Изобретение относится к биотехнологии и может найти применение в медицине, ветеринарии, сельском хозяйстве и пищевой промышленности.
Известно, что начало 70-х годов ознаменовалось наступлением эры генетической инженерии. В основе современной промышленной биотехнологии лежит возможность направленного конструирования генетического материала, введения его в живые клетки и реализация с их помощью содержащейся в нем генетической информации. Возможность введения любого сегмента ДНК в клетки позволяет создавать промышленные микроорганизмы, способные, например, синтезировать ценнейшие белки. Так, первая описанная рекомбинация ДНК состояла из фрагмента ДНК вируса ОВ40 и бактериофага Pdvga1 с галактозным опероном Escherichia Coli (I. D.A. Jackson, R.H.Symons, P.Berg. Proc. Nat. Acad, Sei, USA, 1972, v.69, p.2904).
E.coli излюбленный объект подавляющего большинства работ по клонированию и генетической инженерии. Из других бактерий следует отметить Bacillus subtilis грамположительный, непатогенный, непаразитический микроорганизм, а также штаммы Pseudomonas, Streptomyces и т.д.
В настоящее время E.coli считается малоудобным продуцентом гетеролитических белков. Причем клетки E.coli не обладают биохимическим механизмом, способным осуществить сплайсинг РНК. Представляются перспективными нитчатые грибы, дрожжи, клетки насекомых и млекопитающих. Возможно, что клетки животных в ближайшее время могут найти широкое применение для получения рекомбинантных белков, моноклональных антител, вирусных вакцин, факторов иммунорегуляции, гормонов и ферментов (A.T.Bull. Proc. 4-th Europ. Congress Biotechnol. 1987, vol 4, p. 189-202).
В генетической инженерии существует несколько возможных подходов к осуществлению эффективной экспрессии чужеродных генов у микроорганизмов. Идеальным представляется химический или микробиологический синтез эукариотического гена либо внедрение чужеродного гена внутрь структурного гена бактериальной или дрожжевой клетки, получение структурного гена с использованием обратной транскрипции соответствующей мРНК, не содержащего ни одного интрона с нуклеотидной последовательностью, кодирующей первичную структуру этого эукаротического белка.
Однако этот ген, включенный в E.coli, нормально не экспрессируется, т. е. синтезируемый полипептид будет резко отличаться от нормального эукаротического белка.
Что касается проблемы синтеза гена, то сегодня она представляет пока трудноразрешимую задачу, которая лишь несколько облегчается при использовании соответствующих наборов олигонуклеотидов с частично перекрывающимися последовательностями, способными к самосборке и синтезу двухцепочной ДНК (гена).
Второй подход метод прямой экспрессии основан на стыковке структурного гена в области инициации трансляции с образованием гибридного сайта узнавания рибосомами. Учитывая, что процесс отщепления сигнальных последовательностей в бактериальной клетке не всегда возможен, в конструкцию перед структурным геном вставляется кодон инициации трансляции (метиониновый кодон).
Согласно третьей схеме чужеродный ген встраивается за первым геном оперона таким образом, что сайт инициации трансляции чужеродного гена перекрывается на один нуклеотид с сайтом терминации трансляции первого гена оперона.
Не останавливаясь подробно на тонкостях по клонированию гена и экспрессии соответствующего белка методами технологии рекомбинантных ДНК, отметим, что клонирование фермента/ов чужеродной ДНК является первым этапом ее конструирования, а структурная модификация клетки реципиента, обеспечивающая эффективную экспрессию чужеродного/ых гена/ов у микроорганизмов относится к второму этапу конструирования. Третий этап конструирования это собственно сам подбор клетки хозяина.
Последовательность этапов работы по клонированию гена и экспрессии соответствующего белка методами технологии рекомбинантных ДНК можно представить в виде следующих этапов конструирования (Бэйли Дж. Оллис Д. Основы биохимической инженерии. М. Мир, 1989, т.1, с. 440-462):
получение фрагмента (чужеродного) ДНК;
введение чужеродного фрагмента ДНК в плазмидный вектор;
трансформация;
обнаружение необходимого клона;
выращивание культуры и выделение плазмид;
определение нуклеотидной последовательности клонированного фрагмента ДНК;
конструирование и построение плазмиды для экспрессии;
трансформация;
обнаружение необходимого клона;
выращивание культуры и выделение плазмид;
проверка нуклеотидной последовательности ДНК;
трансформация;
выращивание культуры для производства белка.
Таким образом, из приведенного базисного материала можно заключить, что современная генетическая инженерия и биотехнология трудоемка и многостадийна.
В настоящее время в отраслях сельского хозяйства, в пищевой промышленности и медицине наблюдается тенденция к сдерживанию развития биотехнологии в целом и генетической инженерии в частности (Biofutur, 1989, New Sci. 1990, p. 40). В ряде стран (Германия, Швейцария и некоторые др.) запрещено использование продуктов биотехнологии (например, D. Mackenzie New Sci. 1990, N 1707, p. 28). В генетической инженерии бактерии E.coli являются не только малоудобными продуцентами гетеролитических белков, но и таят в себе летальную опасность для человека, так как содержат элементы неожиданной неопластической трансформации клеток.
Методы технологии рекомбинантных ДНК позволяют получать микроорганизмы с выдающимися признаками, но они малоудобны или неприемлемы для многоклеточных организмов (Баев А.А. Генетическая инженерия. Журнал Всесоюзного химического общества им. Д.И.Менделеева, 1984, т.XXIX, N 2, с. 4-6).
Важно подчеркнуть, что выдающиеся признаки новых штаммов микроорганизмов не закрепляются в последующих поколениях. Частично это объяснимо, во-первых, потерей плазмид микроорганизмами и, во-вторых, вследствие структурных перестроек генетического материала рекомбинантных плазмид.
Для получения генетического материала с полезными свойствами известно применение факторов, вызывающих мутирование генома клетки.
Так, известны физические факторы (например, все виды ионизирующих излучений), вызывающие мутирование генома клетки (Большая мед. энциклопедия. М. Сов. энциклопедия, 1981, т. 16, с.26. Мутагены).
Однако эффект различных ионизирующих излучений принципиально одинаков: образование ионов в облучаемой популяции клеток является началом сложной цепи физико-химических процессов, приводящих к грубому нарушению хода биохимических реакций и в конечном итоге к возникновению генных мутаций и структурных перестроек хромосом. Направленность процессов, вызываемых различными видами ионизирующих излучений, непредсказуема и неуправляема, а вызванные ими мутации не закрепляются в последующих поколениях.
Известны химические средства, вызывающие мутацию (там же).
Однако подавляющее большинство из них обладает канцерогенной активностью, что делает опасной работу с ними. К тому же мутации, вызываемые известными средствами, не закрепляются в последующих поколениях.
На примере штаммов E.coli показана возможность направленной мутации генома бактерии, обеспечивающей, например, утилизацию лактозы салицина и других субстратов (B. G. Hall, L.L.Parker, P.W.Betts et al. Genetics, vol. 121, N 3, p.423; J.A.Heinemann, G.F.Sprague. -Nature, 1989, vol.340, N 6230, p.205).
Но даже если направленное мутирование и описано для E.coli, то оно не описано для эукариот и тем более для высших организмов.
Известны соли N-замещенных 3-оксипиридиниев (авт.св. N 993609, 1982; Tn. Zinoke, G.Muhlhausen. Berichte der deutschen-chemischen gasellschaft, 1905, yd.38, Bd. IV, S.3824).
Техническим результатом, достигнутым при реализации настоящего изобретения, является обнаружение группы веществ с принципиально новым свойством, ранее не описанным ни для каких известных соединений.
Такими соединениями, согласно изобретению, являются соли N-замещенных 3-оксипиридиниев, которые рекомендуются к применению в качестве средства, вызывающего наследуемое и закрепляемое в потомстве направленное мутирование генома клетки одноклеточных и многоклеточных организмов.
Впервые установлено, что существуют такие соединения, использование которых позволяет вызывать направленные полезные мутации, которые наследуются и закрепляются в потомстве.
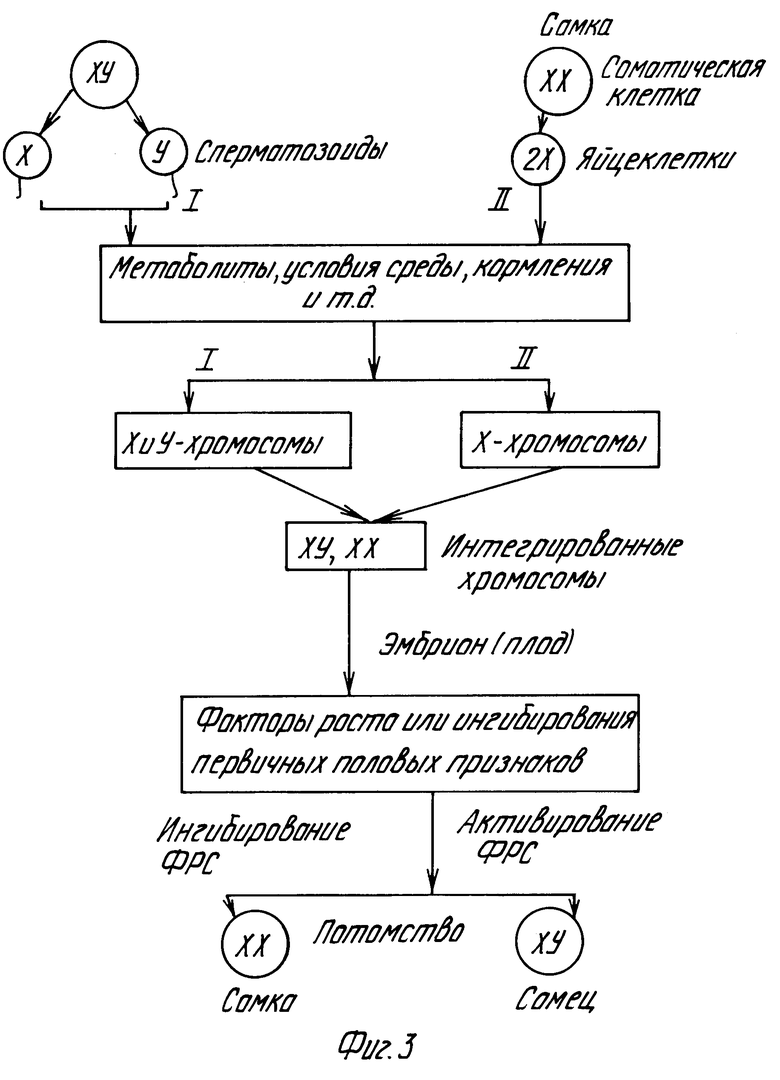
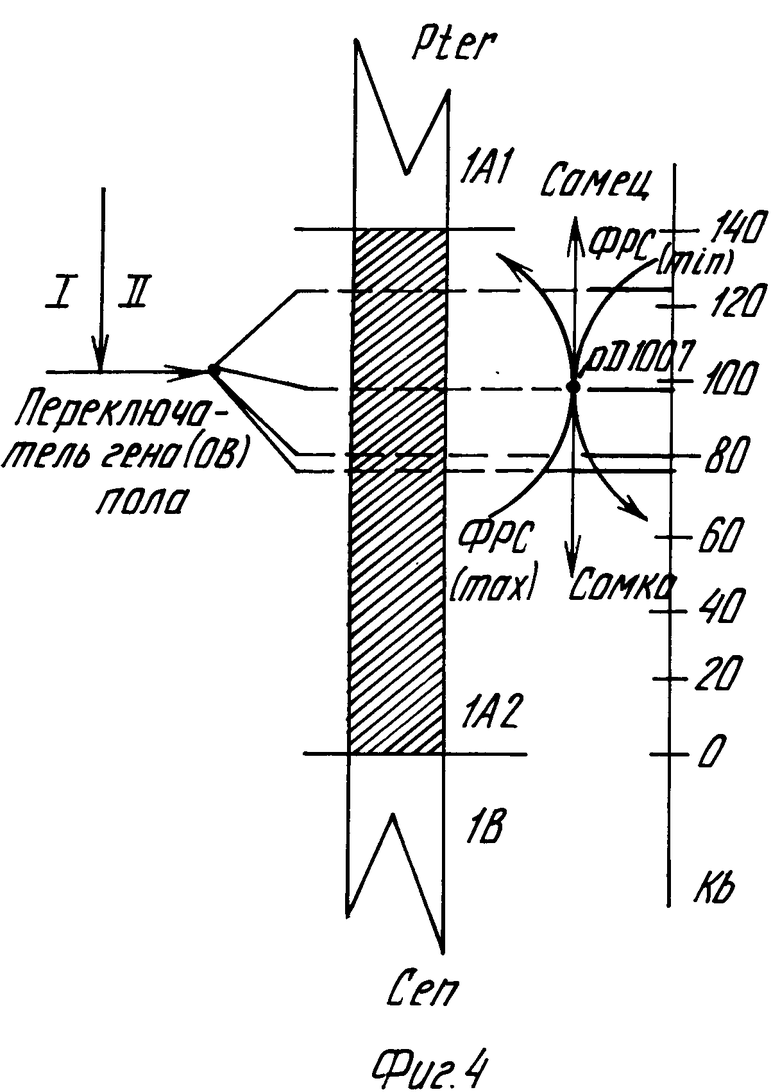
На фиг.1 графически представлена зависимость включения [3H]-тимидина (а) и [3H]-уридина (б) в клетках линии карциномы яичника человека от концентрации перхлоратов N-фенил-3-оксипиридиния (1), N-оксифенил-3-оксипиридиния (2) и N-толил-3-оксипиридиния (3); на фиг.2,а - диаграмма влияния перхлората N-фенил-3-оксипиридиния на рост дрожжевых клеток C.tropicalis штамм СК-4; на фиг. 2,б графически результаты изучения роста колонии коммерческих дрожжевых клеток и популяции C.tropicalis штамм CR-4 в условиях жесткой селекции на крахмальной среде без препаратов (контроль 1, 3) и под влиянием перхлоратов N-оксифенил-3-оксипиридиния (2) и N-толил-3-оксипиридиния (4); на фиг.3 схема формирования признака пола у плода при копуляционном размножении; на фиг. 4 принципиальная схема влияния перхлоратов N-фенил-3-оксипиридиния и N-оксифенил-3-оксипиридиния на переключатель гена/ов пола в локусе pД 1007Х или Y-хромосоме человека с эволюционно устойчивыми сегментами 77, 80, 97 и 124 Kb в интервале 1А2, равном примерно 140 Kb или 0,2% от Y-хромосомы.
Пример 1. Изучали действие перхлоратов N-фенил-, N-оксифенил- и N-толил-3-оксипиридиниев на опухолевые клетки линий карциномы яичника человека CaOγ Скорость синтеза ДНК и РНК оценивали по включению [3H]-тимидина и [3H]-уридина в указанные клетки. Оценку цитостатического действия препарата проводили радиометрическим способом (Добрынин Я.В. Стеняева Т.И. Кондратьева А.Н. Проблемы химиотерапии злокачественных опухолей. М. 1974, с.175).
Результаты приведены на фиг.1 и свидетельствуют о зависимости синтеза ДНК и РНК (включения [3H]-тимидина и [3H]-уридина) от заявленных соединений и от их структуры.
Например, под влиянием N-толил-3-оксипиридиния в два раза возрастает синтез ДНК при дозе препарата 100 мкг/мл (фиг.1). Этот эффект действия препарата на синтез ДНК сохраняется и при более низких концентрациях. Препарат N-фенил-3-оксипиридиний при дозе 100 мкг/мл оказывает частичное ингибирующее влияние на скорость включения [3H]-тимидина. Синтез ДНК возрастает при дозах 10,0 и 1,0 мкг/мл указанного препарата. Соединение обладает высокой цитотоксической активностью в изучаемых дозах.
Результаты по зависимости включения [3H]-уридина в клетках линии карциномы яичника человека от концентрации и структуры исследуемых заявленных соединений позволяет выделить данные по N-оксифенил-3-оксипиридинию. Этот препарат практически полностью угнетает синтез ДНК и одновременно значительно стимулирует синтез РНК: включение [3H]-уридина возрастает при дозе 10 мкг/мл на 214% относительно контрольного показателя. В качестве пограничного критерия активности принимают 50%-ное угнетение включения [3H]-тимидина и [3H]-уридина при испытываемой концентрации тимидина и уридина 0,5•10-3 М.
Пример 2. Препараты политенных хромосом (ПХ) из слюнных желез Drosophila melanogaster изготавливали по известной методике (Кикнадзе И.И. Функциональная организация хромосом. Л. Наука, 1972, с.212). Анализ функциональной активности политенных хромосом проводили по физиологической карте.
В выделенных в последнем третьем личиночном возрасте опытных дрозофилл F1- и F5-поколений низкомутантных линий D-32, вскормленных на N-фенил-3-оксипиридинии; в четвертой хромосоме обнаруживаются интенсивно функционирующие участки в третьей области колец Бальбиани (КБ1) и в меньшей степени в четвертой области КБ2 в пределах дисков 4-3А1-6 и 4-4А4-6. Остальные три длинные хромосомы содержат несколько гигантских пуфов с функционированием до 35 дисков и более, фактически переходящие в ядрышки. О степени выраженности признаков можно судить и по следующим показателям: средний диаметр политенной хромосомы опытных дрозофилл в 2 раза больше контрольного. Отношение диаметра пуфа к диаметру хромосомы и длине пуфа у опытных животных соответственно составляет 1,9-2,0 и 0,5-0,8 см против 3,5-1,4 и 2,3-4,6 см в контроле.
ПХ контрольных животных хотя и содержит большое число поперечных дисков и участки, напоминающие пуфы, генетически мало активны из-за относительно больших гетерохроматических участков.
Известно (Кикнадзе И.И. Функциональная организация хромосом. Л. Наука, 1972, с. 212), что интенсивно функционирующие участки в третьей и четвертой областях КБ1 и КБ2 IV хромосомы свидетельствуют об активности образования РКН. В свою очередь результаты исследования остальных данных I-ПI хромосом, содержащих сложные пуфы с функционированием 20-35 дисков, фактически представляющих собой ядрышки, свидетельствуют об интенсивном синтезе рибосомной РНК на ДНК ядрышкового организатора (Стент Г. Келиндар Р. Молекулярная генетика. М. Мир, 1981, с.602).
Образование избыточного пула рРНК (матричного, рибосомного, информационного и т.д.) в клетках, обнаруженное направленное мутирование и устойчивое закрепление новой полезной мутации в поколении подтверждают возможность появления в геноме большей смысловой информации в молекулах рРНК, чем это закодировано в исходной кодирующей цепи ДНК на матрице рРНК по механизму обратной транскрипции. Имеется в виду активация под влиянием исследуемых заявленных соединений ревертазы и РНК зависимой ДНК - полимеразы, обеспечивающих обратный порядок передачи генетической информации, синтез цепи ДНК, комплементарной матрице и РНК.
Пример 3. Исследование влияния N-фенил-3-оксипиридиния на рост Candida tropicalis (16 популяций) проводили по следующей методике: в питательные среды (солодовое сусло или среде Сабуро) в среднем в 100 повторяющихся опытах вносили различные дозы препарата (0,13333; 0,10; 0,0666; 0,03333 и 0,01333 мг/л). Через 15 ч инкубации при 35oC в аэробных условиях содержания в контрольных и опытных группах подсчитывали в камере Горяева число дрожжевых клеток (F1 поколение). С целью выявления мутации и возможного закрепления в поколении признака в новые питательные среды в той же последовательности, что и выше, переносили такое минимальное количество дрожжевых клеток, чтобы оно соответствовало 3-4 клеткам/мл. Далее после инкубации клетки снова переносили на новые питательные среды и т.д. повторяя эту процедуру с числом повторяющихся опытов до 10 поколений (F10).
В другом опыте к 0,5%-ному раствору крахмала и 2 мл 1%-ного раствора пекарских дрожжей вносили 10-8 моль препарата N-фенил- или N-оксифенил-3-оксипиридиния. Контрольный вариант отличался от опытного отсутствием в нем исследуемых препаратов. Через 24 ч инкубации при 20-25oC в аэробных условиях содержания подсчитывали число дрожжевых клеток. Для выявления возможного закрепления мутации в поколении из опытного и контрольного вариантов переносили на новые крахмальные растворы такое количество клеток, чтобы оно соответствовало в опыте 53/мл и в контроле 67/мл.
Подсчет дрожжевых клеток проводили в камере Горяева ежедневно. Морфологию клеток исследовали под обычным биологическим микроскопом.
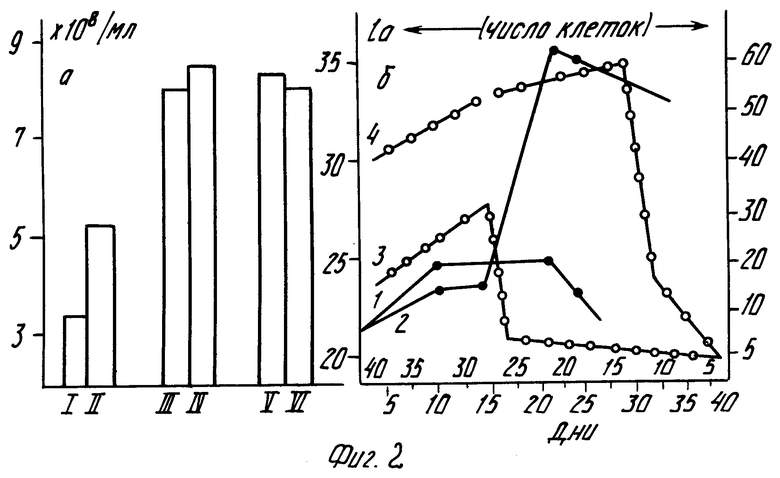
На фиг. 2, а представлены результаты изучения влияния перхлората N-фенил-3-оксипиридиния на рост дрожжевых клеток C.tropicalis штамм СК-4. Действие этого заявленного соединения испытано на 16 популяциях клеток, установлено, что в среднем на 100 исследований введения N-фенил-3-оксипиридиния в питательную среду при оптимальной концентрации 0,1333 мг/л число дрожжевых клеток через 15 ч инкубирования составило в среднем 834 млн/мл и превосходило контрольные популяции, инкубированные так же в пивном сусле без добавления препарата, в 2,49 раз (p < 0,001). При последующем выращивании новых популяций из опытных дрожжевых культур в течение 10 поколений (срок наблюдения) в указанной выше питательной среде без внесения препарата пролиферация дрожжевых культур сохраняется в среднем до 1,5 раза на период F1 F10 поколений.
Каких-либо изменений в морфологии клеток не выявлено. Синтез белка на период F1 F10 поколений соответствует скорости пролиферации.
Результаты влияния солей N-фенил- и N-оксифенил-3-оксипиридиниев на рост колонии коммерческих пекарских дрожжей на крахмальной питательной среде представлены на фиг.2,б. Небольшое число пекарских дрожжей инокулировали в определенные объекты крахмальной лимитирующей среды. На диаграмме обнаруживаются экспоненциальные фазы логарифм-роста, деления клеток с постоянной скоростью. Причем экспоненциальная фаза роста пекарских дрожжей под влиянием N-оксифенил-3-оксипиридиния в течение 30 дней значительно выше аналогичных для контрольных групп фаз роста.
Инокулирование небольшого числа опытных и контрольных эукариот на новые крахмальные среды показало значительно более высокую (на два и более порядков) скорость деления клеток в опыте.
При внесении N-фенил-3-оксипиридиния в среду, содержащую пекарские дрожжи на крахмальном рационе, обнаруживается определенный период лаг-фазы (время наступления деления клеток) с последующим взрывообразным ростом колонии клеток (фиг.2,б: 3 контроль; 4 опыт). Важно отметить, что под влиянием исследуемых заявленных соединений скорость деления опытных эукариот после лаг-фазы взрывообразно возрастает, превосходя контрольный показатель в десятки, сотни и тысячи раз, причем эта высокая скорость пролиферации закреплялась в течение длительного периода времени (60 дней срок наблюдения).
В обычных условиях дрожжевые клетки не утилизируют крахмал (Промышленная микробиология. /Под ред. Н.С.Егорова. М. Высшая школа, 1989, с.416). Значительный взрывообразный рост колонии пекарских дрожжей только на крахмальной среде свидетельствует о направленной мутации генома клеток в сторону метаболизма крахмала. Возможность синтеза гена/ов и их фрагмента/ов при химическом воздействии на клетки создает чрезвычайно перспективную область биохимической инженерии, позволяющую в более значительной степени, чем методом современной генетической инженерии и биотехнологии, заставить клетку синтезировать новые аминокислотные последовательности, крайне важные для нужд энзимологии, а также перспективные в качестве лекарственных средств или компонентов пищевых продуктов.
Известно, что ген Cdc2 клеток дрожжей и клеток человека идентичен и обеспечивает нормальный ход событий клеточного цикла. Активность протеинкиназы C, кодируемая геном Cdc2, в исследуемых условиях метаболизма микроорганизмов, очевидно, не нарушается, что подтверждается тем, что морфология дрожжевых клеток не изменяется и синтез белков на период F1 F10 поколений соответствует скорости пролиферации.
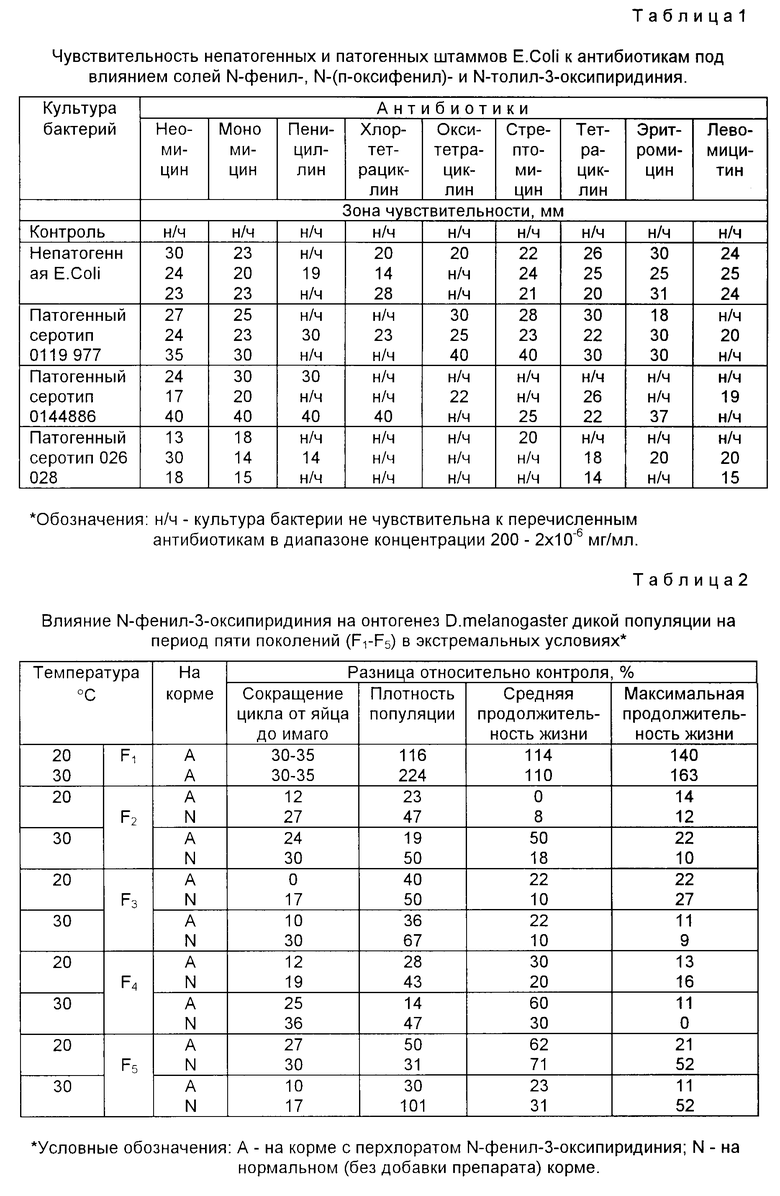
Пример 3. В исследованиях использовали непатогенную культуру кишечной палочки E.coli и три патогенных его серотипа 026; 0119 и 0144, которые были выделены из клинического материала Северо-Осетинской республиканской детской больницы. Все используемые бактерии относились к высокоустойчивым рекомбинантам R.
Влияние исследуемых заявленных соединений на преодоление резистентности инфекционных высокоустойчивых рекомбинантных микроорганизмов E.coli определяли по отношению к следующим антибиотикам неомицину, мономицину, пенициллину, хлортетрациклину, эритромицину, окситетрациклину, стрептомицину, тетрациклину, левомицетину.
Определение чувствительности к антибиотикам проводили по общепринятым методикам: диффузия в агар и серийные разведения в бульоне. Материал равномерно растирали на поверхности среды. Чашки подсушивали 30-40 мин при комнатной температуре, затем на поверхность засеянной среды накладывали диски на равном расстоянии друг от друга и на расстоянии 2 см от края чашек. Чашки с дисками выдерживали 30-40 мин при комнатной температуре и далее 16-18 ч при 36-38oC.
Оценку влияния перхлоратов N-фенил-, N-толил и N-оксифенил-3-оксипиридиниев проводили по диаметру задержки роста колоний вокруг диска, включая и диаметр самого диска. Чем больше зона задержки роста испытуемой культуры, тем выше ее чувствительность к данной концентрации антибиотика: до 10 мм чувствительность принимали за малочувствительную. При испытании препаратов использовали 24-часовую бульонную культуру, для выращивания которой в питательную среду вносили следующие дозы антибиотиков, мг/мл: 200,0; 20,0; 2,0; 0,2; 0,02; 0,002; 0,0002; 0,00002; 0,000002; 0,0000002.
В табл.1 приведены результаты по определению чувствительности непатогенных (I) и патогенных серотипов E.coli 0 119977 (П), 0144886 (III) и 026028 (IV) к антибиотикам. Из приведенных результатов видно, что E.coli резистентна к воздействию всех девяти исследуемых антибиотиков. При внесении в питательную среду N-фенил-3-оксипиридиния преодолевается резистентность непатогенных бактерий к неомицину, мономицину, тетрациклину, стрептомицину, эритромицину, левомицетину и остается полностью устойчивой только к пенициллину.
В присутствии N-фенил-3-оксипиридиния серотип (II) проявляет высокую чувствительность к неомицину, мономицину и тетрациклину, а серотип (III) к неомицину, мономицину и пенициллину. Что касается серотипа (IV), то он оказался малочувствительным ко всем антибиотикам.
Препарат N-толил-3-оксипиридиний малоэффективен для преодоления непатогенной культуры E.coli к воздействию пенициллина, хлортетрациклина и окситетрациклина, но делает ее чувствительной и высокочувствительной к остальным антибиотикам. Серотипы (II IV) проявляют различную чувствительность к антибиотикам под влиянием исследуемых препаратов.
При введении в питательную среду N-оксифенил-3-оксипиридиния серотип (IV) теряет устойчивость к хлортетрациклину, окситетрациклину, а серотип (II) к пенициллину, хлортетрациклину и эритромицину.
Если рассматривать диаметр зон задержки роста культуры, то под влиянием исследуемых заявленных соединений не только преодолевается устойчивость, но и возрастает чувствительность патогенных и непатогенных культур к антибиотикам в 20-40 раз, что ассоциируется с блокированием активности амплифицирующего гена и, следовательно, кодированием гликопротеина Р.
Практическое значение полученных данных было подтверждено в эксперименте на обезьянах макаках-резус Macaca mulatta, болеющих рецидивирующим дезентирийным колитом. Доказан высокий адаптогенный и терапевтический эффект действия N-фенил-3-оксипиридиния (Отчет Института экспериментальной патологии и терапии АМН СССР, 1981).
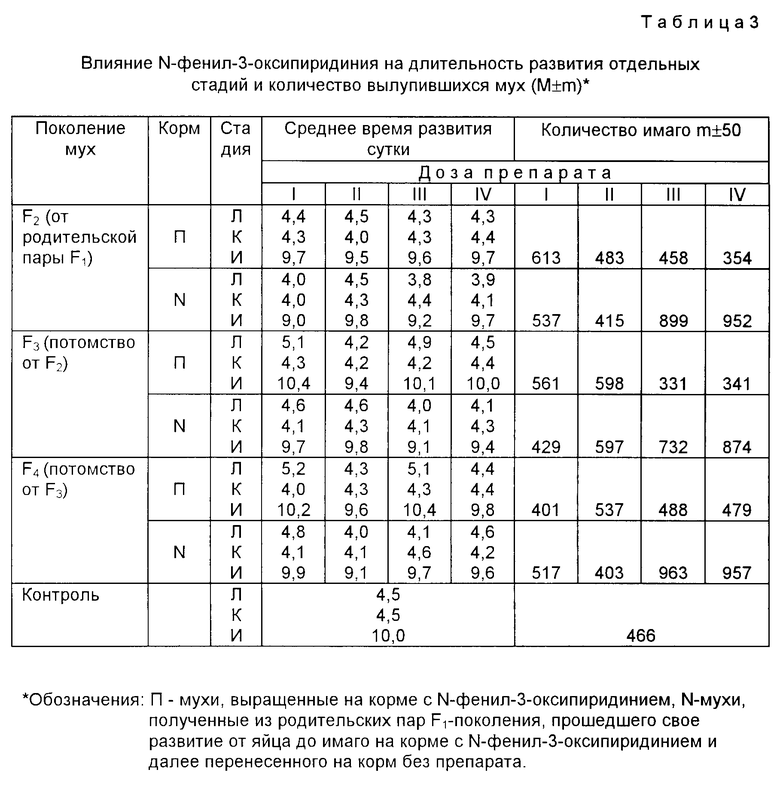
Пример 4. В работе использовали более 10000 особей низкомутантных линий D. melanogaster D-32 и плодовой мушки дикой линии Berlin wild. Виргинных мух D.melanogaster линии D-32 для скрещивания попарно 3q × 3o↗ помещали в отдельные пробирки диаметром 18 мм на 48 ч. Родительскую пару (РР) удаляли, а их потомство проходило свое развитие от яйца до имаго на корме из агара, изюма и манной крупы с различными дозами перхлората N-фенил-3-оксипиридиния. Контрольную группу вскармливали на корме без препарата. Из одновозрастных виргинных мух F1 составляли новые семьи. Одна часть РР1 вскармливалась на корме с различными дозами перхлората N-фенил-3-оксипиридиния (группа I - 0,05 г; группа II 0,10 г; группа III 0,15 г и группа IV на 0,20 г в 100 г корма). Другая часть РР1 была перенесена на корм без препарата N. Мух, полученных от последних РР1, обозначали как F2. По аналогичной схеме были получены F3 и F4 поколений. Мух содержали в термостате при 25oC и каждые 10 дней пересаживали на свежий корм. Ежедневно проводили фиксацию длительности развития стадий времени личинок (Л), куколок (К), имаго (И) и количество вылупившихся мух. Результаты приведены в табл.2.
Исследование картины пуфинга проводили у F1 и F5 поколений личинок третьего, последнего возраста, выползавших из корма и слегка подсохнувших. Влияние условий среды, возрастного фактора и другие параметры были сведены, по возможности, к минимуму.
В другом опыте, отобранные случайным образом виргинные мухи на третий день их имагинальной жизни помещали попарно 3q × 3o↗ в отдельные пробирки для скрещивания на 48 ч, после чего родительскую пару РР удаляли. Новое поколение мух проходило весь цикл развития на среде с различными дозами N-фенил-3-оксипиридиния. Полученных мух обозначали как F1. Из виргинных мух первого F1 поколения также случайным выбором были составлены новые родительские пары. Одну часть РР1 переносили на нормальный, без препарата, корм, а другую на корм с теми же дозами, что и выше, препарата. От F1 таким же образом в зависимости от содержания на нормальном корме (N) или на корме с препаратом (А) получено F2-поколения имаго, а от F2 далее F3 F5-поколений. Через каждые 7-8 дней мух пересаживали на свежий корм. Во всех вариантах опыта использовались виргинные однодневные мухи, которых содержали в термостате при температуре 20±0,1o; 25±0,1o и 30±0,1oC. Учет погибших мух вели ежедневно. Результаты приведены в табл.2.
Минимальная и максимальная деятельность жизни контрольного F1 поколения при 25oC составляло для самок 34-50 и самцов 37-49 дней, а при 20o и 30o 22 ± 1,0 и 10 ± 1,5 дней соответственно (P < 0,001). В работе использовали результаты, в которых наблюдали принципиальное совпадение нескольких повторных опытов.
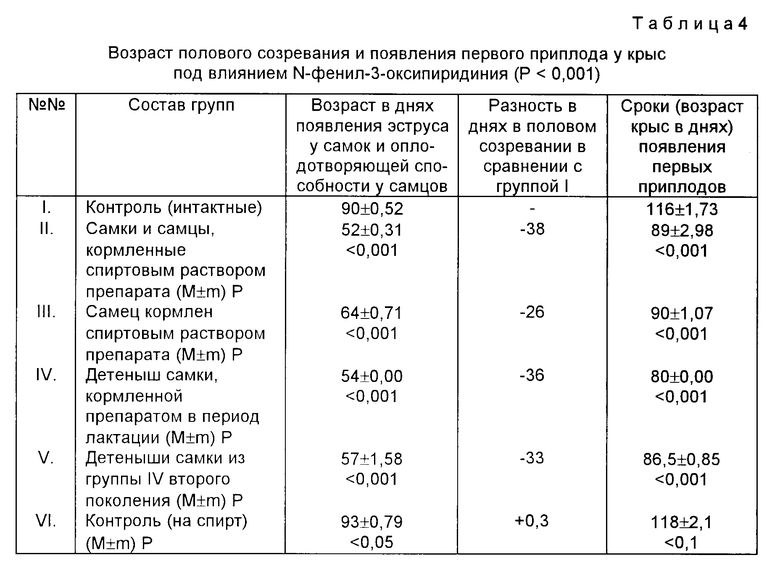
В табл. 3 приведены результаты по изучению влияния перхлората N-фенил-3-оксипиридиния на длительность развития отдельных стадий у дрозофил низкомутантной линии D-32. Можно отметить, что в F3 и F4 поколениях происходит почти полная синхронизация среднего времени развития личиночной стадии, стадии окукливания и имаго у контрольных и опытных мух. Плотность F1 популяции от PP, только однократно вскормленной препаратом, в условиях N содержания (на корме без препарата) возрастает относительно контрольного опыта в два раза, и эта высокая плодовитость наследуется у F1 F5 поколений (срок наблюдения).
В табл.2 приведены результаты выращивания родительских пар D.melanogaster дикой популяции на корме с N-фенил-3-оксипиридинием. Из представленных в табл. 2 данных следует, что в экстремальных условиях их содержания (20oC или 30oC), т.е. в условиях, когда синтез белков кроме белка теплового шока в организме дрозофил полностью почти прекращается, наблюдаются мощные F1 поколения по целому комплексу признаков (P < 0,001): сокращение развития цикла от яйца до имаго на 30-35% увеличение длины тела на 10% плотность популяции возросла в 2-4 раза; средняя и максимальная продолжительность жизни выросла на 110-160% относительно контрольных показателей, закрепляющихся в F1 F5 поколениях (срок наблюдения). Проведенный эксперимент объясним с точки зрения влияния исследуемых заявленных соединений на сигнальные участки генов онтогенеза дрозофил.
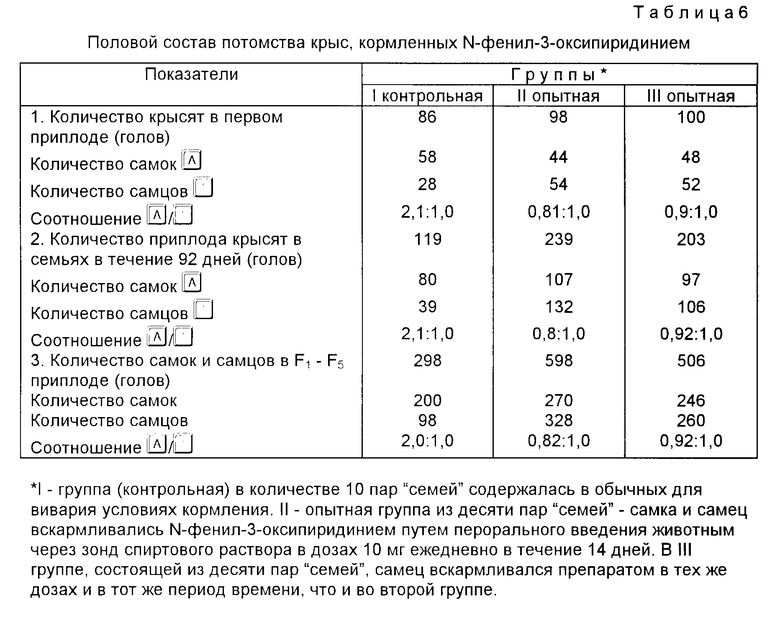
Пример 5. В работе использовали нелинейных крыс, крыс линий Вистар и мышей, а также низкомутантных дрозофил линий D-32. Для исследования влияния солей N-фенил- и N-оксофенил-оксипиридиниев на генеративную активность и возможность искусственного регулирования соотношения полов животных подбирали по принципу пар-аналогов с учетом возраста и живой массы. В опыте использовали 70 половозрелых крыс обоего пола. Все животные были разделены на три группы попарно (самец самка). Составленные родительские пары отсаживались в отдельные клетки, где находились в течение всего эксперимента (до получения от этих пар пяти поколений потомств) в одинаковых условиях обычного кормления, водного режима и содержания в виварии. Животные контрольной группы (1) в количестве десяти пар "семей" спаривались из самцов и самок шестимесячного возраста весом 180-220 г. Животные во второй группе, состоящей из десяти пар "семей", самец и самка шестимесячного возраста и весом 180-220 г за 38-40 дней до спаривания вскармливались N-фенил-3-оксипиридинием путем перорального введения через зонд в форме спиртового раствора в дозах 10 мг ежедневно 1 раз в сутки в течение 15 дней. В третьей группе из восьми родительских пар аналогичного возраста и веса, что и в группах I и II, самцу за 38-40 дней до спаривания вскармливали исследуемый препарат в тех же дозах. Опыты проводили для контрольных и опытных групп животных в апреле мае сентябре, когда соотношение полов по литературным данным (Гамбарян П.П. Дульская Н.М. Крыса. М. Советская наука, 1955, с. 252) составляет самец-самка, 50/50; 42,2/57,0; 45,0/55,0 соответственно.
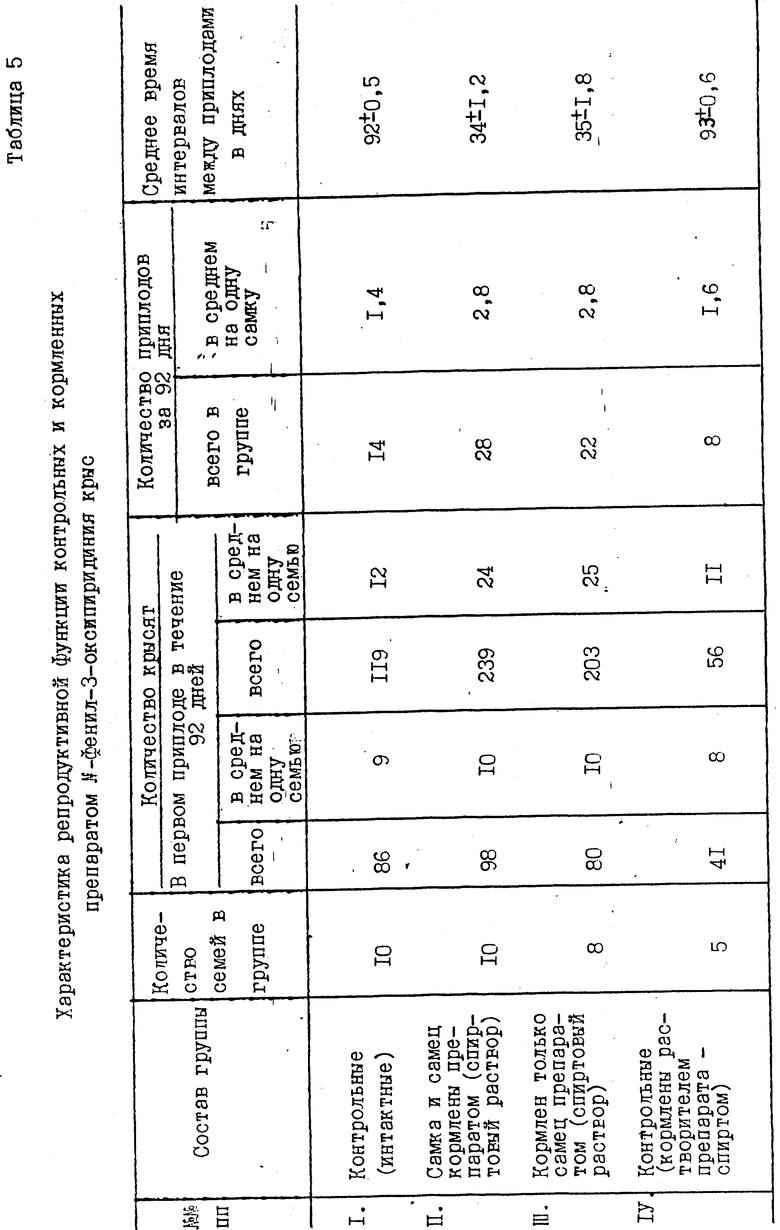
Результаты влияния N-фенил-3-оксипиридиния на половое созревание, появление приплодов, вес семенников и придатков крыс, а также соотношение массы органов к массе тела и смешения соотношения полов представлены в табл.4-8. Из приведенных в табл.4 результатов можно заключить, что у крысят, родившихся от кормленных препаратом родителей, как в группе II, так и в группе III выявлено ускорение истинного полового созревания на 38-26 дней, сроки появления первых приплодов сокращены со 116-118 дней в контроле до 89-80 дней в опытных группах, т.е. на 27-26 дней.
Учет приплода в "семьях" вели в течение 92 дней от момента первого приплода, так как этот срок составляет среднее время интервала между приплодами у контрольных крыс (табл.4). Следует при этом отметить, что в работе пользовались общепринятым в отечественной литературе обозначением плодовитости способности животных регулярно приносить приплод в количестве, свойственном каждому виду (Ветеринарная энциклопедия, т. 4, с.1045, 1973).
За указанный период в десяти контрольных "семьях" (группа I) приплод составил 119 крысят, тогда как в десяти опытных "семьях" (группа II) 239 крысят, в восьми "семьях" группы III 203 крысенка, в пяти "семьях" группы IV 56 крысят. В пересчете на одну пару количество крысят составляет: группа I - 12; группа II 24; группа III 25; группа IV 11 крысят. Из приведенных в табл. 4 данных следует, что популяционный рост идет не за счет увеличения особей в приплоде, а за счет сокращения интервалов между приплодами и увеличения количества приплодов: в группе I (контрольной) среднее время интервалов между приплодами составляет 92 дня (разброс 90-96 дней), группе II 34 дня (разброс 27-45 дней), группе III 36 дней (разброс 28-49 дней) и в группе IV 94 дня (разброс 90-99 дней). Данные по плодовитости и половому составу потомства крыс, потреблявших препарат, представлены в табл.5 и 6.
Увеличение в 2 раза и более плодовитости нелинейных крыс за счет сокращения интервалов между приплодами, а также появление раннего полового созревания под влиянием N-фенил-3-оксипиридиния наблюдали в течение пяти поколений.
За период наблюдения не отмечено заметного старения репродуктивной функции крыс. Физическое развитие крысят, рожденных в опытных семьях, в течение первого месяца постнатальной жизни не отличалось от контрольных: через 24 ч после рождения вес крысят в контрольных семьях составлял 6-8 г; в семьях группы II 6-7 г; группы III 7-9 г; группы IV 7-9 г.
Прибавка веса, раскрытие глаз, приобретение шерстяного покрова у крысят в опытных семьях идет в соответствии с физиологическими нормативами.
Анализ метафазных пластинок костного мозга крыс показал, что хромосомные аберрации отсутствуют, все хромосомы акроцентрические, нормальные, спирализация не нарушена.
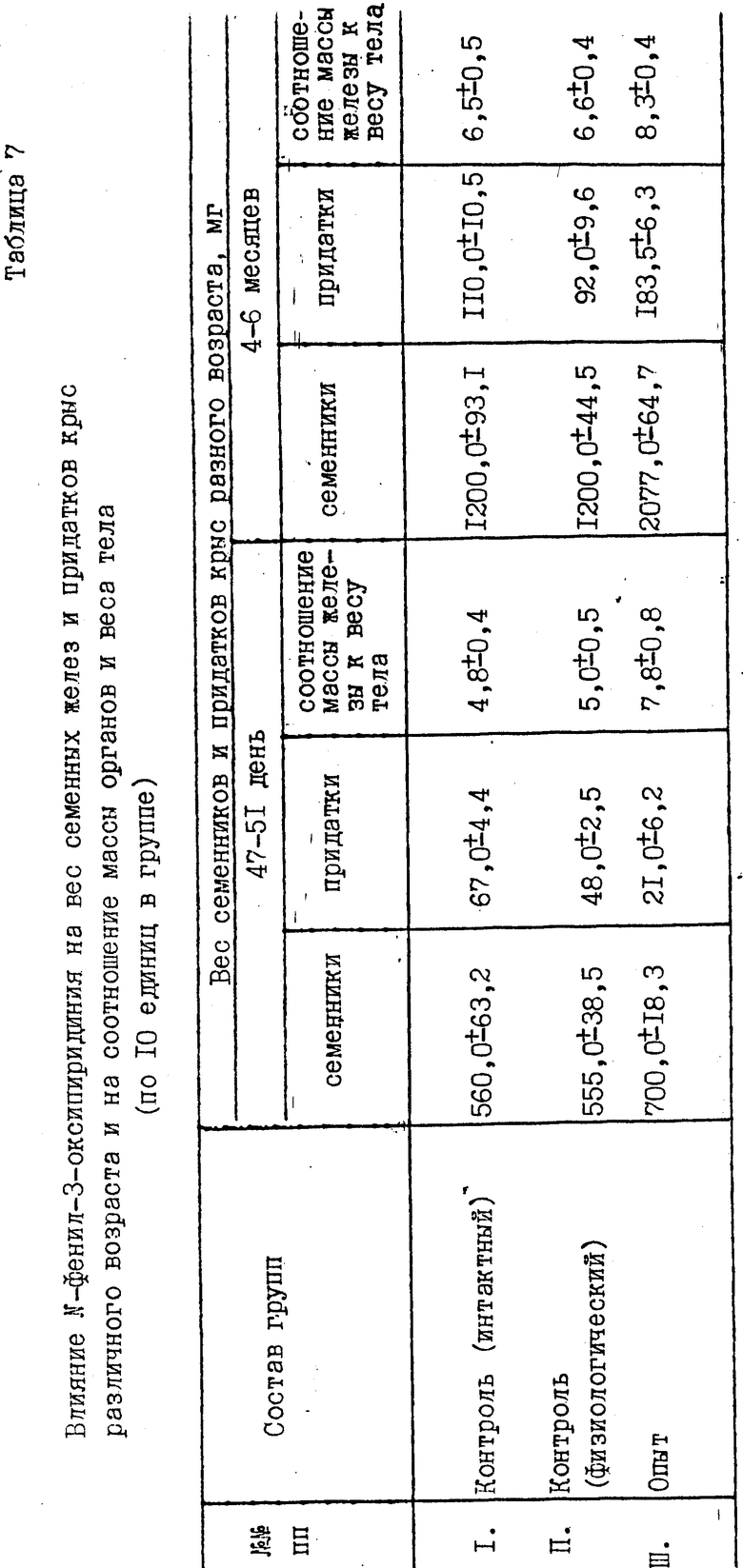
В эксперименте доказано наличие линейной зависимости между влиянием N-фенил-3-оксипиридиния не активацию семенников и их придатков, что проявляется в быстром и интенсивном росте их массы, превышающей вес семенников и придатков у контрольных крыс того же возраста и веса (табл.7), и смещением соотношения полов у крыс в сторону мужских особей (табл.6).
Гистологическое исследование семенников крыс, окрашенных по Фельгену и Браше, показало значительное увеличение толщины стенок извитых канальцев семенников до 250-320 мкм (в контроле 150-200 мкм), увеличение всех слоев сперматогенного эпителия, а также количества активно подвижных сперматозоидов (табл.8).
Клетки Лейдига, продуцирующие мужские половые гормоны и поддерживающие сперматогенез, функционально активны, что выражается в увеличении объемов ядер, их общих размеров по сравнению с контролем. Ядерно-цитоплазматические соотношения в пределах, выявленных у контрольных крыс (табл.8).
Пример 6. В табл.9 представлены результаты по влиянию N-оксифенил-3-оксипиридиния на онтогенез и соотношение полов D.melanogaster низкомутантной линии D-32.
Как известно (Вредные вещества в промышленности. Л. Химия, т.1, 1976), соли метафосфорной кислоты вызывают нарушение в метаболических процессах. Однако в противоположность утвердившимся в физиологии представлениям (D.C. Page, R.Mosher, E.U.Simpson, et al, Cell, 1987, vol 51, p.1091) под влиянием N4PO3 соотношения полов у дрозофил F1 поколений смещается в сторону женских особей. Хотя в F1 поколении соотношение q/o↗ в опыте составляло 1,06:1,0 против 1,30:1,0 в контроле. Двойное действие метафосфата натрия и N-оксифенил-3-оксипиридиния приводит к более высокой частоте расщепления самок 1,60: 1,0.
Таким образом, приведенные в примерах 5 и 6 экспериментальные результаты однозначно показывают, что в зависимости от характера влияния солей N-арил-3-оксипиридиния на генеративные функции на примере крыс и дрозофил происходит различная активация факторов роста первичных половых признаков, кодируемые X- и Y-хромосомами, следовательно, различна и частота расщепления пола в сторону мужских и женских особей (фиг.3). Эти результаты нашли подтверждение в недавней работе (New Sci, 1988, vol.120, N 1635, p.38), где показано, что в эмбриогенезе различия в скорости роста эмбрионов мужского и женского пола определяются Y-хромосомой и, по-видимому, фактором детерминации семенников (фиг.4).
Анализ представленных в примерах данных свидетельствует о высокой эффективности заявленных соединений как средств, вызывающих наследуемое и закрепляемое в потомстве направленное мутирование генома клетки одноклеточных и многоклеточных организмов.
Впервые выявленному воздействию заявленных соединений на геном клетки автор дал название экпресс-морфогенез.
Направленное мутирование и наследование новой мутации в большей ступени достигается коопуляционным (половым) путем, что следует из примеров действия солей N-арил-3-оксипиридиния на различные штаммы кишечной палочки, дрожжевых культур, дрозофил и крыс. Полученные экспериментальные результаты подтверждают предположение, что исследуемые заявленные соединения значительно влияют на процессы "редактирования", т.е. появления большей генетической информации в молекулах РНК, чем это закодировано в исходной ДНК. Тем самым обеспечивается по механизму (обратному общепринятой биологической догме) передача распределения генной активности по схеме, т.е. синтезируется цепь ДНК, комплементарной матрице мРНК:
РНК _→ ДНК _→ синтез белка,
от одного поколения клеток к другому в ходе эмбриогенеза (экспресс-морфологическое наследование признака).
Экспресс-морфогенез в противоположность современной генетической инженерии и биотехнологии позволяет при использовании заявленных соединений проводить изменения генома одноклеточных и многоклеточных организмов и, очевидно, позволяет реализовать огромный потенциал, заложенный эволюцией в организме человека и животных. К ним в первую очередь относятся возможность значительного смещения активной фазы человеческой деятельности до 100 лет и более, успешной борьбы с такими болезнями, как СПИД, онкологические, кардиологические, инфекционные болезни, связанные с нарушением функции эндокринной системы, созданием новых выдающихся штаммов одноклеточных и многоклеточных организмов, закрепление полезной мутации в поколении и т.д.
Экспресс-морфогенез свободен от тех недостатков, которые свойственны современной генетической инженерии и биотехнологии. В то же время в генетической инженерии сложилась парадоксальная ситуация, когда медикаменты биотехнологии для нужд медицины, сельского хозяйства и других областей относительно малоэффективны на практике (моноклональные антитела, гемопоэтические факторы роста, интерфероны, белковые гормоны и т.д.). Мировой товарный рынок биотехнологической продукции значительно растет и составит 13,08 млрд долл. в 1991-1995 гг. а в 1995-2000 гг. 20,12 (расчеты фирмы Business Communications, США). Ожидается, что на долю мирового рынка медикаментов придется 14,086 млрд долл. По прогнозам же экономистов исследовательской фирмы SRI International, США мировой объем продаж терапевтических препаратов, диагностических средств, вакцин, пестицидов и лекарственных средств для сельского хозяйства в 2000 г. составит около 50 млрд долл. из общей продажи продуктов новейшей биотехнологии в объеме 66 млрд долл.
Проведенные расчеты развития мирового рынка продуктов современной генетической инженерии и биотехнологии при очень низкой их эффективности и высокой токсичности подтверждают высокую эффективность от внедрения экспресс-морфогенеза в практику. Возможно, экспресс-морфогенез станет лидером Фонда, по которому будет развиваться мировая биотехнологическая индустрия XXI в. (идея соревновательности для японо-американо-европейских стран), которая была предложена фирмой Cetus,США и поддержана японской фирмой Suntory Ltd.

Использование: в биотехнологии, медицине, ветеринарии, сельском хозяйстве, пищевой промышленности. Сущность изобретения: выявлено, что соли N-замещенных 3-оксипиридиниев могут служить средствами, вызывающими наследуемое и закрепляемое в потомстве направленное мутирование генома клетки одноклеточных и многоклеточных организмов. 4 ил., 8 табл.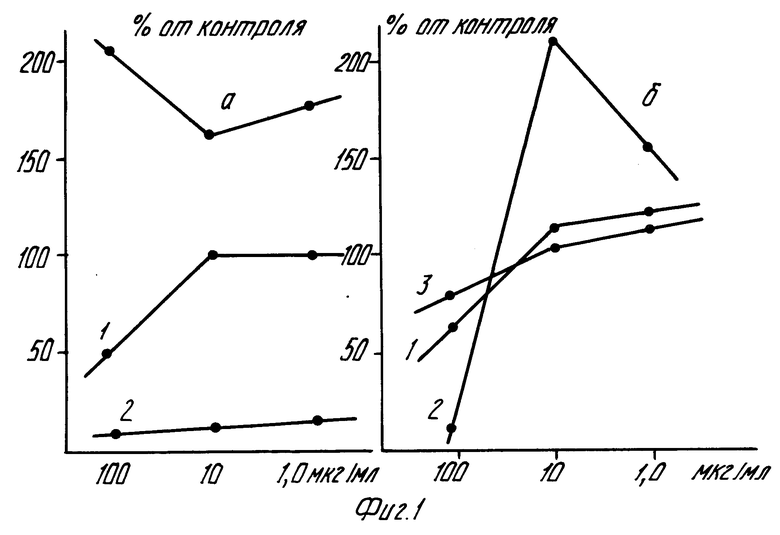
Применение солей N-замещенных 3-оксипиридиниев в качестве средства, вызывающего наследуемое и закрепляемое в потомстве направленное мутирование генома клетки одноклеточных и многоклеточных организмов.
| Th | |||
| Zinoke, G | |||
| Muhlhausen//Berichte der deutschenchemischen gasellschaft | |||
| Дисковая машина для резки капусты и прочих овощей | 1924 |
|
SU1905A1 |
| Дисковая машина для резки капусты и прочих овощей | 1924 |
|
SU1905A1 |
