ОБЛАСТЬ ТЕХНИКИ
Данное раскрытие относится к пептидным иммуногенным конструкциям, нацеленным на ближайший к мембране внеклеточный домен мембраносвязанного IgE (или EMPD IgE), и к их составам в качестве универсальной вакцины для лечения и/или профилактики опосредованных IgE аллергических заболеваний.
УРОВЕНЬ ТЕХНИКИ
Аллергии, также известные как аллергические заболевания, опосредованные иммуноглобулином Е (IgE), представляют собой ряд патологических состояний, вызванных повышенной чувствительностью иммунной системы к чему-либо в окружающей среде, что обычно вызывает небольшие проблемы или вообще не вызывает их у большинства людей. Эти заболевания включают лекарственную и пищевую аллергию, аллергию на укусы насекомых, аллергический ринит (сенная лихорадка), атопический дерматит, аллергическую астму, конъюнктивит, экзему, аллергическую сыпь (крапивницу) и анафилаксию (веб-сайт: en.wikipedia.org/wiki/Allergy). С аллергией часто связаны различные симптомы, которые могут включать покраснение глаз, зудящую сыпь, чихание, насморк, одышку или отек.
Распространенность аллергических заболеваний увеличивается. В начале 20-го века аллергию считали редким заболеванием. Однако с тех пор несколько факторов вызвали резкое увеличение распространенности аллергических заболеваний. Респираторные проявления являются наиболее распространенными, затрагивая до 30% населения в целом. Согласно статистическим данным Всемирной организации здравоохранения (ВОЗ), сотни миллионов людей в мире страдают от ринита и, по оценкам, 235 миллионов человек страдают астмой (веб-сайт: www.who.int/mediacentre/factsheets/fs307/en/index.html). Общественные издержки аллергических заболеваний значительны, главным образом из-за высокой распространенности аллергического риноконъюнктивита и связанной с этим потери продуктивности. Шведское исследование оценило стоимость потери производительности, вызванной ринитом, в 2,7 млрд. евро в год только в одной Швеции, а американское исследование установило, что ринит является наиболее дорогостоящим заболеванием для американских работодателей (Larsen JN, et al., 2016).
Аллергическая реакция - это аномально сильный иммунный ответ, при котором иммунная система борется с предполагаемой угрозой от определенного типа антигена, аллергена, который в противном случае был бы безвредным для организма (веб-сайт: en.wikipedia.org/wiki/Allergen). В частности, аллерген представляет собой антиген, способный стимулировать реакцию гиперчувствительности I типа у склонных к аллергиям индивидуумов посредством ответов IgE. Существует множество источников аллергенов, например, выделения пылевого клеща, пыльца, перхоть домашних животных, определенные продукты питания или химические/физические раздражители. Пищевая аллергия встречается не так часто, как чувствительность к пищевым продуктам, но некоторые продукты, такие как арахис (бобовые), орехи, морепродукты и моллюски, являются причиной серьезной аллергии у многих людей.
Аллергия - это системное иммунологическое заболевание, вызванное примированием адаптивного иммунного ответа на распространенные аллергены. IgE играет центральную роль в опосредовании реакций гиперчувствительности I типа, которые ответственны за возникновение опосредованных IgE аллергических заболеваний. Опосредованные IgE аллергические заболевания характеризуются наличием аллерген-специфических антител IgE и эозинофильным воспалением. Аллергическая реакция является двухфазной, с немедленной реакцией, происходящей в течение нескольких минут после воздействия аллергена, и реакцией поздней фазы, происходящей спустя часы. Немедленная реакция вызвана высвобождением предварительно сформированных медиаторов (например, гистамина, протеаз, хемокинов, гепарина) из базофилов и тучных клеток при перекрестном сшивании IgE, связанного с рецепторами с высокой аффинностью на поверхности клетки. Поздняя фаза аллергической реакции вызвана мобилизацией и привлечением воспалительных клеток, таких как эозинофилы, базофилы, нейтрофилы и мононуклеарные клетки.
Аллергены приводят к повышенным уровням общего свободного иммуноглобулина Е (IgE) в сыворотке и аллерген-специфического IgE у склонных к аллергии людей. Аллерген-специфическая опосредованная IgE гиперчувствительность I типа является ключевой в патогенезе опосредованных IgE аллергических заболеваний (Фиг. 1). IgE сенсибилизирует тучные клетки и базофилы, связываясь с рецептором IgE с высокой аффинностью, FcεRI, на поверхности этих эффекторных клеток. Связывание антигенов с IgE, который уже связан с FcεRI на тучных клетках, вызывает перекрестное сшивание связанного IgE и агрегацию изначально связанного FcεRI. Сшитые рецепторы инициируют каскад сигнальной трансдукции и быструю дегрануляцию. Тучные клетки и базофилы высвобождают накопленный гистамин с последующим синтезом и высвобождением брадикининов, простагландинов, лейкотриенов, цитокинов и других медиаторов воспаления. Они дополнительно привлекают и активируют воспалительные клетки, которые вызывают симптомы аллергии, и активируют биосинтез IgE B-клетками, что способствует повышенной чувствительности. Взаимодействия IgE-FcεRI и дегрануляция являются центральными для аллергических реакций I типа и развития атопической астмы.
Как и другие изотипы иммуноглобулинов, IgE обнаруживается в 2 формах: секретируемая сывороточная форма иммуноглобулина и мембраносвязанная форма (mIgE). Исследования генных сегментов, кодирующих прикрепленный к мембране пептид мышиного и человеческого mIgE, показывают, что разница между mIgE и секретируемым IgE заключается в том, что mIgE содержит три дополнительных области: (1) центральный консервативный участок из 25 гидрофобных незаряженных аминокислотных остатков, который охватывает плазматическую мембрану; (2) С-концевой цитоплазматический хвост; и (3) N-концевой внеклеточный участок, прикрепленный к мембране сегмента mIgE. У человека эпсилон-цепь mIgE на поверхности В-лимфоцитов присутствует в виде как коротких, так и длинных изоформ. Короткая изоформа содержит 15 аминокислот в ближайшем к мембране внеклеточном домене mIgE, называемом EMPD IgE, тогда как длинная изоформа содержит дополнительный сегмент из 52 аминокислотных остатков, что в сумме составляет 67 аминокислот в EMPD. Эти две изоформы генерируются в результате альтернативного сплайсинга между донорным сайтом на 3'-конце экзона CH4 и двумя акцепторными сайтами, разделенными 156 п.н. и расположенными примерно в двух тысячах нуклеотидов в направлении по ходу транскрипции от экзона CH4. Транскрипт длинной формы обнаруживался на уровнях в 100 раз выше уровней обнаружения короткой формы в IgE-продуцирующих миеломных клетках и В-клетках миндалин, обработанных IL-4 с CD40 (Peng et al., 1992 и Zhang et al 1992); тогда как короткая форма была необнаружима на уровнях белка (Peng et al., 1992). EMPD IgE специфичен для формы mIgE и не обнаруживался в секретируемом сывороточном IgE (Фиг. 2).
Современные клинические рекомендации по лечению опосредованных IgE аллергических заболеваний включают сочетание просвещения пациентов, избегания аллергенов, фармакотерапии, иммунотерапии на основе аллергенов и терапевтического таргетирования IgE, но эти варианты лечения имеют свои ограничения. Например, избегание аллергена показано при возможности это сделать, хотя на практике сложно достичь адекватного контроля симптомов с помощью одного только избегания аллергена. Кроме того, хотя безопасные и недорогие лекарственные средства доступны для лечения аллергических симптомов, многие пациенты сообщают о недостаточном контроле симптомов этими препаратами. Важно отметить, что фармакотерапия не влияет на прогрессирование заболевания, и лечение должно проводиться повторно до тех пор, пока преобладают симптомы, что часто означает пожизненное лечение. Только классическая иммунотерапия на основе аллергенов обладает потенциалом модификации заболевания и считается оптимальной стратегией лечения.
Иммунотерапия на основе аллергенов (AIT) включает подкожное введение постепенно увеличивающихся доз аллергена для подавления симптомов при последующем повторном воздействии этого аллергена. Количество аллергена, представляемого иммунной системе на слизистой оболочке в естественных условиях воздействия является относительно низким, но это приводит к эффективной стимуляции аллергического ответа, и симптомы аллергии появляются в течение нескольких минут. В отличие от этого, когда аллерген вводится в качестве иммунотерапии, количество аллергена является относительно высоким, одна доза, вводимая в иммунотерапии, соответствует приблизительно 100-кратному расчетному максимальному годовому потреблению при естественном воздействии. Количественное различие в сочетании с различными путями проникновения в организм оказывает глубокое влияние на иммунную систему, и ее реакция приводит к возникновению иммунологической толерантности к аллергену.
Первоначальная форма введения AIT представляла собой подкожную инъекцию, и этот режим лечения традиционно проводится в две фазы: (1) начальная фаза повышенного дозирования и (2) последующая фаза поддерживающей терапии. Фаза повышенного дозирования представляет собой индивидуальное титрование, при котором увеличивающиеся дозы вводят с целью постепенного наращивания толерантности и тщательной оценки чувствительности пациента. Затем на протяжении фазы поддерживающей терапии пациенту вводится максимальная переносимая доза или максимальная рекомендуемая доза, в зависимости от того, что достигнуто первым.
Считается, что в AIT главную роль играют два механизма: (1) иммунная девиация и (2) индукция регуляторных Т-клеток. Относительный вклад иммунной девиации и регуляторных Т-клеток не установлен, но конечным результатом является снижение и в некоторых случаях - устранение способности вызывать аллергическую реакцию в ответ на воздействие аллергена.
Иммунная девиация - это термин, обозначающий модифицированный иммунологический ответ на воздействие аллергена в ситуации, когда аллерген-специфические T-хелперные клетки типа 1 (Th1) мобилизуются и стимулируются вместо Th2-клеток. Th1-клетки продуцируют гамма-интерферон (IFN-γ), который стимулирует В-клетки продуцировать IgG вместо IgE, а IgG не способен вызывать аллергическую реакцию.
Регуляторные Т-клетки представляют собой разнообразную группу Т-клеток, активных в регуляции иммунных реакций. После проведения AIT было продемонстрировано повышение количества аллерген-специфических CD4+CD25+ регуляторных Т-клеток в периферической крови. Эти клетки продуцируют интерлейкин (IL)-10 и трансформирующий фактор роста (TGF)-β и обладают потенциалом подавлять местные ответы Th2-клеток и перенаправлять переключение классов антител в пользу IgG4 (фактор переключения изотипа IL10) и IgA (фактор переключения изотипа TGF-β). Аллерген-специфические антитела IgG4 прерывают презентацию аллергена Th2-клеткам и, кроме того, блокируют индуцированную аллергеном активацию тучных клеток и базофилов, тем самым значительно ослабляя аллергическую реакцию.
Хотя иммунотерапия на основе антигенов может быть эффективной, все еще существуют серьезные проблемы и неудовлетворенные потребности, связанные с AIT-терапией аллергических заболеваний, опосредованных IgE. Во-первых, все инъекции для AIT делаются в кабинете врача, потому что существует небольшой риск возникновения аллергических реакций, которые могут стать серьезными или даже угрожающими жизни, если их не лечить своевременно и надлежащим образом. Во-вторых, клинически продемонстрирован аспект модификации заболевания с помощью AIT только для нескольких аллергенных продуктов. В-третьих, было описано только несколько структур специфических аллергенов, и определение аллергена главным образом основано на функциональном критерии способности вызывать ответ IgE у восприимчивых людей. Таким образом, аллергены обычно определяются иммунной системой отдельного пациента, и любой иммуногенный белок (антиген) как таковой обладает аллергенным потенциалом, даже если большинство пациентов с аллергией имеют антитела IgE, специфичные для относительно ограниченного количества основных аллергенов. В-четвертых, у каждого пациента есть уникальный паттерн сенсибилизации в отношении эпитопов и молекул аллергенов. В-пятых, все имеющиеся на рынке аллергенные продукты получают путем водной экстракции исходных аллергенных материалов, полученных из натурального сырья, такого как пыльца, культуры клещей домашней пыли, шерсть и/или перхоть животных или яды насекомых, и такое натуральное сырье по своей природе изменчиво по составу. Таким образом, продукты аллергенов, используемые для AIT, не являются общими и отличаются по своему составу, эффективности связывания IgE и степени контроля качества между производителями. Не существует единых действующих международных стандартов. Это означает, что продукты от разных производителей могут работать по-разному у пациентов и, как следствие, клинические результаты не могут быть экстраполированы непосредственно от одного продукта аллергена к другому.
Обновленный обзор иммунотерапии на основе аллергенов: будущее лечения аллергии включено сюда в качестве ссылки (Drug Discovery Today Volume 21, Issue 1, January 2016, Pages 26-37), в нем можно найти все подтверждающие документы для заявлений, сделанных в этом справочном разделе.
В дополнение к AIT было изучено терапевтическое нацеливание молекулы IgE при лечении аллергических заболеваний, опосредованных IgE.
Было показано, что терапевтическое нацеливание моноклонального антитела к IgE на растворимый в сыворотке IgE эффективно при лечении аллергических заболеваний, опосредованных IgE. В настоящее время для лечения персистирующей аллергической астмы средней и тяжелой степени у взрослых и подростков был одобрен препарат омализумаб (XOLAIR®), представляющий собой рекомбинантное гуманизированное моноклональное антитело. Омализумаб останавливает аллергический каскад, связываясь с циркулирующим несвязанным свободным IgE, и предотвращает его связывание с IgE FcεRI на поверхности иммунных эффекторных клеток. Лечение с использованием омализумаба приводит к заметному снижению уровня свободного IgE и снижению регуляции клеточных IgE-рецепторов (Chang et al., 2007). Хотя было показано, что лечение омализумабом является эффективным, оно имеет свои ограничения. В частности, омализумаб способен нейтрализовать свободный IgE в сыворотке, но он не влияет на выработку IgE. Следовательно, для поддержания достаточного подавления сывороточного IgE омализумаб следует вводить часто и постоянно.
Также для лечения опосредованных IgE аллергических заболеваний изучалось терапевтическое воздействие ближайшего к мембране внеклеточного домена мембраносвязанного IgE (EMPD IgE). Сшивание В-клеточного рецептора (BCR) в отсутствие дополнительных костимулирующих сигналов может привести к апоптозу В-клеток. Апоптотическое истощение B-клеток посредством сшивания BCR широко описано для незрелых B-клеток в качестве механизма, посредством которого из репертуара B-клеток удаляются аутореактивные B-клетки. Было показано, что моноклональные антитела к EMPD IgE, такие как 47H4 (Brightbill et al., 2010), 4B12 и 26H2 (Chen et al., 2010), сшивают IgE BCR и вызывают апоптоз экспрессирующих mIgE B-клеток (Фиг. 2). Brightbill et al. также обнаружили, что терапевтическая доставка 47H4 in vivo может снижать установленные IgE-ответы, что наблюдается в моделях инфекции N. brasiliensis и аллергической астмы (Chen et al. 2010). Для истощения В-клеток линии IgE с целью снижения уровня IgE в сыворотке были изучены и идентифицированы определенные антитела и эпитопы для оценки нацеливания на IgE-EMPD, особенно в пределах дополнительной области формы длиной 52 аминокислоты (Chen et al., 2010, Chang et al., 2015, Chen et al., 2002). Одна группа сообщила об использовании ядерного антигена вируса гепатита B (HBcAg) в качестве носителя, содержащего фрагменты EMPD IgE в качестве вставок для индукции образования антител, специфических к EMPD IgE, у мышей BALB/c. Клонированные конструкции спонтанно собираются в вирусоподобные частицы (VPL) с различными фрагментами EMPD IgE, представленными на кончике «шипа» VLP для усиления иммуногенности. Антитела IgG, очищенные из сыворотки иммунизированных мышей, способны вызывать апоптоз экспрессирующих mIgE.FcL клеток Ramos через BCR-зависимый путь каспазы, а также антителозависимую клеточно-опосредованную цитотоксичность (ADCC) в экспрессирующих mIgE.FcL клетках A20 с использованием очищенных мышиных селезеночных NK-клеток в качестве эффекторных клеток (Lin, et al., 2012). Вышеуказанный подход вызвал некоторый интерес к разработке вакцин для лечения аллергии. Однако система экспрессии антигена является громоздкой, большинство генерируемых антител нацелены на VLP-носители, и антигены и система доставки, далеко не оптимальны для эффективной разработки вакцины, применимой для промышленного и клинического применения.
Ввиду вышесказанного существует неудовлетворенная потребность в разработке иммунотерапевтического подхода к лечению и/или профилактике опосредованных IgE аллергических заболеваний, который является аллерген-независимым, способен вызывать высокоспецифичные иммунные ответы против IgE, легко вводится пациентам, способен производится в соответствии с жесткими правилами надлежащей производственной практики (GMP) и является экономически эффективным для применения во всем мире, чтобы заменить вековую практику AIT.
Ссылки:
1. LARSEN, J.N., et al. “Allergy Immunotherapy: The Future of Allergy Treatment”, Drug Discovery Today, 1:26-37 (2016).
2. PENG, C., et al. “A New Isoform of Human Membrane-Bound IgE”, J. Immunol. 148:129-136 (1992).
3. ZHANG, K., et al. “Two unusual forms of human immunoglobulin E encoded by alternative RNA splicing of epsilon heavy chain membrane exons”, J. Exp. Med., 176:233-243 (1992).
4. CHEN, J.B., et al. “Unique epitopes on CɛmX in IgE-B cell receptors are potentially applicable for targeting IgE-committed B cells”, J. Immunol, 184:1748-1756 (2010).
5. LIN, C.J., et al. “CεmX peptide-carrying HBcAg virus-like particles induced antibodies that down-regulate mIgE-B lymphocytes”, Mol. Immunol., 52:190-199 (2012).
6. CHANG, T.W., et al. “C(Epsilon)mX Peptides for Inducing Immune Responses to Human mIgE on B Lymphocytes”, US Patent No. 8,974,794 B2 (2015).
7. CHEN, H.Y., et al. “Monoclonal Antibodies against the CεmX Domain of Human Membrane-Bound IgE and Their Potential Use for Targeting IgE-Expressing B Cells”, Immunol., 128:315-324. (2002).
8. BRIGHTBILL, H.D., et al. “Antibodies specific for a segment of human membrane IgE deplete IgE-producing B cells in humanized mice”, J Clin. Invest., 120:2218-2229 (2010).
9. LU, C.S., et al. “Generating allergen-specific human IgEs for immunoassays by employing human ε gene knockin mice”, Allergy, 70:384-390 (2015).
10. WU, P.C., et al. “The IgE gene in primates exhibits extraordinary evolutionary diversity”, Immunogenetics, 64:279-287 (2012).
11. CHANG, T.W., et al. “Anti-IgE Antibodies for the Treatment of IgE-Mediated Allergic Diseases”, Advances in Immunology, 93:63-119 (2007).
12. TRAGGIAI, E., et al. “An efficient method to make human monoclonal antibodies from memory B cells: potent neutralization of SARS coronavirus”, Nature Medicine, 10: 871-875 (2004).
13. “Asthma Fact Sheet” World Health Organization website, website address: www.who.int/mediacentre/factsheets/fs307/en/index.html (accessed August 18, 2017).
14. “2016 Appendix to GINA Report” Global Initiative For Asthma website, website address: ginasthma.org/wp-content/uploads/2016/04/GINA-Appendix-2016-final.pdf (accessed August 18, 2017).
15. “Allergy” Wikipedia, The Free Encyclopedia, website address: en.wikipedia.org/wiki/Allergy (accessed August 18, 2017).
16. website: en.wikipedia.org/wiki/Allergen)
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к отдельным пептидным иммуногенным конструкциям, нацеленным на ближайший к мембране внеклеточный домен мембраносвязанного IgE (EMPD IgE), для лечения и/или профилактики опосредованных IgE аллергических заболеваний. Данное описание также относится к композициям, содержащим пептидные иммуногенные конструкции, способам получения и применения пептидных иммуногенных конструкций и антител, продуцируемых против пептидных иммуногенных конструкций.
Описанные пептидные иммуногенные конструкции содержат около 20 или более аминокислот. Пептидные иммуногенные конструкции содержат В-клеточный эпитоп из состоящей из 67 аминокислот последовательности полноразмерного EMPD IgE (SEQ ID NO: 1). В-клеточный эпитоп может быть связан с гетерологичным Т-хелперным (Th) эпитопом, полученным из патогенных белков, через необязательный гетерологичный спейсер. Раскрытые пептидные иммуногенные конструкции стимулируют генерацию высокоспецифичных антител, направленных против EMPD IgE, и могут связываться с рекомбинантным белком, содержащим EMPD IgE, EMPD IgE на B-клетках, несущих mIgE, и/или с рекомбинантным растворимым белком EMPD IgE, содержащим Fc-часть человеческого IgG1 и EMPD IgE человеческого мембраносвязанного IgE (обозначается как «γ1-em67»). Описанные пептидные иммуногенные конструкции могут быть использованы в качестве аллерген-независимой, рентабельной, универсальной иммунотерапии для множества пациентов, страдающих от аллергических заболеваний, опосредованных IgE.
Часть В-клеточного эпитопа пептидных иммуногенных конструкций содержит аминокислотные последовательности из полноразмерной последовательности EMPD IgE (SEQ ID NO: 1). В некоторых вариантах осуществления B-клеточный эпитоп содержит последовательность, содержащую внутреннюю внутримолекулярную петлю, образованную эндогенными цистеинами (C18-C39), в соответствии с нумерацией полноразмерной последовательности EMPD IgE (SEQ ID NO: 1). В некоторых конкретных вариантах осуществления В-клеточный эпитоп имеет аминокислотную последовательность EMPD IgE 1-39 (SEQ ID NO: 5), EMPD IgE 7-40 (SEQ ID NO: 6), EMPD IgE 19-38 (SEQ ID NO: 8) или EMPD IgE 1-40 (SEQ ID NO: 9).
Пептидные иммуногенные конструкции согласно данному описанию могут содержать аминокислотную последовательность гетерологичного Th-эпитопа, полученную из патогенного белка (например, SEQ ID NO: 59-87). В некоторых вариантах осуществления гетерологичный Th-эпитоп получен из природных патогенов, таких как токсин дифтерии (SEQ ID NO: 63), Plasmodium Falciparum (SEQ ID NO: 64), токсин холеры (SEQ ID NO: 66). В других вариантах осуществления гетерологичный Th-эпитоп представляет собой идеализированный искусственный Th-эпитоп, полученный из слитого белка вируса кори (MVF 1-5) или поверхностного антигена гепатита B (HBsAg 1-3) в форме одной последовательности или комбинаторных последовательностей (например, SEQ ID NO: 70, 69 и 71).
В некоторых вариантах осуществления пептидные иммуногенные конструкции содержат В-клеточный эпитоп из EMPD IgE, связанный с гетерологичным Т-хелперным (Th) эпитопом через необязательный гетерологичный спейсер. В некоторых вариантах осуществления пептидные иммуногенные конструкции содержат В-клеточную антигенную детерминанту, содержащую более 20 аминокислот из EMPD IgE 1-40 (SEQ ID NO: 9), связанную с гетерологичным Th-эпитопом, полученным из патогенного белка (например, SEQ ID NO: 59-87), через необязательный гетерологичный спейсер. В некоторых вариантах осуществления необязательный гетерологичный спейсер представляет собой молекулу или химическую структуру, способную связывать две аминокислоты и/или пептида вместе. В некоторых вариантах осуществления спейсер представляет собой природную аминокислоту, неприродную аминокислоту или их комбинацию. В конкретных вариантах осуществления пептидные иммуногенные конструкции имеют аминокислотную последовательность SEQ ID NO: 88-95 и 98-124 и 130.
Данное раскрытие также относится к композициям, содержащим пептидную иммуногенную конструкцию EMPD IgE. В некоторых вариантах осуществления описанные композиции содержат более одной пептидной иммуногенной конструкции EMPD IgE. В некоторых вариантах осуществления композиции содержат смесь пептидных иммуногенных конструкций EMPD IgE G1-C39 (например, любую комбинацию SEQ ID NO: 88-95, 98-124 и 130) для охвата обширного генетического фона у субъектов. Композиции, содержащие смесь пептидных иммуногенных конструкций, могут приводить к более высокой процентной доле пациентов с клиническим ответом при иммунизации вакциной для лечения опосредованного IgE аллергического заболевания, по сравнению с композициями, содержащими только одну пептидную иммуногенную конструкцию.
Данное раскрытие также относится к фармацевтическим композициям, включая вакцинные составы, для лечения и/или профилактики аллергических заболеваний, опосредованных IgE. В некоторых вариантах осуществления фармацевтические композиции содержат описанные пептидные иммуногенные конструкции в форме стабилизированного иммуностимулирующего комплекса, образованного посредством электростатических ассоциаций путем смешивания CpG-олигомера с композицией, содержащей пептидный иммуногенный комплекс. Такие стабилизированные иммуностимулирующие комплексы способны дополнительно усиливать иммуногенность пептидных иммуногенных конструкций. В некоторых вариантах осуществления фармацевтические композиции содержат адъюванты, такие как минеральные соли, включая гель на основе квасцов (ALHYDROGEL), фосфат алюминия (ADJUPHOS) или водно-масляные эмульсии, включая MONTANIDE ISA 51 или 720.
Данное раскрытие также относится к антителам, направленным против описанных пептидных иммуногенных конструкций EMPD IgE. В частности, пептидные иммуногенные конструкции согласно данному раскрытию способны стимулировать образование высокоспецифических антител, которые перекрестно реагируют с аминокислотной последовательностью IgE EMPD 1-52 (SEQ ID NO: 2), аминокислотной последовательностью EMPD IgE 1-67 (SEQ ID NO: 1) и их фрагментами (например, SEQ ID NO: 5 и 6) при введении субъекту. Высокоспецифичные антитела, вырабатываемые пептидными иммуногенными конструкциями, перекрестно реагируют с рекомбинантным содержащим IgE белком IgE γ1-em67 и/или EMPD IgE на -В-клетках, несущих мембраносвязанный IgE. Используемые для усиления иммуногенности описанные антитела связываются с EMPD IgE с высокой специфичностью, без особой специфичности, и с любой специфичностью, направленной на гетерологичные Th-эпитопы, что резко контрастирует со стандартным белком или другими биологическими носителями, используемыми для такого усиления пептидной антигенности.
Данное описание также включает способы лечения и/или профилактики опосредованных IgE аллергических заболеваний с использованием описанных пептидных иммуногенных конструкций и/или антител, направленных против пептидных иммуногенных конструкций. В некоторых вариантах осуществления способы лечения и/или профилактики опосредованных IgE аллергических заболеваний включают введение хозяину композиции, содержащей описанную пептидную иммуногенную конструкцию. В некоторых вариантах осуществления композиции, используемые в способах, содержат описанную пептидную иммуногенную конструкцию в форме стабильного иммуностимулирующего комплекса с отрицательно заряженными олигонуклеотидами, такими как CpG-олигомеры, посредством электростатической ассоциации, причем к этим комплексам дополнительно добавляют, необязательно, минеральные соли или масло в качестве адъюванта, для введения субъектам с опосредованными IgE аллергическими заболеваниями. Описанные способы также включают схемы применения, лекарственные формы и способы введения пептидных иммуногенных конструкций хозяину, имеющему риск или страдающему от опосредованных IgE аллергических заболеваний.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
Фигура 1 является иллюстрацией, которая изображает механизм опосредованного IgE пути аллергического заболевания. Наивные зрелые В-клетки начинают с экспрессии мембраносвязанного IgM (mIgE). При обнаружении аллергена эти клетки активируются с помощью родственных T-хелперных клеток (TH), которые обеспечивают В-клетки необходимыми костимулирующими сигналами и цитокинами. Активированные аллерген-специфические B-клетки, которым помогает множество цитокинов, таких как IL-4 и IL-13, становятся IgE-коммитированными B-клетками, экспрессирующими mIgE, посредством рекомбинации с переключением классов. Такие IgE-коммитированные B-клетки окончательно дифференцируются в IgE-секретирующие плазматические клетки. Большинство IgE-секретирующих плазматических клеток недолговечны и мигрируют в область воспаления, а затем отмирают; однако несколько долгоживущих клеток мигрируют в соответствующие ниши в костном мозге. Аллерген-специфический IgE, который секретируется плазматическими клетками, связывается с высокоаффинным рецептором IgE.Fc, FcεRI, на поверхности базофилов крови и тучных клеток ткани. Аллерген-индуцированная агрегация IgE, связанного с FcεRI, стимулирует дегрануляцию базофилов или тучных клеток и высвобождение медиаторов, таких как гистамин, лейкотриены, PGD2, триптаза и различные цитокины, которые запускают немедленную гиперчувствительность и способствуют рекрутированию различных типов клеток, таких как TH2-клетки и эозинофилы.
Фигуры 2А и 2В - иллюстрации, показывающие структурные различия между секретируемым IgE и мембраносвязанным IgE (mIgE) и обоснование истощения B-клеток с mIgE нацеливанием на EMPD IgE.На фигуре 2А показано, что IgE экспрессируется в двух формах: секреторный IgE и мембраносвязанный IgE (mIgE). Секреторный IgE захватывается на клеточной поверхности базофилов и тучных клеток через FcεRI, в то время как mIgE присутствует исключительно на IgE-коммитированных B-клетках как часть B-клеточного рецептора. Ближайший к мембране внеклеточный домен (EMPD) mIgE представляет собой пептидный сегмент из 67 аминокислот (SEQ ID NO: 1) между доменом CH4 и трансмембранной областью, и обнаруживается исключительно в B-клетках с mIgE. Подчеркнутые аминокислоты представляют остатки, которые находятся в короткой изоформе EMPD. Уникальность EMPD IgE обеспечила привлекательный сайт для нацеливания на mIgE и B-клетки с mIgE. На фигуре 2B показан механизм истощения B-клеток с mIgE путем нацеливания на EMPD IgE, который вызывает подавление выработки аллерген-специфического IgE до дифференцировки B-клеток с mIgE в новые IgE-секретирующие плазматические клетки. Существующие плазматические клетки, секретирующие IgE, с их ограниченной продолжительностью жизни в конечном итоге отмирают, что приводит к постепенному снижению общего и аллерген-специфического IgE.
На фигуре 3 представлена блок-схема, идентифицирующая процесс разработки от открытия до коммерциализации (индустриализации) вакцинного состава в соответствии с конкретным вариантом осуществления, раскрытым в данном документе. Настоящее раскрытие включает разработку пептидного иммуногена, разработку пептидной композиции, разработку вакцинного состава, разработку исследования функциональной антигенности in vitro, разработку исследования иммуногенности и эффективности in vivo, а также разработку клинического протокола, как показано в этой таблице. Детальная оценка и анализ каждого из этапов приводит к серии экспериментов, приводящих к коммерциализации безопасного и эффективного вакцинного состава.
На фигуре 4 представлен график, который иллюстрирует кинетику образования антител в течение 8-недельного периода у морских свинок, иммунизированных различными пептидными иммуногенными конструкциями EMPD IgE (SEQ ID NO: 88-94, 96 и 97). Сыворотку разводили от 1:100 до 1:100000 10-кратным серийным разведением. Планшеты для ИФА покрывали пептидом EMPD IgE 1-39 (SEQ ID NO: 5) из расчета 0,5 мкг пептида на лунку. Титр тестируемой сыворотки, выраженный как Log10, рассчитывали с помощью линейного регрессионного анализа A450 с порогом A450, установленным на 0,5.
На фигуре 5 представлен график, который иллюстрирует кривую титрования различных очищенных поликлональных антител к EMPD IgE, полученных от различных иммуногенных конструкций EMPD IgE (SEQ ID NO: 88-94, 96 и 97). Планшеты для ИФА покрывали рекомбинантным, содержащим EMPD IgE (SEQ ID NO: 1) белком γ1-em67 (SEQ ID NO: 1). Поликлональные антитела к EMPD IgE, очищенные из сыворотки морских свинок с помощью хроматографии на основе белка А, разбавляли от 100 мкг/мл до 0,0238 нг/мл путем 4-кратного серийного разведения. EC50 каждого поликлонального антитела к EMPD IgE рассчитывали путем нелинейной регрессии с подбором четырехпараметрической логистической кривой для иммуногенных конструкций SEQ ID NO: 89 и 93, показывающих наилучшую эффективность связывания.
Фигуры с 6А по 6С содержат гистограммы проточной цитометрии, которые иллюстрируют связывание очищенных поликлональных антител из сыворотки морских свинок, собранных у животных, иммунизированных иммуногенными конструкциями EMPD IgE, с клеточной линией клеток Ramos, экспрессирующей либо mIgE.FcL (левая сторона), либо mIgE.FcS (правая сторона). Поликлональные антитела к EMPD IgE, очищенные из сыворотки морских свинок с помощью хроматографии на основе белка А, использовали при 10 мкг/мл с иммуногенными конструкциями, содержащими SEQ ID NO: 88-94, показывающими значительное связывание с B-клетками с mIgEL, по сравнению с иммуногенными конструкциями 96 и 97 с эпитопом EMPD IgE у B-клетки с размерами пептида менее 20 аминокислотных остатков. Фигура 6А содержит гистограммы для поликлональных антител, направленных против иммуногенной конструкции EMPD IgE с SEQ ID NO: 88-90. Фигура 6B содержит гистограммы для поликлональных антител, направленных против иммуногенной конструкции EMPD IgE с SEQ ID NO: 91-93. Фигура 6C содержит гистограммы для поликлональных антител, направленных против иммуногенных конструкций EMPD IgE с SEQ ID NO: 94, 96 и 97.
Фигура 7 представляет собой график, который показывает апоптоз экспрессирующих mIgE.FcL клеток Ramos, индуцированных различными поликлональными антителами к EMPD IgE, направленными против иммуногенных конструкций EMPD IgE (SEQ ID NO: 88-93). Уровень апоптоза выражается в % аннексина V+/PI-. Поликлональные антитела к EMPD IgE, очищенные из сыворотки морских свинок с помощью хроматографии на основе белка А, разбавляли от 1000 до 62,5 нг/мл путем 2-кратного серийного разведения. В качестве положительного контроля использовали гуманизированное моноклональное антитело к IgE, Xolair®. EC50 каждого набора поликлональных антител к EMPD IgE показана под графиком и была рассчитана путем нелинейной регрессии с подбором четырехпараметрической логистической кривой для иммуногенных конструкций, содержащих SEQ ID NO: 88, 90 и 93, показывающих лучшую эффективность индуцирования апоптоза в В-клетках с mIgEL.
Фигура 8 представляет собой столбчатую диаграмму, которая показывает антителозависимую клеточно-опосредованную цитотоксичность (ADCC) экспрессирующих mIgE.FcL клеток Ramos, индуцированных поликлональными антителами к EMPD IgE, направленными на иммуногенные конструкции EMPD IgE (SEQ ID NO: 88-93) при соотношении эффектор/мишень 1/30. Поликлональные антитела к EMPD IgE, очищенные из сыворотки морских свинок с помощью хроматографии на основе белка А, использовали в количестве 10 мкл/мл. В качестве эффекторных клеток использовали клетки селезенки мышей, стимулированные IL-2, а в качестве положительного контроля использовали 5D5, моноклональное антитело мыши к IgE.
Фигура 9 представляет собой схему, которая иллюстрирует картирование эпитопов для анализа специфичности на малом участке с помощью ИФА с использованием перекрывающихся 10-мерных пептидов, покрывающих аминокислоты 9-50 из EMPD IgE иммунной сывороткой морских свинок. Преобладающий эпитоп, распознаваемый антителами в образцах сыворотки, специфичен для области, представляющей петлевую структуру EMPD IgE. Эта схема иллюстрирует внутреннюю петлю, образованную внутримолекулярным дисульфидным мостиком между аминокислотами C18 и C39 в нативном EMPD IgE. Сыворотку разводили 1:1000 для картирования эпитопа. Планшеты для ИФА покрывали 10-мерными пептидами (0,5 мкг пептида на лунку). Реакционноспособные центры были идентифицированы путем исследования картирования эпитопов с использованием образцов иммунных сывороток, собранных у морских свинок, предварительно иммунизированных иммуногенными конструкциями с SEQ ID NO: 88, 89, 93, 96 и 97, и помечены соответствующим образом.
Фигура 10 представляет собой схему, которая иллюстрирует экспериментальную разработку для оценки индуцированных папаином первичных и вторичных иммунных ответов после иммунизации пептидными иммуногенными конструкциями по настоящему изобретению. Гибридных мышей с нокином человеческого IGHE иммунизировали (внутримышечно) пептидной иммуногенной конструкцией EMPD IgE три раза на 0, 3 и 5 неделях. В модели первичного иммунного ответа IgE мышам вводили (подкожно) папаин/TiterMax Gold в неделю 10 и папаин-специфический человеческий IgE (hIgE) измеряли на 12 неделе (показано на Фиг. 13). Для оценки вторичного иммунного ответа IgE мышам подкожно снова подкожно вводили папаин/TiterMax Gold на 16 неделе и измеряли специфический к папаину человеческий IgE (hIgE) на 18 неделе (показано на Фиг. 14). Сыворотку иммунизированных мышей также тестировали на протяжении всего исследования для оценки выработки антител к IgE (Фиг. 11) и изменений в сывороточном IgE (Фиг. 12).
Фигура 11 представляет собой график, который иллюстрирует кинетику выработки антител к IgE в течение 20-недельного периода при реализации схемы эксперимента, описанной на Фиг. 10. В частности, на графике показан образование антител у гибридных мышей с нокином человеческого IGHE (hIGHE x Balb/c, n=8 на группу), иммунизированных внутримышечно иммуногенной конструкцией EMPD IgE (SEQ ID NO: 88 или 93) в указанных дозах (100 мкл на иммунизацию) три раза на 0, 3 и 5 неделях, которым подкожно вводили папаин/TiterMax на 10 и 16 недели. Мышиную сыворотку разводили от 1:100 до 1:(4,19 × 108) 4-кратным серийным разведением. Планшеты для ИФА покрывали рекомбинантным, содержащим EMPD IgE белком γ1-em67. Титр тестируемой сыворотки, выраженный как Log(EC50) коэффициента разведения, рассчитывали с использованием нелинейной регрессии с подгонкой четырехпараметрической логистической кривой.
Фигура 12 представляет собой график, который иллюстрирует изменения сывороточного IgE в течение 20-недельного периода при реализации схемы эксперимента, описанной на Фиг. 10. В частности, на графике показан уровень сывороточного IgE у гибридных мышей с нокином человеческого IGHE (hIGHE x Balb/c, n=8 на группу), внутримышечно иммунизированных иммуногенной конструкцией EMPD IgE (SEQ ID NO: 88 или 93) в указанных дозах (100 мкл на иммунизацию) три раза на 0, 3 и 5 неделях, которым подкожно вводили папаин/TiterMax на 10 и 16 неделях. Сывороточный IgE измеряли с помощью количественного ИФА человеческого IgE (hIgE). Мышиная сыворотка была разведена 1:20. Очищенный из клеток миеломы U266 hIgE использовали для получения стандартной кривой. Концентрация IgE рассчитывалась путем интерполяции A450 к стандартной кривой, полученной путем нелинейной регрессии с подгонкой четырехпараметрической логистической кривой.
Фигура 13 представляет собой график, который показывает подавление выработки папаин-специфического человеческого IgE (hIgE) в первичном ответе IgE, измеренной на 12 неделе, при реализации схемы эксперимента, описанной на Фиг.10. В частности, гибридных мышей с нокином человеческого IGHE (hIGHE x Balb/c, n=8 на группу) иммунизировали внутримышечно иммуногенной конструкцией EMPD IgE (SEQ ID NO: 88 или 93) в указанных дозах (100 мкл на иммунизацию) три раза на 0, 3 и 5 неделе. Сывороточный папаин-специфический hIgE измеряли с помощью количественного ИФА. Мышиная сыворотка была разведена 1:10. Моноклональный химерный папаин-специфичный hIgE использовали для получения стандартной кривой. Концентрацию специфичного для папаина hIgE рассчитывали путем интерполяции A450 к стандартной кривой, полученной путем нелинейной регрессии с подгонкой четырехпараметрической логистической кривой.
Фигура 14 представляет собой подавление выработки папаин-специфического человеческого IgE (hIgE) во вторичном ответе IgE, измеренной на 18 неделе, при реализации схемы эксперимента, описанной на Фиг.10. В частности, гибридных мышей с нокином человеческого IGHE (hIGHE x Balb/c, n=8 на группу) иммунизировали внутримышечно иммуногенной конструкцией EMPD IgE (SEQ ID NO: 88 или 93) в указанных дозах (100 мкл на иммунизацию) три раза на 0, 3 и 5 неделях. Сывороточный папаин-специфический hIgE измеряли с помощью количественного ИФА. Мышиная сыворотка была разведена 1:10. Моноклональный химерный папаин-специфичный hIgE использовали для получения стандартной кривой. Концентрацию специфичного для папаина hIgE рассчитывали путем интерполяции A450 к стандартной кривой, полученной путем нелинейной регрессии с подгонкой четырехпараметрической логистической кривой.
Фигура 15 представляет собой схему, которая иллюстрирует экспериментальную разработку для оценки индуцированных папаином сенсибилизации и вторичного иммунного ответа после иммунизации пептидными иммуногенными конструкциями по настоящему изобретению. Гибридных мышей с нокином человеческого IGHE подкожно (п/к) сенсибилизировали папаином/TiterMax Gold на 0 неделе, а затем иммунизировали внутримышечно пептидной иммуногенной конструкцией EMPD IgE три раза на 3, 6 и 8 неделях. Специфический для папаина вторичный иммунный ответ вызывали путем внутрикожной инъекции папаина в растворе ФСБ в подушечку лапы на 12 неделе. Уровни общего IgE и папаин-специфического IgE/IgG оценивали между 0 и 6 неделями эксперимента (Фиг. 16), а уровень папаин-специфического IgE оценивали на 12, 13 и 14 неделях (Фиг. 17).
Фигура 16 содержит графики, которые показывают результат сенсибилизации папаином всех гибридных мышей с нокином человеческого IGHE при реализации схемы эксперимента, описанной на фигуре 15. Папаин-специфический титр мышиного IgG, экспрессируемый при Log (EC50), и человеческого IgE (нг/мл) определяли в течение 6-недельного периода. Кроме того, общий уровень IgE (нг/мл) был повышен благодаря активации иммунного ответа на данный эпитоп из-за активации иммунного ответа в ходе эксперимента.
Фигура 17 представляет собой график, который показывает подавление выработки папаин-специфического человеческого IgE (hIgE) на 12, 13 и 14 неделях при реализации схемы эксперимента, описанной на Фиг. 15. Конкретно, график показывает специфический для папаина вторичный иммунный ответ у сенсибилизированных гибридных мышей с нокином человеческого IGHE (hIGHE x Balb/c, n=8 на группу), внутримышечно иммунизированных SEQ ID NO: 88 или 93 при 400 мкг/мл (100 мкл на иммунизацию) три раза на 3, 6 и 8 неделях. Сывороточный папаин-специфический hIgE измеряли с использованием количественного ИФА. Мышиная сыворотка была разведена 1:10. Моноклональный химерный папаин-специфический hIgE (нг/мл) использовали для получения стандартной кривой. Концентрацию специфичного для папаина hIgE рассчитывали путем интерполяции A450 к стандартной кривой, полученной путем нелинейной регрессии с подгонкой четырехпараметрической логистической кривой.
Фигуры 18А и 18В представляют собой графики, которые показывают иммуногенность прототипов иммунотерапевтических вакцинных составов от аллергии у яванских макаков, иммунизированных иммуногенной конструкцией EMPD IgE (SEQ ID NO: 88), в четырех дозах по 30, 100, 300 и 1000 мкг на дозу, а также контрольным составом плацебо на 0, 3 и 6 неделях в указанных дозах и анализом на титры антител к EMPD IgE с помощью ИФА. На фигуре 18А показано образование антител у макаков, иммунизированных составом, содержащим Montanide™ ISA 51 и CpG ODN. На фигуре 18В показан ответ антител у макаков, иммунизированных составом, содержащим ADJUPHOS и CpG ODN.
Фигура 19 представляет собой графики, которые иллюстрируют кинетику образования антител в течение 20-недельного периода у яванских макаков (2 самца и 2 самки на группу), иммунизированных внутримышечно иммуногенной конструкцией (SEQ ID NO: 125 или 126) в количестве 300 мкг/мл (500 мкл на иммунизацию) три раза на 0, 3 и 6 неделях. Сыворотку макаков разводили от 1:100 до 1:(4,19 × 108) 4-кратным серийным разведением. Планшеты для ИФА покрывали SEQ ID NO: 5. Титр тестируемой сыворотки, выраженный как Log10 коэффициента разведения, рассчитывали с использованием нелинейной регрессии с подгонкой четырехпараметрической логистической кривой. Установленный порог составлял 2-кратное среднее значение A450 для всех образцов сыворотки при разведении 1:100.
На фигуре 20 показана кинетика образования антител IgG, IgA и IgM в течение 20-недельного периода у яванских макаков (2 самца и 2 самки на группу), иммунизированных внутримышечно иммуногенной конструкцией EMPD IgE (SEQ ID NO: 125 или 126) в количестве 300 мкг/мл (500 мкл на иммунизацию) три раза на 0, 3 и 6 неделях. Сыворотку макаков разводили от 1:100 до 1:(4,19 × 108) 4-кратным серийным разведением. Планшеты для ИФА покрывали пептидом EMPD IgE 1-39 (SEQ ID NO: 5). Титр, выраженный как Log10, рассчитывали путем взаимодействия порога с четырехпараметрической логистической кривой, полученной из данных каждой тестируемой сыворотки. Установленный порог составлял 2-кратное среднее значение A450 для всех образцов сыворотки при разведении 1:100.
Фигура 21 представляет собой графики, которые иллюстрируют изменения уровня IgE в сыворотке в течение 20-недельного периода у яванских макаков (2 самца и 2 самки в группе), внутримышечно иммунизированных иммуногенной конструкцией EMPD IgE (SEQ ID NO: 125 или 126) в количестве 300 мкг/мл (500 мкл на иммунизацию) три раза на 0, 3 и 6 неделях. Уровень сывороточного IgE измеряли в количественном ИФА IgE макака. Сыворотка макака была разведена в 1:20. IgE макаков использовали для получения стандартной кривой. Концентрация IgE рассчитывалась путем интерполяции A450 к стандартной кривой, полученной путем нелинейной регрессии с подгонкой четырехпараметрической логистической кривой. Результат представлял собой среднее значение ± СО. Для определения статистической разницы до 0 недели использовали парный двусторонний t-критерий Стьюдента: *P <0,05, **P <0,01 и ***P <0,001.
ПОДРОБНОЕ ОПИСАНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Настоящее раскрытие направлено на пептидные иммуногенные конструкции, нацеленные на ближайший к мембране внеклеточный домен (EMPD) мембраносвязанного IgE (или EMPD IgE). Данное описание также относится к композициям, содержащим пептидные иммуногенные конструкции, способам получения и применения пептидных иммуногенных конструкций и антител, вырабатываемых хозяином, получивших иммунизацию пептидными иммуногенными конструкциями.
Описанные пептидные иммуногенные конструкции содержат около 20 или более аминокислот. Пептидные иммуногенные конструкции содержат В-клеточный эпитоп из состоящей из 67 аминокислот последовательности полноразмерного EMPD IgE (SEQ ID NO: 1). В-клеточный эпитоп может быть связан с гетерологичным Т-хелперным (Th) эпитопом, полученным из патогенных белков, через необязательный гетерологичный спейсер. Описанные пептидные иммуногенные конструкции стимулируют генерацию высокоспецифичных антител, направленных против EMPD IgE, и могут связываться с рекомбинантным содержащим EMPD IgE белком γ1-em67 и/или с EMPD IgE на B-клетках, несущих mIgE. Описанные пептидные иммуногенные конструкции могут быть использованы в качестве аллерген-независимой, рентабельной, универсальной иммунотерапии для множества пациентов, страдающих от аллергических заболеваний, опосредованных IgE.
Часть В-клеточного эпитопа пептидных иммуногенных конструкций содержит аминокислотные последовательности из полноразмерной последовательности EMPD IgE (SEQ ID NO: 1). В некоторых вариантах осуществления B-клеточный эпитоп содержит последовательность, содержащую внутреннюю внутримолекулярную петлю EMPD IgE, образованную эндогенными цистеинами (C18-C39), в соответствии с нумерацией полноразмерной последовательности EMPD IgE (SEQ ID NO: 1). В некоторых конкретных вариантах осуществления В-клеточный эпитоп имеет аминокислотную последовательность EMPD IgE 1-39 (SEQ ID NO: 5), EMPD IgE 7-40 (SEQ ID NO: 6), EMPD IgE 19-38 (SEQ ID NO: 8) или EMPD IgE 1-40 (SEQ ID NO: 9).
Пептидные иммуногенные конструкции согласно данному описанию могут содержать аминокислотную последовательность гетерологичного Th-эпитопа, полученную из патогенного белка (например, SEQ ID NO: 59-87). В некоторых вариантах осуществления гетерологичный Th-эпитоп получен из природных патогенов, таких как токсин дифтерии (SEQ ID NO: 63), Plasmodium Falciparum (SEQ ID NO: 64), токсин холеры (SEQ ID NO: 66). В других вариантах осуществления гетерологичный Th-эпитоп представляет собой идеализированный искусственный Th-эпитоп, полученный из слитого белка вируса кори (MVF 1-5) или поверхностного антигена гепатита B (HBsAg 1-3) в форме одной последовательности (например, SEQ ID NO: 60, 67, 72 и 73) или комбинаторных последовательностей (например, SEQ ID NO: 70, 69 и 71).
В некоторых вариантах осуществления пептидные иммуногенные конструкции содержат В-клеточный эпитоп из EMPD IgE, связанный с гетерологичным Т-хелперным (Th) эпитопом через необязательный гетерологичный спейсер. Необязательный гетерологичный спейсер может представлять собой молекулу или химическую структуру, способную связывать две аминокислоты и/или пептида вместе. В некоторых вариантах осуществления спейсер представляет собой природную аминокислоту, неприродную аминокислоту или их комбинацию.
В некоторых вариантах осуществления пептидные иммуногенные конструкции содержат В-клеточную антигенную детерминанту, содержащую более 20 аминокислот из EMPD IgE 1-40 (SEQ ID NO: 9), связанную с гетерологичным Th-эпитопом, полученным из патогенного белка (например, SEQ ID NO: 59-87), через необязательный гетерологичный спейсер. В конкретных вариантах осуществления пептидные иммуногенные конструкции имеют аминокислотную последовательность SEQ ID NO: 88-95 и 98-124 и 130.
Данное раскрытие также относится к композициям, содержащим пептидную иммуногенную конструкцию EMPD IgE. В некоторых вариантах осуществления описанные композиции содержат более одной пептидной иммуногенной конструкции EMPD IgE. В некоторых вариантах осуществления композиции содержат смесь пептидных иммуногенных конструкций, содержащих В-клеточную эпитопную часть EMPD IgE-1-39, связанную с различными Th-эпитопами (например, любую комбинацию SEQ ID NO: 98-124) для охвата обширного генетического фона у пациентов. Композиции, содержащие смесь пептидных иммуногенных конструкций, могут приводить к более высокой процентной доле пациентов с клиническим ответом при иммунизации вакциной для лечения опосредованного IgE аллергического заболевания, по сравнению с композициями, содержащими только одну пептидную иммуногенную конструкцию.
Данное раскрытие также относится к фармацевтическим композициям, включая вакцинные составы, для лечения и/или профилактики аллергических заболеваний, опосредованных IgE. В некоторых вариантах осуществления фармацевтические композиции содержат описанные пептидные иммуногенные конструкции в форме стабилизированного иммуностимулирующего комплекса, образованного посредством электростатических ассоциаций путем смешивания CpG-олигомера с композицией, содержащей пептидный иммуногенный комплекс. Такие стабилизированные иммуностимулирующие комплексы способны дополнительно усиливать иммуногенность пептидных иммуногенных конструкций. В некоторых вариантах осуществления фармацевтические композиции содержат адъюванты, такие как минеральные соли, включая гель на основе квасцов (ALHYDROGEL), фосфат алюминия (ADJUPHOS) или водно-масляные эмульсии, включая MONTANIDE ISA 51 или 720.
Данное раскрытие также относится к антителам, направленным против описанных пептидных иммуногенных конструкций EMPD IgE. В частности, пептидные иммуногенные конструкции согласно данному описанию способны стимулировать выработку высокоспецифических антител, которые перекрестно реагируют с В-клеточной эпитопной частью IgE EMPD пептидных иммуногенных конструкций. Используемые для усиления иммуногенности описанные антитела связываются с EMPD IgE с высокой специфичностью, без особой специфичности, и с любой специфичностью, направленной на гетерологичные Th-эпитопы, что резко контрастирует с антителами, выработанными с использованием стандартных белков или других биологических носителей, используемых для такого усиления пептидной антигенности. Таким образом, раскрытые пептидные иммуногенные конструкции способны разрушать иммунную толерантность к аутоантигену с высокой частотой ответа по сравнению с другими пептидными или белковыми иммуногенами.
В некоторых вариантах осуществления антитела направлены против и специфически связываются с аминокислотной последовательностью EMPD IgE 1-52 (SEQ ID NO: 2), аминокислотной последовательностью EMPD IgE 1-67 (SEQ ID NO: 1) и ее фрагментами (например, SEQ ID NO: 5 и 6) при введении таких пептидных иммуногенных конструкций субъекту. Высокоспецифичные антитела, вырабатываемые пептидными иммуногенными конструкциями, перекрестно реагируют с растворимыми содержащими EMPD IgE пептидами и белками, содержащими EMPD IgE гибридными пептидами и белками, γ1-em67, и/или EMPD IgE на несущих связанный с мембраной IgE В-клетках. Полученные антитела способны связываться с рецептором B-клеток IgE (BCR) на B-лимфоцитах, экспрессирующих mIgE, и перекрестно сшиваться с ним. Такое сшивание индуцирует цитолитические эффекты, такие как апоптоз и антитело-зависимая клеточно-опосредованная цитотоксичность (ADCC), что приводит к снижению выработки сывороточного IgE.
Основываясь на своих уникальных характеристиках и свойствах, раскрытые антитела способны обеспечить универсальный иммунотерапевтический подход к лечению опосредованных IgE аллергических заболеваний независимо от вызывающих их аллергенов.
Данное раскрытие также направлено на способы получения раскрытых пептидных иммуногенных конструкций, композиций и антител. Раскрытые способы обеспечивают низкую стоимость изготовления и контроля качества пептидных иммуногенных конструкций и композиций, содержащих конструкции, которые могут быть использованы в способах лечения опосредованных IgE аллергических заболеваний независимо от вызывающих их аллергенов.
Данное описание также включает способы лечения и/или профилактики опосредованных IgE аллергических заболеваний, независимо от вызывающих их аллергенов, с использованием описанных пептидных иммуногенных конструкций и/или антител, направленных против пептидных иммуногенных конструкций. В некоторых вариантах осуществления способы лечения и/или профилактики опосредованных IgE аллергических заболеваний включают введение хозяину композиции, содержащей описанную пептидную иммуногенную конструкцию. В некоторых вариантах осуществления композиции, используемые в способах, содержат описанную пептидную иммуногенную конструкцию в форме стабильного иммуностимулирующего комплекса с отрицательно заряженными олигонуклеотидами, такими как CpG-олигомеры, посредством электростатической ассоциации, к которым может быть дополнительно добавлен адъювант, для введения субъектам с опосредованными IgE аллергическими заболеваниями. Описанные способы также включают схемы применения, лекарственные формы и способы введения пептидных иммуногенных конструкций хозяину, имеющему риск или страдающему от опосредованных IgE аллергических заболеваний.
Заголовки разделов, используемые в данном документе, предназначены только для целей структурирования и не должны рассматриваться как ограничивающие заявленный предмет. Все ссылки или части ссылок, приведенные в данной заявке, специально включены в данный документ посредством ссылки во всей их полноте для любых целей.
Если не указано иное, все технические и научные термины, используемые в данном документе, имеют то же значение, которое обычно понимает средний специалист в данной области техники, к которой относится это изобретение. Формы единственного числа включают ссылки на формы множественного числа, если в контексте явно не указано иное. Подобным образом, слово «или» предназначено для охвата «и», если в контексте явно не указано иное. Следовательно, фраза «содержащий А или В» означает включающий А, или В, или А и В. Кроме того, следует понимать, что все размеры аминокислот и все значения молекулярного веса или молекулярной массы, приведенные для полипептидов, являются приблизительными и представлены для описания. Хотя способы и материалы, подобные или эквивалентные тем, которые описаны в данном документе, могут использоваться на практике или при тестировании описанного способа, подходящие способы и материалы описаны ниже. Все публикации, заявки на патенты, патенты и другие ссылки, упоминаемые в данном документе, включены посредством ссылки во всей своей полноте. В случае конфликта приоритет будет иметь данное описание, включая объяснения терминов. В дополнение, описанные в данном документе материалы, способы и примеры являются исключительно иллюстративными и не подразумевают ограничения.
Пептидная иммуногенная конструкция EMPD IgE
Данное описание относится к пептидным иммуногенным конструкциям, содержащим В-клеточный эпитоп с аминокислотной последовательностью из EMPD IgE, ковалентно связанной с гетерологичным Т-хелперным (Th) эпитопом непосредственно или через необязательный гетерологичный спейсер.
В контексте данного документа фраза «пептидная иммуногенная конструкция EMPD IgE» или «пептидная иммуногенная конструкция» относится к пептиду, содержащему (а) В-клеточный эпитоп, имеющий около 20 или более аминокислотных остатков из последовательности полноразмерного EMPD IgE длиной 67 аминокислот (SEQ ID NO: 1); (b) гетерологичный Th-эпитоп; и (с) необязательный гетерологичный спейсер.
В некоторых вариантах осуществления пептидная иммуногенная конструкция EMPD IgE может быть представлена формулами:
Пептидная иммуногенная конструкция EMPD IgE может быть представлена формулами:
(Th)m-(A)n-(фрагмент EMPD IgE)-X
или
(фрагмент EMPD IgE)-(A)n-(Th)m-X
или
(Th)m-(A)n-(фрагмент EMPD IgE)-(A)n-(Th)m-X,
где
Th представляет собой гетерологичный T-хелперный эпитоп;
А представляет собой гетерологичный спейсер;
(фрагмент EMPD IgE) представляет собой В-клеточный эпитоп, имеющий от около 20 до около 40 аминокислотных остатков из EMPD IgE;
X представляет собой α-COOH или α-CONH2 аминокислоты;
m равно от 1 до около 4; и
n равно от 0 до около 10.
Пептидные иммуногенные конструкции EMPD IgE по настоящему изобретению были сконструированы и отобраны на основании ряда обоснований. Некоторые из этих обоснований включают использование пептидной иммуногенной конструкции EMPD IgE, которая:
i. сама по себе не является иммуногенной, поскольку является собственной молекулой;
ii. может стать иммуногенной при завлечении белка-носителя или сильного Т-хелперного эпитопа;
iii. когда она становится иммуногенной и вводится хозяину:
a. вызывает образование высоких титров антител, направленных против пептидной последовательности EMPD IgE (В-клеточный эпитоп), а не против белка-носителя или Т-хелперного эпитопа;
b. нарушает иммунную толерантность у иммунизированного хозяина и генерирует высокоспецифичные антитела, обладающие перекрестной реактивностью с EMPD IgE (SEQ ID NO: 1), либо в виде рекомбинантного белка, очищенного из экспрессирующих mIgE.FcL клеток CHO, либо на мембране несущей mIgE B-клетки (например, Ramos), трансфицированной рекомбинантной ДНК, кодирующей mIgE.FcL;
c. генерирует высокоспецифичные антитела, способные индуцировать антителозависимую клеточную цитотоксичность (ADCC) и апоптоз экспрессирующих IgE B-лимфоцитов in vitro (пример 6); и
d. вырабатывает высокоспецифичные антитела, способные привести к снижению базального уровня IgE в крови in vivo, а также к значительному снижению антигенспецифического уровня IgE при примировании и повышении при стимуляции аллергеном (примеры 8-11).
Раскрытые пептидные иммуногенные конструкции EMPD IgE и их составы могут эффективно функционировать в качестве вакцин для снижения или устранения опосредованной IgE аллергической патологии у пациентов, страдающих от опосредованных IgE аллергических заболеваний.
Различные компоненты раскрытой пептидной иммуногенной конструкции EMPD IgE описаны в подробностях ниже.
а. В-клеточный эпитоп EMPD IgE
Настоящее раскрытие направлено на новую пептидную композицию для генерации высоких титров поликлональных антител со специфичностью в отношении пептида EMPD IgE с перекрестной реактивностью в отношении мембраносвязанного IgE, экспрессируемого на В-клетках человека, склонных к секреции IgE. Сайт-специфичность пептидной композиции благодаря усилиям, затраченным на рациональный дизайн, минимизирует образование антител, которые направлены на нерелевантные сайты на белках-носителях.
Используемый в данном документе термин «IgE» относится к иммуноглобулину Е в любой форме, включая секретируемый IgE, мембраносвязанный IgE и их фрагменты. Секретируемые и мембраносвязанные формы IgE показаны на Фиг. 2А.
Термин «mIgE» в контексте настоящего описания относится конкретно к мембраносвязанной форме IgE и ее фрагментам. В некоторых вариантах осуществления mIgE представляет собой мембраносвязанную форму IgE у людей, имеющую аминокислотную последовательность, указанную как форма 2 С-области цепи эпсилон Ig человека (фрагмент); номер доступа PH1215. Связанный с мембраной IgE показан на Фиг.2A (справа).
Используемый в данном документе термин «EMPD IgE» относится к ближайшему к мембране внеклеточному домену (EMPD) мембраносвязанного IgE (mIgE) и его фрагментов. EMPD IgE, также называемый CεmX, расположен между доменом CH4 и заякоренным на мембране С-концевым трансмембранным пептидом и обнаруживается исключительно в B-клетках с mIgE. EMPD IgE является результатом альтернативного сплайсинга ε-РНК-транскрипта, расположенного на расстоянии 156 п. н. против хода транскрипции относительно акцепторного сайта сплайсинга, используемого «короткой» изоформой. Полноразмерная «длинная» изоформа EMPD человеческого IgE имеет длину 67 аминокислот (SEQ ID NO: 1), включая 52 аминокислоты (SEQ ID NO: 2), которых нет в «короткой» изоформе. Мембраносвязанный IgE показан на Фиг. 2А с выделенной частью EMPD. Аминокислотная последовательность полноразмерного EMPD IgE (SEQ ID NO: 1) и ее фрагменты (SEQ ID NO: 2-58 и 127) показаны в таблице 1.
EMPD IgE содержит внутримолекулярную петлю между эндогенными цистеинами (C18-C39), нумерация основана на аминокислотной нумерации 67-аминокислотных и 52-аминокислотных последовательностей EMPD IgE (SEQ ID NO: 1 и 2, соответственно). Внутренняя внутримолекулярная петля IgE показана на Фиг. 9.
В-клеточный эпитоп пептидных иммуногенных конструкций EMPD IgE содержит внутримолекулярную петлевую структуру EMPD IgE или ее части. В некоторых вариантах осуществления B-клеточный эпитоп содержит от около 20 до около 40 аминокислот EMPD IgE.
В некоторых вариантах осуществления аминокислотная последовательность В-клеточной эпитопной части пептидной иммуногенной конструкции EMPD IgE содержит от около 20 до около 40 аминокислотных остатков из полноразмерного EMPD IgE (SEQ ID NO: 1). В некоторых вариантах осуществления B-клеточный эпитоп содержит аминокислотную последовательность из внутренней внутримолекулярной петли EMPD IgE, образованной эндогенными цистеинами (C18-C39) в соответствии с нумерацией полноразмерной EMPD IgE (SEQ ID NO: 1). В конкретных вариантах осуществления последовательность В-клеточного эпитопа заканчивается остатком Arg (R) на 38, Cys (C) на 39 или His (H) на 40 на C-конце внутримолекулярной петлевой структуры EMPD IgE.
В некоторых вариантах осуществления B-клеточный эпитоп имеет аминокислотную последовательность EMPD IgE-1-39 (SEQ ID NO: 5), EMPD IgE-7-40 (SEQ ID NO: 6), EMPD IgE-19-38 (SEQ ID NO: 8) или EMPD IgE-1-40 (SEQ ID NO: 9), как показано в таблице 1.
Фрагмент EMPD IgE по настоящему изобретению также включает иммунологически функциональные аналоги или гомологи пептидов EMPD IgE (SEQ ID NO: 5, 6, 8 и 9) и более 20 их аминокислотных фрагментов. Функциональные иммунологические аналоги или гомологи пептида EMPD IgE и его более 20 аминокислотных фрагментов включают варианты, которые сохраняют по существу ту же иммуногенность, что и исходный пептид. Иммунологически функциональные аналоги могут иметь консервативную замену в положении аминокислоты; изменение общего заряда; ковалентную связь к другому фрагменту; или добавления, вставки или делеции аминокислот; и/или любую их комбинацию.
b. Гетерологичные Т-хелперные эпитопы (Th-эпитопы)
Данное описание относится к пептидным иммуногенным конструкциям, содержащим В-клеточный эпитоп из EMPD IgE, ковалентно связанный с гетерологичным Т-хелперным (Th) эпитопом непосредственно или через необязательный гетерологичный спейсер.
Гетерологичный Th-эпитоп в пептидной иммуногенной конструкции EMPD IgE усиливает иммуногенность фрагмента EMPD IgE, что облегчает выработку высоких титров специфических антител, направленных против оптимизированного целевого В-клеточного эпитопа (то есть фрагмента EMPD IgE), посредством рационального дизайна.
Используемый в данном документе термин «гетерологичный» относится к аминокислотной последовательности, которая получена из аминокислотной последовательности, которая не является частью или гомологична последовательности EMPD IgE дикого типа. Таким образом, гетерологичный Th-эпитоп представляет собой Th-эпитоп, полученный из аминокислотной последовательности, которая не обнаружена в природе в EMPD IgE (т.е. Th-эпитоп не является аутологичным по отношению к EMPD IgE). Поскольку Th-эпитоп является гетерологичным по отношению к EMPD IgE, при ковалентной связи гетерологичного Th-эпитопа с фрагментом EMPD IgE природная аминокислотная последовательность EMPD IgE не удлиняется ни в N-концевом, ни в C-концевом направлениях.
Гетерологичный Th-эпитоп согласно данному описанию может представлять собой любой Th-эпитоп, который не имеет аминокислотной последовательности, которая не обнаружена в природе в EMPD IgE. Th-эпитоп также может иметь разнородные мотивы связывания с молекулами ГКГС класса II нескольких видов. В некоторых вариантах осуществления Th-эпитоп содержит множество разнородных мотивов связывания ГКГС класса II для обеспечения максимальной активации Т-хелперов, которая приводит к инициации и регуляции иммунных ответов. Th-эпитоп предпочтительно является иммуномолчащим сам по себе, то есть небольшое количество антител, генерируемых пептидными иммуногенными конструкциями EMPD IgE, если таковые вообще имеются, будет направлено на Th-эпитоп, таким образом обеспечивая очень сфокусированный иммунный ответ, направленный на целевой B-клеточный эпитоп фрагмента EMPD IgE.
Эпитопы согласно данному описанию включают, но не ограничиваются ими, аминокислотные последовательности, полученные из чужеродных патогенов, как показано в таблице 2 (SEQ ID NO: 59-87). Кроме того, Th-эпитопы включают идеализированные искусственные Th-эпитопы и комбинаторные идеализированные искусственные Th-эпитопы (например, SEQ ID NO: 60 и 67-73). Гетерологичные Th-эпитопные пептиды, представленные в виде комбинаторной последовательности (например, SEQ ID NO: 68-71), содержат смесь аминокислотных остатков, представленных в определенных положениях в пределах пептидного остова на основе вариабельных остатков гомологов для этого конкретного пептида. Сборка комбинаторных пептидов может быть синтезирована в одном процессе путем добавления смеси указанных защищенных аминокислот вместо одной конкретной аминокислоты в определенное положение в процессе синтеза. Такие комбинаторные пептиды гетерологичных Th-эпитопов могут обеспечивать широкий охват Th-эпитопов для животных, имеющих разнообразный генетический фон. Типовые комбинаторные последовательности гетерологичных Th-эпитопных пептидов включают SEQ ID NO: 68-71, которые приведены в таблице 2. Th-эпитопные пептиды согласно данному изобретению обеспечивают широкую реактивность и иммуногенность для животных и пациентов из генетически разнообразных популяций.
Пептидные иммуногенные конструкции EMPD IgE, содержащие Th-эпитопы, продуцируются одновременно в одном твердофазном пептидном синтезе в тандеме с фрагментом EMPD IgE. Th-эпитопы также включают иммунологические аналоги Th-эпитопов. Иммунологические аналоги Th включают иммуномодулирующие аналоги, перекрестно-реактивные аналоги и сегменты любого из этих Th-эпитопов, которые достаточны для усиления или стимуляции иммунного ответа на фрагменты EMPD IgE.
Функциональные иммунологические аналоги Th-эпитопных пептидов также эффективны и включены как часть данного изобретения. Функциональные иммунологические аналоги Th могут включать консервативные замены, добавления, делеции и вставки от одного до около пяти аминокислотных остатков в Th-эпитопе, которые по существу не модифицируют Th-стимулирующую функцию Th-эпитопа. Консервативные замены, добавления и вставки могут быть выполнены с природными или неприродными аминокислотами, как описано выше для фрагментов EMPD IgE. Таблица 2 идентифицирует другой вариант функционального аналога для Th-эпитопного пептида. В частности, SEQ ID NO: 60 и 67 для MvF1 и MvF2 Th являются функциональными аналогами SEQ ID NO: 70 и 72 для MvF4 и MvF5 в том смысле, что они отличаются в аминокислотном остове делецией (SEQ ID NO: 60 и 67) или включением (SEQ ID NO: 70 и 72) двух аминокислот, каждая на N- и C-концах. Различия между этими двумя сериями аналогичных последовательностей не влияют на функцию Th-эпитопов, содержащихся в пределах этих последовательностях. Следовательно, функциональные иммунологические аналоги Th включают несколько версий Th-эпитопа, полученного из слитого белка вируса кори MvF1-4 Th (SEQ ID NO: 60, 67, 68, 70, и 72) и из поверхностного белка гепатита HBsAg 1-3 Th (SEQ ID NO: 69, 71, и 73).
Th-эпитоп в пептидной иммуногенной конструкции EMPD IgE может быть ковалентно связан в N- или C-концевой области фрагмента пептида EMPD IgE. В некоторых вариантах осуществления Th-эпитоп ковалентно связан с N-концевой областью фрагмента пептида EMPD IgE. В других вариантах осуществления Th-эпитоп ковалентно связан с С-концевой областью фрагмента пептида EMPD IgE. В некоторых вариантах осуществления более одного Th-эпитопа ковалентно связаны с фрагментом EMPD IgE. Когда более чем один Th-эпитоп связан с фрагментом EMPD IgE, каждый Th-эпитоп может иметь одинаковую аминокислотную последовательность или разные аминокислотные последовательности. Кроме того, когда более чем один Th-эпитоп связан с фрагментом EMPD IgE, Th-эпитопы могут быть расположены в любом порядке. Например, Th-эпитопы могут быть последовательно связаны с N-концевой областью фрагмента EMPD IgE или могут быть последовательно связаны с C-концевой областью фрагмента EMPD IgE, или Th-эпитоп может быть ковалентно связан с N -концевой областью фрагмента EMPD IgE, в то время как отдельный Th-эпитоп ковалентно связан с C-концевой областью фрагмента EMPD IgE. В расположении Th-эпитопов отсутствуют ограничения по отношению к фрагменту EMPD IgE.
В некоторых вариантах осуществления Th-эпитоп непосредственно ковалентно связан с фрагментом EMPD IgE. В других вариантах осуществления Th-эпитоп ковалентно связан с фрагментом EMPD IgE через гетерологичный спейсер, более подробно описанный ниже.
с. Гетерологичный спейсер
Описанные пептидные иммуногенные конструкции EMPD IgE необязательно содержат гетерологичный спейсер, который ковалентно связывает В-клеточный эпитоп из EMPD IgE с гетерологичным Т-хелперным (Th) эпитопом.
Как обсуждалось выше, термин «гетерологичный» относится к аминокислотной последовательности, которая получена из аминокислотной последовательности, которая не является частью или гомологична последовательности EMPD IgE естественного типа. Таким образом, природная аминокислотная последовательность EMPD IgE не удлиняется ни в N-концевом, ни в C-концевом направлениях, когда гетерологичный спейсер ковалентно связан с B-клеточным эпитопом из EMPD IgE, поскольку спейсер является гетерологичным по отношению к последовательности EMPD IgE.
Спейсер представляет собой любую молекулу или химическую структуру, способную связывать две аминокислоты и/или пептида вместе. Спейсер может варьироваться по длине или полярности в зависимости от применения. Спейсер может быть присоединен через амидную или карбоксильную связь, но возможно и через другие функциональные группы. Спейсер может включать химическое соединение, природную аминокислоту или неприродную аминокислоту.
Спейсер может обеспечивать структурные особенности пептидной иммуногенной конструкции EMPD IgE. В структурном отношении спейсер обеспечивает физическое разделение Th-эпитопа и B-клеточного эпитопа фрагмента EMPD IgE. Физическое разделение спейсером может нарушить любые искусственные вторичные структуры, созданные путем присоединения Th-эпитопа к B-клеточному эпитопу. Кроме того, физическое разделение эпитопов при помощи спейсера может устранить интерференции между Th-клеточными и/или B-клеточными ответами. Кроме того, спейсер может быть предназначен для создания или модификации вторичной структуры пептидной иммуногенной конструкции. Например, спейсер может быть сконструирован так, чтобы действовать как гибкий шарнирный участок для усиления разделения Th-эпитопа и B-клеточного эпитопа. Спейсер гибкого шарнирного участка также может обеспечить более эффективные взаимодействия между представленным пептидным иммуногеном и соответствующими Th-клетками и B-клетками для усиления иммунных ответов на Th-эпитоп и B-клеточный эпитоп. Примеры последовательностей, кодирующих гибкие шарнирные участки, находятся в шарнирном участке тяжелой цепи иммуноглобулина, которые часто богаты пролином. Один особенно полезный гибкий шарнирный участок, который можно использовать в качестве спейсера, обеспечивается последовательностью Pro-Pro-Xaa-Pro-Xaa-Pro (SEQ ID NO: 128), где Xaa представляет собой любую аминокислоту, и предпочтительно аспарагиновую кислоту.
Спейсер также может обеспечивать функциональные особенности пептидной иммуногенной конструкции EMPD IgE. Например, спейсер может быть предназначен для изменения общего заряда пептидной иммуногенной конструкции EMPD IgE, который может влиять на растворимость пептидной иммуногенной конструкции. Кроме того, изменение общего заряда пептидной иммуногенной конструкции EMPD IgE может влиять на способность пептидной иммуногенной конструкции связываться с другими соединениями и реактивами. Как более подробно обсуждается ниже, пептидная иммуногенная конструкция EMPD IgE может образовывать стабильный иммуностимулирующий комплекс с высоко заряженным олигонуклеотидом, таким как CpG-олигомеры, посредством электростатической ассоциации. Общий заряд пептидной иммуногенной конструкции EMPD IgE важен для образования этих стабильных иммуностимулирующих комплексов.
Химические соединения, которые можно использовать в качестве спейсера, включают, но не ограничиваются ими, (2-аминоэтокси) уксусную кислоту (AEA), 5-аминовалериановую кислоту (AVA), 6-аминокапроновую кислоту (Ahx), 8-амино-3,6-диоксаоктановую кислоту (AEEA, мини-ПЭГ1), 12-амино-4,7,10-триоксадодекановую кислоту (мини-ПЭГ2), 15-амино-4,7,10,13-тетраоксапента-декановую кислоту (мини-ПЭГ3), триоксатридекан-янтарную кислоту (Ttds), 12-амино-додекановую кислоту, Fmoc-5-амино-3-оксапентановую кислоту (O1Pen) и тому подобное.
Природные аминокислоты включают аланин, аргинин, аспарагин, аспарагиновую кислоту, цистеин, глутаминовую кислоту, глутамин, глицин, гистидин, изолейцин, лейцин, лизин, метионин, фенилаланин, пролин, серин, треонин, триптофан, тирозин и валин.
Неприродные аминокислоты включают, но не ограничиваются ими, ε-N-лизин, β-аланин, орнитин, норлейцин, норвалин, гидроксипролин, тироксин, γ-амино масляную кислоту, гомосерин, цитруллин, аминобензойную кислоту, 6-аминокапроновую кислоту (Aca; 6-аминогексановую кислоту), гидроксипролин, меркаптопропионовую кислоту (MPA), 3-нитро-тирозин, пироглутаминовую кислоту и тому подобное.
Спейсер в пептидной иммуногенной конструкции EMPD IgE может быть ковалентно связан в N- или С-концевой области Th-эпитопа и пептида EMPD IgE. В некоторых вариантах осуществления спейсер ковалентно связан с C-концевой областью Th-эпитопа и с N-концевой областью пептида EMPD IgE. В других вариантах осуществления спейсер ковалентно связан с C-концевой областью пептида EMPD IgE и с N-концевой областью Th-эпитопа. В определенных вариантах осуществления можно использовать более одного спейсера, например, когда в пептидной иммуногенной конструкции присутствует более одного Th-эпитопа. Когда используется более одного спейсера, каждый спейсер может быть таким же, как и другой, или отличаться от него. Кроме того, когда в пептидной иммуногенной конструкции присутствует более одного Th-эпитопа, Th-эпитопы могут быть разделены спейсером, который может быть таким же или отличаться от спейсера, используемого для разделения Th-эпитопа и B-клеточного эпитопа. В расположении спейсера отсутствуют ограничения относительно Th-эпитопа или фрагмента EMPD IgE.
В некоторых вариантах осуществления гетерологичный спейсер представляет собой природную аминокислоту или неприродную аминокислоту. В других вариантах осуществления спейсер содержит более одной природной или неприродной аминокислоты. В конкретных вариантах осуществления спейсер представляет собой Lys-, Gly-, Lys-Lys-Lys-, (α, ε-N)Lys, или ε-N-Lys-Lys-Lys-Lys (SEQ ID NO: 129).
d. Конкретные варианты осуществления пептидных иммуногенных конструкций EMPD IgE
В некоторых вариантах осуществления пептидные иммуногенные конструкции EMPD IgE могут быть представлены следующими формулами:
(Th)m-(A)n-(фрагмент EMPD IgE)-X
или
(фрагмент EMPD IgE)-(A)n-(Th)m-X
или
(Th)m-(A)n-(фрагмент EMPD IgE)-(A)n-(Th)m-X,
где
Th представляет собой гетерологичный T-хелперный эпитоп;
А представляет собой гетерологичный спейсер;
(фрагмент EMPD IgE) представляет собой В-клеточный эпитоп, имеющий от около 20 до около 40 аминокислотных остатков из EMPD IgE;
X представляет собой α-COOH или α-CONH2 аминокислоты;
m равно от 1 до около 4; и
n равно от 0 до около 10.
В конкретных вариантах осуществления гетерологичный Th-эпитоп в пептидной иммуногенной конструкции EMPD IgE имеет аминокислотную последовательность, выбранную из любой из SEQ ID NO: 59-87 и их комбинаций, приведенных в таблице 2. В некоторых вариантах осуществления пептидная иммуногенная конструкция EMPD IgE содержит более одного Th-эпитопа.
В определенных вариантах осуществления необязательный гетерологичный спейсер выбран из любого из Lys-, Gly-, Lys-Lys-Lys-, (α, ε-N)Lys, ε-N-Lys-Lys-Lys-Lys (SEQ ID NO: 129), и их комбинаций. В конкретных вариантах осуществления гетерологичный спейсер представляет собой ε-N-Lys-Lys-Lys-Lys (SEQ ID NO: 129).
В некоторых вариантах осуществления фрагмент EMPD IgE содержит от примерно 20 до примерно 40 аминокислотных остатков из EMPD IgE SEQ ID NO: 1 или 2. В конкретных вариантах осуществления фрагмент EMPD IgE содержит аминокислотную последовательность из внутренней внутримолекулярной петли EMPD IgE, образованной эндогенными цистеинами (C18-C39) в соответствии с нумерацией полноразмерного EMPD IgE (SEQ ID NO: 1). В конкретных вариантах осуществления фрагмент EMPD IgE имеет аминокислотную последовательность EMPD IgE-1-39 (SEQ ID NO: 5), EMPD IgE 7-40 (SEQ ID NO: 6), EMPD IgE 19-38 (SEQ ID NO: 8) или EMPD IgE 1-40 (SEQ ID NO: 9), как показано в таблице 1.
В некоторых вариантах осуществления пептидная иммуногенная конструкция EMPD IgE имеет аминокислотную последовательность, выбранную из любой из SEQ ID NO: 88-130, как показано в таблице 3. В конкретных вариантах осуществления пептидная иммуногенная конструкция EMPD IgE имеет аминокислотную последовательность, выбранную из любой из SEQ ID NO: 88-95, 98-124 и 130.
e. Варианты, гомологи и функциональные аналоги
Также могут быть использованы варианты и аналоги вышеуказанных иммуногенных пептидов, которые индуцируют и/или перекрестно реагируют с антителами к предпочтительным эпитопам EMPD IgE. Аналоги, включая аллельные, видовые и индуцированные варианты, обычно отличаются от природных пептидов в одном, двух или более положениях, часто в силу консервативных замен. Аналоги обычно демонстрируют по меньшей мере 80 или 90% идентичности последовательности с природными пептидами. Некоторые аналоги также включают неприродные аминокислоты или модификации N- или C-концевых аминокислот в одном, двух или более положениях.
Варианты, которые являются функциональными аналогами, могут иметь консервативную замену в положении аминокислоты; изменение общего заряда; ковалентную связь к другому фрагменту; или добавления, вставки или делеции аминокислот; и/или любую их комбинацию.
Консервативные замены - это когда один аминокислотный остаток заменен другим аминокислотным остатком с аналогичными химическими свойствами. Например, неполярные (гидрофобные) аминокислоты включают аланин, лейцин, изолейцин, валин, пролин, фенилаланин, триптофан и метионин; полярные нейтральные аминокислоты включают глицин, серин, треонин, цистеин, тирозин, аспарагин и глутамин; положительно заряженные (основные) аминокислоты включают аргинин, лизин и гистидин; и отрицательно заряженные (кислые) аминокислоты включают аспарагиновую кислоту и глутаминовую кислоту.
В конкретном варианте осуществления функциональный аналог имеет по меньшей мере 50% идентичности с исходной аминокислотной последовательностью. В другом варианте осуществления функциональный аналог имеет по меньшей мере 80% идентичности с исходной аминокислотной последовательностью. В еще одном варианте осуществления функциональный аналог имеет по меньшей мере 85% идентичности с исходной аминокислотной последовательностью. В еще одном варианте осуществления функциональный аналог имеет по меньшей мере 90% идентичности с исходной аминокислотной последовательностью.
Варианты также содержат вариации фосфорилированных остатков. Например, варианты могут содержать различные остатки в пределах пептидов, которые фосфорилированы. Вариантные иммуногенные пептиды EMPD IgE также могут включать псевдофосфорилированные пептиды. Псевдофосфорилированные пептиды получают путем замены одного или более фосфорилированных остатков серина, треонина и тирозина в пептидах EMPD IgE на кислые аминокислотные остатки, такие как глутаминовая кислота и аспарагиновая кислота.
Композиции
Данное описание также обеспечивает композиции, содержащие описанную иммуногенную конструкцию EMPD IgE.
а. Пептидные композиции
Композиции, содержащие описанную пептидную иммуногенную конструкцию EMPD IgE, могут находиться в жидкой или твердой форме. Жидкие композиции могут содержать воду, буферы, растворители, соли и/или любой другой приемлемый реактив, который не изменяет структурные или функциональные свойства пептидной иммуногенной конструкции EMPD IgE. Пептидные композиции могут содержать одну или более описанных пептидных иммуногенных конструкций EMPD.
b. Фармацевтические композиции
Данное описание также относится к фармацевтическим композициям, содержащим описанную пептидную иммуногенную конструкцию EMPD IgE.
Фармацевтические композиции могут содержать носители и/или другие добавки в фармацевтически приемлемой системе доставки. Соответственно, фармацевтические композиции могут содержать фармацевтически эффективное количество пептидной иммуногенной конструкции EMPD IgE вместе с фармацевтически приемлемым носителем, адъювантом и/или другими вспомогательными веществами, такими как разбавители, добавки, стабилизирующие агенты, консерванты, солюбилизирующие агенты, буферы и тому подобное.
Фармацевтические композиции могут содержать один или более адъювантов, которые действуют для ускорения, пролонгации или усиления иммунного ответа на пептидную иммуногенную конструкцию EMPD IgE без какого-либо специфического антигенного действия. Адъюванты, используемые в фармацевтической композиции, могут включать масла, масляные эмульсии, соли алюминия, соли кальция, иммуностимулирующие комплексы, бактериальные и вирусные производные, виросомы, углеводы, цитокины, полимерные микрочастицы. В некоторых вариантах осуществления адъювант может быть выбран из квасцов (калий алюминий фосфат), фосфата алюминия (например, ADJU-PHOS®), гидроксида алюминия (например, ALHYDROGEL®), фосфата кальция, неполного адъюванта Фрейнда (IFA), полного адъюванта Фрейнда, MF59, адъюванта 65, Lipovant, ISCOM, липосина, сапонина, сквалена, L121, Emulsigen®, монофосфориллипида A (MPL), Quil A, QS21, MONTANIDE® ISA 35, ISA 50V, ISA 50V2, ISA 51, ISA 206, ISA 720, липосом, фосфолипидов, пептидогликанов, липополисахаридов (ЛПС), ASO1, ASO2, ASO3, AsO4, AF03, липофильных фосфолипидов (липида А), гамма-инулина, альгаммулина, глюкана, декстранов, глюкоманнанов, галактоманнанрв, леванов, ксиланов, бромида диметилдиоктадециламмония (DDA), а также других адъювантов и эмульгаторов.
В некоторых вариантах осуществления фармацевтическая композиция содержит Montanide™ ISA 51 (масляная адъювантная композиция, состоящая из растительного масла и маннида олеата для получения водно-масляных эмульсий), Tween® 80 (также известный как: Полисорбат 80 или Полиоксиэтилен (20) сорбитан моноолеат), CpG-олигонуклеотид и/или любую их комбинацию. В других вариантах осуществления фармацевтическая композиция представляет собой эмульсию вода-в-масле-в-воде (то есть в/м/в) с Emulsigen или Emulsigen D в качестве адъюванта.
Фармацевтические композиции могут также содержать фармацевтически приемлемые добавки или вспомогательные вещества. Например, фармацевтические композиции могут содержать антиоксиданты, связывающие вещества, буферы, объемообразующие агенты, носители, хелатирующие агенты, красители, разбавители, дезинтегранты, эмульгаторы, наполнители, гелеобразующие агенты, рН-буферные агенты, консерванты, солюбилизирующие агенты, стабилизаторы и тому подобное.
Фармацевтические композиции могут быть составлены в виде составов с немедленным высвобождением или в виде составов с замедленным высвобождением. Кроме того, фармацевтические композиции могут быть составлены для индукции системного или локализованного иммунитета слизистой оболочки путем захвата иммуногена и совместного введения с микрочастицами. Такие системы доставки легко определяются средним специалистом в данной области техники.
Фармацевтические композиции могут быть приготовлены в виде впрыскиваемых лекарственных средств, или в виде жидких растворов, или суспензий. Жидкие носители, содержащие пептидную иммуногенную конструкцию EMPD IgE, также можно приготовить перед инъекцией. Фармацевтическую композицию можно вводить любым подходящим способом применения, например, п/к, в/в, п/о, в/м, интраназально, перорально, подкожно и т.д., и в любом подходящем устройстве доставки. В некоторых вариантах осуществления фармацевтическая композиция составлена для внутривенного, подкожного, интрадермального или внутримышечного введения. Также могут быть приготовлены фармацевтические композиции, подходящие для других способов введения, включая пероральное и интраназальное применение.
Фармацевтические композиции также могут быть составлены в подходящей единичной дозированной форме. В некоторых вариантах осуществления фармацевтическая композиция содержит от около 0,1 мкг до около 1 мг пептидной иммуногенной конструкции EMPD IgE на кг массы тела. Эффективные дозы фармацевтических композиций варьируются в зависимости от многих различных факторов, включая способы введения, целевой участок, физиологическое состояние пациента, является ли пациент человеком или животным, вводятся другие лекарственные средства и является ли лечение профилактическим или терапевтическим. Как правило, пациент представляет собой человека, но также можно лечить отличных от человека млекопитающих, включая трансгенных млекопитающих. При доставке в нескольких дозах фармацевтические композиции могут быть удобно разделены на подходящее количество на единичную дозированную форму. Вводимая доза будет зависеть от возраста, массы и общего состояния здоровья субъекта, что хорошо известно в области терапии.
В некоторых вариантах осуществления фармацевтическая композиция содержит более одной пептидной иммуногенной конструкции EMPD IgE. Фармацевтическая композиция, содержащая смесь более чем одну пептидную иммуногенную конструкцию EMPD IgE, для обеспечения синергетического усиления иммуноэффективности конструкций. Фармацевтические композиции, содержащие более одной пептидной иммуногенной конструкции EMPD IgE, могут быть более эффективными в большей генетической популяции благодаря широкому охвату ГКГС класса II, таким образом обеспечивая улучшенный иммунный ответ на пептидные иммуногенные конструкции EMPD IgE.
В некоторых вариантах осуществления фармацевтическая композиция содержит пептидную иммуногенную конструкцию EMPD IgE, выбранную из SEQ ID NO: 88-95, 98-124 и 130 (таблица 3), а также ее гомологи, аналоги и/или комбинации.
В некоторых вариантах осуществления пептидные иммуногенные конструкции EMPD IgE (SEQ ID NO: 107 и 108) с гетерологичными Th-эпитопами, полученными из MVF и HBsAg в комбинаторной форме (SEQ ID NO: 68 и 69), смешивали в эквимолярном соотношении для использования в вакцинном составе, чтобы обеспечить максимальный охват популяции вакцинируемых хозяев, имеющей обширный генетический фон. В пептидных композициях по настоящему изобретению наблюдалось синергетическое усиление в иммуногенных конструкциях EMPD IgE G1-C39 и A7-C40 (SEQ ID NO: 88-95), и гуморальный ответ, вызванный такими конструкциями (например, SEQ ID NO: 95), был в основном (>90%) сосредоточен на желаемой перекрестной реактивности против пептида В-клеточного эпитопа EMPD IgE (SEQ ID NO: 2) без значительной (или при отсутствии таковой) реактивности, направленной на гетерологичные Th-эпитопы, используемые для усиления иммуногенности (пример 7, таблица 7). Это резко контрастирует с обычным белком, таким как KLH или другие биологические белковые носители, используемым для такого усиления пептидной антигенности.
В других вариантах осуществления фармацевтические композиции, содержащие пептидную композицию, например, смеси пептидных иммуногенных конструкций EMPD IgE в контакте с минеральными солями, включая гель на основе квасцов (ALHYDROGEL) или фосфат алюминия (ADJUPHOS), в качестве адъюванта для образования вакцинного состава в форме суспензии для введения хозяину, получающему вакцину.
Фармацевтические композиции, содержащие пептидную иммуногенную конструкцию EMPD IgE, можно использовать для индукции иммунного ответа и выработки антител у хозяина после введения.
с. Иммуностимулирующие комплексы
Настоящее изобретение также относится к фармацевтическим композициям, содержащим пептидную иммуногенную конструкцию EMPD IgE в форме иммуностимулирующего комплекса с CpG-олигонуклеотидом. Такие иммуностимулирующие комплексы специально приспособлены для действия в качестве адъюванта и стабилизатора пептидного иммуногена. Иммуностимулирующие комплексы находятся в форме частиц, которые могут эффективно представлять пептидный иммуноген EMPD IgE клеткам иммунной системы, вызывая иммунный ответ. Иммуностимулирующие комплексы могут быть составлены в виде суспензии для парентерального введения. Иммуностимулирующие комплексы также могут быть составлены в форме эмульсий вода-в-масле, в виде суспензии в комбинации с минеральной солью или с желирующим полимером in situ для эффективной доставки пептидного иммуногена EMPD IgE к клеткам иммунной системы хозяина после парентерального введения.
Стабилизированный иммуностимулирующий комплекс может быть образован путем комплексирования пептидной иммуногенной конструкции EMPD IgE с анионной молекулой, олигонуклеотидом, полинуклеотидом или их комбинациями посредством электростатической ассоциации. Стабилизированный иммуностимулирующий комплекс может быть включен в фармацевтическую композицию в качестве системы доставки иммуногена.
В некоторых вариантах осуществления пептидная иммуногенная конструкция EMPD IgE сконструирована с возможностью содержания катионной части, которая положительно заряжена при рН в диапазоне от 5,0 до 8,0. Общий заряд катионной части пептидной иммуногенной конструкции EMPD IgE или смеси конструкций рассчитывают, назначая заряд +1 для каждого лизина (K), аргинина (R) или гистидина (H), заряд -1 для каждой аспарагиновой кислоты (D) или глутаминовой кислоты (E) и заряд 0 для другой аминокислоты в пределах последовательности. Заряды суммируются в пределах катионной части пептидной иммуногенной конструкции EMPD IgE и выражаются как общий средний заряд. Подходящий пептидный иммуноген содержит катионную часть с общим средним положительным зарядом +1. Предпочтительно пептидный иммуноген имеет общий положительный заряд в диапазоне, который больше чем +2. В некоторых вариантах осуществления катионная часть пептидной иммуногенной конструкции EMPD IgE представляет собой гетерологичный спейсер. В некоторых вариантах осуществления катионная часть пептидной иммуногенной конструкции EMPD IgE имеет заряд +4, когда последовательность спейсера представляет собой (α, ε-N)Lys, ε-N-Lys-Lys-Lys-Lys (SEQ ID NO: 129).
«Анионная молекула», как описано в данном документе, относится к любой молекуле, которая заряжена отрицательно при рН в диапазоне 5,0-8,0. В определенных вариантах осуществления анионная молекула представляет собой олигомер или полимер. Общий отрицательный заряд на олигомере или полимере рассчитывается путем назначения заряда -1 для каждой фосфодиэфирной или фосфоротиоатной группы в олигомере. Подходящим анионным олигонуклеотидом является одноцепочечная молекула ДНК с 8-64 нуклеотидными основаниями с числом повторов CpG-мотива в диапазоне от 1 до 10. Предпочтительно CpG иммуностимулирующие молекулы одноцепочечной ДНК содержат 18-48 нуклеотидных оснований с числом повторов CpG-мотива в диапазоне от 3 до 8.
Более предпочтительно анионный олигонуклеотид представлен формулой: 5' X1CGX2 3', где C и G являются неметилированными; и X1 выбран из группы, состоящей из A (аденина), G (гуанина) и T (тимина); и Х2 представляет собой С (цитозин) или Т (тимин). Или анионный олигонуклеотид представлен формулой: 5' (X3)2CG(X4)2 3', где C и G являются неметилированными; и X3 выбран из группы, состоящей из A, T или G; и Х4 представляет собой С или Т.
Полученный иммуностимулирующий комплекс находится в форме частиц с размером, обычно находящимся в диапазоне 1-50 мкм и являющимся функцией многих факторов, включая стехиометрию относительного заряда и молекулярную массу взаимодействующих частиц. Иммуностимулирующий комплекс в виде частиц обладает тем преимуществом, что обеспечивает адъювантный эффект и активацию специфических иммунных ответов in vivo. Кроме того, стабилизированный иммуностимулирующий комплекс подходит для приготовления фармацевтических композиций различными способами, включая водно-масляные эмульсии, суспензии минеральных солей и полимерные гели.
Данное раскрытие также относится к фармацевтическим композициям, включая вакцинные составы, для лечения и профилактики аллергических заболеваний, опосредованных IgE. В некоторых вариантах осуществления фармацевтические композиции содержат стабилизированный иммуностимулирующий комплекс, который получают путем смешивания CpG-олигомера с пептидной композицией, содержащей смесь пептидных иммуногенных конструкций EMPD IgE (например, SEQ ID NO: 88-95, 98-124 и 130), посредством электростатической ассоциации для дальнейшего усиления иммуногенности пептида EMPD IgE и выработки антител к пептиду EMPD IgE SEQ ID NO: 1 или 2 для связывания с B-клетками, экспрессирующими mIgE, и индуцирования их антителозависимой клеточной цитотоксичности (ADCC) и апоптоза (пример 6).
В еще других вариантах осуществления фармацевтические композиции содержат смесь пептидных иммуногенных конструкций EMPD IgE (например, любую комбинацию SEQ ID NO: 8-90, 94, 95, 98-124 и 130) в форме стабилизированного иммуностимулирующего комплекса с CpG-олигомерами, которые необязательно смешаны с минеральными солями, включая гель на основе квасцов (ALHYDROGEL) или фосфат алюминия (ADJUPHOS) в качестве адъюванта с высоким фактором безопасности для образования суспензионного вакцинного состава для введения хозяину, получающему вакцину.
Антитела
Данное описание обеспечивает антитела, образованные в ответ на пептидную иммуногенную конструкцию EMPD IgE.
В настоящем изобретении предложены пептидные иммуногенные конструкции EMPD IgE и их составы, экономически выгодные в изготовлении и оптимальные по своей конструкции, которые способны вызывать образование высоких титров антител, нацеленные на мембраносвязанный IgE, способные нарушать иммунную толерантность к аутоантигену с высоким уровнем ответа у хозяина, получившего вакцину. Антитела, генерируемые иммуногенными конструкциями пептидного EMPD IgE, имеют высокую аффинность к белку EMPD IgE в виде растворимого пептида, гибридного белка или IgE, присутствующего в В-клетках, несущих IgE. Полученные антитела способны связывать и перекрестно сшивать BCR IgE на B-лимфоцитах, экспрессирующих mIgE, вызывая цитолитические эффекты, такие как апоптоз и ADCC. Апоптотическое истощение В-лимфоцитов с мембраносвязанным IgE также приводит к снижению выработки сывороточного IgE. Следовательно, нацеливание на человеческие B-клетки с mIgE апоптоз-индуцирующих IgE-EMPD-специфических антител, генерируемых раскрытыми пептидными иммуногенными конструкциями EMPD IgE и их составами, обеспечивает новую терапию и вакцину против опосредованных IgE аллергических заболеваний.
В некоторых вариантах осуществления пептидные иммуногенные конструкции EMPD IgE для вызова антител содержат гибрид пептида EMPD IgE, имеющий В-клеточный эпитоп, содержащий от 20 до 40 аминокислот, покрывающих структуру центральной внутримолекулярной петли, полученных из пептида EMPD IgE (SEQ ID NO: 1) (например, пептиды EMPD IgE из EMPD IgE G1-C39 (SEQ ID NO: 5), EMPD IgE A7-H40 (SEQ ID NO: 6), EMPD IgE H19-R38 (SEQ ID NO: 8) и EMPD IgE G1-H40 (SEQ ID NO: 9)), связанных с гетерологичным Th-эпитопом, полученным из таких патогенных белков как гибридный белок вируса кори (MVF) (SEQ ID NO: 73) и др. (SEQ ID NO: 59-87), через дополнительный спейсер. В-клеточный эпитоп и Th-эпитоп пептидных иммуногенных конструкций EMPD IgE действуют совместно, стимулируя образование высокоспецифичных антител, перекрестно-реактивных с пептидом EMPD IgE 1-52 (SEQ ID NO: 2), белком EMPD IgE 1-67 (SEQ ID NO: 1) либо в виде рекомбинантного белка, содержащего EMPD IgE (например, очищенного из стабильной клеточной линии Flp-In CHO, трансфицированной рекомбинантной ДНК, кодирующей Fc-часть человеческого IgG1 и EMPD IgE мембраносвязанного IgE человека, γ1-em67) или на мембране - клетки, несущей мембраносвязанный IgE, (например, клеточной линии Ramos, трансфицированной рекомбинантной ДНК, кодирующей mIgE.FcL).
Традиционные способы иммуноопотенцирования пептида, такие как химическое связывание с белком-носителем, например, гемоцианином Limpet Keyhole (KLH) или другими белками-носителями, такими как белок дифтерийного анатоксина (DT) и столбнячного анатоксина (TT), обычно приводят к генерации большого количества антител, направленных против белка-носителя. Таким образом, основной недостаток таких вакцин на основе пептидного белка-носителя заключается в том, что большинство (>90%) антител, генерируемых иммуногеном, представляют собой нефункциональные антитела, направленные против белка-носителя KLH, DT или TT, что может привести к подавлению эпитопов.
В отличие от традиционного способа иммунопотенцирования пептида, антитела, генерируемые раскрытыми пептидными иммуногенными конструкциями EMPD IgE, связываются с высокой специфичностью к фрагменту EMPD IgE с незначительным количеством (или при отсутствии таковых) антител, направленных против гетерологичного Th-эпитопа или необязательного гетерологичного спейсера. В частности, поликлональные антитела, образованные у вакцинированных животных, с высокой специфичностью связываются с центральной областью, охватывающей петлевую структуру EMPD IgE, как показано на Фиг. 9.
Способы
Данное описание также направлено на способы получения и применения пептидных иммуногенных конструкций EMPD IgE, композиций и фармацевтических композиций.
а. Способы получения пептидной иммуногенной конструкции EMPD IgE
Пептидные иммуногенные конструкции EMPD IgE согласно данному описанию могут быть получены способами химического синтеза, хорошо известными специалисту в данной области техники (см., например, Fields et al., Chapter 3 in Synthetic Peptides: A User’s Guide, ed. Grant, W. H. Freeman & Co., New York, NY, 1992, p. 77). Пептидные иммуногенные конструкции EMPD IgE могут быть синтезированы с использованием автоматизированных методов твердофазного синтеза Merrifield с α-NH2, защищенным t-Boc- или F-moc-группами, с использованием аминокислот с боковой цепью, например, на модели синтезатора пептидов Applied Biosystems 430А или 431. Приготовление пептидных иммуногенных конструкций EMPD IgE, содержащих пептиды комбинаторной библиотеки для Th-эпитопов, может быть достигнуто путем обеспечения смеси альтернативных аминокислот для связывания в данном вариабельном положении.
После полной сборки желаемой пептидной иммуногенной конструкции EMPD IgE, смолу можно обработать в соответствии со стандартными процедурами для отщепления пептида от смолы, и функциональные группы на боковых цепях аминокислот могут быть разблокированы. Свободный пептид может быть очищен с помощью ВЭЖХ и охарактеризован биохимически, например, при помощи аминокислотного анализа или секвенирования. Способы очистки и определения характеристик пептидов хорошо известны специалисту в данной области техники.
Качество пептидов, получаемых при помощи этого химического процесса, можно контролировать и определять, и в результате можно гарантировать воспроизводимость пептидных иммуногенных конструкций EMPD IgE, иммуногенность и выход. Подробное описание получения пептидной иммуногенной конструкции EMPD IgE посредством твердофазного пептидного синтеза приведено в Примере 1.
Было обнаружено, что диапазон структурной изменчивости, который допускает сохранение предполагаемой иммунологической активности, является гораздо более приемлемым, чем диапазон структурной изменчивости, допускаемый для сохранения специфической активности лекарственного средства с помощью низкомолекулярного лекарственного средства или желаемых активностей и нежелательных токсичностей, обнаруженных в больших молекулах, которые производятся совместно с полученными биологическим лекарственными средствами. Таким образом, аналоги пептидов, или намеренно сконструированные, или неизбежно получаемые в результате ошибок синтетического процесса в виде смеси побочных продуктов делеции последовательности, которые имеют хроматографические и иммунологические свойства, аналогичные предполагаемому пептиду, часто так же эффективны, как очищенный препарат желаемого пептида. Разработанные аналоги и непреднамеренные аналоговые смеси эффективны до тех пор, пока разрабатывается четкая процедура контроля качества для контроля как производственного процесса, так и процесса оценки продукта, чтобы гарантировать воспроизводимость и эффективность конечного продукта, в котором используются эти пептиды.
Пептидные иммуногенные конструкции EMPD IgE также могут быть получены с использованием технологии рекомбинантных ДНК, включая молекулы нуклеиновой кислоты, векторы и/или клетки-хозяева. Таким образом, молекулы нуклеиновой кислоты, кодирующие пептидную иммуногенную конструкцию EMPD IgE и ее иммунологически функциональные аналоги, также включены в данное описание как часть данного изобретения. Подобным образом векторы, включая векторы экспрессии, содержащие молекулы нуклеиновой кислоты, а также клетки-хозяева, содержащие векторы, также включены в данное описание как часть данного изобретения.
Различные иллюстративные варианты осуществления также охватывают способы получения пептидной иммуногенной конструкции EMPD IgE и ее иммунологически функциональных аналогов. Например, способы могут включать стадию инкубации клетки-хозяина, содержащей вектор экспрессии, содержащий молекулу нуклеиновой кислоты, кодирующую пептидную иммуногенную конструкцию EMPD IgE и/или его иммунологически функциональный аналог, в таких условиях, когда экспрессируется пептид и/или аналог. Более длинные синтетические пептидные иммуногены могут быть синтезированы хорошо известными методами рекомбинантных ДНК. Такие методы представлены в известных стандартных руководствах с подробными протоколами. Для конструирования гена, кодирующего пептид согласно изобретению, аминокислотную последовательность подвергают обратной трансляции для получения последовательности нуклеиновой кислоты, кодирующей аминокислотную последовательность, предпочтительно с кодонами, которые являются оптимальными для организма, в котором должен экспрессироваться ген. Затем синтетический ген получают, как правило, путем синтеза олигонуклеотидов, которые кодируют пептид и любые регуляторные элементы, если это необходимо. Синтетический ген вставляют в подходящий клонирующий вектор и трансфицируют в клетку-хозяина. Затем пептид экспрессируется в подходящих условиях, подходящих для выбранной системы экспрессии и хозяина. Пептид очищают и характеризуют при помощи стандартных способов.
b. Способы получения иммуностимулирующих комплексов
Различные иллюстративные варианты осуществления также охватывают способы получения иммуностимулирующих комплексов, содержащих пептидные иммуногенные конструкции EMPD IgE и молекулу CpG-олигодезоксинуклеотида (ODN). Стабилизированные иммуностимулирующие комплексы (ISC) получают из катионной части пептидной иммуногенной конструкции EMPD IgE и полианионной молекулы CpG-ODN. Самособирающаяся система приводится в действие электростатической нейтрализацией заряда. Стехиометрия отношения молярного заряда катионной части пептидной иммуногенной конструкции EMPD IgE к анионному олигомеру определяет степень ассоциации. Нековалентная электростатическая ассоциация пептидной иммуногенной конструкции EMPD IgE и CpG-ODN является полностью воспроизводимым процессом. Агрегаты иммуностимулирующего комплекса пептид/CpG-ODN, которые облегчают презентирование «профессиональными» антигенпрезентирующими клетками (АПК) иммунной системы, таким образом, дополнительно усиливают иммуногенность комплексов. Эти комплексы легко характеризуются для контроля качества при производстве. ISC пептид/CpG хорошо переносятся in vivo. Эта новая система частиц, содержащая CpG ODN и пептидные иммуногенные конструкции, полученные из фрагмента EMPD IgE, была разработана для использования преимуществ общей митогенности в отношении В-клеток, связанной с использованием CpG-ODN, и в то же время для стимуляции сбалансированных ответов типа Th-1/Th-2.
CpG-ODN в описанных фармацевтических композициях на 100% связывается с иммуногеном в процессе, опосредованном электростатической нейтрализацией противоположного заряда, что приводит к образованию частиц микронного размера. Форма частиц позволяет значительно снизить дозировку CpG по сравнению с обычным использованием адъювантов CpG, вероятность неблагоприятных врожденных иммунных ответов и облегчить альтернативные пути процессинга иммуногена, включая антигенпрезентирующие клетки (АПК). Следовательно, такие составы являются концептуально новыми и предлагают потенциальные преимущества, способствуя стимуляции иммунных ответов альтернативными механизмами.
с. Способы получения фармацевтических композиций
Различные иллюстративные варианты осуществления также охватывают фармацевтические композиции, содержащие пептидные иммуногенные конструкции EMPD IgE. В некоторых вариантах осуществления в фармацевтических композициях используются водно-масляные эмульсии и суспензии с минеральными солями.
Чтобы большая популяция могла использовать фармацевтическую композицию, а частью цели введения было предотвращение агрегации EMPD IgE, еще одним важным фактором для рассмотрения становится безопасность. Несмотря на использование водно-масляных эмульсий у людей для многих составов в клинических испытаниях, квасцы остаются основным адъювантом для использования в составах благодаря своей безопасности. Поэтому квасцы или их минеральные соли алюминия фосфата (ADJUPHOS) часто используется в качестве адъювантов при подготовке к клиническому применению.
Другие адъюванты и иммуностимулирующие агенты включают 3-Де-O-ацилированный монофосфориллипид A (MPL) или 3-DMP, полимерные или мономерные аминокислоты, такие как полиглутаминовая кислота или полилизин. Такие адъюванты могут быть использованы с или без других специфических иммуностимулирующих агентов, таких как мурамилпептиды (например, N-ацетилмурамил-L-треонил-D-изоглутамин (thr-MDP), N-ацетилнормурамил-L-аланил-D-изоглутамин (nor-MDP), N-ацетилмурамил-L-аланил-D-изоглутаминил-L-аланин-2- (1'-2'-дипальмитоил-sn-глицеро-3-гидроксифосфорилокси)-этиламин (MTP-PE), N- ацетилглюксаминил-N-ацетилмурамил-L-Al-D-изоглу-L-аланин-дипальмитокси пропиламид (DTP-DPP) Theramide™) или другие компоненты бактериальной клеточной стенки. Водно-масляные эмульсии включают MF59 (см. WO 90/14837, Van Nest et al., которая полностью включена в данный документ посредством ссылки во всей своей полноте), содержащую 5% сквалена, 0,5% Tween 80 и 0,5% Span 85 (необязательно содержащий различные количества MTP-PE), составленные в субмикронные частицы с использованием микрофлюидизатора; SAF, содержащий 10% сквалена, 0,4% Tween 80, 5% плюроник-блокированного полимера L121 и thr-MDP, или микрофлюидизированный в субмикронную эмульсию, или перемешанный на вортексе с образованием эмульсии с более крупным размером частиц; и адъювантную систему Ribi™ (RAS) (Ribi ImmunoChem, Хамильтон, Монтана), содержащую 2% сквалена, 0,2% Tween 80 и один или более компонентов бактериальной клеточной стенки, выбранных из группы, состоящей из монофосфориллипида-A (MPL), трегалозы димиколата (TDM) и скелета клеточной стенки (CWS), предпочтительно MPL+CWS (Detox™). Другие адъюванты включают полный адъювант Фрейнда (CFA), неполный адъювант Фрейнда (IFA) и цитокины, такие как интерлейкины (ИЛ-1, ИЛ-2 и ИЛ-12), макрофагальный колониестимулирующий фактор (М-КСФ), и фактор некроза опухоли (ФНО).
Выбор адъюванта зависит от стабильности иммуногенного состава, содержащего адъювант, пути введения, схемы применения, эффективности адъюванта для вакцинируемых видов, а у людей фармацевтически приемлемый адъювант представляет собой тот, который был одобрен или может быть одобрен для введения человеку соответствующими регулирующими органами. Например, квасцы, MPL или неполный адъювант Фрейнда (Chang et al., Advanced Drug Delivery Reviews 32: 173-186 (1998), который включен в данный документ посредством ссылки во всей своей полноте) отдельно или необязательно все их комбинации подходят для введения человеку.
Композиции могут содержать фармацевтически приемлемые, нетоксичные носители или разбавители, которые определяются как носители, обычно используемые для составления фармацевтических композиций для введения животным или людям. Разбавитель выбирается таким образом, чтобы не влиять на биологическую активность комбинации. Примерами таких разбавителей являются дистиллированная вода, физиологический раствор с фосфатным буфером, растворы Рингера, раствор декстрозы и раствор Хэнка. Кроме того, фармацевтическая композиция или состав может также содержать другие носители, адъюванты или нетоксичные, нетерапевтические, неиммуногенные стабилизаторы и тому подобное.
Фармацевтические композиции также могут включать крупные медленно метаболизируемые макромолекулы, такие как белки, полисахариды, такие как хитозан, полимолочные кислоты, полигликолевые кислоты и сополимеры (например, функционализированная латексом сефароза, агароза, целлюлоза и тому подобное), полимерные аминокислоты, аминокислотные сополимеры, и липидные агрегаты (например, масляные капли или липосомы). Кроме того, эти носители могут функционировать в качестве иммуностимулирующих агентов (то есть адъювантов).
Фармацевтические композиции согласно данному изобретению могут дополнительно содержать подходящий носитель для доставки. Подходящие носители для доставки включают, но не ограничиваются ими, вирусы, бактерии, биоразлагаемые микросферы, микрочастицы, наночастицы, липосомы, минипеллеты коллагена и кохлеаты.
d. Способы с использованием фармацевтических композиций
Данное раскрытие также включает способы применения фармацевтических композиций, содержащих пептидные иммуногенные конструкции EMPD IgE.
В некоторых вариантах осуществления фармацевтические композиции, содержащие пептидные иммуногенные конструкции EMPD IgE, могут применяться для лечения и/или профилактики опосредованных иммуноглобулином E (IgE) аллергических заболеваний, включая, но не ограничиваясь ими, аллергию на лекарства, пищу и укусы насекомых, аллергический ринит (сенная лихорадка), атопический дерматит, аллергическую астму, конъюнктивит, экзему, аллергическую сыпь (крапивницу) и анафилаксию.
В некоторых вариантах осуществления способы включают введение фармацевтической композиции, содержащей фармакологически эффективное количество иммуногенной конструкции пептида EMPD IgE, хозяину, нуждающемуся в этом. В некоторых вариантах осуществления способы включают введение фармацевтической композиции, содержащей фармакологически эффективное количество пептидной иммуногенной конструкции EMPD IgE, теплокровному животному (например, людям, яванским макакам, мышам) для выявления высокоспецифичных антител, перекрестно-реактивных с пептидом EMPD IgE 1-52 (SEQ ID NO: 2), белком EMPD IgE 1-67 (SEQ ID NO: 1) либо в виде рекомбинантного белка, содержащего EMPD IgE, (например, очищенного из стабильной клеточной линии Flp-In CHO, трансфицированной рекомбинантной ДНК, кодирующей Fc-часть человеческого IgG1 и EMPD IgE мембраносвязанного IgE человека, γ1-em67) или на мембране клетки, несущей мембраносвязанный IgE,(например, клеточной линии Ramos, трансфицированной рекомбинантной ДНК, кодирующей mIgE.FcL).
В определенных вариантах осуществления фармацевтические композиции, содержащие пептидные иммуногенные конструкции EMPD IgE, можно использовать для лечения и/или профилактики опосредованных IgE заболеваний путем образования антител, направленных против EMPD IgE. Такие антитела способны (a) связываться с mIgE-экспрессирующими В-клетками и индуцировать антителозависимую клеточную цитотоксичность (ADCC) и апоптоз; (b) приводить к снижению in vivo базального уровня IgE в крови; (c) приводить к снижению in vivo антигенспецифического уровня IgE в крови; и (d) уменьшать или устранять IgE-опосредованную аллергическую патологию у пациентов, страдающих от опосредованных IgE аллергических заболеваний.
e. Функциональные анализы in vitro и доказательство эффективности концептуальных исследований in vivo
Антитела, продуцируемые пептидными иммуногенными конструкциями EMPD IgE, могут применяться в функциональных анализах in vitro. Такие функциональные анализы включают, но не ограничиваются ими:
(a) связывание in vitro с пептидом EMPD IgE 1-52 (SEQ ID NO: 1) в виде рекомбинантного белка, очищенного из клеток CHO, экспрессирующих mIgE.FcL (пример 3);
(b) связывание in vitro с клеткой, несущей мембраносвязанный IgE, из линии В-клеток Ramos, которая была трансфицирована рекомбинантной ДНК, кодирующей IgE.FcL (пример 3);
(c) антителозависимая клеточная цитотоксичность (ADCC) in vitro (пример 6);
(d) индукция in vitro апоптоза IgE-несущих В-лимфоцитов (пример 6);
(e) доказательство эффективности in vivo, демонстрирующее снижение базального уровня IgE в крови вакцинированных хозяев (примеры 8-10);
(f) доказательство эффективности in vivo, демонстрирующее снижение антиген-специфического уровня IgE при примировании и повышение при стимуляции аллергеном (примеры 8-10).
В соответствии с данным раскрытием пептидные иммуногенные конструкции EMPD IgE и их составы могут эффективно функционировать в качестве вакцин для снижения или устранения опосредованной IgE аллергической патологии у пациентов, страдающих от опосредованных IgE аллергических заболеваний.
Конкретные варианты осуществления изобретения
(1) Пептидная иммуногенная конструкция EMPD IgE, представленная формулами:
(Th)m-(A)n-(фрагмент EMPD IgE)-X
или
(фрагмент EMPD IgE)-(A)n-(Th)m-X
или
(Th)m-(A)n-(фрагмент EMPD IgE)-(A)n-(Th)m-X,
где
Th представляет собой гетерологичный T-хелперный эпитоп;
А представляет собой гетерологичный спейсер;
(фрагмент EMPD IgE) представляет собой В-клеточный эпитоп, содержащий от около 20 до около 40 аминокислотных остатков из центральной внутримолекулярной петли EMPD IgE;
X представляет собой α-COOH или α-CONH2 аминокислоты;
m равно от 1 до около 4; и
n равно от 0 до около 10.
(2) Пептидная иммуногенная конструкция EMPD IgE в соответствии с (1), отличающаяся тем, что фрагмент EMPD IgE выбран из группы, состоящей из SEQ ID NO: 5, 6, 8 и 9.
(3) Пептидная иммуногенная конструкция EMPD IgE в соответствии с любым из (1) или (2), отличающаяся тем, что Th-эпитоп выбран из группы, состоящей из SEQ ID NO: 59-87.
(4) Пептидная иммуногенная конструкция EMPD IgE в соответствии с (1), отличающаяся тем, что пептидная иммуногенная конструкция выбрана из группы, состоящей из SEQ ID NO: 88-95, 98-124 и 130.
(5) Пептидная иммуногенная конструкция EMPD IgE, содержащая:
В-клеточный эпитоп, содержащий от около 20 до около 40 аминокислотных остатков из последовательности EMPD IgE в соответствии с SEQ ID NO: 1 или SEQ ID NO: 2;
Т-хелперный эпитоп, содержащий аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 59-87; и
необязательный гетерологичный спейсер, выбранный из группы, состоящей из аминокислоты, Lys-, Gly-, Lys-Lys-Lys-, (α, ε-N)Lys, и ε-N-Lys-Lys-Lys-Lys (SEQ ID NO: 129),
при этом В-клеточный эпитоп ковалентно связан с Т-хелперным эпитопом напрямую или через необязательный гетерологичный спейсер.
(6) Пептидная иммуногенная конструкция EMPD IgE согласно (5), отличающаяся тем, что В-клеточный эпитоп выбран из группы, состоящей из SEQ ID NO: 5, 6, 8 и 9.
(7) Пептидная иммуногенная конструкция EMPD IgE согласно (5), отличающаяся тем, что Т-хелперный эпитоп выбран из группы, состоящей из SEQ ID NO: 59-87.
(8) Пептидная иммуногенная конструкция EMPD IgE согласно (5), отличающаяся тем, что необязательный гетерологичный спейсер представляет собой (α, ε-N)Lys или ε-N-Lys-Lys-Lys-Lys (SEQ ID NO: 129).
(9) Пептидная иммуногенная конструкция EMPD IgE согласно (5), отличающаяся тем, что Т-хелперный эпитоп ковалентно связан с N-концом В-клеточного эпитопа.
(10) Пептидная иммуногенная конструкция EMPD IgE согласно (5), отличающаяся тем, что Т-хелперный эпитоп ковалентно связан с N-концом В-клеточного эпитопа через необязательный гетерологичный спейсер.
(11) Композиция, содержащая пептидную иммуногенную конструкцию в соответствии с любым из (1)-(10).
(12) Фармацевтическая композиция, содержащая:
а. пептидную иммуногенную конструкцию в соответствии с любым из (1)-(10); и
b. фармацевтически приемлемый носитель для доставки и/или адъювант.
(13) Фармацевтическая композиция согласно (12), отличающаяся тем, что
а. пептидная иммуногенная конструкция EMPD IgE выбрана из группы, состоящей из SEQ ID NO: 88-95 и 98-124 и 130; и
b. пептидная иммуногенная конструкция EMPD IgE смешивается с CpG-олигодезоксинуклеотидом (ODN) для образования стабилизированного иммуностимулирующего комплекса.
(14) Выделенное антитело или его эпитопсвязывающий фрагмент, которые специфически связываются с В-клеточным эпитопом пептидной иммуногенной конструкции EMPD IgE в соответствии с любым из (1)-(10).
(15) Выделенное антитело или его эпитопсвязывающий фрагмент в соответствии с (14) связаны с пептидной иммуногенной конструкцией EMPD IgE.
(16) Выделенное антитело или его эпитопсвязывающий фрагмент, которые специфически связываются с В-клеточным эпитопом пептидной иммуногенной конструкции EMPD IgE в соответствии с любым из (1)-(10).
(17) Композиция, содержащая выделенное антитело или его эпитопсвязывающий фрагмент в соответствии с любым из (14)-(16).
Подробное описание используемых процедур приведено в следующих примерах. Следующие примеры служат для иллюстрации настоящего изобретения и не должны использоваться для ограничения объема изобретения.
Пример 1
СИНТЕЗ ПЕПТИДОВ, РОДСТВЕННЫХ EMPD IgE, И ИЗГОТОВЛЕНИЕ ИХ СОСТАВОВ
а. Синтез пептидов, родственных EMPD IgE
Описаны способы синтеза сконструированных пептидов, родственных EMPD IgE, которые были включены в процесс разработки пептидных иммуногенных конструкций EMPD IgE. Пептиды были синтезированы в небольших количествах, которые достаточны для серологических анализов, лабораторных поисковых и экспериментальных исследований, а также в значительных количествах (килограммы), которые применимы для промышленного/коммерческого изготовления фармацевтических композиций. Большой репертуар связанных с EMPD IgE антигенных пептидов, имеющих последовательности длиной от около 20 до 70 аминокислот, был разработан для скрининга и выбора наиболее оптимальных пептидных конструкций для применения в эффективной вакцине от аллергии, опосредованной IgE.
Типичные полноразмерные EMPD IgE 1-67 (SEQ ID NO: 1), EMPD IgE 1-52 (SEQ ID NO: 2) и сегменты EMPD IgE, включая EMPD IgE 1-17 (SEQ ID NO: 7), EMPD IgE 19-38 (SEQ ID NO: 8) и различные 10-мерные пептиды (SEQ ID NO: 10-58), используемые для картирования эпитопов в различных серологических анализах, показаны в таблице 1 (SEQ ID NO: 1-58).
Выбранные пептиды В-клеточного эпитопа EMPD IgE были превращены в иммуногенные конструкции пептида EMPD IgE путем синтетического связывания с тщательно сконструированным эпитопом Т-хелперных клеток (Th), полученным из белков патогенов, включая гибридный белок вируса кори (MVF), белок поверхностного антигена гепатита B (HBsAg), вирус гриппа, Clostridum tetani и вирус Эпштейна-Барра (EBV), как показано в таблице 2 (SEQ ID NO: 59-87). Th-эпитопы использовали или в одной последовательности (SEQ ID NO: 59-67 и 72-87), или в комбинаторной библиотеке (SEQ ID NO: 68-71) для усиления иммуногенности соответствующих им пептидных иммуногенных конструкций EMPD IgE.
В таблице 3 указаны типовые пептидные иммуногенные конструкции EMPD IgE (SEQ ID NO: 88-124 и 130), выбранные из более 100 пептидных конструкций.
Все пептиды, используемые для исследований иммуногенности или связанных с ними серологических тестов для обнаружения и/или измерения антител к EMPD IgE, синтезировали в небольшом масштабе с использованием химии F-moc с помощью пептидных синтезаторов Applied BioSystems Models 430A, 431 и/или 433. Каждый пептид получали при помощи отдельного синтеза на твердофазной подложке с F-moc-защитой на N-конце и защитными группами трифункциональных аминокислот боковой цепи. Полученные пептиды отщепляли от твердой подложки, а защитные группы боковых цепей удаляли при помощи 90% трифторуксусной кислотой (TFA). Препараты на основе синтетических пептидов оценивали с помощью время-пролетной масс-спектрометрии с лазерной десорбцией/ионизацией с использованием матрицы (MALDI-TOF) для гарантии правильного содержания аминокислот. Каждый синтетический пептид также оценивали с помощью обращенно-фазовой ВЭЖХ (ОФ-ВЭЖХ), чтобы подтвердить профиль синтеза и концентрацию препарата. Несмотря на строгий контроль процесса синтеза (включая поэтапный контроль эффективности сочетания), пептидные аналоги также образовывались из-за нежелательных явлений во время циклов элонгации, включая вставку аминокислот, делецию, замену и преждевременную терминацию. Таким образом, синтезированные препараты обычно включали множество пептидных аналогов наряду с целевым пептидом.
Несмотря на включение таких непреднамеренных пептидных аналогов, полученные синтезированные пептидные препараты, тем не менее, были пригодны для применения в иммунологических целях, включая иммунодиагностику (в качестве антигенов для захвата антител) и фармацевтических композициях (в качестве пептидных иммуногенов). Как правило, такие пептидные аналоги, или специально сконструированные, или полученные путем синтеза в виде смеси побочных продуктов, часто столь же эффективны, как и очищенный препарат желаемого пептида, при условии, что разработана четкая процедура контроля качества для контроля как процесса производства, так и процесса оценки продукта, чтобы гарантировать воспроизводимость и эффективность конечного продукта, в котором используются эти пептиды. Крупномасштабный синтез пептидов в количествах от нескольких сотен грамм до килограммов проводили на специализированном автоматизированном синтезаторе пептидов UBI2003 или подобном в масштабе от 15 ммоль до 50 ммоль.
Для активных ингредиентов, которые использовались в конечной фармацевтической композиции для клинических исследований, родственные EMPD IgE пептидные конструкции очищали при помощи препаративной ОФ-ВЭЖХ в условиях низкого градиента элюирования и характеризуют при помощи масс-спектрометрии MALDI-TOF, аминокислотного анализа и ОФ-ВЭЖХ в отношении чистоты и идентичности.
b. Изготовление композиций, содержащих пептидные иммуногенные конструкции EMPD IgE
Составы были изготовлены с использованием эмульсий вода-в-масле и в суспензии с минеральными солями. Поскольку фармацевтическая композиция была разработана для использования обширной популяцией, а частью цели введения была профилактика, еще одним важным фактором для рассмотрения становится безопасность. Несмотря на использование водно-масляных эмульсий для многих фармацевтических композиций для введения людям в клинических испытаниях, квасцы остаются основным адъювантом для использования в составах благодаря своей безопасности. Поэтому квасцы или их минеральная соль ADJUPHOS (фосфат алюминия) часто используется в качестве адъювантов при подготовке к клиническому применению.
Вкратце, составы, указанные в каждой из исследовательских групп, описанных ниже, в общем, содержали все типы разработок пептидных иммуногенных конструкций EMPD IgE. Более 100 разработок пептидных иммуногенных конструкций EMPD IgE были первоначально оценены на морских свинках для определения относительной иммуногенности с соответствующим пептидом EMPD IgE, представляющим пептид В-клеточного эпитопа иммуногена, а также для оценки серологической перекрестной реактивности среди различных гомологичных пептидов с помощью анализов ИФА с использованием планшетов, покрытых различными пептидами, выбранными из пептидов с последовательностью SEQ ID NO: 1-126.
Пептидные иммуногенные конструкции EMPD IgE получали (i) в эмульсии вода-в-масле с Seppic Montanide™ ISA 51 в качестве масла, одобренного для использования у человека или (ii) в смеси с минеральными солями ADJUPHOS (фосфат алюминия) или ALHYDROGEL (квасцы), в различных количествах пептидных конструкций, как указано. Композиции, как правило, готовили путем растворения пептидных иммуногенных конструкций EMPD IgE в воде в концентрации примерно от 20 до 800 мкг/мл и смешивали с Montanide™ ISA 51 в эмульсию вода-в-масле (1:1 по объему) или с минеральными солями или ALHYDROGEL (квасцы) (1:1 по объему). Композиции выдерживали при комнатной температуре в течение примерно 30 минут и перемешивали на вихревой мешалке в течение примерно 10-15 секунд до иммунизации. Некоторых животных иммунизировали 2-3 дозами определенной композиции, вводимыми в моменты времени 0 (основной) и через 3 недели после первичной иммунизации (нпи) (бустер), необязательно через 5 или 6 нпи в качестве второго бустера внутримышечным путем. Затем таких иммунизированных животных тестировали выбранным пептидом В-клеточного эпитопа для оценки иммуногенности различных пептидных иммуногенных конструкций EMPD IgE, присутствующих в препарате, а также их перекрестной реактивности с соответствующими целевыми белками или пептидами. Такие пептидные иммуногенные конструкции EMPD IgE, показавшие сильную иммуногенность при первоначальном скрининге у морских свинок, затем дополнительно тестировали на приматах в составах в виде эмульсии вода-в-масле, в составах на основе минеральных солей и в составах на основе квасцов для определения схем применения в течение определенного периода, как продиктовано протоколами иммунизации.
Только наиболее перспективные пептидные иммуногенные конструкции EMPD IgE были дополнительно подвергнуты тщательной оценке перед включением в окончательные составы для исследований иммуногенности, длительности, токсичности и эффективности в доклинических исследованиях в соответствии с НЛП при подготовке к подаче заявки на применение нового исследуемого лекарственного средства и проведению клинических испытаний у пациентов с заболеваниями, опосредованными IgE.
ПРИМЕР 2
СЕРОЛОГИЧЕСКИЕ АНАЛИЗЫ И РЕАГЕНТЫ
Серологические анализы и реагенты для оценки функциональной иммуногенности синтетических пептидных конструкций и их составов подробно описаны ниже.
а. Тесты ИФА EMPD IgE 1-52, EMPD IgE 1-39, EMPD IgE 1-17, EMPD IgE 19-38, EMPD IgE 7-40 на основе пептидов для анализа специфичности антител
Тесты ИФА для оценки образцов иммунной сыворотки, описанные в следующих примерах, были разработаны и описаны ниже. Лунки 96-луночных планшетов покрывали индивидуально в течение 1 часа при 37°С 100 мкл целевого пептида EMPD IgE 1-52, EMPD IgE 1-39, EMPD IgE 1-17, EMPD IgE 19-38, EMPD IgE 7-40 и т.д. (например, SEQ ID NO: 2 и 5-8) в концентрации 2 мкг/мл (если не указано иное) в 10 мМ буфера NaHCO3, рН 9,5 (если не указано иное).
Лунки, покрытые пептидом, инкубировали с 250 мкл 3% по массе желатина с ФСБ при 37°С в течение 1 часа для блокирования сайтов неспецифического связывания белка, затем три раза промывали ФСБ, содержащим 0,05% TWEEN® 20, и сушили. Анализируемые сыворотки разбавляли 1:20 (если не указано иное) ФСБ, содержащим 20% по объему нормальной козьей сыворотки, 1% по массе желатина и 0,05% по объему TWEEN® 20. В каждую лунку добавляли сто микролитров (100 мкл) разведенных образцов (например, сыворотки, плазмы) и оставляли реагировать в течение 60 минут при 37°С. Затем лунки шесть раз промывали 0,05% по объему TWEEN® 20 в ФСБ для удаления несвязанных антител. Конъюгированные с пероксидазой хрена (ПХ) специфичные для вида (например, мыши, морской свинки или человека) козьи анти-IgG, IgA или IgM использовали в качестве меченого индикатора связывания с комплексом антитело/пептидный антиген, образованным в положительных лунках. Сто микролитров меченного пероксидазой козьего анти-IgG в предварительно титрованном оптимальном разведении и в 1% по объему нормальной козьей сыворотки с 0,05% по объему TWEEN® 20 в ФСБ добавляли в каждую лунку и инкубировали при 37°С еще 30 минут. Лунки промывали шесть раз 0,05% по объему TWEEN® 20 в ФСБ для удаления несвязанного антитела и реагировали с 100 мкл смеси субстратов, содержащей 0,04% по массе 3’,3’,5’,5’-тетраметилбензидина (TMB) и 0,12% по объему перекиси водорода в натриево-цитратном буфере в течение еще 15 минут. Данную субстратную смесь использовали для обнаружения метки пероксидазы путем образования окрашенного продукта. Реакции останавливали добавлением 100 мкл 1,0 М H2SO4 и определяли поглощение при 450 нм (A450). Для определения титров антител вакцинированных животных, получавших различные вакцинные составы на основе пептидов EMPD IgE, 10-кратные серийные разведения сывороток от 1:100 до 1:10000 или 4-кратные серийные разведения сывороток от 1:100 до 1:4,19 × 108, и титр исследуемой сыворотки, выраженный как Log10, рассчитывали с помощью линейного регрессионного анализа A450 с порогом A450, установленным на 0,5.
b. Оценка реактивности антител в отношении Th-пептида с помощью тестов ИФА на основе Th-пептидов
Лунки 96-луночных планшетов для ИФА покрывали индивидуально в течение 1 часа при 37°C 100 мкл Th-пептида в количестве 2 мкг/мл (если не указано иное) в 10 мМ буфера NaHCO3, pH 9,5 (если не указано иное) в процессе проведения аналогичного теста ИФА, выполняемого в соответствии с описанным выше. Для определения титров антител вакцинированных животных, получавших различные вакцинные составы на основе пептидов EMPD IgE, 10-кратные серийные разведения сывороток от 1:100 до 1:10000 и титр исследуемой сыворотки, выраженный как Log10, рассчитывали с помощью линейного регрессионного анализа A450 с порогом A450, установленным на 0,5.
с. Оценка анализа специфичности на малом участке и картирования эпитопа в отношении EMPD IgE и IgE CH4 (человеческого IgE CH4 к мембране) с помощью тестов ИФА на основе 10-мерного пептидного кластера В-клеточного эпитопа
Точный анализ специфичности антител к EMPD IgE у иммунизированных хозяев определяли путем картирования эпитопов. Вкратце, лунки 96-луночных планшетов покрывали отдельными 10-мерными пептидами EMPD IgE (SEQ ID NO: 10-58) в количестве 0,5 мкг на 0,1 мл на лунку, а затем 100 мкл образцов сыворотки (разведение 1:100 в ФСБ) инкубировали в лунках планшета с 10-мерным пептидом в двух экземплярах, в точности выполняя стадии теста антител ИФА, описанные выше. В-клеточный эпитоп пептидной иммуногенной конструкции EMPD IgE и соответствующие анализы специфичности на малом участке антител к EMPD IgE из иммунной сыворотки иммунизированных хозяев тестировали с соответствующим пептидом EMPD IgE 1-52 без спейсера или последовательностями Th (SEQ ID NO: 1) или с неродственным контрольным пептидом для подтверждения дополнительной реактивности и специфичности.
d. Оценка иммуногенности
Образцы сыворотки от субъектов-животных или людей или от животных до и после иммунизации собирали в соответствии с протоколами экспериментальной вакцинации и нагревали при 56°С в течение 30 минут для инактивации факторов комплемента сыворотки. После введения вакцинных составов собирали образцы крови в соответствии с протоколами и оценивали их иммуногенность в отношении специфических целевых сайтов. Было проведено тестирование серийно разведенных сывороток и положительные титры были выражены как Log10 кратности разведения. Иммуногенность конкретного вакцинного состава оценивали по его способности приводить к образованию высоких титров антител, направленных против желаемой последовательности В-клеточного эпитопа в целевом антигене, сохраняя при этом низкую или незначительную реакционную способность антител по отношению к последовательности эпитопа Т-хелперных клеток, используемой для обеспечения усиления желаемых ответов B-клеток.
e. Иммуноанализ для оценки уровня человеческого IgE в сыворотке мыши
Уровни человеческого IgE у мышей с нокином линии huIGHE измеряли с помощью сэндвич-ИФА с использованием анти-человеческого IgE, HP6061 (Abcam), в качестве захватывающего антитела, и меченного биотином анти-человеческого IgE, HP6029 (Abcam), в качестве детектирующего антитела. Вкратце, HP6061 иммобилизовали на 96-луночных планшетах в количестве 100 нг/лунку в покрывающем буфере (15 мМ Na2CO3, 35 мМ NaHCO3, pH 9,6) и инкубировали при 4°C в течение ночи. Покрытые лунки блокировали аналитическим разбавителем (0,5% BSA, 0,05% TWEEN-20, 0,02% ProClin 300 в ФСБ) в количестве 200 мкл/лунку при комнатной температуре в течение 1 часа. Планшеты промывали 3 раза промывочным буфером (ФСБ с 0,05% TWEEN-20) в количестве 200 мкл/лунку. Очищенный IgE U266 использовали для получения стандартной кривой (диапазон от 0 до 800 нг/мл при 2-кратном серийном разведении) в аналитическом разбавителе с 5% сывороткой мыши. В покрытые лунки добавляли 50 мкл разведенных сывороток (1:20) и стандартов. Инкубацию проводили при комнатной температуре в течение 1 часа. Содержимое все лунок аспирировали, а лунки промывали 6 раз 200 мкл/лунку промывочного буфера. Захваченный человеческий IgE инкубировали с 100 мкл раствора детектирующего антитела (50 нг/мл биотина, меченного HP6029 в разбавителе для анализа) при комнатной температуре в течение 1 часа. Затем связанный биотин-HP6029 детектировали с использованием полиПХ-стрептавидина (разведение 1:10000, Thermo Pierce) в течение 1 часа (100 мкл/лунку). Содержимое все лунок аспирировали, а лунки промывали 6 раз 200 мкл/лунку промывочного буфера. Наконец, лунки обрабатывали 100 мкл/лунку субстрата NeA-Blue TMB (Clinical Scientific Products), и реакцию останавливали добавлением 100 мкл/лунку 1 М H2SO4. Стандартная кривая была создана с помощью программного обеспечения SoftMax Pro (Molecular Devices) для создания подгонки четырехпараметрической логистической кривой и была использована для расчета концентраций IgE во всех тестируемых образцах. Для сравнения данных применяли t-критерий Стьюдента использованием программного обеспечения Prism.
f. Иммуноанализ для оценки уровня папаин-специфического IgE в мышиной сыворотке
Папаин-специфический IgE, вырабатываемый у мышей с нокином линии huIGHE, измеряли прямым ИФА с использованием папаина в качестве материала покрытия и меченного биотином IgE человека, HP6029 (Abcam), в качестве детектирующего антитела. Вкратце, папаин иммобилизовали на 96-луночных планшетах в количестве 500 нг/лунку в покрывающем буфере (15 мМ Na2CO3, 35 мМ NaHCO3, pH 9,6) и инкубировали при 4°C в течение ночи. Покрытые лунки блокировали аналитическим разбавителем (0,5% BSA, 0,05% TWEEN-20, 0,02% ProClin 300 в ФСБ) в количестве 200 мкл/лунку при комнатной температуре в течение 1 часа. Планшеты промывали 3 раза промывочным буфером (ФСБ с 0,05% TWEEN-20) в количестве 200 мкл/лунку. Моноклональный специфичный к химерному папаину человеческий IgE (AllerMAbs Co., Ltd.) использовали для получения стандартной кривой (диапазон от 0 до 30 нг/мл при 2-кратном серийном разведении) в аналитическом разбавителе с 10% сывороткой мыши. В покрытые лунки добавляли 50 мкл разведенных сывороток (1:10) и стандартов. Инкубацию проводили при комнатной температуре в течение 1 часа. Содержимое все лунок аспирировали, а лунки промывали 6 раз 200 мкл/лунку промывочного буфера. Захваченный человеческий IgE инкубировали с 100 мкл раствора детектирующего антитела (50 нг/мл биотина, меченного HP6029 в разбавителе для анализа) при комнатной температуре в течение 1 часа. Затем связанный биотин-HP6029 детектировали с использованием полиПХ-стрептавидина (разведение 1:10000, Thermo Pierce) в течение 1 часа (100 мкл/лунку). Содержимое все лунок аспирировали, а лунки промывали 6 раз 200 мкл/лунку промывочного буфера. Наконец, лунки обрабатывали 100 мкл/лунку субстрата NeA-Blue TMB (Clinical Science Products), и реакцию останавливали добавлением 100 мкл/лунку 1 М H2SO4. Стандартная кривая была создана с помощью программного обеспечения SoftMax Pro (Molecular Devices) для создания подгонки четырехпараметрической логистической кривой и была использована для расчета концентраций папаин-специфического IgE во всех тестируемых образцах. Для сравнения данных применяли t-критерий Стьюдента использованием программного обеспечения Prism.
g. Иммуноанализ для оценки уровня IgE в сыворотке макаков
Уровни IgE макака у яванских макаков измеряли с помощью сэндвич-ИФА с использованием анти-человеческого IgE, MB10-5C4 (Miltenyi Biotec), в качестве захватывающего антитела, и меченного биотином поликлонального анти-макакового IgE (Alpha Diagnostic International Inc.) в качестве детектирующего антитела. Вкратце, MB10-5C4 иммобилизовали на 96-луночных планшетах в количестве 100 нг/лунку в покрывающем буфере (15 мМ Na2CO3, 35 мМ NaHCO3, pH 9,6) и инкубировали при 4°C в течение ночи. Покрытые лунки блокировали аналитическим разбавителем (0,5% BSA, 0,05% TWEEN-20, 0,02% ProClin 300 в ФСБ) в количестве 200 мкл/лунку при комнатной температуре в течение 1 часа. Планшеты промывали 3 раза промывочным буфером (ФСБ с 0,05% TWEEN-20) в количестве 200 мкл/лунку. Очищенный IgE макака использовали для получения стандартной кривой (диапазон от 0 до 10000 нг/мл при 2-кратном серийном разведении) в аналитическом разбавителе с 10% сывороткой макака. В покрытые лунки добавляли 100 мкл разведенных сывороток (1:10) и стандартов. Инкубацию проводили при комнатной температуре в течение 1 часа. Содержимое все лунок аспирировали, а лунки промывали 6 раз 200 мкл/лунку промывочного буфера. Захваченный человеческий IgE инкубировали с 100 мкл раствора детектирующего антитела (50 нг/мл биотина, меченного HP6029 в разбавителе для анализа) при комнатной температуре в течение 1 часа. Затем связанный биотин-HP6029 детектировали с использованием полиПХ-стрептавидина (разведение 1:10000, Thermo Pierce) в течение 1 часа (100 мкл/лунку). Содержимое все лунок аспирировали, а лунки промывали 6 раз 200 мкл/лунку промывочного буфера. Наконец, лунки обрабатывали 100 мкл/лунку субстрата NeA-Blue TMB (Clinical Science Products), и реакцию останавливали добавлением 100 мкл/лунку 1 М H2SO4. Стандартная кривая была создана с помощью программного обеспечения SoftMax Pro (Molecular Devices) для создания подгонки четырехпараметрической логистической кривой и была использована для расчета концентраций IgE во всех тестируемых образцах. Для сравнения данных применяли t-критерий Стьюдента использованием программного обеспечения Prism.
ПРИМЕР 3
ОЦЕНКА ФУНКЦИОНАЛЬНЫХ СВОЙСТВ АНТИТЕЛ, ОБРАЗОВАННЫХ В ОТВЕТ НА ПЕПТИДНЫЕ ИММУНОГЕННЫЕ КОНСТРУКЦИИ EMPD IGE И ИХ СОСТАВЫ У ЖИВОТНЫХ
Иммунные сыворотки или очищенные антитела к EMPD IgE от иммунизированных хозяев тестировали на их способность связываться с рекомбинантным растворимым белком EMPD IgE и клеточными линиями Ramos, которые были трансфицированы либо рекомбинантной ДНК, кодирующей mIgE.FcL (CH2-CM, включая EMPD), либо mIgE.FcS (CH2-CM без EMPD).
а. Клетки
Клеточная линия Ramos была приобретена в Американской коллекции типовых культур (ATCC, Манассас, Вирджиния) и выращена в среде RPMI 1640 (Invitrogen, Карлсбад, Калифорния) с добавлением 10% инактивированной нагреванием ФБС (Invitrogen), 4 мМ L-глютамина, 25 мМ HEPES и 1 мМ пирувата натрия (Invitrogen; полная среда RPMI). Клетки Ramos трансфицировали рекомбинантной ДНК, кодирующей mIgE.FcL. или mIgE.FcS. Клетки Ramos, экспрессирующие mIgE.FcL, трансфицировали сегментом ДНК, кодирующим сегмент длинной изоформы цепи ε mIgE, расположенной от CH2 до цитоплазматического пептида, включая EMPD. Клетки Ramos, экспрессирующие mIgE.FcS, трансфицировали сегментом ДНК, кодирующим сегмент короткой или обычной изоформы цепи ε mIgE, расположенной от CH2 до цитоплазматического пептида, исключая EMPD. Стабильные трансфектанты клеток Ramos, экспрессирующих mIgE.FcL или mIgE.FcS, выдерживали в полной среде RMPI 1640 с добавлением 400 мг/мл зеоцина (Invitrogen).
b. Приготовление рекомбинантного растворимого белка EMPD IgE для тестирования ИФА
Клетки Flp-In CHO, экспрессирующие рекомбинантный растворимый белок EMPD IgE, трансфицировали сегментом ДНК, кодирующим Fc-часть человеческого IgG1 и EMPD IgE человеческого мембраносвязанного IgE, γ1-em67. Стабильные Flp-In CHO-трансфектанты выдерживали в среде IMDM (Invitrogen, Carlsbad, CA) с добавлением 10% инактивированной нагреванием ФБС (Invitrogen), 4 мМ L-глутамина, 25 мМ HEPES и 1 мМ пирувата натрия (Invitrogen; полная среда IMDM). Белок γ1-em67 очищали из культуральной среды с использованием протеин-А-сефарозы (GE Healthcare) в соответствии с инструкциями производителя.
с. Очистка поликлональных антител от иммунных сывороток
Поликлональные IgG из различных иммунных сывороток очищали с использованием протеин-А-сефарозы (GE Healthcare) в соответствии с инструкциями производителя.
d. Связывание поликлональных антител с растворимым белком EMPD IgE или mIgE.FcL на В-клетках
Очищенные поликлональные антитела каждого из иммунизированных животных исследовали на их относительную активность по связыванию с (а) рекомбинантным белком γ1-em67 (описанным выше) с помощью ИФА или (b) клетками Ramos, экспрессирующими mIgE.FcL, с помощью анализа на проточном цитофлуорометре. 96-луночный микропланшет был произведен компанией Nalge NUNC International, с плоским дном (№ кат. 442404) для оптического считывания и с V-образным дном для клеточной инкубации (№ кат. 249570). Оптическую плотность считывали на считывающем устройстве для микропланшетов VersaMax (Molecular Devices). Флуоресцентные красители обнаруживали на цитометре BD FACSCanto II (DB Biosciences); и результирующие данные были получены с помощью соответствующего программного обеспечения FACSDiva. Данные связывания из ИФА и FACS были импортированы в программное обеспечение Prism 6 для количественного анализа. Более конкретно, на микропланшетах с V-образным дном добавляли аликвоты 2 × 105 клеток в 0,1 мл на лунку, центрифугировали и удаляли жидкость. Клетки инкубировали на льду в течение 1 часа с аликвотами 100 мкл образца антитела в различных концентрациях. Клетки промывали один раз, центрифугировали при 300 g в течение 5 минут и окрашивали на льду в течение 30 минут 100 мкл козьего F(ab)2, специфичного для Fc антивидового IgG, конъюгированного с FITC (250 нг/мл). Клетки промывали один раз и жидкость удаляли после центрифугирования. На каждую лунку добавляли аликвоты по 200 мкл связывающего буфера и переносили в микропробирки для разведения для проточной цитометрии. Интенсивность связывания (среднее геометрическое значение интенсивности флуоресценции, GeoMFI), основанную на входе 10000 клеток на образец, считывали на FACS.
e. Анализы апоптоза
Клетки Ramos, стабильно экспрессирующие mIgE.FcL (5 × 105 клеток/мл), инкубировали с очищенными иммунными или контрольными антителами в полной среде RPMI 1640 в течение 1 часа при 37°C. Затем клетки обрабатывали вторичным антителом, козьим F(ab')2, специфичным для Fc IgG морской свинки (Jackson ImmunoResearch Laboratories, Вест Гроув., Пенсильвания), в конечной концентрации 10 мкг/мл, и инкубировали еще 24 ч при 37°С. Степень апоптоза клеток анализировали следующим образом. Для анализов с аннексином V экспозицию фосфатидилсерина (PS) измеряли путем ресуспендирования клеток в окрашивающем растворе в течение 15 минут в темноте при комнатной температуре. Раствор для окрашивания содержал меченый FITC аннексин V (Biovision, Маунтин-Вью, Калифорния), разведенный 1/200, и 2,5 мкг/мл иодида пропидия (ИП) в буфере с 10 мМ HEPES/NaOH (pH 7,4), 140 мМ NaCl и 5 мМ CaCl2. Окрашенные клетки анализировали на цитометре FACSCanto II (BD Biosciences, Сан-Хосе, Калифорния). При анализе точечной диаграммы был получен процент апоптотических клеток, определяемых как положительные в отношении аннексина V и отрицательные в отношении ИП.
f. Анализ антителозависимой клеточной цитотоксичности (ADCC)
Из селезенки мышей Balb/c (самка в возрасте от 6 до 8 недель) выделяли лимфоциты селезенки путем повторного гипотонического шока эритроцитов с использованием буфер для лизиса ККТ (Thermo Fisher Scientific Inc.). После удаления эритроцитов лимфоциты селезенки культивировали в количестве 3 × 106 клеток/мл в течение 3 дней в полной среде RPMI, дополненной 50 мкМ 2-ME и 100 ед/мл рекомбинантного человеческого IL-2 (PeproTech, Inc). Клетки Ramos, экспрессирующие mIgE.FcL (клетки-мишени), метили CFSE (Invitrogen) в ФСБ/0,1% БСА в течение 10 минут при 37°C. После трех промываний холодной полной средой RPMI 1640 клетки доводили до количества 105 клеток/мл. Аликвоты из 20000 меченых клеток в 200 мкл полной среды RPMI покрывали очищенными поликлональными антителами IgG соответствующих иммунных сывороток в количестве 10 мкг/мл в течение 30 минут при 37°С, а затем объединяли с активированными IL-2 лимфоцитами селезенки (эффекторные клетки) в соотношении E/T=30. После 24-часовой инкубации все клетки окрашивали 7-аминоактиномицином D (7-AAD, Invitrogen) в количестве 2,5 мкг/мл в течение 15 минут на льду, а затем анализировали на цитометре Becton Dickinson FACS Canto II (BD Biosciences). Живые клетки-мишени были определены в анализе точечных диаграмм как положительные на CFSE и отрицательные на 7-AAD. Процент лизированных целевых клеток при данном соотношении E/T составил: 100 × [(процент живых целевых клеток в независимом от антитела контроле - процент живых целевых клеток в образце) / процент живых целевых клеток в независимом от антитела контроле].
ПРИМЕР 4
ЖИВОТНЫЕ, ИСПОЛЬЗУЕМЫЕ В ИССЛЕДОВАНИЯХ БЕЗОПАСНОСТИ, ИММУНОГЕННОСТИ, ТОКСИЧНОСТИ И ЭФФЕКТИВНОСТИ
а. Морские свинки:
Исследования иммуногенности были проведены на половозрелых, наивных взрослых самцах и самках морских свинок линии Дункан-Хартли (массой 300-350 г). В экспериментах использовали не менее 3 морских свинок на группу. Протоколы с участием морских свинок линии Дункан-Хартли (в возрасте 8-12 недель; Covance Research Laboratories, Денвер, Пенсильвания, США) были выполнены в соответствии с утвержденными заявками IACUC в виварии, с которым был заключен соответствующий контракт, а также в компании United Biomedical, Inc. (UBI), выступающей в качестве спонсора.
b. Яванские макаки:
Исследования иммуногенности и токсичности при повторных дозах у взрослых самцов и самок обезьян (Macaca flavicularis, приблизительно 4 года; Joinn Laboratories, Сучжоу, Китай) проводились в соответствии с утвержденными заявками IACUC в виварии, с которым был заключен соответствующий контракт, а также в компании UBI, выступающей в качестве спонсора.
c. Мыши с нокином hIGHE:
Штамм мыши, у которого ген IGHG1 заменен геном IGHE человека путем нацеливания на гомологичный ген в генетическом фоне C57BL/B6, экспрессирует секреторный и мембраносвязанный IgE человека (Lu, el al., 2015). Мыши HIGHE экспрессируют человеческий IgE под регуляторным контролем мышиных транскрипционных элементов IGHG1 и экспрессируют мембраносвязанный человеческий IgE посредством альтернативного сплайсинга РНК человеческих регуляторных элементов. Сывороточный IgE был обнаружен в кровотоке уже в возрасте от 8 до 10 недель. В модели профилактики первичного иммунного ответа/иммунологической памяти и в терапевтической модели сенсибилизации/вторичного иммунного ответа использовались молодое потомство смешанной линии HIGHE x Balb/c (возраст 10 ~ 12 недель). Оба исследования проводились в соответствии с утвержденными заявками IACUC в виварии, с которым был заключен соответствующий контракт (Национальный исследовательский институт здравоохранения, Тайвань), а также в UBI, выступающей в качестве спонсора.
Наблюдали эффекты внутримышечных вакцинаций в течение 16-недельного периода для оценки образования антител с помощью ИФА-анализа сывороточного IgE человека и для доказательства снижения уровней общего IgE в сыворотке, а также антиген-специфического IgE при стимуляции антигеном.
Перед иммунизацией образцы сыворотки отдельных животных тестировали на наличие сывороточного IgE человека в соответствии со способами, описанными выше в этом примере. Каждое животное иммунизировали пептидными иммуногенными конструкциями EMPD IgE на дозу вакцинных составов в зависимости от вида и протокола.
ПРИМЕР 5
ВАКЦИННЫЕ СОСТАВЫ ДЛЯ ОЦЕНКИ ИММУНОГЕННОСТИ ПЕПТИДНЫХ КОНСТРУКЦИЙ EMPD IgE У МОРСКИХ СВИНОК, ТРАНСГЕННЫХ МЫШЕЙ И ЯВАНСКИХ МАКАКОВ ДЛЯ ЗАКЛЮЧИТЕЛЬНОГО ВЫБОРА ПРОДУКТА
Фармацевтические композиции и вакцинные составы, используемые в каждом эксперименте, более подробно описаны ниже. Вкратце, составы, указанные в каждой из групп исследования, в основном содержали в все типы разработок пептидных иммуногенных конструкций EMPD IgE со связью сегмента пептида EMPD IgE посредством различного типа спейсеров (например, εK или KKK для повышения растворимости пептидной конструкции), и вариации универсальных эпитопов Т-хелперных клеток, включая два набора искусственных эпитопов Т-хелперных клеток, полученных из гибридного белка вируса кори и поверхностного антигена гепатита В с сегментами пептида EMPD IgE, связанным на N-конце разработанных пептидных конструкций. Более 100 разработанных пептидных конструкций EMPD IgE первоначально были оценены на морских свинках на относительную иммуногенность с EMPD IgE 1-52 и, кроме того, на перекрестную реактивность с мембранным IgE на В-клетках, несущих IgE, (клеточная линия Ramos). Пептидные конструкции EMPD IgE получали в эмульсии вода-в-масле с Seppic Montanide™ ISA 51 в качестве одобренного масла для использования в вакцинах для людей или смешивали с минеральными солями ADJUPHOS или ALHYDROGEL (квасцы) с использованием различных количеств пептидных конструкций в соответствии с описанным в данном документе. Вакцины обычно изготавливали путем растворения пептидных конструкций EMPD IgE в воде в концентрации от 20 до 800 мкг/мл и смешивали с Montanide™ ISA 51 в виде эмульсий вода-в-масле (1:1 по объему) или минеральными солями ADJUPHOS или ALHYDROGEL (квасцы) (1:1 по объему). Вакцинные составы выдерживали при комнатной температуре в течение примерно 30 минут и перемешивали на вихревой мешалке в течение примерно 10-15 секунд до иммунизации.
Животных иммунизировали 2-3 дозами конкретного вакцинного состава, которые вводили в моменты времени 0 (первичное) и через 3 недели после начальной иммунизации (нпи) (бустер), необязательно через 5 или 6 нпи в качестве второго бустера, внутримышечно. Затем этих иммунизированных животных тестировали для оценки иммуногенности различных синтетических пептидных иммуногенов EMPD IgE, присутствующих в вакцинном составе, а также их перекрестной реактивности с EMPD IgE 1-52. Пептидные иммуногены EMPD IgE, показавшие сильную иммуногенность при первоначальном скрининге у морских свинок, затем дополнительно тестировали на макаках в виде эмульсии вода-в-масле, в составах на основе минеральных солей и составах на основе квасцов у макак для определения схем применения в течение определенного периода, как продиктовано протоколами иммунизации.
Только наиболее многообещающие кандидаты из пептидных иммуногенов EMPD IgE были дополнительно оценены в отношении способности иммунных сывороток опосредовать ADCC, апоптоз B-клеток, несущих mIgE, и - у трансгенных мышей с нокином и у макак - на их способность нарушать толерантность у тех же видов с собственным антигеном EMPD IgE перед включением в окончательные вакцинные составы для исследований иммуногенности, длительности, токсичности и доказательной эффективности в соответствии с правилами надлежащей лабораторной практики при подготовке к подаче заявки на новое лекарственное средство для проведения исследований и клинических испытаний у пациентов с аллергическими заболеваниями, опосредованными IgE.
ПРИМЕР 6
ОБОСНОВАНИЕ КОНСТРУКЦИИ, СКРИНИНГ, ИДЕНТИФИКАЦИЯ, ОЦЕНКА ФУНКЦИОНАЛЬНЫХ СВОЙСТВ И ОПТИМИЗАЦИЯ МУЛЬТИКОМПОНЕНТНЫХ ВАКЦИННЫХ СОСТАВОВ, СОДЕРЖАЩИХ ПЕПТИДНЫЕ ИММУНОГЕННЫЕ КОНСТРУКЦИИ EMPD IgE 1-39, ДЛЯ ЛЕЧЕНИЯ ОПОСРЕДОВАННЫХ IgE АЛЛЕРГИЧЕСКИХ ЗАБОЛЕВАНИЙ
Фигуры 2А и 2Виллюстрируют обоснование истощения B-клеток с mIgE путем нацеливания на EMPD IgE. IgE экспрессируются в двух формах: секреторный IgE и мембраносвязанный IgE (mIgE) (Фигура 2А, слева и справа, соответственно). Секреторный IgE захватывается на клеточной поверхности базофилов и тучных клеток через FcRI, в то время как mIgE присутствует исключительно на IgE-коммитированных B-клетках как часть B-клеточного рецептора (BCR). Ближайший к мембране внеклеточный домен (EMPD) mIgE представляет собой пептидный сегмент из 67 аминокислот (SEQ ID NO: 1) между доменом CH4 и трансмембранной областью, обнаруживаемый исключительно в B-клетках с mIgE. Уникальность EMPD IgE обеспечила привлекательный сайт для нацеливания на mIgE-несущие В-клетки. Истощение -B-клеток с mIgE путем нацеливания на EMPD IgE позволяет подавлять аллерген-специфическую выработку IgE до их дифференцировки в новые IgE-секретирующие плазматические клетки (Фигура 2B). Существующие плазматические клетки, секретирующие IgE, с их ограниченной продолжительностью жизни будут постепенно отмирать, что приведет к постепенному снижению общего и аллерген-специфического IgE.
На фигуре 3 представлена блок-схема, идентифицирующая процесс разработки от открытия до коммерциализации (индустриализации) вакцинного состава в соответствии с конкретным вариантом осуществления, раскрытым в данном документе Настоящее раскрытие включает конструкцию пептидного иммуногена, конструкцию пептидной композиции, конструкцию вакцинного состава, конструкцию функциональной антигенности in vitro, конструкцию исследования иммуногенности и эффективности in vivo, а также дизайн клинического протокола, как кратко изложено ниже. Подробная оценка для каждого из этапов неожиданно приводит к серии экспериментов, приводящих к окончательной коммерциализации безопасного и эффективного вакцинного состава.
Краткое резюме выполненных шагов приведено далее:
а. История разработки
Каждая пептидная иммуногенная конструкция или иммунотерапевтический продукт требует особого подхода к разработке, основанного на специфическом механизме заболевания и целевых белках, необходимых для вмешательства. Мишень, моделируемая в таких процессах разработки, может включать клеточные белки, вовлеченные в путь заболевания, или инфекционный агент, в который могут быть вовлечены несколько белков патогена. Процесс от исследования до коммерциализации очень длительный, обычно требуется одно или более десятилетий.
После выбора целевой молекулы требуется обширный процесс серологической проверки. Идентификация В-клеточных и Т-клеточных эпитопов и функциональных сайтов в целевой молекуле, подвергаемой вмешательству, очень важна для разработки иммуногенной конструкции. После распознавания В-клеточного эпитопа, являющегося мишенью, проводят последовательные пилотные исследования иммуногенности на небольших животных, включающих различные носители Т-хелпера (белки-носители или подходящие Т-хелперные пептиды), для оценки функциональных свойств антител, вызванных фармацевтическими композициями на основе сконструированных пептидов. Такое серологическое применение затем проверяют на животных целевых видов для дальнейшей оценки иммуногенности и функциональных свойств вызванных антител. Все исследования проводятся в нескольких параллельных группах с использованием сывороток, взятых у иммунизированных хозяев для оценки. Также для дальнейшей проверки иммуногенности и направления разработок проводят ранние исследования иммуногенности у целевых видов или, в случае фармацевтических композиций, предназначенных для человека, у приматов, не являющихся человеком. Затем целевые пептиды готовят в различных смесях для оценки тонких различий в функциональных свойствах, связанных с соответствующими взаимодействиями между пептидными конструкциями, при использовании в комбинациях для приготовления соответствующих составов препаратов. После дополнительной оценки устанавливают конечные пептидные конструкции, пептидные композиции и их составы, а также соответствующие физические параметры составов, что дает возможность запустить процесс разработки конечного продукта.
b. Разработка и валидация полученных из EMPD IgE пептидных иммуногенных конструкций для фармацевтических композиций с потенциалом лечения пациентов с IgE-опосредованным аллергическим заболеванием
Для создания наиболее сильных пептидных конструкций для включения в фармацевтические композиции проводили исследования иммуногенности на морских свинках, для чего в пептидные иммуногенные конструкции EMPD IgE вносили обширный репертуар пептидов В-клеточного эпитопа EMPD IgE (SEQ ID NO: 5-8) (таблица 1) и универсальных Т-хелперных эпитопов, полученных из различных патогенов, или дополнительно сконструированные искусственные Т-хелперные эпитопы (SEQ ID NO: 59-87) (таблица 2).
i) Выбор EMPD IgE G1-H40 в качестве целевой области для конструирования иммуногена.
IgE экспрессируются в двух формах: секреторный IgE и мембраносвязанный IgE (mIgE). Секреторный IgE захватывается на клеточной поверхности базофилов и тучных клеток через FcRI, в то время как mIgE присутствует исключительно на IgE-коммитированных B-клетках как часть B-клеточного рецептора (BCR). Ближайший к мембране полноразмерный внеклеточный домен (EMPD) mIgE представляет собой пептидный сегмент из 67 аминокислот (SEQ ID NO: 1) между доменом CH4 и трансмембранной областью, обнаруживаемый исключительно в B-клетках с mIgE. Существующие плазматические клетки, секретирующие IgE, с их ограниченной продолжительностью жизни будут постепенно отмирать, что приведет к постепенному снижению общего и аллерген-специфического IgE. Из многих пептидных иммуногенных конструкций, протестированных на иммуногенность у морских свинок, были получены данные серии пептидных иммуногенных конструкций, полученных из EMPD IgE (SEQ ID NO: 88-93), включающих репрезентативные пептиды В-клеточного эпитопа EMPD IgE, полученные из EMPD IgE 1-52 (SEQ ID NO: 2) и типичный Th-эпитопный пептид UBITh®1 (SEQ ID NO: 72), которые были использованы для тестирования иммуногенности на морских свинках с использованием пептида EMPD IgE 1-39 (SEQ ID NO: 5) для покрытия планшетов, как показано в таблице 4. Высокая иммуногенность была обнаружена у шести пептидных иммуногенных конструкций EMPD IgE с тремя ориентациями, в которых пептид Th-эпитопа UBITh®1 был связан с пептидом В-клеточного эпитопа EMPD IgE на С-конце (SEQ ID NO: 88 и 91) или на N-конце (SEQ ID NO: 89, 90 и 92), или и на С-, и на N-конце (SEQ ID NO: 93) пептида В-клеточного эпитопа EMPD IgE посредством спейсера εК. Для обеспечения высокой иммуногенности при разработке конструкции также использовали пептидные иммуногенные конструкции EMPD IgE, содержащие более длинный линкер εK-KKK (SEQ ID NO: 129) (например, SEQ ID NO: 94-97).
Более короткие фрагменты пептидов В-клеточного эпитопа EMPD IgE (SEQ ID NO: 7 и 8) также потенцировали пептидом Т-хелперного эпитопа, таким как UBITh®1 (SEQ ID NO: 96 и 97, соответственно), для повышения иммуногенности. Однако эти конструкции вырабатывают антитела, обладающие слабой способностью связываться с EMPD IgE (например, в отношении более длинных фрагментов с SEQ ID NO: 5, 6, 7 и 8), как показано в таблице 5. Конструкции SEQ ID NO: 96 и 97 также вырабатывали антитела, имеющие крайне ограниченные профили связывания, как показано в исследованиях линейного картирования эпитопов, где реактивность показали только 10-мерные пептиды с SEQ ID NO: 25 (аминокислоты 8-17) и SEQ ID NO: 39 (аминокислоты 22-31), соответственно. Кроме того, было обнаружено, что выработанные антитела конструкций SEQ ID NO: 96 и 97 обладают минимальным эффектом связывания mIgE-B-клеток (как показано на Фиг. 6C), что является необходимым условием для индукции апоптоза mIgE-B-клеток.
ii) Ранжирование гетерологичных Т-хелперных эпитопов, полученных из патогенов, и их включение в иммуногенные конструкции пептида EMPD IgE для усиления иммуногенности выбранного пептида В-клеточного эпитопа EMPD IgE.
В таблице 2 перечислено в общей сложности 29 гетерологичных Th-эпитопов (SEQ ID NO: 59-87), которые были протестированы на их относительную активность в отношении усиления иммуногенности В-клеточных эпитопов на животных нескольких видов: мышах, крысах, морских свинках, бабуинах, макаках и т.д.
В ходе исследования иммуногенности на морских свинках было проведено репрезентативное исследование иммуногенных конструкций пептида EMPD IgE, содержащих пептид В-клеточного эпитопа EMPD IgE 1-39 (SEQ ID NO: 5), связанный через спейсер εK с отдельными универсальными T-хелперными эпитопами (SEQ ID NO: 88, 98-124, и 130) для ранжирования относительной эффективности соответствующих гетерологичных Т-хелперных эпитопов, как показано в таблице 6. Из-за способности некоторых Th-эпитопов значительно усиливать B-эпитоп, результаты, полученные через 3 недели после первоначальной иммунизации (3 нпи) после единственной иммунизации морских свинок, были использованы для ранжирования 29 различных иммуногенных пептидных конструкций EMPD IgE. Хотя все выбранные Th-эпитопы обладали способностью усиливать иммуногенность пептида EMPD IgE B-эпитопа, было обнаружено, что наиболее сильной конструкцией является конструкция SEQ ID NO: 88, а наиболее слабой является конструкция SEQ ID NO: 101.
Тщательная калибровка иммуногенности для всех без исключения пептидных иммуногенных конструкций EMPD IgE у разных видов, включая приматов, обеспечивала окончательный выбор Th-пептида и успех в разработке окончательных вакцинных составов.
iii) Оценка иммуногенности пептидных иммуногенных конструкций EMPD IgE G1-C39 по реакционной способности соответствующих антител с соответствующими полноразмерными и EMPD IgE G1-C39 пептидами.
Фигура 4 иллюстрирует кинетику образования антител в течение 8-недельного периода у морских свинок, иммунизированных различными иммуногенными пептидными конструкциями EMPD IgE (SEQ ID NO: 88-94, 96 и 97). Сыворотку разводили от 1:100 до 1:10000000 посредством 10-кратного серийного разведения. Планшеты для ИФА покрывали пептидом EMPD IgE 1-39 (SEQ ID NO: 5) из расчета 0,5 мкг пептида на лунку. Титр тестируемой сыворотки, выраженный как Log10, рассчитывали с помощью линейного регрессионного анализа А450 нм с порогом А450, установленным на 0,5.
Фиг. 5 иллюстрирует кривую титрования различных очищенных поликлональных антител к EMPD IgE, полученных при анализе различных иммуногенных конструкций EMPD IgE (SEQ ID NO: 88-94, 96 и 97). Планшеты для ИФА покрывали содержащим EMPD IgE рекомбинантным белком γ1-em67, который содержит последовательность SEQ ID NO: 1. Поликлональные антитела к EMPD IgE, очищенные из сыворотки морской свинки с помощью хроматографии на основе белка А, разбавляли от 100 мкг/мл до 0,0238 нг/мл 4-кратным серийным разведением. EC50 каждого препарата поликлональных антител к EMPD IgE рассчитывали путем нелинейной регрессии с четырехпараметрической логистической кривой.
Как показано на Фиг.4 и 5, пептидные иммуногенные конструкции с SEQ ID NO: 88-94, которые были выбраны из многих других конструкций, продемонстрировали высокую иммуногенность с титрами Log10 в основном выше 4. Более точные измерения EC50 с использованием очищенных антител из каждой группы, как показано на Фиг. 5, соответствуют образцам от 0,02111 до 0,08892 мкг/мл с пептидными иммуногенными конструкциями EMPD IgE, содержащими более длинные пептиды В-клеточных эпитопов EMPD IgE 1-39 и EMPD IgE 7-40 (SEQ ID NO: 88-94), которые демонстрировали значительно более высокую иммуногенность по сравнению с пептидными иммуногенными конструкциями, содержащими более короткие пептиды В-клеточных эпитопов EMPD IgE (<20 остатков в длину), такие как EMPD IgE 1-17 или EMPD IgE 19-38 (SEQ ID NO: 96 и 97).
Пептидные иммуногенные конструкции EMPD IgE, содержащие более длинный спейсер между В-клеточными и Th-клеточными эпитопами, также значительно снижали иммуногенность, как в случае конструкции пептида В-клеточного эпитопа EMPD IgE (SEQ ID NO: 94 против 89 для соответствующих EC50 0,08892 против 0,02368). Как было обнаружено, среди пептидов В-клеточного эпитопа длиной более 20 остатков оптимальным по конструкции является EMPD IgE 1-39 и, таким образом, именно он будет наиболее часто использоваться в следующих примерах в качестве пептида В-клеточного эпитопа в типичных конструкциях пептидного иммуногена для проведения дальнейшей оценки посредством различных функциональных анализов in vitro и оценки эффективности in vivo.
iv) Оценка иммуногенности пептидных иммуногенных конструкций EMPD IgE в отношении связывания антител с IgE-несущими В-клетками, которые направляют дифференцировку В-клеток для секреции IgE.
Фиг.6А-6С иллюстрируют связывание с В-клетками, происходящими из клеточных линий Ramos, которые экспрессируют mIgE.FcL или mIgE.FcS (способ цитофлуорографического окрашивания см. в Примере 2), очищенными поликлональными антителами из сыворотки морской свинки, собранными у каждой группы животных, иммунизированных иммуногенными конструкциями EMPD IgE (SEQ ID NO: 88-97). Поликлональные антитела к EMPD IgE, очищенные из сыворотки морских свинок с помощью хроматографии на основе белка А, использовали в количестве 10 мкг/мл.
Низкое фоновое связывание с mIgE.FC клеток линии Ramos, в которых отсутствует фрагмент EMPD целевой последовательности, было обнаружено у очищенных антител всех групп, что свидетельствует о высокой специфичности данного анализа связывания клеток. Как показано на Фиг. 6А-6С, антитела, полученные у животных, иммунизированных пептидными иммуногенными конструкциями EMPD IgE с SEQ ID NO: 88-94, 96 и 97, имели различную аффинность связывания с mIgE.FCL клеток линии Ramos. В частности, антитела, образованные в ответ на пептидные иммуногенные конструкции EMPD IgE, содержащие длинный пептид В-клеточного эпитопа EMPD IgE 1-39 (SEQ ID NO: 88-94), продемонстрировали более высокое связывание mIgE у mIgE.FCL клеток линии Ramos по сравнению с антителами, сгенерированными для пептидных иммуногенных конструкций EMPD IgE, содержащие пептиды В-клеточного эпитопа с менее чем 20 остатками (SEQ ID NO: 96-97). Кроме того, антитела, образованные в ответ на пептидную иммуногенную конструкцию EMPD IgE, имеющую более короткий спейсер (εK), показали более высокое связывание mIgE у mIgE.FCL клеток линии Ramos по сравнению с конструкциями, имеющими более длинный спейсер (εK-KKK; SEQ ID NO: 129), что можно увидеть, сравнив результаты SEQ ID NO: 89 на фиг. 6A с результатами SEQ ID NO: 94 на Фиг. 6C.
Отрицательная корреляция была обнаружена между значениями EC50 группы поликлональных антител и процентом (%) положительного связывания IgE-несущих клеток. Процент связывания IgE-несущих В-клеток является важным функциональным параметром для оценки как перекрестной реактивности антител, так и функциональной эффективности иммуногенности пептидной иммуногенной конструкции EMPD IgE.
v) Оценка иммуногенности пептидных иммуногенных конструкций EMPD IgE по их способности генерировать антитела с высокой эффективностью в отношении индуцирования апоптоза несущих IgE В-клеток, которые направляют дифференцировку В-клеток для секреции IgE.
На фигуре 7 показано, что апоптоз был индуцирован различными препаратами поликлональных антител к EMPD IgE, вырабатываемых иммуногенными конструкциями EMPD IgE (SEQ ID NO: 88-93), в дозозависимом режиме на поверхности экспрессирующих mIgE.FcL клеток линии Ramos. Поликлональные антитела к EMPD IgE, очищенные из сыворотки морских свинок с помощью хроматографии на основе белка А, разбавляли от 1000 до 62,5 нг/мл путем 2-кратного серийного разведения. В качестве положительного контроля использовали гуманизированное моноклональное антитело к IgE, Xolair®. EC50 каждого набора поликлональных антител к EMPD IgE рассчитывали путем нелинейной регрессии с четырехпараметрической логистической кривой.
Как показано на Фиг. 7, хотя Xolair® (моноклональное антитело к IgE, нацеленное на связывающий Fc-рецептор домен CH3 молекулы IgE) показало наименьшее значение EC50 (121,4 нг/мл), указывающее на наивысшую эффективность, его присутствие в сыворотке нейтрализует возможность индуцирования апоптоза из-за нейтрализации сывороточным IgE. Среди отобранных протестированных пептидных иммуногенных конструкций EMPD IgE (SEQ ID NO: 88-93) наблюдались значения ЕС50 от 277,5 до 536 нг/мл, свидетельствующие о высокой эффективности индуцирования апоптоза без интерференции с сывороточным IgE из-за связывания с уникальной последовательностью целевой области EMPD. Такие В-клетки, несущие IgE, представляют собой клетки, которые направляют дифференцировку В-клеток для секреции IgE.
Индукция апоптоза таких клеток путем иммунизации хозяина пептидными иммуногенными конструкциями EMPD IgE (например, SEQ ID NO: 88-93) может вызвать подавление синтеза IgE, что приводит к снижению в сыворотке количества IgE, основной причины аллергических заболеваний.
vi) Оценка иммуногенности пептидных иммуногенных конструкций EMPD IgE в отношении их способности генерировать антитела с высокой эффективностью инициации антителозависимой клеточно-опосредованной цитотоксичности (ADCC) IgE-несущих B-клеток, которые направляют дифференцировку B-клеток для секреции IgE.
На Фиг. 8 показано, что различные препараты поликлональных антител к EMPD IgE, генерируемых различными иммуногенными конструкциями EMPD IgE (SEQ ID NO: 88-93), способны индуцировать ADCC в отношении экспрессирующих mIgE.FcL клеток линии Ramos при соотношении эффектор/мишень=30. Поликлональные антитела к EMPD IgE, очищенные из сыворотки морских свинок с помощью хроматографии на основе белка А, использовали в количестве 10 мкг/мл. В качестве эффекторных клеток использовали стимулированные IL-2 клетки мышиной селезенки. В качестве положительного контроля использовали секрецию 5D5, мышиного моноклонального антитела к IgE.
Как показано на Фиг. 8, все пептидные иммуногенные конструкции с длинными пептидами В-клеточного эпитопа, имеющими более 20 аминокислотных остатков (SEQ ID NO: 88-93), индуцировали ADCC B-клеток, несущих IgE, которые направляют дифференцировку B-клеток для секреции IgE, что указывает на снижение и подавление уровней IgE в сыворотке при иммунизации хозяина такими пептидными иммуногенными конструкциями.
vii) Расширение охвата ГКГС с использованием полученных из EMPD IgE пептидных иммуногенных конструкций с различными универсальными T-хелперными эпитопами.
При разработке фармацевтической композиции для лечения пациентов с различным генетическим фоном важно учитывать, что конструкция должна охватывать большинство популяций с разнообразным генетическим фоном. Поскольку разнородные Т-хелперные эпитопы, полученные из MVF или HBsAg, представляют собой один из наиболее эффективных способов для обеспечения такого усиления иммуногенности, можно было бы создать комбинацию пептидных конструкций, содержащих такие два Т-хелперных эпитопа для обеспечения синергетического эффекта иммуногенности. Можно ожидать, что смесь двух пептидных иммуногенных конструкций с одинаковыми В-клеточными эпитопами будет вызывать значительный иммунный ответ по сравнению с таковым, вызванным отдельной соответствующей пептидной конструкцией.
viii) Картирование эпитопа для анализа специфичности на малом участке иммунной сыворотки (9 нпи), выработанной различными пептидными иммуногенными конструкциями EMPD IgE
Разработка вакцины EMPD IgE, содержащей пептидную иммуногенную конструкцию EMPD IgE, была сфокусирована на особой петлевидной структуре C18-C39 в средней области EMPD IgE в качестве функциональной и структурной мишени. Эта конструкция на основе структуры направлена на использования структуры нативной внеклеточной петли в качестве иммуногенной мишени.
Четыре репрезентативных пептидных фрагмента EMPD IgE: 1-17 (SEQ ID NO: 7), 19-38 (SEQ ID NO: 8), 1-39 (SEQ ID NO: 5) и 7-40 (SEQ ID NO: 6), были использованы для конструирования пептидов В-клеточных эпитопов , которые были связаны с UBITh®1 (SEQ ID NO: 72) или UBITh®2 (SEQ ID NO: 73) на N- или С-конце пептидов В-клеточных эпитопов с образованием прототипов иммуногенных пептидов. Для образования пептидных иммуногенных конструкций, показанных в таблице 3 (SEQ ID NO: 88-97), связь между B-клеточным и Th-эпитопами обеспечивалась посредством линкера εK или спейсера εK-KKK (SEQ ID NO: 129). Все пептидные фрагменты с аминокислотами (ак) 1-39 и 7-40 были сконструированы с циклической структурой с ограниченным циклом C18-C39.
Реактивность антител к гипериммунным сывороткам, полученным от морских свинок, иммунизированных соответствующими им пептидными иммуногенными конструкциями (SEQ ID NO: 96, 97, 88, 89, 93) оценивали с помощью ИФА-тестов с использованием отдельных пептидов В-клеточного эпитопа EMPD IgE 1-17 (SEQ ID NO: 7), 19-38 (SEQ ID NO: 8), 1-39 (SEQ ID NO: 5) и 7-40 (SEQ ID NO: 6) для покрытия планшетов. Результаты показали, что конструкции SEQ ID NO: 88, 89 и 93 индуцировали высокие титры антител против всех четырех пептидов В-клеточного эпитопа EMPD IgE, в то время как антисыворотка морской свинки, индуцированная пептидными иммуногенными конструкциями EMPD IgE 1-17 (SEQ ID NO: 96) и IgE -EMPD 19-38 (SEQ ID NO: 97), показывала реакционную способность только в отношении антител с соответствующими им пептидами эпитопа B-клеток (например, SEQ ID NO: 7 или 8), но не имела перекрестной реактивности с пептидами В-клеточного эпитопа EMPD IgE (SEQ ID NO: 8 или 7) из несоответствующего соседнего эпитопа, что свидетельствует о высокой специфичности иммуногенности, то есть, сконструированные иммуногенные конструкции способны вызывать образование специфических антител для реакции с соответствующими доменами В-клеточных эпитопов EMPD IgE (таблица 5).
В исследовании картирования эпитопа на малом участке (таблица 5) для локализации сайтов связывания антитела со специфическими остатками в пределах целевой области было синтезировано 50 перекрывающихся 10-мерных пептидов (SEQ ID NO: 10-58), которые охватывают первые 8 аминокислот C-конца IgE CH4 и от 1 до 50 областей аминокислотной последовательности EMPD IgE. Эти 10-мерные пептиды по-отдельности наносили на лунки 96-луночного планшета для микротитрования в качестве твердофазных иммуноабсорбентов. Объединенные образцы антисыворотки морской свинки в разведении 1:100 в буфере для разведения образца добавляли в лунки планшета, покрытые 10-мерными пептидами, в количестве 2,0 мкг/мл с последующим инкубированием в течение одного часа при температуре 37°С. После промывки лунок планшета буфером для промывания добавляли конъюгированный с пероксидазой хрена rProtein A/G и инкубировали в течение 30 мин. После повторного промывания ФСБ, в лунки добавляли субстрат для измерения оптической плотности при 450 нм на считывателе для планшетов для ИФА при анализе образцов в двух экземплярах. Связывание выработанной иммуногенными пептидами EMPD IgE иммунной сыворотки с соответствующими лунками, покрытыми пептидами В-клеточного эпитопа EMPD IgE, представляет собой максимальный сигнал связывания антител.
Результаты картирования эпитопа на малом участке показали, что объединенные сыворотки морских свинок из пептидной иммуногенной конструкции, полученной из EMPD IgE 19-38 (SEQ ID NO: 97, с нециклизованным линейным В-клеточным эпитопом, H19-R38) распознают пептид EMPD IgE 22-31 (SEQ ID NO: 39), расположенный в средней области EMPD IgE. Они также реагировали с пептидом EMPD IgE 19-38 (SEQ ID NO: 8), 1-39 (SEQ ID NO: 5) и 7-40 (SEQ ID NO: 6), но не с 1-17 (SEQ ID NO: 7).
Антисыворотки, индуцированные EMPD IgE 1-17 (SEQ ID NO: 96, нециклизованный линейный В-клеточный эпитоп, G1-L17), распознавали пептид EMPD IgE 8-17 (SEQ ID NO: 25) в N-концевой области EMPD IgE, как и EMPD IgE 1-17 (SEQ ID NO: 7), 1-39 (SEQ ID NO: 5) и 7-40 (SEQ ID NO: 6), но не 19-38 (SEQ ID NO: 8). Антисыворотки, индуцированные иммуногенными конструкциями EMPD IgE1-17 (SEQ ID NO: 96) и EMPD IgE 19-38 (SEQ ID NO: 97), не имели перекрестной реактивности.
Интересно, что две аналогичные иммуногенные конструкции EMPD IgE 1-39 с эпитопом UBITh®1, связанным с пептидом В-клеточного эпитопа EMPD IgE на N-конце (SEQ ID NO: 89) или на С-конце (SEQ ID NO: 88), приводили к выработке иммунных сывороток, распознающих отчетливо различную картину связывания эпитопа. Иммунные сыворотки, индуцированные иммуногенной конструкцией (SEQ ID NO: 89, UBTh1, расположенный на N-конце пептида), сильно реагировали только с одним 10-мерным пептидом (SEQ ID NO: 47) на аминокислотах 30-39, рядом с С-концевой областью EMPD IgE. Они также слабо реагировали с EMPD IgE 9-18 в N-концевой области (SEQ ID NO: 27).
Однако в случае другой иммуногенной конструкции EMPD IgE 1-39 (SEQ ID NO: 88, UBTh1, расположенной на С-конце) индуцированная антисыворотка сильно реагировала с тремя прерывистыми линейными эпитопами, представленными EMPD IgE 9-19 (SEQ ID NO: 27 и 28); EMPD IgE 19-31 (SEQ ID NO: 37, 38, 39 и 40) и EMPD IgE 30-43 (SEQ ID NO: 48, 49, 50, 51 и 52). Пептидная иммуногенная конструкция (SEQ ID NO: 88) продемонстрировала гораздо более сильную иммуногенность и прореагировала с гораздо более обширной поверхностью по сравнению с пептидной иммуногенной конструкцией EMPD IgE (SEQ ID NO: 89). Это обнаружение гораздо более сильной иммуногенности, связанной с пептидной иммуногенной конструкцией (SEQ ID NO: 88), также показало гораздо более сильную активность ADCC и апоптоза в экспрессирующих IgE лимфоцитах по сравнению с пептидной иммуногенной конструкцией EMPD IgE (SEQ ID NO: 89).
Что касается иммуногенной конструкции EMPD IgE 7-40 (SEQ ID NO: 93, с двумя UBTh1, расположенными на N- и С-концах пептида В-клеточного эпитопа EMPD IgE), индуцированная ей антисыворотка распознала две основные антигенные области, сходные с распознаваемыми EMPD IgE 1-39 в сильном реагировании с пептидами в области EMPD IgE 18-33, покрытой пептидами SEQ ID NO: 35-41; EMPD IgE 29-43 (SEQ ID NO: 45-51). Иммуногенная конструкция EMPD IgE 7-40 (SEQ ID NO: 93) имеет антигенные детерминантные области, сходные с таковыми у иммуногенной конструкции EMPD IgE 1-39 (SEQ ID NO: 88, UBTh1 в С-конце пептида В-клеточного эпитопа EMPD IgE). Наибольшую эффективность среди всех разработанных конструкций пептидного иммуногена показала EMPD IgE 1-39 (SEQ ID NO: 88), что подтверждено несколькими функциональными анализами, которые коррелировали с результатами, представленными картированием эпитопа на малом участке, в том, что данная иммуногенная конструкция вырабатывала высокосвязывающие поликлональные антитела, распознающие обширную поверхность, покрытую тремя или более пептидами В-клеточного эпитопа на белке внеклеточной мембраны EMPD IgE. Область эпитопа EMPD IgE 30-43 (SEQ ID NO: 47-51) представляет собой очень важную область В-клеточного эпитопа, расположенную в С-концевой области петлевой структуры с близостью к базальной мембране В-клеток, несущих IgE, восприимчивой к опосредованному антителами апоптозу и ADCC. Кроме того, циклизованная петлевая структура представляет собой иммуногенную конструкцию лучшего качества по сравнению с ее нециклическим аналогом (Таблица 5, сравнение SEQ ID NO: 88 с SEQ ID NO: 88, нециклизован).
Таким образом, сконструированные синтетические пептидные иммуногенные конструкции EMPD IgE (EMPD IgE 1-39, SEQ ID NO: 88) и (EMPD IgE 7-40, SEQ ID NO: 93), обе с пептидом В-клеточного эпитопа, представленного петлевой структурой внутри EMPD IgE, связанного с пептидом эпитопа UBITh®1, который индуцировал устойчивый иммунный ответ, генерирующий поликлональные антитела, нацеленные на новую эпитопную область (аминокислоты 29-43) на белке EMPD IgE, имеющем непосредственную близость к клеточной мембране из-за его расположения в С-концевой области структуры центральной петли, позволяют сшивать как можно больше мембранного IgE для индуцирования ADCC и апоптоза для истощения IgE-экспрессирующих B-лимфоцитов (таблица 5).
ПРИМЕР 7
БЫЛО ОБНАРУЖЕНО, ЧТО СФОКУСИРОВАННЫЙ ГУМОРАЛЬНЫЙ ОТВЕТ НАПРАВЛЕН ИСКЛЮЧИТЕЛЬНО НА КЛЕТОЧНЫЙ ЭПИТОП ПЕПТИДНОЙ ИММУНОГЕННОЙ КОНСТРУКЦИИ IgE EMPD
Хорошо известно, что все белки-носители (например, Keyhole Limpet Hemocyanin (KLH) или другие белки-носители, такие как белки дифтерийного анатоксина (DT) и столбнячного анатоксина (TT)), используемые для усиления иммунного ответа, направленного против целевого пептида В-клеточного эпитопа путем химического конъюгирования такого пептида В-клеточного эпитопа с соответствующим белком-носителем, будет вызывать образование более 90% антител, направленных против потенцирующего белка-носителя, и менее 10% антител, направленных против целевого эпитопа В-клеток в иммунизированных хозяевах.
Следовательно, представляет интерес оценка специфичности антител, образуемых в ответ на пептидные иммуногенные конструкции EMPD IgE по настоящему изобретению. Для оценки иммуногенности была изготовлена репрезентативная пептидная иммуногенная конструкция EMPD IgE (SEQ ID NO: 94) с В-клеточным эпитопом (SEQ ID NO: 5), связанная через спейсерную последовательность εK-KKK (SEQ ID NO: 129) с гетерологичным Т-клеточным эпитопом UBITh®1 (SEQ ID NO: 72). На планшеты наносили UBITh®1 (T-хелперный пептид, используемый для иммунопотенцирования B-эпитопа), и для испытания на перекрестную реактивность с пептидом UBITh®1, используемым для иммунопотенцирования, использовали сыворотку иммунизированных морских свинок. В таблице 7 показано, что антитела, полученные из иммуногенной пептидной конструкции EMPD IgE (SEQ ID NO: 94), обладали высокой иммуногенностью по отношению к соответствующему целевому B-эпитопу EMPD IgE (SEQ ID NO: 5); при этом было обнаружено, что большинство (если не все) иммунных сывороток не реагируют на пептид UBITh®1 (SEQ ID NO: 72).
Таким образом, конструкция иммуногена, содержащая целевой В-клеточный эпитоп, связанный с тщательно отобранным Т-хелперным эпитопом, приводит к генерации сфокусированного и чистого иммунного ответа, который вызывает образование антител, нацеленных только на В-клеточный эпитоп EMPD IgE, а не на Th-эпитоп, используемый для усиления иммунного ответа. Что касается разработки фармацевтической композиции, то чем более специфичен иммунный ответ, вызываемый иммуногеном, тем выше профиль безопасности, который он обеспечивает для композиции. Таким образом, пептидная иммуногенная конструкция EMPD IgE по настоящему изобретению является высокоспецифичной и в то же время высокоэффективной в отношении ее мишени.
ПРИМЕР 8
ВЛИЯНИЕ ИММУНОТЕРАПЕВТИЧЕСКОЙ ВАКЦИНЫ ОТ АЛЛЕРГИИ В ПРОФИЛАКТИЧЕСКИХ И ТЕРАПЕВТИЧЕСКИХ МОДЕЛЯХ НА УРОВНИ IgE В СЫВОРОТКЕ У ГЕНЕТИЧЕСКИ МОДИФИЦИРОВАННЫХ МЫШЕЙ С НОКИНОМ
Пептидные иммуногенные конструкции EMPD IgE SEQ ID NO: 88 и SEQ ID NO: 93 были отобраны для оценки выработки IgE в первичном ответе и вторичном ответе/анамнестической реакции в доказательстве концептуального исследования с использованием генетически модифицированных мышей, экспрессирующих IgE человека, потомства F1 мышей с нокином Balb/c x HuIGHE.
Первоначально иммунизации прайм-бустом пептидного иммуногена EMPD IgE проводили перед сенсибилизацией аллергеном (например, папаином). Восемь животных (n=8) были отнесены к каждой из шести групп лечения и одной группе плацебо, всего 7 групп. Для каждого из двух пептидных иммуногенов были назначены три подгруппы с различными дозами, получавшие 9 (низкий уровень), 18 (средний уровень) и 40 (высокий уровень) мкг/дозу внутримышечно на 0, 3 и 5 неделях. Были составлены пептидные иммуногены с адъювантами ISA 51VG и CpG для усиления иммунных ответов. Мышам в группе плацебо внутримышечно вводили только носитель состава раствора на 0, 3 и 5 неделях. На 10 и 16 неделях все подгруппы животных, включая группу плацебо, сенсибилизировали 50 мкг папаина с адъювантом TiterMax Gold подкожным путем (Фиг.10).
Образование антител (титры IgG против гибридного белка γ1-εm67) определяли анализом ИФА до двум пептидными иммуногенами, как описано в Примере 2. У всех мышей развивались устойчивые ответы антител в шести получавших препарат группах, иммунизированных различными дозами на 0, 3. 5 неделях. Данные ИФА показывают, что образцы сыворотки всех экспериментальных групп с высокими титрами антител специфически связываются с рекомбинантным гибридным белком γ1-εm67 начиная на 3 неделе, и демонстрируют такие же высокие титры до 20 недели (фигура 11). Напротив, мыши в группе плацебо не генерировали специфических антител против рекомбинантного гибридного белка γ1-εm67. Данные результаты показывают, что все получавшие препарат группы генерировали антитела к EMPD IgE с потенциальной способностью нацеливаться на IgE-экспрессирующие В-лимфоциты, что приводит к ингибированию выработки IgE. Фигура 11 иллюстрирует, что высокие титры антител сохранялись на протяжении всего периода исследования 20 недель. Эти результаты также указывают на то, что пептидные иммуногены (SEQ ID NO: 88 и SEQ ID NO: 93) являются сильно иммуногенными в отношении индуцирования специфического иммунного ответа, генерирующего высокие титры антител к EMPD IgE даже в группе с низкой дозой (9 мкг/доза) каждый из двух вакцинных иммуногенов.
Эффекты иммунизации на выработку IgE в первичных и вторичных ответах у иммунизированных мышей были исследованы путем измерения базовых уровней IgE в сыворотке и аллерген-специфических уровней IgE с использованием процедур анализа, описанных в примере 2. Базовые уровни IgE в сыворотке крови до и после вакцинации показаны на фигуре 12, где явно продемонстрировано, что базальные уровни IgE в сыворотке мыши во всех получавших препарат группах постепенно снижались по сравнению с соответствующим моментом времени в группе плацебо. На 10 неделе, до сенсибилизации, базальные уровни IgE снизились до самого низкого уровня во всех шести получавших препарат группах, в то время как базовые сывороточные уровни IgE в группе плацебо существенно не изменились. Этот результат указывает на то, что на 10 неделе базовая выработка IgE во всех получавших препарат группах было значительно подавлена у животных, получавших вакцинный состав, содержащий пептидную иммуногенную конструкцию EMPD IgE (либо SEQ ID NO: 88, либо 93), по сравнению с группой плацебо. Фигура 12 также показывает, что базовые уровни IgE в сыворотке во всех получавших препарат группах подавлялись в течение 20-недельного периода исследования, даже после двух аллергенных сенсибилизаций на 10 и 16 неделях.
Аллерген-специфическая выработка IgE, индуцированная папаином после первичной сенсибилизации на неделе 12 и вторичной сенсибилизации на неделе 18 показана на фигурах 13 и 14, соответственно. Эти результаты показывают, что обе пептидные иммуногенные конструкции (SEQ ID NO: 88 и 93) были способны значительно подавлять специфичные для папаина уровни IgE после первичной и вторичной сенсибилизации аллергеном по сравнению с группой плацебо. Не было обнаружено существенных различий между тремя уровнями доз ни в одном из двух пептидных иммуногенов. В этом исследовании после генерации аллерген-специфического IgE при первичном ответе на неделе 12 пептидный иммуноген (SEQ ID NO: 88) показывал немного большую эффективность по сравнению с (SEQ ID NO: 93), как показано на Фиг. 13. Обе пептидные иммуногенныех конструкции EMPD IgE показали значительное подавление аллерген-специфической выработки IgE по сравнению с группой плацебо как в первичном ответе, так и во вторичном ответе (Фиг. 13 и 14).
Для дальнейшего изучения потенциального терапевтического эффекта иммуногенных пептидных конструкций EMPD IgE (SEQ ID NO: 88 и SEQ ID NO: 93), направленных на экспрессирующую IgE B-клетку для подавления выработки IgE и лечения IgE-опосредованного аллергического заболевания, был разработан дополнительный протокол, предназначенный для оценки влияния данных двух пептидных иммуногенных конструкций на модели сенсибилизации/вторичного иммунного ответа у мышей с нокином HuIGHE. В каждую из двух групп лечения было определено по 6 животных (N=6), а в группу плацебо назначили четырех животных (N=4), в общей сложности три группы. Мыши во всех группах получали сенсибилизацию дважды, до и после пептидной вакцинации, соответственно, 50 мкг папаина/адъюванта TiterMax Gold подкожным путем на неделе 0 и в подушечки лап?ы на неделе 12. Режим прайм-буст иммунизации 40 мкг/0,1 мл состава, содержащего одну из двух пептидных иммуногенных конструкций, на неделях 3, 6 и 8 оценивали в обеих получавших препарат группах, причем группе плацебо вводили только адъювантный носитель (см.Фиг. 15).
Результаты показали, что все группы, включая плацебо, сенсибилизированные папаином, вызывали образование высоких титров папаин-специфических IgG во всех трех группах после недели 2 и титры оставались высокими до недели 6 (последний наблюдаемый период времени). Общий сывороточный IgE и папаин-специфические уровни IgE достигли наивысших уровней на неделе 2, а затем постепенно снижались, начиная с 3 недели, и возвращались к исходному уровню на 6 неделе (Фиг. 16).
Вакцинация с использованием иммуногена пептида EMPD IgE приводит к выработке высоких титров антител, которые специфически распознают гибридный белок γ1-εm67. После трех инъекций пептидных иммуногенов на 3, 6 и 8 неделях титры выработанных антител IgG (антител к γ1-εm67 или к EMPD IgE) постоянно повышались и достигли наивысшего уровня на 10 неделе (данные не показаны). На 12 неделе после второй сенсибилизации папаином ни в одной из двух получавших препарат групп не наблюдалось повышенных уровней IgE, специфичных для папаина; однако специфичный для папаина уровень IgE заметно повысился в группе плацебо на 12 и 13 неделях, а затем снизился до более низкого уровня на 14 неделе (данные не показаны).
Результаты исследования показывают, что пептидные иммуногены EMPD IgE SEQ ID NO: 88 и 93 способны индуцировать специфический гуморальный иммунный ответ для предотвращения повторного пролиферативного ответа В-клеток памяти in vivo, который полностью блокировал специфическую для папаина выработку IgE, вызванную повторным введением папаина на 12 неделе по сравнению с группой плацебо (Фиг. 17). В целом, данное исследование указывает на то, что раскрытые пептидные иммуногены EMPD IgE индуцируют образование антител, которые не только ингибируют выработку аллерген-специфического сывороточного IgE при первичной сенсибилизации, но также устойчиво подавляют выработанный аллерген-специфический сывороточный IgE при вторичной аллергенной стимуляции. Такой результат исследования демонстрирует, что раскрытое изобретение обеспечивает потенциальную эффективную терапевтическую вакцину для лечения опосредованных IgE аллергических заболеваний, таких как астма. После тщательного изучения терапевтической эффективности ослабления аллерген-специфического IgE был сделан вывод, что оба пептидных иммуногена (SEQ ID NO: 88 и SEQ ID NO: 93) демонстрируют сходную эффективность в ингибировании аллерген-специфической выработки IgE.
ПРИМЕР 9
ИССЛЕДОВАНИЯ ДОЗИРОВАНИЯ И СОСТАВА ПОСРЕДСТВОМ ОЦЕНКИ ИММУНОГЕННОСТИ ПРОТОТИПА СОСТАВОВ ИММУНОТЕРАПЕВТИЧЕСКОЙ ВАКЦИНЫ ОТ АЛЛЕРГИИ У ЯВАНСКИХ МАКАКОВ
а. Конечная цель:
Цель этого исследования состояла в оценке влияния на иммуногенность внутримышечной иммунизации выбранной пептидной иммуногенной конструкцией EMPD IgE SEQ ID NO: 88 в течение 20-недельного периода. В качестве животной модели для оценки иммуногенности прототипа вакцинного состава EMPD IgE и схема применения до проведения исследований на людях были выбраны яванские макаки. Репрезентативная пептидная иммуногенная конструкция (SEQ ID NO: 88) была составлена в виде двух широко используемых составов. В первом составе пептидная иммуногенная конструкция EMPD IgE SEQ ID NO: 88 была превращена в стабилизированный иммуностимулирующий комплекс с CpG перед изготовлением эмульсии вода-в-масле в смеси с Montanide™ ISA51 (часть A исследования). Во втором составе пептидная иммуногенная конструкция EMPD IgE SEQ ID NO: 88 была превращена в стабилизированный иммуностимулирующий комплекс с CpG перед изготовлением состава в виде суспензии с ADJUPHOS (часть B исследования). В данном всеобъемлющем исследовании иммуногенности были оценены четыре дозировки каждого из составов от 30 мкг, 100 мкг, 300 мкг до 1000 мкг на дозу.
b. Краткое содержание протокола
Для оценки влияния пептидных иммуногенов EMPD IgE на иммуногенность и уровень IgE в сыворотке макак были отобраны взрослые яванские макаки массой 2,5-4,0 кг. Всего 20 макак были разделены на 10 групп: животным контрольной группы, получающим плацебо (n=2) вводили только адъювант (Montanide™ ISA 51 плюс олигодезоксинуклеотид CpG) или (ADJUPHOS плюс олигодезоксинуклеотид CpG). Экспериментальным животным вводили пептидный иммуноген EMPD IgE (SEQ ID NO: 88) в дозах 30, 100, 300 и 1000 мкг на группу (всего 500 мкл вакцины на животное; n=2 на группу, 1 самец и 1 самка). Всего было проведено три внутримышечные иммунизации на 0, 3 и 6 неделях. У всех макаков контролировали иммуногенность и уровень IgE в сыворотке на 0, 3, 6, 8, 10, 12, 14, 16, 20 и 24 неделях.
с. Определение титров антител к EMPD IgE
У всех животных отбирали кровь на 0, 3, 6, 8, 10, 12, 14, 16, 20 и 24 неделях. При каждом взятии крови от нее отделяли сыворотку для определения титра антител к EMPD IgE с использованием ИФА γ1-cyno em67. У животных, получавших плацебо, титры антител к EMPD IgE практически не определялись (Фиг. 18A и 18B). Однако все животные, получившие три иммунизации, показали наличие детектируемых титров антител IgG к эпитопам B-клеток EMPD IgE с пиковыми титрами, полученными на недели с 8 по 12. Такая специфическая реактивность поддерживалась в течение всего 24-недельного исследования дозозависимым образом (Фиг. 18А и 18В). Все животные показали высокие антисывороточные титры прости рекомбинантного белка γ1-cyno εm67, и наиболее оптимальной дозой для обоих составов оказалась доза 300 мкг в 0,5 мл на дозу, поскольку доза композиции 1000 мкг приведет к избытку иммуногенного пептида по сравнению с адъювантами ISA51V или ADJUPHOS в соответствующих составах. Результаты показывают, что составы, содержащие ISA51/CpG ODN, имели более высокие результаты при оценке иммуногенности по сравнению с составами с ADJUPHOS/CpG ODN. Образование антител на В-клеточный эпитоп EMPD IgE усиливалось вместе с каждым бустом иммуногенного пептида, и затем титры антител к EMPD IgE постепенно снижались с течением времени. Значительно высокие антисывороточные титры сохранялись в течение 24 недель исследования (Фиг. 18А и 18В).
ПРИМЕР 10
ДОКАЗАТЕЛЬСТВО ЭФФЕКТИВНОСТИ С СУРРОГАТНОЙ ИММУНОТЕРАПЕВТИЧЕСКОЙ ВАКЦИНОЙ ОТ АЛЛЕРГИИ У ЯВАНСКИХ МАКАКОВ
а. Конечная цель:
Цель данного исследования состояла в оценке влияния внутримышечной иммунизации иммуногенными пептидами EMPD IgE, последовательность которых была получена из собственных mIgE, в течение 20-недельного периода на иммуногенность и уровень IgE в сыворотке у яванских макаков, которые являются животной моделью с выработкой IgE, схожей с такой выработкой у человека. EMPD IgE mIgE у приматов, не являющихся человеком (например, у мартышкообразных, капуцинообразных и человекоподобных обезьян), эволюционно консервативен и подобные последовательности EMPD IgE не были обнаружены у других видов (например, грызунов, кроликов и собак). Аминокислотная последовательность EMPD IgE яванского макака (SEQ ID NO: 127) имеет высокую идентичность последовательности (90%) с последовательностью EMPD IgE человека (SEQ ID NO: 2).
b. Краткое содержание протокола
Для оценки влияния пептидных иммуногенов EMPD IgE на иммуногенность и уровень IgE в сыворотке макак были отобраны взрослые яванские макаки массой 2,5-4,0 кг. Всего 12 макак были разделены на 3 группы: животным контрольной группы, получавшим плацебо (n=4, 2 самца и 2 самки), вводили только адъювант (Montanide™ ISA 51 плюс олигодезоксинуклеотид CpG); экспериментальным животным вводили иммуногенные пептиды EMPD IgE (SEQ ID NO: 125 или 126) в дозе 300 мкг (общий объем вакцины - 500 мкл на животное; n=4 на группу, 2 самца и 2 самки). Всего было проведено три внутримышечных иммунизации на 0, 3 и 6 неделях. У всех макаков контролировали иммуногенность и уровень IgE в сыворотке на 0, 3, 6, 8, 10, 12, 14, 16, 18 и 20 неделях.
с. Определение титров антител к EMPD IgE
У всех животных отбирали кровь на 0, 3, 6, 8, 10, 12, 14, 16, 18 и 20 неделях. При каждом взятии крови от нее отделяли сыворотку для определения титра антител к EMPD IgE с использованием ИФА γ1-cyno em67. У животных, получавших плацебо, титры антител к EMPD IgE не определялись (Фиг. 19). Однако все животные, получившие три иммунизации SEQ ID NO: 125 или SEQ ID NO: 126, показали наличие детектируемых титров антител IgG к эпитопам B-клеток EMPD IgE с пиковыми титрами, полученными на недели с 8 по 12 (Фиг. 19). Такая специфическая реактивность сохранялась в течение 20 недель исследования. Кроме того, у всех иммунизированных животных развились специфические титры антител IgM и IgA в течение 20-недельного периода (Фиг. 20).
d. Измерение уровня сывороточного IgE
У всех животных отбирали кровь на 0, 3, 6, 8, 10, 12, 14, 16, 18 и 20 неделях. В каждом образце крови отделяли сыворотку для измерения сывороточного IgE с использованием количественного ИФА IgE у макаки. Базовые прижизненные уровни IgE в группе плацебо были изменчивыми. Наблюдаемое снижение IgE в группе, которой вводили SEQ ID NO: 125, было статистически значимым, в то время как в группе, которой вводили SEQ ID NO: 126, также наблюдалась тенденция снижения IgE в течение наблюдаемого периода (Фиг. 19).
e. Результаты
Было оценено влияние пептидных иммуногенов EMPD IgE в суррогатной модели для оценки иммуногенности против собственных mIgE и концентрации IgE в сыворотке в образцах сыворотки взрослых яванских макаков. В данном исследовании обоснованности концепции четырем животным в каждой группе вводили смесь CpG ODN и Montanide™ ISA 51 в качестве плацебо-контроля и 8 взрослых яванских макаков (n=4 на группу) иммунизировали на 0, 3 и 6 неделях дозой 300 мкг иммуногенных пептидов макак EMPD IgE: SEQ ID NO: 125 или 126, в комплексе с запатентованными иммуностимулирующими комплексами (ISC) с олигодезоксинуклеотидами CpG (CpG ODN) и в составе с адъювантом Montanide™ ISA 51. Последовательности SEQ ID NO: 125 и 126 являются аналогами иммуногенов EMPD IgE человека, SEQ ID NO: 93 и 88 соответственно. Два полученных из макака иммуногена с составом ОДН CpG/Montanide™ ISA 51 приводили к значительному образованию антител к EMPD IgE-IgG у всех животных (Фиг. 19). Кроме того, у всех животных выработались антитела IgM и IgA против EMPD IgE (Фиг. 20). Наблюдалась также тенденция к снижению уровня IgE в базовой сыворотке макак (Фиг. 21). Не было отмечено побочных реакций в месте инъекции. Исследование продемонстрировало, что синтетические иммуногенные пептиды EMPD IgE (SEQ ID NO: 125 и 126), последовательность которых получена из собственных последовательностей, с усиленным эпитопом T-клеток UBITh®, способны вызывать значительное образование антител к EMPD IgE, приводящее к подавлению выработки IgE и показывающие тенденции к снижению базового уровня IgE у макаков.
f. Выводы
Яванским макакам в течение 20 недель 3 раза внутримышечно вводили иммуногенные пептиды EMPD IgE (SEQ ID NO: 125 и 126) или плацебо-контроль. Животные показывали хорошую общую переносимость и проявляли иммунную толерантность. Все иммунизированные макаки развивали транзиторные специфические IgM-антитела вместе с развитием сильных и устойчивых титров антител IgG (до 105) и IgA (до 104) к соответствующему В-клеточному эпитопу конструкций EMPD IgE. У всех субъектов, ответивших на лечение, наблюдались пониженные уровни базального IgE. Эти результаты подтверждают механизм действия антитела к EMPD IgE, в котором антитела нацелены на В-клетки, экспрессирующие мембраносвязанный IgE, что приводит к подавлению выработки IgE в дальнейшем.
Таблица 1
Пептид EMPD IgE и его фрагменты, используемые в серологических анализах
Таблица 1 (продолжение)
ID
NO:
Таблица 2
Выбранные универсальные T-хелперные эпитопы для использования в конструировании пептидных иммуногенных конструкций, полученных из EMPD IgE
NO:
T RT TR T
RRRIKII RII I L IR
VRVV VV V I V
F FF FF F V F
F
T RT TR
FFLL L ITTI
Таблица 3
Повышение иммуногенности пептидных фрагментов EMPD IgE посредством полученных из белка патогена Th-эпитопов, включая универсальный искусственный Th-эпитоп, для индукции образования специфических антител в разработке иммуногенных пептидных конструкций EMPD IgE
NO:
T RT TR T
RRRIKII RII I L IR
VRVV VV V I V
F FF FF F V F
F
T RT TR
FFLL L ITTI
Таблица 3 (продолжение)
NO:
*Пептиды циклизуются цистеиновыми дисульфидными связями, цистеины подчеркнуты.
Таблица 4
Оценка иммуногенности иммуногенных пептидных конструкций, полученных из EMPD IgE, на морских свинках
конструкции EMPD IgE, собранной на разные нпи
IgEEMPD (1-39)
IgEEMPD (1-39)-εK-UBITh1
IgEEMPD (7-40)
IgEEMPD (7-40)-εK-UBITh1
a Группа 1-6: вакцинированы на 0, 2, 4, 6, 8 и 10 нпи; взятие крови на 0, 4, 6, 8, 10, 12 и 14 нпи
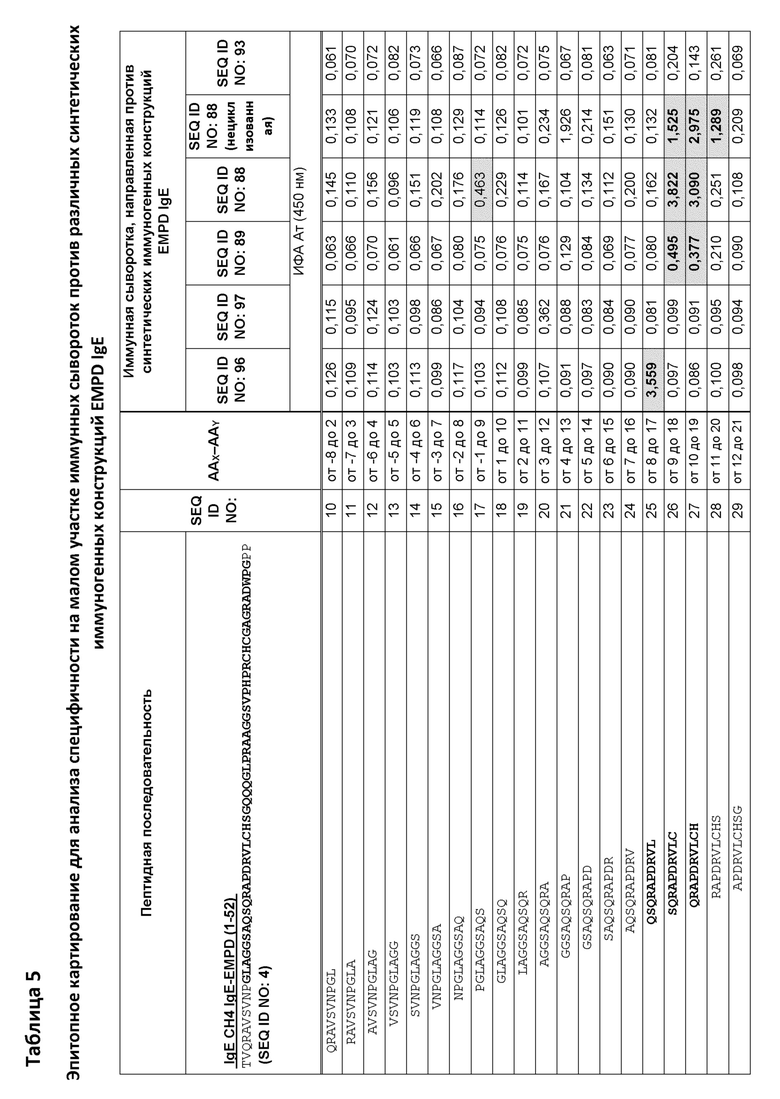
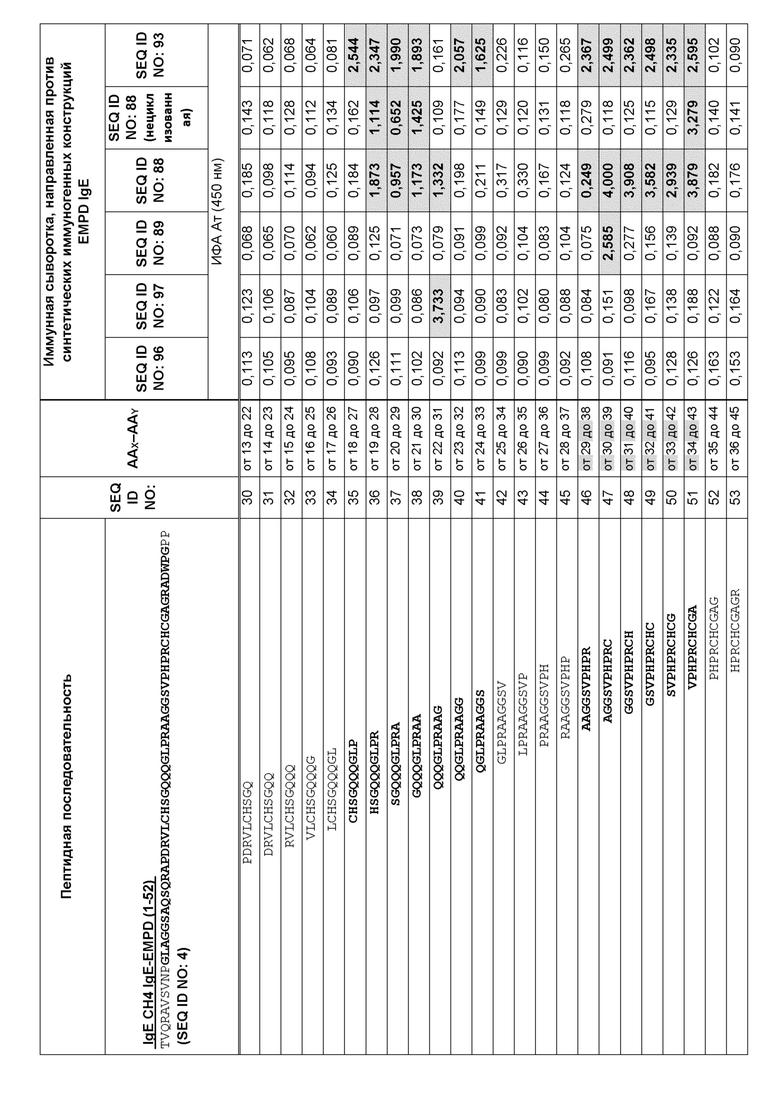
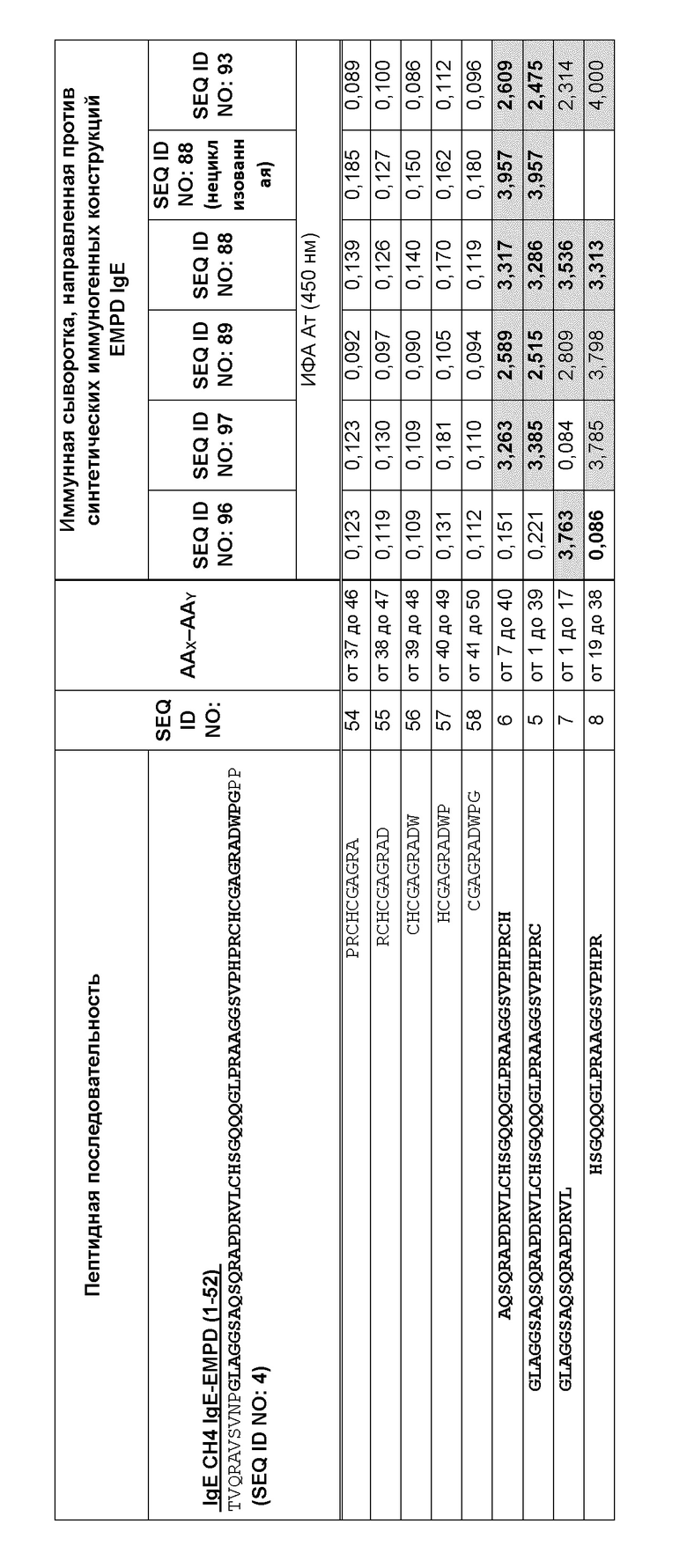
Таблица 6
Ранжирование усиливающей способности В-клеточного эпитопа EMPD IgE в отношении полученных из патогена Th-эпитопов посредством оценки иммуногенности у морских свинок
ИФА титр Log10
ИФА титр Log10
Таблица 6 (продолжение)
ИФА титр Log10
ИФА титр Log10
Таблица 7
Оценка иммуногенности иммунных сывороток морской свинки, направленной против Th-пептида репрезентативной пептидной иммуногенной конструкции EMPD IgE (SEQ ID NO: 94) при примировании (0 нпи) и повторном бусте (3 и 6 нпи)
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> United Biomedical, Inc.
Wang, Chang Yi
Lin, Feng
Chen, Jun Bo
<120> ПЕПТИДНЫЕ ИММУНОГЕНЫ И СОДЕРЖАЩИЕ ИХ СОСТАВЫ, НАЦЕЛЕННЫЕ НА
МЕМБРАНОСВЯЗАННЫЕ IgE ДЛЯ ЛЕЧЕНИЯ ОПОСРЕДОВАННЫХ IgE АЛЛЕРГИЧЕСКИХ ЗАБОЛЕВАНИЙ
<130> 2038-WO
<140> TBD
<141> 31.12.2017
<160> 130
<170> PatentIn версия 3.5
<210> 1
<211> 67
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(67)
<223> IgE-EMPD 1-67
<400> 1
Gly Leu Ala Gly Gly Ser Ala Gln Ser Gln Arg Ala Pro Asp Arg Val
1 5 10 15
Leu Cys His Ser Gly Gln Gln Gln Gly Leu Pro Arg Ala Ala Gly Gly
20 25 30
Ser Val Pro His Pro Arg Cys His Cys Gly Ala Gly Arg Ala Asp Trp
35 40 45
Pro Gly Pro Pro Glu Leu Asp Val Cys Val Glu Glu Ala Glu Gly Glu
50 55 60
Ala Pro Trp
65
<210> 2
<211> 52
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(52)
<223> IgE-EMPD 1-52
<400> 2
Gly Leu Ala Gly Gly Ser Ala Gln Ser Gln Arg Ala Pro Asp Arg Val
1 5 10 15
Leu Cys His Ser Gly Gln Gln Gln Gly Leu Pro Arg Ala Ala Gly Gly
20 25 30
Ser Val Pro His Pro Arg Cys His Cys Gly Ala Gly Arg Ala Asp Trp
35 40 45
Pro Gly Pro Pro
50
<210> 3
<211> 77
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(76)
<223> IgE-EMPD -10-66
<400> 3
Thr Val Gln Arg Ala Val Ser Val Asn Pro Gly Leu Ala Gly Gly Ser
1 5 10 15
Ala Gln Ser Gln Arg Ala Pro Asp Arg Val Leu Cys His Ser Gly Gln
20 25 30
Gln Gln Gly Leu Pro Arg Ala Ala Gly Gly Ser Val Pro His Pro Arg
35 40 45
Cys His Cys Gly Ala Gly Arg Ala Asp Trp Pro Gly Pro Pro Glu Leu
50 55 60
Asp Val Cys Val Glu Glu Ala Glu Gly Glu Ala Pro Trp
65 70 75
<210> 4
<211> 62
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(62)
<223> IgE-EMPD -10-52
<400> 4
Thr Val Gln Arg Ala Val Ser Val Asn Pro Gly Leu Ala Gly Gly Ser
1 5 10 15
Ala Gln Ser Gln Arg Ala Pro Asp Arg Val Leu Cys His Ser Gly Gln
20 25 30
Gln Gln Gly Leu Pro Arg Ala Ala Gly Gly Ser Val Pro His Pro Arg
35 40 45
Cys His Cys Gly Ala Gly Arg Ala Asp Trp Pro Gly Pro Pro
50 55 60
<210> 5
<211> 39
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(39)
<223> IgE-EMPD 1-39
<400> 5
Gly Leu Ala Gly Gly Ser Ala Gln Ser Gln Arg Ala Pro Asp Arg Val
1 5 10 15
Leu Cys His Ser Gly Gln Gln Gln Gly Leu Pro Arg Ala Ala Gly Gly
20 25 30
Ser Val Pro His Pro Arg Cys
35
<210> 6
<211> 34
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(34)
<223> IgE-EMPD 7-40
<400> 6
Ala Gln Ser Gln Arg Ala Pro Asp Arg Val Leu Cys His Ser Gly Gln
1 5 10 15
Gln Gln Gly Leu Pro Arg Ala Ala Gly Gly Ser Val Pro His Pro Arg
20 25 30
Cys His
<210> 7
<211> 17
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(17)
<223> IgE-EMPD 1-17
<400> 7
Gly Leu Ala Gly Gly Ser Ala Gln Ser Gln Arg Ala Pro Asp Arg Val
1 5 10 15
Leu
<210> 8
<211> 20
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(20)
<223> IgE-EMPD 19-38
<400> 8
His Ser Gly Gln Gln Gln Gly Leu Pro Arg Ala Ala Gly Gly Ser Val
1 5 10 15
Pro His Pro Arg
20
<210> 9
<211> 10
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(10)
<223> IgE-EMPD -9-1
<400> 9
Val Gln Arg Ala Val Ser Val Asn Pro Gly
1 5 10
<210> 10
<211> 10
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(10)
<223> IgE-EMPD -8-2
<400> 10
Gln Arg Ala Val Ser Val Asn Pro Gly Leu
1 5 10
<210> 11
<211> 10
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(10)
<223> IgE-EMPD -7-3
<400> 11
Arg Ala Val Ser Val Asn Pro Gly Leu Ala
1 5 10
<210> 12
<211> 10
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(10)
<223> IgE-EMPD -6-4
<400> 12
Ala Val Ser Val Asn Pro Gly Leu Ala Gly
1 5 10
<210> 13
<211> 10
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(10)
<223> IgEEMPD -5-5
<400> 13
Val Ser Val Asn Pro Gly Leu Ala Gly Gly
1 5 10
<210> 14
<211> 10
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(10)
<223> IgE-EMPD -4-6
<400> 14
Ser Val Asn Pro Gly Leu Ala Gly Gly Ser
1 5 10
<210> 15
<211> 10
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(10)
<223> IgE-EMPD -3-7
<400> 15
Val Asn Pro Gly Leu Ala Gly Gly Ser Ala
1 5 10
<210> 16
<211> 10
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(10)
<223> IgE-EMPD -2-8
<400> 16
Asn Pro Gly Leu Ala Gly Gly Ser Ala Gln
1 5 10
<210> 17
<211> 10
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(10)
<223> IgE-EMPD -1-9
<400> 17
Pro Gly Leu Ala Gly Gly Ser Ala Gln Ser
1 5 10
<210> 18
<211> 10
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(10)
<223> IgE-EMPD 1-10
<400> 18
Gly Leu Ala Gly Gly Ser Ala Gln Ser Gln
1 5 10
<210> 19
<211> 10
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(10)
<223> IgE-EMPD 2-11
<400> 19
Leu Ala Gly Gly Ser Ala Gln Ser Gln Arg
1 5 10
<210> 20
<211> 10
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(10)
<223> IgE-EMPD 3-12
<400> 20
Ala Gly Gly Ser Ala Gln Ser Gln Arg Ala
1 5 10
<210> 21
<211> 10
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(10)
<223> IgE-EMPD 4-13
<400> 21
Gly Gly Ser Ala Gln Ser Gln Arg Ala Pro
1 5 10
<210> 22
<211> 10
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(10)
<223> IgE-EMPD 5-14
<400> 22
Gly Ser Ala Gln Ser Gln Arg Ala Pro Asp
1 5 10
<210> 23
<211> 10
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(10)
<223> IgE-EMPD 6-15
<400> 23
Ser Ala Gln Ser Gln Arg Ala Pro Asp Arg
1 5 10
<210> 24
<211> 10
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(10)
<223> IgE-EMPD 7-16
<400> 24
Ala Gln Ser Gln Arg Ala Pro Asp Arg Val
1 5 10
<210> 25
<211> 10
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(10)
<223> IgE-EMPD 8-17
<400> 25
Gln Ser Gln Arg Ala Pro Asp Arg Val Leu
1 5 10
<210> 26
<211> 10
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(10)
<223> IgE-EMPD 9-18
<400> 26
Ser Gln Arg Ala Pro Asp Arg Val Leu Cys
1 5 10
<210> 27
<211> 10
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(10)
<223> IgEEMPD 10-19
<400> 27
Gln Arg Ala Pro Asp Arg Val Leu Cys His
1 5 10
<210> 28
<211> 10
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(10)
<223> IgE-EMPD 11-20
<400> 28
Arg Ala Pro Asp Arg Val Leu Cys His Ser
1 5 10
<210> 29
<211> 10
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(10)
<223> IgE-EMPD 12-21
<400> 29
Ala Pro Asp Arg Val Leu Cys His Ser Gly
1 5 10
<210> 30
<211> 10
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(10)
<223> IgE-EMPD 13-22
<400> 30
Pro Asp Arg Val Leu Cys His Ser Gly Gln
1 5 10
<210> 31
<211> 10
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(10)
<223> IgE-EMPD 14-23
<400> 31
Asp Arg Val Leu Cys His Ser Gly Gln Gln
1 5 10
<210> 32
<211> 10
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(10)
<223> IgE-EMPD 15-24
<400> 32
Arg Val Leu Cys His Ser Gly Gln Gln Gln
1 5 10
<210> 33
<211> 10
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(10)
<223> IgE-EMPD 16-25
<400> 33
Val Leu Cys His Ser Gly Gln Gln Gln Gly
1 5 10
<210> 34
<211> 10
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(10)
<223> IgEEMPD 17-26
<400> 34
Leu Cys His Ser Gly Gln Gln Gln Gly Leu
1 5 10
<210> 35
<211> 10
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(10)
<223> IgE-EMPD 18-27
<400> 35
Cys His Ser Gly Gln Gln Gln Gly Leu Pro
1 5 10
<210> 36
<211> 10
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(10)
<223> IgE-EMPD 19-28
<400> 36
His Ser Gly Gln Gln Gln Gly Leu Pro Arg
1 5 10
<210> 37
<211> 10
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(10)
<223> IgE-EMPD 20-29
<400> 37
Ser Gly Gln Gln Gln Gly Leu Pro Arg Ala
1 5 10
<210> 38
<211> 10
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(10)
<223> IgE-EMPD 21-30
<400> 38
Gly Gln Gln Gln Gly Leu Pro Arg Ala Ala
1 5 10
<210> 39
<211> 10
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(10)
<223> IgE-EMPD 22-31
<400> 39
Gln Gln Gln Gly Leu Pro Arg Ala Ala Gly
1 5 10
<210> 40
<211> 10
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(10)
<223> IgE-EMPD 23-32
<400> 40
Gln Gln Gly Leu Pro Arg Ala Ala Gly Gly
1 5 10
<210> 41
<211> 10
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(10)
<223> IgE-EMPD 24-33
<400> 41
Gln Gly Leu Pro Arg Ala Ala Gly Gly Ser
1 5 10
<210> 42
<211> 10
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(10)
<223> IgE-EMPD 25-34
<400> 42
Gly Leu Pro Arg Ala Ala Gly Gly Ser Val
1 5 10
<210> 43
<211> 10
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(10)
<223> IgE-EMPD 26-35
<400> 43
Leu Pro Arg Ala Ala Gly Gly Ser Val Pro
1 5 10
<210> 44
<211> 10
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(10)
<223> IgE-EMPD 27-36
<400> 44
Pro Arg Ala Ala Gly Gly Ser Val Pro His
1 5 10
<210> 45
<211> 10
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(10)
<223> IgEEMPD 28-37
<400> 45
Arg Ala Ala Gly Gly Ser Val Pro His Pro
1 5 10
<210> 46
<211> 10
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(10)
<223> IgE-EMPD 29-38
<400> 46
Ala Ala Gly Gly Ser Val Pro His Pro Arg
1 5 10
<210> 47
<211> 10
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(10)
<223> IgE-EMPD 30-39
<400> 47
Ala Gly Gly Ser Val Pro His Pro Arg Cys
1 5 10
<210> 48
<211> 10
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(10)
<223> IgE-EMPD 31-40
<400> 48
Gly Gly Ser Val Pro His Pro Arg Cys His
1 5 10
<210> 49
<211> 10
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(10)
<223> IgE-EMPD 32-41
<400> 49
Gly Ser Val Pro His Pro Arg Cys His Cys
1 5 10
<210> 50
<211> 10
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(10)
<223> IgE-EMPD 33-42
<400> 50
Ser Val Pro His Pro Arg Cys His Cys Gly
1 5 10
<210> 51
<211> 10
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(10)
<223> IgE-EMPD 34-43
<400> 51
Val Pro His Pro Arg Cys His Cys Gly Ala
1 5 10
<210> 52
<211> 10
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(10)
<223> IgE-EMPD 35-44
<400> 52
Pro His Pro Arg Cys His Cys Gly Ala Gly
1 5 10
<210> 53
<211> 10
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(10)
<223> IgE-EMPD 36-45
<400> 53
His Pro Arg Cys His Cys Gly Ala Gly Arg
1 5 10
<210> 54
<211> 10
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(10)
<223> IgE-EMPD 37-46
<400> 54
Pro Arg Cys His Cys Gly Ala Gly Arg Ala
1 5 10
<210> 55
<211> 10
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(10)
<223> IgE-EMPD 38-47
<400> 55
Arg Cys His Cys Gly Ala Gly Arg Ala Asp
1 5 10
<210> 56
<211> 10
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(10)
<223> IgE-EMPD 39-48
<400> 56
Cys His Cys Gly Ala Gly Arg Ala Asp Trp
1 5 10
<210> 57
<211> 10
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(10)
<223> IgEEMPD 40-49
<400> 57
His Cys Gly Ala Gly Arg Ala Asp Trp Pro
1 5 10
<210> 58
<211> 10
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(10)
<223> IgE-EMPD 41-50
<400> 58
Cys Gly Ala Gly Arg Ala Asp Trp Pro Gly
1 5 10
<210> 59
<211> 17
<212> Белок
<213> Clostridium tetani
<220>
<221> ПЕПТИД
<222> (1)..(17)
<223> Clostridium tetani 1 Th
<400> 59
Lys Lys Gln Tyr Ile Lys Ala Asn Ser Lys Phe Ile Gly Ile Thr Glu
1 5 10 15
Leu
<210> 60
<211> 15
<212> Белок
<213> вирус кори
<220>
<221> ПЕПТИД
<222> (1)..(15)
<223> MvF1 Th
<400> 60
Leu Ser Glu Ile Lys Gly Val Ile Val His Arg Leu Glu Gly Val
1 5 10 15
<210> 61
<211> 24
<212> Белок
<213> Bordetella pertussis
<220>
<221> ПЕПТИД
<222> (1)..(24)
<223> Bordetella pertussis Th
<400> 61
Gly Ala Tyr Ala Arg Cys Pro Asn Gly Thr Arg Ala Leu Thr Val Ala
1 5 10 15
Glu Leu Arg Gly Asn Ala Glu Leu
20
<210> 62
<211> 17
<212> Белок
<213> Clostridium tetani
<220>
<221> ПЕПТИД
<222> (1)..(17)
<223> Clostridium tetani 2 Th
<400> 62
Trp Val Arg Asp Ile Ile Asp Asp Phe Thr Asn Glu Ser Ser Gln Lys
1 5 10 15
Thr
<210> 63
<211> 23
<212> Белок
<213> diphtheria bacilli
<220>
<221> ПЕПТИД
<222> (1)..(23)
<223> Diphtheria Th
<400> 63
Asp Ser Glu Thr Ala Asp Asn Leu Glu Lys Thr Val Ala Ala Leu Ser
1 5 10 15
Ile Leu Pro Gly His Gly Cys
20
<210> 64
<211> 21
<212> Белок
<213> Plasmodium falciparum
<220>
<221> ПЕПТИД
<222> (1)..(21)
<223> Plasmodium falciparum Th
<400> 64
Asp His Glu Lys Lys His Ala Lys Met Glu Lys Ala Ser Ser Val Phe
1 5 10 15
Asn Val Val Asn Ser
20
<210> 65
<211> 17
<212> Белок
<213> Schistosoma mansoni
<220>
<221> ПЕПТИД
<222> (1)..(17)
<223> Schistosoma mansoni Th
<400> 65
Lys Trp Phe Lys Thr Asn Ala Pro Asn Gly Val Asp Glu Lys His Arg
1 5 10 15
His
<210> 66
<211> 25
<212> Белок
<213> холерный токсин
<220>
<221> ПЕПТИД
<222> (1)..(25)
<223> холерный токсин Th
<400> 66
Ala Leu Asn Ile Trp Asp Arg Phe Asp Val Phe Cys Thr Leu Gly Ala
1 5 10 15
Thr Thr Gly Tyr Leu Lys Gly Asn Ser
20 25
<210> 67
<211> 15
<212> Белок
<213> вирус кори
<220>
<221> ПЕПТИД
<222> (1)..(15)
<223> MvF 2 Th
<400> 67
Ile Ser Glu Ile Lys Gly Val Ile Val His Lys Ile Glu Gly Ile
1 5 10 15
<210> 68
<211> 22
<212> Белок
<213> вирус кори
<220>
<221> ПЕПТИД
<222> (1)..(22)
<223> KKKMvF 3 Th
<220>
<221> SITE
<222> (7)..(7)
<223> S или T
<220>
<221> SITE
<222> (10)..(10)
<223> K или R
<220>
<221> SITE
<222> (11)..(11)
<223> G или T
<220>
<221> SITE
<222> (15)..(15)
<223> H или T
<220>
<221> SITE
<222> (16)..(16)
<223> K или R
<220>
<221> SITE
<222> (19)..(19)
<223> G или T
<400> 68
Lys Lys Lys Ile Ser Ile Xaa Glu Ile Xaa Xaa Val Ile Val Xaa Xaa
1 5 10 15
Ile Glu Xaa Ile Leu Phe
20
<210> 69
<211> 18
<212> Белок
<213> вирус гепатита B
<220>
<221> ПЕПТИД
<222> (1)..(18)
<223> HBsAg 1 Th
<220>
<221> SITE
<222> (1)..(1)
<223> K или R
<220>
<221> SITE
<222> (2)..(2)
<223> K или R
<220>
<221> SITE
<222> (3)..(3)
<223> K или R
<220>
<221> SITE
<222> (4)..(4)
<223> L или I или V или F
<220>
<221> SITE
<222> (5)..(5)
<223> F или K или R
<220>
<221> SITE
<222> (6)..(6)
<223> L или I или V или F
<220>
<221> SITE
<222> (7)..(7)
<223> L или I или V или F
<220>
<221> SITE
<222> (9)..(9)
<223> K или R
<220>
<221> SITE
<222> (10)..(10)
<223> L или I или V или F
<220>
<221> SITE
<222> (11)..(11)
<223> L или I или V или F
<220>
<221> SITE
<222> (13)..(13)
<223> L или I или V или F
<220>
<221> SITE
<222> (15)..(15)
<223> Q или L или I или V или F
<220>
<221> SITE
<222> (17)..(17)
<223> L или I или V или F
<220>
<221> SITE
<222> (18)..(18)
<223> D или R
<400> 69
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Thr Xaa Xaa Xaa Thr Xaa Pro Xaa Ser
1 5 10 15
Xaa Xaa
<210> 70
<211> 19
<212> Белок
<213> вирус кори
<220>
<221> ПЕПТИД
<222> (1)..(19)
<223> MvF 4 Th
<220>
<221> SITE
<222> (4)..(4)
<223> S или T
<220>
<221> SITE
<222> (7)..(7)
<223> K или R
<220>
<221> SITE
<222> (8)..(8)
<223> G или T
<220>
<221> SITE
<222> (12)..(12)
<223> H или T
<220>
<221> SITE
<222> (13)..(13)
<223> K или R
<400> 70
Ile Ser Ile Xaa Glu Ile Xaa Xaa Val Ile Val Xaa Xaa Ile Glu Thr
1 5 10 15
Ile Leu Phe
<210> 71
<211> 18
<212> Белок
<213> вирус гепатита B
<220>
<221> ПЕПТИД
<222> (1)..(18)
<223> HBsAg 2 Th
<220>
<221> SITE
<222> (4)..(4)
<223> I или F
<220>
<221> SITE
<222> (5)..(5)
<223> I или F
<220>
<221> SITE
<222> (6)..(6)
<223> T или L
<220>
<221> SITE
<222> (7)..(7)
<223> I или L
<220>
<221> SITE
<222> (11)..(11)
<223> I или L
<220>
<221> SITE
<222> (14)..(14)
<223> P или I
<220>
<221> SITE
<222> (15)..(15)
<223> Q или T
<220>
<221> SITE
<222> (16)..(16)
<223> S или T
<220>
<221> SITE
<222> (17)..(17)
<223> L или I
<400> 71
Lys Lys Lys Xaa Xaa Xaa Xaa Thr Arg Ile Xaa Thr Ile Xaa Xaa Xaa
1 5 10 15
Xaa Asp
<210> 72
<211> 19
<212> Белок
<213> вирус кори
<220>
<221> ПЕПТИД
<222> (1)..(19)
<223> MvF 5 Th
<400> 72
Ile Ser Ile Thr Glu Ile Lys Gly Val Ile Val His Arg Ile Glu Thr
1 5 10 15
Ile Leu Phe
<210> 73
<211> 18
<212> Белок
<213> вирус гепатита B
<220>
<221> ПЕПТИД
<222> (1)..(18)
<223> HBsAg 3 Th
<400> 73
Lys Lys Lys Ile Ile Thr Ile Thr Arg Ile Ile Thr Ile Ile Thr Thr
1 5 10 15
Ile Asp
<210> 74
<211> 11
<212> Белок
<213> вирус гриппа
<220>
<221> ПЕПТИД
<222> (1)..(11)
<223> матричный белок гриппа 1 _1 Th
<220>
<221> ПЕПТИД
<222> (1)..(11)
<223> матричный белок гриппа 1_1 Th
<400> 74
Phe Val Phe Thr Leu Thr Val Pro Ser Glu Arg
1 5 10
<210> 75
<211> 15
<212> Белок
<213> вирус гриппа
<220>
<221> ПЕПТИД
<222> (1)..(15)
<223> матричный белок гриппа 1_2 Th
<400> 75
Ser Gly Pro Leu Lys Ala Glu Ile Ala Gln Arg Leu Glu Asp Val
1 5 10 15
<210> 76
<211> 9
<212> Белок
<213> вирус гриппа
<220>
<221> ПЕПТИД
<222> (1)..(9)
<223> неструктурный белок гриппа 1 Th
<400> 76
Asp Arg Leu Arg Arg Asp Gln Lys Ser
5
<210> 77
<211> 19
<212> Белок
<213> вирус Эпштейна-Барр
<220>
<221> ПЕПТИД
<222> (1)..(19)
<223> EBV BHRF1 Th
<400> 77
Ala Gly Leu Thr Leu Ser Leu Leu Val Ile Cys Ser Tyr Leu Phe Ile
1 5 10 15
Ser Arg Gly
<210> 78
<211> 15
<212> Белок
<213> Clostridium tetani
<220>
<221> ПЕПТИД
<222> (1)..(15)
<223> Clostridium tetani TT1 Th
<400> 78
Gln Tyr Ile Lys Ala Asn Ser Lys Phe Ile Gly Ile Thr Glu Leu
1 5 10 15
<210> 79
<211> 20
<212> Белок
<213> вирус Эпштейна-Барр
<220>
<221> ПЕПТИД
<222> (1)..(20)
<223> EBV EBNA-1 Th
<400> 79
Pro Gly Pro Leu Arg Glu Ser Ile Val Cys Tyr Phe Met Val Phe Leu
1 5 10 15
Gln Thr His Ile
20
<210> 80
<211> 21
<212> Белок
<213> Clostridium tetani
<220>
<221> ПЕПТИД
<222> (1)..(21)
<223> Clostridium tetani TT2 Th
<400> 80
Phe Asn Asn Phe Thr Val Ser Phe Trp Leu Arg Val Pro Lys Val Ser
1 5 10 15
Ala Ser His Leu Glu
20
<210> 81
<211> 16
<212> Белок
<213> Clostridium tetani
<220>
<221> ПЕПТИД
<222> (1)..(16)
<223> Clostridium tetani TT3 Th
<400> 81
Lys Phe Ile Ile Lys Arg Tyr Thr Pro Asn Asn Glu Ile Asp Ser Phe
1 5 10 15
<210> 82
<211> 16
<212> Белок
<213> Clostridium tetani
<220>
<221> ПЕПТИД
<222> (1)..(16)
<223> Clostridium tetani TT4 Th
<400> 82
Val Ser Ile Asp Lys Phe Arg Ile Phe Cys Lys Ala Leu Asn Pro Lys
1 5 10 15
<210> 83
<211> 18
<212> Белок
<213> вирус Эпштейна-Барр
<220>
<221> ПЕПТИД
<222> (1)..(18)
<223> EBV CP Th
<400> 83
Val Pro Gly Leu Tyr Ser Pro Cys Arg Ala Phe Phe Asn Lys Glu Glu
1 5 10 15
Leu Leu
<210> 84
<211> 14
<212> Белок
<213> цитомегаловирус человека
<220>
<221> ПЕПТИД
<222> (1)..(14)
<223> HCMV IE1 Th
<400> 84
Asp Lys Arg Glu Met Trp Met Ala Cys Ile Lys Glu Leu His
1 5 10
<210> 85
<211> 15
<212> Белок
<213> вирус Эпштейна-Барр
<220>
<221> ПЕПТИД
<222> (1)..(15)
<223> EBV GP340 Th
<400> 85
Thr Gly His Gly Ala Arg Thr Ser Thr Glu Pro Thr Thr Asp Tyr
1 5 10 15
<210> 86
<211> 13
<212> Белок
<213> вирус Эпштейна-Барр
<220>
<221> ПЕПТИД
<222> (1)..(13)
<223> EBV BPLF1 Th
<400> 86
Lys Glu Leu Lys Arg Gln Tyr Glu Lys Lys Leu Arg Gln
1 5 10
<210> 87
<211> 11
<212> Белок
<213> вирус Эпштейна-Барр
<220>
<221> ПЕПТИД
<222> (1)..(11)
<223> EBV EBNA-2 Th
<400> 87
Thr Val Phe Tyr Asn Ile Pro Pro Met Pro Leu
1 5 10
<210> 88
<211> 59
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(39)
<223> IgE-EMPD 1-39
<220>
<221> SITE
<222> (40)..(40)
<223> эпсилон K в качестве спейсера
<220>
<221> ПЕПТИД
<222> (41)..(59)
<223> UBITh(R)1
<400> 88
Gly Leu Ala Gly Gly Ser Ala Gln Ser Gln Arg Ala Pro Asp Arg Val
1 5 10 15
Leu Cys His Ser Gly Gln Gln Gln Gly Leu Pro Arg Ala Ala Gly Gly
20 25 30
Ser Val Pro His Pro Arg Cys Lys Ile Ser Ile Thr Glu Ile Lys Gly
35 40 45
Val Ile Val His Arg Ile Glu Thr Ile Leu Phe
50 55
<210> 89
<211> 59
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(19)
<223> UBITh(R)1
<220>
<221> SITE
<222> (20)..(20)
<223> эпсилон K в качестве спейсера
<220>
<221> ПЕПТИД
<222> (21)..(59)
<223> IgE-EMPD 1-39
<400> 89
Ile Ser Ile Thr Glu Ile Lys Gly Val Ile Val His Arg Ile Glu Thr
1 5 10 15
Ile Leu Phe Lys Gly Leu Ala Gly Gly Ser Ala Gln Ser Gln Arg Ala
20 25 30
Pro Asp Arg Val Leu Cys His Ser Gly Gln Gln Gln Gly Leu Pro Arg
35 40 45
Ala Ala Gly Gly Ser Val Pro His Pro Arg Cys
50 55
<210> 90
<211> 79
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(19)
<223> UBITh(R)1
<220>
<221> SITE
<222> (20)..(20)
<223> эпсилон K в качестве спейсера
<220>
<221> ПЕПТИД
<222> (21)..(59)
<223> IgE-EMPD 1-39
<220>
<221> SITE
<222> (60)..(60)
<223> эпсилон K в качестве спейсера
<220>
<221> ПЕПТИД
<222> (61)..(79)
<223> UBITh(R)1
<400> 90
Ile Ser Ile Thr Glu Ile Lys Gly Val Ile Val His Arg Ile Glu Thr
1 5 10 15
Ile Leu Phe Lys Gly Leu Ala Gly Gly Ser Ala Gln Ser Gln Arg Ala
20 25 30
Pro Asp Arg Val Leu Cys His Ser Gly Gln Gln Gln Gly Leu Pro Arg
35 40 45
Ala Ala Gly Gly Ser Val Pro His Pro Arg Cys Lys Ile Ser Ile Thr
50 55 60
Glu Ile Lys Gly Val Ile Val His Arg Ile Glu Thr Ile Leu Phe
65 70 75
<210> 91
<211> 54
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(34)
<223> IgE-EMPD 7-40
<220>
<221> SITE
<222> (35)..(35)
<223> эпсилон K в качестве спейсера
<220>
<221> ПЕПТИД
<222> (36)..(54)
<223> UBITh(R)1
<400> 91
Ala Gln Ser Gln Arg Ala Pro Asp Arg Val Leu Cys His Ser Gly Gln
1 5 10 15
Gln Gln Gly Leu Pro Arg Ala Ala Gly Gly Ser Val Pro His Pro Arg
20 25 30
Cys His Lys Ile Ser Ile Thr Glu Ile Lys Gly Val Ile Val His Arg
35 40 45
Ile Glu Thr Ile Leu Phe
50
<210> 92
<211> 54
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(19)
<223> UBITh(R)1
<220>
<221> SITE
<222> (20)..(20)
<223> эпсилон K в качестве спейсера
<220>
<221> ПЕПТИД
<222> (21)..(54)
<223> IgE-EMPD 7-40
<400> 92
Ile Ser Ile Thr Glu Ile Lys Gly Val Ile Val His Arg Ile Glu Thr
1 5 10 15
Ile Leu Phe Lys Ala Gln Ser Gln Arg Ala Pro Asp Arg Val Leu Cys
20 25 30
His Ser Gly Gln Gln Gln Gly Leu Pro Arg Ala Ala Gly Gly Ser Val
35 40 45
Pro His Pro Arg Cys His
50
<210> 93
<211> 74
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(19)
<223> UBITh(R)1
<220>
<221> SITE
<222> (20)..(20)
<223> эпсилон K в качестве спейсера
<220>
<221> ПЕПТИД
<222> (21)..(54)
<223> IgE-EMPD 7-40
<220>
<221> SITE
<222> (55)..(55)
<223> эпсилон K в качестве спейсера
<220>
<221> ПЕПТИД
<222> (56)..(74)
<223> UBITh(R)1
<400> 93
Ile Ser Ile Thr Glu Ile Lys Gly Val Ile Val His Arg Ile Glu Thr
1 5 10 15
Ile Leu Phe Lys Ala Gln Ser Gln Arg Ala Pro Asp Arg Val Leu Cys
20 25 30
His Ser Gly Gln Gln Gln Gly Leu Pro Arg Ala Ala Gly Gly Ser Val
35 40 45
Pro His Pro Arg Cys His Lys Ile Ser Ile Thr Glu Ile Lys Gly Val
50 55 60
Ile Val His Arg Ile Glu Thr Ile Leu Phe
65 70
<210> 94
<211> 62
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(19)
<223> UBITh(R)1
<220>
<221> ПЕПТИД
<222> (20)..(23)
<223> эпсилон K-KKK как спейсер
<220>
<221> SITE
<222> (20)..(20)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (24)..(62)
<223> IgE-EMPD 1-39
<400> 94
Ile Ser Ile Thr Glu Ile Lys Gly Val Ile Val His Arg Ile Glu Thr
1 5 10 15
Ile Leu Phe Lys Lys Lys Lys Gly Leu Ala Gly Gly Ser Ala Gln Ser
20 25 30
Gln Arg Ala Pro Asp Arg Val Leu Cys His Ser Gly Gln Gln Gln Gly
35 40 45
Leu Pro Arg Ala Ala Gly Gly Ser Val Pro His Pro Arg Cys
50 55 60
<210> 95
<211> 61
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(18)
<223> UBITh(R)2
<220>
<221> ПЕПТИД
<222> (19)..(22)
<223> эпсилон K-KKK как спейсер
<220>
<221> SITE
<222> (19)..(19)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (23)..(61)
<223> IgE-EMPD 1-39
<400> 95
Lys Lys Lys Ile Ile Thr Ile Thr Arg Ile Ile Thr Ile Ile Thr Thr
1 5 10 15
Ile Asp Lys Lys Lys Lys Gly Leu Ala Gly Gly Ser Ala Gln Ser Gln
20 25 30
Arg Ala Pro Asp Arg Val Leu Cys His Ser Gly Gln Gln Gln Gly Leu
35 40 45
Pro Arg Ala Ala Gly Gly Ser Val Pro His Pro Arg Cys
50 55 60
<210> 96
<211> 40
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(19)
<223> UBITh(R)1
<220>
<221> ПЕПТИД
<222> (20)..(23)
<223> эпсилон K-KKK как спейсер
<220>
<221> SITE
<222> (20)..(20)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (24)..(40)
<223> IgE-EMPD 1-17
<400> 96
Ile Ser Ile Thr Glu Ile Lys Gly Val Ile Val His Arg Ile Glu Thr
1 5 10 15
Ile Leu Phe Lys Lys Lys Lys Gly Leu Ala Gly Gly Ser Ala Gln Ser
20 25 30
Gln Arg Ala Pro Asp Arg Val Leu
35 40
<210> 97
<211> 43
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(19)
<223> UBITh(R)1
<220>
<221> ПЕПТИД
<222> (20)..(23)
<223> эпсилон K-KKK как спейсер
<220>
<221> SITE
<222> (20)..(20)
<223> эпсилон-K
<220>
<221> ПЕПТИД
<222> (24)..(43)
<223> IgE-EMPD 19-38
<400> 97
Ile Ser Ile Thr Glu Ile Lys Gly Val Ile Val His Arg Ile Glu Thr
1 5 10 15
Ile Leu Phe Lys Lys Lys Lys His Ser Gly Gln Gln Gln Gly Leu Pro
20 25 30
Arg Ala Ala Gly Gly Ser Val Pro His Pro Arg
35 40
<210> 98
<211> 57
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(17)
<223> Clostridium tetani 1 Th
<220>
<221> SITE
<222> (18)..(18)
<223> эпсилон K в качестве спейсера
<220>
<221> ПЕПТИД
<222> (19)..(57)
<223> IgE-EMPD 1-39
<400> 98
Lys Lys Gln Tyr Ile Lys Ala Asn Ser Lys Phe Ile Gly Ile Thr Glu
1 5 10 15
Leu Lys Gly Leu Ala Gly Gly Ser Ala Gln Ser Gln Arg Ala Pro Asp
20 25 30
Arg Val Leu Cys His Ser Gly Gln Gln Gln Gly Leu Pro Arg Ala Ala
35 40 45
Gly Gly Ser Val Pro His Pro Arg Cys
50 55
<210> 99
<211> 55
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(15)
<223> MvF1 Th
<220>
<221> SITE
<222> (16)..(16)
<223> эпсилон K в качестве спейсера
<220>
<221> ПЕПТИД
<222> (17)..(55)
<223> IgE-EMPD 1-39
<400> 99
Leu Ser Glu Ile Lys Gly Val Ile Val His Arg Leu Glu Gly Val Lys
1 5 10 15
Gly Leu Ala Gly Gly Ser Ala Gln Ser Gln Arg Ala Pro Asp Arg Val
20 25 30
Leu Cys His Ser Gly Gln Gln Gln Gly Leu Pro Arg Ala Ala Gly Gly
35 40 45
Ser Val Pro His Pro Arg Cys
50 55
<210> 100
<211> 64
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(24)
<223> Bordetella pertussis Th
<220>
<221> SITE
<222> (25)..(25)
<223> эпсилон K в качестве спейсера
<220>
<221> ПЕПТИД
<222> (26)..(64)
<223> IgE-EMPD 1-39
<400> 100
Gly Ala Tyr Ala Arg Cys Pro Asn Gly Thr Arg Ala Leu Thr Val Ala
1 5 10 15
Glu Leu Arg Gly Asn Ala Glu Leu Lys Gly Leu Ala Gly Gly Ser Ala
20 25 30
Gln Ser Gln Arg Ala Pro Asp Arg Val Leu Cys His Ser Gly Gln Gln
35 40 45
Gln Gly Leu Pro Arg Ala Ala Gly Gly Ser Val Pro His Pro Arg Cys
50 55 60
<210> 101
<211> 57
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(17)
<223> Clostridium tetani 2 Th
<220>
<221> SITE
<222> (18)..(18)
<223> эпсилон K в качестве спейсера
<220>
<221> ПЕПТИД
<222> (19)..(57)
<223> IgE-EMPD 1-39
<400> 101
Trp Val Arg Asp Ile Ile Asp Asp Phe Thr Asn Glu Ser Ser Gln Lys
1 5 10 15
Thr Lys Gly Leu Ala Gly Gly Ser Ala Gln Ser Gln Arg Ala Pro Asp
20 25 30
Arg Val Leu Cys His Ser Gly Gln Gln Gln Gly Leu Pro Arg Ala Ala
35 40 45
Gly Gly Ser Val Pro His Pro Arg Cys
50 55
<210> 102
<211> 63
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(23)
<223> Diphtheria Th
<220>
<221> SITE
<222> (24)..(24)
<223> эпсилон K в качестве спейсера
<220>
<221> ПЕПТИД
<222> (25)..(63)
<223> IgE-EMPD 1-39
<400> 102
Asp Ser Glu Thr Ala Asp Asn Leu Glu Lys Thr Val Ala Ala Leu Ser
1 5 10 15
Ile Leu Pro Gly His Gly Cys Lys Gly Leu Ala Gly Gly Ser Ala Gln
20 25 30
Ser Gln Arg Ala Pro Asp Arg Val Leu Cys His Ser Gly Gln Gln Gln
35 40 45
Gly Leu Pro Arg Ala Ala Gly Gly Ser Val Pro His Pro Arg Cys
50 55 60
<210> 103
<211> 61
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(21)
<223> Plasmodium falciparum Th
<220>
<221> SITE
<222> (22)..(22)
<223> эпсилон K в качестве спейсера
<220>
<221> ПЕПТИД
<222> (23)..(61)
<223> IgE-EMPD 1-39
<400> 103
Asp His Glu Lys Lys His Ala Lys Met Glu Lys Ala Ser Ser Val Phe
1 5 10 15
Asn Val Val Asn Ser Lys Gly Leu Ala Gly Gly Ser Ala Gln Ser Gln
20 25 30
Arg Ala Pro Asp Arg Val Leu Cys His Ser Gly Gln Gln Gln Gly Leu
35 40 45
Pro Arg Ala Ala Gly Gly Ser Val Pro His Pro Arg Cys
50 55 60
<210> 104
<211> 57
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(17)
<223> Schistosoma mansoni Th
<220>
<221> SITE
<222> (18)..(18)
<223> эпсилон K в качестве спейсера
<220>
<221> ПЕПТИД
<222> (19)..(57)
<223> IgE-EMPD 1-39
<400> 104
Lys Trp Phe Lys Thr Asn Ala Pro Asn Gly Val Asp Glu Lys His Arg
1 5 10 15
His Lys Gly Leu Ala Gly Gly Ser Ala Gln Ser Gln Arg Ala Pro Asp
20 25 30
Arg Val Leu Cys His Ser Gly Gln Gln Gln Gly Leu Pro Arg Ala Ala
35 40 45
Gly Gly Ser Val Pro His Pro Arg Cys
50 55
<210> 105
<211> 65
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(25)
<223> холерный токсин Th
<220>
<221> SITE
<222> (26)..(26)
<223> эпсилон K в качестве спейсера
<220>
<221> ПЕПТИД
<222> (27)..(65)
<223> IgE-EMPD 1-39
<400> 105
Ala Leu Asn Ile Trp Asp Arg Phe Asp Val Phe Cys Thr Leu Gly Ala
1 5 10 15
Thr Thr Gly Tyr Leu Lys Gly Asn Ser Lys Gly Leu Ala Gly Gly Ser
20 25 30
Ala Gln Ser Gln Arg Ala Pro Asp Arg Val Leu Cys His Ser Gly Gln
35 40 45
Gln Gln Gly Leu Pro Arg Ala Ala Gly Gly Ser Val Pro His Pro Arg
50 55 60
Cys
65
<210> 106
<211> 55
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(15)
<223> MvF 2 Th
<220>
<221> SITE
<222> (16)..(16)
<223> эпсилон K в качестве спейсера
<220>
<221> ПЕПТИД
<222> (17)..(55)
<223> IgE-EMPD 1-39
<400> 106
Ile Ser Glu Ile Lys Gly Val Ile Val His Lys Ile Glu Gly Ile Lys
1 5 10 15
Gly Leu Ala Gly Gly Ser Ala Gln Ser Gln Arg Ala Pro Asp Arg Val
20 25 30
Leu Cys His Ser Gly Gln Gln Gln Gly Leu Pro Arg Ala Ala Gly Gly
35 40 45
Ser Val Pro His Pro Arg Cys
50 55
<210> 107
<211> 62
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(22)
<223> KKKMvF 3 Th
<220>
<221> SITE
<222> (7)..(7)
<223> S или T
<220>
<221> SITE
<222> (10)..(10)
<223> K или R
<220>
<221> SITE
<222> (11)..(11)
<223> G или T
<220>
<221> SITE
<222> (15)..(15)
<223> H или T
<220>
<221> SITE
<222> (16)..(16)
<223> K или R
<220>
<221> SITE
<222> (19)..(19)
<223> G или T
<220>
<221> SITE
<222> (23)..(23)
<223> эпсилон K в качестве спейсера
<220>
<221> ПЕПТИД
<222> (24)..(62)
<223> IgE-EMPD 1-39
<400> 107
Lys Lys Lys Ile Ser Ile Xaa Glu Ile Xaa Xaa Val Ile Val Xaa Xaa
1 5 10 15
Ile Glu Xaa Ile Leu Phe Lys Gly Leu Ala Gly Gly Ser Ala Gln Ser
20 25 30
Gln Arg Ala Pro Asp Arg Val Leu Cys His Ser Gly Gln Gln Gln Gly
35 40 45
Leu Pro Arg Ala Ala Gly Gly Ser Val Pro His Pro Arg Cys
50 55 60
<210> 108
<211> 58
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(18)
<223> HBsAg 1 Th
<220>
<221> SITE
<222> (1)..(1)
<223> K или R
<220>
<221> SITE
<222> (2)..(2)
<223> K или R
<220>
<221> SITE
<222> (3)..(3)
<223> K или R
<220>
<221> SITE
<222> (4)..(4)
<223> L или I или V или F
<220>
<221> SITE
<222> (5)..(5)
<223> F или K или R
<220>
<221> SITE
<222> (6)..(6)
<223> L или I или V или F
<220>
<221> SITE
<222> (7)..(7)
<223> L или I или V или F
<220>
<221> SITE
<222> (9)..(9)
<223> K или R
<220>
<221> SITE
<222> (10)..(10)
<223> L или I или V или F
<220>
<221> SITE
<222> (11)..(11)
<223> L или I или V или F
<220>
<221> SITE
<222> (13)..(13)
<223> L или I или V или F
<220>
<221> SITE
<222> (15)..(15)
<223> Q или L или I или V или F
<220>
<221> SITE
<222> (17)..(17)
<223> L или I или V или F
<220>
<221> SITE
<222> (18)..(18)
<223> D или R
<220>
<221> SITE
<222> (19)..(19)
<223> эпсилон K в качестве спейсера
<220>
<221> ПЕПТИД
<222> (20)..(58)
<223> IgE-EMPD 1-39
<400> 108
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Thr Xaa Xaa Xaa Thr Xaa Pro Xaa Ser
1 5 10 15
Xaa Xaa Lys Gly Leu Ala Gly Gly Ser Ala Gln Ser Gln Arg Ala Pro
20 25 30
Asp Arg Val Leu Cys His Ser Gly Gln Gln Gln Gly Leu Pro Arg Ala
35 40 45
Ala Gly Gly Ser Val Pro His Pro Arg Cys
50 55
<210> 109
<211> 59
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(19)
<223> MvF 4 Th
<220>
<221> SITE
<222> (4)..(4)
<223> S или T
<220>
<221> SITE
<222> (7)..(7)
<223> K или R
<220>
<221> SITE
<222> (8)..(8)
<223> G или T
<220>
<221> SITE
<222> (12)..(12)
<223> H или T
<220>
<221> SITE
<222> (13)..(13)
<223> K или R
<220>
<221> SITE
<222> (20)..(20)
<223> эпсилон K в качестве спейсера
<220>
<221> ПЕПТИД
<222> (21)..(59)
<223> IgE-EMPD 1-39
<400> 109
Ile Ser Ile Xaa Glu Ile Xaa Xaa Val Ile Val Xaa Xaa Ile Glu Thr
1 5 10 15
Ile Leu Phe Lys Gly Leu Ala Gly Gly Ser Ala Gln Ser Gln Arg Ala
20 25 30
Pro Asp Arg Val Leu Cys His Ser Gly Gln Gln Gln Gly Leu Pro Arg
35 40 45
Ala Ala Gly Gly Ser Val Pro His Pro Arg Cys
50 55
<210> 110
<211> 58
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(18)
<223> HBsAg 2 Th
<220>
<221> SITE
<222> (4)..(4)
<223> I или F
<220>
<221> SITE
<222> (5)..(5)
<223> I или F
<220>
<221> SITE
<222> (6)..(6)
<223> T или L
<220>
<221> SITE
<222> (7)..(7)
<223> I или L
<220>
<221> SITE
<222> (11)..(11)
<223> I или L
<220>
<221> SITE
<222> (14)..(14)
<223> P или I
<220>
<221> SITE
<222> (15)..(15)
<223> Q или T
<220>
<221> SITE
<222> (16)..(16)
<223> S или T
<220>
<221> SITE
<222> (17)..(17)
<223> L или I
<220>
<221> SITE
<222> (19)..(19)
<223> эпсилон K в качестве спейсера
<220>
<221> ПЕПТИД
<222> (20)..(58)
<223> IgE-EMPD 1-39
<400> 110
Lys Lys Lys Xaa Xaa Xaa Xaa Thr Arg Ile Xaa Thr Ile Xaa Xaa Xaa
1 5 10 15
Xaa Asp Lys Gly Leu Ala Gly Gly Ser Ala Gln Ser Gln Arg Ala Pro
20 25 30
Asp Arg Val Leu Cys His Ser Gly Gln Gln Gln Gly Leu Pro Arg Ala
35 40 45
Ala Gly Gly Ser Val Pro His Pro Arg Cys
50 55
<210> 111
<211> 51
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(11)
<223> Th Гриппа MP1_1
<220>
<221> SITE
<222> (12)..(12)
<223> эпсилон K в качестве спейсера
<220>
<221> ПЕПТИД
<222> (13)..(51)
<223> IgE-EMPD 1-39
<400> 111
Phe Val Phe Thr Leu Thr Val Pro Ser Glu Arg Lys Gly Leu Ala Gly
1 5 10 15
Gly Ser Ala Gln Ser Gln Arg Ala Pro Asp Arg Val Leu Cys His Ser
20 25 30
Gly Gln Gln Gln Gly Leu Pro Arg Ala Ala Gly Gly Ser Val Pro His
35 40 45
Pro Arg Cys
50
<210> 112
<211> 55
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(15)
<223> Th Гриппа MP1_2
<220>
<221> SITE
<222> (16)..(16)
<223> эпсилон K в качестве спейсера
<220>
<221> ПЕПТИД
<222> (17)..(55)
<223> IgE-EMPD 1-39
<400> 112
Ser Gly Pro Leu Lys Ala Glu Ile Ala Gln Arg Leu Glu Asp Val Lys
1 5 10 15
Gly Leu Ala Gly Gly Ser Ala Gln Ser Gln Arg Ala Pro Asp Arg Val
20 25 30
Leu Cys His Ser Gly Gln Gln Gln Gly Leu Pro Arg Ala Ala Gly Gly
35 40 45
Ser Val Pro His Pro Arg Cys
50 55
<210> 113
<211> 49
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(9)
<223> Th Гриппа NSP1
<220>
<221> SITE
<222> (10)..(10)
<223> эпсилон K в качестве спейсера
<220>
<221> ПЕПТИД
<222> (11)..(49)
<223> IgE-EMPD 1-39
<400> 113
Asp Arg Leu Arg Arg Asp Gln Lys Ser Lys Gly Leu Ala Gly Gly Ser
1 5 10 15
Ala Gln Ser Gln Arg Ala Pro Asp Arg Val Leu Cys His Ser Gly Gln
20 25 30
Gln Gln Gly Leu Pro Arg Ala Ala Gly Gly Ser Val Pro His Pro Arg
35 40 45
Cys
<210> 114
<211> 59
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(19)
<223> EBV BHRF1 Th
<220>
<221> SITE
<222> (20)..(20)
<223> эпсилон K в качестве спейсера
<220>
<221> ПЕПТИД
<222> (21)..(59)
<223> IgE-EMPD 1-39
<400> 114
Ala Gly Leu Thr Leu Ser Leu Leu Val Ile Cys Ser Tyr Leu Phe Ile
1 5 10 15
Ser Arg Gly Lys Gly Leu Ala Gly Gly Ser Ala Gln Ser Gln Arg Ala
20 25 30
Pro Asp Arg Val Leu Cys His Ser Gly Gln Gln Gln Gly Leu Pro Arg
35 40 45
Ala Ala Gly Gly Ser Val Pro His Pro Arg Cys
50 55
<210> 115
<211> 55
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(15)
<223> Clostridium tetani TT1 Th
<220>
<221> SITE
<222> (16)..(16)
<223> эпсилон K в качестве спейсера
<220>
<221> ПЕПТИД
<222> (17)..(55)
<223> IgE-EMPD 1-39
<400> 115
Gln Tyr Ile Lys Ala Asn Ser Lys Phe Ile Gly Ile Thr Glu Leu Lys
1 5 10 15
Gly Leu Ala Gly Gly Ser Ala Gln Ser Gln Arg Ala Pro Asp Arg Val
20 25 30
Leu Cys His Ser Gly Gln Gln Gln Gly Leu Pro Arg Ala Ala Gly Gly
35 40 45
Ser Val Pro His Pro Arg Cys
50 55
<210> 116
<211> 60
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(20)
<223> EBV EBNA-1 Th
<220>
<221> SITE
<222> (21)..(21)
<223> эпсилон K в качестве спейсера
<220>
<221> ПЕПТИД
<222> (22)..(60)
<223> IgE-EMPD 1-39
<400> 116
Pro Gly Pro Leu Arg Glu Ser Ile Val Cys Tyr Phe Met Val Phe Leu
1 5 10 15
Gln Thr His Ile Lys Gly Leu Ala Gly Gly Ser Ala Gln Ser Gln Arg
20 25 30
Ala Pro Asp Arg Val Leu Cys His Ser Gly Gln Gln Gln Gly Leu Pro
35 40 45
Arg Ala Ala Gly Gly Ser Val Pro His Pro Arg Cys
50 55 60
<210> 117
<211> 61
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(21)
<223> Clostridium tetani TT2 Th
<220>
<221> SITE
<222> (22)..(22)
<223> эпсилон K в качестве спейсера
<220>
<221> ПЕПТИД
<222> (23)..(61)
<223> IgE-EMPD 1-39
<400> 117
Phe Asn Asn Phe Thr Val Ser Phe Trp Leu Arg Val Pro Lys Val Ser
1 5 10 15
Ala Ser His Leu Glu Lys Gly Leu Ala Gly Gly Ser Ala Gln Ser Gln
20 25 30
Arg Ala Pro Asp Arg Val Leu Cys His Ser Gly Gln Gln Gln Gly Leu
35 40 45
Pro Arg Ala Ala Gly Gly Ser Val Pro His Pro Arg Cys
50 55 60
<210> 118
<211> 56
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(16)
<223> Clostridium tetani TT3 Th
<220>
<221> SITE
<222> (17)..(17)
<223> эпсилон K в качестве спейсера
<220>
<221> ПЕПТИД
<222> (18)..(56)
<223> IgE-EMPD 1-39
<400> 118
Lys Phe Ile Ile Lys Arg Tyr Thr Pro Asn Asn Glu Ile Asp Ser Phe
1 5 10 15
Lys Gly Leu Ala Gly Gly Ser Ala Gln Ser Gln Arg Ala Pro Asp Arg
20 25 30
Val Leu Cys His Ser Gly Gln Gln Gln Gly Leu Pro Arg Ala Ala Gly
35 40 45
Gly Ser Val Pro His Pro Arg Cys
50 55
<210> 119
<211> 56
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(16)
<223> Clostridium tetani TT4 Th
<220>
<221> SITE
<222> (17)..(17)
<223> эпсилон K в качестве спейсера
<220>
<221> ПЕПТИД
<222> (18)..(56)
<223> IgE-EMPD 1-39
<400> 119
Val Ser Ile Asp Lys Phe Arg Ile Phe Cys Lys Ala Leu Asn Pro Lys
1 5 10 15
Lys Gly Leu Ala Gly Gly Ser Ala Gln Ser Gln Arg Ala Pro Asp Arg
20 25 30
Val Leu Cys His Ser Gly Gln Gln Gln Gly Leu Pro Arg Ala Ala Gly
35 40 45
Gly Ser Val Pro His Pro Arg Cys
50 55
<210> 120
<211> 58
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(18)
<223> EBV CP Th
<220>
<221> SITE
<222> (19)..(19)
<223> эпсилон K в качестве спейсера
<220>
<221> ПЕПТИД
<222> (20)..(58)
<223> IgE-EMPD 1-39
<400> 120
Val Pro Gly Leu Tyr Ser Pro Cys Arg Ala Phe Phe Asn Lys Glu Glu
1 5 10 15
Leu Leu Lys Gly Leu Ala Gly Gly Ser Ala Gln Ser Gln Arg Ala Pro
20 25 30
Asp Arg Val Leu Cys His Ser Gly Gln Gln Gln Gly Leu Pro Arg Ala
35 40 45
Ala Gly Gly Ser Val Pro His Pro Arg Cys
50 55
<210> 121
<211> 54
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(14)
<223> HCMV IE1 Th
<220>
<221> SITE
<222> (15)..(15)
<223> эпсилон K в качестве спейсера
<220>
<221> ПЕПТИД
<222> (16)..(54)
<223> IgE-EMPD 1-39
<400> 121
Asp Lys Arg Glu Met Trp Met Ala Cys Ile Lys Glu Leu His Lys Gly
1 5 10 15
Leu Ala Gly Gly Ser Ala Gln Ser Gln Arg Ala Pro Asp Arg Val Leu
20 25 30
Cys His Ser Gly Gln Gln Gln Gly Leu Pro Arg Ala Ala Gly Gly Ser
35 40 45
Val Pro His Pro Arg Cys
50
<210> 122
<211> 55
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(15)
<223> EBV GP340 Th
<220>
<221> SITE
<222> (16)..(16)
<223> эпсилон K в качестве спейсера
<220>
<221> ПЕПТИД
<222> (17)..(55)
<223> IgE-EMPD 1-39
<400> 122
Thr Gly His Gly Ala Arg Thr Ser Thr Glu Pro Thr Thr Asp Tyr Lys
1 5 10 15
Gly Leu Ala Gly Gly Ser Ala Gln Ser Gln Arg Ala Pro Asp Arg Val
20 25 30
Leu Cys His Ser Gly Gln Gln Gln Gly Leu Pro Arg Ala Ala Gly Gly
35 40 45
Ser Val Pro His Pro Arg Cys
50 55
<210> 123
<211> 53
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(13)
<223> EBV BPLF1 Th
<220>
<221> ПЕПТИД
<222> (14)..(14)
<223> эпсилон K в качестве спейсера
<220>
<221> ПЕПТИД
<222> (15)..(53)
<223> IgE-EMPD 1-39
<400> 123
Lys Glu Leu Lys Arg Gln Tyr Glu Lys Lys Leu Arg Gln Lys Gly Leu
1 5 10 15
Ala Gly Gly Ser Ala Gln Ser Gln Arg Ala Pro Asp Arg Val Leu Cys
20 25 30
His Ser Gly Gln Gln Gln Gly Leu Pro Arg Ala Ala Gly Gly Ser Val
35 40 45
Pro His Pro Arg Cys
50
<210> 124
<211> 51
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(11)
<223> EBV EBNA-2 Th
<220>
<221> SITE
<222> (12)..(12)
<223> эпсилон K в качестве спейсера
<220>
<221> ПЕПТИД
<222> (13)..(51)
<223> IgE-EMPD 1-39
<400> 124
Thr Val Phe Tyr Asn Ile Pro Pro Met Pro Leu Lys Gly Leu Ala Gly
1 5 10 15
Gly Ser Ala Gln Ser Gln Arg Ala Pro Asp Arg Val Leu Cys His Ser
20 25 30
Gly Gln Gln Gln Gly Leu Pro Arg Ala Ala Gly Gly Ser Val Pro His
35 40 45
Pro Arg Cys
50
<210> 125
<211> 74
<212> Белок
<213> Macaca fascicularis
<220>
<221> ПЕПТИД
<222> (1)..(19)
<223> UBITh(R)1
<220>
<221> SITE
<222> (20)..(20)
<223> эпсилон K в качестве спейсера
<220>
<221> ПЕПТИД
<222> (21)..(54)
<223> IgE-EMPD 7-40 (Macaca fascicularis)
<220>
<221> SITE
<222> (55)..(55)
<223> эпсилон K в качестве спейсера
<220>
<221> ПЕПТИД
<222> (56)..(74)
<223> UBITh(R)1
<400> 125
Ile Ser Ile Thr Glu Ile Lys Gly Val Ile Val His Arg Ile Glu Thr
1 5 10 15
Ile Leu Phe Lys Ala Gln Ser Gln Arg Ala Pro Asp Arg Val Leu Cys
20 25 30
His Ser Glu Gln Gln Gln Gly Leu Pro Arg Ala Ala Arg Gly Ser Val
35 40 45
Pro Asp His Arg Cys His Lys Ile Ser Ile Thr Glu Ile Lys Gly Val
50 55 60
Ile Val His Arg Ile Glu Thr Ile Leu Phe
65 70
<210> 126
<211> 59
<212> Белок
<213> Macaca fascicularis
<220>
<221> ПЕПТИД
<222> (1)..(39)
<223> IgE-EMPD 1-39 (Macaca fascicularis)
<220>
<221> SITE
<222> (40)..(40)
<223> эпсилон K в качестве спейсера
<220>
<221> ПЕПТИД
<222> (41)..(59)
<223> UBITh(R)1
<400> 126
Gly Leu Ala Gly Gly Ser Ala Gln Ser Gln Arg Ala Pro Asp Arg Val
1 5 10 15
Leu Cys His Ser Glu Gln Gln Gln Gly Leu Pro Arg Ala Ala Arg Gly
20 25 30
Ser Val Pro Asp His Arg Cys Lys Ile Ser Ile Thr Glu Ile Lys Gly
35 40 45
Val Ile Val His Arg Ile Glu Thr Ile Leu Phe
50 55
<210> 127
<211> 52
<212> Белок
<213> Macaca fascicularis
<220>
<221> ПЕПТИД
<222> (1)..(52)
<223> IgE-EMPD 1-52 (Macaca fascicularis)
<400> 127
Gly Leu Ala Gly Gly Ser Ala Gln Ser Gln Arg Ala Pro Asp Arg Val
1 5 10 15
Leu Cys His Ser Glu Gln Gln Gln Gly Leu Pro Arg Ala Ala Arg Gly
20 25 30
Ser Val Pro Asp His Arg Cys His Cys Gly Ala Gly Arg Ala Asp Trp
35 40 45
Pro Gly Leu Pro
50
<210> 128
<211> 6
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(6)
<223> Спейсер 1
<400> 128
Pro Pro Xaa Pro Xaa Pro
1 5
<210> 129
<211> 4
<212> Белок
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(4)
<223> Спейсер 2
<220>
<221> SITE
<222> (1)..(1)
<223> эпсилон K
<400> 129
Lys Lys Lys Lys
1
<210> 130
<211> 58
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический пептид
<220>
<221> SITE
<222> (1)..(19)
<223> UBITh(R)2
<220>
<221> SITE
<222> (20)..(20)
<223> эпсилон K в качестве спейсера
<220>
<221> SITE
<222> (21)..(58)
<223> IgEEMPD 1-39
<400> 130
Lys Lys Lys Ile Ile Thr Ile Thr Arg Ile Ile Thr Ile Ile Thr Thr
1 5 10 15
Ile Asp Lys Gly Leu Ala Gly Gly Ser Ala Gln Ser Gln Arg Ala Pro
20 25 30
Asp Arg Val Leu Cys His Ser Gly Gln Gln Gln Gly Leu Pro Arg Ala
35 40 45
Ala Gly Gly Ser Val Pro His Pro Arg Cys
50 55
<---
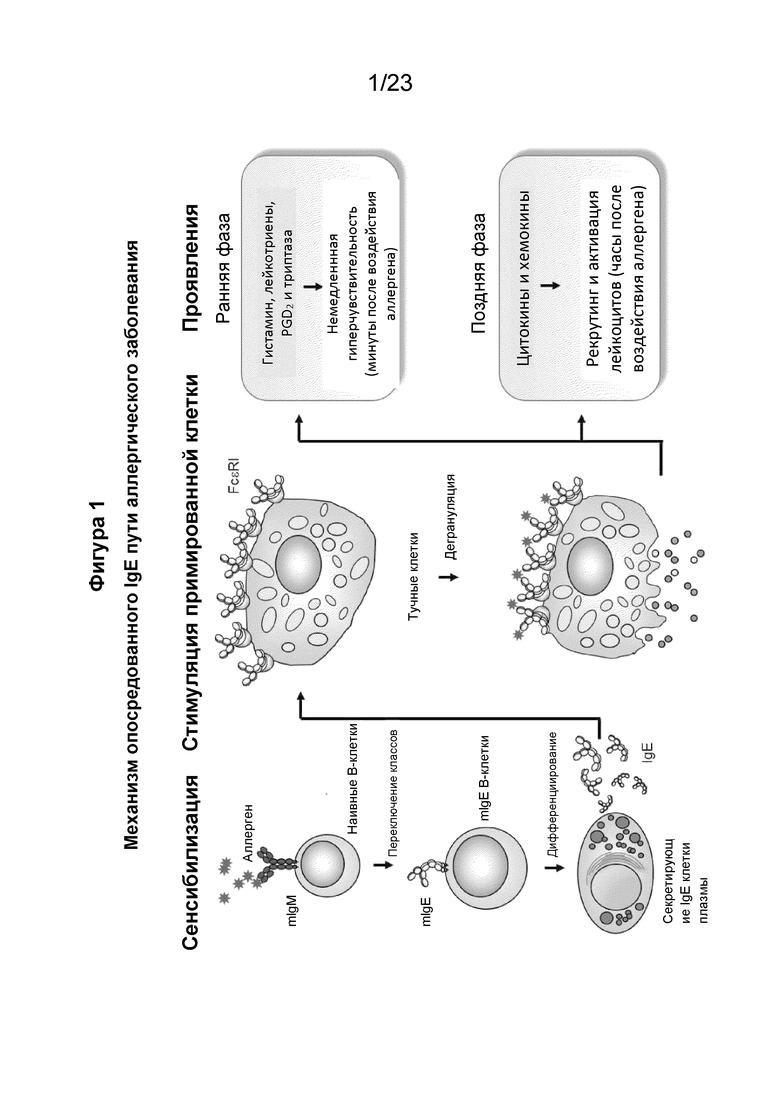
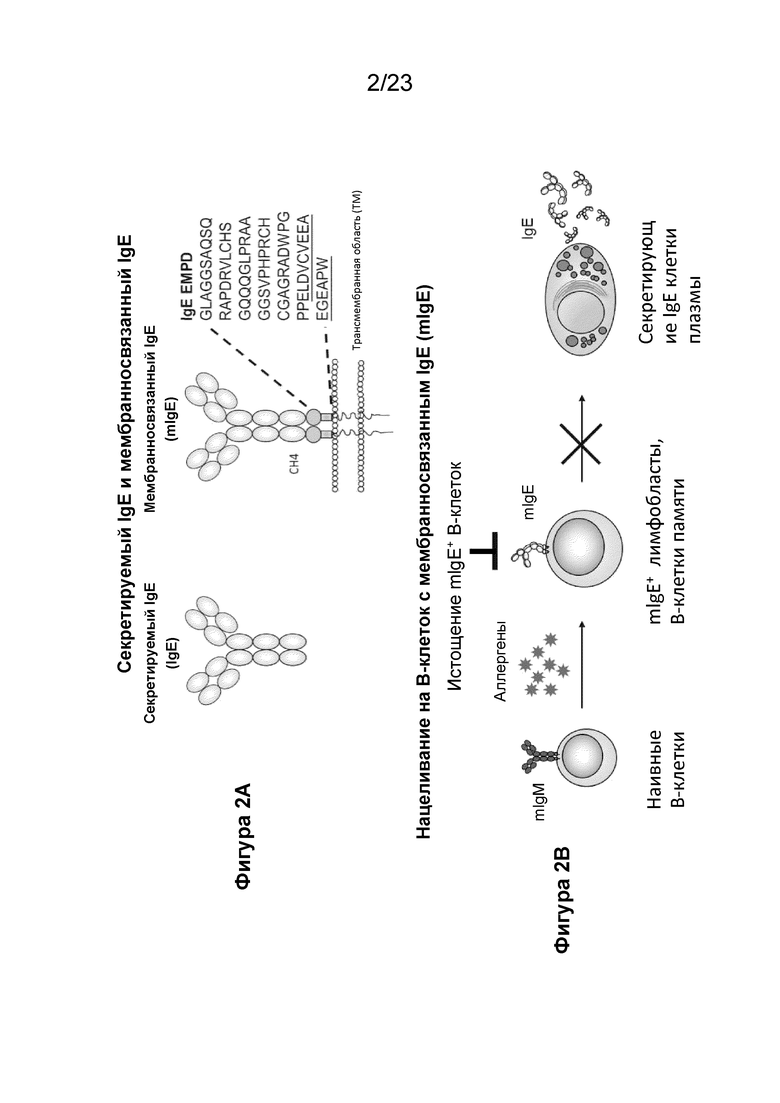

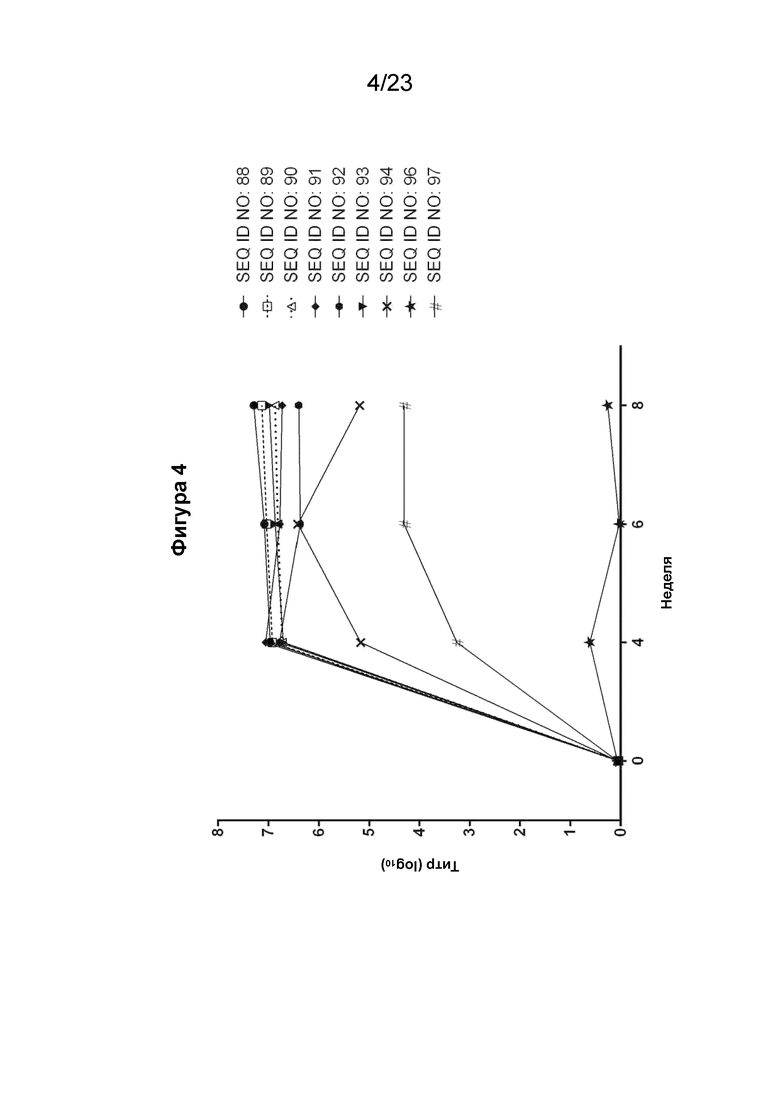
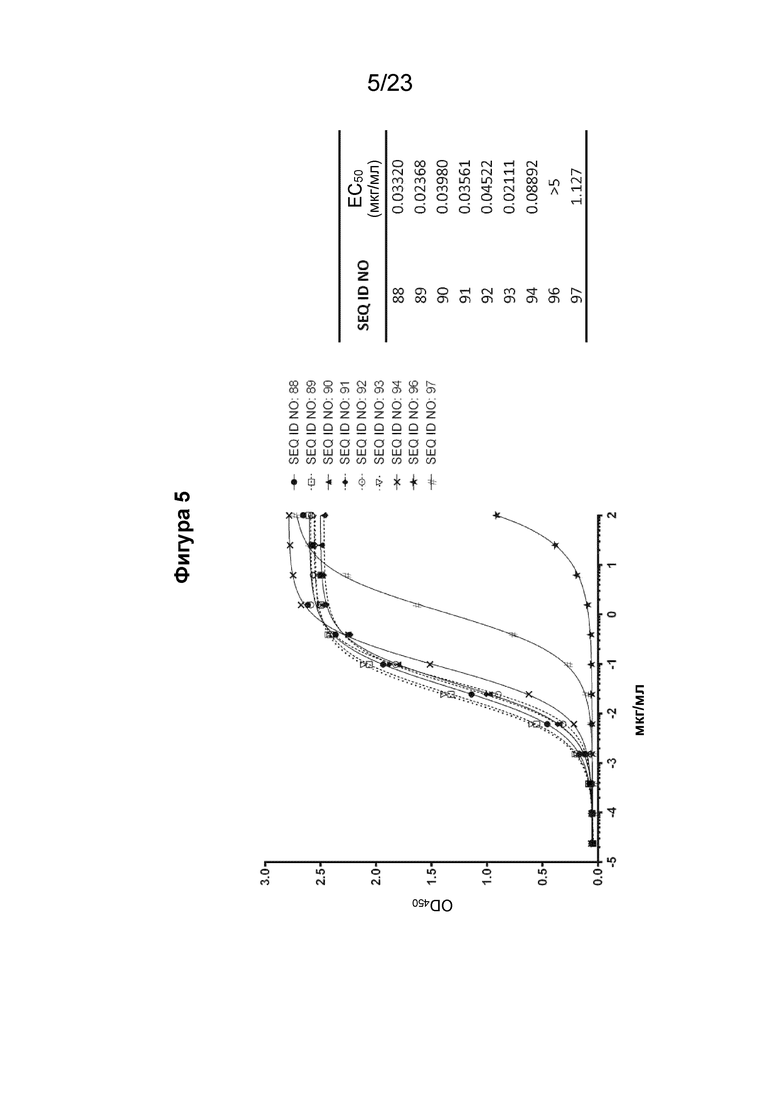
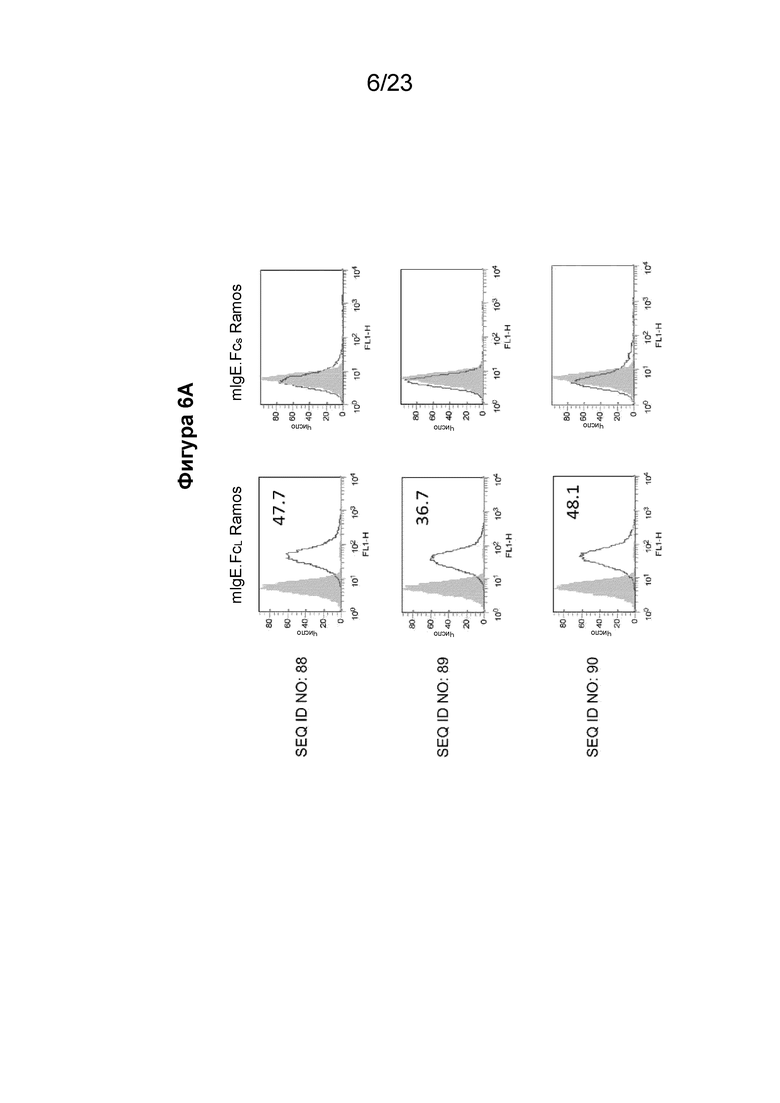
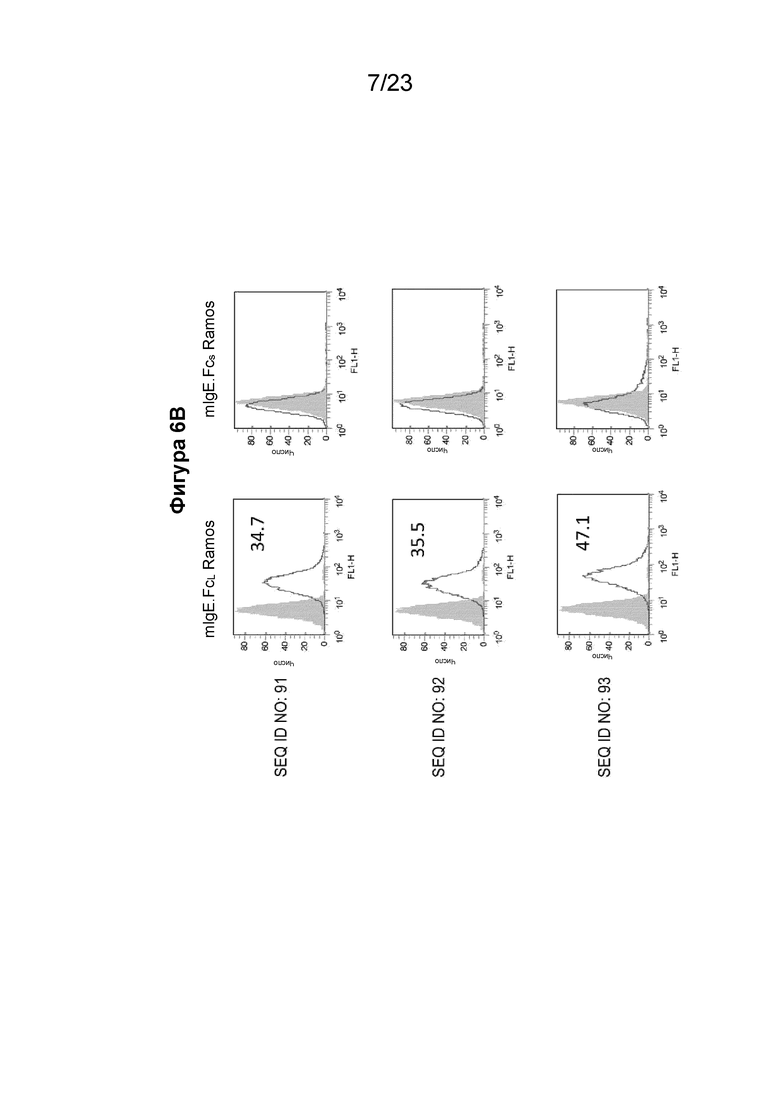
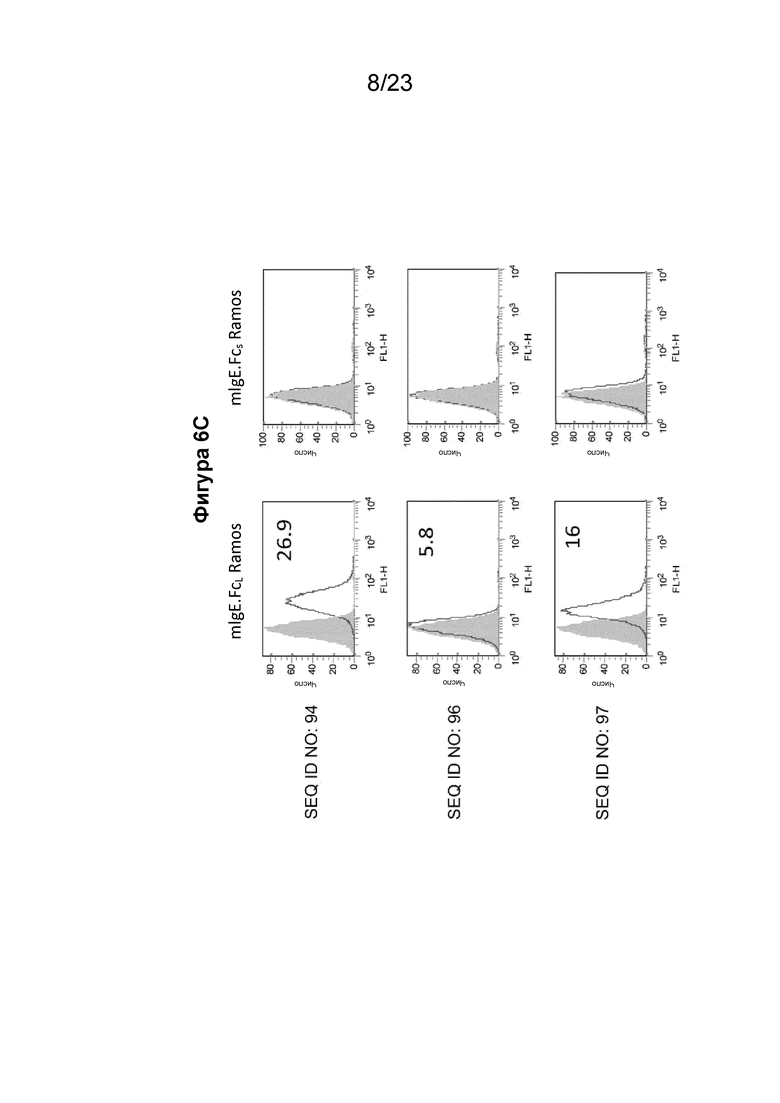
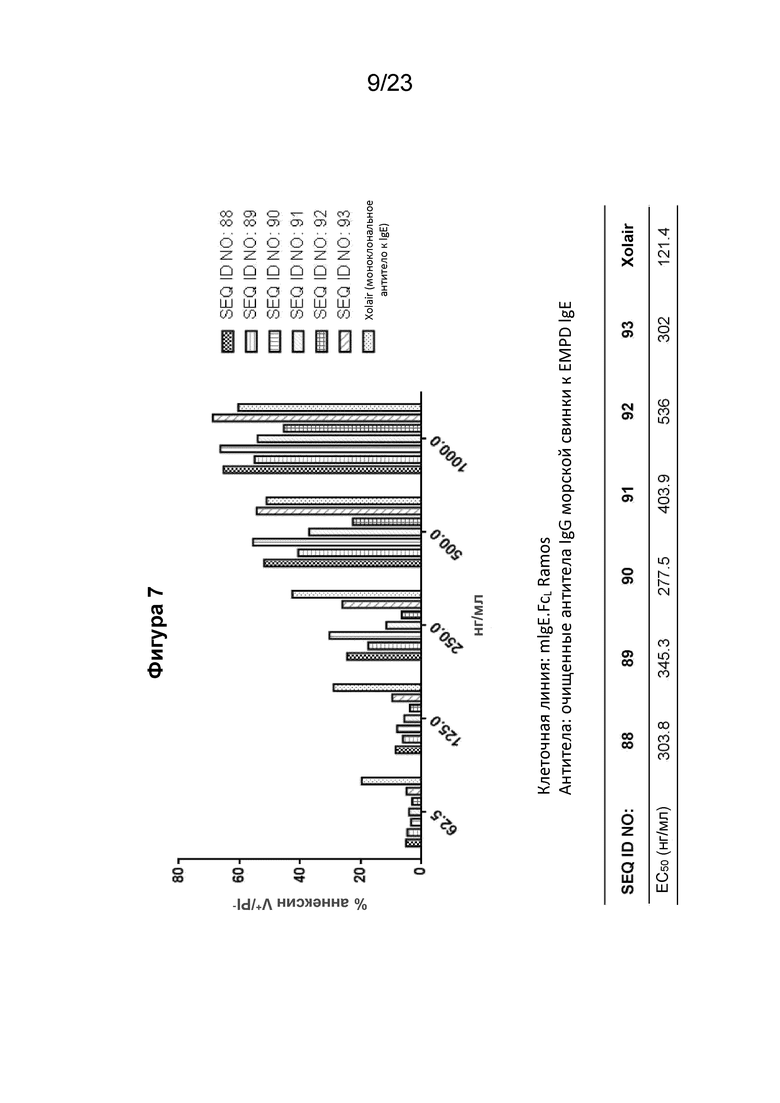
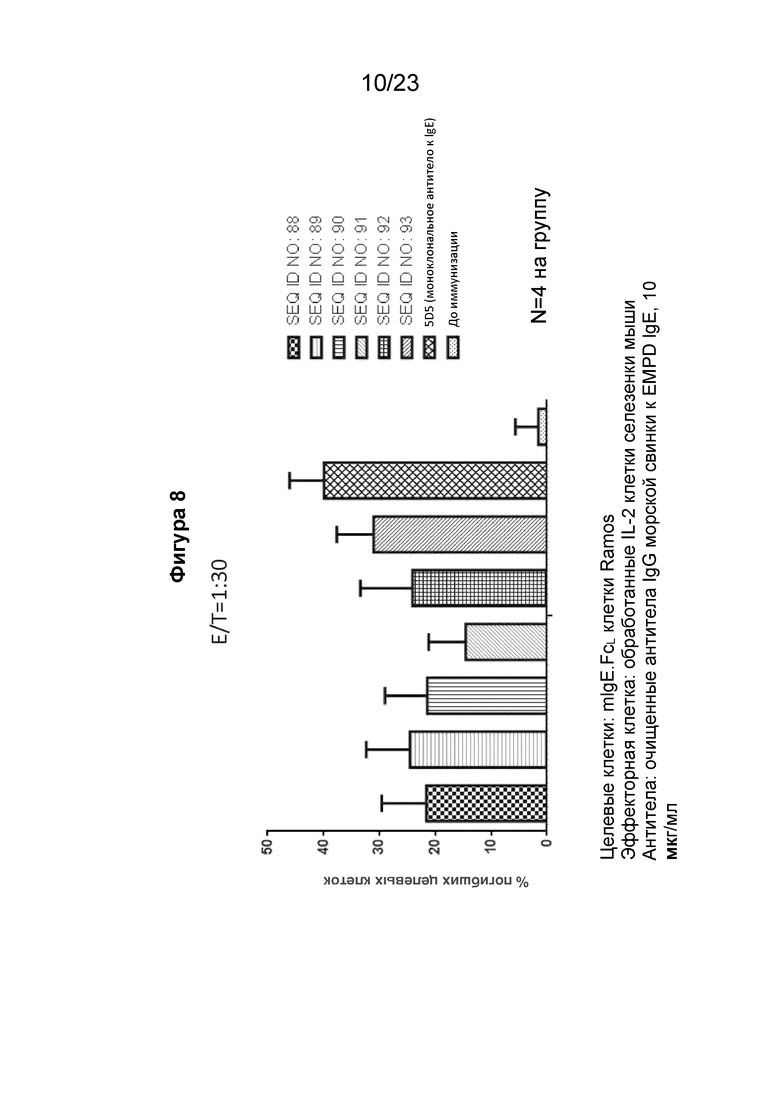
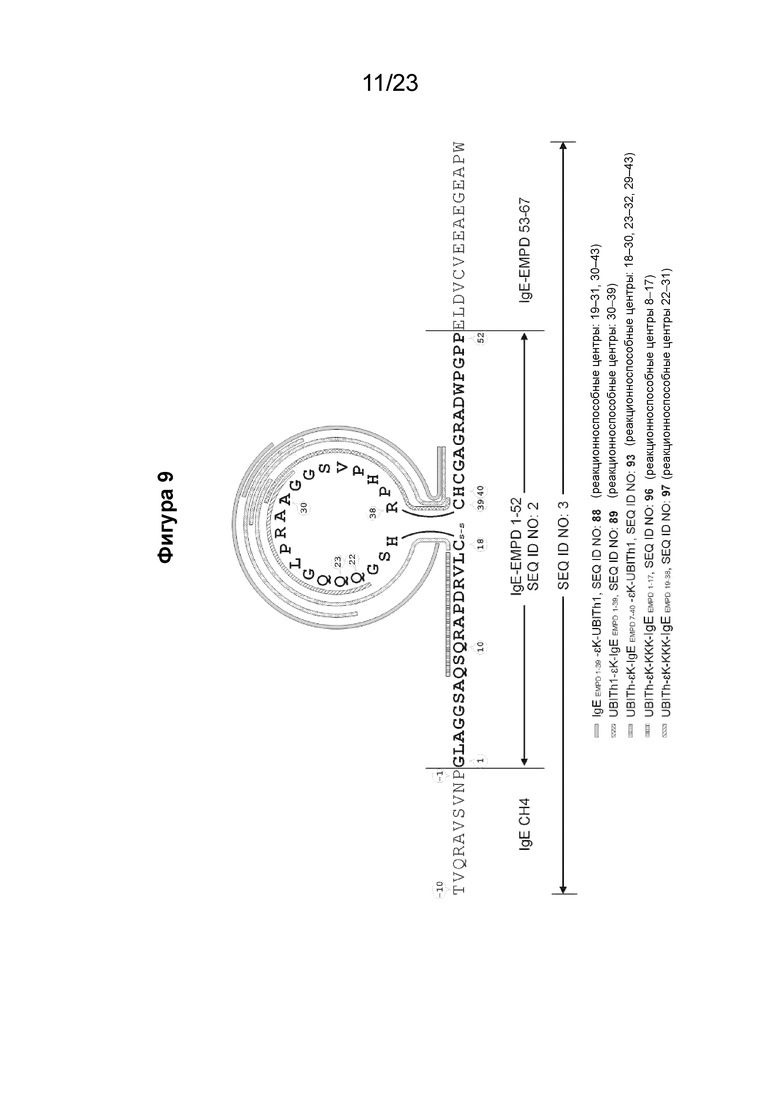
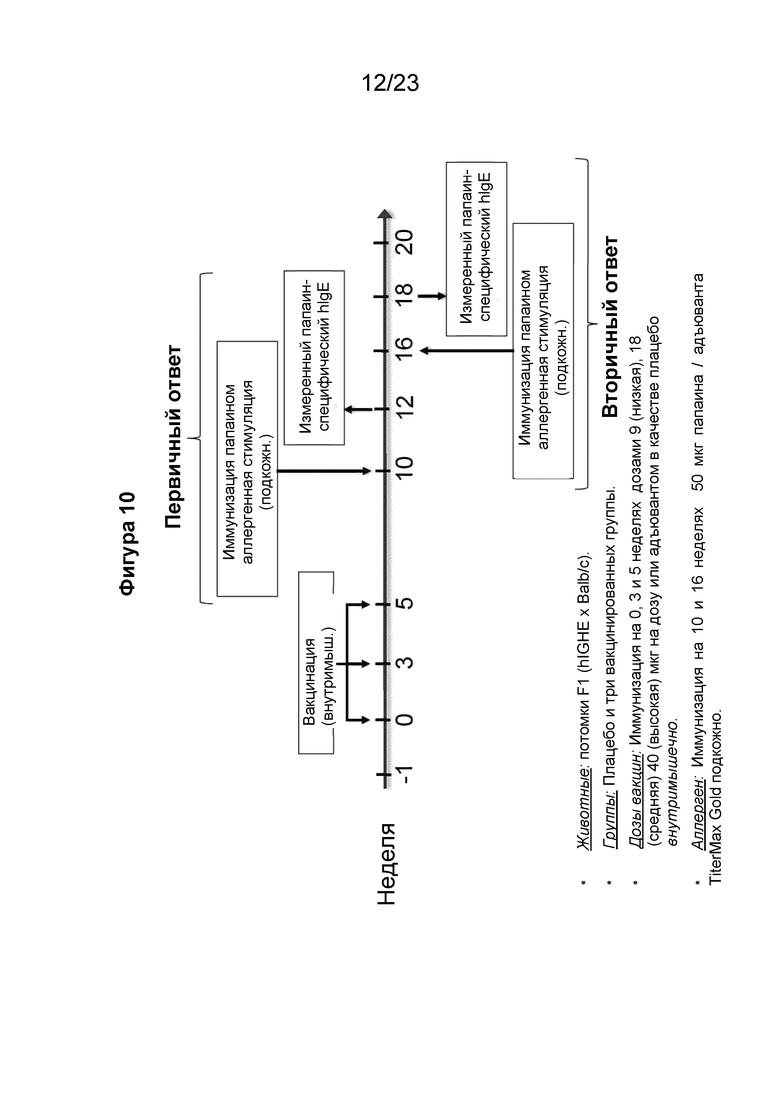
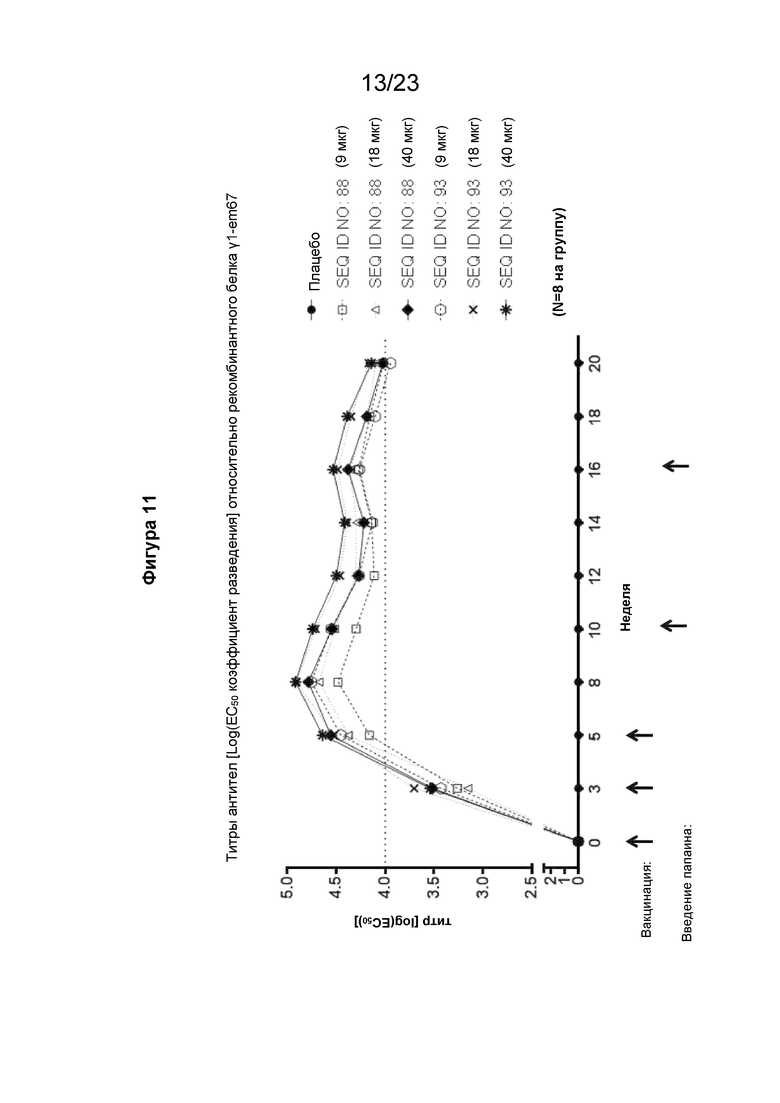
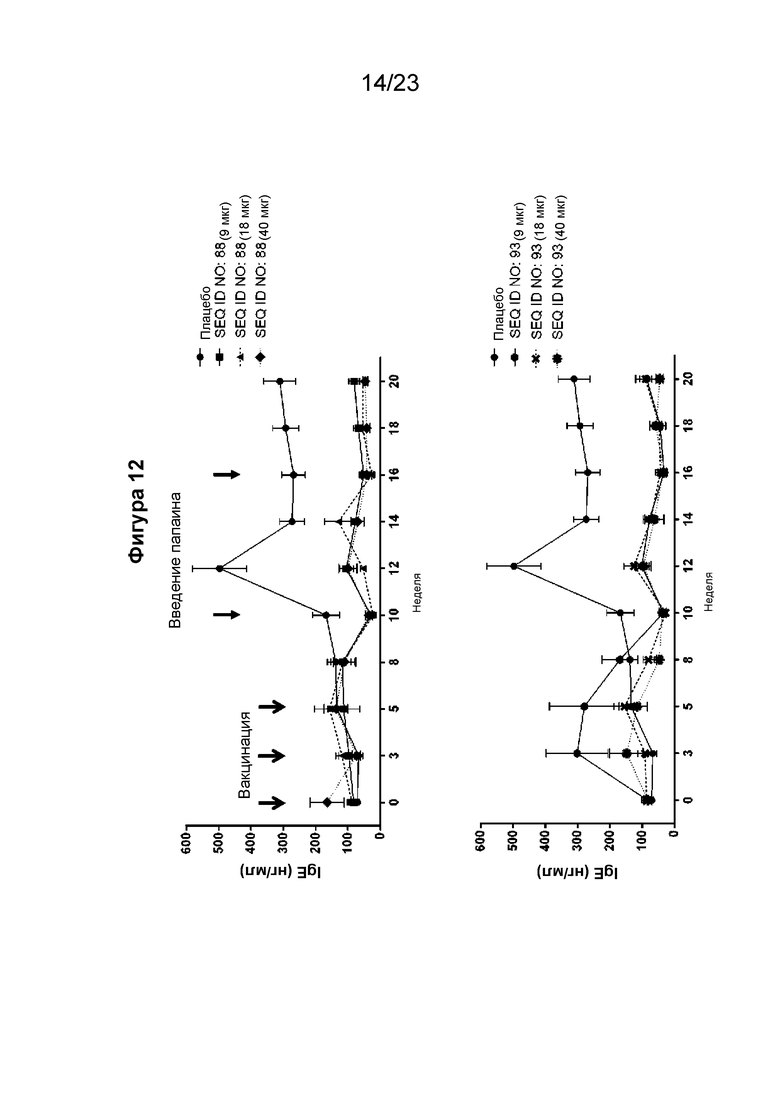
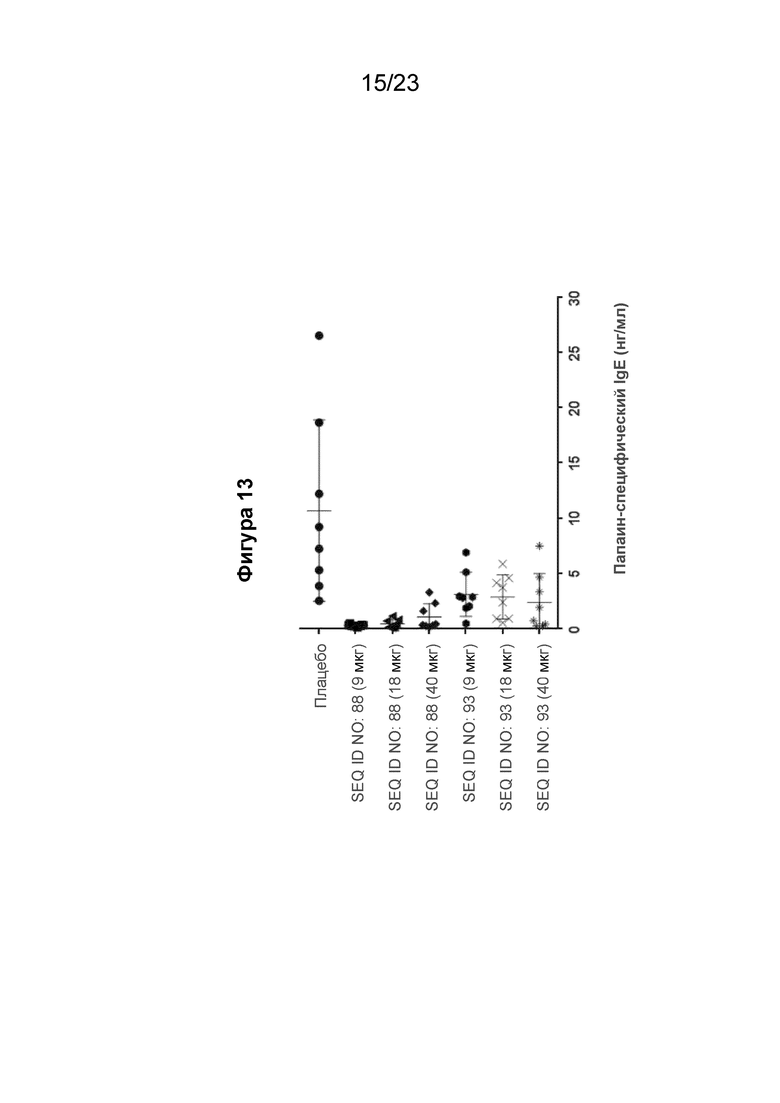
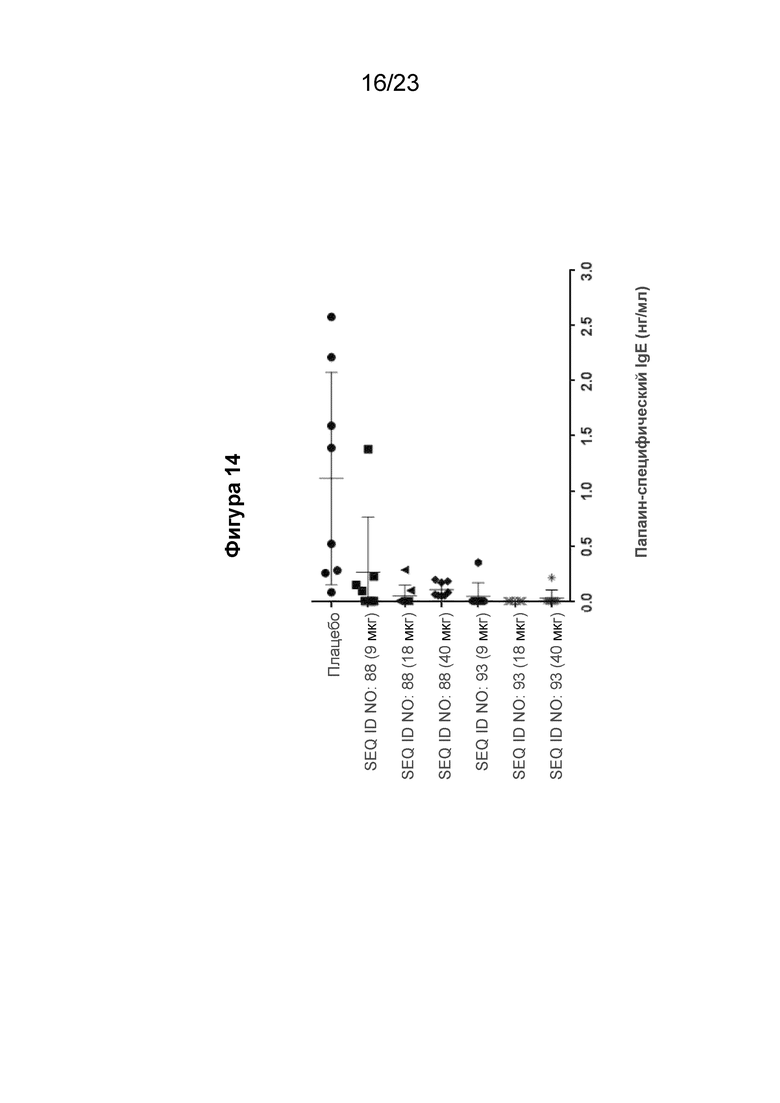
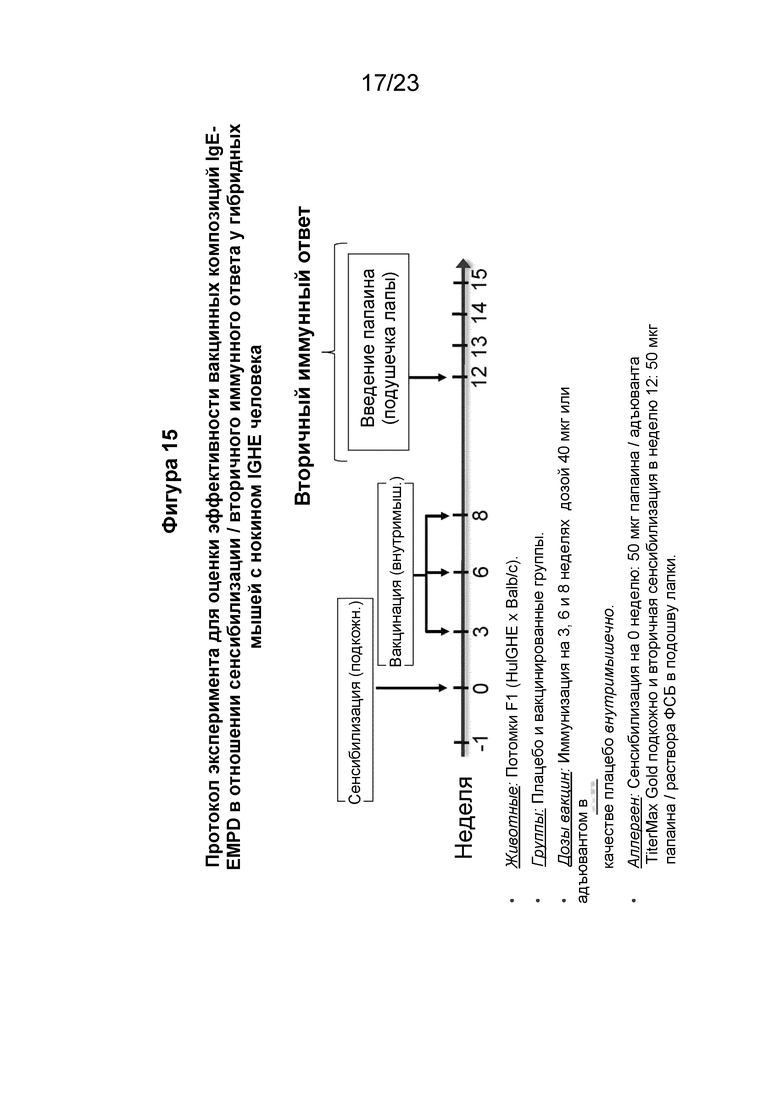
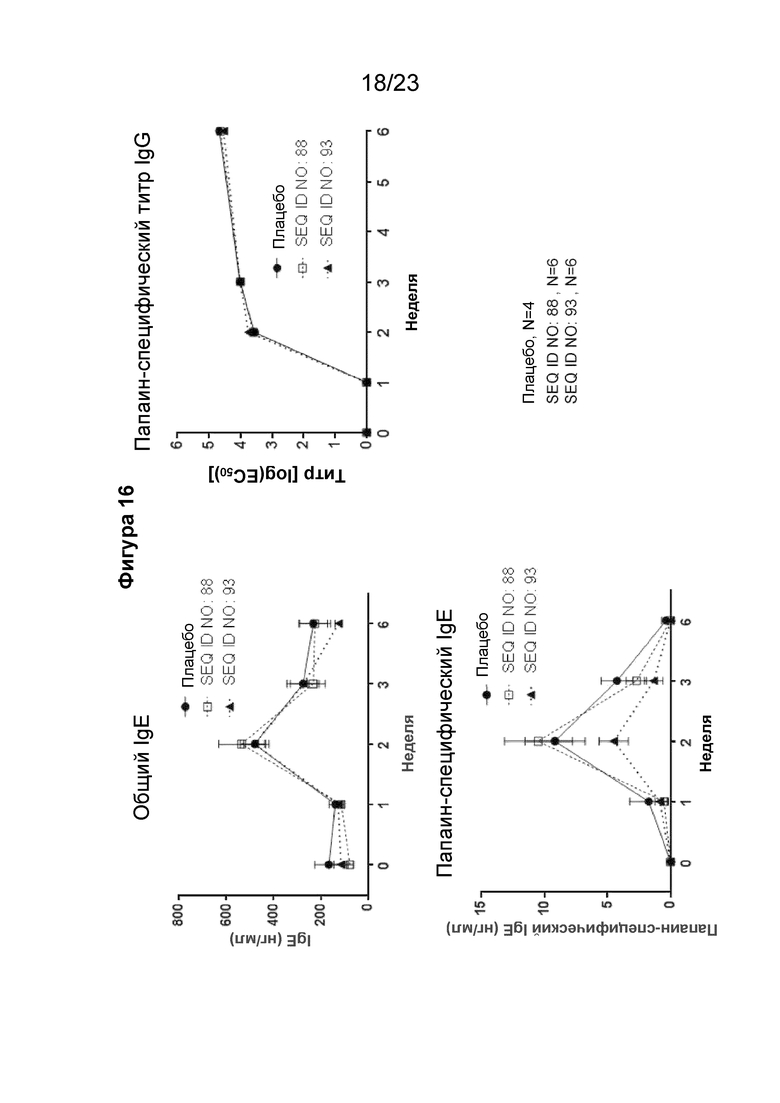
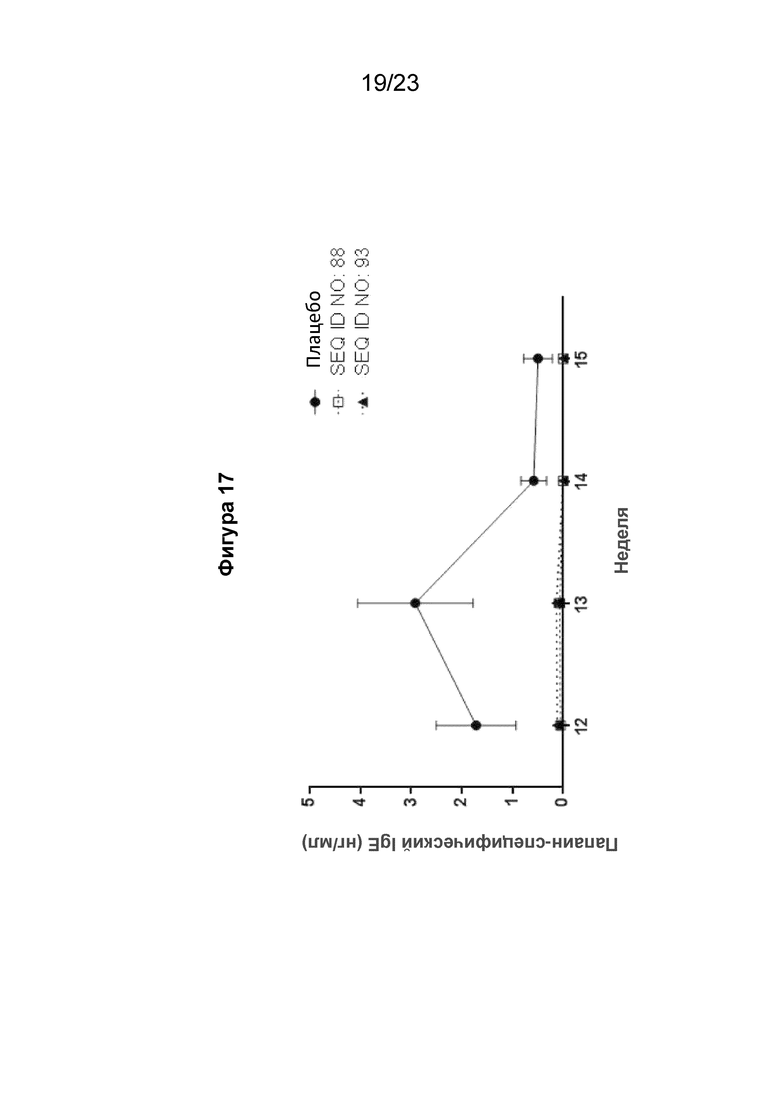
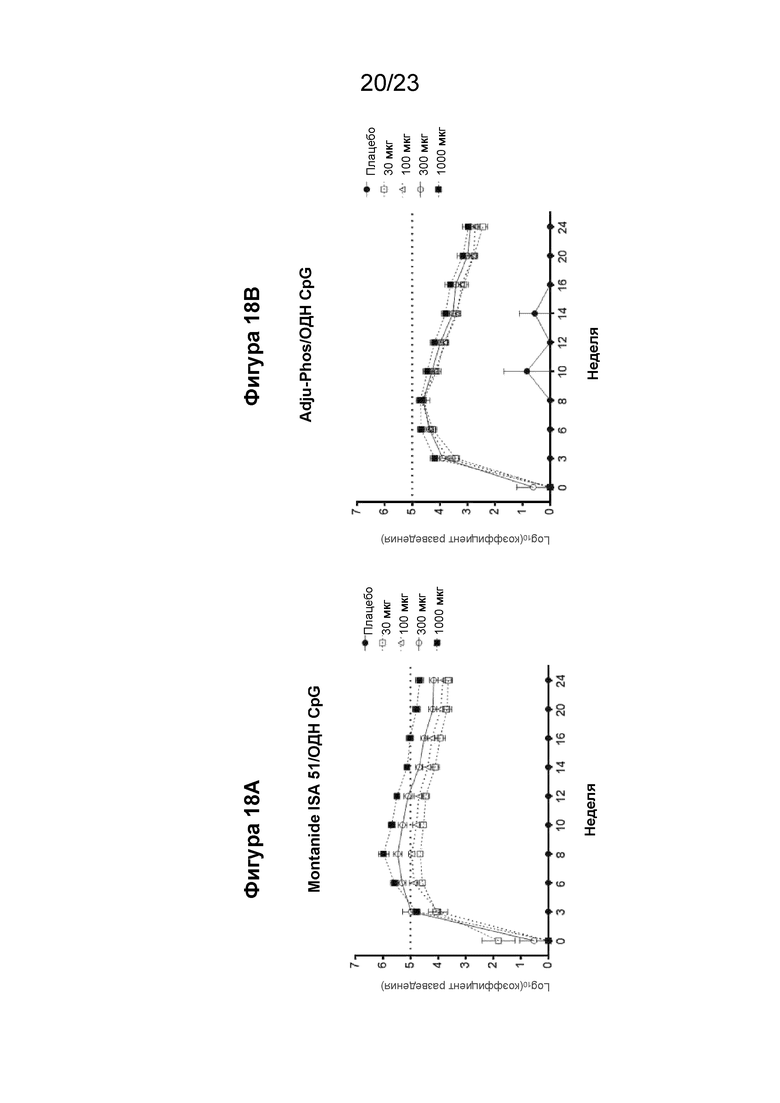
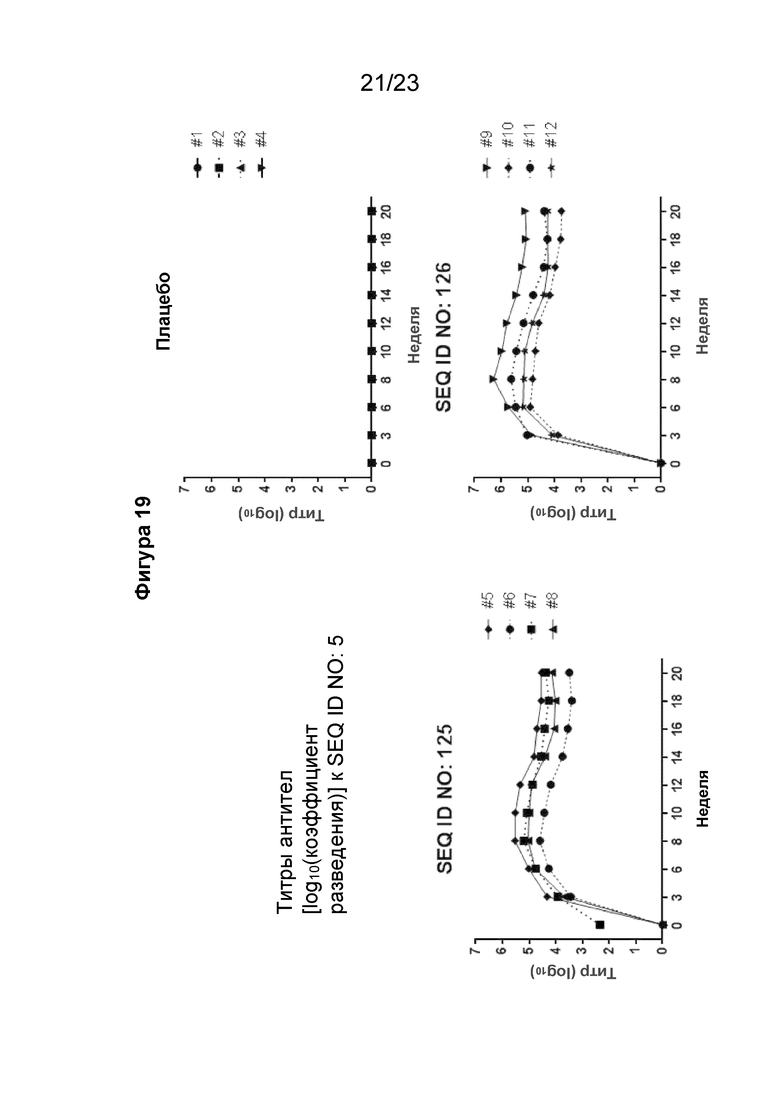
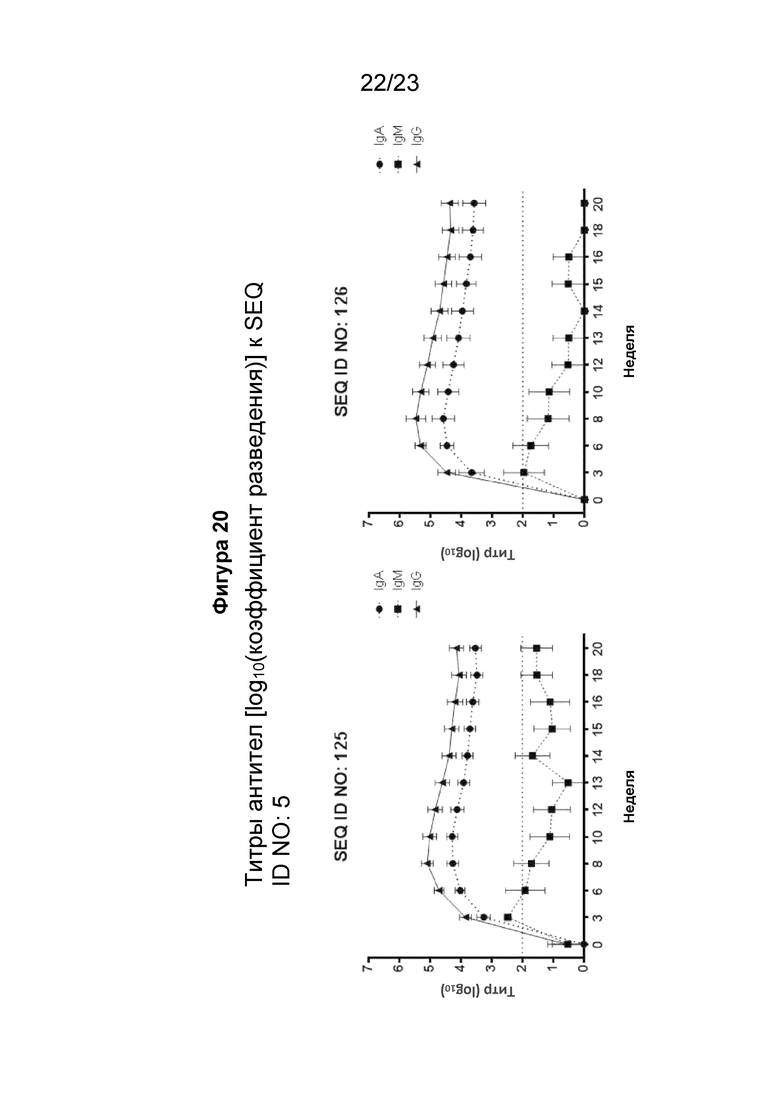
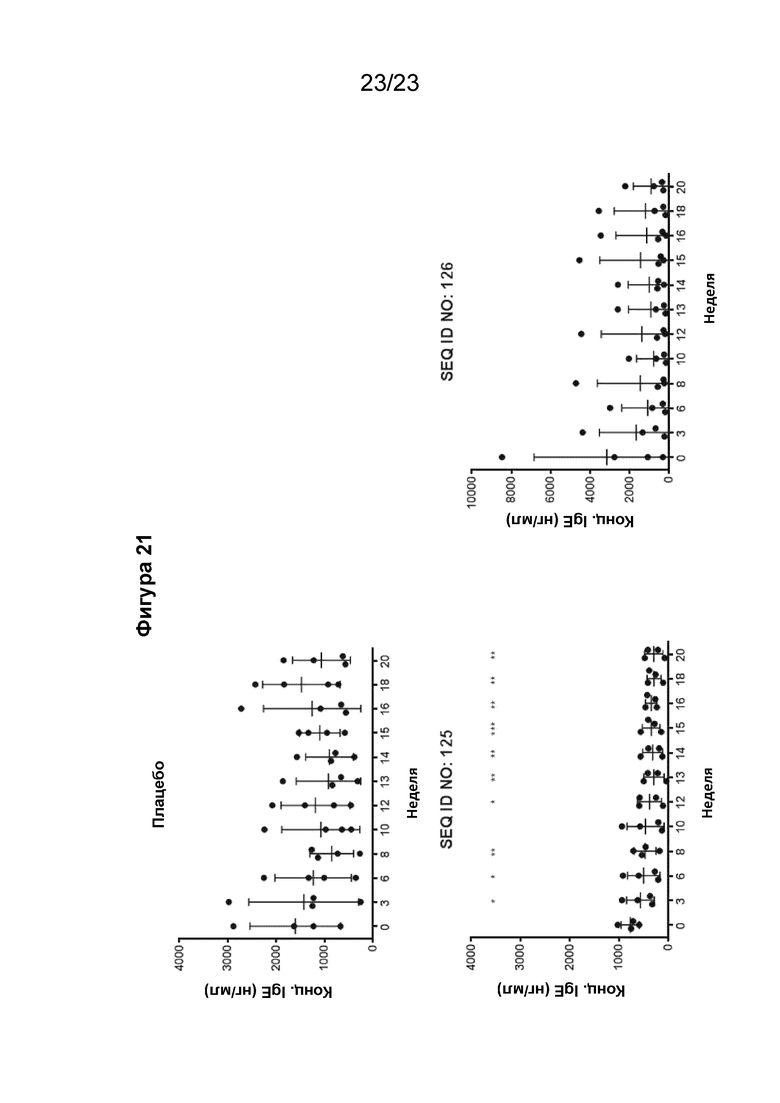
Изобретение относится к области биотехнологии, конкретно к пептидной иммуногенной конструкции EMPD IgE, и может быть использовано в медицине. Пептидная иммуногенная конструкция EMPD IgE с SEQ ID NO: 88-95, 98-124 или 130 может быть использована для эффективного лечения опосредованного IgE аллергического заболевания. 2 н. и 1 з.п. ф-лы, 21 ил., 7 табл., 10 пр.
1. Пептидная иммуногенная конструкция EMPD IgE, выбранная из группы, состоящей из SEQ ID NO: 88-95, 98-124 и 130.
2. Фармацевтическая композиция для лечения опосредованного IgE аллергического заболевания, содержащая:
a. эффективное количество пептидной иммуногенной конструкции EMPD IgE по п. 1; и
b. фармацевтически приемлемый носитель для доставки и/или адъювант.
3. Фармацевтическая композиция по п. 2, где пептидная иммуногенная конструкция EMPD IgE смешивается с CpG-олигодезоксинуклеотидом (ODN) для образования стабилизированного иммуностимулирующего комплекса.
| WO 1996012740, 02.05.1996 | |||
| PAKULA A.A | |||
| et al., Genetic analysis of protein stability and function, Annual review of genetics, 1989, V | |||
| Прибор для равномерного смешения зерна и одновременного отбирания нескольких одинаковых по объему проб | 1921 |
|
SU23A1 |
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| SCHUMACHER T | |||
| N | |||
| M | |||
| et al., Identification of D-peptide ligands through mirror-image phage display, Science, 1996, V | |||
| Искроудержатель для паровозов | 1920 |
|
SU271A1 |
| MULLER S | |||
| et al., Spliceosomal peptide P140 for | |||

