактериального штамма и терапевтические МВ, продуцированные пробиотическим бактериальным штаммом или другим пробиотическим бактериальным штаммом. Пробиотический бактериальный штамм и другой пробиотический бактериальный штамм выбраны из группы, состоящей из штамма Lactobacillus, штамма Bifidobacterium и их комбинации.
В одном варианте осуществления пробиотический бактериальный штамм или другой пробиотический бактериальный штамм подверглись индуцирующей биотической обработке согласно вариантам осуществления изобретения. Следовательно, в одном варианте осуществления терапевтические МВ в пробиотической композиции были получены способом получения терапевтических МВ в соответствии с вариантами осуществления изобретения. Следовательно, в одном варианте осуществления пробиотическая композиция содержит бактерии пробиотического бактериального штамма и терапевтические микровезикулы, продуцированные пробиотическим бактериальным штаммом или другим пробиотическим бактериальным штаммом, подверганием бактерий пробиотического бактериального штамма или другого пробиотического бактериального штамма индуцирующей биотической обработке во время культивирования для индукции продуцирования бактериями терапевтических микровезикул. Пробиотический бактериальный штамм и другой пробиотический бактериальный штамм выбран из группы, состоящей из штамма Lactobacillus, штамма Bifidobacterium и их комбинации. Индуцирующая биотическая обработка выбрана из группы, состоящей из сокультивирования бактерий с бактериями другого бактериального штамма, культивирования бактерий в присутствии кондиционированной среды после культивирования бактерий другого бактериального штамма и их комбинации. Другой бактериальный штамм представляет собой штамм Bifidobacterium, и другой бактериальный штамм отличается от пробиотического бактериального штамма и другого пробиотического бактериального штамма.
В одном варианте осуществления терапевтические МВ выделяют из бактерий пробиотического бактериального штамма, такого как описан выше, подверганием бактерий индуцирующей обработке во время культивирования и последующим выделением терапевтических МВ из культуральной среды. Затем выделенные терапевтические МВ могут быть добавлены к выделенным бактериям того же пробиотического бактериального штамма, который использовался для получения терапевтических МВ. В этом случае пробиотическая композиция содержит смесь выделенных бактерий пробиотического бактериального штамма и терапевтических МВ, выделенных из бактерий пробиотического бактериального штамма. В другом варианте осуществления пробиотическая композиция содержит бактерии первого пробиотического бактериального штамма и терапевтические МВ, выделенные из бактерий второго, другого пробиотического бактериального штамма. В данном последнем случае можно сочетать свойства или характеристики различных пробиотических бактериальных штаммов смешиванием выделенных бактерий пробиотического бактериального штамма с терапевтическими МВ, выделенными из бактерий другого пробиотического бактериального штамма. Также возможно обеспечить пробиотическую композицию, содержащую бактерии первого пробиотического бактериального штамма и терапевтические МВ из бактерий первого пробиотического бактериального штамма и терапевтические МВ, выделенные из бактерий второго, другого пробиотического бактериального штамма. В еще одном варианте осуществления пробиотическая композиция включает смесь, по меньшей мере, одного бактериального штамма и терапевтических МВ, по меньшей мере, из того же бактериального штамма или продуцированных другим бактериальным штаммом.
Бактерии, содержащиеся в пробиотической композиции согласно вышеизложенному, предпочтительно находятся в пробиотической композиции в высушенном виде, например, высушенные замораживанием или лиофилизированные, высушенные распылительной сушкой или высушенные распылительной лиофильной сушкой или высушенные вакуумной сушкой бактерии.
Как описано выше, пробиотический бактериальный штамм предпочтительно представляет собой пробиотический бактериальный штамм, продуцирующий молочную кислоту (такой как Lactobacillus), такой как пробиотический штамм Lactobacillus reuteri, и более предпочтительно L. reuteri DSM 17938 и/или L. reuteri DSM 32846.
Экспериментальные данные, представленные в настоящем документе, показывают, что терапевтические МВ, полученные и выделенные из пробиотических бактериальных штаммов, не только способны воспроизводить положительные эффекты на моторику желудочно-кишечного тракта и передачу болевых сигналов, бактерий, продуцирующих пробиотические МВ. Важно отметить, что эти терапевтические МВ были фактически более эффективными, чем сами пробиотические бактерии, о чем свидетельствует более быстрое начало благоприятного действия терапевтических МВ по сравнению с самими бактериями. Экспериментальные данные, представленные здесь, также показывают, что терапевтические МВ, выделенные из пробиотического бактериального штамма, обладают иммуностимулирующим действием. МВ были способны снижать уровень определенных цитокинов, связанных с аутоиммунными заболеваниями, и также было показано, что они защищают целостность эпителиального барьера. Экспериментальные данные показали, что терапевтические МВ согласно вариантам осуществления изобретения можно использовать для подавления, лечения или профилактики различных заболеваний или расстройств, которые, как было показано ранее, подавляются, лечатся или профилактируются с использованием пробиотических бактерий. Таким же образом можно ожидать, что терапевтические МВ при введении млекопитающему будут вызывать аналогичные общие и/или специфические эффекты и, возможно, также улучшенные эффекты по сравнению с пробиотическими бактериями при введении субъекту, такому как любое млекопитающее. Экспериментальные данные, представленные в настоящем документе, также показывают, что продукцию МВ можно индуцировать биотической обработкой во время культивирования, например, добавлением супернатанта, полученного при культивировании другого бактериального штамма, к пробиотическому бактериальному штамму или добавлением бактериальных клеток из другого бактериального штамма к пробиотическому бактериальному штамму во время культивирования (так называемое сокультивирование). Такая индуцирующая биотическая обработка генерирует более эффективные или сильные МВ при оценке на различных моделях по сравнению с МВ из неиндуцированных или нестимулированных бактериальных препаратов.
Более быстрое начало благоприятного действия, наблюдаемое для терапевтических МВ, можно использовать для пробиотической композиции согласно вариантам осуществления изобретения для достижения пролонгированного терапевтического эффекта при введении млекопитающему. Таким образом, терапевтические МВ в пробиотической композиции вызывают или генерируют ранний эффект у млекопитающего за счет более быстрого начала их действия, тогда как более поздний, но обычно пролонгированный эффект вызывается или генерируется бактериями пробиотического бактериального штамма, содержащегося в пробиотической композиции. Кроме того, терапевтические МВ также могут оказывать повышенный терапевтический эффект по сравнению с терапевтическим эффектом бактерий пробиотического бактериального штамма. Это означает, что пробиотическая композиция согласно вариантам осуществления изобретения обеспечивает значительно повышенные терапевтические эффекты у млекопитающего по сравнению с просто введенными пробиотическими бактериями.
Другими словами, композиция, содержащая как пробиотические бактериальные клетки, так и терапевтические МВ, обеспечивает преимущества по сравнению с композициями либо только с пробиотическими бактериальными клетками, либо только с МВ. В композиции, содержащей как пробиотические бактериальные клетки, так и терапевтические МВ, более быстрое начало благоприятного действия, наблюдаемое с терапевтическими МВ, сочетается с пролонгированным действием пробиотических бактериальных клеток с улучшением пробиотической композиции для быстрого начала и пролонгированного терапевтического при введении субъекту, такому как млекопитающее.
Следовательно, аспект вариантов осуществления изобретения относится к пробиотической композиции, содержащей быстродействующий компонент в форме терапевтических МВ из бактерий пробиотического бактериального штамма. Пробиотический бактериальный штамм выбран из группы, состоящей из штамма Lactobacillus, штамма Bifidobacterium и их комбинации. Пробиотическая композиция также содержит медленнодействующий или пролонгированный компонент в форме бактерий штамма пробиотических бактерий, продуцирующих МВ, или другого штамма пробиотических бактерий. Другой пробиотический бактериальный штамм выбран из группы, состоящей из штамма Lactobacillus, штамма Bifidobacterium и их комбинации. Таким образом, быстродействующий компонент и медленнодействующий компонент вместе обеспечивают повышенный, с ранним началом и продолжительный терапевтический эффект при введении субъекту.
В одном варианте осуществления пробиотический бактериальный штамм или другой пробиотический бактериальный штамм подвергается индуцирующей биотической обработке согласно вариантам осуществления изобретения. Следовательно, в одном варианте осуществления терапевтические МВ в быстродействующем компоненте пробиотической композиции были получены способом получения терапевтических МВ в соответствии с вариантами осуществления изобретения. Следовательно, в одном варианте осуществления пробиотическая композиция содержит быстродействующий компонент в форме терапевтических микровезикул, продуцированных бактериями пробиотического бактериального штамма, подверганием бактерий индуцирующей биотической обработке во время культивирования для индукции продукции терапевтических микровезикул бактериями. Пробиотический бактериальный штамм выбран из группы, состоящей из штамма Lactobacillus, штамма Bifidobacterium и их комбинации. Индуцирующая биотическая обработка выбрана из группы, состоящей из сокультивирования бактерий с бактериями другого бактериального штамма, культивирования бактерий в присутствии кондиционированной среды из бактерий другого бактериального штамма и их комбинации. Другой бактериальный штамм представляет собой штамм Bifidobacterium, и другой бактериальный штамм отличается от пробиотического бактериального штамма. Пробиотическая композиция также содержит медленнодействующий компонент в виде бактерий пробиотического бактериального штамма или другого пробиотического бактериального штамма. Другой пробиотический бактериальный штамм выбран из группы, состоящей из штамма Lactobacillus, штамма Bifidobacterium и их комбинации. Другой пробиотический бактериальный штамм отличается от другого штамма бактерий. Быстродействующий компонент и медленнодействующий компонент вместе обеспечивают пролонгированный терапевтический эффект при введении субъекту.
В одном варианте осуществления быстродействующий компонент имеет более раннее начало проявления терапевтического эффекта у субъекта по сравнению с медленнодействующим компонентом. Следовательно, быстрые и медленные по отношению к быстродействующему компоненту и медленнодействующему компоненту определяют относительное начало терапевтического эффекта, индуцированного этими компонентами. Другими словами, быстродействующий компонент является более быстрым или более быстродействующим компонентом по сравнению с медленнодействующим компонентом, который можно рассматривать как более медленный или более медленнодействующий компонент по сравнению с быстродействующим компонентом с точки зрения индукции терапевтического эффекта у субъекта.
В одном варианте осуществления быстродействующий компонент находится в форме выделенных терапевтических МВ из бактерий пробиотического бактериального штамма.
Обсуждение, представленное выше в отношении использования одного и того же пробиотического бактериального штамма или разных пробиотических бактериальных штаммов для терапевтических МВ и бактерий, также применимо к данному варианту осуществления.
Пробиотические композиции согласно вариантам осуществления изобретения можно использовать в качестве лекарственного средства и, в частности, использовать для лечения желудочно-кишечного расстройства.
Аспект вариантов осуществления определяет пробиотическую композицию, содержащую бактерии пробиотического бактериального штамма и терапевтические МВ, продуцированные пробиотическим бактериальным штаммом или другим бактериальным штаммом, для применения в качестве лекарственного средства и, в частности, для применения в лечении колик. Аспект вариантов осуществления изобретения также определяет пробиотическую композицию, содержащую вышеописанные быстродействующий компонент и медленнодействующий компонент, для применения в качестве лекарственного средства и, в частности, для применения в лечении колик.
В одном варианте осуществления колика представляет собой младенческую колику (также называемую коликой у младенцев).
Другие аспекты вариантов осуществления изобретения определяют пробиотическую композицию, содержащую бактерии пробиотического бактериального штамма и терапевтические МВ, продуцированные пробиотическим бактериальным штаммом или другим бактериальным штаммом, или пробиотическую композицию, содержащую быстродействующий компонент и медленнодействующий компонент, для применения в лечении заболевания, выбранного из группы, состоящей из желудочно-кишечного расстройства или заболевания у младенцев или детей, желудочно-кишечного болевого расстройства, заболевания, связанного с потерей костной массы, заболевания пародонта и их комбинации.
Терапевтические МВ могут быть продуцированы бактериями того же пробиотического бактериального штамма, которые включены в пробиотическую композицию. Альтернативно или дополнительно терапевтические МВ могут быть продуцированы бактериями другого пробиотического бактериального штамма, чем бактерии, включенные в пробиотическую композицию.
В одном варианте осуществления бактерии пробиотического бактериального штамма в пробиотической композиции подвергались индуцирующей биотической обработке согласно вариантам осуществления.
Терапевтические МВ, выделенные из бактерий пробиотического бактериального штамма, можно использовать в лечении колик, таких как младенческие колики. Пробиотический бактериальный штамм выбран из группы, состоящей из штамма Lactobacillus, штамма Bifidobacterium и их комбинации.
Варианты осуществления также относятся к терапевтическим МВ, выделенным из бактерий пробиотического бактериального штамма, которые можно использовать в лечении заболевания, выбранного из группы, состоящей из желудочно-кишечного расстройства или заболевания у младенцев или детей, желудочно-кишечного болевого расстройства, заболевания, связанного с потерей костной массы, заболевания пародонта, и их комбинации. Пробиотический бактериальный штамм выбран из группы, состоящей из штамма Lactobacillus, штамма Bifidobacterium и их комбинации.
Терапевтические МВ предпочтительно получены из штамма Lactobacillus reuteri и даже более предпочтительно из Lactobacillus reuteri DSM 17938 и/или Lactobacillus reuteri DSM 32846.
Выделенные терапевтические МВ или терапевтические МВ в пробиотической композиции предпочтительно были продуцированы бактериями пробиотического бактериального штамма, подвергнутыми индуцирующей биотической обработке, как раскрыто в данном документе. Вариант осуществления относится к терапевтическим микровезикулам, выделенным из бактерий пробиотического бактериального штамма, подвергнутым индуцирующей биотической обработке во время культивирования для индукции продуцирования терапевтических микровезикул бактериями. Пробиотический бактериальный штамм выбран из группы, состоящей из штамма Lactobacillus, штамма Bifidobacterium и их комбинации. Индуцирующая биотическая обработка выбрана из группы, состоящей из сокультивирования бактерий с бактериями другого бактериального штамма, культивирования бактерий в присутствии кондиционированной среды после культивирования бактерий другого бактериального штамма и их комбинации. Другой бактериальный штамм представляет собой штамм Bifidobacterium, и другой бактериальный штамм отличается от пробиотического бактериального штамма.
В одном варианте осуществления изобретения пробиотическая композиция и/или терапевтические МВ могут альтернативно использоваться для лечения желудочно-кишечного расстройства. Желудочно-кишечное расстройство предпочтительно представляет собой функциональное желудочно-кишечное расстройство, выбранное из группы, состоящей из функционального расстройства пищевода, такого как функциональная изжога, функциональная боль в груди пищеводного происхождения, функциональная дисфагия и «глобус»; функционального гастродуоденального расстройства, такого как функциональная диспепсия, аэрофагия, неуточненная чрезмерная отрыжка, хроническая идиопатическая тошнота, функциональная рвота, синдром циклической рвоты и синдром руминации; функционального расстройства кишечника, такого как синдром раздраженного кишечника (IBS), функциональный запор, функциональная диарея и неуточненное функциональное расстройство кишечника; синдрома функциональной абдоминальной боли, такого как функциональная абдоминальная боль (FAP), функциональное расстройство желчного пузыря и сфинктера Одди, например, функциональное расстройство желчного пузыря, функциональное расстройство сфинктера Одди билиарного типа и функциональное расстройство сфинктера Одди панкреатического типа; функционального аноректального расстройства, такого как функциональное недержание кала, функциональная аноректальная боль и функциональное расстройство дефекации; функционального расстройства желудочно-кишечного тракта у детей, такого как срыгивание у младенцев, синдром руминации у младенцев, синдром циклической рвоты у младенцев, функциональная диарея, младенческая дисхезия и функциональный запор.
В конкретном варианте осуществления желудочно-кишечное расстройство выбрано из группы, состоящей из расстройства моторики желудочно-кишечного тракта, желудочно-кишечной боли, колик, синдрома раздраженного кишечника и запора.
В одном варианте осуществления желудочно-кишечное расстройство или заболевание у младенцев или детей представляет собой желудочно-кишечное расстройство или заболевание у младенцев, такое как функциональное желудочно-кишечное расстройство или заболевание у младенцев. В конкретном варианте осуществления желудочно-кишечное расстройство или заболевание у младенцев выбрано из группы, состоящей из расстройства моторики желудочно-кишечного тракта у младенцев, желудочно-кишечной боли у младенцев, младенческих колик, синдрома раздраженного кишечника у младенцев, пищевой непереносимости у младенцев, запоров у младенцев, диареи у младенцев, регургитации у младенцев, синдрома руминации у младецев, младенческой дисхезии, функционального запора у младенцев и их комбинации.
В еще одном конкретном варианте осуществления желудочно-кишечное расстройство или заболевание у младенцев выбрано из группы, состоящей из младенческих колик или пищевой непереносимости у младенцев и их комбинации.
В еще одном конкретном варианте осуществления желудочно-кишечное расстройство или заболевание у младенцев представляет собой расстройство моторики желудочно-кишечного тракта у младенцев, предпочтительно запор у младенцев и/или диарею у младенцев и их комбинацию.
В еще одном конкретном варианте осуществления желудочно-кишечное расстройство или заболевание у младенцев представляет собой расстройство моторики желудочно-кишечного тракта у младенцев и/или младенческие колики.
В одном варианте осуществления желудочно-кишечное расстройство или заболевание у младенцев или детей представляет собой желудочно-кишечное расстройство или заболевание у детей, такое как функциональное желудочно-кишечное расстройство или заболевание у детей.
В конкретном варианте осуществления желудочно-кишечное расстройство или заболевание у детей выбрано из группы, состоящей из детской регургитации, детского синдрома руминации, функциональной диареи у детей, детской дискезии, функционального запора у детей и их комбинации.
В еще одном конкретном варианте осуществления желудочно-кишечное расстройство у детей выбрано из группы, состоящей из срыгивания у детей, детской дисхезии и их комбинации.
В одном варианте осуществления желудочно-кишечное болевое расстройство выбрано из группы, состоящей из функциональной абдоминальной боли (FAP), абдоминальной боли в виде колик, частой рецидивирующей абдоминальной боли (FRAP) и их комбинации.
В одном варианте осуществления заболевание, связанное с потерей костной массы, выбрано из группы, состоящей из остеопороза, остеопении и их комбинации.
В одном варианте осуществления пробиотические композиции предназначены для применения в лечении остеопороза или остеопении.
В одном варианте осуществления заболевание пародонта выбрано из группы, состоящей из пародонтита, гингивита и их комбинации.
В еще одном варианте осуществления пробиотические композиции предназначены для применения в лечении пародонтита.
В дополнительном конкретном варианте осуществления расстройство моторики желудочно-кишечного тракта выбрано из группы, состоящей из вздутия живота, рецидивирующей непроходимости, абдоминальной боли в виде колик, запора, гастроэзофагеальной рефлюксной болезни, трудноизлечимой рецидивирующей рвоты, диареи, воспалительного заболевания кишечника (IBD), недержания кала, частой рецидивирующей абдоминальной боли (FRAP), срыгивания или непереносимости пищи.
Варианты осуществления также относятся к применению пробиотической композиции или терапевтических МВ, выделенных из пробиотического бактериального штамма, в качестве лекарственного средства и для производства лекарственного средства для лечения желудочно-кишечного расстройства.
Варианты осуществления дополнительно включают способ подавления, лечения или профилактики желудочно-кишечного расстройства. Способ включает введение пробиотической композиции или терапевтических МВ, выделенных из пробиотического бактериального штамма, субъекту для подавления, лечения или профилактики желудочно-кишечного расстройства.
Экспериментальные данные, представленные в настоящем документе, также показывают, что терапевтические МВ, выделенные из пробиотического бактериального штамма, обладают иммуностимулирующим действием и способны подавлять секрецию IFN-γ и IL-17A. Также наблюдались эффекты повышения секреции IL-6. Таким образом, такие терапевтические МВ можно использовать в качестве модуляторов иммунитета человека.
Пробиотические композиции по настоящему изобретению и/или терапевтические МВ по настоящему изобретению можно использовать для подавления или снижения уровня цитокинов IFN-γ/или IL-17A при введении субъекту. Следовательно, пробиотические композиции и/или терапевтические МВ можно использовать в подавлении, лечении или профилактике заболеваний, характеризующихся аберрантной экспрессией IFN-γ и/или IL-17A, включая воспалительные, аутовоспалительные и аутоиммунные заболевания. В конкретном варианте воспалительное, аутовоспалительное и аутоиммунное заболевание выбрано из группы, состоящей из SLE, MCTD, RA, SS, DM, SSc, MS, псориаза, потери костной массы, остеопороза, остеопении, пародонтита, гингивита, саркопении, кахексии, недоедания и аллергии, такой как AD, AR и астма, а также пищевой непереносимости и аллергии.
Пробиотические композиции по изобретению и/или терапевтические МВ по изобретению можно использовать для повышения секреции цитокина IL-6 при введении субъекту. IL-6 представляет собой плейотропный цитокин, выполняющий множество функций в организме. Большинство таких функций и процессов связаны с воспалительными реакциями, однако эти функции и реакции важны, например, для защиты от патогенов. Однако провоспалительные процессы в организме должны быть хорошо сбалансированы, и было описано, что L. reuteri DSM 17938 увеличивает количество регуляторных Т-клеток (например, Liu Y., Fatheree N.Y., Dingle B.M., Tran D.Q., & Rhoads J.M. (2013). Lactobacillus reuteri DSM 17938 изменяет частоту Foxp3+ регуляторных Т-клеток в кишечнике и брызжеечном лимфатическом узле при экспериментальном некротическом энтероколите. PloS One, 8 (2), e56547. Http://doi.org/10.1371/journal.pone.0056547). Повышенная экспрессия IL-6 в сочетании с более высокой частотой регуляторных Т-клеток может представлять собой способ сделать иммунную систему более «настороженной» и усилить защиту от инфекций без увеличения риска развития воспаления. Следовательно, пробиотические композиции и/или терапевтические МВ можно использовать для балансирования противовоспалительных и провоспалительных процессов иммунной системы.
Энтеротоксигенная Escherichia coli (ETEC) представляет собой тип E. coli и одну из основных бактериальных причин диареи в развивающихся странах, а также самую частую причину «диареи путешественников». По оценкам, примерно 157000 смертей происходят каждый год, в основном среди детей, в результате ETEC. Основными отличительными признаками ETEC являются экспрессия одного или более энтеротоксинов и наличие фимбрий, которые используются для прикрепления к клеткам кишечника хозяина.
Экспериментальные данные, представленные здесь, показывают, что терапевтические МВ, выделенные из пробиотического бактериального штамма, защищают целостность эпителиального барьера от разрушительного воздействия ETEC, что является моделью для изучения эффектов на проницаемость эпителия. Основная функция кишечного барьера заключается в регуляции всасывания питательных веществ, электролитов и воды из просвета в кровоток и, с другой стороны, в предотвращении прохождения патогенных микроорганизмов и токсичных веществ из просвета в кровоток, таким образом, неповрежденный барьер является важным для обеспечения здорового состояния. Нарушение кишечного барьера связано с некоторыми различными заболеваниями и состояниями, такими как синдром раздраженного кишечника (IBS), болезнь Крона, депрессия, расстройства аутистического спектра, дивертикулярная болезнь, пародонтит, остеопения и остеопороз.
Также сообщалось, что изменения кишечного барьера могут быть основной причиной некоторых кахектических состояний (Bindels et al. (2018)).
Целостность эпителиального барьера важна не только в кишечнике, неповрежденный барьер важен также, например, в ротовой полости. Эпителий десен представляет собой первую линию защиты ротовой полости от микробов. В случае нарушения их целостности бактерии коллективно получают доступ к подлежащей соединительной ткани, что может привести к воспалению и разрушению аттачментов зубов (DiRienzo (2014)). Следовательно, пробиотические композиции и/или терапевтические МВ можно использовать в подавлении, лечении или профилактике заболеваний или состояний, вызывающих нарушение целостности эпителиального барьера, т. е. дисфункции эпителиального барьера, включая IBS, болезнь Крона, кахексию, остеопению, остеопороз, гингивит, пародонтит, депрессию, расстройства аутистического спектра, дивертикулярную болезнь, и/или в подавлении, лечении или профилактике инфекции ETEC, а также в подавлении, лечении или профилактике диареи и/или «диареи путешественников», вызванной ETEC или другими патогенными бактериями.
Более быстрое начало лечебного или пробиотического эффекта, наблюдаемое у терапевтических МВ по сравнению с пробиотическими бактериями, может быть пригодным в лечении субъектов, страдающих заболеванием или расстройством, таким как желудочно-кишечное расстройство, воспалительное, аутовоспалительное или аутоиммунное заболевание, дисфункция эпителиального барьера, инфекция ETEC, диарея и/или «диарея путешественников». Это означает, что при введении терапевтических МВ или пробиотической композиции, содержащей такие терапевтические МВ, может быть получено более быстрое начало терапевтического эффекта, такого как лечебный или пробиотический, по сравнению с введением просто пробиотических бактерий. Различное время начала проявления лечебных или пробиотических эффектов, наблюдаемых между терапевтическими МВ и пробиотическими бактериями, также можно использовать, как упомянуто выше, для достижения пролонгированного лечебного или пробиотического эффекта у субъекта. Таким образом, терапевтические МВ, введенные субъекту, будут способствовать быстрому или немедленному терапевтическому эффекту у субъекта, тогда как пробиотические бактерии, введенные субъекту отдельно или вместе с терапевтическими МВ, будут способствовать более медленному или отсроченному терапевтическому эффекту у субъекта. Как следствие, посредством введения субъекту терапевтических МВ и пробиотических бактерий, отдельно или вместе, можно получить пролонгированный лечебный или пробиотический эффект, т. е. как немедленный, так и отсроченный.
Подходящий способ введения и формуляции пробиотической композиции или терапевтических МВ можно выбрать в зависимости от заболевания или расстройства. Предпочтительным способом введения является пероральный. Другие способы введения включают интраназальное, внутриглазное, местное или некоторые другие формы местного введения в кожу, прямую кишку, нос, глаза, влагалище или десны или внутривенную, подкожную или внутримышечную инъекцию.
Соответствующие дозы пробиотической композиции или терапевтических МВ, как здесь определено, можно легко выбрать в зависимости от заболевания или расстройства, подлежащего лечению, способа введения и рассматриваемого состава. Например, дозировка и режим введения выбираются для гарантии того, что терапевтические МВ или пробиотическая композиция, введенные субъекту в соответствии с настоящим изобретением, могут привести к желаемым терапевтическим эффектам, профилактическим эффектам или пользе для здоровья. Таким образом, предпочтительно дозировка представляет собой терапевтически или профилактически эффективную дозу, которая подходит для типа субъекта и заболевания или расстройства, которое лечат. Например, можно использовать суточные дозы в целом от 104 до 1010, например от 105 до 109, или от 106 до 108, или от 108 до 1010 КОЕ бактерий. Предпочтительная суточная доза в целом составляет примерно 108 КОЕ, например, от 107 до 109 или от 108 до 109 КОЕ бактерий. Например, можно использовать суточные дозы от 104 до 1014, например от 105 до 1013, или от 106 до 1012, или от 108 до 1012, или от 1010 до, 1012, или от 1010 до 1014 общего количества терапевтических МВ. Предпочтительная суточная доза составляет примерно 1010 общего количества терапевтических МВ, например, от 109 до 1011 или от 1010 до 1011 терапевтических МВ. Другая предпочтительная суточная доза составляет примерно 109 общего количества терапевтических МВ, например от 108 до 1010 терапевтических МВ. Другая предпочтительная суточная доза составляет примерно 108 общего количества терапевтических МВ, например от 107 до 109 терапевтических МВ. Другим примером может быть использование МВ, продуцированных фиксированным числом бактерий, например 108 или 109 КОЕ бактерий.
Настоящее изобретение также относится к способам подавления, лечения или профилактики колик, в частности младенческих колик, и/или заболевания, выбранного из группы, состоящей из желудочно-кишечного расстройства или заболевания у младенцев или детей, желудочно-кишечного болевого расстройства, заболевания, связанного с потерей костной массы, пародонтоза и их комбинации у субъекта. Способ включает введение субъекту пробиотической композиции и/или терапевтических МВ по изобретению.
Подавление заболевания или расстройства в контексте настоящего описания включает отсрочку начала развития заболевания или расстройства или симптома, ассоциированного с заболеванием или расстройством.
Субъектом предпочтительно является млекопитающее и более предпочтительно человек.
Примеры, приведенные здесь, описывают протоколы получения и выделения терапевтических МВ из хорошо изученного бактериального штамма L. reuteri DSM 17938 с целью последующего исследования и использования пробиотического действия пробиотического бактериального штамма и терапевтических МВ на его основе. Пробиотический эффект выделенных МВ сравнивали с действием целых бактерий. Было обнаружено, что МВ способны воспроизводить полезные эффекты целых бактерий на модели ex vivo, используемых для изучения перистальтики желудочно-кишечного тракта и на модели in vitro, используемых для изучения передачи болевых сигналов. Удивительно, но МВ не только воспроизводили бактериальный пробиотический эффект, но и были даже более эффективными по сравнению с целыми бактериями, о чем свидетельствует их способность действовать быстрее на модели передачи нервных сигналов, что приводило к более раннему началу проявления наблюдаемого эффекта. Для еще большего усиления результатов, был исследован другой штамм L. reuteri (L. reuteri DSM 32846), который также показал аналогичные эффекты. Кроме того, эффект L. reuteri DSM 17938 и L. reuteri DSM 32846 может быть дополнительно улучшен, если бактерии подвергаются индуцирующей обработке, включая эксперименты по сокультивированию с некоторыми другими бактериальными штаммами или с кондиционированной средой из других бактериальных штаммов, которые усиливают способность бактерий продуцировать и выделять МВ.
Для исследования терапевтических МВ, полученных из штаммов пробиотических бактерий, и их эффекта на различных физиологических моделях выделенные фракции МВ выделяли культивированием бактерий с последующим выделением высвобожденных МВ. Авторы изобретения определили усовершенствованные способы культивирования пробиотических бактерий для повышения продукции МВ, которые сохраняют, и также демонстрируют повышенную биологическую активность.
Примеры
Пример 1 - Изменение ферментативной активности, ассоциированное с продукцией МВ
Lactobacillus reuteri DSM 17938 культивировали и подвергали различным индуцирующим обработкам на определенные временные точки. Ответ на эти индуцирующие обработки определяли с использованием ферментативного анализа и сравнивали с ответом, полученным с контрольными обработками. Индуцирующие обработки включали индукцию стресса у бактерий во время их фазы роста, и затем влияние на ферментативную активность измеряли в кондиционированной бактериальной среде.
Материалы и методы
Культивирование/сбор образцов
L. reuteri DSM 17938 инокулировали из замороженного стокового материала в 25 мл среды де Мана-Рогоза-Шарпа (MRS) в нормальных условиях культивирования, т. е. культивировали в анаэробных условиях при 37°C в течение ночи. Затем бактерии (20 мл) реинокулировали в 200 мл MRS и подвергали их различным индуцирующим обработкам, как более подробно описано ниже. Образцы бактерий центрифугировали при 5000 × g в течение 10 мин, супернатанты переносили в другую пробирку, затем центрифугировали при 10000 × g в течение 10 мин. Супернатанты фильтровали через фильтр с размером пор 0,45 мкм и хранили на льду перед дальнейшим центрифугированием с использованием ультрацентрифуги при 32000 об/мин при 4°C в течение 3 ч (Beckman SW 32 Ti Rotor, Swing bucket, пробирки емкостью 30 мл). Супернатанты отбрасывали (осторожно сливали с помощью пипетки). Осадки осторожно ресуспендировали в среде для ресуспендирования (забуференный фосфатом физиологический раствор (PBS)). Объем ресуспендирования варьировался от 100 до 300 мкл в зависимости от размера осадка. Образцы разделяли на аликвотные порции и хранили при -70°C.
Обработка кислородом
Повышенные концентрации кислорода (окислительный стресс) моделировали интенсивным встряхиванием бактериальной культуры. Моделирование окислительного стресса непрерывно проводили при встряхивании в течение 24 ч.
Высокотемпературная обработка
Индуцированный температурой стресс воспроизводили повышением температуры с Т=37°С до Т=50°С на временной точке, когда оптическая плотность (OD) достигала значения оптической плотности примерно 1,6, и выдерживали при Т=50°С в течение 20 мин. После такой высокотемпературной обработки бактерии культивировали в нормальных условиях при Т=37°С. Общее время культивирования составляло 24 ч.
Обработка изменением pH
Стресс, индуцированный pH, воспроизводили понижением значения pH с pH 6,5 до pH 2 на временной точке, когда оптическая плотность (OD) достигала поглощения примерно 1,6. Сдвиг pH получали спин-центрифугированием бактериальных клеток и добавлением искусственного желудочного сока к бактериальному осадку для достижения pH 2. Пониженное значение pH сохраняли в течение 10 мин перед добавлением супернатанта обратно к бактериальным клеткам для нормализации pH до 6,5. Время культивирования варьировалось за счет того, что были взяты два разных образца. Они были названы по разному, как показано на фиг. 1. pH: образец был отобран через 24 ч культивирования; pH+желудочный сок: образец был отобран сразу после 10 мин индукции желудочным соком.
Обработка сокультивированием
В данной обработке L. reuteri DSM 17938 культивировали совместно с Bifidobacterium longum DSM 32947 и B. longum DSM 32948 на среде SIM (имитированная кишечная среда, состав, описанный ниже) добавлением супернатанта, т. е. кондиционированной среды, полученной при культивировании B. longum DSM 32947 или B. longum DSM 32948. В качестве контроля L. reuteri DSM 17938 культивировали на среде SIM.
Таблица 1
Состав SIM (имитированная кишечная среда)
pH доводили до 6,8; автоклавировали при 121°C в течение 15 мин. Перед посевом добавляли стерильный фильтрованный сахар и растворы акцепторов электронов. Конечные концентрации: по 15 мМ каждого. Сахар: галактоолигосахариды (GOS) или глюкоза. Акцептор электронов: цитрат, 1,2-пропандиол или фруктоза.
Ферментативная активность
Образцы, полученные после индуцирующих обработок, описанных выше, размораживали и затем тестировали в анализе активности 5'-нуклеотидазы с использованием набора для определения активности 5'-нуклеотидазы Crystal Chem (Crystal Chem, Elk Grove Village, IL, США). Вкратце, процедуру проводили в две стадии. Сначала к образцам супернатанта добавляли реагент 1 (CC1), содержащий АМФ, для превращения АМФ в аденозин под действием любого фермента 5'-нуклеотидазы, присутствующего в образцах супернатанта. Аденозин далее гидролизовался до инозина и гипоксантина компонентами реагента 1. На второй стадии добавляли реагент 2 (CC2) для превращения гипоксантина в мочевую кислоту и гидроксид водорода, который использовался для получения хинонового красителя, кинетическое измерение которого проводили при 550 нм на спектрофотометре. Активность 5'-нуклеотидазы в образцах определяли расчетом изменения оптической плотности в период между 3 и 5 мин и сравнением со значением, полученным с образцом калибратора.
Результаты
Как следует из результатов, представленных на фиг.1, активность 5'-нуклеотидазы увеличивалась за счет обработки кислородом (DSM17938+O2) и сокультивирования L. reuteri DSM 17938 и B. longum DSM 32947 или B. longum DSM. 32948 на среде SIM (DSM17938+DSM32947 на среде SIM или DSM17938+DSM32948 на среде SIM).
Другие индуцирующие обработки не вызывали какого-либо повышения активности 5'-нуклеотидазы по сравнению с контрольным культивированием L. reuteri DSM 17938 (DSM17938 на среде MRS или DSM17938 на среде SIM).
Пример 2 - Специфические условия культивирования ассоциированы с повышенной продукцией МВ
Lactobacillus reuteri DSM 17938 культивировали и подвергали различным индуцирующим обработкам. Количество МВ, продуцированных в результате этих индуцирующих обработок, определяли с использованием анализа траекторий наночастиц (NTA), и результаты сравнивали с ответом, полученным при контрольных обработках. Индуцирующие обработки включали обработку кислородом и обработку сахарозой, и затем измеряли влияние на продукцию МВ в бактериальной кондиционированной среде.
Материалы и методы
Обработка кислородом
L. reuteri DSM 17938 культивировали в нормальных условиях культивирования, т. е. культивировали в анаэробных условиях на среде де Мана-Рогоза-Шарпа (MRS) при 37°C во флаконе/колбе. Обработку кислородом проводили интенсивным встряхиванием бактериальной культуры в течение 24 ч. В качестве контроля L. reuteri DSM 17938 культивировали в нормальных условиях культивирования, т. е. культивировали в анаэробных условиях в среде MRS при 37°C во флаконе/колбе без повышения концентрации кислорода (без обработки кислородом).
Обработка сахарозой
L. reuteri DSM 17938 культивировали в нормальных условиях культивирования, т. е. культивировали в анаэробных условиях в среде Lactobacillus Carrying Medium (LCM) при 37°C во флаконе/колбе. Стресс индуцировали добавлением сахарозы (конечная концентрация 2% в LCM) к бактериальной культуре в начале ферментации. Общее время культивирования составляло 24 ч. В качестве контроля L. reuteri DSM 17938 культивировали в нормальных условиях культивирования с добавлением 2% глюкозы вместо 2% сахарозы.
Анализ траекторий наночастиц (NTA)
Физико-химические характеристики МВ исследовали с использованием NTA. МВ соответственно разбавляли PBS, не содержащем частиц (фильтровали через 0,02 мкм фильтр), с получением концентрации в пределах рекомендованного диапазона измерений (1-10×108 частиц/мл) и непосредственно отслеживали траектории с использованием системы NanoSight NS300 (технология NanoSightTM, Malvern, Великобритания). Анализ проводили на следующем инструментальном устройстве:
1. Загрузка образца: загрузка образца шприцевым насосом в верхнюю пластину с уплотнительным кольцом, которая была установлена на лазерном модуле (лазерный луч 488 нм).
2. Образец для измерения: после оптимизации изображения собирали видеозаписи при 25°C с помощью высокочувствительной sCMOS-камеры и анализировали с использованием программного обеспечения NTA (версия 3.2) после захвата в режиме управления событиями (3 видеозаписи по 90 с на измерение) с использованием скорости шприцевого насоса, равной 50.
3. Анализ образцов: образцы собирали и анализировали с применением оптимизированных для прибора настроек, что обеспечивает наилучшую визуализацию частиц за счет применения программных настроек (уровень камеры, фокус и порог обнаружения) для оптимизации результатов анализа по отношению к различным образцам. Дополнительные настройки, такие как размывание, минимальная длина траектории и минимальный ожидаемый размер, были установлены на «автоматический» режим, и вязкость на 0,890 сП. Программное обеспечение NTA было оптимизировано и затем отслеживало каждую частицу по кадрам, и ее броуновское движение отслеживалось и измерялось от кадра к кадру путем захвата видеофайла. Программа отслеживала множество частиц по отдельности и с помощью уравнения Стокса-Эйнштейна рассчитывала их гидродинамические диаметры. Было записано много видеозаписей продолжительностью 90 с, генерируя повторные гистограммы, которые были усреднены для каждого образца.
Результаты
Обработка кислородом и добавление сахарозы приводили к повышению продукции МВ по сравнению с соответствующими контролями, см. таблицу 2 ниже.
Таблица 2
Продукция МВ
Пример 3 - Выделенные бактериальные микровезикулы воспроизводят действие бактерий на перистальтику кишечника
Материалы и методы
Животные
Взрослых мышей-самцов Swiss Webster (6-8 недель) получали из питомника Charles River Laboratories (Wilmington, MA, США). Животных содержали по 4-5 особей в клетке с 12-ч циклом свет/темнота и корм и воду давали ad libitum. Последующие процедуры проводили in vitro после эвтаназии смещением шейных позвонков в соответствии с Этическим советом по исследованиям на животных McMaster (AREB) (разрешение 16-08-30).
Регистрация флотации тканей в ванне
Регистрацию флотации тканей в ванне выполняли, как описано в Wu et al. (2013). Извлекали отрезки тощей и ободочной кишки длиной не менее 4 см и помещали в ванну емкостью 20 мл для флотации тканей, заполненную насыщенным кислородом раствором Кребса при 34°C. Оральный конец отрезков канюлировали, и содержимое вымывали из просвета перфузией под действием силы тяжести газированным углекислым газом раствором Кребсом с использованием бутылей Мариотта. После очистки анальный конец отрезков канюлировали силиконовой трубкой для оттока. Внутрипросветный компартмент перфузировали раствором Кребса с комнатной температурой со скоростью 5 мл/мин. Серозный компартмент перфузировали нагретым до 34°C газированным углекислым газом раствором Кребсом со скоростью 2 мл/мин. Насыщенный кислородом раствор Кребса состоял из (ммоль л-1): 118 NaCl, 4,8 KCl, 25 NaHCO3, 1,0 NaH2PO4, 1,2 MgSO4, 11,1 глюкозы и 2,5 CaCl2, барботированных газообразным углекислым газом (95% O2 и 5% CO2). Перед записью внутрипросветное давление доводили до 2-3 гПа увеличением и уменьшением высоты впускных и выпускных трубок. Обработки проводили открытием и закрытием соответствующих запорных кранов для остановки внутрипросветного потока раствора Кребса и началом потока бактерий. L. reuteri DSM 17938 использовали в концентрации 8-log колониеобразующих единиц (КОЕ)/мл. Кондиционированную среду, полученную при культивировании L. reuteri DSM 17938, микровезикулы, продуцированные L. reuteri DSM 17938, и кондиционированную среду с удаленными микровезикулами применяли в концентрациях, равных концентрации целых бактерий.
Регистрация видеозаписей
Видеозаписи регистрировали с помощью веб-камеры JVC, размещенной на 7 см выше отрезка ткани. Видеоклипы записывали и сохраняли в формате файла MOV с частотой кадров 10 кадров в секунду и соотношением сторон 4:3 с использованием программы NCH Debut Video Capture. Продолжительность записи варьировалась от 20 мин до 40 мин. С помощью VideoPad Video Editor видеозаписи были увеличены до 4 см с использованием принудительного соотношения сторон 4:3. Видеозаписи были преобразованы в черно-белые с помощью настройки цветовых кривых и применения двухтонального цветового фильтра. Черно-белые видеозаписи экспортировались со скоростью 10 кадров в секунду при разрешении 400 × 300 пикселей.
Анализ
Весь процесс получения, обработки и анализа карт пространственно-временного диаметра выполняли, как описано в публикации Wu et al. (2013). Видеозаписи анализировали с помощью StMap plagin для программного обеспечения NIH Image J. Используя обычную процедуру определения границ, диаметр каждой позиции по всей кишке был представлен в виде значения оттенка от 0 до 255. Сокращения кишечника, где диаметр был меньше, приближаются к значению оттенка 0 и представлены в виде более темных черных областей. Области расширения или расслабления достигают значения оттенка 255 и являются белыми. Программа создает пространственно-временную карту на протяжении всей видеозаписи. На карте чередуются темные и светлые оттенки в зависимости от положения вдоль кишечника, времени и диаметра. Пространственно-временная карта проходит от орального к анальному концу по вертикальной оси и во времени по горизонтальной оси. Скорость распространения сократительных комплексов (PCC) определяли измерением наклона больших темных сокращений. Частоту PPC определяли измерением количества сокращений между интервалами. Амплитуду измеряли как высоту (диаметр кишечника) на максимальных сокращениях.
Бактерии
L. reuteri DSM 17938 из стокового материала культивировали в среде де Мана-Рогоза-Шарпа (MRS), собирали через 48-72 ч, промывали забуференным фосфатом физиологическим раствором (PBS) и хранили при -20°C в аликвотах по 1,1 мл при 1×1010 КОЕ/мл, и выделяли их микровезикулы, как описано ниже.
МВ выделяли из бульонной культуры L. reuteri DSM 17938 (48-72 ч). После центрифугирования при 600 × g в течение 30 мин супернатанты фильтровали через фильтр с размером пор 0,22 мкм, дважды промывали PBS с центрифугированием при 100000 × g при 4°C, ресуспендировали в стерильном PBS, в объеме, соответствующем объему исходной культуры L. reuteri DSM 17938, и хранили при -80°C в аликвотах по 0,5 мл при 1×1012 КОЕ/мл. МВ определяли количественно по числу жизнеспособных бактерий в культуре, и также стандартизировали по содержанию белка (последовательно 5-8 мг/мл белка, 25-60 нг/мл ДНК и 18-30 нг/мл РНК; n=10) измеряли NanoDrop ND-1000 (NanoDrop Technologies, Wilmington, DE, США). Если не указано иное, то во всех экспериментах использовали препараты МВ в количестве, эквивалентном 1010 КОЕ/мл.
Бактерии разводили до концентрации 8-log КОЕ/мл для использования в исследовании.
Результаты
L. reuteri DSM 17938 и продукты их культивирования вводили интралюминально на препаратах мышиной тощей и ободочной кишки in vitro, для определения того, могут ли микровезикулы, продуцированные L. reuteri DSM 17938, воспроизвести действие родительских бактерий на перистальтику кишечника. Кондиционированную среду (CM) определяли как питательную среду (бульон), в которой культивировали бактерии L. reuteri DSM 17938. Бактерии отделяли от CM центрифугированием, и оставшуюся CM вводили интралюминально на кишечных препаратах in vitro, как описано выше. Микровезикулы (МВ) выделяли центрифугированием из L. reuteri DSM 17938, культивированных в течение 72 ч, затем ресуспендировали в растворе Кребса. Оставшуюся кондиционированную среду после удаления микровезикул и бактерий (CM-МВ) затем вводили интралюминально в ткани. В качестве отрицательного контроля отдельно применяли питательную среду, используемую для культивирования бактерий (бульон). Эффект от этих обработок сравнивали с контрольным раствором Кребса и измеряли по трем параметрам распространения сократительных комплексов (PCC) в отрезках кишечника: скорости, частоте и амплитуде.
Результаты достоверно воспроизводились при 24-ч приготовлении.
L. reuteri DSM 17938 и его продукты снижали перистальтику тонкого кишечника
Скорость PCC в тощей кишке
L. reuteri DSM 17938, CM и МВ в одинаковой степени снижали скорость PCC в тощей кишке. L. reuteri DSM 17938 достоверно снижали скорость PCC в тощей кишке на 34% при интралюминарном введении (p=0,0067, n=20) (фиг. 2A). CM воспроизводила эффект родительских бактерий и снижала скорость PCC на 29% (p=0,0107, n=28) (фиг. 2B). В качестве отрицательного контроля бульон, используемый в качестве среды для культивирования бактерий, тестировали независимо, и он оказывал незначительное влияние на скорость PCC в тощей кишке (снижение на 5%) по сравнению с контрольным раствором Кребса (p=0,0877, n=20) (фиг. 2C). Микровезикулы, выделенные из 72-ч культуры, достоверно снижали скорость PCC в тощей кишке на 19% (p=0,0002, n=20) (фиг. 2D). CM-МВ не изменяла скорость PCC в тощей кишки при введении в просвет (p=0,5203, n=20) (фиг. 2E).
Частота PCC в тощей кишке
Снижение частоты PCC в тощей кишке также индуцировалось под действием L. reuteri DSM 17938, CM и МВ. L. reuteri DSM 17938 достоверно снижали частоту PCC на 26% в тощей кишке (p=0,0482, n=20) (фиг. 3A). Аналогично CM уменьшила частоту PCC на 21% (p=0,0139, n=28) (фиг. 3B). Бульон не вызывал значительного изменения частоты PCC в тощей кишке (снижение на 6%) при введении в просвет (p=0,2424, n=20) (фиг. 3C). Микровезикулы достоверно снижали частоту PCC в тощей кишке на 26% по сравнению с бактериями (p=0,0004, n=20) (фиг. 3D). На частоту PCC в тощей кишки введение CM-МВ в просвет не оказало значительного влияния (p=0,3408, n=20) (фиг. 3E).
Амплитуда PCC в тощей кишке
Амплитуда PCC в тощей кишке не претерпела достоверных изменений ни в одной из групп обработки, за исключением микровезикул. Микровезикулы уменьшали амплитуду PCC в тощей кишке на 17% (p=0,0453, n=20) (фиг. 4D), несмотря на то, что данный эффект отсутствовал в исследовании с L. reuteri DSM 17938 или CM (p=0,3917, n=20 и p=0,1989, n=28 соответственно, фиг. 4A и 4B). Бульон и CM-МВ не изменяли амплитуду PCC в тощей кишке (p=0,8472 и p=0,5627, n=20) (фиг. 4C и 4E).
L. reuteri DSM-17938 и продукты на их основе увеличивали показатели моторики ободочной кишки
Скорость PCC в ободочной кишке
Сократительную подвижность ободочной кишки стимулировали добавлением L. reuteri DSM 17938, CM или МВ. L. reuteri DSM 17938 достоверно увеличивали скорость сокращений PCC в ободочной кишке на 65% (p=0,0004, n=20) (фиг. 5A). Данный эффект воспроизводился CM, которая достоверно увеличила скорость PCC на 72% в ободочной кишке (p=0,0021, n=28) (фиг. 5B). Бульон продолжал оказывать незначительное влияние на перистальтику кишечника, увеличивая скорость PCC в ободочной кишке на 8%, но не в пределах диапазона значимости 0,05 (p=0,1861, n=20) (фиг. 5C). Микровезикулы достоверно увеличивали скорость PCC в ободочной кишке на 24% (p=0,0051, n=20), но в меньшей степени, чем продуцированные L. reuteri DSM 17938 и CM (фиг. 5D). CM-МВ, введенная интралюминарно, не вызывала достоверного изменения скорости PCC в ободочной кишке (p=0,6475, n=20) (фиг. 5E).
Частота PCC в ободочной кишке
L. reuteri DSM 17938, CM и МВ все стимулировали перистальтику ободочной кишки за счет увеличения частоты сокращений PCC. L. reuteri DSM 17938 достоверно увеличивали частоту PCC в ободочной кишке на 30% (p=0,0231, n=20) (фиг. 6A). Аналогично CM значительно увеличила частоту PCC на 31% в ободочной кишке (p=0,0073, n=28) (фиг. 6B). Бульон увеличивал частоту PCC в ободочной кишке всего на 4%, и недостоверно (p=0,7219, n=20) (фиг. 6C). Аналогично тому, что было отмечено в отношении скорости PCC, микровезикулы увеличивали частоту PCC в ободочной кишке на 18% (p=0,0424, n=20); (фиг. 6D). CM-МВ не оказывала достоверного влияния на частоту PCC в ободочной кишке (p=0,3298, n=20) (фиг. 6E).
Амплитуда PCC в ободочной кишке
На амплитуду PCC в ободочной кишке достоверного влияния не оказали L. reuteri DSM 17938 или любая другая группа обработки (фиг. 7A-E).
Заключение
L. reuteri DSM 17938 оказывали региональное воздействие на перистальтику кишечника; имело место снижение скорости PCC в тощей кишке и повышение скорости PCC в ободочной кишке и частоты сокращений. Результаты настоящего исследования показали, что как микровезикулы, так и кондиционированная среда воспроизводят эффект L. reuteri DSM 17938 на перистальтику кишечника как в тонком, так и в толстом кишечнике. Кроме того, эти результаты не наблюдались при применении кондиционированной среды после удаления микровезикул (CM-МВ). Все результаты представлены на фиг.8.
Полученные результаты демонстрируют роль микровезикул в передаче пробиотических сигналов Lactobacillus с организмом-хозяином и их механизм действия в системе микробиом-кишечник-мозг. Данные показывают, что микровезикулы, продуцированные или высвобождаемые бактериями, ответственны за изменения перистальтики кишечника, вызванные L. reuteri DSM 17938.
Пример 4 - Выделенные бактериальные микровезикулы воспроизводят действие бактерий на передачу болевых сигналов
Эффект микровезикул, выделенных из культуральной среды L. reuteri DSM 17938, на передачу болевых сигналов исследовали с использованием TrpV1-экспрессирующих клеток Jurkat в присутствии 10 мкМ капсаицина.
Материалы и методы
Культура клеток
Клетки Jurkat (клон E6-1 (ATCC® TIB-152TM), ATCC) суспендировали в 2% фетальной телячьей сыворотке (FBS) в среде, разработанной Roswell Park Memorial Institute (RPMI) при титре примерно 5 ×106 клеток/мл в общем объеме 20 мл.
L. reuteri DSM 17938 культивировали, собирали и хранили, как описано в примере 3 выше. Выделение микровезикул из L. reuteri DSM 17938 также проводили, как описано в примере 3 выше (48 ч). Бактерии разводили в бульоне MRS до конечного титра 1010 КОЕ/мл и хранили замороженными при -80°C до использования в экспериментах.
Ратиометрический поток кальция, измеренный методом проточной цитометрии
50 мкг каждого из двух красителей, Fluo-3 AM (F1242; Sigma) и Fura Red AM (F3021; Sigma) растворяли в 100 мкл 0,1% плуроновой кислоты (PLURONIC® F127, растворенный в диметилсульфоксиде (DMSO)). Затем 50 мкл раствора Fluo-3 и 100 мкл раствора Fura Red добавляли к 20 мл клеток Jurkat, в результате чего соотношение Fluo-3 и Fura Red составляло 1:2,5. Затем клетки инкубировали при 37°C в течение 1 ч и промывали PBS (центрифугирование при 300 × g в течение 10 мин). Затем клетки ресуспендировали в среде Игла, модифицированной по способу Дульбекко (DMEM) или среде RPMI 1640, содержащей 2% FBS.
В день постановки эксперимента бактерии оттаивали, трижды промывали PBS, и затем добавляли к культурам клеток. Альтернативно, микровезикулы, выделенные из бактерий (как описано в примере 3), оттаивали, промывали и добавляли к культурам клеток. Инкубацию продолжали при 37°C в течение еще 1 ч. Затем клеточную суспензию центрифугировали, как описано выше, и ресуспендировали в PBS, содержащем 1,25 мМ Ca2+.
Все эксперименты проводили на проточном цитометре BD FACSCelesta (BD Bioscience, Mississauga, Канада), снабженном следующими лазерами: синий лазер, излучающий на длине волны 488 нм, красный лазер, излучающий на длине волны 640 нм, и фиолетовый лазер, излучающий на длине волны 406 нм. Калибровку проводили с использованием гранул BD PMT (BD, Mississauga, Канада). Компенсации выполнялись с помощью одноцветных компенсационных гранул BD.
Для эксперимента с FACS 1 мл клеточной суспензии переносили в пробирку емкостью 5 мл с клеточным ситом (Falcon 352235) и вращали в течение 1 мин перед анализом. Капсаицин готовили из 100 мМ стокового раствора и разбавляли до 100 мкМ в PBS, содержащем кальций и магний.
Фон, соответствующий неспецифическому потоку кальция, регистрировали в течение 30 с. Все образцы собирали в течение фиксированного времени (30 или 60 с) с постоянной скоростью потока (количество клеток/секунду). 100 мкл раствора капсаицина (с конечной концентрацией 10 мкМ) добавляли к клеточной суспензии непосредственно перед регистрацией с использованием FACSCelesta.
Регистрация была непрерывной со скоростью 400-600 событий/секунду в течение 30-60 секунд в целом.
Fluo-3 и Fura Red возбуждали при 488 нм, с детектированием эмиссии Fluo-3 при 575 нм, и детектированием эмиссии Fura Red при 610 нм. Данные собирали в виде гистограмм, отображающих соотношение флуоресценции фиолетовой к синей Fluo-3 в зависимости от времени и флуоресценции Fura Red в зависимости от времени.
Ратиометрический анализ Fluo-3/Fura Red проводили при возбуждении синим лазером (488 нм). Эмиссию регистрировали двумя различными наборами фильтров: увеличение эмиссии отслеживали с помощью фиолетового лазера (610/20 нм), и уменьшение эмиссии регистрировалось с помощью синего лазера (575/25 нм). Ратиометрическое соотношение Fluo-3/Fura Red было рассчитано как возрастающий сигнал, стимулированный фиолетовым лазером, по сравнению с снижающимся сигналом, стимулированным синим лазером (406 нм/488 нм), с использованием инструмента Kinetics в программном обеспечении FlowJo (Tree Star Inc., OR, США).
Результаты
Капсаицин сам по себе или МВ, выделенные из другого бактериального штамма, JB-1 (МВ-JB-1), использовали в качестве контроля. Показателем активации капсаицином (фиг. 9) было проникновение кальция в клетки Jurkat, что приводило к увеличению соотношения FLuo-3/Fura Red. Капсаицин сам по себе в количестве 10 мкМ индуцировал поступление кальция в клетки Jurkat, ответ, который достоверно блокировался при использовании МВ, выделенных из L. reuteri DSM 17938 (DSM-МВ) в высокой концентрации, соответствующей L. reuteri DSM 17938 при 1011 КОЕ/мл. Более низкие концентрации МВ из L. reuteri DSM 17938, соответствующие L. reuteri DSM 17938 при 109-1010 КОЕ/мл, имели аналогичный, но несколько более низкий эффект, тогда как МВ из L. rhamnosus JB-1 при высокой концентрации, соответствующей JB-1 при концентрации 1011 КОЕ/мл не оказывали достоверного влияния на ответ на обработку капсаицином.
Пример 5 - Начало действия на спинномозговой нерв выделенных бактериальных микровезикул происходит быстрее, чем начало действия целых бактерий
Авторы настоящего изобретения неожиданно обнаружили, что микровезикулы, полученные из Lactobacillus reuteri DSM 17938, не только воспроизводят действие целых бактерий на возбуждение брыжеечного нерва, но и что МВ способны оказывать усиленный эффект по сравнению с целыми бактериями. Данный результат был получен анализом времени, которое проходит от начала обработки до максимального ответа (начало полного эффекта) при использовании микровезикул, выделенных из L. reuteri DSM17938, по сравнению с использованием целых бактерий.
Материалы и методы
L. reuteri DSM 17938 культивировали, собирали и хранили, как описано в примере 3 выше. Выделение и получение микровезикул из L. reuteri DSM 17938 выполняли, как описано в примере 3 выше.
Регистрацию возбуждения брыжеечного нерва в тощей кишке выполняли, как описано ранее (Perez-Burgos et al. (2013); Perez-Burgos et al. (2015)). Вкратце, иссекали отрезок мышиной тощей кишки длиной 3 см и помещали на покрытую агаром чашку Петри, заполненную насыщенным кислородом раствором Кребса и блокатором кальциевых каналов L-типа, никардипином (3 мкМ). Содержимое просвета тощей кишки вымывали с использованием раствора Кребса, и оральный и анальный концы канюлировали силиконовой трубкой для обеспечения введения материалов для обработки в отрезок тощей кишки. Пучок брыжеечных нервов осторожно изолировали от отрезка тощей кишки, осторожно соскребая прикрепленную брыжейку тонкими щипцами. Затем чашку, содержащую препарат нервов, устанавливали на предметный столик микроскопа и непрерывно перфузировали теплым насыщенным кислородом раствором Кребса с помощью насоса.
Обнаженный нервный пучок отсасывали с помощью стеклянной микропипетки, прикрепленной к держателю электродов с накладным зажимом. Мультиэлементную электрическую активность регистрировали от нервного пучка с помощью усилителя Multi-Clamp 700B и преобразователя сигналов Digidata 1440A. Контрольные периоды регистрировались по 15-30 мин во время люминальной перфузии раствором Кребса. Люминальные L. reuteri DSM 17938 или микровезикулы применяли сразу после контроля в течение 20-30 мин.
Мультиэлементную электрическую активность анализировали для одноэлементной активности с использованием метода главных компонентов (PCA) и анализа формы волны в программе Dataview (Heitler (2007)). Время до максимального ответа измеряли с момента начала обработки до момента, когда был виден ответ (изменение скорости возбуждения).
Результаты
МВ из L. reuteri (МВ) были способны инициировать ответную реакцию возбуждения нерва быстрее, что приводило к более раннему началу действия по сравнению с целыми бактериями (DSM), как показано на фиг. 10.
Пример 6 - протокол культивирования и выделения
Типичная схема получения МВ включает следующие стадии: культивирование, удаление интактных бактерий и выделение МВ из культурального фильтрата и, необязательно, предварительное концентрирование и очистку. Выбор конкретных методов зависит от многих факторов, например, количества материала, предназначенного для обработки, и требуемой чистоты в соответствии с последующими применениями.
Условия культивирования
Авторы изобретения определили параметры культивирования бактериальных штаммов, приводящие к продукции терапевтических МВ с повышенным эффектом, установленным на предпочтительных моделях перистальтики желудочно-кишечного тракта и желудочно-кишечной боли. L. reuteri DSM 17938 культивировали в нормальных условиях культивирования, т. е. в анаэробных условиях культивирования в среде де Мана-Рогоза-Шарпа (MRS) при 37°C в течение 24 ч во флаконе/колбе с добавлением 2% сахарозы в качестве индуцирующего фактора.
Условия выделения
Приготовление бактериальных МВ включало вышеописанные определенные стадии культивирования бактерий с последующим выделением МВ. Для получения фракций МВ с высоким числом, высокой чистотой и с сохраненным биологическим эффектом бактериальный супернатант центрифугировали при 4000 об/мин в течение 20 мин (высокоскоростная центрифуга Beckman с ротором с фиксированным углом JA-18) с последующим центрифугированием в течение секунды при 10000 × g в течение 20 мин, и затем фильтровали через фильтр с размером пор 0,45 мкм. На этих первых стадиях удалялись живые бактерии и крупный дебрис. Затем осадок отбрасывали, и супернатант, содержащий МВ, наносили на «подушку» из сахарозы и ультрацентрифугировали при 118000 × g при 4°C в течение 20,5 ч. Наконец, осадок промывали и центрифугировали при 118000 × g, при 4°C в течение ночи с получением терапевтических МВ в виде осадка.
Пример 7 - Иммунная стимуляция микровезикулами (МВ), полученными из Lactobacillus reuteri DSM 17938
В данном примере было показано, что МВ, полученные из бесклеточного супернатанта (CFS) Lactobacillus reuteri DSM 17938, обладают иммуностимулирующей активностью и подавляющей активностью для интерферона гамма (IFN-γ), показывая, что внеклеточные МВ, полученные из кишечных бактерий, могут быть важными модуляторами иммунитета человека.
Материал и методы
Этическое заявление и выделение мононуклеарных клеток периферической крови
В данное исследование включали здоровых анонимных взрослых добровольцев (в возрасте 18-65), которое было одобрено Региональным комитетом по этике Каролинского института, Стокгольм, Швеция {Dnr 04-106/1 и 2014/2052-32}. Все участники исследования давали информированное письменное согласие. Образцы венозной крови собирали в гепаринизированные пробирки (вакуумные контейнеры BD Biosciences Pharmingen) и разбавляли клеточной культуральной средой RPMI-1640 с добавлением 20 мМ HEPES (HyClone Laboratories, Inc.). Затем мононуклеарные клетки периферической крови (PBMC) выделяли градиентным разделением в системе Ficoll-Hypaque (GE Healthcare Bio-Sciences AB). PBMC промывали RPMI-1640, ресуспендировали в замораживающей среде, содержащей 40% RPMI-1640, 50% эмбриональной телячьей сыворотки и 10% ДМСО, постепенно замораживали в морозильном контейнере (Mr Frosty, Nalgene Cryo 1°C; Nalge Co.) и хранили в жидком азоте.
Стимуляция PBMC in vitro
РВМС размораживали, промывали и оценивали жизнеспособность окрашиванием трипановым синим с последующим подсчетом с использованием светового микроскопа при увеличении 40×. Клетки ресуспендировали в среде для культивирования клеток (RPMI-1640 с добавлением HEPES (20 мМ), пенициллина (100 Е/мл), стрептомицина (100 мкг/мл), L-глутамина (2 мМ) (все производства HyClone Laboratories, Inc.) и 10% фетальной телячьей сыворотки (Gibco by Life Technologies)) с конечным титром 1×106 клеток/мл. Клетки высевали в планшеты с плоским дном для культивирования клеток и инкубировали при 37°C в атмосфере 5% CO2. Бесклеточный супернатант Staphylococcus aureus (CFS) использовали в качестве стимулятора в концентрации 2,5% (об./об.), и выделенные МВ из Lactobacillus reuteri DSM 17938 добавляли к PBMC при соотношении МВ к клеткам 500:1, 100: 1 и 20:1.
Выделение микровезикул
Бактериальные клетки Lactobacillus reuteri DSM 17938 культивировали в среде де Мана-Рогоза-Шарпа (Oxoid) в течение 24 ч при 37°C. Бактериальные клетки удаляли из культурального бульона центрифугированием при 5000 × g в течение 10 мин при 4°C с последующим центрифугированием при 10000 × g в течение 10 мин при 4°C. Затем супернатанты фильтровали с использованием фильтра с размером пор 0,45 мкм (Millipore). Бесклеточные супернатанты концентрировали с использованием фильтрующего устройства Amicon Ultra с отсечением по молекулярной массе 100 кДа, которое удаляет белки и другие молекулы с молекулярной массой ниже 100 кДа. Супернатанты наносили на верхнюю часть 12% сахарозной «подушки» с 50 мМ Трис-буфером, pH 7,2, в объемном соотношении 5:1, и центрифугировали на ультрацентрифуге Beckman coulter Optima L-80XP (Beckman coulter, США) при 118000 × g при 4ºC в течение 3 ч. Супернатанты отбрасывали, осадок ресуспендировали в буфере PBS и ультрацентрифугировали второй раз (118000 × g при 4°C в течение 3 ч). Затем осадки растворяли в PBS, разделяли на аликвотные порции и хранили при -70°C.
Экспериментальная процедура
МВ, выделенные из Lactobacillus reuteri DSM 17938, добавляли к PBMC при соотношении МВ к клеткам 500:1, 100:1 и 20:1 и инкубировали в течение 48 ч. Супернатанты клеточных культур собирали и анализировали на индукцию цитокинов с помощью ELISA.
ELISA
Уровни секретированных цитокинов IL-1ra (R&D Systems-BioTechne), IL-1β, IL-6, IL-10, IL-17A и IFN-γ (MabTech AB) измеряли в супернатантах клеточных культур с использованием наборов для сэндвич-ELISA в соответствии с инструкциями производителя. Поглощение измеряли при длине волны 405 нм с использованием микропланшетного ридера (Molecular Devices Corp.), и результаты анализировали с использованием программы SoftMax Pro 5.2 rev C (Molecular Devices Corp.).
Статистика
Все статистические тесты выполняли с использованием программного обеспечения GraphPad Prism (GraphPad Software). Все данные считались непараметрическими, в результате чего использовали множественные сравнения Данна или t-критерии Манна-Уитни. Различия считались значимыми, если p < 0,05, и также использовали следующие уровни значимости * p <0,05; ** р <0,01.
Результаты
МВ, выделенные из Lactobacillus reuteri DSM 17938, четко индуцировали продукцию как IL-6, так и IL-10 в зависимости от концентрации, в то время как индукции IFN-γ или IL-17A не было обнаружено (фиг. 11A). Более того, добавление выделенных МВ к PBMC, стимулированным S. aureus, достоверно снижало секрецию IFN-γ и IL-17A в такой же степени, как и фракция с высоким содержанием МВ (фиг. 11B).
Пример 8 - Микровезикулы (МВ), полученные из Lactobacillus reuteri, защищают целостность эпителиального барьера от разрушительного воздействия энтеротоксигенных Escherichia coli
Материал и методы
Выделение внеклеточных микровезикул (МВ)
Бактериальные клетки Lactobacillus reuteri DSM 17938 культивировали в среде де Мана-Рогоза-Шарпа, собирали через 24 ч и удаляли из культурального бульона центрифугированием при 5000 × g в течение 10 мин при 4°C с последующим центрифугированием при 10000 × g в течение 10 мин при 4°C, после чего любые оставшиеся клетки удаляли из супернатанта фильтрованием с использованием фильтра с размером пор 0,45 мкм. Супернатанты концентрировали с использованием фильтрующего устройства Amicon Ultra (100 кДа), которое удаляет белки и другие молекулы с молекулярной массой ниже 100 кДа. Супернатанты центрифугировали на ультрацентрифуге Beckman coulter Optima L-80XP (Beckman coulter, США) при 118000 × g при 4°C в течение 3 ч. Супернатанты отбрасывали, осадки ресуспендировали в буфере PBS и ультрацентрифугировали второй раз (118000 × g при 4°C в течение 3 ч). Затем осадки растворяли в PBS, распределяли на аликвотные порции и хранили при -70°C.
Кишечная проницаемость in vitro (сокультуры клеток Caco-2/HT29)
Культура эпителиальных клеток (Caco-2/HT29)
Клетки Caco-2 и HT29 культивировали по отдельности в колбах для тканевых культур в среде Игла, модифицированной по способу Дульбекко (DMEM) с добавлением 10% фетальной бычьей сыворотки, 1% заменимых аминокислот и 1% пенициллина и стрептомицина, при температуре ниже 37°C в атмосфере с 5% CO2 и относительной влажности 90%. Клетки Caco-2 и HT29 культивировали в колбах для тканевых культур емкостью 25 см2 и разделяли при достижении 80-90% слияния с использованием 0,25% раствора трипсина и 0,02% раствора этилендиаминтетрауксусной кислоты (EDTA). Клетки высевали из расчета 6×104 клеток на колбу емкостью 25 см2.
Сокультуры клеток
Клетки Caco-2 и HT29 высевали в апикальную камеру вставок Transwell (мембранные фильтры Transwell-COL, покрытые коллагеном) в соотношении 9:1 и культивировали в 12-луночных планшетах Transwell с конечной плотностью 1×105 клеток/см2 в каждой вставке. Клетки поддерживали в тех же условиях и позволяли расти в течение 21 суток, меняя среду (0,5 мл на апикальной стороне и 1,5 мл на базолатеральной стороне) через день, чтобы клетки могли дифференцироваться.
Целостность клеточного слоя
Целостность клеточного слоя определяли с использованием двух методов: трансэпителиального электрического сопротивления (TEER) и определения проницаемости для флуоресцеин изотиоцианат-декстрана (FITC-декстран).
Целостность клеточного монослоя во время экспериментов определяли измерением трансэпителиального электрического сопротивления (TEER) с использованием системы электрического сопротивления Millicell (Millipore, Darmstadt, Германия). Для определения значений TEER в каждой лунке были выбраны три разные области, и конечными результатами были средние значения. Для исследования проницаемости использовали значения TEER выше 250 см2.
Высеянные клетки Caco-2/HT29 предварительно обрабатывали живыми клетками Lactobacillus reuteri DSM 17938 при множественности бактерий 100 (MOB) или внеклеточными микровезикулами (МВ) из клеток Lactobacillus reuteri DSM 17938 при множественности МВ 200 в течение 6 ч перед провокационным инфицированием ETEC (патогенная энтеротоксигенная E. coli, известная своим разрушительным действием на целостность эпителия) при множественности инфекции 100 (MOI) в течение дополнительных 6 ч. TEER измеряли до предварительной обработки и инфицировании ETEC, после чего измеряли через каждый час в течение всего периода провокационной обработки. Для количественной оценки параклеточной проницаемости монослоев 1 мг/мл 4 кДа флуоресцеин изотиоцианат-декстрана (FITC-декстран; Sigma) добавляли на апикальной стороне вставок в начале инфицирования ЕТЕС. Образцы из базолатерального компартмента отбирали через 6 ч инкубации. Затем диффундированный флуоресцентный индикатор анализировали в трех повторностях с помощью флуорометрии (возбуждение, 485 нм; эмиссия, 520 нм) с использованием ридера для микропланшетов FLUOstar Omega (BMG Labtech, Ortenberg, Германия).
Результаты
Инфицирование ETEC вызывало снижение TEER. МВ, полученные из L. reuteri, и бактериальные клетки были частично способны защищать эпителиальный монослой от данной обработки (фиг. 12). Через 6 ч после инфицирования ETEC снижение TEER для группы ETEC достигло 35%, и предварительная обработка бактериальными клетками L. reuteri и МВ показала достоверно более высокое значение TEER по сравнению с группой, обработанной ETEC.
Защитный эффект МВ, полученных из L. reuteri, и бактериальных клеток против повреждения ETEC монослоя также был очевиден по результатам экспериментов с утечкой FITC-декстрана (фиг. 12). Предварительная обработка монослоев МВ, полученных из L. reuteri, и бактериальных клеток уменьшала утечку FITC-декстрана по сравнению с группой ETEC.
Таким образом, как МВ, полученные из L. reuteri, так и бактериальные клетки показали защитный эффект от вызванного ЕТЕС повреждения монослоев сокультур Caco-2/HT29.
Пример 9 - Изменение ферментативной активности, связанное с продукцией МВ
Lactobacillus reuteri DSM 17938 и Lactobacillus reuteri DSM 32846 культивировали и подвергали серии индуцирующих биотических обработок. Ответ, выражающийся в изменениях продукции МВ на эти индуцирующие обработки, определяли с помощью ферментативного анализа и сравнивали с ответом, полученным для контролей, т. е. бактериальных культур без индуцирующих обработок. Затем измеряли ферментативную активность в кондиционированной бактериальной среде.
Материалы и методы
Культивирование и биотическая обработка
L. reuteri DSM 17938 или L. reuteri DSM 32846 инокулировали из замороженного стокового материала в 25 мл среды Мана-Рогоза-Шарпа (MRS) в нормальных условиях культивирования, т. е. культивировали в анаэробных условиях при 37°C в течение ночи. Затем бактерии (40 мл) реинокулировали в 400 мл SIM вместе с супернатантом (4%) из других бактериальных культур или с добавлением других бактериальных клеток (25%, отмытых и суспендированных в PBS), и затем культивировали в течение еще 48 ч. Исследованные образцы бактерий представлены в таблице ниже.
Таблица 3
Суммарные данные по бактериальным образцам и способам обработки
Bifidobacterium longum ATCC BAA-999 и Lactobacillus paracasei LMG-P-17806 являются коммерчески доступными бактериальными штаммами и депонированы в АТСС (Американская коллекция типовых культур) и Бельгийских скоординированных коллекциях микроорганизмов, лаборатория микробиологии, соответственно.
Экспериментальный дизайн относится к пробиотическим бактериальным штаммам и различным биотическим обработкам и был разработан для имитации реальной масштабированной ситуации в производственных условиях или для имитации ситуации в желудочно-кишечном тракте человека. 4% супернатант после биотических обработок во время культивирования был выбран в качестве релевантной концентрации, достаточной для возможного проявления эффекта, с одной стороны, но с другой стороны, без излишнего добавления, когда может иметь место риск наличия компонентов от индуцирующих биотических бактерий в виде части конечного продукта. Была выбрана более высокая концентрация, 25%, клеток после биотической обработки, чтобы имитировать эффект, который будет иметь место локально в желудочно-кишечном тракте человека, если два (или более) бактериальных штамма вводить вместе в виде одной комбинированной композиции.
Отбор образцов
Бактериальные образцы сначала центрифугировали при 5000 × g в течение 10 мин, после чего супернатанты переносили в другую пробирку, и затем центрифугировали второй раз при 10000 × g в течение 10 мин для удаления бактериальных клеток и дебриса бактериальных клеток. Супернатанты, теперь содержащие МВ, фильтровали через фильтр с размером пор 0,45 мкм и хранили на льду до дальнейшего центрифугирования с использованием ультрацентрифуги при 32000 об/мин при 4°C в течение 3 ч (Beckman SW 32 Ti Rotor, Swing bucket, пробирки емкостью 30 мл). Супернатанты отбрасывали (осторожно сливали с помощью пипетки). Осадки, содержащие МВ, осторожно ресуспендировали в среде для ресуспендирования (забуференный фосфатом физиологический раствор (PBS)) и снова центрифугировали при 32000 об/мин при 4°C, смывая остатки культуральной среды. Объем ресуспендирования варьировался от 100 до 300 мкл в зависимости от размера осадка. Образцы разделяли на аликвотные порции и хранили при -70°C.
Ферментативная активность
Активность фермента 5'-нуклеотидазы использовали в качестве показателя для количественной оценки изменений числа и/или активности МВ, продуцированных с использованием различных индуцирующих биотических обработок. Образцы, полученные после индуцирующих обработок, описанных описанных выше, размораживали и затем тестировали в анализе активности 5'-нуклеотидазы с использованием набора для анализа 5'-нуклеотидазы Crystal Chem (Crystal Chem, Elk Grove Village, IL, США). Вкратце, процедуру проводили в две стадии. Сначала к образцам супернатанта добавляли реагент 1 (CC1), содержащий АМФ, для превращения АМФ в аденозин под действием любого фермента 5'-нуклеотидазы, присутствующего в образцах супернатанта. Аденозин далее гидролизовался до инозина и гипоксантина компонентами реагента 1. На второй стадии добавляли реагент 2 (CC2) для превращения гипоксантина в мочевую кислоту и гидроксид водорода, который использовался для получения хинонового красителя, кинетическое измерение которого проводили при 550 нм на спектрофотометре. Активность 5'-нуклеотидазы в образцах определяли расчетом изменения оптической плотности в период между 3 и 5 мин и сравнением со значением, полученным с образцом калибратора.
Результаты
На фиг. 13 показана 5'-нуклеотидазная активность в образцах МВ, полученных из L. reuteri DSM 17938 (для контрольных и обработанных образцов). Как следует по данным фигуры, активность 5'-нуклеотидазы была выше в образцах, полученных в результате культивирования L. reuteri DSM 17938 с добавлением 4% супернатанта из B. longum ATCC BAA-999 или B. longum DSM 32947 на среде SIM (DSM 17938+4% DSM 32947 sup. или DSM 17938+4% ATCC BAA-999 sup.) по сравнению с L. reuteri DSM 17938 на среде SIM (контроль) или L. reuteri DSM 17938 с 4% супернатанта L. paracasei LMG-P-17806 (DSM 17938 в SIM или DSM 17938+4% LMG-P-17806 sup.). Эффект индуцирующей биотической обработки на активность 5'-нуклеотидазы был наиболее выражен, когда L. reuteri DSM 17938 культивировали с супернатантом B. longum DSM 32947, но эффект также получали с супернатантом B. longum ATCC BAA-999. Показатели оптической плотности (OD) для каждого образца свидетельствуют о том, что относительное число клеток достоверно не изменилось между обработками.
Результаты, представленные на фиг. 14, аналогичны представленным на фиг. 13, но были нормализованы по отношению к активности 5'-нуклеотидазы и оптической плотности DSM 17938 на среде SIM, что упрощает сравнение кратных изменений в 5'-нуклеотидазной активности между разными экспериментами.
Результаты на фиг.15 (нормализованные данные) дополнительно показывают, что активность 5'-нуклеотидазы увеличивалась после индуцирующей биотической обработки по сравнению с контрольными образцами (т.е. DSM 17938 на среде SIM) при скультивировании L. reuteri DSM 17938 с 25% клетками B. longum DSM 32947 в среде SIM. Показатели OD для этих образцов иллюстрируют относительную разницу в количестве клеток, полученную при этих различных биотических обработках (т. е. повышенный показатель для образцов после индуцирующей биотической обработки за счет более высокого общего количества бактериальных клеток).
На фиг.16 приведены нормализованные значения активности 5'-нуклеотидазы, полученные для контрольных и обработанных образцов с использованием L. reuteri DSM 32846. Как следует из графика, ферментативная активность увеличивалась при культивировании L. reuteri DSM 32846 с добавлением 4% супернатанта B. longum DSM 32947 в среде SIM (DSM 32846+4% DSM 32947 sup.) По сравнению с контрольными L. reuteri DSM 32846 в среде SIM (DSM 32846 в SIM) показатели OD для этих образцов иллюстрируют относительную разницу в количестве клеток, полученную с помощью этих различных биотических обработок.
Пример 10 - Повышенный антагонистический эффект Lactobacillus reuteri DSM 17938 на TrpV1-опосредованную передачу болевых сигналов после сокультивирования с Bifidobacterium longum DSM 32947 или Bifidobacterium longum ATCC BAA-999
Влияние биотических обработок на продукцию МВ Lactobacillus reuteri DSM 17938 исследовали на модели стимуляции электрического поля (EFS) in vitro с использованием TrpV1-экспрессирующих нейронов, полученных из ганглиев задних спинальных корешков крыс (DRG) и платформы Cellectricon Cellaxess Elektra. L. reuteri DSM 17938 культивировали в течение 48 ч либо в качестве контроля в искусственной кишечной среде (DSM 17938, контрольная SIM в течение 48 ч), либо с использованием индуцирующей биотической обработки, состоящей из сокультивирования с 25% бактериальных клеток любого из двух Bifidobacterium longum DSM 32947 или ATCC BAA-999 в среде SIM в течение 48 ч (состав среды SIM можно найти в примере 1). МВ выделяли из различных бактериальных препаратов, как описано в примере 6. Первичные культуры нейронов DRG крыс культивировали в течение 48 ч в 384-луночных планшетах вместе с фактором роста нервов (NGF) для имитации периферической сенсибилизации. Затем оценивали антагонистический эффект полученных препаратов МВ на вызванную капсаицином активацию TrpV1. В день постановки эксперимента культуры DRG окрашивали индикатором Ca2+ (Ca5; набор для скрининга без для отмывки) для визуализации переходных процессов кальция, вызванных капсаицином, специфическим агонистом TrpV1. Сначала в отдельных экспериментах EFS определяли значение ЕС50 капсаицина, и затем эту концентрацию добавляли ко всем культурам DRG для индукции активации TrpV1. Затем оценивали влияние препаратов МВ на активацию TrpV1 в формате зависимости доза-ответ (шесть концентраций тестировали в трех повторностях в каждом планшете) с начальной концентрацией 1:10 от исходной концентрации МВ и с использованием стадий разбавления 1:3. Культуры DRG инкубировали с препаратами МВ в течение 1 ч перед проведением эксперимента EFS. Планшеты помещали на платформу Cellaxess, и использовали серию протоколов EFS. Данные протоколы EFS включали последовательности импульсов для регистрации изменений возбудимости, которые происходят в культурах DRG в результате инкубации с препаратами МВ. Затем анализировали эффект препаратов МВ на вызванную капсаицином активацию TrpV1, и значения ЕС50 определяли для каждого препарата (ЕС50 обычно описывается как половина максимальной эффективной концентрации и относится к концентрации вещества, которая вызывает половинный ответ между базовым уровнем и максимум после указанного времени действия). В данном случае EC50 представляет собой % от основной концентрации МВ, экстрагированных из 400 мл жидкой бактериальной культуры). Эксперимент проводили дважды и рассчитывали среднее значение ЕС50. Значение ЕС50 в контрольном эксперименте составляло 7,4, тогда как значения ЕС50 для обеих индуцирующих биотических обработок были намного ниже (1,8 для DSM 17938+DSM 32947; 2,7 для DSM 17938+ATCC BAA-999). Полученные результаты показывают, что индуцирующие биотические обработки усиливают антагонистический эффект препарата МВ на передачу сигналов TrpV1, т.е. меньшее количество препаратов индуцированных МВ требуется для ингибирования передачи сигналов TrpV1, индуцированных капсаицином, по сравнению с контрольным препаратом МВ. В данном случае следует указать важный аспект, заключающийся в том, что контроль (DSM 17938 SIM 48 ч) ранее показал ингибирующий эффект на передачу сигналов TrpV1 (см. пример 4) и, следовательно, может считаться положительным контролем. Суммируя полученные данные, можно заключить, что в результате подвергания бактерий индуцирующей биотической обработке во время культивирования в соответствии с изобретением, у бактерий имеет место индукция продуцирования терапевтических МВ. Это, в свою очередь, повышало ингибирующий/блокирующий эффект на активацию TrpV1, вызванную капсаицином.
Таблица 4
Значение EC50 у бактерий для ингибирования TrpV1
Значения ЕС50 представлены в виде % от исходной концентрации МВ, экстрагированных из 400 мл жидкой бактериальной культуры, и показывают усиленный антагонистический эффект препаратов МВ из DSM 17938 на TrpV1 в результате индуцирующей биотической обработки.
Пример 11 - Изменения иммунной модуляции в результате биотической обработки во время культивирования
В данном примере было показано, что МВ, полученные из бесклеточного супернатанта (CFS) Lactobacillus reuteri DSM 32846, обладают более сильным иммуномодулирующим эффектом (повышение IL-6) по сравнению с МВ, полученными из бесклеточного супернатанта (CFS) Lactobacillus reuteri DSM 17938. Также было показано, что иммуномодулирующий эффект МВ, полученных из бесклеточного супернатанта (CFS) Lactobacillus reuteri DSM 32846 или Lactobacillus reuteri DSM 17938, усиливается после биотической обработки во время культивирования.
Материал и методы
Этическое заявление и выделение мононуклеарных клеток периферической крови
В данное исследование включали здоровых анонимных взрослых добровольцев (в возрасте 18-65), которое было одобрено Региональным комитетом по этике Каролинского института, Стокгольм, Швеция {Dnr 04-106/1 и 2014/2052-32}. Все участники исследования давали информированное письменное согласие. Образцы венозной крови собирали в гепаринизированные пробирки (вакуумные контейнеры BD Biosciences Pharmingen) и разбавляли клеточной кульутральной средой RPMI-1640 с добавлением 20 мМ HEPES (HyClone Laboratories, Inc.). Затем мононуклеарные клетки периферической крови (PBMC) выделяли градиентным разделением в системе Ficoll-Hypaque (GE Healthcare Bio-Sciences AB). PBMC промывали RPMI-1640, ресуспендировали в среде для замораживания, содержащей 40% RPMI-1640, 50% эмбриональной телячьей сыворотки и 10% ДМСО, постепенно замораживали в морозильном контейнере (Mr Frosty, Nalgene Cryo 1°C; Nalge Co.) и хранили в жидком азоте.
Стимуляция PBMC in vitro
РВМС размораживали, промывали и оценивали жизнеспособность окрашиванием трипановым синим с последующим подсчетом с использованием светового микроскопа при увеличении 40×. Клетки ресуспендировали в среде для культивирования клеток (RPMI-1640 с добавлением HEPES (20 мМ), пенициллина (100 Е/мл), стрептомицина (100 мкг/мл), L-глутамина (2 мМ) (все производства HyClone Laboratories, Inc.) и 10% фетальной телячьей сыворотки (Gibco by Life Technologies)) с конечным титром 1×106 клеток/мл. Клетки высевали в планшеты с плоским дном для культивирования клеток и инкубировали при 37°C в атмосфере 5% CO2. Выделенные МВ из различных бактериальных препаратов, как более детально описано ниже, добавляли к PBMC при соотношении МВ к клеткам 500:1.
Выделение микровезикул
Бактериальные клетки Lactobacillus reuteri DSM 17938 или DSM 32846 культивировали в среде SIM (имитированная кишечная среда, состав приведен в примере 1) в течение 48 ч при 37°C. Для экспериментов, в которых изучали различные виды биотической обработки, бактериальные клетки Lactobacillus reuteri DSM 17938 или DSM 32846 культивировали в присутствии супернатантов отдельно выращенных Bifidobacterium longum DSM 32947 или Bifidobacterium longum ATCC BAA-999 в среде SIM в течение 48 ч при 37°C. Также бактериальные клетки Lactobacillus reuteri DSM 17938 культивировали в присутствии 25% бактериальных клеток Bifidobacterium longum DSM 32947.
Бактериальные клетки удаляли из культурального бульона центрифугированием при 5000 × g в течение 10 мин при 4°C с последующим повторным центрифугированием при 10000 × g в течение 10 мин при 4°C. Затем супернатанты фильтровали с использованием фильтра с размером пор 0,45 мкм (Millipore). Бесклеточные супернатанты концентрировали с использованием фильтрующего устройства Amicon Ultra с отсечением по молекулярной массе 100 кДа, которое удаляет белки и другие молекулы с молекулярной массой ниже 100 кДа. Супернатанты центрифугировали на ультрацентрифуге Beckman Optima L-80XP (Beckman coulter, США) при 118000 × g при 4°C в течение 3 ч. Супернатанты отбрасывали, осадок ресуспендировали в буфере PBS и ультрацентрифугировали второй раз (118000 × g при 4°C в течение 3 ч). Затем осадки растворяли в Neurobasal A+добавка B27+Glutamax, разделяли на аликвотные порции и хранили при -70°C.
Экспериментальная процедура
Выделенные МВ из различных бактериальных препаратов, описанных выше, добавляли к PBMC при соотношении МВ к клеткам 500:1 и инкубировали в течение 48 ч. Супернатанты клеточных культур собирали и анализировали на индукцию цитокинов с использованием ELISA.
ELISA
Уровни секретированного цитокина IL-6 измеряли в супернатантах клеточных культур с использованием наборов для сэндвич-ELISA в соответствии с инструкциями производителя (MabTech AB). Поглощение измеряли при длине волны 405 нм с использованием ридера для микропланшетов (Molecular Devices Corp.), и результаты анализировали с помощью программного обеспечения SoftMax Pro 5.2 rev C (Molecular Devices Corp.).
Результаты
Микровезикулы из Lactobacillus reuteri DSM 32846 были более эффективны в индукции секреции IL-6 по сравнению с микровезикулами, выделенными из Lactobacillus reuteri DSM 17938 (фиг. 17). Более того, МВ, выделенные из обоих штаммов Lactobacillus reuteri после сокультивирования со штаммом Bifidobacterium longum (как в виде 4% супернатанта, так и 25% клеток), увеличивали индуцированную продукцию IL-6 по сравнению с контролем (фиг. 18, 19 и 20). Полученные результаты показывают, что выделенные МВ более эффективны после биотической обработки во время культивирования.
Пример 12 - Микровезикулы (МВ), полученные из Lactobacillus reuteri, защищают целостность эпителиального барьера от разрушительного воздействия энтеротоксигенной Escherichia coli
Материал и методы
Выделение внеклеточных микровезикул (МВ)
Бактериальные клетки Lactobacillus reuteri DSM 17938 культивировали в среде де Мана-Рогоза-Шарпа, собирали через 24 ч и удаляли из культурального бульона центрифугированием при 5000 × g в течение 10 мин при 4°C с последующим центрифугированием при 10000 × g в течение 10 мин при 4°C, после чего любые оставшиеся клетки удаляли из супернатанта фильтрованием с использованием фильтра с размером пор 0,45 мкм. Супернатанты концентрировали с использованием фильтрующего устройства Amicon Ultra (100 кДа), которое удаляет белки и другие молекулы с молекулярной массой нижк 100 кДа. Супернатанты центрифугировали на ультрацентрифуге Beckman coulter Optima L-80XP (Beckman coulter, США) при 118000 × g при 4°C в течение 3 ч. Супернатанты отбрасывали, осадки ресуспендировали в буфере PBS и ультрацентрифугировали второй раз (118000 × g при 4°C в течение 3 ч). Затем осадки растворяли в PBS, распределяли на аликвотные порции и хранили при -70°C.
Кишечная проницаемость in vitro (сокультуры клеток Caco-2/HT29)
Культура эпителиальных клеток (Caco-2/HT29)
Клетки Caco-2 и HT29 культивировали по отдельности в колбах для тканевых культур в среде Игла, модифицированной по способу Дульбекко (DMEM) с добавлением 10% фетальной бычьей сыворотки, 1% заменимых аминокислот и 1% пенициллина и стрептомицина, при температуре ниже 37°C в атмосфере с 5% CO2 и относительной влажности 90%. Клетки Caco-2 и HT29 культивировали в колбах для тканевых культур емкостью 25 см2 и разделяли при достижении 80-90% слияния с использованием 0,25% раствора трипсина и 0,02% раствора этилендиаминтетрауксусной кислоты (EDTA). Клетки высевали из расчета 6×104 клеток на колбу емкостью 25 см2.
Сокультуры клеток
Клетки Caco-2 и HT29 высевали в апикальную камеру вставок Transwell (мембранные фильтры Transwell-COL, покрытые коллагеном) в соотношении 9:1 и культивировали в 12-луночных планшетах Transwell с конечной плотностью 1×105 клеток/см2 в каждой вставке. Клетки поддерживали в тех же условиях и позволяли расти в течение 21 суток, меняя среду (0,5 мл на апикальной стороне и 1,5 мл на базолатеральной стороне) через день, чтобы клетки могли дифференцироваться.
Целостность клеточного слоя
Целостность клеточного слоя определяли с использованием двух методов: трансэпителиального электрического сопротивления (TEER) и определения проницаемости для флуоресцеин изотиоцианат-декстрана (FITC-декстран).
Целостность клеточного монослоя во время экспериментов определяли измерением трансэпителиального электрического сопротивления (TEER) с использованием системы электрического сопротивления Millicell (Millipore, Darmstadt, Германия). Для определения значений TEER в каждой лунке были выбраны три разные области, и конечными результатами были средние значения. Для исследования проницаемости использовали значения TEER выше 250 см2.
Высеянные клетки Caco-2/HT29 предварительно обрабатывали живыми клетками Lactobacillus reuteri DSM 17938 при множественности бактерий 100 (MOB) или внеклеточными микровезикулами (МВ) из клеток Lactobacillus reuteri DSM 17938 при множественности МВ 200 в течение 6 ч перед провокационным инфицированием ETEC (патогенная энтеротоксигенная E. coli, известная своим разрушительным действием на целостность эпителия) при множественности инфекции 100 (MOI) в течение дополнительных 6 ч. TEER измеряли до предварительной обработки и инфицирования ETEC, после чего измеряли через каждый час в течение всего периода инфицирования. Для количественной оценки параклеточной проницаемости монослоев 1 мг/мл 4 кДа флуоресцеин изотиоцианат-декстрана (FITC-декстран; Sigma) добавляли к апикальной стороне вставок в начале инфицирования ЕТЕС. Образцы из базолатерального компартмента отбирали через 6 ч инкубации. Затем диффундированный флуоресцентный индикатор анализировали в трех повторностях с помощью флуорометрии (возбуждение, 485 нм; эмиссия, 520 нм) с использованием ридера для микропланшетов FLUOstar Omega (BMG Labtech, Ortenberg, Германия).
Результаты
Инфицирование ETEC вызывало снижение TEER, что свидетельствует о том, что МВ, полученные из L. reuteri DSM 32846, частично способны защищать монослой эпителия от такого инфицирования (фиг. 21). Через 6 ч после инфицирования ETEC снижение TEER для группы ETEC достигло 27%, тогда как предварительная обработка МВ, полученными из L. reuteri DSM 32846, с 10, 50, 100 и 200 MOM, и показала значительно более высокий TEER по сравнению с группой, обработанной ETEC. Необработанные клетки оставались на уровне примерно 90%.
Данный защитный эффект МВ, полученных из L. reuteri DSM 32846, против повреждения ETEC монослоя был также очевиден в экспериментах с утечкой FITC-декстрана (фиг. 21). Предварительная обработка монослоев производными L. reuteri DSM 32846 МВ 10, 50, 100 и 200 MOM уменьшала утечку FITC-декстрана по сравнению с группой ETEC.
Таким образом, МВ, полученные из L. reuteri DSM 32846, показали защитный эффект от вызванного ETEC повреждения монослоев совместных культур Caco-2/HT29.
На фиг. 22 приведено сравнение защитного эффекта МВ, полученных из L. reuteri DSM 32846, как показано в эксперименте с утечкой FITC-декстрана, с эффектом, полученным с МВ, полученными из L. reuteri DSM 17938. Предварительная обработка монослоев эпителиальных клеток МВ, полученными из L. reuteri DSM 32846, уменьшала утечку FITC-декстрана более эффективно, особенно в более низких концентрациях МВ, по сравнению с МВ, полученными из L. reuteri DSM 17938. Данный факт свидетельствует о том, что МВ, полученные из L. reuteri DSM 32846, более эффективны, чем МВ, полученные из L. reuteri DSM 17938, в защите целостности эпителиального барьера.
Варианты осуществления, описанные выше, следует понимать в качестве нескольких иллюстративных примеров настоящего изобретения. Специалистам в данной области техники будет понятно, что в варианты осуществления могут быть внесены различные модификации, комбинации и изменения, не выходящие за рамки объема настоящего изобретения. В частности, различные частичные решения в различных вариантах осуществления можно объединить в других конфигурациях, где это технически возможно. Однако объем настоящего изобретения определяется прилагаемой формулой изобретения.
Ссылки
Abusleme L. et al. IL-17; overview and role in oral immunity and microbiome. Oral Dis., 23 (7): 854-865 (2017).
Bindels L. et al. Increased gut permeability in cancer cachexia: mechanisms and clinical relevance. Oncoraget., Vol. 9, (No.26), pp: 18224-18238 (2018).
DiRienzo J. Breaking the Gingival Epithelial Barrier: Role of the Aggregatibacter actinomycetemcomitans Cytolethal Distending Toxin in Oral Infectious Disease. Cells, 3, 476-499 (2014).
Funauchi M. et al. Serum level of interferon-gamma in autoimmune diseases, Tohoku J. Exp. Med., 164 (4): 259-267 (1991).
Gu Z. W. et al. Neutralization of interleukion-17 suppresses allergic rhinitis symptoms by downregulating Th2 and Th17 responses and upregulating the Treg response. Oncoraget., Vol. 8, (No.14), pp: 22361-22369 (2017).
Heitler, W.J. DataView: A Tutorial Tool for Data Analysis. Template-based Spike Sorting and Frequency Analysis. J. Undergrad. Neurosci. Educ., 6, A1-7 (2007).
Klonowska J. et al. New Cytokines in the Pathogenesis of Atopic Dermatitis - New Therapeutic Targets. Int. J. Mol. Sci., 19, 3086 (2018).
Koga C. et al. Possible Pathogenic Role of Th17 Cells for Atopic Dermatitis. Journal of Investigative Dermatology, 128, 2625-2630, (2008).
Lee, Youngkyun. The role of interleukin-17 in bone metabolism and inflammatory skeletal diseases. BMB Reports, 46 (1): 479-483 (2013).
Perez-Burgos A. et al. Psychoactive bacteria Lactobacillus rhamnosus (JB-1) elicits rapid frequency facilitation in vagal afferents. Am. J. Physiol. Gastrointest. Liver Physiol., 304, G211-G220 (2013).
Perez-Burgos A. et al. Transient receptor potential vanilloid 1 channel in rodents is a major target for antinociceptive effect of the probiotic L. reuteri DSM 17938. J. Physiol., 17, n/a-n/a (2015).
Pollard K. M et al. Interferon-γ and systemic autoimmunity, Discov. Med., 16 (87), 123-131 (2013).
Tachibana K. et al. IL-17 and VEGF are increased and correlated to systemic inflammation, immune suppression, and malnutrition in patients with breast cancer. International Journal of Immunopathology and Pharmacology. Vol., 15 (3) 219-228 (2017).
Wang Y-H. et al. The cytokine family and their role in allergic inflammation. Curr Opin Immunol., 20 (6): 697-702 (2008).
Wu R. Y. et al. Spatiotemporal maps reveal regional differences in the effects on gut motility for Lactobacillus reuteri and rhamnosus strains. Neurogastroenterol. Motil., 25, e205-e214 (2013).
Zbikowska-Gotz M. et al. Expression of IL17A concentration and effector functions of peripheral blood neutrophils in food allergy hypersensitivity patients. International Journal of Immunopathology and Pharmacology, Vol. 29 (1) 90-98, (2015).
Zhang J. et al. Changes of serum cytokines-related Th1/Th2/Th17 concentration in patients with postmenopausal osteoporosis. Gynecological Endrocrinology, 31:3, 183-190 (2015).
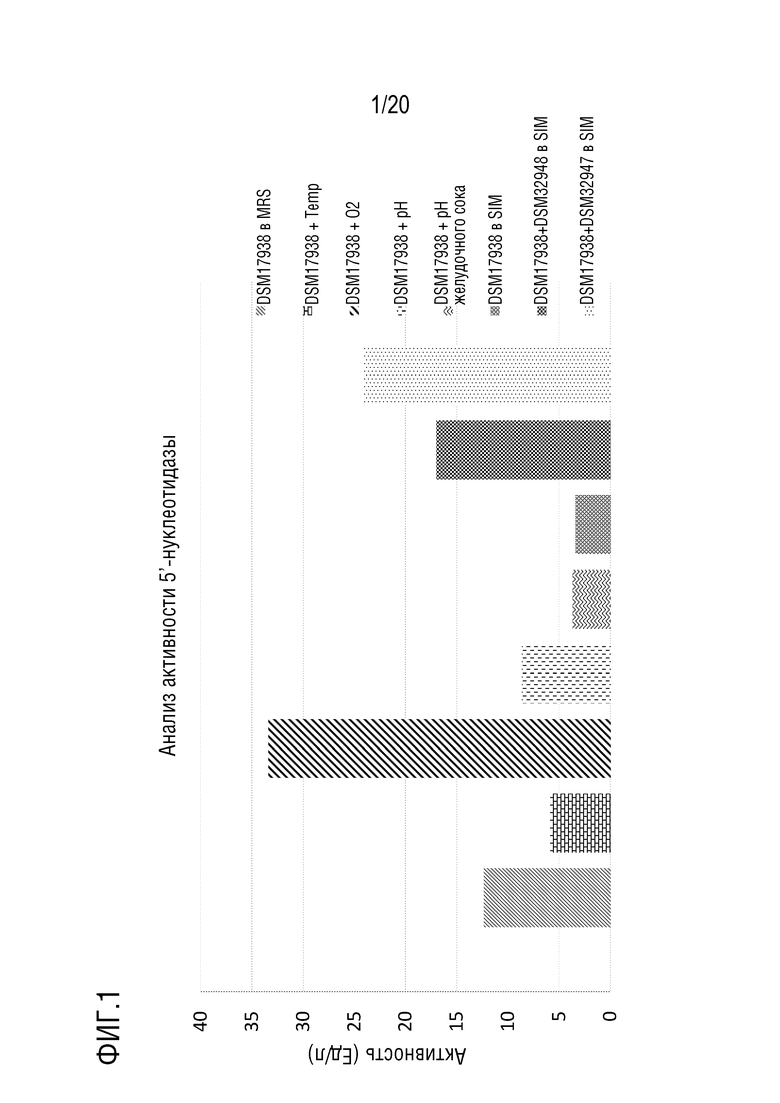
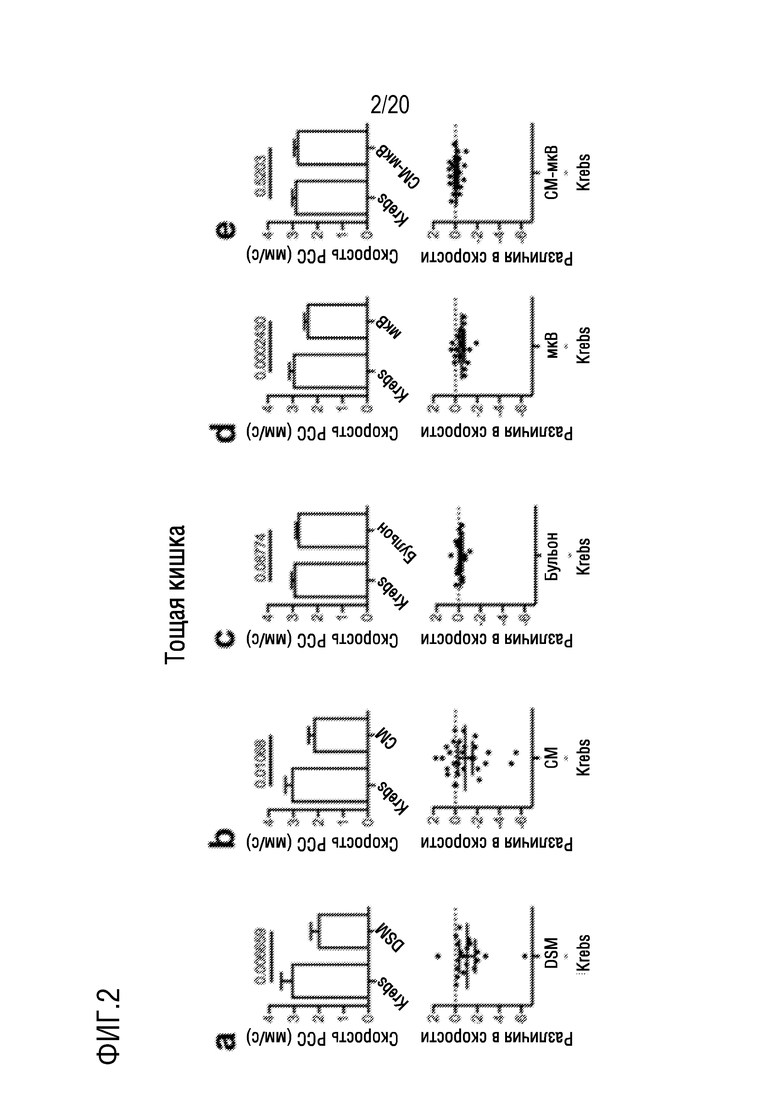
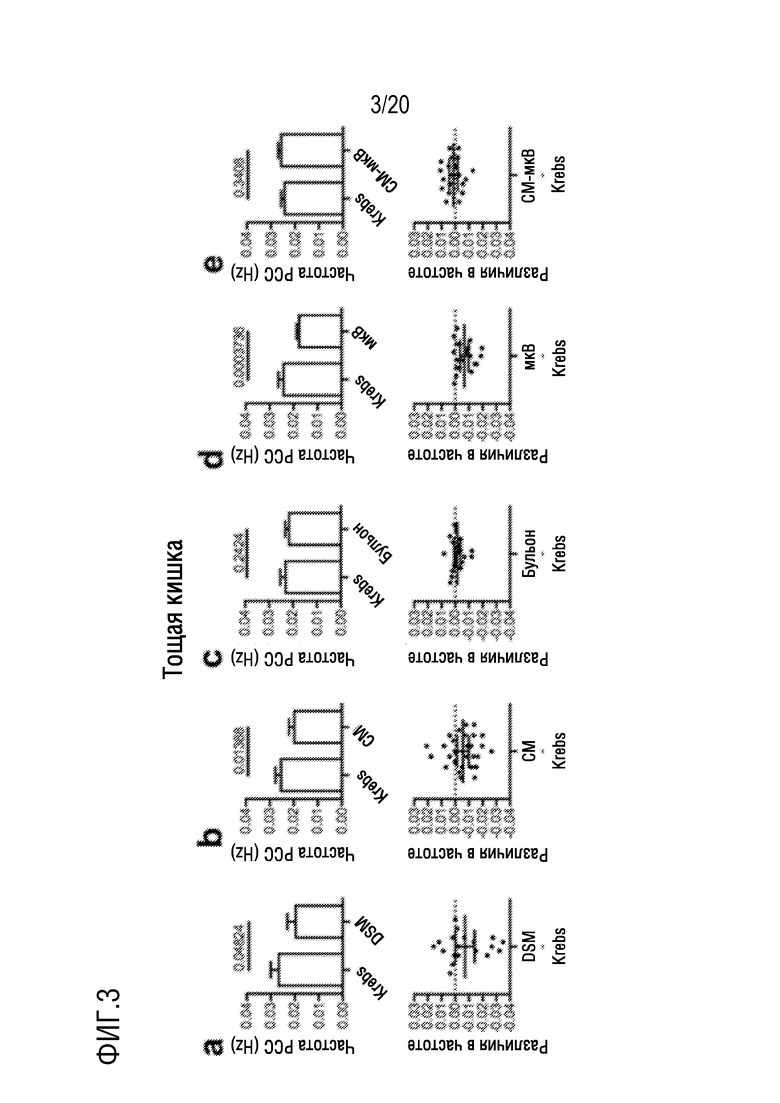
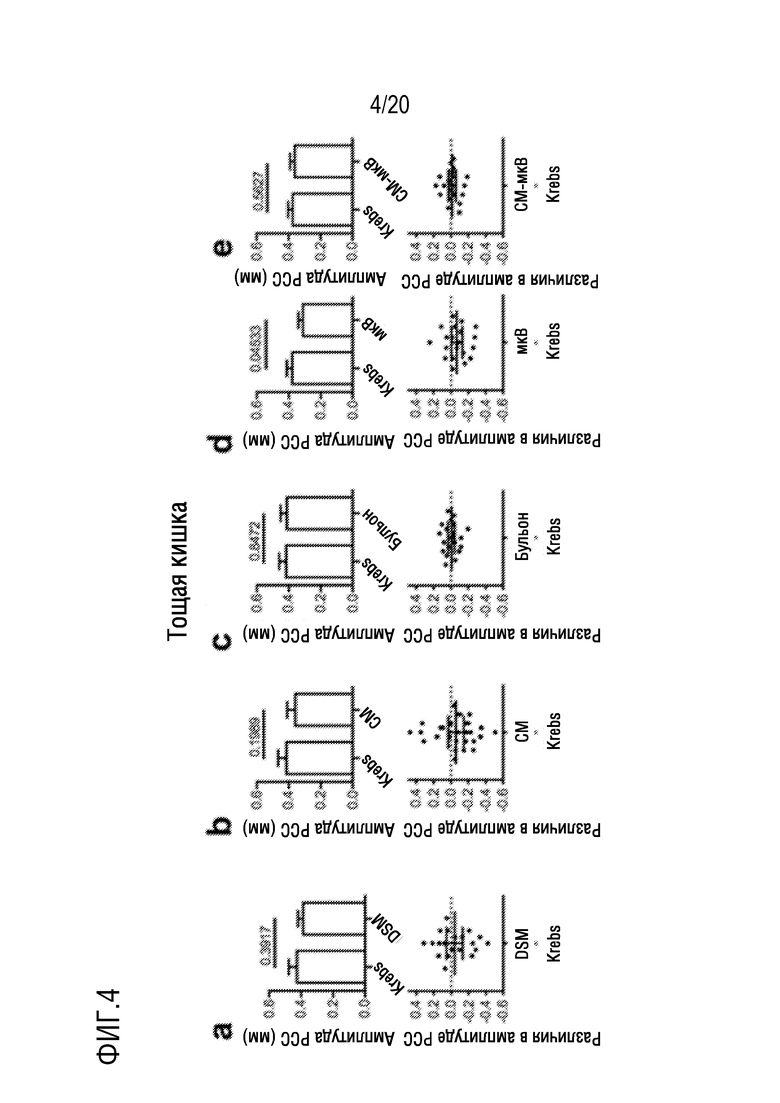
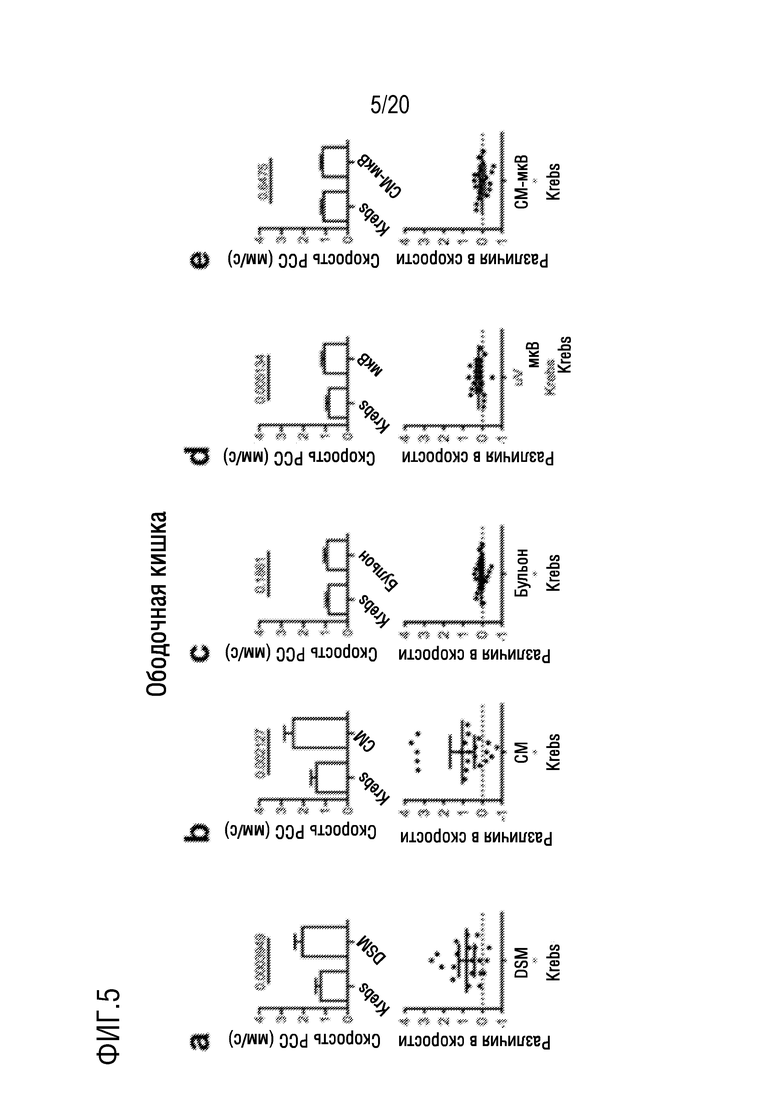
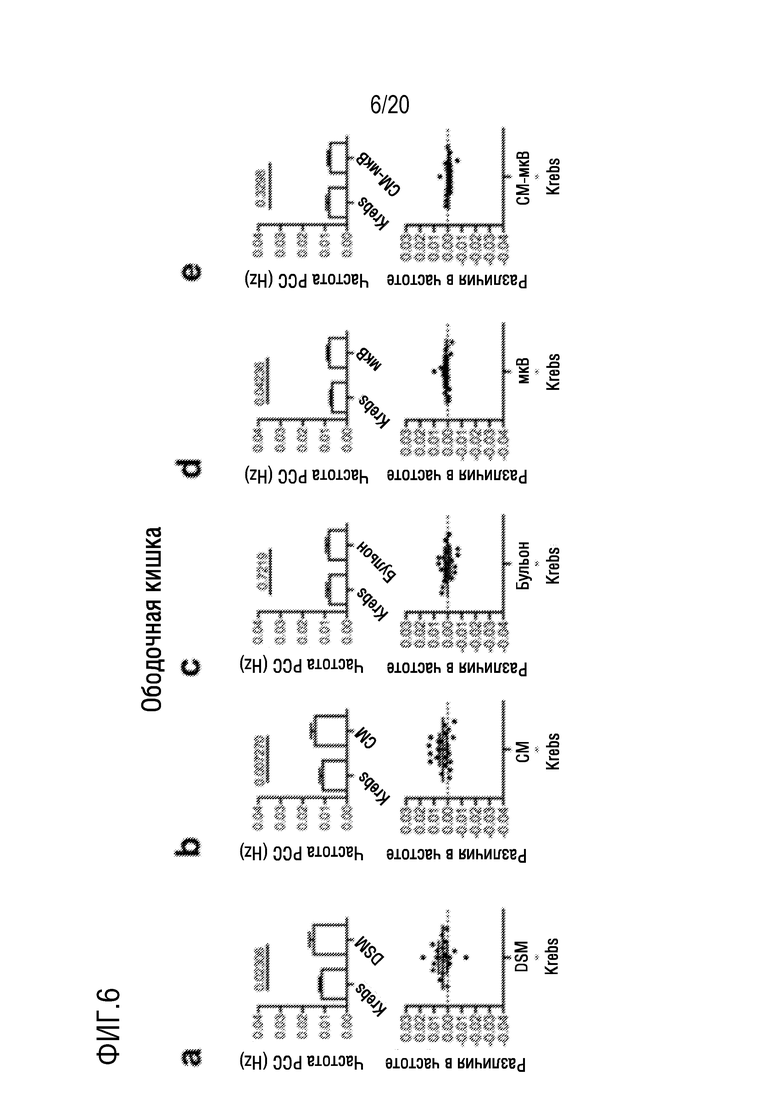
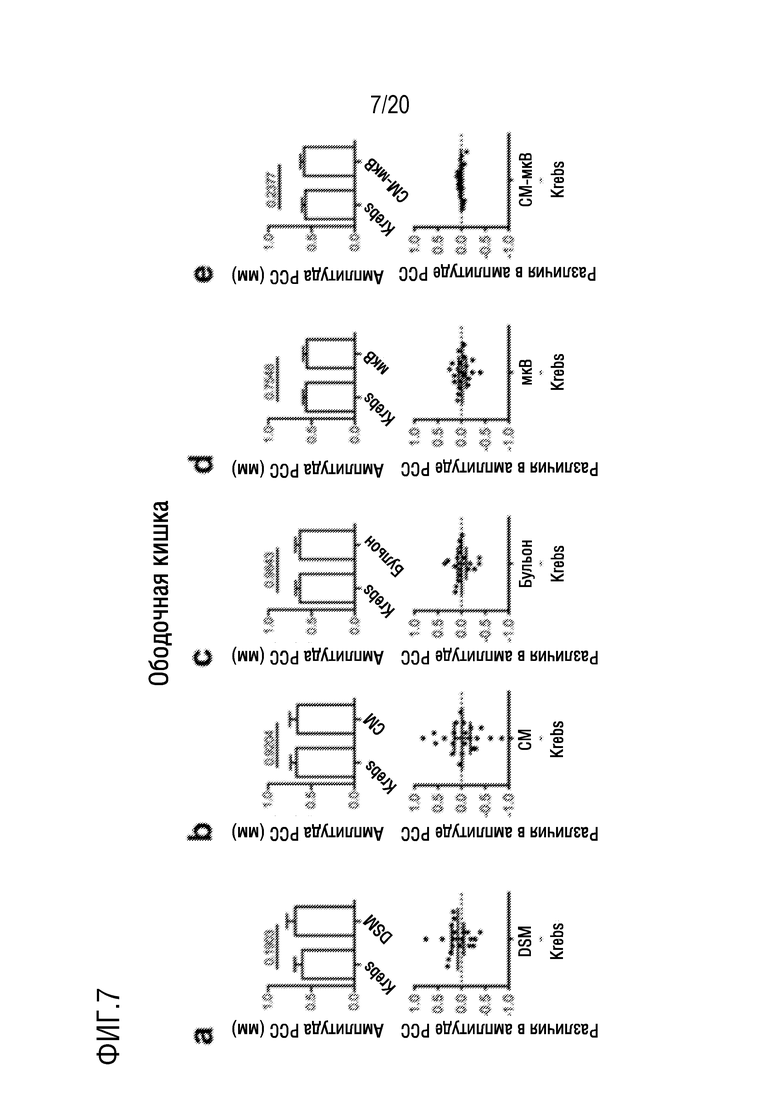
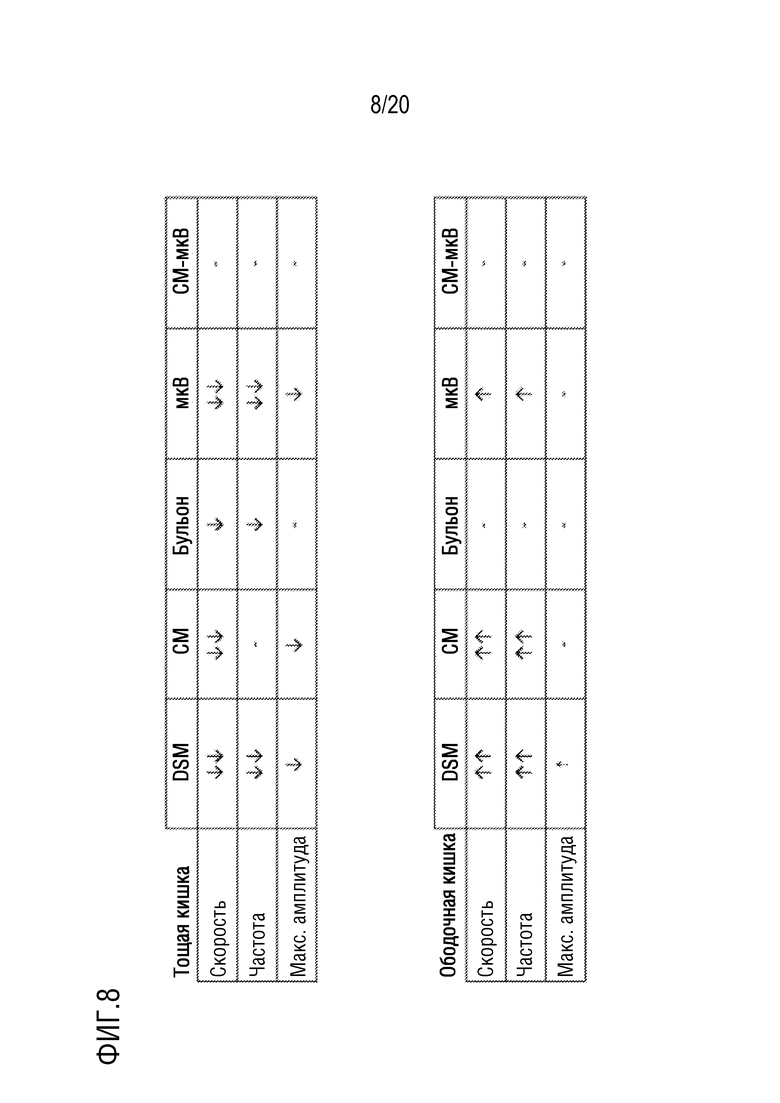
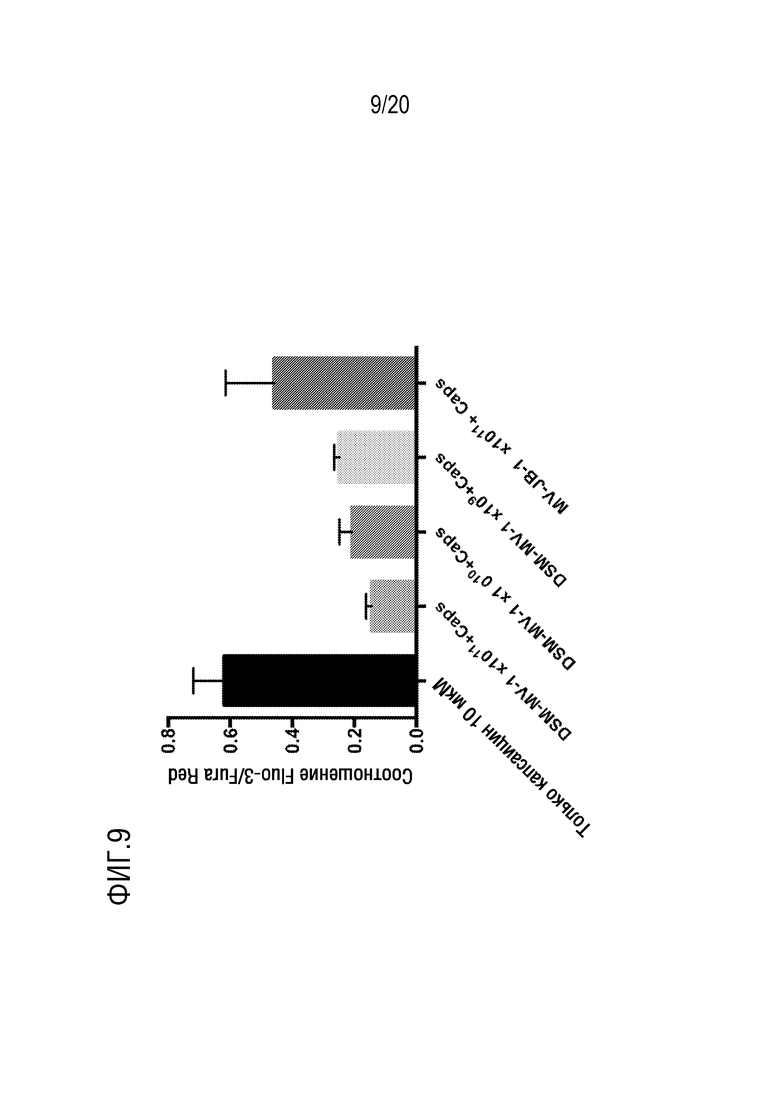
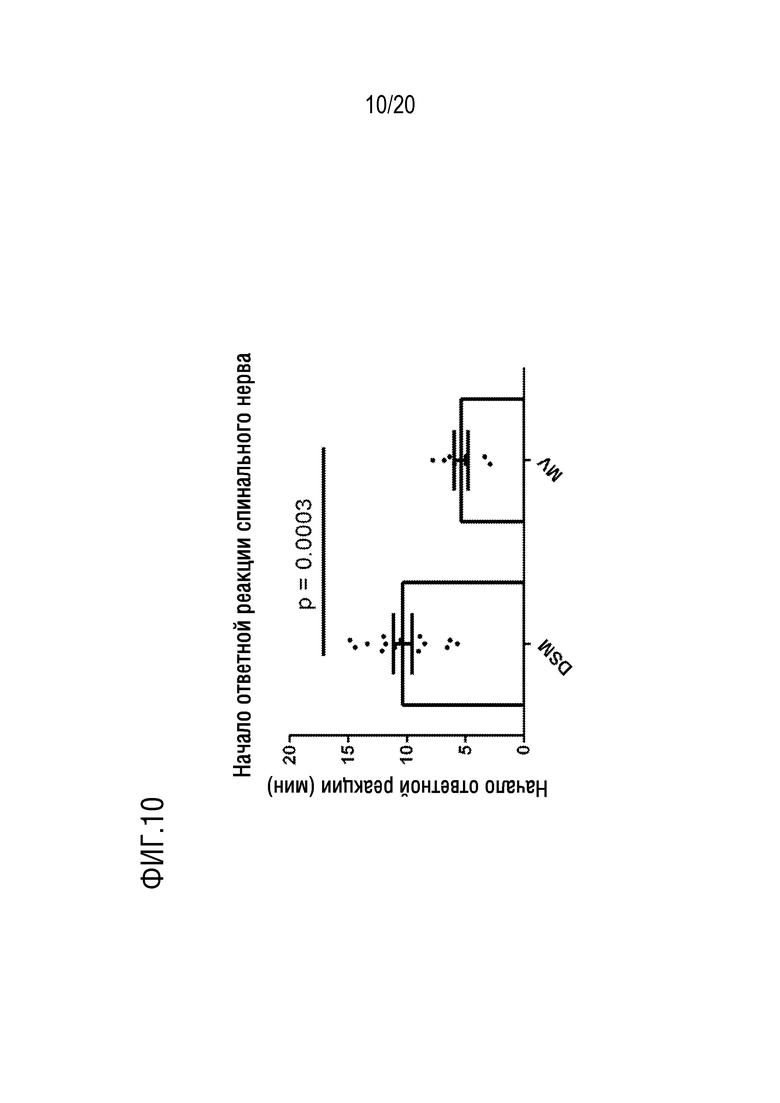
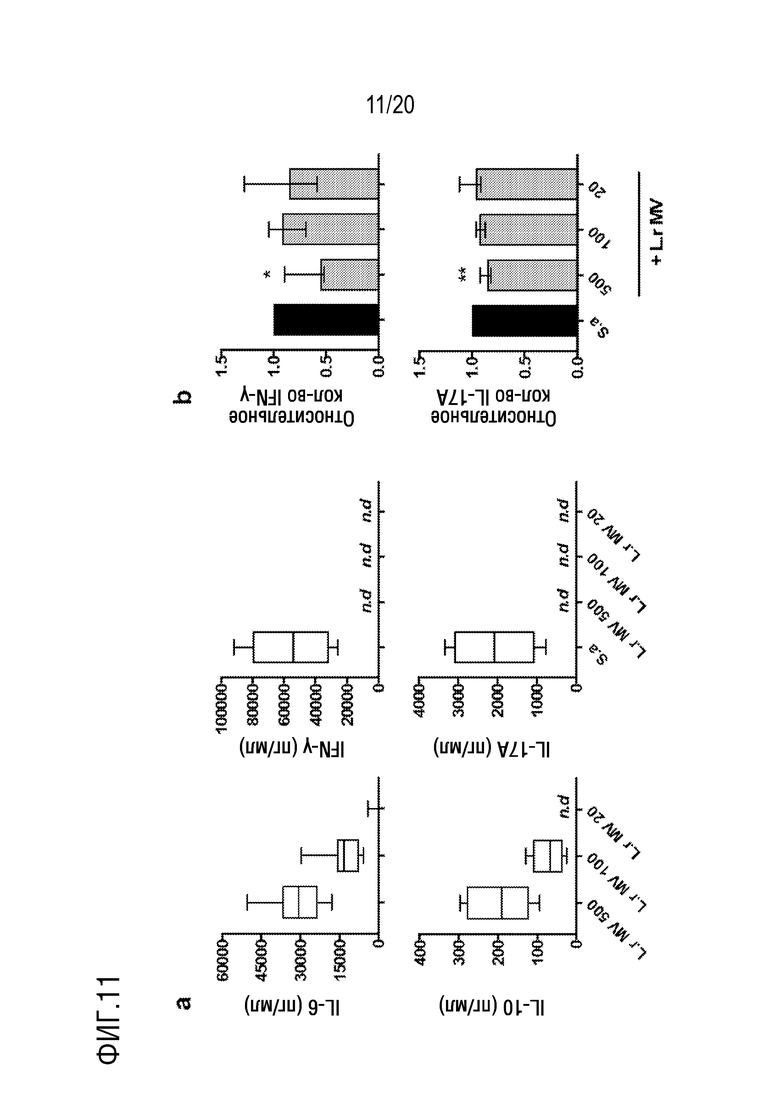
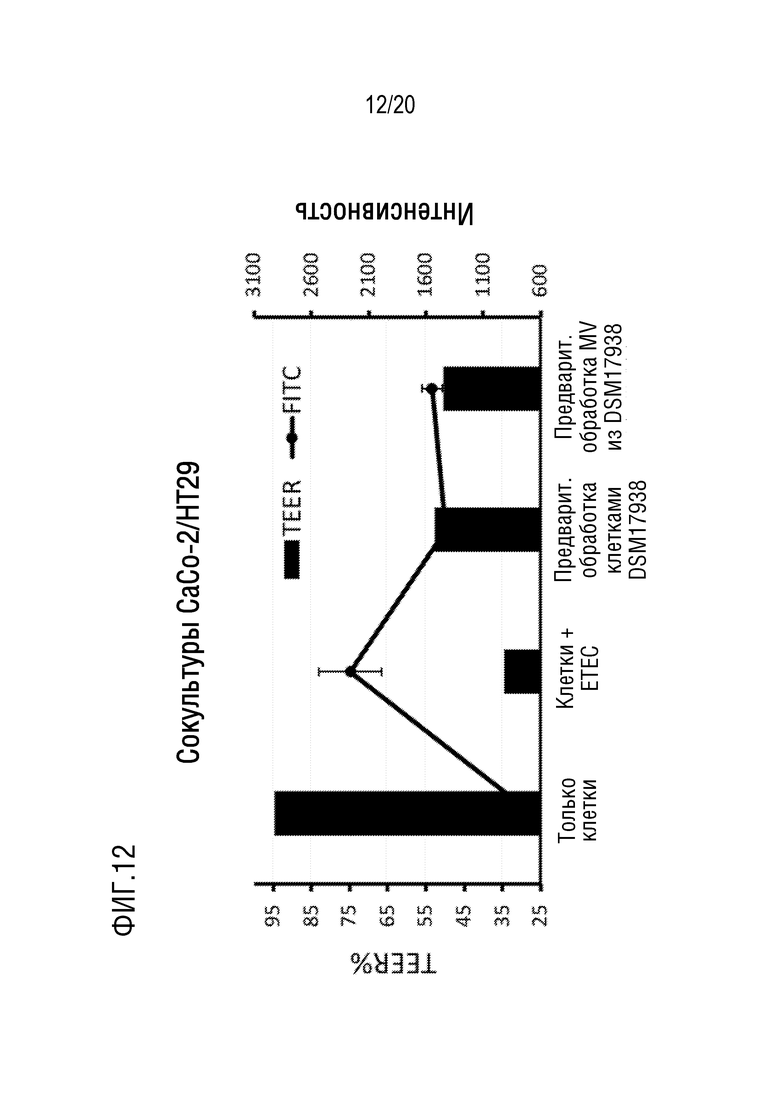
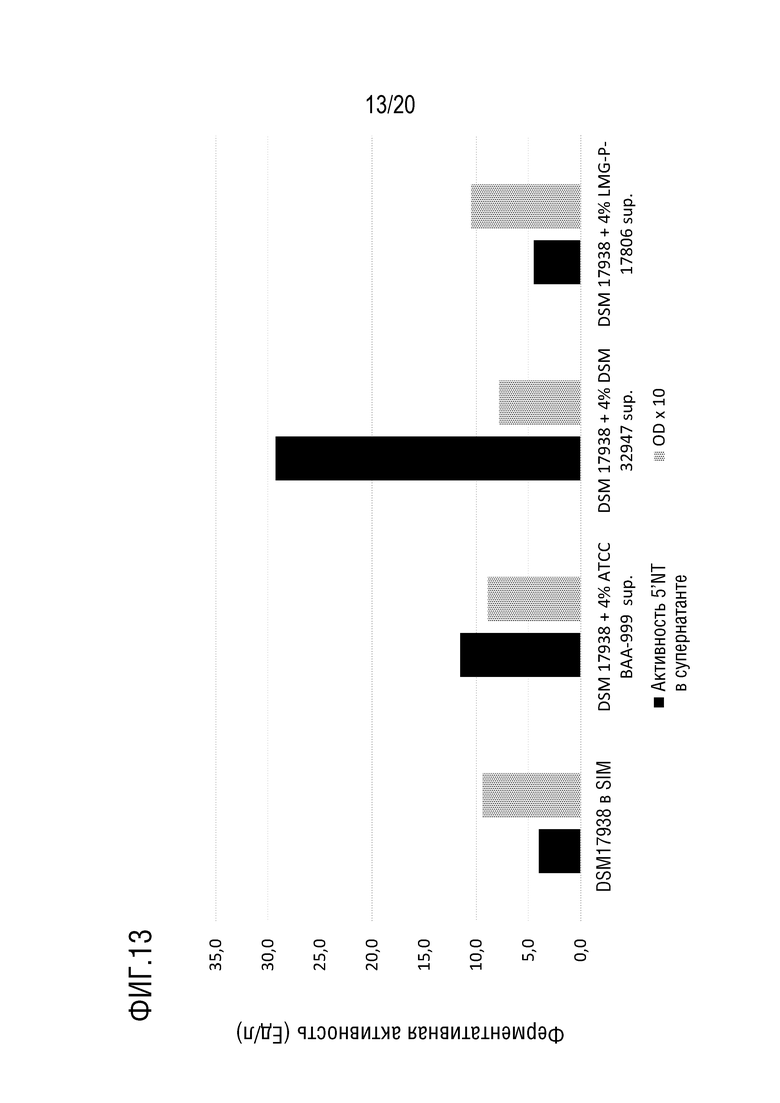
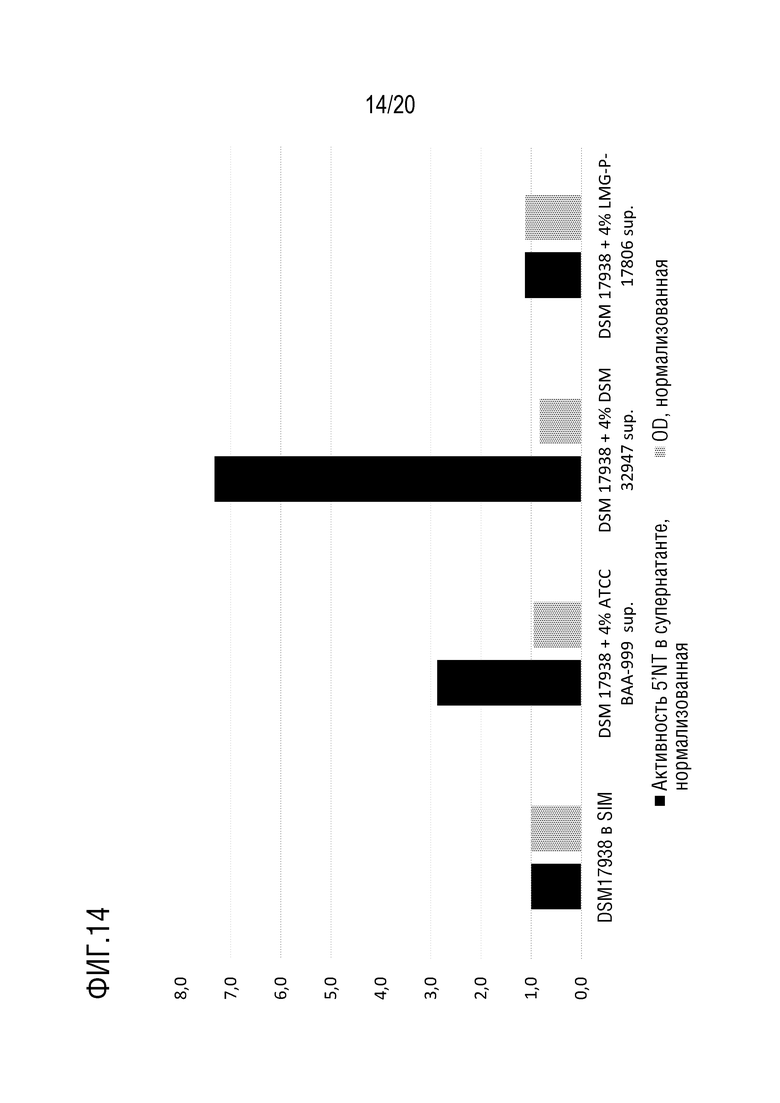
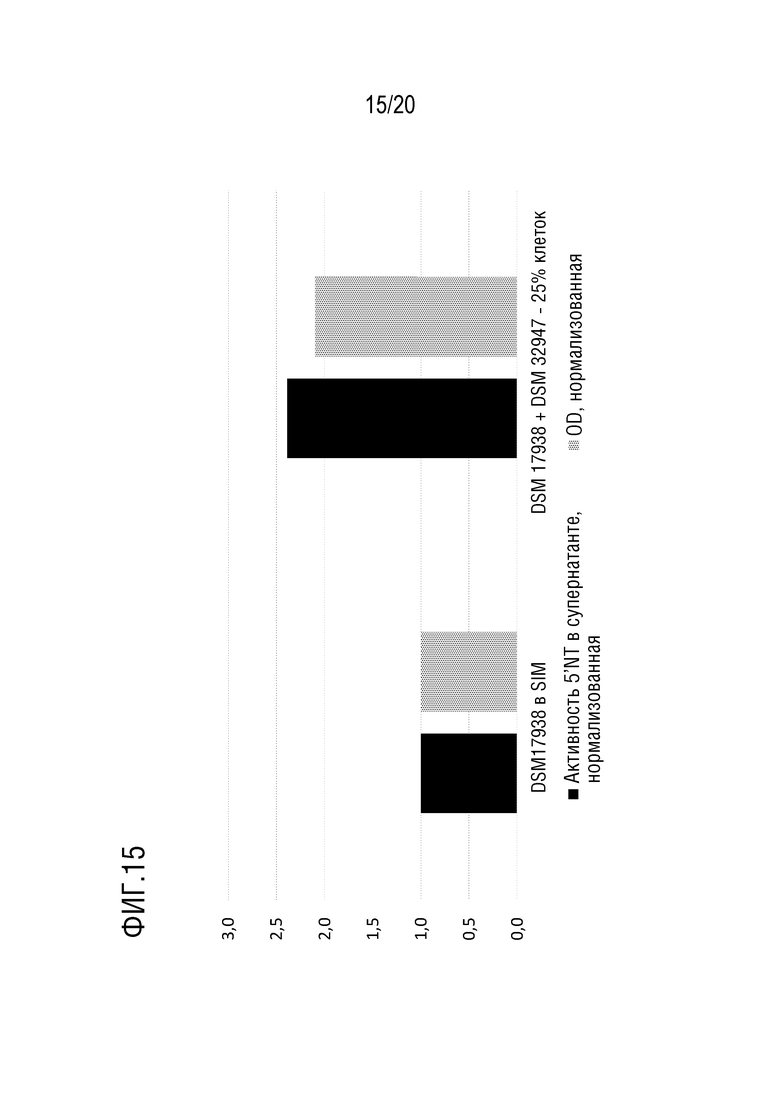
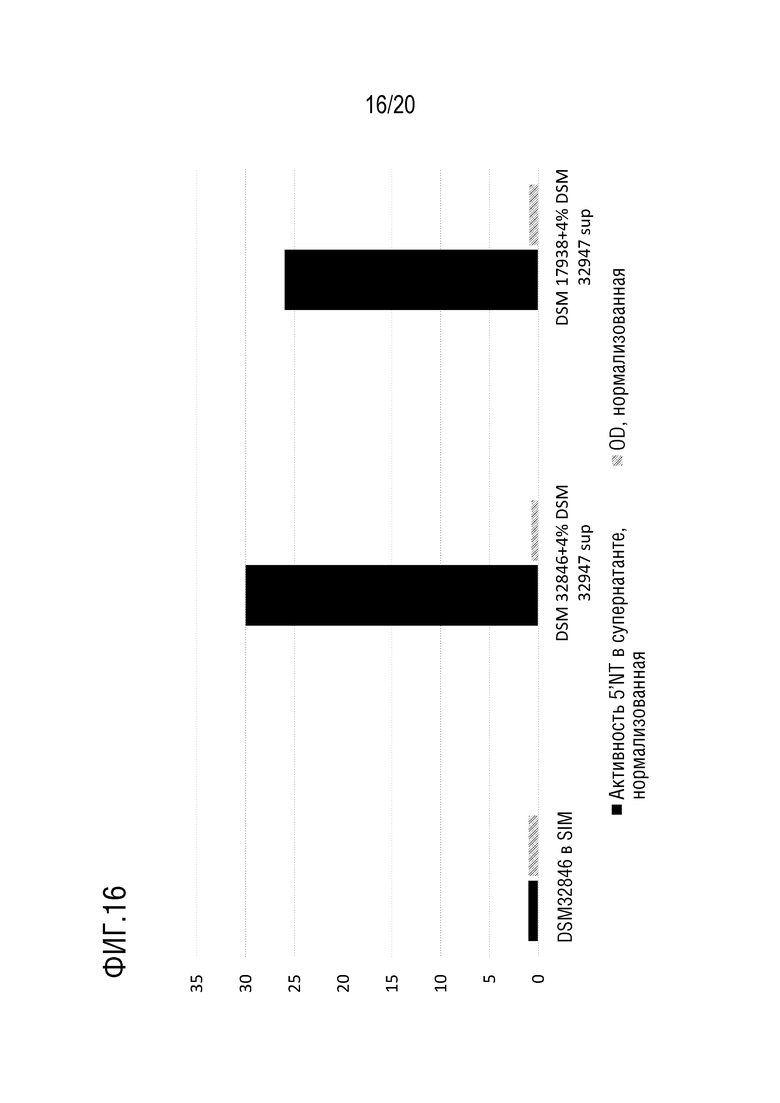
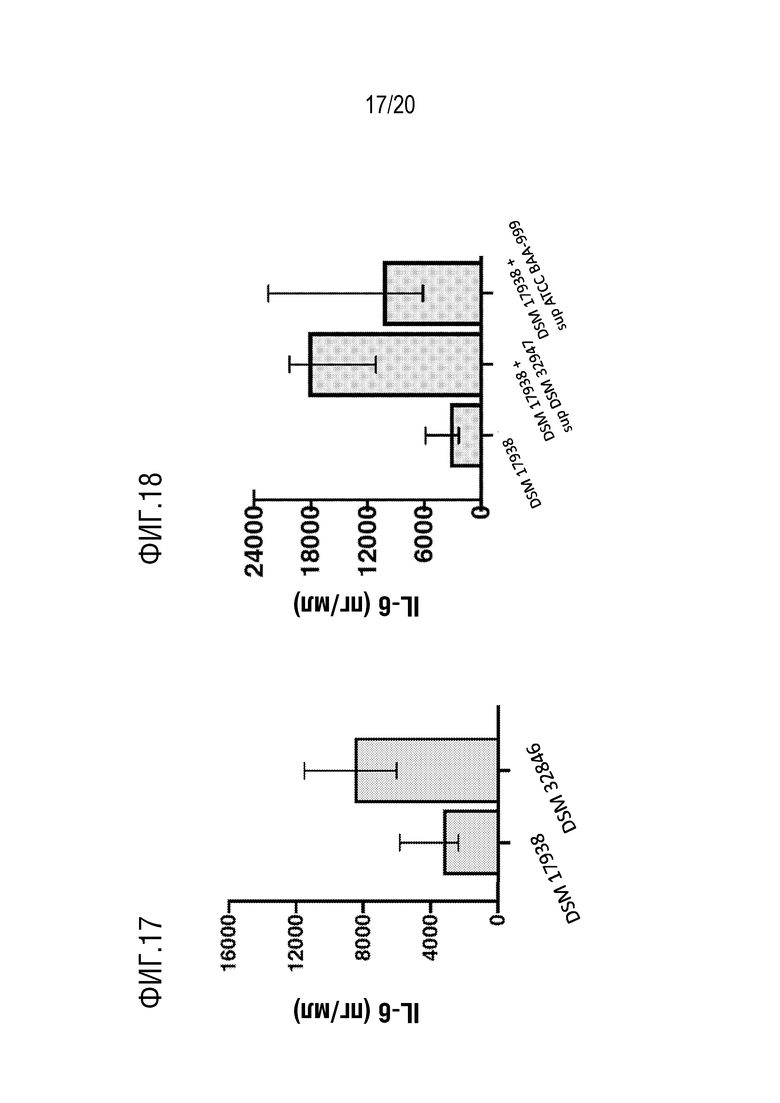
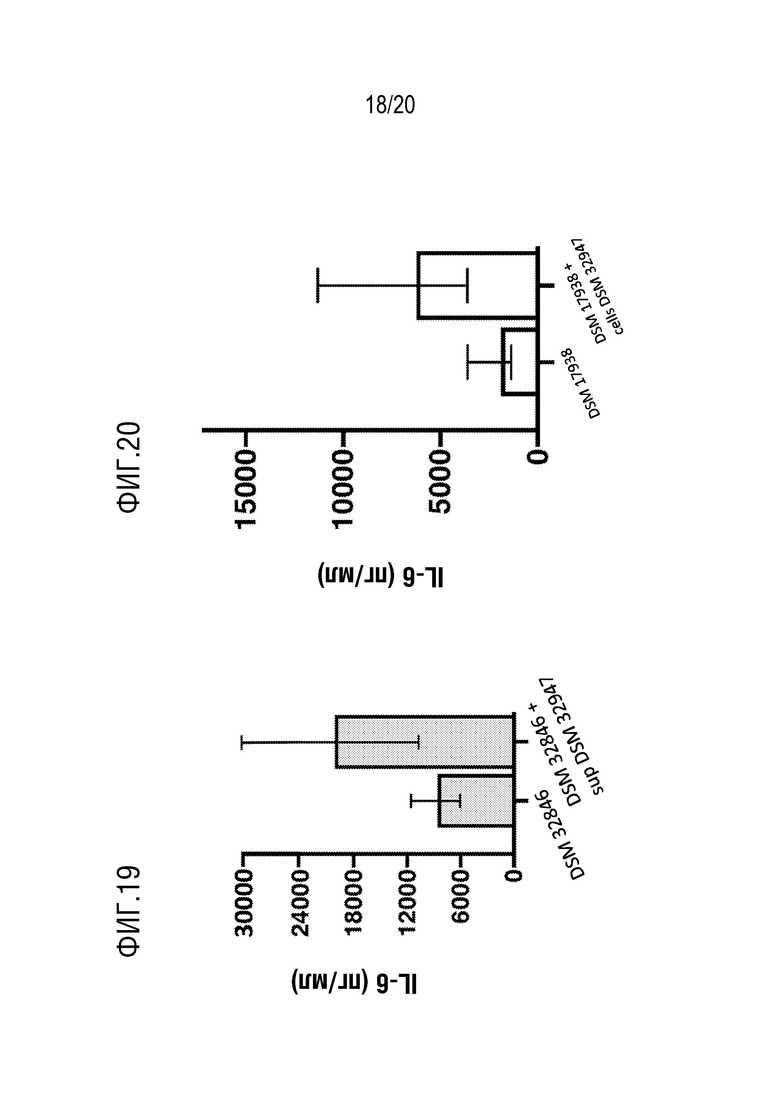
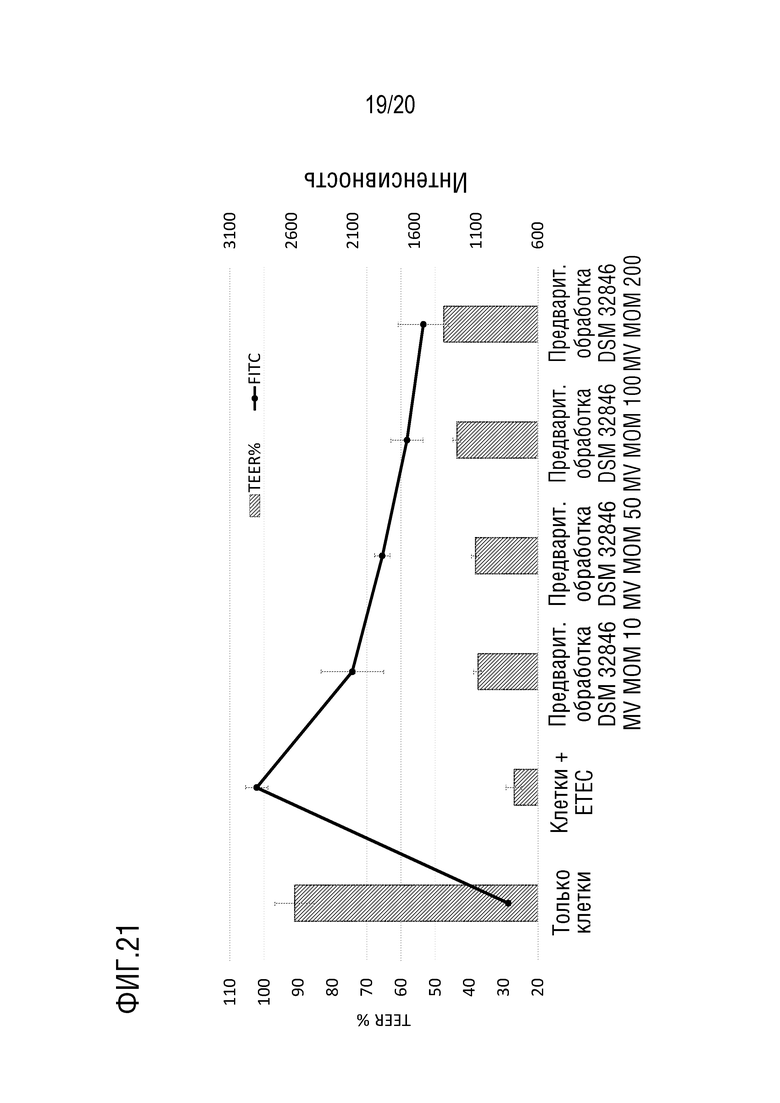
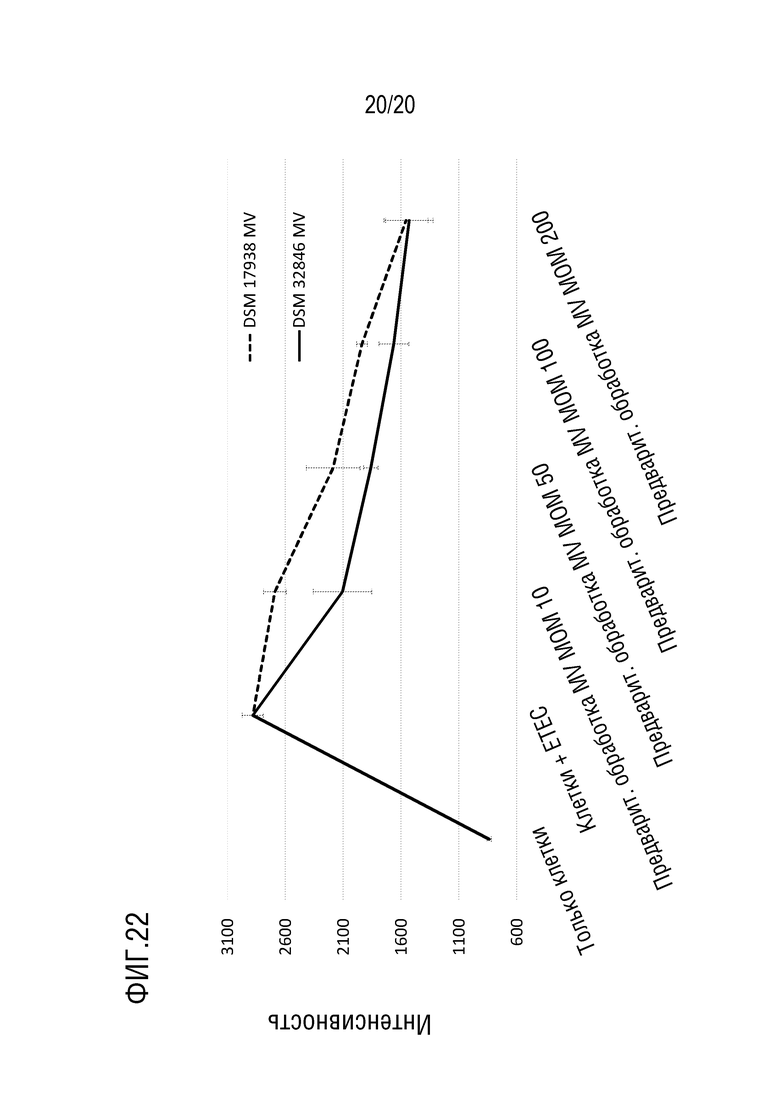
Группа изобретений относится к новому штамму и содержащей его композиции. Предложен бактериальный штамм Bifidobacterium longum DSM 32947 для индуцирующей биотической обработки пробиотических бактерий. Группа изобретений также относится к пробиотической композиции, содержащей бактерии штамма Lactobacillus и бактерии штамма Bifidobacterium longum DSM 32947 или кондиционированную среду после культивирования указанного штамма B. longum. Изобретение обеспечивает повышение продукции бактериями терапевтических микровезикул, которые демонстрируют повышенную биологическую активность. 2 н. и 2 з.п. ф-лы, 22 ил., 4 табл., 12 пр.
1. Бактериальный штамм для индуцирующей биотической обработки пробиотических бактерий, где бактериальный штамм представляет собой Bifidobacterium longum DSM 32947.
2. Бактериальный штамм по п. 1, где бактериальный штамм находится в высушенной или лиофилизированной форме.
3. Пробиотическая композиция, содержащая:
бактерии штамма Lactobacillus; и
бактерии штамма Bifidobacterium longum DSM 32947 или кондиционированную среду после культивирования указанного штамма B. longum.
4. Пробиотическая композиция по п. 3, где штамм Lactobacillus представляет собой штамм L. reuteri, предпочтительно штамм L. reuteri, выбранный из группы, состоящей из L. reuteri DSM 17938, L. reuteri DSM 32846 и их комбинации.
| WO 2013098032 A1, 04.07.2013 | |||
| Способ получения цианистых соединений | 1924 |
|
SU2018A1 |
| EP 3212208 B1, 03.03.2021 | |||
| ПРОБИОТИКИ ДЛЯ УВЕЛИЧЕНИЯ СЕКРЕЦИИ IgA У МЛАДЕНЦЕВ, РОЖДЕННЫХ ПОСРЕДСТВОМ КЕСАРЕВА СЕЧЕНИЯ | 2009 |
|
RU2496505C2 |
| SZAJEWSKA H | |||
| et al | |||
| Приспособление для уменьшения шума при клепальных работах | 1929 |
|
SU17938A1 |
| The Journal of Pediatrics, v | |||
| Деревянное стыковое скрепление | 1920 |
|
SU162A1 |
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |

