Изобретение относится к применению полипептида для определения способности фермента модулировать уровень фосфорилирования полипептида. Другие аспекты изобретения относятся к способу определения такой активности, а также к способу идентификации веществ, которые модифицируют эту способность фермента.
Инсулин представляет собой белковый гормон, который влияет на большое число путей роста и обмена веществ за счет того, что он связывается с рецептором инсулина и таким образом активирует его внутреннюю тирозинкиназу. Это событие приводит к фосфорилированию множества белков, которые могут связываться с рецептором инсулина (IR) по специфическим остаткам тирозина. К таким образом фосфорилированным белкам относится также семейство белков субстрата рецептора инсулина (IRS).
Субстратом рецептора инсулина 1 (IRS-1) является клеточный белок, который может фосфорилироваться множеством протеинкиназ (тирозинспецифичные или серин-/треонинспецифичные протеинкиназы) по остаткам тирозина, и/или серина, и/или треонина. При этом, в зависимости от фермента, вероятно специфически фосфорилируются различные остатки тирозина или серина/треонина. Кроме фосфорилирования посредством тирозинкиназ, как, например, в случае рецептора инсулина (White, 2002), рецептора IGF-1 (White, 2002) или JAK 1/2 (Thirone et al., 1999), IRS-1, как известно, фосфорилируется также серин-/тирозинкиназами, например, киназами из семейства РКС (Schmitz-Peiffer, 2002), комплекс ингибитор-каппа-В-киназа (Gao et al., 2002), с-Jun NH(2)-концевая киназа (JNK, Aguirre et al., 2000), протеинкиназа А (Sun et al., 1991), активированная митогеном протеинкиназа (Mothe et al., 1996), протеинкиназа В (Paz et al., 1999), казеинкиназа (Tanasijevic et al., 1993), гликогенсинтазакиназа-бета (Eldar-Finkelmann et al., 1997), АМР-активированная киназа (Jacobsen et al., 2001) или фосфоинозитол-3-киназа (Freund et al., 1995). Молекулы IRS являются ключевыми молекулами пути сигнальной трансдукции инсулина и играют центральную роль в поддерживании клеточных функций, таких как рост, выживание и обмен веществ. Фосфорилированные IRS-белки при этом служат в качестве “стыковочных” белков с множеством сайтов стыковки рецептора инсулина и комплексной сети внутриклеточных сигнальных молекул с так называемыми 2 гомологичными доменами комплекса узнавания сигнала (SRC) (SH2-домены). Активация этих Sh2-доменов белков при этом активирует определенные сигнальные каскады, что снова приводит к активации различных эффекторов, которые располагаются дальше вниз в сигнальном каскаде, что в конечном счете приводит к опосредованию сигнала инсулина в разветвленной серии других внутриклеточных сигнальных каскадов (в отношении обзора см. White, 2002).
IRS относится к группе фосфопротеинов массой 160-185 кДа, которые служат в качестве субстрата рецептора инсулина. Известны четыре члена семейства IRS (IRS-1, IRS-2, IRS-3 и IRS-4). Они различаются по распределению в тканях, субклеточной локализации, специфичной в отношении развития экспрессии, типу связывания с рецептором инсулина и роду SH2-белков, с которыми они взаимодействуют. Четыре члена семейства IRS по своей основной белковой структуре очень подобны: все обладают амино(N)-концевым плекстрин-гомологичным доменом (РН-домен), который связывается с мембранными фосфолипидами, фосфотирозинсвязывающим доменом (РТВ-домен), который карбоксильным (С) концом прямо присоединяется к РН-домену и опосредует распознавание последовательности Asp-Pro-Glu фосфотирозина (NPEpY), которая локализована в околомембранной области бета-субъединицы рецептора инсулина. Далее, они обладают несколько менее сильно консервативной С-концевой частью, которая обладает различными потенциальными мотивами фосфорилирования тирозина, с которыми могут связываться особые содержащие SH2-домены белки.
IRS-1 содержит 21 возможных сайта фосфорилирования тирозина, из которых некоторые локализованы в звеньях аминокислотных последовательностей, которые могут связываться с содержащими SH2-домены белками. IRS-1, далее, содержит 30 потенциальных сайтов фосфорилирования серина/треонина в мотивах, которые могут распознаваться различными киназами, такими как, например, киназы из семейства РКС (Schmitz-Peiffer, 2002), комплекс ингибитор-каппа-В-киназа (Gao et al., 2002), с-Jun NH(2)-концевая киназа (JNK, Aguirre et al., 2000), протеинкиназа А (Sun et al., 1991), активированная митогеном протеинкиназа (Mothe et al., 1996), протеинкиназа В (Paz et al., 1999), казеинкиназа (Tanasijevic et al., 1993), гликогенсинтазакиназа-бета (Eldar-Finkelmann et al., 1997), АМР-активированная киназа (Jacobsen et al., 2001) или фосфоинозитол-3-киназа (PI3-киназа, Freund et al., 1995). Ингибирующие воздействия на сигнальный путь рецептора инсулина могут по меньшей мере частично объясняться недавно обнаруженной ролью фосфорилирования серина/треонина IRS-1, которую связывают с ухудшением взаимодействия с рецептором инсулина и/или уменьшением фосфорилирования тирозина IRS-1 и/или с ухудшением взаимодействия с последующими сигнальными белками, которые могут связываться с тирозинфосфорилированным IRS-1 (в отношении обзора см. White, 2002). Для различных киназ, как, например, киназы из семейства РКС (Schmitz-Peiffer, 2002), комплекс ингибитор-каппа-В-киназа (Gao et al., 2002), с-Jun NH(2)-концевая киназа (JNK, Aguirre et al., 2000), протеинкиназа А (Sun et al., 1991), активированная митогеном протеинкиназа (Mothe et al., 1996), протеинкиназа В (Paz et al., 1999), казеинкиназа (Tanasijevic et al., 1993), гликогенсинтазакиназа-бета (Eldar-Finkelmann et al., 1997), АМР-активированная киназа (Jacobsen et al., 2001) или фосфоинозитол-3-киназа (PI3-киназа, Freund et al., 1995), до сих пор смогли показать, что in vitro они прямо фосфорилируют IRS-1. При этом в каждом случае повышенная киназная активность в интактных клетках ингибирует активность пути сигнальной трансдукции инсулина. Далее, in vitro фосфорилирование IRS-1 по остаткам серина/треонина, согласно некоторым исследованиям, находится в прямой связи с уменьшенным фосфорилированием тирозина за счет рецептора инсулина (Le Marchand-Brustel, 1999)). Последовательности IRS-1, -2, -3 и -4 общедоступны. Кодирующие полинуклеотидные последовательности и относящиеся к ним белковые последовательности этих генов в базе данных по нуклеотидам NCBI находятся под номерами NM_005544 (IRS-1 hs), XM:007095 (IRS-2 hs), NM:03274 (IRS-3 rat), NM:003604 (IRS-4 hs). NCBI означает Международный информационный центр по биотехнологии (почтовый адрес: National Center for Biotechnology Information, National Library of Medicine, Building 38A, Bethesda, MD 20894, USA; Web-Adress: www.ncbi.nhm.nih.gov). Клонирование IRS-1-гена описано, в частности, Araki et al., 1993 и Siemeister et al., 1996; клонирование IRS-2, -3 и -4 описано Araki et al., 1994, Lavan et al., 1997а и Lavan et al., 1997b.
Для определения способности и измерения активности различных киназ в случае фосфорилирования IRS-1 в уровне техники известны различные способы, которые базируются либо на радиоактивных способах обнаружения (например, перенос от радиактивно меченого фосфата на субстрат), либо на нерадиоактивных способах обнаружения.
Так, известно определение фосфорилирования IRS-1 по всей длине IRS-1-белка, его фрагментов или пептидов, которые обладают еще по меньшей мере одним сайтом фосфорилирования, с помощью способа, в случае которого путем инкубации с радиоактивно меченым АТФ и тестируемой киназой, в зависимости от способности киназы фосфорилировать IRS-1, радиоактивные фосфатные остатки переносятся на IRS-1. Затем осуществляют хроматографическое или электрофоретическое выделение IRS-1 и обнаружение количества перенесенного фосфата путем проточной сцинтилляции или авторадиографии (как, например, описывается для полного белка IRS-1 и гликогенсинтазакиназы-3-бета согласно Eldar-Finkelman et al., 1997, для фрагмента IRS-1 (аминокислоты 516-777) и рецептора инсулина, рецептора IGF-1 или рекомбинантной киназы рецептора инсулина согласно Siemeister et al., 1995, или пептида IRS-1 (аминокислоты 601-616) с помощью клеточных лизатов, которые содержат активированную протеинкиназу из семейства РКС, согласно De Fea et al., 1997). Далее, согласно Siemeister et al., 1995, способность фрагментов IRS-1, например, фрагмента IRS-1 (аминокислоты 516-777) и рецептора инсулина фосфорилировать рецептор IGF-1 или рекомбинантную киназу рецептора инсулина посредством инкубации с радиактивно меченым АТФ (аденозин-трифосфат), нанесения по каплям субстрата на положительно заряженную мембрану (нитроцеллюлоза или подобный материал), промывки и обнаружения связанного радиоактивно меченого субстрата с помощью авторадиографии или измерения радиоактивного излучения.
Инкубация биотинилированного пептида IRS-1 (аминокислоты 601-616) с радиоактивно меченой АТФ, нанесение по каплям субстрата на покрытую стрептавидином мембрану, промывка и обнаружение связанного радиоактивно меченого субстрата с помощью авторадиографии или измерения радиоактивного излучения - это другой способ определения способности киназ фосфорилировать IRS-1 (см. De Fea et al., 1997).
Недостаток вышеописанных радиоактивных способов тестирования очевиден, так как обращение с радиоактивностью вносит существенную опасность, является очень дорогостоящим и, таким образом, мало пригодно в особенности для высокопроизводительных способов (HTS-способы).
Недостаток вышеописанных основанных на использовании короткоцепочечных пептидов способов состоит в том, что эти пептиды обладают неблагоприятными кинетическими константами (Vmax, Km) и к тому же пространственная структура пептидов сильно отличается от физиологических ферментных субстратов. Это проявляется, во-первых, в полностью отличающейся конформации, так что отсутствуют определенные биологические пространства, которые придают специфичность взаимодействию фермент-субстрат, следствием чего является либо отсутствующее распознавание (и, таким образом, модификация), либо неспецифическое распознавание (и, таким образом, модификация), и в конечном счете приходят к неправильным результатам. К тому же пептиды из-за их короткой цепи имеют только одно или небольшое количество сайтов фосфорилирования, так что для исследования модификации за счет фосфорилирования определенного субстрата посредством различных ферментов требуются разные пептиды. Также следствием этого являются опять высокие стоимости и только относительная применимость для способов в рамках HTS.
Задача изобретения поэтому состоит в нахождении варианта определения активности фосфорилирующих и/или дефосфорилирующих белок ферментов, который не обладал бы вышеуказанными недостатками.
Эта задача решается путем применения полипептида (Def GGs для пептида) для определения способности фермента, его функционального фрагмента или производного модулировать уровень фосфорилирования полипептида, отличающегося тем, что полипептид является биотинилированным.
Изобретение основано на результатах исследований, в которых было неожиданно обнаружено, что также в случае полипептидов или белков полной длины не возникает какого-либо стерического затруднения в отношении связывания биотина благодаря стрептавидину и биотинилирование также не интерферирует с фосфорилированием исследуемого субстрата с помощью киназ.
Под термином “полипептид” в рамках настоящего изобретения подразумевают молекулу, содержащую связанные пептидными связями аминокислоты, которая включает по меньшей мере 50 линейно связанных таким образом аминокислот. Более короткие молекулы такого рода обозначают как пептиды. Название “белок” относится к молекулам, которые включают по меньшей мере одну полипептидную цепь, однако, также могут состоять из нескольких, ассоциированных друг с другом или связанных полипептидных цепей. Термин “белок” охватывает, таким образом, и название “полипептид”.
Согласно предпочтительному варианту осуществления различных аспектов настоящего изобретения полипептид обладает длиной из 50 аминокислот и более, предпочтительно, 50-300 аминокислот.
Согласно дальнейшему предпочтительному варианту осуществления различных аспектов изобретения полипептид имеет массу 1 кДа и более, предпочтительно, 1-100 кДа и, особенно предпочтительно, 10-50 кДа.
В качестве субстрата фермента понимают любую имеющуюся молекулу, которая пригодна для модификации посредством фермента. Природными субстратами в рамках настоящего изобретения являются молекулы, которые имеют структуру такого рода, которую они имеют в физиологическом или патологическом окружении в естественном состоянии, и способны подвергаться модификации соответствующим ферментом.
Модуляцией уровня фосфорилирования с помощью фермента называют тип модификации субстрата посредством фермента, в случае которого по меньшей мере одна фосфатная группа переносится на субстрат или удаляется из него. Существенные согласно настоящему изобретению ферменты обладают поэтому способностью катализировать ту и/или другую реакцию. Они, таким образом, по меньшей мере обладают этой способностью киназ и/или фосфатаз, сверх того, однако, обладают еще другими ферментными свойствами (например, свойства протеаз и т.д.). Различные категории ферментов и их свойства в достаточной степени известны специалисту.
Функциональным фрагментом фермента является в данном случае любой фрагмент фермента (следовательно, уменьшенная, соответственно, укороченная по отношению к имеющейся естественной форме молекула), который еще обладает способностью модулировать уровень фосфорилирования по меньшей мере одного полипептида. Термин “функциональное производное” фермента при этом охватывает любой тип модификации фермента по отношению к имеющейся в естественном состоянии форме, который не представляет собой никакой укороченной формы, причем производное фермента еще обладает способностью модулировать уровень фосфорилирования по меньшей мере одного полипептида. Настоящее изобретения относится при этом также к функциональным производным фрагментов ферментов, которые могут модулировать уровень фосфорилирования по меньшей мере одного полипептида.
Определение способности фермента модулировать уровень фосфорилирования полипептида при этом можно осуществлять как качественно, так и также количественно (следовательно, в виде измерения с определением количества).
Предлагаемое согласно изобретению применение обладает тем преимуществом, что таким образом достигаемые результаты на основании длины используемых субстратов являются более информативными, так как они охватывают третичную структуру, которая больше соответствует физиологическим данным. Сверх того, используемые полипептиды, в противоположность известным из уровня техники пептидам, обладают хорошими кинетическими константами (например, в случае IRS-1: Km составляет 19 мкМ, по сравнению с пептидами: >200 мкМ; см. Siemeister et al., 1995) и при анализе субстратов с несколькими сайтами фосфорилирования необходим только один субстрат, с помощью которого можно определять, например, способность также различных ферментов.
Предпочтительный вариант осуществления изобретения относится к применению, при котором определяют способность фермента фосфорилировать полипептид.
К особенно целесообразным видам ферментов с киназной активностью для различных аспектов настоящего изобретения относятся серин-/треонин- или тирозинкиназы. Особенно пригодные примеры киназ включают, в частности, рецептор инсулина, рецептор IGF-1, trK-рецептор, EGF-рецептор, казеинкиназу II, члены семейства протеинкиназы С, протеинкиназу В/Akt, активированную митогеном протеинкиназу (МАР-киназа), GSK-3 бета, ERK 1/2, IKK-бета-киназу, АМР-киназу, PI3-киназу или JNK. Сверх того, предлагаемое согласно изобретению применение также пригодно для определения способности фермента дефосфорилировать полипептид.
Следующий аспект изобретения относится к способу определения способности фермента, его функционального фрагмента или производного модулировать уровень фосфорилирования биотинилированного полипептида. К пригодным способам определения уровня фосфорилирования биотинилированных полипептидов относятся, например, способы, которые, как известно, пригодны для определения уровня фосфорилирования пептидов с короткой цепью. Они известны специалисту.
Согласно предпочтительному варианту осуществления предлагаемый согласно изобретению способ представляет собой способ, в случае которого способность фермента, его функционального фрагмента или производного фосфорилировать полипептид определяют путем осуществления следующих стадий:
а) введение в контакт фермента или его функционального фрагмента или производного с биотинилированным полипептидом и инициация реакции фосфорилирования в пригодной реакционной смеси;
b) введение в контакт реакционной смеси со связанным с носителем средством, которое способно связывать биотинилированный полипептид;
с) определение уровня фосфорилирования, связанного со средством полипептида.
Другой предпочтительный вариант осуществления изобретения относится к способу определения способности фермента, его функционального фрагмента или производного дефосфорилировать полипептид путем осуществления следующих стадий:
а) введение в контакт фермента или его функционального фрагмента или производного с биотинилированным полипептидом, который обладает по меньшей мере одним фосфатным остатком, и инициация реакции фосфорилирования в пригодной реакционной смеси;
b) введение в контакт реакционной смеси со связанным с носителем средством, которое способно связывать биотинилированный полипептид;
с) определение уровня фосфорилирования, связанного со средством полипептида.
Средством при этом может быть любой род молекулы или надмолекулярного объединения (например, тело или приспособление), который пригоден для связывания биотинилированного полипептида. Связывание при этом может происходить с биотиновой частью или с самим полипептидом, причем в случае связывания с самим полипептидом предпочтительно связывание, зависящее от уровня фосфорилирования (например, связывание только в фосфорилированном или нефосфорилированном состоянии в отношении отдельных или нескольких сайтов фосфорилирования). К предпочтительным вариантам осуществления в отношении средства относятся поэтому стрептавидин или специфичные к фосфору антитела (т.е., антитела, которые могут распознавать фосфорилирование определенных остатков в полипептиде и специфически связываться с фосфорилированным там полипептидом).
Используемая в рамках различных аспектов изобретения реакционная смесь при этом может быть биохимического (то есть, in vitro) или клеточного типа. Состав биохимических смесей зависит при этом от “потребностей” исследуемого фермента, причем пригодные компоненты и составы, например, АТФ, буфер для установления желательного значения рН среды и желательная концентрация соли для обеспечения активности фермента, однако, известны компетентному специалисту. В случае биохимических смесей фермент или полипептид могут находиться в виде рекомбинантной и/или в виде частично или полностью очищенной из природных источников молекулы и/или в форме экстрактов из биологического материала, в особенности экстрактов из клеток или тканей.
Биологический материал, в частности, может включать: клетки ткани или органа (такого как, например, головной мозг, кровь, печень, селезенка, почки, сердце, кровеносные сосуды), предпочтительно клетки позвоночного животного, включая человека, или клетки из клеточной культуры. Используемые в рамках изобретения клетки включают при этом все виды клеток, например, эукариотические или прокариотические одноклеточные организмы (как бактерии, например, E.coli, или дрожжи, например, S.pombe или s.cerevisiae), или клеточные линии, которые происходят от многоклеточных организмов (как, например, HeLa, COS, NIH-3T3, CHO и т.д.), причем предпочтительны клеточные линии млекопитающих. Клетки тканевой системы или органа позвоночного животного, включая человека, можно получать посредством общепринятых технологий, как взятие крови, пункция ткани или оперативная технология. Получение такого рода рекомбинантных молекул, очистка естественно имеющихся молекул из клеток или тканей и получение экстрактов из клеток или тканей в достаточной степени известно специалисту (см. также нижеприведенные примеры стандартной литературы).
Клеточные системы, пригодные для применения в рамках различных аспектов, также известны специалисту и предпочтительно включают изолированные клетки, которые первоначально происходят от тканевых систем (предпочтительно, от позвоночных животных, особенно предпочтительно, от млекопитающих и, в особенности, от человека), особенно предпочтительно в форме культивированных клеточных линий; они включают, далее, одноклеточные живые организмы (эукариоты или прокариоты), как, например, клетки дрожжей или бактерий, в особенности в форме культивированных штаммов.
Носителями могут быть любые виды молекул или надмолекулярных объединений (как, например, тела или приспособления), которые пригодны для удаления из реакционной смеси пептида, связанного с ними за счет взаимодействия биотин-стрептавидин, или его маркировки. Пригодными приспособлениями являются, например, мембраны, пластинки или тела самой различной формы (обозначаемые в данном случае как гранулы) из различных материалов, которые в достаточной степени известны в уровне техники. Тип носителя зависит при этом от цели способа (например, диагностика, обнаружение биологически активного вещества или обнаружение нового компонента взаимодействия) и рода детектирования, при этом выбор пригодных носителей находится в компетенции профессионального знания.
Согласно одному варианту осуществления предлагаемого в изобретении способа к реакционной смеси добавляют радиоактивно меченый γ32Р-АТФ и определение уровня фосфорилирования осуществляют путем измерения остающейся на носителе, предпочтительно мембране или пластинке, радиоактивности после осуществления по меньшей мере одной стадии промывки. Таким образом, можно просто удалять не связанные со стрептавидином компоненты реакционной смеси, включая свободную радиоактивность, так что уровень фосфорилирования полипептида можно просто определять при помощи иммобилизированной на носителе радиоактивности. В качестве средства для связывания биотинилированного полипептида в этом случае особенно пригоден стрептавидин.
Согласно другому предпочтительному варианту осуществления предлагаемого в изобретении способа к реакционной смеси добавляют антитело (BSP), которое способно специфически связываться с фосфорилированным полипептидом. Антитело при этом как само по себе может представлять собой средство, так и также может дополнительно добавляться к средству (причем оно тогда предпочтительно не представляет собой никакого специфичного к фосфору антитела и особенно предпочтительно является стрептавидином). Определение уровня фосфорилирования в этом случае осуществляют путем определения количества связанного с полипептидом антитела. Пригодные меры для маркировки и детекции антитела известны специалисту. Так, с одной стороны, можно использовать пригодным образом меченые первичные антитела, которые прямо детектируются, или используются пригодным образом меченые вторичные антитела, направленные против части FC (иммуноглобулиновый фрагмент) первичного антитела, что повышает специфичность детекции.
Понятие “антитело” при этом включает как моноклональные антитела, так и также поликлональные антисыворотки, рекомбинантно полученные антитела и рекомбинантно полученные одноцепочечные антитела. Выбор и получение такого рода антител находятся в компетенции профессионального знания, к тому же, далее, следует указать на нижеприведенную стандартную литературу. Также пригодные маркировки такого рода антител известны в уровне техники и включают, например, ферментные маркировки, как CIP (телячья кишечная фосфатаза) или HRP (пероксидаза хрена), флуорецирующие молекулы, которые при возбуждении за счет облучения светом определенной длины волны генерируют детектируемый сигнал, как техасский красный, Су3, FITC (флуоресцеин-изотиоцианат), или известные флуоресцирующие белки. Выбор пригодных маркировок также соответствует профессиональному знанию. Пригодные маркированные или немаркированные первичные и вторичные антитела, а также их получение представляют собой известный уровень техники, сверх того, такого рода антитела выпускаются различными производителями. Первичные и вторичные антитела выпускаются, например, фирмами Becton Dickinson, Pharmacia или Santa Cruz Biotech.
Согласно предпочтительному варианту осуществления вышеуказанного способа определение количества связанного с полипептидом антитела проводят путем определения количества антитела, остающегося на носителе, предпочтительно, мембране или пластинке, после осуществления по меньшей мере одной стадии промывки.
Согласно другому предпочтительному варианту осуществления предлагаемого в изобретении способа, связанный со средством носитель представляет собой первый носитель, который включает первый генератор сигнала и связывает полипептид со вторым носителем, который включает второй генератор сигнала, причем два генератора сигнала способны генерировать детектируемый сигнал, если они находятся в непосредственной близости друг к другу, и определение уровня фосфорилирования осуществляют посредством определения, генерируется ли детектируемый сигнал. Предпочтительно, согласно данному контексту, носители представляют собой гранулы. Предпочтительно, согласно данному контексту, средство представляет собой специфичное к фосфору антитело. Носитель при этом прямо или непрямо может быть связан с антителом, предпочтительно, непрямо, через протеин А, который связан с носителем. Второй носитель при этом прямо или непрямо может быть связан с полипептидом, предпочтительно, непрямо, через биотиновую часть биотинилированного полипептида; это осуществляют предпочтительно через связанный с носителем стрептавидин.
Генератор сигнала при этом может быть любого рода средством или молекулой, которое(ая) пригодно(на) для генерации детектируемых сигналов; примеры включают флуорофоры, которые после возбуждения за счет воздействия энергии эмиттируют свет, который детектируется прямо или после усиления сигнала с помощью пригодных средств, которые известны в уровне техники. Генератор сигнала при этом в рамках настоящего изобретения выбирают так, что сигнал генерируется только тогда, когда происходит прямое взаимодействие средства (следовательно, предпочтительно, специфичного к фосфору антитела) с полипептидом. Пригодные носители и генераторы сигналов известны (например, в форме ALPHAScreen™ или LANCE™, Perkin-Elmer Life Sciences; HTRF™, CIS Bio International). При этом для генерации сигнала решающей является непосредственная близость носителей друг к другу. Поэтому очень неожиданно оказалось, что этот вид способа пригоден для использования в сочетании с полипептидами, хотя они отчетливо больше по величине, чем используемые в уровне техники пептиды.
Полипептид в рамках различных объектов настоящего изобретения при этом предпочтительно представляет собой природный субстрат фермента, предпочтительно, неукороченной длины.
Особенно пригодные полипептиды включают любые субстраты киназы рецептора инсулина. Особенно предпочтительны при этом полипептиды семейства субстратов рецептора инсулина (IRS), предпочтительно, IRS-1, -2, -3 или -4 и, особенно предпочтительно, IRS-1 или его функциональные фрагменты или производные; следовательно, фрагменты или производные (соответственно, производные фрагментов), которые обладают способностью к фосфорилированию с помощью рецептора инсулина. Далее, предпочтительно, когда IRS представляет собой человеческий IRS. Далее, в рамках различных аспектов настоящего изобретения, предпочтительно использование IRS-1, в особенности, человеческого IRS-1 с последовательностью согласно идентифицированной последовательности №1, особенно предпочтительно, человеческого IRS-1, кодируемого последовательностью согласно идентифицированной последовательности №2. Вышеуказанные полипептиды при этом особенно пригодны для определения способности рецептора инсулина их фосфорилировать. Предпочтительным фрагментом IRS-1 является полипептид с аминокислотной последовательностью согласно идентифицированной последовательности №3.
Различные аспекты изобретения можно применять в различных областях. Целесообразным является их использование, в частности, при идентификации веществ, которые модифицируют способность фермента или его функционального фрагмента или производного модулировать уровень фосфорилирования полипептида. Пригодные аналитические способы или системы, так называемые количественные анализы, с помощью которых измеряют активность или концентрацию, соответственно, количество определенных молекул-мишеней организма (так называемые “мишени”, в данном случае уровень фосфорилирования полипептида), в качестве параметров эффективности потенциальных биологически активных веществ, известны в уровне техники. Они могут представлять собой, например, количественные анализы in vitro, следовательно, биохимические количественные анализы при использовании выделенных или частично выделенных компонентов, образующих реакционную смесь, с помощью которых можно определять эффективность потенциальных биологически активных веществ. Сверх того, это могут быть также клеточные тест-системы (количественные анализы), с помощью которых можно определять активность протеина-мишени (следовательно, в данном случае фермента) и эффективность потенциальных биологически активных веществ в отношении активности этой молекулы-мишени в клеточном окружении.
Количественный анализ при этом представляет собой любой тип аналитических методов, с помощью которых можно следить за биологическим процессом. При этом обычно регулируют протекания молекулярных процессов и сигнальные каскады, части физиологических путей обмена веществ и механизмы регуляции, а также патологические состояния, обнаруживаемые в клеточных или биохимических системах. Фармакологическую активность биологически активного вещества затем можно определять по его способности опосредовать эти пути и механизмы.
Для использования в рамках обнаружения биологически активного вещества, в особенности высокопроизводительного скрининга биологически активных веществ, количественный анализ должен быть воспроизводимым и предпочтительно также стандартизируемым и достоверным (следовательно, малочувствительным к внешним влияниям). Количественный анализ должен быть пригоден предпочтительно для высокопроизводительного скрининга химических веществ в отношении их способности воздействовать на активность молекул-мишеней. Тип количественного анализа при этом зависит, в частности, от рода используемой молекулы-мишени (например, точный тип или род биохимической основной молекулы, как, например, полипептид или полинуклеотид) и “read out”, то есть, параметров, с помощью которых определяют активность молекулы-мишени.
В уровне техники известны различные типы количественных анализов и большей частью также тест-системы для них выпускаются промышленными производителями.
Пригодные для определения взаимодействия двух компонентов связывания анализы включают, например, радиоизотопный или флуоресцентный количественные анализы, например, анализ методом поляризации флуоресценции, тест-системы для которых выпускаются, например, фирмами Panvera, Perkin-Elmer Life Sciences (NEN, LANCE™, AlphaScreen™) или CIS Bio International (HTRF™). Дальнейшие примеры количественных анализов включают клеточные анализы, в случае которых клеточная линия, по желанию, стабильно (индуцируемо или конститутивно; хромосомно или эписомно) или нестабильно экспрессирует рекомбинантный протеин. Эти анализы включают, например, анализы репортер-гена, в случае которых определяют регуляцию определенного промотора или регуляцию пути сигнальной трансдукции или члена каскада сигнальной трансдукции по активности репортер-фермента, экспрессия которого находится под контролем соответствующего промотора. Для этого типа анализа необходимо генерировать рекомбинантную клеточную линию, которая экспрессирует репортер-ген под контролем определенного промотора, который сам исследуют или который регулируется исследуемым каскадом сигнальной трансдукции. Пригодные репортер-ферменты общеизвестны компетентному специалисту и включают люциферазу светляков, люциферазу Renilla (обе выпускаются, например, фирмой Packard Reagents), β-галактозидазу и т.д. Выбор пригодных клеточных линий известен компетентному специалисту и зависит, в частности, от цели анализа или “read out”. Как правило, это клеточные линии, которые можно просто культивировать и трансфицировать, такие, как, например, HeLa, COS, CHO или NIH-3T3 клетки.
Для определения фосфорилирования белка, соответственно, киназной активности пригодны, например, анализ методом поляризации флуоресценции, например, тест-система для которого выпускается фирмой Panvera; анализ методом гомогенной флуоресценции с разрешением по времени (HTRF™, Cis Bio International) или LANCE™ (Perkin-Elmer Life Sciences) или анализ методом гомогенной усиленной проксимальной люминесценции (ALPHAScreen™ фирмы Perkin-Elmer Life Sciences).
Особенно предпочтительное определение, в рамках настоящего изобретения, киназной активности с помощью тест-системы ALPHAScreen™ фирмы Perkin-Elmer Life Sciences осуществляют, например, тем, что с помощью исследуемой киназы фосфорилируют биотинилированный пептид в биохимической смеси в присутствии АТФ. Фосфорилированный пептид затем связывается посредством специфичного к фосфору антитела, с которым связаны конъюгированные с белком А или снабженные пригодными вторичными антителами гранулы акцептора. В той же самой смеси находятся связанные со стрептавидином гранулы донора, которые связываются с биотиновой частью пептида. За счет связывания с пептидом гранулы акцептора и донора находятся в непосредственной близости, что вызывает каскад химических реакций, которые генерируют очень усиленный детектируемый сигнал люминесценции: посредством лазерного возбуждения в грануле донора возбуждается светочувствительность, причем кислород окружающей среды переходит в синглетное состояние. Синглетный кислород затем диффундирует в гранулу акцептора, где он возбуждает тиоксеновое производное, которое, таким образом, вызывает хемилюминесценцию с длиной волны 370 нм, которая, со своей стороны, возбуждает другие флуорофоры в грануле акцептора, люминесцируя свет с длинами волн от 520 нм до 620 нм. Так как возбуждение флуорофоров за счет синглетного кислорода происходит только тогда, когда гранулы донора и акцептора находятся в тесной близости, только тогда генерируются детектируемые сигналы.
Другие виды анализов и другие виды “read outs” также в достаточной степени известны специалисту.
При этом особенно предпочтительно применение в форме высокопроизводительных способов (HTS, высокопроизводительный скрининг), благодаря которым можно анализировать за самое короткое время большое количество веществ.
В зависимости от постановки задачи, модификация модулирования при этом может означать ингибирование или активацию модулирования с помощью фермента. Тип модификации включает при этом любые возможные влияния, которые в конечном счете воздействуют на катализируемый ферментом уровень фосфорилирования полипептида, как модификация взаимодействия субстрат-фермент или модификация каталитической активности фермента, однако, также (предпочтительно, в случае анализа с помощью клеточных реакционных смесей) модификация экспрессии фермента и т.д.
Следующий аспект изобретения относится к способу идентификации веществ, которые модифицируют способность фермента или его функционального фрагмента или производного модулировать степень фосфорилирования полипептида, путем осуществления следующих стадий:
а) определение способности фермента, или его функционального фрагмента, или производного модулировать уровень фосфорилирования полипептида согласно предлагаемому в изобретении вышеуказанному способу, причем в реакционную смесь не добавляют тестируемое вещество;
b) определение способности фермента, или его функционального фрагмента, или производного модулировать уровень фосфорилирования полипептида по вышеописанному способу согласно изобретению, причем в реакционную смесь добавляют тестируемое вещество;
с) сравнение способности по п. а) с таковой по п. b).
Предлагаемые согласно изобретению способы при этом в особенности пригодны для идентификации фармакологически активных веществ для лечения инсулиннезависимого диабета (NIDDM), в онкологии (IGFRK) или для лечения воспалительных процессов (IKK-киназа).
Изобретение подробнее поясняется ниже с помощью различных фигур и примеров, которые не ограничивают объема охраны изобретения.
Пример 1
Используемый фрагмент IRS-1
Для демонстрации возможности биотинилировать полипептидный субстрат из сигнального пути инсулина и использовать его для предлагаемых согласно изобретению применений и способов, был выбран большой фрагмент длиной 262 аминокислоты из человеческого IRS-1 (аа516-аа777), который содержит центральные потенциальные сайты фосфорилирования тирозина (выделено жирным шрифтом), как также серина (подчеркнуто) (Siemeister et al., J. Biol. Chem., 1995). Фрагмент содержит пять потенциальных сайтов фосфорилирования тирозина, которые подчеркнуты на фигуре 3 и представлены ниже вместе с их мотивом, выделенным жирным шрифтом:


Серины 612, 632, 662 и 731, которые представляют собой четыре возможных сайта фосфорилирования серинкиназ в YMXMSP-мотивах, локализованы вблизи сайтов фосфорилирования тирозина рецептора инсулина, которые расположены в сайтах связывания SH2-доменов. Мутация этих остатков серина до аланина приводит к повышению опосредованной IRS-1 активности фосфатидилинозитолтрифосфаткиназы (PI3K) (Mothe et al., 1996), что указывает на то, что ей придана ингибирующая функция. Однако нельзя исключать, что имеются еще другие сайты фосфорилирования серина, которые в настоящее время еще неизвестны.
Пример 2
Клонирование и биотинилирование hlRS-1-p30
Для исследования домены длиной 262 аминокислоты D516-P777 (hlRS-1-p30) человеческого IRS-1 сначала экспрессировали в E. coli, как описывается Siemeister et al., 1995. Экспрессирующие векторы при этом получали путем инсерции полинуклеотида с последовательностью согласно идентифицированной последовательности №10 (кДНК-последовательность hlRS-1-p30) в плазмиду pET3d (выпускается под регистрационным номером 69421 фирмой Novagen) обычными способами. Для этого сначала “пустой” вектор рестриктировали с помощью ферментов NcoI (выпускается фирмой Roche Diagnostics GmbH, Маннхайм, под регистрационным номером 835315) и BamHI (выпускается фирмой Roche Diagnostics GmbH, Маннхайм, под регистрационным номером 656275) в стандартных условиях и очищали при использовании вращающихся колонок (выпускаются фирмой Quiagen, Hilden, под регистрационным номером 28104).
Биотинилирование осуществляли при этом, в соответствии с инструкцией коммерческого поставщика ферментов, Дармштадт, Германия, посредством общепринятых технологий. Экспрессию фрагмента рецептора инсулина hlRS-1-p30 осуществляли при этом, как описывается Siemeister et al., 1995. Для проверки результатов экспрессии получали белковые экстракты из E.coli (штамм: E.coli BL21, поставляется фирмой Novagen под регистрационным номером 69451-3), разделяли путем электрофореза в полиакриламидном геле с додецилсульфатом натрия в стандартных условиях (см., например, нижеуказанную стандартную литературу) и получали в стандартных условиях при окрашивании с помощью раствора для окрашивания с Coomassie (см., например, нижеуказанную стандартную литературу). Очистку hlRS-1-p30 осуществляли также согласно Siemeister и др. Биотинилирование hlRS-1-30 осуществляли ферментативно при использовании трансглутаминазы.
Пример 3
ALPHAScreen™: Фосфорилирование биотинилированного фрагмента IRS-1 c помощью очищенного путем аффинной хроматографии с использованием лектина пшеничных зародышей рецептора инсулина из печени крыс.
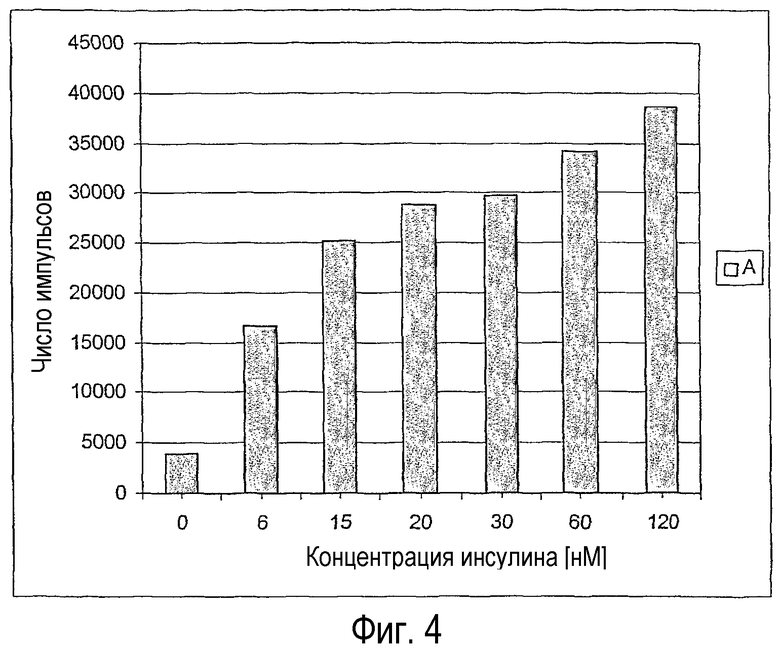
В эксперименте, результаты которого представлены на фигуре 4, очищенный путем аффинной хроматографии с использованием лектина пшеничных зародышей рецептор инсулина из печени крыс (WGA-IR, SEQUACC номер NP_058767 или выпускаемый фирмой Sigma под регистрационным номером 70543) инкубировали с различными концентрациями человеческого инсулина (например, выпускаемый фирмой Sigma под регистрационным номером I-9266) и 85 нМ биотинилированного фрагмента IRS в течение 10 минут при температуре 4°С в 50 мМ Трис-буфере, рН 7,4, 8 мМ MgCl2, 2 мМ MnCl2, с последующей инкубацией после добавки АТФ (конечная концентрация 50 мкМ) в течение 30 минут при температуре 30°С. Реакцию затем прекращали путем добавки этилендиаминтетрауксусной кислоты до конечной концентрации 20 мМ и фосфорилирование IRS-1 определяли путем использования непосредственно связанного с акцептором специфичного к p-Tyr антитела (поставляется фирмой Perkin-Elmer Life Sciences под регистрационным номером 6760601С), как видно из представленного на фигуре 4 “Readout”. С помощью этого способа можно определять EC50 (концентрация, вызывающая 50%-ный эффект) для инсулина до 10 нМ.
Пример 4
ALPHAScreen™: Фосфорилирование биотинилированного фрагмента IRS-1 c помощью РКС и рекомбинантной киназы рецептора инсулина
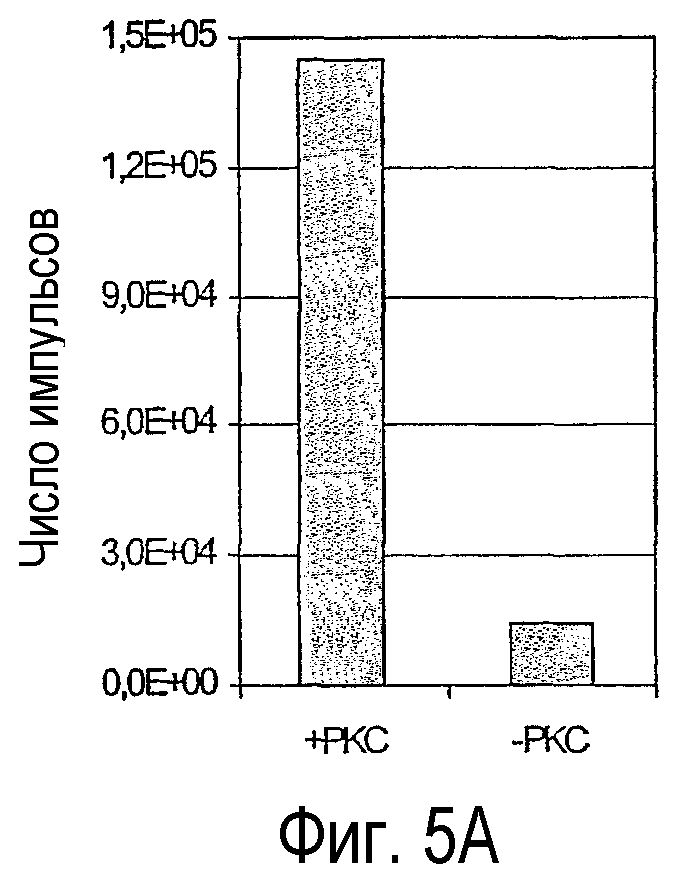
ALPHAScreen™ фирмы Perkin-Elmer Life Sciences позволяет осуществлять детекцию взаимодействия между фосфорилированным фрагментом IRS-1 и антителами, которые распознают фосфорилированные остатки серина или тирозина (p-Ser/p-Tyr-антитела). Биотинилированный IRS-1 при этом связывается с стрептавидин-донором и антителом с помощью связанного с акцептором белка А или пригодного, связанного с акцептором вторичного антитела. Когда происходит взаимодействие, акцептор достигает и остается в непосредственной близости от донора, так что синглетные атомы кислорода, которые генерируются донором, путем диффузии могут достигать хемилюминесцентных групп в грануле акцептора, следствием чего в конечном счете является эмиссия детектируемого света.
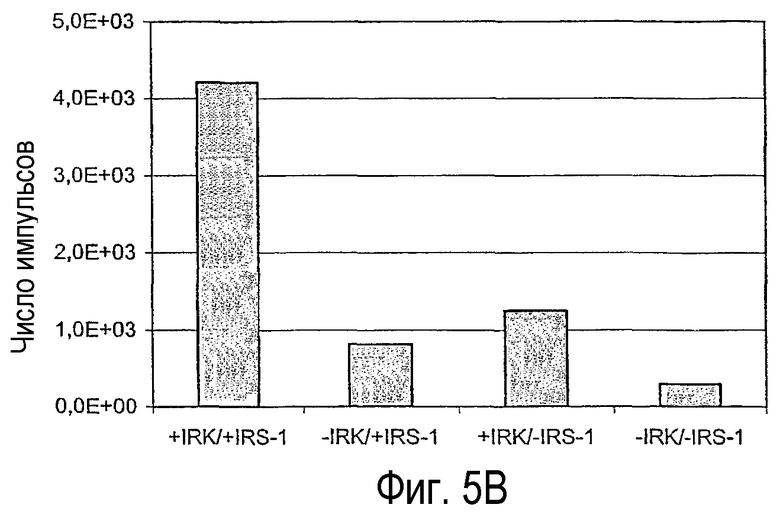
Интенсивности света, генерированные согласно вышеуказанному анализу и представленные в виде гистограмм на фигуре 5А и 5В (так называемые “Readout”), после инкубации в течение 30 минут IRS-1 с протеинкиназой С и АТФ и последующего добавления p-Ser-антител (поставляется фирмой Biosource, Бельгия, под регистрационным номером 44-550) и дальнейшей инкубации в течение 120 минут, детектируют путем измерения с помощью Perkin-Elmer Fusion или AlphaQuest Instrument и количественно определяют. Сравнение генерированных интенсивностей света в присутствии и в отсутствие РКС представлено на фигуре 11А. В эксперименте, результат которого представлен на фигуре 11В, рекомбинантную киназу рецептора инсулина (IRK, аминокислоты 941-1343, NCBI, номер доступа NM_000208) активируют путем инкубации с полилизином в течение 10 минут при температуре 30°С в реакционном буфере: 50 мМ Трис-буфер, рН7,4, 8 мМ MgCl2, 50 мкМ АТФ, и затем добавляют субстрат IRS к IRK с последующей инкубацией в течение 30 минут при температуре 30°С. Фосфорилирование IRS-1 определяют путем использования непосредственно связанного с акцептором специфичного к p-Tyr антитела (поставляется фирмой Perkin-Elmer Life Sciences под регистрационным номером 6760601С), в результате чего получают представленный на фигуре 11В “Readout”.
Благодаря вышеуказанным исследованиям впервые можно было продемонстрировать, что биотинилированные полипептиды можно фосфорилировать с помощью киназ. Это продемонстрировано с помощью фрагмента hlRS-1 массой 28 кДа, который в биотинилированном состоянии может фосфорилироваться с помощью серинкиназы РКСδ и тирозинкиназы рецептора инсулина. Детекция с помощью специфичных к фосфору антител при этом также оказалось успешным, причем стерическое затруднение за счет величины полипептида в сочетании с биотиновым остатком не мешают реакции детектирования. Благодаря этому можно создать, базируясь на принципе ALPHA-Screens™, гомогенную систему анализа, с помощью которой можно определять уровень фосфорилирования полипептидов при использовании возможных за счет биотинилирования технологий очистки и детектирования. Этот принцип анализа, согласно данному контексту, впервые применен к белковому фрагменту величины полипептида (точнее, 28 кДа). Это позволяет осуществлять усовершенствованные поиски фармакологически активных веществ, которые взаимодействуют со сложной системой фосфорилирования и дефосфорилирования клетки (диагноз зависимых от фосфорилирования заболеваний), индентификацию новых протеинкиназ для специальных полипептидов в больших, даже структурно интактных физиологических субстратах, что существенно повышает специфичность фосфорилирований, лежащих в основе этих исследований, и, таким образом, информативность полученных данных. В используемой согласно данному контексту системе анализа “Readout” к тому же является не радиоактивным, а люминесцентным, что представляет собой преимущество в отношении использования в способах высокопроизводительного скрининга (HTS). Представленный согласно данному контексту количественный анализ, таким образом, можно использовать для HTS любых ферментов, модулирующих уровень фосфорилирования полипептидов и белков, как киназы и фосфатазы, для идентификации нового рода биологически активных веществ или верификации известных биологически активных веществ. Точно также он пригоден для других способов, как вышеуказанные способы поиска новых ферментов, которые фосфорилируют определенные полипептиды, так, например, новых фосфорилирующих IRS-1 киназ в полных клеточных лизатах.
Описание фигур
Фигура 1
Белковые последовательности IRS-1-IRS-4 (идентифицированные последовательности №1-4). Номера доступа последовательностей (NCBI, база данных для протеинов) четырех членов семейства представляют собой: NM_005544 (IRS-1 hs);:M:007095 (IRS-2 hs); NM:032074 (IRS-3 rat); NM_003604 (IRS-4 hs).
Фигура 2
Кодирующие ДНК-последовательности IRS-1-IRS-4 (идентифицированные последовательности №5-8). Номера доступа последовательностей (NCBI, база данных для нуклеотидов) четырех членов семейства представляют собой: NM_005544 (IRS-1 hs);:M:007095 (IRS-2 hs); NM:032074 (IRS-3 rat); NM_003604 (IRS-4 hs).
Фигура 3
Включающий 262 аминокислоты домен белка IRS-1 (hlRS-1-p30), который используют для настоящих исследований. Серины 612, 632, 662 и 731 подчеркнуты. Мотивы фосфорилирования тирозина YXXM выделены жирным шрифтом.
Фигура 4
Результаты ALPHAScreens при применении рецептора инсулина, очищенного аффинной хроматографией с использованием лектина пшеничных зародышей.
Фигура 5
Результаты ALPHAScreens™.
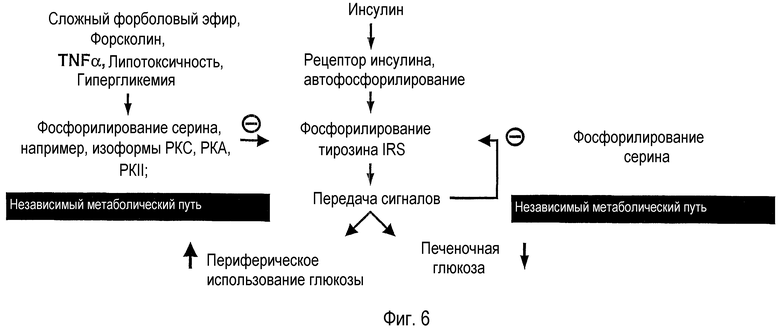
Фигура 6
Наглядное представление взаимодействий рецептора инсулина, IRS-1 и серинкиназ.
Фигура 7
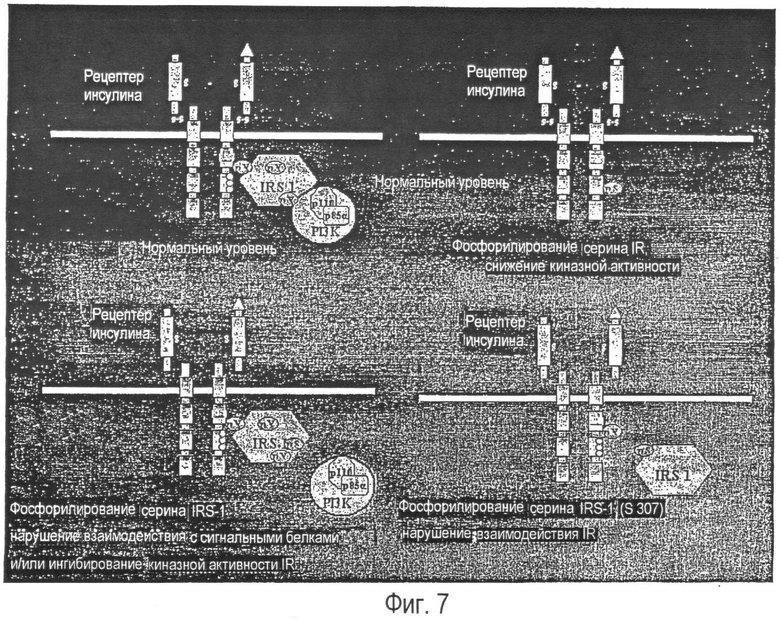
Наглядное представление возможных молекулярных механизмов активного в отношении ингибирования фосфорилирования серина.





Материалы и методы
Если не указано иное, способы представляют собой известные компетентному специалисту стандартные способы и сведения о них можно получить из вышеуказанной литературы, в особенности литературы по стандартным способам (в настоящем тексте также называют как стандартная литература).
Сокращения
Для аминокислот (в общем, также сокращенно как АА или AS) используют трех- или однобуквенные коды (см. также указанную стандартную литературу); для нуклеотидов используют общеупотребительные однобуквенные сокращения (см. также указанную стандартную литературу).

























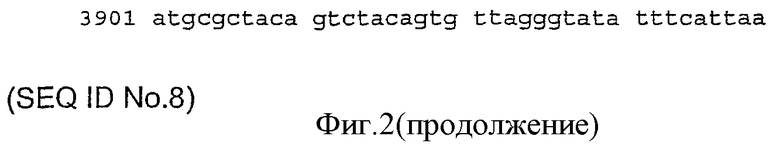

Изобретение относится к области биохимии и может быть использовано при поиске фармакологически активных соединений, обладающих белок-фосфорилирующей активностью или способностью модифицировать указанную активность. Предложен способ определения способности фермента изменять уровень фосфорилирования полипептида, предусматривающий проведение реакции исследуемого фермента с субстратом, в качестве которого используют биотинилированный фрагмент от 516 до 777 остатка человеческого субстрата рецептора инсулина 1 (hIRS-1-р30), связывание продукта реакции с иммобилизованным на носителе стрептавидином и определение уровня его фосфорилирования с помощью антител, специфичных в отношении фосфорилированных остатков полипептида. Способ по изобретению позволяет идентифицировать тирозиновые и сериновые протеинкиназы и может быть положен в основу тест-системы для поиска новых модуляторов их активности. 2 н. и 7 з.п. ф-лы, 7 ил.
1. Способ определения способности фермента модулировать уровень фосфорилирования биотинилированного полипептида по тирозину или серину с использованием стадий:
a) контактирование фермента с биотинилированным фрагментом 516-777 человеческого IRS-1 (hIRS-1-р30) и инициация реакции фосфорилирования в подходящей реакционной смеси,
b) контактирование этой реакционной смеси с иммобилизованным на носителе средством, которое обнаруживает способность связываться с hIRS-1-р30,
c) определение уровня фосфорилирования, связанного с этим средством hIRS-1-р30,
отличающийся тем, что к реакционной смеси добавляют антитело, которое способно специфически связываться с фосфорилированным hIRS-1-р30, и определение уровня фосфорилирования осуществляют путем определения количества связанного с hIRS-1-р30 антитела.
2. Способ по п.1, отличающийся тем, что носителем является мембрана или пластинка.
3. Способ по п.1, отличающийся тем, что средством является стрептавидин.
4. Способ по п.3, отличающийся тем, что связанный с иммобилизованным средством носитель является первым носителем, который содержит в себе первый генератор сигнала, а антитело связано со вторым носителем, который содержит второй генератор сигнала, причем оба генератора сигнала способны генерировать детектируемый сигнал, если они находятся в непосредственной близости друг к другу, и определение уровня фосфорилирования осуществляется посредством определения, генерируется ли сигнал.
5. Способ по п.1, отличающийся тем, что фермент является тирозинкиназой.
6. Способ по п.1, отличающийся тем, что фермент является одной из следующих киназ или их функциональным фрагментом или производным: рецептора инсулина, рецептора IGF-1, trK-рецептора, EGF-рецептора, казеинкиназы II, протеинкиназы С, протеинкиназы B/Akt, активируемой митогеном протеинкиназы (МАР-киназы), GSK-3, ERK или JNK.
7. Способ по п.1 для идентификации веществ, которые модифицируют способность фермента модулировать уровень фосфорилирования полипептида.
8. Способ идентификации веществ, которые модифицируют способность фермента или его функциональных фрагментов или производных модулировать уровень фосфорилирования полипептида, с использованием стадий:
a) определение способности фермента модулировать уровень фосфорилирования полипептида согласно способу по одному из пп.1-4,
b) определение способности фермента модулировать уровень фосфорилирования полипептида согласно способу по одному из пп.1-4, где к реакционной смеси добавляют тестируемое вещество, с) сравнение результата измерения стадии а) с результатом измерения стадии b).
9. Способ по п.7 для идентификации фармакологически активных веществ в целях лечения инсулинзависимого сахарного диабета (NIDDM).
| US 5759787 (TULARIK INC.), 02.06.1998 | |||
| BOGE A., ROTH R., Anal | |||
| Biochem., 1995, 231 (2), 323-332 | |||
| SIEMEISTER ET AL., J | |||
| Biol | |||
| Chem., 1995, 270(9), 4870-4874 | |||
| FREUD G | |||
| ET AL., Biochem | |||
| Biophys | |||
| Res | |||
| Commun., 1995, 206(1), 272-278. |

