Настоящее изобретение относится к вакцине, предназначенной для облегчения симптомов или предупреждения индукции опосредуемых IgE аллергических реакций. Хотя изобретение в основном относится к вакцине для млекопитающих, предпочтительное его осуществление относится к вакцине для людей и поэтому изобретение будет описываться ниже в основном относительно вакцины, используемой для людей.
Предпосылки создания изобретения
IgE (иммуноглобулин E) является, несмотря на его очень низкие концентрации в человеческой плазме в норме (10-400 нг/мл), главной причиной гиперчувствительности, обнаруженной в человеческой популяции. Это свойство связано с его взаимодействием с рецепторами с высоким сродством к IgE на тучных клетках и базофильных лейкоцитах.
Перекрестное связывание двух IgE рецепторов на поверхности этих типов клеток путем связывания аллергеном инициирует выделение ряда физиологически активных веществ, таких как гистамин, ФАТ (фактор активации тромбоцитов), гепарин, хемотаксические факторы для эозинофильных и нейтрофильных гранулоцитов, лейкотриены и тромбоксаны. Это те медиаторы, которые вызывают непосредственные симптомы опосредуемых IgE аллергических реакций (гиперчувствительности типа I). Болезненные состояния, относящиеся к этой группе, включают большинство типов астмы, аллергии к меху, пыльцевой аллергии, многие типы пищевых аллергий и некоторые типы экземы.
Рецепторы с высоким сродством к IgE охарактеризованы как на белковом, так и на генном уровне у мышей, крыс и человека Kinet et al. 1987; Shimizu et al. 1988; Tepler et al. 1989; Blank et al. 1989; Kinet et al 1989). Эти рецепторы возможно присутствуют только на тучных клетках и базофильных лейкоцитах в нашем организме. Рецептор является комплексом из трех различных субъединиц, так называемых α, β и γ цепей. Экстраклеточно главным образом локализуется α цепь, которая взаимодействует с молекулой IgE.
Детальные исследования, касающиеся области цепи эпсилон молекулы IgE, взаимодействующей с рецепторами с высоким сродством к IgE, показали, что область из 76 аминокислот на границе между доменами CH2 и CH3 (CH = константным доменам в тяжелой цепи) имеет решающее значение для взаимодействия между молекулой IgE и рецептором с высоким сродством к ней.
Этот пептид, как было показано, in vitro способен подавлять взаимодействие между природным IgE и рецептором с высоким сродством к нему в молярном соотношении около 1: 1 при сравнении с целой областью CH2-CH3-CH4 (Helm et al. , 1988). Как было также показано, пептид способен подавлять опосредуемую IgE реакцию воспалительной гиперемии при стимуляции аллергеном. Однако в этом случае концентрация примерно в 10 раз больше концентрации, которая с природным IgE обеспечивает такой же ингибирующий эффект (Helm et al., 1989).
Когда молекула IgE связывается со своим рецептором, определенные области цепи эпсилон будут блокированы от взаимодействия с другими молекулами, такими как, например, антитела, направленные против эпитопов в той же самой области молекулы IgE. Затем антитело IgE может связаться только с или антителом IgE, направленным против области CH2-CH3 молекулы IgE, или с рецептором и, таким образом, никогда с обеими этими молекулами одновременно. Антитела, которые связываются с эпитопами за пределами области, непосредственно взаимодействующей с рецептором, будут в противоположность упомянутым ранее давать повышение перекрестного связывания молекул IgE, которые связаны с поверхностью тучных клеток. В этом случае будут присутствовать очень сильное выделение гранул и анафилактический шок у субъекта, которому введены такие антитела. Антитела, связывающиеся с частью, связывающейся с рецептором, напротив, будут неспособны к перекрестному связыванию этих рецепторов, и никакой немедленной реакции не возникает, кроме эффекта более продолжительного снижения концентрации свободно циркулирующего IgE. Это, возможно, предотвратит выделение гранул тем, что никаких IgE антител не присутствует больше в плазме крови субъекта.
Эти анти-IgE антитела будут, возможно, также более долговременно устранять продуцирующую IgE популяцию B-клеток, что увеличивает возможность получить более продолжительную супрессию синтеза IgE. Во время периодов сильной экспозиции с пыльцой в этом случае антитела будут связываться и полностью устранять всю совокупность IgE, которые являются причиной сильной воспалительной реакции у субъектов с аллергией на пыльцу. Ряд наблюдений показывает, что не страдающие аллергией субъекты имеют относительно высокие концентрации эндогенных анти-IgE антител, которые, как полагают, обладают сходным подавляющим аллергию эффектом.
Действие вакцины, соответствующей этому изобретению, основано на ее способности индуцировать иммунный ответ на собственные IgE организма, которое благодаря этому будет предотвращать связывание IgE антител с этими рецепторами. Вследствие этого будет предотвращаться выделение индуцирующих аллергию веществ, хранящихся в тучных клетках.
Предшествующий уровень знаний
A. Связывающиеся в рецептором пептиды и другие рецепторные антагонисты
В настоящее время в мире несколько групп исследователей работают над получением коротких пептидов или других молекул со способностью связываться с IgE рецептором и таким путем предотвращать связывание антиген-специфичного IgE. Затем такие вещества, как ожидается, можно будет использовать в качестве лекарств для лечения аллергий.
Проблема в этом случае состоит в больших трудностях при получении молекулы, имеющей силу связывания с рецептором, соответствующую очень сильному взаимодействию между природной молекулой IgE и ее рецептором. Чтобы получить эффективный препарат, возможно, нужно работать с веществами, необратимо связывающимися с рецептором. Однако такие вещества являются относительно токсичными, так как они могут связываться ковалентно и блокировать другие структурно сходные молекулы в организме. В этом контексте представляет интерес то, что α цепь IgE рецепторов принадлежит к более обширному семейству генов, куда включены, между прочим, несколько из различных IgE рецепторов. Эти рецепторы абсолютно необходимы для защиты организма от, между прочим, и бактериальных инфекций. Молекулы с повышенной реактивной способностью для ковалентного связывания, кроме того, часто являются относительно нестабильными и поэтому они, вероятно, должны вводиться несколько раз в день и к тому же в относительно высоких концентрациях, чтобы сделать возможной полную блокировку постоянно обновляемой совокупности IgE рецепторов на тучных клетках и базофильных лейкоцитах.
B. Анти-IgE моноклональные антитела для лечения аллергии
Биотехнологическая компания в США работает по модели, включающей продукцию у мышей моноклональных антител, направленных против связывающейся с IgE рецептором области молекулы человеческого IgE. Эти антитела затем "гуманизируются" с помощью генной инженерии так, что константные области мышиных моноклональных антител замещаются соответствующими человеческими областями. И далее эти антитела должны получаться в чистом виде в большом масштабе, чтобы использоваться для инъекций. Гуманизация используется для снижения иммунной реакции организма против вышеназванных антител, которые в противном случае после второй или третьей инъекции вызовут массивное образование иммунных комплексов, которое может привести к непосредственно угрожающим жизни осложнениям.
Для лечения субъектов с аллергией к пыльце, вероятно, будет необходимо вводить эти антитела один или несколько раз в неделю в течение периодов высокой концентрации пыльцы. Проблема при инъекциях моноклональных антител состоит в том, что они, даже если они "гуманизированы", будут содержать высокие концентрации новых эпитопов, действию которых организм никогда ранее не подвергался, и, вероятно, относительно быстро появятся высокие титры антител еще и против вышеназванных моноклональных антител. Это, вероятно, вызовет таким же образом, как и в отношении мышиных моноклональных антител, угрожающие жизни осложнения при образовании иммунных комплексов, так как моноклональные антитела все еще имеют "структурные" области из мышиной V-области. Чтобы избежать этих осложнений, возможно, будет необходимо использовать очень большой список гуманизированных антител, которые должны последовательно чередоваться во время курса лечения. Даже если они вводятся очень правильно, это, однако, все время будет сопровождаться риском, что и тогда до какой-то степени будут образовываться иммунные комплексы.
C. Пептидные вакцины (CH4-ε)
Группа исследователей, руководимая доктором Stanworth, работала в соответствии со стратегией, включающей использование человеческого пептида, соединенного с гетерологичным несущим белком для иммунизации кроликов или крыс. В экспериментах, описанных им, использовали гетерологичную антисыворотку от кролика, чтобы лечить крыс. Они не смогли определить какой-либо иммунный ответ с помощью ELISA у кроликов, которым вводили антиген, что показывает, что иммунный ответ даже на невидоспецифический пептид является настолько слабым, что он не определяется одним из наиболее чувствительных методов, известных в настоящее время (Stanworth et al. 1990). Вышеназванная группа использует короткие пептиды из области, которая находится явно вне части, связывающейся с рецептором (CH2-CH3) молекулы IgE, что тем самым явно отличается от идеи настоящего изобретения, которая будет раскрыта детально ниже. Основной недостаток при использовании коротких пептидов состоит в том, что у них почти полностью отсутствует способность образовывать стабильную вторичную структуру, которая, вероятно, также является одной из причин очень слабого иммунного ответа в модельной системе доктора Stanworth.
Предмет изобретения
Предметом изобретения является получение вакцины, предназначенной для облегчения симптомов или снижения индукции опосредуемых IgE аллергических реакций у млекопитающих, включая человека, вакцины, которая индуцирует иммунный ответ против собственного IgE организма и тем самым будет предупреждать связывание антител IgE с рецепторами IgE, посредством чего будет предотвращаться выделение индуцируемых аллергию веществ, накапливаемых в тучных клетках.
Краткое изложение изобретения
Вышеуказанная цель достигается согласно изобретению с помощью вакцины, которая отличается тем, что содержит белок, имеющий полную аминокислотную последовательность константных доменов CH2-CH3 эпсилон-цепи молекулы IgE рассматриваемого вида млекопитающих или структурно стабильную единицу вышеназванной аминокислотной последовательности, содержащей более 12 аминокислот в их подлинной или в мутантной или в полимеризованной форме, причем белок по желанию соединяется с одним или более гетерологичных белков-носителей и по желанию содержит адъювант.
Изобретение также относится к способу получения вакцины и применения полной аминокислотной последовательности константных доменов CH2-CH3 эпсилон-цепи молекулы IgE вида млекопитающих или структурно стабильной единицы вышеназванной аминокислотной последовательности, содержащей более 12 аминокислот в ее подлинной или в мутантной или полимеризованной форме для получения вакцины против аллергических реакций, опосредуемых IgE, у рассматриваемых видов млекопитающих.
Краткое описание чертежей
На прилагаемых чертежах показано следующее.
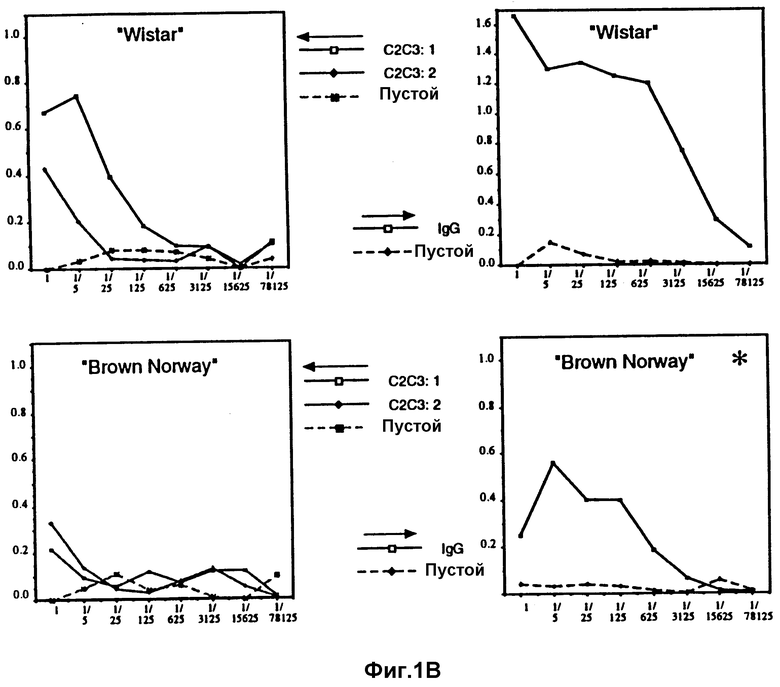
Фиг. 1А и 1В показывают гуморальный иммунный ответ (количество анти-IgE антител) у набора из четырех различных линий крыс по определению с помощью ELISA титров антител против природного IgE человека (в качестве контроля).
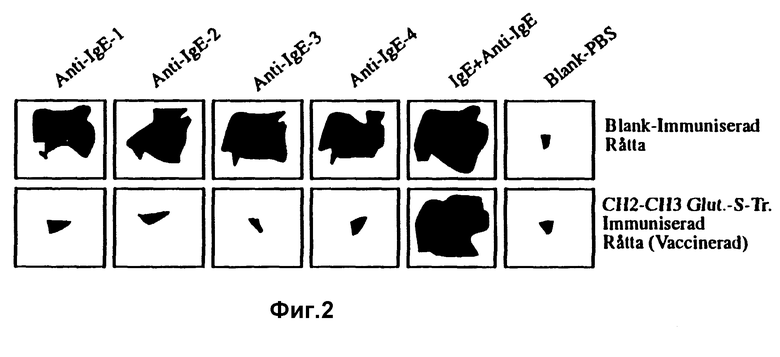
Фиг. 2 показывает подавление опосредуемой IgE воспалительной реакции у вакцинированной крысы в сравнении с иммунизированной плацебо крысой в ответ на инъекцию поликлональной анти-IgE антисыворотки.
Описание изобретения
Определение, что аминокислотная последовательность (полная последовательность или ее части) константных доменов CH2-CH3 присутствует в "мутантной" форме, означает, что аминокислоты в последовательности могут быть изменены путем прямого обмена или путем делеций и вставок соответственно. Затем, "слегка" измененная форма означает, что глазная часть аминокислотной последовательности все еще является последовательностью белка в его подлинной форме и что тем самым эффект является почти тем же самым или, возможно, немного сниженным относительно исходной последовательности; таким образом, белок, имеющий "слегка" измененную аминокислотную последовательность, является все еще допустимым для организма и должен поэтому, подобно его первоначальной форме, соединяться в соответствии с изобретением с гетерологичным белком-носителем, чтобы действовать как антиген.
Под "сильно" измененной формой подразумевается, что замена, делеция и/или вставка аминокислот включает такую интерференцию первоначальной аминокислотной последовательности, что она сама функционирует в качестве антигена, так как создаются новые T-клеточные эпитопы путем произведенного изменения, в отношении которых организм не является толерантным; в этом случае нет необходимости соединять белок с гетерологичным белком-носителем.
Под "полимеризованной" формой подразумевается, что аминокислотная последовательность (целая последовательность или ее часть) белка была полимеризована до формы, содержащей две или более повторяющихся ее единиц; с помощью этой процедуры могут генерироваться новые T-клеточные апитопы на границе между отдельными единицами, давая возможность этой полимеризованной форме белка также функционировать в качество антигена самой по себе, тем самым нет необходимости какого-либо соединения с гетерологичным белком-носителем.
Предпочтительное в настоящее время осуществление изобретения состоит в использовании белка, имеющего аминокислотную последовательность (полную последовательность или ее часть) константных доменов CH2-CH3 эпсилон-цепи молекулы IgE в ее подлинной форме, причем белок соединен с одним или более гетерологичных белков-носителей, и поэтому изобретение будет описано ниже относительно этого конкретного осуществления.
Изобретение основано на полностью новой идее решить проблемы, связанные с прототипными способами иммунизации против аллергических реакций, опосредуемых IgE, тем что вакцина этого изобретения обладает способностью вызывать иммунный ответ против собственного IgE организма.
Путем приведения в действие пусковых механизмов организма для продукции им самим поликлонального анти IgE ответа получаются антитела, которые полностью видоспецифичны и благодаря этому значительно менее иммуногенны. Эти антитела будут затем связывать совокупность свободного IgE, циркулирующего в организме, и тем самым предотвращать связывание с IgE рецептором. Тот факт, что иммунный ответ является поликлональным и тем самым число молекул одного и того же идиотипа очень низко, приведет к почти полному устранению проблемы антиидиотипической реакции.
Тот факт, что подход, описанный здесь, не предпринимался в прототипе, возможно, связан с проблемой, как получить сильный IgE-ответ против собственного IgE организма, так как это является молекулой, к которой организм был толерантен с рождения и, таким образом, не реагирует против нее иммунологически. Чтобы решить эту проблему, в соответствии с изобретением здесь используется свойство иммунной системы, которое едва известно, но которое уникальным путем может решить проблемы, которые с большой вероятностью повлияют на проекты по моноклональным антителам. Путем соединения CH2-CH3 области (белок) с невидоспецифическим белком толерантность иммунной системы к собственному IgE организма обходится. Это приводит к восстановлению не ставшей толерантной популяции T-клеток, которая в норме приводила бы только к гуморальному иммунному ответу против чужеродной молекулы, выбранной в качестве носителя, но которая также будет способствовать B-клеткам, продуцирующим антитела против видоспецифических молекул. Этот эффект получается путем соединения области CH2-CH3 непосредственно с белком-носителем.
Последствия этого феномена почти полностью игнорировались в иммунологическом сообществе, чем можно объяснить, почему другая группа исследователей во всем мире не пробовала подобного подхода. Изобретатель детально проанализировал возможность использования такого феномена, так как было выполнено очень немного таких исследований или они отсутствовали. В проведенных исследованиях, включающих в себя соединение пептидов, использовались человеческие пептиды для инъекции кроликам, мышам или крысам, а не пептиды от тех же самых видов животных, и причиной этого является доминирующее мнение, что нельзя получить сильный иммунный ответ против видоспецифических молекул.
Чтобы подтвердить, согласно изобретению, что сильный иммунный ответ против видоспецифического IgE может быть получен, были выполнены эксперименты, в которых набор линий крыс иммунизировали синтезированным белком, содержащим молекулу носителя, соединенного с целыми CH2-CH3 доменами IgE крысы (пример 2). У этих крыс был получен (через только 4 недели) сильный гуморальный иммунный ответ против природного IgE крыс, т.е. формы IgE, циркулирующего в плазме крысы. Этот гуморальный ответ имеет степень, которая только незначительно ниже, чем уровень (гуморального ответа), который при определении по ELISA получается против полностью невидоспецифического белка, в этом случае человеческого IgE (фиг. 1).
Причина, почему изобретатель, в отличие от доктора Stanworth, получает такой сильный иммунный ответ, состоит, вероятно, в том, что в изобретении используются области, большие по размеру, чем пептиды, имеющие длину только около 10 аминокислот, т.е. в этом случае вплоть до целых доменов CH2-CH3. Это является причиной того, почему получается значительно большее число эпитопов, против которых могут образовываться антитела, и, кроме того, что эти эпитопы присутствуют в той же самой конфигурации, что и у природной молекулы IgE.
Под "гетерологичным белком-носителем" здесь подразумевается любой невидоспецифичный белок, который скорее не обладает очень высокой гомологией по отношению к соответствующему белку вида, для которого белок должен использоваться в качестве носителя. Однако нужно избегать таких белков, которые обычно не находятся в нашем окружении, так как очень сильная иммунная реакции может вызвать проблемы, если мы часто контактируем с этим белком.
Путем соединения небольших пептидов с гетерологичным белком-носителем обычно получается только относительно слабый иммунный ответ против очень ограниченной области молекулы. Кроме того, пептиды до очень большой степени дают повышение иммунного ответа, который действует только против пептида, а не против соответствующей области природного белка.
Важно получить очень сильный иммунный ответ, при котором, кроме того, распознается природный IgE, так как константа связывания для взаимодействия между молекулой IgE и высокоаффинным рецептором очень высока и находится в пределах от 1•10-8 до 2,6•10-1 или выше (Froese, 1980). Путем получения после иммунизации поликлонального ответа несколько различных антител, направленных против различных эпитопов в области CH2-CH3, будут способны связываться одновременно с тем же самым IgE антителом. Это обеспечивает получение очень высокой общей силы связывания для свободного IgE. Тем самым анти-IgE антитела значительно легче, по сравнению со случаем моноклональных антител или антител, образованных против коротких пептидов, будут способны конкурировать за свободный IgE иммунизированного субъекта. Это имеет большое значение, так как взаимодействие между IgE и высокоаффинным рецептором является очень сильным. Идея изобретения по использованию целых доменов или их структурно стабильных частей поэтому влечет за собой очень большое преимущество и полностью новую концепцию по сравнению с ранее описанными пептидными методами.
Гуморальный ответ против пептидов часто имеет низкое сродство или не имеет его к соответствующей аминокислотной области природного белка, которая означает, что гуморальный ответ, полученный против целых доменов или структурно стабильных единиц доменов влечет за собой решающее различие по сравнению с предыдущими подходами в этой области.
При использовании только области CH2-CH3 (а не, как ранее описано, гетерологичных CH4 пептидов) молекулы IgE организм после иммунизации будет продуцировать почти исключительно антитела против области молекулы IgE, взаимодействующей с рецептором IgE. Теоретически существует риск, что антитела, направленные против N-концевой части CH2 домена или C-кольцевой части CH3-домена, могут вызвать анафилактический шок у млекопитающих, у которых эти антитела образуются. Однако, вариабельная область антитела соответствует размеру целого домена и поэтому в большинстве случаев будет получаться пространственное препятствие для связывания рецептора даже с такими антителами. Кроме того, антитела к большому числу эпитопов будут продуцироваться постоянно, пока нарастает иммунный ответ, и поэтому возможность, что только одно из этих очень немногочисленных антител отдельно свяжется с молекулой IgE, вероятно, очень низка. Однако ряд испытаний на животных показал, что такие эффекты иммунизации не могут быть обнаружены (сравните пример 2 ниже). У крыс, имеющих высокое содержание анти-CH2-CH3 антител в крови, не показано какой-либо тенденции к симптомам какого-либо анафилактического шока. С точки зрения здоровья из нельзя отличить от животных, иммунизированных только адъювантом (без антигена), или от животных, иммунизированных человеческим IgE. Это показывает, что у испытуемых животных отсутствуют определяемые отрицательные эффекты этой иммунизации, даже если животные имеют очень высокие титры анти-IgE. К тому же у этих крыс не проявляется каких-либо определяемых отрицательных эффектов этого лечения, что указывает на то, что этот метод лечения с большой вероятностью сможет также применяться без осложнений для лечения людей. Кроме того, эти данные убедительно показывают, что направленная на IgE-рецепторы совокупность IgE у этих крыс фактически отсутствует. Иными словами, сильная анафилактическая реакция анти-IgE антител, которые способны связываться с ранее упомянутыми наружными областями CH2 и CH3 доменов, должна индуцироваться, даже если титр антител относительно низок для таких антител. Это происходит из-за того, что тучные клетки очень чувствительны даже к очень низким концентрациям перекрестно связывающихся антител. В предварительных исследованиях было показано, что в ответ на иммунизацию животные, у которых присутствуют высокие анти-IgE титры, облают сильно сниженной тенденцией выделения гранул их тучными клетками после провокации анти-IgE антителами (пример 3; фиг. 2). Изобретатель в настоящее время проводит значительно более широкое исследование, включающее лечение крыс, у которых смоделирована сильная аллергия к куриному овальбумину. Результаты испытаний, выполненных до сих пор на крысах, являются очень обещающими в нескольких аспектах, так как они показывают, что получается сильный иммунный ответ, который не обладает отрицательными эффектами, и, наконец, что получается сильно сниженное выделение гранул у этих крыс при добавлении поликлонального активатора.
Вакцина этого типа может встретиться с определенными трудностями. Одним из факторов, сильно влияющих на ее возможности, является концентрация вещества, которое желательно удалить из организма. Высокие концентрации в этом случае представляют большие трудности. Если в этой ситуации выбран любой из других иммуноглобулиновых изотипов, проблема должна была бы быть даже большей из-за обычно значительно более высоких концентраций в плазме этих антител.
Из-за его низких уровней в плазме IgE в этом смысле является идеальным. Уровни в плазме у нормальной популяции субъектов без аллергии составляют от 10 до 400 нг/мл. Это количество соответствует менее чем 0,01% общего количества иммуноглобулина в нашей крови. Эти уровни часто немного увеличены у субъектов с аллергией, но очень редко превышают 5 мкг/мл. В качестве сравнения здесь можно упомянуть некоторые испытания, выполненные на мышах, в которых путем использования антител, направленных против одной из легких цепей иммуноглобулинов, были получены животные, у которых почти полностью отсутствовал этот тип иммуноглобулинов. В этих испытаниях, путем введения моноклональных антител против к-цепи иммуноглобулина мышей, была получена соответствующая примерно 95% общего количества иммуноглобулина у мыши, почти полная потеря этих антител в крови мыши (Weiss et al, 1984). Оставшиеся антитела, обнаруженные у мышей, являются почти на 100% Igλ типа (первоначальная концентрация Igλ составляет только примерно 5%). Это показывает, что даже в случае значительно более высоких концентраций вещества, которое нужно удалить из организма, существует возможность почти полностью удалить указанные вещества из кровообращения.
Другое возможное осложнение состоит в том, что ничего не известно о долговременных эффектах индукции сильного аутоиммунитета. Однако преимущество состоит в том, что в этом случае используются T-клетки, направленные против невидоспецифических молекул, для генерации иммунного ответа против видоспецифического антигена. Если программа вакцинации прерывается, гуморальный ответ против собственного IgE организма будет медленно снижаться до неопределяемых уровней после примерно месяца. Это происходит, если вакцинация не усилена CH2-CH3, соединенными в значительной степени такой же молекулой-носителем, которая использована при первоначальной вакцинации. Этот феномен был показан во многих детальных исследованиях на мышах, в которых было показано, что вторичный иммунный ответ против гаптенов, соединенных с различными носителями, целиком зависит от молекулы носителя, выбранной первоначально. Если какой-либо субъект по какой-то невероятной непредвиденной причине реагировал бы отрицательно на эту иммунизацию, иммунизация может быть прервана, и титры антител будут, возможно, достигать неопределяемых уровней в пределах нескольких месяцев. Это сходно с тем, что было бы в случае введения моноклональных антител, направленных против собственно IgE организма.
Объединенный белок, используемый в качестве главного компонента противоаллергической вакцины, может с технической точки зрения получаться двумя различными путями. Один включает продукцию уже объединенного белка, так называемого слитого белка, в прокариотических или эукариотических клетках-хозяевах. В качестве прокариотических хозяев обычно используется бактерия E. coli, в то же время в качестве эукариотических хозяев могут использоваться ряд различных систем, таких как клетки дрожжей или линии клеток. Клеточные линии могут происходить начиная от насекомых и кончая человеком. Однако человеческих клеток обычно избегают для клинического применения, так как всегда существует риск загрязнения человеческими вирусами в клеточных культурах. Второй метод основан на прямом химическом связывании белка-носителя и активного видоспецифического компонента, в этом случае, целой или части области CH2-CH3 молекулы IgE. Кроме того, белки получаются отдельно. Этим методом обычно является один из используемых в иммунизации синтетическими пептидами, так же как и в иммунизации небольшими гаптенами (из веществ, которые не являются белками) и основывается на химической активации белка-носителя, например, CNBr, и затем перемешивании активированного носителя с пептидом или фрагментом белка, которые хотят соединить вместе. Эти два компонента затем соединяются ковалентно друг с другом.
Иммунизация выполняется путем получения смеси растворимой или аггрегированной белковой вакцины с потенцирующим иммунный ответ веществом (адъювант), которая затем вводится подкожно, внутрибрюшинно или внутримышечно. При исследованиях на крысах до сих пор вакцину вводили подкожно или внутрибрюшинно. В этих испытаниях использовались количества антигена примерно 100 мкг на крысу и случай иммунизации, и при этих концентрациях был получен очень сильный иммунный ответ у ряда различных линий крыс (фиг. 1). У людей будут использоваться в первую очередь относительно слабые нетоксичные адъюванты, такие как Alum, или в качестве альтернативы инъекции больших количеств аггрегированного слитого белка без добавления адъюванта. Аггрегированный слитый белок предназначен для увеличения иммуногенности белка.
Кроме того, у людей будут использоваться, вероятно, значительно большие количества антигена, возможно порядка 100-500 мг чистого белка. С технической точки зрения это не представляет каких-либо больших проблем, так как возможно получение очень больших количеств этого слитого белка в очень чистой форме и по цене, которая не слишком отпугивает. При настоящей ситуации существует ряд вариантов соединенных белков при маломасштабном производстве для человеческих вакцин, так же как и для крысиной вакцины. Однако для анализа человеческой вакцины ожидаются результаты очень широкомасштабного изучения, которое предпринято в настоящее время на различных линиях крыс.
Чтобы получить сильный иммунный ответ, первоначально повторные инъекции производятся с примерно 3-недельным интервалом. С этого времени будет, возможно, необходимо проводить иммунизации только за несколько недель перед каждым периодом пыльцы для субъектов с аллергией к пыльце, чтобы активизировать предыдущий иммунный ответ и резко усилить этот ответ перед новым периодом высокого риска.
К тому же, путем использования ряда гетерологичных молекул-носителей, с которым были соединены видоспецифические белки или фрагменты белков, процентное число T-клеток будет увеличиваться. Эти клетки помогут B-клеткам, которые продуцируют антитела, направленные против, как в этом случае, области CH2-CH3 молекулы человеческого IgE. При этом усовершенствовании условий иммунизации ожидается, что будет возможно снизить количество антигена, которое необходимо применить для иммунизации при сохранении иммунизирующего действия. Этот последний подход не будет представлять насущного интереса до тех пор, пока не будут проведены клинические испытания у людей, при которых не могут использоваться сильные адъюванты, и поэтому должны использоваться все возможные способы для усиления иммуногенности вакцины.
Как упомянуто выше, целью изобретения является, в первую очередь, получение вакцины для людей. Однако изобретение включает также вакцины для млекопитающих, которые экономически важно вакцинировать против аллергических реакций, опосредуемых IgE. Примерами таких видов являются собаки, лошади и свиньи.
Изобретение будет проиллюстрировано, кроме того, нижеследующими конкретными рабочими примерами.
Пример 1. Получение препарата слитого белка области CH2-CH3 эпсилон-цепи IgE человека и крысы.
В этом примере была использована система, в которой видоспецифичный белок и белок-носитель продуцируются бактериями, в этом случае E.coli, в соединенной форме. Посредством метода PCR (полимеразной цепной реакции) последовательности кДНК для CH2-CH3 областей, как человеческой, так и крысы, эпсилон-цепи IgE были клонированы и лигированы в коммерчески доступный вектор для продукции слитого белка в бактериальных хозяевах. Использованный вектор был представителем так называемых p EX векторов формы 1, 2 или 3 с разными рамками считывания для лигирования фрагментов кДНК (Smith and Johnson, 1988). Этот тип векторов в этом случае, как было показано, дает высокий выход чистого слитого белка для прямой иммунизации. В этом векторном семействе целая кодирующая область для 26KD клютатион-S-трансферазы (sj 26) из паразитического червя Schistosoma japoncium клонирована после сильного и индуцибельного бактериального промотора. Этот промотор, так называемый тас-промотор, негативно регулируется ас-репрессором. Чтобы получить большие количества белка, подавление промотора ослабляется посредством IPTG (изопропил -β- D-тиогалактозидом). После лигирования CH2-CH3 фрагмента в вектор в C-концевой части sj 26 гена этот вектор трансформируется в штамм E. coli для продукции слитого белка. Ночная культура этой новой бактерии, содержащей вектор, в который был лигирован желаемый фрагмент, разбавляется в отношении 1: 10 средой роста бактерий и ей дают подрасти еще в течение 2 ч. Затем добавляется IPTG до 100 мкМ и культура инкубируется при интенсивном встряхивании в течение последующих 4 ч. Затем бактерии отделяются центрифугированием и клеточный осадок промывается 3 раза PBS. После отмывания клетки суспендируют в PBS + 1% Triton X-100 оch, обрабатываются ультразвуком в течение 5х15 с, чтобы разрушить клеточные стенки бактерий для освобождения белка из клеток. Было показано, что как в случае крысиного, так и человеческого CH2-CH3 слитых белков, белок осаждается внутриклеточно в виде кристаллов и поэтому должен переводиться в растворимое состояние с помощью раствора, содержащего 8 М мочевину. Затем человеческий белок может диализироваться против чистого PBS и стать полностью растворимым. В настоящее время продолжается работа по получению протокола для крупномасштабной очистки до чистоты почти 100% для человеческого слитого белка. Однако CH2-CH3 слитый белок крыс более нерастворим и большая часть белка осаждается уже после диализа в течение от получаса до часа. В последующих примерах использовался препарат слитого белка крыс CH2-CH3, имеющий чистоту примерно 50%. Эти препараты использовались для изучения возможностей блокирования сильной опосредуемой IgE воспалительной реакции у крыс. Остальные 50% белка в препарате состоят из различных загрязняющих бактериальных белков, причем один белок составляет не более 10% от общего белка.
Пример 2. Иммунизация крыс - определение иммунного ответа.
Иммунизация или вакцинация выполняется посредством препарата слитого белка из примера 1 в смеси с потенцирующим иммунный ответ веществом (адъювантом) для получения вакцины. Изучаемым крысам вакцину вводили подкожно или внутрибрюшинно вместе со смесью растворимого и аггрегированного белка. В этих испытаниях использовались количества антигена около 100 мкг на крысу в 0,2 мл полного адъюванта Фрейда и неполного адъюванта соответственно на иммунизацию и крысу. При этих концентрациях и адъювантах были получены очень сильные иммунные ответы у ряда линий крыс.
На фиг. 1А и В представлено испытание на четырех различных линиях крыс по три крысы на линию. Титры антител против природного IgE определяли с помощью ELISA. Это исследование выполнялось таким образом, что природный IgE в покрывающем буфере (5 мкг/мл) использовался для сенсибилизации плат при ELISA. Затем последовательные разведения (1/5) сыворотки крыс от разных испытуемых животных испытывали на реакцию окрашивания в ELISA. Значения поглощения при 400 нм отмечены по оси Y на фиг. 1, а различные разведения 1/5 с увеличением разведений вправо на фиг. 1 отмечены по осям X.
Анализы проводили на четырех разных линиях крыс (Lewis, Sprague Dawley, Wistar and Brown Norwey) в левой части на их способность реагировать на CH2-CH3 вакцину, а в правой части на человеческий IgG (в качестве контроля). Использованной вакциной является CH2-CH3 крыс, который у крыс полностью соответствует человеческой вакцине. Этих крыс иммунизировали только дважды; для начала одной вакцинацией с полным адъювантом Фрейда и раствором белка и затем второй вакцинацией через три недели тем же самым раствором белка в неполном адъюванте Фрейда. Через одну неделю после второй вакцинации у крыс брали пробы крови. Содержание анти-IgE антител в крови затем определяли по ELISA. Как ясно можно видеть из фигуры, три линии реагировали на вакцину очень хорошо, тогда как четвертая линия является так называемым "неотвечающим", который не является настолько необычным, если используются, как в этом случае, конгенные штаммы. Это означает, что эта самая линия крыс не может давать этот антиген для иммунной системы и что поэтому было бы необходимо использовать другой гетерологичный белок-носитель у этой линии крыс, чтобы получить желаемый эффект вакцины против аллергии.
Как можно видеть из этих первоначальных определений по ELISA, получается очень сильный иммунный ответ против только двух доменов IgE крысы, которые должны сравниваться с только слегка более сильной реакцией, полученной против человеческого IgG, который к тому же имеет размер, соответствующий четырем доменам. У этих крыс, показавших очень высокие титры анти-I-антител, не обнаружено каких-либо отрицательных симптомов. Практически их нельзя отличить по любым критериям от крыс, иммунизированных только чистым PBS с адъювантом Фрейда (контроли, отмеченные "пустой" на фиг. 1).
Пример 3. Иммунизация крыс - подавление опосредуемой IgE воспалительной реакции.
Также были выполнены исследования, направленные на оценку способности вакцины согласно изобретению подавлять сильную опосредованную IgE воспалительную реакцию. В качестве системы для исследования использовался тот факт, что анти-IgE антитела обладают способностью перекрестно связываться с IgE антителами, связанными с тучными клетками кожи, и индуцировать, путем своей способности перекрестно связывать эти молекулы IgE, сильное выделение гранул и тем самым вызывать сильную воспалительную реакцию в месте, куда инъекцировали антитела (в этом случае, кожи). В этом примере использовалась поликлональная анти-IgE антисыворотка, направленная против целой константной области IgE. Поэтому эта сыворотка должна содержать большие количества перекрестно-связывающих антител, которые также подтверждаются результатами. Сила воспаления затем измерялась посредством цветной реакции. Проницаемость и, соответственно, просачивание из крови различных белков крови сильно повышалось в области, где вызывалось воспаление. Чем сильнее воспаление, тем большая синяя зона получается, если вводится 1%-ный раствор синего Эванса в кровь испытуемых крыс за два часа до считывания размера синей зоны под кожей испытуемых животных. Проба, которая схематически показана на фиг. 2, выполнялась таким образом, что производилось четыре инъекции, по 50 мкл каждая, концентрированного раствора поликлональной анти-IgE антисыворотки под кожу вакцинированной крысы и иммунизированной плацебо крысы за два часа до удаления кожи и измерения зон воспаления. В качестве контрольных инъекций производились инъекции IgE + анти-IgE на одно пятно на крысу и только PBS на одно пятно. Два типичных примера этих крыс показаны на фигуре, в которых одну из крыс иммунизировали вакциной CH2-CH3 в адъюванте Фрейда и другую контрольную крысу иммунизировали PBS в адъюванте Фрейда. Зоны контролей IgE + анти-IgE имеют очень сходный размер для двух крыс, тогда как зоны для инъекций только анти-IgE антител были уменьшены до около нуля для вакцинированной крысы. Одни анти-IgE антитела вызывали большие синие зоны у крысы, иммунизированной плацебо (в качестве контроля). Это показывает, что у вакцинированной крысы, возможно, полностью отсутствуют IgE антитела на поверхности ее тучных клеток, что находится в полном согласии с результатом, ожидавшимся от иммунизаций, когда у этих крыс были обнаружены высокие концентрации эндогенных анти-IgE антител. Однако у них не отсутствуют тучные клетки, так как возможно сохранить нормальную синюю зону путем добавления экзогенного IgE вместе с анти-IgE антителами и тем самым опять связать рецепторы тучных клеток на этих тучных клетках, которые, возможно, первоначально были свободны от IgE.
В настоящее время выполнена работа с очень обещающими результатами на новой модели на крысах, во-первых, вызывался очень сильный ответ IgE у крыс, Wistar, которые давали хороший ответ на крысиную вакцину, соответствующую изобретению (сравните фиг. 1). Иммунизация крыс выполнена специфическим антигеном, которым в этом случае является овальбумин вместе с токсином рицином, согласно вновь разработанному протоколу доктора Kemeny (Diar-Sancher and Kemeny 1991). Через несколько недель у этих крыс получается очень сильный IgE ответ на овальбумин и, кроме того, этот иммунный ответ является относительно длительным. Ряд крыс проанализировали на их способность давать сильную воспалительную реакцию в коже после инъекции 50 мкл раствора, содержащего 5 мг/мл овальбумина. Это давало синие зоны, которые примерно в два раза больше, чем ранее упомянутые положительные контроли с IgE + анти-IgE, что показывает, что эти крысы являются чрезвычайно аллергичными на овальбумин. Этот тип крыс в настоящее время используется для изучения возможности блокирования аллергических реакций в коже и, кроме того, для изучения эффектов на бронхостеноз и другие типичные симптомы, связанные с аллергией.
Ниже будет следовать в четырех пунктах краткое изложение существенных различий между настоящим изобретением и известными.
1. Изобретение фокусируется единственно на доменах молекулы IgE, которые непосредственно включаются во взаимодействие с рецептором IgE, и не направлено, как в предыдущих исследованиях, на области в невзаимодействующих с рецепторами C4-доменах.
2. Вакцина согласно изобретению содержит видоспецифические белковые фрагменты, соединенные с одним или более гетерологичных носителей, которая делает возможным создание сильного аутоиммунного ответа, в отличие от более ранних исследований, в которых использовались эпсилон-пептиды из другого вида, чем тот, которому инъецируется соединенный пептид, т.е. человеческие пептиды - мыши, крысе или кролику,
3. В противоположность проектам по моноклональным антителам, имеющим место в некоторых лабораториях мира, изобретение основывается на генерации поликлонального иммунного ответа на видоспецифический белок, который значительно повышает возможности получить успешный результат, в то время как осложнения, связанные с иммунными комплексами, приводящие к угрожающим жизни воспалительным реакциям, будут, наиболее вероятно, исключены.
4. Наиболее важное различие состоит в том, что согласно изобретению используются целые домены или структурно стабильные их части (имеющие более 12 аминокислот) в качестве вакцины. Тем самым при испытании в системах на животных получается сильный иммунный поликлональный ответ против природного IgE, что является очень важным свойством вакцины этого типа. Этот эффект получается уже после нескольких недель иммунизации, что означает очень большой прогресс по сравнению с предыдущими методами с использованием коротких синтетических пептидов, разработанными в этой области.
| название | год | авторы | номер документа |
|---|---|---|---|
| УСОВЕРШЕНСТВОВАННЫЕ ВАКЦИНЫ | 1999 |
|
RU2214279C2 |
| АПОПТОТИЧЕСКИЕ АНТИТЕЛА ПРОТИВ IgE | 2008 |
|
RU2500686C2 |
| ПЕПТИДНЫЕ ИММУНОГЕНЫ И СОДЕРЖАЩИЕ ИХ СОСТАВЫ, НАЦЕЛЕННЫЕ НА МЕМБРАНОСВЯЗАННЫЕ IgE ДЛЯ ЛЕЧЕНИЯ ОПОСРЕДОВАННЫХ IgE АЛЛЕРГИЧЕСКИХ ЗАБОЛЕВАНИЙ | 2017 |
|
RU2769983C2 |
| ВАКЦИНА ПРОТИВ ПЕПТИДА CH3 IGE | 2009 |
|
RU2495049C2 |
| ПРОФИЛАКТИЧЕСКАЯ ПРОТИВОРАКОВАЯ ВАКЦИНА | 2004 |
|
RU2385163C2 |
| ПРОФИЛАКТИЧЕСКАЯ ПРОТИВОРАКОВАЯ ВАКЦИНА | 2004 |
|
RU2556128C2 |
| СПОСОБ ОПРЕДЕЛЕНИЯ B-ЛИМФОЦИТОВ, ПРОДУЦИРУЮЩИХ IGE | 1988 |
|
RU2047177C1 |
| СПОСОБЫ ПОВЫШЕНИЯ ЭФФЕКТИВНОСТИ ВАКЦИНЫ ПУТЕМ ВВЕДЕНИЯ АНТАГОНИСТА IL-4R | 2017 |
|
RU2753869C2 |
| МЕЗОТЕЛИН-НАЦЕЛЕННЫЕ ВАКЦИНЫ ОТ РАКА И ИХ ПРИМЕНЕНИЕ | 2018 |
|
RU2751252C1 |
| КОНЪЮГАТ АНГИОТЕНЗИНОВОЙ ПЕПТИДНОЙ СОСТАВЛЯЮЩЕЙ С НОСИТЕЛЕМ, КОМПОЗИЦИЯ ВАКЦИНЫ, СПОСОБ ИММУНИЗАЦИИ ЖИВОТНОГО И СПОСОБ ЛЕЧЕНИЯ ИЛИ ПРОФИЛАКТИКИ ФИЗИЧЕСКОГО НАРУШЕНИЯ, АССОЦИИРОВАННОГО С АКТИВИРУЕМОЙ РЕНИНОМ АНГИОТЕНЗИНОВОЙ СИСТЕМОЙ | 2002 |
|
RU2350622C2 |
Вакцина против опосредуемых IqE аллергических реакций, которая содержит белок, имеющий полную аминокислотную последовательность константных доменов СН2 - СН3эпсилон-цепи молекулы IqE или структурно стабильную единицу указанной аминокислотной последовательности, причем белок произвольно соединен с одним или более гетерологичных белков-носителей и возможно содержит адъювант. Вакцина вводится с адъювантом или без него для повышения концентрации эндогенных анти-IqE-тел в плазме субъекта с аллергией. На практике вакцина может использоваться против всех типов опосредуемых IqE аллергий, так как антитела не зависят от антигенной специфичности молекулы IqE, а будут снижать всю совокупность IqE субъекта. 3 с. и 9 з.п.ф-лы, 2 ил.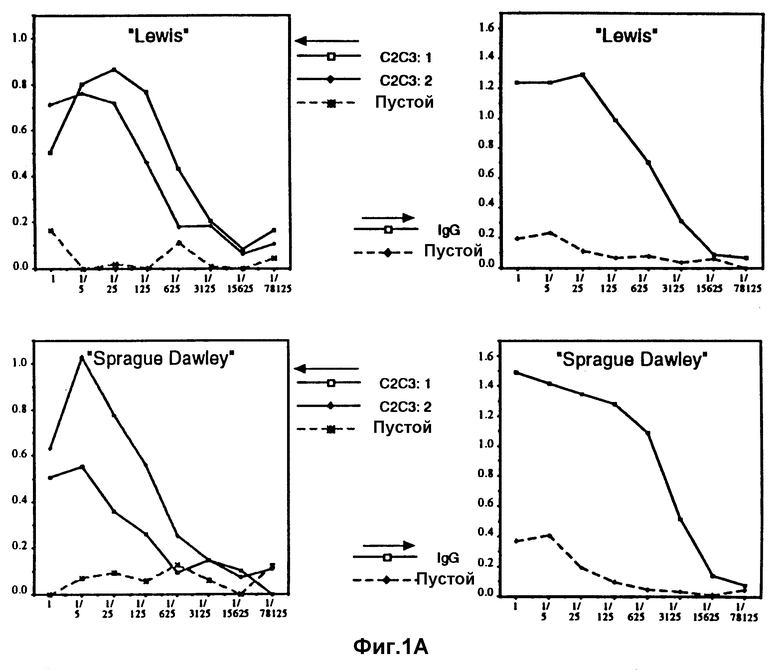
| WO 8904834 A1 (Research Corporation Limited) 01.06.89, C 07 K 7/00 | |||
| EP 0403312 A1 (National Research Development Corp.) 19.12.90, A 61 K 39/00. |

