Изобретение разработано на средства National Institutes of Health (Grant N GE000918). Правительство Соединенных Штатов может иметь определенные права на это изобретение.
Изобретение относится к способам и композициям для улучшенного биологического контроля над насекомыми -вредителями. Более конкретно, данное изобретение относится к использованию палочковидного вируса с измененным геном, продукт которого эффективен в отношении регулирования роста и развития насекомых. Данное изобретение относится также собственно к генетически видоизмененным палочковидным вирусам и способам их получения.
Интерес к биологическому способу борьбы с насекомыми-вредителями обусловлен недостатками традиционных химических пестицидов. Химические пестициды, как правило, воздействуют как на вредные, так и на полезные виды. Имеется тенденция, что насекомые -вредители приобретают устойчивость к подобным химикалиям, так что быстро может развиться популяция устойчивых к таким пестицидам насекомых. Далее, остатки химических веществ наносят урон окружающей среде и, потенциально, здоровью людей. Биологический контроль представляет собой альтернативный способ воздействия на насекомых, который позволяет уменьшить зависимость от химических пестицидов.
Основная стратегия биологического контроля включает использование встречающихся в природе организмов, патогенных по отношению к насекомым (энтомопатогены), и разведение более устойчивых по отношению к насекомым -вредителям культур. Подход к решению этих задач включает идентификацию и характеристику генов насекомых или продуктов генов, которые могут служить подходящей основой для получения агентов контроля, идентификацию и эксплуатацию ранее не использовавшихся микроорганизмов (сюда относится также модифицирование существующих в природе непатогенных микроорганизмов с целью превращения их в патогенные по отношению к насекомым), модифицирование и улучшение используемых в настоящее время энтомопатогенов, а также создание методами генной инженерии культур, обладающих большей устойчивостью к насекомым-вредителям.
К энтомопатогенам, предложенным в качестве биологических пестицидов, относятся вирусы, вызывающие естественные эпизоотии в популяциях насекомых (Heimpel A.M. Japan seminar on microbial control, 1968, p. 51-62, Биологические средства защиты растений, М. 1974, 335-345), в том числе Бакуловирусы. Палочковидные вирусы (бакуловирусы) представляют большую группу вирусов, инфицирующих только членистоногих насекомых (Miller L.K. (1981), в Genetic Engineering in the Plant Sciences, N. Panopoulous, (ed.) Praeger Publ. New York, pp. 203-224; Carstens. (1980) Trends in Biochemical Science 52 107-110; Harrap Payne (1979) в Advances in Virus Research. v. 25, Lawfer et al. (eds. ) Academic Press N.Y. p. 273-355). Многие бакуловирусы инфицируют насекомых, являющихся вредителями промышленно важных сельскохозяйственных и лесных культур. Такие бакуловирусы потенциально представляют собой ценные агенты биологического контроля. Агентством США по охране окружающей среды (U. S. Environmental Protection Agency) рекомендовано к использованию в качестве инсектицидов четыре палочковидных вируса. Одним из преимуществ бакуловирусов как биологических пестицидов является их специфичность по отношению к насекомому-хозяину. Бакуловирусы как группа инфицируют только членистоногих, а каждый индивидуальный штамм палочковидного вируса обычно инфицирует только один или несколько видов насекомых. Таким образом, они не представляют угрозы ни для окружающей среды, ни для человека и могут быть использованы без вреда для полезных видов насекомых.
Группа бакуловирусов включает подгруппы вирусов ядерного полиэдроза (ЯПВ), вирусов ядерного гранулеза (ГВ) и палочковидных вирусов, не образующих тела включения. У окклюдирующих форм палочковидных вирусов вирионы (заключенные в оболочку нуклеокапсиды) помещаются внутри кристаллической белковой матрицы. Такая структура, так называемое тело включения или тело "вкрапления" это форма нахождения вируса в природе вне организма, которая обуславливает передачу инфекции от организма к организму. Отличительной чертой вирусов ЯП является то, что в каждом теле включения содержаться много вирионов. Тела включения ЯП вирусов обладают относительно большими размерами (до 5 мкм). Тела включения ГВ мельче и содержат по одному вириону. У обеих форм кристаллическая белковая матрица тел включения состоит главным образом из одного полипептида от 25000 до 33000 дальтон, называемого полиэдрином или соответственно, гранулином. Палочковидные вирусы неокклюдирующих форм не производят белок полиэдрин или гранулин и не образуют тел включения.
В природе инфекция инициируется, когда насекомое глотает загрязненную частицами палочковидных вирусов пищу, как правило, в виде тел включения. В щелочной среде среднего кишечника насекомого тела включения диссоциируют, высвобождая обособленные вирусные частицы, которые затем проникают в выстилающие стенки кишечника эпителиальные клетки. Внутри клетки-хозяина палочковидный вирус мигрирует к ядру, где производит процесс репликации. Первоначально в инфицированной клетке синтезируются некоторые специфичные вирусные белки путем транскрипции и трансляции так называемых "ранних генов". Эти белки необходимы, кроме других функций, для репликации вирусной ДНК, которая начинается 4-6 ч спустя после проникновения вируса в клетку. Интенсивная репликация вирусной ДНК продолжается примерно в течение 12 ч после заражения (post-infection pi). Примерно через 8-10 ч pi инфицированная клетка производит большие количества так называемых "поздних продуктов вирусных генов". К ним относятся компоненты нуклеокапсида, окружающего вирусную ДНК в процессе образования дочерних вирусных частиц. Дочерние вирусные частицы начинают образовываться примерно через 12 ч pi. Первоначально дочерние вирусы мигрируют к клеточной мембране, где, после выхода на поверхность клетки, они приобретают оболочку. Подобный неокклюдированный вирус может затем инфицировать другие клетки насекомого -хозяина. Синтез полиэдрина начинается через 12-18 ч после заражения и к 24 ч pi он быстро нарастает до очень высокого уровня. В это время наблюдается снижение количества вышедших из клетки вирусных частиц, и затем дочерний вирус образует тела включения. Образование тел включения продолжается до гибели или распада клетки. Некоторые палочковидные вирусы инфицируют все ткани насекомого -хозяина, так что по завершении процесса развития инфекции все насекомое превращается жидкость, при этом выделяется большое количество тел включения, которые далее распространяют инфекцию на другие особи. (Обзор приводится в The Biology of Baculoviruses, v.I and II Granados and Federici (eds), CRC Press, Boca Ration, Florida, 1986).
Одним из существенных недостатков применения палочковидных вирусов в качестве пестицидов является продолжительный период времени между моментом, когда насекомое заглатывает вирус, и его смертью. В течение этого времени вредитель продолжает питаться и вредит урожаю. В связи с этим определяющим представляется уменьшение времени питания насекомого до минимума, т.е. создание пестицида, снижающего питание насекомого до наступления гибели. Желательно также, чтобы смерть насекомого наступала быстрее. Такие результаты могут быть получены путем воздействия на вирусный геном, осуществить которое в частности позволяют методы генной инженерии (EP 0225777A1, C 12 N 15/00).
Данное изобретение выявляет ген палочковидного вируса и продукт этого гена, влияющий на рост, развитие или поведение насекомых. Оно обеспечивает способы использования этого гена и продукта этого гена, а также способы инактивации этого гена или продукта этого гена в целях контроля над насекомыми. В предпочтительном варианте воплощения используют ген egt, который кодирует экдистероид UDP глюкозил-трансферазу (EGT) вируса AcMNPV. Экспрессия гена egt палочковидных вирусов вызывает образование EGT, которая инактивирует гормоны, обслуживающие линьку насекомых, предотвращая этим линьку или окукливание личинки. Инактивирование гена egt палочковидных вирусов обеспечивает нормальное протекание линьки и окукливания личинки.
Предлагаемое изобретение включает целый ряд агентов контроля над насекомыми с использованием генов egt и продуктов функционирования этих генов. Эти агенты контроля либо вызывают у насекомого-вредителя несоответствующий синтез белка EGT, либо подавляют нормальное функционирование или проявление белка EGT, так что нормальное развитие насекомого нарушается.
Предпочтительно, чтобы организм, содержащий ген egt, представлял собой специфичный для данного насекомого вирус и чтобы ген egt вируса был инактивирован, что обеспечило бы увеличение продолжительности развития инфицированного этим вирусом насекомого -хозяина. Длительное развитие насекомого -хозяина связано с изменением его поведения, в частности с понижением питания, и следующим за этим снижением роста насекомого и более быстрым наступлением смерти. Данное изобретение включает также способ производства улучшенного вирусного пестицида. Кроме того, оно включает способ контроля над насекомыми-вредителями, который заключается в том, что насекомое -вредитель подвергается воздействию улучшенного вирусного пестицида.
Предлагаемые данным изобретением генетически измененные вирусы являются более эффективными пестицидами по сравнению с использовавшимися до сих пор вирусами. У палочковидных вирусов, таких, например, как вирус ядерного полиэдроза Autographa californica (AcMNPV) или вирус ядерного полиэдроза Orgyia pseudotsugata (OpMNPV), естественно проявляется ген egt. Экспрессия его увеличивает период времени, в продолжение которого инфицированная личинка питается, не страдая от вирусной инфекции. Данное изобретение включает инактивирование гена egt в геноме вируса. Ген egt можно инактивировать введением на его место или внутрь него другого гена, например, гена-маркера, кодирующего синтез β -галактозидазы. Должно быть ясно, что для нарушения гена egt может использоваться любая последовательность ДНК, поскольку она нарушает проявление кодирующей последовательности egt. Можно, напротив, удалить из генома все части гена egt с помощью делеции соответствующего кодирующего сегмента ДНК или удалить либо изменить регуляторные элементы генома, контролирующие экспрессию гена egt. Палочковидные вирусы с делециями, инактивирующими ген egt, могут быть также получены последовательным пропусканием вируса через культуру клеток насекомого. Полученные при этом вирусы с делециями имеют следующее преимущество: они не содержат чужеродной ДНК и отличаются от вирусов дикого типа только отсутствием функционального гена egt. Подобные модифицированные бакуловирусы эффективнее в качестве агентов контроля над вредителями, чем те, которые применяются в настоящее время. Как простые делеционные мутанты делеции, так и пестициды, полученные путем генной инженерии, они должны быть приемлемы для регулирующих ведомств (например, U.S. EPA), поскольку в них не содержится негомологичная ДНК. Палочковидные вирусы, в которых не проявляется функциональный ген egt, дополнительно могут быть видоизменены за счет введения отличных от egt генов, которые могут воздействовать на развитие насекомого, повышая этим эффективность таких вирусов как агентов контроля насекомых.
Данное изобретение включает также способ контролирования развития насекомых-вредителей путем инфицирования личинок насекомых мутантным вирусом, в котором интактный ген egt отсутствует или не способен производить функциональный продукт EGT. Личинки, инфицированные мутантным вирусом, стремятся линять и окукливаться, и, следовательно, период их питания сокращается по сравнению с личинками, инфицированными диким типом или другими известными в настоящее время вирусами. Зараженные мутантным вирусом личинки также умирают быстрее, чем инфицированные диким типом или другими применяемыми в настоящее время вирусами.
Целью данного изобретения является создание рекомбинантного вируса, который является более эффективным по сравнению с вирусами дикого типа и другими формами, применяемыми в настоящее время. Примером данного изобретения является рекомбинантный вирус AcMNPV, обозначаемый как VEGTZ, в котором часть гена egt заменена на lacZ бактериальный ген, кодирующий образование b
галактозидазы. Как понятно специалистам в данной области, может быть заменена любая последовательность ДНК, инактивирующая ген egt. Примером реализации данного изобретения является также рекомбинантный палочковидный вирус, обозначаемый vEGTDEL в котором часть гена egt удалена. Примером осуществления данного изобретения является пестицид на основе палочковидного вируса с удаленным геном egt. Сюда относятся также палочковидные вирусы, у которых к инактивации гена egt привели мутации, произошедшие естественным путем.
Дополнительной целью данного изобретения является создание с помощью генной инженерии вируса, который не только представляет собой эффективный пестицид, но одновременно и безвреден для окружающей среды.
Еще один предмет изобретения рекомбинантный вирусный пестицид, в котором отсутствует функциональный ген egt и проявляется второй ген, влияющий на развитие насекомых, причем в естественном виде этот второй ген не присутствует в организме. Указанный второй ген кодирует синтез влияющего на метаморфоз продукта. Таким продуктом гена может быть гормон насекомых, влияющий на развитие насекомого, или фермент, который инактивирует регулирующий метаморфоз гормон. Особые примеры включают проторакотропный гормон, гормон вылупления и эстеразу ювенильного гормона. Если требуется с целью получения агента контроля ввести в вирус насекомых кодирующие эти белки гены, то этот вирус любо не должен содержать гена egt, либо ген egt в нем должен быть инактивирован.
Далее целью данного изобретения является создание инсектицидных композиций, пригодных для использования в сельском хозяйстве. В состав таких композиций входят, как известно, приемлемый в сельскохозяйственном смысле носитель и вирус насекомых, например, палочковидный вирус, предварительно модифицированный генетически таким образом, что ген egt указанного вируса инактивирован. Такие egt-бакуловирусы могут далее быть подвергнуты генетическим изменениям, вызывающим экспрессию гетерологичного гена, который кодирует гормон насекомых, влияющий на метаморфоз, или фермент, инактивирующий такой оказывающий влияние на метаморфоз гормон насекомых. К гетерологичным генам, продукты которых влияют на метаморфоз, относятся, но не исчерпывают их, проторакотропный гормон, гормон вылупления и эстераза ювенильного гормона. В предпочтительном исполнении инсектицидные композиции, включающие генетически измененные палочковидные вирусы, предназначены для распыления.
Любая из вышеупомянутых инсектицидных композиций может также включать ингредиенты, стимулирующие питание насекомых. Инсектицидные композиции по данному изобретению насекомое-вредитель может глотать при нанесении его на растения, и насекомые -вредители, чувствительные к активному агенту данной композиции, будут демонстрировать пониженное питание и погибнут.
Также целью данного изобретения является обеспечение модифицированного биологического пестицида, который ингибирует проявление гена egt или функционирование продукта этого гена.
На фиг. 1 схематически представлен геном AcMNPV с указанием локализации гена egt. Геном AcMNPV представлен в единицах карты и в виде карты рестрикции EcoR1 и Hind III.
На фиг. 2 схематически представлены результаты исследования последовательности гена egt и анализа этой последовательности. На фиг. 1A изображена рестрикционная карта данной области генома. На фиг. 1B показаны результаты компьютерного анализа последовательности на наличие открытых рамок считывания. Вертикальными линиями показаны терминирующие кодоны, являющиеся сигналами остановки синтеза полипептидной цепи. Последовательность "транслируется" во всех трех потенциальных открытых рамках считывания в каждой нити ДНК (1, 2, 3, 1', 2', 3'). Соответствующая egt открытая рамка считывания (2) обозначена как EGT.
На фиг. 3 представлены схематически структуры областей гена egt вируса AcMNPV (A) и рекомбинантных вирусов vEGTZ (B) и vEGTDEL (C). Заштрихованный прямоугольник представляет собой ген lacZ.
На фиг. 4 схематически представлены электрофорез в геле агарозы и анализ пятен по Сузерну, проводимые для идентификации гена egt палочковидного вируса OpMNPV.
На фиг. 5 схематически представлена карта рестрикции вирусного генома OpMNPV.
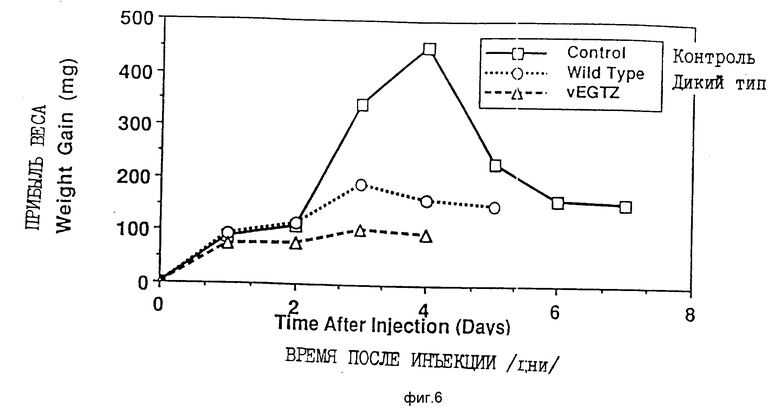
На фиг. 6 представлен график прибыли веса контрольных неинфицированных личинок или личинок четвертого возраста после заражения вирусами AcMNPV дикого типа или vEGTZ.
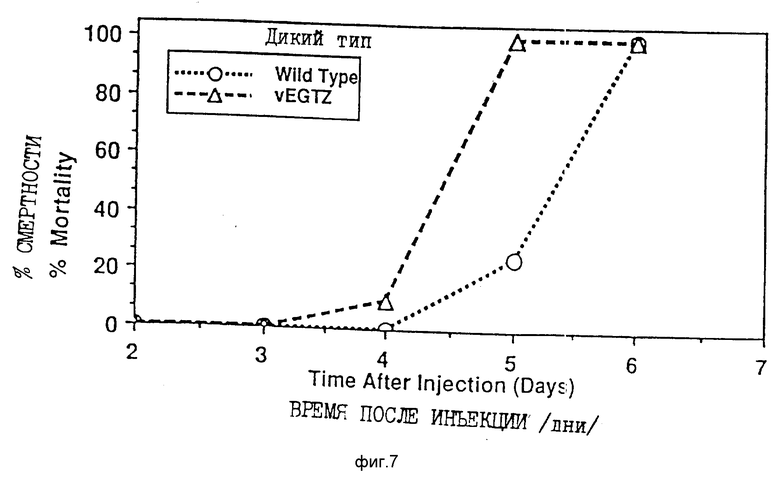
На фиг. 7 приведен график смертности личинок после заражения личинок 4-го возраста вирусами AcMNPV дикого типа или vEGTZ.
На фиг. 8 представлен график прибыли веса после инфицирования личинок 5-го возраста вирусами AcMNPV дикого типа или vEGTZ.
На фиг. 9 приведен график смертности личинок после инфицирования личинок 5-го возраста вирусами дикого типа AcMNPV или vEGTZ.
На фиг. 10 приведен график смертности личинок после инфицирования личинок первого возраста вирусами дикого типа AcMNPV или vEGTZ с концентрацией полиэдрических тел включения (ПТВ) 4,8 х 10. ПТВ/мл.
На фиг. 11 приведен график смертности личинок после инфицирования личинок первого возраста вирусами дикого типа AcMNPV или vEGTZ с концентрацией 2,4 х 10 ПТВ/мл.
Детальное описание вариантов исполнения изобретения.
Чешуекрылые насекомые в процессе развития от яйца до взрослой особи претерпевают хорошо охарактеризованную последовательность превращений (детальное описание см. Comprehensive Insect Physiology, Biochemistry and Pharmacology, v. 7, 8, Kerkut and Gilbert (eds), Pergamon Press, Oxford, 1984). После выхода из яйца у личинки насекомого начинается период интенсивного питания, в течение которого она будет несколько раз линять, что обеспечивает ей непрерывный рост. Промежутки времени между последовательными линьками называются возрастными (личиночными) стадиями. По завершении периода роста личинка окукливается и в конце концов превращается во взрослое насекомое. Процессы линьки и окукливания (называемые собирательно метаморфозом) регулируются совместным действием гормонов нескольких различных групп. Первоначальным стимулом является выделение определенными клетками мозга проторакотропного гормона (PTTH). Это стимулирует секрецию проторакальными железами экдистероидов, которые часто называют гормонами линьки насекомых. Присутствие ювенильного гормона обеспечивает линьку личинки, тогда как в его отсутствии личинка будет окукливаться. Гормон вылупления также представляется важным для осуществления некоторых связанных с метаморфозом поведенческих изменений.
Палочковидный вирус AcMNPV, используемый в качестве модельной системы для многих исследований палочковидных вирусов, совершенно неожиданным образом вмешивается в описанный выше процесс развития насекомого. Инфицированные AcMNPV личинки насекомых не способны более линять или окукливаться в связи с тем, что AcMNPV направляет синтез фермента, известного как экдистероид UDP - глюкозилтрансфераза (EGT), который избирательно инактивирует экдистероиды насекомых.
Авторами данного изобретения идентифицирован кодирующий EGT ген, он простирается от 8,4 до 9,6 единиц карты генома AcMNPV (фиг. 1 и 2). Как показано на фиг. 1C, содержащие ген egt фрагменты вирусной ДНК были клонированы в плазмиды рис. 19, Bluescript M13+ и Bluescript M13-.
На фиг. 2 показаны карты рестрикции области egt генома и компьютерный анализ на наличие в данной области открытых рамок считывания. Только рамка 2 содержит относительно длинную открытую последовательность считывания, которая была идентифицирована как область кодирования гена egt.
На фиг. 12 и 13 приведена последовательность нуклеотидов гена egt и выведенная из нее последовательность из 506 аминокислот. Кодирующая последовательность гена egt простирается примерно от нуклеотида 149 до нуклеотида 1670.
В предпочтительном воплощении данного изобретения ген egt бакуловируса AcMNPV инактивируется путем замещения его части на бактериальную последовательность, кодирующую синтез b -галактозидазы. Такой рекомбинантный бакуловирус был обозначен vEGTZ. Во втором варианте предпочтительного воплощения, как показано на фиг. 3, часть гена egt вируса AcMNPV удаляется без замены. Сравнение белков, синтезируемых при инфицировании AcMNPV дикого типа и vEGTZ выявило, что белок EGT представляет собой пептидную цепь с молекулярной массой 60 КДа, выделяемую инфицированными клетками. Альтернативным механизмом инактивации гена egt вируса насекомых является внедрение гена, который кодирует синтез влияющего на метаморфоз гормона или инактивирующего такой гормон фермента, и который проявляется в инфицированной указанным вирусом насекомых клетке.
Обследования Базы данных Генобанка выявило, что от 21 до 22% последовательности аминокислот egt гомологично UDP -глюкозилтрансферазам млекопитающих. Была выявлена также гомология с UDP-глюкозилтрансферазой растений. Сопоставление последовательности аминокислот egt и некоторых этих ферментов показано на фиг. 14.
Фиг. 14 иллюстрирует сопоставление последовательности аминокислот egt и ряда UDP -глюкозилтрансфераз других биологических видов. Предсказанная последовательность аминокислот egt сравнивается с человеческой (HUMUDPGAT) (Jackson et al. (1987) Biochem. J. 242:581; мышиной (MUSUDPGAT) Kimura and Owens (1987) Eur. J. Biochem. 168:515) и крысиной (RATUDPGAT) (Mackenzie (1987) J. Bicl Chem. 262:9744) UDP -глюкозилтрансферазой кукурузы (ZMAYUDPGT) (Ralston et al. (1988) Genetics 119:185) с помощью алгоритма FASTER (Zipman and Pearson (1985), предлагаемого International Biotechnologies Inc. Буквы верхнего регистра указывают точное совпадение: буквы нижнего регистра определяют замещения, часто встречающиеся среди родственных белков (Dayhoff (1978) Atlas of Protein Sequence and Structure, National Biomedical Research Foundation, v. 5, Supplement 3, Silver Spring, MD); точки обозначают редко встречающиеся замещения, знак дефиса указывает пропуски в последовательности, знак вставки приводится в местах делеции аминокислот из последовательности. В vEGTZ и vEGTDEL аминокислоты гена egt в промежутке между стрелками удалены. Гомологичность гена AcMNPV известным трасферазе UDP-клюкозы и UDP-глюкоронозилтрасферазе подтверждает идентификацию этой последовательности AcMNPV как кодирующей последовательности гена egt.
У млекопитающих UDP -глюкоронозилтрансферазы катализируют перенос глюкуроновой кислоты к целому ряду как экзогенных так и эндогенных липофильных субстратов (смотри обзор в Glucuronidation of Drugs and Other Compounds, Dutton (ed. ) CRC Press, Boca Raton, Florida, 1986). Эта реакция присоединения играет существенную роль в детоксикации и безопасном выведении определенных лекарственных препаратов и канцерогенов. Кроме того, через соединения с глюкуроновой кислотой протекают процессы нормального метаболизма и размещения различных эндогенных соединений, таких, как билирубин и стероидные гормоны. Имеющиеся по системам насекомых данные указывают, что подобные реакции присоединения сахаров включают перенос глюкозы, а не глюкороновой кислоты (обзор смотри Smith (1977) в Drug Metabolism From Microbe to Man, Parke and Smith (eds), Taylor and Francis Ltd, London, p. 219-232). Как и у млекопитающих, у насекомых в подобные реакции присоединения вступают разнообразные экзогенные и эндогенные соединения.
Авторы данного изобретения показали, что EGT-белок вируса AcMNPV представляет собой UDP -глюкоронозилтрансферазу, которая избирательно соединяет глюкозу с экдистероидами, такими как экдизон, 20-гидроксиэкдизон и макистерон A (табл.1). Ни лизаты, ни внеклеточная среда неинфицированных клеток или инфицированных vEGTZ клеток не модифицируют экдизон. Большая часть активной экдистероид -глюкозидтрансферазы, вырабатываемой инфицированными AcMNPV клетками, выделяется в межклеточную среду; только относительно низкий уровень ее активности наблюдается в лизатах инфицированных AcMNPV клеток.
С использованием гена egt вируса AcMNPV в качестве метки ген egt был идентифицирован, как показано на фиг. 4, и в другом палочковидном вирусе, а именно, в вирусе ядерного полиэдроза Orgyia pseudotsugata (OpMNPV). Для специалистов должны быть понятны преимущества, предоставляемые этим открытием, то есть что ген egt любого палочковидного вируса, другого вируса насекомых или самого насекомого может быть локализован, охарактеризован и выделен аналогичным способом. Гены egt, нуклеотидная последовательность которых не менее чем на 70% гомологична последовательности приведенной на фиг.13, считаются эквивалентными этой последовательности, при условии, что гомологичные гены кодируют фермент, представляющий собой экдистероид-UDP -глюкоронозилтрансферазу.
Функциональными эквивалентами гена egt являются эквиваленты, которые также катализируют инактивацию экдистероидов, таких как экдизон, путем переноса половины глюкозы от UDP-глюкозы к экдистероиду (экдистероидам). Такие функциональные egt-эквиваленты могут быть идентифицированы описанными здесь методами.
Путем локализации, идентификации и выделения гена egt специалист может, используя положения данного открытия и известные методики, инактивировать этот ген и получить агент, обеспечивающий более эффективный контроль над развитием и ростом насекомого.
Сравнивая свойства vEGTZ со свойствами AcMNPV дикого типа (wt), авторы данного изобретения показали, что экспрессия гена egt предотвращает линьку или окукливание насекомого. Инфицированные egt AcMNPV насекомые не линяют и не окукливаются, тогда как насекомые, инфицированные vEGTZ, линяют и стремятся окукливаться (табл.2).
За счет подавления линьки и окукливания инфицирование вирусом AcMNPV дикого типа действительно может удлинить период питания личинки. Личинки, зараженные вирусом дикого типа в начале пятой возрастной стадии (последняя возрастная стадия), продолжают питаться до момента гибели, который наступает на 5 или 6 день после заражения. Однако неинфицированные личинки прекращают питаться, готовясь к окукливанию, через 2-3 дня после вступления в пятую возрастную стадию (фиг. 8). Аналогичный эффект наблюдается при обследовании личинок более ранних возрастных стадий. Во время линьки неинфицированные личинки прекращают питаться на период около 24 ч, тогда как зараженные wt AcMNPV личинки не линяют и, соответственно, продолжают питаться (фиг. 6).
Рекомбинантные палочковидные вирусы с отсутствующим функциональным геном egt не увеличивают продолжительность периода питания личинки. Таким образом, личинки, инфицированные vEGTZ в начальный период пятой возрастной стадии, прекращают питаться через два дня после заражения и готовятся к окукливанию (фиг. 8). Однако они не окукливаются и вместо этого погибают от вирусной инфекции даже быстрее, как показано на фиг. 9, чем личинки, инфицированные вирусом дикого типа. Аналогичным образом, личинки, инфицированные вирусом в ранней четвертой возрастной стадии, прекращают питаться спустя два дня после инфицирования и умирают затем быстрее, чем зараженные вирусом дикого типа (фиг. 6 и 7). То, что палочковидный вирус отсутствующим функциональным геном egt убивает насекомых быстрее, наиболее ярко видно, когда вирусом vEGTZ инфицируются только что вышедшие из яиц личинки первой возрастной стадии (фиг. 10 и 11). Зараженные vEGTZ личинки умирают от вирусной инфекции на 3-4 дня раньше по сравнению с теми, которые были заражены wt AcMNPV. Следовательно, рекомбинантные палочковидные вирусы с отсутствующим функциональным геном egt значительно более эффективны как агенты контроля развития насекомых, чем палочковидные вирусы дикого типа. Для специалистов в данной области должны быть очевидны преимущества данного открытия, заключающегося в том, что ген egt может быть сделан нефункциональным в любом палочковидном вирусе или вирусе насекомых любым известным способом.
Эффект, описанный выше и в последующих примерах, на полях будет еще более сильным. Инфицированные vEGTZ личинки испытывают затруднения при линьке, которая все-таки происходит только в тщательно контролируемых лабораторных условиях. В условиях отсутствия жесткого контроля температуры и освещенности многие личинки не могут завершить линьку. Такие насекомые не начинают снова питаться и вскоре после этого погибают.
Несмотря на то, что несколько сокращается продолжительность времени, в течение которого потомство вируса может накапливаться в личинке, зараженной палочковидным вирусом, у которого отсутствует функциональный ген egt, и питание инфицированного насекомого снижается, наблюдается значительное продуцирование дочернего вируса. Количество вируса в расчете на одну личинку, полученное после заражения vEGTZ личинок поздних возрастных стадий, составляет примерно половину от полученного при заражении вирусом дикого типа. Этого достаточно для обеспечения переноса вируса в поле и эффективного в смысле стоимости получения больших количеств вирусных частиц.
В другом варианте воплощения данного изобретения вирус насекомых с отсутствующим функциональным геном egt модифицируют с помощью генной инженерии таким образом, что его эффективность как агента биологического контроля повышается за счет введения второго гена, продукт которого оказывает влияние на развитие насекомого.
Ген, кодирующий PTTH (пептидный гормон) может быть введен в геном вируса с отсутствующим геном egt и экспрессия PTTH при этом будет достаточно высока, чтобы оказывать влияние на процессы линьки. У инфицированных таким вирусом личинок в высшей степени нарушен гормональный контроль развития. Такие насекомые быстро заболевают, что влечет за собой ущербное развитие и рост, снижение питания и ускорение смерти.
Гормон вылупления также представляет собой мелкий пептидный гормон, ген которого может быть введен с помощью известных методов в вирусный геном с нефункциональным геном egt. В связи с тем, что гормоном вылупления определяются многие связанные с метаморфозом поведенческие изменения, насекомое, зараженное вирусом Egt-, продуцирующим этот гормон в достаточно больших количествах, будет демонстрировать аномальное поведение и/или развитие, например, пониженное питание.
Первичным регулятором активности ювенильного гормона в насекомом является фермент эстераза, инактивирующая ювенильный гормон. Рекомбинантный вирус, лишенный функционального egt и продуцирующий достаточно высокий уровень эстеразы ювенильного гормона, может оказывать неблагоприятное воздействие на поведение и/или развитие насекомого.
Важно отметить, что, хотя все вышеупомянутые гены могут быть введены в геном вирусов дикого типа, нельзя ожидать, что в этом случае они в значительной степени будут влиять на поведение насекомого, поскольку экспрессия гена egt вирусами дикого типа инактивирует экдистероидные гормоны линьки и это препятствует, несмотря на продуцирование других гормонов, нормальному протеканию метаморфоза. Таким образом, успешная стратегия должна включать, как описано в данном изобретении, генерирование вирусов, предназначенных для изменения протекания метаморфоза в зависимости от предшествующего инактивирования гена egt.
В данном изобретении агент контроля над ростом и развитием насекомых представляет собой композицию или активный ингредиент композиции, которая оказывает неблагоприятное воздействие на насекомое-вредитель. Реакцией на агент контроля насекомых является снижение питания, нарушается нормальный метаморфоз, в результате чего смерть насекомого становится неизбежной. Активным ингредиентом композиции по данному изобретению является вирус насекомых, в частности бакуловирус, генетически измененный с целью инактивации гена, кодирующего синтез, обуславливающего модифицирование экдистероида фермента или вирус насекомых, подвергнутый дальнейшим генетическим изменениям, результатом которых является экспрессия гетерологичного гена, оказывающего влияние на метаморфоз.
Инсектицидные композиции, годные для нанесения на растения с целью контроля насекомых-вредителей, содержат, кроме активного ингредиента, приемлемый в сельскохозяйственном отношении носитель. Нанесение инсектицидной композиции данного изобретения может предохранить растения от насекомых -вредителей за счет снижения питания и гибели восприимчивых насекомых.
Специалисты знают, каким образом выбрать активное начало композиции, а именно вирус, подходящий для определенного насекомого-вредителя.
Для специалистов ясно, что насекомые-вредители могут быть подвергнуты воздействию контролирующего агента данного изобретения любым традиционным способом, включая глотание, вдыхание или прямой контакт.
Описанные в данном изобретении подвергнутые генетическим изменениям палочковидные вирусы найдут основное применение именно как компоненты сельскохозяйственных композиций, предназначенных для нанесения на растения с биологического контроля насекомых, являющихся вредителями растений. Известны разнообразные способы приготовления таких приемлемых для сельского хозяйства композиций.
Концентрация основного ингредиента, которая требуется для получения эффективных инсектицидных композиций, применяемых в сельском хозяйстве для защиты растений, зависит от типа организма, используемой мутации и от формулы композиции. Как понятно специалисту, эффективные значения концентраций контролирующего агента в инсектицидной композиции можно легко определить экспериментально. Например, концентрацию вируса, дающую инсектицидный эффект, можно легко определить описанными в любом из примеров VI-XI способами.
Сельскохозяйственные композиции должны подходить для применения в сельском хозяйстве и хорошо диспергироваться в поле. В общем случае, компоненты композиции не должны быть фитотоксичными и наносить вред всему сообществу окклюдирующих вирусов. Нанесение на листья не должно приносить вреда листьям или повреждать их. Кроме соответствующих твердых или, что является более предпочтительным, жидких носителей, сельскохозяйственные композиции могут включать компоненты, повышающие липучесть или адгезию, эмульгаторы или увлажнители, но не компоненты, снижающие питание насекомых или функционирование вируса. Может оказаться также желательным введение компонентов, предохраняющих контролирующий агент от инактивации под действием ультрафиолетового излучения. Сельскохозяйственные композиции для контроля насекомых-вредителей могут также включать агенты, стимулирующие питание насекомых.
Имеются обзоры, описывающие способы нанесения агентов биологического контроля насекомых и применение их в сельском хозяйстве. Смотри, например, Couch and Ignoffo (1981) в Microbial Control of Pests and Plant Disease 1970-1980, Burges (ed) chapter 34, p. 621-634; Corke and Rishbeth, ibid, chapter 39, p. 717-732; Brockwell (1980) в Methods for Evaluating Nitrogen Fixation. Bergersen (ed) p. 417-488; Burton (1982) в Biological Nitrogen Fixation Technology for Tropical Agriculture, Graham and Harris (eds) p. 105-114; и Roughley (1982) ibid, p. 115-127; The Biology of Baculoviruses, v. II Supra.
Данное изобретение иллюстрируется приводимыми ниже примерами, которые ни в коей мере нельзя рассматривать как ограничения области данного изобретения. Ясно, что специалистам могут представиться различные варианты конкретного воплощения, модификации и эквиваленты изобретения, не выходящие за пределы идеи данного изобретения и/или прилагаемой формулы изобретения.
Пример 1.
Положение гена egt в геноме AcMNPV показано на фиг. 1, над картой генома AcMNPV показан масштаб в единицах карты. Для определения нуклеотидной последовательности этого типа и соседних с ним областей, что позволило бы осуществить последующие генетические манипуляции, необходимо вначале клонировать некоторые фрагменты ДНК, фланкирующие данную область, в плазмидных векторах (стандартные методики клонирования смотри: T. Maniatis et al. (1982) в Molecular Cloning: A Laboratory Manual, Cold Sprihg Harbor Laboratory, Cold Spring Harbor, N.Y.). На панели A фиг. 1 показана линейная карта генома AcMNPV после расщепления его рестриктазами Eco RI и Hind III. На панели B представлена в увеличенном масштабе область генома от 7,6 до 11,1 единиц карты, показывающая положение гена egt. В качестве исходного AcMNPV использовался штамм L1 (Lee and Miller (1978) J. Virol. 27:754). Подвергаемые клонированию фрагменты ДНК и названия полученных плазмид показаны на фиг. 1. Панель C. Фрагмент 1, который занимает область от сайта PstI, расположенного в точке 7, 6, у.е. до сайта Dam HI (11,1 у.е.), клонируется в плазмидный вектор pUC19; оба фрагмента 2 и 3 [от PstI (7,6 у.е.) до Eco RI (8,65 у.е.) и от Eco RI (8,65 у. е.) до SalI (10,5 у.е.), соответственно] клонируются в вектора Bluescript M13+ и Bluescript M13- (Stratagene, San Diego, California). Фрагмент 5 [расположен от Bst EII сайта (8,35 у.е.) до Bst EI сайта (8,7 у.е.)] клонируется в Bluescript M13+
Затем генерируется большое количество субклонов плазмид BCPSE и BCES (Henikoff (1984) Gene 28:351). Эти подклоны отличаются последовательно увеличивающимися делециями вирусных вставок, так что они содержат различные количества вирусной ДНК, от менее чем 50 пар оснований для полного вирусного фрагмента. Многие такие субклоны и плазмиды BCB затем наращиваются (Sanger et al. (1977) Proc. Natl. Acad. Sci. USA 74:5463), так что в обоих направлениях восстанавливается последовательность целого гена egt. Затем полученная нуклеотидная последовательность анализируется на предмет наличия открытых рамок считывания, которые могут кодировать протеин, с помощью компьютерных программ Pustell and Kefatas (1984), Nucl. Acids Res 12:643-655 и Devereaux et al. (1984) Nucl. Acids Res 12:387-396. Такой анализ показывает, что ген egt кодирует протеин, состоящий из 506 аминокислот. Последовательность нуклеотидов гена egt и предсказанная аминокислотная последовательность продукта этого гена приведена на фиг.13.
Пример 2
Для того, чтобы сконструировать рекомбинантные вирусы, неспособные к экспрессии функционального гена egt, необходимо произвести дальнейшие манипуляции с описанными в примере 1 клонами плазмид. Плазмиду pUCBCPsB расщепляют с помощью эндонуклеаз рестрикции Eco RI и Xba I (сайты рестрикции внутри гена egt, см. фиг. 3) и отбрасывают маленький фрагмент. Ген lacZ Escherichia coli, отщепленный от pSKS 104 (Casadaban et al. (1983) Methods Enzymbol. 100:293-303) с помощью рестриктаз Eco RI и Aha III, внедряется между сайтами Eco RI и Xba I после того, как свободные концы Xba I заполняются с помощью ДНК-полимеразы фага T4. Полученную плазмиду обозначают pEGTZ. В этой плазмиде внедренный ген lac Z находится в пределах рамки вместе с предшествующей кодирующей последовательностью egt. Плазмиду pEGTDEL, напротив, конструируют простым связыванием сайтов Eco RI и Xba I (после того, как концы обоих сайтов заполняются) не вставляя между ними какой-либо нуклеотидной последовательности. Все вирусы являются производными от исходного штамма L-I AcMNPV (Lee and Miller (1978) Supra), очищаются через бляшки и размножаются на культуре линии клеток Spodoptera frugiperda IPLBST 21 (SF-клетки) Vaughn et al. (1977) in vitro 13:213-217) с использованием описанных ранее методов (Lee and Miller (1978); Miller et al. (1986) Genetic Engineering, Principles and Methods, v. 8 (eds. J. Sitlow and A. Hollaender), Plenum Press, N.Y. 277-298, 1986).
Затем плазмиду pEGTZ переносят вместе с ДНК AcMNPV дикого типа в SF -клетки, как описано в Miller et al. (1986) Supra. Эта процедура обеспечивает гомологичную рекомбинацию между последовательностями вирусной и плазмидной ДНК, что приводит к замещению гена вируса egt на связку генов плазмиды egt - lac Z. Поскольку остающаяся кодирующая последовательность egt находится в рамке считывания вместе с последовательностью lac Z, такой рекомбинантный вирус будет продуцировать слитый белок, представляющий собой первые 84 аминокислоты egt, соединенные с b -галактозидазой. Рекомбинантный вирус, обозначенный vEGTZ, поддается идентификации, так как экспрессия b -галактозидазы приводит к образованию голубых вирусных бляшек в присутствии хромогенного индикатора, такого, как 5-бром-4-хлор-3-индолил- b -D-галактопиранозида (X-gal). Диаграмма гена egt вируса vEGTZ приведена на панели B фиг. 2.
Рекомбинантный вирус vEGTDEL получают перенесением плазмиды pEGTDEL и ДНК из вируса vEGTZ в SF клетки. Гомологичная рекомбинация приводит к замещению в vEGTZ генной связки egt-lac Z на ген egt с делецией. Рекомбинантный вирус vEGTDEL идентифицируют по его неспособности образовывать голубые бляшки в присутствии X-gal. Структура гена egt вируса vEGTDEL показана на фиг. 3, C.
Пример 3.
Продукт гена egt идентифицируют путем сравнения белков, синтезируемых вирусом AcMNPV дикого типа (который продуцирует EGT) и вирусами vEGTZ или vEGTDEL (не способны продуцировать EGT). SF-клетки инфицируют либо AcMNPV дикого типа, либо vEGTZ с множественностью заражений (MOI) 20, как описано O'Reilli and Miller (1990) J. Virol. 64:1321-1328. Инфицированные клетки также анализируют. Спустя 6 ч после заражения клетки инкубируют в течение 1 ч в присутствии [35 S]-метионина с целью внесения радиоактивных меток во все белки. Клетки затем подвергают лизису и отделяют белки электрофорезом в геле SDS -полиакриламида (SDS-PAGE)(Laemmli et al. (1970) Nature 227:680-685). Все секретированные клетками белки также собираются и анализируются. После проведения SDS-PAGE меченные радиоизотопами белки определяются с помощью авторадиографии. Инфицированные wt AcMNPV клетки секретировали белок с молекулярной массой 60 KDA, чего не произошло в случае инфицированных vEGTZ или неинфицированных клеток. Этот белок однако не определяется в лизатах клеток, инфицированных диким типом AcMNPV, что доказывает его секрецию клетками. Эти данные показывают, что продуктом гена egt является синтезируемый 60 KDA протеин, что хорошо согласуется с данными по последовательности нуклеотидов и аминокислот.
Пример 4
Ферментивную активность белка EGT определяли путем сравнения SF-клеток, инфицированных wt AcMNPV или vEGTZ.
SF-клетки, инфицированные wt AcMNPV или vEGTZ описаны в примере 3. Через 12 ч "pi" клетки и межклеточную среду собрали и обработали по отдельности. Параллельно обрабатывали неинфицированные клетки. Клеточный лизат или межклеточную среду инкубировали в присутствии 1 мМ UDP-глюкозы и 0,25 m Ci [-3H] экдизона, как описано у O'Reilly and Miller (1989) Science 245:1110-1112.
Активность экдистероид-UDP -глюкозилтрансферазы в клеточных лизатах или межклеточной жидкости будет катализировать перенос глюкозы от UDP -глюкозы к экдизону с образованием соединения (конъюгата) экдизон-глюкоза. Экдизон и экдизон-глюкоза разделяются тонкослойной хроматографией на силикагеле (Bansal and Gessner (1988) Anal. Biochem. 109-321) и визуализируются с помощью авторадиографии Комплекс экдизон-глюкоза (G) образуется только при тестировании инфицированных AcMNPV клеточных лизатов или межклеточной жидкости. Его не наблюдают при использовании неинфицированных или инфицированных vEGTZ клеток или межклеточной жидкости, что доказывает, что активность обусловлена экспрессией egt. Большая часть активности сосредоточена в межклеточной среде, что находится в соответствии с обсуждаемыми в примере 3 данными. Доказательство присоединения глюкозы к экдизону получают при тестировании wt-инфицированных лизатов описанным выше способом, за исключением того, что UDP-глюкозу заменяют на меченную радиоактивными изотопами UDP [U-14C] -глюкозу. Реакции, протекающие с участием немеченой UDP-глюкозы или меченной UDP [U-14C]-глюкозы, с присутствующим в обеих реакциях [3H]-экдизоном, и подсчет импульсов образующихся соединений показывает, что может быть определен и 3H из экдинона, и 14 C из глюкозы. Эти данные доказывают, что продуктом гена egt является UDP -глюкозилтрансфераза, которая является катализатором переноса глюкозы от UDP -глюкозы к экдизону.
Далее проводили эксперименты с целью более тщательного определения субстратной специфичности EGT. В этих экспериментах инкубировали различные субстраты (1 мМ) в присутствии межклеточной среды, извлеченной из инфицированных wt AcMNPV клеток, в которых наблюдалась довольно высокая активность egt. Для обеспечения уверенности в том, что все наблюдаемые случаи образования соединения обусловлены EGT, каждый субстрат инкубировали также со средой, извлеченной из неинфицированных и инфицированных vEGTZ клеток, которые не инфицированы продуцировать EGT. В каждом случае в реакцию вводили 0,05 m Ci UDP-[U-14C] -глюкозы. Все соединения, выступающие по отношению к EGT как субстраты, должны присоединять глюкозу, они поддаются определению, поскольку глюкоза помечена радиоизотопами.
Субстраты инкубируют в присутствии среды, полученной из соответствующим образом инфицированных клеток и 0,05 m CiUDP-[U-14C]-глюкозы. Количество перенесенной глюкозы вычисляется, исходя из числа импульсов от соответствующих областей хроматографических пластинок.
Для дальнейшего контроля вводили UDP-[U-14C]-глюкуроновую кислоту в отдельные системы реакций со средами, полученными из инфицированных вирусом дикого типа клеток, в целях демонстрации того, что глюкуроновая кислота в этих реакциях не переносится. Полученные данные представлены в табл. 1. В качестве субстратов идентифицированы только экдизон, 20-гидроксиэкдизон и макистерон A (все экдистероиды). В средах, извлеченных из ложно инфицированных или инфицированных vEGTZ клеток, конъюгаты не обнаружены, что подтверждает обусловленность наблюдаемой активности экспрессией egt. При использовании UDP-[U-14C]-глюкуроновой кислоты конъюгаты не наблюдаются, что является демонстрацией того, что глюкуроновая кислота не переносится с помощью EGT.
Пример 5.
Для того, чтобы получить доказательства наличия также и у других бакуловирусов генов, гомологичных в значительной степени гену egt AcMNPV, отделили ДНК вируса OpMNPV и подвергли ее по отдельности воздействию эндонуклеаз рестрикции Eco RI, Bam HI и Hind III. Эти ферменты расщепляют вирусную ДНК на несколько сегментов различного размера, положение которых в геноме OpMNPV уже известно (Leisy et al. (1984) J. Virol. 52:699). Затем провели гибридизацию по Саузерну описанным T.Maniatis et al. (1982) Supra способом. Вырезали внутренний сегмент гена egt AcMNPV из плазмиды BCES с помощью ферментов Eco RI и Xba I (фиг. 1). Этот фрагмент пометили радиоактивным изотопом 32P и использовали как средство идентификации любых родственных последовательностей в геноме OpMNPV. Ясно, что в соответствующих условиях фрагмент ДНК присоединит другой фрагмент, в составе которого имеется аналогичная или идентичная последовательность. Таким образом, на нейлоновой мембране происходит присоединение метки-гена egt AcMNPV к любому фрагменту ДНК OpMNPV, в котором содержатся родственные последовательности ДНК. Место присоединения метки можно визуализировать с помощью экспозиции мембраны рентгеновским лучам. Вначале происходит гибридизация метки egt с нейлоновой мембраной в нежестких условиях (1 М хлорида натрия, 0,3 М цитрата натрия, 5% сульфата декстрана, 5 х раствор Denhardt'a, 0,25% SDS при 37oC). Это допускает гибридизацию метки с относительно удаленными родственными последовательностями. Далее постепенно увеличивают жесткость гибридизации, поднимая температуру гибридизации или вводя в раствор гибридизации формамид, до тех пор, пока не возникнут специфические гибридизационные связи. Гибридизация, показанная на фиг. 4, протекает в следующих условиях: 1 М хлорида натрия, 0,3 М цитрата натрия, 5% сульфата декстрана, 5 х раствор Denhardt'a, 0,25% SDS; температура 68oC, продолжительность 15 ч. Затем мембрану дважды промывают, каждый раз в течение 15 мин, в растворе 0,3 М хлорида натрия, 0,1 М цитрата натрия и 0,1% SDS при 68oC. На фиг. 4 показано, что метка egt AcMNPV присоединяется к определенным фрагментам ДНК OpMNPV, а именно, к фрагменту B Eco RI, фрагменту A Bam HI и к фрагментам N и S Hind III. Отметим, что ген egt занимает в геноме OpMNPV такое же относительное положение, что и ген AcMNPV (фиг. 5). Аналогичная пропись используется для идентификации egt -гомологичных генов у других энтомопатогенов. Понятно, что для подтверждения EGT-активности в случае сильно расходящихся последовательностей потребуется метод рестрикции, описанный в примере 4. Тогда для выделения гена, кодирующего фермент EGT, потребуется молекулярно-генетический анализ, методология которого известна.
Для обнаружения гена, кодирующего EGT в других организмах используется, в отличие от вышеописанного, специфический способ тестирования на активность UDP -глюкозилтрансферазы, описанной в примере 4. Этот тест используется для определения энзима в процессе очистки стандартными биологическими методами. После очистки определяется частичная аминокислотная последовательность белка EGT. Полученную информацию используют для создания олигонуклеотидной метки, с помощью которой затем определяют положение гена EGT в геноме.
Пример 6.
Титр экдистероида в гемолимфе изменяется циклически, регулируя превращения и личинка-личинка, и личинка -куколка, и, поскольку предполагается, что присоединение глюкозы инактивирует экдистероиды (Warren et al. (1986) J. Liq. Chromatogr. 9: 1759; Thompson et al. (1987) Arch. Insect. Biochem. Physiol. 4:1; Thompson et al. (1988) Arch. Insect. Biochem. Physiol. 7:157), то представляется возможной взаимосвязь нарушения процесса нормального развития насекомого при инфицировании вирусом AcMNPV с экспрессией egt. Нарушение развития демонстрируют следующим образом: только что перенесшие линьку личинки четвертого возраста S. frugiperda инфицировали вирусами wt AcMNPV или vEGTZ путем впрыскивания и ежедневно наблюдали за возможными изменениями их развития. Для отрицательного контроля одной группе личинок ввели жидкость культуры ткани. Результаты эксперимента приведены в табл. 2. Все личинки, которым ввели культуру ткани (ложное инфицирование), как и ожидалось, перелиняли, превратившись в личинки пятого возраста. Из шестнадцати личинок, инфицированных вирусом дикого типа, подобная трансформация наблюдалась только у одной. Напротив, все личинки, инфицированные мутантным вирусом vEGTZ, перелиняли из четвертой в пятую возрастную стадию. Таким образом, экспрессия гена egt вируса wt AcMNPV явно и специфично подавляет линьку насекомого-хозяина. Впоследствии обе инфицированные группы личинок погибли от вирусной инфекции, что доказывает, что нарушение функции egt вируса vEGTZ не препятствует ему убивать хозяина.
Личинкам S. frugiperda четвертого возраста ввели 1•105 pfu wt AcMNPV или vEGTZ в 5 mл Ложно-инфицированным личинкам ввели 5 μл жидкой культуры ткани. Каждая группа состояла из 16 личинок, которые содержались в следующих условиях: искусственная диета (R.L. Burton (1969) ARS publication p. 33-134), температура 28oC с циклом "свет: темнота" 14:10 ч. Личинки каждый день осматривались на предмет линьки, и на 7 -й день регистрировалась смертность.
При инъекции вируса личинкам ранней пятой стадии получены аналогичные результаты. Ни одна из только что перелинявших личинок пятого возраста, инфицированных вирусом дикого типа, не обнаружила каких-либо признаков окукливания, тогда как большинства инфицированных vEGTZ личинок проявили некоторые поведенческие отклонения (прекращение питания, беспокойство и плетение кокона), что является показателем приближения превращения из личинки в куколку. Однако все личинки, которым была введена вирусная инфекция, погибли до окукливания. Эти данные показывают, что инфицирование AcMNPV препятствует линьке или окукливанию личинок. Они показывают также, что подобное нарушение развития насекомых обусловлено экспрессией гена egt.
Пример 7.
Биоанализ in vivo AcMNPV дикого типа (wt) и vEGTZ выявил, что экспрессия гена egt увеличивает время, в течение которого личинка питается, и что нарушение функции egt улучшает характеристики вируса как пестицида. При проведении данных исследований личинкам S. frugiperda делали инъекции либо wt AcMNPV, либо vEGTZ на ранней 4-й стадии. Для сравнения контрольным личинкам вводили среду культуры ткани, которая не содержала вируса. У личинок каждый день проверяли прибыль веса, признаки линьки или окукливания и смертность. На фиг. 6 и 7 соответственно приведен средний дневной привес личинок различных групп, а также процент смертности. Контрольные личинки в течение первых двух дней показали умеренный рост. В течение второго дня все они перелиняли в пятую стадию. Далее личинки резко выросли за два дня, после чего прекратили питаться и приготовились к окукливанию. Только одна из 16 личинок, инфицированных wt AcMNPV, перелиняла, вместо чего остальные в течение трех дней после заражения личинки непрерывно росли. На этой стадии они заболевают, но вплоть до 5 дня ни одна личинка не погибла. К 6-му дню погибли все инфицированные wt AcMNPV личинки. Напротив, все личинки, инфицированные vEGTZ, перенесли линьку из четвертой в пятую возрастную стадию и в течение этого периода времени они не питались. Этим объясняется отсутствие привеса с 1-го по 2-й день. После линьки они возобновляют питание, но к 3-му дню начинают проявлять признаки болезни. Четыре дня спустя после заражения они начинают умирать и к 5-му дню все погибают. Личинки, инфицированные производным от AcMNPV вирусом, у которого отсутствует функциональный ген egt, меньше питаются и погибают после заражения быстрее, по сравнению с личинками, инфицированными диким типом AcMNPV.
При инфицировании личинок 5-й возрастной стадии эти явления наблюдаются еще более четко (фиг. 8 и 9). Как и ожидалось, контрольные личинки за два дня значительно выросли, после чего прекратили питаться, готовясь к окукливанию. Прекращение питания сопровождается резким падением веса. Личинки, инфицированные wt AcMNPV, не проявили признаков прекращения питания, они продолжали питаться и прибавлять в весе в течение еще двух дней, а затем начали проявляться признаки заболевания. Смертность до седьмого дня после заражения не была полной.
Можно сделать следующий вывод. Личинки, инфицированные vEGTZ, как и контрольные, питаются только в течение первых двух дней после заражения. Затем наблюдается, поскольку они готовятся к окукливанию, резкая потеря веса. Однако, ни одна из этих личинок не окукливается; к третьему дню наблюдается признаки болезни, а через шесть дней после заражения все они погибают.
Пример 8.
Сравнивается воздействие заражения vEGTZ и wt AcMNPV на только что вылупившиеся личинки S. frugiperda первого возраста. Новорожденным личинкам S. frugiperda дают корм, в котором содержатся в различных количествах полиэ - дрические тела включения vEGTZ или wt AcMNPV, и ежедневно наблюдают смертность. На фиг. 10 и 11 приведены результаты, полученные для двух различных доз. Можно видеть, что в обоих случаях у инфицированных vEGTZ личинок значительная степень смертности наступает намного раньше по сравнению с личинками, инфицированными вирусом дикого типа. Вообще, личинки, зараженные рекомбинантным вирусом, погибают на три -четыре дня раньше, чем зараженные wt вирусом. Этот результат является дополнительным доказательством того, что бакуловирусы, содержащие инактивированные гены egt, лучше действуют в качестве биологических пестицидов, чем бакуловирусы дикого типа с интактными генами egt.
Пример 9.
Для конструирования рекомбинантных вирусов, с экспрессией проторакотропного гормона (PTTH), ген PTTH клонируют в плазмидный вектор переноса pEV mod XIV, который описан в U.S. Patent Application Serial N 07/353,847 от 17 мая 1989 г. заявка включена здесь в качестве ссылки. Эта плазмида содержит восходящую и нисходящую последовательности гена полиэдрина AcMNPV, который является медиатором гомологической рекомбинации экспрессируемого гена PTTH с геном вируса EGT-. Сайт множественного клонирования располагается у обычного сайта инициирования трансляции полиэдрина, вниз от LS XIV модифицированного промотора полиэдрина (Rankin et al. Gene 70:39 (1988). Ген PTTH Bombix mori выделяется из плазмиды pBc22k-C19 (Kawakami et al. Science 247:1333 (1990) с помощью рестрикционного фермента Hind III. Связующие концы Hind III заполняются с помощью ДНК-полимеразы фага T4, а затем плазмиду расщепляют с помощью Eco RI. Плазмиду pEV mod XIV расщепляют с помощью KpnI. "Липкий" конец Kpn I удаляют ДНК-полимеразой фага T4 и расщепляют плазмиду с помощью Eco RI. Фрагмент, в котором содержится ген PTTH, клонируют через эти сайты в плазмиду переноса. В полученной рекомбинантной плазмиде, обозначаемой pEVPTTH, ген PTTH расположен сразу после LS XIV модифицированного промотора полиэдрина.
Рекомбинантные вирусы с экспрессией PTTH получают путем контрасфекции pEVPTTH в SF -клетки с ДНК вирусов wt AcMNPV или vEGTDEL. Рекомбинация между последовательностями, фланкирующими полиэдрин в вирусной ДНК, и последовательностями, фланкирующими ген PTTH в плазмиде pEVPTTH, приводит к замещению гена полиэдрина на ген PTTH. Вирусы, в геноме которых имела место такая рекомбинация, идентифицируют по отрицательному по окклюзии фенотипу (Miller et al. в Genetic Engineering, Principles and Methods. v. 8, J. Setlow and A. Hollaender, eds. Plenum Press N.Y. 1986, p. 277-298). Соответствующие рекомбинантные вирусы обозначаются vEGTPTTH и vWTPTTH.
Аналогичным образом получают рекомбинантные вирусы с экспрессией эстеразы ювенильного гормона. Сначала вырезают ген эстеразы ювенильного гормона Heliothis virescens Hanzlik et al. (1989) J. Biol Chem. 264:12419) из плазмиды pCHE16B (Hammock et al. (1990) Nature 344:458) посредством расщепления Eco RI и Kpn I. Плазмиду переноса pEV mod XIV также отщепляют посредством Eco RI и Kpn I и фрагмент, включающий ген эстеразы ювенильного гормона внедряют между этими сайтами. Таким образом, pEVJHE включает последовательности гена полиэдрина, фланкирующие ген эстеразы ювенильного гормона. Рекомбинантные вирусы EGT-JHE и vWTJHE получают контрансфекцией pEVJHE с ДНК vEGTZ или wt AcMNPV соответственно и отбором бляшек без тел включения.
Для конструирования рекомбинантных вирусов с экспрессией гормона вылупления вырезают ген гормона вылупления Manduca sexta из плазмиды pF 5-3 (Horodyski et al. (1989) Proc. Natl. Acad. Sci. USA 86:8123) путем расщепления Eco RI и Hpal.pEV mod XIV расщепляют с помощью KpnI, свободный конец KpnI удаляют посредством ДНК полимеразы фага T4, а затем расщепляют с помощью Eco RI. Внедрение фрагмента, содержащего ген гормона вылупления, в плазмиду перемещения дает рекомбинант pEVEH, в котором ген гормона вылупления располагается вниз от промотора LS XIV модифицирования полиэдрина. Контрансфекция этой плазмиды с ДНК vEGTZ или vDA26Z позволяет выделить соответственно рекомбинантные вирусы vEGT-EH и vDA26Z EH. vDA26Z представляет собой рекомбинантный вирус со вставкой гена lac Z E.coli внутри гена DA26 (O'Reilly et al. J. Gene Virol. in Press). Функция гена DA26 неизвестна, а по фенотипу, что касается метаморфоза, vDA26Z представляет собой дикий тип. Из -за присутствия гена lac Z производные vDA26Z дают голубые бляшки в присутствии X-gal.
Пример 10.
Чтобы оценить in vivo эффект таких рекомбинантных вирусов, личинкам S. frugiperda ввели 2•105 pfu вирусов vEGTDEL, vEGT-PTTH, vwTPTTH или одновременно vEGT-PTTH и vEGT- JHE (по 1•105 pfu каждого); личинки находились в поздней 3-й возрастной стадии или в ранней 4-й. При введении всех EGT- -вирусов инфицированные насекомые перенесли линьку и пережили 4-ю стадию, а к 3-му дню обнаружили соскальзывание головной капсулы, что указывает на приближающуюся линьку. После инфицирования одним vEGT- PTTH или совместно vEGT-PTTH и vEGT- JHE экспрессия PTTH приводит к быстрому заболеванию насекомых, а к 4-му дню наблюдается значительная смертность. Напротив, насекомые, инфицированные vEGTDEL, вплоть до 5-го дня не обнаруживают значительной смертности. Как и ожидалось, насекомые, инфицированные вирусом vWPTTH, никогда не проявляют признаков перехода на 5-ю стадию. Соскальзывания головной капсулы не наблюдается и до 6-го дня не замечено значительной смертности. Таким образом, экспрессия PTTH в отсутствие EGT приближает на 1 день по сравнению с инфекцией vEGTDEL срок гибели насекомых в результате заражения. Эти данные демонстрируют улучшение пестицидных свойств бакуловирусов за счет экспрессии генов, влияющих на метаморфоз насекомых, и показывают, что эффективность такой стратегии возможна только при инактивировании гена egt (табл. 3).
Пример 11.
Генерирование положительных по признаку образования тел включения вирусов с экспрессией гетерологичных белков, влияющих на развитие насекомых, может осуществляться по следующей схеме. Во-первых, конструируется отрицательный по признаку окклюзии вирус с экспрессией β -галактозидазы. Ген lac Z E. coli, кодирующий фермент b -галактозидазу, клонируют в вектор переноса pSyn VI-, описанный в U.S. Patent Application Serial N 07/353,847 от 17 мая 1989 г. Вектор pSyn VI- включает последовательности, расположенные в геноме AcMNPV по восходящей и по нисходящей от гена полиэдрина, причем сайт множественного клонирования расположен непосредственно по нисходящей от синтетического модифицированного промотора полиэдрина. Ген lac Z вырезают из плазмиды PSKS 105 (Casadaban et al. 1983) supra посредством расщепления Pst I. Выступающий конец удаляется с помощью нуклеазы mung bean, и плазмиду расщепляют посредством Sst II. pSyn VI- расщепляют по сайтам Eco RV и Sst II и клонируют ген lac Z в эти сайты. Полученная плазмида обозначается pSyn VI- gal, в ней ген lac Z располагается непосредственно по нисходящей от синтетического модифицированного промотора полиэдрина. pSyn VI- gal переносится в SF-клетки с ДНК вируса vEGTDEL, после чего отделяются отрицательные по признаку окклюзии бляшки с экспрессией b -галактозидазы, которые представляют собой вирус vEGT- Syn VI- gal.
Для построения положительного по признаку образования тел включения рекомбинантного вируса, у которого имеет место экспрессия PTTH, ген PTTH клонируют в вектор переноса pSp XIVVI+X3.
При конструировании pSp XIVVI+X3 сначала строится промежуточная плазмида pSp XIVVI+. Плазмида pSp LSXIVVI+CAT (описанная в заявке на патент США серийный номер 07/353,847 от 17 мая 1989 г.) разрезается по BgLII и концы заполняются ДНК-полимеразой. Плазмиду pSynVI + wtp (описана в заявке на патент США серийный номер 07/353,847 от 17 мая 1989 г.) разрезают по EcoRV и SacI и маленький фрагмент EcoRV SacI очищают. Фрагменты двух плазмид связывают и отбирают pSp XIVVI+CAT. Плазмида pSp XIVVI + идентична плазмиде pSpLSVIVVI+CAT за исключением того, что сайт множественного клонирования в промежутке от сайта BgLII до сайта SacI совпадает с pSynVI+wtp. Для конструирования сайта множественного клонирования V3 (описано в заявке на патент США серийный номер 07/353,847 от 17 мая 1989 г.) между сайтами EcoRV и Sacl плазмиды pSp XIVVI+ вводится полилинкер. Последовательность сайта множественного клонирования в pSp XIVVI+X3 от сайта BgLII до сайта SacI такова AGATCATC GAATTCTCGAG CTGCAGATCT GTCGACCCGGG AATAAA GAGCTC.
EcoRV/Bgl EcoRI-XhoI PstI-BglI SalI-SmaI poly A SacI.
Эта плазмида содержит интактный ген полиэдрина под контролем промотора полиэдрина дикого типа. Синтетический модифицированный промотор полиэдрина расположен по восходящей от гена полиэдрина и с противоположной ориентацией. Положение сайта множественного клонирования обеспечивает возможность внедрения генов, экспрессия которых должна осуществляться под контролем синтетического модифицированного промотора полиэдрина. Ген PTTH вырезают из плазмиды pBc22k (Kawasaki et al. (1990) supra) путем расщепления по Hind III. Свободный конец Hind III заполняют ДНК-полимеразой фага T4 и расщепляют плазмиду по EcoRI. pSp XIVVI+X3 расщепляют по EcoRI и SmaI и клонируют ген PTTH в эти сайты, что дает плазмиду pSpPTTH. Ген PTTH в этой плазмиде расположен по нисходящей ветви от синтетического промотора модифицированного полиэдрина, но по соседству и с противоположной ориентацией относительно промотора полиэдрина дикого типа и кодирующих последовательностей. Оба гена, ген PTTH и полиэдрина, фланкируются и по восходящей, и по нисходящей, последовательностями гена полиэдрина генома AcMNPV.
pSp PTTH переносится в SF-клетки совместно с ДНК vEGT- SynVI- gal. Рекомбинация между восходящими и нисходящими последовательностями полиэдрина в вирусной ДНК и последовательностями, фланкирующими гены PTTH и полиэдрина в pSp PTTH, приводит к замещению гена lac Z EGT-SynVI- gal на гены PTTH и полиэдрина из pSp PTTH. Рекомбинантный вирус EGT- SpPTTH идентифицируют по фенотипу, отрицательному по признаку b -галактозидазы и положительному по признаку окклюзии.
Для конструирования положительного по признаку окклюзии вируса с экспрессией экстеразы ювенильного гормона вырезали ген эстеразы ювенильного гормона из плазмиды pJHE16B (Hammock et al. (1990) supra) посредством расщепления по KpnI, удаления свободного конца посредством ДНК-полимеразы фага T4 и рассекания по EcoRI. Затем клонировали ген экстеразы ювенильного гормона в плазмиду pSp XIVVI+X3, которую расщепили по EcoRI и SmaI. Полученную плазмиду, pSp JHE, подвергают контрансфекции с ДНК vEGT- SynVI- gal, получая рекомбинантный вирус vEGT- Sp JHE. У такого положительного по окклюзии вируса экспрессия эстеразы ювенильного гормона происходит под контролем синтетического промотора модифицированного полиэдрина.
Аналогичным образом, вырезают ген гормона вылупления из плазмиды pF 5-3 (Horodyski et al. (1990) supra) посредством рассекания по EcoRI и Hpal и клонирования в pSp XIVVI+X3, которая затем расщепляется по EcoRI и SmaI. В результате контрансфекции полученной плазмиды pSpEH, с ДНК vEGT- SynVI- gal получают рекомбинантный вирус vEGT- pEH.
Положительные по признаку образования тел включения EGT--вирусы подвергают далее генетическим изменениям, вызывающим экспрессию белка, который влияет на метаморфоз насекомых (пептидный гормон насекомых или фермент, инактивирующий гормоны насекомых), в результате чего они снижают питание и ускоряют гибель инфицированных личинок насекомых эффективнее по сравнению с бакуловирусами дикого типа или с бакуловирусами, генетические изменения в которых вызвали только инактивирование гена egt.
Поскольку регуляторы роста и развития насекомых примера 11 образуют тела включения, эти вирусы могут вводиться в эффективные инсектицидные, приемлемые для сельского хозяйства композиции, которые можно наносить на инфицированные посевы. Глотание таких окклюдированных вирусных частиц насекомыми приведет к распространению вирусов по полям и, тем самым, к распространению контролирующего агента. Инфекция вызовет гибель насекомых и, следовательно, защитит посевы от насекомых-вредителей.
Должно быть понятно, что вышесказанное относится только к особым, предпочтительным примерам воплощения данного изобретения, и что многочисленные модификации или изменения могут соответствовать идее и охвату изобретения, как и постулируется в прилагаемой формуле изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| ФРАГМЕНТ РЕКОМБИНАНТНОЙ ДНК (ВАРИАНТЫ) И РЕКОМБИНАНТНЫЙ БАКУЛОВИРУСНЫЙ ВЕКТОР ДЛЯ ЭКСПРЕССИИ НЕЙРОПАРАЛИТИЧЕСКОГО ТОКСИНА (ВАРИАНТЫ) | 1991 |
|
RU2130968C1 |
| РЕКОМБИНАНТНЫЙ СТАБИЛЬНЫЙ ОККЛЮДИРОВАННЫЙ ВИРУС ЯДЕРНОГО ПОЛИЭДРОЗА (ВАРИАНТЫ), ИНСЕКТИЦИДНАЯ КОМПОЗИЦИЯ И СПОСОБ БОРЬБЫ С НАСЕКОМЫМИ НА УЧАСТКЕ | 1992 |
|
RU2156300C2 |
| ИНСЕКТИЦИДНАЯ СМЕСЬ ДЛЯ РЕГУЛИРОВАНИЯ ЧИСЛЕННОСТИ НАСЕКОМЫХ И СПОСОБ РЕГУЛИРОВАНИЯ ЧИСЛЕННОСТИ НАСЕКОМЫХ | 1995 |
|
RU2200394C2 |
| ВЕКТОРЫ | 2009 |
|
RU2569183C2 |
| ВЕКТОР ДЛЯ ПЕРЕНОСА И ВАКЦИНА ПРОТИВ ТУБЕРКУЛЕЗА | 2007 |
|
RU2453603C2 |
| СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО ВИРУСНОГО ВЕКТОРА APMV-8 | 2010 |
|
RU2592544C2 |
| ФАРМАЦЕВТИЧЕСКИЕ КОМПОЗИЦИИ, СОЕДИНЕНИЯ, СПОСОБЫ ПОЛУЧЕНИЯ СОЕДИНЕНИЙ | 1995 |
|
RU2168995C2 |
| ЭФФЕКТИВНЫЕ ВИРУСОПОДОБНЫЕ ЧАСТИЦЫ (VLPS) ГРИППА | 2004 |
|
RU2369405C2 |
| L-НУКЛЕОЗИДЫ, ОБЛАДАЮЩИЕ АНТИ-HBV ИЛИ АНТИ-EBV АКТИВНОСТЬЮ, СПОСОБ ИНГИБИРОВАНИЯ HBV ИЛИ EBV ИНФЕКЦИИ | 1995 |
|
RU2171809C2 |
| БАКУЛОВИРУСНЫЙ ВЕКТОР И ЕГО ПРИМЕНЕНИЕ | 2007 |
|
RU2588464C2 |




Использование: биотехнология, в частности генноинженерные объекты для биологической защиты растений от вредителей. Сущность: предложена группа изобретений, которая включает способ получения генетически модифицированного вируса ядерного полиэдроза Autographa californica, предусматривающий воздействие на геном вируса путем инактивации гена, кодирующего фермент экдистероид UDP - глюкозил-трансферазу. Полученный рекомбинантный вирус, проявляет большую эффективность в инсектицидных композициях по сравнению с вирусами дикого типа или вирусами, применяемыми в настоящее время. Следующим объектом изобретения в предложенной группе является способ борьбы с насекомыми, который включает прием нанесения генетически модифицированного вируса на растения. 4 с. и 7 з. п. ф-лы, 14 ил, 3 табл.
| Биологические средства защиты растений | |||
| - М.: Колос, 1974, с | |||
| Способ получения коричневых сернистых красителей | 1922 |
|
SU335A1 |
| ЕР, заявка, 0225777, кл | |||
| Способ гальванического снятия позолоты с серебряных изделий без заметного изменения их формы | 1923 |
|
SU12A1 |

