Изобретение относится к анализам и, в частности к катализируемому осаждению репортера через активированный конъюгат для усиления детектируемого сигнала, вследствие чего улучшается детекция и/или количество аналита в образце.
Введение иммунодиагностических анализов в 1960-х и 1970-х годах значительно увеличило количество аналитов, поддающееся точному и тщательному измерению.
Радиоиммуноанализы (RIAS) и иммунорадиометрические анализы (IRMA) используют радиоизотопное мечение либо антитела, либо конкурирующего антигена для измерения аналита. Системы детекции, основанные на ферментах или флуоресцентных метках, были развиты как альтернативные системам изотопной детекции. Анализы, основанные на ферментах, оказались более чувствительными, более быстро осуществимыми, менее зависимыми от дорогого, сложного оборудования.
Необходимость диагностических анализов, имеющих более простой объем, с повышенной чувствительностью, с меньшей зависимостью от сложного дорогого оборудования побудила исследователей попытаться вовлечь каталитическую способность ферментов, чтобы развивать эти новые анализы.
D. L. Bates, Trends in Biotechnology, p. 204-209, V.5, N 7 (1987) описывает диагностику, которая использует метод энзимной амплификации для развития более чувствительных и простых иммуноанализов. В этом методе фермент вторичной системы связывается с ферментом первичной метки, например первичный энзим можно связать каталитически с дополнительной системой, такой как субстратный цикл или каскадная реакция с участием энзимов. Таким образом, суть энзимной амплификации в соответствии с Bates заключается в соединении каталитических процессов, где фермент модулируется действием второго фермента либо посредством прямой модификации, либо при взаимодействии с продуктом этого контролирующего фермента.
US Patent 4668621 описывает анализ энзим-связанного коагулята (ELCA) для совершенствования иммуноанализа с амплификацией, используя свертывающийся каскад для усиления детекции иммунных комплексов. Этот процесс включает формирование сгустка, обусловленное тромбином активируемое образованием фибрина из нерастворимого фибриногена и меченого растворимого фибриногена. Амплификация количества лиганда-репортера, прикрепленного к твердому носителю, получается только комбинированием использования свертывающегося фактора конъюгатов с последующими каскадными реакциями коагуляции. Один из недостатков этой системы то, что она может использоваться только для измерения присутствия факторов, которые модулируют активность одного или более факторов свертывания крови. Другой недостаток заключается в том, что первичный фермент, тромбин, нельзя иммобилизовать или связать с репортером, или членом группы специфических связывающих пар.
US Patent 4463090, вышедший в авторстве Harris 31 июля, 1984, описывает иммуноанализ с использованием каскадной амплификации, требующий комбинации по крайней мере двух последовательных катализаторов, где первый фермент активирует второй фермент, который в свою очередь действует на субстрат.
Еще одна система амплификации описана в US Patent 4598042, вышедшем в авторстве Self 1 июля, 1986, и UK Patent Application N 2059421, которая опубликована 23 апреля, 1981, которые раскрывают иммуноанализ, использующий ферментную метку для образования прямым или непрямым способом вещества, которое способно влиять на каталитический процесс без истощения его в процессе катализа. Более подробно, система первичного фермента дает или удаляет вещество, способное модулировать систему вторичного фермента, которая приводит к амплификации. Эти ферментные системы используют неконъюгировавшие ферменты, чтобы избежать тенденцию инактивировать определенные энзимы при конъюгации.
European Patent Application Publication N 123265, который опубликован 31 октября, 1984, описывает еще один иммуноанализ каскадной амплификацией, где зимоген-родственный энзим связывают с последовательностью каскадной реакции зимоген-энзим, чтобы получить множественные стадии амплификации в получении материала детектируемого маркера, используемого для оценки количества аналита.
European Patent Application Publication N 144744, опубликованный 19 июня, 1985 описывает анализ с использованием специфического связывания, основанный на энзимной каскадной амплификации, где меченый компонент, используемый для детектируемого реагента, является участником или модулятором энзимной каскадной реакции, где первый фермент действует на первый субстрат, чтобы получить второй фермент. Может следовать производство второго фермента или второй фермент может действовать на второй субстрат для получения третьего фермента.
Подобным образом US Patent 4318980, выданный Богуславскому и др. 9 марта, 1982, описывает анализ на гетерогенное специфическое связывание, используя конъюгат, сформированный из специфической связывающейся последовательности, соединенной с реагентом. Способность реагента участвовать в контролировании реакции детектирования присутствия аналита изменяется в присутствии лиганда в среде. Таким образом, конъюгант в его свободном состоянии более активный в контролировании реакции, чем в связанном виде.
Анализ на гетерогенное специфическое связывание, используя энзимную аплификацию, описан в British Patent Application N 1401297, который опубликовано 30 июля, 1975 и US Patent 4376825, выданный Рубинштейну и др. 15 марта, 1983. Амплификация достигается связыванием компаунда, который надо анализировать, или его аналога с ферментом. Полученный энзим-связанный лиганд конкурирует со свободным лигандом за специфические местоположения рецептора. Когда энзим-связанный лиганд замещается свободным лигандом, фермент тогда свободен для реагирования с большим количеством молекул субстрата и можно измерить концентрацию остающегося субстрата или продукта. Заявка РСТ International Publication No Wo 81/00 725, которая опубликована 19 марта, 1981, описывает метод определения субстрата в образце, который включает превращение субстрата в продукт на первой стадии последовательности циклической реакции и превращение продукта обратно в субстрат на второй реакционной стадии последовательности циклической реакции. По крайней мере одна из первой и второй реакционных стадий катализируется ферментом.
РСТ Application, имеющая International Publication Number Wo 84/02 193, которая опубликована 7 июня, 1984, описывает иммуноанализ с хромогенным носителем, где аналит контактирует с меченным ферментом антителом, в котором сигнал, даваемый реакцией фермента со своим субстратом, концентрируется на активном носителе.
European Patent Application Publication N 181762, опубликованное 21 мая, 1986, описывает метод определения энзиматической активности в жидком образце путем агглютинации или ингибирования частиц агглютинации.
Субстрат /кофактор циклизация другой пример амплификации, которая основана на включении в цикл кофактора или субстрата, который образован первичной ферментной меткой. Первичный фермент превращает первичный субстрат в активную форму, которую можно циклизовать двумя ферментами цикла амплификации. Эти два фермента поставляются в высоких концентрациях и уравновешиваются для превращения высокими концентрациями субстрата, но сохраняются до тех пор, пока образуется циклический субстрат. Продукт первичного фермента является каталитическим активатором цикла амплификации, который реагирует пропорционально концентрации субстрата и, следовательно, концентрации энзимной метки.
В начале шестидесятых Lowry и др. Journal of Biological Chemistry, p. 2746 2755, V. 236, N 10 (октябрь 1961) описал измерение пиримидиновых нуклеотидов путем энзиматической циклизации, в которой коэнзим, который надо было определить, использовали для амплификации энзиматической дисмутации между двумя субстратами.
Более сложная система циклизации субстрата описана US Patent 4745054, выданном Rabin и др. 17 мая, 1988. Система Rabin включает использование маленького энзиматически инактивированного пептидного фрагмента фермента в качестве метки и конъюгированного с комплементарным фрагментом для формирования фермента, который катализирует первичную реакцию, чей продукт является или приводит к необходимому для второго фермента коэнзима или простетической группе, которые катализируют вторичную реакцию, приводящую к детектируемому результату, указывающему на наличие аналита.
Vary и др. Clinical Chemistry, р. 1696 1701, V.32, (1986) описывает метод амплификации, подходящий для нуклеиновых кислот. Это стандартная реакция замещения, которая использует уникальную способность какого-либо полинуклеотида действовать как метка субстрата, которая может освобождаться фосфорилазой.
Изобретение касается способа обнаружения или определения количества аналита, согласно которому в анализе используется аналитзависимая ферментная активирующая система, включающая по крайней мере один фермент, взаимодействующий с конъюгатом, состоящим из детектируемо меченого субстрата, специфичного для ферментной системы, для формирования активированного конъюгата, который осаждается субстратом где бы ни был иммобилизован, по крайней мере один рецептор для активированного конъюгата, причем указанный рецептор не реагирует с аналитзависимой активирующей системой, а осажденные детектируемые метки непосредственно или посредственно вызывают сигнал, который можно детектировать или количественно определить.
Причем, аналитзависимая ферментная активирующая система имеет по крайней мере один фермент, выбранный из группы, состоящей из оксидоредуктаз, гидролаз, лиаз, трансфераз, изомераз или лигаз.
Целесообразно, чтобы фермент был выбран из группы, состоящей из пероксидазы, глюкозооксидазы, фосфатазы, эстеразы и глюкозидазы, предпочтительно из проксидазы хрена, глюкозооксидазы, щелочной фосфатазы, эстеразы печени свиньи и бета-галактозидазы; наиболее предпочтительно, когда фермент является пероксидазой хрена.
Согласно заявленному способу конъюгат выбирают из группы, состоящей из биотин-тирамина, n-гидроксифенилпропионил-биоцитина или флюоресцинтирамина или N -(4'''-гидроксифенетил)-6-(фенокси-(4'-азо-2''- бензойной кислоты)) гексамида, а также конъюгатом является 6-(фенокси- (4'-азо-2''-карбоксиэтилфенил) гексаноил)-щелочная фосфотаза.
В заявленном способе конъюгат реагирует с детектируемо меченным антителом; более предпочтительно конъюгат реагирует с детектируемо меченным стрептавидином.
По способу согласно изобретению осуществляют иммобилизацию аналитзависимой ферментной активирующей системы, которая катализирует осаждение фермента путем активации конъюгата, состоящего из меченного ферментом специфического субстрата для энзимной системы, причем данный конъюгат реагирует с аналитзависимой ферментной активирующей системой для формирования активированного конъюгата, который выпадает в осадок, где бы ни был иммобилизован рецептор для активированного конъюгата.
В другом варианте согласно заявленному изобретению способ обнаружения или определения количества аналита заключается в том, что в анализе используется аналитзависимая ферментная активирующая система, реагирующая с иммобилизованным аналитом и включающая по крайней мере один фермент, взаимодействующий с конъюгатом, состоящим из детектируемо меченного субстрата, специфичного для ферментной системы, для формирования активированного конъюгата, который осаждается субстратом где бы ни был иммобилизован, по крайней мере один рецептор для активированного конъюгата, причем указанный рецептор не реагирует с аналитзависимой активирующей системой, а осажденные детектируемые метки непосредственно или посредственно вызывают сигнал, который можно детектировать или количественно определить.
В указанном варианте аналитзависимая ферментная активирующая система имеет по крайней мере один фермент, выбранный из группы, состоящей из оксидоредуктаз, гидролаз, лиаз, трансфераз, изомераз или лигаз.
И в этом случае фермент выбран из группы, состоящей из пероксидазы, глюкозооксидазы, фосфатазы, эстеразы и глюкозидазы; фермент может быть также выбран из пероксидазы хрена, глюкозооксидазы, щелочной фосфатазы, эстеразы печени свиньи и бета-галактозидазы.
Предпочтительно, когда фермент является пероксидазой хрена.
По способу согласно вышеуказанному варианту конъюгат выбирают из группы, состоящей из биотин-тирамина, n-гидроксифенилпропионил-биоцитина, или флюоресцин-тирамина, или N-(4'''-гидроксифенетил)-6-(фенокси-(4'-азо- 2''-бензойной кислоты))гексамида.
При этом конъюгат реагирует с детектируемо меченным членом группы специфических связывающих пар; или конъюгат реагирует с детектируемо меченным антителом, а анализ является иммуноанализом.
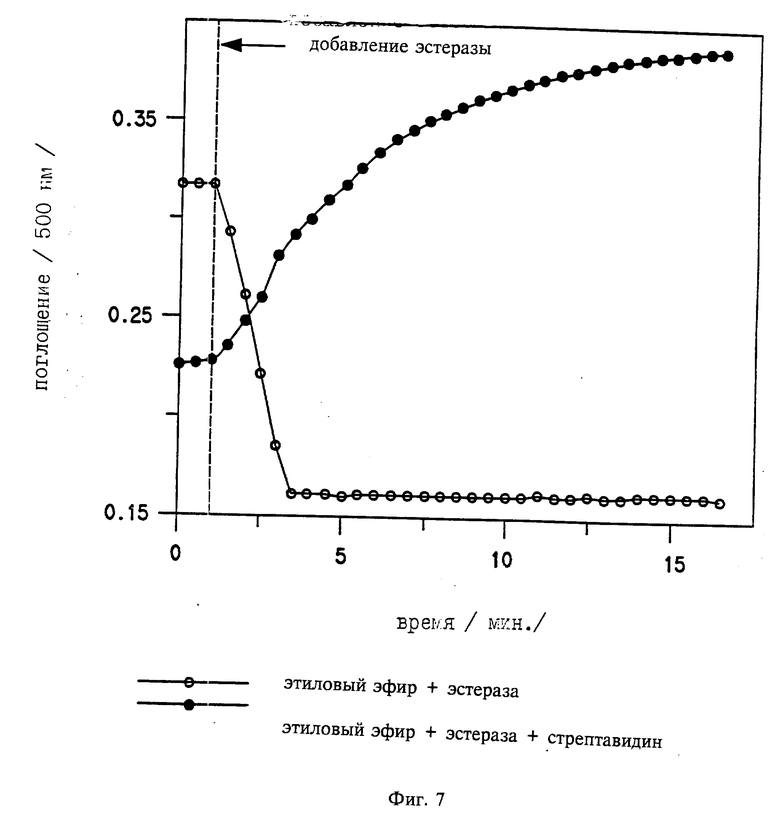
На фиг. 1 приведен график, сравнивающий результаты HSY антигенного анализа, проводимого с и без катализируемого осаждения репортера; на фиг. 2 - график, сравнивающий результаты анализа с использованием HIV p 24 ядра антигена, используя концентрации конъюгата 0,2; 0,4 и 0,8 мкл/мл (Amp. 1,2 и 3 соответственно). "HRP" представляет "неамплифицированный" анализ, где антитело-детектор непосредственно мечено HRP. "Биотин" указывает на еще один "неамплифицированный" анализ, где антитело-детектор конъюгировано с биотином и детектировалось с HRP меченным стрептавидином; на фиг. 3 график анализа IgG мыши, проведенного, используя HRP ADEAS для катализа осаждения биотин-тирамина, который детектировали со стрептавидин HRP (HRP Amp. HRP) или со стрептавидин AР (HRP Amp. AР). Анализ также проводили, используя только HRP меченный детектор-антитело или AP меченное антитело-детектор; на фиг. 4 два графика, сравнивающих результаты, полученные с Du Pont HIV p 24 антиген ELISA, сделанным с и без использования катализируемого осаждения репортера для амплификации сигнала репортера. на фиг. 5 график, сравнивающий результаты анализа IgG мыши без катализируемого осаждения репортера (HRP) и с катализируемым осаждением репортера (HRP- β -Gal); фиг. 6 описывает приготовление этил 2-(4'-гидроксифенилазо)бензоат- 6-щелочной фосфатазы (НЕЕ-6-AР), синтез которой описан в примере 10. фиг. 7 иллюстрирует катализируемое эстеразой превращение НЕЕ в 2- (4'-гидроксифенилазо) бензойную кислоту (HABA) стрептавидиновый комплекс.
Термин аналитзависимая ферментная активирующая система (ADEAS) относится к ферментной системе (i), в которой по крайней мере один фермент связан любым образом, известным для специалистов в этой области, с членом группы специфических связывающих пар, или (ii) ферменту, который не надо связывать с членом группы специфических связывающих пар, когда он является аналитом. Этот фермент либо сам, либо вместе с вторым ферментом катализирует образование активированного конъюгата, который затем осаждается, где бы ни был иммобилизован рецептор активированного конъюгата.
Термин амплификация в том виде, как здесь использован, обозначает амплификацию сигнала репортера, обусловленного осаждением конъюгата, активированного ADEAS.
Термин конъюгат обозначает детектируемый меченый субстрат, специфичный для ADEAS, как если бы это был, одноферментный или мультиферментый ADEAS. Субстрат должен иметь по меньшей мере один компонент, но не ограничиваться им. Например, субстрат может состоять из двух компонентов. Один компонент содержит связывающий сайт для рецептора и мечен для детекции. Другой компонент составная часть, которая предотвращает или мешает связыванию с рецептором до тех пор, пока ADEAS подготавливает конъюгат, как обсуждается ниже. Другой пример конъюгата биотин-тирамин, где тирамин является субстратной частью и биотин образует детектируемую метку, как описано ниже. Ниже также описаны более подробно конъюгаты.
Термин детектируемо меченый означает, что субстрат может быть связан или с репортером, или с немеченым первым членом из группы специфических связывающих пар при условии, что репортер предоставляет другую половину субстрата. Когда субстрат после осаждения связывается с немеченым членом группы специфических связывающих пар, комплекс субстрат-специфических связывающихся партнеров реагирует с вторым членом группы специфических связывающихся пар, который присоединен к репортеру. Альтернативно, комплекс субстрат- специфических связывающихся партнеров может предварительно реагировать с детектируемо меченым другим членом группы специфически связывающихся пар перед осаждением.
Термин осаждение означает непосредственное связывание активированного конъюгата с рецептором, которое происходит либо из-за формирования ковалентной связи, либо при специфическом взаимодействии связывающихся пар.
Термин рецептор обозначает сайт, который будет связываться с активированным конъюгатом либо посредством формирования ковалентной связи, либо специфического взаимодействия связывающихся пар.
Термин активированный конъюгат обозначает, что данный конъюгат подготовлен ADEAS для связывания с рецептором.
Одна из уникальных особенностей этого изобретения заключается в том, что аналитзависимая ферментная активирующая система, которая катализирует осаждение конъюгата путем превращения субстратной части конъюгата в активированную форму, осаждается куда бы ни был иммобилизован специфический рецептор для активированного конъюгата. ADEAS не использует ферментные каскадные реакции или энзимный цикл для влияния на амплификацию, скорее она использует либо один энзим, либо комбинацию энзимов для активации конъюгата. Осаждение конъюгата происходит только, если аналит или аналитзависимая ферментная активирующая система, которая может быть та же, если аналит-энзим, например, при детекции такого энзима, как щелочная фосфотаза, или какого другого, иммобилизован, и рецептор иммобилизован, чтобы связать активированный конъюгат. Таким образом, ADEAS, конъюгат и рецептор выбраны, чтобы сформировать оперативное трио.
Одно воплощение одноферментной ADEAS системы, прилагаемой к препровождаемому иммуноанализу по типу сэндвича: тестируемый образец, содержащий аналит, реагирует с иммобилизованным захватывающим реагентом, таким как антитело; избыток реагента отмывается; иммобилизованный захваченный комплекс антитело-аналит реагирует с ADEAS, таким как второе антитело, специфичное к аналиту, который связан с ферментом, например пероксидазой хрена (HRP), щелочной фосфатазой (AP) и т.д.
ADEAS связывает только, если аналит связан с захватывающим реагентом. Иначе реагенты смываются. Связывание энзима со специфическим связывающим партнером не влияет на энзимную способность реагировать с субстратной частью конъюгата. Когда конъюгат, такой как биотин тирамин или HABA-тираминовый аналог (например, N (4'''-гидроксифенетил)-6- (фенокси-(4'-азо-2''-бензойной кислоты)) гексамид) добавляют к иммобилизованному захватывающему комплексу антитело-аналит-второе антитело-энзим, фермент реагирует с субстратной частью конъюгата, например с тираминовой частью конъюгата, превращая ее в активную форму, которая будет связываться с иммобилизованным рецептором, который является либо эндогенным, либо экзогенным в данной системе анализа.
Количество выпавшего конъюгата будет функцией иммобилизованного ADEAS. Осажденный конъюгат, такой как биотин тирамин или HABA тираминовый аналог, можно затем обнаружить путем реагирования со стрептавидин-HRP и ортофенилендиамином. Термин HABA-тираминовый аналог означает незамещенный или замещенный HABA, связанный или не связанный с гидроксифенилсодержащим соединением, таким как тирамин. Если конъюгатом является флуоресцеин-тирамин, в этом случае осажденный конъюгат может детектироваться непосредственно или вслед за реакцией с меченным антифлуоресцеин антителом.
Таким образом, ADEAS используется для катализирования выпадения детектируемо меченого субстрата (конъюгата), чтобы дать дополнительный сигнал. ADEAS детектируется непосредственно как часть общего сигнала, когда ферментный компонент ADEAS тот же, что и фермент, использованный в качестве репортера. Фиг. 3 иллюстрирует эту ситуацию, так же как и ситуацию в которой ADEAS ферментный компонент и фермент-репортер разные и поэтому ADEAS ферментный компонент не детектируется непосредственно как часть суммарного сигнала.
Иммуноанализ с использованием мультиферментного ADEAS включает в себя подобный подход. В вышеуказанном примере ADEAS может быть антителом, связанным с ферментом, таким как AP. В добавок к иммобилизованному захватывающему комплексу антитело-аналит-второе антитело-AP, второй фермент, такой как HRP, можно было бы иммобилизовать на носителе. Конъюгатом может быть детектируемо меченый фенилфосфат, который не может реагировать с HRP до тех пор, пока его не дефосфарилировали. AP дефосфарилирует фенол, который затем освобождается для реагирования с иммобилизованным HRP, чтобы сформировать активированный фенольный конъюгат, который осаждает где бы ни были иммобилизованы рецепторы. После удаления избытка реагента выпавший репортер детектируют и оценивают количественно. Альтернативно HRP может быть связан с вторым антителом и AP может быть иммобилизован на поверхности носителя.
Изобретение является удивительным и неожиданным, потому что амплификацию сигнала репортера получают через выпавший активированный конъюгат без использования каскадных механизмов или ферментного цикла. ADEAS реагирует с конъюгатом с формированием активированного конъюгата, который будет связываться с иммобилизованным репортером, специфичным для активированного конъюгата. Количества рецептора и активированного конъюгата берутся в избытке к количеству иммобилизованного ADEAS.
В выборе ADEAS руководствуются способностью определенного фермента или ферментов превращать конъюгат в активированную форму, которая будет связываться с иммобилизованным рецептором, эндогенным либо экзогенным. Таким образом, детальное значение каталитических свойств каждого специфического фермента нужно для того, чтобы правильно подобрать субстрат и рецептор. Другие важные факторы включают наличие данного фермента или ферментов, сравнительная легкость и трудность связываться с членом группы специфических связывающихся пар, стабильность этого фермента или ферментов, также как и стабильность конъюгата и рецептора. В некоторых случаях ADEAS можно купить (в зависимости от объема анализа).
Энзимы, подходящие для использования в ADEAS, включают гидролазы, лиазы, оксидоредуктазы, трансферазы, изомеразы и лигазы. Можно упомянуть и пероксидазу, глюкозооксидазу, фосфатазу, эстеразу и глюкозидазу. Специфические примеры включают щелочную фосфатазу, липазы, бета-галактозидазу, пероксидазу хрена и эстеразу из печени свиньи.
Члены группы специфических связывающих пар, подходящие для использования согласно изобретению, могут быть иммунного или неиммунного типа. Иммунные специфические связывающие пары служат примером систем антиген/антитело или систем гаптен/антигаптен. Антитело поликлональное, моноклональное или иммунореактивный фрагмент его, как член группы связывающих пар, может продуцироваться привычными методами, знакомыми специалистам в этой области. Термины иммунореактивный фрагмент антитела или иммунореактивный фрагмент обозначают фрагменты, которые содержат связывающую область антитела. Такие фрагменты могут быть Fab-типа, фрагментами, которые определены как фрагменты без Fc части, например, Fab, Fab' и F(ab')2 фрагменты, или могут быть так называемыми фрагментами "полумолекул", полученными путем восстановительного расщепления дисульфидных связей, соединяющих компоненты тяжелой цепи интактного антитела. Если антигенный член специфических связывающих пар неиммуногенный, например гаптен, он может быть ковалентно связан с белком-носителем, чтобы сделать его иммуногенным.
Неимунные связывающие пары включают системы, где два компонента разделяют естественное сродство друг с другом, но не являются антителами. Примеры неиммунных связывающих пар: биотин-авидин или биотин-стрептавидин, фолиевая кислота-фолят связывающий белок, комплементарные пробы нуклеиновых кислот и т. д. Включены также неиммунные связывающие пары, которые формируют ковалентную связь друг с другом, но не являются антителами. Примеры ковалентно связывающихся пар включают сульфгидрильные реактивные группы, такие как малеимиды и галоацетилпроизводные, и аминные реактивные группы, такие как изотиоцианаты, эфиры сукцинимидина и сульфонилгалоидные соединения и т.д.
Подходящие подложки, используемые в анализах, включают синтетические подложки из полимеров, такие как полистирол, полипропилен, замещенный полистирол, например, аминированный или карбоксилированный полистирол, полиакриламиды, полиамиды, поливинилхлорид и т.д. стеклошарики, агароза, нитроцеллюлоза и т.д.
Другой важный компонент изобретения конъюгат. т.е. детектируемо меченый субстрат, который должен быть специфичным для ADEAS. Как оговорено выше, когда конъюгат реагирует с ADEAS, фермент или ферменты катализируют образование активированного конъюгата, который связывает рецептор, где бы он ни был иммобилизован, экзогенный ли или эндогенный. Иммобилизованный экзогенный рецептор означает рецептор, который не возникает в процессе анализа. Он может быть иммобилизован на поверхности носителя перед добавлением конъюгата в реакционную смесь. Эндогенный рецептор означает рецептор, который образуется в процессе этого анализа и не требует иммобилизации перед добавлением конъюгата, потому что рецептор иммобилизован в системе анализа.
Например, когда HRP ADEAS (HRP, связанный с членом группы специфических связывающих пар) реагирует с конъюгатом, содержащим фенольный субстрат, образуется активированный фенольный субстрат. Считается, что активированный фенольный субстрат связывается с электрон- избыточными остатками, такими как тирозин и триптофан, присутствующими в белках на твердом носителе. Однако, если используется какой-либо другой конъюгат, такой как меченый 3-метил-2-бензотиазолин гидразин (MBTH), который обсуждается ниже, рецептор, такой как 3-(диметиламино) бензойная кислота (DMAB), должен быть иммобилизован перед добавлением конъюгата.
Другое воплощение включает реагирование конъюгата, который фосфорилирует с помощью ADEAS. Активированный (фосфорилированный) конъюгат способен затем реагировать с антителом, специфичным для активированного конъюгата.
В еще одном варианте с ADEAS может реагировать конъюгат, включающий компонент, который затем в активированной форме будет связываться с рецептором и который связывается с компонентом, имеющим тиольную реактивную группу, такую как малеимид. Осажденная малеимидная часть может затем детектироваться реакцией с сульфогидрилсодержащим репортером, который может быть эндогенным для репортера, например бета-галактозидаза, или сульфгидрильные группы могут быть добавлены к репортерам, таким как HRP или AP, используя меркаптидные реагенты, такие как N-сукцинимидил-S-ацетилтиоацетат (SATA), S-ацетилмеркаптосукцинангидрид (SAMSA) или сукцинимидил-3-(ацетилтио) пропионат (SATP).
Альтернативно субстрат можно связать с защищенной сульфогидрилсодержащей группой и это может быть использовано как конъюгат. После связывания с рецептором можно удалить защиту, используя общепринятые технологии, известные специалистам в этой области. Детекция может проводиться, используя репортер, имеющий тиоловую реактивную группу, такую как малеимид-HRP или иодоацетил-HRP.
Еще одна альтернатива использовать конъюгат, где субстрат имеет два компонента, как описано выше, детектируемо меченый первый компонент, который будет связываться с рецептором после того, как второй компонент активирован и удален с помощью ADEAS. Примером этого является любая маленькая органическая молекула, такая как 2-(4'-гидроксифенилазо)-бензойная кислота (HABA), которая связывается специфически с авидином и стрептавидином. Термины авидин и стрептавидин используются здесь взаимозаменяемо. HABA можно детектируемо пометить, используя любой из рецепторов, описанных ниже, например радиоизотопы, ферменты и т.д. Например, щелочную фосфатазу можно конъюгировать с HABA, используя технологии, хорошо известные специалистам с спейсером или без него, с функциональной группой HABA. Пример такой функциональной группы является 4'-гидроксильный остаток. Однако HABA можно модифицировать, чтобы завладеть вторым компонентом, который предотвращает связывание до тех пор, пока он не удален ADEAS. Например, этерификация HABA с этанолом дает этиловый эфир HABA, который не связывается со стрептавидином. Детектируемо меченые этиловые эфиры HABA не будут связываться со стрептавидином до тех пор, пока не гидролизована соответствующей карбоновой кислотой эфирная группа. Гидролиз можно проводить, используя фермент, такой как эстераза, например эстераза из печени свиньи. Таким образом, детектируемо меченные эфиры HABA такие, как 6-(фенокси-(4'-азо-2''-карбоксиэтилфенил))-гексаноил-щелочная фосфатаза, можно осаждать, используя ADEAS, имеющий подходящую эстеразу, которая будет гидролизовать эфир, чтобы осуществить связывание детектируемо меченого HABA со стрептавидином (т. е. экзогенным рецептором), который иммобилизован на поверхности носителя. Соединения (компаунды) формулы
где R1 R8 одинаковые или разные и выбираются из группы, состоящей из прямой или разветвленной цепи алкильных групп, имеющих 1-4 атома углерода, F, Cl, Br или J;
z фенил или нафтил;
n 1 19;
х N, О, S;4 R9 может быть H или прямая, или разветвленная цепь алкильных групп, имеющая 1-4 атома углерода, может быть использована для синтеза конъюгатов типа HABA, как описано в примерах ниже.
Термин конъюгат типа HABA означает HABA производные: (i), которые могут быть замещенными или незамещенными и связанными через спейсер с репортером, и (ii), которые содержат ADEAS активированную часть, чтобы предохранить конъюгат от связывания со стрептавидином до тех пор, пока он не активирован или удален ADEAS. Синтез такого компаунда проиллюстрирован на фиг. 6 так же, как в примере 10. Нижеописанный подход модификации, чтобы синтезировать любой из этих компаундов, может быть использован специалистами в этой области. Другие комбинации небольшая органическая молекула/рецептор являются подходящими, чтобы использовать данное изобретение в приложении к гаптены/антитела, сахара и олигосахариды/лектины, биотин и красители/авидин и/или стрептавидин.
Как показано в табл 1, подходит большое количество рецепторов. Выбор рецептора будет зависеть от выбранного конъюгата.
Оптимальная концентрация конъюгата определяется в соответствии с процедурой, описанной в примере 1. Оптимальные концентрации будут варьировать в зависимости от фермента, использованного в ADEAS, и субстрата, выбранного, чтобы получить конъюгат.
Конъюгат можно синтезировать, используя общепринятые процедуры связывания и мочения. Выбор субстрата будет зависеть от выбранной ADEAS. Повторяясь, следует сказать, что детальные знания требуются, касательно каталитических свойств каждого специфического энзима, для того, чтобы определить правильно полезный синтетический субстрат и, если необходимо, рецептор.
Широкое разнообразие репортеров имеется в наличии для связывания данного субстрата, чтобы получить конъюгат или связать его с членом группы специфических связывающих пар. Как обсуждалось выше, репортер подводит другую часть (конъюгата) к субстрату. Репортерами могут быть радиоактивный изотоп, такой как 125J, энзимы, флюорогенные, химическилюминесцентные, электрохимические или магнетические материалы. Изнутри меченные репортеры (например, тритий или другие такие радионуклиды), которые не вводят другую часть в дополнение к субстрату, не рассматриваются при осуществлении данного изобретения.
Примеры ферментов-репортеров, которые можно использовать в практике данного изобретения, включают гидролазы, лиазы, оксидоредуктазы, трансферазы, изомеразы и лигазы, более предпочтительные фосфатазы, эстеразы, глюкозидазы и пероксидазы. Можно упомянуть пероксидазу. глюкозоксидазу, фосфатазу, эктеразу и глюкозидазу. Специфические примеры включают щелочную фосфатазу, липазы, бета-галактозидазу и эстеразу из печени свиньи. Как было замечено выше, если фермент использовался в качестве репортера, он может быть тем же или отличным от фермента или ферментов, используемых в ADEAS. Данное изобретение можно использовать для катализа выпадения радиоизотопно- меченого конъюгата или энзиммеченого конъюгата и т.д.
Другое воплощение иммуноанализа по типу сэндвича, описанное выше, включает реагирование комплекса захватывающее антитело-аналит-второе антитело с ADEAS, состоящим из анти-антитела, связанного с ферментом, таким как HRP или AP. Анти-антитело связывается с эпитопом второго антитела.
Это изобретение не ограничивается иммуноанализом по типу сэндвича. Оно применимо к широкому разнообразию видов анализа, например гибридизации нуклеиновых кислот, таких как для РНК, так и для ДНК.
В табл. 1 представлены для дальнейшей иллюстрации данного изобретения примеры одно- и мультиферментных ADEAS, конъюгатов, рецепторов и типов рецепторов.
В AP/HRP мультиферментом ADEAS, описанном выше, конъюгат должен быть фосфорилирован перед тем, как он будет реагировать с HRP; в β -gal/HRP мультиферментном ADEAS конъюгат должен быть дегликозилирован перед реагированием с HRP.
Для тех, кто является специалистом в этой области должно быть ясно, что большое количество вариаций возможно и все эти вариации попадают в пределы изобретения.
Следующие примеры предназначены для иллюстрации данного изобретения. Если не оговорено особо, использовали по 100 мкл каждого реагента. Одно исключение, когда использовали 200 мкл блокирующего буфера.
Пример 1. Приготовление конъюгатов и подбор оптимальной концентрации конъюгата.
Пара-гидроксифенилпропионил биоцитин (НРРВ) готовили смешиванием раствора n-гидроксифенилпропионовой кислоты-N-гидроксисукцинимида эфира (50 мг [0,2 мМ] /2 мл диметилсульфоксида) с биоцитином (70,75 мг 0,2 мМ/2 мл 0,1 М NaHCO3) в течение ночи при комнатной температуре (RT). Биотин-тирамин (ВТ) готовили смешиванием раствора тирамина (40 мг [0,3 мМ]/1 мл диметилсульфоксида) со сложным эфиром биотин-N- гидроксисукцинимида (100 мг [0,3 мМ]/1 мл диметилсульфоксида) в течение ночи при RT. Использовали полученные растворы HPPB и ВТ. Подсчитанные концентрации были 26 мг/мл для HPPB и 55 мг/мл для ВТ.
Полистироловые EIA стрипы (NUNC) были покрыты поликлональными анти-антителами к вирусу простого герпеса (HSV) (Dako, Carpenteria, CA) в 0,1 М карбонатном буфере pH 9,6 в течение ночи при 4oC и затем блокированны 2% бычьего сывороточного альбумина (BSA) в карбонатном буфере, затем отмыты 10 мМ буференной фосфатной солью, 0,05% Твин 20, pH 7,4 (PBST). Разведение HSV антигена в 1% BSA 10 мМ буферной фосфатой солью, 0,05% Твин 20, pH 7,4 (BSA
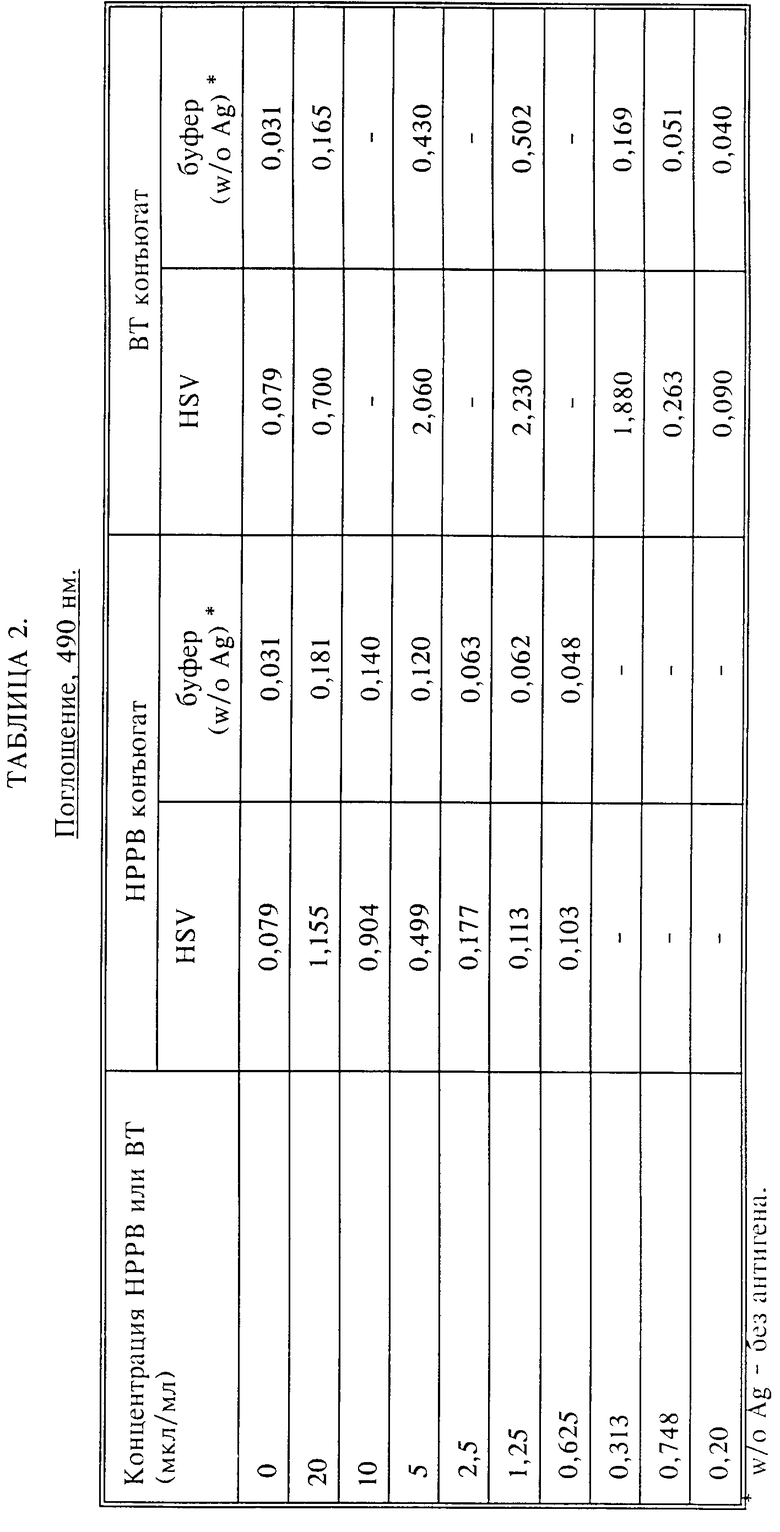
PBST) или буфере без антигена инкубировали 1 ч при 37oC. Разведение было достаточным для того, чтобы получить оптимальные плотности в пределах, указанных в табл. 1. Его отмывали с PBST. Аналит- зависимая ферментная активированная система состояла из HRP, связанного с анти-HSN (HRP ADEAS), которая была закуплена у Dako. HRP ADEAS добавляли и инкубировали 30 мин при RT и отмывали PBST. Варьирующие концентрации HPPB или ВТ, как изложено в таб. 1 ниже, добавляли в 50 мМ трис-HCl, 0,01% H2O2, pH 8,0, в течение 15 мин при RT. После отмывание с PBST добавляли стрептавидин-HRP и инкубировали 15 мин при RT для реакции с осажденными биотинами. Плашку затем отмывали PBST. HRP субстрат, о-фенилендиамин (OPD) добавляли, инкубировали 30 мин при RT и останавливали реакцию 4 H H2SO4. Оптические плотности при 490 нм записывали на счетчике микротитрационных планшетов.
Результаты представлены в табл. 2. Колонка 1 отражает варьирующие концентрации в мкл/мл HPPB или ВТ. Колонки 2 и 3 отражают оптические плотности, записанные как функция HPPB концентрации. Колонки 4 и 5 отражают результаты, полученные при использовании ВТ.
HPPB и ВТ превращали в активированные формы с помощью HRP ADEAS. Катализируемое осаждение репортера достигалось без иммобилизации рецептора.
В выборе оптимальной концентрации необходимо смотреть как на величину амплификации сигнала, так и на соотношение сигнала к шуму. Учитывая это, оптимальная концентрация HPPB была 20 мкл/мл (приблизительно 0,5 мг/мл) и ВТ приблизительно 0,3 мкл/мл (приблизительно 16 мкг/мл).
Пример 2. Амплификация сигнала детектора в HSV анализе, используя катализируемое осаждение репортера.
Покрытие анти-HSV EIA стрипы готовили, как описано в примере 1. Разведение 1: 100 HSV антигена готовили и делали четыре серийных разведения. Эти разведения HSV инкубировали в течение 2 ч при 37oC с анти-HSV покрытыми EIA стрипами. Избыток реагента отмывали PBST. ADEAS был тот же, что в примере 1. Он был добавлен в покрытие анти-HSV EIA стрипы, содержащие комплекс анти-HSV-HSV и инкубировали 30 мин при RT и затем отмывали PBST. 20 мкл/мл HPPB конъюгата, как определено в примере 1, добавляли в 50 Мм трис-HCl, 0,01% H2О2, pH 8,0 и инкубировали 15 мин при RT и затем отмывали PBST. Выпавшие биотины реагировали со стрептавидин-HRP (SA-HRP) в течение 15 мин при RT. Его отмывали PBST. Субстрат, ОРД, добавляли и инкубировали 30 мин при RT, реакцию останавливали 4 H H2SO4 и записывали поглощение при 490 нм на счетчике микротитрационных плашек.
Анализы без амплификации проводились, в которых:
а) не были использованы ни HPPB, ни SA-HRP,
б) HPPB использовали без SA-HRP,
в) SA-HRP использовали без HPPB.
Результаты.
Результаты, показанные на фиг. 1, демонстрируют, что:
а) получено катализируемое осаждение репортера,
б) как конъюгат, так и SA-HRP необходимы для детекции, потому что конъюгат содержал немеченый член группы специфических связывающих пар.
Результаты анализа без амплификации (нет HPPB, нет SA-HRP) не наблюдали. Результаты по другим анализам не наблюдали, потому что дополнительные наблюдения перекроются с результатами без амплификации, уже наблюдаемыми.
Пример 3. Амплификация сигнала детектора в HIV p 24 анализе, используя катализируемое осаждение репортера: эффект концентрации конъюгата.
Полистероловые EIA стрипы (NUNC) покрывали кроличьими анти-HIV p 24 антителами в 0,1 М карбонатном буфере, pH 9,6, в течение ночи при 4oC и блокировали реакцию 2% BSA в карбонатном буфере, за чем следовала отмывка в PBST. NIV антиген инкубировали 2 ч при 37oC (концентрации указаны на фиг. 2). Плашку затем отмывали PBST. Кроличий анти-HIV p 24-HRP аналитзависимая ферментная активированная система была затем проинкубирована в течение 2 ч при 37oC и отмыта с PBST. Различные концентрации ВТ конъюгата (0,2; 0,4 и 0,8 мкл/мл) в 0,1 М боратном буфере, 0,01% H2O2, pH 8,5, инкубировали 15 мин при RT, затем отмывли с PBST. После стрептавидин-HRP инкубировали 15 мин при RT.
Для сравнения использовали биотинилированное анти-HIV p 24 антитело, его детектировали со стрептавидин-HRP. ОРД добавляли и инкубировали с ним 30 мин, реакцию останавливали 4 H H2SO4 и записывали оптические плотности при 490 нм на счетчике микротитрационных плашек.
Результаты.
Результаты показаны на фиг. 2, где Amp. 1, Amp. 2 и Amp. 3 относятся к концентрациям ВТ 0,2; 0,4 и 0,8 мкл/мл соответственно. Разные уровни амплификации достигались, используя катализируемое осаждение репортера, зависящее от концентраций конъюгата.
Фиг. 2 также представляет результаты для анализа без амплификации, используя систему детекции биотинилированное антитело/SA-HRP (биотин) и анализ без амплификации, где анти-HIV p 24 детектор был непосредственно мечен HRP. Полученные результаты, используя анти-HIV p 24 HRP детектор, были плохими по сравнению со значительным увеличением сигнала детектора, полученного при использовании катализируемого осаждения репортера.
В зависимости от концентрации конъюгата, сигнал, как он ни хорош, и более интенсивный, так и те, что получены с биотинилированным антителом, были получены, используя метод катализируемого осаждения репортера настоящего изобретения. Наилучшие результаты получены, используя концентрацию конъюгата около оптимальной, как и было определено в примере 1.
Пример 4. Приготовление и характеристика биотин тирамина.
Приготовление биотин тирамина: раствор биотин-N-гидроксисукцинимида, 170 мг (0,5 мМ) и тирамина (перекристаллизованного из воды) 68,5 мг (0,5 мМ) в 25 мл диметилформамида обрабатывали 10 мл 1 М триэтиламмония бикарбоната, pH 7,5 и затем прогревали при 50oC 3 ч.
Выделение: раствор концентрировали до сухого состояния на роторном испарителе и осадок перекристаллизовывали из воды с выходом 72%
Характеристика: точка плавления определена 192-193oC.
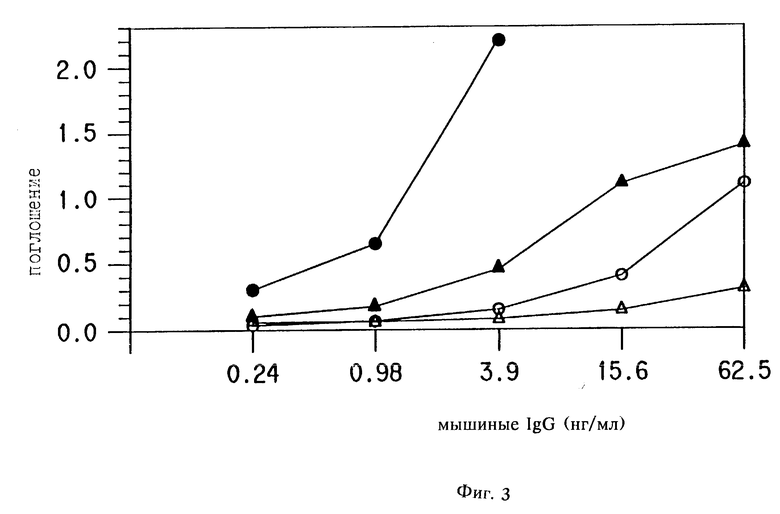
Пример 5. Амплификация сигнала детектора в анализе с использованием мыши, используя катализируемое осаждение репортера.
Полистироловые EIA стрипы (NUNC) покрывали анти-мышиными IgG (Fc фрагмент специфический) антителами козы (ICN) в 0,1 М карбонатном буфере, pH 9,6; в течение ночи при RT. Затем их блокировали 2% BSA в карбонатном буфере и отмывали PBST. Разведения мышиных IgG в BSA-PBST инкубировали на стенках в течение 1 ч при 37oC, после чего отмывали PBST.
Концентрации представлены на фиг. 3. Козлиные анти-мышиные IgG-HRP (HRP ADEAS) и козлиную антимышиную IgG-щелочную фосфатазу (AP ADEAS) (Boehringer Mannehim) разводили, как рекомендовано изготовителем, и инкубировали 1 час при 37oC. Анализы проводились с и без катализируемого осаждения репортера. AP ADEAS не использовали для катализа осаждения репортера в этом эксперименте.
Для катализируемого осаждения репортера, используя HRP ADEAS 1 мг/мл стоковый раствор биотин тирамина (как описано в примере 4) в диметилсульфоксиде, приготавливали и потом добавляли к 0,1 М боратному буферу, pH 8,5; 0,01% H2О2, в 10 мкл/мл (10 мкг/мл биотин триамина) и инкубировали 15 мин при RT. Плашку затем отмывали PBST. Стрептавидин-HRP (для HRP-Amp. HRP) или стрептавидин-щелочную фосфатазу (для HRP -Amp Alk Phos) инкубировали 15 мин при RT, и плашку отмывали PBST. После добавления OPD (для HRP) или п-нитрофенилфосфата (для AP) совершали спектрофотометрическое определение (инкубация после добавления 15 мин при RT). Реакции останавливали добавлением 4 H H2SO4 (HRP/OPD) или 1 N NaOH (Alk Phos/pNPP). Оптические плотности при 490 нм для HRP/OPD и 405 нм для Alk Phos/pNPP записывали на счетчике микротитрационных плашек.
Результаты.
Результаты показаны на фиг. 3. Как видно, можно достигать амплифицированного сигнала с сопутствующим ограничением низкой детекции путем допущения HRP ADEAS катализировать выпадение активированного ВТ конъюгата, за которым следует детекция со стрептавидином, связанным с HRP или AP. Этот пример показывает, что если репортер является ферментом, он может быть тем же или отличным от фермента, использованного в ADEAS.
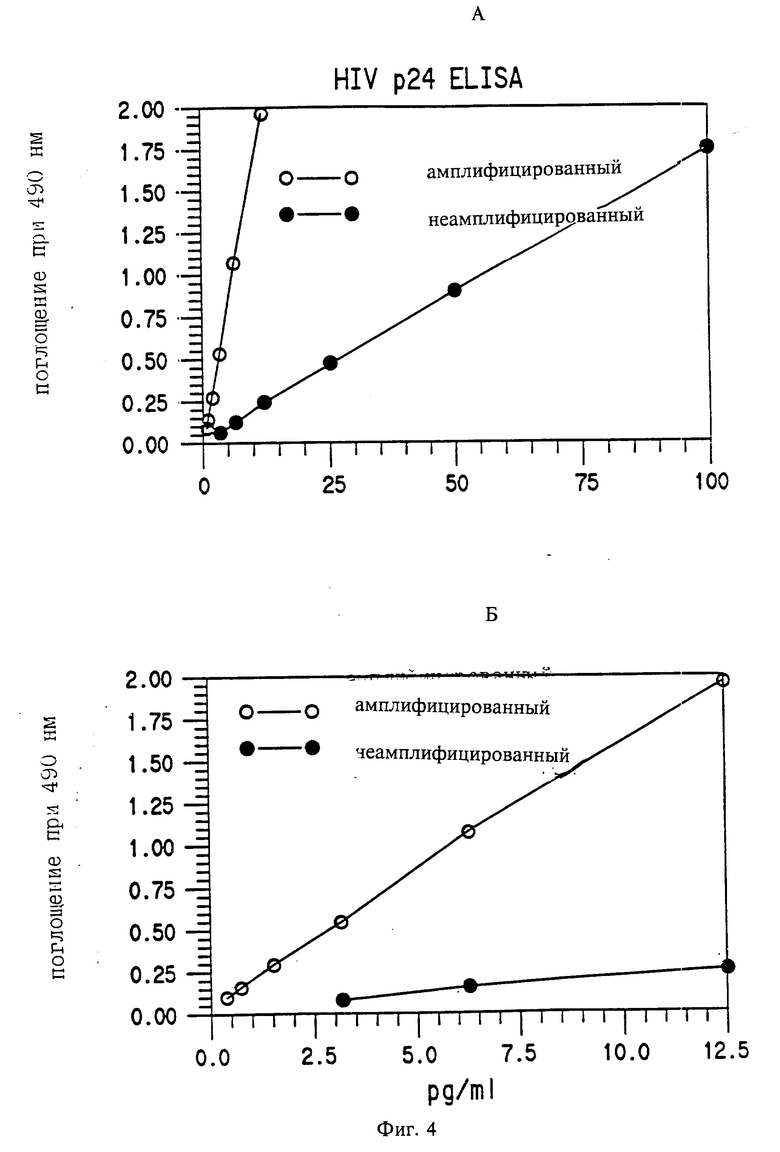
Пример 6. Амплификация сигнала детектора в анализе на HIV p 24, которая использует систему детекции биотинилированное антитело-детектор/стрептавидин
HRP.
Du Pont HIV p 24 антиген ELISA (номер в каталоге NEK 060) была модифицирована для катализируемого осаждения репортера следующим образом: SA-HRP был использован в концентрации 1/4, указанной в руководствах. За этим следовало 15 мин RT инкубация с биотин тирамином, 10 мкг/мл в 0,1 М борате; 0,01% H2О2, pH 8,5 буфере (как в примере 5). Вслед за отмывкой в PBST SA-HRP в концентрации 1/16 инкубировали 15 мин при RT. В завершение добавляли ОРД как по руководству к "киту" (набору реагентов). За исключением продления стандартных концентраций до 0,39 пг/мл, других изменений сделано не было.
Результаты.
Результаты показаны на фиг. 4. Этот эксперимент продемонстрировал, что можно амплифицировать сигнал, образованный системой биотинилированное антитело/SA-HRP, используя катализируемое осаждение репортера. Так как концентрация SA-HRP для обоих инкубаций была гораздо меньше, чем для анализа без амплификации, ясно, что возросший сигнал являлся характерной чертой для осаждения репортера, а не для удвоенной инкубации.
Пример 7. Осаждение репортера на мембранах.
Нитроцеллюлоза (Schleicher Schuell, BA85) покрывалась антигеном и затем блокировалась 1% BSA, 1% обезжиренное сухое молоко в PBS буфере в течение ночи. Эти мембраны инкубировали 1 ч при RT с аналитзависимой ферментной активирующей системой, описанной в примере 1. Затем мембраны инкубировали с биотин тирамином (из примера 1) в 2 мкл/10 мл 50 мМ трис-НС1; 0,01% H2О2, pH 8,0 буфере 15 мин при RT, за которой следовала инкубация со стрептавидин-щелочной фосфатазой 15 мин при RT. Проводились контроли там, где биотин тирамин инкубировали без стрептавидин-щелочной фосфатазы и стрептавидин-щелочная фосфатаза инкубировалась без биотин тирамина. Визуальное наблюдение (визуализацию) выпавшей щелочной фосфатазы усиливали добавлением BCIP/NBT (Kirkegaard Perry). BCIP это 5-бром-4-хлор-индоксил фосфат и NBT это 2,2'-ди-(п- нитрофенил)-5,5'-дифенил-3,3'-(3,3'-диметокси-4,4'-дифенилен)- дитетразолия хлорид. Визуализацию связанного анти-HSV-HRP конъюгата облегчали добавлением диаминобензидино (DAB).
Результаты.
Добавление DAB давало наблюдаемые коричневые пятна там, где находится на нитроцеллюлозной мембране HSV антиген. Добавление BCIP/NBT давало наблюдаемые голубые пятна там, где находился HSV антиген, когда с этой мембраной инкубировали биотин тирамин и стрептавидин-щелочную фосфатазу. Это показывает, что осаждение щелочной фосфатазы обусловлено HRP активацией этого биотин тираминового конъюгата, за которым следует определение стрептавидин-щелочной фосфатазы.
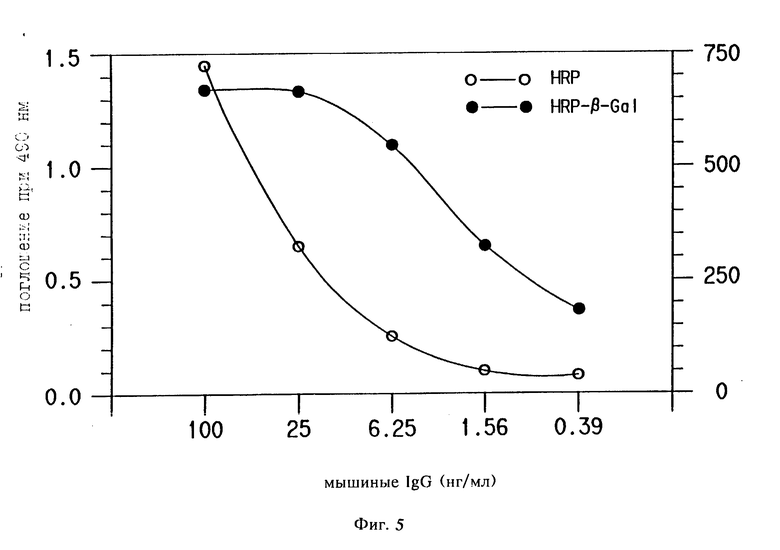
Пример 8. Осаждение бета-галактозидазы пероксидазой хрена и детекция методом флуоресценции.
Полистироловые EIA стрипы (NUNC) покрывали козлиными антимышиными IgG (к специфическому Fc фрагменту) антителами (ICN) в 0,1 М карбонатном буфере, pH 9,6; в течение ночи при RT. Затем они блокировались 2% BSA в карбонатном буфере и отмывались PBST. Концентрации мышиных IgG в BSA- PBST, как изложено на фиг. 5, инкубировались 1 ч при 37oC, после чего следовало отмывание с PBST. Козлиные антимышиные IgG-HRP (ADEAS), закупленные у Boehringer Mannheim, разводили как рекомендовано производителем и инкубировали 1 ч при 37oC. Плашку затем отмывали PBST. 1 мг/мл стоковый раствор ВТ конъюгата (как описано в примере 4) в диметилсульфоксиде приготавливали и потом добавляли к 0,1 М боратному; 0,01% H2О2, pH 8,5 буферу в 10 мкл/мл (10 мкг/мл биотин тирамина). Эту смесь добавляли в плашку и инкубировали 15 мин при RT и затем отмывали с PBST. Стрептавидин-бета галактозидазу (Bethesda Research Labs) добавляли и инкубировали 15 мин при RT. Этот анализ также проводили без катализируемого осаждения репортера, т.е. без добавления ВТ. Калориметрическое определение при анализе без амплификации совершали после инкубации с OPD (для HRP) в течение 15 мин при RT. Флуоресцентную детекцию при анализе с амплификацией совершали после добавления 4-метиламбеллиферил бета-D-галактозида (MUG) (для HRP-бета Gal). Оптические плотности при 490 нм записывались для HRP/OPD на счетчике микротитрационных плашек. Флуоресценция для HRP-бета Gal/MUG записывалась на флуоресцентном счетчике для микротитрационных плашек (Dunatech Laboratories).
Результаты.
Результаты показаны на фиг. 5. Флуоресцентный сигнал был обусловлен количественным осаждением биотин тирамина с помощью ADEAS, за которым следовала инкубация со стрептавидин бета-галактозидазой.
Пример 9. Амплификация в мембранном анализе.
Флуоресцеин-тирамин (FT) готовили следующим образом: растворы 46,6 мг 5-(и 6)-карбоксифлюоресцеинсукцинимидил сложного эфира и 14,6 мг тирамина в 0,3 мл диметилсульфоксида были приготовлены. Конъюгат образовывался до конца смешиванием 0,25 мл каждого раствора в течение ночи при RT. Этот раствор использовался в готовом виде.
На три нитроцеллюлозных (Schieicher Schuell, BA 83) полоски было накапано 1 мкл мышиных IgG в 10 мкг/мл и два серийных разведения в PBS. Эти мембраны блокировали 5%-ным обезжиренным сухим молоком в PBST в течение 30 мин и затем отмывали три раза в PBST. Козлиный антимышиный IgG-HRP конъюгат (Boehringer Mannehim), разбавленный 1/2000 в 1% BSA-PBST, инкубировали 30 мин при RT и отмывали мембраны три раза в PBST. Третью мембрану инкубировали с FT в 20 мкг/мл в 0,1 М борате; 0,01% H2О2, pH 8,5 буфере 15 мин при RT и отмывали три раза в PBST. Потом вторую и третью мембраны инкубировали с антифлуоресцеин антителом (Chernicon), которое было сконъюгировано с HRP (с помощью SMCC метода Ishikawa, G. и др. S Immuncassay, 4, 209-327, 1983), разведенного в 1% BSA-PBST. Визуализацию всех трех полос облегчали добавлением диаминобензидина на 5 мин.
Результаты.
Можно было видеть на первых двух полосках три пятна, указывающих предел детекции 2,5 мкг/мл, и что анти-флуоресцеин-HRP конъюгат не вносил вклад в дополнительный сигнал. На третьей полоске можно было видеть шесть пятен, указывающие предел детекции 313 нг/мл. Использование катализируемого осаждения репортера метода амплификации данного изобретения улучшило предел детекции этого анализа в восемь раз над пределом анализа без амплификации.
Пример 10. Синтез конъюгата, HEE-6-AP, из (N-сукцинимидил)-6-(фенокси-(4'-азо- 2''-карбоксиэтилфенил))гексаната, HEE-6-NHS, и щелочной фосфатазы (AP).
Следующая схема реакции проиллюстрирована на фиг. 6: этил-2-(4'-гидроксифенилазо) бензоат (НЕЕ) готовили из 2-(4'-гидроксифенилазо) бензойной кислоты (HABA, (1) безводного этанола и каталитического количества ацетилхлорида. Этиловый сложный эфир (НЕЕ) реагировал с третбутил-6-иодогексанатом и гидридом натрия, используя основную процедуру, о которой сообщалось Castellanos и др. Tetrahedron, p. 1691- 1696, V. 37, (1981) для получения третбутил-6-(фенокси-(4'-азо-2''- карбоксиэтилфенил)) гексаноат (HEE-6-t-Bu, (3)). Третбутиловый сложный эфир гидролизовали обработкой трифтбруксусной кислотой, используя основную процедуру, о которой сообщено Bryan и др. Journal of the American Chemical Society, p. 2353-2355, v. 99, (1977), чтобы получить 6-(фенокси-(4'-азо-2''-карбоксиэтилфенил)-гексановую кислоту (НЕЕ-6-Н). (N- сукцинимидил)-6-(фенокси-(4'-азо-2''-карбоксиэтилфенил)гексаноат (HEE-6-NHS, (5)) готовили из НЕЕ-6-Н, N-гидроксисукцинимида и дициклогексилкарбодиимида в THF, используя основную процедуру, о которой сообщалось Bryan и др. Macromolecular Chemie, p. 2375-2382, v. 186, (1985). Эфир NHS (5) растворяют в минимальном объеме DMSO и добавляют к уравновешенному буфером раствору щелочной фосфатазы (например, из кишечника теленка), используя основную процедуру, о которой сообщалось О' Sullivan и др. Methods in Enzymology, p. 147-166, v. 73, (1981), чтобы дать конъюгат (НЕЕ-6-АР, (6).
Пример 11. Синтез HABA-тираминконъюгата (H-T) из (N-сукцинимидил)-6-(фенокси- (4'-азо-2''-карбоксиэтилфенил)) гексаноата, (HEE-6-NHS, (5)) и тирамина.
NHS сложный эфир (5) (1 мМ), описанный в примере 10, и тирамин (1 ммоль) растворяли в DMF (5 мл) и перемешивали при RT в течение 48 ч. Раствор выпаривали до сухого состояния под вакуумом. Осадок суспендировали в H2О (pH 8,0; 50 мл) и добавляли эстеразу из печени свиньи, pH раствора поддерживалось добавлением 0,1 н NaOH, насколько требовалось. Раствор выпаривали до сухого состояния после 24 ч. Полученный H-T конъюгат выделяли хроматографией (силикагель, хлороформ/метанол).
Пример 12. Амплификация сигнала детектора в анализе с мышиными IgG, используя эстеразу из печени свиньи (PLE), катализирующую осаждение репортер - энзим.
Микротитрационные полоски плашек (Nunc) покрываются смесью козлиных антимышиных IgG (специфичных для Fc фрагмента) антител (ICN) и стрептавидина (Scripps Laboratories) в 0,1 М карбонатном буфере, pH 9,6 в течение ночи при RT. Затем их блокировали 2% BSA в PBS и отмывали PBST. Разведения мышиных IgG (0-100 нг/мл) в BSA-PBST инкубировали в лунках 1 ч при 37oC, после чего отмывали PBST. Приготавливали препарат козлиных антимышиных IgG-PLE (PLE ADEAS) (0,75 мг/мл) основным методом, описанным Hashida и др. Jornal of Applied Biochemistry, v.6, p. 56-63, 1984, и разводили 1:100 1:2000 фосфатным буфером (0,1 М; pH 8,0; 0,2 BSA), инкубировали 1 ч при 37oC и отмывали с PBST. НЕЕ-6-Ар (1 мг/мл) (приготовленный, как описано в примере 10), добавляли в лунку микротитрационной плашки и инкубировали по крайней мере 15 мин при 37oC. Затем плашку отмывали PBST. Спектрофотометрическое определение достигалось после добавления п-нитрофенил фосфата. Реакции останавливали добавлением 1 н NaOH. Оптические плотности при 405 нм записывались на счетчике микротитрационных плашек. Амплификация сигнала детектора в этом образце происходит от катализируемого осаждения репортера-энзима. т.е. PLE катализирует выпадение HEE-6-AP, где рецептор, стрептавидин иммобилизованы на поверхности микротитрационной плашки.
Пример 13. Демонстрация модулируемого ферментом связывания блокированного связующего компонента.
Суспензию эстеразы из печени свиньи добавили (10 мкд. 2860 ед/мл, Cat. N E 3 128, Sigma Chemical, St. Louis, MO) к раствору этил-2-(4'- гидроксилфенилазо)бензоата (НЕЕ) (0,25 мМ) и стрептавидина (0,2 мг/мл) в фосфатном буфере (0,1 М, pH 8,0; 2 мл). Второе разведение, идентичное первому, готовили то, которое не содержало стрептавидина. Поглощение этих двух растворов измеряли при 500 нм как функцию времени. Фиг. 7 показывает поглощение раствора, который не содержал стрептавидина, снижающегося с течением времени, указывая на гидролиз НЕЕ в 2-(4'- гидроксифенилазо) бензойную кислоту (HABA). Поглощение раствора, который содержал стрептавидин, увеличивалось с течением времени, указывая на образование комплекса HABA-стрептавидин, который, как известно, имеет сильное поглощение при 500 нм.
| название | год | авторы | номер документа |
|---|---|---|---|
| Способ конкурентного иммуноферментного анализа для детекции аналита в образце с помощью мультимодальных наночастиц | 2023 |
|
RU2836697C1 |
| СПОСОБ БЕЗЭЛЕКТРОЛИЗНОГО НАНЕСЕНИЯ МЕТАЛЛИЧЕСКОГО ПОКРЫТИЯ НА ПОВЕРХНОСТЬ ИЗДЕЛИЯ ИЗ АРОМАТИЧЕСКОГО ПОЛИАМИДА И ИЗДЕЛИЕ ИЗ АРОМАТИЧЕСКОГО ПОЛИАМИДА С МЕТАЛЛИЧЕСКИМ ПОКРЫТИЕМ | 1992 |
|
RU2104328C1 |
| Способ синтеза функциональных углеродных квантовых точек | 2023 |
|
RU2824005C1 |
| СПОСОБ УСИЛЕНИЯ ЭФФЕКТА ДИСПЕРСНЫХ ПОЛЕЗНЫХ АГЕНТОВ | 2006 |
|
RU2404740C2 |
| ПРИМЕНЕНИЕ БИОТИНИЛИРОВАННОГО ПОЛИПЕПТИДА ДЛЯ ОПРЕДЕЛЕНИЯ АКТИВНОСТИ БЕЛОК-ФОСФОРИЛИРУЮЩИХ ФЕРМЕНТОВ | 2004 |
|
RU2395813C2 |
| ПРОИЗВОДНЫЕ БОРСОДЕРЖАЩИХ ПЕПТИДОВ | 1991 |
|
RU2017749C1 |
| ФОТОАКТИВИРУЕМОЕ ХИМИЧЕСКОЕ ОБЕСЦВЕЧИВАНИЕ КРАСИТЕЛЕЙ | 2012 |
|
RU2623880C2 |
| СПОСОБ ИММУНОФЕРМЕНТНОГО ОПРЕДЕЛЕНИЯ АНТИГЕНОВ | 2005 |
|
RU2303783C1 |
| Способ биоимиджинга | 2023 |
|
RU2838135C1 |
| Способ детекции аналита в образце с помощью иммунохроматографии | 2023 |
|
RU2835200C1 |



Использование: в области медицины, в частности в методах катализа осаждения репортера, чтобы улучшить детекцию или количественное определение аналита в образце путем амплификации сигнала детектора. Сущность изобретения: проводят иммобилизацию аналит-зависимой ферментной активирующей системы с катализируемым осаждением репортера посредством активации конъюгата, состоящего из детектируемо меченого субстрата, специфичного для этой ферментной системы, названный конъюгат реагирует с аналит-зависимой ферментной активирующей системой, формируя активированный конъюгат, который затем выпадает где бы ни был иммобилизован рецептор для активированного конъюгата, названный рецептор не реагирует с аналит-зависимой ферментной активирующей системой. Другое воплощение этого изобретения относится к анализу для детекции или определения количества наличия или отсутствия аналита в образце, используя катализируемое осаждение репортера для амплификации сигнала репортера. Способ позволяет улучшить предел детекции этого анализа. 2 с. и 17 з.п. ф-лы, 2 табл., 7 ил.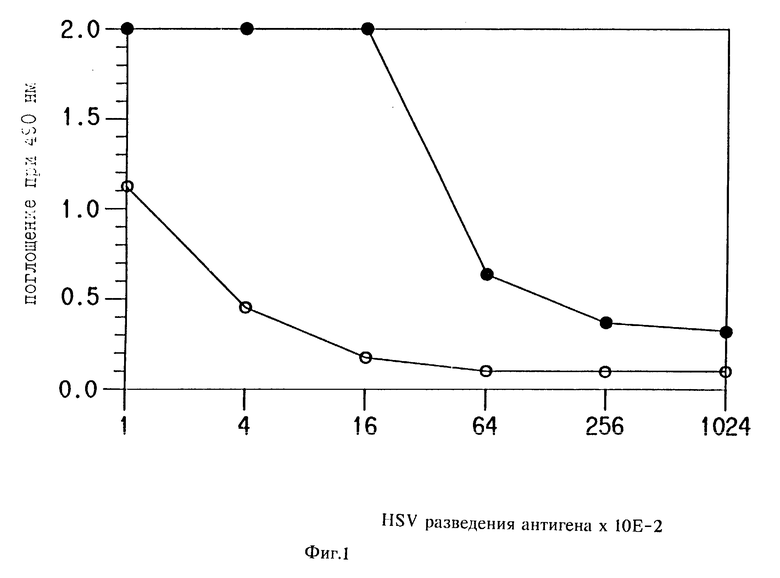
Приоритет по пунктам:
29.09.89 по пп.1 и 2 (за исключением признаков, относящихся к использованию трансфераз, изомераз и лигаз), п. 3 (за исключением признака, относящегося к использованию эстеразы), п. 4 (за исключением признака, относящегося к использованию экстераза печени свиньи), пп. 5 и 6 (за исключением признаков, относящихся к использованию флюоресцин-тирамина, N-(4'''-гидроксифенетил)-6-(фенокси-4'- азо-2''-бензойной кислоты))гексамида), пп. 8 13 (за исключением признака, относящегося к использованию эстеразы), п. 14 (за исключением признака, относящегося к использованию эстеразы печени свиньи), пп. 15 и 16 (за исключением признаков, относящихся к использованию флюоресцин-тирамина, N-(4'''-гидроксифенетил)-6-(фенокси-(4'-азо-2''- бензойной кислоты))гексамида), п. 19;
20.03.90 по п. 2 (признаки относящиеся к использованию трансфераз, изомераз и лигаз), п. 3 (признак, относящийся к использованию эстеразы), п. 4 (признак, относящийся к использованию эстеразы печени свиньи), п. 6 (признаки, относящиеся к использованию флюоресцин-тирамина, N-(4'''-гидроксифенетил)-6-(фенокси-(4'-азо-2''-бензойной кислоты))гексамида), пп. 7 и 13 (признак, относящийся к использованию эстеразы), п. 14 (признак, относящийся к использованию эстеразы печени свиньи), п. 16 (признаки, относящиеся к использованию флюоресцин-тирамина, N-(4'''- гидроксифенетил)-6-(фенокси-(4'-азо-2''-бензойной кислоты))гексамида), пп. 17 и 18.
| EP, патент, 0123265, кл | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
