Предполагаемое изобретение относится к способам гидролиза растворимых и нерастворимых биологических субстратов с целью получения аминокислот, углеводов и других продуктов, и может быть использовано в химической, медицинской, микробиологической, пищевой промышленностях, а также для чистки бытовых и промышленных сточных вод.
Известен способ гидролиза сахара и аналогичных соединений протеиназой /1/, который осуществляется путем передавливания раствора белка или полисахаридов через ячейку ультрафильтрации, к мембране которой присоединен фермент.
Недостатком известного способа является низкая скорость гидролиза, т.к. взаимодействие с субстратом происходит только на относительно небольшой поверхности мембраны. Недостаток способа также заключается в невозможности гидролиза крупных объемов высокомолекулярных субстратов ввиду того, что гидролиз таких субстратов, из-за крупности их молекул, возможен только с одной стороны поверхности мембраны.
Известен способ гидролиза, осуществляемый с помощью только одного нативного фермента /2/.
Однако известно, что для осуществления полного гидролиза нативным ферментом, например, гидролиза дрожжей, нативного фермента оризин ПК требуется большое количество около 1,5% мас. к сухому весу дрожжей /2/. При этом фермент необратимо теряется, расход его велик, продукт загрязняется энзимом и продуктами его распада, а интенсивность гидролиза только одним нативным ферментом низка.
Известен наиболее близкий предлагаемому способ /3/ колоночного гидролиза субстратов с применением иммобилизованного на пористых матрицах фермента, с целью получения растворов аминокислот.
Особенностью известного способа является невозможность проведения гидролиза высокомолекулярных белков и недостаточная скорость, интенсивность процесса.
Целью предлагаемого изобретения является интенсификация процесса гидролиза, повышение выхода готового продукта, расширение функциональных границ способа за счет возможности использования в качестве сырья субстратов с неограниченной молекулярной массой. Целью также является исключения загрязнения готового продукта нативным ферментом и более эффективное использование последнего.
Поставленная цель достигается тем, что в способе гидролиза биологических субстратов путем их контакта с иммобилизованным на пористых матрицах ферментом, согласно изобретению, в объем обрабатываемого сырья дополнительно водят нативный фермент в количестве 0,001 0,1% мас. от массы гидролизуемого субстрата, и модификатор, придающий молекулам субстрата электрический заряд, а гидролиз ведут при pH среды, поддерживаемом в интервале величин между изоэлектрическими точками нативного фермента и субстрата. Для гидролиза используют матрицы, средний размер пор которых превышает средний размер молекул нативного фермента, при этом суспензию или раствор субстрата с находящимися в них нативным ферментом, последовательно обрабатывают сначала на матрицах, диаметры пор которых меньше размеров молекул нативного фермента, а затем на матрицах с диаметрами их пор, превышающими размеры молекул нативного фермента. Субстрат с ферментом помещают в электрическое поле с напряженность, от 3 до 25 в/см для создания направленной электромиграции молекул субстрата к поверхности иммобилизированного фермента и электромиграции нативного фермента от поверхности иммобилизованного. При гидролизе белковых субстратов в качестве модификатора используют преимущественно фосфорную кислоту или ее соли в количестве 0,1 1,4 мас. от массы обрабатываемого субстрата. При гидролизе углеводных субстратов в качестве модификатора преимущественно используют борную кислоту или бораты в количестве 0,2 3% мас. от массы субстрата.
Сопоставительный анализ заявленного решения с прототипом показывает, что заявляемый способ отличается от известного тем, что в гидролизуемый субстрат одновременно с иммобилизованным ферментом вводят нативный фермент, а также определенное количество модификатора с целью, придания молекулам субстрата электрического заряда, противоположного по знаку заряду молекул нативного фермента. При этом гидролиз ведут в направленном электрическом поле с определенной напряженностью, и при определенных пределах величин pH реакционной среды. Кроме того, для процесса используют носители (матрицы) иммобилизованного фермента с определенными диаметрами пор. Таким образом, заявляемый способ соответствует критерию "новизна".
Анализ других известных способов гидролиза биологических субстратов не позволил выявить в них совокупность признаков, отличающую заявляемое решение от прототипа. Сумма известных и отличительных существенных признаков в предложенном способе дает положительный эффект, позволяющий достичь поставленной цели. Таким образом, по мнению автора, предложенное решение соответствует критерию "существенные отличия".
Суть предложенного способа состоит в том, что в раствор или суспензию с гидролизуемым белковым или углеводным субстратом, перед их гидролизом (или одновременно с началом гидролиза) иммобилизованным ферментом, вводят суперкаталическое количество нативного фермента. Нативный фермент, взаимодействуя с субстратом, подвергает высокомолекулярный субстрат предварительному гидролизу, расщепляя молекулы субстрата на части. При этом с объеме реакционной среды происходит некоторое снижение молекулярной массы субстрата и последний становится пригодным для последующего полного гидролиза иммобилизованным ферментом. Сам по себе полный гидролиз субстратов только с помощью нативного фермента представляет собой весьма долгий процесс, длящийся до нескольких суток. Гидролиз же высокомолекулярных соединений, особенно нерастворимых, невозможен в полном объеме иммобилизованным ферментом, т.к. процесс выходит через некоторое время на плато и в обрабатываемом растворе остаются непрогидролизованные молекулы субстрата с большим молекулярным весом.
Нативный фермент не только подготавливает высокомолекулярные соединения к гидролизу на матрицах путем предварительного разрушения массивных молекул субстрата в растворе, но и является катализатором гидролиза субстрата иммобилизованным ферментом, значительно ускоряя своим присутствием в растворе этот процесс. Замечено, что присутствие суперкаталического количества нативного фермента в растворе /суспензии/ является достаточным для катализа процесса полного гидролиза субстрата иммобилизованным ферментом.
Молекулы нативного фермента, в отличие от молекул субстрата, имеют заряд. Для создания взаимного притяжения молекул субстрата и нативного фермента необходимо придать молекулам субстрата противоположный заряд. Это дает убыстрение объемной скорости взаимодействия нативного фермента обрабатываемым субстратом за счет взаимопритяжения между их молекулами.
Молекулы нативного и иммобилизованного ферментов, обладая одинаковыми по знаку зарядами, отталкиваются друг от друга. Однако взаимодействие некоторого количества их молекул происходит, что приводит к разрушению молекул нативного фермента. Чтобы как можно полнее избежать потерь нативного фермента, а также одновременно увеличить скорость поступления фермента к субстрату, в объеме гидролизера организуют направленное электрическое поле с определенной напряженностью. При наличии такого поля повышается скорость электромиграции молекул субстрата к молекулам нативного фермента, что увеличивает объемную скорость расщепления высокомолекулярного субстрата нативным ферментом, а значит и скорость полного гидролиза субстрата.
Одновременно с этим, наличие направленного электрического поля полностью исключает контакт молекул нативного фермента с иммобилизованным ферментом за счет электромиграции молекул нативного фермента от поверхности иммобилизованного. Этим молекулы нативного фермента предохраняются от разрушения и функционируют в растворе продолжительное время. Кроме этого, достигается дополнительный эффект снижение вероятности попадания нативного фермента в готовый продукт, т.к. происходит электромиграция молекул фермента в противоположную от выходного патрубка гидролизера сторону.
Загрязнение готового продукта нативным ферментом полностью исключается применением в гидролизере гранульных матриц с определенным диаметром пор. Верхний слой матриц в колонке может иметь поры, диаметр которых не превышает среднего размера молекул нативного фермента, и поэтому нативный фермент в растворе может присутствовать в этой зоне колонки. При дальнейшем опускании в колонке обрабатываемого раствора /суспензии/, раствор попадает в зону размещения матриц с порами, диаметры которых больше размера молекул нативного фермента. Если, невзирая на наличие направленного электрического поля, оттягивающего нативный фермент вверх на колонки, какое-то количество фермента "проскочит" в нижний уровень колонки, то его молекулы попадают в крупные поры матриц и разрушаются иммобилизованным ферментом.
Чтобы не нейтрализовать заряды молекул субстрата или нативного фермента, гидролиз ведут при pH среды, величина которого находится в интервале между изоэлектрическими точками нативного фермента и субстрата.
Осуществление предложенного способа поясняется с помощью устройства, представленного на чертеже. Устройство содержит реакционную колонку 1, заполненную пористыми матрицами 2 с иммобилизованным на их порах ферментом. Матрицы 2 могут быть выполнены в виде гранул силикагеля. Верхний уровень колонки 1, например, до середины, заполнен матрицами 2 с порами, диаметр которых не превышает размер молекул нативного фермента, а нижний уровень колонки 1 заполнен матрицами 2, имеющими поры, превышающими по диаметру средний размер молекул нативного фермента. Колонка 1 имеет входную верхнюю и выходную нижнюю горловины. Колонка 1 помещена внутрь емкости 3 и сообщена своей верхней горловиной с полостью емкости 3, а нижней горловиной с камерой 4 для отбора готового продукта. Емкость 3 снабжена патрубком 6 для подвода раствора или суспензии субстрата, а камера 4 патрубком 7 для отвода готового продукта. Объем емкости 3 больше объема колонки 1 в 1,5oC6 раз. Внутри емкости 3 установлен электрод 8, а в камере 4 установлен электрод 9. Электроды 8 и 9 подключены к источнику тока. Электрод 8 в несколько раз больше по площади электрода 9.
Предложенный способ реализуется следующим способом. В емкость 3 через патрубок 8 подают борный раствор или суспензию обрабатываемого биологического субстрата. Субстрат проникает в толщу матриц 2 в колонке 1, где подвергается полному гидролизу. Одновременно с этим в емкость 3 на устройства 5 непременно поступает необходимое количество нативного фермента (0,001 0,1% мас. от массы гидролизуемого субстрата). Кроме того, в емкость 3 добавляется модификатор для придания молекулам субстрата электрического заряда, противоположного по знаку молекул нативного фермента. Если гидролизуется белковый субстрат, то в качестве модификатора применяют преимущественно фосфорную кислоту или ее соли в количестве 0,1 1,4 мас. от массы гидролизуемого субстрата. При гидролизе углеводных субстратов в качестве модификатора преимущественно используют борную кислоту или бораты в количестве 0,2 3 мас. от массы субстрата.
Имеющие противоположные по знаку заряды, молекулы субстрата и нативного фермента притягиваются друг к другу, и молекулы субстрата расщепляются ферментом. Высокомолекулярный субстрат становится низкомолекулярным, пригодным для гидролиза иммобилизованным ферментом. При этом pH реакционной среды не должно совпадать по значению с изоэлектрическими точками субстрата и нативного фермента, чтобы не нейтрализовать их заряды и действие электрического поля, которое поддерживается с напряженностью 3-25 в/см электродами 8 и 9.
Присутствие нативного фермента является катализатором гидролиза субстрата иммобилизованным ферментом. Вышеуказанное количество нативного фермента в субстрате достаточно для катализирования полного гидролиза субстрата на иммобилизированном ферменте.
При контакте молекул нативного фермента с иммобилизированным, молекулы нативного фермента разрушаются и его количество в растворе субстрата может снизиться. Однако, благодаря одинаковому знаку заряда молекул этих ферментов, они взаимоотталкиваются, и, кроме того, электрическое поле вызывает электромиграцию молекул нативного фермента от иммобилизованного фермента, что снижает вероятность их контактирования.
Для полного исключения попадания нативного фермента в готовый продукт при выходе последнего из патрубка 7 камеры 4, предусмотрено увеличение диаметра пор матриц 2, расположенных в нижней половине колонки 1. Если нативный фермент достигает в колонке 1 зоны с крупнопористыми матрицами 2, фермент адсорбируется в порах и разрушается иммобилизованным ферментом. Находясь же в зоне мелкопористых матриц, нативный фермент не соприкасается с иммобилизованным, а свободно расщепляет здесь молекулы субстрата, являясь одновременно, как указано выше, и катализатором для гидролиза иммобилизованным ферментом. Таким образом, нативный фермент действует во всем объеме емкости 3 и в части объема колонки 1.
Примеры конкретной реализации предлагаемого способа приведены ниже.
Пример 1 Контрольный.
Гидролиз проводят без участия нативного фермента. В колбу загружают 17 мас. ч. желатина, 1 мас.ч. иммобилизованного по ионно-координационному механизму фермента Протосубтилина на 1,2 мас.ч. матрицы силикагеля с порами 1000 А и 100 мас.ч. воды. Гидролиз проводили при 30oC, при непрерывном перемешивании со скоростью 60 об/мин. мешалки. Полный гидролиз не осуществляется даже за 100 ч. гидролиз при pH 7,2 кислотности, выходит на плато уже за 130 мин. но даже за 100 ч. не превышает 27%
Пример 2.
Пример осуществляли по примеру 1, но дополнительно вводят нативный фермент Протосубтилин, в количестве 0,01 мас.ч. Время полного гидролиза (99%) составляет 32 мин. Кислотность среды 7,2.
Пример 3 Контрольный.
Все осуществляется по примеру 2, но без иммобилизованного фермента, время полного гидролиза, на 99% составляет 57 ч.
Пример 4.
Осуществляли по примеру 2, но загружали нативного фермента /Протосубтилина/ всего 0,00001 мас.ч. Гидролизат с конверсией 99% получают за 30 ч.
Пример 5.
В колонну, снабженную устройством для ввода нативного фермента, выполненной в виде капельной воронки, устройством для отвода гидролизата и устройства для ввода субстрата, помещают 100 мас. ч. трипсина, изоэлектрическая точка 4,2, иммобилизованного на матрице силикагеля с порами, размером 56А /5,6 нм/, с содержанием белка 31% В колонну подают раствор желатина, с изоэлектрической точкой 7,24 с концентрацией 10% раствор нативного Дрожжелитина /изоэлектрическая точка 4,8/ со средним размером молекул 27 А /2,7 нм/, так, чтобы концентрация желатина в растворе составляла 8% мас. а Дрожжелитина 0,008% мас. при pH 6,3 боратного буфера с концентрацией 1 моль/л. Раствор подавали с линейной скоростью 1 см/мин при высоте слоя иммобилизованного фермента в 10 см. Получают гидролизат с содержанием аминокислот 8% мас. с конверсией 99,9% не содержащего нативного фермента. Время контакта 10 минут. За десять минут конверсия составляет 99,9%
Пример 6.
Осуществляют согласно Примеру 5, но используют желатин с концентрацией 12% мас. перед колонкой помещают предколонку с размером пор в силикагеле 18 А /1,8 нм/. Получают раствор аминокислот с концентрацией 10% мас. не содержащей нативный Дрожжелитин, конверсией 100% время контакта 11 мин.
Пример 7.
По примеру 5, но вводят электроды и создают напряженность электрического поля 3 В/см. Проводят гидролиз крахмала с концентрацией его в растворе 8% мас. на иммобилизованной и нативной амилазе с концентрацией амилазы на иммобилизованном ферменте в матрице силикагеля 235 мг/г и концентрацией нативной амилазы в растворе 0,0001 мас. Изоэлектрическая точка амилазы 5,3. В субстрат вводят 0,2% мас. боратного буфера поддерживающего кислотность субстрата 3,4. Время контакта 20 мин. конверсия 99% нативный фермент в гидролизате отсутствует, пока есть напряжение, при отключении напряжения проникает нативная амилаза с концентрацией 0,0001% мас.
Пример 8.
По примеру 7, но создают напряженность электрического поля 25 В/см. Гидролизуют яичный альбумин /изоэлектрическая точка 6,2/ в присутствии фосфатного буфера с концентрацией 0,1% мас. при концентрации белка 10% мас. при pH 5,3. Концентрация нативного Дрожжелитина 0,0001% его изоэлектрическая точка 3,8. В этом примере использован иммобилизованный трипсин по примеру 5. При времени контакта в 60 мин, получают конверсию 100% концентрации аминокислот 10% полном отсутствии нативного фермента.
Пример 9 Контрольный.
По примеру 8, но без напряжения на электродах. При этом на выходе из колонки появляется нативный фермент в количестве 0,00008% мас. по активности.
Пример 10.
По примеру 7, но модификатор вводят в количестве 3% мас. от массы субстрата. Время контакта 20 мин. конверсия 99,9% нативный фермент отсутствует.
Пример 11.
По Примеру 8, но модификатор вводят в количестве 1,4 мас. Через 50 минут контакта выход составил 99,6% мас. концентрация аминокислот 10% мас. нативный фермент в гидролизате отсутствует.
При концентрации модификатора меньше указанных минимальных пределов, заряд молекул фермента и субстрата недостаточен для электромиграции. При большей начинает уменьшаться активность нативного и иммобилизованного ферментов. При напряженности электрического поля менее 3 В/см скорость электромиграции недостаточна для эффективного предотвращения диффузии нативного фермента и ускоренной диффузии субстрата к иммобилизованному ферменту. При большей, чем 25 В/см, напряженности, раствор сильно перегревается и за счет электролиза теряется активность фермента.
Предложенный способ гидролиза биологических субстратов интенсифицирует процесс гидролиза, повышает качество и чистоту получаемого продукта, позволяет использовать в качестве обрабатываемого сырья не только низкомолекулярные, но и высокомолекулярные белковые и полисахаридные субстраты. Исключается потеря нативного фермента и увеличивается выход готового продукта. Сокращаются потери на закупку дорогостоящего оборудования. Очистка сточных вод, содержащих отходы с утилизацией аминокислот, позволит улучшить экологическую ситуацию и получить дополнительный экономический эффект.
Источники информации.
1. Пат. N 79-06616 Нидерланды, МКИ C 12 P 12/02, 1980 г.
2. Неклюдов А.Д. Навашин С.М. "Получение белковых гидролизатов с заданными свойствами", журнал "Прикладная биохимия и микробиология", 1985, т.21, N1, с. 3-17.
3. Менеке Г. Полянский Д. "Реакционоспособные носители для иммобилизации биокатализаторов" в сб. Синтез и химические превращения полимеров, Л. ЛГУ, 1986 г. с. 45-75 (прототип).
| название | год | авторы | номер документа |
|---|---|---|---|
| Способ получения силикагеля | 1989 |
|
SU1773868A1 |
| СПОСОБ ПОЛУЧЕНИЯ ГИДРОЛИЗАТОВ И ИСПОЛЬЗОВАНИЕ ИХ ПРИ ИЗГОТОВЛЕНИИ ПИТАТЕЛЬНЫХ СРЕД ДЛЯ КУЛЬТИВИРОВАНИЯ МИКРООРГАНИЗМОВ | 1996 |
|
RU2103345C1 |
| Фармацевтическая субстанция для лечения инфицированных ран различного генеза | 2018 |
|
RU2687102C1 |
| Способ изомеризации глюкозы во фруктозу | 1983 |
|
SU1523056A3 |
| ПОЛУЧЕНИЕ ИММУНОГЛОБУЛИНОВ С ПОНИЖЕННЫМ СОДЕРЖАНИЕМ ТРОМБОГЕННЫХ АГЕНТОВ И ИММУНОГЛОБУЛИНОВАЯ КОМПОЗИЦИЯ | 2012 |
|
RU2627162C2 |
| СПОСОБ ПОЛУЧЕНИЯ ИММОБИЛИЗОВАННОГО ФЕРМЕНТНОГО ПРЕПАРАТА НА ОСНОВЕ БРОМЕЛАЙНА, ГИАЛУРОНОВОЙ КИСЛОТЫ И ПОЛИСАХАРИДОВ, МОДИФИЦИРОВАННЫХ ВИНИЛОВЫМИ МОНОМЕРАМИ | 2020 |
|
RU2750377C1 |
| Способ получения сиропа, содержащего глюкозу и фруктозу | 1982 |
|
SU1449014A3 |
| СПОСОБ ПОЛУЧЕНИЯ ГЕТЕРОГЕННОГО ПРЕПАРАТА БРОМЕЛАЙНА, КОВАЛЕНТНО СВЯЗАННОГО С МАТРИЦЕЙ ХИТОЗАНА | 2018 |
|
RU2711786C1 |
| Способ получения гидролизата молочных белков | 1986 |
|
SU1472041A1 |
| Способ получения фумаровой кислоты | 2020 |
|
RU2748229C1 |
Область использования: изобретение относится к способам гидролиза высокомолекулярных биологических субстратов и может быть использовано в химической, медицинской, микробиологической, пищевой промышленности, а также для очистки бытовых и промышленных сточных вод. Сущность изобретения. Гидролиз биологических субстратов проводят путем их контакта с иммобилизованным на пористых носителях-матрицах ферментом. В реакционную среду дополнительно вводят нативный фермент и модификатор для придания молекулам субстрата электрического заряда. Реакционную среду с субстратом и ферментами помещают в направленное электрическое поле, а гидролиз ведут при pH среды, величина которого поддерживается не равной величинам изоэлектрических точек субстрата и нативного фермента. Для гидролиза используют матрицы с определенными диаметрами пор. 6 з.п.ф-лы, 1 ил.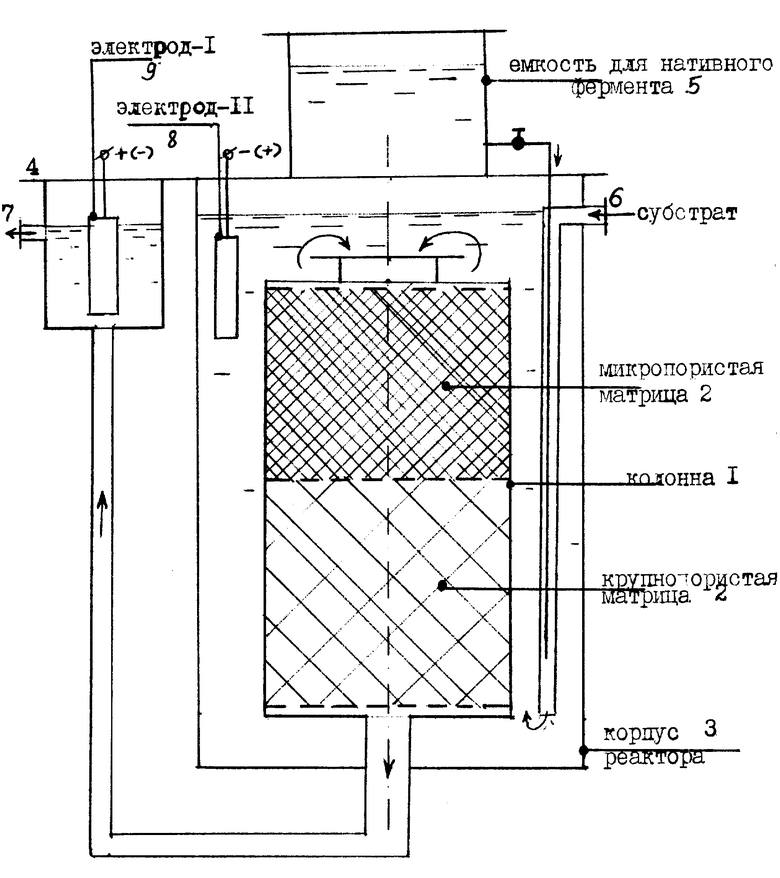
| Менеке Г., Полянский Д | |||
| Реакционноспособные носители для иммобилизации биокатализаторов | |||
| Сб | |||
| Синтез и химические превращения полимеров | |||
| - Л.: ЛГУ, 1986, с.45 - 75. |
