Изобретение относится к биотехнологии.
Известен метод получения гранулоцитарного колониестимулирующего фактора (Г-КСФ) из таких источников, как кровь, где он в норме присутствует. Этот метод трудоемок и дорог в связи с низкой концентрацией биологически активных веществ и ограниченным количеством исходного сырья (Werner RG, Berthold W; Purification of proteins produced by biotechnological process. Arzneimittelforschung 1988 Mar; 38(3):422-8). Другой опасностью этого метода является возможность присутствия в конечной продукции инфекционных для человека агентов.
Другим источником полезных биологически активных веществ являются получение их с использованием рекомбинантных организмов в биоферментерах (Am J Hosp Pharm 1989 Sep; 46(9): 1834-44). Получение белков таким методом сопряжено с определенными проблемами. Система синтеза прокариот не может правильно провести посттрансляционный процессинг белков - фолдинг, модификации белков, такие как гликозилирование, силиконирование и т.д. Показано, что для функциональной активности Г-КСФ важно гликозилирование. Гликозилированная форма Г-КСФ в 2 раза превосходит по способности стимулировать рост колоний гранулоцитов и в 20 раз быстрее приводит к положительному эффекту по сравнению с существующим коммерческим негликозилированным рекомбинантным препаратом Filgrastim (Nissen с. Glycosilation of recombinant human granulocyte colony stimulating factor for stability and potency. Eur. J. Cancer, 1994, 30A, Suppl 3: S12-4). Используя некоторые процессы, можно для отдельных веществ увеличить выход биологически активного белка, например, инсулин, проурокиназа. Однако для большинства веществ этот способ неприменим. Лучшими результатами отличается наработка белка в культуре клеток млекопитающих. Все необходимые модификации нормально происходят, но концентрация искомого белка низка, а сам процесс культивирования дорогостоящ и требует высокого технологического обеспечения.
Известен метод получения биологически активных веществ с использованием трансгенных млекопитающих животных, таких как кролики, овцы, крупный рогатый скот, с секрецией этих веществ в кровяное русло. Этим достигается практически полная идентичность получаемых веществ нативным. Ограничением метода является значительная стоимость исходного сырья и невысокая концентрация в нем продуцируемых веществ, а также возможность неблагоприятного влияния получаемого белка на организм трансгенного животного (И.Л. Гольдман с соавт. Трансгенные сельскохозяйственные животные: экспрессия чужеродных генов. Биотехнология, 1996, N 9, с. 3-23).
Известен метод получения трансгенных животных, экспрессирующих альфа-1-антитрипсин, фактор IX (Э.Д. Кларк, Р. Лейз, ППЛ Терапьютикс Лтд., "Способ получения трансгенной овцы", патент РФ Ru #2085587 C1, 1986) и гамма-интерферон человека (Лагутин О. В. с сотр, "Гибридный ген BLG-HIFN-G для экспрессии гамма-интерферона человека в молочной железе трансгенного животного", патент РФ N 2084525 C1, 1997) в молочной железе лактирующей самки.
Авторами данного изобретения предложен способ получения трансгенных животных, предусматривающий конструирование генетической конструкции на основе плазмидной ДНК, кодирующей слитный белок hG-CSF под контролем промоторов генов белков молока, обеспечивающих секрецию рекомбинантного белка в молочной железе лактирующих самок, микроинъекцию фрагмента в пронуклеус оплодотворенной яйцеклетки и пересадки яйцеклетки в половые пути псевдобеременной самки. Этот метод обеспечивает высокую идентичность получаемого белка нативному, а также высокую концентрацию вещества в исходном сырье.
Человеческий гранулоцитарный колониестимулирующий фактор (Г-КСФ) вызывает стимуляцию образования нейтрофилов и широко применяется в комбинации с химиотерапией при лечении онкологических больных, больных нейтрофилией и анемией.
В настоящее время за рубежом рекомбинантный Г-КСФ получают из бактериальных клеток в виде препарата Нейпоген и Filgrastim. Использование прокариотов для получения рекомбинантных белков, как известно, не обеспечивает ряд посттрансляционных модификаций, в частности, гликозилирование, фолдинг белка и отщепление лишнего метионина с N-конца белка, что приводит к снижению биологической активности получаемого препарата. К тому же его стоимость очень высокая. Рекомбинантные белки, получаемые из трансгенных эукариотических организмов, например, путем направленной экспрессии в молоко трансгенных животных, в частности кроликов, лишены этих недостатков. Использование трансгенных животных позволяет с минимальными затратами получать достаточное количество препарата в максимально сжатые сроки. Концентрация искомого белка в молоке может достигать 5-15 г/л, что составляет до 50% общего белка молока. С этой целью авторами патента созданы оригинальные генные конструкции Г-КСФ человека на основе имеющейся геномной копии ДНК, обеспечивающие эффективную секрецию Г-КСФ с молоком трансгенных животных. Получены трансгенные кролики на основе методов, разработанных в Научно-производственном биотехнологическом центре по животноводству РАСХН.
Созданный авторами гибридный ген для экспрессии Г-КСФ в молоко трансгенных животных характеризуются следующими признаками:
- обеспечивают экспрессию человеческого белка Г-КСФ в молочной железе трансгенных животных;
- состоят из:
- фрагмента Kpn 1 и Cla 1 размером 0,9 тыс. п.н. 5'-фланкирующей области гена бета-казеина быка, содержащей промотор гена BLG (бета-лактоглобулин быка);
- фрагмента размером 1,5 тыс.п.н., содержащего геномную копию гена Г-КСФ человека;
- фрагмента размером 0,35 тыс.п.н., содержащий 3'-фланкирующую область гена бета-казеина быка;
- участка полилинкера, содержащего уникальные сайты узнавания эндонуклеазами рестрикции EcoR I, Hind III, Sal I, BamH I, Xba I, Not 1. Схема вектора гибридного гена hGM-l приведена на фиг .1. Последовательность гибридного гена приведена на фиг. 3.
Создание гибридного гена Г-КСФ человека и получение трансгенных животных иллюстрируется следующими примерами:
ПРИМЕР 1.
Создание гибридного гена h-GM-1.
Схема конструкции приведена на фиг. 1.
Из вектора pbBLG-3, содержащего геномную копию гена бета-лактоглобулина быка с фланкирующими его 5' и 3' последовательностями, по уникальным сайтам рестриктаз Kpn 1 и Cla 1 выделяют фрагмент 5'-фланкирующей области, содержащей промотор гена BLG и его сигнальный пептид. Из этого же вектора методом полимеразной цепной реакции со специфическими праймерами был выделен 3'-фланкирующий участок гена BLG.
5'-фрагмент был переклонирован в сайт рестриктазы Cla 1 в вектор phGCSF2, содержащий геномную копию гена Г-КСФ человека, перед ATG сайтом гена Г-КСФ. Правильность клонирования и сохранность рамки считывания проверяли методом секвенирования фрагмента конструкции, амплифицированного по специфическим праймерам CCT GCA GAG CTC AGA AGC GTG и AGG CGG CTC TCC CAT CCT GGG. Полученную конструкцию назвали p5LG-GCSF.
3'-фрагмент был переклонирован в сайт рестриктазы Xho 1 в вектор p5LG-GCSF, после стоп кодона кодирующей части гена Г-КСФ. Полученный вектор был проверен на правильность кодирующей последовательности методом секвенирования ключевых участков гибридного гена. Полученный вектор был назван phGM-1.
При подготовке фрагмента к микроинъекции его вырезали из 20 мкг векторной ДНК phGM-1 эндонуклеазами Крп 1 и Not 1. Затем выделяли фрагмент из смеси методом фракционирования в агарозном геле и очищали фенол-хлороформным методом. Окончательную очистку проводили на наборе Genomic DNA Purification Kit (Promega). Окончательно растворяли фрагмент в буфере 0,1хТЕ (1 мМ Tris-CI, 0,1 мМ EDTA, pH 8,0).
ПРИМЕР 2.
Получение трансгенных кроликов
В качестве доноров зигот используют половозрелых самок кроликов породы шиншилла в возрасте 5-7 месяцев. Для вызывания суперовуляции у доноров применяют гонадотропин сыворотки жеребых кобыл (Сергон, Чехия). Каждому донору инъецируют 100 ME сергона, через 48-72 часа самку спаривают с самцом и индуцируют овуляцию внутривенным введением 200 ME хорионического гонадотропина (Московский эндокринный завод). Для извлечения зигот у самок-доноров используют оперативное промывание яйцеводов in vivo (Adams C.E. Egg transfer in the rabbit. Mammalian egg transfer. Веса Ration: CRC Press, 1982. P. 29-48). В качестве анестетиков используют кетамин 5%, ромпун 2%. Наркотизированное животное фиксируют на операционном столе и выбривают шерсть в области брюха. Репродуктивный тракт обнажают через разрез по белой линии живота. В воронку яйцевода вводят полиэтиленовый катетер, в стенке рога матки металлической канюлей со стилетом делают отверстие, затем в просвет матки вводят теплую манипуляционную среду и направляют ее ток через соединение матки с фаллопиевыми трубами в яйцевод.
Поиск оплодотворенных яйцеклеток (зигот) осуществляли на бинокулярной лупе Nikon при увеличении 40х.
Зиготы с визуализированными пронуклеусами инъецировали 1-2 пл раствора ДНК в один пронуклеус (Brem G. et al. Production of transgenic, mice, rabbits and pigs by microinjection into pronuclei// Zuchthyg, 1985, v. 20, P. 251-252; Hammer R.E. et al. Production of transgenic rabbits, sheep and pigs by microinjection. Nature. 1985, V. 315, P. 680-683). Микроинъекции проводят в специальной камере, заполненной культуральной средой (PBS с добавлением 5% фетальной сыворотки). Камера состоит из двух параллельно расположенных силиконизированных покровных стекол, между которыми находится капля манипуляционной среды в виде столбика. Все остальное пространство заполняют минеральным маслом (Sigma, d=0,84 г/мл). Предназначенные для инъецирования зиготы помещают в столбик манипуляционной среды, удерживают на стеклянной пипетке и оценивают пронуклеусы с использованием инвертированного микроскопа Axiovert 35 при увеличении 400х. Раствор ДНК инъецируют с помощью микроинъекционной иглы, вытянутой из капиллярной трубочки с микрофиламентом (тонкостенное тугоплавкое стекло, диаметр 1 мм) на вертикальном пулере (НПО "Биоприбор" РАН).
Микроинъекционные иглы заполняли раствором ДНК, используя действие капиллярных сил, затем устанавливали в манипуляторе. Положение обоих микроинструментов (фиксирующей пипетки и микроинъекционной иглы) регулируют с помощью микроманипуляторов (НПО "Биоприбор" РАН). Микроинъекционную иглу с инъецируемым растворам ДНК соединяют силиконовой трубочкой с микроинъектором. Об успехе микроинъекции судили по увеличению объема пронуклеуса. Инъецированные зиготы инкубируют 30-60 мин, чтобы удалить поврежденные. К дегенерированным относили зиготы с фрагментированной цитоплазмой и с поврежденной цитоплазматической мембраной. Зиготы, имеющие нормальный внешний вид, пересаживают реципиентным крольчихам, у которых половой цикл синхронизирован с циклом доноров. Самок-реципиентов спаривали с вазоэктомированным самцом и вводили хорионический гонадотропин в то же время, когда спаривали донорную самку с полноценным самцом. Возможна подготовка реципиента непосредственно перед трансплантацией эмбрионов. Эмбрионы трансплантировали в яйцеводы самок-реципиентов хирургическим способом. Для пересадки зигот используют стерильные катетеры для пересадки эмбрионов (Биомедикол, Россия). Реципиенту через воронку яйцевода вводили катетер, содержащий манипуляционную среду с зиготами. Каждому реципиенту пересаживали 15-20 зигот, распределяя их поровну между яйцеводами.
После трансплантации эмбрионов, каждая крольчиха-реципиент находится в отдельной клетке на весь срок беременности. В течение 7-10 дней проводится санация операционного шва. На 14-16 дни после трансплантации эмбрионов методом пальпации устанавливают наличие плодов у реципиентов. Беременных реципиентов переводят на рацион кормления для сухральных и лактирующих самок. Окрол ожидают на 29-30 дни после трансплантации эмбрионов. В случае задержки окролов на следующий день после ожидаемой даты с целью стимуляции родов внутривенно инъецируют 3 ME окситоцина. Вели четкий учет числа родившихся крольчат. Через 5-7 дней всех крольчат метили и одновременно брали образцы ткани уха для анализа ДНК на интеграцию трансгена.
Наличие интеграции определяли методом полимеразной цепной реакции (ПЦР) со специфическими праймерами GCA CAG CCT GTA GGT GGC ACA и CCT GCA GAG CTC AGA AGC GTG. Положительные по ПЦР пробы подтверждали методом блот-гибридизации (фиг. 2).
В результате проведенных исследований было получено 2 живых трансгенных кролика (N 7 и N 25), а также 3 мертворожденных.
Список литературы.
И.Л. Гольдман с соавт. Трансгенные сельскохозяйственные животные: экспрессия чужеродных генов. Биотехнология, 1996, N 9, с. 3-23
Hammer R. E. et al. Production of transgenic rabbits, sheep and pigs by microinjection. Nature. 1985, V. 315, P.680-683.
Э.Д. Кларк, P. Лейз, ППЛ Терапьютикс Лтд., "Способ получения трансгенной овцы", патент России Ru #2085587 C1, 1986.
Nissen с. Glycosilation of recombinant human granulocyte colony stimulating factor for stability and potency. Eur. J. Cancer, 1994, 30A, Suppl 3: S12-4.
Adams C.E. Egg transfer in the rabbit. Mammalian egg transfer. Веса Ration: CRC Press, 1982. P.29-48.
Werner RG, Berthold W; Purification of proteins produced by biotechnological process. Arzneimittelforschung 1988 Mar; 38 (3):422-8.
Am J Hosp Pharm 1989 Sep; 46(9):1834-44.
Brem G. et al. Production of transgenic mice, rabbits and pigs by microinjection into pronuclei// Zuchthyg, 1985, v.20, P. 251-252.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ ТРАНСГЕННЫХ КРОЛИКОВ, ПРОДУЦИРУЮЩИХ БЕЛКИ В МОЛОЧНУЮ ЖЕЛЕЗУ | 2007 |
|
RU2402211C2 |
| СПОСОБ ПОЛУЧЕНИЯ ЧЕЛОВЕЧЕСКОГО ГРАНУЛОЦИТАРНОГО КОЛОНИЕСТИМУЛИРУЮЩЕГО ФАКТОРА | 2002 |
|
RU2207373C1 |
| ГЕННО-ИНЖЕНЕРНАЯ КОНСТРУКЦИЯ pGoatcasGCSF, ОБЕСПЕЧИВАЮЩАЯ ПРОДУКЦИЮ ГРАНУЛОЦИТ-КОЛОНИЕСТИМУЛИРУЮЩЕГО ФАКТОРА ЧЕЛОВЕКА В МОЛОКО ТРАНСГЕННЫХ ЖИВОТНЫХ | 2010 |
|
RU2422529C1 |
| ГИБРИДНЫЙ ГЕН BLG-HIFN-G ДЛЯ ЭКСПРЕССИИ ГАММА-ИНТЕРФЕРОНА ЧЕЛОВЕКА В МОЛОЧНОЙ ЖЕЛЕЗЕ ТРАНСГЕННОГО ЖИВОТНОГО (ВАРИАНТЫ) | 1994 |
|
RU2084526C1 |
| Способ получения кролика с нокаутом гена CSN2 | 2023 |
|
RU2834016C1 |
| ТРАНСГЕН ДЛЯ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО ПОЛИПЕПТИДА В МОЛОКЕ ТРАНСГЕННЫХ КОРОВ, СПОСОБ ПОЛУЧЕНИЯ ТРАНСГЕННОЙ КОРОВЫ (ВАРИАНТЫ), МОЛОКО ОТ ТРАНСГЕННОЙ КОРОВЫ, ПИЩЕВОЙ СОСТАВ | 1990 |
|
RU2095414C1 |
| СПОСОБ ПОЛУЧЕНИЯ ТРАНСГЕННЫХ КУР ОПОСРЕДОВАННЫМ ПЕРЕНОСОМ ГЕНА ПУТЕМ ОБЫЧНОГО ИСКУССТВЕННОГО ОСЕМЕНЕНИЯ | 2011 |
|
RU2517731C2 |
| Генетическая конструкция рБТШ70 для экспрессии основного человеческого стресс белка в молоке трансгенных животных | 2015 |
|
RU2644663C2 |
| ОСНОВА МИКРОИНЪЕКЦИОННОГО БУФЕРА ДЛЯ ПОЛУЧЕНИЯ ТРАНСГЕННЫХ ЖИВОТНЫХ | 1999 |
|
RU2160569C1 |
| Способ получения мышиной модели для изучения синдрома Леша-Нихена путем внесения делеции p.Val8del в ген hprt1 | 2021 |
|
RU2768048C1 |



Человеческий гранулоцитарный колониестимулирующий фактор (Г-КСФ) получают путем экспрессии рекомбинантных белков в молочной железе трансгенных животных. Для получения белка Г-КСФ человека используют молоко трансгенных животных. Созданы оригинальные генные конструкции Г-КСФ человека на основе имеющейся геномной копии ДНК и регуляторных участков генов белков молока, обеспечивающие эффективную секрецию Г-КСФ с молоком трансгенных животных. Изобретение может быть использовано в иммунологии. 2 c.п. ф-лы, 3 ил.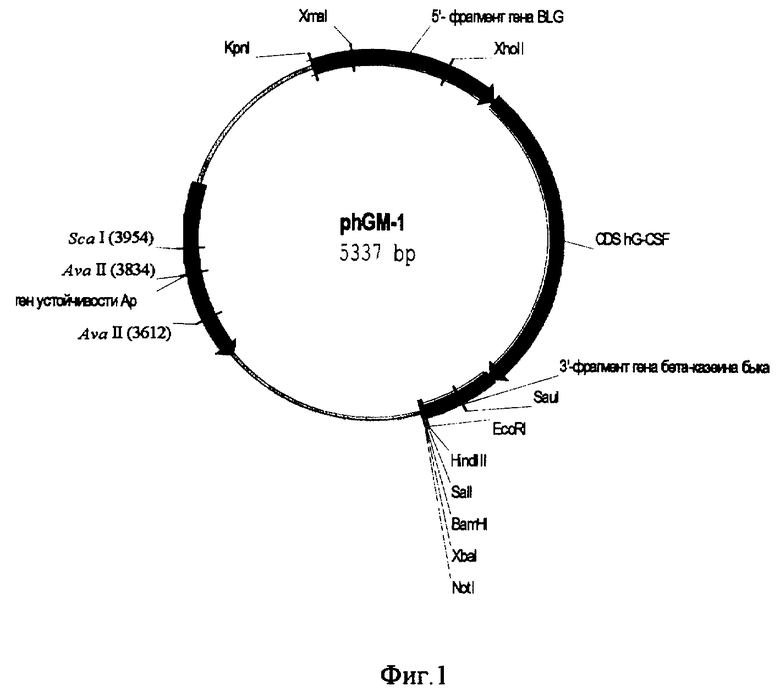
| СПОСОБ ПОЛУЧЕНИЯ ТРАНСГЕННЫХ ЖИВОТНЫХ | 1996 |
|
RU2108714C1 |
