Изобретение относится к способу получения трансгенных свиней, в организме которых образуются полные антитела заданной специфичности.
Для экспрессии чужеродных ДНК-последовательностей в организме животных можно ввести нужную ДНК в мужской пронуклеус оплодотворенных яйцеклеток путем микроинъекции, затем имплантировать яйцеклетки в яйцевод соответствующих животных и разводить потомков, в организме которых экспрессируется введенный генетический материал. Осуществлено выращивание мышей, несущих генную мутацию, путем ретровирусного инфицирования эмбрионов или переноса генетически измененных исходных клеток в бластоциты (Janisch, R. Science 240, 1468 (1988); Palmiter R. D. und Brinster, Ann. Rev. Genet. 20, 465 (1986); R.L. Brinster, und R. D. Palmiter, Harvey Lectures, Series 80. (Liss, New York, 1986), pp. 1 38).
Описано также и введение чужеродного генетического материала в зародышевые пути различных видов животных. Так, например, кур (Bosselman et al. Science 243, 533 (1989), рыб (Brem, G. Brenig, B. Hoerstgen-Schwark, G.U. Winnacker, E.L.Aquaculture, 68, 209 (1988), кроликов (R.G. Hammer et al. Nature 315, 680 (1985); G.Brem. et al. Zuchthygiene 20, 251 (1985), овец (A.J. Clark et al. Bio/Technology 7, 487 (1989); T. Simons et al. Bio/Technology 6, 179 (1988); C.E. Rexroad, Jr. et al. Mol. Reprod. Dev. 1, 164 (1989) и свиней (G. Brem et al. Zuchthygiene 20, 251 (1985); K.M. Ebert et al. Mol. Endocrinol 2, 277 (1988); Brem G. Brenig, B. Muller, M. Kransslich, H. und Winnacker, E.L. Oce. Publ. Br. Soc.Anim.Prod. 12 (1988) 15-31).
У кроликов активировали c-myc-ген под контролем регуляторных последовательностей иммуноглобулинового гена, в результате чего у трансгенных животных был вызван лейкоз (Knight, K.L. Spicker-Potal, H. Kazdin, D.Oi, V.T. Proc. Nat. Acad. Sci. USA 85 (1988) 3130-3134).
Получены также и трансгенные кролики с активным геном соматотропина человека (Ениколопов и др. Докл. АН СССР, 299 (1988) 1246-1249). Описана и экспрессия "фактора выделения гормона роста человека" у трансгенных кроликов (Катарян и др. Докл. АН СССР, 305 (1989) 726-728).
Показана возможность экспрессии гормона роста крупного рогатого скота у трансгенных свиней и исследован эффект данного гормона на рост трансгенных животных (J.Anim. Sci. 66, (Suppl.1) 267 (1988).
Кроме того, получены трансгенные свиньи, в организме которых экспрессируется поверхностный антиген вируса гепатита "В" человека (Докл. АН СССР, 306 (1989), 206-209.
Гены легких и тяжелых цепей антител с определенной специфичностью (направленной против нитрофенола, тринитрофенола, фосфорилхолина) были выделены из соответствующих гибридомных линий и введены в зародышевый путь мышей. Гены легкой и тяжелой цепей экспрессировались в организме трансгенных животных с образованием рекомбинированных антител (Rusconi S. und Kohler, G. Nature 314, 330 (1985); Grosschedl, R. Weaver, D. Baltimore, D. und Constantini, F. cell 38 (1984), 647; v. Storb et al. J. Exp. Med. 164 (1984), 627; Ritchie, K.A. Brinster, R.L. und Storb U. Nature 312, 517 (1984); Weaver, D. Constantini, T. Imanishikari, T. und Baltimore, D. Cell 42, 117 (1985); Iglesias, A. Lamers, M. und Kohler, G. Nature 330, 482 (1987); Nenberger, M. S. Caskey, H. M. Petersson, M. Williams, T. Surani, M.A. Nature 338, 350 (1989).
Как указано в некоторых из приведенных источников информации, в зародышевый путь мышей вводили гены только для легкой или только для тяжелой цепи антитела. В этих исследованиях обнаружилось, что перестроенный "трансген" подавляет перестройку эндогенных иммуноглобулиновых генов. Была показана лишь небольшая степень экспрессии введенных генов антител (около 5-10 мг на мл сыворотки) в организме трансгенных мышей. У других видов животных не показана экспрессия иммуноглобулиновых генов после введения в зародышевые пути.
Задача изобретения заключается в разработке способа, с помощью которого можно получать большое количество антител из организма трансгенных животных.
Сущность изобретения заключается в получении трансгенных свиней, когда путем микроинъекции вводят одну или несколько заданных последовательностей ДНК, кодирующих нужные антитела, в мужской пронуклеус оплодотворенной яйцеклетки свиньи, имплантируют яйцеклетки в яйцевод свиньи, выращивают потомков и затем обычным образом выделяют антитела. При этом используемые для микроинъекции последовательности ДНК свободны от бактериальных чужеродных последовательностей.
Для экспрессии антител в лимфоидных клетках преимущественно используют иммуноглобулиновый промотор и элементы-усилители (энхансеры). Гены антител субклонируют в прокариотический вектор и размножают в подходящих клетках-хозяевах. Соответствующие гены после расщепления вектора рестрикционными эндонуклеазами освобождают от прокариотических последовательностей и инъецируют в оплодотверенные яйцеклетки. При этом можно применять циклические или линеаризованные молекулы ДНК. После делеции интрон-последовательностей (одной или нескольких) должна осуществиться экспрессия генов антител в организме трансгенных животных. При использовании комплементарной ДНК (кДНК) для экспрессии легких и тяжелых цепей антител можно клонировать иммуноглобулиновый промотор ген и последовательности энхансеров в экспрессионных векторах. Возможно также слияние геномных фрагментов с кДНК (Gillies, S.D. et al. Bio/Technology 7 (1989) 799-804; Orlandi et al. Proc. Nat. Acad. of Sci. USA 86 (1989) 3833-3837).
Другой способ усиления экспрессии генов антител заключается в применении регуляторных элементов, расположенных в некодирующих участках генов антител. Так, для генов легкой k-цепи lg человека недавно был описан расположенный в 3'-нетранслируемой области энхансер (Meyer, K.B. и Nenberger M.S.EMBO. J. (1989) 1959-1965). Для гена λ1 мыши также описан энхансер, расположенный в 3'-нетранслируемой области (Bich Thuy, L. und Qucen, C. Nucl. Acids Res. 17 (1989) 5307-5321).
При осуществлении изобретения полные гены для легкой и тяжелой цепей антитела вводят в оплодотворенные яйцеклетки свиньи. При этом возможна экспрессия антител всех изотипов. Предпочтительно использовать γ- и k- цепи. Антитела могут иметь любое происхождение. Особенно предпочтительна экспрессия антител человека. Кодирующие антитела человека гены могут быть получены известными способами, из соответствующих гибридомных линий. Кроме того, изобретение обеспечивает получение химерных антител, гетеробиспецифических антител, фрагментов антител, слитых антител, а также антитела с измененными свойствами, например обладающих повышенной активностью к связыванию комплемента, усиленными цитолитическими свойствами и т.д.
Благодаря предлагаемому способу можно "иммунизировать" животных. Гены антител против определенных патогенных организмов, в частности вызывающих у свиней опасные заболевания, например инфлюэнцу или чуму, можно ввести в зародышевый путь этих животных. Затем при выращивании потомков можно получить животных, имеющих сопротивляемость данным инфекционным заболеваниям.
Благодаря заявленному способу возможно обеспечение секреции терапевтически значимых белков в молоко.
Гены, кодирующие DlgA, можно слить с генами других белков, например фактора свертывания крови VIII, активатора плазминогена тканевого типа и т.д. Слитые гены можно ввести в зародышевый путь животных и получить потомков, в молоко которых секретируется гибридный белок.
На основании изложенного выше становится ясно, что изобретение относится к способу получения трансгенных свиней, согласно которому в мужской пронуклеус оплодотворенной яйцеклетки свиньи вводят одну или несколько ДНК-последовательностей, кодирующих антитело, свободных от чужеродных последовательностей, имплантируют яйцеклетки в яйцевод самки и выращивают потомков. При этом можно использовать, например, ДНК- последовательности, кодирующие антитела, направленных против возбудителей опасных заболеваний. Подобные заболевания известны специалистам, как и вызывающие их патогены. Таким образом можно получить, например, трансгенное животное, которое обладает иммунитетом против определенного заболевания.
На чертеже представлены гены антитела, применяемые для микроинъекции.
Пример 1. Получение ДНК- последовательностей антитела
Исходными векторами являются плазмиды pBMSI (DSM 5229) и pBMS2 (DSM 5230), описанные в патенте EP-A 0388964 (Способ получения гетеробиспецифических антител). Векторы содержат k-ген и g1-ген антитела мыши в различной ориентации. Данные гены были выделены из клеток гибридомы, которая секретирует lgG1-антитело, обозначенное A20/44 (Sablitzky, F. Wildner, G. und Rajewsky, K: EMBO J.4 (1985) 345-350; Sablitzky, F. Weisbaum, D. und Rajewsky, K. EMBO J.4 (1985) 3435-3437; Kocks, C. und Rajewsky, K. Proc. Nat. Acad. Sci. USA 85 (1988) 8206-8210) A20/44 представляет собой анти-идиотипическое антитело, направленное против специфического для гаптена NP/4-гидрокси-3-нитрофенилацетат/антитела. Последнее антитело имеет легкую λ-цепь и g2a-тяжелую цепь (lgG2a).
Описанные в патенте EP-A 0388964 векторы содержат k-ген в виде 5,5 кв Sal l-фрагмента, γ1-ген в виде 9,25 кв Sal l-фрагмента. Кроме того, векторы содержат гены неофосфотрансферазы и дигидрофолатредуктазы мыши под контролем раннего промотора SV pBMSI и pBMS2 (содержащие по 3 сайта рестрикции для Sal l), частично расщепляют с помощью Sal l, обрабатывают S1 (1 мкг частично расщепленной Sal l, инкубируют с 10 единицами S1-нуклеазы в течение 60 мин при 30oС в 50 ммоль/л ацетата натрия, pH 4,5; 1 ммоль/л ZnSO4 и 0,5 мас. глицерина), выделяют линеаризированную плазмиду из низкоплавкого геля агарозы и концы лигируют Т4-лигазой. Лигированные конструкции трансинфицируют в штамм НВ101 кишечной палочки E.coli и устойчивые к ампициллину (50 мкг/мл) колонии изолируют на агаровых пластинках. Путем рестрикционного эндонуклеазного анализа определяют те плазмиды, у которых вследствие вышеописанной манипуляции делетирован Sal l-сайт, расположенный между k-геном и γ1-геном. Полученные плазмиды обозначают как pBMSI (ΔSal) и pBMS2 (DSal).
PBMSI (DSal) и PBMS2 (DSal) расщепляют с помощью Sal l и фрагмент Sal l, содержащий k-ген и g1-ген антитела, изолируют с помощью низкоплавкого геля агарозы и затем растворяют в 10 мМ TPIS. HCl, pH 7,5, 0,25 ЭДТК.
Таким образом, гены антитела отделяют от последовательностей вектора для последующего эксперимента по микроинъекции (см. чертеж).
Пример 2. Получение трансгенных свиней
Получение трансгенных млекопитающих включает в себя приготовление инъецируемого раствора ДНК, получение оплодотверенных яйцеклеток и эмбрионов, микроинъекцию раствора ДНК в пронуклеусы и ядра, перенос инъецированных зигот синхронизированным животным-реципиентам и исследование родившихся животных на интеграцию. При этом для отдельных видов млекопитающих, например мыши, кролика и свиньи, следует учитывать некоторые обусловленные видом различия при подготовке животных-переносчиков и животных-реципиентов, при получении и переносе эмбрионов, а также при микроинъекции.
1. Приготовление инъецируемого раствора ДНК
После изолирования фрагмента ДНК и определения содержания ДНК раствор ДНК разбавляют трис-буферным раствором так, чтобы в пиколитре раствора содержалось до 1000 копий генного конструктора. Все применяемые для микроинъекции растворы ДНК должны быть тщательно очищены во избежания закупорки инъекционных пипеток.
2. Получение эмбрионов
Для получения эмбрионов свиньи можно использовать препубертатных молодых самок, имеющих вес 60 90 кг. В день 0 животных-доноров переводят в другие станки и вечером вводят 1250 IE PSMG. Через 72 ч после этого генерируют овуляцию с помощью 750 IE HCG. Через 24-36 ч после введения HCG животных осеменяют. Эмбрионы получают через 24-27 ч после оплодотворения. Для хирургического получения эмбрионов животных подвергают наркозу с помощью 160 мг азаперона (стресснила) и 400 мг метомидатгидрохлорида. После подготовки поля операции кожу открывают разрезом около 10 см длиной по срединной линии на высоте последних двух пар сосков и выкладывают матку, яйцевод и яичник. Матку перфорируют тупым инструментом так, что в полость можно ввести и закрепить стеклянную канюлю длиной около 5 см. В яйцевод через воронку фибрии вводят и закрепляют изогнутую глазную канюлю длиной около 8 см, которая соединена с резиновым шлангом длиной около 30 см. Яйцевод прополаскивают 50 мл раствора PBS. Промывочную жидкость сливают в чашку Петри и исследуют на эмбрионы. Эмбрионы можно получать после убоя животных-доноров.
3. Микроинъекция раствора ДНК
Для микроинъекции применяют инверсионный микроскоп, два микроманипулятора и инъекционный аппарат. На одном манипуляторе закрепляют фиксирующую пипетку, с помощью которой эмбрион может фиксироваться под действием пониженного давления. На втором микроманипуляторе закрепляют и соединяют с инъекционным аппаратом наполненную раствором ДНК инъекционную пипетку. Острие инъекционной пипетки имеет диаметр 1 2 мкм. Для микроинъекции острие пипетки вводят через (Zona Pellucida), клеточную мембрану и ядерную мембрану в полость ядра и оставляют здесь около 1 2 пиколитра раствора ДНК. Увеличение объема пронуклеуса свидетельствует об успешной микроинъекции. Микроинъекцию ядер эмбрионов осуществляют и на стадии двух клеток.
В противоположность мыши или кролику у большинства видов сельскохозяйственных животных (свинья, крупный рогатый скот) яйцеклетки и эмбрионы необходимо предварительно обработать, чтобы сделать видимыми пронуклеусы и клеточные ядра (центрифугирование при 15000 g в течение 3 5 мин). Микроинъекцию осуществляют в капле среды на покровном стекле или в так называемой инъекционной камере. После микроинъекции яйцеклетки или эмбрионы культивируют до переноса.
4. Перенос микроинъецированных яйцеклеток
В качестве животных-реципиентов также можно использовать препубертатных молодых свиней. Через 12 ч после свиней-доноров свиньям-реципиентам вводят 750 межд.ед. /IE/ PMSG и еще через 72 ч 750 межд.ед. /IE/ HCG. Эта 12-часовая асинхронность между животными-донорами и животными-реципиентами увеличивает шансы выживания эмбрионов после переноса. Через 135-136 ч после начала программы (час 0 инъекция PMSG животным-донорам) производят перенос эмбрионов. Все инъецированные эмбрионы (35-45 штук) переносят хирургическим путем в яйцевод свиньи-реципиента. Эмбрионы равномерно распределяются на обоих маточных рогах. Предложенный способ позволяет осуществлять бескровный перенос эмбрионов в матку свиньи. Рождение молодых животных происходит через 113-116 дней после переноса.
5. Исследование на интеграцию
Для исследования инъецированных ДНК на интеграцию необходимо получить у родившихся животных пробы ткани (ткань хвоста, кровь или биопсия). Из этих проб ткани изолируют высокомолекулярную геномную ДНК. Для доказательства интеграции проводят анализы.
Положительных животных (животные с интегрированными генетическими конструкциями) выращивают и по достижении племенной зрелости спаривают с нетрансгенными половыми партнерами. Родившихся от этого спаривания потомков исследуют на то, унаследовали ли они трансген от трансгенного родителя. Путем спаривания гемизиготных трансгенных животных выводят затем гомозиготных трансгенных F2-потомков.
Для исследования продукции антител у трансгенных животных после взятия крови получают сыворотку. Пробу крови отбирают из яремной вены (Vena jugularis) или других доступных вен свиньи при убое животных.
Пример 3. Определение титра антител в сыворотке трансгенных свиней.
Метод определения титра антител описан в патенте EP-A-0388964. На пластинки для микротитрования наносят антитела против гаптена NP/4-гидрокси-3-нитро-фенилацетата/. Эти антитела относятся к изотипу lgG 2a (легкая цепь- λ, тяжелая цепь- g 2а). Буферный раствор для нанесения состоит из 0,2 ммоль/л карбоната/бикарбоната, pH 9,4. Через 2 ч пластинки инкубируют с буферным раствором для нанесения следующего слоя (50 ммоль/л HEPES, 0,15 моль/л NaCl, 1% кротеин C, pH 7,0). Все реакции проводят при комнатной температуре со встряхиванием. Градуировочную кривую строят при использовании содержащего антитело A 20/44/lgG1/ основного раствора (ряд разбавлений). Эталонные пробы и сыворотки разбавляют с буферным раствором для инкубирования (50 ммоль/л HERESa 0,15 моль/л NaCl, 1% кротеина C, pH 7,0 0,2 моль/л динатрийтартрата 0,75% полиэтиленгликоля /PEG/, 0,5% плюроника® F 68, 0,75% PEG 40.000, 0,01% фенола) и выдерживают в течение 2 ч при комнатной температуре. После отсасывания из лунок раствора и двукратного промывания их инкубационным буфером в лунки на два часа вносят конъюгат Fab фрагментов антитела, направленного против NP, с пероксидазой (POD). Предварительно конъюгат разбавляют в инкубационном буферном растворе до 150 мЕ/мл POD- активности. После удаления из лунок конъюгата и трехкратного отмывания их специальным буферным раствором (50 ммоль/л HEPESa, 0,15 ммоль/л NaCl, 0,1 плюроника® F 68 pH 7,0) лунки в течение 60 мин инкубируют с ABTS®/2,2'азино-ди-3-этилбензтиазолин-сульфонатом в качестве субстрата. Поглощение определяют с помощью фотометра при 405 нм против 490 нм. Концентрацию антител определяют по стандартной градуировочной кривой. Она составляет 1000 мкг/мл сыворотки.
Пример 4. Характеристика антител, содержащихся в сыворотке свиньи
а) Изоэлектрическое фокусирование и иммунофиксация
Изоэлектрическое фокусирование проводят в геле (Pharmachia) по указаниям изготовителя. Сыворотки трансгенной и контрольной свиньи разбавляют в соотношении 1 1000 и наносят на гель в неденатурирующих условиях. Градиент рН в геле (рН 5 8) визуализируют с помощью стандартного белка (Pharmachia). Время разгонки геля составляет 30 мин. Затем в течение 45 мин его инкубируют со 100 мкг поликлонального антитела (овцы к Fc γ мыши) в объеме 150 мкл и накрывают полосками ацетатцеллюлозы. Нереагирующие белки удаляют отмыванием в буферном растворе, содержащем 50 ммоль/л фосфата калия, 0,15 моль/л NaCl, 0,05% твина®, pH 7,2, при встряхивании в течение ночи. В заключение иммунокомплексы выявляют при окрашивании серебром (Pharmachia). В качестве контроля используют антитело А 20/44, полученное из асцитной жидкости.
В сыворотке трансгенной свиньи идентифицируют те же самые цепи, что и у очищенного антитела 20/44. В сыворотке контрольного животного эти цепи не обнаруживаются.
б) Выделение антитела 20/44 из сыворотки трансгенной свиньи и его очистка
А20/44 является анти- идиотипическим антителом, направленным к антителу (IgG 2a), которое связывается с гаптеном NP. Последнее выделяют из асцитной жидкости и связывают с активированной бромистым цианом сефарозой по предписанию изготовителя. Антитело А20/44 адсорбируется на полученной таким образом колонке после нанесения на нее 100 мл сыворотки трансгенной свиньи, затем его элюируют и идентифицируют. Оно по всем своим свойствам соответствует выделенному из асцитной жидкости антителу А20/44.
Пример 5. Плазмиды рГСНIМ и pkappa CHIM, описанные в европейской патентной заявке 0460674, содержат последовательности ДНК, кодирующие легкие (k) или тяжелые (γ) цепи моноклонального антитела С10/179, направленного против a-цепочки рецептора интерлейкина-2. Фрагменты ДНК, кодирующие цепи антител, выделяют с помощью соответствующих рестрикционных ферментов из плазмид и отделяют от векторных последовательностей при использовании низкоплавкого геля агарозы. Затем фрагменты ДНК, кодирующие цепи антител, отделяют обычным путем от векторных последовательностей в геле агарозы и добавляют к 10 мМ/л Трис-HCl, pH 7,5, 0,25 мМ/л ЕДТА.
Полученные таким образом фрагменты ДНК, так же, как и в примере 2, вводят микроинъекцией в зародышевый путь свиней. Полученные таким образом трансгенные животные вырабатывают антитела, соответствующие антителам С10/179.
Пример 6. Плазмиды pUHW, СД 4ГI (DSM 6155) и pUHW СД4 (DSM 6156), описанные в европейской патентной заявке 0481502, содержат последовательности ДНК, кодирующие легкие (k) или тяжелые (g) цепи моноклонального антитела 15 I/P3/ 14, направленного против антигена СД 4. Фрагменты ДНК этих плазмид, кодирующие цепи антител, выделяют с помощью соответствующих рестрикционных ферментов из плазмид, отделяют полностью от векторных последовательностей с помощью низкоплавкого геля агарозы и после выделения из геля переносят в соответствующий буфер.
Полученные таким образом молекулы ДНК так же, как в примере 2, вводят микроинъекцией в зародышевый путь свиней. Полученные таким образом трансгенные животные вырабатывают антитела, соответствующие антителам 15-I/P3/14.
Пример 7. Плазмиды P 16031 и P 16026, описанные в немецкой патентной заявке P 4142077.2, содержат последовательности ДНК, кодирующие моноклональные антитела А23А41 (DSM ACC 2015), направленные против b-цепочки рецептора интерлейкина-2. Фрагменты ДНК этих плазмид, кодирующие цепи антител, выделяют из плазмид с помощью соответствующих рестрикционных ферментов и полностью отделяют от векторных последовательностей, а затем переносят в соответствующий буфер.
Выделенные таким образом фрагменты ДНК, так же, как и в примере 2, вводят микроинъекцией в зародышевый путь свиней. Полученные таким образом животные вырабатывают антитела, соответствующие моноклональным антителам А23А41.
Таким образом, заявленный способ позволяет получать трансгенных свиней, в организме которых образуются антитела заданой специфичности. Титр антител в сыворотке животных составляет около 1000 мкг/мл.
| название | год | авторы | номер документа |
|---|---|---|---|
| ТРАНСГЕНЕЗ У МЛЕКОПИТАЮЩИХ ПУТЕМ ИНТРАЦИТОПЛАЗМАТИЧЕСКОЙ ИНЪЕКЦИИ СПЕРМЫ | 1999 |
|
RU2267270C2 |
| СПОСОБ ПОЛУЧЕНИЯ ЭМБРИОНОВ ЖИВОТНЫХ И СПОСОБ ВЫРАЩИВАНИЯ ЖИВОТНОГО ИЗ ЭМБРИОНОВ | 1998 |
|
RU2216592C2 |
| ТРАНСГЕН ДЛЯ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО ПОЛИПЕПТИДА В МОЛОКЕ ТРАНСГЕННЫХ КОРОВ, СПОСОБ ПОЛУЧЕНИЯ ТРАНСГЕННОЙ КОРОВЫ (ВАРИАНТЫ), МОЛОКО ОТ ТРАНСГЕННОЙ КОРОВЫ, ПИЩЕВОЙ СОСТАВ | 1990 |
|
RU2095414C1 |
| СПОСОБ ПОЛУЧЕНИЯ ТРАНСГЕННОГО ЖИВОТНОГО, ЭКСПРЕССИРУЮЩЕГО В МОЛОЧНОЙ ЖЕЛЕЗЕ ГРАНУЛОЦИТАРНЫЙ КОЛОНИЕСТИМУЛИРУЮЩИЙ ФАКТОР ЧЕЛОВЕКА И ГИБРИДНЫЙ ГЕН h-GM-1 ДЛЯ ОСУЩЕСТВЛЕНИЯ СПОСОБА | 1999 |
|
RU2157846C1 |
| СПОСОБ ПОЛУЧЕНИЯ ТРАНСГЕННОЙ ОВЦЫ | 1987 |
|
RU2085587C1 |
| РЕКОМБИНАНТНЫЙ ВЕКТОР, ОБЕСПЕЧИВАЮЩИЙ СИНТЕЗ ФИЗИОЛОГИЧЕСКИ АКТИВНОГО ВЕЩЕСТВА (ВАРИАНТЫ), СОДЕРЖАЩИЙ ПРОМОТОРНЫЙ САЙТ ГЕНА β-КАЗЕИНА КОРЕЙСКИХ МЕСТНЫХ КОЗЛОВ, СПОСОБ ПОЛУЧЕНИЯ ФИЗИОЛОГИЧЕСКИ АКТИВНОГО ВЕЩЕСТВА (ВАРИАНТЫ) | 1998 |
|
RU2191826C2 |
| МОЛЕКУЛА ДНК, ВЕКТОР ЭКСПРЕССИИ | 1993 |
|
RU2128708C1 |
| ВАРИАНТЫ СТИМУЛИРУЕМОЙ СОЛЯМИ ЖЕЛЧИ ЛИПАЗЫ, КОДИРУЮЩИЕ ИХ МОЛЕКУЛЫ ДНК И ТРАНСГЕННЫЕ МЛЕКОПИТАЮЩИЕ, НЕ ПРИНАДЛЕЖАЩИЕ К ЧЕЛОВЕКУ | 1994 |
|
RU2219239C2 |
| СПОСОБ ТРАНСФОРМАЦИИ ГЕНОМА ПТИЦЫ | 1988 |
|
RU2022014C1 |
| Способ получения мышиной модели для изучения синдрома Леша-Нихена путем внесения делеции p.Val8del в ген hprt1 | 2021 |
|
RU2768048C1 |
Использование: при получении диагностических и профилактических препаратов на основе антител. Сущность изобретения: в мужской пронуклеус оплодотворенной яйцеклетки свиньи вводят ДНК-последовательности, кодирующие легкие и тяжелые цепи полного антитела, свободные от чужеродных бактериальных последовательностей, имплантируют яйцеклетку в яйцевод самки свиньи и выращивают потомство. 1 ил.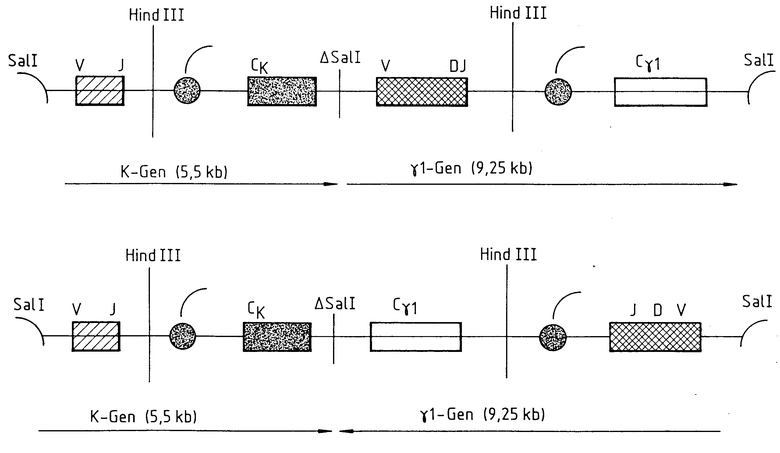
Способ получения трансгенных свиней, в организме которых образуются полные антитела определенной специфичности, заключающийся в том, что в мужской пронуклеус оплодотворенной яйцеклетки свиньи вводят ДНК-последовательности, кодирующие легкие и тяжелые цепи полного антитела, свободные от чужеродных бактериальных последовательностей, имплантируют яйцеклетку в яйцевод самки свиньи и выращивают потомство.

