Область техники
Изобретение относится к смесям инсектицидов, предназначенным для регулирования численности насекомых и включающим смеси генетически модифицированных вирусов насекомых с химическими и биологическими инсектицидами, для обеспечения более эффективного регулирования численности насекомых.
Уровень техники
В настоящее время существуют различные способы борьбы с насекомыми-вредителями и несколько подходов к контролю за насекомыми-вредителями, поражающими важные для народного хозяйства культуры. Широко используются химические инсектициды, однако их применение вызывает серьезные возражения по следующим причинам: одновременно с хозяйственно-опасными видами насекомых-вредителей химические инсектициды могут уничтожать полезные виды насекомых. Постепенно насекомые приобретают резистентность к таким препаратам, что требует разработки новых категорий инсектицидов. Эти препараты могут не разрушаться в окружающей среде в течение длительного периода времени с момента их применения, что приводит к ее загрязнению.
С целью сокращения использования химических инсектицидов для регулирования численности насекомых на личиночных стадиях используют видоспецифичные вирусы. Видоспецифичные вирусы включают как ДНК-, так и РНК-вирусы. ДНК-вирусы включают энтомопоксвирусы (Entomopoxvirus) и бакуловирусы (Baculoviridae), включающие вирусы ядерного полиэдроза (ВЯП), гранулеза (ВГ) и бакуловирины (Bacuiovirinae), подгруппа бакуловирусов, не образующих включений (НОВ). РНК-вирусы включают тогавирусы, флавивирусы, пикорнавирусы, вирусы цитоплазматических полиэдрозов (ВЦП) и т.п. Подсемейство вирусов с двухцепочечной ДНК Eubaculovirinae включает два рода. вирусов: ядерного полиэдроза (ВЯП) и гранулеза (ВГ), наиболее часто используемых в программах биологической борьбы с насекомыми-вредителями, поскольку они в своем жизненном цикле образуют в тканях насекомых характерные тельца-включения (ТВ).
Вирус ядерного полиэдроза поражают: Lymantria dispar (непарный шелкопряд) ВЯП, Autographa californica (пяденица люцерновая калифорнийская) МВЯП, Syngrapha falcifera (совка-пяденица) ВЯП, Spodopfera litturalis ВЯП, Spodoptera frugiperda ВЯП, Spodoptera exigua ВЯП, Heliothis armigera ВЯП, Mamestra brassicae (капустная совка) ВЯП, Choristoneura fumiferana (сливовая плодожорка) ВЯП, Trichoplusia ni (совка ни) ВЯП, Helicoverpa zea ВЯП и т.д.
Примерами ВГ могут служить Cydia pomonella ВГ (яблонная плодожорка ВГ), Pieris brassicae ВГ, Trichoplusia ni ВГ и т.д. Примерами НОВ могут служить Orcytes rhinoceros НОВ и Heliothis zea НОВ. Примерами вирусов энтомопокса (ВЭП) могут служить Melolontha melonotha ВЭП, Amsacta moorei ВЭП, Locustа migratoria (саранча) ВЭП, Melanoplus sanguinipes ВЭП, Schistocerca gregaria ВЭП, Aedes aegypti ВЭП, Chironomus luridus ВЭП и т.д.
Более 400 изолятов бакуловируса было описано как присутствующие у беспозвоночных. Вирус множественного ядерного полиэдроза Autographa californica (АсМВЯП) является прототипом для всего семейства вирусов Baculoviridae и имеет широкий спектр хозяев. АсМВЯП был первоначально изолирован из Autographa californica (A cal.), взрослая стадия которой является ночной бабочкой)), известной под названием alfalfa looper. Этот вирус поражает насекомых 12 семейств и более чем 30 видов из отряда Lepidoptera. Нет данных о том, что он эффективно поражает какие-либо виды, не принадлежащие к данному отряду.
Жизненный цикл бакуловирусов, на примере АсМВЯП, включает две стадии. Каждая из стадий представлена определенной формой вируса: внеклеточные вирусные частицы(ВКВ), которые не заключены в оболочку, и заключенные в оболочку вирусные частицы. Внеклеточные и заключенные в оболочку вирусные частицы имеют одинаковый геном, но разные биологические свойства. Репродуцирование каждой из двух форм вируса регулируется отдельным набором вирусных генов, некоторые из которых уникальны для каждой формы.
Во встречающейся в природе поражающей насекомых форме вируса многочисленные вирусные частицы заключены в матрикс, образованный белком паракристаллином, и образуют так называемые тельца-включения (ТВ), которые также носят название полиэдральных телец-включений (ПТВ). Белок полиэдрин, имеющий молекулярную массу 29 кД, является основным структурным белком вирусных оболочек, кодируемым вирусным геномом. (Подобно этому вирусы гранулеза образуют включения, состоящие преимущественно из гранулина, а не полиэдрина). Заключенные в оболочку вирусные частицы являются важной частью естественного жизненного цикла бакуловирусов, обеспечивая возможность горизонтальной (от насекомого к насекомому) передачи между поражаемыми видами насекомых. В окружающей среде поражаемые насекомые (обычно на личиночной стадии) поглощают заключенные в оболочку вирусы из зараженного источника пищи, например растения. Кристаллические оболочки растворяются в кишечнике поражаемых насекомых с образованием инфицирующих вирусных частиц. Эти вирусные частицы проникают и реплицируются в клетках ткани кишечника.
Считается, что вирусные частицы проникают в клетку путем эндоцитоза или диффузии, и что ДНК вируса освобождается от оболочки в области ядерной поры или в ядре. Репликация вирусной ДНК обнаруживается спустя шесть часов. Через 10-12 часов после инфицирования ("п.и".) вторичная инфекция распространяется на другие ткани насекомого путем отпочковывания внеклеточных вирусов (ВКВ) с поверхности клетки. ВКВ форма вируса отвечает за передачу вируса от клетки к клетке в пределах отдельного пораженного насекомого, а также за передачу инфекции в культуре клеток.
Позднее в цикле инфицирования через 12 часов п.и. в пораженных клетках можно обнаружить белок полиэдрин. Не раньше, чем через 18-24 часа п.и., происходит сборка белка полиэдрина и вирусные частицы заключаются в белковые оболочки. Заключенные в оболочки вирусы собираются в больших количествах, и через 4-5 дней происходит лизис клетки. Эти заключенные в оболочки из полиэдрина вирусы не играют активной роли в распространении инфекции в личинке. ВКВ распространяются в пораженной личинке, что приводит к ее гибели. После гибели пораженной личинки миллионы заключенных в оболочку вирусов остаются в разлагающейся ткани, в то время как ВКВ разрушаются. Когда другая личинка поглощает заключенный в оболочку из полиэдрина вирус, например, путем поедания зараженных растений или других пищевых материалов, цикл повторяется.
Итак, заключенная в оболочку форма вируса отвечает за первоначальное поражение насекомого через кишечник, а также за стабильность вируса в окружающей среде. ПТВ слабо поражают клетки при введении путем инъекции, но очень активны при оральном введении. Не заключенная в оболочку форма вируса (т.е. ВКВ) отвечает за распространение вируса внутри организма и передачу инфекции от клетки к клетке в культуре. ВКВ эффективно поражают клетки в культуре или внутри насекомого при инъекции, но неэффективны при оральном введении.
Эти вирусы насекомых не патогенны для позвоночных и растений. К тому же бакуловирусы, как правило, имеют узкий круг хозяев. Многие штаммы поражают только один или несколько видов.
Использование бакуловирусов в качестве биоинсектицидов многообещающе. Одним из препятствий для их широкого использования в сельском хозяйстве является промежуток времени между первичным заражением насекомого и его гибелью. Этот промежуток может варьировать от нескольких дней до нескольких недель. В течение этого периода личинка продолжает питаться, принося вред растению. Несколько исследователей пытались преодолеть этот недостаток путем внедрения гетерологичного гена в геном вируса, чтобы экспрессировать контролирующее или модифицирующее вещество насекомого, например токсин, нейропептид, гормон или фермент.
Тем не менее до настоящего времени такие генетически модифицированные вирусы не использовали в комбинации с химическими инсектицидами как часть интегрированного механизма контроля численности вредителей. Описывалось применение комбинации вирусов дикого типа насекомых и химических инсектицидов, но их результаты не были оптимальными из-за ограничений, присущих вирусам дикого типа (библиография, пункты 1-5). Исследователи также пытались регулировать численность насекомых при помощи других биологических агентов, таких как бактерии (например, Bacillus thuringiensis), грибы, простейшие и нематоды, взятые отдельно или в комбинации с вирусами насекомых или химическими инсектицидами, но они также не добились оптимальных результатов (2, 3, 5, 6). Поэтому существует необходимость разработки комбинаций химических инсектицидов и генетически модифицированных вирусов насекомых, которые обеспечат использование преимуществ обоих компонентов и снизят число используемых ядохимикатов и время уничтожения, по сравнению с наблюдаемым для вирусов дикого типа, путем использования вирусов, полученных методами генной инженерии.
Сущность изобретения
Задачей данного изобретения является создание инсектицидных смесей для использования против насекомых из отряда Lepidoptera, состоящих из смесей генетически модифицированных вирусов насекомых с химическими и биологическими инсектицидами, для более эффективного регулирования численности хозяйственно-опасных видов насекомых. Генетическая модификация вирусов включает внедрение гена, экспрессирующего контролирующее или модифицирующее вещество насекомого, например токсин, нейропептид или гормон, или фермент. Генетическая модификация вируса также включает в себя делецию в гене.
Данное изобретение представляет инсектицидные смеси, включающие:
(а) эффективное количество химического инсектицида, отобранного из класса веществ, состоящего из пиретроидов, арилпирролов, диацилгидразинов и формамидинов; и
(б) эффективное количество генетически модифицированного вируса множественного ядерного полиэдроза Autographa californica ("АсМВЯП"), который содержит: (i) внедренный ген, экспрессирующий внутренний токсин (AaTH) Autographa californica, или (ii) делецию в гене, кодирующем экдистероид UDP-глюкозил трансферазу ("ЭГТ") АсМВЯП,
при этом указанные смеси используются для борьбы с насекомыми Lepidoptera при использовании их для регулирования численности совки Heliothis zea, а химический инсектицид является формамидином, генетически модифицированный вирус содержит внедренный ген, экспрессирующий AaTH.
В одном из воплощений данное изобретение представляет инсектицидные смеси для использования против Heliothis virescens, включающие:
(а) эффективное количество химического инсектицида, отобранного из класса веществ, состоящего из пиретроидов и арилпирролов; и
(б) эффективное количество генетически модифицированного АсМВЯП, который содержит: (i) внедренный ген, экспрессирующий AaTH, или (ii) делецию в гене, кодирующем ЭГТ АсМВЯП.
В другом варианте данное изобретение представляет инсектицидные смеси для использования против Heliothis zea, включающие:
(а) эффективное количество химического инсектицида, отобранного из класса веществ, состоящего из арилпирролов и диацилгидразинов; и
(б) эффективное количество генетически модифицированного АсМВЯП, который содержит: (i) внедренный ген, экспрессирующий АаТН, или (ii) делецию в гене, кодирующем ЭГТ АсМВЯП.
В еще одном варианте данное изобретение представляет инсектицидные смеси для использования против Heliothis zea, включающие:
(а) эффективное количество химического инсектицида, отобранного из класса веществ, состоящего из формамидинов; и
(б) эффективное количество генетически модифицированного АсМВЯП, который содержит внедренный ген, экспрессирующий АаТН.
Данное изобретение также представляет способ регулирования численности насекомых из отряда Lepidoptera, включающий в себя нанесение на указанных насекомых или культурные растения, которыми указанные насекомые питаются, описанных выше инсектицидных смесей.
Краткое описание фиг.1 и 2
Фиг. 1 представляет собой графическое изображение данных, представленных в таблице 13, т.е. процент смертности насекомых на 1, 4 и 10 дни для первых трех обработок, приведенных в таблице 13, за исключением данных для "Необработанного контроля" из таблицы 13, которые не отражены на фиг.1.
Фиг. 2 представляет собой графическое изображение данных, представленных в таблице 14, т.е. процент смертности насекомых на 1, 4 и 10 день для первых трех обработок, приведенный в таблице 14, за исключением данных для "Необработанного контроля" из таблицы 14, которые не отражены на фиг. 2. "АсМВЯП с инсертированным АаТН" в таблице 14 - это то же, что "rNPV" на фиг. 2.
Подробное описание изобретения
Насекомые в процессе развития от яйца до взрослой особи (имаго) проходят несколько хорошо изученных стадий. После вылупления из яйца личинка насекомого, которую у Lepidoptera называют гусеницей, вступает в период усиленного питания. В течение этого времени она несколько раз линяет, чтобы обеспечить возможность непрерывного роста. Промежутки между последовательными линьками называются возрастами. В конце периода роста гусеница превращается в куколку и выходит из нее взрослым насекомым.
Задача данного изобретения - повышение эффективности борьбы с насекомыми-вредителями в течение личиночного развития. Отряд Lepidoptera, к которому относятся известные вредители сельскохозяйственных культур, включает семейства: Noctuidae, Notodontidae, Arctiidae, Pyralidae, Plutellidae, Pieridae и Geometridae.
Для определения того, обеспечивает ли инсектицидная смесь эффективный контроль за численностью вредителей, существует два критерия.
Первый - это количество уничтоженных в течение определенного периода времени гусениц (личинок). Он называется "% смертности".
Другой - скорость уничтожения. Даже если % смертности в течение всего периода питания гусениц не повышен, преимущественным является уничтожение большего количества гусениц на ранних стадиях, т.к. при этом время, на протяжении которого личинки питаются, меньше, и, соответственно, меньший вред наносится культурному растению.
Таким образом, тестируемая смесь имеет преимущества перед существующими смесями, если при ее применении увеличивается процент смертности и скорость уничтожения насекомых.
Смесь генетически модифицированных вирусов насекомых с химическими или биологическими инсектицидами называется "синергической", если смертность при ее применении превышает сумму смертностей для ее компонентов, применяемых отдельно; "аддитивной", если смертность для смеси равна сумме смертностей для компонентов, применяемых отдельно; "субаддитивной", если смертность для смеси превышает смертность для любого из ее компонентов, применяемого отдельно, но меньше суммы смертностей для отдельных компонентов; "антагонистической", если смертность для смеси меньше смертности для любого из компонентов, применяемого отдельно.
Выигрыш можно получить, если смеси являются синергическими или аддитивными. Даже если смесь является аддитивной, путем сокращения дозы одного или обоих компонентов по сравнению с дозой для самостоятельного применения, затраты сокращаются, а также получается экологический выигрыш, например сокращается количество химического инсектицида, что приводит к снижению персистентности и образования устойчивости.
Применение инсектицидной смеси имеет преимущества, если оно обеспечивает улучшенный контроль за одним из или одновременно пермиссивными и полупермиссивными насекомыми. Пермиссивное насекомое, как правило, в 100-1000 раз более чувствительно к действию вируса или химического инсектицида, чем полупермиссивное насекомое. Например, вид tobacco budworm (H.virescens), пермиссивный по отношению к АсМВЯП, a cotton bollworm (H.zea) - полупермиссивный по отношению к АсМВЯП.
Дополнительным преимуществом данного изобретения является то, что мишенями для смеси химического инсектицида и вируса насекомого может быть большее количество насекомых, чем в случае самостоятельного использования каждого отдельного компонента. И химический инсектицид, и вирус насекомого имеют ограниченный круг хозяев. Смесь же благодаря присутствию обоих компонентов может иметь более широкий круг хозяев. В то же время этот эффект достигается не за счет взаимодействия между компонентами инсектицидной смеси.
Для регулирования численности насекомых-вредителей используется широкий круг классов веществ. Ниже будет представлено перечисление этих классов и описание механизма их действия.
Пиретроиды - это соединения, связывающиеся с белком натриевого канала, вследствие чего происходит изменение потенциала действия на мембране аксона. В свою очередь это нарушает нормальное функционирование нервной системы насекомого. Из пиретроидов используют циперметрин (α-циано-3-феноксибензил-цис/транс-3-(2,2-дихлорвинил)-2,2-диметилциклопропанкарбоксилат; FMC, Inc.). PERMETHRINTM (3-феноксибензил-цис/транс-3-(2,2-дихлорвинил)-2,2-диметилциклопропанкарбоксилат; корпорации Coulston International), фенвалерат (α-циано-3-феноксибензил-2-(4-хлорфенил)-3-метилбутират) и циалотрин (α-циан-3-феноксибензил-3-(2-хлоро-3,3,3-трифторо-проп-1-енил)-диметилциклопропанкарбоксилат).
Формамидины - это соединения, имеющие несколько механизмов действия, включая связывание с рецептором октопамина (нейрогормон/нейротрансмиттер) и действие в качестве его антагониста, усиление образования сАМП и индукция изменений в поведении или ингибирование смешанной функции моноамин оксидаз. Из формамидинов используют Амитраз (N'-(2,4-диметилфенил)-N-[[(2,4-диметилфенил)имино] метил] -N-метилметанимидамид; NOR-AM, Sobering AG) и хлордимеформ (N'-(4-хлоро-О-толил)-N,N-диметилформамидин).
Арилпирролы являются митохондриальными токсинами, летальный эффект которых состоит в нарушении процесса окислительного фосфорилирования. Из арилпирролов используют 4-бром-2-(р-хлорфенил)-1-(этоксиметил)-5-(трифторметил)-пиррол-3-карбонитрил (патент США 5310938) и соединения, описанные в патенте США 5010098.
Диацилгиразины - это нестероидные регуляторы роста насекомых, основной механизм действия которых состоит в том, что они являются антагонистами экдизона. Из диацилгидразинов используют дибензоил-t-бутилгидразин (изготовление которого описано в патенте США 5300688) и MIMICTM (3,5-диметилбензойная кислота 1-(1,1-диметилэтил)-2-(4-этилбензоил) гидразид; корп. Rohm & Haas).
Циклодиены связываются с рецепторной субъединицей комплекс GABA. Из циклодиенов используют эндосульфан (6,7,8,9,10,10-гексахлоро-1,5,5,6,9,9-гексагидро-6,9-метано-2,4,3-бензодиоксатиепин 3-оксид; Hoechst).
Карбаматы действуют как ингибиторы холинэстеразы. Из карбаматов используют тиодикарб (диметил-N,N-тиобис метилимино)карбонилокси)-бис(этанимидотиоат); Rhone-Poulenc) и метомил (S-метил N-[(метил-карбамоил)окси] тиоацетимидат).
Органофосфаты действуют как ингибиторы холинэстеразы. Из органофосфатов используют профенофос(О-4-бромо-2-хлорфенил O-этил S-пропил фосфоротиоат; Ciba-Geigy), малатион (O,O-диметил фосфородитиоат диэтил меркаптосукцината), сульпрофос (O-этил О-[4-(метилтио) фенил] S-пропил фосфородитиоата) и диметоат (О,O-диметил (S-метил-карбаметил)-фосфородитиоат).
Пиразолы являются ингибиторами митохондриального дыхания путем специфического действия на Комплекс I электронно-транспортной системы. Из пиразолов используют тебуфенпирад (N-(4-t-бутилбензил)-4-хлоро-3-этил-1-метилпира-зол-5-карбоксамид; Mitsubishi Kasei, American Cyanamid Company) и соединения, описанные в опубликованном европейском патенте 289879.
Нитрогуанидины препятствуют связыванию ацетилхолина с определенными ацетилхолиновыми рецепторами на постсинаптической мембране; путем связывания с самими рецепторами эти соединения нарушают передачу нервного импульса. Из нитрогуанидинов используют имидаклоприд (1-[(6-хлоро-3-пиридинил)метил]-N-нитро-2-имидазолидинимин; Вауеr) и его производные.
Милбемицины сначала связываются с сайтом на комплексе GABA рецептор/хлорид-ионный канал, а затем вызывают паралич и гибель насекомого путем ингибирования передачи сигнала в нейромышечном соединении. Из милбемицинов используют абамектин (смесь авермектинов, содержащая >80% авермектина В1а и <20% авермектина Blb; Merck, Sharp & Dohme).
Бензоилфенилуреаты являются регуляторами роста насекомых, которые нарушают синтез хитина, таким образом нарушая процесс образования кутикулы во время линьки насекомого. Из бензоилфенилуреата используют дифторбензурон (1-(4-хлорофенил)-3-(2,6-дифторбензоил) мочевина; корпорации Uniroyal Chemical Co.).
Амидингидразоны являются ингибиторами митохондриального дыхания путем ингибирования транспорта электронов на комплексе II. Из амидингидразонов используют гидраметилнон (тетрагидро-5,5-диметил-2(1Н)-пиримидинон [3-[4-(трифторметил)фенил] -1-[2-[4-(трифторметил) фенил] этенил]-2-пропенилдиен; American Cyanamid Company).
Для специалиста в данной области будет понятно, что известны другие примеры вышеуказанных классов химических веществ, сведения о которых могут быть получены у коммерческих поставщиков или из патентной или научной литературы.
В соответствии с настоящим изобретением инсектицидная смесь включает в себя химический инсектицид (или биологический инсектицид, как описано ниже) и генетически модифицированный вирус насекомого.
В одном из вариантов данного изобретения генетическая модификация вируса насекомого включает инсертирование в какое-нибудь подходящее место в геноме вируса гена, экспрессирующего контролирующее или модифицирующее вещество насекомого. Это вещество может быть, например, токсином, нейропептидом, гормоном или ферментом. Экспрессируемое таким образом вещество повышает биоинсектицидное действие вируса.
К таким токсинам относятся видоспецифичный токсин АаТН скорпиона Androctonus auatralls (7), токсин чесоточного клеща вида Pyemotes tritici (8), токсин Bacillus thuringiensis (9, 10) и токсин, выделенный из паучьего яда (11). Примерами таких нейропептидов или гормонов могут служить гормон эклозион (12), проторацикотропный гормон (ПТТГ), адипокинетический гормон, диуретический гормон и проктолин (13). Примером таких ферментов может служить эстераза ювенильного гормона (ЭЮГ) (14).
Для описания данного изобретения приводится пример с использованием генетически модифицированного АсМВЯП, содержащего инсертированный ген, экспрессирующий АаТН. Исходным для генетической модификации является штамм дикого типа, обозначаемый Е2 (АТСС VR-1344). Внедряемым токсином является АаТН, получаемый из яда северо-африканского скорпиона Androctonus austral is Hector. Этот токсин состоит из 70 аминокислот и связывается с натриевыми каналами насекомых, вызывая паралич при попадании на гусеницу в количестве от нанограмма до микрограмма. Так как АаТН не связывается с натриевыми каналами млекопитающих, он может быть использован в качестве биоинсектицида, не принося вреда здоровью человека.
Регион перед кодирующим регионом гена АаТН содержит сигнальную последовательность, которая отвечает за секрецию АаТН за пределы клетки. Сигнальная последовательность направляет токсин по секреторным путям на поверхность клетки, где токсин секретируется за пределы клетки. Во время транспортировки ферменты удаляют сигнальную последовательность, оставляя активный АаТН.
Было обнаружено, что гетерологичные сигнальные последовательности полезны для экспрессии и секреции токсинов насекомых, например АаТН (15). Наиболее предпочтительной гетерологичной сигнальной последовательностью является кутикулярная сигнальная последовательность Drosophila melanogaster (для белка экзоскелета), которая секретирует большое количество связанных с нею зрелых белков. В свою очередь используется последовательность ДНК, кодирующая кутикулярную сигнальную последовательность, с оптимизацией кодонов. Вырожденность генетического кода позволяет менять последовательность нуклеотидов, получая полипептид, имеющий такую же последовательность аминокислот, как и полипептид, кодируемый нативной последовательностью ДНК. Процедура, известная как оптимизация кодонов, обеспечивает наличие средств для создания такой измененной ДНК, последовательность которой отражает частоту кодонов, характерную для организма насекомого-хозяина. В данном изобретении для создания последовательностей ДНК с оптимизацией кодонов, кодирующих кутикулярную сигнальную последовательность и АаТН, использованы таблицы кодонов для Drosophila melanogaster.
Дополнительным средством повышения уровня экспрессии АаТН является использование "раннего" промотора АсМВЯП DA26. Этот промотор инсертируют перед последовательностью ДНК, кодирующую кутикулярную сигнальную последовательность и АаТН, с оптимизированными кодонами.
Образцы генетически модифицированного АсМВЯП Е2 штамма, содержащие промотор DA26 и ДНК с оптимизацией кодонов, кодирующую кутикулярную сигнальную последовательность и АаТН, синтезируют в соответствии с процедурой, описанной в патентной заявке США 08/070164.
Образцы полученного вирусного соединения, обозначенного АС 1001, сданы на хранение в Американскую Коллекцию Типовых Культур и зарегистрированы в АКТК под номером VR-2404. Другие соединения, использующие последовательности ДНК дикого типа (АаТН), другие гетерологичные сигнальные последовательности и другие промоторы могут быть созданы специалистами в данной области при помощи обычных методик.
Повышение эффективности действия вируса насекомого путем генетической модификации вируса также может состоять во внесении делеции в ген. Примером может служить делеция в гене, кодирующем экдистероид UDP-глюкозил трансферазу ("ЭГТ"). Миллер и др. описали создание таких ЭГТ- штаммов вирусов (16). В частности, Миллер описал создание АсМВЯП ЭГТ- штамма.
Экспрессия egt гена приводит к выработке ЭГТ. ЭГТ инактивирует гормон линьки насекомых (экдизон), в результате чего гусеница не линяет и не окукливается. Когда ген egt инактивирован, например, путем выработки ЭГТ- штамма, линька и окукливание гусеницы, пораженной вирусом, могут продолжаться. В свою очередь это продолжающееся развитие насекомого приводит к таким выгодным для защиты растений результатам, как сокращение питания, замедление роста и более быстрой гибели. Причиной этого является то, что ЭГТ- вирус не может блокировать линьку и окукливание насекомого, а вместе с ними и прекращение питания. Следовательно, насекомые, пораженные ЭГТ- вирусом, гораздо более склонны к более ранней гибели при переходе к линьке в инфицированном состоянии, чем насекомые, пораженные вирусом дикого типа (ЭРГ+). Таким образом, инфицирование насекомых ЭГТ- штаммами более эффективно, чем инфицирование вирусом дикого типа по показателю значения LT50 (время, необходимое для гибели половины насекомых из группы после инфицирования вирусом).
Ген egt инактивируют путем его замены или инсертирования в него другого гена, например невирусного маркерного гена β-галактозидазы. Для нарушения egt гена можно использовать любую последовательность ДНК, которая нарушает экспрессию кодирующей последовательности egt. С другой стороны, все делеции или часть последовательности egt гена можно удалить из генома путем делеции или внесения мутации в определенном участке кодирующей последовательности. Кроме того, можно изменить или удалить регуляторную часть генома, контролирующую экспрессию egt гена. Результатом таких изменений будет отсутствие экспрессии egt гена. Делеции, инактивирующие egt ген, могут быть также получены путем серийных пассажей вируса в насекомых или в культуре клеток насекомого. Все эти инсертирования, делеции или мутации осуществляют при помощи обычных средств. Получаемые путем делеции вирусы имеют то преимущество, что они не содержат чужеродной ДНК и отличаются от вирусов дикого типа только тем, что не имеют функционального egt гена.
Миллер привел в качестве примера АсМВЯП ЭГТ- вируса рекомбинант, обозначенный vEGTDEL, в котором часть egt гена была удалена. Миллер получил vEGTDEL путем котрансфекции в SF клетки плазмиды pEGTDEL (которая является продуктом расщепления плазмиды, содержащей ген egt с EcoRI и XbaI, с удалением части гена) и ДНК из вируса vEGTZ (содержащего lacZ ген, инсертированный без сдвига рамки считывания по отношению к предшествующей кодирующей последовательности egt). Результатом гомологичной рекомбинации является замена сцепленных генов egt-lacZ в vEGTZ на ген egt с удаленным участком из pEGTDEL, что дает рекомбинантный вирус vEGTDEL, являющийся ЭГТ-.
Миллер использовал штамм АсМВЯП, обозначенный L1, который является клональным изолятом первоначально изолированного штамма дикого типа (АТСС VR-1345). Позже был изолирован и охарактеризован штамм АсМВЯП, обозначенный V8. Образцы штамма V8 были приняты на хранение Американской Коллекцией Типовых Культур, 12301 Parklawn Drive, Rockville, Maryland 20852, U.S.A., и были зарегистрированы в АКТК под номером [VR 24651. Описанная Миллером методика создания L1 ЭГТ- применима к созданию штамма V8 ЭГТ-.
Для приготовления смесей в соответствии с данным изобретением используется обычная технология разработки рецептуры, известная специалистам в данной области. Смеси готовят в форме смачиваемых порошков, гранул, суспензий, эмульсий, растворов, растворов для аэрозолей, приманок и других распространенных инсектицидных препаратов.
Смеси часто включают в себя инертный носитель, которым может служить жидкость, например вода, спирт, углеводороды или другие органические растворители, или минеральное, животное или растительное масло, или порошок, например тальк, глина, силикат.
Инсектицидные смеси, в соответствии с данным изобретением, применяются при помощи обычной техники, известной специалистам в данной области: введение вируса в организм насекомого с пищей и путем прямого контакта, при вдыхании (путем разбрызгивания или опыливания растений, которыми питаются насекомые).
Инсектицидные смеси применяют несколькими способами. Вирус и химический инсектицид применяют одновременно в виде одной дозированной формы или одновременно двух дозированных форм. Если используют две дозированные формы, их упаковывают отдельно и потом смешивают, при необходимости в присутствии растворителя, для получения конечной смеси. Альтернативно, вирус или химический инсектицид может быть применен первым для подавления насекомого, а потом применен второй компонент.
Инсектицидные смеси, в соответствии с данным изобретением, применяют в дозе от 2.4•108-2.4•1012 ПТВ/гектар генетически модифицированного вируса и 0.001-1.0 кг/гектар химического инсектицида. Эти нормы представляют дозы, установленные для каждого отдельного компонента, с возможными, благодаря комбинации инсектицидных смесей, в соответствии с данным изобретением, сокращениями.
Концентрации каждого из активных компонентов, необходимы для получения оптимально эффективных инсектицидных смесей для защиты растений, зависят от типа организма, модификаций химического инсектицида и вируса насекомых, используемых в рецептуре смеси. Эти концентрации определяют специалисты в данной области.
В качестве альтернативы химическим инсектицидам с вирусами насекомых комбинируют агенты биологического контроля. Агенты биологического контроля включают бактерии, например Bacillus thuringiensis, которые можно получить в Abbott Laboratories как XENTARITM и DIPELTM 2X. Другие агенты биологического контроля включают простейших, например, Nosema polyvora, M. grandis и Bracon mellitor (5). Кроме них агентами биологического контроля являются энтомопатогенные грибы (5) и нематоды. Нематод применяют в жидких составах или диспергированными в геле, где они находятся в состоянии покоя до момента применения.
Для более глубокого понимания данного изобретения приведены следующие примеры. (Примеры имеют исключительно иллюстративный характер и не должны истолковываться, как ограничивающие диапазон изобретения.)
Примеры
Пример 1
Техника биологических опытов
Используемой в этих примерах методикой биологических опытов является метод нанесения агентов на поверхность питательной среды. Опыты проводят следующим образом. Используются насекомые Н. virescens (табачная листовертка-почкоед) и Heliothis. zea (коробочный червь хлопчатника). Гусениц выращивают на питательной среде, включающей семена сои, зародышей пшеницы и агар (среда Stoneville), полученной от USDA Insectary Labs, Stoneville MS. Каждую партию насекомых содержали при 28oС и при постоянном люминесцентном освещении. Все опыты проводят на среде Stoneville, с гусеницами второго возраста (четырехдневные Н. virescens и трехдневные Н. zea].
Каждая кювета для опытов (корпорации C-D International, Pitman, New Jersey) содержит 32 отдельных ячейки, размером 4•4 см2, содержащих 5 мл среды Stoneville. После обработки среды и заражения насекомых кюветы накрывают прозрачными крышками, смазанными по краям клеем, с вентиляцией (корпорации C-D International). Прозрачные крышки позволяют без затруднений проводить наблюдения.
Для log-пробит анализа (корпорация HRO Group) из исходных ацетоноводных растворов вируса готовят серию разведении: воду дважды дистиллируют. Разведение проводят с шагом от 1•108 до 1•101 ПТВ/мл, в зависимости от изучаемого вида. При необходимости, путем центрифугирования растворы вирусов концентрируют. Растворы химических инсектицидов готовят в различных концентрациях, измеряемых в частях на миллион (ррm), исходя из массы инсектицида на объем растворителя.
На поверхность искусственной среды, после ее остывания, наносят пипеткой 0.4 мл раствора в ацетоне:воде (60:40) одного из следующих компонентов: раствор вируса, раствор химического инсектицида, раствор вируса плюс раствор химического инсектицида или необработанный раствор. Для растворов вируса разведение колеблется от 1•108 до 1•101 ПТВ/мл при 10-кратном разведении, в зависимости от изучаемого вида насекомого. Концентрация используемого химического инсектицида колеблется от 1000 ppm до 0.1 ppm, в зависимости от типа химического инсектицида и вида изучаемого насекомого. Каждый изучаемый раствор проверяют на 32 гусеницах в 3-4-кратной повторности. Применяемые растворы равномерно распределяют по поверхности среды путем вращения кюветы, растворам дают высохнуть в вытяжном шкафу. После высыхания в каждую ячейку помещают одну гусеницу и дают ей возможность питаться в течение 8-10 дней. Н. virescens питаются в течение 8 дней; H.zea 12 дней. Кюветы для опытов содержат при температуре 28oС при непрерывном люминесцентном освещении в течение всего периода изучения. Наблюдения проводят раз в день, чтобы заметить время начала заражения. В каждом наблюдении гусеницу считают мертвой, если она не двигается, даже после встряхивания кюветы со средой, или если ее тело разжижается. Значения LD50 и LD20 для вируса и химического вещества (концентрации, при которых наблюдается 50% и 20% смертность) рассчитывают на основании 3-4 повторностей. Статистическую обработку результатов проводят при помощи SAS log-пробит анализа, определяя значение доза-смертность на 8 и 10 день после обработки. После определения пробит-значений проводят опыты отдельно с химическими инсектицидами, используя предварительно определенные дозы LD50 и LD20, отдельно с вирусами и с LD20 и LD50 дозами, и со всеми возможными комбинациями вирусов с химическими инсектицидами, используя тот же метод нанесения их на поверхность питательной среды. "LD20" - это доза, при применении которой происходит гибель 20% гусениц, "LC50" - доза, при применении которой происходит гибель 50% гусениц.
В таблицах использованы следующие обозначения и сокращения. Концентрация ПТВ/мл обозначена буквой Е, где "Е" - это экспонента. Например, 5Е4 соответствует 5•104. "ДПО" соответствует дню (дням) после обработки. В этих таблицах АсМВЯП "с инсертированным АаТН" - это генетически модифицированный штамм Е2, содержащий промотор DA26 и последовательность ДНК с оптимизированными кодонами, кодирующую кутикулярную сигнальную последовательность и АаТН.
О повышении эффективности борьбы с вредителями при применении смесей, содержащих комбинацию генетически модифицированного вируса насекомых и химического инсектицида, свидетельствовало или увеличение смертности, или скорости уничтожения насекомых, или обоих эффектов одновременно. Примеры 2-5 представляют результаты экспериментов с Helicoverpa zea; примеры 6-8 представляют результаты экспериментов с Heliothis virescens.
Пример 2
Комбинация формамидина амитраза с генетически модифицированными вирусами насекомых
В первом эксперименте формамидин амитраз изучают в комбинации с вирусом насекомых АсМВЯП, модифицированным генетически и содержащим АаТН или ЭГТ-.
Результаты представлены в таблицах 1 и 2.
Результаты приведенных в таблице 1 и 2 данных свидетельствуют о том, что амитраз в концентрации 100 ppm синергирует биоактивность АсМВЯП "с инсертированным АаТН" для гусениц H.zea. Синергизм вышеуказанного вируса в некоторой степени зависит от дозы, поскольку комбинации этого рекомбинантного вируса с амитразом в концентрации 1000 ppm оказывают скорее аддитивный, чем синергический эффект на H.zea. С другой стороны, амитраз не оказывает значительного влияния на биоактивность АсМВЯП "с удаленным ЭГТ" на гусениц Н.zеа. Гибель гусениц при использовании комбинации формамидин/вирус с "удаленным ЭГТ" несколько меньше, чем при применении комбинации вирус "с удаленным ЭГТ".
Пример 3
Комбинация арилпиррола с генетически модифицированными вирусами насекомых
В этом эксперименте арилпиррол 4-бром-2-(р-хлорфенил)-1-(этоксиметил)-5-(трифторметил)-пиррол-3-карбонитрил изучали в комбинации с вирусом насекомых АсМВЯП, который модифицирован генетически таким образом, что содержит или АаТН, или ЭГТ-. Результаты представлены в таблицах 3 и 4.
Результаты приведенных в таблицах 3 и 4 данных показывают, что арилпиррол 4-бром-2-(р-хлорфенил)-1-(этоксиметил)-5-(трифторметил)-пиррол-3-карбонитрил в комбинации с АсМВЯП "с инсертированным АаТН" значительно повышает скорость уничтожения гусениц Н. zea, что видно из анализа данных, полученных на третий день после обработки. В то же время на пятый и восьмой дни после обработки гибель гусениц при применении комбинации арилпиррол/рекомбинантный вирус аддитивная (или чуть меньше, чем аддитивная).
Арилпиррол не оказывает заметного влияния на смертность гусениц H.zea второго возраста при использовании его в комбинации с АсМВЯП-V8 "с делецированным ЭГТ". Тем не менее полученные результаты (гибель гусениц на 3 день после обработки) показывают, что арилпиррол несколько повышает скорость уничтожения гусениц Н. zea при использовании вируса "с делецированным ЭГТ".
Пример 4
Комбинация диацилгидразина с генетически модифицированными вирусами насекомых
В следующем эксперименте диацилгидразин дибензоил-t-бутилгидразин изучался в комбинации с вирусом насекомых АсМВЯП, который модифицирован генетически таким образом, что содержит или АаТН, или ЭГТ-. Результаты представлены в таблицах 5 и 6.
Результаты, представленные в таблицах 5 и 6, свидетельствуют, что диацилгидразин дибензоил-t-бутилгидразин при использовании его в комбинации с АсМВЯП "с инсертированным АаТН" увеличивает скорость уничтожения гусениц Н. zea, что видно из анализа данных, полученных на 3-й день после инфицирования.
Диацилгидразин также значительно увеличивает скорость уничтожения гусениц Н. zea при использовании его в смеси с вирусом АсМВЯП "с делецированным ЭГТ" [исходя из данных, полученных на 3-й день после обработки].
Пример 5
Комбинация бензоилфенилуреата с генетически модифицированными вирусами насекомых.
В следующем эксперименте бензоилфенилуреат дифторбензурон изучается в комбинации с вирусом насекомых АсМВЯП, который модифицирован генетически таким образом, что содержит или АаТН, или является ЭГТ-. Результаты представлены в таблицах 7 и 8.
Результаты, представленные в таблицах 7 и 8, показывают, что добавление дибензофенилуреата дифторбензурона не повышает инсектицидного действия АсМВЯП-Е2 "с инсертированным АаТН" для гусениц Н. zea, более того, смертность гусениц Н. zea при использовании этой комбинации меньше аддитивной.
Бензофенилуреат также не повышает эффективности действия АсМВЯП-Е2 "с делецированным ЭГТ" на гусениц Н. zea, более того, смертность гусениц Н. zea при применении этой комбинации меньше аддитивной.
Пример 6
Комбинация пиретроида с диким типом или генетически модифицированным вирусом насекомых.
В следующем эксперименте (с использованием гусениц Н. zea второго возраста) пиретроид циперметрин изучают в комбинации с вирусом насекомых АсМВЯП, который принадлежит дикому типу или модифицирован генетически таким образом, что содержит или АаТН, или является ЭГТ-. Результаты представлены в таблицах 9-14.
В таблице 9 отображены данные, полученные при использовании комбинации циперметрина со штаммом АсМВЯП-Е2 дикого типа. В этой комбинации используются дозы, вызывающие при применении каждого компонента отдельно смертность 20% гусениц (LD20).
Результаты, представленные в таблице 9 свидетельствуют, что при использовании комбинации циперметрина с вирусом АсМВЯП "дикого типа" явления синергизма не наблюдается (смертность гусениц при использовании комбинации и отдельных компонентов существенно не различалась). Аналогичные результаты были получены для Аспирота (1).
В таблице 10 представлены данные, полученные при использовании комбинации циперметрина и штамма АсМВЯП V8 ЭГТ-. В этой комбинации используются дозы, вызывающие при применении каждого компонента отдельно смертность 20% гусениц (LD20).
Результаты, представленные в таблице 10, свидетельствуют, что при использовании комбинации циперметрина с АсМВЯП " с делецированным ЭГТ" наблюдается явление синергизма в отличие от комбинации циперметрина с АсМВЯП "дикого типа".
Таблица 11 отображает данные для комбинации циперметрина и штамма АсМВЯП Е2 "с инсертированным АаТН". В этой комбинации используются дозы, вызывающие при применении каждого компонента смертность 20% гусениц (LD20).
Результаты, представленные в таблице 11, свидетельствуют, что при использовании комбинации циперметрина с АсМВЯП " с инсертированным АаТН" наблюдается, по сравнению с комбинацией циперметрина с АсМВЯП "дикого типа", увеличение скорости уничтожения гусениц. Так, смертность гусениц на 1 и 4 дни после обработки при использовании циперметрина с АсМВЯП "дикого типа" составляло соответственно 5 и 31%, а при использовании циперметрина с АсМВЯП "с инсертированным АаТН" соответственно 22 и 38%.
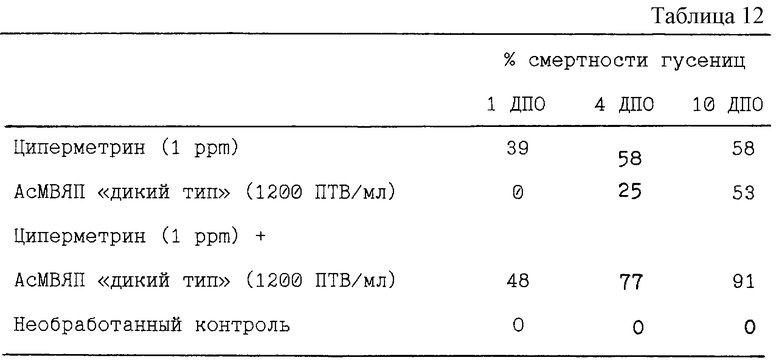
Таблица 12 отображает данные для комбинации циперметрина и штамма АсМВЯП Е2 "дикого типа". В этой комбинации используются дозы, вызывающие при применении каждого отдельного компонента смертность 50% гусениц (LD50).
Результаты, представленные в таблице 12, свидетельствуют, что при использовании циперметрина с АсМВЯП "дикого типа" синергизм наблюдается лишь на 10 день после обработки гусениц.
В таблице 13 отображены данные о смертности гусениц при использовании комбинации циперметрина и штамма АсМВЯП V8 ЭГТ-. В этой комбинации применены дозы, вызывающие при использовании отдельно циперметрина смертность 20% гусениц (LD20), а для штамма АсМВЯП V8 ЭГТ- - смертность 50% гусениц (L50).
Результаты, представленные в таблице 13, свидетельствуют, что при использовании комбинации циперметрина с АсМВЯП "с делецированным ЭГТ" смертность гусениц больше, чем при использовании каждого компонента этой комбинации отдельно, что свидетельствует о синергизме, в отличие от наблюдений при применении комбинации циперметрина с вирусом "дикого типа". При этом при применении циперметрина с генетически модифицированным вирусом используется низкая доза концентрации циперметрина.
В таблице 14 отображены данные о смертности гусениц при использовании комбинации циперметрина и штамма АсМВЯП Е2 "с инсертированным АаТН". В этой комбинации используются дозы, вызывающие при применении каждого компонента отдельно гибель 50% гусениц (LC50).
Результаты, представленные в таблице 14, свидетельствуют, что при использовании комбинации циперметрина с АсМВЯП "с инсертированным АаТН" смертность гусениц на 4 и 10 дни после обработки была выше, чем при использовании отдельно каждого компонента, что свидетельствует о синергизме между примененными компонентами, в отличие от отсутствия синергизма при применении комбинации циперметрина с АсМВЯП "дикого типа".
Итак, комбинация циперметрина с вирусом, который модифицирован генетически таким образом, что содержит или АаТН, или является ЭГТ-, более эффективна для борьбы с гусеницами Н.zea, чем комбинация циперметрина и вируса "дикого типа". Эти результаты нельзя предсказать, исходя из полученных ранее данных для комбинаций вирусов "дикого типа" с пиретроидами (1).
Пример 7
Комбинация диацилгидразина с вирусом насекомых "дикого типа" или с модифицированным генетически
В следующем эксперименте (с Н. virescens третьего возраста) диацилгидразин дибензоил-t-бутилгидразин изучается в комбинации с вирусом насекомых АсМВЯП, который принадлежит к "дикому типу" или модифицирован генетически таким образом, что является ЭГТ- (штамм L1). Результаты представлены в таблицах 15-16. В этих комбинациях используются более низкие дозы, чем в примере 6.
В таблице 15 отображены данные о смертности гусениц при использовании комбинации диацилгидразина и штамма АсМВЯП L1 "дикого типа".
Синергизм для этой комбинации наблюдается на 4 и 10 дни после обработки.
В таблице 16 отображены данные о смертности гусениц при использовании комбинации диацилгидразина и генетически модифицированного АсМВЯП ЭГТ- (штамм L1).
Результаты свидетельствуют, что использование этой комбинации приводило, по сравнению с применением отдельных компонентов, к небольшому увеличению смертности на 4 день после обработки.
Пример 8
Комбинация арилпиррола с вирусом насекомых "дикого типа" или модифицированными генетически
В следующем эксперименте (с гусеницами Н. virescens второго возраста) арилпиррол 4-бром-2-(р-хлорфенил)-1-(этоксиметил)-5-трифторметил)-пиррол-3-карбонитрил изучается в комбинации с вирусом насекомых АсМВЯП, который принадлежит "дикому типу" или модифицирован генетически таким образом, что содержит АаТН или является V8 ЭГТ-. Результаты представлены в таблицах 17-19. В таблице 17 отображены данные для комбинации диацилгидразина и штамма АсМВЯП Е2 "дикого типа". В этой комбинации используются дозы, вызывающие при применении каждого агента отдельно гибель 20% насекомых (LD20).
Анализ данных о смертности гусениц в таблице 17 свидетельствует, что при использовании комбинации арилпиррола с АсМВЯП "дикого типа" явление синергизма наблюдали на 4 и 10 дни после обработки.
В таблице 18 отображены данные о смертности гусениц при использовании комбинации арилпиррола с генетически модифицированным АсМВЯП ЭГТ- (штамм V8).
Анализ приведенных в таблице 18 данных свидетельствует об увеличении скорости уничтожения гусениц при использовании арилпиррола с АсМВЯП "с делецированным ЭГТ" по сравнению с комбинацией арилпиррола с АсМВЯП "дикого типа" (таблица 17), о чем свидетельствует смертность гусениц на 1-й день после обработки, составившая соответственно 33% и 2%.
В таблице 19 отображены данные о смертности гусениц при использовании комбинации арилпиррола с генетически модифицированным штаммом АсМВЯП Е2 с инсертированным АаТН.
Анализ приведенных в таблицах 19 и 17 данных свидетельствует, что применение комбинации арилпиррола с АсМВЯП "с инсертированным АаТН" увеличивало, по сравнению с комбинацией арилпиррола с АсМВЯП "дикого типа", скорость уничтожения гусениц, которая составила на 1-й и 4-й дни после обработки 39 и 69% (таблица 19) и 2 и 19% (таблица 17).
Итак, на основе всех опытов можно сказать, что комбинация арилпиррола 4-бром-2-(р-хлорфенил)-1-(этоксиметил)-5-(трифторметил)-пиррол-3-карбонитрила с вирусом модифицированного генетически таким образом, что он содержит АаТН или является ЭГТ-, более эффективна, чем комбинация арилпиррола с вирусом "дикого типа".
Источники информации
1. Аспирот Дж. и др., патент США 4668511.
2. Мохамед А. И. и др., Environ. Entomology, 12, 478-481 (1983).
3. Мохамед А.И. и др., Environ. Entomology, 12, 1403-1405 (1983).
4. Величкова-Кожухарова М. и др., Rasteniev'dni Nauki, 25, 80-86 (1988).
5. Жакс Р.П. и др. "Совместимость патогенов с другими методами контроля численности вредителей и с различными культурами", глава 38, страницы 695-715.
6. Джирвлайт Дж. Б.Ф. и др., Med. Fac. Landbouww. Rijksuniv. Gent. 56, 305-311 (1991).
7. 3лоткин Э. и др., Toxicon, 9, 1-8 (1971).
8. Томальский М.Д. и др., патент США 5266317.
9. Мартенc Дж. У. М. и др., App.& Envir. Microbiology, 56, 2764-2770 (1990).
10. Федеричи Б.A., In Vitro, 28, 50A (1992).
11. Джексон Дж. Р. X. и др., патент США 4925664.
12. Элдридж Р. и др., Insect Biochem.. 21, 341-351 (1992).
13. Мэнн Дж. Дж. и др. J. Agric. Food Chem., 37, 271-278 (1989).
14. Хэммок Б. Д. и др. Nature, 344, 458-461 (1990).
15. Патентная заявка США, серийный номер 08/009, 265, зарегистрирована 25 января 1993.
16. Миллер Л. К. и др. Международная патентная заявка WO 91/00014.















Изобретение относится к области биотехнологии. Инсектицидные композиции включают смеси генетически модифицированных вирусов насекомых с химическими и биологическими инсектицидами. Генетически модифицированный вирус содержит инсертированный ген, который экспрессирует насекомое, или модифицирующее вещество, например токсин, нейропептид, гормон или энзим. Генетически модифицированный вирус также содержит делецию в гене. Инсектицидная смесь обладает высокой биологической активностью. 2 с. и 3 з.п. ф-лы, 19 табл., 2 ил.
| Огнетушитель | 0 |
|
SU91A1 |
| ТАТУИРОВОЧНАЯ МАШИНА | 2013 |
|
RU2532522C1 |
| US 4668511 А, 05.05.1987. | |||

