1
Изобретение относится к селекции растений и может быть использовано в сельском хозяйстве.
Цель изобретения - снижение трудоемкости, расширение генофонда фертильной свеклы и повышение выхода особей на фертильной основе.
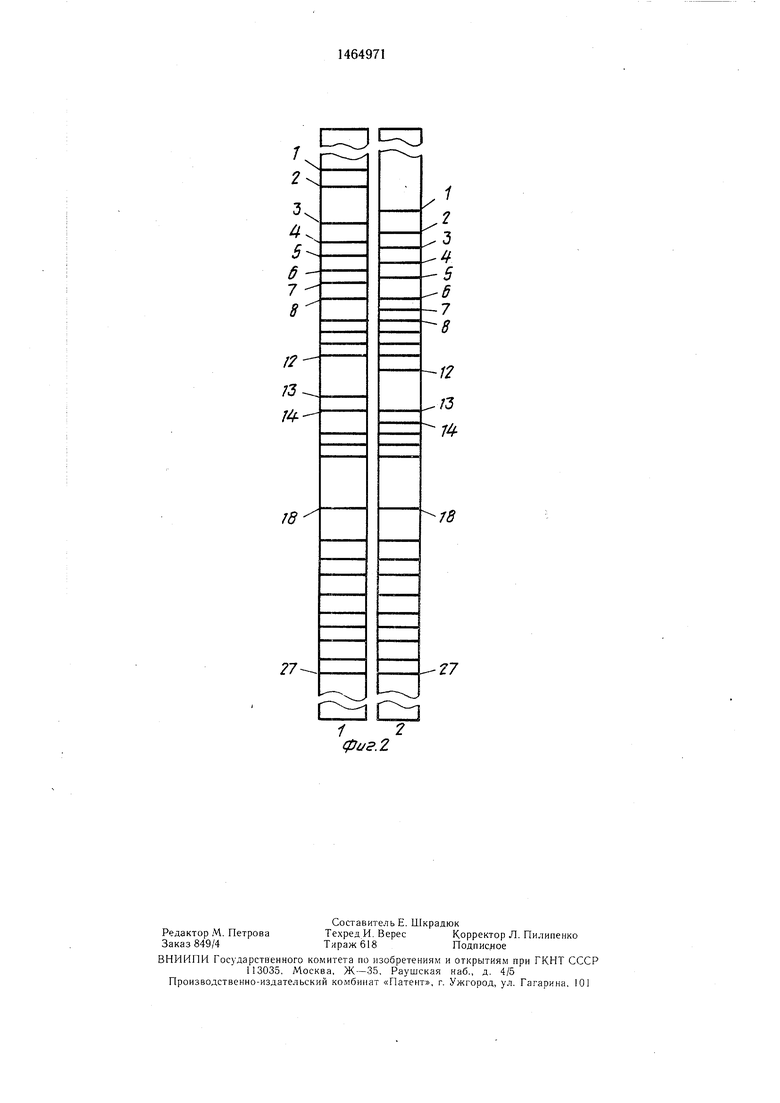
На фиг. 1 представлены схемы спектров митохондриальной ДНК корнеплодов сахарной свеклы фертильной линии ИМ; на фиг. 2 - схемы спектров в сравнении корнеплода сахарной свеклы опылителя О-типа БУО-1 и сахарной свеклы со стандартной цитоплазмой БУО N-2.
Способ осуществляют следующим образом.
Берут корнеплоды исходной популяции фертильной сахарной свеклы. Количество корнеплодов в пробе для отбора одного источника новой цитоплазмы в зависимости от исходной популяции может составлять до 150 шт. В каждом корнеплоде определяют
особенности генов цитоплазмы. Для определения этих особенностей используют метод рестриктного анализа митохондриальной ДНК, причем из гомогенизированных тканей корнеплодов сахарной свеклы выделяют ДНК митохондрий, которую гидролизуют с помощью фермента, подвергают электрофорезу, гель-электрофорез фотографируют в ультрафиолетовом свете. Полученные рестриктные спектры ДНК сравнивают со стан дартным спектром исходной популяции. На основании такого сравнения отбирают корнеплоды с мутацией цитоплазмы, которые идентифицируют между собой.
Для повыщения выхода корнеплодов- мутантов, отличающихся по цитоплазме, отбор их ведут среди опылителей О-типа.
Положительный эффект предлагаемого способа заключается в том, что он позволяет расширить генофонд и направленно вовлечь в селекционный процесс растения с разной цитоплазмой для создания линий и гибридов
05
со
фертильной сахарной свеклы с новыми свой- ciaaMH, определяемыми генетическими раз- лр1чиями цитоплазмы.
I Отбор мутантов среди опылителей О-типа дает возможность повысить вероятност встречи с растениями, отличающимися цитоплазмой, и, следовательно, сократить /в пределах до 150 раз) затраты времени, труда и средств на отбор каждого источника новой цитоплазмы по сравнению с отбором его из оэычной популяции культуры свеклы.
Пример. В качестве исходных популяций с;1харной свеклы взяли фертильные линии - Ивановская многосемянная (ИМ) и сорт Белоцерковская односемянная (БЦО). Кро- мЬ того, испытанию подвергли корнеплоды сахарной свеклы - опылителей О-типа (за- к|)епителей стерильности), сорта Белоцер- к|)вская односемянная (БЦО), линий Ива- н|)вская односемянная (ИО 117, ИО 194, 3X4 ГЦ 83), Ялтушковская односемян- н|1я (ЯО 4128, ЯО 4130) и др.
I В указанных корнеплодах фертильной сахарной свеклы определили биохимические оаобенности генов цитоплазмы, для чего ис|пользовали рестриктный метод анализа м||1тохондриальной ДНК- Ткани корнеплодов (|о 50 г) гомогенизировали в буфере (А), содержащем 50 мМ трис; 3 мМ ЭДТА; 330 мМ сорбита; 0,1% В А; 0,01 мМ меркап- тоэтанола; рН 8,0.
; Затем выделили из го.могенизированных тЦаней корнеплодов митохондрии. Массу О11жали через фильтр. Гомогенат центрифугировали при 3200 об/мин и температуре
0;С в течение 10 мин. Супернатант понторно центрифугировали при 10000 об/мин и тем- пфатуре 0°С в течение 15 мин.
Осадок митохондрий суспендировали в 10 мл буфера (А). Митохондрии обработали ДИКазой для удаления примесей ядерной и Хлоропластной ДНК. К суспендированному ос1адку добавили 10 мМ MgCb и 250 мкг ДНКазы. Гидролиз провели при температуре в течение 30 мин.
Действие фермента прекратили путем добавления 3-кратного объе.ма буфера (Б), содержащего 25 .мМ ЭДТА; 0,6 мМ сахарозы 50 мМ трис - НС1; рН 8,0.
После этого митохондрии осадили путем центрифугирования при 10000 об/мин в те- че ние 15 мин. Осадок митохондрий суспендировали в 10 мл буфера (Б).
Затем суспендированный осадок наслоили на ступенчатый градиент сахарозы: 45, 36 и 30%-ной сахарозы соответственно по 5, 10 и 10 мл.
Градиент сахарозы с наслоенным осадком митохондрий центрифугировали при 2СЮОО об/мин и температуре 4°С в течение
1ч. Очищенные митохондрии собрали в слое между 45 и 36%-ной сахарозой.
ДНК из очищенных митохондрий выделили следующим образом. В митохондрии
добавили 1 мл раствора .проназы концентрацией 6 мг/мл и суспендировали их. Затем добавили 1,2 мл 10%-него раствора лаурил- саркозилата натрия, перемешивая, довели объем смеси до 6 мл буфером, содержащим 50 мМ трис, 20 мМ ЭДТА, рН 8,0. Инкубировали смесь при комнатной температуре в течение 1 ч. В полученный лизат влили 2 мл 4 М хлористого цезия. После этого ли- зат центрифугировали при 2000 об/мин в течение 30 мин.
К надосадочной жидкости добавили 3,2 г
сухого хлористого цезия, 100 мкл раствора
бромистого этидия концентрацией 10 мг/мл
и подслоили 3 мл 6 М раствора хлористого
5 цезия.
Градиент центрифугировали при 25000 об/мин в течение 12 ч.
Слой митохондрий отобрали в ультрафиолетовом свете, развели в три раза водой и центрифугировали при 35000 об/мин в те- чение 3 ч.
Полученный осадок ДНК растворили в 100 мкл воды, добавили к раствору 2-кратный объем этанола и центрифугировали при 15000 об/мин в течение 1 мин. 5 Осадок ДНК промыли 2 раза 70%-ным этанолом, выгушили и растворили в 100 мкл воды.
Гидролиз ДНК провели с помощью фермента Ват HI в 50 мкл реакционной среды,
0 содержащей 10 мМ трис НС, рН 7,6; 60 мМ NaCl; 10 мМ 1 мМ меркаптоэтанола и 2 мкг митохондриальной ДНК.
После инкубации длительностью 3 ч при температуре 37°С фрагменты подвергли электрофорезу в 0,8%;-ном агарозном геле
5 в течение 16 ч.
Гель-электрофорез ДНК митохондрий
фотографировали в ультрафиолетовом свете.
Таким образом, получили рестриктный
спектр митохондриальной ДНК по каждому
Q корнеплоду отобранных для исследования растений сахарной свеклы.
Рестриктные спектры митохондриальной ДНК корнеплодов опылителей О-типа сравнили с тождественными между собой спектрами стандартной цитоплазмы исходных
5 популяций, из которых получен тот или иной опылитель. В результате сравнения установили, что сорт БЦО со стандартной цитоплазмой отличается от всех других изученных популяций фертильной сахарной свеклы, стандартная цитоплазма которых выще,
0 тождественна.
На основании сравнения установили закономерность, которая находит выражение в том, что каждый опылитель О-типа является мутантом по цитоплазме в отнощенин к исходной популяции, на основе которой
- он создан.
Как видно из материалов, представленных на фиг. 2, спектр митохондриальной ДНК опылителя О-типа БЦО (фиг. 2.1) отличается от спектра цитоплазмы исходной популяции (фиг. 2.2) наличием в нем фраг- метов ДНК - 1-7, 13, отсутствующих в смежном (контрольном) спектре, который, с другой стороны, имеет свои отличительные фрагменты ДНК - 1-5, 7, 12, 14.
По сравнению с отбором фертильных растений-мутантов из исходных популяций сахарной свеклы по известному способу выход источников новой цитоплазмы увеличивается многократно, поскольку каждый опылитель О-типа является мутантом по цитоплазме в отношении популяции, на основе которой он создан.
Предлагаемый способ позволяет выявлять, отбирать и идентифицировать на генетической основе фертильные растения сахар0
5
ной свеклы как среди ее популяций, так и среди опылителей О-типа.
Формула изобретения
1.Способ получения исходного селекционного материала сахарной свеклы, включающий отбор из популяции растений с модифицированной цитоплазмой, отличающийся тем, что, с целью снижения трудоемкости, расширения генофонда фертильной свеклы и выхода особей на фертильной основе, отбор растений с модифицированной цитоплазмой ведут среди закрепителей стерильности ЦМС-форм.
2.Способ по п. 1, отличающийся тем, что модификацию цитоплазмы определяют методом рестриктного анализа митохондриаль- ной ДНК корнеплодов.


| название | год | авторы | номер документа |
|---|---|---|---|
| Способ отбора опылителей О-типа у сахарной свеклы | 1987 |
|
SU1507262A1 |
| Способ отбора форм сахарной свеклы с цитоплазматической мужской стерильностью | 1986 |
|
SU1546020A1 |
| СПОСОБ СОЗДАНИЯ РАЗДЕЛЬНОПЛОДНЫХ МУЖСКО-СТЕРИЛЬНЫХ ГИБРИДОВ САХАРНОЙ СВЕКЛЫ | 1996 |
|
RU2107430C1 |
| СПОСОБ ИДЕНТИФИКАЦИИ СТЕРИЛЬНОСТИ/ФЕРТИЛЬНОСТИ ПОДСОЛНЕЧНИКА | 2012 |
|
RU2524135C2 |
| Способ анализа митохондриальной ДНК растений | 1990 |
|
SU1759334A1 |
| Способ получения гибридных семян свеклы и способ выращивания тетраплоидных форм свеклы для получения гибридных семян | 1987 |
|
SU1584836A1 |
| ФРАГМЕНТ ДНК, ДЕТЕРМИНИРУЮЩИЙ СТЕРИЛЬНОСТЬ OQURA И СООБЩАЮЩИЙ ЦИТОПЛАЗМИТИЧЕСКУЮ МУЖСКУЮ СТЕРИЛЬНОСТЬ ПРИ НАЛИЧИИ ЕГО В МИТОХОНДРИАЛЬНОМ ГЕНОМЕ РАСТЕНИЙ И ПРОБА ДНК ДЛЯ ГИБРИДИЗАЦИИ С ДАННЫМ ФРАГМЕНТОМ | 1991 |
|
RU2117704C1 |
| СПОСОБ ПОЛУЧЕНИЯ РАСТЕНИЯ BRASSICA OLERACEA | 1997 |
|
RU2142013C1 |
| Способ получения односемянных самофертильных опылителей "о"-типа у сахарной свеклы | 1981 |
|
SU997634A1 |
| ПОЛУЧЕНИЕ СЕМЯН ГИБРИДА Lactuca sativa | 2008 |
|
RU2593940C2 |
Изобретение относится к селекции растений и может быть использовано в сельском хозяйстве. Цель изобретения - снижение трудоемкости, расширение генофонда фертильной свеклы и повышение выхода особей на фертильной основе. Из гомогенизированных тканей корнеплодов сахарной свеклы выделяют ДНК митохондрий, которую гид- ролизуют с помощью фермента и подвергают электрофорезу. Полученные рестриктные спектры ДНК сравнивают со стандартным спектром исходной популяции. На основании сравнения отбирают корнеплоды с мутацией цитоплазмы. Для анализа выделяют ДНК из закрепителей стерильности ЦМС-форм. 1 3. п. ф-лы, 2 ил. § (Л
CZI3
czD ci:
2
3
J2 -75
.16 -17
J3 -20
23
г4
.28 28
фиг.1
QnCrj
/2
ф1/г.2
| Quetier F | |||
| Udel F | |||
| Heterogeneous population of mitohondrial DNA molecules in higher plants.-Nature, 1977, v | |||
| Способ изготовления гибких труб для проведения жидкостей (пожарных рукавов и т.п.) | 1921 |
|
SU268A1 |
| Станок для нарезания зубьев на гребнях | 1921 |
|
SU365A1 |
